
ワクチン
この発明はC型肝炎ウイルス(HCV)感染症ならびにこれに関連する症候および疾患の治療および予防に有用な方法および組成物に関する。特に、この発明はHCVタンパク質をコードするポリヌクレオチド配列を含むDNAワクチン、および本発明のワクチンを投与することを含むHCVに感染した個体の治療方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、C型肝炎ウイルス(HCV)感染、ならびにそれに関連する症候および疾患の治療および予防に有用な方法および組成物に関する。特に、本発明は、HCVタンパク質をコードするポリヌクレオチド配列を含むDNAワクチン、ならびに、本発明のワクチンの投与を含む、HCVに感染した個体の治療方法に関する。
【背景技術】
【0002】
HCVは、輸血後および市中獲得型非A非B型肝炎の主要な原因として近年確認された。約1億7千万人がHCVに慢性的に感染しており、有病率は1〜10%である。有病率が1.8%の米国における医療費は20億ドルであると推定される。40〜60%の肝疾患はHCVによるものであり、30%の英国移植患者はHCV感染のためである。HCVは、初め無症状の感染であり、患者の90%以上が慢性疾患を発症する。この疾患過程は、典型的に、慢性活動性肝炎(70%)、線維症、肝硬変(40%)から肝細胞癌(60%)に移行する。感染から肝硬変までの期間の中央値は20年であり、肝細胞癌までは、20年である(Lauer G.およびWalker B. 2001, N. Engl J. Med 345, 41, Cohen J. 2001, Science 285 (5424) 26)。
【0003】
HCVの治療を改善することが強く求められる。現在、利用可能な小分子複製阻害剤は存在しない。リボビリンとPEG化インターフェロンからなる現在の最良の標準治療が、HCV感染治療の主軸を表している。しかし、現行の治療法が持続的な効果を達成する能力は依然として最適以下である(6ヶ月までの間、全体で50%有効率だが、遺伝子型1bの場合、有効率はそれより低い(27%))。この治療はまた、不快な副作用も伴う。これにより、特に、治療を始めて6ヶ月が過ぎると、有効率が急激に下降する。
【0004】
いくつかの研究から、個々のHVCタンパク質は正常なマウス(DNAによる免疫を受けたものを含む)において免疫原性であることがわかっている。数種のHCVワクチンが、予防または治療のいずれかを目的として現在臨床試験中である。最も進んでいるのは、E1またはE2エンベロープタンパク質を用いたChironおよびInnogeneticsによるものでフェーズ2にある。Transvaxによるエピトープワクチンもフェーズ2にある。また、各種の送達系(DNAを含む)による、コアおよび非構造抗原由来の配列を用いる数種のワクチンが、前臨床開発段階にある。
【0005】
HCVはフラビウイルス科のプラス鎖RNAウイルスであり、そのゲノムは長さが9.4 kbで、1つのオープンリーディングフレームを有する。HCVゲノムは、単一のポリタンパク質として翻訳され、これが、宿主およびウイルスプロテアーゼによりプロセシングされて、構造タンパク質(コア、エンベロープE1およびE2、ならびにp7)と、様々な酵素活性を有する6つの非構造タンパク質を産生する。1b遺伝子型の一例であるHCV J4L6分離物のゲノムは、登録番号AF054247(Yanagi, M., St Claire,M., Shapiro,M., Emerson,S.U., Purcell,R.H.およびBukh,J. “Transcripts of a chimeric cDNA clone of hepatitis C virus genotype 1b are infectious in vivo”. Virology 244 (1), 161−172 (1998))として見出されているが、これを図1に示す。
【0006】
エンベロープタンパク質によって、標的細胞の認識、これへの結合および進入が可能になる。ウイルス複製に関与する主要な非構造タンパク質としては、NS2(Zn依存性メタロプロテイナーゼ)、NS3(セリンプロテアーゼ/ヘリカーゼ)、NS4A(プロテアーゼ補因子)、NS4B、NS5AおよびNS5B(RNAポリメラーゼ)が挙げられる(Bartenschlager BおよびLohmann V. 2000. Replication of hepatitis C virus. J. Gen Virol 81, 1631)。
【0007】
HCVポリタンパク質の構造は、次のように示すことができる(数字は、各タンパク質の第1アミノ酸の位置を示す;J4L6分離物の完全ポリタンパク質は、長さが3010アミノ酸である)。
【化1】

【0008】
ウイルスの突然変異率は高く、保存および非保存領域のヌクレオチド配列に基づき、少なくとも6つの主要遺伝子型が明らかにされている。しかし、単一の患者から単離したHCVは、常に近縁のゲノムもしくは準種の混合物として提示されるため、さらなる不均質性が存在する。
【0009】
HCVゲノムは高度の遺伝子多様性を示し、これは、6つの主要な遺伝子型(1a、1b、2、3、4、5および6)に分類される。遺伝子型1a、1b、2および3は、欧州、北および南アメリカ、アジア、中国、日本およびオーストラリアで最も優勢である。遺伝子型4および5はアフリカで、また遺伝子型6は東南アジアでそれぞれ優勢である。
【0010】
HCV感染の治療法の改善と共に、多数のHCV遺伝子型を治療できる多様な能力の治療法も強く求められている。本発明の第1の態様では、様々な遺伝子型に対する保護が多様な新規ワクチン製剤を提供する。
【0011】
1種以上のHCVタンパク質をコードするポリヌクレオチドを含むHCVワクチンが記載されている。N3をコードするプラスミドDNAまたはセムリキフォレストウイルスベクターを含むワクチンがBrinsterらにより記載されている(2002, Journal of General Virology, 83, 369−381)。NS5BをコードするポリヌクレオチドワクチンがWO99/51781に開示されている。HCV E1、E1+E2融合物、NS5A およびNS5Bタンパク質をコードするコドン最適化遺伝子、ならびに、それを含むワクチンが、WO97/47358に記載されている。WO 01/04149は、コア、NS3、NS4もしくはNS5Aに由来する、HCVエピトープのモザイクをコードするポリペプチドまたはポリヌクレオチドを開示している。ワクチンに有用な融合タンパク質(NS3、NS4、NS5AおよびNS5Bを含む)、ならびにこのような融合タンパク質をコードするDNAがWO 01/30812に記載されている;随意に、上記融合タンパク質はコアタンパク質の断片を含むと言われている。WO 03/031588には、HCVタンパク質NS3−NS4A−NS4B−NS5A−NS5Bをコードするアデノウイルスベクターが記載され、これは、ワクチンとしての使用に適している。
【0012】
「非処理の」コアタンパク質と非構造タンパク質を含むポリペプチドを含むワクチンがWO 96/37606に記載されている。
【発明の開示】
【課題を解決するための手段】
【0013】
本発明は、HCVタンパク質であるコア、NS3、NS4BおよびNS5Bをコードするポリヌクレオチドワクチンの提供に関する。本発明のポリヌクレオチドワクチンは、NS4A HCVタンパク質および/またはNS5Aタンパク質をコードしない。また、本発明のポリヌクレオチドワクチンは、コア、NS3、NS4BおよびNS5B HCVタンパク質をコードし、それ以外のHCVタンパク質はコードしないのが好ましい。本発明はまた、医療における、および、HCV感染の治療または予防のための医薬品の製造における、前記抗原をコードするポリヌクレオチドワクチンの使用を提供する。
【0014】
本発明のワクチンに使用するポリヌクレオチド配列は、好ましくは、DNA配列である。
【0015】
HCVタンパク質をコードするポリヌクレオチドは、様々な組合せまたは立体配置のものでありうる。例えば、タンパク質は、個々のタンパク質、もしくは融合タンパク質として発現されてもよい。融合物(DNAまたはタンパク質レベルのいずれでもよい)の例として、NS4BおよびNS5Bのアミノ酸配列を含む、もしくはコードする単一ポリペプチドまたはポリヌクレオチドからなる二重融合物(NS4B−NS5B)、NS3−NS4B−NS5Bのアミノ酸配列を含む、もしくはコードする三重融合物、あるいは、本発明の4つの抗原すべての融合物(mCore−NS3−NS4B−NS5B)が挙げられる。
【0016】
本発明の好ましい融合物は、NS4BとNS5B(NS4B−NS5BまたはNS5B−NS4B);ならびに、コアとNS3(NS3−コアまたはコア−NS3)の二重融合物をコードするポリヌクレオチドである。好ましい三重融合物は、NS3−NS4B−NS5Bのアミノ酸配列をコードするポリヌクレオチドである。
【0017】
単一の抗原または融合タンパク質をコードする本発明のポリヌクレオチドは、単一のまたは複数の発現ベクター中に存在することができる。各抗原をコードするポリヌクレオチドが、同じ発現ベクターまたはプラスミド中に存在することが好ましい。これに関連して、HCVタンパク質をコードするポリヌクレオチドは、単一発現カセットに存在するか、もしくはシリーズ発現カセットに複数存在してもよい。
【0018】
他のHCVタンパク質の発現を最適化するために、HCVコアタンパク質をコードするポリヌクレオチドは、少なくとも1つの他のHCVタンパク質をコードするポリヌクレオチドを含む発現カセットの下流にある発現カセットに存在するのが好ましい。HCVコアタンパク質は、NS5Bをコードするポリヌクレオチドを含む発現カセットの下流にある発現カセットに存在するのが好ましい。
【0019】
本発明のオリゴヌクレオチドワクチンによりコードされるポリペプチドは、全長アミノ酸配列を含むか、あるいは、このポリペプチドは、全長タンパク質より短くてもよく、その際、該ポリペプチドは、短くなった遺伝子の発現産物が、全長タンパク質と交差反応する免疫応答を生じさせることを可能にするのに十分な比率の全長ポリヌクレオチド配列を含む。例えば、本発明のポリヌクレオチドは、本来の配列の領域が欠失した切断型HCVタンパク質であるHCVタンパク質の断片をコードすることができ、その際、最終断片は、本来の全長アミノ酸配列の90%以下を含むが、本来の配列の70%以下もしくは50%以下であってもよい。別の言い方をすれば、長さが少なくとも8、例えば、8〜10アミノ酸、あるいは長さが最高20、50、60、70、80、100、150もしくは200アミノ酸までの断片をコードするポリヌクレオチドは、コードされたオリゴまたはポリペプチドがHCV抗原性を示す限り、本発明の範囲内にあると考えられる。特に、しかし、限定するわけではないが、本発明のこの態様は、ポリヌクレオチドが完全なHCVタンパク質配列の断片をコードし、該タンパク質の1以上の別々のエピトープを呈示しうる場合の状況も含む。
【0020】
本発明の好ましいワクチンでは、HCVポリペプチドの少なくとも1つ、好ましいすべてを(末端)切断(truncation)または突然変異により不活性化させる。例えば、NS3のヘリカーゼおよびプロテアーゼ活性は、遺伝子の突然変異により低減または破壊するのが好ましい。発現させたポリペプチドのNS5Bポリメラーゼ活性は、突然変異により低減または破壊するのが好ましい。発現させたポリペプチドのNS4B活性は、突然変異により低減または破壊するのが好ましい。発現させたポリペプチドのコアタンパク質の活性は、切断または突然変異により低減または破壊するのが好ましい。この意味での突然変異には、上記ポリペプチドをコードするポリヌクレオチドに対する付加、欠失、置換もしくは転位事象が含まれる。これ以外にも、全長配列を2つ以上の別々の部分に発現させてもよい。
【0021】
HCVポリペプチドNS3およびNS5Bの機能構造および酵素機能については当分野で記載されている。
【0022】
NS5Bは、RNA依存性RNAポリメラーゼとして記載されている(Qinら、2001, Hepatology, 33, pp 728−737;Lohmannら、2000, Journal of Viral Hepatitis;Lohmannら、1997, Nov., Journal of Virology, 8416−8428;De Francescoら、2000, Seminars in Liver Disease, 20(1), 69−83)。NS5Bポリペプチドは、4つの機能性モチーフA、B、CおよびDを有するものとして記載されている。
【0023】
本発明のポリヌクレオチドワクチンによりコードされるNS5Bポリペプチドを突然変異させることにより、RNA依存性RNAポリメラーゼ活性を低減または除去するのが好ましい。好ましくは、上記ポリペプチドを突然変異させることにより、NS5BのモチーフAを破壊するが、例えば、2639位のアスパラギン酸(D)をグリシン(G)に置換する;または2644位のアスパラギン酸(D)をグリシン(G)に置換するなどがある。ワクチンポリヌクレオチドによりコードされるNS5Bポリペプチドは、これら両方のアスパラギン酸突然変異を含むのが好ましい。
【0024】
好ましくは、コードされたNS5Bポリペプチドは、そのモチーフCでの破壊を含む。例えば、非変異アスパラギン酸残基であるD2737をH、NまたはEに置換する突然変異によってNS5Bの完全な不活性化が起こる。
【0025】
好ましくは、本発明のDNAワクチンによりコードされるNS5Bは、モチーフA突然変異を含むが、随意にモチーフC突然変異を含んでもよい。モチーフAでの好ましい突然変異には、アスパラギン酸(D)2639のグリシンへの置換、およびアスパラギン酸(D)2644のグリシンへの置換が含まれる。後述の実施例1に記載するように、さらに別のコンセンサス突然変異が存在してもよい。
【0026】
NS3は、プロテアーゼとヘリカーゼ活性の両方を持つものとして記載されている。本発明のDNAワクチンによりコードされるNS3ポリペプチドを突然変異させることにより、NS3のプロテアーゼおよびヘリカーゼ活性の両方を破壊するのが好ましい。NS3のプロテアーゼ活性は、H−1083、D−1107およびS−1165の「触媒三つ組(catalytic triad)」に結合していることがわかっている。本発明のワクチンによりコードされるNS3は、触媒三つ組残基に突然変異を含むのが好ましく、最も好ましくは、NS3はセリン1165からバリンへの単一の点突然変異を含む(De Francesco, R., Pessi, a およびSteinkuhler C. 1998. The hepatitis C Virus NS3 proteinase : structure and function of a zinc containing proteinase. Anti− Viral Therapy 3, 1−18)。
【0027】
NS3の構造および機能は次のように表すことができる:
【化2】
【0028】
NS3のヘリカーゼ活性については、I、II、IIIおよびIVの4つの重要なモチーフが確認されている。本発明のDNAワクチンによりコードされるNS3は、これらモチーフの少なくとも1つに対する破壊的突然変異を含むのが好ましい。アスパラギン酸1316からグルタミンへの置換が存在するのが最も好ましい(Paolini, C, Lahm A, De Francesco RおよびGallinari P 2000, Mutational analysis of hepatitis C virus NS3−associated helicase. J.Gen Virol. 81, 1649)。これらの最も好ましいNS3突然変異、S−1165VもしくはD1316Qのいずれも、公知または予測されるT細胞エピトープ内に存在しない。
【0029】
最も好ましくは、本発明のDNAワクチンによりコードされるNS3ポリペプチドは、セリン(S)1165からバリン(V)およびアスパラギン酸(D)1316からグルタミン(Q)への突然変異を含む。加えて、後述の実施例1に記載するように、1以上のコンセンサス突然変異が存在してもよい。
【0030】
HCVコアタンパク質の生物学的機能は複雑であり、個々の点突然変異と相関しない(McLauchlan J. 2000. Properties of the hepatitis C virus core protein: a structural protein that modulates cellular processes. J of Viral Hepatitis 7, 2−4)。コアがリンホトキシンβ受容体と直接的に相互作用し、NFκBおよびPKR経路とも相互作用することができ、かつ細胞の生存およびアポトーシスにも影響を与えることができることが証明されている。コアを発現する組換えワクシニア構築物が、in vivoでコアをより有毒にするワクシニアに対する細胞応答を阻害することが見出された。
【0031】
感染中にコアタンパク質は宿主細胞のプロテアーゼによってウイルスポリタンパク質から2ヶ所で切断される。第1の切断は191位で生じ、E1のN末端を生成する。第2の切断が生じる残基は正確に位置付けられておらず、174〜191位のアミノ酸に存在し、それによって約17アミノ酸の長さの短いコアペプチド配列を遊離させる(McLauchlan J. (2000) J. Viral Hepatitis. 7, 2−14; YasuiK, Lau JYN, Mizokami M.,ら, J. Virol 1998. 72 6048−6055)。
【0032】
本発明のワクチンに使用するコアポリペプチドは全長あるいは切断型の形態のいずれかである。コアポリペプチドは全長であってもよいが、その配列を再配列することによりコアタンパク質のあらゆる活性を破壊(又は阻害)する。コアポリペプチドを少なくとも2つの断片に分割してもよく、コアアミノ酸66〜191とそれに続くアミノ酸1〜65からなるポリペプチド、あるいはコアアミノ酸105〜191とそれに続くコアアミノ酸1〜104からなるポリペプチドを形成することが最も好ましい。
【0033】
同一の細胞における他のHCVタンパク質の産生に与えるコアの負の作用を最小にするために、使用されるコアタンパク質が切断型タンパク質であることが最も好ましい。本発明の好適な態様では、コードされるコアタンパク質は、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断される。最も好ましくは、産生されるタンパク質の配列がコアの第2切断から生じる天然の遊離C末端ペプチド配列を欠くように;より好ましくはこのタンパク質が少なくとも最後の10アミノ酸を欠くように、好ましくは少なくとも最後の15アミノ酸を欠くように、より好ましくは最後の20アミノ酸を欠くように、より好ましくは最後の26アミノ酸を欠くように、そして最も好ましくは最後の40アミノ酸を欠くように、コアタンパク質がカルボキシ末端から切断される。本発明における使用に適したコアをコードする最も好適なポリヌクレオチドは、1〜171位、1〜165位、1〜151位のアミノ酸を含有する切断型コアをコードするものである。本発明の使用に適したコアをコードするポリヌクレオチドが、1〜151位のアミノ酸の切断型コアタンパク質をコードするものであるのが最も好ましい。実施例1で説明するような1以上のコンセンサス突然変異が存在してもよい。
【0034】
本発明のポリヌクレオチドによりコードされる好ましいNS4Bポリペプチドは、HCV分離物と遺伝子型の間で超可変の領域を除去するようなN末端切断を含む。好ましくは、NS4Bポリペプチドは、N末端から30〜100アミノ酸、さらに好ましくは40〜80アミノ酸の欠失、最も好ましくは最初のN末端の48アミノ酸の欠失を含む(J4L6分離物の場合、これは、アミノ酸1760までの切断に相当し、これは、NS4Bの最初の48アミノ酸の消失である;その他のHCV分離物における同等の切断も本発明に含まれる)。さらに、NS4B配列を2つ以上の断片に分割し、野生型分子に認められるものとは違う順序で配置したNS4Bの配列を有するポリペプチドに発現させてもよい。
【0035】
本発明のワクチンに存在するポリヌクレオチドは、HCVウイルスに認められる天然ヌクレオチド配列を含んでもよいが、ヌクレオチド配列のコドンは、哺乳動物細胞での発現のために最適化されているものが好ましい。
【0036】
コドンの最適化に加えて、HCVコア、NS3、NS4BおよびNS5Bをコードする本発明のポリヌクレオチドでのコドン使用頻度を変更することにより、希有コドンが濃縮クラスターに現れるのではなく、反対に、ポリヌクレオチド配列全体にわたって比較的均等な間隔をおいて存在するか、あるいはコドン最適化遺伝子から排除されるようにする。
【0037】
DNAコードは4文字(A、T、CおよびG)を有し、これらを用いて、3文字の「コドン」を綴るが、このコドンは、生物の遺伝子においてコードされたタンパク質のアミノ酸を表すものである。DNA分子に沿ったコドンの直鎖状配列をこれら遺伝子によりコードされたタンパク質中のアミノの直鎖状配列に翻訳する。コードは高度に縮重しており、61個のコドンが20個の天然アミノ酸をコードし、3個のコドンが「停止」シグナルを表す。従って、ほとんどのアミノ酸は2個以上のコドンによりコードされる。実際には、数個が、4個以上の異なるコドンによりコードされる。
【0038】
所与のアミノ酸をコードするのに1個以上のコドンが利用可能である場合には、生物のコドン使用頻度パターンは極めて非ランダムである。種によってコドン選択に偏りがあり、さらにコドンの使用は、単一の種においても、高および低レベルで発現する遺伝子間では著しく異なる可能性がある。この偏りは、ウイルス、植物、細菌および哺乳動物細胞で異なり、いくつかの種は、他の種よりランダムコドン選択から離れた強い偏りを示す。例えば、ヒトおよびその他の哺乳動物は、特定の細菌またはウイルスと比較してあまり強く偏っていない。これらの理由から、大腸菌に発現させた哺乳動物遺伝子、または哺乳動物細胞に発現させたウイルス遺伝子は、効率的な発現にとって不適切なコドン分布を有する。しかし、大腸菌発現に適したコドン使用頻度パターンを有する遺伝子をヒトに効率的に発現させることもできる。発現を起こそうとする宿主に稀に観察されるように、異種DNA配列にコドンのクラスターが存在すると、その宿主での異種発現レベルが低いことが推定される。
【0039】
宿主に希有なものから宿主が優先するものへコドンを変えて(「コドン最適化」)、異種発現レベルを増強するいくつかの例があり、例えば、BPV(ウシパピローマウイルス)後期遺伝子L1およびL2は、哺乳動物コドン使用頻度パターンについて最適化され、これは、哺乳動物(Cos−1)細胞培養物における野生型HPV配列に対する発現レベルを増強することがわかっている(Zhouら、J. Virol 1999. 73, 4972−4982)。この研究では、哺乳動物と比べてBPVで2倍以上の頻度で生じた(使用頻度比>2)すべてのBPVコドンと、使用頻度比が>1.5のほとんどのコドンを、優先的に用いられる哺乳動物コドンで保存的に置換した。WO97/31115、WO97/48370およびWO98/34640(Merck & Co., Inc.)において、HIV遺伝子またはそのセグメントのコドン最適化により、最適化を意図した宿主哺乳動物においてコドン最適化した配列をDNAワクチンとして用いると、タンパク質発現が増大し、免疫原性が向上することがわかっている。これらの文献では、各ウイルスコドンを、意図する宿主の最適コドンで保存的に置換するため、配列は、全体が最適化コドンから構成される(ただし、これが、不要な制限部位、イントロンスプライス部位などを導入する場合を除く)。
【0040】
「コドン使用頻度パターン」という用語は、上記のヌクレオチド配列、遺伝子もしくは遺伝子のクラス(例えば、高度に発現した哺乳動物遺伝子)におけるすべてのコドンについての平均頻度を意味する。哺乳動物(ヒトを含む)についてのコドン使用頻度パターンを文献にみいだすことができる(例えば、Nakamuraら、Nucleic Acids Research 1996、24:214−215参照)。
【0041】
本発明のポリヌクレオチドでは、コドン使用頻度パターンは、HCVの典型的なものから、標的生物、例えば、大腸菌または哺乳動物、特にヒトのコドン出現偏りをより忠実に表すものに変更するのが好ましい。「コドン使用頻度係数」すなわちコドン適合指数(Sharp PM. Li WH. Nucleic Acids Research. 15(3):1281−95, 1987)は、所与のポリヌクレオチド配列のコドン使用頻度パターンが、標的種のパターンにどれくらい密に類似しているかの尺度である。61個のコドンの各々についてのコドン出現頻度(選択したクラスの遺伝子について1000コドン当たりの出現数として表される)を20個の天然アミノ酸の各々について標準化して、各アミノ酸について最も使用頻度の高いコドンの値を1に設定し、これより少ない共通コドンの頻度が、0〜1の間になるように比例的に定められる。すなわち、61個のコドンの各々が、標的種の高度発現遺伝子について1以下の値を割り当てられる。これを優先値(W)と呼ぶ。その種の高度発現遺伝子に対する特定のポリヌクレオチドのコドン使用頻度係数を算出するため、特定のポリヌクレオチドの各コドンについて定めた値を記録し、これらすべての値の相乗平均を求める(コドンの総数でこれらの値の自然対数の和を割り算し、真数(anti−log)を求める)。係数は0から1の間の値になり、係数が高いほど、そのポリヌクレオチドにおいて使用頻度の高いコドンが多い。ポリヌクレオチド配列のコドン使用頻度係数が1の場合には、全コドンが、標的種の高度発現遺伝子について「最も使用頻度の高い」コドンである。
【0042】
