
一本鎖抗体の製造方法
【課題】 抗体をその結合性や抗原特異性を維持させたまま、非常に簡便に絹糸から調製する方法を提供すること。
【解決手段】 抗体として一本鎖抗体(scFv)を選択し、このscFvをフィブロインタンパク質と融合させて絹糸に発現させ、これにより得られた絹糸に対し精錬せずに溶解と再構成の処理を行うことにより、結合性及び抗原特異性を高く保持した一本鎖抗体を製造できることを見出した。
【解決手段】 抗体として一本鎖抗体(scFv)を選択し、このscFvをフィブロインタンパク質と融合させて絹糸に発現させ、これにより得られた絹糸に対し精錬せずに溶解と再構成の処理を行うことにより、結合性及び抗原特異性を高く保持した一本鎖抗体を製造できることを見出した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一本鎖抗体(scFv)の製造方法に関し、より詳しくは、scFvとフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解し、再構成処理を施すことを特徴とする、scFvの製造方法に関する。
【背景技術】
【0002】
抗体は、抗原と結合するドメイン(Fab)と免疫グロブリン受容体に結合するドメイン(Fc)より構成され、Fabドメインを介して病原体の抗原に結合し、それを排除して中和する免疫調節作用を発揮する。そして、その作用の根幹は、ペプチド、核酸、糖鎖、糖脂質、低分子化合物等の種々様々な抗原に対して特異的に結合できることにある。そのため、抗体は、前記免疫調節作用を目的とした医薬品(抗体医薬)のみならず、ウェスタンブロッティング、免疫沈降(プルダウンアッセイ)、フローサイトメトリー、免疫組織染色等の各種検出・精製方法に適したツールとしても利用され、開発が進められており、これらの市場は急速に拡大している。
【0003】
通常、抗体は、検出・精製の対象とする抗原をウサギやマウス等の免疫動物に接種し、該動物の免疫系を刺激した後、その動物の血清(ポリクローナル抗体)を回収することで得ることができる。また、このように免疫系を刺激した動物から脾臓細胞等の抗体産生細胞を回収し、該抗体産生細胞とミエローマ細胞とを融合し、得られたハイブリドーマを選択することにより所望の抗原のみを検出・精製することのできる抗体(モノクローナル抗体)を作製することもできる。しかしながら、かかる動物や動物細胞を利用した方法には、活性を有する抗体を安定的に取得できる反面、大量生産に適していない、生産コストが非常に高くなるといった欠点がある。
【0004】
これに対し、所望の抗体をコードする遺伝子をクローニングすることにより、タンパク質の大量生産に適した大腸菌等で抗体を生産することも試みられている。しかし、大腸菌で生産した場合には、得られる抗体がしばしば不溶化することが知られており、その精製にあたっては、ドデシル硫酸ナトリウム(SDS)等の変性剤でタンパク質を可溶化させる工程やその変性剤を取り除く透析等の工程が必要となる。さらにそのような多くの工程を経て得られた抗体は概して活性が低くなる傾向にあるため、大腸菌等を利用した方法には、活性を有する抗体を大量に生産しにくいという欠点がある。
【0005】
一方、カイコは蛹化する際に自身を保護する目的で繭を作り出す。この繭はフィブロイン(Fibroin)という繊維構造とそれを取り巻くセリシンとの2種の構造からなる絹糸から作られている構造体である。絹糸は、およそ1.5kmに達する単繊維として存在するため、人類はこれをほどいて繊維とし、衣類等に利用してきた。近年では、絹糸を構成するするフィブロイン及びセリシンをそれぞれ加工することによって、パウダー(特許文献1)やフィルム(特許文献2及び3)、及びスポンジ様構造体(特許文献4)として利用することも可能となってきた。
【0006】
2000年にカイコの遺伝子組換え技術が確立され(非特許文献1)、外来遺伝子を導入することによりカイコのシルクタンパク質を改変し、物性の向上や、機能性の付加を実現した新しいシルクを設計・創出することが可能となった。現在では目的のタンパク質を絹タンパク質の主成分であるフィブロイン層とセリシン層のどちらにでも発現させることが可能となり、これまでにフィブロインと蛍光タンパク質やサイトカイン等との融合タンパク質を発現する組換えカイコや(非特許文献2〜4)、またセリシン層にヒト血清アルブミンや抗体分子を発現する組換えカイコの作出に成功し(非特許文献5及び6)、蛍光絹糸としての利用や発現産物を絹糸から分離・精製して利用する試みが行われており、前述の抗体のみならず、機能的なタンパク質の生産の場としてカイコは注目を集めている。
【0007】
例えば、飯塚らは、マウスモノクローナル抗体のH鎖及びL鎖を中部絹糸腺細胞で発現させ、セリシン層にそれら抗体分子を分泌する組換えカイコを作製した。そして、該組換えカイコが作出した絹糸を3M 尿素(Urea)を含む50mM Tris−HCl(pH7.4)溶液で処理し、抗体分子を含んだセリシン溶液を透析後、プロテインGセファロースを用いてアフィニティー精製することにより、抗原特異的な結合活性を有する抗体分子を得ている(非特許文献6)。
【0008】
また、外来タンパク質をフィブロインに発現させ、活性をもつ該タンパク質を調製するためには、一般的に絹糸を高濃度の臭化リチウム溶液やチオシアン酸リチウム溶液、又は塩化カルシウム/エタノール溶液で一度完全に溶解し、得られたシルク複合体から目的の外来タンパク質をプロテアーゼや還元剤を用いて遊離させ、再びリフォールディングすることが必要である。実際、日野らは、フィブロインL鎖にヒトbFGFを融合させた組換えタンパク質を絹糸に発現させ、絹糸を精練し、セリシン成分を除去した後、高濃度のチオシアン酸リチウムに溶解し、2−メルカプトエタノールを用いて内在性フィブロインから遊離させ、さらに透析を施し、変性剤の濃度を徐々に下げていくことで、活性のあるbFGFを獲得している(非特許文献4)。
【0009】
このように、外来タンパク質を発現させた絹糸から活性を有する該タンパク質を調製するためには、アフィニティー精製、プロテアーゼ処理、還元剤の添加及び精練等を行い、外来タンパク質から内在性タンパク質を遊離・除去する工程が必要であった。また、絹糸から外来タンパク質を抽出するためには、変性剤(尿素、臭化リチウム溶液、塩化カルシウム/エタノール溶液、チオシアン酸リチウム、塩酸グアニジン、SDS等)にて該タンパク質を変性させ可溶化する工程が必要となる。さらに、変性して可溶化させたタンパク質から変性剤を取り除き、該タンパク質が活性を有するようにするために、リフォールデイング処理を施す工程を要する。しかしながら、かかる多工程を経て調製されたタンパク質、特に抗体は概してその活性が低くなる傾向にあった。
【0010】
そこで、絹糸から抗体等の外来タンパク質をその活性を維持させたまま、非常に簡便に調製する方法の開発が望まれているが、未だ実用化されていないのが現状である。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】特開2004−123683号公報
【特許文献2】特開2008−173312号公報
【特許文献3】特開平9−192210号公報
【特許文献4】特開2002−186847号公報
【非特許文献】
【0012】
【非特許文献1】Tamura,T.ら、Nat.Biotechnol.、2000年、18巻、81〜84ページ
【非特許文献2】Inoue,S.ら、Insect Biochem.Mol.Biol.、2005年、35巻、51〜59ページ
【非特許文献3】Kojima,K.ら、Biosci.Biotechnol.Biochem.、2007年、71巻、2943〜2951ページ
【非特許文献4】Hino,R.ら、Biomaterials.、2006年、27巻、5715〜5724ページ
【非特許文献5】Ogawa,S.ら、J.Biotechnol.、2007年、128巻、531〜544ページ
【非特許文献6】Iizuka,M.ら、FEBS J、2009年、276巻、5806〜5820ページ
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、前記従来技術の有する課題に鑑みてなされたものであり、絹糸から抗体をその結合性や抗原特異性を維持させたまま、内在性タンパク質を遊離・除去する工程等を要せずに非常に簡便に調製することを可能とする、抗体の製造方法を提供することを目的とする。
【課題を解決するための手段】
【0014】
本発明者らは、前記目的を達成すべく鋭意研究を重ねた結果、抗体として一本鎖抗体(scFv)を選択し、このscFvをフィブロインタンパク質と融合させて絹糸に発現させ、これにより得られた絹糸に対し、精錬せずに溶解と再構成の処理を行うことにより、結合性及び抗原特異性を高く保持した一本鎖抗体を製造できることを見出し、本発明を完成するに至った。
【0015】
本発明は、より詳しくは、下記を提供するものである。
<1> 一本鎖抗体(scFV)とフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解する工程と、該工程で得られた前記融合タンパク質の溶解液に再構成処理を施す工程と、を含むことを特徴とする、scFVの製造方法。
<2> 分子内又は分子間の水素結合を切断する性質を有する溶液で絹糸を溶解する、<1>に記載の製造方法。
<3> 前記再構成処理を施した融合タンパク質の溶解液を凍結乾燥して粉砕する工程をさらに含むことを特徴とする、<1>又は<2>に記載の製造方法。
<4> 前記フィブロインタンパク質が、フィブロインL鎖及びフィブロインH鎖からなる群から選択される少なくとも一のタンパク質であることを特徴とする、<1>〜<3>のうちのいずれか一に記載の製造方法。
<5> 前記再構成処理が透析処理であることを特徴とする、<1>〜<4>のうちのいずれか一に記載の製造方法。
【発明の効果】
【0016】
本発明によれば、絹糸から一本鎖抗体をその結合性や抗原特異性を維持させたまま、非常に簡便に調製することが可能となる。
【図面の簡単な説明】
【0017】
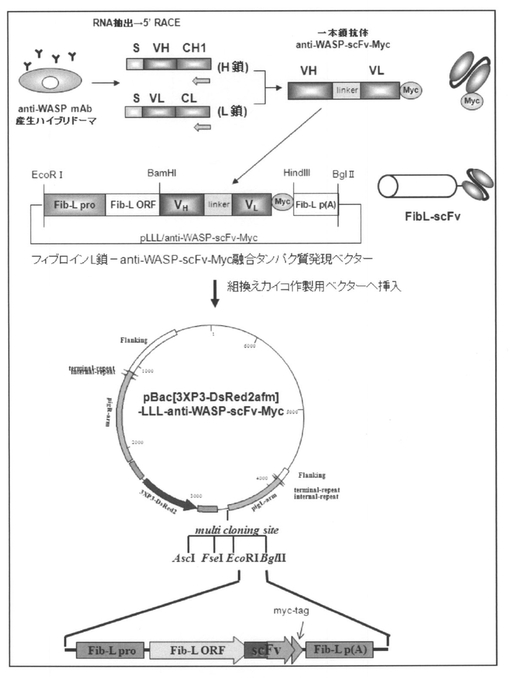
【図1】抗WASPモノクローナル抗体(anti−WASP mAb)を基に一本鎖抗体(抗WASP−scFv(anti−WASP−scFv−Myc))を得た過程と、フィブロインL鎖−抗WASP−scFv(フィブロインL鎖−anti−WASP−scFv−Myc融合タンパク質)をカイコが産生する絹糸に発現させるためのベクター(pBac[3XP3−DsRed2afm]−LLL−anti−WASP−scFv−Myc)のコンストラクトとを示す、概要図である。
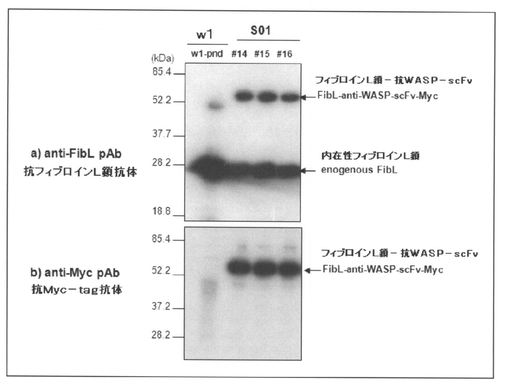
【図2】組換えカイコ(S01)産生繭におけるフィブロインL鎖−抗WASP−scFvの発現を、ウェスタンブロッティングによって検出した結果を示す図である。なお、図中の「w1」は非組換え体カイコ(w1−pnd、陰性対照)産生繭における結果であることを示し、「♯14〜16」はS01系統内の異なる個体由来の繭における結果であることを示す。また、a)は抗フィブロインL鎖抗体(anti−FibL pAb)を用いたウェスタンブロッティングの結果を示し、b)は抗Myc−tag抗体(anti−Myc pAb)を用いたウェスタンブロッティングの結果を示す。
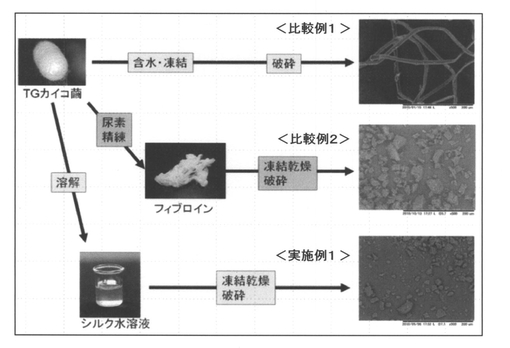
【図3】組換えカイコ(S01)産生繭(TGカイコ繭)から融合タンパク質 フィブロインL鎖−抗WASP−scFvを有するシルクパウダーを調製するための3種類の方法(実施例1、比較例1〜2)の概略、並びにこれらの方法によって得られたシルクパウダーを電子顕微鏡で観察した結果を示す写真である。
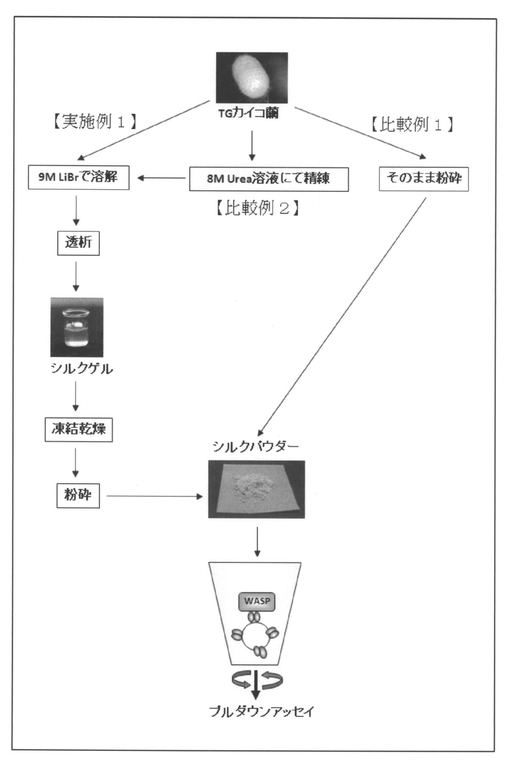
【図4】組換えカイコ(S01)産生繭(TGカイコ繭)から融合タンパク質 フィブロインL鎖−抗WASP−scFvを有するシルクパウダーを調製するための3種類の方法(実施例1、比較例1〜2)の概略、及びこれらの方法によって得られたシルクパウダーと抗WASP−scFvの抗原であるWASPタンパク質との複合体形成を調べるためのプルダウンアッセイの概略を示す図である。
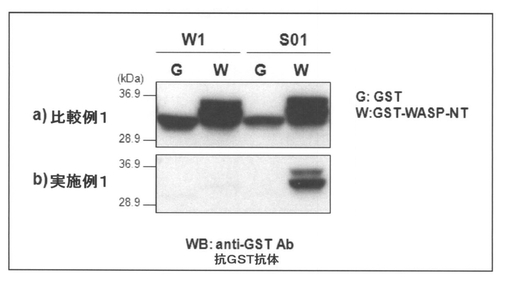
【図5】抗GST抗体を用いたウェスタンブロッティングによって分析した、図4に示したプルダウンアッセイの結果を示す図である。なお、図中の「W1」は非組換え体カイコ(w1−pnd、陰性対照)産生繭における結果であることを示し、「W」及び「G」は「WASPタンパク質(GST−WASP−NT)」及び「GSTタンパク質(陰性対照、GST)」を用いたプルダウンアッセイの結果であることを各々示す。また、a)は比較例1の方法によって得られたシルクパウダーの分析結果であることを示し、b)は実施例1の方法によって得られたシルクパウダーの分析結果であることを示す。
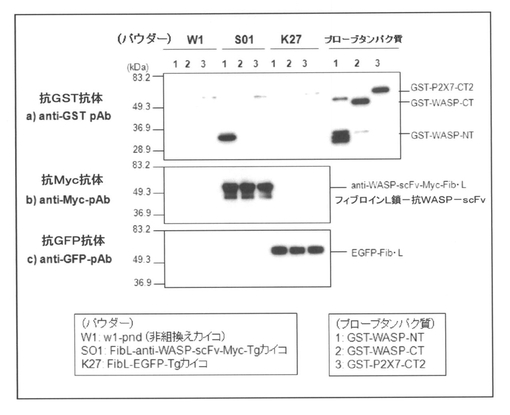
【図6】本発明(実施例1)の方法によって得られたシルクパウダーと、各種プローブタンパク質との複合体形成をプルダウンアッセイ並びにウェスタンブロッティングによって分析した結果を示す図である。
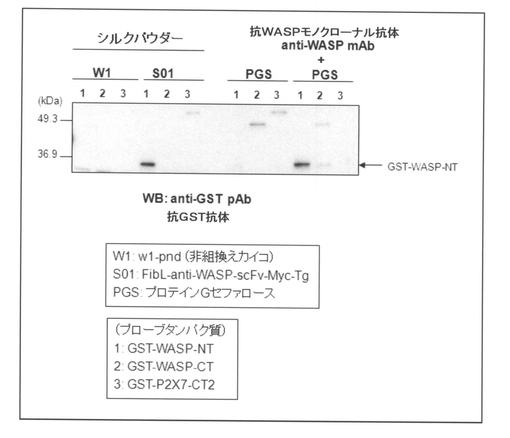
【図7】比較例2に記載の方法によって得られたシルクパウダーと、各種プローブタンパク質との複合体形成をプルダウンアッセイ並びにウェスタンブロッティングによって分析した結果を示す図である。
【図8】本発明(実施例1)の方法によって得られたフィブロインL鎖−抗WASP−scFvを有するシルクパウダーと、抗WASP−scFvの元になった抗WASPモノクローナル抗体とを、結合性及び抗原特異性の点において、各種プローブタンパク質を用いたプルダウンアッセイ並びにウェスタンブロッティングによって分析して比較した結果を示す図である。
【図9】抗WASPモノクローナル抗体(anti−WASP mAb)を基に一本鎖抗体(抗WASP−scFv(anti−WASP−scFv−Myc))を得た過程と、フィブロインH鎖−抗WASP−scFv(フィブロインH鎖−anti−WASP−scFv−Myc融合タンパク質)をカイコが産生する絹糸に発現させるためのベクター(pBac[3XP3−DsRed2afm]−HC−anti−WASP−scFv−Myc)のコンストラクトとを示す、概要図である。
