
ワクチン
本発明は、ナイセリアワクチン組成物、それらの製造、及びかかる組成物の医薬における使用の分野に関する。より具体的には、本発明は、ナイセリア(特に髄膜炎菌)の外膜小胞(すなわちブレブ)ワクチンの製造により好適な、新規な操作された髄膜炎菌の作製方法に関する。また、ヒト被験者における使用により安全でありかつより有効である新規なLOSサブユニット又は髄膜炎菌外膜小胞(すなわちブレブ)ワクチンの使用に基づく有利な方法及びワクチン製品を記載する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ナイセリアワクチン組成物、それらの製造、及びかかる組成物の医薬における使用の分野に関する。より具体的には、ナイセリア(特に髄膜炎菌)の外膜小胞(すなわち、ブレブ)ワクチンの製造により適した新規な操作された髄膜炎菌株の作製方法に関する。新規のLOSサブユニット又は髄膜炎菌外膜小胞(すなわち、ブレブ)ワクチンの使用に基づく有利な方法及びワクチン製品もまた記載しており、これはヒト被験者における使用のために安全性及び有効性をより高めたものである。
【背景技術】
【0002】
髄膜炎菌(Neisseria meningitidis)は、しばしばヒト上気道より単離されるグラム陰性細菌である。この細菌は、菌血症及び髄膜炎等の重篤な侵襲性細菌性疾患の原因である。髄膜炎菌疾患の発生数は、地理的、季節的及び年次的な差異を示す(Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。この細菌は通常、その莢膜多糖による血清群に従って分類される。
【0003】
温暖な国々で最も多い疾患は、血清群B株によるものであり、発症率は総人口の1〜10人/100,000人/年の間で変動し、それより高い値に達する場合もある(Kaczmarski, E.B. (1997), Commun. Dis. Rep. Rev. 7: R55-9, 1995;Scholten, R.J.P.M., Bijlmer, H.A., Poolman, J.T.ら Clin. Infect. Dis. 16:237-246, 1993;Cruz, C., Pavez, G., Aguilar, E.,ら Epidemiol. Infect. 105:119-126, 1990)。
【0004】
血清群A髄膜炎菌が優位を占める流行の大部分は中央アフリカで発生し、その発症率が1000人/100,000人/年のレベルにまで達する場合もある(Schwartz, B., Moore, P.S., Broome, C.V. Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。全体として、髄膜炎菌疾患のほとんど全ての症例は、血清群A、B、C、W−135及びYの髄膜炎菌によって引き起こされ、四価のA、C、W−135、Y莢膜多糖ワクチンが利用可能である(Armand, J., Arminjon, F., Mynard, M.C., Lafaix, C., J. Biol. Stand. 10:335-339, 1982)。
【0005】
髄膜炎菌感染の頻度は、過去数十年、多くのヨーロッパの国々において増加してきている。これは、社会的活動の場(例えば、水泳プール、劇場等)の増加による伝染の増加に起因してきた。いくつかの標準的な抗生物質に対して感受性が低い又は耐性の髄膜炎菌株を単離することは、もはや珍しいことではない。この現象は、新たな抗菌剤、ワクチン、薬剤スクリーニング法、及びこれらの生物についての診断検査に対する、未だ満たされていない医学的ニーズ及び需要を生み出した。
【0006】
利用可能な多糖ワクチンは、現在、それらをキャリアタンパク質に化学的にコンジュゲートする方法によって改良が行われている(Lieberman, J.M., Chiu, S.S.Wong, V.K.らJAMA 275:1499-1503, 1996)。
【0007】
しかしながら、血清群Bワクチンは利用可能な状況にない。血清群B莢膜多糖は、免疫原性を示さないことが分かってきたが、それは恐らくそれが宿主成分との構造類似性を有するためであろう(Wyle, F.A., Artenstein, M.S., Brandt, M.L.ら J. Infect. Dis. 126:514-522, 1972;Finne, J.M., Leinonen, M., Makela, P.M. Lancet ii.:355-357, 1983)。従って、外膜小胞(すなわちブレブ)又はそれらから得た精製タンパク質成分から、血清群Bワクチンの開発を試みることに焦点を当てた努力がされてきた。
【0008】
ワクチン開発のための別の髄膜炎菌抗原は、髄膜炎菌のリポオリゴ糖(LOS)である。これらは外膜に結合した糖脂質であり、O側鎖を欠損しているという点で腸内細菌科(Enterobacteriaceae)細菌のリポ多糖(LPS)と異なり、従ってLPSのラフ型と類似している(GriffissらRev Infect Dis 1988, 10:S287-295)。LOSのオリゴ糖部分の異質性は、異なる髄膜炎菌株における構造的な及び抗原性の多様性を生み出す(GriffissらInf. Immun. 1987, 55:1792-1800)。このことは、かかる株を12種類の免疫型にさらに分類するために用いられている(ScholtanらJ Med Microbiol 1994, 41:236-243)。免疫型の決定は、通常、既知の免疫型のLOSに対して生成された吸着ポリクローナル抗体を用いるオクタロニー法によって行われる(Poolman JT, Hopman CTP及びZanen HC, FEMS Microbiol Letters (1982) 13: 339-348)。免疫型L3、L7及びL9は、免疫学的に同一であり、かつ構造的に類似(若しくは全く同一)であり、従ってL3,7,9(又は、本明細書において総称的に「L3」)と呼ばれている。髄膜炎菌のLOS L3,7,9(L3)、L2及びL5は、シアル化によって、又はシチジン5'−一リン酸−N−アセチルノイラミン酸の付加によって修飾することができる。L2、L4及びL6 LOSは免疫学的に区別することができるが、しかしそれらは構造的に類似しており、本明細書でL2に言及する場合にはL4又はL6のいずれかと任意に置換することができ、これは本発明の範囲内となる。LOSに対する抗体は、実験用ラットにおいて感染に対し防御し、また髄膜炎菌に感染した小児において殺菌活性に寄与することが示されている(GriffissらJ Infect Dis 1984, 150:71-79)。
【0009】
しかしながら髄膜炎菌ワクチンにおけるLOSの使用に伴う問題は、その毒性(そのリピドA部分による毒性)にある。
【0010】
またLOSは髄膜炎菌のブレブの表面に存在する。髄膜炎菌の外膜小胞(すなわちブレブ)に基づいたワクチンの開発に焦点を当てて、長年にわたる努力がされてきた(de Moraes, J.C., Perkins, B., Camargo, M.C.らLancet 340:1074-1078, 1992、Bjune, G., Hoiby, E.A. Gronnesby, J.K.ら338:1093-1096, 1991)。そのようなワクチンは、宿主に投与した場合に防御免疫応答を誘発することのできる、正確に折り畳まれた立体構造をとったいくつかの完全な外膜タンパク質を含むという利点を有する。さらに、ナイセリア株(髄膜炎菌血清群B、すなわちmenBを含む)は、工業的規模での製造を可能にするために十分な量の外膜ブレブを分泌する。しかしながら、ブレブは大抵、細菌細胞の0.5%界面活性剤(例えばデオキシコレート)抽出を含む方法により調製される(例えばEP11243)。それは上記のようなLOSの毒性(内毒素とも呼ばれる)のために望ましいが、ワクチンからほとんどのLOS抗原を除去する作用もまたある。
【0011】
ワクチン抗原としてのLOSの使用に伴うさらなる問題は、多様な糖鎖構造を有する12のLPS免疫型が存在することである(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021;Mol Microbiol 2002, 43:931-43)。ある免疫型に対する抗体は、異なる免疫型を認識することができない。LOS免疫型のオリゴ糖部分の共通「コア」領域を生成することに努力が集中されてきたが(例えばWO94/08021号)、改変されたLOSに対して生起された抗体の殺菌活性は失われる。従って、ワクチンは、有効であるために異なる免疫型の多くのLOS成分を有する必要があるだろう。
【0012】
ヒトワクチンにおける抗原としてのLOS(LPS又はリポ多糖としても知られる)の使用に伴うさらなる問題が存在し、それはすなわちそれらがヒト糖鎖構造(例えば、ヒト赤血球細胞上の)に類似した糖鎖構造を担持しているため、それらの使用に伴う安全性の問題が突きつけられることである。さらにLOS構造の改変は、LOS抗原の殺菌効果の構造的感受性のため、問題を含んでいる。
【0013】
WO2004/014417号は、これらの問題についてのいくつかの解決法を記載している。
【発明の概要】
【0014】
本発明者はさらに以下の点を見出した:
−LOS内部コアの修飾(decoration)は、免疫型の殺菌性エピトープの確定に重要である、
−L2 LOSをヘプトースIIに結合したGlcNAc残基においてO−アセチル化修飾する酵素が見いだされ、それをコードする遺伝子がOac1である、
−O−アセチル化されているL3免疫型LOSが見出され(以前に報告されていない)、これはL3株に広範囲に存在すると考えられる、
−従来のL3 LOSに対する抗体はO−アセチル化L3株を死滅させることができるが、この殺菌は従来のL3株に対してそれほど有効ではない。
【0015】
上記知見により、本発明者は、ナイセリアワクチン製剤におけるO−アセチル化L3 LOSの使用を検討するに至った。
【0016】
発明の概要
従って、本発明の一態様においては、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているナイセリア株由来のL3 LOSを含む免疫原性組成物を提供する。ナイセリア株は、天然にそのようなLOSを産生するもの(例えばNZ124株)であってもよいし、又は機能的oac1遺伝子の挿入によって作出されたもの(以下参照)であってもよい。本発明の目的のため、L3V免疫型は、L2免疫型とより免疫学的に類似しているため、L3株の分類ではない。
【0017】
L3 LOSは、以下の構造を有しうる:
【化1】
【0018】
〔式中、
R1=
【化2】
【0019】
であり、
R2=PEAであり、
R3=Hであり、
R4=OAcであり、
R5=H、PEA、又はGlyである〕。
【0020】
上記の用語はLOS分野の標準的な略語であり、例えばGlcはグルコース(又はD−グルコピラノース)を表し、KDOは2−ケト−3−デオキシオクトネートを表し、HepはL−グリセロ−D−マンノ−ヘプトースを表し、GlcNAcはN−アセチルグルコサミンを表し、Galはガラクトースを表し、NeuNacはシアル酸を表し、OAcはO−アセチルを表し、PEAはホスホエタノールアミン(又は2−アミノエチルホスフェート)を表し、Glyはグリシンを表す、などである。
【0021】
本明細書において「ナイセリア(neisserial)」は、いかなる場合にも、髄膜炎菌(N. meningitidis)、例えば血清群A、B、C、W135及びYを示す。また、本発明のLOSを産生しうる他の任意の株(例えば淋菌又はナイセリア・ラクタミカ(Neisseria lactamica))を示すこともある。
【0022】
本発明のL3(L2/L4/L10)LOSはそれぞれL3(L2/L4/L10)免疫型のLOSを包含するが、これに必ずしも限られない場合がある(例えば本発明のL2 LOSは、L3免疫型のものとして現在知られているL3V LOSを含むが、L3V LOSを有する株は、L2免疫型を有するLOSに対して生成された血清によって死滅する。下記参照)。よって、本発明の「LOS」、「L2 LOS」、「L3 LOS」、「L4 LOS」及び「L10」LOSという用語の機能的な解釈は、この広い意味で解釈されるべきである。例えば、本発明のL3 LOSを含む免疫原性組成物は、L3免疫型の株を死滅する抗体を誘発することができるなどである。
【0023】
本発明の免疫原性組成物はさらに、ナイセリア株由来のL2及び/又はL10及び/又はL4 LOSを含んでもよい。本発明のL2、L3、L4又はL10 LOSは、髄膜炎菌A株、B株、C株、W135株又はY株から単離することができる。
【0024】
本発明の免疫原性組成物はさらに、以下の構造を有するL2 LOS:
【化3】
【0025】
〔式中、
R1=
【化4】
【0026】
であり、
R2=PEA又はGlcであり、
R3=PEAであり、
R4=H又はOAcであり、
R5=H、PEA、又はGlyである〕
を含んでもよい。
【0027】
本発明の免疫原性組成物はさらに、以下の構造を有するL10 LOS:
【化5】
【0028】
〔式中、
R1=ヘキソース−ヘキソース−であり、
R2=H又はPEAであり、
R3=PEAであり、
R4=OAcであり、
R5=H又はGlyである〕
を含んでもよい。
【0029】
「R1=ヘキソース−ヘキソース−」は、Gal−Gal−、Gal−Glc−、Glc−Gal−、又はGlc−Glc−残基、例えば:
R1=
【化6】
【0030】
を包含する。
【0031】
本発明のL10 LOSにおけるR1末端ヘキソースは、シアル化されていてもよいし、されていなくてもよい(シアル化されている場合には、これは場合によりα−Neu5Ac−(2→3)基を介していてもよい)。
【0032】
本発明の免疫原性組成物はさらに、以下の構造を有するL4 LOS:
【化7】
【0033】
〔L4 LOSについて、式中、
R1=
【化8】
【0034】
であり、
R2=Hであり、
R3=PEAであり、
R4=OAcであり、
R5=Glyである〕
を含んでもよい。
【0035】
あるいは、本発明のL4 LOSは、R4=H及び/又はR5=Hであってもよい。
【0036】
本明細書における「本発明の1以上のLOS」若しくは「本発明のLOS」又は類似の文言は、本発明のL2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSを意味する。本明細書における「本発明のブレブ調製物」又は類似の文言は、本発明のL2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSを有する本発明の1又はそれ以上のブレブを意味する。本発明の細菌株は、それから本発明の1以上のLOSを単離することができるものである。
【0037】
本発明の1以上のLOSは、タンパク質キャリアとコンジュゲートされていてもよく、そのタンパク質キャリアは、Tヘルパーエピトープ源、例えば破傷風トキソイド、ジフテリアトキソイド、CRM197、又は髄膜炎菌ブレブ上の外膜タンパク質などである(下記参照)。その1以上は、化学的に(下記参照)又は遺伝的に(例えばmsbB(−)及び/又はhtrB(−)であるナイセリア株から単離された場合。下記参照)、無毒化されたそのリピドA部分を有するものである。
【0038】
本発明の1以上のLOSは、精製LOS調製物として、リポソーム調製物(典型的には精製LOSを含む)として、又はブレブ調製物(下記参照)として、免疫原性組成物中に存在しうる。
【0039】
本発明のブレブ調製物(複数でもよい)は、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離することができる。それらの各ナイセリア株は、それが鉄を利用可能な条件下、又は鉄除去の条件下(例えば、デスフェラールなどの鉄キレート剤を添加する。下記参照)で培養されたものであってもよい。本発明のブレブ調製物は、莢膜多糖を合成することができないナイセリア株から単離されてもよい。例えば、その株は、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、又は欠失している(すなわち、その遺伝子からの機能的発現がない)ものである。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、それが由来する株はそれぞれの株において同じ莢膜多糖遺伝子の発現が下方制御されていることが好ましい。ナイセリア株は、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(すなわち、その遺伝子からの機能的発現がない)ものであってもよい。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、ブレブが由来する株はそれぞれの株において同じリピドA遺伝子(複数でもよい)の発現が下方制御されていることが好ましい。ナイセリア株は、それが由来する天然株と比較して、外膜タンパク質遺伝子porA、porB、opA、opC、pilC、lbpA又はfrpBの1又はそれ以上の発現が下方制御されている、好ましくは欠失している(すなわち、その株の外膜上に遺伝子産物の発現がない)ものであってもよい。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、これらの株は各ナイセリア株において同じ外膜タンパク質遺伝子(複数でもよい)の発現が下方制御されていることが好ましい。ナイセリア株は、外膜タンパク質抗原NspA、TbpA low、TbpA high、Hsf、Hap、OMP85、PilQ、NadA、GNA1870、MltAの1又はそれ以上の発現が上方制御されているものであってもよい。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、それが由来する株はそれぞれの株において1又はそれ以上の異なる外膜タンパク質抗原の発現が上方制御されていることが好ましい。
【0040】
さらに、有効量の本発明の免疫原性組成物及び薬学的に許容される担体を含むワクチン組成物を提供する。ワクチンは、アジュバント、例えば水酸化アルミニウム又はリン酸アルミニウム、をさらに含んでもよい。ワクチンは、以下の株:髄膜炎菌血清群A、髄膜炎菌血清群C、髄膜炎菌血清群W−135、髄膜炎菌血清群Y、及びインフルエンザ菌(H. influenzae)b型に由来する、1又はそれ以上のコンジュゲートした莢膜多糖又はオリゴ糖をさらに含んでもよい。
【0041】
以下のリスト:髄膜炎菌血清群A、B、C、W135及びYから選択される1又はそれ以上によって引き起こされる疾患の予防又は治療のための医薬の製造における、本発明の免疫原性組成物又は本発明のワクチンの使用もまた提供する。さらに、以下のリスト:髄膜炎菌血清群A、B、C、W135及びYから選択される1又はそれ以上によって引き起こされる疾患の予防又は治療方法であって、本発明の免疫原性組成物又は本発明のワクチンをその必要があるヒト患者に投与するステップを含む方法を提供する。
【0042】
本発明の免疫原性組成物又はワクチンの製造方法であって、L3 LOSを単離するステップ、場合によりそれを必要に応じて単離されたL2及び/又はL10 LOSと組み合わせるステップ、並びに薬学的に許容される賦形剤と共にL3 LOSを製剤化するステップを含む方法をさらに提供する。
【0043】
髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、本発明の免疫原性組成物又はワクチンの使用も提供する。髄膜炎菌免疫型L3疾患の予防又は治療方法であって、その必要があるヒト患者に、有効量の本発明の免疫原性組成物又はワクチンを投与するステップを含む方法である。髄膜炎菌免疫型L3疾患は、そのヘプトースII残基に結合したGlcNAc残基においてO−アセチル化されているLOS、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないLOS、又はそのヘプトースII残基に結合したGlcNac残基において部分的にO−アセチル化されておりかつ部分的にO−アセチル化されていないLOS、のいずれかのLOSを有する株によって引き起こされるものである。
【0044】
本発明のさらなる態様において、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株により引き起こされる髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の使用を提供する。また、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株によって引き起こされる髄膜炎菌免疫型L3疾患を予防又は治療する方法であって、その必要があるヒト患者に、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の有効量を投与するステップを含む方法を提供する。
【0045】
上記使用又は方法において、免疫原性組成物は、以下の構造を有するL3 LOS:
【化9】
【0046】
〔式中、
R1=
【化10】
【0047】
であり、
R2=PEAであり、
R3=Hであり、
R4=Hであり、
R5=H、PEA、又はGlyである。〕
を含むことができる。
【0048】
本発明のLOSについて本明細書中で説明するように、免疫原性組成物中のL3 LOSは、タンパク質キャリアとコンジュゲートしていてもよいし(さらなる説明については上記及び下記参照)、無毒化リピドA部分、例えばmsbB(−)ナイセリア株から単離されたLOSと一致する第2(secondary)アシル鎖を欠損している(及び/又は、LOSを以下に記載するリピドA無毒化ペプチドと複合体化することによる)もの、を含んでもよいし、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として免疫原性組成物中に存在してもよい。ブレブ調製物の場合、ブレブ調製物は、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものとすることができる。ナイセリア株は、莢膜多糖を合成することができないものであってもよい、例えば、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものとすることができる。ナイセリア株は、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものとすることができる。
【0049】
本発明のまた別の態様において、通常はそのLOSのヘプトースII残基に結合したGlcNAc残基をO−アセチル化するナイセリア株のLOSを脱O−アセチル化する方法であって、oac1遺伝子の機能的発現を破壊することによって、機能的Oac1を発現することができないようにするステップを含む方法を提供する。
【0050】
本明細書の全体においてoac1とは、LOSのHepII−GlcNacをOAc基によって誘導体化する反応の触媒に関与するナイセリアの遺伝子を意味する。この活性を有する遺伝子配列及びオープンリーディングフレームは、例えば図3Dに示されている。この配列から、あらゆるナイセリアoac1配列を見出すことができる(例えば、図3Dに示されるオープンリーディングフレームと少なくとも70、80、90、95又は99%の配列同一性を有するオープンリーディングフレーム)。本発明のさらなる態様において、本発明のoac1操作方法を実施するために使用することができる、図3Dからの10、15、20、30、35又はそれ以上の連続するヌクレオチドのプライマー配列を提供する。本発明のさらなる態様において、グラム陰性細菌LOSのHepIIに結合したGlcNAc残基をO−アセチル化する方法であって、非(又は部分的)O−アセチル化LOSと単離されたOac1酵素(これは公知の組換え技術によって調製することができる)とを混合するステップを含む方法を提供する。
【0051】
本発明のさらなる態様において、通常はそのLOSのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されている又は部分的にのみO−アセチル化されているナイセリア株のLOSをO−アセチル化する又はさらにO−アセチル化する方法であって、ナイセリア株においてoac1遺伝子の機能的発現を増大するステップを含む方法を提供する。これは、oac1遺伝子の機能的発現を増大するステップが、oac1遺伝子の機能的コピー(これはさらなる機能的コピーであってもよい)(例えば、場合によりその天然のプロモーター配列を含んでいてもよい図3Dのオープンリーディングフレーム)をナイセリア株に導入することによって行うように、実施することができる。あるいは、又はそれに加えて、oac1遺伝子の機能的発現を増大するステップは、存在する非機能的遺伝子のポリGフレーム変動領域(poly-G phase variable region)(後述するようにoac1遺伝子にはそのような領域が2つある)を、oac1遺伝子の機能的発現のためにフレーム内となるようにすることによって行うことができる。これは、公知の突然変異誘発技術によって行うことができる。ポリGフレーム変動領域は、通常は、活性を有する遺伝子のATG開始コドンから1136〜1140位のヌクレオチドの間にある(そして不活性遺伝子においてはフレーム外である1140以上にまで及ぶことがある)。2つ目のポリG領域は、開始コドンから354位に見出されている。
【0052】
さらに、ナイセリア株において機能的oac1発現のフレーム変動性を少なくする方法であって、oac1遺伝子のポリGフレーム変動領域(の一方又は両方)を、より少ないGヌクレオチドを有するコドンを用いて同じアミノ酸がコードされるように、変化させるステップを含む方法を提供する。例えば、ATG開始コドンから1135〜1137位のヌクレオチドによってコードされるアルギニン残基のコドンを、AGGからCGT、CGC、CGA又はAGAに変化させてもよいし、並びに/あるいは、ATG開始コドンから1138〜1140位のヌクレオチドによってコードされるグリシン残基のコドンを、GGGからGGT、GGA又はGGCに変化させてもよい。
【0053】
上記のoac1操作方法は、L2又はL3免疫型のナイセリア株に対して行うことができる。さらに、本発明のoac1操作方法を実施するステップ、得られるナイセリア株からLOSを単離するステップ、及び薬学的に許容される担体と共に有効量のLOSを製剤化するステップを含む、免疫原性組成物の製造方法を提供する。また、本明細書の全体に記載されているように、LOSをタンパク質キャリアにコンジュゲートしてもよいし、そのリピドA部分を、例えばmsbB(−)ナイセリア株からLOSを単離する及び/又は後述するようにリピドA無毒化ペプチドと複合体化することにより、無毒化してもよい。LOSは、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として単離されたものとすることができる。ブレブ調製物の場合には、ブレブ調製物を、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものとすることができる。ナイセリア株は、莢膜多糖を合成することができないものであってよく、例えば、ナイセリア株は、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものであってよい。ナイセリア株は、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものであってもよい。
【0054】
本明細書における「ナイセリア」は、いかなる場合にも、髄膜炎菌(N. meningitidis)、例えば血清群A、B、C、W135及びYを意味する。
【図面の簡単な説明】
【0055】
【図1】L1〜L8免疫型の構造によって示される、髄膜炎菌LOS構造の共通置換パターンである(Kahlerら Glycobiology 2005 15:409-419 / 2006 JBC 281:19939-19948より転載)。L7免疫型構造(示さない)は、L3免疫型構造の非シアル化形態である。保存されている内部コア領域をR1〜R5として示す可変付属基と共に示す。α鎖(R1)の組成は、lgtA−Eトランスフェラーゼ、及び末端α−Neu5Ac(シアル酸)基を付加するシアリルトランスフェラーゼをコードするlstのフレーム変動的(phase-variable)発現によって制御される。内部コアへのグリシンの結合は第2のHep残基上の7位を介することに留意されたい。この略図に示されているKDO残基に結合する、全ての免疫型の内部コアに存在することが知られているさらなるKDO残基は示されていないことに留意されたい。
【図2A】質量分析によって決定された、種々のL3髄膜炎菌株のLOS構造の概略である。興味深いことに、H44/76株由来血清によって殺菌される傾向が低いL3株のいくつかはO−アセチル化されている。
【図2B−1】種々の髄膜炎菌免疫型の共通LOS構造である。LOS構造の特定の部分を形成する酵素をコードする既知の遺伝子の名称を示している。
【図2B−2】図2Bの続き
【図3A】Z2491株由来の髄膜炎菌LOS O−アセチル化遺伝子NMA2202を示す。オープンリーディングフレーム中の5つのG(大文字)の配列によってオープンリーディングフレームがフレーム内となることに留意されたい。
【図3B−1】MC58株由来の髄膜炎菌LOS O−アセチル化遺伝子NMB0285を示す。オープンリーディングフレームを大文字で、周囲の配列(例えばプロモーター配列)を小文字で示す。オープンリーディングフレーム中の6つのG(下線)の配列によってオープンリーディングフレームがフレーム外となることに留意されたい。
【図3B−2】図3Bの続き
【図3C】760676株由来の髄膜炎菌LOS O−アセチル化遺伝子(NMB0285等価物)を示す。オープンリーディングフレームを大文字で、周囲の配列(例えばプロモーター配列)を小文字で示す。オープンリーディングフレーム中の5つのG(下線)の配列によってオープンリーディングフレームがフレーム内となることに留意されたい。
【図3D】(MenB、L3)NZ124株由来の髄膜炎菌LOS O−アセチル化遺伝子(NMB0285等価物)を示す。オープンリーディングフレームを大文字で、周囲の配列(例えばプロモーター配列)を小文字で示す。オープンリーディングフレーム中の5つのG(下線)の配列によってオープンリーディングフレームがフレーム内となることに留意されたい。
【図4A】(A)LOS6275からの内部コアオリゴ糖(ES−);(B)LOS6275からの内部コアオリゴ糖(MS/MS ES+m/z 1803.6)。3位及び6位におけるPEAに連結したHepIIと、O−アセチル化された内部コアを示す構造の概略を示す。
【図4B】(A)LOS6275からの内部コアオリゴ糖(ES−);(B)LOS6275からの内部コアオリゴ糖(MS/MS ES+m/z 1803.6)。3位及び6位におけるPEAに連結したHepIIと、O−アセチル化された内部コアを示す構造の概略を示す。
【図4C】(C)MenC C11株からの内部コアオリゴ糖(ES−)を示す。
【図5】L2由来OMVワクチンによる交差反応性殺菌抗体の誘導に対するL2 LOSのシアル化の影響を示す。SBA力価(50%死滅のGMT)及びセロコンバージョン(%)を示す。
【図6】MenA 3125のL10 LOSの質量分析構造解析を示す。
【発明を実施するための形態】
【0056】
発明の説明
本明細書中に記載される刊行物及び特許、又は特許出願の主題又はそこに開示される情報は、参照により本明細書に組み入れる。
【0057】
「リポオリゴ糖」(又は「LOS」)に関する言及は、「リポ多糖」又は「LPS」にも言及するものとする。
【0058】
本明細書中の用語「含む」(「comprising」「comprise」「comprises」)は、全ての場合に、任意に用語「からなる」(「consisting of」「consist of」「consist of」)に各々置き換えることができるものと発明者は意図している。
【0059】
本発明者らは、LOSオリゴ糖構造の短縮が殺菌性免疫応答を誘発し得るエピトープの減少を導くことを見出した。あるいは、本発明者らは、ワクチン製剤においてLOSを最も有効に使用するためには、LOSオリゴ糖構造をできるだけ保持するべきであるが、たった2種類(又は3若しくは4種類)のLOS抗原の組み合わせによって広範に有効なナイセリアの(好ましくは髄膜炎菌の)ワクチンを生成し得ることを見出した。本発明の第1の態様は、ナイセリア(好ましくは髄膜炎菌)の免疫型L2のLOS及び免疫型L3のLOS(又は、本発明のL2及びL3 LOS、そして場合により本発明のL10及び/若しくはL4 LOS)を含む、ナイセリア(好ましくは髄膜炎菌又は髄膜炎菌B型)疾患の予防又は治療のための免疫原性組成物である。LOSは、公知の精製法のいずれかにより単離してもよいし、あるいは例えばL2及びL3ナイセリア株由来の少なくとも2種類の外膜小胞(すなわちブレブ)調製物中に存在するものでもよい。ブレブ調製物から、毒性を有し緩く保持されたLOSを除去するが、ブレブ内に組み込まれたLOS抗原は高レベルで保持するようにするために、ブレブを低濃度の界面活性剤、すなわち0〜0.3%、好ましくは0.05〜0.2%、最も好ましくは約0.1%の、好ましくはデオキシコレート(すなわちDOC)、を用いて抽出することが好ましい。LOS抗原のそのような組み合わせ(特にブレブワクチン中の)は、驚くべきことに90%を超える髄膜炎菌株に対して有効であるという点で有利である。
【0060】
本発明者らはまた、本発明の上記ブレブ免疫原性組成物、及び実際にはあらゆるナイセリア(好ましくは淋菌又は髄膜炎菌)由来のブレブ免疫原性組成物が、免疫優性外膜タンパク質の特定の組み合わせの発現が下方制御(好ましくは欠失)される場合に、それらの表面上の防御抗原(LOSを含む)の効果が増強されることを見出した。従って、本発明のさらなる態様は、天然の非改変株と比較して、発現が下方制御された、好ましくは欠失した、以下の外膜タンパク質:PorA、PorB、OpA、OpC又はPilC、を2つ以上有するナイセリア株由来の、本発明の1以上のナイセリアブレブ調製物である。好ましくは、PorAとOpA、PorAとOpC、OpAとOpC、又はPorAとOpAとOpCが下方制御されているか又は欠失している。FrpBの発現の下方制御(好ましくは欠失)はまた、交差防御抗原の効果の増強において有利であり、特に、鉄制限状態で増殖させたナイセリア株から作製したブレブ調製物中において有利であることが示されてきた。従って、この突然変異を有する株由来の本発明のナイセリアブレブは、上記の1又はそれ以上の下方制御とFrpB下方制御との組み合わせから得られるブレブとしての、本発明のさらなる実施形態である。好ましくは、PorAが下方制御される場合にはPorBは下方制御されるべきではなく、またその逆も同様である。
【0061】
上記突然変異は、本発明のブレブ免疫原性組成物が由来することになるあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしながらL2又はL3免疫型ナイセリア(好ましくは髄膜炎菌、最も好ましくはmenB)株を用いることが好ましく、典型的には本明細書中上記の低DOC%抽出法で抽出する。好ましくは、本発明のブレブ免疫原性組成物は、L2及びL3ブレブを両方含み、その少なくとも一方(好ましくは両方)が上記の免疫優性外膜タンパク質(又はOMP)の組み合わせを欠損している。それらの遺伝子を下方制御するための技術はWO01/09350号(参照により本明細書に組み入れられる)に記載されている。4つの異なるOpa遺伝子が髄膜炎菌ゲノム中に存在することが知られており(Ahoら1991 Mol. Microbiol. 5:1429-37)、ゆえにOpaの発現が下方制御されていると言われる場合、このことは好ましくは髄膜炎菌に存在する1、2、3つの遺伝子、又は(好ましくは)4つ全ての遺伝子がそのように下方制御されていることを意味する。そのような下方制御は、WO01/09350号に記載のように遺伝学的に達成するか、又は、容易に発見される天然の安定した髄膜炎菌株(Opa座位からの発現がないか若しくは低いもの)を探索することによって達成し得る。そのような株は、Poolmanら(1985 J. Med. Micro. 19:203-209)に記載される技術を用いて見つけ出すことができ、その際、Opa−である細胞は、プレート上又は顕微鏡下において細胞の外見を観察することによって見出されうるOpa発現細胞とは異なる表現型を有する。一度見つけられれば、Opaの欠失を確立するための発酵操作の後に、細胞内容物についてウエスタンブロットを行うことによって、株が安定的にOpa−であることを示すことができる。
【0062】
上記LOS免疫原性組成物の安全性
ヒトスフィンゴ糖脂質に存在するラクト−N−ネオテトラオースオリゴ糖基(Galβ1−4GlcNAcβ1−3Galβ1−4Glcβ1−、図1)と類似した構造が存在することから、L3又はL2 LOSに対して生起された抗体の安全性が問われてきている。多くの人々が残存量のL3 LOSを含むデオキシコレート抽出小胞ワクチンで安全にワクチン接種されてきているが(G. Bjuneら, Lancet(1991), 338, 1093-1096、GVG. Sierraら, NIPH ann(1991), 14, 195-210)、LOSが本明細書に記載されるように抗原として保持される場合、LOS糖鎖構造の末端部分の欠失が、ヒト組織の表面に提示される構造との抗LOS免疫応答の交差反応の阻止において有利であることが、本発明者らによって発見された。好ましい実施形態において、lgtB遺伝子の不活性化が末端ガラクトース残基及びシアル酸が欠損した中間体LOS構造を生じる(図1及び2を参照、突然変異はL2及びL3及びL4 LOSに4GlcNAcβ1−3Galβ1−4Glcβ1−構造を残存させる)。そのような中間体は、L3及び/又はL2(及び/又はL4)LOS株において獲得され得る。別の及びあまり好ましくないLOSの(短い)バージョンは、lgtE遺伝子のスイッチオフによって得ることができる。さらに別の及びあまり好ましくない型のLOSは、lgtA遺伝子のスイッチオフによって得ることができる。そのようなlgtA−突然変異を選択する場合、非免疫原性L1免疫型の形成を阻止するためにlgtC発現をスイッチオフすることがまた好ましい。
【0063】
lgtB−突然変異が最も好ましいのは、これが殺菌(及びさらに交差反応性殺菌)抗体反応を依然として誘導し得るLOS防御性オリゴ糖エピトープを保持したままで安全性の課題を解決するために最適な末端切断物であることを本発明者らが発見したためである。
【0064】
