
ルミネセンス発生及びルミネセンス非発生多重アッセイ
【課題】ルミネセンス発生/ルミネセンス非発生多重アッセイにおける酵素媒介反応に関する少なくとも1種の分子の存在又は量を検出する方法の提供。
【解決手段】第1の酵素媒介反応に関する第1の分子、及び第2の酵素媒介反応に関する第2の分子の存在又は量を検出する方法であって、a)サンプルを、前記第1の酵素により媒介された反応が、ルミネセンス発生生成物を生成し、前記第2の酵素により媒介された反応が、ルミネセンス非発生生成物を生成する前記第1の反応及び前記第2の反応に関する反応混合物に接触させること;及びb)前記サンプルにおける前記第1の分子及び前記第2の分子の存在又は量を検出することを含む方法。
【解決手段】第1の酵素媒介反応に関する第1の分子、及び第2の酵素媒介反応に関する第2の分子の存在又は量を検出する方法であって、a)サンプルを、前記第1の酵素により媒介された反応が、ルミネセンス発生生成物を生成し、前記第2の酵素により媒介された反応が、ルミネセンス非発生生成物を生成する前記第1の反応及び前記第2の反応に関する反応混合物に接触させること;及びb)前記サンプルにおける前記第1の分子及び前記第2の分子の存在又は量を検出することを含む方法。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、参照として本明細書に組み込まれる、2004年1月22日出願の米国特許出願番号第10/762,836号の出願日の利益を主張する。
【背景技術】
【0002】
ルミネセンスは、ルシフェラーゼに媒介された酸化反応の結果として、ある一定の生物において産生される。極めて多種多様の種のルシフェラーゼ遺伝子、特に、フォティヌス・ピラリス(Photinus pyralis)及びフォツリス・ペンシルバニカ(Photuris pennsylvanica)(北米の蛍)、ピロフォルス・プラギオフタラムス(Pyrophorus plagiophthalamus)(ジャマイカのコメツキムシ)、レニラ・レニフォルミス(Renilla reniformis)(シーパンジー(sea pansy))、及びいくつかの細菌(例えば、キセノルハブデュス・ルミネセンス(Xenorhabdus luminescens)及びビブリオ(Vibrio)種)のルシフェラーゼ遺伝子は、極めて一般的なルミネセンスレポーター遺伝子である。蛍のルシフェラーゼはまた、ATP濃度を測定するための一般的なレポーターであり、その役割で、バイオマスの検出に広く用いられている。他の酵素によっても、それらを一定の合成基質、例えば、アルカリホスファターゼ及びアダマンチルジオキセタンホスフェート、又はセイヨウワサビペルオキシダーゼ及びルミノールと混合すると、ルミネセンスが産生される。
【0003】
ルシフェラーゼ遺伝子は、その非放射性、感度、及びルミネセンスアッセイの極めて直線的範囲のため、遺伝子レポーターとして広く用いられている。例えば、わずか10−20モルの蛍ルシフェラーゼを検出することができる。その結果、遺伝子活性のルシフェラーゼアッセイは、原核細胞及び真核細胞双方の培養、遺伝子組換えの植物及び動物、及び無細胞発現系など、事実上あらゆる生物学的実験システムに用いられている。同様に、ATP濃度測定に用いられるルシフェラーゼアッセイは、高感度であり、10−16モル以下の検出が可能である。
【0004】
ルシフェラーゼは、酵素特異的な基質、例えば、ルシフェリン類の酸化によって光を発生させることができる。蛍ルシフェラーゼ及び他の全てのカブトムシルシフェラーゼでは、発光は、マグネシウムイオン、酸素、及びATPの存在下で生じる。ウミシイタケ(Renilla)ルシフェラーゼなどの花虫類動物のルシフェラーゼでは、基質のコエレントラジンと共に、酸素のみを必要とする。一般に、遺伝子活性を測定するためのルミネセンスアッセイにおいて、反応基質及び他のルミネセンス活性化試薬が、レポーター酵素を発現すると推測される生物系に導入される。次いで、ルミネセンスが生じた場合は、それを、ルミノメータ又は任意の好適な放射エネルギー測定装置を用いて測定する。該アッセイは、極めて迅速で高感度であり、放射性試薬を必要とせずに、迅速に容易に遺伝子発現データを提供する。
【0005】
ルシフェラーゼ類、例えば、蛍ルシフェラーゼ、ウミシイタケ(Renilla)ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、ベータガラクトシダーゼ(lacZ)、ベータグルクロニダーゼ(GUS)及び分泌アルカリホスファターゼ(SEAP)などの種々のホスファターゼ類、並びに遺伝子活性のコレポーターとして組み合わせて用いられてきたウテロフェリン(Uf:酸ホスファターゼ)は、多数のレポーターの1つである。二重酵素レポーター系は、単一系内の2種の個別のレポーター酵素の使用、発現、及び測定に関する。遺伝子レポーティングにおいて、二重レポーターアッセイは、2種の異なるレポーター遺伝子を同時に発現するように遺伝子操作される個体の細胞又は細胞集団(培養物中に分散された細胞、分離組織、又は動物全体など)におけるアッセイにとって特に有用である。多くの場合、1つの遺伝子活性が、特定の実験条件の影響をレポートする一方、第2のレポーター遺伝子の活性は、全ての組の実験値を正規化できる内部対照を提供する。二重酵素レポーター法は、実験レポーター酵素及び対照レポーター酵素をコードする独立した遺伝子材料の同時翻訳、又は結合転写及び結合翻訳のために誘導された細胞溶解物などの無細胞再構成系と共に使用することもできる。同様に、単一サンプル内の実験値及び対照値双方の二重レポーティングのために、免疫アッセイをデザインできる。
【0006】
二重酵素レポーターアッセイの実施はいずれも、構成要素である酵素の化学の特性及びそれらの各々で生じたデータセットに関連する能力に基づく。種々のレポーター酵素の酵素動態、アッセイ化学及び温置要件の不同性により、2つのレポーター酵素を組み合わせて単一の管又はウェルの二重レポーターアッセイフォーマットに組み込むことは複雑になり得る。二重レポーターアッセイ組み込みへの1つの方法は、特許文献1に記載されており、ここでは、カブトムシ及びウミシイタケ(Renilla)ルシフェラーゼアッセイ用の特定の一般的又は特異的消光剤が開示され、蛍ルシフェラーゼのルミネセンス、次いでウミシイタケ(Renilla)ルシフェラーゼのルミネセンスを連続的に判定するための典型的な二重レポーターアッセイが示されている。同様に、特許文献2には、いくつかの二重レポーターアッセイ系が開示されている。特許文献1に開示された二重レポーター系のように、ルミネセンスは、特許文献2における2つの反応の各々の測定可能な生成物である。異なった基質だけでなく、異なった検出法を組み込むレポーターアッセイの二重化法は、非特許文献1及び非特許文献2に記載されている。例えば、Liuらは、同一生物中で、ルシフェラーゼ活性及びGFP活性を報告しており、ここで、酵素活性は、段階的方法で、ルミネセンス検出及び蛍光検出によって判定される(非特許文献1)。
【0007】
また、レポーター類は、細胞内又は上澄み液内の分子の存在又は活性を検出するためにも有用である。例えば、プロテアーゼは、血液凝固、炎症、再生、線維素溶解、及び免疫応答におけるタンパク質のターンオーバーなどの多様な生理学的過程に関与する重要な酵素の大群を構成している。多くの疾患状態は、特定のプロテアーゼ類及びそれらの阻害剤の活性変化によって引き起こされるか、又はそれを特徴とする。研究において、又は臨床の場において、これらのプロテアーゼを測定する能力は、疾患状態の研究、治療及び管理にとって重要である。例えば、カスパーゼ−3及びカスパーゼ−7は、システインアスパルチル特異的プロテアーゼ(アスパルテート特異的システインプロテアーゼ、「ASCP」としても知られている)ファミリーのメンバーであり、哺乳動物細胞の細胞死において主要エフェクターの役割を演じている(非特許文献3、非特許文献4、非特許文献5及び非特許文献6)。
【0008】
しかし、プロテアーゼを、それらの天然基質を用いてアッセイすることは容易ではない。さらに、現在利用可能な多くの合成基質は、高価で感度が低く、非選択的である。
【0009】
プロテアーゼを測定するために、多数の色素産生及び蛍光産生基質が用いられてきており(非特許文献7及び非特許文献8)、修飾ルシフェリン類が蛍光指示薬の代替物として提供されている(特許文献3及び特許文献4)。代理基質として、ヒドロラーゼに対する認識部位を有する修飾ルシフェリン類を用いる方法は、Miska及びGeigerによって初めて記載され(非特許文献9)、この方法では、修飾ルシフェリンを、一定の期間ヒドロラーゼと共に温置し、次いで、該混合物のアリコートをルシフェラーゼを含有する溶液に移し入れることによって、不均一アッセイが行われた。Masuda−Nishimuraらは、β−ガラクトシダーゼ基質修飾ルシフェリンを用いた単一管(均一)アッセイの使用を報告している(非特許文献10)。
【0010】
酵素反応の蛍光若しくはルミネセンスの基質又は生成物が、タンパク質アッセイの多重化に用いられている。例えば、2つ以上の異なったサイトカインを検出するために、15個までの異なったサイトカインに対するリガンドを有する蛍光ビーズが用いられ(非特許文献11)、アルカリホスファターゼ及び一定のプロテアーゼを検出するために、蛍光ジホスフェート及びカゼインBODIPY−FLが用いられた(非特許文献12)。
【0011】
しかし、異なった検出方法を用いて2種以上のタンパク質を検出するために必要なのは、改善されたアッセイ、例えば、均一アッセイである。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】米国特許出願第5,744,320号
【特許文献2】米国特許出願第6,586,196号
【特許文献3】米国特許出願第5,035,999号
【特許文献4】米国特許出願第5,098,828号
【非特許文献】
【0013】
【非特許文献1】Liu et al., Luminescence, 15: 45 (2000).
【非特許文献2】Oazi et al., Luminescence, 17:106 (2002).
【非特許文献3】Thronberry et al., Nature, 356:768 (1992).
【非特許文献4】Nicholson et al., Nature, 376: 37 (1995).
【非特許文献5】Tewari et al., Cell, 81: 801 (1995).
【非特許文献6】Fernandes-Alnemri et al. , PNAS USA, 93: 7464 (1996).
【非特許文献7】Monsees et al. , Anal. Biochem., 221 : 329 (1994).
【非特許文献8】Monsees et al. , J. Biolum. Chemilum., 10: 213 (1995).
【非特許文献9】Miska and Geiger, J. Clin. Chem. Clin. Biochem., 25: 23 (1989).
【非特許文献10】Masuda-Nishimura et al., Lett. Appl. Microbio., 30: 130 (2000).
【非特許文献11】DeJager et al., Clin. Diag. Lab. Immunol., 10: 133 (2003).
【非特許文献12】Nolkrantz et al. , Anal. Chem., 7: 4300 (2002).
【発明の概要】
【課題を解決するための手段】
【0014】
本発明は、サンプル中、ATPなどの酵素反応に関する補因子、分子に結合する及び/又は分子の立体配置を変化させるタンパク質(ペプチド又はポリペプチド)、例えば、ペプチド基質又はポリペプチド基質を修飾するか又は開裂させるタンパク質、又はタンパク質に結合するか又はタンパク質によって変化する分子などの1つ又は複数の部分の量(例えば、活性)又は存在を検出するための、ルミネセンス非発生、例えば、蛍光又は比色アッセイ、及びルミネセンス発生アッセイの、例えば、同一ウェル内での多重化を提供する。本明細書に用いられる「ルミネセンス発生アッセイ」は、第1の分子、例えば、第1の酵素に対するペプチド基質又はポリペプチド基質、第1の分子と適切な(第1の)タンパク質との間の反応の生成物、及び/又は別のタンパク質と第1の反応の生成物との間の反応生成物がルミネセンスを発生する反応を含む。したがって、ルミネセンス発生アッセイでは、反応の補因子、タンパク質に結合する、及び/又はタンパク質によって変化する分子、又はタンパク質、の量又は存在を、直接又は間接に検出、例えば測定できる。例えば、一実施形態において、ATP濃度を検出するために、ルミネセンス発生アッセイにおいて、カブトムシルシフェラーゼ及び適切なルシフェリン基質を使用でき、一方、他の実施形態において、プロテアーゼを検出するために、ルミネセンス発生アッセイにおいて、ルシフェラーゼが存在する場合は、プロテアーゼ認識部位を含有するように修飾されている(例えば、共有結合を介して修飾されている)ルシフェラーゼに対する基質を使用できる。ルミネセンス発生アッセイとしては、限定はしないが、ルシフェラーゼ、β−ガラクトシダーゼ、β−グルクロニダーゼ、β−ラクタマーゼ、プロテアーゼ、アルカリホスファターゼ、又はペルオキシダーゼを使用又は検出するもの、及び対応する好適な基質、例えば、ルシフェリンの修飾体、コエレンテラジン、ルミノール、ペプチド又はポリペプチド、ジオキセタン、ジオキセタノン及び関連アクリジニウムエステルを含む化学発光及び生物発光アッセイが挙げられる。本明細書に用いられる「ルミネセンス発生アッセイ試薬」は、基質、並びに補因子(1種又は複数種)又はタンパク質、例えば、ルミネセンス発生反応に関する酵素などの他の分子(1種又は複数種)を含む。一実施形態において、ルミネセンス発生アッセイ試薬は、Z−DEVD−アミノルシフェリン、Z−LETD−アミノルシフェリン、Z−LEHD−アミノルシフェリンであり得るか、又は他の基質、例えば、アミノルシフェリン、ジヒドロルシフェリン、ルシフェリン6’メチルエーテル、又はルシフェリン6’クロロエチルエーテルに結合したペプチド又はポリペプチド基質であり得る。ルミネセンス発生アッセイは、ルミネセンス発生反応が、対応するルミネセンス非発生アッセイよりも少なくとも1%、例えば、少なくとも10%、多くの光を発生するものである。
【0015】
「ルミネセンス非発生アッセイ」は、第1の分子、例えば、タンパク質(ペプチド又はポリペプチド)、第1の分子と好適な(第1の)タンパク質(ペプチド又はポリペプチド)との間の反応の(第1の)生成物、又は別のタンパク質と第1の生成物との間の反応生成物がルミネセンスを発生しないが、検出可能であり得る反応を含む。例えば、該基質及び/又は生成物(1つ又は複数)は、該反応の補因子、該分子又は該分子と相互作用するタンパク質の量又は存在を、直接又は間接に測定する蛍光アッセイ又は比色アッセイを用いて検出される。例えば、酵素が基質と反応し、蛍光体が一定の波長又は一定の波長の範囲の光に接触し(曝露され)た後だけに一定の波長の光を放出する蛍光体を含有するように、該酵素に対する該基質を修飾できる。例えば、(Z−DEVD)2−ローダミン−110は、カスパーゼに対する基質であり、カスパーゼによるその基質の開裂は、ローダミン−110の蛍光によってモニターできる。本明細書に用いられる「蛍光発生アッセイ試薬」は、蛍光発生反応のために、基質、並びに補因子(1種又は複数種)又は他の分子(1種又は複数種)、例えば、タンパク質を含む。ルミネセンス非発生アッセイは、ルミネセンス非発生反応が、対応するルミネセンス発生アッセイのルミネセンスシグナルの約10%未満、例えば、約1%未満又はそれより小さい値未満を発生するものである。
【0016】
一実施形態において、本発明のアッセイに用いられる分子、例えば、タンパク質と結合及び/又はタンパク質によって変化する分子としては、レポーター分子、すなわち、例えば、1つ又は複数のその後の反応の後に、検出可能又は検出能力のある分子を含有するように修飾されている分子が挙げられる。例えば、一実施形態において、本発明のルミネセンス発生アッセイに用いられる基質としては、検出される酵素に対する基質であって、ルミネセンス発生反応の基質に共有結合している基質が挙げられ、一方、他の実施形態において、蛍光発生アッセイに用いられる基質としては、検出される酵素に対する基質であって、1つ又は複数の蛍光体に共有結合している基質を挙げることができる。いくつかの実施形態において、タンパク質に結合している及び/又はタンパク質によって変化している分子はレポーター分子を含有しない。
【0017】
本明細書に記載されているように、サンプル中の2つ以上のプロテアーゼの量又は存在が、少なくとも2つの異なった基質を用いて検出されたが、その1つは、ルミネセンス読み取りを有し、その1つ又は複数は蛍光読み取りを有した。例えば、低濃度の細胞プロテアーゼの検出は、より高感度のルミネセンス法、例えば、基質、Z−LETD−アミノルシフェリンによるカスパーゼ−8の検出、引き続いて、他の基質を用いた他のプロテアーゼの検出、例えば、(Z−DEVD)2−ローダミン−110によるカスパーゼ−3の検出を用いて達成された。このように、このアッセイは、蛍光発生試薬及びルシフェラーゼに媒介されたルミネセンス反応の感度双方の強力を組み合わせている。さらに、驚くべきことに、蛍光特性を有し、ルミネセンスアッセイにしばしば比較的大量に存在する分子、ルシフェリンの存在によって、蛍光/ルミネセンス組合せアッセイにおいて、有意な妨害は生じなかった。さらに、驚くべきことに、2種のカスパーゼ及びルシフェラーゼは、カスパーゼ−8の基質、(Z−LETD−アミノルシフェリン)、並びにカスパーゼ−3の2種の基質、(Z−DEVD)2−ローダミン−110及びAc−DEVD−AMCを含む同一反応混合物中で検出された。このように、本発明は、多重アッセイに使用される分子、例えば、蛍光発生アッセイの基質と組み合わせるルミネセンス発生アッセイの基質において、より多くの適応性を提供する。さらに、2つの酵素媒介反応が適合的な試薬条件を有すれば、該アッセイは一段階アッセイであり得る。
【0018】
したがって、本発明のルミネセンス発生/ルミネセンス非発生の組合せアッセイフォーマットにより、1種又は複数のペプチド又はポリペプチド、例えば、酵素、ペプチド(1種又は複数種)又はポリペプチド(1種又は複数種)に結合している、及び/又はそれらによって変化している1種又は複数の分子、例えば、各酵素に対するペプチド基質又はポリペプチド基質、及び/又は各アッセイに関する1種又は複数の補因子、又はそれらの組合せに関するアッセイの多重化が可能になる。したがって、一実施形態において、本発明は、第1の酵素媒介反応に関する第1の分子の存在又は量及び第2の酵素媒介反応に関する第2の分子の存在又は量を検出する方法を提供する。該方法は、第1及び/又は第2の分子を有すると推測されるサンプルを、第1及び/又は第2の分子を欠いている第1及び第2の酵素媒介反応に関する反応混合物と接触させることを含む。次いで、第1及び第2の分子の存在又は量が検出される。ルミネセンスアッセイを含む多重化を用いることによって、ルミネセンスアッセイを用いて検出される分子に対する感度の増加が提供される。一実施形態において、第1の酵素に媒介された反応によりルミネセンス発生生成物が得られ、一方、第2の酵素に媒介された反応によりルミネセンス非発生生成物が得られる。一実施形態において、アッセイの1つが内部対照を提供するものを含むルミネセンス発生/蛍光発生の組合せアッセイが提供される。本明細書に記載されたアッセイは、レポーターアッセイ、核酸ベースアッセイ又は免疫学ベースアッセイ及び他の非関連酵素アッセイなどの他のアッセイと共に使用できる。
【0019】
また、本発明は、サンプル中の少なくとも1種の分子の活性又は存在を測定する方法を提供する。該方法は、酵素媒介反応に関する少なくとも1種の分子を含有し得るサンプル、例えば、該酵素を含有し得るサンプルを提供し、該サンプルを、該酵素媒介反応に関する該分子を欠いている反応混合物、例えば、該酵素に対する基質を含有する反応混合物と接触させて該分子の存在又は量がルミネセンス発生アッセイによって検出できる反応混合物を得ることを含む。一実施形態において、該サンプル及び/又は反応混合物はまた、第2の酵素媒介反応に関する分子の存在又は量がルミネセンス非発生アッセイによって検出できる第2の酵素媒介反応に関する分子を検出する試薬と接触させる。
【0020】
一実施形態において、本発明は、サンプル中の第1の酵素によって媒介された反応に関するその第1の酵素及び/又は補因子の存在又は量を検出する方法を提供する。該方法は、該サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、反応混合物を得ることを含む。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。例えば、それらは同じ基質を実質的に認識しない。すなわち、それらは同じ基質に結合しないか、又はそれらが同じ基質に結合し反応するとしても、該酵素の一方は、他方の酵素と同じ程度(効率)で基質と反応しない。すなわち、双方の酵素に対する基質が存在する場合、該酵素の一方は、他方の酵素に対する基質と実質的に反応しない。本明細書に用いられる第2の酵素に対する基質と実質的に反応しない酵素(第1の酵素)としては、第2の酵素並びに第1の酵素に対する基質及び第2の酵素に対する基質を等しい量で有する反応液中で、第1の酵素と第1の酵素に対する基質との間の反応に比較して、第2の酵素に対する基質との交差反応が、25%以下、例えば、15%以下、10%以下又は5%以下の酵素が挙げられる。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との生成物との間の反応生成物はルミネセンスを発生する。第2の基質、第2の基質と第2の酵素との間の(第2の)反応生成物、及び/又は他の酵素と第2の生成物との間の反応生成物はルミネセンスを発生しないが、検出可能である。第1の酵素及び/又は補因子の存在又は量が検出又は測定される。一実施形態において、第2の酵素に媒介される反応に関する第2の酵素及び/又は補因子の存在又は量もまた、検出又は測定される。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。検出される酵素は、天然酵素又は組換え酵素、例えば、融合タンパク質などであり得る。同様に、該サンプルに加えられる最適な酵素(1種又は複数種)は、天然酵素又は組換え酵素であり得る。
【0021】
他の実施形態において、本発明は、サンプル中の第1の酵素及び/又はその第1の酵素に媒介される反応に関する補因子の存在又は量を検出する方法を提供する。該方法は、該サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、場合によっては、少なくとも第1の酵素と第2の酵素とが同じではない反応混合物を得ることを含む。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との間の反応生成物との間の反応生成物はルミネセンスを発生しないが、検出可能である。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は他の酵素と第2の生成物との間の反応生成物はルミネセンスを発生する。第1の酵素及び/又は補因子の存在又は量が検出又は測定される。一実施形態において、第2の酵素の存在又は量もまた、検出又は測定される。反応混合物に検出又は使用される酵素は、天然酵素又は組換え酵素であり得る。
【0022】
さらに、第1の酵素に媒介される反応に関する該酵素又は補因子を検出するために、該酵素に媒介されるルミネセンス反応をアッセイする方法が提供される。該方法は、サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、反応混合物を得ることを含み、ここでの第1の酵素と第2の酵素とは同じではない。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との生成物との間の生成物はルミネセンスを発生する。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は第2の生成物と他の酵素との間の反応生成物はルミネセンスを発生しないが、検出可能である。次いで、ルミネセンスが検出される。該方法はさらに、第2の酵素の存在又は量を、例えば、ルミネセンス非発生の基質又は生成物(1つ又は複数)の存在又は量を検出することによって検出することを含む。一実施形態において、第2の酵素は第1の基質と結合又は反応しないが、他の実施形態においては、第1の酵素は第2の基質と結合又は反応しない。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。該反応混合物中に検出又は使用される酵素は、天然酵素又は組換え酵素であり得る。
【0023】
また、第1の酵素に媒介される反応に関する該酵素又は補因子を検出するために、該酵素に媒介されるルミネセンス反応をアッセイする方法が提供される。該方法は、サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び第3の酵素に接触させて、反応混合物を得ることを含む。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との生成物との生成物はルミネセンスを発生しないが、検出可能である。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は第2の生成物と他の酵素との間の反応生成物はルミネセンスを発生する。次いで、ルミネセンスが検出される。該方法はさらに、第1の酵素又は第1の酵素と第1の基質との生成物の存在又は量を検出することを含み得る。一実施形態において、第2の酵素は第1の基質と実質的に結合又は反応しないが、他の実施形態においては、第1の酵素は第2の基質と実質的に結合又は反応しない。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。該反応混合物中に検出又は使用される酵素は、天然酵素又は組換え酵素、例えば融合タンパク質などであり得る。
【0024】
さらに、サンプル中の少なくとも2種の分子の存在又は量を検出するための方法が提供される。該方法は、サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、反応混合物を得ることを含み、ここで、少なくとも第1の酵素と第2の酵素とは同じではない。第1の酵素と第1の基質との間の反応、又は第3の酵素と、第1の基質と第1の酵素との間の反応生成物との反応により、ルミネセンスを発生する生成物が得られる。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は第2の生成物と他の酵素との間の反応生成物はルミネセンスを発生しない。次いで、第1の酵素及び第2の酵素及び/又は補因子(1つ又は複数)の存在又は量が検出される。一実施形態において、第1の酵素及び/又は補因子を検出するためにルミネセンスが用いられ、他の少なくとも1種の酵素及び/又は補因子を検出するために蛍光又は比色が用いられる。一実施形態において、2つの異なる酵素に対する基質がサンプルに同時に組み合わされて反応混合物が得られる。該基質の1種と該酵素の1種との間の反応は直接又は間接にルミネセンスシグナルを発生し、一方、他の基質と酵素との間の反応は、直接又は間接に蛍光シグナルを発生する。温置期間後、1種の酵素及び/又は補因子の存在又は量を検出するために蛍光シグナルが用いられ、他の酵素及び/又は補因子の存在又は量を検出するためにルミネセンスシグナルが用いられる。特定の緩衝液条件は、検出中の酵素及び/又は補因子によって変化し得、インビトロアッセイ、例えば、酵素アッセイの業者により決定できる。或いは、該アッセイは、第1のアッセイと第2のアッセイとの間で試薬調整をした2段階アッセイであり得る。例えば、試薬調整としては、第1の反応液に対する消光剤、及び/又は第2の反応液に対する増強剤の添加を挙げることができる。
【0025】
一実施形態において、第1の酵素に媒介された反応に関する第1の酵素又は補因子、並びに第2の酵素に媒介された反応に関する第2の酵素又は補因子を検出するために、サンプルを、第1の基質及び第2の基質に同時に接触させる。他の実施形態において、サンプルを、第1の基質の前に第2の基質に接触させるか、又は、第2の基質の前に第1の基質に接触させる。一実施形態において、第3の又は異なる酵素を、1種又は複数の基質と共に、1種又は複数の基質の前に、又は1種又は複数の基質の後に添加できる。
【0026】
一実施形態において、第1の酵素に媒介された反応に関する第1の酵素又は補因子、並びに第2の酵素に媒介された反応に関する第2の酵素又は補因子を検出するために、サンプルを、第1の酵素及び第2の酵素に同時に接触させる。他の実施形態において、サンプルを、第1の酵素の前に第2の酵素に接触させるか、又は、第2の酵素の前に第1の酵素に接触させる。
【0027】
一実施形態において、第1の酵素に媒介された反応に関する第1の酵素又は補因子、並びに第2の酵素に媒介された反応に関する第2の基質又は補因子を検出するために、サンプルを、第1の基質及び第2の酵素に同時に接触させる。他の実施形態において、サンプルを、第1の基質の前に第2の酵素に接触させるか、又は、第2の酵素の前に第1の基質に接触させる。一実施形態において、第1の酵素に媒介された反応に関する第1の基質又は補因子、並びに第2の酵素に媒介された反応に関する第2の酵素又は補因子を検出するために、サンプルを、第1の酵素及び第2の基質に同時に接触させる。他の実施形態において、サンプルを、第1の酵素の前に第2の基質に接触させるか、又は、第2の基質の前に第1の酵素に接触させる。
【0028】
本発明の方法に用いられるサンプルは、細胞溶解物、インビトロ転写/翻訳反応物、細胞培養物の上澄み液、生理学的体液サンプル、例えば、血液、血漿、血清、脳脊髄液、涙液又は尿のサンプルであり得、無処置細胞を含み得る。該細胞、細胞溶解物、又は上澄み液は、原核細胞又は真核細胞から得ることができる。
【0029】
また、本発明は、第1及び第2のタンパク質、例えば、酵素、又は補因子の少なくとも1種によって媒介される反応、例えば、場合によっては、前記反応の1つを消光、又は前記反応の1つを増強/促進することのない同時的反応、又は連続的反応に関する前記タンパク質の存在又は量の同時的又は連続的検出を提供する。一実施形態において、第1の基質と第2の基質がサンプルに同時に加えられ、第1の酵素及び/又は補因子の量又は存在が、第2の酵素及び/又は補因子の量又は存在が検出されるより前に検出される。他の実施形態において、第1の基質と第2の基質がサンプルに同時に加えられ、第2の酵素及び/又は補因子の量又は存在が、第1の酵素及び/又は補因子の量又は存在が検出されるより前に検出される。或いは、第1の基質と第2の基質がサンプルに同時に加えられ、第1及び第2の酵素及び/又は補因子の存在又は量が同時に検出される。酵素及び/又は補因子の存在又は量が、単一の反応において検出されること、例えば、全ての反応が単一の容器、例えば、ウェル内で実施されることが好ましい。
【0030】
他の実施形態において、本発明は、蛍光タンパク質、例えば、緑色蛍光タンパク質の発現と関連させて、酵素媒介反応に関する分子の存在又は量を検出する方法を提供する。例えば、蛍光タンパク質、又は脱ハロゲン化酵素など、蛍光を発するように細胞内で標識化できるタンパク質を、一時的に又は安定して発現する細胞を、蛍光発生アッセイにより蛍光タンパク質の存在又は量をアッセイできると共に、酵素によって媒介される反応に関する、細胞内に存在するか又は細胞によって分泌される少なくとも1つの追加分子、例えば、酵素、基質又は補因子に関して、ルミネセンス発生アッセイによってアッセイできる。一実施形態において、異なる分子の存在又は量もまた、例えば、ルミネセンス非発生アッセイにおいて検出又は測定される。次いで、該分子(1種又は複数種)の存在又は量を、蛍光タンパク質から作製されたデータを用いて正規化できる。
【0031】
したがって、本発明は、第1の酵素によって媒介された反応に関する分子の存在又は量を検出する方法を提供する。該方法は、蛍光タンパク質を発現する細胞を含むサンプルを、該分子を欠いている第1の酵素、場合によっては、第2の酵素に関する反応混合物と接触させることを含む。第1の酵素によって媒介された反応からルミネセンス発生生成物が得られる。次いで、該分子の存在又は量及び蛍光タンパク質の存在又は量が検出される。
【0032】
一実施形態において、異なった特徴、例えば、異なった色を有する生成物を生成するルミネセンス発生及び/又は蛍光発生アッセイでは、さらなる多重化(すなわち、他の基質によって)を用いてもよい。例えば、さらなる多重化は、異なるルシフェラーゼベースの反応又は基質によって放出された異なる色の使用、又は異なる励起/発光スペクトルによる蛍光発生アッセイを含み得る。
【0033】
また、本発明は、細胞集団、例えば、細胞培養物集団における生細胞及び/又は死細胞の存在又は数を測定する方法を提供する。該方法は、細胞膜の透過性及び完全性に関連したタンパク質分解活性の差に基づいている。該方法の利点としては、高感度、簡便性、下流多重化適用及び集団応答の正規化に関するアッセイ読み取りの適応性が挙げられる。生存度の測定値差は、1つのプロテアーゼ基質の相対的不透過性及び細胞環境から放出されるプロテアーゼの他のプロテアーゼ基質に対する酵素活性の低さに基づいている。この実施形態において有用な基質としては、N末端又はC末端でブロックされている基質を含む、エキソプロテアーゼ又はエンドプロテアーゼに対する基質が挙げられる。一実施形態において、少なくとも2種の異なる蛍光発生基質(蛍光発生アッセイ試薬)が用いられ、その1つは、実質的に細胞不透過性であり、細胞外環境で活性であるプロテアーゼ、例えば、偏在性の保存放出プロテアーゼに特異的である。他の基質は、実質的に細胞透過性であり、生細胞内で活性であるが細胞外環境に存在している場合は実質的に非活性である細胞内プロテアーゼに特異的である。実質的に細胞不透過性のプロテアーゼ基質は、死細胞に関するアッセイにおけるエンドポイントを測定するために一般に用いられる時間、例えば、サンプルへのプロテアーゼ基質の添加後、5時間未満、4時間未満、3時間未満、2時間未満又は1.5時間未満の時間中に、生細胞において検出できないものである。実質的に細胞透過性のプロテアーゼ基質は、生細胞に関するアッセイにおけるエンドポイントを測定するために一般に用いられる時間、例えば、サンプルへの基質添加後、5分超、15分超、30分超、60分超、又は120分超の時間に、生細胞に入るものである。ある条件下で実質的に非活性なプロテアーゼは、そのプロテアーゼの最適活性の約10%未満を有するものである。一実施形態において、実質的に細胞不透過性の蛍光発生基質としては、トリ又はテトラペプチド基質が挙げられる。一実施形態において、実質的に細胞透過性の蛍光発生基質としては、アミノ酸、又はジ若しくはトリペプチド基質が挙げられる。サンプルは2種の基質に接触させるが、該サンプルは、場合によっては、細胞溶解現象を生じる、又は細胞溶解物を生じることが意図されない(一般に非破壊的)量の1つ又は複数の試験条件又は試薬によって処理される。2種の蛍光発生基質における蛍光体は、異なるスペクトルを有し、蛍光発生基質に接触させたサンプルから得られた相対的蛍光単位(RFLU)は、該サンプル中の生細胞及び死細胞の測定を考慮に入れている。
【0034】
他の実施形態において、サンプル中の生細胞及び死細胞の存在又は量を検出又は測定するために、蛍光発生プロテアーゼ基質及びルミネセンス発生プロテアーゼ基質が用いられる。1つの基質は、実質的に細胞不透過性であり、細胞外環境で活性であるプロテアーゼに特異的であり、他の基質は、実質的に細胞透過性であり、生細胞内で活性であるが細胞外環境に存在している場合は実質的に非活性である細胞内プロテアーゼに特異的である。一実施形態において、実質的に細胞不透過性のプロテアーゼ基質としては、トリ又はテトラペプチド基質が挙げられる。一実施形態において、実質的に細胞透過性のプロテアーゼ基質としては、アミノ酸、又はジ若しくはトリペプチド基質が挙げられる。サンプルは、例えば、細胞溶解現象を生じる条件の不在下で、2種の基質に接触させ、プロテアーゼ開裂(RFLU及びRLU)から生じる蛍光発生生成物及びルミネセンス発生生成物の検出は、該サンプル中の生細胞及び死細胞の測定を考慮に入れている。例えば、細胞溶解現象を生じさせないルシフェラーゼ検出試薬を、ルミネセンス発生基質に接触させたウェルに加えることができる。
【0035】
放出され保持されたプロテアーゼ活性のスペクトルの異なるシグナルを、例えば、蛍光光度計又は蛍光光度計/ルミノメータ機器を用いて測定できる。このような測定は反比例し、したがって相補的である。データの正規化、対照化、及び改善に、生存度及び細胞毒性アッセイを使用できる。
【0036】
本明細書において、Ala−Ala−Phe−AMC(放出プロテアーゼ基質)及びGly−Phe−AFC(生細胞保持プロテアーゼ)は、生細胞及び細胞溶解物の%混合物と組み合わされた。細胞生存度は、凍結/解凍の繰返し、界面活性剤処理により、並びにアポトーシスを誘導する薬剤(例えば、スタウロスポリン、rTRAIL、及び抗Fas mAb)により損なわれた。さらに、細胞生存度のこれらの測定はまた、他の細胞生存度測定(CellTiter−Glo(商標)、CellTiter−Blue(商標)、又はCytoTox−ONE(商標))、又はアポトーシス的細胞毒性の特異的測定(Caspase−Glo(商標)3/7、8、9又はApo−ONE(商標))のいずれかと多重化した。やはり本明細書に記載されているように、他の基質、例えば、ローダミン−110を含有する基質などの他の蛍光発生基質又はアミノルシフェリンベースの基質などのルミネセンス発生基質を、プロテアーゼ放出アッセイ及び/又はプロテアーゼ保持アッセイに使用できる。
【0037】
したがって、本明細書に記載された生細胞及び/又は死細胞アッセイの使用は、細胞の健康状態の逆測定及び相補的測定を提供し、細胞溶解段階を必要とせずに、条件の変化、例えば、化合物による処理の影響を検出するために使用できる。さらに、基質(1種又は複数種)の色は無視し得るか又は非内在的であるため、ペアードエンドポイントアッセイに見られるシグナル消光効果はない。さらに、プロテアーゼ放出及び保持双方の活性成分は実用的な感度を有し(10,000細胞/ウェルの生存度において<2〜5%差の検出)これらの感度を15分という短さで得ることができる。また、ウェル用量を大きく変化させることなく、基質(1種又は複数種)を細胞ウェルに混合でき、次の段階のエンドポイント化学の適応性が増加する。例えば、dsDNA割込み記号又は他の好適なエンドポイントと結合させた生/死アッセイの使用は、死細胞%対対照、生細胞%対対照、細胞総数対対照、及び/又は細胞死の機構(カスパーゼ活性化)又は他のエンドポイントレポーターアッセイ測定値の検出又は測定のために使用できる。
【0038】
したがって、本発明により、例えば、同一ウェル内で、個々の非破壊的蛍光発生若しくは非破壊的ルミネセンス発生プロテアーゼベースアッセイ、又は非破壊的蛍光発生及び/又は非破壊的ルミネセンス発生プロテアーゼベース生/死細胞アッセイの多重化、或いは非破壊的蛍光発生プロテアーゼベースアッセイと他のアッセイとの組合せが提供される。一実施形態において、生/死細胞アッセイにおいて検出又は測定されるプロテアーゼは同じではなく、例えば、実質的に同じ基質を認識することはない。すなわち、それらは同じ基質に結合しないか、又はそれらが同じ基質に結合し反応するとしても、それらプロテアーゼの一方は、他方のプロテアーゼと同程度(効率)に基質と反応することはない。すなわち、双方のプロテアーゼに対する基質が存在する場合、それらプロテアーゼの一方は、他方のプロテアーゼに対する基質と実質的に反応しない。
【0039】
したがって、本発明は、サンプル中の生細胞及び/又は死細胞を検出する方法を提供する。該方法は、サンプルを、第1のプロテアーゼに対する基質及び第2のプロテアーゼに対する基質と接触させることを含む。一方のプロテアーゼによって媒介される一方の基質を用いる反応によって、蛍光発生生成物が生じ、他方のプロテアーゼによって媒介される他方の基質を用いる反応によって、ルミネセンス発生生成物又は蛍光発生生成物が生じる。一方の基質は実質的に細胞透過性であり、他方の基質は実質的に細胞不透過性である。次いで、サンプル中の蛍光及び/又はルミネセンスが検出測定され、それによって次に、サンプル中の生細胞及び/又は死細胞の数又は存在が検出又は測定される。
【0040】
一実施形態において、2種の異なるプロテアーゼに対する基質が同時にサンプルと組み合わされる。他の実施形態において、サンプルを、第1の基質の前に第2の基質と接触させるか、又は第2の基質の前に第1の基質と接触させる。一実施形態において、該アッセイは、場合によっては、第1のアッセイと第2のアッセイとの間で試薬調整をした2段階アッセイであり得る。本発明はまた、第1及び第2のプロテアーゼの存在又は量の同時的又は連続的検出を提供する。一実施形態において、第1及び第2の基質をサンプルに同時に加え、一方のプロテアーゼの量又は存在を、他方のプロテアーゼの量又は存在を検出する前に検出する。或いは、第1及び第2の基質をサンプルに同時に加え、第1及び第2のプロテアーゼの存在又は量を同時に検出する。プロテアーゼの存在又は量は、単一の反応において検出されること、例えば、全ての反応が単一の容器、例えば、ウェル内で実施されることが好ましい。
