
ロタウイルス感染阻害活性を有する新規糖タンパク質
【課題】本発明では、新たなロタウイルス感染防御用組成物並びにロタウイルス感染防御に有用な食品、医薬組成物及び飼料を提供すること。
【解決手段】プロテオースペプトンを硫安沈殿と遠心分離処理を組み合わせた方法で処理して得られる分画物から得られる、ロタウイルス感染阻害活性に優れた新規糖タンパク質。
【解決手段】プロテオースペプトンを硫安沈殿と遠心分離処理を組み合わせた方法で処理して得られる分画物から得られる、ロタウイルス感染阻害活性に優れた新規糖タンパク質。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ロタウイルス感染防御用組成物とその製造法並びにそれを含むロタウイルス感染防御に有用な飲食品、医薬組成物及び家畜飼料に関する。
【背景技術】
【0002】
ヒトロタウイルスを原因とする乳幼児冬季下痢症は主に2歳以下の乳幼児に、発熱、嘔吐、下痢及び脱水症状を引き起こす疾患である。我が国では、灰白色の便の性状から白痢と呼ばれたが、ヒトロタウイルス感染症であることが明らかにされている。先進国での疫学調査から、衛生状態の改善はロタウイルスの有病率を減少させることはできないとされ、その対策の一つとして世界的なロタウイルスワクチン開発が進められている。
【0003】
RRV-TV(rhesus rotavirus tetravalent)ワクチンは、先進国と発展途上国の両方でロタウイルスによる重症の下痢症の予防に高い効果を示し、1998年8月、米国食品医薬局はRRV-TVワクチンを世界で初めてロタウイルスワクチンとして認可した。しかしながら、腸重積症が副反応であることが米国疾病防疫センターから報告され、RRV-TVワクチンの投与は中止された(非特許文献1)。
【0004】
一方、ロタウイルス感染を阻害する食品成分あるいは組成物として例えば、乳ムチン(非特許文献2)、ウシ乳清(以下、ホエイともいう)由来の高分子糖タンパク質混合物(特許文献1)などが提案されている。また、本発明者らもホエイの精密濾過処理の保持液又はその乾燥物に抗ロタウイルス感染阻害活性を持つことを過去に報告している(特許文献2)。前者ではヒトロタウイルスの最小阻害濃度が5μg/ml、後者では11μg/mlと強力なヒトロタウイルス感染阻害活性を見出しているが、最小阻害濃度だけでなく、感染阻害率の点でも優れたロタウイルス感染防御用組成物の登場が望まれている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開平8−99896号公報
【特許文献2】国際公開第2004/080475号パンフレット
【非特許文献】
【0006】
【非特許文献1】石田真一他、「ロタウイルスワクチン」小児科診療 Vol.63, 2000年, p.1045-1049
【非特許文献2】Yolken, R.Y. et al., 「Human Milk Mucin Inhibits Rotavirus Replication and Prevents Experimental Gastroenteritis」 J Clin. Invest., Vol.90, 1992年, p.1984-1991
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、新たなロタウイルス感染防御用組成物並びにロタウイルス感染防御に有用な飲食品、医薬組成物及び飼料を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく検討した結果、ウシ乳清中に見出される糖タンパク質に強力なロタウイルス感染阻害活性を見出した。すなわち、本発明は以下の通りである。
[1] ウシ乳清中に見出され、SDS-PAGEで測定される分子量が15〜20kDa、等電点pIが6〜9であり、N末端から5残基のアミノ酸配列が(1)Ser-Ser-Arg-Gln-Proであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
[2] ウシ乳清中に見出され、SDS-PAGEで測定される分子量が14〜19kDa、等電点pIが6〜9であり、N末端から7残基のアミノ酸配列が(2)Ile-Leu-Lys-Glu-Lys-His-Leuであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
これら[1]及び[2]の糖タンパク質は、ウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画することによって得られるものであることが好ましい。
[3] 前記[1]又は[2]記載の糖タンパク質を含むことを特徴とするロタウイルス感染防御用組成物。
[4] 前記[3]記載のロタウイルス感染防御用組成物を含む食品。
[5] 乳児用食品、幼児用食品、授乳婦用食品、高齢者用食品、病者用食品、保健機能食品、サプリメントからなる群より選択される前記[4]記載の食品。
[6] 前記[3]記載のロタウイルス感染防御用組成物を含む医薬組成物。
[7] 前記[3]記載のロタウイルス感染防御用組成物を含む家畜飼料。
[8] ウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画することを特徴とする、前記[1]又は[2]記載の糖タンパク質の製造法。
【発明の効果】
【0009】
本発明によりロタウイルス感染阻害活性を有する新規糖タンパク質が提供される。
【図面の簡単な説明】
【0010】
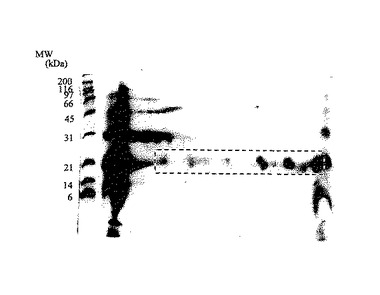
【図1】実施例1に従って精製したタンパク質の二次元電気泳動パターンを示した。ゲルはCBB染色を行った。図の左側の数字は標準タンパク質(BIO-RAD社製、商品名:SDS-PAGEスタンダード Broad)の分子量をkDaの単位で示したものである。
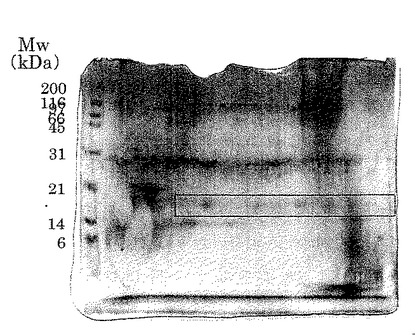
【図2】実施例2に従って精製したタンパク質の二次元電気泳動パターンを示した。ゲルはCBB染色を行った。図の左側の数字は図1と同様の標準タンパク質の分子量をkDaの単位で示したものである。
【図3】実施例1により得られたタンパク質スポット群A、B、C、D、E、Fを示す。
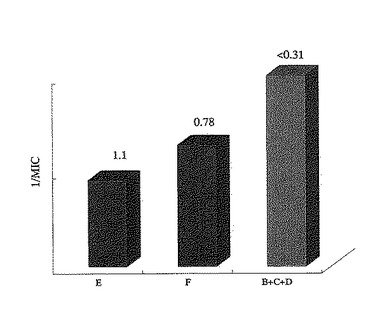
【図4】実施例4における各試料のウイルス感染を阻害する最小阻害濃度(MIC)を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。
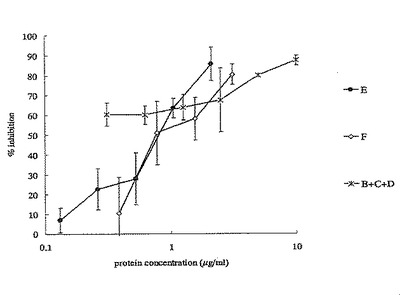
【図5】実施例4における各試料のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。縦軸はロタウイルス感染阻害率(%)を示す。図中の誤差棒は標準偏差を示す。
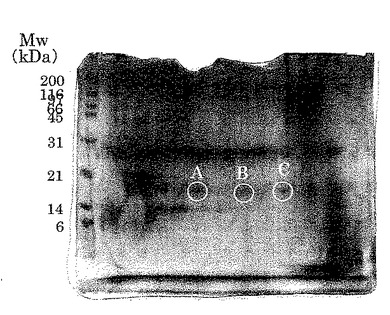
【図6】実施例2により得られたタンパク質スポット群A、B、Cを示す。
【図7】実施例5における各試料のウイルス感染を阻害する最小阻害濃度(MIC)を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。<0.56のMICを示したロタミルクは対照サンプルである。
【図8】実施例5における各試料のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。縦軸はロタウイルス感染阻害率(%)を示す。
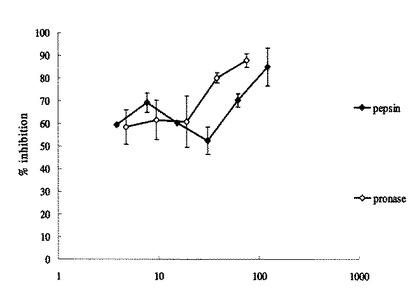
【図9】実施例6における各試料(ペプシン分解物及びプロナーゼ分解物)のウイルス感染を阻害する最小阻害濃度を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。左がペプシン分解物、右がプロナーゼ分解物の試料を示す。
【図10】実施例6における各試料(ペプシン分解物及びプロナーゼ分解物)のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。
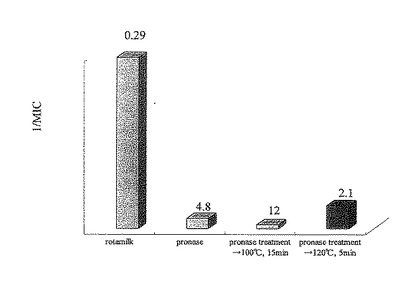
【図11】実施例6における各試料(プロナーゼ分解物(100℃で15分間の加熱処理)、プロナーゼ分解物(120℃で5分間の加熱処理)、ロタミルク)のウイルス感染を阻害する最小阻害濃度(MIC)を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。
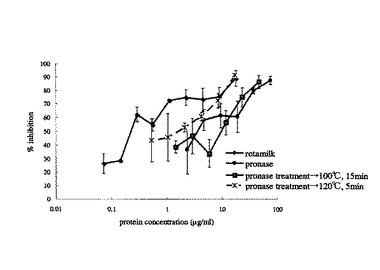
【図12】実施例6における各試料(プロナーゼ分解物(100℃で15分間の加熱処理)、プロナーゼ分解物(120℃で5分間の加熱処理)、ロタミルク)のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。図中の誤差棒は標準偏差を示す。
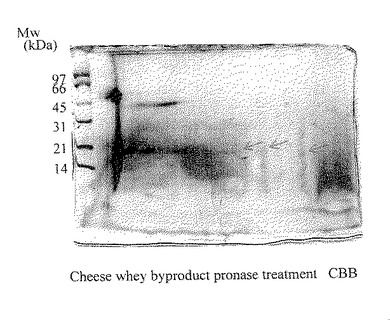
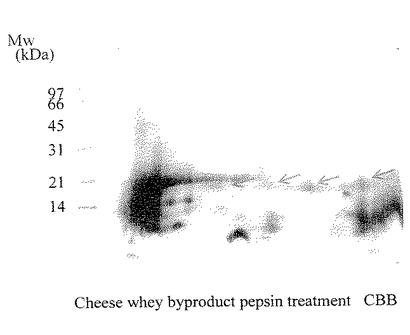
【図13】実施例6により得られたチーズホエイ副産物のプロナーゼ分解物の二次元電気泳動パターンを示した。ゲルはCBB染色を行い、スポット群は矢印で示した。
【図14】実施例6により得られたチーズホエイ副産物のペプシン分解物の二次元電気泳動パターンを示した。ゲルはCBB染色を行い、スポット群は矢印で示した。
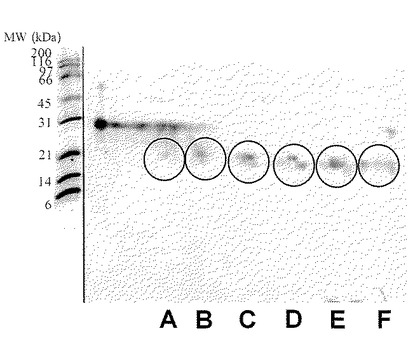
【図15】実施例1に従って精製したプロテオースペプトン画分の二次元電気泳動パターンを示した(弱酸性側からスポット群A、B、C、D、E、Fとする)。PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行い、スポット群は円で示した。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
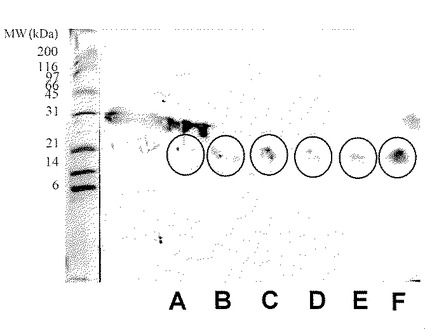
【図16】実施例1に従って精製した硫安画分の二次元電気泳動パターンを示した(弱酸性側からスポット群A、B、C、D、E、Fとする)。PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行い、スポット群は円で示した。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
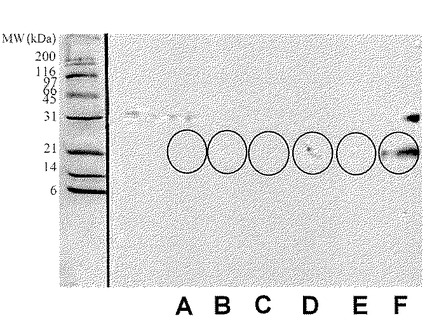
【図17】実施例3で酵素分解物の調製に用いた乳清タンパク質の二次元電気泳動パターンを示した(弱酸性側からスポット群A、B、C、D、E、Fとする)。PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行い、スポット群は円で示した。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
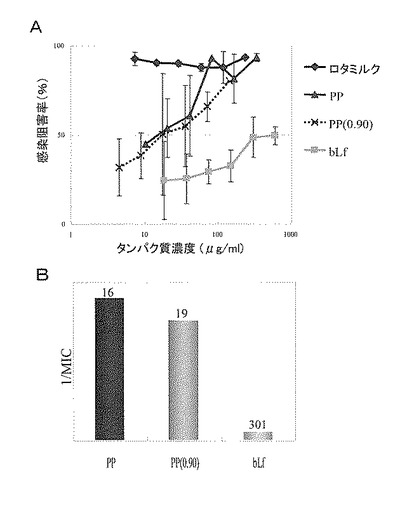
【図18】プロテオースペプトン(PP)画分及び実施例8に従って精製したPP(0.9)画分のロタウイルス感染阻害活性を示す図である。Aは、各サンプルのロタウイルス感染阻害活性を示すグラフ、Bは、各サンプルについて最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。対照サンプルとして、ヒトロタウイルスで過免疫した乳牛の初乳中の抗体を含むロタミルク、及び陰性対照としてのウシラクトフォリン(bLf)を用いた。
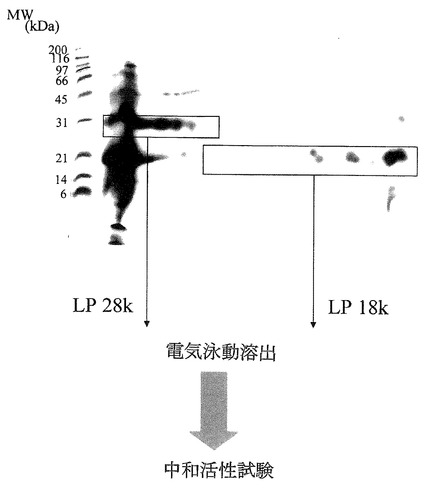
【図19】実施例8に従って精製したPP(0.9)画分の二次元電気泳動パターンとその後の実験の流れを示した。
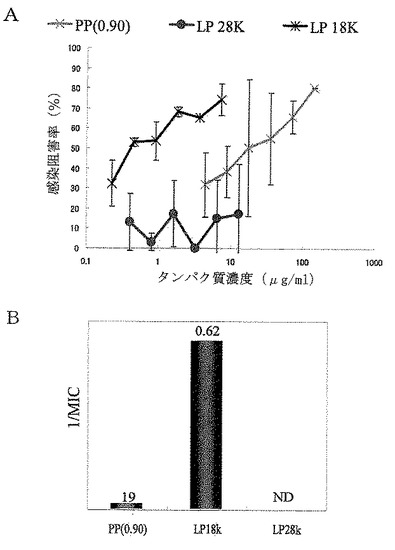
【図20】実施例8に従って精製したPP(0.9)画分並びにその画分から分離した約18kDaのタンパク質(LP 18K)及び約28kDaのタンパク質(LP 28K)のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。AのグラフにおけるX軸(横軸)のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
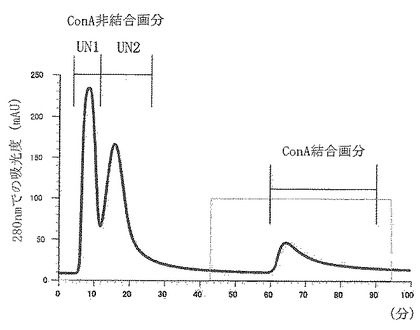
【図21】実施例8に従って調製したプロテオースペプトン画分(PP画分)の、ConAアフィニティークロマトグラフィーを使用した溶出パターンを示した。
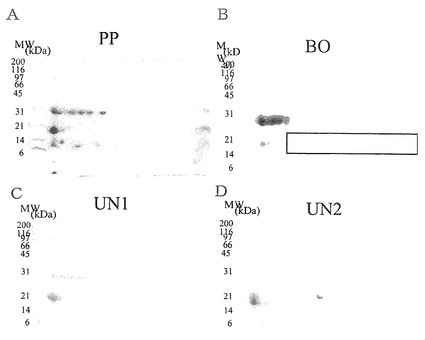
【図22】ConAアフィニティークロマトグラフィーによりPP画分から得られた各画分の二次元電気泳動パターンを示した。AはPP画分、BはBO画分、CはUN1画分、DはUN2画分の結果である。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
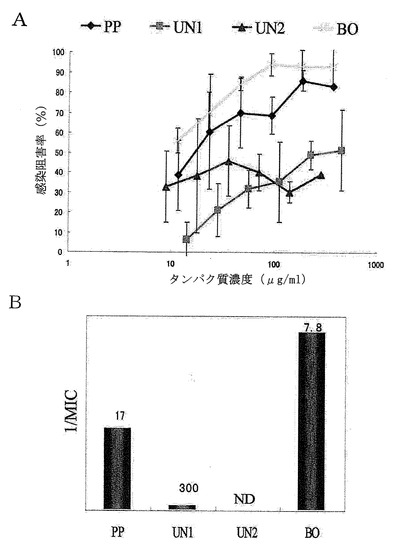
【図23】ConAアフィニティークロマトグラフィーによりPP画分から得られた各画分のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。Aのグラフにおける横軸のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
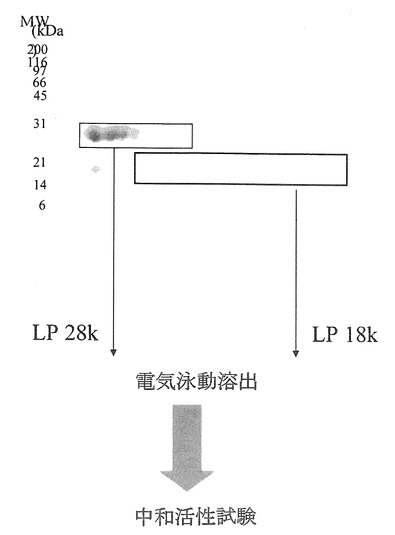
【図24】ConA結合画分であるBO画分の二次元電気泳動パターンとその後の実験の流れを示した。
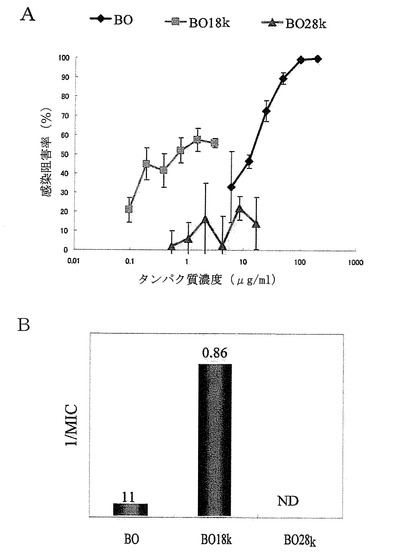
【図25】ConA結合画分であるBO画分並びにその画分から分離した約18kDaのタンパク質(LP 18Kに相当)及び約28kDaのタンパク質(LP 28Kに相当)のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。AのグラフにおけるX軸(横軸)のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
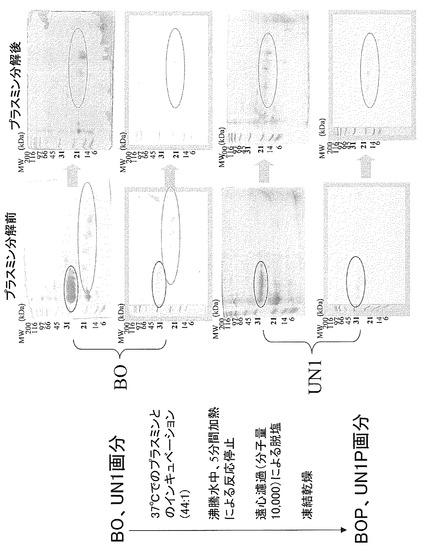
【図26】ConA結合画分であるBO画分及びConA非結合画分であるUN1画分とそれらのプラスミン処理物が示す二次元電気泳動パターンを示した。ゲルはCBB染色を行った。また、PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行った。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
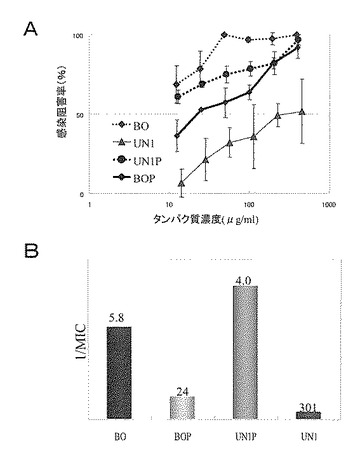
【図27】ConA結合画分であるBO画分及びConA非結合画分であるUN1画分とそれらのプラスミン処理物のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。AのグラフにおけるX軸(横軸)のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
【発明を実施するための形態】
【0011】
以下、本発明を詳細に説明する。本発明は、ウシ乳清中に見出され、下記特徴を有する新規糖タンパク質に関する。
【0012】
ウシ乳清中に見出され、SDS-PAGEで測定される分子量が15〜20kDa、等電点pIが6〜9であり、N末端から5残基のアミノ酸配列が(1)Ser-Ser-Arg-Gln-Proであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
【0013】
ウシ乳清中に見出され、SDS-PAGEで測定される分子量が14〜19kDa、等電点pIが6〜9であり、N末端から7残基のアミノ酸配列が(2)Ile-Leu-Lys-Glu-Lys-His-Leuであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
【0014】
次いで製造法を述べるが、本発明は、これに限定されるものではない。