従って、本発明の上記のL2及び/又はL3調製物(あるいは上述した本発明の1以上のLOS)(精製されたもの、又は単離したブレブ状のもの)あるいは一般的な本発明の髄膜炎菌のブレブ調製物(特にL2及び/又はL3)は、lgtB、lgtA又はlgtE遺伝子由来の機能的遺伝子産物の発現が永続的に下方制御されるように、好ましくは遺伝子のスイッチオフにより、最も好ましくは当該遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により、遺伝子操作したナイセリア株(好ましくは髄膜炎菌株)から、有利に得られる。
【0065】
好ましくは、本発明のナイセリア株は、莢膜多糖の合成に欠損を有する。
【0066】
本発明の上記ブレブ調製物を髄膜炎菌B株から得る場合、莢膜多糖(ヒト様糖鎖構造も含む)もまた除去されることが特に好ましい。これを達成するために多くの遺伝子がスイッチオフされ得るが、本発明者らは、siaD遺伝子由来の機能的遺伝子産物の発現を永続的に下方制御(すなわちα−2−8ポリシアリルトランスフェラーゼの活性を下方制御)するように、ブレブ産生株を、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により、遺伝的に操作することが好ましいということを有利に示してきた。そのような不活性化はWO01/09350号に記載されている。siaD(synDとしても知られる)突然変異が、莢膜多糖からヒト類似エピトープを除去し得る多くの突然変異の中で最も有利であるのは、それが、LOSの防御エピトープの生合成に影響を及ぼさない最良の突然変異の1つであり、そのため、LOSを防御抗原として最終的に用いることを目的とし、かつ細菌の増殖に対して最小限の影響を与える方法において有利であるためである。従って、本発明の好ましい態様は、lgtE− siaD−、lgtA− siaD−又は好ましくはlgtB− siaD−髄膜炎菌B突然変異株由来のブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0067】
siaD−突然変異は上記の理由のために好ましいが、髄膜炎菌B(又は一般的に髄膜炎菌)の莢膜多糖合成をスイッチオフする他の突然変異を用いることができる。従って、ブレブ産生株を、以下の遺伝子:ctrA、ctrB、ctrC、ctrD、synA(synX及びsiaAと同等)、synB(siaBと同等)又はsynC(siaCと同等)遺伝子、の1つ以上に由来する機能的遺伝子産物の発現を永続的に下方制御するように、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により、遺伝的に操作してもよい。lgtE−突然変異はこれらの突然変異の1以上と組み合わせてもよい。好ましくは、lgtB−突然変異はこれらの突然変異のうち1つ以上と組み合わされる。従って、本発明のさらなる態様は、髄膜炎菌B(又は一般的に髄膜炎菌)の突然変異株のそのような組み合わせから得られる上記ブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0068】
lgtB及びlgtEなどの様々なlgt遺伝子を含むナイセリア遺伝子座、並びにその配列は、当技術分野において公知である(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021及びそれに引用されている参考文献;J. Exp. Med. 180:2181-2190(1994);WO96/10086号を参照)。
【0069】
最終産物として本発明の全長(非末端切断型)LOSを使用する場合、LOSがシアル化されていないことが望ましいが、そのようなLOSは、最も危険な侵襲性髄膜炎菌B株(これも同様にシアル化されていない)に対する免疫応答を生じることがあるためである。そのような場合、synA(synX及びsiaAと同等)、synB(siaBと同等)又はsynC(siaCと同等)遺伝子が欠失した莢膜陰性株の使用が有利であり、それはまたそのような突然変異によってmenB LOSがシアル化され得ないようにするためである。本発明の一実施形態において、lst遺伝子(Gilbertら, JBC 1996, 271:28271-6)を機能的に不活性とする(例えば、その遺伝子の欠失若しくは破壊、又は発現の低減により)か、あるいはその遺伝子が天然に破壊されている株を本発明のために選択する(免疫型のL3ファミリーについては、そのような欠損株はL7免疫型と称されることが多い)。lstは、末端シアル酸をLOSα鎖に付加するα−2,3−シアリルトランスフェラーゼである(しかし、シアル酸産生には影響を及ぼさない)。一実施形態において、本発明の株はlst(−)かつsiaD(−)である。
【0070】
上記突然変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしL2又はL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、典型的には本明細書に記載した低DOC%抽出法によって抽出されたものを使用するのが好ましい。好ましくは、本発明のブレブ免疫原性組成物が、L2及びL3ブレブの両方を含み、かつ、少なくとも一方(好ましくは双方)が上記遺伝子の発現が欠損した株に由来する。
【0071】
LOSの毒性
本発明の上記の精製LOS又はブレブ免疫原性組成物はまた、それらが由来する細菌性産生株における特定の遺伝子の発現の下方制御によって、毒性をより少なくすることができる。そのような無毒化は、天然のOMVによる鼻腔内免疫には不必要であるだろうが(J.J Drabickら, Vaccine(2000), 18, 160-172)、非経口ワクチン接種における無毒化には有用であろう。好ましくは、本発明の精製LOS又はブレブ免疫原性組成物は、リピドA生合成に関与する遺伝子(特にリピドAへの第2アシル鎖(secondary acyl chain)の付加に関与する遺伝子)の突然変異/改変/不活性化により(特にmsbB及び/又はhtrB遺伝子由来の機能的遺伝子産物の発現の下方制御により、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により)、ナイセリア産生株を遺伝的に操作することによって無毒化される。あるいはまた(又はさらに)、精製LOS又はブレブ免疫原性組成物は、以下の遺伝子:pmrA、pmrB、pmrE及びpmrFのうちの1つ以上の遺伝子が(より強力なプロモーターの導入、又は遺伝子のさらなるコピーの組み込みによって)上方制御されるように遺伝的に改変されたナイセリア株から得ることができる。あるいはまた(又はさらに)、精製LOS又はブレブ免疫原性組成物は、ポリミキシンBの機能的等価物である非毒性ペプチド[リピドAに対して高いアフィニティの有する分子]を組成物に添加することにより無毒化され得る(以下参照、すなわち、本発明の1以上のLOSが、LOSの毒性を低減させるために好適なリピドA結合ペプチド、例えばSAEP2又はSAEPIIと複合体化されている)。
【0072】
上記無毒化法並びに関連プロモーター/遺伝子配列、並びに上方制御方法及び下方制御方法については、より詳細についてWO01/09350号を参照されたい。またナイセリアmsbB及びhtrB遺伝子は各々lpxL1及びlpxL2とも呼ばれ(WO00/26384号参照)、これらの遺伝子の欠失突然変異は、msbB−突然変異LOSが野生型と比較して1つの第2アシル鎖を欠損する(並びに4つの第1アシル鎖(primary acyl chain)及び1つの第2アシル鎖を保持する)こと、そしてhtrB−突然変異LOSが第2アシル鎖の両方を欠損することの、表現型としての特徴を有する。そのような突然変異は好ましくは、ブレブに対する無毒化LOSの最適な提示を確実にするため、又は無毒化サブユニットLOSの精製を助けるために、ナイセリア産生株が莢膜多糖を欠損していること(上記参照)を確実にする突然変異と組み合わせて用いられる。
【0073】
本発明の組成物中に使用することができるポリミキシンB(本発明のLOSの毒性を低減させるために好適なリピドA結合ペプチド)の機能的等価物である非毒性ペプチドのさらなる詳細、特にペプチドSAEP2(2つのシステインがジスルフィド架橋を形成するKTKCKFLKKC配列を有する)及びSAEPII(WO2006/108586号の請求項1〜10に記載されているペプチド二量体)の使用については、WO93/14115号、WO95/03327号、WO2006/108586号、Velucchiら(1997)J Endotoxin Res 4:1-12及びEP976402を参照されたい。本明細書におけるこのようなリピドA結合ペプチドは、上記引用した特許出願の特許請求の範囲又は実施例に記載されるペプチドの具体的な又は一般的な式のいずれかを指す。
【0074】
本明細書において「機能的遺伝子産物の発現の下方制御」は、対象の遺伝子のプロモーター又はオープンリーディングフレームに対して付加、欠失又は置換が為され、それにより全遺伝子産物の生合成活性が低減(60、70、80、90、95%若しくは最も好ましくは100%低減)されることを意味する。明らかにフレームシフトする突然変異を導入してもよいし、又はより弱いプロモーターを置換してもよいが、最も好ましくは、オープンリーディングフレーム及び/又はプロモーターのほとんど又は全てを、(活性のある)遺伝子産物の永続的な下方制御を確実にするために欠失させる(WO01/09350号に記載)。
【0075】
上記変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有用であるが、L2又はL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株を用いることが好ましく、典型的には本明細書に記載の低DOC%抽出法で抽出されることが好ましい。本発明のブレブ免疫原性組成物は、L2及びL3ブレブの両方(又は本発明のブレブ調製物)を含み、その少なくとも一方(及び好ましくは双方)が上記遺伝子の発現を欠損している株に由来することが好ましい。
【0076】
本発明のさらなる態様は、本発明のLOS又はブレブ免疫原性調製物が由来する上記の遺伝的に改変されたナイセリア(好ましくは髄膜炎菌又は淋菌又は髄膜炎菌B)株を含む。
【0077】
本発明のLOS又はLOS含有ブレブ調製物
本発明のさらなる態様は、本発明のナイセリア株から単離されるLOS調製物(特に上述した任意のもの)である。好ましくは、単離されたLOS(又はLOS含有ブレブ)は、L2又はL3免疫型であり、好ましくは本発明の免疫原性組成物は本発明のL2及びL3 LOS(又はブレブ)調製物を両方含む。
【0078】
そのような調製物はまた、上記LOS(精製されているものであれブレブ調製物に存在しているものであれ)のオリゴ糖部分をT細胞エピトープ源を含むキャリアにコンジュゲートする(従ってLOSをより良好な(T依存的)免疫原とする)ことにより改良することができる。あるいはまた(又はさらに)本発明の精製LOS調製物は、当技術分野で公知のリポソーム製剤中にそれを存在させることにより、より良好な抗原となり得る(例えばWO96/40063号及びそこに引用される参考文献を参照)。
【0079】
細菌からLOSを単離する方法は当技術分野で周知である(例えばWesphal及びJannの温水−フェノール法(Meth. Carbo. Chem. 1965, 5:83-91)を参照)。またGalanosら1969, Eur J Biochem 9:245-249及びWuら1987, Anal Bio Chem 160:281-289も参照されたい。単離されたLOSをコンジュゲートするための技術もまた公知である(例えばEP941738(参照により本明細書に組み入れる)を参照)。
【0080】
本発明の目的のため、「T細胞エピトープ源を含むキャリア」とは、通常、ペプチド、又は好ましくはポリペプチド若しくはタンパク質である。コンジュゲーション技術は当技術分野で周知である。典型的なキャリアとしては、非定型インフルエンザ菌(Haemophilus influenzae)由来のプロテインD、破傷風毒素、ジフテリア毒素、CRM197又は(特にナイセリア又は髄膜炎菌の)ブレブ調製物に存在する外膜タンパク質が挙げられる。
【0081】
本発明の好ましい単離されたLOS組成物としては、各LOSのオリゴ糖部分がT細胞エピトープ源を含むキャリアに任意にコンジュゲートされていてもよいL2及びL3単離LOSを含有する組成物、lgtB−髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープ源を含むキャリアに任意にコンジュゲートされていてもよいL2又はL3 LOSを含有する組成物、並びに最も好ましくは、lgtB−髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープ源を含むキャリアに任意にコンジュゲートされていてもよいL2及びL3単離LOSを含有する組成物である。
【0082】
好ましくは、本発明のLOS組成物(又は本発明の1以上のLOS)は無毒化されている。これは、分子からアシル鎖を除去する(しかし分子の防御効果は減少する可能性がある)ヒドラジン又はアルカリ加水分解化学処理の公知の技術によって行うことができるが、好ましくはhtrB−若しくはmsbB−髄膜炎菌突然変異株(上記のような、特に莢膜多糖欠損株)からLOSを単離することにより、又はポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高いアフィニティを有する分子)、特にSAEP2又はSAEPII(上記)の、組成物への添加により行われ得る。
【0083】
本発明のLOSは、単離された状態で(通常、リピドA部分が無傷のままであればミセル形態で)投与することができるし、又はリポソーム中に入れて投与することもできる。そのような場合、外膜タンパク質はリポソームに添加してもよく、そしてLOSを、オリゴ糖がT依存的抗原となるようにリポソーム内部でそのような外膜タンパク質にコンジュゲートしてもよい。これは、後述するブレブ内LOS架橋について記載されるものと同様の化学法によって行われ得る。
【0084】
LOSのオリゴ糖部分の、ブレブの表面上に存在する外膜タンパク質とのブレブ内架橋(コンジュゲーション)
LOS(特に本発明のLOS)がブレブ製剤中に存在する場合、LOSは好ましくは、ブレブ調製物上にも存在する1又はそれ以上の外膜タンパク質(例えば髄膜炎菌におけるPorA又はPorB)へのLOSのコンジュゲーションを可能にする方法により、in situにおいてコンジュゲートされる。従って、本発明のさらなる態様は、その外膜中に、LOSとコンジュゲートされた外膜タンパク質が組み込まれるグラム陰性細菌株由来のブレブ調製物(本発明の1以上のブレブ調製物)である。LOSはコンジュゲートのためにブレブ調製物に添加してもよいが、LOSはブレブ調製物の表面に天然に存在することが好ましい。
【0085】
この方法は、ブレブ製剤中のLOS抗原の安定性及び/又は免疫原性(T細胞の助けを提供する)、及び/又は抗原性を、外膜の表面上の天然環境にあるLOSのように有利に増強し、それゆえその最も防御的な立体構造をとったT非依存的オリゴ糖免疫原に対し、T細胞の助けを与える。さらに、ブレブ内でのLOSのコンジュゲーションは、LOSの無毒化を生じ得る(理論に縛られることは望まないが、コンジュゲートされると、リピドA部分はより安定的に外膜中に埋没し、従って毒性を生じるように利用されにくくなると考えられる)。従って、上記のhtrB−若しくはmsbB−変異株からのブレブの単離による無毒化法、又はポリミキシンBの機能的等価物である非毒性ペプチドの組成物への添加による上記の無毒化法は、必要ではないこともある(しかしさらなる安全のために組み合わせて添加され得る)。
【0086】
本発明のコンジュゲート化ブレブ調製物は、典型的には、ブレブ中のLOSの毒性が同量の全くコンジュゲートしていないLOSを有する同じブレブと比較して低減されている。LOS毒性は当業者であれば容易に測定することができ、例えばヨーロッパ薬局方におけるLOSウサギ発熱アッセイ(pyrogenicity assay)を用いることができる(WO2004/014417号の実施例7参照)。
【0087】
本発明のコンジュゲート化ブレブ調製物は、コンジュゲートしたLOSが宿主内における免疫応答の誘発に適した立体構造を有し、それに由来する血清がコンジュゲートしていないLOS(好ましくはブレブ調製物が作製された由来の細菌上に存在し、最も好ましくはSBAアッセイにおける殺菌性で存在する)との反応性を有する(結合可能である)ため、有利である。
【0088】
ナイセリアブレブをLOSにコンジュゲートする場合、及び本明細書に記載するように、ブレブが1以上の免疫優性外膜タンパク質が下方制御された株由来である場合、好ましくは、PorAが下方制御されている場合にはPorBは下方制御されるべきではなく、かつその逆も同様である。このことによって、LOSの大部分を主要な外膜タンパク質と架橋することが可能になり、従ってブレブに存在する交差防御性の微量外膜抗原に対するコンジュゲートのあらゆる効果が最小化される。
【0089】
特に、本発明者らは、ブレブに存在するLOSがブレブ内で外膜タンパク質(これもまたブレブに存在する)にコンジュゲートしているブレブを含む組成物が、そのブレブが由来する生物によって引き起こされる疾患の治療又は予防のためのワクチンの基礎を形成しうることを見出した。ここでそのようなワクチンは毒性が低減しており(好ましくは実質的に無毒性であり)、及び/又はその天然環境におけるLOSに対するT依存的殺菌反応を誘導することが可能である。
【0090】
従って、本発明はそのようなブレブ内LOSコンジュゲート化ブレブ調製物をさらに提供する。「ブレブ内」とは、ブレブ中に天然に存在するLOSが同じブレブ上に存在する外膜タンパク質とコンジュゲートしていることを意味する。
【0091】
そのようなブレブ調製物は、対象の細菌より単離することができ(WO01/09350号参照)、次にLOSのオリゴ糖部分の基(例えばNH2又はCOOH)をブレブ外膜タンパク質の基(例えばNH2又はCOOH)に結合するための、既知のコンジュゲーション化学法に供すればよい。グルタルアルデヒド、ホルムアルデヒド又はグルタルアルデヒド/ホルムアルデヒド混合物を使用した架橋技術を用いることができるが、EDAC又はEDAC/NHS等のより選択的な化学法を用いるのが好ましい(J.V. Staros,R.W. Wright及びD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)並びにBioconjugates Techniques. Greg T. Hermanson(1996)pp173-176)。本発明において使用することができるLOS及びタンパク質分子間の共有結合を生じることの可能な他のコンジュゲーション化学法又は処理は、EP941738に記載されている。
【0092】
好ましくは、ブレブ調製物は莢膜多糖の非存在下でコンジュゲートされる。ブレブは莢膜多糖を産生しない株(天然の若しくは突然変異によるもの)から単離することができ、又は大半の混入莢膜多糖から精製(60、70、80、90若しくは99%以上が除去された)、好ましくは全ての混入莢膜多糖から精製することができる。この方法において、ブレブ内LOSコンジュゲーション反応はさらに非常に効果的である。
【0093】
好ましくはブレブ内に存在するLOSの5、10、20、30、40、50、60、70、80、90又は95%を超える割合が架橋/コンジュゲートされる。
【0094】
好ましくは、本発明のブレブは、精製LOSを標準物質として使用したSDS−PAGE電気泳動後の銀染色によって測定した場合に(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)、ブレブのLOS含量が、3〜30、5〜25、10〜25、15〜22、そして最も好ましくはおよそ又は正確に20%LOS含量となるように調製されたものである。髄膜炎菌ブレブにおける20%LOSを0.1%低DOC抽出法で達成することができ、これは緩く保持されたLOS分子を除去することができるが、抗原の大部分は保存するものである。
【0095】
ブレブ内コンジュゲート化ブレブが髄膜炎菌由来である場合、それが由来する株が莢膜多糖を産生することができない突然変異株(例えば上記の突然変異株の1つ、特にsiaD−)であることが好ましい。髄膜炎菌疾患に対して有効な免疫原性組成物がL2及びL3ブレブの双方を含むこともまた好ましく、ここでL2及びL3 LOSは双方ともにブレブ外膜タンパク質にコンジュゲートしている。さらに、ブレブ内コンジュゲート化ブレブ中のLOS構造がlgtB−髄膜炎菌株由来のものと一致していることが好ましい。最も好ましい免疫原性組成物は以下のブレブ内コンジュゲート化ブレブを含む:莢膜多糖を産生することができずlgtB−である、L2若しくはL3突然変異髄膜炎菌株に由来するもの、莢膜多糖を産生することができない突然変異髄膜炎菌株由来のL2及びL3ブレブを含むもの、lgtB−である突然変異髄膜炎菌株由来のL2及びL3ブレブを含むもの、又は最も好ましくは莢膜多糖を産生することができずlgtB−である突然変異髄膜炎菌株由来のL2及びL3ブレブを含むもの。
【0096】
本発明に使用することができる典型的なL3髄膜炎菌株は、H44/76 menB株である。典型的なL2株は、B16B6 menB株又は39E髄膜炎菌C型株又は760676株である。典型的なL10株は、3125 menA株である。L4株は、C19 MenC株である。
【0097】
上述のように、本発明のブレブはコンジュゲーションの作用によってある程度無毒化されるので、さらなる無毒化を必要としないが、しかし、さらなる無毒化法をさらなる安全性のために用いてもよい。それは例えばhtrB−若しくはmsbB−である髄膜炎菌株由来のブレブを使用することによるか、又はポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高いアフィニティを有する分子)(好ましくはSEAP2又はSEAPII)をブレブ組成物に(上記のように)添加することによる。このように、LOSの(特にブレブ内の)コンジュゲーションは、驚くべきことに同量のコンジュゲートしていないLOSを含む調製物と比較して低いLOS毒性を示す。従って、(特に髄膜炎菌の)ブレブの無毒化のための一般的な方法は、さらにLOSのブレブ外膜タンパク質へのブレブ内コンジュゲーションによって提供され、そしてLOSの無毒化のための方法もまた、ブレブ外膜タンパク質へLOSをコンジュゲートする手段によって提供される。
【0098】
上記の方法において、髄膜炎菌ブレブ、及びブレブを含む免疫原性組成物が提供され、これらは、毒性が低減しており(好ましくは実質的に無毒性であり)、自己免疫の問題がなく、T依存的特徴を持ち、その天然環境に存在し、そして潜在的に90%を超える髄膜炎菌株に対する殺菌性抗体反応(L2+L3組成物の場合)を誘導することのできる、重要な抗原としてのLOSを有する。
【0099】
1以上のMenA、C、Y又はW莢膜多糖又はオリゴ糖(好ましくは少なくともMenC、又はMenA及びMenC、又はMenC及びMenY)を、本発明のブレブの外膜タンパク質上に、同様にコンジュゲートすることができる。これはLOS架橋と同じ反応において行うことができるが、別々の(好ましくは後の)反応において行うことが好ましい。
【0100】
最適なブレブ内LOSコンジュゲーションの方法は、本発明のさらなる態様である。かかる方法は、(好ましくは、本明細書に記載されるように低パーセンテージのDOCを使用して)グラム陰性細菌からブレブを単離するステップ、ブレブに存在するLOSを(好ましくはそのオリゴ糖部分を介して)同じブレブに存在する外膜タンパク質にコンジュゲートするのに適した化学法を行うステップ、ブレブ内コンジュゲート化ブレブ調製物を単離するステップ、並びに任意により、同じ方法によって作製されるが異なるLOS免疫型を有する、さらなるブレブ内コンジュゲート化ブレブ調製物とともにブレブ内コンジュゲート化ブレブ調製物を製剤化するステップ(好ましくはL2及びL3ナイセリア/髄膜炎菌ブレブを混合するステップ)、及び/又はワクチン組成物を製造するために薬学的に許容される賦形剤とともにブレブ調製物を製剤化するステップを含むべきである。
【0101】
ブレブ内コンジュゲーションは、好ましくは以下の方法のステップの1若しくは2つ又は3つ全てを含むべきである:コンジュゲーションpHはpH7.0を上回り、好ましくはpH7.5以上(最も好ましくはpH9以下)であるべきである;1〜5%、好ましくは2〜4%、最も好ましくは約3%ショ糖の条件が反応中維持されるべきである;コンジュゲーション反応におけるNaClは最小限にするべきであり、好ましくは0.1M、0.05M、0.01M、0.005M、0.001M以下であり、全く存在しないことが最も好ましい。これら全ての方法の特徴は、コンジュゲーション過程を通してブレブを安定にかつ溶液中に維持することを確実にするものである。
【0102】
EDAC/NHSコンジュゲーション法はブレブ内コンジュゲーションのために好ましい方法である。EDAC/NHSは、非常に高度な架橋が可能であるがゆえに濾過性に不利な影響を及ぼすホルムアルデヒドより好ましい。EDACはカルボン酸(LOS中のKDO等)と反応して活性エステル中間体を生成する。アミン求核試薬(PorB等の外膜タンパク質中のリジン等)存在下において、アミド結合はイソ尿素副生成物の放出を伴って形成される。しかしながら、EDAC仲介反応の効率は、スルホNHSエステル中間体の形成により増大し得る。スルホNHSエステルは、EDACのみとカルボキシレートとの反応より形成される活性エステルより長く水性溶液中に残存する。従って、この2段階の方法を用いて高収率のアミド結合形成を実現し得る。EDAC/NHSコンジュゲーションは、J.V. Staros,R.W. Wright及びD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)並びにBioconjugates Techniques. Greg T. Hermanson(1996)pp173-176に記載されている。好ましくは、0.01〜5mg EDAC/mgブレブ、より好ましくは0.05〜1mg EDAC/mgブレブを反応に使用する。使用するEDACの量はサンプル中に存在するLOSの量に依存し、このLOSの量はまたブレブを抽出するために使用するデオキシコレート(DOC)%に依存する。低パーセンテージ(%)DOC(例えば0.1%)においては多量のEDAC(1mg/mg及びそれ以上)が使用されるが、しかし、高%DOC(例えば0.5%)においては少量のEDAC(0.025〜0.1mg/mg)が、過剰なブレブ内架橋を回避するために使用される。
【0103】
本発明の好ましい方法は、従って、EDAC/NHS存在下において、pHがpH7.0からpH9.0の間(好ましくは約pH7.5)で、1〜5%(好ましくは約3%)ショ糖中で、かつ任意に(上記のように)実質的にNaClを含まない条件において、ブレブをコンジュゲートするステップ、並びに反応混合物からコンジュゲート化ブレブを単離するステップを含む、ブレブ内コンジュゲート化LOS(好ましくは髄膜炎菌のもの)を製造する方法である。
【0104】
この反応に続いて、反応時間の経過に伴いブレブ中のLOSの割合の増加に応じてLOS分子量が増加することを示すために、抗LOS(例えば抗L2又は抗L3)モノクローナル抗体(mAb)を使用して反応混合物をウェスタン分離ゲルに供することができる。
【0105】
収量99%のブレブがそのような技術を用いて回収され得る。
【0106】
EDACは、LOSのT依存的免疫原性を改善するために十分な、LOSをOMPに架橋する点で優れたブレブ内架橋剤であることが見出されたが、しかしそれほど高度にそれを架橋しなかったのは、乏しい濾過性、凝集及びブレブ間架橋等の問題が発生したためであった。生じたブレブの形態は、コンジュゲートしていないブレブのものと類似している(電子顕微鏡による)。さらに上記のプロトコルは、過度に高度の架橋(ブレブの表面上に天然に存在する防御OMP(例えばTbpA又はHsf)の免疫原性を低減し得る)が生じることを回避した。
【0107】
ブレブ単離のための技術
本発明の外膜小胞(OMV又はブレブ)は、多くの公知の技術により単離することができる(Fredriksenら, NIPH Annals(1991), 14, 67-79、Zollingerら, J. Clin Invest(1979), 63, 836-848、Saundersら, Infect Immun(1999), 67, 113-119、J.J. Drabickら, Vaccine(1999), 18, 160-172)。これらは2つの主要な群に分類される。すなわち、デオキシコレート(約0.5%)を使用して髄膜炎菌からブレブを抽出する技術と、低レベルのデオキシコレート(DOC)を使用するか又は全くデオキシコレートを使用しない技術である。DOCを使用しない方法では、ブレブはOMV中に高レベルのLOSを維持するという興味深い特徴を有する。このことはLOSが防御抗原であるワクチンにおいて有利である。DOC抽出ブレブと比較して、DOCを使用しない方法により得られるOMV中のL3抗原(Ag)の濃度は約10倍高い。界面活性剤を用いない(好ましくはDOCを用いない)ブレブ調製法は、この理由により本発明の方法の目的にとって好ましいが、低レベルの界面活性剤(好ましくはDOC)を含むバッファーでの抽出もまた、ブレブ内で強固に相互作用するLOSの大半を残す一方、より毒性があり緩く保持されたLOSを除去するステップであるという点で有利であり得る。典型的には、0〜0.5%、好ましくは0.02〜0.4%、0.04〜3%又は0.06〜2%界面活性剤(好ましくはDOC)がブレブ抽出に使用され、より好ましくは0.08〜0.15%、最も好ましくはおよそ又は正確に0.1%を用いて、最適な量のLOSをブレブ中に安定に存在させて得る。LOSが1以上の上記の方法により無毒化されている場合には、DOCを用いない(又は低DOC〜0.3%DOC若しくはそれ以下の)抽出法が特に好ましい。
【0108】
本発明の全ての実施形態におけるブレブのLOS含量は、精製LOSを標準物質として使用したSDS−PAGE電気泳動後の銀染色によって測定した場合に(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)、3〜30、5〜25、10〜25、15〜22%であることが好ましく、最も好ましくはおよそ又は正確に20%LOS含量である。Nmen L3 LOSをこの方法における標準物質として使用すると、一般的に、0.1%DOCで抽出されたNmen L3免疫型ブレブ中のLOS含量は約20%LOSであり、0.2%DOCで抽出した場合は約15%LOS、0.3%DOCでは約10%LOS、そして0.5%DOCでは約5%LOSである。
【0109】
ブレブの作製は、細胞又は細胞片からブレブを分離するための適当な技術のいずれかを用いて(例えば低速遠心分離により)行うことができる。ブレブ調製物は、超遠心分離を使用して(ブレブをペレット化する)、あるいはFrasch et al. ("Outer membrane protein vesicle vaccines for meningococcal disease" Methods in Molecular Medicine, vol 66, Meningococcal Vaccines: Methods and Protocols 2001 pp81-107 (A.J. Pollard及びM.C. Maiden編, Humana Press Totowa, NJ))に記載されているように超遠心分離及び/又はダイアフィルトレーション(diafiltration)のより穏やかな技術を用いて、さらに精製することができる。
【0110】
ワクチン組成物
本発明の免疫原性組成物は、薬学的に許容される賦形剤の添加によってワクチン組成物として容易に製剤化することができる。
【0111】
さらに、上記の本発明の精製LOS(好ましくはL2若しくはL3)を単離するステップ又は上記の本発明の単離ブレブ(好ましくはL2若しくはL3免疫型を有する)を生成するステップ、及びそのLOS又はブレブを薬学的に許容される賦形剤とともに製剤化するステップを含む、本発明のナイセリア(好ましくは髄膜炎菌)免疫原性組成物又はワクチンの製造方法を提供する。好ましくは、本発明の免疫型L2及びL3の両方の精製LOS、又は本発明の免疫型L2及びL3の両方のブレブ、又はL2の精製LOS及びL3のブレブ(若しくはその逆)を、混合ステップで混合する。好ましくは、本発明の精製LOS又はブレブは、単離された後に上述のようにコンジュゲートされる。さらなるリポソーム製剤化ステップを精製LOSに対して追加してもよい(当技術分野で公知の技術を用いる。例えばWO96/40063号及びそこに引用される参考文献を参照のこと)。好ましくは、ブレブ調製物は低濃度のDOC(又はDOCなし)での抽出によって(上記のように)単離される。
【0112】
このようなL2及びL3の組み合わせ方法は、ほとんど全ての髄膜炎菌B株に対して有効なワクチンをもたらすことができる。
【0113】
上記の免疫原性組成物(又は方法)は、血清群A、C、Y又はW由来の1以上(2、3又は4)の髄膜炎菌多糖又はオリゴ糖(上記のような、そのままの又はT細胞エピトープを含むキャリアにコンジュゲートしたもの)が該組成物に添加されたものであってもよい。好ましくは、少なくともCが添加(最も好ましくはコンジュゲート)されたものであり、より好ましくはA及びC又はY及びCが添加(好ましくは全てコンジュゲート化)されたものであり、最も好ましくはA、C、Y及びWが添加(好ましくは全てコンジュゲート化)されたものである。有利には、コンジュゲート化されたインフルエンザ菌B型莢膜多糖又はオリゴ糖も上記の組成物中に含有させて、広範な髄膜炎ワクチンを作製する。
【0114】
好ましくは、特にWO94/08021号において個別に具体的に示した組成物からなる又はそれを含む組成物は、本発明において特許請求の範囲に含められていない。場合により、US2006/0047106に具体的に特定されている組成物からなる又はそれを含む組成物は、本発明において特許請求の範囲に含められていない。
【0115】
本発明のワクチン製剤
本発明の免疫原性組成物は、適切なアジュバントとともに製剤化することで本発明のワクチン組成物を作製することができる。
【0116】
適切なアジュバントとしては、水酸化アルミニウムゲル(ミョウバン)又はリン酸アルミニウム(好ましくは水酸化アルミニウム)等のアルミニウム塩があるが、しかしカルシウム(特に炭酸カルシウム)、鉄又は亜鉛の塩でもよく、あるいはアシル化チロシン、若しくはアシル化糖、カチオン若しくはアニオン誘導体化多糖、又はポリホスファゼンの不溶性懸濁液でもよい。
【0117】
添加され得る適切なTh1アジュバント系には、モノホスホリルリピドA、特に3−脱−O−アシル化モノホスホリルリピドA(又は他のLPSの非毒性誘導体)、及びアルミニウム塩(好ましくはリン酸アルミニウム)とモノホスホリルリピドA、好ましくは3−脱−O−アシル化モノホスホリルリピドA(3D−MPL)(又は非毒性LPS誘導体)との組み合わせが含まれる。増強系は、モノホスホリルリピドAとサポニン誘導体の組み合わせ、特にWO94/00153号に記載されるようなQS21(若しくは他のサポニン)と3D−MPL(若しくは非毒性LPS誘導体)の組み合わせ、又はWO96/33739号に記載されるような、QS21(若しくはサポニン)がコレステロールによりクエンチされているより反応原性の低い組成物を用いる。水中油型エマルジョン中のQS21、3D−MPL及びトコフェロールを含む特に強力なアジュバント製剤は、WO95/17210号に記載されており、これは添加され得る好ましい製剤である。添加され得る他のアジュバントは、サポニン、より好ましくはQS21及び/又は水中油型エマルジョン及びトコフェロールを含む。オリゴヌクレオチドを含む非メチル化CpG(WO96/02555号)もまた添加され得る。
【0118】
ワクチン調製物は一般的にVaccine Design(「The subunit and adjuvant approach」(Powell M.F.及びNewman M.J.編)(1995)Plenum Press New York)に記載されている。
【0119】
免疫防御用量のワクチンが、全身経路又は粘膜経路によって投与され得る。これらの投与には、筋肉内、腹腔内、皮内若しくは皮下経路による注入、又は口腔/消化器(好ましくは鼻腔内投与)、呼吸器、尿生殖路への粘膜投与による注入が挙げられる。典型的には、各ワクチン用量中のブレブの量は、典型的な被ワクチン接種者において著しく有害な副作用を引き起こすことなく免疫防御反応を誘導する量が選択される。そのような量は、どの特定の免疫原が使用され、かつそれをどのように提示するかによって変動する。一般的に、各用量は、本発明の各ブレブ又はLOSを1〜100μg、好ましくは5〜50μg、最も典型的には5〜25μgの範囲内で含むことが予想される。
【0120】
本発明のブレブ免疫原性組成物に対するさらなる改良
本発明の上記のブレブ組成物は、それらが由来するナイセリア株(淋菌及び好ましくは髄膜炎菌、最も好ましくは髄膜炎菌Bを含む)が、ゲノム中への遺伝子のさらなるコピーを挿入することにより、又は既存の遺伝子の上流により強力なプロモーターを導入することにより、又は非改変株と比較して1.2、1.5、2、3、5若しくは10倍を超える抗原レベルを生じるよう改変された株を誘導することが可能なWO01/09350号に記載のあらゆる他の方法により、上方制御された1又はそれ以上の下記遺伝子(防御抗原をコードする遺伝子)を有する場合、本発明のワクチンにおける有効性がさらに改良され得る。すなわち、NspA(WO96/29412号)、Hsf又はその末端切断物(WO99/31132号及びWO01/55182号、NhhAとしても知られる)、Hap(PCT/EP99/02766号)、OMP85(WO00/23595号)、PilQ(PCT/EP99/03603号)、PldA(PCT/EP99/06718号)、FrpB(WO96/31618号)、TbpA(WO92/03467号、米国特許第5912336号、WO93/06861号及びEP586266)、TbpB(WO93/06861号及びEP586266)、NadA(Comanducciら J. Exp. Med. 2002 195:1445-1454、NMB 1994)、FrpA/FrpC、又は5以上の反復配列を含むこれらの抗原間に共通の部分(WO92/01460号;Thompsonら,(1993) J. Bacteriol. 175:811-818; Thompsonら,(1993) Infect. Immun. 61:2906-2911)、LbpA、LbpB(PCT/EP98/05117号)、FhaB(WO98/02547号、配列番号38(ヌクレオチド3083〜9025))、HasR(PCT/EP99/05989号)、lipo02(PCT/EP99/08315号)、Tbp2(WO99/57280号;NMB0460)、MltA(WO99/57280号;NMB0033)、TspA(WO00/03003号)、TspB(WO00/03003号)、ctrA(PCT/EP00/00135号)、MafA(NMB0652)、MafB(NMB0643)、Omp26(NMB0181)、アドヘシンX(NMB0315)、アドヘシンY(NMB0995)、アドヘシンZ(NMB1119)及びOstA(NMB0280)である。NMB配列の例は、www.neisseria.orgのデータベースで見出すことができる。本明細書でHsfについて言及する場合、この用語はあらゆる場合においてHsf末端切断物(特にWO01/55182号に開示されたもの)と置換可能である。
