
ワクチン
【課題】ナイセリアワクチン組成物、それらの製造、およびかかる組成物の医薬における使用の方法を提供する。
【解決手段】ナイセリアの、特に髄膜炎菌の外膜小胞(ブレブ)ワクチンの産生により適した新規の操作された髄膜炎菌株を製造する方法に関する。新規のLOSサブユニットまたは髄膜炎菌の外膜小胞(ブレブ)ワクチンの使用に基づく有利な方法およびワクチン製品もまた記載しており、これはヒト被験体における使用のために安全性をより高めたものである。特に、遺伝子の下方制御の組み合わせが記載され、その組み合わせとしてはPorAおよびOpA、PorAおよびOpC、OpAおよびOpC、ならびにPorAおよびOpAおよびOpC等が挙げられる。莢膜多糖を欠損した髄膜炎菌突然変異体由来のブレブワクチンについて、LOSが重要な抗原として保持されることになるブレブ調製物を製造する有利な方法として、さらに記載される。
【解決手段】ナイセリアの、特に髄膜炎菌の外膜小胞(ブレブ)ワクチンの産生により適した新規の操作された髄膜炎菌株を製造する方法に関する。新規のLOSサブユニットまたは髄膜炎菌の外膜小胞(ブレブ)ワクチンの使用に基づく有利な方法およびワクチン製品もまた記載しており、これはヒト被験体における使用のために安全性をより高めたものである。特に、遺伝子の下方制御の組み合わせが記載され、その組み合わせとしてはPorAおよびOpA、PorAおよびOpC、OpAおよびOpC、ならびにPorAおよびOpAおよびOpC等が挙げられる。莢膜多糖を欠損した髄膜炎菌突然変異体由来のブレブワクチンについて、LOSが重要な抗原として保持されることになるブレブ調製物を製造する有利な方法として、さらに記載される。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、ナイセリアワクチン組成物、それらの製造、およびかかる組成物の医薬における使用の分野に関する。より具体的には、ナイセリアの、特に髄膜炎菌の外膜小胞(すなわち、ブレブ)ワクチンの産生により適した新規の操作された髄膜炎菌株を製造する方法に関する。新規のLOSサブユニットまたは髄膜炎菌の外膜小胞(すなわち、ブレブ)ワクチンの使用に基づく有利な方法およびワクチン製品もまた記載しており、これはヒト被験体における使用のために安全性および有効性をより高めたものである。
【背景技術】
【0002】
発明の背景
ナイセリア・メニンジティディス(Neisseria meningitidis)(髄膜炎菌)は、しばしばヒト上気道より単離されるグラム陰性細菌である。この細菌は、菌血症および髄膜炎等の重篤な侵襲性細菌性疾患の原因である。髄膜炎菌疾患の発生数は、地理的、季節的および年次的な差異を示す(Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。この菌は通常、その莢膜多糖による血清型に従って分類される。
【0003】
温暖な国々で最も多い疾患は、血清型B株によるものであり、発症率は総人口の1〜10/100,000/年の間で変動し、それより高い値に達する場合もある(Kaczmarski, E.B. (1997), Commun. Dis. Rep. Rev. 7: R55-9, 1995、Scholten, R.J.P.M., Bijlmer, H.A., Poolman, J.T.ら Clin. Infect. Dis. 16:237-246, 1993、Cruz, C., Pavez, G., Aguilar, E.,ら Epidemiol. Infect. 105:119-126, 1990)。
【0004】
血清型A髄膜炎菌が優位を占める流行の大部分は中央アフリカで発生し、その発症率が1000/100,000/年のレベルにまで達する場合もある(Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。全体として、髄膜炎菌疾患のほとんど全ての症例は、血清型A、B、C、W-135およびYの髄膜炎菌によって引き起こされ、四価のA、C、W-135、Y莢膜多糖体ワクチンが利用可能である(Armand, J., Arminjon, F., Mynard, M.C., Lafaix, C. J. Biol. Stand. 10:335-339, 1982)。
【0005】
ナイセリア・メニンジティディス感染の頻度は、過去数十年、多くのヨーロッパの国々において増加してきている。これは、社会的活動の場(例えば、水泳プール、劇場等)の増加による伝染の増加に起因してきた。いくつかの標準的な抗生物質に対して感受性が低い、または耐性のナイセリア・メニンジティディス株を単離することは、もはや珍しいことではない。この現象は、新たな抗菌剤、ワクチン、薬剤スクリーニング法、およびこれらの生物についての診断試験に対する、未だ満たされていない医学的ニーズおよび需要を生み出した。
【0006】
利用可能な多糖ワクチンは、現在、それらを担体タンパク質に化学的にコンジュゲートする方法によって改良が行われている(Lieberman, J.M., Chiu, S.S.Wong, V.K.らJAMA 275:1499-1503, 1996)。
【0007】
しかしながら、血清型Bワクチンは利用可能な状況にない。血清型B莢膜多糖は、免疫原性を示さないことが分かってきた−それは恐らくそれが宿主成分との構造類似性を有するためであろう(Wyle, F.A., Artenstein, M.S., Brandt, M.L.ら, J. Infect. Dis. 126:514-522, 1972、Finne, J.M., Leinonen, M., Makela, P.M. Lancet ii.:355-357, 1983)。従って、外膜小胞(もしくはブレブ)またはそれらから得た精製タンパク質構成成分から、血清型Bワクチンの開発を試みることに焦点を当てた努力がされてきた。
【0008】
ワクチン開発のための別の髄膜炎菌抗原は、髄膜炎菌のリポオリゴ糖(LOS)である。これらは糖脂質に結合した外膜であり、O側鎖が欠如しているという点で腸内細菌科(Enterobacteriaceae)細菌のリポ多糖(LPS)と異なり、従ってLPSのラフ型と類似している(GriffissらRev Infect Dis 1988, 10:S287-295)。LOSのオリゴ糖部分における不均一性は、異なる髄膜炎菌株間における構造的なおよび抗原性の多様性を生み出す(GriffissらInf. Immun. 1987, 55:1792-1800)。このことは、かかる株を12種類の免疫型にさらに分類するために用いられてきた(ScholtanらJ Med Microbiol 1994, 41:236-243)。免疫型L3、L7およびL9は、免疫学的に同一であり、かつ構造的に類似(もしくは全く同一)しており、従ってL3,7,9(または、本明細書において、一般的に「L3」)と呼ばれてきた。髄膜炎菌のLOS L3,7,9(L3)、L2およびL5は、シアリル化によって、またはシチジン5'-一リン酸-N-アセチルノイラミン酸の付加によって修飾することができる。L2、L4およびL6 LOSは免疫学的に区別することができるが、しかしそれらは構造的に類似しており、本明細書でL2に言及する場合にはL4またはL6のいずれかと任意に置換することができ、これは本発明の範囲内となる。LOSに対する抗体は、実験用ラットにおいて感染に対し防御し、またナイセリア・メニンジティディスに感染した小児において殺菌活性に寄与することが示されている(GriffissらJ Infect Dis 1984, 150:71-79)。
【0009】
しかしながら髄膜炎菌ワクチンにおけるLOSの使用に伴う問題は、その毒性(そのリピドA部分による毒性)にある。
【0010】
またLOSは髄膜炎菌のブレブの表面に存在する。髄膜炎菌の外膜小胞(またはブレブ)に基づいたワクチンの開発に焦点を当てて、長年にわたる努力がされてきた(de Moraes, J.C., Perkins, B., Camargo, M.C.らLancet 340:1074-1078, 1992、Bjune, G., Hoiby, E.A. Gronnesby, J.K.ら338:1093-1096, 1991)。そのようなワクチンは、宿主に投与した場合に防御免疫反応を誘発することのできる、正確に折り畳まれた立体構造をとったいくつかの完全な外膜タンパク質を含むという利点を有する。さらに、ナイセリア株(ナイセリア・メニンジティディス血清型B、すなわちmenBを含む)は、工業的規模での製造を可能にするために十分な量の外膜ブレブを排出する。しかしながら、ブレブは大抵、細菌細胞(例えばEP 11243)の0.5%界面活性剤(例えばデオキシコレート)抽出を含む方法により調製される。それは上記のようなLOSの毒性(内毒素とも呼ばれる)のために望まれるが、しかし、ワクチンからほとんどのLOS抗原を除去する効果もまた有する。
【0011】
ワクチン抗原としてのLOSの使用に伴うさらなる問題は、多様な幅を持った糖鎖構造を有する12のLPS免疫型が存在することである(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021、Mol Microbiol 2002, 43:931-43)。ある免疫型に対する抗体は、異なる免疫型を認識することができない。LOS免疫型のオリゴ糖部分の共通「コア」領域を生成することに努力が集中されてきたが(例えばWO 94/08021)、改変されたLOSに対して生じた抗体の殺菌活性は失われる。従って、ワクチンは、有効であるために異なる免疫型の多くのLOS成分を有する必要があるだろう。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】国際公開WO 94/08021
【非特許文献】
【0013】
【非特許文献1】Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989
【非特許文献2】Kaczmarski, E.B. (1997), Commun. Dis. Rep. Rev. 7: R55-9, 1995
【非特許文献3】Scholten, R.J.P.M., Bijlmer, H.A., Poolman, J.T.ら Clin. Infect. Dis. 16:237-246, 1993
【非特許文献4】Cruz, C., Pavez, G., Aguilar, E.,ら Epidemiol. Infect. 105:119-126, 1990
【非特許文献5】Armand, J., Arminjon, F., Mynard, M.C., Lafaix, C. J. Biol. Stand. 10:335-339, 1982
【非特許文献6】Lieberman, J.M., Chiu, S.S.Wong, V.K.らJAMA 275:1499-1503, 1996
【非特許文献7】Wyle, F.A., Artenstein, M.S., Brandt, M.L.ら, J. Infect. Dis. 126:514-522, 1972
【非特許文献8】Finne, J.M., Leinonen, M., Makela, P.M. Lancet ii.:355-357, 1983
【非特許文献9】GriffissらRev Infect Dis 1988, 10:S287-295
【非特許文献10】Griffissら, Inf. Immun. 1987, 55:1792-1800
【非特許文献11】Scholtanら, J Med Microbiol 1994, 41:236-243
【非特許文献12】GriffissらJ Infect Dis 1984, 150:71-79
【非特許文献13】de Moraes, J.C., Perkins, B., Camargo, M.C.ら, Lancet 340:1074-1078, 1992
【非特許文献14】Bjune, G., Hoiby, E.A. Gronnesby, J.K.ら338:1093-1096, 1991
【非特許文献15】M.P.Jenningsら, Microbiology 1999, 145, 3013-3021
【非特許文献16】Mol Microbiol 2002, 43:931-43
【発明の概要】
【発明が解決しようとする課題】
【0014】
ヒトワクチンにおける抗原としてのLOS(LPSまたはリポ多糖としても知られる)の使用に伴うさらなる問題が存在し、それはすなわちそれらがヒト糖鎖構造(例えば、ヒト赤血球細胞上の)に類似した糖鎖構造を担持しているため、それらの使用に伴う安全性の問題が突きつけられることである。さらにLOS構造の改変は、LOS抗原の殺菌効果の構造的感受性のため、問題を含んでいる。
【課題を解決するための手段】
【0015】
本発明は、1以上の上記の問題を改善するための方法を提供し、かつ防御抗原としての髄膜炎菌LOS(特に、外膜小胞上に提示される場合)に基づいた新規のワクチンの製造方法を提供する。
【図面の簡単な説明】
【0016】
【図1】図1はL3およびL2免疫型LOSの模式図を示す。
【図2】図2はL3およびL1免疫型LOSの模式図を示す。
【図3】図3は抗L3 LOS ELISA、血清殺菌アッセイ、およびLOS枯渇実験の結果を示す。
【図4】図4はMAb 1B2-1B7およびMab L3.7.9を用いたELISAの結果を示す。
【図5】図5はマウス血清におけるブレブの抗原性を示す。
【図6】図6は抗LNnT ELISAの結果を示す。
【発明を実施するための形態】
【0017】
発明の説明
本明細書中に記載される刊行物および特許、または特許出願の主題またはそこに開示される情報は、参照により本明細書に組み入れる。
【0018】
「リポオリゴ糖」(または「LOS」)に関する言及は、「リポ多糖」または「LPS」にも言及するものとしてみなされる。
【0019】
本明細書中の用語「含む」(「comprising」「comprise」「comprises」)は、全ての場合に、任意に用語「からなる」(「consisting of」「consist of」「consist of」)に各々置き換えることができるものと発明者は意図している。
【0020】
本発明者らは、LOSオリゴ糖構造の短縮が殺菌性免疫反応を誘発し得るエピトープの減少を導くことを見出した。あるいは、本発明者らは、ワクチン製剤においてLOSを最も有効に使用するためには、LOSオリゴ糖構造をできるだけ保持するべきであるが、しかしたった2種類のLOS抗原の組み合わせによって広範に有効なナイセリアの(好ましくは髄膜炎菌の)ワクチンを生成し得ることを見出した。本発明の第1の態様は、ナイセリア(好ましくは髄膜炎菌)の免疫型L2のLOSおよび免疫型L3のLOSを含む、ナイセリア(好ましくは髄膜炎菌もしくは髄膜炎菌B型)疾患の予防または治療のための免疫原性組成物である。LOSは公知の精製法のいずれかにより単離することができ、またはL2およびL3ナイセリア株由来の少なくとも2種類の外膜小胞(もしくはブレブ)調製物中に存在し得る。ブレブ調製物から、毒性を有し緩く保持されたLOSを除去するが、ブレブ内に組み込まれたLOS抗原は高レベルで保持するようにするために、ブレブを低濃度の界面活性剤、すなわち0〜0.3%、好ましくは0.05〜0.2%、最も好ましくは約0.1%の、好ましくはデオキシコレート(DOC)、を用いて抽出することが好ましい。LOS抗原のそのような組み合わせ(特にブレブワクチン中の)は、驚くべきことに90%を超えるナイセリア・メニンジティディス株に対して効果的であるという点で有利である。
【0021】
本発明者らはまた、本発明の上記ブレブ免疫原性組成物、および実際にはあらゆるナイセリア(好ましくは淋菌または髄膜炎菌)由来のブレブ免疫原性組成物が、免疫優性外膜タンパク質の特定の組み合わせの発現が下方制御(好ましくは欠失)される場合に、それらの表面上の防御抗原(LOSを含む)の効果が増強されることを見出した。従って、本発明の第2の態様は、天然の非改変株と比較して発現が下方制御された、好ましくは欠失した以下の外膜タンパク質:PorA、PorB、OpA、OpCまたはPilC、を2つ以上有するナイセリア株由来のナイセリアブレブ調製物である。好ましくは、PorAとOpA、PorAとOpC、OpAとOpC、またはPorAとOpAとOpCが下方制御されているかまたは欠失している。FrpBの発現の下方制御(好ましくは欠失)はまた、交差防御抗原の効果の増強において有利であり、特に、鉄制限状態で増殖させたナイセリア株から作製したブレブ調製物中において有利であることが示されてきた。従って、この突然変異を有する株由来のナイセリアブレブは、上記の1以上の下方制御とFrpB下方制御との組み合わせから得られるブレブとしての、本発明のさらなる実施形態である。好ましくは、PorAが下方制御される場合にはPorBは下方制御されるべきではなく、またその逆も同様である。
【0022】
上記突然変異は、ブレブ免疫原性組成物が由来することになるあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしながらL2またはL3免疫型ナイセリア(好ましくは髄膜炎菌、最も好ましくはmenB)株を用いることが好ましく、典型的には上記の低DOC%抽出法で抽出することが好ましい。好ましくは、本発明のブレブ免疫原性組成物は、L2およびL3ブレブを両方含み、かつその少なくとも一方(好ましくは両方)が上記の免疫優性外膜タンパク質(またはOMP)の組み合わせを欠損していることが好ましい。それらの遺伝子を下方制御するための技術はWO 01/09350(参照により本明細書に組み入れられる)に記載されている。4つの異なるOpa遺伝子が髄膜炎菌ゲノム中に存在することが知られており(Ahoら1991 Mol. Microbiol. 5:1429-37)、ゆえにOpaの発現が下方制御されていると言われる場合、このことは好ましくは髄膜炎菌に存在する1、2、もしくは3つの遺伝子または(好ましくは)4つ全ての遺伝子がそのように下方制御されていることを意味する。そのような下方制御は、WO 01/09350に記載のように遺伝学的に達成するか、または、容易に発見される天然の安定した髄膜炎菌株(Opa座位からの発現がないかもしくは低いもの)を探索することによって達成し得る。そのような株は、Poolmanら(1985 J. Med. Micro. 19:203-209)に記載される技術を用いて見つけ出すことができ、その際、Opa-である細胞は、プレート上または顕微鏡下において細胞の外見を観察することによって見出されうるOpa発現細胞とは異なる表現型を有する。一度見つけられれば、Opaの欠失を確立するための発酵操作の後に、細胞内容物についてウエスタンブロットを行うことによって、株が安定的にOpa-であることを示すことができる。
【0023】
上記LOS免疫原性組成物の安全性
ヒトスフィンゴ糖脂質に存在するラクト-N-ネオテトラオースオリゴ糖基(Galβ1-4GlcNAcβ1-3Galβ1-4Glcβ1-、図1)と類似した構造が存在することから、L3またはL2 LOSに対して生じた抗体の安全性が問われてきている。多くの人々が残存量のL3 LOSを含むデオキシコレート抽出小胞ワクチンで安全にワクチン接種されてきているが(G. Bjuneら, Lancet(1991), 338, 1093-1096、GVG. Sierraら, NIPH ann(1991), 14, 195-210)、LOSが本明細書に記載されるように抗原として保持される場合、LOS糖鎖構造の末端部分の欠失が、ヒト組織の表面に提示される構造との抗LOS免疫反応の交差反応の阻止において有利であることが、本発明者らによって発見された。好ましい実施形態において、lgtB遺伝子の不活性化が末端ガラクトース残基およびシアル酸が欠損した中間体LOS構造を生じる(図1および2を参照、突然変異はL2およびL3 LOSに4GlcNAcβ1-3Galβ1-4Glcβ1-構造を残存させる)。そのような中間体は、L3および/またはL2 LOS株において獲得され得る。別のおよびあまり好ましくないLOSの(短い)バージョンは、lgtE遺伝子のスイッチオフによって得られうる。さらに別のおよびあまり好ましくない型のLOSは、lgtA遺伝子のスイッチオフによって得られうる。そのようなlgtA-突然変異を選択する場合、非免疫原性L1免疫型の形成を阻止するためにlgtC発現をスイッチオフすることがまた好ましい。
【0024】
lgtB-突然変異が最も好ましいのは、これが殺菌性(およびさらに交差殺菌性の)抗体反応を依然として誘導し得るLOS防御オリゴ糖エピトープを保持したままで安全性の課題を解決するために最適な末端切断物であることを本発明者らが発見したためである。
【0025】
従って、本発明の上記のL2および/またはL3調製物(精製されたもの、または単離したブレブ状のもの)あるいは一般的な髄膜炎菌のブレブ調製物(特にL2および/またはL3)は、lgtB、lgtAまたはlgtE遺伝子由来の機能的遺伝子産物の発現が永続的に下方制御されるように、好ましくは遺伝子のスイッチオフにより、最も好ましくは当該遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により遺伝子操作したナイセリア株(好ましくは髄膜炎菌株)から、有利に得られる。
【0026】
好ましくは、本発明のナイセリア株は、莢膜多糖の合成に欠損を有する。
【0027】
本発明の上記ブレブ調製物を髄膜炎菌B株から得る場合、莢膜多糖(ヒト様糖鎖構造も含む)もまた除去されることが特に好ましい。これを達成するために多くの遺伝子がスイッチオフされ得るが、本発明者らは、siaD遺伝子由来の機能的遺伝子産物の発現を永続的に下方制御(例えばα-2-8ポリシアル酸転移酵素の活性を下方制御)するように、ブレブ産生株を、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により、遺伝的に操作することが好ましいということを有利に示してきた。そのような不活性化はWO 01/09350に記載されている。siaD(synDとしても知られる)突然変異が、莢膜多糖からヒト類似エピトープを除去し得る多くの突然変異の中で最も有利であるのは、それが、LOSの防御エピトープの生合成に影響を及ぼさない最良の突然変異の1つであり、そのため、このようにLOSを防御抗原として最終的に用いることを目的とし、かつ細菌の増殖に対して最小限の影響を与える方法において有利であるためである。従って、本発明の好ましい態様は、lgtE- siaD-、lgtA- siaD-または好ましくはlgtB- siaD-髄膜炎菌B突然変異株由来の上記ブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0028】
siaD-突然変異は上記の理由のために好ましいが、髄膜炎菌B(または一般的に髄膜炎菌)の莢膜多糖合成をスイッチオフする他の突然変異が用いられ得る。従って、ブレブ産生株を、以下の遺伝子の1つ以上に由来する機能的遺伝子産物の発現を永続的に下方制御するように、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により、遺伝的に操作してもよい:ctrA、ctrB、ctrC、ctrD、synA(synXおよびsiaAと同等)、synB(siaBと同等)またはsynC(siaCと同等)遺伝子。lgtE-突然変異はこれらの突然変異の1以上と組み合わせてもよい。好ましくは、lgtB-突然変異はこれらの突然変異のうち1つ以上と併用される。本発明のさらなる態様は、従って、髄膜炎菌B(または一般的に髄膜炎菌)の突然変異株のそのような組み合わせから得られる上記ブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0029】
lgtBおよびlgtEを包含する、様々なlgt遺伝子を含むナイセリア遺伝子座、ならびにその塩基配列は、当技術分野において公知である(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021および以下に引用する参考文献、J. Exp. Med. 180:2181-2190(1994)、WO 96/10086を参照)。
【0030】
最終産物として完全長(非末端切断型)LOSを使用する場合、LOSがシアリル化されていないことが望ましく、そのようなLOSは、最も危険な侵襲性髄膜炎菌B株(これも同様にシアリル化されていない)に対する免疫反応を生じる。そのような場合、synA(synXおよびsiaAと同等)、synB(siaBと同等)またはsynC(siaCと同等)遺伝子が欠失した莢膜陰性株の使用が有利であり、それはまたそのような突然変異によってmenB LOSがシアリル化され得ないようにするためである。
【0031】
上記突然変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしL2またはL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、典型的には本明細書に記載した低DOC%抽出法によって抽出されたものを使用するのが好ましい。好ましくは、本発明のブレブ免疫原性組成物が、L2およびL3ブレブの両方を含み、かつ、そのうちの少なくとも一方(および好ましくは双方)が上記遺伝子の発現が欠損した株に由来する。
【0032】
LOSの毒性
本発明の上記の精製LOSまたはブレブ免疫原性組成物はまた、それらが由来する細菌性産生株における特定の遺伝子の発現の下方制御によって、毒性をより少なくすることができる。そのような無毒化は、天然のOMVによる鼻腔内免疫化には不必要であるだろうが(J.J Drabickら, Vaccine(2000), 18, 160-172)、非経口ワクチン接種における無毒化には有用であろう。好ましくは、本発明の精製LOSまたはブレブ免疫原性組成物であるLOSは、リピドA生合成に関与する遺伝子(特にリピドAへの第2アシル鎖の付加に関与する遺伝子)の突然変異/修飾/不活性化により(特にmsbBおよび/またはhtrB遺伝子由来の機能的遺伝子産物の発現の下方制御により、および好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により、ナイセリア産生株を遺伝的に操作することによって無毒化される。あるいはまた(またはさらに)、精製LOSまたはブレブ免疫原性組成物は、遺伝的に改変して以下の遺伝子:pmrA、pmrB、pmrEおよびpmrFのうちの1つ以上の遺伝子が(より強力なプロモーターの導入、もしくは遺伝子の余分なコピーの組み込みによって)上方制御されるようにしたナイセリア株から得ることができる。あるいはまた(またはさらに)、精製LOSまたはブレブ免疫原性組成物は、ポリミキシンBの機能的等価物である非毒性ペプチド[リピドAに対して高い親和性の有する分子]を組成物に添加することにより無毒化され得る。