本発明は、HCVコア、NS3、NS4BもしくはNS5Bアミノ酸配列をコードするポリヌクレオチド配列であって、該ポリヌクレオチド配列のコドン使用頻度パターンが、高度に発現した哺乳動物遺伝子のものに類似している、上記ポリヌクレオチドを提供する。好ましくは、上記ポリヌクレオチド配列はDNA配列である。上記ポリヌクレオチド配列のコドン使用頻度パターンは、高度に発現したヒト遺伝子のものに類似しているのが望ましい。
【0043】
HCVコア(1−191)をコードするコドン最適化ポリヌクレオチド配列を図2に示す。S1165VおよびD1316Qポリペプチド突然変異を含むHCV NS3をコードするコドン最適化ポリヌクレオチド配列を図3に示す。ポリペプチドのN末端1−48切断を含むHCV NS4Bをコードするコドン最適化ポリヌクレオチドを図4に示す。D2639GおよびD2644Gポリペプチド突然変異を含むHCV NS5Bをコードするコドン最適化ポリヌクレオチド配列を図5に示す。
【0044】
従って、HCVコア、NS3、NS4BもしくはNS5Bアミノ酸配列を一緒にコードする複数のコドンを含み、本発明のワクチンを形成する合成遺伝子であって、上記アミノ酸配列をコードするのに用いられる可能なコドンの選択を最適哺乳動物コドン使用頻度に類似するように変更することにより、上記合成遺伝子におけるコドン使用頻度が、C型肝炎ウイルス遺伝子のそれより、高度に発現した哺乳動物遺伝子の使用頻度に密に類似するようにする、上記合成遺伝子が提供される。コドン使用頻度パターンは、高度に発現したヒト遺伝子のものと実質的に同じであるのが好ましい。「天然の」HCVコア、NS3、NS4BおよびNS5B配列のコドン使用頻度については分析がなされている。HCVタンパク質のコドン使用頻度係数は、コア(0.487)、NS3(0.482)、NS4B(0.481)およびNS5B(0.459)である。本発明のポリヌクレオチドは、一般に、高度に発現したヒト遺伝子について、0.5以上、好ましくは0.6以上、最も好ましくは0.7以上であるが、1以下のコドン使用頻度係数(すでに定義した通り)を有する。ポリヌクレオチドはまた、高度に発現した大腸菌遺伝子について0.5以上、好ましくは0.6以上、最も好ましくは0.7以上のコドン使用頻度係数を有する。
【0045】
コドン最適化のほかにも、合成遺伝子を突然変異させることにより、希有コドンのクラスターの出現を排除する。これは2つの方法のいずれかにより達成することができる。これを達成する好ましい方法は、遺伝子配列から希有コドンを排除するものである。希有コドンを定義する一つの方法によれば、希有コドンは、標的生物の高度発現遺伝子において、特定のアミノ酸に用いられるコドンの<20%を占めるコドン、好ましくは、特定のアミノ酸について<10%を占めるコドンである。あるいは、標的生物の高度発現遺伝子において、相対同義コドン使用頻度(RSCU)値が<0.3、または好ましくは<0.2のコドンとして希有コドンを定義することもできる。RSCU値は、観察されたコドン数を、該アミノ酸の全コドンが同等に高頻度で用いられた場合に予想される数で割って得られる数値である。希有コドンの適切な定義は当業者に明らかであろう。
【0046】
あるいは、HCVコア、NS3、NS4BおよびNS5Bポリヌクレオチドを最適化することにより、濃縮領域に存在する希有の非最適コドンのクラスター形成を防止する。従って、個々の希有コドン、例えば、RSCUが<0.4(さらに好ましくは<0.3)がポリヌクレオチド全体に均等な間隔をおいて存在するように、ポリヌクレオチドを最適化する。
【0047】
ウエスタンブロットで評価されるように、コドンが最適化された突然変異コア、NS3およびNS5Bの発現レベルは野生型と比べて増加することが示されている。切断型のコドン最適化NS4BはNS5Bとの融合物として発現し、この融合物は十分に発現する。
【0048】
本発明のワクチンは、HCVポリペプチドの個々の発現を指令するベクターを含む。あるいは、HCVポリペプチドを1以上の融合タンパク質として発現させてもよい。
【0049】
本発明の好ましいワクチンは、タンパク質またはポリヌクレオチドレベルのいずれかで四重融合物を含み、そのような融合物として以下のものが挙げられる:
HCV組合せ1: HCV500
【化3】
【0050】
HCV組合せ2: HCV510
【化4】
【0051】
HCV組合せ3: HCV520
【化5】
【0052】
HCV組合せ4: HCV530
【化6】
【0053】
HCV組合せ5: HCV501
【化7】
【0054】
HCV組合せ6: HCV502
【化8】
【0055】
HCV組合せ7:
【化9】
【0056】
他の好適な融合物はHCV組合せ1、2および3に類似しているが、ここで、コアタンパク質は切断型コアタンパク質、典型的にはコア1−151である。本発明の別の好ましいワクチンを以下に挙げるが、これらは、同じプラスミド内の様々な発現カセットに存在するポリヌクレオチド二重および三重融合物を含み、各カセットは、プロモーターユニット(例:HCMV IE)の独立した制御下にある(矢印で示す)。
【0057】
このような二重プロモーター構築物は、同じ細胞内で4つのタンパク質抗原の発現を2つの別個のタンパク質として駆動する(矢印で示す)。
【化10】
【0058】
好適な構築物はHCV組合せ7、9、11、または12である。特に好適なものは7と11である。
【0059】
本発明の別の態様では、ポリヌクレオチドワクチンは場合によりコアタンパク質をコードするポリヌクレオチドを含まない。例えば、本発明のこの態様における好適なポリヌクレオチドは下記のものを含む。
【化11】
【0060】
上記HCV組合せ8〜19について、用いた用語、例えば、(コアNS3)+(NS4B5B)は、各々が個別のプロモーターにより独立に制御される2つの発現カセット、この例の場合、コアNS3二重融合タンパク質をコードする1つの発現カセットと、NS4B−NS5B二重融合タンパク質をコードする別のカセットを含むポリヌクレオチドベクターの開示を意味する。各HCV組合せ8〜19も同様に解釈すべきである。
【0061】
前記HCV組合せ1〜19は、HCVタンパク質、ポリタンパク質融合物、もしくはポリヌクレオチドの相対配向を開示する。また、前記HCV組合せ1〜19のすべては、構成タンパク質の活性を除去するのに好ましい突然変異または切断の各々と一緒に開示されることが具体的に記載される。例えば、組合せ1〜19の好ましい変異体(別途記載のない限り)は、コア(1−191(生物学的活性を不能にする2以上の断片への分割配列を除く)または好ましくは、その切断型1−151または1−165もしくは1−171で存在するコア);NS3 1027−1657(ヘリカーゼ(アスパラギン酸1316からグルタミン)およびプロテアーゼ(セリン1165からバリン)活性を不活性化するための突然変異;ポリメラーゼ活性を不活性化させるためのNS5B 2420−3010(アスパラギン酸2639からグリシンおよびアスパラギン酸2644からグリシンへの突然変異、モチーフA);ならびに、NS4B 1712−1972(随意に1760−1972に切断して、N−末端高度可変断片を除去する)を含む。
【0062】
本発明は、前述したような新規のDNAワクチンとポリペプチドを提供する。また、本発明により、記載したポリペプチドの類似体、ならびにそれらを含むDNAワクチンが提供される。
【0063】
用語「類似体」とは、本発明の別のポリペプチドと同じアミノ酸をコードするが、遺伝子コードの重複性を通して、異なるヌクレオチド配列を有するが、同じコドン使用頻度パターンを維持する、例えば、同じコドン使用頻度係数、または他方のポリヌクレオチドのそれの0.1以内、好ましくは、0.05以内のコドン使用頻度係数を有するポリヌクレオチドを意味する。
【0064】
HCVポリヌクレオチド配列は、様々なHCV遺伝子型、株もしくは分離物のいずれに由来するものでもよい。HCV分離物は、1以上のサブタイプを含む以下の6つの主要遺伝子型:HCV 1(1a、1bもしくは1c)、HCV 2(2a、2bもしくは2c)、HCV 3(3a、3b、10a)、HCV 4(4a)、HCV 5(5a)ならびにHCV 6(6a, 6b, 7b, 8b, 9aおよび11a)に分類することができる(Simmonds, J. Gen. Virol., 2001, 693−712)。本発明に関して、各HCVタンパク質は、同じHCV遺伝子型またはサブタイプ、あるいは、HCV遺伝子型またはサブタイプのいずれかの組合せに由来するものでよく、さらには、HCVタンパク質を用いてもよい。遺伝子は、1b遺伝子型、例えば、感染性クローンJ4L6(登録番号AF0542478、図1を参照)に由来するのが好ましい。
【0065】
配列決定されている具体的株として、HCV−J(Katoら、1990, PNAS, USA, 87;9724−9528)およびBK(Takamizawaら、1991, J.Virol. 65:1105−1113)が挙げられる。
【0066】
本発明のポリヌクレオチドは、コードされたタンパク質の発現(in vitro、in vivoもしくはex vivoのいずれで起こってもよい)による生産に有益である。従って、上記ヌクレオチドは、例えば、収率を高めるための組換えタンパク質合成に関与する、あるいは、DNAワクチン接種法に使用される治療剤として独立に用いることも可能である。本発明のポリヌクレオチドをin vitroもしくはex vivoでコードしたタンパク質の生産に用いる場合には、例えば、細胞培養物中の細胞を改変して、発現させようとするポリヌクレオチドを含むようにする。このような細胞として、一過性の、または好ましくは安定な哺乳動物細胞系が挙げられる。本発明のポリタンパク質をコードするベクターの挿入により改変することができる細胞の特定例としては、HEK293T、CHO、HeLa、293およびCOS細胞が挙げられる。選択する細胞系は、安定しているだけでなく、ポリタンパク質の成熟グリコシル化と細胞表面発現を可能にするものが好ましい。発現は、形質転換した卵母細胞で達成することができる。ポリペプチドは、トランスジェニック非ヒト動物、好ましくはマウスの細胞において、本発明のポリヌクレオチドから発現させることができる。本発明のポリヌクレオチドからポリペプチドを発現させるトランスジェニック非ヒト動物も本発明の範囲に含まれる。
【0067】
本発明は、本発明のヌクレオチド配列を含む発現ベクターを包含する。このような発現ベクターは、分子生物学の分野では日常的に構築されており、例えば、プラスミドDNAと好適な開始剤、プロモーター、エンハンサーおよびその他のエレメント(例えば、タンパク質発現に必要で、そのために正しい配向で配置されるポリアデニル化シグナルなど)を含むことができる。その他の好適なベクターは、当業者には明らかであろう。これに関するさらに別の例については、Sambrook ら、Molecular Cloning: a Laboratory Manual. 2nd Edition. CSH Laboratory Press.(1989)を参照されたい。
【0068】
好ましくは、本発明のポリヌクレオチド、またはベクターにおいて本発明で使用するためのポリヌクレオチドは、宿主細胞によるコード配列の発現を達成することが可能な制御配列に機能しうる形で連結されている。すなわち、このベクターは発現ベクターである。「機能しうる形で連結されている」という用語は、記載した要素が、意図される様式でそれらが機能できるような関係にある並置を意味する。コード配列に「機能しうる形で連結された」調節配列(例えば、プロモーターなど)は、調節配列と適合する条件下でコード配列の発現が達成されるように配置される。
【0069】
発現カセットは、目的の配列または遺伝子の発現を指令することができる集合体である。発現カセットは、制御エレメント、例えば、目的の遺伝子に機能しうる形で連結されたプロモーターなどを含む。
【0070】
上記ベクターは、例えば、複製起点を備えるプラスミド、人工染色体(例:BAC、PAC、YAC)、ウイルスもしくはファージベクターでよく、随意に、ポリヌクレオチドの発現用のプロモーター、また随意にプロモーターのレギュレータでもよい。ベクターは、1つ以上の選択マーカー遺伝子を含み、例えば、細菌プラスミドの場合には、アンピシリンまたはカナマイシン耐性遺伝子を、あるいは、真菌ベクターの場合には耐性遺伝子を含む。ベクターをin vitroで用いて、例えば、DNAまたはRNAを産生することもできるし、あるいはベクターを用いて宿主細胞、例えば、哺乳動物宿主細胞をトランスフェクションまたは形質転換することにより、例えば、ベクターがコードしたタンパク質を生産することも可能である。また、ベクターを改変して、DNAワクチン接種法または遺伝子治療法にin vivoで用いることも可能である。
【0071】
プロモーターおよびその他の発現調節シグナルは、発現が設計される宿主細胞と適合するように選択することができる。例えば、哺乳動物プロモーターとして、カドミウムのような重金属に応答して誘導することができるメタロチオネインプロモーター、およびβアクチンプロモーターが挙げられる。また、SV40ラージT抗原プロモーター、ヒトサイトメガロウイルス(CMV)即時型(IE)プロモーター、ラウス肉腫ウイルスLTRプロモーター、アデノウイルスプロモーター、もしくはHPVプロモーター、特にHPV上流調節領域(URR)を用いてもよい。これらプロモーターはすべて周知であり、当分野において容易に入手可能である。
【0072】
好適なウイルスベクターの例として、単純ヘルペスウイルスベクター、ワクシニアもしくはαウイルスベクターおよびレトロウイルス、例えば、レンチウイルス、アデノウイルスおよびアデノ随伴ウイルスが挙げられる。これらのウイルスを用いた遺伝子移入の方法は当業者には知られている。例えば、レトロウイルスベクターを用いて、本発明のポリヌクレオチドを宿主ゲノムに安定に組み込むことができるが、このような組換えは好ましくない。対照的に、複製能欠損アデノウイルスベクターは、エピソームのままであるため、一過的発現を可能にする。例えば、サブユニットワクチンとして、または免疫アッセイで使用するために、昆虫細胞(例えば、バキュロウイルスベクター)、ヒト細胞もしくは細菌での発現を駆動することができるベクターを用いて、本発明のポリヌクレオチドがコードした多量のHCVタンパク質を生産することができる。
【0073】
別の態様では、本発明は、本明細書に記載したポリヌクレオチドを含む医薬組成物を提供する。好ましくは、この組成物は、本発明の第2の態様に従うDNAベクターを含む。好ましい実施形態では、上記組成物は、HCVアミノ酸配列をコードするポリヌクレオチド配列をコードするベクターを含むDNAでコーティングした複数の粒子、好ましくは金粒子を含み、上記ポリヌクレオチド配列のコドン使用頻度パターンは、高度に発現させた哺乳動物遺伝子、特にヒト遺伝子のパターンに類似している。別の実施形態では、前記組成物は、医薬的に許容されうる賦形剤と、本発明の第2の態様のDNAベクターを含む。前記組成物はまた、アジュバントも含む。
【0074】
DNAワクチンは、液体ワクチンの筋肉への間隙投与(WO90/11092)または筋内注射以外の機構により送達することができる。例えば、皮膚への送達は、免疫機構が、感染に対する障壁となる組織(例えば、皮膚や粘膜など)において極めて活発であることから有利である。皮膚への送達は、注射、ジェット式注射器(皮膚、または筋肉などの下層組織に高圧下で液体を強制的に送り出す)もしくはパーティクルボンバードメント(この場合、上皮を貫通するのに十分な密度の粒子にDNAをコーティングしうる)(米国特許第5371015号)により実施することができる。例えば、Haynesら、J. Biotechnology 44: 37−42(1996)に記載されているように、ヌクレオチド配列をプラスミドに組み込み、これを金ビーズにコーティングした後、これらビーズを高圧下で表皮に投与する。皮膚へのこれら粒子の投入により、表皮細胞および表皮ランゲルハンス(Langerhan)細胞両方の直接トランスフェクションが達成される。ランゲルハンス細胞は、抗原提示細胞(APC)であり、DNAを取り込み、コードされたペプチドを発現し、これらをプロセッシングして細胞表面MHCタンパク質に展示する。トランスフェクションされたランゲルハンス細胞は、リンパ節に移動し、そこで、展示された抗原断片をリンパ球に提示して、免疫応答を引き起こす。粒子が媒介する皮膚への送達により免疫応答を誘発するには、非常に少量のDNA(1μg以下、往々にして0.5μg以下)でよいが、これは、直接筋内注射後に免疫応答を生じさせる場合、ミリグラム量のDNAを必要とすることが知られているのとは対照的である。
【0075】
本発明のポリヌクレオチドを治療薬、例えば、DNAワクチン接種に使用する場合には、ワクチン接種しようとする哺乳動物、例えば、ヒトに核酸を投与する。RNAまたはDNA、好ましくはDNAのような核酸は、前記のようなベクターの形態で提供され、これを該哺乳動物の細胞に発現させることができる。ポリヌクレオチドは、利用可能な方法のどれで投与してもよい。例えば、核酸は、針注射、好ましくは、皮内、皮下もしくは筋内注射により投与する。あるいは、粒子媒介DNA送達(PMDD)のような核酸送達装置を用いて、核酸を皮膚内に直接送達することもできる。この方法では、不活性粒子(金ビーズなど)を核酸でコーティングし、例えば、投射装置からの高圧下放出により、粒子が受容体(例:皮膚)の表面を貫通するのに十分な速度で加速する。(本発明の核酸分子でコーティングした粒子は、このような粒子を装填した送達装置と同様に、本発明の範囲に含まれる)。組成物は、0.5〜5μm、好ましくは約2μmの平均直径を有する金粒子を含むのが望ましい。好ましい実施形態では、コーティングした金ビーズをカートリッジとして機能する管に装填し、各カートリッジが、1カートリッジ当たり0.1〜5μg、好ましくは約0.5μgのDNAをコーティングした0.1〜1mg、好ましくは約0.1 mgの金を含むようにする。
【0076】
本発明の別の態様によれば、本明細書に記載したようなポリヌクレオチド配列を含む宿主細胞が提供される。宿主細胞は、細菌、例えば、大腸菌、哺乳動物、例えば、ヒトでもよいし、あるいは昆虫細胞でもよい。本発明のベクターを含む哺乳動物細胞は、in vitroでトランスフェクションした培養細胞でもよいし、あるいは、哺乳動物へのベクターの投与により哺乳動物細胞をin vivoでトランスフェクションすることもできる。
【0077】
別の態様では、本発明は、前記のような医薬組成物を製造する方法であって、野生型HCVヌクレオチド配列のコドン使用頻度パターンを変更するか、またはポリヌクレオチド配列を合成により形成して、高度発現哺乳動物遺伝子に類似したコドン使用頻度パターンを有し、かつ野生型HCVアミノ酸配列、もしくはポリペプチドの1以上の天然の機能を不活性化するのに十分なアミノ酸変化を有する野生型配列を含む突然変異HCVアミノ酸配列、をコードする配列を製造するステップを含む、上記方法を提供する。
【0078】
また、HCV感染の治療または予防における、本明細書に記載したポリヌクレオチドまたはワクチンの使用も提供する。
【0079】
患者に裸のポリヌクレオチドまたはベクターを導入する好適な方法として、好適なビヒクルを用いた局所適用がある。例えば、鼻内、経口、膣内もしくは直腸内投与により、核酸を皮膚、または粘膜表面に局所投与してもよい。裸のポリヌクレオチドまたはベクターは、リン酸緩衝食塩水(PBS)のような医薬的に許容されうる賦形剤と一緒に存在してもよい。DNA取込みは、ブピバカインのような促進剤を別々に、またはDNA製剤に含有させて用いることにより、さらに容易にすることができる。核酸を受容者に直接投与する別の方法としては、超音波、電気刺激、エレクトロポレーションおよびマイクロシーディング(米国特許第5,697,901号に記載されている)が挙げられる。
【0080】
核酸構築物の取込みは、複数の周知のトランスフェクション法、例えば、トランスフェクション剤の使用を含む方法により増強することができる。これらの薬剤の例としては、カチオン剤、例えば、リン酸カルシウムおよびDEAE−デキストラン、ならびに、リポフェクタント、例えば、リポフェクタムおよびトランスフェクタムが挙げられる。投与しようとする核酸の用量は変更することができる。典型的に核酸は、粒子媒介遺伝子送達の場合には、1pg〜1mg、好ましくは1pg〜10μg核酸の範囲、またその他の経路の場合には、10μg〜1mgの範囲の量を投与する。
【0081】
また、本発明の核酸配列は、遺伝子治療に有用な専用送達ベクターにより投与することもできる。遺伝子治療法については、例えば、Vermeら、Nature 1997, 389:239−242に記載されている。ウイルスおよび非ウイルスベクター系のいずれを用いてもよい。ウイルス系としては、レトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、カナリア痘およびワクシニア−ウイルスベースの系が挙げられる。好ましいアデノウイルスベクターは、ヒト以外の霊長類に由来するものである。特に、米国特許第6083716号に記載されているPan 9(C68)、WO03/046124に記載されているPan5、6 もしくは7が挙げられる。
【0082】
非ウイルスベースの系としては、核酸、ミクロスフィアカプセル化法(ポリ(ラクチド−コ−グリコリド)と、リポソームベースの系の直接投与が挙げられる。最初のワクチン接種後にブースター注射を実施するのが望ましい場合には、ウイルスおよび非ウイルス送達系を組み合わせてもよく、例えば、プラスミドのような非ウイルスベクターを用いた最初の「初回感作(プライム)」DNAワクチン接種の後、ウイルスベクターまたは非ウイルスベースの系を用いた1回以上の「追加免疫(ブースト)」ワクチン接種を行なう。プライム−ブーストプロトコルは、アジュバント中のタンパク質で初回感作し、本発明のポリヌクレオチドをコードするDNAまたはウイルスベクターで追加免疫するという利点もある。あるいは、タンパク質を基剤とするワクチンをブースターとして用いてもよい。タンパク質ワクチンは、DNA/ウイルスベクターのワクチンが含むすべての抗原を含むのが好ましい。しかし、タンパク質は個別の形態でも、ポリタンパク質の形態をしていてもよい。
【0083】
本発明の核酸配列は、形質転換した細胞により投与することもできる。このような細胞として、被験者から採取した細胞がある。本発明の裸のポリヌクレオチドまたはベクターをこのような細胞にin vitroで導入し、その後、形質転換した細胞を被験者に戻すことができる。本発明のポリヌクレオチドは、相同組換え事象により細胞にすでに存在する核酸に組み込むことができる。所望であれば、形質転換した細胞をin vitroで増殖させ、こうして得られた1個以上の細胞を本発明で用いてもよい。公知の手術またはマイクロサージャリー法(例:移植、マイクロインジェクションなど)により患者の適切な部位に細胞を送達することができる。
【0084】
好適な細胞としては、抗原提示細胞(APC)があり、例えば、樹状細胞、マクロファージ、B細胞、単球、ならびに有効なAPCに工学的に操作されてもよいその他の細胞が挙げられる。このような細胞は、遺伝子的に改変することにより、抗原提示能力を増大する、T細胞応答の活性化および/または維持を改善する、それ自体で抗腫瘍、例えば抗子宮頸部癌効果を有する、ならびに/あるいは受容体(すなわち、対合HLAハプロタイプ)と免疫学的に適合性となるようにすることもできるが、必ずしも必要なわけではない。APCは一般に、生物由来の多様な体液および器官(腫瘍および腫瘍周辺組織を含む)のいずれから単離されてもよく、オートロガス、同種異系、同系もしくは異種細胞のいずれでもよい。
【0085】
本発明の特定の好ましい実施形態では、樹状細胞またはその前駆細胞を抗原提示細胞として用い、in vitroで形質転換させてから患者に戻すか、あるいは、例えば、粒子媒介DNA送達により、ワクチン中の被送達ヌクレオチドのin vivo標的として用いる。樹状細胞は高い能力を有するAPCであり(Banchereau およびSteinman, Nature 392:245−251, 1998)、予防または治療のための抗腫瘍免疫を誘発する生理学的アジュバントとして有効であることが示されている(TimmermanおよびLevy, Ann. Rev. Med. 50:507−529, 1999)。一般に、樹状細胞は、その典型的な形状(in situで星形、in virto視覚化可能な顕著な細胞質突起(樹状突起)を有する)と、高い効率で抗原を取り込み、プロセッシングおよび提示する能力、ナイーブT細胞応答を活性化する能力に基づいて、確認することができる。勿論、樹状細胞を工学的に操作して、in vivoまたはex vivoで樹状細胞に通常認められない特定の細胞−表面受容体またはリガンド、例えば、本発明の構築物にコードされた抗原を発現させることも可能であり、このような改変された樹状細胞も本発明によって意図される。樹状細胞に代わるものとして、分泌小胞抗原負荷樹状細胞(secreted vesicles antigen−loaded dendritic cells)(エキソソームと称する)をワクチンの中で用いてもよい(Zitvogelら, Nature Med. 4:594−600, 1998参照のこと)。
【0086】
樹状細胞および前駆細胞は、末梢血、骨髄、腫瘍浸潤細胞、腫瘍周辺組織−浸潤細胞、リンパ節、脾臓、皮膚、臍帯血もしくはその他の好適な組織または体液から取得することができる。例えば、樹状細胞は、GM−CSF、IL−4、IL−13および/またはTNFのようなサイトカインの組合せを末梢血液から収集した単球の培養物に添加することにより、ex vivoで分化させることができる。あるいは、GM−CSF、IL−3、TNF、CD40リガンド、リポ多糖LPS、flt3リガンド(プロフェショナル抗原提示細胞、特に樹状細胞の産生に重要なサイトカイン)および/または樹状細胞の分化、成熟および増殖を誘発するその他の化合物の組合せを培養培地に添加することにより、末梢血、臍帯血もしくは骨髄から収集したCD34陽性細胞を樹状細胞に分化させることができる。
