
リン酸化−脱リン酸化反応検出用基質群およびそれを用いた検出方法
【課題】プロテインキナーゼまたはプロテインホスファターゼの反応性を簡便に測定可能な手段の提供。
【解決手段】プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換してなる人工ペプチド基質を少なくとも1種含有するプロテインキナーゼの基質群、前記基質群を固定してなるペプチドアレイ、前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、リン酸化および/または脱リン酸化反応を検出する方法、前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、プロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法、ならびに前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、プロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法。
【解決手段】プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換してなる人工ペプチド基質を少なくとも1種含有するプロテインキナーゼの基質群、前記基質群を固定してなるペプチドアレイ、前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、リン酸化および/または脱リン酸化反応を検出する方法、前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、プロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法、ならびに前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、プロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、リン酸化−脱リン酸化反応検出用基質群およびそれを用いた検出方法に関する。詳しくは、本発明は、Non-RI法でリン酸化−脱リン酸化反応を高感度に検出するペプチド性基質群およびそれを用いたリン酸化−脱リン酸化反応の検出方法に関する。
【背景技術】
【0002】
生体内のタンパク質は、様々な生理的条件下で生化学的修飾を受けることによってその性質を変化させている。リン酸化は、タンパク質の主要な翻訳後修飾反応の1つであり、その反応は瞬時かつ可逆的におこるため、タンパク質の活性/不活性をコントロールする基本的な素過程の1つとして、生体内で重要な役割を担っていることが推測される。実際、真核細胞においては細胞内タンパク質の多くはリン酸化されており、多くのシグナル伝達系において、タンパク質リン酸化が情報伝達の方法として広く用いられていることが知られている。
【0003】
一般に、タンパク質がリン酸化反応、あるいは脱リン酸化反応を受けると、タンパク質を構成するアミノ酸残基の局所的な極性に変化が生じることにより、結果としてタンパク質の高次構造に基づく機能や活性度に変化を生じることにより、タンパク質の活性/不活性がコントロールされていると考えられている。このように、細胞内情報伝達に深く関わっているタンパク質リン酸化酵素の働きを調べることは、細胞機能を解析する上で大変重要である。さらに、医薬品探索研究やポストゲノム研究として注目されているプロテオームの研究にも関連し大変興味深い。プロテオーム解析のツールとして、タンパク質またはペプチドを基盤上に固定するマイクロアレイ技術が知られている(特許文献1および2、非特許文献1〜6)。
【0004】
タンパク質リン酸化酵素(プロテインキナーゼ)は、タンパク質の特定の部位をリン酸化してその機能を変える働きをもち、その基質であるタンパク質のセリンまたはスレオニン残基をリン酸化する「セリン/スレオニンキナーゼ」と、チロシン残基をリン酸化する「チロシンキナーゼ」の2つに大別される。ここで、「チロシンキナーゼ」は細胞膜内あるいは近傍に存在し情報伝達の最上流に位置し細胞外の環境変化を直接検知する場合が多く、「セリン/スレオニンキナーゼ」は上流からの信号を受けて逐次的に情報伝達を行い、細胞機能の直接的な調節に関与する場合が多い。
【0005】
プロテインキナーゼは、細胞外部からの刺激を細胞内部に伝達し、これらの刺激に対する細胞の応答に関与する。このように、プロテインキナーゼはリン酸化を通して、代謝、増殖、分化、情報伝達、運動などの多彩な細胞機能の調節を担っている。細胞内でのプロテインキナーゼは特定のタンパク質を基質として認識することで特異性の高いリン酸化反応を進行させ、これらリン酸化反応の逐次的反応の組み合わせとして、細胞外の環境変化や薬剤に対する応答として信号を伝達し細胞機能を調節している。遺伝子配列の解析から、約800種類のプロテインキナーゼが存在して細胞内のシグナル伝達に関与していると予想されている。タンパク質リン酸化の異常は、癌、糖尿病、関節炎などの様々な疾患の原因または結果と考えられている。
【0006】
一方、プロテインホスファターゼは、リン酸化されたタンパク質を基質として脱リン酸化を触媒する酵素であり、タンパク質リン酸化の可逆性を保障している。プロテインキナーゼは、それ自体がキナーゼの基質でもあり、スレオニン、セリン(ホスホリラーゼなど)、チロシン(srcなどガン遺伝子関連のタンパク質に例が多い)などのいずれかの残基がリン酸化されたプロテインキナーゼは、プロテインホスファターゼによって再び元の酵素に戻り、次の情報による活性化に備える。シグナル伝達経路の共通の特徴として、その反応の一過性がある。この一過性は、シグナル伝達分子の量や活性化状態の一過性の反映である。タンパク質のリン酸化は、ある場合には活性化を、ある場合には不活性化を引き起こす。実際、タンパク質の脱リン酸化反応が積極的なスイッチオンの役割を担っている場合も知られている。
【0007】
プロテインキナーゼは、その基質特異性が高いことが特徴であるが、基質タンパク質の被リン酸化部位を含むアミノ酸配列からなるペプチド性部分を抽出したペプチドもまた基質としてリン酸化を受けることが広く知られている。そのため、プロテインキナーゼの反応性を調べる際には、ペプチド性基質を用いて調べることが可能である。
【0008】
前記「プロテインキナーゼの反応性」は、その適当な基質を用いてリン酸化反応を行わせ、生成物であるリン酸化基質やADPの増加、あるいは出発物である基質やATPの減少を何らかの方法で計測することで判定が可能である。今日では、タンパク質リン酸化は、放射性核種32Pの取り込みを直接測定する方法か、または抗リン酸化タンパク質抗体を用いる方法が一般的である。
【0009】
最近では、基質に含まれるリン酸化セリン残基、リン酸化スレオニン残基またはリン酸化チロシン残基を、抗体および表面プラズモン共鳴によって検出する方法(特許文献3)や、適当なキレート性検出試薬および表面プラズモン共鳴によって検出する方法(特許文献4)が注目されている。これらの方法では、放射性物質を取り扱う必要がなく操作性にも優れているが、抗体や検出試薬の認識能力に強く依存し、これらの認識能力を基とした間接的な検出と測定を行うことになる。そのため、認識能力の優れた抗体や検出試薬の開発が重要かつ不可欠となる。
【0010】
チロシン残基のリン酸化の検出に関しては、タンパク質またはペプチド配列内のリン酸化チロシン残基を幅広く認識する多くの優れた抗リン酸化チロシン抗体が知られており、市販されている。そのため、それらの抗体を用いることでチロシンキナーゼの反応性は比較的容易に測定可能である。一方、セリン/スレオニンキナーゼの場合は、幅広くかつ一般的にリン酸化セリンあるいはリン酸化スレオニン残基を認識可能な優れた抗体は知られていない。そのため、抗体を使ってセリン/スレオニンキナーゼの反応性を調べる場合、特定のリン酸化タンパク質あるいはリン酸化ペプチドを特異的に認識する抗体をその都度作製して用いなければならず、セリン/スレオニンキナーゼの反応性を抗体により一般的かつ簡便に検出し測定する方法はほとんど知られていない。
【特許文献1】WO 02/083933 A2
【特許文献2】WO 02/083884 A2
【特許文献3】特開2004−283114号公報
【特許文献4】特開2004−309303号公報
【非特許文献1】M. Bettencourt-Dias, R. Giet, R. Sinka, A. Mazumdar, W. G. Lock, F. Balloux, P. J. Zafiropoulos, S. Yamaguchi, S. Winter, R. W. Carthew, M. Cooper, D. Jones, L. Frenz, D. M. Glover. Nature. 2004, 432, 980-7.
【非特許文献2】G. Manning, D. B. Whyte, R. Martnez, T. Hunter, S. Sudarsanam. Science. 2002, 298, 1912-1934.
【非特許文献3】S. J. Lee, S. Y. Lee. Anal. Biochem. 2004, 330, 311-316.
【非特許文献4】R. C. Panicker, X. Huang, S. Q. Yao. Comb. Chem. High Throughput Screen. 2004, 7, 547-56.
【非特許文献5】D. S. Y. Yeo, R. C. Panicker, L. P. Tan, S. Q. Yao. Comb. Chem. High Throughput Screen. 2004, 7, 213-221.
【非特許文献6】B. T. Houseman, J. H. Huh, S. J. Kron, M. Mrksich. Nature Biotech. 2002. 20. 270-274.
【発明の開示】
【発明が解決しようとする課題】
【0011】
上述の理由から、現在においては、セリン/スレオニンキナーゼのアッセイに関しては、放射性核種の32Pの取り込みを直接測定する方法が一般的となっているが、実験施設や作業環境に制約があり、また実験者の放射能被爆の問題もある為、あまり大規模なハイスループット(high throughput)解析には適さないという問題がある。したがって、大規模なプロテオーム解析を実施して疾患とタンパク質リン酸化カスケードの関連を調べる上で、放射化標識されたATP等を用いないNon-RI法で、タンパク質のリン酸化/脱リン酸化を高感度に検出するアッセイを可能にすることが重要な課題となっている。
本発明の目的は、プロテインキナーゼまたはプロテインホスファターゼの反応性を簡便に測定可能な手段の提供である。
【課題を解決するための手段】
【0012】
本発明者らは、上記問題点に鑑み、鋭意検討した結果、プロテインキナーゼまたはプロテインホスファターゼの基質として以下に示すような人工ペプチドを用いることを見出し、本発明を完成するに至った。即ち、本願発明は、以下に示す通りである。
【0013】
〔1〕 プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換してなる人工ペプチド基質を少なくとも1種含有するプロテインキナーゼの基質群。
〔2〕 プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をリン酸化スレオニン残基に置換してなる人工ペプチド基質を少なくとも1種さらに含有する、前記〔1〕に記載の基質群。
〔3〕 前記人工ペプチド基質が5〜30のアミノ酸残基を有するものである、前記〔1〕または〔2〕に記載の基質群。
〔4〕 前記プロテインキナーゼがセリン/スレオニンキナーゼである前記〔1〕〜〔3〕いずれかに記載の基質群。
〔5〕 前記セリン/スレオニンキナーゼがcAMP依存性プロテインキナーゼAである、前記〔4〕に記載の基質群。
〔6〕 前記セリン/スレオニンキナーゼがプロテインキナーゼCである、前記〔4〕に記載の基質群。
〔7〕 前記セリン/スレオニンキナーゼがAMP活性化プロテインキナーゼである、前記〔4〕に記載の基質群。
〔8〕 前記人工ペプチド基質が配列番号11〜299に記載のアミノ酸配列群から選択されるアミノ酸配列を有するものである前記〔1〕〜〔7〕いずれかに記載の基質群。
〔9〕 前記〔1〕〜〔8〕いずれかに記載の基質群を固定してなるペプチドアレイ。
〔10〕 前記〔9〕に記載のペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、リン酸化および/または脱リン酸化反応を検出する方法。
〔11〕 前記抗リン酸化スレオニン抗体がポリクローナル抗体である、前記〔10〕に記載の方法。
〔12〕 下記工程:
被験者由来の生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分を調製する工程;
前記画分と前記〔9〕に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を、対照における検出結果と比較し、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する工程
を含むプロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法。
〔13〕 前記疾患が癌、悪性腫瘍、糖尿病、糖尿病性合併症、代謝性疾患、自己免疫疾患、免疫異常疾患、免疫不全、神経変性疾患、虚血性疾患、内分泌疾患、肝疾患または骨疾患である前記〔12〕に記載の診断方法。
〔14〕 下記工程:
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、前記〔9〕に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を被検物質を含まない試料における検出結果と比較し、被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する工程
を含む、プロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法。
【発明の効果】
【0014】
本発明のプロテインキナーゼの基質群によると、適切に設計されたアミノ酸配列に基づいて合成した人工基質を含むことから、Non-RI法で高感度にタンパク質のリン酸化または脱リン酸化を測定可能な一連の基質を提供することが可能となり、さらにコンビナトリアルケミストリーの技術を応用すれば、ペプチド基質ライブラリーを作製することができるため、プロテインキナーゼおよび/またはプロテインホスファターゼのハイスループット解析に供することができる。
本発明のペプチドアレイおよびそれを用いたリン酸化および/または脱リン酸化の検出方法によると、プロテオーム解析の手段として、ハイスループット解析が可能であるとともにリン酸化と脱リン酸化を1つのアレイで同時に検出することも可能である。
本発明の診断方法によると、セリン/スレオニンキナーゼによるリン酸化および/またはプロテインホスファターゼによる脱リン酸化を高感度かつ迅速に検出することが可能であるので、かかるリン酸化および/または脱リン酸化が関与する疾患の有無を迅速に判定することできる。
本発明のスクリーニング方法によると、セリン/スレオニンキナーゼによるリン酸化またはプロテインホスファターゼによる脱リン酸化を高感度かつ迅速に検出することが可能であるので、プロテインキナーゼまたはプロテインホスファターゼ活性を促進または阻害する物質を効率よく選択することができる。
【発明を実施するための最良の形態】
【0015】
本発明においてプロテインキナーゼとは、大別してセリン/スレオニンキナーゼとチロシンキナーゼとを含む概念であるが、以下、これまで有用なキナーゼ活性の測定方法が開発されていなかったセリン/スレオニンキナーゼを中心に述べる。
【0016】
セリン/スレオニンキナーゼとしては、アミノ酸配列中のセリンまたはスレオニンをリン酸化する酵素であれば特に限定されるものではないが、例えば、cAMP依存性プロテインキナーゼ(PKA)、cGMP依存性プロテインキナーゼ(PKG)、カルシウム依存性プロテインキナーゼ、AMP活性化プロテインキナーゼ(AMPK)、二本鎖RNA依存性プロテインキナーゼ、カゼインキナーゼ、リボソームタンパク質S6キナーゼ、Rafキナーゼ、サイクリン依存性キナーゼ(CDK)などがあげられ、前記カルシウム依存性プロテインキナーゼとしては、ホスホリラーゼキナーゼ、ミオシン軽鎖キナーゼ、カルモデュリン依存性プロテインキナーゼ、リン脂質依存性プロテインキナーゼ(プロテインキナーゼC:PKC)などがあげられる。これらの中でも、細胞内における役割が解明されつつあり、疾患との関連が示唆されているPKA、AMPK、PKCなどが好ましい。
【0017】
本発明においてプロテインホスファターゼとは、リン酸化セリン/スレオニン残基、リン酸化チロシン残基またはリン酸化セリン/スレオニン/チロシン残基のリン酸エステル結合を加水分解する酵素をいい、好ましくはリン酸化セリン/スレオニン残基のリン酸エステル結合を加水分解する酵素である。具体的には、リン酸化されたセリン/スレオニン残基に特異的なホスファターゼとしては、PP1、PP2A、PP2BおよびPP2Cなどのホスファターゼがあげられる。これらは、それぞれ基質特異性や阻害剤に対する感受性、金属イオン要求性の違いによって分類されている。PP2Cは単量体酵素であるが、PP1、PP2AおよびPP2Bは、調節サブユニットと活性サブユニットで構成される複合体を形成し、細胞内での局在性は、その調節サブユニットとの相互作用により調節されていると言われている。タイプ2Bは、一般に、カルシニューリンともよばれている。一方、リン酸化されたチロシン残基を特異的に脱リン酸化するホスファターゼとしては、細胞膜貫通ドメインを持つ受容体型と、それらをもたない細胞質型が存在し、それぞれに分子構造や活性調節機構を異にする多様な分子種を含んでいる。またリン酸化されたセリン/スレオニン/チロシン残基を脱リン酸化するホスファターゼとしても、基質特異性を異にする複数の分子種が存在することが知られている。
【0018】
本発明において基質とは、プロテインキナーゼが認識するアミノ酸配列を含有するペプチドまたはタンパク質をいう。前記ペプチドまたはタンパク質を構成するアミノ酸としては、天然アミノ酸として知られるαアミノ酸、βアミノ酸もしくはγアミノ酸に加えて、それらを化学的に修飾したもの、さらにはアミノ酸のアミノ基または水酸基とカルボキシル基やスルフォニル基やフォスフォリル基等を介してペプチド様化学結合を行うことが可能な分子量約800以下の化学構造を有する部分化学構造体もその範囲に含まれる。
【0019】
前記基質としては、人工的に適切なアミノ酸配列を設計、合成可能で、かつ、保存性および操作性に優れているペプチド基質が好ましい。
【0020】
本発明において天然に存在する基質とは、プロテインキナーゼの基質として知られているタンパク質を構成するアミノ酸配列の全部または一部からなるものをいい、セリン/スレオニン残基、好ましくはセリン/スレオニンキナーゼが認識してリン酸化可能なセリン/スレオニン残基を1つまたは複数含むアミノ酸配列からなるものである。天然に存在する基質におけるアミノ酸残基の数としては、5〜タンパク質の全長のアミノ酸残基の数であり、好ましくは5〜30、より好ましくは10〜20である。
【0021】
本発明において人工ペプチド基質とは、前記天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換したものをいい、好ましくはセリン/スレオニンキナーゼが認識してリン酸化可能なセリン残基をスレオニン残基またはリン酸化スレオニン残基で置換したものである。人工ペプチド基質は、酵素の特異性や検出の容易性を担保するためには、セリン/スレオニンキナーゼが認識可能なセリン残基をスレオニン残基(またはリン酸化されたスレオニン残基)に置換した部位を1つ含むものであることが好ましい。人工ペプチド基質中のアミノ酸残基の数としては、セリン/スレオニンキナーゼが認識できる程度のペプチド長となれば特に限定されるものではないが、通常5〜500、好ましくは5〜30、より好ましくは10〜20である。
【0022】
なお、前記人工ペプチド基質は、前記天然に存在する基質から設計されたものであるから、天然に存在する基質中のセリン/スレオニンキナーゼが認識できる程度のアミノ酸残基からなるペプチド部分を含有していればよく、人工ペプチド基質および天然に存在する基質のアミノ酸残基の数が互いに一致する必要はない。
【0023】
本発明のプロテインキナーゼの基質群は、前記人工ペプチド基質を少なくとも1種含有することを特徴とする。「セリン/スレオニンキナーゼ」は、タンパク質性またはペプチド性基質のセリンおよび/またはスレオニン残基をリン酸化するが、セリン残基とスレオニン残基ではそれらの反応性に若干の差はあるものの共にリン酸化を受けるものと考えられる。そこで、本発明では、セリン/スレオニンキナーゼのリン酸化能を調べる目的のためにセリン/スレオニンキナーゼの基質として被リン酸化セリン残基をスレオニン残基に置換した人工ペプチド基質を用いる。あるいは、プロテインホスファターゼの脱リン酸化能を調べる目的のために、前記被リン酸化セリン残基をリン酸化スレオニン残基に置換した人工ペプチド基質を用いてもよい。
【0024】
本発明の基質群に含まれる基質の数は、前記人工ペプチド基質を少なくとも1種含有していれば特に限定されるものではないが、好ましくは、1〜100000、より好ましくは2〜50000、さらにより好ましくは10〜5000である。かかる範囲内の基質群は、ペプチドアレイまたはペプチドチップとして、1個の担体上に固定化することが可能である。
【0025】
本発明の基質群は、そのすべてが前記人工ペプチド基質であってもよく、前記人工ペプチド基質と天然に存在する基質とを含んでいてもよく、本発明の検出方法等に用いる場合に、陽性対照および/または陰性対照となる人工ペプチド基質または天然の基質を含んでいてもよい。陽性対照および陰性対照となる人工ペプチド基質には、セリン残基をリン酸化スレオニン残基またはアラニン残基に置換した人工ペプチド基質も含まれる。
【0026】
人工ペプチド基質の具体的な配列は、配列番号11〜299に記載のアミノ酸配列群から選択されるアミノ酸配列が好ましい。かかるアミノ酸配列を全部用いてもよく、あるいは、対象とするプロテインキナーゼに応じて適宜組み合わせて用いてもよい。
【0027】
本発明の人工ペプチド基質は、天然に存在する基質に含まれるアミノ酸配列中の被リン酸化セリン残基をスレオニン残基またはリン酸化スレオニン残基に置換することによって調製することができる。以下、かかる人工ペプチド基質配列の設計と調製方法について説明する。
【0028】
基質配列設計の方法としては、ゲノム情報またはタンパク質情報からセリン/スレオニンキナーゼの基質となるタンパク質の被リン酸化部位を含むペプチド断片を予測し抽出する。抽出したペプチド断片を、セリン/スレオニンキナーゼのペプチド性基質配列として既に知られているものをも含め、「ペプチド基質の基本配列」と定義して用いる。
【0029】
次に、前記ペプチド基質の基本配列の被リン酸化部位(セリン残基)を全てスレオニン残基に置換し、その他のアミノ酸残基は基本配列と同様にしてもよいし、適当な方法で一部のアミノ酸残基をさらに他のアミノ酸残基に置換してもよい。さらなる置換は、セリン/スレオニンキナーゼの人工ペプチド基質の製造、固定化等に有利な置換基を導入するために行われる。このように人工的に配列設計されたペプチドを、基質群(集合体あるいはペプチド基質ライブラリー)として供する。配列設計するペプチド配列は、化学合成の効率と活性発現を担保するために、40アミノ酸残基以下とすることが好ましい。
【0030】
前記ライブラリーには、被リン酸化部位をリン酸化スレオニンに置換した基質を少なくとも1種含有してもよい。リン酸化スレオニン置換した基質は、リン酸化の陽性対照となるばかりでなく、プロテインホスファターゼの基質ともなり、リン酸化/脱リン酸化を同時に検出することができるので好ましい。
【0031】
「配列設計されたペプチド性基質の集合体(ペプチド性基質ライブラリー)」は、一般的な化学的ペプチド合成方法(例えば、Fmoc法での固相合成等)により化学合成されて供与される。ここで重要な点は、天然に存在する被リン酸化部位としてのセリン残基を敢えてスレオニン残基に置換して配列設計し、化学合成することである。
【0032】
上記の方法によって設計し化学合成した人工ペプチド基質を含有する基質群を、溶液状態または「固相状態」でセリン/スレオニンキナーゼによるリン酸化反応の基質として用いる。キナーゼ反応の生成物であるペプチド性リン酸化基質を抗リン酸化スレオニン抗体で認識させ検出することで、セリン/スレオニンキナーゼの種々の条件下での反応性を検出し測定することができる。逆に、ホスファターゼ反応の生成物である脱リン酸化基質が抗体で検出されなくなることで、プロテインホスファターゼの種々の条件下での反応性、好ましくはセリン/スレオニンキナーゼとの競合性を検出し測定することができる。
【0033】
ここでいう「固相状態」とは、ペプチド性基質を種々の固相担体に直接またはリンカーと呼ばれる適当な化学構造体を介して結合した状態をいう。固相担体としては、有機高分子もしくは無機高分子材料(プラスチック等)、ガラス、金属等で作製された平板、試験管(生化学分野で一般的にプレートと呼称されるものを含む)、薄膜、微粒子等を指す。
【0034】
「リンカー」としては、特に限定されるものではないが、例えば、酸素、窒素、硫黄またはリン原子を含む鎖状または環状構造を含む炭化水素基などがあげられる。
【0035】
「固相担体とペプチド性基質との直接またはリンカーを介した結合」としては、共有結合、静電的結合、水素結合、疎水結合、吸着、接着等が含まれる。当然ながら、生化学分野で96穴プレート、384穴プレート、1536穴プレートと一般的に呼ばれている実験器材にペプチド性基質を結合させたものやスライドガラスや金薄膜上に微細格子状にペプチド性基質を結合させたペプチドアレイも本発明における「固相状態」の範疇に含まれる。
【0036】
本発明においては、本発明の基質群を固定してなるペプチドアレイが好ましく用いられる。
【0037】