本発明における新規糖タンパク質(以下、本発明物質ということもある)は、ウシプロテオースペプトン溶液に硫安を加え、溶液の飽和度が55〜90%のときに得られる沈殿物に見出すことができる。あるいは、本発明における新規糖タンパク質は、ウシプロテオースペプトン溶液を直接分画することによっても得ることができる。プロテオースペプトンは、95℃で20分間の加熱処理を行った脱脂乳をpH 4.6にしたときに沈殿しない耐熱性の乳清タンパク質画分である(「ミルク総合辞典」朝倉書店 1992年 p.37)。本発明物質は、ウシ乳清を加熱処理して得られる可溶性画分、いわゆるウシプロテオースペプトン画分を硫安沈殿と遠心分離処理を組み合わせた方法で処理して得られる分画物に見出され、ロタウイルスの感染阻害活性が非常に高いことが特徴である。上記の本発明の糖タンパク質は、それぞれ、ラクトフォリン(プロテオースペプトン画分に含まれる耐熱性と可溶性(水溶性)を有する糖タンパク質)を構成するタンパク質成分(ラクトフォリンコンポーネントとも呼ばれる)の一つである。ラクトフォリンは、プロテオースペプトン画分の一分画であるプロテオースペプトン3画分中に見出されたことから、プロテオースペプトン3又はプロテオースペプトンコンポーネント3とも呼ばれる。
【0015】
乳清はスイートホエイ(sweet whey)及び酸ホエイ(acid whey)に分類され、ウシの各泌乳期の乳もしくはこれらの濃縮物又は乾燥物(以下、これらをまとめて乳ということもある)を原料として常法により調製することができる。スイートホエイは熟成型チーズの副産物であり、pHが5.9〜6.3で甘味がある。一方、酸ホエイは非熟成型(フレッシュ型)チーズの製造で得られ、通常のpHが4.4〜4.6で酸味がある。また、カゼインの製造時に得られるホエイも酸ホエイである。本発明の乳清にはスイートホエイ及び酸ホエイの両者を含む。
【0016】
本発明では、乳清そのものに加え、乳清を精密濾過処理、限外濾過処理、ダイアフィルトレーション処理などを行うことで、乳清中のタンパク質を濃縮した乳清タンパク質濃縮物あるいはその精製物を用いることができる。市販品に、乳清タンパク質濃縮物(WPC)、乳清タンパク質単離物(WPI)などの濃縮物又は乾燥物が知られ、これらをいずれも用いることができる。また、常法に従って調製しても良い。限外濾過処理などにより乳清タンパク質を濃縮する場合には、分画分子量が300万Da以上の細孔径(ポアサイズ)を持つ分離膜を用いることが望ましい。
【0017】
乳清タンパク質濃縮物を精製して用いる場合には例えば、デキストランの分画分子量が4×104〜2×107Daに相当する担体(商品名:Sephacryl S-500 HR、アマシャムファルマシアバイオテク社製など)を充填したゲル濾過カラムクロマトグラフィーに適用し、波長280nmの吸光度を指標に、最初のピークに相当する画分にウイルスの感染阻害活性が確認されている。溶離液にはトリス塩酸、リン酸などのpHが6〜8の公知の緩衝液が用いられ、硫酸ドデシルナトリウム、トリトンX-100などの界面活性剤を解離剤として併用することもできる。
【0018】
次いで乳清そのもの、乳清タンパク質濃縮物あるいはその精製物を加熱処理し、得られた可溶性画分に硫安を加える。そのために、まず、これら原料の濃度を適宜調整した後に70〜125℃で5〜40分、好ましくは80〜95℃で30分程度に相当する加熱処理を行う。遠心分離処理を行い、上清すなわちプロテオースペプトン溶液を採取する。この上清に溶液の飽和度が50〜90%、好ましくは55〜90%となるように硫安を添加し(硫安の添加法は「実験農芸化学 下」朝倉書店 1978年 p.59-60を参照)、硫安沈殿物を回収する(硫酸アンモニウム分画法、硫酸アンモニウム塩析法又は硫酸アンモニウム沈殿法ともいう)。その際に硫安沈殿により生成した上清に硫安を加え、硫安沈殿物を繰り返し回収することも望ましい。
【0019】
本発明では、プロテオースペプトン溶液の硫安沈殿物に代わり、プロテオースペプトン画分そのもの、又は乳清の酵素分解物を用いることができる。また、乳清に限らず、乳清タンパク質濃縮物、その精製物などを酵素分解処理したものも好ましい。酵素としては、ペプシン、プロナーゼ、トリプシン、α-キモトリプシン、パパインなどが挙げられ、各酵素の至適pHにして、30〜60℃で1〜5時間の加水分解処理を行う。このとき、タンパク質:酵素の重量比率は、100:0.1〜2とする。
【0020】
こうして得られるプロテオースペプトン画分、プロテオースペプトン溶液の硫安沈殿画分、あるいは乳清などの酵素分解物を一次元目にpIが5〜8の等電点電気泳動、二次元目に15%のポリアクリルアミド単一ゲルを用いた二次元電気泳動を行うことで、ウイルス感染阻害活性の高いタンパク質群を採取することができる。後述する実施例の通り、乳清を加熱処理して得られるプロテオースペプトン画分を用いた場合には、ポリアクリルアミドゲルから電気泳動溶出した18kDa付近に少なくとも6つのスポットとして検出される。SDS-PAGEによる分子量が15〜20kDa(以下、約18kDaともいう)、等電点pIが6〜9であり、6つのスポットを弱酸性側からA、B、C、D、E、Fとしたときに、単独物ではE及びFのスポットがウイルスの感染を阻害する最小阻害濃度が1.1μg/ml及び0.78μg/mlと非常に強く、混合物ではB+C+Dが0.31μg/ml未満と高い活性を有することが分かった。
【0021】
また、アミノ酸シークエンスによりスポットC、D、E、FのN末端から5残基がSer-Ser-Arg-Gln-Pro(配列番号:1)であることが確認された。糖鎖の有無を調べるため、レクチン染色を行ったところ、RCA120及びConAとの反応性では、6つのスポット全てにおいてポジティブな反応を示した。RCA120は糖鎖にあるβ-ガラクトースを認識し、ConAはマンノースを認識して結合するものであることから、これらタンパク質が糖鎖にβ-ガラクトース及びマンノースを有する糖タンパク質であることが判明した。更にN-アセチル-D-グルコサミンがβ-1、4結合したオリゴ糖などを認識するWGAレクチンにも、6つのスポット全てがポジティブな反応を示した。
【0022】
一方、乳清を限外濾過処理により濃縮した後に、ゲル濾過により得られる画分を加熱処理して得られるプロテオースペプトン画分を用いた場合には、ポリアクリルアミドゲルから電気泳動溶出した16kDa付近に複数のスポットが検出される。弱酸性側からスポットをA、B、Cとしたときに、CのN末端から7残基がIle-Leu-Lys-Glu-Lys-His-Leu(配列番号:2)で、SDS-PAGEによる分子量が14〜19kDa(以下、約16kDaともいう)、等電点pIが6〜9の新規糖タンパク質であることが確認された。Cを含む16kDa付近にスポットされるタンパク質群のウイルスの感染を阻害する最小阻害濃度を測定したところ、0.16μg/ml未満であった。また、レクチン染色により、RCA120レクチンと反応性が見られたことから、約16kDa付近にスポットされるタンパク質群はβ-ガラクトースを有する糖タンパク質であることが示された。
【0023】
本発明に係る糖タンパク質の少なくとも一部は、上記のようにレクチンとの結合性を有することからN型糖鎖(N結合型糖鎖、アスパラギン結合型糖鎖ともいう)を有することが示された。また、本発明に係る糖タンパク質は、上記で耐熱性と可溶性を有する画分に見出されたように、耐熱性があり、可溶性(水溶性)である。従って、本発明の糖タンパク質は、プロテオースペプトン画分などのウシ乳清由来耐熱性可溶画分からアフィニティークロマトグラフィー法などによって得られるレクチン(ConA、RCA120など)結合画分を、電気泳動などの公知のタンパク質分画法によって更に分画し、そこから本発明の糖タンパク質を規定する上記分子量及び等電点を示す画分を採取することによって調製することもできる。
【0024】
本発明の新規糖タンパク質を含む組成物は、食品(とりわけ育児用調製粉乳や仔ウシの飼料)に、その有効量を混合することにより、ヒト乳児、仔ウシ、仔ウマなどにおけるロタウイルスが原因の下痢症を予防又は治療するのに有効であることが期待できる。
【0025】
ロタウイルス感染における食品成分の抗ロタウイルス感染効果を検定・評価する方法は多岐に渡り、万全の検定・評価系はないとされている。これまでに様々な抗ロタウイルス感染効果の検定・評価法が報告されている。例えば、本発明で用いた方法以外に、実験動物(マウス)にロタウイルス及び食品成分を投与し、下痢の発症及び消化管粘膜へのロタウイルスの結合量を測定するなどの検定・評価法が挙げられる(Duffy L.C. Pediatr. Res. 35: 690-695(1994))。本発明は、これらの系を適切に用いて該組成物の抗ロタウイルス感染活性を更に詳細に調べることが可能である。
【0026】
本発明において用いる新規糖タンパク質は、乳由来であることから長い食経験の中で安全性は確立されている。従って、ヒトによる試験で本発明品の有効性及び有効量の確認が可能である。
【0027】
本発明では、上記のような新規糖タンパク質をロタウイルス感染防御用組成物として用いることができる。本発明のロタウイルス感染防御用組成物は、本発明に係る2種の糖タンパク質を両方とも含んでも良いが、いずれか一方のみを含むものでも良い。
【0028】
本発明のロタウイルス感染防御用組成物は、適量を添加することにより、飲食品や医薬組成物にロタウイルス感染防御作用を付与することができる。このロタウイルス感染防御用組成物は、それ自身(液状又は粉末状)又は他の活性物質と共に、あるいは他の薬理学的な活性物質と共に用いることができる。形態は例えば、錠剤もしくは被覆錠、カプセル、タブレット、溶液、シロップ、乳液又は分散性粉末などの任意の形態であって良い。摂取量は年齢、身体状態などに依存して変化するが、1日当たり0.001〜1000mg/kg体重、好ましくは0.001〜200mg/kg体重である。
【0029】
本発明は、ロタウイルス感染防御用組成物を添加した食品にも関する。本明細書において食品とは、液体状、固体状、ゲル状、顆粒状、粉末状、タブレット状などのいずれでも良く、特に限定するものではないが、飲料、食品及び機能性食品を含む。
【0030】
食品として具体的には、牛乳、乳製品、清涼飲料水、菓子類、パン、麺、練り製品、調理加工食品、畜肉加工食品、調味料などを挙げることができる。食品の形態として中でも好ましいのが機能性食品である。本発明の機能性食品は、生体に対して一定の機能性を有する食品を意味し、例えば、特定保健用食品及び栄養機能食品を含む保健機能食品、特別用途食品、栄養補助食品、健康補助食品、サプリメント(例えば、錠剤、被覆錠、糖衣錠、カプセル及び液剤などの各種剤形のもの)及び美容食品などの、いわゆる健康食品全般を含む。また、本発明の機能性食品には、コーデックス(FAO/WHO合同食品規格委員会)の商品規格に基づく健康強調表示(Health claim)が適用される健康食品を含む。
【0031】
本発明の好適な機能性食品として好ましい具体例には、乳児用、幼児用、授乳婦用、高齢者用、病者用などの特別用途食品がある。乳児用には例えば、乳児用調製乳が含まれ、通常の乳幼児用調製乳の原料に添加して、ロタウイルス感染に対する防御能の発達が未熟な乳幼児のための育児用調製粉乳又は液体調製乳に使用できる。育児用調製粉乳には、乳児用調製粉乳、フォローアップミルク、低出生体重児を対象とする低出生体重児用調製粉乳、牛乳アレルギーや乳糖不耐症などの病的状態を有する小児の治療に用いられる各種治療用ミルクなどが含まれる。また、高齢者用には、ロタウイルス感染に対する防御能の低下した高齢者向けに、その有効量を食品に添加してロタウイルス感染防御用食品とすることができる。更に該組成物を家畜用飼料に、その有効量を添加してロタウイルス感染防御用飼料とすることができる。
【0032】
本発明のロタウイルス感染防御用組成物は、当業者が利用可能である任意の適切な方法によって食品に含めれば良い。例えば、ロタウイルス感染防御用組成物を、液体状、ゲル状、固体状、粉末状又は顆粒状にした後に、それを食品に配合する。あるいは、ロタウイルス感染防御用組成物を、食品の原料中に直接で混合又は溶解しても良い。また、食品に塗布、被覆、浸透又は吹き付けても良い。あるいは、飲食品中に均一に分散又は偏在させても良い。また、可食フィルムや食用コーティング剤などで包み込んでも良い。ロタウイルス感染防御用組成物を含む飲食品を更に加工しても良く、そのような加工品も本発明の範囲に含まれる。
【0033】
本発明の食品の製造において、食品に使用される各種添加物を使用しても良い。具体的には、発色剤(亜硝酸ナトリウムなど)、着色料、香料、甘味料、保存料、乳化剤、酸化防止剤、pH調整剤、化学調味料、増粘剤、安定剤、膨張剤、消泡剤、結着剤、栄養強化剤などが挙げられる。
【0034】
本発明のロタウイルス感染防御用組成物を、医薬組成物として使用する場合には、医薬製剤上で許容される担体又は添加物を配合しても良い。具体的には、水、医薬的に許容される有機溶剤、コラーゲン、ポリビニルアルコール、ポリビニルピロリドン、カルボキシビニルポリマー、アルギン酸ナトリウム、水溶性デキストラン、ペクチン、キサンタンガム、アラビアゴム、カゼイン、ゼラチン、寒天、グリセリン、プロピレングリコール、ポリエチレングリコール、ワセリン、パラフィン、マンニトール、ラクトース、ソルビトール、医薬添加物として許容される界面活性剤などが挙げられる。使用される添加物は、製剤の剤形に応じて適宜又は組み合わせて選択される。本発明の医薬組成物は更に他の薬理成分を含有しても良い。そして、錠剤、顆粒剤、散剤、丸剤、カプセル剤などの固形製剤、ジェル剤もしくは液剤、懸濁剤、シロップ剤などの液体製剤などの剤形にして経口的に投与する他に非経口的に投与することもできる。
【0035】
本発明のロタウイルス感染防御用組成物を配合する飼料としては、家畜、家禽、ペットなどの配合飼料又は配合飼料用原料が挙げられる。
【0036】
また、本発明のロタウイルス感染防御用組成物に加え、乳酸菌、ビフィズス菌などの他のロタウイルス感染防御作用を有する組成物を併用しても良い。
【実施例】
【0037】
以下、本発明の効果を試験例及び実施例に基づき具体的に説明するが、本発明は、これらの具体例に限定されるものではない。
【0038】
[試験例1] 本発明における二次元電気泳動法
等電点電気泳動
1.膨潤化
泳動に用いるStripゲルを膨潤させる。
[IPG Ready Strip]
7cm pH 3-10(BIO-RAD社製)
[膨潤用緩衝液]
2.4gの Urea
0.05gの DTT
0.2gの CHAPS
0.01mlの Bio-Lytes
5μl、50mg/mlの Orange G
これに5mlのMilli-Q(Millipore社)で作成した超純水(以下、MilliQともいう)を加え、冷凍保存した。
【0039】
フォーカシングトレイ(一次元目の泳動で用いる電極装置)のスロットに、膨潤緩衝液で希釈した試料の必要量を入れる。Ready Stripマイナス側をピンセットで持ち、ゲルが付着している面を下側にして、プラス側の方から、ゆっくりと膨潤緩衝液に触れさせる。気泡をトラップしないように注意しながら、数回、Stripを前後にスライドさせ、膨潤緩衝液をゲル面に均一に行き渡らせる。1時間程放置して膨潤緩衝液をゲルに吸収させた後に、ミネラルオイル(BIO-RAD社製)を端から、ゆっくり滴下してゲル全面をカバーし、トレイの蓋をする。なお、膨潤する際には、トレイをIEFセル本体にセットし、20℃、active 50 Voltに設定し、12時間程放置する。
【0040】
2.泳動
プログラム、プリンターは以下のように設定し、この条件下で一次元目の泳動を行う。
[プログラム]
step1:250V、15min、20℃
step2:Start 250-End、4,000V、2hr、20℃
ボルトラインピング設定
電流値リミットは、50μA/Strip
step3:4,000V-20,000VH、5hr、20℃
[プリンター]
Voltage Slopeは、LINEARを選択し、穏やかな電圧上昇のもとで泳動し、泳動が終了した後に、Ready Stripに流れたタンパク質が乱れないように、500V/HOLDを設定する。
【0041】
15%のポリアクリルアミド単一ゲルを用いたSDS-PAGE
1.分離ゲルの作成
[A溶液:30%のアクリルアミド溶液]
29.2gのアクリルアミド(モノマー)
0.8gのN,N’-メチレンビスアクリルアミド
純水で100mlとする。
[B溶液:1.5MのTris-HCl緩衝液(pH 8.8)]
18.17gの Tris
0.4gの Sodium dodecyl sulfate(SDS)
純水で溶解し、HClでpHを8.8に調整した後に、最終容量を100mlとする。
[C溶液:10%の過硫酸アンモニウム溶液]
0.1gの過硫酸アンモニウム
純水1mlを加える。
[TEMED]
N,N,N',N',-Tetramethylethylenediamine
[泳動用緩衝液]
14.4gのグリシン
3gのTris
1gのSDS
純水で1000mlとする。
【0042】
15%のアクリルアミド溶液は、A溶液を8ml、B溶液を4ml、C溶液を0.0063ml、TEMEDを0.01ml、純水を4mlで混合して調製した。ゲル板の中央よりピペットマンでゲル調製溶液を注入する。ゲル調製溶液を導入した後に、その上部に純水を静かに注いで空気を遮断し、アクリルアミドをゲル化させる。
【0043】
2.Ready Stripの平衡化とゲル板へのセット
IPG Ready Stripを二次元目の泳動に必要なSDS緩衝液系にするため、平衡化を行う。使用する溶液は、以下の通りである。
