
レポーター酵素変異体相補作用の利用によるGタンパク共役受容体およびオーファン受容体機能の高感度検知のための改良システム
【課題】G蛋白共役受容体(GPCR)活性のアッセイ法、GPCRリガンドのスクリーニング法、Gタンパク質共役受容体キナーゼ(GRK)活性の解明。
【解決手段】GPCR活性をアッセイするICAST技術を拡張する方法。活性型GPCRに対するアレスチンの親和性を高め、あるいは活性型GPCRとアレスチンの結合時間を長くする、既知あるいはオーファンGPCRのオープン・リーディング・フレームへのセロトニン/トレオニンリン酸化部位の設計、Gタンパク質共役受容体キナーゼを制限した状態での活性型GPCRに結合した変異型アレスチンタンパク質の設計、活性型GPCRの親和性を高めた変異型スーパーアレスチンタンパク質の設計。薬物リード化合物の発見、オーファンGPCRのリガンドおよび機能の発見、相補的なICAST酵素フラグメントを利用し、GPCRホモ、ヘテロダイマー形成をモニターするための特異的な方法を含む。
【解決手段】GPCR活性をアッセイするICAST技術を拡張する方法。活性型GPCRに対するアレスチンの親和性を高め、あるいは活性型GPCRとアレスチンの結合時間を長くする、既知あるいはオーファンGPCRのオープン・リーディング・フレームへのセロトニン/トレオニンリン酸化部位の設計、Gタンパク質共役受容体キナーゼを制限した状態での活性型GPCRに結合した変異型アレスチンタンパク質の設計、活性型GPCRの親和性を高めた変異型スーパーアレスチンタンパク質の設計。薬物リード化合物の発見、オーファンGPCRのリガンドおよび機能の発見、相補的なICAST酵素フラグメントを利用し、GPCRホモ、ヘテロダイマー形成をモニターするための特異的な方法を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2000年2月7日に出願された米国仮出願番号第60/180,669号から利益を請求する、2000年9月1日に出願された米国出願番号第09/654,499号の一部継続である。前述の特許出願の全開示は引用により本明細書に編入する。
【0002】
発明の分野
本発明はGタンパク質-共役受容体(GPCR)の活性を検知する手段に関連し、GPCR活性のアッセイ法、GPCRリガンド、アゴニストおよび/またはアンタゴニストのスクリーニング法、オーファンGPCRの自然なリガンドおよび代理リガンドのスクリーニング法、GPCR制御過程の化合物と相互に作用する化合物のスクリーニング法を提供する。
【背景技術】
【0003】
発明の背景
細胞外の多くのシグナル活動はGタンパク質-共役受容体(GPCR)とグアニンヌクレオチド-結合制御タンパク質(Gタンパク質)の相互作用により仲介される。Gタンパク質-仲介信号系は哺乳類や酵母など、多くの多細胞生物において確認されている。GPCRは分岐アミノ酸鎖を有するタンパク質の大きなスーパーファミリーであるが、共通の構造的特徴、特に7つの膜貫通領域(蛇皮状)の存在を共有する。GPCRは細胞外シグナル(神経伝達物質、ホルモン、臭気、光、その他)に応答する。個々のGPCRタイプは特定のシグナル伝達経路を活性化、少なくとも10個の異なる信号伝達経路がGPCRにより活性化されることが知られている。例えばβ2アドレナリン受容体(β2AR)は、原型哺乳GPCRである。β2AR受容体はアゴニスト結合に応答して、Gタンパク質(Gs)を活性化し、それが呼応してアデニレートシクラーゼ活性を刺激し、細胞内に環状アデノシン1リン酸(cAMP)を増加生成せしめる。
【0004】
GPCRの刺激によって生じたシグナル経路と最終的な細胞応答は、特定の受容体の結合対象であるGタンパク質の特定クラスによる。(「Gタンパク質シグナルの多くの顔(The Many Faces of G-Protein Signaling)」、ハム著、ジェイ・バイオル・ケム(J Biol. Chem.)誌1998年273:669-672を参照のこと)
【0005】
例えば、Gタンパク質のGsクラスとの結合は、cAMPの産生と、タンパク質キナーゼAおよびC経路の活性化を誘発し、そこにおけるGタンパク質のGiクラスとの結合がcAMPを抑制する。カルシウム、ホスホリパーゼC、ホスファチジルイノシトール3などのほかのセカンドメッセンジャーの利用も可能である。ゆえに、GPCRシグナルイベントは、これらのセカンドメッセンジャープロダクトの定量化により大部分測定されることとなる。
【0006】
持続的刺激に対する応答の減少は、よく知られている生物学的現象である。多様なGPCRによるシグナルは同一の2段階のメカニズムにより中断されると信じられている。活性化された受容体は先ず、GPCRキナーゼ(GRK)によりリン酸化される。アレスチンタンパク質は活性化およびリン酸化された受容体に結合し、その結果Gタンパク質の相互作用を遮断する。この過程は一般的に「脱感作(desensitization)」と呼ばれ、機能的に多様なGPCRを表すひとつの一般的メカニズムである。アレスチンはまた、異なるGPCRの間でヘテロジニアスの過程である、GPCR内在化および脱感作の抑制における役割を果たす。(ジェイ・バイオル・ケム(J Biol. Chem.)誌1999年274:32248-32257、オークリー(Oakley)他著を参照のこと。)内在化および再感作の過程におけるアレスチンとGPCRの間の相互作用は、特定のGPCRのカルボキシル末端内の特定の配列モティーフにより決定される。カルボキシル末端に3つのセリンまたはスレオニン残基クラスタを持つGPCRのサブセットのみが、リガンド刺激の後に飲食小胞内にアレスチンとともに共輸送することがわかっている。GPCRの脱感作に関わる受容体キナーゼとアレスチンの数はかなり限られている。
【0007】
GPCR生理学の一般的特徴は、受容体リン酸化、飲食、脱リン酸化の過程を通しての受容体の脱感作とリサイクルである。(「Gタンパク質-共役受容体の調節:Gタンパク質-共役受容体キナーゼとアレスチンの役割(G-protein-coupled receptor regulation: role of G-protein-coupled receptor kinases and arrestins)」ファーグソン(Ferguson)et. al. 編集、Can. J. Physiol Pharmacol誌1996年74:1095-1110を参照のこと。)リガンドで占められるGPCRは、セリン/スレオニンキナーゼの2つのファミリー、Gタンパク質-共役受容体キナーゼ(GRK)、タンパク質イドナーゼAやタンパク質キナーゼCなどの、セカンドメッセンジャー依存のタンパク質キナーゼによりリン酸化できる。 キナーゼのいずれかのクラスによるリン酸化は、対応するGタンパク質からの解離により受容体を抑制する作用をする。GRK-リン酸化はまた、受容体の細胞質領域を結合し、受容体の飲食小胞へのクラスタ化を促すアレスチンと呼ばれるタンパク質の一クラスを漸増することにより受容体を抑制する役割も果たす。受容体が一度飲食されると、リソソームに分解されるか、あるいは、リン酸化され、完全機能の受容体として形質膜に再循環される。
【0008】
アレスチンタンパク質の活性化受容体への結合は、ロドプシンからβ2AR、さらに、ニューロテンシン受容体までの様々なGPCRの一般的現象として発表されている。(「Gタンパク質-共役受容体活性検知のためのβアレスチン/緑色蛍光融合タンパク質(GFP)バイオセンサー(A β arrestin/Green Fluorescent Fusion Protein Biosensor for Detecting G-Protein-Coupled Receptor Activation)」バラク(Barak)et. al. 編集、ジェイ・バイオル・ケム(J Biol. Chem.)誌1997年272:27497-500を参照のこと。)その結果、特定GPCRとアレスチンの相互作用の監視は、GPCR活性測定の一般的ツールとして利用できる。同様に、単一Gタンパク質とGRKも様々な受容体と組み合わせることができ(ハム(Hamm)他著(1998年)および「Gタンパク質-共役受容体キナーゼ(G-Protein-Coupled Receptor Kinases)」、ピッチャー(Pitcher)他著、Annu. Rev. Biochem誌1998年67:653-92を参照のこと)、これらタンパク質/タンパク質相互作用も受容体の活性状態を決定するために監視が可能である。
【0009】
GPCRは、血管拡張、心拍数、気管支拡張、内分泌、腸蠕動を含む生命生理学的応答を調節することから、今日使われている多くの治療薬がGPCRをターゲットとしている。(例としてレフコヴィッツ(Lefkowitz)他著、Annu. Rev. Biochem.誌 52:159 (1983年)を参照のこと。)これら治療薬にはこの受容体のリガンドを模倣したものもある。その他の薬剤は、受容体の自発的活性から発生した疾患の場合、受容体を拮抗する作用を持つ。
ヒトゲノムプロジェクトなどの努力は、生理学的役割やリガンドが未知の、新たなGPCR(オーファン受容体)を発見することである。ヒトゲノムには数千のGPCRが存在すると推定されている。
【0010】
刺激薬に対する細胞内活動の監視には、例えば酵素結合免疫吸着検定法(ELISA)、フロレッスンスイメージングプレートリーダー検定法(FLIPRtm、モラキュラー・デバイシーズ社、カリフォルニア州サニーベル)、EVOscreen tm, EVOTEC tm, Evotec Biosystems Gmbh(ドイツ、ハンブルグ)、また、ペンシルバニア州ピッツバーグセロミクス社のCELLOMICS tm 開発技術など、様々な試みがこれまで採用されている。
【0011】
ジャーミノ(Germino)et. al. 編集「インビボタンパク質-タンパク質相互作用のスクリーニング(Screening for in vivo protein-protein interactions)」Natl. Acad. Sci.誌 90(3):933-937 (1993年)は、着目するタンパク質との相互作用を行うタンパク質の単離にインビボ法を開示している。
フィジッキー(Phizicky)他「タンパク質-タンパク質相互作用法:検知および解析方法(Protein-protein interactions: methods for detection and analysis)」Microbiol Rev.誌59(l):94-123 (1995年)は、タンパク質-タンパク質相互作用の研究に使われる生物化学、分子生物学、遺伝法の論評を開示している。
オファーマン他著「Gα15およびGα16が多種の受容体をホスホリパーゼCに結合する(Gα15 and Gα16 Couple a Wide Variety of Receptors to Phospholipase C)」J. Biol. Chem.誌 270(25):15175-15180 (1995年)は、Gα15 およびGα16が多様なGタンパク質-共役受容体により活性化されることを開示している。活性化受容体のGタンパク質個別模様への選択的結合は、正確なシグナルトランスダクションId の達成のための重要な必要条件と考えられている。
バラク(Barak)他著「Gタンパク質-共役受容体検知のためのβアレスチン/緑色蛍光融合タンパク質(GFP)(A β-arrestin/Green Fluorescent Protein Biosensor for Detecting G Protein-coupled Receptor Activation.)」J. Biol. Chem.誌272(44):27497--27500 (1997年)と、米国特許番号第5,891,646号および第6,110,693号は、光学機器を使ってのGPCRの刺激によるタンパク質転座の造影へのβアレスチン/緑色蛍光融合タンパク質(GFP)の利用を開示している。
【発明の概要】
【発明が解決しようとする課題】
【0012】
上記に参照された研究にはそれぞれ以下の例にような欠点がある。
・従来技術の方法では、タンパク質の過剰発現が必要であり、人為結果を招いたり、細胞調節器の均衡が崩れるなどの恐れがある。
・従来の可視化または造影アッセイ法ではスループットが低く、完全な定量化に欠く。従って、スループットの高い薬理/運動学的アッセイ法には不適である。
さらに、従来技術のアッセイ法では、GPCRの隔離を必要とし、細胞内でのGPCRの観察ができない。そのため、GPCR機能の監視のためには方法を改善する必要があった。
【課題を解決するための手段】
【0013】
発明の要約
本発明は米国出願連続番号第09/654,499号における開示に変更を提供するものである。特に本発明は、アッセイ法の感度をさらに強化する下記の発明の側面を変更する方向付けを持つ。変更は、GPCRがリン酸化か非リン酸化に関わらず活性化されたGPCRへの結合を強化する、遺伝的変異されたアレスチンの利用、および、独自の構造的モチーフを有しないオーファン受容体のスクリーニングアッセイを簡易化するセリン/スレオニンクラスター戦略の利用、および、さらに強化された検知を達成するための上記変更をの併用を含む。
【0014】
本発明の最初の側面はGPCR機能を受容体活性化の箇所で近位的に監視し、従って、より少数の競合メカニズムにより、薬剤発見目的に多くの情報を提供せしめる方法である。GPCRの活性化は、受容体と、アレスチン、Gタンパク質、GRK、または、その他のキナーゼなどの、受容体への結合が受容体のアゴニスト居住に依存する、調整化合物の相互作用の出力により測定される。本発明は、変異レポーター酵素の相補によるタンパク質-タンパク質相互作用の検知に関するものである。
活性化GPCRへのアレスチンの結合は、これまで研究されているほとんどのGPCRで実証されている脱感作の第一段階における共通の過程である。変異酵素相補検定(ICASTなど)を通してのGPCRのアレスチンとの相互作用の測定は、多様なGPCRとオーファン受容体に適用されるより一般的なアッセイ技術を提供するものである。
【0015】
本発明のさらなる側面は、変異レポーター酵素への融合タンパク質として、例えばムスカリン、アドレナリン、ドーパミン、アンギオテンシン、エンドセリン様のGPCRを発現せしめる検査細胞を提供し、例えばGタンパク質、アレスチン、GRKなど、相補変異レポーター酵素との融合タンパク質として、GPCR経路内でタンパク質を相互作用させることより、検査状況下でGPCR経路活性化を査定する方法である。検査状況下で、検査細胞がターゲットGPCRの既知のアゴニストに提示されると、GPCRの活性化がレポーター酵素の相補により監視される。増加したレポーター酵素活性は、相互作用するそのタンパク質パートナーとの、GPCRの相互作用を反映する。
【0016】
本発明のさらなる側面は、検査アレスチン、例えばβアレスチンなどの存在下でGPCR経路の活性化を査定する方法である。
本発明のさらなる側面は、検査Gタンパク質の存在下でGPCR経路の活性化を査定する方法である。
本発明のさらなる側面は、検査細胞を検査リガンドに提示した状態でのGPCR経路の活性化を査定する方法である。
本発明のさらなる側面は、セカンドレセプターの検査細胞内の共発現におけるGPCR経路の活性化を査定する方法である。セカンドレセプターは同一GPCRまたはオーファン受容体(ホモダイマー)、異なるGPCRまたはオーファン受容体(ヘテロダイマー)か、あるいは、別のタイプのレセプターがあり得る。
本発明のさらなる側面は、オーファンGPCRのリガンドまたはアゴニストのスクリーニング方法である。リガンドまたはアゴニストは自然または合成ライブラリまたは混合に含まれるか、あるいは、物理的刺激があり得る。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質として、オーファンGPCRを発現させ、さらに、例えば、別のβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチンまたはアレスチンの変異体を発現するものが提供されている。受容体活性時におけるアレスチンのオーファンGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は検査化合物に曝され、レポーター酵素活性の増加がリガンドまたはアゴニストの存在を示す。
【0017】
本発明のさらなる側面は、リン酸化または活性化されたGPCRへの結合能力を知るために、着目するタンパク質、例えばアレスチンタンパク質(または、アレスチンタンパク質の変異体)をスクリーニングする法である。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを発現させ、さらに、例えば、別のβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチン(またはアレスチンの変異体)を含むものが提供されている。受容体活性時におけるアレスチンのGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は周知のGPCRアゴニストに曝され、その後レポーター酵素活性が検知される。レポーター酵素活性の増加が、βアレスチン分子がリン酸化または活性化された検査細胞内のGPCRと結合され得ることを示す。
【0018】
本発明のさらなる側面は、特定のGPCRのアゴニストのスクリーニング方法である。アゴニストは自然または合成ライブラリまたは混合に含まれるか、あるいは、物理的刺激があり得る。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを、さらに、例えば、別のβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチンを発現するものが提供されている。受容体活性時におけるアレスチンのGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は検査化合物に曝され、レポーター酵素活性の増加がアゴニストの存在を示す。検査細胞は既知のGPCR、または、既知の多種GPCRを発現するか、または、未知の多種GPCRをGPCR発現することもある。このGPCRは例えば臭気GPCRまたはβAR GPCRともなり得る。
【0019】
本発明のさらなる側面は、GPCRアンタゴニスト活性の検査化合物のスクリーニング方法である。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを、さらに、例えば、もうひとつのβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチンを発現するものが提供されている。受容体活性時におけるアレスチンのGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は検査化合物に曝され、レポーター酵素活性の増加がアゴニストの存在を示す。細胞はの検査細胞およびGPCRアゴニストに曝され、その後レポーター酵素活性が検知される。検査化合物への提示と同時、または、それに続いてアゴニストへの提示が提起されると、検査化合物への提示後のレポーター酵素活性の増加が、検査化合物がGPCRに対するアンタゴニスト活性を有していることを示す。
【0020】
本発明のさらなる側面は、GPCRへのアゴニスト、アンタゴニスト、リガンドの存在をみるための、試料液のスクリーニング方法である。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを発現し、さらに、例えば、もうひとつのβガラクトシダーゼ変異体など相補変異レポーターとの融合タンパク質としてβアレスチンを含んでいる。検査細胞は試料液に提示され、レポーター酵素活性が検知される。試料液への提示の後に変化したレポーター酵素活性は、試料液が細胞内に発現したGPCRに対しアゴニスト、アンタゴニスト、または、リガンドを含んでいることを示す。
【0021】
本発明のさらなる側面は、GPCRの存在をみるための細胞スクリーニング方法である。この側面に従うと、変異レポーター酵素を含むアレスチン融合タンパク質、および、変異レポーター酵素を含むGPCRの下流信号融合タンパク質がGPCR活性の見地のために採用されている。細胞本側面の変更を、GPCRを含むこれらの細胞の細胞数多性をスクリーニングする方法として用いることもできる。本側面に従うと、レポーター酵素とともに融合タンパク質としてのβアレスチンで構成される共役を含む細胞数多性が提供され、数多細胞をGPCRアゴニストに提示し、レポーター酵素の活性化が検知される。GPCRアゴニストへの提示後のレポーター酵素活性の増加がβアレスチンタンパク質がGPCRに結合していることを示し、その結果、細胞がGPCRアゴニストに応答するGPCRを含んでいることが示される。
【0022】
本発明のさらなる側面は、GPCR仲介のシグナル経路のマッピング方法である。例えば本システムを、c-SRCのβアレスチン1との、GPCR活性化時の相互作用を監視するために利用することも可能である。さらに本システムを、GPCRシグナリング経路と、レセプターチロシンキナーゼやRas/Rafなどのその他経路間のクロストークに関わるタンパク質/タンパク質相互作用を監視するために利用することも可能である。本側面に従うと、検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素を含むGPCRや関連タンパク質を発現し、さらに、例えば、もうひとつのβガラクトシダーゼ変異体など相補変異レポーター酵素を含む融合タンパク質としての、別の経路からのタンパク質を含むものが提供されている。レポーター酵素の増加がタンパク質/タンパク質相互作用を示す。
【0023】
本発明の更なる側面は、アゴニストまたはアンタゴニスト刺激時の、GPCRのホモダイマーまたはヘテロダイマーの監視方法である。GPCRダイマーが生体活動に重要であることは多くの文献証拠で示されている。(アブダーラ(AbdAlla). et al編集、「AT1受容体ヘテロダイマーが示すGタンパク質活性強化および変容受容体金属イオン封鎖(ATI -receptor heterodimers show enhanced G-protein activation and altered receptor sequestration.)Nature誌 407:94-98 (2000年); ボッカート(Bockaert) et al.編集「Gタンパク質-共役受容体の分子操作:進化成功度(Molecular tinkering of G protein-coupled receptors: an evolutionary success.) EMBO J.誌 18:1723-29 (1999年);ジョーダン(Jordan) et al.編集「Gタンパク質-共役受容体ヘテロダイマーによる受容体機能の調節(G-protein-coupled receptor heterodimerization modulates receptor function.)」 Nature誌, 399:697-700 (1999年) は、2つの非機能性オピオイドレセプター、kappaおよびdeltaがヘテロダイマー化し、機能性レセプターを形成することを示している。ゴードン(Gordon) et al.編集「(Dopamine D2 receptor dimers and receptor blocking peptides.)」Bioch. Biophys. Res. Commun.誌 227:200-204 (1996年)は、ドーパミンレセプターD2におけるモノメリックおよびダイメリックフォームに関連する異なる薬理学的特性を示している。D2レセプターは、スピペロンの選択的ターゲットであるモノマーとして、または、ネモナプリドのターゲットであるダイマーフォームとして存在する。ハーバート(Herbert) et al.編集「β2アドレナリンレセプター膜貫通領域から発生するペプチドによる受容体のダイマー化および活性化の阻害(A peptide derived from a β2-adrenergic receptor transmembrane domain inhibits both receptor dimerization and activation.)」J.B.C.誌 271:16384-92 (1996年)は、アゴニスト刺激により、受容体の二量体状態が安定化し、そこにおいて逆拮抗薬がモノメリックフォームに有利に働くことが発見されていることを示している。実際に、同じ研究が、β2アドレナリンレセプターの第六膜貫通領域に対応するペプチドが、受容体のダイマー化と活性化を阻害したことを示している。さらに、アンガース(Angers) et al.編集「生物発光エネルギー伝達を利用した生細胞内β2アドレナリンレセプターダイマー化の検知(Detection of β-2- adrenergic receptor dimerization in living cells using bioluminescence resonance energy transfer)」Proc. Natl. Acad. Sci.誌USA, 97(7)-.3684-3689は、β2アドレナリンレセプターダイマー化研究のためのβ2アドレナリンレセプター融合タンパク質(ルシフェラーゼに融合したβ2アドレナリンレセプターおよび赤色よりの強化GFPに融合したβ2アドレナリンレセプター)の使用を開示している。
【0024】