本発明のリン酸化および/または脱リン酸化反応を検出する方法は、前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする。具体的には、前記ペプチドアレイを用いることで多様なセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼの反応を行い、抗リン酸化スレオニン抗体を用いることで、多様なセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼの反応性を幅広くかつ同時に検出する。
【0038】
前記抗リン酸化スレオニン抗体は、リン酸化スレオニン残基を認識するものであれば特に限定されるものではなく、モノクローナル抗体またはポリクローナル抗体でもよいが、基質の配列に限定されずにリン酸化スレオニン残基を認識する可能性が高いという観点から、ポリクローナル抗体が好ましい。かかる抗体は、公知の方法(Current Protocol in Molecular Biology, Chapter 11.12〜11.13(2000);Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley and Sons. Section 11.4〜11.11)により製造することができる。あるいは、市販の抗体の中から本発明の目的に適うものを適宜選択することもできる。好適な市販品の抗体としては、ポリクローナル抗リン酸化スレオニン抗体(Zymed)などがあげられる。
【0039】
さらに、ポリクローナル抗リン酸化スレオニン抗体を用いれば、リン酸化スレオニン残基を含むペプチド性基質配列のみならずリン酸化セリン残基を含むペプチド性配列をも幅広く一般的に検出することが可能である。リン酸化セリン残基を含むペプチド配列に対する抗リン酸化セリン抗体の親和力は、抗リン酸化スレオニン抗体の親和力よりも低い傾向がある。
【0040】
反応性を検出する「プロテインキナーゼ」としては、生体試料から抽出した画分中に含まれるキナーゼ;遺伝子組み換えにより発現精製されたキナーゼ;プロテインキナーゼ遺伝子からタンパク質として発現されたプロテインキナーゼであって精製されることなく細胞抽出液もしくは細胞破砕液として適当な緩衝溶液に可溶化された状態のキナーゼ;細胞培養時に分子量約1000以下の化学物質(医薬品、生理活性化合物を含む)、ペプチド、核酸、糖鎖等を培地に添加して刺激を与えることで細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液中に含まれるキナーゼ;または種々の機能既知もしくは未知遺伝子を導入することで、細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるキナーゼなどがあげられる。
【0041】
反応性を検出する「プロテインホスファターゼ」としては、生体試料から抽出した画分中に含まれるホスファターゼ;遺伝子組み換えにより発現精製されたホスファターゼ;プロテインホスファターゼ遺伝子からタンパク質として発現されたプロテインホスファターゼであって精製されることなく細胞抽出液もしくは細胞破砕液として適当な緩衝溶液に可溶化された状態のホスファターゼ;細胞培養時に分子量約1000以下の化学物質(医薬品、生理活性化合物を含む)、ペプチド、核酸、糖鎖等を培地に添加して刺激を与えることで細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるホスファターゼ;または種々の機能既知もしくは未知遺伝子を導入することで、細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるホスファターゼなどがあげられる。
【0042】
本発明において、セリン/スレオニンキナーゼの基質配列内のセリン残基とスレオニン残基を互いに置換しあった配列を準備し、それらの反応性の違いを比較検討した結果、セリン/スレオニンキナーゼが基質ペプチド配列を認識する場合には、被リン酸化部位を含むある一定の周辺配列を認識し局所的な被リン酸化部位としてのセリンあるいはスレオニン残基の構造の違いを厳密に区別してはいないものと考えられる。
【0043】
本発明の検出方法は、本発明のペプチドアレイに前記検出対象の「セリン/スレオニンキナーゼ」および/または「プロテインホスファターゼ」を含む溶液を加えて、キナーゼ反応および/またはホスファターゼ反応が進行する条件下でインキュベートする。かかる工程を、以下、接触工程とも称する。前記条件は、検出対象のキナーゼまたはホスファターゼに応じて適宜設定することができるが、通常、0.05〜0.5mMのATP存在下、pH6.5〜7.5、温度30〜40℃で、5分〜24時間である。なお、プロテインキナーゼおよびプロテインフォスファターゼの分子種は、いずれも大変多岐に渡るため、単独の阻害剤としては、それぞれの全ての分子種に対して十分に効果的な阻害効果を示す汎用的なものは見当たらない。したがって、キナーゼ反応のみを進行させる場合は、それぞれ特異性を異にする公知の複数の種類のプロテインホスファターゼ阻害剤を溶液中に共存させることが好ましく、また、ホスファターゼ反応のみを進行させる場合には、それぞれ特異性を異にする公知の複数の種類のキナーゼ阻害剤を溶液中に共存させることが好ましい。また、いずれの反応を行う場合でも、さらに公知のプロテアーゼ阻害剤を溶液中に共存させることが好ましい。
【0044】
前記ホスファターゼ阻害剤としては、例えば、オカダ酸、カリキュリンA、トウトマイシンなどがあげられ、前記キナーゼ阻害剤としては、PKCに比較的特異性の高い Calphostin C、 Hypericin、 Phloretin、 H-7 等;PKAに比較的特異性の高い KT5720 等; PTKに特異性の高い Curcumin、 AG1024 等;カルモジュリンキナーゼに特異性の高い HA1077、 Lavendustin C などがあげられる。前記プロテアーゼ阻害剤としては、PMSF(phenylmethanesulfonyl fluoride)、ロイペプチン(leupeptin)、アプロチニン(aprotinin)、ペプスタチン(pepstatin)、E−64、AEBSF (4-(2-Aminoethyl)-bezenesulfonylfluoride)、DFP(diisopropyl fluorophosphate)、ZPCK(N-carbobenzoxy-L-phenylalanine chloromethyl ketone)などがあげられる。以上は例示的に挙げたにすぎず、今日では、数多くのホスファターゼ阻害剤、キナーゼ阻害剤、プロテアーゼ阻害剤が市販されているので、通常の知識を有する当業者であれば、それらの市販された特異性の異なる複数の試薬を適宜組み合わせてカクテルとして使用することにより、容易に目的を達成することができる。
【0045】
次に、前記キナーゼおよび/またはホスファターゼ反応が終了した後、抗リン酸化スレオニン抗体を用いて、ペプチドアレイ上のリン酸化スレオニンの有無を検出する。
【0046】
検出手段としては、ペプチドアレイ上のリン酸化スレオニンに対する抗リン酸化スレオニン抗体の認識能を検出可能な適当な物理化学量に変換する手段をとる。具体的には、抗体を蛍光標識化することで蛍光強度に変換すること、化学発光色素を発する酵素を結合させた2次抗体(抗リン酸化スレオニン抗体の非可変部位を認識する)を用いて発光強度や吸光強度に変換すること、抗体結合による分子量増加分による表面プラズモン共鳴強度に変換すること等があげられる。
【0047】
例えば、マイクロタイタープレートを用いて作製したプレートアレイでセリン/スレオニンキナーゼの反応性を測定する手段は、「酵素結合抗体法(Enzyme Linked Immunosorbent Assay:ELISA)」である。このELISAを応用した本発明の検出方法を、より詳細に説明する。マイクロタイタープレートとしてアミノコートプレート(住友ベークライト、コーニング製)を用い、リンカー構造としてグルタルアルデヒドを介して本発明のペプチド基質群を固定化する。牛血清アルブミン等でブロッキング操作を行った後、対象とするセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼによるリン酸化および/または脱リン酸化反応を固相上で行う。反応液を除いた後、洗浄操作を繰り返し、1次抗体としてポリクローナル抗リン酸化スレオニン抗体(Zymed)等でリン酸化スレオニン残基を認識させる。続いてホースラディッシュペルオキシダーゼ(HRP)結合性2次抗体等で1次抗体を認識させ、ペルオキシダーゼ反応による発色を測定する。
【0048】
別の態様として、スライドガラスを用いて作製したペプチドアレイでのセリン/スレオニンキナーゼの反応性を測定する手段は、前記ELISAと同様である。ただし、酵素による発色反応を測定するのではなく、蛍光標識化された抗体を用いて蛍光強度を測定することが好ましい。より詳細に説明すると、ガラス表面がアミノ修飾されたDNAマイクロアレイ用スライドガラスを基本基板(例えば、松浪ガラス製高密度アミノ修飾スライドガラス)として用いる。これに対して、両末端にカルボン酸を有する直鎖型リンカーを活性エステルとし、基本基板に作用させる。その結果、ガラス表面に活性エステル基を有するスライドガラスが作製される。一方、ペプチド性基質のN末端にはシステイン残基を導入したものを合成し準備しておく。ペプチド性基質をリン酸緩衝液(pH7.4)に溶解させ、DNAマイクロアレイ作製用アレイヤーで微細格子状に微量分注(約1nL/スポット)する。ガラス表面の活性エステルとペプチド性基質のN末端のシステイン残基が位置選択的に反応し、ペプチド性基質が微細格子状にスライドガラスに固定化される。過剰の溶液を洗い流した後、大部分のガラス表面を占める活性エステルを化学的にブロッキングする。即ち、ブロッキング剤として適度に親水性と親油性とが制御された性質を持ち1級アミノ基を有する化学構造体を作製し、ペプチド性基質が微細格子状に固相化されたスライドガラス表面全体に作用させる。このことで、キナーゼまたはホスファターゼによるリン酸化/脱リン酸化反応と抗体反応が円滑に行われ得るペプチドマイクロアレイが作製される。作製されたマイクロアレイを、プレートアレイと同様に牛血清アルブミン等でブロッキングし、対象とするセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼによるリン酸化/脱リン酸化反応をマイクロアレイ上で行う。反応液を除いた後、洗浄操作を繰り返し、蛍光標識化されたポリクローナル抗リン酸化スレオニン抗体(Zymed)等でリン酸化スレオニン残基を認識させる。過剰の抗体溶液を洗い流し乾燥させた後、DNAマイクロアレイ解析用蛍光スキャナーで蛍光強度を測定することで、対象とするセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼの反応性を調べることが可能となる。
【0049】
本発明の検出方法において、プロテインホスファターゼの反応性は、抗リン酸化スレオニン抗体によって認識され得るリン酸化スレオニン残基に置換したペプチド基質の量を、ホスファターゼの添加前後において比較定量することにより可能である。さらに、例えば、反応液にプロテインキナーゼとプロテインホスファターゼが共存することもあり得るが、この場合は両酵素が共に反応しうる条件下で、酵素間の競合反応を測定することも可能である。その際、それぞれの特異的な配列を有する基質について、前述の特定のキナーゼ阻害剤やホスファターゼ阻害剤を適宜添加した場合と無添加の場合の反応性を比較することによって、それらの酵素間の関係を詳細に解析できる可能性がある。
【0050】
本発明の検出方法により、
(1) 単離精製されたセリン/スレオニンキナーゼのペプチド性基質を見いだすこと
(2) セリン/スレオニンキナーゼを適当な細胞で遺伝子から発現させ、単離精製することなく細胞抽出液のまま使用しペプチド性基質を見いだすこと
(3) 細胞培養の際に薬剤刺激、対象遺伝子の導入に対する細胞の応答や細胞機能変化をリン酸化反応変化として検出可能なペプチド性基質を見いだすこと
が可能である。
【0051】
上記のペプチド性基質を見いだす際に、ペプチドアレイ上の多数のペプチド性基質は、単離精製セリン/スレオニンキナーゼ、遺伝子導入細胞の細胞抽出液、薬剤刺激細胞の細胞抽出液といったリン酸化条件に応答する形でリン酸化反応性を示し、あるパターンを与える。この得られたリン酸化パターンにより元の単離精製セリン/スレオニンキナーゼの機能、遺伝子導入に用いた遺伝子の機能、薬剤刺激をかけた刺激薬剤の機能を定義づけることが可能となる。ここでは、このような機能の定義づけを「リン酸化プロファイリング(Phosphorylation Profiling)」と称する。
【0052】
本発明のプロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法は、下記工程:
被験者由来の生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分を調製する工程;
前記画分と前記ペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出し、対照と比較する工程;ならびに
前記比較結果に基づき、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する工程
を含むことを特徴とする。
【0053】
前記診断対象となる疾患としては、癌、悪性腫瘍、糖尿病、糖尿病性合併症、代謝性疾患、リウマチや関節炎等の自己免疫疾患、免疫不全やアトピー性疾患などの免疫異常疾患、アルツハイマー病、ハンチントン病、プリオン病などの神経変性疾患、肥大性心疾患、脳梗塞、心筋梗塞、肝硬変等の虚血性疾患、内分泌疾患、肝疾患、骨疾患等の様々な疾患があげられるが、これらに限定されない。
【0054】
被験者由来の生体試料としては、特に限定されるものではないが、血液、尿、粘膜、各種組織などがあげられる。
【0055】
前記生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分の調製は、自体公知のタンパク質の精製方法により行うことができる。生体試料が細胞の場合、細胞抽出液、細胞膜画分、細胞質画分、細胞核画分などの調製が可能である。
【0056】
前記画分と前記ペプチドアレイとの接触工程は、前記本発明の検出方法で記載したとおりである。
【0057】
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程は、前記したとおりである。
【0058】
次に、比較工程において、前記検出工程で得られた結果を対照と比較する。ここで、対照としては、正常者由来の生体試料を被験者由来の生体試料の場合と同様にして前記検出工程に供して得られた結果;予め正常範囲となる基準のリン酸化または脱リン酸化を決定しておいた基準などがあげられる。比較結果は、特定の基質のリン酸化または脱リン酸化の有無として得られる。それぞれの基質のリン酸化または脱リン酸化の有無と疾患との関連を調べることにより、疾患の有無で反応性の異なっている特定の基質のアミノ酸配列を同定することが可能となる。そして、タンパク質配列データベースが整備されてきた今日では、それらの配列をクエリーとして既存の配列データベースをホモロジーサーチすることによって、当該配列に相当する部分配列を含むタンパク質を同定し、その実態を解明できる可能性がある。それらの疾患関連タンパク質は、特定のプロテインキナーゼおよび/またはプロテインホスファターゼが作用する標的として機能しており、例えば、治療薬開発におけるスクリーニング系に利用できる可能性がある。
【0059】
最後に、判定工程において、前記比較結果に基づき、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する。ここで、あらかじめ精製されたプロテインキナーゼまたはフォスファターゼによる同一基質群の反応性を確認しておくことで、疾患の有無で反応性が変化しているプロテインキナーゼまたはフォスファターゼを同定することが可能となる場合も有り得る。
【0060】
これまでに特定の疾患と特定のタンパク質のリン酸化との関連については、多くの研究がなされているが、それらを制御するキナーゼやフォスファターゼの実態については、必ずしも明確になっていない場合が多い。例えば、アルツハイマー病においては、tau タンパク質の過剰かつ異常なリン酸化が神経変性の引き金となることを示唆する報告が多いが、実際にどのような制御因子がtau タンパク質のリン酸化に関与しているかについては、諸説あって未だに解明されていない。また、Pin 1タンパク質のリン酸化と神経変性疾患との関連や、p53 や Myc 、EGFRのリン酸化と癌、悪性腫瘍との関連を示唆する報告も多いが、リン酸化の生理的意味については、必ずしもよく理解されてはいない。本発明の方法は、プロテオーム技術の網羅的方法として利用されることにより、これらの疾患特異的な重要な分子メカニズムを解明する上での有力な手段を提供することができる。
【0061】
本発明のプロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法は、下記工程:
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、前記ペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出し、前記検出結果と被検物質を含まない試料における検出結果とを比較する工程;
被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する工程
を含むことを特徴とする。
【0062】
前記被検物質としては、いかなる公知物質および新規物質であってもよく、例えば、核酸、糖質、脂質、タンパク質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分などがあげられる。
【0063】
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料としては、生体試料から抽出した画分中に含まれるキナーゼまたはホスファターゼに被検物質を添加したもの;遺伝子組み換えにより発現精製されたキナーゼまたはホスファターゼに被検物質を添加した試料;プロテインキナーゼまたはプロテインホスファターゼ遺伝子からタンパク質として発現されたキナーゼまたはホスファターゼであって精製されることなく細胞抽出液もしくは細胞破砕液として適当な緩衝溶液に可溶化された状態のキナーゼまたはホスファターゼに被検物質を添加した試料;細胞培養時に被検物質を培地に添加して刺激を与えることで細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液中に含まれるキナーゼまたはホスファターゼ;または種々の機能既知もしくは未知遺伝子を導入することで、細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるキナーゼまたはホスファターゼに被検物質を添加した試料などがあげられる。
【0064】
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、ペプチドアレイとの接触工程は、前記本発明の検出方法で記載したとおりである。
【0065】
プロテインキナーゼ活性を変動させる被検物質のスクリーニング方法の場合、ペプチドアレイにはスレオニン残基を含むペプチド基質を固定させ、陽性対照としてリン酸化スレオニン残基を含むペプチド基質を固定させたものを用いることが好ましい。次いで、検出工程において、接触工程前後のペプチドアレイについて抗リン酸化スレオニン抗体を用いてリン酸化ペプチドを定性的または定量的に検出し、比較工程において、前記検出結果と被検物質を含まない試料における検出結果とを比較する。
【0066】
プロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法の場合、ペプチドアレイにはリン酸化スレオニン残基を含むペプチド基質を固定させ、陽性対照としてスレオニン残基を含むペプチド基質を固定させたものを用いることが好ましい。次いで、検出工程において、接触工程前後のペプチドアレイについて抗リン酸化スレオニン抗体を用いたリン酸化ペプチドの検出を介して、脱リン酸化により検出されなくなったペプチド基質を定性的または定量的に検出し、比較工程において、前記検出結果と被検物質を含まない試料における検出結果とを比較する。
【0067】
最後に、判定工程において、被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する。プロテインキナーゼ活性は、抗リン酸化スレオニン抗体により検出されるペプチド基質の数および検出強度で示すことができる。プロテインホスファターゼ活性は、抗リン酸化スレオニン抗体により検出されなくなったペプチド基質の数および検出強度で示すことができる。
【0068】
前記判定工程において、プロテインキナーゼ活性が促進されている場合、被検物質は、プロテインキナーゼの活性化を介した疾患の治療剤の候補として選択することができる。プロテインキナーゼ活性が阻害されている場合、被検物質は、プロテインキナーゼ活性の抑制化を介した疾患の治療剤の候補として選択することができる。被検物質は、プロテインキナーゼ調節剤または研究用試薬としても有用である。あるいは、前記判定工程において、プロテインホスファターゼ活性が促進されている場合、被検物質は、プロテインホスファターゼの活性化を介した疾患の治療剤の候補として選択することができる。プロテインホスファターゼ活性が阻害されている場合、被検物質は、プロテインホスファターゼ活性の抑制化を介した疾患の治療剤の候補として選択することができる。被検物質は、プロテインホスファターゼ調節剤または研究用試薬としても有用である。
【実施例】
【0069】
次に、実施例により本発明をさらに詳しく説明するが、本発明はこれら実施例に何ら限定されるものではない。なお、本実施例において市販のキットあるいは試薬を用いた部分については、特に断りのない限り添付のプロトコールに従って実験を行った。本実施例で汎用的に使用した試薬、培地、緩衝溶液を以下に記す。
【0070】
ペプチド基質
被リン酸化部位としてスレオニンまたはセリン残基を含みN末端にシステイン残基を含む10から18アミノ酸からなるペプチド配列を通常のFmoc法による化学合成を行い使用した。
以下の10配列は、合成した粗精製品を逆相クロマトグラフにより精製を行い使用した。pS*はリン酸化セリン残基、pT*はリン酸化スレオニン残基を示す。
【0071】
PKA用基質ペプチド
Kemptide(A): CGGLRRAALG-NH2 (配列番号1)
Kemptide(S): CGGLRRASLG-NH2(配列番号2)
Kemptide(T) : CGGLRRATLG-NH2(配列番号3)
Kemptide(pS*): CGGLRRA(pS*)LG-NH2(配列番号4)
Kemptide(pT*) : CGGLRRA(pT*)LG-NH2(配列番号5)
AMPK用基質ペプチド
SAMA: CGGHMRSAMAGLHLVKRR-NH2(配列番号6)
SAMT: CGGHMRSAMTGLHLVKRR-NH2(配列番号7)
PKC用基質ペプチド
IQAA: CGGAAKIQAAFRGHMARKK-NH2(配列番号8)
IQAT: CGGAAKIQATFRGHMARKK-NH2(配列番号9)
IQA(pT*): CGGAAKIQA(pT*)FRGHMARKK-NH2(配列番号10)
【0072】
以下に示す289配列は、合成した粗精製品を精製することなく使用した。pTはリン酸化されていてもよいスレオニン残基を示す。
PT001: CGGHMRSAM(pT)GLHLVKRR (配列番号11)
PT002: CGGAMARAA(pT)AAALARRR (配列番号12)
PT003: CGGHQGKFLQ(pT)FAGSPLY (配列番号13)
PT004: CGGKSGELLA(pT)WAGSPPY (配列番号14)
PT005: CGGKSGEPLS(pT)WAGSPPY (配列番号15)
PT006: CGGQKDKFLQ(pT)FAGSPLY (配列番号16)
PT007: CGGSDGEFLR(pT)SAGSPNY (配列番号17)
PT008: CGGTFGNKLD(pT)FAGSPPY (配列番号18)
PT009: CGGTLGSKLD(pT)FAGSPPY (配列番号19)
PT010: CGGTVGGKLD(pT)FAGSPPY (配列番号20)
PT011: CGGTVGNKLD(pT)FAGSPPY (配列番号21)
PT012: CGGVGDSLLE(pT)SAGSPHY (配列番号22)
PT013: CGGKSAEPLS(pT)WAGSPPY (配列番号23)
PT014: CGGVGDSLFE(pT)SAGSPHY (配列番号24)
PT015: CGGTVGGKID(pT)FAGSPPY (配列番号25)
PT016: CGGKSGEPLS(pT)WAGSPPH (配列番号26)
PT017: CGGTLGSKLD(pT)FAGNPPY (配列番号27)
PT018: CGGTVAGKLD(pT)FAGSPPY (配列番号28)
PT019: CGGVGDSLLE(pT)SAGSPHH (配列番号29)
PT020: CGGVGDSLLE(pT)SAASPHY (配列番号30)
PT021: CGGYQGKFLQ(pT)FAGSPLY (配列番号31)
PT022: CGGKSGELLA(pT)WSGSPPY (配列番号32)
PT023: CGGSDGEFLR(pT)TAGSPNY (配列番号33)
PT024: CGGKSGDPLS(pT)WAGSPPY (配列番号34)
PT025: CGGTFGNKLD(pT)FAGSPPF (配列番号35)
PT026: CGGTVANKLD(pT)FAGSPPY (配列番号36)
PT027: CGGKSGEPLS(pT)WAGTPPY (配列番号37)
PT028: CGGKSGEPLS(pT)LAGSPPY (配列番号38)
PT029: CGGTIGNKLD(pT)FAGSPPY (配列番号39)
PT030: CGGSDGEFLK(pT)SAGSPNY (配列番号40)
PT031: CGGTLGSKLD(pT)FAGSPAY (配列番号41)
PT032: CGGSDGEFLR(pT)SAGSPNF (配列番号42)
PT033: CGGKSGELLA(pT)WAASPPY (配列番号43)