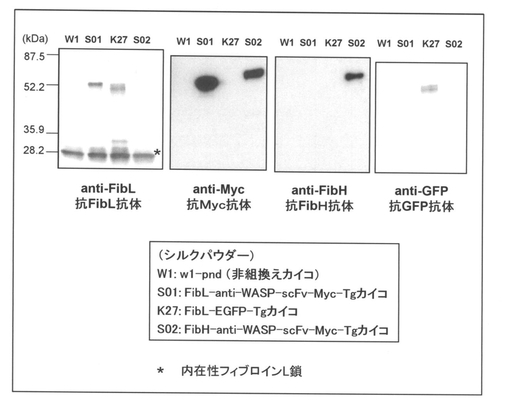
【図10】本発明(実施例2)の方法によって得られた組換えカイコ(S02)産生繭由来シルクパウダーにおけるフィブロインH鎖−抗WASP−scFvの発現を、ウェスタンブロッティングによって検出した結果を示す図である。なお、図中の「*(アスタリスク)」は内在性フィブロインL鎖の位置を示す。
【図11】本発明(実施例2)の方法によって得られたシルクパウダーと、各種プローブタンパク質との複合体形成をプルダウンアッセイ並びにウェスタンブロッティングによって分析した結果を示す図である。
【発明を実施するための形態】
【0018】
本発明は、機能的な一本鎖抗体(scFV)の製造方法であって、scFVとフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解する工程と、該工程で得られた前記融合タンパク質の溶解液に再構成処理を施す工程と、を含むことを特徴とする方法である。
【0019】
本発明において「一本鎖抗体(single chain fragment variable:scFv)」とは、抗体分子の抗原と結合するドメイン(Fab)を構成する重鎖(H鎖)及び軽鎖(L鎖)から可変領域(VH及びVL)を取り出して、リンカーを介して結合させた分子を意味する。本発明にかかる一本鎖抗体には、単一のポリペプチド鎖中に複数のVHとVLとを有している抗体も含まれる。かかる抗体としては、例えば、二つのVHと二つのVLとを前記リンカー等で結合した一本鎖抗体(sc(FV)2)が挙げられる。かかる抗体においては、通常複数のリンカーが必要となるが、全て同じリンカーであってもよく、異なるリンカーであってもよい。また、本発明にかかる一本鎖抗体には、リンカーペプチドの長さを短く(例えば5アミノ酸程度)に設定して、同一ポリペプチド鎖内でのVHとVLとの結合を抑制することにより形成される二量体(ダイアボディ、diabody)も含まれる。
【0020】
本発明にかかる一本鎖抗体におけるリンカーは、その両端に連結されたVH及びVLの発現や、VHとVLとの結合を阻害するものでなければ特に制限されない。リンカーペプチドの長さは、通常、1〜100アミノ酸、好ましくは1〜50アミノ酸、より好ましくは1〜30アミノ酸、特に好ましくは12〜18アミノ酸(例えば15アミノ酸)である。本発明においては、「(GGGGS)×3」という15アミノ酸からなるポリペプチドをリンカーとして好適に用いることができる(なお、GはグリシンをSはセリンを表わす)。
【0021】
一本鎖抗体の調製は、例えば、次のようにして行うことができる。まず、目的の抗原と特異的に結合するモノクローナル抗体を発現しているハイブリドーマよりmRNAを単離し、そのmRNAを用いて作製したcDNAライブラリーから、そのモノクローナル抗体のVH及びVLをコードするcDNAをそれぞれ単離する。次いで、VHをコードするcDNAとVLをコードするcDNAの間にリンカーをコードするDNAを挿入し、遺伝子組換え技術により、一本鎖抗体をコードする組換えDNAを構築する。
【0022】
cDNAライブラリーを作製せずに、mRNAよりRT−PCTを用いて、VH及びVLをコードする遺伝子を直接増幅させ、こうして増幅して得られた断片を用いて、遺伝子組換え技術により、一本鎖抗体をコードするDNAを構築することもできる。
【0023】
本発明にかかる「フィブロインタンパク質」とは、フィブロインL鎖、フィブロインH鎖及びフィブロインヘキサメリン(P25)からなる群から選択される少なくとも一のタンパク質を意味する。フィブロインタンパク質は、フィブロインL鎖とフィブロインH鎖とがジスルフィド結合し、この結合した分子6個に対し1つの割合でフィブロインヘキサメリンが結合し、複合体(フィブロイン複合体)が形成されることが知られている。
【0024】
本発明にかかる「フィブロインL鎖」は、分子量が約3万Daのフィブロインタンパク質であり、典型例として、GenBankアクセッション No.NP_001037488.1で特定されるタンパク質のうちの17〜262番目のアミノ酸配列からなるタンパク質(GenBankアクセッション No.NM_001044023.1で特定されるDNAのうちの90〜827番目の塩基配列からなる遺伝子)が挙げられる。また、このタンパク質は、シグナルペプチド(GenBankアクセッション No.NP_001037488.1で特定されるタンパク質のうちの1〜16番目のアミノ酸配列からなるタンパク質)が前駆体より切断、除去されることにより得られる成熟型であるから、本発明にかかる「フィブロインL鎖」としては、例えば、GenBankアクセッション No.NP_001037488.1で特定されるタンパク質(GenBankアクセッション No.NM_001044023.1で特定される遺伝子)が挙げられる
本発明にかかる「フィブロインH鎖」は、分子量が約35万Daのフィブロインタンパク質であり、典型例として、GenBankアクセッション No.NP_001106733.1で特定されるタンパク質のうちの22〜5263番目のアミノ酸配列からなるタンパク質(GenBankアクセッション No.NM_001113262.1で特定されるDNAのうちの64〜15789番目の塩基配列からなる遺伝子)が挙げられる。また、このタンパク質は、シグナルペプチド(GenBankアクセッション No.NP_001106733.1で特定されるタンパク質のうちの1〜21番目のアミノ酸配列からなるタンパク質)が前駆体より切断、除去されることにより得られる成熟型であるから、本発明にかかる「フィブロインL鎖」としては、例えば、GenBankアクセッション No.NP_001106733.1で特定されるタンパク質(GenBankアクセッション No.NM_001113262.1で特定される遺伝子)が挙げられる
本発明にかかる「フィブロインヘキサメリン」は、分子量が約2.5万Daのフィブロインタンパク質であり、「P25」とも称されるフィブロインタンパク質である。その典型例として、GenBankアクセッション No.NP_001139413.1で特定されるタンパク質のうちの17〜220番目のアミノ酸配列からなるタンパク質(GenBankアクセッション No.NM_001145941.1で特定されるDNAのうちの49〜660番目の塩基配列からなる遺伝子)が挙げられる。また、このタンパク質は、シグナルペプチド(GenBankアクセッション No.NP_001139413.1で特定されるタンパク質のうちの1〜16番目のアミノ酸配列からなるタンパク質)が前駆体より切断、除去されることにより得られる成熟型であるから、本発明にかかる「フィブロインL鎖」としては、例えば、GenBankアクセッション No.NP_001139413.1で特定されるタンパク質(GenBankアクセッション No.NM_001145941.1で特定される遺伝子)が挙げられる
また、タンパク質のアミノ酸配列は、自然界において(すなわち、非人工的に)変異し得る。従って、本発明にかかる「フィブロインタンパク質」には、このような天然の変異体も含まれる。さらに、本発明にかかる「フィブロインタンパク質」としては、前記天然のフィブロインタンパク質やその天然の変異体の全長のみならず、これらの部分的な断片であってもよい。
【0025】
さらに、本発明にかかる「フィブロインタンパク質」としては、本発明にかかる融合タンパク質がフィブロイン複合体の一部になれればよく、前記「フィブロインL鎖」、「フィブロインH鎖」又は「フィブロインヘキサメリン」の部分的断片も本発明にかかる「フィブロインタンパク質」に含まれる。かかる部分的断片としては、例えばフィブロインL鎖−GFP融合タンパク質において、下記部分的断片の有効性が実証されていることから、「フィブロインL鎖」においては、GenBankアクセッション No.NP_001037488.1で特定されるタンパク質のうちの1(メチオニン残基)〜242番目(アラニン残基)のアミノ酸配列からなるタンパク質であることが好ましい。また、「フィブロインH鎖」においては、フィブロインL鎖とヘテロダイマーを形成して(S−S結合を形成して)フィブロインタンパク質として分泌可能なことが確認されたフィブロインH鎖の最小単位であるという観点から、GenBankアクセッション No.NP_001106733.1で特定されるタンパク質のうちの、1(メチオニン残基)〜153番目(アラニン残基)のアミノ酸配列からなるタンパク質及び5205(セリン残基)〜5263番目(システイン残基)のアミノ酸配列からなるタンパク質であることが好ましい。
【0026】
本発明にかかる「一本鎖抗体とフィブロインタンパク質との融合タンパク質」としては、前記一本鎖抗体と、フィブロインL鎖、フィブロインH鎖及びフィブロインヘキサメリンからなる群から選択される少なくとも一のタンパク質とが融合しているタンパク質であればよい。一本鎖抗体が前記フィブロイン複合体の表面上に配置され易く、より高い抗原特異性の一本鎖抗体が得られ易くなるという観点から、前記一本鎖抗体と融合させるフィブロインタンパク質は、フィブロインL鎖及びフィブロインH鎖からなる群から選択される少なくとも一のタンパク質であることが好ましい。
【0027】
フィブロインタンパク質は、一本鎖抗体のN末側、C末側のいずれに融合させてもよく、またN末側及びC末側、両方に融合させてもよい。さらに、直接的に一本鎖抗体に融合させてもよく、リンカーを介して間接的に融合させてもよい。リンカーペプチドの長さは、通常、1〜100アミノ酸、好ましくは1〜50アミノ酸、より好ましくは1〜30アミノ酸である。「一本鎖抗体とフィブロインタンパク質との融合タンパク質」には、また他の機能性タンパク質が融合されていてもよい。この場合、他の機能性タンパク質は、融合タンパク質のN末側、C末側のどちらか一方若しくは両側、又は一本鎖抗体とフィブロインタンパク質との間に、直接的に又は間接的に融合させることができる。他の機能性タンパク質としては特に制限はなく、本発明にかかる融合タンパク質に付与したい機能に応じて適宜選択される。例えば、融合タンパク質に付与したい機能が該融合タンパク質の精製や検出等であれば、緑色蛍光タンパク質(GFP)、ルシフェラーゼタンパク質、Myc−タグ(tag)タンパク質、His−タグタンパク質、ヘマグルチン(HA)−タグタンパク質、FLAG−タグタンパク質(登録商標、Sigma−Aldrich社)、グルタチオン−S−トランスフェラーゼ(GST)タンパク質が挙げられる。
【0028】
前記融合タンパク質の絹糸における発現は、当業者は公知の手法により行うことができる。かかる公知の手法としては、前記融合タンパク質を発現させるためのベクターを構築し、該ベクターをカイコに導入して前記融合タンパク質を発現させる方法が挙げられる。
【0029】
フィブロインタンパク質を合成・分泌するカイコの後部絹糸腺において、前記融合タンパク質を発現させる場合、前記ベクターにおいて、前記融合タンパク質をコードするDNAは、カイコの後部絹糸腺における発現を保証するプロモーターの下流に連結されている必要がある。このようなプロモーターとしては、例えば、フィブロインL鎖遺伝子、フィブロインH鎖遺伝子又はフィブロインヘキサメリン(P25)遺伝子のプロモーターを好適に用いることができる。ベクターは、その他の発現制御領域、エンハンサー、ターミネーター、ポリAシグナル、非翻訳領域(5’−UTR、3’−UTR)等を有していてもよい。
【0030】
また、前記ベクターにおいて、前記融合タンパク質をコードするDNAとその制御領域とを備えた発現カセット以外に、他のタンパク質をコードするDNA及びその発現制御領域を備えていてもよい。かかる他のタンパク質をコードするDNAとしては特に制限はなく、例えば、GFPやDsRed等のレポータータンパク質をコードする遺伝子、薬剤耐性遺伝子が挙げられる。他のタンパク質をコードするDNAの発現制御領域は、配列内リボソーム進入部位(Internal Ribosome Entry Site、IRES)を介して、前記融合タンパク質をコードするDNAのプロモータ−やエンハンサーを共有していてもよく、前記融合タンパク質をコードするDNAの発現制御領域とは異なるプロモーター等を備えていてもよい。発現制御領域としては、例えば、3×P3プロモーター、カイコアクチンプロモーター、ショウジョウバエ由来のHSP70プロモーター、エンハンサー、ターミネーター、ポリAシグナル、非翻訳領域(5’−UTR、3’−UTR)が挙げられる。
【0031】
前記ベクターにおける発現カセット等をカイコの染色体内に挿入することができるベクターとしては、例えば、トランスポゾンを利用したベクターを好適に用いることができる。トランスポゾンを利用したベクターとしては、例えば、piggyBac、Minos、Mariner等のDNAトランスポゾンを利用したベクターが挙げられる。なお、前記ベクターにおいて、かかるDNAトランスポゾンの一対の末端反復配列(逆位末端反復配列)の間に前記発現カセットを配置することにより、転移酵素によって前記ベクターから該末端反復配列に挟まれた前記発現カセットが切り出され、カイコの染色体内の特定の配列(例えば、piggyBacを用いた場合には、TTAA配列)を標的としてランダムに挿入されるようになる。
【0032】
従って、かかるトランスポゾンを利用したベクターを用いる場合には、各トランスポゾンに対応した転移酵素もカイコに導入する必要がある。かかる転移酵素はそのままカイコに導入してもよく、かかる転移酵素をコードするmRNA又はかかる転移酵素をコードするプラスミドベクター(ヘルパープラスミド)を導入してもよい。かかるヘルパープラスミドにおいて、前記転移酵素を発現させるためのプロモーターとしては、例えば、内在性の転移酵素遺伝子のプロモーター、カイコアクチンプロモーター、ショウジョウバエ由来のHSP70プロモーターが挙げられる。
【0033】
本発明において、このように構築した前記融合タンパク質を発現させるためのベクターを導入し、該融合タンパク質が発現している絹糸を産生するカイコを調製する方法としては、例えば、該ベクター等をカイコの卵に注入し、該卵から孵化して成育させ得られたF0カイコを同胞交配又は野生型カイコと交配し、得られるF1カイコから前記融合タンパク質を絹糸に発現するカイコを選択する方法が挙げられる。
【0034】
