
レプリキンペプチドとその使用
【課題】本発明は、迅速な複製に関連するペプチドの新規クラス並びに疾患の診断、予防及び処置におけるそれらの使用を提供する。
【解決手段】特定のモチーフから成るインフルエンザペプチド。
【解決手段】特定のモチーフから成るインフルエンザペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
他の出願に対する相互参照
本出願は、出願US 09/984,057 and US 09/984,056(いずれも2001年10月26日出願)の一部継続出願であるPCT出願PCT/US02/09240(2002年3月26日出願)であり、これは、仮出願60/303,396号(2001年7月9日出願)の優先権を主張する(この主題は、参照することにより本明細書に組み込まれる)。
【0002】
発明の分野
本発明は、構造的な特徴を共有する新たに発見されたクラスのペプチドであるレプリキン類(Replikins)の同定と使用に関する。特に、本発明は、ウイルス、細菌、真菌、癌関連タンパク質、植物および単細胞寄生体中に見いだされたレプリキン、および疾患の治療または予防法の開発における標的としてのこれらの使用に関する。 さらに、本発明は、これらの疾患の検出におけるレプリキンの使用に関する。 また本発明は、食品のために使われる植物の成長を促進するためのレプリキンの使用に関する。
【背景技術】
【0003】
発明の概要と背景
迅速な複製は、いくつかの細菌、ウイルスおよび悪性腫瘍の病原性(virulence)に特徴的であるが、異なった生物の迅速な複製に共通する化学はこれまで開示されていない。本特許出願は、迅速な複製に関連する新しいクラスのタンパク質構造を開示する。レプリキン(Replikin)と称される、迅速な複製に関連する保存された小タンパク質の新しいファミリーは、多数の生物や疾患の迅速な複製を予測または抑制するために、且つ植物と動物の迅速な複製を誘導するために使用される。
【0004】
我々は、レプリキンのためのアルゴリズム検索を作成した。ここで発明されたアルゴリズムの応用において、エピトープの機能(迅速な複製)が明らかになっただけでなく、迅速な複製に関連する機能を有する同族体の全ファミリーが発見され、我々はこれをレプリキン類と名付けた。
【0005】
このアルゴリズムは、以下に基づく:1) 免疫系は、認識の時タンパク質全体ではなく部分を見ているという証拠。タンパク質鎖はまず、それに対する免疫防御を開始する免疫系による外来構造の認識プロセスの一部として、免疫系により小片(しばしば6〜10アミノ酸の長さ)に加水分解される。例えば免疫系は、疾患物質のタンパク質を切り刻んで小さなペプチド配列にし、それらを読むことにより疾患の配列を認識する。この原理は、その構造がいったんわかると、マリグニン(malignin)癌エピトープの同族体を検索するためのアルゴリズムの基礎として使用される;2) 上記の認識(6〜10アミノ酸の長さ)のために免疫系が使用する見かけの「規則」に従って3つのリジン(K's)のうちの2つが8残基離れている、マリグニン癌エピトープの特異的構造;3) マリグニン癌エピトープが、非常に強い抗原(すなわち、強い免疫応答の生成元)であること;10量体ペプチド神経膠腫レプリキン中に3つのリジン(K's)があること、およびK's は、ウイルスの侵入のためのアンカー候補としてしばしばDNAおよびRNAに結合することが証明されたという事実;および4) 8残基離れた2つのK'sの間に、マリグニンエピトープの配列中に1つのヒスチジン(H)が含まれており、これは複製のエネルギーを供給するのに必要な酸化還元系の金属への関連を示唆する。
【0006】
適切に配置された官能基を有する調整された結合ポケットを有する、作り変えた酵素と触媒性抗体は、時にすばらしい効率で多くの化学変換を触媒するのに成功している。特異的でかなり異なる機能を有する2つ以上の異なるタンパク質がある生物により一緒に合成され、次に別々に切断されて「別々の機能をする」ことがことがしばしば認められるように、レプリキン構造はユニークな機能を有するユニークなタンパク質であり、これは、免疫系により別々に認識されるようであり、現在は合理的に作り変えられ、例えば機能性単位を産生するように合成される。
【0007】
プロテオミクスの観点から、新たに決定された神経膠腫ペプチド配列に基づくこの鋳型により、関連する保存された構造と特定の機能(この場合は複製)を有する広いクラスのタンパク質が発見された。疾患の病原性に伴うレプリキン濃度の上昇例は、インフルエンザ、HIV、癌およびトマトリーフカールウイルス(tomato leaf curl virus)を含む疾患で現れる。このクラスの構造は、酵母、藻類、植物、ジェミニカールリーフトマトウイルス(gemini curl leaf tomato virus)、HIVおよび癌のような多様な生物の迅速な複製の現象に関連している。
【0008】
迅速に複製している生物中のレプリキンの存在を検出する以外に我々は、1) レプリキン濃度(100個のアミノ酸あたりのレプリキンの数)、および2) 迅速な複製に依存する特定の機能状態にあるレプリキン組成は、それらが存在する生物の複製速度にレプリキンが定量的ならびに定性的に関係するという知見の基礎を提供する、ことを見いだした。これらの機能的証拠の例には、神経膠芽細胞腫細胞中の迅速な複製と病原性との関係、インフルエンザウイルス中のレプリキンとインフルエンザの汎流行と流行の予測との関係、およびHIV中のレプリキン濃度と迅速な複製との関係がある。
【0009】
迅速な複製におけるレプリキンの役割の最初の機能的基礎は、神経膠腫レプリキン[脳の多形神経膠芽腫(神経膠腫)(250kDの細胞タンパク質)中のマリグニン(Malignin)と呼ぶ10kDのペプチド]の性質中で見いだされた。ほとんどまたはすべてのタイプの細胞で現在使用されている癌の早期段階診断検査により測定すると、抗マリグニン抗体の血清濃度(AMAS)は上昇した。マリグニンは、組織培養においてこのペプチドの発現と抽出可能な膜タンパク質1mgあたりのその濃度が、単位時間当たりの細胞分裂の速度の上昇に伴って上昇したため、この名前が付けられた。細胞数の増加に比例してマリグニンの量が増加するのみでなく、マリグニンの量が増加し、すなわち細胞数はわずかに5倍の増加であったが濃度が10倍増加した。
【0010】
マリグニンタンパク質の構造は、加水分解して質量分析により決定され、これにより新規の16量体のペプチド配列が明らかになった。健常人ゲノムのデータベースでこの16量体のペプチド配列(我々は神経膠腫レプリキンタンパク質(Glioma Replikin protein)と命名した)を検索し、これはこれらのデータベース中に存在しないことを見いだした。
【0011】
従って、一定の要件アルゴリズムを使用して、他の生物で神経膠腫レプリキンタンパク質またはその同族体について検索した。「PubMed」データベース中の4,000を超えるタンパク質配列を検索し、特に迅速な複製に関連するウイルスおよび植物で同族体が見つかった。そのようなレプリキンタンパク質の同族体はしばしば、それらの研究者が「複製タンパク質」と呼ぶタンパク質中に存在した。
【0012】
レプリキン配列の同族体はすべての腫瘍ウイルス(癌を引き起こすウイルス)中、および藻類、植物、真菌、ウイルスおよび細菌の「複製タンパク質」中で見つかった。
【0013】
神経膠腫細胞の迅速な複製中の細胞数と膜タンパク質濃度の5倍の上昇と比較してマリグニンが10倍濃縮されることは、レプリキンと複製との重要な関係を示唆する。神経膠腫レプリキンをin vitroで合成し、合成ワクチンとしてウサギに投与すると、多量の抗マリグニン抗体が産生され、血清中の抗マリグニン抗体(AMAS)の抗原性基礎を厳密に確立し、最初の合成癌ワクチン候補と、他の生物中のレプリキンワクチンのプロトタイプを提供している。
【0014】
レプリキンと複製、および癌レプリキンの特異性の共有に基づく癌レプリキンに対する自然の免疫応答(細胞タイプに限定されない)との関係の証明は、抗マリグニン抗体による免疫の受動的増強と、合成レプリキンワクチンによる能動的増強とを可能にする。
【0015】
癌患者と対照からの8,090の血清試料の研究により、健常人では、癌の頻度が増加するように、抗マリグニン抗体の濃度が年齢とともに増加し、早期悪性腫瘍では細胞のタイプに無関係に2〜3倍増加することが証明された。in vitroではこの抗体は、癌細胞1個当たりピコグラム(フェトモル)の癌細胞に対して細胞障害性であり、in vivoでは抗マリグニン抗体の濃度は、癌患者の生存に定量的に関連する。神経膠腫細胞で証明されるように、細胞は不死化の悪性段階に変換されているのみで停止もしくは休止している癌の段階は、レプリキンの濃度の上昇を特徴とする活動性の生命に危険のある複製状態とは区別される。さらに、癌のウイルス病原性の鍵は、神経膠腫糖タンパク質10Bが、正常な10Bと比較すると、炭水化物残基が50%低下しているという事実にあるかも知れない。この低下は、他の例ではウイルス侵入に関連し、従って、悪性状態への変換の過程としての、神経膠腫細胞の10Bへのウイルスレプリキンの供給のためのウイルスの結合の証拠かも知れない。
【0016】
神経膠腫と関連する癌のレプリキンについて抗マリグニン抗体で証明されるように、多様なクラスのメンバーによる免疫学的特異性の共有は、B細胞とその生成物である抗体が、同様の認識「言語」を使用してレプリキンを認識するかも知れないことを示唆する。レプリキンの発見により、この免疫学的特異性の共有は、以前は困難であると考えられてきたことを説明できるかも知れない:なぜ、抗マリグニン抗体はすべての癌で増加しているのか、およびなぜ癌細胞に細胞障害性であるか、なぜほとんどまたはすべての細胞タイプで癌患者の生存に関係しているのか。すなわち、抗マリグニン抗体は癌レプリキンに対して産生され、これは免疫学的特異性を共有し、細胞タイプではなく迅速な複製の現象に関連している。
【0017】
迅速な複製におけるレプリキンの役割の第2の機能的基礎は、インフルエンザウイルス血球凝集素タンパク質配列についての過去100年間のデータの研究およびインフルエンザの流行または汎流行の疫学研究である。今日まで、インフルエンザウイルスについては血清学的血球凝集素と抗体の分類のみが開示され、株特異的な保存されたペプチド配列は開示されておらず、疫学的に記録された流行病または迅速な複製に関連する株特異的ペプチド配列の濃度と組成の変化については何も開示されていない。
【0018】
4つの主要な株[インフルエンザB、(A)H1N1、(A)H2N2、および(A)H3N2]中のそれぞれの1つで株特異的インフルエンザレプリキン濃度の4〜10倍の増加が見つかり、レプリキン濃度のそのような増加は、1902〜2001年に各株により特異的に引き起こされたインフルエンザ流行に関連していた。次に、これらの濃度の増加は、その消失から1〜64年後の少なくとも1つの特異的レプリキン組成の再出現と、新しい株特異的レプリキン組成の出現によることが証明された。以前は、次のインフルエンザシーズンにどの株が主流になるかを予測するための、またはワクチンのために毎年の全ウイルス株の混合物を作成するための、株特異的化学構造は不明であった。H3N2レプリキン濃度の最近の急激な上昇(1997〜2000)(H3N2の歴史で最大)、および1968年の高死亡率のH3N2汎流行と、1975年と1977年の2つの高死亡率流行で最後に見られた特異的レプリキン組成の再出現(しかし20〜25年は出現しなかった)は一緒に、これから来る流行の警告かも知れない。
【0019】
合成レプリキンは新しいワクチンである。同一の構造が100年間持続し、1〜64年の消失後に再出現する同一の構造で観察されるレプリキン構造の高度の保存は、インフルエンザタンパク質中のアミノ酸のランダム置換による病原性の変化であると以前考えられていたものは、レプリキンの組織化された保存プロセスによる変化である可能性が高い。実際、各アミノ酸のランダム置換が起きると、1年間(100年は言うまでもなく)保存されている平均的長さのインフルエンザレプリキン配列に対する変化は、2の27乗対1のオーダーであると計算される。
【0020】
レプリキンの高度の保存は、インフルエンザウイルスにユニークなものではなく、手足口病ウイルスO型およびHIV、ならびにコムギにも存在する。
【0021】
迅速な複製におけるレプリキンの役割の第3の機能的基礎は、HIV中の迅速な複製に関連することが証明されたレプリキン濃度の増加である。早期感染によくあるHIVの増殖の遅い低力価株(NS1、「Bru」)中のレプリキン濃度は、後期HIV感染によくあるHIVの迅速に増殖している高力価株(SI、「Lai」)中のレプリキン濃度の6分の1であった。
【0022】
レプリキンと迅速な複製との関係についての他の例が示される。例えば、トマトカールリーフジェミニウイルス(これは、トマトの収穫に壊滅的被害を与える)では、DNAに結合することが証明されている「複製タンパク質」の最初の161 アミノ酸がレプリキンを含有する。
【0023】
マラリア(迅速な複製で有名)では、1つのトリパノソーマから数万のトリパノソーマが肝臓中に放出される。そこには、100アミノ酸当たり最大36個の重複レプリキンの濃度を有する複数の新規なほとんど「火炎状」のレプリキン構造が見られる。
【0024】
インフルエンザ流行におけるレプリキン濃度の上昇は、悪性神経膠腫細胞の迅速な複製中の神経膠腫レプリキン濃度の上昇に機能的に匹敵し、HIVおよび多様な範囲の他の生物中の迅速な複製に匹敵する。従ってレプリキンは、異なる生物中の迅速な複製の構造的基礎に関連しており、この一部であるようである。
【0025】
従ってレプリキン濃度と組成は、複製を検出しそのプロセスを制御するための新しい方法を提供し、これは、各生物集団の生存と支配に重要である。迅速な複製に関連するこれらの新しいタンパク質の発見は、新しい機会を与える:1) レプリキンの定性的および定量的測定による病原体の検出、2) 未変性のレプリキンを標的とすることによるおよび合成レプリキンをワクチンとして使用することによる、迅速な複製が重要な要因である広範囲の疾患の制御、および3) 藻類と植物食物の成長を促進するためのレプリキンの使用。
【発明の概要】
【発明が解決しようとする課題】
【0026】
検出や治療が困難であり、有効なワクチンが無い多くの疾患や病原体がある。すなわち、各疾患について、これらの疾患および病原体を検出、治療または予防するための有効な方法を提供する標的を開発するニーズがある。
【課題を解決するための手段】
【0027】
本発明は、レプリキン配列を含むヌクレオチドまたはアミノ酸配列の同定法を提供する。本明細書において本方法は3点認識法と呼ぶ。「3点認識」法を使用することにより、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含むペプチド(レプリキン)(新しいクラスのペプチドを構成する)が、複製、形質転換、または酸化還元機能を有する藻類、酵母、真菌、アメーバ、細菌、植物およびウイルスタンパク質で明らかになった。
【0028】
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成されたペプチドが提供される。このペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を含む。
【0029】
本発明はまた、生体試料または環境試料中の汚染生物の存在を検出する方法であって:
(1) 生体試料または環境試料から核酸を単離する;
(2) レプリキン構造の存在について核酸をスクリーニングする;そして
(3) レプリキン構造の存在を汚染生物の存在と相関させる、
ことを含む方法を提供する。
【0030】
本発明の別の態様において、レプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの実施態様は、出現する生物の株(もしそのような株が出現するなら)中に存在する少なくとも1つのペプチドを含む。
【0031】
本発明はまた、本明細書に記載のレプリキンに特異的に結合する抗体、ならびにレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、レプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0032】
本発明の1つの態様において、他のタンパク質から単離または分離された、ウイルスレプリキン配列を含有する、組換えまたは合成ペプチドまたは他の方法が提供される。ウイルスレプリキンペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸(ウイルスレプリキン)を含む。
【0033】
本発明はまた、生体試料または環境試料中の汚染ウイルスの存在を検出する方法であって:
(1) 生体試料または環境試料から核酸を単離する;
(2) ウイルスレプリキン構造の存在について核酸をスクリーニングする;そして
(3) ウイルスレプリキン構造の存在、その濃度および組成を、汚染ウイルスの存在と相関させる、
ことを含む方法を提供する。
【0034】
本発明の別の態様において、ウイルスレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの実施態様は、出現するウイルスの株(もしそのような株が出現するなら)中に存在する少なくとも1つのペプチドを含む。
【0035】
本発明はまた、本明細書に記載のように、ウイルスレプリキンに特異的に結合する抗体、ならびにウイルスレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、ウイルスレプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0036】
本発明はまた、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を有する、1つ以上の単離されたウイルスペプチドと、医薬として許容される担体とを含む治療用組成物を提供する。
【0037】
本発明の別の態様において、ウイルスレプリキンのmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列は、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸である、アンチセンス核酸分子が提供される。
【0038】
本発明のさらに別の態様において、ウイルスに対する抗体を産生するように被験体の免疫系を刺激する方法であって、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を有する少なくとも1つのウイルスレプリキンペプチドの有効量を投与することを含む方法が提供される。
【0039】
別の態様において、予防または治療用ウイルスワクチンに含有されるウイルスペプチドの選択方法であって:
(1) 複数の該ウイルス株の各株の少なくとも1つの分離株を得る;
(2) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列を、レプリキン配列の存在と濃度について分析する;
(3) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するウイルスの株を同定する;そして
(5) 工程(4)で同定されるウイルスペプチドの株に存在する少なくとも1つのレプリキン配列を、ウイルスワクチン中に含有させるためのペプチドとして選択する、
ことを含む方法が提供される。
【0040】
本発明はまた、予防または治療用ウイルスワクチンの作成方法であって:
(1) ウイルスの株を出現する株として同定する、
(2) 出現する株中に存在する少なくとも1つのレプリキン配列を、ウイルスワクチン製造のためのペプチド鋳型として選択する、
(3) 工程(2)で選択した少なくとも1つのレプリキン配列のアミノ酸配列を有するペプチドを合成する、そして
(4) 治療上有効量の工程(3)のペプチドを医薬として許容される担体および/またはアジュバントと組合わせる、
ことを含む方法が提供される。
【0041】
別の態様において本発明は、診断、予防または治療目的の出現するウイルス株を同定する方法であって:
(1) 複数の該ウイルス株の各株の少なくとも1つの分離株を得る;
(2) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列を、レプリキン配列の存在と濃度について分析する;
(3) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);そして
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するウイルスの株を同定する
ことを含む方法が提供される。
【0042】
本発明のさらに別の態様において、出現するウイルス株のタンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ウイルスワクチンが提供される。
【0043】
また本発明により、必要な患者に、出現するウイルス株のタンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ウイルスワクチンを投与することを含む、ウイルス感染症を予防または治療する方法が提供される。
【0044】
インフルエンザ
インフルエンザは、世界的に重要な急性の呼吸系疾患である。ワクチン接種によりインフルエンザウイルス大流行を抑制しようとする国際的な試みにもかかわらず、インフルエンザ感染症はいまだに、罹患と死亡の重要な原因である。歴史的に不規則で以前は予測できなかった間隔で、世界的なインフルエンザの流行と汎流行が起きており、これはこれからも続くと予測される。罹患率、死亡率、および経済コストの点で、汎流行と流行のインフルエンザの影響は大きい。
【0045】
インフルエンザワクチンは、いまだにインフルエンザウイルスに対する最も有効な防御法であるが、このウイルスの突然変異能力と非ヒト宿主を利用できるために、インフルエンザは今後も出現するかまたは再出現する感染症であるであろう。世界的なインフルエンザ監視は、インフルエンザシーズンの間に、国の中で、国と国の間で、および大陸間で、インフルエンザウイルスが変化することを示している。ウイルスの監視は、抗原性のシフトとドリフトの追跡において重要である。疾患の監視はまた、流行の影響を評価するのに重要である。両方の情報がワクチン組成物の基礎と抗ウイルス剤の正しい使用を提供している。しかし今日まで、増加する数のインフルエンザウイルスの出現する株について毎年事後の血液学的分類があるのみで、近づいてくるインフルエンザの流行または汎流行の指標としての、ウイルスの特異的な化学構造は同定されていない。現在、ある年に活動性、非活動性または流行性としてのインフルエンザウイルスの毎年の分類の唯一の基礎は、ウイルス血球凝集素とノイラミニダーゼタンパク質の活性である。流行または汎流行の定量的警告のために、またはより有効で安全なワクチンを設計するために、使用できるインフルエンザウイルスの化学構造は本出願前には同定されていない。
【0046】
インフルエンザワクチンを毎年投与することとワクチンを投与できる期間が短いことのために、ワクチンの適用範囲を改良するための方策は非常に重要である。
【0047】
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成されたインフルエンザウイルスペプチドが提供される。インフルエンザレプリキンウイルスペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸(インフルエンザレプリキン)を含む。
【0048】
本発明の別の態様において、インフルエンザウイルスのレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのインフルエンザウイルスのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの好適な実施態様において組成物は、インフルエンザウイルスの出現する株中に存在する少なくとも1つのペプチドを含む。
【0049】
本発明はまた、本明細書に記載のように、インフルエンザウイルスのレプリキンに特異的に結合する抗体、ならびにインフルエンザウイルスのレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、インフルエンザレプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0050】
本発明はまた、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する1つ以上の単離されたインフルエンザウイルスペプチドと、医薬として許容される担体とを含む治療用組成物を提供する。
【0051】
本発明の別の態様において、インフルエンザウイルスの血球凝集素レプリキンのmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列が、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を意味する、アンチセンス核酸分子が提供される。
【0052】
本発明のさらに別の態様において、インフルエンザウイルスに対する抗体を産生するように被験体の免疫系を刺激する方法であって、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する少なくとも1つのインフルエンザウイルスのレプリキンペプチドの有効量を投与することを含む方法が提供される。
【0053】
さらに別の態様において、予防または治療用インフルエンザウイルスワクチンに含有されるインフルエンザウイルスペプチドの選択方法であって:
(1) 複数のインフルエンザ株の各株の少なくとも1つの分離株を得る;
(2) レプリキン配列の存在と濃度について、複数のインフルエンザ株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列を分析する;
(3) 複数のインフルエンザ株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株の血球凝集素アミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するインフルエンザウイルス株を同定する;そして
(5) 工程(4)で同定されるインフルエンザウイルス株のペプチドに存在する少なくとも1つのレプリキン配列を、インフルエンザウイルスワクチン中に含有させるためのペプチドとして選択する、
ことを含む方法が提供される。
【0054】
本発明はまた、予防または治療用インフルエンザウイルスワクチンの作成方法であって:
(1) インフルエンザウイルス株を出現する株として同定する、
(2) 出現する株中に存在する少なくとも1つのレプリキン配列を、インフルエンザウイルスワクチン製造のためのペプチド鋳型として選択する、
(3) 工程(2)で選択した少なくとも1つのレプリキン配列のアミノ酸配列を有するペプチドを合成する、そして
(4) 治療上有効量の工程(3)のペプチドを医薬として許容される担体および/またはアジュバントと組合わせる、
ことを含む方法が提供される。
【0055】
別の態様において本発明は、診断、予防または治療目的のインフルエンザウイルスの出現する株を同定する方法であって:
(1) 複数のインフルエンザ株の各株の少なくとも1つの分離株を得る;
(2) レプリキン配列の存在と濃度について、複数のインフルエンザウイルス株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列を分析する;
(3) 複数のインフルエンザウイルス株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株の血球凝集素アミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);そして
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するインフルエンザウイルスの株を同定する
ことを含む方法が提供される。
【0056】
本発明のさらに別の態様において、インフルエンザウイルスの出現する株の血球凝集素タンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用インフルエンザウイルスワクチンが提供される。
【0057】
また本発明により、必要な患者に、インフルエンザウイルスの出現する株のタンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ウイルスワクチンを投与することを含む、インフルエンザウイルス感染を予防または治療する方法が提供される。
【0058】
トリパノソーマ
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成されたトリパノソーマペプチドが提供される。トリパノソーマレプリキンペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基(トリパノソーマレプリキン)を含む、7〜約50個のアミノ酸(トリパノソーマレプリキン)を含む。
【0059】
マラリア
治療が困難で有効なワクチンが存在しない1つのトリパノソーマ疾患はマラリアである。マラリアは、熱帯地域で高い死亡率と、肉体的および経済的苦痛を与える。マラリアは、主に熱帯熱マラリア原虫(Plasmodium falciparum)により引き起こされ、これは、極めて難治性であり、今日までマラリアのワクチンは得られていない。すなわち、有効なマラリアワクチン、およびこの疾患を治療または予防する方法に対するニーズがある。本出願は、そのようなワクチンと治療および予防方法の基礎を提供する。上記したレプリキンウイルスワクチンおよびレプリキンインフルエンザウイルスワクチンの産生およびこれらを用いる治療方法は、レプリキンマラリアワクチンの産生およびこれを用いる治療に応用できる。
【0060】
本発明では、マラリアを予防または治療するためのワクチンと方法が提供される。マラリアワクチンは、少なくとも1つの単離された熱帯熱マラリア原虫レプリキンを含む。本発明はまた、少なくとも1つの単離された熱帯熱マラリア原虫レプリキンを含む予防または治療用ワクチン組成物の有効量を、患者に投与することを含む、マラリアの治療または予防方法を提供する。
【0061】
また本発明により、熱帯熱マラリア原虫のマラリア抗原中に存在するレプリキンに特異的に結合する抗体、抗体カクテル、および抗体を含む組成物が提供される。
【0062】
マラリアのように本発明により治療されるトリパノソーマの別の例である梅毒トレポネーマ(Treponema pallidum)(梅毒)のレプリキンは、梅毒の検出、予防、治療に使用することができる。
【0063】
細菌
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成された細菌ペプチド(細菌レプリキン)が提供される。この細菌ペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸(細菌レプリキン)を含む。US特許出願第10/105,232号(2002年3月26日出願)は、参照することによりその全体(特に限定されないが、細菌配列リストと情報を含む)が本明細書に組み込まれる。
【0064】
本発明はまた、生体試料または環境試料中の汚染細菌の存在を検出する方法であって:
(1) 生体試料または環境試料から核酸を単離する;
(2) レプリキン構造の存在について核酸をスクリーニングする;そして
(3) レプリキン構造の存在を汚染生物の存在と相関させる、
ことを含む方法を提供する。
【0065】
本発明の別の態様において、細菌レプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つの細菌のレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの好適な実施態様は、出現する細菌株(そのような新しい株が出現する場合)中に存在する少なくとも1つの細菌ペプチドを含む。
【0066】
本発明はまた、本明細書に記載のように、細菌のレプリキンに特異的に結合する抗体、ならびに細菌のレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、細菌レプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0067】
本発明はまた、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する1つ以上の単離された細菌ペプチド;および
(4) 医薬として許容される担体
を含む治療用組成物を提供する。
【0068】
本発明の別の態様において、細菌のレプリキンのmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列が、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を意味する、アンチセンス核酸分子が提供される。
【0069】
本発明のさらに別の態様において、細菌に対する抗体を産生するように被験体の免疫系を刺激する方法であって、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する少なくとも1つの細菌のレプリキンペプチドの有効量を投与することを含む方法が提供される。
【0070】
さらに別の態様において、予防または治療用細菌ワクチンに含有される細菌レプリキンペプチドの選択方法であって:
(1) 複数の細菌株の各株の少なくとも1つの分離株を得る;
(2) 細菌レプリキン配列の存在と濃度について、複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列を分析する;
(3) 複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列中の細菌レプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察される細菌レプリキン配列の濃度と比較して、少なくとも2つの時期の細菌レプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内、または迅速に突然変異する細菌ではそれより早い時期である);
(4) 少なくとも2つの時期の間の細菌レプリキン配列濃度の最大の上昇を有する細菌の株を同定する;そして
(5) 工程(4)で同定される細菌ペプチドの株に存在する少なくとも1つの細菌レプリキン配列を、細菌ワクチン中に含有させるためのペプチドとして選択する、
ことを含む方法が提供される。
【0071】
本発明はまた、予防または治療用細菌ワクチンの作成方法であって:
(1) 細菌の株を出現する株として同定する、
(2) 出現する株中に存在する少なくとも1つの細菌レプリキン配列を、細菌ワクチン製造のためのペプチド鋳型として選択する、
(3) 工程(2)で選択した少なくとも1つの細菌レプリキン配列のアミノ酸配列を有するペプチドを合成する、そして
(4) 治療上有効量の工程(3)のペプチドを医薬として許容される担体および/またはアジュバントと組合わせる、
ことを含む方法が提供される。
【0072】
別の態様において本発明は、診断、予防または治療目的の細菌の出現する株を同定する方法であって:
(1) 複数の細菌株の各株の少なくとも1つの分離株を得る;
(2) 細菌レプリキン配列の存在と濃度について、複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列を分析する;
(3) 複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列中の細菌レプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察される細菌レプリキン配列の濃度と比較して、少なくとも2つの時期の細菌レプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);そして
(4) 少なくとも2つの時期の間の細菌レプリキン配列濃度の最大の上昇を有する細菌の株を同定する
ことを含む方法が提供される。
【0073】
本発明のさらに別の態様において、細菌の出現する株のタンパク質中に存在する少なくとも1つの単離された細菌レプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用細菌ワクチンが提供される。
【0074】
細菌(マイコバクテリア(Mycobacteria))として分類される2つの重要な細菌の亜種は、その30sリボゾームタンパク質がC末端レプリキンを有するらい菌(Mycobacterium leprae)と、そのATPaseが3つのレプリキンを有する結核菌(Mycobacterium tuberculosis)である。
らい菌(Mycobacterium leprae)(らい病)の30s リボゾームタンパク質s6中のレプリキンは以下の通りである:
kvmrtdkh
結核菌(Mycobacterium tuberculosis)のATPaseのレプリキンは以下の通りである:
hprpkvaaalkdsyrlk
hprpkvaaalk
ksaqkwpdkflagaaqvah
大腸菌(E. coli)のB-D-ガラクトシダーゼ中のレプリキンは以下の通りである:
hawqhqgktlfisrk
hqgktlfisrk
アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)中のレプリキン:
hsdqqlavmiaakrlddyk
hlldhpasvgqldlramlaveevkidnpvymek
hpasvgqldlramlaveevkidnpvymek
kcvmakncnikcpaglttnqeafngdpralaqylmniah
kncnikcpaglttnqeafngdpralaqylmniah
hhdtysiedlaqlihdakaaruvivk
hdtysiedlaqlihdakaarvrvivk
hdakaarvrvivk
kigqgakpgeggqlpspkvtveiaaarggtpgvelvsppphh
kigqgakpgeggqlpspkvtveiaaarggtpgyelvsppph
kaseitktlasgamshgalvaaaheavahgtnmvggmsnsgeggeh
kaseitktlasgamshgalvaaaheavah
kaseitktlasgamshgalvaaah
kaseitktlasgamsh
kryfpnvktpvggvtfaviaqavadwh
hhiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaekslmk
hhiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaekslmk
hhiaaglgfgasavyplgvqfraeekfgadaokafkrfakaaek
hhiaaglgfgasavyplgvqfraeekfgadadkafkrfak
hhiaaglgfgasavyplgvqfraeekfgadadk
hiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaekslnk
hiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaek
hiaaglgfgasavyplgvqfraeekfgadadkafkrfak
hiaaglgfgasavyplgvqfraeekfgadadk
kfglydaafeksscgvgfitrkdgvqth
【0075】
また本発明により、必要な患者に、細菌の出現する株のタンパク質中に存在する少なくとも1つの単離された細菌レプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ワクチンを投与することを含む、細菌感染症を予防または治療する方法が提供される。
【0076】
真菌
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成された真菌ペプチドが提供される。この真菌ペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基(トリパノソーマレプリキン)を含む、7〜約50個のアミノ酸(真菌レプリキン)を含む。
【0077】
細菌レプリキンワクチンの産生とこれを用いる治療について記載した上記のすべての方法は、真菌レプリキンワクチンの産生とこれを用いる治療に適用される。
【0078】
本発明の別の態様において、真菌のレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つの真菌レプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。
【0079】
本発明はまた、本明細書に記載のように、ウイルスレプリキンに特異的に結合する抗体、ならびにウイルスレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、真菌レプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0080】
本発明はまた、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する1つ以上の単離された真菌ペプチド;および
(4) 医薬として許容される担体
とを含む治療用組成物を提供する。
【0081】
本発明の別の態様において、真菌のレプリキンmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列が、
(1) 第2のリジン残基から6〜10残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する、上記分子が提供される。
【0082】
本発明の別の態様において、真菌のレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。
【0083】
複製の上昇
本発明のさらに別の態様において、生物中で複製機能を有する酵素または他のタンパク質をコードする遺伝子を、少なくとも1つのレプリキン構造を用いて形質転換することを含む、生物の複製速度を上昇させる方法が提供される。
定義
【0084】
本明細書において用語「ペプチド」または「タンパク質」は、1つのアミノ酸のカルボキシル基が別のアミノ酸のアミノ基に結合して、ペプチド結合を形成する2つ以上のアミノ酸の化合物を意味する。用語ペプチドはまた、そのような化合物をコードするアミノ酸配列を意味するのにも使用される。本明細書において「単離された」または「合成された」ペプチドまたはその生物活性のある部分は、そこからペプチドが得られる細胞または組織源からの細胞物質または他の汚染ペプチドを実質的に含まない、または任意の方法により合成される時化学前駆体または他の化学物質を実質的に含まない、または組換え遺伝子技術により合成される時汚染ペプチドを実質的に含まない、ペプチドを意味する。
【0085】
本明細書において、レプリキンペプチドまたはレプリキンタンパク質は、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸配列である。同様にレプリキン配列は、そのようなペプチドまたはタンパク質をコードするアミノ酸配列である。
【0086】
本明細書において、「出現する株」は、他の株の生物中のレプリキンの濃度と比較して、1つ以上のタンパク質配列中のレプリキン配列の濃度が上昇しているとして同定されたウイルス、細菌、真菌、または他の生物の株を意味する。レプリキン濃度の上昇は、例えばインフルエンザウイルスでは、少なくとも6ヶ月の期間、好ましくは少なくとも約1年間、最も好ましくは少なくとも約3年間またはそれ以上の期間起きるが、細菌や他の生物でははるかに短い期間である。
【0087】
本明細書において「突然変異」は、アミノ酸の置換により引き起こされる生物の構造および性質の変化を意味する。これに対して、本明細書において用語「保存」は、置換の欠如のための特定のアミノ酸の保存を意味する。
【図面の簡単な説明】
【0088】
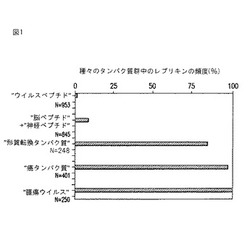
【図1】図1は、種々の生物中のレプリキンの出現頻度を示す棒グラフである。
【図2】図2は、神経膠腫細胞の嫌気的複製中の、総膜タンパク質1mg当たりのマリグニンのパーセントを示すグラフである。
