
レトロウイルス表面糖タンパク質を含む可溶性複合体
【課題】より多くのレトロウイルス表面糖タンパク質エピトープ、例えば、e-gp41分子またはe-gp36を可溶型で提供すること。
【解決手段】非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化され、および形成されるタンパク質− シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびシャペロンを含有する可溶性複合体の製造方法。
【解決手段】非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化され、および形成されるタンパク質− シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびシャペロンを含有する可溶性複合体の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、HIV感染の診断に関する。本発明は特に、可溶性のレトロウイルス表面糖タンパク質−(または膜貫通糖タンパク質)−シャペロン複合体の生成、および特に、好ましくは二重抗原架橋コンセプト(doubleantigen bridge concept)による、イムノアッセイにおけるHIVに対する抗体の検出におけるシャペロン抗原複合体の有利な使用、または免疫原としての使用を教示している。本発明はまた、それぞれ、HIV-1gp41の改変体、またはHIV-2 gp36の改変体、ならびにペプチジル−プロリル−イソメラーゼクラスのシャペロンから選択されたシャペロンを含む可溶性の複合体を開示する。それぞれ、HIV-1gp41の、またはHIV-2 gp36のNヘリックス状ドメインにおいて、特定のアミノ酸置換を含む改変体がまた記載される。
【背景技術】
【0002】
背景
ヒト免疫不全ウイルス(HIV)は、後天性免疫不全症候群の因子であり、通常、その頭文字によって、AIDSと呼ばれる。このウイルスの株には、HIV-1、およびHIV-2と命名された2つの主な株が存在する。HIウイルスは、現在、広範に伝播しており、そして世界的な健康および保健に対して重大な脅威となっており、このせいで、公的な保健機関は、HIVの診断およびAIDSの治療に関して、莫大な金額を費やさせられる。
【0003】
ウイルス伝播の経路の1つは、感染した血液または血液製品の輸血である。実質的に全ての先進工業国、および多くの発展途上国は、現在のところ、このウイルスのさらなる伝播を防ぐためには、全ての献血の強制的な検査を必要とする。血液によるHIV感染を、できるだけ確実に、かつ感染後すぐに診断することが、この分野における、全ての診断方法の課題である。
【0004】
基本的に、以下の3つの異なる診断様式が利用可能である:
(1)ポリメラーゼ連鎖反応(PCR)のような鋭敏な核酸診断手順による、血液由来のウイルスゲノム材料の診断、
(2)血液由来のウイルス抗原の検出、および
(3)体液由来のHIVに対する抗体の検出。
【0005】
HIV感染の経過の間、いくつかの診断上別個で、かつ診断上関連する段階が知られている。感染の初期段階では、HIVウイルス由来のタンパク質またはポリペプチドのみが見出され得る(「ウイルス感染期(viraemicphase)」が、抗HIV抗体はまだ存在しない。その後の、セロコンバージョンと名付けられる段階で、HIV抗原に対する抗体が出現し、一方でウイルス抗原の量(ウイルス量)が低下する。セロコンバージョンの初期段階で形成される抗体のほとんどが、クラスMの免疫グロブリン(IgM)に属する。後に、HIVに対する免疫応答は、クラスGの免疫グロブリン(IgM)に切り換わり、次にこれが、HIVに対する抗体のほとんどを構築する。感染のさらなる経過の間、抗HIV抗体のレベルは、低下し得るが、体液中のウイルス量(ウイルス粒子またはウイルス抗原の存在)は再度増大し得る。HIV感染の存在についてのスクリーニングは、好ましくは、HIV抗原に対する抗体を検出する血清学的アッセイ(時々、HIV抗原の検出と組み合わせて)によって行われる。患者内の免疫応答は、感染の経過中に変化し、そしてまた患者間で異なるので、サブクラスIgM、およびIgGに属する抗HIV抗体を検出する、極めて感受性でかつ信頼のできるイムノアッセイを有することが重要である。HIV感染の検出のための多くの異なるアプローチが記載されている。ウイルスタンパク質に対する抗体の、早期で、信頼でき、かつ鋭敏な検出は、重大でかつ大変重要である。
【0006】
ウイルスタンパク質(しばしば、ウイルス抗原と名付けられる)は、感染の発現時、およびこの疾患の非常に後期段階でのみ検出可能であり得る。従って、p24(HIV-1由来)、またはp26(HIV-2由来)(両方ともウイルスのコアタンパク質である)を測定するアッセイなどの、ウイルス抗原の検出のためのアッセイは、HIV感染を確実に検出するために他の診断手段と組み合わせてのみ用いられ得る。
【0007】
宿主中において抗体形成を誘導し得、それによって診断手順において抗原として用いられ得る、ウイルス抗原の3つのグループが、理論上利用可能である。これらは、エンベロープタンパク質(env遺伝子領域によってコードされる)、ウイルス酵素、または調節性タンパク質(例えば、逆転写酵素、またはインテグラーゼ)(pol遺伝子領域によってコードされる)、および構造的なコアタンパク質(gag遺伝子領域によってコードされる)である。HIV-1、およびHIV-2の両方におけるウイルスエンベロープタンパク質は、ポリペプチド前駆体タンパク質として合成される糖タンパク質である(HIVに関しては、gp160、HIV-2に関してはgp140)。これらの高分子量前駆体は、合成後、切断されて、それぞれ、gp120、およびgp41(HIV-1)、またはgp110、およびgp36(HIV-2)を生じる。これらの大きいポリペプチド(それぞれ、gp120、またはgp110)は、表面サブユニットを形成し、これがルーズな接触を介して、膜を貫通する、さらに小さいポリペプチド(それぞれ、gp41、およびgp36)に会合する。多くの宿主(患者)においては、エンベロープ糖タンパク質が、抗ウイルス免疫応答の好ましい標的である。Ratner,Lら、Nature 313(1985)277-84によって、特にこれらのエンベロープタンパク質の膜貫通(すなわち、それぞれgp41、またはgp36)は、これらのウイルスタンパク質の中でも最も免疫学的な可能性を保有することが実証された。
【0008】
HIウイルスによってコードされるポリペプチドを使用する、例えば、ELISA(酵素結合免疫吸着アッセイ)のようなイムノアッセイ法は、診断、およびスクリーニングにおいては広範に用いられている。ウイルスポリペプチドは、ウイルス材料から直接調製されるか、または組み換えDNA技術を用いるインビトロもしくはインビボの発現系に由来する。抗原生成の両方の方法とも、重大な限界がある。ウイルス調製物由来のポリペプチドは、生存可能なウイルス、または感染性遺伝子材料によって汚染され得、これによって、この材料を用いる人に対して害をもたらす。組み換え体由来材料は、非HIV宿主タンパク質によって汚染され得、これによって、このようなアッセイの特異性の低下、または感度の低下が生じ得る。
【0009】
ウイルス病原体のような、病原性因子に対する抗体の検出において、例えば、US4,945,042号に記載される、二重抗原架橋形式(doubleantigen bridge format)に従って、きわめて高頻度に、かつ極めて有利な抗体検出系が用いられる。この架橋コンセプトによるイムノアッセイは、固相に直接または間接的に結合した抗原、および同じまたは交差反応性の容易に可溶性の抗原(直接または間接的に検出可能である)の使用を必要とする。検討中の抗体は、もし存在するならば、固相結合抗原と、標識された検出抗原との間の架橋を形成する。2つの抗原が、特定の抗体によって架橋される場合にのみ、陽性のシグナルが生じる。
【0010】
抗HIV抗体の検出のための抗原として組換え産生されたgp41を用いるいくつかの試みが記載されている。組換え産生されたgp41は、抗HIV抗体を検出するために用いられ得るが、いくつかの限界を伴う。このようなgp41は、抗HIV抗体を測定するために、単独で、または他のHIV抗原と組み合わせてのいずれかで用いられる。今日、HIV抗原、および/または抗HIV抗体の両方の検出を独立して目的とするアッセイが公知である。WO93/21346においては、HIV-1gp41、およびHIV-2 gp36に対するgp24の抗原および抗体の同時の検出のための「組み合わせ試験(combi-test)」が記載される。このアッセイでは、固相が用いられ、この固相に対して組み換え産生されたgp41が直接コーディングされている。
【0011】
極度に高いpH値または低いpH値の使用が、溶液中でgp41(またはgp36)を保持するための1つの方法であるということもまた十分に確立されている。組み換え産生されたgp41は、ほぼpH3.0未満、またはほぼpH11.0を超えて可溶性であることが公知である。
【0012】
しかし、不幸にも、それぞれ、HIV-1 gp41、およびHIV-2 gp36は両方とも、生理的な緩衝条件下では本質的に不溶性である。
【0013】
一般的なイムノアッセイは、生理的なpHで実施される。生理的な緩衝条件下でのそれらの不溶性に起因して、多くのイムノアッセイにおいては、レトロウイルス表面糖タンパク質抗原は、固相材料上に直接コーティングされて用いられる。しかし、固相材料に対する抗原の直接コーティングは、多くの場合、有害であり、そしてコンフォメーションの変化、分子変性(unfolding)、抗原性の変化、不安定性、およびバックグラウンドの問題などの不利な点を生じる(Butler,J. E.ら、J. Immunol. Methods 150(1992)77-90を参照のこと)。
【0014】
強力なカオトロピック剤、または適切な界面活性剤によって、レトロウイルス表面糖タンパク質(retroviral surface glycoprotein)(rsgp)を可溶性にすることが可能であるが、このような様式で可溶性にされた材料は、診断ツールとしての使用に限定される。
【0015】
生理的な緩衝条件でのレトロウイルス表面糖タンパク質の不溶性によって、さらに、これらのタンパク質には、慣用的な(生物)化学的手順の極めて困難な標的になる。大部分の「標識化学(labeling chemistries)」、すなわち、標識(例えば、ポリペプチドに対するマーカー基)を結合するために用いられる化学的手順は、求核化学に基づいており、従って、pH約6〜pH約8のpH幅に制限されるのではなく、従って、生理的な緩衝条件以上または以下でのみ働く。例えば、「Bioconjugation」(1998)216〜363、M.Aslam、およびA. Dent編,McMillan Reference, Londonの、Aslam, M、およびDent, A., Thepreparation of protein-protein conjugatesに記載されるような、これらの慣用的な手順は、レトロウイルス表面糖タンパク質を可溶化するために必要な極端なpH値(またはSDSのような界面活性剤の存在下)では適切に作用しないか、または実行することが困難であるかのいずれかである。
【0016】
上記で言及したとおり、架橋コンセプトによるイムノアッセイは、病原性生物体と反応性の抗体の検出を目的とする広範な種々の異なるアッセイにおいて利点が証明されている。しかし、その不溶性に起因して、このようなアッセイの設定において、それぞれ、HIV-1のe-gp41分子(すなわち、「糖タンパク質41のエクトドメイン(ectodomain of glycoprotein 41)」、またはe-gp36分子を用いることは、例えば、不可能であった。
【0017】
直接のコーティングの不利な点を補償するために、種々のアッセイが設計されており、このアッセイは、e-gp41抗原を用いることに代わって、合成的に、または組換え産生されたその抗原の部分的配列(多少ともイムノドミナント、いわゆるループ領域にまたがる)を利用する。このようなアッセイの例は、以下に考察する特許文献に示される。
【0018】
gp41の細胞外部分におけるループ領域は、N末端ヘリックス状ドメインを、同様にヘリックス状のC末端ドメインに連結する分子の、非ヘリックス状の先端ヘアピンである。gp41に対して反応性の抗血清の大部分は、この先端ループモチーフに対する抗体を含む。従って、このジスルフィド架橋したヘアピン、またはループ構造は、gp41のイミノドミナント領域を示す。従って、組み換え的に駆動されたgp41に関連する問題を克服するための迂回路の1つは、gp41の部分的配列を表すペプチドの化学的生成である。それぞれgp41、またはgp36が、本発明において言及される場合、いわゆるエクトドメイン(ループ接続されたN-ヘリックス、およびC-ヘリックスを包含するが、N末端融合ペプチド、およびC末端膜貫通セグメントは欠く)として規定されることに注意することが重要である。
【0019】
種々のHIV抗原のペプチド断片が、関連の特許文献(オーストラリア特許出願番号597,884(57733/86)、ならびに米国特許第4,735,896号、および同第4,879,212号)中で開示される。詳細には、これらの3つの明細書は、gp41糖タンパク質の保存されたイムノドミナント領域(HIV-1の主なエンベロープタンパク質のループ領域)を開示する。HIV-2のgp36タンパク質の類似のイムノドミナント領域がまた、合成されている。これらのループ領域に対応するペプチド(このエクトドメインの先端部を構成する)は、HIV-1、およびHIV-2の早期診断を可能にし、そしてアッセイにとって十分だが至適ではない感度および良好な特異性をもたらす。しかし、それらの限界は、特定の患者におけるセロコンバージョンの初日のIgM抗体の検出に関して明らかになる。
【0020】
WO92/22573は、骨格と共通して、すなわち、種々の哺乳動物の免疫不全ウイルスの膜貫通エンベロープタンパク質(例えば、gp41、またはgp36)のイミノドミナント領域と共通して、免疫学的な特性を有するペプチドを開示する。このイムノドミナント領域は、異なる哺乳動物種に由来する免疫不全ウイルス単離体において高度に保存されている、ジスルフィドループを含むことをさらに確認する。
【0021】
EP396559は、HIVの天然に存在するアミノ酸配列に相当するアミノ酸配列を保有する、人工的ペプチドに関する。このエピトープは、やはり、それぞれgp41、またはgp36のループ構造に相当する配列に由来する。それらは、イミノドミナントループの2つのシステイン残基の間の化学的酸化段階によって形成されたジスルフィド結合を含むように、さらに純化されている。
【0022】
しかし、HIV感染した患者の抗HIV抗血清に含まれた抗体のかなり大きい割合が、gp41またはgp36のイムノドミナントループ由来の配列モチーフ、またはその改変体とは反応しない。これらのペプチド抗原は、有利な架橋コンセプトと組み合わせて用いられ得るが、HIVgp41のループ領域の外側のエピトープと反応性の抗体は、検出されない。HIV感染の非常に早期の診断が重要なだけでなく、HIV-1、およびHIV-2のできるだけ多くのサブタイプが検出されることも極めて重要である。エピトープがより多く、特に、rsgpの正確に折り畳まれたコンフォメーションのエピトープが、より多く存在するほど、偽陰性の診断に起因して、感染したサンプルを見逃す可能性は低くなる。
【0023】
従って、レトロウイルス表面糖タンパク質分子の(特に、HIV-1由来のgp41の)より大きい部分を可溶型で提供するために、継続的な努力が行われている。
【0024】
gp41の生物物理学的な、そして生化学的な特性は、過去に広範に研究されてきた。Lu,Mら、Nat. Struct. Biol. 2(1995)1075-82)は、gp41の三量体構造を部分的に解明している。gp41は、生理的な条件下で、不溶性の凝集物を形成するので、検討は、短縮型のエクトドメインgp41に対して限定された。
【0025】
gp41のネイティブな三量体が、3つの平行なN末端中央ヘリックス(これに対してC末端ヘリックスは、逆平行方向に詰める)を含む、6つのヘリックス状バンドルを形成することが、NMR分光法によって最近確認されている(Caffrey,Mら、J Biol Chem 275(2000)19877-82)。
【0026】
gp41の高分子凝集物がまた記載されている。このような凝集物は、gp41のいわゆる先端部ループ領域の相互作用によって形成される可能性が最も高い。
【0027】
タンパク質設計による、標的細胞へのHIV-1進入のインヒビターは、Root, M.J.ら、Science 291(2001)884-8によって開発された。このインヒビターは、gp41由来のN末端ヘリックスドメイン由来の3つのストレッチ、およびこの分子由来のC末端ヘリックスドメインの2つのストレッチを含む。しかし、この遺伝子操作された構築物は、ネイティブな分子の多くのドメイン、および多くの抗原性エピトープを欠き、特にいわゆるループモチーフ(特に免疫原性エピトープを保有することが公知である(上記を参照のこと))を含まない。
【発明の開示】
【発明が解決しようとする課題】
【0028】
従って、できるだけ多くのレトロウイルス表面糖タンパク質エピトープを可溶型で提供するという非常に大きい必要性が依然として存在する。特に種々の治療的な適用、および診断的な適用における使用のために、それぞれ、HIV-1由来のgp41、またはHIV-2由来のgp36を含む、このような可溶性抗原を提供する必要性が存在する。
【0029】
より多くのレトロウイルス表面糖タンパク質エピトープ、またはそれぞれe-gp41分子もしくはe-gp36でさえ、可溶型で提供することが可能であるか否かを検討することが、本発明の課題であった。
【0030】
本発明のさらなる課題は、それぞれgp41、およびgp36の改変体(これは、取り扱うことがより容易であり、そして/または特に、イムノアッセイを実施するために必要とされるような、または免疫に必要とされるような緩衝条件下では、複合体型(この改変体およびペプチジル−プロリル−イソメラーゼクラスのシャペロンのシャペロンを含む)で可溶性である)を提供することが可能であるか否かを検討することであった。
【課題を解決するための手段】
【0031】
本発明の要旨は、
〔1〕非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化され、および形成されるタンパク質− シャペロン複合体
が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびシャペロンを含有する可溶性複合体の製造方法、
〔2〕非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子又はSlyDの結合コンピテント断片とを混合する工程、ここで該タンパク質および該断片の両方が可溶化され、および形成されるタンパク質− 断片複合体が可溶である生理的条件
にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質および断片を含有する可溶性複合体の製造方法、
〔3〕非生理的なバッファー中でレトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここでレトロウイルス表面糖タンパク質およびシャペロンの両方が可溶化され、複合体を形成し、および該複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質−シャペロン複合体の製造方法、
〔4〕レトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを含有してなる可溶性複合体、
〔5〕〔4〕記載の可溶性複合体を含有してなる試薬の組成物
に関する。
【発明の効果】
【0032】
本発明により、より多くのレトロウイルス表面糖タンパク質エピトープ、例えば、e-gp41分子またはe-gp36が可溶型で提供される。
【発明を実施するための最良の形態】
【0033】
古典的な「折り畳みヘルパー(folding helpers)」として知られるシャペロンは、他のタンパク質の折り畳み、および構造的な完全性の維持を補助するポリペプチドである。それらは、インビボ、およびインビトロの両方でポリペプチドの折り畳みを促進する能力を保持する。一般に、折り畳みヘルパーは、折り畳み触媒、およびシャペロンに小分割される。折り畳み触媒は、その触媒性機能に起因してタンパク質折り畳みにおける律速段階を加速する。触媒の例は、以下にさらに記載される。シャペロンは、変性されたポリペプチドまたは部分的に変性されたポリペプチドに結合し、これによってタンパク質を再生することを補助することが公知である。従って、折り畳み触媒とは異なり、シャペロンは、結合機能を発揮するに過ぎない(Buchner,J., Faseb J 10(1996)10-19)。
【0034】
シャペロンは、タンパク質成熟、折り畳み、転位、および分解に関与する、偏在性のストレス誘導性タンパク質である(Gething, M. J.、およびSambrook, J., Nature 355(1992)33-45)。正常な増殖条件下でも存在するが、それらは、ストレス条件下で、豊富に誘導される。このことによって、それらの生理的な機能が、ストレス条件に対処することであるという理論がさらに支持される。
【0035】
現在のところ、いくつかの異なるファミリーのシャペロンが知られている。これらのシャペロンの全てが、折り畳まれていないか、または部分的に折り畳まれていないタンパク質に対して結合する能力によって特徴付けられており、そしてタンパク質の正確な折り畳み、または変性もしくは凝集したタンパク質の除去に関係する生理的機能を有する。
【0036】
シャペロンの十分特徴付けられた例は、タンパク質のいわゆる熱ショックファミリーのメンバーである。これらは、相対的な分子量に従って命名される;例えば、hsp100、hsp90、hsp70、およびhsp60、ならびにいわゆるshsps(小熱ショックタンパク質(smallheat-shock-proteins))(Buchner, J., Faseb J 10(1996)10-19によって、ならびにBeissinger, M.およびBuchner,J., Biol. Chem. 379(1998)245-59によって記載されるような)。
【0037】
折り畳み触媒は、シャペロンと異なり、規定された律速段階を加速することによって折り畳みを補助し、これによって凝集しやすい折り畳み中間体の濃度を低下させる。触媒の1つのクラスである、タンパク質ジスルフィドアイソマー(あるいは、チオール−ジスルフィド−オキシド−レダクターゼと命名された)は、分泌タンパク質におけるジスルフィド結合の形成または再編成を触媒する。グラム陰性の細菌において、周辺質における分泌タンパク質の酸化的折り畳みは、DsbA、DsbB、DsbC、およびDsbDと命名された、タンパク質ジスルフィドイソメラーゼのカスケードによって調整される(Bardwell,J.C., Mol Microbiol 14(1994)199-205、およびMissiakas, D.,ら、Embo J 14(1995)3415-24)。
【0038】
ペプチジルプロリルシス/トランスイソメラーゼ(PPI)と呼ばれる折り畳み酵素の別の重要なクラスは、以下のような異なるメンバーを含む:CypA、PpiD(Dartigalongue,C.、およびRaina, S., Embo J 17(1998)3968-80, FkpA(Danese, P. N.ら、Genes Dev 9(1995)387-98)、誘発因子(triggerfactor)(Crooke, E、およびWickner, W., Proc Natl Acad Sci USA 84(1987)5216-20、ならびにStoller,Gら、Embo J 14(1995)4939-48)、およびSly D(Hottenrott, Sら、J Biol Chem 272(1997)15697-701)。これらのなかでも、FkpA、SlyD、および誘発因子は、配列のアラインメントに基づいて、関連していることが見出されている。
【0039】
ペプチジルプロリルイソメラーゼFkpAは、グラム陰性細菌の周辺質に局在している。このシャペロンが、細菌の外部膜タンパク質の輸送および転位に重要であることが推測されている。Ramm,K.、およびPluckthun, A., J Biol Chem 275(2000)17106-13)によって、FkpAは、タンパク質の正しい折り畳みに対する、その有益な効果を、2つの異なる方法で表すことが示されている。第一に、FkpAは、初期の折り畳み中間体と相互作用し、それによってそれらの凝集を妨げる。第二に、FkpAは、可能性としては、部分的に折り畳まれていない種(それは、凝集型と平衡して存在し得る)に対する結合によっても、不活性なタンパク質を再活性化する能力を有する。
【0040】
いくつかの折り畳みヘルパーは、触媒的に活性なドメイン、およびシャペロン(またはポリペプチド結合)ドメインの両方を含む。代表的な例は、例えば、誘発因子(Zarnt, Tら、J Mol Biol 271(1997)827-37)、Wang, C. C.、およびTsuo, C. L.,Faseb J 7(1993)1515-7)、SurA(Behrensら、EMBO J(2001)20(1)、285-294)、およびDsbA(Frech,C.,ら、Embo J 15(1996)392-98)である。本発明者らの観察によれば、同じモジュール構造が、それぞれ、PPIases FkpA、およびSlyDにおいて現実化されると思われる。
【0041】
異なる独立した系において、シャペロンの強化された発現によって、ポリペプチドの組み換え生成が促進され得ることが実証されている。これに対する例は、WO94/08012に見出され得る。
【0042】
タンパク質の産生の増大は、ポリペプチドコード配列、およびシャペロン配列を含む遺伝子構築物を用いることによって達成され得ることがまた、公知である。例えば、この融合コンセプトは、ヒトのプロインスリン(pro-insulin)遺伝子、およびDsbAを含む遺伝子構築物を用いることによって、Escherichia coliの周辺質におけるヒトのプロインスリンの有意に増大した産生を生じることが示されている(Winter,J.ら、Journal of Biotechnology 84(2000)175-185)。
【0043】
未変性様の折り畳まれたポリペプチドの産生の増大のためにシャペロンを用いるアプローチは主に、結合、およびこれによるシャペロンタンパク質の機能の可溶化に起因している。シャペロンおよび標的タンパク質を含む融合ポリペプチドの組み換え産生後、このシャペロンは習慣的に、得られたポリペプチドから切り離され、所望のポリペプチドが純粋な形態で得られる。対照的に、本発明は、一方では、レトロウイルス表面糖タンパク質と会合している、適切なシャペロンの有益な可溶性効果に基づく。
【0044】
驚くべきことに、折り畳みヘルパー、例えば、ペプチジルプロリルイソメラーゼ(PPI)クラスの多くのメンバー(特にFKBPファミリー由来)が、触媒性活性を示すだけでなく、アミロイド形成性のタンパク質の(またはより一般的に言えば、凝集する傾向にあるタンパク質の)溶解度に対して強烈な有益な効果をもたらすことを、本発明者らは見出した。それらがそのような効果をもたらすのは、このようなタンパク質(そうでなければ(すなわち、非シャペロンの単離型では)凝集しやすい)と可溶性の複合体を形成することによってである。このようなタンパク質(さもなければ、生理的条件下では、ほとんど溶解しないか、または不溶性である)は、一旦それらが、適切なPPIシャペロンと複合して結合すれば、穏やかな生理的条件下では結局、可溶性になる(すなわち、界面活性剤またはカオロトピック剤などの可溶化添加物の必要がない)。このように、本発明者らは、例えば、可溶性のタンパク質−シャペロン複合体(例えば、凝集しやすい標的タンパク質としてHIV-1のgp41タンパク質、ならびに可溶性付与シャペロンとしてFkpA、または他のFKBPを含む)を産生することができた。
【0045】
さらに、本発明者らは、それぞれ、HIV-1 gp41、またはHIV-2 gp36の特定の十分規定された改変体が、PPI-クラスのシャペロンと可溶性の複合体を形成するのに特に適切であることを見出した。
【0046】
gp41、およびFkpAの複合体、またはgp36およびFkpAの複合体は、例えば、生理的条件下で、例えば、容易に可溶性であり、それらを、都合の良いpH範囲で容易に標識することが可能であり、そしてそれらを用いて、HIV(それぞれ、1または2)のそれぞれgp41、またはgp36に対する抗体の検出において、従ってHIV感染の診断において、多大な利益を得ることができる。
【0047】
詳細な説明
本発明は、本質的に不溶性である標的タンパク質、およびペプチジル−プロリル−イソメラーゼクラスのシャペロンを含む可溶性複合体を生成する方法に関し、この方法は、バッファー中でこのタンパク質およびこのシャペロンを混合する工程、ここでこのタンパク質およびシャペロンの両方は、可溶化される、ならびにこのバッファーを生理的な条件に調整する工程(ここで、この形成されたタンパク質−シャペロン複合体は可溶性である)を包含する。
【0048】
本発明による「標的タンパク質」とは、20mMリン酸ナトリウム、および150mM塩化ナトリウムからなるpH7.4の水性緩衝溶液中で本質的に不溶性である任意のタンパク質であってもよい。例えば、好ましい標的タンパク質は、アミロイド形成性タンパク質、アミロイド形成性ウイルスの表面糖タンパク質、レトロウイルス表面糖タンパク質、特にHIV-1gp41、HIV-2 gp36、およびHTLV gp21である。
【0049】
標的ポリペプチドの重要な群の1つは、いわゆるアミロイド形成性タンパク質、またはポリペプチドである。このようなアミロイド形成性タンパク質は、体液または区画中において凝集型で見出されている。周知の例は、血清アミロイドA(sAA)、いわゆるb-A4、またはAb(アルツハイマー病の脳で特徴的なアミロイド沈着を形成することが知られている42または43個のアミノ酸のポリペプチド)、いわゆるプリオンタンパク質(PrPsc型は、BSEまたはクロイツフェルトヤコブ病における凝集において蓄積する)、およびHIV-1gp41のようなレトロウイルス表面糖タンパク質(HAD(HIV関連痴呆)に罹患している患者の脳におけるアミロイド様プラークにおいて見出される)である。好ましい実施形態において、シャペロン、好ましくはPPIシャペロンを用いて、アミロイド形成性タンパク質、およびシャペロンを含む可溶性複合体を形成する。極めて有利になるように、このような複合体を、多くの異なるイムノアッセイ手順において用いてもよい。好ましくは、このような複合体を、二重抗原架橋コンセプトに従って、イムノアッセイにおいて用いる。
【0050】
HIVおよび他のエンベロープウイルス(例えば、HTLV、インフルエンザウイルス、およびエボラウイルス)は、全てが、表面糖タンパク質を発現し、これが細胞付着および膜融合の両方を媒介する。これらの機能を果たすために、これらの表面糖タンパク質の全てが、極端に疎水性のセグメントを含み、これがインビトロでそれらを取り扱うことを困難にさせ、凝集しやすくし、それらを、インビトロの再折り畳みの労力に関して困難な標的にさせる。さらに好ましい実施形態において、本発明は、PPIシャペロン、およびエンベロープウイルスの表面糖タンパク質を含む可溶性複合体に関する。本発明は、PPIシャペロン、およびエンベロープウイルスの表面糖タンパク質を含む複合体を、この表面糖タンパク質に対する抗体の検出のためのイムノアッセイにおいて用いることに特に関する。
【0051】
HADは、HIV感染の周知の合併症である。Caffrey, Mら(前出)によって記載されたように、組織学的な現象学に関して、HADは、クロイツフェルトヤコブ病と呼ばれる海綿状脳症に極めて類似している。クロイツフェルトヤコブ病の病因論は一般に、改変プリオンタンパク質を含むプラークのアミロイド形成性蓄積から生じると考えられる(Prusiner,S. B., Proc Natl Acad Sci USA 95(1998)13363-83)。E-gp41の高分子量凝集物に関与するHADについての類似の病因論は極めて確率が高い。HIV脳症における神経学的な病変が、ビンスヴァンガー病と、病理的特徴および放射線医学的特徴を共有することは注目に値する。
【0052】
本発明による好ましいアミロイド形成性タンパク質は、それぞれ、HIV-1由来のgp41、HIV-2由来のgp36、またはHTLV由来のgp21である。
【0053】
タンパク質は、20mMリン酸ナトリウム(pH7.4)150mM NaClからなるバッファーにおいて、そのタンパク質が、50nM以下の濃度で可溶性である場合に、「本質的に不溶性」であるとみなされる。
【0054】
PPIシャペロンおよび標的タンパク質を含む本発明の複合体は、生理的な緩衝条件下で、例えば、20mMリン酸ナトリウム、pH7.4、150mMNaClからなるバッファーにおいて、このPPIシャペロン複合体中に含まれるこの標的タンパク質が、100nM以上の濃度で可溶性である場合に、「可溶性」であるとみなされる。
【0055】
本発明者らは、バッファー中で標的タンパク質とシャペロンとを混合する工程(ここでは、タンパク質およびシャペロンの両方とも可溶性である)、およびこのバッファーを生理的条件に調整する工程(ここで、この形成されたタンパク質−シャペロン複合体は、可溶性のままである)を包含する方法を開発した。
【0056】
可溶性シャペロン−標的タンパク質複合体の生成は、可溶化バッファー条件から、すなわち、バッファー(ここでは、標的タンパク質およびシャペロンの両方が可溶性である)から開始する。「非生理的な(non-physiological)」、または「可溶化(solubilizing)」バッファーと名付けられ得る、適切なバッファーは、標的タンパク質およびPPIシャペロンの両方が変性されないか、または少なくとも不可逆的に変性されないという要件を満たさなければならない。このような緩衝条件から開始すれば、シャペロンは、標的タンパク質に結合し、そして非生理的な条件から生理的な条件への緩衝条件の変化は、標的タンパク質の沈殿なしに可能である。
【0057】
適切な(非生理的な)バッファー、すなわち、バッファー(ここで、本質的に不溶性である標的タンパク質、およびPPIシャペロンの両方が、可溶性である)は、高いpHもしくは低いpH、または高いカオトロピック塩濃度またはそれらの組み合わせのいずれかを利用する。
【0058】
PPIシャペロンおよび標的タンパク質(これは、本質的に不溶性である)を含む分子内複合体の産生の場合、非生理的なバッファーは、好ましくは、かなり高いpH、またはかなり低いpHを有するバッファーである。好ましくは、このようなバッファーは、高いpH範囲において9〜12のpH、または低いpH範囲においては2〜4.5のpHを有する。
【0059】
PPIシャペロンおよび標的タンパク質(これは、本質的に不溶性である)を含む分子内複合体の産生の場合、可溶化バッファーは、好ましくは、約6のpHにおいて、かなり高い濃度のカオトロピック塩、例えば、6.0M塩化グアニジウムを有するバッファーである。再生の際、標的タンパク質は、その未変性様の構造および分子内複合体型を仮定する。
【0060】
本発明の状況において、生理的なバッファー条件は、必要に応じてこのバッファー中に存在し得る他の非塩成分(例えば、糖、アルコール、界面活性剤)にかかわらず、このような添加物が標的タンパク質およびシャペロンを含む複合体の溶解度を損なわない限り、5.0〜8.5の間のpH値、および総塩濃度500mM未満によって規定される。
【0061】
さらなる好ましい実施形態において、本発明は、可溶性のレトロウイルス表面糖タンパク質−シャペロン複合体を生成する方法に関しており、この方法は以下:バッファー中でレトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼを混合する工程(ここで、レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼの両方は、可溶性であり、複合体を形成する)、ならびにこのバッファーを生理的な条件に調節する工程(ここで、この複合体は可溶性である)を包含する。
【0062】
「レトロウイルス表面糖タンパク質」、または「rsgp」という用語は、本発明において用いる場合、HIV-1のgp41およびHIV-2のgp36、ならびに他の哺乳動物免疫不全ウイルス由来の対応するエンベロープ糖タンパク質を含むはずである。好ましいレトロウイルス表面糖タンパク質は、HIV-1由来のgp41、HIV-2由来のgp36、およびHTLVのgp21である。特に好ましいrsgpは、HIV-1のgp41、およびHIV-2のgp36である。rsgpという用語は、ここで概説されるようにまた、天然に存在するresp、および合成的に操作されたrsgpの改変体を含む。
【0063】