【0121】
Hsf及びTbpA(低若しくは高分子量形態、又は低及び高分子量形態の両方(EP586266))、又はHsf及びOMP85、又はOMP85及びTbpA(低若しくは高分子量形態、又は低及び高分子量形態の両方)、又はNspA及びHsf、又はNspA及びOMP85、又はNspA及びTbpA(低若しくは高分子量形態、又は低及び高分子量形態の両方)の双方が、いずれも上方制御されている場合が特に好ましい。組成物中に2つのブレブが含まれている場合、各ブレブは異なる上方制御を受けていることが好ましい。高及び低分子量のTbpAがどちらも上方制御される場合、これらは2つの形態のTbpAを天然に含む2つの株由来の組成物中に存在する、2つの別々のブレブにおいて上方制御されていることが好ましい。最も好ましくは、かかる2つの株がL2及びL3 LOS免疫型を有することである。TbpAは遺伝子工学的に上方制御させるか、又はナイセリア/髄膜炎菌産生株を鉄制限条件(例えば50〜70μMデスフェラール(メシル酸デフォロキサミン、Sigmaより入手可能)存在下)で増殖させることにより、上方制御させ得る。後者の方法を採用する場合、FrpB遺伝子発現を下方制御(好ましくは欠失)することが好ましいが、これはこの可変性抗原が鉄制限条件下で単離される髄膜炎菌株から単離されたブレブにおいて免疫優性になり得るためである。
【0122】
好ましい実施形態において、本発明の組成物は、好ましくは高分子量TbpA(TbpA High)及びHsfが上方制御されている、lgtB−(又はlst−)莢膜多糖−msbB−株由来のL3ブレブと、低分子量TbpA(TbpA Low)及びOmp85が上方制御されている、lgtB−(又はlst−)莢膜多糖−msbB−株由来のL2ブレブとを含む。より好ましくは、どちらのブレブもさらにPorA及び/又はFrpBの発現、そして任意にOpC及び/又はOpA発現が、下方制御されている。ブレブは、最も好ましくは上記の低DOC法によって単離され、両ブレブ中のLOSは外膜タンパク質とブレブ内架橋されている。
【0123】
ゴースト又は死菌全細胞ワクチン
本発明者らは、ブレブに関わる上記組成物及びワクチンが、(同一の利点を有する)ゴースト又は死菌全細胞調製物及びワクチンに関する方法にまで容易に拡張できることを意図している。グラム陰性細菌株からゴースト調製物(完全なエンベロープを有する空の細胞)を製造する方法は当技術分野で周知である(例えばWO92/01791号を参照)。全細胞を殺滅して、ワクチンに使用するための不活性化細胞調製物を作製する方法もまた周知である。従って、本明細書を通して記載されるブレブを含む組成物及びワクチンは、等価物である本発明のゴースト及び死菌全細胞調製物を含む同じ組成物又はワクチンにも当てはまることを意図している。
【0124】
本発明の組成物に関する血清殺菌アッセイ
血清殺菌アッセイは、本発明の免疫原性組成物中で組み合わせた場合に抗原間の相乗作用的な関係を評価するための好ましい方法である。
【0125】
そのような相乗反応は、抗原の組み合わせによって誘発されるSBAにより、各抗原によって別個に誘発されるSBAと比べて少なくとも50%、2倍、3倍、好ましくは4倍、5倍、6倍、7倍、8倍、9倍、最も好ましくは10倍高いものとして特徴付けられ得る。好ましくはSBAは、抗原が由来するのと同種の株、及びさらに好ましくは一群の異種の株に対して測定される(以下の代表的な群、例えばA−4クラスターに属するBZ10(B:2b:P1.2)、ET−37複合体に属するB16B6(B:2a:P1.2)、及びH44/76(B:15:P1.7,16)を参照)。SBAは、髄膜炎菌ワクチンの有効性を評価するための、最も一般的に受け入れられている免疫学的マーカーである(Perkinsら J Infect Dis. 1998, 177:683-691)。十分なSBAはあらゆる公知の方法により確認することができる。SBAは、動物モデル又はヒト被験体から得た血清を用いて行うことができる。
【0126】
ヒト血清を用いてSBAを行う好ましい方法は以下の通りである。血液サンプルは最初のワクチン接種の前、2回目のワクチン接種の2ヶ月後及び3回目のワクチン接種の1ヶ月後に採取する(3回のワクチン接種は1年以内に行い、通常のヒト第1期ワクチン接種計画では、例えば0、2及び4ヶ月、又は0、1及び6ヶ月の時点で投与される)。そのようなヒト第1期ワクチン接種計画は、1歳未満の幼児(例えばHibワクチン接種を実施するのと同時)に実施することができ、又は2〜4歳児若しくは青年もそのような第1期ワクチン接種計画でワクチン接種することによってSBAを試験することができる。妥当であれば、さらなる血液サンプルは第1期ワクチン接種の6〜12ヶ月後、及び追加免疫投与の1ヶ月後に採取され得る。
【0127】
(2〜4歳児若しくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、本発明の抗原が由来する髄膜炎菌株に対するSBA(抗体希釈率)力価が(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が、被験体の30%を超え、好ましくは40%を超え、より好ましくは50%を超え、最も好ましくは60%を超える場合、SBAは同種の殺菌活性を有する抗原又はブレブ調製物について十分であろう。
【0128】
当然ながら、異種の殺菌活性を有する抗原又はブレブ調製物は、それが由来する髄膜炎菌株に対して同様に十分なSBAを誘導することができるならば、同種の殺菌活性を有するブレブ調製物を構成することもできる。
【0129】
(2〜4歳児若しくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、髄膜炎菌の3種の異種株に対するSBA(抗体希釈率)力価において(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が、被験体の20%を超え、好ましくは30%を超え、より好ましくは35%を超え、最も好ましくは40%を超える場合、SBAは異種の殺菌活性を有する抗原又はブレブ調製物について十分であろう。かかる試験は、異種の殺菌活性を有する抗原又はブレブ調製物が、様々な髄膜炎菌株に対する交差反応性殺菌抗体を誘導し得るか否かの優れた指標である。3種の異種株は、好ましくは互いに異なる電気泳動タイプ(ET)複合体、又は多座配列タイピング(MLST)パターン(Maidenら PNAS USA 1998, 95:3140-5参照)を有し、好ましくはその異種殺菌活性を有する抗原又はブレブ調製物が作製されるか又は由来する起源の株とも異なるべきである。当業者は、髄膜炎菌間で観察される遺伝的多様性、特に、重篤な病苦の原因として認識されているか、及び/又は強い病原性が認識されているMenB系統(Maidenら 前述)である髄膜炎菌B型株において観察される遺伝的多様性を反映した、異なるET−複合体を有する3種の株を決定することは容易にできるであろう。例えば、使用され得る3種の株は以下のものである:A−4クラスターに属するBZ10(B:2b:P1.2)、ET−37複合体に属するB16B6(B:2a:P1.2)、及びET−5複合体に属するH44/76(B:15:P1.7,16)、又は同じET/クラスターに属するあらゆる他の株。そのような株は、かかる株から作製された若しくはそれらに由来する異種殺菌活性を有する抗原又はブレブ調製物を試験するために使用することができ、例えば、ET−5複合体に属する髄膜炎菌CU385株(B:4:P1.15)を使用することができる。使用され得る別のサンプル株は、系統3流行クローン(例えばNZ124(B:4:P1.7,4))由来のものである。別のET−37株は、NGP165(B:2a:P1.2)である。
【0130】
SBA活性を測定するための方法は、当技術分野において公知である。例えば、使用され得る方法がWO99/09176号の実施例10Cに記載されている。一般的には、試験される株の培養物を対数増殖期まで増殖させる(好ましくは、増殖培地にEDDA等の鉄キレート剤を添加することによる、鉄枯渇条件下で)。これを、BSAを加えた培地中(0.3%BSAを含むハンクス培地等)に懸濁し、約20000CFU/mlに調整した標準(working)細胞懸濁液を得てもよい。一連の反応混合物は、試験される血清の一連の2倍希釈物(好ましくは56℃で30分間熱不活性化されたもの)(例えば、50μl/ウェル容量)と試験される20000CFU/mlの髄膜炎菌株の懸濁液(例えば、25μl/ウェル容量)との混合により、作製することができる。反応バイアルをインキュベートし(例えば37℃で15分間)、振とう(例えば210rpmにて)するべきである。最終反応混合物(例えば容量100μl)は、さらに補体源(例えば、最終容量の25%の予備試験済みの仔ウサギ血清、又はヒト血清学のためのヒト血清)を含み、上記のように(例えば37℃で60分間)インキュベートする。滅菌ポリスチレンU底96穴マイクロタイタープレートをこのアッセイに使用できる。マルチチャネルピペットを用いて、各ウェルからアリコート(例えば10μl)を採取し、ミューラーヒントン寒天プレート(好ましくは、1%イソビタレックス(Isovitalex)及び1%熱不活性化ウマ血清を含む)に滴下し、それを(例えば、5%CO2中37℃で18時間)インキュベートしてもよい。好ましくは、個々のコロニーは、1アリコート当たり最大80CFUまで計数することができる。以下の3つの試験サンプルを対照として使用し得る:バッファー+細菌+補体、バッファー+細菌+不活性化補体、血清+細菌+不活性化補体。回帰計算によって、50%の細胞死滅に相当する希釈度の測定値を与えるようにデータを処理するプログラムを用いて、SBA力価が直接算出され得る。
【0131】
本特許明細書中に引用される全ての文献又は特許出願は、参照により本明細書に組み入れられる。
【0132】
実施例
以下の実施例は、特に詳細に記載しない限りは当業者にとって周知でありルーチンである標準的な技術を用いて実施される。本実施例は例示的なものであり、本発明を限定するものではない。
【実施例1】
【0133】
髄膜炎菌(例えば、MenB)の莢膜多糖産生に関わるタンパク質をコードする遺伝子の欠失を説明する例、PorA遺伝子の欠失、髄膜炎菌ブレブの表面上の様々な防御外膜タンパク質の上方制御、免疫優性タンパク質又は生合成酵素の下方制御(例えば、siaA(−)突然変異)、及びブレブを単離する方法は、WO01/09350号に記載されている。さらなる情報が、WO2004/014417号及びWO2004/014418号に記載されている。尚、本明細書におけるNMB及びNMA遺伝子配列の参照は、www.neisseria.orgからアクセス可能な配列の参照番号を指していることに留意されたい。LOS免疫型の通常の構造を示す概略図を図1に示す(Kahlerら、2005 Glycobiology 15:409-419/2006 JBC 281: 19939-19948より)。また、図2Bも参照されたい。
【実施例2】
【0134】
内部コアLOS O−アセチル化:殺菌力価に対する潜在的影響
要約
・MS−MS分析から、NZ124(L3)株の内部コアLOSのN−アセチル−グルコサミン(GlcNAc)はO−アセチル化されていることがわかっている。H44/76株及びM687株、並びにH44/76株由来のL3 lgtB(−)株及びL3 1st(−)ワクチン株(それぞれ、B1854=TrL3及びB1948=L7)についてはそうではない。
・NZ124株は、H44/76株及びM687株と比較して、抗体媒介性補体殺菌に対する抵抗性が高いわけではない。
・露出が乏しい表面エピトープと殺菌抗体との接触性は、NZ124株及びH44/76株で類似しているようである。
・4つの動物種を用いた5つの動物モデルにおいて、どんな製剤を用いても、抗TrL3及び抗L7ブレブ血清は、H44/76株及びM687株と比較して、NZ124株の死滅を媒介する効果が低かった。
・これらの結果は、内部コアLOSのGlcNAcのアセチル化が、抗「非アセチル化LOS」抗体により媒介される死滅の効力を低減する可能性があることを示唆している。
【0135】
概説
MS/MS分析に基づき、髄膜炎菌のL3 LOSの2つの異なる構造を説明する。これら2つの構造は、内部コアのGlcNAc上にアセチル基が存在するかしないかによって区別される(図2A)。文献には、この追加アセチル基なしでL3構造が記載されている。
【0136】
TrL3及びL7ブレブの内部コアLOSのGlcNAcはアセチル化されていない。このことは、野生型H44/76株及びM97250687(M687)株についても同様である。対照的に、野生型NZ124株は、アセチル化GlcNAcを有する。これら3つの野生型血清群B髄膜炎菌株はL3としての免疫型を有する。NZ124は、1998年に分離されたニュージーランド流行性菌株であり、ニュージーランド環境科学研究院(New Zealang Institute of Environmental Science and Research;ニュージーランド、ウェリントン)から入手可能である。
【0137】
TrL3又はL7ブレブで免疫したマウス由来の血清は、H44/76株及びM687株に対する高レベルの殺菌抗体を含んでいる。しかし、これらの血清は、NZ124株に対しては効果が劣る。実際、NZ124株について抗L7/TrL3ブレブ血清を用いて測定した殺菌力価は、H44/76株及びM687株について測定した力価より3〜20倍低い。
【0138】
NZ124株に対して測定された、低い殺菌抗体力価は、以下に挙げる少なくとも3つの仮定により説明することができる:
・この株は本来、抗体及び補体が媒介する殺菌に対して、H44/76株及びM687株より抵抗性が高い可能性がある;
・この株では、LOSエピトープは殺菌抗体との接触性が低い可能性がある;
・内部コアLOSのアセチル化は、抗「非アセチル化」LOS抗体が媒介する殺菌の効力にマイナスの影響を与える可能性がある。
【0139】
結果
NZ124株は、抗体媒介性殺菌に対し、より抵抗性なのか?
この疑問に答えるために、本発明者らは、様々なPorA+ブレブワクチンで免疫したマウス由来の血清をSBAにおいて分析した。抗血清を相同及び異種PorA株に対しSBAで試験した。表1に示す結果は、2つの実験から得られたものである。
【0140】
P1.7,16ブレブでマウスを免疫すると、相同PorA P1.7,16株の死滅を媒介することができる殺菌抗体の産生が誘導された(H44/76に対して力価1/2300)が、異種PorA株(M687及びNZ124)に対してはそうではなかった。相同PorA株に対する防御殺菌応答を誘導することだけができるP1.19,15ワクチンについても同様の観察結果が得られた(M687に対して力価1/900)。P1.7,4 PorAを含むワクチンで免疫したマウスは、NZ124株に対する高レベルの殺菌抗体(力価1/6200)を有したが、H44/76株に対してはそうではなかった。
【表1】
【0141】
これらの実験において、NZ124株に対して最も高い殺菌抗体力価が測定されたが、これは、この株が、抗体及び補体媒介性殺菌に対し、H44/76株及びM687株より抵抗性が高くはないことを示唆している。
【0142】
NZ124株のLOSエピトープは抗体に十分接触可能なのか?
特に、NspAループは、例えば、PorAのVR1及びVR2ループと比較して、比較的小さいことから、莢膜の大きさがNspA表面エピトープと抗体との接触性を制限する可能性があることが以前から示唆されてきた。実際、莢膜の大きさと、抗NspA血清が補体媒介性殺菌を誘導する能力との間には関係が証明されている(Moeら、1999 I&I 167:5664-75)。
【0143】
髄膜炎菌のLOSは短い糖鎖を有するため、抗体によるその防御エピトープとの接触性は、莢膜の厚さのため損なわれる可能性がある。このような作用機構が、NZ124株に対する抗LOS抗体で測定された低い殺菌力価を説明できるかどうかを決定するために、NZ124を含む異なる3つのMenB株に対して抗NspA MAb(MAb Me−7)を用いて、殺菌アッセイを実施した。
【0144】
1/2560を超える殺菌力価により示されるように、補体の存在下で、H44/76株及びNZ124株は、Mab Me−7によって容易に死滅した。対照的に、M687株は、抗NspA MAb媒介の死滅に対してより抵抗性が高く、わずか1/347の力価が測定された。
【0145】
結論として、防御NspAエピトープの接触性は、H44/76株及びNZ124株で類似している。従って、NZ124株死滅の媒介に関して抗TrL3ブレブ抗体の効力が低いのは、防御LOSエピトープの接触性が低いからではないと仮定することができる。
【0146】
内部コアLOSのGlcNAcのアセチル化は抗「非アセチル化」LOS殺菌抗体の効力を低減するのか?
マウス、ウサギ、モルモット、仔ラット及び成体ラットを、porA KOブレブで免疫した。実験の大部分は、ペンタ−アセチル化(penta-acylated)リピドA LOSを産生する株から取得したブレブ(msbB突然変異、B1853、B1854及びB1948ブレブ)を用いて行ったが、少数の実験では、ヘキサ−アセチル化(hexa-acylated)LOS含有ブレブ(B1820ブレブ)を用いて実施した。アルミニウム塩(Al(OH)3若しくはAlPO4)を用いて、又は用いずに(非吸収製剤)、またCpGを用いて、様々な製剤を試験した。H44/76株、M687株及びNZ124株に対する仔ウサギ補体を用いて、抗ブレブ血清をSBAにおいて試験した。
【0147】
TrL3 V1820ブレブで免疫した動物由来の血清で測定した殺菌力価を表2に示す。動物種にかかわらず、NZ124株で得られた殺菌力価は、H44/76株及びM687株で得られた殺菌力価より低い。
【表2】
【0148】
また、TrL3 msbB KOブレブ(B1853若しくはB1854)、又はL7 msbB KOブレブ(B1948)での免疫も、NZ124株よりH44/76株及びM687株に対して高い血清殺菌力価を誘導する。これは、試験した5種の異なる動物モデル(マウス、仔ラット及び成体ラット(表3a)、モルモット及びウサギ(表3b))のすべてにおいて、用いた製剤にかかわらず観察される。
【0149】
結論として、「非アセチル化LOS」を含むブレブで動物を免疫すると、「非アセチル化株」(H44/76及びM687)よりも低く「アセチル化株」(NZ124)に対して殺菌抗体応答が誘発される。
【表3】
【0150】
論考、結論及び展望
TrL3又はL7ブレブでマウスを免疫すると、H44/76株及びM687株の死滅を媒介する高レベルの殺菌抗体が誘発された。マウス血清を用いて測定したSBA力価は、H44/76株及びM687株に対するものと比較してNZ124株に対して3〜20倍低かったことから、上記抗体は、NZ124株に対しては殺菌力が低い。
【0151】
予備臨床データによれば、NZ124株が、H44/76株及びM687株より、抗体−補体媒介性殺菌に対する抵抗性が高いわけではないことが示唆されている。加えて、NZ124の表面露出が乏しい防御エピトープは、H44/76株の表面上の類似エピトープよりも、抗体への接触性が低いわけではないことを示唆する証拠もある。この知見を確認するために、電子顕微鏡検査及び/又はフローサイトメトリー分析を実施することができる。
【0152】
NZ124株のLOSとH44/76株及びM687株のLOSとの違いの1つは、NZ124 LOSにしか観察されない内部コアのGlcNAcのアセチル化である。この違いにより、H44/76株及びM687株と比較して、NZ124株の死滅の媒介において抗TrL3ブレブ血清及び抗L7ブレブ血清の効力が低いことの説明が可能である。確かに、防御糖エピトープのアセチル化が、抗体によるこのようなエピトープの認識にプラスに又はマイナスに影響しうることは知られている。しかし、ある動物種について記載された防御エピトープの免疫原性/抗原性に対するアセチル化/非アセチル化の影響が、他の動物種及びヒトに常に観察されるわけではない。
【0153】
免疫原性に対する糖エピトープのアセチル化の影響は、「動物種」に応じて、しかしまた抗原に応じても変動する。それにもかかわらず、MenBの場合には、試験した5種の動物モデルにおいて同様の観察結果が得られている。4種の異なる動物種に共通するこれらの観察結果から、「非アセチル化LOS」での免疫により誘導された殺菌抗体は、NZ124株のような「アセチル化」株の死滅の媒介において効力がより低いことが示唆される。
【0154】
この仮定を確認するために、第2のL3アセチル化株を用いた血清殺菌アッセイを実施することにする(BZ10株)。さらには、アセチル化H44/76株及び脱アセチル化NZ124株などの遺伝子改変株を用いたSBAの実施も計画する。その結果に基づき、新規L3ブレブ(アセチル化GlcNAcを含む)を評価することができるだろう。
【実施例3】
【0155】
内部コアLOSのO−アセチル化:LOS内部コアのO−アセチル化のためのナイセリア遺伝子
α鎖の糖組成に加えて、ヘプトースIIの「修飾(decoration)」は、LOS免疫原性に影響を与えるようである。PEAの数及び位置、3位におけるグルコースの存在、7位におけるグリシンの存在、並びにGlcNAcのO−アセチル化が、交差防御の重要な決定因子であるようだ。
【0156】
lpt3遺伝子(MacKinnonら、2002 Mol Microbiol. 43: 931-943)は、ヘプトースIIの3位にPEAを付加する酵素を発現する。この遺伝子(NMB2010)はフレーム変動性(phase variable)ではない。
【0157】
lgtG遺伝子は、ヘプトースIIの3位にグルコースを付加する酵素である。この遺伝子は、フレーム変動性である(WO04/015099号参照)。この遺伝子は、多数の髄膜炎菌株において、単独で、又はlpt6と一緒に欠失している。
【0158】
lpt6遺伝子(Wrightら、2004 J Bact. 186: 6970-6982)は、ヘプトースIIの6位にPEAを付加する酵素を発現する。この遺伝子(NMA0408)は、多数の髄膜炎菌株において、単独で、又はlgtGと一緒に欠失している。この遺伝子は、フレーム変動性ではない。lpt6及びlgtGは、lgt3と称される染色体上の同じ領域に位置する。
【0159】
ヘプトースIIの7位にPEAを付加する酵素は不明である。
ヘプトースIIの7位にグリシンを付加する酵素は不明である。
GlcNAcにO−アセチルを付加する酵素は、本試験の時点まで不明であった。
【0160】
O−アセチル化遺伝子の同定
・翻訳されたナイセリアゲノム(ナイセリア・ゴノローエ(N.gonorrhoeae)、髄膜炎菌MenB MC58、MenC FAM18及びMenA Z2491)上のOafAタンパク質(サルモネラ由来;ヘモフィルスO−アセチラーゼHi0391+Hi0392と相同)を用いたBLAST研究後、ナイセリアにoac1(MC58遺伝子NMB0285に代表される)とoac2(MC58遺伝子NMB1836に代表される)と称される2つの遺伝子ファミリーが見出され、oac2よりoac1ファミリーの方がOafAに近縁であった。
・両遺伝子ともにフレーム変動性である:すなわち、ORFにポリG区間(stretch)が存在する。
・MC58(非O−アセチル化LOS)において、oac1遺伝子(NMB0285)はフレーム外(out of phase)であるが、oac2(NMB1836)はフレーム内(in phase)である。
・試験した各株について、oac1及びoac2に対応するPCR産物を取得した。
・ポリG区間による機能性を調べるためにポリG区間を配列決定した。
【0161】
SBA髄膜炎菌株におけるNMB0285及びNMB1836遺伝子の存在及び機能性:MS−MS(質量分光法)を用いたARDにより得られた分析データ。分子生物学により決定した遺伝子の存在/機能性。かっこ内の数値は遺伝子ORFにおけるポリG区間内のGの数である。
【0162】
【0163】
結論
・NMB0285遺伝子の機能性と、MS−MSによるO−アセチル基の検出との間に完全な相関がある(16/16)。
・NMB1836遺伝子の機能性と、MS−MSにより得られたデータとの間には相関がない(7/16)。
・ナイセリアにおけるO−アセチル化遺伝子はNMB0285であるという強力な証拠を得た。
【0164】
図3Bは、MC58株由来の髄膜炎菌LOS O−アセチル化遺伝子NMB0285(oac1)(H44/76株オープンリーディングフレーム遺伝子配列と100%同一)を示す。大文字でオープンリーディングフレームを、小文字で周囲配列(例えば、プロモーター配列)を示す。オープンリーディングフレーム中の6つのGからなる配列(下線部)によってこのオープンリーディングフレームがフレーム外となることに留意されたい。図3Aには、Z2491株由来の等価遺伝子NMA2202(oac1)を示す。オープンリーディングフレーム(大文字)中の5つのGからなる配列によってこのオープンリーディングフレームがフレーム内となることに留意されたい。
【0165】
1136-GGG GGG ATA TTG AA-1150 MC58
1136-GGG GGA TAT TGA A-1149 Z2491(NMA2202)
【0166】
また、760676株の場合もフレーム内となる(図3C)[H44/76及び760676のoac1のオープンリーディングフレーム遺伝子配列同士は97%の同一性]。
【0167】
W3193株では、前述した領域に5つのGを有するにもかかわらず、LOSはアセチル化されていなかった。これは、ORFのヌクレオチド354で開始する遺伝子における別のポリG区域(tract)によるものであることがわかった。MC58株、760676株及びNZ124株の場合、この位置で、遺伝子に4つのGが存在し(フレーム内、活性遺伝子)、W3193株には3つのGが見とめられた(フレーム外であり、その脱O−アセチル化状態を説明している)。
【0168】
NZ124におけるO−アセチル化遺伝子の不活性化
NZ124、すなわち、O−アセチル化され(活性コンホメーションのoac1による)、H44/76(非アセチル化)由来のブレブにより誘導された血清に対してより抵抗性が高いL3株において、等価のNMB0285遺伝子(oac1)をノックアウトした。質量分光分析によりO−アセチラーゼ遺伝子の不活性化が初めて確認されたため、KO突然変異株をSBA分析に用いて、ワクチン交差防御におけるLOS O−アセチル化の関与をさらに詳しく調べることにする。
【0169】
・NMB0285(NZ124)遺伝子座を配列決定した(図4D)ところ、等価のMC58のNMB0285遺伝子座と98.6%の同一性を有した(NZ124において「フレーム内(IN PHASE)」のGの数の確認)。停止コドンの後に、IS1106に類似した別の配列があり、これは、MC58及び760676にも存在したが、NZ124には存在しなかった。
・O−アセチル化遺伝子の5’及び3’フランキング領域に対応するNZ124組換え5’及び3’領域を含むNMB0285 KOプラスミドを構築した(pMG−T−easyベクター)。NMB0285遺伝子の代わりにKanRマーカーを導入した。このプラスミドはpRIT 15574と称する。
・NZ124のoac1 KO株を構築した(これに由来するLOSは脱O−アセチル化されている)ところ、oac1はNmenのLOS O−アセチラーゼであることがさらに示された。
【0170】
新規のL3由来株におけるO−アセチル化遺伝子のオン
H44/76株由来のブレブはL3免疫型(非アセチル化)である。該ブレブは、L3 O−アセチル化株(例えば、NZ124、BZ10)に対する殺菌抗体を誘導する能力が低減しているため、機能的O−アセチル化遺伝子(oac1)をH44/76由来のブレブ産生株に再導入することが提案される。
【0171】
戦略:
・NZ124遺伝子(O−アセチル化オン株)によるH44/76のNMB0285遺伝子の置換(遺伝子オフ)。
・NZ124プロモーター及び上流調節配列を含有させた(NZ124オープンリーディングフレーム及びNZ124の448bp上流を挿入した)。
・挿入は、センス配向のエリスロマイシン耐性遺伝子を用いて行なった。
・ワクチン異種防御効力に関するブレブL3 LOS O−アセチル化の重要性を試験した(前述の方法で改変したH44/76ブレブによるNZ124の殺菌改善があったか否かを観察するため)。このL3ブレブとO−アセチル化L2株760676由来のブレブの組合せも試験することにより、異種殺菌抗体産生について調べることにする。
【0172】
結果
・前記のように実施したところ、質量分光測定により、改変H44/76株から得られたLOSがHepIIにおいてO−アセチル化されていることがわかった。
【実施例4】
【0173】
オクタロニー法(免疫型決定)、MS/MS分析及び分子生物学分析による6275株及びC11株のLOSの特性決定
要約
・特異的なポリクローナル抗体を用いた免疫拡散法(オクタロニー法)により、6725株及びC11株の免疫型を決定した。MS/MS分析により内部コアLOS組成を決定した。PCR及び配列決定により、LOS内部コア修飾に関与する酵素をコードする遺伝子を分析した。
・MS/MS分析によれば、6275株はHepIIに2個のPEA残基を有する。この株はL3株として免疫型決定されたが、通常のL3株には(HepIIの3位に)1個のPEAしかない。
・MS/MSにより、C11株の内部コアLOSの2つの異なる組成が観察された。一方の集団はHepIIに1個のPEA残基を含むのに対し、他方の集団は2個のPEA残基を含む。この株は、抗L2血清と非常に弱い反応を有するL3株として免疫型決定した。
【0174】
概説
髄膜炎菌LOSの内部コアLOS組成は、これまでの記載より複雑であると思われる。近年まで、内部コアLOSは、HepIIにPEAを含まないか、又は3若しくは6(7)位に1個だけPEA残基を含むと提唱されていた。しかし、近年、3位及び6(又は7)位に2個のPEAを含む新規の内部コアLOSが記載された。この新しいLOS構造は、これまでL3として免疫型決定された株由来のものと記載されていることから、この新規のLOS構造は、L3変異体にちなんで「L3v」と名付けられた。
【0175】
L3v構造の発見前に、LOSの免疫型決定に基づく疫学データから、約70%の侵襲性血清群B株はL3であり(3位にPEAを含む)、残りの株の大部分はL2(6位にPEAを含む)であることが示されている。
【0176】
一連のL3及びL2株、並びにL3由来のブレブ又はL2由来のブレブのいずれかで免疫した動物から得た血清を用いて得られた殺菌データに基づき、本発明者らは、抗L3由来の血清のみがL3株の補体死滅を媒介することができ、また、抗L2血清のみがL2株を死滅させることができると結論付けた。しかし、興味深いことに、2つの新しいL3v株について得られた殺菌データは、本発明者らの以前の結論と一致していない。確かに、これら2つの株の補体死滅は、抗L2由来の血清により媒介されるのであり、抗L3血清によってではない。これら2つの「非定型」株は血清群Bの6275株と血清群Cの標準株C11である。
【0177】
免疫型と殺菌結果との間で相違する結果を理解するために、6275株及びC11株の内部コア組成をMS/MSにより決定した。さらに、オクタロニー法(特異的ポリクローナル血清を用いた免疫拡散法)を用いて、これら株の免疫型決定を実施した。また、分子生物学的方法を用いて、機能的lpt3、lpt6及びlgtG遺伝子の存在を分析した。これらの遺伝子は、HepIIで、それぞれ、3位のPEA、6位のPEA、及び3位のグルコースの付加に関与する酵素をコードする。
【0178】
結果
1.MS/MS分析による内部コア組成
MS/MS分析(図4参照)により、以下のことが明らかになった:
・両方の株において、α鎖LOSは、末端シアル酸基を有するL2及びL3について記載された典型的LNnT四糖である。
・6275株の内部コアLOSは、ほぼ確実に3位及び6(7)位に2つのPEA残基を有するが、NMR分析により確認する必要がある。
・C11株は、2つの異なる内部コア:
−1個のPEAを含む(HepIIでのその位置は明確にされていないが、HepIIのグリシンに対し弱いシグナルが認められることから、7位のPEAは排除される)第1集団と、
−2個のPEAを含む(このHepIIでもグリシンが検出されたため、ほぼ確実に3位及び6位に含む)第2集団
から構成されていた。
・いずれの株についても、HepIIにグルコースは検出されない。
【0179】
2.LOS免疫型決定
一連の特異的抗血清を用いて、免疫型決定を実施した。L3及びL2抗血清以外の抗血清(L1、L4、L5…)で得られた結果は、6275株及びC11株に関して陰性であったため、L3抗血清及びL2抗血清を用いて得られた結果のみを以下に記載する。これらの実験では、GSK Bioで現在用いられている6275株及びC11株(GSK6275−1&2、及びGSK C11)を、アムステルダム大学で20年以上フリーザー内に保存された類似株(Zol 6275株及びRIV C11株)と比較した。
【0180】
2つの6275株(Zol 6275とGSK6275−1&2)は免疫型決定アッセイにおいて類似した挙動を有する。両者は抗L3血清と強力に反応するが、抗L2血清とはそうではなかった。
【0181】
GSK C11株とRIV C11株も同じ結果を示す。いずれの株も、抗L3血清について陽性であり、抗L2血清については弱い陽性である。
【0182】
C11株と抗L2血清を用いて得られた極めて弱い沈降を確認するため、1980年にL3,2として予め型決定された5人の患者の分離菌と組み合わせて、新たな免疫拡散実験を実施した。C11の結果は、抗L2血清との極めて弱い沈降を再度示し、また5種の病原菌株の型決定を確認した。
【0183】
【0184】
3.分子特性決定
3位にPEAを付加する酵素が同定されており、これは、lpt3遺伝子によりコードされる。HepIIの3位のPEAは、超毒性髄膜炎菌株の70%に検出され、Wright及びその共同研究者らは、分析した株の86%においてPCRによりlpt3遺伝子を検出した。この遺伝子は、フレーム変動により調節されず、様々な血清群C及びA株において部分的に欠失していることがわかった。3位へのPEAの付加は、同じ位置でのグルコースの付加と競合していると仮定されている。関与する酵素は、lgtG遺伝子によりコードされ(WO94/015099号参照)、ORF内のポリG区域でのフレーム変動により調節される。Moxon研究所の仮定は、lgtGのORFが存在し、かつフレーム内であれば、機能的酵素が産生され、3位にグルコースが付加されるため、この位置にPEAは付加されないというものである。
【0185】
6位にPEAを付加する酵素をコードする遺伝子は、lgtG遺伝子と並んで位置するlpt6である。この遺伝子は、フレーム変動による調節を受けやすい領域は含んでおらず、髄膜炎菌の48%に検出された(Wrightら、2004)。HepIIの7位にグリシンを付加する酵素をコードする遺伝子は不明である。
【0186】
menB 6275株及びmenC C11株を、対応するL3及びL2標準株と並行して分析した。PCR増幅及び配列決定実験を実施した:
・lpt3遺伝子の完全コピーの存在を評価しながら(PCR)、lgtG遺伝子の機能性についての研究を続行した(PCRによる存在、ORF内のポリC区間の配列)
・lpt6遺伝子の存在をPCRにより評価した。遺伝子の完全コピーが存在すれば、この株が6位にPEAを有するLOSを含むことが推定される。
【0187】
【0188】
これらのデータによれば、menB 6275株及びmenC C11株はL2免疫型に関係しているようである。しかし、MS/MS分析は、いずれの株についても、2個のPEA基を示し、グルコースは示していない。*は、lgtG遺伝子がフレーム変動領域に11(活性遺伝子に見とめられる通常の数)ではなく14の連続したCヌクレオチドを有することを示す。これは、オープンリーディングフレームがフレーム内であり、従って、機能的タンパク質を産生しうることを意味するが、別のプロリン残基の付加により、このタンパク質構造、従って、その機能が破壊される可能性があることも考えられる。
【0189】
4.要約
次の表は、多様な方法を用いた様々な髄膜炎菌株の特性決定を要約したものである。
【0190】
【0191】
論考
内部コアLOS組成の多様性は、これまで記載されているより複雑である。初めに、HepIIに1個のPEA基を含まない又は含む(3位又は6/7位のいずれか)株が記載されたが、近年、2個のPEA基を含む株が記載された。このような株は、例えば、血清群Bの6275株及び血清群CのC11株である。
【0192】
驚くべきことに、6275株及びC11株で得られた免疫型決定結果と血清殺菌結果は一致しない。確かに、これらの2株はL3株として型決定されるが、その死滅は、抗L2由来のブレブ血清により媒介されるのであり、抗L3由来のブレブ血清によってではない(次の実施例を参照)。
【0193】
これら株の関連について、また、2個のPEA基の存在が、継続的なin vitro継代培養による研究室の人工産物でありうるのか否かについて疑問が生じた。本発明者らは、6275株及びC11株の様々なシード(seeds)を比較する機会を得た。各株について、現在本発明者らが用いたシードを、20年以上保存されていた古いシード(アムステルダム大学)と比較した。各株について、2つのシードはオクタロニーアッセイにおいて同じ挙動を示したが、これは、少なくとも過去20年間変化がなかったことを示唆する。それでも、2個のPEA残基の存在を確認するために、最も古いシードの内部コアLOS組成を決定する必要がある。
【0194】