前記ベクター等のカイコの卵への注入は、当業者に公知の手法を適宜選択して行うことができる。かかる公知の手法としては、例えば、DNA注入用の管を用いてカイコの発生初期卵に直接注入(マイクロインジェクション)する方法や、針やレーザー等を利用して物理的又は化学的に卵殻に穴を空けた後に、該穴から前記ベクター等を注入する方法が挙げられる。また、かかる前記ベクターをカイコの卵の注入する際に、前記転移酵素や前記ヘルパープラスミドを併せて注入してもよい。
【0035】
F1カイコから前記融合タンパク質を絹糸に発現するカイコを選択する方法としては、例えば、前記発現ベクターとしてGFPやDsRed等の蛍光タンパク質をコードする遺伝子を備えているものを用いた場合には、F1世代の卵や幼虫に励起光を照射することにより卵や幼虫から発せられる蛍光を指標に選択する方法が挙げられる。特に、3×P3プロモーターによって発現が制御されている蛍光タンパク質を利用した場合には、3×P3プロモーターは神経系統(特に眼)での遺伝子発現を促す作用を有するため、F1世代の卵や幼虫の神経系統や眼において蛍光を発する個体を選択することにより、前記融合タンパク質が発現している絹糸を産生するカイコを選択することができる。
【0036】
このように調製した前記カイコから、一本鎖抗体とフィブロインタンパク質との融合タンパク質を発現させた絹糸を調製する方法としては、例えば、カイコの絹糸腺内腔内に蓄積された絹糸を回収する方法、カイコが吐糸した絹糸を回収する方法、またはカイコが形成した繭を回収する方法が挙げられる。
【0037】
なお、後部絹糸腺で合成されたフィブロイン繊維は、中部絹糸腺に送られて濃縮され、さらに、ここで合成されているセリシンに包まれることになる。そして、このようにして合成された絹糸(液状絹糸)は吐糸口につながる前部絹糸腺に送られることになる。従って、絹糸腺内腔内に蓄積された絹糸(液状絹)は、絹糸腺をカイコから採取し、回収した絹糸腺から絹糸腺を構成する細胞(絹糸腺細胞)を除去することにより調製することができる。絹糸腺細胞の除去する方法としては、例えば、絹糸腺を純水に浸漬し、絹糸腺細胞をふやけさせた後に該細胞を取り除く方法、抽出した絹糸腺を30〜100%エタノールに浸漬し、フィブロイン及びセリシンを固定化した後に、絹糸腺細胞を除去する方法が挙げられる。
【0038】
本発明においては、こうして調製した、一本鎖抗体とフィブロインタンパク質との融合タンパク質を発現させた絹糸を溶解する。「絹糸」は、カイコの後部絹糸腺で合成・分泌される3種のフィブロインタンパク質からなるフィブロイン繊維と、該繊維とを包むセリシン層とからなる動物繊維である。また、本発明において溶解する対象となる「絹糸」は、セリシン層が除去(精錬)されていないことが必要である。
【0039】
一本鎖抗体とフィブロインタンパク質との融合タンパク質を発現させた絹糸の溶解においては、分子内又は分子間の水素結合を切断する性質を有する溶液を好適に用いることができる。
【0040】
かかる溶液としては、例えば、臭化リチウム水溶液、塩化カルシウム水溶液、塩化エチレンジアミン水溶液、チオシアン酸ナトリウム水溶液、チオシアン酸リチウム水溶液、硝酸マグネシウム水溶液、塩化カルシウム/エタノール、MMNO(N−メチルモルホリン N−オキシド)、塩化リチウム/N,N−DMAc(N,N−ジメチルアセトアミド)、ヘキサフルオロイソプロパノール、硝酸カルシウム/メタノール、又はこれらの混合溶液が挙げられる。これら水溶液の中では、溶解中にタンパク質分子の分解が確認されず、溶解時に加熱の必要がなく、また溶解にかかるコストが低いという観点から、臭化リチウム水溶液を本発明の方法において用いることが好ましい。
【0041】
また、分子内又は分子間の水素結合を切断する性質を有する溶液のpHとしては特に制限はないが、pH2〜12の溶液であることが好ましい。さらに、前記絹糸の溶解に用いられる溶液の濃度としては、通常8〜10M、好ましくは9〜9.3Mである。また、前記絹糸の溶解は、20〜100℃で10分〜12時間かけて攪拌しながら行うことが好ましく、9M 臭化リチウム水溶液を用いて室温(20〜30℃)にて4時間程度攪拌しながら行うことがより好ましい。
【0042】
本発明においては、こうして得られた前記融合タンパク質の溶解液に再構成処理を施す。かかる再構成処理としては、溶解により変性した前記融合タンパク質を脱塩することによって非変性状態に戻し、且つ前記融合タンパク質中のフィブロインタンパク質が内在性の他のフィブロインタンパク質(フィブロインL鎖、フィブロインH鎖、フィブロインヘキサメリン)、さらにはセリシンと複合体を構成し、フィブロインタンパク質が繊維化(ゲル化)できる処理であれば特に制限はない。かかる脱塩は、例えば、透析処理、限外濾過処理、ゲル濾過クロマトグラフィー処理、脱塩カラム処理、希釈処理、およびこれらの組合せにより行うことができる。これらの中では、フィブロインタンパク質の濃度を維持し、一定の性質のゲル化フィブロインを安価に、大量に得ることが容易であるという観点から、透析処理が好ましい。また、本発明にかかる再構成処理は、前記融合タンパク質の変性、分解、凝集等が生じにくいという観点から、4〜60℃の条件下で実施することが好ましい。
【0043】
「透析処理」においては、例えば、セルロース製の透析膜、多孔性のポリテトラフルオロエチレン製の透析膜、セルロースアセテート製の透析膜、セルロースナイトレート製の透析膜、再生セルロース透析膜、電解透析膜又は逆浸透膜を用いることができる。これら膜を用いて区画した前記融合タンパク質の溶解液を、任意の量の溶媒(例えば、精製水(RO水、Elix水等)、緩衝液)に1〜24時間浸漬させる工程をシルクがゲル化するまで繰り返すことにより、透析処理を実施することができる。また、かかる緩衝液としてはその種類、pH等においても特に制限はなく、例えば、リン酸緩衝液、Tris−HCl緩衝液、ハンクス液が挙げられる。これらの中では、再生セルロース透析膜を用いて区画した前記融合タンパク質の溶解液(シルク溶液)を、該溶液に対して100〜500倍量の純水(Elix)に12時間浸漬させる工程を5〜10回繰り返すことが好ましい。
【0044】
なお、かかる透析処理によって、通常、前記融合タンパク質の溶解液はゲル状(シルクゲル)となるが、ゲル化しない場合は、透析膜のまま空中に放置してタンパク質を濃縮させ、pH3〜7の緩衝液に対して透析し、又は1〜50%の水溶性有機溶剤(エタノール、メタノール、2−プロパノール、DMSO,グリセリンなど)若しくはpH5程度のバッファー(酢酸ナトリウム等)を加えることにより、ゲル化させることが出来る
また、本発明においては、一度再構成した抗体分子の機能(立体構造)を保持したままパウダー化できるという観点から、前記再構成処理を施した融合タンパク質の溶解液をさらに凍結乾燥して粉砕することが好ましく、また、前記再構成処理を施した融合タンパク質の溶解液をスプレードライしてパウダー化することも好ましい。さらに、前記再構成処理を施した融合タンパク質の溶解液を−20℃に一晩、次いで−80℃に1時間以上おいて凍結させ、さらに凍結乾燥機を用いて48時間凍結乾燥させた後、得られた乾燥シルクをワンダーブレンダーで粉砕し、一辺が1〜40μmのブロック状にパウダー化することがより好ましい。
【0045】
こうして再構成された絹糸においては、「機能的な一本鎖抗体」が発現している。ここで「機能的な一本鎖抗体」とは、抗原に対する結合性を有している一本鎖抗体を意味する。本発明により製造される一本鎖抗体は、その元になったモノクローナル抗体(親抗体)と比較して、少なくとも10%以上(好ましくは、20%以上、30%以上、40%以上、50%以上、60%以上、70%以上、80%以上、90%以上)の結合性を有する。結合性は、免疫学的手法を用いて得られる測定値(例えば、後述の図8に示すウェスタンブロッティング解析によって得られるバンドの濃さ)によって評価することができる。
【0046】
また、本発明によれば、その元になったモノクローナル抗体(親抗体)と同程度の結合特異性を有している「機能的な一本鎖抗体」を製造することができる。結合特異性は、免疫学的手法(例えば、ウェスタンブロッティング解析)を実施した際のバックグラウンドを指標に、評価することができる。すなわち、後述の図5に示すように、バックグラウンドが少ないほど特異性が高いと評価することができる。
【0047】
また、本発明の方法において、前記融合タンパク質の溶解の際又は前記再構成処理の際に、前記融合タンパク質の溶解液又は前記融合タンパク質を含むシルクゲルに酸化鉄ナノ粒子を添加してもよい。このように調製された酸化鉄ナノ粒子を含むシルクゲルを凍結乾燥等して粉砕することにより、酸化鉄ナノ粒子を含むシルクパウダーを得ることができ、該シルクパウダーは、磁気ビーズ抗体として目的とする抗原、ウィルス及び菌体を分離する用途等に好適に用いることができる。
【実施例】
【0048】
以下、実施例及び比較例に基づいて本発明をより具体的に説明するが、本発明は以下の実施例に限定されるものではない。
【0049】
(調製例1)
<フィブロインL鎖−抗WASP−scFv融合タンパク質を発現する遺伝子組換えカイコベクターの構築>
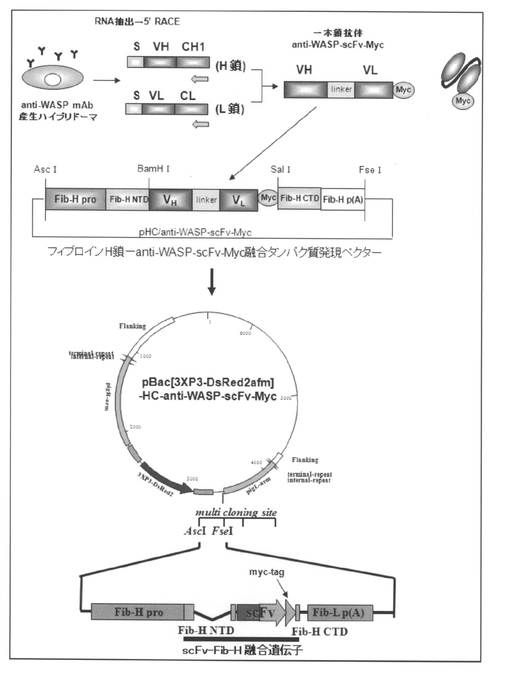
先ず、Sato,M.ら、FEBS J.、2005年、272巻、6131〜6144ページの記載に沿って、マウス ウィスコット−アルドリッチ症候群タンパク質(Wiskott−Aldrich syndrome protein、WASP)のN末端領域のEna/VASP homology 1(EVH1)ドメインに特異的なモノクローナル抗体を産生するハイブリドーマから抗体遺伝子をクローニングし、H鎖及びL鎖の可変領域(VH、VL)をフレキシブルなリンカー配列(アミノ酸配列:GGGGS×3)でつないだ一本鎖抗体(抗WASP−scFv)を構築した。次いで、得られたscFvのC末側に発現チェック用のMyc−tag(アミノ酸配列:EQKLISEEDL)を付加した。そして、この抗WASP−scFv−Myc融合タンパク質をコードするDNAフラグメントをpLLLベクターのBamHI−HindIIIサイトへ挿入し、フィブロインL鎖と抗WASP−scFv−Mycとの融合タンパク質を発現するDNAコンストラクトを作製した。さらに、この融合タンパク質(以下「フィブロインL鎖−抗WASP−scFv」とも称する)の発現ユニットを遺伝子組換えカイコ用ベクターpBac[3XP3−DsRed2afm]のEcoRI−BglIIサイトに挿入し、目的タンパク質を発現する組換えカイコ用プラスミドベクターpBac[3XP3−DsRed2afm]−LLL−anti−WASP−scFv−Mycを構築した(図1 参照)。なお、pLLLベクターについては非特許文献2の記載を、pBac[3XP3−DsRed2afm]については非特許文献1の記載を参照のこと。また、図1に記載のEcoRIサイトからBglIIサイトまでの塩基配列を配列表の配列番号:1にて示し、また該塩基配列がコードするアミノ酸配列を配列表の配列番号:2にて示す。なお、配列番号:1に記載の塩基配列において、1〜6bpはEcoRIサイトであり、7〜677bpはFib−L Pro(フィブロインL鎖遺伝子のプロモーター配列)であり、678〜1402bpはFib−L ORF(フィブロインL鎖の1位(メチオニン残基)〜242位(アラニン残基)からなるタンパク質をコードする塩基配列)であり、1403〜1772bpはVH(抗WASP−scFvのVHをコードする塩基配列)であり、1773〜1817bpはlinker(「フレキシブルなリンカー配列(アミノ酸配列:GGGGS×3)」をコードする塩基配列)であり、1818〜2165bpはVL(抗WASP−scFvのVLをコードする塩基配列)であり、2172〜2201bpはmyc−tag(Myc−tagをコードする塩基配列)であり、2205〜2210bpはHidIIIサイトであり、2211〜2678bpはFib−Lp(A)(フィブロインL鎖遺伝子の3’UTR及びポリAシグナル)であり、2679〜2684bpはBglIIサイトである。
【0050】
(調製例2)
<遺伝子組換えカイコの作出>
次に、構築したプラスミドpBac[3XP3−DsRed2afm]−LLL−anti−WASP−scFv−Mycと、piggyBacヘルパープラスミドpHA3PIGのDNA溶液(DNAの濃度が0.2μg/μlになるよう0.5mM リン酸緩衝液(pH7.0),5mM KClにて調製)とを、産卵後4〜10時間のカイコ(品種:w1−pnd)の受精卵794個にマイクロインジェクションした。
【0051】
なお、piggyBacヘルパープラスミドpHA3PIGについては非特許文献1の記載を参照のこと、また該ヘルパープラスミドには、カイコアクチンA3プロモーターで制御したpiggyBacの転移酵素遺伝子がクローニングされている。そのため、前記2種類のプラスミドDNAをカイコ受精卵に注入すると、ヘルパープラスミドから転移酵素が転写・翻訳され、この働きでベクタープラスミドから末端反復配列に挟まれたフラグメントが切り出され、ゲノム中のTTAA配列を標的としてランダムに挿入されるようになる。
【0052】
次いで、前記2種類のプラスミドDNAを注入した受精卵を25℃でインキュベーションして、幼虫を孵化させた。孵化した655頭の幼虫を人工飼料(日本農産工業)で飼育し、成虫に育てた。得られた成虫を兄妹交配して、162組の受精卵を得た。これらを産卵後6〜10日目に蛍光顕微鏡下で観察し、胚の神経系統で赤色蛍光タンパク質の発現が認められる個体すなわち目的の組換えカイコを識別した。そして、得られた遺伝子組換えカイコ個体(以下「S01系統」又は「S01」とも称する)飼育を続けた。
【0053】
(調製例3)
<組換えカイコ産生繭におけるフィブロインL鎖−抗WASP−scFvの発現>
前記の通りにして得られた「S01」等由来の繭を2mm角程度に切り、繭20mgに対して9M 臭化リチウム(LiBr)溶液1mLを加え、ローテーターで、室温、3時間撹拌して完全に溶解した。そして、10,000×gで20分間(室温)遠心後、上清を回収し、水で2倍希釈した後、等量の2×SDSサンプルバッファーを加えて、65℃で30分間インキュベートした。調製したサンプルをSDS−PAGEにて分離し、PVDFメンブレンにトランスファーした後、抗フィブロインL鎖抗体(anti−FibL pAb、本発明者小島作製)および抗Myc−tag抗体(anti−Myc pAb、MBL社製)と室温で1時間反応後、HRP−標識抗ウサギ抗体と室温で1時間反応させ、ECL試薬(GEヘルスケア社製)を用いて検出した。得られた結果を図2に示す。
【0054】
