
ワクチン
【課題】細菌、とりわけ微生物Photobacterium damselae subsp.piscicidaによる感染に対して、魚類を予防および/または治療処理するワクチン組成物の提供。
【解決手段】特定量の鉄を含む培地で細菌細胞を培養した、Photobacterium damselaeの培養物物由来の生物材料を含むワクチン組成物。培養物を、該成分が生産された後に好ましくはホルマリンで処理することによって、使用前に該微生物を殺菌するために処理する。宿主細胞への侵入に関与し、かつ過剰の鉄を含む培地中で培養物を増殖させたときに多量に生成する該成分中の2成分は、魚類に注入したときに、魚類細胞内への微生物の侵入を防止する抗体の生成を誘発する。これらの抗体は、微生物による感染から魚類を保護する。
【解決手段】特定量の鉄を含む培地で細菌細胞を培養した、Photobacterium damselaeの培養物物由来の生物材料を含むワクチン組成物。培養物を、該成分が生産された後に好ましくはホルマリンで処理することによって、使用前に該微生物を殺菌するために処理する。宿主細胞への侵入に関与し、かつ過剰の鉄を含む培地中で培養物を増殖させたときに多量に生成する該成分中の2成分は、魚類に注入したときに、魚類細胞内への微生物の侵入を防止する抗体の生成を誘発する。これらの抗体は、微生物による感染から魚類を保護する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、細菌による感染に対して魚類を予防処理するため、とりわけPhotobacterium damselae subsp.piscicida等の細菌による感染から魚類を保護するための、ワクチンおよび医薬として使用する新規な組成物に関する。
【背景技術】
【0002】
旧名Photobacterium damsela subsp.piscicidaのPhotobacterium damselae subsp.piscicidaは、Pasteurella piscicidaから再区分され、小サブユニット16sリボソームRNAの相同性に基づいて、vibrionaceae科に割当て直された(Gaulthier等(1995)、Int.J.Syst.Bacteriol.45:139−144)。P.damselaeは、仮性結核と普通呼ばれる重篤な致死性疾患を暖水海水魚に引き起こし、主要な器官や組織に感染して、筋肉組織中に特徴的な潰瘍または偽結核結節を生じる。
【0003】
抗生物質耐性の出現、ならびに、感染したとき魚は摂食を止め、したがって抗生物質入り餌を消費しないことが、化学療法の失敗原因と示唆されてきた。その上、宿主内コロニー形成中に、細胞内に生存する時期が存在することを示唆する証拠が益々増えている(Barnes等(1999)、Microbiology 145:483−494)。したがって、免疫による予防が、この疾患を抑制する最も望ましい手段と思われる。
【0004】
ホルマリン殺菌細菌(バクテリン)に基づく様々なワクチン接種戦略が提案されてきたが、これらの多くは再現性のある保護効果を生じることができなかった(Romalde & Magarinos(1997) Fish Vaccinology,Karger,Basel,Switzerland,pp167−177)。これは、感染剤に対する宿主の低い抗体反応性を反映するものと思われる(Arijo等、(1997) Fish Shellfish Immunol.8:63−72)。その上、これらの製剤中に含まれる保護抗原については殆ど何もわかっていない。鉄制限外膜蛋白質(IROMP)に基づく製剤および細胞外生成物の豊富なバクテリン(ECP)が記載されてきた(Magarinos等、(1994)Bull.Eur.Ass.Fish Pathol.14:120−122)が、これらは商品として無効であった。
【0005】
細菌が絶対的に鉄を必要とし、P.damselaeも例外でないことは、確立された事実である。培地中に十分な量を含めないか、またはその利用率を減少させるキレート剤を含めることにより、鉄が制限的である条件下で、様々な高親和性鉄取込み系が発現することが示された(Magarinos等、(1994)Appl.Environ.Microbiol.60:2990−2998)。これらは、シデロフォア(siderophore)および鉄制限外膜蛋白質(IROMP)を含む。Aeromonas salmonicida(Hirst & Ellis(1994) Fish Shellfish Immunol.4:29−45)、Pasteurella haemolytica(英国特許明細書第8805253号 W Donachie、英国)等の他の細菌種では、ワクチン中の保護抗原としてIROMPを開発して成功した。しかし、これらが、P.damselaeにおいて保護作用を示すとは認められない(Romalde & Magari−os(1997) Fish Vaccinology,Karger,Basel,Switzerland,pp167−177)。この保護作用の欠如が、P.damselaeによる感染の戦略を反映するものと思われる。鉄制限下で、P.damselaeは、プロテアーゼ活性の増加(Bakopolous(1997) J.Fish Dis.20:297−305)および莢膜多糖(capsular polysaccharide)の増加を起こす。莢膜多糖は、血清殺菌力に対する耐性を増加させ、宿主マクロファージによる食作用を防止する(Arijo等、(1998) Fish Shellfish Immunol.8:63−72)。このことは、マクロファージの呼吸バースト中に生成する反応性酸素種による殺菌に対する適応性を欠いた結果として(Barnes等、(1999)Microbiology 145:483−495)、P.damselaeが食細胞による攻撃に耐性を示さないことから(Skarmeta等、(1995)Dis.Aquat.Org.23:51−57)、明らかである。しかし、赤血球の溶解により増加した蛋白分解活性が、ヘム形態の鉄を遊離させることができる。ヘムは莢膜によって吸収され(Ana Do Vale,Pers.Comm.)、その鉄はIROMPによって膜を通過する。したがって、鉄取込みの第1段階として作用する莢膜多糖のために、罹病過程の間にIROMPが露出することはない。鉄濃度が増加するにつれ、莢膜生産は減少し、IROMPは不活性化され、蛋白分解活性は検出不能となる(Bakopolous(1997)J.Fish Dis.20:297−305)。この結果、P.damselaeは、抗体、マクロファージのいずれの攻撃も受け易くなると思われる。しかし、この時点でその細菌は、非食細胞性魚類細胞に接着し(Yoshida等、(1997) J.Fish Dis.20:77−88)、侵入する(Magari−os等、(1996)FEMS Microbiology Lett. 138:29−34)ことができ、したがって抗体や食細胞による攻撃を回避する。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】英国特許第8805253号明細書
【非特許文献】
【0007】
【非特許文献1】Gaulthier等(1995)、Int.J.Syst.Bacteriol.45:139−144
【非特許文献2】Romalde & Magarinos(1997) Fish Vaccinology,Karger,Basel,Switzerland,pp167−177
【非特許文献3】Arijo等、(1997) Fish Shellfish Immunol.8:63−72
【非特許文献4】Magarinos等、(1994)Bull.Eur.Ass.Fish Pathol.14:120−122
【非特許文献5】Magarinos等、(1994)Appl.Environ.Microbiol.60:2990−2998
【非特許文献6】Hirst & Ellis(1994) Fish Shellfish Immunol.4:29−45
【非特許文献7】Bakopolous(1997) J.Fish Dis.20:297−305