PT034: CGGTVGNKLD(pT)LAGSPPY (配列番号44)
PT035: CGGKSGEPLT(pT)WAGSPPY (配列番号45)
PT036: CGGHQGKFLQ(pT)FAGSPLH (配列番号46)
PT037: CGGTVGGKLD(pT)FAASPPY (配列番号47)
PT038: CGGHQGKFLQ(pT)IAGSPLY (配列番号48)
PT039: CGGKSAELLA(pT)WAGSPPY (配列番号49)
PT040: CGGKTGELLA(pT)WAGSPPY (配列番号50)
PT041: CGGQKDKFLQ(pT)FAGSALY (配列番号51)
PT042: CGGQKDKFIQ(pT)FAGSPLY (配列番号52)
PT043: CGGTVGGKLD(pT)FAGNPPY (配列番号53)
PT044: CGGTLGGKLD(pT)FAGSPPY (配列番号54)
PT045: CGGHQGKLLQ(pT)FAGSPLY (配列番号55)
PT046: CGGKSGEPLS(pT)WAGSPPF (配列番号56)
PT047: CGGTFGNKLD(pT)LAGSPPY (配列番号57)
PT048: CGGVGDSLLE(pT)SAGSPYY (配列番号58)
PT049: CGGQKDRFLQ(pT)FAGSPLY (配列番号59)
PT050: CGGVGDSLLE(pT)SAGSPHF (配列番号60)
PT051: CGGTVGGKLD(pT)FAGSPAY (配列番号61)
PT052: CGGHQGKFLQ(pT)FAGSPFY (配列番号62)
PT053: CGGSVGNKLD(pT)FAGSPPY (配列番号63)
PT054: CGGTVGGKLD(pT)FAGSAPY (配列番号64)
PT055: CGGVGDSLVE(pT)SAGSPHY (配列番号65)
PT056: CGGKSGEPLS(pT)WAASPPY (配列番号66)
PT057: CGGTLGSKLE(pT)FAGSPPY (配列番号67)
PT058: CGGSDGEFLR(pT)SAGSANY (配列番号68)
PT059: CGGSDGELLR(pT)SAGSPNY (配列番号69)
PT060: CGGRSGEPLS(pT)WAGSPPY (配列番号70)
PT061: CGGHQGKFLQ(pT)FAGSPIY (配列番号71)
PT062: CGGTFGNKLD(pT)FAGSAPY (配列番号72)
PT063: CGGQKDKFLQ(pT)FAGSPVY (配列番号73)
PT064: CGGKSGELLA(pT)LAGSPPY (配列番号74)
PT065: CGGKTGEPLS(pT)WAGSPPY (配列番号75)
PT066: CGGTVGGKLD(pT)FAGTPPY (配列番号76)
PT067: CGGSDGEFLR(pT)SAGTPNY (配列番号77)
PT068: CGGQKDKFLQ(pT)FSGSPLY (配列番号78)
PT069: CGGIGDSLLE(pT)SAGSPHY (配列番号79)
PT070: CGGQKDKFFQ(pT)FAGSPLY (配列番号80)
PT071: CGGVGDSLLE(pT)TAGSPHY (配列番号81)
PT072: CGGTFGNKLD(pT)FAGSPAY (配列番号82)
PT073: CGGVGDSLLE(pT)SAGSAHY (配列番号83)
PT074: CGGTLGSKLD(pT)FAGSAPY (配列番号84)
PT075: CGGAGDSLLE(pT)SAGSPHY (配列番号85)
PT076: CGGQKDKFLQ(pT)FAGSPFY (配列番号86)
PT077: CGGVGDSILE(pT)SAGSPHY (配列番号87)
PT078: CGGQKDKILQ(pT)FAGSPLY (配列番号88)
PT079: CGGLGDSLLE(pT)SAGSPHY (配列番号89)
PT080: CGGQKDKFLQ(pT)LAGSPLY (配列番号90)
PT081: CGGQKDKFLQ(pT)FAASPLY (配列番号91)
PT082: CGGHQGKFLQ(pT)FAGSPVY (配列番号92)
PT083: CGGQKDKFLQ(pT)FAGSPLH (配列番号93)
PT084: CGGVGESLLE(pT)SAGSPHY (配列番号94)
PT085: CGGVGDSLLE(pT)SAGSPRY (配列番号95)
PT086: CGGSDGEFLR(pT)SAGSPNH (配列番号96)
PT087: CGGHQGKILQ(pT)FAGSPLY (配列番号97)
PT088: CGGSEGEFLR(pT)SAGSPNY (配列番号98)
PT089: CGGKSGELVA(pT)WAGSPPY (配列番号99)
PT090: CGGHQGKFLQ(pT)FAGSPLF (配列番号100)
PT091: CGGQKDKFVQ(pT)FAGSPLY (配列番号101)
PT092: CGGKSGDLLA(pT)WAGSPPY (配列番号102)
PT093: CGGSDAEFLR(pT)SAGSPNY (配列番号103)
PT094: CGGVGDSLLD(pT)SAGSPHY (配列番号104)
PT095: CGGSDGEFLR(pT)SAASPNY (配列番号105)
PT096: CGGTFGNKLE(pT)FAGSPPY (配列番号106)
PT097: CGGQRDKFLQ(pT)FAGSPLY (配列番号107)
PT098: CGGHQAKFLQ(pT)FAGSPLY (配列番号108)
PT099: CGGQKDKFLQ(pT)FAGNPLY (配列番号109)
PT100: CGGQKDKFLQ(pT)FAGTPLY (配列番号110)
PT101: CGGQKEKFLQ(pT)FAGSPLY (配列番号111)
PT102: CGGHQGKFLQ(pT)FAGNPLY (配列番号112)
PT103: CGGSDGEFVR(pT)SAGSPNY (配列番号113)
PT104: CGGHQGKFVQ(pT)FAGSPLY (配列番号114)
PT105: CGGKSGEPLS(pT)WAGSPAY (配列番号115)
PT106: CGGVGDNLLE(pT)SAGSPHY (配列番号116)
PT107: CGGKSGEPVS(pT)WAGSPPY (配列番号117)
PT108: CGGVADSLLE(pT)SAGSPHY (配列番号118)
PT109: CGGVGDSLLE(pT)SAGTPHY (配列番号119)
PT110: CGGKSGELLV(pT)WAGSPPY (配列番号120)
PT111: CGGVGDSVLE(pT)SAGSPHY (配列番号121)
PT112: CGGHQGKFLQ(pT)FAGSALY (配列番号122)
PT113: CGGHQGKFLQ(pT)LAGSPLY (配列番号123)
PT114: CGGQKDKFLQ(pT)IAGSPLY (配列番号124)
PT115: CGGHQGKFLQ(pT)FAGTPLY (配列番号125)
PT116: CGGHQGKFLQ(pT)FSGSPLY (配列番号126)
PT117: CGGKSGELLA(pT)WAGTPPY (配列番号127)
PT118: CGGKNGELLA(pT)WAGSPPY (配列番号128)
PT119: CGGTVGGKLE(pT)FAGSPPY (配列番号129)
PT120: CGGKSGEPLS(pT)WSGSPPY (配列番号130)
PT121: CGGKSGEPLS(pT)WAGSAPY (配列番号131)
PT122: CGGKSGELLA(pT)WAGSPPF (配列番号132)
PT123: CGGQKDKLLQ(pT)FAGSPLY (配列番号133)
PT124: CGGQKDKFLQ(pT)FAGSPLF (配列番号134)
PT125: CGGHQGRFLQ(pT)FAGSPLY (配列番号135)
PT126: CGGHQGKFLQ(pT)FAASPLY (配列番号136)
PT127: CGGQKDKFLQ(pT)FAGSPIY (配列番号137)
PT128: CGGKSGELLA(pT)WAGSPAY (配列番号138)
PT129: CGGVGDSLLE(pT)SSGSPHY (配列番号139)
PT130: CGGKSGELLA(pT)WAGSAPY (配列番号140)
PT131: CGGHQGKFLQ(pT)SAGSPHY (配列番号141)
PT132: CGGKSGELLA(pT)FAGSPLY (配列番号142)
PT133: CGGTVGNKLD(pT)SAGSPHY (配列番号143)
PT134: CGGKSGEPLS(pT)SAGSPHY (配列番号144)
PT135: CGGVGDSLLE(pT)SAGSPNY (配列番号145)
PT136: CGGLALHIRS(pT)WSGLHLV (配列番号146)
PT137: CGGLHIRSSW(pT)GLHLVKQ (配列番号147)
PT138: CGGLEMVKAA(pT)ETNLPLL (配列番号148)
PT139: CGGHNMLDDS(pT)DPILTTI (配列番号149)
PT140: CGGYRYPRPA(pT)VPPSPSL (配列番号150)
PT141: CGGPSLSRHS(pT)PHQSEDE (配列番号151)
PT142: CGGSSGSPAN(pT)FHFKEAW (配列番号152)
PT143: CGGSPRVRTL(pT)GSRPPLL (配列番号153)
PT144: CGGNLMLDRL(pT)GKILHID (配列番号154)
PT145: CGGVTHTFSG(pT)IEYMAPE (配列番号155)
PT146: CGGVTHTFAG(pT)IEYMAPE (配列番号156)
PT147: CGGSQFDSKF(pT)RQTPVDS (配列番号157)
PT148: CGGNQVFLGF(pT)YVAPSVL (配列番号158)
PT149: CGGEPMRRSV(pT)EAALAQP (配列番号159)
PT150: CGGWTETKKQ(pT)FKQTGEF (配列番号160)
PT151: CGGEPLERRL(pT)LVPDSEQ (配列番号161)
PT152: CGGRVPTMRP(pT)MSGLHLV (配列番号162)
PT153: CGGPTMRPSM(pT)GLHLVKR (配列番号163)
PT154: CGGKTPNLRM(pT)EKKSSWA (配列番号164)
PT155: CGGLNVESVS(pT)HRERSED (配列番号165)
PT156: CGGLNVESVA(pT)HRERSED (配列番号166)
PT157: CGGNPLMRRN(pT)VTPLASP (配列番号167)
PT158: CGGPLASPEP(pT)KKPRINS (配列番号168)
PT159: CGGEPSRAPA(pT)LLPPAPE (配列番号169)
PT160: CGGYYSSIKR(pT)GSQAHEQ (配列番号170)
PT161: CGGWRRKRKE(pT)SNTDSAG (配列番号171)
PT162: CGGVEDPPAP(pT)EPVAVEQ (配列番号172)
PT163: CGGPQLLRLL(pT)TLAEEPR (配列番号173)
PT164: CGGTSRIRTQ(pT)FSLQERQ (配列番号174)
PT165: CGGHFAIAAD(pT)EAEQDSW (配列番号175)
PT166: CGGSGDYMPM(pT)PKSVSAP (配列番号176)
PT167: CGGQHLRLST(pT)SGRLLYA (配列番号177)
PT168: CGGPLGSGES(pT)STRRSSE (配列番号178)
PT169: CGGQHLRLST(pT)SGRILYA (配列番号179)
PT170: CGGTSRIRTQ(pT)ISLQERQ (配列番号180)
PT171: CGGNKRSRTR(pT)DTYSAGQ (配列番号181)
PT172: CGGDSLPSSP(pT)TATPHSQ (配列番号182)
PT173: CGGSHMVHNR(pT)KINLQDL (配列番号183)
PT174: CGGRSRTRTD(pT)YSAGQSI (配列番号184)
PT175: CGGPTMRPSM(pT)GLHLVRR (配列番号185)
PT176: CGGLHIRSSW(pT)GLHLLKQ (配列番号186)
PT177: CGGLHIRSSW(pT)GLYLVKQ (配列番号187)
PT178: CGGQHLRVST(pT)SGRLLYA (配列番号188)
PT179: CGGDTLPSSP(pT)SATPHSQ (配列番号189)
PT180: CGGTSRIRTQ(pT)FSFQERQ (配列番号190)
PT181: CGGNPLMRRN(pT)VTPLANP (配列番号191)
PT182: CGGSHMFHNR(pT)KINLQDL (配列番号192)
PT183: CGGLHIRSSW(pT)GLHVVKQ (配列番号193)
PT184: CGGRSRTRTD(pT)YTAGQSV (配列番号194)
PT185: CGGQHLRLST(pT)SGRLIYA (配列番号195)
PT186: CGGQHLRLST(pT)SGRLLYG (配列番号196)
PT187: CGGPTMKPSM(pT)GLHLVKR (配列番号197)
PT188: CGGTSHIRTQ(pT)FSLQERQ (配列番号198)
PT189: CGGLHIRSSW(pT)GLHLVRQ (配列番号199)
PT190: CGGESLPSSP(pT)SATPHSQ (配列番号200)
PT191: CGGTSRIRTQ(pT)FTLQERQ (配列番号201)
PT192: CGGSHMIHNR(pT)KINIQDL (配列番号202)
PT193: CGGNPLMRRN(pT)VTPLGSP (配列番号203)
PT194: CGGQHLRLTT(pT)SGRLLYA (配列番号204)
PT195: CGGNPFMRRN(pT)VTPLASP (配列番号205)
PT196: CGGDSIPSSP(pT)SATPHSQ (配列番号206)
PT197: CGGSHMIHNR(pT)KVNLQDL (配列番号207)
PT198: CGGATMRPSM(pT)GLHLVKR (配列番号208)
PT199: CGGLHIRSSW(pT)ALHLVKQ (配列番号209)
PT200: CGGQRLRLST(pT)SGRLLYA (配列番号210)
PT201: CGGNPVMRRN(pT)VTPLASP (配列番号211)
PT202: CGGNPLMRRN(pT)VNPLASP (配列番号212)
PT203: CGGQHLRLST(pT)SGRLVYA (配列番号213)
PT204: CGGQHLRLST(pT)SGRVLYA (配列番号214)
PT205: CGGNPLMRRN(pT)VTPIASP (配列番号215)
PT206: CGGNPLLRRN(pT)VTPLASP (配列番号216)
PT207: CGGPSMRPSM(pT)GLHLVKR (配列番号217)
PT208: CGGPTMRPSM(pT)GFHLVKR (配列番号218)
PT209: CGGLHIRTSW(pT)GLHLVKQ (配列番号219)
PT210: CGGDSVPSSP(pT)SATPHSQ (配列番号220)
PT211: CGGNPLMRRN(pT)ATPLASP (配列番号221)
PT212: CGGQHLRLST(pT)TGRLLYA (配列番号222)
PT213: CGGDSLPSSP(pT)SATAHSQ (配列番号223)
PT214: CGGPTMRPSL(pT)GLHLVKR (配列番号224)
PT215: CGGPTMRASM(pT)GLHLVKR (配列番号225)
PT216: CGGQHFRLST(pT)SGRLLYA (配列番号226)
PT217: CGGDSLPSSP(pT)SATPHNQ (配列番号227)
PT218: CGGLHIRSSW(pT)GLHLAKQ (配列番号228)
PT219: CGGLHIRSNW(pT)GLHLVKQ (配列番号229)
PT220: CGGQYLRLST(pT)SGRLLYA (配列番号230)
PT221: CGGQHLHLST(pT)SGRLLYA (配列番号231)
PT222: CGGTSRIRTQ(pT)FSLQDRQ (配列番号232)
PT223: CGGSHMIHNK(pT)KINLQDL (配列番号233)
PT224: CGGLRIRSSW(pT)GLHLVKQ (配列番号234)
PT225: CGGSHMIHNR(pT)RINLQDL (配列番号235)
PT226: CGGVHIRSSW(pT)GLHLVKQ (配列番号236)
PT227: CGGLHIRSSW(pT)GLRLVKQ (配列番号237)
PT228: CGGNPLMRRN(pT)VSPLASP (配列番号238)
PT229: CGGQHLRLST(pT)SGRLLHA (配列番号239)
PT230: CGGQHLRLSS(pT)SGRLLYA (配列番号240)
PT231: CGGNKRSRTK(pT)DSYSAGQ (配列番号241)
PT232: CGGQHIRLST(pT)SGRLLYA (配列番号242)
PT233: CGGNPIMRRN(pT)VTPLASP (配列番号243)
PT234: CGGQHLRLST(pT)SGRFLYA (配列番号244)
PT235: CGGSHMIHNR(pT)KINVQDL (配列番号245)
PT236: CGGDSLPSSP(pT)SATPYSQ (配列番号246)
PT237: CGGSYMIHNR(pT)KINLQDL (配列番号247)
PT238: CGGQHLRFST(pT)SGRLLYA (配列番号248)
PT239: CGGTSRIRTQ(pT)FSLQEHQ (配列番号249)
PT240: CGGNKRSRTR(pT)DSYSGGQ (配列番号250)
PT241: CGGEAMRRSV(pT)EAALAQP (配列番号251)
PT242: CGGPTMRPSM(pT)GLRLVKR (配列番号252)
PT243: CGGQHLRLST(pT)SGRLLFA (配列番号253)
PT244: CGGDSLPTSP(pT)SATPHSQ (配列番号254)
PT245: CGGTHMIHNR(pT)KINLQDL (配列番号255)
PT246: CGGRSRTRTD(pT)YSAGQSL (配列番号256)
PT247: CGGQHLRLST(pT)SGHLLYA (配列番号257)
PT248: CGGLHVRSSW(pT)GLHLVKQ (配列番号258)
PT249: CGGLHIRSSW(pT)GLHLIKQ (配列番号259)
PT250: CGGLHIRSSW(pT)GFHLVKQ (配列番号260)
PT251: CGGSHMIHNR(pT)KINFQDL (配列番号261)
PT252: CGGPTMRPSM(pT)GLHLVKK (配列番号262)
PT253: CGGPTMRPSM(pT)GVHLVKR (配列番号263)
PT254: CGGTNRLRSE(pT)IAFIEEN (配列番号264)
PT255: CGGPTLRPSM(pT)GLHLVKR (配列番号265)
PT256: CGGPTMRPNM(pT)GLHLVKR (配列番号266)
PT257: CGGQHVRLST(pT)SGRLLYA (配列番号267)
PT258: CGGLHIKSSW(pT)GLHLVKQ (配列番号268)
PT259: CGGLHIRSSW(pT)GVHLVKQ (配列番号269)
PT260: CGGDSLPSSP(pT)SATPHTQ (配列番号270)
PT261: CGGDSLPSSA(pT)SATPHSQ (配列番号271)
PT262: CGGLHIRSSL(pT)GLHLVKQ (配列番号272)
PT263: CGGNPLMRRN(pT)VTPLVSP (配列番号273)
PT264: CGGLHIRSSW(pT)GLHIVKQ (配列番号274)
PT265: CGGLHIRSTW(pT)GLHLVKQ (配列番号275)
PT266: CGGPTMRPSM(pT)GIHLVKR (配列番号276)
PT267: CGGPTMRPSM(pT)GLHVVKR (配列番号277)
PT268: CGGQHLRIST(pT)SGRLLYA (配列番号278)
PT269: CGGQHLRLST(pT)SARLLYA (配列番号279)
PT270: CGGPTMRPSM(pT)GLHLAKR (配列番号280)
PT271: CGGDSFPSSP(pT)SATPHSQ (配列番号281)
PT272: CGGPTMRPSM(pT)GLHLLKR (配列番号282)
PT273: CGGLYIRSSW(pT)GLHLVKQ (配列番号283)
PT274: CGGPNMRPSM(pT)GLHLVKR (配列番号284)
PT275: CGGQHLRLST(pT)SGRLFYA (配列番号285)
PT276: CGGFHIRSSW(pT)GLHLVKQ (配列番号286)
PT277: CGGLRRA(pT)LG (配列番号287)
PT278: CGGRPRAA(pT)F (配列番号288)
PT279: CGGFKK(pT)FKL (配列番号289)
PT280: CGGTIYRRG(pT)RRWRKL (配列番号290)
PT281: CGGQKRPRRKD(pT)P (配列番号291)
PT282: CGGRKR(pT)RAE (配列番号292)
PT283: CGGAPGLI(pT)PGGPP (配列番号293)
PT284: CGGKKALHRQE(pT)VDAL (配列番号294)
PT285: CGGMHRQE(pT)VDCLK (配列番号295)
PT286: CGGHA(pT)PPKKKRK (配列番号296)
PT287: CGGPK(pT)PKKAKKL (配列番号297)
PT288: CGGRRADD(pT)DDDDD (配列番号298)
PT289: CGGRRRDDD(pT)DDD (配列番号299)
【0073】
TBS−T緩衝溶液:
0.05%のTween20を含むトリス塩酸緩衝生理食塩水(pH7.5)を使用した。
ブロッキング緩衝溶液:
3%牛血清アルブミンと0.05%のTween20を含むトリス塩酸緩衝生理食塩水(pH7.5)を使用した。
抗リン酸化セリン抗体緩衝溶液:
市販品(後述)をブロッキング緩衝溶液で500倍希釈して使用した。
抗リン酸化スレオニン抗体緩衝溶液:
市販品(後述)をブロッキング緩衝溶液で500倍希釈して使用した。
ホースラディッシュペルオキシダーゼ(HRP)標識抗マウス抗体緩衝溶液:
市販品(Zymed)をブロッキング緩衝溶液で5000倍希釈して使用した。
ホースラディッシュペルオキシダーゼ(HRP)標識抗ウサギ抗体緩衝溶液:
市販品(Zymed)をブロッキング緩衝溶液で1000倍希釈して使用した。
PKA酵素:市販品(Promega, Madison, WI、またはUpstate Biotechnology, Lake Placid, NY)を用いた。
PKC酵素:市販品(Upstate)を用いた。
AMPK酵素:市販品(Upstate)を用いた。
PKA反応緩衝溶液:以下の組成の緩衝溶液を用いた。
40 mM Tris/HCl (pH 7.2)
15 mM MgCl2
0.2 mM ATP
AMPK反応緩衝溶液:以下の組成の緩衝溶液を用いた。
40 mM HEPES (pH 7.4)
80 mM NaCl
5 mM Mg(OAc)2
1 mM DTT
0.2 mM 5’-AMP
0.2 mM ATP
PKC反応緩衝溶液(全亜型共通):以下の組成の緩衝溶液を用いた。
20 mM MOPS (pH 7.2)
25 mM β-glycerophosphate
5 mM EGTA
0.2 mM DTT
1 mM CaCl2
20 mM MgCl2
1/5 Lipid Activator (Upstate)
0.2 mM ATP
カルシウムとマグネシウムを含まないリン酸緩衝生理食塩水(PBS−):
塩化ナトリウム80g、塩化カリウム2g、リン酸水素ナトリウム11.5g、リン酸二水素カリウム2gを精製水1Lに溶解したものを10倍希釈しオートクレーブで滅菌したものを用いた。
細胞剥離液:
トリプシン−EDTA液(GIBCO)を滅菌PBS−で10倍希釈して用いた。
HEK293細胞生育培地:
500 mLのDulbeco’s リン酸緩衝生理食塩水(Sigma)に50 mLの牛胎児血清(56℃で30分間加熱し非働化したもの)と5 mLのペニシリン/ストレプトマイシン(10000 units/ml、10000 mg/ml)加えたものを用いた。
PKAプラスミド:
市販品(Stratagene)を使用した。
GFPプラスミド:
市販品(CloneTech)を使用した。
プラスミド溶液:
プラスミド4μgを250μLのOptiMem I (Invitrogen)と250μLのLipofectamine 2000 (Invitrogen)の混合液に溶解させたものを細胞培養用6穴プレートの1穴あたり用いた。
HEK293細胞抽出用緩衝液:以下の組成の緩衝溶液を用いた。
20 mM Tris/HCl (pH 7.2)
0.5 % TritonX-100
150 mM NaCl
20 mM β-glycerophosphate
2 mM EGTA
5 mM EDTA
1 mM Na3VO4(sodium orthovanadate)
0.2 mM PMSF
5 mg/ml Aprotinin
3 mg/ml Pepstatin A
L6細胞生育培地:
500 mLのαMEM(Sigma)に50 mLの牛胎児血清(56℃で30分間加熱し非働化したもの)、5 mLのペニシリン/ストレトマイシン(10000 units/ml、10000 mg/ml)と10 mLの200 mMグルタミンを加えたものを用いた。
L6細胞分化培地:
500 mLのαMEM(Sigma)に10 mLの牛胎児血清、5 mLのペニシリン/ストレプトマイシン(10000 units/ml、10000 mg/ml)混合液と10 mLの200 mMグルタミンを加えたものを用いた。
L6細胞抽出用緩衝液: 以下の組成の緩衝溶液を用いた。
30 mM HEPES (pH 7.4)
2.5 mM EGTA