細胞性理学および薬理学的意味合いにおけるGPCRダイマー化は、本発明に従って監視が可能である。例えばβガラクトシダーゼ相補は、例えばGPCR1ΔαやGPCR2Δω などのβガラクトシダーゼ変異酵素のGPCR融合タンパク質を共発現する検査細胞内で測定可能である(図27を参照されたい)。本側面に従うと、GPCRまたはオーファン受容体の単量体と二量体間の相互変換が変異レポーター酵素相補により測定可能である。図27は、βガラクトシダーゼのΔα体を有する融合タンパク質(例えばGPCR1Δα)としてGPCRまたはオーファン受容体を、また、同じGPCRまたはオーファン受容体をβガラクトシダーゼのΔω体を有する融合タンパク質(例えばGPCR1Δω)として共発現している検査細胞を示している。GPCRホモダイマーの形成は活性酵素の形成により反転され、これはGal-Screen(tm)アッセイ法などの酵素活性アッセイで測ることができる。同様に、2つの個別GPCR間か、2つのオーファン受容体間、または、1つの既知GPCRと1つのオーファン受容体間のヘテロダイマー化を例えばGPCR1ΔαやGPCR2Δωなどの融合タンパク質を共発現する検査細胞内で測ることもできる。βガラクトシダーゼ活性の増加が2つの受容体がヘテロダイマーを形成できることを示している。
【0025】
本発明の更なる側面は、アゴニストまたはアンタゴニスト治療影響下で、GPCRの単量体および二量体相互変換を監視する方法である。検査受容体は、同じGPCRまたはオーファン受容体(ホモダイマー)間、または、2つの個別GPCRあるいはオーファン受容体(ヘテロダイマー)間に置くことができる。化合物を使った治療後のβガラクトシダーゼ活性の増加は、その化合物が受容体の二量体に結合および/またはそれを安定化することを意味している。化合物を使った治療後のβガラクトシダーゼ活性の減少は、その化合物が受容体の単量体に結合および/またはそれを安定化することを意味している。
本発明のさらなる側面は、GPCRアゴニストに対するGPCRの応答をみるための細胞スクリーニング方法である。細胞はGPCR経路内の下流方向に相互作用するタンパク質パートナーを含むものが提供される。タンパク質パートナーは、酵素を相補しながら変異体との融合タンパク質として発現され、GPCR活性監視に使われる。検査細胞はGPCRアゴニストに提示され、その後レポーター酵素の酵素的活性が検知される。レポーター酵素活性の増加は、細胞がアゴニストに応答するGPCRを含んでいることを示す。
【0026】
本発明は、GPCRシグナリング内のタンパク質/タンパク質相互作用を監視するための独自開発技術(ICAST(tm)、Intercistronic Complementation Analysis Screening Technology、Gal-Screen(tm)等)の併用に関連する。米国出願番号第09/654,499号で開示されているごとく、本発明の方法は、結果的に2つの非活性化βガラクトシダーゼ変異体(それぞれがGPCRやアレスチンなどの2つの相互作用ターゲットタンパク質ペアの1つに融合)の利用に関与するICAST(tm)の使用と一部関連している。活性化βガラクトシダーゼ群の形成は、ターゲットタンパク質の相互作用により促される。本システムでは、βガラクトシダーゼ活性化は例えば、直接細胞溶解とβガラクトシダーゼレポーター酵素の高速超高感度化学発光法検知とを組み合わせるGal-Screen(tm)アッセイシステム等を使って検知できる。このシステムは、GPCR活性の出力として照度計内の測定に例えばGalacton-Star(r) chemiluminescent substrate を使っている。
【0027】
図23は、本発明の方法に相補技術を利用した例を表す概略図である。図23は、アレスチン分子と活性化GPCRまたはオーファン受容体の融合パートナー間で、特定の相互作用により強制的に結合された際に活性化する、2つの非活性βガラクトシダーゼ変異体を示している。このアッセイ技術は、オーファン受容体を釣り餌としてリガンドを同定する(「脱オーファン」と呼ばれるプロセス)ハイスループットスクリーニングアッセイに有効である。図28で示されている通り、オーファン受容体のβガラクトシダーゼ融合タンパク質(例としてGPCRorphanΔα)はβアレスチンの融合タンパク質(例としてβ-ArrΔw)を伴う検査細胞内に共発現される。検査細胞の被験が天然または合成化合物に対して行われる場合は、βガラクトシダーゼ活性の増加は、化合物がこのGPCRの自然の、または、代理のリガンドであることを意味する。同じアッセイシステムは新規GPCRの薬剤リード発見に使うことも可能である。治療後の検査細胞内におけるβガラクトシダーゼ活性の増加は、化合物のアゴニスト作用を示している。治療後の検査細胞内におけるβガラクトシダーゼ活性の減少は、化合物のアンタゴニスト作用または逆アゴニスト作用を示している。さらに、本発明の方法は、Gタンパク質、GRK、プロトオンコジーンc-Srcなどのその他下流シグナリング化合物を介したGPCR仲介シグナリング経路の監視にも使うことが可能である。
【0028】
本発明は、シグナリング経路のマッピングにICAST(tm) タンパク質/タンパク質相互作用スクリーニングを使用して、一部達成されている。本技術は多様な機能を備える、多様な既知/未知のGPCRに適用可能である。それらは下記を含むがこれに限定されないGPCRのサブファミリーを含む。
(a) アミン様リガンド-アセチルコリンムスカリン様レセプター(M1-M5)、αおよびβアドレノセプター、ドーパミンレセプター(D1、D2、D3、D4)、ヒスタミンレセプター(H1およびH2)、オクトパミンレセプター、セロトニンレセプター(5HT1、5HT2、5HT4、5HT5、5HT6、5HT7)に結合するレセプター;
(b)ペプチドリガンド-アンギオテンシンレセプター、ボンベシンレセプター、ブラジキニンレセプター、C-Cケモキネレセプター(CCR1 - CCR8およびCCR10)、C-X-Xタイプケモキネレセプター(CXC-R5)、コレシストキニンタイプAレセプター、CCKタイプレセプター、エンドセリンレセプター、ニューロテンシンレセプター、FMLP-関連レセプター、ソマトスタチンレセプター(タイプ1-5)、オピオイドレセプター(タイプD、K、M、X)に結合するレセプター;
(c) ホルモンタンパク質-卵胞刺激ホルモンレセプター、甲状腺刺激レセプター、ルトロピン-絨毛性性腺刺激ホルモンレセプターに結合するレセプター;
(d)神経伝達物質-サブスタンスPレセプター、サブスタンスKレセプター、神経ペプチドYレセプターに結合するレセプター;
(e) 嗅受レセプター-嗅受タイプ1-11、味覚レセプター、臭覚レセプター;
(f) プロスタノイドレセプター、プロスタグランジンE2(EP1-EP4サブタイプ) 、プロスタサイクリン、トロンボキサン;
(g) メタボトロピックサブスタンス-メタボトロピックグルタミングループI-IIIレセプターと結合するレセプター
(h) 光などの物理的刺激に応答するレセプターおよび味覚、嗅覚などの化学的刺激に応答するレセプター;
(i) オーファンGPCR-レセプターの自然なリガンドは未定義。
【0029】
本発明と組み合わせたICAST(tm)技術の使用は、GPCRスクリーニングプロセスに多くの利点を提供するものである。それらには、細胞以下のすべての部分-膜、サイトソル、核とのタンパク質相互作用を監視する能力;タンパク質の過剰発現の必要なく、生理学的により関連の深いモデルを達成する能力;レセプター結合のための機能的アッセイを達成し、高い情報内容を提供する能力、が含まれる。
【図面の簡単な説明】
【0030】
【図1】β2アドレナリン受容体(β2AR)およびβ-アレスチン-2(βArr2)のC2クローン中細胞発現レベルβ-ガラクトシダーゼ(β-gal)融合タンパク質の定量は、標準的なELISAアッセイにより、抗β-gal抗体と精製したβ-galタンパク質の滴定曲線を用いて行った。図1Aでは、(発現ベクターpICAST ALC中の)β2AR-βgalΔαクローン発現レベルを示している。図1Bには、クローン9-3、-7、-9、-10、-19、-24の発現ベクターpICAST OMC4あるいはクローン12-4、-9、-16、-18、-22、-24の発現ベクターpICAST OMN4中のβArr2-βgalΔωの発現レベルを示している。
【図2】β2AR受容体の活性は、アゴニストで刺激されるcAMP産生により測定した。pICAST ALC β2AR(クローン5)を発現したC2細胞あるいは親細胞には、濃度を増加させながら(-)イソプロテレノールと0.1mM IBMX処理を行った。cAMPレベルの量はpmol/wellとして示した。
【図3】活性化β2AR受容体とアレスチンの相互作用は、β-ガラクトシダーゼの相補性により測定することができる。図3Aには、(発現ベクターpICAST ALCおよびpICAST OMC、同一のコロニーから単離されたクローン(43-1、43-2、43-7、43-8)中の)β2AR-βgalΔαを発現したC2(発現ベクターpICAST ALC中のβ2ARのみ、あるいはβ2AR-βgalΔαおよびβArr2-βgalΔωを共発現し、二重に形質導入されたC2プール)について、アゴニストである(-)イソプロテレノール刺激に反応して活性化されたβ-ガラクトシダーゼの時間経過を示している。図3Bには、(発現ベクターpICAST ALC中の)β2AR-βgalΔαのみを発現したC2細胞、および(発現ベクターpICAST ALCおよびpICAST OMC中の)β2AR-βgalΔαおよびβArr1-βgalΔωを共発現したC2クローンについて、アゴニストである(-)イソプロテレノール刺激に反応して活性化されたβ-ガラクトシダーゼの時間経過を示している。
【図4】β2ARとアレスチンの相互作用に対するアゴニスト用量反応は、β-ガラクトシダーゼの相補性により測定することができる。図4Aには、β2AR-βgalΔαおよびβArr2-βgalΔω融合体を共発現したC2細胞中の、アゴニスト(-)イソプロテレノールおよびプロカテロールに対する用量反応を示している。図4Bには、β2AR-βgalΔαおよびβArr1-βgalΔω融合体を共発現したC2細胞中の、アゴニスト(-)イソプロテレノールおよびプロカテロールに対する用量反応を示している。
【図5】受容体活性のアンタゴニストによる抑制は、β2AR-βgalΔαおよびβArr-βgalΔωを共発現した細胞中のβ-ガラクトシダーゼの相補性により測定することができる。図5Aには、アゴニスト(-)イソプロテレノールをインキュベート後、β2AR-βgalΔαおよびβArr2-βgalΔω融合体を共発現したC2クローン中の、アドレナリン拮抗薬ICI-118,551およびプロプラノロールによるβ-ガラクトシダーゼの特異的抑制を示している。図5Bには、アゴニスト(-)イソプロテレノール存在下、β2AR-βgalΔαおよびβArr1-βgalΔω融合体を共発現したC2クローン中の、アドレナリン拮抗薬ICI-118,551およびプロプラノロールによるβ-ガラクトシダーゼの特異的抑制を示している。
【図6】アデノシン受容体A2aを発現したC2細胞は、アゴニスト(CGS-21680)の処理に反応してcAMPを誘導することが示されている。A2aR-βgalΔαおよびβArr1-βgalΔωを共発現したC2親細胞とC2細胞は、プールあるいは特定のクローン(47-2, 47-13)として用い、アゴニストで誘導されるcAMP反応(pmol/well)を測定した。
【図7】ドパミン受容体D1(D1-βgalΔα)およびβ-アレスチン-2(βArr2-βgalΔω)を共発現したC2細胞のアゴニストで刺激されたcAMP反応。このアッセイでは、βArr2-βgalΔω(Arr2のみ)を発現したクローンをネガティブコントロールとして用いた。βArr2-βgalΔωおよびDl-βgalΔαを発現した細胞は、アゴニスト処理(3-ヒドロキシトラミン塩酸塩、3μM)に反応した。D1(PIC2)あるいはD1(PIC3)はそれぞれ、発現ベクターpICAST ALC2あるいはpICAST ALC4でD1を発現した。
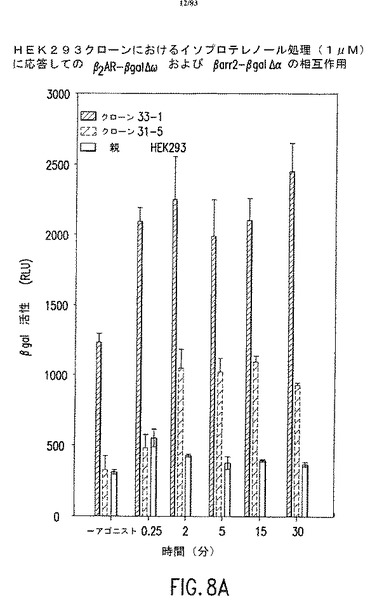
【図8】種々の哺乳類細胞株を用いて、GPCRとアレスチンの相互作用をモニターするための、安定した細胞を作成することができる。図8A、図8B、図8Cには、例として、β-ガラクトシダーゼ変異株のアドレナリン受容体β2ARおよびアレスチン融合タンパク質を共発現した、HEK 293、CHO、およびCHW細胞株を示している。β-ガラクトシダーゼ活性を利用し、アゴニストにより誘導されたβ2ARとアレスチンタンパク質の相互作用をモニターした。
【図9】βgalの相補性を用いて、β2アドレナリン受容体のホモダイマー形成をモニターすることができる。図9Aには、β2AR-βgalΔαおよびβ2AR-βgalΔωを共発現したHEK 293中のβ-ガラクトシダーゼ活性を示している。図9Bには、β2AR-βgalΔαおよびβ2AR-βgalΔωを共発現したHEK 293クローン中のアゴニスト(-)イソプロテレノールに対するcAMPの反応を示している。このアッセイでは、HEK293親細胞をネガティブコントロールとして含めている。
【図10A】pICAST ALC:標的タンパク質のC末端に融合させてβ-galΔαを発現させるためのベクター。このベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、NeoR、ネオマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図10B】pICAST ALCのヌクレオチド配列。
【図11A】pICAST ALN:標的タンパク質のN末端に融合させてβ-galΔαを発現させるためのベクター。このベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、NeoR、ネオマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColE1ori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図11B】pICAST ALNのヌクレオチド配列。
【図12A】pICAST OMC:標的タンパク質のC末端に融合させてβ-galΔωを発現させるためのベクター。このベクターは、MCS、β-galΔω骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、Hygro、ハイグロマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図12B】pICAST OMCのヌクレオチド配列。
【図13A】pICAST OMN:標的タンパク質のN末端に融合させてβ-galΔωを発現させるためのベクター。このベクターは、MCS、β-galΔω骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、Hygro、ハイグロマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図13B】pICAST OMNのヌクレオチド配列。
【図14】pICAST ALC βArr2:β―アレスチン―2のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトβ―アレスチン―2(遺伝子バンク受入番号:NM_004313)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図15】pICAST OMC βArr2:β―アレスチン―2のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトβ―アレスチン―2(遺伝子バンク受入番号:NM_004313)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図16】pICAST ALC βArr1:β―アレスチン―1のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトβ―アレスチン―1(遺伝子バンク受入番号:NM_004041)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図17】pICAST OMC βArr1:β―アレスチン―1のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトβ―アレスチン―1(遺伝子バンク受入番号:NM_004041)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図18】pICAST ALC β2AR:β2アドレナリン受容体のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトβ2アドレナリン受容体(遺伝子バンク受入番号:NM_000024)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図19】pICAST OMC β2AR:β2アドレナリン受容体のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトβ2アドレナリン受容体(遺伝子バンク受入番号:NM_000024)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図20】pICAST ALC A2aR:アデノシン2a受容体のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトアデノシン2a受容体(遺伝子バンク受入番号:NM_000675)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図21】pICAST OMC A2aR:アデノシン2a受容体のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトアデノシン2a受容体(遺伝子バンク受入番号:NM_000675)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図22】pICAST ALC D1:ドパミンD1受容体のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトドパミンD1受容体(遺伝子バンク受入番号:X58987)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図23】本発明の方法で用いられている相補技術の図解。図23では、対応する融合パートナー、GPCRおよびβ-アレスチンが相互作用すると活性化する、2つの不活性な変異した受容体酵素を示している。
【図24】β-galΔαと融合させ、セロトニン/トレオニンのアミノ酸配列を挿入してGPCRを発現させるためのベクター。既知あるいはオーファンGPCRのオープン・リーディング・フレームを設計し、SSS(セロトニン、セロトニン、セロトニン)などのセロトニン/トレオニン配列をC末端に追加している。設計したGPCRは、pICAST ALCベクターのβ-galΔα骨格にクローニングしている。pICAST ALCベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、NeoR、ネオマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図25】β-galΔωと融合させ、変異型(R170E)β-アレスチン2を発現させるためのベクター。アルギニン170をグルタミン酸に変換するポイントミューテーションを行うため、β-アレスチン2のオープン・リーディング・フレームを設計している。変異型β-アレスチン2は、pICAST OMCベクターのβ-galΔω骨格中にクローニングしている。pICAST OMCベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、Hygro、ハイグロマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coli中の成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図26】リン酸化非感受性変異株R170E β-アレスチン2Δωが、アゴニストの活性化に反応してβ2ARΔαに結合。β2ARΔα C2親細胞株は、変異型R170E β-アレスチン2Δωと形質導入した。2構造を共発現したクローン個体は、96ウェルプレートに10,000 cells/wellにて播種し、指示された期間、10μM (-)イソプロテレノール、0.3mMアスコルビン酸を処理した。β-ガラクトシダーゼ活性は、Tropix Gal-ScreenTMアッセイ系の基質(Applied Biosystems社)を加えて測定し、発光はTropix IR717TM照度計(Applied Biosystems社)を用いて測定した。処理は3回行った。比較のため、β2ARΔαおよび野生型β-アレスチン2Δωを共発現したクローン細胞株(43-8)を10,000 cells/wellにて播種し、同様のアゴニスト処理を行った。(-)イソプロテレノール処理後の時間(分)はX軸に示され、β-ガラクトシダーゼ活性は相対的な光ユニット(relative light units: RLU)としてY軸に示されている。
【図27】GPCRのダイマー形成は、β-ガラクトシダーゼの相補性により測定した。GPCRのホモ、ヘテロダイマー形成をモニターする上での、本発明の利用を図解している。1つ目のGPCRは1つの酵素補部フラグメントに融合させ、2つ目のGPCRは2つ目の酵素補部フラグメントに融合させている。2つのGPCRの相互作用は、活性酵素複合体を生成する酵素フラグメントの相補性によりモニターしている(β-ガラクトシダーゼ活性など)。GPCRのホモ、ヘテロダイマー形成は、リガンド、アゴニスト、インバースアゴニスト、アンタゴニストの有無に関わらず、モニターすることができる。
【図28】ICASTTM系における、変異型β-ガラクトシダーゼの相補性によるオーファン受容体のリガンドスクリーニング(ligand fishing)。オーファンGPCR用のligand fishingおよびアゴニスト/アンタゴニストスクリーニングに対する、本発明利用の図解。例として、2種類のβ-gal融合タンパク質、GPCRorphan-ΔαおよびアレスチンΔωを発現したテスト細胞に、天然あるいは合成の化合物ライブラリ、組織抽出物、培養細胞のならし培地から採取したサンプルを処理している。処理後にβ-gal活性が上昇したことから、テストサンプル中のリガンドにより、オーファン受容体が活性化されたことが示されている。β-gal活性の上昇値は、β-アレスチンと活性化したGPCRオーファン受容体との相互作用を反映している。従って、テスト受容体の同系リガンドあるいは代理リガンドが示されている。
【発明を実施するための形態】
【0031】
好適な態様の詳細な説明
本発明はGPCR機能と経路を探索する方法を提供する。Gタンパク質-共役スーパーファミリーは、新規受容体としての急速な拡大を続け、cDNAライブラリまたはゲノムDNAの自動配列をとおして発見される。ヒトゲノムには数千のGPCRが存在すると推定されている。クローン化されているのはその一部のみで、リガンドと結び付けられている数はさらに少ない。これら、または新規発見されるオーファン受容体をそれぞれの同族リガンドや生理学的機能と結び付ける方法は、生物学的/生物医学的研究の大きな課題である。オーファン受容体の同定は一般的に個別化されたアッセイと機能に関する推測を必要とする。本発明は、検査GPCRまたはオーファン受容体とβアレスチン間の活性依存タンパク質/タンパク質相互作用を利用した、受容体の活性化の監視によるGPCR機能の探索に関連する。この特定のタンパク質/タンパク質相互作用は、本明細書に開示されている変異体酵素相補技術により測られる。このアッセイシステムは、その他シグナリングイベントの既存の知識の有無に関わらず、実行できるため、必要な推定を限定するものである。これは感度が高く、高速に、簡易に実行できるものであり、これら受容体の大部分が共通のメカニズムにより脱感受されているため、ほとんどすべてのGPCRに適用できる。
【0032】
本発明は、GPCR経路内のタンパク質/タンパク質相互作用監視のための完全なアッセイシステムを提供するものである。本発明は相補技術、ICAST (tm)(Intercistronic Complementation Analysis Screening Technology、1998年4月1日に申請された米国特許係属中申請番号第053,614号で、その全開示は引用により本明細書に編入する)を採用しているICAST(tm) テクノロジーは、着目するタンパク質に融合されたレポーター酵素の2つの変異体の使用に関連する。着目するタンパク質が相互に作用しない場合、レポーター酵素は不活性のままである。着目するタンパク質が相互に作用しない場合、レポーター酵素は不活性状態のままである。本発明の態様によると、βガラクトシダーゼの活性は、化学発光法サブストレートであるGalacton-Star(r)に関連する、Advanced Discovery Sciences(tm)による開発のGalScreen(tm)アッセイシステムで検知が可能である。GalScreen(tm)アッセイシステムおよびGalacton-Star(r)化学発光法サブストレートは米国特許番号第5,851,771号、第5,538,847号、第5,326,882号、第5,145,772、第4,978,614号、第4,931,569号で開示されており、その全開示は引用により本明細書に編入する。本発明は高速、非放射性アッセイフォーマットである、GPCR結合アッセイを含む一連のアッセイを提供するものである。本発明の方法は、緑色蛍光融合タンパク質(GFP)を含む融合としてのGPCRコンポーネントの顕微鏡的画像化や分光法に依存する、米国特許番号第5,891,646号および第6,110,693号、また、前記アンガース(Angers)et al.編集で開示の発明をさらに発展させたものである。米国特許番号第5,891,646号および第6,110,693号とAngers et alでの分光法ベースの技術は、ロースループットおよび完全定量化の不足により制限を受ける。