【非特許文献8】Barnes等、(1999)Microbiology 145:483−495
【非特許文献9】Skarmeta等、(1995)Dis.Aquat.Org.23:51−57
【非特許文献10】Yoshida等、(1997) J.Fish Dis.20:77−88
【非特許文献11】Magari−os等、(1996)FEMS Microbiology Lett. 138:29−34
【発明の概要】
【0008】
本発明の一目的は、Photobacterium damselae等の細菌による感染から、魚類を保護するワクチンを提供することである。
【0009】
本発明の他の目的は、このようなワクチンを生産する方法を提供することである。
【0010】
本発明によれば、その細菌の通常の増殖に必要な量より過剰の鉄を含む培地で、その細菌細胞を培養したことを特徴とするPhotobacterium damselaeの培養物由来の生物材料を含む、ワクチンおよび/または治療組成物が提供される。
【0011】
「過剰の鉄を含む」により、培地は、標準トリプトンダイズブロス(Oxoid)中の鉄量の少なくとも2倍を含むべきである。
【0012】
培地が0.5μMから1mMの鉄を含むのが好ましい。
培地が10μMから500μMの鉄を含むのがより好ましい。
培地が25μMから500μMの鉄を含むのが最も好ましい。
【0013】
普通、宿主細胞への侵入に関与する外膜蛋白質またはinvasinまたはadhesinは、通常培地での量より多量に発現する。
【0014】
「通常培地での量より多量に発現する」により、該蛋白またはinvasinまたはadhesinは、通常の発現量の少なくとも2倍で発現する。通常の発現量は、標準トリプトンダイズブロス(Oxoid)中での発現量である。
【0015】
普通、invasinと血清学的に関連する細胞外蛋白質は、通常培地中より多量に発現する。
【0016】
普通、55Kda細胞外蛋白複合体および/または97Kda外膜蛋白質は、通常培地中より多量に発現する。
【0017】
本発明は、ワクチンとして、またはワクチン製剤中に使用する、記載のような精製蛋白質も提供する。
【0018】
本発明は、該精製蛋白質に対する抗体も提供する。
【0019】
更に、本発明は、培地からの鉄の直線的取込みがもはや起こらなくなるまで細胞に鉄を担持させるように、十分な鉄を含んだ培地中で細胞を培養する生産方法を提供する。
【0020】
細胞が鉄で飽和されるのが好ましい。
鉄を第二鉄塩の形態で培地に供給するのが好ましい。
培養後、細菌を不活性化してしまうのが好ましい。
ホルムアルデヒド組成物を使用して、不活性化を行うのが好ましい。
【0021】
更に、本発明は、培養細菌の細胞または細胞膜または細胞外生成物を提供する。
その生物材料は、生理学的に許容し得る担体中で提供されるのが好ましい。
【0022】
本発明の組成物は、免疫反応を増強するのに適したアジュバントを含むのが好ましい。
【0023】
したがって、ワクチン組成物を、細菌、とりわけ微生物Photobacterium damselae subsp.piscicidaによる感染に対して、魚類を予防および/または治療処理するために提供する。
【0024】
該組成物は、該微生物の培養によって生成する成分を含む。成分が生成した後、好ましくはホルマリンで処理することによって、使用前に微生物を殺すために、培養物を処理する。
【0025】
宿主細胞への侵入に関与し、かつ過剰の鉄を含む培地中で培養物を増殖させたときに多量に生成する成分中の2成分は、魚類に注入したときに、魚類細胞内への微生物の侵入を防止する抗体の生成を誘発する。
これらの抗体は、微生物による感染から魚類を保護する。
したがって、本発明は抗体も提供する。
【0026】
特定の一実施形態では、その生物材料は、英国、アバディーンのN.C.I.M.B.に2000年8月4日に登録番号41062で寄託されたPhotobacterium damselae MT1415株の培養物から誘導される。
【0027】
本発明者は、鉄の豊富な条件下で外膜中に発現する蛋白質(invasinまたはadhesin)が、シーバスの繊維芽細胞によるP.damselaeのインターナリゼーションに関係していると決定した。この蛋白質を特異的に結合するレクチンは、P.damselaeの侵入能力を阻害するが、一方、その蛋白質を結合しないレクチンは、シーバスの幼若繊維芽細胞系(SBL)への侵入を顕著には阻害しない。その上、シーバス中でこの蛋白質に対して生じた抗体は、魚類上皮細胞(EPC)による該細菌のインターナリゼーションを防止する。塩化第二鉄の形態で第二鉄を増殖培地に添加することにより、この蛋白質の発現量を増加させることができる。例えば、100μMの塩化第二鉄を添加すると、その蛋白質の発現が少なくとも2.6倍増加する。このような鉄の補充下でP.damselaeを増殖させることにより生産したバクテリンは、ニジマスおよび幼若なタイの臨床的パスツレラ症に対して保護作用を示す。
【0028】
invasinが増加するように、細菌細胞による鉄の取込みを起こす任意の形態で、鉄を培地に供給してよい。他の培養条件もこの蛋白の発現を増加させ得る。例えば、十分な鉄の存在下で嫌気的条件におかれると、より多くの該蛋白が発現し得る。しかし、より高い細胞密度は、酸素含量を維持するために連続的振盪または他の通気法で、細菌を22℃から35℃で増殖させることにより実現される。
【0029】
本発明のワクチン細胞は、任意の標準的方法によって不活性化されるが、ホルムアルデヒドを使用して不活性化するのが都合良い。アジュバントの添加、細胞濃度、許容し得る担体中への再懸濁等のワクチンの更なる調製によって、追加の保護が得られよう。
【0030】
本発明のワクチンおよびその生産方法を、単に実例として以下の非制限的な実施例と図を参考にして、これから説明する。本請求項の範囲に入る更なる実施形態は、これらの実例に鑑みて、当分野の技術者に明らかである。
【図面の簡単な説明】
【0031】
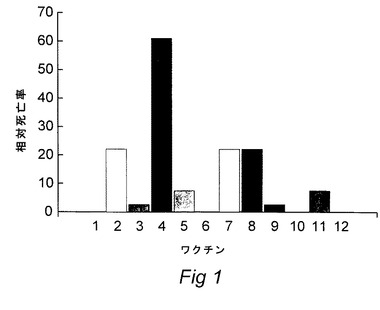
【図1】Photobacterium属の微生物で免疫試験をするニジマスのワクチン処理に対する生存率パーセント(RPS)のヒストグラムを示す図である。
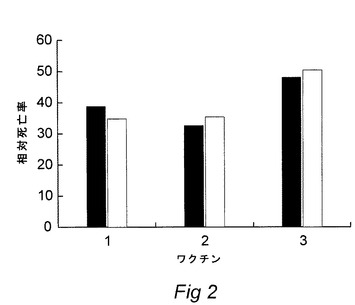
【図2】Photobacterium属の微生物で免疫試験をするヨーロッパヘダイ(Sparus aurata)のワクチン処理に対する生存率パーセント(RPS)のヒストグラムを示す図である。
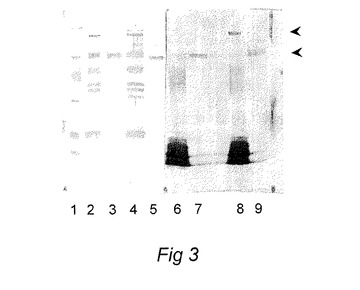
【図3】Aurodye(レーン1〜5の蛋白に対して)およびシーバス抗鉄補充P.damselae血清(レーン6〜9)で染色したP.damselae外膜蛋白質(OMP、レーン2、4、6、8)および細胞外生成物(ECP、レーン3、5、7、9)のウエスタンブロットを示し、97Kda蛋白(レーン8)および55Kda蛋白複合体(レーン7、9)を明確に示す図である。
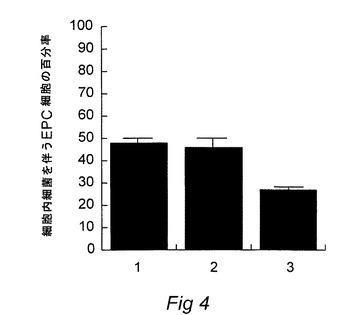
【図4】2)正常シーバス血清、または3)シーバス抗P.damselaeinvasin抗血清で予備インキュベートした、少なくとも1つの細胞内P.damselaeを有するEPC細胞のパーセンテージを、1)対照と共に示す図である。
【図5】様々なレクチンとインキュベートしたP.damselaeによるEPC/SBL細胞の侵入を、レクチンと予備インキュベートしていない対照に対して示す図である。
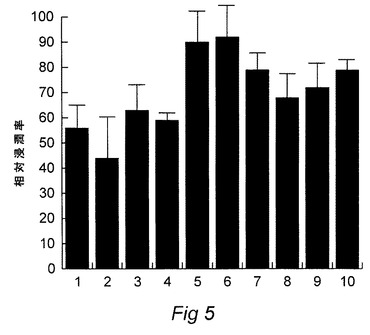
【図6】レクチン、シーバス抗P.damselae抗血清およびウサギ抗55KDa複合体血清をプローブとした、鉄補充条件下で生産したP.damselaeOMPのウエスタンブロットを示す図である。各レーンは、1)ウサギ抗55KDa複合体抗体、2)バス抗MT1415抗体、3)ビオチニル化dolichos biflorusアグルチニン、4)ビオチニル化コンカナバリンAレクチン、5)Sigmaビオチニル化分子量マーカー。
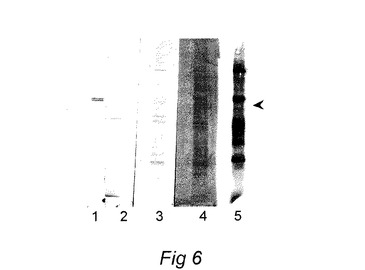
