
ワクチンにおいて使用するための組換えタンパク質、前記タンパク質に対する抗体、ならびに該タンパク質を含む診断法および療法
本発明は、タンパク質および/またはその断片および誘導体、ならびにワクチンとしての、およびバイオテクノロジー法におけるその使用に関する。ワクチンには、特に、ウシにおける趾皮膚炎から単離されるトレポネーマ属(Treponema)種における免疫原性タンパク質が含まれる。本発明はさらに、前記タンパク質またはその断片に対して作製される抗体、ならびに抗体がウシにおける趾皮膚炎の徴候として検出される診断法における前記タンパク質の使用にも関する。
なし
なし
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、タンパク質および/またはその断片および誘導体、ならびにワクチンとしての、およびバイオテクノロジー法におけるその使用に関する。ワクチンには、特に、ウシにおける趾皮膚炎から単離されるトレポネーマ属(Treponema)種における免疫原性タンパク質が含まれる。本発明はさらに、前記タンパク質またはその断片に対して作製される抗体、ならびに抗体がウシにおける趾皮膚炎の徴候として検出される診断法における前記タンパク質の使用にも関する。
【背景技術】
【0002】
趾皮膚炎(DD)は、ウシにおける、最も一般的には集約的酪農生産において見られる、跛行を引き起こす感染性鉤爪疾患である。該疾患は、1974年、イタリアで最初に記載された。スウェーデンでは、最近、DDの最初の群れが記載された(HillstroemおよびBergsten、2005)が、以前には、散発的な非定型の症例しか報告されてきていない(Manskeら、2002)。湿った/汚い鉤爪環境およびDDの発生の間に強い関連があり(Rodriguez−Lainzら、1996)、例えば通路(alleys)上の糞便および尿の集積が典型的な衛生上の問題である小部屋(cubicle)系において強い関連がある。動物福祉の問題であることに加えて、ミルク産生減少および体重喪失による経済的な損失が、DDに関連付けられている(Losinger、2006)。
【0003】
DD病変を抗生物質で処置すると迅速な反応があることから、細菌が原因であることが強く裏付けられる。異なる属の多くの細菌、例えばトレポネーマ属、フゾバクテリウム属(Fusobacterium)、ディケロバクター属(Dichelobacter)、プレボテラ属(Prevotella)、およびポルフィロモナス属(Porphyromonas)がDD病変から単離され、そして原因が多微生物性であるとしばしば論じられる。しかし、DDの病因において、トレポネーマ属種が主因である強い状況証拠がある。早くは1964年に、ウシの「腐蹄」徴候の異なる変種由来のスメアにおいて、スピロヘータ(spirochete)が観察された(Guptaら、1964)。スピロヘータの別の初期観察は、1988年に行われ、これは、英国においてDDが記載された最初の例であった(BloweyおよびSharp、1988)。DDからの最初のスピロヘータ培養が1995年に報告された(Walkerら、1995)。DD病変由来の組織プレパラートにおいて、トレポネーマが表皮のより深部の層に浸潤していることが見出されている(Moterら、1998)。さらに、感染したウシにおいて、トレポネーマ属種に対する体液性免疫反応が立証されてきている(Walkerら、1997; Trottら、2003)。DD病変からの新鮮な剥離物の接種を通じた疾患の実験的伝染の成功が1996年に記載された(ReadおよびWalker、1996)。接種1〜2週間後に、スピロヘータが組織に浸潤することもまた、組織病理によって確認された(Readら、1998)。
【0004】
トレポネーマ属のいくつかの系統型が同じ病変中に存在する可能性もある。異なる系統型が同じ動物から単離されてきており(Walkerら、1995; Evansら、2008)、そして16S rRNA遺伝子のクローニングおよび配列決定によって、5つの異なる系統型が4頭のウシ由来のプール試料中で同定された(Choiら、1997)。DD病変由来の生検に対する蛍光in situハイブリダイゼーションによって、皮層における分布が系統型間で異なることもまた立証されてきている(Moterら、1998)。トレポネーマ・ファゲデニス(Treponema phagedenis)様系統型は主に、角質層および有棘層に位置した。いくつかの系統型は、いまだに培養されたとは報告されていない。最近、いくつかの研究において、トレポネーマ・ファゲデニス様系統型は、DDの発病における重要な病原体であることが示されてきている(Klitgaardら、2008、Nordhoffら、2008、Yanoら、2009)。
【0005】
DDが蔓延している国々では、抗生物質を含有するフットバスがしばしば用いられる。これらのフットバスは、迅速に糞便および汚れで汚染され、そしてしたがって、抗生物質耐性細菌の巨大選択培養物として機能する。スウェーデンにおいてはテトラサイクリンが用いられるが、個々の動物の局所治療に関してのみであり、これは、群れのレベルでは、硫酸銅を含むフットバスが推奨されているためである。
【0006】
現在まで、DDに関する商業的なワクチンまたは血清学的試験は利用可能でない。トレポネーマ属種に対する体液性反応は、DDに罹患したウシにおいて示されてきており、そして研究において、全細胞溶解物ELISA研究に用いられている(Demirkanら、1999、Trottら、2003、Vinkら、2009、Walkerら、1997)。Novartisは、2000年代初期に数年間、USA市場用に全細胞溶解物DDワクチン(TrepShield)を製造した(Berryら、2004、Keilら、2002)。
【0007】
ワクチン開発のための技術および戦略は、とりわけ、Vaccine Design: Innovative Approaches and Novel Strategies(Caister Academic Press, 2011)およびVaccines: From Concept to Clinic: A Guide to the Development and Clinical Testing of Vaccines for Human Use(Informa Healthcare, 1998)に記載される。ワクチンとしての組換えタンパク質の使用は、Erdileら、1997に記載される。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】HillstroemおよびBergsten、2005
【非特許文献2】Manskeら、2002
【非特許文献3】Rodriguez−Lainzら、1996
【非特許文献4】Losinger、2006
【非特許文献5】Guptaら、1964
【非特許文献6】BloweyおよびSharp、1988
【非特許文献7】Walkerら、1995
【非特許文献8】Moterら、1998
【非特許文献9】Walkerら、1997
【非特許文献10】Trottら、2003
【非特許文献11】ReadおよびWalker、1996
【非特許文献12】Readら、1998
【非特許文献13】Evansら、2008
【非特許文献14】Choiら、1997
【非特許文献15】Klitgaardら、2008
【非特許文献16】Nordhoffら、2008
【非特許文献17】Yanoら、2009
【非特許文献18】Demirkanら、1999
【非特許文献19】Vinkら、2009
【非特許文献20】Berryら、2004
【非特許文献21】Keilら、2002
【非特許文献22】Vaccine Design: Innovative Approaches and Novel Strategies(Caister Academic Press, 2011)
【非特許文献23】Vaccines: From Concept to Clinic: A Guide to the Development and Clinical Testing of Vaccines for Human Use(Informa Healthcare, 1998)
【非特許文献24】Erdileら、1997
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、動物における皮膚炎、特に反芻動物における趾皮膚炎の効率的な診断法および該皮膚炎に対する免疫保護、ならびに前記目的のための製品を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明は、ウシにおいて趾皮膚炎から単離されるトレポネーマ属種における免疫原性タンパク質、そしてより具体的には組換えタンパク質に関する。
第一の側面において、本発明は、以下にさらに定義するように、それぞれ配列番号2、配列番号4または配列番号6記載のアミノ酸配列を持つ単離トレポネーマ・ファゲデニス様タンパク質、TmpA、Ttm、およびPrrAに関し、トレポネーマ属種に対する免疫反応を誘導可能なその断片および誘導体に関し、そして前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能な断片および誘導体に関する。
【0011】
本発明の1つの態様において、トレポネーマ属タンパク質、ならびにその断片および誘導体は、組換え的に産生される。
1つの側面において、本発明は、本発明記載のタンパク質、断片および誘導体をコードする核酸分子に関する。
【0012】
本発明はまた、獣医学的医薬品における、特に、趾皮膚炎の防止のためのワクチンとしての、前記タンパク質、その断片および誘導体の使用にも関する。
さらなる側面において、本発明は、趾皮膚炎に対する保護のための獣医学的ワクチンであって、1またはそれより多い前記組換えタンパク質および/またはその活性断片、ならびに慣用的でそして適切なアジュバントを含む、前記ワクチンを提供する。こうしたワクチンには、さらに、より広い免疫反応のためのワクチンにおいて、他のトレポネーマ属免疫原、または異なるトレポネーマ属種の全細胞溶解物がさらに含まれてもよいし、または含まれなくてもよい。
【0013】
別の側面にしたがって、本発明はまた、動物における趾皮膚炎の防止のための方法であって、前記ワクチンを投与する必要がある動物に前記ワクチンを投与する工程を含む、前記方法にも関する。
【0014】
さらにさらなる側面にしたがって、前記組換えタンパク質および/または活性断片が、前記試料中のトレポネーマ属種に対する抗体の存在を検出するのに用いられる、試料中のトレポネーマ属種に対する抗体の存在を検出する方法を提供する。
【0015】
さらにさらなる側面にしたがって、前記組換えタンパク質および/または活性断片が、動物中のトレポネーマ属種に対する抗体の存在を検出するのに用いられる、動物における趾皮膚炎の診断のための方法を提供する。
【0016】
前記検出法または診断法の1つの態様において、前記組換えタンパク質および/または活性断片が、ELISA(酵素連結免疫吸着アッセイ)法において用いられる。
1つの側面において、本発明は、前記免疫原性タンパク質、あるいはその免疫原性誘導体または断片に対して作製される抗体に関する。こうした抗体は、受動免疫による、トレポネーマ属種によって引き起こされる疾患の治療に有用であり、そしてまたトレポネーマ属種細菌の免疫磁気分離などの多様な実験法においてもまた有用である。
