
ワクチンおよび免疫方法
【課題】 プリオンの感染予防に有効である、新規なワクチンおよび免疫方法を提供する。
【解決手段】 異種動物由来、好ましくは異種哺乳動物由来、さらに好ましくはマウス、ハムスター、ウシ、ヒツジ、ヒト由来のプリオン蛋白をワクチンの有効成分とする。特に、抗体のエピトープ(抗原決定基)に相当する中和活性部位を含む、組み替えプリオン蛋白が好ましく用いられる。
【解決手段】 異種動物由来、好ましくは異種哺乳動物由来、さらに好ましくはマウス、ハムスター、ウシ、ヒツジ、ヒト由来のプリオン蛋白をワクチンの有効成分とする。特に、抗体のエピトープ(抗原決定基)に相当する中和活性部位を含む、組み替えプリオン蛋白が好ましく用いられる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なワクチンに関する。また、本発明は、このワクチンを使用する新規な免疫方法に関する。
【背景技術】
【0002】
伝達性海綿状脳症、所謂プリオン病の予防は大きな社会的問題である。伝達性海綿状脳症は、ヒトに於けるクロイツフェルト−ヤコブ病(CJD)及び動物に於けるスクレイピー及び牛海綿状脳症(BSE)を含め、異常な感染性物質、所謂プリオンが感染する悪性の神経退行性疾患群である。疫学的な調査からスクレイピープリオンはヒトに感染しないと言われている。事実、異種間におけるプリオンの最初の感染において、潜伏期間の延長或いは非伝達性が実験的に観察されている。異種間のプリオン非伝達は、「種間バリアー」と呼ばれてきた。
【0003】
しかし、BSEプリオンは、汚染された食品を介して、ヒトに経口伝染し、若い人に新しい変異型のCJDを発症させることが、最近の多くの証拠から明らかになってきた。また、動物に於ける異なるタイプの病気、すなわち慢性消耗性疾患(CWD)が、北米に生息するミュールジカ及びヘラジカの中で急激に広がり、このCWDプリオンがヒトに伝染する心配から、深刻な問題を引き起こしている。最近、輸血がヒトでのプリオン伝染の危険因子で、輸血を受けた患者がその後CJDを発症する可能性が報告された。しかしながら、この様なプリオン感染に対する予防法は未だ開発されていない。
【0004】
免疫の中で、これまで試みられた予防法は、受動免疫とワクチンに分けられる。
受動免疫としては、ポリクローナル抗体及びモノクローナル抗体の投与で、プリオン感染症が予防できる可能性は示唆されている。プリオンは、主にPrPScと表記されるプロテナーゼK(PK)−抵抗性、アミロイド形成性である異常型アイソフォームのプリオン蛋白から構成されていると考えられている。これは、未だ解明されていない翻訳後修飾を介して、正常細胞アイソフォームのプリオン蛋白(PrPC)が構造変換され、生成されると考えられている。PrPCは神経細胞に最とも多く発現する、グリコシルフォスファチジルイノシトール(GPI)に固着する膜性糖蛋白である。したがって、PrPがプリオンに対する予防方法開発のためのターゲット分子と考えるのは大変合理的である。Gabizon等はPrPに対するポリクローナル抗体はハムスタープリオンの感染性を約100倍減少できると報告した(非特許文献1参照。)。最近、Heppner等は抗PrPモノクローナル抗体6H4を遺伝子導入で発現させたマウスは、マウススクレーピーRMLプリオンの腹腔内接種後の発症に抵抗性を示すことを報告した(非特許文献2参照。)。また、White等は他の2種の抗PrPモノクローナル抗体、ICSM18及び35がマウスでの受動免疫によるプリオン感染を予防できると報告した(非特許文献3参照。)。抗PrP抗体によりプリオン感染が有効的に予防できることから、PrPに対する能動的な免疫すなわちワクチン化がプリオン伝染の予防法として可能性があると思われる。しかし、内因性PrPCに対する宿主免疫寛容は、プリオンに対する能動免疫療法の開発にとって大きな障害であった。
【0005】
一方、ワクチンは、大きくプリオン蛋白および融合プリオン蛋白によるワクチン化の2つに分けられる。マウス組換えプリオン蛋白で免疫(ワクチン化)したマウスに、マウススクレーピー139Aを接種・感染させたモデルにおいて、潜伏期間の延長が報告されている(非特許文献4参照。)。これに反し、マウス組換えプリオン蛋白(23−231)で免疫したマウスに、マウススクレーピーRMLプリオンを接種・感染させたモデルでは、発症予防効果がないと報告されている(非特許文献5参照。)。
【非特許文献1】Ruth Gabizon, Proc. Natl. Acad. Sci., 1988, 85, 6617-6621
【非特許文献2】Frank L.. Heppner, Science, 2001, 294, 178-182
【非特許文献3】Anthony R. White, Nature, 2003, 422, 80-83
【非特許文献4】Einar M. Sigurdsson, Amer. J. Pathol., 2002, 161, 13-17
【非特許文献5】Magdalini Polymenidou, Proc. Natl. Acad. Sci., 2004, 101, 14670-14676
【発明の開示】
【発明が解決しようとする課題】
【0006】
しかしながら、発明者が同様な実験を福岡−1プリオン接種マウスモデルで行い、マウス組換えプリオン蛋白の効果を試験したが、その有効性を見出せなかった。現時点で、マウスプリオン蛋白(同種プリオン蛋白)をマウスに免疫し、プリオン感染が予防できるかどうか明確な解答はなく、むしろ否定的と考えるべきであろう。
このため、これらの課題を解決する、新規なワクチンおよびこのワクチンを使用する新規な免疫方法の開発が望まれている。
【0007】
本発明は、このような課題に鑑みてなされたものであり、新規なワクチンを提供することを目的とする。
また、本発明は、このワクチンを使用する新規な免疫方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
上記課題を解決し、本発明の目的を達成するため、本発明のワクチンは、異種動物由来のプリオン蛋白またはその擬態分子を含有する。
【0009】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0010】
本発明のワクチンは、異種動物由来のプリオンの中和活性部位を含有する。
【0011】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0012】
本発明のワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有する。
【0013】
ここで、限定されるわけではないが、哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属することが好ましい。
【0014】
本発明のワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトである。
【0015】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種動物由来のプリオン蛋白またはその擬態分子を含有することを特徴とする。
【0016】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0017】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種動物由来のプリオンの中和活性部位を含有することを特徴とする。
【0018】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0019】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有することを特徴とする。
【0020】
ここで、限定されるわけではないが、哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属することが好ましい。
【0021】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有し、前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトであることを特徴とする。
【発明の効果】
【0022】
本発明は、以下に記載されるような効果を奏する。
【0023】
本発明は、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規なワクチンを提供することができる。
【0024】
本発明は、異種動物由来のプリオンの中和活性部位を含有するので、新規なワクチンを提供することができる。
【0025】
本発明は、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規なワクチンを提供することができる。
【0026】
本発明は、異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規なワクチンを提供することができる。
【0027】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規な免疫方法を提供することができる。
【0028】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオンの中和活性部位を含有するので、新規な免疫方法を提供することができる。
【0029】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規な免疫方法を提供することができる。
【0030】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有し、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規な免疫方法を提供することができる。
【発明を実施するための最良の形態】
【0031】
以下、ワクチンおよび免疫方法にかかる発明を実施するための最良の形態について説明する。
【0032】
微生物と宿主抗原間に存在する分子擬態は、自己抗体及び/或いは自己反応性T細胞産生を介する自己免疫疾患の引き金に関し、良く知られた仮説的な作用機序である。この仮説は、微生物と宿主抗原の間で同じアミノ酸配列すなわち相同性を持つことが分子擬態の初期過程において必須であると言う考えに基づく。PrPは哺乳類では非常に保存された分子で、アミノ酸配列及びその3次構造において著しい類似性を保っている。したがって、異種PrPsが宿主PrPの擬態的に作用し、プリオン蛋白に対する免疫寛容を解除でき、抗PrP抗体を発現しうると、発明者は考えた。
