
ワクチン
【課題】新規多糖抗原ワクチン提供。
【解決手段】本発明により、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)由来のタンパク質、及び場合によりTh1誘導性アジュバントと配合された、ニューモコッカル多糖抗原、典型的には、ニューモコッカル多糖コンジュゲート抗原を含むワクチンが提供される。
【解決手段】本発明により、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)由来のタンパク質、及び場合によりTh1誘導性アジュバントと配合された、ニューモコッカル多糖抗原、典型的には、ニューモコッカル多糖コンジュゲート抗原を含むワクチンが提供される。
【発明の詳細な説明】
【技術分野】
【0001】
産業上の利用分野
本発明は、細菌多糖抗原ワクチン、それらの製造および医療におけるこのような多糖の使用に関する。
【0002】
特に本発明は、以下の3つの相互関連局面に関する:A−肺炎球菌多糖抗原、典型的には肺炎連鎖球菌(Streptococcus pneumoniae)からのタンパク質抗原および任意にTh1誘導性アジュバントを用いて処方された肺炎球菌多糖複合体抗原を含むワクチン;B−Th1アジュバントで活性調節された特異的有益肺炎球菌多糖複合体;ならびにC−概してインフルエンザ菌からのプロテインDと連結された細菌多糖複合体。
【背景技術】
【0003】
発明の背景
肺炎連鎖球菌は、少なからぬ罹患率および死亡率(特に若年および老齢者における)に関与するグラム陽性細菌であって、侵襲性疾患、例えば肺炎、菌血症および髄膜炎、ならびに集落形成に関連した疾患、例えば急性中耳炎を引き起こす。60歳より上の人々に関する米国での肺炎球菌性肺炎率は、100,000人に3〜8人であると見積もられる。症例の20%において、これは菌血症およびその他の症状発現、例えば髄膜炎を引き起こし、死亡率は、抗生物質治療を用いた場合でも30%近くである。
【0004】
肺炎球菌は、血清型特異性を付与する化学的結合多糖に被包されている。肺炎球菌の90の既知の血清型が存在し、莢膜は、補体から細菌の内面を防御するだけでなく、それ自体免疫原性に乏しいので、肺炎球菌に関する素因ビルレンス決定基である。多糖はT非依存性抗原であり、T細胞と相互作用するためにMHC分子上で処理加工または提示され得ない。しかしながら、それらは、B細胞上の表面受容体の架橋を包含する代替的メカニズムにより免疫系を刺激し得る。
【0005】
侵襲性肺炎球菌性疾患は莢膜に特異的な抗体と最も強く相関し、防御は血清型特異的である、ということがいくつかの実験で示された。
【0006】
多糖抗原ベースのワクチンは、当業界で周知である。ヒト使用に関して登録されている4つとしては、腸チフス菌のVi多糖、インフルエンザ菌からのPRP多糖、血清型A、C、W135およびYから成る四価髄膜炎菌ワクチン、ならびに血清型1、2、3、4、5、6B、7F、8、9N、9V、10A、11A、12F、14、15B、17F、18C、19A、19F、20、22F、23F、および33に対応する多糖から成る23価肺炎球菌ワクチン(肺炎球菌血液単離物の少なくとも90%を占める)が挙げられる。
【0007】
後者3つのワクチンは、呼吸器感染を引き起こして乳児における重度罹患率および死亡率を生じる細菌に対する防御を付与するが、これらのワクチンは2歳未満の小児における使用に関しては、この年齢群では免疫原性が不十分であるために、未だ認可されていない[Peltola et al.(1984), N. Engl. J. Med. 310:1561-1566]。肺炎連鎖球菌は、乳児および幼児における侵襲性細菌性疾患および中耳炎の最も一般的な原因である。同様に、老齢者は肺炎球菌ワクチンに対して不十分な応答を増し[Roghmann et al.(1987), J. Gerontol.42:265-270]、それゆえこの集団における細菌性肺炎の発生数が増大する[Verghese and Berk, (1983)Medicine(Baltimore)62:271-285]。
【0008】
乳児における免疫原性のこの欠如を克服するよう意図された戦略としては、大型免疫原性タンパク質との多糖の連結が挙げられるが、これは、バイスタンダーT細胞援助を提供し、それが連結される多糖抗原に対する免疫記憶を誘導する。肺炎球菌糖タンパク質複合体ワクチンは、種々の年齢群における安全性、免疫原性および効能に関して一般に評価されつつある。
【0009】
A)肺炎球菌多糖ワクチン
23価非複合肺炎球菌ワクチンは、0%から81%までの臨床的効能の広範な変異を示している(Fedson et al.(1994)Arch Intern Med. 154:2531-2535)。効能は、免疫感作されつつある危険群、例えば老齢者、ホジキン病、脾摘出、鎌状赤血球症および無γグロブリン血症患者に(Fine et al.(1994)Arch Intern Med. 154:2666-2677)、そして疾患発現にも関連していると思われる。23価ワクチンは、肺炎球菌性肺炎(いくつかの高危険群、例えば老齢者において)および中耳炎に対する防御を実証しない。
【0010】
したがって、改良型肺炎球菌ワクチン組成物、特に老齢者および幼児における肺炎球菌性疾患(特に肺炎)の予防または改善により有効であるものが必要とされている。
【発明の概要】
【0011】
本発明は、このような改良型ワクチンを提供する。
【0012】
B)選定肺炎球菌多糖複合体+3D−MPL組成物
商業化非複合肺炎球菌ワクチンの防御効力がワクチン接種時に誘導される抗体の濃度に多少関連するということは、一般に容認される。実際、23の多糖が、専ら各構成成分多糖の免疫原性に関して商業的許可を容認された(Ed. Williams et al. New York Academy of Sciences 1995 pp. 241-249)。したがって、肺炎球菌多糖に対する抗体応答のさらなる強化は、多糖または多糖複合体の初回注射に対する防御レベルの抗体を有して応答する乳児および老齢者のパーセンテージを増大し得たし、肺炎連鎖球菌により引き起こされる感染に対する防御免疫を誘導するのに必要な注射の投与量および回数を低減し得た。
【0013】
20世紀初頭以来、研究者達は、ワクチン組成物におけるそれらの免疫原性を改善するために抗原に付加され得る厖大な数の化合物を用いて実験してきた[M.F. Powell & M.J. Newman, Plenum Press, NY, "Vaccine Design-the Subunit and Adjuvant Approach"(1995)Chapter 7"A Compendium of Vaccine Ajuvants and Excipients"で再検討された]。多くは非常に効率的であるが、しかしヒトワクチン組成物におけるそれらの使用を妨げる有意の局所性および全身性悪反応を引き起こす。1926年に最初に記載されたアルミニウムベースのアジュバント(例えばアルム、水酸化アルミニウムまたはリン酸アルミニウム)は、依然として米国で認可されたヒトワクチン中に用いられる唯一の免疫アジュバントである。
【0014】
アルミニウムベースのアジュバントは、それが誘導する「デポー製剤作用」を介して働く担体類のアジュバントの例である。抗原はその表面に吸着され、組成物が注入された場合、アジュバントおよび抗原は血流中に直ちに消散されない。その代わりに、組成物は注入の局所環境中に存続し、より顕著な免疫応答が結果として生じる。このような担体アジュバントは、例えばいくつかの多糖抗原を分解する傾向がある抗原を安定化するのに適しているという付加的な既知の利点を有する。
【0015】
3D−MPLは、非担体アジュバントの一例である。そのフルネームは3−O−デアシル化モノホスホリル脂質A(または3デ−O−アシル化ものホスホリル脂質Aまたは3−O−デスアシル−4’モノホスホリル脂質A)であり、還元末端グルコサミンの位置3がデ−O−アシル化されることを示すために3D−MPLと呼ばれる。その調製のためには、英国特許出願第2220211号を参照されたい。化学的には、それは3−デアシル化モノホスホリル脂質Aの4、5または6アシル化鎖との混合物である。それは最初は、脂質Aの4’−モノホスホリル誘導体(MPL)を3−O−デアシル化するための方法が、免疫刺激活性の変化を伴わずにさらなる弱毒化毒性を有する分子をもたらした1990年代初頭に製造された。
【0016】
3D−MPLは、単独で、または優先的にはデポー製剤型担体アジュバント、例えば水酸化アルミニウム、リン酸アルミニウムまたは水中油型エマルションと組合せて、アジュバントとして用いられてきた。この溶暗組成物中では、抗原および3D−MPLは同一粒子構造中に含入されて、抗原性および免疫刺激性シグナルのより効率的送達を可能にする。複数の研究が、3D−MPLはアルム吸着化抗原の免疫原性をさらに増強し得る、ということを示している[Thoelen et al. Vaccine(1998)16:708-14; EP 689454-B1]。このような組合せも、吸着傾向がある抗原に関しては当業界で選択される(例えば、細菌多糖複合体)が、この場合、アルムへの吸着は、抗原を安定化する傾向がある。沈降アルミニウムベースのアジュバントは、それらは認可ヒトワクチンに一般に用いられる唯一のアジュバントであるので、主として用いられる。したがって、アルミニウムベースのアジュバントと組合せて3D−MPLを含有するワクチンは、それらの易開発性およびマーカー上への導入速度のために、当業界で好まれる。
【0017】
MPL(非3−デアシル化)は、いくつかの一価多糖複合体ワクチン抗原に関するアジュバントとして評価されてきた。生理食塩水中のMPLの同時注入は、4つの一価多糖複合体:肺炎球菌PS6B破傷風毒素、肺炎球菌PS 12−ジフテリア毒素、ならびに緑膿菌外毒素Aに連結された5型黄色ブドウ球菌および8型黄色ブドウ球菌に対する血清抗体応答を増強する[Schneerson et al. J. Immunology(1991)147:2136-2140]。応答増強は、抗原特異的であると教示された。水中油型エマルション中のMPL(担体型アジュバント)は、同一粒子構造中のMPLおよび抗原の存在のために、生理食塩水中のMPLの作用を一貫して増強し、そして他の多糖複合体ワクチンの最適供給のために選択さるべきアジュバント系であると考えられた。
【0018】
Devi等[Infect. Immun.(1991)59:3700-7]は、クリプトコックス属のCryptococcus neoformans莢膜多糖のTT複合体に対するネズミ抗体応答に及ぼす生理食塩水中のMPL(非3−デアシル化)のアジュバント作用を評価した。MPLが複合体と同時に用いられた場合、PSに対するIgM−およびIgG−特異的応答の両方において限界増大のみが認められた。しかしながら、MPLは、複合体の2日後に投与された場合、非常に大きい効果を示した。特に乳児において、抗原と比較してMPLの投与の遅延をようする免疫感作計画を用いることの実用性は疑問である。多糖および多糖−タンパク質複合体に関するMPLのアジュバント効果は、組成物依存性であると思われる。さらに、適切な徐放性送達系(例えば、担体アジュバント使用)中のMPLの混入は、より永続的なアジュバント効果を提供し、調時および遅延投与の問題を免れる。
【0019】
要するに、当業界の状態は、特に多糖または多糖複合体抗原に関しては、MPLまたは3D−MPLがアジュバントとして用いられる場合、その免疫刺激作用を最大にするために、それは、担体アジュバント(例えば、アルミニウムベースのアジュバント)とともに有益に用いられる、ということを教示している。
【0020】
意外なことに、ある種の肺炎球菌多糖複合体に関しては、ワクチン組成物の免疫原性は、担体アジュバント(例えばアルミニウムベースのアジュバント)とともに用いられる3D−MPLよりむしろ3D−MPL単独を用いて抗原が処方される場合に有意に大きい、ということを本発明は見出した。さらに、観察された改善は、用いた3D−MPLの濃度とは、そして特定の複合体が一価組成物中に存在するか否か、あるいはそれらが併用されて多価組成物を形成するか否かとは無関係である。
【0021】
C)細菌多糖−プロテインD複合体
前記のように、多糖抗原ベースのワクチンは、当業界で周知である。前記の認可多糖ワクチンは、異なる実証済み臨床効能を有する。Vi多糖ワクチンは、培養確証腸チフスの防止に際して55%〜77%の効能を有すると概算されている(Plotkin and Cam,(1995)Arch Intern Med 155:2293-99)。髄膜炎菌C多糖ワクチンは、流行条件下で79%の効能を有することが示された(De Wals P, et al.(1996)Bull World Health Organ. 74:407-411)。23価肺炎球菌ワクチンは、0%〜81%の臨床効能の広範な変動を示した(Fedson et al.(1994)Arch Intern Med.154:2531-2535)。前記のように、肺炎球菌ワクチンの防御効能は、ワクチン接種時に誘導される抗体の濃度に多少関連する、ということが容認される。
【0022】
ワクチン接種への多糖アプローチに関連した問題の1つは、多糖それ自体が貧免疫原性であるという事実である。免疫原性のこの欠如を克服するよう意図された戦略としては、大型高免疫原性タンパク質との多糖の連結が挙げられるが、これは、バイスタンダーT細胞援助を提供する。
【0023】
多糖免疫原の産生のために今日一般に用いられるこれらの高免疫原性担体の例としては、ジフテリア毒素(CTまたはCRM197突然変異体)、破傷風毒素(TT)、カギアナカサガイヘモシアニン(KLH)およびツベルクリンの精製タンパク質誘導体(PPD)が挙げられる。
【0024】
一般的に用いられる担体に関連した問題
多数の問題が、例えばGMP複合体の産生におけるならびに複合体の免疫学的特徴における、これらの一般的に用いられる担体の各々に関連している。
【0025】
これらの担体の一般的使用および抗多糖抗体応答の誘導におけるそれらの成功例にもかかわらず、それらはいくつかの欠点に関連する。例えば、抗原特異的免疫応答は、担体(この場合、破傷風毒素)に対して向けられる先在する抗体の存在により抑制され得る(エピトープ抑制)ということが知られている(Di John et al;(1989)Lancet, 2:1415-8)。集団全般では、人々はこれらの抗原でルーチンにワクチン接種されるので、非常に高パーセンテージの人々がDTおよびTTの両方に対する先在免疫を有する。英国では、例えば95%の小児がDTおよびTTの両方を含むDTPワクチンを接種されている。他の著者等は、動物モデルにおけるペプチドワクチンに対するエピトープ抑制の問題を記載している(Sad et al, Immunology, 1991; 74:223-227; Schutze et al, J. Immunol. 135:4, 1985; 2319-2322)。
【0026】
さらに、定期的追加免疫を要するワクチンに関しては、高免疫原性担体、例えばTTおよびDTの使用は、数回注射後の多糖抗体応答を抑制すると思われる。これらの多数回予防接種は、遅延型過応答性(DTH)のような望ましくない反応によっても成し遂げられ得る。
【0027】
KLHは、効力のある免疫原として既知であり、ヒト臨床試験におけるIgEペプチドのための担体としてすでに用いられている。しかしながら、いくつかの悪反応(DTH様反応またはIgE感作)ならびに抗体に対する抗体応答が観察されている。
【0028】
したがって、多糖ベースのワクチンのための担体タンパク質の選択は、すべての患者において作用する(広範なMHC認識)担体に対する必要性、高レベルの抗多糖抗体応答の誘導および担体に対する低抗体応答の間の平衡を必要とする。
【0029】
したがって、多糖ベースのワクチンのために予め用いられる担体は、多数の欠点を有する。これは、種々の多糖抗原に関して同一担体が用いられる場合に、エピトープ抑制が特に問題となる組合せワクチンにおいて特にそうである。WO 98/51339では、組合せワクチン中の多担体は、この作用を乗り越えようとするために用いられた。
【0030】
本発明は、前記の欠点を蒙らない多糖/多ペプチドベースの免疫原性複合体の調製に際して用いるための新規の担体を提供する。
【0031】
本発明は、ワクチンを含めた多糖ベースの免疫原性組成物のための担体として、インフルエンザ菌からのプロテインD(EP 0 594 610 B1)またはその断片を提供する。この担体の使用は、組合せワクチンにおいて特に有益である。
【0032】
発明の要約
A)肺炎球菌多糖ワクチン
したがって、本発明は、少なくとも1つの肺炎連鎖球菌多糖抗原(好ましくは複合化)および肺炎連鎖球菌タンパク質抗原またはその免疫学的機能等価物を、任意にTh1アジュバント(Th1免疫応答を誘導するアジュバント)とともに含むワクチンを提供する。好ましくは、肺炎球菌タンパク質およびTh1アジュバントがともに含まれる。本発明の組成物は、老齢者肺炎の治療に特に適している。
【0033】
肺炎球菌多糖ワクチン(複合化または非複合化)は、この疾患の発生数が非常に高い老齢者集団における肺炎に対して防御し得ない。肺炎球菌に対する重要な防衛メカニズムはオプソニン食菌作用(肺炎球菌多糖に対する抗体の産生により引き起こされるヒトB細胞/好中球媒介性事象。細菌は最終的には食菌されるようになる)であるが、しかしながら、関与するオプソニン性メカニズムの一部、即ちPMN(多形核球)によるスーパーオキシド産生、その他の反応性酸素種産生、PMNの可動化、PMNのアポトーシス、PMNの変形能が、老齢者では損傷される。
【0034】
普通に容認された教義と反対に、正常レベルの抗莢膜多糖抗体は、肺炎球菌が宿主細胞に侵襲して免疫系のこの部門を回避し得るので、細菌の完全クリアランスには有効でない可能性がある。
【0035】
意外にも、免疫系の体液性部門(B細胞媒介性免疫)の他に、免疫系の細胞媒介性部門(例えば、T細胞媒介性免疫)を同時刺激することにより、宿主からの肺炎球菌のクリアランスを増強し得る相乗作用(または協同作用)を結果として生じる、ということを本発明は見出した。これは、概して肺炎球菌感染の予防(または治療)に役立つ発見であるが、しかし多糖ベースのワクチンが効能を示さない老齢者における肺炎の予防(または治療)に特に重要である。
【0036】
肺炎球菌多糖(好ましくは複合化)が肺炎球菌タンパク質(好ましくは肺炎球菌の表面で発現され、あるいは分泌または放出されるタンパク質で、これは感染哺乳類細胞の表面で暮らすIIおよびMHCクラスIの情況においてプロセッシングされ、提示され得る)とともに投与される場合、免疫系の両部門はこのように共働し得る、ということを本発明は見出した。肺炎球菌タンパク質はそれ自体で細胞媒介性免疫の引き金となり得るが、しかし、ワクチン処方物中のTh1誘導性アジュバントの存在が免疫系のこの部門の役に立ち、そして意外にも、免疫系の両部門間の相乗作用をさらに増強する、ということを本発明人等は見出した。
【0037】
B)選定肺炎球菌多糖複合体+3D−MPL組成物
したがって、本発明は、3D−MPLで活性調節され、実質的にアルミニウムベースのアジュバントを欠く1つ又はそれ以上の肺炎球菌多糖複合体を含む抗原性組成物も提供するが、この場合、少なくとも1つの肺炎球菌多糖複合体は、アルミニウムベースのアジュバントとともに3D−MPLを含む組成物と比較して、3D−MPLを含む組成物中で有意により免疫原性である。
【0038】
提供される好ましい実施態様は、1つ又はそれ以上の以下の肺炎球菌莢膜多糖:血清型4、6B、18C、19Fおよび23Fの複合体を含む抗原性組成物である。このような組成物中では、多糖は各々、意外にも、3D−MPLおよびアルミニウムベースのアジュバントを含む組成物と比較して、3D−MPL単独を含む組成物中でより免疫原性である。
【0039】
したがって、本発明の一実施態様においては、免疫原性タンパク質および3D−MPLアジュバントに連結された肺炎連鎖球菌莢膜多糖血清型4、6B、18C、19Fまたは23Fを含む抗原性組成物が提供され、この場合、組成物は実質的にはアルミニウムベースのアジュバントを欠く。
【0040】
第二の実施態様では、本発明は、アルミニウムベースのアジュバントを実質的に欠き、そして血清型4、血清型6B、血清型18C、血清型19Fおよび血清型23Fから成る群から選択される3D−MPLおよび2つまたはそれ以上の肺炎球菌多糖複合体を含む組合せ抗原性組成物を提供する。
【0041】
C)細菌多糖−プロテインD複合体
したがって、本発明は、インフルエンザ菌からのプロテインDまたはそのプロテインD断片に連結された病原性細菌から得られる多糖抗原を含む多糖複合体抗原を提供する。さらに、本発明は、1つ又はそれ以上の多糖抗原がプロテインDに連結される多価ワクチン組成物を提供する。
【図面の簡単な説明】
【0042】
【図1A】マウスにおけるニューモリジンに対する血清IgG。
【図1B】ニューモリジンで免疫化されたマウスにおける血清溶解阻害力価。
【図1C】血清型6Bによる鼻内感染後6時間目におけるOF1マウスにおけるニューモコッカル肺コロニー形成。
【発明を実施するための形態】
【0043】
発明の詳細な説明
A)肺炎球菌多糖ワクチン
本発明は、特に老齢者(および/または乳児および幼児)の肺炎球菌感染の予防または改善のための改良型ワクチンを提供する。
【0044】
本発明の情況では、患者は、年齢が55歳以上、典型的には60歳以上、さらに一般的には65歳以上であれば、老齢者であると考えられる。
【0045】
したがって、本発明の一実施態様では、少なくとも1つの肺炎連鎖球菌多糖抗原および少なくとも1つの肺炎連鎖球菌タンパク質抗原を含む、老齢者(および/または小児、乳児(Infants)および幼児)において用いるのに適したワクチン組成物が提供される。
【0046】
第二の好ましい実施態様では、本発明は、少なくとも1つの肺炎連鎖球菌多糖抗原および少なくとも1つの肺炎連鎖球菌タンパク質抗原およびTh1アジュバントを含む(老齢者における肺炎の予防に適した)ワクチンを提供する。
【0047】
このようなワクチンは、例えば乳児または幼児のような集団の他の高度危険群における肺炎球菌感染(例えば中耳炎)を治療するのにも有用であることが意図される。
【0048】
第三の実施態様では、肺炎球菌多糖抗原およびTh1アジュバントを含むワクチン組成物が提供される。
【0049】
本発明の肺炎連鎖球菌多糖抗原
典型的には、本発明の肺炎連鎖球菌ワクチンは、多糖抗原(好ましくは複合化)を含み、この場合、多糖は少なくとも4つの血清型の肺炎球菌から得られる。好ましくは、4つの血清型としては、6B、14、19Fおよび23Fが挙げられる。さらに好ましくは少なくとも7つの血清型、例えば4、6B、9V、14、18C、19Fおよび23Fから得られるものが組成物中に含まれる。さらに好ましくは、少なくとも11の血清型が組成物に含まれ、例えば一実施態様における組成物は、血清型1、3、4、5、6B、7F、9V、14、18C、19Fおよび23Fから得られる莢膜多糖(好ましくは複合化(conjugated))を含む。本発明の好ましい実施態様では、少なくとも13の多糖抗原(好ましくは複合化)が含まれるが、しかしさらなる多糖抗原、例えば23価(例えば血清型1、2、3、4、5、6B、7F、8、9N、9V、10A、11A、12F、14、15B、17F、18C、19A、19F、20、22F、23Fおよび33F)も本発明により意図される。
【0050】
老齢者予防接種(例えば、肺炎予防のための)に関しては、血清型8および12F(そして最も好ましくはさらに15および22)を前記の11価抗原性組成物に含入して、15価ワクチンを生成するのが有益であるが、一方、乳児または幼児用(中耳炎がより問題である)には、血清型6Aおよび19Aが含入されて、13価ワクチンを生成するのが有益である。
【0051】
老齢者(+55歳)集団における肺炎ならびに乳児(18ヶ月まで)および幼児(典型的には18ヶ月〜5歳)における中耳炎の予防/改善のためには、本明細書に記載したような多価肺炎連鎖球菌多糖を肺炎連鎖球菌タンパク質またはその免疫学的機能等価物と併用することが、本発明の好ましい実施態様である。
【0052】
本発明の肺炎球菌タンパク質
本発明の目的のために、「免疫学的機能等価物」とは、本発明のタンパク質からの少なくとも1つの防御的エピトープを含むタンパク質のペプチドと定義される。このようなエピトープは、特徴的には、表面露呈され、高度保存され、そして宿主における殺細菌性抗体応答を引き出し、あるいは毒性作用を阻止し得る。好ましくは、機能的等価物は、本発明のタンパク質からの少なくとも15、好ましくは30またはそれ以上の連続アミノ酸を有する。最も好ましくは、タンパク質の断片、欠失、例えばその膜貫通欠失変異体(即ち、タンパク質の細胞外ドメインの使用)、融合、化学的または遺伝的脱毒素化誘導体などが用いられ得るが、但し、それらはネイティブタンパク質と同一免疫応答を実質的に生じ得る。
【0053】
本発明の好ましいタンパク質は、肺炎球菌の外表面上に露呈される肺炎球菌タンパク質(肺炎球菌の少なくとも一部のライフサイクル中に宿主の免疫系により認識され得る)であるか、あるいは肺炎球菌により分泌または放出されるタンパク質である。最も好ましくは、タンパク質は毒素、粘着素、2−成分シグナル変換体または肺炎連鎖球菌のリポタンパク質、あるいはそれらの免疫学的機能等価物である。
【0054】
このような組合せワクチン中に含入される特に好ましいタンパク質としては以下のものが挙げられるが、これらに限定されない:ニューモリジン(好ましくは、化学処理または突然変異により脱毒素化)[Mitchell et al. Nucleic Acids Res. 1990 Jul 11;18(3):4010 "Comparison of pneumolysin genes and proteins from Steptococcus pneumoniae types 1 and 2.", Mitchell et al. Biochim Biophys Acta 1989 Jan 23; 1007(1):67-72 "Expression of the pneumolysin gene in Escherichia coli: rapid purification and biological properties.", WO 96/05859(A. Cyanamid), WO 90/06951(Paton et al), WO 99/03884(NAVA)];PspAおよびその膜貫通欠失変異体(米国特許第5804193号−Briles等);PspCおよびその膜貫通欠失変異体(WO 97/09994−Briles等);PsaAおよびその膜貫通欠失変異体(Berry & Paton, Infect Immun 1996 Dec; 64(12):5255-62 "Sequence heterogeneity of PsaA, a 37-kilodalton putative adhesin essential for virulence of Streptococcus pneumoniae");肺炎球菌コリン結合タンパク質およびその膜貫通欠失変異体;CbpAおよびその膜貫通欠失変異体(WO 97/41151; WO 99/51266);グリセルアルデヒド−3−ホスフェート−デヒドロゲナーゼ(Infect. Immun. 1996, 64:3544);HSP70(WO 96/40928);PcpA(Sanchez-Beato et al. FEMS Microbiol Lett 1998, 164:207-14);M様タンパク質(SB特許出願EP 0837130);および粘着素18627(SB特許出願EP 0834568)。
【0055】
本発明に用いられるタンパク質は、好ましくは、ニューモリジン群PsaA、PspA、PspC、CbpA、あるいは2またはそれ以上のこのようなタンパク質の組合せから選択される。本発明は、このようなタンパク質の免疫学的機能等価物も包含する(前記)。
【0056】
組成物中では、タンパク質は、肺炎球菌性疾患に対するT細胞媒介性応答を誘導するのに役立ち、特に肺炎に対する防御に必要とされ、これは肺炎球菌による侵襲を阻止するために、そしてオプソニン食菌作用を刺激するために免疫系の体液性部門と協同する。
【0057】
タンパク質抗原を含入することのさらなる利点は、オプソニン食菌作用過程のためのさらなる抗原の提示、および細菌付着の抑制(粘着素(adhesin)が用いられる場合)または毒素の中和(毒素が用いられる場合)である。
【0058】
本発明の一実施態様によれば、少なくとも4つの血清型、好ましくは少なくとも7つの血清型、さらに好ましくは少なくとも11の血清型から得られる多糖抗原、ならびに少なくとも1つの、しかし好ましくは2つの肺炎連鎖球菌タンパク質を含む肺炎球菌多糖複合体ワクチンを包含する肺炎連鎖球菌ワクチンが提供される。好ましくは、タンパク質の1つは、ニューモリジン(Pneumolysin)あるいはPsaAまたはPspAまたはCbpA(最も好ましくは脱毒素化ニューモリジン)である。好ましい組合せは、少なくともニューモリジンまたはその誘導体およびPspAを含有する。
【0059】
前記のように、予防接種への多糖アプローチに関連した問題は、多糖それ自体が貧免疫原性であるという事実である。これを克服するために、多糖は、バイスタンダーT細胞援助を提供するタンパク質担体に連結され得る。したがって、本発明に利用される多糖はこのようなタンパク質担体に結合されるのが好ましい。多糖免疫原の産生のために現在一般的に用いられるこのような担体の例としては、ジフテリアおよび破傷風毒素(それぞれDT、DT CRM197およびTT)、カギアナカサガイヘモシアニン(KLH)、髄膜炎菌からのOMPC、ならびにツベルクリンの精製タンパク質誘導体(PPD)が挙げられる。
【0060】
しかしながら多数の問題が、これらの一般的に用いられる担体の各々に関連している(前記の「一般的に用いられる担体に関連した問題」の節参照)。
【0061】
本発明は、好ましい実施態様において、これらの欠点を蒙らない多糖ベースの免疫原構築物の調製に際して用いるための新規の担体を提供する。肺炎球菌多糖ベースの免疫原性組成物(またはワクチン)のための好ましい担体は、インフルエンザ菌からのプロテインD(EP 594610-B)またはその断片である。用いるのに適した断片としては、T−ヘルパーエピトープを包含する断片が挙げられる。特に、プロテインD断片は、好ましくはタンパク質のN末端1/3を含有する。
【0062】
肺炎球菌多糖のためのさらに好ましい担体は、肺炎球菌タンパク質それ自体である(「本発明の肺炎球菌タンパク質」の節に前記)。
【0063】
本発明のワクチンは、好ましくは活性調節される。適切なアジュバントとしては、水酸化アルミニウムゲル(アルム)またはリン酸アルミニウムのようなアルミニウム塩が挙げられるが、しかしカルシウム、鉄または亜鉛の塩でもあり得るし、あるいはアシル化チロシンまたはアシル化糖、陽イオン的または陰イオン的誘導化多糖またはポリホスファゼンの不溶性懸濁液であり得る。
【0064】
アジュバントは、免疫応答の細胞媒介性部門を活性調節するためにTH1型の応答の優先的誘導物質であるよう選択されるのが好ましい。
【0065】
本発明のTH1アジュバント
高レベルのTh1型サイトカインは、所定の抗原に対する細胞媒介性免疫応答の誘導を援助する傾向があるが、一方、高レベルのTh2型サイトカインは、抗原に対する体液性免疫応答の誘導を援助する傾向がある。