文献によれば、約70%の侵襲性髄膜炎菌株がL3である。L3v株で得られたオクタロニー結果は、この方法がL3株とL3v株を区別しないことを示唆するものであった。従って、両試験において「真のL3」株の数を過大評価する可能性がある。lpt3及びlpt6遺伝子は、HepIIのそれぞれ3位及び6位におけるPEA基付加に寄与している。約36%の循環性株が両方の遺伝子を含み、50%がlpt3のみを、また12%がlpt6のみを有する(Wright JCら、2004)。従って、L3として型決定されたとしても、潜在的に36%の株がL3vでありうる。しかし、様々な内部コアLOS構造に特異的な一連のMAbを用いて得られた近年の疫学データは、2%以下の株がHepIIに2個のPEA基を有することを示唆していた(Gidney MAJら、Infect Immun. 2004 72:559-69)。これら2つの試験の差異(36%対2%)は、6位に1個のPEA基を有する株のヒト補体に対する高い感受性により説明することができる(Ram Sら、J Biol Chem. 2003 278:50853-62)。以上すべてのデータを考え合わせると、侵襲性株の大部分が「真のL3」であることを示唆している。
【0195】
lgtGとlpt−3はHepIIのO−3位について競合し、PEA残基よりもGlc残基の付加に前記の偏りが生じることが示唆されている(Wright JCら、2004)。この仮定は、機能的lgtG遺伝子の存在下でも、6275株及びC11株の内部コアLOSに2つのPEAが存在する(フレーム変動領域において14のCヌクレオチドを含むフレーム内lgtG遺伝子が活性ではない限り。前記参照)ことより証明されるように、普遍的な法則ではないと考えられる。Moxon研究所の仮定に加えて、NmB株のLOS内部コア構造の調節/組成に関与する別の系が近年記載された。この系は、MisR/MisS二成分調節系(Tzeng YLら、J Biol Chem. 2004 279: 35053-62)である。結論として、内部コアLOSの組成に関与する機構は複合的であるようであり、十分に解明されていない。
【0196】
患者から単離した5つの株は、抗L3血清との優勢な沈降だけではなく、抗L2血清との弱い沈降も示すことから、意外なLOS免疫型を呈示した。これはまた、抗L2血清との沈降が非常に弱いにもかかわらず、C11株についても同様である。C11株の内部コアLOSのMS/MS分析も、L3及びL2 LOSの共発現と部分的に一致する2つの異なる内部コア組成を示している。免疫沈降により同定されたL3 LOSは、実際にはL3v LOS(2個のPEA残基を含む)であると考えられるが、L2 LOSは、C11株に明らかに機能的なlgtG遺伝子が検出されているため、特定のLOS分子には存在するはずの検出可能なGlcが存在しなくても、6位に1個のPEAを有する(NMR分析により確認する必要がある)内部コアLOSに関連しているだろう。それでもやはり、これら患者分離菌の1以上の内部コアLOSについて分析(MS/MS、分子特性決定及びSBA)を実施し、L3及びL2株がL3v及びL2 LOSを共発現することを確認する必要がある。
【0197】
結論として、その内部コアに2個のPEAを有する株はL3株として免疫型決定されるが、この免疫型決定は、SBAにおける生物学的反応性、並びに「機能的」lgtG、lpt3及びlpt6遺伝子の存在の分析と一致しない。本発明者のデータは、これらL3v株が恐らくL3株に由来するものではなく(lpt6遺伝子の存在のため)、従って、その名称を改めるべきであることを示している。
【実施例5】
【0198】
髄膜炎菌ワクチン:LOS濃縮ブレブをベースとするワクチンの二価組成物
【0199】
デスフェラール(desferal)を用いて又は用いずに実施した培養からのブレブの作製
・デスフェラールの存在下で、B1854、B1948、B1971、B1984及びB1987ブレブを培養物から作製した。
・B1900ブレブは、デスフェラールを用いずに、培養物から作製した。
【0200】
方法
動物実験の手順:
30匹のマウスの群を筋内経路(IM)によって3回、0、21及び28日目にOMV(約15〜20%の無毒化LOSを含む)で免疫した。毎回の接種材料は、5μg(タンパク質含量)の非吸着OMVからなるものであった。OMVは、莢膜多糖及びPorAが下方制御され、LOSが無毒化される(msbB突然変異)ように操作した髄膜炎菌(Nmen)株から作製した。産生株(前記表を参照)は、遺伝子改変された野生型H44/76株(この場合、これらはL7 LOS又はTrL3 LOSのいずれかを発現した)あるいは遺伝子改変された野生型760676株(この場合、これらは、シアル化された若しくはシアル化されていないL2 LOS、又はTrL2 LOSを発現した)のいずれかに由来するものであった。前述した培養条件下で増殖させた株からブレブを単離した。42日目に、一連の髄膜炎菌株(表1参照)を用いた血清殺菌アッセイ(SBA)による分析のために血液サンプルを採取した。プールした血液サンプル又は個々の血清(1群につき10〜30の血清)のいずれかについてSBAを実施した。
【0201】
仔ラット実験を以下のように実施した。20匹の7日齢ラットの群をIM経路によって(0、14、28及び63日目に)免疫した。毎回の接種材料は、10μg(タンパク質含量)の非吸着ブレブからなるものであった。4回目の注射から14日後に血液サンプルを採取した。
【0202】
モルモット実験を次のように実施した。20匹のモルモットの群をIM経路によって(0、14、28日目に)免疫した。毎回の接種材料は、20μg(タンパク質含量)の非吸着ブレブからなるものであった。3回目の注射から14日後に血液サンプルを採取した。
【0203】
ウサギ実験を次のように実施した。5匹のニュージーランドシロウサギの群をIM経路によって(0、21、42日目に)免疫した。毎回の接種材料は、20μg(タンパク質含量)の非吸着ブレブからなるものであった。3回目の注射から14日後に血液サンプルを採取した。
【0204】
デスフェラールを含む液体培養物を用いたSBA:
37℃+5%CO2にて、MH+1%Polyvitex+1%ウマ血清ペトリ皿上で髄膜炎菌株を一晩培養した。これらを振盪しながら、37℃で50μMのデスフェラール(鉄キレート剤)を添加した液体TSB培地において470nmでのODが0.5に達するまで3時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10又は1/50希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈(8希釈)した。細菌をPBS−グルコース0.1%で希釈することにより、5300CFU/mlを達成し、この希釈物18.8μlを血清希釈物に添加した。また、ウサギ補体(6.2μl)も各ウェルに添加した。振盪しながら37℃で75分インキュベートした後、50μlのMH+0.9%寒天をウェルに添加し、約30分後に50μlのPBS+0.9%寒天を添加した。37℃+CO2で、マイクロプレートを一晩インキュベートした。CFUを計数し、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0205】
寒天培養物(デスフェラールを含まない)を用いたSBA:
37℃+5%CO2にて、BHI+1%ウマ血清ペトリ皿上で髄膜炎菌株を一晩培養した。これらを、37℃+5%CO2で、BHI+1%ウマ血清を用いて4時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈(8希釈)した。細菌をPBS−グルコース0.1%で希釈することにより、6400CFU/mlを達成し、この希釈物12.5μlを血清希釈物に添加した。また、ウサギ補体(12.5μl)も各ウェルに添加した。振盪しながら37℃で75分インキュベートした後、50μlのTSB+0.9%寒天をウェルに添加し、約30分後に50μlのPBS+0.9%寒天を添加した。35℃又は37℃+CO2で、マイクロプレートを一晩インキュベートした。CFUを計数し、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0206】
様々な免疫型の内部コアLOS組成
・L3=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに別のアセチルはない。
・“L3”=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・L2=HepII(ほぼ確実に6位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・変異体=HepII(ほぼ確実に3位及び6位)に2個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する;しかし、W3193株においてはHepIIにOacは一切存在しないことに留意されたい。
・L10及びL11=構造は決定されていない。
・L3、“L3”、L2及び変異体は、LNnT四糖(シアル化又は非シアル化)を含む。
【0207】
結果
以下の表1に要約した結果及び図5から、以下のことがわかる:
・L2由来のブレブは、NmenB L2株に対する殺菌抗体を誘導するが、NmenB L3株(M97250687を除く)に対するものは誘導しない。これに対し、L3由来のブレブは、NmenB L3及び“L3”株に対して、また、血清群Aの3125株(L10)及び血清群YのM01.0240539株(表ではM01.539)に対して防御応答を誘導するが、L2株に対しては誘導しない。質量分光法による3125のL10 LOSの構造を図6に示す。
・加えて、L2由来のブレブは、変異型LOSを保有する株(NmenB 6275株及びNmenC C11株)に対して防御を誘導し、また、血清群C(C19)、血清群Y(S1975)、血清群A(F8238、L11)及び血清群W−135(S4383株及び3193株)由来の他の株に対しても誘導する。
【0208】
・L2及びL3株由来の濃縮LOSブレブをベースとする二価ワクチンは、試験したすべての株(どの血清群及びLOS免疫型でも)に対して防御応答(殺菌抗体)を誘導する(L2 NS及びTrL3 OMVを用いた二価ワクチンは試験しなければならない)。
・非末端切断型L2 LOSは、lgtB突然変異体(=TrL2)より優れた「交差防御」を誘導する。これは、L3由来のブレブ(TrL3=L7)については、そうではない。
・非シアル化L2ブレブは、シアル化L2ブレブと類似レベルの殺菌抗体を誘導する(少なくとも6275株に対して)。
・SBAにおいて対象とした(ターゲティング)株によれば、二価ワクチンについて妨害が観察されたり、されなかったりするが、これは、試験した株のすべてに対して最も優れた効力(SBA力価)を確実にするためには、二価製剤/組成物の微調整が必要であることを示している。
・内部コアの修飾は、交差反応性殺菌抗体の誘導に大きな影響を与える。
【0209】
【0210】
表2の結果から、以下のことがわかる:
・非血清群B株及び6275株に対し、L2由来のブレブ(末端シアル酸を含む又は含まないL2、及びTrL2)により付与される交差防御は、モルモット、ウサギ及び仔ラットにおいても観察される。
・一般に、L2由来のブレブは、M97250687(表ではM687)を除いて、L3株の補体死滅を媒介することができる有意なレベルの殺菌抗体を誘導することができない(仔ラット及びウサギ血清を用いたH44/76株に関するデータは、陰性血清について測定した活性のバックグランドにより、信頼性に欠けるため、反復しなければならない)。
・モルモットでは、TrL2ブレブは、L2 lst−ブレブ及びL2ブレブより高レベルの殺菌抗体を誘導するようであるが、仔ラット、マウス及びウサギでは、L2≧L2 lst−≧TrL2の順位が観察される。
・これらの結果から、殺菌抗体の誘導を誘発するためにL2 LOSのシアル化は必要ではないこと、また、TrL2ブレブ(lgtB突然変異)も、殺菌抗体の産生を誘発することができる(TrL3 LOSと同様に)ことがわかる。
【0211】
【0212】
結論:L2及びL3ベースのLOSワクチンを組み合わせると、Men A、B、C、W135及びY株を死滅させうる抗体を誘発することができることが初めて証明された。L2ベースのワクチンは、L2株、変異(L3v)株及びL11株を死滅させることができる。また、L3ベースのワクチンはL3株、“L3”株(効力は劣る)、及びL10株を死滅させることができる。末端切断型(lgtB(−))及び全長(又はlst(−))α鎖は殺菌することができるが、末端切断型はL3ワクチンに有益であり、全長(又はlst(−))はL2ワクチンに有益であるようだ。これら2つのLOSベースのワクチンを組み合わせると、髄膜炎菌に対するワクチンとして非常に効力があることがわかる。
【実施例6】
【0213】
髄膜炎菌ワクチン:二価OMV(L7由来及びL2由来)はABCWY株に対して交差防御を誘導する
方法
動物実験の手順:
−30匹のマウスの群を筋内経路(IM)によって3回、0、21及び28日目に、二価OMV製剤(約15〜20%の無毒化LOSを含む、L3由来のOMV+L2由来のOMV)で免疫した。毎回の接種材料は、0.8μg+0.8μg(LOS含量)の非吸着OMVからなるものであった。OMVは、莢膜多糖及びPorAが下方制御され、LOSが無毒化される(msbB突然変異)ように操作した髄膜炎菌(Nmen)株から作製した。産生株は、遺伝子改変された野生型H44/76株(L3)又は遺伝子改変された野生型760676株(L2)のいずれかに由来するものであった(遺伝子改変及び培養条件を記載する以下の表を参照)。42日目に、血清群A、B、C、W135(W)及びY由来の一連の22種の髄膜炎菌株を用いた血清殺菌アッセイ(SBA)による分析のために血液サンプルを採取した。プールした血液サンプルについてSBAを実施した。血清は実験20060425及び20060426から得たものである。
【0214】
−以下のように仔ラット実験を実施した。7日齢のラット(1群につきn=20)をIM経路によって(0、14、28及び63日目に)免疫した。毎回の接種材料は、1.6μg+1.6μg(LOS含量)の非吸着OMVからなるものであった。4回目の注射から14日後に血液サンプルを採取し、プールした(実験20060484)。
【0215】
−モルモット実験を次のように実施した。20匹のモルモットの群をIM経路によって(0、14、28日目に)免疫した。毎回の接種材料は、3.2μg+3.2μg(LOS含量)の非吸着OMVからなるものであった。3回目の注射から14日後に血液サンプルを採取し、プールした(実験20060487)。
【0216】
−ウサギ実験を次のように実施した。5匹のニュージーランドシロウサギの群をIM経路によって(0、21、42日目に)免疫した。毎回の接種材料は、3.2μg+3.2μg(LOS含量)の非吸着OMVからなるものであった。最初の注射の前及び3回目の注射から14日後に血液サンプルを採取し、これらをSBAで個々に試験した(実験20060486)。
【0217】
【0218】
デスフェラールを用いて又は用いずに実施した培養からのOMVの作製
デスフェラールを用いて実施した培養物からB1854及びB1948 OMVを作製した。
【0219】
デスフェラールを用いずに実施した培養物からB2071及びB2084 OMVを作製した。
【0220】
SBA:
37℃+5%CO2にて、髄膜炎菌株をペトリ皿上で一晩培養した。次に、37℃+5%CO2にて、デスフェラール(鉄キレート剤)を含む又は含まないペトリ皿上で、上記株を4時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10又は1/50希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈した。次に、細菌(PBS−グルコース0.1%で希釈することにより、1ウェル当たり約100〜150CFU/mlを達成する)と、仔ウサギ補体(マイクロウェル中の最終濃度:12.5%v/v)との混合物25μlを血清希釈物に添加した。振盪しながら37℃で75分インキュベートした後、2層の寒天(0.9%)をウェルに添加した。35℃又は37℃+CO2で、マイクロプレートを一晩インキュベートした後、CFUを計数し、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0221】
様々な免疫型の内部コアLOS組成
・L3=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに別のアセチルはない。
・“L3”=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・L2=HepII(ほぼ確実に6位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・変異体=HepII(ほぼ確実に3位及び6位)に2個のPEAを有し、一般に内部コアGlcNAcに1個の別のアセチルを有する。
・L10及びL11=構造は完全には決定されていない。
・L3、“L3”、L2及び変異体は、LNnT四糖(シアル化、又は非シアル化)を含む。
【0222】
結果
殺菌力価の4倍増加(対照動物由来のプール血清サンプル、又は比較物としてのワクチン前血清サンプルを用いた)に基づき、得られた結果(次の表を参照)から以下のことがわかる:
・L7+NSL2 OMV又はTrL3+TrL2 OMVのいずれかを含む非吸着二価製剤は、血清群A、B、C、W及びYに属する髄膜炎菌株に対する交差防御を付与することができる。
・この交差防御は、マウスだけではなく、仔ラット、モルモット及びウサギでも観察される。
・しかし、上記製剤は、L4 LOS免疫型を発現する株に対しては防御応答を誘発することができない。これらの実験において、血清はL10株に対して殺菌性ではない。
・L3又は“L3”又は変異体又はL2 LOSを発現する株のほとんどは、マウス、仔ラット及びモルモット血清により死滅する。これらの株に対し、交差防御の割合(%)は90%近い。
・交差防御は、ウサギにおいて、より低いようである。
【0223】
結論
L3及びL2髄膜炎菌株由来のOMNをベースとする二価ワクチンは、L3又は“L3”又は変異体(L3v)又はL2 LOSのいずれかを発現する髄膜炎菌株に対する防御を付与することができる。この防御は所与の血清群に制限されるわけではない。
【0224】
【実施例7】
【0225】
L7 OMVの免疫原性に対するOAcの影響
方法
動物実験の手順:
−30匹のマウスの群を筋内(IM)経路によって3回、0、21及び28日目にOMV(約15〜20%の無毒化LOSを含む)で免疫した。毎回の接種材料は、0.8μg(LOS含量)の非吸着OMVからなるものであった。OMVは、莢膜多糖及びPorAが下方制御され、LOSが無毒化される(msbB突然変異)ように操作した髄膜炎菌株から作製した。産生株は、遺伝子改変された野生型H44/76株に由来するものであった(遺伝子改変及び培養条件を記載する以下の表を参照)。42日目に、髄膜炎菌株を用いた血清殺菌アッセイ(SBA)による分析のために血液サンプルを採取した。個々の血清についてSBAを実施した。血清は実験20060634から得たものである。
【0226】
−モルモット実験を次のように実施した。20匹のモルモットの群をIM経路によって(0、14、28日目に)免疫した。毎回の接種材料は、3.2μg(LOS含量)の非吸着OMVからなるものであった。3回目の注射から14日後に血液サンプルを採取した(実験20060636)。
【0227】
【0228】
SBA:
37℃+5%CO2にて、髄膜炎菌株(OAc−であるH44/76及びM97250687と、OAc+であるNZ124)をペトリ皿上で一晩培養した。次に、37℃+5%CO2にて、デスフェラール(鉄キレート剤)を含むペトリ皿上で、上記株を4時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10又は1/50希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈した。次に、細菌(PBS−グルコース0.1%で希釈することにより、1ウェル当たり約100〜150CFU/mlを達成する)と、仔ウサギ補体(マイクロウェル中の最終濃度:12.5%v/v)との混合物25μlを血清希釈物に添加した。振盪しながら37℃で75分インキュベートした後、2層の寒天(0.9%)をウェルに添加した。35℃又は37℃+CO2で、マイクロプレートを一晩インキュベートした。CFUを計数した後、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0229】
結果
マウスの場合、L7 OAc+OMVはL7 OAc−OMVより免疫原性が高い(以下の表を参照)。確かに、L7 OAc+OMVで免疫したマウス由来の血清は、L7 OAc−OMVで免疫したマウス由来の血清より高い補体媒介殺菌活性を示す。これは、OAc−及びOAc+野生型株に対して観察される。しかし、有意な差異はOAc−株(H44/76及びM97205687)に対してしか観察されない。
【0230】
【0231】
モルモットの場合には、SBAにおいてOMVの両方の型がOAc+及びOAc−野生型株に対して同様の免疫原性を示す(以下の表参照)。
【0232】
【0233】
結論
内部コアGlcNAcのO−アセチル化は、マウスにおいてOMVの免疫原性(SBA力価)を増強するが、モルモットではそうではないことが証明された。
【技術分野】
【0001】
本発明は、ナイセリアワクチン組成物、それらの製造、及びかかる組成物の医薬における使用の分野に関する。より具体的には、ナイセリア(特に髄膜炎菌)の外膜小胞(すなわち、ブレブ)ワクチンの製造により適した新規な操作された髄膜炎菌株の作製方法に関する。新規のLOSサブユニット又は髄膜炎菌外膜小胞(すなわち、ブレブ)ワクチンの使用に基づく有利な方法及びワクチン製品もまた記載しており、これはヒト被験者における使用のために安全性及び有効性をより高めたものである。
【背景技術】
【0002】
髄膜炎菌(Neisseria meningitidis)は、しばしばヒト上気道より単離されるグラム陰性細菌である。この細菌は、菌血症及び髄膜炎等の重篤な侵襲性細菌性疾患の原因である。髄膜炎菌疾患の発生数は、地理的、季節的及び年次的な差異を示す(Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。この細菌は通常、その莢膜多糖による血清群に従って分類される。
【0003】
温暖な国々で最も多い疾患は、血清群B株によるものであり、発症率は総人口の1〜10人/100,000人/年の間で変動し、それより高い値に達する場合もある(Kaczmarski, E.B. (1997), Commun. Dis. Rep. Rev. 7: R55-9, 1995;Scholten, R.J.P.M., Bijlmer, H.A., Poolman, J.T.ら Clin. Infect. Dis. 16:237-246, 1993;Cruz, C., Pavez, G., Aguilar, E.,ら Epidemiol. Infect. 105:119-126, 1990)。
【0004】
血清群A髄膜炎菌が優位を占める流行の大部分は中央アフリカで発生し、その発症率が1000人/100,000人/年のレベルにまで達する場合もある(Schwartz, B., Moore, P.S., Broome, C.V. Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。全体として、髄膜炎菌疾患のほとんど全ての症例は、血清群A、B、C、W−135及びYの髄膜炎菌によって引き起こされ、四価のA、C、W−135、Y莢膜多糖ワクチンが利用可能である(Armand, J., Arminjon, F., Mynard, M.C., Lafaix, C., J. Biol. Stand. 10:335-339, 1982)。
【0005】
髄膜炎菌感染の頻度は、過去数十年、多くのヨーロッパの国々において増加してきている。これは、社会的活動の場(例えば、水泳プール、劇場等)の増加による伝染の増加に起因してきた。いくつかの標準的な抗生物質に対して感受性が低い又は耐性の髄膜炎菌株を単離することは、もはや珍しいことではない。この現象は、新たな抗菌剤、ワクチン、薬剤スクリーニング法、及びこれらの生物についての診断検査に対する、未だ満たされていない医学的ニーズ及び需要を生み出した。
【0006】
利用可能な多糖ワクチンは、現在、それらをキャリアタンパク質に化学的にコンジュゲートする方法によって改良が行われている(Lieberman, J.M., Chiu, S.S.Wong, V.K.らJAMA 275:1499-1503, 1996)。
【0007】
しかしながら、血清群Bワクチンは利用可能な状況にない。血清群B莢膜多糖は、免疫原性を示さないことが分かってきたが、それは恐らくそれが宿主成分との構造類似性を有するためであろう(Wyle, F.A., Artenstein, M.S., Brandt, M.L.ら J. Infect. Dis. 126:514-522, 1972;Finne, J.M., Leinonen, M., Makela, P.M. Lancet ii.:355-357, 1983)。従って、外膜小胞(すなわちブレブ)又はそれらから得た精製タンパク質成分から、血清群Bワクチンの開発を試みることに焦点を当てた努力がされてきた。
【0008】
ワクチン開発のための別の髄膜炎菌抗原は、髄膜炎菌のリポオリゴ糖(LOS)である。これらは外膜に結合した糖脂質であり、O側鎖を欠損しているという点で腸内細菌科(Enterobacteriaceae)細菌のリポ多糖(LPS)と異なり、従ってLPSのラフ型と類似している(GriffissらRev Infect Dis 1988, 10:S287-295)。LOSのオリゴ糖部分の異質性は、異なる髄膜炎菌株における構造的な及び抗原性の多様性を生み出す(GriffissらInf. Immun. 1987, 55:1792-1800)。このことは、かかる株を12種類の免疫型にさらに分類するために用いられている(ScholtanらJ Med Microbiol 1994, 41:236-243)。免疫型の決定は、通常、既知の免疫型のLOSに対して生成された吸着ポリクローナル抗体を用いるオクタロニー法によって行われる(Poolman JT, Hopman CTP及びZanen HC, FEMS Microbiol Letters (1982) 13: 339-348)。免疫型L3、L7及びL9は、免疫学的に同一であり、かつ構造的に類似(若しくは全く同一)であり、従ってL3,7,9(又は、本明細書において総称的に「L3」)と呼ばれている。髄膜炎菌のLOS L3,7,9(L3)、L2及びL5は、シアル化によって、又はシチジン5'−一リン酸−N−アセチルノイラミン酸の付加によって修飾することができる。L2、L4及びL6 LOSは免疫学的に区別することができるが、しかしそれらは構造的に類似しており、本明細書でL2に言及する場合にはL4又はL6のいずれかと任意に置換することができ、これは本発明の範囲内となる。LOSに対する抗体は、実験用ラットにおいて感染に対し防御し、また髄膜炎菌に感染した小児において殺菌活性に寄与することが示されている(GriffissらJ Infect Dis 1984, 150:71-79)。
【0009】
しかしながら髄膜炎菌ワクチンにおけるLOSの使用に伴う問題は、その毒性(そのリピドA部分による毒性)にある。
【0010】
またLOSは髄膜炎菌のブレブの表面に存在する。髄膜炎菌の外膜小胞(すなわちブレブ)に基づいたワクチンの開発に焦点を当てて、長年にわたる努力がされてきた(de Moraes, J.C., Perkins, B., Camargo, M.C.らLancet 340:1074-1078, 1992、Bjune, G., Hoiby, E.A. Gronnesby, J.K.ら338:1093-1096, 1991)。そのようなワクチンは、宿主に投与した場合に防御免疫応答を誘発することのできる、正確に折り畳まれた立体構造をとったいくつかの完全な外膜タンパク質を含むという利点を有する。さらに、ナイセリア株(髄膜炎菌血清群B、すなわちmenBを含む)は、工業的規模での製造を可能にするために十分な量の外膜ブレブを分泌する。しかしながら、ブレブは大抵、細菌細胞の0.5%界面活性剤(例えばデオキシコレート)抽出を含む方法により調製される(例えばEP11243)。それは上記のようなLOSの毒性(内毒素とも呼ばれる)のために望ましいが、ワクチンからほとんどのLOS抗原を除去する作用もまたある。
【0011】
ワクチン抗原としてのLOSの使用に伴うさらなる問題は、多様な糖鎖構造を有する12のLPS免疫型が存在することである(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021;Mol Microbiol 2002, 43:931-43)。ある免疫型に対する抗体は、異なる免疫型を認識することができない。LOS免疫型のオリゴ糖部分の共通「コア」領域を生成することに努力が集中されてきたが(例えばWO94/08021号)、改変されたLOSに対して生起された抗体の殺菌活性は失われる。従って、ワクチンは、有効であるために異なる免疫型の多くのLOS成分を有する必要があるだろう。
【0012】
ヒトワクチンにおける抗原としてのLOS(LPS又はリポ多糖としても知られる)の使用に伴うさらなる問題が存在し、それはすなわちそれらがヒト糖鎖構造(例えば、ヒト赤血球細胞上の)に類似した糖鎖構造を担持しているため、それらの使用に伴う安全性の問題が突きつけられることである。さらにLOS構造の改変は、LOS抗原の殺菌効果の構造的感受性のため、問題を含んでいる。
【0013】
WO2004/014417号は、これらの問題についてのいくつかの解決法を記載している。
【発明の概要】
【0014】
本発明者はさらに以下の点を見出した:
−LOS内部コアの修飾(decoration)は、免疫型の殺菌性エピトープの確定に重要である、
−L2 LOSをヘプトースIIに結合したGlcNAc残基においてO−アセチル化修飾する酵素が見いだされ、それをコードする遺伝子がOac1である、
−O−アセチル化されているL3免疫型LOSが見出され(以前に報告されていない)、これはL3株に広範囲に存在すると考えられる、
−従来のL3 LOSに対する抗体はO−アセチル化L3株を死滅させることができるが、この殺菌は従来のL3株に対してそれほど有効ではない。
【0015】
上記知見により、本発明者は、ナイセリアワクチン製剤におけるO−アセチル化L3 LOSの使用を検討するに至った。
【0016】
発明の概要
従って、本発明の一態様においては、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているナイセリア株由来のL3 LOSを含む免疫原性組成物を提供する。ナイセリア株は、天然にそのようなLOSを産生するもの(例えばNZ124株)であってもよいし、又は機能的oac1遺伝子の挿入によって作出されたもの(以下参照)であってもよい。本発明の目的のため、L3V免疫型は、L2免疫型とより免疫学的に類似しているため、L3株の分類ではない。
【0017】
L3 LOSは、以下の構造を有しうる:
【化1】
【0018】
〔式中、
R1=
【化2】
【0019】
であり、
R2=PEAであり、
R3=Hであり、
R4=OAcであり、
R5=H、PEA、又はGlyである〕。
【0020】
上記の用語はLOS分野の標準的な略語であり、例えばGlcはグルコース(又はD−グルコピラノース)を表し、KDOは2−ケト−3−デオキシオクトネートを表し、HepはL−グリセロ−D−マンノ−ヘプトースを表し、GlcNAcはN−アセチルグルコサミンを表し、Galはガラクトースを表し、NeuNacはシアル酸を表し、OAcはO−アセチルを表し、PEAはホスホエタノールアミン(又は2−アミノエチルホスフェート)を表し、Glyはグリシンを表す、などである。
【0021】
本明細書において「ナイセリア(neisserial)」は、いかなる場合にも、髄膜炎菌(N. meningitidis)、例えば血清群A、B、C、W135及びYを示す。また、本発明のLOSを産生しうる他の任意の株(例えば淋菌又はナイセリア・ラクタミカ(Neisseria lactamica))を示すこともある。
【0022】
本発明のL3(L2/L4/L10)LOSはそれぞれL3(L2/L4/L10)免疫型のLOSを包含するが、これに必ずしも限られない場合がある(例えば本発明のL2 LOSは、L3免疫型のものとして現在知られているL3V LOSを含むが、L3V LOSを有する株は、L2免疫型を有するLOSに対して生成された血清によって死滅する。下記参照)。よって、本発明の「LOS」、「L2 LOS」、「L3 LOS」、「L4 LOS」及び「L10」LOSという用語の機能的な解釈は、この広い意味で解釈されるべきである。例えば、本発明のL3 LOSを含む免疫原性組成物は、L3免疫型の株を死滅する抗体を誘発することができるなどである。
【0023】
本発明の免疫原性組成物はさらに、ナイセリア株由来のL2及び/又はL10及び/又はL4 LOSを含んでもよい。本発明のL2、L3、L4又はL10 LOSは、髄膜炎菌A株、B株、C株、W135株又はY株から単離することができる。
【0024】
本発明の免疫原性組成物はさらに、以下の構造を有するL2 LOS:
【化3】
【0025】
〔式中、
R1=
【化4】
【0026】
であり、
R2=PEA又はGlcであり、
R3=PEAであり、
R4=H又はOAcであり、
R5=H、PEA、又はGlyである〕
を含んでもよい。
【0027】
本発明の免疫原性組成物はさらに、以下の構造を有するL10 LOS:
【化5】
【0028】
〔式中、
R1=ヘキソース−ヘキソース−であり、
R2=H又はPEAであり、
R3=PEAであり、
R4=OAcであり、
R5=H又はGlyである〕
を含んでもよい。
【0029】
「R1=ヘキソース−ヘキソース−」は、Gal−Gal−、Gal−Glc−、Glc−Gal−、又はGlc−Glc−残基、例えば:
R1=
【化6】
【0030】
を包含する。
【0031】
本発明のL10 LOSにおけるR1末端ヘキソースは、シアル化されていてもよいし、されていなくてもよい(シアル化されている場合には、これは場合によりα−Neu5Ac−(2→3)基を介していてもよい)。
【0032】
本発明の免疫原性組成物はさらに、以下の構造を有するL4 LOS:
【化7】
【0033】
〔L4 LOSについて、式中、
R1=
【化8】
【0034】
であり、
R2=Hであり、
R3=PEAであり、
R4=OAcであり、
R5=Glyである〕
を含んでもよい。
【0035】
あるいは、本発明のL4 LOSは、R4=H及び/又はR5=Hであってもよい。
【0036】
本明細書における「本発明の1以上のLOS」若しくは「本発明のLOS」又は類似の文言は、本発明のL2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSを意味する。本明細書における「本発明のブレブ調製物」又は類似の文言は、本発明のL2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSを有する本発明の1又はそれ以上のブレブを意味する。本発明の細菌株は、それから本発明の1以上のLOSを単離することができるものである。
【0037】
本発明の1以上のLOSは、タンパク質キャリアとコンジュゲートされていてもよく、そのタンパク質キャリアは、Tヘルパーエピトープ源、例えば破傷風トキソイド、ジフテリアトキソイド、CRM197、又は髄膜炎菌ブレブ上の外膜タンパク質などである(下記参照)。その1以上は、化学的に(下記参照)又は遺伝的に(例えばmsbB(−)及び/又はhtrB(−)であるナイセリア株から単離された場合。下記参照)、無毒化されたそのリピドA部分を有するものである。
【0038】
本発明の1以上のLOSは、精製LOS調製物として、リポソーム調製物(典型的には精製LOSを含む)として、又はブレブ調製物(下記参照)として、免疫原性組成物中に存在しうる。
【0039】
本発明のブレブ調製物(複数でもよい)は、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離することができる。それらの各ナイセリア株は、それが鉄を利用可能な条件下、又は鉄除去の条件下(例えば、デスフェラールなどの鉄キレート剤を添加する。下記参照)で培養されたものであってもよい。本発明のブレブ調製物は、莢膜多糖を合成することができないナイセリア株から単離されてもよい。例えば、その株は、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、又は欠失している(すなわち、その遺伝子からの機能的発現がない)ものである。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、それが由来する株はそれぞれの株において同じ莢膜多糖遺伝子の発現が下方制御されていることが好ましい。ナイセリア株は、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(すなわち、その遺伝子からの機能的発現がない)ものであってもよい。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、ブレブが由来する株はそれぞれの株において同じリピドA遺伝子(複数でもよい)の発現が下方制御されていることが好ましい。ナイセリア株は、それが由来する天然株と比較して、外膜タンパク質遺伝子porA、porB、opA、opC、pilC、lbpA又はfrpBの1又はそれ以上の発現が下方制御されている、好ましくは欠失している(すなわち、その株の外膜上に遺伝子産物の発現がない)ものであってもよい。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、これらの株は各ナイセリア株において同じ外膜タンパク質遺伝子(複数でもよい)の発現が下方制御されていることが好ましい。ナイセリア株は、外膜タンパク質抗原NspA、TbpA low、TbpA high、Hsf、Hap、OMP85、PilQ、NadA、GNA1870、MltAの1又はそれ以上の発現が上方制御されているものであってもよい。L2及びL3ブレブの両方が存在する(あるいは2以上の本発明のブレブ調製物が存在する)場合、それが由来する株はそれぞれの株において1又はそれ以上の異なる外膜タンパク質抗原の発現が上方制御されていることが好ましい。