【0033】
上記無毒化法ならびに当該プロモーター/遺伝子配列、ならびに上方制御方法および下方制御方法については、より詳細についてWO 01/09350を参照せよ。またナイセリアmsbBおよびhtrB遺伝子は各々lpxL1およびlpxL2とも呼ばれ(WO 00/26384を参照)、これらの遺伝子の欠失突然変異は、msbB-突然変異LOSが野生型と比較して1つの第2アシル鎖を欠損する(ならびに4つの第1および1つの第2アシル鎖を保持する)こと、およびhtrB-突然変異LOSが第2アシル鎖の両方を欠損することによって、表現型として特徴付けられる。そのような突然変異は好ましくは、ブレブ上への無毒化されたLOSの最適な提示を確実にするため、または無毒化サブユニットLOSの精製を助けるために、ナイセリア産生株が莢膜多糖を欠損していること(上記参照)を確実にする突然変異と組み合わせて用いられる。本発明の組成物中に使用され得るポリミキシンBの機能的等価物である非毒性ペプチドのさらなる詳細、特にペプチドSAEP 2(2つのシステインがジスルフィド架橋を形成するKTKCKFLKKC配列を有する)の使用については、WO 93/14115、WO 95/03327、Velucchiら(1997)J Endotoxin Res 4:1-12およびEP 976402を参照せよ。
【0034】
本明細書において「機能的遺伝子産物の発現の下方制御」は、問題の遺伝子のプロモーターまたはオープンリーディングフレームに対して付加、欠失または置換が為され、それにより全遺伝子産物の生合成活性が低減(60、70、80、90、95もしくは最も好ましくは100%低減)されることを意味する。明らかにフレームシフト突然変異が導入され得、またはより弱いプロモーターが置換され得るが、しかし、最も好ましくは、オープンリーディングフレームおよび/またはプロモーターのほとんどまたは全てを、(活性のある)遺伝子産物の永続的な下方制御を確実にするために欠失させる(WO 01/09350に記載)。
【0035】
上記変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有用であるが、L2またはL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株を用いることが好ましく、典型的には本明細書に記載の低DOC%抽出法で抽出されることが好ましい。本発明のブレブ免疫原性組成物は、L2およびL3ブレブの両方を含み、その少なくとも一方(および好ましくは双方)が上記遺伝子の発現を欠損している株に由来することが好ましい。
【0036】
本発明のさらなる態様は、本発明のLOSまたはブレブ免疫原性調製物が由来する上記の遺伝的に改変されたナイセリア(好ましくは髄膜炎菌または淋菌または髄膜炎菌B)株を含む。
【0037】
本発明のLOSまたはLOS含有ブレブ調製物
本発明のさらなる態様は、本発明のナイセリア株から単離されるLOS調製物(特に上述した任意のもの)である。好ましくは、単離されたLOS(またはLOS含有ブレブ)は、L2またはL3免疫型であり、好ましくは本発明の免疫原性組成物は本発明のL2およびL3 LOS(またはブレブ)調製物を両方含む。
【0038】
そのような調製物はまた、上記LOS(精製されているものであれブレブ調製物に存在しているものであれ)のオリゴ糖部分をT細胞エピトープの供給源を含む担体にコンジュゲートする(従ってLOSをよりよい(T依存的)免疫原とする)ことにより改良され得る。あるいはまた(またはさらに)本発明の精製LOSは、当技術分野で公知のリポソーム製剤中にそれを提示することにより、よりよい抗原となり得る(例えばWO 96/40063およびそこに引用される参考文献を参照)。
【0039】
細菌からLOSを単離する方法は当技術分野で周知である(例えばWesphalおよびJannの温水-フェノール法(Meth. Carbo. Chem. 1965, 5:83-91)を参照)。またGalanosら1969, Eur J Biochem 9:245-249およびWuら1987, Anal Bio Chem 160:281-289も参照せよ。単離したLOSをコンジュゲートするための技術もまた公知である(例えばEP 941738(参照により本明細書に組み入れる)を参照)。
【0040】
本発明の目的のため、「T細胞エピトープの供給源を含む担体」とは、通常、ペプチド、または好ましくはポリペプチドもしくはタンパク質である。コンジュゲーション技術は当技術分野で周知である。典型的な担体は非定型インフルエンザ菌(Haemophilus influenzae)由来のプロテインD、破傷風毒素、ジフテリア毒素、CRM197または(特にナイセリアまたは髄膜炎菌の)ブレブ調製物に存在する外膜タンパク質を含む。
【0041】
本発明の好ましい単離されたLOS組成物としては、各LOSのオリゴ糖部分がT細胞エピトープの供給源を含む担体に任意にコンジュゲートしたL2およびL3単離LOSを含有する組成物、lgtB-髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープの供給源を含む担体に任意にコンジュゲートしているL2またはL3 LOSを含む組成物、ならびに最も好ましくは、lgtB-髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープの供給源を含む担体に任意にコンジュゲートしているL2およびL3単離LOSを含有する組成物である。
【0042】
好ましくは、本発明のLOS組成物は無毒化されている。これは、分子からアシル鎖を除去する(しかし分子の防御効果は減少し得る)ヒドラジンもしくはアルカリ加水分解化学処理の公知の技術によって行われ得るが、しかし、好ましくはhtrB-もしくはmsbB-髄膜炎菌突然変異体(上記のような、特に莢膜多糖欠損株)からLOSを単離することにより、またはポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高い親和性を有する分子)、特にSAEP 2(上記)の、組成物への添加により行われ得る。
【0043】
本発明のLOSは単離された状態で(通常、リピドA部分が無傷のままであればミセル形態で)投与してもよく、またはリポソーム中に入れて投与してもよい。そのような場合、外膜タンパク質はリポソームに添加してもよく、またLOSを、オリゴ糖がT依存的抗原となるようにリポソーム内部でそのような外膜タンパク質にコンジュゲートしてもよい。これは、下記のブレブ内LOS架橋について記載されるものと類似した化学法によって行われ得る。
【0044】
ブレブの表面上に存在する外膜タンパク質へのLOSのオリゴ糖部分のブレブ内架橋(コンジュゲーション)
LOS(特に本発明のLOS)がブレブ製剤中に存在する場合、LOSは好ましくは、ブレブ調製物上にも存在する1以上の外膜タンパク質(例えば髄膜炎菌におけるPorAまたはPorB)へのLOSのコンジュゲーションを可能にする方法により、in situにおいてコンジュゲートされる。従って、本発明のさらなる態様は、その外膜中に、LOSにコンジュゲートした外膜タンパク質が組み込まれるグラム陰性細菌株由来のブレブ調製物である。LOSはコンジュゲートするためにブレブ調製物に添加してもよいが、LOSはブレブ調製物の表面に天然に存在することが好ましい。
【0045】
この方法は、ブレブ製剤中のLOS抗原の安定性および/または免疫原性(T細胞の助けを提供する)、および/または抗原性を、外膜の表面上の天然環境にあるLOSのように有利に増強し、すなわちその最も防御的な構造をとったT非依存的オリゴ糖免疫原に対し、T細胞の助けを与える。さらに、ブレブ内でのLOSのコンジュゲーションは、LOSの無毒化を生じ得る(理論に縛られることは望まないが、コンジュゲートされると、リピドA部分はより安定的に外膜中に埋没され、従って毒性を生じるように利用されにくいものと思われる)。従って、上記のhtrB-もしくはmsbB-変異体からのブレブの単離についての無毒化法、またはポリミキシンBの機能的等価物である非毒性ペプチドの組成物への添加による上記の無毒化法は、必要でないかも知れない(しかしさらなる安全のために組み合わせて添加され得る)。
【0046】
本発明のコンジュゲート化ブレブ調製物は、典型的には、ブレブ中のLOSの毒性が同量の全くコンジュゲートしていないLOSを有する同じブレブと比較して低減されている。LOS毒性は当業者によって容易に測定され得、それには例えばヨーロッパ薬局方におけるLOSウサギ発熱アッセイ(pyrogenicity assay)を用いることができる(実施例7参照)。
【0047】
本発明のコンジュゲート化ブレブ調製物は、コンジュゲートしたLOSが宿主内における免疫反応の誘発に適した立体構造を有し、それに由来する血清がコンジュゲートしていないLOS(好ましくはブレブ調製物が作製された由来の細菌上に存在し、最も好ましくはSBAアッセイにおける殺菌様式で存在する)との反応性を有する(結合可能である)ため、有利である。
【0048】
ナイセリアブレブがLOSにコンジュゲートする場合、および本明細書に記載するように、ブレブが1以上の免疫優性外膜タンパク質が下方制御された株由来である場合、好ましくは、PorAが下方制御されている場合にはPorBは下方制御されるべきではなく、かつその逆も同様である。このことによって、LOSの大部分を主要な外膜タンパク質と架橋することが可能になり、従ってブレブに存在する交差防御性の微量外膜抗原に対するコンジュゲートのあらゆる効果が最小化される。
【0049】
特に、本発明者らは、ブレブに存在するLOSがブレブ内様式で外膜タンパク質(これもまたブレブに存在する)にコンジュゲートしたブレブを含む組成物が、ブレブが由来する生物によって引き起こされる疾患の治療または予防のためのワクチンの基礎を形成しうることを見出した。ここでそのようなワクチンは毒性が低減しており(好ましくは実質的に無毒性)、および/またはその天然環境におけるLOSに対するT依存的殺菌反応を誘導することが可能である。
【0050】
従って、本発明はそのようなブレブ内LOSコンジュゲート化ブレブ調製物をさらに提供する。「ブレブ内」とは、ブレブ中に天然に存在するLOSが同じブレブ上に存在する外膜タンパク質とコンジュゲートしていることを意味する。好ましくは、ブレブはブレブを産生し得るあらゆるグラム陰性生物(WO 01/09350参照)から得られるものであり、かかるグラム陰性生物は好ましくはモラクセラ・カタラーリス(Moraxella catarrhalis)、非定型インフルエンザ菌またはナイセリア(最も好ましくは髄膜炎菌)である。
【0051】
そのようなブレブ調製物は、問題の細菌より単離することができ(WO 01/09350参照)、次にLOSのオリゴ糖部分の基(例えばNH2またはCOOH)をブレブ外膜タンパク質上の基(例えばNH2またはCOOH)に結合するための、既知のコンジュゲーション化学法に供すればよい。グルタルアルデヒド、ホルムアルデヒドまたはグルタルアルデヒド/ホルムアルデヒド混合物を使用した架橋技術を用いることができるが、EDACまたはEDAC/NHS等のより選択的な化学法を用いるのが好ましい(J.V. Staros,R.W. WrightおよびD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)ならびにBioconjugates Techniques. Greg T Hermanson(1996)pp173-176)。本発明に使用できるLOSおよびタンパク質分子間の共有結合を生じることの可能な他のコンジュゲーション化学法または処理は、EP 941738に記載される。
【0052】
好ましくは、ブレブ調製物は莢膜多糖の非存在下でコンジュゲートされる。ブレブは莢膜多糖を産生しない株(天然のもしくは突然変異によるもの)から単離することができ、または大半の混入莢膜多糖(60、70、80、90もしくは99%以上が除去された)および好ましくは全ての混入莢膜多糖から精製することができる。この方法において、ブレブ内LOSコンジュゲーション反応はさらに非常に効果的である。
【0053】
好ましくはブレブ内に存在するLOSの5、10、20、30、40、50、60、70、80、90または95%を超える割合が架橋/コンジュゲートされる。
【0054】
好ましくは、本発明のブレブは、精製LOSを標準物質として使用したSDS-PAGE電気泳動後の銀染色によって測定したブレブのLOS含量が、3〜30、5〜25、10〜25、15〜22、および最も好ましくはおよそまたは正確に20%LOS含量となるように調製された(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)。髄膜炎菌ブレブにおいて20%LOSを0.1%低DOC抽出で達成することができ、これは緩く保持されたLOS分子を除去することができるが、抗原の大部分は保存する。
【0055】
ブレブ内コンジュゲート化ブレブが髄膜炎菌由来である場合、由来する株が莢膜多糖を産生することができない突然変異株(例えば上記の突然変異株の1つ、特にsiaD-)であることが好ましい。髄膜炎菌疾患に対して有効な免疫原性組成物がL2およびL3ブレブの双方を含むこともまた好ましく、ここでL2およびL3 LOSは双方ともにブレブ外膜タンパク質にコンジュゲートしている。さらに、ブレブ内コンジュゲート化ブレブ内のLOS構造がlgtB-髄膜炎菌株由来のものと一致していることが好ましい。最も好ましい免疫原性組成物は以下のブレブ内-コンジュゲート化ブレブを含む:莢膜多糖を産生することができず、lgtB-である、L2もしくはL3突然変異髄膜炎菌株に由来するもの、莢膜多糖を産生することができない突然変異髄膜炎菌株由来のL2およびL3ブレブを含むもの、lgtB-である突然変異髄膜炎菌株由来のL2およびL3ブレブを含むもの、または最も好ましくは莢膜多糖を産生することができず、lgtB-である突然変異髄膜炎菌株由来のL2およびL3ブレブを含むもの。
【0056】
本発明に使用され得る典型的なL3髄膜炎菌株は、H44/76 menB株である。典型的なL2株は、B16B6 menB株または39E髄膜炎菌C型株または760676株である。
【0057】
上述のように、本発明のブレブはコンジュゲーションの作用によってある程度無毒化されるので、さらなる無毒化を必要としないが、しかし、さらなる無毒化法をさらなる安全性のために用いてもよい。それは例えばhtrB-もしくはmsbB-である髄膜炎菌株由来のブレブを使用することによるか、またはポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高い親和性を有する分子)(好ましくはSEAP 2)をブレブ組成物に(上記のように)添加することによる。LOSの(特にブレブ内様式における)コンジュゲーションは、このように、驚いたことに同量のコンジュゲートしていないLOSを含む調製物と比較して、LOSの低い毒性を示す。従って、(特に髄膜炎菌の)ブレブの無毒化のための一般的な方法は、さらにLOSのブレブ外膜タンパク質へのブレブ内コンジュゲーションによって提供され、かつLOSの無毒化のための方法もまた、ブレブ外膜タンパク質へLOSをコンジュゲートする手段によって提供される。
【0058】
上記の方法において、髄膜炎菌ブレブ、およびブレブを含む免疫原性組成物が提供され、これらは、毒性が低減しており(かつ好ましくは実質的に無毒性)、自己免疫の問題がなく、T依存的特徴を持ち、その天然環境に存在し、かつ潜在的に90%を超える髄膜炎菌株に対する殺菌性抗体反応(L2+L3組成物の場合)を誘導することのできる、重要な抗原としてのLOSを有する。
【0059】
1以上のMen A、C、YもしくはW莢膜多糖またはオリゴ糖(好ましくは少なくともMen C、またはMen AおよびMen C、またはMen CおよびMen Y)もまた、本発明のブレブの外膜タンパク質上に、同様にコンジュゲートされ得る。これはLOS架橋と同じ反応において行われ得るが、別々の(好ましくは後の)反応において行われることが好ましい。
【0060】
最適なブレブ内LOSコンジュゲーションの方法は、本発明のさらなる態様である。かかる方法は、(好ましくは、本明細書に記載されるように低パーセンテージのDOCを使用して)グラム陰性細菌からブレブを単離するステップ、ブレブに存在するLOSを(好ましくはそのオリゴ糖部分を介して)同じブレブに存在する外膜タンパク質にコンジュゲートするのに適した化学法を行うステップ、ブレブ内コンジュゲート化ブレブ調製物を単離するステップ、ならびに同じ方法によって作製されるが異なるLOS免疫型を有する、さらなるブレブ内コンジュゲート化ブレブ調製物を有するブレブ内コンジュゲートブレブ調製物を任意に製剤化するステップ(好ましくはL2およびL3ナイセリア/髄膜炎菌ブレブを混合する)、および/またはワクチン組成物を製造するために製薬上許容される賦形剤とともにブレブ調製物を製剤化するステップを組み込むべきである。
【0061】
ブレブ内コンジュゲーションは、好ましくは以下の方法のステップのうち1もしくは2工程または3工程全てを組み込むべきである:コンジュゲーションpHはpH 7.0を上回り、好ましくはpH 7.5以上(最も好ましくはpH 9以下)であるべきである、1〜5%、好ましくは2〜4%、最も好ましくは約3%ショ糖の条件が反応中維持されるべきである、コンジュゲーション反応におけるNaClは最小限にするべきであり、好ましくは0.1M、0.05M、0.01M、0.005M、0.001M以下であり、全く存在しないことが最も好ましい。これら全ての方法的特徴は、コンジュゲーション過程を通してブレブを安定にかつ溶液中に維持することを確実にするものである。
【0062】
EDAC/NHSコンジュゲーション法はブレブ内コンジュゲーションのために好ましい方法である。EDAC/NHSは、非常に高度な架橋が可能であるがゆえに濾過性に不利な影響を及ぼすホルムアルデヒドより好ましい。EDACはカルボン酸(LOS中のKDO等)と反応して活性エステル中間体を生成する。アミン求核試薬(PorB等の外膜タンパク質中のリジン等)存在下において、アミド結合はイソ尿素副生成物の放出を伴って形成される。しかしながら、EDAC仲介反応の効果は、スルホNHSエステル中間体の形成により増加し得る。スルホNHSエステルは、EDACのみとカルボキシレートとの反応より形成される活性エステルより長く水性溶液中に残存する。従って、この2段階の方法を用いて高収率のアミド結合形成を実現し得る。EDAC/NHSコンジュゲーションは、J.V. Staros,R.W. WrightおよびD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)ならびにBioconjugates Techniques. Greg T Hermanson(1996)pp173-176に記載される。好ましくは、0.01〜5 mg EDAC/mgブレブ、より好ましくは0.05〜1 mg EDAC/mgブレブを反応に使用する。使用するEDACの量はサンプル中に存在するLOSの量に依存し、このLOSの量はまたブレブを抽出するために使用するデオキシコレート(DOC)%に依存する。低パーセンテージ(%)DOC(例えば0.1%)においては多量のEDAC(1mg/mgおよびそれ以上)が使用されるが、しかし、高%DOC(例えば0.5%)においては少量のEDAC(0.025〜0.1mg/mg)が、過剰なブレブ内架橋を回避するために使用される。
【0063】
本発明の好ましい方法は、従って、EDAC/NHS存在下において、pHがpH7.0からpH9.0の間(好ましくは約pH7.5)で、1〜5%(好ましくは約3%)ショ糖中で、かつ任意に(上記のように)実質的にNaClを含まない条件において、ブレブをコンジュゲートし、反応混合物よりコンジュゲート化ブレブを単離するステップを含む、ブレブ内コンジュゲート化LOS(好ましくは髄膜炎菌の)を製造する方法である。
【0064】
この反応に続いて、反応時間の経過に伴いブレブ中のLOSの割合の増加に応じてLOS分子量が増加することを示すために、抗LOS(例えば抗L2または抗L3)モノクローナル抗体(mAb)を使用して反応混合物をウェスタン分離ゲルに供することができる。
【0065】
収量99%のブレブがそのような技術を用いて回収され得る。
【0066】
EDACは、LOSのT依存的免疫原性を改良するために十分な、LOSをOMPに架橋する点で優れたブレブ内架橋剤であることが見出されたが、しかしそれほど高度にそれを架橋しなかったのは、乏しい濾過性、凝集およびブレブ内架橋等の問題が発生したためであった。生じたブレブの形態は、コンジュゲートしていないブレブのものと類似している(電子顕微鏡による)。さらに上記のプロトコルは、過度に高度の架橋(ブレブの表面上に天然に存在する防御OMP(例えばTbpAまたはHsf)の免疫原性を低減し得る)が生じることを回避した。
【0067】
ブレブ単離のための技術
本発明の外膜小胞(OMVまたはブレブ)は、多くの公知の技術により単離され得る(Fredriksenら, NIPH Annals(1991), 14, 67-79、Zollingerら, J. Clin Invest(1979), 63, 836-848、Saundersら, Infect Immun(1999), 67, 113-119、J.J. Drabickら, Vaccine(1999), 18, 160-172)。これらは2つの主要な群に分類される。すなわち、デオキシコレート(約0.5%)を使用して髄膜炎菌由来のブレブを抽出するために技術と、低レベルのデオキシコレート(DOC)を使用するかまたは全くデオキシコレートを使用しない技術である。DOC非含有法では、ブレブはOMV中に高レベルのLOSを維持するという興味深い特徴を有する。このことはLOSが防御抗原であるワクチンにおいて有利である。DOC抽出ブレブと比較して、DOC非含有法により得られるOMV中のL3抗原(Ag)の濃度は約10倍高い。界面活性剤を用いない(好ましくはDOCを用いない)ブレブ調製法は、この理由により本発明の方法の目的にとって好ましいが、低レベルの界面活性剤(好ましくはDOC)を含むバッファーでの抽出もまた、ブレブ内で強固に相互作用するLOSの大半を残留する一方、より毒性があり緩く保持されたLOSを除去するステップであるという点で有利であり得る。典型的には、0〜0.5%および0.02〜0.4%、0.04〜3%または0.06〜2%界面活性剤(好ましくはDOC)がブレブ抽出に使用され、より好ましくは0.08〜0.15%、および最も好ましくはおよそまたは正確に0.1%を用いて、最適な量のLOSをブレブ中に安定に存在させて得る。LOSが1以上の上記の方法により無毒化されている場合には、DOC非含有(または低DOC〜0.3%DOCもしくはそれ以下の)抽出法が特に好ましい。
【0068】
本発明の全ての実施形態におけるブレブのLOS含量は、精製LOSを標準物質として使用したSDS-PAGE電気泳動後の銀染色によって測定した場合に(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)、3〜30、5〜25、10〜25、15〜22%であることが好ましく、かつ最も好ましくはおよそまたは正確に20%LOS含量である。Nmen L3 LOSをこの方法における標準物質として使用すると、一般的に、0.1%DOCで抽出されたNmen L3免疫型ブレブ中のLOS含量は約20%LOSであり、0.2%DOCで抽出した場合は約15%LOS、0.3%DOCでは約10%LOS、および0.5%DOCでは約5%LOSである。
【0069】
ワクチン組成物
本発明の免疫原性組成物は、製薬上許容される賦形剤の添加によってワクチン組成物として容易に製剤化され得る。
【0070】
さらに、上記のような本発明の精製LOS(好ましくはL2もしくはL3)を単離するステップまたは上記のような本発明の単離ブレブ(好ましくはL2もしくはL3免疫型を有する)を生成するステップ、およびそのLOSまたはブレブを製薬上許容される賦形剤とともに製剤化するステップを含む、本発明のナイセリア(好ましくは髄膜炎菌)免疫原性組成物またはワクチンの製造方法が提供される。好ましくは、本発明の両方の免疫型L2およびL3の精製LOS、または本発明の両方の免疫型L2およびL3のブレブ、またはL2の精製LOSおよびL3のブレブ(もしくはその逆)を、混合ステップで混合する。好ましくは、本発明の精製LOSまたはブレブは、単離された後に上記のようにコンジュゲートされている。さらなるリポソーム製剤化ステップもまた精製LOSに追加され得る(当技術分野で公知の技術を用いて。例えばWO 96/40063およびそこに引用される参考文献を参照せよ)。好ましくは、ブレブ調製物は低濃度のDOC(またはDOCなし)での抽出によって(上記のように)単離される。
【0071】
このようなL2およびL3の組み合わせ方法は、ほとんど全ての髄膜炎菌B株に対して有効なワクチンをもたらすことができる。
【0072】
上記の免疫原性組成物(または方法)は、血清型A、C、YまたはW由来の1以上(2、3もしくは4)の髄膜炎菌多糖またはオリゴ糖(上記のような、そのままのもしくはT細胞エピトープを含む担体にコンジュゲートしたもの)が組成物に添加されたものであり得る。好ましくは、少なくともCが添加(最も好ましくはコンジュゲート)されたものであり、より好ましくはAおよびCまたはYおよびCが添加(好ましくは全てコンジュゲート化)されたものであり、最も好ましくはA、C、YおよびWが添加(好ましくは全てコンジュゲート化)されたものである。有利には、コンジュゲート化されたインフルエンザ菌B型莢膜多糖またはオリゴ糖はまた、上記の組成物中に含有させて、広範な髄膜炎ワクチンを作製する。
【0073】
好ましくは、特にWO 94/08021において個別に具体的に示した組成物からなるまたはそれを含む組成物は、本発明において特許請求の範囲に含められていない。
【0074】
本発明のワクチン製剤
本発明の免疫原性組成物は、適切なアジュバントとともに製剤化することで本発明のワクチン組成物を作製することができる。
【0075】
適切なアジュバントは、水酸化アルミニウムゲル(ミョウバン)またはリン酸アルミニウム(好ましくは水酸化アルミニウム)等のアルミニウム塩を含むが、しかしカルシウム塩(特に炭酸カルシウム)、鉄または亜鉛でもよく、あるいはアシル化チロシン、アシル化糖、カチオンもしくはアニオン誘導体化多糖、またはポリホスファゼンの不溶性懸濁液でもよい。
【0076】