3.0 mM EDTA
70 mM KCl
0.1 % Nonident P-40
20 mM β-glycerophosphate
20 mM NaF
2 mM NaPPi (sodium pyrophosphate)
1 mM Na3VO4(sodium orthovanadate)
0.2 mM PMSF
0.01 mM Pepstatin A
【0074】
実施例1:マイクロタイタープレートでの抗体認識反応(1)
1%グルタルアルデヒドの50mM炭酸ナトリウム緩衝液(pH9.5)を96穴アミノプレート(住友ベークライトまたはコーニング製)に50μLずつ注ぎ入れ、37℃で2時間反応させ、プレート表面を活性化させた。反応液を取り除き、精製水で洗浄した。ペプチド基質を50mM炭酸ナトリウム緩衝液(pH9.5)に溶解させ(ペプチド濃度1μM〜20μM)、活性化させた96穴マイクロタイタープレートの各穴に50μLずつ注ぎ入れた。37℃で2時間反応させた後、反応液を除いた。TBS−T緩衝溶液と精製水で順次洗浄することでペプチド基質を固定化した。ペプチド基質を固定化させたマイクロタイタープレートを、ブロッキング緩衝溶液を用いて30℃で1時間ブロッキング処理した。TBS−T緩衝溶液で3回洗浄した後、抗リン酸化セリン抗体緩衝溶液または抗リン酸化スレオニン抗体緩衝溶液を1穴あたり50μLずつ用いて30℃で1〜2時間抗体認識反応を行った。反応液を除いた後、TBS−T緩衝溶液で3回洗浄した。続いて、HRP標識抗マウス緩衝溶液あるいは抗ウサギ抗体緩衝溶液で認識反応させた。認識反応は1穴あたり50μLずつ用いて30℃で1時間行い、反応液を除いた後、TBS−T緩衝溶液で4回洗浄した。TMB(3,3’,5,5’−テトラメチルベンジジン)溶液を100μL加えいれ30℃で約10分間反応させた。2N硫酸水溶液で反応を停止した後、吸光光度計で450nmでの吸光度を定量した(図1および2、表1)。その結果、下記抗体16がリン酸化セリンおよびリン酸化スレオニンの両方を認識することがわかった。
【0075】
評価した抗リン酸化セリン抗体10種類と抗リン酸化スレオニン抗体6種類を以下に示す。
抗体1.マウスモノクローナル抗リン酸化セリン抗体:
クローン7F12、Alexis Biochemicals (San Diego, CA)
抗体2.マウスモノクローナル抗リン酸化セリン抗体:
クローン16B4、Alexis Biochemicals (San Diego, CA)
抗体3.マウスモノクローナル抗リン酸化セリン抗体:
クローンQ5、Qiagen (Valencia, CA)
抗体4.マウスモノクローナル抗リン酸化セリン抗体:
クローンab6639、Abcam (Cambridge, UK)
抗体5.マウスモノクローナル抗リン酸化セリン抗体:
クローンPSR-45、Sigma (St. Lois, MO)
抗体6.マウスモノクローナル抗リン酸化セリン抗体:
クローン608、Zymed (South San Francisco, CA)
抗体7.マウスモノクローナル抗リン酸化セリン抗体:
クローンIC8、Calbiochem (San Diego, CA)
抗体8.マウスモノクローナル抗リン酸化セリン抗体:
クローン4A3、Calbiochem (San Diego, CA)
抗体9.マウスモノクローナル抗リン酸化セリン抗体:
クローン4A9、Calbiochem (San Diego, CA)
抗体10.ウサギポリクローナル抗リン酸化セリン抗体:
Zymed (South San Francisco, CA)
抗体11.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンPTHR-1E11、nanoTools (Germany)
抗体12.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンPTHR-4D11、nanoTools (Germany)
抗体13.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンPTHR-14B3、nanoTools (Germany)
抗体14.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンQ7、Qiagen (Valencia, CA)
抗体15.ウサギポリクローナル抗リン酸化スレオニン抗体:
CHEMICON (Temecula, CA)
抗体16.ウサギポリクローナル抗リン酸化スレオニン抗体:
Zymed (South San Francisco, CA)
【0076】
【表1−1】

【0077】
【表1−2】
【0078】
実施例2:マイクロタイタープレートでの抗体認識反応(2)
実施例1で見出した抗体16について、N末端にシステイン残基を有しリン酸化スレオニン残基を含む10から18アミノ酸からなる289種類のペプチド配列(PT001〜PT289)に対する認識能を調べた。
抗体認識能は、実施例1に示すように1次抗体としての抗リン酸化スレオニン抗体(抗体16)と2次抗体としての抗ウサギ抗体とを組み合わせて調べた。しかし、1次抗体としての抗リン酸化スレオニン抗体(抗体16)の機能をより詳細に調べる目的で289ペプチドの全配列について以下の2つの条件で抗体認識反応を比較した。さらに実験に用いる各プレートには実施例1で用いたKemptide(A)、Kemptide(T)、Kemptide(pT*)を標準サンプルとして常に固相化させ比較検討した(表2)。
【0079】
条件1:1次抗体(−)、二次抗体(+)
ペプチド性基質を実施例1と同様の方法で固定化させた96穴マイクロタイタープレートをブロッキング緩衝溶液を用いて30℃で1時間ブロッキング処理を行った。TBS−T緩衝溶液で3回洗浄した後、HRP標識抗ウサギ抗体緩衝溶液で認識反応させた。認識反応は30℃で1時間行い、反応液を除いた後、TBS−T緩衝溶液で4回洗浄した。TMB溶液を100μL加えいれ30℃で約10分間反応させた。2N硫酸水溶液で反応を停止した後、吸光光度計で450nmでの吸光度を定量した。
【0080】
条件2:1次抗体(+)、二次抗体(+)
ペプチド性基質を実施例1と同様の方法で固定化させた96穴マイクロタイタープレートをブロッキング緩衝溶液を用いて30℃で1時間ブロッキング処理を行った。TBS−T緩衝溶液で3回洗浄した後、抗体16の抗リン酸化スレオニン抗体緩衝溶液により30℃で1〜2時間反応させた。反応液を除いた後、TBS−T緩衝溶液で3回洗浄した。続いて、HRP標識抗ウサギ抗体緩衝溶液で認識反応させた。認識反応は30℃で1時間行い、反応液を除いた後、TBS−T緩衝溶液で4回洗浄した。TMB溶液を100μL加えいれ30℃で約10分間反応させた。2N硫酸水溶液で反応を停止した後、吸光光度計で450nmでの吸光度を定量した。
【0081】
【表2−1】
【0082】
【表2−2】
【0083】
【表2−3】
【0084】
【表2−4】
【0085】
【表2−5】
【0086】
【表2−6】
【0087】
【表2−7】
【0088】
実施例3:プレートでの精製PKAでのリン酸化検出
PKA用基質ペプチドを実施例1と同様の方法で96穴マイクロタイタープレートに固定化し、1穴あたり1Uまたは150 mUのPKA(Promega, Madison, WIまたはUpstate)を含むPKA反応緩衝溶液を1穴あたり50μLずつ用いて30℃でリン酸化反応を行い経時変化を追った。反応溶液を除いた後、TBS−T緩衝溶液と精製水で洗浄した。実施例1に示す方法で、生成物であるリン酸化ペプチドを抗体16を用いて測定した(図3−1、3−2)。
図3−1、3−2より、PKAは基質ペプチド(配列番号2および3)中のセリンおよびスレオニン残基を経時的にリン酸化し、抗体16は、リン酸化セリンおよびリン酸化スレオニンを同程度に認識することがわかった。
【0089】
実施例4:プレートでの精製AMPKでのリン酸化検出
AMPK用基質ペプチドを実施例1に示した方法で96穴マイクロタイタープレートに固定化し、1穴あたり10 mUのAMPK(Upstate)を含むAMPK反応緩衝溶液を1穴あたり50μLずつ用いて30℃でリン酸化反応を行い経時変化を測定した。反応溶液を除いた後、TBS−T緩衝溶液と精製水で洗浄した。さらに、実施例1に示す方法で、生成物であるリン酸化ペプチドを測定した。AMPK反応緩衝溶液からATPとAMPを除いた場合、AMPだけ除いた場合のリン酸化反応も同時に測定した(図4)。
図4より、AMPKは基質ペプチドSAMT(配列番号7)を経時的にリン酸化し、抗体16を用いて当該リン酸化を検出することにより、ATPの存在下よりもAMPとATPの共存下の方がリン酸化の程度が強いことがわかった。
【0090】
実施例5:プレートでの精製PKCでのリン酸化検出
PKC用基質ペプチドを実施例1と同様の方法で96穴マイクロタイタープレートに固定化し、各穴あたり6.3 mUのPKC(Upstate)を含むPKC反応緩衝溶液を1穴あたり50μLずつ用いて30℃で1時間リン酸化反応を行った。用いたPKCは、PKCα、PKCβII、PKCγ、PKCδ、PKCε、PKCη、PKCθおよびPKCζであった。反応溶液を除いた後、TBS−T緩衝溶液と精製水で洗浄した。実施例1に示す方法により、生成物であるリン酸化ペプチドを抗体16を用いて測定した(図5)。
図5より、用いたPKCはすべて基質ペプチドIQAT(配列番号9)をATP存在下でリン酸化することがわかった。
【0091】
実施例6:HEK293細胞でPKAとGFP遺伝子を発現させた細胞抽出液でのリン酸化反応の検出
50mLの滅菌済み試験管にHEK293細胞を入れ、37℃に温めたHEK293細胞生育培地20mLをゆっくりと加え懸濁した。毎分1000回転で5分間遠心分離して細胞を沈殿させ上清を捨てた。HEK293細胞生育培地15mLを加えて細胞を再懸濁した。得られた懸濁液を細胞培養用F75フラスコ(コーニング)に移し、二酸化炭素培養装置内で37℃で培養した。細胞が約80%コンフルエント状態に達したら、次のように継代を行った。フラスコ内の培地を捨て、10mLのPBS−で2回洗浄した。約10mLの細胞剥離液で細胞をはがした。HEK293細胞生育培地で細胞を回収し、毎分1000回転で5分間遠心分離し、細胞を沈殿させ上清を捨てた。HEK293細胞生育培地を加えて細胞を再懸濁した。得られた細胞懸濁液の一部を使い、新しい細胞培養用F75フラスコで同じように増殖させた。必要量の細胞数を得るために継代を繰り返した。継代を繰り返して得たHEK293細胞を300,000/mlの密度で細胞培養用6穴プレート(SUMILON MS-8496F5)に播き、1穴あたりHEK293細胞生育培地で約2mLにし、PKAもしくはGFP遺伝子のプラスミド溶液を加え入れ、37℃で約4時間遺伝子導入処理を行った。4時間後、継代作業と同じように新しいHEK293細胞生育培地に交換し、コンフルエント状態に達するまで細胞を増殖させた。培地を取り除き、冷やしたPBS−で2回洗浄した。PBS−EDTA緩衝液で細胞を回収した。毎分3000回転で5分間遠心分離した後上清を除き、HEK293細胞抽出用緩衝液を加え、4℃で細胞成分を溶解させた。得られた溶解液を毎分15,000回転で遠心分離し、上清を細胞抽出液として得た。得られた細胞抽出液を各穴あたり0.03μgのタンパク量になるようにPKA反応緩衝液50μLに混合した溶液を用いて、実施例3と同様に、PKA用基質ペプチドを固定化した96穴マイクロタイタープレートでリン酸化反応を30℃で1時間行い、実施例1と同様にして、リン酸化ペプチドを抗体16により検出した(図6)。
図6より、本発明のペプチドアレイを用いて、細胞抽出液中に含まれるPKA活性を測定できることがわかった。
【0092】
実施例7:L6培養細胞の細胞抽出液でのリン酸化反応の検出
50mLの滅菌済み試験管にL6細胞を入れ、37℃に温めたL6細胞生育培地20 mLをゆっくりと加え、懸濁した。毎分1000回転で5分間遠心分離して細胞を沈殿させ、上清を捨てた。L6細胞生育培地15mLを加えて細胞を再懸濁した。得られた懸濁液を細胞培養用F75フラスコ(コーニング)に移し、二酸化炭素培養装置内で37℃で培養した。約80%コンフルエント状態で、以下のように継代を行った。フラスコ内の培地を捨て、10mLのPBS−で2回洗浄した。約10mLの細胞剥離液で室温下処理し細胞をはがした。L6細胞生育培地で細胞を回収し、毎分1000回転で5分間遠心分離して細胞を沈殿させ、上清を捨てた。L6細胞生育培地を加えて細胞を再懸濁した。得られた細胞懸濁液の一部を使い、新しい細胞培養用F75フラスコで同じように増殖させた。継代を繰り返し、必要量の細胞数に達したら分化を行った。継代の際と同じようにして得た所定の密度のL6細胞生育培地で懸濁した細胞を細胞培養用6穴プレート(SUMILON MS-8496F5)に100μLずつ分注して培養した。コンフルエント状態にして培地を完全に除き、L6細胞分化培地100μLに置換した。そのまま1〜3日間培養し、筋芽が筋管に分化したのを確認した後、90μLのL6細胞分化培地に交換した。AICAR(5-amino-4- imidazole carboxamide)とメトフォルミンを所定の10倍濃度にL6細胞分化培地で希釈し、各ウェルに10μLずつ添加し2時間インキュベートした。2時間後、培地を取り除き、冷やしたPBS−で2回洗浄した。各ウェルあたり50μLのL6細胞抽出用緩衝液を加え、4℃で攪拌しながら細胞成分を抽出した。抽出物を毎分5000回転で遠心分離し、上清を得た。作製した細胞抽出液を各穴あたり0.1μgのタンパク量になるようにAMPK反応緩衝液50μLに混合した溶液を用いて、実施例4と同様に、AMPK用基質ペプチドを固定化した96穴マイクロタイタープレートでリン酸化反応を30℃で1時間行い、実施例1と同様にしてリン酸化ペプチドを抗体16により検出した(図7)。
図7より、本発明のペプチドアレイを用いて、AICARまたはメトフォルミンで誘導したAMPK活性を測定できることがわかった。
【0093】
実施例8:Cy3標識化抗体16の緩衝溶液の調製
50mM炭酸ナトリウム緩衝液(pH9.5)で2倍に希釈した抗体16をCy3標識キット(アマシャムバイオサイエンス:コード番号PA33001)を用いて添付手順書に従ってCy3標識化した。得られたCy3標識化抗体16をブロッキング緩衝溶液で1000倍に希釈しCy3標識化抗体16の緩衝溶液とした。
【0094】
実施例9:スライドガラス表面に固定化したペプチド基質を用いてのリン酸化検出
高密度化アミノ基導入コートDNAマイクロアレイ用スライドガラス(松浪ガラス:品番SDM0011)に4−マレイミドブチル酸のN−スクシイミドエステル(CAS No. 80307-12-6: Fluka)の100mMのDMF溶液を室温下12時間反応させた後、反応液をDMFで洗い流した。さらに、DMF、精製水、エタノールを用いて順に2回ずつ洗浄し、減圧室温下乾燥した。得られたスライドガラスにDNAマイクロアレイ作製用アレイヤー(Genetic Microsystems:GMS417 Arrayer)を用いて0.38μm間隔でPKA用基質ペプチドの100μMリン酸緩衝生理食塩水溶液(pH7.2)を分注し固定化した。固定化したスライドガラスを室温下3時間静置した後、水、DMF、水、エタノールで順に2回ずつ洗浄し、減圧室温下乾燥した。乾燥したスライドガラスをブロッキング緩衝溶液で室温下1時間ブロッキング処理した後、実施例3および実施例6と同様の方法でスライドガラス上でリン酸化反応を行った。PKAを使った場合は、スライドガラス1枚あたり150mUの酵素量を、PKAおよびGFP発現細胞抽出液を使った場合は、スライドガラス1枚あたり0.1μgのタンパク質量を用いた。30℃で1時間反応させた後、スライドガラスをTBS−T緩衝溶液および精製水で2回ずつ洗浄し、再度ブロッキング緩衝溶液で室温下1時間ブロッキング処理した。実施例8で作製したCy3標識化抗体16の緩衝溶液をスライドガラスに30℃で1時間作用させた。TBS−T緩衝溶液で2回と精製水で5回洗浄を行った後、毎分500回転の遠心操作を行い乾燥させた。得られたスライドガラスはDNAマイクロアレイ用蛍光スキャナー(ScanArray 5000: GSI Lumonics)を用いて、励起波長(543nm)検出波長(570nm)で蛍光イメージング画像を取得した(図8)。
図8の写真は、左からそれぞれ、キナーゼを加えないコントロール〔Kinase(-)〕、PKA酵素を使った場合〔PKA(+)〕、PKA発現細胞抽出液を使った場合〔PKA Lysate(+)〕、GFP発現細胞抽出液を使った場合〔GFP Lysate(+)〕の結果を示す。マイクロタイタープレートを用いて作製したプレートアレイでの結果と同様に、スライドガラス上に作製したマイクロアレイ上でも、PKA遺伝子を発現させた未精製の細胞抽出液を作用させることによって、精製されたPKA酵素を作用させた場合と同様に、PKAによるリン酸化反応が円滑に進行している状態が検出できた。
【産業上の利用可能性】
【0095】
本発明は、これまで標準とされてきた放射化標識されたATPを用いたリン酸化検出方法に置き換わる新しいリン酸化検出方法ならびにその実施に使用される基質群およびペプチドアレイを提供する。さらに、より優れたポリクローナル抗リン酸化スレオニン抗体を作製することで、より高感度なリン酸化検出方法、ならびにこれに基づく診断方法やスクリーニング方法をも提供することが可能となる。
【図面の簡単な説明】
【0096】
【図1】リン酸化セリン残基の抗体認識反応を450nmでの吸光度により定量 (n=1)した図である。抗体1〜抗体16は、それぞれ、[表1−1]の各抗体を意味する。
【図2】リン酸化スレオニン残基の抗体認識反応を450nmでの吸光度により定量 (n=1)した図である。抗体11〜抗体16は、それぞれ、[表1−2]の各抗体を意味する。
【図3−1】PKAによるセリン残基のリン酸化の抗体16を使った検出を示す図である。PKA 1U/well、30℃で反応A450nm (n=3)*、 * 平均値±標準誤差(n=3)
【図3−2】PKAによるスレオニン残基のリン酸化の抗体16を使った検出を示す図である。PKA 1U/well、30℃で反応A450nm (n=3)*、 * 平均値±標準偏差(n=3)
【図4】AMPKによるスレオニン残基のリン酸化の抗体16を使った検出を示す図である。AMPK 10 mU/well、30℃で反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図5】PKCによるスレオニン残基のリン酸化の抗体16を使った検出を示す図である。PKC 6.3 mU/well、30℃で1時間反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図6】HEK293細胞でPKAとGFP遺伝子を発現させた細胞抽出液でのリン酸化反応の検出を示す図である。タンパク量0.03μg/well、30℃で1時間反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図7】L6細胞でのメトフォルミンとAICAR刺激細胞抽出液でのリン酸化反応の検出を示す図である。 タンパク量0.1μg/well、30℃で1時間反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図8】スライドガラス表面に固定化したペプチドアレイを用いて、リン酸化反応を蛍光イメージング画像で示した図である。 写真は、左からそれぞれ、キナーゼを加えないコントロール〔Kinase(-)〕、PKA酵素を使った場合〔PKA(+)〕、PKA発現細胞抽出液を使った場合〔PKA Lysate(+)〕、GFP発現細胞抽出液を使った場合〔GFP Lysate(+)〕の結果を示す。
【技術分野】
【0001】
本発明は、リン酸化−脱リン酸化反応検出用基質群およびそれを用いた検出方法に関する。詳しくは、本発明は、Non-RI法でリン酸化−脱リン酸化反応を高感度に検出するペプチド性基質群およびそれを用いたリン酸化−脱リン酸化反応の検出方法に関する。
【背景技術】
【0002】
生体内のタンパク質は、様々な生理的条件下で生化学的修飾を受けることによってその性質を変化させている。リン酸化は、タンパク質の主要な翻訳後修飾反応の1つであり、その反応は瞬時かつ可逆的におこるため、タンパク質の活性/不活性をコントロールする基本的な素過程の1つとして、生体内で重要な役割を担っていることが推測される。実際、真核細胞においては細胞内タンパク質の多くはリン酸化されており、多くのシグナル伝達系において、タンパク質リン酸化が情報伝達の方法として広く用いられていることが知られている。
【0003】
一般に、タンパク質がリン酸化反応、あるいは脱リン酸化反応を受けると、タンパク質を構成するアミノ酸残基の局所的な極性に変化が生じることにより、結果としてタンパク質の高次構造に基づく機能や活性度に変化を生じることにより、タンパク質の活性/不活性がコントロールされていると考えられている。このように、細胞内情報伝達に深く関わっているタンパク質リン酸化酵素の働きを調べることは、細胞機能を解析する上で大変重要である。さらに、医薬品探索研究やポストゲノム研究として注目されているプロテオームの研究にも関連し大変興味深い。プロテオーム解析のツールとして、タンパク質またはペプチドを基盤上に固定するマイクロアレイ技術が知られている(特許文献1および2、非特許文献1〜6)。
【0004】
タンパク質リン酸化酵素(プロテインキナーゼ)は、タンパク質の特定の部位をリン酸化してその機能を変える働きをもち、その基質であるタンパク質のセリンまたはスレオニン残基をリン酸化する「セリン/スレオニンキナーゼ」と、チロシン残基をリン酸化する「チロシンキナーゼ」の2つに大別される。ここで、「チロシンキナーゼ」は細胞膜内あるいは近傍に存在し情報伝達の最上流に位置し細胞外の環境変化を直接検知する場合が多く、「セリン/スレオニンキナーゼ」は上流からの信号を受けて逐次的に情報伝達を行い、細胞機能の直接的な調節に関与する場合が多い。
【0005】
プロテインキナーゼは、細胞外部からの刺激を細胞内部に伝達し、これらの刺激に対する細胞の応答に関与する。このように、プロテインキナーゼはリン酸化を通して、代謝、増殖、分化、情報伝達、運動などの多彩な細胞機能の調節を担っている。細胞内でのプロテインキナーゼは特定のタンパク質を基質として認識することで特異性の高いリン酸化反応を進行させ、これらリン酸化反応の逐次的反応の組み合わせとして、細胞外の環境変化や薬剤に対する応答として信号を伝達し細胞機能を調節している。遺伝子配列の解析から、約800種類のプロテインキナーゼが存在して細胞内のシグナル伝達に関与していると予想されている。タンパク質リン酸化の異常は、癌、糖尿病、関節炎などの様々な疾患の原因または結果と考えられている。
【0006】
一方、プロテインホスファターゼは、リン酸化されたタンパク質を基質として脱リン酸化を触媒する酵素であり、タンパク質リン酸化の可逆性を保障している。プロテインキナーゼは、それ自体がキナーゼの基質でもあり、スレオニン、セリン(ホスホリラーゼなど)、チロシン(srcなどガン遺伝子関連のタンパク質に例が多い)などのいずれかの残基がリン酸化されたプロテインキナーゼは、プロテインホスファターゼによって再び元の酵素に戻り、次の情報による活性化に備える。シグナル伝達経路の共通の特徴として、その反応の一過性がある。この一過性は、シグナル伝達分子の量や活性化状態の一過性の反映である。タンパク質のリン酸化は、ある場合には活性化を、ある場合には不活性化を引き起こす。実際、タンパク質の脱リン酸化反応が積極的なスイッチオンの役割を担っている場合も知られている。
【0007】
プロテインキナーゼは、その基質特異性が高いことが特徴であるが、基質タンパク質の被リン酸化部位を含むアミノ酸配列からなるペプチド性部分を抽出したペプチドもまた基質としてリン酸化を受けることが広く知られている。そのため、プロテインキナーゼの反応性を調べる際には、ペプチド性基質を用いて調べることが可能である。
【0008】
前記「プロテインキナーゼの反応性」は、その適当な基質を用いてリン酸化反応を行わせ、生成物であるリン酸化基質やADPの増加、あるいは出発物である基質やATPの減少を何らかの方法で計測することで判定が可能である。今日では、タンパク質リン酸化は、放射性核種32Pの取り込みを直接測定する方法か、または抗リン酸化タンパク質抗体を用いる方法が一般的である。
【0009】
最近では、基質に含まれるリン酸化セリン残基、リン酸化スレオニン残基またはリン酸化チロシン残基を、抗体および表面プラズモン共鳴によって検出する方法(特許文献3)や、適当なキレート性検出試薬および表面プラズモン共鳴によって検出する方法(特許文献4)が注目されている。これらの方法では、放射性物質を取り扱う必要がなく操作性にも優れているが、抗体や検出試薬の認識能力に強く依存し、これらの認識能力を基とした間接的な検出と測定を行うことになる。そのため、認識能力の優れた抗体や検出試薬の開発が重要かつ不可欠となる。
【0010】
チロシン残基のリン酸化の検出に関しては、タンパク質またはペプチド配列内のリン酸化チロシン残基を幅広く認識する多くの優れた抗リン酸化チロシン抗体が知られており、市販されている。そのため、それらの抗体を用いることでチロシンキナーゼの反応性は比較的容易に測定可能である。一方、セリン/スレオニンキナーゼの場合は、幅広くかつ一般的にリン酸化セリンあるいはリン酸化スレオニン残基を認識可能な優れた抗体は知られていない。そのため、抗体を使ってセリン/スレオニンキナーゼの反応性を調べる場合、特定のリン酸化タンパク質あるいはリン酸化ペプチドを特異的に認識する抗体をその都度作製して用いなければならず、セリン/スレオニンキナーゼの反応性を抗体により一般的かつ簡便に検出し測定する方法はほとんど知られていない。
【特許文献1】WO 02/083933 A2
【特許文献2】WO 02/083884 A2
【特許文献3】特開2004−283114号公報
【特許文献4】特開2004−309303号公報
【非特許文献1】M. Bettencourt-Dias, R. Giet, R. Sinka, A. Mazumdar, W. G. Lock, F. Balloux, P. J. Zafiropoulos, S. Yamaguchi, S. Winter, R. W. Carthew, M. Cooper, D. Jones, L. Frenz, D. M. Glover. Nature. 2004, 432, 980-7.