【図3】図3は、リコグニン(recognin)16量体への暴露に応答して産生される抗マリグニン抗体の量を示す棒グラフである。
【図4A】図4Aは、普通の蛍光灯で撮った血液スメアの写真である。
【図4B】図4Bは、2つの白血病細胞の存在を例示する普通の蛍光灯で撮った血液スメアの写真である。
【図4C】図4Cは、抗マリグニン抗体の存在下の神経膠腫細胞の密な層の写真である。
【図4D】図4Dは、抗マリグニン抗体の添加の30分後に撮った、図4Cの細胞の層の写真である。
【図4E】図4Eは、抗マリグニン抗体の添加の45分後に撮った、図4Cの細胞の層の写真である。
【図4F】図4Fは、抗マリグニン抗体による小細胞肺癌細胞の増殖のin vitroでの阻害を示す棒グラフである。
【図5】図5は、手術前および後の良性または悪性乳癌患者の血清中に存在する抗マリグニン抗体の量のプロットである。
【図6】図6は、レプリキン配列を同定する3点認識法を実施するのにコンピューターが使用される、本発明の実施態様を示すボックス図である。
【図7】図7は、1918年から2001年まで毎年、インフルエンザBとインフルエンザA株(H1N1)の血球凝集素で観察されたレプリキンの濃度を示すグラフである。
【図8】図8は、1950年から2001年まで毎年、インフルエンザA株(H2N2とH3N2)、ならびに成分レプリキンにより規定された出現する株の血球凝集素で観察されたレプリキンの濃度を示すグラフである。
【発明を実施するための形態】
【0089】
発明の詳細な説明
迅速な複製の現象に関連した小ペプチドの新しいファミリー(ここで、レプリキンと呼ぶ)の同定は、試料中の病原体の検出のための、および治療法の開発(ワクチン開発を含む)のための標的となる。一般に、このファミリーのペプチドの知識および同定は、レプリキンを有する任意の生物の有効な治療法やワクチンの開発を可能にする。このファミリーのペプチドの同定はまた、ウイルスの同定およびウイルスワクチンの開発を可能にする。
【0090】
例えば、このファミリーのペプチドの同定は、インフルエンザウイルスの検出を提供し、インフルエンザ治療法の新しい標的を提供する。このファミリーのペプチドの同定はまた、例えばマラリアの検出を提供し、マラリアワクチンの開発のための新しい標的を提供する。このファミリーのペプチドの同定により提供されるさらなる例には、感染症レプリキン、癌免疫レプリキンおよび構造タンパク質レプリキンの検出がある。
【0091】
迅速な複製は、いくつかの細菌、ウイルスおよび悪性腫瘍の病原性に特徴的であるが、異なった生物の迅速な複製に共通する化学はこれまで開示されていない。我々は、迅速な複製に関連する保存された小タンパク質の新しいファミリーを見いだし、これをレプリキンを名付けた。そのようなレプリキンは、有効な検出法と治療法を開発するための新しい標的を与える。見いだされた最初のレプリキンは、神経膠腫レプリキンであり、これは、マリグニン(Malignin)と呼ぶ脳の多形神経膠芽腫(神経膠腫)細胞タンパク質中で同定された。
【0092】
マリグニンの加水分解と質量スペクトル測定により、神経膠腫レプリキンを含有する新規の16量体のペプチド配列が明らかになった。このレプリキンは、健常人ゲノムのデータベース中には見つからず、従って体外の何らかの起源に由来するものと思われる。
【0093】
我々は、神経膠腫レプリキンまたはその同族体を検索するためのアルゴリズムを作成した。同族体は4,000を越えるタンパク質配列に共通するものではなかったが、驚くべきことに、すべての腫瘍ウイルス、および藻類、植物、真菌、ウイルスおよび細菌の複製タンパク質中に存在した。
【0094】
我々は、1) レプリキン濃度(100アミノ酸当たりのレプリキンの数)、および2) レプリキン組成の両方が、迅速な複製の機能的現象に相関することを確認した。これらの関係は、レプリキンが複製速度に定量的および定性的に関係することを決定するための機能的基礎を提供する。
【0095】
迅速な複製へのレプリキンの役割の最初の機能的基礎は、神経膠腫複製で見られる。神経膠腫細胞の迅速な複製中で細胞数と膜タンパク質との5倍の増加に比較して、神経膠腫マリグニンが10倍濃縮されている事実は、レプリキンと複製の基本的な関係を示唆する。神経膠腫レプリキンをin vitroで合成し、合成ワクチンとしてウサギにに投与すると、多量の抗マリグニン抗体が産生された。これは、血清中の抗マリグニン抗体(AMAS)試験の抗原性基礎を確立し、他の生物中の最初の合成癌ワクチン候補およびレプリキンワクチンの原型を提供する。癌レプリキンと迅速な複製の共有される特異性に基づく、レプリキンと複製との天然の免疫関係、および癌レプリキンに対するこの天然の免疫応答(これは、細胞タイプに無関係である)の証明により、抗マリグニン抗体による免疫の受動的増強と合成レプリキンによる能動免疫増強の両方が可能である。
【0096】
抗マリグニン抗体の存在と患者の生存との関係が、癌患者の8,090個の血清試料の研究により証明された。この試験は、集団中の癌の発症率のように、抗マリグニン抗体の濃度が年齢とともに上昇すること、および細胞タイプに無関係に早期悪性腫瘍でさらに2〜3倍増加することを証明した。in vitroでは、抗マリグニン抗体は、癌細胞当たりピコグラム(フェムトモル)の癌細胞に細胞障害性であり、in vivoでは抗マリグニン抗体の濃度は、癌患者の生存に定量的に関連する。神経膠腫細胞で証明されるように、細胞は不死化の悪性段階に変換されるのみで停止もしくは休止している段階は、レプリキンの濃度の上昇を特徴とするより活発な生命に危険のある複製段階(これは、レプリキンの濃度の上昇が特徴である)とは区別される。さらに癌のウイルス病原性への鍵は、神経膠腫糖タンパク質10Bは、正常な10Bと比較すると、炭水化物残基が50%低下しているという事実にあるかも知れない。この低下は、他の例のウイルス侵入に関連し、従って、悪性段階への変換の工程として神経膠腫細胞の10Bへのウイルスレプリキンの供給のための、ウイルスの結合の証拠かも知れない。
【0097】
過去100年間のインフルエンザウイルス血球凝集素のタンパク質配列とインフルエンザ流行についての我々の研究は、レプリキンと迅速な複製との関係の第2の機能的基礎を提供した。今日まで、インフルエンザについては血清学的血球凝集素と抗体の分類のみが開示され、株特異的保存ペプチド配列は開示されていない。さらに、疫学的に記録された流行病または迅速な複製と相関する株特異的ペプチド配列の濃度と組成の変化は、記載されていない。この研究では、4つの主要な株(インフルエンザB、(A)H1N1、(A)H2N2、および(A)143N2)中のそれぞれの1つで株特異的インフルエンザレプリキン濃度の4〜10倍の増加が、1902〜2001年に各株により引き起こされたインフルエンザ流行に関連していることが証明されている。
【0098】
次に我々は、これらの濃度の増加は、その消失から1〜64年後の少なくとも1つの特異的レプリキン組成の再出現と、新しい株特異的レプリキン組成の出現によることを証明した。以前は、次のインフルエンザシーズにどの株が主流になるかを予測するための、またはワクチンのためのウイルス全体の株の毎年の混合物を作成するための、株特異的化学構造は不明であった。H3N2レプリキン濃度の最近の急激な上昇(1997〜2000)(H3N2の歴史で最大)および1968年の高死亡率のH3N2汎流行と1975年と1977年の2つの高死亡率流行で最後に見られた特異的レプリキン組成の再出現(しかし20〜25年は出現しなかった)は一緒に、これから来る流行を警告しているかも知れない。同じ構造が100年間持続するか、または1〜64年の欠如後に再出現する、観察されるレプリキン構造の高度の保存は、インフルエンザタンパク質中のアミノ酸のランダム置換による変化であると以前考えられていたものは、レプリキンの保存の組織化されたプロセスによる変化である可能性が高いことを示す。
【0099】
レプリキンの保存は、インフルエンザウイルスにユニークなものではなく、他の供給源、例えば手足口病ウイルスO型およびHIV、ならびにコムギでも見られた。
【0100】
迅速な複製におけるレプリキンの役割の第3の機能的基礎は、HIV中の迅速な複製の上昇に見られる。レプリキン濃度は、HIV中の迅速な複製に関連することが証明された。我々は、増殖の遅い低力価HIV株(NS1、「Bru」)中のレプリキン濃度は、迅速に増殖している高力価HIV株(SI、「Lai」)(後期HIV感染によくある)中のレプリキン濃度の6分の1であることを見いだした。
【0101】
さらなる例は、レプリキンと迅速な複製との関係を示す。トマトカールリーフジェミニウイルス(これは、トマトの収穫に壊滅的被害を与える)の「複製タンパク質」では、DNAに結合することが証明されているの最初の161 アミノ酸がレプリキンを含有することが証明された。マラリア(迅速な複製で有名)では、1つのトリパノソーマから数万のトリパノソーマが肝臓中に放出されると、100アミノ酸当たり最大36個の重複レプリキンの濃度の、複数の新規なほとんど「火炎状」のレプリキン構造が見られた。
【0102】
ある構造の保存は、その構造が攻撃と破壊または刺激のための安定な不変の標的となるかどうかにとって決定的に重要である。ある構造がその生物の基本的な生存機構と何らかの意味で関係している時は、その構造は保存される傾向にある。変化する構造は、変わりやすい標的となり、これは、攻撃物を避けるための良好な方法(例えば、以前の構造に対して特異的に作成された抗体)であり、従って修飾型に対しては無効である。この方法が、例えばインフルエンザウイルスにより利用され、以前のワクチンは、現在の病原性ウイルスに対して全く無効となることがある。
治療のための安定な標的としてのレプリキン
【0103】
細菌とHIVともに、レプリキンおよび非レプリキンアミノ酸を有する。例えば最近HIVでは、突然変異により薬剤耐性が9%から13%まで上昇しており、これは、非レプリキンアミノ酸の置換である(本明細書に記載のHIVのTATタンパク質の詳細な分析を参照されたい)。細菌では「耐性株」の出現は、同じ機構による。しかし我々は、レプリキン構造が、非レプリキンアミノ酸と同じ程度までには突然変異も変化もしないことを見いだした(また、本明細書に記載のレプリキンの手足口病ウイルスの説明も参照されたい)。非レプリキン構造とは反対にレプリキン構造は、保存され、従って治療の新しい恒常的標的となる。
【0104】
生存機能にあまりにも密接に関連したいくつかの構造は、絶えず変化することはできない。レプリキン構造の必須成分はヒスチジン(h)であり、これはしばしば、酸化還元酵素の金属基に結合し、おそらく複製に必要なエネルギーの源であることが知られており、このヒスチジン構造は変化しないため、この構造は、破壊または刺激のさらに魅力的な標的である。
【0105】
プロテオミクス的観点から、新に決定された神経膠腫ペプチド配列に基づく鋳型の本発明者らの構築により、関連する保存構造と特定の機能(この場合は複製)を有する広いクラスのタンパク質が発見された。疾患の病原性とのレプリキン濃度の上昇の例には、インフルエンザ、HIV、癌およびトマトリーフカールウイルスがある。この新たに認識されたクラスの構造は、酵母、藻類、植物、ジェミニカールリーフトマトウイルス、HIVおよび癌のような多様な生物の迅速な複製の現象に関連する。
【0106】
レプリキン濃度と組成は、複製(これは、各生物集団の生存と支配にとって重要である)のプロセスを検出し制御するための新しい定量的方法を提供する。神経膠腫と関連する癌のレプリキンについての抗マリグニン抗体で示されるように、多様なクラスのメンバーによる免疫学的特異性の共有は、B細胞とその生成物である抗体が、同様の認識言語を使用してレプリキンを認識することを示唆する。
【0107】
癌レプリキンのペプチド配列またはレプリキンを含有するペプチド配列(すなわち、神経膠腫ペプチドkagvaflhkkの同族体)の例は、特に限定されないが、肺、脳、肝臓、軟組織、唾液腺、鼻咽腔、食堂、胃、結腸、直腸、胆嚢、乳房、前立腺、子宮、子宮頸部、膀胱、眼、黒色腫、リンパ腫、白血病、および腎臓の癌に見いだされる。
【0108】
レプリキンは、1) レプリキンの定性的および定量的測定による病原体の検出;2) 未変性のレプリキンを標的化とすることにより、および合成レプリキンをワクチンとして使用することにより、迅速な複製が重要な要因である広範囲の疾患の治療と制御;および3) 藻類および植物性食物の増殖速度の上昇の促進、を提供する。
【0109】
同定された最初のレプリキン配列は、脳癌タンパク質マリグニン中に見いだされた癌細胞レプリキンであり、これは、多形神経膠芽腫(神経膠腫)細胞の迅速な嫌気的複製中に10倍濃縮されていることが証明された(図2)。マリグニンは、多形神経膠芽腫(神経膠腫)細胞からin vivoおよびin vitroで単離された250 kDaの糖タンパク質10Bの10 KDa部分である。マリグニンの加水分解と質量スペクトル測定により、16量体のペプチド配列、ykagvaflhkkndide(配列番号4)が明らかになり、これは本明細書において神経膠腫レプリキンと呼び、より短いペプチドkagvaflhkk(配列番号1)を含み、この両方とも正常ヒトゲノムには存在しない。
【表1】

【0110】
16量体の神経膠腫レプリキンを合成し、合成ワクチンとしてウサギに注射すると、多量の抗マリグニン抗体が産生された(Bogochら、Cancer Detection and Prevention, 26 (増刊 1):402(2002))。in vivoの血清中の抗マリグニン抗体の濃度は、癌患者の生存に定量的に関連することが証明されている(Bogochら、Protides of Biological Fluids, 31:739-747 (1984))。in vitro抗マリグニン抗体は、癌細胞当たりピコグラム(フェムトモル)濃度で癌細胞に細胞障害性であることが証明されている(Bogochら、Cancer Detection and Prevention, 26 (増刊 1):402(2002))。
【0111】
本発明者らが行った研究は、神経膠腫レプリキンが健常人ゲノム中に存在しないことを示した。従って、種々の生物の公表された配列の分析により、レプリキン配列の起源と同族体候補の検索を行った。
【0112】
16量体の神経膠腫レプリキン配列を鋳型として使用し、そして認識プロテオミクスシステムを構築して、いくつかの異なる生物のタンパク質のアミノ酸配列を視覚的にスキャンすることにより、新しいクラスのペプチド(レプリキン)が同定された。本発明は、レプリキン配列を含むヌクレオチド配列またはアミノ酸配列の同定法を提供する。この方法は本明細書において、3点認識法と呼ぶ。3点認識法は:(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸のペプチド(レプリキン)を含む。これらのペプチドまたはタンパク質は、複製、形質転換、または酸化還元機能を有する藻類、酵母、真菌、アメーバ、細菌、植物、ウイルス、および癌タンパク質を含む種の新しいクラスのペプチドを構成する。レプリキンペプチドは、より大きい「複製」および「形質転換」タンパク質(その研究者らによりそう記載された、表2を参照)および癌細胞で濃縮されていることがわかった。マリグニン16量体ペプチドと同一の配列は見つかっていない。
【表2】
【表3】
【0113】
レプリキンとしてのまたはレプリキンを含有するもの(すなわち、神経膠腫ペプチド kagvaflhkkの同族体)としてのアミノ酸配列の同定には、以下の3つの要件が満たされなければならない。3点認識システムでは、7〜約50個の残基のアミノ酸配列以内に配列は3つの要素を有する:(1) 別のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジンの組成。
【0114】
レプリキン配列を含有するタンパク質配列について、ナショナルライブラリーオブメディシン(National library of Medicine)のキーワード「PubMed」記述子を使用して、データベースを検索した。4,000を越えるタンパク質配列を、同族体について視覚的に調べた。PubMedで分類されたタンパク質の各群内のすべての個々のタンパク質の配列を、上記の3つの要件を満たすペプチドについて視覚的に調べた。全体に「ウイルスペプチド」(1.5%(N=953))、および悪性形質転換または複製に関連するとされていない他のペプチド、例えば「脳ペプチド」および「神経ペプチド」(まとめて8.5%)(N=845)で、低率の同族体が観察された。しかし驚くべきことに、同族体は大きな「複製タンパク質」中でより高頻度に同定され、これらは細菌、藻類およびウイルスで複製の確立された機能を有するとして同定された。さらに驚くべきことは、「腫瘍ウイルス」(N=250)の100%、「癌タンパク質」(N=401)の97%、および「形質転換ウイルス」(N=248)の85%に存在するという知見である。これらの結果は、細胞タイプに無関係に癌の病原性に共通の性質があることを示唆し、発癌性でのウイルスの役割(すなわち休止状態であるが形質転換された状態から、より病原性の活性な複製状態への細胞の変換)を示唆する。
【0115】
3点認識法により定義される以下のアミノ酸配列 kagvaflhkk の同族体は、特に限定されないが、アデノウイルス(adenovirus)、レンチウイルス(lentivirus)、a-ウイルス(a-virus)、レトロウイルス(retrovirus)、アデノ関連ウイルス(andenoassociated virus)、ヒト免疫不全症ウイルス(human immunodeficiency virus)、肝炎ウイルス(hepatitis virus)、インフルエンザウイルス(influenza virus)、トウモロコシストリークウイルス(maize streak virus)、ヘルペスウイルス(herpes virus)、ウシヘルペスウイルス(bovine herpes virus)、ネコ免疫不全症ウイルス(feline immunodeficiency virus)、手足口病ウイルス(foot and mouth disease virus)、天然痘ウイルス(small pox virus)、ラウス肉腫ウイルス(rous sarcome virus)、神経芽腫RASウイルス癌遺伝子(neuroblastoma RAS viral oncogene)、ポリアマウイルス(plyamavirus)、ヒトパピローマウイルス(human papilloma virus)、骨髄性単球性腫瘍ウイルス(myelomonocytic tumor virus)、マウス急性白血病(murine acute leukemia)、T細胞リンパ栄養性ウイルス(T-cell lymphotropic virus)、およびトマトリーフカールウイルス(tomato leaf curl virus)のようなウイルスまたはウイルスペプチド中に存在する。
【0116】
レプリキンは、特に限定されないが、アセトバクター(Acetobacter)、アクロモバクター(Achromobacter)、アクチノミセス(Actinomyces)、アエロバクター(Aerobacter)、アルカリゲネス(Alcaligenes)、アルスロバクター(Arthrobacter)、アゾトバクター(Azotobacter)、バチルス(Bacillus)、ブレビバクテリウム(Brevibacterium)、チャイニア(Chainia)、クロストリディウム(Clostridium)、コリネバクテリウム(Corynebacterium)、エルウィニア(Erwinia)、エシェリキア(Escheria)、レブシエラ(Lebsiella)、ラクトバチルス(Lactobacillus)、ヘモフィルス(Haemophilus)、フラボバクテリウム(Flavobacterium)、メチロモナス(Methylomonas)、ミクロコッカス(Micrococcus)、マイコバクテリウム(Mycobacterium)、ミクロノムスポラ(Micronomspora)、マイコプラズマ(Mycoplasma)、ナイセリア(Neisseria)、ノカルジア(Nocardia)、プロテウス(Proteus)、シュードモナス(Pseudomonas)、リゾビウム(Rhizobium)、サルモネラ(Salmonella)、セラチア(Serratia)、スタフィロコッカス(Staphylococcus)、連鎖球菌(Streptocossus)、ストレプトミセス(Streptomyces)、ストレプトスポランギウム(Streptosporangium)、ストレプトビルチシリウム(Streptovirticillium)、ビブリオ(Vibrio)、ペプチド、およびキサントモナス(Xanthomas)のような細菌に存在する。
【0117】
レプリキンは、特に限定されないが、ペニシリウム(Penicillium)、ディセウラ(Diseula)、オフィストーマ・ノボウリム(Ophiostoma novo-ulim)、ミコフィコフタ(Mycophycophta)、フィトフトラ・インフェスタンス(Phytophthora infestans)、アブシジア(Absidia)、アスペルギルス(Aspergillus)、カンジダ(Candida)、セファロスポリウム(Cephalosporium)、フサリウム(Fusarium)、ハンゼヌラ(Hansenula)、ムコール(Mucor)、パエシロミセス(Paecilomyces)、ピヒア(Pichia)、リゾプス(Rhizopus)、トルロプシス(Torulopsis)、トリコデルマ(Trichoderma)、およびエリシフェ(Erysiphe)のような真菌中に存在する。
【0118】
レプリキンは、特に限定されないが、サッカロミセス(Saccharomyces)、クリプトコッカス(Cryptococcus)(クリプトコッカス‐ネオフォルマンス(Cryptococcus neoformas)を含む)、シゾサッカロミセス(Schizosaccharomyces)、およびオリザ(Oryza)のような酵母中に存在する。
【0119】
レプリキンは、特に限定されないが、カルドフェラ(Caldophera)、イソレピスプロリフェラ(Isolepisprolifera)、ツノマタ(Chondrus)、グラシラリア(Gracilaria)、ゲリジウム(Gelidium)、カウレルパ(Caulerpa)、ラウレンシア(Laurencia)、クラドフェキサ(Cladophexa)、ホンダワラ(Sargassum)、ペニシロス(Penicillos)、サボテングサ(Halimeda)、コンブ(Laminaria)、ヒバマタ(Fucus)、アスコフィルム(Ascophyllum)、ワカメ(Undari)、ダルス(Rhodymenia)、マクロシスティス(Macrocystis)、キリンサイ(Eucheuma)、フササイミ(Ahnfeltia)、プテロクラシア(Pteroclasia)のような藻類中に存在する。
【0120】
レプリキンは、特に限定されないが、エントアメーバ(Entamoeba)(エントアメーバ・インバデンス(Entamoeba invadens)を含む)、アメービダエ(Amoebidae)、アカントアメーバ(Acanthamoeba)およびネグレリア(Naegleria)のようなアメーバ中に存在する。
【0121】
レプリキンは、特に限定されないが、アラビドプシス(Arabidopsis)、コムギ(wheat)、イネ(rice)、およびトウモロコシ(maize)のような植物中に存在する。
【0122】
補助的規格
レプリキンのサブタイプの分類を可能にするために、基本的な「3点認識」要件に追加のまたは「補助的規格」を加える:(a) 例えば、癌細胞タンパク質(例えば、形質転換タンパク質 P21B(K-RAS2B)、肺、表2、配列番号89)中の隣接するジ−およびポリリジンの共通の存在のような、およびTOLL様受容体のような構造に基づくか、または (b) 既存のATPase、チロシンキナーゼ、または表2に見られるような酸化還元活性のような、機能に基づく。
【0123】
機能性誘導体
本明細書に記載のレプリキンの「機能性誘導体」は、レプリキンに対して特異的な抗体と少なくとも免疫学的交差反応性の部分を保持する、レプリキンの断片、変種、類似体、または化学誘導体である。レプリキンペプチドの断片は、分子の任意のサブセットである。変種ペプチドは、例えば当該分野で公知の方法を使用して直接化学合成により作成される。レプリキンの類似体は、全タンパク質またはその断片に実質的に類似の非天然のタンパク質である。レプリキンの化学誘導体は、通常はペプチドの一部またはペプチド断片ではない追加の化学残基を含有する。
【0124】
図2で明らかなように、細胞複製の速度が上昇する嫌気的呼吸中に、マリグニンは濃縮される。すなわち、細胞数や総膜タンパク質の増加に比例して増加するのみではなく、休止状態の3%から出発して総膜タンパク質の30%に達するまで、濃度が10倍も濃縮される。神経膠腫細胞複製に伴うレプリキン濃度の顕著な上昇のこの明瞭な証明は、種々の生物で3点認識法により同定されたレプリキンの存在を示すかまたはこれに一致する。例えばレプリキンは、「サッカロミセス・セレビッシェ(Saccharomyces cerevisiae)複製結合タンパク質」(配列番号2)(hsikrelgiifdk);「トウモロコシストリークウイルスの複製関連タンパク質A」(配列番号8)(kyivcareahk)と(配列番号9)(kekkpskdeimrdiish);「黄色ブドウ球菌(Staphylococcus aureus)の複製関連タンパク質」(配列番号10)(kkektthnk);「ウシヘルペスウイルス4のDNA複製タンパク質」(配列番号11)(hkinitngqk);および「シチメンチョウ(Mealigrid)ヘルペスウイルス1複製結合タンパク質」(配列番号12)(hkdlyrllmk)のようなタンパク質中で同定された。トマトリーフカールジェミニウイルスの以前の研究は、ウイルス蓄積の制御が、ウイルス複製中のリーフDNAおよび他の複製タンパク質分子への、そのウイルスの「複製タンパク質」のアミノ酸1〜160の結合が関与らしいことを示す。この配列の分析は、この「複製タンパク質」のアミノ酸1〜163が、5つのレプリキン、すなわち(配列番号13)kfrinaknyfltyph、(配列番号14)knletpvnklfiricrefh、(配列番号15)hpniqaaksstdvk、(配列番号16)ksstdvkaymdkdgdvldh、および(配列番号17)kasalnilrekapkdfvlqfh を含有することを示した。
【0125】
表2は、レプリキン含有タンパク質がまた、しばしば酸化還元機能、およびタンパク質合成もしくは伸長、ならびに細胞複製に関係していることを示す。金属ベースの酸化還元機能との関連、嫌気的複製中のレプリキン含有神経膠腫マリグニン濃度の濃縮、および低濃度(ピコグラム/細胞)での抗マリグニンの細胞障害性(図4c〜f)はすべて、レプリキンが中枢呼吸生存機能に関連し、非レプリキンアミノ酸に特徴的な突然変異をあまり受けないことを示唆する。
【0126】
特に興味深いことに、調べたインフルエンザウイルスの個々の株のほとんどすべての血球凝集素タンパク質中で、100アミノ酸当たり少なくとも1つのレプリキンが存在することが観察された。インフルエンザウイルスの4つの主要な株(インフルエンザB、H1N1、H2N2、およびH3N2)のそれぞれの分離株の血球凝集素タンパク質中に存在することが観察されたレプリキン配列(各年についてそのアミノ酸配列データは入手できる(1902〜2001))を、下記の表3、4、5および6に示す。
【表4】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
【0127】
観察されたレプリキンの濃度とタイプ(すなわち、観察されたレプリキン組成)の両方は、インフルエンザの汎流行と流行に関連することがわかった。インフルエンザウイルス中のレプリキンの濃度を、過去1世紀(すなわち、1900年〜2001年)にわたって世界中でヒトおよび動物宿主から単離されたインフルエンザ株について、ナショナルライブラリーオブメディシン(National library of Medicine)「PubMed」データベースで公表された血球凝集素アミノ酸配列を視覚的にスキャンして調べた。次に、各株についてこれらのレプリキン濃度(100アミノ酸当たりのレプリキンの数、平均±SD)をプロットした。
【0128】
レプリキンの濃度は、インフルエンザの汎流行と流行の発生に直接関係することがわかった。インフルエンザB血球凝集素およびインフルエンザA株H1N1中のレプリキン濃度を図7に示し、他の2つの共通のインフルエンザウイルスA株(H2N2とH3N2)中のレプリキン濃度を図8(H2N2、H3N2)に示す。図8のデータはまた、その構成レプリキンにより規定されるようにインフルエンザウイルスの出現する新しい株を示す(H3N2(R))。
【0129】
各インフルエンザA株は、1つの汎流行に関与している(それぞれ、1918年、1957年、および1968年)。図7と8のデータは、調べた4つの共通のインフルエンザウイルスのすべての分離株のインフルエンザ血球凝集素タンパク質のそれぞれに、100アミノ酸当たり少なくとも1つのレプリキンが存在することを示し、生存レベルの複製の維持におけるレプリキンの機能を示唆している。1990年代には、H3N2の衰退中に多くのH3N2分離株にはレプリキンが存在しなかったが、H3N2分離株には高濃度の新しいレプリキンが現れ、これはH3N2(R)株の出現を規定する。
【0130】
レプリキン濃度のいくつかの性質は、すべての4つのインフルエンザウイルス株に共通であることが図7と8でわかる。まず濃度は、何年にもわたって循環性であり、上昇と低下の1回のサイクルは2〜30年の期間にわたって起きる。この上昇と低下は、血球凝集素とノイラミニダーゼ分類により、個々のインフルエンザウイルス株支配の既知の漸増‐漸減に一致する。第2に、汎流行に関与することがすでに証明されている各インフルエンザウイルス株のピークのレプリキン濃度が、汎流行の各3年間に具体的にかつ個々に関連することが観察された。例えば1918年の汎流行(ここでは、インフルエンザウイルス株H1N1が関与することが証明された)については、H1N1でレプリキンのピーク濃度が独立にあり(P1);1957年の汎流行(ここでは、インフルエンザウイルス株H2N2が関与することが証明された)については、H2N2でレプリキンのピーク濃度があり(P2);1968年の汎流行(ここでは、インフルエンザウイルス株H3N2が関与することが証明された)については、H3N2でレプリキンのピーク濃度がある(P3)ことが証明された。第3に、上記の各3年間の汎流行直後の年には、特異的なレプリキン濃度は顕著に低下し、これはおそらく各ケースに広く分布した免疫を反映している。すなわち、この汎流行後の低下は、関与していた汎流行(P1)直後のH1N1に特異的であり、その時のすべての株の一般的な性質ではない。インフルエンザB中のレプリキン濃度の増加は、H1N1中のレプリキン濃度の低下と同時に繰り返し起き、例えば1951年のEB1と1976年のEB2は両方とも、最も高い死亡率を有するインフルエンザB流行に関連していた。(Stuart-Harrisら、Edward Arnold Ltd. (1985))。第4に、第2のピーク濃度(これは、第1のピーク濃度上昇を越えた)は、それぞれ3つの汎流行の15年後に起き、この第2のピークは:1918年の汎流行の15年後のH1N1「流行」年(E1);1957年の汎流行の8年後のH2N2「流行」年(E2);1968年の汎流行の7年後のH3N2「流行」年(E3)の流行を伴った。特異的レプリキンのこれらの第2のピーク濃度は、株の回復を反映するかも知れない。第5に、各株の特異的レプリキン濃度のピークはしばしば、1つまたは両方の他の株のレプリキン濃度の低下に関連するようであり、宿主部位について株の間の競合を示唆する。第6に、35年(H2N2)〜60年(インフルエンザB)の期間にわたって各株のレプリキン濃度が低下する全体的傾向があるようである。これは、インフルエンザワクチンの一般的な使用の前に、1901年〜1964年のインフルエンザBの場合に明らかであったため、この低下は、ワクチンの影響とは言えないようである。インフルエンザBの場合、低下からのレプリキンの回復は1965年から起きるように見えるが、レプリキン濃度は1997年〜2000年に再度低下した(図7)。これは、最近の分離株中のインフルエンザBの低頻度に相関する。H1N1レプリキン濃度は、H1N1株の再出現と流行とともに1978〜1979年にピークをうち(図7)、次にH1N1流行とともに1996年にピークをうった(図7)。H1N1レプリキン濃度はまた、1997〜2000年に低下し、これらの年に得られた分離株では、H1N1株の存在は低下していた。H2N2レプリキンについては、36年間の低下からの回復は起きず(図8)、これは、最近の分離株からのH2N2の欠如に相関する。H3N2については、多くの分離株のレプリキン濃度は、1996〜2000年の間にゼロまで低下したが、他のH3N2分離株は、レプリキン濃度の大きな鋭い上昇を示した。これは、H3N2の亜株の出現を示す(これは本明細書においてH3N2(R)と呼ぶ)。
【0131】
図7と8は、レプリキン濃度がピークに達する前に、しばしば1〜3年の段階的上昇が観察されることを示す。この段階的上昇は、流行の発生(これは、レプリキンピークと同時に起きる)に先行する。すなわち、特定の株の濃度の段階的上昇は、その株が流行または汎流行を引き起こす可能性の最も高い候補であるというシグナルである。
【0132】
現在、インフルエンザウイルスのH3N2(R)株中のレプリキン濃度は増加している(図8、1997〜2000年)。H3N2レプリキン濃度の3つの類似の以前のピーク上昇は、1968年のH3N2ベースの汎流行(図8)、および1972と1975年のH3N2ベースの流行(図8)で起きたことがわかる。これらの汎流行および流行のそれぞれは、過度の死亡率に関連していた(Ailingら、Am J. Epidemiol., 113(1):30-43 (1981))。従って、1997〜2000年のH3N2レプリキンのH3N2(R)亜種の濃度の急激な上昇は、統計的に大きな流行または汎流行が近づく早期警告である。H3N2流行は2000年にロシアで起き(図8、E4)、2001年12月のCDC報告は、現在H3N2が世界中で最も頻繁に単離されるインフルエンザウイルス株であるとしている(罹患率と死亡率ウィークリーレポート(MMWR)、疾病管理センター(Centers for Disease Control);50(48):1084-68 (Dec.7, 2001)。
【0133】
インフルエンザウイルスの汎流行または流行の各場合に、新しいレプリキンが出現する。ある特定の分離株の特定の血球凝集素で2つの同じレプリキンは観察されていない。新しいレプリキンの出現が、別の動物または鳥類プールからの移行に対して、どの程度が突然変異であるかは不明である。ある場合には、毎年1つ以上の元々のレプリキン構造が保存され、同時に新しいレプリキンが出現する。例えば、インフルエンザウイルスB血球凝集素では、1919〜2001年に5つのレプリキンが絶えず保存され、一方同じ時期に26個のレプリキンが行き来した(一部は数年間欠如の後に再出現した)。特定のレプリキン構造の消失と再出現は、レプリキンが、新規突然変異ではなく別のウイルス宿主プールから戻ることを示唆する。
【0134】
H1N1レプリキンの場合、1918年の汎流行に関連するP1ピークで存在した2つのレプリキンは、1933年の回復E1ピーク中には存在しなかった(これは12の新しいレプリキンを含有する)。従って、絶えず保存されるレプリキンは、単独または組合せでも、ワクチンの最適な選択である。しかし、1年間の濃度上昇に伴う最近出現しているレプリキンでさえ持続し、さらに1年以上上昇し濃度ピークに達して流行を起こし、従って早期警告となり、合成レプリキンを用いてワクチン接種すべき時期を提供する(例えば、1990年代初期のH1N1、図7)。
【0135】
図7と8のデータは、インフルエンザタンパク質配列中の特定のレプリキンの存在と濃度と、インフルエンザの汎流行および流行の発生との、直接の関係を示す。すなわち、レプリキンの存在と濃度についてのインフルエンザウイルス血球凝集素タンパク質配列の解析は、インフルエンザ汎流行および/または流行の予測変数、ならびにインフルエンザワクチン形成の標的を提供する。
【0136】
インフルエンザウイルスB株中のレプリキンの組成:この株で同定された全部で26個のレプリキンのうち、以下の10個のレプリキンは、1902〜2001年に調べたすべてのインフルエンザB分離株中に存在する。重複するレプリキン配列を別々にリストする。リジンとヒスチジンは太字で示して、「3点認識」と一致する同族関係を示す。
kshfanlk(配列番号104)
kshfanlkgtk(配列番号105)
kshfanlkgtktrgklcpk(配列番号106)
hekygglnk(配列番号107)
hekygglnksk(配列番号108)
hekygglnkskpyytgehak(配列番号10)
hakaigncpiwvk(配列番号110)
hakaigncpiwvkktplklangtk(配列番号111)
hakaigncpiwvktplklangtkyrppak(配列番号112)
hakaigncpiwvktplklangtkyrppakllk(配列番号113)
【0137】
表3と4は、H1N1レプリキンと比較して、インフルエンザB血球凝集素のレプリキン構造にはるかに大きな安定性があるらしいことを示す。インフルエンザBは、どの汎流行にも関与せず、動物または鳥類宿主も無いようである(Stuart-Harrisら、Edward Arnold Ltd. (1985))。
【0138】
インフルエンザH1N1レプリキン:1918年(株が最初に出現し、その年の汎流行を引き起こした)から2000年までの配列が入手できるすべてH1N1分離株には、ただ1つのレプリキン「hp(v/i)tigecpkyv(r/k)(s/t)(t/a)k」のみが存在する(表4)。(「(v/i)」は、アミノ酸vまたはiが異なる年に同じ位置に存在することを示す)。H1N1はただ1つの持続性のレプリキンを含有するが、H1N1は、インフルエンザBより豊富なようである。H1N1について82年間で95の異なるレプリキン構造があり、インフルエンザB分離株では100年間で31の異なるレプリキンがある(表4)。新しいレプリキン構造の数の上昇は流行の年に見られ(表3、4、5および6)、総レプリキン濃度の上昇と相関する(図7と8)。
【0139】
インフルエンザH2N2レプリキン:インフルエンザH2N2は、1957年のヒト汎流行に関与した。1957年についてその株で同定された20個のレプリキンのうち3つは、PubMedで1995年までに試験が可能であったH2N2分離株のそれぞれで保存されていた(表5)。
ha(k/q/m)(d/n)ilekthngk(配列番号232)
ha(k/q/m)(d/n)ilekthngklc(k/r)(配列番号233)
kgsnyp(v/i)ak(g/r)synntsgeqmliiwq(v/i)h(配列番号238)
【0140】
しかしH1N1と比較して、1961年に始まるH2N2では13個のみの追加のレプリキンが見つかっている。新しいレプリキンの出現が少ないことは、何年間にもわたる分離株中のH2N2レプリキンの濃度の低下とH2N2分離株の出現の低下と相関する。
【0141】
インフルエンザH3N2レプリキン:インフルエンザH3N2は、1968年のヒト汎流行に関与していた。1968年に現れた5つのレプリキンは1977年後に消失したが、1990年代に再出現した(表6)。22年間持続した唯一のレプリキン構造はhcd(g/q)f(q/r)nekwdlf(v/i)er(s/t)kであり、これは1977年に最初に出現し、1998年まで持続した。1990年代半ばの12個の新しいH3N2レプリキンの出現(表6)は、同時期のレプリキン濃度の上昇(表8)と相関し、これらの分離株の一部からのすべてのレプリキンの同時消失とともに、最近の分離株のH3N2株の流行(図8)に相関し、これは新しい亜種H3N2(R)の出現を示唆する。
【0142】
図1と2は、インフルエンザ流行と汎流行が、インフルエンザウイルスのレプリキン濃度の上昇と相関し、これは、その消失の1〜59年後の少なくとも1つのレプリキンの再出現によることを示す。また、A株のみでは、新しい株特異的レプリキン組成の出現がある(表4〜6)。単一のタンパク質内の個々のレプリキンの繰り返しによるレプリキン濃度の上昇は、インフルエンザウイルスで起きるのはなく他の生物で見られるようである。
【0143】
異なるインフルエンザ株の活性の変化は、インフルエンザ血球凝集素の配列変化に関連すると考えられており、これはさらに、2つのあまり理解されていないプロセスの1つにより行われる置換の生成物である:i) 抗原ドリフト(血球凝集素分子の一連の点突然変異の蓄積によると考えられる)、またはii) 抗原シフト(ここでは、変化が大きいため、ヒトと非ヒト宿主のウイルスの間で遺伝的組換えが起きる)。まず現在のデータは、異なるインフルエンザ株の活性の変化が、非特異的配列変化に関連するよりも、流行に関連する株特異的レプリキンの濃度の上昇および複製の株特異的上昇に基づくか、またはこれに関連することを示唆する。さらに、どの配列変化が「ドリフト」または「シフト」によるのか、およびどれが、保存、宿主中の保存、および再出現によるのかを理解するために、データを調べた。データは、レプリキン濃度中の流行に関連した上昇が、血球凝集素についての既存のレプリキンの複製によるのではなく、その消失の1〜59年後の少なくとも1つのレプリキン組成の再出現、およびA株のみでは、新しい株特異的レプリキン組成の出現によることを示す(表3〜6)。すなわち、1951〜1977年のインフルエンザB流行におけるレプリキン濃度の上昇は、流行の年の新しいレプリキン組成の出現には関連せず、それ以前の年に出現し次に消失したレプリキン組成の再出現にのみ関連している(表3)。これに対して、A株については、以前消失したウイルスレプリキンの再出現以外に、新しい組成が現れる(例えば、1996年の流行の年のH1N1では、以前の6つのレプリキンの再出現以外に、10個の新しい組成が現れた)。A株のみ(インフルエンザBではそうではない)が非ヒト動物および鳥類宿主に接近できるため、「3点認識」の基本的要件以外に新しい組成に似ていないようである既存のヒトレプリキンの突然変異からではなく、おそらく非ヒト宿主からまったく新しい組成が得られる(表2〜5)。Bと比較した場合のH1N1の多産性と、汎流行はB株ではなく3つのA株によってのみ引き起こされているという事実の両方とも、本ウイルス宿主から新しいレプリキン組成を受けるヒトA株の能力の関数かも知れない。
【0144】
あるレプリキンは1年のみ現れ、消失し、今日まで再出現していない(表3〜6)。他のレプリキンは1〜81年消失し、同じレプリキン配列が再出現する。主要なレプリキン「k」および「h」アミノ酸、およびこれらの間のスペースは、以下の株特異的レプリキンについて(インフルエンザBの10個、H1N1の単一のレプリキン、およびH2N3の単一のレプリキン)、ならびに消失後の同一のレプリキンの再出現について、表23-6に示すように、何年にもわたって特定のレプリキンが絶えず存在するために保存される。残りの血球凝集素配列におけるレプリキン構造の中または外の両方の他のアミノ酸の顕著な代替または置換活性にもかかわらず、インフルエンザレプリキンのヒスチジン(h)は決して置換されず、そしてリジン(k)はまれに置換されるようである。この保存の例は、1918〜2000年の間一定のH1N1レプリキン「hp(v/i)tigecpkyv(r/k)(s/t)(t/a)k」(配列番号135)、1975〜1998年の間一定のH3N2レプリキン「hcd(g/q)f(q,r)nekwdlf(v/i)er(s/t)k」(配列番号277)、および1975年に初めて出現し25年間消失し、次に2000年に再出現したH3N2レプリキン「hqn(s/e)(e/q)g(t/s)g(q/y)aad(l/q)kstq(a/n)a(i/l)d(q/g)I(n/t)(g/n)k,(l/v)n(r/s)vi(e/c)k」(配列番号276)中に見られる。多くのアミノ酸が置換されたが、基本的なレプリキン構造(約50アミノ酸以下の中の、6〜10残基離れた2つのリジン、1つのヒスチジン、少なくとも6%のリジン)は保存された。
【0145】
全くランダムな置換では、これらのH1N1とH3N2レプリキンの持続も、1902年から2001年のインフルエンザBの10個のレプリキン構造の持続も、1919年の18量体のレプリキンの74年間の消失後の1993年の再出現も、不可能であったであろう。ランダムな置換ではなく、この一定性は、秩序だった制御されたプロセス、または少なくともレプリキン残基を保護して、これらが何らかの方法により固定または結合されることを示唆する(リジンはおそらく核酸に結合し、ヒスチジンはおそらく呼吸系酸化還元酵素に結合する)。この保存を制御する機構は現在のところ不明である。