添加され得る適切なTh 1アジュバント系には、モノホスホリルリピドA、特に3-de-O-アシル化モノホスホリルリピドA(または他のLPSの非毒性誘導体)、およびアルミニウム塩(好ましくはリン酸アルミニウム)とモノホスホリルリピドA、好ましくは3-de-O-アシル化モノホスホリルリピドA(3D-MPL)(または非毒性LPS誘導体)との組み合わせを包含する。増強系は、モノホスホリルリピドAとサポニン誘導体の組み合わせ、特にWO 94/00153に記載されるようなQS21(もしくは他のサポニン)と3D-MPL(もしくは非毒性LPS誘導体)の組み合わせ、またはWO96/33739に記載されるような、QS21(もしくはサポニン)がコレステロールによりクエンチングされているより反応原性の低い組成物を用いる。水中油型乳濁液中のQS21、3D-MPLおよびトコフェロールを含む特に強力なアジュバント製剤は、WO 95/17210に記載されており、これは添加され得る好ましい製剤である。添加され得る他のアジュバントは、サポニン、より好ましくはQS21および/または水中油型乳濁液ならびにトコフェロールを含む。オリゴヌクレオチドを含む非メチル化CpG(WO 96/02555)もまた添加され得る。
【0077】
ワクチン調製物は一般的にVaccine Design(「The subunit and sdjuvant approach」(Powell M.F.およびNewman M.J.編)(1995)Plenum Press New York)に記載されている。
【0078】
免疫防御用量のワクチンが、全身経路または粘膜経路を介して投与され得る。これらの投与には、筋肉内、腹腔内、皮内もしくは皮下経路を通じた注入、または口腔/消化器(好ましくは鼻腔内投与)、呼吸器、尿生殖路への粘膜投与を介した注入が挙げられる。典型的には、各ワクチン用量中のブレブの量は、典型的な被ワクチン接種者において著しく有害な副作用を引き起こすことなく免疫防御反応を誘導する量が選択される。そのような量は、どの特定の免疫原が使用され、かつそれをどのように提示するかによって変動する。一般的に、各用量は各ブレブを1〜100μg、好ましくは5〜50μgおよび最も典型的には5〜25μgの範囲内で含むことが期待される。
【0079】
本発明のブレブ免疫原性組成物に対するさらなる改良
本発明の上記のブレブ組成物は、それらが由来するナイセリア株(淋菌および好ましくは髄膜炎菌、最も好ましくは髄膜炎菌Bを含む)が、ゲノム中への遺伝子のさらなるコピーを挿入することにより、または既存の遺伝子の上流により強力なプロモーターを導入することにより、または非改変株と比較して1.2、1.5、2、3、5もしくは10倍を超える抗原レベルを生じるよう改変された株を誘導することが可能なWO 01/09350に記載のあらゆる他の方法により上方制御された1以上の下記遺伝子(防御抗原をコードする)を有する場合、本発明のワクチンにおける有効性がさらに改良され得る:NspA(WO 96/29412)、Hsfまたはその末端切断物(WO 99/31132およびWO 01/55182、NhhAとしても知られる)、Hap(PCT/EP99/02766)、OMP85(WO 00/23595)、PilQ(PCT/EP99/03603)、PldA(PCT/EP99/06718)、FrpB(WO 96/31618)、TbpA(WO 92/03467、US5912336、WO 93/06861およびEP586266)、TbpB(WO 93/06861およびEP586266)、NadA(Comanducciら J. Exp. Med. 2002 195:1445-1454、NMB 1994)、FrpA/FrpC、または5以上の反復配列を含むこれらの抗原間に共通の部分(WO 92/01460; Thompsonら,(1993) J. Bacteriol. 175:811-818; Thompsonら,(1993) Infect. Immun. 61:2906-2911)、LbpA、LbpB(PCT/EP98/05117)、FhaB(WO 98/02547 配列番号38(ヌクレオチド3083-9025))、HasR(PCT/EP99/05989)、lipo02(PCT/EP99/08315)、Tbp2(WO 99/57280; NMB 0460)、MltA(WO 99/57280; NMB 0033)、TspA(WO 00/03003)、TspB(WO 00/03003)、ctrA(PCT/EP00/00135)、MafA(NMB 0652)、MafB(NMB 0643)、Omp26(NMB 0181)、アドヘシンX(NMB 0315)、アドヘシンY(NMB 0995)、アドヘシンZ(NMB 1119)およびOstA(NMB 0280)。NMB配列の例は、www.neisseria.orgのデータベースで見出すことができる。本明細書でHsfについて言及する場合、この用語はあらゆる場合においてHsf末端切断物と置換可能である(特にWO 01/55182に開示されたものにおいて)。
【0080】
HsfおよびTbpA(低もしくは高分子量形態、または低および高分子量形態の両方(EP 586266))、またはHsfおよびOMP85、またはOMP85およびTbpA(低もしくは高分子量形態、または低および高分子量形態の両方)、またはNspAおよびHsf、またはNspAおよびOMP85、またはNspAおよびTbpA(低もしくは高分子量形態、または低および高分子量形態の両方)の双方が、いずれも上方制御されている場合が特に好ましい。組成物中に2つのブレブが含まれている場合、各ブレブは異なる上方制御を受けていることが好ましい。高および低分子量のTbpAがどちらも上方制御される場合、これらは2つの形態のTbpAを天然において含む2つの株由来の組成物中に存在する、2つの別々のブレブにおいて上方制御されていることが好ましい。最も好ましくは、かかる2つの株がL2およびL3 LOS免疫型を有することである。TbpAは遺伝子工学的に上方制御させるか、またはナイセリア/髄膜炎菌産生株を鉄制限状態(例えば50〜70μMデスフェラル(メシル酸デフォロキサミン、Sigmaより入手可能)存在下)で増殖させることにより、上方制御させ得る。後者の方法を採用する場合、FrpB遺伝子発現を下方制御(好ましくは欠失)することが好ましいが、これはこの可変性抗原が鉄制限状態で単離される髄膜炎菌株より単離されたブレブにおいて免疫優性になり得るためである。
【0081】
好ましい実施形態において、本発明の組成物は、好ましくは高分子量TbpAおよびHsfが上方制御されている、lgtB-莢膜多糖-msbB-株由来のL3ブレブと、低分子量TbpAおよびOmp85が上方制御されている、lgtB-莢膜多糖-msbB-株由来のL2ブレブとを含む。より好ましくは、どちらのブレブもさらにPorAおよび/またはFrpBの発現、ならびに任意にOpCおよび/またはOpA発現が、下方制御されている。ブレブは最も好ましくは上記の低DOC法によって単離され、両ブレブ中のLOSは外膜タンパク質にブレブ内架橋をしている。
【0082】
ゴーストまたは死菌全細胞ワクチン
本発明者らは、ブレブに関わる上記組成物およびワクチンが、(同一の利点を有する)ゴーストまたは死菌全細胞調製物およびワクチンに関する方法にまで容易に拡張できることを意図している。グラム陰性株からゴースト調製物(完全なエンペロープを有する空の細胞)を製造する方法は当技術分野で周知である(例えばWO 92/01791を参照)。全細胞を殺滅して、ワクチンに使用するための不活性化細胞調製物を作製する方法もまた周知である。従って、本明細書を通して記載されるブレブを含む組成物およびワクチンは、等価物である本発明のゴーストおよび死菌全細胞調製物を含む同じ組成物またはワクチンにも当てはまることを意図している。
【0083】
本発明の組成物における血清殺菌アッセイ
血清殺菌アッセイは、本発明の免疫原性組成物中で組み合わせた場合に抗原間の相乗作用的な関係を評価するための好ましい方法である。
【0084】
そのような相乗反応は、抗原の組み合わせによって誘発されるSBAにより、各抗原によって別個に誘発されるSBAと比べて少なくとも50%、2倍、3倍、好ましくは4倍、5倍、6倍、7倍、8倍、9倍および最も好ましくは10倍高いものとして特徴付けられ得る。好ましくはSBAは、抗原が由来するのと同種の株、およびさらに好ましくは一群の異種の株に対して測定される(以下の代表的な群、例えばA-4クラスターに属するBZ10(B:2b:P1.2)、ET-37複合体に属するB16B6(B:2a:P1.2)、およびH44/76(B:15:P1.7,16)を参照)。SBAは、髄膜炎菌ワクチンの有効性を評価するための、最も一般的に受け入れられている免疫学的マーカーである(Perkinsら J Infect Dis. 1998, 177:683-691)。十分なSBAはあらゆる公知の方法により確認することができる。SBAは動物モデルまたはヒト被験体から得た血清を用いて行うことができる。
【0085】
ヒト血清を用いてSBAを行う好ましい方法は以下の通りである。血液サンプルは最初のワクチン接種の前、2回目のワクチン接種の2ヶ月後および3回目のワクチン接種の1ヶ月後に採取する(3回のワクチン接種は1年以内に行い、通常のヒト第1期ワクチン接種計画では、例えば0、2および4ヶ月、または0、1および6ヶ月の時点で投与される)。そのようなヒト第1期ワクチン接種計画は、1歳未満の幼児(例えばHibワクチン接種を同時に実施する)に実施することができ、または2〜4歳児もしくは青年もそのような第1期ワクチン接種計画でワクチン接種することによってSBAを試験することができる。妥当であれば、さらなる血液サンプルは初回ワクチン接種の6〜12ヶ月後、および追加免疫投与の1ヶ月後に採取され得る。
【0086】
(2〜4歳児もしくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、本発明の抗原が由来する髄膜炎菌株に対するSBA(抗体希釈)力価が(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が被験体の30%を超え、好ましくは40%を超え、より好ましくは50%を超え、最も好ましくは60%を超える場合、SBAは同種の殺菌活性を有する抗原またはブレブ調製物について十分であろう。
【0087】
当然ながら、異種の殺菌活性を有する抗原またはブレブ調製物は、それが由来する髄膜炎菌株に対して同様に十分なSBAを誘導することができるならば、同種の殺菌活性を有するブレブ調製物を構成することもできる。
【0088】
(2〜4歳児もしくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、髄膜炎菌の3種の異種株に対するSBA(抗体希釈)力価において(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が、被験体の20%を超え、好ましくは30%を超え、より好ましくは35%を超え、最も好ましくは40%を超える場合、SBAは異種の殺菌活性を有する抗原またはブレブ調製物について十分であろう。かかる試験は、異種の殺菌活性を有する抗原またはブレブ調製物が、様々な髄膜炎菌株に対する交差殺菌抗体を誘導し得るか否かの優れた指標である。3種の異種株は好ましくは互いに異なる電気泳動タイプ(ET)複合体、または多座配列タイピング(MLST)パターン(Maidenら PNAS USA 1998, 95:3140-5参照)を有し、さらに好ましくはその異種殺菌活性を有する抗原またはブレブ調製物が作製されるかまたは由来する起源の株とも異なるべきである。当業者は、髄膜炎菌において観察される遺伝的多様性、特に、重篤な病苦の原因として認識されているか、および/または強い病原性が認識されているMenB系統(Maidenら 前述)である髄膜炎菌B型株において観察される遺伝的多様性を反映した、異なるET-複合体を有する3種の株を容易に決定することは容易にできるであろう。例えば、使用され得る3種の株は以下のものである:A-4クラスターに属するBZ10(B:2b:P1.2)、ET-37複合体に属するB16B6(B:2a:P1.2)、およびET-5複合体に属するH44/76(B:15:P1.7,16)、または同じET/クラスターに属するあらゆる他の株。そのような株は、かかる株から作製されたもしくはそれらに由来する異種殺菌活性を有する抗原またはブレブ調製物を試験するために使用してもよく、例えば、ET-5複合体に属する髄膜炎菌株CU385(B:4:P1.15)を使用してもよい。使用され得る別のサンプル株は、系統3流行クローン(例えばNZ124(B:4:P1.7,4))由来のものである。別のET-37株は、NGP165(B:2a:P1.2)である。
【0089】
SBA活性を測定するための方法は、当技術分野において知られている。例えば、使用され得る方法がWO 99/09176の実施例10Cに記載されている。一般的には、試験される株の培養物を対数増殖期まで増殖させる(好ましくは、鉄枯渇条件下で、増殖培地にEDDA等の鉄キレート剤を添加することによる)。これを、BSAを加えた培地中(0.3%BSAを含むハンクス培地等)に懸濁し、約20000 CFU/mlに調整した作業用の細胞懸濁液を得てもよい。一連の反応混合物は、試験される血清の一連の2倍希釈物(好ましくは56℃で30分間熱不活性化されたもの)(例えば、50μl/ウェル容量)と試験される20000 CFU/mlの髄膜炎菌株の懸濁液(例えば、25μl/ウェル容量)との混合により、作製することができる。反応バイアルをインキュベートし(例えば37℃で15分間)、振とう(例えば210 rpmで)するべきである。最終反応混合物(例えば容量100μl)は、さらに補体源(例えば、最終容量の25%の予備試験済みの仔ウサギ血清、またはヒト血清学のためのヒト血清)を含み、上記のように(例えば37℃で60分間)インキュベートする。滅菌ポリスチレンU底96穴マイクロタイタープレートをこのアッセイに使用できる。マルチチャネルピペットを用いて、各ウェルからアリコート(例えば10μl)を採取し、ミューラーヒントン寒天プレート(好ましくは、1%イソビタレックス(Isovitalex)および1%熱不活性化ウマ血清を含む)に滴下し、それを(例えば、5%CO2中37℃で18時間)インキュベートしてもよい。好ましくは、個々のコロニーは、1アリコート当たり最大80 CFUまで計数できる。以下の3つの試験サンプルが対照として使用され得る:バッファー+細菌+補体、バッファー+細菌+不活性化補体、血清+細菌+不活性化補体。回帰計算によって、50%の細胞死滅に相当する希釈度の測定値を与えるようにデータを処理するプログラムを用いて、SBA力価が直接算出され得る。
【0090】
本特許明細書中に引用される全ての文献または特許出願は、参照により本明細書に組み入れられる。
【実施例】
【0091】
以下の実施例は、特に詳細に記載しない限りは当業者にとって周知でありルーチンである標準的な技術を用いて行われる。本実施例は例示的なものであり、本発明を限定するものではない。
【0092】
実施例1:
髄膜炎菌BのB莢膜多糖産生に関わるタンパク質をコードする遺伝子の欠失、PorA遺伝子の欠失、髄膜炎菌ブレブの表面上の様々な防御外膜タンパク質の上方制御、免疫優性タンパク質または生合成酵素の下方制御、およびブレブを単離する方法を記載する例は、WO 01/09350に記載される。
【0093】
実施例2:LOS:重要な交差防御抗原
潜在的な交差防御抗原としてのLOSの役割を評価するために、H44/76野生型(WT)の髄膜炎菌B株(L3 LOSを発現する)および「galE-様LOS」(lgtE- LOSに関しては短い構造を有する)を発現する改変H44/76株を使用し、2つの異なる方法に従ってブレブを生成した。第1の方法では高レベルのLOSを含むブレブを得るために0.1%DOCを使用し、第2の方法では生じたブレブ中に低レベルのLOSを得るために0.5%DOCを使用した。
【0094】
マウスに、1投与量当たりAl3+塩(リン酸ルミニウム)に吸着させたブレブ5μgおよび3D-MPLを、IM経路によって3回(0、21および28日目)注入した。3回目の注入の14日後に血液サンプルを採取した。
【0095】
抗L3 LOS ELISAは、プールした血清について精製L3 LOSを使用して行った。図3Aの結果は、0.1%DOC法がマウスにおいて抗LOS反応を誘発することのできるブレブを産生したことを明白に示す。これは、galE- LOSおよびL3 LOSが抗体の産生を誘導することができることを証明する。一方、0.5%DOCはそれがブレブワクチンにおいて重要な抗原として作用するためには過剰なLOSを抽出した。
【0096】
血清殺菌アッセイ
SBAは、個々の血清について異なるNmenB株を使用して行った:同種のWT H44/76株、PorA(-)H44/76株、および2つの異種の株(血清亜型に基づくもの)Cu385およびNZ124。これら4つの株はL3 LOSを発現する。5番目の株も加えられた。H44/76と比較して、この株(B16B6)はPorAに対してだけでなくLOS(これは免疫型L2株である)に対しても異種性である。
【0097】
図3Bの結果は、交差殺菌反応がL3株のみに対してであり、それはDOC 0.1% WTブレブを用いた場合のみ認められることを示す。DOC 0.1%galE-ブレブおよびDOC 0.5%WTブレブについては交差殺菌反応が観察されない。さらに、PorA抗体によって誘導される殺菌反応が血清型依存的であることは周知である。このことはまたDOC 0.5%WTブレブまたはgalE-ブレブを用いたこの実験において、そしてまたPorA(-)H44/76株を用いて行ったSBAデータでも観察される。
【0098】
これらの結果全ては、L3 LOSを高いパーセンテージで含むブレブによって誘導される交差殺菌反応が、LOS抗原に対する抗体(Abs)の産生によるものであることを示唆する。
【0099】
L3 LOSのみ(galE- LOSではない)が殺菌抗体の産生を誘発することができる。ELISAにおいて、DOC 0.1%galE-ブレブを用いて良好な抗LOS反応が観察されるが、この反応には生物学的な関連性がない(SBAではない)。
【0100】
さらに、この反応は、抗L3 LOS AbsがL2株ではなくL3株のみを死滅させるようなLOS免疫型特異的であるようでもあり、これはまた最適なワクチンが理想的には最適な適用範囲でL3およびL2 LOSを含むべきであることを示している。
【0101】
枯渇実験
WT DOC 0.1%ブレブにより誘導される反応が主として抗LOS抗体によることを示すために、血清プールを異なる濃度の精製L3 LOSを用いて枯渇させた。枯渇後、血清を、同種WT H44/76株に対する殺菌アッセイにおいて使用した。
【0102】
DOC 0.1%WTブレブに対して生じた血清を用いて得られた結果(図3C参照)は明白な投与範囲阻害を示し、これはこの調製物によって誘導される抗体の大半がLOSに対するものであることを示している(これはPorA(-)H44/76株を用いて得られたSBA結果を確認するものである)。反対に、WT DOC 0.5%によって誘導される反応は、PorA(-)H44/76株を用いて行ったSBAによって示され、またLOS枯渇によって示唆されるように、LOSに対するものではない
この結果は以下のL2 LOSについてのものと類似する。
【0103】
実施例3:L3および中間体(lgtB-)DOC非含有ブレブ(非無毒化LOS)を用いた実験は交差殺菌抗体を誘導した
使用したMC58髄膜炎菌誘導株はB:P1.7.16、opc-、siaD-である。この株を遺伝学的に改変して、L3(2G2株)または中間体エピトープ(2G EcoN1b-1株、2G2と同様であるがさらにlgtB-)またはLPSの短いバージョン(lgtE-であるC6株)のいずれかを発現させた。OMVを、通常の高(0.5%)DOC法またはDOC非含有法のどちらかに従って生成した。
【0104】
マウス(1群につき10匹)を筋肉内経路によって3回、0、20および28日目に免疫した。これらにはAl(OH)3上に製剤化されたブレブ1または10μg(タンパク質含量)を与えた。血液サンプルは、28日(Post II)および42日目(Post III)に採取した。
【0105】
殺菌アッセイは、プールされた血清について、同種の株(MC58およびH44/76)ならびに2種の異種の株(M97250687およびM9725078)を使用して、仔ウサギ血清を外来性補体源として用いて行った。
【0106】
以下の表は結果を要約する(50%死滅に対する殺菌力価)。
【表1】

【0107】
明らかに、L3(2g2)または中間体(2gecon1b-1)エピトープの存在は交差殺菌抗体を誘導し、一方、末端切断されたLPS株(C6)由来のブレブは低レベルの交差反応抗体を誘導する。これは特に1μgのOMVを注入した場合に示された。
【0108】
さらに、DOCで精製したOMVについて示されるように、ブレブのLPS含量の低下が交差殺菌抗体の誘導を低減する。増加したLPSとは別に、DOC非含有ブレブはまた、リポタンパク質のようなOMVとゆるやかに相互作用するいくつかのタンパク質を有利に保持することはできる。
【0109】
実施例4:L3 LOSおよび外膜タンパク質のブレブ内架橋
使用したMenBブレブは、SiaD-(従って莢膜多糖を発現しない)およびPorA-であるH44/76株(LOS免疫型L3)から得た。2種の異なる株を使用した:完全なL3(B1717株、siad(-) PorA(-) 完全なL3)および末端切断型L3(B1727株、siad(-) PorA(-) lgtB(-) TrL3)。
【0110】
EDAC/NHSコンジュゲーション法を公知の方法に従って用いて、ブレブ内でLOSとOMPを架橋し、LOSのオリゴ糖成分をT依存性抗原とした(EDAC/NHSは、非常に高度に架橋するがゆえに濾過性に不利な影響を与えることが分かったホルムアルデヒドよりも好ましい)。EDACはカルボン酸と反応して活性エステル中間体を生成する。アミン求核試薬の存在下において、アミド結合がイソ尿素副生成物の放出とともに形成される。しかしながら、EDAC仲介反応の効率はスルホNHSエステル中間体の形成を通して増加し得る。スルホNHSエステルは、EDACのみがカルボキシレートと反応することより形成される活性エステルより長く水性溶液中に存続する。従って、この2段階法を用いて高生成のアミド結合形成を実現し得る。EDAC/NHSコンジュゲーションは、J.V. Staros,R.W. WrightおよびD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)ならびにBioconjugates Techniques. Greg T Hermanson(1996)pp173-176に記載されている。
【0111】
反応混合物は、1.5 mgスルホNHSおよび5 mg EDACを3%ショ糖(ブレブの安定性のため)中に1mL容量で含んだ。ブレブは0.025mg EDAC/mgブレブの比率で存在した。ブレブは2 mg/mlの濃度で存在し、pHをHCl 0.1NまたはNaOH 0.1Nで7.5に調整した。
【0112】
反応を室温で4時間そのままにし、その混合物を3%ショ糖を含む2 mMリン酸バッファー、pH 7.5に対して透析した。該混合物を次にSterivex G10 0.22μmで濾過した。99%の収量のブレブが回収された。
【0113】
この反応に続いて、抗L3モノクローナル抗体(mAb)を使用したウエスタンブロットを行うことができた。その反応を通じ、ゲル上で低分子量LOSはより微弱になり、かつ新たなより高分子量のバンドが現れる。より高分子量のこのバンドが優勢と見られ、コンジュゲート化LOSの大部分がPorBに共有結合するようになることを示していると思われる。
【0114】
EDACはLOSをOMPに不可逆的に架橋する点で優れたブレブ内架橋剤であり、かつLOSのT依存的免疫原性を改良するために十分であることが見出されたが、しかしそれほど高度にそれを架橋しなかったのは、乏しい濾過性、凝集およびブレブ内架橋等の問題が発生したためであった。生じたブレブの形態は、コンジュゲートしていないブレブのものと類似している(電子顕微鏡の観察による)。さらに上記のプロトコルは、過度に高度の架橋(ブレブの表面上に天然に存在する防御OMP(例えばTbpA)の免疫原性を低減し得る)が生じることを回避した。
【0115】
実施例5:L3および末端切断型(中間体、lgtB-)L3は、末端切断型(中間体 lgtB-; TrL3)L3 LOSを認識する殺菌抗体(Ab)の産生を誘導できる
OMV(ブレブ)は、MenB株H44/76 siaD- PorA- L3から、またはH44/76 siad- porA- TrL3から産生された。2つの異なる抽出を行い、使用したDOCの割合を0.1または0.5%のいずれかとした。2つの異なるアジュバント製剤:Al(OH)3、またはリン酸アルミニウム+3D-MPLもまた、評価した。マウス(OF1メスマウス、6〜8週齢、1群につき30匹)にIM経路によって3回(0、21および28日目)注入した(5μgブレブ/注入)。SBAを、PostII(28日)およびPostIII(42日目)血清(プールされた血清または個々の血清)について収集した。
【0116】
50%細胞死滅に関する幾何平均力価およびプール血清力価は、0.5%DOC抽出物と比較して0.1%DOCで抽出したブレブによって誘導された血清の方がより高かった。これは後者と比較して前者のブレブにおいて2.5倍量のLOSが存在する傾向があるという事実によっておそらく説明される。完全なL3 LOSまたは末端切断型L3 LOSを含むブレブを用いて誘導される血清のSBAの間に顕著な差異はなかった。ブレブがリン酸アルミニウム+3D-MPLによりアジュバント作用を受ける場合には、水酸化アルミニウムと比較してSBAが増加する。
【0117】
血清枯渇実験もまた行った。1 mg/mL精製L3またはtrL3 LOSを使用して血清を枯渇させ、次にこれらの枯渇血清においてSBAを行った。この結果は、血清のtrL3 LOS前処理によって殺菌Ab(抗L3抗体を含む)をほとんど完全に枯渇させることができ、また血清のL3 LOS前処理によって殺菌Ab(抗trL3抗体を含む)をほとんど完全に枯渇させることができることを示した。抗L3殺菌抗体(Ab)はこのようにtrL3 LOSと反応することができ、抗trL3殺菌抗体(Ab)はこのようにL3 LOSと反応することができる。さらに、L3およびtrL3 LOSの双方に存在するLOS構造に対する殺菌Abの特異性がこのように証明された。
【0118】
結論として、本発明者らは(OMVにおける)TrL3構造が、L3株に対する殺菌Abの産生を誘導することができることを証明した。枯渇実験との組み合わせにおいて、本発明者らはTrL3およびL3 LOSが免疫学的基礎に基づき非常に近い構造であり、L3株を死滅させることが可能な抗体を産出するためにtrL3を使用することができることを示した。
【0119】