【非特許文献2】G. Manning, D. B. Whyte, R. Martnez, T. Hunter, S. Sudarsanam. Science. 2002, 298, 1912-1934.
【非特許文献3】S. J. Lee, S. Y. Lee. Anal. Biochem. 2004, 330, 311-316.
【非特許文献4】R. C. Panicker, X. Huang, S. Q. Yao. Comb. Chem. High Throughput Screen. 2004, 7, 547-56.
【非特許文献5】D. S. Y. Yeo, R. C. Panicker, L. P. Tan, S. Q. Yao. Comb. Chem. High Throughput Screen. 2004, 7, 213-221.
【非特許文献6】B. T. Houseman, J. H. Huh, S. J. Kron, M. Mrksich. Nature Biotech. 2002. 20. 270-274.
【発明の開示】
【発明が解決しようとする課題】
【0011】
上述の理由から、現在においては、セリン/スレオニンキナーゼのアッセイに関しては、放射性核種の32Pの取り込みを直接測定する方法が一般的となっているが、実験施設や作業環境に制約があり、また実験者の放射能被爆の問題もある為、あまり大規模なハイスループット(high throughput)解析には適さないという問題がある。したがって、大規模なプロテオーム解析を実施して疾患とタンパク質リン酸化カスケードの関連を調べる上で、放射化標識されたATP等を用いないNon-RI法で、タンパク質のリン酸化/脱リン酸化を高感度に検出するアッセイを可能にすることが重要な課題となっている。
本発明の目的は、プロテインキナーゼまたはプロテインホスファターゼの反応性を簡便に測定可能な手段の提供である。
【課題を解決するための手段】
【0012】
本発明者らは、上記問題点に鑑み、鋭意検討した結果、プロテインキナーゼまたはプロテインホスファターゼの基質として以下に示すような人工ペプチドを用いることを見出し、本発明を完成するに至った。即ち、本願発明は、以下に示す通りである。
【0013】
〔1〕 プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換してなる人工ペプチド基質を少なくとも1種含有するプロテインキナーゼの基質群。
〔2〕 プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をリン酸化スレオニン残基に置換してなる人工ペプチド基質を少なくとも1種さらに含有する、前記〔1〕に記載の基質群。
〔3〕 前記人工ペプチド基質が5〜30のアミノ酸残基を有するものである、前記〔1〕または〔2〕に記載の基質群。
〔4〕 前記プロテインキナーゼがセリン/スレオニンキナーゼである前記〔1〕〜〔3〕いずれかに記載の基質群。
〔5〕 前記セリン/スレオニンキナーゼがcAMP依存性プロテインキナーゼAである、前記〔4〕に記載の基質群。
〔6〕 前記セリン/スレオニンキナーゼがプロテインキナーゼCである、前記〔4〕に記載の基質群。
〔7〕 前記セリン/スレオニンキナーゼがAMP活性化プロテインキナーゼである、前記〔4〕に記載の基質群。
〔8〕 前記人工ペプチド基質が配列番号11〜299に記載のアミノ酸配列群から選択されるアミノ酸配列を有するものである前記〔1〕〜〔7〕いずれかに記載の基質群。
〔9〕 前記〔1〕〜〔8〕いずれかに記載の基質群を固定してなるペプチドアレイ。
〔10〕 前記〔9〕に記載のペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、リン酸化および/または脱リン酸化反応を検出する方法。
〔11〕 前記抗リン酸化スレオニン抗体がポリクローナル抗体である、前記〔10〕に記載の方法。
〔12〕 下記工程:
被験者由来の生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分を調製する工程;
前記画分と前記〔9〕に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を、対照における検出結果と比較し、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する工程
を含むプロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法。
〔13〕 前記疾患が癌、悪性腫瘍、糖尿病、糖尿病性合併症、代謝性疾患、自己免疫疾患、免疫異常疾患、免疫不全、神経変性疾患、虚血性疾患、内分泌疾患、肝疾患または骨疾患である前記〔12〕に記載の診断方法。
〔14〕 下記工程:
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、前記〔9〕に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を被検物質を含まない試料における検出結果と比較し、被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する工程
を含む、プロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法。
【発明の効果】
【0014】
本発明のプロテインキナーゼの基質群によると、適切に設計されたアミノ酸配列に基づいて合成した人工基質を含むことから、Non-RI法で高感度にタンパク質のリン酸化または脱リン酸化を測定可能な一連の基質を提供することが可能となり、さらにコンビナトリアルケミストリーの技術を応用すれば、ペプチド基質ライブラリーを作製することができるため、プロテインキナーゼおよび/またはプロテインホスファターゼのハイスループット解析に供することができる。
本発明のペプチドアレイおよびそれを用いたリン酸化および/または脱リン酸化の検出方法によると、プロテオーム解析の手段として、ハイスループット解析が可能であるとともにリン酸化と脱リン酸化を1つのアレイで同時に検出することも可能である。
本発明の診断方法によると、セリン/スレオニンキナーゼによるリン酸化および/またはプロテインホスファターゼによる脱リン酸化を高感度かつ迅速に検出することが可能であるので、かかるリン酸化および/または脱リン酸化が関与する疾患の有無を迅速に判定することできる。
本発明のスクリーニング方法によると、セリン/スレオニンキナーゼによるリン酸化またはプロテインホスファターゼによる脱リン酸化を高感度かつ迅速に検出することが可能であるので、プロテインキナーゼまたはプロテインホスファターゼ活性を促進または阻害する物質を効率よく選択することができる。
【発明を実施するための最良の形態】
【0015】
本発明においてプロテインキナーゼとは、大別してセリン/スレオニンキナーゼとチロシンキナーゼとを含む概念であるが、以下、これまで有用なキナーゼ活性の測定方法が開発されていなかったセリン/スレオニンキナーゼを中心に述べる。
【0016】
セリン/スレオニンキナーゼとしては、アミノ酸配列中のセリンまたはスレオニンをリン酸化する酵素であれば特に限定されるものではないが、例えば、cAMP依存性プロテインキナーゼ(PKA)、cGMP依存性プロテインキナーゼ(PKG)、カルシウム依存性プロテインキナーゼ、AMP活性化プロテインキナーゼ(AMPK)、二本鎖RNA依存性プロテインキナーゼ、カゼインキナーゼ、リボソームタンパク質S6キナーゼ、Rafキナーゼ、サイクリン依存性キナーゼ(CDK)などがあげられ、前記カルシウム依存性プロテインキナーゼとしては、ホスホリラーゼキナーゼ、ミオシン軽鎖キナーゼ、カルモデュリン依存性プロテインキナーゼ、リン脂質依存性プロテインキナーゼ(プロテインキナーゼC:PKC)などがあげられる。これらの中でも、細胞内における役割が解明されつつあり、疾患との関連が示唆されているPKA、AMPK、PKCなどが好ましい。
【0017】
本発明においてプロテインホスファターゼとは、リン酸化セリン/スレオニン残基、リン酸化チロシン残基またはリン酸化セリン/スレオニン/チロシン残基のリン酸エステル結合を加水分解する酵素をいい、好ましくはリン酸化セリン/スレオニン残基のリン酸エステル結合を加水分解する酵素である。具体的には、リン酸化されたセリン/スレオニン残基に特異的なホスファターゼとしては、PP1、PP2A、PP2BおよびPP2Cなどのホスファターゼがあげられる。これらは、それぞれ基質特異性や阻害剤に対する感受性、金属イオン要求性の違いによって分類されている。PP2Cは単量体酵素であるが、PP1、PP2AおよびPP2Bは、調節サブユニットと活性サブユニットで構成される複合体を形成し、細胞内での局在性は、その調節サブユニットとの相互作用により調節されていると言われている。タイプ2Bは、一般に、カルシニューリンともよばれている。一方、リン酸化されたチロシン残基を特異的に脱リン酸化するホスファターゼとしては、細胞膜貫通ドメインを持つ受容体型と、それらをもたない細胞質型が存在し、それぞれに分子構造や活性調節機構を異にする多様な分子種を含んでいる。またリン酸化されたセリン/スレオニン/チロシン残基を脱リン酸化するホスファターゼとしても、基質特異性を異にする複数の分子種が存在することが知られている。
【0018】
本発明において基質とは、プロテインキナーゼが認識するアミノ酸配列を含有するペプチドまたはタンパク質をいう。前記ペプチドまたはタンパク質を構成するアミノ酸としては、天然アミノ酸として知られるαアミノ酸、βアミノ酸もしくはγアミノ酸に加えて、それらを化学的に修飾したもの、さらにはアミノ酸のアミノ基または水酸基とカルボキシル基やスルフォニル基やフォスフォリル基等を介してペプチド様化学結合を行うことが可能な分子量約800以下の化学構造を有する部分化学構造体もその範囲に含まれる。
【0019】
前記基質としては、人工的に適切なアミノ酸配列を設計、合成可能で、かつ、保存性および操作性に優れているペプチド基質が好ましい。
【0020】
本発明において天然に存在する基質とは、プロテインキナーゼの基質として知られているタンパク質を構成するアミノ酸配列の全部または一部からなるものをいい、セリン/スレオニン残基、好ましくはセリン/スレオニンキナーゼが認識してリン酸化可能なセリン/スレオニン残基を1つまたは複数含むアミノ酸配列からなるものである。天然に存在する基質におけるアミノ酸残基の数としては、5〜タンパク質の全長のアミノ酸残基の数であり、好ましくは5〜30、より好ましくは10〜20である。
【0021】
本発明において人工ペプチド基質とは、前記天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換したものをいい、好ましくはセリン/スレオニンキナーゼが認識してリン酸化可能なセリン残基をスレオニン残基またはリン酸化スレオニン残基で置換したものである。人工ペプチド基質は、酵素の特異性や検出の容易性を担保するためには、セリン/スレオニンキナーゼが認識可能なセリン残基をスレオニン残基(またはリン酸化されたスレオニン残基)に置換した部位を1つ含むものであることが好ましい。人工ペプチド基質中のアミノ酸残基の数としては、セリン/スレオニンキナーゼが認識できる程度のペプチド長となれば特に限定されるものではないが、通常5〜500、好ましくは5〜30、より好ましくは10〜20である。
【0022】
なお、前記人工ペプチド基質は、前記天然に存在する基質から設計されたものであるから、天然に存在する基質中のセリン/スレオニンキナーゼが認識できる程度のアミノ酸残基からなるペプチド部分を含有していればよく、人工ペプチド基質および天然に存在する基質のアミノ酸残基の数が互いに一致する必要はない。
【0023】
本発明のプロテインキナーゼの基質群は、前記人工ペプチド基質を少なくとも1種含有することを特徴とする。「セリン/スレオニンキナーゼ」は、タンパク質性またはペプチド性基質のセリンおよび/またはスレオニン残基をリン酸化するが、セリン残基とスレオニン残基ではそれらの反応性に若干の差はあるものの共にリン酸化を受けるものと考えられる。そこで、本発明では、セリン/スレオニンキナーゼのリン酸化能を調べる目的のためにセリン/スレオニンキナーゼの基質として被リン酸化セリン残基をスレオニン残基に置換した人工ペプチド基質を用いる。あるいは、プロテインホスファターゼの脱リン酸化能を調べる目的のために、前記被リン酸化セリン残基をリン酸化スレオニン残基に置換した人工ペプチド基質を用いてもよい。
【0024】
本発明の基質群に含まれる基質の数は、前記人工ペプチド基質を少なくとも1種含有していれば特に限定されるものではないが、好ましくは、1〜100000、より好ましくは2〜50000、さらにより好ましくは10〜5000である。かかる範囲内の基質群は、ペプチドアレイまたはペプチドチップとして、1個の担体上に固定化することが可能である。
【0025】
本発明の基質群は、そのすべてが前記人工ペプチド基質であってもよく、前記人工ペプチド基質と天然に存在する基質とを含んでいてもよく、本発明の検出方法等に用いる場合に、陽性対照および/または陰性対照となる人工ペプチド基質または天然の基質を含んでいてもよい。陽性対照および陰性対照となる人工ペプチド基質には、セリン残基をリン酸化スレオニン残基またはアラニン残基に置換した人工ペプチド基質も含まれる。
【0026】
人工ペプチド基質の具体的な配列は、配列番号11〜299に記載のアミノ酸配列群から選択されるアミノ酸配列が好ましい。かかるアミノ酸配列を全部用いてもよく、あるいは、対象とするプロテインキナーゼに応じて適宜組み合わせて用いてもよい。
【0027】
本発明の人工ペプチド基質は、天然に存在する基質に含まれるアミノ酸配列中の被リン酸化セリン残基をスレオニン残基またはリン酸化スレオニン残基に置換することによって調製することができる。以下、かかる人工ペプチド基質配列の設計と調製方法について説明する。
【0028】
基質配列設計の方法としては、ゲノム情報またはタンパク質情報からセリン/スレオニンキナーゼの基質となるタンパク質の被リン酸化部位を含むペプチド断片を予測し抽出する。抽出したペプチド断片を、セリン/スレオニンキナーゼのペプチド性基質配列として既に知られているものをも含め、「ペプチド基質の基本配列」と定義して用いる。
【0029】
次に、前記ペプチド基質の基本配列の被リン酸化部位(セリン残基)を全てスレオニン残基に置換し、その他のアミノ酸残基は基本配列と同様にしてもよいし、適当な方法で一部のアミノ酸残基をさらに他のアミノ酸残基に置換してもよい。さらなる置換は、セリン/スレオニンキナーゼの人工ペプチド基質の製造、固定化等に有利な置換基を導入するために行われる。このように人工的に配列設計されたペプチドを、基質群(集合体あるいはペプチド基質ライブラリー)として供する。配列設計するペプチド配列は、化学合成の効率と活性発現を担保するために、40アミノ酸残基以下とすることが好ましい。
【0030】
前記ライブラリーには、被リン酸化部位をリン酸化スレオニンに置換した基質を少なくとも1種含有してもよい。リン酸化スレオニン置換した基質は、リン酸化の陽性対照となるばかりでなく、プロテインホスファターゼの基質ともなり、リン酸化/脱リン酸化を同時に検出することができるので好ましい。
【0031】
「配列設計されたペプチド性基質の集合体(ペプチド性基質ライブラリー)」は、一般的な化学的ペプチド合成方法(例えば、Fmoc法での固相合成等)により化学合成されて供与される。ここで重要な点は、天然に存在する被リン酸化部位としてのセリン残基を敢えてスレオニン残基に置換して配列設計し、化学合成することである。
【0032】
上記の方法によって設計し化学合成した人工ペプチド基質を含有する基質群を、溶液状態または「固相状態」でセリン/スレオニンキナーゼによるリン酸化反応の基質として用いる。キナーゼ反応の生成物であるペプチド性リン酸化基質を抗リン酸化スレオニン抗体で認識させ検出することで、セリン/スレオニンキナーゼの種々の条件下での反応性を検出し測定することができる。逆に、ホスファターゼ反応の生成物である脱リン酸化基質が抗体で検出されなくなることで、プロテインホスファターゼの種々の条件下での反応性、好ましくはセリン/スレオニンキナーゼとの競合性を検出し測定することができる。
【0033】
ここでいう「固相状態」とは、ペプチド性基質を種々の固相担体に直接またはリンカーと呼ばれる適当な化学構造体を介して結合した状態をいう。固相担体としては、有機高分子もしくは無機高分子材料(プラスチック等)、ガラス、金属等で作製された平板、試験管(生化学分野で一般的にプレートと呼称されるものを含む)、薄膜、微粒子等を指す。
【0034】
「リンカー」としては、特に限定されるものではないが、例えば、酸素、窒素、硫黄またはリン原子を含む鎖状または環状構造を含む炭化水素基などがあげられる。
【0035】
「固相担体とペプチド性基質との直接またはリンカーを介した結合」としては、共有結合、静電的結合、水素結合、疎水結合、吸着、接着等が含まれる。当然ながら、生化学分野で96穴プレート、384穴プレート、1536穴プレートと一般的に呼ばれている実験器材にペプチド性基質を結合させたものやスライドガラスや金薄膜上に微細格子状にペプチド性基質を結合させたペプチドアレイも本発明における「固相状態」の範疇に含まれる。
【0036】
本発明においては、本発明の基質群を固定してなるペプチドアレイが好ましく用いられる。
【0037】
本発明のリン酸化および/または脱リン酸化反応を検出する方法は、前記ペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする。具体的には、前記ペプチドアレイを用いることで多様なセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼの反応を行い、抗リン酸化スレオニン抗体を用いることで、多様なセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼの反応性を幅広くかつ同時に検出する。
【0038】
前記抗リン酸化スレオニン抗体は、リン酸化スレオニン残基を認識するものであれば特に限定されるものではなく、モノクローナル抗体またはポリクローナル抗体でもよいが、基質の配列に限定されずにリン酸化スレオニン残基を認識する可能性が高いという観点から、ポリクローナル抗体が好ましい。かかる抗体は、公知の方法(Current Protocol in Molecular Biology, Chapter 11.12〜11.13(2000);Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley and Sons. Section 11.4〜11.11)により製造することができる。あるいは、市販の抗体の中から本発明の目的に適うものを適宜選択することもできる。好適な市販品の抗体としては、ポリクローナル抗リン酸化スレオニン抗体(Zymed)などがあげられる。
【0039】
さらに、ポリクローナル抗リン酸化スレオニン抗体を用いれば、リン酸化スレオニン残基を含むペプチド性基質配列のみならずリン酸化セリン残基を含むペプチド性配列をも幅広く一般的に検出することが可能である。リン酸化セリン残基を含むペプチド配列に対する抗リン酸化セリン抗体の親和力は、抗リン酸化スレオニン抗体の親和力よりも低い傾向がある。
【0040】
反応性を検出する「プロテインキナーゼ」としては、生体試料から抽出した画分中に含まれるキナーゼ;遺伝子組み換えにより発現精製されたキナーゼ;プロテインキナーゼ遺伝子からタンパク質として発現されたプロテインキナーゼであって精製されることなく細胞抽出液もしくは細胞破砕液として適当な緩衝溶液に可溶化された状態のキナーゼ;細胞培養時に分子量約1000以下の化学物質(医薬品、生理活性化合物を含む)、ペプチド、核酸、糖鎖等を培地に添加して刺激を与えることで細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液中に含まれるキナーゼ;または種々の機能既知もしくは未知遺伝子を導入することで、細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるキナーゼなどがあげられる。
【0041】
反応性を検出する「プロテインホスファターゼ」としては、生体試料から抽出した画分中に含まれるホスファターゼ;遺伝子組み換えにより発現精製されたホスファターゼ;プロテインホスファターゼ遺伝子からタンパク質として発現されたプロテインホスファターゼであって精製されることなく細胞抽出液もしくは細胞破砕液として適当な緩衝溶液に可溶化された状態のホスファターゼ;細胞培養時に分子量約1000以下の化学物質(医薬品、生理活性化合物を含む)、ペプチド、核酸、糖鎖等を培地に添加して刺激を与えることで細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるホスファターゼ;または種々の機能既知もしくは未知遺伝子を導入することで、細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるホスファターゼなどがあげられる。
【0042】
本発明において、セリン/スレオニンキナーゼの基質配列内のセリン残基とスレオニン残基を互いに置換しあった配列を準備し、それらの反応性の違いを比較検討した結果、セリン/スレオニンキナーゼが基質ペプチド配列を認識する場合には、被リン酸化部位を含むある一定の周辺配列を認識し局所的な被リン酸化部位としてのセリンあるいはスレオニン残基の構造の違いを厳密に区別してはいないものと考えられる。
【0043】
本発明の検出方法は、本発明のペプチドアレイに前記検出対象の「セリン/スレオニンキナーゼ」および/または「プロテインホスファターゼ」を含む溶液を加えて、キナーゼ反応および/またはホスファターゼ反応が進行する条件下でインキュベートする。かかる工程を、以下、接触工程とも称する。前記条件は、検出対象のキナーゼまたはホスファターゼに応じて適宜設定することができるが、通常、0.05〜0.5mMのATP存在下、pH6.5〜7.5、温度30〜40℃で、5分〜24時間である。なお、プロテインキナーゼおよびプロテインフォスファターゼの分子種は、いずれも大変多岐に渡るため、単独の阻害剤としては、それぞれの全ての分子種に対して十分に効果的な阻害効果を示す汎用的なものは見当たらない。したがって、キナーゼ反応のみを進行させる場合は、それぞれ特異性を異にする公知の複数の種類のプロテインホスファターゼ阻害剤を溶液中に共存させることが好ましく、また、ホスファターゼ反応のみを進行させる場合には、それぞれ特異性を異にする公知の複数の種類のキナーゼ阻害剤を溶液中に共存させることが好ましい。また、いずれの反応を行う場合でも、さらに公知のプロテアーゼ阻害剤を溶液中に共存させることが好ましい。
【0044】
前記ホスファターゼ阻害剤としては、例えば、オカダ酸、カリキュリンA、トウトマイシンなどがあげられ、前記キナーゼ阻害剤としては、PKCに比較的特異性の高い Calphostin C、 Hypericin、 Phloretin、 H-7 等;PKAに比較的特異性の高い KT5720 等; PTKに特異性の高い Curcumin、 AG1024 等;カルモジュリンキナーゼに特異性の高い HA1077、 Lavendustin C などがあげられる。前記プロテアーゼ阻害剤としては、PMSF(phenylmethanesulfonyl fluoride)、ロイペプチン(leupeptin)、アプロチニン(aprotinin)、ペプスタチン(pepstatin)、E−64、AEBSF (4-(2-Aminoethyl)-bezenesulfonylfluoride)、DFP(diisopropyl fluorophosphate)、ZPCK(N-carbobenzoxy-L-phenylalanine chloromethyl ketone)などがあげられる。以上は例示的に挙げたにすぎず、今日では、数多くのホスファターゼ阻害剤、キナーゼ阻害剤、プロテアーゼ阻害剤が市販されているので、通常の知識を有する当業者であれば、それらの市販された特異性の異なる複数の試薬を適宜組み合わせてカクテルとして使用することにより、容易に目的を達成することができる。
【0045】
次に、前記キナーゼおよび/またはホスファターゼ反応が終了した後、抗リン酸化スレオニン抗体を用いて、ペプチドアレイ上のリン酸化スレオニンの有無を検出する。
【0046】