【図面の簡単な説明】
【0017】
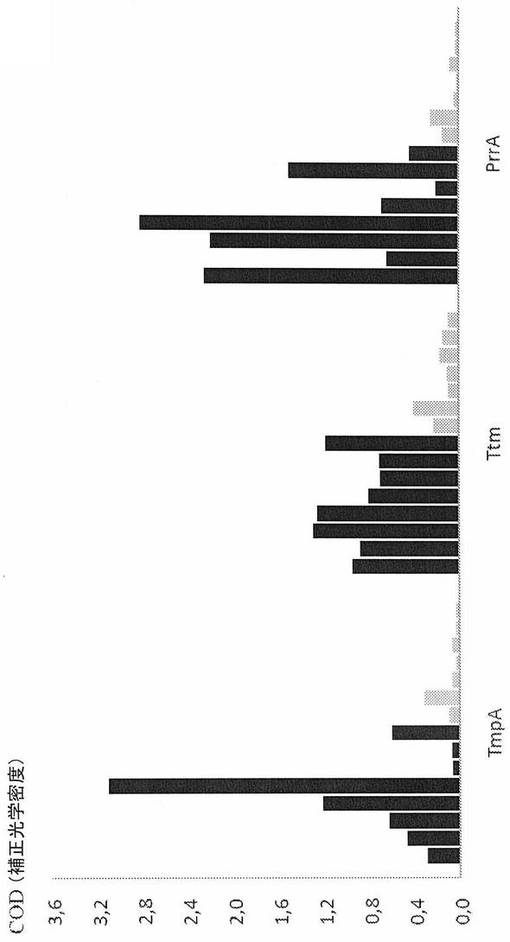
【図1A】トレポネーマ・ファゲデニス様株V1免疫原性タンパク質TmpA、Ttm、およびPrrAを抗原として用いた酵素連結免疫吸着アッセイ。該アッセイは、急性趾皮膚炎に罹患した8頭の乳牛(黒いバー)、趾皮膚炎の既知の病歴がない2頭の乳牛および6〜7ヶ月齢の5頭の子ウシ(灰色のバー)由来の血清を用いて行われた。西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート化ウサギ抗ウシIgG抗体(Sigma)(A)またはモノクローナル22:26抗ウシIgG−HRP抗体(Svanova Biotech AB)(B)を二次抗体として用いた。補正光学密度(COD)を450nmで測定した。
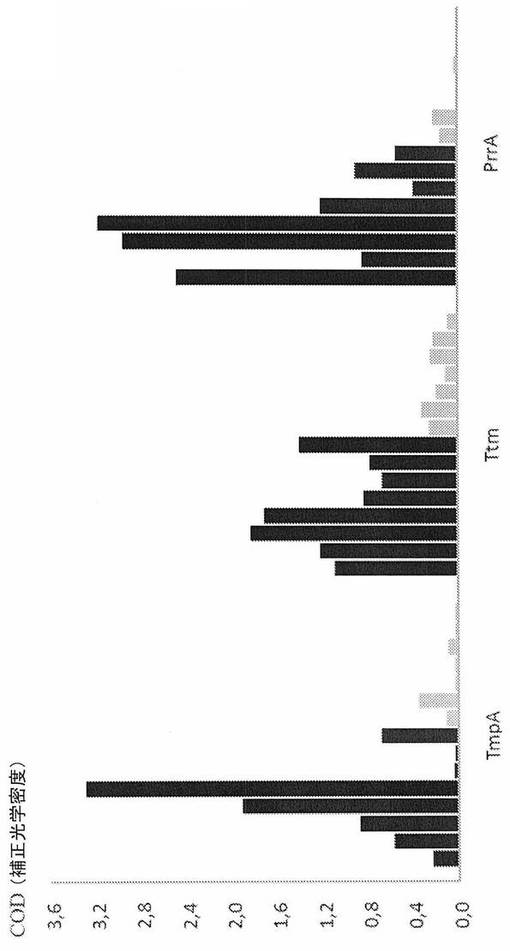
【図1B】トレポネーマ・ファゲデニス様株V1免疫原性タンパク質TmpA、Ttm、およびPrrAを抗原として用いた酵素連結免疫吸着アッセイ。該アッセイは、急性趾皮膚炎に罹患した8頭の乳牛(黒いバー)、趾皮膚炎の既知の病歴がない2頭の乳牛および6〜7ヶ月齢の5頭の子ウシ(灰色のバー)由来の血清を用いて行われた。西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート化ウサギ抗ウシIgG抗体(Sigma)(A)またはモノクローナル22:26抗ウシIgG−HRP抗体(Svanova Biotech AB)(B)を二次抗体として用いた。補正光学密度(COD)を450nmで測定した。
【発明を実施するための形態】
【0018】
定義
「免疫原性物質」または「免疫原」は、場合によってアジュバントと組み合わされて、患者に投与された際に、それ自体に対する免疫学的反応を誘導可能である。
【0019】
「活性断片」または「活性誘導体」は、本明細書において、場合によってアジュバントと組み合わされて、患者に投与された際に、前記天然免疫原性物質に対する免疫学的反応を誘導可能な、天然免疫原性物質の断片または誘導体である。活性断片または誘導体は、少なくとも1つの「エピトープ」または「抗原決定基」を含むかまたは模倣する。
【0020】
「結合性断片」または「結合性誘導体」は、本明細書において、前記天然免疫原性物質に対する免疫反応において、被験体によって産生される抗体に免疫特異的に結合可能な、天然免疫原性物質の断片または誘導体である。結合性断片または誘導体は、少なくとも1つの「エピトープ」または「抗原決定基」を含むかまたは模倣する。
【0021】
タンパク質の「誘導体」は、元来のタンパク質に対する実質的な配列相同性を示すタンパク質であってもよい。配列相同性は、アミノ酸配列の50%同一性またはそれより大きく、例えば65%、80%、85%、90%、95%または99%同一性であってもよい。置換されるアミノ酸は好ましくは保存性置換である。置換されるアミノ酸は、天然または非天然アミノ酸であってもよい。
【0022】
用語「エピトープ」または「抗原決定基」は、Bおよび/またはT細胞が反応する抗原上の部位を指す。B細胞エピトープは、隣接するアミノ酸、またはタンパク質の三次フォールディングによって並置される非隣接アミノ酸の両方から形成可能である。隣接アミノ酸から形成されるエピトープは、典型的には、変性溶媒への曝露に際して保持されるが、三次フォールディングによって形成されるエピトープは、典型的には、変性溶媒での処理に際して失われる。エピトープには、典型的には、ユニークな空間コンホメーションにある、少なくとも3、そしてより一般的には、少なくとも5または8〜10アミノ酸が含まれる。エピトープの空間コンホメーションを決定する方法には、例えば、X線結晶学および二次元核磁気共鳴が含まれる。例えば、Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, Glenn E. Morris監修(1996)を参照されたい。
【0023】
同じエピトープを認識する抗体は、1つの抗体が、ターゲット免疫原、あるいはその断片または誘導体に対する別の抗体の結合を遮断する能力を示す、単純なイムノアッセイにおいて、同定可能である。
【0024】
用語「抗体」は、損なわれていない(intact)抗体またはその結合性断片を指す。抗体は、完全抗体分子(ポリクローナル、モノクローナルまたはキメラを含む)を含んでもよいし、またはその抗原結合性断片を含んでもよい。抗体断片には、F(ab’)2、Fab、Fab’、Fv、Fc、およびFd断片が含まれ、そして単一ドメイン抗体、一本鎖抗体、マキシボディ、ミニボディ、イントラボディ、ディアボディ、トリアボディ、テトラボディ、v−NARおよびビス−scFv内に取り込まれていてもよい(例えばHollingerおよびHudson, 2005, Nature Biotechnology, 23, 9, 1126−1136を参照されたい)。
【0025】
配列表
【0026】
【表1】

【0027】
詳細な説明
本発明の側面および態様のさらなる概要
1つの側面において、本発明は、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能な断片または誘導体に関する。前記タンパク質、断片または誘導体は、獣医学的医薬品において使用されてもよく、例えば、トレポネーマ属種によって引き起こされる疾患、例えば趾皮膚炎の防止において使用されてもよい。
【0028】
別の側面において、本発明は、トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体を、被験体に投与する工程を含む、前記方法に関する。前記方法は、趾皮膚炎などの疾患のために使用されてもよい。
【0029】
さらなる側面において、本発明は、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、および場合による薬学的に許容されうるアジュバント、キャリアーおよび/または希釈剤を含む、薬学的組成物に関する。
【0030】
さらに別の側面において、本発明は、試料において、トレポネーマ属種由来のタンパク質に対する抗体の存在を検出するための方法であって:
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する
工程を含む、前記方法に関する。
【0031】
本発明のさらなる側面は、トレポネーマ属種によって引き起こされる疾患のin vitro診断のための方法であって:
−被験体由来の体液または組織の試料を得て;
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する;
ここで、前記タンパク質、断片または誘導体に結合する抗体の存在が、トレポネーマ属種によって引き起こされる疾患の指標となる
工程を含む、前記方法に関する。
【0032】
前記方法は、趾皮膚炎などの疾患のために使用されてもよい。
本発明のさらなる側面は、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質に特異的に結合する抗体、またはその結合性断片に関する。
【0033】
本発明の別の側面は、トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、前記抗体を被験体に投与する工程を含む、前記方法に関する。趾皮膚炎はこうした疾患の一例である。
【0034】
本発明のさらなる側面は、試料からトレポネーマ属細菌を分離するための方法であって:
−固相に結合した前記抗体と、前記試料を接触させ;
−前記抗体が、前記トレポネーマ属細菌中のトレポネーマ属タンパク質に結合するのを可能にし;そして
−前記試料から前記固相を分離し
それによって、前記試料から前記トレポネーマ属細菌を分離する
工程を含む、前記方法に関する。
【0035】
前記方法において、分離は、例えば免疫磁気分離によって達成可能である。
本発明の別の側面は、本発明記載のタンパク質、断片または誘導体をコードする核酸分子に関する。1つの態様において、タンパク質をコードする核酸分子は、配列番号1、配列番号3、配列番号5およびその一部からなる群より選択されるヌクレオチド配列を有する。
【0036】
本発明のさらなる側面は、前記核酸分子、および場合によって宿主細胞における発現のための制御配列を含む、ベクターに関する。
本発明の別の側面は、前記ベクターを含むトランスジェニック宿主細胞に関する。
【0037】
本発明はまた、本発明記載のタンパク質、断片または誘導体を産生するための方法であって:
−適切な培地中で本発明記載の宿主細胞を培養し;そして
−前記培地から前記タンパク質、断片または誘導体を単離する
工程を含む、前記方法にも関する。
【0038】
詳細
ショットガン・ファージディスプレイを用いて、T.ファゲデニスに緊密に関連するDDトレポネーマ属系統型の単離体(V1)における3つの免疫原性タンパク質を同定した。この系統型は、DDの発病における重要な病原体であることがいくつかの研究において示されてきている(Klitgaardら、2008、Nordhoffら、2008、Yanoら、2009)。ファージ・ライブラリーは、生きた細菌で免疫したウサギ由来の抗体に対して選択された。
【0039】
梅毒トレポネーマ亜種パリダム(T. pallidum subsp. pallidum)のよく性質決定された免疫原性タンパク質TmpAに対する相同体、ならびにいかなる既知のスピロヘータ・タンパク質にも相同性を持たない2つのタンパク質が同定された。454 SequencingTMによって生成されたV1のゲノム配列から、これらのタンパク質の完全アミノ酸配列を予測した。3つの特異的免疫原性タンパク質、およびそのアミノ酸配列は、配列番号2、4および6に記載される。
【0040】
本発明の第一の側面記載のタンパク質、断片および誘導体は、T.ファゲデニス株V1に緊密に関連するトレポネーマ属系統型の培養物から単離されるか、または好ましくは以下に記載するように組換え的に産生されてもよい。
【0041】
ウェスタンブロットを行って、免疫したウサギ由来の抗体ならびに天然感染したウシ由来の抗体の両方が、組換え的に産生されたTmpA相同体およびTtm断片に結合することを示した。
【0042】
パイロットELISA実験を行い、そしてDDに罹患したおよびしていないウシ由来の血清間で、吸光度における相違が記録されてきている(表1)。単一抗原(TmpA相同体またはTtm断片)を用いると、健康群および感染群の結果間にはわずかな重複しかなく、そして2つの抗原の組み合わせを用いると、重複はなかった。
【0043】
本発明記載の免疫原性タンパク質、およびその活性断片を、T.ファゲデニスに緊密に関連するトレポネーマ属系統型または他のトレポネーマ属種によって少なくとも部分的に引き起こされる疾患に対するワクチンにおいて使用してもよい。
【0044】