【0146】
レプリキン構造の保存
レプリキン構造が保存されるかまたは広範な自然の突然変異を受けるかどうかを、手足口病ウイルス(FMDV)(これらのウイルスのタンパク質の突然変異は、何十年も世界的に記録されている)の種々の分離株のタンパク質配列をスキャンすることにより調べた。FMDV分離株のタンパク質配列を、全レプリキンと、特定のレプリキンで観察された成分レプリキンアミノ酸残基のそれぞれとの存在について、視覚的に調べた。
【0147】
隣接するアミノ酸で起きるように時間とともに広範な置換を受けるというより、レプリキン構造を構成するアミノ酸はほとんどまたは全く置換されず、レプリキン構造は保存される。
【0148】
例えば、FMDV O型のタンパク質VP1では、レプリキン(配列番号3)「hkqkivapvk」は、PubMedで報告された236分離株のうち78%で保存されており、各アミノ酸は、以下のように個々の分離株で保存されていることがわかった:his、95.6%;lys、91.8%;gln、92.3%;lys、84.1%;ile、90.7%;val、91.8%;ala、97.3%;pro、96.2%;ala、75.4%;およびlys、88.4%。高率の保存は、レプリキン構造の構造的および機能的安定性を示唆し、治療の一定の標的となる。
【0149】
同様に、HIVの異なる分離株中でそのレプリキンについて配列保存がみつかった(例えばHIV 1型の(配列番号5)「kcfncgkegh」または(配列番号6)「kvylawvpahk」、およびHIV 2型の(配列番号7)「kcwncgkegh」(表2))。配列保存のさらなる例は、HIV tatタンパク質中にあり、例えば(配列番号613)「hclvckqkkglgisygrkk」であり、ここで主要なリジンとヒスチジンアミノ酸は保存されている(表7を参照)。
【0150】
同様に、配列保存は植物(例えばコムギ、例えばコムギユビキチン(ubiquitin)活性化酵素E(配列番号614〜616))中で観察された。コムギ中のレプリキンは、本明細書に記載のような植物増殖の刺激の信頼できる標的となった。他の保存例は、神経膠腫細胞の10年以上の組織培養にわたっての連続的世代でのマリグニンの持続的な存在、および米国、英国、ヨーロッパおよびアジアからの8,090のヒト血清からの免疫吸着により単離された抗マリグニン抗体に対する神経膠腫タンパク質の親和力の継続により見られる(例えば、図5および米国特許第6,242,578 B1号)。
【0151】
同様に保存は、HIVの分離株のトランスアクチベーター(Tat)タンパク質中で観察された。Tat(トランスアクチベーター)タンパク質はレンチウイルスの転写を制御する早期RNA結合タンパク質である。これらのタンパク質は、すべての公知のレンチウイルス(例えば、ヒト免疫不全症ウイルス(HIV))のライフサイクルの必須成分である。Tatは、トランス活性化応答配列(TAR)RNAに結合することにより作用し、転写開始および/またはLTRプロモーターからの伸長を活性化する転写レギュレータータンパク質である。HIVはtat無しでは複製できないが、この化学基礎は不明である。HIV tatタンパク質配列の89〜102残基中に、我々は、他の生物の迅速な複製に関連するレプリキンを見いだした。このレプリキンのアミノ酸配列は「hclvcfqkkglgisygrkk」である。実際我々は、すべてのHIV tatタンパク質中にこのレプリキンが存在することを見いだした。表8に示すようにいくつかのtatアミノ酸は、別のアミノ酸によりしばしば置換される(最も頻繁なアミノ酸の下に小さいフォントで示す(表7)、優性のレプリキン「hclvcfqkkglgisygrkk」の保存パーセント)。これらの置換は、個々のアミノ酸のほとんどで現れた。しかしレプリキン配列内の主要なリジンとヒスチジンアミノ酸(これは、レプリキン構造を規定する)は、配列中で100%保存される;一方、他のアミノ酸の他の場所(レプリキンの内および外)で置換が一般的であり、これらの主要なヒスチジンアミノ酸上で置換は存在しない。
【0152】
表7に示すように、tatタンパク質アミノ酸配列中でリジンが置換されないことではない。表の左側から、すぐ隣接する配列中であるがレプリキン配列の外の最初のリジン、およびレプリキン内の配列中の第2のリジン(k)(しかし、レプリキン形成にとって必須ではないという点で「余分」である)は両方とも、しばしば置換される。しかし3番目、4番目および5番目のリジンおよびカッコ中の1つのヒスチジン(これらは一緒にレプリキン構造を構成する)は、決して置換されない。すなわちこれらの主要なアミノ酸配列は、100%保存されている。インフルエンザウイルスレプリキンの場合に観察されるように、ランダムな置換は、この選択的置換と選択的な非置換が偶然起きることは、可能にしないであろう。
【表16】
【0153】
レプリキン構造の保存は、レプリキン構造が、保存されなければならないHIVウイルスの特定の生存機能を有し、犠牲になって抗体および他の「攻撃」を避けるためにウイルス「防御」手段のアミノ酸置換をすることはできないことを示唆する。これらの「防御」機能は必須ではないが、HIV複製のウイルス生存機能と「競合」することはできない。
【0154】
さらなる保存が、HIVの異なる分離株中で、そのレプリキン、例えばHIV 1型の「kcfncgkegh」(配列番号5)または「kvylawvpahk」(配列番号6)、およびHIV 2型の「kcwnegkegh」(配列番号7)中で観察された。
【0155】
FMVDおよびHIVレプリキン中で観察されたこの高い保存率は、インフルエンザレプリキンのレプリキン中で観察される保存もまた、ウイルスレプリキンの一般的性質であることを示唆する。この保存は、例えば、神経膠腫レプリキンについて例示されるようにインフルエンザ、FMVDまたはHIVのような特異的レプリキンを使用する破壊、または刺激にとって、一定の信頼できる標的としている。
【0156】
同様に、ウイルス(インフルエンザウイルス、FMDV、およびHIVを含む)(ここでは、レプリキンの高率の保存が、ウイルスレプリキンの一般的性質であり、レプリキンを、破壊または刺激の一定の信頼できる標的としていることを示唆する)中の例に提供されるように、レプリキン構造の保存は植物でも起きる。例えば、コムギ植物では、レプリキンが保存され、刺激の信頼できる標的となる。コムギ植物のユビキチン(ubiquitin)活性化酵素Eで保存されたレプリキンの例には以下がある:
E3 hkdrltkkvvdiarevakvdvpeyrrh(配列番号614)
E2 hkerldrkvvdvarevakvevpsyrrh(配列番号615)
E1 hkerldrkvvdvarevakmevpsyrrh(配列番号616)
【0157】
HIV tatタンパク質中の保存と同様に、コムギ植物のユビキチン(ubiquitin)活性化酵素E中のレプリキンは保存されている。HIV tatタンパク質と同様に、コムギ植物のユビキチン(ubiquitin)活性化酵素E中のレプリキン変種型に隣接するアミノ酸の置換(「*」で示す)は一般的である。しかしレプリキン構造を形成する主要なkおよびhアミノ酸は変化しないが、(左側の最初のkから)わずかに5アミノ酸の「必須ではない」kは置換される。
【0158】
抗レプリキン抗体
抗レプリキン抗体は、レプリキンに対する抗体である。抗レプリキン抗体についてのデータもまた、レプリキンクラスの統一性を支持する。抗レプリキン抗体応答は、固定化されたマリグニンに対する血清抗マリグニン抗体の免疫吸着により定量されている(米国特許第5,866,690号中の方法を参照)。16量体のペプチドの合成物(その配列はマリグニンから得られた)をウサギに投与することによる抗マリグニン抗体の多量の産生は、炭水化物又は他の基を産生せず、この免疫応答の充分な基礎となる(図3)。16量体ペプチドは、IgM型およびIgG型の抗体の両方を産生した。抗マリグニン抗体は、感染後約15年〜25年後に出現する肝癌の普通に観察されるはるかに前に、感染の最初の5年のB型肝炎とC型肝炎の米国とアジアの79症例のうちの37%で血清中の濃度が上昇していた。感染性肝炎とHIV感染症の両方に関して、形質転換された細胞は、ウイルスの安全な避難所であり、細胞寿命が延長し、ウイルスの追い立てを逃れ、その結果ウイルスは抗ウイルス治療法に接することがない。
【0159】
レプリキンの投与は免疫系を刺激して、細胞障害作用を有する抗体を生じるため、ある期間中に最も濃縮されることが観察されている特定のインフルエンザウイルスレプリキンまたはレプリキン群に基づくペプチドワクチンは、あるインフルエンザシーズンに大流行を引き起こす可能性が最も高いインフルエンザの特定の株(例えば、出現する株または再出現する株)に対する防御となる。例えば、毎年または2年ごとにインフルエンザウイルス血球凝集素のアミノ酸配列を分析すると、その年の特異的に標的化されたインフルエンザワクチンを調製するのに有用なデータが提供される。そのような分析は、地域ごとにまたは任意の所望の時期に行われて、その結果世界中の異なる地域で出現する株を検出することができ、各地域の特異的に標的化されたワクチンを調製することができる。
【0160】
インフルエンザ
現在、国際的なWHOおよびCDC会議で、ウイルス製剤は年に2回変更される。ワクチン製剤は、世界のある地域のインフルエンザウイルス株の最も最近の優勢の血清学的証拠に基づいている。しかし本発明以前には、インフルエンザウイルス株に特異的なアミノ酸配列の変化と、インフルエンザの流行または汎流行の発生との相関はなかった。
【0161】
インフルエンザウイルスタンパク質中の特異的レプリキンの観察とその濃度は、インフルエンザの汎流行および流行の最初の特異的な定量的早期化学相関現象であり、世界の特定の地域のインフルエンザウイルスの流行している出現性または再出現性株を処理するために具体的に作成されるインフルエンザウイルスの産生とタイミング良い投与を提供する。インフルエンザウイルスの分離株のタンパク質配列(例えば、血球凝集素タンパク質配列)を、レプリキンの存在、濃度および/または保存について分析することにより、インフルエンザウイルスの汎流行と流行を予測することができる。さらに、ある時期(例えば約1年〜約3年)にウイルス分離株で最も豊富であるかまたは上昇していることがわかったレプリキン配列に基づいてインフルエンザペプチドワクチンを投与することにより、インフルエンザのそのような大流行の深刻さを大きく減少させることができる。
【0162】
本発明のインフルエンザペプチドワクチンは、インフルエンザウイルス株で観察される単一のレプリキンペプチド配列を含有するか、または複数のレプリキン配列を含有する。好ましくはペプチドワクチンは、ある期間に濃度が上昇していることが証明されており少なくともその期間は保存されているレプリキン配列に基づく。しかしワクチンは、新しいレプリキンペプチドとともに保存されたレプリキンペプチドを含むか、または新しいレプリキンペプチド配列に基づいてもよい。レプリキンペプチドは、任意の方法(化学合成または組換え遺伝子技術を含む)により合成することができ、非レプリキン配列を含有してもよいが、レプリキン配列のみを含有するペプチドに基づくワクチンが好ましい。好ましくは本発明のワクチン組成物はまた、医薬として許容される担体および/またはアジュバントを含有する。
【0163】
本発明のインフルエンザワクチンは、単独でまたは抗ウイルス剤[例えば、ガンシクロビル;インターフェロン;インターロイキン;M2インヒビター(例えば、アマンタジン、リマンタジン);ノイラミニダーゼインヒビター(例えば、ザナミビルおよびオセルタミビル)等]との組合せで、ならびに抗ウイルス剤の組合せで投与することができる。
【0164】
マラリア中のレプリキンデコイ
メロゾイト表面および/または寄生菌含有空胞内に位置する熱帯熱マラリア原虫のマラリア抗原の1次構造の分析は、この生物もまたインフルエンザウイルスのように、多数のレプリキンを含有することを明らかにした(表8)。しかし、熱帯熱マラリア原虫とインフルエンザウイルス分離株中のレプリキンの観察にはいくつかの差がある。例えば熱帯熱マラリア原虫はいくつかの部分的レプリキン(ここでは「レプリキンデコイ(Replikin decoy)」と呼ぶ)を含有する。これらのデコイの構造は、多数のリジン残基を含有するが、レプリキン構造に必要なヒスチジンは欠如している。具体的にはこれらのデコイは、真のマラリアリコグニン(recognins)のように、重複して6〜10残基離れて多くのリジンを含有するが、ヒスチジン残基は無い。デコイ構造は、抗マラリア抗体または他の物質が、ヒスチジン(これは、呼吸系酵素中のレプリキンのようにレプリキン構造中に存在し、トリパノソーマの破壊を引き起こし得る)に結合するのではなく、リジンを含有するあまり重要ではない構造(すなわち、レプリキンデコイ)に結合する機会を最大にしていると考えられる。例えば、レプリキン構造に対する特異性を持って侵入する抗体は、レプリキンデコイ構造に結合して、真のレプリキン構造をそのまま残すかも知れない。
【0165】
従って、マラリアの抗レプリキン治療には、2つの相が必要である(2重治療):i) タンパク質分解酵素を用いてレプリキンデコイを切断し、特異的抗レプリキン治療の「安全な通過」を可能にする、予備処理;およびii) 特異抗体を用いる、または合成マラリアレプリキンワクチンより発生する細胞性免疫による、またはマラリアレプリキンを標的とする有機的手段による、マラリアレプリキンの攻撃。
【0166】
レプリキン構造の繰り返しと重複
熱帯熱マラリア原虫で見られるもう1つの差は、インフルエンザウイルスでは観察されなかった、単一タンパク質中の個々のレプリキン構造の頻繁な繰り返しである。繰り返しは、(a) レプリキン間のリジン残基の共有、および(b) 別のレプリキン配列内のレプリキン配列の一部の繰り返し、により起きる。
【0167】
インフルエンザウイルス分離株と熱帯熱マラリア原虫で観察されるレプリキン構造の第3の大きな差は、マラリアタンパク質中のレプリキン構造の顕著な重複である[例えば、配列番号393の39アミノ酸配列内の9つの重複するレプリキン(レプリキン濃度=23.1/100アミノ酸);および配列番号467の41アミノ酸中の15の重複するレプリキン(レプリキン濃度=36.6/100アミノ酸)]。これらの重複レプリキン構造の両方とも、血液段階のトロフォゾイトとシゾントで起きる。これに対してインフルエンザウイルスレプリキンは、タンパク質中により分散しており、最大のレプリキン濃度は約7.5/100アミノ酸であり(図7);重複するレプリキンを有することが観察されたトマトリーフカールジェミニウイルスも、わずかに約3.1/100アミノ酸である。
【0168】
多数のリジンのこの機構はまた、癌タンパク質(例えば、胃ガン形質転換タンパク質)のレプリキンktkkgnrvsptmkvth(配列番号88)、および肺の形質転換タンパク質P21B(K-RAS 2B)のレプリキンkhkekmskdgkkkkkks(配列番号89)でも見られる。
【0169】
高レプリキン濃度と迅速な複製の関係はまた、HIV分離株の分析によっても確認される。HIVの増殖の遅い低力価株(NSI、「Bru」、これは早期HIV感染によくある)は、100アミノ酸当たり1.1(±1.6)レプリキンのレプリキン濃度であり、一方HIVの迅速に増殖する高力価株(S1、「Lai」、これは後期HIV感染によくある)は、100アミノ酸残基当たり6.8(±2.7)レプリキンのレプリキン濃度である。
【0170】
マラリア、インフルエンザウイルスおよび癌細胞中の重複レプリキンの高濃度は、マラリア微生物の有名な高および迅速複製能力に一致する。マラリア中の多数の重複レプリキンはまた、この微生物が宿主に殺到しその免疫系を混乱させて、こうして間違った抗体が作成され永続される機会を大きくして、主要なマラリア抗原が攻撃されないようにする。
【0171】
例えばインフルエンザウイルスの場合のように、マラリア微生物中に存在するレプリキン構造に基づくペプチドワクチンは、マラリアを予防および/または治療するための有効な手段となる。マラリアに対するワクチン接種は、熱帯熱マラリア原虫で観察される1つのレプリキン構造またはその混合物を含有する組成物を投与することにより行われる。さらに、マラリアレプリキンに対する抗体を作成し、受動免疫のためにまたはマラリア検出のために投与することができる。
【0172】
表8は、いくつかの熱帯熱マラリア原虫レプリキン配列のリストを提供する。これは完全なものではないことを理解されたい。この微生物の異なる分離株は、他のレプリキン構造を含むこともある。
【表17】
【表18】
【表19】
【表20】
【表21】
【表22】
【0173】
構造タンパク質中のレプリキン
いくつかの構造タンパク質はレプリキン構造を含むことも測定されている。構造タンパク質は、組織および臓器の支持に関与する分子であり、例えば皮膚や結合組織および膜構造物中のコラーゲン、および例えば脳中のアミロイドA4前駆体タンパク質(APP)である。これらのタンパク質の過剰産生は疾患に関係し、特に皮膚のコラーゲンの過剰産生の場合の強皮症(表9)、および脳のAPPの過剰産生の場合のアルツハイマー病がある(表10)。
【0174】
強皮症と悪性腫瘍の関連は、近年議論の的である。これまでの報告で、いくつかの関連機構が示唆されている。最近の長期研究は、強皮症と悪性腫瘍の関係の上昇を示唆する。しかし基本的な機構はいまだに不明である(Wenzel, J. Eur. J. Dermatol. 2002 May-Jun;12(3):296-300)。
【0175】
強皮症中のタンパク質の過剰産生に関連するいくつかのタンパク質は、レプリキン構造を含有することがわかっている。すなわちこれらは、過剰コラーゲン産生の阻害または停止のための、まだ認識されていない標的のさらなる例である。表9は、強皮症中のタンパク質と関連するレプリキンのリストである。
【0176】
APPタンパク質は、アミロイドベータA4タンパク質の供給源であり、これは、過剰にあると脳中の細胞外スペース中でプラークを形成し、アルツハイマー病の神経細胞喪失に関連する毒性作用を示す。今日までの多くの試験は、A4の過剰沈着を防ぐことができないことに注目してきたが、これは、廃棄物の除去問題というより、実際は、前駆体タンパク質APPの過剰産生の問題であるかも知れない。APP中のレプリキンの高濃度(100アミノ酸当たり3.3レプリキン)は、過剰産生がアルツハイマー病の原因であることを強く示唆する(表10)。従って、表10中に含まれるレプリキンは、神経膠腫レプリキンで詳述されるものと同じ方法により阻止または阻害することができる。
【表23】
【表24】
【表25】
【0177】
受動免疫
本発明の別の実施態様において、単離されたレプリキンペプチドは抗体を作成するのに使用され、この抗体は例えば個体の受動免疫を与えるのに使用される。将来のインフルエンザ感染の原因である可能性が最も高いと本発明の方法により同定されたインフルエンザ株に対する受動免疫は、インフルエンザウイルスの同定された株のレプリキン配列に対する抗体を、必要な患者に投与することにより得られる。同様に、マラリアに対する受動免疫は、熱帯熱マラリア原虫レプリキンに対する抗体を投与することにより得られる。
【0178】
当該分野で公知の種々の方法を使用して、レプリキン配列に対する抗体を産生してもよい。そのような抗体には、特に限定されないが、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、ヒト化抗体、1本鎖抗体、Fab断片およびFab発現ライブラリーにより産生される断片がある。細胞障害剤に連結した抗体も作成される。抗体はまた、抗ウイルス剤と組合せて投与してもよい。さらに、異なるレプリキンに対する抗体の組合せを、抗体カクテルとして投与してもよい。
【0179】
抗体産生のために、レプリキンペプチドまたはレプリキンペプチドの組合せを、特に限定されないがウサギ、マウス、ラット、およびより大きな哺乳動物に注射することにより、種々の宿主動物または植物を免疫してもよい。
【0180】
レプリキンに対するモノクローナル抗体は、抗体分子を産生する任意の方法を使用して調製される。これらには、特に限定されないが、元々はKohlerとMilstein(Nature, 256:495-497 (1975))により開示されたハイブリドーマ法、ヒトB細胞ハイブリドーマ法(Kosborら、1983, Immunology Today, 4:72)、およびEBV ハイブリドーマ法(Coleら、モノクローナル抗体と癌療法(Monoclonal Antibodies and Cancer Therapy), Alan R. Liss, Inc., pp. 77-96)がある。さらに、キメラ抗体の産生のために開発された方法(Morrisonら、Proc. Natl. Acad. Sci. USA, 81:6851-6855)、または他の方法も使用される。あるいは、1本鎖抗体の産生について開示された方法(US4,946,778)を、レプリキン特異的1本鎖抗体を産生するように適応させてもよい。
【0181】
本発明の特に有用な抗体は、インフルエンザウイルスのペプチドおよび/またはポリペプチド中に含有されるレプリキン配列に、特異的に結合するものである。例えば、インフルエンザウイルスの出現性または再出現性株中に存在することが観察されている任意のペプチドに対する抗体およびそのような抗体の組合せは、インフルエンザの治療および/または予防に有用である。同様に、マラリア抗原上に存在する任意のレプリキンに対する抗体およびそのような抗体の組合せは、マラリアの予防と治療に有用である。
【0182】
レプリキンに対する結合部位を含有する抗体断片は、公知の方法により作成される。例えばそのような断片には、特に限定されないが、抗体分子のペプシン消化により産生することができるF(ab')2 断片、およびF(ab')2 断片のジスルフィド結合を還元することにより作成することができるFab断片がある。あるいは、Fab発現ライブラリーを作成(Huseら、1989, Science, 246:1275-1281)して、所望の特異性を有するモノクローナルFab断片の迅速かつ容易な同定が可能になる。
【0183】
癌細胞のタイプに無関係にヒト悪性腫瘍では抗マリグニン抗体の濃度が上昇している(図5)という事実、およびこの抗体は細胞タイプに無関係に癌細胞に結合するという事実は、本明細書においてほとんどの悪性腫瘍中に存在することがわかったレプリキン構造の存在により説明される(図1と表2)。集団での研究は、健常成人では年齢とともに抗マリグニン抗体の濃度が上昇し、これは、癌の頻度が上昇するように、ハイリスク家族では特にそうである。早期悪性腫瘍で起きるさらなる2倍またはそれ以上の抗体の上昇は、1〜10mmのサイズの乳癌で97%の特異性で独立に確認されている。in vivoで癌細胞中に選択的に局在化することが証明いる抗体は、組織学的には正常な細胞には結合しないが、in vitroで形質転換された細胞に選択的に結合(図4a、b)し、かつこれに細胞障害性が高い(図c-f)。これらの例では、いくつかのタイプの細胞(すなわち脳神経膠腫、造血細胞(白血病)、および肺の小細胞癌)により同じ抗体が結合されるため、悪性のレプリキンクラスの統一性も示される。
【0184】
抗マリグニンは、良性の増殖といっしょには上昇しないが、in vivoで乳房での悪性形質転換と複製によってのみ特異的に上昇し、悪性細胞を排除すると高値から正常値に戻る(図5)。抗マリグニン抗体濃度は、癌患者の生存に定量的に関連すること、すなわち抗体が多いほど生存期間が長いことが証明されている。まとめるとこれらの結果は、抗レプリキン抗体が、細胞の形質転換と複製の制御機構の一部かも知れないことを示唆する。この免疫応答の増強は、ワクチンとして合成レプリキンを用いて能動的に、または抗レプリキン抗体の投与により受動的に、またはレプリキンを特異的に標的とするように同様に設計された非免疫ベースの有機物質(例えば、炭水化物、脂質など)の導入により、複製を制御するのに有用であるかも知れない。
【0185】
本発明の別の実施態様において、1つ以上のレプリキンに暴露された個体から得られた1つ以上のレプリキンに対する抗体を含有する免疫血清を使用して、別の個体または動物で受動免疫が導入される。免疫血清は、静脈経由で、治療の必要な被験体に投与される。受動免疫はまた、1つ以上のレプリキンに対するあらかじめ形成された抗体を用いて受容者に注射することにより行われる。受動免疫は、感染性生物に暴露された個体に対して即時的防御を与えるのに使用してもよい。免疫血清またはあらかじめ形成された抗体の投与はルーチンの手技であり、熟練者は容易に、血清または抗体の量を確認して所望の作用を達成することができる。
【0186】
合成レプリキンワクチン(能動免疫)
神経膠腫レプリキン(配列番号1)「kagvaflhkk」またはC型肝炎レプリキン(配列番号18)「hyppkpgcivpak」、またはHIVレプリキン、例えば(配列番号5)「kcfncgkegh」もしくは(配列番号6)「kvylawvpahk」のようなレプリキンに基づく合成レプリキンワクチン、または好ましくは、ある期間の保存されたおよび/または出現するかまたは再出現するレプリキンに基づくインフルエンザワクチンを使用して、抗体濃度を増強して、各ウイルスで感染された細胞を溶解し、ウイルスを細胞外に放出させると、ここで化学処理が有効となる。同様に、例えば、メロゾイト表面上のもしくはパラジトフォラス(parasitophorous)小腔内の熱帯熱マラリア原虫マラリア抗原で観察されるレプリキンに基づくマラリアワクチンを使用して、マラリアに対する細胞障害性抗体を作成することができる。
【0187】
リコグニンおよび/またはレプリキンペプチドを被験体に投与して、被験体の免疫系を誘導して抗レプリキン抗体を産生してもよい。一般に0.5〜約2mg用量、好ましくは1mg用量の各ペプチドを被験体に投与して、免疫応答を誘導する。以後の用量は、所望であれば投与される。
【0188】
レプリキン配列構造は、複製機能に関連している。すなわち、本発明のレプリキンが、例えば診断的同定、複製の促進、または複製の阻害もしくは攻撃を目的としてレプリキンを含有する配列を標的とするのに使用されるかどうかは、レプリキンの構造−機能相関が基本的である。
【0189】
レプリキン断片を認識しこれに結合する抗体を誘導して細胞の破壊を引き起こしたい時は、特異的なレプリキン構造のみを利用することが好ましい。より大きなタンパク質配列が「複製関連機能」を有するとして当該分野で公知であるが、より大きなタンパク質を使用するワクチンは、しばしば成功しなかったかまたはあまり有効ではなかった。
【0190】
本発明者らは1つの理論に拘泥されるつもりはないが、本明細書に記載の研究は、先行技術のワクチンがより大きなタンパク質配列の使用に基づくため、有効ではないことを示唆する。より大きなタンパク質配列は必ず、1つ以上のエピトープ(特異抗体形成を誘導することができる独立した抗原配列)を有し、レプリキン構造は通常、これらのエピトープ候補の1つを含む。より大きなタンパク質内の他のエピトープの存在は、レプリキン抗原を先制攻撃する無関係の抗原刺激で免疫系に「殺到」(この抗原優先の公知の現象の考察については、Webster, R.G., J. Immunol., 97(2):177-183 (1966);およびWebsterら、J. Infect Dis., 134:48-58, 1976;Klenermanら、Nature 394:421-422 (1998)を参照されたい)して、レプリキンに対する抗体の充分な形成を妨害し、こうして、免疫系により提示され認識される第1のペプチドエピトープが多くなり、他のペプチドエピトープが同時に存在してもこれに対する抗体が作成される。これは、ワクチン製剤では、レプリキンが先制攻撃されずに免疫記憶に入れられるように、生物からの他のエピトープを提示する前に、免疫系に一定のレプリキンペプチドを最初に提示することが重要である別の理由である。
【0191】
非レプリキンエピトープに対する抗体の生成は、細胞への結合を可能にするが、必ずしも細胞を破壊はしない。マラリアタンパク質のC末端上の構造的「デコイ」の存在は、デコイエピトープがリジン残基を有するがヒスチジン残基は無いため、他のエピトープが、有効な抗レプリキン抗体の結合を妨害する能力の、他の側面である。すなわちデコイエピトープは抗レプリキン抗体に結合するが、その抗体をヒスチジン結合呼吸系酵素からは離して維持する。従って治療は2段階で最も有効となる:1) デコイを加水分解するプロテアーゼ、次に;2) 抗レプリキン抗体または他の抗レプリキン物質。
【0192】
「外来」タンパク質に対する抗体産生の過程では、タンパク質がまず小断片に加水分解されることは、当該分野で公知である。通常、約6〜10アミノ酸を含有する断片が抗体生成のために選択される。すなわち、タンパク質の加水分解によりレプリキン含有断片が得られない場合、抗レプリキン抗体は産生されないであろう。この点で、リジン残基は膜に結合することが知られているため、6〜10アミノ酸離れて位置するリジン残基をレプリキンが含有することは興味深い。
【0193】
さらにレプリキン配列は、少なくとも1つのヒスチジン残基を含有する。ヒスチジンはしばしば、酸化還元中心への結合に関与する。すなわち、レプリキン配列を特異的に認識する抗体は、レプリキンが位置している細胞を不活性化または破壊する可能性が高く(抗マリグニン抗体で見られるように)、これはおそらく、ピコグラム/細胞で活性のある、これまで記載された最も細胞障害性が高い抗癌抗体である。
【0194】
疾患を引き起こす物質の特定のタンパク質抗原に対して作成したワクチンが、疾患に対する防御を与えるのに充分に有効ではない(例えば、VP1タンパク質またはVP1タンパク質の大きなセグメントに対して開発された手足口病ワクチン)理由の1つは、最適な抗体が産生されていないこと、すなわち、レプリキンに対する抗体が産生されていない可能性があることである。レプリキンは産生されていない。すなわち、より大きなタンパク質断片中に存在するレプリキン以外のエピトープが、上記の抗原優先の現象に従って妨害するか、および/または抗体を産生するためのプロセシングのために、より大きなタンパク質配列のより小さい配列への加水分解が、存在するレプリキン構造の完全性を失わせるために、レプリキンが2つに切断されるか、および/またはヒスチジン残基が加水分解プロセシングで失われる。本研究は、有効なワクチンが産生されるためには、レプリキン配列がワクチンとして使用されて他のエピトープは使用されないことであることを示唆する。例えば、本発明のワクチンは、3点認識システムにより同定されたレプリキンペプチドの任意の1つを使用して作成することができる。
【0195】
特に好適なペプチド、例えばインフルエンザワクチンは、1年以上、好ましくは約3年以上の期間にわたって保存されることが証明されているか、および/または、他のインフルエンザウイルス株(例えば、出現する株)中のレプリキン濃度と比較してレプリキン濃度が最も上昇していることが証明されている、インフルエンザウイルスの株中に存在するペプチドを含む。レプリキン濃度の上昇はおそらく、少なくとも約6ヶ月から1年、好ましくは少なくとも約2年以上、および最も好ましくは約3年以上の期間にわたって起きる。インフルエンザウイルスワクチンで使用するために好適なレプリキンペプチドは、1年以上血球凝集素アミノ酸配列から欠如の後に「再出現」することが観察されているレプリキンである。
【0196】
本発明のレプリキンペプチドは、単独または種々の組合せで、被験体に、好ましくは静脈内または筋肉内注射により、被験体の免疫系を刺激してペプチドに対する抗体を産生させるために投与される。一般にペプチドの用量は、約0.1μg〜約10mg、好ましくは約10μg〜約1mg、および最も好ましくは約50μg〜約500μgの範囲である。熟練者は、有効な免疫応答を生成するのに必要な用量および投与の回数を、容易に決定することができるであろう。
レプリキンワクチンに対する早期応答の定量的測定
【0197】
レプリキンワクチンに対する数日または数週間の早期特異抗体の応答を定量的に測定できる能力は、数ヶ月または数年後にのみ測定できる他のワクチンに対して大きな実際的利点である。
【0198】
アジュバント
宿主の種に依存して、免疫応答を増強するのに種々のアジュバントが使用され、特に限定されないが、フロインド(完全アジュバントまたは不完全アジュバント)、ミネラルゲル(例えば水酸化アルミニウム)、界面活性物質(例えば、リソレシチン)、プルロニックポリオール、ポリアニオン、ペプチド、油エマルジョン、キーキンペットヘモシアニン、ジニトロフェノール、および有用な可能性のあるヒトアジュバント(例えば、BCGまたはコリネバクテリウム・パルブム(Corynebacterium parvum))がある。
【0199】
レプリキンヌクレオチド配列
レプリキンDNAまたはRNAは、ウイルス、細菌または他のレプリキンをコードする物質による感染に起因する疾患の診断に多くの用途がある。例えば、レプリキンヌクレオチド配列は、生検組織または血液の、インサイチューハイブリダイゼーション測定法を含むハイブリダイゼーション測定法(例えば、サザンまたはノーザン分析)で、例えば組織試料または環境試料中の特定の生物の存在を診断するために使用される。本発明はまた、目的の特定の病原体中に存在する特定のレプリキンに特異的な抗体を含有するか、または特定のレプリキンに特異的にハイブリダイズする核酸分子(センスまたはアンチセンス)、および随時種々の緩衝液および/または診断に必要な試薬を含有するキットを包含する。
【0200】
また本発明の範囲には、アンチセンスRNAおよびDNA分子を含むオリゴヌクレオチド配列、およびレプリキンもしくはリコグニン含有mRNAの翻訳を阻害するように機能するリボザイムも含まれる。アンチセンスRNAおよびDNA分子およびリボザイムの両方とも、当該分野で公知の任意の方法により調製される。アンチセンス分子は、被験体への供給のために広範囲のベクター中に取り込むことができる。熟練者は、投与の最適経路を容易に決定することができるが、一般的には静脈内または筋肉内投与がルーチン法である。投与量もまた、確認することができる。
【0201】
特に好適なアンチセンス核酸分子は、例えばインフルエンザウイルスポリペプチドをコードするmRNA中に含有されるレプリキン配列に相補的なものであり、ここでレプリキン配列は、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基、を含む7〜約50個のアミノ酸を含む。さらに好ましくは、遺伝子のコード鎖中に存在するレプリキンまたはインフルエンザウイルス血球凝集素タンパク質をコードするmRNAに相補的なアンチセンス核酸分子であり、ここでアンチセンス核酸分子は、6ヶ月から1年以上の期間にわたって保存されることが証明されているかまたは他のインフルエンザウイルス株中のレプリキン濃度と比較してレプリキン濃度が上昇していることが証明されているインフルエンザウイルスの株中に存在する、レプリキンをコードするヌクレオチド配列に相補的である。レプリキン濃度の上昇は好ましくは、少なくとも6ヶ月、好ましくは約1年、最も好ましくは約2〜3年またはそれ以上の期間にわたって存在する。
【0202】
同様に、mRNAに相補的なアンチセンス核酸分子は、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む7〜約50個のアミノ酸のレプリキン配列を含む細菌レプリキンをコードするmRNAに相補的なものである。より好適なものは、遺伝子のコード鎖または細菌のタンパク質をコードするmRNAに相補的なアンチセンス核酸分子である。
【0203】
診断応用
珪藻プランクトン、手足口病ウイルス、トマトリーフカールジェミニウイルス、B型肝炎ウイルスとC型肝炎ウイルス、HIV、インフルエンザウイルスおよび悪性腫瘍細胞のような生物にとって、同定された成分レプリキンはワクチンとして有用であり、診断目的にも有用に標的化される。例えば、輸血用に集めた血液は、汚染生物に特異的であることが証明されたレプリキンの存在についてスクリーニングすることにより、生物(例えばHIV)の汚染についてスクリーニングされる。また、特定の病原性生物に特異的なレプリキン構造のスクリーニングにより、体組織またはその環境中の生物の診断用検出が可能になる。
【0204】
増殖のレプリキン刺激
本発明の別の実施態様において、細胞、組織または臓器の複製速度を上昇させるためにレプリキン構造が使用される。複製の速度および収率を上昇させることが他の細胞、組織または生物について当該分野で知られている細胞分裂への、適切な刺激有りまたは無しで、より迅速に複製することが好ましい他の細胞、組織または臓器にとって特異的なレプリキン構造の添加により、複製速度を上昇させる方法が知られている。これは、例えば当該分野で公知の方法により、少なくとも1つのレプリキン構造を有する生物中で複製機能を有するタンパク質または酵素をコードするかまたはこれに関連する遺伝子を修飾もしくは形質転換することにより行われる。
【0205】
本発明の別の態様において、レプリキン構造は生物の複製を上昇させるために使用される。本発明は、例えばインフルエンザウイルスにおいて、流行に関連する複製の上昇がレプリキン濃度の上昇に関連することを示す。この上昇は、1) 前の年には存在したが、次に1年以上消失している、特定のレプリキン構造の再出現;および/または2) 新しいレプリキン組成の出現、による。さらにマラリアレプリキンでは、単一のタンパク質中で同じレプリキンの繰り返しが起きる。
【0206】
すなわち本発明は、生物の複製を上昇させるための方法と組成物を与える。同様に、異なる生物のレプリキンを任意の生物の複製を阻害するように標的化することができる方法で、レプリキンを使用して任意の生物の複製を上昇させることができる。例えば、イネ、トウモロコシ、およびコムギ穀物の生産(これらは、世界の大人口を養うのに決定的に重要である)は、例えば任意の特定の株のイネの濃度(100アミノ酸残基当たりのレプリキンの数)を上昇させることにより改良することができる。
【0207】
例えば、イネ(Oryza sativa)株では、未成熟の種子から単離されたカタラーゼは、タンパク質の491アミノ酸配列内に以下の異なるレプリキンを含有することが観察された:
kfpdvihafkpnprsh (配列番号638)
kfpdvihafk (配列番号639)
karyvkfhwk (配列番号640)
hpkvspelraiwvnylsqedeslgvkianlnvk (配列番号641)
katihkqndtk (配列番号642)
happtpitprpvvgrrqkatihkqndfk (配列番号643)
kfrpsssfdtkttttnagapvwndnealtvgprgpilledyhliekvah (配列番号644)
kfrpsssfdtkttttnagapvwndnealtvgprgpilledyn (配列番号645)
【0208】
すなわち、当該分野で公知の組換え遺伝子クローニング法を使用することにより、生物(例えば、食用穀物植物)中のレプリキン構造の濃度を上昇させることができ、これは、生物の複製の上昇を促進するであろう。例えば、当該分野で公知の方法によりイネ(Oryza sativa)カタラーゼ遺伝子中に上記で同定したレプリキンのような追加のレプリキン配列を挿入することは、この生物の複製を促進するであろう。
【0209】
同様に、イネ(Oryza sativa)(日本の栽培品種)のNBS-LRRタンパク質中に以下のレプリキンが見いだされた:
kvkahfqkh (配列番号647)
kvkahfqk (配列番号648)
kdyeidkddlih (配列番号648)
hmkqcfafcavfpk (配列番号650)
hvfwelvwrsffqnvkqigsifqrkvyrygqsdvttskihdlmhdlavh (配列番号651)
kqigsifqrkvrygpsdvttskihdlmhdlavh (配列番号652)
kqigsifqrkvyrygpsdvttskihdlmh (配列番号653)
kqigsifqrkvyrygqsdvttskih (配列番号654)
【0210】
さらに、アスパラギン酸プロテイナーゼオリザシン1前駆体タンパク質について、以下のレプリキンが見いだされた:
khgvsagik (SEQ ID N0.655)
htvfdygkmrvgfak (配列番号657)
hsryksgqsstyqkngk (配列番号658)
【0211】
同様に、イネ(Oryza sativa)(インディカ品種群)のMADS−ボックスタンパク質FDRMADS3転写因子中で、以下のレプリキンが見いだされた:
kqeamvlkqeinllqkglryiygnraneh (配列番号659)
kqeinllqkglryiygnraneh (配列番号660)
kskegmlkaaneilqekiveqnglidvgmmvadqqngh (配列番号661)
kaaneilqekiveqnglidvgmmvadqqngh (配列番号662)
【0212】
同様に、ロニメイズ(LONI MAIZE)(ATP結合性酸化還元関連ヒドロラーゼ;セリンプロテアーゼ;マルチ遺伝子ファミリー;ミトコンドリア)について、以下のレプリキンが見いだされた:
kvlaahrygik (配列番号663)
klkiamkhliprvleqh (配列番号664)
klkiamkh (配列番号665)
ktslassiakalnrkfirislggvkdeadirgh (配列番号666)
kalnrkfirislggvkdeadirgh (配列番号667)
kfirislggvkdeadirgh (配列番号668)
kvrlskatelvdrhlqsilvaekitqkvegqlsksqk (配列番号669)
hlqsilvaekitqkvegglsksqk (配列番号670)
kvrlskatelvdrh (配列番号671)
kvggsavesskqdtkngkepihwhskgvaaralh (配列番号672)
kvggsavesskqdtkngkepihwh (配列番号673)
kvggsavesskqdtkngkepih (配列番号674)
kqdtkngkepihwhskgvaaralh (配列番号675)
kqdtkngkepih (配列番号676)
【0213】
同様に、グリセロアルデヒド-3-ホスフェート脱水素酵素A、葉緑体前駆体について、以下のレプリキンが見いだされた:
hrdlrraraaalnivptstgaakavslvlpnlk (配列番号677)
kvlddqkfgiikgtmttth (配列番号678)
hiqagakkvlitapgk (配列番号679)
hgrgdaspldviaindtggvkqashllk (配列番号680)
kqashllk (配列番号71O)
【0214】
さらに、見いだされたさび耐性様タンパク質RP1-4(トウモロコシ(Zea mays)の例は、以下のレプリキンを含む:
kvrrvlskdysslkqlmtlmmdddiskhlqiiesgleeredkvwmkeniik (配列番号681)
kvrrvlskdysslkqlmtlmmdddiskh (配列番号682)
hlqiiesgleeredkvwmkeniik (配列番号683)
hdlreniimkaddlask (配列番号684)
hvqnlenvigkdealask (配列番号685)
kkqgyelrqlkdlnelggslh (配列番号686)
kqgyelrqlkdlnelggslh (配列番号687)
klylksrlkelilewssengmdamnilh (配列番号688)
hlqllqlngmverlpnkvcnlsklrylrgykdqipnigk (配列番号689)
hlqllqlngmverlpnkvcnlskrylrgyk (配列番号690)
hlqllqlngmverlpnkvcnlsk (配列番号691)
hnsnklpksvgelk (配列番号692)
klpkvgelkh (配列番号693)
hlsvrvesmqkhkeiiyk (配列番号694)
khkeiiyk (配列番号695)
klrdilqesqkfllvldlalfkh (配列番号696)
hafsgaeikdqllrmklqdtaeeiakrlgqcplaakvlgsrmcrrk (配列番号697)
hafsgaeikdqllrmk (配列番号698)
klqdtaeeiakrlgqclaakvlgsrmcrrkdiaewkaadvwfeksh (配列番号699)
kvlgsrmcrrkdiaewkaadvwfeksh (配列番号700)
kdiaewkaadvwfeksh (配列番号701)
kaadvwfeksh (配列番号702)
hvptttslptskvfgrnsdrdrivkfllgktttaeasstk (配列番号703)
kailteakqlrdllglph (配列番号704)