【0041】
本発明は、サンプル中の生細胞を検出する方法を提供する。該方法は、サンプルを、プロテアソーム、アミノペプチダーゼ又はカテプシンに関連したプロテアーゼに対する、蛍光を発生する実質的に細胞透過性の基質に接触させ、サンプル中の蛍光を検出又は測定し、それによってサンプル中の生細胞の数又は存在を検出又は測定することを含む。
【0042】
また、サンプル中の死細胞を検出する方法が提供される。該方法は、サンプルを、蛍光発生又はルミネセンス発生細胞不透過性基質であるトリペプチジルペプチダーゼ、カルパイン又はキモトリプシンに接触させ、サンプル中の蛍光又はルミネセンスを検出又は測定し、それによってサンプル中の死細胞の数又は存在を検出又は測定することを含む。
【0043】
また、本発明のアッセイに使用される1種又は複数の試薬を含むキットが提供される。一実施形態において、本発明は、サンプル中の生細胞及び/又は死細胞の検出に有用なキットを提供する。例えば、本発明は、第1のプロテアーゼに対する第1の、蛍光を発生する、又はルミネセンスを発生する実質的に細胞不透過性の基質及び第2のプロテアーゼに対する第2の、蛍光を発生する実質的に細胞不透過性の基質を有する組成物;並びにサンプル中の生細胞及び/又は死細胞を検出するための該組成物の使用について使用者に指示する説明書を含むキットを提供する。一実施形態において、該組成物は溶液、例えば、溶媒、例えば、有機溶媒中、該基質が0.005Mから約1.0M、例えば、0.05Mから約0.2Mで存在する溶液である。他の実施形態において、本発明は、Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、又はそれらの組合せを含んでなる組成物を含んでなるキットを含む。一実施形態において、該組成物は溶液、例えば、該基質が0.005Mから約1.0M、例えば、0.05Mから約0.2Mで存在する溶液である。
【0044】
該アッセイはまた、薬物発見ツールとしての使用法を有する。多くの薬物試験化合物は蛍光/ルミネセンス多重アッセイを妨害し得る蛍光特性を有する。本発明は、間違った結果を検出するためのアッセイを提供する。本明細書に記載されているように、カスパーゼ−3に対する同じコンセンサス基質配列が、異なるスペクトル読取りを有する、例えば、2種は蛍光読取り、1種はルミネセンス読取りを有する異なるレポーター分子に結合した。カスパーゼ−3及びルシフェラーゼが、カスパーゼ−3阻害剤又はルシフェラーゼ阻害剤の存在下又は不在下でアッセイされた。このデータは、3種のレポーター分子間にはほとんど妨害がなく、間違った結果を抑えるための正規化アッセイにルシフェラーゼを使用し得ることを示した。
【0045】
したがって、酵素のモジュレーター、例えば阻害剤の存在又は量を、本発明の多重アッセイ、例えば、蛍光発生/ルミネセンス発生組合せアッセイを用いて検出できる。一実施形態において、該方法は、第1の酵素に対するルミネセンス非発生基質、第1の酵素に対する第2の基質、ルミネセンス発生アッセイに関する第2の酵素、及び試験試薬を含む反応混合物を提供することを含む。第2の基質と第1の酵素との間ではなく、非ルミネセンス発生基質と、第1の酵素との間の反応により、非ルミネセンス発生生成物が得られ、第2の基質と第1の酵素との間の反応により、第2の酵素に対する基質、例えば、ルシフェラーゼに対する基質が得られる。第2の酵素に対する基質と第2の酵素との間の反応により、ルミネセンス発生生成物が得られる。ルミネセンス発生生成物とルミネセンス非発生生成物の存在又は量を、試験反応物及び対照反応物において比較する。これら2つの結果の比較により、該ルミネセンス発生アッセイに関する酵素に及ぼすモジュレーターの影響が示され、それによって間違った結果を排除することができる。
【図面の簡単な説明】
【0046】
【図1A】蛍光発生アッセイ試薬及びルミネセンス発生アッセイ試薬双方の存在下、カスパーゼ−3及びカスパーゼ−8の酵素活性を測定する多重アッセイ。相対的光単位(RLU)対時間を示す図である。
【図1B】蛍光発生アッセイ試薬及びルミネセンス発生アッセイ試薬双方の存在下、カスパーゼ−3及びカスパーゼ−8の酵素活性を測定する多重アッセイ。経時的な相対的蛍光単位(RFU)を示す図である。
【図2A】カスパーゼ−3及びカスパーゼ−8の多重アッセイ。AMCに関するシグナル対背景蛍光を示す図である。
【図2B】カスパーゼ−3及びカスパーゼ−8の多重アッセイ。ローダミン−110に関するシグナル対背景蛍光を示す図である。
【図2C】カスパーゼ−3及びカスパーゼ−8の多重アッセイ。ルミネセンスのシグナル対背景蛍光を示す図である。
【図3A】カスパーゼ−3、カスパーゼ−8、及びトリプシンの活性を測定する三重アッセイ。ローダミン−110に関するRFUを示す図である。
【図3B】カスパーゼ−3、カスパーゼ−8、及びトリプシンの活性を測定する三重アッセイ。AMCに関するRFUを示す図である。
【図3C】カスパーゼ−3、カスパーゼ−8、及びトリプシンの活性を測定する三重アッセイ。RLUを示す図である。
【図4A】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。1/2時間及び18時間におけるRLUを示す図である。
【図4B】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。2時間及び18時間におけるRFUを示す図である。
【図4C】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。1/2時間及び18時間におけるRLUを示す図である。
【図4D】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。2時間及び18時間におけるRFUを示す図である。
【図5A】ルシフェリンの励起及び発光のスペクトルを示す図である。
【図5B】アミノルシフェリンの励起及び発光のスペクトルを示す図である。
【図5C】Z−LETD−アミノルシフェリンの励起及び発光のスペクトルを示す図である。
【図6】カスパーゼ−3アッセイにおいて3つのチャネル;ローダミン−110、AMC及びルミネセンスにおけるシグナル対背景比を示す図である。
【図7A】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対ATP濃度を示す図である。
【図7B】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図7C】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対ATP濃度を示す図である。
【図7D】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図8A】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ−3濃度を示す図である。
【図8B】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図8C】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ−3濃度を示す図である。
【図8D】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図9A】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ濃度を示す図である。
【図9B】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対PKA濃度を示す図である。
【図9C】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ濃度を示す図である。
【図9D】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対PKA濃度を示す図である。
【図10A】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対スタウロスポリン濃度を示す図である。
【図10B】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対カスパーゼ−3濃度を示す図である。
【図10C】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対スタウロスポリン濃度を示す図である。
【図10D】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対カスパーゼ−3濃度を示す図である。
【図11A】RFLU対界面活性剤又は媒体及びプロテアーゼ放出アッセイ試薬で処理されたHL−60細胞数のプロットを示す図である。
【図11B】蛍光(AMC)プロテアーゼ放出アッセイの感度を示す図である。
【図12A】プロテアーゼ放出アッセイ試薬で処理した生存ジャーカット細胞のパーセントに対するRFLUのプロットを示す図である。
【図12B】蛍光((Ala−Ala−Phe)2−R110)プロテアーゼ放出アッセイの感度を示す図である。
【図13A】ルミネセンスプロテアーゼ放出アッセイ試薬で処理した生存ジャーカット細胞のパーセントに対するルミネセンスのプロットを示す図である。
【図13B】ルミネセンスプロテアーゼ放出アッセイ試薬で処理した生存ジャーカット細胞のパーセントに対するシグナル対ノイズ比のプロットを示す図である。
【図14A】異なるプロテアーゼ放出アッセイ試薬で処理し、超音波処理の有無によるHL−60細胞数に対するRLUのプロットを示す図である。
【図14B】種々の基質によるルミネセンスプロテアーゼアッセイの感度を示す図である。
【図15A】プロテアーゼ放出(細胞死)アッセイ(AN32504)及びCytoTox−ONE(商標)アッセイにおけるスタウロスポリン曝露時間に対するシグナル対背景比のプロットを示す図である。
【図15B】RFLU対スタウロスポリン曝露時間対LDH放出のプロットを示す図である。
【図16】細胞溶解の有無及び異なるpHにおけるプロテアーゼ放出アッセイ試薬で処理したHL−60細胞数に対するRFLUのプロットを示す図である。
【図17】Ala−Ala−Phe−アミノルシフェリンと接触させ、異なる細胞溶解に供されたHL−60細胞からのプロテアーゼ放出を示す図である。
【図18】プロテアーゼ保持アッセイ試薬対rTRAIL濃度に関するATPルミネセンスとRFLUのプロットを示す図である。
【図19A】3種の異なるプロテアーゼ保持アッセイ試薬で処理したジャーカット細胞数に対するRFLUのプロットを示す図である。
【図19B】プロテアーゼ保持アッセイにおける3種の異なる基質の感度を示す図である。
【図20】サポニン処理後に放出されたプロテアーゼについてのプロテアーゼ活性の半減期を示す図である。
【図21A】プロテアーゼベースの生/死細胞アッセイを示す図である。
【図21B】生、死又は生/死細胞アッセイによる連続多重適用におけるATP生存度アッセイを示す図である。
【図21C】蛍光生又は死アッセイ及びルミネセンスATPアッセイの連続多重を示す図である。
【図22A】SK−MEL−28細胞におけるイオノマイシン又はスタウロスポリンの濃度増加に対する生細胞RFLU及びカスパーゼ3/7活性に関するRFLUを示す図である。
【図22B】ACHN細胞におけるスタウロスポリンの濃度増加に対する生細胞RFLU及びカスパーゼ3/7活性に関するRLUを示す図である。
【図23】タキシフェン処理の時間に対する死HeLa細胞ルミネセンス及び生HeLa細胞蛍光のプロットを示す図である。
【図24A】タモキシフェンで処理され、PicoGreen(商標)で着色されたHeLa細胞によるプロテアーゼベースの生/死アッセイを示す図である。
【図24B】イオノマイシンで処理され、PicoGreen(商標)で着色されたHep2G細胞によるプロテアーゼベースの生/死アッセイを示す図である。
【発明を実施するための形態】
【0047】
本発明は、タンパク質(ペプチド又はポリペプチド)、例えば、反応に関する酵素、又は基質若しくは補因子などの1種又は複数の部分を検出するために、タンパク質に結合する、及び/又はタンパク質によって変化している少なくとも2種の異なる分子が、反応混合物中に同時に又は連続的に提供される多重アッセイ法を提供する。例えば少なくとも1種の酵素基質を、該基質と該酵素間との反応生成物へと変換する上で効果的な条件下、1つ又は複数の酵素媒介反応が実施される。反応混合物中の各分子、例えば、該反応における基質、又は生成物は、他の分子(1種又は複数種)又は生成物(1種又は複数種)とは異なる特性を有することが好ましく、一実施形態において、少なくとも1種の分子は、直接的又は間接的に検出可能なシグナルを生成できるレポーター分子を含む。生じたシグナルは、検出する分子の存在又は量に関連する。一実施形態において、該方法は、各基質を対応する生成物へと変換する上で効果的な条件下、少なくとも2種の異なる酵素基質の存在下、2種以上の酵素反応を実施することを含み、ここで、各反応の少なくとも基質又は生成物、及び/又は生成物の1種と、第3の、例えば異なる酵素との間の反応の生成物が、他方の基質(1種又は複数種)及び/又は生成物(1種又は複数種)とは異なった検出可能な特性、例えば異なる光学的特性を有する。該反応を、同時に又は連続的に実施した後、該反応(1つ又は複数)の1種又は複数の基質、又は1種又は複数の生成物の存在又は量を、検出又は測定する。これから、対応する酵素(1種又は複数種)及び/又は補因子の存在又は量を測定できる。
【0048】
このように、2つの一般的タイプの多重アッセイが考慮されている。第1のアッセイでは、複数の部分、例えば、酵素媒介反応に関する1種又は複数の酵素、1種又は複数の基質及び/又は1種又は複数の補因子を、同じ反応混合物中でアッセイする。各酵素は、基質の少なくとも1種を対応する生成物に変換することができ、ここで、該基質(1種又は複数種)及び/又は対応する生成物(1種又は複数種)、又は対応する生成物の1種と他の酵素との間の反応生成物(1種又は複数種)は、基質及び/又は生成物が同じ反応混合物中に存在する場合に、それらを個別に検出することを可能にする異なる検出可能な特性を有する。本発明のアッセイに関して、分子を加える順序は、変化し得る。このように、個々の反応は、同時に又は連続的に、開始及び/又は実施できる。連続的に開始され、実施される場合、検出可能な異なる特性は、異なる検出方法、及び/又は反応条件、例えば試薬濃度、温度又は追加の試薬の調整を必要とすることがある。例えば、反応の間に、消光剤又は増強剤を添加できる(例えば、その開示が、参照として、本明細書に具体的に組み込まれている米国特許出願第5,774,320号及び第6,586,196号を参照)。好ましい一実施形態において、2つ以上の反応が単一の反応混合物中で同時に実施され、ここで、各々の酵素が反応混合物中の基質の1種を生成物に変換する上で有効である。この実施形態は、例えば、細胞、細胞溶解物又は細胞上澄み液における少なくとも2種の異なる酵素及び/又は補因子の存在又は量を測定するために使用できる。また、該反応は、1種又は複数の試験試薬、例えば、酵素阻害剤又は酵素活性化剤、及び/又は異なる濃度の阻害剤、活性化剤、又は基質を含有し得る。
【0049】
場合によっては、該アッセイは、均一アッセイとして使用される。例えば、1種又は複数の基質及び追加成分を混合してから、該混合物をサンプルに加える。試薬の追加移入なしに、結果を読み取ることができる。
【0050】
第2のアッセイタイプでは、2つ以上の酵素媒介反応がタンデムに実施される。同じ時間に、又は異なる時間に、別個の反応が実施できる。該反応は、同一の又は異なる1種又は複数の酵素、同一の又は異なる1種又は複数の試験試薬、例えば、酵素阻害剤又は酵素活性化剤、及び/又は異なる濃度の阻害剤、活性化剤、又は基質を含有し得る。一実施形態において、各反応混合物は、生成物に変換できる少なくとも2種の基質を含有し、該基質(1種又は複数種)及び/又は対応する生成物(1種又は複数種)、及び/又は酵素/基質対の一方の生成物と異なる酵素との間の反応生成物(1種又は複数種)は、検出可能な異なる特性を有する。
【0051】
したがって、本発明のアッセイは、サンプル、例えば、真核細胞、例えば、酵母、鳥類、植物、昆虫の細胞、又は、限定はしないが、ヒト、サル、ネズミ、イヌ、ウシ、ウマ、ネコ、ヒツジ、ヤギ又はブタの細胞などの哺乳動物の細胞、又は原核細胞、又は2つ以上の異なる生物からの細胞、又は細胞溶解物又はその上澄み液を含むサンプル中の複数の酵素又は補因子の検出を可能にする。該細胞は、組換え法によって遺伝子操作されていない場合もあり得るし(非組換え細胞)、又は組換えDNAにより一時的に形質移入されている、及び/又はそのゲノムが組換えDNAによって安定して増加している、又はそのゲノムが遺伝子を破壊、例えば、プロモーター、イントロン又はオープンリーディングフレームを破壊する、又は1つのDNA断片を他のものと置換する組換え細胞の場合もあり得る。組換えDNA又はDNA断片置換は、本発明の方法によって検出される分子、検出される分子の濃度又は活性を変化させる部分、及び/又は該分子の濃度又は活性を変化させる分子又は部分に関連のない遺伝子産物をコードし得る。
【0052】
一実施形態において、本発明による方法は、細胞のアリコート又はその溶解物などの単一サンプル中の酵素を含む複数の部分を同時に又は連続的に検出するための迅速で高感度の方法を提供する。一実施形態において、該方法は、ルミネセンス発生アッセイにおける第1の酵素、基質又は補因子の存在又は量(活性)の定量化、及び蛍光発生アッセイなどのルミネセンス非発生アッセイにおける第2の酵素、基質又は補因子の存在又は量の定量化を含む。一実施形態において、試薬、例えば、各反応の基質は一緒に加えてもよいし、連続的に加えてもよい。他の実施形態において、該方法は、蛍光発生アッセイにおける第1の酵素、基質又は補因子の存在又は量の定量化、並びにルミネセンス発生アッセイにおける第2の酵素、基質又は補因子の存在又は量の定量化を含む。したがって、他の実施形態において、該方法は、ルミネセンス発生アッセイにおける補因子の存在又は量の定量化、並びにルミネセンス非発生アッセイにおける異なる分子の定量化を含む。さらに他の実施形態において、該方法は、ルミネセンス非発生アッセイにおける補因子の存在又は量の定量化、並びにルミネセンス発生アッセイにおける異なる分子の定量化を含む。ルミネセンス発生シグナル又はルミネセンス非発生シグナルの強度はそれぞれの分子の存在又は量の関数である。
【0053】
本発明はさらに、1種又は複数のエキソプロテアーゼ及び/又はエンドプロテアーゼに対する1種又は複数の基質が、細胞溶解に供さないサンプルなどのサンプルに提供される個別的な多重アッセイ法を提供し、該基質は、該サンプル中の生細胞及び/又は死細胞の数又は存在を検出又は測定する上で有用である。
【0054】
一実施形態において、本発明は、単一の細胞アリコート又はその溶解物中の1種又は複数の酵素の存在又は量を測定する方法に関する。一実施形態において、少なくとも酵素の1種は、内在性酵素である。例えば、一実施形態において、本発明は、原核細胞又は真核細胞からの精製調製物、細胞溶解物、又は培養真核細胞、例えば、哺乳動物細胞などの細胞の上澄み液などの、プロテアーゼ及び他の酵素を含む調製物中の少なくとも1種のプロテアーゼ、及び場合によっては他の酵素の活性をモニタリングするための改善された高感度の方法を提供する。分泌プロテアーゼ及び細胞内プロテアーゼなどの種々の細胞位置に存在する酵素では、各酵素に対する基質を、無処理細胞と共にウェルに加えることができる。分泌プロテアーゼの存在又は量は、細胞溶解後など、細胞内プロテアーゼの検出前に検出でき、例えば、細胞内プロテアーゼの検出は、分泌プロテアーゼと同じ容器、例えば、同じウェルにおいて行われる。一実施形態において、細胞内プロテアーゼに対する細胞不透過性基質及び分泌若しくは放出プロテアーゼに対する基質を、細胞を含むサンプルに加え、次いで、該細胞を、場合によっては溶解させる。分泌若しくは放出プロテアーゼの検出は、細胞溶解の前でもよいし、後でもよい。他の実施形態において、細胞内酵素又は分泌若しくは放出プロテアーゼに対する細胞不透過性基質及び第2の細胞内酵素に対する細胞透過性基質を、細胞を含むサンプルに加える。第2の細胞内酵素及び分泌若しくは放出プロテアーゼの存在は、溶解せずに検出できる。さらに他の実施形態において、分泌若しくは放出プロテアーゼ、細胞内酵素(細胞透過性基質又は細胞非透過性基質を用いて)、DNA若しくはATPなどの別の分子、又は第2の酵素、例えば細胞内酵素(細胞透過性基質又は細胞非透過性基質を用いて)を検出するために、三重アッセイが実施される。一実施形態において、分泌若しくは放出タンパク質は、蛍光、ルミネセンス又は吸光分光分析を用いて検出される。
【0055】
当該方法は、任意の酵素又は酵素の任意の組などの任意の分子を検出するために使用できる。該方法に用いられる酵素、検出される酵素又は基質若しくは補因子を検出するために有用な酵素は、組換え酵素及び内在性(天然)酵素などの酵素の任意の組合せから選択できる。一実施形態において、検出される全ての酵素が内在性酵素である。他の実施形態において、検出される2種の酵素は内在性酵素であり、もう一方の酵素は組換え酵素である。他の実施形態において、1種の酵素が内在性酵素であり、もう一方の酵素は組換え酵素である。通常の当業者に明白な他の組合せが、本明細書の教示による当該アッセイ及び方法に使用できる。該酵素としては、限定はしないが、プロテアーゼ、ホスファターゼ、ペルオキシダーゼ、スルファターゼ、ペプチダーゼ、及びグリコシダーゼが挙げられる。該酵素は、触媒反応の性質に基づいた、限定はしないが、ヒドロラーゼ、オキシドレダクターゼ、リアーゼ、トランスフェラーゼ、イソメラーゼ、リガーゼ、又はシンターゼなどの種々の群に由来し得るか、又は該酵素は、少なくとも酵素の1種が、他の酵素の少なくとも1種と部分的に重なる、又は好ましくは実質的に異なる基質特異性を有する限り、同じ群に由来するものでもよい。生理学的重要性を有する酵素のクラスは特に興味深い。これらの酵素としては、タンパクキナーゼ、ペプチダーゼ、エステラーゼ、タンパクホスファターゼ、イソメラーゼ、グリコシラーゼ、シンセターゼ、プロテアーゼ、デヒドロゲナーゼ、オキシダーゼ、レダクターゼ、メチラーゼなどが挙げられる。興味深い酵素としては、有機、無機双方のエステルの生成又は加水分解、グリコシル化、及びアミドの加水分解に関与するものが挙げられる。いずれのクラスにおいても、キナーゼ類のように、さらなる細分があり得、キナーゼは、ペプチド及びタンパク質におけるセリン、トレオニン及び/又はチロシン残基のリン酸化に特異的であり得る。したがって、該酵素は、例えば、環式ヌクレオチド調節タンパクキナーゼ、タンパクキナーゼC、Ca2+/CaMによって調節されるキナーゼ、サイクリン依存キナーゼ、ERK/MAPキナーゼ、及びタンパク質−チロシンキナーゼなどのキナーゼの種々の機能群のキナーゼであり得る。キナーゼは、ERKキナーゼ、S6キナーゼ、IRキナーゼ、P38キナーゼ、及びAbIキナーゼなどのオリゴペプチド基質をリン酸化する上で有用な、シグナル伝達経路におけるタンパクキナーゼ酵素であり得る。これらに対する基質としては、オリゴペプチド基質が挙げられる。興味深い他のキナーゼとしては、例えば、Srcキナーゼ、JNK、MAPキナーゼ、サイクリン依存キナーゼ、P53キナーゼ、血小板由来成長因子受容体、上皮成長因子受容体、及びMEKを挙げることができる。
【0056】
特に、本発明に有用な酵素としては、酵素活性を示す任意のタンパク質、例えばリパーゼ、ホスホリパーゼ、スルファターゼ、ウレアーゼ、ペプチダーゼ、プロテアーゼ及び酸ホスファターゼ、グルコシダーゼ、グルクロニダーゼ、ガラクトシダーゼ、カルボキシルエステラーゼ、及びルシフェラーゼなどのエステラーゼが挙げられる。一実施形態において、酵素の1種は、加水分解性酵素である。他の実施形態において、少なくとも酵素の2種が加水分解性酵素である。加水分解性酵素の例としては、アルカリ及び酸ホスファターゼ、エステラーゼ、デカルボキシラーゼ、ホスホリパーゼD、P−キシロシダーゼ、β−D−フコシダーゼ、チオグルコシダーゼ、β−D−ガラクトシダーゼ、α−D−ガラクトシダーゼ、α−D−グルコシダーゼ、β−D−グルコシダーゼ、β−D−グルクロニダーゼ、α−D−マンノシダーゼ、β−D−マンノシダーゼ、β−D−フルクトフラノシダーゼ、及びβ−D−グルコシデュロナーゼが挙げられる。
【0057】
任意の具体的な酵素媒介反応に関する基質又は補因子は、当業者に知られている。いくつかのプロテアーゼに関する典型的な開裂部位は、表1に記載されている。
【0058】
【表1】

【0059】
アルカリホスファターゼに対して、基質は、3−(2’−スピロアダマンタン)−4−メトキシ−4−(3’’−ホスホリルオキシ)フェニル−1,2−ジオキセタン、二ナトリウム塩、又は3−(4−メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]デカン]−4−イル]フェニルリン酸二ナトリウム、又は2−クロロ−5−(4−メトキシスピロ{1,2−ジオキセタン−3,2’−(5’−クロロ)−トリシクロ{3.3.1.13,7]デカン}−4−イル)−1−フェニルリン酸二ナトリウム又は2−クロロ−5−(4−メトキシスピロ{1,2−ジオキセタン−3,2’−トリシクロ[3.3.1.13,7]デカン}−4−イル)−1−フェンジルリン酸二ナトリウム(それぞれ、AMPPD、CSPD、CDP−Star(登録商標))及びADP−Star(商標)などのリン酸含有ジオキセタンを含むことが好ましい。
【0060】
β−ガラクトシダーゼに対して、基質は、ガラクトシダーゼ開裂可能基又はガラクトピラノシド基を含有するジオキセタンを含むことが好ましい。アッセイにおけるルミネセンスは、ジオキセタン基質の糖部分の酵素的開裂から生じる。このような基質の例としては、3−(2’−スピロアダマンタン)−4−メトキシ−4−(3’’−β−D−ガラクトピラノシル)フェニル−1,2−ジオキセタン(AMPGD)、3−(4−メトキシスピロ[1,2−ジオキセタン−3,2’−(5’−クロロ)トリシクロ[3.3.1.13,7]−デカン]−4−イル−フェニル−β−D−ガラクトピラノシド(Galacton(登録商標))、5−クロロ−3−(メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)トリシクロ[3.3.13,7]−デカン]−4−イル−フェニル−β−D−ガラクトピラノシド(Galacton Plus(登録商標))、及び2−クロロ−5−(4−メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]−デカン]−4−イル)フェニル−β−D−ガラクトピラノシド(Galacton−Star(登録商標))が挙げられる。
【0061】
β−グルクロニダーゼ及びβ−グルコシダーゼに関するアッセイにおいて、基質としては、グルクロニド、例えば、3−(4−メトキシスピロ{1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]デカン}−4−イル)フェニル−β−D−グルクロン酸ナトリウム(Glucuron(商標))などのβ−グルクロニダーゼ開裂基を含有するジオキセタンが挙げられる。カルボキシルエステラーゼに関するアッセイにおいて、基質としては、ジオキセタンに結合した好適なエステル基が挙げられる。プロテアーゼ及びホスホリパーゼに関するアッセイにおいて、基質としては、ジオキセタンに結合した好適な酵素開裂基が挙げられる。
【0062】
アッセイにおける各酵素に対する基質は、異なっていることが好ましい。1種のジオキセタン含有基質を含むアッセイでは、該基質は、場合によっては、置換又は非置換アダマンチル基、置換又は非置換であり得るY基及び酵素開裂基を含有する。好ましいジオキセタンの例としては、上記に記載したもの、例えば、Galacton(登録商標)、Galacton Plus(登録商標)、CDP−Star(登録商標)、Glucuron(商標)、AMPPD、Galacton−Star(登録商標)、及びADP−Star(商標)、と称されるもの、並びに3−(4−メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]デカン}−4−イル)フェニル−β−D−グルコピラノシド(Glucon(商標))、CSPD、3−クロロ−5−(4−メトキシスピロ{1,2−ジオキセタン−3,2’−(5’−クロロ)−トリシクロ{3.3.1.13,7]デカン)−4−イル)−1−フェニルリン酸二ナトリウム(CDP)が挙げられる。
【0063】
検出される少なくとも1種の酵素に対する基質は、レポーター分子を含有するように修飾されることが好ましい。レポーター分子は、その分子に結合した基質、該酵素と該基質との間の反応から生じる生成物、又はその生成物と他の酵素との間の反応生成物が、別個に検出される、好ましくは定量的に検出されることを可能にする任意の分子である。レポーター分子としては、限定はしないが、蛍光体などの光学的分子、吸収性有色粒子又は色素、放射標識、好適な反応成分の存在下、検出反応を触媒する上で有効な触媒的部分などの酵素、他のサブユニット(1つ又は複数)又は断片(1つ又は複数)と結合した際に機能的な酵素のサブユニット又は断片、又は引き続く反応に関する基質、例えば、その反応の生成物が検出可能である基質が挙げられる。本明細書に用いられる「蛍光体」には、ある波長領域でエネルギーを吸収し、該吸収領域以外の波長領域でエネルギーを放出することのできる分子が含まれる。用語の「励起波長」とは、蛍光体がエネルギーを吸収する波長領域を言う。用語の「発光波長」とは、蛍光体がエネルギー又は蛍光を放出する波長領域を言う。
【0064】
一実施形態において、レポーター分子は蛍光を発する。蛍光発生物の一群は、キサンテン色素であり、これには、フルオレセイン、ローザミン及びローダミンが含まれる。これらの化合物は、フェニル基上に置換基を有した商品として入手でき、結合のための部位として、又は結合官能基として使用できる。例えば、アミノ置換及びイソチオシアネート置換したフルオレセイン化合物を入手できる。
【0065】
蛍光性化合物の他の群は、アルファ位又はベータ位、通常はアルファ位にアミノ基を有するナフチルアミン類である。ナフチルアミノ化合物の中には、1−ジメチルアミノナフチル−5−スルフォネート、スルホン酸1−アニリノ−8−ナフタレン及びスルホン酸2−p−トルイジニル−6−ナフタレンが含まれる。いくつかのナフタレン化合物は、タンパク質に対していくらか非特異的な結合が見られるため、それらの使用には、タンパク質の量が最少化されているアッセイ培地を用いる必要がある。他の蛍光発生物は、窒素含有マクロ環を含む多座リガンドであり、これらは、パイ電子を有する共役環系を有する。これらのマクロ環は、場合によっては、架橋炭素上、又は窒素上の置換などの置換がされていてもよい。好適なマクロ環としては、ポルフィリン、アザポルフィリン、コリン、サフィリン及びポルフィセン、並びに、広く非局在化している電子を含有する他のマクロ環様の誘導体が挙げられる。アザポルフィリン誘導体としては、フタロシアニン、ベンゾトリアザポルフィリン及びナフタロシアン並びにそれらの誘導体が挙げられる。
【0066】
いくつかの例では、蛍光融合タンパク質、例えば、ポリペプチド基質に融合させた、緑色、赤色又は青色蛍光タンパク質又は他の蛍光タンパク質を使用できる。他の実施形態において、蛍光タンパク質は、それ自体、加水分解性酵素に対する基質であり得る。「蛍光タンパク質」は、完全長蛍光タンパク質又はそれの蛍光断片である。
【0067】
本発明に使用される化学的蛍光体の非限定的リストを、それらの励起波長及び発光波長と共に、表2に示してある。励起値及び発光値は、pH、緩衝系又は溶媒などの反応条件に依存して変化できる。
【0068】
【表2】
【0069】
一実施形態において、酵素の1種は、アミノ修飾ルシフェリン又はそのカルボキシ保護誘導体を含む基質を用いて検出され、前記修飾は、該酵素に対する基質を含む。一実施形態において、前記修飾は、プロテアーゼに対する認識部位を含む1つ又は複数のアミノ酸残基である。一実施形態において、該基質は、ペプチド結合を介して、アミノルシフェリン又はそのカルボキシ修飾誘導体のアミノ基に共有結合している。一実施形態において、ペプチド又はタンパク質基質のN末端は、例えば、アミノ末端保護基を用いて、アミノペプチダーゼによる分解を防ぐように修飾される。適切な酵素又は補因子の不在下では、このような基質及びルシフェラーゼを含む混合物は、最少のアミノルシフェリンが存在するため、最少の光を生じる。適切な酵素の存在下では、該基質及びアミノルシフェリンとの結合を該酵素によって開裂することができ、ルシフェラーゼの基質であるアミノルシフェリンが生じる。したがって、ルシフェラーゼ、例えば、天然、組換え又は変異ルシフェラーゼ、及びいずれかの補因子並びに適切な反応条件の存在下、該酵素の存在又は活性に比例する光が生じる。
【0070】
一実施形態において、酵素の1種は、蛍光体を含む基質を用いて検出される。一実施形態において、該基質は、プロテアーゼに対する認識部位を含む1つ又は複数のアミノ酸残基を含む。一実施形態において、該基質は、1種又は複数の蛍光体に共有結合している。適切な酵素又は補因子の不在下では、基質−蛍光体共役体の特性が変化して、該蛍光体単独に比較して該共役体の蛍光特性の変化、例えば低下が生じるように、該蛍光体の蛍光特性が、例えば、消光基の近接によって消光されるに従い、そのような基質を含む混合物は、その発光波長において最小の光を発生する。適切な酵素の存在下、共役体の開裂によって蛍光体が生じる。他の実施形態において、開裂前の共役体は蛍光性であるが、酵素による開裂後、生成物(1つ又は複数)はスペクトルを変化させた。
【0071】
一実施形態において、少なくとも2つの反応に関する条件は適合性である。例えば、少なくとも2種の酵素の条件、好ましくは3種以上の酵素、例えば、4種以上の酵素の条件は適合性である。類似した酵素の群は、pHやイオン強度などの反応条件に一般に適合性であるが、比較的低質量濃度を有するアッセイ成分、例えば、補因子に関係する補因子要件、金属イオン要件などは共通である必要はない。共通の条件としては、各酵素が反応の経過中、測定可能な比率を提供し、一般に、各酵素が他の酵素(1種又は複数種)に対して、加えられた成分から有意な妨害なしに、特定の基質に対する最大回転率の少なくとも約10%、通常は少なくとも約20%、好ましくは少なくとも約50%の回転率を有するような条件が挙げられる。
【0072】
或いは、双方の反応に関する基質が存在しているが、1つの反応に関する条件は、他の反応に適合性でない場合があり得る。このような実施形態において、1種の酵素は活性であるが、その基質と反応できない。例えば、2つの反応に関する条件が適合性でない一実施形態において、個々の酵素反応アッセイは、連続的に及び/又は個別の反応混合物において実施される。酵素アッセイ後、該反応混合物(又はその一部)を他の反応と組み合わせ得る。各々、個別の反応混合物が、1種又は複数の酵素及び1種又は複数の基質を含有し得る。その最も単純な形態では、アッセイする単一の酵素及びその酵素に対する単一の基質が、各反応混合物中にある。該反応に使用される基質の組は、単一反応多重アッセイに必要とされるものと同じ一般的特性を有する。すなわち、各基質及び/又は対応する生成物は独特の特性を有し、それらを互いに区別することを可能にする。
【0073】
該反応における分子の検出順序は変化し得る。一実施形態において、反応が同時に開始されるか否かに関わりなく、ルミネセンス発生アッセイによって検出される分子が検出され、次いで非ルミネセンス発生アッセイによって検出される分子が検出される。或いは、反応が同時に開始されるか否かに関わりなく、非ルミネセンス発生アッセイによって検出される分子が検出され、次いでルミネセンス発生アッセイによって検出される分子が検出される。他の実施形態において、2種以上の分子の存在又は量が本質的に同時に検出される。一実施形態において、検出される1種の分子の存在又は活性を、第2の分子の存在又は活性を検出する前に、例えば、第1のシグナルが、例えば少なくとも50%減少するまで待つことによって、又は第1の反応に対する消光剤を加えることによって、実質的に減少させる。したがって、いくつかの実施形態において、1種又は複数の反応を、検出前に、例えば、該反応に関する酵素を阻害することによって終了させる。1つのアッセイによって生じたシグナルが、他の少なくとも1つのアッセイによって生じたシグナルの定量化を実質的に妨害しないことが好ましい。
【0074】
また、本発明は、無処置細胞、細胞溶解物、例えば、少なくとも部分的に精製された溶解物、又は細胞上澄み液を含むサンプルなどのサンプル中の1種又は複数のペプチド又はタンパク質、ペプチド又はタンパク質に結合する及び/又はペプチド又はタンパク質によって変化する分子、又は補因子の存在又は活性を検出するためのキットを提供する。このようなキットは、少なくとも1種の酵素に対する基質などの、少なくとも1種のペプチド及び/又はタンパク質、ペプチド及び/又はタンパク質に結合した、及び/又はペプチド及び/又はタンパク質によって変化した分子、又は補因子を定量化するための少なくとも1種の試薬を含む。
【0075】
本発明を、以下の非限定的実施例によって、さらに説明する。すべての実施例に関し、好適な制御反応は、当業者によって容易にデザインされる。
【実施例】
【0076】
(実施例I)
蛍光/ルミネセンス多重アッセイ
A.単一ウェル、多重アッセイにおけるカスパーゼ−3及びカスパーゼ−8の測定
Caspase−Glo(商標)8 Reagent(Caspase−Glo(商標)8 Assay System、Promega社)を、均一ルミネセンス発生カスパーゼ−8及びルミネセンス非発生カスパーゼ−3酵素アッセイの多重化を可能にする能力に関して評価した。Caspase−Glo(商標)8 Reagentは、Caspase−Glo(商標)8 Buffer及びルミネセンス発生基質であるZ−LETD−アミノルシフェリンからなる。図1Aにおけるルミネセンス発生アッセイに関して、多重化されたルミネセンス発生及びルミネセンス非発生アッセイの実行可能性を実証するために、Caspase−Glo(商標)8 Reagent(菱形)又はカスパーゼ−3に対する50μMの蛍光発生基質、(Z−DEVD)2−ローダミン−110(四角形)も含有するCaspase−Glo(商標)8 Reagentのいずれかを用いた。図1Bにおける蛍光発生アッセイに関して、50μM (Z−DEVD)2−ローダミン−110及び10mM DTT(四角形)又は50μM (Z−DEVD)2−ローダミン−110及びZ−LETD−アミノルシフェリン(四角形)のいずれかを含有するCaspase−Glo(商標)8 Bufferを用いた。
【0077】
カスパーゼ−8酵素、カスパーゼ−3酵素、及びカスパーゼ−8とカスパーゼ−3酵素との組合せ(Biomol Research Laboratories)の希釈液を、100単位/mlの最終濃度に、RPMI1640(Sigma社)中で調製した。100μlのカスパーゼ−8希釈液、カスパーゼ−8希釈液とカスパーゼ−3希釈液との混合液、又はカスパーゼ−3希釈液を、96ウェルプレートの個別のウェルに加えた。50μM (Z−DEVD)2−ローダミン−110を有する又は有さない100μlのCaspase−Glo(商標)8 Reagent(図1A)、又はZ−LETD−アミノルシフェリンを有する又は有さない、(Z−DEVD)2−ローダミン−110及びDTTを添加した100μlのCaspase−Glo(商標)8 Bufferを、200μl/ウェルの最終容量に達するように加えた。該反応プレートを室温で、プレートシェーカー上、少なくとも10分間温置した。
【0078】
温置後、DYNEX Laboratories MLX(商標)プレートルミノメータを用いて相対的ルミネセンスを測定し、485EX/530EMフィルターセットを装備したCytoFluorII蛍光プレート読取り装置により相対的蛍光を測定した。
【0079】
結果
図1に、単一ウェル中、2種のプロテアーゼ酵素に関する蛍光及びルミネセンスの同時測定が示されている。図1Aに見られる通り、カスパーゼ−8(四角形)に関するルミネセンス発生アッセイにおいて、カスパーゼ−3及びその蛍光発生基質である(Z−DEVD)2−ローダミン−110の存在は、ルミネセンス反応を大きく変化させることはない。同様に、カスパーゼ−3(四角形)に関する蛍光発生アッセイにおいて、カスパーゼ−8及びそのルミネセンス発生基質であるZ−LETD−アミノルシフェリンの存在は、カスパーゼ−3に関する蛍光発生アッセイに影響を与えない。
【0080】
B.カスパーゼ−3及びカスパーゼ−8多重アッセイに関する背景測定