特定の態様において、完全タンパク質がワクチンにおいて用いられる。特定の態様において、適切なエピトープを含む断片のみが用いられる。特定の態様において、1またはそれより多いタンパク質の1またはそれより多いエピトープが単一の分子に組み合わされ、そしてワクチンにおいて用いられる。組換えタンパク質、その誘導体または断片は、したがって、単独で、または異なる組み合わせで、または結合性エピトープの融合タンパク質として用いられうる。
【実施例】
【0045】
方法
ヘルパーファージ、細菌株、増殖条件、およびDNA技術
ファージR408(Promega)をヘルパー・ファージとして用いた。ファージまたはファージミドが関与するすべての実験において、大腸菌(Escherichia coli)TG1(Δ(lac−proAB)Δ(mcrB−hsdSM)5(rK−mK−)thi−1 supE[F’traD36 proAB lacIqZΔM15]; Stratagene)を宿主として用い、そしてLuria−Bertaniブロス(LB)中、またはLuria−Bertani寒天(LA)上で増殖させた。適切な場合、50μg/mlアンピシリン(C16H18N3O4SNa、Roche)を添加した。インキュベーションは37℃であった。ファージライブラリーの構築のため、トレポネーマ属種株V1由来の染色体DNAを用いた。嫌気性ジャー中、振盪装置(90rpm)上で、FABGS(1リットルあたり2.0g D−グルコースおよび25%ウシ胎児血清、S 0115、Biochrom AG、ドイツを含む、偏好性嫌気性細菌ブロス、LAB 71、LabM、英国ランカシャー州)を含むフラスコ中、37℃でインキュベーションし、トレポネーマ属株を増殖させた。ブロス培養物を等張生理食塩水(pH6.3)で3回洗浄し、その後、リン酸緩衝生理食塩水(PBS、pH7.3)で1回洗浄した。慣用的なフェノール−クロロホルム抽出によってトレポネーマ属DNAを調製した。制限酵素および修飾酵素は、MBI Fermentas AB由来であり、そしてこれらを製造者の指示にしたがって用いた。QIAprepTM Miniprep(QIAGEN)を用いて、プラスミドを調製した。
【0046】
免疫およびポリクローナル抗体の精製
研究のこの部分は、ウプサラにおける動物実験に関する倫理委員会によって認可された(C 300/8)。ニュージーランドホワイト・ウサギを、トレポネーマ属種株V1の生菌培養物で皮下免疫した。2回洗浄し、そして0.5ml体積の等張生理食塩水中に分配したおよそ109細菌の用量を20日おいて2回注射した。最初の免疫の38日後の最終出血由来の血清を、この研究のための抗体(IgG)の精製に用いた。0.45μmシリンジフィルターを通じて10mlの血清を滅菌ろ過し、そして5ml HiTrapTMプロテインG HPカラム(GE Healthcare)上に適用した。Ab緩衝液キット(GE Healthcare)を用いて、製造者の指示にしたがって、ウサギIgGを精製した。Zebaスピン脱塩カラム(Pierce)を用いて、精製抗体の溶出物を脱塩し、そしてマイナス20℃、PBS中で保存した。
【0047】
トレポネーマ属種ファージディスプレイライブラリーの構築および結合性ファージの選択(パニング)
pG8SAETファージミドベクター中で、ファージライブラリーを構築した。大部分の断片が長さ0.4〜1.5kbになるまで、トレポネーマ属種株V1染色体DNAを超音波処理によって断片化した。T4 DNAポリメラーゼおよびT4 DNAキナーゼ処理によって、断片を平滑端化し、そして次いで、Ready−To−GoTM T4 DNAリガーゼ試験管(GE Healthcare)を用いて、SnaB1消化しそして脱リン酸化したファージミドベクターpG8SAET内に連結した。連結した物質を大腸菌TG1細胞に電気形質転換し(2.5kV、25μF、360Ω)、ヘルパーファージを用いて感染させ、そしてファージ粒子を増殖させることによって、最終ライブラリーを生成した。この方法は、ユニークなクローンと見なされる、4x107形質転換体を生じ、14のランダムに選択されたクローンに対するコロニーPCRによって決定すると、このうち86%が挿入物を所持した。最終ライブラリーは、1mlあたり、1x1011コロニー形成単位の力価を有した。
【0048】
ウサギ抗トレポネーマ属種株V1 IgGに対するパニングによって、免疫原性ポリペプチドをディスプレイするファージを単離した。3回のパニング実験を行った。マイクロウェル(MaxiSorpTM、Nalge Nunc International)を、200μlの50mM炭酸ナトリウム、pH9.5中、10μgの濃度のZymed組換えプロテインG(Invitrogen)でコーティングした。その後、0.05% Tween20を含むリン酸緩衝生理食塩水pH7.4(PBS−T)でウェルをブロッキングした。ウサギ抗トレポネーマ属種株V1 IgGを200μl PBSまたは100μl PBS+100μl未精製大腸菌溶解物(ブロッキング用)中、85または215μgの濃度で添加した。洗浄後、200μlのファージライブラリーを添加した。ウェルを室温で3時間または2℃で一晩インキュベーションし、その後、これらを25回洗浄した後、50mMクエン酸Na/140mM NaCl pH2.0を添加することによって、ファージを溶出させた。2M Tris緩衝液pH8.0で溶出物を直ちに中和し、そしてアンピシリンを含むLAプレート(LAamp)上にプレーティングした大腸菌TG1を感染させるのに用いた。1つのパニング実験において、ウェルに添加したTG1細胞を、結合したファージによって直接感染させることによってもまた、溶出を行った。一晩インキュベーションした後、コロニーを計数し、そして100のコロニーをLAampプレートに移した。次いで、マウス抗E−タグ抗体(GE Healthcare)および西洋ワサビ・ペルオキシダーゼ標識した二次ヒツジ抗マウス抗体(GE Healthcare)を用いたE−タグ発現のスクリーニングのために、これらのコロニーをニトロセルロース・フィルターにトランスファーした。残ったコロニーをプレートから洗い流し、そしてヘルパーファージで重感染させて、濃縮ライブラリー/ファージストックを作製し、同じプロトコルにしたがって、これを二次濃縮サイクル(再パニング)において用いた。全部で2回の再パニングを行った。プラスミド調製およびプライマーSAsekv(5’−TAT CTG GTG GCG TAA CAC CTG CT−3’、配列番号17)を用いた挿入物の配列決定のため、200を超えるE−タグ陽性コロニーを選択した。ウプサラ・ゲノム・センターで、3730xl DNA分析装置(Applied Biosystems)上、プラスミドDNAを配列決定し、そしてCLC Main Workbenchソフトウェア(CLC bio)で分析した。挿入物の分析によって、3つの異なる遺伝子から、それぞれ、9、9、および8の重複する部分配列が明らかになった。
【0049】
ゲノム配列決定および配列分析
トレポネーマ属種株V1の染色体DNAを配列決定し、そしてロングリードGS FLXチタン化学反応および454新規アセンブリー装置、Newbler(454 Life Sciences、米国コネチカット州ブランフォード)とともに、ゲノム配列決定装置FLX系を用いて、KTH王立工科大学のKTHゲノムセンター、スウェーデン・ストックホルムでアセンブリーした。CLC Genomics Workbench 3(CLC bio)を用いて読み取り配列のさらなる新規アセンブリーを行い、そしてさらなる配列編集のため、CLC Main Workbench 5(CLC bio)を用いた。
【0050】
ゲノム配列を用いて、3つの免疫原性タンパク質の全長オープンリーディングフレームおよび対応するアミノ酸配列を予測した。National Center for Biotechnology InformationのBLASTアルゴリズムを用いて、相同性検索を行った。シグナルペプチドの予測には、グラム陽性データを含むSignalP 3.0サーバーを用いた。1つのタンパク質は、Setubalら、2006にしたがって、リポタンパク質と予測された。
【0051】
タンパク質発現および精製のためのクローンの構築
グラム陰性細菌用のプロトコルにしたがって、DNeasy血液および組織キット(QIAGEN)を用いて、PCR用のゲノム・トレポネーマ属種株V1 DNAを調製した。MgSO4を含む10X Pfu緩衝剤(Fermentas)5μl、各0.2mMのデオキシヌクレオチド、表2に示すような0.2μMの順方向および逆方向プライマー、1.25U Pfu DNAポリメラーゼ(Fermentas)および50ngゲノムDNAの50μl反応混合物を調製した。熱サイクリング条件は、95℃1分間、30周期の95℃30秒間、50℃30秒間および72℃3分間、ならびに72℃5分間の最終伸長であった。アガロースゲル電気泳動によってPCR産物を分析し、そしてillustra GFX PCR DNAおよびゲルバンド精製キット(GE Healthcare)を用いて精製した。製造者の指示(Fast digest、Fermentas)にしたがって、精製アンプリコンを、BamHIおよびXhoIまたはNdeIおよびSapIのいずれかで消化し、そして先に記載するように精製した。消化アンプリコンを、ReadyToGo T4DNAリガーゼ(GE Healthcare)を用いて、それぞれのベクター、BamHIおよびXhoIで消化したpGEX−6P−1(バルクGST精製モジュール、GE Healthcare)またはNdeIおよびSapIで消化したpTXB1(IMPACTTMキット、New England BioLabs)内に連結した。連結した材料を、コンピテント大腸菌株BL21(DE3)(GST)またはER2566(IMPACT)内に電気形質転換し、そしてアンピシリン(最終濃度50μg/ml)を補充したLA上にスプレッドした。ベクター配列決定プライマーを用いたPCRによって、多くのコロニー中の挿入物の存在を分析した。DNA配列決定によって、正しいサイズの挿入物を持つクローンをさらに分析した。
【0052】
組換え免疫原性トレポネーマ属タンパク質の産生
組換え免疫原性トレポネーマ属タンパク質の産生のため、製造者の指示にしたがって、商業的に入手可能なタンパク質発現および精製系、例えばバルクGST精製モジュール(GE Healthcare)またはIMPACTTMキット(New England BioLabs)を用いた。アンピシリン(最終濃度50μg/ml)を補充したLB培地中、37℃で、組換えクローンを増殖させた。光学密度(OD600nm)〜0.6の際、増殖培地にIPTG(最終濃度0.3mM)を補充し、そして増殖温度を20℃にシフトした。一晩インキュベーションした後、細胞を採取し、そして緩衝液[20mM Tris−HCl(pH8.0)、500mM NaCl、0.1mM EDTA、および0.05%(v/v)TWEEN20]中に再懸濁し、そして凍結融解によって溶解した。遠心分離後、上清を滅菌ろ過し、そしてキチンカラム上に適用した。同じ緩衝液を用いてカラムを徹底的に洗浄し、そして続いて切断緩衝液[20mM Tris−HCl(pH8.0)、50mM NaCl、0.1mM EDTA、および30mMジチオスレイトール(DTT)]で処理した。抗原を含有する溶出試料を、リン酸緩衝生理食塩水[PBS; 137mM NaCl、2.7mM KCl、10mM Na2HPO4、1.4mM KH2PO4(pH7.4)]に対して透析した。