kakaksgkgpllredessstattvmkpfh (配列番号705)
ksphrgkleswlrrlkeafydaedlldeh (配列番号706)
ksphrgkleswlrrlk (配列番号707)
hrgkleswlrrlk (配列番号708)
ksphrgk (配列番号709)
【0215】
前述のように、コムギユビキチン(ubiquitin)活性化酵素E(配列番号614〜616))中のレプリキンは保存されている。このレプリキン構造の保存は、植物成長の刺激のための信頼できる標的となる。
【0216】
レプリキンと酸化還元酵素との密接な関係もまた、コムギのこの構造に明瞭に示される。すなわちこのコムギユビキチン(ubiquitin)活性化酵素Eは、まずそのカルボキシ末端グリシン残基をATPでアデニル化し、次にこの残基をE1(配列番号614)中のシステイン残基の側鎖に連結して、ユビキチン−E1チオールエステルと遊離のAMPを産生することにより、ユビキチンを活性化する。
【0217】
コムギレプリキンと酸化還元酵素との関係のさらなる例はまた、Chinese spring wheat Chloroplast Triticum aestivamから単離されたPSABコムギタンパク質, フォトシステムI P700 クロロフィルA アポタンパク質 A2 (PsaB) (PSI-B) 中に見いだされる。このタンパク質は以下のように機能する:PsaAとPsaBは9700(フォトシステムI P700の1次電子供与者)ならびに電子アクセプターA0、A1およびFXに結合する。PSIは、プラストシアニン/サイトクローム c6-フェレドキシン酸化還元酵素として機能する。補助因子P700はクロロフィルAダイマーであり、A0はクロロフィルAであり、A1はフィロキノンであり、FXは4Fe-4S鉄−イオウ中心である。サブユニットA psaA/Sヘテロダイマーは、P700クロロフィル特殊対に結合し、次に電子アクセプターに結合する。高等植物と藻類のPSI反応中心は、1つの少なくとも11個のサブユニットからなる。これは、クロロプラストチラコイド膜の重要な膜タンパク質である。「h」が結合する4Fe-4S鉄−イオウ「中心」は決定的に重要である;従ってレプリキン構造中の「h」は重要である。細菌レプリキンの次に、これらのコムギレプリキンと植物レプリキンは、複製および複製のために必要なエネルギー源に対するレプリキン構造の重要性の、最も初歩的な進化的例示である。この基礎的関係は、複製に関して藻類、ウイルスレプリキン、細菌、癌細胞、およびおそらくすべての細胞まで続く。
【0218】
レプリキンのさらなる例は、PSABコムギタンパク質中に見いだされ、これはコムギの成長に決定的に重要である。これらには以下を含む:
hlqpkwkpslswfknaesrlnhh (配列番号617)
hlqpkwkpslswfk (配列番号618)
kwkyslswfknaesrlnhh (配列番号619)
kwkpslswfknaesrlnh (配列番号620)
kpslswfknaesrlnhh (配列番号621)
kpslswfknaesrlnh (配列番号622)
hhaialglhtttlilvkgaldargsklmpdkk (配列番号623)
haialglhtttlilvkgaldargsklmpdkk (配列番号624)
hhaialglhtttlilvkgaldargsk (配列番号625)
haialglhtttlilvkgaldargsk (配列番号626)
htttlilvkgaldargsklmpdkk (SEQ ID N0.627)
htttlilvkgaldargsklmpdk (配列番号628)
htttlilvkgaldargsk (配列番号629)
【0219】
コムギレプリキンと酸化還元との関係のさらなる例は、PSAA_コムギフォトシステムI 9700 クロロフィルA アポタンパク質 A1中に提供され、これは以下を含む:
hhhlaiailfliaghmyrtnwgighglkdileahkgpftgqghk (配列番号630)
hhlaiailfliaghmyrtnwgighglkdileahkgpftgqghk (配列番号631)
hlaiailfliaghmyrtnwgighglkdileahkgpftgqghk (配列番号632)
hmyrtnwgighglkdileahkgpftgqghk (配列番号633)
hglkdileahkgpftgqghk (配列番号634)
hdileahkgpftgqghk (配列番号635)
hkgpftgqghk (配列番号636)
kgpftgqgnk (配列番号637)
レプリキンを同定するためのコンピューターソフトウェア
【0220】
本発明はまた、アミノ酸または核酸配列中のレプリキン配列の同定法を提供する。本発明の3点認識法の開発において、4000を越える配列の視覚的スキャンを行った。しかし、ヌクレオチドおよび/またはアミノ酸配列を含むデータバンクはまた、3点認識要件を満足する配列の存在について、コンピューターによりスキャンすることができる。
【0221】
本発明の別の実施態様において、本明細書に記載の3点認識法はコンピューターにより行われる。図6は、本発明の前記実施態様で使用できるコンピューターのブロック図である。コンピューターは、プロセッサー、入力/出力装置、および前記実施態様の3点認識法である実施可能なプログラム命令を保存するメモリーを含む。メモリーは、静的メモリー、揮発性メモリーおよび/または不揮発性メモリーがある。静的メモリーは、磁気、または電気または光学保存媒体で提供されるリードオンリーメモリー(「ROM」)でもよい。揮発性メモリーは、ランダムアクセスメモリー(「RAM」)でもよく、キャッシュとしてプロセッサー内に取り込まれるか、または別の集積回路としてプロセッサーから外部に提供される。不揮発性メモリーは、電気、磁気または光学保存媒体でもよい。
【0222】
プロテオミクス的観点から、新しい神経膠腫ペプチド配列に基づく「3点認識」鋳型の構築により、関連構造および機能を有する生物学的に広いクラスのタンパク質が直接同定された。3点認識法の実施は、「キーワード」検索の使用による同定に似ているが、典型的な配列ホモジネート検索やアミノ酸のヌクレオチド特定におけるようにキーワード「kagvaflhkk」(配列番号1)の正確なスペリングを使用する代わりに、「3点認識」パラメータで制限したキーワードの抽出が使用される。この制限抽出(単一の比較的短いアミノ酸配列から得られるが)により、同じ規格で規定される構造を有するタンパク質のクラスが同定される。構造以外に特定の機能(この場合形質転換と複製)がまた、露出されたクラスのメンバーに共有されることは、これらの構造と機能が関連していることを示唆する。すなわち、この新たに同定された短いペプチド配列から、分子認識「言語」が考案され、これは従来は開示されていない。さらに、ここで癌レプリキンについて示されるように、クラスの多様なメンバーによる免疫学的特異性の共有は、B細胞とその生成物である抗体が、同様の認識言語でレプリキンを認識することを示唆する。
T細胞プライマーの他の用途
【0223】
「3点認識」は、特定のクラスのタンパク質を特定するプロテオミクス法であるため、他のペプチドについて3つ以上の異なる認識点を使用することは、他のタンパク質クラスに関する有用な情報を与えるであろう。さらに「3点認識」法は、他のリコグニン、例えば生物のリポ多糖のTOLL「innate」認識に応用される。3点認識法はまた、共有結合した有機分子の他の有用な化合物(他の共有結合したアミノ酸、ヌクレオチド、炭水化物、脂質またはこれらの組合せを含む)を同定するように修飾される。本発明のこの実施態様において配列は、3つ以上の所望の構造的特徴を有するサブ配列についてスクリーニングされる。共有結合したアミノ酸、脂質または炭水化物からなる化合物のスクリーニングにおいて、7〜約50の共有結合した単位のサブ配列は、(1) 最初のアミノ酸、炭水化物または脂質残基の2番目から7〜10残基に位置する少なくとも1つの最初のアミノ酸、炭水化物または脂質残基を含有し;(2) 少なくとも1つの第2のアミノ酸、炭水化物または脂質残基をコードし;および(3) 最初のアミノ酸、炭水化物または脂質残基の少なくとも6%を含有する。ヌクレオチド配列のスクリーニングの場合、約21〜約150ヌクレオチドのサブ配列は、(1) 最初のアミノ酸残基をコードする第2のコドンから18〜30ヌクレオチド以内に位置する最初のアミノ酸をコードする少なくとも1つのコドン;(2) 少なくとも1つの第2のアミノ酸残基を含有し;および(3) 最初のアミノ酸残基の少なくとも6%をコードする。
【0224】
本発明のいくつかの実施態様を、本明細書に記載のように具体的に例示する。しかし、本発明の修飾や変更態様が、本発明の精神と範囲を逸脱することなく、上記教示および特許請求の範囲に包含されることは理解されるであろう。
実施例1
レプリキンの抽出、単離および同定法と、レプリキン含有生物を標的化、標識または破壊するためのレプリキンの使用
a) 藻類
【0225】
以下の藻類は、バミューダ海地域から採取し、同日抽出したかまたは−20℃に凍結し、翌日抽出した。藻類を低温室(0〜5℃)で、中性緩衝液、例えば0.005Mのリン酸緩衝液(pH7)100cc中で1グラムのアリコートでWaringブレンダー中で15分ホモジナイズし、3000rpmで遠心分離し、溶媒を留去して上清を濃縮し、低温でリン酸緩衝液で透析して、約15mlの容量を得た。この抽出溶液の容量を記録し、タンパク質分析のためにアリコートを取り、残りを分画して、pKが1〜4の範囲のタンパク質画分を得た。
【0226】
好適な分画法は以下のようにクロマトグラフィーである:抽出溶液を低温室(4℃)で、0.005Mリン酸緩衝液で平衡化しておいたDEAEセルロース(Cellex-D)カラム2.5×11.0cmで分画した。段階的溶出溶媒の変化は以下の溶液を用いて作成する:
溶液1 − 4.04gのNaH2PO4と0.5gのNaH2PO4を15リットルの蒸留水に溶解する(0.005モル、pH7);
溶液2 − 8.57gのNaH2PO4を2,480mlの蒸留水に溶解する;
溶液3 − 17.1gのNaH2PO4を2480mlの蒸留水に溶解する(0.05モル、pH4.7);
溶液4 − 59.65gのNaH2PO4を2470mlの蒸留水に溶解する(0.175モル);
溶液5 − 101.6gのNaH2PO4を2455mlの蒸留水に溶解する(pH4.3);
溶液6 − 340.2gのNaH2PO4を2465mlの蒸留水に溶解する(1.0モル、pX-i 4.1);
溶液7 − 283.63gの80%リン酸(H3PO4)を2460mlの蒸留水に作成する(1.0モル、pH1.0)。
【0227】
抽出溶液(6〜10ml容量)をカラムに流し、溶液1を重層し、300mlの溶液1の容器を取り付け、重力でカラムに滴下させる。3mlアリコートの溶出液を集め、溶液1で除去すべきすべてのタンパク質がカラムから除去されるまで、OD280でタンパク質含量を分析する。次にカラムに溶液2を加え、続いて溶液3、4、5、6および7を、各溶液で除去すべきすべてのタンパク質がカラムから除去されるまで、加える。溶液7からの溶出液を集め、リン酸緩衝液で透析し、透析外液と透析内液の両方のタンパク質含量を測定し、両方をゲル電気泳動で分析する。溶液7で、分子量が3,000〜25,000ダルトンのペプチドまたはタンパク質の1つまたは2つのバンドが得られる。例えば藻類カウレルパ・メキシカナ(Caulerpa mexicana)、ラウレンシア・オブツラ(Laurencia obtura)、クラドフェキサ・プロリフェラ(Cladophexa prolifera)、サルガスム・ナタンス(Sargassum natans)、カウレルパ・ヴェルチシラータ(Caulerpa verticillata)、ハリメダ・ツナ(Halimeda tuna)、およびペニシロス・カピタツス(Penicillos capitatus)は、上記のように抽出と処理した後、溶液7溶出液で、この分子量範囲で汚染物質の無い鋭いペプチドバンドを示した。これらの溶液7タンパク質またはこれらの溶出バンドを加水分解し、アミノ酸組成を測定する。こうして得られたペプチド(リジン組成が6%またはそれ以上)はレプリキン前駆体である。これらのレプリキンペプチド前駆体を次に、アミノ酸配列について測定し、米国特許第6,242,578 B1号に詳述されているように、加水分解と質量スペクトル法によりレプリキンを測定する。「3点認識」法により規定される基準を満足するものをレプリキンとして同定する。この方法はまた、酵母、細菌および任意の植物レプリキンを得るのに応用できる。
b) ウイルス
【0228】
藻類についての上記a)と同じ抽出およびカラムクロマトグラフィー分離法を使用して、ウイルスに感染された細胞中のレプリキンを単離し同定する。
c) in vivoおよびin vitro組織培養での腫瘍細胞
【0229】
藻類についての上記a)と同じ抽出およびカラムクロマトグラフィー分離法を使用して、腫瘍細胞中のレプリキンを単離し同定する。例えば、上記a)のように処理した、悪性脳腫瘍から単離したアストロシチン(Astrocytin)のレプリキン前駆体、神経膠芽細胞腫腫瘍細胞から単離したマリグニン(Aglyco 1OB)、組織培養のMCF7乳癌細胞、組織培養のP3Jリンパ腫細胞はそれぞれ、リジン含量が9.1%、6.7%、6.7%、および6.5%のレプリキン前駆体を与えた。実施例10(米国特許第6,242,578 B1号)に記載されているように、Aglyco 1OBの加水分解と質量スペクトル法により、アミノ酸配列 ykagvaflhkkndiide 16量体レプリキンが得られた。
実施例2:
【0230】
レプリキンの診断的用途の例として:Aglyco 1OBまたは16量体レプリキンを抗原として使用して、米国特許第6,242,578 B1号の図2、3、4および7に記載のように、診断目的に血清中に存在する対応する抗体の量を定量する。
【0231】
標識、栄養または破壊目的にレプリキンに結合する物質の産生の例として:16量体レプリキンをウサギに注射して16量体レプリキンに対する特異抗体を産生することが、米国特許第6,242,578 B1号の実施例6と図9Aおよび9Bに記載されている。
【0232】
レプリキンを標識する物質の使用の例として:16量体レプリキンに対する抗体を使用して、このレプリキンを含有する特異的細胞を標識することが、米国特許第6,242,578 B1号の図5と実施例6に記載されている。
【0233】
レプリキンを破壊する物質の使用の例として:16量体レプリキンに対する抗体を使用して、このレプリキンを含有する特異的細胞を阻害または破壊することが、米国特許第6,242,578 B1号の図6に記載されている。
実施例3
【0234】
レプリキンの存在と濃度についてのインフルエンザウイルス血球凝集素タンパク質またはノイラミニダーゼタンパク質の分離株の配列データの分析は、配列の視覚的スキャニングまたは本明細書に記載の3点認識システムに基づくコンピュータープログラムを使用して行う。インフルエンザウイルスの分離株が得られ、インフルエンザ血球凝集素および/またはノイラミニダーゼタンパク質のアミノ酸配列は、当該分野で公知の任意の方法(例えば血球凝集素またはノイラミニダーゼ遺伝子を配列決定してそこからタンパク質配列を得る)により得られる。各分離株中の新しいレプリキンの存在、経時的なレプリキンの保存、およびレプリキンの濃度について、配列をスキャンする。レプリキン配列と濃度を、先に得られた(例えば約6ヶ月〜約3年前)分離株から得られたアミノ酸配列と比較すると、来るインフルエンザシーズンにインフルエンザの原因となる可能性の高い株の出現を予測するのに使用され、季節的インフルエンザペプチドワクチンまたは核酸ベースのワクチンの基礎を形成するデータが得られる。濃度の上昇(特に、約6ヶ月〜約3年またはそれ以上の期間のインフルエンザウイルスのある株でのレプリキンの濃度の段階的上昇)の観察は、将来のインフルエンザ流行または汎流行の原因となる可能性のある株の出現の予測因子である。
【0235】
出現する株で観察されたレプリキンに基づきペプチドワクチンまたは核酸ベースのワクチンが作成される。出現する株は、この期間の血球凝集素および/またはノイラミニダーゼ配列内のレプリキン配列濃度の最大の上昇を有するインフルエンザウイルスの株として同定される。好ましくはペプチドまたは核酸ワクチンは、出現する株で保存されていることが観察されている任意のレプリキン配列に基づくかまたはこれを含む。保存されているレプリキンは、好ましくは約2年または好ましくはそれ以上の期間、血球凝集素またはノイラミニダーゼタンパク質配列中に存在するレプリキン配列である。ワクチンは、出現する株で同定されるレプリキン配列の任意の組合せを含んでよい。
【0236】
ワクチン産生のために、有効なワクチンのために有用であると同定されるレプリキンペプチドは、任意の方法(化学合成および分子生物学法、例えばクローニング、宿主細胞中での発現、およびそこからの精製を含む)により合成される。ペプチドは好ましくは、医薬として許容される担体と、これに対する治療抗体を誘導することが決定されている量で混合される。一般に投与量は、約0.1μg〜約10mgである。
【0237】
インフルエンザワクチンは好ましくは、「インフルエンザシーズン」が始まる前に必要な患者に投与される。インフルエンザシーズンは一般に、10月の後半から始まり4月の後半まで続く。しかしワクチンは、年の任意の時期に投与される。好ましくはインフルエンザワクチンは年に1回投与され、存在すること、好ましくはインフルエンザウイルスの出現する株で保存されていることが、観察されたレプリキン配列に基づく。インフルエンザワクチンに含めるのに好適な別のレプリキンは、1年以上欠如していた後に再出現したインフルエンザ株中であることが証明されているレプリキンである。
実施例4
【0238】
レプリキンの存在と濃度についての熱帯熱マラリア原虫抗原の分離株の配列データの分析は、配列の視覚的スキャニングまたは本明細書に記載の3点認識法に基づくコンピュータープログラムを使用して行う。熱帯熱マラリア原虫の分離株、およびタンパク質のアミノ酸配列は、当該分野で公知の任意の方法、例えば遺伝子を配列決定してそこからタンパク質配列を得ることにより得られる。配列を、各分離株中のレプリキンの存在、経時的なレプリキンの保存、およびレプリキンの濃度について配列をスキャンする。この情報は、抗マラリアペプチドワクチンまたは核酸ベースのワクチンの基礎を形成するのに使用されるデータを提供する。
【0239】
マラリアを引き起こす生物で観察されたレプリキンに基づくペプチドワクチンまたは核酸ベースのワクチンが作成される。好ましくはペプチドまたは核酸ワクチンは、生物の表面抗原上に存在することが観察されている任意のレプリキン配列に基づくかまたはこれを含む。ワクチンは、マラリアを引き起こす株で同定されるレプリキン配列の任意の組合せを含んでよい。
【0240】
ワクチン産生のために、有効なワクチンのために有用であると同定されるレプリキンペプチドは、任意の方法(化学合成および分子生物学法、例えばクローニング、宿主細胞中での発現、およびそこからの精製を含む)により合成される。ペプチドは好ましくは、医薬として許容される担体と、これに対する治療抗体を誘導することが決定されている量で混合される。一般に投与量は、約0.1μg〜約10mgである。
【0241】
マラリアワクチンは好ましくは、年の任意の時期に必要な患者に投与され、特に熱帯地方への旅行の前に投与される。
【0242】
他の実施態様は、遺伝子のコード鎖に相補的なアンチセンス核酸分子、または生物(特に限定されないが、ウイルス、トリパノソーマ、細菌、真菌、藻類、アメーバ、および植物を含む)中のレプリキンについてのmRNAをコードする生物を含み、ここで該アンチセンス核酸分子は、レプリキン含有生物のヌクレオチド配列に相補的である。
【技術分野】
【0001】
他の出願に対する相互参照
本出願は、出願US 09/984,057 and US 09/984,056(いずれも2001年10月26日出願)の一部継続出願であるPCT出願PCT/US02/09240(2002年3月26日出願)であり、これは、仮出願60/303,396号(2001年7月9日出願)の優先権を主張する(この主題は、参照することにより本明細書に組み込まれる)。
【0002】
発明の分野
本発明は、構造的な特徴を共有する新たに発見されたクラスのペプチドであるレプリキン類(Replikins)の同定と使用に関する。特に、本発明は、ウイルス、細菌、真菌、癌関連タンパク質、植物および単細胞寄生体中に見いだされたレプリキン、および疾患の治療または予防法の開発における標的としてのこれらの使用に関する。 さらに、本発明は、これらの疾患の検出におけるレプリキンの使用に関する。 また本発明は、食品のために使われる植物の成長を促進するためのレプリキンの使用に関する。
【背景技術】
【0003】
発明の概要と背景
迅速な複製は、いくつかの細菌、ウイルスおよび悪性腫瘍の病原性(virulence)に特徴的であるが、異なった生物の迅速な複製に共通する化学はこれまで開示されていない。本特許出願は、迅速な複製に関連する新しいクラスのタンパク質構造を開示する。レプリキン(Replikin)と称される、迅速な複製に関連する保存された小タンパク質の新しいファミリーは、多数の生物や疾患の迅速な複製を予測または抑制するために、且つ植物と動物の迅速な複製を誘導するために使用される。
【0004】
我々は、レプリキンのためのアルゴリズム検索を作成した。ここで発明されたアルゴリズムの応用において、エピトープの機能(迅速な複製)が明らかになっただけでなく、迅速な複製に関連する機能を有する同族体の全ファミリーが発見され、我々はこれをレプリキン類と名付けた。
【0005】
このアルゴリズムは、以下に基づく:1) 免疫系は、認識の時タンパク質全体ではなく部分を見ているという証拠。タンパク質鎖はまず、それに対する免疫防御を開始する免疫系による外来構造の認識プロセスの一部として、免疫系により小片(しばしば6〜10アミノ酸の長さ)に加水分解される。例えば免疫系は、疾患物質のタンパク質を切り刻んで小さなペプチド配列にし、それらを読むことにより疾患の配列を認識する。この原理は、その構造がいったんわかると、マリグニン(malignin)癌エピトープの同族体を検索するためのアルゴリズムの基礎として使用される;2) 上記の認識(6〜10アミノ酸の長さ)のために免疫系が使用する見かけの「規則」に従って3つのリジン(K's)のうちの2つが8残基離れている、マリグニン癌エピトープの特異的構造;3) マリグニン癌エピトープが、非常に強い抗原(すなわち、強い免疫応答の生成元)であること;10量体ペプチド神経膠腫レプリキン中に3つのリジン(K's)があること、およびK's は、ウイルスの侵入のためのアンカー候補としてしばしばDNAおよびRNAに結合することが証明されたという事実;および4) 8残基離れた2つのK'sの間に、マリグニンエピトープの配列中に1つのヒスチジン(H)が含まれており、これは複製のエネルギーを供給するのに必要な酸化還元系の金属への関連を示唆する。
【0006】
適切に配置された官能基を有する調整された結合ポケットを有する、作り変えた酵素と触媒性抗体は、時にすばらしい効率で多くの化学変換を触媒するのに成功している。特異的でかなり異なる機能を有する2つ以上の異なるタンパク質がある生物により一緒に合成され、次に別々に切断されて「別々の機能をする」ことがことがしばしば認められるように、レプリキン構造はユニークな機能を有するユニークなタンパク質であり、これは、免疫系により別々に認識されるようであり、現在は合理的に作り変えられ、例えば機能性単位を産生するように合成される。
【0007】
プロテオミクスの観点から、新たに決定された神経膠腫ペプチド配列に基づくこの鋳型により、関連する保存された構造と特定の機能(この場合は複製)を有する広いクラスのタンパク質が発見された。疾患の病原性に伴うレプリキン濃度の上昇例は、インフルエンザ、HIV、癌およびトマトリーフカールウイルス(tomato leaf curl virus)を含む疾患で現れる。このクラスの構造は、酵母、藻類、植物、ジェミニカールリーフトマトウイルス(gemini curl leaf tomato virus)、HIVおよび癌のような多様な生物の迅速な複製の現象に関連している。
【0008】
迅速に複製している生物中のレプリキンの存在を検出する以外に我々は、1) レプリキン濃度(100個のアミノ酸あたりのレプリキンの数)、および2) 迅速な複製に依存する特定の機能状態にあるレプリキン組成は、それらが存在する生物の複製速度にレプリキンが定量的ならびに定性的に関係するという知見の基礎を提供する、ことを見いだした。これらの機能的証拠の例には、神経膠芽細胞腫細胞中の迅速な複製と病原性との関係、インフルエンザウイルス中のレプリキンとインフルエンザの汎流行と流行の予測との関係、およびHIV中のレプリキン濃度と迅速な複製との関係がある。
【0009】
迅速な複製におけるレプリキンの役割の最初の機能的基礎は、神経膠腫レプリキン[脳の多形神経膠芽腫(神経膠腫)(250kDの細胞タンパク質)中のマリグニン(Malignin)と呼ぶ10kDのペプチド]の性質中で見いだされた。ほとんどまたはすべてのタイプの細胞で現在使用されている癌の早期段階診断検査により測定すると、抗マリグニン抗体の血清濃度(AMAS)は上昇した。マリグニンは、組織培養においてこのペプチドの発現と抽出可能な膜タンパク質1mgあたりのその濃度が、単位時間当たりの細胞分裂の速度の上昇に伴って上昇したため、この名前が付けられた。細胞数の増加に比例してマリグニンの量が増加するのみでなく、マリグニンの量が増加し、すなわち細胞数はわずかに5倍の増加であったが濃度が10倍増加した。
【0010】
マリグニンタンパク質の構造は、加水分解して質量分析により決定され、これにより新規の16量体のペプチド配列が明らかになった。健常人ゲノムのデータベースでこの16量体のペプチド配列(我々は神経膠腫レプリキンタンパク質(Glioma Replikin protein)と命名した)を検索し、これはこれらのデータベース中に存在しないことを見いだした。
【0011】
従って、一定の要件アルゴリズムを使用して、他の生物で神経膠腫レプリキンタンパク質またはその同族体について検索した。「PubMed」データベース中の4,000を超えるタンパク質配列を検索し、特に迅速な複製に関連するウイルスおよび植物で同族体が見つかった。そのようなレプリキンタンパク質の同族体はしばしば、それらの研究者が「複製タンパク質」と呼ぶタンパク質中に存在した。
【0012】
レプリキン配列の同族体はすべての腫瘍ウイルス(癌を引き起こすウイルス)中、および藻類、植物、真菌、ウイルスおよび細菌の「複製タンパク質」中で見つかった。
【0013】
神経膠腫細胞の迅速な複製中の細胞数と膜タンパク質濃度の5倍の上昇と比較してマリグニンが10倍濃縮されることは、レプリキンと複製との重要な関係を示唆する。神経膠腫レプリキンをin vitroで合成し、合成ワクチンとしてウサギに投与すると、多量の抗マリグニン抗体が産生され、血清中の抗マリグニン抗体(AMAS)の抗原性基礎を厳密に確立し、最初の合成癌ワクチン候補と、他の生物中のレプリキンワクチンのプロトタイプを提供している。
【0014】
レプリキンと複製、および癌レプリキンの特異性の共有に基づく癌レプリキンに対する自然の免疫応答(細胞タイプに限定されない)との関係の証明は、抗マリグニン抗体による免疫の受動的増強と、合成レプリキンワクチンによる能動的増強とを可能にする。
【0015】
癌患者と対照からの8,090の血清試料の研究により、健常人では、癌の頻度が増加するように、抗マリグニン抗体の濃度が年齢とともに増加し、早期悪性腫瘍では細胞のタイプに無関係に2〜3倍増加することが証明された。in vitroではこの抗体は、癌細胞1個当たりピコグラム(フェトモル)の癌細胞に対して細胞障害性であり、in vivoでは抗マリグニン抗体の濃度は、癌患者の生存に定量的に関連する。神経膠腫細胞で証明されるように、細胞は不死化の悪性段階に変換されているのみで停止もしくは休止している癌の段階は、レプリキンの濃度の上昇を特徴とする活動性の生命に危険のある複製状態とは区別される。さらに、癌のウイルス病原性の鍵は、神経膠腫糖タンパク質10Bが、正常な10Bと比較すると、炭水化物残基が50%低下しているという事実にあるかも知れない。この低下は、他の例ではウイルス侵入に関連し、従って、悪性状態への変換の過程としての、神経膠腫細胞の10Bへのウイルスレプリキンの供給のためのウイルスの結合の証拠かも知れない。
【0016】
神経膠腫と関連する癌のレプリキンについて抗マリグニン抗体で証明されるように、多様なクラスのメンバーによる免疫学的特異性の共有は、B細胞とその生成物である抗体が、同様の認識「言語」を使用してレプリキンを認識するかも知れないことを示唆する。レプリキンの発見により、この免疫学的特異性の共有は、以前は困難であると考えられてきたことを説明できるかも知れない:なぜ、抗マリグニン抗体はすべての癌で増加しているのか、およびなぜ癌細胞に細胞障害性であるか、なぜほとんどまたはすべての細胞タイプで癌患者の生存に関係しているのか。すなわち、抗マリグニン抗体は癌レプリキンに対して産生され、これは免疫学的特異性を共有し、細胞タイプではなく迅速な複製の現象に関連している。
【0017】
迅速な複製におけるレプリキンの役割の第2の機能的基礎は、インフルエンザウイルス血球凝集素タンパク質配列についての過去100年間のデータの研究およびインフルエンザの流行または汎流行の疫学研究である。今日まで、インフルエンザウイルスについては血清学的血球凝集素と抗体の分類のみが開示され、株特異的な保存されたペプチド配列は開示されておらず、疫学的に記録された流行病または迅速な複製に関連する株特異的ペプチド配列の濃度と組成の変化については何も開示されていない。
【0018】
4つの主要な株[インフルエンザB、(A)H1N1、(A)H2N2、および(A)H3N2]中のそれぞれの1つで株特異的インフルエンザレプリキン濃度の4〜10倍の増加が見つかり、レプリキン濃度のそのような増加は、1902〜2001年に各株により特異的に引き起こされたインフルエンザ流行に関連していた。次に、これらの濃度の増加は、その消失から1〜64年後の少なくとも1つの特異的レプリキン組成の再出現と、新しい株特異的レプリキン組成の出現によることが証明された。以前は、次のインフルエンザシーズンにどの株が主流になるかを予測するための、またはワクチンのために毎年の全ウイルス株の混合物を作成するための、株特異的化学構造は不明であった。H3N2レプリキン濃度の最近の急激な上昇(1997〜2000)(H3N2の歴史で最大)、および1968年の高死亡率のH3N2汎流行と、1975年と1977年の2つの高死亡率流行で最後に見られた特異的レプリキン組成の再出現(しかし20〜25年は出現しなかった)は一緒に、これから来る流行の警告かも知れない。
【0019】
合成レプリキンは新しいワクチンである。同一の構造が100年間持続し、1〜64年の消失後に再出現する同一の構造で観察されるレプリキン構造の高度の保存は、インフルエンザタンパク質中のアミノ酸のランダム置換による病原性の変化であると以前考えられていたものは、レプリキンの組織化された保存プロセスによる変化である可能性が高い。実際、各アミノ酸のランダム置換が起きると、1年間(100年は言うまでもなく)保存されている平均的長さのインフルエンザレプリキン配列に対する変化は、2の27乗対1のオーダーであると計算される。
【0020】
レプリキンの高度の保存は、インフルエンザウイルスにユニークなものではなく、手足口病ウイルスO型およびHIV、ならびにコムギにも存在する。
【0021】
迅速な複製におけるレプリキンの役割の第3の機能的基礎は、HIV中の迅速な複製に関連することが証明されたレプリキン濃度の増加である。早期感染によくあるHIVの増殖の遅い低力価株(NS1、「Bru」)中のレプリキン濃度は、後期HIV感染によくあるHIVの迅速に増殖している高力価株(SI、「Lai」)中のレプリキン濃度の6分の1であった。
【0022】
レプリキンと迅速な複製との関係についての他の例が示される。例えば、トマトカールリーフジェミニウイルス(これは、トマトの収穫に壊滅的被害を与える)では、DNAに結合することが証明されている「複製タンパク質」の最初の161 アミノ酸がレプリキンを含有する。
【0023】
マラリア(迅速な複製で有名)では、1つのトリパノソーマから数万のトリパノソーマが肝臓中に放出される。そこには、100アミノ酸当たり最大36個の重複レプリキンの濃度を有する複数の新規なほとんど「火炎状」のレプリキン構造が見られる。
【0024】
インフルエンザ流行におけるレプリキン濃度の上昇は、悪性神経膠腫細胞の迅速な複製中の神経膠腫レプリキン濃度の上昇に機能的に匹敵し、HIVおよび多様な範囲の他の生物中の迅速な複製に匹敵する。従ってレプリキンは、異なる生物中の迅速な複製の構造的基礎に関連しており、この一部であるようである。
【0025】
従ってレプリキン濃度と組成は、複製を検出しそのプロセスを制御するための新しい方法を提供し、これは、各生物集団の生存と支配に重要である。迅速な複製に関連するこれらの新しいタンパク質の発見は、新しい機会を与える:1) レプリキンの定性的および定量的測定による病原体の検出、2) 未変性のレプリキンを標的とすることによるおよび合成レプリキンをワクチンとして使用することによる、迅速な複製が重要な要因である広範囲の疾患の制御、および3) 藻類と植物食物の成長を促進するためのレプリキンの使用。
【発明の概要】
【発明が解決しようとする課題】
【0026】
検出や治療が困難であり、有効なワクチンが無い多くの疾患や病原体がある。すなわち、各疾患について、これらの疾患および病原体を検出、治療または予防するための有効な方法を提供する標的を開発するニーズがある。
【課題を解決するための手段】
【0027】
本発明は、レプリキン配列を含むヌクレオチドまたはアミノ酸配列の同定法を提供する。本明細書において本方法は3点認識法と呼ぶ。「3点認識」法を使用することにより、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含むペプチド(レプリキン)(新しいクラスのペプチドを構成する)が、複製、形質転換、または酸化還元機能を有する藻類、酵母、真菌、アメーバ、細菌、植物およびウイルスタンパク質で明らかになった。
【0028】
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成されたペプチドが提供される。このペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を含む。
【0029】
本発明はまた、生体試料または環境試料中の汚染生物の存在を検出する方法であって:
(1) 生体試料または環境試料から核酸を単離する;
(2) レプリキン構造の存在について核酸をスクリーニングする;そして
(3) レプリキン構造の存在を汚染生物の存在と相関させる、
ことを含む方法を提供する。
【0030】
本発明の別の態様において、レプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの実施態様は、出現する生物の株(もしそのような株が出現するなら)中に存在する少なくとも1つのペプチドを含む。
【0031】
本発明はまた、本明細書に記載のレプリキンに特異的に結合する抗体、ならびにレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、レプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0032】
本発明の1つの態様において、他のタンパク質から単離または分離された、ウイルスレプリキン配列を含有する、組換えまたは合成ペプチドまたは他の方法が提供される。ウイルスレプリキンペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸(ウイルスレプリキン)を含む。
【0033】
本発明はまた、生体試料または環境試料中の汚染ウイルスの存在を検出する方法であって:
(1) 生体試料または環境試料から核酸を単離する;
(2) ウイルスレプリキン構造の存在について核酸をスクリーニングする;そして
(3) ウイルスレプリキン構造の存在、その濃度および組成を、汚染ウイルスの存在と相関させる、
ことを含む方法を提供する。
【0034】
本発明の別の態様において、ウイルスレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの実施態様は、出現するウイルスの株(もしそのような株が出現するなら)中に存在する少なくとも1つのペプチドを含む。
【0035】
本発明はまた、本明細書に記載のように、ウイルスレプリキンに特異的に結合する抗体、ならびにウイルスレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、ウイルスレプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0036】
本発明はまた、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を有する、1つ以上の単離されたウイルスペプチドと、医薬として許容される担体とを含む治療用組成物を提供する。
【0037】
本発明の別の態様において、ウイルスレプリキンのmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列は、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸である、アンチセンス核酸分子が提供される。
【0038】
本発明のさらに別の態様において、ウイルスに対する抗体を産生するように被験体の免疫系を刺激する方法であって、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を有する少なくとも1つのウイルスレプリキンペプチドの有効量を投与することを含む方法が提供される。
【0039】
別の態様において、予防または治療用ウイルスワクチンに含有されるウイルスペプチドの選択方法であって:
(1) 複数の該ウイルス株の各株の少なくとも1つの分離株を得る;
(2) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列を、レプリキン配列の存在と濃度について分析する;
(3) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するウイルスの株を同定する;そして
(5) 工程(4)で同定されるウイルスペプチドの株に存在する少なくとも1つのレプリキン配列を、ウイルスワクチン中に含有させるためのペプチドとして選択する、
ことを含む方法が提供される。
【0040】
本発明はまた、予防または治療用ウイルスワクチンの作成方法であって:
(1) ウイルスの株を出現する株として同定する、
(2) 出現する株中に存在する少なくとも1つのレプリキン配列を、ウイルスワクチン製造のためのペプチド鋳型として選択する、
(3) 工程(2)で選択した少なくとも1つのレプリキン配列のアミノ酸配列を有するペプチドを合成する、そして
(4) 治療上有効量の工程(3)のペプチドを医薬として許容される担体および/またはアジュバントと組合わせる、
ことを含む方法が提供される。
【0041】
別の態様において本発明は、診断、予防または治療目的の出現するウイルス株を同定する方法であって:
(1) 複数の該ウイルス株の各株の少なくとも1つの分離株を得る;
(2) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列を、レプリキン配列の存在と濃度について分析する;
(3) 複数のウイルス株の各株の少なくとも1つの分離株のアミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);そして
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するウイルスの株を同定する
ことを含む方法が提供される。
【0042】
本発明のさらに別の態様において、出現するウイルス株のタンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ウイルスワクチンが提供される。
【0043】
また本発明により、必要な患者に、出現するウイルス株のタンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ウイルスワクチンを投与することを含む、ウイルス感染症を予防または治療する方法が提供される。
【0044】
インフルエンザ
インフルエンザは、世界的に重要な急性の呼吸系疾患である。ワクチン接種によりインフルエンザウイルス大流行を抑制しようとする国際的な試みにもかかわらず、インフルエンザ感染症はいまだに、罹患と死亡の重要な原因である。歴史的に不規則で以前は予測できなかった間隔で、世界的なインフルエンザの流行と汎流行が起きており、これはこれからも続くと予測される。罹患率、死亡率、および経済コストの点で、汎流行と流行のインフルエンザの影響は大きい。
【0045】
インフルエンザワクチンは、いまだにインフルエンザウイルスに対する最も有効な防御法であるが、このウイルスの突然変異能力と非ヒト宿主を利用できるために、インフルエンザは今後も出現するかまたは再出現する感染症であるであろう。世界的なインフルエンザ監視は、インフルエンザシーズンの間に、国の中で、国と国の間で、および大陸間で、インフルエンザウイルスが変化することを示している。ウイルスの監視は、抗原性のシフトとドリフトの追跡において重要である。疾患の監視はまた、流行の影響を評価するのに重要である。両方の情報がワクチン組成物の基礎と抗ウイルス剤の正しい使用を提供している。しかし今日まで、増加する数のインフルエンザウイルスの出現する株について毎年事後の血液学的分類があるのみで、近づいてくるインフルエンザの流行または汎流行の指標としての、ウイルスの特異的な化学構造は同定されていない。現在、ある年に活動性、非活動性または流行性としてのインフルエンザウイルスの毎年の分類の唯一の基礎は、ウイルス血球凝集素とノイラミニダーゼタンパク質の活性である。流行または汎流行の定量的警告のために、またはより有効で安全なワクチンを設計するために、使用できるインフルエンザウイルスの化学構造は本出願前には同定されていない。
【0046】
インフルエンザワクチンを毎年投与することとワクチンを投与できる期間が短いことのために、ワクチンの適用範囲を改良するための方策は非常に重要である。
【0047】
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成されたインフルエンザウイルスペプチドが提供される。インフルエンザレプリキンウイルスペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸(インフルエンザレプリキン)を含む。
【0048】
本発明の別の態様において、インフルエンザウイルスのレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのインフルエンザウイルスのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの好適な実施態様において組成物は、インフルエンザウイルスの出現する株中に存在する少なくとも1つのペプチドを含む。
【0049】
本発明はまた、本明細書に記載のように、インフルエンザウイルスのレプリキンに特異的に結合する抗体、ならびにインフルエンザウイルスのレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、インフルエンザレプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0050】
本発明はまた、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する1つ以上の単離されたインフルエンザウイルスペプチドと、医薬として許容される担体とを含む治療用組成物を提供する。
【0051】
本発明の別の態様において、インフルエンザウイルスの血球凝集素レプリキンのmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列が、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸を意味する、アンチセンス核酸分子が提供される。
【0052】