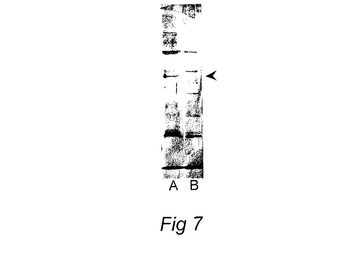
【図7】鉄補充培養物(ワクチンA)と比較して、鉄枯渇培養物(ワクチンB)中に55KDa複合体がないことを示すワクチン製剤から得た外膜蛋白質の、シーバス抗Photobacterium damselae抗体で染色したウエスタンブロットを示す図である。
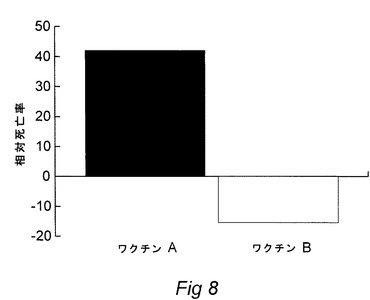
【図8】A)55KDa複合体および97KDa OMPを発現するワクチン、またはB)55KDa複合体、97KDa OMPのいずれも発現しないワクチンで接種した後、Photobacterium damselae subsp.piscicidaで免疫試験をしたシーバスDicentrarclus Labraxの生存率パーセント(RPS)を示す図である(図7も参照されたい)。
【実施例】
【0032】
実施例1:invasin発現P.damselaeワクチンの製造
トリプトン大豆ブロス(オキソイド)を30g/lで蒸留水中で調製し、塩化ナトリウムを三角フラスコ中で20g/lで添加したが、その際、フラスコの容量は十分なエアレーションが可能であるように媒体容量の5倍であった。121℃で15分間のオートクレーブ処理による滅菌の後に、塩化第二鉄を、蒸留水中100ミリモルの無菌ストックからの100マイクロモルの最終濃度に添加した。ブロスを25℃に予備加温し、これに、フォトバクテリウム damselae(MT1415株、アバディーン(英国)のN.C.I.M.Bに2000年8月4日に受入番号41062番で寄託されているようなカプセルポジティブビルレント分離菌)の、塩2%を含有する一晩トリプトン大豆ブロス培養1/10000容量を播種した。インキュベーションを140rpmでの振盪下に25℃で40時間継続した。インキュベーションの後に、ホルマリン0.5v/v%(遊離ホルムアルデヒド0.2%)の添加により培養を不活化し、ブロスを25℃で24時間放置して、完全に不活化した。最終ワクチン製剤の光学濃度を求め、ワクチンを必要になるまで4℃で貯蔵した。
【0033】
投与のために、この細胞懸濁液をいくつの方法のうちの1つで投与した。
【0034】
25℃の真水に入れたニジマス(10〜15g)にMS222(sigma)で麻酔をかけた。麻酔をかけられた魚に適切なワクチン製剤(100μl)または対照製剤または対照としての無菌リン酸緩衝生理食塩水を腹腔内注射により播種した。免疫化後少なくとも500度日後に魚にビルレントP.damselae108cfuの腹腔内注射でチャレンジした。死亡率を毎日記録し、対照魚に比較してのRPS(相対生存百分率)を求めた。結果を下記の図1に示したが、この際、本発明で使用したRPSをいくつかの対照ワクチンと比較している;1 対照ワクチン、2 対照ワクチン、3 鉄制限ワクチン、4 鉄補足ワクチン(本発明のワクチン)、5 標準トリプトン大豆ブロス(TSB)ワクチン、6〜12 対照ワクチン。
【0035】
もしくは、アメリカチヌ(0.3g)を本発明または対照ワクチンの10倍希釈液中で一回60秒浸漬することにより免疫化した。ワクチン接種後500日の後の魚をビルレントP.damselae約105cfu/mlを含有する懸濁液に25℃で1時間浸漬することによりチャレンジした。死亡率を上記と同様に記録した。結果を図2に示した;1 鉄制限ワクチン、2 TSBワクチン、3 鉄補足ワクチン(本発明)。
【0036】
防御抗原の同定
腹腔内注射によりスズキ(Dicentrarchus labrax)を免疫化するために、上記と同様に調製されたワクチンを使用した。フロインド不完全アジュバントを1:1の比率で同時に投与した。魚を25℃の海水に20日間保持し、その後、これらに2回目の同じ用量を与えた。さらに20日の後に魚から採血し、血液を血餅にし、赤血球を遠心分離により除去して、血清を集めた。血清をリン酸緩衝生理食塩水に対して透析し、−80℃で凍結貯蔵した。
【0037】
外膜タンパク質(OMP)および細胞外生成物(ECP)を鉄制限下に、標準TSM中で、または鉄補足下に培養したP.damselaeから調製した。鉄制限のために2,2−ジピリジル(100マイクロモル)をトリプトン大豆ブロスに添加して、P.damselaeの接種前に鉄をキレート化した。次いでインキュベーションを前記と同様に実施した。沈殿、続いて内膜のサルコシル可溶化により、OMPを調製した(Hancock & Poxton(1988)Bacterial Cell Surface Techniques,John Wiley & Sons,Chichester(英国))。細胞外生成物をブロス培養上澄み液から回収した。
【0038】
非還元条件下にECPおよびOMP(同じタンパク質濃度)をSDS−PAGEゲル上に泳動させ、PVDF膜上にブロッティングした。膜を抗鉄補足P.damselaeバス血清で、続いて、抗バス免疫グロブリンマウスモノクローナル抗体で(Santos 他(1997)Fish.Shellfish Immunol.7:175−191)、続いて抗マウスヤギ血清アルカリフォスファターゼ複合体で調べた。バンドを視覚化するために膜を、ニトロブルーテトラゾリウムおよび5−ブロモ−4−クロロ−3−インドリルフォスフェートからなる基質中でインキュベートした。結果を図3および図7に示している。鉄含有培養からのOMP調製物では澄明なバンドが97Kdaマーカー付近で視認されたが、これは鉄制限下では存在しない(図3および7)。デンシトメトリーを走査することにより、標準TSM培養に対して鉄補足培養からのOMPでは少なくともその2.6倍以上濃縮されていて、鉄制限培養では検出されなかったことがこのバンドで決定された。鉄含有媒体中で培養した細胞からのECPでは、2つのバンドが明らかであり、それらのうち、移動度がおよそ97Kda付近のバンドは、OMP調製物で認められたバンドと同一であると確認された。もう一方のバンドはより低分子量で、55KDaマーカー付近ではあるが、それよりも下に存在した。これらのバンドのどちらも鉄不足培地中で培養された細胞からのOMPSまたはECP中では検出されなかった。
【0039】
鉄補足P.damselaeに対する抗血清による魚類上皮細胞の浸潤の抑制
Bandin 他(1995)(Dis Aquat.Org.23:221〜227)によって記載された蛍光標識直接カウント法によって、魚類上皮細胞(EPC)に浸潤するP.damselaeの能力を決定した。フルオレセインイソチオシアネート(FITC)(0.1mg/ml)を使用してP.damselae細胞を1時間標識した。次いでP.damselaeをPBS中で十分に洗い、109細胞/mlの密度に再懸濁した。細胞懸濁液を分け、1つのアリコットを前記の抗鉄補足P.damselaeスズキ抗血清を加熱することにより調製された熱不活化免疫血清と共に45℃で15分間、インキュベーションし、第2のアリコットは熱不活化正常スズキ血清と共にインキュベーションした。血清処理されたP.damselae細胞のアリコット(10μl)をG−MEM1ml中の6×105EPC細胞に添加し、25℃で2時間、接触および浸潤させた。PBS中で洗うことにより外部細菌を除去し、EPCを臭化エチジウムで対比染色した。アリコット(10μl)をガラススライドに置き、ガラスカバーで覆い、その後、蛍光顕微鏡検査法により分析した。少なくとも100場での直接カウントにより、内在化したバクテリアの割合を決定した。実験を4回、反復した。結果を図4に示す:1 対照(血清を伴わないP.damselae)、2 熱不活化正常スズキ血清と共にインキュベートしたP.damselae、3 熱不活化抗血清と共にインキュベートしたP.damselae(スズキvsP.damselae)。
【0040】
レクチンを使用しての、スズキ細胞中へのP.damselae内在化に随伴するタンパク質の同定