【0053】
GST−グルタチオン・アフィニティ系において、上記方法にしたがって、増殖、誘導および採取後、最終濃度0.1%(v/v)TWEEN20を補充したPBS(PBST)中に大腸菌細胞を懸濁したら、凍結融解によって細胞を溶解した。遠心分離後、上清を滅菌ろ過し、そしてグルタチオン−セファロース・ビーズでバッチ精製した。PBSTを用いて徹底的に洗浄した後、グルタチオン溶出緩衝液で融合タンパク質を溶出させるか、またはscissorプロテアーゼで処理して、生じたタンパク質を放出させた。最後に、分光光度測定を用いて、得られた抗原の量を決定し、そしてSDS−PAGEクーマシー染色によって品質を分析した。最後にタンパク質を−20℃で保存した。
【0054】
酵素連結免疫吸着アッセイ(ELISA)
ELISAにおいて、全細胞溶解物に比較して、大腸菌で組換え的に産生された精製タンパク質を用いる利点は、大規模産生の可能性があり、そして試験が最適化されることである(タンパク質の異なる組み合わせを用いると、しばしば、特異性がより高くなるため、バックグラウンドがより低くなる)。趾皮膚炎(DD)に罹患したおよびしていないウシ由来の血清、ならびに免疫したウサギ(上記)由来の血清を用いて、最初のELISA試験を行った。マイクロプレート(PolySorpTM、Nalge Nunc International)を、100μlの50mM炭酸ナトリウム、pH9.5中、それぞれ2.5μgおよび/または0.31μgの濃度で、組換えTmpA相同体および/または組換えテール・テープ測定タンパク質断片で、2℃で一晩、コーティングした。0.05%Tween20を含む400μlのリン酸緩衝生理食塩水、pH7.4(PBS−T)で、ウェルを2回洗浄した。その後、PBS−Tで、室温で1時間、ウェルをブロッキングした。100μlの血清またはPBS−Tを各ウェルに添加した。各血清の4つの希釈、1:25、1:50、1:100、および1:200を用いた。マイクロプレートを37℃で1時間インキュベーションし、そして次いで、400μl PBS−Tで3回洗浄した。それぞれ1:4000および1:500希釈した西洋ワサビ・ペルオキシダーゼ・コンジュゲート化ブタ抗ウサギ(Dako)およびウサギ抗ウシ(Dako)抗体を適切なウェルに添加し、そしてプレートを37℃で1時間インキュベーションした。400μl PBS−Tでウェルを3回洗浄した後、1:20で混合した20mMテトラメチルベンジジン(TMB)と0.1Mクエン酸カリウム/H2O2(230μl/l)pH4.25からなる100μl溶液を添加した。プレートを室温で10分間インキュベーションした。反応を停止するため、50μlの10%硫酸を添加した。光学密度(OD)を450nmで測定し、そして読み取り値を試料緩衝液ブランクに対して補正した。
【0055】
抗体を用いた免疫磁気分離
トレポネーマ属種は、増殖のために複雑な培地および嫌気性環境を必要とする偏好性生物である。さらに、ウシ鉤爪由来の試料は、培養を汚染させる大量の他の細菌を有する。培養およびDNA単離のためにトレポネーマ属種を濃縮しそして精製するオプションは、免疫磁気分離である(Demirkanら、1999、Demirkanら、2001、Choiら、1996)。例えば前記免疫原性タンパク質、その誘導体または活性断片に対して作製されたウサギ抗体とカップリングされた抗ウサギIgGで共有的にコーティングされた免疫磁気ビーズを、T.ファゲデニスに緊密に関連するDDトレポネーマ属系統型の特異的分離に用いてもよい。
【0056】
ワクチン
免疫系を刺激して抗体を産生させる細菌タンパク質はまた、ワクチン開発にも使用可能である。組換えタンパク質を免疫刺激複合体(ISCOM)および/または全細胞溶解物と組み合わせて、動物における免疫反応を、そしてしたがって、疾患原因物質(単数または複数)に対する保護を増加させることも可能である。
【0057】
行ったさらなる実験
酵素連結免疫吸着アッセイ(ELISA)
材料および方法:Tpl株V1が単離された群れからの、急性DDに罹患した8頭の乳牛、既知のDDの病歴がない別の群れからの2頭の乳牛、および6〜7ヶ月齢の5頭の子ウシ由来の血清を用いてアッセイを行った。趾皮膚炎診断は、視覚的検査によって行われた。マイクロプレート(PolySorpTM、Nalge Nunc International)を、100μlの50mM炭酸ナトリウム、pH9.5中、1μg/ml TmpA、0.8μg/ml Ttmまたは0.02μg/ml PrrAの濃度の組換えタンパク質で、2℃で一晩コーティングした。PBS−Tでウェルを2回洗浄し、そしてPBS−Tで、室温で1時間、ブロッキングした。PBS−T中で1:100希釈した171の100μl血清を各ウェルに添加した。マイクロプレートを37℃で1時間インキュベーションし、そして次いでPBS−Tで洗浄した。1:8000希釈した西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート化ウサギ抗ウシIgG抗体(Sigma)または1:4000希釈したモノクローナル22:26抗ウシIgG−HRP抗体(Svanova Biotech AB)をウェルに添加し、そして37℃で1時間、プレートをインキュベーションした。PBS−Tでウェルを3回洗浄し、その後、0.1Mクエン酸カリウムpH4.25中の1mMテトラメチルベンジジンおよび0.006%H2O2からなる100μl溶液を添加した。プレートを室温で10分間インキュベーションした。反応を停止するため、50μlの10%硫酸を添加した。光学密度を450nmで測定し、そして読み取り値から試料緩衝液ブランクの光学密度を減じて、補正光学密度(COD)を生じた。
【0058】
結果:Tpl株V1で同定された3つの免疫原性タンパク質を組換え的に産生した;予測されるリポタンパク質シグナルペプチドのシステイン残基に対してaa+1から停止コドン前の最後のaaまでの全長成熟タンパク質としてのPrrA(aa:s22〜251)、予測されるリポタンパク質シグナルペプチドのシステイン残基に対してaa+7から停止コドン前の最後のaaまでのTmpA(aa:s29〜344)、およびパニング実験由来の重複Ttm配列のコンセンサス配列を構成するaa:sであるaa:s689〜970を含む部分的ポリペプチドとしてのTtm。これらのタンパク質/ポリペプチドを、間接ELISAにおける抗原として用い、ここでDDに罹患したおよびしていないウシ由来の血清試料を、抗原に対する抗体の存在に関して分析した。異なる濃度の抗原、血清、および二次抗体をパイロット実験において試験した(データ未提示)。DDに罹患したおよびしていないウシの間の最適な区別を達成する条件を、最終実験で用いた。TmpA抗原に関して、DDに罹患したウシ由来の3つの試料に関する光学密度は、臨床的に健康な269頭のウシ由来の試料に関する最高値よりも低く、一方、TtmおよびPrrAを用いた試験は、1つを除きすべての場合で識別力があった(図1AおよびB)。
【0059】
【表2】
【0060】
表1.趾皮膚炎(DD)に罹患したおよびしていないウシ由来の血清の最初のELISA試験。450nmで光学密度(OD)を測定し、そして試料緩衝液ブランクに対して読み取り値を補正した。血清を1:100に希釈し、TmpAの濃度は2.5μg/ウェルであり、そして推定上のテール・テープ測定タンパク質断片の濃度は0.31μg/ウェルであった。
【0061】
【表3】
【0062】
表2:デオキシオリゴリボヌクレオチド
参考文献
【0063】
【化1−1】
【0064】
【化1−2】
【0065】
【化1−3】
【技術分野】
【0001】
本発明は、タンパク質および/またはその断片および誘導体、ならびにワクチンとしての、およびバイオテクノロジー法におけるその使用に関する。ワクチンには、特に、ウシにおける趾皮膚炎から単離されるトレポネーマ属(Treponema)種における免疫原性タンパク質が含まれる。本発明はさらに、前記タンパク質またはその断片に対して作製される抗体、ならびに抗体がウシにおける趾皮膚炎の徴候として検出される診断法における前記タンパク質の使用にも関する。
【背景技術】
【0002】
趾皮膚炎(DD)は、ウシにおける、最も一般的には集約的酪農生産において見られる、跛行を引き起こす感染性鉤爪疾患である。該疾患は、1974年、イタリアで最初に記載された。スウェーデンでは、最近、DDの最初の群れが記載された(HillstroemおよびBergsten、2005)が、以前には、散発的な非定型の症例しか報告されてきていない(Manskeら、2002)。湿った/汚い鉤爪環境およびDDの発生の間に強い関連があり(Rodriguez−Lainzら、1996)、例えば通路(alleys)上の糞便および尿の集積が典型的な衛生上の問題である小部屋(cubicle)系において強い関連がある。動物福祉の問題であることに加えて、ミルク産生減少および体重喪失による経済的な損失が、DDに関連付けられている(Losinger、2006)。
【0003】
DD病変を抗生物質で処置すると迅速な反応があることから、細菌が原因であることが強く裏付けられる。異なる属の多くの細菌、例えばトレポネーマ属、フゾバクテリウム属(Fusobacterium)、ディケロバクター属(Dichelobacter)、プレボテラ属(Prevotella)、およびポルフィロモナス属(Porphyromonas)がDD病変から単離され、そして原因が多微生物性であるとしばしば論じられる。しかし、DDの病因において、トレポネーマ属種が主因である強い状況証拠がある。早くは1964年に、ウシの「腐蹄」徴候の異なる変種由来のスメアにおいて、スピロヘータ(spirochete)が観察された(Guptaら、1964)。スピロヘータの別の初期観察は、1988年に行われ、これは、英国においてDDが記載された最初の例であった(BloweyおよびSharp、1988)。DDからの最初のスピロヘータ培養が1995年に報告された(Walkerら、1995)。DD病変由来の組織プレパラートにおいて、トレポネーマが表皮のより深部の層に浸潤していることが見出されている(Moterら、1998)。さらに、感染したウシにおいて、トレポネーマ属種に対する体液性免疫反応が立証されてきている(Walkerら、1997; Trottら、2003)。DD病変からの新鮮な剥離物の接種を通じた疾患の実験的伝染の成功が1996年に記載された(ReadおよびWalker、1996)。接種1〜2週間後に、スピロヘータが組織に浸潤することもまた、組織病理によって確認された(Readら、1998)。
【0004】
トレポネーマ属のいくつかの系統型が同じ病変中に存在する可能性もある。異なる系統型が同じ動物から単離されてきており(Walkerら、1995; Evansら、2008)、そして16S rRNA遺伝子のクローニングおよび配列決定によって、5つの異なる系統型が4頭のウシ由来のプール試料中で同定された(Choiら、1997)。DD病変由来の生検に対する蛍光in situハイブリダイゼーションによって、皮層における分布が系統型間で異なることもまた立証されてきている(Moterら、1998)。トレポネーマ・ファゲデニス(Treponema phagedenis)様系統型は主に、角質層および有棘層に位置した。いくつかの系統型は、いまだに培養されたとは報告されていない。最近、いくつかの研究において、トレポネーマ・ファゲデニス様系統型は、DDの発病における重要な病原体であることが示されてきている(Klitgaardら、2008、Nordhoffら、2008、Yanoら、2009)。