[平衡化緩衝液1]
3.6g/10mlの Urea
0.2g/10mlの SDS
2.5ml/10ml、1.5Mの Tris-HCl
2ml/10mlの Glycerol
200mg/10mlの DTT
以上、各試薬10mlを合わせて50mlとする。
[平衡化緩衝液2]
3.6g/10mlの Urea
0.2g/10mlの SDS
2.5ml/10ml、1.5Mの Tris-HCl
2ml/10mlの Glycerol
250mg/10mlの Iodoacetamide
以上、各試薬10mlを合わせて50mlとする。
【0044】
等電点電気泳動が終了したStripゲルを、ゲル面を上にして膨潤トレイに入れ、4mlの平衡化緩衝液1を注ぎ、静かに振とうしながら、10分間平衡化する。更に平衡化緩衝液2で同様にして、10分間平衡化する。作成したゲルのインナーガラスプレートを手前に向けて置き、Stripのプラスチックフィルムをアウターガラスに付け、Stripの端から気泡が入らないように、ゆっくりウェルに入れる。その上を電子レンジで溶解して人肌程度に冷ました1%のアガロースゲルで固定化し、泳動槽にセットして泳動用緩衝液を注ぐ。
[泳動条件]
200V、30mAの定電圧、定電流で泳動を行う。
【0045】
3.ポリアクリルアミドゲルの染色
CBB染色
電気泳動後のタンパク質染色にはCBB染色を用いる。使用する溶液は、以下の通りである。
[CBB染色]
2.5gの CBB
454mlのメタノール
75mlの 酢酸
これに純水を加えて1000mlとし、3回濾過する。
[脱色液]
250mlのメタノール
375mlの酢酸
純水で5000mlとする。
【0046】
ゲルを染色液に約20分間浸した後に、脱色液でバックグラウンドの色が抜けるまで脱色する。
【0047】
[試験例2] ポリアクリルアミドゲルからのタンパク質の電気泳動溶出
ポリアクリルアミドゲルからのタンパク質の電気泳動溶出は、Centrilutor micro-electroeluter(Millipore社製)を用いて行うことができる。キャップと底に穴(孔)が空いた0.5mlのサンプルチューブに切り出したポリアクリルアミドゲル細片を入れる。下部のバッファーチェンバーに溶出緩衝液(25mMのTris、192mMのグリシン、0.02%のSDS、20%のメタノール)を注ぎ、上部のバッファーチェンバーラックの穴(孔)に分画分子量が10000Daのセントリコン装置を取り付ける。使用しない穴(孔)はストッパーで閉め、下部のバッファーチェンバーに上部のバッファーチェンバーをのせる。上段のバッファーチェンバーを溶出緩衝液で満たし、セントリコン装置の下部に気泡がないか確認する。気泡がある場合には、シリンジを用いて除去する。それぞれのセントリコン装置のサンプルリザーバーにゲル端片の入ったサンプルチューブを差し込む。溶出機に安全カバーを着け、電極をコンセントに差し込み、200Vで4時間溶出を行う。
【0048】
[試験例3] タンパク質濃度の測定
タンパク質の定量にはBradford法に従う。Protein assay reagent(BIO-RAD社製)を使用し、ウシ血清アルブミンを標準溶液として測定する。各試料は、PBS(−)[Ca及びMgイオンを除去したDulbecco's Phosphate Buffered Saline]で希釈し、マイクロプレートに10μlずつ注ぐ。5倍に希釈した200μlのBradford試薬を加えてピペッティングし、595nmにおける吸光度を測定する。
【0049】
[試験例4] 抗ロタウイルス活性の測定法(ロタウイルス中和活性試験)
ウシ乳由来組成物の加熱処理物を試料として、ヒトロタウイルスMO株の感染阻害活性を以下のように測定することができる。
(1)培養細胞の調製
ウイルスの増殖及び抗ロタウイルス活性の測定に、37℃で7日間培養した単層のアカゲザル腎臓由来株化細胞MA104細胞(J.Clin.Microbiol.Oct. 1982, 727-730)を用いる。継代後の4日間は、10%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地中で培養し、継代して5日目に2%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地に交換する。
(2)ウイルスの調製
ウイルスには、ヒトロタウイルスMO株(血清型3)(J.Clin.Microbiol.Oct.1982,727-730)を用いる。あらかじめ20μg/mlのトリプシンにより、37℃で30分間混合し、M.O.I(Multiplicity Of Infection;細胞1個当たりの接種ウイルス数)=1となるように希釈し、これを0.11%のウシ血清アルブミン、ジエチルアミノエチル、デキストランを加えたHanks液で3回洗浄した単層細胞上に接種する。37℃で60分間吸着させた後に、ウイルスを除去し、前記Hanks液で洗浄した後に、0.5μg/mlのトリプシンを含む2%のウシ胎児血清、5%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地を加え、37℃で回転培養する。大部分の細胞が壁面から剥離したら、その培養液を回収し、遠心分離処理(3000rpm、10分間)を行い、その上清をストックウイルスとして分注し、使用時まで−80℃で保存する。
(3)ウイルスの感染
ウイルスの接種は同時接種法で行う。すなわち、PBS(−)[Ca及びMgイオンを除去したDulbecco's Phosphate Buffered Saline]で溶解した試料原液を10%ウシ胎児血清、10%トリプトース・ホスフェイト・ブロスを含むイーグルMEN培地で段階希釈し、50μlの希釈試料(試料溶液は0.45μmのフィルターで濾過滅菌)を0.5mlのエッペンドルフチューブに分注する。次いで105〜106FCFU(Fluorescent Cell Focus Forming Unit)/mlのヒトロタウイルスMO株液の0.4mlを、20μg/mlのトリプシン溶液の0.4mlにより、37℃で30分間の処理を行い、10%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地で20〜100倍に希釈した後に、各エッペンドルフチューブに50μlずつ加え、37℃で1時間培養した。空試験用(試料無添加時)には、前記希釈試料の50μlに代えて、10%ウシ胎児血清、10%トリプトース・ホスフェイト・ブロスを含むイーグルMEN培地を用いる。
【0050】
培養が終了した後に、アカゲザル腎臓由来のMA-104細胞の細胞浮遊液をウイルス−試料混合液が入った各エッペンドルフチューブに100μlずつ加え、24ウェルスライドグラス上に各20μlずつ分注して、37℃で45〜65時間培養する。
【0051】
前記細胞浮遊液は、37℃で7日間培養した単層のアカゲザル腎臓由来のMA-104細胞を、0.025%のEDTAを添加した0.125%のトリプシン溶液で剥離し、更に遠心分離処理して上清を取り除いた後に、2×105/mlとなるように培養液に加えることで得られる。そして、培養液には10%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地を用いる。
【0052】
培養が終了した後に、アセトンで固定し、ヒトロタウイルスMO株感染細胞数を間接蛍光抗体法で検出する。なお、一次抗体として鳩ロタウイルスPO-13株のVP6を特異的に認識する抗ロタウイルスモノクローナル抗体(Minamoto,N. et.al.,Epidem,Inf.,100,481-492(1988))及び二次抗体として蛍光標識したヤギ抗マウスIgG血清(Organon Teknika-Cappel,NC,U.S.A.)を用いる。
【0053】
増殖阻害活性の評価は、100×(1−(試料添加時の感染細胞数)/(試料無添加時の感染細胞数))の値を阻害率として行う。
【0054】
また、前記値が50%以上のものをポジティブとして判断し、活性が認められた最も低い試料濃度を最小阻害濃度(MIC)として活性を数値化する。
【0055】
[実施例1] 乳清からの抗ロタウイルス感染組成物の調製
1.プロテオースペプトン画分の調製
搾乳した未加熱の10リットルの生乳(原乳)を5℃に冷却し、遠心分離処理(3000rpm、20分)で乳脂肪を除去し、脱脂乳を調製した。pHを4.6に調整した後に、沈殿したカゼインを遠心分離処理(20℃、3000rpm、15分)で除去し、乳清画分(酸ホエイ)を得た。次いで、酸ホエイを95℃で30分間の加熱処理を行い、30℃まで冷却した後に、変性したタンパク質を遠心分離処理(5000G、20℃、30分間)で除去し、プロテオースペプトン画分を得た。
【0056】
2.硫安画分の調製
前記プロテオースペプトン画分に100%の飽和硫安溶液を55%の飽和度になるまで加えた。数時間放置した後に、遠心分離処理(7000G、20℃、30分間)を行った。上清を採取し、90%の飽和度になるまで、更に100%の飽和硫安溶液を加えた。スターラーで撹拌しながら、一晩放置した後に、遠心分離処理(7000G、20℃、30分間)を行い、生じた沈殿を脱イオン蒸留水に溶解し、48時間透析を行った。
【0057】
3.二次元電気泳動による精製
透析した後に得られた溶液を試料として二次元電気泳動を行った。試験例1記載の条件で一次元目にはpIが5〜8の等電点電気泳動、二次元目には15%のポリアクリルアミド単一ゲルを用いたSDS-PAGEを行った。そして、ポリアクリルアミドゲル上に認められた等電点pIが6〜9で分子量約18kDa付近のタンパク質スポット群を、試験例2に従って電気泳動溶出し、採取した(図1)。
【0058】
[実施例2] 乳清タンパク質濃縮物からの抗ロタウイルス感染組成物の調製
実施例1に従って酸ホエイを調製し、pHを6.0とした。0.15MのNaCl及び0.02%のNaN3を含む10リットルのリン酸緩衝液(pH 6.0)を用い、分画分子量が300万Daの分離膜で限外濾過処理を行った。その保持液を採取し、常法により凍結乾燥することで、500mgの乳清タンパク質濃縮物の粉体を得た。この粉末を溶出緩衝液で濃度を9mg/mlに調整し、Sephacryl S-500(アマシャムファルマシアバイオテク社製)を充填したカラム(5×60cm、ゲル容積1200ml)に30mlをアプライした。なお、溶出緩衝液には、0.15MのNaCl、1mMのNa2EDTA、0.02%のNaN3を含む0.05MのTris-HCl緩衝液を用い、280nmの吸光度における最初のピークに相当する分画を採取した。その後に実施例1と同様にして、画分を加熱処理した後に硫安沈殿を行い、生じた沈殿を脱イオン蒸留水に溶解し、48時間透析を行った。
【0059】
透析した後に得られた溶液を試料として実施例1と同様の方法で二次元電気泳動を行い、等電点pIが6〜9で約16kDa付近に認められたタンパク質スポット群を採取した(図2)。
【0060】
[実施例3] 抗ロタウイルス活性を有する乳清タンパク質分解物の調製
搾乳した未加熱の100 kgの生乳(原乳)を63℃で30分間の加熱処理を行い、常法に従ってゴーダチーズを調製し、生成したホエイ(pH 6.0)を分離した。このホエイを加温し、遠心分離処理(3000G、20分間、40℃)を行い、脱脂ホエイを得た。この脱脂ホエイを国際公開第2004/080475号の実施例1に記載された方法に従い、精密濾過処理し、保持液を回収した。更に得られた保持液に脱イオン水を添加し、精密濾過処理を3回繰り返した(ダイアフィルトレーション処理)。最後に得られた保持液はスプレードライ法により乾燥した。次いで乾燥試料の2%の水溶液を調製し、そのpHを1NのHCl及び1NのNaOHでpHを3.0に調整した。55℃に加温してから乾燥試料重量の0.5%に当たるペプシン(Sigma社製)を添加し、3時間の酵素処理を行った。酵素処理液は沸騰水浴中で5分間の加熱処理を行うことで酵素を失活させ、その後に凍結乾燥することで抗ロタウイルス活性の試験試料(ペプシン分解物)を得た。全く同様にして、2%の水溶液をpHで8.0に調整し、ペプシンの代わりにプロナーゼ(Merck社製)を用いることでプロナーゼ分解物を得た。
【0061】
[実施例4] 抗ロタウイルス活性成分の分画1
1.レクチン染色
実施例1で確認された約18kDaのスポットについてレクチン染色を行うため、実施例1に従って二次元電気泳動を行った後に、イモビロンPVDF膜に転写した。転写した後に、CBB染色を行い、次いでレクチン染色をした。
【0062】
まず、ろ紙、スポンジパットを転写緩衝液(25mMのTris、192mMのグリシン、0.02%のSDS、20%のメタノール)に15分間浸し、平衡化させた。イモビロンPVDF膜メタノールに1分間浸し、脱イオン蒸留水で洗浄した後に、転写緩衝液に入れ、振とうした。ゲルをイモビロンPVDF膜に密着させ、ろ紙、スポンジパット、ホルダーに挟み、緩衝液を満たした電気泳動槽中にゲルをマイナス側にして垂直に固定し、30Vで一晩通電した。転写した後のPVDF膜にCBB染色液を加え、染色の度合いに応じて約10分間振とう染色を行った。染色した後に、脱色の度合いに応じてCBB脱色液で10〜15分間脱色し、脱イオン蒸留水で洗浄し、風乾した。更に緩衝液(0.9%のNaCl、0.05%のTween20、10mMのTris-HCl、pH 7.4)にペルオキシダーゼを結合させた5μl/10mlのレクチンを加え、PVDF膜上のタンパク質に対し、1時間振とう反応を行った。反応が終了した後に洗浄し、15mMのリン酸緩衝液(pH 6.8)で平衡化を行って発色液で発色させた。レクチンにはRCA-120、ConA、WGAを用いた。
【0063】
実施例1で確認された約18kDaのスポットに対し、弱酸性側からA、B、C、D、E、Fとしたところ(図3)、RCA-120及びConAレクチンとの反応性は、6つすべてのスポットにおいてポジティブな反応であった。従って、約18kDaのタンパク質はβ-ガラクトース及びマンノースを構成単糖として持つことが示された。また、WGAレクチンとの反応性でも、全てのスポットにおいてポジティブな反応を示した。WGAレクチンによって認識される糖鎖構造はN-アセチル-D-グルコサミンがβ-1,4結合したオリゴ糖、バイセクティングN-アセチル-D-グルコサミンを有する直鎖型の糖鎖構造、N-アセチルノイラミン酸が挙げられる。このように親和性を示す糖鎖構造がいくつか報告されているため、WGAレクチンによる染色からは、どのような糖鎖構造を持つか特定はできないが、3種のレクチン染色から、約18kDaタンパク質スポットは共通した糖鎖構造を持つことが示された。また、各種レクチン染色において、pIの高いスポットの方が、よりポジティブに染色していたことから、スポットによって修飾を受けている糖鎖の量が異なっていることが示唆された。
【0064】
2.N末端アミノ酸配列の特定
実施例1で採取したスポット群のうち、スポットC、D、E、FのN末端から5残基を気相シークエンス(綱沢進 他、続生化学実験講座 1 タンパク質の化学(上)、東京化学同人、347-373(1987))(Procese 49-HT プロテインシークエンサー)により測定したところ、これらのN末端配列5残基がプロテオースペプトン3(Esben S.S.et.al.,J Dairy Res. 60 535-542(1993))のアミノ酸54位から58位の配列(Ser-Ser-Arg-Gln-Pro)と一致する新規な糖タンパク質であることが確認された。
【0065】
3.抗ロタウイルス活性の測定
更にB+C+Dからなるスポット群、スポットE及びスポットFの抗ロタウイルス活性を測定した。以下に手順を示す。
【0066】
二次元電気泳動を行ったゲルから、試験例2に従ってタンパク質を電気泳動溶出することで試料を分離した。試料原液に含まれるタンパク質濃度を測定後(試験例3参照)、この原液を6段階希釈し、1試料につき計6つずつ試料を用意した。そして、これら試料のウイルス増殖阻害活性を測定した(試験例4参照)。
【0067】
その結果、ウイルスの感染を阻害する最小阻害濃度は、スポット群(B+C+D)が0.31μg/ml未満、スポットEが1.1μg/ml、スポットFが0.78μg/mlであった(図4)。なお、スポット群(B+C+D)は、低いタンパク質濃度でも高い感染阻害率をし、タンパク質の濃度が高くなるにつれ、100%に近い感染阻害率を示すことが分かった(図5)。
【0068】
[実施例5] 抗ロタウイルス活性成分の分画2
1.レクチン染色
実施例4と同様にして、実施例2で確認された約16kDaのスポットについてRCA-120を用いたレクチン染色を行った。その結果、レクチンと反応性が見られたことから、約16kDaのタンパク質スポット群は、β-ガラクトースを持つことが示された。
【0069】
2.N末端アミノ酸配列の特定