【0033】
我々は、異種なマムスター、ヒツジ、ウシ、及びヒトの組み換えPrPsがマウスで強い抗原性を示し、マウスPrPと交差反応する抗体を効率的に産生することを、ここで示す。これに対し、マウスの組み換えPrPはマウスで非常に弱い抗体反応しか引き起こさなかった。また、ウシ及びヒツジで免疫したマウスは、非免疫マウスに比較し、マウスプリオンの腹腔内接種による発病の潜伏期間を延長させたが、マウスPrPではこの様な作用は認められなかった。これらの結果から、分子擬態に基づく新規プリオンワクシンの開発における実験的な基礎が出来たと考える。
【0034】
本発明のワクチンについて説明する。
【0035】
本発明のワクチンは、異種動物(異種哺乳動物)由来のプリオン蛋白またはその擬態分子を含有する。また、本発明のワクチンは、異種動物(異種哺乳動物)由来のプリオンの中和活性部位を含有する。
【0036】
動物としては、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属するものを採用することができる。
【0037】
哺乳動物としては、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属するものを採用することができる。
【0038】
哺乳動物は具体的に、マウス、ハムスター、ウシ、ヒツジ、またはヒトを採用することができる。哺乳動物は、これらに限定されるものではない。このほか哺乳動物としては、鹿、ネコ、犬、サル、トラ、ピューマなどを採用することができる。
【0039】
異種とは、免疫をされる個体と異なる種のことである。
【0040】
同種とは、免疫をされる個体と同じ種のことである。
【0041】
プリオン蛋白には、正常型プリオン蛋白のほかに異常型プリオン蛋白を変性したものを含む。異常型プリオン蛋白を変性したものとは、蛋白変性剤や加熱等により変性させた異常型プリオン蛋白である。
【0042】
プリオンの中和活性部位とは、プリオン感染を抑制できる予防免疫を誘導するプリオン蛋白の部位である。
【0043】
プリオン蛋白の擬態分子とは、プリオン蛋白と同じあるいは同様なアミノ酸配列を有する分子のことである。
【0044】
プリオン蛋白、プリオン蛋白の擬態分子、およびプリオンの中和活性部位は、形質転換した大腸菌を培養することにより作製することができる。プリオン蛋白、プリオン蛋白の擬態分子、およびプリオンの中和活性部位の作製方法は、これに限定されるものではない。このほか作製方法としては、真核細胞、人工的に合成などを採用することができる。
【0045】
本発明の免疫方法について説明する。本発明の免疫方法は、ワクチンを接種する方法である。
【0046】
免疫において、プリオン蛋白等の摂取量は1〜1000μgの範囲内にあることが好ましい。
接種の回数は1〜10回の範囲内にあることが好ましい。
接種の間隔は1〜5週間の範囲内にあることが好ましい。
【0047】
接種方法としては、腹腔内接種、表皮接種、皮内接種、皮下注射、静脈内注射、筋肉内注射、経口接種、経鼻接種、気管内投与などを採用することができる。
【0048】
以上のことから、本発明を実施するための最良の形態によれば、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規なワクチンを提供することができる。
【0049】
また、本発明を実施するための最良の形態によれば、異種動物由来のプリオンの中和活性部位を含有するので、新規なワクチンを提供することができる。
【0050】
また、本発明を実施するための最良の形態によれば、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規なワクチンを提供することができる。
【0051】
また、本発明を実施するための最良の形態によれば、異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規なワクチンを提供することができる。
【0052】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規な免疫方法を提供することができる。
【0053】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオンの中和活性部位を含有するので、新規な免疫方法を提供することができる。
【0054】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規な免疫方法を提供することができる。
【0055】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有し、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規な免疫方法を提供することができる。
【0056】
なお、本発明は上述の発明を実施するための最良の形態に限らず本発明の要旨を逸脱することなくその他種々の構成を採り得ることはもちろんである。
【実施例】
【0057】
つぎに、本発明にかかる実施例について具体的に説明する。ただし、本発明はこれら実施例に限定されるものではないことはもちろんである。
【0058】
本実施例の方法について説明する。
【0059】
組み換えプリオン蛋白の免疫原の発現及び精製
マウス、ハムスター、ウシ、ヒツジ、或いはヒト、全長組み換えプリオン蛋白(図1〜5参照)を発現させるために、マウスプリオン蛋白23−231残基(GenBank、注文番号M13685)、ハムスタープリオン蛋白23−232残基(GenBank、注文番号M14054)、ヒツジプリオン蛋白25−234残基(GenBank、注文番号U67922)、ウシプリオン蛋白25−242残基(GenBank、注文番号AJ298878)、及びヒトプリオン蛋白残基(GenBank、注文番号M13899)に相当するDNAフラグメントを、表1に示す様に、相当するプライマーペアーを用い、ポリメレース連鎖反応(PCR)で増幅させた。これらのPCR産物の配列確認後に、これらのフラグメントはBamHI及びHindIIIを用い消化し、pQE30ベクター(QIAGEN、Hilden、独国)の中へ挿入し、pQE30−マウスプリオン蛋白、pQE30−ハムスタープリオン蛋白、pQE30−ヒツジプリオン蛋白、pQE30−ウシプリオン蛋白、pQE30−ヒトプリオン蛋白を作製した。pQE30ベクターはN末端に6×His標識を持つ蛋白を生成するように開発されている。
【0060】
【表1】

【0061】
大腸菌(M15)は、各プラスミドにより新たに形質転換し、1mMイソプロピルチオ−β−ガラクトシド(IPTG)を含有するLB培地で培養し、5,000×g、20分間遠心分離した。この様にして集められた細胞はデオキシリボヌクラーゼI存在下、CelLytic B Bacterial Cell Lysis/Extraction Reagent(Sigma−Aldrich Co. St Louis,米国)用いて溶解させ、その溶解液を25,000×g、10分間遠心分離した。生成した沈査は0.2mg/mlのリゾチームを含む10mlのCelLytic B Bacterial Cell Lysis/Extraction Reagent中に懸濁させ、室温(RT)で15分間、時々攪拌しながらインキュベートした。1:10に希釈したCelLytic B Bacterial Cell Lysis/Extraction Reagentを加え、懸濁液の容量を増やし、25,000×g、10分間遠心分離した。生成した沈査を40mlの1:10希釈CelLytic B Bacterial Cell Lysis/Extraction Reagentで3回洗浄し、溶解緩衝液中(8M 尿素、10mM Tris−HCl、pH8.0、100mM Na2HPO4)に懸濁させた。次いで、QIAGEN社のプロトコールで推薦されている様に、変性条件化でNi−NTAカラム(QIAGEN)を用い精製した。
【0062】
6×His標識を含まないマウス組み換えプリオン蛋白の精製
6×His標識を含まないマウス組み換えプリオン蛋白を発現させるために、マウスプリオン蛋白23−231に相当するDNAフラグメントを適切なプライマーペアー(表1)を用い、PCRで増幅させた。配列の確認後、このフラグメントをNdeI及びBamHI用い消化し、pET11a−マウスプリオン蛋白を作製するために、pET11aベクター(Novagen,Inc.、WI、米国)の中へ挿入した。大腸菌(BL21)をpET11a−マウスプリオン蛋白で形質変換させ、1mMのIPTGを含むLB培地中で培養した。その細胞を5,000×gで20分間遠心分離し集め、300μg/mlのリゾチームを含有する緩衝液((50mM Tris−HCl,pH8,1mM EDTA, 100mM NaCl,1mM PMSF)に懸濁させた。室温で20分間保温後、80mgのデオキシコール酸を加え、更に20分間保温した。次いで、ゲノムDNAをデオキシリボヌクラーゼIを用い、室温で30分間消化した。その消化液を25,000×g、20分間遠心分離し、その残渣を緩衝液中(8M 尿素,50mM Tris−HCl, 1mM EDTA,pH8)に懸濁させた。この抽出液をCM−セファローズカラム(Amersham Pharmacia Biotech AB,Uppsala,スエーデン)にかけ、組み換えプリオン蛋白を、上記緩衝液による直線的食塩勾配(0〜500mM)を用い溶出した。
【0063】
GST−融合プリオン蛋白の発現と精製