実施例6:TrL3は、完全なL3構造による自己免疫の潜在的問題を解決する
L3およびtrL3構造が、防御抗体に関して免疫学的に非常に近い関係である場合、L3(およびL2)LOSに付随する可能性としての自己免疫の問題に関して、構造(ラクト-N-ネオテトラオース部分による)の間に何らかの差異はあるのだろうか。本発明者らは、寒冷凝集素がtrL3 LOSを認識することが可能か否かに注目することによってこの問題に取り組んできた。
【0120】
モノクローナル抗体(MAb)1B2-1B7(J Bio Chem 256(1981) 10967〜10972およびATCC受託番号TIB-189)は、低温でヒト成人赤血球(RBC)を凝集し、かつLNnT(ラクト-N-ネオテトラオース)と反応することが知られている。これは典型的な寒冷凝集素である。
【0121】
このモノクローナル抗体は、以下の実験において、L3髄膜炎菌株を死滅させることのできるMabL3.7.9モノクローナル抗体とともに使用された。
【0122】
これら2つのmAbは、ポリ-L-リジンでプレコート(1μg/ml、37℃で2時間)し、次に精製L3または精製TrL3 LOSでコート(5μg、4℃で一晩)したマイクロプレートを用いたELISAにおいて、使用された。かかるプレートを次にBSAで飽和させた(1%、室温で30分間)。その後、2種の抗体の各々を用いて標準ELISAを行った。
【0123】
結果(図4)は、Mab L379がL3およびTrL3と反応するが(図4B)、しかし1B2-1B7はL3 LOSに対してのみ反応する(図4A)ことを明らかに示す。従って、TrL3は、LNnT四糖を含む構造(L3 LOSおよびヒト赤血球等)に対して反応する寒冷凝集素によっては認識されないと言うことができる。
【0124】
TrL3 LOSは、このように防御エピトープを保持するには十分に長く、しかしヒト自己免疫の影響を示す可能性があるエピトープを喪失するには十分に短いという最適な特徴を有する。
【0125】
このことが本特許出願において提案する末端切断型L2(lgtB-)LOS構造の場合も同様であるはずがないことを示唆する根拠はない。
【0126】
実施例7:B1820 DOC 0.1%ブレブの発熱性/抗原性における架橋の影響
ブレブ(B1820株由来[siaD(-)PorA(-)FrpB(-)であり、上方制御された末端切断型Hsfを有するH44/76から得られ、lgtB(-)突然変異株であることによりL3が末端切断されており、デスフェラル(desferral)存在下で培養され、ブレブはDOC 0.1%を用いて抽出される])は、異なる濃度のEDAC(EDACがより多く存在するほど、より多くのブレブが架橋される)を使用して架橋された。架橋は、ブレブの無菌的濾過によって示されるとおりブレブ内でなされる。
【0127】
精製Nmen L3 LOSを標準物質として用いたSDS-PAGE電気泳動後の銀染色によって測定されるブレブ中のLOS含量は18%である(Tsai, J. Biol. Standardization(1986) 14:25〜33)。一般的に、0.1%DOCで抽出されるブレブ中のLOS含量は約20%LOS、0.2%DOCでは約15%LOS、0.3%DOCでは約10%LOS、そして0.5%DOCでは約5%LOSである。一般的に、10%のコンジュゲートしていないLOSまたはそれ以上を含むブレブは、許容できない発熱性を有する。
【0128】
ウサギにおける発熱性
2種の製剤(Al(OH)3またはAlPO4に吸着したブレブ)を試験し、発熱試験においてウサギにヨーロッパ薬局方に記載されるようにして500ng/kgをIV経路によって投与した。
【0129】
この結果は、ブレブの発熱性に対するブレブ内架橋のプラスの影響を(以下の表に)明確に示す。同じバッチのブレブを対照として使用し、あるいは異なるEDAC濃度を用いて架橋した。ブレブがより多く架橋される(EDACがより多い)ほど、その発熱性は低くなった。このことは2種の異なる製剤について観察された。
【表2】
【0130】
架橋したブレブの抗原性
上記ブレブ(非吸着)の抗原性を評価して、この架橋がブレブの抗原性に対して何らかの影響を有するか否かを調べた。ブレブの異なる調製物(架橋されているまたはされていないもの)をマイクロプレート上にコートした(10μg/ml、4℃で一晩)。洗浄および飽和の後、MAb L379の連続希釈物またはB1820 DOC 0.1%または0.5%で免疫したマウス由来の血清を該プレートに添加した(振盪しながら室温で30分間)。コートしたブレブに対する抗体の固定は、ビオチンと結合した抗マウスIgを用い、次にストレプトアビジンペルオキシダーゼ複合体を使用し、その後OPDおよびH2O2を用いた発色によって、示した。各マイクロウエルの吸光度はマイクロプレートリーダーを用いて測定した。
【0131】
この結果は、MAb L379(L3 LOSに対してだけでなく、TrL3 LOS(lgtB-突然変異体)とも反応でき、L3株に対して殺菌性である)は、未処理の(コンジュゲートしていない)B1820ブレブおよび異なる架橋ブレブを(どのようなEDAC濃度を使用しても)同じように認識することを示す。図5Aを参照せよ。EDAC 0.2および1で得たより高い反応は、これらのEDAC濃度でのブレブにおけるLOSのよりよい固定化、または少なくとも架橋ブレブにおけるLOSのよりよい安定性を反映し得たものであった。
【0132】
これらブレブの抗原性を評価するためにマウス血清もまた使用した。2種の異なる血清を使用した;第1はB1820 DOC 0.5%ブレブ(8%以下の低LOS含量を有し、主として抗タンパク質抗体を誘導するブレブ)を用いて免疫したマウスより得た。第2の血清はB1820 DOC 0.1%ブレブ(15%以上のLOS含量を有し、主としてLOSに対する交差殺菌Abを誘導するブレブ)を用いて免疫したマウスより得た。L379 MAbを用いて観察されるように、これら2種の血清で得た結果(各々図5Bおよび5C)は、未処理の(コンジュゲートしていない)ブレブおよび架橋ブレブの間に(どのようなEDAC濃度を使用しても)いかなる差異も示さなかった。
【0133】
結論として、LOSの抗原性は架橋によって影響されず、かつブレブのその「広範な」抗原性もまたEDAC処理によっては改変されなかったものと思われる。(高濃度のEDACでの)架橋が重要な防御抗原の免疫原性を損なわないことを確認するために、マウスにおける免疫原性実験が進行中である。しかしながら、架橋がDOC 0.5%抽出ブレブに対してEDAC 0.025を用いて為された場合に示される予備的な結果(実施例8)は、、EDAC処理後のこれらブレブの免疫原性における増加を示す。
【0134】
実施例8:架橋ブレブの免疫原性(EDAC 0.025mg化学法)
この実験において、ブレブはB1727株より産生された。この株は遺伝的に改変されたH44/76株であり、これはsiaD(-)PorA(-)trL3(lgtB-)HsfでありそしてTbpAが上方制御されている。ブレブは0.5%DOCを用いて抽出した。マウスは3回(0、21および28日目)、IM経路により免疫した。1回の注入で、それらマウスにはAl(OH)3に吸着したブレブ5μgを投与した。
【0135】
血清殺菌アッセイはH44/76株に対して、3回目の注入の14日後に採取した個々の血清について行った。その結果は、EDAC処理が応答者の数(100を超えるSBA力価を示すマウスの数)にプラスの影響を与えたことを示す。EDAC処理ブレブに対しては応答者が37%、非改変ブレブでは応答者は17%だけであった。
【0136】
製剤中の3D-MPLの欠如、および0.5%DOC抽出後のブレブ調製物における比較的低い割合のLOSは、低い反応性を説明する。
【表3】
【0137】
また抗Hsf ELISAを行い、架橋がこのタンパク質の免疫原性に影響を及ぼすか否かを調べた。(プールした血清について得た)その結果は、架橋がIgG抗Hsf反応に影響を及ぼさないことを示す。IgMは検出されなかった。
【表4】
【0138】
実施例9:TrL3 LOSデータ
以下の実験を評価した:
- LNnT(ラクト-N-ネオテトラオース)と反応することができる抗体の誘導に対するTrL3(lgtB(-)L3 LOS)の影響、
- 上記構築物の殺菌抗体の誘導。
【0139】
ブレブは2種の遺伝的に改変したH44/76株より作製した。どちらもsiaD(-)PorA(-)であるがしかし、一方はWT L3 LOSを産生し、もう一方はTrL3 LOS(lgtB(-))であった。これらのブレブは2つの異なる方法に従って作製され、高LOS含量(約18%、DOC 0.1%抽出を使用)または低LOS含量(5%近く、DOC 0.5%抽出を使用)を有するようにした。
【0140】
マウスは、3回(0、21および28日に)、IM経路によって、3D-MPLとともにまたはそれなしで、Al(OH)3に吸着したブレブ5μg(注入1回当たり)で免疫した。
【0141】
抗LNnT ELISA
方法:マイクロプレートは、スペーサー(ADH)を介してヒト血清アルブミンにコンジュゲートしたLNnTでコートした(PBS中1ml当たり5μgのコンジュゲート、マイクロウエル当たり100μl)。4℃で一晩インキュベートした後、プレートをPBS-BSA 1%で洗浄し飽和させた(室温で40分)。洗浄の後、PBS 0.2%、BSA 0.05%、Tween20中への連続希釈液を添加した(室温で30分)。IgGのLNnTへの固定は、ペルオキシダーゼ(Jackson)に結合した抗マウスIgGによって、続いてOPDAおよびH2O2によるインキュベーションによって明らかにされた。
【0142】
結果:陽性対照は1B2-1B7 MAbである。このMAbはLNnTおよびL3LOSと反応し(しかしTrL3 LOSとは反応しない)(上記実施例を参照)、かつヒト赤血球を凝集させる。陰性対照(-)はアジュバントのみで免疫したマウス由来の血清である。
【0143】
結果(図6)は、高LOS含量を有するL3ブレブ(DOC 0.1%)のみがLNnTと反応可能なIgGの産生を誘導することを明白に示す。同様のLOS含量を有するTrL3ブレブは、LNnTに対するIgGの産生を誘導せず、低量のLOSを含む双方のブレブ調製物(DOC 0.5%)が誘導しないのと同様である。
【0144】
H44/76株についてのSBA
H44/76株に対するSBAアッセイを、3回目の注入の14日後に採取した個々の血清について実施した。以下の結果はtrL3(lgtB(-))LOSブレブがL3 LOSと同等のレベルで殺菌抗体を誘導することを明白に示す(GMTの他、>1/100のSBA力価を有するマウスの数(=SC)を参照)。
【表5】
【0145】
実施例10:FrpBノックアウト
以下のデータは2つの前臨床試験の概要である。
【0146】
これらの実験において、2種の遺伝的に改変されたH44/76株を使用して、0.1%DOCを用いてブレブを生成した。この方法によって得たブレブは20%近いLOS含量を有する。
【0147】
2種のH44/76株は以下の通りであった:
- B1733:siaD(-) PorA(-) Tr(末端切断型)Hsfの上方制御 lgtB(-)
- B1820:siaD(-) PorA(-) TrHsfの上方制御 lgtB(-) FrpB(-)。
【0148】
LbpA/B、TbpA/B、FrpB(B1733中)等の鉄依存性タンパク質の産生を上方制御するように、株をデスフェラル存在下で増殖させた後にブレブを生成した。
【0149】
これらの異なるブレブ調製物はAl(OH)3に吸着させ、マウスにIM経路にて3週間隔で2回注入した。血液サンプルは2回目の投与の7日後に採取した。マウスには1回の注入当たり5μgのブレブを投与した。
【0150】
SBA結果
殺菌アッセイは3つのL3株について行った(同種の野生型株H44/76および2つの異種のL3株:NZ124およびM97250687)。この結果は、FrpB(-)(ノックアウト)(B1820)ブレブが、FrpB(+)ブレブ(B1733)よりも優れた異種交差殺菌反応(高い力価およびより優れた血清転換SC)を誘導することを明白に示す。同種反応は、FrpB欠失により低下するにも関わらず、十分なままである。
【0151】
これらのデータは、FrpBが、ブレブによって誘発される免疫反応において主要な駆動因子ではあるが、この外膜タンパク質は可変性が高いため、このタンパク質に対する抗体は同種株の死滅を誘導することだけしかできないことを示している。従って、ブレブ産生株におけるFrpBの欠失は、産生されたブレブワクチンの適用範囲を改善するための有利な手段である。
【表6】
【0152】
実施例11:ブレブの発熱性に対するmsbB(lpxL1)突然変異の影響
2つのNmenB株をこの評価に使用した:
- galE(-)である(従って莢膜多糖を生成できない)対照株。
【0153】
- msbB突然変異株:これはgalE(-)およびmsbB(-)である。
【0154】
これら2種の株から0.1%のDOCを使用してブレブを生成し、OMV(ブレブ)中のLOS含量が15%を超えるようにした。上記の実施例で述べたように、LOS含量が10%を超えるブレブ調製物は発熱性という観点から十分でなく、ヨーロッパ薬局方のウサギ発熱性アッセイに適合しない。
【0155】
上記のブレブを、ウサギにおける発熱性アッセイ(500ngのブレブ/kgをIV経路で注入)のためにAl(OH)3に対して製剤化した(50μgのOMV/500μgのAl3+塩)。
【0156】
以下の結果は、msbBの欠失(特に莢膜多糖を産生できない株において)により、15%を超えるLOS含量であっても、ウサギにおいて非発熱性のブレブの産生が可能になることを明白に示す。
【表7】
【0157】
結論:
lgtB(-)およびmsbB(-)突然変異を有する髄膜炎菌に由来し、かつより低い(例えば0.1%)デオキシコレート濃度で抽出されたL3およびL2ブレブを含む組成物は、髄膜炎菌Bに対する効果的で安全なワクチンに対する強力な基礎を提供する。ブレブ産生株は理想的には莢膜多糖合成欠損性であり、かつ該ブレブはブレブ内で外膜タンパク質に架橋しているLOSを有する。PorA(-)およびFrpB(-)のいずれかまたは両方が、交差殺菌効果を改良する上でさらに有用であり、Hsfおよび/またはTbpA抗原の上方制御も同様に有用である。
【技術分野】
【0001】
発明の分野
本発明は、ナイセリアワクチン組成物、それらの製造、およびかかる組成物の医薬における使用の分野に関する。より具体的には、ナイセリアの、特に髄膜炎菌の外膜小胞(すなわち、ブレブ)ワクチンの産生により適した新規の操作された髄膜炎菌株を製造する方法に関する。新規のLOSサブユニットまたは髄膜炎菌の外膜小胞(すなわち、ブレブ)ワクチンの使用に基づく有利な方法およびワクチン製品もまた記載しており、これはヒト被験体における使用のために安全性および有効性をより高めたものである。
【背景技術】
【0002】
発明の背景
ナイセリア・メニンジティディス(Neisseria meningitidis)(髄膜炎菌)は、しばしばヒト上気道より単離されるグラム陰性細菌である。この細菌は、菌血症および髄膜炎等の重篤な侵襲性細菌性疾患の原因である。髄膜炎菌疾患の発生数は、地理的、季節的および年次的な差異を示す(Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。この菌は通常、その莢膜多糖による血清型に従って分類される。
【0003】
温暖な国々で最も多い疾患は、血清型B株によるものであり、発症率は総人口の1〜10/100,000/年の間で変動し、それより高い値に達する場合もある(Kaczmarski, E.B. (1997), Commun. Dis. Rep. Rev. 7: R55-9, 1995、Scholten, R.J.P.M., Bijlmer, H.A., Poolman, J.T.ら Clin. Infect. Dis. 16:237-246, 1993、Cruz, C., Pavez, G., Aguilar, E.,ら Epidemiol. Infect. 105:119-126, 1990)。
【0004】
血清型A髄膜炎菌が優位を占める流行の大部分は中央アフリカで発生し、その発症率が1000/100,000/年のレベルにまで達する場合もある(Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。全体として、髄膜炎菌疾患のほとんど全ての症例は、血清型A、B、C、W-135およびYの髄膜炎菌によって引き起こされ、四価のA、C、W-135、Y莢膜多糖体ワクチンが利用可能である(Armand, J., Arminjon, F., Mynard, M.C., Lafaix, C. J. Biol. Stand. 10:335-339, 1982)。
【0005】
ナイセリア・メニンジティディス感染の頻度は、過去数十年、多くのヨーロッパの国々において増加してきている。これは、社会的活動の場(例えば、水泳プール、劇場等)の増加による伝染の増加に起因してきた。いくつかの標準的な抗生物質に対して感受性が低い、または耐性のナイセリア・メニンジティディス株を単離することは、もはや珍しいことではない。この現象は、新たな抗菌剤、ワクチン、薬剤スクリーニング法、およびこれらの生物についての診断試験に対する、未だ満たされていない医学的ニーズおよび需要を生み出した。
【0006】
利用可能な多糖ワクチンは、現在、それらを担体タンパク質に化学的にコンジュゲートする方法によって改良が行われている(Lieberman, J.M., Chiu, S.S.Wong, V.K.らJAMA 275:1499-1503, 1996)。
【0007】
しかしながら、血清型Bワクチンは利用可能な状況にない。血清型B莢膜多糖は、免疫原性を示さないことが分かってきた−それは恐らくそれが宿主成分との構造類似性を有するためであろう(Wyle, F.A., Artenstein, M.S., Brandt, M.L.ら, J. Infect. Dis. 126:514-522, 1972、Finne, J.M., Leinonen, M., Makela, P.M. Lancet ii.:355-357, 1983)。従って、外膜小胞(もしくはブレブ)またはそれらから得た精製タンパク質構成成分から、血清型Bワクチンの開発を試みることに焦点を当てた努力がされてきた。
【0008】
ワクチン開発のための別の髄膜炎菌抗原は、髄膜炎菌のリポオリゴ糖(LOS)である。これらは糖脂質に結合した外膜であり、O側鎖が欠如しているという点で腸内細菌科(Enterobacteriaceae)細菌のリポ多糖(LPS)と異なり、従ってLPSのラフ型と類似している(GriffissらRev Infect Dis 1988, 10:S287-295)。LOSのオリゴ糖部分における不均一性は、異なる髄膜炎菌株間における構造的なおよび抗原性の多様性を生み出す(GriffissらInf. Immun. 1987, 55:1792-1800)。このことは、かかる株を12種類の免疫型にさらに分類するために用いられてきた(ScholtanらJ Med Microbiol 1994, 41:236-243)。免疫型L3、L7およびL9は、免疫学的に同一であり、かつ構造的に類似(もしくは全く同一)しており、従ってL3,7,9(または、本明細書において、一般的に「L3」)と呼ばれてきた。髄膜炎菌のLOS L3,7,9(L3)、L2およびL5は、シアリル化によって、またはシチジン5'-一リン酸-N-アセチルノイラミン酸の付加によって修飾することができる。L2、L4およびL6 LOSは免疫学的に区別することができるが、しかしそれらは構造的に類似しており、本明細書でL2に言及する場合にはL4またはL6のいずれかと任意に置換することができ、これは本発明の範囲内となる。LOSに対する抗体は、実験用ラットにおいて感染に対し防御し、またナイセリア・メニンジティディスに感染した小児において殺菌活性に寄与することが示されている(GriffissらJ Infect Dis 1984, 150:71-79)。
【0009】
しかしながら髄膜炎菌ワクチンにおけるLOSの使用に伴う問題は、その毒性(そのリピドA部分による毒性)にある。
【0010】
またLOSは髄膜炎菌のブレブの表面に存在する。髄膜炎菌の外膜小胞(またはブレブ)に基づいたワクチンの開発に焦点を当てて、長年にわたる努力がされてきた(de Moraes, J.C., Perkins, B., Camargo, M.C.らLancet 340:1074-1078, 1992、Bjune, G., Hoiby, E.A. Gronnesby, J.K.ら338:1093-1096, 1991)。そのようなワクチンは、宿主に投与した場合に防御免疫反応を誘発することのできる、正確に折り畳まれた立体構造をとったいくつかの完全な外膜タンパク質を含むという利点を有する。さらに、ナイセリア株(ナイセリア・メニンジティディス血清型B、すなわちmenBを含む)は、工業的規模での製造を可能にするために十分な量の外膜ブレブを排出する。しかしながら、ブレブは大抵、細菌細胞(例えばEP 11243)の0.5%界面活性剤(例えばデオキシコレート)抽出を含む方法により調製される。それは上記のようなLOSの毒性(内毒素とも呼ばれる)のために望まれるが、しかし、ワクチンからほとんどのLOS抗原を除去する効果もまた有する。
【0011】
ワクチン抗原としてのLOSの使用に伴うさらなる問題は、多様な幅を持った糖鎖構造を有する12のLPS免疫型が存在することである(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021、Mol Microbiol 2002, 43:931-43)。ある免疫型に対する抗体は、異なる免疫型を認識することができない。LOS免疫型のオリゴ糖部分の共通「コア」領域を生成することに努力が集中されてきたが(例えばWO 94/08021)、改変されたLOSに対して生じた抗体の殺菌活性は失われる。従って、ワクチンは、有効であるために異なる免疫型の多くのLOS成分を有する必要があるだろう。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】国際公開WO 94/08021
【非特許文献】
【0013】
【非特許文献1】Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989
【非特許文献2】Kaczmarski, E.B. (1997), Commun. Dis. Rep. Rev. 7: R55-9, 1995
【非特許文献3】Scholten, R.J.P.M., Bijlmer, H.A., Poolman, J.T.ら Clin. Infect. Dis. 16:237-246, 1993
【非特許文献4】Cruz, C., Pavez, G., Aguilar, E.,ら Epidemiol. Infect. 105:119-126, 1990
【非特許文献5】Armand, J., Arminjon, F., Mynard, M.C., Lafaix, C. J. Biol. Stand. 10:335-339, 1982
【非特許文献6】Lieberman, J.M., Chiu, S.S.Wong, V.K.らJAMA 275:1499-1503, 1996
【非特許文献7】Wyle, F.A., Artenstein, M.S., Brandt, M.L.ら, J. Infect. Dis. 126:514-522, 1972
【非特許文献8】Finne, J.M., Leinonen, M., Makela, P.M. Lancet ii.:355-357, 1983
【非特許文献9】GriffissらRev Infect Dis 1988, 10:S287-295
【非特許文献10】Griffissら, Inf. Immun. 1987, 55:1792-1800
【非特許文献11】Scholtanら, J Med Microbiol 1994, 41:236-243
【非特許文献12】GriffissらJ Infect Dis 1984, 150:71-79
【非特許文献13】de Moraes, J.C., Perkins, B., Camargo, M.C.ら, Lancet 340:1074-1078, 1992
【非特許文献14】Bjune, G., Hoiby, E.A. Gronnesby, J.K.ら338:1093-1096, 1991
【非特許文献15】M.P.Jenningsら, Microbiology 1999, 145, 3013-3021
【非特許文献16】Mol Microbiol 2002, 43:931-43
【発明の概要】
【発明が解決しようとする課題】
【0014】
ヒトワクチンにおける抗原としてのLOS(LPSまたはリポ多糖としても知られる)の使用に伴うさらなる問題が存在し、それはすなわちそれらがヒト糖鎖構造(例えば、ヒト赤血球細胞上の)に類似した糖鎖構造を担持しているため、それらの使用に伴う安全性の問題が突きつけられることである。さらにLOS構造の改変は、LOS抗原の殺菌効果の構造的感受性のため、問題を含んでいる。
【課題を解決するための手段】
【0015】
本発明は、1以上の上記の問題を改善するための方法を提供し、かつ防御抗原としての髄膜炎菌LOS(特に、外膜小胞上に提示される場合)に基づいた新規のワクチンの製造方法を提供する。
【図面の簡単な説明】
【0016】
【図1】図1はL3およびL2免疫型LOSの模式図を示す。
【図2】図2はL3およびL1免疫型LOSの模式図を示す。
【図3】図3は抗L3 LOS ELISA、血清殺菌アッセイ、およびLOS枯渇実験の結果を示す。
【図4】図4はMAb 1B2-1B7およびMab L3.7.9を用いたELISAの結果を示す。
【図5】図5はマウス血清におけるブレブの抗原性を示す。
【図6】図6は抗LNnT ELISAの結果を示す。
【発明を実施するための形態】
【0017】
発明の説明
本明細書中に記載される刊行物および特許、または特許出願の主題またはそこに開示される情報は、参照により本明細書に組み入れる。
【0018】
「リポオリゴ糖」(または「LOS」)に関する言及は、「リポ多糖」または「LPS」にも言及するものとしてみなされる。
【0019】
本明細書中の用語「含む」(「comprising」「comprise」「comprises」)は、全ての場合に、任意に用語「からなる」(「consisting of」「consist of」「consist of」)に各々置き換えることができるものと発明者は意図している。
【0020】