【0040】
さらに、有効量の本発明の免疫原性組成物及び薬学的に許容される担体を含むワクチン組成物を提供する。ワクチンは、アジュバント、例えば水酸化アルミニウム又はリン酸アルミニウム、をさらに含んでもよい。ワクチンは、以下の株:髄膜炎菌血清群A、髄膜炎菌血清群C、髄膜炎菌血清群W−135、髄膜炎菌血清群Y、及びインフルエンザ菌(H. influenzae)b型に由来する、1又はそれ以上のコンジュゲートした莢膜多糖又はオリゴ糖をさらに含んでもよい。
【0041】
以下のリスト:髄膜炎菌血清群A、B、C、W135及びYから選択される1又はそれ以上によって引き起こされる疾患の予防又は治療のための医薬の製造における、本発明の免疫原性組成物又は本発明のワクチンの使用もまた提供する。さらに、以下のリスト:髄膜炎菌血清群A、B、C、W135及びYから選択される1又はそれ以上によって引き起こされる疾患の予防又は治療方法であって、本発明の免疫原性組成物又は本発明のワクチンをその必要があるヒト患者に投与するステップを含む方法を提供する。
【0042】
本発明の免疫原性組成物又はワクチンの製造方法であって、L3 LOSを単離するステップ、場合によりそれを必要に応じて単離されたL2及び/又はL10 LOSと組み合わせるステップ、並びに薬学的に許容される賦形剤と共にL3 LOSを製剤化するステップを含む方法をさらに提供する。
【0043】
髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、本発明の免疫原性組成物又はワクチンの使用も提供する。髄膜炎菌免疫型L3疾患の予防又は治療方法であって、その必要があるヒト患者に、有効量の本発明の免疫原性組成物又はワクチンを投与するステップを含む方法である。髄膜炎菌免疫型L3疾患は、そのヘプトースII残基に結合したGlcNAc残基においてO−アセチル化されているLOS、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないLOS、又はそのヘプトースII残基に結合したGlcNac残基において部分的にO−アセチル化されておりかつ部分的にO−アセチル化されていないLOS、のいずれかのLOSを有する株によって引き起こされるものである。
【0044】
本発明のさらなる態様において、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株により引き起こされる髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の使用を提供する。また、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株によって引き起こされる髄膜炎菌免疫型L3疾患を予防又は治療する方法であって、その必要があるヒト患者に、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の有効量を投与するステップを含む方法を提供する。
【0045】
上記使用又は方法において、免疫原性組成物は、以下の構造を有するL3 LOS:
【化9】
【0046】
〔式中、
R1=
【化10】
【0047】
であり、
R2=PEAであり、
R3=Hであり、
R4=Hであり、
R5=H、PEA、又はGlyである。〕
を含むことができる。
【0048】
本発明のLOSについて本明細書中で説明するように、免疫原性組成物中のL3 LOSは、タンパク質キャリアとコンジュゲートしていてもよいし(さらなる説明については上記及び下記参照)、無毒化リピドA部分、例えばmsbB(−)ナイセリア株から単離されたLOSと一致する第2(secondary)アシル鎖を欠損している(及び/又は、LOSを以下に記載するリピドA無毒化ペプチドと複合体化することによる)もの、を含んでもよいし、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として免疫原性組成物中に存在してもよい。ブレブ調製物の場合、ブレブ調製物は、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものとすることができる。ナイセリア株は、莢膜多糖を合成することができないものであってもよい、例えば、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものとすることができる。ナイセリア株は、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものとすることができる。
【0049】
本発明のまた別の態様において、通常はそのLOSのヘプトースII残基に結合したGlcNAc残基をO−アセチル化するナイセリア株のLOSを脱O−アセチル化する方法であって、oac1遺伝子の機能的発現を破壊することによって、機能的Oac1を発現することができないようにするステップを含む方法を提供する。
【0050】
本明細書の全体においてoac1とは、LOSのHepII−GlcNacをOAc基によって誘導体化する反応の触媒に関与するナイセリアの遺伝子を意味する。この活性を有する遺伝子配列及びオープンリーディングフレームは、例えば図3Dに示されている。この配列から、あらゆるナイセリアoac1配列を見出すことができる(例えば、図3Dに示されるオープンリーディングフレームと少なくとも70、80、90、95又は99%の配列同一性を有するオープンリーディングフレーム)。本発明のさらなる態様において、本発明のoac1操作方法を実施するために使用することができる、図3Dからの10、15、20、30、35又はそれ以上の連続するヌクレオチドのプライマー配列を提供する。本発明のさらなる態様において、グラム陰性細菌LOSのHepIIに結合したGlcNAc残基をO−アセチル化する方法であって、非(又は部分的)O−アセチル化LOSと単離されたOac1酵素(これは公知の組換え技術によって調製することができる)とを混合するステップを含む方法を提供する。
【0051】
本発明のさらなる態様において、通常はそのLOSのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されている又は部分的にのみO−アセチル化されているナイセリア株のLOSをO−アセチル化する又はさらにO−アセチル化する方法であって、ナイセリア株においてoac1遺伝子の機能的発現を増大するステップを含む方法を提供する。これは、oac1遺伝子の機能的発現を増大するステップが、oac1遺伝子の機能的コピー(これはさらなる機能的コピーであってもよい)(例えば、場合によりその天然のプロモーター配列を含んでいてもよい図3Dのオープンリーディングフレーム)をナイセリア株に導入することによって行うように、実施することができる。あるいは、又はそれに加えて、oac1遺伝子の機能的発現を増大するステップは、存在する非機能的遺伝子のポリGフレーム変動領域(poly-G phase variable region)(後述するようにoac1遺伝子にはそのような領域が2つある)を、oac1遺伝子の機能的発現のためにフレーム内となるようにすることによって行うことができる。これは、公知の突然変異誘発技術によって行うことができる。ポリGフレーム変動領域は、通常は、活性を有する遺伝子のATG開始コドンから1136〜1140位のヌクレオチドの間にある(そして不活性遺伝子においてはフレーム外である1140以上にまで及ぶことがある)。2つ目のポリG領域は、開始コドンから354位に見出されている。
【0052】
さらに、ナイセリア株において機能的oac1発現のフレーム変動性を少なくする方法であって、oac1遺伝子のポリGフレーム変動領域(の一方又は両方)を、より少ないGヌクレオチドを有するコドンを用いて同じアミノ酸がコードされるように、変化させるステップを含む方法を提供する。例えば、ATG開始コドンから1135〜1137位のヌクレオチドによってコードされるアルギニン残基のコドンを、AGGからCGT、CGC、CGA又はAGAに変化させてもよいし、並びに/あるいは、ATG開始コドンから1138〜1140位のヌクレオチドによってコードされるグリシン残基のコドンを、GGGからGGT、GGA又はGGCに変化させてもよい。
【0053】
上記のoac1操作方法は、L2又はL3免疫型のナイセリア株に対して行うことができる。さらに、本発明のoac1操作方法を実施するステップ、得られるナイセリア株からLOSを単離するステップ、及び薬学的に許容される担体と共に有効量のLOSを製剤化するステップを含む、免疫原性組成物の製造方法を提供する。また、本明細書の全体に記載されているように、LOSをタンパク質キャリアにコンジュゲートしてもよいし、そのリピドA部分を、例えばmsbB(−)ナイセリア株からLOSを単離する及び/又は後述するようにリピドA無毒化ペプチドと複合体化することにより、無毒化してもよい。LOSは、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として単離されたものとすることができる。ブレブ調製物の場合には、ブレブ調製物を、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものとすることができる。ナイセリア株は、莢膜多糖を合成することができないものであってよく、例えば、ナイセリア株は、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものであってよい。ナイセリア株は、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ものであってもよい。
【0054】
本明細書における「ナイセリア」は、いかなる場合にも、髄膜炎菌(N. meningitidis)、例えば血清群A、B、C、W135及びYを意味する。
【図面の簡単な説明】
【0055】
【図1】L1〜L8免疫型の構造によって示される、髄膜炎菌LOS構造の共通置換パターンである(Kahlerら Glycobiology 2005 15:409-419 / 2006 JBC 281:19939-19948より転載)。L7免疫型構造(示さない)は、L3免疫型構造の非シアル化形態である。保存されている内部コア領域をR1〜R5として示す可変付属基と共に示す。α鎖(R1)の組成は、lgtA−Eトランスフェラーゼ、及び末端α−Neu5Ac(シアル酸)基を付加するシアリルトランスフェラーゼをコードするlstのフレーム変動的(phase-variable)発現によって制御される。内部コアへのグリシンの結合は第2のHep残基上の7位を介することに留意されたい。この略図に示されているKDO残基に結合する、全ての免疫型の内部コアに存在することが知られているさらなるKDO残基は示されていないことに留意されたい。
【図2A】質量分析によって決定された、種々のL3髄膜炎菌株のLOS構造の概略である。興味深いことに、H44/76株由来血清によって殺菌される傾向が低いL3株のいくつかはO−アセチル化されている。
【図2B−1】種々の髄膜炎菌免疫型の共通LOS構造である。LOS構造の特定の部分を形成する酵素をコードする既知の遺伝子の名称を示している。
【図2B−2】図2Bの続き
【図3A】Z2491株由来の髄膜炎菌LOS O−アセチル化遺伝子NMA2202を示す。オープンリーディングフレーム中の5つのG(大文字)の配列によってオープンリーディングフレームがフレーム内となることに留意されたい。
【図3B−1】MC58株由来の髄膜炎菌LOS O−アセチル化遺伝子NMB0285を示す。オープンリーディングフレームを大文字で、周囲の配列(例えばプロモーター配列)を小文字で示す。オープンリーディングフレーム中の6つのG(下線)の配列によってオープンリーディングフレームがフレーム外となることに留意されたい。
【図3B−2】図3Bの続き
【図3C】760676株由来の髄膜炎菌LOS O−アセチル化遺伝子(NMB0285等価物)を示す。オープンリーディングフレームを大文字で、周囲の配列(例えばプロモーター配列)を小文字で示す。オープンリーディングフレーム中の5つのG(下線)の配列によってオープンリーディングフレームがフレーム内となることに留意されたい。
【図3D】(MenB、L3)NZ124株由来の髄膜炎菌LOS O−アセチル化遺伝子(NMB0285等価物)を示す。オープンリーディングフレームを大文字で、周囲の配列(例えばプロモーター配列)を小文字で示す。オープンリーディングフレーム中の5つのG(下線)の配列によってオープンリーディングフレームがフレーム内となることに留意されたい。
【図4A】(A)LOS6275からの内部コアオリゴ糖(ES−);(B)LOS6275からの内部コアオリゴ糖(MS/MS ES+m/z 1803.6)。3位及び6位におけるPEAに連結したHepIIと、O−アセチル化された内部コアを示す構造の概略を示す。
【図4B】(A)LOS6275からの内部コアオリゴ糖(ES−);(B)LOS6275からの内部コアオリゴ糖(MS/MS ES+m/z 1803.6)。3位及び6位におけるPEAに連結したHepIIと、O−アセチル化された内部コアを示す構造の概略を示す。
【図4C】(C)MenC C11株からの内部コアオリゴ糖(ES−)を示す。
【図5】L2由来OMVワクチンによる交差反応性殺菌抗体の誘導に対するL2 LOSのシアル化の影響を示す。SBA力価(50%死滅のGMT)及びセロコンバージョン(%)を示す。
【図6】MenA 3125のL10 LOSの質量分析構造解析を示す。
【発明を実施するための形態】
【0056】
発明の説明
本明細書中に記載される刊行物及び特許、又は特許出願の主題又はそこに開示される情報は、参照により本明細書に組み入れる。
【0057】
「リポオリゴ糖」(又は「LOS」)に関する言及は、「リポ多糖」又は「LPS」にも言及するものとする。
【0058】
本明細書中の用語「含む」(「comprising」「comprise」「comprises」)は、全ての場合に、任意に用語「からなる」(「consisting of」「consist of」「consist of」)に各々置き換えることができるものと発明者は意図している。
【0059】
本発明者らは、LOSオリゴ糖構造の短縮が殺菌性免疫応答を誘発し得るエピトープの減少を導くことを見出した。あるいは、本発明者らは、ワクチン製剤においてLOSを最も有効に使用するためには、LOSオリゴ糖構造をできるだけ保持するべきであるが、たった2種類(又は3若しくは4種類)のLOS抗原の組み合わせによって広範に有効なナイセリアの(好ましくは髄膜炎菌の)ワクチンを生成し得ることを見出した。本発明の第1の態様は、ナイセリア(好ましくは髄膜炎菌)の免疫型L2のLOS及び免疫型L3のLOS(又は、本発明のL2及びL3 LOS、そして場合により本発明のL10及び/若しくはL4 LOS)を含む、ナイセリア(好ましくは髄膜炎菌又は髄膜炎菌B型)疾患の予防又は治療のための免疫原性組成物である。LOSは、公知の精製法のいずれかにより単離してもよいし、あるいは例えばL2及びL3ナイセリア株由来の少なくとも2種類の外膜小胞(すなわちブレブ)調製物中に存在するものでもよい。ブレブ調製物から、毒性を有し緩く保持されたLOSを除去するが、ブレブ内に組み込まれたLOS抗原は高レベルで保持するようにするために、ブレブを低濃度の界面活性剤、すなわち0〜0.3%、好ましくは0.05〜0.2%、最も好ましくは約0.1%の、好ましくはデオキシコレート(すなわちDOC)、を用いて抽出することが好ましい。LOS抗原のそのような組み合わせ(特にブレブワクチン中の)は、驚くべきことに90%を超える髄膜炎菌株に対して有効であるという点で有利である。
【0060】
本発明者らはまた、本発明の上記ブレブ免疫原性組成物、及び実際にはあらゆるナイセリア(好ましくは淋菌又は髄膜炎菌)由来のブレブ免疫原性組成物が、免疫優性外膜タンパク質の特定の組み合わせの発現が下方制御(好ましくは欠失)される場合に、それらの表面上の防御抗原(LOSを含む)の効果が増強されることを見出した。従って、本発明のさらなる態様は、天然の非改変株と比較して、発現が下方制御された、好ましくは欠失した、以下の外膜タンパク質:PorA、PorB、OpA、OpC又はPilC、を2つ以上有するナイセリア株由来の、本発明の1以上のナイセリアブレブ調製物である。好ましくは、PorAとOpA、PorAとOpC、OpAとOpC、又はPorAとOpAとOpCが下方制御されているか又は欠失している。FrpBの発現の下方制御(好ましくは欠失)はまた、交差防御抗原の効果の増強において有利であり、特に、鉄制限状態で増殖させたナイセリア株から作製したブレブ調製物中において有利であることが示されてきた。従って、この突然変異を有する株由来の本発明のナイセリアブレブは、上記の1又はそれ以上の下方制御とFrpB下方制御との組み合わせから得られるブレブとしての、本発明のさらなる実施形態である。好ましくは、PorAが下方制御される場合にはPorBは下方制御されるべきではなく、またその逆も同様である。
【0061】
上記突然変異は、本発明のブレブ免疫原性組成物が由来することになるあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしながらL2又はL3免疫型ナイセリア(好ましくは髄膜炎菌、最も好ましくはmenB)株を用いることが好ましく、典型的には本明細書中上記の低DOC%抽出法で抽出する。好ましくは、本発明のブレブ免疫原性組成物は、L2及びL3ブレブを両方含み、その少なくとも一方(好ましくは両方)が上記の免疫優性外膜タンパク質(又はOMP)の組み合わせを欠損している。それらの遺伝子を下方制御するための技術はWO01/09350号(参照により本明細書に組み入れられる)に記載されている。4つの異なるOpa遺伝子が髄膜炎菌ゲノム中に存在することが知られており(Ahoら1991 Mol. Microbiol. 5:1429-37)、ゆえにOpaの発現が下方制御されていると言われる場合、このことは好ましくは髄膜炎菌に存在する1、2、3つの遺伝子、又は(好ましくは)4つ全ての遺伝子がそのように下方制御されていることを意味する。そのような下方制御は、WO01/09350号に記載のように遺伝学的に達成するか、又は、容易に発見される天然の安定した髄膜炎菌株(Opa座位からの発現がないか若しくは低いもの)を探索することによって達成し得る。そのような株は、Poolmanら(1985 J. Med. Micro. 19:203-209)に記載される技術を用いて見つけ出すことができ、その際、Opa−である細胞は、プレート上又は顕微鏡下において細胞の外見を観察することによって見出されうるOpa発現細胞とは異なる表現型を有する。一度見つけられれば、Opaの欠失を確立するための発酵操作の後に、細胞内容物についてウエスタンブロットを行うことによって、株が安定的にOpa−であることを示すことができる。
【0062】
上記LOS免疫原性組成物の安全性
ヒトスフィンゴ糖脂質に存在するラクト−N−ネオテトラオースオリゴ糖基(Galβ1−4GlcNAcβ1−3Galβ1−4Glcβ1−、図1)と類似した構造が存在することから、L3又はL2 LOSに対して生起された抗体の安全性が問われてきている。多くの人々が残存量のL3 LOSを含むデオキシコレート抽出小胞ワクチンで安全にワクチン接種されてきているが(G. Bjuneら, Lancet(1991), 338, 1093-1096、GVG. Sierraら, NIPH ann(1991), 14, 195-210)、LOSが本明細書に記載されるように抗原として保持される場合、LOS糖鎖構造の末端部分の欠失が、ヒト組織の表面に提示される構造との抗LOS免疫応答の交差反応の阻止において有利であることが、本発明者らによって発見された。好ましい実施形態において、lgtB遺伝子の不活性化が末端ガラクトース残基及びシアル酸が欠損した中間体LOS構造を生じる(図1及び2を参照、突然変異はL2及びL3及びL4 LOSに4GlcNAcβ1−3Galβ1−4Glcβ1−構造を残存させる)。そのような中間体は、L3及び/又はL2(及び/又はL4)LOS株において獲得され得る。別の及びあまり好ましくないLOSの(短い)バージョンは、lgtE遺伝子のスイッチオフによって得ることができる。さらに別の及びあまり好ましくない型のLOSは、lgtA遺伝子のスイッチオフによって得ることができる。そのようなlgtA−突然変異を選択する場合、非免疫原性L1免疫型の形成を阻止するためにlgtC発現をスイッチオフすることがまた好ましい。
【0063】
lgtB−突然変異が最も好ましいのは、これが殺菌(及びさらに交差反応性殺菌)抗体反応を依然として誘導し得るLOS防御性オリゴ糖エピトープを保持したままで安全性の課題を解決するために最適な末端切断物であることを本発明者らが発見したためである。
【0064】
従って、本発明の上記のL2及び/又はL3調製物(あるいは上述した本発明の1以上のLOS)(精製されたもの、又は単離したブレブ状のもの)あるいは一般的な本発明の髄膜炎菌のブレブ調製物(特にL2及び/又はL3)は、lgtB、lgtA又はlgtE遺伝子由来の機能的遺伝子産物の発現が永続的に下方制御されるように、好ましくは遺伝子のスイッチオフにより、最も好ましくは当該遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により、遺伝子操作したナイセリア株(好ましくは髄膜炎菌株)から、有利に得られる。
【0065】
好ましくは、本発明のナイセリア株は、莢膜多糖の合成に欠損を有する。
【0066】
本発明の上記ブレブ調製物を髄膜炎菌B株から得る場合、莢膜多糖(ヒト様糖鎖構造も含む)もまた除去されることが特に好ましい。これを達成するために多くの遺伝子がスイッチオフされ得るが、本発明者らは、siaD遺伝子由来の機能的遺伝子産物の発現を永続的に下方制御(すなわちα−2−8ポリシアリルトランスフェラーゼの活性を下方制御)するように、ブレブ産生株を、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により、遺伝的に操作することが好ましいということを有利に示してきた。そのような不活性化はWO01/09350号に記載されている。siaD(synDとしても知られる)突然変異が、莢膜多糖からヒト類似エピトープを除去し得る多くの突然変異の中で最も有利であるのは、それが、LOSの防御エピトープの生合成に影響を及ぼさない最良の突然変異の1つであり、そのため、LOSを防御抗原として最終的に用いることを目的とし、かつ細菌の増殖に対して最小限の影響を与える方法において有利であるためである。従って、本発明の好ましい態様は、lgtE− siaD−、lgtA− siaD−又は好ましくはlgtB− siaD−髄膜炎菌B突然変異株由来のブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0067】
siaD−突然変異は上記の理由のために好ましいが、髄膜炎菌B(又は一般的に髄膜炎菌)の莢膜多糖合成をスイッチオフする他の突然変異を用いることができる。従って、ブレブ産生株を、以下の遺伝子:ctrA、ctrB、ctrC、ctrD、synA(synX及びsiaAと同等)、synB(siaBと同等)又はsynC(siaCと同等)遺伝子、の1つ以上に由来する機能的遺伝子産物の発現を永続的に下方制御するように、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により、遺伝的に操作してもよい。lgtE−突然変異はこれらの突然変異の1以上と組み合わせてもよい。好ましくは、lgtB−突然変異はこれらの突然変異のうち1つ以上と組み合わされる。従って、本発明のさらなる態様は、髄膜炎菌B(又は一般的に髄膜炎菌)の突然変異株のそのような組み合わせから得られる上記ブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0068】
lgtB及びlgtEなどの様々なlgt遺伝子を含むナイセリア遺伝子座、並びにその配列は、当技術分野において公知である(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021及びそれに引用されている参考文献;J. Exp. Med. 180:2181-2190(1994);WO96/10086号を参照)。
【0069】
最終産物として本発明の全長(非末端切断型)LOSを使用する場合、LOSがシアル化されていないことが望ましいが、そのようなLOSは、最も危険な侵襲性髄膜炎菌B株(これも同様にシアル化されていない)に対する免疫応答を生じることがあるためである。そのような場合、synA(synX及びsiaAと同等)、synB(siaBと同等)又はsynC(siaCと同等)遺伝子が欠失した莢膜陰性株の使用が有利であり、それはまたそのような突然変異によってmenB LOSがシアル化され得ないようにするためである。本発明の一実施形態において、lst遺伝子(Gilbertら, JBC 1996, 271:28271-6)を機能的に不活性とする(例えば、その遺伝子の欠失若しくは破壊、又は発現の低減により)か、あるいはその遺伝子が天然に破壊されている株を本発明のために選択する(免疫型のL3ファミリーについては、そのような欠損株はL7免疫型と称されることが多い)。lstは、末端シアル酸をLOSα鎖に付加するα−2,3−シアリルトランスフェラーゼである(しかし、シアル酸産生には影響を及ぼさない)。一実施形態において、本発明の株はlst(−)かつsiaD(−)である。
【0070】
上記突然変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしL2又はL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、典型的には本明細書に記載した低DOC%抽出法によって抽出されたものを使用するのが好ましい。好ましくは、本発明のブレブ免疫原性組成物が、L2及びL3ブレブの両方を含み、かつ、少なくとも一方(好ましくは双方)が上記遺伝子の発現が欠損した株に由来する。
【0071】
LOSの毒性
本発明の上記の精製LOS又はブレブ免疫原性組成物はまた、それらが由来する細菌性産生株における特定の遺伝子の発現の下方制御によって、毒性をより少なくすることができる。そのような無毒化は、天然のOMVによる鼻腔内免疫には不必要であるだろうが(J.J Drabickら, Vaccine(2000), 18, 160-172)、非経口ワクチン接種における無毒化には有用であろう。好ましくは、本発明の精製LOS又はブレブ免疫原性組成物は、リピドA生合成に関与する遺伝子(特にリピドAへの第2アシル鎖(secondary acyl chain)の付加に関与する遺伝子)の突然変異/改変/不活性化により(特にmsbB及び/又はhtrB遺伝子由来の機能的遺伝子産物の発現の下方制御により、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーター及び/又はオープンリーディングフレームの全体又は一部の欠失により)、ナイセリア産生株を遺伝的に操作することによって無毒化される。あるいはまた(又はさらに)、精製LOS又はブレブ免疫原性組成物は、以下の遺伝子:pmrA、pmrB、pmrE及びpmrFのうちの1つ以上の遺伝子が(より強力なプロモーターの導入、又は遺伝子のさらなるコピーの組み込みによって)上方制御されるように遺伝的に改変されたナイセリア株から得ることができる。あるいはまた(又はさらに)、精製LOS又はブレブ免疫原性組成物は、ポリミキシンBの機能的等価物である非毒性ペプチド[リピドAに対して高いアフィニティの有する分子]を組成物に添加することにより無毒化され得る(以下参照、すなわち、本発明の1以上のLOSが、LOSの毒性を低減させるために好適なリピドA結合ペプチド、例えばSAEP2又はSAEPIIと複合体化されている)。
【0072】
上記無毒化法並びに関連プロモーター/遺伝子配列、並びに上方制御方法及び下方制御方法については、より詳細についてWO01/09350号を参照されたい。またナイセリアmsbB及びhtrB遺伝子は各々lpxL1及びlpxL2とも呼ばれ(WO00/26384号参照)、これらの遺伝子の欠失突然変異は、msbB−突然変異LOSが野生型と比較して1つの第2アシル鎖を欠損する(並びに4つの第1アシル鎖(primary acyl chain)及び1つの第2アシル鎖を保持する)こと、そしてhtrB−突然変異LOSが第2アシル鎖の両方を欠損することの、表現型としての特徴を有する。そのような突然変異は好ましくは、ブレブに対する無毒化LOSの最適な提示を確実にするため、又は無毒化サブユニットLOSの精製を助けるために、ナイセリア産生株が莢膜多糖を欠損していること(上記参照)を確実にする突然変異と組み合わせて用いられる。
【0073】
本発明の組成物中に使用することができるポリミキシンB(本発明のLOSの毒性を低減させるために好適なリピドA結合ペプチド)の機能的等価物である非毒性ペプチドのさらなる詳細、特にペプチドSAEP2(2つのシステインがジスルフィド架橋を形成するKTKCKFLKKC配列を有する)及びSAEPII(WO2006/108586号の請求項1〜10に記載されているペプチド二量体)の使用については、WO93/14115号、WO95/03327号、WO2006/108586号、Velucchiら(1997)J Endotoxin Res 4:1-12及びEP976402を参照されたい。本明細書におけるこのようなリピドA結合ペプチドは、上記引用した特許出願の特許請求の範囲又は実施例に記載されるペプチドの具体的な又は一般的な式のいずれかを指す。
【0074】
本明細書において「機能的遺伝子産物の発現の下方制御」は、対象の遺伝子のプロモーター又はオープンリーディングフレームに対して付加、欠失又は置換が為され、それにより全遺伝子産物の生合成活性が低減(60、70、80、90、95%若しくは最も好ましくは100%低減)されることを意味する。明らかにフレームシフトする突然変異を導入してもよいし、又はより弱いプロモーターを置換してもよいが、最も好ましくは、オープンリーディングフレーム及び/又はプロモーターのほとんど又は全てを、(活性のある)遺伝子産物の永続的な下方制御を確実にするために欠失させる(WO01/09350号に記載)。
【0075】
上記変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有用であるが、L2又はL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株を用いることが好ましく、典型的には本明細書に記載の低DOC%抽出法で抽出されることが好ましい。本発明のブレブ免疫原性組成物は、L2及びL3ブレブの両方(又は本発明のブレブ調製物)を含み、その少なくとも一方(及び好ましくは双方)が上記遺伝子の発現を欠損している株に由来することが好ましい。
【0076】
本発明のさらなる態様は、本発明のLOS又はブレブ免疫原性調製物が由来する上記の遺伝的に改変されたナイセリア(好ましくは髄膜炎菌又は淋菌又は髄膜炎菌B)株を含む。
【0077】
本発明のLOS又はLOS含有ブレブ調製物
本発明のさらなる態様は、本発明のナイセリア株から単離されるLOS調製物(特に上述した任意のもの)である。好ましくは、単離されたLOS(又はLOS含有ブレブ)は、L2又はL3免疫型であり、好ましくは本発明の免疫原性組成物は本発明のL2及びL3 LOS(又はブレブ)調製物を両方含む。
【0078】
そのような調製物はまた、上記LOS(精製されているものであれブレブ調製物に存在しているものであれ)のオリゴ糖部分をT細胞エピトープ源を含むキャリアにコンジュゲートする(従ってLOSをより良好な(T依存的)免疫原とする)ことにより改良することができる。あるいはまた(又はさらに)本発明の精製LOS調製物は、当技術分野で公知のリポソーム製剤中にそれを存在させることにより、より良好な抗原となり得る(例えばWO96/40063号及びそこに引用される参考文献を参照)。
【0079】
細菌からLOSを単離する方法は当技術分野で周知である(例えばWesphal及びJannの温水−フェノール法(Meth. Carbo. Chem. 1965, 5:83-91)を参照)。またGalanosら1969, Eur J Biochem 9:245-249及びWuら1987, Anal Bio Chem 160:281-289も参照されたい。単離されたLOSをコンジュゲートするための技術もまた公知である(例えばEP941738(参照により本明細書に組み入れる)を参照)。
【0080】
本発明の目的のため、「T細胞エピトープ源を含むキャリア」とは、通常、ペプチド、又は好ましくはポリペプチド若しくはタンパク質である。コンジュゲーション技術は当技術分野で周知である。典型的なキャリアとしては、非定型インフルエンザ菌(Haemophilus influenzae)由来のプロテインD、破傷風毒素、ジフテリア毒素、CRM197又は(特にナイセリア又は髄膜炎菌の)ブレブ調製物に存在する外膜タンパク質が挙げられる。
【0081】
本発明の好ましい単離されたLOS組成物としては、各LOSのオリゴ糖部分がT細胞エピトープ源を含むキャリアに任意にコンジュゲートされていてもよいL2及びL3単離LOSを含有する組成物、lgtB−髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープ源を含むキャリアに任意にコンジュゲートされていてもよいL2又はL3 LOSを含有する組成物、並びに最も好ましくは、lgtB−髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープ源を含むキャリアに任意にコンジュゲートされていてもよいL2及びL3単離LOSを含有する組成物である。
【0082】
好ましくは、本発明のLOS組成物(又は本発明の1以上のLOS)は無毒化されている。これは、分子からアシル鎖を除去する(しかし分子の防御効果は減少する可能性がある)ヒドラジン又はアルカリ加水分解化学処理の公知の技術によって行うことができるが、好ましくはhtrB−若しくはmsbB−髄膜炎菌突然変異株(上記のような、特に莢膜多糖欠損株)からLOSを単離することにより、又はポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高いアフィニティを有する分子)、特にSAEP2又はSAEPII(上記)の、組成物への添加により行われ得る。
【0083】
本発明のLOSは、単離された状態で(通常、リピドA部分が無傷のままであればミセル形態で)投与することができるし、又はリポソーム中に入れて投与することもできる。そのような場合、外膜タンパク質はリポソームに添加してもよく、そしてLOSを、オリゴ糖がT依存的抗原となるようにリポソーム内部でそのような外膜タンパク質にコンジュゲートしてもよい。これは、後述するブレブ内LOS架橋について記載されるものと同様の化学法によって行われ得る。
【0084】
LOSのオリゴ糖部分の、ブレブの表面上に存在する外膜タンパク質とのブレブ内架橋(コンジュゲーション)
LOS(特に本発明のLOS)がブレブ製剤中に存在する場合、LOSは好ましくは、ブレブ調製物上にも存在する1又はそれ以上の外膜タンパク質(例えば髄膜炎菌におけるPorA又はPorB)へのLOSのコンジュゲーションを可能にする方法により、in situにおいてコンジュゲートされる。従って、本発明のさらなる態様は、その外膜中に、LOSとコンジュゲートされた外膜タンパク質が組み込まれるグラム陰性細菌株由来のブレブ調製物(本発明の1以上のブレブ調製物)である。LOSはコンジュゲートのためにブレブ調製物に添加してもよいが、LOSはブレブ調製物の表面に天然に存在することが好ましい。
【0085】
この方法は、ブレブ製剤中のLOS抗原の安定性及び/又は免疫原性(T細胞の助けを提供する)、及び/又は抗原性を、外膜の表面上の天然環境にあるLOSのように有利に増強し、それゆえその最も防御的な立体構造をとったT非依存的オリゴ糖免疫原に対し、T細胞の助けを与える。さらに、ブレブ内でのLOSのコンジュゲーションは、LOSの無毒化を生じ得る(理論に縛られることは望まないが、コンジュゲートされると、リピドA部分はより安定的に外膜中に埋没し、従って毒性を生じるように利用されにくくなると考えられる)。従って、上記のhtrB−若しくはmsbB−変異株からのブレブの単離による無毒化法、又はポリミキシンBの機能的等価物である非毒性ペプチドの組成物への添加による上記の無毒化法は、必要ではないこともある(しかしさらなる安全のために組み合わせて添加され得る)。
【0086】
本発明のコンジュゲート化ブレブ調製物は、典型的には、ブレブ中のLOSの毒性が同量の全くコンジュゲートしていないLOSを有する同じブレブと比較して低減されている。LOS毒性は当業者であれば容易に測定することができ、例えばヨーロッパ薬局方におけるLOSウサギ発熱アッセイ(pyrogenicity assay)を用いることができる(WO2004/014417号の実施例7参照)。
【0087】
本発明のコンジュゲート化ブレブ調製物は、コンジュゲートしたLOSが宿主内における免疫応答の誘発に適した立体構造を有し、それに由来する血清がコンジュゲートしていないLOS(好ましくはブレブ調製物が作製された由来の細菌上に存在し、最も好ましくはSBAアッセイにおける殺菌性で存在する)との反応性を有する(結合可能である)ため、有利である。
【0088】
ナイセリアブレブをLOSにコンジュゲートする場合、及び本明細書に記載するように、ブレブが1以上の免疫優性外膜タンパク質が下方制御された株由来である場合、好ましくは、PorAが下方制御されている場合にはPorBは下方制御されるべきではなく、かつその逆も同様である。このことによって、LOSの大部分を主要な外膜タンパク質と架橋することが可能になり、従ってブレブに存在する交差防御性の微量外膜抗原に対するコンジュゲートのあらゆる効果が最小化される。
【0089】
特に、本発明者らは、ブレブに存在するLOSがブレブ内で外膜タンパク質(これもまたブレブに存在する)にコンジュゲートしているブレブを含む組成物が、そのブレブが由来する生物によって引き起こされる疾患の治療又は予防のためのワクチンの基礎を形成しうることを見出した。ここでそのようなワクチンは毒性が低減しており(好ましくは実質的に無毒性であり)、及び/又はその天然環境におけるLOSに対するT依存的殺菌反応を誘導することが可能である。
【0090】
従って、本発明はそのようなブレブ内LOSコンジュゲート化ブレブ調製物をさらに提供する。「ブレブ内」とは、ブレブ中に天然に存在するLOSが同じブレブ上に存在する外膜タンパク質とコンジュゲートしていることを意味する。