以前の研究により、魚類細胞中へのP.damselaeの内在化に伴う実体は糖蛋白質でありうるということが提示されている(Magari−os 他(1996)FEMS Microbiol Lett.138:29〜34)。糖蛋白質の炭水化物側鎖は特異的に特定のレクチンに結合しうる。レクチンは特定の糖の配置に対して高い特異的な親和性を有する植物抽出物である。異なる種類のレクチンと共にP.damselaeをインキュベーションし、次いでスズキ幼生線維芽細胞(SBL)に浸潤するその能力を決定し、浸潤を抑制しうるレクチンおよび抑制しえないレクチンを同定した。次いで、浸潤に伴う糖蛋白質をビオチン化レクチンを用いてのウエスタンブロットでプローブすることにより同定することができた。
【0041】
FITCで標識し、洗浄し、前記と同様に109cfu/mlの密度に再懸濁したP.damselaeを100μg/mlの濃度の様々なレクチン(Vector Laboratories)と共に1時間インキュベーションした。インキュベーションの後に、細胞を十分に洗い、浸潤アッセイを前記と同様に実施した。結果を図5に示す:P.damselaeを次と共にインキュベーションした;1 Sophora japonicaグルチニン、2 コンカナバリンAアグルチニン、3 Lens culinarisアグルチニン、4 Griffonia simplicifoliaアグルチニン、5 スクシニル化小麦胚芽アグルチニン、6 ドリコスアグルチニン、7 ピーナッツアグルチニン、8 大豆アグルチニン、9 Ulex europaeusアグルチニン、10 小麦胚芽アグルチニン。
【0042】
P.damselaeによる浸潤を強力に抑制しうる2種のレクチン、Sophora japonicaグルチニン(SJA)およびConAアグルチニン(ConA)を選択した。浸潤を抑制しなかった1種のレクチン、ドリコスアグルチニン(DBA)も選択した。これらのレクチン(Vector Laboratories)のビオチン化製剤をP.damselaeからのSDS−PAGE分離OMPのウエスタンブロットを染色するために使用した。結果を図6に示した。
【0043】
全てのレクチンがいくつかの炭水化物/糖蛋白質を染色した。しかしながら、1つの領域のみがConAによって染色され、DBAによっては染色されなかった。この領域は2つの蛋白質バンドのコンプレックスからなり、非限定的条件下に近似的分子量55Kdaを有し、鉄飽和条件下に培養されたP.damselaeからのOMP製剤中でのみ検出され、鉄制限条件下に培養されたP.damselaeからの製剤では検出されなかった。さらに、P.damselaeからのOMPのウエスタンブロットを切り出し、ConAレクチンおよび抗鉄補足P.damselaeスズキ抗血清で染色すると、通常のバンド、55Kdaコンプレックスが両方の方法で染色された。レーン1は抗55KDaコンプレックスウサギでプローブされたOMPを示し、レーン2は浸潤抑制研究で使用された抗フォトバクテリウムdamsealeMT1415スズキ抗血清でプローブされたOMPを示す。レーン3はドリコアグルチニンでプローブされたOMPを示し、レーン4はコンカナバリンAレクチンでプローブされたOMPを示し、レーン5はSigmaビオチン化分子量マーカー(SDS−6B)を示す。
【0044】
97KDaOMPと55KDaECPとの間の血清学的関連性
97KDaOMPおよび55KDaECPを慎重にポリアクリルアミドゲルから切除し、ホモジナイズし、スズキに注入した。30日後、血清を集め、OMPおよびECPのウエスタンブロットをプローブするために使用した。97KDaOMPに対してスズキで生じた抗血清を55KDaと交叉反応させた。同様に、55KDaECPに対してスズキで生じた抗体も、KDaOMPと交叉反応させた。本発明者は55KDaECPは97KDaOMPの分泌バージョンの1つであると提案する。
【0045】
精製およびN末端配列
55KDa蛋白質の後続精製および配列決定はこの領域の3個の蛋白質を明らかにした。主要な抗原蛋白質はN−末端ブロックされていて、転写後のグリコシレーションと矛盾せず、したがって配列を求めることはできないが、このフラクションは強い赤血球凝集活性を有し、内在化での概然的波及を示す。第2のたんぱく質はβ−1,4N−アセチルムラミダーゼ、他のバクテリアに対する防御に対して100%相同性を有するN−末端アミノ酸配列を与えた:AMKRHGLDNYRGYSLGNWVC。
【0046】
第3のたんぱく質はデアミナーゼまたは異化作用性デヒドラターゼのフラグメントでありうる:NVVLHGDNFDSTXVXVKAV。
【0047】
スズキ(Dicentrarchus labrax)での攻撃研究での、55KDaコンプレックスを発現するワクチンと発現しないワクチンとの防護効力の比較
次の研究を独立して、CEFAS Weymouth Laboratoryで研究プロトコルP0075、参照99008で行った。
【0048】
55KDaたんぱく質コンプレックスを発現するワクチンを次のように調製した:2.5lエルレンマイヤーフラスコ中塩化第二鉄200マイクロモルを含有するトリプトン大豆ブロス500ml+NaCl2%(TSB2)にフォトバクテリウムdamseale subsp.piscicida MT1415の18時間TSB2培養の接種物0.01v/v%を接種した。この培養を後期指数成長段階まで(約40時間)140rpmの振盪下に24℃で成長させた。生じた培養をホルマリン(最終濃度0.2%)で不活化し、フェニルメチルスルホニルフルオリド(PMSF)を添加して、イソプロパノール中の100ミリモル原液から100マイクロモルの最終濃度にして、プロテアーゼを不活化した。
【0049】
55KDaコンプレックスの発現が完全に抑制されるワクチンを製造するために、フォトバクテリウムdamselae subsp.piscicida分離菌MT1415を、2,2ジピリジル、鉄キレート化剤100マイクロモルを含有するTSB2中で2回、18時間継代培養した。生じた完全に鉄を消尽した培養をワクチン培養のための接種材料(0.5v/v%)として使用したが、これを、2.5lエルレンマイヤーフラスコ中の2,2ジピリジル100マイクロモルを含有するTSB2 500ml中、140rpmでの振盪下に、遅い指数成長段階まで(約48時間)、24℃で成長させた。この製剤からの55Daコンプレックスの不在が、重複培養から調製された外膜たんぱく質のウエスタンブロットにより確認された(図7参照)。
【技術分野】
【0001】
本発明は、細菌による感染に対して魚類を予防処理するため、とりわけPhotobacterium damselae subsp.piscicida等の細菌による感染から魚類を保護するための、ワクチンおよび医薬として使用する新規な組成物に関する。
【背景技術】
【0002】
旧名Photobacterium damsela subsp.piscicidaのPhotobacterium damselae subsp.piscicidaは、Pasteurella piscicidaから再区分され、小サブユニット16sリボソームRNAの相同性に基づいて、vibrionaceae科に割当て直された(Gaulthier等(1995)、Int.J.Syst.Bacteriol.45:139−144)。P.damselaeは、仮性結核と普通呼ばれる重篤な致死性疾患を暖水海水魚に引き起こし、主要な器官や組織に感染して、筋肉組織中に特徴的な潰瘍または偽結核結節を生じる。
【0003】
抗生物質耐性の出現、ならびに、感染したとき魚は摂食を止め、したがって抗生物質入り餌を消費しないことが、化学療法の失敗原因と示唆されてきた。その上、宿主内コロニー形成中に、細胞内に生存する時期が存在することを示唆する証拠が益々増えている(Barnes等(1999)、Microbiology 145:483−494)。したがって、免疫による予防が、この疾患を抑制する最も望ましい手段と思われる。
【0004】
ホルマリン殺菌細菌(バクテリン)に基づく様々なワクチン接種戦略が提案されてきたが、これらの多くは再現性のある保護効果を生じることができなかった(Romalde & Magarinos(1997) Fish Vaccinology,Karger,Basel,Switzerland,pp167−177)。これは、感染剤に対する宿主の低い抗体反応性を反映するものと思われる(Arijo等、(1997) Fish Shellfish Immunol.8:63−72)。その上、これらの製剤中に含まれる保護抗原については殆ど何もわかっていない。鉄制限外膜蛋白質(IROMP)に基づく製剤および細胞外生成物の豊富なバクテリン(ECP)が記載されてきた(Magarinos等、(1994)Bull.Eur.Ass.Fish Pathol.14:120−122)が、これらは商品として無効であった。