【0066】
Th1およびTh2型免疫応答の区別は絶対的であるというわけではない、ということを思い起こすことは重要である。実際は、個体は主としてTh1または主としてTh2であると説明される免疫応答を支持する。しかしながら、MosmannとCoffiman(Mosmann, T.R. and Coffiman, R.L.(1989)TH1 and TH2 cell: different patterns of lymphokine secretion lead to different functional properties. Annual Review of Immunology, 7, p145-173)によりネズミCD4+ve T細胞クローンで説明されたことに関してサイトカイン族を考えるのがしばしば便利である。伝統的に、Th1型応答は、Tリンパ球によるINF−γおよびIL−2サイトカインの産生に関連する。Th1型免疫応答の誘導にしばしば直接関連するその他のサイトカインは、IL−12のように、T細胞により産生されない。これに対比して、Th2型応答は、Il−4、IL−5、IL−6、IL−10の分泌に関連する。主要Th1応答を促す適切なアジュバント系としては、モノホスホリル脂質A、ならびにモノホスホリル脂質A、好ましくは3−デ−O−アシル化モノホスホリル脂質A(3D−MPL)とアルミニウム塩の組合せが挙げられる。
【0067】
増強系は、モノホスホリル脂質Aおよびサポニン誘導体の組合せ、特にWO 94/00153に開示されたようなQS21および3D−MPLの組合せ、またはWO 96/33739に開示されているようにQS21がコレステロールで抑制される低反応原性組成物を包含する。
【0068】
水中油型エマルション中にQS21、3D−MPLおよびトコフェロールを包含する特に強力なアジュバント処方物は、WO 95/17210に記載されており、好ましい処方物である。
【0069】
好ましくは、ワクチンは、サポニンを、さらに好ましくはQS21を付加的に含む。処方物は、水中油型エマルションおよびトコフェロールも含み得る(WO 95/17210)。
【0070】
本発明は、ワクチン処方物の製造方法であって、本発明のタンパク質を製薬上許容可能な賦形剤、例えば3D−MPLと一緒に混合することを包含する方法も提供する。
【0071】
オリゴヌクレオチドを含有する非メチル化CpG(WO 96/02555)もTH応答の選択的誘導物質であり、本発明に用いるのに適している。
【0072】
本発明の特に好ましい組成物は、1つ又はそれ以上の複合肺炎球菌多糖、1つ又はそれ以上の肺炎球菌タンパク質およびTh1アジュバントを含む。肺炎球菌タンパク質による細胞媒介性応答の誘導(前記)および免疫系の両部門の間の協同は、このようなアジュバントを用いて活性調節されて、概して肺炎球菌性疾患に対する、そして重要なのは老齢者における肺炎球菌性肺炎に対して特に有効なワクチンを生じる。
【0073】
本発明のさらなる局面において、医療に用いるための本明細書中に記載したような免疫原またはワクチンが提供される。
【0074】
本発明のさらに別の局面では、不応答者の集団内で多糖抗原に対する体液性抗体応答を血清変換または誘導し得る、肺炎球菌多糖複合体およびTh1アジュバント(好ましくは3D−MPL)を含む組成物が提供される。
【0075】
集団の10〜30%は、多糖免疫感作に対する不応答者(ワクチン中の50%より多い血清型に応答しない)であることが知られている(Konradsen et al., Scand. J. Immun 40:251(1994); Rodriguez et al., JID, 173:1347(1996))。これも、複合化ワクチンに関する症例であり得る(Musher et al. Clin. Inf. Dis. 27:1487(1998))。これは、高危険域の集団(乳児、幼児および老齢者)に関しては特に重大である。
【0076】
複合肺炎球菌多糖(これは特定の集団において低応答傾向がある)のTh1アジュバント(前記の「本発明のTh1アジュバント」参照)との組合せは、意外にもこの不応答性を克服し得る、ということを本発明は見出した。好ましくは3D−MPLが用いられるべきであり、最も好ましくは3D−MPLはアルミニウムベースのアジュバントを欠く(依然として良好な応答を提供する)。したがって本発明はこのような組成物を提供し、さらにこのような組成物を投与することにより肺炎球菌多糖に対する不応答者を治療する方法を提供し、そしてその上さらに、多糖抗原に不応答性である個体における肺炎球菌性疾患に対する治療(または防御)における複合肺炎球菌多糖抗原を含む薬剤の製造に際してのTh1アジュバントの使用を提供する。
【0077】
一実施態様では、老齢者における肺炎の予防または改善方法であって、肺炎連鎖球菌多糖抗原およびTh1アジュバントまたは肺炎球菌タンパク質(好ましくは両方)を含む本明細書に記載したような安全且つ有効量のワクチンを前記の老齢患者に投与することを包含する方法がある。
【0078】
さらに別の実施態様では、乳児または幼児における中耳炎の予防または改善方法であって、肺炎連鎖球菌多糖抗原および肺炎連鎖球菌タンパク質抗原またはTh1アジュバント(好ましくは両方)を含む安全且つ有効量のワクチンを前記の乳児または幼児に投与することを包含する方法が提供される。
【0079】
好ましくは、前記のような本発明の方法においては、多糖抗原は多糖タンパク質複合体として存在する。
【0080】
本発明のワクチン製剤
本発明のワクチン製剤は、全身または粘膜経路により前記のワクチンを投与することにより、感染に感受性の哺乳類を防御または治療するために用いられ得る。これらの投与は、筋内、腹腔内、皮内または皮下経路による、あるいは口腔/栄養管、気道、尿生殖路への粘膜性糖よによる注入を含み得る。肺炎または中耳炎の治療のためのワクチンの鼻腔内投与は好ましい(肺炎球菌の鼻咽頭運搬はより有効に予防され得るので、したがって、その最初期段階で感染を減衰し得る)。
【0081】
各ワクチン中の複合抗原の量は、典型的ワクチンにおいて有意の副作用を伴わずに免疫防御応答を誘導する量として選択される。このような量は、どの特異的免疫原が用いられるか、そしてそれがどのように存在するかによって変わる。一般的に、各用量は、0.1〜100μg、好ましくは0.1〜50μg、好ましくは0.1〜10μgの多糖を含むと予測され、1〜5μgが最も好ましい範囲である。
【0082】
ワクチン中のタンパク質抗原の含量は、典型的には1〜100μg、好ましくは5〜50μg、最も典型的には5〜25μgの範囲である。
【0083】
特定のワクチンに関する構成成分の最適量は、被験者における適切な免疫応答の観察を含めた標準試験により確かめられ得る。初回予防接種後、適当な間隔で1回または数回の追加免疫感作を被験者に施し得る。
【0084】
ワクチン製剤は、Vaccine Design("The subunit and adjuvant approach")(eds Powell M.F. & Newman M.J.(1995)Plenum Press New York)に一般的に記載されている。リポソーム内封入は、米国特許第4,235,877号(Fullerton)に記載されている。
【0085】
B)選定肺炎球菌多糖複合体+3D−MPL組成物
本発明の目的のために、「本発明の肺炎球菌多糖複合体」という用語は、アルミニウムベースのアジュバントとともに3D−MPLを含む組成物と比較して、3D−MPLを含む組成物中でより免疫原性である肺炎連鎖球菌莢膜多糖の複合体(例えば、血清型4、血清型6B、血清型18C、血清型19Fおよび血清型23Fの複合体)を記述する。
【0086】
本発明の目的のために、「アルミニウムベースのアジュバントを実質的に欠く」という用語は、付加的アルミニウムベースのアジュバントを伴わずに3D−MPLを含む等価組成物と比較した場合の本発明の肺炎球菌多糖複合体の免疫原性の任意の低減を引き起こスクリーニング十分なアルミニウムベースのアジュバント(例えば水酸化アルミニウムおよび、特にリン酸アルミニウム)を含有しない組成物を記述する。好ましくは、抗原性組成物は、本質的に3D−MPLから成るアジュバントを含有すべきである。用量当たりで付加されるアルミニウムベースのアジュバントの量は、好ましくは50μg未満、さらに好ましくは30μg未満、さらに好ましくは10μg未満であるべきであり、最も好ましくは本発明の抗原性組成物に付加されるアルミニウムベースのアジュバントは存在しない。
【0087】
本発明の目的のために、アルミニウムベースのアジュバントとともに3D−MPLを含む組成物と比較して、3D−MPLを含む組成物中で肺炎球菌多糖複合体が十分により免疫原性であるか否かの決定は、実施例2に記載されているように確定されるべきである。3D−MPL単独を含む場合に組成物が有意により免疫原性であるか否かの指標として、3D−MPL単独を含む組成物対リン酸アルミニウムアジュバントとともに3D−MPLを含む等価組成物間のGMC IgG濃度(実施例2で確定されるような)の比は、2より大きい、好ましくは5より大きい、さらに好ましくは7より大きい、さらに好ましくは9より大きい、最も好ましくは14より大きい値であるべきである。
【0088】
予防接種への多糖アプローチに関連した問題の1つは、多糖それ自体が貧免疫原性であるという事実である。免疫原性のこの欠如を克服するよう意図された戦略としては、大型免疫原性タンパク質担体との多糖の結合(連結(conjugating))が挙げられるが、これは、バイスタンダーT細胞援助を提供する。本発明の肺炎球菌多糖がバイスタンダーT細胞援助を提供するタンパク質担体に結合されるのが好ましい。用いられ得るこのような担体の例としては、ジフテリア、ジフテリア突然変異体および破傷風毒素(それぞれDT、DT CRM197およびTT)、カギアナカサガイヘモシアニン(KLH)、ツベルクリンの精製タンパク質誘導体(PPD)ならびに髄膜炎菌からのOMPCが挙げられる。
【0089】
最も好ましくは、インフルエンザ菌からのプロテインD(EP 0 594 610-B)またはその断片(節C参照)は、本発明の肺炎球菌多糖のための免疫原性タンパク質担体として用いられる。
【0090】
一実施態様では、本発明の抗原性組成物は、免疫原性タンパク質に連結され、3D−MPLアジュバントを処方された肺炎球菌多糖血清型(PS)4を含むが、この場合、組成物はアルミニウムベースのアジュバントを実質的に欠く。さらなる実施態様では、抗原性組成物は、免疫原性タンパク質に連結され、3D−MPLアジュバントを処方されたそれぞれPS6B、18C、19Fまたは23Fを含むが、この場合、組成物はアルミニウムベースのアジュバントを実質的に欠く。
【0091】
本発明のさらに別の実施態様では、3D−MPLアジュバントを処方されたPS4、PS6B、PS18C、PS19FおよびPS23F群からの2つまたはそれ以上の肺炎球菌多糖複合体を含む組合せ抗原性組成物が提供されるが、この場合、組成物はアルミニウムベースのアジュバントを実質的に欠く。
【0092】
本発明の肺炎球菌多糖複合体の免疫原性は、他の肺炎球菌多糖複合体との組合せにより有意に影響されない(実施例3)。したがって、本発明の好ましい局面は、本発明の1つ又はそれ以上の肺炎球菌多糖複合体を、1つ又はそれ以上のさらに別の肺炎球菌多糖複合体と組合せて含む組合せ抗原性組成物を提供するが、この場合、組成物は3D−MPLアジュバントを処方されるが、しかしアルミニウムベースのアジュバントを実質的に欠いている。
【0093】
本発明のさらなる好ましい実施態様では、少なくとも1つの、好ましくは2、3、4または5つすべてのPS4、6B、18C、19Fまたは23F肺炎球菌多糖複合体を、さらに3D−MPLアジュバントを処方されるがしかしアルミニウムベースのアジュバントを実質的に欠いているその他の肺炎球菌多糖複合体の任意の組合せを含有する組合せ抗原性組成物が提供される。
【0094】
典型的には、本発明の肺炎連鎖球菌組合せ抗原性組成物は多糖複合体抗原を含むが、この場合、多糖は、少なくとも4つ、7つ、11、13、15または23の血清型から得られる(治療される疾患による血清型の好ましい組合せに関する前記の「本発明の肺炎連鎖球菌多糖抗原」参照)。
【0095】
本発明の抗原性組成物は、好ましくは、特に老齢者および乳児および幼児における肺炎球菌感染を予防(または治療)するためのワクチン組成物として用いられる。
【0096】
本発明のさらなる実施態様は、薬剤中に用いるための前記の抗原性組成物の提供;肺炎連鎖球菌莢膜多糖複合体に対する免疫応答の誘導方法であって、安全且つ有効量の前記の抗原性組成物の1つを患者に投与する過程を含む方法;ならびに肺炎球菌性疾患の予防(または治療)のための薬剤の製造における前記の抗原性組成物の1つの使用を包含する。
【0097】
老齢者(+55歳)集団における肺炎ならびに乳児(18ヶ月まで)および幼児(典型的には18ヶ月〜5歳)における中耳炎の予防/改善のためには、本明細書に記載したように処方される多価肺炎連鎖球菌多糖複合体を肺炎連鎖球菌タンパク質またはその免疫学的機能等価物と併用することが、本発明のさらなる好ましい実施態様である。好ましいタンパク質/タンパク質組合せに関しては、前節「本発明の肺炎球菌タンパク質」を参照されたい。
【0098】
好ましくは、本明細書中に前記した抗原性組成物(およびワクチン)は、それらが用いられるまで凍結乾燥され、使用時点で、それらは希釈剤を用いて即座に再構築される。さらに好ましくは、それらは3D−MPLの存在下で凍結乾燥され、食塩溶液で即座に再構築される。
【0099】
組成物を凍結乾燥すると、より安定な組成物が結果的に生じる(例えば、それは多糖抗原の分解を防止する)。その過程は、意外にも、肺炎球菌多糖に対するより高い抗体力価にも関与する。これは、PS6B複合体に関して特に有意であることが示されている。したがって、本発明の別の局面は、3D−MPLで活性調節され、アルミニウムベースのアジュバントを実質的に欠くPS6B複合体を含む凍結乾燥抗原性組成物である。
【0100】
ワクチンの調製に関しては、前記の「本発明のワクチン製剤」の節を参照されたい。
【0101】
C)細菌多糖−プロテインD複合体
組合せワクチンに対する傾向は、レシピエントへの不快を軽減し、予定を立て易くし、レジメンの完了を保証するという利点を有するが、しかしワクチンの効能を低減するという付随する危険性も存在する(担体タンパク質の過剰使用によるエピトープ抑制における考察に関する前記を参照)。したがって、ある集団の要求を満たし、さらにそれらの構成成分間の免疫原性干渉を示さないワクチン組合せを製造することは有益である。これらの利点は、乳児、幼児または老齢者のような高危険群への組合せワクチンの投与に対して特別な利点を有する本発明の免疫原性組成物(またはワクチン)により実現され得る。
【0102】
本発明は、多糖ベースの免疫原性組成物、例えばワクチンのための担体として、インフルエンザ菌からのプロテインDまたはそのだPWを提供する。使用に適した断片としては、Tヘルパーエピトープを包含する断片が挙げられる。特に、プロテインD断片は、好ましくはタンパク質のN末端1/3を含有する。
【0103】
プロテインDは、インフルエンザ菌からのIgD結合タンパク質であり(EP 0 594 610 B1)、潜在的免疫原である。
【0104】
本発明により意図されるプロテインDに連結される多糖としては、腸チフス菌Salmonella typhiに対するVi多糖抗原、髄膜炎菌多糖(A、C、W135およびY型、ならびにB群髄膜炎菌の多糖および修飾多糖を含む)、黄色ブドウ球菌からの多糖、連鎖球菌属のStreptococcus agalactiaeからの多糖、肺炎連鎖球菌からの多糖、ミコバクテリウム属、例えばヒト結核菌Mycobacterium tuberculosisからの多糖(例えば、マンノホスホイニシチドトレハロース、ミコール酸、マンノース面冠アラビノマンナン、それからの莢膜およびアラビノガラクタンス)、クリプトコックス属のCryptococcus neoformansからの多糖、非分類可能インフルエンザ菌のリポ多糖、b型インフルエンザ菌からの莢膜多糖、モラクセラ属のMoraxella catharralisのリポ多糖、ゾンネ赤痢菌のリポ多糖、クルーズトリパノソーマのリポペプチドホスホグリカン(LPPG)、癌関連ガングリオシドGD3、GD2、腫瘍関連ムチン、特にT−F抗原およびシアリルT−F抗原、ならびにT−F抗原に構造的に関連するHIV関連多糖が挙げられるが、これらに限定されない。
【0105】
多糖は、既知の方法により、担体タンパク質に結合され得る(例えば、Likhite(米国特許第4,372,945号)およびArmor等(米国特許第4,474,757号)による)。好ましくは、CDAP複合が実行される(WO 95/08348)。
【0106】
CDAPでは、シアニル化試薬1−シアノ−ジメチルアミノピリジニウムテトラフルオロボレート(CDAP)は、好ましくは多糖−タンパク質複合体の合成のために用いられる。シアニル化反応は、アルカリ性感受性多糖の加水分解を回避する相対的に低刺激性条件下で実施され得る。この合成は、担体タンパク質との直接結合を可能にする。
【0107】
多糖は、水または食塩溶液中に可溶化される。CDAPは、アセトニトリル中に溶解されて、多糖溶液に直接付加される。CDAPは多糖のヒドロキシル基と反応して、シアネートエステルを生成する。活性化工程後、担体タンパク質が付加される。しリンのアミノ基は、活性化多糖と反応して、イソウレア共有結合を生成する。
【0108】
カップリング反応後、大余分量のグリシンが次に付加されて、残留活性化官能基をクエンチングさせる。次に生成物をゲル浸透に通して見反応担体タンパク質および残留試薬を除去する。したがって、本発明は、多糖プロテインD複合体の製造方法であって、多糖を活性化し、多糖をプロテインDと結合する工程を包含する方法を提供する。
【0109】
本発明の好ましい実施態様では、肺炎連鎖球菌感染の予防のための免疫原性組成物(またはワクチン)処方物が提供される。
【0110】
肺炎球菌が肺、脳脊髄液および血液に拡がるメカニズムは、十分には分かっていない。正常肺胞に達する細菌の増殖は、それらの相対的乾燥により、そして肺胞マクロファージの食菌活性により抑制される。これらの等位防衛のあらゆる解剖学的または生理学的変化は、感染に対する肺の感受性を増大する傾向がある。肺炎連鎖球菌の細胞壁は、肺の肺胞における炎症性応答の発生において重要な役割を有する(Gillespie et al.(1997), I&I 65:3936)。
【0111】
典型的には、本発明の肺炎連鎖球菌ワクチンはプロテインD多糖複合体を含むが、この場合、多糖は少なくとも4、7、11、13、15または23の血清型から得られる。治療される疾患による血清型の好ましい組合せに関する前記の「本発明の肺炎連鎖球菌多糖抗原」を参照されたい。
【0112】
本発明のさらなる実施態様では、特に血清型A、C、W-135およびYから、髄膜炎菌ワクチンが提供される。髄膜炎菌は、細菌性髄膜炎の最も重要な原因の1つである。これらの生物の炭水化物莢膜は、ビルレンス決定基および防御抗体のための標的として作用する。それにもかかわらず、炭水化物は、低年齢小児において貧免疫原性であることが周知である。本発明は、これらの多糖に特に適したタンパク質担体であるプロテインDを提供するが、これは、多糖抗原特異的B細胞増殖および成熟、ならびに免疫記憶の誘導を補佐するためにT細胞応答を活性化し得るT細胞エピトープを提供する。
【0113】
本発明の代替的実施態様では、b型インフルエンザ菌の莢膜多糖(PRP)−プロテインD複合体が提供される。
【0114】
本発明は、一連の異なる病原体に対する防御を提供する組合せワクチンも意図する。プロテインD担体は、意外にも、多重多糖抗原が連結された組合せワクチン中の担体として有用である。前記のように、エピトープ抑制は、各多糖に関して同一担体が用いられる場合に起こると思われる。WO 98/51339は、DTに関する組成物中のある比率の多糖とTTに関する残りの比率のものを連結することによりこの妨害を最小限にしようとするための組成物を示した。
【0115】
意外なことに、組合せワクチンにおけるこのようなエピトープ抑制作用を最小限にするのにプロテインDは特に適していることを、本発明は見出した。組合せにおける1つ又はそれ以上の多糖は、プロテインDと連結されるのが有益であり、好ましくは、抗原はすべて、このような組合せワクチン内でプロテインDに連結される。
【0116】
好ましい組合せとしては、髄膜炎菌CおよびY(そして好ましくはA)に対する防御をもたらすワクチンが挙げられるが、この場合、1つ又はそれ以上の血清型YおよびC(そして最も好ましくはA)からの多糖抗原はプロテインDに結合される。
【0117】
インフルエンザ菌多糖ベースのワクチン(好ましくはTT、DTまたはCRM197と、あるいは最も好ましくはプロテインDと連結されたPRP)は、前記の組合せワクチンを用いて処方され得る。
【0118】
多数の小児科ワクチンは、目下、小児が受ける注射の回数を低減するために組合せワクチンとして投与されている。したがって、小児科ワクチンに関しては、他の抗原は、本発明のワクチンを用いて処方され得る。例えば本発明のワクチンは、ジフテリア毒素(DT)、破傷風毒素(TT)および百日咳構成成分[典型的には脱毒素価百日咳毒素(PT)、ならびに任意のペルタクチン(PRN)および/またはアグルチニン1+2を有する糸状赤血球凝集素(FHA)]を含む周知の「三価」組合せワクチン、例えばDT、TT、PT、FHAおよびPRN抗原を含有する市販のワクチンINFANRIX-DTPa(SmithKlineBeecham Biologicals)を用いて、あるいは、例えばTritanrixとしてこれもSmithKlineBeecham Biologicalsにより市販されているような全細胞百日咳菌構成成分を用いて処方され、あるいは別々に、しかし同時に投与され得る。組合せワクチンは、その他の抗原、例えばB型肝炎表面抗原(HBsAg)、ポリオウイルス抗原(例えば、不活性化三価ポリオウイルス−IPV)、モラクセラ属のMoraxella catarrhalis外膜タンパク質、非分類可能インフルエンザ菌タンパク質、B型髄膜炎菌外膜タンパク質も含み得る。
【0119】
組合せワクチン(特に中耳炎の予防用)中に含入され得る好ましいMoraxella catarrhalisタンパク質抗原の例を以下に示す:OMP106[WO 97/41731(Antex)およびWO 96/34960(PMC)];OMP21;LbpAおよびLbpB[WO 98/55606(PMC)];TbpAおよびTbpB[WO 97/13785およびWO 97/32980(PMC)];CopB[Helminen ME, et al.(1993)Infect. Immun. 61:2003-2010];UspA1/2[WO 93/03761(テキサス大学)];ならびにOmpCD。組合せワクチン(特に中耳炎予防用)中に含入され得る非分類可能インフルエンザ菌抗原の例としては以下のものが挙げられる:フィンブリンタンパク質[(米国特許第5766608号-Ohio State Research Foundation)]およびそれからのペプチドを含む融合物[例えば、LB1(f)ペプチド融合物;米国特許第5843464号(OSU)またはWO 99/64067];OMP26[WO 97/01638(Cortecs)];P6[EP 281673(ニューヨーク州立大学)];TbpAおよびTbpB;Hia;Hmw1、2;Hap;ならびにD15。
【0120】
本発明により意図される好ましい小児科ワクチンを以下に示す:
a)C型髄膜炎菌多糖複合体およびb型インフルエンザ菌多糖複合体。任意にAおよび/またはY型髄膜炎菌多糖複合体を有するが、但し、少なくとも1つの、好ましくはすべての多糖抗原がプロテインDに連結される。
【0121】
b)DT、TT、百日咳構成成分(望ましいPT、FHAおよびPRN)、B型肝炎表面抗原およびIPV(不活性化三価ポリオウイルスワクチン)を有するa)のワクチン。
【0122】
c)プロテインDに連結された肺炎連鎖球菌多糖抗原。
【0123】
d)モラクセラ属のMoraxella catarrhalisおよび/または非分類可能インフルエンザ菌からの1つ又はそれ以上の抗原を有するc)のワクチン。
【0124】
前記の組合せワクチンはすべて、担体としてのプロテインDの含入により利益を蒙り得る。明らかに、組合せワクチン(例えばエピトープ抑制を克服するために)中に包含される担体が多いほど、最終ワクチンはより高価で複雑になる。したがって、プロテインDに連結される組合せワクチンの多糖抗原のすべてのまたは大多数を有ると、かなりの利点を提供する。
【0125】
老齢者(+55歳)集団における肺炎ならびに乳児または幼児における中耳炎の予防のためには、本明細書に記載したような多価肺炎球菌多糖−プロテインD抗原を肺炎連鎖球菌タンパク質またはその免疫学的機能等価物と併用することが、本発明の好ましい実施態様である。このような組合せ中に含入され得る好ましいタンパク質/タンパク質組合せに関しては、前記の「本発明の肺炎球菌タンパク質」の節を参照されたい。
【0126】
したがって、本発明は、肺炎連鎖球菌多糖−プロテインD複合体および肺炎連鎖球菌タンパク質抗原を含む免疫原性組成物を提供する。
【0127】
本発明の多糖−プロテインD複合体抗原は、好ましくは、本発明のワクチン処方物中で活性調節される。適切なアジュバントとしては、水酸化アルミニウムゲル(アルム)またはリン酸アルミニウムのようなアルミニウム塩が挙げられるが、しかしカルシウム、鉄または亜鉛の塩でもあり得るし、あるいはアシル化チロシンまたはアシル化糖、陽イオン的または陰イオン的誘導化多糖またはポリホスファゼンの不溶性懸濁液であり得る。
【0128】
老齢者ワクチンに関しては、アジュバントはTH1型の応答の望ましい誘導物質であるよう選択されるのが好ましい。
【0129】
特定のTh1アジュバントに関しては、前記の「本発明のTh1アジュバント」を参照されたい。
【0130】
本発明のさらなる局面では、医療に用いるための本明細書中に記載したような免疫原またはワクチンが提供される。
【0131】
ワクチン製剤/複合体の投与に関しては、前記の「本発明のワクチン製剤」を参照されたい。
【0132】
プロテインDは、基本的には、非分類可能インフルエンザ菌(ntHi)に対するB細胞媒介性防御を生じ得る免疫原であるので、中耳炎に対するワクチン中にも有益に用いられる。ntHiは、宿主細胞を侵襲し、タンパク質抗原により誘導されるB細胞媒介性作用を回避する。中耳炎ワクチンのための抗原としてのプロテインDの有効性(それ自体または多糖のための担体としての)を増大する方法を、本発明は意外にも見出した。これは、免疫系の細胞媒介性部門がプロテインDに対して最適化されるよう、強力なTh1応答が被験者において誘発されるようにプロテインDを活性調節することによりなされる。これは、意外にも、投与直前に再構築される、プロテインDおよびTh1アジュバント(好ましくは3D−MPL)を含む凍結乾燥化組成物を用いて成し遂げられる。したがって、本発明は、このような組成物、このような組成物の製造方法(プロテインDおよびTh1アジュバントを含む混合物を凍結乾燥することによる)、ならびに中耳炎の治療におけるこのような組成物の使用も提供する。
【0133】
広義では、本発明は、Th1アジュバント(「本発明のTh1アジュバント」参照)、好ましくは3D−MPLの存在下での免疫原の凍結乾燥は、一般に、免疫原に対するTh1免疫応答を増大する、ということを意図する。したがって、本発明は、より強力なTh1免疫応答が必要とされるあらゆる免疫原に適用可能である。このような免疫原は、細菌、ウイルスおよび腫瘍タンパク質抗原、ならびに自己タンパク質およびペプチドを包含する。
【実施例】
【0134】
実施例
実施例で本発明を説明するが、本発明はそれらに限定されない。
【0135】
実施例1
肺炎連鎖球菌莢膜多糖:
11価候補ワクチンとしては、本質的にはEP 72513に記載されたように作製された莢膜多糖血清型1、3、4、5、6B、7F、9V、14、18C、19Fおよび23Fが挙げられる。各多糖は、CDAP化学作用を用いて活性化され、誘導されて(WO 95/08348)、タンパク質担体と連結される。多糖はすべて、血清型3(その粘度を減少させるためにサイズ低減される)以外は、それらのネイティブ形態で連結される。
【0136】
タンパク質担体:
選択されるタンパク質担体は、大腸菌中で発現された非分類可能インフルエンザ菌からの組換え体プロテインD(PD)である。
【0137】
プロテインDの発現
インフルエンザ菌プロテインD
プロテインD発現のための遺伝子構築
出発物質
プロテインDコードDNA
プロテインDは、すべての血清型および非分類可能菌株のインフルエンザ菌の間で高度に保存される。全プロテインD遺伝子をコードするDNA配列を含有するベクターpHIC348は、A. Forsgren博士(Department of Medical Microbiology, University of Lund, Malmo General Hospital, Malmo, Sweden)から入手した。プロテインDのDNA配列は、Janson et al.(1991)Infect. Immun. 59:119-125により発表されている。
【0138】
発現ベクターpMG1
発現ベクターpMG1は、外来挿入遺伝子の転写および翻訳のためのバクテリオファージλ由来対照素子が導入されたpBR322(Gross et al., 1985)の誘導体である(Shatzman et al., 1983)。さらに、アンピシリン耐性遺伝子をカナマイシン耐性遺伝子と交換した。
【0139】
大腸菌AR58菌株
SA500誘導体(galE::TN10, λKil-cI857△H1)上で予め増殖させたP1ファージストックを用いたN99の形質導入により、大腸菌AR58菌株を生成した。N99およびSA500は、Martin Rosenberg博士の実験室(National Institute of Health)から得られた大腸菌K12菌株である。
【0140】
発現ベクターpMG1
プロテインDの製造のために、そのタンパク質をコードするDNAを発現ベクターpMG 1中でクローン化した。このプラスミドは、λファージDNAからのシグナルを利用して挿入外来遺伝子の転写および翻訳を駆動する。ベクターは、プロモーターPL、オペレーターOLおよび2つの利用部位(NutLおよびNutR)を含有して、Nタンパク質が提供された場合の転写性極性作用を軽減する(Gross et al., 1985)。PLプロモーターを含有するベクターを大腸菌溶原性宿主中に°にして、プラスミドDNAを安定化する。溶原性宿主菌株は、ゲノム中に組み込まれる複製欠陥λファージDNAを含有する(Shatzman et al., 1983)。染色体λファージDNAは、ベクターのOLリプレッサーと結合し、PLプロモーターとのRNAポリメラーゼの結合を阻止し、それにより挿入遺伝子の転写を阻止するcIリプレッサータンパク質の合成を指図する。発現菌株AR58のcI遺伝子は、PL特異的転写が温度シフトにより調節され得るように、即ち、培養温度の増大がリプレッサーを不活性化し、外来タンパク質の合成が開始されるように、温度感受性突然変異体を含有する。この発現系は、特に細胞に対して毒性であり得るものの外来タンパク質の合成の制御を可能にする(Shimataka & Rosenberg, 1981 )。