【0005】
DDが蔓延している国々では、抗生物質を含有するフットバスがしばしば用いられる。これらのフットバスは、迅速に糞便および汚れで汚染され、そしてしたがって、抗生物質耐性細菌の巨大選択培養物として機能する。スウェーデンにおいてはテトラサイクリンが用いられるが、個々の動物の局所治療に関してのみであり、これは、群れのレベルでは、硫酸銅を含むフットバスが推奨されているためである。
【0006】
現在まで、DDに関する商業的なワクチンまたは血清学的試験は利用可能でない。トレポネーマ属種に対する体液性反応は、DDに罹患したウシにおいて示されてきており、そして研究において、全細胞溶解物ELISA研究に用いられている(Demirkanら、1999、Trottら、2003、Vinkら、2009、Walkerら、1997)。Novartisは、2000年代初期に数年間、USA市場用に全細胞溶解物DDワクチン(TrepShield)を製造した(Berryら、2004、Keilら、2002)。
【0007】
ワクチン開発のための技術および戦略は、とりわけ、Vaccine Design: Innovative Approaches and Novel Strategies(Caister Academic Press, 2011)およびVaccines: From Concept to Clinic: A Guide to the Development and Clinical Testing of Vaccines for Human Use(Informa Healthcare, 1998)に記載される。ワクチンとしての組換えタンパク質の使用は、Erdileら、1997に記載される。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】HillstroemおよびBergsten、2005
【非特許文献2】Manskeら、2002
【非特許文献3】Rodriguez−Lainzら、1996
【非特許文献4】Losinger、2006
【非特許文献5】Guptaら、1964
【非特許文献6】BloweyおよびSharp、1988
【非特許文献7】Walkerら、1995
【非特許文献8】Moterら、1998
【非特許文献9】Walkerら、1997
【非特許文献10】Trottら、2003
【非特許文献11】ReadおよびWalker、1996
【非特許文献12】Readら、1998
【非特許文献13】Evansら、2008
【非特許文献14】Choiら、1997
【非特許文献15】Klitgaardら、2008
【非特許文献16】Nordhoffら、2008
【非特許文献17】Yanoら、2009
【非特許文献18】Demirkanら、1999
【非特許文献19】Vinkら、2009
【非特許文献20】Berryら、2004
【非特許文献21】Keilら、2002
【非特許文献22】Vaccine Design: Innovative Approaches and Novel Strategies(Caister Academic Press, 2011)
【非特許文献23】Vaccines: From Concept to Clinic: A Guide to the Development and Clinical Testing of Vaccines for Human Use(Informa Healthcare, 1998)
【非特許文献24】Erdileら、1997
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、動物における皮膚炎、特に反芻動物における趾皮膚炎の効率的な診断法および該皮膚炎に対する免疫保護、ならびに前記目的のための製品を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明は、ウシにおいて趾皮膚炎から単離されるトレポネーマ属種における免疫原性タンパク質、そしてより具体的には組換えタンパク質に関する。
第一の側面において、本発明は、以下にさらに定義するように、それぞれ配列番号2、配列番号4または配列番号6記載のアミノ酸配列を持つ単離トレポネーマ・ファゲデニス様タンパク質、TmpA、Ttm、およびPrrAに関し、トレポネーマ属種に対する免疫反応を誘導可能なその断片および誘導体に関し、そして前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能な断片および誘導体に関する。
【0011】
本発明の1つの態様において、トレポネーマ属タンパク質、ならびにその断片および誘導体は、組換え的に産生される。
1つの側面において、本発明は、本発明記載のタンパク質、断片および誘導体をコードする核酸分子に関する。
【0012】
本発明はまた、獣医学的医薬品における、特に、趾皮膚炎の防止のためのワクチンとしての、前記タンパク質、その断片および誘導体の使用にも関する。
さらなる側面において、本発明は、趾皮膚炎に対する保護のための獣医学的ワクチンであって、1またはそれより多い前記組換えタンパク質および/またはその活性断片、ならびに慣用的でそして適切なアジュバントを含む、前記ワクチンを提供する。こうしたワクチンには、さらに、より広い免疫反応のためのワクチンにおいて、他のトレポネーマ属免疫原、または異なるトレポネーマ属種の全細胞溶解物がさらに含まれてもよいし、または含まれなくてもよい。
【0013】
別の側面にしたがって、本発明はまた、動物における趾皮膚炎の防止のための方法であって、前記ワクチンを投与する必要がある動物に前記ワクチンを投与する工程を含む、前記方法にも関する。
【0014】
さらにさらなる側面にしたがって、前記組換えタンパク質および/または活性断片が、前記試料中のトレポネーマ属種に対する抗体の存在を検出するのに用いられる、試料中のトレポネーマ属種に対する抗体の存在を検出する方法を提供する。
【0015】
さらにさらなる側面にしたがって、前記組換えタンパク質および/または活性断片が、動物中のトレポネーマ属種に対する抗体の存在を検出するのに用いられる、動物における趾皮膚炎の診断のための方法を提供する。
【0016】
前記検出法または診断法の1つの態様において、前記組換えタンパク質および/または活性断片が、ELISA(酵素連結免疫吸着アッセイ)法において用いられる。
1つの側面において、本発明は、前記免疫原性タンパク質、あるいはその免疫原性誘導体または断片に対して作製される抗体に関する。こうした抗体は、受動免疫による、トレポネーマ属種によって引き起こされる疾患の治療に有用であり、そしてまたトレポネーマ属種細菌の免疫磁気分離などの多様な実験法においてもまた有用である。
【図面の簡単な説明】
【0017】
【図1A】トレポネーマ・ファゲデニス様株V1免疫原性タンパク質TmpA、Ttm、およびPrrAを抗原として用いた酵素連結免疫吸着アッセイ。該アッセイは、急性趾皮膚炎に罹患した8頭の乳牛(黒いバー)、趾皮膚炎の既知の病歴がない2頭の乳牛および6〜7ヶ月齢の5頭の子ウシ(灰色のバー)由来の血清を用いて行われた。西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート化ウサギ抗ウシIgG抗体(Sigma)(A)またはモノクローナル22:26抗ウシIgG−HRP抗体(Svanova Biotech AB)(B)を二次抗体として用いた。補正光学密度(COD)を450nmで測定した。
【図1B】トレポネーマ・ファゲデニス様株V1免疫原性タンパク質TmpA、Ttm、およびPrrAを抗原として用いた酵素連結免疫吸着アッセイ。該アッセイは、急性趾皮膚炎に罹患した8頭の乳牛(黒いバー)、趾皮膚炎の既知の病歴がない2頭の乳牛および6〜7ヶ月齢の5頭の子ウシ(灰色のバー)由来の血清を用いて行われた。西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート化ウサギ抗ウシIgG抗体(Sigma)(A)またはモノクローナル22:26抗ウシIgG−HRP抗体(Svanova Biotech AB)(B)を二次抗体として用いた。補正光学密度(COD)を450nmで測定した。
【発明を実施するための形態】
【0018】
定義
「免疫原性物質」または「免疫原」は、場合によってアジュバントと組み合わされて、患者に投与された際に、それ自体に対する免疫学的反応を誘導可能である。
【0019】
「活性断片」または「活性誘導体」は、本明細書において、場合によってアジュバントと組み合わされて、患者に投与された際に、前記天然免疫原性物質に対する免疫学的反応を誘導可能な、天然免疫原性物質の断片または誘導体である。活性断片または誘導体は、少なくとも1つの「エピトープ」または「抗原決定基」を含むかまたは模倣する。
【0020】
「結合性断片」または「結合性誘導体」は、本明細書において、前記天然免疫原性物質に対する免疫反応において、被験体によって産生される抗体に免疫特異的に結合可能な、天然免疫原性物質の断片または誘導体である。結合性断片または誘導体は、少なくとも1つの「エピトープ」または「抗原決定基」を含むかまたは模倣する。
【0021】
タンパク質の「誘導体」は、元来のタンパク質に対する実質的な配列相同性を示すタンパク質であってもよい。配列相同性は、アミノ酸配列の50%同一性またはそれより大きく、例えば65%、80%、85%、90%、95%または99%同一性であってもよい。置換されるアミノ酸は好ましくは保存性置換である。置換されるアミノ酸は、天然または非天然アミノ酸であってもよい。
【0022】
用語「エピトープ」または「抗原決定基」は、Bおよび/またはT細胞が反応する抗原上の部位を指す。B細胞エピトープは、隣接するアミノ酸、またはタンパク質の三次フォールディングによって並置される非隣接アミノ酸の両方から形成可能である。隣接アミノ酸から形成されるエピトープは、典型的には、変性溶媒への曝露に際して保持されるが、三次フォールディングによって形成されるエピトープは、典型的には、変性溶媒での処理に際して失われる。エピトープには、典型的には、ユニークな空間コンホメーションにある、少なくとも3、そしてより一般的には、少なくとも5または8〜10アミノ酸が含まれる。エピトープの空間コンホメーションを決定する方法には、例えば、X線結晶学および二次元核磁気共鳴が含まれる。例えば、Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, Glenn E. Morris監修(1996)を参照されたい。
【0023】
同じエピトープを認識する抗体は、1つの抗体が、ターゲット免疫原、あるいはその断片または誘導体に対する別の抗体の結合を遮断する能力を示す、単純なイムノアッセイにおいて、同定可能である。
【0024】