【0033】
Advanced Discovery Sciences(tm) テクノロジーと本発明のアッセイシステムを組み合わせることにより、ハイスループットスクリーニング(HTS)が可能なGPCR経路探索用高感度細胞ベースアッセイ法が実現される。本発明と併用が可能なAdvanced DiscoverySciences(tm)開発のテクノロジーには、Gal-Screen(tm)(上記記述を参照されたい)アッセイシステムおよびcANP-Screen(tm)イミュノアッセイシステム等がある。cAMP-Screen(tm)イミュノアッセイシステムは、細胞溶解液中のcAMP濃度を超高感度で測定する。 cAMP Screen(tm)アッセイは、超高感度化学発光アルカリフォスタファーゼ(AP)サブストレートCSPD(r) (disodium 3-(4- methoxyspiro {1,2-dioxetane-3,2'-(5'-chloro) tricyclo 3.3. 1.1.3,7}decan-4-yl phenyl phosphate)およびSapphire-II(tm) 発光エンハンサーを利用している。
【0034】
高スループットアッセイのタンパク質/タンパク質相互作用監視に使われるyeast-based-two-hybrid アッセイとは違い、本発明では(1)哺乳動物細胞、植物細胞、大腸菌などの原生細胞、また、酵母、粘菌変形菌(Dictyostelium)昆虫などの無脊椎源の細胞、など種々の細胞を利用でき、(2)例えばヌクレアスなどの限定された細胞限局化ではなく、受容体ターゲット部位の膜における、あるいは、下流ターゲットタンパク質の部位におけるシトソル内の相互作用を検知できる他、(3)転写活性化などの間接的出力に依存しない。従って、本発明はより高い生理学的関連性と、偽陽性の減化を提供するものである。
【0035】
本発明は、発明GPCRアッセイ法の感度を強化するために、前記記載の米国特許出願番号第053,614号開示の態様に改変を加えて開発されている。態様に従うと、本発明はGPCR-アレスチンの相互作用を強化/延長化して検知の信頼性をより高めるため、セリン/スレオニンクラスターの利用を伴うものである。これらクラスターは、これらのシークエンスモチーフを持たないオーファン受容体または既知GPCRに利用できる。このシークエンスを受容体のc-末端に付加し、例えばβガラクトシダーゼ変異体を伴うターゲットタンパク質の融合タンパク質からのβガラクトシダーゼ相補など、GPCRへのアレスチン結合を出力することにより、受容体の活性化が簡易に検知できる。
【0036】
別の態様に従うと、本発明は、特定GRKの活性化によるリン酸化の必要を回避する目的で、活性時にGPCRのc-末端上または細胞間ループ上のアレスチンポイントミューテーションを利用する。態様はi) 特定のGPCRまたはオーファン受容体と同族のGRKが未知であり、また、 ii) 着目する(または検査下の)受容体の特定GRKが、受容体活性化アッセイに使われるホスト細胞内に不存在または低活性である可能性を含む。
【0037】
別の態様に従うと、本発明は、活性化されたGPCRへのアレスチンの結合効果を強化し、GPCR脱感作中、GPCR/アレスチン群を安定化する目的でのスーパーアレスチンの利用を伴う。本態様はGPCRが通常、脱感作後急速に再感作する、ICAST/GPCR態様の信頼性を増すために利用できる。
【実施例】
【0038】
これら方法を各、以下に述べる。
本発明を、限定されない以下の実施例で説明する。
実施例:
本発明のひとつの態様によると、GPCR活性化はリガンド-活性化GPCRへのアレスチンの結合監視を通して測定できる。本アッセイシステムでは、例えばβアドレナリン受容体(β2AR)などのGPCRおよび例えばβアレスチンなどのアレスチンは、例えばβガラクトシダーゼ(β-gal)などのレポーター酵素の変異体を伴う融合タンパク質と同じ細胞内に共発現される。図23で示されている通り、β2ARはβ-gal変異体のΔαを伴う融合タンパク質(β2ARΔα)として、βアレスチンはβ-gal変異体のΔωを伴う融合タンパク質(β-ArrΔω)として発現する。当初、例えばGPCRの膜とアレスチンのシトソルなど、2つの異なる区画内の静止(または非刺激性)細胞内に存在する、これら2つの融合タンパク質は、活性のβガラクトシダーゼ酵素を形成することはできない。かかる細胞がアゴニストまたはリガンドにより治療されると、リガンド居住の活性化受容体は、アレスチンにとっての高親和結合部位となる。活性化GPCR、β2ARΔα、アレスチンβ-ArrΔω間の相互作用がβ-gal変異体相補を誘導する。酵素活性化は、分割に際して比色、蛍光、化学発光(例えばGal-Screen(tm)アッセイシステム等)法により測定可能な生成物を解離する、酵素基質を使って測定できる。
【0039】
実験プロトコル
1. まず始めに、選択可能なレトロウイルスベクターpICAST ALC(図18で説明)およびpICAST OMC(図15で説明)に、β2ARΔαおよびβArr2Δωの発現ベクターを組み込んだ。
2. 第二に、2つの発現体を、C2Cl2心筋細胞、あるいはCOS-7、CHO、A431、HEK 293、CHWなどの他の哺乳類細胞株に形質導入した。抗生物質を選択した後、2種類の融合タンパク質両方を適切なレベルで発現している、安定なクローンを選択した。
3. 最後に、β2ARΔαおよびβArr2Δωの両方を発現している細胞の反応性を、アゴニスト/リガンドで刺激されるβ-ガラクトシダーゼ活性により測定した。3種類の細胞サンプルを、96ウェル培養プレートのウェルに、100マイクロリットル、10,000細胞にて播種した。細胞は24時間培養し、アッセイを行った。アゴニストのアッセイ(図3および4)については、(-)イソプロテレノール、プロカテロール、ドブタミン、L-L-フェニレフリンなど、アゴニストの濃度を変えて、60分、37℃にて細胞を処理した。誘導したβ-ガラクトシダーゼの活性は、Tropix Gal-ScreenTMアッセイ系の基質(Applied Biosystems社)を加えて測定し、発光はTropix TR717TM照度計(Applied Biosystems社)を用いて測定した。アンタゴニストのアッセイ(図5)については、まず調整したばかりの培地に、血清なし、ICI-118,551あるいはプロプラノロール存在下で10分間細胞をインキュベートし、次に10マイクロモル濃度の(-)イソプロテレノールを加えた。
【0040】
セリン/スレオニンクラスターストラテジー
背景
βアレスチンの構造-機能研究によると、βアレスチンのアミノ末端内の大領域(活性化認識領域と呼ばれる)がGPCRのアゴニスト活性化状況を認識する。βアレスチンにおけるこの領域は、正電荷を帯びたリン酸化認識領域(phosphorylation-recognition domain) と呼ばれる小領域(ネット電荷+7で約20個のアミノ酸)も含み、これはGRK-リン酸化されたGPCRのカルボキシル末端と相互作用しているように見られる。
【0041】
GPCRはβアレスチンへの親和性により2つのクラスに分けられる。オークリー(Oakley) et al., 編集「クラスリン仲介飲食作用中のβアレスチンのGタンパク質-共役受容体との結合による受容体再感作のプロフィールの検知(Association of β-Arrestin with G Protein-Coupled Receptors During Clathrin-Mediated Endocytosis Dictates the Profile of Receptor Resensitization.)」J. Biol. Chem.誌、274(45):32248-32257 (1999年)。この分類の根底にある分子決定因子が、受容体のカルボキシル端子に位置する特定セリンまたはスレオニン残基内に存在しているようである。カルボキシル末端に、離散したセリンおよびスレオニン残基のみしか備えていない受容体クラスは、低い親和性でβアレスチンと結合し、内在化時にβアレスチンから解離するのに対し、カルボキシル端子内のセリン/スレオニンクラスター(連続する3つ、または、4つのうち3つの配列を持つセリンまたはスレオニン残基として定義される)を含む受容体クラスは、活性化およびリン酸化時、高い親和性でβアレスチンと結合し、受容体の内在化後もβアレスチンと結合を維持する。バソプレシンV2レセプター(Oakley, et al)、ニューロテンシンレセプター1、アンジオテンシンIIレセプタータイプ1A(ツァン(Zhang) et al編集「Gタンパク質-共役受容体/βアレスチンエンドサイトーシス群の細胞輸送(Cellular Trafficking of G Protein- Coupled Receptor/β-Arrestin Endocytic Complexes.)」J. Biol. Chem.誌、274(16):10999-11006 (1999年))など、カルボキシル末端内にこのようなセリン/スレオニンクラスターを1つあるいは複数持ついくつかの既知のGPCRは、βアレスチンを高い親和性で結合することが示されている。
【0042】
実施例
本発明の態様によると、セリン/スレオニンクラスターストラテジーは、独自にこういった構造的モチーフを持たないオーファン受容体のスクリーニングアッセイ法の簡易化に利用できる。オーファン受容体は連鎖配列により簡単に分類化ができる。セリン/スレオニンクラスターを備えないオーファン受容体を、それぞれ、受容体のカルボキシル末端に1つあるいは複数のセリン/スレオニンクラスターを誘導すべく改変された発現ベクターにクローニングする。(図24)セリン/スレオニンクラスターが、強力なイオン相互作用を通して、活性化およびリン酸化された受容体(負電荷)とβアレスチン(リン酸化認識領域中に正電荷)との間の受容体活性化依存相互作用を強化させる。オーファン受容体末端の修飾により、このように、受容体活性化の検知信頼性が強化される。
【0043】
実験プロトコル
1. まず始めに、セロトニン/トレオニンクラスターのない、オーファン受容体のオープン・リーディング・フレーム(ORF)を、図10Aで説明したpICAST ALCなどの修飾した発現ベクターにクローニングする。この修飾したpICAST ALCには、1つ以上のセロトニン/トレオニンクラスター(例えばSSSあるいはSST)のコード配列が、オーファン受容体のORF挿入部位下流に位置している(図24)。
2. 第二に、キメラオーファン受容体であるORForphan-(SSS)n-Δαを、図15で説明したβArr2Δωなどのβ-アレスチンキメラを持つ哺乳類細胞に共発現させる。
3. 第三に、この細胞にアゴニストまたはリガンドを処理し、リン酸化セリンクラスターを持つ活性型受容体を、β-gal相補性の値が強いシグナルを示す高親和性β-アレスチンに結合させる。
【0044】
このアッセイは、オーファン受容体の機能活性化を測定できる高感度の方法であり、オーファン受容体の天然リガンドあるいは代理リガンドのスクリーニングに用いることができる。このプロセスは、新しいGPCRのデオーファニング(de-orphaning)あるいはターゲット発見と呼ばれる(図28)。さらに、このアッセイは、GPCRのリード発見に用いる潜在的なアゴニストおよびアンタゴニストのスクリーニングにも有用である。
【0045】
GPCRリン酸化の存在時、非存在時における強化アレスチン結合
背景
これまでGタンパク質-共役受容体キナーゼ(GRK)の6つの異なるクラスが特定され、さらにこれら各クラスが複数のmultiple variantとして発現することが報告されている。クループニック(Krupnick) et al.編集「Gタンパク質-共役受容体調節におけるレセプタキナーゼおよびアレスチンの役割(The role of receptor kinases and arrestins in G protein-coupled receptor regulation.)」Ann. Rev. Pharmacol. Toxicol.誌、38:289-319 (1998)。細胞株は多くが種々のGRKを発現するが、あるGPCRのリン酸化に必要な特定のGRKが、組換GPCRおよびアレスチン発現に使われる細胞株に、常に存在するとは限らない。これは、同族GRKが未知である可能性が高い、オーファン受容体を使う態様で特に問題となる。その他の場合では、組換え発現作業に使われる細胞株が必要なGRKを備えていたとしても、GRKの発現率が低い可能性がある。このような危険性を回避するために、活性化GPCRに特異的に結合されているがGRKリン酸化の必要のない、遺伝子的に改変されたアレスチンが採用されている。
【0046】
アレスチン上の変異誘発に関する研究は、リン酸化認識領域におけるポイントミューテーション、特にArg175(視覚系アレスチンの)を逆の電荷を帯びたグルタミンなどの残基へと変換する変異(R175E変異)は、GPCRに特異的に結合するアレスチンを生むが、リン酸化の必要なくそれを行うことを示している。
【0047】
多くの研究により、アレスチンがGPCRへの親和性の低い非活性状態中に存在するとの仮説を導いている。GPCRの活性化とリン酸化の両方がひとたび起こると、GPCR C末端のリン酸化領域がアレスチンのリン酸化認識領域と相互作用し、アレスチンの構造変化、引いては、活性化認識領域の提示と活性化/リン酸化された受容体への結合が導かれる。ヴィシュニヴェツキー(Vishnivetskiy) et al.編集「ロドプシンのリン酸化過程に対するアレスチンの応答(How does arrestin respond to the phosphorylated state of rhodopsin?)」 J. Biol. Chem.誌 274(17):11451-11454 (1999年);グレヴィッチ(Gurevich) et al.編集「Gタンパク質-共役受容体とのアレスチンの相互作用:野生/変異型アレスチンとロドプシンβ2アドレナリン/m2ムスカリン様アセチルコリン受容体の直接結合研究(Arrestin interactions with G protein-coupled receptors. Direct binding studies of wild-type and mutant arrestins with rhodopsin, beta 2-adrenergic and m2 muscarinic cholinergic receptors.)」J. Biol. Chem.誌270(2):720-731, (1995年);グレヴィッチ(Gurevich) et al.編集「視覚系アレスチンによるリン酸化認識と高親和性結合部位へのアレスチンの転位メカニズム(Mechanism of phosphorylation-recognition by visual arrestin and the transition of arrestin into a high affinity binding site.)」Mol. Pharmacol.誌51(l):161-169 (1997年);コーヴァー(Kovoor) et al.編集「細胞内に構成活性性を有するリン酸化非依存性βアレスチン変異体のターゲット構成(Targeted construction of phosphorylation-independent beta-arrestin mutants with constitutive activity in cells.)」J. Biol. Chem.誌274(11):6831-6834 (1999年)。結論的には、非活性化、非活性化/リン酸化、活性化/非リン酸化、リン酸化された視覚系/非視覚系GPCRとの単一変異株、二重変異株、欠失、キメラアレスチンの結合研究のすべてが本モデルを論証してるのである。
【0048】
実施例
変異レポータータンパク質に融合されたアレスチンのリン酸化非感作変異体を、リン酸化非依存の様態で活性化されたGPCRと結合すべく産生することが可能である。この考え方の証明として、本発明に従い、βアレスチン2、R170E βアレスチン2のポイントミューテーションが産生され、β2AR との相互作用が分析された。
【0049】
実験プロトコル:
1) まず始めに、Arg 170をGluに変換するなど、β-アレスチン2を変異させた。この変異は、視覚系アレスチンのR175E変異と同等である。変異型β-アレスチン2のオープン・リーディング・フレームを、pICAST OMC発現ベクターのΔω-β-ガラクトシダーゼフレームにクローニングし、修飾発現ベクターR170E β-アレスチン2を作成した(図25)。
2) 第二に、R170E β-アレスチン2発現体をC2C12心筋細胞株に形質導入した。C2C12心筋細胞株は、図18、米国出願シリアル番号09/654,499に説明した通り、Δα-β-ガラクトシダーゼに融合させてβ2ARを発現させるために設計したものである。抗生物質を選択した後、2種類の融合タンパク質両方を発現しているクローンを採取した。
3)最後に、R170E β-アレスチン2Δωおよびβ2ARΔαの両方を発現しているこの細胞集団の反応性を、図26に示されている通り、アゴニスト/リガンドで刺激されるβ-ガラクトシダーゼ活性により測定した。β2ARΔαおよび野生型β-アレスチン2Δωを共発現しているC2Cl2クローン43-8(図26)を、対照として用いた。3種類の細胞サンプルを、96ウェル培養プレートのウェルに、100マイクロリットル、10,000細胞にて播種した。細胞は24時間培養し、アッセイを行った。図26のアゴニストアッセイについては、細胞を0.3mMアスコルビン酸で安定化させた10μm (-)イソプロテレノールにより、37℃、0、5、10、15、30、45あるいは60分間、処理した。誘導したβ-ガラクトシダーゼの活性は、Tropix Gal-ScreenTMアッセイ系の基質(Applied Biosystems社)を加えて測定し、発光はTropix TR717TM照度計(Applied Biosystems社)を用いて測定した。図26に示した通り、変異型アレスチンは、アゴニスト依存的にβ2ARと相互作用し、野生型アレスチンと同等であった。
4) リン酸化非感受性のアレスチンを用いた応用に拡張するため、C2C12、CHO、HEK 293などの細胞株をR170E β-アレスチン2Δω構造を発現させる開発を行う。アゴニストおよびアンタゴニストスクリーニング用の細胞株を開発するため、これらの細胞株を用いて、Δα-β-ガラクトシダーゼに融合させて、オーファンあるいは既知のGPCRを形質導入することができる。
【0050】
スーパーアレスチンの開発
背景
アレスチン経路によるGPCRシグナリングのアティニュエーションは、細胞または微生物が刺激に対し過剰に反応を防ぐ作用をする。同時に、アレスチン経路はGPCRを、一時的に非活性化し、すばやく再感作して新規刺激に対する感作を可能にするために再循環する作用も持つ。ダウンレギュレーション過程では、アレスチンとエンドサイトーシスとを結合しながら、受容体のリン酸化が行われる。脱感作された受容体のエンドサイトーシスに続き、受容体はライソソームにディグレードするか、または、再感作され、膜に戻される。再感作により、アレスチンが受容体から解離、リン酸化が行われ、膜へと戻る。活性化に伴いGPCRがたどる実際の経路は、生物学的機能と生体のニーズに拠る。ダウンレギュレーション経路の必要の可能性を含むこれら経路の多様性により、活性化したGPCRへのアレスチン親和性は受容体により異なる。そのため、自然が提供するものよりも高い、活性化されたGPCRへの親和性と結合活性を備えたスーパーアレスチンの精製は極めて有益である。
【0051】
アレスチンの変異、欠失、キメラ研究は、これまで、GPCRリン酸化状態に応答する分子の調節スイッチの解明に集中してきたが、アレスチンの改変組換えが活性化リン酸化されたGPCRへの分子内の結合強化をもたらしている。Arg175の、ヒスチジン、タイロシン、フェニルアラニン、スレオニンへの変換は、野生型アレスチンまたはR175Eアレスチンよりも、リン酸化、活性化されたロドプシンへの相当量に高い結合量を導いているが、これら変異体は活性化、非リン酸化された受容体には低い結合となっている。(ビレヴィッチ(Gurevich) et al.編集(1997年))。さらに、バリン170のアラニンへの変換は、R175E変異の構成効果を増加させているが、野生型アレスチンの活性化/リン酸化ロドプシンとの相互作用量もほとんど倍化させている。(グレヴィッチ(Gurevich) et al.編集(1997年))。
【0052】
βアレスチン1のアミノ酸382での切形は、R169E(アレスチンR175Eと同等)と野生型βアレスチンのいずれもの、活性化または活性化/リン酸化受容体への結合をそれぞれ強化していることが報告されている。(コーヴァー(Kovoor) et al.編集)。視覚系アレスチンの機能領域がβアレスチン1のそれとスワップされている、キメラアレスチンの場合、活性化/リン酸化されたGPCRへの結合親和性が変化することが報告されている。(グレヴィッチ(Gurevich) et al.編集(1995年))。これらキメラのうち、視覚系アレスチン極性N末端を伴うβアレスチン1などのいくつかのキメラが、野生型βアレスチン1よりも、リン酸化/活性化されたGPCRに特異的に結合を増加させていることを示している。(ブレクイヴィッチ(Gurevich) et al編集 (1995年))。前記に説明の、活性化GPCRへのアレスチン親和性を強化する改変は、アレスチン/GPCR群が安定化および/または長命化することから、リン酸化か非リン酸化かに関わらず、βガラクトシダーゼ活性のノイズへのシグナルも強化させる可能性がある。活性化されたGPCRとより高い親和性を持つ変異体アレスチンを利用することにより、GPCRターゲットの発明技術の改善が、受容体/リガンドの生物学を妥協させることなく、実現される。
【0053】
さらに、この「スーパーアレスチン」アプローチとアレスチンポイントミューテーションの利用とを組み合わせて、GRKの必要の有無に関わらずノイズへのシグナルを強化させることもできる。
【0054】
実施例
変異レポーターと融合されたアレスチン変異体を、活性化GPCRへのアレスチンの結合強化目的のため産生、結果、検知の感度を強化する。
【0055】
実験プロトコル
1) まず始めに、変異型β-アレスチン2構造に、R170E/T/Y/あるいはH、V165Aを組み込み、視覚系アレスチンのaa. 1-43とa.a. 1-47を置換、あるいはC末端を欠失させるか、これらの変質を組み合わせて行う。図25に示したR170E β-アレスチン2変異のクローニングと同様に、変異型β-アレスチン2のオープン・リーディング・フレームを、pICAST OMC発現ベクターのΔω-β-ガラクトシダーゼ骨格にクローニングする。
2) 第二に、変異型発現体を、Δα-β-ガラクトシダーゼに融合させるとβ2ARを発現するように設計したC2Cl2心筋細胞株に形質導入する。抗生物質を選択した後、2種類の融合タンパク質両方を発現しているクローン個体を採取する。野生型およびR170E β-アレスチン2の構造を形質導入し、対照の基準クローンを作成する。
3) 第三に、β-アレスチン2Δω(変異型あるいは野生型)およびβ2ARΔαの両方を発現した細胞集団の反応性を、アゴニスト/リガンドで刺激されるβ-ガラクトシダーゼ活性により測定する。
4) 次に、野生型β-アレスチン2Δωと比較し、アゴニストアッセイでノイズ比よりも有意に高いシグナルを示した変異型(スーパー)β-アレスチン2Δω構造を選択する。この構造を用い、既知あるいはオーファンGPCRに形質導入することができ、「スーパー」β-アレスチン2Δωを発現している安定な細胞株を開発する。スーパーβ-アレスチン2Δωを用い、ICAST/GPCRの応用に関し、ノイズに対するシグナルを増加させることができ、リードおよびリガンド発見のためのスクリーニング効果を改善することができる。
【0056】
スーパーアレスチンを用い、GPCRの脱感作中、活性型GPCRおよび安定したGPCR/アレスチン複合体へのアレスチンの結合効率を向上させる。この応用技術を用いることで、通常はGPCRが脱感作後、急速に再感作されるケースで、ICAST/GPCR技術の信頼性を高めることができる。
【0057】
本発明のアッセイ、応用と調整法は、包括的にも、特定の例を用いても説明してある。例は、限定的なものとして用いているわけではない。他の代替アイデンティティ、性質、アッセイは、発明力を駆使しなくとも、この分野における普通の技術の中で見出されることであろう。後述する特許請求の範囲に関する詳述で除外されていない限り、そのような修正は本発明の範囲内に収まるものである。
【図1A】