【0005】
細菌が絶対的に鉄を必要とし、P.damselaeも例外でないことは、確立された事実である。培地中に十分な量を含めないか、またはその利用率を減少させるキレート剤を含めることにより、鉄が制限的である条件下で、様々な高親和性鉄取込み系が発現することが示された(Magarinos等、(1994)Appl.Environ.Microbiol.60:2990−2998)。これらは、シデロフォア(siderophore)および鉄制限外膜蛋白質(IROMP)を含む。Aeromonas salmonicida(Hirst & Ellis(1994) Fish Shellfish Immunol.4:29−45)、Pasteurella haemolytica(英国特許明細書第8805253号 W Donachie、英国)等の他の細菌種では、ワクチン中の保護抗原としてIROMPを開発して成功した。しかし、これらが、P.damselaeにおいて保護作用を示すとは認められない(Romalde & Magari−os(1997) Fish Vaccinology,Karger,Basel,Switzerland,pp167−177)。この保護作用の欠如が、P.damselaeによる感染の戦略を反映するものと思われる。鉄制限下で、P.damselaeは、プロテアーゼ活性の増加(Bakopolous(1997) J.Fish Dis.20:297−305)および莢膜多糖(capsular polysaccharide)の増加を起こす。莢膜多糖は、血清殺菌力に対する耐性を増加させ、宿主マクロファージによる食作用を防止する(Arijo等、(1998) Fish Shellfish Immunol.8:63−72)。このことは、マクロファージの呼吸バースト中に生成する反応性酸素種による殺菌に対する適応性を欠いた結果として(Barnes等、(1999)Microbiology 145:483−495)、P.damselaeが食細胞による攻撃に耐性を示さないことから(Skarmeta等、(1995)Dis.Aquat.Org.23:51−57)、明らかである。しかし、赤血球の溶解により増加した蛋白分解活性が、ヘム形態の鉄を遊離させることができる。ヘムは莢膜によって吸収され(Ana Do Vale,Pers.Comm.)、その鉄はIROMPによって膜を通過する。したがって、鉄取込みの第1段階として作用する莢膜多糖のために、罹病過程の間にIROMPが露出することはない。鉄濃度が増加するにつれ、莢膜生産は減少し、IROMPは不活性化され、蛋白分解活性は検出不能となる(Bakopolous(1997)J.Fish Dis.20:297−305)。この結果、P.damselaeは、抗体、マクロファージのいずれの攻撃も受け易くなると思われる。しかし、この時点でその細菌は、非食細胞性魚類細胞に接着し(Yoshida等、(1997) J.Fish Dis.20:77−88)、侵入する(Magari−os等、(1996)FEMS Microbiology Lett. 138:29−34)ことができ、したがって抗体や食細胞による攻撃を回避する。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】英国特許第8805253号明細書
【非特許文献】
【0007】
【非特許文献1】Gaulthier等(1995)、Int.J.Syst.Bacteriol.45:139−144
【非特許文献2】Romalde & Magarinos(1997) Fish Vaccinology,Karger,Basel,Switzerland,pp167−177
【非特許文献3】Arijo等、(1997) Fish Shellfish Immunol.8:63−72
【非特許文献4】Magarinos等、(1994)Bull.Eur.Ass.Fish Pathol.14:120−122
【非特許文献5】Magarinos等、(1994)Appl.Environ.Microbiol.60:2990−2998
【非特許文献6】Hirst & Ellis(1994) Fish Shellfish Immunol.4:29−45
【非特許文献7】Bakopolous(1997) J.Fish Dis.20:297−305
【非特許文献8】Barnes等、(1999)Microbiology 145:483−495
【非特許文献9】Skarmeta等、(1995)Dis.Aquat.Org.23:51−57
【非特許文献10】Yoshida等、(1997) J.Fish Dis.20:77−88
【非特許文献11】Magari−os等、(1996)FEMS Microbiology Lett. 138:29−34
【発明の概要】
【0008】
本発明の一目的は、Photobacterium damselae等の細菌による感染から、魚類を保護するワクチンを提供することである。
【0009】
本発明の他の目的は、このようなワクチンを生産する方法を提供することである。
【0010】
本発明によれば、その細菌の通常の増殖に必要な量より過剰の鉄を含む培地で、その細菌細胞を培養したことを特徴とするPhotobacterium damselaeの培養物由来の生物材料を含む、ワクチンおよび/または治療組成物が提供される。
【0011】
「過剰の鉄を含む」により、培地は、標準トリプトンダイズブロス(Oxoid)中の鉄量の少なくとも2倍を含むべきである。
【0012】
培地が0.5μMから1mMの鉄を含むのが好ましい。
培地が10μMから500μMの鉄を含むのがより好ましい。
培地が25μMから500μMの鉄を含むのが最も好ましい。
【0013】
普通、宿主細胞への侵入に関与する外膜蛋白質またはinvasinまたはadhesinは、通常培地での量より多量に発現する。
【0014】
「通常培地での量より多量に発現する」により、該蛋白またはinvasinまたはadhesinは、通常の発現量の少なくとも2倍で発現する。通常の発現量は、標準トリプトンダイズブロス(Oxoid)中での発現量である。
【0015】
普通、invasinと血清学的に関連する細胞外蛋白質は、通常培地中より多量に発現する。
【0016】
普通、55Kda細胞外蛋白複合体および/または97Kda外膜蛋白質は、通常培地中より多量に発現する。
【0017】
本発明は、ワクチンとして、またはワクチン製剤中に使用する、記載のような精製蛋白質も提供する。
【0018】
本発明は、該精製蛋白質に対する抗体も提供する。
【0019】
更に、本発明は、培地からの鉄の直線的取込みがもはや起こらなくなるまで細胞に鉄を担持させるように、十分な鉄を含んだ培地中で細胞を培養する生産方法を提供する。
【0020】
細胞が鉄で飽和されるのが好ましい。
鉄を第二鉄塩の形態で培地に供給するのが好ましい。
培養後、細菌を不活性化してしまうのが好ましい。
ホルムアルデヒド組成物を使用して、不活性化を行うのが好ましい。
【0021】
更に、本発明は、培養細菌の細胞または細胞膜または細胞外生成物を提供する。
その生物材料は、生理学的に許容し得る担体中で提供されるのが好ましい。
【0022】
本発明の組成物は、免疫反応を増強するのに適したアジュバントを含むのが好ましい。
【0023】
したがって、ワクチン組成物を、細菌、とりわけ微生物Photobacterium damselae subsp.piscicidaによる感染に対して、魚類を予防および/または治療処理するために提供する。
【0024】
該組成物は、該微生物の培養によって生成する成分を含む。成分が生成した後、好ましくはホルマリンで処理することによって、使用前に微生物を殺すために、培養物を処理する。
【0025】