用語「抗体」は、損なわれていない(intact)抗体またはその結合性断片を指す。抗体は、完全抗体分子(ポリクローナル、モノクローナルまたはキメラを含む)を含んでもよいし、またはその抗原結合性断片を含んでもよい。抗体断片には、F(ab’)2、Fab、Fab’、Fv、Fc、およびFd断片が含まれ、そして単一ドメイン抗体、一本鎖抗体、マキシボディ、ミニボディ、イントラボディ、ディアボディ、トリアボディ、テトラボディ、v−NARおよびビス−scFv内に取り込まれていてもよい(例えばHollingerおよびHudson, 2005, Nature Biotechnology, 23, 9, 1126−1136を参照されたい)。
【0025】
配列表
【0026】
【表1】
【0027】
詳細な説明
本発明の側面および態様のさらなる概要
1つの側面において、本発明は、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能な断片または誘導体に関する。前記タンパク質、断片または誘導体は、獣医学的医薬品において使用されてもよく、例えば、トレポネーマ属種によって引き起こされる疾患、例えば趾皮膚炎の防止において使用されてもよい。
【0028】
別の側面において、本発明は、トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体を、被験体に投与する工程を含む、前記方法に関する。前記方法は、趾皮膚炎などの疾患のために使用されてもよい。
【0029】
さらなる側面において、本発明は、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、および場合による薬学的に許容されうるアジュバント、キャリアーおよび/または希釈剤を含む、薬学的組成物に関する。
【0030】
さらに別の側面において、本発明は、試料において、トレポネーマ属種由来のタンパク質に対する抗体の存在を検出するための方法であって:
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する
工程を含む、前記方法に関する。
【0031】
本発明のさらなる側面は、トレポネーマ属種によって引き起こされる疾患のin vitro診断のための方法であって:
−被験体由来の体液または組織の試料を得て;
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する;
ここで、前記タンパク質、断片または誘導体に結合する抗体の存在が、トレポネーマ属種によって引き起こされる疾患の指標となる
工程を含む、前記方法に関する。
【0032】
前記方法は、趾皮膚炎などの疾患のために使用されてもよい。
本発明のさらなる側面は、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質に特異的に結合する抗体、またはその結合性断片に関する。
【0033】
本発明の別の側面は、トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、前記抗体を被験体に投与する工程を含む、前記方法に関する。趾皮膚炎はこうした疾患の一例である。
【0034】
本発明のさらなる側面は、試料からトレポネーマ属細菌を分離するための方法であって:
−固相に結合した前記抗体と、前記試料を接触させ;
−前記抗体が、前記トレポネーマ属細菌中のトレポネーマ属タンパク質に結合するのを可能にし;そして
−前記試料から前記固相を分離し
それによって、前記試料から前記トレポネーマ属細菌を分離する
工程を含む、前記方法に関する。
【0035】
前記方法において、分離は、例えば免疫磁気分離によって達成可能である。
本発明の別の側面は、本発明記載のタンパク質、断片または誘導体をコードする核酸分子に関する。1つの態様において、タンパク質をコードする核酸分子は、配列番号1、配列番号3、配列番号5およびその一部からなる群より選択されるヌクレオチド配列を有する。
【0036】
本発明のさらなる側面は、前記核酸分子、および場合によって宿主細胞における発現のための制御配列を含む、ベクターに関する。
本発明の別の側面は、前記ベクターを含むトランスジェニック宿主細胞に関する。
【0037】
本発明はまた、本発明記載のタンパク質、断片または誘導体を産生するための方法であって:
−適切な培地中で本発明記載の宿主細胞を培養し;そして
−前記培地から前記タンパク質、断片または誘導体を単離する
工程を含む、前記方法にも関する。
【0038】
詳細
ショットガン・ファージディスプレイを用いて、T.ファゲデニスに緊密に関連するDDトレポネーマ属系統型の単離体(V1)における3つの免疫原性タンパク質を同定した。この系統型は、DDの発病における重要な病原体であることがいくつかの研究において示されてきている(Klitgaardら、2008、Nordhoffら、2008、Yanoら、2009)。ファージ・ライブラリーは、生きた細菌で免疫したウサギ由来の抗体に対して選択された。
【0039】
梅毒トレポネーマ亜種パリダム(T. pallidum subsp. pallidum)のよく性質決定された免疫原性タンパク質TmpAに対する相同体、ならびにいかなる既知のスピロヘータ・タンパク質にも相同性を持たない2つのタンパク質が同定された。454 SequencingTMによって生成されたV1のゲノム配列から、これらのタンパク質の完全アミノ酸配列を予測した。3つの特異的免疫原性タンパク質、およびそのアミノ酸配列は、配列番号2、4および6に記載される。
【0040】
本発明の第一の側面記載のタンパク質、断片および誘導体は、T.ファゲデニス株V1に緊密に関連するトレポネーマ属系統型の培養物から単離されるか、または好ましくは以下に記載するように組換え的に産生されてもよい。
【0041】
ウェスタンブロットを行って、免疫したウサギ由来の抗体ならびに天然感染したウシ由来の抗体の両方が、組換え的に産生されたTmpA相同体およびTtm断片に結合することを示した。
【0042】
パイロットELISA実験を行い、そしてDDに罹患したおよびしていないウシ由来の血清間で、吸光度における相違が記録されてきている(表1)。単一抗原(TmpA相同体またはTtm断片)を用いると、健康群および感染群の結果間にはわずかな重複しかなく、そして2つの抗原の組み合わせを用いると、重複はなかった。
【0043】
本発明記載の免疫原性タンパク質、およびその活性断片を、T.ファゲデニスに緊密に関連するトレポネーマ属系統型または他のトレポネーマ属種によって少なくとも部分的に引き起こされる疾患に対するワクチンにおいて使用してもよい。
【0044】
特定の態様において、完全タンパク質がワクチンにおいて用いられる。特定の態様において、適切なエピトープを含む断片のみが用いられる。特定の態様において、1またはそれより多いタンパク質の1またはそれより多いエピトープが単一の分子に組み合わされ、そしてワクチンにおいて用いられる。組換えタンパク質、その誘導体または断片は、したがって、単独で、または異なる組み合わせで、または結合性エピトープの融合タンパク質として用いられうる。
【実施例】
【0045】
方法
ヘルパーファージ、細菌株、増殖条件、およびDNA技術
ファージR408(Promega)をヘルパー・ファージとして用いた。ファージまたはファージミドが関与するすべての実験において、大腸菌(Escherichia coli)TG1(Δ(lac−proAB)Δ(mcrB−hsdSM)5(rK−mK−)thi−1 supE[F’traD36 proAB lacIqZΔM15]; Stratagene)を宿主として用い、そしてLuria−Bertaniブロス(LB)中、またはLuria−Bertani寒天(LA)上で増殖させた。適切な場合、50μg/mlアンピシリン(C16H18N3O4SNa、Roche)を添加した。インキュベーションは37℃であった。ファージライブラリーの構築のため、トレポネーマ属種株V1由来の染色体DNAを用いた。嫌気性ジャー中、振盪装置(90rpm)上で、FABGS(1リットルあたり2.0g D−グルコースおよび25%ウシ胎児血清、S 0115、Biochrom AG、ドイツを含む、偏好性嫌気性細菌ブロス、LAB 71、LabM、英国ランカシャー州)を含むフラスコ中、37℃でインキュベーションし、トレポネーマ属株を増殖させた。ブロス培養物を等張生理食塩水(pH6.3)で3回洗浄し、その後、リン酸緩衝生理食塩水(PBS、pH7.3)で1回洗浄した。慣用的なフェノール−クロロホルム抽出によってトレポネーマ属DNAを調製した。制限酵素および修飾酵素は、MBI Fermentas AB由来であり、そしてこれらを製造者の指示にしたがって用いた。QIAprepTM Miniprep(QIAGEN)を用いて、プラスミドを調製した。
【0046】
免疫およびポリクローナル抗体の精製
研究のこの部分は、ウプサラにおける動物実験に関する倫理委員会によって認可された(C 300/8)。ニュージーランドホワイト・ウサギを、トレポネーマ属種株V1の生菌培養物で皮下免疫した。2回洗浄し、そして0.5ml体積の等張生理食塩水中に分配したおよそ109細菌の用量を20日おいて2回注射した。最初の免疫の38日後の最終出血由来の血清を、この研究のための抗体(IgG)の精製に用いた。0.45μmシリンジフィルターを通じて10mlの血清を滅菌ろ過し、そして5ml HiTrapTMプロテインG HPカラム(GE Healthcare)上に適用した。Ab緩衝液キット(GE Healthcare)を用いて、製造者の指示にしたがって、ウサギIgGを精製した。Zebaスピン脱塩カラム(Pierce)を用いて、精製抗体の溶出物を脱塩し、そしてマイナス20℃、PBS中で保存した。
【0047】
トレポネーマ属種ファージディスプレイライブラリーの構築および結合性ファージの選択(パニング)
pG8SAETファージミドベクター中で、ファージライブラリーを構築した。大部分の断片が長さ0.4〜1.5kbになるまで、トレポネーマ属種株V1染色体DNAを超音波処理によって断片化した。T4 DNAポリメラーゼおよびT4 DNAキナーゼ処理によって、断片を平滑端化し、そして次いで、Ready−To−GoTM T4 DNAリガーゼ試験管(GE Healthcare)を用いて、SnaB1消化しそして脱リン酸化したファージミドベクターpG8SAET内に連結した。連結した物質を大腸菌TG1細胞に電気形質転換し(2.5kV、25μF、360Ω)、ヘルパーファージを用いて感染させ、そしてファージ粒子を増殖させることによって、最終ライブラリーを生成した。この方法は、ユニークなクローンと見なされる、4x107形質転換体を生じ、14のランダムに選択されたクローンに対するコロニーPCRによって決定すると、このうち86%が挿入物を所持した。最終ライブラリーは、1mlあたり、1x1011コロニー形成単位の力価を有した。
【0048】