【図1B】
【技術分野】
【0001】
本出願は、2000年2月7日に出願された米国仮出願番号第60/180,669号から利益を請求する、2000年9月1日に出願された米国出願番号第09/654,499号の一部継続である。前述の特許出願の全開示は引用により本明細書に編入する。
【0002】
発明の分野
本発明はGタンパク質-共役受容体(GPCR)の活性を検知する手段に関連し、GPCR活性のアッセイ法、GPCRリガンド、アゴニストおよび/またはアンタゴニストのスクリーニング法、オーファンGPCRの自然なリガンドおよび代理リガンドのスクリーニング法、GPCR制御過程の化合物と相互に作用する化合物のスクリーニング法を提供する。
【背景技術】
【0003】
発明の背景
細胞外の多くのシグナル活動はGタンパク質-共役受容体(GPCR)とグアニンヌクレオチド-結合制御タンパク質(Gタンパク質)の相互作用により仲介される。Gタンパク質-仲介信号系は哺乳類や酵母など、多くの多細胞生物において確認されている。GPCRは分岐アミノ酸鎖を有するタンパク質の大きなスーパーファミリーであるが、共通の構造的特徴、特に7つの膜貫通領域(蛇皮状)の存在を共有する。GPCRは細胞外シグナル(神経伝達物質、ホルモン、臭気、光、その他)に応答する。個々のGPCRタイプは特定のシグナル伝達経路を活性化、少なくとも10個の異なる信号伝達経路がGPCRにより活性化されることが知られている。例えばβ2アドレナリン受容体(β2AR)は、原型哺乳GPCRである。β2AR受容体はアゴニスト結合に応答して、Gタンパク質(Gs)を活性化し、それが呼応してアデニレートシクラーゼ活性を刺激し、細胞内に環状アデノシン1リン酸(cAMP)を増加生成せしめる。
【0004】
GPCRの刺激によって生じたシグナル経路と最終的な細胞応答は、特定の受容体の結合対象であるGタンパク質の特定クラスによる。(「Gタンパク質シグナルの多くの顔(The Many Faces of G-Protein Signaling)」、ハム著、ジェイ・バイオル・ケム(J Biol. Chem.)誌1998年273:669-672を参照のこと)
【0005】
例えば、Gタンパク質のGsクラスとの結合は、cAMPの産生と、タンパク質キナーゼAおよびC経路の活性化を誘発し、そこにおけるGタンパク質のGiクラスとの結合がcAMPを抑制する。カルシウム、ホスホリパーゼC、ホスファチジルイノシトール3などのほかのセカンドメッセンジャーの利用も可能である。ゆえに、GPCRシグナルイベントは、これらのセカンドメッセンジャープロダクトの定量化により大部分測定されることとなる。
【0006】
持続的刺激に対する応答の減少は、よく知られている生物学的現象である。多様なGPCRによるシグナルは同一の2段階のメカニズムにより中断されると信じられている。活性化された受容体は先ず、GPCRキナーゼ(GRK)によりリン酸化される。アレスチンタンパク質は活性化およびリン酸化された受容体に結合し、その結果Gタンパク質の相互作用を遮断する。この過程は一般的に「脱感作(desensitization)」と呼ばれ、機能的に多様なGPCRを表すひとつの一般的メカニズムである。アレスチンはまた、異なるGPCRの間でヘテロジニアスの過程である、GPCR内在化および脱感作の抑制における役割を果たす。(ジェイ・バイオル・ケム(J Biol. Chem.)誌1999年274:32248-32257、オークリー(Oakley)他著を参照のこと。)内在化および再感作の過程におけるアレスチンとGPCRの間の相互作用は、特定のGPCRのカルボキシル末端内の特定の配列モティーフにより決定される。カルボキシル末端に3つのセリンまたはスレオニン残基クラスタを持つGPCRのサブセットのみが、リガンド刺激の後に飲食小胞内にアレスチンとともに共輸送することがわかっている。GPCRの脱感作に関わる受容体キナーゼとアレスチンの数はかなり限られている。
【0007】
GPCR生理学の一般的特徴は、受容体リン酸化、飲食、脱リン酸化の過程を通しての受容体の脱感作とリサイクルである。(「Gタンパク質-共役受容体の調節:Gタンパク質-共役受容体キナーゼとアレスチンの役割(G-protein-coupled receptor regulation: role of G-protein-coupled receptor kinases and arrestins)」ファーグソン(Ferguson)et. al. 編集、Can. J. Physiol Pharmacol誌1996年74:1095-1110を参照のこと。)リガンドで占められるGPCRは、セリン/スレオニンキナーゼの2つのファミリー、Gタンパク質-共役受容体キナーゼ(GRK)、タンパク質イドナーゼAやタンパク質キナーゼCなどの、セカンドメッセンジャー依存のタンパク質キナーゼによりリン酸化できる。 キナーゼのいずれかのクラスによるリン酸化は、対応するGタンパク質からの解離により受容体を抑制する作用をする。GRK-リン酸化はまた、受容体の細胞質領域を結合し、受容体の飲食小胞へのクラスタ化を促すアレスチンと呼ばれるタンパク質の一クラスを漸増することにより受容体を抑制する役割も果たす。受容体が一度飲食されると、リソソームに分解されるか、あるいは、リン酸化され、完全機能の受容体として形質膜に再循環される。
【0008】
アレスチンタンパク質の活性化受容体への結合は、ロドプシンからβ2AR、さらに、ニューロテンシン受容体までの様々なGPCRの一般的現象として発表されている。(「Gタンパク質-共役受容体活性検知のためのβアレスチン/緑色蛍光融合タンパク質(GFP)バイオセンサー(A β arrestin/Green Fluorescent Fusion Protein Biosensor for Detecting G-Protein-Coupled Receptor Activation)」バラク(Barak)et. al. 編集、ジェイ・バイオル・ケム(J Biol. Chem.)誌1997年272:27497-500を参照のこと。)その結果、特定GPCRとアレスチンの相互作用の監視は、GPCR活性測定の一般的ツールとして利用できる。同様に、単一Gタンパク質とGRKも様々な受容体と組み合わせることができ(ハム(Hamm)他著(1998年)および「Gタンパク質-共役受容体キナーゼ(G-Protein-Coupled Receptor Kinases)」、ピッチャー(Pitcher)他著、Annu. Rev. Biochem誌1998年67:653-92を参照のこと)、これらタンパク質/タンパク質相互作用も受容体の活性状態を決定するために監視が可能である。
【0009】
GPCRは、血管拡張、心拍数、気管支拡張、内分泌、腸蠕動を含む生命生理学的応答を調節することから、今日使われている多くの治療薬がGPCRをターゲットとしている。(例としてレフコヴィッツ(Lefkowitz)他著、Annu. Rev. Biochem.誌 52:159 (1983年)を参照のこと。)これら治療薬にはこの受容体のリガンドを模倣したものもある。その他の薬剤は、受容体の自発的活性から発生した疾患の場合、受容体を拮抗する作用を持つ。
ヒトゲノムプロジェクトなどの努力は、生理学的役割やリガンドが未知の、新たなGPCR(オーファン受容体)を発見することである。ヒトゲノムには数千のGPCRが存在すると推定されている。
【0010】
刺激薬に対する細胞内活動の監視には、例えば酵素結合免疫吸着検定法(ELISA)、フロレッスンスイメージングプレートリーダー検定法(FLIPRtm、モラキュラー・デバイシーズ社、カリフォルニア州サニーベル)、EVOscreen tm, EVOTEC tm, Evotec Biosystems Gmbh(ドイツ、ハンブルグ)、また、ペンシルバニア州ピッツバーグセロミクス社のCELLOMICS tm 開発技術など、様々な試みがこれまで採用されている。
【0011】
ジャーミノ(Germino)et. al. 編集「インビボタンパク質-タンパク質相互作用のスクリーニング(Screening for in vivo protein-protein interactions)」Natl. Acad. Sci.誌 90(3):933-937 (1993年)は、着目するタンパク質との相互作用を行うタンパク質の単離にインビボ法を開示している。
フィジッキー(Phizicky)他「タンパク質-タンパク質相互作用法:検知および解析方法(Protein-protein interactions: methods for detection and analysis)」Microbiol Rev.誌59(l):94-123 (1995年)は、タンパク質-タンパク質相互作用の研究に使われる生物化学、分子生物学、遺伝法の論評を開示している。
オファーマン他著「Gα15およびGα16が多種の受容体をホスホリパーゼCに結合する(Gα15 and Gα16 Couple a Wide Variety of Receptors to Phospholipase C)」J. Biol. Chem.誌 270(25):15175-15180 (1995年)は、Gα15 およびGα16が多様なGタンパク質-共役受容体により活性化されることを開示している。活性化受容体のGタンパク質個別模様への選択的結合は、正確なシグナルトランスダクションId の達成のための重要な必要条件と考えられている。
バラク(Barak)他著「Gタンパク質-共役受容体検知のためのβアレスチン/緑色蛍光融合タンパク質(GFP)(A β-arrestin/Green Fluorescent Protein Biosensor for Detecting G Protein-coupled Receptor Activation.)」J. Biol. Chem.誌272(44):27497--27500 (1997年)と、米国特許番号第5,891,646号および第6,110,693号は、光学機器を使ってのGPCRの刺激によるタンパク質転座の造影へのβアレスチン/緑色蛍光融合タンパク質(GFP)の利用を開示している。
【発明の概要】
【発明が解決しようとする課題】
【0012】
上記に参照された研究にはそれぞれ以下の例にような欠点がある。
・従来技術の方法では、タンパク質の過剰発現が必要であり、人為結果を招いたり、細胞調節器の均衡が崩れるなどの恐れがある。
・従来の可視化または造影アッセイ法ではスループットが低く、完全な定量化に欠く。従って、スループットの高い薬理/運動学的アッセイ法には不適である。
さらに、従来技術のアッセイ法では、GPCRの隔離を必要とし、細胞内でのGPCRの観察ができない。そのため、GPCR機能の監視のためには方法を改善する必要があった。
【課題を解決するための手段】
【0013】
発明の要約
本発明は米国出願連続番号第09/654,499号における開示に変更を提供するものである。特に本発明は、アッセイ法の感度をさらに強化する下記の発明の側面を変更する方向付けを持つ。変更は、GPCRがリン酸化か非リン酸化に関わらず活性化されたGPCRへの結合を強化する、遺伝的変異されたアレスチンの利用、および、独自の構造的モチーフを有しないオーファン受容体のスクリーニングアッセイを簡易化するセリン/スレオニンクラスター戦略の利用、および、さらに強化された検知を達成するための上記変更をの併用を含む。
【0014】
本発明の最初の側面はGPCR機能を受容体活性化の箇所で近位的に監視し、従って、より少数の競合メカニズムにより、薬剤発見目的に多くの情報を提供せしめる方法である。GPCRの活性化は、受容体と、アレスチン、Gタンパク質、GRK、または、その他のキナーゼなどの、受容体への結合が受容体のアゴニスト居住に依存する、調整化合物の相互作用の出力により測定される。本発明は、変異レポーター酵素の相補によるタンパク質-タンパク質相互作用の検知に関するものである。
活性化GPCRへのアレスチンの結合は、これまで研究されているほとんどのGPCRで実証されている脱感作の第一段階における共通の過程である。変異酵素相補検定(ICASTなど)を通してのGPCRのアレスチンとの相互作用の測定は、多様なGPCRとオーファン受容体に適用されるより一般的なアッセイ技術を提供するものである。
【0015】
本発明のさらなる側面は、変異レポーター酵素への融合タンパク質として、例えばムスカリン、アドレナリン、ドーパミン、アンギオテンシン、エンドセリン様のGPCRを発現せしめる検査細胞を提供し、例えばGタンパク質、アレスチン、GRKなど、相補変異レポーター酵素との融合タンパク質として、GPCR経路内でタンパク質を相互作用させることより、検査状況下でGPCR経路活性化を査定する方法である。検査状況下で、検査細胞がターゲットGPCRの既知のアゴニストに提示されると、GPCRの活性化がレポーター酵素の相補により監視される。増加したレポーター酵素活性は、相互作用するそのタンパク質パートナーとの、GPCRの相互作用を反映する。
【0016】
本発明のさらなる側面は、検査アレスチン、例えばβアレスチンなどの存在下でGPCR経路の活性化を査定する方法である。
本発明のさらなる側面は、検査Gタンパク質の存在下でGPCR経路の活性化を査定する方法である。
本発明のさらなる側面は、検査細胞を検査リガンドに提示した状態でのGPCR経路の活性化を査定する方法である。
本発明のさらなる側面は、セカンドレセプターの検査細胞内の共発現におけるGPCR経路の活性化を査定する方法である。セカンドレセプターは同一GPCRまたはオーファン受容体(ホモダイマー)、異なるGPCRまたはオーファン受容体(ヘテロダイマー)か、あるいは、別のタイプのレセプターがあり得る。
本発明のさらなる側面は、オーファンGPCRのリガンドまたはアゴニストのスクリーニング方法である。リガンドまたはアゴニストは自然または合成ライブラリまたは混合に含まれるか、あるいは、物理的刺激があり得る。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質として、オーファンGPCRを発現させ、さらに、例えば、別のβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチンまたはアレスチンの変異体を発現するものが提供されている。受容体活性時におけるアレスチンのオーファンGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は検査化合物に曝され、レポーター酵素活性の増加がリガンドまたはアゴニストの存在を示す。
【0017】
本発明のさらなる側面は、リン酸化または活性化されたGPCRへの結合能力を知るために、着目するタンパク質、例えばアレスチンタンパク質(または、アレスチンタンパク質の変異体)をスクリーニングする法である。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを発現させ、さらに、例えば、別のβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチン(またはアレスチンの変異体)を含むものが提供されている。受容体活性時におけるアレスチンのGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は周知のGPCRアゴニストに曝され、その後レポーター酵素活性が検知される。レポーター酵素活性の増加が、βアレスチン分子がリン酸化または活性化された検査細胞内のGPCRと結合され得ることを示す。
【0018】
本発明のさらなる側面は、特定のGPCRのアゴニストのスクリーニング方法である。アゴニストは自然または合成ライブラリまたは混合に含まれるか、あるいは、物理的刺激があり得る。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを、さらに、例えば、別のβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチンを発現するものが提供されている。受容体活性時におけるアレスチンのGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は検査化合物に曝され、レポーター酵素活性の増加がアゴニストの存在を示す。検査細胞は既知のGPCR、または、既知の多種GPCRを発現するか、または、未知の多種GPCRをGPCR発現することもある。このGPCRは例えば臭気GPCRまたはβAR GPCRともなり得る。
【0019】
本発明のさらなる側面は、GPCRアンタゴニスト活性の検査化合物のスクリーニング方法である。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを、さらに、例えば、もうひとつのβガラクトシダーゼ変異体など相補変異レポーター酵素との融合タンパク質としてアレスチンを発現するものが提供されている。受容体活性時におけるアレスチンのGPCRとの相互作用は、相補されたレポーター酵素の酵素的活性により測られる。検査細胞は検査化合物に曝され、レポーター酵素活性の増加がアゴニストの存在を示す。細胞はの検査細胞およびGPCRアゴニストに曝され、その後レポーター酵素活性が検知される。検査化合物への提示と同時、または、それに続いてアゴニストへの提示が提起されると、検査化合物への提示後のレポーター酵素活性の増加が、検査化合物がGPCRに対するアンタゴニスト活性を有していることを示す。
【0020】
本発明のさらなる側面は、GPCRへのアゴニスト、アンタゴニスト、リガンドの存在をみるための、試料液のスクリーニング方法である。検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素との融合タンパク質としてGPCRを発現し、さらに、例えば、もうひとつのβガラクトシダーゼ変異体など相補変異レポーターとの融合タンパク質としてβアレスチンを含んでいる。検査細胞は試料液に提示され、レポーター酵素活性が検知される。試料液への提示の後に変化したレポーター酵素活性は、試料液が細胞内に発現したGPCRに対しアゴニスト、アンタゴニスト、または、リガンドを含んでいることを示す。
【0021】
本発明のさらなる側面は、GPCRの存在をみるための細胞スクリーニング方法である。この側面に従うと、変異レポーター酵素を含むアレスチン融合タンパク質、および、変異レポーター酵素を含むGPCRの下流信号融合タンパク質がGPCR活性の見地のために採用されている。細胞本側面の変更を、GPCRを含むこれらの細胞の細胞数多性をスクリーニングする方法として用いることもできる。本側面に従うと、レポーター酵素とともに融合タンパク質としてのβアレスチンで構成される共役を含む細胞数多性が提供され、数多細胞をGPCRアゴニストに提示し、レポーター酵素の活性化が検知される。GPCRアゴニストへの提示後のレポーター酵素活性の増加がβアレスチンタンパク質がGPCRに結合していることを示し、その結果、細胞がGPCRアゴニストに応答するGPCRを含んでいることが示される。
【0022】
本発明のさらなる側面は、GPCR仲介のシグナル経路のマッピング方法である。例えば本システムを、c-SRCのβアレスチン1との、GPCR活性化時の相互作用を監視するために利用することも可能である。さらに本システムを、GPCRシグナリング経路と、レセプターチロシンキナーゼやRas/Rafなどのその他経路間のクロストークに関わるタンパク質/タンパク質相互作用を監視するために利用することも可能である。本側面に従うと、検査細胞は、例えばβガラクトシダーゼ変異体など変異レポーター酵素を含むGPCRや関連タンパク質を発現し、さらに、例えば、もうひとつのβガラクトシダーゼ変異体など相補変異レポーター酵素を含む融合タンパク質としての、別の経路からのタンパク質を含むものが提供されている。レポーター酵素の増加がタンパク質/タンパク質相互作用を示す。
【0023】
本発明の更なる側面は、アゴニストまたはアンタゴニスト刺激時の、GPCRのホモダイマーまたはヘテロダイマーの監視方法である。GPCRダイマーが生体活動に重要であることは多くの文献証拠で示されている。(アブダーラ(AbdAlla). et al編集、「AT1受容体ヘテロダイマーが示すGタンパク質活性強化および変容受容体金属イオン封鎖(ATI -receptor heterodimers show enhanced G-protein activation and altered receptor sequestration.)Nature誌 407:94-98 (2000年); ボッカート(Bockaert) et al.編集「Gタンパク質-共役受容体の分子操作:進化成功度(Molecular tinkering of G protein-coupled receptors: an evolutionary success.) EMBO J.誌 18:1723-29 (1999年);ジョーダン(Jordan) et al.編集「Gタンパク質-共役受容体ヘテロダイマーによる受容体機能の調節(G-protein-coupled receptor heterodimerization modulates receptor function.)」 Nature誌, 399:697-700 (1999年) は、2つの非機能性オピオイドレセプター、kappaおよびdeltaがヘテロダイマー化し、機能性レセプターを形成することを示している。ゴードン(Gordon) et al.編集「(Dopamine D2 receptor dimers and receptor blocking peptides.)」Bioch. Biophys. Res. Commun.誌 227:200-204 (1996年)は、ドーパミンレセプターD2におけるモノメリックおよびダイメリックフォームに関連する異なる薬理学的特性を示している。D2レセプターは、スピペロンの選択的ターゲットであるモノマーとして、または、ネモナプリドのターゲットであるダイマーフォームとして存在する。ハーバート(Herbert) et al.編集「β2アドレナリンレセプター膜貫通領域から発生するペプチドによる受容体のダイマー化および活性化の阻害(A peptide derived from a β2-adrenergic receptor transmembrane domain inhibits both receptor dimerization and activation.)」J.B.C.誌 271:16384-92 (1996年)は、アゴニスト刺激により、受容体の二量体状態が安定化し、そこにおいて逆拮抗薬がモノメリックフォームに有利に働くことが発見されていることを示している。実際に、同じ研究が、β2アドレナリンレセプターの第六膜貫通領域に対応するペプチドが、受容体のダイマー化と活性化を阻害したことを示している。さらに、アンガース(Angers) et al.編集「生物発光エネルギー伝達を利用した生細胞内β2アドレナリンレセプターダイマー化の検知(Detection of β-2- adrenergic receptor dimerization in living cells using bioluminescence resonance energy transfer)」Proc. Natl. Acad. Sci.誌USA, 97(7)-.3684-3689は、β2アドレナリンレセプターダイマー化研究のためのβ2アドレナリンレセプター融合タンパク質(ルシフェラーゼに融合したβ2アドレナリンレセプターおよび赤色よりの強化GFPに融合したβ2アドレナリンレセプター)の使用を開示している。
【0024】