宿主細胞への侵入に関与し、かつ過剰の鉄を含む培地中で培養物を増殖させたときに多量に生成する成分中の2成分は、魚類に注入したときに、魚類細胞内への微生物の侵入を防止する抗体の生成を誘発する。
これらの抗体は、微生物による感染から魚類を保護する。
したがって、本発明は抗体も提供する。
【0026】
特定の一実施形態では、その生物材料は、英国、アバディーンのN.C.I.M.B.に2000年8月4日に登録番号41062で寄託されたPhotobacterium damselae MT1415株の培養物から誘導される。
【0027】
本発明者は、鉄の豊富な条件下で外膜中に発現する蛋白質(invasinまたはadhesin)が、シーバスの繊維芽細胞によるP.damselaeのインターナリゼーションに関係していると決定した。この蛋白質を特異的に結合するレクチンは、P.damselaeの侵入能力を阻害するが、一方、その蛋白質を結合しないレクチンは、シーバスの幼若繊維芽細胞系(SBL)への侵入を顕著には阻害しない。その上、シーバス中でこの蛋白質に対して生じた抗体は、魚類上皮細胞(EPC)による該細菌のインターナリゼーションを防止する。塩化第二鉄の形態で第二鉄を増殖培地に添加することにより、この蛋白質の発現量を増加させることができる。例えば、100μMの塩化第二鉄を添加すると、その蛋白質の発現が少なくとも2.6倍増加する。このような鉄の補充下でP.damselaeを増殖させることにより生産したバクテリンは、ニジマスおよび幼若なタイの臨床的パスツレラ症に対して保護作用を示す。
【0028】
invasinが増加するように、細菌細胞による鉄の取込みを起こす任意の形態で、鉄を培地に供給してよい。他の培養条件もこの蛋白の発現を増加させ得る。例えば、十分な鉄の存在下で嫌気的条件におかれると、より多くの該蛋白が発現し得る。しかし、より高い細胞密度は、酸素含量を維持するために連続的振盪または他の通気法で、細菌を22℃から35℃で増殖させることにより実現される。
【0029】
本発明のワクチン細胞は、任意の標準的方法によって不活性化されるが、ホルムアルデヒドを使用して不活性化するのが都合良い。アジュバントの添加、細胞濃度、許容し得る担体中への再懸濁等のワクチンの更なる調製によって、追加の保護が得られよう。
【0030】
本発明のワクチンおよびその生産方法を、単に実例として以下の非制限的な実施例と図を参考にして、これから説明する。本請求項の範囲に入る更なる実施形態は、これらの実例に鑑みて、当分野の技術者に明らかである。
【図面の簡単な説明】
【0031】
【図1】Photobacterium属の微生物で免疫試験をするニジマスのワクチン処理に対する生存率パーセント(RPS)のヒストグラムを示す図である。
【図2】Photobacterium属の微生物で免疫試験をするヨーロッパヘダイ(Sparus aurata)のワクチン処理に対する生存率パーセント(RPS)のヒストグラムを示す図である。
【図3】Aurodye(レーン1〜5の蛋白に対して)およびシーバス抗鉄補充P.damselae血清(レーン6〜9)で染色したP.damselae外膜蛋白質(OMP、レーン2、4、6、8)および細胞外生成物(ECP、レーン3、5、7、9)のウエスタンブロットを示し、97Kda蛋白(レーン8)および55Kda蛋白複合体(レーン7、9)を明確に示す図である。
【図4】2)正常シーバス血清、または3)シーバス抗P.damselaeinvasin抗血清で予備インキュベートした、少なくとも1つの細胞内P.damselaeを有するEPC細胞のパーセンテージを、1)対照と共に示す図である。
【図5】様々なレクチンとインキュベートしたP.damselaeによるEPC/SBL細胞の侵入を、レクチンと予備インキュベートしていない対照に対して示す図である。
【図6】レクチン、シーバス抗P.damselae抗血清およびウサギ抗55KDa複合体血清をプローブとした、鉄補充条件下で生産したP.damselaeOMPのウエスタンブロットを示す図である。各レーンは、1)ウサギ抗55KDa複合体抗体、2)バス抗MT1415抗体、3)ビオチニル化dolichos biflorusアグルチニン、4)ビオチニル化コンカナバリンAレクチン、5)Sigmaビオチニル化分子量マーカー。
【図7】鉄補充培養物(ワクチンA)と比較して、鉄枯渇培養物(ワクチンB)中に55KDa複合体がないことを示すワクチン製剤から得た外膜蛋白質の、シーバス抗Photobacterium damselae抗体で染色したウエスタンブロットを示す図である。
【図8】A)55KDa複合体および97KDa OMPを発現するワクチン、またはB)55KDa複合体、97KDa OMPのいずれも発現しないワクチンで接種した後、Photobacterium damselae subsp.piscicidaで免疫試験をしたシーバスDicentrarclus Labraxの生存率パーセント(RPS)を示す図である(図7も参照されたい)。
【実施例】
【0032】
実施例1:invasin発現P.damselaeワクチンの製造
トリプトン大豆ブロス(オキソイド)を30g/lで蒸留水中で調製し、塩化ナトリウムを三角フラスコ中で20g/lで添加したが、その際、フラスコの容量は十分なエアレーションが可能であるように媒体容量の5倍であった。121℃で15分間のオートクレーブ処理による滅菌の後に、塩化第二鉄を、蒸留水中100ミリモルの無菌ストックからの100マイクロモルの最終濃度に添加した。ブロスを25℃に予備加温し、これに、フォトバクテリウム damselae(MT1415株、アバディーン(英国)のN.C.I.M.Bに2000年8月4日に受入番号41062番で寄託されているようなカプセルポジティブビルレント分離菌)の、塩2%を含有する一晩トリプトン大豆ブロス培養1/10000容量を播種した。インキュベーションを140rpmでの振盪下に25℃で40時間継続した。インキュベーションの後に、ホルマリン0.5v/v%(遊離ホルムアルデヒド0.2%)の添加により培養を不活化し、ブロスを25℃で24時間放置して、完全に不活化した。最終ワクチン製剤の光学濃度を求め、ワクチンを必要になるまで4℃で貯蔵した。
【0033】
投与のために、この細胞懸濁液をいくつの方法のうちの1つで投与した。
【0034】
25℃の真水に入れたニジマス(10〜15g)にMS222(sigma)で麻酔をかけた。麻酔をかけられた魚に適切なワクチン製剤(100μl)または対照製剤または対照としての無菌リン酸緩衝生理食塩水を腹腔内注射により播種した。免疫化後少なくとも500度日後に魚にビルレントP.damselae108cfuの腹腔内注射でチャレンジした。死亡率を毎日記録し、対照魚に比較してのRPS(相対生存百分率)を求めた。結果を下記の図1に示したが、この際、本発明で使用したRPSをいくつかの対照ワクチンと比較している;1 対照ワクチン、2 対照ワクチン、3 鉄制限ワクチン、4 鉄補足ワクチン(本発明のワクチン)、5 標準トリプトン大豆ブロス(TSB)ワクチン、6〜12 対照ワクチン。
【0035】
もしくは、アメリカチヌ(0.3g)を本発明または対照ワクチンの10倍希釈液中で一回60秒浸漬することにより免疫化した。ワクチン接種後500日の後の魚をビルレントP.damselae約105cfu/mlを含有する懸濁液に25℃で1時間浸漬することによりチャレンジした。死亡率を上記と同様に記録した。結果を図2に示した;1 鉄制限ワクチン、2 TSBワクチン、3 鉄補足ワクチン(本発明)。
【0036】
防御抗原の同定
腹腔内注射によりスズキ(Dicentrarchus labrax)を免疫化するために、上記と同様に調製されたワクチンを使用した。フロインド不完全アジュバントを1:1の比率で同時に投与した。魚を25℃の海水に20日間保持し、その後、これらに2回目の同じ用量を与えた。さらに20日の後に魚から採血し、血液を血餅にし、赤血球を遠心分離により除去して、血清を集めた。血清をリン酸緩衝生理食塩水に対して透析し、−80℃で凍結貯蔵した。
【0037】