【0141】
大腸菌AR58菌株
プロテインD担体の産生に用いられるAR58溶原性大腸菌株は、標準NIH大腸菌K12株N99の誘導体(F-su-galK2、lacZ-thr-)である。それは、欠陥溶原性λファージ(galE::TN10、λKil-cI857△H1)を含有する。Kil-表現型は、宿主高分子合成の遮断を阻止する。cI857突然変異は、cIリプレッサーに温度感受性損傷を付与する。△H1欠失は、λファージ正当オペロンおよび宿主bio、uvr3およびchlA遺伝子座を除去する。SA500誘導体(galE::TN10, λKil-cI857△H1)上で予め増殖させたP1ファージストックを用いたN99の形質導入により、大腸菌AR58菌株を生成した。隣接galE遺伝子中のエト等サイクリン耐性をコードするTN10トランスポゾンの存在によりテトラサイクリンを用いて、欠陥リソゲンのN99中への導入を選択した。
【0142】
ベクターpMGMDPPrDの構築
インフルエンザウイルスの非構造S1タンパク質をコードする遺伝子を含有するpMG1ベクター(pMGNSI)を用いて、pMGMDPPrDを構築した。それぞれ5‘および3’末端にNcoIおよびXbaI制限部位を含有するPCRプライマーを用いて、pHIC348ベクター(Janson et al. 1991)からのPCRにより、プロテインD遺伝子を増幅した。次に、 NcoIおよびXbaI間のpMGNS1中にNcoI/XbaI断片を導入し、したがってNS1タンパク質のN末端81アミノ酸を含有する融合タンパク質を、その後、PDタンパク質を作製した。このベクターを、pMGNS1PrDと名付けた。
【0143】
前記の構築物に基づいて、プロテインD発現のための最終構築物を生成した。pMGNS1PrDからBamHI/BamHI断片を除去した。このDNA加水分解は、最初の3つのN末端残基以外のNS1コード領域を除去する。ベクターの再結紮時に、その後のN末端アミノ酸配列を有する融合タンパク質をコードする遺伝子が生成された。
‐‐‐‐MDP SSHSSNMANT‐‐‐‐
NS1 プロテインD
プロテインDは、普通は脂質鎖が結合されるリーダーペプチドまたはN末端システインを含有しない。したがって、そのタンパク質は、ペリプラズムに排出されないし、脂質化もされず、可溶性形態で細胞質中に残存する。
【0144】
37℃での熱ショックにより、AR58宿主菌株中に最終構築物pMG−MDPPrDを導入した。カナマイシンの存在下で、プラスミド含有細菌を選択した。選定エンドヌクレアーゼによる単離プラスミドDNAの消化によって、プロテインDコードDNA挿入物の存在を実証した。組換え体大腸菌株をECD4と呼ぶ。
【0145】
プロテインDの発現は、λPLプロモーター/OLオペレーターの制御下にある。宿主菌株AR58は、OLとの結合により低温でλPLからの発現を遮断するゲノム内の温度感受性cI遺伝子を含有する。温度を上げると、cIはOLから放出され、プロテインDが発現される。発酵終了時に、細胞を濃縮し、凍結する。
【0146】
収穫細胞からの抽出およびプロテインDの精製を、以下のように実施した。凍結細胞培養ペレットを解氷し、細胞崩壊溶液(クエン酸塩緩衝液、pH6.0)中に再懸濁して、最終OD650=60とする。P=1000 barで高圧ホモジナイザーに懸濁液を2回通す。遠心分離により細胞培養ホモジネートを清澄化し、濾過により細胞破砕屑を除去する。一次精製過程において、濾過溶解物を陽イオン交換クロマトグラフィーカラム(SP Sepharose Fast Flow)に適用する。PDはイオン相互作用によりゲルマトリックスと結合するが、それを、溶離緩衝液のイオン強度の段階的増大により溶離する。
【0147】
二次精製過程では、不純物を陰イオン交換マトリックス(Q Sepharose Fast Flow)上に保持する。PDはゲル上に結合せず、フロースルーで収集され得る。
【0148】
両カラムクロマトグラフィー過程においては、ODにより分画収集をモニタリングする。精製プロテインDを含有する陰イオン交換カラムクロマトグラフィーのフロースルーを、限外濾過により濃縮する。
【0149】
プロテインD含有限外濾過保持物を最後に0.2μm膜に通す。
【0150】
化学的性質:
活性化およびカップリング化学作用:
活性化およびカップリング条件は、各多糖に特異的である。これらを表1に示す。注射のために、ネイティブ多糖(PS3以外)を2 MのNaCl中または水中に溶解した。すべての血清型に関して、最適多糖濃度を評価した。
【0151】
アセトニトリル中の100 mg/mlストック溶液から、CDAP(CDAP/PS比0.75 mg/mg PS)を多糖溶液に付加した。1.5分後、0.2 Mのトリエチルアミンを付加して、特異的活性化pHを得た。25℃で2分間、このpHで多糖の活性化を実施した。プロテインD(量は初期PS/PD比によっている)を活性化多糖に付加し、特異的pHで1時間、カップリング反応を実施した。次に、25℃で30分間そして4℃で一夜、グリシンを用いて反応を止めた。
【0152】
0.2 MNaClで平衡させたセファクリル500HRゲル濾過カラムを用いてゲル濾過により、複合体を精製した。
【0153】
溶離分画の炭水化物およびタンパク質の含量を確定した。複合体をプールし、0.22μm滅菌膜上で滅菌濾過した。複合体調製物中のPS/タンパク質比を確定した。
【0154】
特性化:
各複合体を特性化して、表2に記載した内訳に合した。レソルシノール検定により多糖含量(μg/ml)を、Lowry検定によりタンパク質含量(μg/ml)を測定した。その濃度比により、最終PS/PD比(w/w)を確定する。
【0155】
残留DMAP含量(ng/μgPS):
CDAPによる多糖の活性化は、多糖中にシアネート基を導入し、DMAP(4−ジメチルアミノ−ピリジン)を遊離する。SBで開発された特異的検定により、残留DMAP含量を確定した。
【0156】
遊離多糖含量(%):
α−PD抗体および飽和硫酸アンモニウムを用いたインキュベーションおよびその後の遠心分離後に得られた上清に関して、4℃に保持するかまたは37℃で7日間貯蔵した複合体の遊離多糖含量を確定した。
【0157】
上清中の遊離多糖の定量のために、α−PS/α−PS ELISAを用いた。α−PD/αPSELISAにより、複合体の非存在も制御した。遊離多糖の量の低減は、改良型複合ワクチンをもたらした。
【0158】
抗原性:
同一複合体上の抗原性を、サンドイッチ型ELISAで分析したが、この場合、抗体の捕獲および検出は、それぞれα−PSおよびα−PDであった。
【0159】
遊離タンパク質含量(%):
試料のSDS処理による方法を用いて、「遊離」残留プロテインDのレベルを確定した。0.1%SDSの存在下で100℃で10分間、複合体を加熱し、SEC−HPLCゲル濾過カラム(TSK3000−PWXL)上に注入した。プロテインDは二量体であるので、SDSでの構造の解離により「遊離」プロテインDのレベルを過剰刺激する危険性がある。
【0160】
分子サイズ(Kav):
SEC−HPLCゲル濾過カラム(TSK5000−PWXL)上で、分子サイズ分類を実施した。
【0161】
安定性:
4℃に保持するかまたは37℃で7日間貯蔵した複合体に関して、 HPLC−SECゲル濾過カラム(TSK6000−PWXL)上で安定性を測定した。
【0162】
11価特性化を表2に示す。
【0163】
タンパク質複合体をリン酸アルミニウム上に吸着させて、プールし、最終ワクチンを生成し得る。
【0164】
結論:
有望なワクチンの構成成分であることが示されている免疫原性複合体を生成した。最良品質最終複合肺炎球菌多糖生成物に関する最適CDAP条件を、11価物の各々に関して発見した。前記の改良型(最適化)CDAP法により得られるこれらの肺炎球菌多糖の複合体は(担体タンパク質にかかわらず、しかし好ましくはプロテインD)、したがって、本発明のさらなる局面である。
【0165】
実施例2−乳児ラットにおける11価肺炎球菌PS−PD複合体ワクチンの免疫原性に及ぼす先進アジュバントの作用の研究
0.1μgの各多糖の用量の11価肺炎球菌PS−PD複合体ワクチン(実施例1の方法にしたがって製造)で、以下のアジュバント処方物:なし、AlPO4、3D−MPL、AlPO4上の3D−MPLを用いて、乳児ラットを免疫感作した。
【0166】
3D−MPLのみを有する処方物は、11の抗原のうち5つに関して、他の処方物より統計学的に(そして意外にも)より免疫原性(最大GMC IgG)であった。
【0167】
オプソニン食菌作用は、GMC結果を確証した。
【0168】
材料と方法
免疫感作プロトコール
乳児OFAラットを異なる母親に対して無作為化し、それらが初回免疫感作を施されたときは7日齢であった。それらには、14および28日後にさらに2回の免疫感作を施した。56日目に(第3回免疫感作後28日目)、採血を実施した。ワクチンを皮下注射した。ラットは、10匹/ワクチン群であった。
【0169】
プロテインD上に以下の多糖血清型:1、3、4、5、6B、7F、9V、14、18C、19F、23Fを連結させたものを含む11価肺炎球菌複合ワクチンで、ラットを免疫感作した。
【0170】
処方物
異なる先進アジュバントの作用を調べるために、複合体の用量を0.1μgの各多糖に一定に保持し、アジュバントAlPO4および3D−MPLを、アジュバントを全く含まないものを含めて、異なる用量および組合せで処方した。これらを参考のために表3に数値で列挙する。
【0171】
AlPO4上の吸着
以下の手法により、濃縮吸着一価物を調製した。50μgのAlPO4(pH5.1)を5μgの複合多糖と2時間混合した。pHをpH5.1に調整し、混合物をさらに16時間放置した。1500mMのNaClを付加して、塩濃度を150mMにした。5分後、5 mg/mlの2−フェノキシエタノールを付加した。さらに30分後、pHを6.1に調整して、4℃で3日以上放置した。
【0172】
希釈剤の調製
NaCl150mM/5 mg/mLフェノキシエタノール中で3つの希釈剤を調製した。
【0173】
A:AlPO4、1mg/mlで。
【0174】
B:AlPO4上の3D−MPL、それぞれ1000および250μg/ml。重量比は、3D−MPL/AlPO4=5/20。
【0175】
C:AlPO4上の3D−MPL、それぞれ1000および561μg/ml。重量比は、3D−MPL/AlPO4=50/89。
【0176】
吸着11価物の調製
11の濃縮吸着PS−PD一価物を適切な比率で混合した。AlPO4の補体を希釈剤Aとして付加した。必要な場合、水性溶液(非吸着価。下記の方法1参照)として、あるいは希釈剤BまたはC(AlPO4上に2用量で吸着された3D−MPL。下記参照)として、3D−MPLを付加した。
【0177】
方法1
3D−MPLを、水性懸濁液として併合吸着複合体に付加した。それを室温で10分間、11価物に混合して、投与まで4℃で貯蔵した。
【0178】
方法2
3D−MPLをAlPO4上に予備吸着させた後、併合吸着複合体に付加した(希釈剤BおよびC)。1 mlの希釈剤を調製するために、3D−MPL(250または561 μg)の水性懸濁液を、室温で5分間、150 mMのNaCl、pH6.3中の1 mgのAlPO4と混合した。この溶液をNaCl、pH6.1/フェノキシ中に稀釈し、4℃で一夜インキュベートした。
【0179】
非吸着化11価物の調製
11のPS−PD複合体を混合し、正しい比率で150 mMのNaCl、pH6.1、フェノキシ中に稀釈した。必要な場合には、3D−MPLを溶液(非吸着化)として付加した。
【0180】
すべての注射用の処方物は、初回投与の18日前に調製した。
【0181】
ELISA
ヒト血清中の肺炎連鎖球菌莢膜多糖に対するIgG抗体の定量のためのELISA手法に関してWHOワークショップから得られたプロトコールを用いて、ELISAを実施し、ラットIgGを測定した。基本的に、精製莢膜多糖は微小滴定プレート上に直接被覆される。血清試料を、すべての肺炎球菌に共通の、そして開示(EP 72513 B1)にしたがって精製される肺炎球菌多糖中に焼く0.5%で存在する細胞壁多糖(物質C)とともに予備インキュベートする。Jackson ImmunoLaboratories Inc.試薬を用いて、結合ネズミIgGを検出した。滴定曲線を、数学論理的対数方程式によりモデル化された内部標準(モノクローナル抗体)に参照した。SoftMax Proソフトウェアを用いて計算を実施した。これらの結果に関する最大絶対誤差は、2の因数内であると予測した。相対誤差は30%未満である。
【0182】
オプソニン食菌作用
精製ヒトPMNおよび嬰児ウサギ補体を用いたCDCプロトコール(分化HL60細胞を用いた肺炎連鎖球菌オプソニン食菌作用。バージョン11)を用いて、血清型3、6B、7F、14、19Fおよび23Fに関してオプソニン力価を決定した。修正は社内肺炎球菌株の使用を含み、食菌性HL60細胞を精製ヒト好中球PMNに置き換えた(これらの食菌性細胞間には高度の相関が認められる)。さらに、3 mmガラスビーズを微小滴定ウエルに付加して、混合を増大し、これは、400であるべきと推奨された食細胞:細菌比の低減を可能にした。
【0183】
結果
IgG濃度
すべての血清型およびPDに関して確定されたゲノム平均IgG濃度を表4〜10に示す。血清型6B、14、19Fおよび23Fに関しては、4価処方物を用いて得られた前の結果を比較のために含める。
【0184】
最高IgG濃度を、表4〜10で強調した。3D−MPL組成物体3D−MPL/AlPO4組成物に関する統計的p値は、表11に示してある。アジュバント処方物4(高用量3D−MPLを有する非吸着化複合体)は、11例中9例で最高GMCを示す。5/11例では、低用量のMPLは、第二の最大免疫原性物である。さらに、アジュバント活性化は、全血清型に関して用量の修正によるより高いGMCを示し(データは示されていない)、これは血清型4、6B、7F、18Cおよび23Fに関しては統計学的に有意である(p<0.05。95%CIから)。
【0185】
オプソニン食菌作用
プール化血清に関するオプソニン食菌作用結果を、血清型3、6B、7F、14、19Fおよび23Fに関して表4〜8に示す。ほとんどの部分に関して、これらのオプソニン力価は、GMC IgGを確証する。実際、IgG濃度との相関は、血清型6B、19F、23Fに関しては85%より大きい(データは示されていない)。血清型3に関しては、3D−MPL群のみがオプソニン活性を閾値より上に誘導した、ということに留意することは重要である。
【0186】
結論
この実験では、3D−MPL単独の使用は最高IgG濃度を誘導する、ということは予測されなかった。
【0187】
アジュバントを修正することにより得られた最大GMCを、PS用量を修正することにより得られた最大GMCと比較し、3D−MPLが、5/11血清型で有意に高い応答を誘導し得た、ということを見出した。
【0188】
表11は、3D−MPLおよび3D−MPL/AlPO4組成物を比較した場合(処方過程および3D−MPLの用量を比較)、3D−MPL+AlPO4よりむしろ3D−MPLだけを用いて処方した場合、免疫原性に関して、多糖複合体のうちの5つが有意に改良されるということを示す:PS4、PS6B、PS18C、PS19FおよびPS23F。
【0189】
実施例3−成体ラットにおけるPS4、PS6B、PS18C、PS19FおよびPS23F複合体の免疫原性に及ぼす組合せの作用の研究
別々に、または多価組成物(4−、5−、7−または10価物)中に併合した肺炎球菌多糖−プロテインD複合体ワクチンを用いて、成体ラットを免疫感作した。ラット10匹の群を28日置いて2回免疫感作して、28日目および42日目(2回目投与の14日後)に被験血液を採取した。
【0190】
肺炎球菌多糖に対するIgG抗体に関して、ELISAにより血清を検査した。ELISAにより測定した場合、複合体はすべて、特異的IgG抗体を誘導した。表12は、2回目投与後14日目にIgG濃度により測定した場合の、成体ラットにおけるそれらの免疫原性に及ぼす1価PS6B、PS18C、PS19FおよびPS23Fの組合せの作用を示す。
【0191】
すべての試料に関して統計分析を実施して、組合せ時の抗体濃度の差が有意であるか否かを確定した。多価ワクチン中の血清型PS6B、PS18C、PS19FおよびPS23FプロテインD複合体の組合せはいずれも、それらの免疫原性を有意に変えなかった。
【0192】
【表1】

【0193】
【表2】
【0194】
【表3】
【0195】
【表4】
【0196】
【表5】
【0197】
【表6】
【0198】
【表7】
【0199】
【表8】
【0200】
【表9】
【0201】
【表10】
【0202】
【表11】
【0203】
【表12】
【0204】
実施例4−マウスにおける肺炎球菌肺コロニー形成に対するPD−複合11価多糖ワクチンの防御的有効性に及ぼすニューモリジンおよび3D−MPLの付加の有益な影響
免疫学的解読
ニューモリジン特異的血清IgGのELISA投与
PBS中に稀釈した100μl/ウエルの2μg/ml組換え体ネイティブニューモリジン(PLY)を用いて、37℃で2時間、Maxisorp Nuncイムノプレートを被覆した。プレートを0.9%NaCl、0.05%トゥイーン20緩衝液で3回洗浄した。次に、標準曲線として付加された(670 ng/mlIgGで出発)抗PLY血清参照の連続2倍稀釈液(PBS/0.05%トゥイーン20、100μl/ウエル)および血清試料(1/10稀釈液で出発)を、撹拌しながら20℃で30分間インキュベートした。前記と同様に洗浄後、 PBS/0.05%トゥイーン20中に5000x稀釈したペルオキシダーゼ−複合ヤギ抗マウスIgG(Jackson)を、撹拌しながら20℃で30分間インキュベートした(100μl/ウエル)。洗浄後、100μl/ウエルの再ベール化緩衝液(100mM、pH4.5クエン酸塩緩衝液中OPDA0.4 mg/mlおよびH2O20.05%)とともに、室温で15分間、プレートをインキュベートした。50μl/ウエルの1 NHClを付加することにより、再ベール化を停止した。Emaxイムノリーダー(Molecular Devices)を用いて、490および620 nmで光学濃度を読み取った。SoftMaxProソフトウェアを用いた4パラメーター数学法により、抗体力価を算定した。
【0205】
溶血抑制
ニューモリジン(PLY)溶血活性を抑制する血清抗体の能力を測定するために、この検定を実行した。コレステロール(PLYとの相互作用に感受性)を排除するために、血清試料に以下の処置を2回施した:それらを1等容量のクロロホルムと混合し、次に撹拌しながら45分間インキュベートした。1000 rpmで10分間遠心分離後に上清を収集した。コレステロール消去血清を、96ウエル微小プレート(Nunc)中で稀釈した(1 mMジチオトレイトール、0.01%BSA、15 mMトリス、150 mMNaCl、pH7.5中の連続2倍稀釈液)。4 HU(溶血単位)のPLYを含有する50μl溶液を各ウエル中に付加し、37℃で15分間インキュベートした。次に、100μlのヒツジ赤血球(1%溶液)を、37℃で30分間、付加した。1000 rpmで10分間遠心分離後、上清(150μl)を収集し、405 nmでの光学密度読み取りのために別の96ウエル微小プレート中に入れた。結果は、中点稀釈力価として表した。
【0206】
ニューモリジン化学的脱毒素化
組換え体ネイティブニューモリジン(PLY)を50 mMリン酸塩、500 mMNaCl、pH7.6緩衝液に対して透析した。時々撹拌しながら、39.5℃で以下の全工程を実行した。1日目に、10%トゥイーン80(1/250 v/v)、57.4 mMのN−アセチルトリプトファン、pH7.6(3/100 v/v)、リン酸塩緩衝液中の2.2 Mグリシン(1/100 v/v)およびリン酸塩緩衝液中の10%ホルムアルデヒド(3/100 v/v)をPLY溶液中に付加した。2および3日目に、10%ホルムアミドを、それぞれ3/100および2/100 v/v比で再び付加した。時々撹拌しながら、7日目まで、39.5℃でインキュベーションを持続した。最後に、50 mMリン酸塩、500 mMNaCl、pH7.6緩衝液に対してPLYを透析した。溶血検定において、PLYの完全不活性化を実証した。
【0207】
OF1マウスにおける肺炎球菌鼻腔内チャレンジ
5.105 CFUのマウス適合化肺炎連鎖球菌血清型6Bを、麻酔下で7週齢OF1雌マウスに鼻腔内接種した。接種後6時間目に肺を切除し、Todd Hewithブロス(THB、Gibco)培地中で均質化した(Ultramax, 24000 rpm、4℃)。肺ホモジネートの連続10倍稀釈液を、37℃で一夜、酵母菌抽出物補充THB寒天を含入するペトリ皿上にプレート化した。肺炎球菌肺感染をCFU数/マウスとして確定し、対数計量平均として表した。検出限界は2.14 log CFU/マウスであった。
【0208】
実施例4A.抗ニューモリジン免疫応答に及ぼす3D−MPLアジュバント作用
本実施例では、ネイティブ組換え体ニューモリジン(PLY、J. Paton, Children's Hospital, North Adelaide, Australiaにより提供された)およびその化学的脱毒素化対応物(DPLY)に対する免疫応答に及ぼす3D−MPLアジュバント活性化の影響を、我々は評価した。前記と同様に化学的脱毒素化を実行した。
【0209】
0、14および21日目に、A:100μgのAlPO;またはB:100μgのAlPO +5μgの3D−MPL(3デ−O−アシル化モノホスホリル脂質A、供給元Ribi Immunochem)中に含入された1μgのPLYまたはDPLYを用いて、10匹の雌6週齢Balb/cマウスの群を筋肉内免疫感作した。図1Aおよび1Bは、第3回血清後に測定したELISA IgGおよび溶血抑制力価(HLI)
どの抗原でも、3D−MPL補充処方物を接種した動物において最良免疫応答を誘導した。興味深いことに、DPLYは、AlPO4+3D−MPLとともに投与した場合、PLYと同様に免疫原性であったが、一方、ALPO4処方物中では免疫原であった。これは、脱毒素化ニューモリジンに対する抗体応答を改良する3D−MPLの有益な能力を示した。
【0210】
ニューモリジンを含有する組成物では、突然変異的脱毒素化ニューモリジンよりむしろ化学的脱毒素化ニューモリジンを用いるのが望ましい。これは、今日までに得られた脱毒素化突然変異体が依然として残留毒素活性を有するためであり、化学的脱毒素化ニューモリジンは残留毒素活性を有さない。したがって、本発明の別の局面では、概して、ワクチン中に用いるために化学的脱毒素化されたニューモリジンを含む組成物は、Th1アジュバント、好ましくは3D−MPLで活性調節されるべきである、と考えられる。本発明は、このような組成物を提供する。組成物にTh1アジュバント(好ましくは3D−MPL)を付加する過程を包含する免疫原性組成物内の化学的脱毒素化ニューモリジンの免疫応答の増大方法も意図される。
【0211】
実施例4B.血清型6Bを鼻腔内チャレンジさせたOF1マウスにおける肺炎球菌肺コロニー形成に対するPD−複合11価多糖ワクチンの防御的有効性に及ぼすニューモリジンおよび3D−MPLアジュバントの弱毒化突然変異体の付加の有益な影響
本実施例では、11価多糖−プロテインD複合体、弱毒化突然変異体ニューモリジン抗原(PdB、WO 90/06951)およびAlPO4+3D−MPLアジュバントを含有するワクチンの予防的効能を、古典的AlPO4吸着11価多糖−プロテインD複合体処方物と比較して、我々は評価した。
【0212】
0および14日目に、A:50μgのAlPO;B:0.1μgPS/血清型のPD複合11価多糖ワクチン+50μgのAlPO4;またはC:0.1μgPS/血清型のPD複合11価多糖ワクチン+10μgのPdB(J. Paton, Children's Hospital, North Adelaide, Australia提供)+50μgのAlPO4+5μgの3D−MPL(供給元Ribi Immunochem)を含有する処方物を用いて、12匹の雌4週齢OF1マウスの群を皮下免疫感作した。21日目に、前記と同様にチャレンジを実行した。
【0213】
図1Cに示したように、PdBを補充し、AlPO4+MPLで活性調節した11価多糖複合ワクチンにより、非常に有意の防御(p<0.007)が付与された(黒棒は算術平均を表す)。これに反して、11価多糖複合体/AlPO4処方物で免疫感作した動物においては、有意の防御は観察されなかった。この結果は、ニューモリジン抗原(弱毒化されていても)および3D−MPLアジュバントの付加が肺炎に対する11価多糖複合ワクチンの有効性を増大することを立証した。
【0214】
実施例4C.実施例4Bに示した防御の免疫相関物
弱毒価突然変異体ニューモリジン(PdB)および3D−MPLを補充した11価多糖複合体ワクチンにより、実施例4Bで付与された防御の免疫相関物を確立するために、多糖6BおよびPdBに対するプレチャレンジ血清学的抗体応答を前記と同様に測定した。
【0215】
次に、抗体力価を、チャレンジ後6時間目に収集した対応する動物の肺で測定された細菌コロニー数と比較した。Log/Log線状回帰に関してR2を算定した。
【0216】
算定R2は、抗PdBおよび抗6B抗体応答に関してそれぞれ0.18および0.02であった。これは、体液性免疫応答と両抗原に対する防御との間の相関の非存在を示した。抗6B抗体力価は、11価複合体ワクチン(GMT=0.318 ng/ml)で免疫感作した群またはPdDおよび3D−MPLを補充した同一ワクチン(GMT=0.458 ng/ml)で免疫感作した群において有意に異ならなかった。したがって、処方物Cで観察された防御改善は、単に多糖6Bに対する高抗体応答によるものではなかった。
【0217】
合わせて考えると、防御は体液性免疫応答単独により媒介されるだけでなく、むしろ3D−MPLの存在下でPdBにより誘導された細胞媒介性免疫によっても媒介された、ということを結果は示唆する。これは、最適防御のための免疫系の両部門を対等にするよう、肺炎球菌多糖複合体ワクチン中のタンパク質抗原(単数または複数)および効力のあるアジュバント(単数または複数)の付加に対してさらなる支持を与えた。
【0218】
実施例5−ニューモリジンで能動的免疫感作され、肺炎球菌PSに対する抗体で受動的免疫感作されたマウスにおける免疫系の両部門の共働
実施例5A−肺炎に対して防御する受動的投与抗6B多糖(抗PS)抗体の濃度を見出す
方法
ワクチン群:下記に詳述した群により(マウス計64匹)、マウス16匹の4群を、−1日目に100μlの非稀釈ラット抗多糖抗血清で受動的免疫感作(腹腔内)させた。
【0219】
【表13】
【0220】
動物:Charles River, Canadaからの体重約35 gの64匹の雄CD−1マウス(約10週齢)。
【0221】
麻酔:イソフルラン(3%)+O2(1 L/分)でマウスを麻酔した。
【0222】
生物:肺炎連鎖球菌N1387(血清型6)を、5%ウマ血液を補充したトリプチカーゼ大豆寒天プレート(TSA)から収穫し、PBS6 ml中に懸濁した。感染直前に、1 ml細菌懸濁液を9 mlの冷却溶融栄養寒天(BBL)中に稀釈して、41℃に保持した。マウスに容量50 ul中の約6.0 log10 cfu/マウスを投与した。
【0223】
感染:0日目に、前記のようにマウスを麻酔して、非外科的気管内挿管による気管支内点滴注入により、肺炎連鎖球菌N1387(50μl冷却細菌懸濁液)を感染させた。この方法は、WoodnutとBerry(Antimicrob. Ag. Chemotherap. 43:29(1999))により記載された。
【0224】
試料:感染後3日目に、マウス8匹/群をCO2過剰投与により屠殺して、肺を切除し、1 mlPBS中で均質化した。10倍連続稀釈液をPBS中で調製して、生育可能細菌数を数え上げた。5%ウマ血液を補充したTSAプレート上に3通り、試料を接種(20μl)し、評価前に37℃で一夜インキュベートした。さらに別の組のマウスを7日目に屠殺して、前記と同様に試料採取した。
【0225】
結果:
【0226】
【表14】
【0227】
結論:概して、あらゆる処置群から単離した細菌数に有意差は認められなかった。これは、測定可能防御が5μg/ml以下の濃度の抗多糖によりもたらされたことを示す。
【0228】
これは、いくつかのヒト臨床試験で観察されるものと同様であり、即ち、抗多糖体はいくつかの集団においては肺炎球菌性肺炎に対して防御するのに不十分である。
【0229】
実施例5B−アジュバントを用いた場合と用いない場合とのPly(ニューモリジン)の能動的投与および亜最適抗PS抗体との共働作用によりもたらされる肺炎からの防御を確定する
方法
動物:6週齢時体重約20 g、10週齢時約38 gの、Charles River, St. Quebec, Constant, Canadaからの128匹の雄CD−1マウス(免疫感作時6週齢、感染時10週齢)。
【0230】
免疫感作:マウス16匹の6群を、-22日目および-14日目に皮下注射により下記に詳述するワクチン100 ulで免疫感作した(マウス計128匹)。PdB(WO 90/06951)は、オーストラリアのJames Paton博士の御好意により入手した。3D−MPLは、Ribi/Corixaから入手した。
【0231】
-1日目に、特定群(下記の表参照)を、濃度4.26μg/ml(4 mlの5μg/ml+1.3 mlの2μg/ml)のマウス抗多糖抗体で受動的免疫感作(腹腔内100 μl)した。
【0232】
【表15】
【0233】
感染:0日目に、マウスを麻酔した(3%イソフルラン+1 L/分O2)。肺炎連鎖球菌N1387(血清型6)の増殖物を、5%ウマ血液を補充したトリプチカーゼ大豆寒天プレート(TSA)から収穫し、PBS6 ml中に懸濁することにより細菌接種物を調製した。感染直前に、10倍稀釈液(1 ml+9 ml)を冷却溶融栄養寒天(41℃に保持)中に調製した。気管内挿管による気管支内点滴注入によりマウスを感染させて、容量50 ul中の約6.0 log10 cfu/マウスを投与した。この方法は、WoodnutとBerry(Antimicrob. Ag. Chemotherap. 43:29(1999))により記載されていた。
【0234】
試料:感染後72時間目に、マウス8匹/群をCO2過剰投与により屠殺して、肺を切除し、1 mlPBS中で均質化した。10倍連続稀釈液をPBS中で調製して、生育可能細菌数を数え上げた。5%ウマ血液を補充したTSAプレート上に3通り、試料を接種(20μl)し、評価前に37℃で一夜インキュベートした。さらに別の組のマウスを感染後8日目に屠殺して、前記と同様に試料採取した。
【0235】
データ分析
処置の比較のための結果測定値は、感染後3および7日目の肺中の細菌数であった。結果は、群平均+標準偏差として示される。スチューデントt検定を用いて統計分析を実施したが、この場合、<0.05のP値を、有意であるとみなした。
【0236】
結果:
感染後72時間
群1−4からの細菌数は、群1−3の場合よりも有意に低かった(p<0.05)。
【0237】
群1−4からの細菌数は、群1−5の場合よりも有意に低かった(p<0.05)。
【0238】
感染後168時間
全群の細菌数は、8日目では3日目より約2 log低かったが、これは感染が消散したことを示す。