それぞれgp41またはgp36のNヘリックス状ドメイン内のアミノ酸の特定の十分規定された置換が、それぞれgp41またはgp39の野生型配列を有するポリペプチドに比べて、これらの分子の全体的特性においてさらなる利点を生じることが見出されている。これらの改変体は、本発明に従って好ましい実施形態を示す。特に、Leu555、Leu566、Ile573、およびIle580の位置(ここで、これらの位置は、HIV-1gp41野生型配列(配列番号1)から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された1つ以上の位置において、少なくとも1つのアミノ酸置換、および多くとも4つのアミノ酸置換を含み、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられる、HIV-1gp41の改変体、あるいは、Leu554、Leu565、およびVal579の位置(ここで、これらの位置は、HIV-2 gp36野生型配列(配列番号2)から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された位置において、少なくとも1つのアミノ酸置換、および多くとも3つのアミノ酸置換を含み、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられるHIV-2gp36の改変体が、当該分野で公知である問題を少なくとも部分的に解決するために適切である。
【0064】
それぞれgp41またはgp36の新規な改変体は、凝集する傾向が低いほど、野生型配列のそれらの対応するポリペプチドに比べて、溶解度が向上し、取り扱いがより簡単になる。生理的緩衝条件下で、gp41またはgp36を可溶型で含む試薬を提供するための労力が一旦払われれば、溶解度の改善は、特に明白になる。ペプチジル−プロリル−イソメラーゼ(PPI)クラスのシャペロンから選択されたシャペロンによって付与された効果を有する新規な改変体の好適な特性を合わせることが特に有利であると証明されている。従って、本発明は、この改変体およびペプチジル−プロリル−イソメラーゼクラスのシャペロンのシャペロンを含む可溶性複合体の生成における、本発明において記載されたようなgp41の改変体、および/またはgp36の改変体の使用にさらに関する。
【0065】
改変HIV糖タンパク質、およびPPIクラスのシャペロンを含む可溶性複合体は好ましくは、それぞれ、改変HIVgp41、またはHIV-2 gp36、ならびにPPIクラスシャペロンの両方を含む単一の組み換えタンパク質から得られる。従って、好ましい実施形態は、本発明に記載されるようなHIV-1gp41、またはHIV-2 gp36の改変体、およびペプチジル−プロリル−イソメラーゼクラスのシャペロンから選択されたシャペロンを含む組み換えタンパク質である。
【0066】
gp41、またはgp36の新規な改変体が、野生型ポリペプチドよりも、取り扱いが容易であるという事実によって、それらの改変体は、免疫原としての使用、または抗原としての使用のような種々の目的に理想的にされる。好ましい実施形態において、本発明は、イムノアッセイにおける、本発明によるgp41、および/もしくはgp36の改変体の、またはPPIシャペロンおよびこのような改変体を含む複合体の、例えば、単一の組み換えタンパク質としての使用に関する。最も興味深いことには、レトロウイルス表面糖タンパク質、およびPPIシャペロンの両方を含む融合タンパク質は、適切な条件下で可溶化かつ再生され得、そしてHIVイムノアッセイにおいて簡便な標識および信頼性のある検出を可能にする可溶性の分子内rsgp-シャペロン複合体を形成することが見出されている。
【0067】
レトロウイルス表面糖タンパク質と、シャペロンとの間の可溶性複合体は、二重抗原架橋コンセプトに従って、抗体の検出のためのイムノアッセイにおいて非常に有利になるように用いることができる。
【0068】
HIV-1由来のgp41、またはHIV-2由来のgp36を含むrsgpシャペロン複合体は、感染の早期段階におけるHIVに対する抗体の検出に特に有利である。可溶性のシャペロン−gp41複合体、または可溶性のシャペロン−gp36複合体を用いれば、好ましくは架橋コンセプトに従って、イムノアッセイを実施することが可能であり、これによって、体液サンプル中におけるHIVに対する抗体の鋭敏かつ早期の検出が可能になる。
【0069】
シャペロンが、他の不溶性タンパク質と複合体を形成し得るという事実をまた用いて、イムノアッセイ(好ましくは、架橋コンセプトに従うイムノアッセイ)をさらに大幅に改善するという、大きい利点をえることができる。この架橋コンセプトによって、第一の抗原としてのシャペロン−抗原複合体(ほとんどが固相側でのいわゆる捕捉抗原)、および第二のシャペロン−抗原複合体(ほとんどが、検出側での検出抗原)の使用が可能になる。シャペロン反応性抗体の結合によって生じたバックグラウンド反応の問題を最小化するために、固相側について第一のシャペロン、そして異なる種由来の検出側について第二のシャペロンを利用することによって、このような架橋アッセイをさらに有利に改変することができる。
【0070】
ここで、標識されたシャペロン−抗原複合体を使用する、架橋コンセプトに従うイムノアッセイを実施することが可能である。シャペロン−抗原複合体(ここでは、このシャペロンのみが標識されており、これによって、この抗原は、このような標識によって改変もされず、負の影響もされない(例えば、高次構造に関して)ことが確実にされる)を生成することがまた可能である。
【0071】
化学的カップリングの様式およびストラテジーは、必要に応じて選択され得る。ポリペプチドの場合、-SH、-NH2、または-COO-残基、ならびにチロシンのOH基、ヒスチジンのイミダゾール基、またはトリプトファンの複素環式イミノ基を標的するカップリング化学は、すぐ手が届くものである。いくつかの適切なカップリング化学が、これらの官能基の各々について公知である(Aslam,M、およびDent, A.,前出)。慣用的なタンパク質カップリング化学は、作用する緩衝条件下、例えば、約5〜8.5のpH範囲内でタンパク質が可溶であることを必要とする。例えば、gp41が、このpH範囲内では、例えばSDSによって変性されない限り可溶ではないので、未変性様に折り畳まれたgp41は、これまでは、化学的カップリングを受けやすくはなかった。本明細書において本発明者らが記載する、gp41−シャペロン複合体は、用いられる検出様式にかかわらず、イムノアッセイのための可溶性の標識されたHIVエンベロープタンパク質を生成する、都合のよい手段を提供する。
【0072】
好ましい実施形態において、本発明は、可溶性のrsgp−シャペロン複合体の生成のためのプロセスに関し、このプロセスは、可溶化されたレトロウイルス表面糖タンパク質、およびペプチジルプロリルイソメラーゼクラスから選択されたシャペロンを、非生理的緩衝条件下で混合する工程、およびその後にこのバッファーを生理的条件に調整し、それによって分子内複合体を形成する工程を包含する。
【0073】
シャペロンおよびレトロウイルス表面糖タンパク質は、別々のポリペプチドとして用いることができるだけではない。本発明者らは、驚くべきことに、両方のタンパク質を共有結合させることが有利であるということを観察した。このような共有結合は、従来の架橋手順によって可能である;しかし、好ましくは、共有結合は、レトロウイルス表面糖タンパク質およびシャペロンを含む組み換えポリペプチドを産生することによって達成される。
【0074】
さらに好ましい実施形態において、本発明は、可溶性のrsgp-シャペロン複合体の生成のためのプロセスに関する。このプロセスは、適切な緩衝条件下で、共有結合したレトロウイルス表面糖タンパク質、およびペプチジルイソメラーゼクラスから選択されたシャペロンタンパク質を含むタンパク質を可溶化する工程、そしてその後にこのバッファーを生理的条件に調節する工程を包含する。これによよって、分子内複合体が得られる。
【0075】
本発明は、ペプチジルプロリルシス/トランスイソメラーゼ(PPI)と命名された折り畳みヘルパーのクラスに由来するシャペロンの使用を教示する(Dartigalongue,C.、およびRaina(前出)を参照のこと)。このファミリーの周知の例は、以下のように呼ばれるメンバーである:CypA、PpiD(Dartigalongue,C.およびRaina, S., Embo J 17(1998)3968-80;Schmid, F. X., Molecular chaperones inthe life cycle of proteins(1998)361-389, 編集A. L. Fink、およびY. Goto, Marcel DeckerIn., New York),FkpA(Danese, P. N.ら., Genes Dev 9(1995)387-98)、および誘発因子(Crooke,E.、およびWickner, W., Proc Natl Acad Sci USA 84(1987)5216-20;Stoller, G.ら、Embo J14(1995)4939-48)。このペプチジルプロリルイソメラーゼは、以下の3つのファミリーに小分割される:パルビュリネス(parvulines)ファミリー(Schmid,F. X.(前出),Rahfeld, J. U.ら、FEBS Lett 352(1994)180-4)、サイクロフィリンファミリー(Fischer, Gら、Nature337(1989)476-8)、およびFKBPファミリー(Lane, W. S.ら、J Protein Chem 10(1991)151-160)。FKBPファミリーは、興味深い生化学的特徴を示す。なぜなら、そのメンバーは、マクロライド(例えば、FK506、およびラパマイシン)に結合する能力によってもともと同定されたからである(Kay,J. E., Biochem J 314(1996)361-85)。
【0076】
プロリルイソメラーゼは、異なる機能の異なるサブユニットまたはモジュール、例えば、触媒活性を示すモジュール、およびシャペロンもしくは結合活性を示すモジュールを含んでもよい。FKBPファミリーのこのようなモジュールメンバーは、FkpA(Ramm, K.およびPluckthun, A., J Biol.Chem275(2000)17106-13)、SlyD(Hottenrott, S.ら、J. Biol Chem 272(1997)15697-701)、および誘発因子(Scholz,Cら、Embo J 16(1997)54-8)である。好ましい実施形態において、本発明は、可溶性複合体であって、レトロウイルス表面糖タンパク質、およびペプチジルプロリルイソメラーゼクラスの折り畳み触媒から選択されたシャペロンを含む複合体に関する。
【0077】
当然ながら、本発明は、ペプチジルプロリルイソメラーゼクラスの特別に言及されたメンバーの使用に対して制限されるものではなく、また、同じクラスから生じるが、異なる種の細菌から派生するシャペロンを用いて実施することもできる。PPIクラスのシャペロンのFKBPファミリーの好ましいメンバーが用いられる。
【0078】
さらなる実施形態において、真核生物由来の相同体を使用することが好ましく、そしてヒト起源のPPIaseを使用することが極めて好ましい。なぜなら、これらのPPIaseは、ヒト血清由来の抗体によって認識されないはずであり、従って、血清学的アッセイ(すなわち、ヒト抗体の検出に基づくアッセイ)においては妨害しないはずであるからである。
【0079】
分子シャペロンの完全な配列を常に使用する必要はないということがまた周知でありかつ理解される。必要な能力および機能を依然として保有するシャペロンの機能的断片(いわゆる、モジュール)がまた、用いられ得る(WO98/13496を参照のこと)。
【0080】
例えば、FkpAは、細菌のサイトゾルにおける不活性前駆体分子として合成されて、細胞質膜を横切って転位される、周辺質PPIである。FkpAの活性型(成熟FkpA、または周辺質FkpA)は、シグナル配列(アミノ酸1〜25)を欠き、従って前駆体分子のアミノ酸26〜270を含む。FkpAに関する関連の配列情報は、登録番号P45523として、公的なデータベースから、例えば、「SWISS-PROT」から容易に入手可能である。
【0081】
FlpAに密接に関連した、いわゆるSlyDは、触媒性およびシャペロン機能を担う構造化されたN末端ドメインから、そしてほとんど構造化されていないC末端(例外的にヒスチジン残基およびシステイン残基に富む)からなる(Hottenrott,S.ら、J. Biol Chem 272(1997)15697-701)。本発明者らは、アミノ酸1-165を含むSlyDのC末端短縮改変体が、gp41、およびgp36においてその可溶性機能を効率的に発揮することを見出した。野生型SlyDにおいてとは異なり、ジスルフィドシャッフリングを損なう危険は、用いられた短縮型SlyD改変体(1-165)においては、首尾よく回避される。
【0082】
上記で考察されたシャペロンの改変体であって、1つまたはいくつかのアミノ酸置換または欠失を保有する改変体がまた、本発明に従うプロセスを実施するために用いられてもよい。
【0083】
別の供給源に由来する適切なシャペロン、およびシャペロンの適切な断片または変異体が、本実施例に記載のような手順を用いることによって容易に選択され得る。それらは、可溶性のrsgp-シャペロン複合体を生成するために、遊離型で、またはレトロウイルス表面糖タンパク質に共有結合されてのいずれかで用いられ得る。本発明による好ましい実施形態において、結合にコンピテントなPPIaseシャペロンは、レトロウイルス表面糖タンパク質に組み換え的に結合されて、細菌のサイトゾル中でこの遺伝子産物の高い発現を生じる。本発明において言及されるような結合コンピテント(bindingcompetent)なPPIaseは、その触媒性のPPIase活性にかかわらず、伸長したポリペプチド基質に対する結合(すなわち、基質結合、またはシャペロンモチーフ)を媒介する、少なくとも機能的な単位を含む。
【0084】
本発明者らは、また、PPIクラスの折り畳み触媒に属さないいくつかのシャペロンが、レトロウイルス表面糖タンパク質と可溶性の複合体を形成し得るということを観察した。従って、本発明に従うさらなる好ましい実施形態は、Skp(OmpHとしても公知;Missiakas,D.ら、Mol Microbiol 21(1996)871-84)と、レトロウイルス表面糖タンパク質との間の可溶性複合体である。しかし、さらに好ましい実施形態は、レトロウイルス表面糖タンパク質、およびGroEL、またはその部分を含む可溶性複合体である。Skpに対して相同であるシャペロンがまた用いられ得る。
【0085】
モジュラーPPIが、変性されたかまたは部分的に変性されたタンパク質に対して優先的に結合することは公知である(例えば、Scholzら、前出)。PPIaseは、ここで、タンパク質の折り畳みを触媒するだけでなく、このようなタンパク質と安定な複合体を形成し、それによって、溶解性が得られるという顕著な特性を有することが見出されている。驚くべきことに、研究されたPPIase(例えば、TF、SlyD、およびFkpA)は、未変性様の折り畳まれたレトロウイルス表面糖タンパク質に結合し、それによって例えば、可溶化する。本発明による「未変性様の(native-like)」、または「未変性様に折り畳まれた(native-likefolded)」gp41とは、遠UV-CDによって評価される場合、二次構造における高ヘリックス含量によって、および近UV-CDによって評価される場合、三次接触によって、その両方で特徴付けられる、これは、それぞれ図1Bおよび5において示されるとおり、代表的な「gp41-サイン(signature)」に反映される。さらに、本発明による、「未変性様の(native-like)」gp41のUVスペクトルは、320nmより高い波長で有意な吸収(これは、凝集物のような迷光粒子を暗示する)を示さない。
【0086】
モデル生体分子間、例えば、抗体と抗原との間の複合体形成について大量の情報がある(概説については、Braden, B. C.、およびPoljak, R. J., Faseb J 9(1995)9-16を参照のこと)。通常、複合体の形成、および解離は平行に生じ、この複合体および結合パートナーは、自由平衡において共存する。同様に、本発明に記載されるようなPPIシャペロンとアミロイド形成性タンパク質との間の複合体についても同じことが真であると考えられる。
【0087】
本発明に記載されるように、複合体の形成は、特に重要な特性である。なぜなら、PPIシャペロンとタンパク質(これは、例えば、生理的な緩衝条件下で、本質的に不溶性である)との間の複合体は、例えば、生理的緩衝条件下で容易に可溶であることが見出されているからである。生理的条件下で可溶性である抗原は、診断適用においては多くの利点を有する。それらは、例えば、標準物質として直接用いられてもよい。さらに、それらは、適切なマーカーに、または適切な結合基に複合体化されてもよい。
【0088】
上記で考察されたように、HIV-2由来のgp36は、類似の機能(すなわち、膜融合、およびウイルス侵入)を果たし、そしてHIV-1由来のgp41と同様の診断適合性である。レトロウイルス表面糖タンパク質のプロトタイプの例としてHIV-1のgp41を用いるこの適用において、多くの技術的問題が考察される。単に明確化のために、この考察および説明は、主にHIV-1のgp41に集中する。しかし、同様の考慮が、他のレトロウイルス表面糖タンパク質、特にHIV-2由来のgp36、およびHTLV由来のgp21にあてはまる。
【0089】
HIV-1、またはHIV-2の天然に存在する単離体は、もともと単離され、かつ記載されたアミノ酸配列の改変体を含み得ることが公知である。哺乳動物の免疫不全rsgpのこのような天然に存在する改変体、および合成的に操作された改変体はまた、本発明の範囲内である。
【0090】
本発明は、好ましい実施形態において、regp、またはヒト免疫不全ウイルスウイルス(HIV)の膜貫通糖タンパク質の改変体に関する。HIV-1gp41、またはHIV-2 gp36のNヘリックス状ドメインにおける特定のアミノ酸置換を含む改変体が開示される。
【0091】
NヘリックスおよびCヘリックス状ドメイン(ヘリックス対ヘリックスの接触に関与する)の両方のアミノ酸位置は、HIV-1gp41の文献から公知であり、そしてHIV-2相同性gp36に対して外挿され得る。これらの位置を突然変異させることは、特に、この改変体およびPPIシャペロンドメインを含む融合タンパク質の状況では、それぞれgp41、またはgp36の特性に影響することが見出されている。
【0092】
gp41ロイシンジッパーのヘリックス状ホイール突出部における「a」、および「d」のアミノ酸位置は、本発明による改変体を作成するための好ましい標的である。「a」位置におけるアミノ酸残基(Chan,D. Cら、Cell 89(1997)263-73による付番)は、Q552、I559、L566、I573、およびI580である;各々の「d」位置におけるアミノ酸残基は、I548、I555、Q562、T569、およびL576である。
【0093】
このジッパーモチーフのヘリックス状の完全性を損なうことなく、溶解度を改善するために、変異位置は、2つ以上のヘリックスターンによってお互いから離されることが好ましい。この必要条件は、例えば、保存的な「a」残基であるQ552、I559、L566、およびI573の置換によって、ならびに例えば、保存的な「d」残基であるI548、L555、Q562、およびT569の置換によって満たされる。言い換えれば、突然変異された残基は、少なくとも6つの野生型アミノ酸残基によってお互いから離され、従って正確に7個のモチーフが続く。置換位置がお互いから2つ以上のヘリックスターンによって離されるという前述の状況下で改変体内の「a」、および「d」残基の両方を変異することがまた可能である。
【0094】
同様に、HIV-2のgp36エクトドメインにおける変更は、驚くべきことに、SlyDまたはFkpAに対して融合された場合、容易に溶解する組み換えタンパク質を生じることが見出された。ここで、「a」位置は、Q551、V558、L565、T572、V579であり、そして「d」位置は、I547、L554、Q561、T568、およびL575である。
【0095】
好ましくは、それぞれ、HIV-1 gp41の位置Q552、I559、L566、I573、I580、I548、L555、Q562、T569、およびL576、HIV-2gp36の位置Q551、V558、L565、T572、V579、I547、L554、Q561、T568、およびL575を含む位置の群から選択される1〜6個のアミノ酸が、より小さいか、またはより親水性のアミノ酸によって置換される。
【0096】
好ましくは、置換対象のアミノ酸位置は、それぞれHIV-1のQ552、I559、L566、I573、およびI580からなる位置の群、ならびにHIV-2gp36のL554、Q561、T568、およびL575からなる位置の群より選択される。
【0097】
好ましい実施形態において、本発明は、HIV-1 gp41の改変体であって、Leu555、Leu566、Ile573、およびIle580の位置(ここで、これらの位置は、配列番号1に記載されるgp41野生型配列から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された位置において、少なくとも1つのアミノ酸置換、および多くとも4つのアミノ酸置換を含む改変体であって、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられる、改変体、に関する。
【0098】
本発明のこの好ましい実施形態は、gp41野生型配列の対応するポリペプチドに比べて、有意な改善を示す野生型gp41の改変体が提供され得るという驚くべき知見に基づく。本発明の改変体をもたらすアミノ酸置換は、Chan,D. C.ら、Cell 89(1997)263-73から公知であって、配列番号1に示されるような、アミノ酸組成およびgp41野生型配列の番号付けに基づいて記載される。
【0099】
明白に、本発明に記載されるアミノ酸置換基はまた、他の公知のgp41、およびまだ未同定のHIV-1単離物の内部の対応する配列位置でアミノ酸を置換するために用いられ得る。「ある位置に対応する(correspondingto a position)」という用語は、HIV-1単離体、およびその改変体が見出され得るか、または生成され得るが、これがさらなるアミノ酸を含んでいるか、またはアミノ酸を欠失しており、それに基づいて、配列番号1に対する配列アラインメントで、対応する配列位置または配列モチーフについての種々の絶対数が得られるということを示すために用いられる。
【0100】
配列番号1の野生型配列とのgp41配列のマルチアラインメントおよび比較を、GCGPackage Version 10.2(Genetics Computer Group,Inc)のPileUPプログラムを用いて実施する。PileUPは、Feng,D. F.Doolittle, R. F., J Mol Evol 25(1987)351-60のプログレッシブアラインメント法の簡便法を用いてマルチ配列アラインメントを作成し、そして同一、類似、または異なるアミノ酸残基についてのスコアリングマトリックスが適宜規定される。このプロセスは、2つの整列された配列のクラスターを生成する、2つのほとんど類似の配列のペアワイズアラインメントで開始する。次いで、このクラスターは、次の最も関連した配列に対して、または整列された配列のクラスターに対して整列され得る。配列の2つのクラスターは、2つの個々の配列のペアワイズアラインメントの単純な伸長によって整列され得る。最終アラインメントは、全ての配列が、最終ペアワイズアラインメントに含まれるまで、一連のプログレッシブ、ペアワイズアラインメント(漸増的に異なる配列およびクラスターを含む)によって達成される。従って、新規なHIV-1単離物のアミノ酸位置、または操作されたgp41のアミノ酸位置(野生型配列の555、566、573、および580の位置に相当する)は、容易に配置される。
【0101】
本発明によるHIV-1 gp41ポリペプチドの好ましい改変体は、その改変体が、555位にアミノ酸置換基を含むという点で特徴付けられ、ここでLeu555は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0102】
本発明によるHIV-1 gp41ポリペプチドのさらなる好ましい改変体は、その改変体が、566位にアミノ酸換基を含むという点で特徴付けられ、ここでLeu566は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0103】
本発明によるHIV-1 gp41ポリペプチドのさらなる好ましい改変体は、その改変体が、573位にアミノ酸換基を含むという点で特徴付けられ、ここでIle573は、セリンによって、またはトレオニンによって置換され、このセリンによる置換が、最も好ましい置換である。
【0104】
本発明によるHIV-1 gp41ポリペプチドのさらなる好ましい改変体は、その改変体が、580位にアミノ酸換基を含むという点で特徴付けられ、ここでIle580は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0105】
本発明はまた、HIV-2 gp36の改変体であって、Leu554、Leu565、およびVal579の位置(ここで、これらの位置は、HIV-2gp36野生型配列(配列番号2)から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された1つの位置において、少なくとも1つのアミノ酸置換、および多くとも3つのアミノ酸置換を含む改変体であって、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられる改変体、に関する。
【0106】
この番号付けは、Guyader, M.ら、Nature 326(1987)662-9によって公開された野生型配列(配列番号2)に従う。この配列から公知の位置に相当するgp36内のアミノ酸位置は、gp41について上記されたように決定される。
【0107】
本発明によるHIV-2 gp36ポリペプチドの好ましい改変体は、その改変体が、554位にアミノ酸置換基を含むという点で特徴付けられ、ここでLeu554は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0108】
本発明によるHIV-2 gp36ポリペプチドのさらなる好ましい改変体は、その改変体が、565位にアミノ酸換基を含むという点で特徴付けられ、ここでLeu565は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0109】
本発明によるHIV-2 gp36ポリペプチドの好ましい改変体は、その改変体が、579位にアミノ酸換基を含むという点で特徴付けられ、ここでVal579は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。好ましい実施形態において、それぞれ改変体gp41、または改変体gp36は、上記のようにこのアミノ酸位置のうち2つで置換を含む。3つのアミノ酸置換を含む改変体がまた好ましい。なおさらなる好ましい実施形態において、改変体gp41は、上記に詳細に考察される4つのアミノ酸位置で置換を含む。
【0110】
好ましい実施形態において、それぞれ、gp41、またはgp36(すなわち、融合ペプチド、および膜貫通セグメントを欠くエクトドメイン)の完全な配列、または対応する哺乳動物免疫不全ウイルスエンベロープタンパク質(例えば、HTLV由来のgp21)の完全な配列を用いて、PPIシャペロンとの複合体を形成する。レトロウイルス表面糖タンパク質の断片(例えば、Luら(前出)によって記載されたHIV-1由来のgp41についての断片)を使用することもまた考えられる。このような断片は好ましくは、gp41の細胞外部分のC末端ヘリックス、およびN末端ヘリックスを含む。
【0111】
アミノ酸535位〜681位を含む診断上関連するgp41(Chan, D. C.ら、Cell89(1997)263-73による命名法)は、標準的な手順による組み換え技術によって生成され得る。Luら(前出)の図1に記載されるとおり、別の興味深いgp41分子は、gp160前駆体分子のアミノ酸540〜669にまたがる。
【0112】
レトロウイルス表面糖タンパク質についての代表的な例として、HIVの小エンベロープタンパク質は、取り扱いが非常に困難であり、かつ非常に異例な特性を示す。既に言及したとおり、e-gp41分子の最も重要な特徴の1つは、生理的な緩衝条件でのその不溶性である。組み換え生成されたgp41は、可溶性であり、かつpH3.0および低イオン強度において未変性様の構造を示す。しかし、このpHでさえ、バッファー中の塩濃度には敏感なままである。用いられる塩によって、100〜500mMを超える塩の存在下でgp41は、沈殿する。以下により詳細に記載されるとおり、これは、カオトロピック剤によって(再度)可溶化(変性型で)され得る。
【0113】
生理的な緩衝条件は、通常、動物の血漿または血清に見出される塩およびpHの条件に相当すると理解されており、そして約7.4のpH値および約150mMの塩濃度によって規定される。本発明によるrsgp-シャペロン複合体は、これらの緩衝条件下で容易に可溶である。そこに存在するrsgpは、免疫学的に活性であり、従って、未変性様の構造に仕向ける。適切な界面活性剤の非存在下で、または適切な界面活性剤による事前処理なしには、gp41は、生理的な緩衝条件(例えば、20mMリン酸ナトリウム、pH7.4、150mMNaCl)下で、本質的に不溶性であるが、本発明によって記載された複合体は、適切なプロトコールに従った再折り畳み後に、容易に可溶性である。gp41エクトドメインは、本発明の複合体に含まれる場合、少なくとも100nMの濃度、好ましくは、1μM以上の濃度、最も好ましくは、10μM以上の濃度で可溶性である。従って、溶解度は、ナノモル未満の濃度から約マイクロモル濃度へ実質的に増大する。
【0114】
本発明の範囲のさらに良好な理解のために、可溶化および再生のために適用された緩衝条件は、必要に応じてかつ適切に改変されてもよく、そして本発明の過度の限定と理解されるべきではなく、本発明は広範な範囲の緩衝条件にわたって首尾よく実行されるということを強調することが必要である。
【0115】
生理的バッファーの全体的塩濃度は、シャペロン−gp41複合体が解離されず、そしてgp41が溶液中に留まることに注意を払いさえすれば、重要ではない。好ましくは、生理的バッファーは、少なくとも10mMの緩衝系、および多くとも200mMを含む。バッファー構成成分の残りは、もしあれば、有意な緩衝能力のない塩、例えば塩化ナトリウムであり得る。この生理的バッファーは、好ましくは、20〜500mM、より好ましくは50〜300mM、そして最も好ましくは100〜200mMの塩濃度を有する。
【0116】
本発明によるプロセスにおいて、この生理的なバッファーは、5.0〜8.5の範囲のpH値を有するように変化されてもよい;より好ましくは、このようなバッファーの範囲は、pH5.5〜pH8.3である。なおより好ましくは、このような生理的バッファー条件は、上記で示されるような塩濃度、および6.0〜8.0のpH値によって規定される;最も好ましくは、このような生理的バッファーのpHは、6.5〜7.8である。
【0117】
本発明において記載されるプロセスに従って、レトロウイルス表面糖タンパク質は、非生理的な緩衝条件下で可溶化され、シャペロンが添加され(または、共有結合したさらなるタンパク質ドメインとして既に存在する)、そして可溶化されたレトロウイルス表面糖タンパク質およびシャペロンを含む混合物が次に、生理的緩衝条件に調節される。レトロウイルス表面糖タンパク質単独では、そうされれば、自然に沈殿するが、それは、上記のプロセスでは驚くべきことに溶液中にとどまる。この重要な知見は、レトロウイルス表面糖タンパク質と、シャペロンとの間の複合体の形成に起因する可能性が最も高い。
【0118】
E. coliにおけるgp41の組み換え生成の場合、組み換え生成されたgp41は、封入体の形態において得られる。この材料は、高度にカオトロピックな試薬、例えば、7.0Mグアニジウムチオシアネートを用いて可溶化される。gp41ポリペプチドはこれらの条件下でほとんど構造化されていない。適切な工程においてこのバッファーを30mMギ酸(pH3.0)に変えることによって、溶液中のgp41は、その未変性様の全ヘリックス状構造として認知されるものの形態になる。タンパク質の正しい折り畳みまたは不正確な折り畳みの状態をモニターするための1つの簡易な方法は、アミド領域(185-250nm)、および芳香族領域(260-320nm)における対応するCDスペクトルを分析することである。それに加えて、迷光粒子(凝集物のような)についての情報は、標準的なUVスペクトルから容易に得られ得る。
【0119】
ここで強調するのが重要なことは、本発明によるレトロウイルス表面糖タンパク質−シャペロン複合体内のレトロウイルス表面糖タンパク質は、未変性様折り畳みと考えられるものの形態をとるという事実である。これに反して、レトロウイルス表面糖タンパク質(カオトロピック剤によって天然のpHで可溶化されている)は、ほとんど構造化されておらず、従って規則正しい高次構造(コンフォメーション)エピトープを失っている。代替的に界面活性剤を用いることによって、レトロウイルス表面糖タンパク質を可溶化することがまた可能である。例えば、ドデシル硫酸ナトリウム(SDS)は、gp41を可溶化するために首尾よく用いられている。しかし、このような「SDS−可溶化材料(SDS-solubilizedmaterial)」は、例えば、gp41に対する抗体の検出のためのイムノアッセイにおける使用のための、選り抜きの材料ではない。さらに、(上記で考察したように)、このようなイムノアッセイは、好ましくはまた、gp41の高次構造(コンフォメーション)エピトープに対する抗体を検出する。そして界面活性剤が、高次構造(コンフォメーション)エピトープを部分的に消滅させることは排除できない。
【0120】
好ましくは、本発明によるrsgp-シャペロン複合体は、未変性様折り畳みされているrsgpによって特徴づけられる。このような複合体内の、この未変性様折り畳みされたrsgpは、例えば、必要な免疫学的または物理的特徴を示す。
【0121】
未変性様折り畳みを、好ましくは、近UV CD分光法によって評価するが、この方法は、緻密な球状タンパク質内の三次接触について報告する。gp41は、約3.0のpHで、そして低イオン強度の塩濃度で容易に可溶性であることが公知である。近UVCDデータによって、このようなバッファー条件下では、gp41は、未変性様折り畳みされた球状タンパク質の代表的なサインをともなう特徴的な楕円率シグナルを示すことが実証される。図5に示されるとおり、gp41、およびFkpAを含む融合ペプチドのgp41部分は、酸性バッファー中のこの代表的な近UVCDスペクトルを示す。生理的緩衝条件下で、本発明による可溶性複合体の近UV CDスペクトルは、正確に折り畳まれたシャペロンのスペクトルと、未変性様折り畳みされたgp41のスペクトルの両方から構成される。これは、図7のFkpA-gp41融合タンパク質について示される。
【0122】
本発明による好ましい実施形態において、gp41-シャペロン複合体中の未変性様折り畳みを、近UVCDを分析することによって評価する。この近UV CDは、分子gp41、およびシャペロンの両方が未変性様折り畳みされていることを実証するために用いられることが好ましい。
【0123】
可溶性シャペロン−gp41複合体の生成は、非生理的緩衝条件から開始する。遊離のシャペロンと遊離の標的タンパク質(例えば、HIV-1由来のgp41)との間の複合体形成の場合、「非生理的」バッファーは、以下の2つの要件を満たさなければならない:(a)gp41が、未変性様の酸性構造で存在すること、および(b)PPIシャペロンが、少なくとも部分的に機能的(すなわち結合コンピテント)であること。このような緩衝条件から開始して、シャペロンは、アミロイド形成性タンパク質に結合し、そして非生理的条件から、幾分生理的な条件までの緩衝条件の変化が、アミロイド形成性タンパク質の沈殿なしに、可能である。
【0124】
シャペロンは、通常変性されたタンパク質に結合して、それらに作用し、それによってそれらの正確な(再)折り畳みを容易にするが、本発明が基づく状況は著しく異なる。適切な非生理的緩衝条件下で可溶化されたgp41は、未変性様型で存在すると考えられる(図1A、および1B、ならびに図5を参照のこと)。シャペロン機能の慣習的な見解とは異なり、本発明の方法では、シャペロンは、未変性様折り畳みタンパク質に結合して、緩衝条件(この緩衝条件下ではgp41はさもなければ不溶性であって沈殿する)において、このタンパク質を安定化するようである。
【0125】
本発明による好ましい実施形態において、PPIシャペロンは、FkpA、SlyD、および誘発因子からなる群より選択される。
【0126】
特にFkpA、またはSlyDがgp41の溶解度を改善して、それとともにかなり安定な複合体を形成することが見出されている。従って、さらに好ましい実施形態は、このシャペロンが、FkpA、およびSlyDを含む群から選択されるという点で特徴付けられる。最も好ましいのは、シャペロンFkpAである。