次いで、約16kDaのタンパク質スポット群を弱酸性側からA、B、Cとし(図6)、実施例2で採取した、これらのスポット群においてスポットCのN末端から7残基をアミノ酸シークエンス(Procese 49-HT プロテインシークエンサー)により測定した。その結果、N末端配列7残基がプロテオースペプトン3(Esben S.S.et.al.,J Dairy Res. 60 535-542(1993))のアミノ酸69位から75位の配列(Ile-Leu-Lys-Glu-Lys-His-Leu)と一致する新規な糖タンパク質であることが確認された。
【0070】
3.抗ロタウイルス活性の測定
実施例2で採取したスポット群のうち、A、B、Cを含むスポット群(A+B+C)の抗ロタウイルス活性を測定した。ロタミルク(EBINA,T.et.al.,J.Med.Virol.38 117-123(1992))をポジティブコントロールとした。また、実施例2における二次元電気泳動の代わりに一次元電気泳動を実施し、CBB染色で約18kDaの位置に認められたタンパク質(heated F1)を比較試料とした。なお、その際に一次元電気泳動のゲルは、試験例1の15%のポリアクリルアミド単一ゲルを用いた。試料の採取では、電気泳動により分離されたタンパク質を試験例2に従って電気泳動溶出することで得た。抗ロタウイルス活性は実施例4と同様にして測定した。
【0071】
その結果、A、B、Cを含むスポット群は、ウイルスの感染を阻害する最小阻害濃度が0.16μg/ml未満と最も高く(図7)、感染阻害率の濃度依存性においてはタンパク質濃度が高濃度になるにつれ、感染阻害率が100%に近付くことが分かった(図8)。
【0072】
[実施例6] 酵素分解物の抗ロタウイルス活性の測定
1.抗ロタウイルス活性の測定1
実施例3で得られたペプシン分解物及びプロナーゼ分解物を試料とし、実施例4と同様にして抗ロタウイルス活性を測定した。
【0073】
各試料の最小阻害濃度は、ペプシン分解物が4μg/ml、プロナーゼ分解物が5μg/mlであった(図9)。また、感染率の濃度依存性の結果は図10に示す通りであり、ペプシン分解物に比べ、プロナーゼ分解物は低濃度で、より高い感染阻害率を示すことが分かった。
【0074】
2.抗ロタウイルス活性の測定2
一方、プロナーゼ分解物については実施例3の試料に加えて、水溶液の状態で更に100℃で15分間あるいは120℃で5分間の加熱処理を行ったものを調製し、凍結乾燥して試料とした。これら2つの試料の抗ロタウイルス活性を、ロタミルク(EBINA,T.et.al.,J.Med.Virol.38 117-123(1992))を対照として実施例4と同様にして測定した。
【0075】
各試料の最小阻害濃度は、12μg/mlのプロナーゼ分解物(100℃で15分間の加熱処理)、2.1μg/mlのプロナーゼ分解物(120℃で5分間の加熱処理)、0.29/mlのロタミルクであった(図11)。また、感染率の濃度依存性は、図12のようになった。
【0076】
3.タンパク質の電気泳動溶出
前記プロナーゼ分解物及びペプシン分解物を試料とし、実施例1と同様の方法で二次元電気泳動を行った。その結果、プロナーゼ分解物は約18kDaに3つのスポットが認められた(図13)。また、ペプシン分解物は約18kDaに4つのスポットが認められた(図14)。
【0077】
以上より、ホエイの精密濾過処理の保持液又はその乾燥物の酵素分解物にも抗ロタウイルス感染阻害活性が認められ、その活性には、約18kDaのタンパク質が関与していることが示された。
【0078】
[実施例7] 抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体との反応性
実施例1や実施例3で確認された約18kDaのスポットに関する情報を得るため、抗ラクトフォリン(プロテオースペプトン3)のモノクローナル抗体染色を行った。抗体染色は、実施例1のプロテオースペプトン画分、その硫安画分及び実施例3で酵素分解物の調製に用いた乳清タンパク質の3つの試料で検討した。
【0079】
実施例1に従って二次元電気泳動を行った後に、実施例4のレクチン染色の場合と同様にして、イモビロンPVDF膜に転写した。転写した後に、一次抗体には抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体1C10(青木らの方法により作製(青木直人 他、ミルクサイエンス、54(2)、pp.83-84(2005)))、二次抗体にはペルオキシダーゼ標識した抗マウスIgG抗体(MP Biomedicals Inc.-Cappel Products)を用い、それぞれイモビロンPVDF膜と反応させた。最後にペルオキシダーゼの活性染色をDAB(ジアミノベンチジン;和光純薬工業)の基質溶液で行った。
【0080】
得られた抗体染色の結果を図15、図16、図17に示した(いずれも弱酸性側からスポット群A、B、C、D、E、Fとする)。プロテオースペプトン画分、その硫安画分及び実施例3で酵素分解物の調製に用いた乳清タンパク質の3つの全ての試料において、約18kDaのスポットが認められ、実施例4でウイルスの感染を阻害する活性が最も高いスポットであることが判明したFに相当する箇所が共通して抗体染色されていた。この結果は、FのN末端配列5残基がプロテオースペプトン3(Esben S.S.et.al.,J Dairy Res. 60 535-542(1993))のアミノ酸54位から58位の配列(Ser-Ser-Arg-Gln-Pro)と一致したことを支持するとともに、抗ロタウイルス活性にプロテオースペプトン3由来のタンパク質が関与することを改めて示した。
【0081】
[実施例8] プロテオースペプトン画分の硫安沈殿画分中の分子量が約28kDaと約18kDaのタンパク質における抗ロタウイルス活性の測定
95℃で30分間の加熱処理を行った脱脂乳に酸添加を行い、カゼイン及び変性タンパク質を沈殿除去して、プロテオースペプトン画分(以下、PP画分ともいう)を調製した。PP画分に、100%飽和硫安溶液を55%の飽和度になるまで加え、遠心分離処理を行い、上清を採取した。更に100%飽和硫安溶液を90%の飽和度になるまで加え、一晩放置した後に、遠心分離処理を行い、生じた沈殿物を採取した。次いで、この沈殿物を脱イオン蒸留水に溶解し、透析を行った。得られた溶液は、プロテオースペプトン画分由来の硫安飽和55%可溶かつ90%不溶の画分であり、以下「PP(0.90)画分」と称した。調製したPP画分とPP(0.90)画分については、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。その活性の評価は、感染阻害率の濃度依存性及び最小阻害濃度(MIC)に基づいて行った。PP画分及びPP(0.90)画分は共に感染阻害活性を示したが、PP画分の最小阻害濃度(MIC)は16μg/ml、PP(0.90)画分の最小阻害濃度(MIC)は19μg/mlであり、活性の強さには大きな違いは認められなかった(図18)。
【0082】
このPP画分及びPP(0.90)画分を膨潤用緩衝液に溶解して、3mg/ml濃度の溶液を調製した。その溶液の125μlを試料とし、試験例1記載の条件で、一次元目にpIが5〜8の等電点電気泳動に供し、二次元目に15%のポリアクリルアミドゲルを用いたSDS-PAGEに供した後に、実施例7に従って、抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体(1C10)を一次抗体として使用するウエスタンブロット解析を行った。その結果、いずれの画分についても、分子量が約28kDaのスポット群及び約18kDaのスポット群に染色反応が認められた(図19)。実施例4の「2.N末端アミノ酸配列の特定」に記載されている通り、本願発明に係る約18kDaのタンパク質は、そのN末端配列がプロテオースペプトン画分中のプロテオースペプトン3の内部配列と一致したことから、本実施例で検出された約28kDaのタンパク質が牛乳に内在するプロテアーゼの一種であるプラスミンによって分解される際に生成したものと考えられた。
【0083】
SDS-PAGEで分画したPP(0.9)画分からは、実施例4に従って約18kDaと約28kDaのタンパク質を個々に電気泳動溶出し(切り出したゲルから低電圧200Vにて4時間反応)、試料として調製した(図19)。溶出した約28kDaのタンパク質の試料はLP 28K、約18kDaのタンパク質の試料はLP 18Kと称した。次いで、3つの試料(LP 28K、LP 18K、PP(0.9)画分)について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0084】
その結果を図20のA及びBに示す。図20に示される通り、約18kDaのタンパク質(LP 18K)の感染阻害率は、調べた中の、どのタンパク質濃度でも高く、かつタンパク質濃度に依存して上昇した。また、約18kDaのタンパク質については最小阻害濃度(MIC)も0.62μg/mlであり、極めて強い活性が示された。一方、約28kDaのタンパク質(LP 28K)は、調べた範囲のタンパク質濃度では十分な感染阻害率を示すことができず、最小阻害濃度から見てもほとんど活性が認められなかった。更に約18kDaのタンパク質の結果をPP(0.9)画分と比較すると、例えば、10μg/mlのタンパク質濃度での感染阻害率がPP(0.9)画分の約2倍であり、最小阻害濃度もPP(0.9)画分の30倍以上低いという優れた結果が示された。
【0085】
[実施例9] ConA結合画分が示す感染阻害活性
実施例8に従って調製したプロテオースペプトン画分(PP画分)を、コンカナバリンA(Concanavalin A; ConA)アフィニティーカラムを使用したアフィニティークロマトグラフィーにかけて分画した。以下の組成のバッファーを使用し、流速は0.7ml/分とした。
・結合バッファー: 5mMのCH3COONa, 1mMのCaCl2, 1mMのMgCl2, 1mMのMnCl2, 1MのNaCl, 0.02%のNaN3(pH 6.8)
・溶出バッファー: 5mMのCH3COONa, 1mMのCaCl2, 1mMのMgCl2, 1mMのMnCl2, 1MのNaCl, 0.5Mのメチル-α-D-グルコピラノシド, 0.02%のNaN3 (pH 6.8)
【0086】
分画の結果を図21に示す。図21中で100 mAUを示す横線の範囲が溶出バッファーを適用した部分である。図21に示される通り、この分画の結果、ConAに結合しなかった2つの画分(非結合画分UN1及びUN2)と、ConAに結合した画分(BO画分)とが得られた。
【0087】
PP画分、BO画分、UN1画分、UN2画分のそれぞれについて膨潤用緩衝液に溶解して、3mg/ml濃度の溶液を調製した。その溶液の125μlを試料とし、試験例1記載の条件で、一次元目にpIが5〜8の等電点電気泳動に供し、二次元目に15%のポリアクリルアミドゲルを用いたSDS-PAGEに供し、電気泳動が完了した後に、CBB染色を行った(図22)。図22に示されるように、BO画分の二次元電気泳動の結果は、実施例1〜7に記載する硫安画分、乳清タンパク質濃縮物、乳清タンパク質分解物、実施例8に記載するPP(0.9)画分の二次元電気泳動の結果と良く似ていた。
【0088】
また、上記で得られた各画分について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0089】
その結果を図23のA及びBに示す。図23に示される通り、BO画分の感染阻害率は、調べた中の、どのタンパク質濃度でもPP画分よりも高く、かつタンパク質濃度に依存して上昇した。BO画分の最小阻害濃度(MIC)も7.8μg/mlであり、MICが17μg/mlであったPP画分と比較して強い活性が示された。すなわち、ConA結合性を示す画分をPP画分から精製することで、より強いロタウイルス感染阻害活性を示すBO画分を得ることができた。一方、UN1画分及びUN2画分は、調べた範囲のタンパク質濃度では十分な感染阻害率を示さず、最小阻害濃度から見てもほとんど活性が認められなかった。
【0090】
ConAはN型糖鎖と結合する性質を有すること(詳細は実施例4を参照)及びプロテオースペプトン画分に含まれるラクトフォリンにはN型糖鎖を持つ種類と持たない種類があることから、ウイルス感染阻害活性が、少なくとも、N型糖鎖を持つプロテオースペプトン画分由来タンパク質(BO画分)に認められることが確認できた。
【0091】
BO画分については、SDS-PAGEで分画した後に、実施例8と同様にして、約18kDaと約28kDaのタンパク質を実施例4に従って個々に電気泳動溶出し(切り出したゲルから低電圧200Vにて4時間反応)、試料として調製した(図24)。溶出した約28kDaのタンパク質の試料はLP 28K、約18kDaのタンパク質の試料はLP 18Kに相当する。3つの試料(LP 28K、LP 18K、BO画分)について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0092】
その結果を図25のA及びBに示す。図25に示される通り、約18kDaのタンパク質の感染阻害率は、調べた中の、どのタンパク質濃度でも高く、かつタンパク質濃度にほぼ依存して上昇した。また、約18kDaのタンパク質については最小阻害濃度(MIC)も0.86であり、BO画分と比較しても強い活性が示された。すなわち、ConA結合性を示すBO画分から更に約18kDaのタンパク質を分画することにより、更に強いロタウイルス感染阻害活性を示す画分を得ることができた。一方、約28kDaのタンパク質は、調べた範囲のタンパク質濃度では十分な感染阻害率を示すことができず、最小阻害濃度から見てもほとんど活性が認められなかった。このため、乳清タンパク質由来のウイルス感染阻害活性は、少なくとも、プロテオースペプトン画分由来のN型糖鎖を有するBO画分中の約18kDaのタンパク質に認められることが示された。以上の結果から、BO画分中のこの約18kDaのConA結合タンパク質が、実施例1〜8において分離したプロテオースペプトン3(ラクトフォリン)由来の約18kDaの乳清タンパク質であることが確認された。
【0093】
[実施例10] ConA非結合画分のプラスミン処理物が示す感染阻害活性
実施例9に従ってプロテオースペプトン画分(PP画分)から調製したConA結合のBO画分又はConA非結合のUN1画分をプラスミンにより処理した。BO画分又はUN1画分のタンパク質濃度を3mg/mlに調整し、これらにプラスミン溶液(1mg/ml、ヒト血清由来、Sigma社製)を44:1となるように加え、混合液を37℃で5分間保持した。次いで、混合液を沸騰水中で5分間加熱し、プラスミンを失活させた後に、分画分子量が10000Daの透析膜を用いて脱塩処理を行い、分解した後のサンプルをそれぞれBOP画分とUN1P画分とした。
【0094】
BOP画分とUN1P画分のそれぞれについて膨潤用緩衝液に溶解して、3mg/ml濃度の溶液を調製した。その溶液の125μlを試料とし、試験例1記載の条件で、一次元目にpIが5〜8の等電点電気泳動に供し、二次元目に15%のポリアクリルアミドゲルを用いたSDS-PAGEに供し、電気泳動が完了した後に、CBB染色及び実施例7と同様の方法で抗体染色を行った(図26)。図26に示されるように、BOP画分とUN1P画分は共に分解する前に確認されるLP28kが、分解した後にはなくなり、LP18kのみとなっていた。また、分解する前と比較して、分解した後に現れたLP18kのスポット群は、やや酸性側に認められた。この変化は、プラスミンによる分解が一部28kDaから18kDaに留まらず、更に進行して修飾糖鎖の一部が失われてしまったことに起因する可能性が考えられた。
【0095】
また、上記で得られた各画分について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0096】
その結果を図27のA及びBに示す。図27に示される通り、BO画分よりも強い活性は得られないが、BOP画分とUN1P画分は共に感染阻害活性が認められた。ConA非結合のUN1P画分にも高い感染阻害活性が認められたことから、感染阻害活性LP18kの持つ活性にN型糖鎖は必ずしも必要では無いことが示唆された。
【産業上の利用可能性】
【0097】
本発明により、新たなロタウイルス感染防御作用を有する組成物が提供される。該組成物は、その有効量を食品や医薬組成物、家畜飼料の原料として利用することができ、極めて有用である。
【技術分野】
【0001】
本発明は、ロタウイルス感染防御用組成物とその製造法並びにそれを含むロタウイルス感染防御に有用な飲食品、医薬組成物及び家畜飼料に関する。
【背景技術】
【0002】
ヒトロタウイルスを原因とする乳幼児冬季下痢症は主に2歳以下の乳幼児に、発熱、嘔吐、下痢及び脱水症状を引き起こす疾患である。我が国では、灰白色の便の性状から白痢と呼ばれたが、ヒトロタウイルス感染症であることが明らかにされている。先進国での疫学調査から、衛生状態の改善はロタウイルスの有病率を減少させることはできないとされ、その対策の一つとして世界的なロタウイルスワクチン開発が進められている。
【0003】