図2に示した結果から明らかなように、抗フィブロインL鎖抗体を用いたウェスタンブロットにおいて、遺伝子組換えカイコ S01由来の繭では、内在性(天然)のフィブロインL鎖(27kDa)と共に抗WASP−scFv−Mycが融合した約60kDaのタンパク質(フィブロインL鎖−抗WASP−scFv)が強く検出された(図2のa 参照)。さらに、抗Myc−tag抗体を用いたウェスタンブロットにおいても同様に60kDaのタンパク質が強く検出されたことから(図2のb 参照)、今回作出した組換えカイコが産生した絹糸において、目的とする融合タンパク質(フィブロインL鎖−抗WASP−scFv)が非常に多く発現していることが確かめられた。
【0055】
このようにして得られた遺伝子組換えカイコ S01由来の繭から以下に示す3種類の方法にて、融合タンパク質 フィブロインL鎖−抗WASP−scFvを調製した(図3及び図4 参照)。
【0056】
(実施例1)
<未精練シルクパウダーの調製方法>
非組換え体であるw1−pnd(以下「w1」とも称する)由来、S01由来及びフィブロインL鎖−緑色蛍光タンパク質(EGFP)の融合タンパク質(EGFP−Fib−L)を発現する組換えカイコ(未発表、以下「K27系統」又は「K27」とも称する)由来の繭をそれぞれ3g用意し、裁断した後9M LiBr溶液30mLに浸し、4時間撹拌して完全に溶解した。得られたシルク溶液を透析膜(再生セルロース膜、製品名:Spectra/Por、MWCO 6−8000)に入れ、精製水(RO水)8Lに対して12時間×6回透析を行った。透析により得られたシルクゲルを−20℃に一晩、次いで−80℃に1時間以上おいて凍結させ、さらに凍結乾燥機を用いて48時間凍結乾燥した。得られた乾燥シルクを粉砕器(製品名:ワンダーブレンダー、大阪ケミカル株式会社製)で粉砕し、未精練シルクパウダーとした。
【0057】
(比較例1)
(繭をそのまま粉砕してパウダーにする方法)
w1由来及びS01由来の繭それぞれ3gをRO水に1日浸し、−80℃で凍結・乾燥後、そのままワンダーブレンダーを用いて粉砕した。
【0058】
(比較例2)
<精練シルクパウダーの調製方法>
w1由来、S01由来及びK27由来の繭をそれぞれ4g用意し、8M Urea,0.5% 2−メルカプトエタノールに浸して80℃で30分間撹拌してセリシンを除去した(尿素精練)。そして、w1由来、S01由来及びK27由来の未精錬の繭3gの代わりに得られた精練シルク(フィブロインのみを含む)3gを用いた以外は、実施例1と同様の操作によりシルクパウダー(精練シルクパウダー)を作製した。
【0059】
このように異なる調製方法(実施例1、比較例1〜2)によって調製されたシルクパウダーについて、下記の方法にて評価した。
【0060】
<電子顕微鏡による観察>
実施例1及び比較例1〜2で得られたシルクパウダーの形状に関して電子顕微鏡観察したところ、比較例1で得られた粉砕しただけのパウダーはまだ繊維状の構造をとっているが(直径10〜15μm、長さ数mm)、実施例1及び比較例2で得られた、一度溶解し、凍結乾燥・粉砕したパウダーは一辺が1〜40μmのブロック状の構造をとっていることが確認された(図3 参照)。
【0061】
<シルクパウダーの抗原特異的な結合解析>
実施例1及び比較例2にて調製したシルクパウダーと、抗原であるGST−WASP−NT及びコントロールであるGSTとを各々反応させ、その後プルダウンアッセイにより、フィブロインL鎖−抗WASP−scFvを有するシルクパウダーと抗原との複合体が形成されているかを調べた(図4 参照)。すなわち、各方法にて調製したシルクパウダーをそれぞれ10mgずつ量り取り、1.5mLチューブに入れ、20%エタノール溶液1mLを加えてよく懸濁し、分散させた。分散後、遠心して上清のエタノール溶液を捨て、続けてブロッキング液(製品名:ブロッキングワン、ナカライテスク社製)1mLを加えて室温、90分間ローテーターにて撹拌しながらインキュベートし、非特異的な結合を抑える操作(ブロッキング)を施した。ブロッキング後、大腸菌で発現させてアフィニティー精製した組換えタンパク質GST、GST−WASP−NT(抗WASP−scFvの抗原)をプローブタンパク質(それぞれ20μg/mLの濃度になるようブロッキング液にて調製)として、シルクパウダーと共に室温で60分間ローテーターにて撹拌しながらインキュベートした。次いで、TBST溶液(10 mM Tris−HCl,pH8.0,0.15M NaCl,0.05% Tween−20)で5回洗浄することにより、プルダウンアッセイを行った(図4 参照)。得られたサンプルに2×SDSサンプルバッファーを加えて100℃で10分間ボイルした。そして、ボイルしたサンプルをSDS−PAGEした後、PVDFメンブレンにトランスファーして、抗GST抗体を用いてウェスタンブロットを行った。得られた結果を図5に示す。
【0062】
図5に示した結果から明らかなように、繭をそのまま粉砕したシルクパウダー(比較例1)においては、w1由来及びS01由来のパウダー共に非特異的なプローブタンパク質の結合が見られたが(図5のa 参照)、実施例1で得られたシルクパウダーでは、非特異的な結合が抑制され、S01由来のシルクパウダーのみに抗原であるGST−WASP−NTとの特異的な結合が検出された(図5のb 参照)。従って、本発明の方法によって調製された一本鎖抗体は、抗原への結合活性が維持されており、しかも非特異的な結合が抑制されていることが明らかになった。さらに、本発明の方法によって調製された一本鎖抗体は、融合しているフィブロインL鎖を介して、内在性の他のフィブロインタンパク質(フィブロインL鎖、フィブロインH鎖、フィブロインヘキサメリン)及びセリシンと複合体を再構成することによりシルクパウダーの表面上に発現していることが示唆された。しかも、前述の通り、一本鎖抗体の活性をシルクパウダーの表面上で再現でき、シルクパウダーを担体として利用することもできるため、本発明の方法によって調製された一本鎖抗体を有するシルクパウダーはそのままプルダウンアッセイに供することができることも明らかになった。
【0063】
<シルクパウダーの抗原特異的な結合活性の評価>
実施例1で得られたシルクパウダーに関して、さらに抗原特異性を評価するために、抗原であるGST−WASP−NT、抗原部位を欠損させたWASPとGSTとの融合タンパク質であるGST−WASP−CT、さらにWASPと全く相同性のないマウスの免疫系細胞で発現しているP2X7レセプターの細胞内ドメインとGSTとの融合タンパク質GST−P2X7−CT2の3種類を大腸菌で発現させてアフィニティー精製し、プローブタンパク質(それぞれ20μg/mLになるようブロッキング液にて調製)として用意した。そして、前記同様に、w1由来、S01由来、K27由来のシルクパウダーと反応させた後、プルダウンアッセイ、並びに抗Myc抗体及び抗GFP抗体を用いたウェスタンブロットにより抗原−抗体反応による複合体を形成しているかを確かめた。得られた結果を図6に示す。
【0064】
また、比較例2で得られたシルクパウダーに関しても、前記実施例1と同様に、抗原特異性を評価するために、抗原であるGST−WASP−NT、GST−WASP−CT又はGST−P2X7−CT2の3種類のプローブタンパク質を用いたプルダウンアッセイにて抗原−抗体反応による複合体を形成しているかを確かめた。得られた結果を図7に示す。
【0065】
図6に示した結果から明らかなように、実施例1で得られたシルクパウダーにおいては、S01由来のシルクパウダーのみに抗原であるGST−WASP−NTとの強い結合が確認され、他のプローブタンパク質との特異的な結合はほとんど見られなかった(図6のa 参照)。さらに抗Myc抗体及び抗GFP抗体を用いたウェスタンブロットにより、結合解析に用いたそれぞれのシルクパウダーの性質及び量に関してほぼ同等であることが確認された(図6のb及びc 参照)。従って、本発明の方法によって調製された、シルクパウダー上の一本鎖抗体は、優れた抗原特異性を維持して、抗体分子として機能していることが確かめられた。
【0066】
一方、図7に示した結果から明らかなように、比較例2で得られたシルクパウダーにおいては、w1由来、S01由来及びK27由来のシルクパウダーのいずれも、GST−WASP−NT、GST−WASP−CT及びGST−P2X7−CT2と結合していた。従って、精錬した繭をLiBr溶液に溶解し、透析して調製したシルクパウダーでは、非特異的結合が強く、抗体分子として機能しているかどうか評価することが出来なかった。
【0067】
<抗WASP−EVH1モノクローナル抗体を用いた免疫沈降>
抗WASP−scFvの親抗体である完全長の抗WASPモノクローナル抗体(anti−WASP mAb)とプロテインGセファロースとの複合体を用いたプルダウンアッセイを行い、実施例1で得られた抗WASP−scFvにおいて確認された抗体活性(抗原特異性や結合性)と比較した。すなわち、組換えタンパク質GST−WASP−NT、GST−WASP−CT及びGST−P2X7−CT2をそれぞれPBSバッファーで20μg/mLに調整し、プロテインGセファロース(GEヘルスケア社製)40μLを加え、4℃で60分間ローテーターにてインキュベートして、プロテインGセファロースに非特異的に吸着する成分を取り除いた(プレクリアー)。そして、10,000×gで10分間遠心した後、上清を回収し、そこに抗WASP−scFvの親抗体である抗WASP−EVH1モノクローナル抗体5μgとプロテインGセファロース40μLとを加え、4℃で3時間ローテーターにてインキュベートした。次いで、PBSで5回洗浄した後、2×SDSサンプルバッファーを加えて100℃で10分間ボイルした。このようにして得られたサンプルをSDS−PAGEした後、PVDFメンブレンにトランスファーして、抗GST抗体を用いてウェスタンブロットを行った。得られた結果を図8に示す。
【0068】
図8に示した結果から明らかなように、実施例1で得られた抗WASP−scFvの抗体活性は、親抗体である抗WASPモノクローナル抗体とほぼ同等であることが確認された。従って、本発明の方法によって調製された一本鎖抗体は、元としたモノクローナル抗体と同等である程、抗原への特異性及び結合性が高く維持されていることが明らかになった。
【0069】
(調製例4)
<フィブロインH鎖−抗WASP−scFv融合タンパク質を発現する遺伝子組換えカイコベクターの構築>
フィブロインL鎖の時と同様に、抗WASP−scFv−MycのDNAコンストラクトをpHCベクターのBamHI−SalIサイトへ挿入し、フィブロインH鎖のN末側の一部(NTD)とC末側の一部(CTD)の間に抗WASP−scFv−Mycが挟まれたような形状で融合タンパク質を発現するDNAコンストラクトを作製した。さらにこの融合タンパク質の発現ユニットを遺伝子組換えカイコ用ベクターpBac[3XP3−Dsred2afm]のAscI−FseIサイトに挿入し、目的タンパク質を発現する組換えカイコ用プラスミドベクターpBac[3XP3−DsRed2afm]−HC−anti−WASP−scFv−Mycを構築した(図9 参照)。なお、pHCベクターについては非特許文献3の記載を、pBac[3XP3−DsRed2afm]については非特許文献1の記載を参照のこと。また、図9に記載のAscIサイトからFseIサイトまでの塩基配列を配列表の配列番号:3にて示し、該塩基配列がコードするアミノ酸配列を配列表の配列番号:4にて示す。なお、配列番号:3に記載の塩基配列において、1〜8bpはAscIサイトであり、9〜1133bpはFib−H Pro(フィブロインH鎖遺伝子のプロモーター配列)であり、1134〜1175bpはFib−H NTD(N−terminal domain(N末端領域))の第1エクソン(フィブロインH鎖の1位(メチオニン残基)〜14位(グルタミン残基)からなるタンパク質をコードする塩基配列)であり、1176〜2144bpはフィブロインH鎖遺伝子の第1イントロンであり、2145〜2561bpはFib−H NTDの第2エクソン(フィブロインH鎖の15位(チロシン残基)〜153位(アラニン残基)からなるタンパク質をコードする塩基配列)であり、2562〜2567bpはBamHIサイトであり、2568〜2936bpはVH(抗WASP−scFvのVHをコードする塩基配列)であり、2937〜2981bpはlinker(「フレキシブルなリンカー配列(アミノ酸配列:GGGGS×3)」をコードする塩基配列)であり、2982〜3329bpはVL(抗WASP−scFvのVLをコードする塩基配列)であり、3336〜3365bpはmyc−tag(Myc−tagをコードする塩基配列)であり、3366〜3371bpはSalIサイトであり、3372〜3551bpはFib−H CTD(C−terminal domain(C末端領域))(フィブロインH鎖の5205位(セリン残基)〜5263位(システイン残基)からなるタンパク質をコードする塩基配列)であり、3552〜3851bpはFib−Hp(A)(フィブロインH鎖遺伝子の3’UTR及びポリAシグナル)であり、3852〜3859bpはFseIサイトである。
【0070】
(調製例5)
<遺伝子組換えカイコの作出>
次に、構築したプラスミドpBac[3XP3−DsRed2afm]−HC−anti−WASP−scFv−Mycと、piggyBacヘルパープラスミドpHA3PIGのDNA溶液(DNAの濃度が0.2μg/μlになるよう、0.5mM リン酸緩衝液(pH7.0),5mM KClにて調製)を産卵後4〜10時間のカイコ(品種:w1−pnd)の受精卵396個にマイクロインジェクションした。これを25℃でインキュベーションして幼虫を孵化させた。孵化した126頭の幼虫を人工飼料(日本農産工業)で飼育し、成虫に育てた。得られた成虫を兄妹交配して、85組の受精卵を得た。これらを産卵6〜10日目に蛍光顕微鏡下で観察し、胚の神経系統で赤色蛍光タンパク質の発現が認められる個体すなわち目的の組換えカイコを識別した。得られた遺伝子組換えカイコ個体を、S02系統又はS02と呼称し、飼育を続けた。
【0071】
(実施例2)
<未精練シルクパウダーの調製方法>
実施例1に記載の方法(フィブロインL鎖とscFvとの融合タンパク質において抗体活性が保持されていたパウダー化の方法)に従って、未精練のS02由来の繭からシルクパウダーを調製した。
【0072】
<組換えカイコ産生繭におけるフィブロインH鎖−抗WASP−scFvの発現>