【0087】
APCは一般に、抗原のHCVアミノ酸配列をコードするポリヌクレオチド、例えば、本発明で考慮されるコドン最適化ポリヌクレオチドでトランスフェクションすることもできる。このようなトランスフェクションは、ex vivoで行うことができ、トランスフェクションしたそのような細胞を含む組成物またはワクチンは、本明細書に記載するように、治療の目的で用いることができる。あるいは、樹状、またはその他の抗原提示細胞をターゲッティングする遺伝子送達ビヒクルを患者に投与し、in vivoでトランスフェクションを起こすことも可能である。樹状細胞のin vivoおよびex vivoトランスフェクションは、例えば、WO 97/24447に記載されているような当分野では公知の方法のいずれか、またはMahviら、Immunology and cell Biology 75:456−460, 1997に記載された粒子媒介方法を用いて、一般に実施することができる。
【0088】
本発明のワクチンおよび医薬組成物は、抗ウイルス剤、例えば、αインターフェロン、好ましくは、PEG化αインターフェロンおよびリボバリンと組合せて用いてもよい。ワクチンおよび医薬組成物は、単位用量または複数回用量の容器、例えば、密封したアンプルまたはバイアルに収容することができる。このような容器は、気密シールすることにより、使用まで製剤の滅菌を保存するのが好ましい。一般に、製剤は、油性または水性ビヒクル中の懸濁液、溶液もしくは乳濁液として保存することができる。これ以外にも、凍結乾燥した状態でワクチンまたは医薬組成物を保存し、使用の直前に滅菌液体担体を添加するだけにしてもよい。粒子媒介送達による投与を意図する場合のヌクレオチド配列含有ワクチンは、圧縮ガス送達装置に用いるのに適したカートリッジの形態をしていてもよく、その際、カートリッジは、中空の管からなり、その内面は、随意に医薬的に許容されうる他の成分が存在してもよいワクチンヌクレオチド配列を担持する粒子でコーティングされている。
【0089】
本発明の医薬組成物は、アジュバント化合物、もしくはDNAによりコードされるタンパク質により誘発される免疫応答をモジュレートまたは増強するのに役立つその他の物質を含んでもよい。これらは、抗原とは別に、または抗原との融合物としてDNAによりコードされるか、あるいは、製剤の非DNAエレメントとして含有させてもよい。本発明の製剤に含有させることができるアジュバントタイプの物質の例として、ユビキチン、リソソーム結合膜タンパク質(LAMP)、B型肝炎ウイルスコア抗原、flt3−リガンドおよびその他のサイトカイン(例:IFN−γおよびGMCSF)が挙げられる。
【0090】
上記以外の好適なアジュバントは、以下に挙げる市販のもの、例えば、フロイント不完全アジュバントおよび完全アジュバント(Difco Laboratories, デトロイト、MI);イミキモド(Imiquimod)(3M, St. Paul, MN);レシミキモド(3M, セントポール、MN);メルクアジュバント(Merck Adjuvant)65(Merck and Company, Inc., ローウェイ、NJ);アルミニウム塩、例えば、水酸化アルミニウムゲル(alum)またはリン酸アルミニウム;カルシウム、鉄もしくは亜鉛の塩;アシル化チロシンの不溶性懸濁液;アシル化糖;カチオンまたはアニオン的に誘導体化した多糖;ポリホスファゼン;生物分解性ミクロスフェア;モノホスホリル脂質AおよびクイルAである。サイトカイン、例えば、GM−CSFもしくはインターロイキン−2、−7もしくは−12もアジュバントとして用いることができる。
【0091】
本発明の製剤では、アジュバント組成物が、主にTh1型の免疫応答を誘発するのが好ましい。従って、アジュバントは、DNAがコードした抗原に応答して生じた免疫応答を主にTh2型から主にTh1型の応答へとモジュレートする上で役立つものである。高レベルのTh−1型サイトカイン(例:IFN−、TFN、IL−2およびIL−12)は、投与された抗原に対する細胞媒介免疫応答の誘発を有利にする傾向がある。応答が主にTh1型である好ましい実施形態では、Th1型サイトカインのレベルは、Th2型サイトカインのレベルより大幅に増大するであろう。これらのサイトカインのレベルは標準的アッセイを用いて、容易に評価することができる。サイトカインのファミリーについては、Mosmann and Coffman, Ann. Rev. Immunol. 7:145−173, 1989を参照されたい。
【0092】
従って、Th1型が優勢な応答を誘発するための使用に適したアジュバントとしては、例えば、モノホスホリル脂質A、好ましくは3−デ−O−アシル化モノホスホリル脂質A(3D−MPL)とアルミニウム塩との組合せが挙げられる。Th1型免疫応答を優先的に誘発するその他の公知のアジュバントとして、CpG含有オリゴヌクレオチドがある。このオリゴヌクレオチドは、CpGジヌクレオチドがメチル化されていないことを特徴とする。このようなオリゴヌクレオチドはよく知られており、例えば、WO96/02555に記載されている。免疫刺激性DNA配列は、例えば、Satoら、Science 273:352, 1996にも記載されている。CpG含有オリゴヌクレオチドは、同じまたは異なるポリヌクレオチド構築物におけるHCV抗原から個別にコードされたものでもよいし、もしくは、例えば、該抗原との融合物として、それらにすぐ隣接していてもよい。あるいはまた、CpG含有オリゴヌクレオチドを個別に、すなわち、コードされた抗原を含む組成物の一部としてではなく、投与してもよい。CpGオリゴヌクレオチドは、単独でも、他のアジュバントと組み合わせて用いてもよい。例えば、増強された系は、CpG含有オリゴヌクレオチドとサポニン誘導体の組合せ、特に、WO 00/09159およびWO 00/62800に記載されているようなCpGとQS21の組合せを含む。この製剤は、さらに水中油形乳濁液および/またはトコフェロールを含むのが好ましい。
【0093】
別の好ましいアジュバントは、サポニン、好ましくはQS21(Aquila Biopharmaceuticals Inc., フラミンガム、MA)であり、これは単独でも、他のアジュバントと組み合わせて用いてもよい。例えば、増強された系は、モノホスホリル脂質Aとサポニン誘導体の組合せ、例えば、WO 94/00153に記載されているQS21と3D−MPLの組合せ、またはWO 96/33739に記載されているように、QS21をコレステロールでクエンチングした、反応性(reactogenic)の低い組成物を含む。別の好ましい製剤は、水中油形乳濁液およびトコフェロールを含む。QS21、3D−MPLおよび水中油形乳濁液中のトコフェロールを含む、特に効力のあるアジュバント製剤については、WO 95/17210に記載されている。
【0094】
別の好ましいアジュバントとして、モンタニドISA 720(Seppic、フランス)、SAF(Chiron、カリフォルニア、米国)、ISCOMS(CSL)、MF−59(Chiron)、デトクス(Detox)(Ribi、ハミルトン、MT)、RC−529(Corixa、ハミルトン、MT)ならびに他のアミノアルキルグルコサミニド4−ホスフェート(AGP)が挙げられる。
【0095】
ワクチンがアジュバントを含む場合、ワクチン製剤を2部に分けて投与してもよい。例えば、抗原をコードするヌクレオチド構築物を含む製剤の部分を、例えば、皮下または筋内注射、あるいは経皮粒子媒介送達により最初に投与してから、アジュバントを含む製剤の部分をその直後、またはワクチン分野に精通した医師には明らかな適当な時間をおいて、投与することができる。このような状況で、アジュバントは、抗原製剤と同じ経路、または別の経路のいずれにより投与してもよい。別の実施形態では、製剤のアジュバント部分を抗原部分の前に投与する。一実施形態では、アジュバントは局所製剤として、抗原をコードするヌクレオチド配列の粒子媒介送達の部位で、その粒子媒介送達の前または後のいずれかに、皮膚に適用する。
【0096】
好ましくは、本発明のDNAワクチンは、有効な免疫応答、典型的には、HCV抗原に対するCD4+およびCD8+免疫を刺激する。好ましくは、広範囲のエピトープに対する免疫を刺激する。治療に用いる場合には、ワクチン接種の後、肝線維症および/または炎症を軽減するのが好ましい。
【0097】
本明細書で用いる「含む」という用語は、非制限的意味で用いることを意図しており、その他の要素の存在も排除されない。しかし、「含んでいる」という用語は、その排他的意味で理解できるように意図することもあり、その場合、「構成される」または「からなる」と同等である。本発明を以下の実施例により説明するが、これらに限定されるわけではない。
【実施例】
【0098】
実施例1 抗原パネルに導入した突然変異
1)コンセンサス突然変異
すべての既知HCV分離物の全ゲノム配列を比較した。J4L6ポリタンパク質内の特定の位置を、他の大部分のHCV分離物から、特異である/逸脱するものとして確認した。特に重要なのは、これらの位置が、1a、2、3群、およびその他の群(この群では、さもなくば、同等の位置に1または2個の別のアミノ酸残基が占めていた。)に及ぶ関連1b群の分離物全体のより一致性の高い残基から逸脱することが認められたことである。選択したコンセンサス突然変異で、既知CD4またはCD8エピトープを妨害するものはない。NS3における2つの変更により、実際に免疫優性HLA−B35制限CD8エピトープを修復する[イソロイシン(I)1365からバリン(V)、およびグリシン(G)1366からアラニン(A)]。
【0099】
不用な可変性のために、NS4Bの最初の51アミノ酸を除去した。
【0100】
コア:
アラニン(A)52からトレオニン(T)へ。
【0101】
NS3:
バリン(V)1040からロイシン(L)へ。
【0102】
ロイシン(L)1106からグルタミン(Q)へ。
【0103】
セリン(S)1124からトレオニン(T)へ。
【0104】
バリン(V)1179からイソロイシン(I)へ。
【0105】
トレオニン(T)1215からセリン(S)へ。
【0106】
グリシン(G)1289からアラニン(A)へ。
【0107】
セリン(S)1290からプロリン(P)へ。
【0108】
イソロイシン(I)1365からバリン(V)へ。
【0109】
グリシン(G)1366からアラニン(A)へ。
【0110】
トレオニン(T)1408からセリン(S)へ。
【0111】
プロリン(P)1428からトレオニン(T)へ。
【0112】
イソロイシン(I)1429からセリン(S)へ。
【0113】
イソロイシン(I)1636からトレオニン(T)へ。
【0114】
NS4B:
フェニルアラニン(F)1760でORFを開始。
【0115】
NS5B:
イソロイシン(I)2824からバリン(V)へ。
【0116】
トレオニン(T)2892からセリン(S)へ。
【0117】
トレオニン(T)2918からバリン(V)へ。
【0118】
N.B.:番号付けは、J4L6分離物のポリタンパク質における位置に従う。
【0119】
実施例2 プラスミドDNAワクチンの構築
HCVコア、NS3、切断型(truncated)NS4B、およびNS5Bをコードするポリヌクレオチド配列に対し、SynGene 2eソフトウエアを用い、哺乳動物コドン使用頻度についてコドン最適化を実施した。コドン使用頻度係数は、各ポリヌクレオチドについて0.7以上まで改善された。コドン最適化、酵素ノックアウト突然変異、ならびにコンセンサス突然変異を組み込んだ新しいポリヌクレオチド配列各々のセンスおよびアンチセンス鎖を40〜60ヌクレオチドの領域(20ヌクレオチドのオーバーラップを含む)に分割した。これらの領域を商業的に合成し、オリゴアッセンブリィPCR法によりポリヌクレオチドを製造した。
【0120】
クローニングに用いられるユニークな制限エンドヌクレアーゼ部位を例示する、各ポリヌクレオチドの外側正方向および逆方向PCRプライマーの概略を以下に記載する。
【化12】
【0121】
単一抗原をコードするポリヌクレオチドはすべて、Not IおよびBamHIユニーククローニング部位を介して哺乳動物発現ベクターp7313ieにクローン化した(図7参照)。
【0122】
コードされたポリタンパク質は、以下の通りであった(突然変異とコドン最適化を含む)。
【化13】
【化14】
【化15】
【化16】
【0123】
実施例3 免疫応答アッセイ
C57BLまたはBALB/cマウスは、p7313ベクターにおいて個別に発現させた4つのHCV抗原のWTまたはコドン最適化+突然変異形態で免疫した。1.0μg/カートリッジの標準用量を含むPMIDによりマウスを免疫し、21日目に追加免疫(追加免疫1)、49日目に再度追加免疫(追加免疫2)した。脾細胞を個々のマウスから採取し、様々なHCV抗原製剤を用いたELISPOTで再刺激した。IL2およびIFNγ応答の両方を測定した。免疫応答を測定するのに用いた試薬は、ミクロゲン(Mikrogen)、ワクシニア−コアおよびワクシニアNS3−5(社内の遺伝子型1b)からの精製HCVコア、NS3、NS4およびNS5B(遺伝子型1b)タンパク質であった。
【0124】
HCVコア:
WT全長(FL−1−191)または切断型(TR1−115)コアで免疫したC57BLマウスをHCVコアタンパク質で再刺激したところ、精製コアタンパク質を用いて良好な応答が観察された(図8)。
【0125】
HCV NS3:
p7313 WTと、PMIDを用いてコドン最適化したNS3とでマウスを免疫した。NS3タンパク質とワクシニア3−5の両方を用いたC57Blマウスにおいて、免疫および1回の追加免疫後、NS3に対する良好な応答が実証され、ELISPOTにより応答を読み出した。IL2およびIFNγ応答の両方を検出した。この実験では、野生型と、コドン最適化した(co + m)形態の構築物との間に有意な差は観察されなかった(図9)。しかし、一過的トランスフェクション後のin vitro発現には、野生型とコドン最適化構築物との間に差が認められた。DNA用量が低い構築物、または一次応答を比較する実験から、プラスミドの効力の差が明らかになると考えられる。
【0126】
HCV NS4B:
BALB/cマウスのPMID免疫後に、全長WT p7313 NS4Bに対する応答を観察した。NS4Bタンパク質およびワクシニア3−5のいずれかでin vitro再刺激後に、IL2およびIFNγ ELISPOT応答の両方を観察した(図10)。
【0127】
NS4Bタンパク質をN末端で切断することにより、高度可変領域を除去したが、N末端領域に対して利用可能な抗血清が産生されていたため、in vitroトランスフェクション実験後に、タンパク質の発現を検出することはできなかった。この領域の発現を確認するために、これをNS5Bタンパク質と融合させた。近年の実験から、NS4Bタンパク質とNS3−5ワクシニアを用いて、切断型NS4Bタンパク質(単独またはNS5Bとの融合物として)に対し免疫応答を検出できることが確認されている。WTおよびコドン最適化NS4Bに対して良好な応答を観察した。
【0128】
HCV NS5B:
WTおよびコドン最適化(co+M)配列を用いた免疫後に、PMID後のNS5Bに対する免疫応答を調べた。NS3タンパク質とワクシニア3−5の両方を用いたC57BLマウスにおいて、免疫および1回の追加免疫後、NS5Bに対する良好な応答が実証され、ELISPOTにより応答を読み出した。NS3を用いた場合と同様、この実験でも、WTとco+m形態の構築物との間に、免疫応答の差は認められなかった(図11)。
【0129】
実施例4 HCVポリタンパク質の発現
4つの選択したHCV抗原コア、NS3、NS4BおよびNS5Bをp7313ieにフォーマット化することにより、単一の融合ポリタンパク質として発現させた。これらの抗原を以下に示すように様々な構築物において異なる順序で発現させた。(4つの抗原の各々がアミノ末端の位置を順に占めるように、単一ポリタンパク質の発現をコードする構築パネルを設計し、この重要な位置で1つの抗原の存在が他の抗原に対し、発現のレベルを有意に改善するのか、あるいは低減させるのかをモニターした。)さらに、コアタンパク質を2つの断片、すなわち、コア66−191>1−65と105−191>1−104を介して再配置した、2つの構築物を製造した。
【化17】
【0130】
標準的な製造者のプロトコルに従い、リポフェクタミン2000トランスフェクション試薬(Invitrogen/Life Technologies)を用いて、標準化量のDNAをHEK 293T細胞にトランスフェクションした。トランスフェクションから24時間後に細胞を回収し、MOPSまたはMES既製バッファー(Invitrogen/Life Technologies)のいずれかで前形成したNuPAGE 4−12%Bis−Trisを用いて、ポリアクリルアミドゲル電気泳動を実施した。分離したタンパク質をPVDF膜にブロッティングし、NS5B全タンパク質に対して産生したウサギ抗血清を用いて、タンパク質発現をモニターした。二次プローブは、西洋ワサビペルオキシダーゼ(hrp)に結合させた抗ウサギ免疫グロブリン抗血清であり、続いて、ECL試薬(Amersham Biosciences)を用いて、化学発光検出を実施した。
【0131】
この発現実験の結果を図12に示す。これらの結果から、ポリタンパク質はすべて、同様の程度まで発現するが、NS5Bを発現する単一抗原に認められるものよりは低いレベルである。HCV500の分子量がやや低いのは、N末端位置からのHCVコアの切断のためである。HCV502は、クローニングの誤りによりこの実験では検出されなかった。別のクローンを用いた反復実験では、HCV502の発現レベルは他のポリタンパク質と類似していた。
【0132】
実施例5 HCVポリタンパク質に対する免疫応答の検出
ポリタンパク質の各々をコードするDNA(1μg)を含むPMIDによりC57BLマウスを免疫し、実施例4に記載したようにそれから3週間後に追加免疫した。ELISPOT、またはHCV抗原に対する細胞内サイトカインの産生を用いて、追加免疫から7日後に免疫応答をモニタリングした。
【0133】
HCV遺伝子産物に対するT細胞応答についてのELISPOTアッセイ:
脾細胞の調製:
追加免疫から7日後に、免疫動物から脾臓を取得した。脾臓をスライドガラスの間で粉砕して処理することにより、細胞懸濁液を生成した。塩化アンモニウム処理で赤血球を溶解させた後、細胞破片を除去することにより、脾細胞の微細な懸濁液を得た。ELISPOTアッセイに用いるため、マウスが一次免疫しか受けていない場合は4×106/ml、またマウスが追加免疫を受けている場合には2×106/mlの濃度で細胞をRPMI完全培地に再懸濁させた。
【0134】
ELISPOTアッセイ:
15μg/ml(PBS中)のラット抗マウスIFNγまたはラット抗マウスIL−2(Phamingen)でプレートを被覆した。プレートは+4℃で一晩被覆した。使用前に、プレートをPBSで3回洗浄した。4×105細胞/ウェルで脾細胞をプレートに添加した。組換えHCV抗原をミクロゲン(Mikrogen)から取得し、1μg/mlで用いた。1〜10μMの最終濃度でペプチドをアッセイに用いることにより、CD4またはCD8応答を測定した。これらのペプチドは、Genemed Synthesisから取得した。各ウェルの合計容量は200μlであった。抗原で刺激した細胞を含むプレートを、加湿した37℃のインキュベーター内で16時間インキュベートした。いくつかの実験では、NS3−5またはワクシニア野生型を発現する組換えワクシニアに感染させた細胞をELISPOTアッセイで抗原として用いた。
【0135】
ELISPOTアッセイプレートの展開:
水で1回(細胞の溶解を確実にするため1分浸ける)、次にPBSで3回洗浄することにより、プレートから細胞を除去した。ビオチン結合ラット抗マウスIFNγまたはIL−2(Phamingen)を1μg/mlでPBSに添加した。プレートを室温で攪拌しながら2時間インキュベートした。次に、プレートをPBSで3回洗浄した後、ストレプトアビジンアルカリホスファターゼ(Caltag)を1/1000希釈で添加した。PBSで3回の洗浄後、BCICP基質(Biorad)を用いた15〜45分のインキュベーションにより、スポットが明らかになった。水を用いて基質を洗浄してから、プレートを乾燥させた。イメージ分析装置を用いて、スポットを数えた。
【0136】
ペプチド刺激に応答するT細胞からのIFNγおよびIL−2産生を検出するためのフローサイトメトリー:
試験管1本当たり約3×106個の脾細胞に等分し、回転することによりペレットとした。上清を除去し、サンプルを激しく攪拌し、ペレットを壊した。0.5μgの抗CD28+0.5μgの抗CD49d(Phamingen)を各試験管に添加し、室温で10分放置することによりインキュベートした。適切な試験管に1mlの培地を添加したが、これは培地だけか、もしくはHCV抗原を含む培地であった。次にサンプルを加熱水浴中に37℃で1時間インキュベートした。10μg/mlのブレフェルジンAを各試験管に添加し、37℃でのインキュベーションをさらに5時間継続した。プログラム化した水浴を6℃に戻し、その温度で一晩維持した。
【0137】
抗マウスCD4−CyChrome(Phamingen)と抗マウスCD8ビオチン(Immnotech)でサンプルを染色した。サンプルを洗浄し、ストレプトアビジン−ECDで染色した。サンプルを洗浄し、「イントラプレップ膜透過試薬(Intraprep Permeabilization Reagent)」キット(Immnotech)から100μgの固定液を15分室温で添加した。洗浄後、イントラプレップキットからの100μgの膜透過試薬を、抗IFN−γ−PE+抗IL−2−FITCを含む各サンプルに添加した。サンプルを室温で15分インキュベートした後、洗浄した。サンプルを0.5 mlバッファー中に再懸濁させ、フローサイトメーターで分析した。
【0138】
1サンプル当たり合計500,000個の細胞を収集してから、CD4およびCD8細胞をゲーティングすることにより、刺激に応答してIFN−γおよび/またはIL−2を分泌する細胞の集団を測定した。
【0139】
これらの結果から、様々な順序でコア、NS3、NS4BおよびNS5Bをコードするポリタンパク質はすべてNS3(すなわち、HCV 500、510、520、530)に対する免疫応答を刺激できることがわかった。IL2(図13A)およびIFN−γ(図13B)ELISPOTによりモニタリングしたところ、NS3タンパク質に対する応答は、HCVポリタンパク質(HCV 500、510、520および530)の各々で類似していた。
【0140】
応答細胞の表現型をICSによりさらに詳細に分析した。良好なCD4+T細胞応答が免疫優性NS3 CD4特異的ペプチドに対して誘発され、これは、HCV 500、510、520および530間で類似していた。
【表1】
【0141】
抗原の存在または非存在下で6時間、最後の4時間はブレフェルジンAの存在下で、脾細胞の刺激後、IFN−γ特異的なT細胞応答が検出された。CD4またはCD8T細胞上でのゲーティングおよびIFNγ FITCでの染色により、IFNgが検出された。
【0142】
また、HCV 500、510、520および530での免疫後、免疫優性NS3特異的ペプチドに対する強いCD8応答が生じ、CD8+細胞の2.5〜6%の頻度に達した。
【0143】
HCV 500、510、520および530での免疫によって、NS4BおよびNS5B抗原の両方に対するCD4およびCD8応答も検出されたが、CD8応答は、後の単一抗原での免疫後に比べ、ポリタンパク質に対して弱かった。
【表2】
【0144】
抗原の存在または非存在下で6時間、最後の4時間はブレフェルジンAの存在下で、脾細胞の刺激後、IFN−γ特異的なT細胞応答が検出された。CD4またはCD8T細胞上でのゲーティングおよびIFNγ FITCでの染色により、IFNgが検出された。
【表3】
【0145】
抗原の存在または非存在下で6時間、最後の4時間はブレフェルジンAの存在下で、脾細胞の刺激後、IFN−γ特異的なT細胞応答が検出された。CD4またはCD8T細胞上でのゲーティングおよびIFNγ FITCでの染色により、IFNgが検出された。
【0146】
用いたペプチドは以下の配列を有する。
【化18】
【0147】
内因的にプロセッシングされた抗原の認識:
HCVポリタンパク質でのPMID免疫が、内因的にプロセッシングされた抗原を認識することができる応答を誘発したか否かを決定するために、NS3−5を発現するワクシニア組換えウイルスに感染させた標的細胞をELISPOTアッセイで刺激因子として用いた。結果から、500、510、520および530での免疫後、良好なIL2およびIFNγ ELISPOT応答が検出されたことがわかる(図14)。
【0148】
HCVポリタンパク質での免疫は機能的CTL活性を誘導する:
NS3だけ、HCV500、510および520をコードする0.01μgのDNAでC57BLマウスを免疫した。初回免疫および1回の追加免疫後、各グループからの脾細胞をNS3 CD8ペプチドおよびIL2で5日間in vitroで再刺激した。同じペプチドを用いてパルスしたEL4細胞に対するCTL活性を測定した。すべての構築物で免疫したマウスが、このアッセイにおいて同様の死滅レベルを示した。
【0149】