細胞性理学および薬理学的意味合いにおけるGPCRダイマー化は、本発明に従って監視が可能である。例えばβガラクトシダーゼ相補は、例えばGPCR1ΔαやGPCR2Δω などのβガラクトシダーゼ変異酵素のGPCR融合タンパク質を共発現する検査細胞内で測定可能である(図27を参照されたい)。本側面に従うと、GPCRまたはオーファン受容体の単量体と二量体間の相互変換が変異レポーター酵素相補により測定可能である。図27は、βガラクトシダーゼのΔα体を有する融合タンパク質(例えばGPCR1Δα)としてGPCRまたはオーファン受容体を、また、同じGPCRまたはオーファン受容体をβガラクトシダーゼのΔω体を有する融合タンパク質(例えばGPCR1Δω)として共発現している検査細胞を示している。GPCRホモダイマーの形成は活性酵素の形成により反転され、これはGal-Screen(tm)アッセイ法などの酵素活性アッセイで測ることができる。同様に、2つの個別GPCR間か、2つのオーファン受容体間、または、1つの既知GPCRと1つのオーファン受容体間のヘテロダイマー化を例えばGPCR1ΔαやGPCR2Δωなどの融合タンパク質を共発現する検査細胞内で測ることもできる。βガラクトシダーゼ活性の増加が2つの受容体がヘテロダイマーを形成できることを示している。
【0025】
本発明の更なる側面は、アゴニストまたはアンタゴニスト治療影響下で、GPCRの単量体および二量体相互変換を監視する方法である。検査受容体は、同じGPCRまたはオーファン受容体(ホモダイマー)間、または、2つの個別GPCRあるいはオーファン受容体(ヘテロダイマー)間に置くことができる。化合物を使った治療後のβガラクトシダーゼ活性の増加は、その化合物が受容体の二量体に結合および/またはそれを安定化することを意味している。化合物を使った治療後のβガラクトシダーゼ活性の減少は、その化合物が受容体の単量体に結合および/またはそれを安定化することを意味している。
本発明のさらなる側面は、GPCRアゴニストに対するGPCRの応答をみるための細胞スクリーニング方法である。細胞はGPCR経路内の下流方向に相互作用するタンパク質パートナーを含むものが提供される。タンパク質パートナーは、酵素を相補しながら変異体との融合タンパク質として発現され、GPCR活性監視に使われる。検査細胞はGPCRアゴニストに提示され、その後レポーター酵素の酵素的活性が検知される。レポーター酵素活性の増加は、細胞がアゴニストに応答するGPCRを含んでいることを示す。
【0026】
本発明は、GPCRシグナリング内のタンパク質/タンパク質相互作用を監視するための独自開発技術(ICAST(tm)、Intercistronic Complementation Analysis Screening Technology、Gal-Screen(tm)等)の併用に関連する。米国出願番号第09/654,499号で開示されているごとく、本発明の方法は、結果的に2つの非活性化βガラクトシダーゼ変異体(それぞれがGPCRやアレスチンなどの2つの相互作用ターゲットタンパク質ペアの1つに融合)の利用に関与するICAST(tm)の使用と一部関連している。活性化βガラクトシダーゼ群の形成は、ターゲットタンパク質の相互作用により促される。本システムでは、βガラクトシダーゼ活性化は例えば、直接細胞溶解とβガラクトシダーゼレポーター酵素の高速超高感度化学発光法検知とを組み合わせるGal-Screen(tm)アッセイシステム等を使って検知できる。このシステムは、GPCR活性の出力として照度計内の測定に例えばGalacton-Star(r) chemiluminescent substrate を使っている。
【0027】
図23は、本発明の方法に相補技術を利用した例を表す概略図である。図23は、アレスチン分子と活性化GPCRまたはオーファン受容体の融合パートナー間で、特定の相互作用により強制的に結合された際に活性化する、2つの非活性βガラクトシダーゼ変異体を示している。このアッセイ技術は、オーファン受容体を釣り餌としてリガンドを同定する(「脱オーファン」と呼ばれるプロセス)ハイスループットスクリーニングアッセイに有効である。図28で示されている通り、オーファン受容体のβガラクトシダーゼ融合タンパク質(例としてGPCRorphanΔα)はβアレスチンの融合タンパク質(例としてβ-ArrΔw)を伴う検査細胞内に共発現される。検査細胞の被験が天然または合成化合物に対して行われる場合は、βガラクトシダーゼ活性の増加は、化合物がこのGPCRの自然の、または、代理のリガンドであることを意味する。同じアッセイシステムは新規GPCRの薬剤リード発見に使うことも可能である。治療後の検査細胞内におけるβガラクトシダーゼ活性の増加は、化合物のアゴニスト作用を示している。治療後の検査細胞内におけるβガラクトシダーゼ活性の減少は、化合物のアンタゴニスト作用または逆アゴニスト作用を示している。さらに、本発明の方法は、Gタンパク質、GRK、プロトオンコジーンc-Srcなどのその他下流シグナリング化合物を介したGPCR仲介シグナリング経路の監視にも使うことが可能である。
【0028】
本発明は、シグナリング経路のマッピングにICAST(tm) タンパク質/タンパク質相互作用スクリーニングを使用して、一部達成されている。本技術は多様な機能を備える、多様な既知/未知のGPCRに適用可能である。それらは下記を含むがこれに限定されないGPCRのサブファミリーを含む。
(a) アミン様リガンド-アセチルコリンムスカリン様レセプター(M1-M5)、αおよびβアドレノセプター、ドーパミンレセプター(D1、D2、D3、D4)、ヒスタミンレセプター(H1およびH2)、オクトパミンレセプター、セロトニンレセプター(5HT1、5HT2、5HT4、5HT5、5HT6、5HT7)に結合するレセプター;
(b)ペプチドリガンド-アンギオテンシンレセプター、ボンベシンレセプター、ブラジキニンレセプター、C-Cケモキネレセプター(CCR1 - CCR8およびCCR10)、C-X-Xタイプケモキネレセプター(CXC-R5)、コレシストキニンタイプAレセプター、CCKタイプレセプター、エンドセリンレセプター、ニューロテンシンレセプター、FMLP-関連レセプター、ソマトスタチンレセプター(タイプ1-5)、オピオイドレセプター(タイプD、K、M、X)に結合するレセプター;
(c) ホルモンタンパク質-卵胞刺激ホルモンレセプター、甲状腺刺激レセプター、ルトロピン-絨毛性性腺刺激ホルモンレセプターに結合するレセプター;
(d)神経伝達物質-サブスタンスPレセプター、サブスタンスKレセプター、神経ペプチドYレセプターに結合するレセプター;
(e) 嗅受レセプター-嗅受タイプ1-11、味覚レセプター、臭覚レセプター;
(f) プロスタノイドレセプター、プロスタグランジンE2(EP1-EP4サブタイプ) 、プロスタサイクリン、トロンボキサン;
(g) メタボトロピックサブスタンス-メタボトロピックグルタミングループI-IIIレセプターと結合するレセプター
(h) 光などの物理的刺激に応答するレセプターおよび味覚、嗅覚などの化学的刺激に応答するレセプター;
(i) オーファンGPCR-レセプターの自然なリガンドは未定義。
【0029】
本発明と組み合わせたICAST(tm)技術の使用は、GPCRスクリーニングプロセスに多くの利点を提供するものである。それらには、細胞以下のすべての部分-膜、サイトソル、核とのタンパク質相互作用を監視する能力;タンパク質の過剰発現の必要なく、生理学的により関連の深いモデルを達成する能力;レセプター結合のための機能的アッセイを達成し、高い情報内容を提供する能力、が含まれる。
【図面の簡単な説明】
【0030】
【図1】β2アドレナリン受容体(β2AR)およびβ-アレスチン-2(βArr2)のC2クローン中細胞発現レベルβ-ガラクトシダーゼ(β-gal)融合タンパク質の定量は、標準的なELISAアッセイにより、抗β-gal抗体と精製したβ-galタンパク質の滴定曲線を用いて行った。図1Aでは、(発現ベクターpICAST ALC中の)β2AR-βgalΔαクローン発現レベルを示している。図1Bには、クローン9-3、-7、-9、-10、-19、-24の発現ベクターpICAST OMC4あるいはクローン12-4、-9、-16、-18、-22、-24の発現ベクターpICAST OMN4中のβArr2-βgalΔωの発現レベルを示している。
【図2】β2AR受容体の活性は、アゴニストで刺激されるcAMP産生により測定した。pICAST ALC β2AR(クローン5)を発現したC2細胞あるいは親細胞には、濃度を増加させながら(-)イソプロテレノールと0.1mM IBMX処理を行った。cAMPレベルの量はpmol/wellとして示した。
【図3】活性化β2AR受容体とアレスチンの相互作用は、β-ガラクトシダーゼの相補性により測定することができる。図3Aには、(発現ベクターpICAST ALCおよびpICAST OMC、同一のコロニーから単離されたクローン(43-1、43-2、43-7、43-8)中の)β2AR-βgalΔαを発現したC2(発現ベクターpICAST ALC中のβ2ARのみ、あるいはβ2AR-βgalΔαおよびβArr2-βgalΔωを共発現し、二重に形質導入されたC2プール)について、アゴニストである(-)イソプロテレノール刺激に反応して活性化されたβ-ガラクトシダーゼの時間経過を示している。図3Bには、(発現ベクターpICAST ALC中の)β2AR-βgalΔαのみを発現したC2細胞、および(発現ベクターpICAST ALCおよびpICAST OMC中の)β2AR-βgalΔαおよびβArr1-βgalΔωを共発現したC2クローンについて、アゴニストである(-)イソプロテレノール刺激に反応して活性化されたβ-ガラクトシダーゼの時間経過を示している。
【図4】β2ARとアレスチンの相互作用に対するアゴニスト用量反応は、β-ガラクトシダーゼの相補性により測定することができる。図4Aには、β2AR-βgalΔαおよびβArr2-βgalΔω融合体を共発現したC2細胞中の、アゴニスト(-)イソプロテレノールおよびプロカテロールに対する用量反応を示している。図4Bには、β2AR-βgalΔαおよびβArr1-βgalΔω融合体を共発現したC2細胞中の、アゴニスト(-)イソプロテレノールおよびプロカテロールに対する用量反応を示している。
【図5】受容体活性のアンタゴニストによる抑制は、β2AR-βgalΔαおよびβArr-βgalΔωを共発現した細胞中のβ-ガラクトシダーゼの相補性により測定することができる。図5Aには、アゴニスト(-)イソプロテレノールをインキュベート後、β2AR-βgalΔαおよびβArr2-βgalΔω融合体を共発現したC2クローン中の、アドレナリン拮抗薬ICI-118,551およびプロプラノロールによるβ-ガラクトシダーゼの特異的抑制を示している。図5Bには、アゴニスト(-)イソプロテレノール存在下、β2AR-βgalΔαおよびβArr1-βgalΔω融合体を共発現したC2クローン中の、アドレナリン拮抗薬ICI-118,551およびプロプラノロールによるβ-ガラクトシダーゼの特異的抑制を示している。
【図6】アデノシン受容体A2aを発現したC2細胞は、アゴニスト(CGS-21680)の処理に反応してcAMPを誘導することが示されている。A2aR-βgalΔαおよびβArr1-βgalΔωを共発現したC2親細胞とC2細胞は、プールあるいは特定のクローン(47-2, 47-13)として用い、アゴニストで誘導されるcAMP反応(pmol/well)を測定した。
【図7】ドパミン受容体D1(D1-βgalΔα)およびβ-アレスチン-2(βArr2-βgalΔω)を共発現したC2細胞のアゴニストで刺激されたcAMP反応。このアッセイでは、βArr2-βgalΔω(Arr2のみ)を発現したクローンをネガティブコントロールとして用いた。βArr2-βgalΔωおよびDl-βgalΔαを発現した細胞は、アゴニスト処理(3-ヒドロキシトラミン塩酸塩、3μM)に反応した。D1(PIC2)あるいはD1(PIC3)はそれぞれ、発現ベクターpICAST ALC2あるいはpICAST ALC4でD1を発現した。
【図8】種々の哺乳類細胞株を用いて、GPCRとアレスチンの相互作用をモニターするための、安定した細胞を作成することができる。図8A、図8B、図8Cには、例として、β-ガラクトシダーゼ変異株のアドレナリン受容体β2ARおよびアレスチン融合タンパク質を共発現した、HEK 293、CHO、およびCHW細胞株を示している。β-ガラクトシダーゼ活性を利用し、アゴニストにより誘導されたβ2ARとアレスチンタンパク質の相互作用をモニターした。
【図9】βgalの相補性を用いて、β2アドレナリン受容体のホモダイマー形成をモニターすることができる。図9Aには、β2AR-βgalΔαおよびβ2AR-βgalΔωを共発現したHEK 293中のβ-ガラクトシダーゼ活性を示している。図9Bには、β2AR-βgalΔαおよびβ2AR-βgalΔωを共発現したHEK 293クローン中のアゴニスト(-)イソプロテレノールに対するcAMPの反応を示している。このアッセイでは、HEK293親細胞をネガティブコントロールとして含めている。
【図10A】pICAST ALC:標的タンパク質のC末端に融合させてβ-galΔαを発現させるためのベクター。このベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、NeoR、ネオマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図10B】pICAST ALCのヌクレオチド配列。
【図11A】pICAST ALN:標的タンパク質のN末端に融合させてβ-galΔαを発現させるためのベクター。このベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、NeoR、ネオマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColE1ori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図11B】pICAST ALNのヌクレオチド配列。
【図12A】pICAST OMC:標的タンパク質のC末端に融合させてβ-galΔωを発現させるためのベクター。このベクターは、MCS、β-galΔω骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、Hygro、ハイグロマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図12B】pICAST OMCのヌクレオチド配列。
【図13A】pICAST OMN:標的タンパク質のN末端に融合させてβ-galΔωを発現させるためのベクター。このベクターは、MCS、β-galΔω骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、Hygro、ハイグロマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図13B】pICAST OMNのヌクレオチド配列。
【図14】pICAST ALC βArr2:β―アレスチン―2のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトβ―アレスチン―2(遺伝子バンク受入番号:NM_004313)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図15】pICAST OMC βArr2:β―アレスチン―2のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトβ―アレスチン―2(遺伝子バンク受入番号:NM_004313)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図16】pICAST ALC βArr1:β―アレスチン―1のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトβ―アレスチン―1(遺伝子バンク受入番号:NM_004041)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図17】pICAST OMC βArr1:β―アレスチン―1のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトβ―アレスチン―1(遺伝子バンク受入番号:NM_004041)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図18】pICAST ALC β2AR:β2アドレナリン受容体のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトβ2アドレナリン受容体(遺伝子バンク受入番号:NM_000024)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図19】pICAST OMC β2AR:β2アドレナリン受容体のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトβ2アドレナリン受容体(遺伝子バンク受入番号:NM_000024)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図20】pICAST ALC A2aR:アデノシン2a受容体のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトアデノシン2a受容体(遺伝子バンク受入番号:NM_000675)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図21】pICAST OMC A2aR:アデノシン2a受容体のC末端に融合させてβ-galΔωを発現させるためのベクター。ヒトアデノシン2a受容体(遺伝子バンク受入番号:NM_000675)のコード配列は、pICAST OMCベクターのβ-galΔω骨格にクローニングした。
【図22】pICAST ALC D1:ドパミンD1受容体のC末端に融合させてβ-galΔαを発現させるためのベクター。ヒトドパミンD1受容体(遺伝子バンク受入番号:X58987)のコード配列は、pICAST ALCベクターのβ-galΔα骨格にクローニングした。
【図23】本発明の方法で用いられている相補技術の図解。図23では、対応する融合パートナー、GPCRおよびβ-アレスチンが相互作用すると活性化する、2つの不活性な変異した受容体酵素を示している。
【図24】β-galΔαと融合させ、セロトニン/トレオニンのアミノ酸配列を挿入してGPCRを発現させるためのベクター。既知あるいはオーファンGPCRのオープン・リーディング・フレームを設計し、SSS(セロトニン、セロトニン、セロトニン)などのセロトニン/トレオニン配列をC末端に追加している。設計したGPCRは、pICAST ALCベクターのβ-galΔα骨格にクローニングしている。pICAST ALCベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、NeoR、ネオマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coliの成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図25】β-galΔωと融合させ、変異型(R170E)β-アレスチン2を発現させるためのベクター。アルギニン170をグルタミン酸に変換するポイントミューテーションを行うため、β-アレスチン2のオープン・リーディング・フレームを設計している。変異型β-アレスチン2は、pICAST OMCベクターのβ-galΔω骨格中にクローニングしている。pICAST OMCベクターは、MCS、β-galΔα骨格の中で標的タンパク質をクローニングするための複数のクローニング部位、GSリンカー、(GGGGS)n、Hygro、ハイグロマイシン耐性遺伝子、IRES、リボソーム内部進入部位、ColElori、E.coli中の成長複製点、5'MoMuLV LTRおよび3’MoMuLV LTR、ウイルスプロモーター、Moloney Murine白血病ウイルス由来のポリアデニル化シグナルなどの機能を持つ。
【図26】リン酸化非感受性変異株R170E β-アレスチン2Δωが、アゴニストの活性化に反応してβ2ARΔαに結合。β2ARΔα C2親細胞株は、変異型R170E β-アレスチン2Δωと形質導入した。2構造を共発現したクローン個体は、96ウェルプレートに10,000 cells/wellにて播種し、指示された期間、10μM (-)イソプロテレノール、0.3mMアスコルビン酸を処理した。β-ガラクトシダーゼ活性は、Tropix Gal-ScreenTMアッセイ系の基質(Applied Biosystems社)を加えて測定し、発光はTropix IR717TM照度計(Applied Biosystems社)を用いて測定した。処理は3回行った。比較のため、β2ARΔαおよび野生型β-アレスチン2Δωを共発現したクローン細胞株(43-8)を10,000 cells/wellにて播種し、同様のアゴニスト処理を行った。(-)イソプロテレノール処理後の時間(分)はX軸に示され、β-ガラクトシダーゼ活性は相対的な光ユニット(relative light units: RLU)としてY軸に示されている。
【図27】GPCRのダイマー形成は、β-ガラクトシダーゼの相補性により測定した。GPCRのホモ、ヘテロダイマー形成をモニターする上での、本発明の利用を図解している。1つ目のGPCRは1つの酵素補部フラグメントに融合させ、2つ目のGPCRは2つ目の酵素補部フラグメントに融合させている。2つのGPCRの相互作用は、活性酵素複合体を生成する酵素フラグメントの相補性によりモニターしている(β-ガラクトシダーゼ活性など)。GPCRのホモ、ヘテロダイマー形成は、リガンド、アゴニスト、インバースアゴニスト、アンタゴニストの有無に関わらず、モニターすることができる。
【図28】ICASTTM系における、変異型β-ガラクトシダーゼの相補性によるオーファン受容体のリガンドスクリーニング(ligand fishing)。オーファンGPCR用のligand fishingおよびアゴニスト/アンタゴニストスクリーニングに対する、本発明利用の図解。例として、2種類のβ-gal融合タンパク質、GPCRorphan-ΔαおよびアレスチンΔωを発現したテスト細胞に、天然あるいは合成の化合物ライブラリ、組織抽出物、培養細胞のならし培地から採取したサンプルを処理している。処理後にβ-gal活性が上昇したことから、テストサンプル中のリガンドにより、オーファン受容体が活性化されたことが示されている。β-gal活性の上昇値は、β-アレスチンと活性化したGPCRオーファン受容体との相互作用を反映している。従って、テスト受容体の同系リガンドあるいは代理リガンドが示されている。
【発明を実施するための形態】
【0031】
好適な態様の詳細な説明