【0127】
さらに上記で記載したように、またシャペロンの断片は、所望の機能をもたらすために用いられ得る。モジュールシャペロンの場合、FKBPと同様に、触媒性モジュール、および結合モジュールを含み、このような断片は、結合ドメインを少なくとも含むこと、またはこれらの断片が、本質的にこの結合ドメインに匹敵する機能を少なくとも示すことが好ましい。
【0128】
FKBP12は、FKBPファミリーのヒトメンバーであり、そしてPPIaseの触媒性イソメラーゼドメインから本質的になる。FKBP12は、さらなるポリペプチド結合ドメインを欠くので、これは、FKBPファミリーの他のメンバーに比べた場合、折り畳まれていないか、または部分的に折り畳まれたタンパク質基質に対して、有意に低下した結合親和性を示す。FKBP12の変性および再折り畳みは、可逆プロセスであることが示されている(Egan,D. A.ら、Biochemistry 32(1993)1920-7;Scholz, C.ら、J Biol Chem 271(1996)12703-7)。本発明者らは、FkpA(25-270)、およびSlyD(1-165)の再折り畳み、および変性が、いずれも可逆であり、従ってここに記載されたプロセスの中心的な必要条件を満たすということを見出す。
【0129】
好ましい実施形態において、本発明は、gp41、および(FKBPファミリーから選択された)シャペロンを含む可溶性複合体に関する。
【0130】
上記のように、このような可溶性複合体であって、HIV-1由来のgp41、または別の哺乳動物免疫不全ウイルス由来の相同体を含む複合体は、PPIシャペロン(例えば、組み換え技術によって生成された)、および組み換え生成されたgp41を混合することによって容易に調製され得る。次いで、この複合体は、2つの独立した分子の間で、すなわち、分子間で形成される。
【0131】
複合体形成は、動的なプロセスであって、ここで解離および再会合が並行して生じる。これは、例えば、FkpAとgp41との間の、分子間、および分子内(例えば、融合構築物内)の会合の両方について真である。gp41は、生理的バッファーから直ちに、かつ定量的に沈殿するので、両方のパートナーの濃度を、選択して、これによって非臨界濃度、または非凝集濃度の遊離型gp41しか存在しないこと、および大量のgp41がgp41−シャペロン複合体の型で結合されかつ安定化されることを確実にしなければならない。
【0132】
用いたシャペロンによって、gp41分子に比較して、モル基準で、少なくとも2倍多いシャペロンを混合することが必要であることが見出されている。好ましい実施形態においては、従って、本発明は、gp41、およびシャペロン(好ましくはFkpA)の混合物を含む試薬に関する。好ましくは、このような混合物は、gp41に比べた場合、FkpAをモル過剰で含む。FkpAより3〜10倍存在することが好ましい。FkpA対gp41の最も好ましいモル比は、4〜6である。
【0133】
例えば、共有結合した、少なくとも1つのrsgpドメインおよび少なくとも1つのPPI-シャペロンドメインを含むタンパク質の異なるドメインの間の分子内複合体の形成が、例えば、安定性および生成の容易さに関して、さらなる有利な効果をもたらすということがまた見出されている。例えば、両方のドメインが共有結合されている場合、1:1(rsgp対シャペロン)の比が、可溶性複合体を形成するのに十分であることが見出されている。
【0134】
レトロウイルス表面糖タンパク質およびシャペロンを組み換え的に連結された形態で含む可溶性複合体は、本発明によるさらなる好ましい実施形態を表す。このような組み換えポリペプチドに含まれる最も好ましいrsgpは、HIV-1由来のgp41、およびHIV-2由来のgp36である。
【0135】
少なくとも1つのrsgpドメイン、および少なくとも1つのPPIシャペロンドメインを含む組み換えタンパク質について、非生理的緩衝条件から、生理的緩衝条件への移行は、異なる方法で達成され得る。gp41とFkpAとの間の可溶性分子内複合体は、非生理的緩衝条件を、透析、急速希釈、またはマトリックス補助再折り畳みによって生理的緩衝条件に調節することによって容易に得られる。可溶性gp41シャペロン複合体を含む混合物は、改変のために直接用いられ得る。
【0136】
本発明による可溶性複合体(例えば、gp41およびPPIシャペロンを含む)はまた、組み換え技術によって得られた両方のタンパク質ドメイン(gp41、およびシャペロン)を含む1つのポリペプチドから開始して生成されてもよい。gp41−シャペロン複合体は、その点で、事実上、分子内である。好ましくは、本発明による組み換えポリペプチドは、gp41とシャペロン、またはgp36とシャペロンを含む。なおさらに好ましい実施形態では、本発明は、組み換えタンパク質であって、少なくとも1つのrsgpドメイン、および少なくとも2つのPPI−シャペロンドメインを含む組み換えタンパク質に関する。1つのrsgpドメイン、および2つのPPIシャペロンを含む組み換えポリペプチドがまた好ましい。
【0137】
標準的な分子生理的技術を適用して、本発明による可溶性gp41−シャペロン複合体を得るために用いた組み換えポリペプチドを発現する。好ましくは、このシャペロン遺伝子は、発現ベクター(これは、gp41およびシャペロンの両方についての遺伝情報、ならびに必要に応じて、適切なペプチド性リンカー配列についての遺伝情報も含む)中に、標的タンパク質遺伝子の上流にインフレームで位置する。このような組み換え融合タンパク質の大規模な生成のために好ましい宿主は、E.coliである。
【0138】
好ましい実施形態において、本発明は、可溶性複合体であって、それぞれgp41、またはgp36、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されたシャペロンを含む複合体に関する。この可溶性複合体が、分子内複合体、であって、好ましくは、gp41、またはgp36、およびPPIシャペロンを含む組み換えポリペプチド内の分子内複合体であることがなお、さらに好ましい。最も好ましくは、組み換えポリペプチドのPPIシャペロン部分は、いずれの輸送シグナルペプチド(対応する前駆体分子の)も欠き、そして成熟PPIシャペロンに相当する。好ましい実施形態において、この組み換えタンパク質は、機能的なシグナル配列を欠くので、この遺伝子産物は、細菌のサイトゾル中に蓄積する。
【0139】
組換え産生されたFkpA-gp41に含まれるgp41の著しい特徴は、「非シャペロン(unchaperoned)」gp41エクトドメインに比較した場合の異例な溶解度である。「カオトロピック材料(chaotropicmaterial)」(すなわち、6.0-7.0M GuHCl中のFkpA-gp41)は、異なる方法で再折り畳みされ得、全てが熱力学的に安定、かつ可溶性の未変性様形態になることは興味深い。再折り畳みは、透析および急速希釈の両方によって、ならびに再生サイズ排除クロマトグラフィー、またはマトリックス補助再折り畳みによって高収率で達成される。これらの知見によって、この共有結合型において、gp41-FkpA融合ポリペプチドが、準安定なタンパク質よりも熱力学的にかなり安定であることが示唆される。
【0140】
組み換えFkpA-gp41ポリペプチドは、異なる折り畳み要件を有する2つのタンパク質ドメインを含む。精製プロトコールは、初期の変性工程を含むので、シャロンの折り畳みが可逆であることは必須である。実際、共有結合したタンパク質複合体内でのFkpAおよびgp41の両方の可逆的かつ独立した再折り畳みについて、説得力のある分光学的な証拠が存在する。C末端切断型SlyDの再折り畳みは、いずれにせよ可逆であることが見出されている。
【0141】
レトロウイルス表面糖タンパク質、およびシャペロンを含む組み換えポリペプチドであって、さらにこれらの2つのポリペプチドドメインの間に適切なペプチドリンカー配列を含む、ポリペプチドがまた好ましい。このようなペプチドリンカー配列は、用いられたrsgpおよびシャペロンドメインの最適の分子内会合を確実にするように選択される。好ましくは、このようなリンカー配列は、約20アミノ酸長であり、そして可変性(flexibility)および溶解度の両方を支持するアミノ酸(例えば、グリシン、およびセリンなど)を含む。好ましくは、このリンカーは、10〜50アミノ酸長である。より好ましくは、この長さは、12〜40アミノ酸であり、そして最も好ましくは、このリンカーは、15〜35アミノ酸を含む。Rsgpおよびシャペロンの両方とも、常に密接に近接している(例えば、適切なリンカーによって互いに支持される)。好ましい実施形態において、この組み換えポリペプチドは、可変性のリンカーを介してその標的タンパク質に連結された成熟FkpAを含む。このことによって、データが示すとおり、さらなる安定化効果がもたらされる。
【0142】
驚くべきことに、gp41は、PPIシャペロンとgp41との間の分子内複合体の一部として、可溶性であり、かつ安定であることが見出されている。同じことが、PPIシャペロンとgp36、またはPPIシャペロンとgp21(HTLV由来)を含む分子内複合体についても真実である。このような複合体におけるgp41の改善された安定性は、さらなる利点をもたらす。例えば、完全に再生された組み換えgp41−シャペロン分子を非常に簡単に獲得することが可能である。組み換えタンパク質は、カオトロピック剤(例えば、塩化グアニジウム)を用いた処理によって最初に可溶化される。可溶化された材料をゲル濾過カラム(適切な生理的バッファーを用いて平衡化された)に単に通過させることによって、共有結合したタンパク質ドメインを含む完全に再生されたタンパク質を得ることができる(実施例2.3、ならびに図7および8を参照)のこと)。
【0143】
可溶性分子内gp41−シャペロン複合体は、なお、さらなる顕著な利点を示す:これは、界面活性剤の変性作用に対してかなり安定である。この効果は、この融合タンパク質が、それぞれ2つのシャペロンおよび1つのgp41、または1つのgp36を含む場合、なおさらに明白になる。ほとんどのイムノアッセイは、非特異的な結合によって生じる問題を低減し、そして少なくとも部分的に回避するために、界面活性剤の存在下で実施される。HIV診断の場合、前述の理由のせいで、ただしさらにウイルス粒子を分解しかつ破壊して、これによってウイルス抗原(gp24など)の検出を容易にするために、かなり強力な界面活性剤が用いられる。
【0144】
SDS(ドデシル硫酸ナトリウム)によって可溶化された、組み換え生成されたgp41エクトドメインは、例えば、抗HIV抗体、またはp24抗原の検出において、慣用的に用いられるアッセイバッファーにおいて免疫反応性ではない(図9を参照のこと)。しかし、同じ緩衝条件下では、gp41(本発明による、PPIシャペロンとの分子内複合体の一部である)は、強力に免疫反応性である。図9から判明するとおり、同じアッセイ条件下で、かつ同じ患者血清を用いて、この材料は、未変性様可溶性gp41(さらに、試験された界面活性剤の存在下で安定である)の存在によってのみ説明され得る、優れた競合曲線を生じる。
【0145】
これは、本発明による、この複合体の非常に重要な特徴であって、この可溶性rsgp−シャペロン複合体内のrsgpは、生理的な緩衝条件、例えば、20mMリン酸、150mM塩化ナトリウムバッファー中で、pH7.4において、未変性性折り畳みされる。これは治療適用のため、そして診断適用のための大きな利点である。好ましい実施形態において、本発明は、生理的緩衝条件下で可溶である試薬の組成物であって、分子内複合体または分子間複合体(これは、レトロウイルス表面糖タンパク質、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されるシャペロンを含む)を含む、組成物に関する。
【0146】
従って、HIV-1由来の未変性様折り畳みgp41、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されるシャペロンを含む可溶性複合体は、本発明の非常に好ましい実施形態を表す。
【0147】
従って、HIV-2由来の未変性様折り畳みgp36、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されるシャペロンを含む可溶性複合体はまた、本発明の非常に好ましい実施形態を表す。
【0148】
治療に関して、それぞれ、「可溶性、かつ未変性様折り畳みされた(soluble andnative-like folded)」gp41またはgp36を提供することによってなされる進歩は、かなり明白である。例えば、gp41は、生理的緩衝条件下で、初めて、この時、利用可能となる。
【0149】
好ましい実施形態において、記載されるような可溶性複合体は、医薬として用いるための試薬の組成物を生成するために用いられる。試薬の組成物は、生理的に許容され得る賦形剤、ならびに必要に応じて、適切な添加物、および/または従来の補助物質とともに、gp41-シャペロン複合体を含む。
【0150】
gp41の7残基反復か、またはgp41C末端ヘリックスに由来するペプチドが、抗ウイルス活性を保有することは公知である(Wild,C.ら、Proc Natl Acad Sci USA 89(1992)10537-41)。それらは、gp41のいわゆる「ヘアピン中間体(hairpin−intermediate)」と特異的に相互作用することによってウイルス侵入を邪魔する(概説については、Doms,R. W.、およびMoore, J. P., J Cell Biol 151(2000)F9-14を参照のこと)。本発明者らは、本発明によるrsgp-シャペロン複合体が抗ウイルス活性を示すことを見出した。gp41−シャペロン複合体、もしくはgp36−シャペロン複合体、または両方を、第一の好ましい治療適用において治療上有効な用量で含有する、試薬の組成物を用いて、宿主生物体内のHIV進入、およびHIV伝播を防止する(「ウイルス侵入阻害(virusentry inhibition)」)。
【0151】
これは、哺乳動物における免疫応答を誘発するための組成物などを使用するための、gp41−シャペロン複合体を含む試薬の組成物のさらなる好ましい治療適用を表す。記載された複合体は、公知の他のいずれのHIV免疫原よりも多いgp41エピトープを利用できる(例えば、Rootら(前出)を参照のこと)。従って、この新規な免疫原は、かなり広範な免疫応答を誘導することが期待される。
【0152】
診断手順に関して、本発明による可溶性rsgp−シャペロン複合体の明白な利点は、例えば、レトロウイルス表面糖タンパク質(例えば、生理的緩衝条件下でのgp41)の安定性の増大、および/または診断感度の増大、および/または高次構造(コンフォメーション)的なエピトープの存在数の増大、および/または正確に折り畳まれたrsgp(gp41など)を容易に標識する能力である。
【0153】
周知の標識は、固相結合基などの、マーカー基、またはエフェクター基である。標識された可溶性rsgp−シャペロン複合体は、本発明によるさらなる好ましい実施形態を表す。
【0154】
標識基は、以下から選択されてもよい:任意の公知の検出可能マーカー基(例えば、色素)、蛍光標識基(例えば、化学発光基)、例えば、アクリジニウムエステル、またはジオキセタン、または蛍光色素、例えば、フルオレセイン、クマリン、ローダミン、オキサジン、レゾルフィン、シアニン、およびそれらの誘導体。標識基の他の例は、蛍光金属錯体(例えば、ルテニウム、またはユーロピウム錯体)、例えば、ELISAのために、またはCEDIAのために用いられるような酵素(Cloned Enzyme Donor Immunoassay、例えば、EP-A-0061 888)、ならびに放射性同位体。
【0155】
エフェクター基は、例えば、バイオアフィン(bioaffine)結合対の1つのパートナーを含む。アッセイを実施する間、エフェクター基は、バイオアフィン結合対の他のパートナーと特異的に、好ましくは非共有結合的に相互作用する。適切な結合対の例は、以下である:ハプテン、または抗原/抗体、ビオチン、またはビオチン類似体(例えば、アミノビオチン)、イミノビオチン、またはデスチオビオチン/アビジン、またはストレプトアビジン、糖/レクチン、核酸、または核酸類似体/相補的核酸、およびレセプター/リガンド、例えば、ステロイドホルモンレセプター/ステロイドホルモン。好ましい結合対のメンバーは、ハプテン、抗原、およびホルモンを含む。特に好ましいのは、ハプテン様ジゴキシン、およびビオチン、ならびにそれらの類似体である。
【0156】
rsgpおよびPPIシャペロンを含む可溶性複合体は、好ましくはrsgpに対する抗体の検出のためのイムノアッセイにおいて用いられる。好ましくは、gp41−シャペロン複合体、および/またはgp36−シャペロン複合体が用いられる。非常に好ましい実施形態において、gp41およびPPIシャペロンを含む標識された可溶性複合体は、gp41に対する抗体の検出のためのイムノアッセイにおいて用いられる。最も好ましくは、標識された複合体は、PPIシャペロンおよびgp41を含む組み換えポリペプチド内の分子内複合体である。
【0157】
イムノアッセイは、当業者に周知である。このようなアッセイを実施するための方法、ならびに実際の適用および手順は、関連の教科書にまとめられている。関連の教科書の例は、「Practice and theory of enzyme immunoassays」(1990)221-278、編集、R. H.Burdon、およびv. P. H. Knippenberg, Elsevier, Amsterdam中の、Tijssen, P., Preparationof enzyme-antibody or other enzyme-macromolecule conjugates、および「Methods inenzymology」(1980)編集、S.P.Colowick, N. O. Caplan、およびS. P., Academic Press中のTijssenの種々の巻であって、免疫学的検出法を取り扱う、特に70、73、74、84、92、および121巻である。
【0158】
新規な可溶性rsgp−PPIシャペロン複合体を用いて、検出の様式(例えば、放射性同位体アッセイ、酵素イムノアッセイ、電気化学発光アッセイなど)、またはアッセイ原理(例えば、テストストリップアッセイ、サンドイッチアッセイ、またはホモジニアスアッセイなど)とは独立して、抗HIV抗体の検出のためのアッセイを改善し得る。
【0159】
HIV感染の確実で、かつ鋭敏な早期検出のために、体液サンプル中のウイルス抗原および抗ウイルス抗体の両方を測定することが必須である。本発明による可溶性複合体は、生理的緩衝条件において、抗gp41抗体、および/または抗gp36抗体の検出を可能にする。抗gp41抗体、および/または抗gp36抗体の検出は、このような併用型HIV検出システムの有益な部分である。従って、好ましい実施形態において、本発明は、HIV検出システムであって、gp41、および/またはgp36シャペロン複合体の使用に基づく抗gp41抗体、および/または抗gp36抗体の検出を包含するシステムに関する。最も好ましくは、このような複合体に基づく、抗gp41抗体、および/または抗gp36抗体の検出は、HIV抗原、好ましくはp24抗原の検出とともに実施される。
【0160】
先行技術から公知のとおり、細菌、真菌、またはウイルスのような感染因子に対する抗体は、好ましくは、二重抗原架橋コンセプトに従って実施されるアッセイによって検出される(時には、このアッセイコンセプト(double antigen bridge concept)はまた、二重抗原架橋コンセプトと名付けられる、なぜなら2つの抗原が抗体によって架橋されるからである)。このようなアッセイにおいては、抗体が、所定の抗原の少なくとも2つの異なる分子に、その2つ(IgG、IgA、IgE)、または10(IgM)のパラトープ(抗原結合部位)で結合する能力が必要とされ、かつ用いられる。
【0161】
この架橋コンセプトに従う体液からの抗体の検出は、多くの異なるアッセイ設定で実施され得る。簡易な設定は、固相に対する抗原の直接コーティング、および同じ抗原の標識型での使用を含む。適切なアッセイ条件下で、サンプル中の抗体は、固相結合抗原と標識抗原との間の架橋を形成する。従って、検討中の抗体がサンプル中に存在する場合のみ、架橋が形成されて、シグナルが検出され得る。
【0162】
「固相抗原(solid phase antigen)」、および「検出抗原(detectionantigen)」の基本的な構造は、好ましくは、同じである。例えば、1または数個のエピトープを含むポリペプチドを用いて、固相を直接または間接的にコーティングしてもよいが、標識またはマーカーに結合した同じ合成ポリペプチドが、検出抗原として用いられる。類似であるが、異なる抗原であって、二重抗原架橋アッセイにおいて免疫学的に交差反応性である抗原を使用することも可能である。このようなアッセイを実施するための必須の要件は、関連のエピトープ(単数または複数)が、両方の抗原上に存在することである。もちろん、二重抗原架橋アッセイ形式の多くの改変型が存在する。このような改変型は、例えば、固相に対する抗原の間接的なコーティングを含む。好ましくは、特定の結合対、最も好ましくは、ビオチン−ストレプトアビジン(またはビオチン−アビジン)系が、固相に対して抗原を間接的に結合するために用いられる。一方では、このようなシステムにおける検出のために用いられる抗原は、マーカー(例えば、放射性同位体、酵素、蛍光分子など)を直接有していなくてもよいが、それよりむしろ、例えば、ハプテン(例えば、ジゴキシン)を担持することによって、間接的に検出され得る。次いで、このような間接的な検出は、例えば、標識された抗ジゴキシン抗体によって実施され得る。
【0163】
好ましい実施形態において、本発明は、第一のシャペロン−抗原複合体を含む第一の抗原、および第二のシャペロン−抗原複合体を含む第二の抗原を含む、二重抗原架橋コンセプトに従うイムノアッセイに関する。
【0164】
さらなる好ましい実施形態において、本発明は、二重抗原架橋コンセプトに従うイムノアッセイであって、第一のシャペロン−抗原複合体が、捕捉抗原として用いられ、そして第二のシャペロン−抗原複合体が検出抗原として用いられることを特徴とするイムノアッセイに関する。
【0165】
本発明において記載されたようなシャペロン−抗原複合体は、種々のポリペプチド(さもなければ取り扱いが困難である)の溶解性をもたらすだけでなく、この二重抗原架橋コンセプトに従って、極めて有利なイムノアッセイを可能にする。
【0166】
この二重抗原架橋コンセプトによる、このようなイムノアッセイの特に魅力的な特徴は、それぞれ、固相結合抗原との複合体形成のため、および検出抗原との複合体形成のために、異なるシャペロンを用いることが可能となるということである。アッセイのこのような改変によって、非特異的結合の問題をさらに改善する。サンプル中の抗体(シャペロンに対して反応性であり、従って偽陽性のシグナルを生じ得る)は、異なるシャペロンを用いてそれぞれ、固相抗原、および検出抗原と複合体化する場合、架橋を形成しない。従って、本発明のこの様式においては、非特異的結合に起因する陽性シグナルの可能性は、大きく低下する。従って、好ましい実施形態において、本発明は、二重抗原架橋コンセプトに従うイムノアッセイであって、第一および第二のシャペロン−抗原複合体の第一シャペロンおよび第二シャペロンが、お互いに異なることを特徴とする、イムノアッセイに関する。
【0167】
最もよく特徴付けられているシャペロンのほとんどは、Escherichia coliから単離されており、生物工学の研究において広範に用いられる。Escherichiacoliは、広範に分布した細菌種であるため、多くの哺乳動物は、この細菌に由来するタンパク質に対する抗体を発現している。このような抗体によって生じる偽陽性反応の可能性を減少させるため、別個の細菌種(好ましくは好熱性の菌種)由来の少なくとも1つのPPIシャペロンを使用することが好ましい。好ましくは、このシャペロンは、特に、Thermatogamaritima、Aquifex aeolicus、およびThermus thermophilusを含む細菌の群の、好極限性細菌由来である。
【0168】
イムノアッセイ一般における、そして好ましくは架橋コンセプトに従うイムノアッセイにおけるシャペロン−抗原複合体の使用はまた、このような複合体のシャペロンを誘導体化する可能性を提供し、そして抗原自体の改変を必要としない。第二の化学部分によるポリペプチドの改変、例えば、その分子に対する標識のカップリングは、このポリペプチドにマイナスに影響するリスクを含むということが一般に認められる。例えば、検討中のエピトープが、損なわれ得るか、または非特異的な結合が、このような標識によって生じ得る。本発明に従って、シャペロン−抗原複合体内のシャペロンを特異的に誘導体化することが、ここで、可能である。
【0169】
好ましい実施形態において、二重抗原架橋コンセプトによるイムノアッセイは、捕捉抗原として用いられる第一のシャペロン−抗原複合体が、固相結合基を含むことをさらなる特徴とする。
【0170】
さらなる好ましい実施形態において、架橋コンセプトによるイムノアッセイが実施される。このイムノアッセイは、検出抗原として用いられる第二のシャペロン−抗原複合体がマーカー基を含むことをさらなる特徴とする。
【0171】
別の実施形態において、rsgpおよびPPIシャペロンを含む可溶性複合体(例えば、gp41−シャペロン複合体、および/またはgp36−シャペロン複合体)はまた、被験体(例えば、ヒトまたは非ヒト動物)における免疫応答を惹起するために用いられ得る。この可溶性複合体は、被験体に対して組成物(例えば、この組成物は、賦形剤、または担体を含んでもよい)として投与され得る。このような組成物はまた、アジュバントを含んでもよい。従来のアジュバントの例としては、フロイントの不完全アジュバント、フロイントの完全アジュバント、Merck65、AS-2、ミョウバン、リン酸アルミニウム、鉱物ゲル(例えば、水酸化アルミニウム)、および表面活性物質(例えばリゾレシチン、プルロニックポリオール類、ポリアニオン、ペプチド、油状エマルジョン、スカシガイヘモシアニン、およびジニトロフェノール)が挙げられるがこれらに限定されない。他の有用なアジュバントとしては、細菌莢膜多糖類、デキストラン、IL-12、GM-CSF、CD40リガンド、IFN-g、IL-1、IL-2、IL-3、IL-4、IL-10、IL-13、IL-18、または任意のサイトカインもしくは細菌のDNA断片が挙げられるがこれらに限定されない。
【0172】
1用量(投与)の可溶性複合体組成物が与えられてもよい。しかし、最初の投与後に、ブースト投与(例えば、1回、2回、3回、それ以上)が続けられてもよい。被験体に投与された用量の回数は、被験体の可溶性複合体組成物に対する応答性にある程度依存する。本発明の範囲内で、適切な投薬の回数としては、可溶性複合体に対して動物を免疫するために必要な任意の回数が挙げられる。
【0173】
適切な複合体組成物の第二の投与(ブースター)は、最初の投与後、約7日〜1年間の間に与えられ得る。最初の投与と2回目の投与との間の時間は、14日〜6ヶ月、21日〜3ヶ月、しばしば、最初の投与後、約28日〜2ヶ月の間であり得る。第3回目の投与(2回目のブースター)は、最初の投与後、約14日〜10年の間に、例えば、最初の投与後、約14日〜3年間の間に、しばしば、約21日〜1年の間に、非常にしばしば、最初の投与後、約28日〜6ヶ月の間に与えられてもよい。その後のブースターは、2週間間隔、または1ヶ月、3ヶ月、もしくは6ヶ月から10年間の間隔で投与されてもよい。
【0174】
典型的には、抗原に対して動物を免疫するのに十分である可溶性複合体の量(すなわち、「免疫学的に有効な用量」、または「治療上有効な用量」)が、被験体に投与される。「免疫学的に有効な量」を達成するのに十分な量は、被験体の体重および健康の全身的状態に、ならびに処方医または資格のある他の人員の判断に、ある程度依存する。
【0175】
可溶性複合体の有効用量は、免疫応答の誘導を達成するために動物モデルにおいて処方され得る;このようなデータを用いて、動物モデルに基づいてヒトに対する投与を容易に最適化し得る。用量は典型的には、約1fg〜約100μgの間、しばしば、約1pg〜約100μgの間、それよりしばしば、約1ng〜約50μgの間、そして通常、約100ng〜約50μgの間である。いくつかの実施形態において、用量は、被験体の体重1kgあたり、約1fg〜約100μgの間、しばしば被験体の体重1kgあたり、約1pg〜約100μgの間、それよりしばしば、約1ng〜約50μgの間、そして通常、約100ng〜約50μgの間である。
【0176】
本発明の可溶性複合体含有組成物は、種々の方法、および種々の形式で投与され得る。可溶性複合体組成物は、担体、および賦形剤(例えば、バッファー、炭水化物、マンニトール、タンパク質、ポリペプチド、またはアミノ酸(例えば、グリシン)、抗酸化剤、静菌剤、キレート剤、懸濁剤、増粘剤、および/または保存剤);水、油、生理食塩水、水性デキストロース、およびグリセロール溶液、適切な生理的条件に必要な他の薬学的に許容され得る補助物質(例えば、緩衝化剤、張度調節剤、湿潤剤)などを含んでもよい。従来のアジュバントもまた、この組成物中に組み込まれてもよい。
【0177】
任意の適切な担体が、本発明の組成物を投与するために用いられ得るが、担体のタイプは、投与の様式によって異なる。化合物はまた、リポソーム内にカプセル化され得る。例えば、Ticeら、米国特許5,942,252,1999に記載されるような生分解性マイクロスフェアは、ある場合には担体として都合がよい。
【0178】
従来の技術(例えば、濾過滅菌)によって達成されるような、この組成物の滅菌が望ましい。得られた水溶液は、そのまま用いるためにパッケージングされてもよいし、凍結乾燥されてもよい。
【0179】
本発明の可溶性複合体組成物は、種々の方法で投与され得る。この方法としては、注射(例えば、皮内、皮下、筋肉内、腹腔内など)による方法、吸入による方法、局所投与による方法、坐剤による方法、経皮パッチを用いることによる方法、または経口による方法が挙げられる。
【0180】
注射によって投与される場合、組成物は、水溶液中に、好ましくは生理的に適合するバッファー(例えば、ハンクス溶液、リンガー溶液、20mMリン酸 150mM塩化ナトリウムバッファー(pH7.4)、または生理的食塩水のバッファー)中に処方され得る。この溶液は、懸濁剤、安定化剤、および/または分散剤のような処方剤(formulatoryagent)を含んでもよい。あるいは、この組成物は、使用前の適切なビヒクル(例えば、滅菌発熱因子無含有水)との構成のために粉末型で存在してもよい。吸入送達型組成物は、適切な噴霧剤(例えば、ジクロロジフルオロメタン、トリクロロフルオロメタン、二酸化炭素、または他の適切なガス)の使用によって、圧縮パックまたはネブライザーからのエアロゾルスプレーとしてであってもよい。圧縮されたエアロゾルの場合、投薬量単位は、一定量を送達するためのバルブを備えることによって決定され得る。インヘイラーまたは吸入器(insufflator)における使用のための例えばゼラチンのカプセル、およびカートリッジが、処方されてもよく、これは、タンパク質および適切な粉末基剤(例えば、ラクトースまたはデンプン)の粉末混合物を含有する。局所投与のために、組成物は、当該分野で周知のとおり、溶液、ゲル、軟膏、クリーム、懸濁液などとして処方されてもよい。いくつかの実施形態では、投与は、経皮パッチによる。坐剤組成物はまた、従来の坐剤基剤を含むように処方されてもよい。
【0181】
投与が経口である場合、組成物は、この組成物と薬学的に許容され得る担体とを合わせることによって容易に処方され得る。固体担体としては、マンニトール、ラクトース、ステアリン酸マグネシウムなどが挙げられる;このような担体は、経口摂取用の、錠剤、丸剤、糖衣錠、カプセル、液体、ゲル、シロップ、スラリー、懸濁物などの形成を可能にする。このような処方物は、粉末、カプセル、および錠剤であってもよい;適切な賦形剤としては、糖、セルロース調製物、顆粒化剤、および結合剤のような増量剤(filler)が挙げられる。
【0182】
ポリクローナル抗体およびモノクローナル抗体(結合断片(例えば、F(ab)2)、および一本鎖型を含む)を生成するための方法は、周知である。しかし、多くの抗原は、十分な抗体応答を誘発できない。1つの実施形態において、本発明の可溶性複合体および抗原を含む組成物が、動物に投与され、これによって動物中の免疫応答が惹起される。ポリクローナル抗体、またはモノクローナル抗体が、標準的な技術によって引き続き調製される。
【0183】
rsgpおよびPPIシャペロンを含む可溶性複合体(例えば、gp41−シャペロン複合体、および/またはgp36−シャペロン複合体)はまた、膜融合を阻害することなどによって、細胞へのウイルス侵入を阻害するために用いられ得る。細胞は、インビトロであっても、インビボであっても、またはエキソビボであってもよい。組成物および投与の方法は、免疫応答を惹起するために用いられた組成物および方法について記載されたものと同様である。細胞へのウイルス侵入阻害が、ワクチン接種を用いて達成されるならば、アジュバントが用いられてもよい。インビトロおよびエキソビボ投与のために、当業者は、細胞、培養条件、および時間制限(もしあれば)に部分的に基づいて適切な方法を選択する。例えば、このような有用な方法の1つは、可溶性複合体を担持するリポソームを処方することである。
【0184】
以下の実施例、参考文献、および図面は、本発明の理解を助けるために提供されるものであって、本発明の真の範囲は、添付の特許請求の範囲に記載される。本発明の趣旨から逸脱することなく、この記載された手順において改変がなされ得ることが理解される。
【実施例】
【0185】
実施例
実施例1 gp41およびPPIシャペロンを含む可溶性分子間複合体の生成
1.1 E.coli FkpAの生成
Bothmann, H.、およびPluckthum, A., J Biol Chem275(2000)17100-5に従って(ただしそれをわずかに改変して)、FkpAをクローニングし、発現させ、そして精製した。保存のために、20mM NaH2PO4/NaOH(pH6.0)、100mMNaClに対してタンパク質溶液を透析して、26mg/ml(1mM)に濃縮した。
【0186】
サイトゾル発現のために、上記の発現ベクターのFkpAコード配列を、シグナルペプチドをコードする配列部分を欠くように、そして代わりに成熟FkpAのコード領域のみを含むように改変した。
【0187】
1.2 gp41(535-681)-His6の生成
gp41(535-681)−His6を、T7プロモーターベースの発現系においてクローニングして発現し、宿主細胞の封入体中に蓄積させた。単離した封入体を、6M塩化グアニジウム中で溶解した。His−タク゛化タンパク質を、Ni−キレートカラム上で精製し、その後に、Sephacryl100上で、6Mグアニジウム中でゲルろ過した。Wingfield, P. T.ら、Protein Sci 6(1997)1653−60に記載のとおり、急速希釈によってタンパク質を再折り畳みした。最終の緩衝条件は、30mMギ酸ナトリウム(pH3.0)であった。折り畳みの状況を、近UVCDおよび遠UV CDを用いて両方の緩衝条件について評価した。図1Aおよび1Bからわかるように、遠UV CDおよび近UV CDの両方のスペクトルによって、gp41がカオトロピック剤の非存在下において、pH3.0でのみ、未変性様折り畳みをとることが示唆される。
【0188】
1.3 E.coli FkpAの存在下における、gp41エクトドメイン(HIV)のpH3.0から生理的pHへのpHシフト
1.3.1 コントロール実験
コントロール実験において、可溶性e-gp41(30mMギ酸塩、pH3.0)を、20mMリン酸ナトリウム(pH7.5)、50mMNaCl, 1mM EDTAの最終緩衝条件中に100倍希釈した。最終タンパク質濃度は、約1μMであった。UVスペクトルを、1分後、および10分後に記録した。非シャペロンのエクトドメインは、中性へのpHシフトの際に、自然に凝集することが、図2におけるUVスペクトルから明白である。図2は、gp41の異例な凝集傾向を強調する意図がある;この分子の自発的な凝集は、上の線によって示される段階をかなり超えて進行する。
【0189】
1.3.2 pH3.0でのFkpAとのgp41のプレインキュベーションによって中性pHにシフトできる
分子シャペロンFkpAの可溶化能力について試験するために、エクトドメインgp41、およびFkpAを、1:2、および1:4の分子比で混合して(約3.5のpHで30mMギ酸塩)、1分間、同時インキュベートさせた。次いで、得られた複合体を、20mMリン酸ナトリウム(pH7.4)、50mMNaCl、1mM EDTAの緩衝条件中への12倍希釈によって、中性のpHにシフトさせた。試験管中における結合パートナーの最終濃度は、それぞれ、1μMのgp41、2μMおよび4μMのFkpAであった。全ての反応を室温で実施した。1分後、および10分後、凝集についてgp41サンプルを試験するために、UVスペクトルを記録した。図3A、および3Bから、FkpAが、用量依存性の様式で、gp41の凝集をかなり低下させることが明白である。Thermatogamaritima由来の誘発因子、およびE. coli由来のC末端切断型SlyDを用いて匹敵するデータが得られている。