ウシプリオン蛋白103−121残基、143−166残基、及び231−242残基、並びにヒトプリオン蛋白91−110残基、132−155残基、及び220−231残基をコード化したDNAフラグメントを適切なプライマーペアー(表1)を用い、PCRで増幅させた。これらのPCR産物の配列を確認後、これらのフラグメントをBamHI及びEcoRIで消化し、pGEX−2TK−boPrP103−121,−boPrP143−166,及び−boPrP231−242並びにpGEX−2TK−huPrP−91−110,−huPrP132−155,及び−huPrP220−231プラスミドを作製するために、pGEX−2TKベクター(Amersham Biosciences Corp,Piscataway,NJ、米国)に挿入した。これらのプラスミドを用い、大腸菌(DH5α)を新たに形質変換させ、100μg/ml ampicilin及び1mM IPTGを含有するLB培地中で培養した。これらの培養液を、5,000×g、20分間、遠心分離した。集められた細胞は、デオキシリボヌクレアーゼI存在下、CelLytic B Bacterial Cell Lysis/Extraction Reagent(Sigma−Aldrich Co.)を用い溶解し、この溶解液を25,000×g、10分間、遠心分離した。その上清に1%グルタチオンセファローズ4B(Amersham Biosciences)を混合し1時間放置した。この混合液を、1M尿素を含むCelLytic B Bacterial Cell Lysis/Extraction Reagent(Sigma−Aldrich Co.)で洗浄し、次いで、GST−融合プリオン蛋白を1M尿素、10mM還元グルタチオン及び50mMTris−HClを含むCelLytic B Bacterial Cell Lysis/Extraction Reagentで溶出した。
【0064】
免疫及び接種
精製した6×His標識を持つ組み換えプリオン蛋白をPBSで透析し、各組み換え蛋白100μgを雌性、8週BALB/cマウス(SLC Japan,静岡、日本)に、2週間間隔で、初回はcomplete Freund’s adjuvant(Difco Laboratories)と共に、2回目以降はincomplete Freund’s adjuvant(Difco Laborato ries)と共に、腹腔内投与し、免疫した。
1%脳ホモジネート(福岡−1プリオン感染マウス)の100ulを、5回免疫の1週間後に、腹腔内に接種した。
抗血清は、最終免疫後1週間に採取し、使用時まで−20℃で保存した。
なお、動物は長崎大学動物実験ガイドラインに基づき、取り扱った。
【0065】
ELISA
96穴ウエル免疫プレート(Nunc)の各ウエルを500ngの6×His標識を含まない精製マウス組み換えプリオン蛋白或いは6×His標識を含む他の組み換えプリオン蛋白で、4℃にて1晩保温し、コートした。次いで、0.05% Tween−20(T−PBS)及び25% Block Ace(Dainihonseiyaku Co.)を含むPBSで、37℃、1時間ブロッッキングした。特異的IgG抗体を検出するために、連続的に10倍希釈した抗血清をウエルに加え、37℃で1時間保温し、非結合抗体をT−PBSで2回洗浄し除去した。免疫複合体は、HRP(Amersham Biosciences),2mM 2,2’−Azino−bis(3−Ethylbenzthiazoline−6−sulfonic acid),及び0.04% H2O2と複合体を形成した第二次ヒツジ抗マウスIgG抗体を用いて検出した。抗プリオン抗体力価は405nmにおける吸光度で表現した。
【0066】
CBB染色
蛋白は、12%SDS−PAG上で泳動し、Quick−CBB(和光)で染色した。
【0067】
本実施例の結果について説明する。
【0068】
ハムスター、ウシ、ヒツジ及びヒトの組み換え異種プリオン蛋白はマウスの抗体反応を強力に促進する
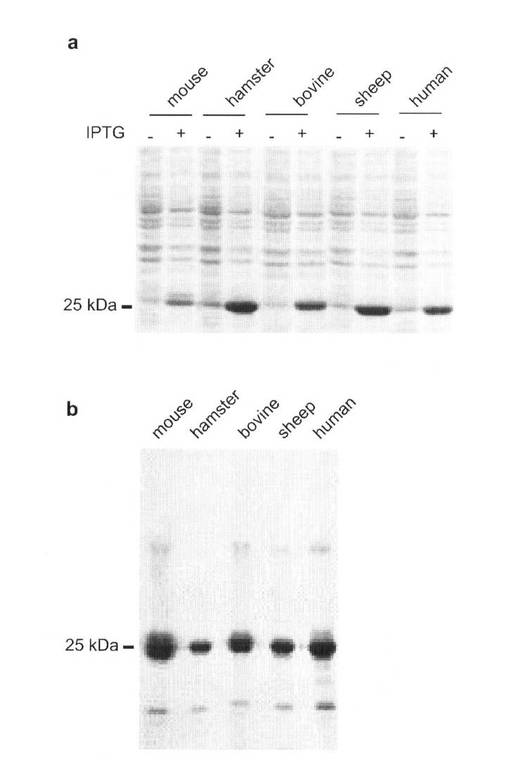
我々は、マウス、ハムスター、ウシ、ヒツジ及びヒトの組み換え異種プリオン蛋白を作製した。これらの組み換え蛋白は、N末端シグナルペプチド及びC末端GPIアンカーシグナルの両方が欠損し、かつN末端を6×His残基で標識したものである。我々はこれらの蛋白を大腸菌で発現させ、その細胞溶解液のナトリウムドデシルサルフェイト−ポリアクリルゲル電気泳動(SDS−PAGE)後、クマシイブリリアントブルー(CBB)染色を行なった。期待する分子量、〜25kD、の組み換えプリオン蛋白が効率的にこれらの細胞に発現された(図6a)。次いで、Ni−NTAアフニティークロマトグラフィーを用い、これらの蛋白を、変性条件下で精製した。各精製フラクションは単量体プリオン蛋白を大量に含み、2量体及び分解プリオン蛋白は少量であった(図6b)。CBB染色では、これらのフラクション中にはその他の蛋白質は検出されなかった(図6b)。
【0069】
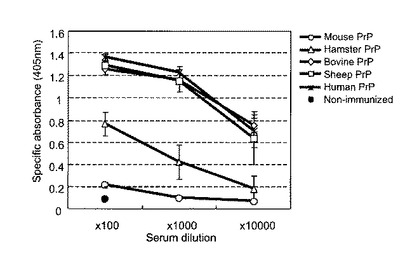
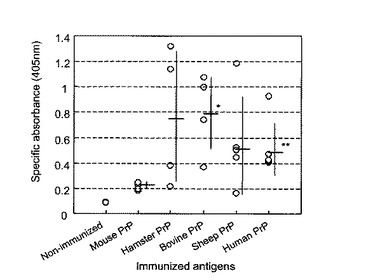
組み換え異種プリオン蛋白がマウスで特異的な抗体反応を起こすかどうかを検討するために、我々はBALB/cマウスに各精製組み換えプリオン蛋白(100μg/マウス)を2週間間隔で腹腔内投与し、免疫した。5回免疫後1週間目に抗血清を採取し、それぞれの免疫した組み換えプリオン蛋白に対する特異的なIgG抗体反応を検出するために、酵素結合免疫吸着検出法(ELISA法)を行なった。マウス組み換えプリオン蛋白で免疫したマウスでは非常に弱い抗体反応しか検出出来なかった。これらのマウスの抗血清は、非免疫マウスに比較し、405nmに於ける光学密度値(OD405)を僅かに上昇させただけであった。これに対し、組み換え異種プリオン蛋白は免疫原性を有し、マウスで非常に強力な抗体反応を示した。抗ハムスター組み換えプリオン蛋白血清は、1:100希釈で、約0.8のOD405を示した(図7)。ウシ、ヒツジ及びヒトの組み換えプリオン蛋白で免疫したマウスの抗血清では、OD405は1.2以上の高値を示した(図7)。
【0070】
抗プリオン蛋白自己抗体は、組み換え異種プリオン蛋白で免疫したマウスで、効率的に作製される
組み換え異種プリオン蛋白がトレランスを解消し、抗プリオン蛋白自己抗体を生成できるかどうかマウスで確認するために、マウス組み換えプリオン蛋白に対して免疫したマウス血清を用いてELISAを行なった。組み換え異種プリオン蛋白は6×His標識を含んでいる。従って、免疫マウスで生成される可能性がある抗体と標識の免疫反応を除去するために、ELISA抗原として標識を含まないマウス組み換えプリオン蛋白を使用した。マウス組み換えプリオン蛋白はマウスにおいてトレラントで、非免疫マウスと比較し、1:100希釈でOD405は僅かに上昇しただけであった(図8)。これに対し、ハムスター、ウシ、ヒツジ及びヒトの組み換えプリオン蛋白で免疫したマウスの抗血清の場合、OD405は、バラツキは見られるが、非免疫マウスと比較し、より高値を示した(図8)。ハムスター或いはヒツジ組み換えプリオン蛋白で免疫したマウスの中でそれぞれ1匹だけが(計2匹)、マウス組み換えプリオン蛋白で免疫したマウスと同程度に、低い抗プリオン自己抗体値を示した(図8)。
【0071】
ウシ及びヒツジ組み換えプリオン蛋白で免疫したマウスでマウスプリオンの減弱が見られたが、マウス組み換えプリオン蛋白では変化が無かった
組み換え異種プリオン蛋白による免疫が、プリオンの感染に対し予防効果を有するかどうか検討するために、マウス、ウシ、ヒツジ組み換えプリオン蛋白で5回免疫後、マウス福岡−1プリオンをBALBマウスに腹腔内投与し、接種(感染)させた。非免疫BALB/cマウスは、接種後293.5±13.4日で発症し、295.0±13.3日で死亡した(表2)。マウス組み換えプリオン蛋白で免疫したマウスは、同様に、接種後269.8±25.6日に発症し、275.0±25.2日で死亡した(表2)。これらのことから、マウス組み換えプリオン蛋白による免疫はマウスプリオンに対して何ら影響を及ぼさない。対照的に、ウシ及びヒツジプリオン蛋白で免疫したマウスは部分的に発症を抑制し、それぞれ接種後332.3±9.0日(p<0.01)及び316.0±7.5日(p<0.05)まで発症を有意に遅延させた(表2)。
【0072】
【表2】
【0073】
抗ウシ及び抗ヒトプリオン抗体は、それぞれ抗BSE及び抗ヒトプリオンの潜在的なエピトープを認識する
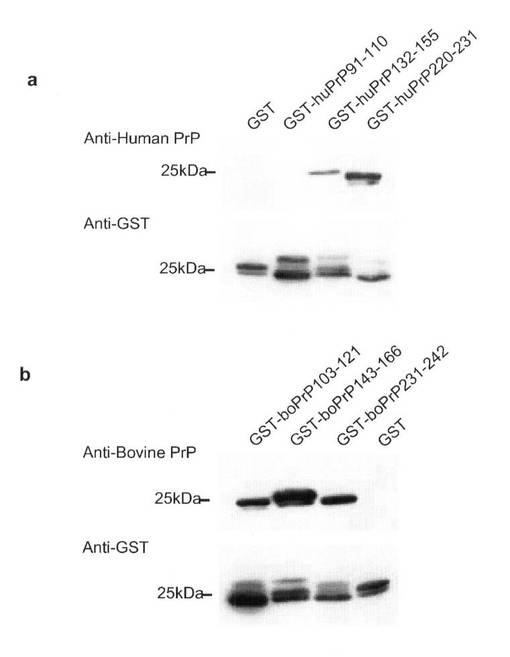
組み換え異種プリオン蛋白による免疫が異種間のプリオン感染に対し予防的かどうか確認するために、ウシ及びヒト組み換えプリオン蛋白に対する産生抗体のエピトープマッピングを行なった。マウスプリオン蛋白の91−110、144−152、及び146−159残基は、それぞれ防御的モノクローナル抗体、ICSN35、6H4及びICSN18の標的である。加えて、プリオン感染神経芽細胞種N2a細胞で異常型プリオン蛋白PrPsc除去能を有する、プリオン蛋白特異的Fabフラグメント、R1及びR2はマウスプリオン蛋白の220−231残基に結合した。これらの結果は、他のプリオン蛋白の相当領域も抗プリオンエピトープである可能性が高いことを強力に示唆している。そこで、これらの相当領域、即ちウシプリオン蛋白103−121、143−166、及び231−242残基、並びにヒトプリオン蛋白91−110、132−155、及び220−231残基を、グルタチオン−S−トランスフェラーゼ(GST)との融合蛋白として、発現させた。これらの融合蛋白を用い、抗ウシ及び抗ヒトプリオン蛋白血清でウエスタンブロッティングを行なった。融合蛋白は、GST単独の場合に比較し、大腸菌の中で、或いはその精製過程で容易に分解された(図9a及び9b)。その理由は現時点では不明である。しかし、各融合蛋白の一部は分解されずに残っていた(図9a及び9b)。GST単独及びGST−ヒトプリオン蛋白91−110は抗ヒトプリオン蛋白血清で検出できなかった(図9a及び9b)。しかし、GST−ヒトプリオン蛋白132−155及び220−231は抗ヒトプリオン蛋白血清で充分検出できた(図9a)。更に、抗ウシプリオン蛋白血清は全てのウシプリオン融合蛋白と反応した(図9b)。これらの結果は、異種プリオン蛋白での免疫がプリオンの異種間の感染に対して、潜在的な抗プリオンエピトープを認識する抗体を誘導することにより、予防的に働く可能性を強く示唆している。
【0074】
本実施例に基づく考察を説明する。
【0075】