本発明者らは、LOSオリゴ糖構造の短縮が殺菌性免疫反応を誘発し得るエピトープの減少を導くことを見出した。あるいは、本発明者らは、ワクチン製剤においてLOSを最も有効に使用するためには、LOSオリゴ糖構造をできるだけ保持するべきであるが、しかしたった2種類のLOS抗原の組み合わせによって広範に有効なナイセリアの(好ましくは髄膜炎菌の)ワクチンを生成し得ることを見出した。本発明の第1の態様は、ナイセリア(好ましくは髄膜炎菌)の免疫型L2のLOSおよび免疫型L3のLOSを含む、ナイセリア(好ましくは髄膜炎菌もしくは髄膜炎菌B型)疾患の予防または治療のための免疫原性組成物である。LOSは公知の精製法のいずれかにより単離することができ、またはL2およびL3ナイセリア株由来の少なくとも2種類の外膜小胞(もしくはブレブ)調製物中に存在し得る。ブレブ調製物から、毒性を有し緩く保持されたLOSを除去するが、ブレブ内に組み込まれたLOS抗原は高レベルで保持するようにするために、ブレブを低濃度の界面活性剤、すなわち0〜0.3%、好ましくは0.05〜0.2%、最も好ましくは約0.1%の、好ましくはデオキシコレート(DOC)、を用いて抽出することが好ましい。LOS抗原のそのような組み合わせ(特にブレブワクチン中の)は、驚くべきことに90%を超えるナイセリア・メニンジティディス株に対して効果的であるという点で有利である。
【0021】
本発明者らはまた、本発明の上記ブレブ免疫原性組成物、および実際にはあらゆるナイセリア(好ましくは淋菌または髄膜炎菌)由来のブレブ免疫原性組成物が、免疫優性外膜タンパク質の特定の組み合わせの発現が下方制御(好ましくは欠失)される場合に、それらの表面上の防御抗原(LOSを含む)の効果が増強されることを見出した。従って、本発明の第2の態様は、天然の非改変株と比較して発現が下方制御された、好ましくは欠失した以下の外膜タンパク質:PorA、PorB、OpA、OpCまたはPilC、を2つ以上有するナイセリア株由来のナイセリアブレブ調製物である。好ましくは、PorAとOpA、PorAとOpC、OpAとOpC、またはPorAとOpAとOpCが下方制御されているかまたは欠失している。FrpBの発現の下方制御(好ましくは欠失)はまた、交差防御抗原の効果の増強において有利であり、特に、鉄制限状態で増殖させたナイセリア株から作製したブレブ調製物中において有利であることが示されてきた。従って、この突然変異を有する株由来のナイセリアブレブは、上記の1以上の下方制御とFrpB下方制御との組み合わせから得られるブレブとしての、本発明のさらなる実施形態である。好ましくは、PorAが下方制御される場合にはPorBは下方制御されるべきではなく、またその逆も同様である。
【0022】
上記突然変異は、ブレブ免疫原性組成物が由来することになるあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしながらL2またはL3免疫型ナイセリア(好ましくは髄膜炎菌、最も好ましくはmenB)株を用いることが好ましく、典型的には上記の低DOC%抽出法で抽出することが好ましい。好ましくは、本発明のブレブ免疫原性組成物は、L2およびL3ブレブを両方含み、かつその少なくとも一方(好ましくは両方)が上記の免疫優性外膜タンパク質(またはOMP)の組み合わせを欠損していることが好ましい。それらの遺伝子を下方制御するための技術はWO 01/09350(参照により本明細書に組み入れられる)に記載されている。4つの異なるOpa遺伝子が髄膜炎菌ゲノム中に存在することが知られており(Ahoら1991 Mol. Microbiol. 5:1429-37)、ゆえにOpaの発現が下方制御されていると言われる場合、このことは好ましくは髄膜炎菌に存在する1、2、もしくは3つの遺伝子または(好ましくは)4つ全ての遺伝子がそのように下方制御されていることを意味する。そのような下方制御は、WO 01/09350に記載のように遺伝学的に達成するか、または、容易に発見される天然の安定した髄膜炎菌株(Opa座位からの発現がないかもしくは低いもの)を探索することによって達成し得る。そのような株は、Poolmanら(1985 J. Med. Micro. 19:203-209)に記載される技術を用いて見つけ出すことができ、その際、Opa-である細胞は、プレート上または顕微鏡下において細胞の外見を観察することによって見出されうるOpa発現細胞とは異なる表現型を有する。一度見つけられれば、Opaの欠失を確立するための発酵操作の後に、細胞内容物についてウエスタンブロットを行うことによって、株が安定的にOpa-であることを示すことができる。
【0023】
上記LOS免疫原性組成物の安全性
ヒトスフィンゴ糖脂質に存在するラクト-N-ネオテトラオースオリゴ糖基(Galβ1-4GlcNAcβ1-3Galβ1-4Glcβ1-、図1)と類似した構造が存在することから、L3またはL2 LOSに対して生じた抗体の安全性が問われてきている。多くの人々が残存量のL3 LOSを含むデオキシコレート抽出小胞ワクチンで安全にワクチン接種されてきているが(G. Bjuneら, Lancet(1991), 338, 1093-1096、GVG. Sierraら, NIPH ann(1991), 14, 195-210)、LOSが本明細書に記載されるように抗原として保持される場合、LOS糖鎖構造の末端部分の欠失が、ヒト組織の表面に提示される構造との抗LOS免疫反応の交差反応の阻止において有利であることが、本発明者らによって発見された。好ましい実施形態において、lgtB遺伝子の不活性化が末端ガラクトース残基およびシアル酸が欠損した中間体LOS構造を生じる(図1および2を参照、突然変異はL2およびL3 LOSに4GlcNAcβ1-3Galβ1-4Glcβ1-構造を残存させる)。そのような中間体は、L3および/またはL2 LOS株において獲得され得る。別のおよびあまり好ましくないLOSの(短い)バージョンは、lgtE遺伝子のスイッチオフによって得られうる。さらに別のおよびあまり好ましくない型のLOSは、lgtA遺伝子のスイッチオフによって得られうる。そのようなlgtA-突然変異を選択する場合、非免疫原性L1免疫型の形成を阻止するためにlgtC発現をスイッチオフすることがまた好ましい。
【0024】
lgtB-突然変異が最も好ましいのは、これが殺菌性(およびさらに交差殺菌性の)抗体反応を依然として誘導し得るLOS防御オリゴ糖エピトープを保持したままで安全性の課題を解決するために最適な末端切断物であることを本発明者らが発見したためである。
【0025】
従って、本発明の上記のL2および/またはL3調製物(精製されたもの、または単離したブレブ状のもの)あるいは一般的な髄膜炎菌のブレブ調製物(特にL2および/またはL3)は、lgtB、lgtAまたはlgtE遺伝子由来の機能的遺伝子産物の発現が永続的に下方制御されるように、好ましくは遺伝子のスイッチオフにより、最も好ましくは当該遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により遺伝子操作したナイセリア株(好ましくは髄膜炎菌株)から、有利に得られる。
【0026】
好ましくは、本発明のナイセリア株は、莢膜多糖の合成に欠損を有する。
【0027】
本発明の上記ブレブ調製物を髄膜炎菌B株から得る場合、莢膜多糖(ヒト様糖鎖構造も含む)もまた除去されることが特に好ましい。これを達成するために多くの遺伝子がスイッチオフされ得るが、本発明者らは、siaD遺伝子由来の機能的遺伝子産物の発現を永続的に下方制御(例えばα-2-8ポリシアル酸転移酵素の活性を下方制御)するように、ブレブ産生株を、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により、遺伝的に操作することが好ましいということを有利に示してきた。そのような不活性化はWO 01/09350に記載されている。siaD(synDとしても知られる)突然変異が、莢膜多糖からヒト類似エピトープを除去し得る多くの突然変異の中で最も有利であるのは、それが、LOSの防御エピトープの生合成に影響を及ぼさない最良の突然変異の1つであり、そのため、このようにLOSを防御抗原として最終的に用いることを目的とし、かつ細菌の増殖に対して最小限の影響を与える方法において有利であるためである。従って、本発明の好ましい態様は、lgtE- siaD-、lgtA- siaD-または好ましくはlgtB- siaD-髄膜炎菌B突然変異株由来の上記ブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0028】
siaD-突然変異は上記の理由のために好ましいが、髄膜炎菌B(または一般的に髄膜炎菌)の莢膜多糖合成をスイッチオフする他の突然変異が用いられ得る。従って、ブレブ産生株を、以下の遺伝子の1つ以上に由来する機能的遺伝子産物の発現を永続的に下方制御するように、好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により、遺伝的に操作してもよい:ctrA、ctrB、ctrC、ctrD、synA(synXおよびsiaAと同等)、synB(siaBと同等)またはsynC(siaCと同等)遺伝子。lgtE-突然変異はこれらの突然変異の1以上と組み合わせてもよい。好ましくは、lgtB-突然変異はこれらの突然変異のうち1つ以上と併用される。本発明のさらなる態様は、従って、髄膜炎菌B(または一般的に髄膜炎菌)の突然変異株のそのような組み合わせから得られる上記ブレブ免疫原性調製物である。かかる株自体が、本発明のさらなる態様である。
【0029】
lgtBおよびlgtEを包含する、様々なlgt遺伝子を含むナイセリア遺伝子座、ならびにその塩基配列は、当技術分野において公知である(M.P.Jenningsら, Microbiology 1999, 145, 3013-3021および以下に引用する参考文献、J. Exp. Med. 180:2181-2190(1994)、WO 96/10086を参照)。
【0030】
最終産物として完全長(非末端切断型)LOSを使用する場合、LOSがシアリル化されていないことが望ましく、そのようなLOSは、最も危険な侵襲性髄膜炎菌B株(これも同様にシアリル化されていない)に対する免疫反応を生じる。そのような場合、synA(synXおよびsiaAと同等)、synB(siaBと同等)またはsynC(siaCと同等)遺伝子が欠失した莢膜陰性株の使用が有利であり、それはまたそのような突然変異によってmenB LOSがシアリル化され得ないようにするためである。
【0031】
上記突然変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有利であるが、しかしL2またはL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、典型的には本明細書に記載した低DOC%抽出法によって抽出されたものを使用するのが好ましい。好ましくは、本発明のブレブ免疫原性組成物が、L2およびL3ブレブの両方を含み、かつ、そのうちの少なくとも一方(および好ましくは双方)が上記遺伝子の発現が欠損した株に由来する。
【0032】
LOSの毒性
本発明の上記の精製LOSまたはブレブ免疫原性組成物はまた、それらが由来する細菌性産生株における特定の遺伝子の発現の下方制御によって、毒性をより少なくすることができる。そのような無毒化は、天然のOMVによる鼻腔内免疫化には不必要であるだろうが(J.J Drabickら, Vaccine(2000), 18, 160-172)、非経口ワクチン接種における無毒化には有用であろう。好ましくは、本発明の精製LOSまたはブレブ免疫原性組成物であるLOSは、リピドA生合成に関与する遺伝子(特にリピドAへの第2アシル鎖の付加に関与する遺伝子)の突然変異/修飾/不活性化により(特にmsbBおよび/またはhtrB遺伝子由来の機能的遺伝子産物の発現の下方制御により、および好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により、ナイセリア産生株を遺伝的に操作することによって無毒化される。あるいはまた(またはさらに)、精製LOSまたはブレブ免疫原性組成物は、遺伝的に改変して以下の遺伝子:pmrA、pmrB、pmrEおよびpmrFのうちの1つ以上の遺伝子が(より強力なプロモーターの導入、もしくは遺伝子の余分なコピーの組み込みによって)上方制御されるようにしたナイセリア株から得ることができる。あるいはまた(またはさらに)、精製LOSまたはブレブ免疫原性組成物は、ポリミキシンBの機能的等価物である非毒性ペプチド[リピドAに対して高い親和性の有する分子]を組成物に添加することにより無毒化され得る。
【0033】
上記無毒化法ならびに当該プロモーター/遺伝子配列、ならびに上方制御方法および下方制御方法については、より詳細についてWO 01/09350を参照せよ。またナイセリアmsbBおよびhtrB遺伝子は各々lpxL1およびlpxL2とも呼ばれ(WO 00/26384を参照)、これらの遺伝子の欠失突然変異は、msbB-突然変異LOSが野生型と比較して1つの第2アシル鎖を欠損する(ならびに4つの第1および1つの第2アシル鎖を保持する)こと、およびhtrB-突然変異LOSが第2アシル鎖の両方を欠損することによって、表現型として特徴付けられる。そのような突然変異は好ましくは、ブレブ上への無毒化されたLOSの最適な提示を確実にするため、または無毒化サブユニットLOSの精製を助けるために、ナイセリア産生株が莢膜多糖を欠損していること(上記参照)を確実にする突然変異と組み合わせて用いられる。本発明の組成物中に使用され得るポリミキシンBの機能的等価物である非毒性ペプチドのさらなる詳細、特にペプチドSAEP 2(2つのシステインがジスルフィド架橋を形成するKTKCKFLKKC配列を有する)の使用については、WO 93/14115、WO 95/03327、Velucchiら(1997)J Endotoxin Res 4:1-12およびEP 976402を参照せよ。
【0034】
本明細書において「機能的遺伝子産物の発現の下方制御」は、問題の遺伝子のプロモーターまたはオープンリーディングフレームに対して付加、欠失または置換が為され、それにより全遺伝子産物の生合成活性が低減(60、70、80、90、95もしくは最も好ましくは100%低減)されることを意味する。明らかにフレームシフト突然変異が導入され得、またはより弱いプロモーターが置換され得るが、しかし、最も好ましくは、オープンリーディングフレームおよび/またはプロモーターのほとんどまたは全てを、(活性のある)遺伝子産物の永続的な下方制御を確実にするために欠失させる(WO 01/09350に記載)。
【0035】
上記変異は、ブレブ免疫原性組成物が由来するあらゆるナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株、特に本明細書に記載する株において有用であるが、L2またはL3免疫型ナイセリア(好ましくは髄膜炎菌の、最も好ましくはmenB)株を用いることが好ましく、典型的には本明細書に記載の低DOC%抽出法で抽出されることが好ましい。本発明のブレブ免疫原性組成物は、L2およびL3ブレブの両方を含み、その少なくとも一方(および好ましくは双方)が上記遺伝子の発現を欠損している株に由来することが好ましい。
【0036】
本発明のさらなる態様は、本発明のLOSまたはブレブ免疫原性調製物が由来する上記の遺伝的に改変されたナイセリア(好ましくは髄膜炎菌または淋菌または髄膜炎菌B)株を含む。
【0037】
本発明のLOSまたはLOS含有ブレブ調製物
本発明のさらなる態様は、本発明のナイセリア株から単離されるLOS調製物(特に上述した任意のもの)である。好ましくは、単離されたLOS(またはLOS含有ブレブ)は、L2またはL3免疫型であり、好ましくは本発明の免疫原性組成物は本発明のL2およびL3 LOS(またはブレブ)調製物を両方含む。
【0038】
そのような調製物はまた、上記LOS(精製されているものであれブレブ調製物に存在しているものであれ)のオリゴ糖部分をT細胞エピトープの供給源を含む担体にコンジュゲートする(従ってLOSをよりよい(T依存的)免疫原とする)ことにより改良され得る。あるいはまた(またはさらに)本発明の精製LOSは、当技術分野で公知のリポソーム製剤中にそれを提示することにより、よりよい抗原となり得る(例えばWO 96/40063およびそこに引用される参考文献を参照)。
【0039】
細菌からLOSを単離する方法は当技術分野で周知である(例えばWesphalおよびJannの温水-フェノール法(Meth. Carbo. Chem. 1965, 5:83-91)を参照)。またGalanosら1969, Eur J Biochem 9:245-249およびWuら1987, Anal Bio Chem 160:281-289も参照せよ。単離したLOSをコンジュゲートするための技術もまた公知である(例えばEP 941738(参照により本明細書に組み入れる)を参照)。
【0040】
本発明の目的のため、「T細胞エピトープの供給源を含む担体」とは、通常、ペプチド、または好ましくはポリペプチドもしくはタンパク質である。コンジュゲーション技術は当技術分野で周知である。典型的な担体は非定型インフルエンザ菌(Haemophilus influenzae)由来のプロテインD、破傷風毒素、ジフテリア毒素、CRM197または(特にナイセリアまたは髄膜炎菌の)ブレブ調製物に存在する外膜タンパク質を含む。
【0041】
本発明の好ましい単離されたLOS組成物としては、各LOSのオリゴ糖部分がT細胞エピトープの供給源を含む担体に任意にコンジュゲートしたL2およびL3単離LOSを含有する組成物、lgtB-髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープの供給源を含む担体に任意にコンジュゲートしているL2またはL3 LOSを含む組成物、ならびに最も好ましくは、lgtB-髄膜炎菌株由来のものに一致する構造を有し、各LOSのオリゴ糖部分がT細胞エピトープの供給源を含む担体に任意にコンジュゲートしているL2およびL3単離LOSを含有する組成物である。
【0042】
好ましくは、本発明のLOS組成物は無毒化されている。これは、分子からアシル鎖を除去する(しかし分子の防御効果は減少し得る)ヒドラジンもしくはアルカリ加水分解化学処理の公知の技術によって行われ得るが、しかし、好ましくはhtrB-もしくはmsbB-髄膜炎菌突然変異体(上記のような、特に莢膜多糖欠損株)からLOSを単離することにより、またはポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高い親和性を有する分子)、特にSAEP 2(上記)の、組成物への添加により行われ得る。
【0043】
本発明のLOSは単離された状態で(通常、リピドA部分が無傷のままであればミセル形態で)投与してもよく、またはリポソーム中に入れて投与してもよい。そのような場合、外膜タンパク質はリポソームに添加してもよく、またLOSを、オリゴ糖がT依存的抗原となるようにリポソーム内部でそのような外膜タンパク質にコンジュゲートしてもよい。これは、下記のブレブ内LOS架橋について記載されるものと類似した化学法によって行われ得る。
【0044】
ブレブの表面上に存在する外膜タンパク質へのLOSのオリゴ糖部分のブレブ内架橋(コンジュゲーション)
LOS(特に本発明のLOS)がブレブ製剤中に存在する場合、LOSは好ましくは、ブレブ調製物上にも存在する1以上の外膜タンパク質(例えば髄膜炎菌におけるPorAまたはPorB)へのLOSのコンジュゲーションを可能にする方法により、in situにおいてコンジュゲートされる。従って、本発明のさらなる態様は、その外膜中に、LOSにコンジュゲートした外膜タンパク質が組み込まれるグラム陰性細菌株由来のブレブ調製物である。LOSはコンジュゲートするためにブレブ調製物に添加してもよいが、LOSはブレブ調製物の表面に天然に存在することが好ましい。
【0045】
この方法は、ブレブ製剤中のLOS抗原の安定性および/または免疫原性(T細胞の助けを提供する)、および/または抗原性を、外膜の表面上の天然環境にあるLOSのように有利に増強し、すなわちその最も防御的な構造をとったT非依存的オリゴ糖免疫原に対し、T細胞の助けを与える。さらに、ブレブ内でのLOSのコンジュゲーションは、LOSの無毒化を生じ得る(理論に縛られることは望まないが、コンジュゲートされると、リピドA部分はより安定的に外膜中に埋没され、従って毒性を生じるように利用されにくいものと思われる)。従って、上記のhtrB-もしくはmsbB-変異体からのブレブの単離についての無毒化法、またはポリミキシンBの機能的等価物である非毒性ペプチドの組成物への添加による上記の無毒化法は、必要でないかも知れない(しかしさらなる安全のために組み合わせて添加され得る)。
【0046】
本発明のコンジュゲート化ブレブ調製物は、典型的には、ブレブ中のLOSの毒性が同量の全くコンジュゲートしていないLOSを有する同じブレブと比較して低減されている。LOS毒性は当業者によって容易に測定され得、それには例えばヨーロッパ薬局方におけるLOSウサギ発熱アッセイ(pyrogenicity assay)を用いることができる(実施例7参照)。
【0047】
本発明のコンジュゲート化ブレブ調製物は、コンジュゲートしたLOSが宿主内における免疫反応の誘発に適した立体構造を有し、それに由来する血清がコンジュゲートしていないLOS(好ましくはブレブ調製物が作製された由来の細菌上に存在し、最も好ましくはSBAアッセイにおける殺菌様式で存在する)との反応性を有する(結合可能である)ため、有利である。
【0048】
ナイセリアブレブがLOSにコンジュゲートする場合、および本明細書に記載するように、ブレブが1以上の免疫優性外膜タンパク質が下方制御された株由来である場合、好ましくは、PorAが下方制御されている場合にはPorBは下方制御されるべきではなく、かつその逆も同様である。このことによって、LOSの大部分を主要な外膜タンパク質と架橋することが可能になり、従ってブレブに存在する交差防御性の微量外膜抗原に対するコンジュゲートのあらゆる効果が最小化される。
【0049】
特に、本発明者らは、ブレブに存在するLOSがブレブ内様式で外膜タンパク質(これもまたブレブに存在する)にコンジュゲートしたブレブを含む組成物が、ブレブが由来する生物によって引き起こされる疾患の治療または予防のためのワクチンの基礎を形成しうることを見出した。ここでそのようなワクチンは毒性が低減しており(好ましくは実質的に無毒性)、および/またはその天然環境におけるLOSに対するT依存的殺菌反応を誘導することが可能である。
【0050】
従って、本発明はそのようなブレブ内LOSコンジュゲート化ブレブ調製物をさらに提供する。「ブレブ内」とは、ブレブ中に天然に存在するLOSが同じブレブ上に存在する外膜タンパク質とコンジュゲートしていることを意味する。好ましくは、ブレブはブレブを産生し得るあらゆるグラム陰性生物(WO 01/09350参照)から得られるものであり、かかるグラム陰性生物は好ましくはモラクセラ・カタラーリス(Moraxella catarrhalis)、非定型インフルエンザ菌またはナイセリア(最も好ましくは髄膜炎菌)である。
【0051】
そのようなブレブ調製物は、問題の細菌より単離することができ(WO 01/09350参照)、次にLOSのオリゴ糖部分の基(例えばNH2またはCOOH)をブレブ外膜タンパク質上の基(例えばNH2またはCOOH)に結合するための、既知のコンジュゲーション化学法に供すればよい。グルタルアルデヒド、ホルムアルデヒドまたはグルタルアルデヒド/ホルムアルデヒド混合物を使用した架橋技術を用いることができるが、EDACまたはEDAC/NHS等のより選択的な化学法を用いるのが好ましい(J.V. Staros,R.W. WrightおよびD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)ならびにBioconjugates Techniques. Greg T Hermanson(1996)pp173-176)。本発明に使用できるLOSおよびタンパク質分子間の共有結合を生じることの可能な他のコンジュゲーション化学法または処理は、EP 941738に記載される。
【0052】
好ましくは、ブレブ調製物は莢膜多糖の非存在下でコンジュゲートされる。ブレブは莢膜多糖を産生しない株(天然のもしくは突然変異によるもの)から単離することができ、または大半の混入莢膜多糖(60、70、80、90もしくは99%以上が除去された)および好ましくは全ての混入莢膜多糖から精製することができる。この方法において、ブレブ内LOSコンジュゲーション反応はさらに非常に効果的である。
【0053】
好ましくはブレブ内に存在するLOSの5、10、20、30、40、50、60、70、80、90または95%を超える割合が架橋/コンジュゲートされる。
【0054】
好ましくは、本発明のブレブは、精製LOSを標準物質として使用したSDS-PAGE電気泳動後の銀染色によって測定したブレブのLOS含量が、3〜30、5〜25、10〜25、15〜22、および最も好ましくはおよそまたは正確に20%LOS含量となるように調製された(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)。髄膜炎菌ブレブにおいて20%LOSを0.1%低DOC抽出で達成することができ、これは緩く保持されたLOS分子を除去することができるが、抗原の大部分は保存する。
【0055】
ブレブ内コンジュゲート化ブレブが髄膜炎菌由来である場合、由来する株が莢膜多糖を産生することができない突然変異株(例えば上記の突然変異株の1つ、特にsiaD-)であることが好ましい。髄膜炎菌疾患に対して有効な免疫原性組成物がL2およびL3ブレブの双方を含むこともまた好ましく、ここでL2およびL3 LOSは双方ともにブレブ外膜タンパク質にコンジュゲートしている。さらに、ブレブ内コンジュゲート化ブレブ内のLOS構造がlgtB-髄膜炎菌株由来のものと一致していることが好ましい。最も好ましい免疫原性組成物は以下のブレブ内-コンジュゲート化ブレブを含む:莢膜多糖を産生することができず、lgtB-である、L2もしくはL3突然変異髄膜炎菌株に由来するもの、莢膜多糖を産生することができない突然変異髄膜炎菌株由来のL2およびL3ブレブを含むもの、lgtB-である突然変異髄膜炎菌株由来のL2およびL3ブレブを含むもの、または最も好ましくは莢膜多糖を産生することができず、lgtB-である突然変異髄膜炎菌株由来のL2およびL3ブレブを含むもの。
【0056】
本発明に使用され得る典型的なL3髄膜炎菌株は、H44/76 menB株である。典型的なL2株は、B16B6 menB株または39E髄膜炎菌C型株または760676株である。
【0057】