実施例1及び2にて得られた未精練シルクパウダー10mgに対して9M臭化リチウム(LiBr)溶液500μLを加え、ローテーターで室温にて3時間撹拌して完全に該シルクパウダーを溶解させた。次いで、10,000xg、室温にて20分間遠心して上清を回収し、水で2倍希釈した後、等量の2xSDSサンプルバッファーを加えて、65℃で30分間インキュベートした。このように調製したサンプルをSDS−PAGEにて分離し、PVDFメンブレンにトランスファーした後、抗フィブロインH鎖(FibH)抗体(本発明者小島作成)、抗Myc−tag抗体(MBL社製)、抗フィブロインL鎖(FibL)抗体又は抗GFP抗体と室温にて1時間反応させた後、HPR−標識抗ウサギ抗体等の前記一次抗体に対応する各二次抗体と室温にて1時間反応させ、ECL試薬(GEヘルスケア社製)を用いて検出した。得られた結果を図10に示す。
【0073】
図10に示した結果から明らかなように、抗Myc抗体及び抗フィブロインH鎖抗体を用いたウェスタンブロットにおいて、約70kDaの大きさに目的とするFibH−anti−WASP−scFv−Myc融合タンパク質がS02由来の未精錬シルクパウダーには発現していることが確認された。なお、内在性のフィブロインH鎖タンパク質は350kDaの巨大なタンパク質なので、本ウェスタンブロット解析では検出されなかった。また、FibH−anti−WASP−scFv−Myc融合タンパク質を構成していないフィブロインL鎖やGFPに対する抗体を用いたウェスタンブロットにおいて、S02由来の未精錬シルクパウダーには特異的なバンドは検出されなかった。
【0074】
<シルクパウダーの抗原特異的な結合解析>
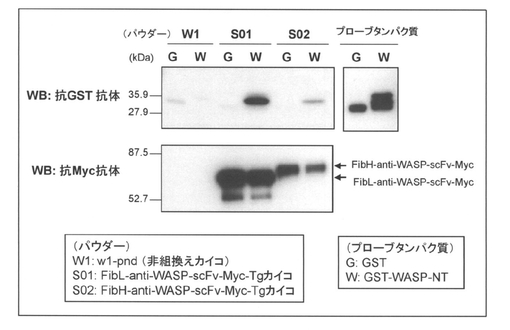
実施例1及び2にて得られた未精練シルクパウダー(W1由来、S01由来、S02由来)を10mgずつ測り取り、1.5mLチューブに入れ、20%エタノール1mLを加えよく懸濁し、分散させた後、遠心して上清のエタノール溶液を捨て、続けて、非特異的な結合を抑えるために、ブロッキング液(製品名:ブロッキングワン、ナカライテスク社製)1mLを加えて室温にて90分間ローテーターを用いて撹拌しながらインキュベートした。ブロッキング後、大腸菌で発現させた組換えタンパク質GST及びGST−WASP−NT(scFvの抗原)をプローブタンパク質(各々20μg/mLになるようブロッキング液にて調製)として、シルクパウダーと室温にて60分間ローテーターを用いて撹拌しながらインキュベートした。次いで、TBST溶液(10mM Tri−HCl、pH8.0、0.15M NaCl、0.05% Tween−20)にて5回洗浄した後、2xSDSサンプルバッファーを加えて100℃にて10分間ボイルした。このように調製したサンプルをSDS−PAGEにて分離した後、PVDFメンブレンにトランスファーして、抗GST抗体、抗Myc抗体を用いてウェスタンブロットを行った。得られた結果を図11に示す。
【0075】
図11に示した結果から明らかなように、S01由来の未精錬シルクパウダーのみならず、S02由来の未精錬シルクパウダーにおいても抗原特異的な結合活性が確認された(図11上部パネル WB:抗GST抗体 参照)。また、かかる結合活性の強さは、各々のパウダーにおける一本鎖抗体(scFv)の発現量に比例していることが示唆された(図11下部パネル WB:抗Myc抗体 参照)。
【0076】
以上の結果から、本発明の方法によって調製されたシルクパウダー上の一本鎖抗体は、フィブロインタンパク質のL鎖及びH鎖のいずれに融合させた形で発現させても、優れた抗原特異性を維持して、抗体分子として機能していることが確かめられた。
【産業上の利用可能性】
【0077】
以上説明したように、本発明によれば、抗体をその結合性や抗原特異性を維持させたまま、非常に簡便に絹糸から調製することが可能となる。しかも、本発明の製造方法によって得られる一本鎖抗体は、該一本鎖抗体の元になったモノクローナル抗体(親抗体)と同程度の極めて高い結合性及び抗原特異性を有しており、さらには該一本鎖抗体に融合しているフィブロインタンパク質等から構成されるシルクパウダーを担体として利用することもできる。したがって、抗体生産・精製・担体への固定化のプロセスが簡略化され、従来の方法と比較して安価な抗体活性を利用したアフィニティー担体を提供することが可能となり、様々な病原体の検出・同定や疾病診断を目的とした新しいバイオマテリアルとしての有効である。
【技術分野】
【0001】
本発明は、一本鎖抗体(scFv)の製造方法に関し、より詳しくは、scFvとフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解し、再構成処理を施すことを特徴とする、scFvの製造方法に関する。
【背景技術】
【0002】
抗体は、抗原と結合するドメイン(Fab)と免疫グロブリン受容体に結合するドメイン(Fc)より構成され、Fabドメインを介して病原体の抗原に結合し、それを排除して中和する免疫調節作用を発揮する。そして、その作用の根幹は、ペプチド、核酸、糖鎖、糖脂質、低分子化合物等の種々様々な抗原に対して特異的に結合できることにある。そのため、抗体は、前記免疫調節作用を目的とした医薬品(抗体医薬)のみならず、ウェスタンブロッティング、免疫沈降(プルダウンアッセイ)、フローサイトメトリー、免疫組織染色等の各種検出・精製方法に適したツールとしても利用され、開発が進められており、これらの市場は急速に拡大している。
【0003】
通常、抗体は、検出・精製の対象とする抗原をウサギやマウス等の免疫動物に接種し、該動物の免疫系を刺激した後、その動物の血清(ポリクローナル抗体)を回収することで得ることができる。また、このように免疫系を刺激した動物から脾臓細胞等の抗体産生細胞を回収し、該抗体産生細胞とミエローマ細胞とを融合し、得られたハイブリドーマを選択することにより所望の抗原のみを検出・精製することのできる抗体(モノクローナル抗体)を作製することもできる。しかしながら、かかる動物や動物細胞を利用した方法には、活性を有する抗体を安定的に取得できる反面、大量生産に適していない、生産コストが非常に高くなるといった欠点がある。
【0004】
これに対し、所望の抗体をコードする遺伝子をクローニングすることにより、タンパク質の大量生産に適した大腸菌等で抗体を生産することも試みられている。しかし、大腸菌で生産した場合には、得られる抗体がしばしば不溶化することが知られており、その精製にあたっては、ドデシル硫酸ナトリウム(SDS)等の変性剤でタンパク質を可溶化させる工程やその変性剤を取り除く透析等の工程が必要となる。さらにそのような多くの工程を経て得られた抗体は概して活性が低くなる傾向にあるため、大腸菌等を利用した方法には、活性を有する抗体を大量に生産しにくいという欠点がある。
【0005】
一方、カイコは蛹化する際に自身を保護する目的で繭を作り出す。この繭はフィブロイン(Fibroin)という繊維構造とそれを取り巻くセリシンとの2種の構造からなる絹糸から作られている構造体である。絹糸は、およそ1.5kmに達する単繊維として存在するため、人類はこれをほどいて繊維とし、衣類等に利用してきた。近年では、絹糸を構成するするフィブロイン及びセリシンをそれぞれ加工することによって、パウダー(特許文献1)やフィルム(特許文献2及び3)、及びスポンジ様構造体(特許文献4)として利用することも可能となってきた。
【0006】
2000年にカイコの遺伝子組換え技術が確立され(非特許文献1)、外来遺伝子を導入することによりカイコのシルクタンパク質を改変し、物性の向上や、機能性の付加を実現した新しいシルクを設計・創出することが可能となった。現在では目的のタンパク質を絹タンパク質の主成分であるフィブロイン層とセリシン層のどちらにでも発現させることが可能となり、これまでにフィブロインと蛍光タンパク質やサイトカイン等との融合タンパク質を発現する組換えカイコや(非特許文献2〜4)、またセリシン層にヒト血清アルブミンや抗体分子を発現する組換えカイコの作出に成功し(非特許文献5及び6)、蛍光絹糸としての利用や発現産物を絹糸から分離・精製して利用する試みが行われており、前述の抗体のみならず、機能的なタンパク質の生産の場としてカイコは注目を集めている。
【0007】
例えば、飯塚らは、マウスモノクローナル抗体のH鎖及びL鎖を中部絹糸腺細胞で発現させ、セリシン層にそれら抗体分子を分泌する組換えカイコを作製した。そして、該組換えカイコが作出した絹糸を3M 尿素(Urea)を含む50mM Tris−HCl(pH7.4)溶液で処理し、抗体分子を含んだセリシン溶液を透析後、プロテインGセファロースを用いてアフィニティー精製することにより、抗原特異的な結合活性を有する抗体分子を得ている(非特許文献6)。
【0008】
また、外来タンパク質をフィブロインに発現させ、活性をもつ該タンパク質を調製するためには、一般的に絹糸を高濃度の臭化リチウム溶液やチオシアン酸リチウム溶液、又は塩化カルシウム/エタノール溶液で一度完全に溶解し、得られたシルク複合体から目的の外来タンパク質をプロテアーゼや還元剤を用いて遊離させ、再びリフォールディングすることが必要である。実際、日野らは、フィブロインL鎖にヒトbFGFを融合させた組換えタンパク質を絹糸に発現させ、絹糸を精練し、セリシン成分を除去した後、高濃度のチオシアン酸リチウムに溶解し、2−メルカプトエタノールを用いて内在性フィブロインから遊離させ、さらに透析を施し、変性剤の濃度を徐々に下げていくことで、活性のあるbFGFを獲得している(非特許文献4)。
【0009】
このように、外来タンパク質を発現させた絹糸から活性を有する該タンパク質を調製するためには、アフィニティー精製、プロテアーゼ処理、還元剤の添加及び精練等を行い、外来タンパク質から内在性タンパク質を遊離・除去する工程が必要であった。また、絹糸から外来タンパク質を抽出するためには、変性剤(尿素、臭化リチウム溶液、塩化カルシウム/エタノール溶液、チオシアン酸リチウム、塩酸グアニジン、SDS等)にて該タンパク質を変性させ可溶化する工程が必要となる。さらに、変性して可溶化させたタンパク質から変性剤を取り除き、該タンパク質が活性を有するようにするために、リフォールデイング処理を施す工程を要する。しかしながら、かかる多工程を経て調製されたタンパク質、特に抗体は概してその活性が低くなる傾向にあった。
【0010】
そこで、絹糸から抗体等の外来タンパク質をその活性を維持させたまま、非常に簡便に調製する方法の開発が望まれているが、未だ実用化されていないのが現状である。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】特開2004−123683号公報
【特許文献2】特開2008−173312号公報
【特許文献3】特開平9−192210号公報
【特許文献4】特開2002−186847号公報
【非特許文献】
【0012】
【非特許文献1】Tamura,T.ら、Nat.Biotechnol.、2000年、18巻、81〜84ページ
【非特許文献2】Inoue,S.ら、Insect Biochem.Mol.Biol.、2005年、35巻、51〜59ページ
【非特許文献3】Kojima,K.ら、Biosci.Biotechnol.Biochem.、2007年、71巻、2943〜2951ページ
【非特許文献4】Hino,R.ら、Biomaterials.、2006年、27巻、5715〜5724ページ
【非特許文献5】Ogawa,S.ら、J.Biotechnol.、2007年、128巻、531〜544ページ
【非特許文献6】Iizuka,M.ら、FEBS J、2009年、276巻、5806〜5820ページ
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、前記従来技術の有する課題に鑑みてなされたものであり、絹糸から抗体をその結合性や抗原特異性を維持させたまま、内在性タンパク質を遊離・除去する工程等を要せずに非常に簡便に調製することを可能とする、抗体の製造方法を提供することを目的とする。
【課題を解決するための手段】
【0014】
本発明者らは、前記目的を達成すべく鋭意研究を重ねた結果、抗体として一本鎖抗体(scFv)を選択し、このscFvをフィブロインタンパク質と融合させて絹糸に発現させ、これにより得られた絹糸に対し、精錬せずに溶解と再構成の処理を行うことにより、結合性及び抗原特異性を高く保持した一本鎖抗体を製造できることを見出し、本発明を完成するに至った。
【0015】
本発明は、より詳しくは、下記を提供するものである。
<1> 一本鎖抗体(scFV)とフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解する工程と、該工程で得られた前記融合タンパク質の溶解液に再構成処理を施す工程と、を含むことを特徴とする、scFVの製造方法。
<2> 分子内又は分子間の水素結合を切断する性質を有する溶液で絹糸を溶解する、<1>に記載の製造方法。
<3> 前記再構成処理を施した融合タンパク質の溶解液を凍結乾燥して粉砕する工程をさらに含むことを特徴とする、<1>又は<2>に記載の製造方法。
<4> 前記フィブロインタンパク質が、フィブロインL鎖及びフィブロインH鎖からなる群から選択される少なくとも一のタンパク質であることを特徴とする、<1>〜<3>のうちのいずれか一に記載の製造方法。
<5> 前記再構成処理が透析処理であることを特徴とする、<1>〜<4>のうちのいずれか一に記載の製造方法。
【発明の効果】
【0016】
本発明によれば、絹糸から一本鎖抗体をその結合性や抗原特異性を維持させたまま、非常に簡便に調製することが可能となる。
【図面の簡単な説明】
【0017】
【図1】抗WASPモノクローナル抗体(anti−WASP mAb)を基に一本鎖抗体(抗WASP−scFv(anti−WASP−scFv−Myc))を得た過程と、フィブロインL鎖−抗WASP−scFv(フィブロインL鎖−anti−WASP−scFv−Myc融合タンパク質)をカイコが産生する絹糸に発現させるためのベクター(pBac[3XP3−DsRed2afm]−LLL−anti−WASP−scFv−Myc)のコンストラクトとを示す、概要図である。
【図2】組換えカイコ(S01)産生繭におけるフィブロインL鎖−抗WASP−scFvの発現を、ウェスタンブロッティングによって検出した結果を示す図である。なお、図中の「w1」は非組換え体カイコ(w1−pnd、陰性対照)産生繭における結果であることを示し、「♯14〜16」はS01系統内の異なる個体由来の繭における結果であることを示す。また、a)は抗フィブロインL鎖抗体(anti−FibL pAb)を用いたウェスタンブロッティングの結果を示し、b)は抗Myc−tag抗体(anti−Myc pAb)を用いたウェスタンブロッティングの結果を示す。