本発明はGPCR機能と経路を探索する方法を提供する。Gタンパク質-共役スーパーファミリーは、新規受容体としての急速な拡大を続け、cDNAライブラリまたはゲノムDNAの自動配列をとおして発見される。ヒトゲノムには数千のGPCRが存在すると推定されている。クローン化されているのはその一部のみで、リガンドと結び付けられている数はさらに少ない。これら、または新規発見されるオーファン受容体をそれぞれの同族リガンドや生理学的機能と結び付ける方法は、生物学的/生物医学的研究の大きな課題である。オーファン受容体の同定は一般的に個別化されたアッセイと機能に関する推測を必要とする。本発明は、検査GPCRまたはオーファン受容体とβアレスチン間の活性依存タンパク質/タンパク質相互作用を利用した、受容体の活性化の監視によるGPCR機能の探索に関連する。この特定のタンパク質/タンパク質相互作用は、本明細書に開示されている変異体酵素相補技術により測られる。このアッセイシステムは、その他シグナリングイベントの既存の知識の有無に関わらず、実行できるため、必要な推定を限定するものである。これは感度が高く、高速に、簡易に実行できるものであり、これら受容体の大部分が共通のメカニズムにより脱感受されているため、ほとんどすべてのGPCRに適用できる。
【0032】
本発明は、GPCR経路内のタンパク質/タンパク質相互作用監視のための完全なアッセイシステムを提供するものである。本発明は相補技術、ICAST (tm)(Intercistronic Complementation Analysis Screening Technology、1998年4月1日に申請された米国特許係属中申請番号第053,614号で、その全開示は引用により本明細書に編入する)を採用しているICAST(tm) テクノロジーは、着目するタンパク質に融合されたレポーター酵素の2つの変異体の使用に関連する。着目するタンパク質が相互に作用しない場合、レポーター酵素は不活性のままである。着目するタンパク質が相互に作用しない場合、レポーター酵素は不活性状態のままである。本発明の態様によると、βガラクトシダーゼの活性は、化学発光法サブストレートであるGalacton-Star(r)に関連する、Advanced Discovery Sciences(tm)による開発のGalScreen(tm)アッセイシステムで検知が可能である。GalScreen(tm)アッセイシステムおよびGalacton-Star(r)化学発光法サブストレートは米国特許番号第5,851,771号、第5,538,847号、第5,326,882号、第5,145,772、第4,978,614号、第4,931,569号で開示されており、その全開示は引用により本明細書に編入する。本発明は高速、非放射性アッセイフォーマットである、GPCR結合アッセイを含む一連のアッセイを提供するものである。本発明の方法は、緑色蛍光融合タンパク質(GFP)を含む融合としてのGPCRコンポーネントの顕微鏡的画像化や分光法に依存する、米国特許番号第5,891,646号および第6,110,693号、また、前記アンガース(Angers)et al.編集で開示の発明をさらに発展させたものである。米国特許番号第5,891,646号および第6,110,693号とAngers et alでの分光法ベースの技術は、ロースループットおよび完全定量化の不足により制限を受ける。
【0033】
Advanced Discovery Sciences(tm) テクノロジーと本発明のアッセイシステムを組み合わせることにより、ハイスループットスクリーニング(HTS)が可能なGPCR経路探索用高感度細胞ベースアッセイ法が実現される。本発明と併用が可能なAdvanced DiscoverySciences(tm)開発のテクノロジーには、Gal-Screen(tm)(上記記述を参照されたい)アッセイシステムおよびcANP-Screen(tm)イミュノアッセイシステム等がある。cAMP-Screen(tm)イミュノアッセイシステムは、細胞溶解液中のcAMP濃度を超高感度で測定する。 cAMP Screen(tm)アッセイは、超高感度化学発光アルカリフォスタファーゼ(AP)サブストレートCSPD(r) (disodium 3-(4- methoxyspiro {1,2-dioxetane-3,2'-(5'-chloro) tricyclo 3.3. 1.1.3,7}decan-4-yl phenyl phosphate)およびSapphire-II(tm) 発光エンハンサーを利用している。
【0034】
高スループットアッセイのタンパク質/タンパク質相互作用監視に使われるyeast-based-two-hybrid アッセイとは違い、本発明では(1)哺乳動物細胞、植物細胞、大腸菌などの原生細胞、また、酵母、粘菌変形菌(Dictyostelium)昆虫などの無脊椎源の細胞、など種々の細胞を利用でき、(2)例えばヌクレアスなどの限定された細胞限局化ではなく、受容体ターゲット部位の膜における、あるいは、下流ターゲットタンパク質の部位におけるシトソル内の相互作用を検知できる他、(3)転写活性化などの間接的出力に依存しない。従って、本発明はより高い生理学的関連性と、偽陽性の減化を提供するものである。
【0035】
本発明は、発明GPCRアッセイ法の感度を強化するために、前記記載の米国特許出願番号第053,614号開示の態様に改変を加えて開発されている。態様に従うと、本発明はGPCR-アレスチンの相互作用を強化/延長化して検知の信頼性をより高めるため、セリン/スレオニンクラスターの利用を伴うものである。これらクラスターは、これらのシークエンスモチーフを持たないオーファン受容体または既知GPCRに利用できる。このシークエンスを受容体のc-末端に付加し、例えばβガラクトシダーゼ変異体を伴うターゲットタンパク質の融合タンパク質からのβガラクトシダーゼ相補など、GPCRへのアレスチン結合を出力することにより、受容体の活性化が簡易に検知できる。
【0036】
別の態様に従うと、本発明は、特定GRKの活性化によるリン酸化の必要を回避する目的で、活性時にGPCRのc-末端上または細胞間ループ上のアレスチンポイントミューテーションを利用する。態様はi) 特定のGPCRまたはオーファン受容体と同族のGRKが未知であり、また、 ii) 着目する(または検査下の)受容体の特定GRKが、受容体活性化アッセイに使われるホスト細胞内に不存在または低活性である可能性を含む。
【0037】
別の態様に従うと、本発明は、活性化されたGPCRへのアレスチンの結合効果を強化し、GPCR脱感作中、GPCR/アレスチン群を安定化する目的でのスーパーアレスチンの利用を伴う。本態様はGPCRが通常、脱感作後急速に再感作する、ICAST/GPCR態様の信頼性を増すために利用できる。
【実施例】
【0038】
これら方法を各、以下に述べる。
本発明を、限定されない以下の実施例で説明する。
実施例:
本発明のひとつの態様によると、GPCR活性化はリガンド-活性化GPCRへのアレスチンの結合監視を通して測定できる。本アッセイシステムでは、例えばβアドレナリン受容体(β2AR)などのGPCRおよび例えばβアレスチンなどのアレスチンは、例えばβガラクトシダーゼ(β-gal)などのレポーター酵素の変異体を伴う融合タンパク質と同じ細胞内に共発現される。図23で示されている通り、β2ARはβ-gal変異体のΔαを伴う融合タンパク質(β2ARΔα)として、βアレスチンはβ-gal変異体のΔωを伴う融合タンパク質(β-ArrΔω)として発現する。当初、例えばGPCRの膜とアレスチンのシトソルなど、2つの異なる区画内の静止(または非刺激性)細胞内に存在する、これら2つの融合タンパク質は、活性のβガラクトシダーゼ酵素を形成することはできない。かかる細胞がアゴニストまたはリガンドにより治療されると、リガンド居住の活性化受容体は、アレスチンにとっての高親和結合部位となる。活性化GPCR、β2ARΔα、アレスチンβ-ArrΔω間の相互作用がβ-gal変異体相補を誘導する。酵素活性化は、分割に際して比色、蛍光、化学発光(例えばGal-Screen(tm)アッセイシステム等)法により測定可能な生成物を解離する、酵素基質を使って測定できる。
【0039】
実験プロトコル
1. まず始めに、選択可能なレトロウイルスベクターpICAST ALC(図18で説明)およびpICAST OMC(図15で説明)に、β2ARΔαおよびβArr2Δωの発現ベクターを組み込んだ。
2. 第二に、2つの発現体を、C2Cl2心筋細胞、あるいはCOS-7、CHO、A431、HEK 293、CHWなどの他の哺乳類細胞株に形質導入した。抗生物質を選択した後、2種類の融合タンパク質両方を適切なレベルで発現している、安定なクローンを選択した。
3. 最後に、β2ARΔαおよびβArr2Δωの両方を発現している細胞の反応性を、アゴニスト/リガンドで刺激されるβ-ガラクトシダーゼ活性により測定した。3種類の細胞サンプルを、96ウェル培養プレートのウェルに、100マイクロリットル、10,000細胞にて播種した。細胞は24時間培養し、アッセイを行った。アゴニストのアッセイ(図3および4)については、(-)イソプロテレノール、プロカテロール、ドブタミン、L-L-フェニレフリンなど、アゴニストの濃度を変えて、60分、37℃にて細胞を処理した。誘導したβ-ガラクトシダーゼの活性は、Tropix Gal-ScreenTMアッセイ系の基質(Applied Biosystems社)を加えて測定し、発光はTropix TR717TM照度計(Applied Biosystems社)を用いて測定した。アンタゴニストのアッセイ(図5)については、まず調整したばかりの培地に、血清なし、ICI-118,551あるいはプロプラノロール存在下で10分間細胞をインキュベートし、次に10マイクロモル濃度の(-)イソプロテレノールを加えた。
【0040】
セリン/スレオニンクラスターストラテジー
背景
βアレスチンの構造-機能研究によると、βアレスチンのアミノ末端内の大領域(活性化認識領域と呼ばれる)がGPCRのアゴニスト活性化状況を認識する。βアレスチンにおけるこの領域は、正電荷を帯びたリン酸化認識領域(phosphorylation-recognition domain) と呼ばれる小領域(ネット電荷+7で約20個のアミノ酸)も含み、これはGRK-リン酸化されたGPCRのカルボキシル末端と相互作用しているように見られる。
【0041】
GPCRはβアレスチンへの親和性により2つのクラスに分けられる。オークリー(Oakley) et al., 編集「クラスリン仲介飲食作用中のβアレスチンのGタンパク質-共役受容体との結合による受容体再感作のプロフィールの検知(Association of β-Arrestin with G Protein-Coupled Receptors During Clathrin-Mediated Endocytosis Dictates the Profile of Receptor Resensitization.)」J. Biol. Chem.誌、274(45):32248-32257 (1999年)。この分類の根底にある分子決定因子が、受容体のカルボキシル端子に位置する特定セリンまたはスレオニン残基内に存在しているようである。カルボキシル末端に、離散したセリンおよびスレオニン残基のみしか備えていない受容体クラスは、低い親和性でβアレスチンと結合し、内在化時にβアレスチンから解離するのに対し、カルボキシル端子内のセリン/スレオニンクラスター(連続する3つ、または、4つのうち3つの配列を持つセリンまたはスレオニン残基として定義される)を含む受容体クラスは、活性化およびリン酸化時、高い親和性でβアレスチンと結合し、受容体の内在化後もβアレスチンと結合を維持する。バソプレシンV2レセプター(Oakley, et al)、ニューロテンシンレセプター1、アンジオテンシンIIレセプタータイプ1A(ツァン(Zhang) et al編集「Gタンパク質-共役受容体/βアレスチンエンドサイトーシス群の細胞輸送(Cellular Trafficking of G Protein- Coupled Receptor/β-Arrestin Endocytic Complexes.)」J. Biol. Chem.誌、274(16):10999-11006 (1999年))など、カルボキシル末端内にこのようなセリン/スレオニンクラスターを1つあるいは複数持ついくつかの既知のGPCRは、βアレスチンを高い親和性で結合することが示されている。
【0042】
実施例
本発明の態様によると、セリン/スレオニンクラスターストラテジーは、独自にこういった構造的モチーフを持たないオーファン受容体のスクリーニングアッセイ法の簡易化に利用できる。オーファン受容体は連鎖配列により簡単に分類化ができる。セリン/スレオニンクラスターを備えないオーファン受容体を、それぞれ、受容体のカルボキシル末端に1つあるいは複数のセリン/スレオニンクラスターを誘導すべく改変された発現ベクターにクローニングする。(図24)セリン/スレオニンクラスターが、強力なイオン相互作用を通して、活性化およびリン酸化された受容体(負電荷)とβアレスチン(リン酸化認識領域中に正電荷)との間の受容体活性化依存相互作用を強化させる。オーファン受容体末端の修飾により、このように、受容体活性化の検知信頼性が強化される。
【0043】
実験プロトコル
1. まず始めに、セロトニン/トレオニンクラスターのない、オーファン受容体のオープン・リーディング・フレーム(ORF)を、図10Aで説明したpICAST ALCなどの修飾した発現ベクターにクローニングする。この修飾したpICAST ALCには、1つ以上のセロトニン/トレオニンクラスター(例えばSSSあるいはSST)のコード配列が、オーファン受容体のORF挿入部位下流に位置している(図24)。
2. 第二に、キメラオーファン受容体であるORForphan-(SSS)n-Δαを、図15で説明したβArr2Δωなどのβ-アレスチンキメラを持つ哺乳類細胞に共発現させる。
3. 第三に、この細胞にアゴニストまたはリガンドを処理し、リン酸化セリンクラスターを持つ活性型受容体を、β-gal相補性の値が強いシグナルを示す高親和性β-アレスチンに結合させる。
【0044】
このアッセイは、オーファン受容体の機能活性化を測定できる高感度の方法であり、オーファン受容体の天然リガンドあるいは代理リガンドのスクリーニングに用いることができる。このプロセスは、新しいGPCRのデオーファニング(de-orphaning)あるいはターゲット発見と呼ばれる(図28)。さらに、このアッセイは、GPCRのリード発見に用いる潜在的なアゴニストおよびアンタゴニストのスクリーニングにも有用である。
【0045】
GPCRリン酸化の存在時、非存在時における強化アレスチン結合
背景
これまでGタンパク質-共役受容体キナーゼ(GRK)の6つの異なるクラスが特定され、さらにこれら各クラスが複数のmultiple variantとして発現することが報告されている。クループニック(Krupnick) et al.編集「Gタンパク質-共役受容体調節におけるレセプタキナーゼおよびアレスチンの役割(The role of receptor kinases and arrestins in G protein-coupled receptor regulation.)」Ann. Rev. Pharmacol. Toxicol.誌、38:289-319 (1998)。細胞株は多くが種々のGRKを発現するが、あるGPCRのリン酸化に必要な特定のGRKが、組換GPCRおよびアレスチン発現に使われる細胞株に、常に存在するとは限らない。これは、同族GRKが未知である可能性が高い、オーファン受容体を使う態様で特に問題となる。その他の場合では、組換え発現作業に使われる細胞株が必要なGRKを備えていたとしても、GRKの発現率が低い可能性がある。このような危険性を回避するために、活性化GPCRに特異的に結合されているがGRKリン酸化の必要のない、遺伝子的に改変されたアレスチンが採用されている。
【0046】
アレスチン上の変異誘発に関する研究は、リン酸化認識領域におけるポイントミューテーション、特にArg175(視覚系アレスチンの)を逆の電荷を帯びたグルタミンなどの残基へと変換する変異(R175E変異)は、GPCRに特異的に結合するアレスチンを生むが、リン酸化の必要なくそれを行うことを示している。
【0047】
多くの研究により、アレスチンがGPCRへの親和性の低い非活性状態中に存在するとの仮説を導いている。GPCRの活性化とリン酸化の両方がひとたび起こると、GPCR C末端のリン酸化領域がアレスチンのリン酸化認識領域と相互作用し、アレスチンの構造変化、引いては、活性化認識領域の提示と活性化/リン酸化された受容体への結合が導かれる。ヴィシュニヴェツキー(Vishnivetskiy) et al.編集「ロドプシンのリン酸化過程に対するアレスチンの応答(How does arrestin respond to the phosphorylated state of rhodopsin?)」 J. Biol. Chem.誌 274(17):11451-11454 (1999年);グレヴィッチ(Gurevich) et al.編集「Gタンパク質-共役受容体とのアレスチンの相互作用:野生/変異型アレスチンとロドプシンβ2アドレナリン/m2ムスカリン様アセチルコリン受容体の直接結合研究(Arrestin interactions with G protein-coupled receptors. Direct binding studies of wild-type and mutant arrestins with rhodopsin, beta 2-adrenergic and m2 muscarinic cholinergic receptors.)」J. Biol. Chem.誌270(2):720-731, (1995年);グレヴィッチ(Gurevich) et al.編集「視覚系アレスチンによるリン酸化認識と高親和性結合部位へのアレスチンの転位メカニズム(Mechanism of phosphorylation-recognition by visual arrestin and the transition of arrestin into a high affinity binding site.)」Mol. Pharmacol.誌51(l):161-169 (1997年);コーヴァー(Kovoor) et al.編集「細胞内に構成活性性を有するリン酸化非依存性βアレスチン変異体のターゲット構成(Targeted construction of phosphorylation-independent beta-arrestin mutants with constitutive activity in cells.)」J. Biol. Chem.誌274(11):6831-6834 (1999年)。結論的には、非活性化、非活性化/リン酸化、活性化/非リン酸化、リン酸化された視覚系/非視覚系GPCRとの単一変異株、二重変異株、欠失、キメラアレスチンの結合研究のすべてが本モデルを論証してるのである。
【0048】
実施例
変異レポータータンパク質に融合されたアレスチンのリン酸化非感作変異体を、リン酸化非依存の様態で活性化されたGPCRと結合すべく産生することが可能である。この考え方の証明として、本発明に従い、βアレスチン2、R170E βアレスチン2のポイントミューテーションが産生され、β2AR との相互作用が分析された。
【0049】
実験プロトコル:
1) まず始めに、Arg 170をGluに変換するなど、β-アレスチン2を変異させた。この変異は、視覚系アレスチンのR175E変異と同等である。変異型β-アレスチン2のオープン・リーディング・フレームを、pICAST OMC発現ベクターのΔω-β-ガラクトシダーゼフレームにクローニングし、修飾発現ベクターR170E β-アレスチン2を作成した(図25)。
2) 第二に、R170E β-アレスチン2発現体をC2C12心筋細胞株に形質導入した。C2C12心筋細胞株は、図18、米国出願シリアル番号09/654,499に説明した通り、Δα-β-ガラクトシダーゼに融合させてβ2ARを発現させるために設計したものである。抗生物質を選択した後、2種類の融合タンパク質両方を発現しているクローンを採取した。
3)最後に、R170E β-アレスチン2Δωおよびβ2ARΔαの両方を発現しているこの細胞集団の反応性を、図26に示されている通り、アゴニスト/リガンドで刺激されるβ-ガラクトシダーゼ活性により測定した。β2ARΔαおよび野生型β-アレスチン2Δωを共発現しているC2Cl2クローン43-8(図26)を、対照として用いた。3種類の細胞サンプルを、96ウェル培養プレートのウェルに、100マイクロリットル、10,000細胞にて播種した。細胞は24時間培養し、アッセイを行った。図26のアゴニストアッセイについては、細胞を0.3mMアスコルビン酸で安定化させた10μm (-)イソプロテレノールにより、37℃、0、5、10、15、30、45あるいは60分間、処理した。誘導したβ-ガラクトシダーゼの活性は、Tropix Gal-ScreenTMアッセイ系の基質(Applied Biosystems社)を加えて測定し、発光はTropix TR717TM照度計(Applied Biosystems社)を用いて測定した。図26に示した通り、変異型アレスチンは、アゴニスト依存的にβ2ARと相互作用し、野生型アレスチンと同等であった。
4) リン酸化非感受性のアレスチンを用いた応用に拡張するため、C2C12、CHO、HEK 293などの細胞株をR170E β-アレスチン2Δω構造を発現させる開発を行う。アゴニストおよびアンタゴニストスクリーニング用の細胞株を開発するため、これらの細胞株を用いて、Δα-β-ガラクトシダーゼに融合させて、オーファンあるいは既知のGPCRを形質導入することができる。
【0050】
スーパーアレスチンの開発
背景
アレスチン経路によるGPCRシグナリングのアティニュエーションは、細胞または微生物が刺激に対し過剰に反応を防ぐ作用をする。同時に、アレスチン経路はGPCRを、一時的に非活性化し、すばやく再感作して新規刺激に対する感作を可能にするために再循環する作用も持つ。ダウンレギュレーション過程では、アレスチンとエンドサイトーシスとを結合しながら、受容体のリン酸化が行われる。脱感作された受容体のエンドサイトーシスに続き、受容体はライソソームにディグレードするか、または、再感作され、膜に戻される。再感作により、アレスチンが受容体から解離、リン酸化が行われ、膜へと戻る。活性化に伴いGPCRがたどる実際の経路は、生物学的機能と生体のニーズに拠る。ダウンレギュレーション経路の必要の可能性を含むこれら経路の多様性により、活性化したGPCRへのアレスチン親和性は受容体により異なる。そのため、自然が提供するものよりも高い、活性化されたGPCRへの親和性と結合活性を備えたスーパーアレスチンの精製は極めて有益である。
【0051】
アレスチンの変異、欠失、キメラ研究は、これまで、GPCRリン酸化状態に応答する分子の調節スイッチの解明に集中してきたが、アレスチンの改変組換えが活性化リン酸化されたGPCRへの分子内の結合強化をもたらしている。Arg175の、ヒスチジン、タイロシン、フェニルアラニン、スレオニンへの変換は、野生型アレスチンまたはR175Eアレスチンよりも、リン酸化、活性化されたロドプシンへの相当量に高い結合量を導いているが、これら変異体は活性化、非リン酸化された受容体には低い結合となっている。(ビレヴィッチ(Gurevich) et al.編集(1997年))。さらに、バリン170のアラニンへの変換は、R175E変異の構成効果を増加させているが、野生型アレスチンの活性化/リン酸化ロドプシンとの相互作用量もほとんど倍化させている。(グレヴィッチ(Gurevich) et al.編集(1997年))。
【0052】