カスパーゼ酵素及びそれらの基質などの種々の濃度のルミネセンス発生試薬及び蛍光発生試薬並びに緩衝液成分を組み合わせて、蛍光及び/又はルミネセンスに対する各構成成分の寄与を確認した。ルミネセンス測定時、蛍光発生基質である(Z−DEVD)2−ローダミン−110は、ローダミンチャネル(485EX/520EM)内でカスパーゼ−3活性を報じ、蛍光発生基質であるAc−DEVD−AMCは、AMCチャネル(360EX/460EM)内でカスパーゼ−3活性を報じるが、Z−LETD−アミノルシフェリン基質は、カスパーゼ−8活性を報じる。表3には、12の異なる反応条件に関する各成分の量(μl)が、各反応条件に関し、合計、約500μlのマスター混合物、又は100μlのマスター混合物/反応(n=4)を生じることが記載されている。「カスパーゼ添加」の列でのこの列の数は過剰に添加されたカスパーゼのタイプを定義するものであって、容量を示しているのではない。
【0081】
【表3】
1.カスパーゼ−8 luc対照
2.カスパーゼ−3ローダミン−110対照
3.カスパーゼ−3AMC対照
4.ローダミン−110を有する多重対照
5.ローダミン−110+阻害剤を有する多重対照
6.AMCを有する多重対照
7.アミノルシフェリン混合物を有するカスパーゼ−3
8.アミノルシフェリン混合物+阻害剤を有するカスパーゼ−3
9.ローダミン−110を有するカスパーゼ−8
10.アミノルシフェリン混合物を有するカスパーゼ−3
11.アミノルシフェリン混合物+阻害剤を有するカスパーゼ−8
12.ローダミン−110を有し、アミノルシフェリン混合物を有さないカスパーゼ−3
【0082】
表IIIの成分を反復ウェルに加え、反応液を室温で2時間温置した。用いられた緩衝液は、Caspase−Glo(商標)8 Assey Systemのものであった。DMSOはSigma−Aldrichから入手し、DTTは、Amrescoから入手した。基質及び阻害剤は、Promega Corpから入手した。
【0083】
相対的ルミネセンスは、DYNEX LaboratoriesのMLX(商標)プレートルミノメータを用いて測定された。蛍光は、ローダミン−110に関しては、485EX/530EMフィルターセット、次いで、AMCチャネルに関しては、360EX/460EMにセットしたCytoFluorII Fluorescentプレート読取り機を用いて測定した。
【0084】
結果
図2A、B、及びCで、全てのカラットは、酵素活性を示す蛍光又はルミネセンスのいずれかが予測される場合を表す。図2Aは、各反応におけるAMC蛍光に関するシグナルを示す。背景以上の蛍光は、Ac−DEVD−AMC及びカスパーゼ−3という適切な基質/酵素の組合せが存在した場合のみ存在した(反応条件3及び6)。図2Bは、各反応におけるローダミン−110蛍光に関するシグナルを示す。背景以上の蛍光は、カスパーゼ−3阻害剤が存在した場合(反応条件5)を除いて、(Z−DEVD)2−ローダミン−110/カスパーゼ−3という基質/酵素の組合せが存在した場合に存在した(反応条件2、4、10、及び12)。背景以上のルミネセンスシグナル(図2C)では、カスパーゼ阻害剤が存在した反応条件(反応条件5、8、及び11)を除いて、Z−LETD−アミノルシフェリン/カスパーゼ−8という適切な基質/酵素の組合せを有した反応が、背景以上のシグナルを示した(反応条件1、4、6、7、9、及び10)。したがって、該データにより、これらの条件下で、背景の蛍光及びルミネセンス測定に及ぼす反応成分の影響は無視できるものだったことが証明された。
【0085】
C.単一ウェル、三重アッセイにおけるカスパーゼ−3、カスパーゼ−8、及びトリプシンの測定
ダルベッコ−リン酸緩衝生理食塩水(Sigma社)中、カスパーゼ−8(150単位/ml、Biomol Research Laboratories)、カスパーゼ−3(Pharmingen社)、トリプシン(Sigma社)、及び3種全ての酵素の組合せの検出可能濃度の希釈液を調製した。100μlの各酵素希釈液を、96ウェルプレートのウェルに加え、100μlの各基質を、単独で、又は適切に組み合わせて対応するウェルに加えた:カスパーゼ−3に対しては、(Z−DEVD)2−ローダミン−110基質、トリプシンに対しては、Z−PRNK−AMC基質(ベータ−トリプターゼに対する基質として、本明細書にその全体が組み込まれている米国特許出願第09/955,639号に記載されているが、トリプシンに対しては、有用性が少ないと認識されている)、及びカスパーゼ−8に対しては、Z−LETD−アミノルシフェリン基質。蛍光アッセイで、基質と共にCaspase−Glo(商標)8緩衝液を用いた場合、10mM DTTを含めた。プレートは、プレートシェーカー上、室温で、少なくとも10分間温置した。
【0086】
温置後、カスパーゼ−8活性に関する相対的ルミネセンスを、BMG Fluorostar(BMG Labtechnologies社)を用いて測定した。相対的蛍光は、Labsystems Fluoroskan Ascent プレート読取り機を用いて測定した。カスパーゼ−3活性には、485EX/527EMのフィルターセットを用いた。トリプシン活性には360EX/460EMのフィルターセットを用いた。
【0087】
結果
図3Aに示されるように、3種の異なる基質及び組み合わされた対応する酵素を有する反応(三重アッセイ)において、カスパーゼ−3の検出に用いられた条件により、対照条件の蛍光以上の比較的高い蛍光が生じた。三重アッセイ(全ての酵素と共に全ての基質)におけるカスパーゼ−3の活性を、カスパーゼ−3単独の活性と比較すると、カスパーゼ−3が同じ反応液中に他の三重酵素反応液と共にあった場合、カスパーゼ−3活性は背景より大きかった。トリプシン(図3B)及びカスパーゼ−8(図3C)に関しても、カスパーゼ−3と同程度ではないにしても、同様な結果が見られた。
【0088】
D.単一ウェル、多重フォーマットにおけるカスパーゼ−3及びβ−ガラクトシダーゼの測定
Beta−Glo(登録商標)緩衝液(Beta−Glo(登録商標)Assay System、Promega社)を用いてBeta−Glo(登録商標)凍結乾燥基質を再構成することにより、又はBeta−Glo(登録商標)緩衝液に(Z−DEVD)2−ローダミン−110(50μM)を加えることにより、又はBeta−Glo(登録商標)緩衝液を用いてBeta−Glo(登録商標)凍結乾燥基質を再構成して(Z−DEVD)2−ローダミン−110(50μM)を加えることにより、試薬を調製した。カスパーゼ−3(2μl/ml、Pharmingen社)、又はβ−ガラクトシダーゼ(0.1μl/ml)、又はカスパーゼ−3及びβ−ガラクトシダーゼを、RPMI1640中で希釈し、100μlを96ウェル白色プレートのウェルに加えた。100μlの適切な試薬を96ウェルプレートのウェルに加え、室温で温置した。DYNEX LaboratoriesのMLX(商標)プレートルミノメータを用いて、30分におけるルミネセンスを測定した。485EX/530EMのフィルターにセットしたCytoFluorII Fluorescentプレート読取り機上で、2時間温置後の蛍光を測定した。増加した蛍光を補正するために、18時間目に、CytoFluorIIフルオロメータ上、異なる増加セッティングで全ての測定を繰り返した。
【0089】
結果
図4A及びCは、β−ガラクトシダーゼに関するルミネセンス発生アッセイが、カスパーゼ−3を測定するための蛍光発生試薬の存在下で機能的であることを証明している。図4B及びDは、カスパーゼ−3を測定するための蛍光発生アッセイが、β−ガラクトシダーゼを測定するためのルミネセンス発生試薬の存在下で機能的であることを証明している。図4B及び4Dに見られるように、背景蛍光に及ぼすルミネセンス発生試薬成分の軽微な影響がある。しかし、ルミネセンスに及ぼす蛍光発生試薬成分の影響はほとんど無い(図4A及び4C)。
【0090】
E.ルミネセンス発生アッセイに関する基質のスペクトル走査
0.1M Tris pH7.3、2mM EDTA、及び10mM MgSO4を含有する緩衝液中、ルシフェリン、アミノルシフェリン、及びZ−LETD−アミノルシフェリンを、およそ2μMに希釈した。1.25mm励起及び発光スリットフィルターを有するSPEX Fluorolog−2分光器上、1nmの波長間隔及び0.2秒の積分時間でサンプルを走査した。全ての走査は、石英キュベットを用いて実施された。
【0091】
結果
ルシフェリン及びアミノルシフェリンに関して、励起が325nmに存在すると、発光は、375nmから750nmで捕捉され、発光が600nmで測定される場合の励起は、280nmから550nmで捕捉された(図5A及び5B)。Z−LETD−アミノルシフェリンでは、励起が325nmに存在すると、発光は、375nmから750nmで捕捉され、発光が525nmで測定される場合の励起は、280nmから500nmで捕捉された(図5C)。興味深いことに、ペプチドがアミノルシフェリンに共役すると(図5C)、共役体の発光ピークは、より短い波長へ青色シフトした。これは予想外のことであり、したがって、特に、アミノルシフェリンと同じ波長領域で発光する蛍光体を用いると、ルミネセンス/蛍光二重測定が可能になる。
【0092】
(実施例II)
偽結果を検出する方法
方法
カスパーゼ−3阻害剤(Ac−DEVD−CHO、10μM)又はルシフェラーゼ阻害剤(Resveratol、5μM)のいずれかと共に、カスパーゼ−3の存在下、Z−DEVD−アミノルシフェリンを含有するCaspase−Glo(商標)3/7試薬(Caspase−Glo(商標)3/7 Assay、Promega社)を、(Z−DEVD)2−ローダミン−110又はAc−DEVD−AMCと組み合わせた。Z−DEVD−アミノルシフェリンのカスパーゼ−3開裂に由来するルミネセンスシグナルは、30分目に読み取り、一方、カスパーゼ−3開裂活性由来の蛍光シグナルは、適切なAMC又はローダミン110フィルターセットを用いて、2時間目に読み取った。
【0093】
結果
ルミネセンスは、背景比に対して最大のシグナルを与え、次いでローダミン−110、次いで、AMCであった(図6)。カスパーゼ−3を検出する3種の基質全てが、知られたカスパーゼ−3阻害剤の添加によって、一貫して影響を受けなかった。このことは、いずれかのアッセイが実施される際に、例えば、可能性のある偽妨害を制御するために、ルミネセンス発生試薬と蛍光発生試薬とを組み合わせることができるということを示唆している。したがって、ある試薬がある一定の酵素の真の阻害剤であるかどうかを判定するために多重シグナルを用いるこができる。
【0094】
(実施例III)
さらなる典型的多重アッセイ
A.単一ウェルフォーマットにおける乳酸デヒドロゲナーゼ(LDH)及びアデノシン三リン酸に関する多重アッセイ
以下の検出試薬を調製した:1)LDH試薬(30mM HEPES、pH7.4、10mM NaCl、20mM MgSO4、250μMレザズリン(Aldrich))を用いて、CytoTox−ONE(商標)Homogeneous Membrane Integrity Assay(Promega社、Technical Bulletin306)からの凍結乾燥基質成分を再構成した;2)ATP試薬(30mM HEPES pH7.4、10mM NaCl、20mM MgSO4)を用いて、CellTiter−Glo(商標)Luminescent Cell Viability Assay(Promega社、Technical Bulletin288)からの凍結乾燥基質成分を再構成した;3)LDH/ATP組合せ試薬(30mM HEPES、pH7.4、10mM NaCl、20mM MgSO4、250μMレザズリン(Aldrich))を用いて、CytoTox−ONE(商標)からの凍結乾燥基質成分を再構成し、次に、これを用いて、CellTiter−Glo(商標)からの凍結乾燥基質成分を再構成した。
【0095】
10mM HEPES pH7.5、0.1%Prionex(PentaPharma社)溶液を用いて、LDH(0、1:8000、1:4000、1:2000、菱形)、ATP(0μM、1.25μM、2.5μM、及び5μM、四角形)、及びLDH/ATPの組合せ(それぞれ、0/0μM、1:8000/1.25μM、1:4000/2.5μM、及び1:2000/5μM、三角形)のサンプル希釈液を作製し、100μlの該希釈液(n=4)を、白色96ウェルプレートのウェルに加えた。該サンプルに適切な検出試薬(100μl)を加え、該プレートを遮光し、30秒間混合し、室温で温置した。温置8分後、フィルターを560EX/590EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。温置30分後に、Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0096】
結果
対照反応(図7C)と比較した場合、ATPに関するルミネセンス発生アッセイに及ぼすLDH及びその蛍光発生検出試薬の軽微な影響があった(図7A);しかし、ATPの検出は、依然として可能であった。LDHに関する蛍光発生アッセイへルミネセンス発生検出試薬を添加しても背景蛍光に影響を与えず(図7B)、対照反応(図7D)に比較して全体的な蛍光は減少したが、LDH活性は、依然として検出可能であった。
【0097】
B.単一ウェルフォーマットにおけるLDH及びカスパーゼ−3に関する多重アッセイ
以下の検出試薬を調製した:1)238μMレザズリンを添加したLDH試薬−Caspase−Glo(商標)3/7緩衝液を用いて、CytoTox−ONE(商標)凍結乾燥基質成分を再構成した;2)カスパーゼ−3試薬−Caspase−Glo(商標)3/7緩衝液を用い、Promega Technical Bulletin322にしたがって、Caspase−Glo(商標)3/7凍結乾燥基質成分を再構成した;3)LDH/カスパーゼ−3組合せ試薬−LDH試薬(上記の通りに調製)を用いて、凍結乾燥Caspase−Glo(商標)3/7基質を再構成した。LDH/カスパーゼ−3組合せ試薬のLDH検出試薬化合物は、Caspase−Glo(商標)3/7凍結乾燥基質中のDTTの存在によって不安定であるため、この試薬は、サンプルへの添加直前に調製した。
【0098】
10mM HEPES pH7.5、0.1% Prionex(PentaPharma社)溶液中で、サンプル希釈液を調製した:LDHの0、1:8000、1:4000、1:2000希釈液(菱形);カスパーゼ−3の0、5、10、及び20U/ml(BIOMOL Laboratories、四角形)、及びLDH/カスパーゼ−3の組合せ(それぞれ、0/0U/ml、1:8000/5U/ml、1:4000/10U/ml、及び1:2000/20U/ml、三角形)。100μlの該希釈液(n=4)を、白色96ウェルプレートに加えた。該サンプルに適切な検出試薬(100μl)を加え、該プレートを遮光し、30秒間混合し、室温で温置した。室温で温置6分後、フィルターを560EX/590EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。温置45分後に、Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0099】
結果
多重反応に蛍光発生LDH検出試薬を加えると、対照反応に比較してルミネセンスの減少があった(それぞれ、図8A及び8C)。図8Aにおける全体的なルミネセンスシグナルの減少に関わらず、蛍光発生LDH検出試薬の存在下、ルミネセンスカスパーゼ−3アッセイは機能的であった。図8Bは、多重反応にルミネセンス発生カスパーゼ−3検出試薬を加えると、対照(図8D)に比較して、蛍光背景の増加があることを示している。;しかし、図8Aは、カスパーゼ−3に関するルミネセンス発生検出試薬の存在下で、LDHに関する蛍光発生アッセイが機能的であることを証明している。背景ルミネセンスに対するLDHの影響はなく(図8C)、また、背景ルミネセンスに対するカスパーゼ−3の影響もなかった(図8D)。
【0100】
C.単一ウェルフォーマットにおけるカスパーゼ−3及びタンパクキナーゼA(PKA)に関する多重アッセイ
以下の検出試薬を調製した:1)PKA試薬−100mMトリス pH7.3、100mM MgCl2、PKAローダミン−110基質(ProFluor(商標)PKA Assay、Promega社、 Technical Bulletin315)の1:1000希釈液、及び400μM ATPを含有する1X反応緩衝液を調製した;2)カスパーゼ−3試薬−100mMトリス pH7.3、100mM MgCl2、150μg/mlの組換え熱安定性ルシフェラーゼ、80μM Z−DEVD−アミノルシフェリン(Promega社)、400μM ATP、100μM DTT(Promega社)、2.5mM CaCl2(Fisher)、40mM MgSO4(Fisher)、及び0.2% Tergitol NP−9(Sigma)を含有する1X反応緩衝液を調製した;3)キナーゼ/カスパーゼ−3組合せ試薬−100mMトリス pH7.3、100mM MgCl2、PKAローダミン−110基質の1:1000希釈液、150μg/mlの組換え熱安定性ルシフェラーゼ、80μM Z−DEVD−アミノルシフェリン、400μM ATP、100μM DTT、2.5mM CaCl2、40mM MgSO4、及び0.2% Tergitol NP−9を含有する1X反応緩衝液を調製した;4)タンパクキナーゼ停止試薬−100mMトリス pH7.3、100mM MgCl2、プロテアーゼ試薬(ProFluor(商標)PKA Assay)の1:50希釈液、30μM スタウロスポリン(BIOMOL Laboratories)を含有する1X停止試薬を調製した。
【0101】
10mM HEPES pH7.5、0.1% Prionex(PentaPharma社)溶液中で、サンプル希釈液を調製し;0、1、2、及び4U/ml PKA(菱形)、0、5、10、及び20U/mlカスパーゼ−3(四角形)、及びPKAとカスパーゼ−3との組合せ(それぞれ、0/0U/ml、1/5U/ml、2/10U/ml、及び4/20U/ml、三角形)、40μlの該希釈液(n=4)を、白色96ウェルプレートに加えた。該サンプルに適切な検出試薬(40μl)を加え、該プレートを遮光し、30秒間混合し、室温で20分間温置した。温置後、キナーゼ試薬単独又はキナーゼ/カスパーゼ−3試薬の組合せのいずれかを含有するウェルに、40μlのタンパクキナーゼ停止試薬を加えた。該プレートをさらに30秒間混合し、遮光し、室温でさらに30分温置した。フィルターを485EX/527EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0102】
結果
蛍光発生PKAアッセイ検出試薬の添加により、PKAは存在するが完全なPKA検出試薬が存在しない対照反応(図9C)に比較して、ルミネセンス背景の増加が生じた。しかし、カスパーゼ−3の活性から生じた反応ルミネセンスは比例して増加し、該条件は、ルミネセンス発生カスパーゼ−3反応それ自体に影響したとは思われなかった。PKAに関する蛍光発生アッセイに、カスパーゼ−3に関する検出試薬を添加すると(図9B)、カスパーゼ−3は存在するが完全なカスパーゼ−3検出試薬が存在しない蛍光対照反応(図9D)に比較して、全体の蛍光が50%超減少した。カスパーゼ−3及びPKA活性は、これらの多重条件を用いて、背景以上を測定できた。
【0103】
D.単一ウェルフォーマットにおけるウミシイタケルシフェラーゼ及びカスパーゼ−3に関する多重アッセイ
以下の検出試薬を調製した:1)ウミシイタケルシフェラーゼに対する細胞透過性修飾セレンテラジン基質であるEnduRen(商標)(Promega社)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェートを添加したF−12組織培養培地中に、600μMに希釈した;2)カスパーゼ−3基質:(Z−DEVD)2−ローダミン−110(Promega社)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェートを添加したF−12組織培養培地中に、250μMに希釈した;3)ルシフェラーゼ/カスパーゼ−3組合せ基質:EnduRen(商標)(600μM)及び(Z−DEVD)2−ローダミン−110(250μM)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェートを添加したF−12組織培養培地中に、希釈した。
【0104】
ウミシイタケルシフェラーゼ(CHO−KlhRL25)を安定して発現するCHO−Kl細胞(ATCC)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェート中で維持し、細胞ベースの実験に用いた。これらの細胞を用いる実験条件には、以下が含まれた:1)スタウロスポリンの添加によるルシフェラーゼ活性レベルの変化、2)カスパーゼ−3酵素の添加と共にスタウロスポリンの添加によるルシフェラーゼ活性レベルの変化、及び3)スタウロスポリンは添加せずにカスパーゼ−3酵素の添加によるルシフェラーゼ活性。
【0105】
CHO−KlhRL25細胞を採収し、96ウェル、透明底、白色壁の組織培養プレート中に20,000細胞/ウェルの密度で塗布し、5%CO2中、37℃で一晩温置した。最終濃度が0、0.5、1、2μMのスタウロスポリン(10μl/ウェル)を適切なウェルに加え、細胞死を開始させ、したがってルシフェラーゼ活性を変化させた。5%CO2中37℃でさらに3.5時間、細胞を温置した。種々の濃度のカスパーゼ−3(BIOMOL Laboratories)を、組織培養培地中、0、5、10、及び20U/mlで、適切なウェルに加えた(10μl/ウェル)。したがって、データポイントに関するスタウロスポリン/カスパーゼ−3組合せ濃度は、それぞれ、0μM/0U/ml、0.5μM/5U/ml、1μM/10U/ml、及び2μM/20U/mlであった。カスパーゼ−3酵素の添加直後、10μl/ウェルのルシフェラーゼ基質、カスパーゼ−3基質、又はルシフェラーゼ/カスパーゼ−3基質のいずれかを、適切なウェルに加えた。検出試薬の添加後、プレートを手短に混合し、5%CO2中37℃で2時間温置した。フィルターを485EX/527EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0106】
結果
これらのアッセイにおいて、ウミシイタケルシフェラーゼの活性を、細胞死に関する内部対照として用いた。したがって、スタウロスポリンの濃度が増加するにつれて、ルシフェラーゼの活性は減少するはずである。図10Aは、カスパーゼ−3基質の添加は、対照反応(図10C)と比較して、ルシフェラーゼ反応に負の影響を与えなかったことを示している。図10Bは、ルシフェラーゼ基質の添加が、図10Dに比較して、全体の蛍光にわずかな増加はあったものの、背景蛍光に影響を与えなかったことを示している。ウミシイタケルシフェラーゼに関するルミネセンス発生アッセイは、カスパーゼ−3又はカスパーゼ−3基質の存在下で十分に機能的であり、カスパーゼ−3に関する蛍光発生アッセイは、わずかに影響を受けただけで、ウミシイタケルシフェラーゼ又はウミシイタケルシフェラーゼ基質の存在下で十分に機能的であった。
【0107】
(実施例IV)
プロテアーゼ保持及び放出細胞生存度多重アッセイ
生細胞及び死細胞アッセイは、特定の化学的、生物学的又は物理的処理に応答した細胞生存度の変化をモニターするために、広く用いられている。生存度と細胞毒性アッセイとは、一般的に逆であり、異なった生物マーカーを測定する。細胞毒性による細胞生存度の一般的変化を評価する方法は、歴史的に外部膜透過性の変化に関連してきた。膜構造の損傷を検出する古典的方法としては、トリパンブルー排除、核酸染色、及び乳酸デヒドロゲナーゼ放出が挙げられる(Rissら、2004年;Myersら、1998年)。細胞機能又は増殖の評価に関するアッセイとしては、トリチウムチミジン取込み、ATP含量、テトラゾリウム色素移行又はフルオレセインジアセテートが挙げられる(Cookら、1989年)。無処置細胞膜は、かさばった荷電分子又はペプチドを、細胞外空間から細胞質内へ進入させないと考えられる。逆に、損傷した膜は、色素又は化合物を細胞内へ、又は細胞内容物を細胞外へ自由に透過させる。この透過現象が、色素標識(「生命」色素、DNA挿入剤、又はエステラーゼ修飾フルオレセイン)並びにLDH放出アッセイの双方にとっての基礎である。細胞生存度を測定する既存の方法は、依然として有用であり、コスト効率の良い適用である一方、それらは多くの技術的又は実用的な欠点を有し、高含量、多重又は高スループットフォーマットにおける利用が制限されている。例えば、LDH放出(CytoTox−ONE(商標))又は色素減少能力(CellTiter−Blue(商標))による細胞膜の完全性の現在の測定は、レザズリン基質の共有及びEx/Emスペクトルの重なりのために、対にする(データ正規化の手段)ことができない。さらに、双方のアッセイに利用される着色レアズリン基質は、他のエンドポイントアッセイ測定(消色)との第2のアッセイシグナルウィンドー強度(及び感度)を制限し、濃度及びフォーマットが、第2のアッセイ試薬ペアリングに関して最適化されない(例えば、容量が限定される)。
【0108】
既存の生/死細胞フォーマットでは、カルボキシフルオレセイン及びエチヂウムホモ二量体が用いられるが、後者は知られた強力な変異誘発因子である。このフォーマットには、洗浄及び細胞培養培地の置換が必要である。さらに、カルボキシフルオレセインは、水性溶液中で自然加水分解を示し、エチヂウムホモ二量体挿入はDNAを染色し、下流のデータ正規化を妨害し得る。
【0109】
培養哺乳動物細胞は、プロテアーゼ類、エステラーゼ類、リパーゼ類、及びヌクレアーゼ類の豊富な環境を含有する。例えば、プロテアーゼの4つの一般的なクラス(アスパラギン酸、システイン、セリン、及び金属依存性)が代表であり、恒常性維持の特異的機能に関連している。これらの細胞質の、リソゾームの、及び膜貫通結合のプロテアーゼは、細胞内タンパク質分解、免疫原性ペプチドの生成、翻訳後修飾、及び細胞分裂に関与している(Tranら、2002年、Constamら、1995年、Vinitskyら、1997年)。これらの酵素の活性は、特殊区画化などの種々の機構によって調節されている(Bondら、1987年)。過剰なストレス、環境の悪状況、又は細胞死プログラムの委任進行に応答して、区画化及び膜完全性の整合喪失が見られる(Syntichakiら、2003年、Haunstetterら、1998年)。したがって、インビトロ細胞モデルにおける細胞培養培地内への安定なタンパク質分解伝達物質の放出は、細胞死の潜在的代理物となる。逆に、保持されたタンパク質分解酵素の細胞酵素学的染色は、細胞健康度の表現型測定にパラレルである。このようなタンパク質分解活性が一緒になって、細胞培養集団、例えば「生/死」アッセイ中の生存細胞又は損傷細胞の相対的数の確認を補助し得る。
【0110】
プロテアーゼベースの生/死細胞アッセイでは、一実施形態において、1つの基質(死細胞に関する)は、細胞質pH、例えば、7.0から7.2において安定で活性であり、スペクトル的に読取り(R/O)の異なる標識を有する、比較的豊富で活性であり保存されたプロテアーゼに対する基質である。この基質の開裂動態は、LDH放出にパラレルであり、活性の条件が毒性物質又は膜変化物質、例えば、塩又はチオール類を含まず、アッセイ時間を高速にすることが好ましい。他の基質(生細胞に関する)は、比較的豊富で保存されたプロテアーゼに対する基質であり、生存細胞に関しては細胞透過性であり、また、該プロテアーゼは、生存細胞の細胞質環境では活性であるが、細胞外環境では不安定である。この基質は、スペクトル的にR/Oの異なる標識を有し、開裂反応は、アッセイ時間を高速にするように進行する。非破壊的アッセイにおける2種の基質の使用により、望ましくない増殖事象を検出でき、また、生存度対細胞毒性の比率は、そのウェルにおける細胞数の変動に独立であるため、異なるスペクトルにおける相補的で独立の代理物を用いることにより、誤った結論及び細胞凝集又はピペット操作エラーによるエラーを減らすことができる。
【0111】
A.AMC若しくはR110蛍光又はアミノルシフェリンルミネセンスレポーターによるプロテアーゼ放出アッセイフォーマット
HL−60細胞を、2倍連続希釈してからトリトンXの添加によって、0.2%最終に細胞溶解させるか、又は媒体添加によって維持した。100mM酢酸ナトリウム、pH4.5中、200μM Ala−Ala−Phe−AMC基質の1/10容量を、溶解物又は細胞に加え、37℃でさらに1時間温置した。次いで、溶解細胞又は生存細胞に関連した蛍光を、CytoFluorIIを用いて、Ex.360 Em.460で測定した。
【0112】
活動的ダブリングを受けているジャーカット細胞を、トリパンブルー排除によって数えると、95%超の生存が判明した。該細胞をRPMI1640+10%FBS中、100,000細胞/mlに調整し、2つのアリコートに分けた。1つのアリコートは、マイクロチップを備えたMisonix3000を、3×5秒パルスの30%出力で用いて超音波処理した。他方の画分は、超音波操作(合計約5分)の間、37℃の水浴中で温置した。次いで、細胞懸濁液及び溶解物画分を比率混合によって、0〜100%の生存度を表す種々の生存度へ混合した。次に、混合した細胞サンプルを、100μl容量で白色壁、透明底の96ウェルプレート(Costar)に加えた。(Ala−Ala−Phe)2−R110を、RPMI−1640中、1000μMni希釈し、1/10容量を該プレートに加えた。該プレートを30分間温置してからCytoFluorIIを用いて、蛍光を、Ex.485 Em.530で測定した。
【0113】
活動的ダブリングを受けているジャーカット細胞を、トリパンブルー排除によって数えると、95%超の生存が判明した。該細胞をRPMI1640+10%FBS中、100,000細胞/mlに調整し、2つのアリコートに分けた。1つのアリコートは、マイクロチップを備えたMisonix3000を、3×5秒パルスの30%出力で用いて超音波処理した。他方の画分は、超音波操作(合計約5分)の間、37℃の水浴中で温置した。次いで、細胞溶液及び溶解物画分を比率混合によって、0〜100%の生存度を表す種々の生存度へ混合した。次に、混合した細胞サンプルを、100μl容量で白色壁、透明底の96ウェルプレート(Costar)に加えた。10mlの10mM Hepes、pH7.5を有するルシフェリン検出試薬ケーク(Promega V859A)の再水和により、ルミネセンス発生プロテアーゼ放出アッセイ試薬を調製し、この試薬に、Ala−Ala−Phe−アミノルシフェリンを添加し、100μMの最終濃度にした。該プレートのウェルに、100μlのルミネセンス発生プロテアーゼ放出アッセイ試薬を加え、BMG FLUOstar Optimaを用いて、動態様式でルミネセンスを測定した。
【0114】
AMC蛍光フォーマットの実際の感度は、CytoTox−ONE(商標)と同等の感度値である、約240個の細胞(図11)であると算出された。R110フォーマット(図12)のアッセイは、同様に鋭敏であり、多重適用のためにさらに別の蛍光体を提供した。特にこれらのアッセイの感度は、下流の多重適用におけるCytoTox−ONE(商標)又は他のレザズリンベースのアッセイの使用の主要な障害である蛍光の消光なしで得られた。ルミネセンスフォーマットの優れた直線性と範囲(図13)により、9800個の生細胞集団中、200個と少ない細胞の統計的検出を可能にした。非溶解性のルミネセンスフォーマットは、細胞毒検出のために別の代替法を提供する。
【0115】
B.種々の酵素標的によるプロテアーゼ放出アッセイのフォーマット
活動的ダブリングHL−60細胞を、100,000個の細胞/mlに調整し、2つのアリコットに分けた。1つのアリコットは、マイクロチップのMisonix3000を用いて3回の5秒パルスの30%出力で超音波処理した。他のアリコットは、37℃で保持した。次に細胞懸濁液及び溶解物を、100μl容量でRPMI 1640+10%FBS中で2倍連続希釈した。培地のみのものは、無細胞対照として寄与した。ルシフェリン検出試薬のケーク(Promega V859A)を、2.0mlの10mM Hepes、pH7.5で再懸濁した。次いでルシフェリン検出試薬を分けて、Z−Leu−Leu−Val−Tyr−アミノルシフェリン又はAla−Ala−Phe−アミノルシフェリンによる1mMを作製した。各試薬を、1/10容量で独立した反復試験のプレートに加え、Me’Courの熱ジャケット水浴ホルダー中37℃で15分間温置させてから、BMG FLUOstar Optimaを用いてルミネセンス測定を行った。
【0116】
Z−LLVY−アミノルシフェリンアッセイは、AAF−アミノルシフェリンシーケンスよりも少ない最適さで実施したが、他のプロテアーゼ類は、完全性の欠いた代用物として使用できることが立証された(図14)。この場合、LLVY活性は、プロテオソームのキモトリプシン活性に起因し得る。
【0117】
C.プロテアーゼ放出の時間経過
HL−60細胞(25,000個/ウェル)を、透明底の白壁で仕切られた96ウェルプレート(Costar)内で5%CO2と共に37℃で7時間の経過にわたり、10μMスタウロスポリン又は適性のDMSO媒体対照により処理した。200μMのAla−Ala−Phe−AMC基質溶液を、100mMのNaアセテート、pH4.5中で作製した。10μl容量の基質(1/10容量のサンプル)をウェルに加えてさらに1時間温置した。「プロテアーゼ放出」活性を、CytoFluor II上Ex.360Em.460で測定した。平行にセットされたウェル内で、CytoTox−ONE(商標)試薬は、膜完全性アッセイ対照として作用した。この試薬を、Ex.560 Em.580で蛍光測定10分前に加えた。
【0118】
細胞透過性の動態学、すなわち、LDHとプロテアーゼ放出は、互いに反映され、細胞集団における二次的壊死の形態学的観察と一致した(図15)。酸性Naアセテート製剤(サンプル中約6.5の最終pH)中アミノペプチダーゼ基質の提示は、潜在的なリソソームプロテアーゼ活性を調整するために実施された。
【0119】
D.プロテアーゼ放出活性のpH要件
プロテアーゼ放出活性のpH要件は、非調整培養培地(水媒体)と比較して、pH2.5、3.5及び4.5に調整された100mM Naアセテートを用いて調査された。Ala−Ala−Phe−AMCを、これらの緩衝液中200μMに加えた。溶液の1/10容量をプレートに加え、軌道振とうにより短時間混合した。このプレートを37℃で40分間温置してから、蛍光を、CytoFluor IIを用いてEx.360Em.460で測定した。
【0120】
pH4.5のNaアセテート1/10容量の添加により、培養培地を約6.5の最終pHに減じた。他のより低いpH溶液/培地の組合せの最終pHは、試験しなかったが、以前の実験により、pH2.5のNaアセテートの1/10容量の添加は、細胞培地のpHを約5.5に減じることを示唆した。非pH調整媒体は、プロテアーゼ放出活性にとって最も好ましいことを示すことが判った(図16)。この活性は、細胞基質のアミノペプチダーゼと一致し、恐らくリソソームプロテアーゼ(カテプシンなど)ではないと思われる。プロテアーゼ放出活性を測定するために、有害な又は潜在的細胞毒性の付加物を必要としないことからこのことは重要である。このことにより、温置時間枠をより柔軟にでき、可能なルミネセンスベースのアッセイをさらに受けられることが考慮に入れられる。
【0121】
E.プロテアーゼ放出酵素の細胞下位置
HL−60細胞を、1ml当り100,000個の細胞に調整し、2つのアリコットに分けた。1つのアリコットは、マイクロチップのMisonix3000を用いて3回の5秒パルスの30%出力で超音波処理した。100μlのこの溶解物(形態学的に確認された)を、透明底96ウェルプレートの複数ウェルに加え、10%FBSを有するRPMI 1640中で2倍連続希釈した。同様に、100μlの非超音波処理の細胞懸濁液を加え、プレートの複数ウェルで連続希釈した。NP−9及びジギトニンを、それぞれ最終0.2%及び30μg/mlで個別のウェルに加えた。未処理の対照は、生細胞及び適性容量の水媒体から構成された。ルシフェリン検出ケーク(Promega V859A)を、2mlの10mM Hepes、pH7.5で再水和し、Ala−Ala−Phe−アミノルシフェリン(Promega)により500μMを作製した。20μlのこのプロルミネセンスプロテアーゼ放出溶液を、全てのウェルに加えて、37℃で15分間温置させてから、BMG FLUOstar Optimaを用いてルミネセンスを測定した。
【0122】
超音波処理及び上記のパラメータと濃度を有するNP−9は、外膜のみならず、リソソーム内容物(カテプシン放出により測定された)も破壊することが知られている(図17)。ジギトニンによる選択的破壊は、リソソーム破壊の証拠のないトリパンブルー染色を考慮に入れている。したがって、活性は、超音波処理又は差別的界面活性の細胞溶解の間で類似していたため、最適pHを考慮して、プロテアーゼ放出アッセイで測定されたプロテアーゼは、恐らく細胞基質性であり、無処置の細胞小器官の外部にあることが推測され得る。
【0123】
F.プロテアーゼ放出又は保持酵素基質の選択性
Ala−Ala−Phe−AMCを、Promegaから入手した。Z−Leu−Leu−Val−Tyr−アミノルシフェリン、Z−Leu−Arg−アミノルシフェリン、Z−Phe−Arg−アミノルシフェリン、Ala−Ala−Phe−アミノルシフェリン、(Ala−Ala−Phe)2−R110、及び(Gly−Phe)2−R110は、Promega Biosciencesにより合成された。Suc−Ala−Ala−Phe−AMC、H−Phe−AMC、H−Tyr−AMC、グルチル−Ala−Ala−Phe−AMC、H−Gly−Phe−AMC、Z−Gly−Ala−Met−AMC、Suc−Leu−Leu−Val−Tyr−AMC、D−Ala−Leu−Lys−AMC、H−Gly−Ala−AMC、H−Gly−Gly−AMC、Suc−Ala−Ala−Phe−AMC、Z−Arg−Leu−Arg−Gly−Gly−AMC、Z−Leu−Arg−Gly−Gly−AMC及びAc−Ala−Ala−Tyr−AMCは、Bachemから供給された。Gly−Phe−AFC、Pro−Phe−Arg−AMC、Gly−Gly−Leu−AMC、及びSer−Tyr−AFCは、Calbiochemから入手した。Z−Phe−Arg−AMC及びSuc−Arg−Pro−Phe−His−Leu−Leu−Val−Tyr−AMCは、Sigmaから購入した。
【0124】
全ての基質は、固有の溶解度によって10mMから100mMでDMSOに可溶性であった。蛍光性基質を、10mM Hepes、pH7.5又は10%血清を有する適性な細胞培地100μMから1mMに希釈し、白壁仕切りの透明底の96ウェルプレート中の溶解細胞(凍結破壊、超音波処理、又は界面活性剤)又は未処理生存細胞に1/10容量で加えた。HL−60又はジャーカットを、それらの容易に操作された懸濁表現型のため実験に交換可能に用いた。プレートを、CytoFluorIIによる蛍光を測定する前に37℃で15〜30分間温置した。
【0125】
ルミネセンス基質を、2mlの10mM Hepes、pH7.5に再懸濁されたルシフェリン検出ケーク(Promega V859A)に500μMで加えた。1/5容量のプロルミネセンス反応混合物を、白壁仕切りの透明底の96ウェルプレート中、溶解細胞(凍結破壊、超音波処理、又は界面活性剤)又は未処理生存細胞に加えた。やはりHL−60又はジャーカットを、実験に交換可能に用いた。プレートを、Caron2050W交換ユニットにより制御されたMeCour’循環熱ブロック内で37℃で温置した。ルミネセンスを、15分から30分の間(シグナル定常状態)で測定した。
【0126】
損傷細胞又は生存細胞内でのプロテアーゼ放出又は保持に関する潜在的基質の優先度を特性化するために、努めて多種多様のタンパク分解基質を調べた(表4を参照)。エンドペプチダーゼ又はエキソペプチダーゼの活性が勝っているかどうかを正確に概説するために、アミノ末端ブロック化基質(Z、Suc−、又はAc−)が選択された。非ブロック化基質(H−など)を、アミノペプチダーゼ活性の寄与を含めるために調べた。このパネルから、少なくとも3つのタンパク分解プロフィルが明らかになった:非ブロック化Ala−Ala−Pheトリペプチドを優先させるアミノペプチダーゼ様活性、ブロック化Leu−Leu−Val−Tyrペプチド類の放出により測定されたプロテオソーマル(キモトリプシン様)活性、及びGly−Phe、Gly−Ala、Phe−、Tyr−又はGly−Gly−Leu基質による極めて不安定な活性。後者の活性は、生存する無処置細胞においてのみ測定できた。さらに重要なことは、これらの活性を検出するためにいくつかの蛍光体又はプロルミネセンス標識が使用でき、最終的には下流の多重化適応性の増大が考慮に入れられることである。
【0127】
【表4】
無しは、対照集団を超える統計的活性が無いことを示す。
(+)から(+++++)は、対照集団を超える多少の活性から強固な活性範囲を示す。
【0128】