RRV-TV(rhesus rotavirus tetravalent)ワクチンは、先進国と発展途上国の両方でロタウイルスによる重症の下痢症の予防に高い効果を示し、1998年8月、米国食品医薬局はRRV-TVワクチンを世界で初めてロタウイルスワクチンとして認可した。しかしながら、腸重積症が副反応であることが米国疾病防疫センターから報告され、RRV-TVワクチンの投与は中止された(非特許文献1)。
【0004】
一方、ロタウイルス感染を阻害する食品成分あるいは組成物として例えば、乳ムチン(非特許文献2)、ウシ乳清(以下、ホエイともいう)由来の高分子糖タンパク質混合物(特許文献1)などが提案されている。また、本発明者らもホエイの精密濾過処理の保持液又はその乾燥物に抗ロタウイルス感染阻害活性を持つことを過去に報告している(特許文献2)。前者ではヒトロタウイルスの最小阻害濃度が5μg/ml、後者では11μg/mlと強力なヒトロタウイルス感染阻害活性を見出しているが、最小阻害濃度だけでなく、感染阻害率の点でも優れたロタウイルス感染防御用組成物の登場が望まれている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開平8−99896号公報
【特許文献2】国際公開第2004/080475号パンフレット
【非特許文献】
【0006】
【非特許文献1】石田真一他、「ロタウイルスワクチン」小児科診療 Vol.63, 2000年, p.1045-1049
【非特許文献2】Yolken, R.Y. et al., 「Human Milk Mucin Inhibits Rotavirus Replication and Prevents Experimental Gastroenteritis」 J Clin. Invest., Vol.90, 1992年, p.1984-1991
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、新たなロタウイルス感染防御用組成物並びにロタウイルス感染防御に有用な飲食品、医薬組成物及び飼料を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく検討した結果、ウシ乳清中に見出される糖タンパク質に強力なロタウイルス感染阻害活性を見出した。すなわち、本発明は以下の通りである。
[1] ウシ乳清中に見出され、SDS-PAGEで測定される分子量が15〜20kDa、等電点pIが6〜9であり、N末端から5残基のアミノ酸配列が(1)Ser-Ser-Arg-Gln-Proであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
[2] ウシ乳清中に見出され、SDS-PAGEで測定される分子量が14〜19kDa、等電点pIが6〜9であり、N末端から7残基のアミノ酸配列が(2)Ile-Leu-Lys-Glu-Lys-His-Leuであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
これら[1]及び[2]の糖タンパク質は、ウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画することによって得られるものであることが好ましい。
[3] 前記[1]又は[2]記載の糖タンパク質を含むことを特徴とするロタウイルス感染防御用組成物。
[4] 前記[3]記載のロタウイルス感染防御用組成物を含む食品。
[5] 乳児用食品、幼児用食品、授乳婦用食品、高齢者用食品、病者用食品、保健機能食品、サプリメントからなる群より選択される前記[4]記載の食品。
[6] 前記[3]記載のロタウイルス感染防御用組成物を含む医薬組成物。
[7] 前記[3]記載のロタウイルス感染防御用組成物を含む家畜飼料。
[8] ウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画することを特徴とする、前記[1]又は[2]記載の糖タンパク質の製造法。
【発明の効果】
【0009】
本発明によりロタウイルス感染阻害活性を有する新規糖タンパク質が提供される。
【図面の簡単な説明】
【0010】
【図1】実施例1に従って精製したタンパク質の二次元電気泳動パターンを示した。ゲルはCBB染色を行った。図の左側の数字は標準タンパク質(BIO-RAD社製、商品名:SDS-PAGEスタンダード Broad)の分子量をkDaの単位で示したものである。
【図2】実施例2に従って精製したタンパク質の二次元電気泳動パターンを示した。ゲルはCBB染色を行った。図の左側の数字は図1と同様の標準タンパク質の分子量をkDaの単位で示したものである。
【図3】実施例1により得られたタンパク質スポット群A、B、C、D、E、Fを示す。
【図4】実施例4における各試料のウイルス感染を阻害する最小阻害濃度(MIC)を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。
【図5】実施例4における各試料のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。縦軸はロタウイルス感染阻害率(%)を示す。図中の誤差棒は標準偏差を示す。
【図6】実施例2により得られたタンパク質スポット群A、B、Cを示す。
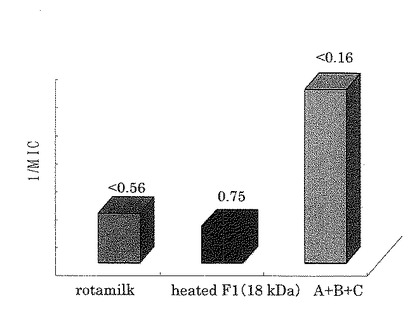
【図7】実施例5における各試料のウイルス感染を阻害する最小阻害濃度(MIC)を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。<0.56のMICを示したロタミルクは対照サンプルである。
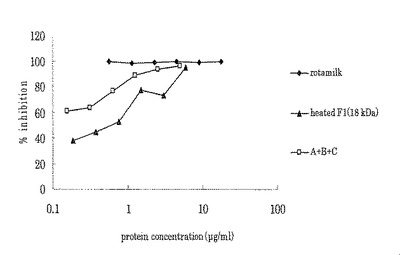
【図8】実施例5における各試料のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。縦軸はロタウイルス感染阻害率(%)を示す。
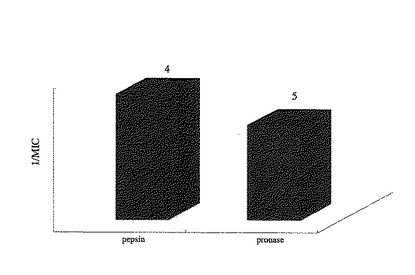
【図9】実施例6における各試料(ペプシン分解物及びプロナーゼ分解物)のウイルス感染を阻害する最小阻害濃度を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。左がペプシン分解物、右がプロナーゼ分解物の試料を示す。
【図10】実施例6における各試料(ペプシン分解物及びプロナーゼ分解物)のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。
【図11】実施例6における各試料(プロナーゼ分解物(100℃で15分間の加熱処理)、プロナーゼ分解物(120℃で5分間の加熱処理)、ロタミルク)のウイルス感染を阻害する最小阻害濃度(MIC)を示したものである。棒グラフはMICの逆数を示し、各グラフの上にはそれぞれのMICを示した。
【図12】実施例6における各試料(プロナーゼ分解物(100℃で15分間の加熱処理)、プロナーゼ分解物(120℃で5分間の加熱処理)、ロタミルク)のウイルス感染阻害率の濃度依存性を示したものである。X軸(横軸)はタンパク質濃度を対数で表示した。図中の誤差棒は標準偏差を示す。
【図13】実施例6により得られたチーズホエイ副産物のプロナーゼ分解物の二次元電気泳動パターンを示した。ゲルはCBB染色を行い、スポット群は矢印で示した。
【図14】実施例6により得られたチーズホエイ副産物のペプシン分解物の二次元電気泳動パターンを示した。ゲルはCBB染色を行い、スポット群は矢印で示した。
【図15】実施例1に従って精製したプロテオースペプトン画分の二次元電気泳動パターンを示した(弱酸性側からスポット群A、B、C、D、E、Fとする)。PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行い、スポット群は円で示した。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
【図16】実施例1に従って精製した硫安画分の二次元電気泳動パターンを示した(弱酸性側からスポット群A、B、C、D、E、Fとする)。PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行い、スポット群は円で示した。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
【図17】実施例3で酵素分解物の調製に用いた乳清タンパク質の二次元電気泳動パターンを示した(弱酸性側からスポット群A、B、C、D、E、Fとする)。PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行い、スポット群は円で示した。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
【図18】プロテオースペプトン(PP)画分及び実施例8に従って精製したPP(0.9)画分のロタウイルス感染阻害活性を示す図である。Aは、各サンプルのロタウイルス感染阻害活性を示すグラフ、Bは、各サンプルについて最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。対照サンプルとして、ヒトロタウイルスで過免疫した乳牛の初乳中の抗体を含むロタミルク、及び陰性対照としてのウシラクトフォリン(bLf)を用いた。
【図19】実施例8に従って精製したPP(0.9)画分の二次元電気泳動パターンとその後の実験の流れを示した。
【図20】実施例8に従って精製したPP(0.9)画分並びにその画分から分離した約18kDaのタンパク質(LP 18K)及び約28kDaのタンパク質(LP 28K)のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。AのグラフにおけるX軸(横軸)のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
【図21】実施例8に従って調製したプロテオースペプトン画分(PP画分)の、ConAアフィニティークロマトグラフィーを使用した溶出パターンを示した。
【図22】ConAアフィニティークロマトグラフィーによりPP画分から得られた各画分の二次元電気泳動パターンを示した。AはPP画分、BはBO画分、CはUN1画分、DはUN2画分の結果である。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
【図23】ConAアフィニティークロマトグラフィーによりPP画分から得られた各画分のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。Aのグラフにおける横軸のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
【図24】ConA結合画分であるBO画分の二次元電気泳動パターンとその後の実験の流れを示した。
【図25】ConA結合画分であるBO画分並びにその画分から分離した約18kDaのタンパク質(LP 18Kに相当)及び約28kDaのタンパク質(LP 28Kに相当)のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。AのグラフにおけるX軸(横軸)のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
【図26】ConA結合画分であるBO画分及びConA非結合画分であるUN1画分とそれらのプラスミン処理物が示す二次元電気泳動パターンを示した。ゲルはCBB染色を行った。また、PVDF膜に転写したタンパク質は抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体による染色を行った。図の左側の数字は標準タンパク質(BIO-RAD社製)の分子量をkDaの単位で示したものである。
【図27】ConA結合画分であるBO画分及びConA非結合画分であるUN1画分とそれらのプラスミン処理物のロタウイルス感染阻害活性を示した。Aは、測定されたロタウイルス感染阻害率とそのタンパク質濃度依存性を示すグラフである。AのグラフにおけるX軸(横軸)のタンパク質濃度は対数表示であり、誤差棒は標準偏差を表す。Bは、最小阻害濃度(MIC)の逆数を縦軸に示し、各棒グラフの上に最小阻害濃度の値を示したグラフである。
【発明を実施するための形態】
【0011】
以下、本発明を詳細に説明する。本発明は、ウシ乳清中に見出され、下記特徴を有する新規糖タンパク質に関する。
【0012】
ウシ乳清中に見出され、SDS-PAGEで測定される分子量が15〜20kDa、等電点pIが6〜9であり、N末端から5残基のアミノ酸配列が(1)Ser-Ser-Arg-Gln-Proであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
【0013】
ウシ乳清中に見出され、SDS-PAGEで測定される分子量が14〜19kDa、等電点pIが6〜9であり、N末端から7残基のアミノ酸配列が(2)Ile-Leu-Lys-Glu-Lys-His-Leuであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質。
【0014】
次いで製造法を述べるが、本発明は、これに限定されるものではない。
本発明における新規糖タンパク質(以下、本発明物質ということもある)は、ウシプロテオースペプトン溶液に硫安を加え、溶液の飽和度が55〜90%のときに得られる沈殿物に見出すことができる。あるいは、本発明における新規糖タンパク質は、ウシプロテオースペプトン溶液を直接分画することによっても得ることができる。プロテオースペプトンは、95℃で20分間の加熱処理を行った脱脂乳をpH 4.6にしたときに沈殿しない耐熱性の乳清タンパク質画分である(「ミルク総合辞典」朝倉書店 1992年 p.37)。本発明物質は、ウシ乳清を加熱処理して得られる可溶性画分、いわゆるウシプロテオースペプトン画分を硫安沈殿と遠心分離処理を組み合わせた方法で処理して得られる分画物に見出され、ロタウイルスの感染阻害活性が非常に高いことが特徴である。上記の本発明の糖タンパク質は、それぞれ、ラクトフォリン(プロテオースペプトン画分に含まれる耐熱性と可溶性(水溶性)を有する糖タンパク質)を構成するタンパク質成分(ラクトフォリンコンポーネントとも呼ばれる)の一つである。ラクトフォリンは、プロテオースペプトン画分の一分画であるプロテオースペプトン3画分中に見出されたことから、プロテオースペプトン3又はプロテオースペプトンコンポーネント3とも呼ばれる。
【0015】
乳清はスイートホエイ(sweet whey)及び酸ホエイ(acid whey)に分類され、ウシの各泌乳期の乳もしくはこれらの濃縮物又は乾燥物(以下、これらをまとめて乳ということもある)を原料として常法により調製することができる。スイートホエイは熟成型チーズの副産物であり、pHが5.9〜6.3で甘味がある。一方、酸ホエイは非熟成型(フレッシュ型)チーズの製造で得られ、通常のpHが4.4〜4.6で酸味がある。また、カゼインの製造時に得られるホエイも酸ホエイである。本発明の乳清にはスイートホエイ及び酸ホエイの両者を含む。
【0016】
本発明では、乳清そのものに加え、乳清を精密濾過処理、限外濾過処理、ダイアフィルトレーション処理などを行うことで、乳清中のタンパク質を濃縮した乳清タンパク質濃縮物あるいはその精製物を用いることができる。市販品に、乳清タンパク質濃縮物(WPC)、乳清タンパク質単離物(WPI)などの濃縮物又は乾燥物が知られ、これらをいずれも用いることができる。また、常法に従って調製しても良い。限外濾過処理などにより乳清タンパク質を濃縮する場合には、分画分子量が300万Da以上の細孔径(ポアサイズ)を持つ分離膜を用いることが望ましい。
【0017】
乳清タンパク質濃縮物を精製して用いる場合には例えば、デキストランの分画分子量が4×104〜2×107Daに相当する担体(商品名:Sephacryl S-500 HR、アマシャムファルマシアバイオテク社製など)を充填したゲル濾過カラムクロマトグラフィーに適用し、波長280nmの吸光度を指標に、最初のピークに相当する画分にウイルスの感染阻害活性が確認されている。溶離液にはトリス塩酸、リン酸などのpHが6〜8の公知の緩衝液が用いられ、硫酸ドデシルナトリウム、トリトンX-100などの界面活性剤を解離剤として併用することもできる。
【0018】
次いで乳清そのもの、乳清タンパク質濃縮物あるいはその精製物を加熱処理し、得られた可溶性画分に硫安を加える。そのために、まず、これら原料の濃度を適宜調整した後に70〜125℃で5〜40分、好ましくは80〜95℃で30分程度に相当する加熱処理を行う。遠心分離処理を行い、上清すなわちプロテオースペプトン溶液を採取する。この上清に溶液の飽和度が50〜90%、好ましくは55〜90%となるように硫安を添加し(硫安の添加法は「実験農芸化学 下」朝倉書店 1978年 p.59-60を参照)、硫安沈殿物を回収する(硫酸アンモニウム分画法、硫酸アンモニウム塩析法又は硫酸アンモニウム沈殿法ともいう)。その際に硫安沈殿により生成した上清に硫安を加え、硫安沈殿物を繰り返し回収することも望ましい。