上述のように、本発明のブレブはコンジュゲーションの作用によってある程度無毒化されるので、さらなる無毒化を必要としないが、しかし、さらなる無毒化法をさらなる安全性のために用いてもよい。それは例えばhtrB-もしくはmsbB-である髄膜炎菌株由来のブレブを使用することによるか、またはポリミキシンBの機能的等価物である非毒性ペプチド(リピドAに対して高い親和性を有する分子)(好ましくはSEAP 2)をブレブ組成物に(上記のように)添加することによる。LOSの(特にブレブ内様式における)コンジュゲーションは、このように、驚いたことに同量のコンジュゲートしていないLOSを含む調製物と比較して、LOSの低い毒性を示す。従って、(特に髄膜炎菌の)ブレブの無毒化のための一般的な方法は、さらにLOSのブレブ外膜タンパク質へのブレブ内コンジュゲーションによって提供され、かつLOSの無毒化のための方法もまた、ブレブ外膜タンパク質へLOSをコンジュゲートする手段によって提供される。
【0058】
上記の方法において、髄膜炎菌ブレブ、およびブレブを含む免疫原性組成物が提供され、これらは、毒性が低減しており(かつ好ましくは実質的に無毒性)、自己免疫の問題がなく、T依存的特徴を持ち、その天然環境に存在し、かつ潜在的に90%を超える髄膜炎菌株に対する殺菌性抗体反応(L2+L3組成物の場合)を誘導することのできる、重要な抗原としてのLOSを有する。
【0059】
1以上のMen A、C、YもしくはW莢膜多糖またはオリゴ糖(好ましくは少なくともMen C、またはMen AおよびMen C、またはMen CおよびMen Y)もまた、本発明のブレブの外膜タンパク質上に、同様にコンジュゲートされ得る。これはLOS架橋と同じ反応において行われ得るが、別々の(好ましくは後の)反応において行われることが好ましい。
【0060】
最適なブレブ内LOSコンジュゲーションの方法は、本発明のさらなる態様である。かかる方法は、(好ましくは、本明細書に記載されるように低パーセンテージのDOCを使用して)グラム陰性細菌からブレブを単離するステップ、ブレブに存在するLOSを(好ましくはそのオリゴ糖部分を介して)同じブレブに存在する外膜タンパク質にコンジュゲートするのに適した化学法を行うステップ、ブレブ内コンジュゲート化ブレブ調製物を単離するステップ、ならびに同じ方法によって作製されるが異なるLOS免疫型を有する、さらなるブレブ内コンジュゲート化ブレブ調製物を有するブレブ内コンジュゲートブレブ調製物を任意に製剤化するステップ(好ましくはL2およびL3ナイセリア/髄膜炎菌ブレブを混合する)、および/またはワクチン組成物を製造するために製薬上許容される賦形剤とともにブレブ調製物を製剤化するステップを組み込むべきである。
【0061】
ブレブ内コンジュゲーションは、好ましくは以下の方法のステップのうち1もしくは2工程または3工程全てを組み込むべきである:コンジュゲーションpHはpH 7.0を上回り、好ましくはpH 7.5以上(最も好ましくはpH 9以下)であるべきである、1〜5%、好ましくは2〜4%、最も好ましくは約3%ショ糖の条件が反応中維持されるべきである、コンジュゲーション反応におけるNaClは最小限にするべきであり、好ましくは0.1M、0.05M、0.01M、0.005M、0.001M以下であり、全く存在しないことが最も好ましい。これら全ての方法的特徴は、コンジュゲーション過程を通してブレブを安定にかつ溶液中に維持することを確実にするものである。
【0062】
EDAC/NHSコンジュゲーション法はブレブ内コンジュゲーションのために好ましい方法である。EDAC/NHSは、非常に高度な架橋が可能であるがゆえに濾過性に不利な影響を及ぼすホルムアルデヒドより好ましい。EDACはカルボン酸(LOS中のKDO等)と反応して活性エステル中間体を生成する。アミン求核試薬(PorB等の外膜タンパク質中のリジン等)存在下において、アミド結合はイソ尿素副生成物の放出を伴って形成される。しかしながら、EDAC仲介反応の効果は、スルホNHSエステル中間体の形成により増加し得る。スルホNHSエステルは、EDACのみとカルボキシレートとの反応より形成される活性エステルより長く水性溶液中に残存する。従って、この2段階の方法を用いて高収率のアミド結合形成を実現し得る。EDAC/NHSコンジュゲーションは、J.V. Staros,R.W. WrightおよびD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)ならびにBioconjugates Techniques. Greg T Hermanson(1996)pp173-176に記載される。好ましくは、0.01〜5 mg EDAC/mgブレブ、より好ましくは0.05〜1 mg EDAC/mgブレブを反応に使用する。使用するEDACの量はサンプル中に存在するLOSの量に依存し、このLOSの量はまたブレブを抽出するために使用するデオキシコレート(DOC)%に依存する。低パーセンテージ(%)DOC(例えば0.1%)においては多量のEDAC(1mg/mgおよびそれ以上)が使用されるが、しかし、高%DOC(例えば0.5%)においては少量のEDAC(0.025〜0.1mg/mg)が、過剰なブレブ内架橋を回避するために使用される。
【0063】
本発明の好ましい方法は、従って、EDAC/NHS存在下において、pHがpH7.0からpH9.0の間(好ましくは約pH7.5)で、1〜5%(好ましくは約3%)ショ糖中で、かつ任意に(上記のように)実質的にNaClを含まない条件において、ブレブをコンジュゲートし、反応混合物よりコンジュゲート化ブレブを単離するステップを含む、ブレブ内コンジュゲート化LOS(好ましくは髄膜炎菌の)を製造する方法である。
【0064】
この反応に続いて、反応時間の経過に伴いブレブ中のLOSの割合の増加に応じてLOS分子量が増加することを示すために、抗LOS(例えば抗L2または抗L3)モノクローナル抗体(mAb)を使用して反応混合物をウェスタン分離ゲルに供することができる。
【0065】
収量99%のブレブがそのような技術を用いて回収され得る。
【0066】
EDACは、LOSのT依存的免疫原性を改良するために十分な、LOSをOMPに架橋する点で優れたブレブ内架橋剤であることが見出されたが、しかしそれほど高度にそれを架橋しなかったのは、乏しい濾過性、凝集およびブレブ内架橋等の問題が発生したためであった。生じたブレブの形態は、コンジュゲートしていないブレブのものと類似している(電子顕微鏡による)。さらに上記のプロトコルは、過度に高度の架橋(ブレブの表面上に天然に存在する防御OMP(例えばTbpAまたはHsf)の免疫原性を低減し得る)が生じることを回避した。
【0067】
ブレブ単離のための技術
本発明の外膜小胞(OMVまたはブレブ)は、多くの公知の技術により単離され得る(Fredriksenら, NIPH Annals(1991), 14, 67-79、Zollingerら, J. Clin Invest(1979), 63, 836-848、Saundersら, Infect Immun(1999), 67, 113-119、J.J. Drabickら, Vaccine(1999), 18, 160-172)。これらは2つの主要な群に分類される。すなわち、デオキシコレート(約0.5%)を使用して髄膜炎菌由来のブレブを抽出するために技術と、低レベルのデオキシコレート(DOC)を使用するかまたは全くデオキシコレートを使用しない技術である。DOC非含有法では、ブレブはOMV中に高レベルのLOSを維持するという興味深い特徴を有する。このことはLOSが防御抗原であるワクチンにおいて有利である。DOC抽出ブレブと比較して、DOC非含有法により得られるOMV中のL3抗原(Ag)の濃度は約10倍高い。界面活性剤を用いない(好ましくはDOCを用いない)ブレブ調製法は、この理由により本発明の方法の目的にとって好ましいが、低レベルの界面活性剤(好ましくはDOC)を含むバッファーでの抽出もまた、ブレブ内で強固に相互作用するLOSの大半を残留する一方、より毒性があり緩く保持されたLOSを除去するステップであるという点で有利であり得る。典型的には、0〜0.5%および0.02〜0.4%、0.04〜3%または0.06〜2%界面活性剤(好ましくはDOC)がブレブ抽出に使用され、より好ましくは0.08〜0.15%、および最も好ましくはおよそまたは正確に0.1%を用いて、最適な量のLOSをブレブ中に安定に存在させて得る。LOSが1以上の上記の方法により無毒化されている場合には、DOC非含有(または低DOC〜0.3%DOCもしくはそれ以下の)抽出法が特に好ましい。
【0068】
本発明の全ての実施形態におけるブレブのLOS含量は、精製LOSを標準物質として使用したSDS-PAGE電気泳動後の銀染色によって測定した場合に(Tsai, J. Biol. Standardization(1986) 14:25-33の方法を参照)、3〜30、5〜25、10〜25、15〜22%であることが好ましく、かつ最も好ましくはおよそまたは正確に20%LOS含量である。Nmen L3 LOSをこの方法における標準物質として使用すると、一般的に、0.1%DOCで抽出されたNmen L3免疫型ブレブ中のLOS含量は約20%LOSであり、0.2%DOCで抽出した場合は約15%LOS、0.3%DOCでは約10%LOS、および0.5%DOCでは約5%LOSである。
【0069】
ワクチン組成物
本発明の免疫原性組成物は、製薬上許容される賦形剤の添加によってワクチン組成物として容易に製剤化され得る。
【0070】
さらに、上記のような本発明の精製LOS(好ましくはL2もしくはL3)を単離するステップまたは上記のような本発明の単離ブレブ(好ましくはL2もしくはL3免疫型を有する)を生成するステップ、およびそのLOSまたはブレブを製薬上許容される賦形剤とともに製剤化するステップを含む、本発明のナイセリア(好ましくは髄膜炎菌)免疫原性組成物またはワクチンの製造方法が提供される。好ましくは、本発明の両方の免疫型L2およびL3の精製LOS、または本発明の両方の免疫型L2およびL3のブレブ、またはL2の精製LOSおよびL3のブレブ(もしくはその逆)を、混合ステップで混合する。好ましくは、本発明の精製LOSまたはブレブは、単離された後に上記のようにコンジュゲートされている。さらなるリポソーム製剤化ステップもまた精製LOSに追加され得る(当技術分野で公知の技術を用いて。例えばWO 96/40063およびそこに引用される参考文献を参照せよ)。好ましくは、ブレブ調製物は低濃度のDOC(またはDOCなし)での抽出によって(上記のように)単離される。
【0071】
このようなL2およびL3の組み合わせ方法は、ほとんど全ての髄膜炎菌B株に対して有効なワクチンをもたらすことができる。
【0072】
上記の免疫原性組成物(または方法)は、血清型A、C、YまたはW由来の1以上(2、3もしくは4)の髄膜炎菌多糖またはオリゴ糖(上記のような、そのままのもしくはT細胞エピトープを含む担体にコンジュゲートしたもの)が組成物に添加されたものであり得る。好ましくは、少なくともCが添加(最も好ましくはコンジュゲート)されたものであり、より好ましくはAおよびCまたはYおよびCが添加(好ましくは全てコンジュゲート化)されたものであり、最も好ましくはA、C、YおよびWが添加(好ましくは全てコンジュゲート化)されたものである。有利には、コンジュゲート化されたインフルエンザ菌B型莢膜多糖またはオリゴ糖はまた、上記の組成物中に含有させて、広範な髄膜炎ワクチンを作製する。
【0073】
好ましくは、特にWO 94/08021において個別に具体的に示した組成物からなるまたはそれを含む組成物は、本発明において特許請求の範囲に含められていない。
【0074】
本発明のワクチン製剤
本発明の免疫原性組成物は、適切なアジュバントとともに製剤化することで本発明のワクチン組成物を作製することができる。
【0075】
適切なアジュバントは、水酸化アルミニウムゲル(ミョウバン)またはリン酸アルミニウム(好ましくは水酸化アルミニウム)等のアルミニウム塩を含むが、しかしカルシウム塩(特に炭酸カルシウム)、鉄または亜鉛でもよく、あるいはアシル化チロシン、アシル化糖、カチオンもしくはアニオン誘導体化多糖、またはポリホスファゼンの不溶性懸濁液でもよい。
【0076】
添加され得る適切なTh 1アジュバント系には、モノホスホリルリピドA、特に3-de-O-アシル化モノホスホリルリピドA(または他のLPSの非毒性誘導体)、およびアルミニウム塩(好ましくはリン酸アルミニウム)とモノホスホリルリピドA、好ましくは3-de-O-アシル化モノホスホリルリピドA(3D-MPL)(または非毒性LPS誘導体)との組み合わせを包含する。増強系は、モノホスホリルリピドAとサポニン誘導体の組み合わせ、特にWO 94/00153に記載されるようなQS21(もしくは他のサポニン)と3D-MPL(もしくは非毒性LPS誘導体)の組み合わせ、またはWO96/33739に記載されるような、QS21(もしくはサポニン)がコレステロールによりクエンチングされているより反応原性の低い組成物を用いる。水中油型乳濁液中のQS21、3D-MPLおよびトコフェロールを含む特に強力なアジュバント製剤は、WO 95/17210に記載されており、これは添加され得る好ましい製剤である。添加され得る他のアジュバントは、サポニン、より好ましくはQS21および/または水中油型乳濁液ならびにトコフェロールを含む。オリゴヌクレオチドを含む非メチル化CpG(WO 96/02555)もまた添加され得る。
【0077】
ワクチン調製物は一般的にVaccine Design(「The subunit and sdjuvant approach」(Powell M.F.およびNewman M.J.編)(1995)Plenum Press New York)に記載されている。
【0078】
免疫防御用量のワクチンが、全身経路または粘膜経路を介して投与され得る。これらの投与には、筋肉内、腹腔内、皮内もしくは皮下経路を通じた注入、または口腔/消化器(好ましくは鼻腔内投与)、呼吸器、尿生殖路への粘膜投与を介した注入が挙げられる。典型的には、各ワクチン用量中のブレブの量は、典型的な被ワクチン接種者において著しく有害な副作用を引き起こすことなく免疫防御反応を誘導する量が選択される。そのような量は、どの特定の免疫原が使用され、かつそれをどのように提示するかによって変動する。一般的に、各用量は各ブレブを1〜100μg、好ましくは5〜50μgおよび最も典型的には5〜25μgの範囲内で含むことが期待される。
【0079】
本発明のブレブ免疫原性組成物に対するさらなる改良
本発明の上記のブレブ組成物は、それらが由来するナイセリア株(淋菌および好ましくは髄膜炎菌、最も好ましくは髄膜炎菌Bを含む)が、ゲノム中への遺伝子のさらなるコピーを挿入することにより、または既存の遺伝子の上流により強力なプロモーターを導入することにより、または非改変株と比較して1.2、1.5、2、3、5もしくは10倍を超える抗原レベルを生じるよう改変された株を誘導することが可能なWO 01/09350に記載のあらゆる他の方法により上方制御された1以上の下記遺伝子(防御抗原をコードする)を有する場合、本発明のワクチンにおける有効性がさらに改良され得る:NspA(WO 96/29412)、Hsfまたはその末端切断物(WO 99/31132およびWO 01/55182、NhhAとしても知られる)、Hap(PCT/EP99/02766)、OMP85(WO 00/23595)、PilQ(PCT/EP99/03603)、PldA(PCT/EP99/06718)、FrpB(WO 96/31618)、TbpA(WO 92/03467、US5912336、WO 93/06861およびEP586266)、TbpB(WO 93/06861およびEP586266)、NadA(Comanducciら J. Exp. Med. 2002 195:1445-1454、NMB 1994)、FrpA/FrpC、または5以上の反復配列を含むこれらの抗原間に共通の部分(WO 92/01460; Thompsonら,(1993) J. Bacteriol. 175:811-818; Thompsonら,(1993) Infect. Immun. 61:2906-2911)、LbpA、LbpB(PCT/EP98/05117)、FhaB(WO 98/02547 配列番号38(ヌクレオチド3083-9025))、HasR(PCT/EP99/05989)、lipo02(PCT/EP99/08315)、Tbp2(WO 99/57280; NMB 0460)、MltA(WO 99/57280; NMB 0033)、TspA(WO 00/03003)、TspB(WO 00/03003)、ctrA(PCT/EP00/00135)、MafA(NMB 0652)、MafB(NMB 0643)、Omp26(NMB 0181)、アドヘシンX(NMB 0315)、アドヘシンY(NMB 0995)、アドヘシンZ(NMB 1119)およびOstA(NMB 0280)。NMB配列の例は、www.neisseria.orgのデータベースで見出すことができる。本明細書でHsfについて言及する場合、この用語はあらゆる場合においてHsf末端切断物と置換可能である(特にWO 01/55182に開示されたものにおいて)。
【0080】
HsfおよびTbpA(低もしくは高分子量形態、または低および高分子量形態の両方(EP 586266))、またはHsfおよびOMP85、またはOMP85およびTbpA(低もしくは高分子量形態、または低および高分子量形態の両方)、またはNspAおよびHsf、またはNspAおよびOMP85、またはNspAおよびTbpA(低もしくは高分子量形態、または低および高分子量形態の両方)の双方が、いずれも上方制御されている場合が特に好ましい。組成物中に2つのブレブが含まれている場合、各ブレブは異なる上方制御を受けていることが好ましい。高および低分子量のTbpAがどちらも上方制御される場合、これらは2つの形態のTbpAを天然において含む2つの株由来の組成物中に存在する、2つの別々のブレブにおいて上方制御されていることが好ましい。最も好ましくは、かかる2つの株がL2およびL3 LOS免疫型を有することである。TbpAは遺伝子工学的に上方制御させるか、またはナイセリア/髄膜炎菌産生株を鉄制限状態(例えば50〜70μMデスフェラル(メシル酸デフォロキサミン、Sigmaより入手可能)存在下)で増殖させることにより、上方制御させ得る。後者の方法を採用する場合、FrpB遺伝子発現を下方制御(好ましくは欠失)することが好ましいが、これはこの可変性抗原が鉄制限状態で単離される髄膜炎菌株より単離されたブレブにおいて免疫優性になり得るためである。
【0081】
好ましい実施形態において、本発明の組成物は、好ましくは高分子量TbpAおよびHsfが上方制御されている、lgtB-莢膜多糖-msbB-株由来のL3ブレブと、低分子量TbpAおよびOmp85が上方制御されている、lgtB-莢膜多糖-msbB-株由来のL2ブレブとを含む。より好ましくは、どちらのブレブもさらにPorAおよび/またはFrpBの発現、ならびに任意にOpCおよび/またはOpA発現が、下方制御されている。ブレブは最も好ましくは上記の低DOC法によって単離され、両ブレブ中のLOSは外膜タンパク質にブレブ内架橋をしている。
【0082】
ゴーストまたは死菌全細胞ワクチン
本発明者らは、ブレブに関わる上記組成物およびワクチンが、(同一の利点を有する)ゴーストまたは死菌全細胞調製物およびワクチンに関する方法にまで容易に拡張できることを意図している。グラム陰性株からゴースト調製物(完全なエンペロープを有する空の細胞)を製造する方法は当技術分野で周知である(例えばWO 92/01791を参照)。全細胞を殺滅して、ワクチンに使用するための不活性化細胞調製物を作製する方法もまた周知である。従って、本明細書を通して記載されるブレブを含む組成物およびワクチンは、等価物である本発明のゴーストおよび死菌全細胞調製物を含む同じ組成物またはワクチンにも当てはまることを意図している。
【0083】
本発明の組成物における血清殺菌アッセイ
血清殺菌アッセイは、本発明の免疫原性組成物中で組み合わせた場合に抗原間の相乗作用的な関係を評価するための好ましい方法である。
【0084】
そのような相乗反応は、抗原の組み合わせによって誘発されるSBAにより、各抗原によって別個に誘発されるSBAと比べて少なくとも50%、2倍、3倍、好ましくは4倍、5倍、6倍、7倍、8倍、9倍および最も好ましくは10倍高いものとして特徴付けられ得る。好ましくはSBAは、抗原が由来するのと同種の株、およびさらに好ましくは一群の異種の株に対して測定される(以下の代表的な群、例えばA-4クラスターに属するBZ10(B:2b:P1.2)、ET-37複合体に属するB16B6(B:2a:P1.2)、およびH44/76(B:15:P1.7,16)を参照)。SBAは、髄膜炎菌ワクチンの有効性を評価するための、最も一般的に受け入れられている免疫学的マーカーである(Perkinsら J Infect Dis. 1998, 177:683-691)。十分なSBAはあらゆる公知の方法により確認することができる。SBAは動物モデルまたはヒト被験体から得た血清を用いて行うことができる。
【0085】
ヒト血清を用いてSBAを行う好ましい方法は以下の通りである。血液サンプルは最初のワクチン接種の前、2回目のワクチン接種の2ヶ月後および3回目のワクチン接種の1ヶ月後に採取する(3回のワクチン接種は1年以内に行い、通常のヒト第1期ワクチン接種計画では、例えば0、2および4ヶ月、または0、1および6ヶ月の時点で投与される)。そのようなヒト第1期ワクチン接種計画は、1歳未満の幼児(例えばHibワクチン接種を同時に実施する)に実施することができ、または2〜4歳児もしくは青年もそのような第1期ワクチン接種計画でワクチン接種することによってSBAを試験することができる。妥当であれば、さらなる血液サンプルは初回ワクチン接種の6〜12ヶ月後、および追加免疫投与の1ヶ月後に採取され得る。
【0086】
(2〜4歳児もしくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、本発明の抗原が由来する髄膜炎菌株に対するSBA(抗体希釈)力価が(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が被験体の30%を超え、好ましくは40%を超え、より好ましくは50%を超え、最も好ましくは60%を超える場合、SBAは同種の殺菌活性を有する抗原またはブレブ調製物について十分であろう。
【0087】
当然ながら、異種の殺菌活性を有する抗原またはブレブ調製物は、それが由来する髄膜炎菌株に対して同様に十分なSBAを誘導することができるならば、同種の殺菌活性を有するブレブ調製物を構成することもできる。
【0088】
(2〜4歳児もしくは青年における、しかし好ましくは生後1年以内の乳児における)(第1期ワクチン接種計画の)3回目のワクチン投与の1ヶ月後に、髄膜炎菌の3種の異種株に対するSBA(抗体希釈)力価において(ワクチン接種前の力価と比較して)4倍に増加する被験体の割合が、被験体の20%を超え、好ましくは30%を超え、より好ましくは35%を超え、最も好ましくは40%を超える場合、SBAは異種の殺菌活性を有する抗原またはブレブ調製物について十分であろう。かかる試験は、異種の殺菌活性を有する抗原またはブレブ調製物が、様々な髄膜炎菌株に対する交差殺菌抗体を誘導し得るか否かの優れた指標である。3種の異種株は好ましくは互いに異なる電気泳動タイプ(ET)複合体、または多座配列タイピング(MLST)パターン(Maidenら PNAS USA 1998, 95:3140-5参照)を有し、さらに好ましくはその異種殺菌活性を有する抗原またはブレブ調製物が作製されるかまたは由来する起源の株とも異なるべきである。当業者は、髄膜炎菌において観察される遺伝的多様性、特に、重篤な病苦の原因として認識されているか、および/または強い病原性が認識されているMenB系統(Maidenら 前述)である髄膜炎菌B型株において観察される遺伝的多様性を反映した、異なるET-複合体を有する3種の株を容易に決定することは容易にできるであろう。例えば、使用され得る3種の株は以下のものである:A-4クラスターに属するBZ10(B:2b:P1.2)、ET-37複合体に属するB16B6(B:2a:P1.2)、およびET-5複合体に属するH44/76(B:15:P1.7,16)、または同じET/クラスターに属するあらゆる他の株。そのような株は、かかる株から作製されたもしくはそれらに由来する異種殺菌活性を有する抗原またはブレブ調製物を試験するために使用してもよく、例えば、ET-5複合体に属する髄膜炎菌株CU385(B:4:P1.15)を使用してもよい。使用され得る別のサンプル株は、系統3流行クローン(例えばNZ124(B:4:P1.7,4))由来のものである。別のET-37株は、NGP165(B:2a:P1.2)である。
【0089】
SBA活性を測定するための方法は、当技術分野において知られている。例えば、使用され得る方法がWO 99/09176の実施例10Cに記載されている。一般的には、試験される株の培養物を対数増殖期まで増殖させる(好ましくは、鉄枯渇条件下で、増殖培地にEDDA等の鉄キレート剤を添加することによる)。これを、BSAを加えた培地中(0.3%BSAを含むハンクス培地等)に懸濁し、約20000 CFU/mlに調整した作業用の細胞懸濁液を得てもよい。一連の反応混合物は、試験される血清の一連の2倍希釈物(好ましくは56℃で30分間熱不活性化されたもの)(例えば、50μl/ウェル容量)と試験される20000 CFU/mlの髄膜炎菌株の懸濁液(例えば、25μl/ウェル容量)との混合により、作製することができる。反応バイアルをインキュベートし(例えば37℃で15分間)、振とう(例えば210 rpmで)するべきである。最終反応混合物(例えば容量100μl)は、さらに補体源(例えば、最終容量の25%の予備試験済みの仔ウサギ血清、またはヒト血清学のためのヒト血清)を含み、上記のように(例えば37℃で60分間)インキュベートする。滅菌ポリスチレンU底96穴マイクロタイタープレートをこのアッセイに使用できる。マルチチャネルピペットを用いて、各ウェルからアリコート(例えば10μl)を採取し、ミューラーヒントン寒天プレート(好ましくは、1%イソビタレックス(Isovitalex)および1%熱不活性化ウマ血清を含む)に滴下し、それを(例えば、5%CO2中37℃で18時間)インキュベートしてもよい。好ましくは、個々のコロニーは、1アリコート当たり最大80 CFUまで計数できる。以下の3つの試験サンプルが対照として使用され得る:バッファー+細菌+補体、バッファー+細菌+不活性化補体、血清+細菌+不活性化補体。回帰計算によって、50%の細胞死滅に相当する希釈度の測定値を与えるようにデータを処理するプログラムを用いて、SBA力価が直接算出され得る。