G.プロテアーゼ保持活性及び生存細胞要件
ジャーカット細胞を、50μl容量で1ウェル当り20,000個の密度で白壁仕切りの透明底の96ウェルプレートに接種した。アポトーシス誘導試剤rTRAIL(BioMol)の連続希釈液を、RPMI1640+10%FBSで500ng/mlから作製し、50μl容量で反復して細胞に加えた。50μl培地の添加は、媒体対照として寄与した。このプレートを、5%CO2と共に37℃で4時間温置した。Gly−Phe−AFCを、RPMI1640中1mMに希釈し、10μ容量で全てのウェルに加えた。次にプレートをMeCour’循環熱ブロックに30分間置いてから、CytoFluor IIによりEx.405Em.530で蛍光を測定した。次に、等しい容量のCellTiter−Glo(商標)試薬をウェルに加え、細胞内に残存するATP含量を、FLUOstar Optimaによるルミネセンス測定により調べた。
【0129】
相対的ATP濃度及びプロテアーゼ保持活性は、事実上多重化でき、最適の保持酵素活性のための細胞生存度又は安定な細胞膜に関する要件を示唆している(図18)。膜完全性の乱れは、保持活性に対して非常に有害であることから、この活性を、集団生存度検出用の「生/死」フォーマットに関する放出活性と結合させることができる。
【0130】
H.種々のペプチド配列及びレポーター類を用いるプロテアーゼ保持アッセイフォーマット
ダブリング活性のジャーカット細胞を、100μl容量で白壁仕切りの透明底の96ウェルプレート内で、RPMI1640+10%FBS中37,500細胞/ウェルから連続希釈した。プレートの半分の細胞を、トリトンXの添加により最終0.2%に細胞溶解した。プレートウェルの他の半分は、適性容量(5μl)の水媒体を受けた。Tyr−AMC、Phe−AMC及びGly−Phe−AFCの全てを、DMSO中100mMに再水和化してから、RPMI1640で1mMに希釈した。希釈基質の1/10容量をウェルに加え、軌道振とうにより短時間混合してから、5%CO2中37℃で1.5時間まで温置した。生じた蛍光を、30分と90分でEx.360 Em.460及びEx.405、Em.500で測定した。
【0131】
生細胞と死細胞とを区別するのに良好に作用するペプチド配列(生細胞によって標的にされ利用される)は、生存細胞の細胞質に恐らく自由に入ることができる非ブロック化モノ−ペプチド基質又はバイ−ペプチド基質のいずれかである(図19)。候補基質の(Gly−Phe)2−R110は、明らかに細胞膜を有効に通過できないか、或いは候補プロテアーゼにより有効に開裂されることはなかった。
【0132】
I.プロテアーゼ放出活性の半減期
ジャーカット細胞を、100μl容量で1ウェル当り20,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種した。サポニン(Sigma)を加え、短時間軌道振とうすることにより最終0.2%濃度(5μl添加)に混合し、8時間の時間経過にわたって毎時間ウェルを反復させた。この同じ時間枠内で等しい容量の10%FBSを有するRPMI1640を対照ウェルに加えた。Ala−Ala−Phe−AMCを、RPMI1640+10%FBSで500μMに希釈し、ウェルに10μl容量で加え、軌道振とうにより短時間混合してから、5%CO2中37℃で1時間温置した。生じた蛍光を、CytoFluor IIで測定した。
【0133】
放出されたプロテアーゼ活性の半減期は、活性減衰を外挿すると10時間近い(図20)。細胞培養溶解物におけるこの延長された活性は、好ましいことにおよそ9時間の推定半減期を有するラクテートデヒドロゲナーゼ(LDH)と同等である。この観察は、処理集団における細胞死に対する代用物であるプロテアーゼ活性に関して重要である。簡単に言うと、シグナル半減期が延長すると、典型的なインビトロプロトコルにおける細胞死の報告におけるアッセイが有する有用性が増加する(活性を減少させて応答を過小評価することなく)。
【0134】
J.プロテアーゼ保持及び放出活性阻害/増強プロフィル
プロマイシン、E−64、フェニルメタンスルホニルフルオリド(PMSF)、アデノシン−5’−トリホスフェート(ATP)、N−(α−ラムノピラノシルオキシヒドロキシホスフィニル)−Leu−Trp二ナトリウム塩(ホスホラミドン)、N−[(2S,3R)−3−アミノ−2−ヒドロキシ−4−フェニルブチリル−L−ロイシン塩酸塩(ベスタチン)、1,10−フェナントロリン、3,4−ジイソクマリン、4−(2−アミノエチル)ベンゼンスルホニルフルオリド(AEBSF)、1,4−ジチオ−DL−トレイトール(DTT)、エデテート二ナトリウム二水和物(EDTA)、イソバレリル−L−バリル−L−バリル−[(3S,4S)−4−アミノ−3−ヒドロキシ−6−メチルヘプタノイル]−L−アラニル[93S,4S]−4−アミノ−3−ヒドロキシ−6−メチルヘプタン酸(ペプスタチンA)、塩化ナトリウム、アプロチニン、N−アセチル−L−ロイシル−L−ロイシル−L−アルギニナルヘミサルフェート塩(ロイペプチン)は全てSigmaから購入した。阻害剤は、溶解物又は生存細胞集団のいずれかに添加するために、Mg++又はCa++(DPBS)の無いダルベッコ燐酸緩衝生理食塩水中、200μM又は200μg/mlの高標的濃度の保存濃度を変えてDMSO中に再懸濁した。DTT、NaCl、EDTA、及びATPもまた、DPBS中に希釈した。全ての化合物を、損傷細胞又は生存細胞と共に37℃で少なくとも30分間(大部分は60分間)温置してから活性を評価した。
【0135】
プロテアーゼ保持アッセイ阻害剤/補助剤の調査は、先に記載されたように、最終100μM濃度でGly−Phe−AFCを用いて、超音波処理、サポニン溶解、又は生存HL−60及び/又はU937に対して実施された。このプロテアーゼ放出アッセイ阻害剤/補助剤の調査は、Ala−Ala−Phe−AMC又は(Ala−Ala−Phe)2−ローダミン110を用いて、超音波処理、サポニン溶解、又は生存HL−60、SK−MEL−28及び/又はU937に対して実施された。
【0136】
種々のクラスの阻害剤又は補助剤の存在下でのプロテアーゼ保持活性プロフィルは、観察された活性の大部分は、アミノペプチダーゼに関するものであることを示している(プロマイシン、EDTA、及びベスタチン感度)(表5及び表6を参照)。この活性は、ATP非依存性並びにDTT非依存性(活性の回復が無い)であり、並びにハライド類(Cl−)に非感受性である。この活性は、システイン又はセリンプロテアーゼクラスの酵素に関連しているように思われる。
【0137】
プロテアーゼ放出活性プロフィルは、トリブシン又はキモトリプシン様活性に選択性を有するものではないが、セリンプロテアーゼ阻害剤に感受性があるように思われる。チオール類(システインクラスに強く示される)にとっての明白な要件はなく、アスパラギン酸及びメタロプロテアーゼの特異的阻害剤は、活性を制御するのに無効である。
【0138】
プロテアーゼ保持活性の原因となる酵素は、生存細胞を必要とし、損傷細胞の外部では検出できない。逆に、プロテアーゼ放出応答を増強させるために混合された補助剤は必要ない。これにより、非毒性、非細胞溶解性フォーマットにおけるアッセイを組み合わせて、それらプロテアーゼ活性の差異に基づいた生細胞と死細胞の検出が考慮に入れられる。
【0139】
【表5】
【0140】
【表6】
【0141】
K.多重プロテアーゼ放出及び保持アッセイ
1.ジャーカット用量応答
ダブリング活性のジャーカット細胞を、50μl容量で1ウェル当り20,000個の密度で96ウェルプレート内に接種した。RPMIアポトーシス誘導リガンド、すなわち、1640中のrTRAILの連続希釈液を、追加の50μl容量中、250ngから244pg/mlの最終濃度で反復ウェルに加えた。RPMIのみのものは、非誘導対照として寄与した。このプレートを、5%CO2中37℃で4時間温置した。Gly−Phe−AFC及びAla−Ala−Phe−AMCを、RPMI中1mMに同時に希釈し、1/10容量でプレートに加え、さらに37℃で30分間温置した。生じた蛍光を、CytoFluor IIを用いてEx 360 Em 460及びEx 405 Em 530で測定した。蛍光測定が完了したら、CellTiter−Glo(登録商標)を等しい添加でウェルに加え、ルミネセンスを、BMG FLUOstar Optimaを用いて測定した。
【0142】
健全細胞の2種の独立した非破壊代用物(プロテアーゼ放出と保持)を、マイクロ−タイタープレートフォーマット内の集団生存度を報告するために多重化した(図21)。生じたデータは、その細胞集団の健全度の逆の目安である。この関係により、対照の使用が考慮に入れられ、正規化レベルが提供される。さらに、生存度の第3の目安(ATP含量)を、妨害又は消光の無い連続多重フォーマット内に加えることができ、データの解釈におけるさらなる信頼が考慮に入れられる。
【0143】
2.SK−MEL−28及びACHN細胞
SK−MEL−28又はACHN細胞を、100μl容量で1ウェル当り10,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種し、5%CO2中37℃で2時間結合させた。結合後、50μlの培地を注意深く除き、MEM+10%FBS中、イオノマイシン又はスタウロスポリンのいずれかの連続希釈で置き換えた。培地のみのものは、対照として寄与した。このプレートを、さらに5時間温置した。Gly−Phe−AFCの1mM溶液をMEM中で作製し、1/10容量でウェルに加えた。生じた蛍光を、CytoFluor IIを用いて測定した。次にCaspase−Glo(商標)3/7試薬を加えて、BMG FLUOstar Optimaを用いて蛍光測定を行った。
【0144】
プロテアーゼ保持基質は、ウェル内の一般的生存度を報告したが、一方、カスパーゼ特異的試薬は、細胞毒の特異的経路を報告した(図22)。この点において、カスパーゼ活性化(したがってアポトーシス誘導)は、SK−MEL−28に対するスタウロスポリンによって明白であるが、一方、イオノマイシンは、壊死タイプのプロフィル開始する。アポトーシスプロフィルはまた、スタウロスポリン処理ACHNによっても観察される。
【0145】
3.HeLa細胞及びタモキシフェン処理
HeLa細胞を、100μl容量で1ウェル当り10,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種し、5%CO2中37℃で2時間結合させた。結合後、0時間、1時間、3時間、5時間、7時間、24時間の曝露時間で50μlの培地を注意深く除き、MEM+10%FBS中、50μMのタモキシフェンで置き換えた。培地のみのものは、対照として寄与した。プロテアーゼ保持及び放出試薬を、2mlの10mM Hepes、pH7.5を有するルシフェリン検出試薬ケークを再水和化することにより調製した。次にこの溶液を、Ala−Ala−Phe−アミノルシフェリン及びGly−Phe−AFC双方により500μMを作製した。この溶液の1/5容量を全てのウェルに加え、Me’Cour熱ユニット内で37℃で15分間温置した。ルミネセンスは、BMG FLUOstar Optimaにより測定し、蛍光は、CytoFluor IIを用いて測定した。
【0146】
本実施例は、混合プラットフォーム(蛍光及びルネセンス)が、プロテアーゼの保持及び放出アッセイの構成において可能であることを立証している(図23)。これらの試薬は、非細胞溶解性で明らかに非毒性であることが顕著であり、それらは、Apo−ONE(商標)アッセイによるカスパーゼ−3/7検出などのスペクトル的に異なる他の下流適用を受けることができることを示唆している。
【0147】
4.DNA染色による生/死プロテアーゼアッセイの利用
HeLa又はHepG2細胞を、100μl容量で1ウェル当り10,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種し、5%CO2中37℃で2時間結合させた。結合後、50μlの培地を注意深く除き、MEM+10%FBS中、タモキシフェン又はイオノマイシンの連続希釈で置き換えた。培地のみのものは、対照として寄与した。化合物との温置を、さらに5時間続けた。プロテアーゼ保持及び放出試薬は、2mlの10mM Hepes、pH7.5を有するルシフェリン検出試薬ケーク(Promega V859A)を再水和化することにより調製した。次にこの溶液を、Ala−Ala−Phe−アミノルシフェリン及びGly−Phe−AFC双方により500μMを作製した。この溶液の1/5容量を全てのウェルに加え、Me’Cour熱ユニット内で37℃で15分間温置した。ルミネセンスは、BMG FLUOstar Optimaにより測定し、蛍光は、CytoFluor IIを用いて測定した。次に残りの生存細胞を、0.4% NP−9界面活性剤の添加により溶解した。軌道振とう機上で短時間混合後、MEM中、PicoGreen(登録商標)(Molecular Probes)の1:20希釈液を、さらに1/10容量で加えた。DNA/色素結合に伴う蛍光は、CytoFluor IIを用いてEx.485 Em.530で測定した。
【0148】
本実験は、さらに2つの接着細胞タイプのスクリーニングができるようにプロテアーゼベースの生存度試験の有用性を拡張するのみならず、DNA染色による「全体」測定を組み込んでいる(図24)。スペクトルの相違及び混合プラットフォームの読取りのため、全ての測定は、非妨害的であり、また、非消光である。
【0149】
考察
益々精巧な細胞モデル系を利用するために、薬物発見及び一次研究双方の努力が継続されている。実験操作後にこれらのインビトロ系における細胞数及び細胞生存度を測定することが絶対に必要であることは、十分に認識されている。この要件は、測定の信頼性を証明するために、又複雑な生物系の背景内でこれらの応答を正規化するために必要である。
【0150】
不幸にして、細胞生存度並びに細胞毒を定義する現代化学は、生物学的疑問の新規な方法論及び技法と歩調を合わせておらず、したがって実験的オプションを限定してきた。例えば、アッセイの多重化、すなわち同じウェル内での組合せアッセイの出現により、アッセイ性能における有意な低減の無い適合性且つスペクトル的に異なるアッセイを組み合わせるための要件が必要となった。この課題は、健全細胞の一般的な補足測定とカスパーゼ活性化又はレポーター遺伝子モジュレーションなどのより具体的な事象とを結合させる点で特に重要である。
【0151】
細胞生存度及び/又は細胞毒レポーターの測定のための前述の方法論は、多くの下流アッセイ適用に適合性である。このことは、多岐にわたる励起及び発光スペクトルをもつ異なる蛍光体により、又はルミネセンスなどの他のレポータープラットフォームを組み込むことにより達成される。これは、細胞溶解なしで恐らく非毒性環境で達成され、エンドポイント判定のためにアッセイウィンドウにおける適合性を考慮に入れられることは注目すべきである。さらに、この技法は、処理能力、小型化及び自動化に適応させる上で十分高感度であり、コスト効率が良い。種々のアッセイにより提供された利点の比較を、表7に示す。
【0152】
【表7】
【0153】
現在までの結論として、哺乳動物プロテアーゼの試験において公表された努力の残りのものは、主として容易に精製されるか、分泌されるか、又は双方であるものを中心に行われてきた。これらの試験から提供された情報は、タンパク分解機構、構造と機能についての洞察を提供しているが、他のプロテアーゼ類については、プロテオミック予測から推測されているもの以外、ほとんど知られていない。簡単に述べると、細胞内プロテアーゼの機能、調節、細胞下分布、存在量、及び重要性に対して多くの研究を実施する必要がある。
【0154】
益々増加している証拠は、多くの細胞基質プロテアーゼ類が、細胞ホメオスタシスの機構に関与していることを示唆している。プロテオソーム類は、細胞質ペプチド類の遊離に関与していることは明白だが、いくつかの知見は、他の保存細胞質プロテアーゼ類に関する役割を示唆している(Vititskyら、1997年;Constamら、1995年)。
【0155】
本明細書に記載された個々のプロテアーゼアッセイ及びプロテアーゼベースの生細胞/死細胞アッセイは、スペクトルの相違性のため多重化に、より適合性であり、アッセイ相補性又は他のエンドポイントアッセイの組合せ、例えば、AMC、AFC、R110、クレシルバイオレット又はルミネセンスの組合せが考慮に入れられ、染色消滅が無く、容量制限が無く、アッセイ化学のレトロエンジニアリングが無く、温置時間が短く、同様な、又はより良好な実用的感度(スクリーニング環境の細胞生存度におけるパーセント変化)であり、DNA結合アッセイによる下流妨害が無く、洗浄又は遠心分離の必要が無く、例えば均一アッセイである。さらに、プロテアーゼベースの生細胞/死細胞アッセイのデータは、比率(細胞毒指標)で用いられる場合、細胞数に係わりなく正規化でき、例えば、DNAインターカレーション(一次的又は二次的壊死の潜在的結果が同定できる)などの他のアッセイと結合される場合、化合物作用の動態における差異の理由を説明するために、細胞/化合物接触ウィンドウを拡張でき、DNAインターカレーション及びカスパーゼ活性により、例えば、ウェル内の不均一集団において細胞サイクルの薬物応答を確認できる。さらに、プロテアーゼ類の基質は、比較的単純な、例えば、ジ又はトリ−ペプチド類であり得、よく知られた化学により、非毒性及び/又は非変異原性で安定な蛍光体又はルミネセンス基質に結合され、種々のフォーマットにおいて、例えば、DMSO中、又は乾燥して提供できる。
(参考文献)
【表8】
【0156】
全ての刊行物、特許及び特許出願は、参照として本明細書に組み込まれている。前述の明細書において、本発明は、ある一定の好ましいその実施形態に関連して記載され、多くの詳細が、説明目的のために記載されているが、本発明は、さらなる実施形態の影響を受け易く、本明細書に記載されたある一定の詳細は、本発明の基本的原則から逸脱することなく、かなり変え得るものであることは当業者にとって明白であろう。
【技術分野】
【0001】
本出願は、参照として本明細書に組み込まれる、2004年1月22日出願の米国特許出願番号第10/762,836号の出願日の利益を主張する。
【背景技術】
【0002】
ルミネセンスは、ルシフェラーゼに媒介された酸化反応の結果として、ある一定の生物において産生される。極めて多種多様の種のルシフェラーゼ遺伝子、特に、フォティヌス・ピラリス(Photinus pyralis)及びフォツリス・ペンシルバニカ(Photuris pennsylvanica)(北米の蛍)、ピロフォルス・プラギオフタラムス(Pyrophorus plagiophthalamus)(ジャマイカのコメツキムシ)、レニラ・レニフォルミス(Renilla reniformis)(シーパンジー(sea pansy))、及びいくつかの細菌(例えば、キセノルハブデュス・ルミネセンス(Xenorhabdus luminescens)及びビブリオ(Vibrio)種)のルシフェラーゼ遺伝子は、極めて一般的なルミネセンスレポーター遺伝子である。蛍のルシフェラーゼはまた、ATP濃度を測定するための一般的なレポーターであり、その役割で、バイオマスの検出に広く用いられている。他の酵素によっても、それらを一定の合成基質、例えば、アルカリホスファターゼ及びアダマンチルジオキセタンホスフェート、又はセイヨウワサビペルオキシダーゼ及びルミノールと混合すると、ルミネセンスが産生される。
【0003】
ルシフェラーゼ遺伝子は、その非放射性、感度、及びルミネセンスアッセイの極めて直線的範囲のため、遺伝子レポーターとして広く用いられている。例えば、わずか10−20モルの蛍ルシフェラーゼを検出することができる。その結果、遺伝子活性のルシフェラーゼアッセイは、原核細胞及び真核細胞双方の培養、遺伝子組換えの植物及び動物、及び無細胞発現系など、事実上あらゆる生物学的実験システムに用いられている。同様に、ATP濃度測定に用いられるルシフェラーゼアッセイは、高感度であり、10−16モル以下の検出が可能である。
【0004】
ルシフェラーゼは、酵素特異的な基質、例えば、ルシフェリン類の酸化によって光を発生させることができる。蛍ルシフェラーゼ及び他の全てのカブトムシルシフェラーゼでは、発光は、マグネシウムイオン、酸素、及びATPの存在下で生じる。ウミシイタケ(Renilla)ルシフェラーゼなどの花虫類動物のルシフェラーゼでは、基質のコエレントラジンと共に、酸素のみを必要とする。一般に、遺伝子活性を測定するためのルミネセンスアッセイにおいて、反応基質及び他のルミネセンス活性化試薬が、レポーター酵素を発現すると推測される生物系に導入される。次いで、ルミネセンスが生じた場合は、それを、ルミノメータ又は任意の好適な放射エネルギー測定装置を用いて測定する。該アッセイは、極めて迅速で高感度であり、放射性試薬を必要とせずに、迅速に容易に遺伝子発現データを提供する。
【0005】
ルシフェラーゼ類、例えば、蛍ルシフェラーゼ、ウミシイタケ(Renilla)ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、ベータガラクトシダーゼ(lacZ)、ベータグルクロニダーゼ(GUS)及び分泌アルカリホスファターゼ(SEAP)などの種々のホスファターゼ類、並びに遺伝子活性のコレポーターとして組み合わせて用いられてきたウテロフェリン(Uf:酸ホスファターゼ)は、多数のレポーターの1つである。二重酵素レポーター系は、単一系内の2種の個別のレポーター酵素の使用、発現、及び測定に関する。遺伝子レポーティングにおいて、二重レポーターアッセイは、2種の異なるレポーター遺伝子を同時に発現するように遺伝子操作される個体の細胞又は細胞集団(培養物中に分散された細胞、分離組織、又は動物全体など)におけるアッセイにとって特に有用である。多くの場合、1つの遺伝子活性が、特定の実験条件の影響をレポートする一方、第2のレポーター遺伝子の活性は、全ての組の実験値を正規化できる内部対照を提供する。二重酵素レポーター法は、実験レポーター酵素及び対照レポーター酵素をコードする独立した遺伝子材料の同時翻訳、又は結合転写及び結合翻訳のために誘導された細胞溶解物などの無細胞再構成系と共に使用することもできる。同様に、単一サンプル内の実験値及び対照値双方の二重レポーティングのために、免疫アッセイをデザインできる。
【0006】
二重酵素レポーターアッセイの実施はいずれも、構成要素である酵素の化学の特性及びそれらの各々で生じたデータセットに関連する能力に基づく。種々のレポーター酵素の酵素動態、アッセイ化学及び温置要件の不同性により、2つのレポーター酵素を組み合わせて単一の管又はウェルの二重レポーターアッセイフォーマットに組み込むことは複雑になり得る。二重レポーターアッセイ組み込みへの1つの方法は、特許文献1に記載されており、ここでは、カブトムシ及びウミシイタケ(Renilla)ルシフェラーゼアッセイ用の特定の一般的又は特異的消光剤が開示され、蛍ルシフェラーゼのルミネセンス、次いでウミシイタケ(Renilla)ルシフェラーゼのルミネセンスを連続的に判定するための典型的な二重レポーターアッセイが示されている。同様に、特許文献2には、いくつかの二重レポーターアッセイ系が開示されている。特許文献1に開示された二重レポーター系のように、ルミネセンスは、特許文献2における2つの反応の各々の測定可能な生成物である。異なった基質だけでなく、異なった検出法を組み込むレポーターアッセイの二重化法は、非特許文献1及び非特許文献2に記載されている。例えば、Liuらは、同一生物中で、ルシフェラーゼ活性及びGFP活性を報告しており、ここで、酵素活性は、段階的方法で、ルミネセンス検出及び蛍光検出によって判定される(非特許文献1)。
【0007】
また、レポーター類は、細胞内又は上澄み液内の分子の存在又は活性を検出するためにも有用である。例えば、プロテアーゼは、血液凝固、炎症、再生、線維素溶解、及び免疫応答におけるタンパク質のターンオーバーなどの多様な生理学的過程に関与する重要な酵素の大群を構成している。多くの疾患状態は、特定のプロテアーゼ類及びそれらの阻害剤の活性変化によって引き起こされるか、又はそれを特徴とする。研究において、又は臨床の場において、これらのプロテアーゼを測定する能力は、疾患状態の研究、治療及び管理にとって重要である。例えば、カスパーゼ−3及びカスパーゼ−7は、システインアスパルチル特異的プロテアーゼ(アスパルテート特異的システインプロテアーゼ、「ASCP」としても知られている)ファミリーのメンバーであり、哺乳動物細胞の細胞死において主要エフェクターの役割を演じている(非特許文献3、非特許文献4、非特許文献5及び非特許文献6)。
【0008】
しかし、プロテアーゼを、それらの天然基質を用いてアッセイすることは容易ではない。さらに、現在利用可能な多くの合成基質は、高価で感度が低く、非選択的である。
【0009】
プロテアーゼを測定するために、多数の色素産生及び蛍光産生基質が用いられてきており(非特許文献7及び非特許文献8)、修飾ルシフェリン類が蛍光指示薬の代替物として提供されている(特許文献3及び特許文献4)。代理基質として、ヒドロラーゼに対する認識部位を有する修飾ルシフェリン類を用いる方法は、Miska及びGeigerによって初めて記載され(非特許文献9)、この方法では、修飾ルシフェリンを、一定の期間ヒドロラーゼと共に温置し、次いで、該混合物のアリコートをルシフェラーゼを含有する溶液に移し入れることによって、不均一アッセイが行われた。Masuda−Nishimuraらは、β−ガラクトシダーゼ基質修飾ルシフェリンを用いた単一管(均一)アッセイの使用を報告している(非特許文献10)。
【0010】
酵素反応の蛍光若しくはルミネセンスの基質又は生成物が、タンパク質アッセイの多重化に用いられている。例えば、2つ以上の異なったサイトカインを検出するために、15個までの異なったサイトカインに対するリガンドを有する蛍光ビーズが用いられ(非特許文献11)、アルカリホスファターゼ及び一定のプロテアーゼを検出するために、蛍光ジホスフェート及びカゼインBODIPY−FLが用いられた(非特許文献12)。
【0011】
しかし、異なった検出方法を用いて2種以上のタンパク質を検出するために必要なのは、改善されたアッセイ、例えば、均一アッセイである。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】米国特許出願第5,744,320号
【特許文献2】米国特許出願第6,586,196号
【特許文献3】米国特許出願第5,035,999号
【特許文献4】米国特許出願第5,098,828号
【非特許文献】
【0013】
【非特許文献1】Liu et al., Luminescence, 15: 45 (2000).
【非特許文献2】Oazi et al., Luminescence, 17:106 (2002).
【非特許文献3】Thronberry et al., Nature, 356:768 (1992).
【非特許文献4】Nicholson et al., Nature, 376: 37 (1995).
【非特許文献5】Tewari et al., Cell, 81: 801 (1995).
【非特許文献6】Fernandes-Alnemri et al. , PNAS USA, 93: 7464 (1996).
【非特許文献7】Monsees et al. , Anal. Biochem., 221 : 329 (1994).
【非特許文献8】Monsees et al. , J. Biolum. Chemilum., 10: 213 (1995).
【非特許文献9】Miska and Geiger, J. Clin. Chem. Clin. Biochem., 25: 23 (1989).
【非特許文献10】Masuda-Nishimura et al., Lett. Appl. Microbio., 30: 130 (2000).
【非特許文献11】DeJager et al., Clin. Diag. Lab. Immunol., 10: 133 (2003).
【非特許文献12】Nolkrantz et al. , Anal. Chem., 7: 4300 (2002).
【発明の概要】
【課題を解決するための手段】
【0014】
本発明は、サンプル中、ATPなどの酵素反応に関する補因子、分子に結合する及び/又は分子の立体配置を変化させるタンパク質(ペプチド又はポリペプチド)、例えば、ペプチド基質又はポリペプチド基質を修飾するか又は開裂させるタンパク質、又はタンパク質に結合するか又はタンパク質によって変化する分子などの1つ又は複数の部分の量(例えば、活性)又は存在を検出するための、ルミネセンス非発生、例えば、蛍光又は比色アッセイ、及びルミネセンス発生アッセイの、例えば、同一ウェル内での多重化を提供する。本明細書に用いられる「ルミネセンス発生アッセイ」は、第1の分子、例えば、第1の酵素に対するペプチド基質又はポリペプチド基質、第1の分子と適切な(第1の)タンパク質との間の反応の生成物、及び/又は別のタンパク質と第1の反応の生成物との間の反応生成物がルミネセンスを発生する反応を含む。したがって、ルミネセンス発生アッセイでは、反応の補因子、タンパク質に結合する、及び/又はタンパク質によって変化する分子、又はタンパク質、の量又は存在を、直接又は間接に検出、例えば測定できる。例えば、一実施形態において、ATP濃度を検出するために、ルミネセンス発生アッセイにおいて、カブトムシルシフェラーゼ及び適切なルシフェリン基質を使用でき、一方、他の実施形態において、プロテアーゼを検出するために、ルミネセンス発生アッセイにおいて、ルシフェラーゼが存在する場合は、プロテアーゼ認識部位を含有するように修飾されている(例えば、共有結合を介して修飾されている)ルシフェラーゼに対する基質を使用できる。ルミネセンス発生アッセイとしては、限定はしないが、ルシフェラーゼ、β−ガラクトシダーゼ、β−グルクロニダーゼ、β−ラクタマーゼ、プロテアーゼ、アルカリホスファターゼ、又はペルオキシダーゼを使用又は検出するもの、及び対応する好適な基質、例えば、ルシフェリンの修飾体、コエレンテラジン、ルミノール、ペプチド又はポリペプチド、ジオキセタン、ジオキセタノン及び関連アクリジニウムエステルを含む化学発光及び生物発光アッセイが挙げられる。本明細書に用いられる「ルミネセンス発生アッセイ試薬」は、基質、並びに補因子(1種又は複数種)又はタンパク質、例えば、ルミネセンス発生反応に関する酵素などの他の分子(1種又は複数種)を含む。一実施形態において、ルミネセンス発生アッセイ試薬は、Z−DEVD−アミノルシフェリン、Z−LETD−アミノルシフェリン、Z−LEHD−アミノルシフェリンであり得るか、又は他の基質、例えば、アミノルシフェリン、ジヒドロルシフェリン、ルシフェリン6’メチルエーテル、又はルシフェリン6’クロロエチルエーテルに結合したペプチド又はポリペプチド基質であり得る。ルミネセンス発生アッセイは、ルミネセンス発生反応が、対応するルミネセンス非発生アッセイよりも少なくとも1%、例えば、少なくとも10%、多くの光を発生するものである。
【0015】
「ルミネセンス非発生アッセイ」は、第1の分子、例えば、タンパク質(ペプチド又はポリペプチド)、第1の分子と好適な(第1の)タンパク質(ペプチド又はポリペプチド)との間の反応の(第1の)生成物、又は別のタンパク質と第1の生成物との間の反応生成物がルミネセンスを発生しないが、検出可能であり得る反応を含む。例えば、該基質及び/又は生成物(1つ又は複数)は、該反応の補因子、該分子又は該分子と相互作用するタンパク質の量又は存在を、直接又は間接に測定する蛍光アッセイ又は比色アッセイを用いて検出される。例えば、酵素が基質と反応し、蛍光体が一定の波長又は一定の波長の範囲の光に接触し(曝露され)た後だけに一定の波長の光を放出する蛍光体を含有するように、該酵素に対する該基質を修飾できる。例えば、(Z−DEVD)2−ローダミン−110は、カスパーゼに対する基質であり、カスパーゼによるその基質の開裂は、ローダミン−110の蛍光によってモニターできる。本明細書に用いられる「蛍光発生アッセイ試薬」は、蛍光発生反応のために、基質、並びに補因子(1種又は複数種)又は他の分子(1種又は複数種)、例えば、タンパク質を含む。ルミネセンス非発生アッセイは、ルミネセンス非発生反応が、対応するルミネセンス発生アッセイのルミネセンスシグナルの約10%未満、例えば、約1%未満又はそれより小さい値未満を発生するものである。
【0016】
一実施形態において、本発明のアッセイに用いられる分子、例えば、タンパク質と結合及び/又はタンパク質によって変化する分子としては、レポーター分子、すなわち、例えば、1つ又は複数のその後の反応の後に、検出可能又は検出能力のある分子を含有するように修飾されている分子が挙げられる。例えば、一実施形態において、本発明のルミネセンス発生アッセイに用いられる基質としては、検出される酵素に対する基質であって、ルミネセンス発生反応の基質に共有結合している基質が挙げられ、一方、他の実施形態において、蛍光発生アッセイに用いられる基質としては、検出される酵素に対する基質であって、1つ又は複数の蛍光体に共有結合している基質を挙げることができる。いくつかの実施形態において、タンパク質に結合している及び/又はタンパク質によって変化している分子はレポーター分子を含有しない。
【0017】
本明細書に記載されているように、サンプル中の2つ以上のプロテアーゼの量又は存在が、少なくとも2つの異なった基質を用いて検出されたが、その1つは、ルミネセンス読み取りを有し、その1つ又は複数は蛍光読み取りを有した。例えば、低濃度の細胞プロテアーゼの検出は、より高感度のルミネセンス法、例えば、基質、Z−LETD−アミノルシフェリンによるカスパーゼ−8の検出、引き続いて、他の基質を用いた他のプロテアーゼの検出、例えば、(Z−DEVD)2−ローダミン−110によるカスパーゼ−3の検出を用いて達成された。このように、このアッセイは、蛍光発生試薬及びルシフェラーゼに媒介されたルミネセンス反応の感度双方の強力を組み合わせている。さらに、驚くべきことに、蛍光特性を有し、ルミネセンスアッセイにしばしば比較的大量に存在する分子、ルシフェリンの存在によって、蛍光/ルミネセンス組合せアッセイにおいて、有意な妨害は生じなかった。さらに、驚くべきことに、2種のカスパーゼ及びルシフェラーゼは、カスパーゼ−8の基質、(Z−LETD−アミノルシフェリン)、並びにカスパーゼ−3の2種の基質、(Z−DEVD)2−ローダミン−110及びAc−DEVD−AMCを含む同一反応混合物中で検出された。このように、本発明は、多重アッセイに使用される分子、例えば、蛍光発生アッセイの基質と組み合わせるルミネセンス発生アッセイの基質において、より多くの適応性を提供する。さらに、2つの酵素媒介反応が適合的な試薬条件を有すれば、該アッセイは一段階アッセイであり得る。
【0018】
したがって、本発明のルミネセンス発生/ルミネセンス非発生の組合せアッセイフォーマットにより、1種又は複数のペプチド又はポリペプチド、例えば、酵素、ペプチド(1種又は複数種)又はポリペプチド(1種又は複数種)に結合している、及び/又はそれらによって変化している1種又は複数の分子、例えば、各酵素に対するペプチド基質又はポリペプチド基質、及び/又は各アッセイに関する1種又は複数の補因子、又はそれらの組合せに関するアッセイの多重化が可能になる。したがって、一実施形態において、本発明は、第1の酵素媒介反応に関する第1の分子の存在又は量及び第2の酵素媒介反応に関する第2の分子の存在又は量を検出する方法を提供する。該方法は、第1及び/又は第2の分子を有すると推測されるサンプルを、第1及び/又は第2の分子を欠いている第1及び第2の酵素媒介反応に関する反応混合物と接触させることを含む。次いで、第1及び第2の分子の存在又は量が検出される。ルミネセンスアッセイを含む多重化を用いることによって、ルミネセンスアッセイを用いて検出される分子に対する感度の増加が提供される。一実施形態において、第1の酵素に媒介された反応によりルミネセンス発生生成物が得られ、一方、第2の酵素に媒介された反応によりルミネセンス非発生生成物が得られる。一実施形態において、アッセイの1つが内部対照を提供するものを含むルミネセンス発生/蛍光発生の組合せアッセイが提供される。本明細書に記載されたアッセイは、レポーターアッセイ、核酸ベースアッセイ又は免疫学ベースアッセイ及び他の非関連酵素アッセイなどの他のアッセイと共に使用できる。
【0019】
また、本発明は、サンプル中の少なくとも1種の分子の活性又は存在を測定する方法を提供する。該方法は、酵素媒介反応に関する少なくとも1種の分子を含有し得るサンプル、例えば、該酵素を含有し得るサンプルを提供し、該サンプルを、該酵素媒介反応に関する該分子を欠いている反応混合物、例えば、該酵素に対する基質を含有する反応混合物と接触させて該分子の存在又は量がルミネセンス発生アッセイによって検出できる反応混合物を得ることを含む。一実施形態において、該サンプル及び/又は反応混合物はまた、第2の酵素媒介反応に関する分子の存在又は量がルミネセンス非発生アッセイによって検出できる第2の酵素媒介反応に関する分子を検出する試薬と接触させる。
【0020】
一実施形態において、本発明は、サンプル中の第1の酵素によって媒介された反応に関するその第1の酵素及び/又は補因子の存在又は量を検出する方法を提供する。該方法は、該サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、反応混合物を得ることを含む。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。例えば、それらは同じ基質を実質的に認識しない。すなわち、それらは同じ基質に結合しないか、又はそれらが同じ基質に結合し反応するとしても、該酵素の一方は、他方の酵素と同じ程度(効率)で基質と反応しない。すなわち、双方の酵素に対する基質が存在する場合、該酵素の一方は、他方の酵素に対する基質と実質的に反応しない。本明細書に用いられる第2の酵素に対する基質と実質的に反応しない酵素(第1の酵素)としては、第2の酵素並びに第1の酵素に対する基質及び第2の酵素に対する基質を等しい量で有する反応液中で、第1の酵素と第1の酵素に対する基質との間の反応に比較して、第2の酵素に対する基質との交差反応が、25%以下、例えば、15%以下、10%以下又は5%以下の酵素が挙げられる。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との生成物との間の反応生成物はルミネセンスを発生する。第2の基質、第2の基質と第2の酵素との間の(第2の)反応生成物、及び/又は他の酵素と第2の生成物との間の反応生成物はルミネセンスを発生しないが、検出可能である。第1の酵素及び/又は補因子の存在又は量が検出又は測定される。一実施形態において、第2の酵素に媒介される反応に関する第2の酵素及び/又は補因子の存在又は量もまた、検出又は測定される。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。検出される酵素は、天然酵素又は組換え酵素、例えば、融合タンパク質などであり得る。同様に、該サンプルに加えられる最適な酵素(1種又は複数種)は、天然酵素又は組換え酵素であり得る。
【0021】
他の実施形態において、本発明は、サンプル中の第1の酵素及び/又はその第1の酵素に媒介される反応に関する補因子の存在又は量を検出する方法を提供する。該方法は、該サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、場合によっては、少なくとも第1の酵素と第2の酵素とが同じではない反応混合物を得ることを含む。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との間の反応生成物との間の反応生成物はルミネセンスを発生しないが、検出可能である。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は他の酵素と第2の生成物との間の反応生成物はルミネセンスを発生する。第1の酵素及び/又は補因子の存在又は量が検出又は測定される。一実施形態において、第2の酵素の存在又は量もまた、検出又は測定される。反応混合物に検出又は使用される酵素は、天然酵素又は組換え酵素であり得る。
【0022】
さらに、第1の酵素に媒介される反応に関する該酵素又は補因子を検出するために、該酵素に媒介されるルミネセンス反応をアッセイする方法が提供される。該方法は、サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、反応混合物を得ることを含み、ここでの第1の酵素と第2の酵素とは同じではない。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との生成物との間の生成物はルミネセンスを発生する。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は第2の生成物と他の酵素との間の反応生成物はルミネセンスを発生しないが、検出可能である。次いで、ルミネセンスが検出される。該方法はさらに、第2の酵素の存在又は量を、例えば、ルミネセンス非発生の基質又は生成物(1つ又は複数)の存在又は量を検出することによって検出することを含む。一実施形態において、第2の酵素は第1の基質と結合又は反応しないが、他の実施形態においては、第1の酵素は第2の基質と結合又は反応しない。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。該反応混合物中に検出又は使用される酵素は、天然酵素又は組換え酵素であり得る。