【0239】
群1−2からの細菌数は、群1−5の場合よりも有意に低かった(p<0.05)。
【0240】
【表16】
【0241】
前記で実証されたように、抗多糖抗体単独(群1−5)は、肺中の肺炎球菌の増殖に対する防御をもたらさない。AlPO4で活性調節したPdBはいずれも防御を付与しないが、しかし8日目に、PdBを3D−MPLと組合せた場合(群1−2)に、防御傾向が認められる。
【0242】
3日目に、群は最も有意に防御し、群1−4は3つの要素、即ちPdB、3D−MPLおよび受動的投与抗多糖抗体をすべて有した。この結論は、死亡率により支持される。群1−4は死亡は2/8だけであったが、これに比して群1−5および1−3に関しては5/10であった。
【0243】
結論:
受動的免疫感作動物を用いて実験を実施したので、ニューモリジンおよびMPLでの能動的免疫感作の共働作用も、多糖抗原に対する抗体レベルの増大によるとは言えない。
【0244】
肺炎球菌多糖に対して動物を受動的免疫感作しただけであったので、8日目までに、このような抗体のレベルは宿主から大いに消散していた。
【0245】
そのような場合でも、肺炎球菌性肺炎に対する有意の防御は、ニューモリジン+3D−MPLで免疫感作した群において、特に濡森+3D−MPL+受動的投与抗多糖抗体で免疫感作した群において認められたが、これは、この組合せの共働作用を示す。
【0246】
能動的(好ましくは、複合多糖を用いて)に抗多糖免疫感作を実行していた場合、B細胞記憶の作用および一定レベルの抗PS抗体が免疫応答協同作用に寄与していたので、作用はさらに顕著であったと思われる(例えば、図1C参照。この場合、多糖およびタンパク質で能動的免疫感作された動物は、チャレンジ後の肺中に細菌が認められなかったことが示された)。
【0247】
実施例6−3D−MPLで活性調節した11価肺炎球菌多糖プロテインD複合体ワクチンの1年齢Balb/Cマウスにおける免疫原性
序論および対象(単数または複数):
肺炎球菌感染に対する防御は、オプソニン食菌作用を介した血清型特異的抗体により媒介される。抗体濃度の増大はより大きい防御を生じ、したがって体液性応答を増大するための方法の発見に多大の努力が費やされてきた、と推測され得る。前臨床試験においてワクチンを連結するために首尾よく適用されてきた一戦略は、免疫刺激性アジュバントの使用である(Poolman et al. 1998, Carbohydrate-Based Bacterial Vaccines. In:Handbook of Experimental Pharmacology eds. P. Perlmann and H. Wigsell. Springer-Verlag, Heidelberg, Dで再検討されている)。
【0248】
この節に提示したデータは、臨床試験を模倣するよう意図されたプロトコールにおける臨床ロットを用いた最新実験結果を示す。
【0249】
プロトコール:
1/10回のヒト投与量の肺炎球菌多糖プロテインD複合体ワクチン、または23価純多糖ワクチンで、1年齢Balb/Cマウスを免疫感作した。用いたワクチンは、それぞれ1 mcg用量の血清型6Bおよび23Fならびに5 mcgの11価複合ワクチンの残りの血清型、0.1 mdg用量の11価複合ワクチン、または5 mcg3D−MPLで活性調節した0.1 mcg用量の11価複合ワクチンに対応する臨床ロットDSP009、DSP013またはDSP014であった。11価複合ワクチンはすべて、50μgのAlPO4でも活性調節した。
【0250】
マウス20匹の群を筋内免疫感作した。下記の表に列挙した群の注射は、0および21日目に実施した。35日目(第2回投与後14日目)に被験血液を採取した。
【0251】
【表17】
【0252】
CDC/WHO合意プロトコール後、即ち細胞壁多糖による血清の中和後、肺炎球菌多糖に対するIgG抗体に関してELISAにより、血清を検査した。血清型特異的IgG1モノクローナル抗体を用いてELISAを較正して、抗体濃度をmcg/mlで得た。
【0253】
UNISTATバージョン5.0βを用いて、比較の統計分析を算定した。log形質転換化IgG濃度に関して、Tukey-HSD法によるANOVAを実施した。Fisherの精度検定を用いて、血清変換の対方式比較を実施した。
【0254】
結果:
第2回免疫感作(投与2)後14日で誘導された11の血清型およびプロテインDに対するGMC IgGおよび95%信頼区間を、下記の表に示す。血清変換率を示すが、この場合、95%信頼区間は算定され得なかった。
【0255】
群1は、普通は動物においてIgMのみを誘導する純多糖による免疫感作の効果を示す。ほとんどのIgGレベルは検出の閾値より下であった。それにもかかわらず、balb/cマウスは少数の肺炎球菌多糖、特に血清型3、19Fおよび14に対してIgGを作り得た。
【0256】
複合体ワクチンによる免疫感作は、23F以外の全血清型に対して高血清変換率でIgG抗体を誘導した。
【0257】
血清型7Fおよび19Fに関してのみ用量依存性応答(群4対群2)を観察したが、しかしこれらの観察は統計学的に有意でなかった。血清型3、6B、7Fおよび19FならびにPDに関して2回の投与後により大きい応答を観察し、これらの観察は、3つの処方物すべてを有する多くの場合に、統計学的に有意であった。
【0258】
最も興味深いのは、3D−MPLの作用である。3D−MPL処方ワクチン(群3b)の2回の投与は、特異的IgGの最高GMCを誘導し、これは、すべての血清型に関して統計的に有意であったが、但し、23Fの場合は、有意に高い血清変換率を示した(p=0.02,群3b対2b。Fisher精度)。
【0259】
【表18】
【0260】
群の定義に関しては、前記の表を参照されたい。
【0261】
結論:
ここに提示したデータは、11価肺炎球菌多糖プロテインD複合体ワクチンへの3D−MPLの付加が、試験したすべての血清型に対する老齢balb/cマウスにおける免疫応答を増大した、ということを実証する。
【0262】
ほとんどの場合、ワクチンの2回投与は、1回投与より高い幾何学平均IgG濃度を誘導した。これは、ヒトにおいてさえ、純多糖ワクチンを用いて観察されないため、T細胞依存性免疫応答および免疫記憶の誘導の指標とみなされる。
【0263】
これらのデータは、アジュバント(好ましくは3D−MPL)で活性調節した複合肺炎球菌多糖を用いるワクチン投与計画を支持し、それにより、少なくとも2回用量の活性調節化ワクチンが、好ましくは1〜12週おきに、最も好ましくは3週間置きに投与される。このような投与計画は、本発明のさらなる局面とみなされる。
【0264】
実験に用いたマウスは、PS23(純または複合)に対して非応答性であった。興味深いことに、用いたワクチン組成物にかかわらず多糖に対する抗体レベルは低いままであったが、しかし3D−MPLをアジュバントとして用いた場合(血清変換は有意に高い)には、より多数のマウスがPS 23に応答した。ワクチン中の肺炎球菌多糖に対する非応答性を軽減するための複合肺炎球菌多糖を含むワクチン組成物中のTh1アジュバント、特に3D−MPLの使用は、本発明のさらに別の局面である。前記の2回投与計画を用いた前記の組成物による非応答性の軽減方法は、さらに別の局面である。
【0265】
実施例7−C型髄膜炎菌多糖−プロテインD複合体(PSC−PD)
A:プロテインDの発現
実施例1と同様。
【0266】
B:多糖Cの製造
C群多糖の供給源は、髄膜炎菌のC11菌株である。これを、古典的発酵技術(EP 72513)を用いて発酵させる。複合過程に用いられる乾燥粉末多糖は、Mencevax(SB Biologicalsも参照)と同一である。
【0267】
C11菌株のアリコートを解氷し、0.1 mlの懸濁液を、酵母菌抽出透析物(10%、v/v)を補充したMueller Hinton培地ペトリ皿上に画線し、水飽和空気インキュベーター中で36℃で23〜25時間インキュベートする。
【0268】
次に、表面増殖物を滅菌発酵培地中に再懸濁して、この懸濁液を、酵母菌抽出透析物(10%、v/v)を補充したMueller Hinton培地および滅菌ビーズガラスを含入する1つのRouxボトル上に接種する。水飽和空気インキュベーター中で36℃で23〜25時間のRouxボトルのインキュベーション後、表面増殖物を10 mlの滅菌発酵培地中に再懸濁して、0.2〜0.3 mlのこの懸濁液を12の他のMueller Hinton培地Rouxボトル上に接種する。
【0269】
水飽和空気インキュベーター中で36℃で23〜25時間のインキュベーション後、表面増殖物を10 mlの滅菌発酵培地中に再懸濁する。細菌懸濁液を円錐フラスコ中にプールする。
【0270】
次にこの懸濁液を、滅菌注射器を用いて無菌的に発酵器に移す。
【0271】
陰圧下で清浄室中に入れた発酵器中で髄膜炎菌の発酵を実施する。発酵は一般に、約1010個の細菌/mlに対応して約10〜12時間後(即ち早期定常期)に完了され、pH増大により検出される。
【0272】
この段階で、全ブロスを熱不活性化(56℃で12分)した後、遠心分離する。不活性化の前後に、ブロスの試料を採取し、Mueller Hinton培地ペトリ皿上に画線する。
【0273】
C:PS精製
精製方法は、全発酵ブロス上で実施される多段階手法である。精製の第一段階では、遠心分離により不活性培養を清澄化して、上清を回収する。
【0274】
多糖精製は、第四級アンモニウム塩(セチルトリメチルアンモニウムブロミド/CTAB、CETAVLON R)による沈降に基づいている。CTABは、それらのpIによって、多糖、核酸およびタンパク質のような多陰イオンと不溶性複合体を形成する。イオン制御条件後、この方法を用いて、不純物(低導電率)または多糖(高導電率)を沈降させ得る。
【0275】
清澄化上清に含有される多糖を、マトリックスとして珪藻土(セライト(商標)545)を用いて沈降させて、異なる沈降/精製中の不溶性不活性塊の形成を回避する。
【0276】
髄膜炎菌多糖Cに関する精製計画:
工程1:セライト(商標)545上でのPSC−CTAB複合体固定および0.05%CTABを用いた洗浄による細胞破砕屑、核酸およびタンパク質の除去。
【0277】
工程2:50%EtOHによるPSの溶離。混濁した、そして不純物およびLPSを含有する第一分画を廃棄する。以後の分画中のPSの存在を、凝集検定により立証する。
【0278】
工程3:セライト(商標)545上でのPSC−CTAB複合体再固定および0.05%CTAB洗浄による小型核酸およびタンパク質の除去。
【0279】
工程4:50%EtOHによるPSの溶離。第一混濁分画を廃棄する。以後の分画中のPSの存在を、凝集検定により立証する。
【0280】
溶離液を濾過し、粗製多糖を含有する濾液を収集する。エタノールを付加して最終濃度を80%とすることにより、多糖を濾液から沈降させる。次に、多糖を白色粉末として回収し、真空乾燥して、-20℃で貯蔵する。
【0281】
D:CDAP複合化(conjugation)
PSCおよびPDの複合化
PSCおよびPDの複合化のためには、古典的CNBr活性化およびスペーサーを介した担体タンパク質との結合よりCDAP複合技法が好まれる。1−シアノ−4−ジメチルアミノピリジニウムテトラフルオロボレート(CDAP)を用いたシアニル化により、多糖を先ず活性化する。CDAPは水溶性シアニル化試薬であって、この場合、シアノ基の求電子性はCNBrの場合を上回って増大され、シアニル化反応を相対的に低刺激性条件下で実施させる。活性化後、多糖は、いかなるスペーサー分子も導入せずに、そのアミノ基により担体タンパク質に直接結合され得る。未反応エステルシアネート基は、グリシンとの広範な反応によりクエンチングされる。複合体ワクチンの調製に関与する工程の総数は低減され、最も重要なのは、免疫原性である可能性を有するスペーサー分子が最終生成物中に存在しないことである。
【0282】
CDAPによる多糖の活性化は、多糖中にシアネート基を導入し、ジメチルアミノピリジン(DMAP)が遊離される。シアネート基は、その後のカップリング手法中にタンパク質中のNH2基と反応して、カルバメートに転換される。
【0283】
PSC活性化およびPSC−PDカップリング
活性化およびカップリングは+25℃で実施する。
【0284】
120 mgのPSを、少なくとも4時間、WFI中に溶解する。
【0285】
CDAP溶液(アセトニトリル中に新たに調製した100 mg/ml)を付加して、CDAP/PS(w/w)比を0.75とする。
【0286】
1分30秒後、トリエチルアミンの付加によりpHを活性化pHまで上げ、PD付加まで安定化させる。
【0287】
3分30秒時点で、NaClを付加して、最終濃度を2 Mとする。
【0288】
4分時点で、精製PDを付加して、PD/PS比を1.5/1とする。pHを直ちにカップリングpH(pH10)に調整する。溶液を、pH調節下で1時間放置する。
【0289】
クエンチング
6 mlの2 Mグリシン溶液をPS/PD/CDAP混合物に付加する。pHをクエンチングpH(pH8.8)に調整する。溶液を作用温度で30分間撹拌した後、連続的にゆっくり撹拌しながら、+2〜8℃で一夜置く。
【0290】
PS−PD精製
濾過(5μm)後、S400HR Sephacrylゲル上でのゲル浸透クロマトグラフィーにより冷室中でPS−PD複合体を精製して、小型分子(DMAPを含む)を除去し、PD:溶離−150 mMNaCl、pH6.5を再複合化して、UV280 nm、pHおよび導電率をモニタリングする。
【0291】
異なる分子サイズの反応構成成分を基礎にして、PS−PD複合体を先ず溶離し、その後遊離PDを、そして最後にDMAPを溶離する。DMAB(PS)およびμBCA(タンパク質)により検出されるような複合体を含有する分画をプールする。プール分画を滅菌濾過(0.2μm)する。
【0292】
E:PSC−PD吸着複合体ワクチンの処方
AlPO4の洗浄
AlPO4上のPSC−PD複合体の吸着を最適化するために、AlPO4を洗浄して、PO43-濃度を低減する。
【0293】
−AlPO4を150mMNaClで洗浄して、遠心分離(4x)する。
【0294】
−次にペレットを150 mMのNaCl中に再懸濁した後、濾過(100μm)する。そして
濾液を加熱滅菌する。
【0295】
この洗浄AlPO4は、WAP(洗浄オートクレーブ処理リン酸塩)と呼ばれる。
【0296】
配合方法
PSC−PD複合体本体をAlPO4 WAP上に吸着させた後、完成品を最終処方する。AlPO4 WAPをPSC−PDとともに室温で5分間撹拌する。pHを5.1に調整し、混合物を室温で18時間撹拌する。NaCl溶液を付加して150 mMとし、混合物を室温で5分間撹拌する。2−フェノキシエタノールを付加して、5 mg/mlとし、混合物を室温で15分間撹拌した後、pH6.1に調整する。
【0297】
最終組成物/用量
PSC−PD: 10μgPS
AlPO4 WAP: 0.25 mgAl3+
NaCl: 150 mM
2−フェノキシエタノール: 2.5 mg
注射用水: 0.5 mlにする
pH: 6.1
F:前臨床情報
マウスにおける他糖複合体の免疫原性
PSC−PD複合体の免疫原性を、6〜8週齢Balb/cマウスで査定した。純(非吸着)複合体またはAlPO4上に吸着した複合体を、一価ワクチンとして注入した。誘導された抗PSC抗体をELISAにより測定し、一方、殺細菌試験により機能性抗体を分析した。両方法は、CDC(Centers for Disease Control and Prevention, Atlanta, USA)プロトコールを基礎にしている。応答対用量およびアジュバント(AlPO4)作用を査定するために実施した2つの異なる実験からの結果を示す。
【0298】
用量範囲実験
この実験では、Balb/CマウスにPSC−PDを2回(2週間間隔)注射した。4つの異なる用量:0.1;0.5;2.5および9.6μg/動物のAlPO4上処方複合体を用いた。14日目(第1回後14日目)、28日目(第2回目後14日目)および42日目(第2回後28日目)に、マウス(10匹/群)を採血した。ELISAにより測定した多糖C特異的抗体の幾何学平均濃度(GMC)を、参照としてIgGを用いて、μgIgG/mlで表した。殺細菌抗体をプール血清に関して測定し、嬰児ウサギ補体の存在下で髄膜炎菌C11菌株を用いて、細菌の50%を殺害し得る稀釈液の逆数として力価を表した。
【0299】
得られた用量−応答曲線は、2.5μgからプラトーを示す。結果は、第1回後14日目と第2回後14日目の間に良好な追加免疫応答が認められるということを示す。第2回後28日目の抗体レベルは、第2回後14日目のレベルと少なくとも同等である。殺細菌抗体力価はELISA濃度と一致し、PSC−PD複合体の免疫原性を確証する。
【0300】
アジュバントの効果
この実験では、AlPO4上に処方された1ロットのPSC−PD複合体を査定し、純(非吸着)複合体を比較のために注射した。2μgの複合体を用いて、皮下経路により、2週間間隔で、マウス10匹/群に2回注射した。14日目(第1回後14日目)、28日目(第2回目後14日目)および42日目(第2回後28日目)にマウスを採血し、そしてELISAおよび機能性抗体力価を測定した(殺細菌検定に関しては第2回後14日目および第2回後28日目のみ)。AlPO4処方物は、非活性調節処方物と比較した場合、10倍までの抗体力価を誘導する。
【0301】
結論:
前記の実験結果から、以下の全般的結論が得られた:
−PSC−PD複合体は、連結された場合に、PSCがT細胞依存性抗原になるということを実証する既往性応答を誘導する。
【0302】
−ELISAにより測定された抗PSC抗体濃度は殺細菌性抗体力価とよく相関し、このことは、PSC−PD複合体により誘導された抗体が髄膜炎菌血清型Cに対して機能的であることを示す。
【0303】
−AlPO4上に吸着された複合体約2.5μgは、マウスにおいて最適抗体応答を引き出すと思われる。
【0304】
−CDAP化学作用は、免疫原性PSC−PD複合体を製造するための適切な方法であると思われる。
【0305】
実施例8−髄膜炎菌血清型A−PD複合体からの多糖の調製
多糖A(PSA)の乾燥粉末を0.2 MNaCl溶液中に1時間溶解して、最終濃度を8 mg/mlとする。次に、HClまたはNaOHを用いてpH値を6に固定し、溶液を25℃に熱調節する。0.75 mgCDAP/mgPSA(調製物を100 mg/mlアセトニトリルとする)をPSA溶液に付加する。pH調節せずに1.5分後、0.2 MNaOHを付加して、pHを10とする。2.5分後、約1のPD/PSA比にしたがって、プロテインD(5 mg/mlに濃縮)を付加する。1時間のカップリング反応時間中は、pHを10に保持する。次に、10 mgのグリシン(2 M、pH9.0)/mgPSAを付加し、25℃で30分間、pH値を9.0に調節する。次に混合物を4℃で一夜保存した後、排除カラムクロマトグラフィー(Sephacryl S400HR, Pharmacia)により精製する。先ず複合体が、その後未反応PDおよび副産物(DMAP、グリシン、塩)が溶離する。複合体を収集し、Sartopore膜(Sartorius)上での0.2μm濾過により滅菌する。
【0306】
実施例9−実施例7および8の生成物のin vitro特性化
【0307】
【表19】
【0308】
in vivo結果
Balb/Cマウスを動物モデルとして用いて、複合体の免疫原性を調べた。複合体をAlPO4またはAl(OH)3上に吸着させる(500μGのAl3+上に10μgのPS)か、あるいは吸着させなかった。マウスに以下のように注射した:2週間間隔で2回注射(2μgPS/注射)。
【0309】
これらの結果から、遊離PSは免疫応答に大いに影響を及ぼす、と先ず結論し得る。10%未満の遊離PSを有する複合体を用いて、より良好な結果を得た。したがって、CDAP法に対する前記の改良は、本発明のさらなる局面である。
【0310】
処方物も重要である。AlPO4は、このモデルにおいて最も適切なアジュバントであると思われる。複合体は、多糖を単独で注射した場合には観察されない追加免疫作用を誘導する。
【0311】
結論
それぞれ1および0.6〜0.7(w/w)の最終PS/タンパク質比を有する髄膜炎菌AおよびCの複合体を得た。遊離PSおよび遊離担体タンパク質は、それぞれ10%および15%以下であった。多糖回収率は、70%より高い。前記の改良型(最適化)CDAP法により得られるPSAおよびPSCの複合体(担体タンパク質にかかわらず、しかし好ましくはプロテインD)は、したがって、本発明のさらなる局面である。
【0312】
実施例10−b型インフルエンザ菌は、2歳以下の小児における髄膜炎の主因の1つである。破傷風毒素上の複合体としてのインフルエンザ菌の莢膜多糖(PRP)が周知である(J. Robbinsにより開発された化学作用により連結される)。CDAPは、改良型化学である。好ましくはPDにPRPを連結するために見出された最適CDAP条件について、以下に説明する。
【0313】
複合化の反応に影響を及ぼすパラメーターを以下に示す:
・多糖の初期濃度(遊離多糖の最終レベルにおよび滅菌濾過工程に二重の影響を及ぼし得る)
・担体タンパク質の初期濃度
・多糖対タンパク質の初期比(遊離多糖の最終レベルにおよび滅菌濾過工程に二重の影響を及ぼし得る)
・用いられるCDAPの量(通常は大余分量で)
・反応温度(多糖の分解、反応の速度論、および反応性基の分解に影響を及ぼし得る)
・活性化およびカップリングのpH
・クエンチングのpH(残留DMAPのレベルに影響を及ぼす)
・活性化、カップリングおよびクエンチングの時間
最終生成物の量を最適化するための3つの最も重要なパラメーターは、多糖/タンパク質の初期比、多糖の初期濃度およびカップリングpHである、ということを本発明人等は見出した。
【0314】
したがって、前記の3つの条件を3つの軸として用いて、反応立方体を設計した。これらの軸に関する中心点(および実験値範囲)は、PS/タンパク質比−1/1(±0.3/1);[PS]=5 mg/ml(±2 mg/ml);およびカップリングpH=8.0(±1.0pH単位)であった。
【0315】
あまり必須でないパラメーターは以下のように固定した:30 mgの多糖を用いた;温度25℃;[CDAP]=0.75 mg/mgPS;pHは0.2 MNaOHで滴定;活性化pH=9.5;活性化温度=1.5分間;カップリング温度=1時間;[タンパク質]=10 mg/ml;クエンチングpH=9.0;クエンチング温度=1時間;溶媒中にPSを溶解する温度=2 MNaCl中で1時間;12 cm/時間で150 mMNaClで用利したSephacryl S-400HR上での精製;ならびに5ml/分でのSARTOLAB P20によるフィルター滅菌。
【0316】
前記の反応立方体内で製品を製造する場合の最適化条件を確立するために考察されたデータを以下に示す:工程データ−濾過後の最大収率、今夕されるタンパク質の最大レベル;ならびに生成物の品質データ−最終PS/タンパク質比、遊離PSレベル、遊離タンパク質レベル、残留DMAP(CDAPの分解生成物)の最小レベル。
【0317】
濾過からの結果
濾過後の結果を左右する因子は、初期[PS]およびカップリングpHおよび初期PS/タンパク質比間の相互作用である。低[PS]では、後者2つの因子との相互作用はほとんど認められず、良好な濾過可能性が常に結果的に生じる(全生成物に関して約95%)。しかしながら、高濃度では、pHおよび初期比が増大すれば、濾過可能性は減少する(高[PS]、最低比、最低pH=99%濾過;しかし高[PS]、最高比およびpH=19%濾過)。
【0318】
タンパク質混入レベル
最終PS/タンパク質比の、初期比に関する比は、カップリングの効率の測定値である。高[PS]では、pHは両比の比に影響しない。しかしながら、初期比は影響する(低初期比で1.75、高初期比で1.26)。低[PS]では、両比の比は、ほとんどの部分に関しては、より低いが、しかしながら、pHは、ここではより大きい影響を有する(低pH、低比=0.96;低pH、高比=0.8;高pH、低比=1.4;および高pH、高比=0.92)。
【0319】
最終PS/タンパク質比
最終比は、初期比および[PS]によっている。大多数のかなり大きい最終比は、組合せ高初期比および高[PS]を用いて得られる。最終比に及ぼすpHの影響は、弱[PS]と同じくらい有意ではない。
【0320】
遊離プロテインDのレベル
最小量の遊離プロテインDは、高pHおよび高[PS]で観察される(0.0に近似のレベル)。高[PS]の作用は、pHが低い場合に特に顕著になる。初期比の上昇は、遊離プロテインDの増大にわずかに寄与する。
【0321】
残留DMAP
初期比は、有意の作用を有さない。これに対比して、DMAPのレベルは、[PS]に伴って増大し、pHが上昇すると低減する。
【0322】
結論
したがって、最も好ましい複合化条件は、以下の通りである:カップリングpH=9.0;[PS]=3 mg/ml;および初期比=1/1。このような条件を用いた場合の、最終生成物の特徴を以下に示す:
【0323】
【表20】
【0324】
前記の改良型(最適化)CDAP法により得られるPRPの複合体は(担体タンパク質にかかわらず、しかし好ましくはプロテインD)、したがって、本発明のさらなる局面である。
【0325】
実施例11:抗原としてのプロテインD−非分類可能インフルエンザ菌に対するその防御効力は3D−MPLを用いてそれを処方することにより以下に改善され得るか
11価肺炎球菌多糖−プロテインD複合体ワクチンを用いて、雌Balb/cマウス(10匹/群)を、20週齢で初回(D0)の免疫感作(筋内)し、2週間後(14日目)に2回目の免疫感作を施した。2回目免疫感作後7日目に、血液を収集した。IgG1、IgG2aおよびIgG2b型抗体の量に関して、プロテインDに対する抗体力価を測定した。
【0326】
室温で15分間撹拌しながら複合体を15.75%のラクトースと併合し、pHを6.1±0.1に調整し、凍結乾燥することにより、凍結乾燥11価ワクチン(AlPO4を含有しない)を調製した(当該サイクルは通常は、-69℃で出発し、漸次3時間に亘って-24℃に調整し、次にこの温度を18時間保持した後、漸次1時間に亘って-16℃に調整し、この温度を6時間保持して、その後漸次3時間に亘って+34℃に調整し、最後にこの温度を9時間保持する)。
【0327】
凍結乾燥物のための処方物および再構築物の組成を表13に示す。
【0328】
Th1型細胞媒介性免疫応答が起きたか否かに関する大多数の特徴的測定値は、IgG2aのレベルと相関することが知られている。データから分かるように、IgG2aの意外にも大きい増大は、プロテインDがTh1アジュバント(この場合は3D−MPL)とともに凍結乾燥された場合に、結果的に生じる。
【0329】
【表21】
【技術分野】
【0001】
産業上の利用分野
本発明は、細菌多糖抗原ワクチン、それらの製造および医療におけるこのような多糖の使用に関する。
【0002】
特に本発明は、以下の3つの相互関連局面に関する:A−肺炎球菌多糖抗原、典型的には肺炎連鎖球菌(Streptococcus pneumoniae)からのタンパク質抗原および任意にTh1誘導性アジュバントを用いて処方された肺炎球菌多糖複合体抗原を含むワクチン;B−Th1アジュバントで活性調節された特異的有益肺炎球菌多糖複合体;ならびにC−概してインフルエンザ菌からのプロテインDと連結された細菌多糖複合体。
【背景技術】
【0003】
発明の背景
肺炎連鎖球菌は、少なからぬ罹患率および死亡率(特に若年および老齢者における)に関与するグラム陽性細菌であって、侵襲性疾患、例えば肺炎、菌血症および髄膜炎、ならびに集落形成に関連した疾患、例えば急性中耳炎を引き起こす。60歳より上の人々に関する米国での肺炎球菌性肺炎率は、100,000人に3〜8人であると見積もられる。症例の20%において、これは菌血症およびその他の症状発現、例えば髄膜炎を引き起こし、死亡率は、抗生物質治療を用いた場合でも30%近くである。
【0004】
肺炎球菌は、血清型特異性を付与する化学的結合多糖に被包されている。肺炎球菌の90の既知の血清型が存在し、莢膜は、補体から細菌の内面を防御するだけでなく、それ自体免疫原性に乏しいので、肺炎球菌に関する素因ビルレンス決定基である。多糖はT非依存性抗原であり、T細胞と相互作用するためにMHC分子上で処理加工または提示され得ない。しかしながら、それらは、B細胞上の表面受容体の架橋を包含する代替的メカニズムにより免疫系を刺激し得る。
【0005】
侵襲性肺炎球菌性疾患は莢膜に特異的な抗体と最も強く相関し、防御は血清型特異的である、ということがいくつかの実験で示された。
【0006】
多糖抗原ベースのワクチンは、当業界で周知である。ヒト使用に関して登録されている4つとしては、腸チフス菌のVi多糖、インフルエンザ菌からのPRP多糖、血清型A、C、W135およびYから成る四価髄膜炎菌ワクチン、ならびに血清型1、2、3、4、5、6B、7F、8、9N、9V、10A、11A、12F、14、15B、17F、18C、19A、19F、20、22F、23F、および33に対応する多糖から成る23価肺炎球菌ワクチン(肺炎球菌血液単離物の少なくとも90%を占める)が挙げられる。
【0007】
後者3つのワクチンは、呼吸器感染を引き起こして乳児における重度罹患率および死亡率を生じる細菌に対する防御を付与するが、これらのワクチンは2歳未満の小児における使用に関しては、この年齢群では免疫原性が不十分であるために、未だ認可されていない[Peltola et al.(1984), N. Engl. J. Med. 310:1561-1566]。肺炎連鎖球菌は、乳児および幼児における侵襲性細菌性疾患および中耳炎の最も一般的な原因である。同様に、老齢者は肺炎球菌ワクチンに対して不十分な応答を増し[Roghmann et al.(1987), J. Gerontol.42:265-270]、それゆえこの集団における細菌性肺炎の発生数が増大する[Verghese and Berk, (1983)Medicine(Baltimore)62:271-285]。
【0008】
乳児における免疫原性のこの欠如を克服するよう意図された戦略としては、大型免疫原性タンパク質との多糖の連結が挙げられるが、これは、バイスタンダーT細胞援助を提供し、それが連結される多糖抗原に対する免疫記憶を誘導する。肺炎球菌糖タンパク質複合体ワクチンは、種々の年齢群における安全性、免疫原性および効能に関して一般に評価されつつある。
【0009】
A)肺炎球菌多糖ワクチン
23価非複合肺炎球菌ワクチンは、0%から81%までの臨床的効能の広範な変異を示している(Fedson et al.(1994)Arch Intern Med. 154:2531-2535)。効能は、免疫感作されつつある危険群、例えば老齢者、ホジキン病、脾摘出、鎌状赤血球症および無γグロブリン血症患者に(Fine et al.(1994)Arch Intern Med. 154:2666-2677)、そして疾患発現にも関連していると思われる。23価ワクチンは、肺炎球菌性肺炎(いくつかの高危険群、例えば老齢者において)および中耳炎に対する防御を実証しない。
【0010】
したがって、改良型肺炎球菌ワクチン組成物、特に老齢者および幼児における肺炎球菌性疾患(特に肺炎)の予防または改善により有効であるものが必要とされている。
【発明の概要】
【0011】