【図3】組換えカイコ(S01)産生繭(TGカイコ繭)から融合タンパク質 フィブロインL鎖−抗WASP−scFvを有するシルクパウダーを調製するための3種類の方法(実施例1、比較例1〜2)の概略、並びにこれらの方法によって得られたシルクパウダーを電子顕微鏡で観察した結果を示す写真である。
【図4】組換えカイコ(S01)産生繭(TGカイコ繭)から融合タンパク質 フィブロインL鎖−抗WASP−scFvを有するシルクパウダーを調製するための3種類の方法(実施例1、比較例1〜2)の概略、及びこれらの方法によって得られたシルクパウダーと抗WASP−scFvの抗原であるWASPタンパク質との複合体形成を調べるためのプルダウンアッセイの概略を示す図である。
【図5】抗GST抗体を用いたウェスタンブロッティングによって分析した、図4に示したプルダウンアッセイの結果を示す図である。なお、図中の「W1」は非組換え体カイコ(w1−pnd、陰性対照)産生繭における結果であることを示し、「W」及び「G」は「WASPタンパク質(GST−WASP−NT)」及び「GSTタンパク質(陰性対照、GST)」を用いたプルダウンアッセイの結果であることを各々示す。また、a)は比較例1の方法によって得られたシルクパウダーの分析結果であることを示し、b)は実施例1の方法によって得られたシルクパウダーの分析結果であることを示す。
【図6】本発明(実施例1)の方法によって得られたシルクパウダーと、各種プローブタンパク質との複合体形成をプルダウンアッセイ並びにウェスタンブロッティングによって分析した結果を示す図である。
【図7】比較例2に記載の方法によって得られたシルクパウダーと、各種プローブタンパク質との複合体形成をプルダウンアッセイ並びにウェスタンブロッティングによって分析した結果を示す図である。
【図8】本発明(実施例1)の方法によって得られたフィブロインL鎖−抗WASP−scFvを有するシルクパウダーと、抗WASP−scFvの元になった抗WASPモノクローナル抗体とを、結合性及び抗原特異性の点において、各種プローブタンパク質を用いたプルダウンアッセイ並びにウェスタンブロッティングによって分析して比較した結果を示す図である。
【図9】抗WASPモノクローナル抗体(anti−WASP mAb)を基に一本鎖抗体(抗WASP−scFv(anti−WASP−scFv−Myc))を得た過程と、フィブロインH鎖−抗WASP−scFv(フィブロインH鎖−anti−WASP−scFv−Myc融合タンパク質)をカイコが産生する絹糸に発現させるためのベクター(pBac[3XP3−DsRed2afm]−HC−anti−WASP−scFv−Myc)のコンストラクトとを示す、概要図である。
【図10】本発明(実施例2)の方法によって得られた組換えカイコ(S02)産生繭由来シルクパウダーにおけるフィブロインH鎖−抗WASP−scFvの発現を、ウェスタンブロッティングによって検出した結果を示す図である。なお、図中の「*(アスタリスク)」は内在性フィブロインL鎖の位置を示す。
【図11】本発明(実施例2)の方法によって得られたシルクパウダーと、各種プローブタンパク質との複合体形成をプルダウンアッセイ並びにウェスタンブロッティングによって分析した結果を示す図である。
【発明を実施するための形態】
【0018】
本発明は、機能的な一本鎖抗体(scFV)の製造方法であって、scFVとフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解する工程と、該工程で得られた前記融合タンパク質の溶解液に再構成処理を施す工程と、を含むことを特徴とする方法である。
【0019】
本発明において「一本鎖抗体(single chain fragment variable:scFv)」とは、抗体分子の抗原と結合するドメイン(Fab)を構成する重鎖(H鎖)及び軽鎖(L鎖)から可変領域(VH及びVL)を取り出して、リンカーを介して結合させた分子を意味する。本発明にかかる一本鎖抗体には、単一のポリペプチド鎖中に複数のVHとVLとを有している抗体も含まれる。かかる抗体としては、例えば、二つのVHと二つのVLとを前記リンカー等で結合した一本鎖抗体(sc(FV)2)が挙げられる。かかる抗体においては、通常複数のリンカーが必要となるが、全て同じリンカーであってもよく、異なるリンカーであってもよい。また、本発明にかかる一本鎖抗体には、リンカーペプチドの長さを短く(例えば5アミノ酸程度)に設定して、同一ポリペプチド鎖内でのVHとVLとの結合を抑制することにより形成される二量体(ダイアボディ、diabody)も含まれる。
【0020】
本発明にかかる一本鎖抗体におけるリンカーは、その両端に連結されたVH及びVLの発現や、VHとVLとの結合を阻害するものでなければ特に制限されない。リンカーペプチドの長さは、通常、1〜100アミノ酸、好ましくは1〜50アミノ酸、より好ましくは1〜30アミノ酸、特に好ましくは12〜18アミノ酸(例えば15アミノ酸)である。本発明においては、「(GGGGS)×3」という15アミノ酸からなるポリペプチドをリンカーとして好適に用いることができる(なお、GはグリシンをSはセリンを表わす)。
【0021】
一本鎖抗体の調製は、例えば、次のようにして行うことができる。まず、目的の抗原と特異的に結合するモノクローナル抗体を発現しているハイブリドーマよりmRNAを単離し、そのmRNAを用いて作製したcDNAライブラリーから、そのモノクローナル抗体のVH及びVLをコードするcDNAをそれぞれ単離する。次いで、VHをコードするcDNAとVLをコードするcDNAの間にリンカーをコードするDNAを挿入し、遺伝子組換え技術により、一本鎖抗体をコードする組換えDNAを構築する。
【0022】
cDNAライブラリーを作製せずに、mRNAよりRT−PCTを用いて、VH及びVLをコードする遺伝子を直接増幅させ、こうして増幅して得られた断片を用いて、遺伝子組換え技術により、一本鎖抗体をコードするDNAを構築することもできる。
【0023】
本発明にかかる「フィブロインタンパク質」とは、フィブロインL鎖、フィブロインH鎖及びフィブロインヘキサメリン(P25)からなる群から選択される少なくとも一のタンパク質を意味する。フィブロインタンパク質は、フィブロインL鎖とフィブロインH鎖とがジスルフィド結合し、この結合した分子6個に対し1つの割合でフィブロインヘキサメリンが結合し、複合体(フィブロイン複合体)が形成されることが知られている。
【0024】
本発明にかかる「フィブロインL鎖」は、分子量が約3万Daのフィブロインタンパク質であり、典型例として、GenBankアクセッション No.NP_001037488.1で特定されるタンパク質のうちの17〜262番目のアミノ酸配列からなるタンパク質(GenBankアクセッション No.NM_001044023.1で特定されるDNAのうちの90〜827番目の塩基配列からなる遺伝子)が挙げられる。また、このタンパク質は、シグナルペプチド(GenBankアクセッション No.NP_001037488.1で特定されるタンパク質のうちの1〜16番目のアミノ酸配列からなるタンパク質)が前駆体より切断、除去されることにより得られる成熟型であるから、本発明にかかる「フィブロインL鎖」としては、例えば、GenBankアクセッション No.NP_001037488.1で特定されるタンパク質(GenBankアクセッション No.NM_001044023.1で特定される遺伝子)が挙げられる
本発明にかかる「フィブロインH鎖」は、分子量が約35万Daのフィブロインタンパク質であり、典型例として、GenBankアクセッション No.NP_001106733.1で特定されるタンパク質のうちの22〜5263番目のアミノ酸配列からなるタンパク質(GenBankアクセッション No.NM_001113262.1で特定されるDNAのうちの64〜15789番目の塩基配列からなる遺伝子)が挙げられる。また、このタンパク質は、シグナルペプチド(GenBankアクセッション No.NP_001106733.1で特定されるタンパク質のうちの1〜21番目のアミノ酸配列からなるタンパク質)が前駆体より切断、除去されることにより得られる成熟型であるから、本発明にかかる「フィブロインL鎖」としては、例えば、GenBankアクセッション No.NP_001106733.1で特定されるタンパク質(GenBankアクセッション No.NM_001113262.1で特定される遺伝子)が挙げられる
本発明にかかる「フィブロインヘキサメリン」は、分子量が約2.5万Daのフィブロインタンパク質であり、「P25」とも称されるフィブロインタンパク質である。その典型例として、GenBankアクセッション No.NP_001139413.1で特定されるタンパク質のうちの17〜220番目のアミノ酸配列からなるタンパク質(GenBankアクセッション No.NM_001145941.1で特定されるDNAのうちの49〜660番目の塩基配列からなる遺伝子)が挙げられる。また、このタンパク質は、シグナルペプチド(GenBankアクセッション No.NP_001139413.1で特定されるタンパク質のうちの1〜16番目のアミノ酸配列からなるタンパク質)が前駆体より切断、除去されることにより得られる成熟型であるから、本発明にかかる「フィブロインL鎖」としては、例えば、GenBankアクセッション No.NP_001139413.1で特定されるタンパク質(GenBankアクセッション No.NM_001145941.1で特定される遺伝子)が挙げられる
また、タンパク質のアミノ酸配列は、自然界において(すなわち、非人工的に)変異し得る。従って、本発明にかかる「フィブロインタンパク質」には、このような天然の変異体も含まれる。さらに、本発明にかかる「フィブロインタンパク質」としては、前記天然のフィブロインタンパク質やその天然の変異体の全長のみならず、これらの部分的な断片であってもよい。
【0025】
さらに、本発明にかかる「フィブロインタンパク質」としては、本発明にかかる融合タンパク質がフィブロイン複合体の一部になれればよく、前記「フィブロインL鎖」、「フィブロインH鎖」又は「フィブロインヘキサメリン」の部分的断片も本発明にかかる「フィブロインタンパク質」に含まれる。かかる部分的断片としては、例えばフィブロインL鎖−GFP融合タンパク質において、下記部分的断片の有効性が実証されていることから、「フィブロインL鎖」においては、GenBankアクセッション No.NP_001037488.1で特定されるタンパク質のうちの1(メチオニン残基)〜242番目(アラニン残基)のアミノ酸配列からなるタンパク質であることが好ましい。また、「フィブロインH鎖」においては、フィブロインL鎖とヘテロダイマーを形成して(S−S結合を形成して)フィブロインタンパク質として分泌可能なことが確認されたフィブロインH鎖の最小単位であるという観点から、GenBankアクセッション No.NP_001106733.1で特定されるタンパク質のうちの、1(メチオニン残基)〜153番目(アラニン残基)のアミノ酸配列からなるタンパク質及び5205(セリン残基)〜5263番目(システイン残基)のアミノ酸配列からなるタンパク質であることが好ましい。
【0026】
本発明にかかる「一本鎖抗体とフィブロインタンパク質との融合タンパク質」としては、前記一本鎖抗体と、フィブロインL鎖、フィブロインH鎖及びフィブロインヘキサメリンからなる群から選択される少なくとも一のタンパク質とが融合しているタンパク質であればよい。一本鎖抗体が前記フィブロイン複合体の表面上に配置され易く、より高い抗原特異性の一本鎖抗体が得られ易くなるという観点から、前記一本鎖抗体と融合させるフィブロインタンパク質は、フィブロインL鎖及びフィブロインH鎖からなる群から選択される少なくとも一のタンパク質であることが好ましい。
【0027】
フィブロインタンパク質は、一本鎖抗体のN末側、C末側のいずれに融合させてもよく、またN末側及びC末側、両方に融合させてもよい。さらに、直接的に一本鎖抗体に融合させてもよく、リンカーを介して間接的に融合させてもよい。リンカーペプチドの長さは、通常、1〜100アミノ酸、好ましくは1〜50アミノ酸、より好ましくは1〜30アミノ酸である。「一本鎖抗体とフィブロインタンパク質との融合タンパク質」には、また他の機能性タンパク質が融合されていてもよい。この場合、他の機能性タンパク質は、融合タンパク質のN末側、C末側のどちらか一方若しくは両側、又は一本鎖抗体とフィブロインタンパク質との間に、直接的に又は間接的に融合させることができる。他の機能性タンパク質としては特に制限はなく、本発明にかかる融合タンパク質に付与したい機能に応じて適宜選択される。例えば、融合タンパク質に付与したい機能が該融合タンパク質の精製や検出等であれば、緑色蛍光タンパク質(GFP)、ルシフェラーゼタンパク質、Myc−タグ(tag)タンパク質、His−タグタンパク質、ヘマグルチン(HA)−タグタンパク質、FLAG−タグタンパク質(登録商標、Sigma−Aldrich社)、グルタチオン−S−トランスフェラーゼ(GST)タンパク質が挙げられる。
【0028】
前記融合タンパク質の絹糸における発現は、当業者は公知の手法により行うことができる。かかる公知の手法としては、前記融合タンパク質を発現させるためのベクターを構築し、該ベクターをカイコに導入して前記融合タンパク質を発現させる方法が挙げられる。
【0029】
フィブロインタンパク質を合成・分泌するカイコの後部絹糸腺において、前記融合タンパク質を発現させる場合、前記ベクターにおいて、前記融合タンパク質をコードするDNAは、カイコの後部絹糸腺における発現を保証するプロモーターの下流に連結されている必要がある。このようなプロモーターとしては、例えば、フィブロインL鎖遺伝子、フィブロインH鎖遺伝子又はフィブロインヘキサメリン(P25)遺伝子のプロモーターを好適に用いることができる。ベクターは、その他の発現制御領域、エンハンサー、ターミネーター、ポリAシグナル、非翻訳領域(5’−UTR、3’−UTR)等を有していてもよい。
【0030】