【0023】
また、第1の酵素に媒介される反応に関する該酵素又は補因子を検出するために、該酵素に媒介されるルミネセンス反応をアッセイする方法が提供される。該方法は、サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び第3の酵素に接触させて、反応混合物を得ることを含む。第1の基質、第1の基質と第1の酵素との間の反応生成物、及び/又は第3の酵素と、第1の酵素と第1の基質との生成物との生成物はルミネセンスを発生しないが、検出可能である。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は第2の生成物と他の酵素との間の反応生成物はルミネセンスを発生する。次いで、ルミネセンスが検出される。該方法はさらに、第1の酵素又は第1の酵素と第1の基質との生成物の存在又は量を検出することを含み得る。一実施形態において、第2の酵素は第1の基質と実質的に結合又は反応しないが、他の実施形態においては、第1の酵素は第2の基質と実質的に結合又は反応しない。一実施形態において、少なくとも第1の酵素と第2の酵素は同じではない。該反応混合物中に検出又は使用される酵素は、天然酵素又は組換え酵素、例えば融合タンパク質などであり得る。
【0024】
さらに、サンプル中の少なくとも2種の分子の存在又は量を検出するための方法が提供される。該方法は、サンプルを、第1の酵素に対する第1の基質、第2の酵素に対する第2の基質、及び場合によっては、第3の酵素に接触させて、反応混合物を得ることを含み、ここで、少なくとも第1の酵素と第2の酵素とは同じではない。第1の酵素と第1の基質との間の反応、又は第3の酵素と、第1の基質と第1の酵素との間の反応生成物との反応により、ルミネセンスを発生する生成物が得られる。第2の基質、第2の基質と第2の酵素との間の第2の反応生成物、及び/又は第2の生成物と他の酵素との間の反応生成物はルミネセンスを発生しない。次いで、第1の酵素及び第2の酵素及び/又は補因子(1つ又は複数)の存在又は量が検出される。一実施形態において、第1の酵素及び/又は補因子を検出するためにルミネセンスが用いられ、他の少なくとも1種の酵素及び/又は補因子を検出するために蛍光又は比色が用いられる。一実施形態において、2つの異なる酵素に対する基質がサンプルに同時に組み合わされて反応混合物が得られる。該基質の1種と該酵素の1種との間の反応は直接又は間接にルミネセンスシグナルを発生し、一方、他の基質と酵素との間の反応は、直接又は間接に蛍光シグナルを発生する。温置期間後、1種の酵素及び/又は補因子の存在又は量を検出するために蛍光シグナルが用いられ、他の酵素及び/又は補因子の存在又は量を検出するためにルミネセンスシグナルが用いられる。特定の緩衝液条件は、検出中の酵素及び/又は補因子によって変化し得、インビトロアッセイ、例えば、酵素アッセイの業者により決定できる。或いは、該アッセイは、第1のアッセイと第2のアッセイとの間で試薬調整をした2段階アッセイであり得る。例えば、試薬調整としては、第1の反応液に対する消光剤、及び/又は第2の反応液に対する増強剤の添加を挙げることができる。
【0025】
一実施形態において、第1の酵素に媒介された反応に関する第1の酵素又は補因子、並びに第2の酵素に媒介された反応に関する第2の酵素又は補因子を検出するために、サンプルを、第1の基質及び第2の基質に同時に接触させる。他の実施形態において、サンプルを、第1の基質の前に第2の基質に接触させるか、又は、第2の基質の前に第1の基質に接触させる。一実施形態において、第3の又は異なる酵素を、1種又は複数の基質と共に、1種又は複数の基質の前に、又は1種又は複数の基質の後に添加できる。
【0026】
一実施形態において、第1の酵素に媒介された反応に関する第1の酵素又は補因子、並びに第2の酵素に媒介された反応に関する第2の酵素又は補因子を検出するために、サンプルを、第1の酵素及び第2の酵素に同時に接触させる。他の実施形態において、サンプルを、第1の酵素の前に第2の酵素に接触させるか、又は、第2の酵素の前に第1の酵素に接触させる。
【0027】
一実施形態において、第1の酵素に媒介された反応に関する第1の酵素又は補因子、並びに第2の酵素に媒介された反応に関する第2の基質又は補因子を検出するために、サンプルを、第1の基質及び第2の酵素に同時に接触させる。他の実施形態において、サンプルを、第1の基質の前に第2の酵素に接触させるか、又は、第2の酵素の前に第1の基質に接触させる。一実施形態において、第1の酵素に媒介された反応に関する第1の基質又は補因子、並びに第2の酵素に媒介された反応に関する第2の酵素又は補因子を検出するために、サンプルを、第1の酵素及び第2の基質に同時に接触させる。他の実施形態において、サンプルを、第1の酵素の前に第2の基質に接触させるか、又は、第2の基質の前に第1の酵素に接触させる。
【0028】
本発明の方法に用いられるサンプルは、細胞溶解物、インビトロ転写/翻訳反応物、細胞培養物の上澄み液、生理学的体液サンプル、例えば、血液、血漿、血清、脳脊髄液、涙液又は尿のサンプルであり得、無処置細胞を含み得る。該細胞、細胞溶解物、又は上澄み液は、原核細胞又は真核細胞から得ることができる。
【0029】
また、本発明は、第1及び第2のタンパク質、例えば、酵素、又は補因子の少なくとも1種によって媒介される反応、例えば、場合によっては、前記反応の1つを消光、又は前記反応の1つを増強/促進することのない同時的反応、又は連続的反応に関する前記タンパク質の存在又は量の同時的又は連続的検出を提供する。一実施形態において、第1の基質と第2の基質がサンプルに同時に加えられ、第1の酵素及び/又は補因子の量又は存在が、第2の酵素及び/又は補因子の量又は存在が検出されるより前に検出される。他の実施形態において、第1の基質と第2の基質がサンプルに同時に加えられ、第2の酵素及び/又は補因子の量又は存在が、第1の酵素及び/又は補因子の量又は存在が検出されるより前に検出される。或いは、第1の基質と第2の基質がサンプルに同時に加えられ、第1及び第2の酵素及び/又は補因子の存在又は量が同時に検出される。酵素及び/又は補因子の存在又は量が、単一の反応において検出されること、例えば、全ての反応が単一の容器、例えば、ウェル内で実施されることが好ましい。
【0030】
他の実施形態において、本発明は、蛍光タンパク質、例えば、緑色蛍光タンパク質の発現と関連させて、酵素媒介反応に関する分子の存在又は量を検出する方法を提供する。例えば、蛍光タンパク質、又は脱ハロゲン化酵素など、蛍光を発するように細胞内で標識化できるタンパク質を、一時的に又は安定して発現する細胞を、蛍光発生アッセイにより蛍光タンパク質の存在又は量をアッセイできると共に、酵素によって媒介される反応に関する、細胞内に存在するか又は細胞によって分泌される少なくとも1つの追加分子、例えば、酵素、基質又は補因子に関して、ルミネセンス発生アッセイによってアッセイできる。一実施形態において、異なる分子の存在又は量もまた、例えば、ルミネセンス非発生アッセイにおいて検出又は測定される。次いで、該分子(1種又は複数種)の存在又は量を、蛍光タンパク質から作製されたデータを用いて正規化できる。
【0031】
したがって、本発明は、第1の酵素によって媒介された反応に関する分子の存在又は量を検出する方法を提供する。該方法は、蛍光タンパク質を発現する細胞を含むサンプルを、該分子を欠いている第1の酵素、場合によっては、第2の酵素に関する反応混合物と接触させることを含む。第1の酵素によって媒介された反応からルミネセンス発生生成物が得られる。次いで、該分子の存在又は量及び蛍光タンパク質の存在又は量が検出される。
【0032】
一実施形態において、異なった特徴、例えば、異なった色を有する生成物を生成するルミネセンス発生及び/又は蛍光発生アッセイでは、さらなる多重化(すなわち、他の基質によって)を用いてもよい。例えば、さらなる多重化は、異なるルシフェラーゼベースの反応又は基質によって放出された異なる色の使用、又は異なる励起/発光スペクトルによる蛍光発生アッセイを含み得る。
【0033】
また、本発明は、細胞集団、例えば、細胞培養物集団における生細胞及び/又は死細胞の存在又は数を測定する方法を提供する。該方法は、細胞膜の透過性及び完全性に関連したタンパク質分解活性の差に基づいている。該方法の利点としては、高感度、簡便性、下流多重化適用及び集団応答の正規化に関するアッセイ読み取りの適応性が挙げられる。生存度の測定値差は、1つのプロテアーゼ基質の相対的不透過性及び細胞環境から放出されるプロテアーゼの他のプロテアーゼ基質に対する酵素活性の低さに基づいている。この実施形態において有用な基質としては、N末端又はC末端でブロックされている基質を含む、エキソプロテアーゼ又はエンドプロテアーゼに対する基質が挙げられる。一実施形態において、少なくとも2種の異なる蛍光発生基質(蛍光発生アッセイ試薬)が用いられ、その1つは、実質的に細胞不透過性であり、細胞外環境で活性であるプロテアーゼ、例えば、偏在性の保存放出プロテアーゼに特異的である。他の基質は、実質的に細胞透過性であり、生細胞内で活性であるが細胞外環境に存在している場合は実質的に非活性である細胞内プロテアーゼに特異的である。実質的に細胞不透過性のプロテアーゼ基質は、死細胞に関するアッセイにおけるエンドポイントを測定するために一般に用いられる時間、例えば、サンプルへのプロテアーゼ基質の添加後、5時間未満、4時間未満、3時間未満、2時間未満又は1.5時間未満の時間中に、生細胞において検出できないものである。実質的に細胞透過性のプロテアーゼ基質は、生細胞に関するアッセイにおけるエンドポイントを測定するために一般に用いられる時間、例えば、サンプルへの基質添加後、5分超、15分超、30分超、60分超、又は120分超の時間に、生細胞に入るものである。ある条件下で実質的に非活性なプロテアーゼは、そのプロテアーゼの最適活性の約10%未満を有するものである。一実施形態において、実質的に細胞不透過性の蛍光発生基質としては、トリ又はテトラペプチド基質が挙げられる。一実施形態において、実質的に細胞透過性の蛍光発生基質としては、アミノ酸、又はジ若しくはトリペプチド基質が挙げられる。サンプルは2種の基質に接触させるが、該サンプルは、場合によっては、細胞溶解現象を生じる、又は細胞溶解物を生じることが意図されない(一般に非破壊的)量の1つ又は複数の試験条件又は試薬によって処理される。2種の蛍光発生基質における蛍光体は、異なるスペクトルを有し、蛍光発生基質に接触させたサンプルから得られた相対的蛍光単位(RFLU)は、該サンプル中の生細胞及び死細胞の測定を考慮に入れている。
【0034】
他の実施形態において、サンプル中の生細胞及び死細胞の存在又は量を検出又は測定するために、蛍光発生プロテアーゼ基質及びルミネセンス発生プロテアーゼ基質が用いられる。1つの基質は、実質的に細胞不透過性であり、細胞外環境で活性であるプロテアーゼに特異的であり、他の基質は、実質的に細胞透過性であり、生細胞内で活性であるが細胞外環境に存在している場合は実質的に非活性である細胞内プロテアーゼに特異的である。一実施形態において、実質的に細胞不透過性のプロテアーゼ基質としては、トリ又はテトラペプチド基質が挙げられる。一実施形態において、実質的に細胞透過性のプロテアーゼ基質としては、アミノ酸、又はジ若しくはトリペプチド基質が挙げられる。サンプルは、例えば、細胞溶解現象を生じる条件の不在下で、2種の基質に接触させ、プロテアーゼ開裂(RFLU及びRLU)から生じる蛍光発生生成物及びルミネセンス発生生成物の検出は、該サンプル中の生細胞及び死細胞の測定を考慮に入れている。例えば、細胞溶解現象を生じさせないルシフェラーゼ検出試薬を、ルミネセンス発生基質に接触させたウェルに加えることができる。
【0035】
放出され保持されたプロテアーゼ活性のスペクトルの異なるシグナルを、例えば、蛍光光度計又は蛍光光度計/ルミノメータ機器を用いて測定できる。このような測定は反比例し、したがって相補的である。データの正規化、対照化、及び改善に、生存度及び細胞毒性アッセイを使用できる。
【0036】
本明細書において、Ala−Ala−Phe−AMC(放出プロテアーゼ基質)及びGly−Phe−AFC(生細胞保持プロテアーゼ)は、生細胞及び細胞溶解物の%混合物と組み合わされた。細胞生存度は、凍結/解凍の繰返し、界面活性剤処理により、並びにアポトーシスを誘導する薬剤(例えば、スタウロスポリン、rTRAIL、及び抗Fas mAb)により損なわれた。さらに、細胞生存度のこれらの測定はまた、他の細胞生存度測定(CellTiter−Glo(商標)、CellTiter−Blue(商標)、又はCytoTox−ONE(商標))、又はアポトーシス的細胞毒性の特異的測定(Caspase−Glo(商標)3/7、8、9又はApo−ONE(商標))のいずれかと多重化した。やはり本明細書に記載されているように、他の基質、例えば、ローダミン−110を含有する基質などの他の蛍光発生基質又はアミノルシフェリンベースの基質などのルミネセンス発生基質を、プロテアーゼ放出アッセイ及び/又はプロテアーゼ保持アッセイに使用できる。
【0037】
したがって、本明細書に記載された生細胞及び/又は死細胞アッセイの使用は、細胞の健康状態の逆測定及び相補的測定を提供し、細胞溶解段階を必要とせずに、条件の変化、例えば、化合物による処理の影響を検出するために使用できる。さらに、基質(1種又は複数種)の色は無視し得るか又は非内在的であるため、ペアードエンドポイントアッセイに見られるシグナル消光効果はない。さらに、プロテアーゼ放出及び保持双方の活性成分は実用的な感度を有し(10,000細胞/ウェルの生存度において<2〜5%差の検出)これらの感度を15分という短さで得ることができる。また、ウェル用量を大きく変化させることなく、基質(1種又は複数種)を細胞ウェルに混合でき、次の段階のエンドポイント化学の適応性が増加する。例えば、dsDNA割込み記号又は他の好適なエンドポイントと結合させた生/死アッセイの使用は、死細胞%対対照、生細胞%対対照、細胞総数対対照、及び/又は細胞死の機構(カスパーゼ活性化)又は他のエンドポイントレポーターアッセイ測定値の検出又は測定のために使用できる。
【0038】
したがって、本発明により、例えば、同一ウェル内で、個々の非破壊的蛍光発生若しくは非破壊的ルミネセンス発生プロテアーゼベースアッセイ、又は非破壊的蛍光発生及び/又は非破壊的ルミネセンス発生プロテアーゼベース生/死細胞アッセイの多重化、或いは非破壊的蛍光発生プロテアーゼベースアッセイと他のアッセイとの組合せが提供される。一実施形態において、生/死細胞アッセイにおいて検出又は測定されるプロテアーゼは同じではなく、例えば、実質的に同じ基質を認識することはない。すなわち、それらは同じ基質に結合しないか、又はそれらが同じ基質に結合し反応するとしても、それらプロテアーゼの一方は、他方のプロテアーゼと同程度(効率)に基質と反応することはない。すなわち、双方のプロテアーゼに対する基質が存在する場合、それらプロテアーゼの一方は、他方のプロテアーゼに対する基質と実質的に反応しない。
【0039】
したがって、本発明は、サンプル中の生細胞及び/又は死細胞を検出する方法を提供する。該方法は、サンプルを、第1のプロテアーゼに対する基質及び第2のプロテアーゼに対する基質と接触させることを含む。一方のプロテアーゼによって媒介される一方の基質を用いる反応によって、蛍光発生生成物が生じ、他方のプロテアーゼによって媒介される他方の基質を用いる反応によって、ルミネセンス発生生成物又は蛍光発生生成物が生じる。一方の基質は実質的に細胞透過性であり、他方の基質は実質的に細胞不透過性である。次いで、サンプル中の蛍光及び/又はルミネセンスが検出測定され、それによって次に、サンプル中の生細胞及び/又は死細胞の数又は存在が検出又は測定される。
【0040】
一実施形態において、2種の異なるプロテアーゼに対する基質が同時にサンプルと組み合わされる。他の実施形態において、サンプルを、第1の基質の前に第2の基質と接触させるか、又は第2の基質の前に第1の基質と接触させる。一実施形態において、該アッセイは、場合によっては、第1のアッセイと第2のアッセイとの間で試薬調整をした2段階アッセイであり得る。本発明はまた、第1及び第2のプロテアーゼの存在又は量の同時的又は連続的検出を提供する。一実施形態において、第1及び第2の基質をサンプルに同時に加え、一方のプロテアーゼの量又は存在を、他方のプロテアーゼの量又は存在を検出する前に検出する。或いは、第1及び第2の基質をサンプルに同時に加え、第1及び第2のプロテアーゼの存在又は量を同時に検出する。プロテアーゼの存在又は量は、単一の反応において検出されること、例えば、全ての反応が単一の容器、例えば、ウェル内で実施されることが好ましい。
【0041】
本発明は、サンプル中の生細胞を検出する方法を提供する。該方法は、サンプルを、プロテアソーム、アミノペプチダーゼ又はカテプシンに関連したプロテアーゼに対する、蛍光を発生する実質的に細胞透過性の基質に接触させ、サンプル中の蛍光を検出又は測定し、それによってサンプル中の生細胞の数又は存在を検出又は測定することを含む。
【0042】
また、サンプル中の死細胞を検出する方法が提供される。該方法は、サンプルを、蛍光発生又はルミネセンス発生細胞不透過性基質であるトリペプチジルペプチダーゼ、カルパイン又はキモトリプシンに接触させ、サンプル中の蛍光又はルミネセンスを検出又は測定し、それによってサンプル中の死細胞の数又は存在を検出又は測定することを含む。
【0043】
また、本発明のアッセイに使用される1種又は複数の試薬を含むキットが提供される。一実施形態において、本発明は、サンプル中の生細胞及び/又は死細胞の検出に有用なキットを提供する。例えば、本発明は、第1のプロテアーゼに対する第1の、蛍光を発生する、又はルミネセンスを発生する実質的に細胞不透過性の基質及び第2のプロテアーゼに対する第2の、蛍光を発生する実質的に細胞不透過性の基質を有する組成物;並びにサンプル中の生細胞及び/又は死細胞を検出するための該組成物の使用について使用者に指示する説明書を含むキットを提供する。一実施形態において、該組成物は溶液、例えば、溶媒、例えば、有機溶媒中、該基質が0.005Mから約1.0M、例えば、0.05Mから約0.2Mで存在する溶液である。他の実施形態において、本発明は、Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、又はそれらの組合せを含んでなる組成物を含んでなるキットを含む。一実施形態において、該組成物は溶液、例えば、該基質が0.005Mから約1.0M、例えば、0.05Mから約0.2Mで存在する溶液である。
【0044】
該アッセイはまた、薬物発見ツールとしての使用法を有する。多くの薬物試験化合物は蛍光/ルミネセンス多重アッセイを妨害し得る蛍光特性を有する。本発明は、間違った結果を検出するためのアッセイを提供する。本明細書に記載されているように、カスパーゼ−3に対する同じコンセンサス基質配列が、異なるスペクトル読取りを有する、例えば、2種は蛍光読取り、1種はルミネセンス読取りを有する異なるレポーター分子に結合した。カスパーゼ−3及びルシフェラーゼが、カスパーゼ−3阻害剤又はルシフェラーゼ阻害剤の存在下又は不在下でアッセイされた。このデータは、3種のレポーター分子間にはほとんど妨害がなく、間違った結果を抑えるための正規化アッセイにルシフェラーゼを使用し得ることを示した。
【0045】
したがって、酵素のモジュレーター、例えば阻害剤の存在又は量を、本発明の多重アッセイ、例えば、蛍光発生/ルミネセンス発生組合せアッセイを用いて検出できる。一実施形態において、該方法は、第1の酵素に対するルミネセンス非発生基質、第1の酵素に対する第2の基質、ルミネセンス発生アッセイに関する第2の酵素、及び試験試薬を含む反応混合物を提供することを含む。第2の基質と第1の酵素との間ではなく、非ルミネセンス発生基質と、第1の酵素との間の反応により、非ルミネセンス発生生成物が得られ、第2の基質と第1の酵素との間の反応により、第2の酵素に対する基質、例えば、ルシフェラーゼに対する基質が得られる。第2の酵素に対する基質と第2の酵素との間の反応により、ルミネセンス発生生成物が得られる。ルミネセンス発生生成物とルミネセンス非発生生成物の存在又は量を、試験反応物及び対照反応物において比較する。これら2つの結果の比較により、該ルミネセンス発生アッセイに関する酵素に及ぼすモジュレーターの影響が示され、それによって間違った結果を排除することができる。
【図面の簡単な説明】
【0046】
【図1A】蛍光発生アッセイ試薬及びルミネセンス発生アッセイ試薬双方の存在下、カスパーゼ−3及びカスパーゼ−8の酵素活性を測定する多重アッセイ。相対的光単位(RLU)対時間を示す図である。
【図1B】蛍光発生アッセイ試薬及びルミネセンス発生アッセイ試薬双方の存在下、カスパーゼ−3及びカスパーゼ−8の酵素活性を測定する多重アッセイ。経時的な相対的蛍光単位(RFU)を示す図である。
【図2A】カスパーゼ−3及びカスパーゼ−8の多重アッセイ。AMCに関するシグナル対背景蛍光を示す図である。
【図2B】カスパーゼ−3及びカスパーゼ−8の多重アッセイ。ローダミン−110に関するシグナル対背景蛍光を示す図である。
【図2C】カスパーゼ−3及びカスパーゼ−8の多重アッセイ。ルミネセンスのシグナル対背景蛍光を示す図である。
【図3A】カスパーゼ−3、カスパーゼ−8、及びトリプシンの活性を測定する三重アッセイ。ローダミン−110に関するRFUを示す図である。
【図3B】カスパーゼ−3、カスパーゼ−8、及びトリプシンの活性を測定する三重アッセイ。AMCに関するRFUを示す図である。
【図3C】カスパーゼ−3、カスパーゼ−8、及びトリプシンの活性を測定する三重アッセイ。RLUを示す図である。
【図4A】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。1/2時間及び18時間におけるRLUを示す図である。
【図4B】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。2時間及び18時間におけるRFUを示す図である。
【図4C】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。1/2時間及び18時間におけるRLUを示す図である。
【図4D】プロテアーゼ(カスパーゼ−3)及び非プロテアーゼ(β−ガラクトシダーゼ)酵素を測定する多重アッセイ。2時間及び18時間におけるRFUを示す図である。
【図5A】ルシフェリンの励起及び発光のスペクトルを示す図である。
【図5B】アミノルシフェリンの励起及び発光のスペクトルを示す図である。
【図5C】Z−LETD−アミノルシフェリンの励起及び発光のスペクトルを示す図である。
【図6】カスパーゼ−3アッセイにおいて3つのチャネル;ローダミン−110、AMC及びルミネセンスにおけるシグナル対背景比を示す図である。
【図7A】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対ATP濃度を示す図である。
【図7B】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図7C】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対ATP濃度を示す図である。
【図7D】ラクテートデヒドロゲナーゼ(LDH)活性及びアデノシントリホスフェート(ATP)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図8A】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ−3濃度を示す図である。
【図8B】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図8C】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ−3濃度を示す図である。
【図8D】LDH及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対LDH希釈を示す図である。
【図9A】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ濃度を示す図である。
【図9B】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対PKA濃度を示す図である。
【図9C】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対カスパーゼ濃度を示す図である。
【図9D】タンパクキナーゼA(PKA)及びカスパーゼ−3を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対PKA濃度を示す図である。
【図10A】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対スタウロスポリン濃度を示す図である。
【図10B】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対カスパーゼ−3濃度を示す図である。
【図10C】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RLU対スタウロスポリン濃度を示す図である。
【図10D】カスパーゼ−3及びウミシイタケルシフェラーゼ(luc)を測定する多重蛍光発生アッセイ及びルミネセンス発生アッセイ。RFU対カスパーゼ−3濃度を示す図である。
【図11A】RFLU対界面活性剤又は媒体及びプロテアーゼ放出アッセイ試薬で処理されたHL−60細胞数のプロットを示す図である。
【図11B】蛍光(AMC)プロテアーゼ放出アッセイの感度を示す図である。
【図12A】プロテアーゼ放出アッセイ試薬で処理した生存ジャーカット細胞のパーセントに対するRFLUのプロットを示す図である。
【図12B】蛍光((Ala−Ala−Phe)2−R110)プロテアーゼ放出アッセイの感度を示す図である。
【図13A】ルミネセンスプロテアーゼ放出アッセイ試薬で処理した生存ジャーカット細胞のパーセントに対するルミネセンスのプロットを示す図である。
【図13B】ルミネセンスプロテアーゼ放出アッセイ試薬で処理した生存ジャーカット細胞のパーセントに対するシグナル対ノイズ比のプロットを示す図である。
【図14A】異なるプロテアーゼ放出アッセイ試薬で処理し、超音波処理の有無によるHL−60細胞数に対するRLUのプロットを示す図である。
【図14B】種々の基質によるルミネセンスプロテアーゼアッセイの感度を示す図である。
【図15A】プロテアーゼ放出(細胞死)アッセイ(AN32504)及びCytoTox−ONE(商標)アッセイにおけるスタウロスポリン曝露時間に対するシグナル対背景比のプロットを示す図である。
【図15B】RFLU対スタウロスポリン曝露時間対LDH放出のプロットを示す図である。
【図16】細胞溶解の有無及び異なるpHにおけるプロテアーゼ放出アッセイ試薬で処理したHL−60細胞数に対するRFLUのプロットを示す図である。
【図17】Ala−Ala−Phe−アミノルシフェリンと接触させ、異なる細胞溶解に供されたHL−60細胞からのプロテアーゼ放出を示す図である。
【図18】プロテアーゼ保持アッセイ試薬対rTRAIL濃度に関するATPルミネセンスとRFLUのプロットを示す図である。
【図19A】3種の異なるプロテアーゼ保持アッセイ試薬で処理したジャーカット細胞数に対するRFLUのプロットを示す図である。
【図19B】プロテアーゼ保持アッセイにおける3種の異なる基質の感度を示す図である。
【図20】サポニン処理後に放出されたプロテアーゼについてのプロテアーゼ活性の半減期を示す図である。
【図21A】プロテアーゼベースの生/死細胞アッセイを示す図である。
【図21B】生、死又は生/死細胞アッセイによる連続多重適用におけるATP生存度アッセイを示す図である。
【図21C】蛍光生又は死アッセイ及びルミネセンスATPアッセイの連続多重を示す図である。
【図22A】SK−MEL−28細胞におけるイオノマイシン又はスタウロスポリンの濃度増加に対する生細胞RFLU及びカスパーゼ3/7活性に関するRFLUを示す図である。
【図22B】ACHN細胞におけるスタウロスポリンの濃度増加に対する生細胞RFLU及びカスパーゼ3/7活性に関するRLUを示す図である。
【図23】タキシフェン処理の時間に対する死HeLa細胞ルミネセンス及び生HeLa細胞蛍光のプロットを示す図である。
【図24A】タモキシフェンで処理され、PicoGreen(商標)で着色されたHeLa細胞によるプロテアーゼベースの生/死アッセイを示す図である。
【図24B】イオノマイシンで処理され、PicoGreen(商標)で着色されたHep2G細胞によるプロテアーゼベースの生/死アッセイを示す図である。
【発明を実施するための形態】
【0047】
本発明は、タンパク質(ペプチド又はポリペプチド)、例えば、反応に関する酵素、又は基質若しくは補因子などの1種又は複数の部分を検出するために、タンパク質に結合する、及び/又はタンパク質によって変化している少なくとも2種の異なる分子が、反応混合物中に同時に又は連続的に提供される多重アッセイ法を提供する。例えば少なくとも1種の酵素基質を、該基質と該酵素間との反応生成物へと変換する上で効果的な条件下、1つ又は複数の酵素媒介反応が実施される。反応混合物中の各分子、例えば、該反応における基質、又は生成物は、他の分子(1種又は複数種)又は生成物(1種又は複数種)とは異なる特性を有することが好ましく、一実施形態において、少なくとも1種の分子は、直接的又は間接的に検出可能なシグナルを生成できるレポーター分子を含む。生じたシグナルは、検出する分子の存在又は量に関連する。一実施形態において、該方法は、各基質を対応する生成物へと変換する上で効果的な条件下、少なくとも2種の異なる酵素基質の存在下、2種以上の酵素反応を実施することを含み、ここで、各反応の少なくとも基質又は生成物、及び/又は生成物の1種と、第3の、例えば異なる酵素との間の反応の生成物が、他方の基質(1種又は複数種)及び/又は生成物(1種又は複数種)とは異なった検出可能な特性、例えば異なる光学的特性を有する。該反応を、同時に又は連続的に実施した後、該反応(1つ又は複数)の1種又は複数の基質、又は1種又は複数の生成物の存在又は量を、検出又は測定する。これから、対応する酵素(1種又は複数種)及び/又は補因子の存在又は量を測定できる。
【0048】
このように、2つの一般的タイプの多重アッセイが考慮されている。第1のアッセイでは、複数の部分、例えば、酵素媒介反応に関する1種又は複数の酵素、1種又は複数の基質及び/又は1種又は複数の補因子を、同じ反応混合物中でアッセイする。各酵素は、基質の少なくとも1種を対応する生成物に変換することができ、ここで、該基質(1種又は複数種)及び/又は対応する生成物(1種又は複数種)、又は対応する生成物の1種と他の酵素との間の反応生成物(1種又は複数種)は、基質及び/又は生成物が同じ反応混合物中に存在する場合に、それらを個別に検出することを可能にする異なる検出可能な特性を有する。本発明のアッセイに関して、分子を加える順序は、変化し得る。このように、個々の反応は、同時に又は連続的に、開始及び/又は実施できる。連続的に開始され、実施される場合、検出可能な異なる特性は、異なる検出方法、及び/又は反応条件、例えば試薬濃度、温度又は追加の試薬の調整を必要とすることがある。例えば、反応の間に、消光剤又は増強剤を添加できる(例えば、その開示が、参照として、本明細書に具体的に組み込まれている米国特許出願第5,774,320号及び第6,586,196号を参照)。好ましい一実施形態において、2つ以上の反応が単一の反応混合物中で同時に実施され、ここで、各々の酵素が反応混合物中の基質の1種を生成物に変換する上で有効である。この実施形態は、例えば、細胞、細胞溶解物又は細胞上澄み液における少なくとも2種の異なる酵素及び/又は補因子の存在又は量を測定するために使用できる。また、該反応は、1種又は複数の試験試薬、例えば、酵素阻害剤又は酵素活性化剤、及び/又は異なる濃度の阻害剤、活性化剤、又は基質を含有し得る。
【0049】
場合によっては、該アッセイは、均一アッセイとして使用される。例えば、1種又は複数の基質及び追加成分を混合してから、該混合物をサンプルに加える。試薬の追加移入なしに、結果を読み取ることができる。
【0050】
第2のアッセイタイプでは、2つ以上の酵素媒介反応がタンデムに実施される。同じ時間に、又は異なる時間に、別個の反応が実施できる。該反応は、同一の又は異なる1種又は複数の酵素、同一の又は異なる1種又は複数の試験試薬、例えば、酵素阻害剤又は酵素活性化剤、及び/又は異なる濃度の阻害剤、活性化剤、又は基質を含有し得る。一実施形態において、各反応混合物は、生成物に変換できる少なくとも2種の基質を含有し、該基質(1種又は複数種)及び/又は対応する生成物(1種又は複数種)、及び/又は酵素/基質対の一方の生成物と異なる酵素との間の反応生成物(1種又は複数種)は、検出可能な異なる特性を有する。
【0051】
したがって、本発明のアッセイは、サンプル、例えば、真核細胞、例えば、酵母、鳥類、植物、昆虫の細胞、又は、限定はしないが、ヒト、サル、ネズミ、イヌ、ウシ、ウマ、ネコ、ヒツジ、ヤギ又はブタの細胞などの哺乳動物の細胞、又は原核細胞、又は2つ以上の異なる生物からの細胞、又は細胞溶解物又はその上澄み液を含むサンプル中の複数の酵素又は補因子の検出を可能にする。該細胞は、組換え法によって遺伝子操作されていない場合もあり得るし(非組換え細胞)、又は組換えDNAにより一時的に形質移入されている、及び/又はそのゲノムが組換えDNAによって安定して増加している、又はそのゲノムが遺伝子を破壊、例えば、プロモーター、イントロン又はオープンリーディングフレームを破壊する、又は1つのDNA断片を他のものと置換する組換え細胞の場合もあり得る。組換えDNA又はDNA断片置換は、本発明の方法によって検出される分子、検出される分子の濃度又は活性を変化させる部分、及び/又は該分子の濃度又は活性を変化させる分子又は部分に関連のない遺伝子産物をコードし得る。
【0052】
一実施形態において、本発明による方法は、細胞のアリコート又はその溶解物などの単一サンプル中の酵素を含む複数の部分を同時に又は連続的に検出するための迅速で高感度の方法を提供する。一実施形態において、該方法は、ルミネセンス発生アッセイにおける第1の酵素、基質又は補因子の存在又は量(活性)の定量化、及び蛍光発生アッセイなどのルミネセンス非発生アッセイにおける第2の酵素、基質又は補因子の存在又は量の定量化を含む。一実施形態において、試薬、例えば、各反応の基質は一緒に加えてもよいし、連続的に加えてもよい。他の実施形態において、該方法は、蛍光発生アッセイにおける第1の酵素、基質又は補因子の存在又は量の定量化、並びにルミネセンス発生アッセイにおける第2の酵素、基質又は補因子の存在又は量の定量化を含む。したがって、他の実施形態において、該方法は、ルミネセンス発生アッセイにおける補因子の存在又は量の定量化、並びにルミネセンス非発生アッセイにおける異なる分子の定量化を含む。さらに他の実施形態において、該方法は、ルミネセンス非発生アッセイにおける補因子の存在又は量の定量化、並びにルミネセンス発生アッセイにおける異なる分子の定量化を含む。ルミネセンス発生シグナル又はルミネセンス非発生シグナルの強度はそれぞれの分子の存在又は量の関数である。
【0053】
本発明はさらに、1種又は複数のエキソプロテアーゼ及び/又はエンドプロテアーゼに対する1種又は複数の基質が、細胞溶解に供さないサンプルなどのサンプルに提供される個別的な多重アッセイ法を提供し、該基質は、該サンプル中の生細胞及び/又は死細胞の数又は存在を検出又は測定する上で有用である。
【0054】
一実施形態において、本発明は、単一の細胞アリコート又はその溶解物中の1種又は複数の酵素の存在又は量を測定する方法に関する。一実施形態において、少なくとも酵素の1種は、内在性酵素である。例えば、一実施形態において、本発明は、原核細胞又は真核細胞からの精製調製物、細胞溶解物、又は培養真核細胞、例えば、哺乳動物細胞などの細胞の上澄み液などの、プロテアーゼ及び他の酵素を含む調製物中の少なくとも1種のプロテアーゼ、及び場合によっては他の酵素の活性をモニタリングするための改善された高感度の方法を提供する。分泌プロテアーゼ及び細胞内プロテアーゼなどの種々の細胞位置に存在する酵素では、各酵素に対する基質を、無処理細胞と共にウェルに加えることができる。分泌プロテアーゼの存在又は量は、細胞溶解後など、細胞内プロテアーゼの検出前に検出でき、例えば、細胞内プロテアーゼの検出は、分泌プロテアーゼと同じ容器、例えば、同じウェルにおいて行われる。一実施形態において、細胞内プロテアーゼに対する細胞不透過性基質及び分泌若しくは放出プロテアーゼに対する基質を、細胞を含むサンプルに加え、次いで、該細胞を、場合によっては溶解させる。分泌若しくは放出プロテアーゼの検出は、細胞溶解の前でもよいし、後でもよい。他の実施形態において、細胞内酵素又は分泌若しくは放出プロテアーゼに対する細胞不透過性基質及び第2の細胞内酵素に対する細胞透過性基質を、細胞を含むサンプルに加える。第2の細胞内酵素及び分泌若しくは放出プロテアーゼの存在は、溶解せずに検出できる。さらに他の実施形態において、分泌若しくは放出プロテアーゼ、細胞内酵素(細胞透過性基質又は細胞非透過性基質を用いて)、DNA若しくはATPなどの別の分子、又は第2の酵素、例えば細胞内酵素(細胞透過性基質又は細胞非透過性基質を用いて)を検出するために、三重アッセイが実施される。一実施形態において、分泌若しくは放出タンパク質は、蛍光、ルミネセンス又は吸光分光分析を用いて検出される。
【0055】
当該方法は、任意の酵素又は酵素の任意の組などの任意の分子を検出するために使用できる。該方法に用いられる酵素、検出される酵素又は基質若しくは補因子を検出するために有用な酵素は、組換え酵素及び内在性(天然)酵素などの酵素の任意の組合せから選択できる。一実施形態において、検出される全ての酵素が内在性酵素である。他の実施形態において、検出される2種の酵素は内在性酵素であり、もう一方の酵素は組換え酵素である。他の実施形態において、1種の酵素が内在性酵素であり、もう一方の酵素は組換え酵素である。通常の当業者に明白な他の組合せが、本明細書の教示による当該アッセイ及び方法に使用できる。該酵素としては、限定はしないが、プロテアーゼ、ホスファターゼ、ペルオキシダーゼ、スルファターゼ、ペプチダーゼ、及びグリコシダーゼが挙げられる。該酵素は、触媒反応の性質に基づいた、限定はしないが、ヒドロラーゼ、オキシドレダクターゼ、リアーゼ、トランスフェラーゼ、イソメラーゼ、リガーゼ、又はシンターゼなどの種々の群に由来し得るか、又は該酵素は、少なくとも酵素の1種が、他の酵素の少なくとも1種と部分的に重なる、又は好ましくは実質的に異なる基質特異性を有する限り、同じ群に由来するものでもよい。生理学的重要性を有する酵素のクラスは特に興味深い。これらの酵素としては、タンパクキナーゼ、ペプチダーゼ、エステラーゼ、タンパクホスファターゼ、イソメラーゼ、グリコシラーゼ、シンセターゼ、プロテアーゼ、デヒドロゲナーゼ、オキシダーゼ、レダクターゼ、メチラーゼなどが挙げられる。興味深い酵素としては、有機、無機双方のエステルの生成又は加水分解、グリコシル化、及びアミドの加水分解に関与するものが挙げられる。いずれのクラスにおいても、キナーゼ類のように、さらなる細分があり得、キナーゼは、ペプチド及びタンパク質におけるセリン、トレオニン及び/又はチロシン残基のリン酸化に特異的であり得る。したがって、該酵素は、例えば、環式ヌクレオチド調節タンパクキナーゼ、タンパクキナーゼC、Ca2+/CaMによって調節されるキナーゼ、サイクリン依存キナーゼ、ERK/MAPキナーゼ、及びタンパク質−チロシンキナーゼなどのキナーゼの種々の機能群のキナーゼであり得る。キナーゼは、ERKキナーゼ、S6キナーゼ、IRキナーゼ、P38キナーゼ、及びAbIキナーゼなどのオリゴペプチド基質をリン酸化する上で有用な、シグナル伝達経路におけるタンパクキナーゼ酵素であり得る。これらに対する基質としては、オリゴペプチド基質が挙げられる。興味深い他のキナーゼとしては、例えば、Srcキナーゼ、JNK、MAPキナーゼ、サイクリン依存キナーゼ、P53キナーゼ、血小板由来成長因子受容体、上皮成長因子受容体、及びMEKを挙げることができる。
【0056】
特に、本発明に有用な酵素としては、酵素活性を示す任意のタンパク質、例えばリパーゼ、ホスホリパーゼ、スルファターゼ、ウレアーゼ、ペプチダーゼ、プロテアーゼ及び酸ホスファターゼ、グルコシダーゼ、グルクロニダーゼ、ガラクトシダーゼ、カルボキシルエステラーゼ、及びルシフェラーゼなどのエステラーゼが挙げられる。一実施形態において、酵素の1種は、加水分解性酵素である。他の実施形態において、少なくとも酵素の2種が加水分解性酵素である。加水分解性酵素の例としては、アルカリ及び酸ホスファターゼ、エステラーゼ、デカルボキシラーゼ、ホスホリパーゼD、P−キシロシダーゼ、β−D−フコシダーゼ、チオグルコシダーゼ、β−D−ガラクトシダーゼ、α−D−ガラクトシダーゼ、α−D−グルコシダーゼ、β−D−グルコシダーゼ、β−D−グルクロニダーゼ、α−D−マンノシダーゼ、β−D−マンノシダーゼ、β−D−フルクトフラノシダーゼ、及びβ−D−グルコシデュロナーゼが挙げられる。