βアレスチン1のアミノ酸382での切形は、R169E(アレスチンR175Eと同等)と野生型βアレスチンのいずれもの、活性化または活性化/リン酸化受容体への結合をそれぞれ強化していることが報告されている。(コーヴァー(Kovoor) et al.編集)。視覚系アレスチンの機能領域がβアレスチン1のそれとスワップされている、キメラアレスチンの場合、活性化/リン酸化されたGPCRへの結合親和性が変化することが報告されている。(グレヴィッチ(Gurevich) et al.編集(1995年))。これらキメラのうち、視覚系アレスチン極性N末端を伴うβアレスチン1などのいくつかのキメラが、野生型βアレスチン1よりも、リン酸化/活性化されたGPCRに特異的に結合を増加させていることを示している。(ブレクイヴィッチ(Gurevich) et al編集 (1995年))。前記に説明の、活性化GPCRへのアレスチン親和性を強化する改変は、アレスチン/GPCR群が安定化および/または長命化することから、リン酸化か非リン酸化かに関わらず、βガラクトシダーゼ活性のノイズへのシグナルも強化させる可能性がある。活性化されたGPCRとより高い親和性を持つ変異体アレスチンを利用することにより、GPCRターゲットの発明技術の改善が、受容体/リガンドの生物学を妥協させることなく、実現される。
【0053】
さらに、この「スーパーアレスチン」アプローチとアレスチンポイントミューテーションの利用とを組み合わせて、GRKの必要の有無に関わらずノイズへのシグナルを強化させることもできる。
【0054】
実施例
変異レポーターと融合されたアレスチン変異体を、活性化GPCRへのアレスチンの結合強化目的のため産生、結果、検知の感度を強化する。
【0055】
実験プロトコル
1) まず始めに、変異型β-アレスチン2構造に、R170E/T/Y/あるいはH、V165Aを組み込み、視覚系アレスチンのaa. 1-43とa.a. 1-47を置換、あるいはC末端を欠失させるか、これらの変質を組み合わせて行う。図25に示したR170E β-アレスチン2変異のクローニングと同様に、変異型β-アレスチン2のオープン・リーディング・フレームを、pICAST OMC発現ベクターのΔω-β-ガラクトシダーゼ骨格にクローニングする。
2) 第二に、変異型発現体を、Δα-β-ガラクトシダーゼに融合させるとβ2ARを発現するように設計したC2Cl2心筋細胞株に形質導入する。抗生物質を選択した後、2種類の融合タンパク質両方を発現しているクローン個体を採取する。野生型およびR170E β-アレスチン2の構造を形質導入し、対照の基準クローンを作成する。
3) 第三に、β-アレスチン2Δω(変異型あるいは野生型)およびβ2ARΔαの両方を発現した細胞集団の反応性を、アゴニスト/リガンドで刺激されるβ-ガラクトシダーゼ活性により測定する。
4) 次に、野生型β-アレスチン2Δωと比較し、アゴニストアッセイでノイズ比よりも有意に高いシグナルを示した変異型(スーパー)β-アレスチン2Δω構造を選択する。この構造を用い、既知あるいはオーファンGPCRに形質導入することができ、「スーパー」β-アレスチン2Δωを発現している安定な細胞株を開発する。スーパーβ-アレスチン2Δωを用い、ICAST/GPCRの応用に関し、ノイズに対するシグナルを増加させることができ、リードおよびリガンド発見のためのスクリーニング効果を改善することができる。
【0056】
スーパーアレスチンを用い、GPCRの脱感作中、活性型GPCRおよび安定したGPCR/アレスチン複合体へのアレスチンの結合効率を向上させる。この応用技術を用いることで、通常はGPCRが脱感作後、急速に再感作されるケースで、ICAST/GPCR技術の信頼性を高めることができる。
【0057】
本発明のアッセイ、応用と調整法は、包括的にも、特定の例を用いても説明してある。例は、限定的なものとして用いているわけではない。他の代替アイデンティティ、性質、アッセイは、発明力を駆使しなくとも、この分野における普通の技術の中で見出されることであろう。後述する特許請求の範囲に関する詳述で除外されていない限り、そのような修正は本発明の範囲内に収まるものである。
【図1A】
【図1B】
【特許請求の範囲】
【請求項1】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1つの変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現する細胞を提供し、
ここで前記細胞は、前記GPCRへのアレスチンの結合を強化させるべく改変されたアレスチンを発現しており、前記アレスチンと前記GPCR間の結合強化が、前記検査条件の前記効果の検知感度を増加させる、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を提示し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項2】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1つの変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現する細胞を提供し、
ここで前記GPCR融合タンパク質が1または複数セットのセリン/スレオニンクラスターを含むべく改変され、前記1または複数セットのセリン/スレオニンクラスターが前記GPCRのアレスチンへの結合を強化させ、前記GPCRと前記アレスチン間で強化された前記結合が前記検査条件の前記効果の検知感度を増加させる、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を暴露し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項3】
生物的に活性のハイブリッドGPCRをコードする配列を含むDNA分子であって、前記ハイブリッドGPCRがレポーター酵素の1つの変異体への融合タンパク質としてのGPCRを構成し、前記ハイブリッドGPCRが前記1または複数セットのセリン/スレオニンクラスターを含むよう改変されており、前記1または複数セットのセリン/スレオニンクラスターが前記ハイブリッドGPCRのアレスチンへの結合を強化させる、DNA分子。
【請求項4】
細胞内へ生物的活性ハイブリッドGPCRの発現を導き得、以下の操作的にリンクした要素:
プロモーター、および
生物的に活性なハイブリッドGPCRをコードする配列を含むDNA分子、を含み、前記ハイブリッドGPCRがレポーター酵素の1つの変異体への融合タンパク質としてのGPCRを構成し、前記ハイブリッドGPCRが前記1または複数セットのセリン/スレオニンクラスターを含むよう改変されており、前記1または複数セットのセリン/スレオニンクラスターが前記ハイブリッドGPCRのアレスチンへの結合を強化させるものであるDNA構築物。
【請求項5】
細胞内へ生物的活性ハイブリッドGPCRの発現を導き得るDNA構築物であって、以下の操作的にリンクした要素:
プロモーター、および、
生物的に活性なハイブリッドGPCRをコードする配列を含むDNA分子、を含み、前記ハイブリッドGPCRがレポーター酵素の1つの変異体への融合タンパク質としてのGPCRを構成し、前記ハイブリッドGPCRが前記1または複数セットのセリン/スレオニンクラスターを含むよう改変されており、前記1または複数セットのセリン/スレオニンクラスターが前記ハイブリッドGPCRのアレスチンへの結合を強化させるものであるDNA構築物によって形質転換された細胞。
【請求項6】
生物的に活性なハイブリッドアレスチンをコードする配列を含み、前記ハイブリッドアレスチンがレポーター酵素の1の変異体への融合タンパク質としてのアレスチンを構成し、前記ハイブリッドアレスチンがGPCRへの前記アレスチンの結合を強化するよう改変されているDNA分子。
【請求項7】
細胞内に生物的に活性なハイブリッドアレスチンの発現を導くことができ、以下の操作的にリンクした要素:
プロモーター、および
生物的に活性なハイブリッドアレスチンをコードする配列を含むDNA分子、を含み、前記ハイブリッドアレスチンがレポーター酵素の1の変異体への融合タンパク質としてのアレスチンを構成し、前記ハイブリッドアレスチンがGPCRへの前記アレスチンの結合を強化させるよう改変されている、DNA構築物。
【請求項8】
細胞内へ生物的に活性なハイブリッドアレスチンの発現を導くことができ、以下の操作的にリンクした要素:
プロモーター、および
生物的に活性なハイブリッドアレスチンをコードする配列を含むDNA分子、を含み、前記ハイブリッドアレスチンがレポーター酵素の1つの変異体への融合タンパク質としてのアレスチンを構成し、前記ハイブリッドアレスチンがGPCRへの前記アレスチンの結合を強化させるよう改変されているDNA構築物により形質転換された細胞。
【請求項9】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1の変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現させる細胞を提供し、
ここで前記細胞はアレスチンをもまた発現し、前記アレスチンがリン酸化認識領域内のポイントミューテーションを誘導することによって、GPCRがアレスチンへ結合する際にGPCRのリン酸化必要であるという条件を取り除き、GPCRがリン酸化されているか否かに関わらず細胞内のGPCRへのアレスチンの結合を可能にしている、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を暴露し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項10】
前記アレスチンが、活性化された非リン酸化GPCRの結合親和性および結合活性からなる群より選択される特性を増強するものである、請求項9記載の方法。
【請求項11】
前記アレスチンがβアレスチン2であり、βアレスチン2がArg169を逆の電荷を有する残基に変換されるよう変異されている、請求項10記載の方法。
【請求項12】
逆の電荷を有する残基が、ヒスチジン、チロシン、フェニルアラニン、スレオニンにより成る群から選択される、請求項11記載の方法。
【請求項13】
前記アレスチンが活性化かつリン酸化されたGPCRの結合親和性および結合活性からなる群から選択される特性を増強する、請求項9記載の方法。
【請求項14】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1つの変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現させる細胞を提供し、
ここで、前記GPCR融合タンパク質が1つあるいは複数のセリン/スレオニンクラスターを含むべく改変され、セリン/スレオニン残基として定義される前記1つあるいは複数のセリン/スレオニンクラスターが前記GPCRのカルボキシル末端内の3つの連続する、あるいは、4つのうち3つの部位を占め、前記1または複数セットのセリン/スレオニンクラスターが前記GPCRのアレスチンへの結合を強化させ、前記GPCRと前記アレスチン間で強化された前記結合が前記検査条件の前記効果の検知感度を増加させるものである、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を提示し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項15】
改変された前記アレスチンが活性化/リン酸化されたGPCRへの結合強化を誘導するものである、請求項1記載の方法。
【請求項16】
改変された前記アレスチンが、ヒスチジン、チロシン、フェニルアラニン、スレオニンにより成るグループから選択されるアミノ酸へのArg169の変換を含む、請求項14記載の方法
【請求項17】
改変された前記アレスチンがVAL170のアラニンへの変換を含む、請求項14記載の方法。
【請求項18】
前記アレスチンがβアレスチン1およびβアレスチン2から成る群から選択され、前記βアレスチン1または前記βアレスチン2がカルボキシル末端の全てまたは一部において前記βアレスチン1または前記βアレスチン2の半分に切形されている、請求項14記載の方法。
【請求項19】
前記βアレスチン1または前記βアレスチン2が前記βアレスチン1または前記βアレスチン2のアミノ酸190から前記βアレスチン1または前記βアレスチン2のカルボキシル末端まで切形されている、請求項18記載の方法。
【請求項20】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系(visual)アレスチンのキメラである、請求項14記載の方法。
【請求項21】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系アレスチンのキメラである、請求項10記載の方法。
【請求項22】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系アレスチンのキメラである、請求項11記載の方法。
【請求項23】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系アレスチンのキメラである、請求項12記載の方法。
【請求項24】
前記アレスチンがβアレスチン1であり、前記βアレスチン2がArg170を逆の電荷を有する残基と変換するよう変異されている、請求項10記載の方法。
【請求項25】
改変された前記アレスチンが、ヒスチジン、チロシン、フェニルアラニン、スレオニンにより成るグループから選択されるアミノ酸へのArg170の変換を含む、請求項14記載の方法。
【請求項1】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1つの変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現する細胞を提供し、
ここで前記細胞は、前記GPCRへのアレスチンの結合を強化させるべく改変されたアレスチンを発現しており、前記アレスチンと前記GPCR間の結合強化が、前記検査条件の前記効果の検知感度を増加させる、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を提示し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項2】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1つの変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現する細胞を提供し、
ここで前記GPCR融合タンパク質が1または複数セットのセリン/スレオニンクラスターを含むべく改変され、前記1または複数セットのセリン/スレオニンクラスターが前記GPCRのアレスチンへの結合を強化させ、前記GPCRと前記アレスチン間で強化された前記結合が前記検査条件の前記効果の検知感度を増加させる、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を暴露し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項3】
生物的に活性のハイブリッドGPCRをコードする配列を含むDNA分子であって、前記ハイブリッドGPCRがレポーター酵素の1つの変異体への融合タンパク質としてのGPCRを構成し、前記ハイブリッドGPCRが前記1または複数セットのセリン/スレオニンクラスターを含むよう改変されており、前記1または複数セットのセリン/スレオニンクラスターが前記ハイブリッドGPCRのアレスチンへの結合を強化させる、DNA分子。
【請求項4】
細胞内へ生物的活性ハイブリッドGPCRの発現を導き得、以下の操作的にリンクした要素:
プロモーター、および
生物的に活性なハイブリッドGPCRをコードする配列を含むDNA分子、を含み、前記ハイブリッドGPCRがレポーター酵素の1つの変異体への融合タンパク質としてのGPCRを構成し、前記ハイブリッドGPCRが前記1または複数セットのセリン/スレオニンクラスターを含むよう改変されており、前記1または複数セットのセリン/スレオニンクラスターが前記ハイブリッドGPCRのアレスチンへの結合を強化させるものであるDNA構築物。
【請求項5】
細胞内へ生物的活性ハイブリッドGPCRの発現を導き得るDNA構築物であって、以下の操作的にリンクした要素:
プロモーター、および、
生物的に活性なハイブリッドGPCRをコードする配列を含むDNA分子、を含み、前記ハイブリッドGPCRがレポーター酵素の1つの変異体への融合タンパク質としてのGPCRを構成し、前記ハイブリッドGPCRが前記1または複数セットのセリン/スレオニンクラスターを含むよう改変されており、前記1または複数セットのセリン/スレオニンクラスターが前記ハイブリッドGPCRのアレスチンへの結合を強化させるものであるDNA構築物によって形質転換された細胞。
【請求項6】
生物的に活性なハイブリッドアレスチンをコードする配列を含み、前記ハイブリッドアレスチンがレポーター酵素の1の変異体への融合タンパク質としてのアレスチンを構成し、前記ハイブリッドアレスチンがGPCRへの前記アレスチンの結合を強化するよう改変されているDNA分子。
【請求項7】
細胞内に生物的に活性なハイブリッドアレスチンの発現を導くことができ、以下の操作的にリンクした要素:
プロモーター、および
生物的に活性なハイブリッドアレスチンをコードする配列を含むDNA分子、を含み、前記ハイブリッドアレスチンがレポーター酵素の1の変異体への融合タンパク質としてのアレスチンを構成し、前記ハイブリッドアレスチンがGPCRへの前記アレスチンの結合を強化させるよう改変されている、DNA構築物。
【請求項8】
細胞内へ生物的に活性なハイブリッドアレスチンの発現を導くことができ、以下の操作的にリンクした要素:
プロモーター、および
生物的に活性なハイブリッドアレスチンをコードする配列を含むDNA分子、を含み、前記ハイブリッドアレスチンがレポーター酵素の1つの変異体への融合タンパク質としてのアレスチンを構成し、前記ハイブリッドアレスチンがGPCRへの前記アレスチンの結合を強化させるよう改変されているDNA構築物により形質転換された細胞。
【請求項9】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1の変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現させる細胞を提供し、
ここで前記細胞はアレスチンをもまた発現し、前記アレスチンがリン酸化認識領域内のポイントミューテーションを誘導することによって、GPCRがアレスチンへ結合する際にGPCRのリン酸化必要であるという条件を取り除き、GPCRがリン酸化されているか否かに関わらず細胞内のGPCRへのアレスチンの結合を可能にしている、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を暴露し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項10】
前記アレスチンが、活性化された非リン酸化GPCRの結合親和性および結合活性からなる群より選択される特性を増強するものである、請求項9記載の方法。
【請求項11】
前記アレスチンがβアレスチン2であり、βアレスチン2がArg169を逆の電荷を有する残基に変換されるよう変異されている、請求項10記載の方法。
【請求項12】
逆の電荷を有する残基が、ヒスチジン、チロシン、フェニルアラニン、スレオニンにより成る群から選択される、請求項11記載の方法。
【請求項13】
前記アレスチンが活性化かつリン酸化されたGPCRの結合親和性および結合活性からなる群から選択される特性を増強する、請求項9記載の方法。
【請求項14】
Gタンパク質-共役受容体(GPCR)経路活性における検査条件の効果の査定方法であって、
a) レポーター酵素の1つの変異体への融合タンパク質としてGPCRを、また、別の酵素変異体への融合として相互作用するタンパク質パートナーを発現させる細胞を提供し、
ここで、前記GPCR融合タンパク質が1つあるいは複数のセリン/スレオニンクラスターを含むべく改変され、セリン/スレオニン残基として定義される前記1つあるいは複数のセリン/スレオニンクラスターが前記GPCRのカルボキシル末端内の3つの連続する、あるいは、4つのうち3つの部位を占め、前記1または複数セットのセリン/スレオニンクラスターが前記GPCRのアレスチンへの結合を強化させ、前記GPCRと前記アレスチン間で強化された前記結合が前記検査条件の前記効果の検知感度を増加させるものである、
b) 前記検査条件の下で前記GPCRのリガンドに対し細胞を提示し、および
c) 前記レポーター酵素の相補により、前記GPCRの活性を監視することを含み、
前記検査条件が不在の場合と比較して細胞内のレポーター酵素活性が増加することは、相互作用するタンパク質パートナーとのGPCRの相互作用が、前記検査条件不在の場合と比較して増加することを示し、また、前記検査条件が不在の場合の発生と比べ細胞内のレポーター酵素活性が減少することが、相互作用するタンパク質パートナーとのGPCRの相互作用が前記検査条件が不在の場合と比べて減少することを示すものである方法。
【請求項15】
改変された前記アレスチンが活性化/リン酸化されたGPCRへの結合強化を誘導するものである、請求項1記載の方法。
【請求項16】
改変された前記アレスチンが、ヒスチジン、チロシン、フェニルアラニン、スレオニンにより成るグループから選択されるアミノ酸へのArg169の変換を含む、請求項14記載の方法
【請求項17】
改変された前記アレスチンがVAL170のアラニンへの変換を含む、請求項14記載の方法。
【請求項18】
前記アレスチンがβアレスチン1およびβアレスチン2から成る群から選択され、前記βアレスチン1または前記βアレスチン2がカルボキシル末端の全てまたは一部において前記βアレスチン1または前記βアレスチン2の半分に切形されている、請求項14記載の方法。
【請求項19】
前記βアレスチン1または前記βアレスチン2が前記βアレスチン1または前記βアレスチン2のアミノ酸190から前記βアレスチン1または前記βアレスチン2のカルボキシル末端まで切形されている、請求項18記載の方法。
【請求項20】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系(visual)アレスチンのキメラである、請求項14記載の方法。
【請求項21】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系アレスチンのキメラである、請求項10記載の方法。
【請求項22】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系アレスチンのキメラである、請求項11記載の方法。
【請求項23】
前記アレスチンがβアレスチン1、βアレスチン2および/または視覚系アレスチンのキメラである、請求項12記載の方法。
【請求項24】
前記アレスチンがβアレスチン1であり、前記βアレスチン2がArg170を逆の電荷を有する残基と変換するよう変異されている、請求項10記載の方法。
【請求項25】
改変された前記アレスチンが、ヒスチジン、チロシン、フェニルアラニン、スレオニンにより成るグループから選択されるアミノ酸へのArg170の変換を含む、請求項14記載の方法。
【図2】