このことから、HCVポリタンパク質によるPMID免疫が機能的CD8応答を誘発できることがわかる。結果を図15に示す。
【0150】
実施例6 二重プロモーター構築物を介したHCV抗原の送達
以下に示す方法を用いて、二重プロモーター構築物を作製した。発現カセット1(アイオワ−レングス(Iowa−length)CMVプロモーター、エクソン1、目的のタンパク質/融合タンパク質をコードする遺伝子、プラス、ウサギグロビンポリ−Aシグナル)を保有する断片をその宿主ベクター、すなわちp7313ieから、ユニーク制限エンドヌクレアーゼ部位ClaIおよびXmnIにより切除した。XmnIは、切除した断片の3−プライム(prime)末端に平滑末端を形成した。
【0151】
受容プラスミドベクターは、発現カセット2を含むp7313ieであった。これは、ユニーク制限エンドヌクレアーゼSse8387Iによる消化後、T4DNAポリメラーゼとインキュベートして、形成された3−プライムオーバーハングを除去することによって、線状分子の5−プライム末端および3−プライム末端の両方を平滑末端にすることにより、作製した。これは、259 bp断片を除去するユニーク制限エンドヌクレアーゼClaIを用いて切断された。
【0152】
ClaI/平滑適合性末端を介して、発現カセット1をp7313ie/発現カセット2にクローン化して、p7313ie/発現カセット1+発現カセット2を作製したが、その際、カセット1はカセット2の上流に位置した。
【0153】
以下のものを含むp7313ieプラスミドを作製した。
【化19】
【0154】
上に示した構築物パネルは完全であり、ウェスタンブロットにより、293T細胞における一過的トランスフェクションからの発現についてモニターした。ウェスタンブロット分析の結果を図16に示す:
レーンの説明:
1.p7313ie/コア
2.p7313ie/NS3
3.p7313ie/NS5B
4.p7313ie/コアNS3
5.p7313ie/NS4B5B
6.p7313ie/NS3コア
7.p7313ie/NS34B5B
8.p7313ie/コアNS3+NS4B5B
9.p7313ie/NS4B5B+コアNS3
10.p7313ie/NS3コア+NS4B5B
11.p7313ie/NS4B5B+NS3コア
12.p7313ie/コア+NS34B5B
13.p7313ie/NS34B5B+コア
構築物の各対は2つの独立した発現カセットを保有する。カセットをベクターに挿入する順序が、いずれかのカセットからの発現に影響を及ぼすとは予想しなかった。しかし、これらの結果から、NS4B5BまたはNS34B5B融合タンパク質の発現に関して、それぞれの発現カセットがコア、NS3コア、もしくはコアNS3カセットの下流に位置するとき、有意に不利であることがわかる。
【0155】
発現レベルは、単一抗原構築物ほど明確ではないが、大きさの有意な増大(175〜228%)のためにある程度の低減が予想され、これは、細胞に送達されるプラスミドのコピー数の減少(同じ質量のDNAに対して約50%以下の減少)を意味する。
【0156】
二重プロモーター構築物により誘発されるin vivo免疫原性:
免疫原性実験のために、4つの抗原すべての最大発現を呈示した3つの二重プロモーター構築物を選択した。これらは、p7313ie NS4B/NS5B+コア/NS3、p7313ie NS4B/NS5B+NS3コアおよびp7313ie NS3/NS4B/NS5B+コアであった。C57BLマウスをPMIDにより1μgのDNAで免疫し、7日後、IL2についてはELISPOTを用いて、優性NS3 CD8T細胞エピトープに対する応答を測定した。結果(図17に示す)から、単一免疫(CD4およびCd8 NS3T細胞特異的なペプチドで刺激した脾細胞)後、すべての二重プロモーター構築物に対して応答が観察されたことがわかる。
【0157】
実施例7 コアの欠失突然変異
コアのORFをコードする多数の遺伝子を、3’末端から1回につき20アミノ酸にわたる領域ずつ徐々に欠失させて作製し、完全に配列決定した。
【表4】
【0158】
図18は、コアの断片をコードする遺伝子の範囲を示すDNAアガロースゲルを表す。これらの構築物の発現を、NS4B5B 融合物(p7313ie/NS4B5B)の発現レベルへのそれらの作用と併せて、293T細胞での共トランスフェクションにより試験した。結果を図19に示す。レーンは以下のようにロードされた。
【表5】
【0159】
コア191、コアΔ15、コア171、コア151、およびコア131の発現は、NS4B5Bの発現の抗NS5B検出後、ウェスタンブロットを抗コアでプローブすると、はっきりと検出される。恐らく、用いたゲル系のサイズキャプチャー(size−capture)制限のために、コアの別の切断形態は検出されていない。
【0160】
結果から、コア191およびコアΔ15の存在下でのNS4B5Bの発現レベルに、両コア種の強い発現にもかかわらず、有意な低減(これは、コア171、そして再度コア151で回復する)が明らかである。この観察は、NS4B5Bで2回、NS3およびNS5Bで1回繰り返した。
【0161】
実施例8 NS3、NS5B、NS4B−NS5B融合物およびNS3−NS4B−NS5B三重融合物の発現に対するコアおよびコア151の影響
実験1−トランスフォーマットでの発現:
コアをトランスで発現させる、すなわち別々のプラスミド上にコードさせた場合に、非構造抗原の発現に対するコア191対コア151の発現の影響を比較してモニタリングするための実験を実施した。実験プロトコルは、実施例7に記載したのと同じであった。簡単にいうと、標準的プロトコル(Invitrogen/Life Technologies)におけるリポフェクタミン2000トランスフェクション試薬を用いて、各々0.5μgの2種のDNAプラスミドベクター(概略を以下の表に示す)をHEK 293T細胞中に共トランスフェクションした。(トランスフェクションおよびウェスタンブロット方法は、実施例4に記載の通り)。
【0162】
結果を図20に示すが、同図では、レーンを以下の表に記載したようにロードしており、ウェスタンブロット分析を実施することにより、主として抗NS3および抗NS5B抗血清を用いて非構造タンパク質の発現、および抗コアを含む同じブロットの二次プローブによりコアの発現を検出した。
【表6】
【0163】
すべてのケースで、コア151と一緒にトランスで産生された場合、非構造タンパク質または融合物(NS3、NS5B、NS4B−NS5B)の量は、コア191と一緒にトランスで発現させたときに産生されるレベルと比較して、有意に増加することが示された。
【0164】
実験2−シスフォーマットでの発現:
コアをシスで発現させる、すなわち、非構造要素と融合した同じプラスミド上にコードさせた場合に、非構造抗原の発現に対するコア191対コア151の発現の影響を比較してモニタリングするための実験を実施した。各ケースで、コア151は、指定した非構造領域とカルボキシ末端融合したコア191に置換した。
【0165】
標準的プロトコル(Invitrogen/Life Technologies)におけるリポフェクタミン2000トランスフェクション試薬を用いて、1μgのDNAプラスミドベクター(概略を以下の表に示す)をHEK 293T細胞中にトランスフェクションさせた(トランスフェクションおよびウェスタンブロット方法は、実施例4に記載の通り)。
【0166】
結果を図21に示す。ウェスタンブロット分析を実施することにより、ゲルAにおいて、主として抗NS3および抗NS5B抗血清を用いて非構造成分の発現、及び抗コアを含む同じブロットの二次プローブによりコアの発現を検出した。レーンは以下の表に記載するようにロードした。
【表7】
【0167】
結果から、抗原がポリタンパク質融合物の形態であるシス様式では、コアの切断により、融合タンパク質の発現が増大することがわかる。
【0168】
NS3に対する免疫応答に対するコア191およびコア151の影響の比較:
C57BLマウスをPMIDにより全DNA1.5μgの2回の注射で免疫した。免疫したグループは、空ベクターp7313ieだけ、p7313ieNS3、p7313ieNS5Bとp7313ieコア191またはp7313ieNS3、p7313ieNS5Bとp7313ieコア151による金ビーズの共コーティングを含んでいた。共コーティングを用いたのは、これによって、すべてのプラスミドが同じ細胞に送達され、この細胞が、前記のin vitro共トランスフェクション実験を模倣すると考えられるためである。初回免疫から14日後、IFNγおよびIL2抗原特異的な応答を測定するための細胞内サイトカイン染色を用いて、NS3からの優性CD8およびCD4T細胞エピトープに対する免疫応答を測定した。結果(図22に示す)から、CD4およびCD8 NS3応答は、コア191と比較してコア151の存在下で約2倍以上高くなったことがわかる。
【0169】
別の実験では、コア191またはコア151のいずれかと一緒に、p7313ieNS3/NS4B/NS5B 三重融合物を発現するプラスミドを共コーティングした金ビーズでC57BLマウスを免疫した。マウスをさらに同じ構築物で追加免疫し、追加免疫から7日後に、応答を測定するために細胞内サイトカイン染色を用いてNS3に対する応答をモニターした。図23に示した結果から、NS3抗原特異的CD4およびCD8応答は、コア191と比較して、コア151の存在下で約2倍高いことがわかる。
【0170】
コアの存在下でのNS3に対する応答を比較するin vivo実験全体は、FLコアと非構造タンパク質の同時送達により非構造抗原の発現が低減する可能性があり、これによって、構築物の免疫原性が低下するというin vitro発現データを支持する。この影響は、C末端40アミノ酸が除去された切断型コアを共コーティングすることにより少なくとも部分的に解消することができる。
【図面の簡単な説明】
【0171】
【図1−1】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−2】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−3】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−4】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−5】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図2】コドン最適化HCVコアポリヌクレオチド。
【図3】コドン最適化HCV NS3ポリヌクレオチド。
【図4】コドン最適化HCV NS4Bポリヌクレオチド。
【図5】コドン最適化HCV NS5Bポリヌクレオチド。
【図6−1】HCV J4L6ゲノム(野生型配列)の翻訳。
【図6−2】HCV J4L6ゲノム(野生型配列)の翻訳。
【図7】p7313−ieの構造。
【図8】コアに対する免疫応答。
【図9】NS3免疫原性。
【図10】NS4Bに対する免疫応答。
【図11】NS5Bに対する免疫応答。
【図12】抗HCV NS5Bを用いるタンパク質発現のモニタリング。
【図13】IL2(A)及びIFN−γ(B) ELISPOTによるNS3タンパク質に対する免疫応答。
【図14】NS3−5を発現するワクシニアウイルスに感染させた脾細胞を刺激因子とするIL2及びIFNγ ELISPOTアッセイ結果。
【図15】HCVポリタンパク質でのPMID免疫による機能的CD8応答の誘発。
【図16】293T細胞での一過的トランスフェクションからの発現を示すウエスタンブロット分析の結果。
【図17】二重プロモーター構築物により誘導されるNS3 T細胞応答の比較。
【図18】コアの断片をコードする遺伝子の範囲を示すDNAアガロースゲル。
【図19】293T細胞での共トランスフェクションとNS4B5B融合物の発現レベル。
【図20】293T細胞での共トランスフェクション後のNS3、NS5B、NS4B5B、およびNS34B5Bの発現に対するコアおよびコア151の影響。
【図21】293T細胞での一過的トランスフェクションにおけるコア151のコア191への置換後の融合タンパク質の発現に対する影響。
【図22】NS3からの優性CD8及びCD4T細胞エピトープに対する免疫応答。
【図23】NS3に対するCD4及びCD8応答。
【技術分野】
【0001】
本発明は、C型肝炎ウイルス(HCV)感染、ならびにそれに関連する症候および疾患の治療および予防に有用な方法および組成物に関する。特に、本発明は、HCVタンパク質をコードするポリヌクレオチド配列を含むDNAワクチン、ならびに、本発明のワクチンの投与を含む、HCVに感染した個体の治療方法に関する。
【背景技術】
【0002】
HCVは、輸血後および市中獲得型非A非B型肝炎の主要な原因として近年確認された。約1億7千万人がHCVに慢性的に感染しており、有病率は1〜10%である。有病率が1.8%の米国における医療費は20億ドルであると推定される。40〜60%の肝疾患はHCVによるものであり、30%の英国移植患者はHCV感染のためである。HCVは、初め無症状の感染であり、患者の90%以上が慢性疾患を発症する。この疾患過程は、典型的に、慢性活動性肝炎(70%)、線維症、肝硬変(40%)から肝細胞癌(60%)に移行する。感染から肝硬変までの期間の中央値は20年であり、肝細胞癌までは、20年である(Lauer G.およびWalker B. 2001, N. Engl J. Med 345, 41, Cohen J. 2001, Science 285 (5424) 26)。
【0003】
HCVの治療を改善することが強く求められる。現在、利用可能な小分子複製阻害剤は存在しない。リボビリンとPEG化インターフェロンからなる現在の最良の標準治療が、HCV感染治療の主軸を表している。しかし、現行の治療法が持続的な効果を達成する能力は依然として最適以下である(6ヶ月までの間、全体で50%有効率だが、遺伝子型1bの場合、有効率はそれより低い(27%))。この治療はまた、不快な副作用も伴う。これにより、特に、治療を始めて6ヶ月が過ぎると、有効率が急激に下降する。
【0004】
いくつかの研究から、個々のHVCタンパク質は正常なマウス(DNAによる免疫を受けたものを含む)において免疫原性であることがわかっている。数種のHCVワクチンが、予防または治療のいずれかを目的として現在臨床試験中である。最も進んでいるのは、E1またはE2エンベロープタンパク質を用いたChironおよびInnogeneticsによるものでフェーズ2にある。Transvaxによるエピトープワクチンもフェーズ2にある。また、各種の送達系(DNAを含む)による、コアおよび非構造抗原由来の配列を用いる数種のワクチンが、前臨床開発段階にある。
【0005】
HCVはフラビウイルス科のプラス鎖RNAウイルスであり、そのゲノムは長さが9.4 kbで、1つのオープンリーディングフレームを有する。HCVゲノムは、単一のポリタンパク質として翻訳され、これが、宿主およびウイルスプロテアーゼによりプロセシングされて、構造タンパク質(コア、エンベロープE1およびE2、ならびにp7)と、様々な酵素活性を有する6つの非構造タンパク質を産生する。1b遺伝子型の一例であるHCV J4L6分離物のゲノムは、登録番号AF054247(Yanagi, M., St Claire,M., Shapiro,M., Emerson,S.U., Purcell,R.H.およびBukh,J. “Transcripts of a chimeric cDNA clone of hepatitis C virus genotype 1b are infectious in vivo”. Virology 244 (1), 161−172 (1998))として見出されているが、これを図1に示す。
【0006】
エンベロープタンパク質によって、標的細胞の認識、これへの結合および進入が可能になる。ウイルス複製に関与する主要な非構造タンパク質としては、NS2(Zn依存性メタロプロテイナーゼ)、NS3(セリンプロテアーゼ/ヘリカーゼ)、NS4A(プロテアーゼ補因子)、NS4B、NS5AおよびNS5B(RNAポリメラーゼ)が挙げられる(Bartenschlager BおよびLohmann V. 2000. Replication of hepatitis C virus. J. Gen Virol 81, 1631)。
【0007】
HCVポリタンパク質の構造は、次のように示すことができる(数字は、各タンパク質の第1アミノ酸の位置を示す;J4L6分離物の完全ポリタンパク質は、長さが3010アミノ酸である)。
【化1】
【0008】
ウイルスの突然変異率は高く、保存および非保存領域のヌクレオチド配列に基づき、少なくとも6つの主要遺伝子型が明らかにされている。しかし、単一の患者から単離したHCVは、常に近縁のゲノムもしくは準種の混合物として提示されるため、さらなる不均質性が存在する。
【0009】
HCVゲノムは高度の遺伝子多様性を示し、これは、6つの主要な遺伝子型(1a、1b、2、3、4、5および6)に分類される。遺伝子型1a、1b、2および3は、欧州、北および南アメリカ、アジア、中国、日本およびオーストラリアで最も優勢である。遺伝子型4および5はアフリカで、また遺伝子型6は東南アジアでそれぞれ優勢である。
【0010】
HCV感染の治療法の改善と共に、多数のHCV遺伝子型を治療できる多様な能力の治療法も強く求められている。本発明の第1の態様では、様々な遺伝子型に対する保護が多様な新規ワクチン製剤を提供する。
【0011】
1種以上のHCVタンパク質をコードするポリヌクレオチドを含むHCVワクチンが記載されている。N3をコードするプラスミドDNAまたはセムリキフォレストウイルスベクターを含むワクチンがBrinsterらにより記載されている(2002, Journal of General Virology, 83, 369−381)。NS5BをコードするポリヌクレオチドワクチンがWO99/51781に開示されている。HCV E1、E1+E2融合物、NS5A およびNS5Bタンパク質をコードするコドン最適化遺伝子、ならびに、それを含むワクチンが、WO97/47358に記載されている。WO 01/04149は、コア、NS3、NS4もしくはNS5Aに由来する、HCVエピトープのモザイクをコードするポリペプチドまたはポリヌクレオチドを開示している。ワクチンに有用な融合タンパク質(NS3、NS4、NS5AおよびNS5Bを含む)、ならびにこのような融合タンパク質をコードするDNAがWO 01/30812に記載されている;随意に、上記融合タンパク質はコアタンパク質の断片を含むと言われている。WO 03/031588には、HCVタンパク質NS3−NS4A−NS4B−NS5A−NS5Bをコードするアデノウイルスベクターが記載され、これは、ワクチンとしての使用に適している。
【0012】
「非処理の」コアタンパク質と非構造タンパク質を含むポリペプチドを含むワクチンがWO 96/37606に記載されている。
【発明の開示】
【課題を解決するための手段】
【0013】
本発明は、HCVタンパク質であるコア、NS3、NS4BおよびNS5Bをコードするポリヌクレオチドワクチンの提供に関する。本発明のポリヌクレオチドワクチンは、NS4A HCVタンパク質および/またはNS5Aタンパク質をコードしない。また、本発明のポリヌクレオチドワクチンは、コア、NS3、NS4BおよびNS5B HCVタンパク質をコードし、それ以外のHCVタンパク質はコードしないのが好ましい。本発明はまた、医療における、および、HCV感染の治療または予防のための医薬品の製造における、前記抗原をコードするポリヌクレオチドワクチンの使用を提供する。
【0014】
本発明のワクチンに使用するポリヌクレオチド配列は、好ましくは、DNA配列である。
【0015】
HCVタンパク質をコードするポリヌクレオチドは、様々な組合せまたは立体配置のものでありうる。例えば、タンパク質は、個々のタンパク質、もしくは融合タンパク質として発現されてもよい。融合物(DNAまたはタンパク質レベルのいずれでもよい)の例として、NS4BおよびNS5Bのアミノ酸配列を含む、もしくはコードする単一ポリペプチドまたはポリヌクレオチドからなる二重融合物(NS4B−NS5B)、NS3−NS4B−NS5Bのアミノ酸配列を含む、もしくはコードする三重融合物、あるいは、本発明の4つの抗原すべての融合物(mCore−NS3−NS4B−NS5B)が挙げられる。
【0016】
本発明の好ましい融合物は、NS4BとNS5B(NS4B−NS5BまたはNS5B−NS4B);ならびに、コアとNS3(NS3−コアまたはコア−NS3)の二重融合物をコードするポリヌクレオチドである。好ましい三重融合物は、NS3−NS4B−NS5Bのアミノ酸配列をコードするポリヌクレオチドである。
【0017】
単一の抗原または融合タンパク質をコードする本発明のポリヌクレオチドは、単一のまたは複数の発現ベクター中に存在することができる。各抗原をコードするポリヌクレオチドが、同じ発現ベクターまたはプラスミド中に存在することが好ましい。これに関連して、HCVタンパク質をコードするポリヌクレオチドは、単一発現カセットに存在するか、もしくはシリーズ発現カセットに複数存在してもよい。
【0018】
他のHCVタンパク質の発現を最適化するために、HCVコアタンパク質をコードするポリヌクレオチドは、少なくとも1つの他のHCVタンパク質をコードするポリヌクレオチドを含む発現カセットの下流にある発現カセットに存在するのが好ましい。HCVコアタンパク質は、NS5Bをコードするポリヌクレオチドを含む発現カセットの下流にある発現カセットに存在するのが好ましい。
【0019】
本発明のオリゴヌクレオチドワクチンによりコードされるポリペプチドは、全長アミノ酸配列を含むか、あるいは、このポリペプチドは、全長タンパク質より短くてもよく、その際、該ポリペプチドは、短くなった遺伝子の発現産物が、全長タンパク質と交差反応する免疫応答を生じさせることを可能にするのに十分な比率の全長ポリヌクレオチド配列を含む。例えば、本発明のポリヌクレオチドは、本来の配列の領域が欠失した切断型HCVタンパク質であるHCVタンパク質の断片をコードすることができ、その際、最終断片は、本来の全長アミノ酸配列の90%以下を含むが、本来の配列の70%以下もしくは50%以下であってもよい。別の言い方をすれば、長さが少なくとも8、例えば、8〜10アミノ酸、あるいは長さが最高20、50、60、70、80、100、150もしくは200アミノ酸までの断片をコードするポリヌクレオチドは、コードされたオリゴまたはポリペプチドがHCV抗原性を示す限り、本発明の範囲内にあると考えられる。特に、しかし、限定するわけではないが、本発明のこの態様は、ポリヌクレオチドが完全なHCVタンパク質配列の断片をコードし、該タンパク質の1以上の別々のエピトープを呈示しうる場合の状況も含む。
【0020】
本発明の好ましいワクチンでは、HCVポリペプチドの少なくとも1つ、好ましいすべてを(末端)切断(truncation)または突然変異により不活性化させる。例えば、NS3のヘリカーゼおよびプロテアーゼ活性は、遺伝子の突然変異により低減または破壊するのが好ましい。発現させたポリペプチドのNS5Bポリメラーゼ活性は、突然変異により低減または破壊するのが好ましい。発現させたポリペプチドのNS4B活性は、突然変異により低減または破壊するのが好ましい。発現させたポリペプチドのコアタンパク質の活性は、切断または突然変異により低減または破壊するのが好ましい。この意味での突然変異には、上記ポリペプチドをコードするポリヌクレオチドに対する付加、欠失、置換もしくは転位事象が含まれる。これ以外にも、全長配列を2つ以上の別々の部分に発現させてもよい。
【0021】
HCVポリペプチドNS3およびNS5Bの機能構造および酵素機能については当分野で記載されている。
【0022】
NS5Bは、RNA依存性RNAポリメラーゼとして記載されている(Qinら、2001, Hepatology, 33, pp 728−737;Lohmannら、2000, Journal of Viral Hepatitis;Lohmannら、1997, Nov., Journal of Virology, 8416−8428;De Francescoら、2000, Seminars in Liver Disease, 20(1), 69−83)。NS5Bポリペプチドは、4つの機能性モチーフA、B、CおよびDを有するものとして記載されている。
【0023】
本発明のポリヌクレオチドワクチンによりコードされるNS5Bポリペプチドを突然変異させることにより、RNA依存性RNAポリメラーゼ活性を低減または除去するのが好ましい。好ましくは、上記ポリペプチドを突然変異させることにより、NS5BのモチーフAを破壊するが、例えば、2639位のアスパラギン酸(D)をグリシン(G)に置換する;または2644位のアスパラギン酸(D)をグリシン(G)に置換するなどがある。ワクチンポリヌクレオチドによりコードされるNS5Bポリペプチドは、これら両方のアスパラギン酸突然変異を含むのが好ましい。
【0024】
好ましくは、コードされたNS5Bポリペプチドは、そのモチーフCでの破壊を含む。例えば、非変異アスパラギン酸残基であるD2737をH、NまたはEに置換する突然変異によってNS5Bの完全な不活性化が起こる。
【0025】
好ましくは、本発明のDNAワクチンによりコードされるNS5Bは、モチーフA突然変異を含むが、随意にモチーフC突然変異を含んでもよい。モチーフAでの好ましい突然変異には、アスパラギン酸(D)2639のグリシンへの置換、およびアスパラギン酸(D)2644のグリシンへの置換が含まれる。後述の実施例1に記載するように、さらに別のコンセンサス突然変異が存在してもよい。
【0026】