【0057】
任意の具体的な酵素媒介反応に関する基質又は補因子は、当業者に知られている。いくつかのプロテアーゼに関する典型的な開裂部位は、表1に記載されている。
【0058】
【表1】
【0059】
アルカリホスファターゼに対して、基質は、3−(2’−スピロアダマンタン)−4−メトキシ−4−(3’’−ホスホリルオキシ)フェニル−1,2−ジオキセタン、二ナトリウム塩、又は3−(4−メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]デカン]−4−イル]フェニルリン酸二ナトリウム、又は2−クロロ−5−(4−メトキシスピロ{1,2−ジオキセタン−3,2’−(5’−クロロ)−トリシクロ{3.3.1.13,7]デカン}−4−イル)−1−フェニルリン酸二ナトリウム又は2−クロロ−5−(4−メトキシスピロ{1,2−ジオキセタン−3,2’−トリシクロ[3.3.1.13,7]デカン}−4−イル)−1−フェンジルリン酸二ナトリウム(それぞれ、AMPPD、CSPD、CDP−Star(登録商標))及びADP−Star(商標)などのリン酸含有ジオキセタンを含むことが好ましい。
【0060】
β−ガラクトシダーゼに対して、基質は、ガラクトシダーゼ開裂可能基又はガラクトピラノシド基を含有するジオキセタンを含むことが好ましい。アッセイにおけるルミネセンスは、ジオキセタン基質の糖部分の酵素的開裂から生じる。このような基質の例としては、3−(2’−スピロアダマンタン)−4−メトキシ−4−(3’’−β−D−ガラクトピラノシル)フェニル−1,2−ジオキセタン(AMPGD)、3−(4−メトキシスピロ[1,2−ジオキセタン−3,2’−(5’−クロロ)トリシクロ[3.3.1.13,7]−デカン]−4−イル−フェニル−β−D−ガラクトピラノシド(Galacton(登録商標))、5−クロロ−3−(メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)トリシクロ[3.3.13,7]−デカン]−4−イル−フェニル−β−D−ガラクトピラノシド(Galacton Plus(登録商標))、及び2−クロロ−5−(4−メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]−デカン]−4−イル)フェニル−β−D−ガラクトピラノシド(Galacton−Star(登録商標))が挙げられる。
【0061】
β−グルクロニダーゼ及びβ−グルコシダーゼに関するアッセイにおいて、基質としては、グルクロニド、例えば、3−(4−メトキシスピロ{1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]デカン}−4−イル)フェニル−β−D−グルクロン酸ナトリウム(Glucuron(商標))などのβ−グルクロニダーゼ開裂基を含有するジオキセタンが挙げられる。カルボキシルエステラーゼに関するアッセイにおいて、基質としては、ジオキセタンに結合した好適なエステル基が挙げられる。プロテアーゼ及びホスホリパーゼに関するアッセイにおいて、基質としては、ジオキセタンに結合した好適な酵素開裂基が挙げられる。
【0062】
アッセイにおける各酵素に対する基質は、異なっていることが好ましい。1種のジオキセタン含有基質を含むアッセイでは、該基質は、場合によっては、置換又は非置換アダマンチル基、置換又は非置換であり得るY基及び酵素開裂基を含有する。好ましいジオキセタンの例としては、上記に記載したもの、例えば、Galacton(登録商標)、Galacton Plus(登録商標)、CDP−Star(登録商標)、Glucuron(商標)、AMPPD、Galacton−Star(登録商標)、及びADP−Star(商標)、と称されるもの、並びに3−(4−メトキシスピロ[1,2−ジオキセタン−3,2’(5’−クロロ)−トリシクロ[3.3.1.13,7]デカン}−4−イル)フェニル−β−D−グルコピラノシド(Glucon(商標))、CSPD、3−クロロ−5−(4−メトキシスピロ{1,2−ジオキセタン−3,2’−(5’−クロロ)−トリシクロ{3.3.1.13,7]デカン)−4−イル)−1−フェニルリン酸二ナトリウム(CDP)が挙げられる。
【0063】
検出される少なくとも1種の酵素に対する基質は、レポーター分子を含有するように修飾されることが好ましい。レポーター分子は、その分子に結合した基質、該酵素と該基質との間の反応から生じる生成物、又はその生成物と他の酵素との間の反応生成物が、別個に検出される、好ましくは定量的に検出されることを可能にする任意の分子である。レポーター分子としては、限定はしないが、蛍光体などの光学的分子、吸収性有色粒子又は色素、放射標識、好適な反応成分の存在下、検出反応を触媒する上で有効な触媒的部分などの酵素、他のサブユニット(1つ又は複数)又は断片(1つ又は複数)と結合した際に機能的な酵素のサブユニット又は断片、又は引き続く反応に関する基質、例えば、その反応の生成物が検出可能である基質が挙げられる。本明細書に用いられる「蛍光体」には、ある波長領域でエネルギーを吸収し、該吸収領域以外の波長領域でエネルギーを放出することのできる分子が含まれる。用語の「励起波長」とは、蛍光体がエネルギーを吸収する波長領域を言う。用語の「発光波長」とは、蛍光体がエネルギー又は蛍光を放出する波長領域を言う。
【0064】
一実施形態において、レポーター分子は蛍光を発する。蛍光発生物の一群は、キサンテン色素であり、これには、フルオレセイン、ローザミン及びローダミンが含まれる。これらの化合物は、フェニル基上に置換基を有した商品として入手でき、結合のための部位として、又は結合官能基として使用できる。例えば、アミノ置換及びイソチオシアネート置換したフルオレセイン化合物を入手できる。
【0065】
蛍光性化合物の他の群は、アルファ位又はベータ位、通常はアルファ位にアミノ基を有するナフチルアミン類である。ナフチルアミノ化合物の中には、1−ジメチルアミノナフチル−5−スルフォネート、スルホン酸1−アニリノ−8−ナフタレン及びスルホン酸2−p−トルイジニル−6−ナフタレンが含まれる。いくつかのナフタレン化合物は、タンパク質に対していくらか非特異的な結合が見られるため、それらの使用には、タンパク質の量が最少化されているアッセイ培地を用いる必要がある。他の蛍光発生物は、窒素含有マクロ環を含む多座リガンドであり、これらは、パイ電子を有する共役環系を有する。これらのマクロ環は、場合によっては、架橋炭素上、又は窒素上の置換などの置換がされていてもよい。好適なマクロ環としては、ポルフィリン、アザポルフィリン、コリン、サフィリン及びポルフィセン、並びに、広く非局在化している電子を含有する他のマクロ環様の誘導体が挙げられる。アザポルフィリン誘導体としては、フタロシアニン、ベンゾトリアザポルフィリン及びナフタロシアン並びにそれらの誘導体が挙げられる。
【0066】
いくつかの例では、蛍光融合タンパク質、例えば、ポリペプチド基質に融合させた、緑色、赤色又は青色蛍光タンパク質又は他の蛍光タンパク質を使用できる。他の実施形態において、蛍光タンパク質は、それ自体、加水分解性酵素に対する基質であり得る。「蛍光タンパク質」は、完全長蛍光タンパク質又はそれの蛍光断片である。
【0067】
本発明に使用される化学的蛍光体の非限定的リストを、それらの励起波長及び発光波長と共に、表2に示してある。励起値及び発光値は、pH、緩衝系又は溶媒などの反応条件に依存して変化できる。
【0068】
【表2】
【0069】
一実施形態において、酵素の1種は、アミノ修飾ルシフェリン又はそのカルボキシ保護誘導体を含む基質を用いて検出され、前記修飾は、該酵素に対する基質を含む。一実施形態において、前記修飾は、プロテアーゼに対する認識部位を含む1つ又は複数のアミノ酸残基である。一実施形態において、該基質は、ペプチド結合を介して、アミノルシフェリン又はそのカルボキシ修飾誘導体のアミノ基に共有結合している。一実施形態において、ペプチド又はタンパク質基質のN末端は、例えば、アミノ末端保護基を用いて、アミノペプチダーゼによる分解を防ぐように修飾される。適切な酵素又は補因子の不在下では、このような基質及びルシフェラーゼを含む混合物は、最少のアミノルシフェリンが存在するため、最少の光を生じる。適切な酵素の存在下では、該基質及びアミノルシフェリンとの結合を該酵素によって開裂することができ、ルシフェラーゼの基質であるアミノルシフェリンが生じる。したがって、ルシフェラーゼ、例えば、天然、組換え又は変異ルシフェラーゼ、及びいずれかの補因子並びに適切な反応条件の存在下、該酵素の存在又は活性に比例する光が生じる。
【0070】
一実施形態において、酵素の1種は、蛍光体を含む基質を用いて検出される。一実施形態において、該基質は、プロテアーゼに対する認識部位を含む1つ又は複数のアミノ酸残基を含む。一実施形態において、該基質は、1種又は複数の蛍光体に共有結合している。適切な酵素又は補因子の不在下では、基質−蛍光体共役体の特性が変化して、該蛍光体単独に比較して該共役体の蛍光特性の変化、例えば低下が生じるように、該蛍光体の蛍光特性が、例えば、消光基の近接によって消光されるに従い、そのような基質を含む混合物は、その発光波長において最小の光を発生する。適切な酵素の存在下、共役体の開裂によって蛍光体が生じる。他の実施形態において、開裂前の共役体は蛍光性であるが、酵素による開裂後、生成物(1つ又は複数)はスペクトルを変化させた。
【0071】
一実施形態において、少なくとも2つの反応に関する条件は適合性である。例えば、少なくとも2種の酵素の条件、好ましくは3種以上の酵素、例えば、4種以上の酵素の条件は適合性である。類似した酵素の群は、pHやイオン強度などの反応条件に一般に適合性であるが、比較的低質量濃度を有するアッセイ成分、例えば、補因子に関係する補因子要件、金属イオン要件などは共通である必要はない。共通の条件としては、各酵素が反応の経過中、測定可能な比率を提供し、一般に、各酵素が他の酵素(1種又は複数種)に対して、加えられた成分から有意な妨害なしに、特定の基質に対する最大回転率の少なくとも約10%、通常は少なくとも約20%、好ましくは少なくとも約50%の回転率を有するような条件が挙げられる。
【0072】
或いは、双方の反応に関する基質が存在しているが、1つの反応に関する条件は、他の反応に適合性でない場合があり得る。このような実施形態において、1種の酵素は活性であるが、その基質と反応できない。例えば、2つの反応に関する条件が適合性でない一実施形態において、個々の酵素反応アッセイは、連続的に及び/又は個別の反応混合物において実施される。酵素アッセイ後、該反応混合物(又はその一部)を他の反応と組み合わせ得る。各々、個別の反応混合物が、1種又は複数の酵素及び1種又は複数の基質を含有し得る。その最も単純な形態では、アッセイする単一の酵素及びその酵素に対する単一の基質が、各反応混合物中にある。該反応に使用される基質の組は、単一反応多重アッセイに必要とされるものと同じ一般的特性を有する。すなわち、各基質及び/又は対応する生成物は独特の特性を有し、それらを互いに区別することを可能にする。
【0073】
該反応における分子の検出順序は変化し得る。一実施形態において、反応が同時に開始されるか否かに関わりなく、ルミネセンス発生アッセイによって検出される分子が検出され、次いで非ルミネセンス発生アッセイによって検出される分子が検出される。或いは、反応が同時に開始されるか否かに関わりなく、非ルミネセンス発生アッセイによって検出される分子が検出され、次いでルミネセンス発生アッセイによって検出される分子が検出される。他の実施形態において、2種以上の分子の存在又は量が本質的に同時に検出される。一実施形態において、検出される1種の分子の存在又は活性を、第2の分子の存在又は活性を検出する前に、例えば、第1のシグナルが、例えば少なくとも50%減少するまで待つことによって、又は第1の反応に対する消光剤を加えることによって、実質的に減少させる。したがって、いくつかの実施形態において、1種又は複数の反応を、検出前に、例えば、該反応に関する酵素を阻害することによって終了させる。1つのアッセイによって生じたシグナルが、他の少なくとも1つのアッセイによって生じたシグナルの定量化を実質的に妨害しないことが好ましい。
【0074】
また、本発明は、無処置細胞、細胞溶解物、例えば、少なくとも部分的に精製された溶解物、又は細胞上澄み液を含むサンプルなどのサンプル中の1種又は複数のペプチド又はタンパク質、ペプチド又はタンパク質に結合する及び/又はペプチド又はタンパク質によって変化する分子、又は補因子の存在又は活性を検出するためのキットを提供する。このようなキットは、少なくとも1種の酵素に対する基質などの、少なくとも1種のペプチド及び/又はタンパク質、ペプチド及び/又はタンパク質に結合した、及び/又はペプチド及び/又はタンパク質によって変化した分子、又は補因子を定量化するための少なくとも1種の試薬を含む。
【0075】
本発明を、以下の非限定的実施例によって、さらに説明する。すべての実施例に関し、好適な制御反応は、当業者によって容易にデザインされる。
【実施例】
【0076】
(実施例I)
蛍光/ルミネセンス多重アッセイ
A.単一ウェル、多重アッセイにおけるカスパーゼ−3及びカスパーゼ−8の測定
Caspase−Glo(商標)8 Reagent(Caspase−Glo(商標)8 Assay System、Promega社)を、均一ルミネセンス発生カスパーゼ−8及びルミネセンス非発生カスパーゼ−3酵素アッセイの多重化を可能にする能力に関して評価した。Caspase−Glo(商標)8 Reagentは、Caspase−Glo(商標)8 Buffer及びルミネセンス発生基質であるZ−LETD−アミノルシフェリンからなる。図1Aにおけるルミネセンス発生アッセイに関して、多重化されたルミネセンス発生及びルミネセンス非発生アッセイの実行可能性を実証するために、Caspase−Glo(商標)8 Reagent(菱形)又はカスパーゼ−3に対する50μMの蛍光発生基質、(Z−DEVD)2−ローダミン−110(四角形)も含有するCaspase−Glo(商標)8 Reagentのいずれかを用いた。図1Bにおける蛍光発生アッセイに関して、50μM (Z−DEVD)2−ローダミン−110及び10mM DTT(四角形)又は50μM (Z−DEVD)2−ローダミン−110及びZ−LETD−アミノルシフェリン(四角形)のいずれかを含有するCaspase−Glo(商標)8 Bufferを用いた。
【0077】
カスパーゼ−8酵素、カスパーゼ−3酵素、及びカスパーゼ−8とカスパーゼ−3酵素との組合せ(Biomol Research Laboratories)の希釈液を、100単位/mlの最終濃度に、RPMI1640(Sigma社)中で調製した。100μlのカスパーゼ−8希釈液、カスパーゼ−8希釈液とカスパーゼ−3希釈液との混合液、又はカスパーゼ−3希釈液を、96ウェルプレートの個別のウェルに加えた。50μM (Z−DEVD)2−ローダミン−110を有する又は有さない100μlのCaspase−Glo(商標)8 Reagent(図1A)、又はZ−LETD−アミノルシフェリンを有する又は有さない、(Z−DEVD)2−ローダミン−110及びDTTを添加した100μlのCaspase−Glo(商標)8 Bufferを、200μl/ウェルの最終容量に達するように加えた。該反応プレートを室温で、プレートシェーカー上、少なくとも10分間温置した。
【0078】
温置後、DYNEX Laboratories MLX(商標)プレートルミノメータを用いて相対的ルミネセンスを測定し、485EX/530EMフィルターセットを装備したCytoFluorII蛍光プレート読取り装置により相対的蛍光を測定した。
【0079】
結果
図1に、単一ウェル中、2種のプロテアーゼ酵素に関する蛍光及びルミネセンスの同時測定が示されている。図1Aに見られる通り、カスパーゼ−8(四角形)に関するルミネセンス発生アッセイにおいて、カスパーゼ−3及びその蛍光発生基質である(Z−DEVD)2−ローダミン−110の存在は、ルミネセンス反応を大きく変化させることはない。同様に、カスパーゼ−3(四角形)に関する蛍光発生アッセイにおいて、カスパーゼ−8及びそのルミネセンス発生基質であるZ−LETD−アミノルシフェリンの存在は、カスパーゼ−3に関する蛍光発生アッセイに影響を与えない。
【0080】
B.カスパーゼ−3及びカスパーゼ−8多重アッセイに関する背景測定
カスパーゼ酵素及びそれらの基質などの種々の濃度のルミネセンス発生試薬及び蛍光発生試薬並びに緩衝液成分を組み合わせて、蛍光及び/又はルミネセンスに対する各構成成分の寄与を確認した。ルミネセンス測定時、蛍光発生基質である(Z−DEVD)2−ローダミン−110は、ローダミンチャネル(485EX/520EM)内でカスパーゼ−3活性を報じ、蛍光発生基質であるAc−DEVD−AMCは、AMCチャネル(360EX/460EM)内でカスパーゼ−3活性を報じるが、Z−LETD−アミノルシフェリン基質は、カスパーゼ−8活性を報じる。表3には、12の異なる反応条件に関する各成分の量(μl)が、各反応条件に関し、合計、約500μlのマスター混合物、又は100μlのマスター混合物/反応(n=4)を生じることが記載されている。「カスパーゼ添加」の列でのこの列の数は過剰に添加されたカスパーゼのタイプを定義するものであって、容量を示しているのではない。
【0081】
【表3】
1.カスパーゼ−8 luc対照
2.カスパーゼ−3ローダミン−110対照
3.カスパーゼ−3AMC対照
4.ローダミン−110を有する多重対照
5.ローダミン−110+阻害剤を有する多重対照
6.AMCを有する多重対照
7.アミノルシフェリン混合物を有するカスパーゼ−3
8.アミノルシフェリン混合物+阻害剤を有するカスパーゼ−3
9.ローダミン−110を有するカスパーゼ−8
10.アミノルシフェリン混合物を有するカスパーゼ−3
11.アミノルシフェリン混合物+阻害剤を有するカスパーゼ−8
12.ローダミン−110を有し、アミノルシフェリン混合物を有さないカスパーゼ−3
【0082】
表IIIの成分を反復ウェルに加え、反応液を室温で2時間温置した。用いられた緩衝液は、Caspase−Glo(商標)8 Assey Systemのものであった。DMSOはSigma−Aldrichから入手し、DTTは、Amrescoから入手した。基質及び阻害剤は、Promega Corpから入手した。
【0083】
相対的ルミネセンスは、DYNEX LaboratoriesのMLX(商標)プレートルミノメータを用いて測定された。蛍光は、ローダミン−110に関しては、485EX/530EMフィルターセット、次いで、AMCチャネルに関しては、360EX/460EMにセットしたCytoFluorII Fluorescentプレート読取り機を用いて測定した。
【0084】
結果
図2A、B、及びCで、全てのカラットは、酵素活性を示す蛍光又はルミネセンスのいずれかが予測される場合を表す。図2Aは、各反応におけるAMC蛍光に関するシグナルを示す。背景以上の蛍光は、Ac−DEVD−AMC及びカスパーゼ−3という適切な基質/酵素の組合せが存在した場合のみ存在した(反応条件3及び6)。図2Bは、各反応におけるローダミン−110蛍光に関するシグナルを示す。背景以上の蛍光は、カスパーゼ−3阻害剤が存在した場合(反応条件5)を除いて、(Z−DEVD)2−ローダミン−110/カスパーゼ−3という基質/酵素の組合せが存在した場合に存在した(反応条件2、4、10、及び12)。背景以上のルミネセンスシグナル(図2C)では、カスパーゼ阻害剤が存在した反応条件(反応条件5、8、及び11)を除いて、Z−LETD−アミノルシフェリン/カスパーゼ−8という適切な基質/酵素の組合せを有した反応が、背景以上のシグナルを示した(反応条件1、4、6、7、9、及び10)。したがって、該データにより、これらの条件下で、背景の蛍光及びルミネセンス測定に及ぼす反応成分の影響は無視できるものだったことが証明された。
【0085】
C.単一ウェル、三重アッセイにおけるカスパーゼ−3、カスパーゼ−8、及びトリプシンの測定
ダルベッコ−リン酸緩衝生理食塩水(Sigma社)中、カスパーゼ−8(150単位/ml、Biomol Research Laboratories)、カスパーゼ−3(Pharmingen社)、トリプシン(Sigma社)、及び3種全ての酵素の組合せの検出可能濃度の希釈液を調製した。100μlの各酵素希釈液を、96ウェルプレートのウェルに加え、100μlの各基質を、単独で、又は適切に組み合わせて対応するウェルに加えた:カスパーゼ−3に対しては、(Z−DEVD)2−ローダミン−110基質、トリプシンに対しては、Z−PRNK−AMC基質(ベータ−トリプターゼに対する基質として、本明細書にその全体が組み込まれている米国特許出願第09/955,639号に記載されているが、トリプシンに対しては、有用性が少ないと認識されている)、及びカスパーゼ−8に対しては、Z−LETD−アミノルシフェリン基質。蛍光アッセイで、基質と共にCaspase−Glo(商標)8緩衝液を用いた場合、10mM DTTを含めた。プレートは、プレートシェーカー上、室温で、少なくとも10分間温置した。
【0086】
温置後、カスパーゼ−8活性に関する相対的ルミネセンスを、BMG Fluorostar(BMG Labtechnologies社)を用いて測定した。相対的蛍光は、Labsystems Fluoroskan Ascent プレート読取り機を用いて測定した。カスパーゼ−3活性には、485EX/527EMのフィルターセットを用いた。トリプシン活性には360EX/460EMのフィルターセットを用いた。
【0087】
結果
図3Aに示されるように、3種の異なる基質及び組み合わされた対応する酵素を有する反応(三重アッセイ)において、カスパーゼ−3の検出に用いられた条件により、対照条件の蛍光以上の比較的高い蛍光が生じた。三重アッセイ(全ての酵素と共に全ての基質)におけるカスパーゼ−3の活性を、カスパーゼ−3単独の活性と比較すると、カスパーゼ−3が同じ反応液中に他の三重酵素反応液と共にあった場合、カスパーゼ−3活性は背景より大きかった。トリプシン(図3B)及びカスパーゼ−8(図3C)に関しても、カスパーゼ−3と同程度ではないにしても、同様な結果が見られた。
【0088】
D.単一ウェル、多重フォーマットにおけるカスパーゼ−3及びβ−ガラクトシダーゼの測定
Beta−Glo(登録商標)緩衝液(Beta−Glo(登録商標)Assay System、Promega社)を用いてBeta−Glo(登録商標)凍結乾燥基質を再構成することにより、又はBeta−Glo(登録商標)緩衝液に(Z−DEVD)2−ローダミン−110(50μM)を加えることにより、又はBeta−Glo(登録商標)緩衝液を用いてBeta−Glo(登録商標)凍結乾燥基質を再構成して(Z−DEVD)2−ローダミン−110(50μM)を加えることにより、試薬を調製した。カスパーゼ−3(2μl/ml、Pharmingen社)、又はβ−ガラクトシダーゼ(0.1μl/ml)、又はカスパーゼ−3及びβ−ガラクトシダーゼを、RPMI1640中で希釈し、100μlを96ウェル白色プレートのウェルに加えた。100μlの適切な試薬を96ウェルプレートのウェルに加え、室温で温置した。DYNEX LaboratoriesのMLX(商標)プレートルミノメータを用いて、30分におけるルミネセンスを測定した。485EX/530EMのフィルターにセットしたCytoFluorII Fluorescentプレート読取り機上で、2時間温置後の蛍光を測定した。増加した蛍光を補正するために、18時間目に、CytoFluorIIフルオロメータ上、異なる増加セッティングで全ての測定を繰り返した。
【0089】
結果
図4A及びCは、β−ガラクトシダーゼに関するルミネセンス発生アッセイが、カスパーゼ−3を測定するための蛍光発生試薬の存在下で機能的であることを証明している。図4B及びDは、カスパーゼ−3を測定するための蛍光発生アッセイが、β−ガラクトシダーゼを測定するためのルミネセンス発生試薬の存在下で機能的であることを証明している。図4B及び4Dに見られるように、背景蛍光に及ぼすルミネセンス発生試薬成分の軽微な影響がある。しかし、ルミネセンスに及ぼす蛍光発生試薬成分の影響はほとんど無い(図4A及び4C)。
【0090】
E.ルミネセンス発生アッセイに関する基質のスペクトル走査
0.1M Tris pH7.3、2mM EDTA、及び10mM MgSO4を含有する緩衝液中、ルシフェリン、アミノルシフェリン、及びZ−LETD−アミノルシフェリンを、およそ2μMに希釈した。1.25mm励起及び発光スリットフィルターを有するSPEX Fluorolog−2分光器上、1nmの波長間隔及び0.2秒の積分時間でサンプルを走査した。全ての走査は、石英キュベットを用いて実施された。
【0091】
結果
ルシフェリン及びアミノルシフェリンに関して、励起が325nmに存在すると、発光は、375nmから750nmで捕捉され、発光が600nmで測定される場合の励起は、280nmから550nmで捕捉された(図5A及び5B)。Z−LETD−アミノルシフェリンでは、励起が325nmに存在すると、発光は、375nmから750nmで捕捉され、発光が525nmで測定される場合の励起は、280nmから500nmで捕捉された(図5C)。興味深いことに、ペプチドがアミノルシフェリンに共役すると(図5C)、共役体の発光ピークは、より短い波長へ青色シフトした。これは予想外のことであり、したがって、特に、アミノルシフェリンと同じ波長領域で発光する蛍光体を用いると、ルミネセンス/蛍光二重測定が可能になる。
【0092】
(実施例II)
偽結果を検出する方法
方法
カスパーゼ−3阻害剤(Ac−DEVD−CHO、10μM)又はルシフェラーゼ阻害剤(Resveratol、5μM)のいずれかと共に、カスパーゼ−3の存在下、Z−DEVD−アミノルシフェリンを含有するCaspase−Glo(商標)3/7試薬(Caspase−Glo(商標)3/7 Assay、Promega社)を、(Z−DEVD)2−ローダミン−110又はAc−DEVD−AMCと組み合わせた。Z−DEVD−アミノルシフェリンのカスパーゼ−3開裂に由来するルミネセンスシグナルは、30分目に読み取り、一方、カスパーゼ−3開裂活性由来の蛍光シグナルは、適切なAMC又はローダミン110フィルターセットを用いて、2時間目に読み取った。
【0093】
結果
ルミネセンスは、背景比に対して最大のシグナルを与え、次いでローダミン−110、次いで、AMCであった(図6)。カスパーゼ−3を検出する3種の基質全てが、知られたカスパーゼ−3阻害剤の添加によって、一貫して影響を受けなかった。このことは、いずれかのアッセイが実施される際に、例えば、可能性のある偽妨害を制御するために、ルミネセンス発生試薬と蛍光発生試薬とを組み合わせることができるということを示唆している。したがって、ある試薬がある一定の酵素の真の阻害剤であるかどうかを判定するために多重シグナルを用いるこができる。
【0094】
(実施例III)
さらなる典型的多重アッセイ
A.単一ウェルフォーマットにおける乳酸デヒドロゲナーゼ(LDH)及びアデノシン三リン酸に関する多重アッセイ
以下の検出試薬を調製した:1)LDH試薬(30mM HEPES、pH7.4、10mM NaCl、20mM MgSO4、250μMレザズリン(Aldrich))を用いて、CytoTox−ONE(商標)Homogeneous Membrane Integrity Assay(Promega社、Technical Bulletin306)からの凍結乾燥基質成分を再構成した;2)ATP試薬(30mM HEPES pH7.4、10mM NaCl、20mM MgSO4)を用いて、CellTiter−Glo(商標)Luminescent Cell Viability Assay(Promega社、Technical Bulletin288)からの凍結乾燥基質成分を再構成した;3)LDH/ATP組合せ試薬(30mM HEPES、pH7.4、10mM NaCl、20mM MgSO4、250μMレザズリン(Aldrich))を用いて、CytoTox−ONE(商標)からの凍結乾燥基質成分を再構成し、次に、これを用いて、CellTiter−Glo(商標)からの凍結乾燥基質成分を再構成した。
【0095】
10mM HEPES pH7.5、0.1%Prionex(PentaPharma社)溶液を用いて、LDH(0、1:8000、1:4000、1:2000、菱形)、ATP(0μM、1.25μM、2.5μM、及び5μM、四角形)、及びLDH/ATPの組合せ(それぞれ、0/0μM、1:8000/1.25μM、1:4000/2.5μM、及び1:2000/5μM、三角形)のサンプル希釈液を作製し、100μlの該希釈液(n=4)を、白色96ウェルプレートのウェルに加えた。該サンプルに適切な検出試薬(100μl)を加え、該プレートを遮光し、30秒間混合し、室温で温置した。温置8分後、フィルターを560EX/590EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。温置30分後に、Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0096】
結果
対照反応(図7C)と比較した場合、ATPに関するルミネセンス発生アッセイに及ぼすLDH及びその蛍光発生検出試薬の軽微な影響があった(図7A);しかし、ATPの検出は、依然として可能であった。LDHに関する蛍光発生アッセイへルミネセンス発生検出試薬を添加しても背景蛍光に影響を与えず(図7B)、対照反応(図7D)に比較して全体的な蛍光は減少したが、LDH活性は、依然として検出可能であった。
【0097】
B.単一ウェルフォーマットにおけるLDH及びカスパーゼ−3に関する多重アッセイ
以下の検出試薬を調製した:1)238μMレザズリンを添加したLDH試薬−Caspase−Glo(商標)3/7緩衝液を用いて、CytoTox−ONE(商標)凍結乾燥基質成分を再構成した;2)カスパーゼ−3試薬−Caspase−Glo(商標)3/7緩衝液を用い、Promega Technical Bulletin322にしたがって、Caspase−Glo(商標)3/7凍結乾燥基質成分を再構成した;3)LDH/カスパーゼ−3組合せ試薬−LDH試薬(上記の通りに調製)を用いて、凍結乾燥Caspase−Glo(商標)3/7基質を再構成した。LDH/カスパーゼ−3組合せ試薬のLDH検出試薬化合物は、Caspase−Glo(商標)3/7凍結乾燥基質中のDTTの存在によって不安定であるため、この試薬は、サンプルへの添加直前に調製した。
【0098】
10mM HEPES pH7.5、0.1% Prionex(PentaPharma社)溶液中で、サンプル希釈液を調製した:LDHの0、1:8000、1:4000、1:2000希釈液(菱形);カスパーゼ−3の0、5、10、及び20U/ml(BIOMOL Laboratories、四角形)、及びLDH/カスパーゼ−3の組合せ(それぞれ、0/0U/ml、1:8000/5U/ml、1:4000/10U/ml、及び1:2000/20U/ml、三角形)。100μlの該希釈液(n=4)を、白色96ウェルプレートに加えた。該サンプルに適切な検出試薬(100μl)を加え、該プレートを遮光し、30秒間混合し、室温で温置した。室温で温置6分後、フィルターを560EX/590EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。温置45分後に、Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0099】
結果
多重反応に蛍光発生LDH検出試薬を加えると、対照反応に比較してルミネセンスの減少があった(それぞれ、図8A及び8C)。図8Aにおける全体的なルミネセンスシグナルの減少に関わらず、蛍光発生LDH検出試薬の存在下、ルミネセンスカスパーゼ−3アッセイは機能的であった。図8Bは、多重反応にルミネセンス発生カスパーゼ−3検出試薬を加えると、対照(図8D)に比較して、蛍光背景の増加があることを示している。;しかし、図8Aは、カスパーゼ−3に関するルミネセンス発生検出試薬の存在下で、LDHに関する蛍光発生アッセイが機能的であることを証明している。背景ルミネセンスに対するLDHの影響はなく(図8C)、また、背景ルミネセンスに対するカスパーゼ−3の影響もなかった(図8D)。
【0100】
C.単一ウェルフォーマットにおけるカスパーゼ−3及びタンパクキナーゼA(PKA)に関する多重アッセイ
以下の検出試薬を調製した:1)PKA試薬−100mMトリス pH7.3、100mM MgCl2、PKAローダミン−110基質(ProFluor(商標)PKA Assay、Promega社、 Technical Bulletin315)の1:1000希釈液、及び400μM ATPを含有する1X反応緩衝液を調製した;2)カスパーゼ−3試薬−100mMトリス pH7.3、100mM MgCl2、150μg/mlの組換え熱安定性ルシフェラーゼ、80μM Z−DEVD−アミノルシフェリン(Promega社)、400μM ATP、100μM DTT(Promega社)、2.5mM CaCl2(Fisher)、40mM MgSO4(Fisher)、及び0.2% Tergitol NP−9(Sigma)を含有する1X反応緩衝液を調製した;3)キナーゼ/カスパーゼ−3組合せ試薬−100mMトリス pH7.3、100mM MgCl2、PKAローダミン−110基質の1:1000希釈液、150μg/mlの組換え熱安定性ルシフェラーゼ、80μM Z−DEVD−アミノルシフェリン、400μM ATP、100μM DTT、2.5mM CaCl2、40mM MgSO4、及び0.2% Tergitol NP−9を含有する1X反応緩衝液を調製した;4)タンパクキナーゼ停止試薬−100mMトリス pH7.3、100mM MgCl2、プロテアーゼ試薬(ProFluor(商標)PKA Assay)の1:50希釈液、30μM スタウロスポリン(BIOMOL Laboratories)を含有する1X停止試薬を調製した。
【0101】
10mM HEPES pH7.5、0.1% Prionex(PentaPharma社)溶液中で、サンプル希釈液を調製し;0、1、2、及び4U/ml PKA(菱形)、0、5、10、及び20U/mlカスパーゼ−3(四角形)、及びPKAとカスパーゼ−3との組合せ(それぞれ、0/0U/ml、1/5U/ml、2/10U/ml、及び4/20U/ml、三角形)、40μlの該希釈液(n=4)を、白色96ウェルプレートに加えた。該サンプルに適切な検出試薬(40μl)を加え、該プレートを遮光し、30秒間混合し、室温で20分間温置した。温置後、キナーゼ試薬単独又はキナーゼ/カスパーゼ−3試薬の組合せのいずれかを含有するウェルに、40μlのタンパクキナーゼ停止試薬を加えた。該プレートをさらに30秒間混合し、遮光し、室温でさらに30分温置した。フィルターを485EX/527EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0102】
結果
蛍光発生PKAアッセイ検出試薬の添加により、PKAは存在するが完全なPKA検出試薬が存在しない対照反応(図9C)に比較して、ルミネセンス背景の増加が生じた。しかし、カスパーゼ−3の活性から生じた反応ルミネセンスは比例して増加し、該条件は、ルミネセンス発生カスパーゼ−3反応それ自体に影響したとは思われなかった。PKAに関する蛍光発生アッセイに、カスパーゼ−3に関する検出試薬を添加すると(図9B)、カスパーゼ−3は存在するが完全なカスパーゼ−3検出試薬が存在しない蛍光対照反応(図9D)に比較して、全体の蛍光が50%超減少した。カスパーゼ−3及びPKA活性は、これらの多重条件を用いて、背景以上を測定できた。
【0103】
D.単一ウェルフォーマットにおけるウミシイタケルシフェラーゼ及びカスパーゼ−3に関する多重アッセイ
以下の検出試薬を調製した:1)ウミシイタケルシフェラーゼに対する細胞透過性修飾セレンテラジン基質であるEnduRen(商標)(Promega社)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェートを添加したF−12組織培養培地中に、600μMに希釈した;2)カスパーゼ−3基質:(Z−DEVD)2−ローダミン−110(Promega社)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェートを添加したF−12組織培養培地中に、250μMに希釈した;3)ルシフェラーゼ/カスパーゼ−3組合せ基質:EnduRen(商標)(600μM)及び(Z−DEVD)2−ローダミン−110(250μM)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェートを添加したF−12組織培養培地中に、希釈した。
【0104】
ウミシイタケルシフェラーゼ(CHO−KlhRL25)を安定して発現するCHO−Kl細胞(ATCC)を、10%ウシ胎仔血清及び500μg/mlのG−418サルフェート中で維持し、細胞ベースの実験に用いた。これらの細胞を用いる実験条件には、以下が含まれた:1)スタウロスポリンの添加によるルシフェラーゼ活性レベルの変化、2)カスパーゼ−3酵素の添加と共にスタウロスポリンの添加によるルシフェラーゼ活性レベルの変化、及び3)スタウロスポリンは添加せずにカスパーゼ−3酵素の添加によるルシフェラーゼ活性。
【0105】
CHO−KlhRL25細胞を採収し、96ウェル、透明底、白色壁の組織培養プレート中に20,000細胞/ウェルの密度で塗布し、5%CO2中、37℃で一晩温置した。最終濃度が0、0.5、1、2μMのスタウロスポリン(10μl/ウェル)を適切なウェルに加え、細胞死を開始させ、したがってルシフェラーゼ活性を変化させた。5%CO2中37℃でさらに3.5時間、細胞を温置した。種々の濃度のカスパーゼ−3(BIOMOL Laboratories)を、組織培養培地中、0、5、10、及び20U/mlで、適切なウェルに加えた(10μl/ウェル)。したがって、データポイントに関するスタウロスポリン/カスパーゼ−3組合せ濃度は、それぞれ、0μM/0U/ml、0.5μM/5U/ml、1μM/10U/ml、及び2μM/20U/mlであった。カスパーゼ−3酵素の添加直後、10μl/ウェルのルシフェラーゼ基質、カスパーゼ−3基質、又はルシフェラーゼ/カスパーゼ−3基質のいずれかを、適切なウェルに加えた。検出試薬の添加後、プレートを手短に混合し、5%CO2中37℃で2時間温置した。フィルターを485EX/527EMにセットしたLabsystems Fluoroskan Ascent プレート読取り機上で蛍光を測定した。Dynex MLXプレートルミノメータを用いて、ルミネセンスを記録した。
【0106】
結果
これらのアッセイにおいて、ウミシイタケルシフェラーゼの活性を、細胞死に関する内部対照として用いた。したがって、スタウロスポリンの濃度が増加するにつれて、ルシフェラーゼの活性は減少するはずである。図10Aは、カスパーゼ−3基質の添加は、対照反応(図10C)と比較して、ルシフェラーゼ反応に負の影響を与えなかったことを示している。図10Bは、ルシフェラーゼ基質の添加が、図10Dに比較して、全体の蛍光にわずかな増加はあったものの、背景蛍光に影響を与えなかったことを示している。ウミシイタケルシフェラーゼに関するルミネセンス発生アッセイは、カスパーゼ−3又はカスパーゼ−3基質の存在下で十分に機能的であり、カスパーゼ−3に関する蛍光発生アッセイは、わずかに影響を受けただけで、ウミシイタケルシフェラーゼ又はウミシイタケルシフェラーゼ基質の存在下で十分に機能的であった。
【0107】
(実施例IV)
プロテアーゼ保持及び放出細胞生存度多重アッセイ
生細胞及び死細胞アッセイは、特定の化学的、生物学的又は物理的処理に応答した細胞生存度の変化をモニターするために、広く用いられている。生存度と細胞毒性アッセイとは、一般的に逆であり、異なった生物マーカーを測定する。細胞毒性による細胞生存度の一般的変化を評価する方法は、歴史的に外部膜透過性の変化に関連してきた。膜構造の損傷を検出する古典的方法としては、トリパンブルー排除、核酸染色、及び乳酸デヒドロゲナーゼ放出が挙げられる(Rissら、2004年;Myersら、1998年)。細胞機能又は増殖の評価に関するアッセイとしては、トリチウムチミジン取込み、ATP含量、テトラゾリウム色素移行又はフルオレセインジアセテートが挙げられる(Cookら、1989年)。無処置細胞膜は、かさばった荷電分子又はペプチドを、細胞外空間から細胞質内へ進入させないと考えられる。逆に、損傷した膜は、色素又は化合物を細胞内へ、又は細胞内容物を細胞外へ自由に透過させる。この透過現象が、色素標識(「生命」色素、DNA挿入剤、又はエステラーゼ修飾フルオレセイン)並びにLDH放出アッセイの双方にとっての基礎である。細胞生存度を測定する既存の方法は、依然として有用であり、コスト効率の良い適用である一方、それらは多くの技術的又は実用的な欠点を有し、高含量、多重又は高スループットフォーマットにおける利用が制限されている。例えば、LDH放出(CytoTox−ONE(商標))又は色素減少能力(CellTiter−Blue(商標))による細胞膜の完全性の現在の測定は、レザズリン基質の共有及びEx/Emスペクトルの重なりのために、対にする(データ正規化の手段)ことができない。さらに、双方のアッセイに利用される着色レアズリン基質は、他のエンドポイントアッセイ測定(消色)との第2のアッセイシグナルウィンドー強度(及び感度)を制限し、濃度及びフォーマットが、第2のアッセイ試薬ペアリングに関して最適化されない(例えば、容量が限定される)。