外膜タンパク質(OMP)および細胞外生成物(ECP)を鉄制限下に、標準TSM中で、または鉄補足下に培養したP.damselaeから調製した。鉄制限のために2,2−ジピリジル(100マイクロモル)をトリプトン大豆ブロスに添加して、P.damselaeの接種前に鉄をキレート化した。次いでインキュベーションを前記と同様に実施した。沈殿、続いて内膜のサルコシル可溶化により、OMPを調製した(Hancock & Poxton(1988)Bacterial Cell Surface Techniques,John Wiley & Sons,Chichester(英国))。細胞外生成物をブロス培養上澄み液から回収した。
【0038】
非還元条件下にECPおよびOMP(同じタンパク質濃度)をSDS−PAGEゲル上に泳動させ、PVDF膜上にブロッティングした。膜を抗鉄補足P.damselaeバス血清で、続いて、抗バス免疫グロブリンマウスモノクローナル抗体で(Santos 他(1997)Fish.Shellfish Immunol.7:175−191)、続いて抗マウスヤギ血清アルカリフォスファターゼ複合体で調べた。バンドを視覚化するために膜を、ニトロブルーテトラゾリウムおよび5−ブロモ−4−クロロ−3−インドリルフォスフェートからなる基質中でインキュベートした。結果を図3および図7に示している。鉄含有培養からのOMP調製物では澄明なバンドが97Kdaマーカー付近で視認されたが、これは鉄制限下では存在しない(図3および7)。デンシトメトリーを走査することにより、標準TSM培養に対して鉄補足培養からのOMPでは少なくともその2.6倍以上濃縮されていて、鉄制限培養では検出されなかったことがこのバンドで決定された。鉄含有媒体中で培養した細胞からのECPでは、2つのバンドが明らかであり、それらのうち、移動度がおよそ97Kda付近のバンドは、OMP調製物で認められたバンドと同一であると確認された。もう一方のバンドはより低分子量で、55KDaマーカー付近ではあるが、それよりも下に存在した。これらのバンドのどちらも鉄不足培地中で培養された細胞からのOMPSまたはECP中では検出されなかった。
【0039】
鉄補足P.damselaeに対する抗血清による魚類上皮細胞の浸潤の抑制
Bandin 他(1995)(Dis Aquat.Org.23:221〜227)によって記載された蛍光標識直接カウント法によって、魚類上皮細胞(EPC)に浸潤するP.damselaeの能力を決定した。フルオレセインイソチオシアネート(FITC)(0.1mg/ml)を使用してP.damselae細胞を1時間標識した。次いでP.damselaeをPBS中で十分に洗い、109細胞/mlの密度に再懸濁した。細胞懸濁液を分け、1つのアリコットを前記の抗鉄補足P.damselaeスズキ抗血清を加熱することにより調製された熱不活化免疫血清と共に45℃で15分間、インキュベーションし、第2のアリコットは熱不活化正常スズキ血清と共にインキュベーションした。血清処理されたP.damselae細胞のアリコット(10μl)をG−MEM1ml中の6×105EPC細胞に添加し、25℃で2時間、接触および浸潤させた。PBS中で洗うことにより外部細菌を除去し、EPCを臭化エチジウムで対比染色した。アリコット(10μl)をガラススライドに置き、ガラスカバーで覆い、その後、蛍光顕微鏡検査法により分析した。少なくとも100場での直接カウントにより、内在化したバクテリアの割合を決定した。実験を4回、反復した。結果を図4に示す:1 対照(血清を伴わないP.damselae)、2 熱不活化正常スズキ血清と共にインキュベートしたP.damselae、3 熱不活化抗血清と共にインキュベートしたP.damselae(スズキvsP.damselae)。
【0040】
レクチンを使用しての、スズキ細胞中へのP.damselae内在化に随伴するタンパク質の同定
以前の研究により、魚類細胞中へのP.damselaeの内在化に伴う実体は糖蛋白質でありうるということが提示されている(Magari−os 他(1996)FEMS Microbiol Lett.138:29〜34)。糖蛋白質の炭水化物側鎖は特異的に特定のレクチンに結合しうる。レクチンは特定の糖の配置に対して高い特異的な親和性を有する植物抽出物である。異なる種類のレクチンと共にP.damselaeをインキュベーションし、次いでスズキ幼生線維芽細胞(SBL)に浸潤するその能力を決定し、浸潤を抑制しうるレクチンおよび抑制しえないレクチンを同定した。次いで、浸潤に伴う糖蛋白質をビオチン化レクチンを用いてのウエスタンブロットでプローブすることにより同定することができた。
【0041】
FITCで標識し、洗浄し、前記と同様に109cfu/mlの密度に再懸濁したP.damselaeを100μg/mlの濃度の様々なレクチン(Vector Laboratories)と共に1時間インキュベーションした。インキュベーションの後に、細胞を十分に洗い、浸潤アッセイを前記と同様に実施した。結果を図5に示す:P.damselaeを次と共にインキュベーションした;1 Sophora japonicaグルチニン、2 コンカナバリンAアグルチニン、3 Lens culinarisアグルチニン、4 Griffonia simplicifoliaアグルチニン、5 スクシニル化小麦胚芽アグルチニン、6 ドリコスアグルチニン、7 ピーナッツアグルチニン、8 大豆アグルチニン、9 Ulex europaeusアグルチニン、10 小麦胚芽アグルチニン。
【0042】
P.damselaeによる浸潤を強力に抑制しうる2種のレクチン、Sophora japonicaグルチニン(SJA)およびConAアグルチニン(ConA)を選択した。浸潤を抑制しなかった1種のレクチン、ドリコスアグルチニン(DBA)も選択した。これらのレクチン(Vector Laboratories)のビオチン化製剤をP.damselaeからのSDS−PAGE分離OMPのウエスタンブロットを染色するために使用した。結果を図6に示した。
【0043】
全てのレクチンがいくつかの炭水化物/糖蛋白質を染色した。しかしながら、1つの領域のみがConAによって染色され、DBAによっては染色されなかった。この領域は2つの蛋白質バンドのコンプレックスからなり、非限定的条件下に近似的分子量55Kdaを有し、鉄飽和条件下に培養されたP.damselaeからのOMP製剤中でのみ検出され、鉄制限条件下に培養されたP.damselaeからの製剤では検出されなかった。さらに、P.damselaeからのOMPのウエスタンブロットを切り出し、ConAレクチンおよび抗鉄補足P.damselaeスズキ抗血清で染色すると、通常のバンド、55Kdaコンプレックスが両方の方法で染色された。レーン1は抗55KDaコンプレックスウサギでプローブされたOMPを示し、レーン2は浸潤抑制研究で使用された抗フォトバクテリウムdamsealeMT1415スズキ抗血清でプローブされたOMPを示す。レーン3はドリコアグルチニンでプローブされたOMPを示し、レーン4はコンカナバリンAレクチンでプローブされたOMPを示し、レーン5はSigmaビオチン化分子量マーカー(SDS−6B)を示す。
【0044】
97KDaOMPと55KDaECPとの間の血清学的関連性
97KDaOMPおよび55KDaECPを慎重にポリアクリルアミドゲルから切除し、ホモジナイズし、スズキに注入した。30日後、血清を集め、OMPおよびECPのウエスタンブロットをプローブするために使用した。97KDaOMPに対してスズキで生じた抗血清を55KDaと交叉反応させた。同様に、55KDaECPに対してスズキで生じた抗体も、KDaOMPと交叉反応させた。本発明者は55KDaECPは97KDaOMPの分泌バージョンの1つであると提案する。
【0045】
精製およびN末端配列
55KDa蛋白質の後続精製および配列決定はこの領域の3個の蛋白質を明らかにした。主要な抗原蛋白質はN−末端ブロックされていて、転写後のグリコシレーションと矛盾せず、したがって配列を求めることはできないが、このフラクションは強い赤血球凝集活性を有し、内在化での概然的波及を示す。第2のたんぱく質はβ−1,4N−アセチルムラミダーゼ、他のバクテリアに対する防御に対して100%相同性を有するN−末端アミノ酸配列を与えた:AMKRHGLDNYRGYSLGNWVC。
【0046】