NS3は、プロテアーゼとヘリカーゼ活性の両方を持つものとして記載されている。本発明のDNAワクチンによりコードされるNS3ポリペプチドを突然変異させることにより、NS3のプロテアーゼおよびヘリカーゼ活性の両方を破壊するのが好ましい。NS3のプロテアーゼ活性は、H−1083、D−1107およびS−1165の「触媒三つ組(catalytic triad)」に結合していることがわかっている。本発明のワクチンによりコードされるNS3は、触媒三つ組残基に突然変異を含むのが好ましく、最も好ましくは、NS3はセリン1165からバリンへの単一の点突然変異を含む(De Francesco, R., Pessi, a およびSteinkuhler C. 1998. The hepatitis C Virus NS3 proteinase : structure and function of a zinc containing proteinase. Anti− Viral Therapy 3, 1−18)。
【0027】
NS3の構造および機能は次のように表すことができる:
【化2】
【0028】
NS3のヘリカーゼ活性については、I、II、IIIおよびIVの4つの重要なモチーフが確認されている。本発明のDNAワクチンによりコードされるNS3は、これらモチーフの少なくとも1つに対する破壊的突然変異を含むのが好ましい。アスパラギン酸1316からグルタミンへの置換が存在するのが最も好ましい(Paolini, C, Lahm A, De Francesco RおよびGallinari P 2000, Mutational analysis of hepatitis C virus NS3−associated helicase. J.Gen Virol. 81, 1649)。これらの最も好ましいNS3突然変異、S−1165VもしくはD1316Qのいずれも、公知または予測されるT細胞エピトープ内に存在しない。
【0029】
最も好ましくは、本発明のDNAワクチンによりコードされるNS3ポリペプチドは、セリン(S)1165からバリン(V)およびアスパラギン酸(D)1316からグルタミン(Q)への突然変異を含む。加えて、後述の実施例1に記載するように、1以上のコンセンサス突然変異が存在してもよい。
【0030】
HCVコアタンパク質の生物学的機能は複雑であり、個々の点突然変異と相関しない(McLauchlan J. 2000. Properties of the hepatitis C virus core protein: a structural protein that modulates cellular processes. J of Viral Hepatitis 7, 2−4)。コアがリンホトキシンβ受容体と直接的に相互作用し、NFκBおよびPKR経路とも相互作用することができ、かつ細胞の生存およびアポトーシスにも影響を与えることができることが証明されている。コアを発現する組換えワクシニア構築物が、in vivoでコアをより有毒にするワクシニアに対する細胞応答を阻害することが見出された。
【0031】
感染中にコアタンパク質は宿主細胞のプロテアーゼによってウイルスポリタンパク質から2ヶ所で切断される。第1の切断は191位で生じ、E1のN末端を生成する。第2の切断が生じる残基は正確に位置付けられておらず、174〜191位のアミノ酸に存在し、それによって約17アミノ酸の長さの短いコアペプチド配列を遊離させる(McLauchlan J. (2000) J. Viral Hepatitis. 7, 2−14; YasuiK, Lau JYN, Mizokami M.,ら, J. Virol 1998. 72 6048−6055)。
【0032】
本発明のワクチンに使用するコアポリペプチドは全長あるいは切断型の形態のいずれかである。コアポリペプチドは全長であってもよいが、その配列を再配列することによりコアタンパク質のあらゆる活性を破壊(又は阻害)する。コアポリペプチドを少なくとも2つの断片に分割してもよく、コアアミノ酸66〜191とそれに続くアミノ酸1〜65からなるポリペプチド、あるいはコアアミノ酸105〜191とそれに続くコアアミノ酸1〜104からなるポリペプチドを形成することが最も好ましい。
【0033】
同一の細胞における他のHCVタンパク質の産生に与えるコアの負の作用を最小にするために、使用されるコアタンパク質が切断型タンパク質であることが最も好ましい。本発明の好適な態様では、コードされるコアタンパク質は、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断される。最も好ましくは、産生されるタンパク質の配列がコアの第2切断から生じる天然の遊離C末端ペプチド配列を欠くように;より好ましくはこのタンパク質が少なくとも最後の10アミノ酸を欠くように、好ましくは少なくとも最後の15アミノ酸を欠くように、より好ましくは最後の20アミノ酸を欠くように、より好ましくは最後の26アミノ酸を欠くように、そして最も好ましくは最後の40アミノ酸を欠くように、コアタンパク質がカルボキシ末端から切断される。本発明における使用に適したコアをコードする最も好適なポリヌクレオチドは、1〜171位、1〜165位、1〜151位のアミノ酸を含有する切断型コアをコードするものである。本発明の使用に適したコアをコードするポリヌクレオチドが、1〜151位のアミノ酸の切断型コアタンパク質をコードするものであるのが最も好ましい。実施例1で説明するような1以上のコンセンサス突然変異が存在してもよい。
【0034】
本発明のポリヌクレオチドによりコードされる好ましいNS4Bポリペプチドは、HCV分離物と遺伝子型の間で超可変の領域を除去するようなN末端切断を含む。好ましくは、NS4Bポリペプチドは、N末端から30〜100アミノ酸、さらに好ましくは40〜80アミノ酸の欠失、最も好ましくは最初のN末端の48アミノ酸の欠失を含む(J4L6分離物の場合、これは、アミノ酸1760までの切断に相当し、これは、NS4Bの最初の48アミノ酸の消失である;その他のHCV分離物における同等の切断も本発明に含まれる)。さらに、NS4B配列を2つ以上の断片に分割し、野生型分子に認められるものとは違う順序で配置したNS4Bの配列を有するポリペプチドに発現させてもよい。
【0035】
本発明のワクチンに存在するポリヌクレオチドは、HCVウイルスに認められる天然ヌクレオチド配列を含んでもよいが、ヌクレオチド配列のコドンは、哺乳動物細胞での発現のために最適化されているものが好ましい。
【0036】
コドンの最適化に加えて、HCVコア、NS3、NS4BおよびNS5Bをコードする本発明のポリヌクレオチドでのコドン使用頻度を変更することにより、希有コドンが濃縮クラスターに現れるのではなく、反対に、ポリヌクレオチド配列全体にわたって比較的均等な間隔をおいて存在するか、あるいはコドン最適化遺伝子から排除されるようにする。
【0037】
DNAコードは4文字(A、T、CおよびG)を有し、これらを用いて、3文字の「コドン」を綴るが、このコドンは、生物の遺伝子においてコードされたタンパク質のアミノ酸を表すものである。DNA分子に沿ったコドンの直鎖状配列をこれら遺伝子によりコードされたタンパク質中のアミノの直鎖状配列に翻訳する。コードは高度に縮重しており、61個のコドンが20個の天然アミノ酸をコードし、3個のコドンが「停止」シグナルを表す。従って、ほとんどのアミノ酸は2個以上のコドンによりコードされる。実際には、数個が、4個以上の異なるコドンによりコードされる。
【0038】
所与のアミノ酸をコードするのに1個以上のコドンが利用可能である場合には、生物のコドン使用頻度パターンは極めて非ランダムである。種によってコドン選択に偏りがあり、さらにコドンの使用は、単一の種においても、高および低レベルで発現する遺伝子間では著しく異なる可能性がある。この偏りは、ウイルス、植物、細菌および哺乳動物細胞で異なり、いくつかの種は、他の種よりランダムコドン選択から離れた強い偏りを示す。例えば、ヒトおよびその他の哺乳動物は、特定の細菌またはウイルスと比較してあまり強く偏っていない。これらの理由から、大腸菌に発現させた哺乳動物遺伝子、または哺乳動物細胞に発現させたウイルス遺伝子は、効率的な発現にとって不適切なコドン分布を有する。しかし、大腸菌発現に適したコドン使用頻度パターンを有する遺伝子をヒトに効率的に発現させることもできる。発現を起こそうとする宿主に稀に観察されるように、異種DNA配列にコドンのクラスターが存在すると、その宿主での異種発現レベルが低いことが推定される。
【0039】
宿主に希有なものから宿主が優先するものへコドンを変えて(「コドン最適化」)、異種発現レベルを増強するいくつかの例があり、例えば、BPV(ウシパピローマウイルス)後期遺伝子L1およびL2は、哺乳動物コドン使用頻度パターンについて最適化され、これは、哺乳動物(Cos−1)細胞培養物における野生型HPV配列に対する発現レベルを増強することがわかっている(Zhouら、J. Virol 1999. 73, 4972−4982)。この研究では、哺乳動物と比べてBPVで2倍以上の頻度で生じた(使用頻度比>2)すべてのBPVコドンと、使用頻度比が>1.5のほとんどのコドンを、優先的に用いられる哺乳動物コドンで保存的に置換した。WO97/31115、WO97/48370およびWO98/34640(Merck & Co., Inc.)において、HIV遺伝子またはそのセグメントのコドン最適化により、最適化を意図した宿主哺乳動物においてコドン最適化した配列をDNAワクチンとして用いると、タンパク質発現が増大し、免疫原性が向上することがわかっている。これらの文献では、各ウイルスコドンを、意図する宿主の最適コドンで保存的に置換するため、配列は、全体が最適化コドンから構成される(ただし、これが、不要な制限部位、イントロンスプライス部位などを導入する場合を除く)。
【0040】
「コドン使用頻度パターン」という用語は、上記のヌクレオチド配列、遺伝子もしくは遺伝子のクラス(例えば、高度に発現した哺乳動物遺伝子)におけるすべてのコドンについての平均頻度を意味する。哺乳動物(ヒトを含む)についてのコドン使用頻度パターンを文献にみいだすことができる(例えば、Nakamuraら、Nucleic Acids Research 1996、24:214−215参照)。
【0041】
本発明のポリヌクレオチドでは、コドン使用頻度パターンは、HCVの典型的なものから、標的生物、例えば、大腸菌または哺乳動物、特にヒトのコドン出現偏りをより忠実に表すものに変更するのが好ましい。「コドン使用頻度係数」すなわちコドン適合指数(Sharp PM. Li WH. Nucleic Acids Research. 15(3):1281−95, 1987)は、所与のポリヌクレオチド配列のコドン使用頻度パターンが、標的種のパターンにどれくらい密に類似しているかの尺度である。61個のコドンの各々についてのコドン出現頻度(選択したクラスの遺伝子について1000コドン当たりの出現数として表される)を20個の天然アミノ酸の各々について標準化して、各アミノ酸について最も使用頻度の高いコドンの値を1に設定し、これより少ない共通コドンの頻度が、0〜1の間になるように比例的に定められる。すなわち、61個のコドンの各々が、標的種の高度発現遺伝子について1以下の値を割り当てられる。これを優先値(W)と呼ぶ。その種の高度発現遺伝子に対する特定のポリヌクレオチドのコドン使用頻度係数を算出するため、特定のポリヌクレオチドの各コドンについて定めた値を記録し、これらすべての値の相乗平均を求める(コドンの総数でこれらの値の自然対数の和を割り算し、真数(anti−log)を求める)。係数は0から1の間の値になり、係数が高いほど、そのポリヌクレオチドにおいて使用頻度の高いコドンが多い。ポリヌクレオチド配列のコドン使用頻度係数が1の場合には、全コドンが、標的種の高度発現遺伝子について「最も使用頻度の高い」コドンである。
【0042】
本発明は、HCVコア、NS3、NS4BもしくはNS5Bアミノ酸配列をコードするポリヌクレオチド配列であって、該ポリヌクレオチド配列のコドン使用頻度パターンが、高度に発現した哺乳動物遺伝子のものに類似している、上記ポリヌクレオチドを提供する。好ましくは、上記ポリヌクレオチド配列はDNA配列である。上記ポリヌクレオチド配列のコドン使用頻度パターンは、高度に発現したヒト遺伝子のものに類似しているのが望ましい。
【0043】
HCVコア(1−191)をコードするコドン最適化ポリヌクレオチド配列を図2に示す。S1165VおよびD1316Qポリペプチド突然変異を含むHCV NS3をコードするコドン最適化ポリヌクレオチド配列を図3に示す。ポリペプチドのN末端1−48切断を含むHCV NS4Bをコードするコドン最適化ポリヌクレオチドを図4に示す。D2639GおよびD2644Gポリペプチド突然変異を含むHCV NS5Bをコードするコドン最適化ポリヌクレオチド配列を図5に示す。
【0044】
従って、HCVコア、NS3、NS4BもしくはNS5Bアミノ酸配列を一緒にコードする複数のコドンを含み、本発明のワクチンを形成する合成遺伝子であって、上記アミノ酸配列をコードするのに用いられる可能なコドンの選択を最適哺乳動物コドン使用頻度に類似するように変更することにより、上記合成遺伝子におけるコドン使用頻度が、C型肝炎ウイルス遺伝子のそれより、高度に発現した哺乳動物遺伝子の使用頻度に密に類似するようにする、上記合成遺伝子が提供される。コドン使用頻度パターンは、高度に発現したヒト遺伝子のものと実質的に同じであるのが好ましい。「天然の」HCVコア、NS3、NS4BおよびNS5B配列のコドン使用頻度については分析がなされている。HCVタンパク質のコドン使用頻度係数は、コア(0.487)、NS3(0.482)、NS4B(0.481)およびNS5B(0.459)である。本発明のポリヌクレオチドは、一般に、高度に発現したヒト遺伝子について、0.5以上、好ましくは0.6以上、最も好ましくは0.7以上であるが、1以下のコドン使用頻度係数(すでに定義した通り)を有する。ポリヌクレオチドはまた、高度に発現した大腸菌遺伝子について0.5以上、好ましくは0.6以上、最も好ましくは0.7以上のコドン使用頻度係数を有する。
【0045】
コドン最適化のほかにも、合成遺伝子を突然変異させることにより、希有コドンのクラスターの出現を排除する。これは2つの方法のいずれかにより達成することができる。これを達成する好ましい方法は、遺伝子配列から希有コドンを排除するものである。希有コドンを定義する一つの方法によれば、希有コドンは、標的生物の高度発現遺伝子において、特定のアミノ酸に用いられるコドンの<20%を占めるコドン、好ましくは、特定のアミノ酸について<10%を占めるコドンである。あるいは、標的生物の高度発現遺伝子において、相対同義コドン使用頻度(RSCU)値が<0.3、または好ましくは<0.2のコドンとして希有コドンを定義することもできる。RSCU値は、観察されたコドン数を、該アミノ酸の全コドンが同等に高頻度で用いられた場合に予想される数で割って得られる数値である。希有コドンの適切な定義は当業者に明らかであろう。
【0046】
あるいは、HCVコア、NS3、NS4BおよびNS5Bポリヌクレオチドを最適化することにより、濃縮領域に存在する希有の非最適コドンのクラスター形成を防止する。従って、個々の希有コドン、例えば、RSCUが<0.4(さらに好ましくは<0.3)がポリヌクレオチド全体に均等な間隔をおいて存在するように、ポリヌクレオチドを最適化する。
【0047】
ウエスタンブロットで評価されるように、コドンが最適化された突然変異コア、NS3およびNS5Bの発現レベルは野生型と比べて増加することが示されている。切断型のコドン最適化NS4BはNS5Bとの融合物として発現し、この融合物は十分に発現する。
【0048】
本発明のワクチンは、HCVポリペプチドの個々の発現を指令するベクターを含む。あるいは、HCVポリペプチドを1以上の融合タンパク質として発現させてもよい。
【0049】
本発明の好ましいワクチンは、タンパク質またはポリヌクレオチドレベルのいずれかで四重融合物を含み、そのような融合物として以下のものが挙げられる:
HCV組合せ1: HCV500
【化3】
【0050】
HCV組合せ2: HCV510
【化4】
【0051】
HCV組合せ3: HCV520
【化5】
【0052】
HCV組合せ4: HCV530
【化6】
【0053】
HCV組合せ5: HCV501
【化7】
【0054】
HCV組合せ6: HCV502
【化8】
【0055】
HCV組合せ7:
【化9】
【0056】
他の好適な融合物はHCV組合せ1、2および3に類似しているが、ここで、コアタンパク質は切断型コアタンパク質、典型的にはコア1−151である。本発明の別の好ましいワクチンを以下に挙げるが、これらは、同じプラスミド内の様々な発現カセットに存在するポリヌクレオチド二重および三重融合物を含み、各カセットは、プロモーターユニット(例:HCMV IE)の独立した制御下にある(矢印で示す)。
【0057】
このような二重プロモーター構築物は、同じ細胞内で4つのタンパク質抗原の発現を2つの別個のタンパク質として駆動する(矢印で示す)。
【化10】
【0058】
好適な構築物はHCV組合せ7、9、11、または12である。特に好適なものは7と11である。
【0059】
本発明の別の態様では、ポリヌクレオチドワクチンは場合によりコアタンパク質をコードするポリヌクレオチドを含まない。例えば、本発明のこの態様における好適なポリヌクレオチドは下記のものを含む。
【化11】
【0060】
上記HCV組合せ8〜19について、用いた用語、例えば、(コアNS3)+(NS4B5B)は、各々が個別のプロモーターにより独立に制御される2つの発現カセット、この例の場合、コアNS3二重融合タンパク質をコードする1つの発現カセットと、NS4B−NS5B二重融合タンパク質をコードする別のカセットを含むポリヌクレオチドベクターの開示を意味する。各HCV組合せ8〜19も同様に解釈すべきである。
【0061】
前記HCV組合せ1〜19は、HCVタンパク質、ポリタンパク質融合物、もしくはポリヌクレオチドの相対配向を開示する。また、前記HCV組合せ1〜19のすべては、構成タンパク質の活性を除去するのに好ましい突然変異または切断の各々と一緒に開示されることが具体的に記載される。例えば、組合せ1〜19の好ましい変異体(別途記載のない限り)は、コア(1−191(生物学的活性を不能にする2以上の断片への分割配列を除く)または好ましくは、その切断型1−151または1−165もしくは1−171で存在するコア);NS3 1027−1657(ヘリカーゼ(アスパラギン酸1316からグルタミン)およびプロテアーゼ(セリン1165からバリン)活性を不活性化するための突然変異;ポリメラーゼ活性を不活性化させるためのNS5B 2420−3010(アスパラギン酸2639からグリシンおよびアスパラギン酸2644からグリシンへの突然変異、モチーフA);ならびに、NS4B 1712−1972(随意に1760−1972に切断して、N−末端高度可変断片を除去する)を含む。
【0062】
本発明は、前述したような新規のDNAワクチンとポリペプチドを提供する。また、本発明により、記載したポリペプチドの類似体、ならびにそれらを含むDNAワクチンが提供される。
【0063】
用語「類似体」とは、本発明の別のポリペプチドと同じアミノ酸をコードするが、遺伝子コードの重複性を通して、異なるヌクレオチド配列を有するが、同じコドン使用頻度パターンを維持する、例えば、同じコドン使用頻度係数、または他方のポリヌクレオチドのそれの0.1以内、好ましくは、0.05以内のコドン使用頻度係数を有するポリヌクレオチドを意味する。
【0064】
HCVポリヌクレオチド配列は、様々なHCV遺伝子型、株もしくは分離物のいずれに由来するものでもよい。HCV分離物は、1以上のサブタイプを含む以下の6つの主要遺伝子型:HCV 1(1a、1bもしくは1c)、HCV 2(2a、2bもしくは2c)、HCV 3(3a、3b、10a)、HCV 4(4a)、HCV 5(5a)ならびにHCV 6(6a, 6b, 7b, 8b, 9aおよび11a)に分類することができる(Simmonds, J. Gen. Virol., 2001, 693−712)。本発明に関して、各HCVタンパク質は、同じHCV遺伝子型またはサブタイプ、あるいは、HCV遺伝子型またはサブタイプのいずれかの組合せに由来するものでよく、さらには、HCVタンパク質を用いてもよい。遺伝子は、1b遺伝子型、例えば、感染性クローンJ4L6(登録番号AF0542478、図1を参照)に由来するのが好ましい。
【0065】
配列決定されている具体的株として、HCV−J(Katoら、1990, PNAS, USA, 87;9724−9528)およびBK(Takamizawaら、1991, J.Virol. 65:1105−1113)が挙げられる。
【0066】
本発明のポリヌクレオチドは、コードされたタンパク質の発現(in vitro、in vivoもしくはex vivoのいずれで起こってもよい)による生産に有益である。従って、上記ヌクレオチドは、例えば、収率を高めるための組換えタンパク質合成に関与する、あるいは、DNAワクチン接種法に使用される治療剤として独立に用いることも可能である。本発明のポリヌクレオチドをin vitroもしくはex vivoでコードしたタンパク質の生産に用いる場合には、例えば、細胞培養物中の細胞を改変して、発現させようとするポリヌクレオチドを含むようにする。このような細胞として、一過性の、または好ましくは安定な哺乳動物細胞系が挙げられる。本発明のポリタンパク質をコードするベクターの挿入により改変することができる細胞の特定例としては、HEK293T、CHO、HeLa、293およびCOS細胞が挙げられる。選択する細胞系は、安定しているだけでなく、ポリタンパク質の成熟グリコシル化と細胞表面発現を可能にするものが好ましい。発現は、形質転換した卵母細胞で達成することができる。ポリペプチドは、トランスジェニック非ヒト動物、好ましくはマウスの細胞において、本発明のポリヌクレオチドから発現させることができる。本発明のポリヌクレオチドからポリペプチドを発現させるトランスジェニック非ヒト動物も本発明の範囲に含まれる。
【0067】
本発明は、本発明のヌクレオチド配列を含む発現ベクターを包含する。このような発現ベクターは、分子生物学の分野では日常的に構築されており、例えば、プラスミドDNAと好適な開始剤、プロモーター、エンハンサーおよびその他のエレメント(例えば、タンパク質発現に必要で、そのために正しい配向で配置されるポリアデニル化シグナルなど)を含むことができる。その他の好適なベクターは、当業者には明らかであろう。これに関するさらに別の例については、Sambrook ら、Molecular Cloning: a Laboratory Manual. 2nd Edition. CSH Laboratory Press.(1989)を参照されたい。
【0068】
好ましくは、本発明のポリヌクレオチド、またはベクターにおいて本発明で使用するためのポリヌクレオチドは、宿主細胞によるコード配列の発現を達成することが可能な制御配列に機能しうる形で連結されている。すなわち、このベクターは発現ベクターである。「機能しうる形で連結されている」という用語は、記載した要素が、意図される様式でそれらが機能できるような関係にある並置を意味する。コード配列に「機能しうる形で連結された」調節配列(例えば、プロモーターなど)は、調節配列と適合する条件下でコード配列の発現が達成されるように配置される。
【0069】
発現カセットは、目的の配列または遺伝子の発現を指令することができる集合体である。発現カセットは、制御エレメント、例えば、目的の遺伝子に機能しうる形で連結されたプロモーターなどを含む。
【0070】
上記ベクターは、例えば、複製起点を備えるプラスミド、人工染色体(例:BAC、PAC、YAC)、ウイルスもしくはファージベクターでよく、随意に、ポリヌクレオチドの発現用のプロモーター、また随意にプロモーターのレギュレータでもよい。ベクターは、1つ以上の選択マーカー遺伝子を含み、例えば、細菌プラスミドの場合には、アンピシリンまたはカナマイシン耐性遺伝子を、あるいは、真菌ベクターの場合には耐性遺伝子を含む。ベクターをin vitroで用いて、例えば、DNAまたはRNAを産生することもできるし、あるいはベクターを用いて宿主細胞、例えば、哺乳動物宿主細胞をトランスフェクションまたは形質転換することにより、例えば、ベクターがコードしたタンパク質を生産することも可能である。また、ベクターを改変して、DNAワクチン接種法または遺伝子治療法にin vivoで用いることも可能である。
【0071】
プロモーターおよびその他の発現調節シグナルは、発現が設計される宿主細胞と適合するように選択することができる。例えば、哺乳動物プロモーターとして、カドミウムのような重金属に応答して誘導することができるメタロチオネインプロモーター、およびβアクチンプロモーターが挙げられる。また、SV40ラージT抗原プロモーター、ヒトサイトメガロウイルス(CMV)即時型(IE)プロモーター、ラウス肉腫ウイルスLTRプロモーター、アデノウイルスプロモーター、もしくはHPVプロモーター、特にHPV上流調節領域(URR)を用いてもよい。これらプロモーターはすべて周知であり、当分野において容易に入手可能である。
【0072】