【0190】
実施例2
共有結合したgp41−FkpAの組み換え生成
2.1 FkpA、およびgp41を含む発現プラスミドの構築
第一の工程において、Stratagene(La Jolla、CA;USA)のQuickChange部位特異的変異誘発キットを用いて、以下のプライマーを用い、実施例1.1のプラスミドから、成熟E.coli FkpAのコード領域における制限部位BamHIを欠失させた:
5’−gcgggtgttccgggtatcccaccgaattc−3’(配列番号3)
5’−gaattcggtgggatacccggaacacccgc−3’(配列番号4)
この構築物を、EcFkpA(DbamHI)[GGGS]3と命名した。
【0191】
第二の工程において、HIV-1エンベロープタンパク質由来のアミノ酸535-681をコードする遺伝子断片を、実施例1.2の構築物から、以下のプライマーを用いて、PCRによって増幅した:
5’−cgggatccggtggcggttcaggcggtggctctggtggcggtacgctg-acggtacaggccag−3’(配列番号5)
5’−ccgctcgaggtaccacagccaatttgttat−3’(配列番号6)
【0192】
この断片を、BamHIおよびXhoI制限部位を用いて、EcFkpA(DbamHI)[GGGS]3中に挿入した。
【0193】
FkpAと、e-gp41との間にグリシン−セリンリンカーのコドンを、FkpAのクローニングのためのリバースプライマーとともに、およびe-gp41のクローニングのためのフォワードプライマーとともに挿入した。
【0194】
得られた構築物を配列決定して、所望のタンパク質をコードすることを見出した。
【0195】
2.2 融合タンパク質の精製
発現プラスミドを保有するE. coli BL21細胞を、0.7のOD600まで増殖させて、37℃の増殖温度で1mMのIPTGを添加することによって、細胞質過剰発現を誘導した。誘導の4時間後、遠心分離によって細胞を収集した(5000gで20分)。50mMリン酸ナトリウムpH7.8、6.0MGuHCl(塩化グアニジウム)、5mMイミダゾール中に、細菌ペレットを再懸濁して、完全に溶かすために室温で(10分)撹拌した。繰り返し遠心分離(SorvallSS34, 20000rpm, 4℃)後、上清を濾過(0.8/0.2μm)して、Ni−NTA−カラム(NTA: ニトリロ三酢酸;Qiagen;Germantown,MD)(溶解バッファー中で事前に平衡化した)にかけた。10カラム容積の溶解バッファーを与えることによって、洗浄工程で、非特異的に結合したタンパク質を取り除いた。最終的に、結合した標的タンパク質を、50mMのリン酸ナトリウム、pH2.5、6.0MGuHClを用いて溶出させて、4ml画分中に収集した。吸光度を280nmで記録した。
【0196】
得られた酸性溶液およびカオトロピック溶液は、さらなる精製工程、またはインビトロ再折り畳み実験のために4℃で保存してもよい。
【0197】
この変性材料で開始して、異なる再折り畳み方法(例えば、透析、急速希釈、再生サイズ排除クロマトグラフィー、またはマトリックス補助再折り畳み)を首尾よく用いかつ実施することができ、その全てが、実質的に同じ未変性様折り畳み、および可溶性タンパク質をもたらす。
【0198】
2.3 透析および急速希釈による再生
上記のように可溶化された材料を、透析によって生理的緩衝条件に移行する。透析チューブの選択された排除値は、4000-6000ダルトンであった。
【0199】
エクトドメイン(共有結合したgp41およびFkpAタンパク質ドメインのgp41部分)の再折り畳みを誘導するために、50mMリン酸ナトリウム、pH2.5、50mM NaCl(塩化ナトリウム)に対する透析によって、溶出したタンパク質から、GuHClを除去した。単離されたエクトドメインは、全てヘリックスであって、この極端なpHで三次接触を形成することが周知である。近UVCDによって、組換え産生されたFkpAを分析する場合、FkpAは、同じ条件下では本質的に未構築であることが見出された。透析によるgp41-FkpAの再折り畳みが、共有結合したgp41およびFkpAタンパク質ドメインを含む、容易に可溶性のタンパク質複合体を生じることは驚くべきことである。UVスペクトル(図4)は、迷光、すなわち、300nmを超える明白な吸収を欠く。迷光は、凝集の指標であり、従って図4に示されるスペクトルは、この再折り畳みされた材料が、有意な量の凝集物を含まないことを意味する。
【0200】
円偏光二色性分光法(CD)は、タンパク質中の二次構造および三次構造の両方を評価するための選り抜きの方法である。芳香族領域における楕円率(260-320nm)は、タンパク質内の三次接触(すなわち、規則的に折り畳まれたタンパク質の球状構造)を報告するが、アミド領域における楕円率は、タンパク質骨格中の規則的な反復性エレメント(すなわち、二次構造)を反映する。
【0201】
図5に示される近UV CDスペクトルによって、エクトドメイン(融合タンパク質の状況において)が、pH2.5で、未変性様三次接触を提示するという、説得力のある証拠が提供される。共有結合したgp41/FkpAタンパク質ドメインのスペクトルは、同一条件下で単離されたエクトドメインのスペクトルとほとんど一致する(データ示さず)。以下の、gp41の典型的なサインが見出された:290nmでの最大楕円率、285nmでの特徴的な肩部分、および260nmでの別の最大値(光学的に活性なジスルフィド結合を反映する)。FkpAは、全ての各々の条件下で、近UVシグナルに貢献しないことに注目することが重要である。実際、pH2.5におけるFkpAの芳香族楕円率は事実上、基準値に等しい(データ示さず)。
【0202】
近UV領域からの結果と一致して、pH2.5の融合構築物の遠UV CDは、ほとんど構造化されたgp41分子を暗示する。220nmおよび208nmの2つの最大値は、全ヘリックス状エクトドメインの典型的サインを形成し、かつそれに相当する(図6)。FkpA-gp41融合ポリペプチドは、急速希釈によって、示された条件(50mMリン酸ナトリウム、pH2.5、50mM NaCl)から、生理的緩衝条件に容易に移行され得る。結論として、近UV CDおよび遠UV CDの両方とも、未変性様構造のgp41は、非常に都合のよい様式で(FkpAも含む融合タンパク質の状況において)利用可能であることを強調する。興味深いことに、本発明者らは、SlyD(1-165)-gp41型の未変性様融合ポリペプチドが、室温での50mMリン酸ナトリウムpH7.4、150mMNaClに対するカオトロピック材料(例えば、7.0M GuHClに溶解された)の透析によって、さらに簡易に得られ得ることを見出す。完全に本発明に従って、例外的に実施された、2つのシャペロン-gp41融合構築物のヌクレオチド配列は、それぞれ、配列番号7および配列番号8に記される。
【0203】
2.4 サイズ排除クロマトグラフィー(SEC)による再生
変性gp41-FkpAポリペプチド(50mMリン酸ナトリウム、pH7.8、7.0MGuHClに溶解した)を、Superdex 200ゲル濾過カラム(20mMリン酸ナトリウム、pH7.4、50mM NaCl、1mM EDTAで平衡化した)にかけた。FkpA-gp41は、本質的に、3つの主な画分:高分子量の要素として、明らかなヘキサマー種として、そして明らかな三量体種として溶出する。この明白な三量体画分を濃縮して、近UVCD測定において、その三次構造について評価した(図7)。
【0204】
得られたグラフは、事実上、オーバーレイの(かぶさった)曲線であるが、これには、キャリアタンパク質FkpA、および標的タンパク質gp41の両方が1:1の比で寄与する。最も幸運なことに、gp41は、中性のpHで三次構造を示し、そして共有結合したシャペロンによって明白に可溶化される。言い換えれば、シャペロンFkpAは、基質として、未変性様構造のエクトドメインgp41を受容し、そして中性の作業pHにおいてこの折り畳み困難なタンパク質を可溶化するようである。従って、診断目的のための多量の可溶性gp41抗原を生成するために重要な要件は満たされる。
【0205】
pH7.4でのFkpA-gp41の遠UV CD(図8)によって、近UV CDは、それぞれFkpAおよびgp41のシグナル寄与の加算性を示すということが確認される。期待通り、このスペクトルは、高度にヘリックス状のgp41エクトドメインによって支配される(それぞれ、220nm、および208nmで最大楕円率)。
【0206】
上述の条件下で、pH7.4で可溶化された、共有結合されたgp41/FkpAタンパク質ドメインを用いて得られたデータによって、FkpAおよびgp41が、このポリペプチド構築物内の独立して折り畳む単位として挙動することが示される。
【0207】
実施例3 イムノアッセイにおいて抗原として用いられる組み換えgp41、および組み換えFkpA-gp41複合体に対する、異なる界面活性剤の効果
3.1 競合型イムノアッセイ
COBAS CORE HIV Combi試験(Roche DiagnosticsGmbH, Germany)は、組み換えgp41の免疫反応性について試験するための従来の手段を提供する。原理的に、このアッセイはまた、HIVのgp41に対する抗体を検出するための二重抗原コンセプトに従って働く。この固相抗原は、直接コーティングされる。検出抗原は、ペルオキシダーゼ標識されたgp41であって、ただし、SDS可溶化されたgp41材料を含む。
【0208】
HIVの検出のためのイムノアッセイにおいて、用いられる試薬は、かなり高濃度の界面活性剤を含むインキュベーションバッファー中で容易に溶解され、かつ安定であることが非常に所望される。このような界面活性剤(例えば、TritonX-100(登録商標)、またはNonident P-40(登録商標))は、ウイルス粒子を破壊するために0.1〜0.2%の濃度で用いられる。
【0209】
実施例1に記載のように生成されたSDS可溶化gp41、およびFkpA-gp41の両方が、COBASCORE HIV Combiアッセイにおいて競合抗原として試験された。それを行うために、市販のインキュベーションバッファーの代わりに、ヒト血清を含まないバッファーマトリックス中に0.1%TritonX100(登録商標)を含むインキュベーションバッファーが用いられる。試験されるべき抗原は、gp41と反応性であることが公知のヒト血清と同時インキュベートされる。
【0210】
gp41−FkpA抗原は、用量依存性の様式において、競合型アッセイ中のシグナルを強力にクエンチするが、SDS可溶化gp41は、本質的に非反応性である(図9)。50%阻害が、0.1μg/ml(2.2nMのモル濃度に相当する)のFkpA−gp41抗原濃度で達成される。
【0211】
試験中で、インタクトなウイルス膜を分解するために、界面活性剤(ヘルパー界面活性剤)として0.1%TritonX-100を含む希釈バッファーを用いた事前処理後に、FkpA−gp41が、優れた免疫反応性を保持することが、際立っている。これは、gp41エクトドメイン単独(SDS中のgp41)に比べて顕著である。gp41エクトドメイン単独では、へルパー界面活性剤の存在下で、免疫活性をほぼ完全に失う(図9)。
【0212】
FkpAが、不十分な結合動態に起因して重要なエピトープをマスクするか、または試験に内在性の界面活性剤TritonX-100が、gp41抗原の凝集を誘導することによって試験能力を破壊することのいずれかが、共有結合されたgp41−FkpA構築物の開発において主要な懸念であった。COBASCOREプラットフォームに対する多くの競合試験の実験結果によって、重要なgp41エピトープが、共有結合されたタンパク質ドメインの状況で、十分に接近可能であるという説得力のある証拠が得られる。さらに、分子内シャペロン-gp41複合体内のgp41の免疫反応性は、TritonX-100のようなヘルパー界面活性剤の存在下で保持される。
【0213】
3.2 電気化学発光アッセイ
二重抗原架橋形式による免疫アッセイは、感染性因子の血清学的な診断において大きな利点があるアッセイである。本発明によるFkpA-gp41は、生理的緩衝条件で可溶性であるので、検出原理として電気化学発光を使用する二重抗原架橋における使用に、この材料が適切であるか否かを検討することができる。
【0214】
ルテニウム標識に対してSDS可溶化gp41を結合させる試みは成功していない。しかし、FkpA-gp41は、生理的緩衝条件下で、容易に可溶であるので、疎水性Ru-標識に対するこの材料の結合は、直接証明された。記載された方法で改変された標的−シャペロン複合体でさえ可溶性のままであることは注目すべきである。Elecsys(登録商標)試験システム(RocheDiagnostics GmbH, Germany)においてアッセイを実施するために、FkpA-gp41を、それぞれビオチン化し、ルテニル化し、そして二重抗原架橋アッセイにおいて、免疫反応性について試験した。
【0215】
いくつかの典型的な抗HIV血清であって、主にIgG(免疫グロブリンG)クラスの抗体を含む血清を試験したところ、共有結合したFkpA−gp41タンパク質ドメインを用いて陽性であった。バックグラウンドのシグナルは、500ng/ml程度の高さの抗原濃度でさえ、内在するガジェット(gadget)バックグラウンドに達することが見出されている。このシグナル対ノイズ比は優れていることがわかった。さらに、キャリアタンパク質(E.coli由来の分子、シャペロンFkpA)が、これらのヒト血清に含まれる抗体の非特異的結合を生じるという証拠はない。
【0216】
上記で考察したように、血清変換の早期検出は、HIVの確実な診断に重要である。感染の経過の間に、IgMクラスの抗体が最初に出現する。非常に早期に、HIV感染を確実に検出するためには、従って、IgMの認識および結合について高いエピトープ密度を有する抗原モジュールを設計することが必須である。実際、FkpA-gp41は、典型的な抗HIV、IgM型血清によって十分に認識される。さらにより重要であるのは、陽性を試験するのが困難であるサンプル(NABI(Miami,Florida)によって供給された9003および4009の血清変換パネル由来のB血清およびC血清など)が、本発明による融合構築物を用いて陽性と試験されたという事実である。これは主要な成果である。なぜなら、ペプチドに基づくgp41抗原は、これらのIgM血清と試験された場合、全く反応性ではないからである。
【0217】
実施例4 可溶性シャペロン-gp41複合体は、ウイルス侵入を阻害する
異なるgp41−シャペロン融合タンパク質を、インビトロアッセイにおいて、HIV-1-媒介膜融合を阻害する能力について評価した。要するに、CD4、CCR5、およびCXCR4を発現するMAGIP4-CCR5レポーター細胞株を、HIV-1株NL4-3を用いて感染させて、Meisterら、Virology(2001)284(2)、287-296に従ってTat依存性b−ガラクトシダーゼ活性について評価した。実際、本発明者らは、nM範囲のIC50値を有する感染の実質的阻害を観察する。例えば、SlyD-gp41(配列番号6を参照のこと)は、IC50< 70nMでもって、ウイルス侵入を有意に阻害する。
【0218】
結論として、それぞれ、HIV-1 gp41、またはHIV-2 gp36を含む可溶性分子内複合体、およびペプチジル−プロリル−イソメラーゼシャペロンは、可溶性および高次構造の統合性(改善された抗HIV抗体試験の設計、および他の市販の適用を可能にする)に関して、著名な特色を保有する。
【0219】
実施例5 gp36エクトドメインに対するシャペロンFkpAの融合によって、インビトロで容易に再折り畳みされ得るサイトゾルポリペプチドが生じる
gp36(gp41のHIV-2ホモログ)を、可溶性かつ免疫学的に活性な形態で得るために、本発明者らは、FF36と名付けられた構築物をクローニングした。この融合ポリペプチドは、2つのFkpA単位、およびgp36単位(各々、可変性のグリシンリッチストレッチによって連結される)を含む。精製を容易にするために、この融合構築物をHis6C末端を用いてタグ化した。本質的に、前述のプロトコールに従って、このタンパク質を精製した:シャペロン溶解後、タンパク質をNi-NTA-カラムに結合させて、50mMリン酸ナトリウム、pH7.8、7.0MGuHClを用いた過剰な洗浄後、pHを下げることによって溶出させた。次いで、50mMリン酸ナトリウム、pH7.8、100mM塩化ナトリウム、1mM EDTAで平衡化したゲル濾過カラムを通過させることによって、溶出したタンパク質を、再折り畳みさせた。この「再生ゲル濾過(renaturinggel filtration)」法から得られたネィティブなタンパク質は、満足な免疫学的かつ分光学的特性を示し(図10を参照のこと)、従って、F41、またはFF41のようなgp41対応物に相当する。本明細書に記載された、この精製、および再折り畳みのプロトコールは、gp36のN末端7残基反復領域において3つの点突然変異を保有するFF36を用いて実施した(配列については、配列番号9を参照のこと)。可溶性タンパク質のより低い収率にもかかわらず、同じプロトコールがまた、wtgp36エクトドメインを含む融合構築物に首尾よく適用された。
【0220】
実施例6 可溶性の免疫反応性FkpA−gp21は、好都合でかつ再現可能な様式で獲得できる
FkpA-gp21-過剰産生E. coli細胞(配列番号10を含む)を、前記のとおり、増殖させ、誘導して、回収した。完全に溶解するために、細胞ペレットを50mMリン酸ナトリウム、pH7.8、7.0MGuHCl中で、室温で1時間撹拌させた。カオトロピック細胞溶解物を、溶解バッファー中で事前に平衡化したNi-NTA-カラム上に付与した。洗浄工程後、pHを低下させることによって、標的タンパク質(C末端Hexa-His-Tagを保有する)を溶出させた。再折り畳みのために、50mMリン酸ナトリウム、pH7.8、7.0M100mM塩化ナトリウム中で事前に平衡化したSuperdex 200ゲル濾過カラムに、GuHCl中FkpA−gp21(50mM、リン酸ナトリウム、pH6.0、7.0MGuHCl 4℃)を通過させた。UVスペクトルによって、FkpA-gp21が、可溶性タンパク質(これは、非シャペロンのgp21とは対照的に、それ以上凝集する傾向がない)として溶出することが実証された(図11/1)。さらに、得られたFkpA−gp21は、競合型COBASCORE実験において評価された場合、優れた免疫学的活性を示す(図11/2)。
【0221】
実施例7:FkpA-gp41に対するさらなるFkpAサブユニットの融合は、gp41エクトドメインの免疫学的特性を改善する
本発明者らは、融合ポリペプチド状況におけるさらなるPPIaseサブユニットが、gp41エクトドメイン−シャペロン複合体の全体的特性を改善し得るか否かという疑問に取り組んだ。このために、本発明者らは、記載されたプロトコールによって、F41(gp41改変体の上流に位置するFkpAドメインの1つ)、およびFF41(gp41改変体の上流に位置する2つのFkpAドメイン)の両方を調製した。次いで、ビオチン化し、かつルテニル化した融合タンパク質をElecsys(登録商標)E2010システムにおいて評価した。この結果は、さらなるシャペロンドメインを含む、FF41構築物の改善された特性を強力に示す。
【0222】
陰性の血清によるバックグラウンドシグナルであって、シグナル対ノイズ比、および力価の低い血清の確実な測定に決定的なバックグラウンドシグナルは、FF41(約1600カウント)を、F41(約3800カウント)と比較した場合、50パーセントよりも低下されることが見出された。
【0223】
【表1】

【0224】
SS41融合タンパク質、すなわち、2つのSlyDドメイン、および1つのgp41ドメイン(シャペロンドメインに対してC末端)を含む融合タンパク質を用いて、同様の陽性の結果が得られている。
【0225】
本発明の態様として、以下のものが挙げられる。
[1]バッファー中で、本質的に不溶性である標的タンパク質とペチジル-プロリル-イソメラーゼクラスシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化される、および形成されるタンパク質-シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびペプチジル-プロリル-イソメラーゼクラスシャペロンを含有する可溶性複合体の製造方法。
[2]生理的バッファーが10〜200mMの濃度のバッファー化合物および20〜500mMの総濃度の塩を含有する[1]記載の方法。
[3]タンパク質が組換え産生される[1]記載の方法。
[4]ペプチジルプロリルイソメラーゼが組換え産生される[1]記載の方法。
[5]タンパク質およびペプチジルプロリルイソメラーゼが組換え産生される[1]記載の方法。
[6]タンパク質がアミロイド形成性タンパク質である[1]記載の方法。
[7]アミロイド形成性タンパク質がレトロウイルス表面糖タンパク質である[6]記載の方法。
[8]タンパク質がHIV-2 gp36またはHIV-1 gp41である[1]記載の方法。
[9]ペプチジルプロリルイソメラーゼがペプチジルプロリルイソメラーゼの結合コンピテント断片である[1]記載の方法。
[10]バッファー中でレトロウイルス表面糖タンパク質とペプチジルプロリルイソメラーゼとを混合する工程、ここでレトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼの両方が可溶化され、複合体を形成する、および複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質-シャペロン複合体の製造方法。
[11]レトロウイルス表面糖タンパク質が組換え産生される[10]記載の方法。
[12]ペプチジルプロリルイソメラーゼが組換え産生される[10]記載の方法。
[13]レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼが組換え産生される[10]記載の方法。
[14]レトロウイルス表面糖タンパク質がHIV gp36またはgp41である[10]記載の方法。
[15]ペプチジルプロリルイソメラーゼが、ペプチジルプロリルイソメラーゼの結合コンピテント断片を含有する[10]記載の方法。
[16]バッファー中でペプチジルプロリルイソメラーゼに共有結合したレトロウイルス表面糖タンパク質を可溶化させる工程、ここでレトロウイルス表面糖タンパク質が可溶化される、およびレトロウイルス表面糖タンパク質-シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質-シャペロン複合体の製造方法。
[17]ペプチジルプロリルイソメラーゼがFKBPシャペロンである[1]、[10]または[16]記載の方法。
[18]FKBPシャペロンがSlyD、FkpAおよびトリガー因子からなる群より選ばれる[17]記載の方法。
[19]レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼを含有してなる可溶性複合体。
[20]レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼシャペロンが共有結合されているレトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼを含有してなる可溶性複合体。
[21]共有結合が化学的共役を含む[20]記載の複合体。
[22]共有結合が組換え結合を含む[21]記載の複合体。
[23]組換え結合がペプチドリンカーを含む[22]記載の複合体。
[24]ペプチドリンカーが少なくとも10アミノ酸を含有してなる[23]記載の複合体。
[25]ペプチドリンカーが少なくとも15アミノ酸を含有してなる[23]記載の複合体。
[26]ペプチドリンカーが多くとも50アミノ酸を含有してなる[23]記載の複合体。
[27]ペプチドリンカーが多くとも40アミノ酸を含有してなる[23]記載の複合体。
[28][19]〜[27]いずれか記載の可溶性複合体を含有してなる試薬の組成物。
[29]サンプルと、表面糖タンパク質とペプチジルプロリルイソメラーゼシャペロンを含有する複合体を含有する組成物とを接触させる工程、および結合抗体を検出する工程を含む、サンプル中のエンベロープウイルス表面糖タンパク質に対する少なくとも1つの抗体を検出する方法。
[30]接触させる工程が、表面糖タンパク質への抗体の結合に適切な条件下で行われる[29]記載の方法。
[31]結合した抗体の検出がサンプル中の抗-ウイルス抗体の存在を示す[30]記載の方法。
[32]第1のシャペロン-抗原複合体を含有する第1の抗原および第2のシャペロン-抗原複合体を含有する第2の抗原を含む二重抗原架橋概念によるイムノアッセイ。
[33]第1のシャペロンおよび第2のシャペロンが1つの種に由来する異なる分子である[32]記載のイムノアッセイ。
[34]第1のシャペロンおよび第2のシャペロンが異なる種に由来する[32]記載のイムノアッセイ。
[35]第1のシャペロンが好熱細菌に由来する[32]〜[34]いずれか記載のイムノアッセイ。
[36]第2のシャペロンが好熱細菌に由来する[32]〜[34]いずれか記載のイムノアッセイ。
[37]第1のシャペロンおよび第2のシャペロンが好熱細菌に由来する[32]または[34]記載のイムノアッセイ。
[38]第1の抗原複合体が固相結合基を含有する[32]記載のイムノアッセイ。
[39]第2の抗原複合体がマーカー基を含有する[32]記載のイムノアッセイ。
[40]可溶性レトロウイルス表面糖タンパク質-シャペロン複合体を含有するワクチンを被験体に注入し、それにより被験体においてレトロウイルス表面糖タンパク質に結合する抗体を誘発する工程を含む、免疫応答の誘発方法。
[41][10]記載の可溶性複合体を細胞に投与することを含む、細胞へのウイルス侵入の阻害方法。
[42]ウイルス侵入の阻害が膜融合を阻害することを含む[41]記載の方法。
[43]生理的バッファーが、10〜200mMの濃度のバッファー化合物および20〜500mMの総濃度の塩を含む[41]記載の方法。
[44]タンパク質が組換え産生される[41]記載の方法。
[45]ペプチジルプロリルイソメラーゼが組換え産生される[41]記載の方法。
[46]タンパク質およびペプチジルプロリルイソメラーゼが組換え産生される[41]記載の方法。
[47]タンパク質がアミロイド形成性タンパク質である[41]記載の方法。
[48]タンパク質がレトロウイルス表面アミロイド形成性糖タンパク質である[41]記載の方法。
[49]タンパク質がHIV-2 gp36またはHIV-1 gp41である[41]記載の方法。
[50]ペプチジルプロリルイソメラーゼがペプチジルプロリルイソメラーゼの結合コンピテント断片である[41]記載の方法。
【0226】
【図面の簡単な説明】
【0227】
【図1】図1Aおよび1Bは、HIV-1エクトドメインgp41(535-681)-His6の遠UVCD(1A)、および近UV CD(1B)を示す。折り畳まれたgp41(太線):緩衝条件(30mM ギ酸ナトリウム,pH3.0)を設定して、gp41の未変性様の全ヘリックス状高次構造(コンフォメーション)を誘導した;変性されたgp41(細線):緩衝条件(50mMリン酸ナトリウムpH3.0,7.0MGuHCl)を設定して、gp41(変性)を完全に変性する。カオトロピック塩の高いモル濃度に起因して、図1A中の参照二色性シグナル(細線)は、215nm未満の波長領域で確実にモニターすることはできない。このスペクトルは、Jasco-720分光偏光計で記録され、ノイズを低下させるために9回平均された。パスレングスは、遠UVCDについては0.2cm(図1A)、そして近UV CDについては0.5cm(図1B)であった。それぞれのタンパク質濃度は、1.5μM、および29μMであった。縦軸の単位は、平均残余楕円率(meanresidue ellipticity)であり、そしてdeg×cm2×dmol-1の次元を有する。
【図2】図2は、生理的バッファー中における「非シャペロン(unchaperoned)」gp41の凝集を示す。3.0から7.5へのpHジャンプ後、1分(下の線)、および10分(上の線)のgp41エクトドメインのUVスペクトルを示す。凝集する分子は、迷光効果をもたらし、そして310nmを超えて明白な吸収を生じる。この図は、gp41の甚大な凝集傾向をまさに実証することを意味する;凝集過程が、上の線によって示された段階で停止しないことは注目に値する。
【図3A】図3Aは、FkpAが中性のpHでgp41エクトドメインを可溶化することを示す。gp41、および成熟FkpAを、低いpHで同時インキュベートして、その後に20mMリン酸ナトリウム、pH7.4;50mMNaCl, 1mM EDTAの最終バッファーにシフトした;1分後、および10分後(それぞれ、下の線、および上の線)、UVスペクトルを記録して、サンプル中の凝集の程度を評価した。図3Aは、2倍モル過剰のシャペロンによる凝集の抑制を示し、図3Bは、4倍過剰の効果を示す。gp41の最終濃度は、約1μMであった。迷光(300nmを超えて明白な吸収をもたらす)は、最小まで低下したので、FkpAは、gp41エクトドメインを用量依存性の様式で効率的に可溶化するという、説得力のある分光光度的な証拠がある。
【図3B】図3Bは、FkpAが中性のpHでgp41エクトドメインを可溶化することを示す。gp41、および成熟FkpAを、低いpHで同時インキュベートして、その後に20mMリン酸ナトリウム、pH7.4;50mMNaCl, 1mM EDTAの最終バッファーにシフトした;1分後、および10分後(それぞれ、下の線、および上の線)、UVスペクトルを記録して、サンプル中の凝集の程度を評価した。図3Aは、2倍モル過剰のシャペロンによる凝集の抑制を示し、図3Bは、4倍過剰の効果を示す。gp41の最終濃度は、約1μMであった。迷光(300nmを超えて明白な吸収をもたらす)は、最小まで低下したので、FkpAは、gp41エクトドメインを用量依存性の様式で効率的に可溶化するという、説得力のある分光光度的な証拠がある。
【図4】図4は、pH2.5でのFkpA-gp41のUVスペクトルを示す。50mMリン酸ナトリウム、pH2.5;50mMNaClに対する透析後の融合ポリペプチドFkpA-gp41のUVスペクトル。驚くべきことに、2つのドメイン構築物は、可溶化カオトロピック剤CuHClの除去後、完全に可溶性のままである。300nmを超える波長で基線ドリフト(baselinedrift)および有意な明白な吸収を生じることが期待される、迷光する凝集物が存在するという証拠はない。
【図5】図5は、pH2.5でのFkpA-gp41の近UV CDスペクトルを示す。20mMリン酸ナトリウム、pH2.5;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、ノイズを低下させるために9回蓄積した。タンパク質濃度は、0.5cmのパスレングスで22.5μMであった。芳香族の楕円率は、gp41の典型的な特徴を示す(基準については、図1Bを参照のこと)。pH2.5では、FkpAは、大部分が構造化されておらず、近UV-CDにおけるシグナルに対して全く寄与しない。
【図6】図6は、pH2.5でのFkpA-gp41の遠UV CDスペクトルを示す。20mMリン酸ナトリウム、pH2.5;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、シグナル対ノイズ比を改善するために9回蓄積した。タンパク質濃度は、0.2cmのパスレングスで2.25μMであった。220および208nmでの最小値は、融合タンパク質の状況では、gp41の大部分のヘリックス構造を暗示する。197nm未満のスペクトルノイズは、高いアミド吸収に起因しており、そして融合タンパク質のいずれの構造的特徴についても報告しない。それにもかかわらず、193nmで典型的なヘリックス最大値が推測され得る。
【図7】図7は、生理的な緩衝条件下でのFkpA-gp41の近UV CDである。20mMリン酸ナトリウム、pH7.4;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、ノイズを低下させるために9回蓄積した。タンパク質濃度は、0.5cmのパスレングスで15.5μMであった。著しいことに、gp41およびFkpAの共有結合したタンパク質ドメインの芳香族楕円率(実線)は、pH3.0での未変性様の全ヘリックス状gp41の寄与(下の破線)、およびpH7.4のFkpAの寄与(上の破線)から相加的に作成される。これによって、キャリアFkpA、および標的gp41(すなわち、2つの別個の機能的な折り畳み単位)が、ポリペプチド融合タンパク質において連結される場合、可逆的かつ半独立的に再折り畳みすることが示される。
【図8】図8は、生理的な緩衝条件下でのFkpA-gp41の遠UV CDを示す。20mMリン酸ナトリウム、pH7.4;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、シグナル対ノイズ比を改善するために9回蓄積した。タンパク質濃度は、0.2cmのパスレングスで1.55μMであった。220および208nmでの強力なシグナルは、それぞれ、融合タンパク質の状況では、gp41の大部分のヘリックス状構造を暗示する。198nm未満のノイズは、高いタンパク質吸収に起因しており、そしてFkpA-gp41のいずれの二次構造的な特性も反映しない。
【図9】図9は、FkpA連結したgp41がHIVアッセイにおいて可溶性であり、かつ高度に免疫反応性であることを示す。FkeA-gp41は、COBASCORE HIV Combiアッセイにおける強力な競合因子である。gp41エクトドメイン単独(白丸)と比較した、希釈バッファー(ヘルパー界面活性剤としてTritonX-100を含有する)を用いた前処理後の可溶性FkpA-gp41ポリペプチド(黒丸)の阻害能力を示す。gp41エクトドメイン(融合タンパク質の分子内複合体内)は、界面活性剤の存在下でさえ高い免疫反応性を保持しているが、一方で裸のエクトドメインは、ほとんど完全に免疫反応性を失っているということが明白である。試験したHIV陽性血清は、1:3000希釈で、内部血清数21284であった。
【図10】図10は、再生ゲル濾過後のFF36のUVスペクトルである。このスペクトルは、gp36融合ペプチドが、実施例の節に記載のとおりの再生ゲル濾過法に従ってSux200カラム上で再折り畳みされた場合に、可溶性であり、かつ凝集しないという有力な証拠を提供する。
【図11】図11(1+2)は、FkpA-gp21が可溶性であり、かつ免疫学的に反応性の融合ポリペプチドであることを示す。再生ゲル濾過後、再折り畳みされたFkpA-gp21融合タンパク質は、高度に可溶性に溶出して、UVスペクトルにおいては凝集傾向を示さない(11/1)。HTLV陽性血清858893-00(1:10希釈)を用いたCOBASCOREにおける競合型の免疫アッセイ実験において評価した場合、FkpA-gp21は、結局、優れた免疫学的特性を保有するようになる(11/2)。
【技術分野】
【0001】
本発明は、HIV感染の診断に関する。本発明は特に、可溶性のレトロウイルス表面糖タンパク質−(または膜貫通糖タンパク質)−シャペロン複合体の生成、および特に、好ましくは二重抗原架橋コンセプト(doubleantigen bridge concept)による、イムノアッセイにおけるHIVに対する抗体の検出におけるシャペロン抗原複合体の有利な使用、または免疫原としての使用を教示している。本発明はまた、それぞれ、HIV-1gp41の改変体、またはHIV-2 gp36の改変体、ならびにペプチジル−プロリル−イソメラーゼクラスのシャペロンから選択されたシャペロンを含む可溶性の複合体を開示する。それぞれ、HIV-1gp41の、またはHIV-2 gp36のNヘリックス状ドメインにおいて、特定のアミノ酸置換を含む改変体がまた記載される。
【背景技術】
【0002】
背景
ヒト免疫不全ウイルス(HIV)は、後天性免疫不全症候群の因子であり、通常、その頭文字によって、AIDSと呼ばれる。このウイルスの株には、HIV-1、およびHIV-2と命名された2つの主な株が存在する。HIウイルスは、現在、広範に伝播しており、そして世界的な健康および保健に対して重大な脅威となっており、このせいで、公的な保健機関は、HIVの診断およびAIDSの治療に関して、莫大な金額を費やさせられる。
【0003】
ウイルス伝播の経路の1つは、感染した血液または血液製品の輸血である。実質的に全ての先進工業国、および多くの発展途上国は、現在のところ、このウイルスのさらなる伝播を防ぐためには、全ての献血の強制的な検査を必要とする。血液によるHIV感染を、できるだけ確実に、かつ感染後すぐに診断することが、この分野における、全ての診断方法の課題である。
【0004】
基本的に、以下の3つの異なる診断様式が利用可能である:
(1)ポリメラーゼ連鎖反応(PCR)のような鋭敏な核酸診断手順による、血液由来のウイルスゲノム材料の診断、
(2)血液由来のウイルス抗原の検出、および
(3)体液由来のHIVに対する抗体の検出。
【0005】