検出手段としては、ペプチドアレイ上のリン酸化スレオニンに対する抗リン酸化スレオニン抗体の認識能を検出可能な適当な物理化学量に変換する手段をとる。具体的には、抗体を蛍光標識化することで蛍光強度に変換すること、化学発光色素を発する酵素を結合させた2次抗体(抗リン酸化スレオニン抗体の非可変部位を認識する)を用いて発光強度や吸光強度に変換すること、抗体結合による分子量増加分による表面プラズモン共鳴強度に変換すること等があげられる。
【0047】
例えば、マイクロタイタープレートを用いて作製したプレートアレイでセリン/スレオニンキナーゼの反応性を測定する手段は、「酵素結合抗体法(Enzyme Linked Immunosorbent Assay:ELISA)」である。このELISAを応用した本発明の検出方法を、より詳細に説明する。マイクロタイタープレートとしてアミノコートプレート(住友ベークライト、コーニング製)を用い、リンカー構造としてグルタルアルデヒドを介して本発明のペプチド基質群を固定化する。牛血清アルブミン等でブロッキング操作を行った後、対象とするセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼによるリン酸化および/または脱リン酸化反応を固相上で行う。反応液を除いた後、洗浄操作を繰り返し、1次抗体としてポリクローナル抗リン酸化スレオニン抗体(Zymed)等でリン酸化スレオニン残基を認識させる。続いてホースラディッシュペルオキシダーゼ(HRP)結合性2次抗体等で1次抗体を認識させ、ペルオキシダーゼ反応による発色を測定する。
【0048】
別の態様として、スライドガラスを用いて作製したペプチドアレイでのセリン/スレオニンキナーゼの反応性を測定する手段は、前記ELISAと同様である。ただし、酵素による発色反応を測定するのではなく、蛍光標識化された抗体を用いて蛍光強度を測定することが好ましい。より詳細に説明すると、ガラス表面がアミノ修飾されたDNAマイクロアレイ用スライドガラスを基本基板(例えば、松浪ガラス製高密度アミノ修飾スライドガラス)として用いる。これに対して、両末端にカルボン酸を有する直鎖型リンカーを活性エステルとし、基本基板に作用させる。その結果、ガラス表面に活性エステル基を有するスライドガラスが作製される。一方、ペプチド性基質のN末端にはシステイン残基を導入したものを合成し準備しておく。ペプチド性基質をリン酸緩衝液(pH7.4)に溶解させ、DNAマイクロアレイ作製用アレイヤーで微細格子状に微量分注(約1nL/スポット)する。ガラス表面の活性エステルとペプチド性基質のN末端のシステイン残基が位置選択的に反応し、ペプチド性基質が微細格子状にスライドガラスに固定化される。過剰の溶液を洗い流した後、大部分のガラス表面を占める活性エステルを化学的にブロッキングする。即ち、ブロッキング剤として適度に親水性と親油性とが制御された性質を持ち1級アミノ基を有する化学構造体を作製し、ペプチド性基質が微細格子状に固相化されたスライドガラス表面全体に作用させる。このことで、キナーゼまたはホスファターゼによるリン酸化/脱リン酸化反応と抗体反応が円滑に行われ得るペプチドマイクロアレイが作製される。作製されたマイクロアレイを、プレートアレイと同様に牛血清アルブミン等でブロッキングし、対象とするセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼによるリン酸化/脱リン酸化反応をマイクロアレイ上で行う。反応液を除いた後、洗浄操作を繰り返し、蛍光標識化されたポリクローナル抗リン酸化スレオニン抗体(Zymed)等でリン酸化スレオニン残基を認識させる。過剰の抗体溶液を洗い流し乾燥させた後、DNAマイクロアレイ解析用蛍光スキャナーで蛍光強度を測定することで、対象とするセリン/スレオニンキナーゼおよび/またはプロテインホスファターゼの反応性を調べることが可能となる。
【0049】
本発明の検出方法において、プロテインホスファターゼの反応性は、抗リン酸化スレオニン抗体によって認識され得るリン酸化スレオニン残基に置換したペプチド基質の量を、ホスファターゼの添加前後において比較定量することにより可能である。さらに、例えば、反応液にプロテインキナーゼとプロテインホスファターゼが共存することもあり得るが、この場合は両酵素が共に反応しうる条件下で、酵素間の競合反応を測定することも可能である。その際、それぞれの特異的な配列を有する基質について、前述の特定のキナーゼ阻害剤やホスファターゼ阻害剤を適宜添加した場合と無添加の場合の反応性を比較することによって、それらの酵素間の関係を詳細に解析できる可能性がある。
【0050】
本発明の検出方法により、
(1) 単離精製されたセリン/スレオニンキナーゼのペプチド性基質を見いだすこと
(2) セリン/スレオニンキナーゼを適当な細胞で遺伝子から発現させ、単離精製することなく細胞抽出液のまま使用しペプチド性基質を見いだすこと
(3) 細胞培養の際に薬剤刺激、対象遺伝子の導入に対する細胞の応答や細胞機能変化をリン酸化反応変化として検出可能なペプチド性基質を見いだすこと
が可能である。
【0051】
上記のペプチド性基質を見いだす際に、ペプチドアレイ上の多数のペプチド性基質は、単離精製セリン/スレオニンキナーゼ、遺伝子導入細胞の細胞抽出液、薬剤刺激細胞の細胞抽出液といったリン酸化条件に応答する形でリン酸化反応性を示し、あるパターンを与える。この得られたリン酸化パターンにより元の単離精製セリン/スレオニンキナーゼの機能、遺伝子導入に用いた遺伝子の機能、薬剤刺激をかけた刺激薬剤の機能を定義づけることが可能となる。ここでは、このような機能の定義づけを「リン酸化プロファイリング(Phosphorylation Profiling)」と称する。
【0052】
本発明のプロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法は、下記工程:
被験者由来の生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分を調製する工程;
前記画分と前記ペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出し、対照と比較する工程;ならびに
前記比較結果に基づき、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する工程
を含むことを特徴とする。
【0053】
前記診断対象となる疾患としては、癌、悪性腫瘍、糖尿病、糖尿病性合併症、代謝性疾患、リウマチや関節炎等の自己免疫疾患、免疫不全やアトピー性疾患などの免疫異常疾患、アルツハイマー病、ハンチントン病、プリオン病などの神経変性疾患、肥大性心疾患、脳梗塞、心筋梗塞、肝硬変等の虚血性疾患、内分泌疾患、肝疾患、骨疾患等の様々な疾患があげられるが、これらに限定されない。
【0054】
被験者由来の生体試料としては、特に限定されるものではないが、血液、尿、粘膜、各種組織などがあげられる。
【0055】
前記生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分の調製は、自体公知のタンパク質の精製方法により行うことができる。生体試料が細胞の場合、細胞抽出液、細胞膜画分、細胞質画分、細胞核画分などの調製が可能である。
【0056】
前記画分と前記ペプチドアレイとの接触工程は、前記本発明の検出方法で記載したとおりである。
【0057】
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程は、前記したとおりである。
【0058】
次に、比較工程において、前記検出工程で得られた結果を対照と比較する。ここで、対照としては、正常者由来の生体試料を被験者由来の生体試料の場合と同様にして前記検出工程に供して得られた結果;予め正常範囲となる基準のリン酸化または脱リン酸化を決定しておいた基準などがあげられる。比較結果は、特定の基質のリン酸化または脱リン酸化の有無として得られる。それぞれの基質のリン酸化または脱リン酸化の有無と疾患との関連を調べることにより、疾患の有無で反応性の異なっている特定の基質のアミノ酸配列を同定することが可能となる。そして、タンパク質配列データベースが整備されてきた今日では、それらの配列をクエリーとして既存の配列データベースをホモロジーサーチすることによって、当該配列に相当する部分配列を含むタンパク質を同定し、その実態を解明できる可能性がある。それらの疾患関連タンパク質は、特定のプロテインキナーゼおよび/またはプロテインホスファターゼが作用する標的として機能しており、例えば、治療薬開発におけるスクリーニング系に利用できる可能性がある。
【0059】
最後に、判定工程において、前記比較結果に基づき、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する。ここで、あらかじめ精製されたプロテインキナーゼまたはフォスファターゼによる同一基質群の反応性を確認しておくことで、疾患の有無で反応性が変化しているプロテインキナーゼまたはフォスファターゼを同定することが可能となる場合も有り得る。
【0060】
これまでに特定の疾患と特定のタンパク質のリン酸化との関連については、多くの研究がなされているが、それらを制御するキナーゼやフォスファターゼの実態については、必ずしも明確になっていない場合が多い。例えば、アルツハイマー病においては、tau タンパク質の過剰かつ異常なリン酸化が神経変性の引き金となることを示唆する報告が多いが、実際にどのような制御因子がtau タンパク質のリン酸化に関与しているかについては、諸説あって未だに解明されていない。また、Pin 1タンパク質のリン酸化と神経変性疾患との関連や、p53 や Myc 、EGFRのリン酸化と癌、悪性腫瘍との関連を示唆する報告も多いが、リン酸化の生理的意味については、必ずしもよく理解されてはいない。本発明の方法は、プロテオーム技術の網羅的方法として利用されることにより、これらの疾患特異的な重要な分子メカニズムを解明する上での有力な手段を提供することができる。
【0061】
本発明のプロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法は、下記工程:
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、前記ペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出し、前記検出結果と被検物質を含まない試料における検出結果とを比較する工程;
被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する工程
を含むことを特徴とする。
【0062】
前記被検物質としては、いかなる公知物質および新規物質であってもよく、例えば、核酸、糖質、脂質、タンパク質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分などがあげられる。
【0063】
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料としては、生体試料から抽出した画分中に含まれるキナーゼまたはホスファターゼに被検物質を添加したもの;遺伝子組み換えにより発現精製されたキナーゼまたはホスファターゼに被検物質を添加した試料;プロテインキナーゼまたはプロテインホスファターゼ遺伝子からタンパク質として発現されたキナーゼまたはホスファターゼであって精製されることなく細胞抽出液もしくは細胞破砕液として適当な緩衝溶液に可溶化された状態のキナーゼまたはホスファターゼに被検物質を添加した試料;細胞培養時に被検物質を培地に添加して刺激を与えることで細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液中に含まれるキナーゼまたはホスファターゼ;または種々の機能既知もしくは未知遺伝子を導入することで、細胞状態を変化させて培養して得られる細胞抽出液もしくは細胞破砕液に含まれるキナーゼまたはホスファターゼに被検物質を添加した試料などがあげられる。
【0064】
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、ペプチドアレイとの接触工程は、前記本発明の検出方法で記載したとおりである。
【0065】
プロテインキナーゼ活性を変動させる被検物質のスクリーニング方法の場合、ペプチドアレイにはスレオニン残基を含むペプチド基質を固定させ、陽性対照としてリン酸化スレオニン残基を含むペプチド基質を固定させたものを用いることが好ましい。次いで、検出工程において、接触工程前後のペプチドアレイについて抗リン酸化スレオニン抗体を用いてリン酸化ペプチドを定性的または定量的に検出し、比較工程において、前記検出結果と被検物質を含まない試料における検出結果とを比較する。
【0066】
プロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法の場合、ペプチドアレイにはリン酸化スレオニン残基を含むペプチド基質を固定させ、陽性対照としてスレオニン残基を含むペプチド基質を固定させたものを用いることが好ましい。次いで、検出工程において、接触工程前後のペプチドアレイについて抗リン酸化スレオニン抗体を用いたリン酸化ペプチドの検出を介して、脱リン酸化により検出されなくなったペプチド基質を定性的または定量的に検出し、比較工程において、前記検出結果と被検物質を含まない試料における検出結果とを比較する。
【0067】
最後に、判定工程において、被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する。プロテインキナーゼ活性は、抗リン酸化スレオニン抗体により検出されるペプチド基質の数および検出強度で示すことができる。プロテインホスファターゼ活性は、抗リン酸化スレオニン抗体により検出されなくなったペプチド基質の数および検出強度で示すことができる。
【0068】
前記判定工程において、プロテインキナーゼ活性が促進されている場合、被検物質は、プロテインキナーゼの活性化を介した疾患の治療剤の候補として選択することができる。プロテインキナーゼ活性が阻害されている場合、被検物質は、プロテインキナーゼ活性の抑制化を介した疾患の治療剤の候補として選択することができる。被検物質は、プロテインキナーゼ調節剤または研究用試薬としても有用である。あるいは、前記判定工程において、プロテインホスファターゼ活性が促進されている場合、被検物質は、プロテインホスファターゼの活性化を介した疾患の治療剤の候補として選択することができる。プロテインホスファターゼ活性が阻害されている場合、被検物質は、プロテインホスファターゼ活性の抑制化を介した疾患の治療剤の候補として選択することができる。被検物質は、プロテインホスファターゼ調節剤または研究用試薬としても有用である。
【実施例】
【0069】
次に、実施例により本発明をさらに詳しく説明するが、本発明はこれら実施例に何ら限定されるものではない。なお、本実施例において市販のキットあるいは試薬を用いた部分については、特に断りのない限り添付のプロトコールに従って実験を行った。本実施例で汎用的に使用した試薬、培地、緩衝溶液を以下に記す。
【0070】
ペプチド基質
被リン酸化部位としてスレオニンまたはセリン残基を含みN末端にシステイン残基を含む10から18アミノ酸からなるペプチド配列を通常のFmoc法による化学合成を行い使用した。
以下の10配列は、合成した粗精製品を逆相クロマトグラフにより精製を行い使用した。pS*はリン酸化セリン残基、pT*はリン酸化スレオニン残基を示す。
【0071】
PKA用基質ペプチド
Kemptide(A): CGGLRRAALG-NH2 (配列番号1)
Kemptide(S): CGGLRRASLG-NH2(配列番号2)
Kemptide(T) : CGGLRRATLG-NH2(配列番号3)
Kemptide(pS*): CGGLRRA(pS*)LG-NH2(配列番号4)
Kemptide(pT*) : CGGLRRA(pT*)LG-NH2(配列番号5)
AMPK用基質ペプチド
SAMA: CGGHMRSAMAGLHLVKRR-NH2(配列番号6)
SAMT: CGGHMRSAMTGLHLVKRR-NH2(配列番号7)
PKC用基質ペプチド
IQAA: CGGAAKIQAAFRGHMARKK-NH2(配列番号8)
IQAT: CGGAAKIQATFRGHMARKK-NH2(配列番号9)
IQA(pT*): CGGAAKIQA(pT*)FRGHMARKK-NH2(配列番号10)
【0072】
以下に示す289配列は、合成した粗精製品を精製することなく使用した。pTはリン酸化されていてもよいスレオニン残基を示す。
PT001: CGGHMRSAM(pT)GLHLVKRR (配列番号11)
PT002: CGGAMARAA(pT)AAALARRR (配列番号12)
PT003: CGGHQGKFLQ(pT)FAGSPLY (配列番号13)
PT004: CGGKSGELLA(pT)WAGSPPY (配列番号14)
PT005: CGGKSGEPLS(pT)WAGSPPY (配列番号15)
PT006: CGGQKDKFLQ(pT)FAGSPLY (配列番号16)
PT007: CGGSDGEFLR(pT)SAGSPNY (配列番号17)
PT008: CGGTFGNKLD(pT)FAGSPPY (配列番号18)
PT009: CGGTLGSKLD(pT)FAGSPPY (配列番号19)
PT010: CGGTVGGKLD(pT)FAGSPPY (配列番号20)
PT011: CGGTVGNKLD(pT)FAGSPPY (配列番号21)
PT012: CGGVGDSLLE(pT)SAGSPHY (配列番号22)
PT013: CGGKSAEPLS(pT)WAGSPPY (配列番号23)
PT014: CGGVGDSLFE(pT)SAGSPHY (配列番号24)
PT015: CGGTVGGKID(pT)FAGSPPY (配列番号25)
PT016: CGGKSGEPLS(pT)WAGSPPH (配列番号26)
PT017: CGGTLGSKLD(pT)FAGNPPY (配列番号27)
PT018: CGGTVAGKLD(pT)FAGSPPY (配列番号28)
PT019: CGGVGDSLLE(pT)SAGSPHH (配列番号29)
PT020: CGGVGDSLLE(pT)SAASPHY (配列番号30)
PT021: CGGYQGKFLQ(pT)FAGSPLY (配列番号31)
PT022: CGGKSGELLA(pT)WSGSPPY (配列番号32)
PT023: CGGSDGEFLR(pT)TAGSPNY (配列番号33)
PT024: CGGKSGDPLS(pT)WAGSPPY (配列番号34)
PT025: CGGTFGNKLD(pT)FAGSPPF (配列番号35)
PT026: CGGTVANKLD(pT)FAGSPPY (配列番号36)
PT027: CGGKSGEPLS(pT)WAGTPPY (配列番号37)
PT028: CGGKSGEPLS(pT)LAGSPPY (配列番号38)
PT029: CGGTIGNKLD(pT)FAGSPPY (配列番号39)
PT030: CGGSDGEFLK(pT)SAGSPNY (配列番号40)
PT031: CGGTLGSKLD(pT)FAGSPAY (配列番号41)
PT032: CGGSDGEFLR(pT)SAGSPNF (配列番号42)
PT033: CGGKSGELLA(pT)WAASPPY (配列番号43)
PT034: CGGTVGNKLD(pT)LAGSPPY (配列番号44)
PT035: CGGKSGEPLT(pT)WAGSPPY (配列番号45)
PT036: CGGHQGKFLQ(pT)FAGSPLH (配列番号46)
PT037: CGGTVGGKLD(pT)FAASPPY (配列番号47)
PT038: CGGHQGKFLQ(pT)IAGSPLY (配列番号48)
PT039: CGGKSAELLA(pT)WAGSPPY (配列番号49)
PT040: CGGKTGELLA(pT)WAGSPPY (配列番号50)
PT041: CGGQKDKFLQ(pT)FAGSALY (配列番号51)
PT042: CGGQKDKFIQ(pT)FAGSPLY (配列番号52)
PT043: CGGTVGGKLD(pT)FAGNPPY (配列番号53)
PT044: CGGTLGGKLD(pT)FAGSPPY (配列番号54)
PT045: CGGHQGKLLQ(pT)FAGSPLY (配列番号55)
PT046: CGGKSGEPLS(pT)WAGSPPF (配列番号56)
PT047: CGGTFGNKLD(pT)LAGSPPY (配列番号57)
PT048: CGGVGDSLLE(pT)SAGSPYY (配列番号58)
PT049: CGGQKDRFLQ(pT)FAGSPLY (配列番号59)
PT050: CGGVGDSLLE(pT)SAGSPHF (配列番号60)
PT051: CGGTVGGKLD(pT)FAGSPAY (配列番号61)
PT052: CGGHQGKFLQ(pT)FAGSPFY (配列番号62)
PT053: CGGSVGNKLD(pT)FAGSPPY (配列番号63)
PT054: CGGTVGGKLD(pT)FAGSAPY (配列番号64)
PT055: CGGVGDSLVE(pT)SAGSPHY (配列番号65)
PT056: CGGKSGEPLS(pT)WAASPPY (配列番号66)
PT057: CGGTLGSKLE(pT)FAGSPPY (配列番号67)
PT058: CGGSDGEFLR(pT)SAGSANY (配列番号68)
PT059: CGGSDGELLR(pT)SAGSPNY (配列番号69)
PT060: CGGRSGEPLS(pT)WAGSPPY (配列番号70)
PT061: CGGHQGKFLQ(pT)FAGSPIY (配列番号71)
PT062: CGGTFGNKLD(pT)FAGSAPY (配列番号72)
PT063: CGGQKDKFLQ(pT)FAGSPVY (配列番号73)
PT064: CGGKSGELLA(pT)LAGSPPY (配列番号74)
PT065: CGGKTGEPLS(pT)WAGSPPY (配列番号75)
PT066: CGGTVGGKLD(pT)FAGTPPY (配列番号76)
PT067: CGGSDGEFLR(pT)SAGTPNY (配列番号77)
PT068: CGGQKDKFLQ(pT)FSGSPLY (配列番号78)
PT069: CGGIGDSLLE(pT)SAGSPHY (配列番号79)
PT070: CGGQKDKFFQ(pT)FAGSPLY (配列番号80)
PT071: CGGVGDSLLE(pT)TAGSPHY (配列番号81)
PT072: CGGTFGNKLD(pT)FAGSPAY (配列番号82)
PT073: CGGVGDSLLE(pT)SAGSAHY (配列番号83)
PT074: CGGTLGSKLD(pT)FAGSAPY (配列番号84)
PT075: CGGAGDSLLE(pT)SAGSPHY (配列番号85)
PT076: CGGQKDKFLQ(pT)FAGSPFY (配列番号86)
PT077: CGGVGDSILE(pT)SAGSPHY (配列番号87)
PT078: CGGQKDKILQ(pT)FAGSPLY (配列番号88)
PT079: CGGLGDSLLE(pT)SAGSPHY (配列番号89)
PT080: CGGQKDKFLQ(pT)LAGSPLY (配列番号90)
PT081: CGGQKDKFLQ(pT)FAASPLY (配列番号91)
PT082: CGGHQGKFLQ(pT)FAGSPVY (配列番号92)
PT083: CGGQKDKFLQ(pT)FAGSPLH (配列番号93)
PT084: CGGVGESLLE(pT)SAGSPHY (配列番号94)
PT085: CGGVGDSLLE(pT)SAGSPRY (配列番号95)
PT086: CGGSDGEFLR(pT)SAGSPNH (配列番号96)
PT087: CGGHQGKILQ(pT)FAGSPLY (配列番号97)
PT088: CGGSEGEFLR(pT)SAGSPNY (配列番号98)
PT089: CGGKSGELVA(pT)WAGSPPY (配列番号99)
PT090: CGGHQGKFLQ(pT)FAGSPLF (配列番号100)
PT091: CGGQKDKFVQ(pT)FAGSPLY (配列番号101)
PT092: CGGKSGDLLA(pT)WAGSPPY (配列番号102)