本研究において、異種PrPsが、分子擬態機序を介してPrPに対する宿主免疫寛容を打破できる可能性を仮説とした。この考えに一致し、異種ハムスター、ウシ、ヒツジ、及びヒト組み換えPrPがマウスで強い抗原性を示し、強力に抗体反応を促進した。一方、同種マウスPrPではその様な作用は見られなかった。ウシ及びヒト組み換えPrPで免疫したマウスは、それぞれ抗BSE及び抗ヒトプリオンに対するエピトープを認識する抗体を誘導した。このことから、異種組み換えPrPsによる免疫は、例えばウシからヒトへの感染の様な、異種間のプリオン感染に対し予防的に作用する可能性が示唆された。更に、異種組み換えPrPで免疫したマウスは、自己免疫疾患に関連する症状を発現することなく、効率的にマウスPrPに対する自己抗体を生成した。また、ヒツジ及びウシ組み換えPrPで免疫したマウスは、福岡−1マウスプリオンを感染させると、有意に延長した発症潜伏期間を示した。これらの結果は、異種組み換えPrPの免疫が同種間のプリオン感染に対しも予防的である可能性を示している。まとめると、異種組み換えPrPによる免疫は別種間及び同種間のプリオン感染に対し予防的な免疫性を発現する可能性がある。
【0076】
ウシ及びヒツジで免疫したマウスにおけるプリオン病の潜伏期間の延長は、それぞれの血清中の抗PrP自己抗体の存在と関連していた。PrPに対する抗体を用いたマウスの受動免疫では、プリオン感染を効率的に阻止することが報告された。更に、抗PrP自己抗体の力価は、マウスプリオンを接種されたマウスにおける症状発現までの時間と良く相関することが報告されている。したがって、異種組み換えPrPで免疫したマウスにおける福岡−1プリオン感染の減弱化は、自己抗体を介する液性免疫に基づくと思われる。しかし、ケイホール リンペット ヘモシアニンとPrP由来ペプチドの複合体でPrP欠損マウスを免疫するとことによって、細胞毒性を示すT細胞様クローンが単利されたと言う報告がある。したがって、細胞性免疫がプリオン感染の予防に関与する可能性も考えられる。
【0077】
以前、Sigurdsson等は、組み換えマウスPrPが抗マウスPrP自己抗体を生成する免疫原性を持ち、マウスA139プリオンで感染後、免疫したマウスでの発症を僅かに遅延させることを報告している[1]。しかし、我々は、マウス組み換えPrPで免疫したマウスにおいて、福岡−1プリオンに対する抗PrP自己抗体の効果的な誘導も、発症予防的な効果も見出せなかった。また、Pollymenidou等は組み換えマウスPrPはマウスで抗PrP自己抗体を誘導出来なかったと報告している[2]。これらの結果は、異種組み換えPrPは、同種PrPより、プリオンに対する防御的免疫を促進する作用が強力である可能性を示している。熱ショック蛋白DnaKとマウスPrPの融合蛋白の投与あるいはCpG−オリゴヌクレオチドとマウスPrPの同時投与はマウスでプリオンに対する免疫原性を高め、PrPに対する自己抗体を産生することが報告されている[3,4]。そこで、異種組み換えPrPの免疫原性がDnakとの融合であるいはCpGゴリゴヌクレオチドとの併用投与により高められるかどうか興味のあるところである。
【0078】
Gilch等は、マウス組み換えPrPでマウスを免疫することにより抗PrP自己抗体を産生出来ると報告している。また、彼等は、組み換えマウスPrP二量体が、プリオンに持続感染しているマウスの神経細胞芽腫瘍N2a細胞のプリオン感染を治療する自己抗体を誘導できるという興味ある報告を出している。しかし、この組み換えPrPはヒトあるいはハムスター由来の3F4エピトープを相当する領域に挿入されているため、マウスPrPと2個のアミノ酸が異なる組み換えPrPとなっている。したがって、この組み換えPrPはマウスで異種PrP様免疫原性を獲得し、そのことにより抗PrP自己抗体を誘導したと考えられる。我々は、単量体異種PrPで免疫したマウスの血清中に、この様な治療効果のある抗体を検出できなかた。したがって、異種組み換えPrPを2量体にすることにより、この様な治療活性を有する抗体を産生するかどうか興味のあるところである。
【0079】
マウス、ハムスター、ウシ、羊、ヒトの組み換えPrPは90%以上のアミノ酸が同じである。ハムスター、ウシ、ヒツジ、ヒト組み換えPrPは、マウスの組み換えPrPと、それぞれ9個、19個、21個、17個のアミノ酸が異なるだけである。したがって、マウスにおける異種PrPによる抗PrP自己抗体の産生は分子擬態機序に基づくと考えるのが妥当であろう。しかし、我々は現時点で他の可能性を否定するものではない。以前、Wucherphennig及びStromingerは、神経変性障害である多発性硬化症の自己抗原である可能性が指摘されている、ミエリン塩基性蛋白(MBP)のある部位に相同性を示す種々のウイルス由来ペプチドが、その強さに差は有るが、MBP特異的なT細胞を刺激出来ることを示した。興味あることに、これらのペプチドの中で、Epstein−Barrウイルス由来のペプチドは、MBPペプチドと同程度或いはそれ以上にT細胞を活性化した。これらの結果を総合的考え、我々の今回の結果は、同種間及び異種間の両プリオン感染に対し予防的である免疫反応を有効的に引き起こす、分子擬態に基づく新しいプリオンワクチン開発の魅力的な可能性を強く示した。
【0080】
以上のことから、本実施例によれば、異種(ハムスター、ウシ、ヒツジ、ヒト)組換えプリオン蛋白でマウスを免疫した場合、免疫したそれぞれの異種プリオン蛋白に対する抗体が産生され、異種組換えプリオン蛋白で免疫することにより、異種間プリオン感染を予防でき、プリオンワクチンの開発に道を開くことができる。
【0081】
この時、マウスの血清中の抗プリオン自己抗体が増加していた。また、異種(ウシ、ヒツジ)組換えプリオン蛋白で免疫したマウスに福岡−1マウスプリオンを感染させたモデルにおいて、有意に潜伏(生存)期間の延長が認められ、感染を予防する効果が示された。このことから、異種(ハムスター、ウシ、ヒツジ、ヒト)組換えプリオン蛋白による免疫は同種間のプリオン感染をも予防・防止できると考えられる。
【0082】
[参考文献]
[1]Einar M. Sigurdsson, Amer. J. Pathol., 2002, 161, 13-17
[2]Magdalini Polymenidou, Proc. Natl. Acad. Sci., 2004, 101, 14670-14676
[3]Michael F. Koller, J. Neuroimmunol., 2002, 132, 113-116
[4]Martine Bruley Rosset, J. Immuno., 2004, 172, 5168-5174
【図面の簡単な説明】
【0083】
【図1】マウスについての、核酸配列とアミノ酸配列である。
【図2】ハムスターについての、核酸配列とアミノ酸配列である。
【図3】ウシについての、核酸配列とアミノ酸配列である。
【図4】ヒツジについての、核酸配列とアミノ酸配列である。
【図5】ヒトについての、核酸配列とアミノ酸配列である。
【図6】(a)IPTG処理の有(+)無(−)における、マウス、ハムスター、ウシ、ヒツジ及びヒトプリオン蛋白を発現した大腸菌のコマシーブリリアントブルー(CBB)染色を示し、(b)精製したマウス、ハムスター、ウシ、ヒツジ及びヒトプリオン蛋白のCBB染色を示す図である。
【図7】マウス、ハムスター、ウシ、ヒツジ及びヒト組み換えプリオン蛋白で免疫したマウスにおける特異的抗体反応を示す図である。5匹のマウスを各精製蛋白で免疫した。抗体力価は405nmにおける特異的な吸光度として表現した。
【図8】マウス、ハムスター、ウシ、ヒツジ及びヒト組み換えプリオン蛋白で免疫したマウスにおける抗マウスプリオン蛋白自己抗体を示す図である。免疫したマウスの各抗血清は1:100に希釈し、マウス組み換えプリオン蛋白に対するELISAを行なった。抗体力価は405nmにおける特異的な吸光度として表現した。図中の横軸及び縦軸は、それぞれ平均及び標準偏差を示す。*,p<0.05;**,p<0.01
【図9】抗ヒト(a)及び抗ウシ(b)プリオン蛋白抗体のエピトープマッピングを示す図である。ヒト及びウシプリオン蛋白のペプチドと融合したGST並びに融合しないGSTを精製し、抗ヒトプリオン蛋白、抗ウシオウリオン蛋白及び抗GST抗体を用いウエスタンブロッティングを行なった。
【技術分野】
【0001】
本発明は、新規なワクチンに関する。また、本発明は、このワクチンを使用する新規な免疫方法に関する。
【背景技術】
【0002】
伝達性海綿状脳症、所謂プリオン病の予防は大きな社会的問題である。伝達性海綿状脳症は、ヒトに於けるクロイツフェルト−ヤコブ病(CJD)及び動物に於けるスクレイピー及び牛海綿状脳症(BSE)を含め、異常な感染性物質、所謂プリオンが感染する悪性の神経退行性疾患群である。疫学的な調査からスクレイピープリオンはヒトに感染しないと言われている。事実、異種間におけるプリオンの最初の感染において、潜伏期間の延長或いは非伝達性が実験的に観察されている。異種間のプリオン非伝達は、「種間バリアー」と呼ばれてきた。
【0003】
しかし、BSEプリオンは、汚染された食品を介して、ヒトに経口伝染し、若い人に新しい変異型のCJDを発症させることが、最近の多くの証拠から明らかになってきた。また、動物に於ける異なるタイプの病気、すなわち慢性消耗性疾患(CWD)が、北米に生息するミュールジカ及びヘラジカの中で急激に広がり、このCWDプリオンがヒトに伝染する心配から、深刻な問題を引き起こしている。最近、輸血がヒトでのプリオン伝染の危険因子で、輸血を受けた患者がその後CJDを発症する可能性が報告された。しかしながら、この様なプリオン感染に対する予防法は未だ開発されていない。
【0004】
免疫の中で、これまで試みられた予防法は、受動免疫とワクチンに分けられる。
受動免疫としては、ポリクローナル抗体及びモノクローナル抗体の投与で、プリオン感染症が予防できる可能性は示唆されている。プリオンは、主にPrPScと表記されるプロテナーゼK(PK)−抵抗性、アミロイド形成性である異常型アイソフォームのプリオン蛋白から構成されていると考えられている。これは、未だ解明されていない翻訳後修飾を介して、正常細胞アイソフォームのプリオン蛋白(PrPC)が構造変換され、生成されると考えられている。PrPCは神経細胞に最とも多く発現する、グリコシルフォスファチジルイノシトール(GPI)に固着する膜性糖蛋白である。したがって、PrPがプリオンに対する予防方法開発のためのターゲット分子と考えるのは大変合理的である。Gabizon等はPrPに対するポリクローナル抗体はハムスタープリオンの感染性を約100倍減少できると報告した(非特許文献1参照。)。最近、Heppner等は抗PrPモノクローナル抗体6H4を遺伝子導入で発現させたマウスは、マウススクレーピーRMLプリオンの腹腔内接種後の発症に抵抗性を示すことを報告した(非特許文献2参照。)。また、White等は他の2種の抗PrPモノクローナル抗体、ICSM18及び35がマウスでの受動免疫によるプリオン感染を予防できると報告した(非特許文献3参照。)。抗PrP抗体によりプリオン感染が有効的に予防できることから、PrPに対する能動的な免疫すなわちワクチン化がプリオン伝染の予防法として可能性があると思われる。しかし、内因性PrPCに対する宿主免疫寛容は、プリオンに対する能動免疫療法の開発にとって大きな障害であった。