HIV感染の経過の間、いくつかの診断上別個で、かつ診断上関連する段階が知られている。感染の初期段階では、HIVウイルス由来のタンパク質またはポリペプチドのみが見出され得る(「ウイルス感染期(viraemicphase)」が、抗HIV抗体はまだ存在しない。その後の、セロコンバージョンと名付けられる段階で、HIV抗原に対する抗体が出現し、一方でウイルス抗原の量(ウイルス量)が低下する。セロコンバージョンの初期段階で形成される抗体のほとんどが、クラスMの免疫グロブリン(IgM)に属する。後に、HIVに対する免疫応答は、クラスGの免疫グロブリン(IgM)に切り換わり、次にこれが、HIVに対する抗体のほとんどを構築する。感染のさらなる経過の間、抗HIV抗体のレベルは、低下し得るが、体液中のウイルス量(ウイルス粒子またはウイルス抗原の存在)は再度増大し得る。HIV感染の存在についてのスクリーニングは、好ましくは、HIV抗原に対する抗体を検出する血清学的アッセイ(時々、HIV抗原の検出と組み合わせて)によって行われる。患者内の免疫応答は、感染の経過中に変化し、そしてまた患者間で異なるので、サブクラスIgM、およびIgGに属する抗HIV抗体を検出する、極めて感受性でかつ信頼のできるイムノアッセイを有することが重要である。HIV感染の検出のための多くの異なるアプローチが記載されている。ウイルスタンパク質に対する抗体の、早期で、信頼でき、かつ鋭敏な検出は、重大でかつ大変重要である。
【0006】
ウイルスタンパク質(しばしば、ウイルス抗原と名付けられる)は、感染の発現時、およびこの疾患の非常に後期段階でのみ検出可能であり得る。従って、p24(HIV-1由来)、またはp26(HIV-2由来)(両方ともウイルスのコアタンパク質である)を測定するアッセイなどの、ウイルス抗原の検出のためのアッセイは、HIV感染を確実に検出するために他の診断手段と組み合わせてのみ用いられ得る。
【0007】
宿主中において抗体形成を誘導し得、それによって診断手順において抗原として用いられ得る、ウイルス抗原の3つのグループが、理論上利用可能である。これらは、エンベロープタンパク質(env遺伝子領域によってコードされる)、ウイルス酵素、または調節性タンパク質(例えば、逆転写酵素、またはインテグラーゼ)(pol遺伝子領域によってコードされる)、および構造的なコアタンパク質(gag遺伝子領域によってコードされる)である。HIV-1、およびHIV-2の両方におけるウイルスエンベロープタンパク質は、ポリペプチド前駆体タンパク質として合成される糖タンパク質である(HIVに関しては、gp160、HIV-2に関してはgp140)。これらの高分子量前駆体は、合成後、切断されて、それぞれ、gp120、およびgp41(HIV-1)、またはgp110、およびgp36(HIV-2)を生じる。これらの大きいポリペプチド(それぞれ、gp120、またはgp110)は、表面サブユニットを形成し、これがルーズな接触を介して、膜を貫通する、さらに小さいポリペプチド(それぞれ、gp41、およびgp36)に会合する。多くの宿主(患者)においては、エンベロープ糖タンパク質が、抗ウイルス免疫応答の好ましい標的である。Ratner,Lら、Nature 313(1985)277-84によって、特にこれらのエンベロープタンパク質の膜貫通(すなわち、それぞれgp41、またはgp36)は、これらのウイルスタンパク質の中でも最も免疫学的な可能性を保有することが実証された。
【0008】
HIウイルスによってコードされるポリペプチドを使用する、例えば、ELISA(酵素結合免疫吸着アッセイ)のようなイムノアッセイ法は、診断、およびスクリーニングにおいては広範に用いられている。ウイルスポリペプチドは、ウイルス材料から直接調製されるか、または組み換えDNA技術を用いるインビトロもしくはインビボの発現系に由来する。抗原生成の両方の方法とも、重大な限界がある。ウイルス調製物由来のポリペプチドは、生存可能なウイルス、または感染性遺伝子材料によって汚染され得、これによって、この材料を用いる人に対して害をもたらす。組み換え体由来材料は、非HIV宿主タンパク質によって汚染され得、これによって、このようなアッセイの特異性の低下、または感度の低下が生じ得る。
【0009】
ウイルス病原体のような、病原性因子に対する抗体の検出において、例えば、US4,945,042号に記載される、二重抗原架橋形式(doubleantigen bridge format)に従って、きわめて高頻度に、かつ極めて有利な抗体検出系が用いられる。この架橋コンセプトによるイムノアッセイは、固相に直接または間接的に結合した抗原、および同じまたは交差反応性の容易に可溶性の抗原(直接または間接的に検出可能である)の使用を必要とする。検討中の抗体は、もし存在するならば、固相結合抗原と、標識された検出抗原との間の架橋を形成する。2つの抗原が、特定の抗体によって架橋される場合にのみ、陽性のシグナルが生じる。
【0010】
抗HIV抗体の検出のための抗原として組換え産生されたgp41を用いるいくつかの試みが記載されている。組換え産生されたgp41は、抗HIV抗体を検出するために用いられ得るが、いくつかの限界を伴う。このようなgp41は、抗HIV抗体を測定するために、単独で、または他のHIV抗原と組み合わせてのいずれかで用いられる。今日、HIV抗原、および/または抗HIV抗体の両方の検出を独立して目的とするアッセイが公知である。WO93/21346においては、HIV-1gp41、およびHIV-2 gp36に対するgp24の抗原および抗体の同時の検出のための「組み合わせ試験(combi-test)」が記載される。このアッセイでは、固相が用いられ、この固相に対して組み換え産生されたgp41が直接コーディングされている。
【0011】
極度に高いpH値または低いpH値の使用が、溶液中でgp41(またはgp36)を保持するための1つの方法であるということもまた十分に確立されている。組み換え産生されたgp41は、ほぼpH3.0未満、またはほぼpH11.0を超えて可溶性であることが公知である。
【0012】
しかし、不幸にも、それぞれ、HIV-1 gp41、およびHIV-2 gp36は両方とも、生理的な緩衝条件下では本質的に不溶性である。
【0013】
一般的なイムノアッセイは、生理的なpHで実施される。生理的な緩衝条件下でのそれらの不溶性に起因して、多くのイムノアッセイにおいては、レトロウイルス表面糖タンパク質抗原は、固相材料上に直接コーティングされて用いられる。しかし、固相材料に対する抗原の直接コーティングは、多くの場合、有害であり、そしてコンフォメーションの変化、分子変性(unfolding)、抗原性の変化、不安定性、およびバックグラウンドの問題などの不利な点を生じる(Butler,J. E.ら、J. Immunol. Methods 150(1992)77-90を参照のこと)。
【0014】
強力なカオトロピック剤、または適切な界面活性剤によって、レトロウイルス表面糖タンパク質(retroviral surface glycoprotein)(rsgp)を可溶性にすることが可能であるが、このような様式で可溶性にされた材料は、診断ツールとしての使用に限定される。
【0015】
生理的な緩衝条件でのレトロウイルス表面糖タンパク質の不溶性によって、さらに、これらのタンパク質には、慣用的な(生物)化学的手順の極めて困難な標的になる。大部分の「標識化学(labeling chemistries)」、すなわち、標識(例えば、ポリペプチドに対するマーカー基)を結合するために用いられる化学的手順は、求核化学に基づいており、従って、pH約6〜pH約8のpH幅に制限されるのではなく、従って、生理的な緩衝条件以上または以下でのみ働く。例えば、「Bioconjugation」(1998)216〜363、M.Aslam、およびA. Dent編,McMillan Reference, Londonの、Aslam, M、およびDent, A., Thepreparation of protein-protein conjugatesに記載されるような、これらの慣用的な手順は、レトロウイルス表面糖タンパク質を可溶化するために必要な極端なpH値(またはSDSのような界面活性剤の存在下)では適切に作用しないか、または実行することが困難であるかのいずれかである。
【0016】
上記で言及したとおり、架橋コンセプトによるイムノアッセイは、病原性生物体と反応性の抗体の検出を目的とする広範な種々の異なるアッセイにおいて利点が証明されている。しかし、その不溶性に起因して、このようなアッセイの設定において、それぞれ、HIV-1のe-gp41分子(すなわち、「糖タンパク質41のエクトドメイン(ectodomain of glycoprotein 41)」、またはe-gp36分子を用いることは、例えば、不可能であった。
【0017】
直接のコーティングの不利な点を補償するために、種々のアッセイが設計されており、このアッセイは、e-gp41抗原を用いることに代わって、合成的に、または組換え産生されたその抗原の部分的配列(多少ともイムノドミナント、いわゆるループ領域にまたがる)を利用する。このようなアッセイの例は、以下に考察する特許文献に示される。
【0018】
gp41の細胞外部分におけるループ領域は、N末端ヘリックス状ドメインを、同様にヘリックス状のC末端ドメインに連結する分子の、非ヘリックス状の先端ヘアピンである。gp41に対して反応性の抗血清の大部分は、この先端ループモチーフに対する抗体を含む。従って、このジスルフィド架橋したヘアピン、またはループ構造は、gp41のイミノドミナント領域を示す。従って、組み換え的に駆動されたgp41に関連する問題を克服するための迂回路の1つは、gp41の部分的配列を表すペプチドの化学的生成である。それぞれgp41、またはgp36が、本発明において言及される場合、いわゆるエクトドメイン(ループ接続されたN-ヘリックス、およびC-ヘリックスを包含するが、N末端融合ペプチド、およびC末端膜貫通セグメントは欠く)として規定されることに注意することが重要である。
【0019】
種々のHIV抗原のペプチド断片が、関連の特許文献(オーストラリア特許出願番号597,884(57733/86)、ならびに米国特許第4,735,896号、および同第4,879,212号)中で開示される。詳細には、これらの3つの明細書は、gp41糖タンパク質の保存されたイムノドミナント領域(HIV-1の主なエンベロープタンパク質のループ領域)を開示する。HIV-2のgp36タンパク質の類似のイムノドミナント領域がまた、合成されている。これらのループ領域に対応するペプチド(このエクトドメインの先端部を構成する)は、HIV-1、およびHIV-2の早期診断を可能にし、そしてアッセイにとって十分だが至適ではない感度および良好な特異性をもたらす。しかし、それらの限界は、特定の患者におけるセロコンバージョンの初日のIgM抗体の検出に関して明らかになる。
【0020】
WO92/22573は、骨格と共通して、すなわち、種々の哺乳動物の免疫不全ウイルスの膜貫通エンベロープタンパク質(例えば、gp41、またはgp36)のイミノドミナント領域と共通して、免疫学的な特性を有するペプチドを開示する。このイムノドミナント領域は、異なる哺乳動物種に由来する免疫不全ウイルス単離体において高度に保存されている、ジスルフィドループを含むことをさらに確認する。
【0021】
EP396559は、HIVの天然に存在するアミノ酸配列に相当するアミノ酸配列を保有する、人工的ペプチドに関する。このエピトープは、やはり、それぞれgp41、またはgp36のループ構造に相当する配列に由来する。それらは、イミノドミナントループの2つのシステイン残基の間の化学的酸化段階によって形成されたジスルフィド結合を含むように、さらに純化されている。
【0022】
しかし、HIV感染した患者の抗HIV抗血清に含まれた抗体のかなり大きい割合が、gp41またはgp36のイムノドミナントループ由来の配列モチーフ、またはその改変体とは反応しない。これらのペプチド抗原は、有利な架橋コンセプトと組み合わせて用いられ得るが、HIVgp41のループ領域の外側のエピトープと反応性の抗体は、検出されない。HIV感染の非常に早期の診断が重要なだけでなく、HIV-1、およびHIV-2のできるだけ多くのサブタイプが検出されることも極めて重要である。エピトープがより多く、特に、rsgpの正確に折り畳まれたコンフォメーションのエピトープが、より多く存在するほど、偽陰性の診断に起因して、感染したサンプルを見逃す可能性は低くなる。
【0023】
従って、レトロウイルス表面糖タンパク質分子の(特に、HIV-1由来のgp41の)より大きい部分を可溶型で提供するために、継続的な努力が行われている。
【0024】
gp41の生物物理学的な、そして生化学的な特性は、過去に広範に研究されてきた。Lu,Mら、Nat. Struct. Biol. 2(1995)1075-82)は、gp41の三量体構造を部分的に解明している。gp41は、生理的な条件下で、不溶性の凝集物を形成するので、検討は、短縮型のエクトドメインgp41に対して限定された。
【0025】
gp41のネイティブな三量体が、3つの平行なN末端中央ヘリックス(これに対してC末端ヘリックスは、逆平行方向に詰める)を含む、6つのヘリックス状バンドルを形成することが、NMR分光法によって最近確認されている(Caffrey,Mら、J Biol Chem 275(2000)19877-82)。
【0026】
gp41の高分子凝集物がまた記載されている。このような凝集物は、gp41のいわゆる先端部ループ領域の相互作用によって形成される可能性が最も高い。
【0027】
タンパク質設計による、標的細胞へのHIV-1進入のインヒビターは、Root, M.J.ら、Science 291(2001)884-8によって開発された。このインヒビターは、gp41由来のN末端ヘリックスドメイン由来の3つのストレッチ、およびこの分子由来のC末端ヘリックスドメインの2つのストレッチを含む。しかし、この遺伝子操作された構築物は、ネイティブな分子の多くのドメイン、および多くの抗原性エピトープを欠き、特にいわゆるループモチーフ(特に免疫原性エピトープを保有することが公知である(上記を参照のこと))を含まない。
【発明の開示】
【発明が解決しようとする課題】
【0028】
従って、できるだけ多くのレトロウイルス表面糖タンパク質エピトープを可溶型で提供するという非常に大きい必要性が依然として存在する。特に種々の治療的な適用、および診断的な適用における使用のために、それぞれ、HIV-1由来のgp41、またはHIV-2由来のgp36を含む、このような可溶性抗原を提供する必要性が存在する。
【0029】
より多くのレトロウイルス表面糖タンパク質エピトープ、またはそれぞれe-gp41分子もしくはe-gp36でさえ、可溶型で提供することが可能であるか否かを検討することが、本発明の課題であった。
【0030】
本発明のさらなる課題は、それぞれgp41、およびgp36の改変体(これは、取り扱うことがより容易であり、そして/または特に、イムノアッセイを実施するために必要とされるような、または免疫に必要とされるような緩衝条件下では、複合体型(この改変体およびペプチジル−プロリル−イソメラーゼクラスのシャペロンのシャペロンを含む)で可溶性である)を提供することが可能であるか否かを検討することであった。
【課題を解決するための手段】
【0031】
本発明の要旨は、
〔1〕非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化され、および形成されるタンパク質− シャペロン複合体
が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびシャペロンを含有する可溶性複合体の製造方法、
〔2〕非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子又はSlyDの結合コンピテント断片とを混合する工程、ここで該タンパク質および該断片の両方が可溶化され、および形成されるタンパク質− 断片複合体が可溶である生理的条件
にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質および断片を含有する可溶性複合体の製造方法、
〔3〕非生理的なバッファー中でレトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここでレトロウイルス表面糖タンパク質およびシャペロンの両方が可溶化され、複合体を形成し、および該複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質−シャペロン複合体の製造方法、
〔4〕レトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを含有してなる可溶性複合体、
〔5〕〔4〕記載の可溶性複合体を含有してなる試薬の組成物
に関する。
【発明の効果】
【0032】
本発明により、より多くのレトロウイルス表面糖タンパク質エピトープ、例えば、e-gp41分子またはe-gp36が可溶型で提供される。
【発明を実施するための最良の形態】
【0033】
古典的な「折り畳みヘルパー(folding helpers)」として知られるシャペロンは、他のタンパク質の折り畳み、および構造的な完全性の維持を補助するポリペプチドである。それらは、インビボ、およびインビトロの両方でポリペプチドの折り畳みを促進する能力を保持する。一般に、折り畳みヘルパーは、折り畳み触媒、およびシャペロンに小分割される。折り畳み触媒は、その触媒性機能に起因してタンパク質折り畳みにおける律速段階を加速する。触媒の例は、以下にさらに記載される。シャペロンは、変性されたポリペプチドまたは部分的に変性されたポリペプチドに結合し、これによってタンパク質を再生することを補助することが公知である。従って、折り畳み触媒とは異なり、シャペロンは、結合機能を発揮するに過ぎない(Buchner,J., Faseb J 10(1996)10-19)。
【0034】
シャペロンは、タンパク質成熟、折り畳み、転位、および分解に関与する、偏在性のストレス誘導性タンパク質である(Gething, M. J.、およびSambrook, J., Nature 355(1992)33-45)。正常な増殖条件下でも存在するが、それらは、ストレス条件下で、豊富に誘導される。このことによって、それらの生理的な機能が、ストレス条件に対処することであるという理論がさらに支持される。
【0035】
現在のところ、いくつかの異なるファミリーのシャペロンが知られている。これらのシャペロンの全てが、折り畳まれていないか、または部分的に折り畳まれていないタンパク質に対して結合する能力によって特徴付けられており、そしてタンパク質の正確な折り畳み、または変性もしくは凝集したタンパク質の除去に関係する生理的機能を有する。
【0036】
シャペロンの十分特徴付けられた例は、タンパク質のいわゆる熱ショックファミリーのメンバーである。これらは、相対的な分子量に従って命名される;例えば、hsp100、hsp90、hsp70、およびhsp60、ならびにいわゆるshsps(小熱ショックタンパク質(smallheat-shock-proteins))(Buchner, J., Faseb J 10(1996)10-19によって、ならびにBeissinger, M.およびBuchner,J., Biol. Chem. 379(1998)245-59によって記載されるような)。
【0037】
折り畳み触媒は、シャペロンと異なり、規定された律速段階を加速することによって折り畳みを補助し、これによって凝集しやすい折り畳み中間体の濃度を低下させる。触媒の1つのクラスである、タンパク質ジスルフィドアイソマー(あるいは、チオール−ジスルフィド−オキシド−レダクターゼと命名された)は、分泌タンパク質におけるジスルフィド結合の形成または再編成を触媒する。グラム陰性の細菌において、周辺質における分泌タンパク質の酸化的折り畳みは、DsbA、DsbB、DsbC、およびDsbDと命名された、タンパク質ジスルフィドイソメラーゼのカスケードによって調整される(Bardwell,J.C., Mol Microbiol 14(1994)199-205、およびMissiakas, D.,ら、Embo J 14(1995)3415-24)。
【0038】
ペプチジルプロリルシス/トランスイソメラーゼ(PPI)と呼ばれる折り畳み酵素の別の重要なクラスは、以下のような異なるメンバーを含む:CypA、PpiD(Dartigalongue,C.、およびRaina, S., Embo J 17(1998)3968-80, FkpA(Danese, P. N.ら、Genes Dev 9(1995)387-98)、誘発因子(triggerfactor)(Crooke, E、およびWickner, W., Proc Natl Acad Sci USA 84(1987)5216-20、ならびにStoller,Gら、Embo J 14(1995)4939-48)、およびSly D(Hottenrott, Sら、J Biol Chem 272(1997)15697-701)。これらのなかでも、FkpA、SlyD、および誘発因子は、配列のアラインメントに基づいて、関連していることが見出されている。
【0039】
ペプチジルプロリルイソメラーゼFkpAは、グラム陰性細菌の周辺質に局在している。このシャペロンが、細菌の外部膜タンパク質の輸送および転位に重要であることが推測されている。Ramm,K.、およびPluckthun, A., J Biol Chem 275(2000)17106-13)によって、FkpAは、タンパク質の正しい折り畳みに対する、その有益な効果を、2つの異なる方法で表すことが示されている。第一に、FkpAは、初期の折り畳み中間体と相互作用し、それによってそれらの凝集を妨げる。第二に、FkpAは、可能性としては、部分的に折り畳まれていない種(それは、凝集型と平衡して存在し得る)に対する結合によっても、不活性なタンパク質を再活性化する能力を有する。
【0040】
いくつかの折り畳みヘルパーは、触媒的に活性なドメイン、およびシャペロン(またはポリペプチド結合)ドメインの両方を含む。代表的な例は、例えば、誘発因子(Zarnt, Tら、J Mol Biol 271(1997)827-37)、Wang, C. C.、およびTsuo, C. L.,Faseb J 7(1993)1515-7)、SurA(Behrensら、EMBO J(2001)20(1)、285-294)、およびDsbA(Frech,C.,ら、Embo J 15(1996)392-98)である。本発明者らの観察によれば、同じモジュール構造が、それぞれ、PPIases FkpA、およびSlyDにおいて現実化されると思われる。
【0041】
異なる独立した系において、シャペロンの強化された発現によって、ポリペプチドの組み換え生成が促進され得ることが実証されている。これに対する例は、WO94/08012に見出され得る。
【0042】
タンパク質の産生の増大は、ポリペプチドコード配列、およびシャペロン配列を含む遺伝子構築物を用いることによって達成され得ることがまた、公知である。例えば、この融合コンセプトは、ヒトのプロインスリン(pro-insulin)遺伝子、およびDsbAを含む遺伝子構築物を用いることによって、Escherichia coliの周辺質におけるヒトのプロインスリンの有意に増大した産生を生じることが示されている(Winter,J.ら、Journal of Biotechnology 84(2000)175-185)。
【0043】
未変性様の折り畳まれたポリペプチドの産生の増大のためにシャペロンを用いるアプローチは主に、結合、およびこれによるシャペロンタンパク質の機能の可溶化に起因している。シャペロンおよび標的タンパク質を含む融合ポリペプチドの組み換え産生後、このシャペロンは習慣的に、得られたポリペプチドから切り離され、所望のポリペプチドが純粋な形態で得られる。対照的に、本発明は、一方では、レトロウイルス表面糖タンパク質と会合している、適切なシャペロンの有益な可溶性効果に基づく。
【0044】
驚くべきことに、折り畳みヘルパー、例えば、ペプチジルプロリルイソメラーゼ(PPI)クラスの多くのメンバー(特にFKBPファミリー由来)が、触媒性活性を示すだけでなく、アミロイド形成性のタンパク質の(またはより一般的に言えば、凝集する傾向にあるタンパク質の)溶解度に対して強烈な有益な効果をもたらすことを、本発明者らは見出した。それらがそのような効果をもたらすのは、このようなタンパク質(そうでなければ(すなわち、非シャペロンの単離型では)凝集しやすい)と可溶性の複合体を形成することによってである。このようなタンパク質(さもなければ、生理的条件下では、ほとんど溶解しないか、または不溶性である)は、一旦それらが、適切なPPIシャペロンと複合して結合すれば、穏やかな生理的条件下では結局、可溶性になる(すなわち、界面活性剤またはカオロトピック剤などの可溶化添加物の必要がない)。このように、本発明者らは、例えば、可溶性のタンパク質−シャペロン複合体(例えば、凝集しやすい標的タンパク質としてHIV-1のgp41タンパク質、ならびに可溶性付与シャペロンとしてFkpA、または他のFKBPを含む)を産生することができた。
【0045】
さらに、本発明者らは、それぞれ、HIV-1 gp41、またはHIV-2 gp36の特定の十分規定された改変体が、PPI-クラスのシャペロンと可溶性の複合体を形成するのに特に適切であることを見出した。
【0046】
gp41、およびFkpAの複合体、またはgp36およびFkpAの複合体は、例えば、生理的条件下で、例えば、容易に可溶性であり、それらを、都合の良いpH範囲で容易に標識することが可能であり、そしてそれらを用いて、HIV(それぞれ、1または2)のそれぞれgp41、またはgp36に対する抗体の検出において、従ってHIV感染の診断において、多大な利益を得ることができる。
【0047】
詳細な説明
本発明は、本質的に不溶性である標的タンパク質、およびペプチジル−プロリル−イソメラーゼクラスのシャペロンを含む可溶性複合体を生成する方法に関し、この方法は、バッファー中でこのタンパク質およびこのシャペロンを混合する工程、ここでこのタンパク質およびシャペロンの両方は、可溶化される、ならびにこのバッファーを生理的な条件に調整する工程(ここで、この形成されたタンパク質−シャペロン複合体は可溶性である)を包含する。
【0048】
本発明による「標的タンパク質」とは、20mMリン酸ナトリウム、および150mM塩化ナトリウムからなるpH7.4の水性緩衝溶液中で本質的に不溶性である任意のタンパク質であってもよい。例えば、好ましい標的タンパク質は、アミロイド形成性タンパク質、アミロイド形成性ウイルスの表面糖タンパク質、レトロウイルス表面糖タンパク質、特にHIV-1gp41、HIV-2 gp36、およびHTLV gp21である。
【0049】
標的ポリペプチドの重要な群の1つは、いわゆるアミロイド形成性タンパク質、またはポリペプチドである。このようなアミロイド形成性タンパク質は、体液または区画中において凝集型で見出されている。周知の例は、血清アミロイドA(sAA)、いわゆるb-A4、またはAb(アルツハイマー病の脳で特徴的なアミロイド沈着を形成することが知られている42または43個のアミノ酸のポリペプチド)、いわゆるプリオンタンパク質(PrPsc型は、BSEまたはクロイツフェルトヤコブ病における凝集において蓄積する)、およびHIV-1gp41のようなレトロウイルス表面糖タンパク質(HAD(HIV関連痴呆)に罹患している患者の脳におけるアミロイド様プラークにおいて見出される)である。好ましい実施形態において、シャペロン、好ましくはPPIシャペロンを用いて、アミロイド形成性タンパク質、およびシャペロンを含む可溶性複合体を形成する。極めて有利になるように、このような複合体を、多くの異なるイムノアッセイ手順において用いてもよい。好ましくは、このような複合体を、二重抗原架橋コンセプトに従って、イムノアッセイにおいて用いる。
【0050】
HIVおよび他のエンベロープウイルス(例えば、HTLV、インフルエンザウイルス、およびエボラウイルス)は、全てが、表面糖タンパク質を発現し、これが細胞付着および膜融合の両方を媒介する。これらの機能を果たすために、これらの表面糖タンパク質の全てが、極端に疎水性のセグメントを含み、これがインビトロでそれらを取り扱うことを困難にさせ、凝集しやすくし、それらを、インビトロの再折り畳みの労力に関して困難な標的にさせる。さらに好ましい実施形態において、本発明は、PPIシャペロン、およびエンベロープウイルスの表面糖タンパク質を含む可溶性複合体に関する。本発明は、PPIシャペロン、およびエンベロープウイルスの表面糖タンパク質を含む複合体を、この表面糖タンパク質に対する抗体の検出のためのイムノアッセイにおいて用いることに特に関する。
【0051】
HADは、HIV感染の周知の合併症である。Caffrey, Mら(前出)によって記載されたように、組織学的な現象学に関して、HADは、クロイツフェルトヤコブ病と呼ばれる海綿状脳症に極めて類似している。クロイツフェルトヤコブ病の病因論は一般に、改変プリオンタンパク質を含むプラークのアミロイド形成性蓄積から生じると考えられる(Prusiner,S. B., Proc Natl Acad Sci USA 95(1998)13363-83)。E-gp41の高分子量凝集物に関与するHADについての類似の病因論は極めて確率が高い。HIV脳症における神経学的な病変が、ビンスヴァンガー病と、病理的特徴および放射線医学的特徴を共有することは注目に値する。
【0052】
本発明による好ましいアミロイド形成性タンパク質は、それぞれ、HIV-1由来のgp41、HIV-2由来のgp36、またはHTLV由来のgp21である。
【0053】
タンパク質は、20mMリン酸ナトリウム(pH7.4)150mM NaClからなるバッファーにおいて、そのタンパク質が、50nM以下の濃度で可溶性である場合に、「本質的に不溶性」であるとみなされる。
【0054】
PPIシャペロンおよび標的タンパク質を含む本発明の複合体は、生理的な緩衝条件下で、例えば、20mMリン酸ナトリウム、pH7.4、150mMNaClからなるバッファーにおいて、このPPIシャペロン複合体中に含まれるこの標的タンパク質が、100nM以上の濃度で可溶性である場合に、「可溶性」であるとみなされる。
【0055】
本発明者らは、バッファー中で標的タンパク質とシャペロンとを混合する工程(ここでは、タンパク質およびシャペロンの両方とも可溶性である)、およびこのバッファーを生理的条件に調整する工程(ここで、この形成されたタンパク質−シャペロン複合体は、可溶性のままである)を包含する方法を開発した。
【0056】
可溶性シャペロン−標的タンパク質複合体の生成は、可溶化バッファー条件から、すなわち、バッファー(ここでは、標的タンパク質およびシャペロンの両方が可溶性である)から開始する。「非生理的な(non-physiological)」、または「可溶化(solubilizing)」バッファーと名付けられ得る、適切なバッファーは、標的タンパク質およびPPIシャペロンの両方が変性されないか、または少なくとも不可逆的に変性されないという要件を満たさなければならない。このような緩衝条件から開始すれば、シャペロンは、標的タンパク質に結合し、そして非生理的な条件から生理的な条件への緩衝条件の変化は、標的タンパク質の沈殿なしに可能である。
【0057】
適切な(非生理的な)バッファー、すなわち、バッファー(ここで、本質的に不溶性である標的タンパク質、およびPPIシャペロンの両方が、可溶性である)は、高いpHもしくは低いpH、または高いカオトロピック塩濃度またはそれらの組み合わせのいずれかを利用する。
【0058】
PPIシャペロンおよび標的タンパク質(これは、本質的に不溶性である)を含む分子内複合体の産生の場合、非生理的なバッファーは、好ましくは、かなり高いpH、またはかなり低いpHを有するバッファーである。好ましくは、このようなバッファーは、高いpH範囲において9〜12のpH、または低いpH範囲においては2〜4.5のpHを有する。
【0059】
PPIシャペロンおよび標的タンパク質(これは、本質的に不溶性である)を含む分子内複合体の産生の場合、可溶化バッファーは、好ましくは、約6のpHにおいて、かなり高い濃度のカオトロピック塩、例えば、6.0M塩化グアニジウムを有するバッファーである。再生の際、標的タンパク質は、その未変性様の構造および分子内複合体型を仮定する。
【0060】
本発明の状況において、生理的なバッファー条件は、必要に応じてこのバッファー中に存在し得る他の非塩成分(例えば、糖、アルコール、界面活性剤)にかかわらず、このような添加物が標的タンパク質およびシャペロンを含む複合体の溶解度を損なわない限り、5.0〜8.5の間のpH値、および総塩濃度500mM未満によって規定される。
【0061】
さらなる好ましい実施形態において、本発明は、可溶性のレトロウイルス表面糖タンパク質−シャペロン複合体を生成する方法に関しており、この方法は以下:バッファー中でレトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼを混合する工程(ここで、レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼの両方は、可溶性であり、複合体を形成する)、ならびにこのバッファーを生理的な条件に調節する工程(ここで、この複合体は可溶性である)を包含する。
【0062】
「レトロウイルス表面糖タンパク質」、または「rsgp」という用語は、本発明において用いる場合、HIV-1のgp41およびHIV-2のgp36、ならびに他の哺乳動物免疫不全ウイルス由来の対応するエンベロープ糖タンパク質を含むはずである。好ましいレトロウイルス表面糖タンパク質は、HIV-1由来のgp41、HIV-2由来のgp36、およびHTLVのgp21である。特に好ましいrsgpは、HIV-1のgp41、およびHIV-2のgp36である。rsgpという用語は、ここで概説されるようにまた、天然に存在するresp、および合成的に操作されたrsgpの改変体を含む。
【0063】
それぞれgp41またはgp36のNヘリックス状ドメイン内のアミノ酸の特定の十分規定された置換が、それぞれgp41またはgp39の野生型配列を有するポリペプチドに比べて、これらの分子の全体的特性においてさらなる利点を生じることが見出されている。これらの改変体は、本発明に従って好ましい実施形態を示す。特に、Leu555、Leu566、Ile573、およびIle580の位置(ここで、これらの位置は、HIV-1gp41野生型配列(配列番号1)から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された1つ以上の位置において、少なくとも1つのアミノ酸置換、および多くとも4つのアミノ酸置換を含み、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられる、HIV-1gp41の改変体、あるいは、Leu554、Leu565、およびVal579の位置(ここで、これらの位置は、HIV-2 gp36野生型配列(配列番号2)から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された位置において、少なくとも1つのアミノ酸置換、および多くとも3つのアミノ酸置換を含み、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられるHIV-2gp36の改変体が、当該分野で公知である問題を少なくとも部分的に解決するために適切である。
【0064】
それぞれgp41またはgp36の新規な改変体は、凝集する傾向が低いほど、野生型配列のそれらの対応するポリペプチドに比べて、溶解度が向上し、取り扱いがより簡単になる。生理的緩衝条件下で、gp41またはgp36を可溶型で含む試薬を提供するための労力が一旦払われれば、溶解度の改善は、特に明白になる。ペプチジル−プロリル−イソメラーゼ(PPI)クラスのシャペロンから選択されたシャペロンによって付与された効果を有する新規な改変体の好適な特性を合わせることが特に有利であると証明されている。従って、本発明は、この改変体およびペプチジル−プロリル−イソメラーゼクラスのシャペロンのシャペロンを含む可溶性複合体の生成における、本発明において記載されたようなgp41の改変体、および/またはgp36の改変体の使用にさらに関する。
【0065】
改変HIV糖タンパク質、およびPPIクラスのシャペロンを含む可溶性複合体は好ましくは、それぞれ、改変HIVgp41、またはHIV-2 gp36、ならびにPPIクラスシャペロンの両方を含む単一の組み換えタンパク質から得られる。従って、好ましい実施形態は、本発明に記載されるようなHIV-1gp41、またはHIV-2 gp36の改変体、およびペプチジル−プロリル−イソメラーゼクラスのシャペロンから選択されたシャペロンを含む組み換えタンパク質である。
【0066】
gp41、またはgp36の新規な改変体が、野生型ポリペプチドよりも、取り扱いが容易であるという事実によって、それらの改変体は、免疫原としての使用、または抗原としての使用のような種々の目的に理想的にされる。好ましい実施形態において、本発明は、イムノアッセイにおける、本発明によるgp41、および/もしくはgp36の改変体の、またはPPIシャペロンおよびこのような改変体を含む複合体の、例えば、単一の組み換えタンパク質としての使用に関する。最も興味深いことには、レトロウイルス表面糖タンパク質、およびPPIシャペロンの両方を含む融合タンパク質は、適切な条件下で可溶化かつ再生され得、そしてHIVイムノアッセイにおいて簡便な標識および信頼性のある検出を可能にする可溶性の分子内rsgp-シャペロン複合体を形成することが見出されている。