【0090】
本特許明細書中に引用される全ての文献または特許出願は、参照により本明細書に組み入れられる。
【実施例】
【0091】
以下の実施例は、特に詳細に記載しない限りは当業者にとって周知でありルーチンである標準的な技術を用いて行われる。本実施例は例示的なものであり、本発明を限定するものではない。
【0092】
実施例1:
髄膜炎菌BのB莢膜多糖産生に関わるタンパク質をコードする遺伝子の欠失、PorA遺伝子の欠失、髄膜炎菌ブレブの表面上の様々な防御外膜タンパク質の上方制御、免疫優性タンパク質または生合成酵素の下方制御、およびブレブを単離する方法を記載する例は、WO 01/09350に記載される。
【0093】
実施例2:LOS:重要な交差防御抗原
潜在的な交差防御抗原としてのLOSの役割を評価するために、H44/76野生型(WT)の髄膜炎菌B株(L3 LOSを発現する)および「galE-様LOS」(lgtE- LOSに関しては短い構造を有する)を発現する改変H44/76株を使用し、2つの異なる方法に従ってブレブを生成した。第1の方法では高レベルのLOSを含むブレブを得るために0.1%DOCを使用し、第2の方法では生じたブレブ中に低レベルのLOSを得るために0.5%DOCを使用した。
【0094】
マウスに、1投与量当たりAl3+塩(リン酸ルミニウム)に吸着させたブレブ5μgおよび3D-MPLを、IM経路によって3回(0、21および28日目)注入した。3回目の注入の14日後に血液サンプルを採取した。
【0095】
抗L3 LOS ELISAは、プールした血清について精製L3 LOSを使用して行った。図3Aの結果は、0.1%DOC法がマウスにおいて抗LOS反応を誘発することのできるブレブを産生したことを明白に示す。これは、galE- LOSおよびL3 LOSが抗体の産生を誘導することができることを証明する。一方、0.5%DOCはそれがブレブワクチンにおいて重要な抗原として作用するためには過剰なLOSを抽出した。
【0096】
血清殺菌アッセイ
SBAは、個々の血清について異なるNmenB株を使用して行った:同種のWT H44/76株、PorA(-)H44/76株、および2つの異種の株(血清亜型に基づくもの)Cu385およびNZ124。これら4つの株はL3 LOSを発現する。5番目の株も加えられた。H44/76と比較して、この株(B16B6)はPorAに対してだけでなくLOS(これは免疫型L2株である)に対しても異種性である。
【0097】
図3Bの結果は、交差殺菌反応がL3株のみに対してであり、それはDOC 0.1% WTブレブを用いた場合のみ認められることを示す。DOC 0.1%galE-ブレブおよびDOC 0.5%WTブレブについては交差殺菌反応が観察されない。さらに、PorA抗体によって誘導される殺菌反応が血清型依存的であることは周知である。このことはまたDOC 0.5%WTブレブまたはgalE-ブレブを用いたこの実験において、そしてまたPorA(-)H44/76株を用いて行ったSBAデータでも観察される。
【0098】
これらの結果全ては、L3 LOSを高いパーセンテージで含むブレブによって誘導される交差殺菌反応が、LOS抗原に対する抗体(Abs)の産生によるものであることを示唆する。
【0099】
L3 LOSのみ(galE- LOSではない)が殺菌抗体の産生を誘発することができる。ELISAにおいて、DOC 0.1%galE-ブレブを用いて良好な抗LOS反応が観察されるが、この反応には生物学的な関連性がない(SBAではない)。
【0100】
さらに、この反応は、抗L3 LOS AbsがL2株ではなくL3株のみを死滅させるようなLOS免疫型特異的であるようでもあり、これはまた最適なワクチンが理想的には最適な適用範囲でL3およびL2 LOSを含むべきであることを示している。
【0101】
枯渇実験
WT DOC 0.1%ブレブにより誘導される反応が主として抗LOS抗体によることを示すために、血清プールを異なる濃度の精製L3 LOSを用いて枯渇させた。枯渇後、血清を、同種WT H44/76株に対する殺菌アッセイにおいて使用した。
【0102】
DOC 0.1%WTブレブに対して生じた血清を用いて得られた結果(図3C参照)は明白な投与範囲阻害を示し、これはこの調製物によって誘導される抗体の大半がLOSに対するものであることを示している(これはPorA(-)H44/76株を用いて得られたSBA結果を確認するものである)。反対に、WT DOC 0.5%によって誘導される反応は、PorA(-)H44/76株を用いて行ったSBAによって示され、またLOS枯渇によって示唆されるように、LOSに対するものではない
この結果は以下のL2 LOSについてのものと類似する。
【0103】
実施例3:L3および中間体(lgtB-)DOC非含有ブレブ(非無毒化LOS)を用いた実験は交差殺菌抗体を誘導した
使用したMC58髄膜炎菌誘導株はB:P1.7.16、opc-、siaD-である。この株を遺伝学的に改変して、L3(2G2株)または中間体エピトープ(2G EcoN1b-1株、2G2と同様であるがさらにlgtB-)またはLPSの短いバージョン(lgtE-であるC6株)のいずれかを発現させた。OMVを、通常の高(0.5%)DOC法またはDOC非含有法のどちらかに従って生成した。
【0104】
マウス(1群につき10匹)を筋肉内経路によって3回、0、20および28日目に免疫した。これらにはAl(OH)3上に製剤化されたブレブ1または10μg(タンパク質含量)を与えた。血液サンプルは、28日(Post II)および42日目(Post III)に採取した。
【0105】
殺菌アッセイは、プールされた血清について、同種の株(MC58およびH44/76)ならびに2種の異種の株(M97250687およびM9725078)を使用して、仔ウサギ血清を外来性補体源として用いて行った。
【0106】
以下の表は結果を要約する(50%死滅に対する殺菌力価)。
【表1】
【0107】
明らかに、L3(2g2)または中間体(2gecon1b-1)エピトープの存在は交差殺菌抗体を誘導し、一方、末端切断されたLPS株(C6)由来のブレブは低レベルの交差反応抗体を誘導する。これは特に1μgのOMVを注入した場合に示された。
【0108】
さらに、DOCで精製したOMVについて示されるように、ブレブのLPS含量の低下が交差殺菌抗体の誘導を低減する。増加したLPSとは別に、DOC非含有ブレブはまた、リポタンパク質のようなOMVとゆるやかに相互作用するいくつかのタンパク質を有利に保持することはできる。
【0109】
実施例4:L3 LOSおよび外膜タンパク質のブレブ内架橋
使用したMenBブレブは、SiaD-(従って莢膜多糖を発現しない)およびPorA-であるH44/76株(LOS免疫型L3)から得た。2種の異なる株を使用した:完全なL3(B1717株、siad(-) PorA(-) 完全なL3)および末端切断型L3(B1727株、siad(-) PorA(-) lgtB(-) TrL3)。
【0110】
EDAC/NHSコンジュゲーション法を公知の方法に従って用いて、ブレブ内でLOSとOMPを架橋し、LOSのオリゴ糖成分をT依存性抗原とした(EDAC/NHSは、非常に高度に架橋するがゆえに濾過性に不利な影響を与えることが分かったホルムアルデヒドよりも好ましい)。EDACはカルボン酸と反応して活性エステル中間体を生成する。アミン求核試薬の存在下において、アミド結合がイソ尿素副生成物の放出とともに形成される。しかしながら、EDAC仲介反応の効率はスルホNHSエステル中間体の形成を通して増加し得る。スルホNHSエステルは、EDACのみがカルボキシレートと反応することより形成される活性エステルより長く水性溶液中に存続する。従って、この2段階法を用いて高生成のアミド結合形成を実現し得る。EDAC/NHSコンジュゲーションは、J.V. Staros,R.W. WrightおよびD.M. Swingle. Enhancement by N-hydroxysuccinimide of water-soluble carbodiimide-mediated coupling reactions. Analytical chemistry 156:220-222(1986)ならびにBioconjugates Techniques. Greg T Hermanson(1996)pp173-176に記載されている。
【0111】
反応混合物は、1.5 mgスルホNHSおよび5 mg EDACを3%ショ糖(ブレブの安定性のため)中に1mL容量で含んだ。ブレブは0.025mg EDAC/mgブレブの比率で存在した。ブレブは2 mg/mlの濃度で存在し、pHをHCl 0.1NまたはNaOH 0.1Nで7.5に調整した。
【0112】
反応を室温で4時間そのままにし、その混合物を3%ショ糖を含む2 mMリン酸バッファー、pH 7.5に対して透析した。該混合物を次にSterivex G10 0.22μmで濾過した。99%の収量のブレブが回収された。
【0113】
この反応に続いて、抗L3モノクローナル抗体(mAb)を使用したウエスタンブロットを行うことができた。その反応を通じ、ゲル上で低分子量LOSはより微弱になり、かつ新たなより高分子量のバンドが現れる。より高分子量のこのバンドが優勢と見られ、コンジュゲート化LOSの大部分がPorBに共有結合するようになることを示していると思われる。
【0114】
EDACはLOSをOMPに不可逆的に架橋する点で優れたブレブ内架橋剤であり、かつLOSのT依存的免疫原性を改良するために十分であることが見出されたが、しかしそれほど高度にそれを架橋しなかったのは、乏しい濾過性、凝集およびブレブ内架橋等の問題が発生したためであった。生じたブレブの形態は、コンジュゲートしていないブレブのものと類似している(電子顕微鏡の観察による)。さらに上記のプロトコルは、過度に高度の架橋(ブレブの表面上に天然に存在する防御OMP(例えばTbpA)の免疫原性を低減し得る)が生じることを回避した。
【0115】
実施例5:L3および末端切断型(中間体、lgtB-)L3は、末端切断型(中間体 lgtB-; TrL3)L3 LOSを認識する殺菌抗体(Ab)の産生を誘導できる
OMV(ブレブ)は、MenB株H44/76 siaD- PorA- L3から、またはH44/76 siad- porA- TrL3から産生された。2つの異なる抽出を行い、使用したDOCの割合を0.1または0.5%のいずれかとした。2つの異なるアジュバント製剤:Al(OH)3、またはリン酸アルミニウム+3D-MPLもまた、評価した。マウス(OF1メスマウス、6〜8週齢、1群につき30匹)にIM経路によって3回(0、21および28日目)注入した(5μgブレブ/注入)。SBAを、PostII(28日)およびPostIII(42日目)血清(プールされた血清または個々の血清)について収集した。
【0116】
50%細胞死滅に関する幾何平均力価およびプール血清力価は、0.5%DOC抽出物と比較して0.1%DOCで抽出したブレブによって誘導された血清の方がより高かった。これは後者と比較して前者のブレブにおいて2.5倍量のLOSが存在する傾向があるという事実によっておそらく説明される。完全なL3 LOSまたは末端切断型L3 LOSを含むブレブを用いて誘導される血清のSBAの間に顕著な差異はなかった。ブレブがリン酸アルミニウム+3D-MPLによりアジュバント作用を受ける場合には、水酸化アルミニウムと比較してSBAが増加する。
【0117】
血清枯渇実験もまた行った。1 mg/mL精製L3またはtrL3 LOSを使用して血清を枯渇させ、次にこれらの枯渇血清においてSBAを行った。この結果は、血清のtrL3 LOS前処理によって殺菌Ab(抗L3抗体を含む)をほとんど完全に枯渇させることができ、また血清のL3 LOS前処理によって殺菌Ab(抗trL3抗体を含む)をほとんど完全に枯渇させることができることを示した。抗L3殺菌抗体(Ab)はこのようにtrL3 LOSと反応することができ、抗trL3殺菌抗体(Ab)はこのようにL3 LOSと反応することができる。さらに、L3およびtrL3 LOSの双方に存在するLOS構造に対する殺菌Abの特異性がこのように証明された。
【0118】
結論として、本発明者らは(OMVにおける)TrL3構造が、L3株に対する殺菌Abの産生を誘導することができることを証明した。枯渇実験との組み合わせにおいて、本発明者らはTrL3およびL3 LOSが免疫学的基礎に基づき非常に近い構造であり、L3株を死滅させることが可能な抗体を産出するためにtrL3を使用することができることを示した。
【0119】
実施例6:TrL3は、完全なL3構造による自己免疫の潜在的問題を解決する
L3およびtrL3構造が、防御抗体に関して免疫学的に非常に近い関係である場合、L3(およびL2)LOSに付随する可能性としての自己免疫の問題に関して、構造(ラクト-N-ネオテトラオース部分による)の間に何らかの差異はあるのだろうか。本発明者らは、寒冷凝集素がtrL3 LOSを認識することが可能か否かに注目することによってこの問題に取り組んできた。
【0120】
モノクローナル抗体(MAb)1B2-1B7(J Bio Chem 256(1981) 10967〜10972およびATCC受託番号TIB-189)は、低温でヒト成人赤血球(RBC)を凝集し、かつLNnT(ラクト-N-ネオテトラオース)と反応することが知られている。これは典型的な寒冷凝集素である。
【0121】
このモノクローナル抗体は、以下の実験において、L3髄膜炎菌株を死滅させることのできるMabL3.7.9モノクローナル抗体とともに使用された。
【0122】
これら2つのmAbは、ポリ-L-リジンでプレコート(1μg/ml、37℃で2時間)し、次に精製L3または精製TrL3 LOSでコート(5μg、4℃で一晩)したマイクロプレートを用いたELISAにおいて、使用された。かかるプレートを次にBSAで飽和させた(1%、室温で30分間)。その後、2種の抗体の各々を用いて標準ELISAを行った。
【0123】
結果(図4)は、Mab L379がL3およびTrL3と反応するが(図4B)、しかし1B2-1B7はL3 LOSに対してのみ反応する(図4A)ことを明らかに示す。従って、TrL3は、LNnT四糖を含む構造(L3 LOSおよびヒト赤血球等)に対して反応する寒冷凝集素によっては認識されないと言うことができる。
【0124】
TrL3 LOSは、このように防御エピトープを保持するには十分に長く、しかしヒト自己免疫の影響を示す可能性があるエピトープを喪失するには十分に短いという最適な特徴を有する。
【0125】
このことが本特許出願において提案する末端切断型L2(lgtB-)LOS構造の場合も同様であるはずがないことを示唆する根拠はない。
【0126】
実施例7:B1820 DOC 0.1%ブレブの発熱性/抗原性における架橋の影響
ブレブ(B1820株由来[siaD(-)PorA(-)FrpB(-)であり、上方制御された末端切断型Hsfを有するH44/76から得られ、lgtB(-)突然変異株であることによりL3が末端切断されており、デスフェラル(desferral)存在下で培養され、ブレブはDOC 0.1%を用いて抽出される])は、異なる濃度のEDAC(EDACがより多く存在するほど、より多くのブレブが架橋される)を使用して架橋された。架橋は、ブレブの無菌的濾過によって示されるとおりブレブ内でなされる。
【0127】
精製Nmen L3 LOSを標準物質として用いたSDS-PAGE電気泳動後の銀染色によって測定されるブレブ中のLOS含量は18%である(Tsai, J. Biol. Standardization(1986) 14:25〜33)。一般的に、0.1%DOCで抽出されるブレブ中のLOS含量は約20%LOS、0.2%DOCでは約15%LOS、0.3%DOCでは約10%LOS、そして0.5%DOCでは約5%LOSである。一般的に、10%のコンジュゲートしていないLOSまたはそれ以上を含むブレブは、許容できない発熱性を有する。
【0128】
ウサギにおける発熱性
2種の製剤(Al(OH)3またはAlPO4に吸着したブレブ)を試験し、発熱試験においてウサギにヨーロッパ薬局方に記載されるようにして500ng/kgをIV経路によって投与した。
【0129】
この結果は、ブレブの発熱性に対するブレブ内架橋のプラスの影響を(以下の表に)明確に示す。同じバッチのブレブを対照として使用し、あるいは異なるEDAC濃度を用いて架橋した。ブレブがより多く架橋される(EDACがより多い)ほど、その発熱性は低くなった。このことは2種の異なる製剤について観察された。
【表2】
【0130】
架橋したブレブの抗原性
上記ブレブ(非吸着)の抗原性を評価して、この架橋がブレブの抗原性に対して何らかの影響を有するか否かを調べた。ブレブの異なる調製物(架橋されているまたはされていないもの)をマイクロプレート上にコートした(10μg/ml、4℃で一晩)。洗浄および飽和の後、MAb L379の連続希釈物またはB1820 DOC 0.1%または0.5%で免疫したマウス由来の血清を該プレートに添加した(振盪しながら室温で30分間)。コートしたブレブに対する抗体の固定は、ビオチンと結合した抗マウスIgを用い、次にストレプトアビジンペルオキシダーゼ複合体を使用し、その後OPDおよびH2O2を用いた発色によって、示した。各マイクロウエルの吸光度はマイクロプレートリーダーを用いて測定した。
【0131】
この結果は、MAb L379(L3 LOSに対してだけでなく、TrL3 LOS(lgtB-突然変異体)とも反応でき、L3株に対して殺菌性である)は、未処理の(コンジュゲートしていない)B1820ブレブおよび異なる架橋ブレブを(どのようなEDAC濃度を使用しても)同じように認識することを示す。図5Aを参照せよ。EDAC 0.2および1で得たより高い反応は、これらのEDAC濃度でのブレブにおけるLOSのよりよい固定化、または少なくとも架橋ブレブにおけるLOSのよりよい安定性を反映し得たものであった。
【0132】
これらブレブの抗原性を評価するためにマウス血清もまた使用した。2種の異なる血清を使用した;第1はB1820 DOC 0.5%ブレブ(8%以下の低LOS含量を有し、主として抗タンパク質抗体を誘導するブレブ)を用いて免疫したマウスより得た。第2の血清はB1820 DOC 0.1%ブレブ(15%以上のLOS含量を有し、主としてLOSに対する交差殺菌Abを誘導するブレブ)を用いて免疫したマウスより得た。L379 MAbを用いて観察されるように、これら2種の血清で得た結果(各々図5Bおよび5C)は、未処理の(コンジュゲートしていない)ブレブおよび架橋ブレブの間に(どのようなEDAC濃度を使用しても)いかなる差異も示さなかった。
【0133】
結論として、LOSの抗原性は架橋によって影響されず、かつブレブのその「広範な」抗原性もまたEDAC処理によっては改変されなかったものと思われる。(高濃度のEDACでの)架橋が重要な防御抗原の免疫原性を損なわないことを確認するために、マウスにおける免疫原性実験が進行中である。しかしながら、架橋がDOC 0.5%抽出ブレブに対してEDAC 0.025を用いて為された場合に示される予備的な結果(実施例8)は、、EDAC処理後のこれらブレブの免疫原性における増加を示す。
【0134】
実施例8:架橋ブレブの免疫原性(EDAC 0.025mg化学法)
この実験において、ブレブはB1727株より産生された。この株は遺伝的に改変されたH44/76株であり、これはsiaD(-)PorA(-)trL3(lgtB-)HsfでありそしてTbpAが上方制御されている。ブレブは0.5%DOCを用いて抽出した。マウスは3回(0、21および28日目)、IM経路により免疫した。1回の注入で、それらマウスにはAl(OH)3に吸着したブレブ5μgを投与した。
【0135】
血清殺菌アッセイはH44/76株に対して、3回目の注入の14日後に採取した個々の血清について行った。その結果は、EDAC処理が応答者の数(100を超えるSBA力価を示すマウスの数)にプラスの影響を与えたことを示す。EDAC処理ブレブに対しては応答者が37%、非改変ブレブでは応答者は17%だけであった。
【0136】
製剤中の3D-MPLの欠如、および0.5%DOC抽出後のブレブ調製物における比較的低い割合のLOSは、低い反応性を説明する。
【表3】
【0137】
また抗Hsf ELISAを行い、架橋がこのタンパク質の免疫原性に影響を及ぼすか否かを調べた。(プールした血清について得た)その結果は、架橋がIgG抗Hsf反応に影響を及ぼさないことを示す。IgMは検出されなかった。
【表4】
【0138】
実施例9:TrL3 LOSデータ
以下の実験を評価した:
- LNnT(ラクト-N-ネオテトラオース)と反応することができる抗体の誘導に対するTrL3(lgtB(-)L3 LOS)の影響、
- 上記構築物の殺菌抗体の誘導。
【0139】
ブレブは2種の遺伝的に改変したH44/76株より作製した。どちらもsiaD(-)PorA(-)であるがしかし、一方はWT L3 LOSを産生し、もう一方はTrL3 LOS(lgtB(-))であった。これらのブレブは2つの異なる方法に従って作製され、高LOS含量(約18%、DOC 0.1%抽出を使用)または低LOS含量(5%近く、DOC 0.5%抽出を使用)を有するようにした。
【0140】
マウスは、3回(0、21および28日に)、IM経路によって、3D-MPLとともにまたはそれなしで、Al(OH)3に吸着したブレブ5μg(注入1回当たり)で免疫した。
【0141】
抗LNnT ELISA
方法:マイクロプレートは、スペーサー(ADH)を介してヒト血清アルブミンにコンジュゲートしたLNnTでコートした(PBS中1ml当たり5μgのコンジュゲート、マイクロウエル当たり100μl)。4℃で一晩インキュベートした後、プレートをPBS-BSA 1%で洗浄し飽和させた(室温で40分)。洗浄の後、PBS 0.2%、BSA 0.05%、Tween20中への連続希釈液を添加した(室温で30分)。IgGのLNnTへの固定は、ペルオキシダーゼ(Jackson)に結合した抗マウスIgGによって、続いてOPDAおよびH2O2によるインキュベーションによって明らかにされた。
【0142】
結果:陽性対照は1B2-1B7 MAbである。このMAbはLNnTおよびL3LOSと反応し(しかしTrL3 LOSとは反応しない)(上記実施例を参照)、かつヒト赤血球を凝集させる。陰性対照(-)はアジュバントのみで免疫したマウス由来の血清である。
【0143】
結果(図6)は、高LOS含量を有するL3ブレブ(DOC 0.1%)のみがLNnTと反応可能なIgGの産生を誘導することを明白に示す。同様のLOS含量を有するTrL3ブレブは、LNnTに対するIgGの産生を誘導せず、低量のLOSを含む双方のブレブ調製物(DOC 0.5%)が誘導しないのと同様である。
【0144】
H44/76株についてのSBA
H44/76株に対するSBAアッセイを、3回目の注入の14日後に採取した個々の血清について実施した。以下の結果はtrL3(lgtB(-))LOSブレブがL3 LOSと同等のレベルで殺菌抗体を誘導することを明白に示す(GMTの他、>1/100のSBA力価を有するマウスの数(=SC)を参照)。
【表5】
【0145】
実施例10:FrpBノックアウト
以下のデータは2つの前臨床試験の概要である。
【0146】
これらの実験において、2種の遺伝的に改変されたH44/76株を使用して、0.1%DOCを用いてブレブを生成した。この方法によって得たブレブは20%近いLOS含量を有する。
【0147】
2種のH44/76株は以下の通りであった:
- B1733:siaD(-) PorA(-) Tr(末端切断型)Hsfの上方制御 lgtB(-)
- B1820:siaD(-) PorA(-) TrHsfの上方制御 lgtB(-) FrpB(-)。
【0148】
LbpA/B、TbpA/B、FrpB(B1733中)等の鉄依存性タンパク質の産生を上方制御するように、株をデスフェラル存在下で増殖させた後にブレブを生成した。
【0149】
これらの異なるブレブ調製物はAl(OH)3に吸着させ、マウスにIM経路にて3週間隔で2回注入した。血液サンプルは2回目の投与の7日後に採取した。マウスには1回の注入当たり5μgのブレブを投与した。
【0150】
SBA結果
殺菌アッセイは3つのL3株について行った(同種の野生型株H44/76および2つの異種のL3株:NZ124およびM97250687)。この結果は、FrpB(-)(ノックアウト)(B1820)ブレブが、FrpB(+)ブレブ(B1733)よりも優れた異種交差殺菌反応(高い力価およびより優れた血清転換SC)を誘導することを明白に示す。同種反応は、FrpB欠失により低下するにも関わらず、十分なままである。
【0151】
これらのデータは、FrpBが、ブレブによって誘発される免疫反応において主要な駆動因子ではあるが、この外膜タンパク質は可変性が高いため、このタンパク質に対する抗体は同種株の死滅を誘導することだけしかできないことを示している。従って、ブレブ産生株におけるFrpBの欠失は、産生されたブレブワクチンの適用範囲を改善するための有利な手段である。
【表6】
【0152】
実施例11:ブレブの発熱性に対するmsbB(lpxL1)突然変異の影響
2つのNmenB株をこの評価に使用した:
- galE(-)である(従って莢膜多糖を生成できない)対照株。
【0153】
- msbB突然変異株:これはgalE(-)およびmsbB(-)である。
【0154】
これら2種の株から0.1%のDOCを使用してブレブを生成し、OMV(ブレブ)中のLOS含量が15%を超えるようにした。上記の実施例で述べたように、LOS含量が10%を超えるブレブ調製物は発熱性という観点から十分でなく、ヨーロッパ薬局方のウサギ発熱性アッセイに適合しない。
【0155】
上記のブレブを、ウサギにおける発熱性アッセイ(500ngのブレブ/kgをIV経路で注入)のためにAl(OH)3に対して製剤化した(50μgのOMV/500μgのAl3+塩)。
【0156】
以下の結果は、msbBの欠失(特に莢膜多糖を産生できない株において)により、15%を超えるLOS含量であっても、ウサギにおいて非発熱性のブレブの産生が可能になることを明白に示す。
【表7】
【0157】
結論:
lgtB(-)およびmsbB(-)突然変異を有する髄膜炎菌に由来し、かつより低い(例えば0.1%)デオキシコレート濃度で抽出されたL3およびL2ブレブを含む組成物は、髄膜炎菌Bに対する効果的で安全なワクチンに対する強力な基礎を提供する。ブレブ産生株は理想的には莢膜多糖合成欠損性であり、かつ該ブレブはブレブ内で外膜タンパク質に架橋しているLOSを有する。PorA(-)およびFrpB(-)のいずれかまたは両方が、交差殺菌効果を改良する上でさらに有用であり、Hsfおよび/またはTbpA抗原の上方制御も同様に有用である。