本発明は、このような改良型ワクチンを提供する。
【0012】
B)選定肺炎球菌多糖複合体+3D−MPL組成物
商業化非複合肺炎球菌ワクチンの防御効力がワクチン接種時に誘導される抗体の濃度に多少関連するということは、一般に容認される。実際、23の多糖が、専ら各構成成分多糖の免疫原性に関して商業的許可を容認された(Ed. Williams et al. New York Academy of Sciences 1995 pp. 241-249)。したがって、肺炎球菌多糖に対する抗体応答のさらなる強化は、多糖または多糖複合体の初回注射に対する防御レベルの抗体を有して応答する乳児および老齢者のパーセンテージを増大し得たし、肺炎連鎖球菌により引き起こされる感染に対する防御免疫を誘導するのに必要な注射の投与量および回数を低減し得た。
【0013】
20世紀初頭以来、研究者達は、ワクチン組成物におけるそれらの免疫原性を改善するために抗原に付加され得る厖大な数の化合物を用いて実験してきた[M.F. Powell & M.J. Newman, Plenum Press, NY, "Vaccine Design-the Subunit and Adjuvant Approach"(1995)Chapter 7"A Compendium of Vaccine Ajuvants and Excipients"で再検討された]。多くは非常に効率的であるが、しかしヒトワクチン組成物におけるそれらの使用を妨げる有意の局所性および全身性悪反応を引き起こす。1926年に最初に記載されたアルミニウムベースのアジュバント(例えばアルム、水酸化アルミニウムまたはリン酸アルミニウム)は、依然として米国で認可されたヒトワクチン中に用いられる唯一の免疫アジュバントである。
【0014】
アルミニウムベースのアジュバントは、それが誘導する「デポー製剤作用」を介して働く担体類のアジュバントの例である。抗原はその表面に吸着され、組成物が注入された場合、アジュバントおよび抗原は血流中に直ちに消散されない。その代わりに、組成物は注入の局所環境中に存続し、より顕著な免疫応答が結果として生じる。このような担体アジュバントは、例えばいくつかの多糖抗原を分解する傾向がある抗原を安定化するのに適しているという付加的な既知の利点を有する。
【0015】
3D−MPLは、非担体アジュバントの一例である。そのフルネームは3−O−デアシル化モノホスホリル脂質A(または3デ−O−アシル化ものホスホリル脂質Aまたは3−O−デスアシル−4’モノホスホリル脂質A)であり、還元末端グルコサミンの位置3がデ−O−アシル化されることを示すために3D−MPLと呼ばれる。その調製のためには、英国特許出願第2220211号を参照されたい。化学的には、それは3−デアシル化モノホスホリル脂質Aの4、5または6アシル化鎖との混合物である。それは最初は、脂質Aの4’−モノホスホリル誘導体(MPL)を3−O−デアシル化するための方法が、免疫刺激活性の変化を伴わずにさらなる弱毒化毒性を有する分子をもたらした1990年代初頭に製造された。
【0016】
3D−MPLは、単独で、または優先的にはデポー製剤型担体アジュバント、例えば水酸化アルミニウム、リン酸アルミニウムまたは水中油型エマルションと組合せて、アジュバントとして用いられてきた。この溶暗組成物中では、抗原および3D−MPLは同一粒子構造中に含入されて、抗原性および免疫刺激性シグナルのより効率的送達を可能にする。複数の研究が、3D−MPLはアルム吸着化抗原の免疫原性をさらに増強し得る、ということを示している[Thoelen et al. Vaccine(1998)16:708-14; EP 689454-B1]。このような組合せも、吸着傾向がある抗原に関しては当業界で選択される(例えば、細菌多糖複合体)が、この場合、アルムへの吸着は、抗原を安定化する傾向がある。沈降アルミニウムベースのアジュバントは、それらは認可ヒトワクチンに一般に用いられる唯一のアジュバントであるので、主として用いられる。したがって、アルミニウムベースのアジュバントと組合せて3D−MPLを含有するワクチンは、それらの易開発性およびマーカー上への導入速度のために、当業界で好まれる。
【0017】
MPL(非3−デアシル化)は、いくつかの一価多糖複合体ワクチン抗原に関するアジュバントとして評価されてきた。生理食塩水中のMPLの同時注入は、4つの一価多糖複合体:肺炎球菌PS6B破傷風毒素、肺炎球菌PS 12−ジフテリア毒素、ならびに緑膿菌外毒素Aに連結された5型黄色ブドウ球菌および8型黄色ブドウ球菌に対する血清抗体応答を増強する[Schneerson et al. J. Immunology(1991)147:2136-2140]。応答増強は、抗原特異的であると教示された。水中油型エマルション中のMPL(担体型アジュバント)は、同一粒子構造中のMPLおよび抗原の存在のために、生理食塩水中のMPLの作用を一貫して増強し、そして他の多糖複合体ワクチンの最適供給のために選択さるべきアジュバント系であると考えられた。
【0018】
Devi等[Infect. Immun.(1991)59:3700-7]は、クリプトコックス属のCryptococcus neoformans莢膜多糖のTT複合体に対するネズミ抗体応答に及ぼす生理食塩水中のMPL(非3−デアシル化)のアジュバント作用を評価した。MPLが複合体と同時に用いられた場合、PSに対するIgM−およびIgG−特異的応答の両方において限界増大のみが認められた。しかしながら、MPLは、複合体の2日後に投与された場合、非常に大きい効果を示した。特に乳児において、抗原と比較してMPLの投与の遅延をようする免疫感作計画を用いることの実用性は疑問である。多糖および多糖−タンパク質複合体に関するMPLのアジュバント効果は、組成物依存性であると思われる。さらに、適切な徐放性送達系(例えば、担体アジュバント使用)中のMPLの混入は、より永続的なアジュバント効果を提供し、調時および遅延投与の問題を免れる。
【0019】
要するに、当業界の状態は、特に多糖または多糖複合体抗原に関しては、MPLまたは3D−MPLがアジュバントとして用いられる場合、その免疫刺激作用を最大にするために、それは、担体アジュバント(例えば、アルミニウムベースのアジュバント)とともに有益に用いられる、ということを教示している。
【0020】
意外なことに、ある種の肺炎球菌多糖複合体に関しては、ワクチン組成物の免疫原性は、担体アジュバント(例えばアルミニウムベースのアジュバント)とともに用いられる3D−MPLよりむしろ3D−MPL単独を用いて抗原が処方される場合に有意に大きい、ということを本発明は見出した。さらに、観察された改善は、用いた3D−MPLの濃度とは、そして特定の複合体が一価組成物中に存在するか否か、あるいはそれらが併用されて多価組成物を形成するか否かとは無関係である。
【0021】
C)細菌多糖−プロテインD複合体
前記のように、多糖抗原ベースのワクチンは、当業界で周知である。前記の認可多糖ワクチンは、異なる実証済み臨床効能を有する。Vi多糖ワクチンは、培養確証腸チフスの防止に際して55%〜77%の効能を有すると概算されている(Plotkin and Cam,(1995)Arch Intern Med 155:2293-99)。髄膜炎菌C多糖ワクチンは、流行条件下で79%の効能を有することが示された(De Wals P, et al.(1996)Bull World Health Organ. 74:407-411)。23価肺炎球菌ワクチンは、0%〜81%の臨床効能の広範な変動を示した(Fedson et al.(1994)Arch Intern Med.154:2531-2535)。前記のように、肺炎球菌ワクチンの防御効能は、ワクチン接種時に誘導される抗体の濃度に多少関連する、ということが容認される。
【0022】
ワクチン接種への多糖アプローチに関連した問題の1つは、多糖それ自体が貧免疫原性であるという事実である。免疫原性のこの欠如を克服するよう意図された戦略としては、大型高免疫原性タンパク質との多糖の連結が挙げられるが、これは、バイスタンダーT細胞援助を提供する。
【0023】
多糖免疫原の産生のために今日一般に用いられるこれらの高免疫原性担体の例としては、ジフテリア毒素(CTまたはCRM197突然変異体)、破傷風毒素(TT)、カギアナカサガイヘモシアニン(KLH)およびツベルクリンの精製タンパク質誘導体(PPD)が挙げられる。
【0024】
一般的に用いられる担体に関連した問題
多数の問題が、例えばGMP複合体の産生におけるならびに複合体の免疫学的特徴における、これらの一般的に用いられる担体の各々に関連している。
【0025】
これらの担体の一般的使用および抗多糖抗体応答の誘導におけるそれらの成功例にもかかわらず、それらはいくつかの欠点に関連する。例えば、抗原特異的免疫応答は、担体(この場合、破傷風毒素)に対して向けられる先在する抗体の存在により抑制され得る(エピトープ抑制)ということが知られている(Di John et al;(1989)Lancet, 2:1415-8)。集団全般では、人々はこれらの抗原でルーチンにワクチン接種されるので、非常に高パーセンテージの人々がDTおよびTTの両方に対する先在免疫を有する。英国では、例えば95%の小児がDTおよびTTの両方を含むDTPワクチンを接種されている。他の著者等は、動物モデルにおけるペプチドワクチンに対するエピトープ抑制の問題を記載している(Sad et al, Immunology, 1991; 74:223-227; Schutze et al, J. Immunol. 135:4, 1985; 2319-2322)。
【0026】
さらに、定期的追加免疫を要するワクチンに関しては、高免疫原性担体、例えばTTおよびDTの使用は、数回注射後の多糖抗体応答を抑制すると思われる。これらの多数回予防接種は、遅延型過応答性(DTH)のような望ましくない反応によっても成し遂げられ得る。
【0027】
KLHは、効力のある免疫原として既知であり、ヒト臨床試験におけるIgEペプチドのための担体としてすでに用いられている。しかしながら、いくつかの悪反応(DTH様反応またはIgE感作)ならびに抗体に対する抗体応答が観察されている。
【0028】
したがって、多糖ベースのワクチンのための担体タンパク質の選択は、すべての患者において作用する(広範なMHC認識)担体に対する必要性、高レベルの抗多糖抗体応答の誘導および担体に対する低抗体応答の間の平衡を必要とする。
【0029】
したがって、多糖ベースのワクチンのために予め用いられる担体は、多数の欠点を有する。これは、種々の多糖抗原に関して同一担体が用いられる場合に、エピトープ抑制が特に問題となる組合せワクチンにおいて特にそうである。WO 98/51339では、組合せワクチン中の多担体は、この作用を乗り越えようとするために用いられた。
【0030】
本発明は、前記の欠点を蒙らない多糖/多ペプチドベースの免疫原性複合体の調製に際して用いるための新規の担体を提供する。
【0031】
本発明は、ワクチンを含めた多糖ベースの免疫原性組成物のための担体として、インフルエンザ菌からのプロテインD(EP 0 594 610 B1)またはその断片を提供する。この担体の使用は、組合せワクチンにおいて特に有益である。
【0032】
発明の要約
A)肺炎球菌多糖ワクチン
したがって、本発明は、少なくとも1つの肺炎連鎖球菌多糖抗原(好ましくは複合化)および肺炎連鎖球菌タンパク質抗原またはその免疫学的機能等価物を、任意にTh1アジュバント(Th1免疫応答を誘導するアジュバント)とともに含むワクチンを提供する。好ましくは、肺炎球菌タンパク質およびTh1アジュバントがともに含まれる。本発明の組成物は、老齢者肺炎の治療に特に適している。
【0033】
肺炎球菌多糖ワクチン(複合化または非複合化)は、この疾患の発生数が非常に高い老齢者集団における肺炎に対して防御し得ない。肺炎球菌に対する重要な防衛メカニズムはオプソニン食菌作用(肺炎球菌多糖に対する抗体の産生により引き起こされるヒトB細胞/好中球媒介性事象。細菌は最終的には食菌されるようになる)であるが、しかしながら、関与するオプソニン性メカニズムの一部、即ちPMN(多形核球)によるスーパーオキシド産生、その他の反応性酸素種産生、PMNの可動化、PMNのアポトーシス、PMNの変形能が、老齢者では損傷される。
【0034】
普通に容認された教義と反対に、正常レベルの抗莢膜多糖抗体は、肺炎球菌が宿主細胞に侵襲して免疫系のこの部門を回避し得るので、細菌の完全クリアランスには有効でない可能性がある。
【0035】
意外にも、免疫系の体液性部門(B細胞媒介性免疫)の他に、免疫系の細胞媒介性部門(例えば、T細胞媒介性免疫)を同時刺激することにより、宿主からの肺炎球菌のクリアランスを増強し得る相乗作用(または協同作用)を結果として生じる、ということを本発明は見出した。これは、概して肺炎球菌感染の予防(または治療)に役立つ発見であるが、しかし多糖ベースのワクチンが効能を示さない老齢者における肺炎の予防(または治療)に特に重要である。
【0036】
肺炎球菌多糖(好ましくは複合化)が肺炎球菌タンパク質(好ましくは肺炎球菌の表面で発現され、あるいは分泌または放出されるタンパク質で、これは感染哺乳類細胞の表面で暮らすIIおよびMHCクラスIの情況においてプロセッシングされ、提示され得る)とともに投与される場合、免疫系の両部門はこのように共働し得る、ということを本発明は見出した。肺炎球菌タンパク質はそれ自体で細胞媒介性免疫の引き金となり得るが、しかし、ワクチン処方物中のTh1誘導性アジュバントの存在が免疫系のこの部門の役に立ち、そして意外にも、免疫系の両部門間の相乗作用をさらに増強する、ということを本発明人等は見出した。
【0037】
B)選定肺炎球菌多糖複合体+3D−MPL組成物
したがって、本発明は、3D−MPLで活性調節され、実質的にアルミニウムベースのアジュバントを欠く1つ又はそれ以上の肺炎球菌多糖複合体を含む抗原性組成物も提供するが、この場合、少なくとも1つの肺炎球菌多糖複合体は、アルミニウムベースのアジュバントとともに3D−MPLを含む組成物と比較して、3D−MPLを含む組成物中で有意により免疫原性である。
【0038】
提供される好ましい実施態様は、1つ又はそれ以上の以下の肺炎球菌莢膜多糖:血清型4、6B、18C、19Fおよび23Fの複合体を含む抗原性組成物である。このような組成物中では、多糖は各々、意外にも、3D−MPLおよびアルミニウムベースのアジュバントを含む組成物と比較して、3D−MPL単独を含む組成物中でより免疫原性である。
【0039】
したがって、本発明の一実施態様においては、免疫原性タンパク質および3D−MPLアジュバントに連結された肺炎連鎖球菌莢膜多糖血清型4、6B、18C、19Fまたは23Fを含む抗原性組成物が提供され、この場合、組成物は実質的にはアルミニウムベースのアジュバントを欠く。
【0040】
第二の実施態様では、本発明は、アルミニウムベースのアジュバントを実質的に欠き、そして血清型4、血清型6B、血清型18C、血清型19Fおよび血清型23Fから成る群から選択される3D−MPLおよび2つまたはそれ以上の肺炎球菌多糖複合体を含む組合せ抗原性組成物を提供する。
【0041】
C)細菌多糖−プロテインD複合体
したがって、本発明は、インフルエンザ菌からのプロテインDまたはそのプロテインD断片に連結された病原性細菌から得られる多糖抗原を含む多糖複合体抗原を提供する。さらに、本発明は、1つ又はそれ以上の多糖抗原がプロテインDに連結される多価ワクチン組成物を提供する。
【図面の簡単な説明】
【0042】
【図1A】マウスにおけるニューモリジンに対する血清IgG。
【図1B】ニューモリジンで免疫化されたマウスにおける血清溶解阻害力価。
【図1C】血清型6Bによる鼻内感染後6時間目におけるOF1マウスにおけるニューモコッカル肺コロニー形成。
【発明を実施するための形態】
【0043】
発明の詳細な説明
A)肺炎球菌多糖ワクチン
本発明は、特に老齢者(および/または乳児および幼児)の肺炎球菌感染の予防または改善のための改良型ワクチンを提供する。
【0044】
本発明の情況では、患者は、年齢が55歳以上、典型的には60歳以上、さらに一般的には65歳以上であれば、老齢者であると考えられる。
【0045】
したがって、本発明の一実施態様では、少なくとも1つの肺炎連鎖球菌多糖抗原および少なくとも1つの肺炎連鎖球菌タンパク質抗原を含む、老齢者(および/または小児、乳児(Infants)および幼児)において用いるのに適したワクチン組成物が提供される。
【0046】
第二の好ましい実施態様では、本発明は、少なくとも1つの肺炎連鎖球菌多糖抗原および少なくとも1つの肺炎連鎖球菌タンパク質抗原およびTh1アジュバントを含む(老齢者における肺炎の予防に適した)ワクチンを提供する。
【0047】
このようなワクチンは、例えば乳児または幼児のような集団の他の高度危険群における肺炎球菌感染(例えば中耳炎)を治療するのにも有用であることが意図される。
【0048】
第三の実施態様では、肺炎球菌多糖抗原およびTh1アジュバントを含むワクチン組成物が提供される。
【0049】
本発明の肺炎連鎖球菌多糖抗原
典型的には、本発明の肺炎連鎖球菌ワクチンは、多糖抗原(好ましくは複合化)を含み、この場合、多糖は少なくとも4つの血清型の肺炎球菌から得られる。好ましくは、4つの血清型としては、6B、14、19Fおよび23Fが挙げられる。さらに好ましくは少なくとも7つの血清型、例えば4、6B、9V、14、18C、19Fおよび23Fから得られるものが組成物中に含まれる。さらに好ましくは、少なくとも11の血清型が組成物に含まれ、例えば一実施態様における組成物は、血清型1、3、4、5、6B、7F、9V、14、18C、19Fおよび23Fから得られる莢膜多糖(好ましくは複合化(conjugated))を含む。本発明の好ましい実施態様では、少なくとも13の多糖抗原(好ましくは複合化)が含まれるが、しかしさらなる多糖抗原、例えば23価(例えば血清型1、2、3、4、5、6B、7F、8、9N、9V、10A、11A、12F、14、15B、17F、18C、19A、19F、20、22F、23Fおよび33F)も本発明により意図される。
【0050】
老齢者予防接種(例えば、肺炎予防のための)に関しては、血清型8および12F(そして最も好ましくはさらに15および22)を前記の11価抗原性組成物に含入して、15価ワクチンを生成するのが有益であるが、一方、乳児または幼児用(中耳炎がより問題である)には、血清型6Aおよび19Aが含入されて、13価ワクチンを生成するのが有益である。
【0051】
老齢者(+55歳)集団における肺炎ならびに乳児(18ヶ月まで)および幼児(典型的には18ヶ月〜5歳)における中耳炎の予防/改善のためには、本明細書に記載したような多価肺炎連鎖球菌多糖を肺炎連鎖球菌タンパク質またはその免疫学的機能等価物と併用することが、本発明の好ましい実施態様である。
【0052】
本発明の肺炎球菌タンパク質
本発明の目的のために、「免疫学的機能等価物」とは、本発明のタンパク質からの少なくとも1つの防御的エピトープを含むタンパク質のペプチドと定義される。このようなエピトープは、特徴的には、表面露呈され、高度保存され、そして宿主における殺細菌性抗体応答を引き出し、あるいは毒性作用を阻止し得る。好ましくは、機能的等価物は、本発明のタンパク質からの少なくとも15、好ましくは30またはそれ以上の連続アミノ酸を有する。最も好ましくは、タンパク質の断片、欠失、例えばその膜貫通欠失変異体(即ち、タンパク質の細胞外ドメインの使用)、融合、化学的または遺伝的脱毒素化誘導体などが用いられ得るが、但し、それらはネイティブタンパク質と同一免疫応答を実質的に生じ得る。
【0053】
本発明の好ましいタンパク質は、肺炎球菌の外表面上に露呈される肺炎球菌タンパク質(肺炎球菌の少なくとも一部のライフサイクル中に宿主の免疫系により認識され得る)であるか、あるいは肺炎球菌により分泌または放出されるタンパク質である。最も好ましくは、タンパク質は毒素、粘着素、2−成分シグナル変換体または肺炎連鎖球菌のリポタンパク質、あるいはそれらの免疫学的機能等価物である。
【0054】
このような組合せワクチン中に含入される特に好ましいタンパク質としては以下のものが挙げられるが、これらに限定されない:ニューモリジン(好ましくは、化学処理または突然変異により脱毒素化)[Mitchell et al. Nucleic Acids Res. 1990 Jul 11;18(3):4010 "Comparison of pneumolysin genes and proteins from Steptococcus pneumoniae types 1 and 2.", Mitchell et al. Biochim Biophys Acta 1989 Jan 23; 1007(1):67-72 "Expression of the pneumolysin gene in Escherichia coli: rapid purification and biological properties.", WO 96/05859(A. Cyanamid), WO 90/06951(Paton et al), WO 99/03884(NAVA)];PspAおよびその膜貫通欠失変異体(米国特許第5804193号−Briles等);PspCおよびその膜貫通欠失変異体(WO 97/09994−Briles等);PsaAおよびその膜貫通欠失変異体(Berry & Paton, Infect Immun 1996 Dec; 64(12):5255-62 "Sequence heterogeneity of PsaA, a 37-kilodalton putative adhesin essential for virulence of Streptococcus pneumoniae");肺炎球菌コリン結合タンパク質およびその膜貫通欠失変異体;CbpAおよびその膜貫通欠失変異体(WO 97/41151; WO 99/51266);グリセルアルデヒド−3−ホスフェート−デヒドロゲナーゼ(Infect. Immun. 1996, 64:3544);HSP70(WO 96/40928);PcpA(Sanchez-Beato et al. FEMS Microbiol Lett 1998, 164:207-14);M様タンパク質(SB特許出願EP 0837130);および粘着素18627(SB特許出願EP 0834568)。
【0055】
本発明に用いられるタンパク質は、好ましくは、ニューモリジン群PsaA、PspA、PspC、CbpA、あるいは2またはそれ以上のこのようなタンパク質の組合せから選択される。本発明は、このようなタンパク質の免疫学的機能等価物も包含する(前記)。
【0056】
組成物中では、タンパク質は、肺炎球菌性疾患に対するT細胞媒介性応答を誘導するのに役立ち、特に肺炎に対する防御に必要とされ、これは肺炎球菌による侵襲を阻止するために、そしてオプソニン食菌作用を刺激するために免疫系の体液性部門と協同する。
【0057】
タンパク質抗原を含入することのさらなる利点は、オプソニン食菌作用過程のためのさらなる抗原の提示、および細菌付着の抑制(粘着素(adhesin)が用いられる場合)または毒素の中和(毒素が用いられる場合)である。
【0058】
本発明の一実施態様によれば、少なくとも4つの血清型、好ましくは少なくとも7つの血清型、さらに好ましくは少なくとも11の血清型から得られる多糖抗原、ならびに少なくとも1つの、しかし好ましくは2つの肺炎連鎖球菌タンパク質を含む肺炎球菌多糖複合体ワクチンを包含する肺炎連鎖球菌ワクチンが提供される。好ましくは、タンパク質の1つは、ニューモリジン(Pneumolysin)あるいはPsaAまたはPspAまたはCbpA(最も好ましくは脱毒素化ニューモリジン)である。好ましい組合せは、少なくともニューモリジンまたはその誘導体およびPspAを含有する。
【0059】
前記のように、予防接種への多糖アプローチに関連した問題は、多糖それ自体が貧免疫原性であるという事実である。これを克服するために、多糖は、バイスタンダーT細胞援助を提供するタンパク質担体に連結され得る。したがって、本発明に利用される多糖はこのようなタンパク質担体に結合されるのが好ましい。多糖免疫原の産生のために現在一般的に用いられるこのような担体の例としては、ジフテリアおよび破傷風毒素(それぞれDT、DT CRM197およびTT)、カギアナカサガイヘモシアニン(KLH)、髄膜炎菌からのOMPC、ならびにツベルクリンの精製タンパク質誘導体(PPD)が挙げられる。
【0060】
しかしながら多数の問題が、これらの一般的に用いられる担体の各々に関連している(前記の「一般的に用いられる担体に関連した問題」の節参照)。
【0061】
本発明は、好ましい実施態様において、これらの欠点を蒙らない多糖ベースの免疫原構築物の調製に際して用いるための新規の担体を提供する。肺炎球菌多糖ベースの免疫原性組成物(またはワクチン)のための好ましい担体は、インフルエンザ菌からのプロテインD(EP 594610-B)またはその断片である。用いるのに適した断片としては、T−ヘルパーエピトープを包含する断片が挙げられる。特に、プロテインD断片は、好ましくはタンパク質のN末端1/3を含有する。
【0062】
肺炎球菌多糖のためのさらに好ましい担体は、肺炎球菌タンパク質それ自体である(「本発明の肺炎球菌タンパク質」の節に前記)。
【0063】
本発明のワクチンは、好ましくは活性調節される。適切なアジュバントとしては、水酸化アルミニウムゲル(アルム)またはリン酸アルミニウムのようなアルミニウム塩が挙げられるが、しかしカルシウム、鉄または亜鉛の塩でもあり得るし、あるいはアシル化チロシンまたはアシル化糖、陽イオン的または陰イオン的誘導化多糖またはポリホスファゼンの不溶性懸濁液であり得る。
【0064】
アジュバントは、免疫応答の細胞媒介性部門を活性調節するためにTH1型の応答の優先的誘導物質であるよう選択されるのが好ましい。
【0065】
本発明のTH1アジュバント
高レベルのTh1型サイトカインは、所定の抗原に対する細胞媒介性免疫応答の誘導を援助する傾向があるが、一方、高レベルのTh2型サイトカインは、抗原に対する体液性免疫応答の誘導を援助する傾向がある。
【0066】
Th1およびTh2型免疫応答の区別は絶対的であるというわけではない、ということを思い起こすことは重要である。実際は、個体は主としてTh1または主としてTh2であると説明される免疫応答を支持する。しかしながら、MosmannとCoffiman(Mosmann, T.R. and Coffiman, R.L.(1989)TH1 and TH2 cell: different patterns of lymphokine secretion lead to different functional properties. Annual Review of Immunology, 7, p145-173)によりネズミCD4+ve T細胞クローンで説明されたことに関してサイトカイン族を考えるのがしばしば便利である。伝統的に、Th1型応答は、Tリンパ球によるINF−γおよびIL−2サイトカインの産生に関連する。Th1型免疫応答の誘導にしばしば直接関連するその他のサイトカインは、IL−12のように、T細胞により産生されない。これに対比して、Th2型応答は、Il−4、IL−5、IL−6、IL−10の分泌に関連する。主要Th1応答を促す適切なアジュバント系としては、モノホスホリル脂質A、ならびにモノホスホリル脂質A、好ましくは3−デ−O−アシル化モノホスホリル脂質A(3D−MPL)とアルミニウム塩の組合せが挙げられる。
【0067】
増強系は、モノホスホリル脂質Aおよびサポニン誘導体の組合せ、特にWO 94/00153に開示されたようなQS21および3D−MPLの組合せ、またはWO 96/33739に開示されているようにQS21がコレステロールで抑制される低反応原性組成物を包含する。
【0068】
水中油型エマルション中にQS21、3D−MPLおよびトコフェロールを包含する特に強力なアジュバント処方物は、WO 95/17210に記載されており、好ましい処方物である。
【0069】
好ましくは、ワクチンは、サポニンを、さらに好ましくはQS21を付加的に含む。処方物は、水中油型エマルションおよびトコフェロールも含み得る(WO 95/17210)。
【0070】
本発明は、ワクチン処方物の製造方法であって、本発明のタンパク質を製薬上許容可能な賦形剤、例えば3D−MPLと一緒に混合することを包含する方法も提供する。
【0071】
オリゴヌクレオチドを含有する非メチル化CpG(WO 96/02555)もTH応答の選択的誘導物質であり、本発明に用いるのに適している。
【0072】
本発明の特に好ましい組成物は、1つ又はそれ以上の複合肺炎球菌多糖、1つ又はそれ以上の肺炎球菌タンパク質およびTh1アジュバントを含む。肺炎球菌タンパク質による細胞媒介性応答の誘導(前記)および免疫系の両部門の間の協同は、このようなアジュバントを用いて活性調節されて、概して肺炎球菌性疾患に対する、そして重要なのは老齢者における肺炎球菌性肺炎に対して特に有効なワクチンを生じる。
【0073】
本発明のさらなる局面において、医療に用いるための本明細書中に記載したような免疫原またはワクチンが提供される。
【0074】
本発明のさらに別の局面では、不応答者の集団内で多糖抗原に対する体液性抗体応答を血清変換または誘導し得る、肺炎球菌多糖複合体およびTh1アジュバント(好ましくは3D−MPL)を含む組成物が提供される。
【0075】
集団の10〜30%は、多糖免疫感作に対する不応答者(ワクチン中の50%より多い血清型に応答しない)であることが知られている(Konradsen et al., Scand. J. Immun 40:251(1994); Rodriguez et al., JID, 173:1347(1996))。これも、複合化ワクチンに関する症例であり得る(Musher et al. Clin. Inf. Dis. 27:1487(1998))。これは、高危険域の集団(乳児、幼児および老齢者)に関しては特に重大である。
【0076】
複合肺炎球菌多糖(これは特定の集団において低応答傾向がある)のTh1アジュバント(前記の「本発明のTh1アジュバント」参照)との組合せは、意外にもこの不応答性を克服し得る、ということを本発明は見出した。好ましくは3D−MPLが用いられるべきであり、最も好ましくは3D−MPLはアルミニウムベースのアジュバントを欠く(依然として良好な応答を提供する)。したがって本発明はこのような組成物を提供し、さらにこのような組成物を投与することにより肺炎球菌多糖に対する不応答者を治療する方法を提供し、そしてその上さらに、多糖抗原に不応答性である個体における肺炎球菌性疾患に対する治療(または防御)における複合肺炎球菌多糖抗原を含む薬剤の製造に際してのTh1アジュバントの使用を提供する。
【0077】
一実施態様では、老齢者における肺炎の予防または改善方法であって、肺炎連鎖球菌多糖抗原およびTh1アジュバントまたは肺炎球菌タンパク質(好ましくは両方)を含む本明細書に記載したような安全且つ有効量のワクチンを前記の老齢患者に投与することを包含する方法がある。
【0078】
さらに別の実施態様では、乳児または幼児における中耳炎の予防または改善方法であって、肺炎連鎖球菌多糖抗原および肺炎連鎖球菌タンパク質抗原またはTh1アジュバント(好ましくは両方)を含む安全且つ有効量のワクチンを前記の乳児または幼児に投与することを包含する方法が提供される。