【0091】
そのようなブレブ調製物は、対象の細菌より単離することができ(WO01/09350号参照)、次にLOSのオリゴ糖部分の基(例えばNH2又はCOOH)をブレブ外膜タンパク質の基(例えばNH2又はCOOH)に結合するための、既知のコンジュゲーション化学法に供すればよい。グルタルアルデヒド、ホルムアルデヒド又はグルタルアルデヒド/ホルムアルデヒド混合物を使用した架橋技術を用いることができるが、EDAC又はEDAC/NHS等のより選択的な化学法を用いるのが好ましい(J.V. Staros,R.W. Wright及びD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)並びにBioconjugates Techniques. Greg T. Hermanson(1996)pp173-176)。本発明において使用することができるLOS及びタンパク質分子間の共有結合を生じることの可能な他のコンジュゲーション化学法又は処理は、EP941738に記載されている。
【0092】
好ましくは、ブレブ調製物は莢膜多糖の非存在下でコンジュゲートされる。ブレブは莢膜多糖を産生しない株(天然の若しくは突然変異によるもの)から単離することができ、又は大半の混入莢膜多糖から精製(60、70、80、90若しくは99%以上が除去された)、好ましくは全ての混入莢膜多糖から精製することができる。この方法において、ブレブ内LOSコンジュゲーション反応はさらに非常に効果的である。
【0093】
好ましくはブレブ内に存在するLOSの5、10、20、30、40、50、60、70、80、90又は95%を超える割合が架橋/コンジュゲートされる。
【0094】
好ましくは、本発明のブレブは、精製LOSを標準物質として使用したSDS−PAGE電気泳動後の銀染色によって測定した場合に(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)、ブレブのLOS含量が、3〜30、5〜25、10〜25、15〜22、そして最も好ましくはおよそ又は正確に20%LOS含量となるように調製されたものである。髄膜炎菌ブレブにおける20%LOSを0.1%低DOC抽出法で達成することができ、これは緩く保持されたLOS分子を除去することができるが、抗原の大部分は保存するものである。
【0095】
ブレブ内コンジュゲート化ブレブが髄膜炎菌由来である場合、それが由来する株が莢膜多糖を産生することができない突然変異株(例えば上記の突然変異株の1つ、特にsiaD−)であることが好ましい。髄膜炎菌疾患に対して有効な免疫原性組成物がL2及びL3ブレブの双方を含むこともまた好ましく、ここでL2及びL3 LOSは双方ともにブレブ外膜タンパク質にコンジュゲートしている。さらに、ブレブ内コンジュゲート化ブレブ中のLOS構造がlgtB−髄膜炎菌株由来のものと一致していることが好ましい。最も好ましい免疫原性組成物は以下のブレブ内コンジュゲート化ブレブを含む:莢膜多糖を産生することができずlgtB−である、L2若しくはL3突然変異髄膜炎菌株に由来するもの、莢膜多糖を産生することができない突然変異髄膜炎菌株由来のL2及びL3ブレブを含むもの、lgtB−である突然変異髄膜炎菌株由来のL2及びL3ブレブを含むもの、又は最も好ましくは莢膜多糖を産生することができずlgtB−である突然変異髄膜炎菌株由来のL2及びL3ブレブを含むもの。
【0096】
本発明に使用することができる典型的なL3髄膜炎菌株は、H44/76 menB株である。典型的なL2株は、B16B6 menB株又は39E髄膜炎菌C型株又は760676株である。典型的なL10株は、3125 menA株である。L4株は、C19 MenC株である。
【0097】
上述のように、本発明のブレブはコンジュゲーションの作用によってある程度無毒化されるので、さらなる無毒化を必要としないが、しかし、さらなる無毒化法をさらなる安全性のために用いてもよい。それは例えばhtrB−若しくはmsbB−である髄膜炎菌株由来のブレブを使用することによるか、又はポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高いアフィニティを有する分子)(好ましくはSEAP2又はSEAPII)をブレブ組成物に(上記のように)添加することによる。このように、LOSの(特にブレブ内の)コンジュゲーションは、驚くべきことに同量のコンジュゲートしていないLOSを含む調製物と比較して低いLOS毒性を示す。従って、(特に髄膜炎菌の)ブレブの無毒化のための一般的な方法は、さらにLOSのブレブ外膜タンパク質へのブレブ内コンジュゲーションによって提供され、そしてLOSの無毒化のための方法もまた、ブレブ外膜タンパク質へLOSをコンジュゲートする手段によって提供される。
【0098】
上記の方法において、髄膜炎菌ブレブ、及びブレブを含む免疫原性組成物が提供され、これらは、毒性が低減しており(好ましくは実質的に無毒性であり)、自己免疫の問題がなく、T依存的特徴を持ち、その天然環境に存在し、そして潜在的に90%を超える髄膜炎菌株に対する殺菌性抗体反応(L2+L3組成物の場合)を誘導することのできる、重要な抗原としてのLOSを有する。
【0099】
1以上のMenA、C、Y又はW莢膜多糖又はオリゴ糖(好ましくは少なくともMenC、又はMenA及びMenC、又はMenC及びMenY)を、本発明のブレブの外膜タンパク質上に、同様にコンジュゲートすることができる。これはLOS架橋と同じ反応において行うことができるが、別々の(好ましくは後の)反応において行うことが好ましい。
【0100】
最適なブレブ内LOSコンジュゲーションの方法は、本発明のさらなる態様である。かかる方法は、(好ましくは、本明細書に記載されるように低パーセンテージのDOCを使用して)グラム陰性細菌からブレブを単離するステップ、ブレブに存在するLOSを(好ましくはそのオリゴ糖部分を介して)同じブレブに存在する外膜タンパク質にコンジュゲートするのに適した化学法を行うステップ、ブレブ内コンジュゲート化ブレブ調製物を単離するステップ、並びに任意により、同じ方法によって作製されるが異なるLOS免疫型を有する、さらなるブレブ内コンジュゲート化ブレブ調製物とともにブレブ内コンジュゲート化ブレブ調製物を製剤化するステップ(好ましくはL2及びL3ナイセリア/髄膜炎菌ブレブを混合するステップ)、及び/又はワクチン組成物を製造するために薬学的に許容される賦形剤とともにブレブ調製物を製剤化するステップを含むべきである。
【0101】
ブレブ内コンジュゲーションは、好ましくは以下の方法のステップの1若しくは2つ又は3つ全てを含むべきである:コンジュゲーションpHはpH7.0を上回り、好ましくはpH7.5以上(最も好ましくはpH9以下)であるべきである;1〜5%、好ましくは2〜4%、最も好ましくは約3%ショ糖の条件が反応中維持されるべきである;コンジュゲーション反応におけるNaClは最小限にするべきであり、好ましくは0.1M、0.05M、0.01M、0.005M、0.001M以下であり、全く存在しないことが最も好ましい。これら全ての方法の特徴は、コンジュゲーション過程を通してブレブを安定にかつ溶液中に維持することを確実にするものである。
【0102】
EDAC/NHSコンジュゲーション法はブレブ内コンジュゲーションのために好ましい方法である。EDAC/NHSは、非常に高度な架橋が可能であるがゆえに濾過性に不利な影響を及ぼすホルムアルデヒドより好ましい。EDACはカルボン酸(LOS中のKDO等)と反応して活性エステル中間体を生成する。アミン求核試薬(PorB等の外膜タンパク質中のリジン等)存在下において、アミド結合はイソ尿素副生成物の放出を伴って形成される。しかしながら、EDAC仲介反応の効率は、スルホNHSエステル中間体の形成により増大し得る。スルホNHSエステルは、EDACのみとカルボキシレートとの反応より形成される活性エステルより長く水性溶液中に残存する。従って、この2段階の方法を用いて高収率のアミド結合形成を実現し得る。EDAC/NHSコンジュゲーションは、J.V. Staros,R.W. Wright及びD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)並びにBioconjugates Techniques. Greg T. Hermanson(1996)pp173-176に記載されている。好ましくは、0.01〜5mg EDAC/mgブレブ、より好ましくは0.05〜1mg EDAC/mgブレブを反応に使用する。使用するEDACの量はサンプル中に存在するLOSの量に依存し、このLOSの量はまたブレブを抽出するために使用するデオキシコレート(DOC)%に依存する。低パーセンテージ(%)DOC(例えば0.1%)においては多量のEDAC(1mg/mg及びそれ以上)が使用されるが、しかし、高%DOC(例えば0.5%)においては少量のEDAC(0.025〜0.1mg/mg)が、過剰なブレブ内架橋を回避するために使用される。
【0103】
本発明の好ましい方法は、従って、EDAC/NHS存在下において、pHがpH7.0からpH9.0の間(好ましくは約pH7.5)で、1〜5%(好ましくは約3%)ショ糖中で、かつ任意に(上記のように)実質的にNaClを含まない条件において、ブレブをコンジュゲートするステップ、並びに反応混合物からコンジュゲート化ブレブを単離するステップを含む、ブレブ内コンジュゲート化LOS(好ましくは髄膜炎菌のもの)を製造する方法である。
【0104】
この反応に続いて、反応時間の経過に伴いブレブ中のLOSの割合の増加に応じてLOS分子量が増加することを示すために、抗LOS(例えば抗L2又は抗L3)モノクローナル抗体(mAb)を使用して反応混合物をウェスタン分離ゲルに供することができる。
【0105】
収量99%のブレブがそのような技術を用いて回収され得る。
【0106】
EDACは、LOSのT依存的免疫原性を改善するために十分な、LOSをOMPに架橋する点で優れたブレブ内架橋剤であることが見出されたが、しかしそれほど高度にそれを架橋しなかったのは、乏しい濾過性、凝集及びブレブ間架橋等の問題が発生したためであった。生じたブレブの形態は、コンジュゲートしていないブレブのものと類似している(電子顕微鏡による)。さらに上記のプロトコルは、過度に高度の架橋(ブレブの表面上に天然に存在する防御OMP(例えばTbpA又はHsf)の免疫原性を低減し得る)が生じることを回避した。
【0107】
ブレブ単離のための技術
本発明の外膜小胞(OMV又はブレブ)は、多くの公知の技術により単離することができる(Fredriksenら, NIPH Annals(1991), 14, 67-79、Zollingerら, J. Clin Invest(1979), 63, 836-848、Saundersら, Infect Immun(1999), 67, 113-119、J.J. Drabickら, Vaccine(1999), 18, 160-172)。これらは2つの主要な群に分類される。すなわち、デオキシコレート(約0.5%)を使用して髄膜炎菌からブレブを抽出する技術と、低レベルのデオキシコレート(DOC)を使用するか又は全くデオキシコレートを使用しない技術である。DOCを使用しない方法では、ブレブはOMV中に高レベルのLOSを維持するという興味深い特徴を有する。このことはLOSが防御抗原であるワクチンにおいて有利である。DOC抽出ブレブと比較して、DOCを使用しない方法により得られるOMV中のL3抗原(Ag)の濃度は約10倍高い。界面活性剤を用いない(好ましくはDOCを用いない)ブレブ調製法は、この理由により本発明の方法の目的にとって好ましいが、低レベルの界面活性剤(好ましくはDOC)を含むバッファーでの抽出もまた、ブレブ内で強固に相互作用するLOSの大半を残す一方、より毒性があり緩く保持されたLOSを除去するステップであるという点で有利であり得る。典型的には、0〜0.5%、好ましくは0.02〜0.4%、0.04〜3%又は0.06〜2%界面活性剤(好ましくはDOC)がブレブ抽出に使用され、より好ましくは0.08〜0.15%、最も好ましくはおよそ又は正確に0.1%を用いて、最適な量のLOSをブレブ中に安定に存在させて得る。LOSが1以上の上記の方法により無毒化されている場合には、DOCを用いない(又は低DOC〜0.3%DOC若しくはそれ以下の)抽出法が特に好ましい。
【0108】
本発明の全ての実施形態におけるブレブのLOS含量は、精製LOSを標準物質として使用したSDS−PAGE電気泳動後の銀染色によって測定した場合に(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)、3〜30、5〜25、10〜25、15〜22%であることが好ましく、最も好ましくはおよそ又は正確に20%LOS含量である。Nmen L3 LOSをこの方法における標準物質として使用すると、一般的に、0.1%DOCで抽出されたNmen L3免疫型ブレブ中のLOS含量は約20%LOSであり、0.2%DOCで抽出した場合は約15%LOS、0.3%DOCでは約10%LOS、そして0.5%DOCでは約5%LOSである。
【0109】
ブレブの作製は、細胞又は細胞片からブレブを分離するための適当な技術のいずれかを用いて(例えば低速遠心分離により)行うことができる。ブレブ調製物は、超遠心分離を使用して(ブレブをペレット化する)、あるいはFrasch et al. ("Outer membrane protein vesicle vaccines for meningococcal disease" Methods in Molecular Medicine, vol 66, Meningococcal Vaccines: Methods and Protocols 2001 pp81-107 (A.J. Pollard及びM.C. Maiden編, Humana Press Totowa, NJ))に記載されているように超遠心分離及び/又はダイアフィルトレーション(diafiltration)のより穏やかな技術を用いて、さらに精製することができる。
【0110】
ワクチン組成物
本発明の免疫原性組成物は、薬学的に許容される賦形剤の添加によってワクチン組成物として容易に製剤化することができる。
【0111】
さらに、上記の本発明の精製LOS(好ましくはL2若しくはL3)を単離するステップ又は上記の本発明の単離ブレブ(好ましくはL2若しくはL3免疫型を有する)を生成するステップ、及びそのLOS又はブレブを薬学的に許容される賦形剤とともに製剤化するステップを含む、本発明のナイセリア(好ましくは髄膜炎菌)免疫原性組成物又はワクチンの製造方法を提供する。好ましくは、本発明の免疫型L2及びL3の両方の精製LOS、又は本発明の免疫型L2及びL3の両方のブレブ、又はL2の精製LOS及びL3のブレブ(若しくはその逆)を、混合ステップで混合する。好ましくは、本発明の精製LOS又はブレブは、単離された後に上述のようにコンジュゲートされる。さらなるリポソーム製剤化ステップを精製LOSに対して追加してもよい(当技術分野で公知の技術を用いる。例えばWO96/40063号及びそこに引用される参考文献を参照のこと)。好ましくは、ブレブ調製物は低濃度のDOC(又はDOCなし)での抽出によって(上記のように)単離される。
【0112】
このようなL2及びL3の組み合わせ方法は、ほとんど全ての髄膜炎菌B株に対して有効なワクチンをもたらすことができる。
【0113】
上記の免疫原性組成物(又は方法)は、血清群A、C、Y又はW由来の1以上(2、3又は4)の髄膜炎菌多糖又はオリゴ糖(上記のような、そのままの又はT細胞エピトープを含むキャリアにコンジュゲートしたもの)が該組成物に添加されたものであってもよい。好ましくは、少なくともCが添加(最も好ましくはコンジュゲート)されたものであり、より好ましくはA及びC又はY及びCが添加(好ましくは全てコンジュゲート化)されたものであり、最も好ましくはA、C、Y及びWが添加(好ましくは全てコンジュゲート化)されたものである。有利には、コンジュゲート化されたインフルエンザ菌B型莢膜多糖又はオリゴ糖も上記の組成物中に含有させて、広範な髄膜炎ワクチンを作製する。
【0114】
好ましくは、特にWO94/08021号において個別に具体的に示した組成物からなる又はそれを含む組成物は、本発明において特許請求の範囲に含められていない。場合により、US2006/0047106に具体的に特定されている組成物からなる又はそれを含む組成物は、本発明において特許請求の範囲に含められていない。
【0115】
本発明のワクチン製剤
本発明の免疫原性組成物は、適切なアジュバントとともに製剤化することで本発明のワクチン組成物を作製することができる。
【0116】
適切なアジュバントとしては、水酸化アルミニウムゲル(ミョウバン)又はリン酸アルミニウム(好ましくは水酸化アルミニウム)等のアルミニウム塩があるが、しかしカルシウム(特に炭酸カルシウム)、鉄又は亜鉛の塩でもよく、あるいはアシル化チロシン、若しくはアシル化糖、カチオン若しくはアニオン誘導体化多糖、又はポリホスファゼンの不溶性懸濁液でもよい。
【0117】
添加され得る適切なTh1アジュバント系には、モノホスホリルリピドA、特に3−脱−O−アシル化モノホスホリルリピドA(又は他のLPSの非毒性誘導体)、及びアルミニウム塩(好ましくはリン酸アルミニウム)とモノホスホリルリピドA、好ましくは3−脱−O−アシル化モノホスホリルリピドA(3D−MPL)(又は非毒性LPS誘導体)との組み合わせが含まれる。増強系は、モノホスホリルリピドAとサポニン誘導体の組み合わせ、特にWO94/00153号に記載されるようなQS21(若しくは他のサポニン)と3D−MPL(若しくは非毒性LPS誘導体)の組み合わせ、又はWO96/33739号に記載されるような、QS21(若しくはサポニン)がコレステロールによりクエンチされているより反応原性の低い組成物を用いる。水中油型エマルジョン中のQS21、3D−MPL及びトコフェロールを含む特に強力なアジュバント製剤は、WO95/17210号に記載されており、これは添加され得る好ましい製剤である。添加され得る他のアジュバントは、サポニン、より好ましくはQS21及び/又は水中油型エマルジョン及びトコフェロールを含む。オリゴヌクレオチドを含む非メチル化CpG(WO96/02555号)もまた添加され得る。
【0118】
ワクチン調製物は一般的にVaccine Design(「The subunit and adjuvant approach」(Powell M.F.及びNewman M.J.編)(1995)Plenum Press New York)に記載されている。
【0119】
免疫防御用量のワクチンが、全身経路又は粘膜経路によって投与され得る。これらの投与には、筋肉内、腹腔内、皮内若しくは皮下経路による注入、又は口腔/消化器(好ましくは鼻腔内投与)、呼吸器、尿生殖路への粘膜投与による注入が挙げられる。典型的には、各ワクチン用量中のブレブの量は、典型的な被ワクチン接種者において著しく有害な副作用を引き起こすことなく免疫防御反応を誘導する量が選択される。そのような量は、どの特定の免疫原が使用され、かつそれをどのように提示するかによって変動する。一般的に、各用量は、本発明の各ブレブ又はLOSを1〜100μg、好ましくは5〜50μg、最も典型的には5〜25μgの範囲内で含むことが予想される。
【0120】
本発明のブレブ免疫原性組成物に対するさらなる改良
本発明の上記のブレブ組成物は、それらが由来するナイセリア株(淋菌及び好ましくは髄膜炎菌、最も好ましくは髄膜炎菌Bを含む)が、ゲノム中への遺伝子のさらなるコピーを挿入することにより、又は既存の遺伝子の上流により強力なプロモーターを導入することにより、又は非改変株と比較して1.2、1.5、2、3、5若しくは10倍を超える抗原レベルを生じるよう改変された株を誘導することが可能なWO01/09350号に記載のあらゆる他の方法により、上方制御された1又はそれ以上の下記遺伝子(防御抗原をコードする遺伝子)を有する場合、本発明のワクチンにおける有効性がさらに改良され得る。すなわち、NspA(WO96/29412号)、Hsf又はその末端切断物(WO99/31132号及びWO01/55182号、NhhAとしても知られる)、Hap(PCT/EP99/02766号)、OMP85(WO00/23595号)、PilQ(PCT/EP99/03603号)、PldA(PCT/EP99/06718号)、FrpB(WO96/31618号)、TbpA(WO92/03467号、米国特許第5912336号、WO93/06861号及びEP586266)、TbpB(WO93/06861号及びEP586266)、NadA(Comanducciら J. Exp. Med. 2002 195:1445-1454、NMB 1994)、FrpA/FrpC、又は5以上の反復配列を含むこれらの抗原間に共通の部分(WO92/01460号;Thompsonら,(1993) J. Bacteriol. 175:811-818; Thompsonら,(1993) Infect. Immun. 61:2906-2911)、LbpA、LbpB(PCT/EP98/05117号)、FhaB(WO98/02547号、配列番号38(ヌクレオチド3083〜9025))、HasR(PCT/EP99/05989号)、lipo02(PCT/EP99/08315号)、Tbp2(WO99/57280号;NMB0460)、MltA(WO99/57280号;NMB0033)、TspA(WO00/03003号)、TspB(WO00/03003号)、ctrA(PCT/EP00/00135号)、MafA(NMB0652)、MafB(NMB0643)、Omp26(NMB0181)、アドヘシンX(NMB0315)、アドヘシンY(NMB0995)、アドヘシンZ(NMB1119)及びOstA(NMB0280)である。NMB配列の例は、www.neisseria.orgのデータベースで見出すことができる。本明細書でHsfについて言及する場合、この用語はあらゆる場合においてHsf末端切断物(特にWO01/55182号に開示されたもの)と置換可能である。
【0121】
Hsf及びTbpA(低若しくは高分子量形態、又は低及び高分子量形態の両方(EP586266))、又はHsf及びOMP85、又はOMP85及びTbpA(低若しくは高分子量形態、又は低及び高分子量形態の両方)、又はNspA及びHsf、又はNspA及びOMP85、又はNspA及びTbpA(低若しくは高分子量形態、又は低及び高分子量形態の両方)の双方が、いずれも上方制御されている場合が特に好ましい。組成物中に2つのブレブが含まれている場合、各ブレブは異なる上方制御を受けていることが好ましい。高及び低分子量のTbpAがどちらも上方制御される場合、これらは2つの形態のTbpAを天然に含む2つの株由来の組成物中に存在する、2つの別々のブレブにおいて上方制御されていることが好ましい。最も好ましくは、かかる2つの株がL2及びL3 LOS免疫型を有することである。TbpAは遺伝子工学的に上方制御させるか、又はナイセリア/髄膜炎菌産生株を鉄制限条件(例えば50〜70μMデスフェラール(メシル酸デフォロキサミン、Sigmaより入手可能)存在下)で増殖させることにより、上方制御させ得る。後者の方法を採用する場合、FrpB遺伝子発現を下方制御(好ましくは欠失)することが好ましいが、これはこの可変性抗原が鉄制限条件下で単離される髄膜炎菌株から単離されたブレブにおいて免疫優性になり得るためである。
【0122】
好ましい実施形態において、本発明の組成物は、好ましくは高分子量TbpA(TbpA High)及びHsfが上方制御されている、lgtB−(又はlst−)莢膜多糖−msbB−株由来のL3ブレブと、低分子量TbpA(TbpA Low)及びOmp85が上方制御されている、lgtB−(又はlst−)莢膜多糖−msbB−株由来のL2ブレブとを含む。より好ましくは、どちらのブレブもさらにPorA及び/又はFrpBの発現、そして任意にOpC及び/又はOpA発現が、下方制御されている。ブレブは、最も好ましくは上記の低DOC法によって単離され、両ブレブ中のLOSは外膜タンパク質とブレブ内架橋されている。
【0123】
ゴースト又は死菌全細胞ワクチン
本発明者らは、ブレブに関わる上記組成物及びワクチンが、(同一の利点を有する)ゴースト又は死菌全細胞調製物及びワクチンに関する方法にまで容易に拡張できることを意図している。グラム陰性細菌株からゴースト調製物(完全なエンベロープを有する空の細胞)を製造する方法は当技術分野で周知である(例えばWO92/01791号を参照)。全細胞を殺滅して、ワクチンに使用するための不活性化細胞調製物を作製する方法もまた周知である。従って、本明細書を通して記載されるブレブを含む組成物及びワクチンは、等価物である本発明のゴースト及び死菌全細胞調製物を含む同じ組成物又はワクチンにも当てはまることを意図している。
【0124】
本発明の組成物に関する血清殺菌アッセイ
血清殺菌アッセイは、本発明の免疫原性組成物中で組み合わせた場合に抗原間の相乗作用的な関係を評価するための好ましい方法である。
【0125】
そのような相乗反応は、抗原の組み合わせによって誘発されるSBAにより、各抗原によって別個に誘発されるSBAと比べて少なくとも50%、2倍、3倍、好ましくは4倍、5倍、6倍、7倍、8倍、9倍、最も好ましくは10倍高いものとして特徴付けられ得る。好ましくはSBAは、抗原が由来するのと同種の株、及びさらに好ましくは一群の異種の株に対して測定される(以下の代表的な群、例えばA−4クラスターに属するBZ10(B:2b:P1.2)、ET−37複合体に属するB16B6(B:2a:P1.2)、及びH44/76(B:15:P1.7,16)を参照)。SBAは、髄膜炎菌ワクチンの有効性を評価するための、最も一般的に受け入れられている免疫学的マーカーである(Perkinsら J Infect Dis. 1998, 177:683-691)。十分なSBAはあらゆる公知の方法により確認することができる。SBAは、動物モデル又はヒト被験体から得た血清を用いて行うことができる。
【0126】
ヒト血清を用いてSBAを行う好ましい方法は以下の通りである。血液サンプルは最初のワクチン接種の前、2回目のワクチン接種の2ヶ月後及び3回目のワクチン接種の1ヶ月後に採取する(3回のワクチン接種は1年以内に行い、通常のヒト第1期ワクチン接種計画では、例えば0、2及び4ヶ月、又は0、1及び6ヶ月の時点で投与される)。そのようなヒト第1期ワクチン接種計画は、1歳未満の幼児(例えばHibワクチン接種を実施するのと同時)に実施することができ、又は2〜4歳児若しくは青年もそのような第1期ワクチン接種計画でワクチン接種することによってSBAを試験することができる。妥当であれば、さらなる血液サンプルは第1期ワクチン接種の6〜12ヶ月後、及び追加免疫投与の1ヶ月後に採取され得る。
【0127】
(2〜4歳児若しくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、本発明の抗原が由来する髄膜炎菌株に対するSBA(抗体希釈率)力価が(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が、被験体の30%を超え、好ましくは40%を超え、より好ましくは50%を超え、最も好ましくは60%を超える場合、SBAは同種の殺菌活性を有する抗原又はブレブ調製物について十分であろう。
【0128】
当然ながら、異種の殺菌活性を有する抗原又はブレブ調製物は、それが由来する髄膜炎菌株に対して同様に十分なSBAを誘導することができるならば、同種の殺菌活性を有するブレブ調製物を構成することもできる。
【0129】
(2〜4歳児若しくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、髄膜炎菌の3種の異種株に対するSBA(抗体希釈率)力価において(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が、被験体の20%を超え、好ましくは30%を超え、より好ましくは35%を超え、最も好ましくは40%を超える場合、SBAは異種の殺菌活性を有する抗原又はブレブ調製物について十分であろう。かかる試験は、異種の殺菌活性を有する抗原又はブレブ調製物が、様々な髄膜炎菌株に対する交差反応性殺菌抗体を誘導し得るか否かの優れた指標である。3種の異種株は、好ましくは互いに異なる電気泳動タイプ(ET)複合体、又は多座配列タイピング(MLST)パターン(Maidenら PNAS USA 1998, 95:3140-5参照)を有し、好ましくはその異種殺菌活性を有する抗原又はブレブ調製物が作製されるか又は由来する起源の株とも異なるべきである。当業者は、髄膜炎菌間で観察される遺伝的多様性、特に、重篤な病苦の原因として認識されているか、及び/又は強い病原性が認識されているMenB系統(Maidenら 前述)である髄膜炎菌B型株において観察される遺伝的多様性を反映した、異なるET−複合体を有する3種の株を決定することは容易にできるであろう。例えば、使用され得る3種の株は以下のものである:A−4クラスターに属するBZ10(B:2b:P1.2)、ET−37複合体に属するB16B6(B:2a:P1.2)、及びET−5複合体に属するH44/76(B:15:P1.7,16)、又は同じET/クラスターに属するあらゆる他の株。そのような株は、かかる株から作製された若しくはそれらに由来する異種殺菌活性を有する抗原又はブレブ調製物を試験するために使用することができ、例えば、ET−5複合体に属する髄膜炎菌CU385株(B:4:P1.15)を使用することができる。使用され得る別のサンプル株は、系統3流行クローン(例えばNZ124(B:4:P1.7,4))由来のものである。別のET−37株は、NGP165(B:2a:P1.2)である。
【0130】
SBA活性を測定するための方法は、当技術分野において公知である。例えば、使用され得る方法がWO99/09176号の実施例10Cに記載されている。一般的には、試験される株の培養物を対数増殖期まで増殖させる(好ましくは、増殖培地にEDDA等の鉄キレート剤を添加することによる、鉄枯渇条件下で)。これを、BSAを加えた培地中(0.3%BSAを含むハンクス培地等)に懸濁し、約20000CFU/mlに調整した標準(working)細胞懸濁液を得てもよい。一連の反応混合物は、試験される血清の一連の2倍希釈物(好ましくは56℃で30分間熱不活性化されたもの)(例えば、50μl/ウェル容量)と試験される20000CFU/mlの髄膜炎菌株の懸濁液(例えば、25μl/ウェル容量)との混合により、作製することができる。反応バイアルをインキュベートし(例えば37℃で15分間)、振とう(例えば210rpmにて)するべきである。最終反応混合物(例えば容量100μl)は、さらに補体源(例えば、最終容量の25%の予備試験済みの仔ウサギ血清、又はヒト血清学のためのヒト血清)を含み、上記のように(例えば37℃で60分間)インキュベートする。滅菌ポリスチレンU底96穴マイクロタイタープレートをこのアッセイに使用できる。マルチチャネルピペットを用いて、各ウェルからアリコート(例えば10μl)を採取し、ミューラーヒントン寒天プレート(好ましくは、1%イソビタレックス(Isovitalex)及び1%熱不活性化ウマ血清を含む)に滴下し、それを(例えば、5%CO2中37℃で18時間)インキュベートしてもよい。好ましくは、個々のコロニーは、1アリコート当たり最大80CFUまで計数することができる。以下の3つの試験サンプルを対照として使用し得る:バッファー+細菌+補体、バッファー+細菌+不活性化補体、血清+細菌+不活性化補体。回帰計算によって、50%の細胞死滅に相当する希釈度の測定値を与えるようにデータを処理するプログラムを用いて、SBA力価が直接算出され得る。
【0131】
本特許明細書中に引用される全ての文献又は特許出願は、参照により本明細書に組み入れられる。
【0132】
実施例
以下の実施例は、特に詳細に記載しない限りは当業者にとって周知でありルーチンである標準的な技術を用いて実施される。本実施例は例示的なものであり、本発明を限定するものではない。
【実施例1】
【0133】
髄膜炎菌(例えば、MenB)の莢膜多糖産生に関わるタンパク質をコードする遺伝子の欠失を説明する例、PorA遺伝子の欠失、髄膜炎菌ブレブの表面上の様々な防御外膜タンパク質の上方制御、免疫優性タンパク質又は生合成酵素の下方制御(例えば、siaA(−)突然変異)、及びブレブを単離する方法は、WO01/09350号に記載されている。さらなる情報が、WO2004/014417号及びWO2004/014418号に記載されている。尚、本明細書におけるNMB及びNMA遺伝子配列の参照は、www.neisseria.orgからアクセス可能な配列の参照番号を指していることに留意されたい。LOS免疫型の通常の構造を示す概略図を図1に示す(Kahlerら、2005 Glycobiology 15:409-419/2006 JBC 281: 19939-19948より)。また、図2Bも参照されたい。
【実施例2】
【0134】
内部コアLOS O−アセチル化:殺菌力価に対する潜在的影響
要約
・MS−MS分析から、NZ124(L3)株の内部コアLOSのN−アセチル−グルコサミン(GlcNAc)はO−アセチル化されていることがわかっている。H44/76株及びM687株、並びにH44/76株由来のL3 lgtB(−)株及びL3 1st(−)ワクチン株(それぞれ、B1854=TrL3及びB1948=L7)についてはそうではない。
・NZ124株は、H44/76株及びM687株と比較して、抗体媒介性補体殺菌に対する抵抗性が高いわけではない。
・露出が乏しい表面エピトープと殺菌抗体との接触性は、NZ124株及びH44/76株で類似しているようである。
・4つの動物種を用いた5つの動物モデルにおいて、どんな製剤を用いても、抗TrL3及び抗L7ブレブ血清は、H44/76株及びM687株と比較して、NZ124株の死滅を媒介する効果が低かった。
・これらの結果は、内部コアLOSのGlcNAcのアセチル化が、抗「非アセチル化LOS」抗体により媒介される死滅の効力を低減する可能性があることを示唆している。
【0135】
概説
MS/MS分析に基づき、髄膜炎菌のL3 LOSの2つの異なる構造を説明する。これら2つの構造は、内部コアのGlcNAc上にアセチル基が存在するかしないかによって区別される(図2A)。文献には、この追加アセチル基なしでL3構造が記載されている。
【0136】
TrL3及びL7ブレブの内部コアLOSのGlcNAcはアセチル化されていない。このことは、野生型H44/76株及びM97250687(M687)株についても同様である。対照的に、野生型NZ124株は、アセチル化GlcNAcを有する。これら3つの野生型血清群B髄膜炎菌株はL3としての免疫型を有する。NZ124は、1998年に分離されたニュージーランド流行性菌株であり、ニュージーランド環境科学研究院(New Zealang Institute of Environmental Science and Research;ニュージーランド、ウェリントン)から入手可能である。
【0137】
TrL3又はL7ブレブで免疫したマウス由来の血清は、H44/76株及びM687株に対する高レベルの殺菌抗体を含んでいる。しかし、これらの血清は、NZ124株に対しては効果が劣る。実際、NZ124株について抗L7/TrL3ブレブ血清を用いて測定した殺菌力価は、H44/76株及びM687株について測定した力価より3〜20倍低い。
【0138】
NZ124株に対して測定された、低い殺菌抗体力価は、以下に挙げる少なくとも3つの仮定により説明することができる:
・この株は本来、抗体及び補体が媒介する殺菌に対して、H44/76株及びM687株より抵抗性が高い可能性がある;
・この株では、LOSエピトープは殺菌抗体との接触性が低い可能性がある;
・内部コアLOSのアセチル化は、抗「非アセチル化」LOS抗体が媒介する殺菌の効力にマイナスの影響を与える可能性がある。
【0139】
結果
NZ124株は、抗体媒介性殺菌に対し、より抵抗性なのか?
この疑問に答えるために、本発明者らは、様々なPorA+ブレブワクチンで免疫したマウス由来の血清をSBAにおいて分析した。抗血清を相同及び異種PorA株に対しSBAで試験した。表1に示す結果は、2つの実験から得られたものである。
【0140】
P1.7,16ブレブでマウスを免疫すると、相同PorA P1.7,16株の死滅を媒介することができる殺菌抗体の産生が誘導された(H44/76に対して力価1/2300)が、異種PorA株(M687及びNZ124)に対してはそうではなかった。相同PorA株に対する防御殺菌応答を誘導することだけができるP1.19,15ワクチンについても同様の観察結果が得られた(M687に対して力価1/900)。P1.7,4 PorAを含むワクチンで免疫したマウスは、NZ124株に対する高レベルの殺菌抗体(力価1/6200)を有したが、H44/76株に対してはそうではなかった。
【表1】
【0141】
これらの実験において、NZ124株に対して最も高い殺菌抗体力価が測定されたが、これは、この株が、抗体及び補体媒介性殺菌に対し、H44/76株及びM687株より抵抗性が高くはないことを示唆している。
【0142】
NZ124株のLOSエピトープは抗体に十分接触可能なのか?
特に、NspAループは、例えば、PorAのVR1及びVR2ループと比較して、比較的小さいことから、莢膜の大きさがNspA表面エピトープと抗体との接触性を制限する可能性があることが以前から示唆されてきた。実際、莢膜の大きさと、抗NspA血清が補体媒介性殺菌を誘導する能力との間には関係が証明されている(Moeら、1999 I&I 167:5664-75)。
【0143】
髄膜炎菌のLOSは短い糖鎖を有するため、抗体によるその防御エピトープとの接触性は、莢膜の厚さのため損なわれる可能性がある。このような作用機構が、NZ124株に対する抗LOS抗体で測定された低い殺菌力価を説明できるかどうかを決定するために、NZ124を含む異なる3つのMenB株に対して抗NspA MAb(MAb Me−7)を用いて、殺菌アッセイを実施した。
【0144】
1/2560を超える殺菌力価により示されるように、補体の存在下で、H44/76株及びNZ124株は、Mab Me−7によって容易に死滅した。対照的に、M687株は、抗NspA MAb媒介の死滅に対してより抵抗性が高く、わずか1/347の力価が測定された。
【0145】
結論として、防御NspAエピトープの接触性は、H44/76株及びNZ124株で類似している。従って、NZ124株死滅の媒介に関して抗TrL3ブレブ抗体の効力が低いのは、防御LOSエピトープの接触性が低いからではないと仮定することができる。
【0146】
内部コアLOSのGlcNAcのアセチル化は抗「非アセチル化」LOS殺菌抗体の効力を低減するのか?