好適なウイルスベクターの例として、単純ヘルペスウイルスベクター、ワクシニアもしくはαウイルスベクターおよびレトロウイルス、例えば、レンチウイルス、アデノウイルスおよびアデノ随伴ウイルスが挙げられる。これらのウイルスを用いた遺伝子移入の方法は当業者には知られている。例えば、レトロウイルスベクターを用いて、本発明のポリヌクレオチドを宿主ゲノムに安定に組み込むことができるが、このような組換えは好ましくない。対照的に、複製能欠損アデノウイルスベクターは、エピソームのままであるため、一過的発現を可能にする。例えば、サブユニットワクチンとして、または免疫アッセイで使用するために、昆虫細胞(例えば、バキュロウイルスベクター)、ヒト細胞もしくは細菌での発現を駆動することができるベクターを用いて、本発明のポリヌクレオチドがコードした多量のHCVタンパク質を生産することができる。
【0073】
別の態様では、本発明は、本明細書に記載したポリヌクレオチドを含む医薬組成物を提供する。好ましくは、この組成物は、本発明の第2の態様に従うDNAベクターを含む。好ましい実施形態では、上記組成物は、HCVアミノ酸配列をコードするポリヌクレオチド配列をコードするベクターを含むDNAでコーティングした複数の粒子、好ましくは金粒子を含み、上記ポリヌクレオチド配列のコドン使用頻度パターンは、高度に発現させた哺乳動物遺伝子、特にヒト遺伝子のパターンに類似している。別の実施形態では、前記組成物は、医薬的に許容されうる賦形剤と、本発明の第2の態様のDNAベクターを含む。前記組成物はまた、アジュバントも含む。
【0074】
DNAワクチンは、液体ワクチンの筋肉への間隙投与(WO90/11092)または筋内注射以外の機構により送達することができる。例えば、皮膚への送達は、免疫機構が、感染に対する障壁となる組織(例えば、皮膚や粘膜など)において極めて活発であることから有利である。皮膚への送達は、注射、ジェット式注射器(皮膚、または筋肉などの下層組織に高圧下で液体を強制的に送り出す)もしくはパーティクルボンバードメント(この場合、上皮を貫通するのに十分な密度の粒子にDNAをコーティングしうる)(米国特許第5371015号)により実施することができる。例えば、Haynesら、J. Biotechnology 44: 37−42(1996)に記載されているように、ヌクレオチド配列をプラスミドに組み込み、これを金ビーズにコーティングした後、これらビーズを高圧下で表皮に投与する。皮膚へのこれら粒子の投入により、表皮細胞および表皮ランゲルハンス(Langerhan)細胞両方の直接トランスフェクションが達成される。ランゲルハンス細胞は、抗原提示細胞(APC)であり、DNAを取り込み、コードされたペプチドを発現し、これらをプロセッシングして細胞表面MHCタンパク質に展示する。トランスフェクションされたランゲルハンス細胞は、リンパ節に移動し、そこで、展示された抗原断片をリンパ球に提示して、免疫応答を引き起こす。粒子が媒介する皮膚への送達により免疫応答を誘発するには、非常に少量のDNA(1μg以下、往々にして0.5μg以下)でよいが、これは、直接筋内注射後に免疫応答を生じさせる場合、ミリグラム量のDNAを必要とすることが知られているのとは対照的である。
【0075】
本発明のポリヌクレオチドを治療薬、例えば、DNAワクチン接種に使用する場合には、ワクチン接種しようとする哺乳動物、例えば、ヒトに核酸を投与する。RNAまたはDNA、好ましくはDNAのような核酸は、前記のようなベクターの形態で提供され、これを該哺乳動物の細胞に発現させることができる。ポリヌクレオチドは、利用可能な方法のどれで投与してもよい。例えば、核酸は、針注射、好ましくは、皮内、皮下もしくは筋内注射により投与する。あるいは、粒子媒介DNA送達(PMDD)のような核酸送達装置を用いて、核酸を皮膚内に直接送達することもできる。この方法では、不活性粒子(金ビーズなど)を核酸でコーティングし、例えば、投射装置からの高圧下放出により、粒子が受容体(例:皮膚)の表面を貫通するのに十分な速度で加速する。(本発明の核酸分子でコーティングした粒子は、このような粒子を装填した送達装置と同様に、本発明の範囲に含まれる)。組成物は、0.5〜5μm、好ましくは約2μmの平均直径を有する金粒子を含むのが望ましい。好ましい実施形態では、コーティングした金ビーズをカートリッジとして機能する管に装填し、各カートリッジが、1カートリッジ当たり0.1〜5μg、好ましくは約0.5μgのDNAをコーティングした0.1〜1mg、好ましくは約0.1 mgの金を含むようにする。
【0076】
本発明の別の態様によれば、本明細書に記載したようなポリヌクレオチド配列を含む宿主細胞が提供される。宿主細胞は、細菌、例えば、大腸菌、哺乳動物、例えば、ヒトでもよいし、あるいは昆虫細胞でもよい。本発明のベクターを含む哺乳動物細胞は、in vitroでトランスフェクションした培養細胞でもよいし、あるいは、哺乳動物へのベクターの投与により哺乳動物細胞をin vivoでトランスフェクションすることもできる。
【0077】
別の態様では、本発明は、前記のような医薬組成物を製造する方法であって、野生型HCVヌクレオチド配列のコドン使用頻度パターンを変更するか、またはポリヌクレオチド配列を合成により形成して、高度発現哺乳動物遺伝子に類似したコドン使用頻度パターンを有し、かつ野生型HCVアミノ酸配列、もしくはポリペプチドの1以上の天然の機能を不活性化するのに十分なアミノ酸変化を有する野生型配列を含む突然変異HCVアミノ酸配列、をコードする配列を製造するステップを含む、上記方法を提供する。
【0078】
また、HCV感染の治療または予防における、本明細書に記載したポリヌクレオチドまたはワクチンの使用も提供する。
【0079】
患者に裸のポリヌクレオチドまたはベクターを導入する好適な方法として、好適なビヒクルを用いた局所適用がある。例えば、鼻内、経口、膣内もしくは直腸内投与により、核酸を皮膚、または粘膜表面に局所投与してもよい。裸のポリヌクレオチドまたはベクターは、リン酸緩衝食塩水(PBS)のような医薬的に許容されうる賦形剤と一緒に存在してもよい。DNA取込みは、ブピバカインのような促進剤を別々に、またはDNA製剤に含有させて用いることにより、さらに容易にすることができる。核酸を受容者に直接投与する別の方法としては、超音波、電気刺激、エレクトロポレーションおよびマイクロシーディング(米国特許第5,697,901号に記載されている)が挙げられる。
【0080】
核酸構築物の取込みは、複数の周知のトランスフェクション法、例えば、トランスフェクション剤の使用を含む方法により増強することができる。これらの薬剤の例としては、カチオン剤、例えば、リン酸カルシウムおよびDEAE−デキストラン、ならびに、リポフェクタント、例えば、リポフェクタムおよびトランスフェクタムが挙げられる。投与しようとする核酸の用量は変更することができる。典型的に核酸は、粒子媒介遺伝子送達の場合には、1pg〜1mg、好ましくは1pg〜10μg核酸の範囲、またその他の経路の場合には、10μg〜1mgの範囲の量を投与する。
【0081】
また、本発明の核酸配列は、遺伝子治療に有用な専用送達ベクターにより投与することもできる。遺伝子治療法については、例えば、Vermeら、Nature 1997, 389:239−242に記載されている。ウイルスおよび非ウイルスベクター系のいずれを用いてもよい。ウイルス系としては、レトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、カナリア痘およびワクシニア−ウイルスベースの系が挙げられる。好ましいアデノウイルスベクターは、ヒト以外の霊長類に由来するものである。特に、米国特許第6083716号に記載されているPan 9(C68)、WO03/046124に記載されているPan5、6 もしくは7が挙げられる。
【0082】
非ウイルスベースの系としては、核酸、ミクロスフィアカプセル化法(ポリ(ラクチド−コ−グリコリド)と、リポソームベースの系の直接投与が挙げられる。最初のワクチン接種後にブースター注射を実施するのが望ましい場合には、ウイルスおよび非ウイルス送達系を組み合わせてもよく、例えば、プラスミドのような非ウイルスベクターを用いた最初の「初回感作(プライム)」DNAワクチン接種の後、ウイルスベクターまたは非ウイルスベースの系を用いた1回以上の「追加免疫(ブースト)」ワクチン接種を行なう。プライム−ブーストプロトコルは、アジュバント中のタンパク質で初回感作し、本発明のポリヌクレオチドをコードするDNAまたはウイルスベクターで追加免疫するという利点もある。あるいは、タンパク質を基剤とするワクチンをブースターとして用いてもよい。タンパク質ワクチンは、DNA/ウイルスベクターのワクチンが含むすべての抗原を含むのが好ましい。しかし、タンパク質は個別の形態でも、ポリタンパク質の形態をしていてもよい。
【0083】
本発明の核酸配列は、形質転換した細胞により投与することもできる。このような細胞として、被験者から採取した細胞がある。本発明の裸のポリヌクレオチドまたはベクターをこのような細胞にin vitroで導入し、その後、形質転換した細胞を被験者に戻すことができる。本発明のポリヌクレオチドは、相同組換え事象により細胞にすでに存在する核酸に組み込むことができる。所望であれば、形質転換した細胞をin vitroで増殖させ、こうして得られた1個以上の細胞を本発明で用いてもよい。公知の手術またはマイクロサージャリー法(例:移植、マイクロインジェクションなど)により患者の適切な部位に細胞を送達することができる。
【0084】
好適な細胞としては、抗原提示細胞(APC)があり、例えば、樹状細胞、マクロファージ、B細胞、単球、ならびに有効なAPCに工学的に操作されてもよいその他の細胞が挙げられる。このような細胞は、遺伝子的に改変することにより、抗原提示能力を増大する、T細胞応答の活性化および/または維持を改善する、それ自体で抗腫瘍、例えば抗子宮頸部癌効果を有する、ならびに/あるいは受容体(すなわち、対合HLAハプロタイプ)と免疫学的に適合性となるようにすることもできるが、必ずしも必要なわけではない。APCは一般に、生物由来の多様な体液および器官(腫瘍および腫瘍周辺組織を含む)のいずれから単離されてもよく、オートロガス、同種異系、同系もしくは異種細胞のいずれでもよい。
【0085】
本発明の特定の好ましい実施形態では、樹状細胞またはその前駆細胞を抗原提示細胞として用い、in vitroで形質転換させてから患者に戻すか、あるいは、例えば、粒子媒介DNA送達により、ワクチン中の被送達ヌクレオチドのin vivo標的として用いる。樹状細胞は高い能力を有するAPCであり(Banchereau およびSteinman, Nature 392:245−251, 1998)、予防または治療のための抗腫瘍免疫を誘発する生理学的アジュバントとして有効であることが示されている(TimmermanおよびLevy, Ann. Rev. Med. 50:507−529, 1999)。一般に、樹状細胞は、その典型的な形状(in situで星形、in virto視覚化可能な顕著な細胞質突起(樹状突起)を有する)と、高い効率で抗原を取り込み、プロセッシングおよび提示する能力、ナイーブT細胞応答を活性化する能力に基づいて、確認することができる。勿論、樹状細胞を工学的に操作して、in vivoまたはex vivoで樹状細胞に通常認められない特定の細胞−表面受容体またはリガンド、例えば、本発明の構築物にコードされた抗原を発現させることも可能であり、このような改変された樹状細胞も本発明によって意図される。樹状細胞に代わるものとして、分泌小胞抗原負荷樹状細胞(secreted vesicles antigen−loaded dendritic cells)(エキソソームと称する)をワクチンの中で用いてもよい(Zitvogelら, Nature Med. 4:594−600, 1998参照のこと)。
【0086】
樹状細胞および前駆細胞は、末梢血、骨髄、腫瘍浸潤細胞、腫瘍周辺組織−浸潤細胞、リンパ節、脾臓、皮膚、臍帯血もしくはその他の好適な組織または体液から取得することができる。例えば、樹状細胞は、GM−CSF、IL−4、IL−13および/またはTNFのようなサイトカインの組合せを末梢血液から収集した単球の培養物に添加することにより、ex vivoで分化させることができる。あるいは、GM−CSF、IL−3、TNF、CD40リガンド、リポ多糖LPS、flt3リガンド(プロフェショナル抗原提示細胞、特に樹状細胞の産生に重要なサイトカイン)および/または樹状細胞の分化、成熟および増殖を誘発するその他の化合物の組合せを培養培地に添加することにより、末梢血、臍帯血もしくは骨髄から収集したCD34陽性細胞を樹状細胞に分化させることができる。
【0087】
APCは一般に、抗原のHCVアミノ酸配列をコードするポリヌクレオチド、例えば、本発明で考慮されるコドン最適化ポリヌクレオチドでトランスフェクションすることもできる。このようなトランスフェクションは、ex vivoで行うことができ、トランスフェクションしたそのような細胞を含む組成物またはワクチンは、本明細書に記載するように、治療の目的で用いることができる。あるいは、樹状、またはその他の抗原提示細胞をターゲッティングする遺伝子送達ビヒクルを患者に投与し、in vivoでトランスフェクションを起こすことも可能である。樹状細胞のin vivoおよびex vivoトランスフェクションは、例えば、WO 97/24447に記載されているような当分野では公知の方法のいずれか、またはMahviら、Immunology and cell Biology 75:456−460, 1997に記載された粒子媒介方法を用いて、一般に実施することができる。
【0088】
本発明のワクチンおよび医薬組成物は、抗ウイルス剤、例えば、αインターフェロン、好ましくは、PEG化αインターフェロンおよびリボバリンと組合せて用いてもよい。ワクチンおよび医薬組成物は、単位用量または複数回用量の容器、例えば、密封したアンプルまたはバイアルに収容することができる。このような容器は、気密シールすることにより、使用まで製剤の滅菌を保存するのが好ましい。一般に、製剤は、油性または水性ビヒクル中の懸濁液、溶液もしくは乳濁液として保存することができる。これ以外にも、凍結乾燥した状態でワクチンまたは医薬組成物を保存し、使用の直前に滅菌液体担体を添加するだけにしてもよい。粒子媒介送達による投与を意図する場合のヌクレオチド配列含有ワクチンは、圧縮ガス送達装置に用いるのに適したカートリッジの形態をしていてもよく、その際、カートリッジは、中空の管からなり、その内面は、随意に医薬的に許容されうる他の成分が存在してもよいワクチンヌクレオチド配列を担持する粒子でコーティングされている。
【0089】
本発明の医薬組成物は、アジュバント化合物、もしくはDNAによりコードされるタンパク質により誘発される免疫応答をモジュレートまたは増強するのに役立つその他の物質を含んでもよい。これらは、抗原とは別に、または抗原との融合物としてDNAによりコードされるか、あるいは、製剤の非DNAエレメントとして含有させてもよい。本発明の製剤に含有させることができるアジュバントタイプの物質の例として、ユビキチン、リソソーム結合膜タンパク質(LAMP)、B型肝炎ウイルスコア抗原、flt3−リガンドおよびその他のサイトカイン(例:IFN−γおよびGMCSF)が挙げられる。
【0090】
上記以外の好適なアジュバントは、以下に挙げる市販のもの、例えば、フロイント不完全アジュバントおよび完全アジュバント(Difco Laboratories, デトロイト、MI);イミキモド(Imiquimod)(3M, St. Paul, MN);レシミキモド(3M, セントポール、MN);メルクアジュバント(Merck Adjuvant)65(Merck and Company, Inc., ローウェイ、NJ);アルミニウム塩、例えば、水酸化アルミニウムゲル(alum)またはリン酸アルミニウム;カルシウム、鉄もしくは亜鉛の塩;アシル化チロシンの不溶性懸濁液;アシル化糖;カチオンまたはアニオン的に誘導体化した多糖;ポリホスファゼン;生物分解性ミクロスフェア;モノホスホリル脂質AおよびクイルAである。サイトカイン、例えば、GM−CSFもしくはインターロイキン−2、−7もしくは−12もアジュバントとして用いることができる。
【0091】
本発明の製剤では、アジュバント組成物が、主にTh1型の免疫応答を誘発するのが好ましい。従って、アジュバントは、DNAがコードした抗原に応答して生じた免疫応答を主にTh2型から主にTh1型の応答へとモジュレートする上で役立つものである。高レベルのTh−1型サイトカイン(例:IFN−、TFN、IL−2およびIL−12)は、投与された抗原に対する細胞媒介免疫応答の誘発を有利にする傾向がある。応答が主にTh1型である好ましい実施形態では、Th1型サイトカインのレベルは、Th2型サイトカインのレベルより大幅に増大するであろう。これらのサイトカインのレベルは標準的アッセイを用いて、容易に評価することができる。サイトカインのファミリーについては、Mosmann and Coffman, Ann. Rev. Immunol. 7:145−173, 1989を参照されたい。
【0092】
従って、Th1型が優勢な応答を誘発するための使用に適したアジュバントとしては、例えば、モノホスホリル脂質A、好ましくは3−デ−O−アシル化モノホスホリル脂質A(3D−MPL)とアルミニウム塩との組合せが挙げられる。Th1型免疫応答を優先的に誘発するその他の公知のアジュバントとして、CpG含有オリゴヌクレオチドがある。このオリゴヌクレオチドは、CpGジヌクレオチドがメチル化されていないことを特徴とする。このようなオリゴヌクレオチドはよく知られており、例えば、WO96/02555に記載されている。免疫刺激性DNA配列は、例えば、Satoら、Science 273:352, 1996にも記載されている。CpG含有オリゴヌクレオチドは、同じまたは異なるポリヌクレオチド構築物におけるHCV抗原から個別にコードされたものでもよいし、もしくは、例えば、該抗原との融合物として、それらにすぐ隣接していてもよい。あるいはまた、CpG含有オリゴヌクレオチドを個別に、すなわち、コードされた抗原を含む組成物の一部としてではなく、投与してもよい。CpGオリゴヌクレオチドは、単独でも、他のアジュバントと組み合わせて用いてもよい。例えば、増強された系は、CpG含有オリゴヌクレオチドとサポニン誘導体の組合せ、特に、WO 00/09159およびWO 00/62800に記載されているようなCpGとQS21の組合せを含む。この製剤は、さらに水中油形乳濁液および/またはトコフェロールを含むのが好ましい。
【0093】
別の好ましいアジュバントは、サポニン、好ましくはQS21(Aquila Biopharmaceuticals Inc., フラミンガム、MA)であり、これは単独でも、他のアジュバントと組み合わせて用いてもよい。例えば、増強された系は、モノホスホリル脂質Aとサポニン誘導体の組合せ、例えば、WO 94/00153に記載されているQS21と3D−MPLの組合せ、またはWO 96/33739に記載されているように、QS21をコレステロールでクエンチングした、反応性(reactogenic)の低い組成物を含む。別の好ましい製剤は、水中油形乳濁液およびトコフェロールを含む。QS21、3D−MPLおよび水中油形乳濁液中のトコフェロールを含む、特に効力のあるアジュバント製剤については、WO 95/17210に記載されている。
【0094】
別の好ましいアジュバントとして、モンタニドISA 720(Seppic、フランス)、SAF(Chiron、カリフォルニア、米国)、ISCOMS(CSL)、MF−59(Chiron)、デトクス(Detox)(Ribi、ハミルトン、MT)、RC−529(Corixa、ハミルトン、MT)ならびに他のアミノアルキルグルコサミニド4−ホスフェート(AGP)が挙げられる。
【0095】
ワクチンがアジュバントを含む場合、ワクチン製剤を2部に分けて投与してもよい。例えば、抗原をコードするヌクレオチド構築物を含む製剤の部分を、例えば、皮下または筋内注射、あるいは経皮粒子媒介送達により最初に投与してから、アジュバントを含む製剤の部分をその直後、またはワクチン分野に精通した医師には明らかな適当な時間をおいて、投与することができる。このような状況で、アジュバントは、抗原製剤と同じ経路、または別の経路のいずれにより投与してもよい。別の実施形態では、製剤のアジュバント部分を抗原部分の前に投与する。一実施形態では、アジュバントは局所製剤として、抗原をコードするヌクレオチド配列の粒子媒介送達の部位で、その粒子媒介送達の前または後のいずれかに、皮膚に適用する。
【0096】
好ましくは、本発明のDNAワクチンは、有効な免疫応答、典型的には、HCV抗原に対するCD4+およびCD8+免疫を刺激する。好ましくは、広範囲のエピトープに対する免疫を刺激する。治療に用いる場合には、ワクチン接種の後、肝線維症および/または炎症を軽減するのが好ましい。
【0097】
本明細書で用いる「含む」という用語は、非制限的意味で用いることを意図しており、その他の要素の存在も排除されない。しかし、「含んでいる」という用語は、その排他的意味で理解できるように意図することもあり、その場合、「構成される」または「からなる」と同等である。本発明を以下の実施例により説明するが、これらに限定されるわけではない。
【実施例】
【0098】
実施例1 抗原パネルに導入した突然変異
1)コンセンサス突然変異
すべての既知HCV分離物の全ゲノム配列を比較した。J4L6ポリタンパク質内の特定の位置を、他の大部分のHCV分離物から、特異である/逸脱するものとして確認した。特に重要なのは、これらの位置が、1a、2、3群、およびその他の群(この群では、さもなくば、同等の位置に1または2個の別のアミノ酸残基が占めていた。)に及ぶ関連1b群の分離物全体のより一致性の高い残基から逸脱することが認められたことである。選択したコンセンサス突然変異で、既知CD4またはCD8エピトープを妨害するものはない。NS3における2つの変更により、実際に免疫優性HLA−B35制限CD8エピトープを修復する[イソロイシン(I)1365からバリン(V)、およびグリシン(G)1366からアラニン(A)]。
【0099】
不用な可変性のために、NS4Bの最初の51アミノ酸を除去した。
【0100】
コア:
アラニン(A)52からトレオニン(T)へ。
【0101】
NS3:
バリン(V)1040からロイシン(L)へ。
【0102】
ロイシン(L)1106からグルタミン(Q)へ。
【0103】
セリン(S)1124からトレオニン(T)へ。
【0104】
バリン(V)1179からイソロイシン(I)へ。
【0105】
トレオニン(T)1215からセリン(S)へ。
【0106】
グリシン(G)1289からアラニン(A)へ。
【0107】
セリン(S)1290からプロリン(P)へ。
【0108】
イソロイシン(I)1365からバリン(V)へ。
【0109】
グリシン(G)1366からアラニン(A)へ。
【0110】
トレオニン(T)1408からセリン(S)へ。
【0111】
プロリン(P)1428からトレオニン(T)へ。
【0112】
イソロイシン(I)1429からセリン(S)へ。
【0113】
イソロイシン(I)1636からトレオニン(T)へ。
【0114】
NS4B:
フェニルアラニン(F)1760でORFを開始。
【0115】
NS5B:
イソロイシン(I)2824からバリン(V)へ。
【0116】
トレオニン(T)2892からセリン(S)へ。
【0117】
トレオニン(T)2918からバリン(V)へ。
【0118】
N.B.:番号付けは、J4L6分離物のポリタンパク質における位置に従う。
【0119】
実施例2 プラスミドDNAワクチンの構築
HCVコア、NS3、切断型(truncated)NS4B、およびNS5Bをコードするポリヌクレオチド配列に対し、SynGene 2eソフトウエアを用い、哺乳動物コドン使用頻度についてコドン最適化を実施した。コドン使用頻度係数は、各ポリヌクレオチドについて0.7以上まで改善された。コドン最適化、酵素ノックアウト突然変異、ならびにコンセンサス突然変異を組み込んだ新しいポリヌクレオチド配列各々のセンスおよびアンチセンス鎖を40〜60ヌクレオチドの領域(20ヌクレオチドのオーバーラップを含む)に分割した。これらの領域を商業的に合成し、オリゴアッセンブリィPCR法によりポリヌクレオチドを製造した。
【0120】
クローニングに用いられるユニークな制限エンドヌクレアーゼ部位を例示する、各ポリヌクレオチドの外側正方向および逆方向PCRプライマーの概略を以下に記載する。
【化12】
【0121】
単一抗原をコードするポリヌクレオチドはすべて、Not IおよびBamHIユニーククローニング部位を介して哺乳動物発現ベクターp7313ieにクローン化した(図7参照)。
【0122】
コードされたポリタンパク質は、以下の通りであった(突然変異とコドン最適化を含む)。
【化13】
【化14】
【化15】
【化16】
【0123】
実施例3 免疫応答アッセイ
C57BLまたはBALB/cマウスは、p7313ベクターにおいて個別に発現させた4つのHCV抗原のWTまたはコドン最適化+突然変異形態で免疫した。1.0μg/カートリッジの標準用量を含むPMIDによりマウスを免疫し、21日目に追加免疫(追加免疫1)、49日目に再度追加免疫(追加免疫2)した。脾細胞を個々のマウスから採取し、様々なHCV抗原製剤を用いたELISPOTで再刺激した。IL2およびIFNγ応答の両方を測定した。免疫応答を測定するのに用いた試薬は、ミクロゲン(Mikrogen)、ワクシニア−コアおよびワクシニアNS3−5(社内の遺伝子型1b)からの精製HCVコア、NS3、NS4およびNS5B(遺伝子型1b)タンパク質であった。
【0124】
HCVコア:
WT全長(FL−1−191)または切断型(TR1−115)コアで免疫したC57BLマウスをHCVコアタンパク質で再刺激したところ、精製コアタンパク質を用いて良好な応答が観察された(図8)。
【0125】
HCV NS3:
p7313 WTと、PMIDを用いてコドン最適化したNS3とでマウスを免疫した。NS3タンパク質とワクシニア3−5の両方を用いたC57Blマウスにおいて、免疫および1回の追加免疫後、NS3に対する良好な応答が実証され、ELISPOTにより応答を読み出した。IL2およびIFNγ応答の両方を検出した。この実験では、野生型と、コドン最適化した(co + m)形態の構築物との間に有意な差は観察されなかった(図9)。しかし、一過的トランスフェクション後のin vitro発現には、野生型とコドン最適化構築物との間に差が認められた。DNA用量が低い構築物、または一次応答を比較する実験から、プラスミドの効力の差が明らかになると考えられる。
【0126】
HCV NS4B:
BALB/cマウスのPMID免疫後に、全長WT p7313 NS4Bに対する応答を観察した。NS4Bタンパク質およびワクシニア3−5のいずれかでin vitro再刺激後に、IL2およびIFNγ ELISPOT応答の両方を観察した(図10)。
【0127】
NS4Bタンパク質をN末端で切断することにより、高度可変領域を除去したが、N末端領域に対して利用可能な抗血清が産生されていたため、in vitroトランスフェクション実験後に、タンパク質の発現を検出することはできなかった。この領域の発現を確認するために、これをNS5Bタンパク質と融合させた。近年の実験から、NS4Bタンパク質とNS3−5ワクシニアを用いて、切断型NS4Bタンパク質(単独またはNS5Bとの融合物として)に対し免疫応答を検出できることが確認されている。WTおよびコドン最適化NS4Bに対して良好な応答を観察した。
【0128】
HCV NS5B:
WTおよびコドン最適化(co+M)配列を用いた免疫後に、PMID後のNS5Bに対する免疫応答を調べた。NS3タンパク質とワクシニア3−5の両方を用いたC57BLマウスにおいて、免疫および1回の追加免疫後、NS5Bに対する良好な応答が実証され、ELISPOTにより応答を読み出した。NS3を用いた場合と同様、この実験でも、WTとco+m形態の構築物との間に、免疫応答の差は認められなかった(図11)。
【0129】
実施例4 HCVポリタンパク質の発現
4つの選択したHCV抗原コア、NS3、NS4BおよびNS5Bをp7313ieにフォーマット化することにより、単一の融合ポリタンパク質として発現させた。これらの抗原を以下に示すように様々な構築物において異なる順序で発現させた。(4つの抗原の各々がアミノ末端の位置を順に占めるように、単一ポリタンパク質の発現をコードする構築パネルを設計し、この重要な位置で1つの抗原の存在が他の抗原に対し、発現のレベルを有意に改善するのか、あるいは低減させるのかをモニターした。)さらに、コアタンパク質を2つの断片、すなわち、コア66−191>1−65と105−191>1−104を介して再配置した、2つの構築物を製造した。
【化17】
【0130】
標準的な製造者のプロトコルに従い、リポフェクタミン2000トランスフェクション試薬(Invitrogen/Life Technologies)を用いて、標準化量のDNAをHEK 293T細胞にトランスフェクションした。トランスフェクションから24時間後に細胞を回収し、MOPSまたはMES既製バッファー(Invitrogen/Life Technologies)のいずれかで前形成したNuPAGE 4−12%Bis−Trisを用いて、ポリアクリルアミドゲル電気泳動を実施した。分離したタンパク質をPVDF膜にブロッティングし、NS5B全タンパク質に対して産生したウサギ抗血清を用いて、タンパク質発現をモニターした。二次プローブは、西洋ワサビペルオキシダーゼ(hrp)に結合させた抗ウサギ免疫グロブリン抗血清であり、続いて、ECL試薬(Amersham Biosciences)を用いて、化学発光検出を実施した。
【0131】
この発現実験の結果を図12に示す。これらの結果から、ポリタンパク質はすべて、同様の程度まで発現するが、NS5Bを発現する単一抗原に認められるものよりは低いレベルである。HCV500の分子量がやや低いのは、N末端位置からのHCVコアの切断のためである。HCV502は、クローニングの誤りによりこの実験では検出されなかった。別のクローンを用いた反復実験では、HCV502の発現レベルは他のポリタンパク質と類似していた。
【0132】
実施例5 HCVポリタンパク質に対する免疫応答の検出
ポリタンパク質の各々をコードするDNA(1μg)を含むPMIDによりC57BLマウスを免疫し、実施例4に記載したようにそれから3週間後に追加免疫した。ELISPOT、またはHCV抗原に対する細胞内サイトカインの産生を用いて、追加免疫から7日後に免疫応答をモニタリングした。
【0133】
HCV遺伝子産物に対するT細胞応答についてのELISPOTアッセイ:
脾細胞の調製:
追加免疫から7日後に、免疫動物から脾臓を取得した。脾臓をスライドガラスの間で粉砕して処理することにより、細胞懸濁液を生成した。塩化アンモニウム処理で赤血球を溶解させた後、細胞破片を除去することにより、脾細胞の微細な懸濁液を得た。ELISPOTアッセイに用いるため、マウスが一次免疫しか受けていない場合は4×106/ml、またマウスが追加免疫を受けている場合には2×106/mlの濃度で細胞をRPMI完全培地に再懸濁させた。
【0134】
ELISPOTアッセイ:
15μg/ml(PBS中)のラット抗マウスIFNγまたはラット抗マウスIL−2(Phamingen)でプレートを被覆した。プレートは+4℃で一晩被覆した。使用前に、プレートをPBSで3回洗浄した。4×105細胞/ウェルで脾細胞をプレートに添加した。組換えHCV抗原をミクロゲン(Mikrogen)から取得し、1μg/mlで用いた。1〜10μMの最終濃度でペプチドをアッセイに用いることにより、CD4またはCD8応答を測定した。これらのペプチドは、Genemed Synthesisから取得した。各ウェルの合計容量は200μlであった。抗原で刺激した細胞を含むプレートを、加湿した37℃のインキュベーター内で16時間インキュベートした。いくつかの実験では、NS3−5またはワクシニア野生型を発現する組換えワクシニアに感染させた細胞をELISPOTアッセイで抗原として用いた。
【0135】
ELISPOTアッセイプレートの展開:
水で1回(細胞の溶解を確実にするため1分浸ける)、次にPBSで3回洗浄することにより、プレートから細胞を除去した。ビオチン結合ラット抗マウスIFNγまたはIL−2(Phamingen)を1μg/mlでPBSに添加した。プレートを室温で攪拌しながら2時間インキュベートした。次に、プレートをPBSで3回洗浄した後、ストレプトアビジンアルカリホスファターゼ(Caltag)を1/1000希釈で添加した。PBSで3回の洗浄後、BCICP基質(Biorad)を用いた15〜45分のインキュベーションにより、スポットが明らかになった。水を用いて基質を洗浄してから、プレートを乾燥させた。イメージ分析装置を用いて、スポットを数えた。
【0136】
ペプチド刺激に応答するT細胞からのIFNγおよびIL−2産生を検出するためのフローサイトメトリー:
試験管1本当たり約3×106個の脾細胞に等分し、回転することによりペレットとした。上清を除去し、サンプルを激しく攪拌し、ペレットを壊した。0.5μgの抗CD28+0.5μgの抗CD49d(Phamingen)を各試験管に添加し、室温で10分放置することによりインキュベートした。適切な試験管に1mlの培地を添加したが、これは培地だけか、もしくはHCV抗原を含む培地であった。次にサンプルを加熱水浴中に37℃で1時間インキュベートした。10μg/mlのブレフェルジンAを各試験管に添加し、37℃でのインキュベーションをさらに5時間継続した。プログラム化した水浴を6℃に戻し、その温度で一晩維持した。
【0137】
抗マウスCD4−CyChrome(Phamingen)と抗マウスCD8ビオチン(Immnotech)でサンプルを染色した。サンプルを洗浄し、ストレプトアビジン−ECDで染色した。サンプルを洗浄し、「イントラプレップ膜透過試薬(Intraprep Permeabilization Reagent)」キット(Immnotech)から100μgの固定液を15分室温で添加した。洗浄後、イントラプレップキットからの100μgの膜透過試薬を、抗IFN−γ−PE+抗IL−2−FITCを含む各サンプルに添加した。サンプルを室温で15分インキュベートした後、洗浄した。サンプルを0.5 mlバッファー中に再懸濁させ、フローサイトメーターで分析した。
【0138】
1サンプル当たり合計500,000個の細胞を収集してから、CD4およびCD8細胞をゲーティングすることにより、刺激に応答してIFN−γおよび/またはIL−2を分泌する細胞の集団を測定した。
【0139】
これらの結果から、様々な順序でコア、NS3、NS4BおよびNS5Bをコードするポリタンパク質はすべてNS3(すなわち、HCV 500、510、520、530)に対する免疫応答を刺激できることがわかった。IL2(図13A)およびIFN−γ(図13B)ELISPOTによりモニタリングしたところ、NS3タンパク質に対する応答は、HCVポリタンパク質(HCV 500、510、520および530)の各々で類似していた。
【0140】
応答細胞の表現型をICSによりさらに詳細に分析した。良好なCD4+T細胞応答が免疫優性NS3 CD4特異的ペプチドに対して誘発され、これは、HCV 500、510、520および530間で類似していた。
【表1】
【0141】
抗原の存在または非存在下で6時間、最後の4時間はブレフェルジンAの存在下で、脾細胞の刺激後、IFN−γ特異的なT細胞応答が検出された。CD4またはCD8T細胞上でのゲーティングおよびIFNγ FITCでの染色により、IFNgが検出された。
【0142】
また、HCV 500、510、520および530での免疫後、免疫優性NS3特異的ペプチドに対する強いCD8応答が生じ、CD8+細胞の2.5〜6%の頻度に達した。
【0143】
HCV 500、510、520および530での免疫によって、NS4BおよびNS5B抗原の両方に対するCD4およびCD8応答も検出されたが、CD8応答は、後の単一抗原での免疫後に比べ、ポリタンパク質に対して弱かった。
【表2】
【0144】
抗原の存在または非存在下で6時間、最後の4時間はブレフェルジンAの存在下で、脾細胞の刺激後、IFN−γ特異的なT細胞応答が検出された。CD4またはCD8T細胞上でのゲーティングおよびIFNγ FITCでの染色により、IFNgが検出された。
【表3】
【0145】
抗原の存在または非存在下で6時間、最後の4時間はブレフェルジンAの存在下で、脾細胞の刺激後、IFN−γ特異的なT細胞応答が検出された。CD4またはCD8T細胞上でのゲーティングおよびIFNγ FITCでの染色により、IFNgが検出された。
【0146】
用いたペプチドは以下の配列を有する。
【化18】
【0147】
内因的にプロセッシングされた抗原の認識:
HCVポリタンパク質でのPMID免疫が、内因的にプロセッシングされた抗原を認識することができる応答を誘発したか否かを決定するために、NS3−5を発現するワクシニア組換えウイルスに感染させた標的細胞をELISPOTアッセイで刺激因子として用いた。結果から、500、510、520および530での免疫後、良好なIL2およびIFNγ ELISPOT応答が検出されたことがわかる(図14)。
【0148】
HCVポリタンパク質での免疫は機能的CTL活性を誘導する:
NS3だけ、HCV500、510および520をコードする0.01μgのDNAでC57BLマウスを免疫した。初回免疫および1回の追加免疫後、各グループからの脾細胞をNS3 CD8ペプチドおよびIL2で5日間in vitroで再刺激した。同じペプチドを用いてパルスしたEL4細胞に対するCTL活性を測定した。すべての構築物で免疫したマウスが、このアッセイにおいて同様の死滅レベルを示した。
【0149】
このことから、HCVポリタンパク質によるPMID免疫が機能的CD8応答を誘発できることがわかる。結果を図15に示す。
【0150】
実施例6 二重プロモーター構築物を介したHCV抗原の送達
以下に示す方法を用いて、二重プロモーター構築物を作製した。発現カセット1(アイオワ−レングス(Iowa−length)CMVプロモーター、エクソン1、目的のタンパク質/融合タンパク質をコードする遺伝子、プラス、ウサギグロビンポリ−Aシグナル)を保有する断片をその宿主ベクター、すなわちp7313ieから、ユニーク制限エンドヌクレアーゼ部位ClaIおよびXmnIにより切除した。XmnIは、切除した断片の3−プライム(prime)末端に平滑末端を形成した。
【0151】
受容プラスミドベクターは、発現カセット2を含むp7313ieであった。これは、ユニーク制限エンドヌクレアーゼSse8387Iによる消化後、T4DNAポリメラーゼとインキュベートして、形成された3−プライムオーバーハングを除去することによって、線状分子の5−プライム末端および3−プライム末端の両方を平滑末端にすることにより、作製した。これは、259 bp断片を除去するユニーク制限エンドヌクレアーゼClaIを用いて切断された。
【0152】
ClaI/平滑適合性末端を介して、発現カセット1をp7313ie/発現カセット2にクローン化して、p7313ie/発現カセット1+発現カセット2を作製したが、その際、カセット1はカセット2の上流に位置した。
【0153】
以下のものを含むp7313ieプラスミドを作製した。
【化19】
【0154】
上に示した構築物パネルは完全であり、ウェスタンブロットにより、293T細胞における一過的トランスフェクションからの発現についてモニターした。ウェスタンブロット分析の結果を図16に示す:
レーンの説明:
1.p7313ie/コア
2.p7313ie/NS3
3.p7313ie/NS5B
4.p7313ie/コアNS3
5.p7313ie/NS4B5B
6.p7313ie/NS3コア
7.p7313ie/NS34B5B
8.p7313ie/コアNS3+NS4B5B
9.p7313ie/NS4B5B+コアNS3
10.p7313ie/NS3コア+NS4B5B
11.p7313ie/NS4B5B+NS3コア
12.p7313ie/コア+NS34B5B
13.p7313ie/NS34B5B+コア
構築物の各対は2つの独立した発現カセットを保有する。カセットをベクターに挿入する順序が、いずれかのカセットからの発現に影響を及ぼすとは予想しなかった。しかし、これらの結果から、NS4B5BまたはNS34B5B融合タンパク質の発現に関して、それぞれの発現カセットがコア、NS3コア、もしくはコアNS3カセットの下流に位置するとき、有意に不利であることがわかる。
【0155】
発現レベルは、単一抗原構築物ほど明確ではないが、大きさの有意な増大(175〜228%)のためにある程度の低減が予想され、これは、細胞に送達されるプラスミドのコピー数の減少(同じ質量のDNAに対して約50%以下の減少)を意味する。
【0156】
二重プロモーター構築物により誘発されるin vivo免疫原性:
免疫原性実験のために、4つの抗原すべての最大発現を呈示した3つの二重プロモーター構築物を選択した。これらは、p7313ie NS4B/NS5B+コア/NS3、p7313ie NS4B/NS5B+NS3コアおよびp7313ie NS3/NS4B/NS5B+コアであった。C57BLマウスをPMIDにより1μgのDNAで免疫し、7日後、IL2についてはELISPOTを用いて、優性NS3 CD8T細胞エピトープに対する応答を測定した。結果(図17に示す)から、単一免疫(CD4およびCd8 NS3T細胞特異的なペプチドで刺激した脾細胞)後、すべての二重プロモーター構築物に対して応答が観察されたことがわかる。
【0157】
実施例7 コアの欠失突然変異
コアのORFをコードする多数の遺伝子を、3’末端から1回につき20アミノ酸にわたる領域ずつ徐々に欠失させて作製し、完全に配列決定した。
【表4】
【0158】
図18は、コアの断片をコードする遺伝子の範囲を示すDNAアガロースゲルを表す。これらの構築物の発現を、NS4B5B 融合物(p7313ie/NS4B5B)の発現レベルへのそれらの作用と併せて、293T細胞での共トランスフェクションにより試験した。結果を図19に示す。レーンは以下のようにロードされた。
【表5】
【0159】
コア191、コアΔ15、コア171、コア151、およびコア131の発現は、NS4B5Bの発現の抗NS5B検出後、ウェスタンブロットを抗コアでプローブすると、はっきりと検出される。恐らく、用いたゲル系のサイズキャプチャー(size−capture)制限のために、コアの別の切断形態は検出されていない。
【0160】
結果から、コア191およびコアΔ15の存在下でのNS4B5Bの発現レベルに、両コア種の強い発現にもかかわらず、有意な低減(これは、コア171、そして再度コア151で回復する)が明らかである。この観察は、NS4B5Bで2回、NS3およびNS5Bで1回繰り返した。
【0161】
実施例8 NS3、NS5B、NS4B−NS5B融合物およびNS3−NS4B−NS5B三重融合物の発現に対するコアおよびコア151の影響
実験1−トランスフォーマットでの発現:
コアをトランスで発現させる、すなわち別々のプラスミド上にコードさせた場合に、非構造抗原の発現に対するコア191対コア151の発現の影響を比較してモニタリングするための実験を実施した。実験プロトコルは、実施例7に記載したのと同じであった。簡単にいうと、標準的プロトコル(Invitrogen/Life Technologies)におけるリポフェクタミン2000トランスフェクション試薬を用いて、各々0.5μgの2種のDNAプラスミドベクター(概略を以下の表に示す)をHEK 293T細胞中に共トランスフェクションした。(トランスフェクションおよびウェスタンブロット方法は、実施例4に記載の通り)。
【0162】
結果を図20に示すが、同図では、レーンを以下の表に記載したようにロードしており、ウェスタンブロット分析を実施することにより、主として抗NS3および抗NS5B抗血清を用いて非構造タンパク質の発現、および抗コアを含む同じブロットの二次プローブによりコアの発現を検出した。
【表6】
【0163】
すべてのケースで、コア151と一緒にトランスで産生された場合、非構造タンパク質または融合物(NS3、NS5B、NS4B−NS5B)の量は、コア191と一緒にトランスで発現させたときに産生されるレベルと比較して、有意に増加することが示された。
【0164】
実験2−シスフォーマットでの発現:
コアをシスで発現させる、すなわち、非構造要素と融合した同じプラスミド上にコードさせた場合に、非構造抗原の発現に対するコア191対コア151の発現の影響を比較してモニタリングするための実験を実施した。各ケースで、コア151は、指定した非構造領域とカルボキシ末端融合したコア191に置換した。
【0165】
標準的プロトコル(Invitrogen/Life Technologies)におけるリポフェクタミン2000トランスフェクション試薬を用いて、1μgのDNAプラスミドベクター(概略を以下の表に示す)をHEK 293T細胞中にトランスフェクションさせた(トランスフェクションおよびウェスタンブロット方法は、実施例4に記載の通り)。
【0166】
結果を図21に示す。ウェスタンブロット分析を実施することにより、ゲルAにおいて、主として抗NS3および抗NS5B抗血清を用いて非構造成分の発現、及び抗コアを含む同じブロットの二次プローブによりコアの発現を検出した。レーンは以下の表に記載するようにロードした。
【表7】
【0167】
結果から、抗原がポリタンパク質融合物の形態であるシス様式では、コアの切断により、融合タンパク質の発現が増大することがわかる。
【0168】
NS3に対する免疫応答に対するコア191およびコア151の影響の比較:
C57BLマウスをPMIDにより全DNA1.5μgの2回の注射で免疫した。免疫したグループは、空ベクターp7313ieだけ、p7313ieNS3、p7313ieNS5Bとp7313ieコア191またはp7313ieNS3、p7313ieNS5Bとp7313ieコア151による金ビーズの共コーティングを含んでいた。共コーティングを用いたのは、これによって、すべてのプラスミドが同じ細胞に送達され、この細胞が、前記のin vitro共トランスフェクション実験を模倣すると考えられるためである。初回免疫から14日後、IFNγおよびIL2抗原特異的な応答を測定するための細胞内サイトカイン染色を用いて、NS3からの優性CD8およびCD4T細胞エピトープに対する免疫応答を測定した。結果(図22に示す)から、CD4およびCD8 NS3応答は、コア191と比較してコア151の存在下で約2倍以上高くなったことがわかる。
【0169】
別の実験では、コア191またはコア151のいずれかと一緒に、p7313ieNS3/NS4B/NS5B 三重融合物を発現するプラスミドを共コーティングした金ビーズでC57BLマウスを免疫した。マウスをさらに同じ構築物で追加免疫し、追加免疫から7日後に、応答を測定するために細胞内サイトカイン染色を用いてNS3に対する応答をモニターした。図23に示した結果から、NS3抗原特異的CD4およびCD8応答は、コア191と比較して、コア151の存在下で約2倍高いことがわかる。
【0170】
コアの存在下でのNS3に対する応答を比較するin vivo実験全体は、FLコアと非構造タンパク質の同時送達により非構造抗原の発現が低減する可能性があり、これによって、構築物の免疫原性が低下するというin vitro発現データを支持する。この影響は、C末端40アミノ酸が除去された切断型コアを共コーティングすることにより少なくとも部分的に解消することができる。
【図面の簡単な説明】
【0171】
【図1−1】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−2】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−3】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−4】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図1−5】HCV J4L6ゲノム野生型cDNA配列(参照登録番号AF054247)。
【図2】コドン最適化HCVコアポリヌクレオチド。
【図3】コドン最適化HCV NS3ポリヌクレオチド。
【図4】コドン最適化HCV NS4Bポリヌクレオチド。
【図5】コドン最適化HCV NS5Bポリヌクレオチド。
【図6−1】HCV J4L6ゲノム(野生型配列)の翻訳。
【図6−2】HCV J4L6ゲノム(野生型配列)の翻訳。
【図7】p7313−ieの構造。
【図8】コアに対する免疫応答。
【図9】NS3免疫原性。
【図10】NS4Bに対する免疫応答。
【図11】NS5Bに対する免疫応答。
【図12】抗HCV NS5Bを用いるタンパク質発現のモニタリング。
【図13】IL2(A)及びIFN−γ(B) ELISPOTによるNS3タンパク質に対する免疫応答。
【図14】NS3−5を発現するワクシニアウイルスに感染させた脾細胞を刺激因子とするIL2及びIFNγ ELISPOTアッセイ結果。
【図15】HCVポリタンパク質でのPMID免疫による機能的CD8応答の誘発。
【図16】293T細胞での一過的トランスフェクションからの発現を示すウエスタンブロット分析の結果。
【図17】二重プロモーター構築物により誘導されるNS3 T細胞応答の比較。
【図18】コアの断片をコードする遺伝子の範囲を示すDNAアガロースゲル。
【図19】293T細胞での共トランスフェクションとNS4B5B融合物の発現レベル。
【図20】293T細胞での共トランスフェクション後のNS3、NS5B、NS4B5B、およびNS34B5Bの発現に対するコアおよびコア151の影響。
【図21】293T細胞での一過的トランスフェクションにおけるコア151のコア191への置換後の融合タンパク質の発現に対する影響。
【図22】NS3からの優性CD8及びCD4T細胞エピトープに対する免疫応答。
【図23】NS3に対するCD4及びCD8応答。
【特許請求の範囲】
【請求項1】
HCVタンパク質コア、NS3、NS4BおよびNS5Bのポリペプチド配列をコードするポリヌクレオチドを含む、医療における使用のためのHCVワクチン。
【請求項2】
前記ポリヌクレオチドが他のHCVタンパク質をコードしない、請求項1に記載のHVCワクチン。
【請求項3】
ポリヌクレオチドが、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断されたコアタンパク質をコードする、請求項1または2に記載のHCVワクチン。
【請求項4】
前記切断型コアタンパク質が、少なくともC末端の10アミノ酸の欠失を有する、請求項3に記載のHCVワクチン。
【請求項5】
前記切断型コアタンパク質がコア1−151配列からなる、請求項4に記載のHCVワクチン。
【請求項6】
前記HCVタンパク質が1以上の前記HCVタンパク質を含有する融合タンパク質の形態で存在する、請求項1に記載のHCVワクチン。
【請求項7】
前記融合タンパク質がNS4BとNS5Bのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項8】
前記融合タンパク質がNS3とコアのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項9】
前記HCVタンパク質が1以上の発現カセット中のポリヌクレオチドによってコードされる、請求項1に記載のHCVワクチン。
【請求項10】
コアタンパク質をコードする前記発現カセットが、少なくとも前記他のHCVタンパク質をコードする発現カセットの下流のシス位置にある、請求項9に記載のHCVワクチン。