【0079】
好ましくは、前記のような本発明の方法においては、多糖抗原は多糖タンパク質複合体として存在する。
【0080】
本発明のワクチン製剤
本発明のワクチン製剤は、全身または粘膜経路により前記のワクチンを投与することにより、感染に感受性の哺乳類を防御または治療するために用いられ得る。これらの投与は、筋内、腹腔内、皮内または皮下経路による、あるいは口腔/栄養管、気道、尿生殖路への粘膜性糖よによる注入を含み得る。肺炎または中耳炎の治療のためのワクチンの鼻腔内投与は好ましい(肺炎球菌の鼻咽頭運搬はより有効に予防され得るので、したがって、その最初期段階で感染を減衰し得る)。
【0081】
各ワクチン中の複合抗原の量は、典型的ワクチンにおいて有意の副作用を伴わずに免疫防御応答を誘導する量として選択される。このような量は、どの特異的免疫原が用いられるか、そしてそれがどのように存在するかによって変わる。一般的に、各用量は、0.1〜100μg、好ましくは0.1〜50μg、好ましくは0.1〜10μgの多糖を含むと予測され、1〜5μgが最も好ましい範囲である。
【0082】
ワクチン中のタンパク質抗原の含量は、典型的には1〜100μg、好ましくは5〜50μg、最も典型的には5〜25μgの範囲である。
【0083】
特定のワクチンに関する構成成分の最適量は、被験者における適切な免疫応答の観察を含めた標準試験により確かめられ得る。初回予防接種後、適当な間隔で1回または数回の追加免疫感作を被験者に施し得る。
【0084】
ワクチン製剤は、Vaccine Design("The subunit and adjuvant approach")(eds Powell M.F. & Newman M.J.(1995)Plenum Press New York)に一般的に記載されている。リポソーム内封入は、米国特許第4,235,877号(Fullerton)に記載されている。
【0085】
B)選定肺炎球菌多糖複合体+3D−MPL組成物
本発明の目的のために、「本発明の肺炎球菌多糖複合体」という用語は、アルミニウムベースのアジュバントとともに3D−MPLを含む組成物と比較して、3D−MPLを含む組成物中でより免疫原性である肺炎連鎖球菌莢膜多糖の複合体(例えば、血清型4、血清型6B、血清型18C、血清型19Fおよび血清型23Fの複合体)を記述する。
【0086】
本発明の目的のために、「アルミニウムベースのアジュバントを実質的に欠く」という用語は、付加的アルミニウムベースのアジュバントを伴わずに3D−MPLを含む等価組成物と比較した場合の本発明の肺炎球菌多糖複合体の免疫原性の任意の低減を引き起こスクリーニング十分なアルミニウムベースのアジュバント(例えば水酸化アルミニウムおよび、特にリン酸アルミニウム)を含有しない組成物を記述する。好ましくは、抗原性組成物は、本質的に3D−MPLから成るアジュバントを含有すべきである。用量当たりで付加されるアルミニウムベースのアジュバントの量は、好ましくは50μg未満、さらに好ましくは30μg未満、さらに好ましくは10μg未満であるべきであり、最も好ましくは本発明の抗原性組成物に付加されるアルミニウムベースのアジュバントは存在しない。
【0087】
本発明の目的のために、アルミニウムベースのアジュバントとともに3D−MPLを含む組成物と比較して、3D−MPLを含む組成物中で肺炎球菌多糖複合体が十分により免疫原性であるか否かの決定は、実施例2に記載されているように確定されるべきである。3D−MPL単独を含む場合に組成物が有意により免疫原性であるか否かの指標として、3D−MPL単独を含む組成物対リン酸アルミニウムアジュバントとともに3D−MPLを含む等価組成物間のGMC IgG濃度(実施例2で確定されるような)の比は、2より大きい、好ましくは5より大きい、さらに好ましくは7より大きい、さらに好ましくは9より大きい、最も好ましくは14より大きい値であるべきである。
【0088】
予防接種への多糖アプローチに関連した問題の1つは、多糖それ自体が貧免疫原性であるという事実である。免疫原性のこの欠如を克服するよう意図された戦略としては、大型免疫原性タンパク質担体との多糖の結合(連結(conjugating))が挙げられるが、これは、バイスタンダーT細胞援助を提供する。本発明の肺炎球菌多糖がバイスタンダーT細胞援助を提供するタンパク質担体に結合されるのが好ましい。用いられ得るこのような担体の例としては、ジフテリア、ジフテリア突然変異体および破傷風毒素(それぞれDT、DT CRM197およびTT)、カギアナカサガイヘモシアニン(KLH)、ツベルクリンの精製タンパク質誘導体(PPD)ならびに髄膜炎菌からのOMPCが挙げられる。
【0089】
最も好ましくは、インフルエンザ菌からのプロテインD(EP 0 594 610-B)またはその断片(節C参照)は、本発明の肺炎球菌多糖のための免疫原性タンパク質担体として用いられる。
【0090】
一実施態様では、本発明の抗原性組成物は、免疫原性タンパク質に連結され、3D−MPLアジュバントを処方された肺炎球菌多糖血清型(PS)4を含むが、この場合、組成物はアルミニウムベースのアジュバントを実質的に欠く。さらなる実施態様では、抗原性組成物は、免疫原性タンパク質に連結され、3D−MPLアジュバントを処方されたそれぞれPS6B、18C、19Fまたは23Fを含むが、この場合、組成物はアルミニウムベースのアジュバントを実質的に欠く。
【0091】
本発明のさらに別の実施態様では、3D−MPLアジュバントを処方されたPS4、PS6B、PS18C、PS19FおよびPS23F群からの2つまたはそれ以上の肺炎球菌多糖複合体を含む組合せ抗原性組成物が提供されるが、この場合、組成物はアルミニウムベースのアジュバントを実質的に欠く。
【0092】
本発明の肺炎球菌多糖複合体の免疫原性は、他の肺炎球菌多糖複合体との組合せにより有意に影響されない(実施例3)。したがって、本発明の好ましい局面は、本発明の1つ又はそれ以上の肺炎球菌多糖複合体を、1つ又はそれ以上のさらに別の肺炎球菌多糖複合体と組合せて含む組合せ抗原性組成物を提供するが、この場合、組成物は3D−MPLアジュバントを処方されるが、しかしアルミニウムベースのアジュバントを実質的に欠いている。
【0093】
本発明のさらなる好ましい実施態様では、少なくとも1つの、好ましくは2、3、4または5つすべてのPS4、6B、18C、19Fまたは23F肺炎球菌多糖複合体を、さらに3D−MPLアジュバントを処方されるがしかしアルミニウムベースのアジュバントを実質的に欠いているその他の肺炎球菌多糖複合体の任意の組合せを含有する組合せ抗原性組成物が提供される。
【0094】
典型的には、本発明の肺炎連鎖球菌組合せ抗原性組成物は多糖複合体抗原を含むが、この場合、多糖は、少なくとも4つ、7つ、11、13、15または23の血清型から得られる(治療される疾患による血清型の好ましい組合せに関する前記の「本発明の肺炎連鎖球菌多糖抗原」参照)。
【0095】
本発明の抗原性組成物は、好ましくは、特に老齢者および乳児および幼児における肺炎球菌感染を予防(または治療)するためのワクチン組成物として用いられる。
【0096】
本発明のさらなる実施態様は、薬剤中に用いるための前記の抗原性組成物の提供;肺炎連鎖球菌莢膜多糖複合体に対する免疫応答の誘導方法であって、安全且つ有効量の前記の抗原性組成物の1つを患者に投与する過程を含む方法;ならびに肺炎球菌性疾患の予防(または治療)のための薬剤の製造における前記の抗原性組成物の1つの使用を包含する。
【0097】
老齢者(+55歳)集団における肺炎ならびに乳児(18ヶ月まで)および幼児(典型的には18ヶ月〜5歳)における中耳炎の予防/改善のためには、本明細書に記載したように処方される多価肺炎連鎖球菌多糖複合体を肺炎連鎖球菌タンパク質またはその免疫学的機能等価物と併用することが、本発明のさらなる好ましい実施態様である。好ましいタンパク質/タンパク質組合せに関しては、前節「本発明の肺炎球菌タンパク質」を参照されたい。
【0098】
好ましくは、本明細書中に前記した抗原性組成物(およびワクチン)は、それらが用いられるまで凍結乾燥され、使用時点で、それらは希釈剤を用いて即座に再構築される。さらに好ましくは、それらは3D−MPLの存在下で凍結乾燥され、食塩溶液で即座に再構築される。
【0099】
組成物を凍結乾燥すると、より安定な組成物が結果的に生じる(例えば、それは多糖抗原の分解を防止する)。その過程は、意外にも、肺炎球菌多糖に対するより高い抗体力価にも関与する。これは、PS6B複合体に関して特に有意であることが示されている。したがって、本発明の別の局面は、3D−MPLで活性調節され、アルミニウムベースのアジュバントを実質的に欠くPS6B複合体を含む凍結乾燥抗原性組成物である。
【0100】
ワクチンの調製に関しては、前記の「本発明のワクチン製剤」の節を参照されたい。
【0101】
C)細菌多糖−プロテインD複合体
組合せワクチンに対する傾向は、レシピエントへの不快を軽減し、予定を立て易くし、レジメンの完了を保証するという利点を有するが、しかしワクチンの効能を低減するという付随する危険性も存在する(担体タンパク質の過剰使用によるエピトープ抑制における考察に関する前記を参照)。したがって、ある集団の要求を満たし、さらにそれらの構成成分間の免疫原性干渉を示さないワクチン組合せを製造することは有益である。これらの利点は、乳児、幼児または老齢者のような高危険群への組合せワクチンの投与に対して特別な利点を有する本発明の免疫原性組成物(またはワクチン)により実現され得る。
【0102】
本発明は、多糖ベースの免疫原性組成物、例えばワクチンのための担体として、インフルエンザ菌からのプロテインDまたはそのだPWを提供する。使用に適した断片としては、Tヘルパーエピトープを包含する断片が挙げられる。特に、プロテインD断片は、好ましくはタンパク質のN末端1/3を含有する。
【0103】
プロテインDは、インフルエンザ菌からのIgD結合タンパク質であり(EP 0 594 610 B1)、潜在的免疫原である。
【0104】
本発明により意図されるプロテインDに連結される多糖としては、腸チフス菌Salmonella typhiに対するVi多糖抗原、髄膜炎菌多糖(A、C、W135およびY型、ならびにB群髄膜炎菌の多糖および修飾多糖を含む)、黄色ブドウ球菌からの多糖、連鎖球菌属のStreptococcus agalactiaeからの多糖、肺炎連鎖球菌からの多糖、ミコバクテリウム属、例えばヒト結核菌Mycobacterium tuberculosisからの多糖(例えば、マンノホスホイニシチドトレハロース、ミコール酸、マンノース面冠アラビノマンナン、それからの莢膜およびアラビノガラクタンス)、クリプトコックス属のCryptococcus neoformansからの多糖、非分類可能インフルエンザ菌のリポ多糖、b型インフルエンザ菌からの莢膜多糖、モラクセラ属のMoraxella catharralisのリポ多糖、ゾンネ赤痢菌のリポ多糖、クルーズトリパノソーマのリポペプチドホスホグリカン(LPPG)、癌関連ガングリオシドGD3、GD2、腫瘍関連ムチン、特にT−F抗原およびシアリルT−F抗原、ならびにT−F抗原に構造的に関連するHIV関連多糖が挙げられるが、これらに限定されない。
【0105】
多糖は、既知の方法により、担体タンパク質に結合され得る(例えば、Likhite(米国特許第4,372,945号)およびArmor等(米国特許第4,474,757号)による)。好ましくは、CDAP複合が実行される(WO 95/08348)。
【0106】
CDAPでは、シアニル化試薬1−シアノ−ジメチルアミノピリジニウムテトラフルオロボレート(CDAP)は、好ましくは多糖−タンパク質複合体の合成のために用いられる。シアニル化反応は、アルカリ性感受性多糖の加水分解を回避する相対的に低刺激性条件下で実施され得る。この合成は、担体タンパク質との直接結合を可能にする。
【0107】
多糖は、水または食塩溶液中に可溶化される。CDAPは、アセトニトリル中に溶解されて、多糖溶液に直接付加される。CDAPは多糖のヒドロキシル基と反応して、シアネートエステルを生成する。活性化工程後、担体タンパク質が付加される。しリンのアミノ基は、活性化多糖と反応して、イソウレア共有結合を生成する。
【0108】
カップリング反応後、大余分量のグリシンが次に付加されて、残留活性化官能基をクエンチングさせる。次に生成物をゲル浸透に通して見反応担体タンパク質および残留試薬を除去する。したがって、本発明は、多糖プロテインD複合体の製造方法であって、多糖を活性化し、多糖をプロテインDと結合する工程を包含する方法を提供する。
【0109】
本発明の好ましい実施態様では、肺炎連鎖球菌感染の予防のための免疫原性組成物(またはワクチン)処方物が提供される。
【0110】
肺炎球菌が肺、脳脊髄液および血液に拡がるメカニズムは、十分には分かっていない。正常肺胞に達する細菌の増殖は、それらの相対的乾燥により、そして肺胞マクロファージの食菌活性により抑制される。これらの等位防衛のあらゆる解剖学的または生理学的変化は、感染に対する肺の感受性を増大する傾向がある。肺炎連鎖球菌の細胞壁は、肺の肺胞における炎症性応答の発生において重要な役割を有する(Gillespie et al.(1997), I&I 65:3936)。
【0111】
典型的には、本発明の肺炎連鎖球菌ワクチンはプロテインD多糖複合体を含むが、この場合、多糖は少なくとも4、7、11、13、15または23の血清型から得られる。治療される疾患による血清型の好ましい組合せに関する前記の「本発明の肺炎連鎖球菌多糖抗原」を参照されたい。
【0112】
本発明のさらなる実施態様では、特に血清型A、C、W-135およびYから、髄膜炎菌ワクチンが提供される。髄膜炎菌は、細菌性髄膜炎の最も重要な原因の1つである。これらの生物の炭水化物莢膜は、ビルレンス決定基および防御抗体のための標的として作用する。それにもかかわらず、炭水化物は、低年齢小児において貧免疫原性であることが周知である。本発明は、これらの多糖に特に適したタンパク質担体であるプロテインDを提供するが、これは、多糖抗原特異的B細胞増殖および成熟、ならびに免疫記憶の誘導を補佐するためにT細胞応答を活性化し得るT細胞エピトープを提供する。
【0113】
本発明の代替的実施態様では、b型インフルエンザ菌の莢膜多糖(PRP)−プロテインD複合体が提供される。
【0114】
本発明は、一連の異なる病原体に対する防御を提供する組合せワクチンも意図する。プロテインD担体は、意外にも、多重多糖抗原が連結された組合せワクチン中の担体として有用である。前記のように、エピトープ抑制は、各多糖に関して同一担体が用いられる場合に起こると思われる。WO 98/51339は、DTに関する組成物中のある比率の多糖とTTに関する残りの比率のものを連結することによりこの妨害を最小限にしようとするための組成物を示した。
【0115】
意外なことに、組合せワクチンにおけるこのようなエピトープ抑制作用を最小限にするのにプロテインDは特に適していることを、本発明は見出した。組合せにおける1つ又はそれ以上の多糖は、プロテインDと連結されるのが有益であり、好ましくは、抗原はすべて、このような組合せワクチン内でプロテインDに連結される。
【0116】
好ましい組合せとしては、髄膜炎菌CおよびY(そして好ましくはA)に対する防御をもたらすワクチンが挙げられるが、この場合、1つ又はそれ以上の血清型YおよびC(そして最も好ましくはA)からの多糖抗原はプロテインDに結合される。
【0117】
インフルエンザ菌多糖ベースのワクチン(好ましくはTT、DTまたはCRM197と、あるいは最も好ましくはプロテインDと連結されたPRP)は、前記の組合せワクチンを用いて処方され得る。
【0118】
多数の小児科ワクチンは、目下、小児が受ける注射の回数を低減するために組合せワクチンとして投与されている。したがって、小児科ワクチンに関しては、他の抗原は、本発明のワクチンを用いて処方され得る。例えば本発明のワクチンは、ジフテリア毒素(DT)、破傷風毒素(TT)および百日咳構成成分[典型的には脱毒素価百日咳毒素(PT)、ならびに任意のペルタクチン(PRN)および/またはアグルチニン1+2を有する糸状赤血球凝集素(FHA)]を含む周知の「三価」組合せワクチン、例えばDT、TT、PT、FHAおよびPRN抗原を含有する市販のワクチンINFANRIX-DTPa(SmithKlineBeecham Biologicals)を用いて、あるいは、例えばTritanrixとしてこれもSmithKlineBeecham Biologicalsにより市販されているような全細胞百日咳菌構成成分を用いて処方され、あるいは別々に、しかし同時に投与され得る。組合せワクチンは、その他の抗原、例えばB型肝炎表面抗原(HBsAg)、ポリオウイルス抗原(例えば、不活性化三価ポリオウイルス−IPV)、モラクセラ属のMoraxella catarrhalis外膜タンパク質、非分類可能インフルエンザ菌タンパク質、B型髄膜炎菌外膜タンパク質も含み得る。
【0119】
組合せワクチン(特に中耳炎の予防用)中に含入され得る好ましいMoraxella catarrhalisタンパク質抗原の例を以下に示す:OMP106[WO 97/41731(Antex)およびWO 96/34960(PMC)];OMP21;LbpAおよびLbpB[WO 98/55606(PMC)];TbpAおよびTbpB[WO 97/13785およびWO 97/32980(PMC)];CopB[Helminen ME, et al.(1993)Infect. Immun. 61:2003-2010];UspA1/2[WO 93/03761(テキサス大学)];ならびにOmpCD。組合せワクチン(特に中耳炎予防用)中に含入され得る非分類可能インフルエンザ菌抗原の例としては以下のものが挙げられる:フィンブリンタンパク質[(米国特許第5766608号-Ohio State Research Foundation)]およびそれからのペプチドを含む融合物[例えば、LB1(f)ペプチド融合物;米国特許第5843464号(OSU)またはWO 99/64067];OMP26[WO 97/01638(Cortecs)];P6[EP 281673(ニューヨーク州立大学)];TbpAおよびTbpB;Hia;Hmw1、2;Hap;ならびにD15。
【0120】
本発明により意図される好ましい小児科ワクチンを以下に示す:
a)C型髄膜炎菌多糖複合体およびb型インフルエンザ菌多糖複合体。任意にAおよび/またはY型髄膜炎菌多糖複合体を有するが、但し、少なくとも1つの、好ましくはすべての多糖抗原がプロテインDに連結される。
【0121】
b)DT、TT、百日咳構成成分(望ましいPT、FHAおよびPRN)、B型肝炎表面抗原およびIPV(不活性化三価ポリオウイルスワクチン)を有するa)のワクチン。
【0122】
c)プロテインDに連結された肺炎連鎖球菌多糖抗原。
【0123】
d)モラクセラ属のMoraxella catarrhalisおよび/または非分類可能インフルエンザ菌からの1つ又はそれ以上の抗原を有するc)のワクチン。
【0124】
前記の組合せワクチンはすべて、担体としてのプロテインDの含入により利益を蒙り得る。明らかに、組合せワクチン(例えばエピトープ抑制を克服するために)中に包含される担体が多いほど、最終ワクチンはより高価で複雑になる。したがって、プロテインDに連結される組合せワクチンの多糖抗原のすべてのまたは大多数を有ると、かなりの利点を提供する。
【0125】
老齢者(+55歳)集団における肺炎ならびに乳児または幼児における中耳炎の予防のためには、本明細書に記載したような多価肺炎球菌多糖−プロテインD抗原を肺炎連鎖球菌タンパク質またはその免疫学的機能等価物と併用することが、本発明の好ましい実施態様である。このような組合せ中に含入され得る好ましいタンパク質/タンパク質組合せに関しては、前記の「本発明の肺炎球菌タンパク質」の節を参照されたい。
【0126】
したがって、本発明は、肺炎連鎖球菌多糖−プロテインD複合体および肺炎連鎖球菌タンパク質抗原を含む免疫原性組成物を提供する。
【0127】
本発明の多糖−プロテインD複合体抗原は、好ましくは、本発明のワクチン処方物中で活性調節される。適切なアジュバントとしては、水酸化アルミニウムゲル(アルム)またはリン酸アルミニウムのようなアルミニウム塩が挙げられるが、しかしカルシウム、鉄または亜鉛の塩でもあり得るし、あるいはアシル化チロシンまたはアシル化糖、陽イオン的または陰イオン的誘導化多糖またはポリホスファゼンの不溶性懸濁液であり得る。
【0128】
老齢者ワクチンに関しては、アジュバントはTH1型の応答の望ましい誘導物質であるよう選択されるのが好ましい。
【0129】
特定のTh1アジュバントに関しては、前記の「本発明のTh1アジュバント」を参照されたい。
【0130】
本発明のさらなる局面では、医療に用いるための本明細書中に記載したような免疫原またはワクチンが提供される。
【0131】
ワクチン製剤/複合体の投与に関しては、前記の「本発明のワクチン製剤」を参照されたい。
【0132】
プロテインDは、基本的には、非分類可能インフルエンザ菌(ntHi)に対するB細胞媒介性防御を生じ得る免疫原であるので、中耳炎に対するワクチン中にも有益に用いられる。ntHiは、宿主細胞を侵襲し、タンパク質抗原により誘導されるB細胞媒介性作用を回避する。中耳炎ワクチンのための抗原としてのプロテインDの有効性(それ自体または多糖のための担体としての)を増大する方法を、本発明は意外にも見出した。これは、免疫系の細胞媒介性部門がプロテインDに対して最適化されるよう、強力なTh1応答が被験者において誘発されるようにプロテインDを活性調節することによりなされる。これは、意外にも、投与直前に再構築される、プロテインDおよびTh1アジュバント(好ましくは3D−MPL)を含む凍結乾燥化組成物を用いて成し遂げられる。したがって、本発明は、このような組成物、このような組成物の製造方法(プロテインDおよびTh1アジュバントを含む混合物を凍結乾燥することによる)、ならびに中耳炎の治療におけるこのような組成物の使用も提供する。
【0133】
広義では、本発明は、Th1アジュバント(「本発明のTh1アジュバント」参照)、好ましくは3D−MPLの存在下での免疫原の凍結乾燥は、一般に、免疫原に対するTh1免疫応答を増大する、ということを意図する。したがって、本発明は、より強力なTh1免疫応答が必要とされるあらゆる免疫原に適用可能である。このような免疫原は、細菌、ウイルスおよび腫瘍タンパク質抗原、ならびに自己タンパク質およびペプチドを包含する。
【実施例】
【0134】
実施例
実施例で本発明を説明するが、本発明はそれらに限定されない。
【0135】
実施例1
肺炎連鎖球菌莢膜多糖:
11価候補ワクチンとしては、本質的にはEP 72513に記載されたように作製された莢膜多糖血清型1、3、4、5、6B、7F、9V、14、18C、19Fおよび23Fが挙げられる。各多糖は、CDAP化学作用を用いて活性化され、誘導されて(WO 95/08348)、タンパク質担体と連結される。多糖はすべて、血清型3(その粘度を減少させるためにサイズ低減される)以外は、それらのネイティブ形態で連結される。
【0136】
タンパク質担体:
選択されるタンパク質担体は、大腸菌中で発現された非分類可能インフルエンザ菌からの組換え体プロテインD(PD)である。
【0137】
プロテインDの発現
インフルエンザ菌プロテインD
プロテインD発現のための遺伝子構築
出発物質
プロテインDコードDNA
プロテインDは、すべての血清型および非分類可能菌株のインフルエンザ菌の間で高度に保存される。全プロテインD遺伝子をコードするDNA配列を含有するベクターpHIC348は、A. Forsgren博士(Department of Medical Microbiology, University of Lund, Malmo General Hospital, Malmo, Sweden)から入手した。プロテインDのDNA配列は、Janson et al.(1991)Infect. Immun. 59:119-125により発表されている。
【0138】
発現ベクターpMG1
発現ベクターpMG1は、外来挿入遺伝子の転写および翻訳のためのバクテリオファージλ由来対照素子が導入されたpBR322(Gross et al., 1985)の誘導体である(Shatzman et al., 1983)。さらに、アンピシリン耐性遺伝子をカナマイシン耐性遺伝子と交換した。
【0139】
大腸菌AR58菌株
SA500誘導体(galE::TN10, λKil-cI857△H1)上で予め増殖させたP1ファージストックを用いたN99の形質導入により、大腸菌AR58菌株を生成した。N99およびSA500は、Martin Rosenberg博士の実験室(National Institute of Health)から得られた大腸菌K12菌株である。
【0140】
発現ベクターpMG1
プロテインDの製造のために、そのタンパク質をコードするDNAを発現ベクターpMG 1中でクローン化した。このプラスミドは、λファージDNAからのシグナルを利用して挿入外来遺伝子の転写および翻訳を駆動する。ベクターは、プロモーターPL、オペレーターOLおよび2つの利用部位(NutLおよびNutR)を含有して、Nタンパク質が提供された場合の転写性極性作用を軽減する(Gross et al., 1985)。PLプロモーターを含有するベクターを大腸菌溶原性宿主中に°にして、プラスミドDNAを安定化する。溶原性宿主菌株は、ゲノム中に組み込まれる複製欠陥λファージDNAを含有する(Shatzman et al., 1983)。染色体λファージDNAは、ベクターのOLリプレッサーと結合し、PLプロモーターとのRNAポリメラーゼの結合を阻止し、それにより挿入遺伝子の転写を阻止するcIリプレッサータンパク質の合成を指図する。発現菌株AR58のcI遺伝子は、PL特異的転写が温度シフトにより調節され得るように、即ち、培養温度の増大がリプレッサーを不活性化し、外来タンパク質の合成が開始されるように、温度感受性突然変異体を含有する。この発現系は、特に細胞に対して毒性であり得るものの外来タンパク質の合成の制御を可能にする(Shimataka & Rosenberg, 1981 )。
【0141】
大腸菌AR58菌株
プロテインD担体の産生に用いられるAR58溶原性大腸菌株は、標準NIH大腸菌K12株N99の誘導体(F-su-galK2、lacZ-thr-)である。それは、欠陥溶原性λファージ(galE::TN10、λKil-cI857△H1)を含有する。Kil-表現型は、宿主高分子合成の遮断を阻止する。cI857突然変異は、cIリプレッサーに温度感受性損傷を付与する。△H1欠失は、λファージ正当オペロンおよび宿主bio、uvr3およびchlA遺伝子座を除去する。SA500誘導体(galE::TN10, λKil-cI857△H1)上で予め増殖させたP1ファージストックを用いたN99の形質導入により、大腸菌AR58菌株を生成した。隣接galE遺伝子中のエト等サイクリン耐性をコードするTN10トランスポゾンの存在によりテトラサイクリンを用いて、欠陥リソゲンのN99中への導入を選択した。
【0142】
ベクターpMGMDPPrDの構築
インフルエンザウイルスの非構造S1タンパク質をコードする遺伝子を含有するpMG1ベクター(pMGNSI)を用いて、pMGMDPPrDを構築した。それぞれ5‘および3’末端にNcoIおよびXbaI制限部位を含有するPCRプライマーを用いて、pHIC348ベクター(Janson et al. 1991)からのPCRにより、プロテインD遺伝子を増幅した。次に、 NcoIおよびXbaI間のpMGNS1中にNcoI/XbaI断片を導入し、したがってNS1タンパク質のN末端81アミノ酸を含有する融合タンパク質を、その後、PDタンパク質を作製した。このベクターを、pMGNS1PrDと名付けた。
【0143】
前記の構築物に基づいて、プロテインD発現のための最終構築物を生成した。pMGNS1PrDからBamHI/BamHI断片を除去した。このDNA加水分解は、最初の3つのN末端残基以外のNS1コード領域を除去する。ベクターの再結紮時に、その後のN末端アミノ酸配列を有する融合タンパク質をコードする遺伝子が生成された。
‐‐‐‐MDP SSHSSNMANT‐‐‐‐
NS1 プロテインD
プロテインDは、普通は脂質鎖が結合されるリーダーペプチドまたはN末端システインを含有しない。したがって、そのタンパク質は、ペリプラズムに排出されないし、脂質化もされず、可溶性形態で細胞質中に残存する。
【0144】
37℃での熱ショックにより、AR58宿主菌株中に最終構築物pMG−MDPPrDを導入した。カナマイシンの存在下で、プラスミド含有細菌を選択した。選定エンドヌクレアーゼによる単離プラスミドDNAの消化によって、プロテインDコードDNA挿入物の存在を実証した。組換え体大腸菌株をECD4と呼ぶ。
【0145】
プロテインDの発現は、λPLプロモーター/OLオペレーターの制御下にある。宿主菌株AR58は、OLとの結合により低温でλPLからの発現を遮断するゲノム内の温度感受性cI遺伝子を含有する。温度を上げると、cIはOLから放出され、プロテインDが発現される。発酵終了時に、細胞を濃縮し、凍結する。
【0146】
収穫細胞からの抽出およびプロテインDの精製を、以下のように実施した。凍結細胞培養ペレットを解氷し、細胞崩壊溶液(クエン酸塩緩衝液、pH6.0)中に再懸濁して、最終OD650=60とする。P=1000 barで高圧ホモジナイザーに懸濁液を2回通す。遠心分離により細胞培養ホモジネートを清澄化し、濾過により細胞破砕屑を除去する。一次精製過程において、濾過溶解物を陽イオン交換クロマトグラフィーカラム(SP Sepharose Fast Flow)に適用する。PDはイオン相互作用によりゲルマトリックスと結合するが、それを、溶離緩衝液のイオン強度の段階的増大により溶離する。
【0147】
二次精製過程では、不純物を陰イオン交換マトリックス(Q Sepharose Fast Flow)上に保持する。PDはゲル上に結合せず、フロースルーで収集され得る。
【0148】
両カラムクロマトグラフィー過程においては、ODにより分画収集をモニタリングする。精製プロテインDを含有する陰イオン交換カラムクロマトグラフィーのフロースルーを、限外濾過により濃縮する。
【0149】
プロテインD含有限外濾過保持物を最後に0.2μm膜に通す。
【0150】
化学的性質:
活性化およびカップリング化学作用:
活性化およびカップリング条件は、各多糖に特異的である。これらを表1に示す。注射のために、ネイティブ多糖(PS3以外)を2 MのNaCl中または水中に溶解した。すべての血清型に関して、最適多糖濃度を評価した。
【0151】
アセトニトリル中の100 mg/mlストック溶液から、CDAP(CDAP/PS比0.75 mg/mg PS)を多糖溶液に付加した。1.5分後、0.2 Mのトリエチルアミンを付加して、特異的活性化pHを得た。25℃で2分間、このpHで多糖の活性化を実施した。プロテインD(量は初期PS/PD比によっている)を活性化多糖に付加し、特異的pHで1時間、カップリング反応を実施した。次に、25℃で30分間そして4℃で一夜、グリシンを用いて反応を止めた。
【0152】
0.2 MNaClで平衡させたセファクリル500HRゲル濾過カラムを用いてゲル濾過により、複合体を精製した。
【0153】