マウス、ウサギ、モルモット、仔ラット及び成体ラットを、porA KOブレブで免疫した。実験の大部分は、ペンタ−アセチル化(penta-acylated)リピドA LOSを産生する株から取得したブレブ(msbB突然変異、B1853、B1854及びB1948ブレブ)を用いて行ったが、少数の実験では、ヘキサ−アセチル化(hexa-acylated)LOS含有ブレブ(B1820ブレブ)を用いて実施した。アルミニウム塩(Al(OH)3若しくはAlPO4)を用いて、又は用いずに(非吸収製剤)、またCpGを用いて、様々な製剤を試験した。H44/76株、M687株及びNZ124株に対する仔ウサギ補体を用いて、抗ブレブ血清をSBAにおいて試験した。
【0147】
TrL3 V1820ブレブで免疫した動物由来の血清で測定した殺菌力価を表2に示す。動物種にかかわらず、NZ124株で得られた殺菌力価は、H44/76株及びM687株で得られた殺菌力価より低い。
【表2】
【0148】
また、TrL3 msbB KOブレブ(B1853若しくはB1854)、又はL7 msbB KOブレブ(B1948)での免疫も、NZ124株よりH44/76株及びM687株に対して高い血清殺菌力価を誘導する。これは、試験した5種の異なる動物モデル(マウス、仔ラット及び成体ラット(表3a)、モルモット及びウサギ(表3b))のすべてにおいて、用いた製剤にかかわらず観察される。
【0149】
結論として、「非アセチル化LOS」を含むブレブで動物を免疫すると、「非アセチル化株」(H44/76及びM687)よりも低く「アセチル化株」(NZ124)に対して殺菌抗体応答が誘発される。
【表3】
【0150】
論考、結論及び展望
TrL3又はL7ブレブでマウスを免疫すると、H44/76株及びM687株の死滅を媒介する高レベルの殺菌抗体が誘発された。マウス血清を用いて測定したSBA力価は、H44/76株及びM687株に対するものと比較してNZ124株に対して3〜20倍低かったことから、上記抗体は、NZ124株に対しては殺菌力が低い。
【0151】
予備臨床データによれば、NZ124株が、H44/76株及びM687株より、抗体−補体媒介性殺菌に対する抵抗性が高いわけではないことが示唆されている。加えて、NZ124の表面露出が乏しい防御エピトープは、H44/76株の表面上の類似エピトープよりも、抗体への接触性が低いわけではないことを示唆する証拠もある。この知見を確認するために、電子顕微鏡検査及び/又はフローサイトメトリー分析を実施することができる。
【0152】
NZ124株のLOSとH44/76株及びM687株のLOSとの違いの1つは、NZ124 LOSにしか観察されない内部コアのGlcNAcのアセチル化である。この違いにより、H44/76株及びM687株と比較して、NZ124株の死滅の媒介において抗TrL3ブレブ血清及び抗L7ブレブ血清の効力が低いことの説明が可能である。確かに、防御糖エピトープのアセチル化が、抗体によるこのようなエピトープの認識にプラスに又はマイナスに影響しうることは知られている。しかし、ある動物種について記載された防御エピトープの免疫原性/抗原性に対するアセチル化/非アセチル化の影響が、他の動物種及びヒトに常に観察されるわけではない。
【0153】
免疫原性に対する糖エピトープのアセチル化の影響は、「動物種」に応じて、しかしまた抗原に応じても変動する。それにもかかわらず、MenBの場合には、試験した5種の動物モデルにおいて同様の観察結果が得られている。4種の異なる動物種に共通するこれらの観察結果から、「非アセチル化LOS」での免疫により誘導された殺菌抗体は、NZ124株のような「アセチル化」株の死滅の媒介において効力がより低いことが示唆される。
【0154】
この仮定を確認するために、第2のL3アセチル化株を用いた血清殺菌アッセイを実施することにする(BZ10株)。さらには、アセチル化H44/76株及び脱アセチル化NZ124株などの遺伝子改変株を用いたSBAの実施も計画する。その結果に基づき、新規L3ブレブ(アセチル化GlcNAcを含む)を評価することができるだろう。
【実施例3】
【0155】
内部コアLOSのO−アセチル化:LOS内部コアのO−アセチル化のためのナイセリア遺伝子
α鎖の糖組成に加えて、ヘプトースIIの「修飾(decoration)」は、LOS免疫原性に影響を与えるようである。PEAの数及び位置、3位におけるグルコースの存在、7位におけるグリシンの存在、並びにGlcNAcのO−アセチル化が、交差防御の重要な決定因子であるようだ。
【0156】
lpt3遺伝子(MacKinnonら、2002 Mol Microbiol. 43: 931-943)は、ヘプトースIIの3位にPEAを付加する酵素を発現する。この遺伝子(NMB2010)はフレーム変動性(phase variable)ではない。
【0157】
lgtG遺伝子は、ヘプトースIIの3位にグルコースを付加する酵素である。この遺伝子は、フレーム変動性である(WO04/015099号参照)。この遺伝子は、多数の髄膜炎菌株において、単独で、又はlpt6と一緒に欠失している。
【0158】
lpt6遺伝子(Wrightら、2004 J Bact. 186: 6970-6982)は、ヘプトースIIの6位にPEAを付加する酵素を発現する。この遺伝子(NMA0408)は、多数の髄膜炎菌株において、単独で、又はlgtGと一緒に欠失している。この遺伝子は、フレーム変動性ではない。lpt6及びlgtGは、lgt3と称される染色体上の同じ領域に位置する。
【0159】
ヘプトースIIの7位にPEAを付加する酵素は不明である。
ヘプトースIIの7位にグリシンを付加する酵素は不明である。
GlcNAcにO−アセチルを付加する酵素は、本試験の時点まで不明であった。
【0160】
O−アセチル化遺伝子の同定
・翻訳されたナイセリアゲノム(ナイセリア・ゴノローエ(N.gonorrhoeae)、髄膜炎菌MenB MC58、MenC FAM18及びMenA Z2491)上のOafAタンパク質(サルモネラ由来;ヘモフィルスO−アセチラーゼHi0391+Hi0392と相同)を用いたBLAST研究後、ナイセリアにoac1(MC58遺伝子NMB0285に代表される)とoac2(MC58遺伝子NMB1836に代表される)と称される2つの遺伝子ファミリーが見出され、oac2よりoac1ファミリーの方がOafAに近縁であった。
・両遺伝子ともにフレーム変動性である:すなわち、ORFにポリG区間(stretch)が存在する。
・MC58(非O−アセチル化LOS)において、oac1遺伝子(NMB0285)はフレーム外(out of phase)であるが、oac2(NMB1836)はフレーム内(in phase)である。
・試験した各株について、oac1及びoac2に対応するPCR産物を取得した。
・ポリG区間による機能性を調べるためにポリG区間を配列決定した。
【0161】
SBA髄膜炎菌株におけるNMB0285及びNMB1836遺伝子の存在及び機能性:MS−MS(質量分光法)を用いたARDにより得られた分析データ。分子生物学により決定した遺伝子の存在/機能性。かっこ内の数値は遺伝子ORFにおけるポリG区間内のGの数である。
【0162】
【0163】
結論
・NMB0285遺伝子の機能性と、MS−MSによるO−アセチル基の検出との間に完全な相関がある(16/16)。
・NMB1836遺伝子の機能性と、MS−MSにより得られたデータとの間には相関がない(7/16)。
・ナイセリアにおけるO−アセチル化遺伝子はNMB0285であるという強力な証拠を得た。
【0164】
図3Bは、MC58株由来の髄膜炎菌LOS O−アセチル化遺伝子NMB0285(oac1)(H44/76株オープンリーディングフレーム遺伝子配列と100%同一)を示す。大文字でオープンリーディングフレームを、小文字で周囲配列(例えば、プロモーター配列)を示す。オープンリーディングフレーム中の6つのGからなる配列(下線部)によってこのオープンリーディングフレームがフレーム外となることに留意されたい。図3Aには、Z2491株由来の等価遺伝子NMA2202(oac1)を示す。オープンリーディングフレーム(大文字)中の5つのGからなる配列によってこのオープンリーディングフレームがフレーム内となることに留意されたい。
【0165】
1136-GGG GGG ATA TTG AA-1150 MC58
1136-GGG GGA TAT TGA A-1149 Z2491(NMA2202)
【0166】
また、760676株の場合もフレーム内となる(図3C)[H44/76及び760676のoac1のオープンリーディングフレーム遺伝子配列同士は97%の同一性]。
【0167】
W3193株では、前述した領域に5つのGを有するにもかかわらず、LOSはアセチル化されていなかった。これは、ORFのヌクレオチド354で開始する遺伝子における別のポリG区域(tract)によるものであることがわかった。MC58株、760676株及びNZ124株の場合、この位置で、遺伝子に4つのGが存在し(フレーム内、活性遺伝子)、W3193株には3つのGが見とめられた(フレーム外であり、その脱O−アセチル化状態を説明している)。
【0168】
NZ124におけるO−アセチル化遺伝子の不活性化
NZ124、すなわち、O−アセチル化され(活性コンホメーションのoac1による)、H44/76(非アセチル化)由来のブレブにより誘導された血清に対してより抵抗性が高いL3株において、等価のNMB0285遺伝子(oac1)をノックアウトした。質量分光分析によりO−アセチラーゼ遺伝子の不活性化が初めて確認されたため、KO突然変異株をSBA分析に用いて、ワクチン交差防御におけるLOS O−アセチル化の関与をさらに詳しく調べることにする。
【0169】
・NMB0285(NZ124)遺伝子座を配列決定した(図4D)ところ、等価のMC58のNMB0285遺伝子座と98.6%の同一性を有した(NZ124において「フレーム内(IN PHASE)」のGの数の確認)。停止コドンの後に、IS1106に類似した別の配列があり、これは、MC58及び760676にも存在したが、NZ124には存在しなかった。
・O−アセチル化遺伝子の5’及び3’フランキング領域に対応するNZ124組換え5’及び3’領域を含むNMB0285 KOプラスミドを構築した(pMG−T−easyベクター)。NMB0285遺伝子の代わりにKanRマーカーを導入した。このプラスミドはpRIT 15574と称する。
・NZ124のoac1 KO株を構築した(これに由来するLOSは脱O−アセチル化されている)ところ、oac1はNmenのLOS O−アセチラーゼであることがさらに示された。
【0170】
新規のL3由来株におけるO−アセチル化遺伝子のオン
H44/76株由来のブレブはL3免疫型(非アセチル化)である。該ブレブは、L3 O−アセチル化株(例えば、NZ124、BZ10)に対する殺菌抗体を誘導する能力が低減しているため、機能的O−アセチル化遺伝子(oac1)をH44/76由来のブレブ産生株に再導入することが提案される。
【0171】
戦略:
・NZ124遺伝子(O−アセチル化オン株)によるH44/76のNMB0285遺伝子の置換(遺伝子オフ)。
・NZ124プロモーター及び上流調節配列を含有させた(NZ124オープンリーディングフレーム及びNZ124の448bp上流を挿入した)。
・挿入は、センス配向のエリスロマイシン耐性遺伝子を用いて行なった。
・ワクチン異種防御効力に関するブレブL3 LOS O−アセチル化の重要性を試験した(前述の方法で改変したH44/76ブレブによるNZ124の殺菌改善があったか否かを観察するため)。このL3ブレブとO−アセチル化L2株760676由来のブレブの組合せも試験することにより、異種殺菌抗体産生について調べることにする。
【0172】
結果
・前記のように実施したところ、質量分光測定により、改変H44/76株から得られたLOSがHepIIにおいてO−アセチル化されていることがわかった。
【実施例4】
【0173】
オクタロニー法(免疫型決定)、MS/MS分析及び分子生物学分析による6275株及びC11株のLOSの特性決定
要約
・特異的なポリクローナル抗体を用いた免疫拡散法(オクタロニー法)により、6725株及びC11株の免疫型を決定した。MS/MS分析により内部コアLOS組成を決定した。PCR及び配列決定により、LOS内部コア修飾に関与する酵素をコードする遺伝子を分析した。
・MS/MS分析によれば、6275株はHepIIに2個のPEA残基を有する。この株はL3株として免疫型決定されたが、通常のL3株には(HepIIの3位に)1個のPEAしかない。
・MS/MSにより、C11株の内部コアLOSの2つの異なる組成が観察された。一方の集団はHepIIに1個のPEA残基を含むのに対し、他方の集団は2個のPEA残基を含む。この株は、抗L2血清と非常に弱い反応を有するL3株として免疫型決定した。
【0174】
概説
髄膜炎菌LOSの内部コアLOS組成は、これまでの記載より複雑であると思われる。近年まで、内部コアLOSは、HepIIにPEAを含まないか、又は3若しくは6(7)位に1個だけPEA残基を含むと提唱されていた。しかし、近年、3位及び6(又は7)位に2個のPEAを含む新規の内部コアLOSが記載された。この新しいLOS構造は、これまでL3として免疫型決定された株由来のものと記載されていることから、この新規のLOS構造は、L3変異体にちなんで「L3v」と名付けられた。
【0175】
L3v構造の発見前に、LOSの免疫型決定に基づく疫学データから、約70%の侵襲性血清群B株はL3であり(3位にPEAを含む)、残りの株の大部分はL2(6位にPEAを含む)であることが示されている。
【0176】
一連のL3及びL2株、並びにL3由来のブレブ又はL2由来のブレブのいずれかで免疫した動物から得た血清を用いて得られた殺菌データに基づき、本発明者らは、抗L3由来の血清のみがL3株の補体死滅を媒介することができ、また、抗L2血清のみがL2株を死滅させることができると結論付けた。しかし、興味深いことに、2つの新しいL3v株について得られた殺菌データは、本発明者らの以前の結論と一致していない。確かに、これら2つの株の補体死滅は、抗L2由来の血清により媒介されるのであり、抗L3血清によってではない。これら2つの「非定型」株は血清群Bの6275株と血清群Cの標準株C11である。
【0177】
免疫型と殺菌結果との間で相違する結果を理解するために、6275株及びC11株の内部コア組成をMS/MSにより決定した。さらに、オクタロニー法(特異的ポリクローナル血清を用いた免疫拡散法)を用いて、これら株の免疫型決定を実施した。また、分子生物学的方法を用いて、機能的lpt3、lpt6及びlgtG遺伝子の存在を分析した。これらの遺伝子は、HepIIで、それぞれ、3位のPEA、6位のPEA、及び3位のグルコースの付加に関与する酵素をコードする。
【0178】
結果
1.MS/MS分析による内部コア組成
MS/MS分析(図4参照)により、以下のことが明らかになった:
・両方の株において、α鎖LOSは、末端シアル酸基を有するL2及びL3について記載された典型的LNnT四糖である。
・6275株の内部コアLOSは、ほぼ確実に3位及び6(7)位に2つのPEA残基を有するが、NMR分析により確認する必要がある。
・C11株は、2つの異なる内部コア:
−1個のPEAを含む(HepIIでのその位置は明確にされていないが、HepIIのグリシンに対し弱いシグナルが認められることから、7位のPEAは排除される)第1集団と、
−2個のPEAを含む(このHepIIでもグリシンが検出されたため、ほぼ確実に3位及び6位に含む)第2集団
から構成されていた。
・いずれの株についても、HepIIにグルコースは検出されない。
【0179】
2.LOS免疫型決定
一連の特異的抗血清を用いて、免疫型決定を実施した。L3及びL2抗血清以外の抗血清(L1、L4、L5…)で得られた結果は、6275株及びC11株に関して陰性であったため、L3抗血清及びL2抗血清を用いて得られた結果のみを以下に記載する。これらの実験では、GSK Bioで現在用いられている6275株及びC11株(GSK6275−1&2、及びGSK C11)を、アムステルダム大学で20年以上フリーザー内に保存された類似株(Zol 6275株及びRIV C11株)と比較した。
【0180】
2つの6275株(Zol 6275とGSK6275−1&2)は免疫型決定アッセイにおいて類似した挙動を有する。両者は抗L3血清と強力に反応するが、抗L2血清とはそうではなかった。
【0181】
GSK C11株とRIV C11株も同じ結果を示す。いずれの株も、抗L3血清について陽性であり、抗L2血清については弱い陽性である。
【0182】
C11株と抗L2血清を用いて得られた極めて弱い沈降を確認するため、1980年にL3,2として予め型決定された5人の患者の分離菌と組み合わせて、新たな免疫拡散実験を実施した。C11の結果は、抗L2血清との極めて弱い沈降を再度示し、また5種の病原菌株の型決定を確認した。
【0183】
【0184】
3.分子特性決定
3位にPEAを付加する酵素が同定されており、これは、lpt3遺伝子によりコードされる。HepIIの3位のPEAは、超毒性髄膜炎菌株の70%に検出され、Wright及びその共同研究者らは、分析した株の86%においてPCRによりlpt3遺伝子を検出した。この遺伝子は、フレーム変動により調節されず、様々な血清群C及びA株において部分的に欠失していることがわかった。3位へのPEAの付加は、同じ位置でのグルコースの付加と競合していると仮定されている。関与する酵素は、lgtG遺伝子によりコードされ(WO94/015099号参照)、ORF内のポリG区域でのフレーム変動により調節される。Moxon研究所の仮定は、lgtGのORFが存在し、かつフレーム内であれば、機能的酵素が産生され、3位にグルコースが付加されるため、この位置にPEAは付加されないというものである。
【0185】
6位にPEAを付加する酵素をコードする遺伝子は、lgtG遺伝子と並んで位置するlpt6である。この遺伝子は、フレーム変動による調節を受けやすい領域は含んでおらず、髄膜炎菌の48%に検出された(Wrightら、2004)。HepIIの7位にグリシンを付加する酵素をコードする遺伝子は不明である。
【0186】
menB 6275株及びmenC C11株を、対応するL3及びL2標準株と並行して分析した。PCR増幅及び配列決定実験を実施した:
・lpt3遺伝子の完全コピーの存在を評価しながら(PCR)、lgtG遺伝子の機能性についての研究を続行した(PCRによる存在、ORF内のポリC区間の配列)
・lpt6遺伝子の存在をPCRにより評価した。遺伝子の完全コピーが存在すれば、この株が6位にPEAを有するLOSを含むことが推定される。
【0187】
【0188】
これらのデータによれば、menB 6275株及びmenC C11株はL2免疫型に関係しているようである。しかし、MS/MS分析は、いずれの株についても、2個のPEA基を示し、グルコースは示していない。*は、lgtG遺伝子がフレーム変動領域に11(活性遺伝子に見とめられる通常の数)ではなく14の連続したCヌクレオチドを有することを示す。これは、オープンリーディングフレームがフレーム内であり、従って、機能的タンパク質を産生しうることを意味するが、別のプロリン残基の付加により、このタンパク質構造、従って、その機能が破壊される可能性があることも考えられる。
【0189】
4.要約
次の表は、多様な方法を用いた様々な髄膜炎菌株の特性決定を要約したものである。
【0190】
【0191】
論考
内部コアLOS組成の多様性は、これまで記載されているより複雑である。初めに、HepIIに1個のPEA基を含まない又は含む(3位又は6/7位のいずれか)株が記載されたが、近年、2個のPEA基を含む株が記載された。このような株は、例えば、血清群Bの6275株及び血清群CのC11株である。
【0192】
驚くべきことに、6275株及びC11株で得られた免疫型決定結果と血清殺菌結果は一致しない。確かに、これらの2株はL3株として型決定されるが、その死滅は、抗L2由来のブレブ血清により媒介されるのであり、抗L3由来のブレブ血清によってではない(次の実施例を参照)。
【0193】
これら株の関連について、また、2個のPEA基の存在が、継続的なin vitro継代培養による研究室の人工産物でありうるのか否かについて疑問が生じた。本発明者らは、6275株及びC11株の様々なシード(seeds)を比較する機会を得た。各株について、現在本発明者らが用いたシードを、20年以上保存されていた古いシード(アムステルダム大学)と比較した。各株について、2つのシードはオクタロニーアッセイにおいて同じ挙動を示したが、これは、少なくとも過去20年間変化がなかったことを示唆する。それでも、2個のPEA残基の存在を確認するために、最も古いシードの内部コアLOS組成を決定する必要がある。
【0194】
文献によれば、約70%の侵襲性髄膜炎菌株がL3である。L3v株で得られたオクタロニー結果は、この方法がL3株とL3v株を区別しないことを示唆するものであった。従って、両試験において「真のL3」株の数を過大評価する可能性がある。lpt3及びlpt6遺伝子は、HepIIのそれぞれ3位及び6位におけるPEA基付加に寄与している。約36%の循環性株が両方の遺伝子を含み、50%がlpt3のみを、また12%がlpt6のみを有する(Wright JCら、2004)。従って、L3として型決定されたとしても、潜在的に36%の株がL3vでありうる。しかし、様々な内部コアLOS構造に特異的な一連のMAbを用いて得られた近年の疫学データは、2%以下の株がHepIIに2個のPEA基を有することを示唆していた(Gidney MAJら、Infect Immun. 2004 72:559-69)。これら2つの試験の差異(36%対2%)は、6位に1個のPEA基を有する株のヒト補体に対する高い感受性により説明することができる(Ram Sら、J Biol Chem. 2003 278:50853-62)。以上すべてのデータを考え合わせると、侵襲性株の大部分が「真のL3」であることを示唆している。
【0195】
lgtGとlpt−3はHepIIのO−3位について競合し、PEA残基よりもGlc残基の付加に前記の偏りが生じることが示唆されている(Wright JCら、2004)。この仮定は、機能的lgtG遺伝子の存在下でも、6275株及びC11株の内部コアLOSに2つのPEAが存在する(フレーム変動領域において14のCヌクレオチドを含むフレーム内lgtG遺伝子が活性ではない限り。前記参照)ことより証明されるように、普遍的な法則ではないと考えられる。Moxon研究所の仮定に加えて、NmB株のLOS内部コア構造の調節/組成に関与する別の系が近年記載された。この系は、MisR/MisS二成分調節系(Tzeng YLら、J Biol Chem. 2004 279: 35053-62)である。結論として、内部コアLOSの組成に関与する機構は複合的であるようであり、十分に解明されていない。
【0196】
患者から単離した5つの株は、抗L3血清との優勢な沈降だけではなく、抗L2血清との弱い沈降も示すことから、意外なLOS免疫型を呈示した。これはまた、抗L2血清との沈降が非常に弱いにもかかわらず、C11株についても同様である。C11株の内部コアLOSのMS/MS分析も、L3及びL2 LOSの共発現と部分的に一致する2つの異なる内部コア組成を示している。免疫沈降により同定されたL3 LOSは、実際にはL3v LOS(2個のPEA残基を含む)であると考えられるが、L2 LOSは、C11株に明らかに機能的なlgtG遺伝子が検出されているため、特定のLOS分子には存在するはずの検出可能なGlcが存在しなくても、6位に1個のPEAを有する(NMR分析により確認する必要がある)内部コアLOSに関連しているだろう。それでもやはり、これら患者分離菌の1以上の内部コアLOSについて分析(MS/MS、分子特性決定及びSBA)を実施し、L3及びL2株がL3v及びL2 LOSを共発現することを確認する必要がある。
【0197】
結論として、その内部コアに2個のPEAを有する株はL3株として免疫型決定されるが、この免疫型決定は、SBAにおける生物学的反応性、並びに「機能的」lgtG、lpt3及びlpt6遺伝子の存在の分析と一致しない。本発明者のデータは、これらL3v株が恐らくL3株に由来するものではなく(lpt6遺伝子の存在のため)、従って、その名称を改めるべきであることを示している。
【実施例5】
【0198】
髄膜炎菌ワクチン:LOS濃縮ブレブをベースとするワクチンの二価組成物
【0199】
デスフェラール(desferal)を用いて又は用いずに実施した培養からのブレブの作製
・デスフェラールの存在下で、B1854、B1948、B1971、B1984及びB1987ブレブを培養物から作製した。
・B1900ブレブは、デスフェラールを用いずに、培養物から作製した。
【0200】
方法
動物実験の手順:
30匹のマウスの群を筋内経路(IM)によって3回、0、21及び28日目にOMV(約15〜20%の無毒化LOSを含む)で免疫した。毎回の接種材料は、5μg(タンパク質含量)の非吸着OMVからなるものであった。OMVは、莢膜多糖及びPorAが下方制御され、LOSが無毒化される(msbB突然変異)ように操作した髄膜炎菌(Nmen)株から作製した。産生株(前記表を参照)は、遺伝子改変された野生型H44/76株(この場合、これらはL7 LOS又はTrL3 LOSのいずれかを発現した)あるいは遺伝子改変された野生型760676株(この場合、これらは、シアル化された若しくはシアル化されていないL2 LOS、又はTrL2 LOSを発現した)のいずれかに由来するものであった。前述した培養条件下で増殖させた株からブレブを単離した。42日目に、一連の髄膜炎菌株(表1参照)を用いた血清殺菌アッセイ(SBA)による分析のために血液サンプルを採取した。プールした血液サンプル又は個々の血清(1群につき10〜30の血清)のいずれかについてSBAを実施した。
【0201】
仔ラット実験を以下のように実施した。20匹の7日齢ラットの群をIM経路によって(0、14、28及び63日目に)免疫した。毎回の接種材料は、10μg(タンパク質含量)の非吸着ブレブからなるものであった。4回目の注射から14日後に血液サンプルを採取した。
【0202】
モルモット実験を次のように実施した。20匹のモルモットの群をIM経路によって(0、14、28日目に)免疫した。毎回の接種材料は、20μg(タンパク質含量)の非吸着ブレブからなるものであった。3回目の注射から14日後に血液サンプルを採取した。
【0203】
ウサギ実験を次のように実施した。5匹のニュージーランドシロウサギの群をIM経路によって(0、21、42日目に)免疫した。毎回の接種材料は、20μg(タンパク質含量)の非吸着ブレブからなるものであった。3回目の注射から14日後に血液サンプルを採取した。
【0204】
デスフェラールを含む液体培養物を用いたSBA:
37℃+5%CO2にて、MH+1%Polyvitex+1%ウマ血清ペトリ皿上で髄膜炎菌株を一晩培養した。これらを振盪しながら、37℃で50μMのデスフェラール(鉄キレート剤)を添加した液体TSB培地において470nmでのODが0.5に達するまで3時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10又は1/50希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈(8希釈)した。細菌をPBS−グルコース0.1%で希釈することにより、5300CFU/mlを達成し、この希釈物18.8μlを血清希釈物に添加した。また、ウサギ補体(6.2μl)も各ウェルに添加した。振盪しながら37℃で75分インキュベートした後、50μlのMH+0.9%寒天をウェルに添加し、約30分後に50μlのPBS+0.9%寒天を添加した。37℃+CO2で、マイクロプレートを一晩インキュベートした。CFUを計数し、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0205】
寒天培養物(デスフェラールを含まない)を用いたSBA:
37℃+5%CO2にて、BHI+1%ウマ血清ペトリ皿上で髄膜炎菌株を一晩培養した。これらを、37℃+5%CO2で、BHI+1%ウマ血清を用いて4時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈(8希釈)した。細菌をPBS−グルコース0.1%で希釈することにより、6400CFU/mlを達成し、この希釈物12.5μlを血清希釈物に添加した。また、ウサギ補体(12.5μl)も各ウェルに添加した。振盪しながら37℃で75分インキュベートした後、50μlのTSB+0.9%寒天をウェルに添加し、約30分後に50μlのPBS+0.9%寒天を添加した。35℃又は37℃+CO2で、マイクロプレートを一晩インキュベートした。CFUを計数し、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0206】
様々な免疫型の内部コアLOS組成
・L3=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに別のアセチルはない。
・“L3”=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・L2=HepII(ほぼ確実に6位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・変異体=HepII(ほぼ確実に3位及び6位)に2個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する;しかし、W3193株においてはHepIIにOacは一切存在しないことに留意されたい。
・L10及びL11=構造は決定されていない。
・L3、“L3”、L2及び変異体は、LNnT四糖(シアル化又は非シアル化)を含む。
【0207】
結果
以下の表1に要約した結果及び図5から、以下のことがわかる:
・L2由来のブレブは、NmenB L2株に対する殺菌抗体を誘導するが、NmenB L3株(M97250687を除く)に対するものは誘導しない。これに対し、L3由来のブレブは、NmenB L3及び“L3”株に対して、また、血清群Aの3125株(L10)及び血清群YのM01.0240539株(表ではM01.539)に対して防御応答を誘導するが、L2株に対しては誘導しない。質量分光法による3125のL10 LOSの構造を図6に示す。
・加えて、L2由来のブレブは、変異型LOSを保有する株(NmenB 6275株及びNmenC C11株)に対して防御を誘導し、また、血清群C(C19)、血清群Y(S1975)、血清群A(F8238、L11)及び血清群W−135(S4383株及び3193株)由来の他の株に対しても誘導する。
【0208】
・L2及びL3株由来の濃縮LOSブレブをベースとする二価ワクチンは、試験したすべての株(どの血清群及びLOS免疫型でも)に対して防御応答(殺菌抗体)を誘導する(L2 NS及びTrL3 OMVを用いた二価ワクチンは試験しなければならない)。
・非末端切断型L2 LOSは、lgtB突然変異体(=TrL2)より優れた「交差防御」を誘導する。これは、L3由来のブレブ(TrL3=L7)については、そうではない。
・非シアル化L2ブレブは、シアル化L2ブレブと類似レベルの殺菌抗体を誘導する(少なくとも6275株に対して)。
・SBAにおいて対象とした(ターゲティング)株によれば、二価ワクチンについて妨害が観察されたり、されなかったりするが、これは、試験した株のすべてに対して最も優れた効力(SBA力価)を確実にするためには、二価製剤/組成物の微調整が必要であることを示している。
・内部コアの修飾は、交差反応性殺菌抗体の誘導に大きな影響を与える。
【0209】
【0210】
表2の結果から、以下のことがわかる:
・非血清群B株及び6275株に対し、L2由来のブレブ(末端シアル酸を含む又は含まないL2、及びTrL2)により付与される交差防御は、モルモット、ウサギ及び仔ラットにおいても観察される。
・一般に、L2由来のブレブは、M97250687(表ではM687)を除いて、L3株の補体死滅を媒介することができる有意なレベルの殺菌抗体を誘導することができない(仔ラット及びウサギ血清を用いたH44/76株に関するデータは、陰性血清について測定した活性のバックグランドにより、信頼性に欠けるため、反復しなければならない)。
・モルモットでは、TrL2ブレブは、L2 lst−ブレブ及びL2ブレブより高レベルの殺菌抗体を誘導するようであるが、仔ラット、マウス及びウサギでは、L2≧L2 lst−≧TrL2の順位が観察される。
・これらの結果から、殺菌抗体の誘導を誘発するためにL2 LOSのシアル化は必要ではないこと、また、TrL2ブレブ(lgtB突然変異)も、殺菌抗体の産生を誘発することができる(TrL3 LOSと同様に)ことがわかる。
【0211】
【0212】
結論:L2及びL3ベースのLOSワクチンを組み合わせると、Men A、B、C、W135及びY株を死滅させうる抗体を誘発することができることが初めて証明された。L2ベースのワクチンは、L2株、変異(L3v)株及びL11株を死滅させることができる。また、L3ベースのワクチンはL3株、“L3”株(効力は劣る)、及びL10株を死滅させることができる。末端切断型(lgtB(−))及び全長(又はlst(−))α鎖は殺菌することができるが、末端切断型はL3ワクチンに有益であり、全長(又はlst(−))はL2ワクチンに有益であるようだ。これら2つのLOSベースのワクチンを組み合わせると、髄膜炎菌に対するワクチンとして非常に効力があることがわかる。
【実施例6】
【0213】
髄膜炎菌ワクチン:二価OMV(L7由来及びL2由来)はABCWY株に対して交差防御を誘導する
方法
動物実験の手順:
−30匹のマウスの群を筋内経路(IM)によって3回、0、21及び28日目に、二価OMV製剤(約15〜20%の無毒化LOSを含む、L3由来のOMV+L2由来のOMV)で免疫した。毎回の接種材料は、0.8μg+0.8μg(LOS含量)の非吸着OMVからなるものであった。OMVは、莢膜多糖及びPorAが下方制御され、LOSが無毒化される(msbB突然変異)ように操作した髄膜炎菌(Nmen)株から作製した。産生株は、遺伝子改変された野生型H44/76株(L3)又は遺伝子改変された野生型760676株(L2)のいずれかに由来するものであった(遺伝子改変及び培養条件を記載する以下の表を参照)。42日目に、血清群A、B、C、W135(W)及びY由来の一連の22種の髄膜炎菌株を用いた血清殺菌アッセイ(SBA)による分析のために血液サンプルを採取した。プールした血液サンプルについてSBAを実施した。血清は実験20060425及び20060426から得たものである。
【0214】
−以下のように仔ラット実験を実施した。7日齢のラット(1群につきn=20)をIM経路によって(0、14、28及び63日目に)免疫した。毎回の接種材料は、1.6μg+1.6μg(LOS含量)の非吸着OMVからなるものであった。4回目の注射から14日後に血液サンプルを採取し、プールした(実験20060484)。
【0215】
−モルモット実験を次のように実施した。20匹のモルモットの群をIM経路によって(0、14、28日目に)免疫した。毎回の接種材料は、3.2μg+3.2μg(LOS含量)の非吸着OMVからなるものであった。3回目の注射から14日後に血液サンプルを採取し、プールした(実験20060487)。
【0216】
−ウサギ実験を次のように実施した。5匹のニュージーランドシロウサギの群をIM経路によって(0、21、42日目に)免疫した。毎回の接種材料は、3.2μg+3.2μg(LOS含量)の非吸着OMVからなるものであった。最初の注射の前及び3回目の注射から14日後に血液サンプルを採取し、これらをSBAで個々に試験した(実験20060486)。
【0217】
【0218】
デスフェラールを用いて又は用いずに実施した培養からのOMVの作製
デスフェラールを用いて実施した培養物からB1854及びB1948 OMVを作製した。
【0219】
デスフェラールを用いずに実施した培養物からB2071及びB2084 OMVを作製した。
【0220】
SBA:
37℃+5%CO2にて、髄膜炎菌株をペトリ皿上で一晩培養した。次に、37℃+5%CO2にて、デスフェラール(鉄キレート剤)を含む又は含まないペトリ皿上で、上記株を4時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10又は1/50希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈した。次に、細菌(PBS−グルコース0.1%で希釈することにより、1ウェル当たり約100〜150CFU/mlを達成する)と、仔ウサギ補体(マイクロウェル中の最終濃度:12.5%v/v)との混合物25μlを血清希釈物に添加した。振盪しながら37℃で75分インキュベートした後、2層の寒天(0.9%)をウェルに添加した。35℃又は37℃+CO2で、マイクロプレートを一晩インキュベートした後、CFUを計数し、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0221】
様々な免疫型の内部コアLOS組成
・L3=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに別のアセチルはない。
・“L3”=HepII(ほぼ確実に3位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・L2=HepII(ほぼ確実に6位)に1個のPEAを有し、内部コアGlcNAcに1個の別のアセチルを有する。
・変異体=HepII(ほぼ確実に3位及び6位)に2個のPEAを有し、一般に内部コアGlcNAcに1個の別のアセチルを有する。
・L10及びL11=構造は完全には決定されていない。
・L3、“L3”、L2及び変異体は、LNnT四糖(シアル化、又は非シアル化)を含む。
【0222】
結果
殺菌力価の4倍増加(対照動物由来のプール血清サンプル、又は比較物としてのワクチン前血清サンプルを用いた)に基づき、得られた結果(次の表を参照)から以下のことがわかる:
・L7+NSL2 OMV又はTrL3+TrL2 OMVのいずれかを含む非吸着二価製剤は、血清群A、B、C、W及びYに属する髄膜炎菌株に対する交差防御を付与することができる。
・この交差防御は、マウスだけではなく、仔ラット、モルモット及びウサギでも観察される。
・しかし、上記製剤は、L4 LOS免疫型を発現する株に対しては防御応答を誘発することができない。これらの実験において、血清はL10株に対して殺菌性ではない。
・L3又は“L3”又は変異体又はL2 LOSを発現する株のほとんどは、マウス、仔ラット及びモルモット血清により死滅する。これらの株に対し、交差防御の割合(%)は90%近い。
・交差防御は、ウサギにおいて、より低いようである。
【0223】
結論
L3及びL2髄膜炎菌株由来のOMNをベースとする二価ワクチンは、L3又は“L3”又は変異体(L3v)又はL2 LOSのいずれかを発現する髄膜炎菌株に対する防御を付与することができる。この防御は所与の血清群に制限されるわけではない。
【0224】
【実施例7】
【0225】
L7 OMVの免疫原性に対するOAcの影響
方法
動物実験の手順:
−30匹のマウスの群を筋内(IM)経路によって3回、0、21及び28日目にOMV(約15〜20%の無毒化LOSを含む)で免疫した。毎回の接種材料は、0.8μg(LOS含量)の非吸着OMVからなるものであった。OMVは、莢膜多糖及びPorAが下方制御され、LOSが無毒化される(msbB突然変異)ように操作した髄膜炎菌株から作製した。産生株は、遺伝子改変された野生型H44/76株に由来するものであった(遺伝子改変及び培養条件を記載する以下の表を参照)。42日目に、髄膜炎菌株を用いた血清殺菌アッセイ(SBA)による分析のために血液サンプルを採取した。個々の血清についてSBAを実施した。血清は実験20060634から得たものである。
【0226】
−モルモット実験を次のように実施した。20匹のモルモットの群をIM経路によって(0、14、28日目に)免疫した。毎回の接種材料は、3.2μg(LOS含量)の非吸着OMVからなるものであった。3回目の注射から14日後に血液サンプルを採取した(実験20060636)。
【0227】
【0228】
SBA:
37℃+5%CO2にて、髄膜炎菌株(OAc−であるH44/76及びM97250687と、OAc+であるNZ124)をペトリ皿上で一晩培養した。次に、37℃+5%CO2にて、デスフェラール(鉄キレート剤)を含むペトリ皿上で、上記株を4時間継代培養した。血清サンプルを56℃で40分かけて不活性化し、次にPBS−グルコース0.1%で1/10又は1/50希釈した後、平底マイクロプレートにおいて25μlの容量で2倍希釈した。次に、細菌(PBS−グルコース0.1%で希釈することにより、1ウェル当たり約100〜150CFU/mlを達成する)と、仔ウサギ補体(マイクロウェル中の最終濃度:12.5%v/v)との混合物25μlを血清希釈物に添加した。振盪しながら37℃で75分インキュベートした後、2層の寒天(0.9%)をウェルに添加した。35℃又は37℃+CO2で、マイクロプレートを一晩インキュベートした。CFUを計数した後、死滅の割合(%)を計算した。SBA力価は、50%の死滅をもたらす希釈度とする。
【0229】
結果
マウスの場合、L7 OAc+OMVはL7 OAc−OMVより免疫原性が高い(以下の表を参照)。確かに、L7 OAc+OMVで免疫したマウス由来の血清は、L7 OAc−OMVで免疫したマウス由来の血清より高い補体媒介殺菌活性を示す。これは、OAc−及びOAc+野生型株に対して観察される。しかし、有意な差異はOAc−株(H44/76及びM97205687)に対してしか観察されない。
【0230】
【0231】
モルモットの場合には、SBAにおいてOMVの両方の型がOAc+及びOAc−野生型株に対して同様の免疫原性を示す(以下の表参照)。
【0232】
【0233】
結論
内部コアGlcNAcのO−アセチル化は、マウスにおいてOMVの免疫原性(SBA力価)を増強するが、モルモットではそうではないことが証明された。
【特許請求の範囲】
【請求項1】
そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているナイセリア株由来のL3 LOSを含む免疫原性組成物。
【請求項2】
L3 LOSが、以下の構造:
【化1】
〔式中、
R1=
【化2】
であり、
R2=PEAであり、
R3=Hであり、
R4=OAcであり、
R5=H、PEA、又はGlyである。〕
を有する、請求項1記載の免疫原性組成物。
【請求項3】
R1=
【化3】
である、請求項2記載の免疫原性組成物。
【請求項4】
R5=H、PEA又はGlyである、請求項2又は3記載の免疫原性組成物。
【請求項5】
ナイセリア株由来のL2 LOS、場合により髄膜炎菌A株、B株、C株、W135株又はY株から単離されたもの、をさらに含む、請求項1〜4のいずれか1項に記載の免疫原性組成物。
【請求項6】
以下の構造を有するL2 LOS:
【化4】
〔L2 LOSについて、式中、
R1=
【化5】
であり、
R2=PEA又はGlcであり、
R3=PEAであり、
R4=H又はOAcであり、
R5=H、PEA、又はGlyである。〕
をさらに含む、請求項1〜5のいずれか1項に記載の免疫原性組成物。
【請求項7】
L2 LOSについて、R1=
【化6】
である、請求項6記載の免疫原性組成物。
【請求項8】
L2 LOSについて、R2=PEA又はGlcである、請求項6又は7記載の免疫原性組成物。