ウサギ抗トレポネーマ属種株V1 IgGに対するパニングによって、免疫原性ポリペプチドをディスプレイするファージを単離した。3回のパニング実験を行った。マイクロウェル(MaxiSorpTM、Nalge Nunc International)を、200μlの50mM炭酸ナトリウム、pH9.5中、10μgの濃度のZymed組換えプロテインG(Invitrogen)でコーティングした。その後、0.05% Tween20を含むリン酸緩衝生理食塩水pH7.4(PBS−T)でウェルをブロッキングした。ウサギ抗トレポネーマ属種株V1 IgGを200μl PBSまたは100μl PBS+100μl未精製大腸菌溶解物(ブロッキング用)中、85または215μgの濃度で添加した。洗浄後、200μlのファージライブラリーを添加した。ウェルを室温で3時間または2℃で一晩インキュベーションし、その後、これらを25回洗浄した後、50mMクエン酸Na/140mM NaCl pH2.0を添加することによって、ファージを溶出させた。2M Tris緩衝液pH8.0で溶出物を直ちに中和し、そしてアンピシリンを含むLAプレート(LAamp)上にプレーティングした大腸菌TG1を感染させるのに用いた。1つのパニング実験において、ウェルに添加したTG1細胞を、結合したファージによって直接感染させることによってもまた、溶出を行った。一晩インキュベーションした後、コロニーを計数し、そして100のコロニーをLAampプレートに移した。次いで、マウス抗E−タグ抗体(GE Healthcare)および西洋ワサビ・ペルオキシダーゼ標識した二次ヒツジ抗マウス抗体(GE Healthcare)を用いたE−タグ発現のスクリーニングのために、これらのコロニーをニトロセルロース・フィルターにトランスファーした。残ったコロニーをプレートから洗い流し、そしてヘルパーファージで重感染させて、濃縮ライブラリー/ファージストックを作製し、同じプロトコルにしたがって、これを二次濃縮サイクル(再パニング)において用いた。全部で2回の再パニングを行った。プラスミド調製およびプライマーSAsekv(5’−TAT CTG GTG GCG TAA CAC CTG CT−3’、配列番号17)を用いた挿入物の配列決定のため、200を超えるE−タグ陽性コロニーを選択した。ウプサラ・ゲノム・センターで、3730xl DNA分析装置(Applied Biosystems)上、プラスミドDNAを配列決定し、そしてCLC Main Workbenchソフトウェア(CLC bio)で分析した。挿入物の分析によって、3つの異なる遺伝子から、それぞれ、9、9、および8の重複する部分配列が明らかになった。
【0049】
ゲノム配列決定および配列分析
トレポネーマ属種株V1の染色体DNAを配列決定し、そしてロングリードGS FLXチタン化学反応および454新規アセンブリー装置、Newbler(454 Life Sciences、米国コネチカット州ブランフォード)とともに、ゲノム配列決定装置FLX系を用いて、KTH王立工科大学のKTHゲノムセンター、スウェーデン・ストックホルムでアセンブリーした。CLC Genomics Workbench 3(CLC bio)を用いて読み取り配列のさらなる新規アセンブリーを行い、そしてさらなる配列編集のため、CLC Main Workbench 5(CLC bio)を用いた。
【0050】
ゲノム配列を用いて、3つの免疫原性タンパク質の全長オープンリーディングフレームおよび対応するアミノ酸配列を予測した。National Center for Biotechnology InformationのBLASTアルゴリズムを用いて、相同性検索を行った。シグナルペプチドの予測には、グラム陽性データを含むSignalP 3.0サーバーを用いた。1つのタンパク質は、Setubalら、2006にしたがって、リポタンパク質と予測された。
【0051】
タンパク質発現および精製のためのクローンの構築
グラム陰性細菌用のプロトコルにしたがって、DNeasy血液および組織キット(QIAGEN)を用いて、PCR用のゲノム・トレポネーマ属種株V1 DNAを調製した。MgSO4を含む10X Pfu緩衝剤(Fermentas)5μl、各0.2mMのデオキシヌクレオチド、表2に示すような0.2μMの順方向および逆方向プライマー、1.25U Pfu DNAポリメラーゼ(Fermentas)および50ngゲノムDNAの50μl反応混合物を調製した。熱サイクリング条件は、95℃1分間、30周期の95℃30秒間、50℃30秒間および72℃3分間、ならびに72℃5分間の最終伸長であった。アガロースゲル電気泳動によってPCR産物を分析し、そしてillustra GFX PCR DNAおよびゲルバンド精製キット(GE Healthcare)を用いて精製した。製造者の指示(Fast digest、Fermentas)にしたがって、精製アンプリコンを、BamHIおよびXhoIまたはNdeIおよびSapIのいずれかで消化し、そして先に記載するように精製した。消化アンプリコンを、ReadyToGo T4DNAリガーゼ(GE Healthcare)を用いて、それぞれのベクター、BamHIおよびXhoIで消化したpGEX−6P−1(バルクGST精製モジュール、GE Healthcare)またはNdeIおよびSapIで消化したpTXB1(IMPACTTMキット、New England BioLabs)内に連結した。連結した材料を、コンピテント大腸菌株BL21(DE3)(GST)またはER2566(IMPACT)内に電気形質転換し、そしてアンピシリン(最終濃度50μg/ml)を補充したLA上にスプレッドした。ベクター配列決定プライマーを用いたPCRによって、多くのコロニー中の挿入物の存在を分析した。DNA配列決定によって、正しいサイズの挿入物を持つクローンをさらに分析した。
【0052】
組換え免疫原性トレポネーマ属タンパク質の産生
組換え免疫原性トレポネーマ属タンパク質の産生のため、製造者の指示にしたがって、商業的に入手可能なタンパク質発現および精製系、例えばバルクGST精製モジュール(GE Healthcare)またはIMPACTTMキット(New England BioLabs)を用いた。アンピシリン(最終濃度50μg/ml)を補充したLB培地中、37℃で、組換えクローンを増殖させた。光学密度(OD600nm)〜0.6の際、増殖培地にIPTG(最終濃度0.3mM)を補充し、そして増殖温度を20℃にシフトした。一晩インキュベーションした後、細胞を採取し、そして緩衝液[20mM Tris−HCl(pH8.0)、500mM NaCl、0.1mM EDTA、および0.05%(v/v)TWEEN20]中に再懸濁し、そして凍結融解によって溶解した。遠心分離後、上清を滅菌ろ過し、そしてキチンカラム上に適用した。同じ緩衝液を用いてカラムを徹底的に洗浄し、そして続いて切断緩衝液[20mM Tris−HCl(pH8.0)、50mM NaCl、0.1mM EDTA、および30mMジチオスレイトール(DTT)]で処理した。抗原を含有する溶出試料を、リン酸緩衝生理食塩水[PBS; 137mM NaCl、2.7mM KCl、10mM Na2HPO4、1.4mM KH2PO4(pH7.4)]に対して透析した。
【0053】
GST−グルタチオン・アフィニティ系において、上記方法にしたがって、増殖、誘導および採取後、最終濃度0.1%(v/v)TWEEN20を補充したPBS(PBST)中に大腸菌細胞を懸濁したら、凍結融解によって細胞を溶解した。遠心分離後、上清を滅菌ろ過し、そしてグルタチオン−セファロース・ビーズでバッチ精製した。PBSTを用いて徹底的に洗浄した後、グルタチオン溶出緩衝液で融合タンパク質を溶出させるか、またはscissorプロテアーゼで処理して、生じたタンパク質を放出させた。最後に、分光光度測定を用いて、得られた抗原の量を決定し、そしてSDS−PAGEクーマシー染色によって品質を分析した。最後にタンパク質を−20℃で保存した。
【0054】
酵素連結免疫吸着アッセイ(ELISA)
ELISAにおいて、全細胞溶解物に比較して、大腸菌で組換え的に産生された精製タンパク質を用いる利点は、大規模産生の可能性があり、そして試験が最適化されることである(タンパク質の異なる組み合わせを用いると、しばしば、特異性がより高くなるため、バックグラウンドがより低くなる)。趾皮膚炎(DD)に罹患したおよびしていないウシ由来の血清、ならびに免疫したウサギ(上記)由来の血清を用いて、最初のELISA試験を行った。マイクロプレート(PolySorpTM、Nalge Nunc International)を、100μlの50mM炭酸ナトリウム、pH9.5中、それぞれ2.5μgおよび/または0.31μgの濃度で、組換えTmpA相同体および/または組換えテール・テープ測定タンパク質断片で、2℃で一晩、コーティングした。0.05%Tween20を含む400μlのリン酸緩衝生理食塩水、pH7.4(PBS−T)で、ウェルを2回洗浄した。その後、PBS−Tで、室温で1時間、ウェルをブロッキングした。100μlの血清またはPBS−Tを各ウェルに添加した。各血清の4つの希釈、1:25、1:50、1:100、および1:200を用いた。マイクロプレートを37℃で1時間インキュベーションし、そして次いで、400μl PBS−Tで3回洗浄した。それぞれ1:4000および1:500希釈した西洋ワサビ・ペルオキシダーゼ・コンジュゲート化ブタ抗ウサギ(Dako)およびウサギ抗ウシ(Dako)抗体を適切なウェルに添加し、そしてプレートを37℃で1時間インキュベーションした。400μl PBS−Tでウェルを3回洗浄した後、1:20で混合した20mMテトラメチルベンジジン(TMB)と0.1Mクエン酸カリウム/H2O2(230μl/l)pH4.25からなる100μl溶液を添加した。プレートを室温で10分間インキュベーションした。反応を停止するため、50μlの10%硫酸を添加した。光学密度(OD)を450nmで測定し、そして読み取り値を試料緩衝液ブランクに対して補正した。
【0055】
抗体を用いた免疫磁気分離
トレポネーマ属種は、増殖のために複雑な培地および嫌気性環境を必要とする偏好性生物である。さらに、ウシ鉤爪由来の試料は、培養を汚染させる大量の他の細菌を有する。培養およびDNA単離のためにトレポネーマ属種を濃縮しそして精製するオプションは、免疫磁気分離である(Demirkanら、1999、Demirkanら、2001、Choiら、1996)。例えば前記免疫原性タンパク質、その誘導体または活性断片に対して作製されたウサギ抗体とカップリングされた抗ウサギIgGで共有的にコーティングされた免疫磁気ビーズを、T.ファゲデニスに緊密に関連するDDトレポネーマ属系統型の特異的分離に用いてもよい。
【0056】
ワクチン
免疫系を刺激して抗体を産生させる細菌タンパク質はまた、ワクチン開発にも使用可能である。組換えタンパク質を免疫刺激複合体(ISCOM)および/または全細胞溶解物と組み合わせて、動物における免疫反応を、そしてしたがって、疾患原因物質(単数または複数)に対する保護を増加させることも可能である。
【0057】
行ったさらなる実験
酵素連結免疫吸着アッセイ(ELISA)