溶離分画の炭水化物およびタンパク質の含量を確定した。複合体をプールし、0.22μm滅菌膜上で滅菌濾過した。複合体調製物中のPS/タンパク質比を確定した。
【0154】
特性化:
各複合体を特性化して、表2に記載した内訳に合した。レソルシノール検定により多糖含量(μg/ml)を、Lowry検定によりタンパク質含量(μg/ml)を測定した。その濃度比により、最終PS/PD比(w/w)を確定する。
【0155】
残留DMAP含量(ng/μgPS):
CDAPによる多糖の活性化は、多糖中にシアネート基を導入し、DMAP(4−ジメチルアミノ−ピリジン)を遊離する。SBで開発された特異的検定により、残留DMAP含量を確定した。
【0156】
遊離多糖含量(%):
α−PD抗体および飽和硫酸アンモニウムを用いたインキュベーションおよびその後の遠心分離後に得られた上清に関して、4℃に保持するかまたは37℃で7日間貯蔵した複合体の遊離多糖含量を確定した。
【0157】
上清中の遊離多糖の定量のために、α−PS/α−PS ELISAを用いた。α−PD/αPSELISAにより、複合体の非存在も制御した。遊離多糖の量の低減は、改良型複合ワクチンをもたらした。
【0158】
抗原性:
同一複合体上の抗原性を、サンドイッチ型ELISAで分析したが、この場合、抗体の捕獲および検出は、それぞれα−PSおよびα−PDであった。
【0159】
遊離タンパク質含量(%):
試料のSDS処理による方法を用いて、「遊離」残留プロテインDのレベルを確定した。0.1%SDSの存在下で100℃で10分間、複合体を加熱し、SEC−HPLCゲル濾過カラム(TSK3000−PWXL)上に注入した。プロテインDは二量体であるので、SDSでの構造の解離により「遊離」プロテインDのレベルを過剰刺激する危険性がある。
【0160】
分子サイズ(Kav):
SEC−HPLCゲル濾過カラム(TSK5000−PWXL)上で、分子サイズ分類を実施した。
【0161】
安定性:
4℃に保持するかまたは37℃で7日間貯蔵した複合体に関して、 HPLC−SECゲル濾過カラム(TSK6000−PWXL)上で安定性を測定した。
【0162】
11価特性化を表2に示す。
【0163】
タンパク質複合体をリン酸アルミニウム上に吸着させて、プールし、最終ワクチンを生成し得る。
【0164】
結論:
有望なワクチンの構成成分であることが示されている免疫原性複合体を生成した。最良品質最終複合肺炎球菌多糖生成物に関する最適CDAP条件を、11価物の各々に関して発見した。前記の改良型(最適化)CDAP法により得られるこれらの肺炎球菌多糖の複合体は(担体タンパク質にかかわらず、しかし好ましくはプロテインD)、したがって、本発明のさらなる局面である。
【0165】
実施例2−乳児ラットにおける11価肺炎球菌PS−PD複合体ワクチンの免疫原性に及ぼす先進アジュバントの作用の研究
0.1μgの各多糖の用量の11価肺炎球菌PS−PD複合体ワクチン(実施例1の方法にしたがって製造)で、以下のアジュバント処方物:なし、AlPO4、3D−MPL、AlPO4上の3D−MPLを用いて、乳児ラットを免疫感作した。
【0166】
3D−MPLのみを有する処方物は、11の抗原のうち5つに関して、他の処方物より統計学的に(そして意外にも)より免疫原性(最大GMC IgG)であった。
【0167】
オプソニン食菌作用は、GMC結果を確証した。
【0168】
材料と方法
免疫感作プロトコール
乳児OFAラットを異なる母親に対して無作為化し、それらが初回免疫感作を施されたときは7日齢であった。それらには、14および28日後にさらに2回の免疫感作を施した。56日目に(第3回免疫感作後28日目)、採血を実施した。ワクチンを皮下注射した。ラットは、10匹/ワクチン群であった。
【0169】
プロテインD上に以下の多糖血清型:1、3、4、5、6B、7F、9V、14、18C、19F、23Fを連結させたものを含む11価肺炎球菌複合ワクチンで、ラットを免疫感作した。
【0170】
処方物
異なる先進アジュバントの作用を調べるために、複合体の用量を0.1μgの各多糖に一定に保持し、アジュバントAlPO4および3D−MPLを、アジュバントを全く含まないものを含めて、異なる用量および組合せで処方した。これらを参考のために表3に数値で列挙する。
【0171】
AlPO4上の吸着
以下の手法により、濃縮吸着一価物を調製した。50μgのAlPO4(pH5.1)を5μgの複合多糖と2時間混合した。pHをpH5.1に調整し、混合物をさらに16時間放置した。1500mMのNaClを付加して、塩濃度を150mMにした。5分後、5 mg/mlの2−フェノキシエタノールを付加した。さらに30分後、pHを6.1に調整して、4℃で3日以上放置した。
【0172】
希釈剤の調製
NaCl150mM/5 mg/mLフェノキシエタノール中で3つの希釈剤を調製した。
【0173】
A:AlPO4、1mg/mlで。
【0174】
B:AlPO4上の3D−MPL、それぞれ1000および250μg/ml。重量比は、3D−MPL/AlPO4=5/20。
【0175】
C:AlPO4上の3D−MPL、それぞれ1000および561μg/ml。重量比は、3D−MPL/AlPO4=50/89。
【0176】
吸着11価物の調製
11の濃縮吸着PS−PD一価物を適切な比率で混合した。AlPO4の補体を希釈剤Aとして付加した。必要な場合、水性溶液(非吸着価。下記の方法1参照)として、あるいは希釈剤BまたはC(AlPO4上に2用量で吸着された3D−MPL。下記参照)として、3D−MPLを付加した。
【0177】
方法1
3D−MPLを、水性懸濁液として併合吸着複合体に付加した。それを室温で10分間、11価物に混合して、投与まで4℃で貯蔵した。
【0178】
方法2
3D−MPLをAlPO4上に予備吸着させた後、併合吸着複合体に付加した(希釈剤BおよびC)。1 mlの希釈剤を調製するために、3D−MPL(250または561 μg)の水性懸濁液を、室温で5分間、150 mMのNaCl、pH6.3中の1 mgのAlPO4と混合した。この溶液をNaCl、pH6.1/フェノキシ中に稀釈し、4℃で一夜インキュベートした。
【0179】
非吸着化11価物の調製
11のPS−PD複合体を混合し、正しい比率で150 mMのNaCl、pH6.1、フェノキシ中に稀釈した。必要な場合には、3D−MPLを溶液(非吸着化)として付加した。
【0180】
すべての注射用の処方物は、初回投与の18日前に調製した。
【0181】
ELISA
ヒト血清中の肺炎連鎖球菌莢膜多糖に対するIgG抗体の定量のためのELISA手法に関してWHOワークショップから得られたプロトコールを用いて、ELISAを実施し、ラットIgGを測定した。基本的に、精製莢膜多糖は微小滴定プレート上に直接被覆される。血清試料を、すべての肺炎球菌に共通の、そして開示(EP 72513 B1)にしたがって精製される肺炎球菌多糖中に焼く0.5%で存在する細胞壁多糖(物質C)とともに予備インキュベートする。Jackson ImmunoLaboratories Inc.試薬を用いて、結合ネズミIgGを検出した。滴定曲線を、数学論理的対数方程式によりモデル化された内部標準(モノクローナル抗体)に参照した。SoftMax Proソフトウェアを用いて計算を実施した。これらの結果に関する最大絶対誤差は、2の因数内であると予測した。相対誤差は30%未満である。
【0182】
オプソニン食菌作用
精製ヒトPMNおよび嬰児ウサギ補体を用いたCDCプロトコール(分化HL60細胞を用いた肺炎連鎖球菌オプソニン食菌作用。バージョン11)を用いて、血清型3、6B、7F、14、19Fおよび23Fに関してオプソニン力価を決定した。修正は社内肺炎球菌株の使用を含み、食菌性HL60細胞を精製ヒト好中球PMNに置き換えた(これらの食菌性細胞間には高度の相関が認められる)。さらに、3 mmガラスビーズを微小滴定ウエルに付加して、混合を増大し、これは、400であるべきと推奨された食細胞:細菌比の低減を可能にした。
【0183】
結果
IgG濃度
すべての血清型およびPDに関して確定されたゲノム平均IgG濃度を表4〜10に示す。血清型6B、14、19Fおよび23Fに関しては、4価処方物を用いて得られた前の結果を比較のために含める。
【0184】
最高IgG濃度を、表4〜10で強調した。3D−MPL組成物体3D−MPL/AlPO4組成物に関する統計的p値は、表11に示してある。アジュバント処方物4(高用量3D−MPLを有する非吸着化複合体)は、11例中9例で最高GMCを示す。5/11例では、低用量のMPLは、第二の最大免疫原性物である。さらに、アジュバント活性化は、全血清型に関して用量の修正によるより高いGMCを示し(データは示されていない)、これは血清型4、6B、7F、18Cおよび23Fに関しては統計学的に有意である(p<0.05。95%CIから)。
【0185】
オプソニン食菌作用
プール化血清に関するオプソニン食菌作用結果を、血清型3、6B、7F、14、19Fおよび23Fに関して表4〜8に示す。ほとんどの部分に関して、これらのオプソニン力価は、GMC IgGを確証する。実際、IgG濃度との相関は、血清型6B、19F、23Fに関しては85%より大きい(データは示されていない)。血清型3に関しては、3D−MPL群のみがオプソニン活性を閾値より上に誘導した、ということに留意することは重要である。
【0186】
結論
この実験では、3D−MPL単独の使用は最高IgG濃度を誘導する、ということは予測されなかった。
【0187】
アジュバントを修正することにより得られた最大GMCを、PS用量を修正することにより得られた最大GMCと比較し、3D−MPLが、5/11血清型で有意に高い応答を誘導し得た、ということを見出した。
【0188】
表11は、3D−MPLおよび3D−MPL/AlPO4組成物を比較した場合(処方過程および3D−MPLの用量を比較)、3D−MPL+AlPO4よりむしろ3D−MPLだけを用いて処方した場合、免疫原性に関して、多糖複合体のうちの5つが有意に改良されるということを示す:PS4、PS6B、PS18C、PS19FおよびPS23F。
【0189】
実施例3−成体ラットにおけるPS4、PS6B、PS18C、PS19FおよびPS23F複合体の免疫原性に及ぼす組合せの作用の研究
別々に、または多価組成物(4−、5−、7−または10価物)中に併合した肺炎球菌多糖−プロテインD複合体ワクチンを用いて、成体ラットを免疫感作した。ラット10匹の群を28日置いて2回免疫感作して、28日目および42日目(2回目投与の14日後)に被験血液を採取した。
【0190】
肺炎球菌多糖に対するIgG抗体に関して、ELISAにより血清を検査した。ELISAにより測定した場合、複合体はすべて、特異的IgG抗体を誘導した。表12は、2回目投与後14日目にIgG濃度により測定した場合の、成体ラットにおけるそれらの免疫原性に及ぼす1価PS6B、PS18C、PS19FおよびPS23Fの組合せの作用を示す。
【0191】
すべての試料に関して統計分析を実施して、組合せ時の抗体濃度の差が有意であるか否かを確定した。多価ワクチン中の血清型PS6B、PS18C、PS19FおよびPS23FプロテインD複合体の組合せはいずれも、それらの免疫原性を有意に変えなかった。
【0192】
【表1】
【0193】
【表2】
【0194】
【表3】
【0195】
【表4】
【0196】
【表5】
【0197】
【表6】
【0198】
【表7】
【0199】
【表8】
【0200】
【表9】
【0201】
【表10】
【0202】
【表11】
【0203】
【表12】
【0204】
実施例4−マウスにおける肺炎球菌肺コロニー形成に対するPD−複合11価多糖ワクチンの防御的有効性に及ぼすニューモリジンおよび3D−MPLの付加の有益な影響
免疫学的解読
ニューモリジン特異的血清IgGのELISA投与
PBS中に稀釈した100μl/ウエルの2μg/ml組換え体ネイティブニューモリジン(PLY)を用いて、37℃で2時間、Maxisorp Nuncイムノプレートを被覆した。プレートを0.9%NaCl、0.05%トゥイーン20緩衝液で3回洗浄した。次に、標準曲線として付加された(670 ng/mlIgGで出発)抗PLY血清参照の連続2倍稀釈液(PBS/0.05%トゥイーン20、100μl/ウエル)および血清試料(1/10稀釈液で出発)を、撹拌しながら20℃で30分間インキュベートした。前記と同様に洗浄後、 PBS/0.05%トゥイーン20中に5000x稀釈したペルオキシダーゼ−複合ヤギ抗マウスIgG(Jackson)を、撹拌しながら20℃で30分間インキュベートした(100μl/ウエル)。洗浄後、100μl/ウエルの再ベール化緩衝液(100mM、pH4.5クエン酸塩緩衝液中OPDA0.4 mg/mlおよびH2O20.05%)とともに、室温で15分間、プレートをインキュベートした。50μl/ウエルの1 NHClを付加することにより、再ベール化を停止した。Emaxイムノリーダー(Molecular Devices)を用いて、490および620 nmで光学濃度を読み取った。SoftMaxProソフトウェアを用いた4パラメーター数学法により、抗体力価を算定した。
【0205】
溶血抑制
ニューモリジン(PLY)溶血活性を抑制する血清抗体の能力を測定するために、この検定を実行した。コレステロール(PLYとの相互作用に感受性)を排除するために、血清試料に以下の処置を2回施した:それらを1等容量のクロロホルムと混合し、次に撹拌しながら45分間インキュベートした。1000 rpmで10分間遠心分離後に上清を収集した。コレステロール消去血清を、96ウエル微小プレート(Nunc)中で稀釈した(1 mMジチオトレイトール、0.01%BSA、15 mMトリス、150 mMNaCl、pH7.5中の連続2倍稀釈液)。4 HU(溶血単位)のPLYを含有する50μl溶液を各ウエル中に付加し、37℃で15分間インキュベートした。次に、100μlのヒツジ赤血球(1%溶液)を、37℃で30分間、付加した。1000 rpmで10分間遠心分離後、上清(150μl)を収集し、405 nmでの光学密度読み取りのために別の96ウエル微小プレート中に入れた。結果は、中点稀釈力価として表した。
【0206】
ニューモリジン化学的脱毒素化
組換え体ネイティブニューモリジン(PLY)を50 mMリン酸塩、500 mMNaCl、pH7.6緩衝液に対して透析した。時々撹拌しながら、39.5℃で以下の全工程を実行した。1日目に、10%トゥイーン80(1/250 v/v)、57.4 mMのN−アセチルトリプトファン、pH7.6(3/100 v/v)、リン酸塩緩衝液中の2.2 Mグリシン(1/100 v/v)およびリン酸塩緩衝液中の10%ホルムアルデヒド(3/100 v/v)をPLY溶液中に付加した。2および3日目に、10%ホルムアミドを、それぞれ3/100および2/100 v/v比で再び付加した。時々撹拌しながら、7日目まで、39.5℃でインキュベーションを持続した。最後に、50 mMリン酸塩、500 mMNaCl、pH7.6緩衝液に対してPLYを透析した。溶血検定において、PLYの完全不活性化を実証した。
【0207】
OF1マウスにおける肺炎球菌鼻腔内チャレンジ
5.105 CFUのマウス適合化肺炎連鎖球菌血清型6Bを、麻酔下で7週齢OF1雌マウスに鼻腔内接種した。接種後6時間目に肺を切除し、Todd Hewithブロス(THB、Gibco)培地中で均質化した(Ultramax, 24000 rpm、4℃)。肺ホモジネートの連続10倍稀釈液を、37℃で一夜、酵母菌抽出物補充THB寒天を含入するペトリ皿上にプレート化した。肺炎球菌肺感染をCFU数/マウスとして確定し、対数計量平均として表した。検出限界は2.14 log CFU/マウスであった。
【0208】
実施例4A.抗ニューモリジン免疫応答に及ぼす3D−MPLアジュバント作用
本実施例では、ネイティブ組換え体ニューモリジン(PLY、J. Paton, Children's Hospital, North Adelaide, Australiaにより提供された)およびその化学的脱毒素化対応物(DPLY)に対する免疫応答に及ぼす3D−MPLアジュバント活性化の影響を、我々は評価した。前記と同様に化学的脱毒素化を実行した。
【0209】
0、14および21日目に、A:100μgのAlPO;またはB:100μgのAlPO +5μgの3D−MPL(3デ−O−アシル化モノホスホリル脂質A、供給元Ribi Immunochem)中に含入された1μgのPLYまたはDPLYを用いて、10匹の雌6週齢Balb/cマウスの群を筋肉内免疫感作した。図1Aおよび1Bは、第3回血清後に測定したELISA IgGおよび溶血抑制力価(HLI)
どの抗原でも、3D−MPL補充処方物を接種した動物において最良免疫応答を誘導した。興味深いことに、DPLYは、AlPO4+3D−MPLとともに投与した場合、PLYと同様に免疫原性であったが、一方、ALPO4処方物中では免疫原であった。これは、脱毒素化ニューモリジンに対する抗体応答を改良する3D−MPLの有益な能力を示した。
【0210】
ニューモリジンを含有する組成物では、突然変異的脱毒素化ニューモリジンよりむしろ化学的脱毒素化ニューモリジンを用いるのが望ましい。これは、今日までに得られた脱毒素化突然変異体が依然として残留毒素活性を有するためであり、化学的脱毒素化ニューモリジンは残留毒素活性を有さない。したがって、本発明の別の局面では、概して、ワクチン中に用いるために化学的脱毒素化されたニューモリジンを含む組成物は、Th1アジュバント、好ましくは3D−MPLで活性調節されるべきである、と考えられる。本発明は、このような組成物を提供する。組成物にTh1アジュバント(好ましくは3D−MPL)を付加する過程を包含する免疫原性組成物内の化学的脱毒素化ニューモリジンの免疫応答の増大方法も意図される。
【0211】
実施例4B.血清型6Bを鼻腔内チャレンジさせたOF1マウスにおける肺炎球菌肺コロニー形成に対するPD−複合11価多糖ワクチンの防御的有効性に及ぼすニューモリジンおよび3D−MPLアジュバントの弱毒化突然変異体の付加の有益な影響
本実施例では、11価多糖−プロテインD複合体、弱毒化突然変異体ニューモリジン抗原(PdB、WO 90/06951)およびAlPO4+3D−MPLアジュバントを含有するワクチンの予防的効能を、古典的AlPO4吸着11価多糖−プロテインD複合体処方物と比較して、我々は評価した。
【0212】
0および14日目に、A:50μgのAlPO;B:0.1μgPS/血清型のPD複合11価多糖ワクチン+50μgのAlPO4;またはC:0.1μgPS/血清型のPD複合11価多糖ワクチン+10μgのPdB(J. Paton, Children's Hospital, North Adelaide, Australia提供)+50μgのAlPO4+5μgの3D−MPL(供給元Ribi Immunochem)を含有する処方物を用いて、12匹の雌4週齢OF1マウスの群を皮下免疫感作した。21日目に、前記と同様にチャレンジを実行した。
【0213】
図1Cに示したように、PdBを補充し、AlPO4+MPLで活性調節した11価多糖複合ワクチンにより、非常に有意の防御(p<0.007)が付与された(黒棒は算術平均を表す)。これに反して、11価多糖複合体/AlPO4処方物で免疫感作した動物においては、有意の防御は観察されなかった。この結果は、ニューモリジン抗原(弱毒化されていても)および3D−MPLアジュバントの付加が肺炎に対する11価多糖複合ワクチンの有効性を増大することを立証した。
【0214】
実施例4C.実施例4Bに示した防御の免疫相関物
弱毒価突然変異体ニューモリジン(PdB)および3D−MPLを補充した11価多糖複合体ワクチンにより、実施例4Bで付与された防御の免疫相関物を確立するために、多糖6BおよびPdBに対するプレチャレンジ血清学的抗体応答を前記と同様に測定した。
【0215】
次に、抗体力価を、チャレンジ後6時間目に収集した対応する動物の肺で測定された細菌コロニー数と比較した。Log/Log線状回帰に関してR2を算定した。
【0216】
算定R2は、抗PdBおよび抗6B抗体応答に関してそれぞれ0.18および0.02であった。これは、体液性免疫応答と両抗原に対する防御との間の相関の非存在を示した。抗6B抗体力価は、11価複合体ワクチン(GMT=0.318 ng/ml)で免疫感作した群またはPdDおよび3D−MPLを補充した同一ワクチン(GMT=0.458 ng/ml)で免疫感作した群において有意に異ならなかった。したがって、処方物Cで観察された防御改善は、単に多糖6Bに対する高抗体応答によるものではなかった。
【0217】
合わせて考えると、防御は体液性免疫応答単独により媒介されるだけでなく、むしろ3D−MPLの存在下でPdBにより誘導された細胞媒介性免疫によっても媒介された、ということを結果は示唆する。これは、最適防御のための免疫系の両部門を対等にするよう、肺炎球菌多糖複合体ワクチン中のタンパク質抗原(単数または複数)および効力のあるアジュバント(単数または複数)の付加に対してさらなる支持を与えた。
【0218】
実施例5−ニューモリジンで能動的免疫感作され、肺炎球菌PSに対する抗体で受動的免疫感作されたマウスにおける免疫系の両部門の共働
実施例5A−肺炎に対して防御する受動的投与抗6B多糖(抗PS)抗体の濃度を見出す
方法
ワクチン群:下記に詳述した群により(マウス計64匹)、マウス16匹の4群を、−1日目に100μlの非稀釈ラット抗多糖抗血清で受動的免疫感作(腹腔内)させた。
【0219】
【表13】
【0220】
動物:Charles River, Canadaからの体重約35 gの64匹の雄CD−1マウス(約10週齢)。
【0221】
麻酔:イソフルラン(3%)+O2(1 L/分)でマウスを麻酔した。
【0222】
生物:肺炎連鎖球菌N1387(血清型6)を、5%ウマ血液を補充したトリプチカーゼ大豆寒天プレート(TSA)から収穫し、PBS6 ml中に懸濁した。感染直前に、1 ml細菌懸濁液を9 mlの冷却溶融栄養寒天(BBL)中に稀釈して、41℃に保持した。マウスに容量50 ul中の約6.0 log10 cfu/マウスを投与した。
【0223】
感染:0日目に、前記のようにマウスを麻酔して、非外科的気管内挿管による気管支内点滴注入により、肺炎連鎖球菌N1387(50μl冷却細菌懸濁液)を感染させた。この方法は、WoodnutとBerry(Antimicrob. Ag. Chemotherap. 43:29(1999))により記載された。
【0224】
試料:感染後3日目に、マウス8匹/群をCO2過剰投与により屠殺して、肺を切除し、1 mlPBS中で均質化した。10倍連続稀釈液をPBS中で調製して、生育可能細菌数を数え上げた。5%ウマ血液を補充したTSAプレート上に3通り、試料を接種(20μl)し、評価前に37℃で一夜インキュベートした。さらに別の組のマウスを7日目に屠殺して、前記と同様に試料採取した。
【0225】
結果:
【0226】
【表14】
【0227】
結論:概して、あらゆる処置群から単離した細菌数に有意差は認められなかった。これは、測定可能防御が5μg/ml以下の濃度の抗多糖によりもたらされたことを示す。
【0228】
これは、いくつかのヒト臨床試験で観察されるものと同様であり、即ち、抗多糖体はいくつかの集団においては肺炎球菌性肺炎に対して防御するのに不十分である。
【0229】
実施例5B−アジュバントを用いた場合と用いない場合とのPly(ニューモリジン)の能動的投与および亜最適抗PS抗体との共働作用によりもたらされる肺炎からの防御を確定する
方法
動物:6週齢時体重約20 g、10週齢時約38 gの、Charles River, St. Quebec, Constant, Canadaからの128匹の雄CD−1マウス(免疫感作時6週齢、感染時10週齢)。
【0230】
免疫感作:マウス16匹の6群を、-22日目および-14日目に皮下注射により下記に詳述するワクチン100 ulで免疫感作した(マウス計128匹)。PdB(WO 90/06951)は、オーストラリアのJames Paton博士の御好意により入手した。3D−MPLは、Ribi/Corixaから入手した。
【0231】
-1日目に、特定群(下記の表参照)を、濃度4.26μg/ml(4 mlの5μg/ml+1.3 mlの2μg/ml)のマウス抗多糖抗体で受動的免疫感作(腹腔内100 μl)した。
【0232】
【表15】
【0233】
感染:0日目に、マウスを麻酔した(3%イソフルラン+1 L/分O2)。肺炎連鎖球菌N1387(血清型6)の増殖物を、5%ウマ血液を補充したトリプチカーゼ大豆寒天プレート(TSA)から収穫し、PBS6 ml中に懸濁することにより細菌接種物を調製した。感染直前に、10倍稀釈液(1 ml+9 ml)を冷却溶融栄養寒天(41℃に保持)中に調製した。気管内挿管による気管支内点滴注入によりマウスを感染させて、容量50 ul中の約6.0 log10 cfu/マウスを投与した。この方法は、WoodnutとBerry(Antimicrob. Ag. Chemotherap. 43:29(1999))により記載されていた。
【0234】
試料:感染後72時間目に、マウス8匹/群をCO2過剰投与により屠殺して、肺を切除し、1 mlPBS中で均質化した。10倍連続稀釈液をPBS中で調製して、生育可能細菌数を数え上げた。5%ウマ血液を補充したTSAプレート上に3通り、試料を接種(20μl)し、評価前に37℃で一夜インキュベートした。さらに別の組のマウスを感染後8日目に屠殺して、前記と同様に試料採取した。
【0235】
データ分析
処置の比較のための結果測定値は、感染後3および7日目の肺中の細菌数であった。結果は、群平均+標準偏差として示される。スチューデントt検定を用いて統計分析を実施したが、この場合、<0.05のP値を、有意であるとみなした。
【0236】
結果:
感染後72時間
群1−4からの細菌数は、群1−3の場合よりも有意に低かった(p<0.05)。
【0237】
群1−4からの細菌数は、群1−5の場合よりも有意に低かった(p<0.05)。
【0238】
感染後168時間
全群の細菌数は、8日目では3日目より約2 log低かったが、これは感染が消散したことを示す。
【0239】
群1−2からの細菌数は、群1−5の場合よりも有意に低かった(p<0.05)。
【0240】
【表16】
【0241】
前記で実証されたように、抗多糖抗体単独(群1−5)は、肺中の肺炎球菌の増殖に対する防御をもたらさない。AlPO4で活性調節したPdBはいずれも防御を付与しないが、しかし8日目に、PdBを3D−MPLと組合せた場合(群1−2)に、防御傾向が認められる。
【0242】
3日目に、群は最も有意に防御し、群1−4は3つの要素、即ちPdB、3D−MPLおよび受動的投与抗多糖抗体をすべて有した。この結論は、死亡率により支持される。群1−4は死亡は2/8だけであったが、これに比して群1−5および1−3に関しては5/10であった。
【0243】
結論:
受動的免疫感作動物を用いて実験を実施したので、ニューモリジンおよびMPLでの能動的免疫感作の共働作用も、多糖抗原に対する抗体レベルの増大によるとは言えない。
【0244】
肺炎球菌多糖に対して動物を受動的免疫感作しただけであったので、8日目までに、このような抗体のレベルは宿主から大いに消散していた。
【0245】
そのような場合でも、肺炎球菌性肺炎に対する有意の防御は、ニューモリジン+3D−MPLで免疫感作した群において、特に濡森+3D−MPL+受動的投与抗多糖抗体で免疫感作した群において認められたが、これは、この組合せの共働作用を示す。
【0246】
能動的(好ましくは、複合多糖を用いて)に抗多糖免疫感作を実行していた場合、B細胞記憶の作用および一定レベルの抗PS抗体が免疫応答協同作用に寄与していたので、作用はさらに顕著であったと思われる(例えば、図1C参照。この場合、多糖およびタンパク質で能動的免疫感作された動物は、チャレンジ後の肺中に細菌が認められなかったことが示された)。
【0247】
実施例6−3D−MPLで活性調節した11価肺炎球菌多糖プロテインD複合体ワクチンの1年齢Balb/Cマウスにおける免疫原性
序論および対象(単数または複数):
肺炎球菌感染に対する防御は、オプソニン食菌作用を介した血清型特異的抗体により媒介される。抗体濃度の増大はより大きい防御を生じ、したがって体液性応答を増大するための方法の発見に多大の努力が費やされてきた、と推測され得る。前臨床試験においてワクチンを連結するために首尾よく適用されてきた一戦略は、免疫刺激性アジュバントの使用である(Poolman et al. 1998, Carbohydrate-Based Bacterial Vaccines. In:Handbook of Experimental Pharmacology eds. P. Perlmann and H. Wigsell. Springer-Verlag, Heidelberg, Dで再検討されている)。
【0248】
この節に提示したデータは、臨床試験を模倣するよう意図されたプロトコールにおける臨床ロットを用いた最新実験結果を示す。
【0249】
プロトコール:
1/10回のヒト投与量の肺炎球菌多糖プロテインD複合体ワクチン、または23価純多糖ワクチンで、1年齢Balb/Cマウスを免疫感作した。用いたワクチンは、それぞれ1 mcg用量の血清型6Bおよび23Fならびに5 mcgの11価複合ワクチンの残りの血清型、0.1 mdg用量の11価複合ワクチン、または5 mcg3D−MPLで活性調節した0.1 mcg用量の11価複合ワクチンに対応する臨床ロットDSP009、DSP013またはDSP014であった。11価複合ワクチンはすべて、50μgのAlPO4でも活性調節した。
【0250】
マウス20匹の群を筋内免疫感作した。下記の表に列挙した群の注射は、0および21日目に実施した。35日目(第2回投与後14日目)に被験血液を採取した。
【0251】
【表17】
【0252】
CDC/WHO合意プロトコール後、即ち細胞壁多糖による血清の中和後、肺炎球菌多糖に対するIgG抗体に関してELISAにより、血清を検査した。血清型特異的IgG1モノクローナル抗体を用いてELISAを較正して、抗体濃度をmcg/mlで得た。
【0253】
UNISTATバージョン5.0βを用いて、比較の統計分析を算定した。log形質転換化IgG濃度に関して、Tukey-HSD法によるANOVAを実施した。Fisherの精度検定を用いて、血清変換の対方式比較を実施した。
【0254】
結果:
第2回免疫感作(投与2)後14日で誘導された11の血清型およびプロテインDに対するGMC IgGおよび95%信頼区間を、下記の表に示す。血清変換率を示すが、この場合、95%信頼区間は算定され得なかった。
【0255】
群1は、普通は動物においてIgMのみを誘導する純多糖による免疫感作の効果を示す。ほとんどのIgGレベルは検出の閾値より下であった。それにもかかわらず、balb/cマウスは少数の肺炎球菌多糖、特に血清型3、19Fおよび14に対してIgGを作り得た。
【0256】
複合体ワクチンによる免疫感作は、23F以外の全血清型に対して高血清変換率でIgG抗体を誘導した。
【0257】
血清型7Fおよび19Fに関してのみ用量依存性応答(群4対群2)を観察したが、しかしこれらの観察は統計学的に有意でなかった。血清型3、6B、7Fおよび19FならびにPDに関して2回の投与後により大きい応答を観察し、これらの観察は、3つの処方物すべてを有する多くの場合に、統計学的に有意であった。
【0258】