第3のたんぱく質はデアミナーゼまたは異化作用性デヒドラターゼのフラグメントでありうる:NVVLHGDNFDSTXVXVKAV。
【0047】
スズキ(Dicentrarchus labrax)での攻撃研究での、55KDaコンプレックスを発現するワクチンと発現しないワクチンとの防護効力の比較
次の研究を独立して、CEFAS Weymouth Laboratoryで研究プロトコルP0075、参照99008で行った。
【0048】
55KDaたんぱく質コンプレックスを発現するワクチンを次のように調製した:2.5lエルレンマイヤーフラスコ中塩化第二鉄200マイクロモルを含有するトリプトン大豆ブロス500ml+NaCl2%(TSB2)にフォトバクテリウムdamseale subsp.piscicida MT1415の18時間TSB2培養の接種物0.01v/v%を接種した。この培養を後期指数成長段階まで(約40時間)140rpmの振盪下に24℃で成長させた。生じた培養をホルマリン(最終濃度0.2%)で不活化し、フェニルメチルスルホニルフルオリド(PMSF)を添加して、イソプロパノール中の100ミリモル原液から100マイクロモルの最終濃度にして、プロテアーゼを不活化した。
【0049】
55KDaコンプレックスの発現が完全に抑制されるワクチンを製造するために、フォトバクテリウムdamselae subsp.piscicida分離菌MT1415を、2,2ジピリジル、鉄キレート化剤100マイクロモルを含有するTSB2中で2回、18時間継代培養した。生じた完全に鉄を消尽した培養をワクチン培養のための接種材料(0.5v/v%)として使用したが、これを、2.5lエルレンマイヤーフラスコ中の2,2ジピリジル100マイクロモルを含有するTSB2 500ml中、140rpmでの振盪下に、遅い指数成長段階まで(約48時間)、24℃で成長させた。この製剤からの55Daコンプレックスの不在が、重複培養から調製された外膜たんぱく質のウエスタンブロットにより確認された(図7参照)。
【特許請求の範囲】
【請求項1】
その細菌の通常の増殖に必要な量より過剰の鉄を含む培地で、その細菌細胞を培養したことを特徴とする、Photobacterium damselaeの培養物から誘導した生物材料を含む組成物。
【請求項2】
生物材料が、宿主細胞への侵入に関与する外膜蛋白、またはinvasinまたはadhesinを含み、かつワクチンの調製に使用する通常の培地中より多量に発現する請求項1に記載の組成物。
【請求項3】
invasinと血清学的に関連する細胞外蛋白が、通常の培養より多量に発現する請求項1または2に記載の組成物。
【請求項4】
培地からの鉄の直線的取込みがもはや起こらなくなるまで細胞に鉄を担持させるように、十分な鉄を含んだ培地中でPhotobacterium damselaeの細菌細胞を培養する段階を含むワクチンの生産方法。
【請求項5】
細胞が鉄で飽和される請求項4に記載の方法。
【請求項6】
鉄が第二鉄塩の形態で培地に供給される請求項4または5に記載の方法。
【請求項7】
培養後、細菌が不活性化される請求項4から6のいずれか一項に記載の方法。
【請求項8】
不活性化がホルムアルデヒド組成物を用いて行われる請求項7に記載の方法。
【請求項9】
その方法の生産物が抗体の生産に使用される請求項4から8のいずれか一項に記載の方法。
【請求項10】
細菌による感染に対して、魚類を予防または治療処理するワクチンの調製における、請求項4から8のいずれか一項に記載の方法により生産される細胞、細胞膜または細胞外生成物の使用。
【請求項11】
細菌による感染に対して、魚類を予防または治療処理するワクチンの調製における、請求項1から3のいずれか一項に記載の組成物の使用。
【請求項12】
ワクチンが生理的に許容し得る担体を含む請求項10または11に記載の使用。
【請求項13】
ワクチンが免疫反応を増強するのに適したアジュバントを含む請求項10から12のいずれか一項に記載の使用。
【請求項14】
細菌、とりわけ微生物Photobacterium damelae subsp.piscicidaによる感染に対して、魚類を予防および/または治療処理するワクチン組成物であって、英国、アバディーンのN.C.I.M.B.に2000年8月4日に登録番号41062で寄託された微生物の培養により生産される成分を含む組成物。
【請求項15】
ワクチンの調製における、英国、アバディーンのN.C.I.M.B.に2000年8月4日に登録番号41062で寄託された細菌の使用。
【請求項16】
請求項9に記載の方法によって生産される抗体。
【請求項1】
その細菌の通常の増殖に必要な量より過剰の鉄を含む培地で、その細菌細胞を培養したことを特徴とする、Photobacterium damselaeの培養物から誘導した生物材料を含む組成物。
【請求項2】
生物材料が、宿主細胞への侵入に関与する外膜蛋白、またはinvasinまたはadhesinを含み、かつワクチンの調製に使用する通常の培地中より多量に発現する請求項1に記載の組成物。
【請求項3】
invasinと血清学的に関連する細胞外蛋白が、通常の培養より多量に発現する請求項1または2に記載の組成物。
【請求項4】
培地からの鉄の直線的取込みがもはや起こらなくなるまで細胞に鉄を担持させるように、十分な鉄を含んだ培地中でPhotobacterium damselaeの細菌細胞を培養する段階を含むワクチンの生産方法。
【請求項5】
細胞が鉄で飽和される請求項4に記載の方法。
【請求項6】
鉄が第二鉄塩の形態で培地に供給される請求項4または5に記載の方法。
【請求項7】
培養後、細菌が不活性化される請求項4から6のいずれか一項に記載の方法。
【請求項8】
不活性化がホルムアルデヒド組成物を用いて行われる請求項7に記載の方法。
【請求項9】
その方法の生産物が抗体の生産に使用される請求項4から8のいずれか一項に記載の方法。
【請求項10】
細菌による感染に対して、魚類を予防または治療処理するワクチンの調製における、請求項4から8のいずれか一項に記載の方法により生産される細胞、細胞膜または細胞外生成物の使用。
【請求項11】
細菌による感染に対して、魚類を予防または治療処理するワクチンの調製における、請求項1から3のいずれか一項に記載の組成物の使用。
【請求項12】
ワクチンが生理的に許容し得る担体を含む請求項10または11に記載の使用。
【請求項13】
ワクチンが免疫反応を増強するのに適したアジュバントを含む請求項10から12のいずれか一項に記載の使用。
【請求項14】
細菌、とりわけ微生物Photobacterium damelae subsp.piscicidaによる感染に対して、魚類を予防および/または治療処理するワクチン組成物であって、英国、アバディーンのN.C.I.M.B.に2000年8月4日に登録番号41062で寄託された微生物の培養により生産される成分を含む組成物。
【請求項15】
ワクチンの調製における、英国、アバディーンのN.C.I.M.B.に2000年8月4日に登録番号41062で寄託された細菌の使用。
【請求項16】
請求項9に記載の方法によって生産される抗体。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2011−207888(P2011−207888A)
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−105283(P2011−105283)
【出願日】平成23年5月10日(2011.5.10)
【分割の表示】特願2001−514975(P2001−514975)の分割
【原出願日】平成12年8月7日(2000.8.7)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【出願番号】特願2011−105283(P2011−105283)
【出願日】平成23年5月10日(2011.5.10)
【分割の表示】特願2001−514975(P2001−514975)の分割
【原出願日】平成12年8月7日(2000.8.7)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
[ Back to top ]