【0108】
既存の生/死細胞フォーマットでは、カルボキシフルオレセイン及びエチヂウムホモ二量体が用いられるが、後者は知られた強力な変異誘発因子である。このフォーマットには、洗浄及び細胞培養培地の置換が必要である。さらに、カルボキシフルオレセインは、水性溶液中で自然加水分解を示し、エチヂウムホモ二量体挿入はDNAを染色し、下流のデータ正規化を妨害し得る。
【0109】
培養哺乳動物細胞は、プロテアーゼ類、エステラーゼ類、リパーゼ類、及びヌクレアーゼ類の豊富な環境を含有する。例えば、プロテアーゼの4つの一般的なクラス(アスパラギン酸、システイン、セリン、及び金属依存性)が代表であり、恒常性維持の特異的機能に関連している。これらの細胞質の、リソゾームの、及び膜貫通結合のプロテアーゼは、細胞内タンパク質分解、免疫原性ペプチドの生成、翻訳後修飾、及び細胞分裂に関与している(Tranら、2002年、Constamら、1995年、Vinitskyら、1997年)。これらの酵素の活性は、特殊区画化などの種々の機構によって調節されている(Bondら、1987年)。過剰なストレス、環境の悪状況、又は細胞死プログラムの委任進行に応答して、区画化及び膜完全性の整合喪失が見られる(Syntichakiら、2003年、Haunstetterら、1998年)。したがって、インビトロ細胞モデルにおける細胞培養培地内への安定なタンパク質分解伝達物質の放出は、細胞死の潜在的代理物となる。逆に、保持されたタンパク質分解酵素の細胞酵素学的染色は、細胞健康度の表現型測定にパラレルである。このようなタンパク質分解活性が一緒になって、細胞培養集団、例えば「生/死」アッセイ中の生存細胞又は損傷細胞の相対的数の確認を補助し得る。
【0110】
プロテアーゼベースの生/死細胞アッセイでは、一実施形態において、1つの基質(死細胞に関する)は、細胞質pH、例えば、7.0から7.2において安定で活性であり、スペクトル的に読取り(R/O)の異なる標識を有する、比較的豊富で活性であり保存されたプロテアーゼに対する基質である。この基質の開裂動態は、LDH放出にパラレルであり、活性の条件が毒性物質又は膜変化物質、例えば、塩又はチオール類を含まず、アッセイ時間を高速にすることが好ましい。他の基質(生細胞に関する)は、比較的豊富で保存されたプロテアーゼに対する基質であり、生存細胞に関しては細胞透過性であり、また、該プロテアーゼは、生存細胞の細胞質環境では活性であるが、細胞外環境では不安定である。この基質は、スペクトル的にR/Oの異なる標識を有し、開裂反応は、アッセイ時間を高速にするように進行する。非破壊的アッセイにおける2種の基質の使用により、望ましくない増殖事象を検出でき、また、生存度対細胞毒性の比率は、そのウェルにおける細胞数の変動に独立であるため、異なるスペクトルにおける相補的で独立の代理物を用いることにより、誤った結論及び細胞凝集又はピペット操作エラーによるエラーを減らすことができる。
【0111】
A.AMC若しくはR110蛍光又はアミノルシフェリンルミネセンスレポーターによるプロテアーゼ放出アッセイフォーマット
HL−60細胞を、2倍連続希釈してからトリトンXの添加によって、0.2%最終に細胞溶解させるか、又は媒体添加によって維持した。100mM酢酸ナトリウム、pH4.5中、200μM Ala−Ala−Phe−AMC基質の1/10容量を、溶解物又は細胞に加え、37℃でさらに1時間温置した。次いで、溶解細胞又は生存細胞に関連した蛍光を、CytoFluorIIを用いて、Ex.360 Em.460で測定した。
【0112】
活動的ダブリングを受けているジャーカット細胞を、トリパンブルー排除によって数えると、95%超の生存が判明した。該細胞をRPMI1640+10%FBS中、100,000細胞/mlに調整し、2つのアリコートに分けた。1つのアリコートは、マイクロチップを備えたMisonix3000を、3×5秒パルスの30%出力で用いて超音波処理した。他方の画分は、超音波操作(合計約5分)の間、37℃の水浴中で温置した。次いで、細胞懸濁液及び溶解物画分を比率混合によって、0〜100%の生存度を表す種々の生存度へ混合した。次に、混合した細胞サンプルを、100μl容量で白色壁、透明底の96ウェルプレート(Costar)に加えた。(Ala−Ala−Phe)2−R110を、RPMI−1640中、1000μMni希釈し、1/10容量を該プレートに加えた。該プレートを30分間温置してからCytoFluorIIを用いて、蛍光を、Ex.485 Em.530で測定した。
【0113】
活動的ダブリングを受けているジャーカット細胞を、トリパンブルー排除によって数えると、95%超の生存が判明した。該細胞をRPMI1640+10%FBS中、100,000細胞/mlに調整し、2つのアリコートに分けた。1つのアリコートは、マイクロチップを備えたMisonix3000を、3×5秒パルスの30%出力で用いて超音波処理した。他方の画分は、超音波操作(合計約5分)の間、37℃の水浴中で温置した。次いで、細胞溶液及び溶解物画分を比率混合によって、0〜100%の生存度を表す種々の生存度へ混合した。次に、混合した細胞サンプルを、100μl容量で白色壁、透明底の96ウェルプレート(Costar)に加えた。10mlの10mM Hepes、pH7.5を有するルシフェリン検出試薬ケーク(Promega V859A)の再水和により、ルミネセンス発生プロテアーゼ放出アッセイ試薬を調製し、この試薬に、Ala−Ala−Phe−アミノルシフェリンを添加し、100μMの最終濃度にした。該プレートのウェルに、100μlのルミネセンス発生プロテアーゼ放出アッセイ試薬を加え、BMG FLUOstar Optimaを用いて、動態様式でルミネセンスを測定した。
【0114】
AMC蛍光フォーマットの実際の感度は、CytoTox−ONE(商標)と同等の感度値である、約240個の細胞(図11)であると算出された。R110フォーマット(図12)のアッセイは、同様に鋭敏であり、多重適用のためにさらに別の蛍光体を提供した。特にこれらのアッセイの感度は、下流の多重適用におけるCytoTox−ONE(商標)又は他のレザズリンベースのアッセイの使用の主要な障害である蛍光の消光なしで得られた。ルミネセンスフォーマットの優れた直線性と範囲(図13)により、9800個の生細胞集団中、200個と少ない細胞の統計的検出を可能にした。非溶解性のルミネセンスフォーマットは、細胞毒検出のために別の代替法を提供する。
【0115】
B.種々の酵素標的によるプロテアーゼ放出アッセイのフォーマット
活動的ダブリングHL−60細胞を、100,000個の細胞/mlに調整し、2つのアリコットに分けた。1つのアリコットは、マイクロチップのMisonix3000を用いて3回の5秒パルスの30%出力で超音波処理した。他のアリコットは、37℃で保持した。次に細胞懸濁液及び溶解物を、100μl容量でRPMI 1640+10%FBS中で2倍連続希釈した。培地のみのものは、無細胞対照として寄与した。ルシフェリン検出試薬のケーク(Promega V859A)を、2.0mlの10mM Hepes、pH7.5で再懸濁した。次いでルシフェリン検出試薬を分けて、Z−Leu−Leu−Val−Tyr−アミノルシフェリン又はAla−Ala−Phe−アミノルシフェリンによる1mMを作製した。各試薬を、1/10容量で独立した反復試験のプレートに加え、Me’Courの熱ジャケット水浴ホルダー中37℃で15分間温置させてから、BMG FLUOstar Optimaを用いてルミネセンス測定を行った。
【0116】
Z−LLVY−アミノルシフェリンアッセイは、AAF−アミノルシフェリンシーケンスよりも少ない最適さで実施したが、他のプロテアーゼ類は、完全性の欠いた代用物として使用できることが立証された(図14)。この場合、LLVY活性は、プロテオソームのキモトリプシン活性に起因し得る。
【0117】
C.プロテアーゼ放出の時間経過
HL−60細胞(25,000個/ウェル)を、透明底の白壁で仕切られた96ウェルプレート(Costar)内で5%CO2と共に37℃で7時間の経過にわたり、10μMスタウロスポリン又は適性のDMSO媒体対照により処理した。200μMのAla−Ala−Phe−AMC基質溶液を、100mMのNaアセテート、pH4.5中で作製した。10μl容量の基質(1/10容量のサンプル)をウェルに加えてさらに1時間温置した。「プロテアーゼ放出」活性を、CytoFluor II上Ex.360Em.460で測定した。平行にセットされたウェル内で、CytoTox−ONE(商標)試薬は、膜完全性アッセイ対照として作用した。この試薬を、Ex.560 Em.580で蛍光測定10分前に加えた。
【0118】
細胞透過性の動態学、すなわち、LDHとプロテアーゼ放出は、互いに反映され、細胞集団における二次的壊死の形態学的観察と一致した(図15)。酸性Naアセテート製剤(サンプル中約6.5の最終pH)中アミノペプチダーゼ基質の提示は、潜在的なリソソームプロテアーゼ活性を調整するために実施された。
【0119】
D.プロテアーゼ放出活性のpH要件
プロテアーゼ放出活性のpH要件は、非調整培養培地(水媒体)と比較して、pH2.5、3.5及び4.5に調整された100mM Naアセテートを用いて調査された。Ala−Ala−Phe−AMCを、これらの緩衝液中200μMに加えた。溶液の1/10容量をプレートに加え、軌道振とうにより短時間混合した。このプレートを37℃で40分間温置してから、蛍光を、CytoFluor IIを用いてEx.360Em.460で測定した。
【0120】
pH4.5のNaアセテート1/10容量の添加により、培養培地を約6.5の最終pHに減じた。他のより低いpH溶液/培地の組合せの最終pHは、試験しなかったが、以前の実験により、pH2.5のNaアセテートの1/10容量の添加は、細胞培地のpHを約5.5に減じることを示唆した。非pH調整媒体は、プロテアーゼ放出活性にとって最も好ましいことを示すことが判った(図16)。この活性は、細胞基質のアミノペプチダーゼと一致し、恐らくリソソームプロテアーゼ(カテプシンなど)ではないと思われる。プロテアーゼ放出活性を測定するために、有害な又は潜在的細胞毒性の付加物を必要としないことからこのことは重要である。このことにより、温置時間枠をより柔軟にでき、可能なルミネセンスベースのアッセイをさらに受けられることが考慮に入れられる。
【0121】
E.プロテアーゼ放出酵素の細胞下位置
HL−60細胞を、1ml当り100,000個の細胞に調整し、2つのアリコットに分けた。1つのアリコットは、マイクロチップのMisonix3000を用いて3回の5秒パルスの30%出力で超音波処理した。100μlのこの溶解物(形態学的に確認された)を、透明底96ウェルプレートの複数ウェルに加え、10%FBSを有するRPMI 1640中で2倍連続希釈した。同様に、100μlの非超音波処理の細胞懸濁液を加え、プレートの複数ウェルで連続希釈した。NP−9及びジギトニンを、それぞれ最終0.2%及び30μg/mlで個別のウェルに加えた。未処理の対照は、生細胞及び適性容量の水媒体から構成された。ルシフェリン検出ケーク(Promega V859A)を、2mlの10mM Hepes、pH7.5で再水和し、Ala−Ala−Phe−アミノルシフェリン(Promega)により500μMを作製した。20μlのこのプロルミネセンスプロテアーゼ放出溶液を、全てのウェルに加えて、37℃で15分間温置させてから、BMG FLUOstar Optimaを用いてルミネセンスを測定した。
【0122】
超音波処理及び上記のパラメータと濃度を有するNP−9は、外膜のみならず、リソソーム内容物(カテプシン放出により測定された)も破壊することが知られている(図17)。ジギトニンによる選択的破壊は、リソソーム破壊の証拠のないトリパンブルー染色を考慮に入れている。したがって、活性は、超音波処理又は差別的界面活性の細胞溶解の間で類似していたため、最適pHを考慮して、プロテアーゼ放出アッセイで測定されたプロテアーゼは、恐らく細胞基質性であり、無処置の細胞小器官の外部にあることが推測され得る。
【0123】
F.プロテアーゼ放出又は保持酵素基質の選択性
Ala−Ala−Phe−AMCを、Promegaから入手した。Z−Leu−Leu−Val−Tyr−アミノルシフェリン、Z−Leu−Arg−アミノルシフェリン、Z−Phe−Arg−アミノルシフェリン、Ala−Ala−Phe−アミノルシフェリン、(Ala−Ala−Phe)2−R110、及び(Gly−Phe)2−R110は、Promega Biosciencesにより合成された。Suc−Ala−Ala−Phe−AMC、H−Phe−AMC、H−Tyr−AMC、グルチル−Ala−Ala−Phe−AMC、H−Gly−Phe−AMC、Z−Gly−Ala−Met−AMC、Suc−Leu−Leu−Val−Tyr−AMC、D−Ala−Leu−Lys−AMC、H−Gly−Ala−AMC、H−Gly−Gly−AMC、Suc−Ala−Ala−Phe−AMC、Z−Arg−Leu−Arg−Gly−Gly−AMC、Z−Leu−Arg−Gly−Gly−AMC及びAc−Ala−Ala−Tyr−AMCは、Bachemから供給された。Gly−Phe−AFC、Pro−Phe−Arg−AMC、Gly−Gly−Leu−AMC、及びSer−Tyr−AFCは、Calbiochemから入手した。Z−Phe−Arg−AMC及びSuc−Arg−Pro−Phe−His−Leu−Leu−Val−Tyr−AMCは、Sigmaから購入した。
【0124】
全ての基質は、固有の溶解度によって10mMから100mMでDMSOに可溶性であった。蛍光性基質を、10mM Hepes、pH7.5又は10%血清を有する適性な細胞培地100μMから1mMに希釈し、白壁仕切りの透明底の96ウェルプレート中の溶解細胞(凍結破壊、超音波処理、又は界面活性剤)又は未処理生存細胞に1/10容量で加えた。HL−60又はジャーカットを、それらの容易に操作された懸濁表現型のため実験に交換可能に用いた。プレートを、CytoFluorIIによる蛍光を測定する前に37℃で15〜30分間温置した。
【0125】
ルミネセンス基質を、2mlの10mM Hepes、pH7.5に再懸濁されたルシフェリン検出ケーク(Promega V859A)に500μMで加えた。1/5容量のプロルミネセンス反応混合物を、白壁仕切りの透明底の96ウェルプレート中、溶解細胞(凍結破壊、超音波処理、又は界面活性剤)又は未処理生存細胞に加えた。やはりHL−60又はジャーカットを、実験に交換可能に用いた。プレートを、Caron2050W交換ユニットにより制御されたMeCour’循環熱ブロック内で37℃で温置した。ルミネセンスを、15分から30分の間(シグナル定常状態)で測定した。
【0126】
損傷細胞又は生存細胞内でのプロテアーゼ放出又は保持に関する潜在的基質の優先度を特性化するために、努めて多種多様のタンパク分解基質を調べた(表4を参照)。エンドペプチダーゼ又はエキソペプチダーゼの活性が勝っているかどうかを正確に概説するために、アミノ末端ブロック化基質(Z、Suc−、又はAc−)が選択された。非ブロック化基質(H−など)を、アミノペプチダーゼ活性の寄与を含めるために調べた。このパネルから、少なくとも3つのタンパク分解プロフィルが明らかになった:非ブロック化Ala−Ala−Pheトリペプチドを優先させるアミノペプチダーゼ様活性、ブロック化Leu−Leu−Val−Tyrペプチド類の放出により測定されたプロテオソーマル(キモトリプシン様)活性、及びGly−Phe、Gly−Ala、Phe−、Tyr−又はGly−Gly−Leu基質による極めて不安定な活性。後者の活性は、生存する無処置細胞においてのみ測定できた。さらに重要なことは、これらの活性を検出するためにいくつかの蛍光体又はプロルミネセンス標識が使用でき、最終的には下流の多重化適応性の増大が考慮に入れられることである。
【0127】
【表4】
無しは、対照集団を超える統計的活性が無いことを示す。
(+)から(+++++)は、対照集団を超える多少の活性から強固な活性範囲を示す。
【0128】
G.プロテアーゼ保持活性及び生存細胞要件
ジャーカット細胞を、50μl容量で1ウェル当り20,000個の密度で白壁仕切りの透明底の96ウェルプレートに接種した。アポトーシス誘導試剤rTRAIL(BioMol)の連続希釈液を、RPMI1640+10%FBSで500ng/mlから作製し、50μl容量で反復して細胞に加えた。50μl培地の添加は、媒体対照として寄与した。このプレートを、5%CO2と共に37℃で4時間温置した。Gly−Phe−AFCを、RPMI1640中1mMに希釈し、10μ容量で全てのウェルに加えた。次にプレートをMeCour’循環熱ブロックに30分間置いてから、CytoFluor IIによりEx.405Em.530で蛍光を測定した。次に、等しい容量のCellTiter−Glo(商標)試薬をウェルに加え、細胞内に残存するATP含量を、FLUOstar Optimaによるルミネセンス測定により調べた。
【0129】
相対的ATP濃度及びプロテアーゼ保持活性は、事実上多重化でき、最適の保持酵素活性のための細胞生存度又は安定な細胞膜に関する要件を示唆している(図18)。膜完全性の乱れは、保持活性に対して非常に有害であることから、この活性を、集団生存度検出用の「生/死」フォーマットに関する放出活性と結合させることができる。
【0130】
H.種々のペプチド配列及びレポーター類を用いるプロテアーゼ保持アッセイフォーマット
ダブリング活性のジャーカット細胞を、100μl容量で白壁仕切りの透明底の96ウェルプレート内で、RPMI1640+10%FBS中37,500細胞/ウェルから連続希釈した。プレートの半分の細胞を、トリトンXの添加により最終0.2%に細胞溶解した。プレートウェルの他の半分は、適性容量(5μl)の水媒体を受けた。Tyr−AMC、Phe−AMC及びGly−Phe−AFCの全てを、DMSO中100mMに再水和化してから、RPMI1640で1mMに希釈した。希釈基質の1/10容量をウェルに加え、軌道振とうにより短時間混合してから、5%CO2中37℃で1.5時間まで温置した。生じた蛍光を、30分と90分でEx.360 Em.460及びEx.405、Em.500で測定した。
【0131】
生細胞と死細胞とを区別するのに良好に作用するペプチド配列(生細胞によって標的にされ利用される)は、生存細胞の細胞質に恐らく自由に入ることができる非ブロック化モノ−ペプチド基質又はバイ−ペプチド基質のいずれかである(図19)。候補基質の(Gly−Phe)2−R110は、明らかに細胞膜を有効に通過できないか、或いは候補プロテアーゼにより有効に開裂されることはなかった。
【0132】
I.プロテアーゼ放出活性の半減期
ジャーカット細胞を、100μl容量で1ウェル当り20,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種した。サポニン(Sigma)を加え、短時間軌道振とうすることにより最終0.2%濃度(5μl添加)に混合し、8時間の時間経過にわたって毎時間ウェルを反復させた。この同じ時間枠内で等しい容量の10%FBSを有するRPMI1640を対照ウェルに加えた。Ala−Ala−Phe−AMCを、RPMI1640+10%FBSで500μMに希釈し、ウェルに10μl容量で加え、軌道振とうにより短時間混合してから、5%CO2中37℃で1時間温置した。生じた蛍光を、CytoFluor IIで測定した。
【0133】
放出されたプロテアーゼ活性の半減期は、活性減衰を外挿すると10時間近い(図20)。細胞培養溶解物におけるこの延長された活性は、好ましいことにおよそ9時間の推定半減期を有するラクテートデヒドロゲナーゼ(LDH)と同等である。この観察は、処理集団における細胞死に対する代用物であるプロテアーゼ活性に関して重要である。簡単に言うと、シグナル半減期が延長すると、典型的なインビトロプロトコルにおける細胞死の報告におけるアッセイが有する有用性が増加する(活性を減少させて応答を過小評価することなく)。
【0134】
J.プロテアーゼ保持及び放出活性阻害/増強プロフィル
プロマイシン、E−64、フェニルメタンスルホニルフルオリド(PMSF)、アデノシン−5’−トリホスフェート(ATP)、N−(α−ラムノピラノシルオキシヒドロキシホスフィニル)−Leu−Trp二ナトリウム塩(ホスホラミドン)、N−[(2S,3R)−3−アミノ−2−ヒドロキシ−4−フェニルブチリル−L−ロイシン塩酸塩(ベスタチン)、1,10−フェナントロリン、3,4−ジイソクマリン、4−(2−アミノエチル)ベンゼンスルホニルフルオリド(AEBSF)、1,4−ジチオ−DL−トレイトール(DTT)、エデテート二ナトリウム二水和物(EDTA)、イソバレリル−L−バリル−L−バリル−[(3S,4S)−4−アミノ−3−ヒドロキシ−6−メチルヘプタノイル]−L−アラニル[93S,4S]−4−アミノ−3−ヒドロキシ−6−メチルヘプタン酸(ペプスタチンA)、塩化ナトリウム、アプロチニン、N−アセチル−L−ロイシル−L−ロイシル−L−アルギニナルヘミサルフェート塩(ロイペプチン)は全てSigmaから購入した。阻害剤は、溶解物又は生存細胞集団のいずれかに添加するために、Mg++又はCa++(DPBS)の無いダルベッコ燐酸緩衝生理食塩水中、200μM又は200μg/mlの高標的濃度の保存濃度を変えてDMSO中に再懸濁した。DTT、NaCl、EDTA、及びATPもまた、DPBS中に希釈した。全ての化合物を、損傷細胞又は生存細胞と共に37℃で少なくとも30分間(大部分は60分間)温置してから活性を評価した。
【0135】
プロテアーゼ保持アッセイ阻害剤/補助剤の調査は、先に記載されたように、最終100μM濃度でGly−Phe−AFCを用いて、超音波処理、サポニン溶解、又は生存HL−60及び/又はU937に対して実施された。このプロテアーゼ放出アッセイ阻害剤/補助剤の調査は、Ala−Ala−Phe−AMC又は(Ala−Ala−Phe)2−ローダミン110を用いて、超音波処理、サポニン溶解、又は生存HL−60、SK−MEL−28及び/又はU937に対して実施された。
【0136】
種々のクラスの阻害剤又は補助剤の存在下でのプロテアーゼ保持活性プロフィルは、観察された活性の大部分は、アミノペプチダーゼに関するものであることを示している(プロマイシン、EDTA、及びベスタチン感度)(表5及び表6を参照)。この活性は、ATP非依存性並びにDTT非依存性(活性の回復が無い)であり、並びにハライド類(Cl−)に非感受性である。この活性は、システイン又はセリンプロテアーゼクラスの酵素に関連しているように思われる。
【0137】
プロテアーゼ放出活性プロフィルは、トリブシン又はキモトリプシン様活性に選択性を有するものではないが、セリンプロテアーゼ阻害剤に感受性があるように思われる。チオール類(システインクラスに強く示される)にとっての明白な要件はなく、アスパラギン酸及びメタロプロテアーゼの特異的阻害剤は、活性を制御するのに無効である。
【0138】
プロテアーゼ保持活性の原因となる酵素は、生存細胞を必要とし、損傷細胞の外部では検出できない。逆に、プロテアーゼ放出応答を増強させるために混合された補助剤は必要ない。これにより、非毒性、非細胞溶解性フォーマットにおけるアッセイを組み合わせて、それらプロテアーゼ活性の差異に基づいた生細胞と死細胞の検出が考慮に入れられる。
【0139】
【表5】
【0140】
【表6】
【0141】
K.多重プロテアーゼ放出及び保持アッセイ
1.ジャーカット用量応答
ダブリング活性のジャーカット細胞を、50μl容量で1ウェル当り20,000個の密度で96ウェルプレート内に接種した。RPMIアポトーシス誘導リガンド、すなわち、1640中のrTRAILの連続希釈液を、追加の50μl容量中、250ngから244pg/mlの最終濃度で反復ウェルに加えた。RPMIのみのものは、非誘導対照として寄与した。このプレートを、5%CO2中37℃で4時間温置した。Gly−Phe−AFC及びAla−Ala−Phe−AMCを、RPMI中1mMに同時に希釈し、1/10容量でプレートに加え、さらに37℃で30分間温置した。生じた蛍光を、CytoFluor IIを用いてEx 360 Em 460及びEx 405 Em 530で測定した。蛍光測定が完了したら、CellTiter−Glo(登録商標)を等しい添加でウェルに加え、ルミネセンスを、BMG FLUOstar Optimaを用いて測定した。
【0142】
健全細胞の2種の独立した非破壊代用物(プロテアーゼ放出と保持)を、マイクロ−タイタープレートフォーマット内の集団生存度を報告するために多重化した(図21)。生じたデータは、その細胞集団の健全度の逆の目安である。この関係により、対照の使用が考慮に入れられ、正規化レベルが提供される。さらに、生存度の第3の目安(ATP含量)を、妨害又は消光の無い連続多重フォーマット内に加えることができ、データの解釈におけるさらなる信頼が考慮に入れられる。
【0143】
2.SK−MEL−28及びACHN細胞
SK−MEL−28又はACHN細胞を、100μl容量で1ウェル当り10,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種し、5%CO2中37℃で2時間結合させた。結合後、50μlの培地を注意深く除き、MEM+10%FBS中、イオノマイシン又はスタウロスポリンのいずれかの連続希釈で置き換えた。培地のみのものは、対照として寄与した。このプレートを、さらに5時間温置した。Gly−Phe−AFCの1mM溶液をMEM中で作製し、1/10容量でウェルに加えた。生じた蛍光を、CytoFluor IIを用いて測定した。次にCaspase−Glo(商標)3/7試薬を加えて、BMG FLUOstar Optimaを用いて蛍光測定を行った。
【0144】
プロテアーゼ保持基質は、ウェル内の一般的生存度を報告したが、一方、カスパーゼ特異的試薬は、細胞毒の特異的経路を報告した(図22)。この点において、カスパーゼ活性化(したがってアポトーシス誘導)は、SK−MEL−28に対するスタウロスポリンによって明白であるが、一方、イオノマイシンは、壊死タイプのプロフィル開始する。アポトーシスプロフィルはまた、スタウロスポリン処理ACHNによっても観察される。
【0145】
3.HeLa細胞及びタモキシフェン処理
HeLa細胞を、100μl容量で1ウェル当り10,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種し、5%CO2中37℃で2時間結合させた。結合後、0時間、1時間、3時間、5時間、7時間、24時間の曝露時間で50μlの培地を注意深く除き、MEM+10%FBS中、50μMのタモキシフェンで置き換えた。培地のみのものは、対照として寄与した。プロテアーゼ保持及び放出試薬を、2mlの10mM Hepes、pH7.5を有するルシフェリン検出試薬ケークを再水和化することにより調製した。次にこの溶液を、Ala−Ala−Phe−アミノルシフェリン及びGly−Phe−AFC双方により500μMを作製した。この溶液の1/5容量を全てのウェルに加え、Me’Cour熱ユニット内で37℃で15分間温置した。ルミネセンスは、BMG FLUOstar Optimaにより測定し、蛍光は、CytoFluor IIを用いて測定した。
【0146】
本実施例は、混合プラットフォーム(蛍光及びルネセンス)が、プロテアーゼの保持及び放出アッセイの構成において可能であることを立証している(図23)。これらの試薬は、非細胞溶解性で明らかに非毒性であることが顕著であり、それらは、Apo−ONE(商標)アッセイによるカスパーゼ−3/7検出などのスペクトル的に異なる他の下流適用を受けることができることを示唆している。
【0147】
4.DNA染色による生/死プロテアーゼアッセイの利用
HeLa又はHepG2細胞を、100μl容量で1ウェル当り10,000個の密度で白壁仕切りの透明底の96ウェルプレート内に接種し、5%CO2中37℃で2時間結合させた。結合後、50μlの培地を注意深く除き、MEM+10%FBS中、タモキシフェン又はイオノマイシンの連続希釈で置き換えた。培地のみのものは、対照として寄与した。化合物との温置を、さらに5時間続けた。プロテアーゼ保持及び放出試薬は、2mlの10mM Hepes、pH7.5を有するルシフェリン検出試薬ケーク(Promega V859A)を再水和化することにより調製した。次にこの溶液を、Ala−Ala−Phe−アミノルシフェリン及びGly−Phe−AFC双方により500μMを作製した。この溶液の1/5容量を全てのウェルに加え、Me’Cour熱ユニット内で37℃で15分間温置した。ルミネセンスは、BMG FLUOstar Optimaにより測定し、蛍光は、CytoFluor IIを用いて測定した。次に残りの生存細胞を、0.4% NP−9界面活性剤の添加により溶解した。軌道振とう機上で短時間混合後、MEM中、PicoGreen(登録商標)(Molecular Probes)の1:20希釈液を、さらに1/10容量で加えた。DNA/色素結合に伴う蛍光は、CytoFluor IIを用いてEx.485 Em.530で測定した。
【0148】
本実験は、さらに2つの接着細胞タイプのスクリーニングができるようにプロテアーゼベースの生存度試験の有用性を拡張するのみならず、DNA染色による「全体」測定を組み込んでいる(図24)。スペクトルの相違及び混合プラットフォームの読取りのため、全ての測定は、非妨害的であり、また、非消光である。
【0149】
考察
益々精巧な細胞モデル系を利用するために、薬物発見及び一次研究双方の努力が継続されている。実験操作後にこれらのインビトロ系における細胞数及び細胞生存度を測定することが絶対に必要であることは、十分に認識されている。この要件は、測定の信頼性を証明するために、又複雑な生物系の背景内でこれらの応答を正規化するために必要である。
【0150】
不幸にして、細胞生存度並びに細胞毒を定義する現代化学は、生物学的疑問の新規な方法論及び技法と歩調を合わせておらず、したがって実験的オプションを限定してきた。例えば、アッセイの多重化、すなわち同じウェル内での組合せアッセイの出現により、アッセイ性能における有意な低減の無い適合性且つスペクトル的に異なるアッセイを組み合わせるための要件が必要となった。この課題は、健全細胞の一般的な補足測定とカスパーゼ活性化又はレポーター遺伝子モジュレーションなどのより具体的な事象とを結合させる点で特に重要である。
【0151】
細胞生存度及び/又は細胞毒レポーターの測定のための前述の方法論は、多くの下流アッセイ適用に適合性である。このことは、多岐にわたる励起及び発光スペクトルをもつ異なる蛍光体により、又はルミネセンスなどの他のレポータープラットフォームを組み込むことにより達成される。これは、細胞溶解なしで恐らく非毒性環境で達成され、エンドポイント判定のためにアッセイウィンドウにおける適合性を考慮に入れられることは注目すべきである。さらに、この技法は、処理能力、小型化及び自動化に適応させる上で十分高感度であり、コスト効率が良い。種々のアッセイにより提供された利点の比較を、表7に示す。
【0152】
【表7】
【0153】
現在までの結論として、哺乳動物プロテアーゼの試験において公表された努力の残りのものは、主として容易に精製されるか、分泌されるか、又は双方であるものを中心に行われてきた。これらの試験から提供された情報は、タンパク分解機構、構造と機能についての洞察を提供しているが、他のプロテアーゼ類については、プロテオミック予測から推測されているもの以外、ほとんど知られていない。簡単に述べると、細胞内プロテアーゼの機能、調節、細胞下分布、存在量、及び重要性に対して多くの研究を実施する必要がある。
【0154】
益々増加している証拠は、多くの細胞基質プロテアーゼ類が、細胞ホメオスタシスの機構に関与していることを示唆している。プロテオソーム類は、細胞質ペプチド類の遊離に関与していることは明白だが、いくつかの知見は、他の保存細胞質プロテアーゼ類に関する役割を示唆している(Vititskyら、1997年;Constamら、1995年)。
【0155】
本明細書に記載された個々のプロテアーゼアッセイ及びプロテアーゼベースの生細胞/死細胞アッセイは、スペクトルの相違性のため多重化に、より適合性であり、アッセイ相補性又は他のエンドポイントアッセイの組合せ、例えば、AMC、AFC、R110、クレシルバイオレット又はルミネセンスの組合せが考慮に入れられ、染色消滅が無く、容量制限が無く、アッセイ化学のレトロエンジニアリングが無く、温置時間が短く、同様な、又はより良好な実用的感度(スクリーニング環境の細胞生存度におけるパーセント変化)であり、DNA結合アッセイによる下流妨害が無く、洗浄又は遠心分離の必要が無く、例えば均一アッセイである。さらに、プロテアーゼベースの生細胞/死細胞アッセイのデータは、比率(細胞毒指標)で用いられる場合、細胞数に係わりなく正規化でき、例えば、DNAインターカレーション(一次的又は二次的壊死の潜在的結果が同定できる)などの他のアッセイと結合される場合、化合物作用の動態における差異の理由を説明するために、細胞/化合物接触ウィンドウを拡張でき、DNAインターカレーション及びカスパーゼ活性により、例えば、ウェル内の不均一集団において細胞サイクルの薬物応答を確認できる。さらに、プロテアーゼ類の基質は、比較的単純な、例えば、ジ又はトリ−ペプチド類であり得、よく知られた化学により、非毒性及び/又は非変異原性で安定な蛍光体又はルミネセンス基質に結合され、種々のフォーマットにおいて、例えば、DMSO中、又は乾燥して提供できる。
(参考文献)
【表8】
【0156】
全ての刊行物、特許及び特許出願は、参照として本明細書に組み込まれている。前述の明細書において、本発明は、ある一定の好ましいその実施形態に関連して記載され、多くの詳細が、説明目的のために記載されているが、本発明は、さらなる実施形態の影響を受け易く、本明細書に記載されたある一定の詳細は、本発明の基本的原則から逸脱することなく、かなり変え得るものであることは当業者にとって明白であろう。
【特許請求の範囲】
【請求項1】
第1の酵素媒介反応に関する第1の分子、及び第2の酵素媒介反応に関する第2の分子の存在又は量を検出する方法であって、
a)サンプルを、前記第1の酵素により媒介された反応が、ルミネセンス発生生成物を生成し、前記第2の酵素により媒介された反応が、ルミネセンス非発生生成物を生成する前記第1の反応及び前記第2の反応に関する反応混合物に接触させること;及び
b)前記サンプルにおける前記第1の分子及び前記第2の分子の存在又は量を検出することを含む方法。
【請求項2】
前記第1の分子が、前記第1の酵素媒介反応に関する基質である請求項1に記載の方法。
【請求項3】
前記第2の分子が、前記第2の酵素媒介反応に関する基質である請求項1に記載の方法。
【請求項4】
前記第1の分子が、前記第1の酵素媒介反応に関する酵素である請求項1に記載の方法。
【請求項5】
前記第2の分子が、前記第2の酵素媒介反応に関する酵素である請求項1に記載の方法。
【請求項6】
前記第1の分子が、前記第1の酵素媒介反応に関する補因子である請求項1に記載の方法。
【請求項7】
前記第2の分子が、前記第2の酵素媒介反応に関する補因子である請求項1に記載の方法。
【請求項8】
ルミネセンスが、前記第1の分子を検出するために使用される請求項1に記載の方法。
【請求項9】
蛍光が、前記第2の分子を検出するために使用される請求項1に記載の方法。
【請求項10】
前記第1の分子及び前記第2の分子の存在又は量を、連続的に検出する請求項1に記載の方法。
【請求項11】
前記サンプルが、細胞溶解物である請求項1に記載の方法。
【請求項12】
前記サンプルを、前記第2の反応に関する反応混合物の前に前記第1の反応に関する反応混合物に接触させる請求項1に記載の方法。
【請求項13】
前記サンプルを、前記第1の反応に関する反応混合物の前に前記第2の反応に関する反応混合物に接触させる請求項1に記載の方法。
【請求項14】
前記サンプルを、前記第1の反応及び前記第2の反応に関する反応混合物に同時に接触させる請求項1に記載の方法。
【請求項15】
第1の酵素媒介反応に関する第1の酵素又は補因子の存在又は量を検出する方法であって、
a)サンプルを、第1の基質と前記第1の酵素との間の反応、又は第3の酵素と、前記第1の酵素と前記第1の基質との間の反応生成物との間の反応が、ルミネセンス発生生成物を生成し、第2の基質及び/又は前記第2の基質と第2の酵素との間の反応生成物がルミネセンスを発生しない、前記第1の酵素に対する前記第1の基質と、前記第2の酵素に対する前記第2の基質と、場合によっては前記第3の酵素に接触させること;及び
b)前記第1の酵素により媒介される反応に関する第1の酵素又は補因子の存在又は量を検出すること
を含む方法。
【請求項16】
第1の酵素媒介反応に関する酵素又は補因子の存在又は量を検出するために酵素媒介ルミネセンス反応をアッセイする方法であって、
a)サンプルを、第1の基質と前記第1の酵素との間の反応、又は第3の酵素と、前記第1の酵素と前記第1の基質との間の反応生成物との間の反応が、ルミネセンス発生生成物を生成し、第2の基質及び/又は前記第2の基質と第2の酵素との間の反応生成物がルミネセンスを発生しない、前記第1の酵素に対する前記第1の基質と、前記第2の酵素に対する前記第2の基質と、場合によっては前記第3の酵素に接触させること;及び
b)ルミネセンスを検出すること
を含む方法。
【請求項17】
ルミネセンスが検出される請求項15に記載の方法。
【請求項18】
ルミネセンスが、前記第1の酵素又は補因子の存在下で増加する請求項15又は16に記載の方法。
【請求項19】
ルミネセンスが、前記第1の酵素又は補因子の存在下で減少する請求項15又は16に記載の方法。
【請求項20】
前記第2の酵素の存在又は量を検出することをさらに含む請求項15又は16に記載の方法。
【請求項21】
蛍光が、前記第2の酵素の存在又は量を検出するために使用される請求項20に記載の方法。
【請求項22】
前記第1の酵素又は補因子の存在又は量及び前記第2の酵素の存在又は量が、連続的に検出される請求項20に記載の方法。
【請求項23】
前記第1の酵素又は補因子の存在又は量及び前記第2の酵素の存在又は量が、同時に検出される請求項20に記載の方法。
【請求項24】
前記第2の酵素の存在又は量が、比色検出される請求項20に記載の方法。
【請求項25】
サンプルを、前記第2の基質と前記第2の酵素との間の蛍光発生生成物を生成する反応生成物間の反応に関する第4の酵素と第3の基質に接触させることによって前記第2の酵素の存在又は量が検出される請求項20に記載の方法。
【請求項26】
前記ルミネセンス非発生の第2の基質又は前記第2の基質と前記第2の酵素との間の反応における前記ルミネセンス非発生生成物の存在又は量を検出することをさらに含む請求項15又は16に記載の方法。
【請求項27】
前記第2の酵素が、前記第1の基質と実質的に反応しない請求項15又は16に記載の方法。
【請求項28】
前記第1の酵素が、前記第2の基質と実質的に反応しない請求項15又は16に記載の方法。
【請求項29】
前記第2の基質又は前記第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項15又は16に記載の方法。
【請求項30】
前記第2の基質又は前記第2の基質と前記第2の酵素との反応生成物が、臭化エチジウム、フルオレセイン、Cy3、BODIPY、ロドール、Rox、5−カルボキシフルオレセイン、6−カルボキシフルオレセイン、アントラセン、2−アミノ−4−メトキシナフタレン、フェナレノン、アクリドン、フッ素化キサンテン誘導体、α−ナフトール、β−ナフトール、1−ヒドロキシピレン、クマリン、7−アミノ−4−メチルクマリン(AMC)、7−アミノ−4−トリフルオロメチルクマリン(AFC)、テキサスレッド、テトラメチルローダミン、カルボキシローダミン、又はローダミン、クレシル、ローダミン−110又はレゾルフィンを含む請求項29に記載の方法。
【請求項31】
1種の酵素が、グリコシダーゼ、ホスファターゼ、キナーゼ、デヒドロゲナーゼ、ペルオキシダーゼ、スルファターゼ、ペプチダーゼ、又はヒドロラーゼである請求項15又は16に記載の方法。
【請求項32】
1種の酵素が、プロテアーゼである請求項15又は16に記載の方法。
【請求項33】
1種の酵素が、カスパーゼである請求項32に記載の方法。
【請求項34】
前記カスパーゼが、カスパーゼ−3、カスパーゼ−7、又はカスパーゼ−8を含む請求項33に記載の方法。
【請求項35】
基質の1種が、DEVD、WEHD、LEHD、VEID、VEVD、VEHD、IETD、AEVD、LEXD、VEXD、IEHD、PEHD、ZEVD又はLETDを含む請求項15又は16に記載の方法。