また、前記ベクターにおいて、前記融合タンパク質をコードするDNAとその制御領域とを備えた発現カセット以外に、他のタンパク質をコードするDNA及びその発現制御領域を備えていてもよい。かかる他のタンパク質をコードするDNAとしては特に制限はなく、例えば、GFPやDsRed等のレポータータンパク質をコードする遺伝子、薬剤耐性遺伝子が挙げられる。他のタンパク質をコードするDNAの発現制御領域は、配列内リボソーム進入部位(Internal Ribosome Entry Site、IRES)を介して、前記融合タンパク質をコードするDNAのプロモータ−やエンハンサーを共有していてもよく、前記融合タンパク質をコードするDNAの発現制御領域とは異なるプロモーター等を備えていてもよい。発現制御領域としては、例えば、3×P3プロモーター、カイコアクチンプロモーター、ショウジョウバエ由来のHSP70プロモーター、エンハンサー、ターミネーター、ポリAシグナル、非翻訳領域(5’−UTR、3’−UTR)が挙げられる。
【0031】
前記ベクターにおける発現カセット等をカイコの染色体内に挿入することができるベクターとしては、例えば、トランスポゾンを利用したベクターを好適に用いることができる。トランスポゾンを利用したベクターとしては、例えば、piggyBac、Minos、Mariner等のDNAトランスポゾンを利用したベクターが挙げられる。なお、前記ベクターにおいて、かかるDNAトランスポゾンの一対の末端反復配列(逆位末端反復配列)の間に前記発現カセットを配置することにより、転移酵素によって前記ベクターから該末端反復配列に挟まれた前記発現カセットが切り出され、カイコの染色体内の特定の配列(例えば、piggyBacを用いた場合には、TTAA配列)を標的としてランダムに挿入されるようになる。
【0032】
従って、かかるトランスポゾンを利用したベクターを用いる場合には、各トランスポゾンに対応した転移酵素もカイコに導入する必要がある。かかる転移酵素はそのままカイコに導入してもよく、かかる転移酵素をコードするmRNA又はかかる転移酵素をコードするプラスミドベクター(ヘルパープラスミド)を導入してもよい。かかるヘルパープラスミドにおいて、前記転移酵素を発現させるためのプロモーターとしては、例えば、内在性の転移酵素遺伝子のプロモーター、カイコアクチンプロモーター、ショウジョウバエ由来のHSP70プロモーターが挙げられる。
【0033】
本発明において、このように構築した前記融合タンパク質を発現させるためのベクターを導入し、該融合タンパク質が発現している絹糸を産生するカイコを調製する方法としては、例えば、該ベクター等をカイコの卵に注入し、該卵から孵化して成育させ得られたF0カイコを同胞交配又は野生型カイコと交配し、得られるF1カイコから前記融合タンパク質を絹糸に発現するカイコを選択する方法が挙げられる。
【0034】
前記ベクター等のカイコの卵への注入は、当業者に公知の手法を適宜選択して行うことができる。かかる公知の手法としては、例えば、DNA注入用の管を用いてカイコの発生初期卵に直接注入(マイクロインジェクション)する方法や、針やレーザー等を利用して物理的又は化学的に卵殻に穴を空けた後に、該穴から前記ベクター等を注入する方法が挙げられる。また、かかる前記ベクターをカイコの卵の注入する際に、前記転移酵素や前記ヘルパープラスミドを併せて注入してもよい。
【0035】
F1カイコから前記融合タンパク質を絹糸に発現するカイコを選択する方法としては、例えば、前記発現ベクターとしてGFPやDsRed等の蛍光タンパク質をコードする遺伝子を備えているものを用いた場合には、F1世代の卵や幼虫に励起光を照射することにより卵や幼虫から発せられる蛍光を指標に選択する方法が挙げられる。特に、3×P3プロモーターによって発現が制御されている蛍光タンパク質を利用した場合には、3×P3プロモーターは神経系統(特に眼)での遺伝子発現を促す作用を有するため、F1世代の卵や幼虫の神経系統や眼において蛍光を発する個体を選択することにより、前記融合タンパク質が発現している絹糸を産生するカイコを選択することができる。
【0036】
このように調製した前記カイコから、一本鎖抗体とフィブロインタンパク質との融合タンパク質を発現させた絹糸を調製する方法としては、例えば、カイコの絹糸腺内腔内に蓄積された絹糸を回収する方法、カイコが吐糸した絹糸を回収する方法、またはカイコが形成した繭を回収する方法が挙げられる。
【0037】
なお、後部絹糸腺で合成されたフィブロイン繊維は、中部絹糸腺に送られて濃縮され、さらに、ここで合成されているセリシンに包まれることになる。そして、このようにして合成された絹糸(液状絹糸)は吐糸口につながる前部絹糸腺に送られることになる。従って、絹糸腺内腔内に蓄積された絹糸(液状絹)は、絹糸腺をカイコから採取し、回収した絹糸腺から絹糸腺を構成する細胞(絹糸腺細胞)を除去することにより調製することができる。絹糸腺細胞の除去する方法としては、例えば、絹糸腺を純水に浸漬し、絹糸腺細胞をふやけさせた後に該細胞を取り除く方法、抽出した絹糸腺を30〜100%エタノールに浸漬し、フィブロイン及びセリシンを固定化した後に、絹糸腺細胞を除去する方法が挙げられる。
【0038】
本発明においては、こうして調製した、一本鎖抗体とフィブロインタンパク質との融合タンパク質を発現させた絹糸を溶解する。「絹糸」は、カイコの後部絹糸腺で合成・分泌される3種のフィブロインタンパク質からなるフィブロイン繊維と、該繊維とを包むセリシン層とからなる動物繊維である。また、本発明において溶解する対象となる「絹糸」は、セリシン層が除去(精錬)されていないことが必要である。
【0039】
一本鎖抗体とフィブロインタンパク質との融合タンパク質を発現させた絹糸の溶解においては、分子内又は分子間の水素結合を切断する性質を有する溶液を好適に用いることができる。
【0040】
かかる溶液としては、例えば、臭化リチウム水溶液、塩化カルシウム水溶液、塩化エチレンジアミン水溶液、チオシアン酸ナトリウム水溶液、チオシアン酸リチウム水溶液、硝酸マグネシウム水溶液、塩化カルシウム/エタノール、MMNO(N−メチルモルホリン N−オキシド)、塩化リチウム/N,N−DMAc(N,N−ジメチルアセトアミド)、ヘキサフルオロイソプロパノール、硝酸カルシウム/メタノール、又はこれらの混合溶液が挙げられる。これら水溶液の中では、溶解中にタンパク質分子の分解が確認されず、溶解時に加熱の必要がなく、また溶解にかかるコストが低いという観点から、臭化リチウム水溶液を本発明の方法において用いることが好ましい。
【0041】
また、分子内又は分子間の水素結合を切断する性質を有する溶液のpHとしては特に制限はないが、pH2〜12の溶液であることが好ましい。さらに、前記絹糸の溶解に用いられる溶液の濃度としては、通常8〜10M、好ましくは9〜9.3Mである。また、前記絹糸の溶解は、20〜100℃で10分〜12時間かけて攪拌しながら行うことが好ましく、9M 臭化リチウム水溶液を用いて室温(20〜30℃)にて4時間程度攪拌しながら行うことがより好ましい。
【0042】
本発明においては、こうして得られた前記融合タンパク質の溶解液に再構成処理を施す。かかる再構成処理としては、溶解により変性した前記融合タンパク質を脱塩することによって非変性状態に戻し、且つ前記融合タンパク質中のフィブロインタンパク質が内在性の他のフィブロインタンパク質(フィブロインL鎖、フィブロインH鎖、フィブロインヘキサメリン)、さらにはセリシンと複合体を構成し、フィブロインタンパク質が繊維化(ゲル化)できる処理であれば特に制限はない。かかる脱塩は、例えば、透析処理、限外濾過処理、ゲル濾過クロマトグラフィー処理、脱塩カラム処理、希釈処理、およびこれらの組合せにより行うことができる。これらの中では、フィブロインタンパク質の濃度を維持し、一定の性質のゲル化フィブロインを安価に、大量に得ることが容易であるという観点から、透析処理が好ましい。また、本発明にかかる再構成処理は、前記融合タンパク質の変性、分解、凝集等が生じにくいという観点から、4〜60℃の条件下で実施することが好ましい。
【0043】
「透析処理」においては、例えば、セルロース製の透析膜、多孔性のポリテトラフルオロエチレン製の透析膜、セルロースアセテート製の透析膜、セルロースナイトレート製の透析膜、再生セルロース透析膜、電解透析膜又は逆浸透膜を用いることができる。これら膜を用いて区画した前記融合タンパク質の溶解液を、任意の量の溶媒(例えば、精製水(RO水、Elix水等)、緩衝液)に1〜24時間浸漬させる工程をシルクがゲル化するまで繰り返すことにより、透析処理を実施することができる。また、かかる緩衝液としてはその種類、pH等においても特に制限はなく、例えば、リン酸緩衝液、Tris−HCl緩衝液、ハンクス液が挙げられる。これらの中では、再生セルロース透析膜を用いて区画した前記融合タンパク質の溶解液(シルク溶液)を、該溶液に対して100〜500倍量の純水(Elix)に12時間浸漬させる工程を5〜10回繰り返すことが好ましい。
【0044】
なお、かかる透析処理によって、通常、前記融合タンパク質の溶解液はゲル状(シルクゲル)となるが、ゲル化しない場合は、透析膜のまま空中に放置してタンパク質を濃縮させ、pH3〜7の緩衝液に対して透析し、又は1〜50%の水溶性有機溶剤(エタノール、メタノール、2−プロパノール、DMSO,グリセリンなど)若しくはpH5程度のバッファー(酢酸ナトリウム等)を加えることにより、ゲル化させることが出来る
また、本発明においては、一度再構成した抗体分子の機能(立体構造)を保持したままパウダー化できるという観点から、前記再構成処理を施した融合タンパク質の溶解液をさらに凍結乾燥して粉砕することが好ましく、また、前記再構成処理を施した融合タンパク質の溶解液をスプレードライしてパウダー化することも好ましい。さらに、前記再構成処理を施した融合タンパク質の溶解液を−20℃に一晩、次いで−80℃に1時間以上おいて凍結させ、さらに凍結乾燥機を用いて48時間凍結乾燥させた後、得られた乾燥シルクをワンダーブレンダーで粉砕し、一辺が1〜40μmのブロック状にパウダー化することがより好ましい。
【0045】
こうして再構成された絹糸においては、「機能的な一本鎖抗体」が発現している。ここで「機能的な一本鎖抗体」とは、抗原に対する結合性を有している一本鎖抗体を意味する。本発明により製造される一本鎖抗体は、その元になったモノクローナル抗体(親抗体)と比較して、少なくとも10%以上(好ましくは、20%以上、30%以上、40%以上、50%以上、60%以上、70%以上、80%以上、90%以上)の結合性を有する。結合性は、免疫学的手法を用いて得られる測定値(例えば、後述の図8に示すウェスタンブロッティング解析によって得られるバンドの濃さ)によって評価することができる。
【0046】
また、本発明によれば、その元になったモノクローナル抗体(親抗体)と同程度の結合特異性を有している「機能的な一本鎖抗体」を製造することができる。結合特異性は、免疫学的手法(例えば、ウェスタンブロッティング解析)を実施した際のバックグラウンドを指標に、評価することができる。すなわち、後述の図5に示すように、バックグラウンドが少ないほど特異性が高いと評価することができる。
【0047】
また、本発明の方法において、前記融合タンパク質の溶解の際又は前記再構成処理の際に、前記融合タンパク質の溶解液又は前記融合タンパク質を含むシルクゲルに酸化鉄ナノ粒子を添加してもよい。このように調製された酸化鉄ナノ粒子を含むシルクゲルを凍結乾燥等して粉砕することにより、酸化鉄ナノ粒子を含むシルクパウダーを得ることができ、該シルクパウダーは、磁気ビーズ抗体として目的とする抗原、ウィルス及び菌体を分離する用途等に好適に用いることができる。
【実施例】
【0048】
以下、実施例及び比較例に基づいて本発明をより具体的に説明するが、本発明は以下の実施例に限定されるものではない。
【0049】
(調製例1)
<フィブロインL鎖−抗WASP−scFv融合タンパク質を発現する遺伝子組換えカイコベクターの構築>
先ず、Sato,M.ら、FEBS J.、2005年、272巻、6131〜6144ページの記載に沿って、マウス ウィスコット−アルドリッチ症候群タンパク質(Wiskott−Aldrich syndrome protein、WASP)のN末端領域のEna/VASP homology 1(EVH1)ドメインに特異的なモノクローナル抗体を産生するハイブリドーマから抗体遺伝子をクローニングし、H鎖及びL鎖の可変領域(VH、VL)をフレキシブルなリンカー配列(アミノ酸配列:GGGGS×3)でつないだ一本鎖抗体(抗WASP−scFv)を構築した。次いで、得られたscFvのC末側に発現チェック用のMyc−tag(アミノ酸配列:EQKLISEEDL)を付加した。そして、この抗WASP−scFv−Myc融合タンパク質をコードするDNAフラグメントをpLLLベクターのBamHI−HindIIIサイトへ挿入し、フィブロインL鎖と抗WASP−scFv−Mycとの融合タンパク質を発現するDNAコンストラクトを作製した。さらに、この融合タンパク質(以下「フィブロインL鎖−抗WASP−scFv」とも称する)の発現ユニットを遺伝子組換えカイコ用ベクターpBac[3XP3−DsRed2afm]のEcoRI−BglIIサイトに挿入し、目的タンパク質を発現する組換えカイコ用プラスミドベクターpBac[3XP3−DsRed2afm]−LLL−anti−WASP−scFv−Mycを構築した(図1 参照)。なお、pLLLベクターについては非特許文献2の記載を、pBac[3XP3−DsRed2afm]については非特許文献1の記載を参照のこと。また、図1に記載のEcoRIサイトからBglIIサイトまでの塩基配列を配列表の配列番号:1にて示し、また該塩基配列がコードするアミノ酸配列を配列表の配列番号:2にて示す。なお、配列番号:1に記載の塩基配列において、1〜6bpはEcoRIサイトであり、7〜677bpはFib−L Pro(フィブロインL鎖遺伝子のプロモーター配列)であり、678〜1402bpはFib−L ORF(フィブロインL鎖の1位(メチオニン残基)〜242位(アラニン残基)からなるタンパク質をコードする塩基配列)であり、1403〜1772bpはVH(抗WASP−scFvのVHをコードする塩基配列)であり、1773〜1817bpはlinker(「フレキシブルなリンカー配列(アミノ酸配列:GGGGS×3)」をコードする塩基配列)であり、1818〜2165bpはVL(抗WASP−scFvのVLをコードする塩基配列)であり、2172〜2201bpはmyc−tag(Myc−tagをコードする塩基配列)であり、2205〜2210bpはHidIIIサイトであり、2211〜2678bpはFib−Lp(A)(フィブロインL鎖遺伝子の3’UTR及びポリAシグナル)であり、2679〜2684bpはBglIIサイトである。