【0067】
レトロウイルス表面糖タンパク質と、シャペロンとの間の可溶性複合体は、二重抗原架橋コンセプトに従って、抗体の検出のためのイムノアッセイにおいて非常に有利になるように用いることができる。
【0068】
HIV-1由来のgp41、またはHIV-2由来のgp36を含むrsgpシャペロン複合体は、感染の早期段階におけるHIVに対する抗体の検出に特に有利である。可溶性のシャペロン−gp41複合体、または可溶性のシャペロン−gp36複合体を用いれば、好ましくは架橋コンセプトに従って、イムノアッセイを実施することが可能であり、これによって、体液サンプル中におけるHIVに対する抗体の鋭敏かつ早期の検出が可能になる。
【0069】
シャペロンが、他の不溶性タンパク質と複合体を形成し得るという事実をまた用いて、イムノアッセイ(好ましくは、架橋コンセプトに従うイムノアッセイ)をさらに大幅に改善するという、大きい利点をえることができる。この架橋コンセプトによって、第一の抗原としてのシャペロン−抗原複合体(ほとんどが固相側でのいわゆる捕捉抗原)、および第二のシャペロン−抗原複合体(ほとんどが、検出側での検出抗原)の使用が可能になる。シャペロン反応性抗体の結合によって生じたバックグラウンド反応の問題を最小化するために、固相側について第一のシャペロン、そして異なる種由来の検出側について第二のシャペロンを利用することによって、このような架橋アッセイをさらに有利に改変することができる。
【0070】
ここで、標識されたシャペロン−抗原複合体を使用する、架橋コンセプトに従うイムノアッセイを実施することが可能である。シャペロン−抗原複合体(ここでは、このシャペロンのみが標識されており、これによって、この抗原は、このような標識によって改変もされず、負の影響もされない(例えば、高次構造に関して)ことが確実にされる)を生成することがまた可能である。
【0071】
化学的カップリングの様式およびストラテジーは、必要に応じて選択され得る。ポリペプチドの場合、-SH、-NH2、または-COO-残基、ならびにチロシンのOH基、ヒスチジンのイミダゾール基、またはトリプトファンの複素環式イミノ基を標的するカップリング化学は、すぐ手が届くものである。いくつかの適切なカップリング化学が、これらの官能基の各々について公知である(Aslam,M、およびDent, A.,前出)。慣用的なタンパク質カップリング化学は、作用する緩衝条件下、例えば、約5〜8.5のpH範囲内でタンパク質が可溶であることを必要とする。例えば、gp41が、このpH範囲内では、例えばSDSによって変性されない限り可溶ではないので、未変性様に折り畳まれたgp41は、これまでは、化学的カップリングを受けやすくはなかった。本明細書において本発明者らが記載する、gp41−シャペロン複合体は、用いられる検出様式にかかわらず、イムノアッセイのための可溶性の標識されたHIVエンベロープタンパク質を生成する、都合のよい手段を提供する。
【0072】
好ましい実施形態において、本発明は、可溶性のrsgp−シャペロン複合体の生成のためのプロセスに関し、このプロセスは、可溶化されたレトロウイルス表面糖タンパク質、およびペプチジルプロリルイソメラーゼクラスから選択されたシャペロンを、非生理的緩衝条件下で混合する工程、およびその後にこのバッファーを生理的条件に調整し、それによって分子内複合体を形成する工程を包含する。
【0073】
シャペロンおよびレトロウイルス表面糖タンパク質は、別々のポリペプチドとして用いることができるだけではない。本発明者らは、驚くべきことに、両方のタンパク質を共有結合させることが有利であるということを観察した。このような共有結合は、従来の架橋手順によって可能である;しかし、好ましくは、共有結合は、レトロウイルス表面糖タンパク質およびシャペロンを含む組み換えポリペプチドを産生することによって達成される。
【0074】
さらに好ましい実施形態において、本発明は、可溶性のrsgp-シャペロン複合体の生成のためのプロセスに関する。このプロセスは、適切な緩衝条件下で、共有結合したレトロウイルス表面糖タンパク質、およびペプチジルイソメラーゼクラスから選択されたシャペロンタンパク質を含むタンパク質を可溶化する工程、そしてその後にこのバッファーを生理的条件に調節する工程を包含する。これによよって、分子内複合体が得られる。
【0075】
本発明は、ペプチジルプロリルシス/トランスイソメラーゼ(PPI)と命名された折り畳みヘルパーのクラスに由来するシャペロンの使用を教示する(Dartigalongue,C.、およびRaina(前出)を参照のこと)。このファミリーの周知の例は、以下のように呼ばれるメンバーである:CypA、PpiD(Dartigalongue,C.およびRaina, S., Embo J 17(1998)3968-80;Schmid, F. X., Molecular chaperones inthe life cycle of proteins(1998)361-389, 編集A. L. Fink、およびY. Goto, Marcel DeckerIn., New York),FkpA(Danese, P. N.ら., Genes Dev 9(1995)387-98)、および誘発因子(Crooke,E.、およびWickner, W., Proc Natl Acad Sci USA 84(1987)5216-20;Stoller, G.ら、Embo J14(1995)4939-48)。このペプチジルプロリルイソメラーゼは、以下の3つのファミリーに小分割される:パルビュリネス(parvulines)ファミリー(Schmid,F. X.(前出),Rahfeld, J. U.ら、FEBS Lett 352(1994)180-4)、サイクロフィリンファミリー(Fischer, Gら、Nature337(1989)476-8)、およびFKBPファミリー(Lane, W. S.ら、J Protein Chem 10(1991)151-160)。FKBPファミリーは、興味深い生化学的特徴を示す。なぜなら、そのメンバーは、マクロライド(例えば、FK506、およびラパマイシン)に結合する能力によってもともと同定されたからである(Kay,J. E., Biochem J 314(1996)361-85)。
【0076】
プロリルイソメラーゼは、異なる機能の異なるサブユニットまたはモジュール、例えば、触媒活性を示すモジュール、およびシャペロンもしくは結合活性を示すモジュールを含んでもよい。FKBPファミリーのこのようなモジュールメンバーは、FkpA(Ramm, K.およびPluckthun, A., J Biol.Chem275(2000)17106-13)、SlyD(Hottenrott, S.ら、J. Biol Chem 272(1997)15697-701)、および誘発因子(Scholz,Cら、Embo J 16(1997)54-8)である。好ましい実施形態において、本発明は、可溶性複合体であって、レトロウイルス表面糖タンパク質、およびペプチジルプロリルイソメラーゼクラスの折り畳み触媒から選択されたシャペロンを含む複合体に関する。
【0077】
当然ながら、本発明は、ペプチジルプロリルイソメラーゼクラスの特別に言及されたメンバーの使用に対して制限されるものではなく、また、同じクラスから生じるが、異なる種の細菌から派生するシャペロンを用いて実施することもできる。PPIクラスのシャペロンのFKBPファミリーの好ましいメンバーが用いられる。
【0078】
さらなる実施形態において、真核生物由来の相同体を使用することが好ましく、そしてヒト起源のPPIaseを使用することが極めて好ましい。なぜなら、これらのPPIaseは、ヒト血清由来の抗体によって認識されないはずであり、従って、血清学的アッセイ(すなわち、ヒト抗体の検出に基づくアッセイ)においては妨害しないはずであるからである。
【0079】
分子シャペロンの完全な配列を常に使用する必要はないということがまた周知でありかつ理解される。必要な能力および機能を依然として保有するシャペロンの機能的断片(いわゆる、モジュール)がまた、用いられ得る(WO98/13496を参照のこと)。
【0080】
例えば、FkpAは、細菌のサイトゾルにおける不活性前駆体分子として合成されて、細胞質膜を横切って転位される、周辺質PPIである。FkpAの活性型(成熟FkpA、または周辺質FkpA)は、シグナル配列(アミノ酸1〜25)を欠き、従って前駆体分子のアミノ酸26〜270を含む。FkpAに関する関連の配列情報は、登録番号P45523として、公的なデータベースから、例えば、「SWISS-PROT」から容易に入手可能である。
【0081】
FlpAに密接に関連した、いわゆるSlyDは、触媒性およびシャペロン機能を担う構造化されたN末端ドメインから、そしてほとんど構造化されていないC末端(例外的にヒスチジン残基およびシステイン残基に富む)からなる(Hottenrott,S.ら、J. Biol Chem 272(1997)15697-701)。本発明者らは、アミノ酸1-165を含むSlyDのC末端短縮改変体が、gp41、およびgp36においてその可溶性機能を効率的に発揮することを見出した。野生型SlyDにおいてとは異なり、ジスルフィドシャッフリングを損なう危険は、用いられた短縮型SlyD改変体(1-165)においては、首尾よく回避される。
【0082】
上記で考察されたシャペロンの改変体であって、1つまたはいくつかのアミノ酸置換または欠失を保有する改変体がまた、本発明に従うプロセスを実施するために用いられてもよい。
【0083】
別の供給源に由来する適切なシャペロン、およびシャペロンの適切な断片または変異体が、本実施例に記載のような手順を用いることによって容易に選択され得る。それらは、可溶性のrsgp-シャペロン複合体を生成するために、遊離型で、またはレトロウイルス表面糖タンパク質に共有結合されてのいずれかで用いられ得る。本発明による好ましい実施形態において、結合にコンピテントなPPIaseシャペロンは、レトロウイルス表面糖タンパク質に組み換え的に結合されて、細菌のサイトゾル中でこの遺伝子産物の高い発現を生じる。本発明において言及されるような結合コンピテント(bindingcompetent)なPPIaseは、その触媒性のPPIase活性にかかわらず、伸長したポリペプチド基質に対する結合(すなわち、基質結合、またはシャペロンモチーフ)を媒介する、少なくとも機能的な単位を含む。
【0084】
本発明者らは、また、PPIクラスの折り畳み触媒に属さないいくつかのシャペロンが、レトロウイルス表面糖タンパク質と可溶性の複合体を形成し得るということを観察した。従って、本発明に従うさらなる好ましい実施形態は、Skp(OmpHとしても公知;Missiakas,D.ら、Mol Microbiol 21(1996)871-84)と、レトロウイルス表面糖タンパク質との間の可溶性複合体である。しかし、さらに好ましい実施形態は、レトロウイルス表面糖タンパク質、およびGroEL、またはその部分を含む可溶性複合体である。Skpに対して相同であるシャペロンがまた用いられ得る。
【0085】
モジュラーPPIが、変性されたかまたは部分的に変性されたタンパク質に対して優先的に結合することは公知である(例えば、Scholzら、前出)。PPIaseは、ここで、タンパク質の折り畳みを触媒するだけでなく、このようなタンパク質と安定な複合体を形成し、それによって、溶解性が得られるという顕著な特性を有することが見出されている。驚くべきことに、研究されたPPIase(例えば、TF、SlyD、およびFkpA)は、未変性様の折り畳まれたレトロウイルス表面糖タンパク質に結合し、それによって例えば、可溶化する。本発明による「未変性様の(native-like)」、または「未変性様に折り畳まれた(native-likefolded)」gp41とは、遠UV-CDによって評価される場合、二次構造における高ヘリックス含量によって、および近UV-CDによって評価される場合、三次接触によって、その両方で特徴付けられる、これは、それぞれ図1Bおよび5において示されるとおり、代表的な「gp41-サイン(signature)」に反映される。さらに、本発明による、「未変性様の(native-like)」gp41のUVスペクトルは、320nmより高い波長で有意な吸収(これは、凝集物のような迷光粒子を暗示する)を示さない。
【0086】
モデル生体分子間、例えば、抗体と抗原との間の複合体形成について大量の情報がある(概説については、Braden, B. C.、およびPoljak, R. J., Faseb J 9(1995)9-16を参照のこと)。通常、複合体の形成、および解離は平行に生じ、この複合体および結合パートナーは、自由平衡において共存する。同様に、本発明に記載されるようなPPIシャペロンとアミロイド形成性タンパク質との間の複合体についても同じことが真であると考えられる。
【0087】
本発明に記載されるように、複合体の形成は、特に重要な特性である。なぜなら、PPIシャペロンとタンパク質(これは、例えば、生理的な緩衝条件下で、本質的に不溶性である)との間の複合体は、例えば、生理的緩衝条件下で容易に可溶であることが見出されているからである。生理的条件下で可溶性である抗原は、診断適用においては多くの利点を有する。それらは、例えば、標準物質として直接用いられてもよい。さらに、それらは、適切なマーカーに、または適切な結合基に複合体化されてもよい。
【0088】
上記で考察されたように、HIV-2由来のgp36は、類似の機能(すなわち、膜融合、およびウイルス侵入)を果たし、そしてHIV-1由来のgp41と同様の診断適合性である。レトロウイルス表面糖タンパク質のプロトタイプの例としてHIV-1のgp41を用いるこの適用において、多くの技術的問題が考察される。単に明確化のために、この考察および説明は、主にHIV-1のgp41に集中する。しかし、同様の考慮が、他のレトロウイルス表面糖タンパク質、特にHIV-2由来のgp36、およびHTLV由来のgp21にあてはまる。
【0089】
HIV-1、またはHIV-2の天然に存在する単離体は、もともと単離され、かつ記載されたアミノ酸配列の改変体を含み得ることが公知である。哺乳動物の免疫不全rsgpのこのような天然に存在する改変体、および合成的に操作された改変体はまた、本発明の範囲内である。
【0090】
本発明は、好ましい実施形態において、regp、またはヒト免疫不全ウイルスウイルス(HIV)の膜貫通糖タンパク質の改変体に関する。HIV-1gp41、またはHIV-2 gp36のNヘリックス状ドメインにおける特定のアミノ酸置換を含む改変体が開示される。
【0091】
NヘリックスおよびCヘリックス状ドメイン(ヘリックス対ヘリックスの接触に関与する)の両方のアミノ酸位置は、HIV-1gp41の文献から公知であり、そしてHIV-2相同性gp36に対して外挿され得る。これらの位置を突然変異させることは、特に、この改変体およびPPIシャペロンドメインを含む融合タンパク質の状況では、それぞれgp41、またはgp36の特性に影響することが見出されている。
【0092】
gp41ロイシンジッパーのヘリックス状ホイール突出部における「a」、および「d」のアミノ酸位置は、本発明による改変体を作成するための好ましい標的である。「a」位置におけるアミノ酸残基(Chan,D. Cら、Cell 89(1997)263-73による付番)は、Q552、I559、L566、I573、およびI580である;各々の「d」位置におけるアミノ酸残基は、I548、I555、Q562、T569、およびL576である。
【0093】
このジッパーモチーフのヘリックス状の完全性を損なうことなく、溶解度を改善するために、変異位置は、2つ以上のヘリックスターンによってお互いから離されることが好ましい。この必要条件は、例えば、保存的な「a」残基であるQ552、I559、L566、およびI573の置換によって、ならびに例えば、保存的な「d」残基であるI548、L555、Q562、およびT569の置換によって満たされる。言い換えれば、突然変異された残基は、少なくとも6つの野生型アミノ酸残基によってお互いから離され、従って正確に7個のモチーフが続く。置換位置がお互いから2つ以上のヘリックスターンによって離されるという前述の状況下で改変体内の「a」、および「d」残基の両方を変異することがまた可能である。
【0094】
同様に、HIV-2のgp36エクトドメインにおける変更は、驚くべきことに、SlyDまたはFkpAに対して融合された場合、容易に溶解する組み換えタンパク質を生じることが見出された。ここで、「a」位置は、Q551、V558、L565、T572、V579であり、そして「d」位置は、I547、L554、Q561、T568、およびL575である。
【0095】
好ましくは、それぞれ、HIV-1 gp41の位置Q552、I559、L566、I573、I580、I548、L555、Q562、T569、およびL576、HIV-2gp36の位置Q551、V558、L565、T572、V579、I547、L554、Q561、T568、およびL575を含む位置の群から選択される1〜6個のアミノ酸が、より小さいか、またはより親水性のアミノ酸によって置換される。
【0096】
好ましくは、置換対象のアミノ酸位置は、それぞれHIV-1のQ552、I559、L566、I573、およびI580からなる位置の群、ならびにHIV-2gp36のL554、Q561、T568、およびL575からなる位置の群より選択される。
【0097】
好ましい実施形態において、本発明は、HIV-1 gp41の改変体であって、Leu555、Leu566、Ile573、およびIle580の位置(ここで、これらの位置は、配列番号1に記載されるgp41野生型配列から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された位置において、少なくとも1つのアミノ酸置換、および多くとも4つのアミノ酸置換を含む改変体であって、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられる、改変体、に関する。
【0098】
本発明のこの好ましい実施形態は、gp41野生型配列の対応するポリペプチドに比べて、有意な改善を示す野生型gp41の改変体が提供され得るという驚くべき知見に基づく。本発明の改変体をもたらすアミノ酸置換は、Chan,D. C.ら、Cell 89(1997)263-73から公知であって、配列番号1に示されるような、アミノ酸組成およびgp41野生型配列の番号付けに基づいて記載される。
【0099】
明白に、本発明に記載されるアミノ酸置換基はまた、他の公知のgp41、およびまだ未同定のHIV-1単離物の内部の対応する配列位置でアミノ酸を置換するために用いられ得る。「ある位置に対応する(correspondingto a position)」という用語は、HIV-1単離体、およびその改変体が見出され得るか、または生成され得るが、これがさらなるアミノ酸を含んでいるか、またはアミノ酸を欠失しており、それに基づいて、配列番号1に対する配列アラインメントで、対応する配列位置または配列モチーフについての種々の絶対数が得られるということを示すために用いられる。
【0100】
配列番号1の野生型配列とのgp41配列のマルチアラインメントおよび比較を、GCGPackage Version 10.2(Genetics Computer Group,Inc)のPileUPプログラムを用いて実施する。PileUPは、Feng,D. F.Doolittle, R. F., J Mol Evol 25(1987)351-60のプログレッシブアラインメント法の簡便法を用いてマルチ配列アラインメントを作成し、そして同一、類似、または異なるアミノ酸残基についてのスコアリングマトリックスが適宜規定される。このプロセスは、2つの整列された配列のクラスターを生成する、2つのほとんど類似の配列のペアワイズアラインメントで開始する。次いで、このクラスターは、次の最も関連した配列に対して、または整列された配列のクラスターに対して整列され得る。配列の2つのクラスターは、2つの個々の配列のペアワイズアラインメントの単純な伸長によって整列され得る。最終アラインメントは、全ての配列が、最終ペアワイズアラインメントに含まれるまで、一連のプログレッシブ、ペアワイズアラインメント(漸増的に異なる配列およびクラスターを含む)によって達成される。従って、新規なHIV-1単離物のアミノ酸位置、または操作されたgp41のアミノ酸位置(野生型配列の555、566、573、および580の位置に相当する)は、容易に配置される。
【0101】
本発明によるHIV-1 gp41ポリペプチドの好ましい改変体は、その改変体が、555位にアミノ酸置換基を含むという点で特徴付けられ、ここでLeu555は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0102】
本発明によるHIV-1 gp41ポリペプチドのさらなる好ましい改変体は、その改変体が、566位にアミノ酸換基を含むという点で特徴付けられ、ここでLeu566は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0103】
本発明によるHIV-1 gp41ポリペプチドのさらなる好ましい改変体は、その改変体が、573位にアミノ酸換基を含むという点で特徴付けられ、ここでIle573は、セリンによって、またはトレオニンによって置換され、このセリンによる置換が、最も好ましい置換である。
【0104】
本発明によるHIV-1 gp41ポリペプチドのさらなる好ましい改変体は、その改変体が、580位にアミノ酸換基を含むという点で特徴付けられ、ここでIle580は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0105】
本発明はまた、HIV-2 gp36の改変体であって、Leu554、Leu565、およびVal579の位置(ここで、これらの位置は、HIV-2gp36野生型配列(配列番号2)から公知の位置であるか、またはそれから公知の位置に相当する)の群から選択された1つの位置において、少なくとも1つのアミノ酸置換、および多くとも3つのアミノ酸置換を含む改変体であって、その置換アミノ酸が、それぞれ、そして独立して、セリン、トレオニン、アスパラギン、グルタミン、アスパラギン酸、およびグルタミン酸からなる群より選択されるという点で特徴付けられる改変体、に関する。
【0106】
この番号付けは、Guyader, M.ら、Nature 326(1987)662-9によって公開された野生型配列(配列番号2)に従う。この配列から公知の位置に相当するgp36内のアミノ酸位置は、gp41について上記されたように決定される。
【0107】
本発明によるHIV-2 gp36ポリペプチドの好ましい改変体は、その改変体が、554位にアミノ酸置換基を含むという点で特徴付けられ、ここでLeu554は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0108】
本発明によるHIV-2 gp36ポリペプチドのさらなる好ましい改変体は、その改変体が、565位にアミノ酸換基を含むという点で特徴付けられ、ここでLeu565は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。
【0109】
本発明によるHIV-2 gp36ポリペプチドの好ましい改変体は、その改変体が、579位にアミノ酸換基を含むという点で特徴付けられ、ここでVal579は、アスパラギン酸によって、またはグルタミン酸によって置換され、このグルタミン酸による置換が、最も好ましい置換である。好ましい実施形態において、それぞれ改変体gp41、または改変体gp36は、上記のようにこのアミノ酸位置のうち2つで置換を含む。3つのアミノ酸置換を含む改変体がまた好ましい。なおさらなる好ましい実施形態において、改変体gp41は、上記に詳細に考察される4つのアミノ酸位置で置換を含む。
【0110】
好ましい実施形態において、それぞれ、gp41、またはgp36(すなわち、融合ペプチド、および膜貫通セグメントを欠くエクトドメイン)の完全な配列、または対応する哺乳動物免疫不全ウイルスエンベロープタンパク質(例えば、HTLV由来のgp21)の完全な配列を用いて、PPIシャペロンとの複合体を形成する。レトロウイルス表面糖タンパク質の断片(例えば、Luら(前出)によって記載されたHIV-1由来のgp41についての断片)を使用することもまた考えられる。このような断片は好ましくは、gp41の細胞外部分のC末端ヘリックス、およびN末端ヘリックスを含む。
【0111】
アミノ酸535位〜681位を含む診断上関連するgp41(Chan, D. C.ら、Cell89(1997)263-73による命名法)は、標準的な手順による組み換え技術によって生成され得る。Luら(前出)の図1に記載されるとおり、別の興味深いgp41分子は、gp160前駆体分子のアミノ酸540〜669にまたがる。
【0112】
レトロウイルス表面糖タンパク質についての代表的な例として、HIVの小エンベロープタンパク質は、取り扱いが非常に困難であり、かつ非常に異例な特性を示す。既に言及したとおり、e-gp41分子の最も重要な特徴の1つは、生理的な緩衝条件でのその不溶性である。組み換え生成されたgp41は、可溶性であり、かつpH3.0および低イオン強度において未変性様の構造を示す。しかし、このpHでさえ、バッファー中の塩濃度には敏感なままである。用いられる塩によって、100〜500mMを超える塩の存在下でgp41は、沈殿する。以下により詳細に記載されるとおり、これは、カオトロピック剤によって(再度)可溶化(変性型で)され得る。
【0113】
生理的な緩衝条件は、通常、動物の血漿または血清に見出される塩およびpHの条件に相当すると理解されており、そして約7.4のpH値および約150mMの塩濃度によって規定される。本発明によるrsgp-シャペロン複合体は、これらの緩衝条件下で容易に可溶である。そこに存在するrsgpは、免疫学的に活性であり、従って、未変性様の構造に仕向ける。適切な界面活性剤の非存在下で、または適切な界面活性剤による事前処理なしには、gp41は、生理的な緩衝条件(例えば、20mMリン酸ナトリウム、pH7.4、150mMNaCl)下で、本質的に不溶性であるが、本発明によって記載された複合体は、適切なプロトコールに従った再折り畳み後に、容易に可溶性である。gp41エクトドメインは、本発明の複合体に含まれる場合、少なくとも100nMの濃度、好ましくは、1μM以上の濃度、最も好ましくは、10μM以上の濃度で可溶性である。従って、溶解度は、ナノモル未満の濃度から約マイクロモル濃度へ実質的に増大する。
【0114】
本発明の範囲のさらに良好な理解のために、可溶化および再生のために適用された緩衝条件は、必要に応じてかつ適切に改変されてもよく、そして本発明の過度の限定と理解されるべきではなく、本発明は広範な範囲の緩衝条件にわたって首尾よく実行されるということを強調することが必要である。
【0115】
生理的バッファーの全体的塩濃度は、シャペロン−gp41複合体が解離されず、そしてgp41が溶液中に留まることに注意を払いさえすれば、重要ではない。好ましくは、生理的バッファーは、少なくとも10mMの緩衝系、および多くとも200mMを含む。バッファー構成成分の残りは、もしあれば、有意な緩衝能力のない塩、例えば塩化ナトリウムであり得る。この生理的バッファーは、好ましくは、20〜500mM、より好ましくは50〜300mM、そして最も好ましくは100〜200mMの塩濃度を有する。
【0116】
本発明によるプロセスにおいて、この生理的なバッファーは、5.0〜8.5の範囲のpH値を有するように変化されてもよい;より好ましくは、このようなバッファーの範囲は、pH5.5〜pH8.3である。なおより好ましくは、このような生理的バッファー条件は、上記で示されるような塩濃度、および6.0〜8.0のpH値によって規定される;最も好ましくは、このような生理的バッファーのpHは、6.5〜7.8である。
【0117】
本発明において記載されるプロセスに従って、レトロウイルス表面糖タンパク質は、非生理的な緩衝条件下で可溶化され、シャペロンが添加され(または、共有結合したさらなるタンパク質ドメインとして既に存在する)、そして可溶化されたレトロウイルス表面糖タンパク質およびシャペロンを含む混合物が次に、生理的緩衝条件に調節される。レトロウイルス表面糖タンパク質単独では、そうされれば、自然に沈殿するが、それは、上記のプロセスでは驚くべきことに溶液中にとどまる。この重要な知見は、レトロウイルス表面糖タンパク質と、シャペロンとの間の複合体の形成に起因する可能性が最も高い。
【0118】
E. coliにおけるgp41の組み換え生成の場合、組み換え生成されたgp41は、封入体の形態において得られる。この材料は、高度にカオトロピックな試薬、例えば、7.0Mグアニジウムチオシアネートを用いて可溶化される。gp41ポリペプチドはこれらの条件下でほとんど構造化されていない。適切な工程においてこのバッファーを30mMギ酸(pH3.0)に変えることによって、溶液中のgp41は、その未変性様の全ヘリックス状構造として認知されるものの形態になる。タンパク質の正しい折り畳みまたは不正確な折り畳みの状態をモニターするための1つの簡易な方法は、アミド領域(185-250nm)、および芳香族領域(260-320nm)における対応するCDスペクトルを分析することである。それに加えて、迷光粒子(凝集物のような)についての情報は、標準的なUVスペクトルから容易に得られ得る。
【0119】
ここで強調するのが重要なことは、本発明によるレトロウイルス表面糖タンパク質−シャペロン複合体内のレトロウイルス表面糖タンパク質は、未変性様折り畳みと考えられるものの形態をとるという事実である。これに反して、レトロウイルス表面糖タンパク質(カオトロピック剤によって天然のpHで可溶化されている)は、ほとんど構造化されておらず、従って規則正しい高次構造(コンフォメーション)エピトープを失っている。代替的に界面活性剤を用いることによって、レトロウイルス表面糖タンパク質を可溶化することがまた可能である。例えば、ドデシル硫酸ナトリウム(SDS)は、gp41を可溶化するために首尾よく用いられている。しかし、このような「SDS−可溶化材料(SDS-solubilizedmaterial)」は、例えば、gp41に対する抗体の検出のためのイムノアッセイにおける使用のための、選り抜きの材料ではない。さらに、(上記で考察したように)、このようなイムノアッセイは、好ましくはまた、gp41の高次構造(コンフォメーション)エピトープに対する抗体を検出する。そして界面活性剤が、高次構造(コンフォメーション)エピトープを部分的に消滅させることは排除できない。
【0120】
好ましくは、本発明によるrsgp-シャペロン複合体は、未変性様折り畳みされているrsgpによって特徴づけられる。このような複合体内の、この未変性様折り畳みされたrsgpは、例えば、必要な免疫学的または物理的特徴を示す。
【0121】
未変性様折り畳みを、好ましくは、近UV CD分光法によって評価するが、この方法は、緻密な球状タンパク質内の三次接触について報告する。gp41は、約3.0のpHで、そして低イオン強度の塩濃度で容易に可溶性であることが公知である。近UVCDデータによって、このようなバッファー条件下では、gp41は、未変性様折り畳みされた球状タンパク質の代表的なサインをともなう特徴的な楕円率シグナルを示すことが実証される。図5に示されるとおり、gp41、およびFkpAを含む融合ペプチドのgp41部分は、酸性バッファー中のこの代表的な近UVCDスペクトルを示す。生理的緩衝条件下で、本発明による可溶性複合体の近UV CDスペクトルは、正確に折り畳まれたシャペロンのスペクトルと、未変性様折り畳みされたgp41のスペクトルの両方から構成される。これは、図7のFkpA-gp41融合タンパク質について示される。
【0122】
本発明による好ましい実施形態において、gp41-シャペロン複合体中の未変性様折り畳みを、近UVCDを分析することによって評価する。この近UV CDは、分子gp41、およびシャペロンの両方が未変性様折り畳みされていることを実証するために用いられることが好ましい。
【0123】
可溶性シャペロン−gp41複合体の生成は、非生理的緩衝条件から開始する。遊離のシャペロンと遊離の標的タンパク質(例えば、HIV-1由来のgp41)との間の複合体形成の場合、「非生理的」バッファーは、以下の2つの要件を満たさなければならない:(a)gp41が、未変性様の酸性構造で存在すること、および(b)PPIシャペロンが、少なくとも部分的に機能的(すなわち結合コンピテント)であること。このような緩衝条件から開始して、シャペロンは、アミロイド形成性タンパク質に結合し、そして非生理的条件から、幾分生理的な条件までの緩衝条件の変化が、アミロイド形成性タンパク質の沈殿なしに、可能である。
【0124】
シャペロンは、通常変性されたタンパク質に結合して、それらに作用し、それによってそれらの正確な(再)折り畳みを容易にするが、本発明が基づく状況は著しく異なる。適切な非生理的緩衝条件下で可溶化されたgp41は、未変性様型で存在すると考えられる(図1A、および1B、ならびに図5を参照のこと)。シャペロン機能の慣習的な見解とは異なり、本発明の方法では、シャペロンは、未変性様折り畳みタンパク質に結合して、緩衝条件(この緩衝条件下ではgp41はさもなければ不溶性であって沈殿する)において、このタンパク質を安定化するようである。
【0125】
本発明による好ましい実施形態において、PPIシャペロンは、FkpA、SlyD、および誘発因子からなる群より選択される。
【0126】
特にFkpA、またはSlyDがgp41の溶解度を改善して、それとともにかなり安定な複合体を形成することが見出されている。従って、さらに好ましい実施形態は、このシャペロンが、FkpA、およびSlyDを含む群から選択されるという点で特徴付けられる。最も好ましいのは、シャペロンFkpAである。
【0127】
さらに上記で記載したように、またシャペロンの断片は、所望の機能をもたらすために用いられ得る。モジュールシャペロンの場合、FKBPと同様に、触媒性モジュール、および結合モジュールを含み、このような断片は、結合ドメインを少なくとも含むこと、またはこれらの断片が、本質的にこの結合ドメインに匹敵する機能を少なくとも示すことが好ましい。
【0128】
FKBP12は、FKBPファミリーのヒトメンバーであり、そしてPPIaseの触媒性イソメラーゼドメインから本質的になる。FKBP12は、さらなるポリペプチド結合ドメインを欠くので、これは、FKBPファミリーの他のメンバーに比べた場合、折り畳まれていないか、または部分的に折り畳まれたタンパク質基質に対して、有意に低下した結合親和性を示す。FKBP12の変性および再折り畳みは、可逆プロセスであることが示されている(Egan,D. A.ら、Biochemistry 32(1993)1920-7;Scholz, C.ら、J Biol Chem 271(1996)12703-7)。本発明者らは、FkpA(25-270)、およびSlyD(1-165)の再折り畳み、および変性が、いずれも可逆であり、従ってここに記載されたプロセスの中心的な必要条件を満たすということを見出す。
【0129】
好ましい実施形態において、本発明は、gp41、および(FKBPファミリーから選択された)シャペロンを含む可溶性複合体に関する。
【0130】
上記のように、このような可溶性複合体であって、HIV-1由来のgp41、または別の哺乳動物免疫不全ウイルス由来の相同体を含む複合体は、PPIシャペロン(例えば、組み換え技術によって生成された)、および組み換え生成されたgp41を混合することによって容易に調製され得る。次いで、この複合体は、2つの独立した分子の間で、すなわち、分子間で形成される。
【0131】
複合体形成は、動的なプロセスであって、ここで解離および再会合が並行して生じる。これは、例えば、FkpAとgp41との間の、分子間、および分子内(例えば、融合構築物内)の会合の両方について真である。gp41は、生理的バッファーから直ちに、かつ定量的に沈殿するので、両方のパートナーの濃度を、選択して、これによって非臨界濃度、または非凝集濃度の遊離型gp41しか存在しないこと、および大量のgp41がgp41−シャペロン複合体の型で結合されかつ安定化されることを確実にしなければならない。
【0132】
用いたシャペロンによって、gp41分子に比較して、モル基準で、少なくとも2倍多いシャペロンを混合することが必要であることが見出されている。好ましい実施形態においては、従って、本発明は、gp41、およびシャペロン(好ましくはFkpA)の混合物を含む試薬に関する。好ましくは、このような混合物は、gp41に比べた場合、FkpAをモル過剰で含む。FkpAより3〜10倍存在することが好ましい。FkpA対gp41の最も好ましいモル比は、4〜6である。
【0133】