【請求項36】
基質の1種が、X1−X2−X3−Dを含み、X1がZ、Y、D、L、V、I、A、W又はPであり、X2がV又はEであり、X3が任意のアミノ酸である請求項15又は16に記載の方法。
【請求項37】
基質の1種が、トリプシン又はトリプターゼに対する基質である請求項15又は16に記載の方法。
【請求項38】
1種の酵素が、アルギニン又はリジンを含む基質を開裂する請求項15又は16に記載の方法。
【請求項39】
前記サンプルが、細胞溶解物である請求項15又は16に記載の方法。
【請求項40】
前記サンプルが、細胞溶解前に細胞死誘導剤で処理される細胞サンプルである請求項39に記載の方法。
【請求項41】
前記サンプルが、無処置細胞を含む請求項15又は16に記載の方法。
【請求項42】
前記第3の酵素が、ルシフェラーゼである請求項15又は16に記載の方法。
【請求項43】
前記ルシフェラーゼが、カブトムシのルシフェラーゼである請求項42に記載の方法。
【請求項44】
前記第2の基質が、ラクテートデヒドロゲナーゼに対する基質である請求項15又は16に記載の方法。
【請求項45】
前記サンプルを、前記第2の基質の前に前記第1の基質に接触させる請求項15又は16に記載の方法。
【請求項46】
前記サンプルを、前記第1の基質及び前記第2の基質に同時に接触させる請求項15又は16に記載の方法。
【請求項47】
前記サンプルを、前記第1の基質の前に前記第2の基質に接触させる請求項15又は16に記載の方法。
【請求項48】
前記サンプルを、前記第1の酵素に接触させることによって補因子の存在又は量が検出される請求項15又は16に記載の方法。
【請求項49】
サンプル中少なくとも2種の分子の存在又は量を検出する方法であって、
a)サンプルを、第1の基質と第1の酵素との間の反応、又は第3の酵素及び前記第1の酵素と前記第1の基質との間の反応生成物が、ルミネセンス発生生成物を生成し、第2の基質及び/又は前記第2の基質と第2の酵素との間の反応生成物がルミネセンスを発生せず、前記第1の酵素と第2の酵素は同じではない、前記第1の酵素に対する前記第1の基質と、前記第2の酵素に対する前記第2の基質と、場合によっては前記第3の酵素に接触させること;及び
b)前記第1又は前記第2の酵素により媒介される反応に関する前記第1及び前記第2の酵素又は補因子の存在又は量を検出すること
を含む方法。
【請求項50】
ルミネセンスが、検出される請求項49に記載の方法。
【請求項51】
少なくとも1種の酵素が、プロテアーゼである請求項49に記載の方法。
【請求項52】
前記第2の酵素が、前記第1の基質と実質的に反応しない請求項49に記載の方法。
【請求項53】
前記第1の酵素が、前記第2の基質と実質的に反応しない請求項49に記載の方法。
【請求項54】
前記第2の基質又は前記第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項49に記載の方法。
【請求項55】
蛍光が、前記第2の酵素又は補因子の存在又は量を検出するために使用される請求項49に記載の方法。
【請求項56】
少なくとも1種の酵素が、カスパーゼである請求項49に記載の方法。
【請求項57】
基質の1種が、トリプシン又はトリプターゼに対する基質である請求項49に記載の方法。
【請求項58】
前記サンプルが、細胞溶解物である請求項49に記載の方法。
【請求項59】
前記サンプルが、細胞溶解前に細胞死誘導剤で処理される細胞サンプルである請求項58に記載の方法。
【請求項60】
前記サンプルが、無処置細胞を含む請求項49に記載の方法。
【請求項61】
前記第3の酵素が、ルシフェラーゼである請求項49に記載の方法。
【請求項62】
前記ルシフェラーゼが、カブトムシのルシフェラーゼである請求項61に記載の方法。
【請求項63】
前記第2の基質又は第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項62に記載の方法。
【請求項64】
前記サンプルを、前記第2の基質の前に前記第1の基質に接触させる請求項49に記載の方法。
【請求項65】
前記サンプルを、前記第1の基質の前に前記第2の基質に接触させる請求項49に記載の方法。
【請求項66】
前記サンプルを、前記第1の基質及び前記第2の基質に同時に接触させる請求項49に記載の方法。
【請求項67】
前記第1の酵素又は第1の補因子の存在又は量が、前記第2の酵素又は第2の補因子の存在又は量が検出される前に検出される請求項49に記載の方法。
【請求項68】
前記第2の酵素又は第2の補因子の存在又は量が、前記第1の酵素又は第1の補因子の存在又は量が検出される前に検出される請求項49に記載の方法。
【請求項69】
前記第2の基質又は第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項68に記載の方法。
【請求項70】
前記第2の基質又は前記第2の基質と前記第2の酵素との反応生成物が、臭化エチジウム、フルオレセイン、Cy3、BODIPY、ロドール、Rox、5−カルボキシフルオレセイン、6−カルボキシフルオレセイン、アントラセン、2−アミノ−4−メトキシナフタレン、フェナレノン、アクリドン、フッ素化キサンテン誘導体、α−ナフトール、β−ナフトール、1−ヒドロキシピレン、クマリン、7−アミノ−4−メチルクマリン(AMC)、7−アミノ−4−トリフルオロメチルクマリン(AFC)、テキサスレッド、テトラメチルローダミン、カルボキシローダミン、又はローダミン、クレシル、ローダミン−110又はレゾルフィンを含む請求項69に記載の方法。
【請求項71】
前記補因子が、ATPである請求項15、16又は49に記載の方法。
【請求項72】
前記サンプルを、前記第2の基質と第2の酵素との間の蛍光発生生成物を生成する反応生成物間の反応に関する第4の酵素及び第3の基質に接触させることにより前記第2の酵素の存在又は量が検出される請求項49に記載の方法。
【請求項73】
第1の酵素媒介反応に関する分子の存在又は量を検出する方法であって、
a)蛍光タンパク質を発現する細胞を含むサンプルを、前記第1の酵素により媒介される反応がルミネセンス発生生成物を生成する前記第1の酵素媒介反応に関する反応混合物に接触させること;及び
b)前記分子の存在又は量及びサンプル中の蛍光タンパク質の存在又は量を検出すること
を含む方法。
【請求項74】
前記分子が、基質である請求項73に記載の方法。
【請求項75】
前記分子が、補因子である請求項73に記載の方法。
【請求項76】
前記分子が、酵素である請求項73に記載の方法。
【請求項77】
ルミネセンス非発生基質;及び
ルミネセンス発生基質
を含むキット。
【請求項78】
前記ルミネセンス発生基質とのルミネセンス反応を媒介できる酵素をさらに含む請求項77に記載のキット。
【請求項79】
前記ルミネセンス非発生基質及び前記ルミネセンス発生基質を含む単一反応容器内で、ルミネセンス発生反応及びルミネセンス非発生反応を実施するための使用説明書をさらに含む請求項77に記載のキット。
【請求項80】
ルミネセンス非発生基質;及び
ルミネセンス発生反応を媒介できる酵素
を含むキット。
【請求項81】
前記酵素に対するルミネセンス発生基質をさらに含む請求項80に記載のキット。
【請求項82】
前記ルミネセンス非発生基質が、蛍光発生基質である請求項77又は80に記載のキット。
【請求項83】
前記酵素が、ルシフェラーゼである請求項78又は80に記載のキット。
【請求項84】
前記ルミネセンス非発生基質及び前記酵素を含む単一反応容器内で、ルミネセンス発生反応及びルミネセンス非発生反応を実施するための使用説明書をさらに含む請求項80に記載のキット。
【請求項85】
サンプル中の生細胞及び/又は死細胞を検出する方法であって、
a)サンプルを、第1のプロテアーゼに対する基質及び第2のプロテアーゼに対する基質であって、前記プロテアーゼの一方により媒介される前記基質の一方との反応が、蛍光発生生成物を生成し、また、他方のプロテアーゼにより媒介される他方の基質との反応が、ルミネセンス発生生成物又は蛍光発生生成物を生成し、基質の一方が、実質的に細胞透過性であり、他方の基質が、実質的に細胞不透過性であり、両反応が、蛍光発生生成物を生成する場合、2種の基質上の蛍光体のスペクトルが異なる前記基質に接触させること;及び
b)サンプル中の蛍光及び/又はルミネセンスを検出するか、又は測定することによって、サンプル中の生細胞及び/又は死細胞の数又は存在を検出又は測定すること
を含む方法。
【請求項86】
反応媒介の各プロテアーゼが、蛍光発生生成物を生成する請求項85に記載の方法。
【請求項87】
一方のプロテアーゼにより媒介される反応が、ルミネセンス発生生成物を生成し、他方のプロテアーゼにより媒介される反応が、蛍光発生生成物を生成する請求項85に記載の方法。
【請求項88】
実質的に細胞透過性基質が、プロテアソーム、アミノペプチダーゼ又はカテプシンと関連するプロテアーゼに対する基質である請求項85に記載の方法。
【請求項89】
実質的に細胞不透過性基質が、トリペプチジルペプチダーゼ、カルパイン又はキモトリプシンに対する基質である請求項85に記載の方法。
【請求項90】
前記サンプルが、哺乳動物細胞を含む請求項85に記載の方法。
【請求項91】
前記2種の基質が、前記サンプルとの接触前に組み合わされる請求項85に記載の方法。
【請求項92】
蛍光及びルミネセンスが、連続的に検出又は測定される請求項87に記載の方法。
【請求項93】
各蛍光発生生成物の蛍光が、同時に検出又は測定される請求項86に記載の方法。
【請求項94】
分子の存在又は量を検出又は測定することをさらに含む請求項85に記載の方法。
【請求項95】
前記分子が、DNAである請求項94に記載の方法。
【請求項96】
前記分子が、酵素である請求項94に記載の方法。
【請求項97】
前記分子が、ATPである請求項94に記載の方法。
【請求項98】
前記サンプルを、細胞を溶解する条件に供することをさらに含む請求項85に記載の方法。
【請求項99】
前記サンプルを、前記基質との接触前に1種又は複数の試薬に接触させることをさらに含む請求項85に記載の方法。
【請求項100】
サンプル中の生細胞を検出する方法であって、
a)サンプルを、プロテアソーム、アミノペプチダーゼ又はカテプシンと関連するプロテアーゼに対する蛍光発生の実質的に細胞透過性基質に接触させること;及び
b)サンプル中の蛍光を検出又は測定することによって、サンプル中の生細胞の数又は存在を検出又は測定すること
を含む方法。
【請求項101】
前記基質が、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、Z−Gly−Gly−Leu−AMC、Phe−AMC、又はTyr−AMCである請求項100に記載の方法。
【請求項102】
サンプル中の死細胞を検出する方法であって、
a)サンプルを、トリペプチジルペプチダーゼ、カルパイン又はキモトリプシンに対する蛍光発生又はルミネセンス発生の細胞不透過性基質に接触させること;及び
b)サンプル中の蛍光又はルミネセンスを検出又は測定することによって、サンプル中の死細胞の数又は存在を検出又は測定すること
を含む方法。
【請求項103】
前記基質が、Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Suc−Leu−Leu−Val−Tyr−AMC、又はZ−Leu−Leu−Val−Tyr−アミノルシフェリンである請求項102に記載の方法。
【請求項104】
第1のプロテアーゼに対する第1の蛍光発生又はルミネセンス発生の実質的に細胞不透過性基質及び第2のプロテアーゼに対する第2の蛍光発生の実質的に細胞透過性基質を含む組成物;サンプル中の生細胞及び/又は死細胞を検出する組成物の使用について使用者に指示する使用説明書を含むキット。
【請求項105】
前記第1の基質が、Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Suc−Leu−Leu−Val−Tyr−AMC、又はZ−Leu−Leu−Val−Tyr−アミノルシフェリンである請求項104に記載のキット。
【請求項106】
前記第2の基質が、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、Z−Gly−Gly−Leu−AMC、Phe−AMC、又はTyr−AMCである請求項104に記載のキット。
【請求項107】
ルミネセンス発生反応に関する1種又は複数の試薬をさらに含む請求項104に記載のキット。
【請求項108】
少なくとも1種の試薬が、凍結乾燥されている請求項107に記載のキット。
【請求項109】
前記組成物は、前記基質が有機溶媒中にある溶液である請求項104に記載のキット。
【請求項110】
前記組成物が、凍結乾燥されている請求項109に記載のキット。
【請求項111】
前記組成物は、前記基質濃度が、約0.005Mから1Mの溶液である請求項104に記載のキット。
【請求項112】
Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、又はそれらの任意の組合せを含む組成物を含むキット。
【請求項113】
前記組成物が、溶媒を含む請求項112に記載のキット。
【請求項114】
Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Gly−Phe−AFC、Gly−Phe−AMC、又はGly−Gly−Leu−AMCの濃度が、約0.005Mから1Mである請求項113に記載のキット。
【請求項1】
第1の酵素媒介反応に関する第1の分子、及び第2の酵素媒介反応に関する第2の分子の存在又は量を検出する方法であって、
a)サンプルを、前記第1の酵素により媒介された反応が、ルミネセンス発生生成物を生成し、前記第2の酵素により媒介された反応が、ルミネセンス非発生生成物を生成する前記第1の反応及び前記第2の反応に関する反応混合物に接触させること;及び
b)前記サンプルにおける前記第1の分子及び前記第2の分子の存在又は量を検出することを含む方法。
【請求項2】
前記第1の分子が、前記第1の酵素媒介反応に関する基質である請求項1に記載の方法。
【請求項3】
前記第2の分子が、前記第2の酵素媒介反応に関する基質である請求項1に記載の方法。
【請求項4】
前記第1の分子が、前記第1の酵素媒介反応に関する酵素である請求項1に記載の方法。
【請求項5】
前記第2の分子が、前記第2の酵素媒介反応に関する酵素である請求項1に記載の方法。
【請求項6】
前記第1の分子が、前記第1の酵素媒介反応に関する補因子である請求項1に記載の方法。
【請求項7】
前記第2の分子が、前記第2の酵素媒介反応に関する補因子である請求項1に記載の方法。
【請求項8】
ルミネセンスが、前記第1の分子を検出するために使用される請求項1に記載の方法。
【請求項9】
蛍光が、前記第2の分子を検出するために使用される請求項1に記載の方法。
【請求項10】
前記第1の分子及び前記第2の分子の存在又は量を、連続的に検出する請求項1に記載の方法。
【請求項11】
前記サンプルが、細胞溶解物である請求項1に記載の方法。
【請求項12】
前記サンプルを、前記第2の反応に関する反応混合物の前に前記第1の反応に関する反応混合物に接触させる請求項1に記載の方法。
【請求項13】
前記サンプルを、前記第1の反応に関する反応混合物の前に前記第2の反応に関する反応混合物に接触させる請求項1に記載の方法。
【請求項14】
前記サンプルを、前記第1の反応及び前記第2の反応に関する反応混合物に同時に接触させる請求項1に記載の方法。
【請求項15】
第1の酵素媒介反応に関する第1の酵素又は補因子の存在又は量を検出する方法であって、
a)サンプルを、第1の基質と前記第1の酵素との間の反応、又は第3の酵素と、前記第1の酵素と前記第1の基質との間の反応生成物との間の反応が、ルミネセンス発生生成物を生成し、第2の基質及び/又は前記第2の基質と第2の酵素との間の反応生成物がルミネセンスを発生しない、前記第1の酵素に対する前記第1の基質と、前記第2の酵素に対する前記第2の基質と、場合によっては前記第3の酵素に接触させること;及び
b)前記第1の酵素により媒介される反応に関する第1の酵素又は補因子の存在又は量を検出すること
を含む方法。
【請求項16】
第1の酵素媒介反応に関する酵素又は補因子の存在又は量を検出するために酵素媒介ルミネセンス反応をアッセイする方法であって、
a)サンプルを、第1の基質と前記第1の酵素との間の反応、又は第3の酵素と、前記第1の酵素と前記第1の基質との間の反応生成物との間の反応が、ルミネセンス発生生成物を生成し、第2の基質及び/又は前記第2の基質と第2の酵素との間の反応生成物がルミネセンスを発生しない、前記第1の酵素に対する前記第1の基質と、前記第2の酵素に対する前記第2の基質と、場合によっては前記第3の酵素に接触させること;及び
b)ルミネセンスを検出すること
を含む方法。
【請求項17】
ルミネセンスが検出される請求項15に記載の方法。
【請求項18】
ルミネセンスが、前記第1の酵素又は補因子の存在下で増加する請求項15又は16に記載の方法。
【請求項19】
ルミネセンスが、前記第1の酵素又は補因子の存在下で減少する請求項15又は16に記載の方法。
【請求項20】
前記第2の酵素の存在又は量を検出することをさらに含む請求項15又は16に記載の方法。
【請求項21】
蛍光が、前記第2の酵素の存在又は量を検出するために使用される請求項20に記載の方法。
【請求項22】
前記第1の酵素又は補因子の存在又は量及び前記第2の酵素の存在又は量が、連続的に検出される請求項20に記載の方法。
【請求項23】
前記第1の酵素又は補因子の存在又は量及び前記第2の酵素の存在又は量が、同時に検出される請求項20に記載の方法。
【請求項24】
前記第2の酵素の存在又は量が、比色検出される請求項20に記載の方法。
【請求項25】
サンプルを、前記第2の基質と前記第2の酵素との間の蛍光発生生成物を生成する反応生成物間の反応に関する第4の酵素と第3の基質に接触させることによって前記第2の酵素の存在又は量が検出される請求項20に記載の方法。
【請求項26】
前記ルミネセンス非発生の第2の基質又は前記第2の基質と前記第2の酵素との間の反応における前記ルミネセンス非発生生成物の存在又は量を検出することをさらに含む請求項15又は16に記載の方法。
【請求項27】
前記第2の酵素が、前記第1の基質と実質的に反応しない請求項15又は16に記載の方法。
【請求項28】
前記第1の酵素が、前記第2の基質と実質的に反応しない請求項15又は16に記載の方法。
【請求項29】
前記第2の基質又は前記第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項15又は16に記載の方法。
【請求項30】
前記第2の基質又は前記第2の基質と前記第2の酵素との反応生成物が、臭化エチジウム、フルオレセイン、Cy3、BODIPY、ロドール、Rox、5−カルボキシフルオレセイン、6−カルボキシフルオレセイン、アントラセン、2−アミノ−4−メトキシナフタレン、フェナレノン、アクリドン、フッ素化キサンテン誘導体、α−ナフトール、β−ナフトール、1−ヒドロキシピレン、クマリン、7−アミノ−4−メチルクマリン(AMC)、7−アミノ−4−トリフルオロメチルクマリン(AFC)、テキサスレッド、テトラメチルローダミン、カルボキシローダミン、又はローダミン、クレシル、ローダミン−110又はレゾルフィンを含む請求項29に記載の方法。
【請求項31】
1種の酵素が、グリコシダーゼ、ホスファターゼ、キナーゼ、デヒドロゲナーゼ、ペルオキシダーゼ、スルファターゼ、ペプチダーゼ、又はヒドロラーゼである請求項15又は16に記載の方法。
【請求項32】
1種の酵素が、プロテアーゼである請求項15又は16に記載の方法。
【請求項33】
1種の酵素が、カスパーゼである請求項32に記載の方法。
【請求項34】
前記カスパーゼが、カスパーゼ−3、カスパーゼ−7、又はカスパーゼ−8を含む請求項33に記載の方法。
【請求項35】
基質の1種が、DEVD、WEHD、LEHD、VEID、VEVD、VEHD、IETD、AEVD、LEXD、VEXD、IEHD、PEHD、ZEVD又はLETDを含む請求項15又は16に記載の方法。
【請求項36】
基質の1種が、X1−X2−X3−Dを含み、X1がZ、Y、D、L、V、I、A、W又はPであり、X2がV又はEであり、X3が任意のアミノ酸である請求項15又は16に記載の方法。
【請求項37】
基質の1種が、トリプシン又はトリプターゼに対する基質である請求項15又は16に記載の方法。
【請求項38】
1種の酵素が、アルギニン又はリジンを含む基質を開裂する請求項15又は16に記載の方法。
【請求項39】
前記サンプルが、細胞溶解物である請求項15又は16に記載の方法。
【請求項40】
前記サンプルが、細胞溶解前に細胞死誘導剤で処理される細胞サンプルである請求項39に記載の方法。
【請求項41】
前記サンプルが、無処置細胞を含む請求項15又は16に記載の方法。
【請求項42】
前記第3の酵素が、ルシフェラーゼである請求項15又は16に記載の方法。
【請求項43】
前記ルシフェラーゼが、カブトムシのルシフェラーゼである請求項42に記載の方法。
【請求項44】
前記第2の基質が、ラクテートデヒドロゲナーゼに対する基質である請求項15又は16に記載の方法。
【請求項45】
前記サンプルを、前記第2の基質の前に前記第1の基質に接触させる請求項15又は16に記載の方法。
【請求項46】
前記サンプルを、前記第1の基質及び前記第2の基質に同時に接触させる請求項15又は16に記載の方法。
【請求項47】
前記サンプルを、前記第1の基質の前に前記第2の基質に接触させる請求項15又は16に記載の方法。
【請求項48】
前記サンプルを、前記第1の酵素に接触させることによって補因子の存在又は量が検出される請求項15又は16に記載の方法。
【請求項49】
サンプル中少なくとも2種の分子の存在又は量を検出する方法であって、
a)サンプルを、第1の基質と第1の酵素との間の反応、又は第3の酵素及び前記第1の酵素と前記第1の基質との間の反応生成物が、ルミネセンス発生生成物を生成し、第2の基質及び/又は前記第2の基質と第2の酵素との間の反応生成物がルミネセンスを発生せず、前記第1の酵素と第2の酵素は同じではない、前記第1の酵素に対する前記第1の基質と、前記第2の酵素に対する前記第2の基質と、場合によっては前記第3の酵素に接触させること;及び
b)前記第1又は前記第2の酵素により媒介される反応に関する前記第1及び前記第2の酵素又は補因子の存在又は量を検出すること
を含む方法。
【請求項50】
ルミネセンスが、検出される請求項49に記載の方法。
【請求項51】
少なくとも1種の酵素が、プロテアーゼである請求項49に記載の方法。
【請求項52】
前記第2の酵素が、前記第1の基質と実質的に反応しない請求項49に記載の方法。
【請求項53】
前記第1の酵素が、前記第2の基質と実質的に反応しない請求項49に記載の方法。
【請求項54】
前記第2の基質又は前記第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項49に記載の方法。
【請求項55】
蛍光が、前記第2の酵素又は補因子の存在又は量を検出するために使用される請求項49に記載の方法。
【請求項56】
少なくとも1種の酵素が、カスパーゼである請求項49に記載の方法。
【請求項57】
基質の1種が、トリプシン又はトリプターゼに対する基質である請求項49に記載の方法。
【請求項58】
前記サンプルが、細胞溶解物である請求項49に記載の方法。
【請求項59】
前記サンプルが、細胞溶解前に細胞死誘導剤で処理される細胞サンプルである請求項58に記載の方法。
【請求項60】
前記サンプルが、無処置細胞を含む請求項49に記載の方法。
【請求項61】
前記第3の酵素が、ルシフェラーゼである請求項49に記載の方法。
【請求項62】
前記ルシフェラーゼが、カブトムシのルシフェラーゼである請求項61に記載の方法。
【請求項63】
前記第2の基質又は第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項62に記載の方法。
【請求項64】
前記サンプルを、前記第2の基質の前に前記第1の基質に接触させる請求項49に記載の方法。
【請求項65】
前記サンプルを、前記第1の基質の前に前記第2の基質に接触させる請求項49に記載の方法。
【請求項66】
前記サンプルを、前記第1の基質及び前記第2の基質に同時に接触させる請求項49に記載の方法。
【請求項67】
前記第1の酵素又は第1の補因子の存在又は量が、前記第2の酵素又は第2の補因子の存在又は量が検出される前に検出される請求項49に記載の方法。
【請求項68】
前記第2の酵素又は第2の補因子の存在又は量が、前記第1の酵素又は第1の補因子の存在又は量が検出される前に検出される請求項49に記載の方法。
【請求項69】
前記第2の基質又は第2の基質と前記第2の酵素との間の反応生成物が、蛍光性である請求項68に記載の方法。
【請求項70】
前記第2の基質又は前記第2の基質と前記第2の酵素との反応生成物が、臭化エチジウム、フルオレセイン、Cy3、BODIPY、ロドール、Rox、5−カルボキシフルオレセイン、6−カルボキシフルオレセイン、アントラセン、2−アミノ−4−メトキシナフタレン、フェナレノン、アクリドン、フッ素化キサンテン誘導体、α−ナフトール、β−ナフトール、1−ヒドロキシピレン、クマリン、7−アミノ−4−メチルクマリン(AMC)、7−アミノ−4−トリフルオロメチルクマリン(AFC)、テキサスレッド、テトラメチルローダミン、カルボキシローダミン、又はローダミン、クレシル、ローダミン−110又はレゾルフィンを含む請求項69に記載の方法。
【請求項71】
前記補因子が、ATPである請求項15、16又は49に記載の方法。
【請求項72】
前記サンプルを、前記第2の基質と第2の酵素との間の蛍光発生生成物を生成する反応生成物間の反応に関する第4の酵素及び第3の基質に接触させることにより前記第2の酵素の存在又は量が検出される請求項49に記載の方法。
【請求項73】
第1の酵素媒介反応に関する分子の存在又は量を検出する方法であって、
a)蛍光タンパク質を発現する細胞を含むサンプルを、前記第1の酵素により媒介される反応がルミネセンス発生生成物を生成する前記第1の酵素媒介反応に関する反応混合物に接触させること;及び
b)前記分子の存在又は量及びサンプル中の蛍光タンパク質の存在又は量を検出すること
を含む方法。
【請求項74】
前記分子が、基質である請求項73に記載の方法。
【請求項75】
前記分子が、補因子である請求項73に記載の方法。
【請求項76】
前記分子が、酵素である請求項73に記載の方法。
【請求項77】
ルミネセンス非発生基質;及び
ルミネセンス発生基質
を含むキット。
【請求項78】
前記ルミネセンス発生基質とのルミネセンス反応を媒介できる酵素をさらに含む請求項77に記載のキット。
【請求項79】
前記ルミネセンス非発生基質及び前記ルミネセンス発生基質を含む単一反応容器内で、ルミネセンス発生反応及びルミネセンス非発生反応を実施するための使用説明書をさらに含む請求項77に記載のキット。
【請求項80】
ルミネセンス非発生基質;及び
ルミネセンス発生反応を媒介できる酵素
を含むキット。
【請求項81】
前記酵素に対するルミネセンス発生基質をさらに含む請求項80に記載のキット。
【請求項82】
前記ルミネセンス非発生基質が、蛍光発生基質である請求項77又は80に記載のキット。
【請求項83】
前記酵素が、ルシフェラーゼである請求項78又は80に記載のキット。
【請求項84】
前記ルミネセンス非発生基質及び前記酵素を含む単一反応容器内で、ルミネセンス発生反応及びルミネセンス非発生反応を実施するための使用説明書をさらに含む請求項80に記載のキット。
【請求項85】
サンプル中の生細胞及び/又は死細胞を検出する方法であって、
a)サンプルを、第1のプロテアーゼに対する基質及び第2のプロテアーゼに対する基質であって、前記プロテアーゼの一方により媒介される前記基質の一方との反応が、蛍光発生生成物を生成し、また、他方のプロテアーゼにより媒介される他方の基質との反応が、ルミネセンス発生生成物又は蛍光発生生成物を生成し、基質の一方が、実質的に細胞透過性であり、他方の基質が、実質的に細胞不透過性であり、両反応が、蛍光発生生成物を生成する場合、2種の基質上の蛍光体のスペクトルが異なる前記基質に接触させること;及び
b)サンプル中の蛍光及び/又はルミネセンスを検出するか、又は測定することによって、サンプル中の生細胞及び/又は死細胞の数又は存在を検出又は測定すること
を含む方法。
【請求項86】
反応媒介の各プロテアーゼが、蛍光発生生成物を生成する請求項85に記載の方法。
【請求項87】
一方のプロテアーゼにより媒介される反応が、ルミネセンス発生生成物を生成し、他方のプロテアーゼにより媒介される反応が、蛍光発生生成物を生成する請求項85に記載の方法。
【請求項88】
実質的に細胞透過性基質が、プロテアソーム、アミノペプチダーゼ又はカテプシンと関連するプロテアーゼに対する基質である請求項85に記載の方法。
【請求項89】
実質的に細胞不透過性基質が、トリペプチジルペプチダーゼ、カルパイン又はキモトリプシンに対する基質である請求項85に記載の方法。
【請求項90】
前記サンプルが、哺乳動物細胞を含む請求項85に記載の方法。
【請求項91】
前記2種の基質が、前記サンプルとの接触前に組み合わされる請求項85に記載の方法。
【請求項92】
蛍光及びルミネセンスが、連続的に検出又は測定される請求項87に記載の方法。
【請求項93】
各蛍光発生生成物の蛍光が、同時に検出又は測定される請求項86に記載の方法。
【請求項94】
分子の存在又は量を検出又は測定することをさらに含む請求項85に記載の方法。
【請求項95】
前記分子が、DNAである請求項94に記載の方法。
【請求項96】
前記分子が、酵素である請求項94に記載の方法。
【請求項97】
前記分子が、ATPである請求項94に記載の方法。
【請求項98】
前記サンプルを、細胞を溶解する条件に供することをさらに含む請求項85に記載の方法。
【請求項99】
前記サンプルを、前記基質との接触前に1種又は複数の試薬に接触させることをさらに含む請求項85に記載の方法。
【請求項100】
サンプル中の生細胞を検出する方法であって、
a)サンプルを、プロテアソーム、アミノペプチダーゼ又はカテプシンと関連するプロテアーゼに対する蛍光発生の実質的に細胞透過性基質に接触させること;及び
b)サンプル中の蛍光を検出又は測定することによって、サンプル中の生細胞の数又は存在を検出又は測定すること
を含む方法。
【請求項101】
前記基質が、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、Z−Gly−Gly−Leu−AMC、Phe−AMC、又はTyr−AMCである請求項100に記載の方法。
【請求項102】
サンプル中の死細胞を検出する方法であって、
a)サンプルを、トリペプチジルペプチダーゼ、カルパイン又はキモトリプシンに対する蛍光発生又はルミネセンス発生の細胞不透過性基質に接触させること;及び
b)サンプル中の蛍光又はルミネセンスを検出又は測定することによって、サンプル中の死細胞の数又は存在を検出又は測定すること
を含む方法。
【請求項103】
前記基質が、Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Suc−Leu−Leu−Val−Tyr−AMC、又はZ−Leu−Leu−Val−Tyr−アミノルシフェリンである請求項102に記載の方法。
【請求項104】
第1のプロテアーゼに対する第1の蛍光発生又はルミネセンス発生の実質的に細胞不透過性基質及び第2のプロテアーゼに対する第2の蛍光発生の実質的に細胞透過性基質を含む組成物;サンプル中の生細胞及び/又は死細胞を検出する組成物の使用について使用者に指示する使用説明書を含むキット。
【請求項105】
前記第1の基質が、Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Suc−Leu−Leu−Val−Tyr−AMC、又はZ−Leu−Leu−Val−Tyr−アミノルシフェリンである請求項104に記載のキット。
【請求項106】
前記第2の基質が、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、Z−Gly−Gly−Leu−AMC、Phe−AMC、又はTyr−AMCである請求項104に記載のキット。
【請求項107】
ルミネセンス発生反応に関する1種又は複数の試薬をさらに含む請求項104に記載のキット。
【請求項108】
少なくとも1種の試薬が、凍結乾燥されている請求項107に記載のキット。
【請求項109】
前記組成物は、前記基質が有機溶媒中にある溶液である請求項104に記載のキット。
【請求項110】
前記組成物が、凍結乾燥されている請求項109に記載のキット。
【請求項111】
前記組成物は、前記基質濃度が、約0.005Mから1Mの溶液である請求項104に記載のキット。
【請求項112】
Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Gly−Phe−AFC、Gly−Phe−AMC、Gly−Gly−Leu−AMC、又はそれらの任意の組合せを含む組成物を含むキット。
【請求項113】
前記組成物が、溶媒を含む請求項112に記載のキット。
【請求項114】
Ala−Ala−Phe−AMC、(Ala−Ala−Phe)2−R110、Ala−Ala−Phe−アミノルシフェリン、Gly−Phe−AFC、Gly−Phe−AMC、又はGly−Gly−Leu−AMCの濃度が、約0.005Mから1Mである請求項113に記載のキット。
【図1A】

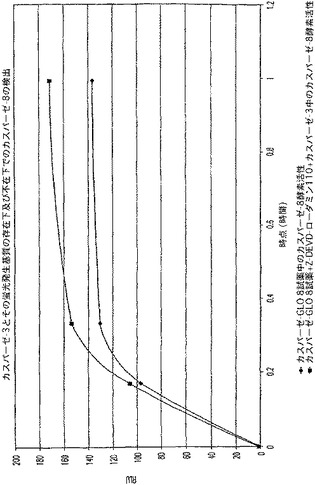
【図1B】
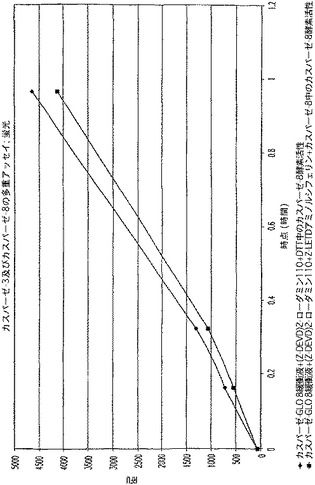
【図2A】
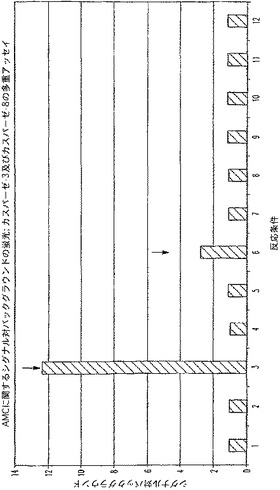
【図2B】

【図2C】
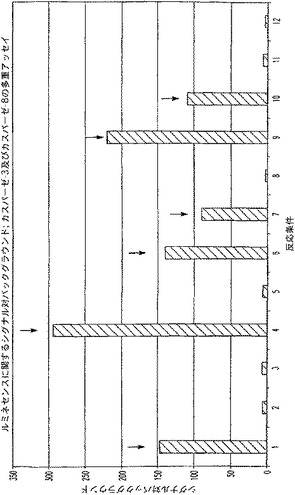
【図3A】
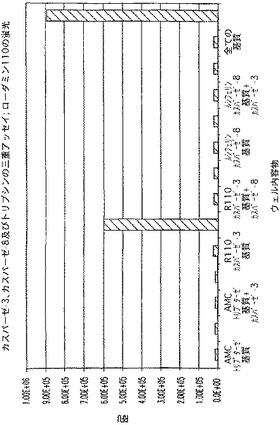
【図3C】
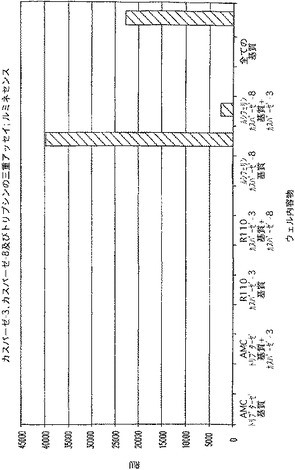
【図4A】
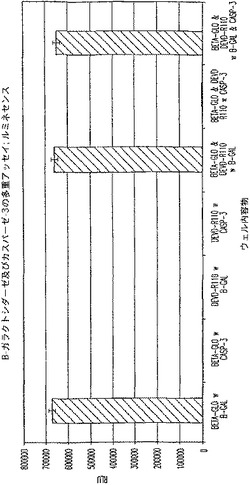
【図4B】
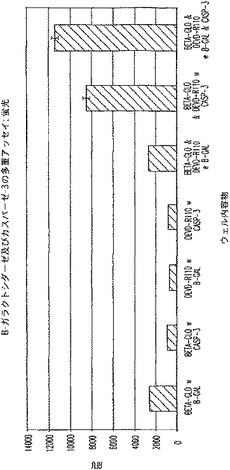
【図4C】
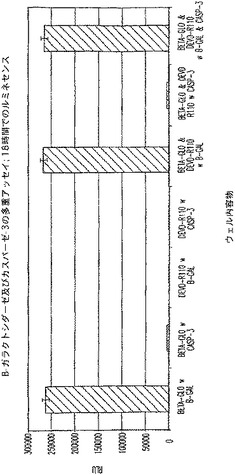
【図4D】

【図5A】
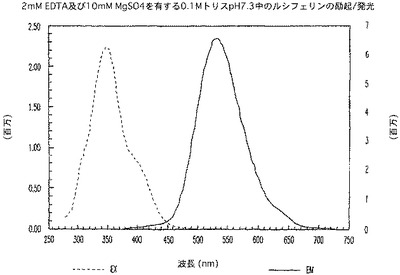
【図5B】
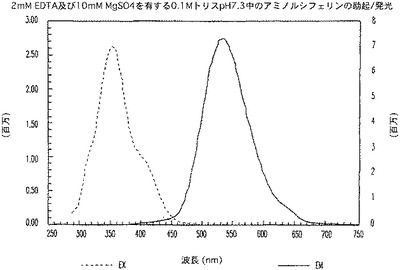
【図5C】
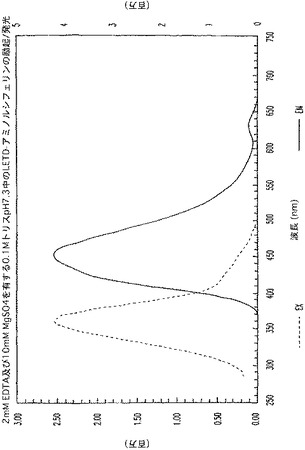
【図7A】
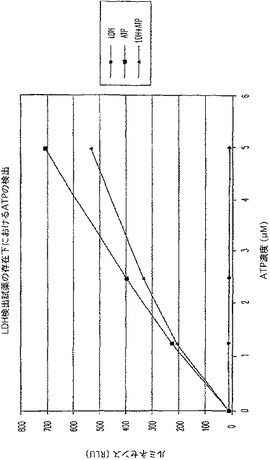
【図7B】
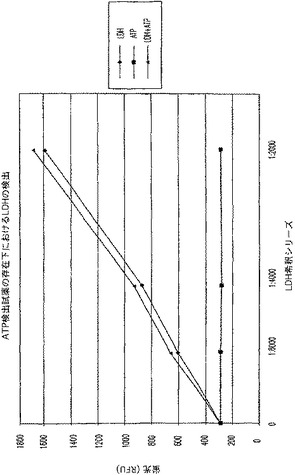
【図7C】
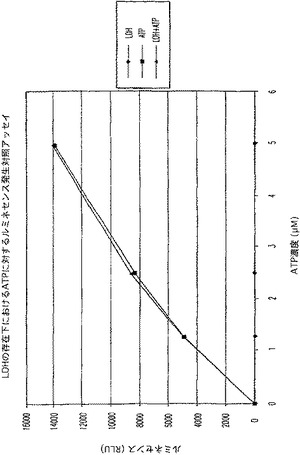
【図7D】
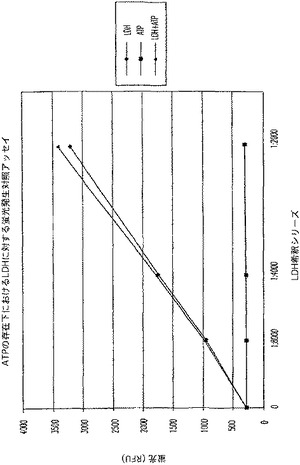
【図8A】
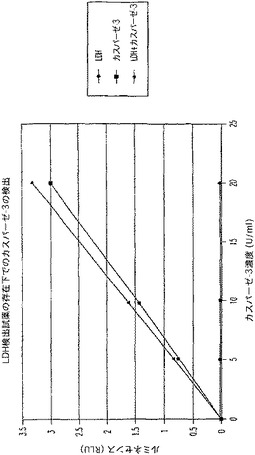
【図8B】
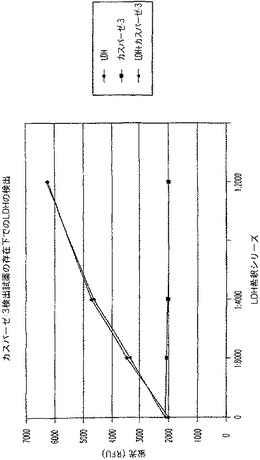
【図8C】
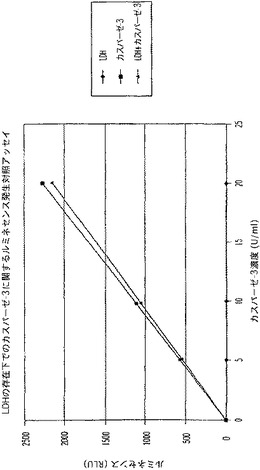
【図8D】
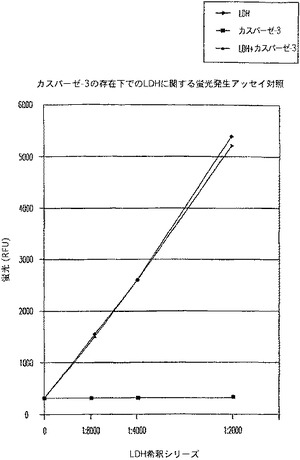
【図9A】
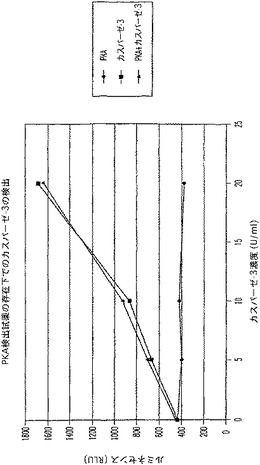
【図9B】
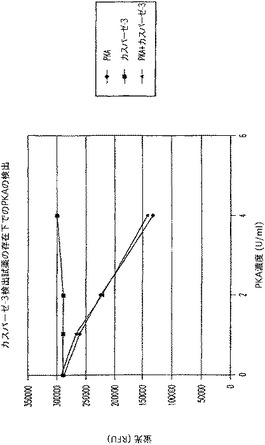
【図9C】
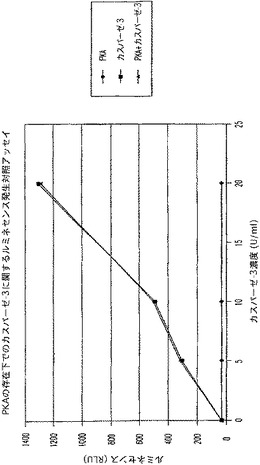
【図9D】
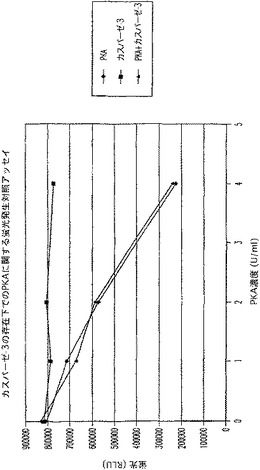
【図10A】
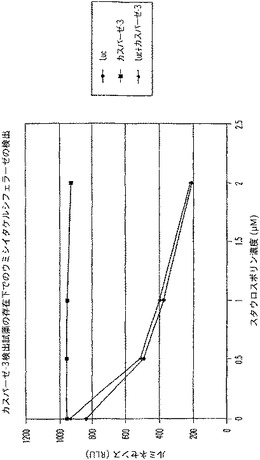
【図10B】
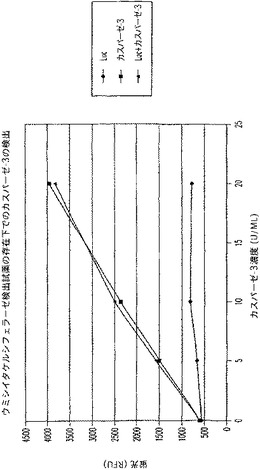
【図10C】
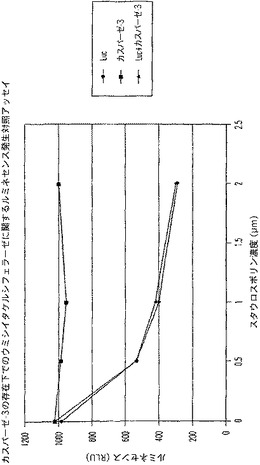
【図10D】
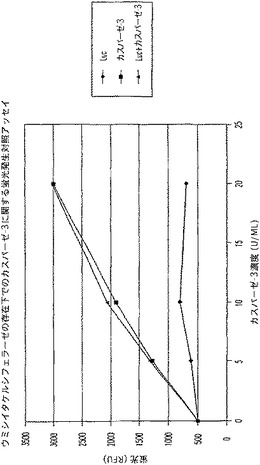
【図11A】
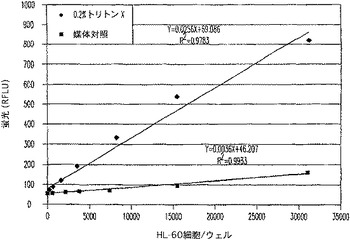
【図11B】
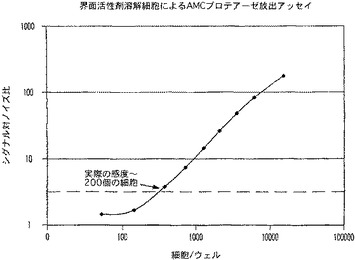
【図12A】
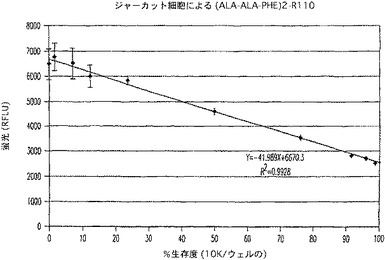
【図12B】
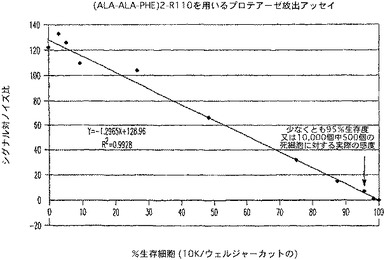
【図13A】
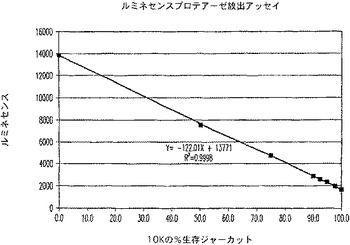
【図13B】
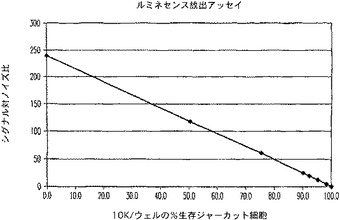
【図14A】
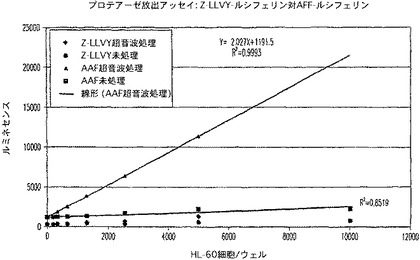
【図14B】

【図15A】
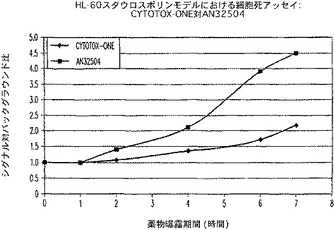
【図15B】
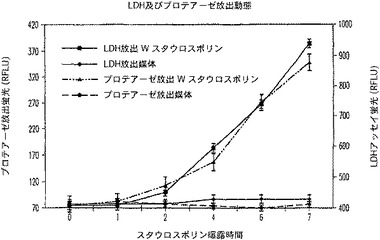
【図16】

【図17】
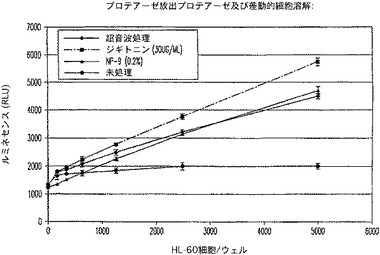
【図18】
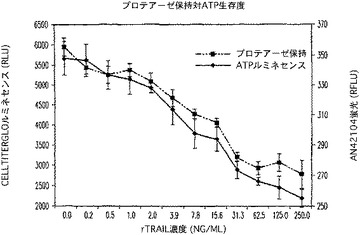
【図19A】
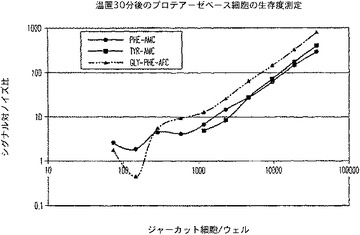
【図19B】
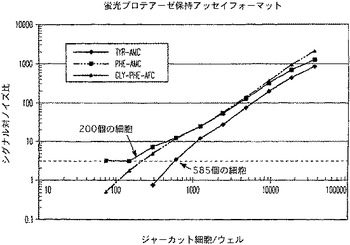
【図20】
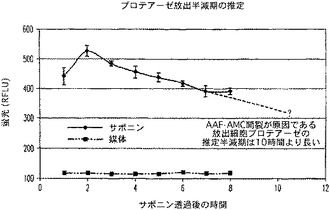
【図21A】
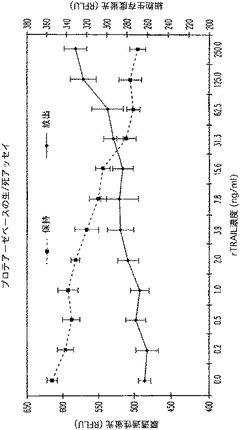
【図21B】
【図21C】
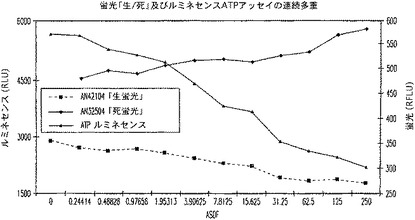
【図22A】
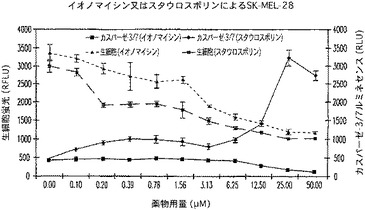
【図22B】
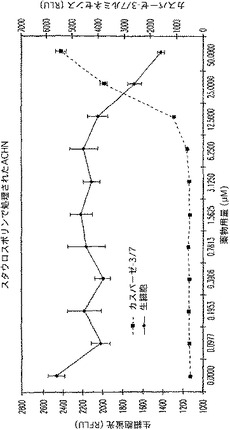
【図23】
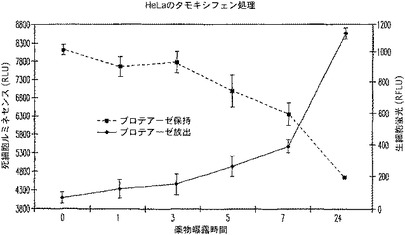
【図24A】
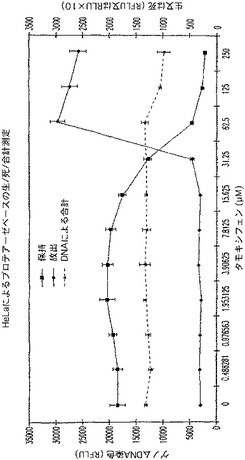
【図24B】
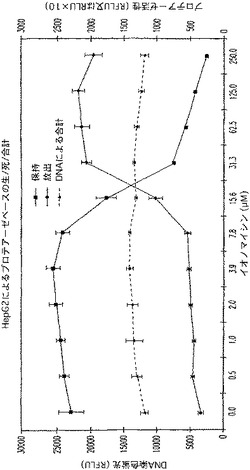
【図3B】

【図6】
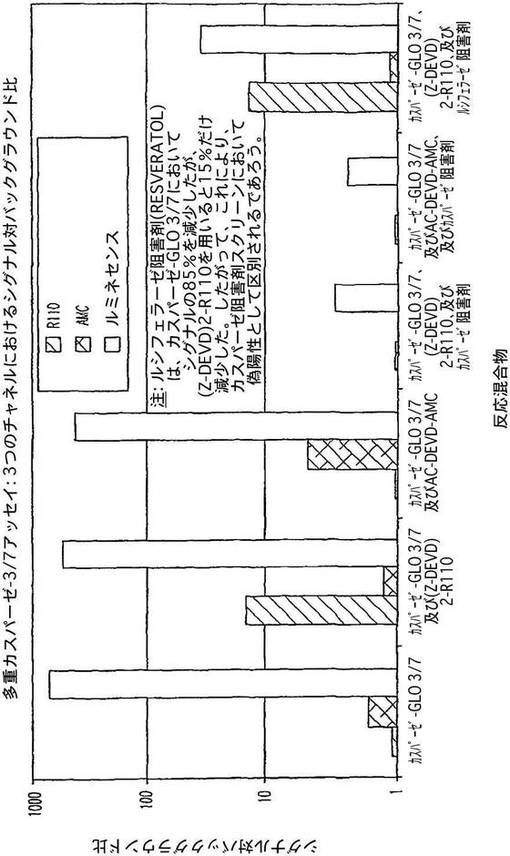
【図1B】
【図2A】
【図2B】
【図2C】
【図3A】
【図3C】
【図4A】
【図4B】
【図4C】
【図4D】
【図5A】
【図5B】
【図5C】
【図7A】
【図7B】
【図7C】
【図7D】
【図8A】
【図8B】
【図8C】
【図8D】
【図9A】
【図9B】
【図9C】
【図9D】
【図10A】
【図10B】
【図10C】
【図10D】
【図11A】
【図11B】
【図12A】
【図12B】
【図13A】
【図13B】
【図14A】
【図14B】
【図15A】
【図15B】
【図16】
【図17】
【図18】
【図19A】
【図19B】
【図20】
【図21A】
【図21B】
【図21C】
【図22A】
【図22B】
【図23】
【図24A】
【図24B】
【図3B】
【図6】
【公開番号】特開2011−188857(P2011−188857A)
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願番号】特願2011−70191(P2011−70191)
【出願日】平成23年3月28日(2011.3.28)
【分割の表示】特願2006−551360(P2006−551360)の分割
【原出願日】平成17年1月24日(2005.1.24)
【出願人】(593089149)プロメガ コーポレイション (57)
【氏名又は名称原語表記】Promega Corporation
【Fターム(参考)】
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願日】平成23年3月28日(2011.3.28)
【分割の表示】特願2006−551360(P2006−551360)の分割
【原出願日】平成17年1月24日(2005.1.24)
【出願人】(593089149)プロメガ コーポレイション (57)
【氏名又は名称原語表記】Promega Corporation
【Fターム(参考)】
[ Back to top ]