【0019】
本発明では、プロテオースペプトン溶液の硫安沈殿物に代わり、プロテオースペプトン画分そのもの、又は乳清の酵素分解物を用いることができる。また、乳清に限らず、乳清タンパク質濃縮物、その精製物などを酵素分解処理したものも好ましい。酵素としては、ペプシン、プロナーゼ、トリプシン、α-キモトリプシン、パパインなどが挙げられ、各酵素の至適pHにして、30〜60℃で1〜5時間の加水分解処理を行う。このとき、タンパク質:酵素の重量比率は、100:0.1〜2とする。
【0020】
こうして得られるプロテオースペプトン画分、プロテオースペプトン溶液の硫安沈殿画分、あるいは乳清などの酵素分解物を一次元目にpIが5〜8の等電点電気泳動、二次元目に15%のポリアクリルアミド単一ゲルを用いた二次元電気泳動を行うことで、ウイルス感染阻害活性の高いタンパク質群を採取することができる。後述する実施例の通り、乳清を加熱処理して得られるプロテオースペプトン画分を用いた場合には、ポリアクリルアミドゲルから電気泳動溶出した18kDa付近に少なくとも6つのスポットとして検出される。SDS-PAGEによる分子量が15〜20kDa(以下、約18kDaともいう)、等電点pIが6〜9であり、6つのスポットを弱酸性側からA、B、C、D、E、Fとしたときに、単独物ではE及びFのスポットがウイルスの感染を阻害する最小阻害濃度が1.1μg/ml及び0.78μg/mlと非常に強く、混合物ではB+C+Dが0.31μg/ml未満と高い活性を有することが分かった。
【0021】
また、アミノ酸シークエンスによりスポットC、D、E、FのN末端から5残基がSer-Ser-Arg-Gln-Pro(配列番号:1)であることが確認された。糖鎖の有無を調べるため、レクチン染色を行ったところ、RCA120及びConAとの反応性では、6つのスポット全てにおいてポジティブな反応を示した。RCA120は糖鎖にあるβ-ガラクトースを認識し、ConAはマンノースを認識して結合するものであることから、これらタンパク質が糖鎖にβ-ガラクトース及びマンノースを有する糖タンパク質であることが判明した。更にN-アセチル-D-グルコサミンがβ-1、4結合したオリゴ糖などを認識するWGAレクチンにも、6つのスポット全てがポジティブな反応を示した。
【0022】
一方、乳清を限外濾過処理により濃縮した後に、ゲル濾過により得られる画分を加熱処理して得られるプロテオースペプトン画分を用いた場合には、ポリアクリルアミドゲルから電気泳動溶出した16kDa付近に複数のスポットが検出される。弱酸性側からスポットをA、B、Cとしたときに、CのN末端から7残基がIle-Leu-Lys-Glu-Lys-His-Leu(配列番号:2)で、SDS-PAGEによる分子量が14〜19kDa(以下、約16kDaともいう)、等電点pIが6〜9の新規糖タンパク質であることが確認された。Cを含む16kDa付近にスポットされるタンパク質群のウイルスの感染を阻害する最小阻害濃度を測定したところ、0.16μg/ml未満であった。また、レクチン染色により、RCA120レクチンと反応性が見られたことから、約16kDa付近にスポットされるタンパク質群はβ-ガラクトースを有する糖タンパク質であることが示された。
【0023】
本発明に係る糖タンパク質の少なくとも一部は、上記のようにレクチンとの結合性を有することからN型糖鎖(N結合型糖鎖、アスパラギン結合型糖鎖ともいう)を有することが示された。また、本発明に係る糖タンパク質は、上記で耐熱性と可溶性を有する画分に見出されたように、耐熱性があり、可溶性(水溶性)である。従って、本発明の糖タンパク質は、プロテオースペプトン画分などのウシ乳清由来耐熱性可溶画分からアフィニティークロマトグラフィー法などによって得られるレクチン(ConA、RCA120など)結合画分を、電気泳動などの公知のタンパク質分画法によって更に分画し、そこから本発明の糖タンパク質を規定する上記分子量及び等電点を示す画分を採取することによって調製することもできる。
【0024】
本発明の新規糖タンパク質を含む組成物は、食品(とりわけ育児用調製粉乳や仔ウシの飼料)に、その有効量を混合することにより、ヒト乳児、仔ウシ、仔ウマなどにおけるロタウイルスが原因の下痢症を予防又は治療するのに有効であることが期待できる。
【0025】
ロタウイルス感染における食品成分の抗ロタウイルス感染効果を検定・評価する方法は多岐に渡り、万全の検定・評価系はないとされている。これまでに様々な抗ロタウイルス感染効果の検定・評価法が報告されている。例えば、本発明で用いた方法以外に、実験動物(マウス)にロタウイルス及び食品成分を投与し、下痢の発症及び消化管粘膜へのロタウイルスの結合量を測定するなどの検定・評価法が挙げられる(Duffy L.C. Pediatr. Res. 35: 690-695(1994))。本発明は、これらの系を適切に用いて該組成物の抗ロタウイルス感染活性を更に詳細に調べることが可能である。
【0026】
本発明において用いる新規糖タンパク質は、乳由来であることから長い食経験の中で安全性は確立されている。従って、ヒトによる試験で本発明品の有効性及び有効量の確認が可能である。
【0027】
本発明では、上記のような新規糖タンパク質をロタウイルス感染防御用組成物として用いることができる。本発明のロタウイルス感染防御用組成物は、本発明に係る2種の糖タンパク質を両方とも含んでも良いが、いずれか一方のみを含むものでも良い。
【0028】
本発明のロタウイルス感染防御用組成物は、適量を添加することにより、飲食品や医薬組成物にロタウイルス感染防御作用を付与することができる。このロタウイルス感染防御用組成物は、それ自身(液状又は粉末状)又は他の活性物質と共に、あるいは他の薬理学的な活性物質と共に用いることができる。形態は例えば、錠剤もしくは被覆錠、カプセル、タブレット、溶液、シロップ、乳液又は分散性粉末などの任意の形態であって良い。摂取量は年齢、身体状態などに依存して変化するが、1日当たり0.001〜1000mg/kg体重、好ましくは0.001〜200mg/kg体重である。
【0029】
本発明は、ロタウイルス感染防御用組成物を添加した食品にも関する。本明細書において食品とは、液体状、固体状、ゲル状、顆粒状、粉末状、タブレット状などのいずれでも良く、特に限定するものではないが、飲料、食品及び機能性食品を含む。
【0030】
食品として具体的には、牛乳、乳製品、清涼飲料水、菓子類、パン、麺、練り製品、調理加工食品、畜肉加工食品、調味料などを挙げることができる。食品の形態として中でも好ましいのが機能性食品である。本発明の機能性食品は、生体に対して一定の機能性を有する食品を意味し、例えば、特定保健用食品及び栄養機能食品を含む保健機能食品、特別用途食品、栄養補助食品、健康補助食品、サプリメント(例えば、錠剤、被覆錠、糖衣錠、カプセル及び液剤などの各種剤形のもの)及び美容食品などの、いわゆる健康食品全般を含む。また、本発明の機能性食品には、コーデックス(FAO/WHO合同食品規格委員会)の商品規格に基づく健康強調表示(Health claim)が適用される健康食品を含む。
【0031】
本発明の好適な機能性食品として好ましい具体例には、乳児用、幼児用、授乳婦用、高齢者用、病者用などの特別用途食品がある。乳児用には例えば、乳児用調製乳が含まれ、通常の乳幼児用調製乳の原料に添加して、ロタウイルス感染に対する防御能の発達が未熟な乳幼児のための育児用調製粉乳又は液体調製乳に使用できる。育児用調製粉乳には、乳児用調製粉乳、フォローアップミルク、低出生体重児を対象とする低出生体重児用調製粉乳、牛乳アレルギーや乳糖不耐症などの病的状態を有する小児の治療に用いられる各種治療用ミルクなどが含まれる。また、高齢者用には、ロタウイルス感染に対する防御能の低下した高齢者向けに、その有効量を食品に添加してロタウイルス感染防御用食品とすることができる。更に該組成物を家畜用飼料に、その有効量を添加してロタウイルス感染防御用飼料とすることができる。
【0032】
本発明のロタウイルス感染防御用組成物は、当業者が利用可能である任意の適切な方法によって食品に含めれば良い。例えば、ロタウイルス感染防御用組成物を、液体状、ゲル状、固体状、粉末状又は顆粒状にした後に、それを食品に配合する。あるいは、ロタウイルス感染防御用組成物を、食品の原料中に直接で混合又は溶解しても良い。また、食品に塗布、被覆、浸透又は吹き付けても良い。あるいは、飲食品中に均一に分散又は偏在させても良い。また、可食フィルムや食用コーティング剤などで包み込んでも良い。ロタウイルス感染防御用組成物を含む飲食品を更に加工しても良く、そのような加工品も本発明の範囲に含まれる。
【0033】
本発明の食品の製造において、食品に使用される各種添加物を使用しても良い。具体的には、発色剤(亜硝酸ナトリウムなど)、着色料、香料、甘味料、保存料、乳化剤、酸化防止剤、pH調整剤、化学調味料、増粘剤、安定剤、膨張剤、消泡剤、結着剤、栄養強化剤などが挙げられる。
【0034】
本発明のロタウイルス感染防御用組成物を、医薬組成物として使用する場合には、医薬製剤上で許容される担体又は添加物を配合しても良い。具体的には、水、医薬的に許容される有機溶剤、コラーゲン、ポリビニルアルコール、ポリビニルピロリドン、カルボキシビニルポリマー、アルギン酸ナトリウム、水溶性デキストラン、ペクチン、キサンタンガム、アラビアゴム、カゼイン、ゼラチン、寒天、グリセリン、プロピレングリコール、ポリエチレングリコール、ワセリン、パラフィン、マンニトール、ラクトース、ソルビトール、医薬添加物として許容される界面活性剤などが挙げられる。使用される添加物は、製剤の剤形に応じて適宜又は組み合わせて選択される。本発明の医薬組成物は更に他の薬理成分を含有しても良い。そして、錠剤、顆粒剤、散剤、丸剤、カプセル剤などの固形製剤、ジェル剤もしくは液剤、懸濁剤、シロップ剤などの液体製剤などの剤形にして経口的に投与する他に非経口的に投与することもできる。
【0035】
本発明のロタウイルス感染防御用組成物を配合する飼料としては、家畜、家禽、ペットなどの配合飼料又は配合飼料用原料が挙げられる。
【0036】
また、本発明のロタウイルス感染防御用組成物に加え、乳酸菌、ビフィズス菌などの他のロタウイルス感染防御作用を有する組成物を併用しても良い。
【実施例】
【0037】
以下、本発明の効果を試験例及び実施例に基づき具体的に説明するが、本発明は、これらの具体例に限定されるものではない。
【0038】
[試験例1] 本発明における二次元電気泳動法
等電点電気泳動
1.膨潤化
泳動に用いるStripゲルを膨潤させる。
[IPG Ready Strip]
7cm pH 3-10(BIO-RAD社製)
[膨潤用緩衝液]
2.4gの Urea
0.05gの DTT
0.2gの CHAPS
0.01mlの Bio-Lytes
5μl、50mg/mlの Orange G
これに5mlのMilli-Q(Millipore社)で作成した超純水(以下、MilliQともいう)を加え、冷凍保存した。
【0039】
フォーカシングトレイ(一次元目の泳動で用いる電極装置)のスロットに、膨潤緩衝液で希釈した試料の必要量を入れる。Ready Stripマイナス側をピンセットで持ち、ゲルが付着している面を下側にして、プラス側の方から、ゆっくりと膨潤緩衝液に触れさせる。気泡をトラップしないように注意しながら、数回、Stripを前後にスライドさせ、膨潤緩衝液をゲル面に均一に行き渡らせる。1時間程放置して膨潤緩衝液をゲルに吸収させた後に、ミネラルオイル(BIO-RAD社製)を端から、ゆっくり滴下してゲル全面をカバーし、トレイの蓋をする。なお、膨潤する際には、トレイをIEFセル本体にセットし、20℃、active 50 Voltに設定し、12時間程放置する。
【0040】
2.泳動
プログラム、プリンターは以下のように設定し、この条件下で一次元目の泳動を行う。
[プログラム]
step1:250V、15min、20℃
step2:Start 250-End、4,000V、2hr、20℃
ボルトラインピング設定
電流値リミットは、50μA/Strip
step3:4,000V-20,000VH、5hr、20℃
[プリンター]
Voltage Slopeは、LINEARを選択し、穏やかな電圧上昇のもとで泳動し、泳動が終了した後に、Ready Stripに流れたタンパク質が乱れないように、500V/HOLDを設定する。
【0041】
15%のポリアクリルアミド単一ゲルを用いたSDS-PAGE
1.分離ゲルの作成
[A溶液:30%のアクリルアミド溶液]
29.2gのアクリルアミド(モノマー)
0.8gのN,N’-メチレンビスアクリルアミド
純水で100mlとする。
[B溶液:1.5MのTris-HCl緩衝液(pH 8.8)]
18.17gの Tris
0.4gの Sodium dodecyl sulfate(SDS)
純水で溶解し、HClでpHを8.8に調整した後に、最終容量を100mlとする。
[C溶液:10%の過硫酸アンモニウム溶液]
0.1gの過硫酸アンモニウム
純水1mlを加える。
[TEMED]
N,N,N',N',-Tetramethylethylenediamine
[泳動用緩衝液]
14.4gのグリシン
3gのTris
1gのSDS
純水で1000mlとする。
【0042】
15%のアクリルアミド溶液は、A溶液を8ml、B溶液を4ml、C溶液を0.0063ml、TEMEDを0.01ml、純水を4mlで混合して調製した。ゲル板の中央よりピペットマンでゲル調製溶液を注入する。ゲル調製溶液を導入した後に、その上部に純水を静かに注いで空気を遮断し、アクリルアミドをゲル化させる。
【0043】
2.Ready Stripの平衡化とゲル板へのセット
IPG Ready Stripを二次元目の泳動に必要なSDS緩衝液系にするため、平衡化を行う。使用する溶液は、以下の通りである。
[平衡化緩衝液1]
3.6g/10mlの Urea
0.2g/10mlの SDS
2.5ml/10ml、1.5Mの Tris-HCl
2ml/10mlの Glycerol
200mg/10mlの DTT
以上、各試薬10mlを合わせて50mlとする。
[平衡化緩衝液2]
3.6g/10mlの Urea
0.2g/10mlの SDS
2.5ml/10ml、1.5Mの Tris-HCl
2ml/10mlの Glycerol
250mg/10mlの Iodoacetamide
以上、各試薬10mlを合わせて50mlとする。
【0044】
等電点電気泳動が終了したStripゲルを、ゲル面を上にして膨潤トレイに入れ、4mlの平衡化緩衝液1を注ぎ、静かに振とうしながら、10分間平衡化する。更に平衡化緩衝液2で同様にして、10分間平衡化する。作成したゲルのインナーガラスプレートを手前に向けて置き、Stripのプラスチックフィルムをアウターガラスに付け、Stripの端から気泡が入らないように、ゆっくりウェルに入れる。その上を電子レンジで溶解して人肌程度に冷ました1%のアガロースゲルで固定化し、泳動槽にセットして泳動用緩衝液を注ぐ。
[泳動条件]
200V、30mAの定電圧、定電流で泳動を行う。
【0045】
3.ポリアクリルアミドゲルの染色
CBB染色
電気泳動後のタンパク質染色にはCBB染色を用いる。使用する溶液は、以下の通りである。
[CBB染色]
2.5gの CBB
454mlのメタノール
75mlの 酢酸
これに純水を加えて1000mlとし、3回濾過する。
[脱色液]
250mlのメタノール
375mlの酢酸
純水で5000mlとする。
【0046】
ゲルを染色液に約20分間浸した後に、脱色液でバックグラウンドの色が抜けるまで脱色する。
【0047】
[試験例2] ポリアクリルアミドゲルからのタンパク質の電気泳動溶出
ポリアクリルアミドゲルからのタンパク質の電気泳動溶出は、Centrilutor micro-electroeluter(Millipore社製)を用いて行うことができる。キャップと底に穴(孔)が空いた0.5mlのサンプルチューブに切り出したポリアクリルアミドゲル細片を入れる。下部のバッファーチェンバーに溶出緩衝液(25mMのTris、192mMのグリシン、0.02%のSDS、20%のメタノール)を注ぎ、上部のバッファーチェンバーラックの穴(孔)に分画分子量が10000Daのセントリコン装置を取り付ける。使用しない穴(孔)はストッパーで閉め、下部のバッファーチェンバーに上部のバッファーチェンバーをのせる。上段のバッファーチェンバーを溶出緩衝液で満たし、セントリコン装置の下部に気泡がないか確認する。気泡がある場合には、シリンジを用いて除去する。それぞれのセントリコン装置のサンプルリザーバーにゲル端片の入ったサンプルチューブを差し込む。溶出機に安全カバーを着け、電極をコンセントに差し込み、200Vで4時間溶出を行う。
【0048】
[試験例3] タンパク質濃度の測定