【0005】
一方、ワクチンは、大きくプリオン蛋白および融合プリオン蛋白によるワクチン化の2つに分けられる。マウス組換えプリオン蛋白で免疫(ワクチン化)したマウスに、マウススクレーピー139Aを接種・感染させたモデルにおいて、潜伏期間の延長が報告されている(非特許文献4参照。)。これに反し、マウス組換えプリオン蛋白(23−231)で免疫したマウスに、マウススクレーピーRMLプリオンを接種・感染させたモデルでは、発症予防効果がないと報告されている(非特許文献5参照。)。
【非特許文献1】Ruth Gabizon, Proc. Natl. Acad. Sci., 1988, 85, 6617-6621
【非特許文献2】Frank L.. Heppner, Science, 2001, 294, 178-182
【非特許文献3】Anthony R. White, Nature, 2003, 422, 80-83
【非特許文献4】Einar M. Sigurdsson, Amer. J. Pathol., 2002, 161, 13-17
【非特許文献5】Magdalini Polymenidou, Proc. Natl. Acad. Sci., 2004, 101, 14670-14676
【発明の開示】
【発明が解決しようとする課題】
【0006】
しかしながら、発明者が同様な実験を福岡−1プリオン接種マウスモデルで行い、マウス組換えプリオン蛋白の効果を試験したが、その有効性を見出せなかった。現時点で、マウスプリオン蛋白(同種プリオン蛋白)をマウスに免疫し、プリオン感染が予防できるかどうか明確な解答はなく、むしろ否定的と考えるべきであろう。
このため、これらの課題を解決する、新規なワクチンおよびこのワクチンを使用する新規な免疫方法の開発が望まれている。
【0007】
本発明は、このような課題に鑑みてなされたものであり、新規なワクチンを提供することを目的とする。
また、本発明は、このワクチンを使用する新規な免疫方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
上記課題を解決し、本発明の目的を達成するため、本発明のワクチンは、異種動物由来のプリオン蛋白またはその擬態分子を含有する。
【0009】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0010】
本発明のワクチンは、異種動物由来のプリオンの中和活性部位を含有する。
【0011】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0012】
本発明のワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有する。
【0013】
ここで、限定されるわけではないが、哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属することが好ましい。
【0014】
本発明のワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトである。
【0015】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種動物由来のプリオン蛋白またはその擬態分子を含有することを特徴とする。
【0016】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0017】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種動物由来のプリオンの中和活性部位を含有することを特徴とする。
【0018】
ここで、限定されるわけではないが、動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属することが好ましい。
【0019】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有することを特徴とする。
【0020】
ここで、限定されるわけではないが、哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属することが好ましい。
【0021】
本発明の免疫方法は、ワクチンを接種する免疫方法において、前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有し、前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトであることを特徴とする。
【発明の効果】
【0022】
本発明は、以下に記載されるような効果を奏する。
【0023】
本発明は、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規なワクチンを提供することができる。
【0024】
本発明は、異種動物由来のプリオンの中和活性部位を含有するので、新規なワクチンを提供することができる。
【0025】
本発明は、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規なワクチンを提供することができる。
【0026】
本発明は、異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規なワクチンを提供することができる。
【0027】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規な免疫方法を提供することができる。
【0028】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオンの中和活性部位を含有するので、新規な免疫方法を提供することができる。
【0029】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規な免疫方法を提供することができる。
【0030】
本発明は、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有し、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規な免疫方法を提供することができる。
【発明を実施するための最良の形態】
【0031】
以下、ワクチンおよび免疫方法にかかる発明を実施するための最良の形態について説明する。
【0032】
微生物と宿主抗原間に存在する分子擬態は、自己抗体及び/或いは自己反応性T細胞産生を介する自己免疫疾患の引き金に関し、良く知られた仮説的な作用機序である。この仮説は、微生物と宿主抗原の間で同じアミノ酸配列すなわち相同性を持つことが分子擬態の初期過程において必須であると言う考えに基づく。PrPは哺乳類では非常に保存された分子で、アミノ酸配列及びその3次構造において著しい類似性を保っている。したがって、異種PrPsが宿主PrPの擬態的に作用し、プリオン蛋白に対する免疫寛容を解除でき、抗PrP抗体を発現しうると、発明者は考えた。
【0033】
我々は、異種なマムスター、ヒツジ、ウシ、及びヒトの組み換えPrPsがマウスで強い抗原性を示し、マウスPrPと交差反応する抗体を効率的に産生することを、ここで示す。これに対し、マウスの組み換えPrPはマウスで非常に弱い抗体反応しか引き起こさなかった。また、ウシ及びヒツジで免疫したマウスは、非免疫マウスに比較し、マウスプリオンの腹腔内接種による発病の潜伏期間を延長させたが、マウスPrPではこの様な作用は認められなかった。これらの結果から、分子擬態に基づく新規プリオンワクシンの開発における実験的な基礎が出来たと考える。
【0034】
本発明のワクチンについて説明する。
【0035】
本発明のワクチンは、異種動物(異種哺乳動物)由来のプリオン蛋白またはその擬態分子を含有する。また、本発明のワクチンは、異種動物(異種哺乳動物)由来のプリオンの中和活性部位を含有する。
【0036】
動物としては、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属するものを採用することができる。
【0037】
哺乳動物としては、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属するものを採用することができる。
【0038】
哺乳動物は具体的に、マウス、ハムスター、ウシ、ヒツジ、またはヒトを採用することができる。哺乳動物は、これらに限定されるものではない。このほか哺乳動物としては、鹿、ネコ、犬、サル、トラ、ピューマなどを採用することができる。
【0039】
異種とは、免疫をされる個体と異なる種のことである。
【0040】
同種とは、免疫をされる個体と同じ種のことである。
【0041】
プリオン蛋白には、正常型プリオン蛋白のほかに異常型プリオン蛋白を変性したものを含む。異常型プリオン蛋白を変性したものとは、蛋白変性剤や加熱等により変性させた異常型プリオン蛋白である。
【0042】
プリオンの中和活性部位とは、プリオン感染を抑制できる予防免疫を誘導するプリオン蛋白の部位である。
【0043】
プリオン蛋白の擬態分子とは、プリオン蛋白と同じあるいは同様なアミノ酸配列を有する分子のことである。
【0044】
プリオン蛋白、プリオン蛋白の擬態分子、およびプリオンの中和活性部位は、形質転換した大腸菌を培養することにより作製することができる。プリオン蛋白、プリオン蛋白の擬態分子、およびプリオンの中和活性部位の作製方法は、これに限定されるものではない。このほか作製方法としては、真核細胞、人工的に合成などを採用することができる。
【0045】
本発明の免疫方法について説明する。本発明の免疫方法は、ワクチンを接種する方法である。
【0046】
免疫において、プリオン蛋白等の摂取量は1〜1000μgの範囲内にあることが好ましい。
接種の回数は1〜10回の範囲内にあることが好ましい。
接種の間隔は1〜5週間の範囲内にあることが好ましい。
【0047】
接種方法としては、腹腔内接種、表皮接種、皮内接種、皮下注射、静脈内注射、筋肉内注射、経口接種、経鼻接種、気管内投与などを採用することができる。