【請求項9】
L2 LOSについて、R5=H、PEA又はGlyである、請求項6〜8のいずれか1項に記載の免疫原性組成物。
【請求項10】
L2 LOSについて、R4=H又はOAcである、請求項6〜9のいずれか1項に記載の免疫原性組成物。
【請求項11】
ナイセリア株由来のL10 LOS、場合により髄膜炎菌A株、B株、C株、W135株又はY株から単離されたもの、をさらに含む、請求項1〜10のいずれか1項に記載の免疫原性組成物。
【請求項12】
以下の構造を有するL10 LOS:
【化7】
〔式中、
R1=ヘキソース−ヘキソース−であり、
R2=H又はPEAであり、
R3=PEAであり、
R4=OAcであり、
R5=H又はGlyである。〕
をさらに含む、請求項1〜11のいずれか1項に記載の免疫原性組成物。
【請求項13】
L10 LOSについて、R1=Gal−Gal−、Gal−Glc−、Glc−Gal−、又はGlc−Glc−である、請求項12記載の免疫原性組成物。
【請求項14】
L10 LOSについて、R1=
【化8】
である、請求項13記載の免疫原性組成物。
【請求項15】
L10 LOSについて、R1の末端ヘキソースが、(場合によりα−Neu5Ac−(2→3)基を介して)シアル化されている、請求項12〜14のいずれか1項に記載の免疫原性組成物。
【請求項16】
L10 LOSについて、R2=H又はPEAである、請求項12〜15のいずれか1項に記載の免疫原性組成物。
【請求項17】
L10 LOSについて、R5=H又はGlyである、請求項12〜16のいずれか1項に記載の免疫原性組成物。
【請求項18】
ナイセリア株由来のL4 LOS、場合により髄膜炎菌A株、B株、C株、W135株又はY株から単離されたもの、をさらに含む、請求項1〜17のいずれか1項に記載の免疫原性組成物。
【請求項19】
以下の構造を有するL4 LOS:
【化9】
〔L4 LOSについて、式中、
R1=
【化10】
であり、
R2=Hであり、
R3=PEAであり、
R4=OAcであり、
R5=Glyである。〕
をさらに含む、請求項1〜18のいずれか1項に記載の免疫原性組成物。
【請求項20】
L4 LOSが、R4=OAcの代わりにHであり、及び/又はR5=Glyの代わりにHである、請求項19記載の免疫原性組成物。
【請求項21】
L3 LOSが髄膜炎菌A株、B株、C株、W135株又はY株から単離されたものである、請求項1〜20のいずれか1項に記載の免疫原性組成物。
【請求項22】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、タンパク質キャリアとコンジュゲートされている、請求項1〜21のいずれか1項に記載の免疫原性組成物。
【請求項23】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、無毒化リピドA部分(例えばmsbB(−)ナイセリア株から単離されたLOSと一致する第2アシル鎖を欠損するもの)を有する、請求項1〜22のいずれか1項に記載の免疫原性組成物。
【請求項24】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、LOSの毒性低減に適したリピドA結合ペプチド(SAEP2又はSAEPIIなど)と複合体化されている、請求項1〜23のいずれか1項に記載の免疫原性組成物。
【請求項25】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、免疫原性組成物中に精製LOS調製物として存在する、請求項1〜24のいずれか1項に記載の免疫原性組成物。
【請求項26】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、免疫原性組成物中にリポソーム調製物として存在する、請求項1〜25のいずれか1項に記載の免疫原性組成物。
【請求項27】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、免疫原性組成物中にブレブ調製物として存在する、請求項1〜26のいずれか1項に記載の免疫原性組成物。
【請求項28】
ブレブ調製物(複数でもよい)が、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものである、請求項27記載の免疫原性組成物。
【請求項29】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSのブレブ調製物(複数でもよい)が、鉄塩の存在下における培養後の各ナイセリア株(複数でもよい)から単離されたものである、請求項27又は28記載の免疫原性組成物。
【請求項30】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSのブレブ調製物(複数でもよい)が、鉄塩の不在下における培養後の各ナイセリア株(複数でもよい)から単離されたものである、請求項27〜29のいずれか1項に記載の免疫原性組成物。
【請求項31】
ブレブ調製物(複数でもよい)が、莢膜多糖を合成することができないナイセリア株(複数でもよい)から単離されたものである、請求項27〜30のいずれか1項に記載の免疫原性組成物。
【請求項32】
ナイセリア株(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失しており(機能的発現がない)、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において同じ莢膜多糖遺伝子の発現が下方制御されている、請求項31記載の免疫原性組成物。
【請求項33】
ブレブ調製物(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ナイセリア株(複数でもよい)から単離されたものであり、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において同じリピドA遺伝子(複数でもよい)の発現が下方制御されている、請求項27〜32のいずれか1項に記載の免疫原性組成物。
【請求項34】
ブレブ調製物(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、外膜タンパク質遺伝子porA、porB、opA、opC、pilC、lbpA又はfrpBの1以上の発現が下方制御されている、好ましくは欠失しているナイセリア株(複数でもよい)から単離されたものであり、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において同じ外膜タンパク質遺伝子(複数でもよい)の発現が下方制御されている、請求項27〜33のいずれか1項に記載の免疫原性組成物。
【請求項35】
鉄塩の存在下で培養されるナイセリア株のいずれか(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、以下の外膜タンパク質遺伝子の組合せ:PorAとOpA、PorAとOpC、OpAとOpC、PorAとOpAとOpC、のいずれかの発現が下方制御されている、好ましくは欠失している、請求項34記載の免疫原性組成物。
【請求項36】
鉄塩の不在下で培養されるナイセリア株のいずれか(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、以下の外膜タンパク質遺伝子の組合せ:PorAとOpA、PorAとOpC、OpAとOpC、PorAとOpAとOpC、PorAとFrpB、OpCとFrpB、OpAとFrpB、PorAとOpA、OpCとFrpB、PorAとLbpA、FrpBとLbpA、PorAとFrpBとLbpAのいずれか、の発現が下方制御されている、好ましくは欠失している、請求項34又は35記載の免疫原性組成物。
【請求項37】
ブレブ調製物(複数でもよい)が、外膜タンパク質抗原NspA、TbpA low、TbpA high、Hsf、Hap、OMP85、PilQ、NadA、GNA1870、MltAの1以上の発現が上方制御されているナイセリア株(複数でもよい)から単離されたものであり、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において1以上の異なる外膜タンパク質抗原の発現が上方制御されている、請求項27〜36のいずれか1項に記載の免疫原性組成物。
【請求項38】
NspAの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含み、好ましくはナイセリア株が鉄塩の存在下で培養されたものである、請求項37記載の免疫原性組成物。
【請求項39】
Hsfの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含む、請求項37又は38記載の免疫原性組成物。
【請求項40】
Tbp highの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含み、好ましくはナイセリア株が鉄塩の不在下で培養されたものである、請求項37〜39のいずれか1項に記載の免疫原性組成物。
【請求項41】
Tbp lowの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含み、好ましくはナイセリア株が鉄塩の不在下で培養されたものである、請求項37〜40のいずれか1項に記載の免疫原性組成物。
【請求項42】
請求項1〜41のいずれか1項に記載の免疫原性組成物の有効量と薬学的に許容される担体又は賦形剤とを含むワクチン組成物。
【請求項43】
アジュバント、例えば水酸化アルミニウム又はリン酸アルミニウム、をさらに含む、請求項42記載のワクチン。
【請求項44】
髄膜炎菌血清群A、髄膜炎菌血清群C、髄膜炎菌血清群W−135、髄膜炎菌血清群Y、及びインフルエンザ菌b型の株に由来する、1以上のコンジュゲートした莢膜多糖又はオリゴ糖をさらに含む、請求項42又は43記載のワクチン。
【請求項45】
髄膜炎菌血清群A、B、C、W135及びYから選択される1以上によって引き起こされる疾患の予防又は治療のための医薬の製造における、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの使用。
【請求項46】
髄膜炎菌血清群A、B、C、W135及びYから選択される1以上によって引き起こされる疾患の予防又は治療方法であって、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの有効量をその必要があるヒト患者に投与するステップを含む方法。
【請求項47】
予防又は治療が髄膜炎菌血清群Aの疾患のものである、請求項45又は46記載の使用又は方法。
【請求項48】
予防又は治療が髄膜炎菌血清群Bの疾患のものである、請求項45〜47のいずれか1項に記載の使用又は方法。
【請求項49】
予防又は治療が髄膜炎菌血清群Cの疾患のものである、請求項45〜48のいずれか1項に記載の使用又は方法。
【請求項50】
予防又は治療が髄膜炎菌血清群W135の疾患のものである、請求項45〜49のいずれか1項に記載の使用又は方法。
【請求項51】
予防又は治療が髄膜炎菌血清群Yの疾患のものである、請求項45〜50のいずれか1項に記載の使用又は方法。
【請求項52】
請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの製造方法であって、L3 LOSを単離するステップ、場合によりそれを必要に応じて単離されたL2及び/又はL10及び/又はL4 LOSと組み合わせるステップ、並びに薬学的に許容される賦形剤と共にL3 LOSを製剤化するステップを含む方法。
【請求項53】
髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの使用。
【請求項54】
髄膜炎菌免疫型L3疾患の予防又は治療方法であって、その必要があるヒト患者に、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの有効量を投与するステップを含む方法。
【請求項55】
髄膜炎菌免疫型L3疾患が、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOS、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないLOS、又はそのヘプトースII残基に結合したGlcNac残基において部分的にO−アセチル化されておりかつ部分的にO−アセチル化されていないLOS、のいずれかのLOSを有する株によって引き起こされる、請求項53又は54記載の使用又は方法。
【請求項56】
そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株により引き起こされる髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の使用。
【請求項57】
そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株によって引き起こされる髄膜炎菌免疫型L3疾患を予防又は治療する方法であって、その必要があるヒト患者に、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の有効量を投与するステップを含む方法。
【請求項58】
免疫原性組成物が、以下の構造を有するL3 LOS:
【化11】
〔式中、
R1=
【化12】
であり、
R2=PEAであり、
R3=Hであり、
R4=Hであり、
R5=H、PEA、又はGlyである。〕
を含む、請求項56又は57記載の使用又は方法。
【請求項59】
R1=
【化13】
である、請求項58記載の使用又は方法。
【請求項60】
R5=H、PEA、又はGlyである、請求項58又は59記載の使用又は方法。
【請求項61】
免疫原性組成物中のL3 LOSがタンパク質キャリアとコンジュゲートされている、請求項56〜60のいずれか1項に記載の使用又は方法。
【請求項62】
免疫原性組成物中のL3 LOSが、無毒化リピドA部分(例えばmsbB(−)ナイセリア株から単離されたLOSと一致する第2アシル鎖を欠損するもの)を含む、請求項56〜61のいずれか1項に記載の使用又は方法。
【請求項63】
L3 LOSが、LOSの毒性低減に適したリピドA結合ペプチド(SAEP2又はSAEPIIなど)との複合体化による無毒化リピドA部分を有する、請求項56〜62のいずれか1項に記載の使用又は方法。
【請求項64】
L3 LOSが、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として免疫原性組成物中に存在する、請求項56〜63のいずれか1項に記載の使用又は方法。
【請求項65】
ブレブ調製物が、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものである、請求項64記載の使用又は方法。
【請求項66】
ブレブ調製物が、莢膜多糖を合成することができないナイセリア株から単離されたものである、請求項64又は65記載の使用又は方法。
【請求項67】
ナイセリア株が、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)、請求項66記載の使用又は方法。
【請求項68】
ブレブ調製物が、それが由来する天然株(複数でもよい)と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ナイセリア株から単離されたものである、請求項64〜67のいずれか1項に記載の使用又は方法。
【請求項69】
通常はそのLOSのヘプトースII残基に結合したGlcNac残基においてO−アセチル化するナイセリア株のLOSを脱O−アセチル化する方法であって、oac1遺伝子の機能的発現を破壊することによって、機能的Oac1を発現することができないようにするステップを含む方法。
【請求項70】
通常はそのLOSのヘプトースII残基に結合したGlcNac残基においてO−アセチル化しない又は部分的にのみO−アセチル化するナイセリア株のLOSをO−アセチル化する又はさらにO−アセチル化する方法であって、oac1遺伝子の機能的発現を増大するステップを含む方法。
【請求項71】
oac1遺伝子の機能的発現を増大するステップが、oac1遺伝子の機能的コピー(例えば、場合により天然のプロモーター配列を含んでいてもよい図3Dのオープンリーディングフレーム)をナイセリア株に導入することによって行われる、請求項70記載の方法。
【請求項72】
oac1遺伝子の機能的発現を増大するステップが、存在する非機能的遺伝子のポリGフレーム変動領域(複数でもよい)を、oac1遺伝子の機能的発現のためにフレーム内となるようにすることによって行われる、請求項70記載の方法。
【請求項73】
ナイセリア株において機能的oac1発現のフレーム変動性を少なくする方法であって、oac1遺伝子のポリGフレーム変動領域(複数でもよい)を、より少ないGヌクレオチドを有するコドンを用いて同じアミノ酸がコードされるように、変化させるステップを含む方法。
【請求項74】
ATG開始コドンから1135〜1137位のヌクレオチドによってコードされるアルギニン残基のコドンを、AGGからCGT、CGC、CGA又はAGAに変化させる、請求項73記載の方法。
【請求項75】
ATG開始コドンから1138〜1140位のヌクレオチドによってコードされるグリシン残基のコドンを、GGGからGGT、GGA又はGGCに変化させる、請求項73又は74記載の方法。
【請求項76】
L2免疫型のナイセリア株に対して行われる、請求項69〜75のいずれか1項に記載の方法。
【請求項77】
L3免疫型のナイセリア株に対して行われる、請求項69〜75のいずれか1項に記載の方法。
【請求項78】
免疫原性組成物の製造方法であって、請求項69〜77のいずれか1項に記載の方法を実施するステップ、得られるナイセリア株からLOSを単離するステップ、及び薬学的に許容される担体又は賦形剤と共に有効量のLOSを製剤化するステップを含む方法。
【請求項79】
単離されたLOSをタンパク質キャリアにコンジュゲートするさらなるステップを含む、請求項78記載の方法。
【請求項80】
単離されたLOSのリピドA部分を無毒化する、例えばmsbB(−)ナイセリア株からLOSを単離することにより無毒化する、さらなるステップを含む、請求項78又は79記載の方法。
【請求項81】
単離されたLOSのリピドA部分が、LOSの毒性低減に適したリピドA結合ペプチド(SAEP2又はSAEPIIなど)との複合体化によって無毒化されている、請求項80記載の方法。
【請求項82】
LOSが、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として単離されている、請求項78〜81のいずれか1項に記載の方法。
【請求項83】
ブレブ調製物が、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものである、請求項82記載の方法。
【請求項84】
ブレブ調製物が、莢膜多糖を合成することができないナイセリア株から単離されたものである、請求項82又は83記載の方法。
【請求項85】
ナイセリア株が、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)、請求項84記載の方法。
【請求項86】
ブレブ調製物が、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ナイセリア株から単離されたものである、請求項82〜85のいずれか1項に記載の方法。
【請求項1】
そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているナイセリア株由来のL3 LOSを含む免疫原性組成物。
【請求項2】
L3 LOSが、以下の構造:
【化1】
〔式中、
R1=
【化2】
であり、
R2=PEAであり、
R3=Hであり、
R4=OAcであり、
R5=H、PEA、又はGlyである。〕
を有する、請求項1記載の免疫原性組成物。
【請求項3】
R1=
【化3】
である、請求項2記載の免疫原性組成物。
【請求項4】
R5=H、PEA又はGlyである、請求項2又は3記載の免疫原性組成物。
【請求項5】
ナイセリア株由来のL2 LOS、場合により髄膜炎菌A株、B株、C株、W135株又はY株から単離されたもの、をさらに含む、請求項1〜4のいずれか1項に記載の免疫原性組成物。
【請求項6】
以下の構造を有するL2 LOS:
【化4】
〔L2 LOSについて、式中、
R1=
【化5】
であり、
R2=PEA又はGlcであり、
R3=PEAであり、
R4=H又はOAcであり、
R5=H、PEA、又はGlyである。〕
をさらに含む、請求項1〜5のいずれか1項に記載の免疫原性組成物。
【請求項7】
L2 LOSについて、R1=
【化6】
である、請求項6記載の免疫原性組成物。
【請求項8】
L2 LOSについて、R2=PEA又はGlcである、請求項6又は7記載の免疫原性組成物。
【請求項9】
L2 LOSについて、R5=H、PEA又はGlyである、請求項6〜8のいずれか1項に記載の免疫原性組成物。
【請求項10】
L2 LOSについて、R4=H又はOAcである、請求項6〜9のいずれか1項に記載の免疫原性組成物。
【請求項11】
ナイセリア株由来のL10 LOS、場合により髄膜炎菌A株、B株、C株、W135株又はY株から単離されたもの、をさらに含む、請求項1〜10のいずれか1項に記載の免疫原性組成物。
【請求項12】
以下の構造を有するL10 LOS:
【化7】
〔式中、
R1=ヘキソース−ヘキソース−であり、
R2=H又はPEAであり、
R3=PEAであり、
R4=OAcであり、
R5=H又はGlyである。〕
をさらに含む、請求項1〜11のいずれか1項に記載の免疫原性組成物。
【請求項13】
L10 LOSについて、R1=Gal−Gal−、Gal−Glc−、Glc−Gal−、又はGlc−Glc−である、請求項12記載の免疫原性組成物。
【請求項14】
L10 LOSについて、R1=
【化8】
である、請求項13記載の免疫原性組成物。
【請求項15】
L10 LOSについて、R1の末端ヘキソースが、(場合によりα−Neu5Ac−(2→3)基を介して)シアル化されている、請求項12〜14のいずれか1項に記載の免疫原性組成物。
【請求項16】
L10 LOSについて、R2=H又はPEAである、請求項12〜15のいずれか1項に記載の免疫原性組成物。
【請求項17】
L10 LOSについて、R5=H又はGlyである、請求項12〜16のいずれか1項に記載の免疫原性組成物。
【請求項18】
ナイセリア株由来のL4 LOS、場合により髄膜炎菌A株、B株、C株、W135株又はY株から単離されたもの、をさらに含む、請求項1〜17のいずれか1項に記載の免疫原性組成物。
【請求項19】
以下の構造を有するL4 LOS:
【化9】
〔L4 LOSについて、式中、
R1=
【化10】
であり、
R2=Hであり、
R3=PEAであり、
R4=OAcであり、
R5=Glyである。〕
をさらに含む、請求項1〜18のいずれか1項に記載の免疫原性組成物。
【請求項20】
L4 LOSが、R4=OAcの代わりにHであり、及び/又はR5=Glyの代わりにHである、請求項19記載の免疫原性組成物。
【請求項21】
L3 LOSが髄膜炎菌A株、B株、C株、W135株又はY株から単離されたものである、請求項1〜20のいずれか1項に記載の免疫原性組成物。
【請求項22】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、タンパク質キャリアとコンジュゲートされている、請求項1〜21のいずれか1項に記載の免疫原性組成物。
【請求項23】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、無毒化リピドA部分(例えばmsbB(−)ナイセリア株から単離されたLOSと一致する第2アシル鎖を欠損するもの)を有する、請求項1〜22のいずれか1項に記載の免疫原性組成物。
【請求項24】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、LOSの毒性低減に適したリピドA結合ペプチド(SAEP2又はSAEPIIなど)と複合体化されている、請求項1〜23のいずれか1項に記載の免疫原性組成物。
【請求項25】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、免疫原性組成物中に精製LOS調製物として存在する、請求項1〜24のいずれか1項に記載の免疫原性組成物。
【請求項26】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、免疫原性組成物中にリポソーム調製物として存在する、請求項1〜25のいずれか1項に記載の免疫原性組成物。
【請求項27】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSが、免疫原性組成物中にブレブ調製物として存在する、請求項1〜26のいずれか1項に記載の免疫原性組成物。
【請求項28】
ブレブ調製物(複数でもよい)が、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものである、請求項27記載の免疫原性組成物。
【請求項29】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSのブレブ調製物(複数でもよい)が、鉄塩の存在下における培養後の各ナイセリア株(複数でもよい)から単離されたものである、請求項27又は28記載の免疫原性組成物。
【請求項30】
L2 LOS、L3 LOS、L10 LOS、L4 LOS、L2及びL3 LOS、L2及びL10 LOS、L3及びL10 LOS、L4及びL2 LOS、L4及びL3 LOS、L4及びL10 LOS、L2及びL3及びL10 LOS、L2及びL3及びL4 LOS、L2及びL4及びL10 LOS、L3及びL4及びL10 LOS、又はL2及びL3及びL10及びL4 LOSのブレブ調製物(複数でもよい)が、鉄塩の不在下における培養後の各ナイセリア株(複数でもよい)から単離されたものである、請求項27〜29のいずれか1項に記載の免疫原性組成物。
【請求項31】
ブレブ調製物(複数でもよい)が、莢膜多糖を合成することができないナイセリア株(複数でもよい)から単離されたものである、請求項27〜30のいずれか1項に記載の免疫原性組成物。
【請求項32】
ナイセリア株(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失しており(機能的発現がない)、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において同じ莢膜多糖遺伝子の発現が下方制御されている、請求項31記載の免疫原性組成物。
【請求項33】
ブレブ調製物(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ナイセリア株(複数でもよい)から単離されたものであり、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において同じリピドA遺伝子(複数でもよい)の発現が下方制御されている、請求項27〜32のいずれか1項に記載の免疫原性組成物。
【請求項34】
ブレブ調製物(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、外膜タンパク質遺伝子porA、porB、opA、opC、pilC、lbpA又はfrpBの1以上の発現が下方制御されている、好ましくは欠失しているナイセリア株(複数でもよい)から単離されたものであり、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において同じ外膜タンパク質遺伝子(複数でもよい)の発現が下方制御されている、請求項27〜33のいずれか1項に記載の免疫原性組成物。
【請求項35】
鉄塩の存在下で培養されるナイセリア株のいずれか(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、以下の外膜タンパク質遺伝子の組合せ:PorAとOpA、PorAとOpC、OpAとOpC、PorAとOpAとOpC、のいずれかの発現が下方制御されている、好ましくは欠失している、請求項34記載の免疫原性組成物。
【請求項36】
鉄塩の不在下で培養されるナイセリア株のいずれか(複数でもよい)が、それが由来する天然株(複数でもよい)と比較して、以下の外膜タンパク質遺伝子の組合せ:PorAとOpA、PorAとOpC、OpAとOpC、PorAとOpAとOpC、PorAとFrpB、OpCとFrpB、OpAとFrpB、PorAとOpA、OpCとFrpB、PorAとLbpA、FrpBとLbpA、PorAとFrpBとLbpAのいずれか、の発現が下方制御されている、好ましくは欠失している、請求項34又は35記載の免疫原性組成物。
【請求項37】
ブレブ調製物(複数でもよい)が、外膜タンパク質抗原NspA、TbpA low、TbpA high、Hsf、Hap、OMP85、PilQ、NadA、GNA1870、MltAの1以上の発現が上方制御されているナイセリア株(複数でもよい)から単離されたものであり、L2及びL3ブレブの両方が存在する場合には、それが由来する株は好ましくはそれぞれの株において1以上の異なる外膜タンパク質抗原の発現が上方制御されている、請求項27〜36のいずれか1項に記載の免疫原性組成物。
【請求項38】
NspAの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含み、好ましくはナイセリア株が鉄塩の存在下で培養されたものである、請求項37記載の免疫原性組成物。
【請求項39】
Hsfの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含む、請求項37又は38記載の免疫原性組成物。
【請求項40】
Tbp highの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含み、好ましくはナイセリア株が鉄塩の不在下で培養されたものである、請求項37〜39のいずれか1項に記載の免疫原性組成物。
【請求項41】
Tbp lowの発現が上方制御されているナイセリア株から単離されたL2及び/又はL3ブレブ調製物を含み、好ましくはナイセリア株が鉄塩の不在下で培養されたものである、請求項37〜40のいずれか1項に記載の免疫原性組成物。
【請求項42】
請求項1〜41のいずれか1項に記載の免疫原性組成物の有効量と薬学的に許容される担体又は賦形剤とを含むワクチン組成物。
【請求項43】
アジュバント、例えば水酸化アルミニウム又はリン酸アルミニウム、をさらに含む、請求項42記載のワクチン。
【請求項44】
髄膜炎菌血清群A、髄膜炎菌血清群C、髄膜炎菌血清群W−135、髄膜炎菌血清群Y、及びインフルエンザ菌b型の株に由来する、1以上のコンジュゲートした莢膜多糖又はオリゴ糖をさらに含む、請求項42又は43記載のワクチン。
【請求項45】
髄膜炎菌血清群A、B、C、W135及びYから選択される1以上によって引き起こされる疾患の予防又は治療のための医薬の製造における、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの使用。
【請求項46】
髄膜炎菌血清群A、B、C、W135及びYから選択される1以上によって引き起こされる疾患の予防又は治療方法であって、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの有効量をその必要があるヒト患者に投与するステップを含む方法。
【請求項47】
予防又は治療が髄膜炎菌血清群Aの疾患のものである、請求項45又は46記載の使用又は方法。
【請求項48】
予防又は治療が髄膜炎菌血清群Bの疾患のものである、請求項45〜47のいずれか1項に記載の使用又は方法。
【請求項49】
予防又は治療が髄膜炎菌血清群Cの疾患のものである、請求項45〜48のいずれか1項に記載の使用又は方法。
【請求項50】
予防又は治療が髄膜炎菌血清群W135の疾患のものである、請求項45〜49のいずれか1項に記載の使用又は方法。
【請求項51】
予防又は治療が髄膜炎菌血清群Yの疾患のものである、請求項45〜50のいずれか1項に記載の使用又は方法。
【請求項52】
請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの製造方法であって、L3 LOSを単離するステップ、場合によりそれを必要に応じて単離されたL2及び/又はL10及び/又はL4 LOSと組み合わせるステップ、並びに薬学的に許容される賦形剤と共にL3 LOSを製剤化するステップを含む方法。
【請求項53】
髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの使用。
【請求項54】
髄膜炎菌免疫型L3疾患の予防又は治療方法であって、その必要があるヒト患者に、請求項1〜41のいずれか1項に記載の免疫原性組成物又は請求項42〜44のいずれか1項に記載のワクチンの有効量を投与するステップを含む方法。
【請求項55】
髄膜炎菌免疫型L3疾患が、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOS、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないLOS、又はそのヘプトースII残基に結合したGlcNac残基において部分的にO−アセチル化されておりかつ部分的にO−アセチル化されていないLOS、のいずれかのLOSを有する株によって引き起こされる、請求項53又は54記載の使用又は方法。
【請求項56】
そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株により引き起こされる髄膜炎菌免疫型L3疾患の予防又は治療のための医薬の製造における、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の使用。
【請求項57】
そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されているLOSを有する株によって引き起こされる髄膜炎菌免疫型L3疾患を予防又は治療する方法であって、その必要があるヒト患者に、そのヘプトースII残基に結合したGlcNac残基においてO−アセチル化されていないL3 LOSを含む免疫原性組成物の有効量を投与するステップを含む方法。
【請求項58】
免疫原性組成物が、以下の構造を有するL3 LOS:
【化11】
〔式中、
R1=
【化12】
であり、
R2=PEAであり、
R3=Hであり、
R4=Hであり、
R5=H、PEA、又はGlyである。〕
を含む、請求項56又は57記載の使用又は方法。
【請求項59】
R1=
【化13】
である、請求項58記載の使用又は方法。
【請求項60】
R5=H、PEA、又はGlyである、請求項58又は59記載の使用又は方法。
【請求項61】
免疫原性組成物中のL3 LOSがタンパク質キャリアとコンジュゲートされている、請求項56〜60のいずれか1項に記載の使用又は方法。
【請求項62】
免疫原性組成物中のL3 LOSが、無毒化リピドA部分(例えばmsbB(−)ナイセリア株から単離されたLOSと一致する第2アシル鎖を欠損するもの)を含む、請求項56〜61のいずれか1項に記載の使用又は方法。
【請求項63】
L3 LOSが、LOSの毒性低減に適したリピドA結合ペプチド(SAEP2又はSAEPIIなど)との複合体化による無毒化リピドA部分を有する、請求項56〜62のいずれか1項に記載の使用又は方法。
【請求項64】
L3 LOSが、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として免疫原性組成物中に存在する、請求項56〜63のいずれか1項に記載の使用又は方法。
【請求項65】
ブレブ調製物が、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものである、請求項64記載の使用又は方法。
【請求項66】
ブレブ調製物が、莢膜多糖を合成することができないナイセリア株から単離されたものである、請求項64又は65記載の使用又は方法。
【請求項67】
ナイセリア株が、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)、請求項66記載の使用又は方法。
【請求項68】
ブレブ調製物が、それが由来する天然株(複数でもよい)と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ナイセリア株から単離されたものである、請求項64〜67のいずれか1項に記載の使用又は方法。
【請求項69】
通常はそのLOSのヘプトースII残基に結合したGlcNac残基においてO−アセチル化するナイセリア株のLOSを脱O−アセチル化する方法であって、oac1遺伝子の機能的発現を破壊することによって、機能的Oac1を発現することができないようにするステップを含む方法。
【請求項70】
通常はそのLOSのヘプトースII残基に結合したGlcNac残基においてO−アセチル化しない又は部分的にのみO−アセチル化するナイセリア株のLOSをO−アセチル化する又はさらにO−アセチル化する方法であって、oac1遺伝子の機能的発現を増大するステップを含む方法。
【請求項71】
oac1遺伝子の機能的発現を増大するステップが、oac1遺伝子の機能的コピー(例えば、場合により天然のプロモーター配列を含んでいてもよい図3Dのオープンリーディングフレーム)をナイセリア株に導入することによって行われる、請求項70記載の方法。
【請求項72】
oac1遺伝子の機能的発現を増大するステップが、存在する非機能的遺伝子のポリGフレーム変動領域(複数でもよい)を、oac1遺伝子の機能的発現のためにフレーム内となるようにすることによって行われる、請求項70記載の方法。
【請求項73】
ナイセリア株において機能的oac1発現のフレーム変動性を少なくする方法であって、oac1遺伝子のポリGフレーム変動領域(複数でもよい)を、より少ないGヌクレオチドを有するコドンを用いて同じアミノ酸がコードされるように、変化させるステップを含む方法。
【請求項74】
ATG開始コドンから1135〜1137位のヌクレオチドによってコードされるアルギニン残基のコドンを、AGGからCGT、CGC、CGA又はAGAに変化させる、請求項73記載の方法。
【請求項75】
ATG開始コドンから1138〜1140位のヌクレオチドによってコードされるグリシン残基のコドンを、GGGからGGT、GGA又はGGCに変化させる、請求項73又は74記載の方法。
【請求項76】
L2免疫型のナイセリア株に対して行われる、請求項69〜75のいずれか1項に記載の方法。
【請求項77】
L3免疫型のナイセリア株に対して行われる、請求項69〜75のいずれか1項に記載の方法。
【請求項78】
免疫原性組成物の製造方法であって、請求項69〜77のいずれか1項に記載の方法を実施するステップ、得られるナイセリア株からLOSを単離するステップ、及び薬学的に許容される担体又は賦形剤と共に有効量のLOSを製剤化するステップを含む方法。
【請求項79】
単離されたLOSをタンパク質キャリアにコンジュゲートするさらなるステップを含む、請求項78記載の方法。
【請求項80】
単離されたLOSのリピドA部分を無毒化する、例えばmsbB(−)ナイセリア株からLOSを単離することにより無毒化する、さらなるステップを含む、請求項78又は79記載の方法。
【請求項81】
単離されたLOSのリピドA部分が、LOSの毒性低減に適したリピドA結合ペプチド(SAEP2又はSAEPIIなど)との複合体化によって無毒化されている、請求項80記載の方法。
【請求項82】
LOSが、精製LOS調製物として、リポソーム調製物として、又はブレブ調製物として単離されている、請求項78〜81のいずれか1項に記載の方法。
【請求項83】
ブレブ調製物が、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15又は0.09〜0.11%の界面活性剤、好ましくはデオキシコーレートを用いる抽出ステップの後に各ナイセリア株から単離されたものである、請求項82記載の方法。
【請求項84】
ブレブ調製物が、莢膜多糖を合成することができないナイセリア株から単離されたものである、請求項82又は83記載の方法。
【請求項85】
ナイセリア株が、それが由来する天然株と比較して、莢膜多糖遺伝子ctrA、ctrB、ctrC、ctrD、synA、synB、synC又は好ましくはsiaDの1つの発現が下方制御されている、好ましくは欠失している(機能的発現がない)、請求項84記載の方法。
【請求項86】
ブレブ調製物が、それが由来する天然株と比較して、リピドA遺伝子msbB又はhtrBの一方又は両方、好ましくは前者、の発現が下方制御されている、好ましくは欠失している(機能的発現がない)ナイセリア株から単離されたものである、請求項82〜85のいずれか1項に記載の方法。
【図1】

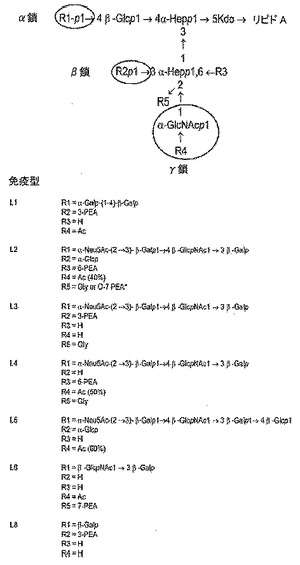
【図2A】
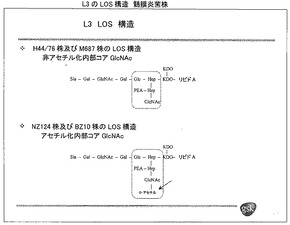
【図2B−1】
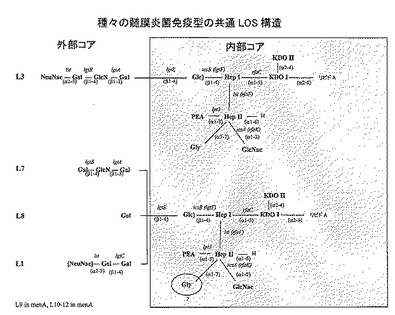
【図2B−2】
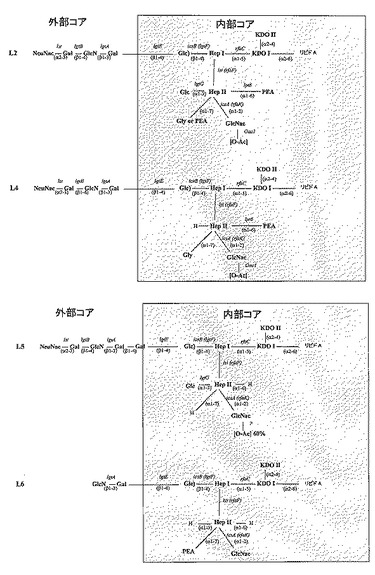
【図3A】

【図3B−1】

【図3B−2】

【図3C】

【図3D】

【図4A】
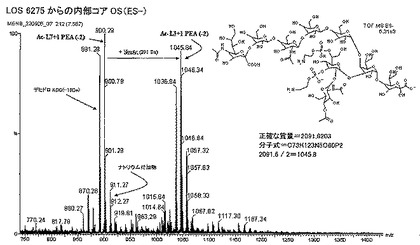
【図4B】
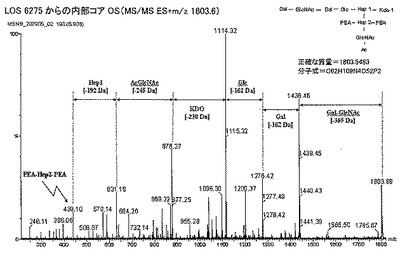
【図4C】
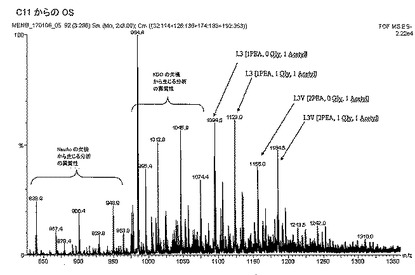
【図5】
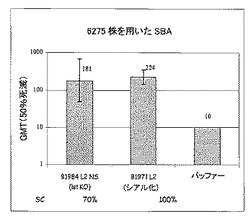
【図6】
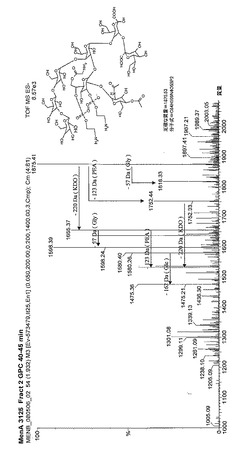
【図2A】
【図2B−1】
【図2B−2】
【図3A】
【図3B−1】
【図3B−2】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【公表番号】特表2009−539922(P2009−539922A)
【公表日】平成21年11月19日(2009.11.19)
【国際特許分類】
【出願番号】特願2009−514767(P2009−514767)
【出願日】平成19年6月8日(2007.6.8)
【国際出願番号】PCT/EP2007/055676
【国際公開番号】WO2007/144316
【国際公開日】平成19年12月21日(2007.12.21)
【出願人】(305060279)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (169)
【Fターム(参考)】
【公表日】平成21年11月19日(2009.11.19)
【国際特許分類】
【出願日】平成19年6月8日(2007.6.8)
【国際出願番号】PCT/EP2007/055676
【国際公開番号】WO2007/144316
【国際公開日】平成19年12月21日(2007.12.21)
【出願人】(305060279)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (169)
【Fターム(参考)】
[ Back to top ]