タンパク質の定量にはBradford法に従う。Protein assay reagent(BIO-RAD社製)を使用し、ウシ血清アルブミンを標準溶液として測定する。各試料は、PBS(−)[Ca及びMgイオンを除去したDulbecco's Phosphate Buffered Saline]で希釈し、マイクロプレートに10μlずつ注ぐ。5倍に希釈した200μlのBradford試薬を加えてピペッティングし、595nmにおける吸光度を測定する。
【0049】
[試験例4] 抗ロタウイルス活性の測定法(ロタウイルス中和活性試験)
ウシ乳由来組成物の加熱処理物を試料として、ヒトロタウイルスMO株の感染阻害活性を以下のように測定することができる。
(1)培養細胞の調製
ウイルスの増殖及び抗ロタウイルス活性の測定に、37℃で7日間培養した単層のアカゲザル腎臓由来株化細胞MA104細胞(J.Clin.Microbiol.Oct. 1982, 727-730)を用いる。継代後の4日間は、10%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地中で培養し、継代して5日目に2%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地に交換する。
(2)ウイルスの調製
ウイルスには、ヒトロタウイルスMO株(血清型3)(J.Clin.Microbiol.Oct.1982,727-730)を用いる。あらかじめ20μg/mlのトリプシンにより、37℃で30分間混合し、M.O.I(Multiplicity Of Infection;細胞1個当たりの接種ウイルス数)=1となるように希釈し、これを0.11%のウシ血清アルブミン、ジエチルアミノエチル、デキストランを加えたHanks液で3回洗浄した単層細胞上に接種する。37℃で60分間吸着させた後に、ウイルスを除去し、前記Hanks液で洗浄した後に、0.5μg/mlのトリプシンを含む2%のウシ胎児血清、5%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地を加え、37℃で回転培養する。大部分の細胞が壁面から剥離したら、その培養液を回収し、遠心分離処理(3000rpm、10分間)を行い、その上清をストックウイルスとして分注し、使用時まで−80℃で保存する。
(3)ウイルスの感染
ウイルスの接種は同時接種法で行う。すなわち、PBS(−)[Ca及びMgイオンを除去したDulbecco's Phosphate Buffered Saline]で溶解した試料原液を10%ウシ胎児血清、10%トリプトース・ホスフェイト・ブロスを含むイーグルMEN培地で段階希釈し、50μlの希釈試料(試料溶液は0.45μmのフィルターで濾過滅菌)を0.5mlのエッペンドルフチューブに分注する。次いで105〜106FCFU(Fluorescent Cell Focus Forming Unit)/mlのヒトロタウイルスMO株液の0.4mlを、20μg/mlのトリプシン溶液の0.4mlにより、37℃で30分間の処理を行い、10%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地で20〜100倍に希釈した後に、各エッペンドルフチューブに50μlずつ加え、37℃で1時間培養した。空試験用(試料無添加時)には、前記希釈試料の50μlに代えて、10%ウシ胎児血清、10%トリプトース・ホスフェイト・ブロスを含むイーグルMEN培地を用いる。
【0050】
培養が終了した後に、アカゲザル腎臓由来のMA-104細胞の細胞浮遊液をウイルス−試料混合液が入った各エッペンドルフチューブに100μlずつ加え、24ウェルスライドグラス上に各20μlずつ分注して、37℃で45〜65時間培養する。
【0051】
前記細胞浮遊液は、37℃で7日間培養した単層のアカゲザル腎臓由来のMA-104細胞を、0.025%のEDTAを添加した0.125%のトリプシン溶液で剥離し、更に遠心分離処理して上清を取り除いた後に、2×105/mlとなるように培養液に加えることで得られる。そして、培養液には10%のウシ胎児血清、10%のトリプトース・ホスフェイト・ブロスを含むイーグルMEN培地を用いる。
【0052】
培養が終了した後に、アセトンで固定し、ヒトロタウイルスMO株感染細胞数を間接蛍光抗体法で検出する。なお、一次抗体として鳩ロタウイルスPO-13株のVP6を特異的に認識する抗ロタウイルスモノクローナル抗体(Minamoto,N. et.al.,Epidem,Inf.,100,481-492(1988))及び二次抗体として蛍光標識したヤギ抗マウスIgG血清(Organon Teknika-Cappel,NC,U.S.A.)を用いる。
【0053】
増殖阻害活性の評価は、100×(1−(試料添加時の感染細胞数)/(試料無添加時の感染細胞数))の値を阻害率として行う。
【0054】
また、前記値が50%以上のものをポジティブとして判断し、活性が認められた最も低い試料濃度を最小阻害濃度(MIC)として活性を数値化する。
【0055】
[実施例1] 乳清からの抗ロタウイルス感染組成物の調製
1.プロテオースペプトン画分の調製
搾乳した未加熱の10リットルの生乳(原乳)を5℃に冷却し、遠心分離処理(3000rpm、20分)で乳脂肪を除去し、脱脂乳を調製した。pHを4.6に調整した後に、沈殿したカゼインを遠心分離処理(20℃、3000rpm、15分)で除去し、乳清画分(酸ホエイ)を得た。次いで、酸ホエイを95℃で30分間の加熱処理を行い、30℃まで冷却した後に、変性したタンパク質を遠心分離処理(5000G、20℃、30分間)で除去し、プロテオースペプトン画分を得た。
【0056】
2.硫安画分の調製
前記プロテオースペプトン画分に100%の飽和硫安溶液を55%の飽和度になるまで加えた。数時間放置した後に、遠心分離処理(7000G、20℃、30分間)を行った。上清を採取し、90%の飽和度になるまで、更に100%の飽和硫安溶液を加えた。スターラーで撹拌しながら、一晩放置した後に、遠心分離処理(7000G、20℃、30分間)を行い、生じた沈殿を脱イオン蒸留水に溶解し、48時間透析を行った。
【0057】
3.二次元電気泳動による精製
透析した後に得られた溶液を試料として二次元電気泳動を行った。試験例1記載の条件で一次元目にはpIが5〜8の等電点電気泳動、二次元目には15%のポリアクリルアミド単一ゲルを用いたSDS-PAGEを行った。そして、ポリアクリルアミドゲル上に認められた等電点pIが6〜9で分子量約18kDa付近のタンパク質スポット群を、試験例2に従って電気泳動溶出し、採取した(図1)。
【0058】
[実施例2] 乳清タンパク質濃縮物からの抗ロタウイルス感染組成物の調製
実施例1に従って酸ホエイを調製し、pHを6.0とした。0.15MのNaCl及び0.02%のNaN3を含む10リットルのリン酸緩衝液(pH 6.0)を用い、分画分子量が300万Daの分離膜で限外濾過処理を行った。その保持液を採取し、常法により凍結乾燥することで、500mgの乳清タンパク質濃縮物の粉体を得た。この粉末を溶出緩衝液で濃度を9mg/mlに調整し、Sephacryl S-500(アマシャムファルマシアバイオテク社製)を充填したカラム(5×60cm、ゲル容積1200ml)に30mlをアプライした。なお、溶出緩衝液には、0.15MのNaCl、1mMのNa2EDTA、0.02%のNaN3を含む0.05MのTris-HCl緩衝液を用い、280nmの吸光度における最初のピークに相当する分画を採取した。その後に実施例1と同様にして、画分を加熱処理した後に硫安沈殿を行い、生じた沈殿を脱イオン蒸留水に溶解し、48時間透析を行った。
【0059】
透析した後に得られた溶液を試料として実施例1と同様の方法で二次元電気泳動を行い、等電点pIが6〜9で約16kDa付近に認められたタンパク質スポット群を採取した(図2)。
【0060】
[実施例3] 抗ロタウイルス活性を有する乳清タンパク質分解物の調製
搾乳した未加熱の100 kgの生乳(原乳)を63℃で30分間の加熱処理を行い、常法に従ってゴーダチーズを調製し、生成したホエイ(pH 6.0)を分離した。このホエイを加温し、遠心分離処理(3000G、20分間、40℃)を行い、脱脂ホエイを得た。この脱脂ホエイを国際公開第2004/080475号の実施例1に記載された方法に従い、精密濾過処理し、保持液を回収した。更に得られた保持液に脱イオン水を添加し、精密濾過処理を3回繰り返した(ダイアフィルトレーション処理)。最後に得られた保持液はスプレードライ法により乾燥した。次いで乾燥試料の2%の水溶液を調製し、そのpHを1NのHCl及び1NのNaOHでpHを3.0に調整した。55℃に加温してから乾燥試料重量の0.5%に当たるペプシン(Sigma社製)を添加し、3時間の酵素処理を行った。酵素処理液は沸騰水浴中で5分間の加熱処理を行うことで酵素を失活させ、その後に凍結乾燥することで抗ロタウイルス活性の試験試料(ペプシン分解物)を得た。全く同様にして、2%の水溶液をpHで8.0に調整し、ペプシンの代わりにプロナーゼ(Merck社製)を用いることでプロナーゼ分解物を得た。
【0061】
[実施例4] 抗ロタウイルス活性成分の分画1
1.レクチン染色
実施例1で確認された約18kDaのスポットについてレクチン染色を行うため、実施例1に従って二次元電気泳動を行った後に、イモビロンPVDF膜に転写した。転写した後に、CBB染色を行い、次いでレクチン染色をした。
【0062】
まず、ろ紙、スポンジパットを転写緩衝液(25mMのTris、192mMのグリシン、0.02%のSDS、20%のメタノール)に15分間浸し、平衡化させた。イモビロンPVDF膜メタノールに1分間浸し、脱イオン蒸留水で洗浄した後に、転写緩衝液に入れ、振とうした。ゲルをイモビロンPVDF膜に密着させ、ろ紙、スポンジパット、ホルダーに挟み、緩衝液を満たした電気泳動槽中にゲルをマイナス側にして垂直に固定し、30Vで一晩通電した。転写した後のPVDF膜にCBB染色液を加え、染色の度合いに応じて約10分間振とう染色を行った。染色した後に、脱色の度合いに応じてCBB脱色液で10〜15分間脱色し、脱イオン蒸留水で洗浄し、風乾した。更に緩衝液(0.9%のNaCl、0.05%のTween20、10mMのTris-HCl、pH 7.4)にペルオキシダーゼを結合させた5μl/10mlのレクチンを加え、PVDF膜上のタンパク質に対し、1時間振とう反応を行った。反応が終了した後に洗浄し、15mMのリン酸緩衝液(pH 6.8)で平衡化を行って発色液で発色させた。レクチンにはRCA-120、ConA、WGAを用いた。
【0063】
実施例1で確認された約18kDaのスポットに対し、弱酸性側からA、B、C、D、E、Fとしたところ(図3)、RCA-120及びConAレクチンとの反応性は、6つすべてのスポットにおいてポジティブな反応であった。従って、約18kDaのタンパク質はβ-ガラクトース及びマンノースを構成単糖として持つことが示された。また、WGAレクチンとの反応性でも、全てのスポットにおいてポジティブな反応を示した。WGAレクチンによって認識される糖鎖構造はN-アセチル-D-グルコサミンがβ-1,4結合したオリゴ糖、バイセクティングN-アセチル-D-グルコサミンを有する直鎖型の糖鎖構造、N-アセチルノイラミン酸が挙げられる。このように親和性を示す糖鎖構造がいくつか報告されているため、WGAレクチンによる染色からは、どのような糖鎖構造を持つか特定はできないが、3種のレクチン染色から、約18kDaタンパク質スポットは共通した糖鎖構造を持つことが示された。また、各種レクチン染色において、pIの高いスポットの方が、よりポジティブに染色していたことから、スポットによって修飾を受けている糖鎖の量が異なっていることが示唆された。
【0064】
2.N末端アミノ酸配列の特定
実施例1で採取したスポット群のうち、スポットC、D、E、FのN末端から5残基を気相シークエンス(綱沢進 他、続生化学実験講座 1 タンパク質の化学(上)、東京化学同人、347-373(1987))(Procese 49-HT プロテインシークエンサー)により測定したところ、これらのN末端配列5残基がプロテオースペプトン3(Esben S.S.et.al.,J Dairy Res. 60 535-542(1993))のアミノ酸54位から58位の配列(Ser-Ser-Arg-Gln-Pro)と一致する新規な糖タンパク質であることが確認された。
【0065】
3.抗ロタウイルス活性の測定
更にB+C+Dからなるスポット群、スポットE及びスポットFの抗ロタウイルス活性を測定した。以下に手順を示す。
【0066】
二次元電気泳動を行ったゲルから、試験例2に従ってタンパク質を電気泳動溶出することで試料を分離した。試料原液に含まれるタンパク質濃度を測定後(試験例3参照)、この原液を6段階希釈し、1試料につき計6つずつ試料を用意した。そして、これら試料のウイルス増殖阻害活性を測定した(試験例4参照)。
【0067】
その結果、ウイルスの感染を阻害する最小阻害濃度は、スポット群(B+C+D)が0.31μg/ml未満、スポットEが1.1μg/ml、スポットFが0.78μg/mlであった(図4)。なお、スポット群(B+C+D)は、低いタンパク質濃度でも高い感染阻害率をし、タンパク質の濃度が高くなるにつれ、100%に近い感染阻害率を示すことが分かった(図5)。
【0068】
[実施例5] 抗ロタウイルス活性成分の分画2
1.レクチン染色
実施例4と同様にして、実施例2で確認された約16kDaのスポットについてRCA-120を用いたレクチン染色を行った。その結果、レクチンと反応性が見られたことから、約16kDaのタンパク質スポット群は、β-ガラクトースを持つことが示された。
【0069】
2.N末端アミノ酸配列の特定
次いで、約16kDaのタンパク質スポット群を弱酸性側からA、B、Cとし(図6)、実施例2で採取した、これらのスポット群においてスポットCのN末端から7残基をアミノ酸シークエンス(Procese 49-HT プロテインシークエンサー)により測定した。その結果、N末端配列7残基がプロテオースペプトン3(Esben S.S.et.al.,J Dairy Res. 60 535-542(1993))のアミノ酸69位から75位の配列(Ile-Leu-Lys-Glu-Lys-His-Leu)と一致する新規な糖タンパク質であることが確認された。
【0070】
3.抗ロタウイルス活性の測定
実施例2で採取したスポット群のうち、A、B、Cを含むスポット群(A+B+C)の抗ロタウイルス活性を測定した。ロタミルク(EBINA,T.et.al.,J.Med.Virol.38 117-123(1992))をポジティブコントロールとした。また、実施例2における二次元電気泳動の代わりに一次元電気泳動を実施し、CBB染色で約18kDaの位置に認められたタンパク質(heated F1)を比較試料とした。なお、その際に一次元電気泳動のゲルは、試験例1の15%のポリアクリルアミド単一ゲルを用いた。試料の採取では、電気泳動により分離されたタンパク質を試験例2に従って電気泳動溶出することで得た。抗ロタウイルス活性は実施例4と同様にして測定した。
【0071】
その結果、A、B、Cを含むスポット群は、ウイルスの感染を阻害する最小阻害濃度が0.16μg/ml未満と最も高く(図7)、感染阻害率の濃度依存性においてはタンパク質濃度が高濃度になるにつれ、感染阻害率が100%に近付くことが分かった(図8)。
【0072】
[実施例6] 酵素分解物の抗ロタウイルス活性の測定
1.抗ロタウイルス活性の測定1
実施例3で得られたペプシン分解物及びプロナーゼ分解物を試料とし、実施例4と同様にして抗ロタウイルス活性を測定した。
【0073】
各試料の最小阻害濃度は、ペプシン分解物が4μg/ml、プロナーゼ分解物が5μg/mlであった(図9)。また、感染率の濃度依存性の結果は図10に示す通りであり、ペプシン分解物に比べ、プロナーゼ分解物は低濃度で、より高い感染阻害率を示すことが分かった。
【0074】
2.抗ロタウイルス活性の測定2
一方、プロナーゼ分解物については実施例3の試料に加えて、水溶液の状態で更に100℃で15分間あるいは120℃で5分間の加熱処理を行ったものを調製し、凍結乾燥して試料とした。これら2つの試料の抗ロタウイルス活性を、ロタミルク(EBINA,T.et.al.,J.Med.Virol.38 117-123(1992))を対照として実施例4と同様にして測定した。
【0075】
各試料の最小阻害濃度は、12μg/mlのプロナーゼ分解物(100℃で15分間の加熱処理)、2.1μg/mlのプロナーゼ分解物(120℃で5分間の加熱処理)、0.29/mlのロタミルクであった(図11)。また、感染率の濃度依存性は、図12のようになった。
【0076】
3.タンパク質の電気泳動溶出
前記プロナーゼ分解物及びペプシン分解物を試料とし、実施例1と同様の方法で二次元電気泳動を行った。その結果、プロナーゼ分解物は約18kDaに3つのスポットが認められた(図13)。また、ペプシン分解物は約18kDaに4つのスポットが認められた(図14)。
【0077】
以上より、ホエイの精密濾過処理の保持液又はその乾燥物の酵素分解物にも抗ロタウイルス感染阻害活性が認められ、その活性には、約18kDaのタンパク質が関与していることが示された。