【請求項11】
コアタンパク質をコードする前記発現カセットが、前記NS5Bタンパク質をコードする発現カセットの下流である、請求項10に記載のHCVワクチン。
【請求項12】
前記HCVタンパク質の少なくとも1つが突然変異により不活性化されている、請求項1に記載のHCVワクチン。
【請求項13】
前記ポリヌクレオチドがモチーフAに突然変異を含むNS5Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項14】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのプロテアーゼ活性が触媒三つ組アミノ酸のいずれかにおける突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項15】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのヘリカーゼ活性がヘリカーゼモチーフI、II、III、もしくはIVの1以上における突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項16】
前記ポリヌクレオチドが、高度可変のN末端領域を除去する切断を含むNS4Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項17】
前記ポリヌクレオチドワクチンがHCV組合せ1〜19のいずれか1つをコードする、請求項1〜16のいずれか1項に記載のHCVワクチン。
【請求項18】
前記ポリヌクレオチドがDNA配列である、請求項1に記載のHCVワクチン。
【請求項19】
前記DNA配列がプラスミドの形態である、請求項18に記載のHCVワクチン。
【請求項20】
前記オリゴヌクレオチドのコドンが哺乳動物細胞での発現のために最適化されている、請求項1〜17のいずれか1項に記載のワクチン。
【請求項21】
請求項1〜17のいずれか1項に記載のワクチンを哺乳動物に投与することを含んでなる、哺乳動物におけるHCV感染症の予防または治療方法。
【請求項22】
請求項1〜17のいずれか1項に記載のポリヌクレオチドワクチンを取得し、金ビーズ上にポリヌクレオチドをコーティングし、該金ビーズを皮膚内に送達することを含む、個体のワクチン接種方法。
【請求項23】
HCV治療用の医薬品の製造における、請求項1〜17のいずれか1項に記載のHCVワクチンの使用。
【特許請求の範囲】
【請求項1】
HCVタンパク質コア、NS3、NS4BおよびNS5Bのポリペプチド配列をコードするポリヌクレオチドを含む医療に使用するためのHCVワクチンであって、該ポリヌクレオチドが他のHCVタンパク質をコードしない、上記ワクチン。
【請求項2】
ポリヌクレオチドが、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断されたコアタンパク質をコードする、請求項1に記載のHCVワクチン。
【請求項3】
前記切断型コアタンパク質が少なくともC末端の10アミノ酸の欠失を有する、請求項2に記載のHCVワクチン。
【請求項4】
前記切断型コアタンパク質がコア1−151配列からなる、請求項3に記載のHCVワクチン。
【請求項5】
前記切断型コアタンパク質がコア1−165配列からなる、請求項3に記載のHCVワクチン。
【請求項6】
前記HCVタンパク質が1以上の前記HCVタンパク質を含有する融合タンパク質の形態で存在する、請求項1に記載のHCVワクチン。
【請求項7】
前記融合タンパク質がNS4BとNS5Bのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項8】
前記融合タンパク質がNS3とコアのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項9】
前記HCVタンパク質が2つ以上の発現カセット中のポリヌクレオチドによってコードされる、請求項1に記載のHCVワクチン。
【請求項10】
コアタンパク質をコードする前記発現カセットが、少なくとも前記他のHCVタンパク質をコードする発現カセットの下流のシス位置にある、請求項9に記載のHCVワクチン。
【請求項11】
コアタンパク質をコードする前記発現カセットが、NS5Bタンパク質をコードする発現カセットの下流にある、請求項10に記載のHCVワクチン。
【請求項12】
前記HCVタンパク質の少なくとも1つが突然変異により不活性化されている、請求項1に記載のHCVワクチン。
【請求項13】
前記ポリヌクレオチドがモチーフAに突然変異を含むNS5Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項14】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのプロテアーゼ活性が触媒三つ組アミノ酸のいずれかにおける突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項15】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのヘリカーゼ活性がヘリカーゼモチーフI、II、IIIもしくはIVの1以上における突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項16】
前記ポリヌクレオチドが、高度可変のN末端領域を除去する切断を含むNS4Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項17】
前記ポリヌクレオチドワクチンがHCV組合せ1〜19のいずれか1つをコードする、請求項1〜16のいずれか1項に記載のHCVワクチン。
【請求項18】
前記ポリヌクレオチドがDNA配列である、請求項1に記載のHCVワクチン。
【請求項19】
前記DNA配列がプラスミドの形態である、請求項18に記載のHCVワクチン。
【請求項20】
前記オリゴヌクレオチドのコドンが哺乳動物細胞での発現のために最適化されている、請求項1〜17のいずれか1項に記載のワクチン。
【請求項21】
請求項1〜17のいずれか1項に記載のワクチンを哺乳動物に投与することを含んでなる、哺乳動物におけるHCV感染症の予防または治療方法。
【請求項22】
請求項1〜17のいずれか1項に記載のポリヌクレオチドワクチンを取得し、金ビーズ上に該ポリヌクレオチドをコーティングし、該金ビーズを皮膚内に送達することを含む、個体のワクチン接種の方法。
【請求項23】
HCV治療用の医薬品の製造における、請求項1〜17のいずれか1項に記載のHCVワクチンの使用。
【特許請求の範囲】
【請求項1】
HCVタンパク質コア、NS3、NS4BおよびNS5Bのポリペプチド配列をコードするポリヌクレオチドを含む医療に使用するためのHCVワクチンであって、該ポリヌクレオチドが他のHCVタンパク質をコードしない、上記ワクチン。
【請求項2】
ポリヌクレオチドが、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断されたコアタンパク質をコードする、請求項1に記載のHCVワクチン。
【請求項3】
前記切断型コアタンパク質が少なくともC末端の10アミノ酸の欠失を有する、請求項2に記載のHCVワクチン。
【請求項4】
前記切断型コアタンパク質がコア1−151配列からなる、請求項3に記載のHCVワクチン。
【請求項5】
前記切断型コアタンパク質がコア1−165配列からなる、請求項3に記載のHCVワクチン。
【請求項6】
前記HCVタンパク質が1以上の前記HCVタンパク質を含有する融合タンパク質の形態で存在する、請求項1に記載のHCVワクチン。
【請求項7】
前記融合タンパク質がNS4BとNS5Bのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項8】
前記融合タンパク質がNS3とコアのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項9】
前記HCVタンパク質が2つ以上の発現カセット中のポリヌクレオチドによってコードされる、請求項1に記載のHCVワクチン。
【請求項10】
コアタンパク質をコードする前記発現カセットが、少なくとも前記他のHCVタンパク質をコードする発現カセットの下流のシス位置にある、請求項9に記載のHCVワクチン。
【請求項11】
コアタンパク質をコードする前記発現カセットが、NS5Bタンパク質をコードする発現カセットの下流にある、請求項10に記載のHCVワクチン。
【請求項12】
前記HCVタンパク質の少なくとも1つが突然変異により不活性化されている、請求項1に記載のHCVワクチン。
【請求項13】
前記ポリヌクレオチドがモチーフAに突然変異を含むNS5Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項14】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのプロテアーゼ活性が触媒三つ組アミノ酸のいずれかにおける突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項15】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのヘリカーゼ活性がヘリカーゼモチーフI、II、IIIもしくはIVの1以上における突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項16】
前記ポリヌクレオチドが、高度可変のN末端領域を除去する切断を含むNS4Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項17】
前記ポリヌクレオチドワクチンがHCV組合せ1〜19のいずれか1つをコードする、請求項1〜16のいずれか1項に記載のHCVワクチン。
【請求項18】
前記ポリヌクレオチドがDNA配列である、請求項1に記載のHCVワクチン。
【請求項19】
前記DNA配列がプラスミドの形態である、請求項18に記載のHCVワクチン。
【請求項20】
前記オリゴヌクレオチドのコドンが哺乳動物細胞での発現のために最適化されている、請求項1〜17のいずれか1項に記載のワクチン。
【請求項21】
HCV治療用の医薬品の製造における、請求項1〜17のいずれか1項に記載のHCVワクチンの使用。
【請求項1】
HCVタンパク質コア、NS3、NS4BおよびNS5Bのポリペプチド配列をコードするポリヌクレオチドを含む、医療における使用のためのHCVワクチン。
【請求項2】
前記ポリヌクレオチドが他のHCVタンパク質をコードしない、請求項1に記載のHVCワクチン。
【請求項3】
ポリヌクレオチドが、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断されたコアタンパク質をコードする、請求項1または2に記載のHCVワクチン。
【請求項4】
前記切断型コアタンパク質が、少なくともC末端の10アミノ酸の欠失を有する、請求項3に記載のHCVワクチン。
【請求項5】
前記切断型コアタンパク質がコア1−151配列からなる、請求項4に記載のHCVワクチン。
【請求項6】
前記HCVタンパク質が1以上の前記HCVタンパク質を含有する融合タンパク質の形態で存在する、請求項1に記載のHCVワクチン。
【請求項7】
前記融合タンパク質がNS4BとNS5Bのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項8】
前記融合タンパク質がNS3とコアのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項9】
前記HCVタンパク質が1以上の発現カセット中のポリヌクレオチドによってコードされる、請求項1に記載のHCVワクチン。
【請求項10】
コアタンパク質をコードする前記発現カセットが、少なくとも前記他のHCVタンパク質をコードする発現カセットの下流のシス位置にある、請求項9に記載のHCVワクチン。
【請求項11】
コアタンパク質をコードする前記発現カセットが、前記NS5Bタンパク質をコードする発現カセットの下流である、請求項10に記載のHCVワクチン。
【請求項12】
前記HCVタンパク質の少なくとも1つが突然変異により不活性化されている、請求項1に記載のHCVワクチン。
【請求項13】
前記ポリヌクレオチドがモチーフAに突然変異を含むNS5Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項14】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのプロテアーゼ活性が触媒三つ組アミノ酸のいずれかにおける突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項15】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのヘリカーゼ活性がヘリカーゼモチーフI、II、III、もしくはIVの1以上における突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項16】
前記ポリヌクレオチドが、高度可変のN末端領域を除去する切断を含むNS4Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項17】
前記ポリヌクレオチドワクチンがHCV組合せ1〜19のいずれか1つをコードする、請求項1〜16のいずれか1項に記載のHCVワクチン。
【請求項18】
前記ポリヌクレオチドがDNA配列である、請求項1に記載のHCVワクチン。
【請求項19】
前記DNA配列がプラスミドの形態である、請求項18に記載のHCVワクチン。
【請求項20】
前記オリゴヌクレオチドのコドンが哺乳動物細胞での発現のために最適化されている、請求項1〜17のいずれか1項に記載のワクチン。
【請求項21】
請求項1〜17のいずれか1項に記載のワクチンを哺乳動物に投与することを含んでなる、哺乳動物におけるHCV感染症の予防または治療方法。
【請求項22】
請求項1〜17のいずれか1項に記載のポリヌクレオチドワクチンを取得し、金ビーズ上にポリヌクレオチドをコーティングし、該金ビーズを皮膚内に送達することを含む、個体のワクチン接種方法。
【請求項23】
HCV治療用の医薬品の製造における、請求項1〜17のいずれか1項に記載のHCVワクチンの使用。
【特許請求の範囲】
【請求項1】
HCVタンパク質コア、NS3、NS4BおよびNS5Bのポリペプチド配列をコードするポリヌクレオチドを含む医療に使用するためのHCVワクチンであって、該ポリヌクレオチドが他のHCVタンパク質をコードしない、上記ワクチン。
【請求項2】
ポリヌクレオチドが、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断されたコアタンパク質をコードする、請求項1に記載のHCVワクチン。
【請求項3】
前記切断型コアタンパク質が少なくともC末端の10アミノ酸の欠失を有する、請求項2に記載のHCVワクチン。
【請求項4】
前記切断型コアタンパク質がコア1−151配列からなる、請求項3に記載のHCVワクチン。
【請求項5】
前記切断型コアタンパク質がコア1−165配列からなる、請求項3に記載のHCVワクチン。
【請求項6】
前記HCVタンパク質が1以上の前記HCVタンパク質を含有する融合タンパク質の形態で存在する、請求項1に記載のHCVワクチン。
【請求項7】
前記融合タンパク質がNS4BとNS5Bのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項8】
前記融合タンパク質がNS3とコアのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項9】
前記HCVタンパク質が2つ以上の発現カセット中のポリヌクレオチドによってコードされる、請求項1に記載のHCVワクチン。
【請求項10】
コアタンパク質をコードする前記発現カセットが、少なくとも前記他のHCVタンパク質をコードする発現カセットの下流のシス位置にある、請求項9に記載のHCVワクチン。
【請求項11】
コアタンパク質をコードする前記発現カセットが、NS5Bタンパク質をコードする発現カセットの下流にある、請求項10に記載のHCVワクチン。
【請求項12】
前記HCVタンパク質の少なくとも1つが突然変異により不活性化されている、請求項1に記載のHCVワクチン。
【請求項13】
前記ポリヌクレオチドがモチーフAに突然変異を含むNS5Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項14】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのプロテアーゼ活性が触媒三つ組アミノ酸のいずれかにおける突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項15】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのヘリカーゼ活性がヘリカーゼモチーフI、II、IIIもしくはIVの1以上における突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項16】
前記ポリヌクレオチドが、高度可変のN末端領域を除去する切断を含むNS4Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項17】
前記ポリヌクレオチドワクチンがHCV組合せ1〜19のいずれか1つをコードする、請求項1〜16のいずれか1項に記載のHCVワクチン。
【請求項18】
前記ポリヌクレオチドがDNA配列である、請求項1に記載のHCVワクチン。
【請求項19】
前記DNA配列がプラスミドの形態である、請求項18に記載のHCVワクチン。
【請求項20】
前記オリゴヌクレオチドのコドンが哺乳動物細胞での発現のために最適化されている、請求項1〜17のいずれか1項に記載のワクチン。
【請求項21】
請求項1〜17のいずれか1項に記載のワクチンを哺乳動物に投与することを含んでなる、哺乳動物におけるHCV感染症の予防または治療方法。
【請求項22】
請求項1〜17のいずれか1項に記載のポリヌクレオチドワクチンを取得し、金ビーズ上に該ポリヌクレオチドをコーティングし、該金ビーズを皮膚内に送達することを含む、個体のワクチン接種の方法。
【請求項23】
HCV治療用の医薬品の製造における、請求項1〜17のいずれか1項に記載のHCVワクチンの使用。
【特許請求の範囲】
【請求項1】
HCVタンパク質コア、NS3、NS4BおよびNS5Bのポリペプチド配列をコードするポリヌクレオチドを含む医療に使用するためのHCVワクチンであって、該ポリヌクレオチドが他のHCVタンパク質をコードしない、上記ワクチン。
【請求項2】
ポリヌクレオチドが、他のHCVタンパク質の発現に与えるコアの抑制作用を低減するのに十分な量でカルボキシ末端から切断されたコアタンパク質をコードする、請求項1に記載のHCVワクチン。
【請求項3】
前記切断型コアタンパク質が少なくともC末端の10アミノ酸の欠失を有する、請求項2に記載のHCVワクチン。
【請求項4】
前記切断型コアタンパク質がコア1−151配列からなる、請求項3に記載のHCVワクチン。
【請求項5】
前記切断型コアタンパク質がコア1−165配列からなる、請求項3に記載のHCVワクチン。
【請求項6】
前記HCVタンパク質が1以上の前記HCVタンパク質を含有する融合タンパク質の形態で存在する、請求項1に記載のHCVワクチン。
【請求項7】
前記融合タンパク質がNS4BとNS5Bのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項8】
前記融合タンパク質がNS3とコアのポリペプチド配列で構成される二重融合物である、請求項6に記載のHCVワクチン。
【請求項9】
前記HCVタンパク質が2つ以上の発現カセット中のポリヌクレオチドによってコードされる、請求項1に記載のHCVワクチン。
【請求項10】
コアタンパク質をコードする前記発現カセットが、少なくとも前記他のHCVタンパク質をコードする発現カセットの下流のシス位置にある、請求項9に記載のHCVワクチン。
【請求項11】
コアタンパク質をコードする前記発現カセットが、NS5Bタンパク質をコードする発現カセットの下流にある、請求項10に記載のHCVワクチン。
【請求項12】
前記HCVタンパク質の少なくとも1つが突然変異により不活性化されている、請求項1に記載のHCVワクチン。
【請求項13】
前記ポリヌクレオチドがモチーフAに突然変異を含むNS5Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項14】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのプロテアーゼ活性が触媒三つ組アミノ酸のいずれかにおける突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項15】
前記ポリヌクレオチドがNS3タンパク質をコードし、そのヘリカーゼ活性がヘリカーゼモチーフI、II、IIIもしくはIVの1以上における突然変異により破壊されている、請求項12に記載のHCVワクチン。
【請求項16】
前記ポリヌクレオチドが、高度可変のN末端領域を除去する切断を含むNS4Bタンパク質をコードする、請求項12に記載のHCVワクチン。
【請求項17】
前記ポリヌクレオチドワクチンがHCV組合せ1〜19のいずれか1つをコードする、請求項1〜16のいずれか1項に記載のHCVワクチン。
【請求項18】
前記ポリヌクレオチドがDNA配列である、請求項1に記載のHCVワクチン。
【請求項19】
前記DNA配列がプラスミドの形態である、請求項18に記載のHCVワクチン。
【請求項20】
前記オリゴヌクレオチドのコドンが哺乳動物細胞での発現のために最適化されている、請求項1〜17のいずれか1項に記載のワクチン。
【請求項21】
HCV治療用の医薬品の製造における、請求項1〜17のいずれか1項に記載のHCVワクチンの使用。
【図1−1】


【図1−2】

【図1−3】

【図1−4】

【図1−5】

【図2】

【図3】

【図4】

【図5】

【図6−1】

【図6−2】

【図7】
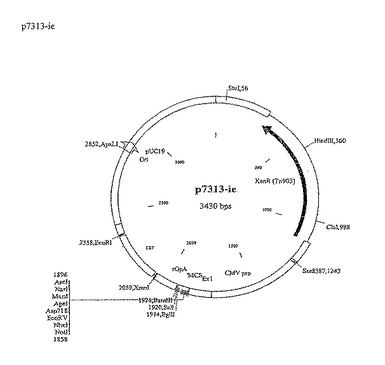
【図8】
【図9】
【図10】
【図11】
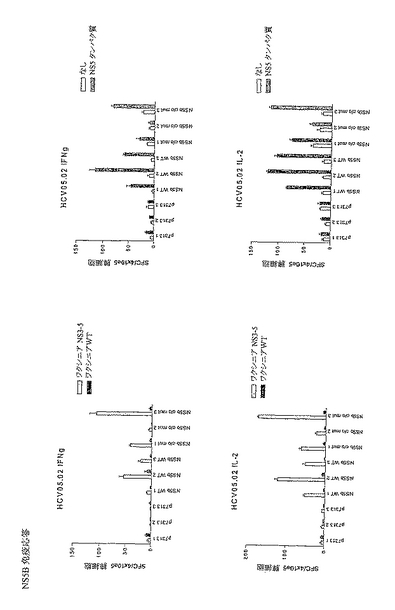
【図12】
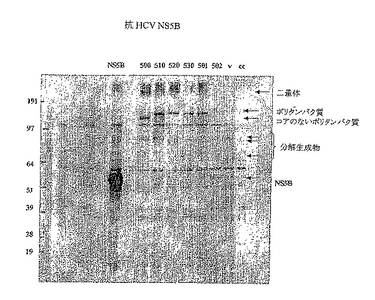
【図13】
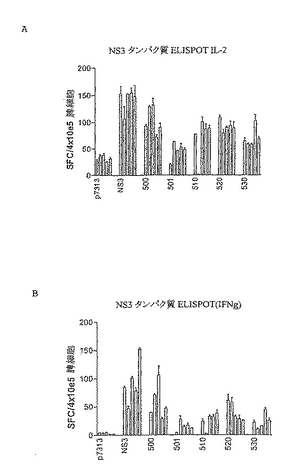
【図14】
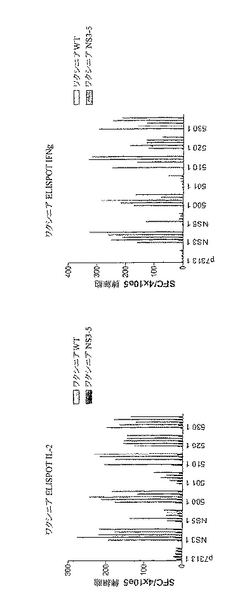
【図15】
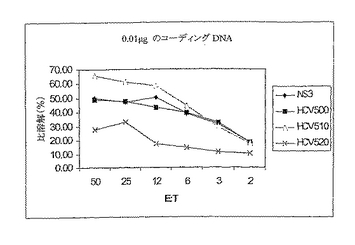
【図16】
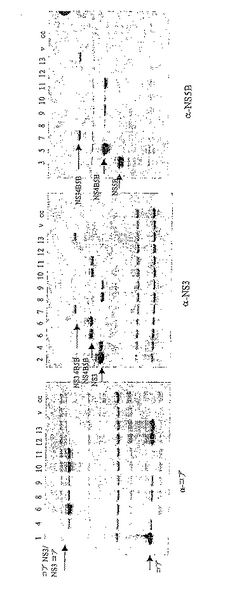
【図17】
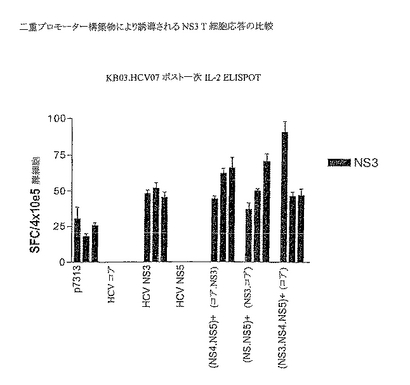
【図18】
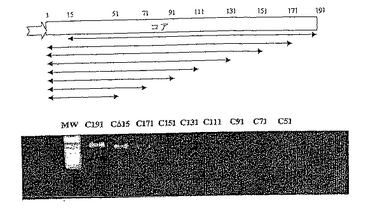
【図19】
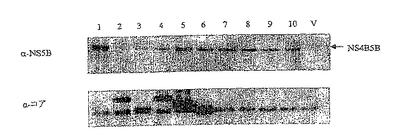
【図20】

【図21】

【図22】
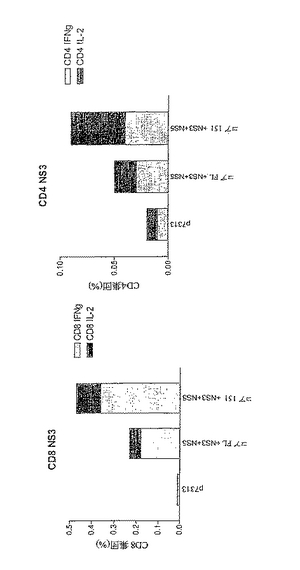
【図23】
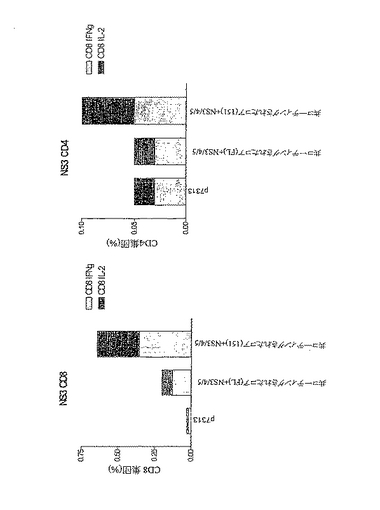
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図2】
【図3】
【図4】
【図5】
【図6−1】
【図6−2】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【公表番号】特表2006−524181(P2006−524181A)
【公表日】平成18年10月26日(2006.10.26)
【国際特許分類】
【出願番号】特願2004−552621(P2004−552621)
【出願日】平成15年11月13日(2003.11.13)
【国際出願番号】PCT/EP2003/012830
【国際公開番号】WO2004/046176
【国際公開日】平成16年6月3日(2004.6.3)
【出願人】(397009934)グラクソ グループ リミテッド (832)
【氏名又は名称原語表記】GLAXO GROUP LIMITED
【住所又は居所原語表記】Glaxo Wellcome House,Berkeley Avenue Greenford,Middlesex UB6 0NN,Great Britain
【Fターム(参考)】
【公表日】平成18年10月26日(2006.10.26)
【国際特許分類】
【出願日】平成15年11月13日(2003.11.13)
【国際出願番号】PCT/EP2003/012830
【国際公開番号】WO2004/046176
【国際公開日】平成16年6月3日(2004.6.3)
【出願人】(397009934)グラクソ グループ リミテッド (832)
【氏名又は名称原語表記】GLAXO GROUP LIMITED
【住所又は居所原語表記】Glaxo Wellcome House,Berkeley Avenue Greenford,Middlesex UB6 0NN,Great Britain
【Fターム(参考)】
[ Back to top ]