本発明のさらに別の態様において、インフルエンザウイルスに対する抗体を産生するように被験体の免疫系を刺激する方法であって、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する少なくとも1つのインフルエンザウイルスのレプリキンペプチドの有効量を投与することを含む方法が提供される。
【0053】
さらに別の態様において、予防または治療用インフルエンザウイルスワクチンに含有されるインフルエンザウイルスペプチドの選択方法であって:
(1) 複数のインフルエンザ株の各株の少なくとも1つの分離株を得る;
(2) レプリキン配列の存在と濃度について、複数のインフルエンザ株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列を分析する;
(3) 複数のインフルエンザ株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株の血球凝集素アミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するインフルエンザウイルス株を同定する;そして
(5) 工程(4)で同定されるインフルエンザウイルス株のペプチドに存在する少なくとも1つのレプリキン配列を、インフルエンザウイルスワクチン中に含有させるためのペプチドとして選択する、
ことを含む方法が提供される。
【0054】
本発明はまた、予防または治療用インフルエンザウイルスワクチンの作成方法であって:
(1) インフルエンザウイルス株を出現する株として同定する、
(2) 出現する株中に存在する少なくとも1つのレプリキン配列を、インフルエンザウイルスワクチン製造のためのペプチド鋳型として選択する、
(3) 工程(2)で選択した少なくとも1つのレプリキン配列のアミノ酸配列を有するペプチドを合成する、そして
(4) 治療上有効量の工程(3)のペプチドを医薬として許容される担体および/またはアジュバントと組合わせる、
ことを含む方法が提供される。
【0055】
別の態様において本発明は、診断、予防または治療目的のインフルエンザウイルスの出現する株を同定する方法であって:
(1) 複数のインフルエンザ株の各株の少なくとも1つの分離株を得る;
(2) レプリキン配列の存在と濃度について、複数のインフルエンザウイルス株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列を分析する;
(3) 複数のインフルエンザウイルス株の各株の少なくとも1つの分離株の血球凝集素アミノ酸配列中のレプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株の血球凝集素アミノ酸配列で観察されるレプリキン配列の濃度と比較して、少なくとも2つの時期のレプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);そして
(4) 少なくとも2つの時期の間のレプリキン配列濃度の最大の上昇を有するインフルエンザウイルスの株を同定する
ことを含む方法が提供される。
【0056】
本発明のさらに別の態様において、インフルエンザウイルスの出現する株の血球凝集素タンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用インフルエンザウイルスワクチンが提供される。
【0057】
また本発明により、必要な患者に、インフルエンザウイルスの出現する株のタンパク質中に存在する少なくとも1つの単離されたレプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ウイルスワクチンを投与することを含む、インフルエンザウイルス感染を予防または治療する方法が提供される。
【0058】
トリパノソーマ
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成されたトリパノソーマペプチドが提供される。トリパノソーマレプリキンペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基(トリパノソーマレプリキン)を含む、7〜約50個のアミノ酸(トリパノソーマレプリキン)を含む。
【0059】
マラリア
治療が困難で有効なワクチンが存在しない1つのトリパノソーマ疾患はマラリアである。マラリアは、熱帯地域で高い死亡率と、肉体的および経済的苦痛を与える。マラリアは、主に熱帯熱マラリア原虫(Plasmodium falciparum)により引き起こされ、これは、極めて難治性であり、今日までマラリアのワクチンは得られていない。すなわち、有効なマラリアワクチン、およびこの疾患を治療または予防する方法に対するニーズがある。本出願は、そのようなワクチンと治療および予防方法の基礎を提供する。上記したレプリキンウイルスワクチンおよびレプリキンインフルエンザウイルスワクチンの産生およびこれらを用いる治療方法は、レプリキンマラリアワクチンの産生およびこれを用いる治療に応用できる。
【0060】
本発明では、マラリアを予防または治療するためのワクチンと方法が提供される。マラリアワクチンは、少なくとも1つの単離された熱帯熱マラリア原虫レプリキンを含む。本発明はまた、少なくとも1つの単離された熱帯熱マラリア原虫レプリキンを含む予防または治療用ワクチン組成物の有効量を、患者に投与することを含む、マラリアの治療または予防方法を提供する。
【0061】
また本発明により、熱帯熱マラリア原虫のマラリア抗原中に存在するレプリキンに特異的に結合する抗体、抗体カクテル、および抗体を含む組成物が提供される。
【0062】
マラリアのように本発明により治療されるトリパノソーマの別の例である梅毒トレポネーマ(Treponema pallidum)(梅毒)のレプリキンは、梅毒の検出、予防、治療に使用することができる。
【0063】
細菌
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成された細菌ペプチド(細菌レプリキン)が提供される。この細菌ペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸(細菌レプリキン)を含む。US特許出願第10/105,232号(2002年3月26日出願)は、参照することによりその全体(特に限定されないが、細菌配列リストと情報を含む)が本明細書に組み込まれる。
【0064】
本発明はまた、生体試料または環境試料中の汚染細菌の存在を検出する方法であって:
(1) 生体試料または環境試料から核酸を単離する;
(2) レプリキン構造の存在について核酸をスクリーニングする;そして
(3) レプリキン構造の存在を汚染生物の存在と相関させる、
ことを含む方法を提供する。
【0065】
本発明の別の態様において、細菌レプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つの細菌のレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。1つの好適な実施態様は、出現する細菌株(そのような新しい株が出現する場合)中に存在する少なくとも1つの細菌ペプチドを含む。
【0066】
本発明はまた、本明細書に記載のように、細菌のレプリキンに特異的に結合する抗体、ならびに細菌のレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、細菌レプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0067】
本発明はまた、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する1つ以上の単離された細菌ペプチド;および
(4) 医薬として許容される担体
を含む治療用組成物を提供する。
【0068】
本発明の別の態様において、細菌のレプリキンのmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列が、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を意味する、アンチセンス核酸分子が提供される。
【0069】
本発明のさらに別の態様において、細菌に対する抗体を産生するように被験体の免疫系を刺激する方法であって、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する少なくとも1つの細菌のレプリキンペプチドの有効量を投与することを含む方法が提供される。
【0070】
さらに別の態様において、予防または治療用細菌ワクチンに含有される細菌レプリキンペプチドの選択方法であって:
(1) 複数の細菌株の各株の少なくとも1つの分離株を得る;
(2) 細菌レプリキン配列の存在と濃度について、複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列を分析する;
(3) 複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列中の細菌レプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察される細菌レプリキン配列の濃度と比較して、少なくとも2つの時期の細菌レプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内、または迅速に突然変異する細菌ではそれより早い時期である);
(4) 少なくとも2つの時期の間の細菌レプリキン配列濃度の最大の上昇を有する細菌の株を同定する;そして
(5) 工程(4)で同定される細菌ペプチドの株に存在する少なくとも1つの細菌レプリキン配列を、細菌ワクチン中に含有させるためのペプチドとして選択する、
ことを含む方法が提供される。
【0071】
本発明はまた、予防または治療用細菌ワクチンの作成方法であって:
(1) 細菌の株を出現する株として同定する、
(2) 出現する株中に存在する少なくとも1つの細菌レプリキン配列を、細菌ワクチン製造のためのペプチド鋳型として選択する、
(3) 工程(2)で選択した少なくとも1つの細菌レプリキン配列のアミノ酸配列を有するペプチドを合成する、そして
(4) 治療上有効量の工程(3)のペプチドを医薬として許容される担体および/またはアジュバントと組合わせる、
ことを含む方法が提供される。
【0072】
別の態様において本発明は、診断、予防または治療目的の細菌の出現する株を同定する方法であって:
(1) 複数の細菌株の各株の少なくとも1つの分離株を得る;
(2) 細菌レプリキン配列の存在と濃度について、複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列を分析する;
(3) 複数の細菌株の各株の少なくとも1つの分離株のアミノ酸配列中の細菌レプリキン配列の濃度を、少なくとも1つのより早い方の時期の各株のアミノ酸配列で観察される細菌レプリキン配列の濃度と比較して、少なくとも2つの時期の細菌レプリキンの濃度を提供する(該少なくとも1つのより早い方の時期は、工程(1)の約6ヶ月〜約3年前以内である);そして
(4) 少なくとも2つの時期の間の細菌レプリキン配列濃度の最大の上昇を有する細菌の株を同定する
ことを含む方法が提供される。
【0073】
本発明のさらに別の態様において、細菌の出現する株のタンパク質中に存在する少なくとも1つの単離された細菌レプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用細菌ワクチンが提供される。
【0074】
細菌(マイコバクテリア(Mycobacteria))として分類される2つの重要な細菌の亜種は、その30sリボゾームタンパク質がC末端レプリキンを有するらい菌(Mycobacterium leprae)と、そのATPaseが3つのレプリキンを有する結核菌(Mycobacterium tuberculosis)である。
らい菌(Mycobacterium leprae)(らい病)の30s リボゾームタンパク質s6中のレプリキンは以下の通りである:
kvmrtdkh
結核菌(Mycobacterium tuberculosis)のATPaseのレプリキンは以下の通りである:
hprpkvaaalkdsyrlk
hprpkvaaalk
ksaqkwpdkflagaaqvah
大腸菌(E. coli)のB-D-ガラクトシダーゼ中のレプリキンは以下の通りである:
hawqhqgktlfisrk
hqgktlfisrk
アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)中のレプリキン:
hsdqqlavmiaakrlddyk
hlldhpasvgqldlramlaveevkidnpvymek
hpasvgqldlramlaveevkidnpvymek
kcvmakncnikcpaglttnqeafngdpralaqylmniah
kncnikcpaglttnqeafngdpralaqylmniah
hhdtysiedlaqlihdakaaruvivk
hdtysiedlaqlihdakaarvrvivk
hdakaarvrvivk
kigqgakpgeggqlpspkvtveiaaarggtpgvelvsppphh
kigqgakpgeggqlpspkvtveiaaarggtpgyelvsppph
kaseitktlasgamshgalvaaaheavahgtnmvggmsnsgeggeh
kaseitktlasgamshgalvaaaheavah
kaseitktlasgamshgalvaaah
kaseitktlasgamsh
kryfpnvktpvggvtfaviaqavadwh
hhiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaekslmk
hhiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaekslmk
hhiaaglgfgasavyplgvqfraeekfgadaokafkrfakaaek
hhiaaglgfgasavyplgvqfraeekfgadadkafkrfak
hhiaaglgfgasavyplgvqfraeekfgadadk
hiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaekslnk
hiaaglgfgasavyplgvqfraeekfgadadkafkrfakaaek
hiaaglgfgasavyplgvqfraeekfgadadkafkrfak
hiaaglgfgasavyplgvqfraeekfgadadk
kfglydaafeksscgvgfitrkdgvqth
【0075】
また本発明により、必要な患者に、細菌の出現する株のタンパク質中に存在する少なくとも1つの単離された細菌レプリキンと、医薬として許容される担体および/またはアジュバントとを含む、予防または治療用ワクチンを投与することを含む、細菌感染症を予防または治療する方法が提供される。
【0076】
真菌
本発明の1つの態様において、レプリキン配列を含有する単離されたかまたは合成された真菌ペプチドが提供される。この真菌ペプチドは、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基(トリパノソーマレプリキン)を含む、7〜約50個のアミノ酸(真菌レプリキン)を含む。
【0077】
細菌レプリキンワクチンの産生とこれを用いる治療について記載した上記のすべての方法は、真菌レプリキンワクチンの産生とこれを用いる治療に適用される。
【0078】
本発明の別の態様において、真菌のレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つの真菌レプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。
【0079】
本発明はまた、本明細書に記載のように、ウイルスレプリキンに特異的に結合する抗体、ならびにウイルスレプリキンに特異的に結合する複数の抗体を含有する抗体カクテルを提供する。本発明のある実施態様において、真菌レプリキンに特異的に結合する抗体と医薬として許容される担体とを含む組成物が提供される。
【0080】
本発明はまた、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する1つ以上の単離された真菌ペプチド;および
(4) 医薬として許容される担体
とを含む治療用組成物を提供する。
【0081】
本発明の別の態様において、真菌のレプリキンmRNA配列に相補的なアンチセンス核酸分子であって、該レプリキンmRNA配列が、
(1) 第2のリジン残基から6〜10残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸を有する、上記分子が提供される。
【0082】
本発明の別の態様において、真菌のレプリキン配列に特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法であって、少なくとも1つのレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む方法が提供される。
【0083】
複製の上昇
本発明のさらに別の態様において、生物中で複製機能を有する酵素または他のタンパク質をコードする遺伝子を、少なくとも1つのレプリキン構造を用いて形質転換することを含む、生物の複製速度を上昇させる方法が提供される。
定義
【0084】
本明細書において用語「ペプチド」または「タンパク質」は、1つのアミノ酸のカルボキシル基が別のアミノ酸のアミノ基に結合して、ペプチド結合を形成する2つ以上のアミノ酸の化合物を意味する。用語ペプチドはまた、そのような化合物をコードするアミノ酸配列を意味するのにも使用される。本明細書において「単離された」または「合成された」ペプチドまたはその生物活性のある部分は、そこからペプチドが得られる細胞または組織源からの細胞物質または他の汚染ペプチドを実質的に含まない、または任意の方法により合成される時化学前駆体または他の化学物質を実質的に含まない、または組換え遺伝子技術により合成される時汚染ペプチドを実質的に含まない、ペプチドを意味する。
【0085】
本明細書において、レプリキンペプチドまたはレプリキンタンパク質は、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;
(3) 少なくとも6%のリジン残基
を含む、7〜約50個のアミノ酸配列である。同様にレプリキン配列は、そのようなペプチドまたはタンパク質をコードするアミノ酸配列である。
【0086】
本明細書において、「出現する株」は、他の株の生物中のレプリキンの濃度と比較して、1つ以上のタンパク質配列中のレプリキン配列の濃度が上昇しているとして同定されたウイルス、細菌、真菌、または他の生物の株を意味する。レプリキン濃度の上昇は、例えばインフルエンザウイルスでは、少なくとも6ヶ月の期間、好ましくは少なくとも約1年間、最も好ましくは少なくとも約3年間またはそれ以上の期間起きるが、細菌や他の生物でははるかに短い期間である。
【0087】
本明細書において「突然変異」は、アミノ酸の置換により引き起こされる生物の構造および性質の変化を意味する。これに対して、本明細書において用語「保存」は、置換の欠如のための特定のアミノ酸の保存を意味する。
【図面の簡単な説明】
【0088】
【図1】図1は、種々の生物中のレプリキンの出現頻度を示す棒グラフである。
【図2】図2は、神経膠腫細胞の嫌気的複製中の、総膜タンパク質1mg当たりのマリグニンのパーセントを示すグラフである。
【図3】図3は、リコグニン(recognin)16量体への暴露に応答して産生される抗マリグニン抗体の量を示す棒グラフである。
【図4A】図4Aは、普通の蛍光灯で撮った血液スメアの写真である。
【図4B】図4Bは、2つの白血病細胞の存在を例示する普通の蛍光灯で撮った血液スメアの写真である。
【図4C】図4Cは、抗マリグニン抗体の存在下の神経膠腫細胞の密な層の写真である。
【図4D】図4Dは、抗マリグニン抗体の添加の30分後に撮った、図4Cの細胞の層の写真である。
【図4E】図4Eは、抗マリグニン抗体の添加の45分後に撮った、図4Cの細胞の層の写真である。
【図4F】図4Fは、抗マリグニン抗体による小細胞肺癌細胞の増殖のin vitroでの阻害を示す棒グラフである。
【図5】図5は、手術前および後の良性または悪性乳癌患者の血清中に存在する抗マリグニン抗体の量のプロットである。
【図6】図6は、レプリキン配列を同定する3点認識法を実施するのにコンピューターが使用される、本発明の実施態様を示すボックス図である。
【図7】図7は、1918年から2001年まで毎年、インフルエンザBとインフルエンザA株(H1N1)の血球凝集素で観察されたレプリキンの濃度を示すグラフである。
【図8】図8は、1950年から2001年まで毎年、インフルエンザA株(H2N2とH3N2)、ならびに成分レプリキンにより規定された出現する株の血球凝集素で観察されたレプリキンの濃度を示すグラフである。
【発明を実施するための形態】
【0089】
発明の詳細な説明
迅速な複製の現象に関連した小ペプチドの新しいファミリー(ここで、レプリキンと呼ぶ)の同定は、試料中の病原体の検出のための、および治療法の開発(ワクチン開発を含む)のための標的となる。一般に、このファミリーのペプチドの知識および同定は、レプリキンを有する任意の生物の有効な治療法やワクチンの開発を可能にする。このファミリーのペプチドの同定はまた、ウイルスの同定およびウイルスワクチンの開発を可能にする。
【0090】
例えば、このファミリーのペプチドの同定は、インフルエンザウイルスの検出を提供し、インフルエンザ治療法の新しい標的を提供する。このファミリーのペプチドの同定はまた、例えばマラリアの検出を提供し、マラリアワクチンの開発のための新しい標的を提供する。このファミリーのペプチドの同定により提供されるさらなる例には、感染症レプリキン、癌免疫レプリキンおよび構造タンパク質レプリキンの検出がある。
【0091】
迅速な複製は、いくつかの細菌、ウイルスおよび悪性腫瘍の病原性に特徴的であるが、異なった生物の迅速な複製に共通する化学はこれまで開示されていない。我々は、迅速な複製に関連する保存された小タンパク質の新しいファミリーを見いだし、これをレプリキンを名付けた。そのようなレプリキンは、有効な検出法と治療法を開発するための新しい標的を与える。見いだされた最初のレプリキンは、神経膠腫レプリキンであり、これは、マリグニン(Malignin)と呼ぶ脳の多形神経膠芽腫(神経膠腫)細胞タンパク質中で同定された。
【0092】
マリグニンの加水分解と質量スペクトル測定により、神経膠腫レプリキンを含有する新規の16量体のペプチド配列が明らかになった。このレプリキンは、健常人ゲノムのデータベース中には見つからず、従って体外の何らかの起源に由来するものと思われる。
【0093】
我々は、神経膠腫レプリキンまたはその同族体を検索するためのアルゴリズムを作成した。同族体は4,000を越えるタンパク質配列に共通するものではなかったが、驚くべきことに、すべての腫瘍ウイルス、および藻類、植物、真菌、ウイルスおよび細菌の複製タンパク質中に存在した。
【0094】
我々は、1) レプリキン濃度(100アミノ酸当たりのレプリキンの数)、および2) レプリキン組成の両方が、迅速な複製の機能的現象に相関することを確認した。これらの関係は、レプリキンが複製速度に定量的および定性的に関係することを決定するための機能的基礎を提供する。
【0095】
迅速な複製へのレプリキンの役割の最初の機能的基礎は、神経膠腫複製で見られる。神経膠腫細胞の迅速な複製中で細胞数と膜タンパク質との5倍の増加に比較して、神経膠腫マリグニンが10倍濃縮されている事実は、レプリキンと複製の基本的な関係を示唆する。神経膠腫レプリキンをin vitroで合成し、合成ワクチンとしてウサギにに投与すると、多量の抗マリグニン抗体が産生された。これは、血清中の抗マリグニン抗体(AMAS)試験の抗原性基礎を確立し、他の生物中の最初の合成癌ワクチン候補およびレプリキンワクチンの原型を提供する。癌レプリキンと迅速な複製の共有される特異性に基づく、レプリキンと複製との天然の免疫関係、および癌レプリキンに対するこの天然の免疫応答(これは、細胞タイプに無関係である)の証明により、抗マリグニン抗体による免疫の受動的増強と合成レプリキンによる能動免疫増強の両方が可能である。
【0096】
抗マリグニン抗体の存在と患者の生存との関係が、癌患者の8,090個の血清試料の研究により証明された。この試験は、集団中の癌の発症率のように、抗マリグニン抗体の濃度が年齢とともに上昇すること、および細胞タイプに無関係に早期悪性腫瘍でさらに2〜3倍増加することを証明した。in vitroでは、抗マリグニン抗体は、癌細胞当たりピコグラム(フェムトモル)の癌細胞に細胞障害性であり、in vivoでは抗マリグニン抗体の濃度は、癌患者の生存に定量的に関連する。神経膠腫細胞で証明されるように、細胞は不死化の悪性段階に変換されるのみで停止もしくは休止している段階は、レプリキンの濃度の上昇を特徴とするより活発な生命に危険のある複製段階(これは、レプリキンの濃度の上昇が特徴である)とは区別される。さらに癌のウイルス病原性への鍵は、神経膠腫糖タンパク質10Bは、正常な10Bと比較すると、炭水化物残基が50%低下しているという事実にあるかも知れない。この低下は、他の例のウイルス侵入に関連し、従って、悪性段階への変換の工程として神経膠腫細胞の10Bへのウイルスレプリキンの供給のための、ウイルスの結合の証拠かも知れない。
【0097】
過去100年間のインフルエンザウイルス血球凝集素のタンパク質配列とインフルエンザ流行についての我々の研究は、レプリキンと迅速な複製との関係の第2の機能的基礎を提供した。今日まで、インフルエンザについては血清学的血球凝集素と抗体の分類のみが開示され、株特異的保存ペプチド配列は開示されていない。さらに、疫学的に記録された流行病または迅速な複製と相関する株特異的ペプチド配列の濃度と組成の変化は、記載されていない。この研究では、4つの主要な株(インフルエンザB、(A)H1N1、(A)H2N2、および(A)143N2)中のそれぞれの1つで株特異的インフルエンザレプリキン濃度の4〜10倍の増加が、1902〜2001年に各株により引き起こされたインフルエンザ流行に関連していることが証明されている。
【0098】
次に我々は、これらの濃度の増加は、その消失から1〜64年後の少なくとも1つの特異的レプリキン組成の再出現と、新しい株特異的レプリキン組成の出現によることを証明した。以前は、次のインフルエンザシーズにどの株が主流になるかを予測するための、またはワクチンのためのウイルス全体の株の毎年の混合物を作成するための、株特異的化学構造は不明であった。H3N2レプリキン濃度の最近の急激な上昇(1997〜2000)(H3N2の歴史で最大)および1968年の高死亡率のH3N2汎流行と1975年と1977年の2つの高死亡率流行で最後に見られた特異的レプリキン組成の再出現(しかし20〜25年は出現しなかった)は一緒に、これから来る流行を警告しているかも知れない。同じ構造が100年間持続するか、または1〜64年の欠如後に再出現する、観察されるレプリキン構造の高度の保存は、インフルエンザタンパク質中のアミノ酸のランダム置換による変化であると以前考えられていたものは、レプリキンの保存の組織化されたプロセスによる変化である可能性が高いことを示す。
【0099】
レプリキンの保存は、インフルエンザウイルスにユニークなものではなく、他の供給源、例えば手足口病ウイルスO型およびHIV、ならびにコムギでも見られた。
【0100】
迅速な複製におけるレプリキンの役割の第3の機能的基礎は、HIV中の迅速な複製の上昇に見られる。レプリキン濃度は、HIV中の迅速な複製に関連することが証明された。我々は、増殖の遅い低力価HIV株(NS1、「Bru」)中のレプリキン濃度は、迅速に増殖している高力価HIV株(SI、「Lai」)(後期HIV感染によくある)中のレプリキン濃度の6分の1であることを見いだした。
【0101】
さらなる例は、レプリキンと迅速な複製との関係を示す。トマトカールリーフジェミニウイルス(これは、トマトの収穫に壊滅的被害を与える)の「複製タンパク質」では、DNAに結合することが証明されているの最初の161 アミノ酸がレプリキンを含有することが証明された。マラリア(迅速な複製で有名)では、1つのトリパノソーマから数万のトリパノソーマが肝臓中に放出されると、100アミノ酸当たり最大36個の重複レプリキンの濃度の、複数の新規なほとんど「火炎状」のレプリキン構造が見られた。
【0102】
ある構造の保存は、その構造が攻撃と破壊または刺激のための安定な不変の標的となるかどうかにとって決定的に重要である。ある構造がその生物の基本的な生存機構と何らかの意味で関係している時は、その構造は保存される傾向にある。変化する構造は、変わりやすい標的となり、これは、攻撃物を避けるための良好な方法(例えば、以前の構造に対して特異的に作成された抗体)であり、従って修飾型に対しては無効である。この方法が、例えばインフルエンザウイルスにより利用され、以前のワクチンは、現在の病原性ウイルスに対して全く無効となることがある。
治療のための安定な標的としてのレプリキン
【0103】
細菌とHIVともに、レプリキンおよび非レプリキンアミノ酸を有する。例えば最近HIVでは、突然変異により薬剤耐性が9%から13%まで上昇しており、これは、非レプリキンアミノ酸の置換である(本明細書に記載のHIVのTATタンパク質の詳細な分析を参照されたい)。細菌では「耐性株」の出現は、同じ機構による。しかし我々は、レプリキン構造が、非レプリキンアミノ酸と同じ程度までには突然変異も変化もしないことを見いだした(また、本明細書に記載のレプリキンの手足口病ウイルスの説明も参照されたい)。非レプリキン構造とは反対にレプリキン構造は、保存され、従って治療の新しい恒常的標的となる。
【0104】
生存機能にあまりにも密接に関連したいくつかの構造は、絶えず変化することはできない。レプリキン構造の必須成分はヒスチジン(h)であり、これはしばしば、酸化還元酵素の金属基に結合し、おそらく複製に必要なエネルギーの源であることが知られており、このヒスチジン構造は変化しないため、この構造は、破壊または刺激のさらに魅力的な標的である。
【0105】
プロテオミクス的観点から、新に決定された神経膠腫ペプチド配列に基づく鋳型の本発明者らの構築により、関連する保存構造と特定の機能(この場合は複製)を有する広いクラスのタンパク質が発見された。疾患の病原性とのレプリキン濃度の上昇の例には、インフルエンザ、HIV、癌およびトマトリーフカールウイルスがある。この新たに認識されたクラスの構造は、酵母、藻類、植物、ジェミニカールリーフトマトウイルス、HIVおよび癌のような多様な生物の迅速な複製の現象に関連する。
【0106】
レプリキン濃度と組成は、複製(これは、各生物集団の生存と支配にとって重要である)のプロセスを検出し制御するための新しい定量的方法を提供する。神経膠腫と関連する癌のレプリキンについての抗マリグニン抗体で示されるように、多様なクラスのメンバーによる免疫学的特異性の共有は、B細胞とその生成物である抗体が、同様の認識言語を使用してレプリキンを認識することを示唆する。
【0107】
癌レプリキンのペプチド配列またはレプリキンを含有するペプチド配列(すなわち、神経膠腫ペプチドkagvaflhkkの同族体)の例は、特に限定されないが、肺、脳、肝臓、軟組織、唾液腺、鼻咽腔、食堂、胃、結腸、直腸、胆嚢、乳房、前立腺、子宮、子宮頸部、膀胱、眼、黒色腫、リンパ腫、白血病、および腎臓の癌に見いだされる。
【0108】
レプリキンは、1) レプリキンの定性的および定量的測定による病原体の検出;2) 未変性のレプリキンを標的化とすることにより、および合成レプリキンをワクチンとして使用することにより、迅速な複製が重要な要因である広範囲の疾患の治療と制御;および3) 藻類および植物性食物の増殖速度の上昇の促進、を提供する。
【0109】
同定された最初のレプリキン配列は、脳癌タンパク質マリグニン中に見いだされた癌細胞レプリキンであり、これは、多形神経膠芽腫(神経膠腫)細胞の迅速な嫌気的複製中に10倍濃縮されていることが証明された(図2)。マリグニンは、多形神経膠芽腫(神経膠腫)細胞からin vivoおよびin vitroで単離された250 kDaの糖タンパク質10Bの10 KDa部分である。マリグニンの加水分解と質量スペクトル測定により、16量体のペプチド配列、ykagvaflhkkndide(配列番号4)が明らかになり、これは本明細書において神経膠腫レプリキンと呼び、より短いペプチドkagvaflhkk(配列番号1)を含み、この両方とも正常ヒトゲノムには存在しない。
【表1】
【0110】
16量体の神経膠腫レプリキンを合成し、合成ワクチンとしてウサギに注射すると、多量の抗マリグニン抗体が産生された(Bogochら、Cancer Detection and Prevention, 26 (増刊 1):402(2002))。in vivoの血清中の抗マリグニン抗体の濃度は、癌患者の生存に定量的に関連することが証明されている(Bogochら、Protides of Biological Fluids, 31:739-747 (1984))。in vitro抗マリグニン抗体は、癌細胞当たりピコグラム(フェムトモル)濃度で癌細胞に細胞障害性であることが証明されている(Bogochら、Cancer Detection and Prevention, 26 (増刊 1):402(2002))。
【0111】
本発明者らが行った研究は、神経膠腫レプリキンが健常人ゲノム中に存在しないことを示した。従って、種々の生物の公表された配列の分析により、レプリキン配列の起源と同族体候補の検索を行った。
【0112】
16量体の神経膠腫レプリキン配列を鋳型として使用し、そして認識プロテオミクスシステムを構築して、いくつかの異なる生物のタンパク質のアミノ酸配列を視覚的にスキャンすることにより、新しいクラスのペプチド(レプリキン)が同定された。本発明は、レプリキン配列を含むヌクレオチド配列またはアミノ酸配列の同定法を提供する。この方法は本明細書において、3点認識法と呼ぶ。3点認識法は:(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む、7〜約50個のアミノ酸のペプチド(レプリキン)を含む。これらのペプチドまたはタンパク質は、複製、形質転換、または酸化還元機能を有する藻類、酵母、真菌、アメーバ、細菌、植物、ウイルス、および癌タンパク質を含む種の新しいクラスのペプチドを構成する。レプリキンペプチドは、より大きい「複製」および「形質転換」タンパク質(その研究者らによりそう記載された、表2を参照)および癌細胞で濃縮されていることがわかった。マリグニン16量体ペプチドと同一の配列は見つかっていない。
【表2】
【表3】
【0113】
レプリキンとしてのまたはレプリキンを含有するもの(すなわち、神経膠腫ペプチド kagvaflhkkの同族体)としてのアミノ酸配列の同定には、以下の3つの要件が満たされなければならない。3点認識システムでは、7〜約50個の残基のアミノ酸配列以内に配列は3つの要素を有する:(1) 別のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジンの組成。
【0114】
レプリキン配列を含有するタンパク質配列について、ナショナルライブラリーオブメディシン(National library of Medicine)のキーワード「PubMed」記述子を使用して、データベースを検索した。4,000を越えるタンパク質配列を、同族体について視覚的に調べた。PubMedで分類されたタンパク質の各群内のすべての個々のタンパク質の配列を、上記の3つの要件を満たすペプチドについて視覚的に調べた。全体に「ウイルスペプチド」(1.5%(N=953))、および悪性形質転換または複製に関連するとされていない他のペプチド、例えば「脳ペプチド」および「神経ペプチド」(まとめて8.5%)(N=845)で、低率の同族体が観察された。しかし驚くべきことに、同族体は大きな「複製タンパク質」中でより高頻度に同定され、これらは細菌、藻類およびウイルスで複製の確立された機能を有するとして同定された。さらに驚くべきことは、「腫瘍ウイルス」(N=250)の100%、「癌タンパク質」(N=401)の97%、および「形質転換ウイルス」(N=248)の85%に存在するという知見である。これらの結果は、細胞タイプに無関係に癌の病原性に共通の性質があることを示唆し、発癌性でのウイルスの役割(すなわち休止状態であるが形質転換された状態から、より病原性の活性な複製状態への細胞の変換)を示唆する。
【0115】
3点認識法により定義される以下のアミノ酸配列 kagvaflhkk の同族体は、特に限定されないが、アデノウイルス(adenovirus)、レンチウイルス(lentivirus)、a-ウイルス(a-virus)、レトロウイルス(retrovirus)、アデノ関連ウイルス(andenoassociated virus)、ヒト免疫不全症ウイルス(human immunodeficiency virus)、肝炎ウイルス(hepatitis virus)、インフルエンザウイルス(influenza virus)、トウモロコシストリークウイルス(maize streak virus)、ヘルペスウイルス(herpes virus)、ウシヘルペスウイルス(bovine herpes virus)、ネコ免疫不全症ウイルス(feline immunodeficiency virus)、手足口病ウイルス(foot and mouth disease virus)、天然痘ウイルス(small pox virus)、ラウス肉腫ウイルス(rous sarcome virus)、神経芽腫RASウイルス癌遺伝子(neuroblastoma RAS viral oncogene)、ポリアマウイルス(plyamavirus)、ヒトパピローマウイルス(human papilloma virus)、骨髄性単球性腫瘍ウイルス(myelomonocytic tumor virus)、マウス急性白血病(murine acute leukemia)、T細胞リンパ栄養性ウイルス(T-cell lymphotropic virus)、およびトマトリーフカールウイルス(tomato leaf curl virus)のようなウイルスまたはウイルスペプチド中に存在する。
【0116】
レプリキンは、特に限定されないが、アセトバクター(Acetobacter)、アクロモバクター(Achromobacter)、アクチノミセス(Actinomyces)、アエロバクター(Aerobacter)、アルカリゲネス(Alcaligenes)、アルスロバクター(Arthrobacter)、アゾトバクター(Azotobacter)、バチルス(Bacillus)、ブレビバクテリウム(Brevibacterium)、チャイニア(Chainia)、クロストリディウム(Clostridium)、コリネバクテリウム(Corynebacterium)、エルウィニア(Erwinia)、エシェリキア(Escheria)、レブシエラ(Lebsiella)、ラクトバチルス(Lactobacillus)、ヘモフィルス(Haemophilus)、フラボバクテリウム(Flavobacterium)、メチロモナス(Methylomonas)、ミクロコッカス(Micrococcus)、マイコバクテリウム(Mycobacterium)、ミクロノムスポラ(Micronomspora)、マイコプラズマ(Mycoplasma)、ナイセリア(Neisseria)、ノカルジア(Nocardia)、プロテウス(Proteus)、シュードモナス(Pseudomonas)、リゾビウム(Rhizobium)、サルモネラ(Salmonella)、セラチア(Serratia)、スタフィロコッカス(Staphylococcus)、連鎖球菌(Streptocossus)、ストレプトミセス(Streptomyces)、ストレプトスポランギウム(Streptosporangium)、ストレプトビルチシリウム(Streptovirticillium)、ビブリオ(Vibrio)、ペプチド、およびキサントモナス(Xanthomas)のような細菌に存在する。
【0117】
レプリキンは、特に限定されないが、ペニシリウム(Penicillium)、ディセウラ(Diseula)、オフィストーマ・ノボウリム(Ophiostoma novo-ulim)、ミコフィコフタ(Mycophycophta)、フィトフトラ・インフェスタンス(Phytophthora infestans)、アブシジア(Absidia)、アスペルギルス(Aspergillus)、カンジダ(Candida)、セファロスポリウム(Cephalosporium)、フサリウム(Fusarium)、ハンゼヌラ(Hansenula)、ムコール(Mucor)、パエシロミセス(Paecilomyces)、ピヒア(Pichia)、リゾプス(Rhizopus)、トルロプシス(Torulopsis)、トリコデルマ(Trichoderma)、およびエリシフェ(Erysiphe)のような真菌中に存在する。
【0118】
レプリキンは、特に限定されないが、サッカロミセス(Saccharomyces)、クリプトコッカス(Cryptococcus)(クリプトコッカス‐ネオフォルマンス(Cryptococcus neoformas)を含む)、シゾサッカロミセス(Schizosaccharomyces)、およびオリザ(Oryza)のような酵母中に存在する。
【0119】
レプリキンは、特に限定されないが、カルドフェラ(Caldophera)、イソレピスプロリフェラ(Isolepisprolifera)、ツノマタ(Chondrus)、グラシラリア(Gracilaria)、ゲリジウム(Gelidium)、カウレルパ(Caulerpa)、ラウレンシア(Laurencia)、クラドフェキサ(Cladophexa)、ホンダワラ(Sargassum)、ペニシロス(Penicillos)、サボテングサ(Halimeda)、コンブ(Laminaria)、ヒバマタ(Fucus)、アスコフィルム(Ascophyllum)、ワカメ(Undari)、ダルス(Rhodymenia)、マクロシスティス(Macrocystis)、キリンサイ(Eucheuma)、フササイミ(Ahnfeltia)、プテロクラシア(Pteroclasia)のような藻類中に存在する。
【0120】
レプリキンは、特に限定されないが、エントアメーバ(Entamoeba)(エントアメーバ・インバデンス(Entamoeba invadens)を含む)、アメービダエ(Amoebidae)、アカントアメーバ(Acanthamoeba)およびネグレリア(Naegleria)のようなアメーバ中に存在する。
【0121】
レプリキンは、特に限定されないが、アラビドプシス(Arabidopsis)、コムギ(wheat)、イネ(rice)、およびトウモロコシ(maize)のような植物中に存在する。
【0122】
補助的規格
レプリキンのサブタイプの分類を可能にするために、基本的な「3点認識」要件に追加のまたは「補助的規格」を加える:(a) 例えば、癌細胞タンパク質(例えば、形質転換タンパク質 P21B(K-RAS2B)、肺、表2、配列番号89)中の隣接するジ−およびポリリジンの共通の存在のような、およびTOLL様受容体のような構造に基づくか、または (b) 既存のATPase、チロシンキナーゼ、または表2に見られるような酸化還元活性のような、機能に基づく。
【0123】
機能性誘導体
本明細書に記載のレプリキンの「機能性誘導体」は、レプリキンに対して特異的な抗体と少なくとも免疫学的交差反応性の部分を保持する、レプリキンの断片、変種、類似体、または化学誘導体である。レプリキンペプチドの断片は、分子の任意のサブセットである。変種ペプチドは、例えば当該分野で公知の方法を使用して直接化学合成により作成される。レプリキンの類似体は、全タンパク質またはその断片に実質的に類似の非天然のタンパク質である。レプリキンの化学誘導体は、通常はペプチドの一部またはペプチド断片ではない追加の化学残基を含有する。
【0124】