【図3A】

【図3B】

【図4A】

【図4B】

【図5A】

【図5B】

【図6】

【図7】

【図8A】
【図8B】

【図8C】

【図9A】

【図9B】

【図10A】

【図10B−1】

【図10B−2】

【図10B−3】

【図10B−4】

【図10B−5】

【図10B−6】

【図10B−7】

【図10B−8】

【図10B−9】

【図10B−10】

【図10B−11】

【図10B−12】

【図10B−13】

【図10B−14】

【図10B−15】

【図11A】

【図11B−1】

【図11B−2】

【図11B−3】

【図11B−4】

【図11B−5】

【図11B−6】

【図11B−7】

【図11B−8】

【図11B−9】

【図11B−10】

【図11B−11】

【図12A】

【図12B−1】

【図12B−2】

【図12B−3】

【図12B−4】

【図12B−5】

【図12B−6】

【図12B−7】

【図12B−8】

【図12B−9】

【図12B−10】

【図12B−11】

【図13A】

【図13B−1】

【図13B−2】

【図13B−3】

【図13B−4】

【図13B−5】

【図13B−6】

【図13B−7】

【図13B−8】

【図13B−9】

【図13B−10】

【図13B−11】

【図14】

【図15】

【図16】

【図17】

【図18】

【図19】

【図20】

【図21】

【図22】

【図23】

【図24】

【図25】

【図26】

【図27】

【図28】

【図3A】
【図3B】
【図4A】
【図4B】
【図5A】
【図5B】
【図6】
【図7】
【図8A】
【図8B】
【図8C】
【図9A】
【図9B】
【図10A】
【図10B−1】
【図10B−2】
【図10B−3】
【図10B−4】
【図10B−5】
【図10B−6】
【図10B−7】
【図10B−8】
【図10B−9】
【図10B−10】
【図10B−11】
【図10B−12】
【図10B−13】
【図10B−14】
【図10B−15】
【図11A】
【図11B−1】
【図11B−2】
【図11B−3】
【図11B−4】
【図11B−5】
【図11B−6】
【図11B−7】
【図11B−8】
【図11B−9】
【図11B−10】
【図11B−11】
【図12A】
【図12B−1】
【図12B−2】
【図12B−3】
【図12B−4】
【図12B−5】
【図12B−6】
【図12B−7】
【図12B−8】
【図12B−9】
【図12B−10】
【図12B−11】
【図13A】
【図13B−1】
【図13B−2】
【図13B−3】
【図13B−4】
【図13B−5】
【図13B−6】
【図13B−7】
【図13B−8】
【図13B−9】
【図13B−10】
【図13B−11】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【公開番号】特開2011−200229(P2011−200229A)
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−88404(P2011−88404)
【出願日】平成23年4月12日(2011.4.12)
【分割の表示】特願2001−558070(P2001−558070)の分割
【原出願日】平成13年2月7日(2001.2.7)
【出願人】(310009775)アプライド バイオシステムズ リミテッド ライアビリティー カンパニー (19)
【Fターム(参考)】
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2011−88404(P2011−88404)
【出願日】平成23年4月12日(2011.4.12)
【分割の表示】特願2001−558070(P2001−558070)の分割
【原出願日】平成13年2月7日(2001.2.7)
【出願人】(310009775)アプライド バイオシステムズ リミテッド ライアビリティー カンパニー (19)
【Fターム(参考)】
[ Back to top ]