最も興味深いのは、3D−MPLの作用である。3D−MPL処方ワクチン(群3b)の2回の投与は、特異的IgGの最高GMCを誘導し、これは、すべての血清型に関して統計的に有意であったが、但し、23Fの場合は、有意に高い血清変換率を示した(p=0.02,群3b対2b。Fisher精度)。
【0259】
【表18】
【0260】
群の定義に関しては、前記の表を参照されたい。
【0261】
結論:
ここに提示したデータは、11価肺炎球菌多糖プロテインD複合体ワクチンへの3D−MPLの付加が、試験したすべての血清型に対する老齢balb/cマウスにおける免疫応答を増大した、ということを実証する。
【0262】
ほとんどの場合、ワクチンの2回投与は、1回投与より高い幾何学平均IgG濃度を誘導した。これは、ヒトにおいてさえ、純多糖ワクチンを用いて観察されないため、T細胞依存性免疫応答および免疫記憶の誘導の指標とみなされる。
【0263】
これらのデータは、アジュバント(好ましくは3D−MPL)で活性調節した複合肺炎球菌多糖を用いるワクチン投与計画を支持し、それにより、少なくとも2回用量の活性調節化ワクチンが、好ましくは1〜12週おきに、最も好ましくは3週間置きに投与される。このような投与計画は、本発明のさらなる局面とみなされる。
【0264】
実験に用いたマウスは、PS23(純または複合)に対して非応答性であった。興味深いことに、用いたワクチン組成物にかかわらず多糖に対する抗体レベルは低いままであったが、しかし3D−MPLをアジュバントとして用いた場合(血清変換は有意に高い)には、より多数のマウスがPS 23に応答した。ワクチン中の肺炎球菌多糖に対する非応答性を軽減するための複合肺炎球菌多糖を含むワクチン組成物中のTh1アジュバント、特に3D−MPLの使用は、本発明のさらに別の局面である。前記の2回投与計画を用いた前記の組成物による非応答性の軽減方法は、さらに別の局面である。
【0265】
実施例7−C型髄膜炎菌多糖−プロテインD複合体(PSC−PD)
A:プロテインDの発現
実施例1と同様。
【0266】
B:多糖Cの製造
C群多糖の供給源は、髄膜炎菌のC11菌株である。これを、古典的発酵技術(EP 72513)を用いて発酵させる。複合過程に用いられる乾燥粉末多糖は、Mencevax(SB Biologicalsも参照)と同一である。
【0267】
C11菌株のアリコートを解氷し、0.1 mlの懸濁液を、酵母菌抽出透析物(10%、v/v)を補充したMueller Hinton培地ペトリ皿上に画線し、水飽和空気インキュベーター中で36℃で23〜25時間インキュベートする。
【0268】
次に、表面増殖物を滅菌発酵培地中に再懸濁して、この懸濁液を、酵母菌抽出透析物(10%、v/v)を補充したMueller Hinton培地および滅菌ビーズガラスを含入する1つのRouxボトル上に接種する。水飽和空気インキュベーター中で36℃で23〜25時間のRouxボトルのインキュベーション後、表面増殖物を10 mlの滅菌発酵培地中に再懸濁して、0.2〜0.3 mlのこの懸濁液を12の他のMueller Hinton培地Rouxボトル上に接種する。
【0269】
水飽和空気インキュベーター中で36℃で23〜25時間のインキュベーション後、表面増殖物を10 mlの滅菌発酵培地中に再懸濁する。細菌懸濁液を円錐フラスコ中にプールする。
【0270】
次にこの懸濁液を、滅菌注射器を用いて無菌的に発酵器に移す。
【0271】
陰圧下で清浄室中に入れた発酵器中で髄膜炎菌の発酵を実施する。発酵は一般に、約1010個の細菌/mlに対応して約10〜12時間後(即ち早期定常期)に完了され、pH増大により検出される。
【0272】
この段階で、全ブロスを熱不活性化(56℃で12分)した後、遠心分離する。不活性化の前後に、ブロスの試料を採取し、Mueller Hinton培地ペトリ皿上に画線する。
【0273】
C:PS精製
精製方法は、全発酵ブロス上で実施される多段階手法である。精製の第一段階では、遠心分離により不活性培養を清澄化して、上清を回収する。
【0274】
多糖精製は、第四級アンモニウム塩(セチルトリメチルアンモニウムブロミド/CTAB、CETAVLON R)による沈降に基づいている。CTABは、それらのpIによって、多糖、核酸およびタンパク質のような多陰イオンと不溶性複合体を形成する。イオン制御条件後、この方法を用いて、不純物(低導電率)または多糖(高導電率)を沈降させ得る。
【0275】
清澄化上清に含有される多糖を、マトリックスとして珪藻土(セライト(商標)545)を用いて沈降させて、異なる沈降/精製中の不溶性不活性塊の形成を回避する。
【0276】
髄膜炎菌多糖Cに関する精製計画:
工程1:セライト(商標)545上でのPSC−CTAB複合体固定および0.05%CTABを用いた洗浄による細胞破砕屑、核酸およびタンパク質の除去。
【0277】
工程2:50%EtOHによるPSの溶離。混濁した、そして不純物およびLPSを含有する第一分画を廃棄する。以後の分画中のPSの存在を、凝集検定により立証する。
【0278】
工程3:セライト(商標)545上でのPSC−CTAB複合体再固定および0.05%CTAB洗浄による小型核酸およびタンパク質の除去。
【0279】
工程4:50%EtOHによるPSの溶離。第一混濁分画を廃棄する。以後の分画中のPSの存在を、凝集検定により立証する。
【0280】
溶離液を濾過し、粗製多糖を含有する濾液を収集する。エタノールを付加して最終濃度を80%とすることにより、多糖を濾液から沈降させる。次に、多糖を白色粉末として回収し、真空乾燥して、-20℃で貯蔵する。
【0281】
D:CDAP複合化(conjugation)
PSCおよびPDの複合化
PSCおよびPDの複合化のためには、古典的CNBr活性化およびスペーサーを介した担体タンパク質との結合よりCDAP複合技法が好まれる。1−シアノ−4−ジメチルアミノピリジニウムテトラフルオロボレート(CDAP)を用いたシアニル化により、多糖を先ず活性化する。CDAPは水溶性シアニル化試薬であって、この場合、シアノ基の求電子性はCNBrの場合を上回って増大され、シアニル化反応を相対的に低刺激性条件下で実施させる。活性化後、多糖は、いかなるスペーサー分子も導入せずに、そのアミノ基により担体タンパク質に直接結合され得る。未反応エステルシアネート基は、グリシンとの広範な反応によりクエンチングされる。複合体ワクチンの調製に関与する工程の総数は低減され、最も重要なのは、免疫原性である可能性を有するスペーサー分子が最終生成物中に存在しないことである。
【0282】
CDAPによる多糖の活性化は、多糖中にシアネート基を導入し、ジメチルアミノピリジン(DMAP)が遊離される。シアネート基は、その後のカップリング手法中にタンパク質中のNH2基と反応して、カルバメートに転換される。
【0283】
PSC活性化およびPSC−PDカップリング
活性化およびカップリングは+25℃で実施する。
【0284】
120 mgのPSを、少なくとも4時間、WFI中に溶解する。
【0285】
CDAP溶液(アセトニトリル中に新たに調製した100 mg/ml)を付加して、CDAP/PS(w/w)比を0.75とする。
【0286】
1分30秒後、トリエチルアミンの付加によりpHを活性化pHまで上げ、PD付加まで安定化させる。
【0287】
3分30秒時点で、NaClを付加して、最終濃度を2 Mとする。
【0288】
4分時点で、精製PDを付加して、PD/PS比を1.5/1とする。pHを直ちにカップリングpH(pH10)に調整する。溶液を、pH調節下で1時間放置する。
【0289】
クエンチング
6 mlの2 Mグリシン溶液をPS/PD/CDAP混合物に付加する。pHをクエンチングpH(pH8.8)に調整する。溶液を作用温度で30分間撹拌した後、連続的にゆっくり撹拌しながら、+2〜8℃で一夜置く。
【0290】
PS−PD精製
濾過(5μm)後、S400HR Sephacrylゲル上でのゲル浸透クロマトグラフィーにより冷室中でPS−PD複合体を精製して、小型分子(DMAPを含む)を除去し、PD:溶離−150 mMNaCl、pH6.5を再複合化して、UV280 nm、pHおよび導電率をモニタリングする。
【0291】
異なる分子サイズの反応構成成分を基礎にして、PS−PD複合体を先ず溶離し、その後遊離PDを、そして最後にDMAPを溶離する。DMAB(PS)およびμBCA(タンパク質)により検出されるような複合体を含有する分画をプールする。プール分画を滅菌濾過(0.2μm)する。
【0292】
E:PSC−PD吸着複合体ワクチンの処方
AlPO4の洗浄
AlPO4上のPSC−PD複合体の吸着を最適化するために、AlPO4を洗浄して、PO43-濃度を低減する。
【0293】
−AlPO4を150mMNaClで洗浄して、遠心分離(4x)する。
【0294】
−次にペレットを150 mMのNaCl中に再懸濁した後、濾過(100μm)する。そして
濾液を加熱滅菌する。
【0295】
この洗浄AlPO4は、WAP(洗浄オートクレーブ処理リン酸塩)と呼ばれる。
【0296】
配合方法
PSC−PD複合体本体をAlPO4 WAP上に吸着させた後、完成品を最終処方する。AlPO4 WAPをPSC−PDとともに室温で5分間撹拌する。pHを5.1に調整し、混合物を室温で18時間撹拌する。NaCl溶液を付加して150 mMとし、混合物を室温で5分間撹拌する。2−フェノキシエタノールを付加して、5 mg/mlとし、混合物を室温で15分間撹拌した後、pH6.1に調整する。
【0297】
最終組成物/用量
PSC−PD: 10μgPS
AlPO4 WAP: 0.25 mgAl3+
NaCl: 150 mM
2−フェノキシエタノール: 2.5 mg
注射用水: 0.5 mlにする
pH: 6.1
F:前臨床情報
マウスにおける他糖複合体の免疫原性
PSC−PD複合体の免疫原性を、6〜8週齢Balb/cマウスで査定した。純(非吸着)複合体またはAlPO4上に吸着した複合体を、一価ワクチンとして注入した。誘導された抗PSC抗体をELISAにより測定し、一方、殺細菌試験により機能性抗体を分析した。両方法は、CDC(Centers for Disease Control and Prevention, Atlanta, USA)プロトコールを基礎にしている。応答対用量およびアジュバント(AlPO4)作用を査定するために実施した2つの異なる実験からの結果を示す。
【0298】
用量範囲実験
この実験では、Balb/CマウスにPSC−PDを2回(2週間間隔)注射した。4つの異なる用量:0.1;0.5;2.5および9.6μg/動物のAlPO4上処方複合体を用いた。14日目(第1回後14日目)、28日目(第2回目後14日目)および42日目(第2回後28日目)に、マウス(10匹/群)を採血した。ELISAにより測定した多糖C特異的抗体の幾何学平均濃度(GMC)を、参照としてIgGを用いて、μgIgG/mlで表した。殺細菌抗体をプール血清に関して測定し、嬰児ウサギ補体の存在下で髄膜炎菌C11菌株を用いて、細菌の50%を殺害し得る稀釈液の逆数として力価を表した。
【0299】
得られた用量−応答曲線は、2.5μgからプラトーを示す。結果は、第1回後14日目と第2回後14日目の間に良好な追加免疫応答が認められるということを示す。第2回後28日目の抗体レベルは、第2回後14日目のレベルと少なくとも同等である。殺細菌抗体力価はELISA濃度と一致し、PSC−PD複合体の免疫原性を確証する。
【0300】
アジュバントの効果
この実験では、AlPO4上に処方された1ロットのPSC−PD複合体を査定し、純(非吸着)複合体を比較のために注射した。2μgの複合体を用いて、皮下経路により、2週間間隔で、マウス10匹/群に2回注射した。14日目(第1回後14日目)、28日目(第2回目後14日目)および42日目(第2回後28日目)にマウスを採血し、そしてELISAおよび機能性抗体力価を測定した(殺細菌検定に関しては第2回後14日目および第2回後28日目のみ)。AlPO4処方物は、非活性調節処方物と比較した場合、10倍までの抗体力価を誘導する。
【0301】
結論:
前記の実験結果から、以下の全般的結論が得られた:
−PSC−PD複合体は、連結された場合に、PSCがT細胞依存性抗原になるということを実証する既往性応答を誘導する。
【0302】
−ELISAにより測定された抗PSC抗体濃度は殺細菌性抗体力価とよく相関し、このことは、PSC−PD複合体により誘導された抗体が髄膜炎菌血清型Cに対して機能的であることを示す。
【0303】
−AlPO4上に吸着された複合体約2.5μgは、マウスにおいて最適抗体応答を引き出すと思われる。
【0304】
−CDAP化学作用は、免疫原性PSC−PD複合体を製造するための適切な方法であると思われる。
【0305】
実施例8−髄膜炎菌血清型A−PD複合体からの多糖の調製
多糖A(PSA)の乾燥粉末を0.2 MNaCl溶液中に1時間溶解して、最終濃度を8 mg/mlとする。次に、HClまたはNaOHを用いてpH値を6に固定し、溶液を25℃に熱調節する。0.75 mgCDAP/mgPSA(調製物を100 mg/mlアセトニトリルとする)をPSA溶液に付加する。pH調節せずに1.5分後、0.2 MNaOHを付加して、pHを10とする。2.5分後、約1のPD/PSA比にしたがって、プロテインD(5 mg/mlに濃縮)を付加する。1時間のカップリング反応時間中は、pHを10に保持する。次に、10 mgのグリシン(2 M、pH9.0)/mgPSAを付加し、25℃で30分間、pH値を9.0に調節する。次に混合物を4℃で一夜保存した後、排除カラムクロマトグラフィー(Sephacryl S400HR, Pharmacia)により精製する。先ず複合体が、その後未反応PDおよび副産物(DMAP、グリシン、塩)が溶離する。複合体を収集し、Sartopore膜(Sartorius)上での0.2μm濾過により滅菌する。
【0306】
実施例9−実施例7および8の生成物のin vitro特性化
【0307】
【表19】
【0308】
in vivo結果
Balb/Cマウスを動物モデルとして用いて、複合体の免疫原性を調べた。複合体をAlPO4またはAl(OH)3上に吸着させる(500μGのAl3+上に10μgのPS)か、あるいは吸着させなかった。マウスに以下のように注射した:2週間間隔で2回注射(2μgPS/注射)。
【0309】
これらの結果から、遊離PSは免疫応答に大いに影響を及ぼす、と先ず結論し得る。10%未満の遊離PSを有する複合体を用いて、より良好な結果を得た。したがって、CDAP法に対する前記の改良は、本発明のさらなる局面である。
【0310】
処方物も重要である。AlPO4は、このモデルにおいて最も適切なアジュバントであると思われる。複合体は、多糖を単独で注射した場合には観察されない追加免疫作用を誘導する。
【0311】
結論
それぞれ1および0.6〜0.7(w/w)の最終PS/タンパク質比を有する髄膜炎菌AおよびCの複合体を得た。遊離PSおよび遊離担体タンパク質は、それぞれ10%および15%以下であった。多糖回収率は、70%より高い。前記の改良型(最適化)CDAP法により得られるPSAおよびPSCの複合体(担体タンパク質にかかわらず、しかし好ましくはプロテインD)は、したがって、本発明のさらなる局面である。
【0312】
実施例10−b型インフルエンザ菌は、2歳以下の小児における髄膜炎の主因の1つである。破傷風毒素上の複合体としてのインフルエンザ菌の莢膜多糖(PRP)が周知である(J. Robbinsにより開発された化学作用により連結される)。CDAPは、改良型化学である。好ましくはPDにPRPを連結するために見出された最適CDAP条件について、以下に説明する。
【0313】
複合化の反応に影響を及ぼすパラメーターを以下に示す:
・多糖の初期濃度(遊離多糖の最終レベルにおよび滅菌濾過工程に二重の影響を及ぼし得る)
・担体タンパク質の初期濃度
・多糖対タンパク質の初期比(遊離多糖の最終レベルにおよび滅菌濾過工程に二重の影響を及ぼし得る)
・用いられるCDAPの量(通常は大余分量で)
・反応温度(多糖の分解、反応の速度論、および反応性基の分解に影響を及ぼし得る)
・活性化およびカップリングのpH
・クエンチングのpH(残留DMAPのレベルに影響を及ぼす)
・活性化、カップリングおよびクエンチングの時間
最終生成物の量を最適化するための3つの最も重要なパラメーターは、多糖/タンパク質の初期比、多糖の初期濃度およびカップリングpHである、ということを本発明人等は見出した。
【0314】
したがって、前記の3つの条件を3つの軸として用いて、反応立方体を設計した。これらの軸に関する中心点(および実験値範囲)は、PS/タンパク質比−1/1(±0.3/1);[PS]=5 mg/ml(±2 mg/ml);およびカップリングpH=8.0(±1.0pH単位)であった。
【0315】
あまり必須でないパラメーターは以下のように固定した:30 mgの多糖を用いた;温度25℃;[CDAP]=0.75 mg/mgPS;pHは0.2 MNaOHで滴定;活性化pH=9.5;活性化温度=1.5分間;カップリング温度=1時間;[タンパク質]=10 mg/ml;クエンチングpH=9.0;クエンチング温度=1時間;溶媒中にPSを溶解する温度=2 MNaCl中で1時間;12 cm/時間で150 mMNaClで用利したSephacryl S-400HR上での精製;ならびに5ml/分でのSARTOLAB P20によるフィルター滅菌。
【0316】
前記の反応立方体内で製品を製造する場合の最適化条件を確立するために考察されたデータを以下に示す:工程データ−濾過後の最大収率、今夕されるタンパク質の最大レベル;ならびに生成物の品質データ−最終PS/タンパク質比、遊離PSレベル、遊離タンパク質レベル、残留DMAP(CDAPの分解生成物)の最小レベル。
【0317】
濾過からの結果
濾過後の結果を左右する因子は、初期[PS]およびカップリングpHおよび初期PS/タンパク質比間の相互作用である。低[PS]では、後者2つの因子との相互作用はほとんど認められず、良好な濾過可能性が常に結果的に生じる(全生成物に関して約95%)。しかしながら、高濃度では、pHおよび初期比が増大すれば、濾過可能性は減少する(高[PS]、最低比、最低pH=99%濾過;しかし高[PS]、最高比およびpH=19%濾過)。
【0318】
タンパク質混入レベル
最終PS/タンパク質比の、初期比に関する比は、カップリングの効率の測定値である。高[PS]では、pHは両比の比に影響しない。しかしながら、初期比は影響する(低初期比で1.75、高初期比で1.26)。低[PS]では、両比の比は、ほとんどの部分に関しては、より低いが、しかしながら、pHは、ここではより大きい影響を有する(低pH、低比=0.96;低pH、高比=0.8;高pH、低比=1.4;および高pH、高比=0.92)。
【0319】
最終PS/タンパク質比
最終比は、初期比および[PS]によっている。大多数のかなり大きい最終比は、組合せ高初期比および高[PS]を用いて得られる。最終比に及ぼすpHの影響は、弱[PS]と同じくらい有意ではない。
【0320】
遊離プロテインDのレベル
最小量の遊離プロテインDは、高pHおよび高[PS]で観察される(0.0に近似のレベル)。高[PS]の作用は、pHが低い場合に特に顕著になる。初期比の上昇は、遊離プロテインDの増大にわずかに寄与する。
【0321】
残留DMAP
初期比は、有意の作用を有さない。これに対比して、DMAPのレベルは、[PS]に伴って増大し、pHが上昇すると低減する。
【0322】
結論
したがって、最も好ましい複合化条件は、以下の通りである:カップリングpH=9.0;[PS]=3 mg/ml;および初期比=1/1。このような条件を用いた場合の、最終生成物の特徴を以下に示す:
【0323】
【表20】
【0324】
前記の改良型(最適化)CDAP法により得られるPRPの複合体は(担体タンパク質にかかわらず、しかし好ましくはプロテインD)、したがって、本発明のさらなる局面である。
【0325】
実施例11:抗原としてのプロテインD−非分類可能インフルエンザ菌に対するその防御効力は3D−MPLを用いてそれを処方することにより以下に改善され得るか
11価肺炎球菌多糖−プロテインD複合体ワクチンを用いて、雌Balb/cマウス(10匹/群)を、20週齢で初回(D0)の免疫感作(筋内)し、2週間後(14日目)に2回目の免疫感作を施した。2回目免疫感作後7日目に、血液を収集した。IgG1、IgG2aおよびIgG2b型抗体の量に関して、プロテインDに対する抗体力価を測定した。
【0326】
室温で15分間撹拌しながら複合体を15.75%のラクトースと併合し、pHを6.1±0.1に調整し、凍結乾燥することにより、凍結乾燥11価ワクチン(AlPO4を含有しない)を調製した(当該サイクルは通常は、-69℃で出発し、漸次3時間に亘って-24℃に調整し、次にこの温度を18時間保持した後、漸次1時間に亘って-16℃に調整し、この温度を6時間保持して、その後漸次3時間に亘って+34℃に調整し、最後にこの温度を9時間保持する)。
【0327】
凍結乾燥物のための処方物および再構築物の組成を表13に示す。
【0328】
Th1型細胞媒介性免疫応答が起きたか否かに関する大多数の特徴的測定値は、IgG2aのレベルと相関することが知られている。データから分かるように、IgG2aの意外にも大きい増大は、プロテインDがTh1アジュバント(この場合は3D−MPL)とともに凍結乾燥された場合に、結果的に生じる。
【0329】
【表21】
【特許請求の範囲】
【請求項1】
少なくとも1のストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)多糖抗原、及び少なくとも1のストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)タンパク質抗原又は免疫学的に機能性のその等価物を含む免疫原性組成物。
【請求項2】
前記タンパク質抗原が、ストレプトコッカス・ニューモニエ又は免疫学的に機能性のその等価物の外表面タンパク質又は分泌タンパク質である、請求項1に記載の免疫原性組成物。
【請求項3】
前記タンパク質抗原が、ストレプトコッカス・ニューモニエ又は免疫学的に機能性のその等価物の毒素、アドヘシン(adhesin)又はリポタンパク質である、請求項1又は2に記載の免疫原性組成物。
【請求項4】
前記タンパク質抗原又は免疫学的に機能性のその等価物が、以下の:ニューモリジン(pneumolysin)、PspA又はそのトランスメンブラン欠失変異体、PspC又はそのトランスメンブラン欠失変異体、PsaA又はそのトランスメンブラン欠失変異体、グリセルアルデヒド−3−ホスフェート・デヒドロゲナーゼ、及びCbpA又はそのトランスメンブラン欠失変異体から成る群から選ばれる、請求項1〜3のいずれかに記載の免疫原性組成物。
【請求項5】
前記多糖抗原が、多糖−タンパク質担体コンジュゲートの形態で提示される、請求項1〜4のいずれかに記載の免疫原性組成物。
【請求項6】
前記担体タンパク質が、以下の:ジフテリア毒素(Diphtheria toxid)、破傷風毒素(Tetanus toxid)、CRM197、キーホール・リンペット・ヘモシアニン(Keyhole Limpet Haemocyanin (KLH))、ツベルクリン(Tuberculin)のタンパク質誘導体(PPD)、及びH.インフルエンザ(H. influenzae)由来のタンパク質から成る群から選ばれる、請求項5に記載の免疫原性組成物。
【請求項7】
前記ワクチンが、異なる血清型からの少なくとも4つのニューモコッカル(pneumococcal)多糖抗原を含む、請求項1〜6のいずれかに記載の免疫原性組成物。
【請求項8】
アジュバントをさらに含む先の請求項のいずれかに記載の免疫原性組成物。
【請求項9】
前記アジュバントが、アルミニウム塩を含む、請求項8に記載の免疫原性組成物。
【請求項10】
前記アジュバントが、TH1応答の優先的誘導物質である、請求項8に記載の免疫原性組成物。
【請求項11】
前記アジュバントが、以下の:3D−MPL、サポニン免疫刺激剤、又は免疫刺激性CpGオリゴヌクレオチドの少なくとも1を含む、請求項10に記載の免疫原性組成物。
【請求項12】
前記アジュバントが、以下の:油/水エマルジョン、リポソーム、及びアルミニウム塩を含む群から選ばれる担体を含む、請求項11に記載の免疫原性組成物。
【請求項13】
医薬として使用される、先の請求項のいずれかに記載の免疫原性組成物。
【請求項14】
請求項1〜12に記載の免疫原性組成物を含むワクチン。
【請求項15】
55歳を超える患者におけるストレプトコッカス・ニューモニエ感染を予防し又は軽減する方法であって、ストレプトコッカス・ニューモニエ多糖及び少なくとも1のストレプトコッカス・ニューモニエ・タンパク質、並びに場合によりTH1誘導性アジュバントを含む有効量のワクチンを投与することを含む、前記方法。
【請求項16】
55歳を超える患者における肺炎の予防のための医薬の製造における、ストレプトコッカス・ニューモニエ・タンパク質抗原と、及び場合によりTH1誘導性アジュバントと併合したニューモコッカル多糖抗原の使用。
【請求項17】
以下のステップ:
1以上のニューモコッカル多糖抗原を選び;
1以上のニューモコッカル・タンパク質抗原を選び;そして
上記多糖抗原及びタンパク質抗原を好適に賦形剤と混合する、
を含む、先の請求項のいずれかに記載の免疫原性組成物の製造方法。
【請求項18】
小児(Infants)における中耳炎の予防又は軽減方法であって、上記小児に、場合によりTH1アジュバントとともに、ストレプトコッカス・ニューモニエ多糖抗原及びストレプトコッカス・ニューモニエ・タンパク質抗原の安全かつ有効な量を投与することを含む、前記方法。
【請求項1】
少なくとも1のストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)多糖抗原、及び少なくとも1のストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)タンパク質抗原又は免疫学的に機能性のその等価物を含む免疫原性組成物。
【請求項2】
前記タンパク質抗原が、ストレプトコッカス・ニューモニエ又は免疫学的に機能性のその等価物の外表面タンパク質又は分泌タンパク質である、請求項1に記載の免疫原性組成物。
【請求項3】
前記タンパク質抗原が、ストレプトコッカス・ニューモニエ又は免疫学的に機能性のその等価物の毒素、アドヘシン(adhesin)又はリポタンパク質である、請求項1又は2に記載の免疫原性組成物。
【請求項4】
前記タンパク質抗原又は免疫学的に機能性のその等価物が、以下の:ニューモリジン(pneumolysin)、PspA又はそのトランスメンブラン欠失変異体、PspC又はそのトランスメンブラン欠失変異体、PsaA又はそのトランスメンブラン欠失変異体、グリセルアルデヒド−3−ホスフェート・デヒドロゲナーゼ、及びCbpA又はそのトランスメンブラン欠失変異体から成る群から選ばれる、請求項1〜3のいずれかに記載の免疫原性組成物。
【請求項5】
前記多糖抗原が、多糖−タンパク質担体コンジュゲートの形態で提示される、請求項1〜4のいずれかに記載の免疫原性組成物。
【請求項6】
前記担体タンパク質が、以下の:ジフテリア毒素(Diphtheria toxid)、破傷風毒素(Tetanus toxid)、CRM197、キーホール・リンペット・ヘモシアニン(Keyhole Limpet Haemocyanin (KLH))、ツベルクリン(Tuberculin)のタンパク質誘導体(PPD)、及びH.インフルエンザ(H. influenzae)由来のタンパク質から成る群から選ばれる、請求項5に記載の免疫原性組成物。
【請求項7】
前記ワクチンが、異なる血清型からの少なくとも4つのニューモコッカル(pneumococcal)多糖抗原を含む、請求項1〜6のいずれかに記載の免疫原性組成物。
【請求項8】
アジュバントをさらに含む先の請求項のいずれかに記載の免疫原性組成物。
【請求項9】
前記アジュバントが、アルミニウム塩を含む、請求項8に記載の免疫原性組成物。
【請求項10】
前記アジュバントが、TH1応答の優先的誘導物質である、請求項8に記載の免疫原性組成物。
【請求項11】
前記アジュバントが、以下の:3D−MPL、サポニン免疫刺激剤、又は免疫刺激性CpGオリゴヌクレオチドの少なくとも1を含む、請求項10に記載の免疫原性組成物。
【請求項12】
前記アジュバントが、以下の:油/水エマルジョン、リポソーム、及びアルミニウム塩を含む群から選ばれる担体を含む、請求項11に記載の免疫原性組成物。
【請求項13】
医薬として使用される、先の請求項のいずれかに記載の免疫原性組成物。
【請求項14】
請求項1〜12に記載の免疫原性組成物を含むワクチン。
【請求項15】
55歳を超える患者におけるストレプトコッカス・ニューモニエ感染を予防し又は軽減する方法であって、ストレプトコッカス・ニューモニエ多糖及び少なくとも1のストレプトコッカス・ニューモニエ・タンパク質、並びに場合によりTH1誘導性アジュバントを含む有効量のワクチンを投与することを含む、前記方法。
【請求項16】
55歳を超える患者における肺炎の予防のための医薬の製造における、ストレプトコッカス・ニューモニエ・タンパク質抗原と、及び場合によりTH1誘導性アジュバントと併合したニューモコッカル多糖抗原の使用。
【請求項17】
以下のステップ:
1以上のニューモコッカル多糖抗原を選び;
1以上のニューモコッカル・タンパク質抗原を選び;そして
上記多糖抗原及びタンパク質抗原を好適に賦形剤と混合する、
を含む、先の請求項のいずれかに記載の免疫原性組成物の製造方法。
【請求項18】
小児(Infants)における中耳炎の予防又は軽減方法であって、上記小児に、場合によりTH1アジュバントとともに、ストレプトコッカス・ニューモニエ多糖抗原及びストレプトコッカス・ニューモニエ・タンパク質抗原の安全かつ有効な量を投与することを含む、前記方法。
【図1A】

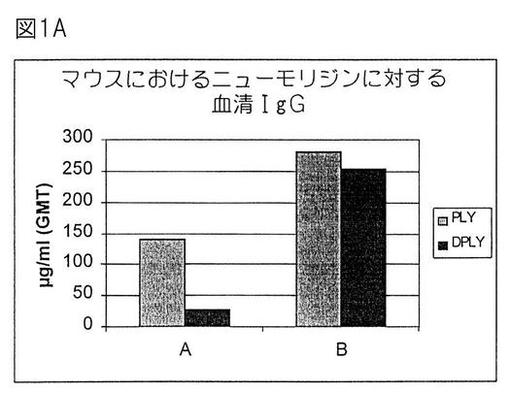
【図1B】
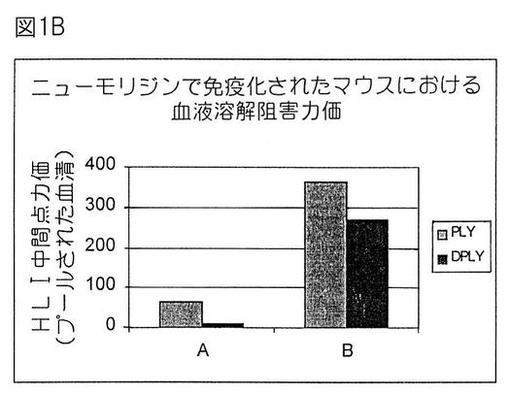
【図1C】
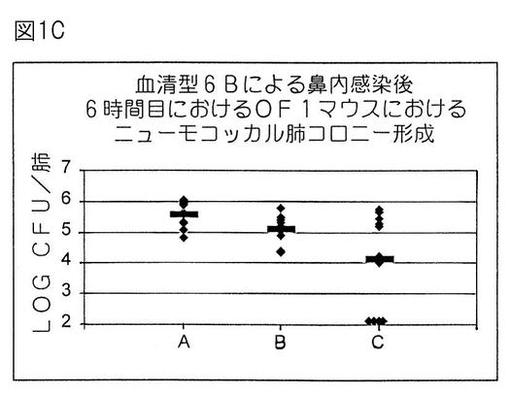
【図1B】
【図1C】
【公開番号】特開2011−57713(P2011−57713A)
【公開日】平成23年3月24日(2011.3.24)
【国際特許分類】
【出願番号】特願2010−288197(P2010−288197)
【出願日】平成22年12月24日(2010.12.24)
【分割の表示】特願2000−606263(P2000−606263)の分割
【原出願日】平成12年3月17日(2000.3.17)
【出願人】(305060279)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (169)
【Fターム(参考)】
【公開日】平成23年3月24日(2011.3.24)
【国際特許分類】
【出願日】平成22年12月24日(2010.12.24)
【分割の表示】特願2000−606263(P2000−606263)の分割
【原出願日】平成12年3月17日(2000.3.17)
【出願人】(305060279)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (169)
【Fターム(参考)】
[ Back to top ]