【0078】
[実施例7] 抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体との反応性
実施例1や実施例3で確認された約18kDaのスポットに関する情報を得るため、抗ラクトフォリン(プロテオースペプトン3)のモノクローナル抗体染色を行った。抗体染色は、実施例1のプロテオースペプトン画分、その硫安画分及び実施例3で酵素分解物の調製に用いた乳清タンパク質の3つの試料で検討した。
【0079】
実施例1に従って二次元電気泳動を行った後に、実施例4のレクチン染色の場合と同様にして、イモビロンPVDF膜に転写した。転写した後に、一次抗体には抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体1C10(青木らの方法により作製(青木直人 他、ミルクサイエンス、54(2)、pp.83-84(2005)))、二次抗体にはペルオキシダーゼ標識した抗マウスIgG抗体(MP Biomedicals Inc.-Cappel Products)を用い、それぞれイモビロンPVDF膜と反応させた。最後にペルオキシダーゼの活性染色をDAB(ジアミノベンチジン;和光純薬工業)の基質溶液で行った。
【0080】
得られた抗体染色の結果を図15、図16、図17に示した(いずれも弱酸性側からスポット群A、B、C、D、E、Fとする)。プロテオースペプトン画分、その硫安画分及び実施例3で酵素分解物の調製に用いた乳清タンパク質の3つの全ての試料において、約18kDaのスポットが認められ、実施例4でウイルスの感染を阻害する活性が最も高いスポットであることが判明したFに相当する箇所が共通して抗体染色されていた。この結果は、FのN末端配列5残基がプロテオースペプトン3(Esben S.S.et.al.,J Dairy Res. 60 535-542(1993))のアミノ酸54位から58位の配列(Ser-Ser-Arg-Gln-Pro)と一致したことを支持するとともに、抗ロタウイルス活性にプロテオースペプトン3由来のタンパク質が関与することを改めて示した。
【0081】
[実施例8] プロテオースペプトン画分の硫安沈殿画分中の分子量が約28kDaと約18kDaのタンパク質における抗ロタウイルス活性の測定
95℃で30分間の加熱処理を行った脱脂乳に酸添加を行い、カゼイン及び変性タンパク質を沈殿除去して、プロテオースペプトン画分(以下、PP画分ともいう)を調製した。PP画分に、100%飽和硫安溶液を55%の飽和度になるまで加え、遠心分離処理を行い、上清を採取した。更に100%飽和硫安溶液を90%の飽和度になるまで加え、一晩放置した後に、遠心分離処理を行い、生じた沈殿物を採取した。次いで、この沈殿物を脱イオン蒸留水に溶解し、透析を行った。得られた溶液は、プロテオースペプトン画分由来の硫安飽和55%可溶かつ90%不溶の画分であり、以下「PP(0.90)画分」と称した。調製したPP画分とPP(0.90)画分については、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。その活性の評価は、感染阻害率の濃度依存性及び最小阻害濃度(MIC)に基づいて行った。PP画分及びPP(0.90)画分は共に感染阻害活性を示したが、PP画分の最小阻害濃度(MIC)は16μg/ml、PP(0.90)画分の最小阻害濃度(MIC)は19μg/mlであり、活性の強さには大きな違いは認められなかった(図18)。
【0082】
このPP画分及びPP(0.90)画分を膨潤用緩衝液に溶解して、3mg/ml濃度の溶液を調製した。その溶液の125μlを試料とし、試験例1記載の条件で、一次元目にpIが5〜8の等電点電気泳動に供し、二次元目に15%のポリアクリルアミドゲルを用いたSDS-PAGEに供した後に、実施例7に従って、抗ラクトフォリン(プロテオースペプトン3)モノクローナル抗体(1C10)を一次抗体として使用するウエスタンブロット解析を行った。その結果、いずれの画分についても、分子量が約28kDaのスポット群及び約18kDaのスポット群に染色反応が認められた(図19)。実施例4の「2.N末端アミノ酸配列の特定」に記載されている通り、本願発明に係る約18kDaのタンパク質は、そのN末端配列がプロテオースペプトン画分中のプロテオースペプトン3の内部配列と一致したことから、本実施例で検出された約28kDaのタンパク質が牛乳に内在するプロテアーゼの一種であるプラスミンによって分解される際に生成したものと考えられた。
【0083】
SDS-PAGEで分画したPP(0.9)画分からは、実施例4に従って約18kDaと約28kDaのタンパク質を個々に電気泳動溶出し(切り出したゲルから低電圧200Vにて4時間反応)、試料として調製した(図19)。溶出した約28kDaのタンパク質の試料はLP 28K、約18kDaのタンパク質の試料はLP 18Kと称した。次いで、3つの試料(LP 28K、LP 18K、PP(0.9)画分)について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0084】
その結果を図20のA及びBに示す。図20に示される通り、約18kDaのタンパク質(LP 18K)の感染阻害率は、調べた中の、どのタンパク質濃度でも高く、かつタンパク質濃度に依存して上昇した。また、約18kDaのタンパク質については最小阻害濃度(MIC)も0.62μg/mlであり、極めて強い活性が示された。一方、約28kDaのタンパク質(LP 28K)は、調べた範囲のタンパク質濃度では十分な感染阻害率を示すことができず、最小阻害濃度から見てもほとんど活性が認められなかった。更に約18kDaのタンパク質の結果をPP(0.9)画分と比較すると、例えば、10μg/mlのタンパク質濃度での感染阻害率がPP(0.9)画分の約2倍であり、最小阻害濃度もPP(0.9)画分の30倍以上低いという優れた結果が示された。
【0085】
[実施例9] ConA結合画分が示す感染阻害活性
実施例8に従って調製したプロテオースペプトン画分(PP画分)を、コンカナバリンA(Concanavalin A; ConA)アフィニティーカラムを使用したアフィニティークロマトグラフィーにかけて分画した。以下の組成のバッファーを使用し、流速は0.7ml/分とした。
・結合バッファー: 5mMのCH3COONa, 1mMのCaCl2, 1mMのMgCl2, 1mMのMnCl2, 1MのNaCl, 0.02%のNaN3(pH 6.8)
・溶出バッファー: 5mMのCH3COONa, 1mMのCaCl2, 1mMのMgCl2, 1mMのMnCl2, 1MのNaCl, 0.5Mのメチル-α-D-グルコピラノシド, 0.02%のNaN3 (pH 6.8)
【0086】
分画の結果を図21に示す。図21中で100 mAUを示す横線の範囲が溶出バッファーを適用した部分である。図21に示される通り、この分画の結果、ConAに結合しなかった2つの画分(非結合画分UN1及びUN2)と、ConAに結合した画分(BO画分)とが得られた。
【0087】
PP画分、BO画分、UN1画分、UN2画分のそれぞれについて膨潤用緩衝液に溶解して、3mg/ml濃度の溶液を調製した。その溶液の125μlを試料とし、試験例1記載の条件で、一次元目にpIが5〜8の等電点電気泳動に供し、二次元目に15%のポリアクリルアミドゲルを用いたSDS-PAGEに供し、電気泳動が完了した後に、CBB染色を行った(図22)。図22に示されるように、BO画分の二次元電気泳動の結果は、実施例1〜7に記載する硫安画分、乳清タンパク質濃縮物、乳清タンパク質分解物、実施例8に記載するPP(0.9)画分の二次元電気泳動の結果と良く似ていた。
【0088】
また、上記で得られた各画分について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0089】
その結果を図23のA及びBに示す。図23に示される通り、BO画分の感染阻害率は、調べた中の、どのタンパク質濃度でもPP画分よりも高く、かつタンパク質濃度に依存して上昇した。BO画分の最小阻害濃度(MIC)も7.8μg/mlであり、MICが17μg/mlであったPP画分と比較して強い活性が示された。すなわち、ConA結合性を示す画分をPP画分から精製することで、より強いロタウイルス感染阻害活性を示すBO画分を得ることができた。一方、UN1画分及びUN2画分は、調べた範囲のタンパク質濃度では十分な感染阻害率を示さず、最小阻害濃度から見てもほとんど活性が認められなかった。
【0090】
ConAはN型糖鎖と結合する性質を有すること(詳細は実施例4を参照)及びプロテオースペプトン画分に含まれるラクトフォリンにはN型糖鎖を持つ種類と持たない種類があることから、ウイルス感染阻害活性が、少なくとも、N型糖鎖を持つプロテオースペプトン画分由来タンパク質(BO画分)に認められることが確認できた。
【0091】
BO画分については、SDS-PAGEで分画した後に、実施例8と同様にして、約18kDaと約28kDaのタンパク質を実施例4に従って個々に電気泳動溶出し(切り出したゲルから低電圧200Vにて4時間反応)、試料として調製した(図24)。溶出した約28kDaのタンパク質の試料はLP 28K、約18kDaのタンパク質の試料はLP 18Kに相当する。3つの試料(LP 28K、LP 18K、BO画分)について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0092】
その結果を図25のA及びBに示す。図25に示される通り、約18kDaのタンパク質の感染阻害率は、調べた中の、どのタンパク質濃度でも高く、かつタンパク質濃度にほぼ依存して上昇した。また、約18kDaのタンパク質については最小阻害濃度(MIC)も0.86であり、BO画分と比較しても強い活性が示された。すなわち、ConA結合性を示すBO画分から更に約18kDaのタンパク質を分画することにより、更に強いロタウイルス感染阻害活性を示す画分を得ることができた。一方、約28kDaのタンパク質は、調べた範囲のタンパク質濃度では十分な感染阻害率を示すことができず、最小阻害濃度から見てもほとんど活性が認められなかった。このため、乳清タンパク質由来のウイルス感染阻害活性は、少なくとも、プロテオースペプトン画分由来のN型糖鎖を有するBO画分中の約18kDaのタンパク質に認められることが示された。以上の結果から、BO画分中のこの約18kDaのConA結合タンパク質が、実施例1〜8において分離したプロテオースペプトン3(ラクトフォリン)由来の約18kDaの乳清タンパク質であることが確認された。
【0093】
[実施例10] ConA非結合画分のプラスミン処理物が示す感染阻害活性
実施例9に従ってプロテオースペプトン画分(PP画分)から調製したConA結合のBO画分又はConA非結合のUN1画分をプラスミンにより処理した。BO画分又はUN1画分のタンパク質濃度を3mg/mlに調整し、これらにプラスミン溶液(1mg/ml、ヒト血清由来、Sigma社製)を44:1となるように加え、混合液を37℃で5分間保持した。次いで、混合液を沸騰水中で5分間加熱し、プラスミンを失活させた後に、分画分子量が10000Daの透析膜を用いて脱塩処理を行い、分解した後のサンプルをそれぞれBOP画分とUN1P画分とした。
【0094】
BOP画分とUN1P画分のそれぞれについて膨潤用緩衝液に溶解して、3mg/ml濃度の溶液を調製した。その溶液の125μlを試料とし、試験例1記載の条件で、一次元目にpIが5〜8の等電点電気泳動に供し、二次元目に15%のポリアクリルアミドゲルを用いたSDS-PAGEに供し、電気泳動が完了した後に、CBB染色及び実施例7と同様の方法で抗体染色を行った(図26)。図26に示されるように、BOP画分とUN1P画分は共に分解する前に確認されるLP28kが、分解した後にはなくなり、LP18kのみとなっていた。また、分解する前と比較して、分解した後に現れたLP18kのスポット群は、やや酸性側に認められた。この変化は、プラスミンによる分解が一部28kDaから18kDaに留まらず、更に進行して修飾糖鎖の一部が失われてしまったことに起因する可能性が考えられた。
【0095】
また、上記で得られた各画分について、試験例4に従うヒトロタウイルス中和活性試験によって、ロタウイルス感染阻害活性を測定した。この中和活性試験では、MA-104細胞に試料とヒトロタウイルスMO株の混合液を同時接種し、45時間感作させ、アセトン固定した後に、間接蛍光抗体法により感染細胞をカウントした。
【0096】
その結果を図27のA及びBに示す。図27に示される通り、BO画分よりも強い活性は得られないが、BOP画分とUN1P画分は共に感染阻害活性が認められた。ConA非結合のUN1P画分にも高い感染阻害活性が認められたことから、感染阻害活性LP18kの持つ活性にN型糖鎖は必ずしも必要では無いことが示唆された。
【産業上の利用可能性】
【0097】
本発明により、新たなロタウイルス感染防御作用を有する組成物が提供される。該組成物は、その有効量を食品や医薬組成物、家畜飼料の原料として利用することができ、極めて有用である。
【特許請求の範囲】
【請求項1】
ウシ乳清中に見出され、SDS-PAGEで測定される分子量が15〜20kDa、等電点pIが6〜9であり、N末端から5残基のアミノ酸配列がSer-Ser-Arg-Gln-Proであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質を含む、ロタウイルス感染防御用組成物。
【請求項2】
前記糖タンパク質が、酸ホエイを加熱処理し、遠心分離により変性タンパク質を除去して得られるウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画し、等電点pIが6〜9で18kDa付近に現れる弱酸性側から2〜4つ目のスポット群、5つめのスポット、又は6つ目のスポットのタンパク質を採取することによって得られる、請求項1記載のロタウイルス感染防御用組成物。
【請求項3】
酸ホエイを加熱処理し、遠心分離により変性タンパク質を除去し、得られるウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画し、等電点pIが6〜9で18kDa付近に現れる弱酸性側から2〜4つ目のスポット群、5つめのスポット、又は6つ目のスポットのタンパク質を採取することを特徴とする請求項1又は2記載のロタウイルス感染防御用組成物の製造法。
【請求項1】
ウシ乳清中に見出され、SDS-PAGEで測定される分子量が15〜20kDa、等電点pIが6〜9であり、N末端から5残基のアミノ酸配列がSer-Ser-Arg-Gln-Proであることを特徴とするロタウイルス感染阻害活性を有する糖タンパク質を含む、ロタウイルス感染防御用組成物。
【請求項2】
前記糖タンパク質が、酸ホエイを加熱処理し、遠心分離により変性タンパク質を除去して得られるウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画し、等電点pIが6〜9で18kDa付近に現れる弱酸性側から2〜4つ目のスポット群、5つめのスポット、又は6つ目のスポットのタンパク質を採取することによって得られる、請求項1記載のロタウイルス感染防御用組成物。
【請求項3】
酸ホエイを加熱処理し、遠心分離により変性タンパク質を除去し、得られるウシプロテオースペプトン溶液に硫安を加え、55〜90%の飽和度で得られる沈殿物を採取し、それを分画し、等電点pIが6〜9で18kDa付近に現れる弱酸性側から2〜4つ目のスポット群、5つめのスポット、又は6つ目のスポットのタンパク質を採取することを特徴とする請求項1又は2記載のロタウイルス感染防御用組成物の製造法。
【図1】


【図2】
【図3】

【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【公開番号】特開2011−254838(P2011−254838A)
【公開日】平成23年12月22日(2011.12.22)
【国際特許分類】
【出願番号】特願2011−186408(P2011−186408)
【出願日】平成23年8月29日(2011.8.29)
【分割の表示】特願2006−39998(P2006−39998)の分割
【原出願日】平成18年2月16日(2006.2.16)
【出願人】(000006138)株式会社明治 (265)
【出願人】(304019399)国立大学法人岐阜大学 (289)
【Fターム(参考)】
【公開日】平成23年12月22日(2011.12.22)
【国際特許分類】
【出願日】平成23年8月29日(2011.8.29)
【分割の表示】特願2006−39998(P2006−39998)の分割
【原出願日】平成18年2月16日(2006.2.16)
【出願人】(000006138)株式会社明治 (265)
【出願人】(304019399)国立大学法人岐阜大学 (289)
【Fターム(参考)】
[ Back to top ]