【0048】
以上のことから、本発明を実施するための最良の形態によれば、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規なワクチンを提供することができる。
【0049】
また、本発明を実施するための最良の形態によれば、異種動物由来のプリオンの中和活性部位を含有するので、新規なワクチンを提供することができる。
【0050】
また、本発明を実施するための最良の形態によれば、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規なワクチンを提供することができる。
【0051】
また、本発明を実施するための最良の形態によれば、異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規なワクチンを提供することができる。
【0052】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオン蛋白またはその擬態分子を含有するので、新規な免疫方法を提供することができる。
【0053】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種動物由来のプリオンの中和活性部位を含有するので、新規な免疫方法を提供することができる。
【0054】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有するので、新規な免疫方法を提供することができる。
【0055】
また、本発明を実施するための最良の形態によれば、ワクチンを接種する免疫方法において、前記ワクチンが、異種の哺乳動物に由来するプリオン蛋白を含有し、前記哺乳動物が、マウス、ハムスター、ウシ、ヒツジ、またはヒトであるので、新規な免疫方法を提供することができる。
【0056】
なお、本発明は上述の発明を実施するための最良の形態に限らず本発明の要旨を逸脱することなくその他種々の構成を採り得ることはもちろんである。
【実施例】
【0057】
つぎに、本発明にかかる実施例について具体的に説明する。ただし、本発明はこれら実施例に限定されるものではないことはもちろんである。
【0058】
本実施例の方法について説明する。
【0059】
組み換えプリオン蛋白の免疫原の発現及び精製
マウス、ハムスター、ウシ、ヒツジ、或いはヒト、全長組み換えプリオン蛋白(図1〜5参照)を発現させるために、マウスプリオン蛋白23−231残基(GenBank、注文番号M13685)、ハムスタープリオン蛋白23−232残基(GenBank、注文番号M14054)、ヒツジプリオン蛋白25−234残基(GenBank、注文番号U67922)、ウシプリオン蛋白25−242残基(GenBank、注文番号AJ298878)、及びヒトプリオン蛋白残基(GenBank、注文番号M13899)に相当するDNAフラグメントを、表1に示す様に、相当するプライマーペアーを用い、ポリメレース連鎖反応(PCR)で増幅させた。これらのPCR産物の配列確認後に、これらのフラグメントはBamHI及びHindIIIを用い消化し、pQE30ベクター(QIAGEN、Hilden、独国)の中へ挿入し、pQE30−マウスプリオン蛋白、pQE30−ハムスタープリオン蛋白、pQE30−ヒツジプリオン蛋白、pQE30−ウシプリオン蛋白、pQE30−ヒトプリオン蛋白を作製した。pQE30ベクターはN末端に6×His標識を持つ蛋白を生成するように開発されている。
【0060】
【表1】
【0061】
大腸菌(M15)は、各プラスミドにより新たに形質転換し、1mMイソプロピルチオ−β−ガラクトシド(IPTG)を含有するLB培地で培養し、5,000×g、20分間遠心分離した。この様にして集められた細胞はデオキシリボヌクラーゼI存在下、CelLytic B Bacterial Cell Lysis/Extraction Reagent(Sigma−Aldrich Co. St Louis,米国)用いて溶解させ、その溶解液を25,000×g、10分間遠心分離した。生成した沈査は0.2mg/mlのリゾチームを含む10mlのCelLytic B Bacterial Cell Lysis/Extraction Reagent中に懸濁させ、室温(RT)で15分間、時々攪拌しながらインキュベートした。1:10に希釈したCelLytic B Bacterial Cell Lysis/Extraction Reagentを加え、懸濁液の容量を増やし、25,000×g、10分間遠心分離した。生成した沈査を40mlの1:10希釈CelLytic B Bacterial Cell Lysis/Extraction Reagentで3回洗浄し、溶解緩衝液中(8M 尿素、10mM Tris−HCl、pH8.0、100mM Na2HPO4)に懸濁させた。次いで、QIAGEN社のプロトコールで推薦されている様に、変性条件化でNi−NTAカラム(QIAGEN)を用い精製した。
【0062】
6×His標識を含まないマウス組み換えプリオン蛋白の精製
6×His標識を含まないマウス組み換えプリオン蛋白を発現させるために、マウスプリオン蛋白23−231に相当するDNAフラグメントを適切なプライマーペアー(表1)を用い、PCRで増幅させた。配列の確認後、このフラグメントをNdeI及びBamHI用い消化し、pET11a−マウスプリオン蛋白を作製するために、pET11aベクター(Novagen,Inc.、WI、米国)の中へ挿入した。大腸菌(BL21)をpET11a−マウスプリオン蛋白で形質変換させ、1mMのIPTGを含むLB培地中で培養した。その細胞を5,000×gで20分間遠心分離し集め、300μg/mlのリゾチームを含有する緩衝液((50mM Tris−HCl,pH8,1mM EDTA, 100mM NaCl,1mM PMSF)に懸濁させた。室温で20分間保温後、80mgのデオキシコール酸を加え、更に20分間保温した。次いで、ゲノムDNAをデオキシリボヌクラーゼIを用い、室温で30分間消化した。その消化液を25,000×g、20分間遠心分離し、その残渣を緩衝液中(8M 尿素,50mM Tris−HCl, 1mM EDTA,pH8)に懸濁させた。この抽出液をCM−セファローズカラム(Amersham Pharmacia Biotech AB,Uppsala,スエーデン)にかけ、組み換えプリオン蛋白を、上記緩衝液による直線的食塩勾配(0〜500mM)を用い溶出した。
【0063】
GST−融合プリオン蛋白の発現と精製
ウシプリオン蛋白103−121残基、143−166残基、及び231−242残基、並びにヒトプリオン蛋白91−110残基、132−155残基、及び220−231残基をコード化したDNAフラグメントを適切なプライマーペアー(表1)を用い、PCRで増幅させた。これらのPCR産物の配列を確認後、これらのフラグメントをBamHI及びEcoRIで消化し、pGEX−2TK−boPrP103−121,−boPrP143−166,及び−boPrP231−242並びにpGEX−2TK−huPrP−91−110,−huPrP132−155,及び−huPrP220−231プラスミドを作製するために、pGEX−2TKベクター(Amersham Biosciences Corp,Piscataway,NJ、米国)に挿入した。これらのプラスミドを用い、大腸菌(DH5α)を新たに形質変換させ、100μg/ml ampicilin及び1mM IPTGを含有するLB培地中で培養した。これらの培養液を、5,000×g、20分間、遠心分離した。集められた細胞は、デオキシリボヌクレアーゼI存在下、CelLytic B Bacterial Cell Lysis/Extraction Reagent(Sigma−Aldrich Co.)を用い溶解し、この溶解液を25,000×g、10分間、遠心分離した。その上清に1%グルタチオンセファローズ4B(Amersham Biosciences)を混合し1時間放置した。この混合液を、1M尿素を含むCelLytic B Bacterial Cell Lysis/Extraction Reagent(Sigma−Aldrich Co.)で洗浄し、次いで、GST−融合プリオン蛋白を1M尿素、10mM還元グルタチオン及び50mMTris−HClを含むCelLytic B Bacterial Cell Lysis/Extraction Reagentで溶出した。
【0064】
免疫及び接種
精製した6×His標識を持つ組み換えプリオン蛋白をPBSで透析し、各組み換え蛋白100μgを雌性、8週BALB/cマウス(SLC Japan,静岡、日本)に、2週間間隔で、初回はcomplete Freund’s adjuvant(Difco Laboratories)と共に、2回目以降はincomplete Freund’s adjuvant(Difco Laborato ries)と共に、腹腔内投与し、免疫した。
1%脳ホモジネート(福岡−1プリオン感染マウス)の100ulを、5回免疫の1週間後に、腹腔内に接種した。
抗血清は、最終免疫後1週間に採取し、使用時まで−20℃で保存した。
なお、動物は長崎大学動物実験ガイドラインに基づき、取り扱った。
【0065】
ELISA
96穴ウエル免疫プレート(Nunc)の各ウエルを500ngの6×His標識を含まない精製マウス組み換えプリオン蛋白或いは6×His標識を含む他の組み換えプリオン蛋白で、4℃にて1晩保温し、コートした。次いで、0.05% Tween−20(T−PBS)及び25% Block Ace(Dainihonseiyaku Co.)を含むPBSで、37℃、1時間ブロッッキングした。特異的IgG抗体を検出するために、連続的に10倍希釈した抗血清をウエルに加え、37℃で1時間保温し、非結合抗体をT−PBSで2回洗浄し除去した。免疫複合体は、HRP(Amersham Biosciences),2mM 2,2’−Azino−bis(3−Ethylbenzthiazoline−6−sulfonic acid),及び0.04% H2O2と複合体を形成した第二次ヒツジ抗マウスIgG抗体を用いて検出した。抗プリオン抗体力価は405nmにおける吸光度で表現した。
【0066】
CBB染色
蛋白は、12%SDS−PAG上で泳動し、Quick−CBB(和光)で染色した。
【0067】
本実施例の結果について説明する。
【0068】