【特許請求の範囲】
【請求項1】
L2 LOS免疫型を有するナイセリア株もしくはL3 LOS免疫型を有するナイセリア株に由来し、ここで該株がlgtB-であるナイセリアブレブ調製物;または、L2 LOS免疫型を有するナイセリア株とL3 LOS免疫型を有するナイセリア株とに由来するブレブの混合物を含み、ここで場合によりその各株がlgtB-であるナイセリアブレブ調製物。
【請求項2】
ナイセリア株が、髄膜炎菌であり、好ましくは血清型Bである、請求項1記載のナイセリアブレブ調製物。
【請求項3】
ナイセリア株が莢膜多糖を合成できないものである、請求項1または2記載のナイセリアブレブ調製物。
【請求項4】
ナイセリア株が、以下の莢膜多糖遺伝子:ctrA、ctrB、ctrC、ctrD、synA、synB、synCまたは好ましくはsiaDのうち1つについて、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において同じ莢膜多糖遺伝子の発現が下方制御されている、請求項3記載のナイセリアブレブ調製物。
【請求項5】
ナイセリア株が、以下のリピドA遺伝子:msbBもしくはhtrBのいずれかまたは両方、好ましくは前者について、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において同じリピドA遺伝子の発現が下方制御されている、請求項1〜4のいずれか1項記載のナイセリアブレブ調製物。
【請求項6】
ナイセリア株が、以下の外膜タンパク質遺伝子:porA、porB、opA、opC、pilCまたはfrpBのうち1以上について、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において同じ外膜タンパク質遺伝子の発現が下方制御されている、請求項1〜5のいずれか1項記載のナイセリアブレブ調製物。
【請求項7】
ナイセリア株が、以下の外膜タンパク質遺伝子の組み合わせ:PorAおよびOpA、PorAおよびOpC、OpAおよびOpC、PorAおよびOpAおよびOpC、PorAおよびFrpB、OpCおよびFrpB、OpAおよびFrpB、PorAおよびOpAおよびOpCおよびFrpB、のいずれかについて、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失している、請求項6記載のナイセリアブレブ調製物。
【請求項8】
ナイセリア株が、以下の外膜タンパク質抗原:NspA、TbpA低、TbpA高、Hsf、Hap、OMP85、PilQ、NadA、LbpA、MltAのうち1以上について、発現が上方制御されているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において1以上の異なる外膜タンパク質抗原の発現が上方制御されている、請求項1〜7のいずれか1項記載のナイセリアブレブ調製物。
【請求項9】
以下の外膜タンパク質:PorA、PorB、OpA、OpC、PilCまたはFrpBのうち2以上について、それが由来する天然株と比較して発現が下方制御されているナイセリア株に由来する、ナイセリアブレブ調製物。
【請求項10】
ナイセリア株が、以下の外膜タンパク質の組み合わせ:PorAおよびOpA、PorAおよびOpC、OpAおよびOpC、PorAおよびOpAおよびOpC、PorAおよびFrpB、OpCおよびFrpB、OpAおよびFrpB、PorAおよびOpAおよびOpCおよびFrpB、のいずれかについて、それが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものである、請求項9記載のナイセリアブレブ調製物。
【請求項11】
請求項1〜10のいずれか1項記載のナイセリアブレブ調製物が由来するナイセリア株。
【請求項12】
免疫型L2および/またはL3のLOSを含む、請求項11記載のナイセリア株より単離されるLOS調製物。
【請求項13】
リポソーム製剤に入れた、請求項12記載のLOS調製物。
【請求項14】
含まれるLOSが、Tヘルパーエピトープの供給源、好ましくはタンパク質もしくは外膜タンパク質に、コンジュゲートしている、請求項1〜10のいずれか1項記載のナイセリアブレブ調製物または請求項12もしくは13記載のLOS調製物。
【請求項15】
ブレブ内架橋の工程により得られる、請求項14記載のナイセリアブレブ調製物。
【請求項16】
請求項1〜10または12〜15のいずれか1項記載のナイセリアブレブ調製物またはLOS調製物と、製薬上許容される賦形剤とを含む、免疫原性組成物またはワクチン。
【請求項17】
アジュバント、好ましくは水酸化アルミニウム、または3D-MPLおよびリン酸アルミニウムをさらに含む請求項16記載のワクチン。
【請求項18】
以下の株:髄膜炎菌血清型A、髄膜炎菌血清型C、髄膜炎菌血清型W-135、髄膜炎菌血清型Yおよびインフルエンザ菌(H. influenzae)b型、に由来する1以上のコンジュゲート化莢膜多糖またはオリゴ糖をさらに含む、請求項16または17記載のワクチン。
【請求項19】
請求項11記載のナイセリア株を培養し、そこからブレブを単離し、任意に、適切である場合にはL2ブレブとL3ブレブを組み合わせ、そしてそれらのブレブを製薬上許容される賦形剤とともに製剤化するステップを含む、請求項16記載のナイセリアブレブ調製物ワクチンの製造方法。
【請求項20】
単離ステップが、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2または0.08〜0.15%デオキシコレート、好ましくはおよそまたは正確に0.1%のデオキシコレートで抽出することにより行われる、請求項19記載の方法。
【請求項21】
LOSにコンジュゲートした外膜タンパク質が外膜に組み込まれているグラム陰性細菌株に由来する、ブレブ調製物。
【請求項22】
コンジュゲート化LOSの10、20、30、40、50、60、70、80、90、95または99%を超える割合が、ブレブの外膜中に組み込まれているリピドA部分、および/またはそれが投与された宿主において毒性を低減または遮断する環境中に組み込まれているリピドA部分を有する、請求項21記載のブレブ調製物。
【請求項23】
ブレブ中のLOSの毒性が、同量のコンジュゲートしていないLOSを有するブレブと比較して低減されている、請求項21または22記載のブレブ調製物。
【請求項24】
コンジュゲート化LOSの10、20、30、40、50、60、70、80、90、95または99%を超える割合が、宿主の免疫系に投与された場合にそれに対する殺菌抗体反応を誘導するのに適した天然の立体構造をとる、請求項21〜23のいずれか1項記載のブレブ調製物。
【請求項25】
コンジュゲート化LOSが、コンジュゲートしていないLOSに対して反応性の宿主における免疫反応の誘導に適した立体構造を有する、請求項21〜24のいずれか1項記載のブレブ調製物。
【請求項26】
外膜タンパク質がLOS分子のオリゴ糖または多糖部分にコンジュゲートしている、請求項21〜25のいずれか1項記載のブレブ調製物。
【請求項27】
外膜タンパク質およびLOS分子が、ブレブが由来するグラム陰性細菌株にとって天然のものである、請求項21〜26のいずれか1項記載のブレブ調製物。
【請求項28】
ブレブ内架橋の工程によって得ることができる、請求項21〜27のいずれか1項記載のブレブ調製物。
【請求項29】
ブレブに存在するLOSの10、20、30、40、50、60、70、80、90または95%を超える割合が、外膜タンパク質に架橋またはコンジュゲートしている、請求項21〜28のいずれか1項記載のブレブ調製物。
【請求項30】
莢膜多糖を産生しないグラム陰性株に由来するか、または莢膜多糖を含まないブレブ調製物に由来する、請求項21〜29のいずれか1項記載のブレブ調製物。
【請求項31】
莢膜多糖が、ブレブ調製物に組み込まれた外膜タンパク質にコンジュゲートしていない、請求項21〜30のいずれか1項記載のブレブ調製物。
【請求項32】
モラクセラ・カタラーリス(Moraxella catarrhalis)または非定型(non typeable)インフルエンザ菌株由来である、請求項21〜31のいずれか1項記載のブレブ調製物。
【請求項33】
ナイセリア株由来、好ましくはナイセリア・メニンジティディス(Neisseria meningitidis)由来である、請求項21〜31のいずれか1項記載のブレブ調製物。
【請求項34】
コンジュゲート化L2 LOS、コンジュゲート化L3 LOS、または、好ましくは少なくとも2つの異なるブレブに別々にコンジュゲートしたコンジュゲート化L2およびL3 LOSの混合物を含む、請求項33記載のブレブ調製物。
【請求項35】
LgtB-株由来であるか、またはLgtB-である株から得られたものと一致するようにLOSが末端切断構造を有する、請求項33または34記載のブレブ調製物。
【請求項36】
htrB-および/またはmsbB-株由来であるか、またはhtrB-および/もしくはmsbB-髄膜炎菌株より単離されたものと一致するようにLOSリピドA部分が第2のアシル鎖を欠失している、請求項32〜35のいずれか1項記載のブレブ調製物。
【請求項37】
請求項21〜36のいずれか1項記載のブレブ調製物と、製薬上許容される賦形剤とを含む、免疫原性組成物またはワクチン。
【請求項38】
アジュバント、好ましくは水酸化アルミニウム、または3D-MPLおよびリン酸アルミニウムをさらに含む、請求項37記載の免疫原性組成物またはワクチン。
【請求項39】
以下の株:髄膜炎菌血清型A、髄膜炎菌血清型C、髄膜炎菌血清型W-135、髄膜炎菌血清型Yおよびインフルエンザ菌b型、に由来する1以上のコンジュゲート化莢膜多糖またはオリゴ糖をさらに含む、請求項37または38記載の免疫原性組成物またはワクチン。
【請求項40】
ブレブ内でコンジュゲートしたブレブ調製物を、グラム陰性細菌株から製造する方法であって、但し、該株の外膜にはLOSにコンジュゲートした外膜タンパク質が組み込まれており、以下のステップを含む方法:
a)該グラム陰性株からブレブを単離するステップ、
b)該ブレブに存在するLOSのオリゴ糖部分を同じブレブに存在する外膜タンパク質にコンジュゲートするのに適した化学法を行うステップ、
c)ブレブ内でコンジュゲートしたブレブ調製物を単離するステップ、および
d)任意に、ブレブ内でコンジュゲートしたブレブ調製物を、同様の方法で作製されるが異なるLOS免疫型を有するブレブ内でコンジュゲートしたさらなるブレブ調製物とともに製剤化し、および/または該ブレブ調製物をワクチン組成物を製造するための製薬上許容される賦形剤とともに製剤化するステップ。
【請求項41】
ステップa)において、ブレブを例えば0〜0.3%、好ましくはおよそまたは正確に0.1%のような低濃度のデオキシコレートを使用して抽出する、請求項40記載の方法。
【請求項42】
ステップb)において、pHを7〜9の間、好ましくはおよそpH 7.5に維持する、請求項40または41記載の方法。
【請求項43】
ステップb)を、1〜5%、好ましくはおよそ3%ショ糖中で行う、請求項40〜42のいずれか1項記載の方法。
【請求項44】
ステップb)を、低NaCl濃度条件において行う、請求項40〜43のいずれか1項記載の方法。
【請求項45】
ステップb)を、EDAC/NHS化学法を用いて行う、請求項40〜44のいずれか1項記載の方法。
【請求項46】
ステップa)において、ブレブを、ナイセリア株、好ましくは髄膜炎菌株、最も好ましくは髄膜炎菌B株より単離する、請求項40〜45のいずれか1項記載の方法。
【請求項47】
前記株が莢膜多糖を作製することができないものであり、好ましくはsiaD-突然変異体である、請求項46記載の方法。
【請求項48】
前記株がlgtB-突然変異体である、請求項46または47記載の方法。
【請求項49】
前記株がmsbB-および/またはhtrB-である、請求項46〜48のいずれか1項記載の方法。
【請求項50】
前記株がL2 LOS免疫型を有する、請求項46〜49のいずれか1項記載の方法。
【請求項51】
前記株がL3 LOS免疫型を有する、請求項46〜50のいずれか1項記載の方法。
【請求項52】
ステップb)において、請求項50に記載の方法により製造されるL2免疫型を有する髄膜炎菌のブレブ内でコンジュゲートしたブレブ調製物を、請求項51に記載の方法により製造されるL3免疫型を有する髄膜炎菌のブレブ内でコンジュゲートしたさらなるブレブ調製物とともに組み合わせる、請求項46〜51のいずれか1項記載の方法。
【請求項1】
L2 LOS免疫型を有するナイセリア株もしくはL3 LOS免疫型を有するナイセリア株に由来し、ここで該株がlgtB-であるナイセリアブレブ調製物;または、L2 LOS免疫型を有するナイセリア株とL3 LOS免疫型を有するナイセリア株とに由来するブレブの混合物を含み、ここで場合によりその各株がlgtB-であるナイセリアブレブ調製物。
【請求項2】
ナイセリア株が、髄膜炎菌であり、好ましくは血清型Bである、請求項1記載のナイセリアブレブ調製物。
【請求項3】
ナイセリア株が莢膜多糖を合成できないものである、請求項1または2記載のナイセリアブレブ調製物。
【請求項4】
ナイセリア株が、以下の莢膜多糖遺伝子:ctrA、ctrB、ctrC、ctrD、synA、synB、synCまたは好ましくはsiaDのうち1つについて、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において同じ莢膜多糖遺伝子の発現が下方制御されている、請求項3記載のナイセリアブレブ調製物。
【請求項5】
ナイセリア株が、以下のリピドA遺伝子:msbBもしくはhtrBのいずれかまたは両方、好ましくは前者について、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において同じリピドA遺伝子の発現が下方制御されている、請求項1〜4のいずれか1項記載のナイセリアブレブ調製物。
【請求項6】
ナイセリア株が、以下の外膜タンパク質遺伝子:porA、porB、opA、opC、pilCまたはfrpBのうち1以上について、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において同じ外膜タンパク質遺伝子の発現が下方制御されている、請求項1〜5のいずれか1項記載のナイセリアブレブ調製物。
【請求項7】
ナイセリア株が、以下の外膜タンパク質遺伝子の組み合わせ:PorAおよびOpA、PorAおよびOpC、OpAおよびOpC、PorAおよびOpAおよびOpC、PorAおよびFrpB、OpCおよびFrpB、OpAおよびFrpB、PorAおよびOpAおよびOpCおよびFrpB、のいずれかについて、それらが由来する天然株と比較して発現が下方制御され、好ましくは欠失している、請求項6記載のナイセリアブレブ調製物。
【請求項8】
ナイセリア株が、以下の外膜タンパク質抗原:NspA、TbpA低、TbpA高、Hsf、Hap、OMP85、PilQ、NadA、LbpA、MltAのうち1以上について、発現が上方制御されているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株は好ましくはそれぞれの株において1以上の異なる外膜タンパク質抗原の発現が上方制御されている、請求項1〜7のいずれか1項記載のナイセリアブレブ調製物。
【請求項9】
以下の外膜タンパク質:PorA、PorB、OpA、OpC、PilCまたはFrpBのうち2以上について、それが由来する天然株と比較して発現が下方制御されているナイセリア株に由来する、ナイセリアブレブ調製物。
【請求項10】
ナイセリア株が、以下の外膜タンパク質の組み合わせ:PorAおよびOpA、PorAおよびOpC、OpAおよびOpC、PorAおよびOpAおよびOpC、PorAおよびFrpB、OpCおよびFrpB、OpAおよびFrpB、PorAおよびOpAおよびOpCおよびFrpB、のいずれかについて、それが由来する天然株と比較して発現が下方制御され、好ましくは欠失しているものである、請求項9記載のナイセリアブレブ調製物。
【請求項11】
請求項1〜10のいずれか1項記載のナイセリアブレブ調製物が由来するナイセリア株。
【請求項12】
免疫型L2および/またはL3のLOSを含む、請求項11記載のナイセリア株より単離されるLOS調製物。
【請求項13】
リポソーム製剤に入れた、請求項12記載のLOS調製物。
【請求項14】
含まれるLOSが、Tヘルパーエピトープの供給源、好ましくはタンパク質もしくは外膜タンパク質に、コンジュゲートしている、請求項1〜10のいずれか1項記載のナイセリアブレブ調製物または請求項12もしくは13記載のLOS調製物。
【請求項15】
ブレブ内架橋の工程により得られる、請求項14記載のナイセリアブレブ調製物。
【請求項16】
請求項1〜10または12〜15のいずれか1項記載のナイセリアブレブ調製物またはLOS調製物と、製薬上許容される賦形剤とを含む、免疫原性組成物またはワクチン。
【請求項17】
アジュバント、好ましくは水酸化アルミニウム、または3D-MPLおよびリン酸アルミニウムをさらに含む請求項16記載のワクチン。
【請求項18】
以下の株:髄膜炎菌血清型A、髄膜炎菌血清型C、髄膜炎菌血清型W-135、髄膜炎菌血清型Yおよびインフルエンザ菌(H. influenzae)b型、に由来する1以上のコンジュゲート化莢膜多糖またはオリゴ糖をさらに含む、請求項16または17記載のワクチン。
【請求項19】
請求項11記載のナイセリア株を培養し、そこからブレブを単離し、任意に、適切である場合にはL2ブレブとL3ブレブを組み合わせ、そしてそれらのブレブを製薬上許容される賦形剤とともに製剤化するステップを含む、請求項16記載のナイセリアブレブ調製物ワクチンの製造方法。
【請求項20】
単離ステップが、0〜0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2または0.08〜0.15%デオキシコレート、好ましくはおよそまたは正確に0.1%のデオキシコレートで抽出することにより行われる、請求項19記載の方法。
【請求項21】
LOSにコンジュゲートした外膜タンパク質が外膜に組み込まれているグラム陰性細菌株に由来する、ブレブ調製物。
【請求項22】
コンジュゲート化LOSの10、20、30、40、50、60、70、80、90、95または99%を超える割合が、ブレブの外膜中に組み込まれているリピドA部分、および/またはそれが投与された宿主において毒性を低減または遮断する環境中に組み込まれているリピドA部分を有する、請求項21記載のブレブ調製物。
【請求項23】
ブレブ中のLOSの毒性が、同量のコンジュゲートしていないLOSを有するブレブと比較して低減されている、請求項21または22記載のブレブ調製物。
【請求項24】
コンジュゲート化LOSの10、20、30、40、50、60、70、80、90、95または99%を超える割合が、宿主の免疫系に投与された場合にそれに対する殺菌抗体反応を誘導するのに適した天然の立体構造をとる、請求項21〜23のいずれか1項記載のブレブ調製物。
【請求項25】
コンジュゲート化LOSが、コンジュゲートしていないLOSに対して反応性の宿主における免疫反応の誘導に適した立体構造を有する、請求項21〜24のいずれか1項記載のブレブ調製物。
【請求項26】
外膜タンパク質がLOS分子のオリゴ糖または多糖部分にコンジュゲートしている、請求項21〜25のいずれか1項記載のブレブ調製物。
【請求項27】
外膜タンパク質およびLOS分子が、ブレブが由来するグラム陰性細菌株にとって天然のものである、請求項21〜26のいずれか1項記載のブレブ調製物。
【請求項28】
ブレブ内架橋の工程によって得ることができる、請求項21〜27のいずれか1項記載のブレブ調製物。
【請求項29】
ブレブに存在するLOSの10、20、30、40、50、60、70、80、90または95%を超える割合が、外膜タンパク質に架橋またはコンジュゲートしている、請求項21〜28のいずれか1項記載のブレブ調製物。
【請求項30】
莢膜多糖を産生しないグラム陰性株に由来するか、または莢膜多糖を含まないブレブ調製物に由来する、請求項21〜29のいずれか1項記載のブレブ調製物。
【請求項31】
莢膜多糖が、ブレブ調製物に組み込まれた外膜タンパク質にコンジュゲートしていない、請求項21〜30のいずれか1項記載のブレブ調製物。
【請求項32】
モラクセラ・カタラーリス(Moraxella catarrhalis)または非定型(non typeable)インフルエンザ菌株由来である、請求項21〜31のいずれか1項記載のブレブ調製物。
【請求項33】
ナイセリア株由来、好ましくはナイセリア・メニンジティディス(Neisseria meningitidis)由来である、請求項21〜31のいずれか1項記載のブレブ調製物。
【請求項34】
コンジュゲート化L2 LOS、コンジュゲート化L3 LOS、または、好ましくは少なくとも2つの異なるブレブに別々にコンジュゲートしたコンジュゲート化L2およびL3 LOSの混合物を含む、請求項33記載のブレブ調製物。
【請求項35】
LgtB-株由来であるか、またはLgtB-である株から得られたものと一致するようにLOSが末端切断構造を有する、請求項33または34記載のブレブ調製物。
【請求項36】
htrB-および/またはmsbB-株由来であるか、またはhtrB-および/もしくはmsbB-髄膜炎菌株より単離されたものと一致するようにLOSリピドA部分が第2のアシル鎖を欠失している、請求項32〜35のいずれか1項記載のブレブ調製物。
【請求項37】
請求項21〜36のいずれか1項記載のブレブ調製物と、製薬上許容される賦形剤とを含む、免疫原性組成物またはワクチン。
【請求項38】
アジュバント、好ましくは水酸化アルミニウム、または3D-MPLおよびリン酸アルミニウムをさらに含む、請求項37記載の免疫原性組成物またはワクチン。
【請求項39】
以下の株:髄膜炎菌血清型A、髄膜炎菌血清型C、髄膜炎菌血清型W-135、髄膜炎菌血清型Yおよびインフルエンザ菌b型、に由来する1以上のコンジュゲート化莢膜多糖またはオリゴ糖をさらに含む、請求項37または38記載の免疫原性組成物またはワクチン。
【請求項40】
ブレブ内でコンジュゲートしたブレブ調製物を、グラム陰性細菌株から製造する方法であって、但し、該株の外膜にはLOSにコンジュゲートした外膜タンパク質が組み込まれており、以下のステップを含む方法:
a)該グラム陰性株からブレブを単離するステップ、
b)該ブレブに存在するLOSのオリゴ糖部分を同じブレブに存在する外膜タンパク質にコンジュゲートするのに適した化学法を行うステップ、
c)ブレブ内でコンジュゲートしたブレブ調製物を単離するステップ、および
d)任意に、ブレブ内でコンジュゲートしたブレブ調製物を、同様の方法で作製されるが異なるLOS免疫型を有するブレブ内でコンジュゲートしたさらなるブレブ調製物とともに製剤化し、および/または該ブレブ調製物をワクチン組成物を製造するための製薬上許容される賦形剤とともに製剤化するステップ。
【請求項41】
ステップa)において、ブレブを例えば0〜0.3%、好ましくはおよそまたは正確に0.1%のような低濃度のデオキシコレートを使用して抽出する、請求項40記載の方法。
【請求項42】
ステップb)において、pHを7〜9の間、好ましくはおよそpH 7.5に維持する、請求項40または41記載の方法。
【請求項43】
ステップb)を、1〜5%、好ましくはおよそ3%ショ糖中で行う、請求項40〜42のいずれか1項記載の方法。
【請求項44】
ステップb)を、低NaCl濃度条件において行う、請求項40〜43のいずれか1項記載の方法。
【請求項45】
ステップb)を、EDAC/NHS化学法を用いて行う、請求項40〜44のいずれか1項記載の方法。
【請求項46】
ステップa)において、ブレブを、ナイセリア株、好ましくは髄膜炎菌株、最も好ましくは髄膜炎菌B株より単離する、請求項40〜45のいずれか1項記載の方法。
【請求項47】
前記株が莢膜多糖を作製することができないものであり、好ましくはsiaD-突然変異体である、請求項46記載の方法。
【請求項48】
前記株がlgtB-突然変異体である、請求項46または47記載の方法。
【請求項49】
前記株がmsbB-および/またはhtrB-である、請求項46〜48のいずれか1項記載の方法。
【請求項50】
前記株がL2 LOS免疫型を有する、請求項46〜49のいずれか1項記載の方法。
【請求項51】
前記株がL3 LOS免疫型を有する、請求項46〜50のいずれか1項記載の方法。
【請求項52】
ステップb)において、請求項50に記載の方法により製造されるL2免疫型を有する髄膜炎菌のブレブ内でコンジュゲートしたブレブ調製物を、請求項51に記載の方法により製造されるL3免疫型を有する髄膜炎菌のブレブ内でコンジュゲートしたさらなるブレブ調製物とともに組み合わせる、請求項46〜51のいずれか1項記載の方法。
【図1】

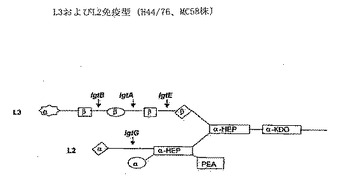
【図2】
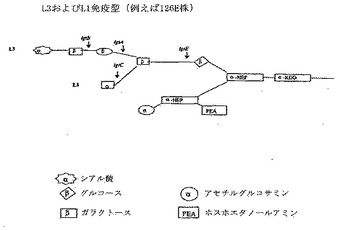
【図3】
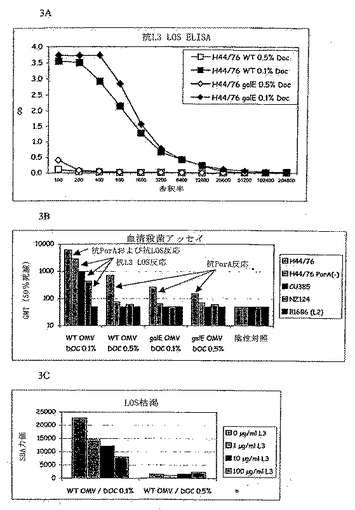
【図4】
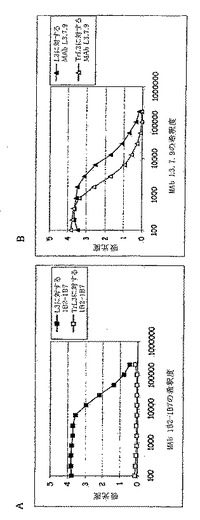
【図5】
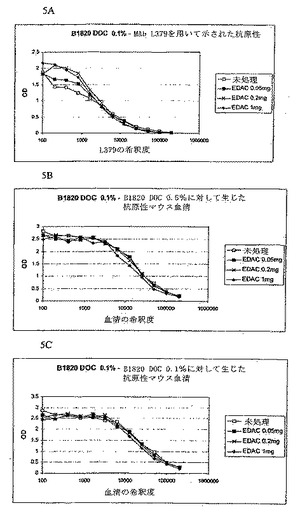
【図6】
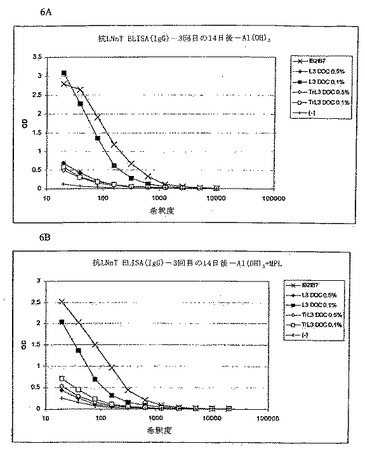
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2011−142916(P2011−142916A)
【公開日】平成23年7月28日(2011.7.28)
【国際特許分類】
【出願番号】特願2011−47056(P2011−47056)
【出願日】平成23年3月4日(2011.3.4)
【分割の表示】特願2005−506112(P2005−506112)の分割
【原出願日】平成15年7月31日(2003.7.31)
【出願人】(305060279)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (169)
【Fターム(参考)】
【公開日】平成23年7月28日(2011.7.28)
【国際特許分類】
【出願日】平成23年3月4日(2011.3.4)
【分割の表示】特願2005−506112(P2005−506112)の分割
【原出願日】平成15年7月31日(2003.7.31)
【出願人】(305060279)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (169)
【Fターム(参考)】
[ Back to top ]