例えば、共有結合した、少なくとも1つのrsgpドメインおよび少なくとも1つのPPI-シャペロンドメインを含むタンパク質の異なるドメインの間の分子内複合体の形成が、例えば、安定性および生成の容易さに関して、さらなる有利な効果をもたらすということがまた見出されている。例えば、両方のドメインが共有結合されている場合、1:1(rsgp対シャペロン)の比が、可溶性複合体を形成するのに十分であることが見出されている。
【0134】
レトロウイルス表面糖タンパク質およびシャペロンを組み換え的に連結された形態で含む可溶性複合体は、本発明によるさらなる好ましい実施形態を表す。このような組み換えポリペプチドに含まれる最も好ましいrsgpは、HIV-1由来のgp41、およびHIV-2由来のgp36である。
【0135】
少なくとも1つのrsgpドメイン、および少なくとも1つのPPIシャペロンドメインを含む組み換えタンパク質について、非生理的緩衝条件から、生理的緩衝条件への移行は、異なる方法で達成され得る。gp41とFkpAとの間の可溶性分子内複合体は、非生理的緩衝条件を、透析、急速希釈、またはマトリックス補助再折り畳みによって生理的緩衝条件に調節することによって容易に得られる。可溶性gp41シャペロン複合体を含む混合物は、改変のために直接用いられ得る。
【0136】
本発明による可溶性複合体(例えば、gp41およびPPIシャペロンを含む)はまた、組み換え技術によって得られた両方のタンパク質ドメイン(gp41、およびシャペロン)を含む1つのポリペプチドから開始して生成されてもよい。gp41−シャペロン複合体は、その点で、事実上、分子内である。好ましくは、本発明による組み換えポリペプチドは、gp41とシャペロン、またはgp36とシャペロンを含む。なおさらに好ましい実施形態では、本発明は、組み換えタンパク質であって、少なくとも1つのrsgpドメイン、および少なくとも2つのPPI−シャペロンドメインを含む組み換えタンパク質に関する。1つのrsgpドメイン、および2つのPPIシャペロンを含む組み換えポリペプチドがまた好ましい。
【0137】
標準的な分子生理的技術を適用して、本発明による可溶性gp41−シャペロン複合体を得るために用いた組み換えポリペプチドを発現する。好ましくは、このシャペロン遺伝子は、発現ベクター(これは、gp41およびシャペロンの両方についての遺伝情報、ならびに必要に応じて、適切なペプチド性リンカー配列についての遺伝情報も含む)中に、標的タンパク質遺伝子の上流にインフレームで位置する。このような組み換え融合タンパク質の大規模な生成のために好ましい宿主は、E.coliである。
【0138】
好ましい実施形態において、本発明は、可溶性複合体であって、それぞれgp41、またはgp36、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されたシャペロンを含む複合体に関する。この可溶性複合体が、分子内複合体、であって、好ましくは、gp41、またはgp36、およびPPIシャペロンを含む組み換えポリペプチド内の分子内複合体であることがなお、さらに好ましい。最も好ましくは、組み換えポリペプチドのPPIシャペロン部分は、いずれの輸送シグナルペプチド(対応する前駆体分子の)も欠き、そして成熟PPIシャペロンに相当する。好ましい実施形態において、この組み換えタンパク質は、機能的なシグナル配列を欠くので、この遺伝子産物は、細菌のサイトゾル中に蓄積する。
【0139】
組換え産生されたFkpA-gp41に含まれるgp41の著しい特徴は、「非シャペロン(unchaperoned)」gp41エクトドメインに比較した場合の異例な溶解度である。「カオトロピック材料(chaotropicmaterial)」(すなわち、6.0-7.0M GuHCl中のFkpA-gp41)は、異なる方法で再折り畳みされ得、全てが熱力学的に安定、かつ可溶性の未変性様形態になることは興味深い。再折り畳みは、透析および急速希釈の両方によって、ならびに再生サイズ排除クロマトグラフィー、またはマトリックス補助再折り畳みによって高収率で達成される。これらの知見によって、この共有結合型において、gp41-FkpA融合ポリペプチドが、準安定なタンパク質よりも熱力学的にかなり安定であることが示唆される。
【0140】
組み換えFkpA-gp41ポリペプチドは、異なる折り畳み要件を有する2つのタンパク質ドメインを含む。精製プロトコールは、初期の変性工程を含むので、シャロンの折り畳みが可逆であることは必須である。実際、共有結合したタンパク質複合体内でのFkpAおよびgp41の両方の可逆的かつ独立した再折り畳みについて、説得力のある分光学的な証拠が存在する。C末端切断型SlyDの再折り畳みは、いずれにせよ可逆であることが見出されている。
【0141】
レトロウイルス表面糖タンパク質、およびシャペロンを含む組み換えポリペプチドであって、さらにこれらの2つのポリペプチドドメインの間に適切なペプチドリンカー配列を含む、ポリペプチドがまた好ましい。このようなペプチドリンカー配列は、用いられたrsgpおよびシャペロンドメインの最適の分子内会合を確実にするように選択される。好ましくは、このようなリンカー配列は、約20アミノ酸長であり、そして可変性(flexibility)および溶解度の両方を支持するアミノ酸(例えば、グリシン、およびセリンなど)を含む。好ましくは、このリンカーは、10〜50アミノ酸長である。より好ましくは、この長さは、12〜40アミノ酸であり、そして最も好ましくは、このリンカーは、15〜35アミノ酸を含む。Rsgpおよびシャペロンの両方とも、常に密接に近接している(例えば、適切なリンカーによって互いに支持される)。好ましい実施形態において、この組み換えポリペプチドは、可変性のリンカーを介してその標的タンパク質に連結された成熟FkpAを含む。このことによって、データが示すとおり、さらなる安定化効果がもたらされる。
【0142】
驚くべきことに、gp41は、PPIシャペロンとgp41との間の分子内複合体の一部として、可溶性であり、かつ安定であることが見出されている。同じことが、PPIシャペロンとgp36、またはPPIシャペロンとgp21(HTLV由来)を含む分子内複合体についても真実である。このような複合体におけるgp41の改善された安定性は、さらなる利点をもたらす。例えば、完全に再生された組み換えgp41−シャペロン分子を非常に簡単に獲得することが可能である。組み換えタンパク質は、カオトロピック剤(例えば、塩化グアニジウム)を用いた処理によって最初に可溶化される。可溶化された材料をゲル濾過カラム(適切な生理的バッファーを用いて平衡化された)に単に通過させることによって、共有結合したタンパク質ドメインを含む完全に再生されたタンパク質を得ることができる(実施例2.3、ならびに図7および8を参照)のこと)。
【0143】
可溶性分子内gp41−シャペロン複合体は、なお、さらなる顕著な利点を示す:これは、界面活性剤の変性作用に対してかなり安定である。この効果は、この融合タンパク質が、それぞれ2つのシャペロンおよび1つのgp41、または1つのgp36を含む場合、なおさらに明白になる。ほとんどのイムノアッセイは、非特異的な結合によって生じる問題を低減し、そして少なくとも部分的に回避するために、界面活性剤の存在下で実施される。HIV診断の場合、前述の理由のせいで、ただしさらにウイルス粒子を分解しかつ破壊して、これによってウイルス抗原(gp24など)の検出を容易にするために、かなり強力な界面活性剤が用いられる。
【0144】
SDS(ドデシル硫酸ナトリウム)によって可溶化された、組み換え生成されたgp41エクトドメインは、例えば、抗HIV抗体、またはp24抗原の検出において、慣用的に用いられるアッセイバッファーにおいて免疫反応性ではない(図9を参照のこと)。しかし、同じ緩衝条件下では、gp41(本発明による、PPIシャペロンとの分子内複合体の一部である)は、強力に免疫反応性である。図9から判明するとおり、同じアッセイ条件下で、かつ同じ患者血清を用いて、この材料は、未変性様可溶性gp41(さらに、試験された界面活性剤の存在下で安定である)の存在によってのみ説明され得る、優れた競合曲線を生じる。
【0145】
これは、本発明による、この複合体の非常に重要な特徴であって、この可溶性rsgp−シャペロン複合体内のrsgpは、生理的な緩衝条件、例えば、20mMリン酸、150mM塩化ナトリウムバッファー中で、pH7.4において、未変性性折り畳みされる。これは治療適用のため、そして診断適用のための大きな利点である。好ましい実施形態において、本発明は、生理的緩衝条件下で可溶である試薬の組成物であって、分子内複合体または分子間複合体(これは、レトロウイルス表面糖タンパク質、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されるシャペロンを含む)を含む、組成物に関する。
【0146】
従って、HIV-1由来の未変性様折り畳みgp41、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されるシャペロンを含む可溶性複合体は、本発明の非常に好ましい実施形態を表す。
【0147】
従って、HIV-2由来の未変性様折り畳みgp36、およびペプチジルプロリルイソメラーゼクラスのシャペロンから選択されるシャペロンを含む可溶性複合体はまた、本発明の非常に好ましい実施形態を表す。
【0148】
治療に関して、それぞれ、「可溶性、かつ未変性様折り畳みされた(soluble andnative-like folded)」gp41またはgp36を提供することによってなされる進歩は、かなり明白である。例えば、gp41は、生理的緩衝条件下で、初めて、この時、利用可能となる。
【0149】
好ましい実施形態において、記載されるような可溶性複合体は、医薬として用いるための試薬の組成物を生成するために用いられる。試薬の組成物は、生理的に許容され得る賦形剤、ならびに必要に応じて、適切な添加物、および/または従来の補助物質とともに、gp41-シャペロン複合体を含む。
【0150】
gp41の7残基反復か、またはgp41C末端ヘリックスに由来するペプチドが、抗ウイルス活性を保有することは公知である(Wild,C.ら、Proc Natl Acad Sci USA 89(1992)10537-41)。それらは、gp41のいわゆる「ヘアピン中間体(hairpin−intermediate)」と特異的に相互作用することによってウイルス侵入を邪魔する(概説については、Doms,R. W.、およびMoore, J. P., J Cell Biol 151(2000)F9-14を参照のこと)。本発明者らは、本発明によるrsgp-シャペロン複合体が抗ウイルス活性を示すことを見出した。gp41−シャペロン複合体、もしくはgp36−シャペロン複合体、または両方を、第一の好ましい治療適用において治療上有効な用量で含有する、試薬の組成物を用いて、宿主生物体内のHIV進入、およびHIV伝播を防止する(「ウイルス侵入阻害(virusentry inhibition)」)。
【0151】
これは、哺乳動物における免疫応答を誘発するための組成物などを使用するための、gp41−シャペロン複合体を含む試薬の組成物のさらなる好ましい治療適用を表す。記載された複合体は、公知の他のいずれのHIV免疫原よりも多いgp41エピトープを利用できる(例えば、Rootら(前出)を参照のこと)。従って、この新規な免疫原は、かなり広範な免疫応答を誘導することが期待される。
【0152】
診断手順に関して、本発明による可溶性rsgp−シャペロン複合体の明白な利点は、例えば、レトロウイルス表面糖タンパク質(例えば、生理的緩衝条件下でのgp41)の安定性の増大、および/または診断感度の増大、および/または高次構造(コンフォメーション)的なエピトープの存在数の増大、および/または正確に折り畳まれたrsgp(gp41など)を容易に標識する能力である。
【0153】
周知の標識は、固相結合基などの、マーカー基、またはエフェクター基である。標識された可溶性rsgp−シャペロン複合体は、本発明によるさらなる好ましい実施形態を表す。
【0154】
標識基は、以下から選択されてもよい:任意の公知の検出可能マーカー基(例えば、色素)、蛍光標識基(例えば、化学発光基)、例えば、アクリジニウムエステル、またはジオキセタン、または蛍光色素、例えば、フルオレセイン、クマリン、ローダミン、オキサジン、レゾルフィン、シアニン、およびそれらの誘導体。標識基の他の例は、蛍光金属錯体(例えば、ルテニウム、またはユーロピウム錯体)、例えば、ELISAのために、またはCEDIAのために用いられるような酵素(Cloned Enzyme Donor Immunoassay、例えば、EP-A-0061 888)、ならびに放射性同位体。
【0155】
エフェクター基は、例えば、バイオアフィン(bioaffine)結合対の1つのパートナーを含む。アッセイを実施する間、エフェクター基は、バイオアフィン結合対の他のパートナーと特異的に、好ましくは非共有結合的に相互作用する。適切な結合対の例は、以下である:ハプテン、または抗原/抗体、ビオチン、またはビオチン類似体(例えば、アミノビオチン)、イミノビオチン、またはデスチオビオチン/アビジン、またはストレプトアビジン、糖/レクチン、核酸、または核酸類似体/相補的核酸、およびレセプター/リガンド、例えば、ステロイドホルモンレセプター/ステロイドホルモン。好ましい結合対のメンバーは、ハプテン、抗原、およびホルモンを含む。特に好ましいのは、ハプテン様ジゴキシン、およびビオチン、ならびにそれらの類似体である。
【0156】
rsgpおよびPPIシャペロンを含む可溶性複合体は、好ましくはrsgpに対する抗体の検出のためのイムノアッセイにおいて用いられる。好ましくは、gp41−シャペロン複合体、および/またはgp36−シャペロン複合体が用いられる。非常に好ましい実施形態において、gp41およびPPIシャペロンを含む標識された可溶性複合体は、gp41に対する抗体の検出のためのイムノアッセイにおいて用いられる。最も好ましくは、標識された複合体は、PPIシャペロンおよびgp41を含む組み換えポリペプチド内の分子内複合体である。
【0157】
イムノアッセイは、当業者に周知である。このようなアッセイを実施するための方法、ならびに実際の適用および手順は、関連の教科書にまとめられている。関連の教科書の例は、「Practice and theory of enzyme immunoassays」(1990)221-278、編集、R. H.Burdon、およびv. P. H. Knippenberg, Elsevier, Amsterdam中の、Tijssen, P., Preparationof enzyme-antibody or other enzyme-macromolecule conjugates、および「Methods inenzymology」(1980)編集、S.P.Colowick, N. O. Caplan、およびS. P., Academic Press中のTijssenの種々の巻であって、免疫学的検出法を取り扱う、特に70、73、74、84、92、および121巻である。
【0158】
新規な可溶性rsgp−PPIシャペロン複合体を用いて、検出の様式(例えば、放射性同位体アッセイ、酵素イムノアッセイ、電気化学発光アッセイなど)、またはアッセイ原理(例えば、テストストリップアッセイ、サンドイッチアッセイ、またはホモジニアスアッセイなど)とは独立して、抗HIV抗体の検出のためのアッセイを改善し得る。
【0159】
HIV感染の確実で、かつ鋭敏な早期検出のために、体液サンプル中のウイルス抗原および抗ウイルス抗体の両方を測定することが必須である。本発明による可溶性複合体は、生理的緩衝条件において、抗gp41抗体、および/または抗gp36抗体の検出を可能にする。抗gp41抗体、および/または抗gp36抗体の検出は、このような併用型HIV検出システムの有益な部分である。従って、好ましい実施形態において、本発明は、HIV検出システムであって、gp41、および/またはgp36シャペロン複合体の使用に基づく抗gp41抗体、および/または抗gp36抗体の検出を包含するシステムに関する。最も好ましくは、このような複合体に基づく、抗gp41抗体、および/または抗gp36抗体の検出は、HIV抗原、好ましくはp24抗原の検出とともに実施される。
【0160】
先行技術から公知のとおり、細菌、真菌、またはウイルスのような感染因子に対する抗体は、好ましくは、二重抗原架橋コンセプトに従って実施されるアッセイによって検出される(時には、このアッセイコンセプト(double antigen bridge concept)はまた、二重抗原架橋コンセプトと名付けられる、なぜなら2つの抗原が抗体によって架橋されるからである)。このようなアッセイにおいては、抗体が、所定の抗原の少なくとも2つの異なる分子に、その2つ(IgG、IgA、IgE)、または10(IgM)のパラトープ(抗原結合部位)で結合する能力が必要とされ、かつ用いられる。
【0161】
この架橋コンセプトに従う体液からの抗体の検出は、多くの異なるアッセイ設定で実施され得る。簡易な設定は、固相に対する抗原の直接コーティング、および同じ抗原の標識型での使用を含む。適切なアッセイ条件下で、サンプル中の抗体は、固相結合抗原と標識抗原との間の架橋を形成する。従って、検討中の抗体がサンプル中に存在する場合のみ、架橋が形成されて、シグナルが検出され得る。
【0162】
「固相抗原(solid phase antigen)」、および「検出抗原(detectionantigen)」の基本的な構造は、好ましくは、同じである。例えば、1または数個のエピトープを含むポリペプチドを用いて、固相を直接または間接的にコーティングしてもよいが、標識またはマーカーに結合した同じ合成ポリペプチドが、検出抗原として用いられる。類似であるが、異なる抗原であって、二重抗原架橋アッセイにおいて免疫学的に交差反応性である抗原を使用することも可能である。このようなアッセイを実施するための必須の要件は、関連のエピトープ(単数または複数)が、両方の抗原上に存在することである。もちろん、二重抗原架橋アッセイ形式の多くの改変型が存在する。このような改変型は、例えば、固相に対する抗原の間接的なコーティングを含む。好ましくは、特定の結合対、最も好ましくは、ビオチン−ストレプトアビジン(またはビオチン−アビジン)系が、固相に対して抗原を間接的に結合するために用いられる。一方では、このようなシステムにおける検出のために用いられる抗原は、マーカー(例えば、放射性同位体、酵素、蛍光分子など)を直接有していなくてもよいが、それよりむしろ、例えば、ハプテン(例えば、ジゴキシン)を担持することによって、間接的に検出され得る。次いで、このような間接的な検出は、例えば、標識された抗ジゴキシン抗体によって実施され得る。
【0163】
好ましい実施形態において、本発明は、第一のシャペロン−抗原複合体を含む第一の抗原、および第二のシャペロン−抗原複合体を含む第二の抗原を含む、二重抗原架橋コンセプトに従うイムノアッセイに関する。
【0164】
さらなる好ましい実施形態において、本発明は、二重抗原架橋コンセプトに従うイムノアッセイであって、第一のシャペロン−抗原複合体が、捕捉抗原として用いられ、そして第二のシャペロン−抗原複合体が検出抗原として用いられることを特徴とするイムノアッセイに関する。
【0165】
本発明において記載されたようなシャペロン−抗原複合体は、種々のポリペプチド(さもなければ取り扱いが困難である)の溶解性をもたらすだけでなく、この二重抗原架橋コンセプトに従って、極めて有利なイムノアッセイを可能にする。
【0166】
この二重抗原架橋コンセプトによる、このようなイムノアッセイの特に魅力的な特徴は、それぞれ、固相結合抗原との複合体形成のため、および検出抗原との複合体形成のために、異なるシャペロンを用いることが可能となるということである。アッセイのこのような改変によって、非特異的結合の問題をさらに改善する。サンプル中の抗体(シャペロンに対して反応性であり、従って偽陽性のシグナルを生じ得る)は、異なるシャペロンを用いてそれぞれ、固相抗原、および検出抗原と複合体化する場合、架橋を形成しない。従って、本発明のこの様式においては、非特異的結合に起因する陽性シグナルの可能性は、大きく低下する。従って、好ましい実施形態において、本発明は、二重抗原架橋コンセプトに従うイムノアッセイであって、第一および第二のシャペロン−抗原複合体の第一シャペロンおよび第二シャペロンが、お互いに異なることを特徴とする、イムノアッセイに関する。
【0167】
最もよく特徴付けられているシャペロンのほとんどは、Escherichia coliから単離されており、生物工学の研究において広範に用いられる。Escherichiacoliは、広範に分布した細菌種であるため、多くの哺乳動物は、この細菌に由来するタンパク質に対する抗体を発現している。このような抗体によって生じる偽陽性反応の可能性を減少させるため、別個の細菌種(好ましくは好熱性の菌種)由来の少なくとも1つのPPIシャペロンを使用することが好ましい。好ましくは、このシャペロンは、特に、Thermatogamaritima、Aquifex aeolicus、およびThermus thermophilusを含む細菌の群の、好極限性細菌由来である。
【0168】
イムノアッセイ一般における、そして好ましくは架橋コンセプトに従うイムノアッセイにおけるシャペロン−抗原複合体の使用はまた、このような複合体のシャペロンを誘導体化する可能性を提供し、そして抗原自体の改変を必要としない。第二の化学部分によるポリペプチドの改変、例えば、その分子に対する標識のカップリングは、このポリペプチドにマイナスに影響するリスクを含むということが一般に認められる。例えば、検討中のエピトープが、損なわれ得るか、または非特異的な結合が、このような標識によって生じ得る。本発明に従って、シャペロン−抗原複合体内のシャペロンを特異的に誘導体化することが、ここで、可能である。
【0169】
好ましい実施形態において、二重抗原架橋コンセプトによるイムノアッセイは、捕捉抗原として用いられる第一のシャペロン−抗原複合体が、固相結合基を含むことをさらなる特徴とする。
【0170】
さらなる好ましい実施形態において、架橋コンセプトによるイムノアッセイが実施される。このイムノアッセイは、検出抗原として用いられる第二のシャペロン−抗原複合体がマーカー基を含むことをさらなる特徴とする。
【0171】
別の実施形態において、rsgpおよびPPIシャペロンを含む可溶性複合体(例えば、gp41−シャペロン複合体、および/またはgp36−シャペロン複合体)はまた、被験体(例えば、ヒトまたは非ヒト動物)における免疫応答を惹起するために用いられ得る。この可溶性複合体は、被験体に対して組成物(例えば、この組成物は、賦形剤、または担体を含んでもよい)として投与され得る。このような組成物はまた、アジュバントを含んでもよい。従来のアジュバントの例としては、フロイントの不完全アジュバント、フロイントの完全アジュバント、Merck65、AS-2、ミョウバン、リン酸アルミニウム、鉱物ゲル(例えば、水酸化アルミニウム)、および表面活性物質(例えばリゾレシチン、プルロニックポリオール類、ポリアニオン、ペプチド、油状エマルジョン、スカシガイヘモシアニン、およびジニトロフェノール)が挙げられるがこれらに限定されない。他の有用なアジュバントとしては、細菌莢膜多糖類、デキストラン、IL-12、GM-CSF、CD40リガンド、IFN-g、IL-1、IL-2、IL-3、IL-4、IL-10、IL-13、IL-18、または任意のサイトカインもしくは細菌のDNA断片が挙げられるがこれらに限定されない。
【0172】
1用量(投与)の可溶性複合体組成物が与えられてもよい。しかし、最初の投与後に、ブースト投与(例えば、1回、2回、3回、それ以上)が続けられてもよい。被験体に投与された用量の回数は、被験体の可溶性複合体組成物に対する応答性にある程度依存する。本発明の範囲内で、適切な投薬の回数としては、可溶性複合体に対して動物を免疫するために必要な任意の回数が挙げられる。
【0173】
適切な複合体組成物の第二の投与(ブースター)は、最初の投与後、約7日〜1年間の間に与えられ得る。最初の投与と2回目の投与との間の時間は、14日〜6ヶ月、21日〜3ヶ月、しばしば、最初の投与後、約28日〜2ヶ月の間であり得る。第3回目の投与(2回目のブースター)は、最初の投与後、約14日〜10年の間に、例えば、最初の投与後、約14日〜3年間の間に、しばしば、約21日〜1年の間に、非常にしばしば、最初の投与後、約28日〜6ヶ月の間に与えられてもよい。その後のブースターは、2週間間隔、または1ヶ月、3ヶ月、もしくは6ヶ月から10年間の間隔で投与されてもよい。
【0174】
典型的には、抗原に対して動物を免疫するのに十分である可溶性複合体の量(すなわち、「免疫学的に有効な用量」、または「治療上有効な用量」)が、被験体に投与される。「免疫学的に有効な量」を達成するのに十分な量は、被験体の体重および健康の全身的状態に、ならびに処方医または資格のある他の人員の判断に、ある程度依存する。
【0175】
可溶性複合体の有効用量は、免疫応答の誘導を達成するために動物モデルにおいて処方され得る;このようなデータを用いて、動物モデルに基づいてヒトに対する投与を容易に最適化し得る。用量は典型的には、約1fg〜約100μgの間、しばしば、約1pg〜約100μgの間、それよりしばしば、約1ng〜約50μgの間、そして通常、約100ng〜約50μgの間である。いくつかの実施形態において、用量は、被験体の体重1kgあたり、約1fg〜約100μgの間、しばしば被験体の体重1kgあたり、約1pg〜約100μgの間、それよりしばしば、約1ng〜約50μgの間、そして通常、約100ng〜約50μgの間である。
【0176】
本発明の可溶性複合体含有組成物は、種々の方法、および種々の形式で投与され得る。可溶性複合体組成物は、担体、および賦形剤(例えば、バッファー、炭水化物、マンニトール、タンパク質、ポリペプチド、またはアミノ酸(例えば、グリシン)、抗酸化剤、静菌剤、キレート剤、懸濁剤、増粘剤、および/または保存剤);水、油、生理食塩水、水性デキストロース、およびグリセロール溶液、適切な生理的条件に必要な他の薬学的に許容され得る補助物質(例えば、緩衝化剤、張度調節剤、湿潤剤)などを含んでもよい。従来のアジュバントもまた、この組成物中に組み込まれてもよい。
【0177】
任意の適切な担体が、本発明の組成物を投与するために用いられ得るが、担体のタイプは、投与の様式によって異なる。化合物はまた、リポソーム内にカプセル化され得る。例えば、Ticeら、米国特許5,942,252,1999に記載されるような生分解性マイクロスフェアは、ある場合には担体として都合がよい。
【0178】
従来の技術(例えば、濾過滅菌)によって達成されるような、この組成物の滅菌が望ましい。得られた水溶液は、そのまま用いるためにパッケージングされてもよいし、凍結乾燥されてもよい。
【0179】
本発明の可溶性複合体組成物は、種々の方法で投与され得る。この方法としては、注射(例えば、皮内、皮下、筋肉内、腹腔内など)による方法、吸入による方法、局所投与による方法、坐剤による方法、経皮パッチを用いることによる方法、または経口による方法が挙げられる。
【0180】
注射によって投与される場合、組成物は、水溶液中に、好ましくは生理的に適合するバッファー(例えば、ハンクス溶液、リンガー溶液、20mMリン酸 150mM塩化ナトリウムバッファー(pH7.4)、または生理的食塩水のバッファー)中に処方され得る。この溶液は、懸濁剤、安定化剤、および/または分散剤のような処方剤(formulatoryagent)を含んでもよい。あるいは、この組成物は、使用前の適切なビヒクル(例えば、滅菌発熱因子無含有水)との構成のために粉末型で存在してもよい。吸入送達型組成物は、適切な噴霧剤(例えば、ジクロロジフルオロメタン、トリクロロフルオロメタン、二酸化炭素、または他の適切なガス)の使用によって、圧縮パックまたはネブライザーからのエアロゾルスプレーとしてであってもよい。圧縮されたエアロゾルの場合、投薬量単位は、一定量を送達するためのバルブを備えることによって決定され得る。インヘイラーまたは吸入器(insufflator)における使用のための例えばゼラチンのカプセル、およびカートリッジが、処方されてもよく、これは、タンパク質および適切な粉末基剤(例えば、ラクトースまたはデンプン)の粉末混合物を含有する。局所投与のために、組成物は、当該分野で周知のとおり、溶液、ゲル、軟膏、クリーム、懸濁液などとして処方されてもよい。いくつかの実施形態では、投与は、経皮パッチによる。坐剤組成物はまた、従来の坐剤基剤を含むように処方されてもよい。
【0181】
投与が経口である場合、組成物は、この組成物と薬学的に許容され得る担体とを合わせることによって容易に処方され得る。固体担体としては、マンニトール、ラクトース、ステアリン酸マグネシウムなどが挙げられる;このような担体は、経口摂取用の、錠剤、丸剤、糖衣錠、カプセル、液体、ゲル、シロップ、スラリー、懸濁物などの形成を可能にする。このような処方物は、粉末、カプセル、および錠剤であってもよい;適切な賦形剤としては、糖、セルロース調製物、顆粒化剤、および結合剤のような増量剤(filler)が挙げられる。
【0182】
ポリクローナル抗体およびモノクローナル抗体(結合断片(例えば、F(ab)2)、および一本鎖型を含む)を生成するための方法は、周知である。しかし、多くの抗原は、十分な抗体応答を誘発できない。1つの実施形態において、本発明の可溶性複合体および抗原を含む組成物が、動物に投与され、これによって動物中の免疫応答が惹起される。ポリクローナル抗体、またはモノクローナル抗体が、標準的な技術によって引き続き調製される。
【0183】
rsgpおよびPPIシャペロンを含む可溶性複合体(例えば、gp41−シャペロン複合体、および/またはgp36−シャペロン複合体)はまた、膜融合を阻害することなどによって、細胞へのウイルス侵入を阻害するために用いられ得る。細胞は、インビトロであっても、インビボであっても、またはエキソビボであってもよい。組成物および投与の方法は、免疫応答を惹起するために用いられた組成物および方法について記載されたものと同様である。細胞へのウイルス侵入阻害が、ワクチン接種を用いて達成されるならば、アジュバントが用いられてもよい。インビトロおよびエキソビボ投与のために、当業者は、細胞、培養条件、および時間制限(もしあれば)に部分的に基づいて適切な方法を選択する。例えば、このような有用な方法の1つは、可溶性複合体を担持するリポソームを処方することである。
【0184】
以下の実施例、参考文献、および図面は、本発明の理解を助けるために提供されるものであって、本発明の真の範囲は、添付の特許請求の範囲に記載される。本発明の趣旨から逸脱することなく、この記載された手順において改変がなされ得ることが理解される。
【実施例】
【0185】
実施例
実施例1 gp41およびPPIシャペロンを含む可溶性分子間複合体の生成
1.1 E.coli FkpAの生成
Bothmann, H.、およびPluckthum, A., J Biol Chem275(2000)17100-5に従って(ただしそれをわずかに改変して)、FkpAをクローニングし、発現させ、そして精製した。保存のために、20mM NaH2PO4/NaOH(pH6.0)、100mMNaClに対してタンパク質溶液を透析して、26mg/ml(1mM)に濃縮した。
【0186】
サイトゾル発現のために、上記の発現ベクターのFkpAコード配列を、シグナルペプチドをコードする配列部分を欠くように、そして代わりに成熟FkpAのコード領域のみを含むように改変した。
【0187】
1.2 gp41(535-681)-His6の生成
gp41(535-681)−His6を、T7プロモーターベースの発現系においてクローニングして発現し、宿主細胞の封入体中に蓄積させた。単離した封入体を、6M塩化グアニジウム中で溶解した。His−タク゛化タンパク質を、Ni−キレートカラム上で精製し、その後に、Sephacryl100上で、6Mグアニジウム中でゲルろ過した。Wingfield, P. T.ら、Protein Sci 6(1997)1653−60に記載のとおり、急速希釈によってタンパク質を再折り畳みした。最終の緩衝条件は、30mMギ酸ナトリウム(pH3.0)であった。折り畳みの状況を、近UVCDおよび遠UV CDを用いて両方の緩衝条件について評価した。図1Aおよび1Bからわかるように、遠UV CDおよび近UV CDの両方のスペクトルによって、gp41がカオトロピック剤の非存在下において、pH3.0でのみ、未変性様折り畳みをとることが示唆される。
【0188】
1.3 E.coli FkpAの存在下における、gp41エクトドメイン(HIV)のpH3.0から生理的pHへのpHシフト
1.3.1 コントロール実験
コントロール実験において、可溶性e-gp41(30mMギ酸塩、pH3.0)を、20mMリン酸ナトリウム(pH7.5)、50mMNaCl, 1mM EDTAの最終緩衝条件中に100倍希釈した。最終タンパク質濃度は、約1μMであった。UVスペクトルを、1分後、および10分後に記録した。非シャペロンのエクトドメインは、中性へのpHシフトの際に、自然に凝集することが、図2におけるUVスペクトルから明白である。図2は、gp41の異例な凝集傾向を強調する意図がある;この分子の自発的な凝集は、上の線によって示される段階をかなり超えて進行する。
【0189】
1.3.2 pH3.0でのFkpAとのgp41のプレインキュベーションによって中性pHにシフトできる
分子シャペロンFkpAの可溶化能力について試験するために、エクトドメインgp41、およびFkpAを、1:2、および1:4の分子比で混合して(約3.5のpHで30mMギ酸塩)、1分間、同時インキュベートさせた。次いで、得られた複合体を、20mMリン酸ナトリウム(pH7.4)、50mMNaCl、1mM EDTAの緩衝条件中への12倍希釈によって、中性のpHにシフトさせた。試験管中における結合パートナーの最終濃度は、それぞれ、1μMのgp41、2μMおよび4μMのFkpAであった。全ての反応を室温で実施した。1分後、および10分後、凝集についてgp41サンプルを試験するために、UVスペクトルを記録した。図3A、および3Bから、FkpAが、用量依存性の様式で、gp41の凝集をかなり低下させることが明白である。Thermatogamaritima由来の誘発因子、およびE. coli由来のC末端切断型SlyDを用いて匹敵するデータが得られている。
【0190】
実施例2
共有結合したgp41−FkpAの組み換え生成
2.1 FkpA、およびgp41を含む発現プラスミドの構築
第一の工程において、Stratagene(La Jolla、CA;USA)のQuickChange部位特異的変異誘発キットを用いて、以下のプライマーを用い、実施例1.1のプラスミドから、成熟E.coli FkpAのコード領域における制限部位BamHIを欠失させた:
5’−gcgggtgttccgggtatcccaccgaattc−3’(配列番号3)
5’−gaattcggtgggatacccggaacacccgc−3’(配列番号4)
この構築物を、EcFkpA(DbamHI)[GGGS]3と命名した。
【0191】
第二の工程において、HIV-1エンベロープタンパク質由来のアミノ酸535-681をコードする遺伝子断片を、実施例1.2の構築物から、以下のプライマーを用いて、PCRによって増幅した:
5’−cgggatccggtggcggttcaggcggtggctctggtggcggtacgctg-acggtacaggccag−3’(配列番号5)
5’−ccgctcgaggtaccacagccaatttgttat−3’(配列番号6)
【0192】
この断片を、BamHIおよびXhoI制限部位を用いて、EcFkpA(DbamHI)[GGGS]3中に挿入した。
【0193】
FkpAと、e-gp41との間にグリシン−セリンリンカーのコドンを、FkpAのクローニングのためのリバースプライマーとともに、およびe-gp41のクローニングのためのフォワードプライマーとともに挿入した。
【0194】
得られた構築物を配列決定して、所望のタンパク質をコードすることを見出した。
【0195】
2.2 融合タンパク質の精製
発現プラスミドを保有するE. coli BL21細胞を、0.7のOD600まで増殖させて、37℃の増殖温度で1mMのIPTGを添加することによって、細胞質過剰発現を誘導した。誘導の4時間後、遠心分離によって細胞を収集した(5000gで20分)。50mMリン酸ナトリウムpH7.8、6.0MGuHCl(塩化グアニジウム)、5mMイミダゾール中に、細菌ペレットを再懸濁して、完全に溶かすために室温で(10分)撹拌した。繰り返し遠心分離(SorvallSS34, 20000rpm, 4℃)後、上清を濾過(0.8/0.2μm)して、Ni−NTA−カラム(NTA: ニトリロ三酢酸;Qiagen;Germantown,MD)(溶解バッファー中で事前に平衡化した)にかけた。10カラム容積の溶解バッファーを与えることによって、洗浄工程で、非特異的に結合したタンパク質を取り除いた。最終的に、結合した標的タンパク質を、50mMのリン酸ナトリウム、pH2.5、6.0MGuHClを用いて溶出させて、4ml画分中に収集した。吸光度を280nmで記録した。
【0196】
得られた酸性溶液およびカオトロピック溶液は、さらなる精製工程、またはインビトロ再折り畳み実験のために4℃で保存してもよい。
【0197】