PT093: CGGSDAEFLR(pT)SAGSPNY (配列番号103)
PT094: CGGVGDSLLD(pT)SAGSPHY (配列番号104)
PT095: CGGSDGEFLR(pT)SAASPNY (配列番号105)
PT096: CGGTFGNKLE(pT)FAGSPPY (配列番号106)
PT097: CGGQRDKFLQ(pT)FAGSPLY (配列番号107)
PT098: CGGHQAKFLQ(pT)FAGSPLY (配列番号108)
PT099: CGGQKDKFLQ(pT)FAGNPLY (配列番号109)
PT100: CGGQKDKFLQ(pT)FAGTPLY (配列番号110)
PT101: CGGQKEKFLQ(pT)FAGSPLY (配列番号111)
PT102: CGGHQGKFLQ(pT)FAGNPLY (配列番号112)
PT103: CGGSDGEFVR(pT)SAGSPNY (配列番号113)
PT104: CGGHQGKFVQ(pT)FAGSPLY (配列番号114)
PT105: CGGKSGEPLS(pT)WAGSPAY (配列番号115)
PT106: CGGVGDNLLE(pT)SAGSPHY (配列番号116)
PT107: CGGKSGEPVS(pT)WAGSPPY (配列番号117)
PT108: CGGVADSLLE(pT)SAGSPHY (配列番号118)
PT109: CGGVGDSLLE(pT)SAGTPHY (配列番号119)
PT110: CGGKSGELLV(pT)WAGSPPY (配列番号120)
PT111: CGGVGDSVLE(pT)SAGSPHY (配列番号121)
PT112: CGGHQGKFLQ(pT)FAGSALY (配列番号122)
PT113: CGGHQGKFLQ(pT)LAGSPLY (配列番号123)
PT114: CGGQKDKFLQ(pT)IAGSPLY (配列番号124)
PT115: CGGHQGKFLQ(pT)FAGTPLY (配列番号125)
PT116: CGGHQGKFLQ(pT)FSGSPLY (配列番号126)
PT117: CGGKSGELLA(pT)WAGTPPY (配列番号127)
PT118: CGGKNGELLA(pT)WAGSPPY (配列番号128)
PT119: CGGTVGGKLE(pT)FAGSPPY (配列番号129)
PT120: CGGKSGEPLS(pT)WSGSPPY (配列番号130)
PT121: CGGKSGEPLS(pT)WAGSAPY (配列番号131)
PT122: CGGKSGELLA(pT)WAGSPPF (配列番号132)
PT123: CGGQKDKLLQ(pT)FAGSPLY (配列番号133)
PT124: CGGQKDKFLQ(pT)FAGSPLF (配列番号134)
PT125: CGGHQGRFLQ(pT)FAGSPLY (配列番号135)
PT126: CGGHQGKFLQ(pT)FAASPLY (配列番号136)
PT127: CGGQKDKFLQ(pT)FAGSPIY (配列番号137)
PT128: CGGKSGELLA(pT)WAGSPAY (配列番号138)
PT129: CGGVGDSLLE(pT)SSGSPHY (配列番号139)
PT130: CGGKSGELLA(pT)WAGSAPY (配列番号140)
PT131: CGGHQGKFLQ(pT)SAGSPHY (配列番号141)
PT132: CGGKSGELLA(pT)FAGSPLY (配列番号142)
PT133: CGGTVGNKLD(pT)SAGSPHY (配列番号143)
PT134: CGGKSGEPLS(pT)SAGSPHY (配列番号144)
PT135: CGGVGDSLLE(pT)SAGSPNY (配列番号145)
PT136: CGGLALHIRS(pT)WSGLHLV (配列番号146)
PT137: CGGLHIRSSW(pT)GLHLVKQ (配列番号147)
PT138: CGGLEMVKAA(pT)ETNLPLL (配列番号148)
PT139: CGGHNMLDDS(pT)DPILTTI (配列番号149)
PT140: CGGYRYPRPA(pT)VPPSPSL (配列番号150)
PT141: CGGPSLSRHS(pT)PHQSEDE (配列番号151)
PT142: CGGSSGSPAN(pT)FHFKEAW (配列番号152)
PT143: CGGSPRVRTL(pT)GSRPPLL (配列番号153)
PT144: CGGNLMLDRL(pT)GKILHID (配列番号154)
PT145: CGGVTHTFSG(pT)IEYMAPE (配列番号155)
PT146: CGGVTHTFAG(pT)IEYMAPE (配列番号156)
PT147: CGGSQFDSKF(pT)RQTPVDS (配列番号157)
PT148: CGGNQVFLGF(pT)YVAPSVL (配列番号158)
PT149: CGGEPMRRSV(pT)EAALAQP (配列番号159)
PT150: CGGWTETKKQ(pT)FKQTGEF (配列番号160)
PT151: CGGEPLERRL(pT)LVPDSEQ (配列番号161)
PT152: CGGRVPTMRP(pT)MSGLHLV (配列番号162)
PT153: CGGPTMRPSM(pT)GLHLVKR (配列番号163)
PT154: CGGKTPNLRM(pT)EKKSSWA (配列番号164)
PT155: CGGLNVESVS(pT)HRERSED (配列番号165)
PT156: CGGLNVESVA(pT)HRERSED (配列番号166)
PT157: CGGNPLMRRN(pT)VTPLASP (配列番号167)
PT158: CGGPLASPEP(pT)KKPRINS (配列番号168)
PT159: CGGEPSRAPA(pT)LLPPAPE (配列番号169)
PT160: CGGYYSSIKR(pT)GSQAHEQ (配列番号170)
PT161: CGGWRRKRKE(pT)SNTDSAG (配列番号171)
PT162: CGGVEDPPAP(pT)EPVAVEQ (配列番号172)
PT163: CGGPQLLRLL(pT)TLAEEPR (配列番号173)
PT164: CGGTSRIRTQ(pT)FSLQERQ (配列番号174)
PT165: CGGHFAIAAD(pT)EAEQDSW (配列番号175)
PT166: CGGSGDYMPM(pT)PKSVSAP (配列番号176)
PT167: CGGQHLRLST(pT)SGRLLYA (配列番号177)
PT168: CGGPLGSGES(pT)STRRSSE (配列番号178)
PT169: CGGQHLRLST(pT)SGRILYA (配列番号179)
PT170: CGGTSRIRTQ(pT)ISLQERQ (配列番号180)
PT171: CGGNKRSRTR(pT)DTYSAGQ (配列番号181)
PT172: CGGDSLPSSP(pT)TATPHSQ (配列番号182)
PT173: CGGSHMVHNR(pT)KINLQDL (配列番号183)
PT174: CGGRSRTRTD(pT)YSAGQSI (配列番号184)
PT175: CGGPTMRPSM(pT)GLHLVRR (配列番号185)
PT176: CGGLHIRSSW(pT)GLHLLKQ (配列番号186)
PT177: CGGLHIRSSW(pT)GLYLVKQ (配列番号187)
PT178: CGGQHLRVST(pT)SGRLLYA (配列番号188)
PT179: CGGDTLPSSP(pT)SATPHSQ (配列番号189)
PT180: CGGTSRIRTQ(pT)FSFQERQ (配列番号190)
PT181: CGGNPLMRRN(pT)VTPLANP (配列番号191)
PT182: CGGSHMFHNR(pT)KINLQDL (配列番号192)
PT183: CGGLHIRSSW(pT)GLHVVKQ (配列番号193)
PT184: CGGRSRTRTD(pT)YTAGQSV (配列番号194)
PT185: CGGQHLRLST(pT)SGRLIYA (配列番号195)
PT186: CGGQHLRLST(pT)SGRLLYG (配列番号196)
PT187: CGGPTMKPSM(pT)GLHLVKR (配列番号197)
PT188: CGGTSHIRTQ(pT)FSLQERQ (配列番号198)
PT189: CGGLHIRSSW(pT)GLHLVRQ (配列番号199)
PT190: CGGESLPSSP(pT)SATPHSQ (配列番号200)
PT191: CGGTSRIRTQ(pT)FTLQERQ (配列番号201)
PT192: CGGSHMIHNR(pT)KINIQDL (配列番号202)
PT193: CGGNPLMRRN(pT)VTPLGSP (配列番号203)
PT194: CGGQHLRLTT(pT)SGRLLYA (配列番号204)
PT195: CGGNPFMRRN(pT)VTPLASP (配列番号205)
PT196: CGGDSIPSSP(pT)SATPHSQ (配列番号206)
PT197: CGGSHMIHNR(pT)KVNLQDL (配列番号207)
PT198: CGGATMRPSM(pT)GLHLVKR (配列番号208)
PT199: CGGLHIRSSW(pT)ALHLVKQ (配列番号209)
PT200: CGGQRLRLST(pT)SGRLLYA (配列番号210)
PT201: CGGNPVMRRN(pT)VTPLASP (配列番号211)
PT202: CGGNPLMRRN(pT)VNPLASP (配列番号212)
PT203: CGGQHLRLST(pT)SGRLVYA (配列番号213)
PT204: CGGQHLRLST(pT)SGRVLYA (配列番号214)
PT205: CGGNPLMRRN(pT)VTPIASP (配列番号215)
PT206: CGGNPLLRRN(pT)VTPLASP (配列番号216)
PT207: CGGPSMRPSM(pT)GLHLVKR (配列番号217)
PT208: CGGPTMRPSM(pT)GFHLVKR (配列番号218)
PT209: CGGLHIRTSW(pT)GLHLVKQ (配列番号219)
PT210: CGGDSVPSSP(pT)SATPHSQ (配列番号220)
PT211: CGGNPLMRRN(pT)ATPLASP (配列番号221)
PT212: CGGQHLRLST(pT)TGRLLYA (配列番号222)
PT213: CGGDSLPSSP(pT)SATAHSQ (配列番号223)
PT214: CGGPTMRPSL(pT)GLHLVKR (配列番号224)
PT215: CGGPTMRASM(pT)GLHLVKR (配列番号225)
PT216: CGGQHFRLST(pT)SGRLLYA (配列番号226)
PT217: CGGDSLPSSP(pT)SATPHNQ (配列番号227)
PT218: CGGLHIRSSW(pT)GLHLAKQ (配列番号228)
PT219: CGGLHIRSNW(pT)GLHLVKQ (配列番号229)
PT220: CGGQYLRLST(pT)SGRLLYA (配列番号230)
PT221: CGGQHLHLST(pT)SGRLLYA (配列番号231)
PT222: CGGTSRIRTQ(pT)FSLQDRQ (配列番号232)
PT223: CGGSHMIHNK(pT)KINLQDL (配列番号233)
PT224: CGGLRIRSSW(pT)GLHLVKQ (配列番号234)
PT225: CGGSHMIHNR(pT)RINLQDL (配列番号235)
PT226: CGGVHIRSSW(pT)GLHLVKQ (配列番号236)
PT227: CGGLHIRSSW(pT)GLRLVKQ (配列番号237)
PT228: CGGNPLMRRN(pT)VSPLASP (配列番号238)
PT229: CGGQHLRLST(pT)SGRLLHA (配列番号239)
PT230: CGGQHLRLSS(pT)SGRLLYA (配列番号240)
PT231: CGGNKRSRTK(pT)DSYSAGQ (配列番号241)
PT232: CGGQHIRLST(pT)SGRLLYA (配列番号242)
PT233: CGGNPIMRRN(pT)VTPLASP (配列番号243)
PT234: CGGQHLRLST(pT)SGRFLYA (配列番号244)
PT235: CGGSHMIHNR(pT)KINVQDL (配列番号245)
PT236: CGGDSLPSSP(pT)SATPYSQ (配列番号246)
PT237: CGGSYMIHNR(pT)KINLQDL (配列番号247)
PT238: CGGQHLRFST(pT)SGRLLYA (配列番号248)
PT239: CGGTSRIRTQ(pT)FSLQEHQ (配列番号249)
PT240: CGGNKRSRTR(pT)DSYSGGQ (配列番号250)
PT241: CGGEAMRRSV(pT)EAALAQP (配列番号251)
PT242: CGGPTMRPSM(pT)GLRLVKR (配列番号252)
PT243: CGGQHLRLST(pT)SGRLLFA (配列番号253)
PT244: CGGDSLPTSP(pT)SATPHSQ (配列番号254)
PT245: CGGTHMIHNR(pT)KINLQDL (配列番号255)
PT246: CGGRSRTRTD(pT)YSAGQSL (配列番号256)
PT247: CGGQHLRLST(pT)SGHLLYA (配列番号257)
PT248: CGGLHVRSSW(pT)GLHLVKQ (配列番号258)
PT249: CGGLHIRSSW(pT)GLHLIKQ (配列番号259)
PT250: CGGLHIRSSW(pT)GFHLVKQ (配列番号260)
PT251: CGGSHMIHNR(pT)KINFQDL (配列番号261)
PT252: CGGPTMRPSM(pT)GLHLVKK (配列番号262)
PT253: CGGPTMRPSM(pT)GVHLVKR (配列番号263)
PT254: CGGTNRLRSE(pT)IAFIEEN (配列番号264)
PT255: CGGPTLRPSM(pT)GLHLVKR (配列番号265)
PT256: CGGPTMRPNM(pT)GLHLVKR (配列番号266)
PT257: CGGQHVRLST(pT)SGRLLYA (配列番号267)
PT258: CGGLHIKSSW(pT)GLHLVKQ (配列番号268)
PT259: CGGLHIRSSW(pT)GVHLVKQ (配列番号269)
PT260: CGGDSLPSSP(pT)SATPHTQ (配列番号270)
PT261: CGGDSLPSSA(pT)SATPHSQ (配列番号271)
PT262: CGGLHIRSSL(pT)GLHLVKQ (配列番号272)
PT263: CGGNPLMRRN(pT)VTPLVSP (配列番号273)
PT264: CGGLHIRSSW(pT)GLHIVKQ (配列番号274)
PT265: CGGLHIRSTW(pT)GLHLVKQ (配列番号275)
PT266: CGGPTMRPSM(pT)GIHLVKR (配列番号276)
PT267: CGGPTMRPSM(pT)GLHVVKR (配列番号277)
PT268: CGGQHLRIST(pT)SGRLLYA (配列番号278)
PT269: CGGQHLRLST(pT)SARLLYA (配列番号279)
PT270: CGGPTMRPSM(pT)GLHLAKR (配列番号280)
PT271: CGGDSFPSSP(pT)SATPHSQ (配列番号281)
PT272: CGGPTMRPSM(pT)GLHLLKR (配列番号282)
PT273: CGGLYIRSSW(pT)GLHLVKQ (配列番号283)
PT274: CGGPNMRPSM(pT)GLHLVKR (配列番号284)
PT275: CGGQHLRLST(pT)SGRLFYA (配列番号285)
PT276: CGGFHIRSSW(pT)GLHLVKQ (配列番号286)
PT277: CGGLRRA(pT)LG (配列番号287)
PT278: CGGRPRAA(pT)F (配列番号288)
PT279: CGGFKK(pT)FKL (配列番号289)
PT280: CGGTIYRRG(pT)RRWRKL (配列番号290)
PT281: CGGQKRPRRKD(pT)P (配列番号291)
PT282: CGGRKR(pT)RAE (配列番号292)
PT283: CGGAPGLI(pT)PGGPP (配列番号293)
PT284: CGGKKALHRQE(pT)VDAL (配列番号294)
PT285: CGGMHRQE(pT)VDCLK (配列番号295)
PT286: CGGHA(pT)PPKKKRK (配列番号296)
PT287: CGGPK(pT)PKKAKKL (配列番号297)
PT288: CGGRRADD(pT)DDDDD (配列番号298)
PT289: CGGRRRDDD(pT)DDD (配列番号299)
【0073】
TBS−T緩衝溶液:
0.05%のTween20を含むトリス塩酸緩衝生理食塩水(pH7.5)を使用した。
ブロッキング緩衝溶液:
3%牛血清アルブミンと0.05%のTween20を含むトリス塩酸緩衝生理食塩水(pH7.5)を使用した。
抗リン酸化セリン抗体緩衝溶液:
市販品(後述)をブロッキング緩衝溶液で500倍希釈して使用した。
抗リン酸化スレオニン抗体緩衝溶液:
市販品(後述)をブロッキング緩衝溶液で500倍希釈して使用した。
ホースラディッシュペルオキシダーゼ(HRP)標識抗マウス抗体緩衝溶液:
市販品(Zymed)をブロッキング緩衝溶液で5000倍希釈して使用した。
ホースラディッシュペルオキシダーゼ(HRP)標識抗ウサギ抗体緩衝溶液:
市販品(Zymed)をブロッキング緩衝溶液で1000倍希釈して使用した。
PKA酵素:市販品(Promega, Madison, WI、またはUpstate Biotechnology, Lake Placid, NY)を用いた。
PKC酵素:市販品(Upstate)を用いた。
AMPK酵素:市販品(Upstate)を用いた。
PKA反応緩衝溶液:以下の組成の緩衝溶液を用いた。
40 mM Tris/HCl (pH 7.2)
15 mM MgCl2
0.2 mM ATP
AMPK反応緩衝溶液:以下の組成の緩衝溶液を用いた。
40 mM HEPES (pH 7.4)
80 mM NaCl
5 mM Mg(OAc)2
1 mM DTT
0.2 mM 5’-AMP
0.2 mM ATP
PKC反応緩衝溶液(全亜型共通):以下の組成の緩衝溶液を用いた。
20 mM MOPS (pH 7.2)
25 mM β-glycerophosphate
5 mM EGTA
0.2 mM DTT
1 mM CaCl2
20 mM MgCl2
1/5 Lipid Activator (Upstate)
0.2 mM ATP
カルシウムとマグネシウムを含まないリン酸緩衝生理食塩水(PBS−):
塩化ナトリウム80g、塩化カリウム2g、リン酸水素ナトリウム11.5g、リン酸二水素カリウム2gを精製水1Lに溶解したものを10倍希釈しオートクレーブで滅菌したものを用いた。
細胞剥離液:
トリプシン−EDTA液(GIBCO)を滅菌PBS−で10倍希釈して用いた。
HEK293細胞生育培地:
500 mLのDulbeco’s リン酸緩衝生理食塩水(Sigma)に50 mLの牛胎児血清(56℃で30分間加熱し非働化したもの)と5 mLのペニシリン/ストレプトマイシン(10000 units/ml、10000 mg/ml)加えたものを用いた。
PKAプラスミド:
市販品(Stratagene)を使用した。
GFPプラスミド:
市販品(CloneTech)を使用した。
プラスミド溶液:
プラスミド4μgを250μLのOptiMem I (Invitrogen)と250μLのLipofectamine 2000 (Invitrogen)の混合液に溶解させたものを細胞培養用6穴プレートの1穴あたり用いた。
HEK293細胞抽出用緩衝液:以下の組成の緩衝溶液を用いた。
20 mM Tris/HCl (pH 7.2)
0.5 % TritonX-100
150 mM NaCl
20 mM β-glycerophosphate
2 mM EGTA
5 mM EDTA
1 mM Na3VO4(sodium orthovanadate)
0.2 mM PMSF
5 mg/ml Aprotinin
3 mg/ml Pepstatin A
L6細胞生育培地:
500 mLのαMEM(Sigma)に50 mLの牛胎児血清(56℃で30分間加熱し非働化したもの)、5 mLのペニシリン/ストレトマイシン(10000 units/ml、10000 mg/ml)と10 mLの200 mMグルタミンを加えたものを用いた。
L6細胞分化培地:
500 mLのαMEM(Sigma)に10 mLの牛胎児血清、5 mLのペニシリン/ストレプトマイシン(10000 units/ml、10000 mg/ml)混合液と10 mLの200 mMグルタミンを加えたものを用いた。
L6細胞抽出用緩衝液: 以下の組成の緩衝溶液を用いた。
30 mM HEPES (pH 7.4)
2.5 mM EGTA
3.0 mM EDTA
70 mM KCl
0.1 % Nonident P-40
20 mM β-glycerophosphate
20 mM NaF
2 mM NaPPi (sodium pyrophosphate)
1 mM Na3VO4(sodium orthovanadate)
0.2 mM PMSF
0.01 mM Pepstatin A
【0074】
実施例1:マイクロタイタープレートでの抗体認識反応(1)
1%グルタルアルデヒドの50mM炭酸ナトリウム緩衝液(pH9.5)を96穴アミノプレート(住友ベークライトまたはコーニング製)に50μLずつ注ぎ入れ、37℃で2時間反応させ、プレート表面を活性化させた。反応液を取り除き、精製水で洗浄した。ペプチド基質を50mM炭酸ナトリウム緩衝液(pH9.5)に溶解させ(ペプチド濃度1μM〜20μM)、活性化させた96穴マイクロタイタープレートの各穴に50μLずつ注ぎ入れた。37℃で2時間反応させた後、反応液を除いた。TBS−T緩衝溶液と精製水で順次洗浄することでペプチド基質を固定化した。ペプチド基質を固定化させたマイクロタイタープレートを、ブロッキング緩衝溶液を用いて30℃で1時間ブロッキング処理した。TBS−T緩衝溶液で3回洗浄した後、抗リン酸化セリン抗体緩衝溶液または抗リン酸化スレオニン抗体緩衝溶液を1穴あたり50μLずつ用いて30℃で1〜2時間抗体認識反応を行った。反応液を除いた後、TBS−T緩衝溶液で3回洗浄した。続いて、HRP標識抗マウス緩衝溶液あるいは抗ウサギ抗体緩衝溶液で認識反応させた。認識反応は1穴あたり50μLずつ用いて30℃で1時間行い、反応液を除いた後、TBS−T緩衝溶液で4回洗浄した。TMB(3,3’,5,5’−テトラメチルベンジジン)溶液を100μL加えいれ30℃で約10分間反応させた。2N硫酸水溶液で反応を停止した後、吸光光度計で450nmでの吸光度を定量した(図1および2、表1)。その結果、下記抗体16がリン酸化セリンおよびリン酸化スレオニンの両方を認識することがわかった。
【0075】
評価した抗リン酸化セリン抗体10種類と抗リン酸化スレオニン抗体6種類を以下に示す。
抗体1.マウスモノクローナル抗リン酸化セリン抗体:
クローン7F12、Alexis Biochemicals (San Diego, CA)
抗体2.マウスモノクローナル抗リン酸化セリン抗体:
クローン16B4、Alexis Biochemicals (San Diego, CA)
抗体3.マウスモノクローナル抗リン酸化セリン抗体:
クローンQ5、Qiagen (Valencia, CA)
抗体4.マウスモノクローナル抗リン酸化セリン抗体:
クローンab6639、Abcam (Cambridge, UK)
抗体5.マウスモノクローナル抗リン酸化セリン抗体:
クローンPSR-45、Sigma (St. Lois, MO)
抗体6.マウスモノクローナル抗リン酸化セリン抗体:
クローン608、Zymed (South San Francisco, CA)
抗体7.マウスモノクローナル抗リン酸化セリン抗体:
クローンIC8、Calbiochem (San Diego, CA)