材料および方法:Tpl株V1が単離された群れからの、急性DDに罹患した8頭の乳牛、既知のDDの病歴がない別の群れからの2頭の乳牛、および6〜7ヶ月齢の5頭の子ウシ由来の血清を用いてアッセイを行った。趾皮膚炎診断は、視覚的検査によって行われた。マイクロプレート(PolySorpTM、Nalge Nunc International)を、100μlの50mM炭酸ナトリウム、pH9.5中、1μg/ml TmpA、0.8μg/ml Ttmまたは0.02μg/ml PrrAの濃度の組換えタンパク質で、2℃で一晩コーティングした。PBS−Tでウェルを2回洗浄し、そしてPBS−Tで、室温で1時間、ブロッキングした。PBS−T中で1:100希釈した171の100μl血清を各ウェルに添加した。マイクロプレートを37℃で1時間インキュベーションし、そして次いでPBS−Tで洗浄した。1:8000希釈した西洋ワサビ・ペルオキシダーゼ(HRP)コンジュゲート化ウサギ抗ウシIgG抗体(Sigma)または1:4000希釈したモノクローナル22:26抗ウシIgG−HRP抗体(Svanova Biotech AB)をウェルに添加し、そして37℃で1時間、プレートをインキュベーションした。PBS−Tでウェルを3回洗浄し、その後、0.1Mクエン酸カリウムpH4.25中の1mMテトラメチルベンジジンおよび0.006%H2O2からなる100μl溶液を添加した。プレートを室温で10分間インキュベーションした。反応を停止するため、50μlの10%硫酸を添加した。光学密度を450nmで測定し、そして読み取り値から試料緩衝液ブランクの光学密度を減じて、補正光学密度(COD)を生じた。
【0058】
結果:Tpl株V1で同定された3つの免疫原性タンパク質を組換え的に産生した;予測されるリポタンパク質シグナルペプチドのシステイン残基に対してaa+1から停止コドン前の最後のaaまでの全長成熟タンパク質としてのPrrA(aa:s22〜251)、予測されるリポタンパク質シグナルペプチドのシステイン残基に対してaa+7から停止コドン前の最後のaaまでのTmpA(aa:s29〜344)、およびパニング実験由来の重複Ttm配列のコンセンサス配列を構成するaa:sであるaa:s689〜970を含む部分的ポリペプチドとしてのTtm。これらのタンパク質/ポリペプチドを、間接ELISAにおける抗原として用い、ここでDDに罹患したおよびしていないウシ由来の血清試料を、抗原に対する抗体の存在に関して分析した。異なる濃度の抗原、血清、および二次抗体をパイロット実験において試験した(データ未提示)。DDに罹患したおよびしていないウシの間の最適な区別を達成する条件を、最終実験で用いた。TmpA抗原に関して、DDに罹患したウシ由来の3つの試料に関する光学密度は、臨床的に健康な269頭のウシ由来の試料に関する最高値よりも低く、一方、TtmおよびPrrAを用いた試験は、1つを除きすべての場合で識別力があった(図1AおよびB)。
【0059】
【表2】
【0060】
表1.趾皮膚炎(DD)に罹患したおよびしていないウシ由来の血清の最初のELISA試験。450nmで光学密度(OD)を測定し、そして試料緩衝液ブランクに対して読み取り値を補正した。血清を1:100に希釈し、TmpAの濃度は2.5μg/ウェルであり、そして推定上のテール・テープ測定タンパク質断片の濃度は0.31μg/ウェルであった。
【0061】
【表3】
【0062】
表2:デオキシオリゴリボヌクレオチド
参考文献
【0063】
【化1−1】
【0064】
【化1−2】
【0065】
【化1−3】
【特許請求の範囲】
【請求項1】
配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能な断片または誘導体。
【請求項2】
獣医学的医薬品において使用するための、請求項1記載のタンパク質、断片または誘導体。
【請求項3】
トレポネーマ属(Treponema)種によって引き起こされる疾患、例えば趾皮膚炎の防止において使用するための、請求項1記載のタンパク質、断片または誘導体。
【請求項4】
トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体を、被験体に投与する工程を含む、前記方法。
【請求項5】
トレポネーマ属種によって引き起こされる疾患が趾皮膚炎である、請求項4記載の方法。
【請求項6】
配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、および場合による薬学的に許容されうるアジュバント、キャリアーおよび/または希釈剤を含む、薬学的組成物。
【請求項7】
試料において、トレポネーマ属種由来のタンパク質に対する抗体の存在を検出するための方法であって:
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する
工程を含む、前記方法。
【請求項8】
トレポネーマ属種によって引き起こされる疾患のin vitro診断のための方法であって:
−被験体由来の体液または組織の試料を得て;
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する;
ここで、前記タンパク質、断片または誘導体に結合する抗体の存在が、トレポネーマ属種によって引き起こされる疾患の指標となる
工程を含む、前記方法。
【請求項9】
前記疾患が趾皮膚炎である、請求項8記載の方法。
【請求項10】
配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質に特異的に結合する抗体、またはその結合性断片。
【請求項11】
トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、請求項10記載の抗体を被験体に投与する工程を含む、前記方法。
【請求項12】
トレポネーマ属種によって引き起こされる疾患が趾皮膚炎である、請求項11記載の方法。
【請求項13】
試料からトレポネーマ属細菌を分離するための方法であって:
−固相に結合した請求項10記載の抗体と、前記試料を接触させ;
−前記抗体が、前記トレポネーマ属細菌中のトレポネーマ属タンパク質に結合するのを可能にし;そして
−前記試料から前記固相を分離し
それによって、前記試料から前記トレポネーマ属細菌を分離する
工程を含む、前記方法。
【請求項14】
前記分離が免疫磁気分離である、請求項13記載の方法。
【請求項15】
請求項1記載のタンパク質、断片または誘導体をコードする核酸分子。
【請求項16】
配列番号1、配列番号3または配列番号5記載のヌクレオチド配列を有する、請求項15記載の核酸分子。
【請求項17】
請求項15または16記載の核酸分子、および場合によって宿主細胞における発現のための制御配列を含む、ベクター。
【請求項18】
請求項17記載のベクターを含むトランスジェニック宿主細胞。
【請求項19】
請求項1記載のタンパク質、断片または誘導体を産生するための方法であって:
−適切な培地中で請求項18記載の宿主細胞を培養し;そして
−前記培地から前記タンパク質、断片または誘導体を単離する
工程を含む、前記方法。
【請求項1】
配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能な断片または誘導体。
【請求項2】
獣医学的医薬品において使用するための、請求項1記載のタンパク質、断片または誘導体。
【請求項3】
トレポネーマ属(Treponema)種によって引き起こされる疾患、例えば趾皮膚炎の防止において使用するための、請求項1記載のタンパク質、断片または誘導体。
【請求項4】
トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体を、被験体に投与する工程を含む、前記方法。
【請求項5】
トレポネーマ属種によって引き起こされる疾患が趾皮膚炎である、請求項4記載の方法。
【請求項6】
配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応を誘導可能なその断片または誘導体、および場合による薬学的に許容されうるアジュバント、キャリアーおよび/または希釈剤を含む、薬学的組成物。
【請求項7】
試料において、トレポネーマ属種由来のタンパク質に対する抗体の存在を検出するための方法であって:
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する
工程を含む、前記方法。
【請求項8】
トレポネーマ属種によって引き起こされる疾患のin vitro診断のための方法であって:
−被験体由来の体液または組織の試料を得て;
−前記試料を、配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質、あるいは前記タンパク質に対する免疫反応において被験体によって産生される抗体に結合可能なその断片または誘導体と接触させ;そして
−前記タンパク質、断片または誘導体に結合する抗体を検出する;
ここで、前記タンパク質、断片または誘導体に結合する抗体の存在が、トレポネーマ属種によって引き起こされる疾患の指標となる
工程を含む、前記方法。
【請求項9】
前記疾患が趾皮膚炎である、請求項8記載の方法。
【請求項10】
配列番号2、配列番号4または配列番号6記載のアミノ酸配列を有する単離タンパク質に特異的に結合する抗体、またはその結合性断片。
【請求項11】
トレポネーマ属種によって引き起こされる疾患の治療または防止のための方法であって、請求項10記載の抗体を被験体に投与する工程を含む、前記方法。
【請求項12】
トレポネーマ属種によって引き起こされる疾患が趾皮膚炎である、請求項11記載の方法。
【請求項13】
試料からトレポネーマ属細菌を分離するための方法であって:
−固相に結合した請求項10記載の抗体と、前記試料を接触させ;
−前記抗体が、前記トレポネーマ属細菌中のトレポネーマ属タンパク質に結合するのを可能にし;そして
−前記試料から前記固相を分離し
それによって、前記試料から前記トレポネーマ属細菌を分離する
工程を含む、前記方法。
【請求項14】
前記分離が免疫磁気分離である、請求項13記載の方法。
【請求項15】
請求項1記載のタンパク質、断片または誘導体をコードする核酸分子。
【請求項16】
配列番号1、配列番号3または配列番号5記載のヌクレオチド配列を有する、請求項15記載の核酸分子。
【請求項17】
請求項15または16記載の核酸分子、および場合によって宿主細胞における発現のための制御配列を含む、ベクター。
【請求項18】
請求項17記載のベクターを含むトランスジェニック宿主細胞。
【請求項19】
請求項1記載のタンパク質、断片または誘導体を産生するための方法であって:
−適切な培地中で請求項18記載の宿主細胞を培養し;そして
−前記培地から前記タンパク質、断片または誘導体を単離する
工程を含む、前記方法。
【図1A】

【図1B】
【図1B】
【公表番号】特表2013−518563(P2013−518563A)
【公表日】平成25年5月23日(2013.5.23)
【国際特許分類】
【出願番号】特願2012−551129(P2012−551129)
【出願日】平成23年1月28日(2011.1.28)
【国際出願番号】PCT/SE2011/050090
【国際公開番号】WO2011/093783
【国際公開日】平成23年8月4日(2011.8.4)
【出願人】(512197342)
【出願人】(512197353)
【Fターム(参考)】
【公表日】平成25年5月23日(2013.5.23)
【国際特許分類】
【出願日】平成23年1月28日(2011.1.28)
【国際出願番号】PCT/SE2011/050090
【国際公開番号】WO2011/093783
【国際公開日】平成23年8月4日(2011.8.4)
【出願人】(512197342)
【出願人】(512197353)
【Fターム(参考)】
[ Back to top ]