この変性材料で開始して、異なる再折り畳み方法(例えば、透析、急速希釈、再生サイズ排除クロマトグラフィー、またはマトリックス補助再折り畳み)を首尾よく用いかつ実施することができ、その全てが、実質的に同じ未変性様折り畳み、および可溶性タンパク質をもたらす。
【0198】
2.3 透析および急速希釈による再生
上記のように可溶化された材料を、透析によって生理的緩衝条件に移行する。透析チューブの選択された排除値は、4000-6000ダルトンであった。
【0199】
エクトドメイン(共有結合したgp41およびFkpAタンパク質ドメインのgp41部分)の再折り畳みを誘導するために、50mMリン酸ナトリウム、pH2.5、50mM NaCl(塩化ナトリウム)に対する透析によって、溶出したタンパク質から、GuHClを除去した。単離されたエクトドメインは、全てヘリックスであって、この極端なpHで三次接触を形成することが周知である。近UVCDによって、組換え産生されたFkpAを分析する場合、FkpAは、同じ条件下では本質的に未構築であることが見出された。透析によるgp41-FkpAの再折り畳みが、共有結合したgp41およびFkpAタンパク質ドメインを含む、容易に可溶性のタンパク質複合体を生じることは驚くべきことである。UVスペクトル(図4)は、迷光、すなわち、300nmを超える明白な吸収を欠く。迷光は、凝集の指標であり、従って図4に示されるスペクトルは、この再折り畳みされた材料が、有意な量の凝集物を含まないことを意味する。
【0200】
円偏光二色性分光法(CD)は、タンパク質中の二次構造および三次構造の両方を評価するための選り抜きの方法である。芳香族領域における楕円率(260-320nm)は、タンパク質内の三次接触(すなわち、規則的に折り畳まれたタンパク質の球状構造)を報告するが、アミド領域における楕円率は、タンパク質骨格中の規則的な反復性エレメント(すなわち、二次構造)を反映する。
【0201】
図5に示される近UV CDスペクトルによって、エクトドメイン(融合タンパク質の状況において)が、pH2.5で、未変性様三次接触を提示するという、説得力のある証拠が提供される。共有結合したgp41/FkpAタンパク質ドメインのスペクトルは、同一条件下で単離されたエクトドメインのスペクトルとほとんど一致する(データ示さず)。以下の、gp41の典型的なサインが見出された:290nmでの最大楕円率、285nmでの特徴的な肩部分、および260nmでの別の最大値(光学的に活性なジスルフィド結合を反映する)。FkpAは、全ての各々の条件下で、近UVシグナルに貢献しないことに注目することが重要である。実際、pH2.5におけるFkpAの芳香族楕円率は事実上、基準値に等しい(データ示さず)。
【0202】
近UV領域からの結果と一致して、pH2.5の融合構築物の遠UV CDは、ほとんど構造化されたgp41分子を暗示する。220nmおよび208nmの2つの最大値は、全ヘリックス状エクトドメインの典型的サインを形成し、かつそれに相当する(図6)。FkpA-gp41融合ポリペプチドは、急速希釈によって、示された条件(50mMリン酸ナトリウム、pH2.5、50mM NaCl)から、生理的緩衝条件に容易に移行され得る。結論として、近UV CDおよび遠UV CDの両方とも、未変性様構造のgp41は、非常に都合のよい様式で(FkpAも含む融合タンパク質の状況において)利用可能であることを強調する。興味深いことに、本発明者らは、SlyD(1-165)-gp41型の未変性様融合ポリペプチドが、室温での50mMリン酸ナトリウムpH7.4、150mMNaClに対するカオトロピック材料(例えば、7.0M GuHClに溶解された)の透析によって、さらに簡易に得られ得ることを見出す。完全に本発明に従って、例外的に実施された、2つのシャペロン-gp41融合構築物のヌクレオチド配列は、それぞれ、配列番号7および配列番号8に記される。
【0203】
2.4 サイズ排除クロマトグラフィー(SEC)による再生
変性gp41-FkpAポリペプチド(50mMリン酸ナトリウム、pH7.8、7.0MGuHClに溶解した)を、Superdex 200ゲル濾過カラム(20mMリン酸ナトリウム、pH7.4、50mM NaCl、1mM EDTAで平衡化した)にかけた。FkpA-gp41は、本質的に、3つの主な画分:高分子量の要素として、明らかなヘキサマー種として、そして明らかな三量体種として溶出する。この明白な三量体画分を濃縮して、近UVCD測定において、その三次構造について評価した(図7)。
【0204】
得られたグラフは、事実上、オーバーレイの(かぶさった)曲線であるが、これには、キャリアタンパク質FkpA、および標的タンパク質gp41の両方が1:1の比で寄与する。最も幸運なことに、gp41は、中性のpHで三次構造を示し、そして共有結合したシャペロンによって明白に可溶化される。言い換えれば、シャペロンFkpAは、基質として、未変性様構造のエクトドメインgp41を受容し、そして中性の作業pHにおいてこの折り畳み困難なタンパク質を可溶化するようである。従って、診断目的のための多量の可溶性gp41抗原を生成するために重要な要件は満たされる。
【0205】
pH7.4でのFkpA-gp41の遠UV CD(図8)によって、近UV CDは、それぞれFkpAおよびgp41のシグナル寄与の加算性を示すということが確認される。期待通り、このスペクトルは、高度にヘリックス状のgp41エクトドメインによって支配される(それぞれ、220nm、および208nmで最大楕円率)。
【0206】
上述の条件下で、pH7.4で可溶化された、共有結合されたgp41/FkpAタンパク質ドメインを用いて得られたデータによって、FkpAおよびgp41が、このポリペプチド構築物内の独立して折り畳む単位として挙動することが示される。
【0207】
実施例3 イムノアッセイにおいて抗原として用いられる組み換えgp41、および組み換えFkpA-gp41複合体に対する、異なる界面活性剤の効果
3.1 競合型イムノアッセイ
COBAS CORE HIV Combi試験(Roche DiagnosticsGmbH, Germany)は、組み換えgp41の免疫反応性について試験するための従来の手段を提供する。原理的に、このアッセイはまた、HIVのgp41に対する抗体を検出するための二重抗原コンセプトに従って働く。この固相抗原は、直接コーティングされる。検出抗原は、ペルオキシダーゼ標識されたgp41であって、ただし、SDS可溶化されたgp41材料を含む。
【0208】
HIVの検出のためのイムノアッセイにおいて、用いられる試薬は、かなり高濃度の界面活性剤を含むインキュベーションバッファー中で容易に溶解され、かつ安定であることが非常に所望される。このような界面活性剤(例えば、TritonX-100(登録商標)、またはNonident P-40(登録商標))は、ウイルス粒子を破壊するために0.1〜0.2%の濃度で用いられる。
【0209】
実施例1に記載のように生成されたSDS可溶化gp41、およびFkpA-gp41の両方が、COBASCORE HIV Combiアッセイにおいて競合抗原として試験された。それを行うために、市販のインキュベーションバッファーの代わりに、ヒト血清を含まないバッファーマトリックス中に0.1%TritonX100(登録商標)を含むインキュベーションバッファーが用いられる。試験されるべき抗原は、gp41と反応性であることが公知のヒト血清と同時インキュベートされる。
【0210】
gp41−FkpA抗原は、用量依存性の様式において、競合型アッセイ中のシグナルを強力にクエンチするが、SDS可溶化gp41は、本質的に非反応性である(図9)。50%阻害が、0.1μg/ml(2.2nMのモル濃度に相当する)のFkpA−gp41抗原濃度で達成される。
【0211】
試験中で、インタクトなウイルス膜を分解するために、界面活性剤(ヘルパー界面活性剤)として0.1%TritonX-100を含む希釈バッファーを用いた事前処理後に、FkpA−gp41が、優れた免疫反応性を保持することが、際立っている。これは、gp41エクトドメイン単独(SDS中のgp41)に比べて顕著である。gp41エクトドメイン単独では、へルパー界面活性剤の存在下で、免疫活性をほぼ完全に失う(図9)。
【0212】
FkpAが、不十分な結合動態に起因して重要なエピトープをマスクするか、または試験に内在性の界面活性剤TritonX-100が、gp41抗原の凝集を誘導することによって試験能力を破壊することのいずれかが、共有結合されたgp41−FkpA構築物の開発において主要な懸念であった。COBASCOREプラットフォームに対する多くの競合試験の実験結果によって、重要なgp41エピトープが、共有結合されたタンパク質ドメインの状況で、十分に接近可能であるという説得力のある証拠が得られる。さらに、分子内シャペロン-gp41複合体内のgp41の免疫反応性は、TritonX-100のようなヘルパー界面活性剤の存在下で保持される。
【0213】
3.2 電気化学発光アッセイ
二重抗原架橋形式による免疫アッセイは、感染性因子の血清学的な診断において大きな利点があるアッセイである。本発明によるFkpA-gp41は、生理的緩衝条件で可溶性であるので、検出原理として電気化学発光を使用する二重抗原架橋における使用に、この材料が適切であるか否かを検討することができる。
【0214】
ルテニウム標識に対してSDS可溶化gp41を結合させる試みは成功していない。しかし、FkpA-gp41は、生理的緩衝条件下で、容易に可溶であるので、疎水性Ru-標識に対するこの材料の結合は、直接証明された。記載された方法で改変された標的−シャペロン複合体でさえ可溶性のままであることは注目すべきである。Elecsys(登録商標)試験システム(RocheDiagnostics GmbH, Germany)においてアッセイを実施するために、FkpA-gp41を、それぞれビオチン化し、ルテニル化し、そして二重抗原架橋アッセイにおいて、免疫反応性について試験した。
【0215】
いくつかの典型的な抗HIV血清であって、主にIgG(免疫グロブリンG)クラスの抗体を含む血清を試験したところ、共有結合したFkpA−gp41タンパク質ドメインを用いて陽性であった。バックグラウンドのシグナルは、500ng/ml程度の高さの抗原濃度でさえ、内在するガジェット(gadget)バックグラウンドに達することが見出されている。このシグナル対ノイズ比は優れていることがわかった。さらに、キャリアタンパク質(E.coli由来の分子、シャペロンFkpA)が、これらのヒト血清に含まれる抗体の非特異的結合を生じるという証拠はない。
【0216】
上記で考察したように、血清変換の早期検出は、HIVの確実な診断に重要である。感染の経過の間に、IgMクラスの抗体が最初に出現する。非常に早期に、HIV感染を確実に検出するためには、従って、IgMの認識および結合について高いエピトープ密度を有する抗原モジュールを設計することが必須である。実際、FkpA-gp41は、典型的な抗HIV、IgM型血清によって十分に認識される。さらにより重要であるのは、陽性を試験するのが困難であるサンプル(NABI(Miami,Florida)によって供給された9003および4009の血清変換パネル由来のB血清およびC血清など)が、本発明による融合構築物を用いて陽性と試験されたという事実である。これは主要な成果である。なぜなら、ペプチドに基づくgp41抗原は、これらのIgM血清と試験された場合、全く反応性ではないからである。
【0217】
実施例4 可溶性シャペロン-gp41複合体は、ウイルス侵入を阻害する
異なるgp41−シャペロン融合タンパク質を、インビトロアッセイにおいて、HIV-1-媒介膜融合を阻害する能力について評価した。要するに、CD4、CCR5、およびCXCR4を発現するMAGIP4-CCR5レポーター細胞株を、HIV-1株NL4-3を用いて感染させて、Meisterら、Virology(2001)284(2)、287-296に従ってTat依存性b−ガラクトシダーゼ活性について評価した。実際、本発明者らは、nM範囲のIC50値を有する感染の実質的阻害を観察する。例えば、SlyD-gp41(配列番号6を参照のこと)は、IC50< 70nMでもって、ウイルス侵入を有意に阻害する。
【0218】
結論として、それぞれ、HIV-1 gp41、またはHIV-2 gp36を含む可溶性分子内複合体、およびペプチジル−プロリル−イソメラーゼシャペロンは、可溶性および高次構造の統合性(改善された抗HIV抗体試験の設計、および他の市販の適用を可能にする)に関して、著名な特色を保有する。
【0219】
実施例5 gp36エクトドメインに対するシャペロンFkpAの融合によって、インビトロで容易に再折り畳みされ得るサイトゾルポリペプチドが生じる
gp36(gp41のHIV-2ホモログ)を、可溶性かつ免疫学的に活性な形態で得るために、本発明者らは、FF36と名付けられた構築物をクローニングした。この融合ポリペプチドは、2つのFkpA単位、およびgp36単位(各々、可変性のグリシンリッチストレッチによって連結される)を含む。精製を容易にするために、この融合構築物をHis6C末端を用いてタグ化した。本質的に、前述のプロトコールに従って、このタンパク質を精製した:シャペロン溶解後、タンパク質をNi-NTA-カラムに結合させて、50mMリン酸ナトリウム、pH7.8、7.0MGuHClを用いた過剰な洗浄後、pHを下げることによって溶出させた。次いで、50mMリン酸ナトリウム、pH7.8、100mM塩化ナトリウム、1mM EDTAで平衡化したゲル濾過カラムを通過させることによって、溶出したタンパク質を、再折り畳みさせた。この「再生ゲル濾過(renaturinggel filtration)」法から得られたネィティブなタンパク質は、満足な免疫学的かつ分光学的特性を示し(図10を参照のこと)、従って、F41、またはFF41のようなgp41対応物に相当する。本明細書に記載された、この精製、および再折り畳みのプロトコールは、gp36のN末端7残基反復領域において3つの点突然変異を保有するFF36を用いて実施した(配列については、配列番号9を参照のこと)。可溶性タンパク質のより低い収率にもかかわらず、同じプロトコールがまた、wtgp36エクトドメインを含む融合構築物に首尾よく適用された。
【0220】
実施例6 可溶性の免疫反応性FkpA−gp21は、好都合でかつ再現可能な様式で獲得できる
FkpA-gp21-過剰産生E. coli細胞(配列番号10を含む)を、前記のとおり、増殖させ、誘導して、回収した。完全に溶解するために、細胞ペレットを50mMリン酸ナトリウム、pH7.8、7.0MGuHCl中で、室温で1時間撹拌させた。カオトロピック細胞溶解物を、溶解バッファー中で事前に平衡化したNi-NTA-カラム上に付与した。洗浄工程後、pHを低下させることによって、標的タンパク質(C末端Hexa-His-Tagを保有する)を溶出させた。再折り畳みのために、50mMリン酸ナトリウム、pH7.8、7.0M100mM塩化ナトリウム中で事前に平衡化したSuperdex 200ゲル濾過カラムに、GuHCl中FkpA−gp21(50mM、リン酸ナトリウム、pH6.0、7.0MGuHCl 4℃)を通過させた。UVスペクトルによって、FkpA-gp21が、可溶性タンパク質(これは、非シャペロンのgp21とは対照的に、それ以上凝集する傾向がない)として溶出することが実証された(図11/1)。さらに、得られたFkpA−gp21は、競合型COBASCORE実験において評価された場合、優れた免疫学的活性を示す(図11/2)。
【0221】
実施例7:FkpA-gp41に対するさらなるFkpAサブユニットの融合は、gp41エクトドメインの免疫学的特性を改善する
本発明者らは、融合ポリペプチド状況におけるさらなるPPIaseサブユニットが、gp41エクトドメイン−シャペロン複合体の全体的特性を改善し得るか否かという疑問に取り組んだ。このために、本発明者らは、記載されたプロトコールによって、F41(gp41改変体の上流に位置するFkpAドメインの1つ)、およびFF41(gp41改変体の上流に位置する2つのFkpAドメイン)の両方を調製した。次いで、ビオチン化し、かつルテニル化した融合タンパク質をElecsys(登録商標)E2010システムにおいて評価した。この結果は、さらなるシャペロンドメインを含む、FF41構築物の改善された特性を強力に示す。
【0222】
陰性の血清によるバックグラウンドシグナルであって、シグナル対ノイズ比、および力価の低い血清の確実な測定に決定的なバックグラウンドシグナルは、FF41(約1600カウント)を、F41(約3800カウント)と比較した場合、50パーセントよりも低下されることが見出された。
【0223】
【表1】
【0224】
SS41融合タンパク質、すなわち、2つのSlyDドメイン、および1つのgp41ドメイン(シャペロンドメインに対してC末端)を含む融合タンパク質を用いて、同様の陽性の結果が得られている。
【0225】
本発明の態様として、以下のものが挙げられる。
[1]バッファー中で、本質的に不溶性である標的タンパク質とペチジル-プロリル-イソメラーゼクラスシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化される、および形成されるタンパク質-シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびペプチジル-プロリル-イソメラーゼクラスシャペロンを含有する可溶性複合体の製造方法。
[2]生理的バッファーが10〜200mMの濃度のバッファー化合物および20〜500mMの総濃度の塩を含有する[1]記載の方法。
[3]タンパク質が組換え産生される[1]記載の方法。
[4]ペプチジルプロリルイソメラーゼが組換え産生される[1]記載の方法。
[5]タンパク質およびペプチジルプロリルイソメラーゼが組換え産生される[1]記載の方法。
[6]タンパク質がアミロイド形成性タンパク質である[1]記載の方法。
[7]アミロイド形成性タンパク質がレトロウイルス表面糖タンパク質である[6]記載の方法。
[8]タンパク質がHIV-2 gp36またはHIV-1 gp41である[1]記載の方法。
[9]ペプチジルプロリルイソメラーゼがペプチジルプロリルイソメラーゼの結合コンピテント断片である[1]記載の方法。
[10]バッファー中でレトロウイルス表面糖タンパク質とペプチジルプロリルイソメラーゼとを混合する工程、ここでレトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼの両方が可溶化され、複合体を形成する、および複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質-シャペロン複合体の製造方法。
[11]レトロウイルス表面糖タンパク質が組換え産生される[10]記載の方法。
[12]ペプチジルプロリルイソメラーゼが組換え産生される[10]記載の方法。
[13]レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼが組換え産生される[10]記載の方法。
[14]レトロウイルス表面糖タンパク質がHIV gp36またはgp41である[10]記載の方法。
[15]ペプチジルプロリルイソメラーゼが、ペプチジルプロリルイソメラーゼの結合コンピテント断片を含有する[10]記載の方法。
[16]バッファー中でペプチジルプロリルイソメラーゼに共有結合したレトロウイルス表面糖タンパク質を可溶化させる工程、ここでレトロウイルス表面糖タンパク質が可溶化される、およびレトロウイルス表面糖タンパク質-シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質-シャペロン複合体の製造方法。
[17]ペプチジルプロリルイソメラーゼがFKBPシャペロンである[1]、[10]または[16]記載の方法。
[18]FKBPシャペロンがSlyD、FkpAおよびトリガー因子からなる群より選ばれる[17]記載の方法。
[19]レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼを含有してなる可溶性複合体。
[20]レトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼシャペロンが共有結合されているレトロウイルス表面糖タンパク質およびペプチジルプロリルイソメラーゼを含有してなる可溶性複合体。
[21]共有結合が化学的共役を含む[20]記載の複合体。
[22]共有結合が組換え結合を含む[21]記載の複合体。
[23]組換え結合がペプチドリンカーを含む[22]記載の複合体。
[24]ペプチドリンカーが少なくとも10アミノ酸を含有してなる[23]記載の複合体。
[25]ペプチドリンカーが少なくとも15アミノ酸を含有してなる[23]記載の複合体。
[26]ペプチドリンカーが多くとも50アミノ酸を含有してなる[23]記載の複合体。
[27]ペプチドリンカーが多くとも40アミノ酸を含有してなる[23]記載の複合体。
[28][19]〜[27]いずれか記載の可溶性複合体を含有してなる試薬の組成物。
[29]サンプルと、表面糖タンパク質とペプチジルプロリルイソメラーゼシャペロンを含有する複合体を含有する組成物とを接触させる工程、および結合抗体を検出する工程を含む、サンプル中のエンベロープウイルス表面糖タンパク質に対する少なくとも1つの抗体を検出する方法。
[30]接触させる工程が、表面糖タンパク質への抗体の結合に適切な条件下で行われる[29]記載の方法。
[31]結合した抗体の検出がサンプル中の抗-ウイルス抗体の存在を示す[30]記載の方法。
[32]第1のシャペロン-抗原複合体を含有する第1の抗原および第2のシャペロン-抗原複合体を含有する第2の抗原を含む二重抗原架橋概念によるイムノアッセイ。
[33]第1のシャペロンおよび第2のシャペロンが1つの種に由来する異なる分子である[32]記載のイムノアッセイ。
[34]第1のシャペロンおよび第2のシャペロンが異なる種に由来する[32]記載のイムノアッセイ。
[35]第1のシャペロンが好熱細菌に由来する[32]〜[34]いずれか記載のイムノアッセイ。
[36]第2のシャペロンが好熱細菌に由来する[32]〜[34]いずれか記載のイムノアッセイ。
[37]第1のシャペロンおよび第2のシャペロンが好熱細菌に由来する[32]または[34]記載のイムノアッセイ。
[38]第1の抗原複合体が固相結合基を含有する[32]記載のイムノアッセイ。
[39]第2の抗原複合体がマーカー基を含有する[32]記載のイムノアッセイ。
[40]可溶性レトロウイルス表面糖タンパク質-シャペロン複合体を含有するワクチンを被験体に注入し、それにより被験体においてレトロウイルス表面糖タンパク質に結合する抗体を誘発する工程を含む、免疫応答の誘発方法。
[41][10]記載の可溶性複合体を細胞に投与することを含む、細胞へのウイルス侵入の阻害方法。
[42]ウイルス侵入の阻害が膜融合を阻害することを含む[41]記載の方法。
[43]生理的バッファーが、10〜200mMの濃度のバッファー化合物および20〜500mMの総濃度の塩を含む[41]記載の方法。
[44]タンパク質が組換え産生される[41]記載の方法。
[45]ペプチジルプロリルイソメラーゼが組換え産生される[41]記載の方法。
[46]タンパク質およびペプチジルプロリルイソメラーゼが組換え産生される[41]記載の方法。
[47]タンパク質がアミロイド形成性タンパク質である[41]記載の方法。
[48]タンパク質がレトロウイルス表面アミロイド形成性糖タンパク質である[41]記載の方法。
[49]タンパク質がHIV-2 gp36またはHIV-1 gp41である[41]記載の方法。
[50]ペプチジルプロリルイソメラーゼがペプチジルプロリルイソメラーゼの結合コンピテント断片である[41]記載の方法。
【0226】
【図面の簡単な説明】
【0227】
【図1】図1Aおよび1Bは、HIV-1エクトドメインgp41(535-681)-His6の遠UVCD(1A)、および近UV CD(1B)を示す。折り畳まれたgp41(太線):緩衝条件(30mM ギ酸ナトリウム,pH3.0)を設定して、gp41の未変性様の全ヘリックス状高次構造(コンフォメーション)を誘導した;変性されたgp41(細線):緩衝条件(50mMリン酸ナトリウムpH3.0,7.0MGuHCl)を設定して、gp41(変性)を完全に変性する。カオトロピック塩の高いモル濃度に起因して、図1A中の参照二色性シグナル(細線)は、215nm未満の波長領域で確実にモニターすることはできない。このスペクトルは、Jasco-720分光偏光計で記録され、ノイズを低下させるために9回平均された。パスレングスは、遠UVCDについては0.2cm(図1A)、そして近UV CDについては0.5cm(図1B)であった。それぞれのタンパク質濃度は、1.5μM、および29μMであった。縦軸の単位は、平均残余楕円率(meanresidue ellipticity)であり、そしてdeg×cm2×dmol-1の次元を有する。
【図2】図2は、生理的バッファー中における「非シャペロン(unchaperoned)」gp41の凝集を示す。3.0から7.5へのpHジャンプ後、1分(下の線)、および10分(上の線)のgp41エクトドメインのUVスペクトルを示す。凝集する分子は、迷光効果をもたらし、そして310nmを超えて明白な吸収を生じる。この図は、gp41の甚大な凝集傾向をまさに実証することを意味する;凝集過程が、上の線によって示された段階で停止しないことは注目に値する。
【図3A】図3Aは、FkpAが中性のpHでgp41エクトドメインを可溶化することを示す。gp41、および成熟FkpAを、低いpHで同時インキュベートして、その後に20mMリン酸ナトリウム、pH7.4;50mMNaCl, 1mM EDTAの最終バッファーにシフトした;1分後、および10分後(それぞれ、下の線、および上の線)、UVスペクトルを記録して、サンプル中の凝集の程度を評価した。図3Aは、2倍モル過剰のシャペロンによる凝集の抑制を示し、図3Bは、4倍過剰の効果を示す。gp41の最終濃度は、約1μMであった。迷光(300nmを超えて明白な吸収をもたらす)は、最小まで低下したので、FkpAは、gp41エクトドメインを用量依存性の様式で効率的に可溶化するという、説得力のある分光光度的な証拠がある。
【図3B】図3Bは、FkpAが中性のpHでgp41エクトドメインを可溶化することを示す。gp41、および成熟FkpAを、低いpHで同時インキュベートして、その後に20mMリン酸ナトリウム、pH7.4;50mMNaCl, 1mM EDTAの最終バッファーにシフトした;1分後、および10分後(それぞれ、下の線、および上の線)、UVスペクトルを記録して、サンプル中の凝集の程度を評価した。図3Aは、2倍モル過剰のシャペロンによる凝集の抑制を示し、図3Bは、4倍過剰の効果を示す。gp41の最終濃度は、約1μMであった。迷光(300nmを超えて明白な吸収をもたらす)は、最小まで低下したので、FkpAは、gp41エクトドメインを用量依存性の様式で効率的に可溶化するという、説得力のある分光光度的な証拠がある。
【図4】図4は、pH2.5でのFkpA-gp41のUVスペクトルを示す。50mMリン酸ナトリウム、pH2.5;50mMNaClに対する透析後の融合ポリペプチドFkpA-gp41のUVスペクトル。驚くべきことに、2つのドメイン構築物は、可溶化カオトロピック剤CuHClの除去後、完全に可溶性のままである。300nmを超える波長で基線ドリフト(baselinedrift)および有意な明白な吸収を生じることが期待される、迷光する凝集物が存在するという証拠はない。
【図5】図5は、pH2.5でのFkpA-gp41の近UV CDスペクトルを示す。20mMリン酸ナトリウム、pH2.5;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、ノイズを低下させるために9回蓄積した。タンパク質濃度は、0.5cmのパスレングスで22.5μMであった。芳香族の楕円率は、gp41の典型的な特徴を示す(基準については、図1Bを参照のこと)。pH2.5では、FkpAは、大部分が構造化されておらず、近UV-CDにおけるシグナルに対して全く寄与しない。
【図6】図6は、pH2.5でのFkpA-gp41の遠UV CDスペクトルを示す。20mMリン酸ナトリウム、pH2.5;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、シグナル対ノイズ比を改善するために9回蓄積した。タンパク質濃度は、0.2cmのパスレングスで2.25μMであった。220および208nmでの最小値は、融合タンパク質の状況では、gp41の大部分のヘリックス構造を暗示する。197nm未満のスペクトルノイズは、高いアミド吸収に起因しており、そして融合タンパク質のいずれの構造的特徴についても報告しない。それにもかかわらず、193nmで典型的なヘリックス最大値が推測され得る。
【図7】図7は、生理的な緩衝条件下でのFkpA-gp41の近UV CDである。20mMリン酸ナトリウム、pH7.4;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、ノイズを低下させるために9回蓄積した。タンパク質濃度は、0.5cmのパスレングスで15.5μMであった。著しいことに、gp41およびFkpAの共有結合したタンパク質ドメインの芳香族楕円率(実線)は、pH3.0での未変性様の全ヘリックス状gp41の寄与(下の破線)、およびpH7.4のFkpAの寄与(上の破線)から相加的に作成される。これによって、キャリアFkpA、および標的gp41(すなわち、2つの別個の機能的な折り畳み単位)が、ポリペプチド融合タンパク質において連結される場合、可逆的かつ半独立的に再折り畳みすることが示される。
【図8】図8は、生理的な緩衝条件下でのFkpA-gp41の遠UV CDを示す。20mMリン酸ナトリウム、pH7.4;50mMNaCl(20℃)中で、Jasco 720分光偏光計においてスペクトルを記録して、シグナル対ノイズ比を改善するために9回蓄積した。タンパク質濃度は、0.2cmのパスレングスで1.55μMであった。220および208nmでの強力なシグナルは、それぞれ、融合タンパク質の状況では、gp41の大部分のヘリックス状構造を暗示する。198nm未満のノイズは、高いタンパク質吸収に起因しており、そしてFkpA-gp41のいずれの二次構造的な特性も反映しない。
【図9】図9は、FkpA連結したgp41がHIVアッセイにおいて可溶性であり、かつ高度に免疫反応性であることを示す。FkeA-gp41は、COBASCORE HIV Combiアッセイにおける強力な競合因子である。gp41エクトドメイン単独(白丸)と比較した、希釈バッファー(ヘルパー界面活性剤としてTritonX-100を含有する)を用いた前処理後の可溶性FkpA-gp41ポリペプチド(黒丸)の阻害能力を示す。gp41エクトドメイン(融合タンパク質の分子内複合体内)は、界面活性剤の存在下でさえ高い免疫反応性を保持しているが、一方で裸のエクトドメインは、ほとんど完全に免疫反応性を失っているということが明白である。試験したHIV陽性血清は、1:3000希釈で、内部血清数21284であった。
【図10】図10は、再生ゲル濾過後のFF36のUVスペクトルである。このスペクトルは、gp36融合ペプチドが、実施例の節に記載のとおりの再生ゲル濾過法に従ってSux200カラム上で再折り畳みされた場合に、可溶性であり、かつ凝集しないという有力な証拠を提供する。
【図11】図11(1+2)は、FkpA-gp21が可溶性であり、かつ免疫学的に反応性の融合ポリペプチドであることを示す。再生ゲル濾過後、再折り畳みされたFkpA-gp21融合タンパク質は、高度に可溶性に溶出して、UVスペクトルにおいては凝集傾向を示さない(11/1)。HTLV陽性血清858893-00(1:10希釈)を用いたCOBASCOREにおける競合型の免疫アッセイ実験において評価した場合、FkpA-gp21は、結局、優れた免疫学的特性を保有するようになる(11/2)。
【特許請求の範囲】
【請求項1】
非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化され、および形成されるタンパク質− シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびシャペロンを含有する可溶性複合体の製造方法。
【請求項2】
非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子又はSlyDの結合コンピテント断片とを混合する工程、ここで該タンパク質および該断片の両方が可溶化され、および形成されるタンパク質− 断片複合体が可溶である生理的条件
にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質および断片を含有する可溶性複合体の製造方法。
【請求項3】
非生理的なバッファー中でレトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここでレトロウイルス表面糖タンパク質およびシャペロンの両方が可溶化され、複合体を形成し、および該複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質−シャペロン複合体の製造方法。
【請求項4】
レトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを含有してなる可溶性複合体。
【請求項5】
請求項4記載の可溶性複合体を含有してなる試薬の組成物。
【請求項1】
非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここで該タンパク質および該シャペロンの両方が可溶化され、および形成されるタンパク質− シャペロン複合体が可溶である生理的条件にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質およびシャペロンを含有する可溶性複合体の製造方法。
【請求項2】
非生理的なバッファー中で、本質的に不溶性である標的タンパク質と、FkpA、誘発因子又はSlyDの結合コンピテント断片とを混合する工程、ここで該タンパク質および該断片の両方が可溶化され、および形成されるタンパク質− 断片複合体が可溶である生理的条件
にバッファーを調整する工程を含む、本質的に不溶性である標的タンパク質および断片を含有する可溶性複合体の製造方法。
【請求項3】
非生理的なバッファー中でレトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを混合する工程、ここでレトロウイルス表面糖タンパク質およびシャペロンの両方が可溶化され、複合体を形成し、および該複合体が可溶である生理的条件にバッファーを調整する工程を含む、可溶性レトロウイルス表面糖タンパク質−シャペロン複合体の製造方法。
【請求項4】
レトロウイルス表面糖タンパク質と、FkpA、誘発因子及びSlyDからなる群より選ばれるシャペロンとを含有してなる可溶性複合体。
【請求項5】
請求項4記載の可溶性複合体を含有してなる試薬の組成物。
【図1】

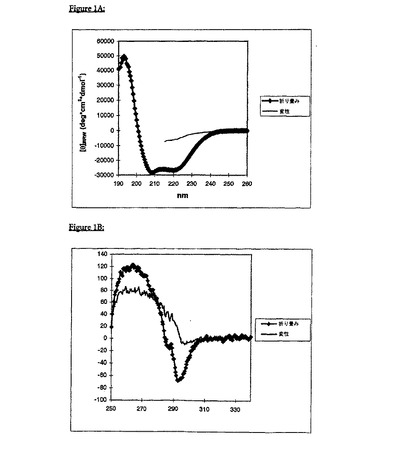
【図2】
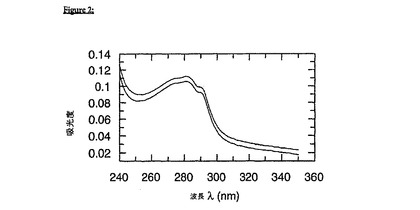
【図3A】
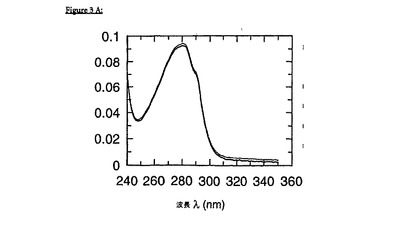
【図3B】
【図4】
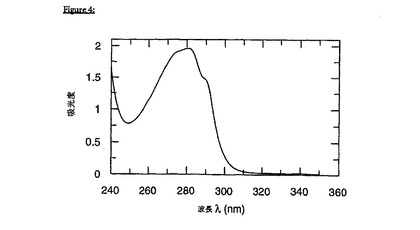
【図5】
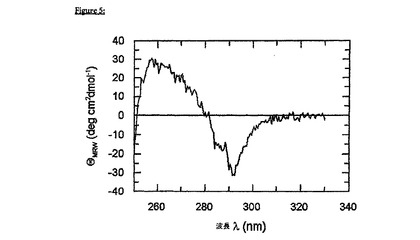
【図6】
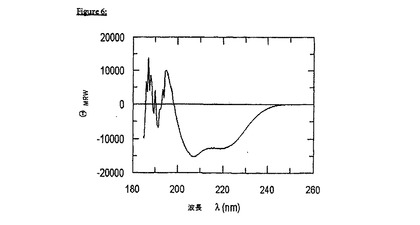
【図7】
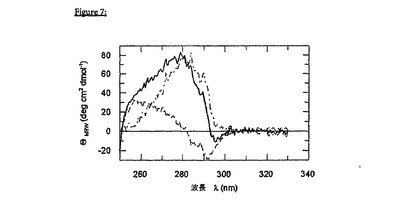
【図8】
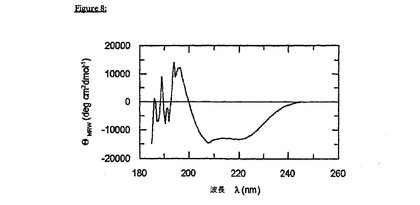
【図9】
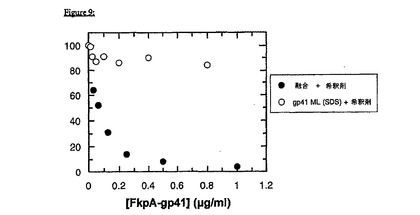
【図10】
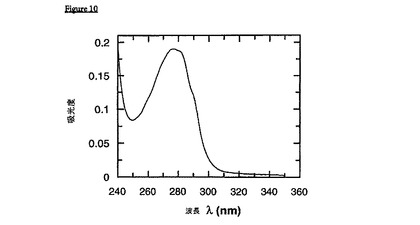
【図11】
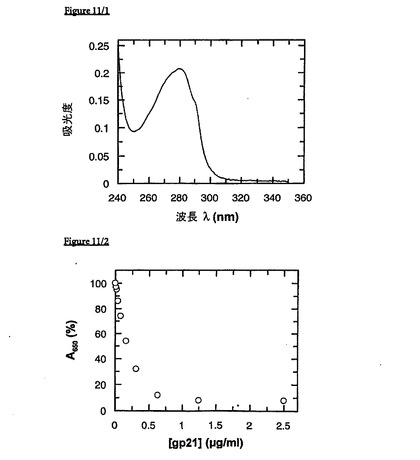
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2009−102320(P2009−102320A)
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願番号】特願2008−286959(P2008−286959)
【出願日】平成20年11月7日(2008.11.7)
【分割の表示】特願2003−507262(P2003−507262)の分割
【原出願日】平成14年6月24日(2002.6.24)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願日】平成20年11月7日(2008.11.7)
【分割の表示】特願2003−507262(P2003−507262)の分割
【原出願日】平成14年6月24日(2002.6.24)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
[ Back to top ]