ハムスター、ウシ、ヒツジ及びヒトの組み換え異種プリオン蛋白はマウスの抗体反応を強力に促進する
我々は、マウス、ハムスター、ウシ、ヒツジ及びヒトの組み換え異種プリオン蛋白を作製した。これらの組み換え蛋白は、N末端シグナルペプチド及びC末端GPIアンカーシグナルの両方が欠損し、かつN末端を6×His残基で標識したものである。我々はこれらの蛋白を大腸菌で発現させ、その細胞溶解液のナトリウムドデシルサルフェイト−ポリアクリルゲル電気泳動(SDS−PAGE)後、クマシイブリリアントブルー(CBB)染色を行なった。期待する分子量、〜25kD、の組み換えプリオン蛋白が効率的にこれらの細胞に発現された(図6a)。次いで、Ni−NTAアフニティークロマトグラフィーを用い、これらの蛋白を、変性条件下で精製した。各精製フラクションは単量体プリオン蛋白を大量に含み、2量体及び分解プリオン蛋白は少量であった(図6b)。CBB染色では、これらのフラクション中にはその他の蛋白質は検出されなかった(図6b)。
【0069】
組み換え異種プリオン蛋白がマウスで特異的な抗体反応を起こすかどうかを検討するために、我々はBALB/cマウスに各精製組み換えプリオン蛋白(100μg/マウス)を2週間間隔で腹腔内投与し、免疫した。5回免疫後1週間目に抗血清を採取し、それぞれの免疫した組み換えプリオン蛋白に対する特異的なIgG抗体反応を検出するために、酵素結合免疫吸着検出法(ELISA法)を行なった。マウス組み換えプリオン蛋白で免疫したマウスでは非常に弱い抗体反応しか検出出来なかった。これらのマウスの抗血清は、非免疫マウスに比較し、405nmに於ける光学密度値(OD405)を僅かに上昇させただけであった。これに対し、組み換え異種プリオン蛋白は免疫原性を有し、マウスで非常に強力な抗体反応を示した。抗ハムスター組み換えプリオン蛋白血清は、1:100希釈で、約0.8のOD405を示した(図7)。ウシ、ヒツジ及びヒトの組み換えプリオン蛋白で免疫したマウスの抗血清では、OD405は1.2以上の高値を示した(図7)。
【0070】
抗プリオン蛋白自己抗体は、組み換え異種プリオン蛋白で免疫したマウスで、効率的に作製される
組み換え異種プリオン蛋白がトレランスを解消し、抗プリオン蛋白自己抗体を生成できるかどうかマウスで確認するために、マウス組み換えプリオン蛋白に対して免疫したマウス血清を用いてELISAを行なった。組み換え異種プリオン蛋白は6×His標識を含んでいる。従って、免疫マウスで生成される可能性がある抗体と標識の免疫反応を除去するために、ELISA抗原として標識を含まないマウス組み換えプリオン蛋白を使用した。マウス組み換えプリオン蛋白はマウスにおいてトレラントで、非免疫マウスと比較し、1:100希釈でOD405は僅かに上昇しただけであった(図8)。これに対し、ハムスター、ウシ、ヒツジ及びヒトの組み換えプリオン蛋白で免疫したマウスの抗血清の場合、OD405は、バラツキは見られるが、非免疫マウスと比較し、より高値を示した(図8)。ハムスター或いはヒツジ組み換えプリオン蛋白で免疫したマウスの中でそれぞれ1匹だけが(計2匹)、マウス組み換えプリオン蛋白で免疫したマウスと同程度に、低い抗プリオン自己抗体値を示した(図8)。
【0071】
ウシ及びヒツジ組み換えプリオン蛋白で免疫したマウスでマウスプリオンの減弱が見られたが、マウス組み換えプリオン蛋白では変化が無かった
組み換え異種プリオン蛋白による免疫が、プリオンの感染に対し予防効果を有するかどうか検討するために、マウス、ウシ、ヒツジ組み換えプリオン蛋白で5回免疫後、マウス福岡−1プリオンをBALBマウスに腹腔内投与し、接種(感染)させた。非免疫BALB/cマウスは、接種後293.5±13.4日で発症し、295.0±13.3日で死亡した(表2)。マウス組み換えプリオン蛋白で免疫したマウスは、同様に、接種後269.8±25.6日に発症し、275.0±25.2日で死亡した(表2)。これらのことから、マウス組み換えプリオン蛋白による免疫はマウスプリオンに対して何ら影響を及ぼさない。対照的に、ウシ及びヒツジプリオン蛋白で免疫したマウスは部分的に発症を抑制し、それぞれ接種後332.3±9.0日(p<0.01)及び316.0±7.5日(p<0.05)まで発症を有意に遅延させた(表2)。
【0072】
【表2】
【0073】
抗ウシ及び抗ヒトプリオン抗体は、それぞれ抗BSE及び抗ヒトプリオンの潜在的なエピトープを認識する
組み換え異種プリオン蛋白による免疫が異種間のプリオン感染に対し予防的かどうか確認するために、ウシ及びヒト組み換えプリオン蛋白に対する産生抗体のエピトープマッピングを行なった。マウスプリオン蛋白の91−110、144−152、及び146−159残基は、それぞれ防御的モノクローナル抗体、ICSN35、6H4及びICSN18の標的である。加えて、プリオン感染神経芽細胞種N2a細胞で異常型プリオン蛋白PrPsc除去能を有する、プリオン蛋白特異的Fabフラグメント、R1及びR2はマウスプリオン蛋白の220−231残基に結合した。これらの結果は、他のプリオン蛋白の相当領域も抗プリオンエピトープである可能性が高いことを強力に示唆している。そこで、これらの相当領域、即ちウシプリオン蛋白103−121、143−166、及び231−242残基、並びにヒトプリオン蛋白91−110、132−155、及び220−231残基を、グルタチオン−S−トランスフェラーゼ(GST)との融合蛋白として、発現させた。これらの融合蛋白を用い、抗ウシ及び抗ヒトプリオン蛋白血清でウエスタンブロッティングを行なった。融合蛋白は、GST単独の場合に比較し、大腸菌の中で、或いはその精製過程で容易に分解された(図9a及び9b)。その理由は現時点では不明である。しかし、各融合蛋白の一部は分解されずに残っていた(図9a及び9b)。GST単独及びGST−ヒトプリオン蛋白91−110は抗ヒトプリオン蛋白血清で検出できなかった(図9a及び9b)。しかし、GST−ヒトプリオン蛋白132−155及び220−231は抗ヒトプリオン蛋白血清で充分検出できた(図9a)。更に、抗ウシプリオン蛋白血清は全てのウシプリオン融合蛋白と反応した(図9b)。これらの結果は、異種プリオン蛋白での免疫がプリオンの異種間の感染に対して、潜在的な抗プリオンエピトープを認識する抗体を誘導することにより、予防的に働く可能性を強く示唆している。
【0074】
本実施例に基づく考察を説明する。
【0075】
本研究において、異種PrPsが、分子擬態機序を介してPrPに対する宿主免疫寛容を打破できる可能性を仮説とした。この考えに一致し、異種ハムスター、ウシ、ヒツジ、及びヒト組み換えPrPがマウスで強い抗原性を示し、強力に抗体反応を促進した。一方、同種マウスPrPではその様な作用は見られなかった。ウシ及びヒト組み換えPrPで免疫したマウスは、それぞれ抗BSE及び抗ヒトプリオンに対するエピトープを認識する抗体を誘導した。このことから、異種組み換えPrPsによる免疫は、例えばウシからヒトへの感染の様な、異種間のプリオン感染に対し予防的に作用する可能性が示唆された。更に、異種組み換えPrPで免疫したマウスは、自己免疫疾患に関連する症状を発現することなく、効率的にマウスPrPに対する自己抗体を生成した。また、ヒツジ及びウシ組み換えPrPで免疫したマウスは、福岡−1マウスプリオンを感染させると、有意に延長した発症潜伏期間を示した。これらの結果は、異種組み換えPrPの免疫が同種間のプリオン感染に対しも予防的である可能性を示している。まとめると、異種組み換えPrPによる免疫は別種間及び同種間のプリオン感染に対し予防的な免疫性を発現する可能性がある。
【0076】
ウシ及びヒツジで免疫したマウスにおけるプリオン病の潜伏期間の延長は、それぞれの血清中の抗PrP自己抗体の存在と関連していた。PrPに対する抗体を用いたマウスの受動免疫では、プリオン感染を効率的に阻止することが報告された。更に、抗PrP自己抗体の力価は、マウスプリオンを接種されたマウスにおける症状発現までの時間と良く相関することが報告されている。したがって、異種組み換えPrPで免疫したマウスにおける福岡−1プリオン感染の減弱化は、自己抗体を介する液性免疫に基づくと思われる。しかし、ケイホール リンペット ヘモシアニンとPrP由来ペプチドの複合体でPrP欠損マウスを免疫するとことによって、細胞毒性を示すT細胞様クローンが単利されたと言う報告がある。したがって、細胞性免疫がプリオン感染の予防に関与する可能性も考えられる。
【0077】
以前、Sigurdsson等は、組み換えマウスPrPが抗マウスPrP自己抗体を生成する免疫原性を持ち、マウスA139プリオンで感染後、免疫したマウスでの発症を僅かに遅延させることを報告している[1]。しかし、我々は、マウス組み換えPrPで免疫したマウスにおいて、福岡−1プリオンに対する抗PrP自己抗体の効果的な誘導も、発症予防的な効果も見出せなかった。また、Pollymenidou等は組み換えマウスPrPはマウスで抗PrP自己抗体を誘導出来なかったと報告している[2]。これらの結果は、異種組み換えPrPは、同種PrPより、プリオンに対する防御的免疫を促進する作用が強力である可能性を示している。熱ショック蛋白DnaKとマウスPrPの融合蛋白の投与あるいはCpG−オリゴヌクレオチドとマウスPrPの同時投与はマウスでプリオンに対する免疫原性を高め、PrPに対する自己抗体を産生することが報告されている[3,4]。そこで、異種組み換えPrPの免疫原性がDnakとの融合であるいはCpGゴリゴヌクレオチドとの併用投与により高められるかどうか興味のあるところである。
【0078】
Gilch等は、マウス組み換えPrPでマウスを免疫することにより抗PrP自己抗体を産生出来ると報告している。また、彼等は、組み換えマウスPrP二量体が、プリオンに持続感染しているマウスの神経細胞芽腫瘍N2a細胞のプリオン感染を治療する自己抗体を誘導できるという興味ある報告を出している。しかし、この組み換えPrPはヒトあるいはハムスター由来の3F4エピトープを相当する領域に挿入されているため、マウスPrPと2個のアミノ酸が異なる組み換えPrPとなっている。したがって、この組み換えPrPはマウスで異種PrP様免疫原性を獲得し、そのことにより抗PrP自己抗体を誘導したと考えられる。我々は、単量体異種PrPで免疫したマウスの血清中に、この様な治療効果のある抗体を検出できなかた。したがって、異種組み換えPrPを2量体にすることにより、この様な治療活性を有する抗体を産生するかどうか興味のあるところである。
【0079】