図2で明らかなように、細胞複製の速度が上昇する嫌気的呼吸中に、マリグニンは濃縮される。すなわち、細胞数や総膜タンパク質の増加に比例して増加するのみではなく、休止状態の3%から出発して総膜タンパク質の30%に達するまで、濃度が10倍も濃縮される。神経膠腫細胞複製に伴うレプリキン濃度の顕著な上昇のこの明瞭な証明は、種々の生物で3点認識法により同定されたレプリキンの存在を示すかまたはこれに一致する。例えばレプリキンは、「サッカロミセス・セレビッシェ(Saccharomyces cerevisiae)複製結合タンパク質」(配列番号2)(hsikrelgiifdk);「トウモロコシストリークウイルスの複製関連タンパク質A」(配列番号8)(kyivcareahk)と(配列番号9)(kekkpskdeimrdiish);「黄色ブドウ球菌(Staphylococcus aureus)の複製関連タンパク質」(配列番号10)(kkektthnk);「ウシヘルペスウイルス4のDNA複製タンパク質」(配列番号11)(hkinitngqk);および「シチメンチョウ(Mealigrid)ヘルペスウイルス1複製結合タンパク質」(配列番号12)(hkdlyrllmk)のようなタンパク質中で同定された。トマトリーフカールジェミニウイルスの以前の研究は、ウイルス蓄積の制御が、ウイルス複製中のリーフDNAおよび他の複製タンパク質分子への、そのウイルスの「複製タンパク質」のアミノ酸1〜160の結合が関与らしいことを示す。この配列の分析は、この「複製タンパク質」のアミノ酸1〜163が、5つのレプリキン、すなわち(配列番号13)kfrinaknyfltyph、(配列番号14)knletpvnklfiricrefh、(配列番号15)hpniqaaksstdvk、(配列番号16)ksstdvkaymdkdgdvldh、および(配列番号17)kasalnilrekapkdfvlqfh を含有することを示した。
【0125】
表2は、レプリキン含有タンパク質がまた、しばしば酸化還元機能、およびタンパク質合成もしくは伸長、ならびに細胞複製に関係していることを示す。金属ベースの酸化還元機能との関連、嫌気的複製中のレプリキン含有神経膠腫マリグニン濃度の濃縮、および低濃度(ピコグラム/細胞)での抗マリグニンの細胞障害性(図4c〜f)はすべて、レプリキンが中枢呼吸生存機能に関連し、非レプリキンアミノ酸に特徴的な突然変異をあまり受けないことを示唆する。
【0126】
特に興味深いことに、調べたインフルエンザウイルスの個々の株のほとんどすべての血球凝集素タンパク質中で、100アミノ酸当たり少なくとも1つのレプリキンが存在することが観察された。インフルエンザウイルスの4つの主要な株(インフルエンザB、H1N1、H2N2、およびH3N2)のそれぞれの分離株の血球凝集素タンパク質中に存在することが観察されたレプリキン配列(各年についてそのアミノ酸配列データは入手できる(1902〜2001))を、下記の表3、4、5および6に示す。
【表4】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
【0127】
観察されたレプリキンの濃度とタイプ(すなわち、観察されたレプリキン組成)の両方は、インフルエンザの汎流行と流行に関連することがわかった。インフルエンザウイルス中のレプリキンの濃度を、過去1世紀(すなわち、1900年〜2001年)にわたって世界中でヒトおよび動物宿主から単離されたインフルエンザ株について、ナショナルライブラリーオブメディシン(National library of Medicine)「PubMed」データベースで公表された血球凝集素アミノ酸配列を視覚的にスキャンして調べた。次に、各株についてこれらのレプリキン濃度(100アミノ酸当たりのレプリキンの数、平均±SD)をプロットした。
【0128】
レプリキンの濃度は、インフルエンザの汎流行と流行の発生に直接関係することがわかった。インフルエンザB血球凝集素およびインフルエンザA株H1N1中のレプリキン濃度を図7に示し、他の2つの共通のインフルエンザウイルスA株(H2N2とH3N2)中のレプリキン濃度を図8(H2N2、H3N2)に示す。図8のデータはまた、その構成レプリキンにより規定されるようにインフルエンザウイルスの出現する新しい株を示す(H3N2(R))。
【0129】
各インフルエンザA株は、1つの汎流行に関与している(それぞれ、1918年、1957年、および1968年)。図7と8のデータは、調べた4つの共通のインフルエンザウイルスのすべての分離株のインフルエンザ血球凝集素タンパク質のそれぞれに、100アミノ酸当たり少なくとも1つのレプリキンが存在することを示し、生存レベルの複製の維持におけるレプリキンの機能を示唆している。1990年代には、H3N2の衰退中に多くのH3N2分離株にはレプリキンが存在しなかったが、H3N2分離株には高濃度の新しいレプリキンが現れ、これはH3N2(R)株の出現を規定する。
【0130】
レプリキン濃度のいくつかの性質は、すべての4つのインフルエンザウイルス株に共通であることが図7と8でわかる。まず濃度は、何年にもわたって循環性であり、上昇と低下の1回のサイクルは2〜30年の期間にわたって起きる。この上昇と低下は、血球凝集素とノイラミニダーゼ分類により、個々のインフルエンザウイルス株支配の既知の漸増‐漸減に一致する。第2に、汎流行に関与することがすでに証明されている各インフルエンザウイルス株のピークのレプリキン濃度が、汎流行の各3年間に具体的にかつ個々に関連することが観察された。例えば1918年の汎流行(ここでは、インフルエンザウイルス株H1N1が関与することが証明された)については、H1N1でレプリキンのピーク濃度が独立にあり(P1);1957年の汎流行(ここでは、インフルエンザウイルス株H2N2が関与することが証明された)については、H2N2でレプリキンのピーク濃度があり(P2);1968年の汎流行(ここでは、インフルエンザウイルス株H3N2が関与することが証明された)については、H3N2でレプリキンのピーク濃度がある(P3)ことが証明された。第3に、上記の各3年間の汎流行直後の年には、特異的なレプリキン濃度は顕著に低下し、これはおそらく各ケースに広く分布した免疫を反映している。すなわち、この汎流行後の低下は、関与していた汎流行(P1)直後のH1N1に特異的であり、その時のすべての株の一般的な性質ではない。インフルエンザB中のレプリキン濃度の増加は、H1N1中のレプリキン濃度の低下と同時に繰り返し起き、例えば1951年のEB1と1976年のEB2は両方とも、最も高い死亡率を有するインフルエンザB流行に関連していた。(Stuart-Harrisら、Edward Arnold Ltd. (1985))。第4に、第2のピーク濃度(これは、第1のピーク濃度上昇を越えた)は、それぞれ3つの汎流行の15年後に起き、この第2のピークは:1918年の汎流行の15年後のH1N1「流行」年(E1);1957年の汎流行の8年後のH2N2「流行」年(E2);1968年の汎流行の7年後のH3N2「流行」年(E3)の流行を伴った。特異的レプリキンのこれらの第2のピーク濃度は、株の回復を反映するかも知れない。第5に、各株の特異的レプリキン濃度のピークはしばしば、1つまたは両方の他の株のレプリキン濃度の低下に関連するようであり、宿主部位について株の間の競合を示唆する。第6に、35年(H2N2)〜60年(インフルエンザB)の期間にわたって各株のレプリキン濃度が低下する全体的傾向があるようである。これは、インフルエンザワクチンの一般的な使用の前に、1901年〜1964年のインフルエンザBの場合に明らかであったため、この低下は、ワクチンの影響とは言えないようである。インフルエンザBの場合、低下からのレプリキンの回復は1965年から起きるように見えるが、レプリキン濃度は1997年〜2000年に再度低下した(図7)。これは、最近の分離株中のインフルエンザBの低頻度に相関する。H1N1レプリキン濃度は、H1N1株の再出現と流行とともに1978〜1979年にピークをうち(図7)、次にH1N1流行とともに1996年にピークをうった(図7)。H1N1レプリキン濃度はまた、1997〜2000年に低下し、これらの年に得られた分離株では、H1N1株の存在は低下していた。H2N2レプリキンについては、36年間の低下からの回復は起きず(図8)、これは、最近の分離株からのH2N2の欠如に相関する。H3N2については、多くの分離株のレプリキン濃度は、1996〜2000年の間にゼロまで低下したが、他のH3N2分離株は、レプリキン濃度の大きな鋭い上昇を示した。これは、H3N2の亜株の出現を示す(これは本明細書においてH3N2(R)と呼ぶ)。
【0131】
図7と8は、レプリキン濃度がピークに達する前に、しばしば1〜3年の段階的上昇が観察されることを示す。この段階的上昇は、流行の発生(これは、レプリキンピークと同時に起きる)に先行する。すなわち、特定の株の濃度の段階的上昇は、その株が流行または汎流行を引き起こす可能性の最も高い候補であるというシグナルである。
【0132】
現在、インフルエンザウイルスのH3N2(R)株中のレプリキン濃度は増加している(図8、1997〜2000年)。H3N2レプリキン濃度の3つの類似の以前のピーク上昇は、1968年のH3N2ベースの汎流行(図8)、および1972と1975年のH3N2ベースの流行(図8)で起きたことがわかる。これらの汎流行および流行のそれぞれは、過度の死亡率に関連していた(Ailingら、Am J. Epidemiol., 113(1):30-43 (1981))。従って、1997〜2000年のH3N2レプリキンのH3N2(R)亜種の濃度の急激な上昇は、統計的に大きな流行または汎流行が近づく早期警告である。H3N2流行は2000年にロシアで起き(図8、E4)、2001年12月のCDC報告は、現在H3N2が世界中で最も頻繁に単離されるインフルエンザウイルス株であるとしている(罹患率と死亡率ウィークリーレポート(MMWR)、疾病管理センター(Centers for Disease Control);50(48):1084-68 (Dec.7, 2001)。
【0133】
インフルエンザウイルスの汎流行または流行の各場合に、新しいレプリキンが出現する。ある特定の分離株の特定の血球凝集素で2つの同じレプリキンは観察されていない。新しいレプリキンの出現が、別の動物または鳥類プールからの移行に対して、どの程度が突然変異であるかは不明である。ある場合には、毎年1つ以上の元々のレプリキン構造が保存され、同時に新しいレプリキンが出現する。例えば、インフルエンザウイルスB血球凝集素では、1919〜2001年に5つのレプリキンが絶えず保存され、一方同じ時期に26個のレプリキンが行き来した(一部は数年間欠如の後に再出現した)。特定のレプリキン構造の消失と再出現は、レプリキンが、新規突然変異ではなく別のウイルス宿主プールから戻ることを示唆する。
【0134】
H1N1レプリキンの場合、1918年の汎流行に関連するP1ピークで存在した2つのレプリキンは、1933年の回復E1ピーク中には存在しなかった(これは12の新しいレプリキンを含有する)。従って、絶えず保存されるレプリキンは、単独または組合せでも、ワクチンの最適な選択である。しかし、1年間の濃度上昇に伴う最近出現しているレプリキンでさえ持続し、さらに1年以上上昇し濃度ピークに達して流行を起こし、従って早期警告となり、合成レプリキンを用いてワクチン接種すべき時期を提供する(例えば、1990年代初期のH1N1、図7)。
【0135】
図7と8のデータは、インフルエンザタンパク質配列中の特定のレプリキンの存在と濃度と、インフルエンザの汎流行および流行の発生との、直接の関係を示す。すなわち、レプリキンの存在と濃度についてのインフルエンザウイルス血球凝集素タンパク質配列の解析は、インフルエンザ汎流行および/または流行の予測変数、ならびにインフルエンザワクチン形成の標的を提供する。
【0136】
インフルエンザウイルスB株中のレプリキンの組成:この株で同定された全部で26個のレプリキンのうち、以下の10個のレプリキンは、1902〜2001年に調べたすべてのインフルエンザB分離株中に存在する。重複するレプリキン配列を別々にリストする。リジンとヒスチジンは太字で示して、「3点認識」と一致する同族関係を示す。
kshfanlk(配列番号104)
kshfanlkgtk(配列番号105)
kshfanlkgtktrgklcpk(配列番号106)
hekygglnk(配列番号107)
hekygglnksk(配列番号108)
hekygglnkskpyytgehak(配列番号10)
hakaigncpiwvk(配列番号110)
hakaigncpiwvkktplklangtk(配列番号111)
hakaigncpiwvktplklangtkyrppak(配列番号112)
hakaigncpiwvktplklangtkyrppakllk(配列番号113)
【0137】
表3と4は、H1N1レプリキンと比較して、インフルエンザB血球凝集素のレプリキン構造にはるかに大きな安定性があるらしいことを示す。インフルエンザBは、どの汎流行にも関与せず、動物または鳥類宿主も無いようである(Stuart-Harrisら、Edward Arnold Ltd. (1985))。
【0138】
インフルエンザH1N1レプリキン:1918年(株が最初に出現し、その年の汎流行を引き起こした)から2000年までの配列が入手できるすべてH1N1分離株には、ただ1つのレプリキン「hp(v/i)tigecpkyv(r/k)(s/t)(t/a)k」のみが存在する(表4)。(「(v/i)」は、アミノ酸vまたはiが異なる年に同じ位置に存在することを示す)。H1N1はただ1つの持続性のレプリキンを含有するが、H1N1は、インフルエンザBより豊富なようである。H1N1について82年間で95の異なるレプリキン構造があり、インフルエンザB分離株では100年間で31の異なるレプリキンがある(表4)。新しいレプリキン構造の数の上昇は流行の年に見られ(表3、4、5および6)、総レプリキン濃度の上昇と相関する(図7と8)。
【0139】
インフルエンザH2N2レプリキン:インフルエンザH2N2は、1957年のヒト汎流行に関与した。1957年についてその株で同定された20個のレプリキンのうち3つは、PubMedで1995年までに試験が可能であったH2N2分離株のそれぞれで保存されていた(表5)。
ha(k/q/m)(d/n)ilekthngk(配列番号232)
ha(k/q/m)(d/n)ilekthngklc(k/r)(配列番号233)
kgsnyp(v/i)ak(g/r)synntsgeqmliiwq(v/i)h(配列番号238)
【0140】
しかしH1N1と比較して、1961年に始まるH2N2では13個のみの追加のレプリキンが見つかっている。新しいレプリキンの出現が少ないことは、何年間にもわたる分離株中のH2N2レプリキンの濃度の低下とH2N2分離株の出現の低下と相関する。
【0141】
インフルエンザH3N2レプリキン:インフルエンザH3N2は、1968年のヒト汎流行に関与していた。1968年に現れた5つのレプリキンは1977年後に消失したが、1990年代に再出現した(表6)。22年間持続した唯一のレプリキン構造はhcd(g/q)f(q/r)nekwdlf(v/i)er(s/t)kであり、これは1977年に最初に出現し、1998年まで持続した。1990年代半ばの12個の新しいH3N2レプリキンの出現(表6)は、同時期のレプリキン濃度の上昇(表8)と相関し、これらの分離株の一部からのすべてのレプリキンの同時消失とともに、最近の分離株のH3N2株の流行(図8)に相関し、これは新しい亜種H3N2(R)の出現を示唆する。
【0142】
図1と2は、インフルエンザ流行と汎流行が、インフルエンザウイルスのレプリキン濃度の上昇と相関し、これは、その消失の1〜59年後の少なくとも1つのレプリキンの再出現によることを示す。また、A株のみでは、新しい株特異的レプリキン組成の出現がある(表4〜6)。単一のタンパク質内の個々のレプリキンの繰り返しによるレプリキン濃度の上昇は、インフルエンザウイルスで起きるのはなく他の生物で見られるようである。
【0143】
異なるインフルエンザ株の活性の変化は、インフルエンザ血球凝集素の配列変化に関連すると考えられており、これはさらに、2つのあまり理解されていないプロセスの1つにより行われる置換の生成物である:i) 抗原ドリフト(血球凝集素分子の一連の点突然変異の蓄積によると考えられる)、またはii) 抗原シフト(ここでは、変化が大きいため、ヒトと非ヒト宿主のウイルスの間で遺伝的組換えが起きる)。まず現在のデータは、異なるインフルエンザ株の活性の変化が、非特異的配列変化に関連するよりも、流行に関連する株特異的レプリキンの濃度の上昇および複製の株特異的上昇に基づくか、またはこれに関連することを示唆する。さらに、どの配列変化が「ドリフト」または「シフト」によるのか、およびどれが、保存、宿主中の保存、および再出現によるのかを理解するために、データを調べた。データは、レプリキン濃度中の流行に関連した上昇が、血球凝集素についての既存のレプリキンの複製によるのではなく、その消失の1〜59年後の少なくとも1つのレプリキン組成の再出現、およびA株のみでは、新しい株特異的レプリキン組成の出現によることを示す(表3〜6)。すなわち、1951〜1977年のインフルエンザB流行におけるレプリキン濃度の上昇は、流行の年の新しいレプリキン組成の出現には関連せず、それ以前の年に出現し次に消失したレプリキン組成の再出現にのみ関連している(表3)。これに対して、A株については、以前消失したウイルスレプリキンの再出現以外に、新しい組成が現れる(例えば、1996年の流行の年のH1N1では、以前の6つのレプリキンの再出現以外に、10個の新しい組成が現れた)。A株のみ(インフルエンザBではそうではない)が非ヒト動物および鳥類宿主に接近できるため、「3点認識」の基本的要件以外に新しい組成に似ていないようである既存のヒトレプリキンの突然変異からではなく、おそらく非ヒト宿主からまったく新しい組成が得られる(表2〜5)。Bと比較した場合のH1N1の多産性と、汎流行はB株ではなく3つのA株によってのみ引き起こされているという事実の両方とも、本ウイルス宿主から新しいレプリキン組成を受けるヒトA株の能力の関数かも知れない。
【0144】
あるレプリキンは1年のみ現れ、消失し、今日まで再出現していない(表3〜6)。他のレプリキンは1〜81年消失し、同じレプリキン配列が再出現する。主要なレプリキン「k」および「h」アミノ酸、およびこれらの間のスペースは、以下の株特異的レプリキンについて(インフルエンザBの10個、H1N1の単一のレプリキン、およびH2N3の単一のレプリキン)、ならびに消失後の同一のレプリキンの再出現について、表23-6に示すように、何年にもわたって特定のレプリキンが絶えず存在するために保存される。残りの血球凝集素配列におけるレプリキン構造の中または外の両方の他のアミノ酸の顕著な代替または置換活性にもかかわらず、インフルエンザレプリキンのヒスチジン(h)は決して置換されず、そしてリジン(k)はまれに置換されるようである。この保存の例は、1918〜2000年の間一定のH1N1レプリキン「hp(v/i)tigecpkyv(r/k)(s/t)(t/a)k」(配列番号135)、1975〜1998年の間一定のH3N2レプリキン「hcd(g/q)f(q,r)nekwdlf(v/i)er(s/t)k」(配列番号277)、および1975年に初めて出現し25年間消失し、次に2000年に再出現したH3N2レプリキン「hqn(s/e)(e/q)g(t/s)g(q/y)aad(l/q)kstq(a/n)a(i/l)d(q/g)I(n/t)(g/n)k,(l/v)n(r/s)vi(e/c)k」(配列番号276)中に見られる。多くのアミノ酸が置換されたが、基本的なレプリキン構造(約50アミノ酸以下の中の、6〜10残基離れた2つのリジン、1つのヒスチジン、少なくとも6%のリジン)は保存された。
【0145】
全くランダムな置換では、これらのH1N1とH3N2レプリキンの持続も、1902年から2001年のインフルエンザBの10個のレプリキン構造の持続も、1919年の18量体のレプリキンの74年間の消失後の1993年の再出現も、不可能であったであろう。ランダムな置換ではなく、この一定性は、秩序だった制御されたプロセス、または少なくともレプリキン残基を保護して、これらが何らかの方法により固定または結合されることを示唆する(リジンはおそらく核酸に結合し、ヒスチジンはおそらく呼吸系酸化還元酵素に結合する)。この保存を制御する機構は現在のところ不明である。
【0146】
レプリキン構造の保存
レプリキン構造が保存されるかまたは広範な自然の突然変異を受けるかどうかを、手足口病ウイルス(FMDV)(これらのウイルスのタンパク質の突然変異は、何十年も世界的に記録されている)の種々の分離株のタンパク質配列をスキャンすることにより調べた。FMDV分離株のタンパク質配列を、全レプリキンと、特定のレプリキンで観察された成分レプリキンアミノ酸残基のそれぞれとの存在について、視覚的に調べた。
【0147】
隣接するアミノ酸で起きるように時間とともに広範な置換を受けるというより、レプリキン構造を構成するアミノ酸はほとんどまたは全く置換されず、レプリキン構造は保存される。
【0148】
例えば、FMDV O型のタンパク質VP1では、レプリキン(配列番号3)「hkqkivapvk」は、PubMedで報告された236分離株のうち78%で保存されており、各アミノ酸は、以下のように個々の分離株で保存されていることがわかった:his、95.6%;lys、91.8%;gln、92.3%;lys、84.1%;ile、90.7%;val、91.8%;ala、97.3%;pro、96.2%;ala、75.4%;およびlys、88.4%。高率の保存は、レプリキン構造の構造的および機能的安定性を示唆し、治療の一定の標的となる。
【0149】
同様に、HIVの異なる分離株中でそのレプリキンについて配列保存がみつかった(例えばHIV 1型の(配列番号5)「kcfncgkegh」または(配列番号6)「kvylawvpahk」、およびHIV 2型の(配列番号7)「kcwncgkegh」(表2))。配列保存のさらなる例は、HIV tatタンパク質中にあり、例えば(配列番号613)「hclvckqkkglgisygrkk」であり、ここで主要なリジンとヒスチジンアミノ酸は保存されている(表7を参照)。
【0150】
同様に、配列保存は植物(例えばコムギ、例えばコムギユビキチン(ubiquitin)活性化酵素E(配列番号614〜616))中で観察された。コムギ中のレプリキンは、本明細書に記載のような植物増殖の刺激の信頼できる標的となった。他の保存例は、神経膠腫細胞の10年以上の組織培養にわたっての連続的世代でのマリグニンの持続的な存在、および米国、英国、ヨーロッパおよびアジアからの8,090のヒト血清からの免疫吸着により単離された抗マリグニン抗体に対する神経膠腫タンパク質の親和力の継続により見られる(例えば、図5および米国特許第6,242,578 B1号)。
【0151】
同様に保存は、HIVの分離株のトランスアクチベーター(Tat)タンパク質中で観察された。Tat(トランスアクチベーター)タンパク質はレンチウイルスの転写を制御する早期RNA結合タンパク質である。これらのタンパク質は、すべての公知のレンチウイルス(例えば、ヒト免疫不全症ウイルス(HIV))のライフサイクルの必須成分である。Tatは、トランス活性化応答配列(TAR)RNAに結合することにより作用し、転写開始および/またはLTRプロモーターからの伸長を活性化する転写レギュレータータンパク質である。HIVはtat無しでは複製できないが、この化学基礎は不明である。HIV tatタンパク質配列の89〜102残基中に、我々は、他の生物の迅速な複製に関連するレプリキンを見いだした。このレプリキンのアミノ酸配列は「hclvcfqkkglgisygrkk」である。実際我々は、すべてのHIV tatタンパク質中にこのレプリキンが存在することを見いだした。表8に示すようにいくつかのtatアミノ酸は、別のアミノ酸によりしばしば置換される(最も頻繁なアミノ酸の下に小さいフォントで示す(表7)、優性のレプリキン「hclvcfqkkglgisygrkk」の保存パーセント)。これらの置換は、個々のアミノ酸のほとんどで現れた。しかしレプリキン配列内の主要なリジンとヒスチジンアミノ酸(これは、レプリキン構造を規定する)は、配列中で100%保存される;一方、他のアミノ酸の他の場所(レプリキンの内および外)で置換が一般的であり、これらの主要なヒスチジンアミノ酸上で置換は存在しない。
【0152】
表7に示すように、tatタンパク質アミノ酸配列中でリジンが置換されないことではない。表の左側から、すぐ隣接する配列中であるがレプリキン配列の外の最初のリジン、およびレプリキン内の配列中の第2のリジン(k)(しかし、レプリキン形成にとって必須ではないという点で「余分」である)は両方とも、しばしば置換される。しかし3番目、4番目および5番目のリジンおよびカッコ中の1つのヒスチジン(これらは一緒にレプリキン構造を構成する)は、決して置換されない。すなわちこれらの主要なアミノ酸配列は、100%保存されている。インフルエンザウイルスレプリキンの場合に観察されるように、ランダムな置換は、この選択的置換と選択的な非置換が偶然起きることは、可能にしないであろう。
【表16】
【0153】
レプリキン構造の保存は、レプリキン構造が、保存されなければならないHIVウイルスの特定の生存機能を有し、犠牲になって抗体および他の「攻撃」を避けるためにウイルス「防御」手段のアミノ酸置換をすることはできないことを示唆する。これらの「防御」機能は必須ではないが、HIV複製のウイルス生存機能と「競合」することはできない。
【0154】
さらなる保存が、HIVの異なる分離株中で、そのレプリキン、例えばHIV 1型の「kcfncgkegh」(配列番号5)または「kvylawvpahk」(配列番号6)、およびHIV 2型の「kcwnegkegh」(配列番号7)中で観察された。
【0155】
FMVDおよびHIVレプリキン中で観察されたこの高い保存率は、インフルエンザレプリキンのレプリキン中で観察される保存もまた、ウイルスレプリキンの一般的性質であることを示唆する。この保存は、例えば、神経膠腫レプリキンについて例示されるようにインフルエンザ、FMVDまたはHIVのような特異的レプリキンを使用する破壊、または刺激にとって、一定の信頼できる標的としている。
【0156】
同様に、ウイルス(インフルエンザウイルス、FMDV、およびHIVを含む)(ここでは、レプリキンの高率の保存が、ウイルスレプリキンの一般的性質であり、レプリキンを、破壊または刺激の一定の信頼できる標的としていることを示唆する)中の例に提供されるように、レプリキン構造の保存は植物でも起きる。例えば、コムギ植物では、レプリキンが保存され、刺激の信頼できる標的となる。コムギ植物のユビキチン(ubiquitin)活性化酵素Eで保存されたレプリキンの例には以下がある:
E3 hkdrltkkvvdiarevakvdvpeyrrh(配列番号614)
E2 hkerldrkvvdvarevakvevpsyrrh(配列番号615)
E1 hkerldrkvvdvarevakmevpsyrrh(配列番号616)
【0157】
HIV tatタンパク質中の保存と同様に、コムギ植物のユビキチン(ubiquitin)活性化酵素E中のレプリキンは保存されている。HIV tatタンパク質と同様に、コムギ植物のユビキチン(ubiquitin)活性化酵素E中のレプリキン変種型に隣接するアミノ酸の置換(「*」で示す)は一般的である。しかしレプリキン構造を形成する主要なkおよびhアミノ酸は変化しないが、(左側の最初のkから)わずかに5アミノ酸の「必須ではない」kは置換される。
【0158】
抗レプリキン抗体
抗レプリキン抗体は、レプリキンに対する抗体である。抗レプリキン抗体についてのデータもまた、レプリキンクラスの統一性を支持する。抗レプリキン抗体応答は、固定化されたマリグニンに対する血清抗マリグニン抗体の免疫吸着により定量されている(米国特許第5,866,690号中の方法を参照)。16量体のペプチドの合成物(その配列はマリグニンから得られた)をウサギに投与することによる抗マリグニン抗体の多量の産生は、炭水化物又は他の基を産生せず、この免疫応答の充分な基礎となる(図3)。16量体ペプチドは、IgM型およびIgG型の抗体の両方を産生した。抗マリグニン抗体は、感染後約15年〜25年後に出現する肝癌の普通に観察されるはるかに前に、感染の最初の5年のB型肝炎とC型肝炎の米国とアジアの79症例のうちの37%で血清中の濃度が上昇していた。感染性肝炎とHIV感染症の両方に関して、形質転換された細胞は、ウイルスの安全な避難所であり、細胞寿命が延長し、ウイルスの追い立てを逃れ、その結果ウイルスは抗ウイルス治療法に接することがない。
【0159】
レプリキンの投与は免疫系を刺激して、細胞障害作用を有する抗体を生じるため、ある期間中に最も濃縮されることが観察されている特定のインフルエンザウイルスレプリキンまたはレプリキン群に基づくペプチドワクチンは、あるインフルエンザシーズンに大流行を引き起こす可能性が最も高いインフルエンザの特定の株(例えば、出現する株または再出現する株)に対する防御となる。例えば、毎年または2年ごとにインフルエンザウイルス血球凝集素のアミノ酸配列を分析すると、その年の特異的に標的化されたインフルエンザワクチンを調製するのに有用なデータが提供される。そのような分析は、地域ごとにまたは任意の所望の時期に行われて、その結果世界中の異なる地域で出現する株を検出することができ、各地域の特異的に標的化されたワクチンを調製することができる。
【0160】
インフルエンザ
現在、国際的なWHOおよびCDC会議で、ウイルス製剤は年に2回変更される。ワクチン製剤は、世界のある地域のインフルエンザウイルス株の最も最近の優勢の血清学的証拠に基づいている。しかし本発明以前には、インフルエンザウイルス株に特異的なアミノ酸配列の変化と、インフルエンザの流行または汎流行の発生との相関はなかった。
【0161】
インフルエンザウイルスタンパク質中の特異的レプリキンの観察とその濃度は、インフルエンザの汎流行および流行の最初の特異的な定量的早期化学相関現象であり、世界の特定の地域のインフルエンザウイルスの流行している出現性または再出現性株を処理するために具体的に作成されるインフルエンザウイルスの産生とタイミング良い投与を提供する。インフルエンザウイルスの分離株のタンパク質配列(例えば、血球凝集素タンパク質配列)を、レプリキンの存在、濃度および/または保存について分析することにより、インフルエンザウイルスの汎流行と流行を予測することができる。さらに、ある時期(例えば約1年〜約3年)にウイルス分離株で最も豊富であるかまたは上昇していることがわかったレプリキン配列に基づいてインフルエンザペプチドワクチンを投与することにより、インフルエンザのそのような大流行の深刻さを大きく減少させることができる。
【0162】
本発明のインフルエンザペプチドワクチンは、インフルエンザウイルス株で観察される単一のレプリキンペプチド配列を含有するか、または複数のレプリキン配列を含有する。好ましくはペプチドワクチンは、ある期間に濃度が上昇していることが証明されており少なくともその期間は保存されているレプリキン配列に基づく。しかしワクチンは、新しいレプリキンペプチドとともに保存されたレプリキンペプチドを含むか、または新しいレプリキンペプチド配列に基づいてもよい。レプリキンペプチドは、任意の方法(化学合成または組換え遺伝子技術を含む)により合成することができ、非レプリキン配列を含有してもよいが、レプリキン配列のみを含有するペプチドに基づくワクチンが好ましい。好ましくは本発明のワクチン組成物はまた、医薬として許容される担体および/またはアジュバントを含有する。
【0163】
本発明のインフルエンザワクチンは、単独でまたは抗ウイルス剤[例えば、ガンシクロビル;インターフェロン;インターロイキン;M2インヒビター(例えば、アマンタジン、リマンタジン);ノイラミニダーゼインヒビター(例えば、ザナミビルおよびオセルタミビル)等]との組合せで、ならびに抗ウイルス剤の組合せで投与することができる。
【0164】
マラリア中のレプリキンデコイ
メロゾイト表面および/または寄生菌含有空胞内に位置する熱帯熱マラリア原虫のマラリア抗原の1次構造の分析は、この生物もまたインフルエンザウイルスのように、多数のレプリキンを含有することを明らかにした(表8)。しかし、熱帯熱マラリア原虫とインフルエンザウイルス分離株中のレプリキンの観察にはいくつかの差がある。例えば熱帯熱マラリア原虫はいくつかの部分的レプリキン(ここでは「レプリキンデコイ(Replikin decoy)」と呼ぶ)を含有する。これらのデコイの構造は、多数のリジン残基を含有するが、レプリキン構造に必要なヒスチジンは欠如している。具体的にはこれらのデコイは、真のマラリアリコグニン(recognins)のように、重複して6〜10残基離れて多くのリジンを含有するが、ヒスチジン残基は無い。デコイ構造は、抗マラリア抗体または他の物質が、ヒスチジン(これは、呼吸系酵素中のレプリキンのようにレプリキン構造中に存在し、トリパノソーマの破壊を引き起こし得る)に結合するのではなく、リジンを含有するあまり重要ではない構造(すなわち、レプリキンデコイ)に結合する機会を最大にしていると考えられる。例えば、レプリキン構造に対する特異性を持って侵入する抗体は、レプリキンデコイ構造に結合して、真のレプリキン構造をそのまま残すかも知れない。
【0165】
従って、マラリアの抗レプリキン治療には、2つの相が必要である(2重治療):i) タンパク質分解酵素を用いてレプリキンデコイを切断し、特異的抗レプリキン治療の「安全な通過」を可能にする、予備処理;およびii) 特異抗体を用いる、または合成マラリアレプリキンワクチンより発生する細胞性免疫による、またはマラリアレプリキンを標的とする有機的手段による、マラリアレプリキンの攻撃。
【0166】
レプリキン構造の繰り返しと重複
熱帯熱マラリア原虫で見られるもう1つの差は、インフルエンザウイルスでは観察されなかった、単一タンパク質中の個々のレプリキン構造の頻繁な繰り返しである。繰り返しは、(a) レプリキン間のリジン残基の共有、および(b) 別のレプリキン配列内のレプリキン配列の一部の繰り返し、により起きる。
【0167】
インフルエンザウイルス分離株と熱帯熱マラリア原虫で観察されるレプリキン構造の第3の大きな差は、マラリアタンパク質中のレプリキン構造の顕著な重複である[例えば、配列番号393の39アミノ酸配列内の9つの重複するレプリキン(レプリキン濃度=23.1/100アミノ酸);および配列番号467の41アミノ酸中の15の重複するレプリキン(レプリキン濃度=36.6/100アミノ酸)]。これらの重複レプリキン構造の両方とも、血液段階のトロフォゾイトとシゾントで起きる。これに対してインフルエンザウイルスレプリキンは、タンパク質中により分散しており、最大のレプリキン濃度は約7.5/100アミノ酸であり(図7);重複するレプリキンを有することが観察されたトマトリーフカールジェミニウイルスも、わずかに約3.1/100アミノ酸である。
【0168】
多数のリジンのこの機構はまた、癌タンパク質(例えば、胃ガン形質転換タンパク質)のレプリキンktkkgnrvsptmkvth(配列番号88)、および肺の形質転換タンパク質P21B(K-RAS 2B)のレプリキンkhkekmskdgkkkkkks(配列番号89)でも見られる。
【0169】
高レプリキン濃度と迅速な複製の関係はまた、HIV分離株の分析によっても確認される。HIVの増殖の遅い低力価株(NSI、「Bru」、これは早期HIV感染によくある)は、100アミノ酸当たり1.1(±1.6)レプリキンのレプリキン濃度であり、一方HIVの迅速に増殖する高力価株(S1、「Lai」、これは後期HIV感染によくある)は、100アミノ酸残基当たり6.8(±2.7)レプリキンのレプリキン濃度である。
【0170】
マラリア、インフルエンザウイルスおよび癌細胞中の重複レプリキンの高濃度は、マラリア微生物の有名な高および迅速複製能力に一致する。マラリア中の多数の重複レプリキンはまた、この微生物が宿主に殺到しその免疫系を混乱させて、こうして間違った抗体が作成され永続される機会を大きくして、主要なマラリア抗原が攻撃されないようにする。
【0171】
例えばインフルエンザウイルスの場合のように、マラリア微生物中に存在するレプリキン構造に基づくペプチドワクチンは、マラリアを予防および/または治療するための有効な手段となる。マラリアに対するワクチン接種は、熱帯熱マラリア原虫で観察される1つのレプリキン構造またはその混合物を含有する組成物を投与することにより行われる。さらに、マラリアレプリキンに対する抗体を作成し、受動免疫のためにまたはマラリア検出のために投与することができる。
【0172】
表8は、いくつかの熱帯熱マラリア原虫レプリキン配列のリストを提供する。これは完全なものではないことを理解されたい。この微生物の異なる分離株は、他のレプリキン構造を含むこともある。
【表17】
【表18】
【表19】
【表20】
【表21】
【表22】
【0173】
構造タンパク質中のレプリキン
いくつかの構造タンパク質はレプリキン構造を含むことも測定されている。構造タンパク質は、組織および臓器の支持に関与する分子であり、例えば皮膚や結合組織および膜構造物中のコラーゲン、および例えば脳中のアミロイドA4前駆体タンパク質(APP)である。これらのタンパク質の過剰産生は疾患に関係し、特に皮膚のコラーゲンの過剰産生の場合の強皮症(表9)、および脳のAPPの過剰産生の場合のアルツハイマー病がある(表10)。
【0174】
強皮症と悪性腫瘍の関連は、近年議論の的である。これまでの報告で、いくつかの関連機構が示唆されている。最近の長期研究は、強皮症と悪性腫瘍の関係の上昇を示唆する。しかし基本的な機構はいまだに不明である(Wenzel, J. Eur. J. Dermatol. 2002 May-Jun;12(3):296-300)。
【0175】
強皮症中のタンパク質の過剰産生に関連するいくつかのタンパク質は、レプリキン構造を含有することがわかっている。すなわちこれらは、過剰コラーゲン産生の阻害または停止のための、まだ認識されていない標的のさらなる例である。表9は、強皮症中のタンパク質と関連するレプリキンのリストである。
【0176】
APPタンパク質は、アミロイドベータA4タンパク質の供給源であり、これは、過剰にあると脳中の細胞外スペース中でプラークを形成し、アルツハイマー病の神経細胞喪失に関連する毒性作用を示す。今日までの多くの試験は、A4の過剰沈着を防ぐことができないことに注目してきたが、これは、廃棄物の除去問題というより、実際は、前駆体タンパク質APPの過剰産生の問題であるかも知れない。APP中のレプリキンの高濃度(100アミノ酸当たり3.3レプリキン)は、過剰産生がアルツハイマー病の原因であることを強く示唆する(表10)。従って、表10中に含まれるレプリキンは、神経膠腫レプリキンで詳述されるものと同じ方法により阻止または阻害することができる。
【表23】
【表24】
【表25】
【0177】
受動免疫
本発明の別の実施態様において、単離されたレプリキンペプチドは抗体を作成するのに使用され、この抗体は例えば個体の受動免疫を与えるのに使用される。将来のインフルエンザ感染の原因である可能性が最も高いと本発明の方法により同定されたインフルエンザ株に対する受動免疫は、インフルエンザウイルスの同定された株のレプリキン配列に対する抗体を、必要な患者に投与することにより得られる。同様に、マラリアに対する受動免疫は、熱帯熱マラリア原虫レプリキンに対する抗体を投与することにより得られる。
【0178】
当該分野で公知の種々の方法を使用して、レプリキン配列に対する抗体を産生してもよい。そのような抗体には、特に限定されないが、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、ヒト化抗体、1本鎖抗体、Fab断片およびFab発現ライブラリーにより産生される断片がある。細胞障害剤に連結した抗体も作成される。抗体はまた、抗ウイルス剤と組合せて投与してもよい。さらに、異なるレプリキンに対する抗体の組合せを、抗体カクテルとして投与してもよい。
【0179】
抗体産生のために、レプリキンペプチドまたはレプリキンペプチドの組合せを、特に限定されないがウサギ、マウス、ラット、およびより大きな哺乳動物に注射することにより、種々の宿主動物または植物を免疫してもよい。
【0180】
レプリキンに対するモノクローナル抗体は、抗体分子を産生する任意の方法を使用して調製される。これらには、特に限定されないが、元々はKohlerとMilstein(Nature, 256:495-497 (1975))により開示されたハイブリドーマ法、ヒトB細胞ハイブリドーマ法(Kosborら、1983, Immunology Today, 4:72)、およびEBV ハイブリドーマ法(Coleら、モノクローナル抗体と癌療法(Monoclonal Antibodies and Cancer Therapy), Alan R. Liss, Inc., pp. 77-96)がある。さらに、キメラ抗体の産生のために開発された方法(Morrisonら、Proc. Natl. Acad. Sci. USA, 81:6851-6855)、または他の方法も使用される。あるいは、1本鎖抗体の産生について開示された方法(US4,946,778)を、レプリキン特異的1本鎖抗体を産生するように適応させてもよい。
【0181】
本発明の特に有用な抗体は、インフルエンザウイルスのペプチドおよび/またはポリペプチド中に含有されるレプリキン配列に、特異的に結合するものである。例えば、インフルエンザウイルスの出現性または再出現性株中に存在することが観察されている任意のペプチドに対する抗体およびそのような抗体の組合せは、インフルエンザの治療および/または予防に有用である。同様に、マラリア抗原上に存在する任意のレプリキンに対する抗体およびそのような抗体の組合せは、マラリアの予防と治療に有用である。
【0182】
レプリキンに対する結合部位を含有する抗体断片は、公知の方法により作成される。例えばそのような断片には、特に限定されないが、抗体分子のペプシン消化により産生することができるF(ab')2 断片、およびF(ab')2 断片のジスルフィド結合を還元することにより作成することができるFab断片がある。あるいは、Fab発現ライブラリーを作成(Huseら、1989, Science, 246:1275-1281)して、所望の特異性を有するモノクローナルFab断片の迅速かつ容易な同定が可能になる。
【0183】
癌細胞のタイプに無関係にヒト悪性腫瘍では抗マリグニン抗体の濃度が上昇している(図5)という事実、およびこの抗体は細胞タイプに無関係に癌細胞に結合するという事実は、本明細書においてほとんどの悪性腫瘍中に存在することがわかったレプリキン構造の存在により説明される(図1と表2)。集団での研究は、健常成人では年齢とともに抗マリグニン抗体の濃度が上昇し、これは、癌の頻度が上昇するように、ハイリスク家族では特にそうである。早期悪性腫瘍で起きるさらなる2倍またはそれ以上の抗体の上昇は、1〜10mmのサイズの乳癌で97%の特異性で独立に確認されている。in vivoで癌細胞中に選択的に局在化することが証明いる抗体は、組織学的には正常な細胞には結合しないが、in vitroで形質転換された細胞に選択的に結合(図4a、b)し、かつこれに細胞障害性が高い(図c-f)。これらの例では、いくつかのタイプの細胞(すなわち脳神経膠腫、造血細胞(白血病)、および肺の小細胞癌)により同じ抗体が結合されるため、悪性のレプリキンクラスの統一性も示される。
【0184】
抗マリグニンは、良性の増殖といっしょには上昇しないが、in vivoで乳房での悪性形質転換と複製によってのみ特異的に上昇し、悪性細胞を排除すると高値から正常値に戻る(図5)。抗マリグニン抗体濃度は、癌患者の生存に定量的に関連すること、すなわち抗体が多いほど生存期間が長いことが証明されている。まとめるとこれらの結果は、抗レプリキン抗体が、細胞の形質転換と複製の制御機構の一部かも知れないことを示唆する。この免疫応答の増強は、ワクチンとして合成レプリキンを用いて能動的に、または抗レプリキン抗体の投与により受動的に、またはレプリキンを特異的に標的とするように同様に設計された非免疫ベースの有機物質(例えば、炭水化物、脂質など)の導入により、複製を制御するのに有用であるかも知れない。
【0185】
本発明の別の実施態様において、1つ以上のレプリキンに暴露された個体から得られた1つ以上のレプリキンに対する抗体を含有する免疫血清を使用して、別の個体または動物で受動免疫が導入される。免疫血清は、静脈経由で、治療の必要な被験体に投与される。受動免疫はまた、1つ以上のレプリキンに対するあらかじめ形成された抗体を用いて受容者に注射することにより行われる。受動免疫は、感染性生物に暴露された個体に対して即時的防御を与えるのに使用してもよい。免疫血清またはあらかじめ形成された抗体の投与はルーチンの手技であり、熟練者は容易に、血清または抗体の量を確認して所望の作用を達成することができる。
【0186】
合成レプリキンワクチン(能動免疫)