抗体8.マウスモノクローナル抗リン酸化セリン抗体:
クローン4A3、Calbiochem (San Diego, CA)
抗体9.マウスモノクローナル抗リン酸化セリン抗体:
クローン4A9、Calbiochem (San Diego, CA)
抗体10.ウサギポリクローナル抗リン酸化セリン抗体:
Zymed (South San Francisco, CA)
抗体11.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンPTHR-1E11、nanoTools (Germany)
抗体12.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンPTHR-4D11、nanoTools (Germany)
抗体13.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンPTHR-14B3、nanoTools (Germany)
抗体14.マウスモノクローナル抗リン酸化スレオニン抗体:
クローンQ7、Qiagen (Valencia, CA)
抗体15.ウサギポリクローナル抗リン酸化スレオニン抗体:
CHEMICON (Temecula, CA)
抗体16.ウサギポリクローナル抗リン酸化スレオニン抗体:
Zymed (South San Francisco, CA)
【0076】
【表1−1】
【0077】
【表1−2】
【0078】
実施例2:マイクロタイタープレートでの抗体認識反応(2)
実施例1で見出した抗体16について、N末端にシステイン残基を有しリン酸化スレオニン残基を含む10から18アミノ酸からなる289種類のペプチド配列(PT001〜PT289)に対する認識能を調べた。
抗体認識能は、実施例1に示すように1次抗体としての抗リン酸化スレオニン抗体(抗体16)と2次抗体としての抗ウサギ抗体とを組み合わせて調べた。しかし、1次抗体としての抗リン酸化スレオニン抗体(抗体16)の機能をより詳細に調べる目的で289ペプチドの全配列について以下の2つの条件で抗体認識反応を比較した。さらに実験に用いる各プレートには実施例1で用いたKemptide(A)、Kemptide(T)、Kemptide(pT*)を標準サンプルとして常に固相化させ比較検討した(表2)。
【0079】
条件1:1次抗体(−)、二次抗体(+)
ペプチド性基質を実施例1と同様の方法で固定化させた96穴マイクロタイタープレートをブロッキング緩衝溶液を用いて30℃で1時間ブロッキング処理を行った。TBS−T緩衝溶液で3回洗浄した後、HRP標識抗ウサギ抗体緩衝溶液で認識反応させた。認識反応は30℃で1時間行い、反応液を除いた後、TBS−T緩衝溶液で4回洗浄した。TMB溶液を100μL加えいれ30℃で約10分間反応させた。2N硫酸水溶液で反応を停止した後、吸光光度計で450nmでの吸光度を定量した。
【0080】
条件2:1次抗体(+)、二次抗体(+)
ペプチド性基質を実施例1と同様の方法で固定化させた96穴マイクロタイタープレートをブロッキング緩衝溶液を用いて30℃で1時間ブロッキング処理を行った。TBS−T緩衝溶液で3回洗浄した後、抗体16の抗リン酸化スレオニン抗体緩衝溶液により30℃で1〜2時間反応させた。反応液を除いた後、TBS−T緩衝溶液で3回洗浄した。続いて、HRP標識抗ウサギ抗体緩衝溶液で認識反応させた。認識反応は30℃で1時間行い、反応液を除いた後、TBS−T緩衝溶液で4回洗浄した。TMB溶液を100μL加えいれ30℃で約10分間反応させた。2N硫酸水溶液で反応を停止した後、吸光光度計で450nmでの吸光度を定量した。
【0081】
【表2−1】
【0082】
【表2−2】
【0083】
【表2−3】
【0084】
【表2−4】
【0085】
【表2−5】
【0086】
【表2−6】
【0087】
【表2−7】
【0088】
実施例3:プレートでの精製PKAでのリン酸化検出
PKA用基質ペプチドを実施例1と同様の方法で96穴マイクロタイタープレートに固定化し、1穴あたり1Uまたは150 mUのPKA(Promega, Madison, WIまたはUpstate)を含むPKA反応緩衝溶液を1穴あたり50μLずつ用いて30℃でリン酸化反応を行い経時変化を追った。反応溶液を除いた後、TBS−T緩衝溶液と精製水で洗浄した。実施例1に示す方法で、生成物であるリン酸化ペプチドを抗体16を用いて測定した(図3−1、3−2)。
図3−1、3−2より、PKAは基質ペプチド(配列番号2および3)中のセリンおよびスレオニン残基を経時的にリン酸化し、抗体16は、リン酸化セリンおよびリン酸化スレオニンを同程度に認識することがわかった。
【0089】
実施例4:プレートでの精製AMPKでのリン酸化検出
AMPK用基質ペプチドを実施例1に示した方法で96穴マイクロタイタープレートに固定化し、1穴あたり10 mUのAMPK(Upstate)を含むAMPK反応緩衝溶液を1穴あたり50μLずつ用いて30℃でリン酸化反応を行い経時変化を測定した。反応溶液を除いた後、TBS−T緩衝溶液と精製水で洗浄した。さらに、実施例1に示す方法で、生成物であるリン酸化ペプチドを測定した。AMPK反応緩衝溶液からATPとAMPを除いた場合、AMPだけ除いた場合のリン酸化反応も同時に測定した(図4)。
図4より、AMPKは基質ペプチドSAMT(配列番号7)を経時的にリン酸化し、抗体16を用いて当該リン酸化を検出することにより、ATPの存在下よりもAMPとATPの共存下の方がリン酸化の程度が強いことがわかった。
【0090】
実施例5:プレートでの精製PKCでのリン酸化検出
PKC用基質ペプチドを実施例1と同様の方法で96穴マイクロタイタープレートに固定化し、各穴あたり6.3 mUのPKC(Upstate)を含むPKC反応緩衝溶液を1穴あたり50μLずつ用いて30℃で1時間リン酸化反応を行った。用いたPKCは、PKCα、PKCβII、PKCγ、PKCδ、PKCε、PKCη、PKCθおよびPKCζであった。反応溶液を除いた後、TBS−T緩衝溶液と精製水で洗浄した。実施例1に示す方法により、生成物であるリン酸化ペプチドを抗体16を用いて測定した(図5)。
図5より、用いたPKCはすべて基質ペプチドIQAT(配列番号9)をATP存在下でリン酸化することがわかった。
【0091】
実施例6:HEK293細胞でPKAとGFP遺伝子を発現させた細胞抽出液でのリン酸化反応の検出
50mLの滅菌済み試験管にHEK293細胞を入れ、37℃に温めたHEK293細胞生育培地20mLをゆっくりと加え懸濁した。毎分1000回転で5分間遠心分離して細胞を沈殿させ上清を捨てた。HEK293細胞生育培地15mLを加えて細胞を再懸濁した。得られた懸濁液を細胞培養用F75フラスコ(コーニング)に移し、二酸化炭素培養装置内で37℃で培養した。細胞が約80%コンフルエント状態に達したら、次のように継代を行った。フラスコ内の培地を捨て、10mLのPBS−で2回洗浄した。約10mLの細胞剥離液で細胞をはがした。HEK293細胞生育培地で細胞を回収し、毎分1000回転で5分間遠心分離し、細胞を沈殿させ上清を捨てた。HEK293細胞生育培地を加えて細胞を再懸濁した。得られた細胞懸濁液の一部を使い、新しい細胞培養用F75フラスコで同じように増殖させた。必要量の細胞数を得るために継代を繰り返した。継代を繰り返して得たHEK293細胞を300,000/mlの密度で細胞培養用6穴プレート(SUMILON MS-8496F5)に播き、1穴あたりHEK293細胞生育培地で約2mLにし、PKAもしくはGFP遺伝子のプラスミド溶液を加え入れ、37℃で約4時間遺伝子導入処理を行った。4時間後、継代作業と同じように新しいHEK293細胞生育培地に交換し、コンフルエント状態に達するまで細胞を増殖させた。培地を取り除き、冷やしたPBS−で2回洗浄した。PBS−EDTA緩衝液で細胞を回収した。毎分3000回転で5分間遠心分離した後上清を除き、HEK293細胞抽出用緩衝液を加え、4℃で細胞成分を溶解させた。得られた溶解液を毎分15,000回転で遠心分離し、上清を細胞抽出液として得た。得られた細胞抽出液を各穴あたり0.03μgのタンパク量になるようにPKA反応緩衝液50μLに混合した溶液を用いて、実施例3と同様に、PKA用基質ペプチドを固定化した96穴マイクロタイタープレートでリン酸化反応を30℃で1時間行い、実施例1と同様にして、リン酸化ペプチドを抗体16により検出した(図6)。
図6より、本発明のペプチドアレイを用いて、細胞抽出液中に含まれるPKA活性を測定できることがわかった。
【0092】
実施例7:L6培養細胞の細胞抽出液でのリン酸化反応の検出
50mLの滅菌済み試験管にL6細胞を入れ、37℃に温めたL6細胞生育培地20 mLをゆっくりと加え、懸濁した。毎分1000回転で5分間遠心分離して細胞を沈殿させ、上清を捨てた。L6細胞生育培地15mLを加えて細胞を再懸濁した。得られた懸濁液を細胞培養用F75フラスコ(コーニング)に移し、二酸化炭素培養装置内で37℃で培養した。約80%コンフルエント状態で、以下のように継代を行った。フラスコ内の培地を捨て、10mLのPBS−で2回洗浄した。約10mLの細胞剥離液で室温下処理し細胞をはがした。L6細胞生育培地で細胞を回収し、毎分1000回転で5分間遠心分離して細胞を沈殿させ、上清を捨てた。L6細胞生育培地を加えて細胞を再懸濁した。得られた細胞懸濁液の一部を使い、新しい細胞培養用F75フラスコで同じように増殖させた。継代を繰り返し、必要量の細胞数に達したら分化を行った。継代の際と同じようにして得た所定の密度のL6細胞生育培地で懸濁した細胞を細胞培養用6穴プレート(SUMILON MS-8496F5)に100μLずつ分注して培養した。コンフルエント状態にして培地を完全に除き、L6細胞分化培地100μLに置換した。そのまま1〜3日間培養し、筋芽が筋管に分化したのを確認した後、90μLのL6細胞分化培地に交換した。AICAR(5-amino-4- imidazole carboxamide)とメトフォルミンを所定の10倍濃度にL6細胞分化培地で希釈し、各ウェルに10μLずつ添加し2時間インキュベートした。2時間後、培地を取り除き、冷やしたPBS−で2回洗浄した。各ウェルあたり50μLのL6細胞抽出用緩衝液を加え、4℃で攪拌しながら細胞成分を抽出した。抽出物を毎分5000回転で遠心分離し、上清を得た。作製した細胞抽出液を各穴あたり0.1μgのタンパク量になるようにAMPK反応緩衝液50μLに混合した溶液を用いて、実施例4と同様に、AMPK用基質ペプチドを固定化した96穴マイクロタイタープレートでリン酸化反応を30℃で1時間行い、実施例1と同様にしてリン酸化ペプチドを抗体16により検出した(図7)。
図7より、本発明のペプチドアレイを用いて、AICARまたはメトフォルミンで誘導したAMPK活性を測定できることがわかった。
【0093】
実施例8:Cy3標識化抗体16の緩衝溶液の調製
50mM炭酸ナトリウム緩衝液(pH9.5)で2倍に希釈した抗体16をCy3標識キット(アマシャムバイオサイエンス:コード番号PA33001)を用いて添付手順書に従ってCy3標識化した。得られたCy3標識化抗体16をブロッキング緩衝溶液で1000倍に希釈しCy3標識化抗体16の緩衝溶液とした。
【0094】
実施例9:スライドガラス表面に固定化したペプチド基質を用いてのリン酸化検出
高密度化アミノ基導入コートDNAマイクロアレイ用スライドガラス(松浪ガラス:品番SDM0011)に4−マレイミドブチル酸のN−スクシイミドエステル(CAS No. 80307-12-6: Fluka)の100mMのDMF溶液を室温下12時間反応させた後、反応液をDMFで洗い流した。さらに、DMF、精製水、エタノールを用いて順に2回ずつ洗浄し、減圧室温下乾燥した。得られたスライドガラスにDNAマイクロアレイ作製用アレイヤー(Genetic Microsystems:GMS417 Arrayer)を用いて0.38μm間隔でPKA用基質ペプチドの100μMリン酸緩衝生理食塩水溶液(pH7.2)を分注し固定化した。固定化したスライドガラスを室温下3時間静置した後、水、DMF、水、エタノールで順に2回ずつ洗浄し、減圧室温下乾燥した。乾燥したスライドガラスをブロッキング緩衝溶液で室温下1時間ブロッキング処理した後、実施例3および実施例6と同様の方法でスライドガラス上でリン酸化反応を行った。PKAを使った場合は、スライドガラス1枚あたり150mUの酵素量を、PKAおよびGFP発現細胞抽出液を使った場合は、スライドガラス1枚あたり0.1μgのタンパク質量を用いた。30℃で1時間反応させた後、スライドガラスをTBS−T緩衝溶液および精製水で2回ずつ洗浄し、再度ブロッキング緩衝溶液で室温下1時間ブロッキング処理した。実施例8で作製したCy3標識化抗体16の緩衝溶液をスライドガラスに30℃で1時間作用させた。TBS−T緩衝溶液で2回と精製水で5回洗浄を行った後、毎分500回転の遠心操作を行い乾燥させた。得られたスライドガラスはDNAマイクロアレイ用蛍光スキャナー(ScanArray 5000: GSI Lumonics)を用いて、励起波長(543nm)検出波長(570nm)で蛍光イメージング画像を取得した(図8)。
図8の写真は、左からそれぞれ、キナーゼを加えないコントロール〔Kinase(-)〕、PKA酵素を使った場合〔PKA(+)〕、PKA発現細胞抽出液を使った場合〔PKA Lysate(+)〕、GFP発現細胞抽出液を使った場合〔GFP Lysate(+)〕の結果を示す。マイクロタイタープレートを用いて作製したプレートアレイでの結果と同様に、スライドガラス上に作製したマイクロアレイ上でも、PKA遺伝子を発現させた未精製の細胞抽出液を作用させることによって、精製されたPKA酵素を作用させた場合と同様に、PKAによるリン酸化反応が円滑に進行している状態が検出できた。
【産業上の利用可能性】
【0095】
本発明は、これまで標準とされてきた放射化標識されたATPを用いたリン酸化検出方法に置き換わる新しいリン酸化検出方法ならびにその実施に使用される基質群およびペプチドアレイを提供する。さらに、より優れたポリクローナル抗リン酸化スレオニン抗体を作製することで、より高感度なリン酸化検出方法、ならびにこれに基づく診断方法やスクリーニング方法をも提供することが可能となる。
【図面の簡単な説明】
【0096】
【図1】リン酸化セリン残基の抗体認識反応を450nmでの吸光度により定量 (n=1)した図である。抗体1〜抗体16は、それぞれ、[表1−1]の各抗体を意味する。
【図2】リン酸化スレオニン残基の抗体認識反応を450nmでの吸光度により定量 (n=1)した図である。抗体11〜抗体16は、それぞれ、[表1−2]の各抗体を意味する。
【図3−1】PKAによるセリン残基のリン酸化の抗体16を使った検出を示す図である。PKA 1U/well、30℃で反応A450nm (n=3)*、 * 平均値±標準誤差(n=3)
【図3−2】PKAによるスレオニン残基のリン酸化の抗体16を使った検出を示す図である。PKA 1U/well、30℃で反応A450nm (n=3)*、 * 平均値±標準偏差(n=3)
【図4】AMPKによるスレオニン残基のリン酸化の抗体16を使った検出を示す図である。AMPK 10 mU/well、30℃で反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図5】PKCによるスレオニン残基のリン酸化の抗体16を使った検出を示す図である。PKC 6.3 mU/well、30℃で1時間反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図6】HEK293細胞でPKAとGFP遺伝子を発現させた細胞抽出液でのリン酸化反応の検出を示す図である。タンパク量0.03μg/well、30℃で1時間反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図7】L6細胞でのメトフォルミンとAICAR刺激細胞抽出液でのリン酸化反応の検出を示す図である。 タンパク量0.1μg/well、30℃で1時間反応A450nm (n=3)*, * 平均値±標準偏差(n=3)
【図8】スライドガラス表面に固定化したペプチドアレイを用いて、リン酸化反応を蛍光イメージング画像で示した図である。 写真は、左からそれぞれ、キナーゼを加えないコントロール〔Kinase(-)〕、PKA酵素を使った場合〔PKA(+)〕、PKA発現細胞抽出液を使った場合〔PKA Lysate(+)〕、GFP発現細胞抽出液を使った場合〔GFP Lysate(+)〕の結果を示す。
【特許請求の範囲】
【請求項1】
プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換してなる人工ペプチド基質を少なくとも1種含有するプロテインキナーゼの基質群。
【請求項2】
プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をリン酸化スレオニン残基に置換してなる人工ペプチド基質を少なくとも1種さらに含有する、請求項1に記載の基質群。
【請求項3】
前記人工ペプチド基質が5〜30のアミノ酸残基を有するものである、請求項1または2に記載の基質群。
【請求項4】
前記プロテインキナーゼがセリン/スレオニンキナーゼである請求項1〜3いずれかに記載の基質群。
【請求項5】
前記セリン/スレオニンキナーゼがcAMP依存性プロテインキナーゼAである、請求項4に記載の基質群。
【請求項6】
前記セリン/スレオニンキナーゼがプロテインキナーゼCである、請求項4に記載の基質群。
【請求項7】
前記セリン/スレオニンキナーゼがAMP活性化プロテインキナーゼである、請求項4に記載の基質群。
【請求項8】
前記人工ペプチド基質が配列番号11〜299に記載のアミノ酸配列群から選択されるアミノ酸配列を有するものである請求項1〜7いずれかに記載の基質群。
【請求項9】
請求項1〜8いずれかに記載の基質群を固定してなるペプチドアレイ。
【請求項10】
請求項9に記載のペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、リン酸化および/または脱リン酸化反応を検出する方法。
【請求項11】
前記抗リン酸化スレオニン抗体がポリクローナル抗体である、請求項10に記載の方法。
【請求項12】
下記工程:
被験者由来の生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分を調製する工程;
前記画分と請求項9に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を、対照における検出結果と比較し、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する工程
を含むプロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法。
【請求項13】
前記疾患が癌、悪性腫瘍、糖尿病、糖尿病性合併症、代謝性疾患、自己免疫疾患、免疫異常疾患、免疫不全、神経変性疾患、虚血性疾患、内分泌疾患、肝疾患または骨疾患である請求項12に記載の診断方法。
【請求項14】
下記工程:
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、請求項9に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を被検物質を含まない試料における検出結果と比較し、被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する工程
を含む、プロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法。
【請求項1】
プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をスレオニン残基に置換してなる人工ペプチド基質を少なくとも1種含有するプロテインキナーゼの基質群。
【請求項2】
プロテインキナーゼの天然に存在する基質に含まれるアミノ酸配列中のセリン残基をリン酸化スレオニン残基に置換してなる人工ペプチド基質を少なくとも1種さらに含有する、請求項1に記載の基質群。
【請求項3】
前記人工ペプチド基質が5〜30のアミノ酸残基を有するものである、請求項1または2に記載の基質群。
【請求項4】
前記プロテインキナーゼがセリン/スレオニンキナーゼである請求項1〜3いずれかに記載の基質群。
【請求項5】
前記セリン/スレオニンキナーゼがcAMP依存性プロテインキナーゼAである、請求項4に記載の基質群。
【請求項6】
前記セリン/スレオニンキナーゼがプロテインキナーゼCである、請求項4に記載の基質群。
【請求項7】
前記セリン/スレオニンキナーゼがAMP活性化プロテインキナーゼである、請求項4に記載の基質群。
【請求項8】
前記人工ペプチド基質が配列番号11〜299に記載のアミノ酸配列群から選択されるアミノ酸配列を有するものである請求項1〜7いずれかに記載の基質群。
【請求項9】
請求項1〜8いずれかに記載の基質群を固定してなるペプチドアレイ。
【請求項10】
請求項9に記載のペプチドアレイおよび抗リン酸化スレオニン抗体を用いることを特徴とする、リン酸化および/または脱リン酸化反応を検出する方法。
【請求項11】
前記抗リン酸化スレオニン抗体がポリクローナル抗体である、請求項10に記載の方法。
【請求項12】
下記工程:
被験者由来の生体試料からプロテインキナーゼおよび/またはプロテインホスファターゼを含む画分を調製する工程;
前記画分と請求項9に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を、対照における検出結果と比較し、疾患の有無に応じて変化するプロテインキナーゼおよび/またはプロテインホスファターゼ活性を判定する工程
を含むプロテインキナーゼおよび/またはプロテインホスファターゼの活性化および/または不活化に基づく疾患の診断方法。
【請求項13】
前記疾患が癌、悪性腫瘍、糖尿病、糖尿病性合併症、代謝性疾患、自己免疫疾患、免疫異常疾患、免疫不全、神経変性疾患、虚血性疾患、内分泌疾患、肝疾患または骨疾患である請求項12に記載の診断方法。
【請求項14】
下記工程:
プロテインキナーゼまたはプロテインホスファターゼおよび被検物質を含む試料と、請求項9に記載のペプチドアレイとを接触させる工程;
前記接触工程前後のペプチドアレイについて、抗リン酸化スレオニン抗体を用いてリン酸化ペプチドおよび/または脱リン酸化ペプチドを検出する工程;ならびに
前記検出結果を被検物質を含まない試料における検出結果と比較し、被検物質の有無により変化するプロテインキナーゼまたはプロテインホスファターゼ活性を判定する工程
を含む、プロテインキナーゼまたはプロテインホスファターゼ活性を変動させる被検物質のスクリーニング方法。
【図1】

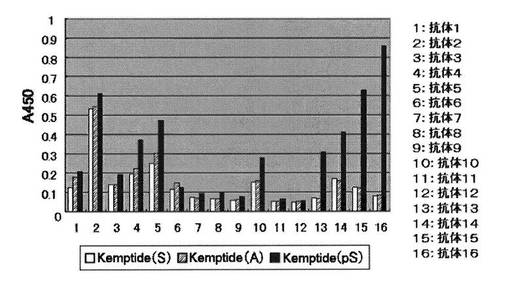
【図2】
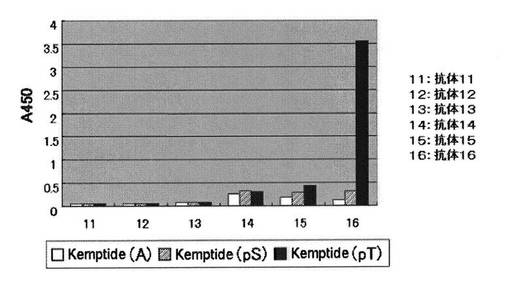
【図3−1】
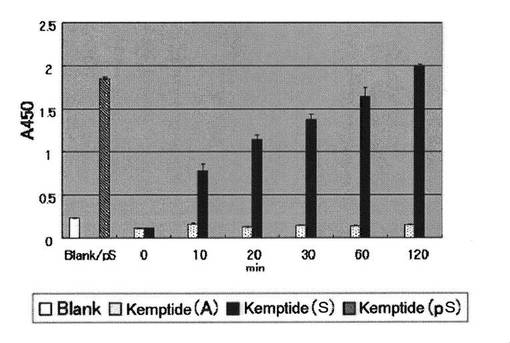
【図3−2】
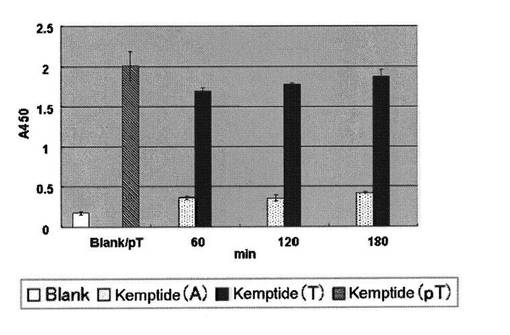
【図4】
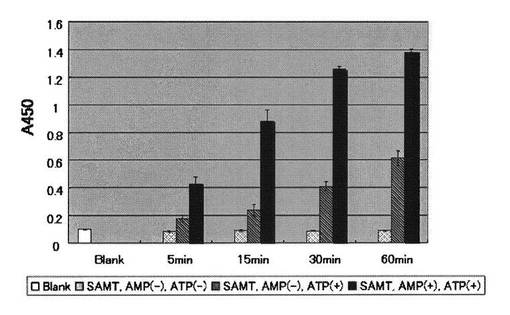
【図5】
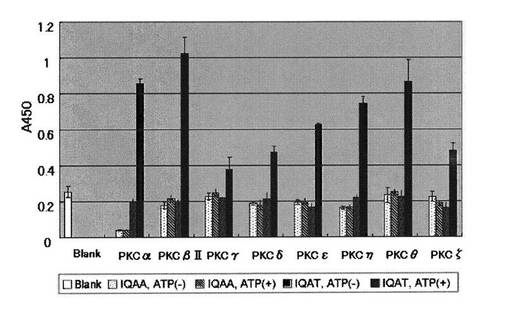
【図6】
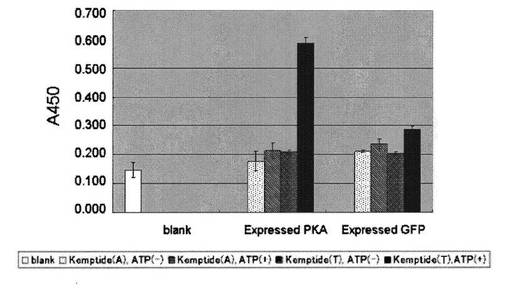
【図7】
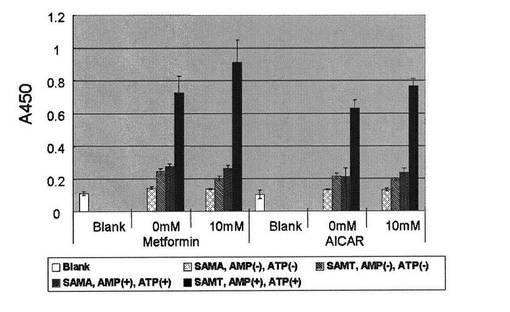
【図8】
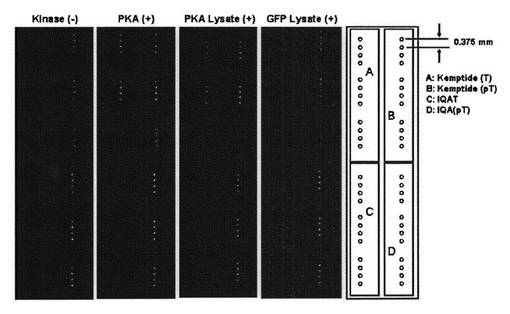
【図2】
【図3−1】
【図3−2】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2008−104356(P2008−104356A)
【公開日】平成20年5月8日(2008.5.8)
【国際特許分類】
【出願番号】特願2005−29690(P2005−29690)
【出願日】平成17年2月4日(2005.2.4)
【出願人】(000006677)アステラス製薬株式会社 (274)
【Fターム(参考)】
【公開日】平成20年5月8日(2008.5.8)
【国際特許分類】
【出願日】平成17年2月4日(2005.2.4)
【出願人】(000006677)アステラス製薬株式会社 (274)
【Fターム(参考)】
[ Back to top ]