マウス、ハムスター、ウシ、羊、ヒトの組み換えPrPは90%以上のアミノ酸が同じである。ハムスター、ウシ、ヒツジ、ヒト組み換えPrPは、マウスの組み換えPrPと、それぞれ9個、19個、21個、17個のアミノ酸が異なるだけである。したがって、マウスにおける異種PrPによる抗PrP自己抗体の産生は分子擬態機序に基づくと考えるのが妥当であろう。しかし、我々は現時点で他の可能性を否定するものではない。以前、Wucherphennig及びStromingerは、神経変性障害である多発性硬化症の自己抗原である可能性が指摘されている、ミエリン塩基性蛋白(MBP)のある部位に相同性を示す種々のウイルス由来ペプチドが、その強さに差は有るが、MBP特異的なT細胞を刺激出来ることを示した。興味あることに、これらのペプチドの中で、Epstein−Barrウイルス由来のペプチドは、MBPペプチドと同程度或いはそれ以上にT細胞を活性化した。これらの結果を総合的考え、我々の今回の結果は、同種間及び異種間の両プリオン感染に対し予防的である免疫反応を有効的に引き起こす、分子擬態に基づく新しいプリオンワクチン開発の魅力的な可能性を強く示した。
【0080】
以上のことから、本実施例によれば、異種(ハムスター、ウシ、ヒツジ、ヒト)組換えプリオン蛋白でマウスを免疫した場合、免疫したそれぞれの異種プリオン蛋白に対する抗体が産生され、異種組換えプリオン蛋白で免疫することにより、異種間プリオン感染を予防でき、プリオンワクチンの開発に道を開くことができる。
【0081】
この時、マウスの血清中の抗プリオン自己抗体が増加していた。また、異種(ウシ、ヒツジ)組換えプリオン蛋白で免疫したマウスに福岡−1マウスプリオンを感染させたモデルにおいて、有意に潜伏(生存)期間の延長が認められ、感染を予防する効果が示された。このことから、異種(ハムスター、ウシ、ヒツジ、ヒト)組換えプリオン蛋白による免疫は同種間のプリオン感染をも予防・防止できると考えられる。
【0082】
[参考文献]
[1]Einar M. Sigurdsson, Amer. J. Pathol., 2002, 161, 13-17
[2]Magdalini Polymenidou, Proc. Natl. Acad. Sci., 2004, 101, 14670-14676
[3]Michael F. Koller, J. Neuroimmunol., 2002, 132, 113-116
[4]Martine Bruley Rosset, J. Immuno., 2004, 172, 5168-5174
【図面の簡単な説明】
【0083】
【図1】マウスについての、核酸配列とアミノ酸配列である。
【図2】ハムスターについての、核酸配列とアミノ酸配列である。
【図3】ウシについての、核酸配列とアミノ酸配列である。
【図4】ヒツジについての、核酸配列とアミノ酸配列である。
【図5】ヒトについての、核酸配列とアミノ酸配列である。
【図6】(a)IPTG処理の有(+)無(−)における、マウス、ハムスター、ウシ、ヒツジ及びヒトプリオン蛋白を発現した大腸菌のコマシーブリリアントブルー(CBB)染色を示し、(b)精製したマウス、ハムスター、ウシ、ヒツジ及びヒトプリオン蛋白のCBB染色を示す図である。
【図7】マウス、ハムスター、ウシ、ヒツジ及びヒト組み換えプリオン蛋白で免疫したマウスにおける特異的抗体反応を示す図である。5匹のマウスを各精製蛋白で免疫した。抗体力価は405nmにおける特異的な吸光度として表現した。
【図8】マウス、ハムスター、ウシ、ヒツジ及びヒト組み換えプリオン蛋白で免疫したマウスにおける抗マウスプリオン蛋白自己抗体を示す図である。免疫したマウスの各抗血清は1:100に希釈し、マウス組み換えプリオン蛋白に対するELISAを行なった。抗体力価は405nmにおける特異的な吸光度として表現した。図中の横軸及び縦軸は、それぞれ平均及び標準偏差を示す。*,p<0.05;**,p<0.01
【図9】抗ヒト(a)及び抗ウシ(b)プリオン蛋白抗体のエピトープマッピングを示す図である。ヒト及びウシプリオン蛋白のペプチドと融合したGST並びに融合しないGSTを精製し、抗ヒトプリオン蛋白、抗ウシオウリオン蛋白及び抗GST抗体を用いウエスタンブロッティングを行なった。
【特許請求の範囲】
【請求項1】
異種動物由来のプリオン蛋白またはその擬態分子を含有するワクチン。
【請求項2】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
請求項1記載のワクチン。
【請求項3】
異種動物由来のプリオンの中和活性部位を含有するワクチン。
【請求項4】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
請求項3記載のワクチン。
【請求項5】
異種の哺乳動物に由来するプリオン蛋白を含有するワクチン。
【請求項6】
哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属する
請求項5記載のワクチン。
【請求項7】
異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、
前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトである
ワクチン。
【請求項8】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種動物由来のプリオン蛋白またはその擬態分子を含有する
ことを特徴とする免疫方法。
【請求項9】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
ことを特徴とする請求項8記載の免疫方法。
【請求項10】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種動物由来のプリオンの中和活性部位を含有する
ことを特徴とする免疫方法。
【請求項11】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
ことを特徴とする請求項10記載の免疫方法。
【請求項12】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有する
ことを特徴とする免疫方法。
【請求項13】
哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属する
ことを特徴とする請求項12記載の免疫方法。
【請求項14】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有し、
前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトである
ことを特徴とする免疫方法。
【請求項1】
異種動物由来のプリオン蛋白またはその擬態分子を含有するワクチン。
【請求項2】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
請求項1記載のワクチン。
【請求項3】
異種動物由来のプリオンの中和活性部位を含有するワクチン。
【請求項4】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
請求項3記載のワクチン。
【請求項5】
異種の哺乳動物に由来するプリオン蛋白を含有するワクチン。
【請求項6】
哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属する
請求項5記載のワクチン。
【請求項7】
異種の哺乳動物に由来するプリオン蛋白を含有するワクチンであって、
前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトである
ワクチン。
【請求項8】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種動物由来のプリオン蛋白またはその擬態分子を含有する
ことを特徴とする免疫方法。
【請求項9】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
ことを特徴とする請求項8記載の免疫方法。
【請求項10】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種動物由来のプリオンの中和活性部位を含有する
ことを特徴とする免疫方法。
【請求項11】
動物は、脊椎動物門、原索動物門、棘皮動物門、半索動物門、毛顎動物門、有鬚動物門 、触手動物門、節足動物門、舌形動物門、緩歩動物門、有爪動物門、星口動物門、ゆむし動物門、環形動物門、軟体動物門、曲形動物門、紐形動物門、扁形動物門、袋形動物門、有櫛動物門、刺胞動物門、または海綿動物門に属する
ことを特徴とする請求項10記載の免疫方法。
【請求項12】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有する
ことを特徴とする免疫方法。
【請求項13】
哺乳動物は、カモノハシ目、フクロネズミ目、アリクイ目、モグラ目、ツパイ目、ヒヨケザル目、コウモリ目、サル目、ネコ目、ゾウ目、ジュゴン目、ウマ目、イワダヌキ目、ツチブタ目、ウシ目、クジラ目、センザンコウ目、ネズミ目、ウサギ目、またはハネジネズミ目に属する
ことを特徴とする請求項12記載の免疫方法。
【請求項14】
ワクチンを接種する免疫方法において、
前記ワクチンは、異種の哺乳動物に由来するプリオン蛋白を含有し、
前記哺乳動物は、マウス、ハムスター、ウシ、ヒツジ、またはヒトである
ことを特徴とする免疫方法。
【図1】


【図2】

【図3】

【図4】

【図5】

【図7】
【図8】
【図6】
【図9】
【図2】
【図3】
【図4】
【図5】
【図7】
【図8】
【図6】
【図9】
【公開番号】特開2008−201674(P2008−201674A)
【公開日】平成20年9月4日(2008.9.4)
【国際特許分類】
【出願番号】特願2005−157931(P2005−157931)
【出願日】平成17年5月30日(2005.5.30)
【出願人】(504205521)国立大学法人 長崎大学 (226)
【Fターム(参考)】
【公開日】平成20年9月4日(2008.9.4)
【国際特許分類】
【出願日】平成17年5月30日(2005.5.30)
【出願人】(504205521)国立大学法人 長崎大学 (226)
【Fターム(参考)】
[ Back to top ]