神経膠腫レプリキン(配列番号1)「kagvaflhkk」またはC型肝炎レプリキン(配列番号18)「hyppkpgcivpak」、またはHIVレプリキン、例えば(配列番号5)「kcfncgkegh」もしくは(配列番号6)「kvylawvpahk」のようなレプリキンに基づく合成レプリキンワクチン、または好ましくは、ある期間の保存されたおよび/または出現するかまたは再出現するレプリキンに基づくインフルエンザワクチンを使用して、抗体濃度を増強して、各ウイルスで感染された細胞を溶解し、ウイルスを細胞外に放出させると、ここで化学処理が有効となる。同様に、例えば、メロゾイト表面上のもしくはパラジトフォラス(parasitophorous)小腔内の熱帯熱マラリア原虫マラリア抗原で観察されるレプリキンに基づくマラリアワクチンを使用して、マラリアに対する細胞障害性抗体を作成することができる。
【0187】
リコグニンおよび/またはレプリキンペプチドを被験体に投与して、被験体の免疫系を誘導して抗レプリキン抗体を産生してもよい。一般に0.5〜約2mg用量、好ましくは1mg用量の各ペプチドを被験体に投与して、免疫応答を誘導する。以後の用量は、所望であれば投与される。
【0188】
レプリキン配列構造は、複製機能に関連している。すなわち、本発明のレプリキンが、例えば診断的同定、複製の促進、または複製の阻害もしくは攻撃を目的としてレプリキンを含有する配列を標的とするのに使用されるかどうかは、レプリキンの構造−機能相関が基本的である。
【0189】
レプリキン断片を認識しこれに結合する抗体を誘導して細胞の破壊を引き起こしたい時は、特異的なレプリキン構造のみを利用することが好ましい。より大きなタンパク質配列が「複製関連機能」を有するとして当該分野で公知であるが、より大きなタンパク質を使用するワクチンは、しばしば成功しなかったかまたはあまり有効ではなかった。
【0190】
本発明者らは1つの理論に拘泥されるつもりはないが、本明細書に記載の研究は、先行技術のワクチンがより大きなタンパク質配列の使用に基づくため、有効ではないことを示唆する。より大きなタンパク質配列は必ず、1つ以上のエピトープ(特異抗体形成を誘導することができる独立した抗原配列)を有し、レプリキン構造は通常、これらのエピトープ候補の1つを含む。より大きなタンパク質内の他のエピトープの存在は、レプリキン抗原を先制攻撃する無関係の抗原刺激で免疫系に「殺到」(この抗原優先の公知の現象の考察については、Webster, R.G., J. Immunol., 97(2):177-183 (1966);およびWebsterら、J. Infect Dis., 134:48-58, 1976;Klenermanら、Nature 394:421-422 (1998)を参照されたい)して、レプリキンに対する抗体の充分な形成を妨害し、こうして、免疫系により提示され認識される第1のペプチドエピトープが多くなり、他のペプチドエピトープが同時に存在してもこれに対する抗体が作成される。これは、ワクチン製剤では、レプリキンが先制攻撃されずに免疫記憶に入れられるように、生物からの他のエピトープを提示する前に、免疫系に一定のレプリキンペプチドを最初に提示することが重要である別の理由である。
【0191】
非レプリキンエピトープに対する抗体の生成は、細胞への結合を可能にするが、必ずしも細胞を破壊はしない。マラリアタンパク質のC末端上の構造的「デコイ」の存在は、デコイエピトープがリジン残基を有するがヒスチジン残基は無いため、他のエピトープが、有効な抗レプリキン抗体の結合を妨害する能力の、他の側面である。すなわちデコイエピトープは抗レプリキン抗体に結合するが、その抗体をヒスチジン結合呼吸系酵素からは離して維持する。従って治療は2段階で最も有効となる:1) デコイを加水分解するプロテアーゼ、次に;2) 抗レプリキン抗体または他の抗レプリキン物質。
【0192】
「外来」タンパク質に対する抗体産生の過程では、タンパク質がまず小断片に加水分解されることは、当該分野で公知である。通常、約6〜10アミノ酸を含有する断片が抗体生成のために選択される。すなわち、タンパク質の加水分解によりレプリキン含有断片が得られない場合、抗レプリキン抗体は産生されないであろう。この点で、リジン残基は膜に結合することが知られているため、6〜10アミノ酸離れて位置するリジン残基をレプリキンが含有することは興味深い。
【0193】
さらにレプリキン配列は、少なくとも1つのヒスチジン残基を含有する。ヒスチジンはしばしば、酸化還元中心への結合に関与する。すなわち、レプリキン配列を特異的に認識する抗体は、レプリキンが位置している細胞を不活性化または破壊する可能性が高く(抗マリグニン抗体で見られるように)、これはおそらく、ピコグラム/細胞で活性のある、これまで記載された最も細胞障害性が高い抗癌抗体である。
【0194】
疾患を引き起こす物質の特定のタンパク質抗原に対して作成したワクチンが、疾患に対する防御を与えるのに充分に有効ではない(例えば、VP1タンパク質またはVP1タンパク質の大きなセグメントに対して開発された手足口病ワクチン)理由の1つは、最適な抗体が産生されていないこと、すなわち、レプリキンに対する抗体が産生されていない可能性があることである。レプリキンは産生されていない。すなわち、より大きなタンパク質断片中に存在するレプリキン以外のエピトープが、上記の抗原優先の現象に従って妨害するか、および/または抗体を産生するためのプロセシングのために、より大きなタンパク質配列のより小さい配列への加水分解が、存在するレプリキン構造の完全性を失わせるために、レプリキンが2つに切断されるか、および/またはヒスチジン残基が加水分解プロセシングで失われる。本研究は、有効なワクチンが産生されるためには、レプリキン配列がワクチンとして使用されて他のエピトープは使用されないことであることを示唆する。例えば、本発明のワクチンは、3点認識システムにより同定されたレプリキンペプチドの任意の1つを使用して作成することができる。
【0195】
特に好適なペプチド、例えばインフルエンザワクチンは、1年以上、好ましくは約3年以上の期間にわたって保存されることが証明されているか、および/または、他のインフルエンザウイルス株(例えば、出現する株)中のレプリキン濃度と比較してレプリキン濃度が最も上昇していることが証明されている、インフルエンザウイルスの株中に存在するペプチドを含む。レプリキン濃度の上昇はおそらく、少なくとも約6ヶ月から1年、好ましくは少なくとも約2年以上、および最も好ましくは約3年以上の期間にわたって起きる。インフルエンザウイルスワクチンで使用するために好適なレプリキンペプチドは、1年以上血球凝集素アミノ酸配列から欠如の後に「再出現」することが観察されているレプリキンである。
【0196】
本発明のレプリキンペプチドは、単独または種々の組合せで、被験体に、好ましくは静脈内または筋肉内注射により、被験体の免疫系を刺激してペプチドに対する抗体を産生させるために投与される。一般にペプチドの用量は、約0.1μg〜約10mg、好ましくは約10μg〜約1mg、および最も好ましくは約50μg〜約500μgの範囲である。熟練者は、有効な免疫応答を生成するのに必要な用量および投与の回数を、容易に決定することができるであろう。
レプリキンワクチンに対する早期応答の定量的測定
【0197】
レプリキンワクチンに対する数日または数週間の早期特異抗体の応答を定量的に測定できる能力は、数ヶ月または数年後にのみ測定できる他のワクチンに対して大きな実際的利点である。
【0198】
アジュバント
宿主の種に依存して、免疫応答を増強するのに種々のアジュバントが使用され、特に限定されないが、フロインド(完全アジュバントまたは不完全アジュバント)、ミネラルゲル(例えば水酸化アルミニウム)、界面活性物質(例えば、リソレシチン)、プルロニックポリオール、ポリアニオン、ペプチド、油エマルジョン、キーキンペットヘモシアニン、ジニトロフェノール、および有用な可能性のあるヒトアジュバント(例えば、BCGまたはコリネバクテリウム・パルブム(Corynebacterium parvum))がある。
【0199】
レプリキンヌクレオチド配列
レプリキンDNAまたはRNAは、ウイルス、細菌または他のレプリキンをコードする物質による感染に起因する疾患の診断に多くの用途がある。例えば、レプリキンヌクレオチド配列は、生検組織または血液の、インサイチューハイブリダイゼーション測定法を含むハイブリダイゼーション測定法(例えば、サザンまたはノーザン分析)で、例えば組織試料または環境試料中の特定の生物の存在を診断するために使用される。本発明はまた、目的の特定の病原体中に存在する特定のレプリキンに特異的な抗体を含有するか、または特定のレプリキンに特異的にハイブリダイズする核酸分子(センスまたはアンチセンス)、および随時種々の緩衝液および/または診断に必要な試薬を含有するキットを包含する。
【0200】
また本発明の範囲には、アンチセンスRNAおよびDNA分子を含むオリゴヌクレオチド配列、およびレプリキンもしくはリコグニン含有mRNAの翻訳を阻害するように機能するリボザイムも含まれる。アンチセンスRNAおよびDNA分子およびリボザイムの両方とも、当該分野で公知の任意の方法により調製される。アンチセンス分子は、被験体への供給のために広範囲のベクター中に取り込むことができる。熟練者は、投与の最適経路を容易に決定することができるが、一般的には静脈内または筋肉内投与がルーチン法である。投与量もまた、確認することができる。
【0201】
特に好適なアンチセンス核酸分子は、例えばインフルエンザウイルスポリペプチドをコードするmRNA中に含有されるレプリキン配列に相補的なものであり、ここでレプリキン配列は、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基、を含む7〜約50個のアミノ酸を含む。さらに好ましくは、遺伝子のコード鎖中に存在するレプリキンまたはインフルエンザウイルス血球凝集素タンパク質をコードするmRNAに相補的なアンチセンス核酸分子であり、ここでアンチセンス核酸分子は、6ヶ月から1年以上の期間にわたって保存されることが証明されているかまたは他のインフルエンザウイルス株中のレプリキン濃度と比較してレプリキン濃度が上昇していることが証明されているインフルエンザウイルスの株中に存在する、レプリキンをコードするヌクレオチド配列に相補的である。レプリキン濃度の上昇は好ましくは、少なくとも6ヶ月、好ましくは約1年、最も好ましくは約2〜3年またはそれ以上の期間にわたって存在する。
【0202】
同様に、mRNAに相補的なアンチセンス核酸分子は、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;(2) 少なくとも1つのヒスチジン残基;および(3) 少なくとも6%のリジン残基を含む7〜約50個のアミノ酸のレプリキン配列を含む細菌レプリキンをコードするmRNAに相補的なものである。より好適なものは、遺伝子のコード鎖または細菌のタンパク質をコードするmRNAに相補的なアンチセンス核酸分子である。
【0203】
診断応用
珪藻プランクトン、手足口病ウイルス、トマトリーフカールジェミニウイルス、B型肝炎ウイルスとC型肝炎ウイルス、HIV、インフルエンザウイルスおよび悪性腫瘍細胞のような生物にとって、同定された成分レプリキンはワクチンとして有用であり、診断目的にも有用に標的化される。例えば、輸血用に集めた血液は、汚染生物に特異的であることが証明されたレプリキンの存在についてスクリーニングすることにより、生物(例えばHIV)の汚染についてスクリーニングされる。また、特定の病原性生物に特異的なレプリキン構造のスクリーニングにより、体組織またはその環境中の生物の診断用検出が可能になる。
【0204】
増殖のレプリキン刺激
本発明の別の実施態様において、細胞、組織または臓器の複製速度を上昇させるためにレプリキン構造が使用される。複製の速度および収率を上昇させることが他の細胞、組織または生物について当該分野で知られている細胞分裂への、適切な刺激有りまたは無しで、より迅速に複製することが好ましい他の細胞、組織または臓器にとって特異的なレプリキン構造の添加により、複製速度を上昇させる方法が知られている。これは、例えば当該分野で公知の方法により、少なくとも1つのレプリキン構造を有する生物中で複製機能を有するタンパク質または酵素をコードするかまたはこれに関連する遺伝子を修飾もしくは形質転換することにより行われる。
【0205】
本発明の別の態様において、レプリキン構造は生物の複製を上昇させるために使用される。本発明は、例えばインフルエンザウイルスにおいて、流行に関連する複製の上昇がレプリキン濃度の上昇に関連することを示す。この上昇は、1) 前の年には存在したが、次に1年以上消失している、特定のレプリキン構造の再出現;および/または2) 新しいレプリキン組成の出現、による。さらにマラリアレプリキンでは、単一のタンパク質中で同じレプリキンの繰り返しが起きる。
【0206】
すなわち本発明は、生物の複製を上昇させるための方法と組成物を与える。同様に、異なる生物のレプリキンを任意の生物の複製を阻害するように標的化することができる方法で、レプリキンを使用して任意の生物の複製を上昇させることができる。例えば、イネ、トウモロコシ、およびコムギ穀物の生産(これらは、世界の大人口を養うのに決定的に重要である)は、例えば任意の特定の株のイネの濃度(100アミノ酸残基当たりのレプリキンの数)を上昇させることにより改良することができる。
【0207】
例えば、イネ(Oryza sativa)株では、未成熟の種子から単離されたカタラーゼは、タンパク質の491アミノ酸配列内に以下の異なるレプリキンを含有することが観察された:
kfpdvihafkpnprsh (配列番号638)
kfpdvihafk (配列番号639)
karyvkfhwk (配列番号640)
hpkvspelraiwvnylsqedeslgvkianlnvk (配列番号641)
katihkqndtk (配列番号642)
happtpitprpvvgrrqkatihkqndfk (配列番号643)
kfrpsssfdtkttttnagapvwndnealtvgprgpilledyhliekvah (配列番号644)
kfrpsssfdtkttttnagapvwndnealtvgprgpilledyn (配列番号645)
【0208】
すなわち、当該分野で公知の組換え遺伝子クローニング法を使用することにより、生物(例えば、食用穀物植物)中のレプリキン構造の濃度を上昇させることができ、これは、生物の複製の上昇を促進するであろう。例えば、当該分野で公知の方法によりイネ(Oryza sativa)カタラーゼ遺伝子中に上記で同定したレプリキンのような追加のレプリキン配列を挿入することは、この生物の複製を促進するであろう。
【0209】
同様に、イネ(Oryza sativa)(日本の栽培品種)のNBS-LRRタンパク質中に以下のレプリキンが見いだされた:
kvkahfqkh (配列番号647)
kvkahfqk (配列番号648)
kdyeidkddlih (配列番号648)
hmkqcfafcavfpk (配列番号650)
hvfwelvwrsffqnvkqigsifqrkvyrygqsdvttskihdlmhdlavh (配列番号651)
kqigsifqrkvrygpsdvttskihdlmhdlavh (配列番号652)
kqigsifqrkvyrygpsdvttskihdlmh (配列番号653)
kqigsifqrkvyrygqsdvttskih (配列番号654)
【0210】
さらに、アスパラギン酸プロテイナーゼオリザシン1前駆体タンパク質について、以下のレプリキンが見いだされた:
khgvsagik (SEQ ID N0.655)
htvfdygkmrvgfak (配列番号657)
hsryksgqsstyqkngk (配列番号658)
【0211】
同様に、イネ(Oryza sativa)(インディカ品種群)のMADS−ボックスタンパク質FDRMADS3転写因子中で、以下のレプリキンが見いだされた:
kqeamvlkqeinllqkglryiygnraneh (配列番号659)
kqeinllqkglryiygnraneh (配列番号660)
kskegmlkaaneilqekiveqnglidvgmmvadqqngh (配列番号661)
kaaneilqekiveqnglidvgmmvadqqngh (配列番号662)
【0212】
同様に、ロニメイズ(LONI MAIZE)(ATP結合性酸化還元関連ヒドロラーゼ;セリンプロテアーゼ;マルチ遺伝子ファミリー;ミトコンドリア)について、以下のレプリキンが見いだされた:
kvlaahrygik (配列番号663)
klkiamkhliprvleqh (配列番号664)
klkiamkh (配列番号665)
ktslassiakalnrkfirislggvkdeadirgh (配列番号666)
kalnrkfirislggvkdeadirgh (配列番号667)
kfirislggvkdeadirgh (配列番号668)
kvrlskatelvdrhlqsilvaekitqkvegqlsksqk (配列番号669)
hlqsilvaekitqkvegglsksqk (配列番号670)
kvrlskatelvdrh (配列番号671)
kvggsavesskqdtkngkepihwhskgvaaralh (配列番号672)
kvggsavesskqdtkngkepihwh (配列番号673)
kvggsavesskqdtkngkepih (配列番号674)
kqdtkngkepihwhskgvaaralh (配列番号675)
kqdtkngkepih (配列番号676)
【0213】
同様に、グリセロアルデヒド-3-ホスフェート脱水素酵素A、葉緑体前駆体について、以下のレプリキンが見いだされた:
hrdlrraraaalnivptstgaakavslvlpnlk (配列番号677)
kvlddqkfgiikgtmttth (配列番号678)
hiqagakkvlitapgk (配列番号679)
hgrgdaspldviaindtggvkqashllk (配列番号680)
kqashllk (配列番号71O)
【0214】
さらに、見いだされたさび耐性様タンパク質RP1-4(トウモロコシ(Zea mays)の例は、以下のレプリキンを含む:
kvrrvlskdysslkqlmtlmmdddiskhlqiiesgleeredkvwmkeniik (配列番号681)
kvrrvlskdysslkqlmtlmmdddiskh (配列番号682)
hlqiiesgleeredkvwmkeniik (配列番号683)
hdlreniimkaddlask (配列番号684)
hvqnlenvigkdealask (配列番号685)
kkqgyelrqlkdlnelggslh (配列番号686)
kqgyelrqlkdlnelggslh (配列番号687)
klylksrlkelilewssengmdamnilh (配列番号688)
hlqllqlngmverlpnkvcnlsklrylrgykdqipnigk (配列番号689)
hlqllqlngmverlpnkvcnlskrylrgyk (配列番号690)
hlqllqlngmverlpnkvcnlsk (配列番号691)
hnsnklpksvgelk (配列番号692)
klpkvgelkh (配列番号693)
hlsvrvesmqkhkeiiyk (配列番号694)
khkeiiyk (配列番号695)
klrdilqesqkfllvldlalfkh (配列番号696)
hafsgaeikdqllrmklqdtaeeiakrlgqcplaakvlgsrmcrrk (配列番号697)
hafsgaeikdqllrmk (配列番号698)
klqdtaeeiakrlgqclaakvlgsrmcrrkdiaewkaadvwfeksh (配列番号699)
kvlgsrmcrrkdiaewkaadvwfeksh (配列番号700)
kdiaewkaadvwfeksh (配列番号701)
kaadvwfeksh (配列番号702)
hvptttslptskvfgrnsdrdrivkfllgktttaeasstk (配列番号703)
kailteakqlrdllglph (配列番号704)
kakaksgkgpllredessstattvmkpfh (配列番号705)
ksphrgkleswlrrlkeafydaedlldeh (配列番号706)
ksphrgkleswlrrlk (配列番号707)
hrgkleswlrrlk (配列番号708)
ksphrgk (配列番号709)
【0215】
前述のように、コムギユビキチン(ubiquitin)活性化酵素E(配列番号614〜616))中のレプリキンは保存されている。このレプリキン構造の保存は、植物成長の刺激のための信頼できる標的となる。
【0216】
レプリキンと酸化還元酵素との密接な関係もまた、コムギのこの構造に明瞭に示される。すなわちこのコムギユビキチン(ubiquitin)活性化酵素Eは、まずそのカルボキシ末端グリシン残基をATPでアデニル化し、次にこの残基をE1(配列番号614)中のシステイン残基の側鎖に連結して、ユビキチン−E1チオールエステルと遊離のAMPを産生することにより、ユビキチンを活性化する。
【0217】
コムギレプリキンと酸化還元酵素との関係のさらなる例はまた、Chinese spring wheat Chloroplast Triticum aestivamから単離されたPSABコムギタンパク質, フォトシステムI P700 クロロフィルA アポタンパク質 A2 (PsaB) (PSI-B) 中に見いだされる。このタンパク質は以下のように機能する:PsaAとPsaBは9700(フォトシステムI P700の1次電子供与者)ならびに電子アクセプターA0、A1およびFXに結合する。PSIは、プラストシアニン/サイトクローム c6-フェレドキシン酸化還元酵素として機能する。補助因子P700はクロロフィルAダイマーであり、A0はクロロフィルAであり、A1はフィロキノンであり、FXは4Fe-4S鉄−イオウ中心である。サブユニットA psaA/Sヘテロダイマーは、P700クロロフィル特殊対に結合し、次に電子アクセプターに結合する。高等植物と藻類のPSI反応中心は、1つの少なくとも11個のサブユニットからなる。これは、クロロプラストチラコイド膜の重要な膜タンパク質である。「h」が結合する4Fe-4S鉄−イオウ「中心」は決定的に重要である;従ってレプリキン構造中の「h」は重要である。細菌レプリキンの次に、これらのコムギレプリキンと植物レプリキンは、複製および複製のために必要なエネルギー源に対するレプリキン構造の重要性の、最も初歩的な進化的例示である。この基礎的関係は、複製に関して藻類、ウイルスレプリキン、細菌、癌細胞、およびおそらくすべての細胞まで続く。
【0218】
レプリキンのさらなる例は、PSABコムギタンパク質中に見いだされ、これはコムギの成長に決定的に重要である。これらには以下を含む:
hlqpkwkpslswfknaesrlnhh (配列番号617)
hlqpkwkpslswfk (配列番号618)
kwkyslswfknaesrlnhh (配列番号619)
kwkpslswfknaesrlnh (配列番号620)
kpslswfknaesrlnhh (配列番号621)
kpslswfknaesrlnh (配列番号622)
hhaialglhtttlilvkgaldargsklmpdkk (配列番号623)
haialglhtttlilvkgaldargsklmpdkk (配列番号624)
hhaialglhtttlilvkgaldargsk (配列番号625)
haialglhtttlilvkgaldargsk (配列番号626)
htttlilvkgaldargsklmpdkk (SEQ ID N0.627)
htttlilvkgaldargsklmpdk (配列番号628)
htttlilvkgaldargsk (配列番号629)
【0219】
コムギレプリキンと酸化還元との関係のさらなる例は、PSAA_コムギフォトシステムI 9700 クロロフィルA アポタンパク質 A1中に提供され、これは以下を含む:
hhhlaiailfliaghmyrtnwgighglkdileahkgpftgqghk (配列番号630)
hhlaiailfliaghmyrtnwgighglkdileahkgpftgqghk (配列番号631)
hlaiailfliaghmyrtnwgighglkdileahkgpftgqghk (配列番号632)
hmyrtnwgighglkdileahkgpftgqghk (配列番号633)
hglkdileahkgpftgqghk (配列番号634)
hdileahkgpftgqghk (配列番号635)
hkgpftgqghk (配列番号636)
kgpftgqgnk (配列番号637)
レプリキンを同定するためのコンピューターソフトウェア
【0220】
本発明はまた、アミノ酸または核酸配列中のレプリキン配列の同定法を提供する。本発明の3点認識法の開発において、4000を越える配列の視覚的スキャンを行った。しかし、ヌクレオチドおよび/またはアミノ酸配列を含むデータバンクはまた、3点認識要件を満足する配列の存在について、コンピューターによりスキャンすることができる。
【0221】
本発明の別の実施態様において、本明細書に記載の3点認識法はコンピューターにより行われる。図6は、本発明の前記実施態様で使用できるコンピューターのブロック図である。コンピューターは、プロセッサー、入力/出力装置、および前記実施態様の3点認識法である実施可能なプログラム命令を保存するメモリーを含む。メモリーは、静的メモリー、揮発性メモリーおよび/または不揮発性メモリーがある。静的メモリーは、磁気、または電気または光学保存媒体で提供されるリードオンリーメモリー(「ROM」)でもよい。揮発性メモリーは、ランダムアクセスメモリー(「RAM」)でもよく、キャッシュとしてプロセッサー内に取り込まれるか、または別の集積回路としてプロセッサーから外部に提供される。不揮発性メモリーは、電気、磁気または光学保存媒体でもよい。
【0222】
プロテオミクス的観点から、新しい神経膠腫ペプチド配列に基づく「3点認識」鋳型の構築により、関連構造および機能を有する生物学的に広いクラスのタンパク質が直接同定された。3点認識法の実施は、「キーワード」検索の使用による同定に似ているが、典型的な配列ホモジネート検索やアミノ酸のヌクレオチド特定におけるようにキーワード「kagvaflhkk」(配列番号1)の正確なスペリングを使用する代わりに、「3点認識」パラメータで制限したキーワードの抽出が使用される。この制限抽出(単一の比較的短いアミノ酸配列から得られるが)により、同じ規格で規定される構造を有するタンパク質のクラスが同定される。構造以外に特定の機能(この場合形質転換と複製)がまた、露出されたクラスのメンバーに共有されることは、これらの構造と機能が関連していることを示唆する。すなわち、この新たに同定された短いペプチド配列から、分子認識「言語」が考案され、これは従来は開示されていない。さらに、ここで癌レプリキンについて示されるように、クラスの多様なメンバーによる免疫学的特異性の共有は、B細胞とその生成物である抗体が、同様の認識言語でレプリキンを認識することを示唆する。
T細胞プライマーの他の用途
【0223】
「3点認識」は、特定のクラスのタンパク質を特定するプロテオミクス法であるため、他のペプチドについて3つ以上の異なる認識点を使用することは、他のタンパク質クラスに関する有用な情報を与えるであろう。さらに「3点認識」法は、他のリコグニン、例えば生物のリポ多糖のTOLL「innate」認識に応用される。3点認識法はまた、共有結合した有機分子の他の有用な化合物(他の共有結合したアミノ酸、ヌクレオチド、炭水化物、脂質またはこれらの組合せを含む)を同定するように修飾される。本発明のこの実施態様において配列は、3つ以上の所望の構造的特徴を有するサブ配列についてスクリーニングされる。共有結合したアミノ酸、脂質または炭水化物からなる化合物のスクリーニングにおいて、7〜約50の共有結合した単位のサブ配列は、(1) 最初のアミノ酸、炭水化物または脂質残基の2番目から7〜10残基に位置する少なくとも1つの最初のアミノ酸、炭水化物または脂質残基を含有し;(2) 少なくとも1つの第2のアミノ酸、炭水化物または脂質残基をコードし;および(3) 最初のアミノ酸、炭水化物または脂質残基の少なくとも6%を含有する。ヌクレオチド配列のスクリーニングの場合、約21〜約150ヌクレオチドのサブ配列は、(1) 最初のアミノ酸残基をコードする第2のコドンから18〜30ヌクレオチド以内に位置する最初のアミノ酸をコードする少なくとも1つのコドン;(2) 少なくとも1つの第2のアミノ酸残基を含有し;および(3) 最初のアミノ酸残基の少なくとも6%をコードする。
【0224】
本発明のいくつかの実施態様を、本明細書に記載のように具体的に例示する。しかし、本発明の修飾や変更態様が、本発明の精神と範囲を逸脱することなく、上記教示および特許請求の範囲に包含されることは理解されるであろう。
実施例1
レプリキンの抽出、単離および同定法と、レプリキン含有生物を標的化、標識または破壊するためのレプリキンの使用
a) 藻類
【0225】
以下の藻類は、バミューダ海地域から採取し、同日抽出したかまたは−20℃に凍結し、翌日抽出した。藻類を低温室(0〜5℃)で、中性緩衝液、例えば0.005Mのリン酸緩衝液(pH7)100cc中で1グラムのアリコートでWaringブレンダー中で15分ホモジナイズし、3000rpmで遠心分離し、溶媒を留去して上清を濃縮し、低温でリン酸緩衝液で透析して、約15mlの容量を得た。この抽出溶液の容量を記録し、タンパク質分析のためにアリコートを取り、残りを分画して、pKが1〜4の範囲のタンパク質画分を得た。
【0226】
好適な分画法は以下のようにクロマトグラフィーである:抽出溶液を低温室(4℃)で、0.005Mリン酸緩衝液で平衡化しておいたDEAEセルロース(Cellex-D)カラム2.5×11.0cmで分画した。段階的溶出溶媒の変化は以下の溶液を用いて作成する:
溶液1 − 4.04gのNaH2PO4と0.5gのNaH2PO4を15リットルの蒸留水に溶解する(0.005モル、pH7);
溶液2 − 8.57gのNaH2PO4を2,480mlの蒸留水に溶解する;
溶液3 − 17.1gのNaH2PO4を2480mlの蒸留水に溶解する(0.05モル、pH4.7);
溶液4 − 59.65gのNaH2PO4を2470mlの蒸留水に溶解する(0.175モル);
溶液5 − 101.6gのNaH2PO4を2455mlの蒸留水に溶解する(pH4.3);
溶液6 − 340.2gのNaH2PO4を2465mlの蒸留水に溶解する(1.0モル、pX-i 4.1);
溶液7 − 283.63gの80%リン酸(H3PO4)を2460mlの蒸留水に作成する(1.0モル、pH1.0)。
【0227】
抽出溶液(6〜10ml容量)をカラムに流し、溶液1を重層し、300mlの溶液1の容器を取り付け、重力でカラムに滴下させる。3mlアリコートの溶出液を集め、溶液1で除去すべきすべてのタンパク質がカラムから除去されるまで、OD280でタンパク質含量を分析する。次にカラムに溶液2を加え、続いて溶液3、4、5、6および7を、各溶液で除去すべきすべてのタンパク質がカラムから除去されるまで、加える。溶液7からの溶出液を集め、リン酸緩衝液で透析し、透析外液と透析内液の両方のタンパク質含量を測定し、両方をゲル電気泳動で分析する。溶液7で、分子量が3,000〜25,000ダルトンのペプチドまたはタンパク質の1つまたは2つのバンドが得られる。例えば藻類カウレルパ・メキシカナ(Caulerpa mexicana)、ラウレンシア・オブツラ(Laurencia obtura)、クラドフェキサ・プロリフェラ(Cladophexa prolifera)、サルガスム・ナタンス(Sargassum natans)、カウレルパ・ヴェルチシラータ(Caulerpa verticillata)、ハリメダ・ツナ(Halimeda tuna)、およびペニシロス・カピタツス(Penicillos capitatus)は、上記のように抽出と処理した後、溶液7溶出液で、この分子量範囲で汚染物質の無い鋭いペプチドバンドを示した。これらの溶液7タンパク質またはこれらの溶出バンドを加水分解し、アミノ酸組成を測定する。こうして得られたペプチド(リジン組成が6%またはそれ以上)はレプリキン前駆体である。これらのレプリキンペプチド前駆体を次に、アミノ酸配列について測定し、米国特許第6,242,578 B1号に詳述されているように、加水分解と質量スペクトル法によりレプリキンを測定する。「3点認識」法により規定される基準を満足するものをレプリキンとして同定する。この方法はまた、酵母、細菌および任意の植物レプリキンを得るのに応用できる。
b) ウイルス
【0228】
藻類についての上記a)と同じ抽出およびカラムクロマトグラフィー分離法を使用して、ウイルスに感染された細胞中のレプリキンを単離し同定する。
c) in vivoおよびin vitro組織培養での腫瘍細胞
【0229】
藻類についての上記a)と同じ抽出およびカラムクロマトグラフィー分離法を使用して、腫瘍細胞中のレプリキンを単離し同定する。例えば、上記a)のように処理した、悪性脳腫瘍から単離したアストロシチン(Astrocytin)のレプリキン前駆体、神経膠芽細胞腫腫瘍細胞から単離したマリグニン(Aglyco 1OB)、組織培養のMCF7乳癌細胞、組織培養のP3Jリンパ腫細胞はそれぞれ、リジン含量が9.1%、6.7%、6.7%、および6.5%のレプリキン前駆体を与えた。実施例10(米国特許第6,242,578 B1号)に記載されているように、Aglyco 1OBの加水分解と質量スペクトル法により、アミノ酸配列 ykagvaflhkkndiide 16量体レプリキンが得られた。
実施例2:
【0230】
レプリキンの診断的用途の例として:Aglyco 1OBまたは16量体レプリキンを抗原として使用して、米国特許第6,242,578 B1号の図2、3、4および7に記載のように、診断目的に血清中に存在する対応する抗体の量を定量する。
【0231】
標識、栄養または破壊目的にレプリキンに結合する物質の産生の例として:16量体レプリキンをウサギに注射して16量体レプリキンに対する特異抗体を産生することが、米国特許第6,242,578 B1号の実施例6と図9Aおよび9Bに記載されている。
【0232】
レプリキンを標識する物質の使用の例として:16量体レプリキンに対する抗体を使用して、このレプリキンを含有する特異的細胞を標識することが、米国特許第6,242,578 B1号の図5と実施例6に記載されている。
【0233】
レプリキンを破壊する物質の使用の例として:16量体レプリキンに対する抗体を使用して、このレプリキンを含有する特異的細胞を阻害または破壊することが、米国特許第6,242,578 B1号の図6に記載されている。
実施例3
【0234】
レプリキンの存在と濃度についてのインフルエンザウイルス血球凝集素タンパク質またはノイラミニダーゼタンパク質の分離株の配列データの分析は、配列の視覚的スキャニングまたは本明細書に記載の3点認識システムに基づくコンピュータープログラムを使用して行う。インフルエンザウイルスの分離株が得られ、インフルエンザ血球凝集素および/またはノイラミニダーゼタンパク質のアミノ酸配列は、当該分野で公知の任意の方法(例えば血球凝集素またはノイラミニダーゼ遺伝子を配列決定してそこからタンパク質配列を得る)により得られる。各分離株中の新しいレプリキンの存在、経時的なレプリキンの保存、およびレプリキンの濃度について、配列をスキャンする。レプリキン配列と濃度を、先に得られた(例えば約6ヶ月〜約3年前)分離株から得られたアミノ酸配列と比較すると、来るインフルエンザシーズンにインフルエンザの原因となる可能性の高い株の出現を予測するのに使用され、季節的インフルエンザペプチドワクチンまたは核酸ベースのワクチンの基礎を形成するデータが得られる。濃度の上昇(特に、約6ヶ月〜約3年またはそれ以上の期間のインフルエンザウイルスのある株でのレプリキンの濃度の段階的上昇)の観察は、将来のインフルエンザ流行または汎流行の原因となる可能性のある株の出現の予測因子である。
【0235】
出現する株で観察されたレプリキンに基づきペプチドワクチンまたは核酸ベースのワクチンが作成される。出現する株は、この期間の血球凝集素および/またはノイラミニダーゼ配列内のレプリキン配列濃度の最大の上昇を有するインフルエンザウイルスの株として同定される。好ましくはペプチドまたは核酸ワクチンは、出現する株で保存されていることが観察されている任意のレプリキン配列に基づくかまたはこれを含む。保存されているレプリキンは、好ましくは約2年または好ましくはそれ以上の期間、血球凝集素またはノイラミニダーゼタンパク質配列中に存在するレプリキン配列である。ワクチンは、出現する株で同定されるレプリキン配列の任意の組合せを含んでよい。
【0236】
ワクチン産生のために、有効なワクチンのために有用であると同定されるレプリキンペプチドは、任意の方法(化学合成および分子生物学法、例えばクローニング、宿主細胞中での発現、およびそこからの精製を含む)により合成される。ペプチドは好ましくは、医薬として許容される担体と、これに対する治療抗体を誘導することが決定されている量で混合される。一般に投与量は、約0.1μg〜約10mgである。
【0237】
インフルエンザワクチンは好ましくは、「インフルエンザシーズン」が始まる前に必要な患者に投与される。インフルエンザシーズンは一般に、10月の後半から始まり4月の後半まで続く。しかしワクチンは、年の任意の時期に投与される。好ましくはインフルエンザワクチンは年に1回投与され、存在すること、好ましくはインフルエンザウイルスの出現する株で保存されていることが、観察されたレプリキン配列に基づく。インフルエンザワクチンに含めるのに好適な別のレプリキンは、1年以上欠如していた後に再出現したインフルエンザ株中であることが証明されているレプリキンである。
実施例4
【0238】
レプリキンの存在と濃度についての熱帯熱マラリア原虫抗原の分離株の配列データの分析は、配列の視覚的スキャニングまたは本明細書に記載の3点認識法に基づくコンピュータープログラムを使用して行う。熱帯熱マラリア原虫の分離株、およびタンパク質のアミノ酸配列は、当該分野で公知の任意の方法、例えば遺伝子を配列決定してそこからタンパク質配列を得ることにより得られる。配列を、各分離株中のレプリキンの存在、経時的なレプリキンの保存、およびレプリキンの濃度について配列をスキャンする。この情報は、抗マラリアペプチドワクチンまたは核酸ベースのワクチンの基礎を形成するのに使用されるデータを提供する。
【0239】
マラリアを引き起こす生物で観察されたレプリキンに基づくペプチドワクチンまたは核酸ベースのワクチンが作成される。好ましくはペプチドまたは核酸ワクチンは、生物の表面抗原上に存在することが観察されている任意のレプリキン配列に基づくかまたはこれを含む。ワクチンは、マラリアを引き起こす株で同定されるレプリキン配列の任意の組合せを含んでよい。
【0240】
ワクチン産生のために、有効なワクチンのために有用であると同定されるレプリキンペプチドは、任意の方法(化学合成および分子生物学法、例えばクローニング、宿主細胞中での発現、およびそこからの精製を含む)により合成される。ペプチドは好ましくは、医薬として許容される担体と、これに対する治療抗体を誘導することが決定されている量で混合される。一般に投与量は、約0.1μg〜約10mgである。
【0241】
マラリアワクチンは好ましくは、年の任意の時期に必要な患者に投与され、特に熱帯地方への旅行の前に投与される。
【0242】
他の実施態様は、遺伝子のコード鎖に相補的なアンチセンス核酸分子、または生物(特に限定されないが、ウイルス、トリパノソーマ、細菌、真菌、藻類、アメーバ、および植物を含む)中のレプリキンについてのmRNAをコードする生物を含み、ここで該アンチセンス核酸分子は、レプリキン含有生物のヌクレオチド配列に相補的である。
【特許請求の範囲】
【請求項1】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成されたペプチド。
【請求項2】
該ペプチドアミノ酸配列は7〜25個のアミノ酸残基を含む、請求項1の単離されたかまたは合成されたペプチド。
【請求項3】
該ペプチドアミノ酸配列は7〜16個のアミノ酸残基を含む、請求項1の単離されたかまたは合成されたペプチド。
【請求項4】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成されたウイルスペプチド。
【請求項5】
インフルエンザウイルスペプチドである、請求項4のペプチド。
【請求項6】
ヒト免疫不全症ウイルス(HIV)ペプチドである請求項4のペプチド。
【請求項7】
該ウイルスペプチドは肝炎ウイルスペプチドである、請求項4のペプチド。
【請求項8】
該ウイルスペプチドはトウモロコシストリークウイルスペプチドである、請求項4のペプチド。
【請求項9】
該ウイルスペプチドはヘルペスウイルスペプチドである、請求項4のペプチド。
【請求項10】
該ウイルスペプチドはウシヘルペスウイルスペプチドである、請求項4のペプチド。
【請求項11】
該ウイルスペプチドはミアリグリッドヘルペスウイルスペプチドである、請求項4のペプチド。
【請求項12】
該ウイルスペプチドはネコ免疫不全症ウイルスペプチドである、請求項4のペプチド。
【請求項13】
該ウイルスペプチドは手足口病ウイルスペプチドである、請求項4のペプチド。
【請求項14】
該ウイルスペプチドはラウス肉腫ウイルスペプチドである、請求項4のペプチド。
【請求項15】
該ウイルスペプチドは鳥類肉腫ウイルスペプチドである、請求項4のペプチド。
【請求項16】
該ウイルスペプチドは神経芽腫RASウイルス癌遺伝子ペプチドである、請求項4のペプチド。
【請求項17】
該ウイルスペプチドはポリオーマウイルスペプチドである、請求項4のペプチド。
【請求項18】
該ウイルスペプチドはシンドビスペプチドである、請求項4のペプチド。
【請求項19】
該ウイルスペプチドはヒトパピローマウイルスペプチドである、請求項4のペプチド。
【請求項20】
該ウイルスペプチドは骨髄性単球性腫瘍ウイルスペプチドである、請求項4のペプチド。
【請求項21】
該ウイルスペプチドはマウス急性白血病ペプチドである、請求項4のペプチド。
【請求項22】
該ウイルスペプチドはヒトT細胞リンパ栄養性ウイルスペプチドである、請求項4のペプチド。
【請求項23】
該ウイルスペプチドはトマトリーフカールウイルスペプチドである、請求項4のペプチド。
【請求項24】
該HIVウイルスペプチドはHIVトランスアクチベーターペプチドである、請求項6のペプチド。
【請求項25】
該ウイルスペプチドは、配列番号613に記載の配列を有する、請求項24のペプチド。
【請求項26】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む単離された細菌ペプチド。
【請求項27】
該細菌ペプチドはヘリコバクターペプチドである、請求項26のペプチド。
【請求項28】
該ヘリコバクターペプチドはヘリコバクターピロリペプチドである、請求項27のペプチド。
【請求項29】
該細菌ペプチドはスタフィロコッカス(Staphylococcus)ペプチドである、請求項26のペプチド。
【請求項30】
該スタフィロコッカス(Staphylococcus)ペプチドはスタフィロコッカス・アウレウス(Staphylococcus aureus)ペプチドである、請求項29のペプチド。
【請求項31】
該細菌ペプチドはレジオネラ(Legionella)ペプチドである、請求項26のペプチド。
【請求項32】
該レジオネラ(Legionella)ペプチドはレジオネラ・ニューモフィリア(Legionella pneumophilia)ペプチドである、請求項31のペプチド。
【請求項33】
該細菌ペプチドはアセトバクター(Acetobacter)ペプチドである、請求項26のペプチド。
【請求項34】
該細菌ペプチドはアクチノミセス(Actinomyces)ペプチドである、請求項26のペプチド。
【請求項35】
該細菌ペプチドはアエロバクター(Aerobacter)ペプチドである、請求項26のペプチド。
【請求項36】
該細菌ペプチドはアルスロバクター(Arthrobacter)ペプチドである、請求項26のペプチド。
【請求項37】
該細菌ペプチドはアゾトバクター(Azotobacter)ペプチドである、請求項26のペプチド。
【請求項38】
該細菌ペプチドはバチルス(Bacillus)ペプチドである、請求項26のペプチド。
【請求項39】
該細菌ペプチドはブレビバクテリウム(Brevibacterium)である、請求項26のペプチド。
【請求項40】
該細菌ペプチドはクロストリディウム(Clostridium)ペプチドである、請求項26のペプチド。
【請求項41】
該細菌ペプチドはコリネバクテリウム(Corynebacterium)ペプチドである、請求項26のペプチド。
【請求項42】
該細菌ペプチドはエルウィニア(Erwinia)ペプチドである、請求項26のペプチド。
【請求項43】
該細菌ペプチドはエシェリキア(Escheria)ペプチドである、請求項26のペプチド。
【請求項44】
該細菌ペプチドはクレブシエラ(Klebsiella)ペプチドである、請求項26のペプチド。
【請求項45】
該細菌ペプチドはラクトバチルス(Lactobacillus)ペプチドである、請求項26のペプチド。
【請求項46】
該細菌ペプチドはヘモフィルス(Haemophilus)ペプチドである、請求項26のペプチド。
【請求項47】
該細菌ペプチドはフラボバクテリウム(Flavobacterium)ペプチドである、請求項26のペプチド。
【請求項48】
該細菌ペプチドはメチロモナス(Methylomonas)ペプチドである、請求項26のペプチド。
【請求項49】
該細菌ペプチドはミクロコッカス(Micrococcus)ペプチドである、請求項26のペプチド。
【請求項50】
該細菌ペプチドはマイコバクテリウム(Mycobacterium)ペプチドである、請求項26のペプチド。
【請求項51】
該細菌ペプチドはミクロノムスポラ(Micronomspora)ペプチドである、請求項26のペプチド。
【請求項52】
該細菌ペプチドはマイコプラズマ(Mycoplasma)ペプチドである、請求項26のペプチド。
【請求項53】
該細菌ペプチドはナイセリア(Neisseria)ペプチドである、請求項26のペプチド。
【請求項54】
該細菌ペプチドはノカルジア(Nocardia)ペプチドである、請求項26のペプチド。
【請求項55】
該細菌ペプチドはプロテウス(Proteus)ペプチドである、請求項26のペプチド。
【請求項56】
該細菌ペプチドはシュードモナス(Pseudomonas)ペプチドである、請求項26のペプチド。
【請求項57】
該細菌ペプチドはリゾビウム(Rhizobium)ペプチドである、請求項26のペプチド。
【請求項58】
該細菌ペプチドはサルモネラ(Salmonella)ペプチドである、請求項26のペプチド。
【請求項59】
該細菌ペプチドはセラチア(Serratia)ペプチドである、請求項26のペプチド。
【請求項60】
該細菌ペプチドはスタフィロコッカス(Staphylococcus)ペプチドである、請求項26のペプチド。
【請求項61】
該細菌ペプチドは連鎖球菌(Streptocossus)ペプチドである、請求項26のペプチド。
【請求項62】
該細菌ペプチドはストレプトミセス(Streptomyces)ペプチドである、請求項26のペプチド。
【請求項63】
該細菌ペプチドはストレプトスポラングルム(Streptosporangrum)ペプチドである、請求項26のペプチド。
【請求項64】
該細菌ペプチドはストレプトビルチシリウム(Streptovirticillium)ペプチドである、請求項26のペプチド。
【請求項65】
該細菌ペプチドはビブリオ(Vibrio)ペプチドである、請求項26のペプチド。
【請求項66】
該細菌ペプチドはキサントモナス(Xanthomas)ペプチドである、請求項26のペプチド。
【請求項67】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された真菌ペプチド。
【請求項68】
該真菌ペプチドはペニシリウム(Penicillium)ペプチドである、請求項67のペプチド。
【請求項69】
該真菌ペプチドはディセウラ(Diseula)ペプチドである、請求項67のペプチド。
【請求項70】
該真菌ペプチドはオフィストーマ(Ophiostoma)ペプチドである、請求項67のペプチド。
【請求項71】
該オフィストーマ(Ophiostoma)ペプチドはオフィストーマ・ノボウルミ(Ophiostoma novo-ulmi)ペプチドである、請求項70のペプチド。
【請求項72】
該真菌ペプチドはカンジダ(Candida)ペプチドである、請求項67のペプチド。
【請求項73】
該真菌ペプチドはアブシジア(Absidia)ペプチドである、請求項67のペプチド。
【請求項74】
該真菌ペプチドはアスペルギルス(Aspergillus)ペプチドである、請求項67のペプチド。
【請求項75】
該真菌ペプチドはセファロスポリウム(Cephalosporium)ペプチドである、請求項67のペプチド。
【請求項76】
該真菌ペプチドはフサリウム(Fusarium)ペプチドである、請求項67のペプチド。
【請求項77】
該真菌ペプチドはハンゼヌラ(Hansenula)ペプチドである、請求項67のペプチド。
【請求項78】
該真菌ペプチドはムコール(Mucor)ペプチドである、請求項67のペプチド。
【請求項79】
該真菌ペプチドはパエシロミセス(Paecilomyces)ペプチドである、請求項67のペプチド。
【請求項80】
該ペニシリウム(Penicillium)ペプチドはペニシリウム・マルネッフェイ(Penicillium marneffei)ペプチドである、請求項68のペプチド。
【請求項81】
該真菌ペプチドはピヒア(Pichia)ペプチドである、請求項67のペプチド。
【請求項82】
該真菌ペプチドはリゾプス(Rhizopus)ペプチドである、請求項67のペプチド。
【請求項83】
該真菌ペプチドはサッカロミセス(Saccharomyces)ペプチドである、請求項67のペプチド。
【請求項84】
該真菌ペプチドはトルロプシス(Torulopsis)ペプチドである、請求項67のペプチド。
【請求項85】
該真菌ペプチドはトリコデルマ(Trichoderma)ペプチドである、請求項67のペプチド。
【請求項86】
該真菌ペプチドはエリシフェ(Erysiphe)ペプチドである、請求項67のペプチド。
【請求項87】
該真菌ペプチドはフィトフトラ・インフェスタンス(Phytophthora infestans)ペプチドである、請求項67のペプチド。
【請求項88】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された藻類ペプチド。
【請求項89】
該藻類ペプチドはカルドフェラ(Caldophera)ペプチドである、請求項88のペプチド。
【請求項90】
該藻類ペプチドはイソレピスプロリフェラ(Isolepisprolifera)ペプチドである、請求項88のペプチド。
【請求項91】
該藻類ペプチドはポルフィラ(Porphyra)ペプチドである、請求項88のペプチド。
【請求項92】
該藻類ペプチドはツノマタ(Chondrus)ペプチドである、請求項88のペプチド。
【請求項93】
該藻類ペプチドはグラシラリア(Gracilaria)ペプチドである、請求項88のペプチド。
【請求項94】
該藻類ペプチドはゲリジウム(Gelidium)ペプチドである、請求項88のペプチド。
【請求項95】
該藻類ペプチドはカウレルパ(Caulerpa)ペプチドである、請求項88のペプチド。
【請求項96】
該藻類ペプチドはラウレンシア(Laurencia)ペプチドである、請求項88のペプチド。
【請求項97】
該藻類ペプチドはクラドフェキサ(Cladophexa)ペプチドである、請求項88のペプチド。
【請求項98】
該藻類ペプチドはホンダワラ(Sargassum)ペプチドである、請求項88のペプチド。
【請求項99】
該藻類ペプチドはペニシロス(Penicillos)ペプチドである、請求項88のペプチド。
【請求項100】
該藻類ペプチドはサボテングサ(Halimeda)ペプチドである、請求項88のペプチド。
【請求項101】
該藻類ペプチドはコンブ(Laminaria)ペプチドである、請求項88のペプチド。
【請求項102】
該藻類ペプチドはヒバマタ(Fucus)ペプチドである、請求項88のペプチド。
【請求項103】
該藻類ペプチドはアスコフィルム(Ascophyllum)ペプチドである、請求項88のペプチド。
【請求項104】
該藻類ペプチドはワカメ(Undari)ペプチドである、請求項88のペプチド。
【請求項105】
該藻類ペプチドはダルス(Rhodymenia)ペプチドである、請求項88のペプチド。
【請求項106】
該藻類ペプチドはマクロシスティス(Macrocystis)ペプチドである、請求項88のペプチド。
【請求項107】
該藻類ペプチドはキリンサイ(Eucheuma)ペプチドである、請求項88のペプチド。
【請求項108】
該藻類ペプチドはフササイミ(Ahnfeltia)ペプチドである、請求項88のペプチド。
【請求項109】
該藻類ペプチドはプテロクラシア(Pteroclasia)ペプチドである、請求項88のペプチド。
【請求項110】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された酵母ペプチド。
【請求項111】
該酵母ペプチドはサッカロミセス(Saccharomyces)ペプチドである、請求項110のペプチド。
【請求項112】
該酵母ペプチドはクリプトコッカス(Cryptococcus)ペプチドである、請求項110のペプチド。
【請求項113】
該クリプトコッカス(Cryptococcus)ペプチドはクリプトコッカス‐ネオフォルマンス(Cryptococcus neoformas)ペプチドである、請求項110のペプチド。
【請求項114】
該酵母ペプチドはシゾサッカロミセス(Schizosaccharomyces)ペプチドである、請求項110のペプチド。
【請求項115】
該酵母ペプチドはオリザ(Oryza)ペプチドである、請求項110のペプチド。
【請求項116】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成されたアメーバペプチド。
【請求項117】
該アメーバペプチドはエントアメーバ(Entamoeba)ペプチドである、請求項116のペプチド。
【請求項118】
該アメーバペプチドはアメービダエ(Amoebidae)ペプチドである、請求項116のペプチド。
【請求項119】
該アメーバペプチドはアカントアメーバ(Acanthamoeba)ペプチドである、請求項116のペプチド。
【請求項120】
該アメーバペプチドはネグレリア(Naegleria)ペプチドである、請求項116のペプチド。
【請求項121】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された植物ペプチド。
【請求項122】
該植物ペプチドはコムギペプチドである、請求項121のペプチド。
【請求項123】
該植物ペプチドはイネペプチドである、請求項121のペプチド。
【請求項124】
該植物ペプチドはトウモロコシペプチドである、請求項121のペプチド。
【請求項125】
該植物ペプチドはユビキチン(ubiquitin)活性化酵素ペプチドである、請求項122のペプチド。
【請求項126】
該植物ペプチドはPsaBコムギペプチドである、請求項125のペプチド。
【請求項127】
該植物ペプチドはPsaAコムギペプチドである、請求項125のペプチド。
【請求項128】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された構造複製関連ペプチド。
【請求項129】
該構造複製関連ペプチドはPMC1ペプチドである、請求項128のペプチド。
【請求項130】
該構造複製関連ペプチドは核小体強皮症抗原ペプチドである、請求項128のペプチド。
【請求項131】
該構造複製関連ペプチドはフィブリラリン(fibrillarin)ペプチドである、請求項128のペプチド。
【請求項132】
該構造複製関連ペプチドはSPOPペプチドである、請求項128のペプチド。
【請求項133】
該構造複製関連ペプチドはセントロメアプロテインCペプチドである、請求項128のペプチド。
【請求項134】
該構造複製関連ペプチドはCTCBF、KU抗原ペプチドである、請求項128のペプチド。
【請求項135】
該構造複製関連ペプチドはATPシンターゼペプチドである、請求項128のペプチド。
【請求項136】
該構造複製関連ペプチドはFBRL核ペプチドである、請求項128のペプチド。
【請求項137】
該構造複製関連ペプチドはHP1Hs−アルファペプチドである、請求項128のペプチド。
【請求項138】
該構造複製関連ペプチドはPM/Sc1核小体ペプチドである、請求項128のペプチド。
【請求項139】
該構造複製関連ペプチドはアミロイドベータA4前駆体ペプチドである、請求項128のペプチド。
【請求項140】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された腫瘍ウイルス関連ペプチド。
【請求項141】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された形質転換関連ペプチド。
【請求項142】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された癌細胞関連ペプチド。
【請求項143】
生物中で複製機能を有する酵素または他のタンパク質をコードする遺伝子を、少なくとも1つのレプリキン構造を用いて形質転換することを含む、生物の複製速度を上昇させる方法。
【請求項144】
生物は食用植物である、請求項143の方法。
【請求項145】
生物はイネ植物である、請求項144の方法。
【請求項146】
生物はコムギ植物である、請求項144の方法。
【請求項147】
生物はトウモロコシ植物である、請求項144の方法。
【請求項148】
7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含むウイルスペプチド配列に特異的に結合する抗体。
【請求項149】
請求項148の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項150】
7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む細菌ペプチド配列に特異的に結合する抗体。
【請求項151】
請求項150の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項152】
7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む真菌ペプチド配列に特異的に結合する抗体。
【請求項153】
請求項152の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項154】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を有する酵母ペプチドまたはポリペプチド配列に特異的に結合する抗体。
【請求項155】
請求項154の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項156】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を有するアメーバペプチドまたはポリペプチド配列に特異的に結合する抗体。
【請求項157】
請求項156の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項158】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含むウイルスペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項159】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む細菌ペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項160】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む真菌ペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項161】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む酵母ペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項162】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含むアメーバペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項163】
少なくとも1つのウイルスレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む、ウイルスレプリキンに特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法。
【請求項164】
細菌レプリキン配列を特異的に標的とする少なくとも1つの非免疫ベースの有機物質を含む組成物の有効量を被験体に投与することを含む、ウイルスレプリキンに特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法。
【請求項165】
物質は核酸である、請求項164の方法。
【請求項166】
核酸はアンチセンス形態である、請求項165の方法。
【請求項167】
投与量は、約1mg剤形として投与される、請求項164の方法。
【請求項168】
(1) 少なくとも1つのタンパク質分解酵素の有効量を投与し;そして
(2) 7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、少なくとも1つの単離された熱帯熱マラリア原虫レプリキンペプチドに特異的な少なくとも1つの抗体の有効量を投与する、ことを含むマラリアを治療するための2重治療法。
【請求項169】
熱帯熱マラリア原虫レプリキンに特異的な複数の抗体が投与される、請求項168の方法。
【請求項170】
(1) 有効量の少なくとも1つのタンパク質分解酵素を投与し;そして
(2) 7〜約50個のアミノ酸を有し、且つ、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む少なくとも1つの単離された熱帯熱マラリア原虫レプリキンペプチドを含む有効量の予防または治療用ワクチンを投与する、ことを含むマラリアの治療方法。
【請求項171】
ワクチンは、熱帯熱マラリア原虫に特異的な複数のレプリキンを含む、請求項170の方法。
【請求項172】
ワクチンは重複するレプリキンを含む、請求項170の方法。
【請求項173】
レプリキンペプチドは、7〜約50個のアミノ酸からなり、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、「3点認識法」と呼ぶレプリキンペプチド構造を認識する方法。
【請求項174】
ウイルスペプチドは天然痘ウイルスである、請求項4のペプチド。
【請求項175】
バチルス(Bacillus)ペプチドは(Bacillus anthracis)ペプチドである、請求項38のペプチド。
【請求項1】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成されたペプチド。
【請求項2】
該ペプチドアミノ酸配列は7〜25個のアミノ酸残基を含む、請求項1の単離されたかまたは合成されたペプチド。
【請求項3】
該ペプチドアミノ酸配列は7〜16個のアミノ酸残基を含む、請求項1の単離されたかまたは合成されたペプチド。
【請求項4】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成されたウイルスペプチド。
【請求項5】
インフルエンザウイルスペプチドである、請求項4のペプチド。
【請求項6】
ヒト免疫不全症ウイルス(HIV)ペプチドである請求項4のペプチド。
【請求項7】
該ウイルスペプチドは肝炎ウイルスペプチドである、請求項4のペプチド。
【請求項8】
該ウイルスペプチドはトウモロコシストリークウイルスペプチドである、請求項4のペプチド。
【請求項9】
該ウイルスペプチドはヘルペスウイルスペプチドである、請求項4のペプチド。
【請求項10】
該ウイルスペプチドはウシヘルペスウイルスペプチドである、請求項4のペプチド。
【請求項11】
該ウイルスペプチドはミアリグリッドヘルペスウイルスペプチドである、請求項4のペプチド。
【請求項12】
該ウイルスペプチドはネコ免疫不全症ウイルスペプチドである、請求項4のペプチド。
【請求項13】
該ウイルスペプチドは手足口病ウイルスペプチドである、請求項4のペプチド。
【請求項14】
該ウイルスペプチドはラウス肉腫ウイルスペプチドである、請求項4のペプチド。
【請求項15】
該ウイルスペプチドは鳥類肉腫ウイルスペプチドである、請求項4のペプチド。
【請求項16】
該ウイルスペプチドは神経芽腫RASウイルス癌遺伝子ペプチドである、請求項4のペプチド。
【請求項17】
該ウイルスペプチドはポリオーマウイルスペプチドである、請求項4のペプチド。
【請求項18】
該ウイルスペプチドはシンドビスペプチドである、請求項4のペプチド。
【請求項19】
該ウイルスペプチドはヒトパピローマウイルスペプチドである、請求項4のペプチド。
【請求項20】
該ウイルスペプチドは骨髄性単球性腫瘍ウイルスペプチドである、請求項4のペプチド。
【請求項21】
該ウイルスペプチドはマウス急性白血病ペプチドである、請求項4のペプチド。
【請求項22】
該ウイルスペプチドはヒトT細胞リンパ栄養性ウイルスペプチドである、請求項4のペプチド。
【請求項23】
該ウイルスペプチドはトマトリーフカールウイルスペプチドである、請求項4のペプチド。
【請求項24】
該HIVウイルスペプチドはHIVトランスアクチベーターペプチドである、請求項6のペプチド。
【請求項25】
該ウイルスペプチドは、配列番号613に記載の配列を有する、請求項24のペプチド。
【請求項26】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む単離された細菌ペプチド。
【請求項27】
該細菌ペプチドはヘリコバクターペプチドである、請求項26のペプチド。
【請求項28】
該ヘリコバクターペプチドはヘリコバクターピロリペプチドである、請求項27のペプチド。
【請求項29】
該細菌ペプチドはスタフィロコッカス(Staphylococcus)ペプチドである、請求項26のペプチド。
【請求項30】
該スタフィロコッカス(Staphylococcus)ペプチドはスタフィロコッカス・アウレウス(Staphylococcus aureus)ペプチドである、請求項29のペプチド。
【請求項31】
該細菌ペプチドはレジオネラ(Legionella)ペプチドである、請求項26のペプチド。
【請求項32】
該レジオネラ(Legionella)ペプチドはレジオネラ・ニューモフィリア(Legionella pneumophilia)ペプチドである、請求項31のペプチド。
【請求項33】
該細菌ペプチドはアセトバクター(Acetobacter)ペプチドである、請求項26のペプチド。
【請求項34】
該細菌ペプチドはアクチノミセス(Actinomyces)ペプチドである、請求項26のペプチド。
【請求項35】
該細菌ペプチドはアエロバクター(Aerobacter)ペプチドである、請求項26のペプチド。
【請求項36】
該細菌ペプチドはアルスロバクター(Arthrobacter)ペプチドである、請求項26のペプチド。
【請求項37】
該細菌ペプチドはアゾトバクター(Azotobacter)ペプチドである、請求項26のペプチド。
【請求項38】
該細菌ペプチドはバチルス(Bacillus)ペプチドである、請求項26のペプチド。
【請求項39】
該細菌ペプチドはブレビバクテリウム(Brevibacterium)である、請求項26のペプチド。
【請求項40】
該細菌ペプチドはクロストリディウム(Clostridium)ペプチドである、請求項26のペプチド。
【請求項41】
該細菌ペプチドはコリネバクテリウム(Corynebacterium)ペプチドである、請求項26のペプチド。
【請求項42】
該細菌ペプチドはエルウィニア(Erwinia)ペプチドである、請求項26のペプチド。
【請求項43】
該細菌ペプチドはエシェリキア(Escheria)ペプチドである、請求項26のペプチド。
【請求項44】
該細菌ペプチドはクレブシエラ(Klebsiella)ペプチドである、請求項26のペプチド。
【請求項45】
該細菌ペプチドはラクトバチルス(Lactobacillus)ペプチドである、請求項26のペプチド。
【請求項46】
該細菌ペプチドはヘモフィルス(Haemophilus)ペプチドである、請求項26のペプチド。
【請求項47】
該細菌ペプチドはフラボバクテリウム(Flavobacterium)ペプチドである、請求項26のペプチド。
【請求項48】
該細菌ペプチドはメチロモナス(Methylomonas)ペプチドである、請求項26のペプチド。
【請求項49】
該細菌ペプチドはミクロコッカス(Micrococcus)ペプチドである、請求項26のペプチド。
【請求項50】
該細菌ペプチドはマイコバクテリウム(Mycobacterium)ペプチドである、請求項26のペプチド。
【請求項51】
該細菌ペプチドはミクロノムスポラ(Micronomspora)ペプチドである、請求項26のペプチド。
【請求項52】
該細菌ペプチドはマイコプラズマ(Mycoplasma)ペプチドである、請求項26のペプチド。
【請求項53】
該細菌ペプチドはナイセリア(Neisseria)ペプチドである、請求項26のペプチド。
【請求項54】
該細菌ペプチドはノカルジア(Nocardia)ペプチドである、請求項26のペプチド。
【請求項55】
該細菌ペプチドはプロテウス(Proteus)ペプチドである、請求項26のペプチド。
【請求項56】
該細菌ペプチドはシュードモナス(Pseudomonas)ペプチドである、請求項26のペプチド。
【請求項57】
該細菌ペプチドはリゾビウム(Rhizobium)ペプチドである、請求項26のペプチド。
【請求項58】
該細菌ペプチドはサルモネラ(Salmonella)ペプチドである、請求項26のペプチド。
【請求項59】
該細菌ペプチドはセラチア(Serratia)ペプチドである、請求項26のペプチド。
【請求項60】
該細菌ペプチドはスタフィロコッカス(Staphylococcus)ペプチドである、請求項26のペプチド。
【請求項61】
該細菌ペプチドは連鎖球菌(Streptocossus)ペプチドである、請求項26のペプチド。
【請求項62】
該細菌ペプチドはストレプトミセス(Streptomyces)ペプチドである、請求項26のペプチド。
【請求項63】
該細菌ペプチドはストレプトスポラングルム(Streptosporangrum)ペプチドである、請求項26のペプチド。
【請求項64】
該細菌ペプチドはストレプトビルチシリウム(Streptovirticillium)ペプチドである、請求項26のペプチド。
【請求項65】
該細菌ペプチドはビブリオ(Vibrio)ペプチドである、請求項26のペプチド。
【請求項66】
該細菌ペプチドはキサントモナス(Xanthomas)ペプチドである、請求項26のペプチド。
【請求項67】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された真菌ペプチド。
【請求項68】
該真菌ペプチドはペニシリウム(Penicillium)ペプチドである、請求項67のペプチド。
【請求項69】
該真菌ペプチドはディセウラ(Diseula)ペプチドである、請求項67のペプチド。
【請求項70】
該真菌ペプチドはオフィストーマ(Ophiostoma)ペプチドである、請求項67のペプチド。
【請求項71】
該オフィストーマ(Ophiostoma)ペプチドはオフィストーマ・ノボウルミ(Ophiostoma novo-ulmi)ペプチドである、請求項70のペプチド。
【請求項72】
該真菌ペプチドはカンジダ(Candida)ペプチドである、請求項67のペプチド。
【請求項73】
該真菌ペプチドはアブシジア(Absidia)ペプチドである、請求項67のペプチド。
【請求項74】
該真菌ペプチドはアスペルギルス(Aspergillus)ペプチドである、請求項67のペプチド。
【請求項75】
該真菌ペプチドはセファロスポリウム(Cephalosporium)ペプチドである、請求項67のペプチド。
【請求項76】
該真菌ペプチドはフサリウム(Fusarium)ペプチドである、請求項67のペプチド。
【請求項77】
該真菌ペプチドはハンゼヌラ(Hansenula)ペプチドである、請求項67のペプチド。
【請求項78】
該真菌ペプチドはムコール(Mucor)ペプチドである、請求項67のペプチド。
【請求項79】
該真菌ペプチドはパエシロミセス(Paecilomyces)ペプチドである、請求項67のペプチド。
【請求項80】
該ペニシリウム(Penicillium)ペプチドはペニシリウム・マルネッフェイ(Penicillium marneffei)ペプチドである、請求項68のペプチド。
【請求項81】
該真菌ペプチドはピヒア(Pichia)ペプチドである、請求項67のペプチド。
【請求項82】
該真菌ペプチドはリゾプス(Rhizopus)ペプチドである、請求項67のペプチド。
【請求項83】
該真菌ペプチドはサッカロミセス(Saccharomyces)ペプチドである、請求項67のペプチド。
【請求項84】
該真菌ペプチドはトルロプシス(Torulopsis)ペプチドである、請求項67のペプチド。
【請求項85】
該真菌ペプチドはトリコデルマ(Trichoderma)ペプチドである、請求項67のペプチド。
【請求項86】
該真菌ペプチドはエリシフェ(Erysiphe)ペプチドである、請求項67のペプチド。
【請求項87】
該真菌ペプチドはフィトフトラ・インフェスタンス(Phytophthora infestans)ペプチドである、請求項67のペプチド。
【請求項88】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された藻類ペプチド。
【請求項89】
該藻類ペプチドはカルドフェラ(Caldophera)ペプチドである、請求項88のペプチド。
【請求項90】
該藻類ペプチドはイソレピスプロリフェラ(Isolepisprolifera)ペプチドである、請求項88のペプチド。
【請求項91】
該藻類ペプチドはポルフィラ(Porphyra)ペプチドである、請求項88のペプチド。
【請求項92】
該藻類ペプチドはツノマタ(Chondrus)ペプチドである、請求項88のペプチド。
【請求項93】
該藻類ペプチドはグラシラリア(Gracilaria)ペプチドである、請求項88のペプチド。
【請求項94】
該藻類ペプチドはゲリジウム(Gelidium)ペプチドである、請求項88のペプチド。
【請求項95】
該藻類ペプチドはカウレルパ(Caulerpa)ペプチドである、請求項88のペプチド。
【請求項96】
該藻類ペプチドはラウレンシア(Laurencia)ペプチドである、請求項88のペプチド。
【請求項97】
該藻類ペプチドはクラドフェキサ(Cladophexa)ペプチドである、請求項88のペプチド。
【請求項98】
該藻類ペプチドはホンダワラ(Sargassum)ペプチドである、請求項88のペプチド。
【請求項99】
該藻類ペプチドはペニシロス(Penicillos)ペプチドである、請求項88のペプチド。
【請求項100】
該藻類ペプチドはサボテングサ(Halimeda)ペプチドである、請求項88のペプチド。
【請求項101】
該藻類ペプチドはコンブ(Laminaria)ペプチドである、請求項88のペプチド。
【請求項102】
該藻類ペプチドはヒバマタ(Fucus)ペプチドである、請求項88のペプチド。
【請求項103】
該藻類ペプチドはアスコフィルム(Ascophyllum)ペプチドである、請求項88のペプチド。
【請求項104】
該藻類ペプチドはワカメ(Undari)ペプチドである、請求項88のペプチド。
【請求項105】
該藻類ペプチドはダルス(Rhodymenia)ペプチドである、請求項88のペプチド。
【請求項106】
該藻類ペプチドはマクロシスティス(Macrocystis)ペプチドである、請求項88のペプチド。
【請求項107】
該藻類ペプチドはキリンサイ(Eucheuma)ペプチドである、請求項88のペプチド。
【請求項108】
該藻類ペプチドはフササイミ(Ahnfeltia)ペプチドである、請求項88のペプチド。
【請求項109】
該藻類ペプチドはプテロクラシア(Pteroclasia)ペプチドである、請求項88のペプチド。
【請求項110】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された酵母ペプチド。
【請求項111】
該酵母ペプチドはサッカロミセス(Saccharomyces)ペプチドである、請求項110のペプチド。
【請求項112】
該酵母ペプチドはクリプトコッカス(Cryptococcus)ペプチドである、請求項110のペプチド。
【請求項113】
該クリプトコッカス(Cryptococcus)ペプチドはクリプトコッカス‐ネオフォルマンス(Cryptococcus neoformas)ペプチドである、請求項110のペプチド。
【請求項114】
該酵母ペプチドはシゾサッカロミセス(Schizosaccharomyces)ペプチドである、請求項110のペプチド。
【請求項115】
該酵母ペプチドはオリザ(Oryza)ペプチドである、請求項110のペプチド。
【請求項116】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成されたアメーバペプチド。
【請求項117】
該アメーバペプチドはエントアメーバ(Entamoeba)ペプチドである、請求項116のペプチド。
【請求項118】
該アメーバペプチドはアメービダエ(Amoebidae)ペプチドである、請求項116のペプチド。
【請求項119】
該アメーバペプチドはアカントアメーバ(Acanthamoeba)ペプチドである、請求項116のペプチド。
【請求項120】
該アメーバペプチドはネグレリア(Naegleria)ペプチドである、請求項116のペプチド。
【請求項121】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された植物ペプチド。
【請求項122】
該植物ペプチドはコムギペプチドである、請求項121のペプチド。
【請求項123】
該植物ペプチドはイネペプチドである、請求項121のペプチド。
【請求項124】
該植物ペプチドはトウモロコシペプチドである、請求項121のペプチド。
【請求項125】
該植物ペプチドはユビキチン(ubiquitin)活性化酵素ペプチドである、請求項122のペプチド。
【請求項126】
該植物ペプチドはPsaBコムギペプチドである、請求項125のペプチド。
【請求項127】
該植物ペプチドはPsaAコムギペプチドである、請求項125のペプチド。
【請求項128】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された構造複製関連ペプチド。
【請求項129】
該構造複製関連ペプチドはPMC1ペプチドである、請求項128のペプチド。
【請求項130】
該構造複製関連ペプチドは核小体強皮症抗原ペプチドである、請求項128のペプチド。
【請求項131】
該構造複製関連ペプチドはフィブリラリン(fibrillarin)ペプチドである、請求項128のペプチド。
【請求項132】
該構造複製関連ペプチドはSPOPペプチドである、請求項128のペプチド。
【請求項133】
該構造複製関連ペプチドはセントロメアプロテインCペプチドである、請求項128のペプチド。
【請求項134】
該構造複製関連ペプチドはCTCBF、KU抗原ペプチドである、請求項128のペプチド。
【請求項135】
該構造複製関連ペプチドはATPシンターゼペプチドである、請求項128のペプチド。
【請求項136】
該構造複製関連ペプチドはFBRL核ペプチドである、請求項128のペプチド。
【請求項137】
該構造複製関連ペプチドはHP1Hs−アルファペプチドである、請求項128のペプチド。
【請求項138】
該構造複製関連ペプチドはPM/Sc1核小体ペプチドである、請求項128のペプチド。
【請求項139】
該構造複製関連ペプチドはアミロイドベータA4前駆体ペプチドである、請求項128のペプチド。
【請求項140】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された腫瘍ウイルス関連ペプチド。
【請求項141】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された形質転換関連ペプチド。
【請求項142】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を含む、単離されたかまたは合成された癌細胞関連ペプチド。
【請求項143】
生物中で複製機能を有する酵素または他のタンパク質をコードする遺伝子を、少なくとも1つのレプリキン構造を用いて形質転換することを含む、生物の複製速度を上昇させる方法。
【請求項144】
生物は食用植物である、請求項143の方法。
【請求項145】
生物はイネ植物である、請求項144の方法。
【請求項146】
生物はコムギ植物である、請求項144の方法。
【請求項147】
生物はトウモロコシ植物である、請求項144の方法。
【請求項148】
7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含むウイルスペプチド配列に特異的に結合する抗体。
【請求項149】
請求項148の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項150】
7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む細菌ペプチド配列に特異的に結合する抗体。
【請求項151】
請求項150の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項152】
7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む真菌ペプチド配列に特異的に結合する抗体。
【請求項153】
請求項152の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項154】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を有する酵母ペプチドまたはポリペプチド配列に特異的に結合する抗体。
【請求項155】
請求項154の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項156】
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む7〜約50個のアミノ酸を有するアメーバペプチドまたはポリペプチド配列に特異的に結合する抗体。
【請求項157】
請求項156の抗体と医薬として許容される担体またはアジュバントとを含む組成物。
【請求項158】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含むウイルスペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項159】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む細菌ペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項160】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む真菌ペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項161】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む酵母ペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項162】
各抗体は:
7〜約50個のアミノ酸を含んで成り、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含むアメーバペプチド配列に特異的に結合する、複数の抗体を含む抗体カクテル。
【請求項163】
少なくとも1つのウイルスレプリキンペプチドを含む組成物の有効量を被験体に投与することを含む、ウイルスレプリキンに特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法。
【請求項164】
細菌レプリキン配列を特異的に標的とする少なくとも1つの非免疫ベースの有機物質を含む組成物の有効量を被験体に投与することを含む、ウイルスレプリキンに特異的に結合する抗体を産生するように被験体の免疫系を刺激する方法。
【請求項165】
物質は核酸である、請求項164の方法。
【請求項166】
核酸はアンチセンス形態である、請求項165の方法。
【請求項167】
投与量は、約1mg剤形として投与される、請求項164の方法。
【請求項168】
(1) 少なくとも1つのタンパク質分解酵素の有効量を投与し;そして
(2) 7〜約50個のアミノ酸を有し、且つ
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、少なくとも1つの単離された熱帯熱マラリア原虫レプリキンペプチドに特異的な少なくとも1つの抗体の有効量を投与する、ことを含むマラリアを治療するための2重治療法。
【請求項169】
熱帯熱マラリア原虫レプリキンに特異的な複数の抗体が投与される、請求項168の方法。
【請求項170】
(1) 有効量の少なくとも1つのタンパク質分解酵素を投与し;そして
(2) 7〜約50個のアミノ酸を有し、且つ、(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む少なくとも1つの単離された熱帯熱マラリア原虫レプリキンペプチドを含む有効量の予防または治療用ワクチンを投与する、ことを含むマラリアの治療方法。
【請求項171】
ワクチンは、熱帯熱マラリア原虫に特異的な複数のレプリキンを含む、請求項170の方法。
【請求項172】
ワクチンは重複するレプリキンを含む、請求項170の方法。
【請求項173】
レプリキンペプチドは、7〜約50個のアミノ酸からなり、
(1) 第2のリジン残基から6〜10アミノ酸残基の位置にある少なくとも1つのリジン残基;
(2) 少なくとも1つのヒスチジン残基;および
(3) 少なくとも6%のリジン残基
を含む、「3点認識法」と呼ぶレプリキンペプチド構造を認識する方法。
【請求項174】
ウイルスペプチドは天然痘ウイルスである、請求項4のペプチド。
【請求項175】
バチルス(Bacillus)ペプチドは(Bacillus anthracis)ペプチドである、請求項38のペプチド。
【図1】

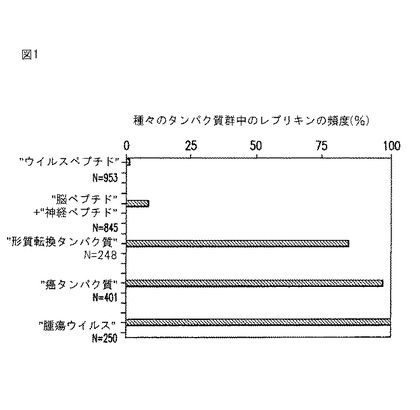
【図2】
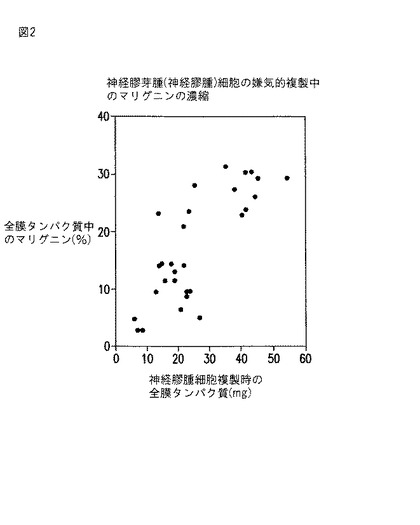
【図3】

【図4F】
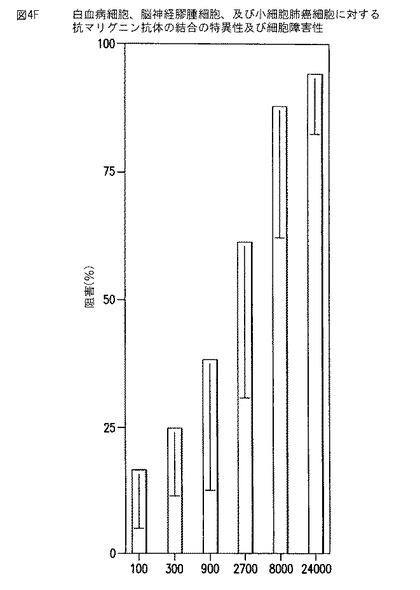
【図5】

【図6】
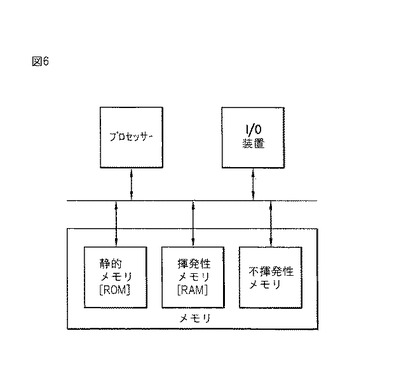
【図7】
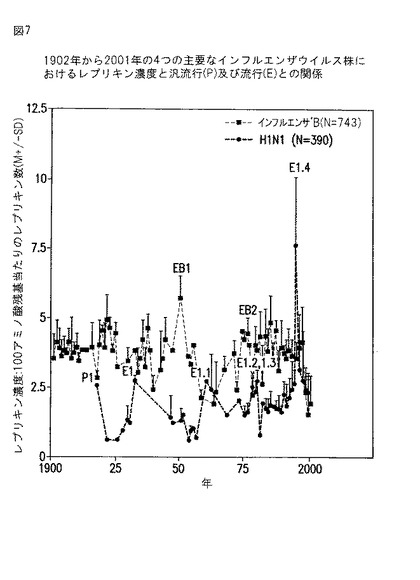
【図8】
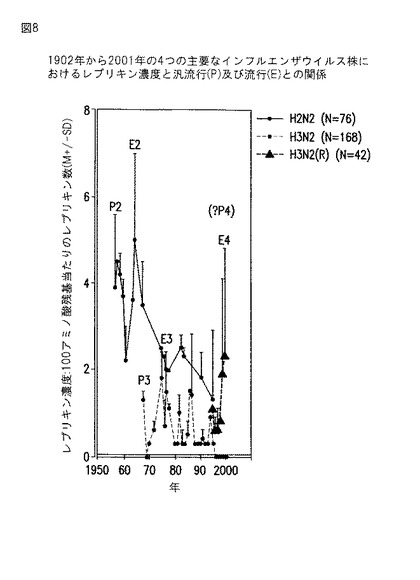
【図4A】
【図4B】

【図4C】

【図4D】
【図4E】
【図2】
【図3】
【図4F】
【図5】
【図6】
【図7】
【図8】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【公開番号】特開2011−157368(P2011−157368A)
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願番号】特願2011−57736(P2011−57736)
【出願日】平成23年3月16日(2011.3.16)
【分割の表示】特願2009−24307(P2009−24307)の分割
【原出願日】平成14年7月9日(2002.7.9)
【出願人】(501141781)
【出願人】(501141792)
【Fターム(参考)】
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願日】平成23年3月16日(2011.3.16)
【分割の表示】特願2009−24307(P2009−24307)の分割
【原出願日】平成14年7月9日(2002.7.9)
【出願人】(501141781)
【出願人】(501141792)
【Fターム(参考)】
[ Back to top ]