【0050】
(調製例2)
<遺伝子組換えカイコの作出>
次に、構築したプラスミドpBac[3XP3−DsRed2afm]−LLL−anti−WASP−scFv−Mycと、piggyBacヘルパープラスミドpHA3PIGのDNA溶液(DNAの濃度が0.2μg/μlになるよう0.5mM リン酸緩衝液(pH7.0),5mM KClにて調製)とを、産卵後4〜10時間のカイコ(品種:w1−pnd)の受精卵794個にマイクロインジェクションした。
【0051】
なお、piggyBacヘルパープラスミドpHA3PIGについては非特許文献1の記載を参照のこと、また該ヘルパープラスミドには、カイコアクチンA3プロモーターで制御したpiggyBacの転移酵素遺伝子がクローニングされている。そのため、前記2種類のプラスミドDNAをカイコ受精卵に注入すると、ヘルパープラスミドから転移酵素が転写・翻訳され、この働きでベクタープラスミドから末端反復配列に挟まれたフラグメントが切り出され、ゲノム中のTTAA配列を標的としてランダムに挿入されるようになる。
【0052】
次いで、前記2種類のプラスミドDNAを注入した受精卵を25℃でインキュベーションして、幼虫を孵化させた。孵化した655頭の幼虫を人工飼料(日本農産工業)で飼育し、成虫に育てた。得られた成虫を兄妹交配して、162組の受精卵を得た。これらを産卵後6〜10日目に蛍光顕微鏡下で観察し、胚の神経系統で赤色蛍光タンパク質の発現が認められる個体すなわち目的の組換えカイコを識別した。そして、得られた遺伝子組換えカイコ個体(以下「S01系統」又は「S01」とも称する)飼育を続けた。
【0053】
(調製例3)
<組換えカイコ産生繭におけるフィブロインL鎖−抗WASP−scFvの発現>
前記の通りにして得られた「S01」等由来の繭を2mm角程度に切り、繭20mgに対して9M 臭化リチウム(LiBr)溶液1mLを加え、ローテーターで、室温、3時間撹拌して完全に溶解した。そして、10,000×gで20分間(室温)遠心後、上清を回収し、水で2倍希釈した後、等量の2×SDSサンプルバッファーを加えて、65℃で30分間インキュベートした。調製したサンプルをSDS−PAGEにて分離し、PVDFメンブレンにトランスファーした後、抗フィブロインL鎖抗体(anti−FibL pAb、本発明者小島作製)および抗Myc−tag抗体(anti−Myc pAb、MBL社製)と室温で1時間反応後、HRP−標識抗ウサギ抗体と室温で1時間反応させ、ECL試薬(GEヘルスケア社製)を用いて検出した。得られた結果を図2に示す。
【0054】
図2に示した結果から明らかなように、抗フィブロインL鎖抗体を用いたウェスタンブロットにおいて、遺伝子組換えカイコ S01由来の繭では、内在性(天然)のフィブロインL鎖(27kDa)と共に抗WASP−scFv−Mycが融合した約60kDaのタンパク質(フィブロインL鎖−抗WASP−scFv)が強く検出された(図2のa 参照)。さらに、抗Myc−tag抗体を用いたウェスタンブロットにおいても同様に60kDaのタンパク質が強く検出されたことから(図2のb 参照)、今回作出した組換えカイコが産生した絹糸において、目的とする融合タンパク質(フィブロインL鎖−抗WASP−scFv)が非常に多く発現していることが確かめられた。
【0055】
このようにして得られた遺伝子組換えカイコ S01由来の繭から以下に示す3種類の方法にて、融合タンパク質 フィブロインL鎖−抗WASP−scFvを調製した(図3及び図4 参照)。
【0056】
(実施例1)
<未精練シルクパウダーの調製方法>
非組換え体であるw1−pnd(以下「w1」とも称する)由来、S01由来及びフィブロインL鎖−緑色蛍光タンパク質(EGFP)の融合タンパク質(EGFP−Fib−L)を発現する組換えカイコ(未発表、以下「K27系統」又は「K27」とも称する)由来の繭をそれぞれ3g用意し、裁断した後9M LiBr溶液30mLに浸し、4時間撹拌して完全に溶解した。得られたシルク溶液を透析膜(再生セルロース膜、製品名:Spectra/Por、MWCO 6−8000)に入れ、精製水(RO水)8Lに対して12時間×6回透析を行った。透析により得られたシルクゲルを−20℃に一晩、次いで−80℃に1時間以上おいて凍結させ、さらに凍結乾燥機を用いて48時間凍結乾燥した。得られた乾燥シルクを粉砕器(製品名:ワンダーブレンダー、大阪ケミカル株式会社製)で粉砕し、未精練シルクパウダーとした。
【0057】
(比較例1)
(繭をそのまま粉砕してパウダーにする方法)
w1由来及びS01由来の繭それぞれ3gをRO水に1日浸し、−80℃で凍結・乾燥後、そのままワンダーブレンダーを用いて粉砕した。
【0058】
(比較例2)
<精練シルクパウダーの調製方法>
w1由来、S01由来及びK27由来の繭をそれぞれ4g用意し、8M Urea,0.5% 2−メルカプトエタノールに浸して80℃で30分間撹拌してセリシンを除去した(尿素精練)。そして、w1由来、S01由来及びK27由来の未精錬の繭3gの代わりに得られた精練シルク(フィブロインのみを含む)3gを用いた以外は、実施例1と同様の操作によりシルクパウダー(精練シルクパウダー)を作製した。
【0059】
このように異なる調製方法(実施例1、比較例1〜2)によって調製されたシルクパウダーについて、下記の方法にて評価した。
【0060】
<電子顕微鏡による観察>
実施例1及び比較例1〜2で得られたシルクパウダーの形状に関して電子顕微鏡観察したところ、比較例1で得られた粉砕しただけのパウダーはまだ繊維状の構造をとっているが(直径10〜15μm、長さ数mm)、実施例1及び比較例2で得られた、一度溶解し、凍結乾燥・粉砕したパウダーは一辺が1〜40μmのブロック状の構造をとっていることが確認された(図3 参照)。
【0061】
<シルクパウダーの抗原特異的な結合解析>
実施例1及び比較例2にて調製したシルクパウダーと、抗原であるGST−WASP−NT及びコントロールであるGSTとを各々反応させ、その後プルダウンアッセイにより、フィブロインL鎖−抗WASP−scFvを有するシルクパウダーと抗原との複合体が形成されているかを調べた(図4 参照)。すなわち、各方法にて調製したシルクパウダーをそれぞれ10mgずつ量り取り、1.5mLチューブに入れ、20%エタノール溶液1mLを加えてよく懸濁し、分散させた。分散後、遠心して上清のエタノール溶液を捨て、続けてブロッキング液(製品名:ブロッキングワン、ナカライテスク社製)1mLを加えて室温、90分間ローテーターにて撹拌しながらインキュベートし、非特異的な結合を抑える操作(ブロッキング)を施した。ブロッキング後、大腸菌で発現させてアフィニティー精製した組換えタンパク質GST、GST−WASP−NT(抗WASP−scFvの抗原)をプローブタンパク質(それぞれ20μg/mLの濃度になるようブロッキング液にて調製)として、シルクパウダーと共に室温で60分間ローテーターにて撹拌しながらインキュベートした。次いで、TBST溶液(10 mM Tris−HCl,pH8.0,0.15M NaCl,0.05% Tween−20)で5回洗浄することにより、プルダウンアッセイを行った(図4 参照)。得られたサンプルに2×SDSサンプルバッファーを加えて100℃で10分間ボイルした。そして、ボイルしたサンプルをSDS−PAGEした後、PVDFメンブレンにトランスファーして、抗GST抗体を用いてウェスタンブロットを行った。得られた結果を図5に示す。
【0062】
図5に示した結果から明らかなように、繭をそのまま粉砕したシルクパウダー(比較例1)においては、w1由来及びS01由来のパウダー共に非特異的なプローブタンパク質の結合が見られたが(図5のa 参照)、実施例1で得られたシルクパウダーでは、非特異的な結合が抑制され、S01由来のシルクパウダーのみに抗原であるGST−WASP−NTとの特異的な結合が検出された(図5のb 参照)。従って、本発明の方法によって調製された一本鎖抗体は、抗原への結合活性が維持されており、しかも非特異的な結合が抑制されていることが明らかになった。さらに、本発明の方法によって調製された一本鎖抗体は、融合しているフィブロインL鎖を介して、内在性の他のフィブロインタンパク質(フィブロインL鎖、フィブロインH鎖、フィブロインヘキサメリン)及びセリシンと複合体を再構成することによりシルクパウダーの表面上に発現していることが示唆された。しかも、前述の通り、一本鎖抗体の活性をシルクパウダーの表面上で再現でき、シルクパウダーを担体として利用することもできるため、本発明の方法によって調製された一本鎖抗体を有するシルクパウダーはそのままプルダウンアッセイに供することができることも明らかになった。
【0063】
<シルクパウダーの抗原特異的な結合活性の評価>
実施例1で得られたシルクパウダーに関して、さらに抗原特異性を評価するために、抗原であるGST−WASP−NT、抗原部位を欠損させたWASPとGSTとの融合タンパク質であるGST−WASP−CT、さらにWASPと全く相同性のないマウスの免疫系細胞で発現しているP2X7レセプターの細胞内ドメインとGSTとの融合タンパク質GST−P2X7−CT2の3種類を大腸菌で発現させてアフィニティー精製し、プローブタンパク質(それぞれ20μg/mLになるようブロッキング液にて調製)として用意した。そして、前記同様に、w1由来、S01由来、K27由来のシルクパウダーと反応させた後、プルダウンアッセイ、並びに抗Myc抗体及び抗GFP抗体を用いたウェスタンブロットにより抗原−抗体反応による複合体を形成しているかを確かめた。得られた結果を図6に示す。
【0064】
また、比較例2で得られたシルクパウダーに関しても、前記実施例1と同様に、抗原特異性を評価するために、抗原であるGST−WASP−NT、GST−WASP−CT又はGST−P2X7−CT2の3種類のプローブタンパク質を用いたプルダウンアッセイにて抗原−抗体反応による複合体を形成しているかを確かめた。得られた結果を図7に示す。
【0065】
図6に示した結果から明らかなように、実施例1で得られたシルクパウダーにおいては、S01由来のシルクパウダーのみに抗原であるGST−WASP−NTとの強い結合が確認され、他のプローブタンパク質との特異的な結合はほとんど見られなかった(図6のa 参照)。さらに抗Myc抗体及び抗GFP抗体を用いたウェスタンブロットにより、結合解析に用いたそれぞれのシルクパウダーの性質及び量に関してほぼ同等であることが確認された(図6のb及びc 参照)。従って、本発明の方法によって調製された、シルクパウダー上の一本鎖抗体は、優れた抗原特異性を維持して、抗体分子として機能していることが確かめられた。
【0066】
一方、図7に示した結果から明らかなように、比較例2で得られたシルクパウダーにおいては、w1由来、S01由来及びK27由来のシルクパウダーのいずれも、GST−WASP−NT、GST−WASP−CT及びGST−P2X7−CT2と結合していた。従って、精錬した繭をLiBr溶液に溶解し、透析して調製したシルクパウダーでは、非特異的結合が強く、抗体分子として機能しているかどうか評価することが出来なかった。
【0067】
<抗WASP−EVH1モノクローナル抗体を用いた免疫沈降>
抗WASP−scFvの親抗体である完全長の抗WASPモノクローナル抗体(anti−WASP mAb)とプロテインGセファロースとの複合体を用いたプルダウンアッセイを行い、実施例1で得られた抗WASP−scFvにおいて確認された抗体活性(抗原特異性や結合性)と比較した。すなわち、組換えタンパク質GST−WASP−NT、GST−WASP−CT及びGST−P2X7−CT2をそれぞれPBSバッファーで20μg/mLに調整し、プロテインGセファロース(GEヘルスケア社製)40μLを加え、4℃で60分間ローテーターにてインキュベートして、プロテインGセファロースに非特異的に吸着する成分を取り除いた(プレクリアー)。そして、10,000×gで10分間遠心した後、上清を回収し、そこに抗WASP−scFvの親抗体である抗WASP−EVH1モノクローナル抗体5μgとプロテインGセファロース40μLとを加え、4℃で3時間ローテーターにてインキュベートした。次いで、PBSで5回洗浄した後、2×SDSサンプルバッファーを加えて100℃で10分間ボイルした。このようにして得られたサンプルをSDS−PAGEした後、PVDFメンブレンにトランスファーして、抗GST抗体を用いてウェスタンブロットを行った。得られた結果を図8に示す。
【0068】
図8に示した結果から明らかなように、実施例1で得られた抗WASP−scFvの抗体活性は、親抗体である抗WASPモノクローナル抗体とほぼ同等であることが確認された。従って、本発明の方法によって調製された一本鎖抗体は、元としたモノクローナル抗体と同等である程、抗原への特異性及び結合性が高く維持されていることが明らかになった。
【0069】
(調製例4)
<フィブロインH鎖−抗WASP−scFv融合タンパク質を発現する遺伝子組換えカイコベクターの構築>
フィブロインL鎖の時と同様に、抗WASP−scFv−MycのDNAコンストラクトをpHCベクターのBamHI−SalIサイトへ挿入し、フィブロインH鎖のN末側の一部(NTD)とC末側の一部(CTD)の間に抗WASP−scFv−Mycが挟まれたような形状で融合タンパク質を発現するDNAコンストラクトを作製した。さらにこの融合タンパク質の発現ユニットを遺伝子組換えカイコ用ベクターpBac[3XP3−Dsred2afm]のAscI−FseIサイトに挿入し、目的タンパク質を発現する組換えカイコ用プラスミドベクターpBac[3XP3−DsRed2afm]−HC−anti−WASP−scFv−Mycを構築した(図9 参照)。なお、pHCベクターについては非特許文献3の記載を、pBac[3XP3−DsRed2afm]については非特許文献1の記載を参照のこと。また、図9に記載のAscIサイトからFseIサイトまでの塩基配列を配列表の配列番号:3にて示し、該塩基配列がコードするアミノ酸配列を配列表の配列番号:4にて示す。なお、配列番号:3に記載の塩基配列において、1〜8bpはAscIサイトであり、9〜1133bpはFib−H Pro(フィブロインH鎖遺伝子のプロモーター配列)であり、1134〜1175bpはFib−H NTD(N−terminal domain(N末端領域))の第1エクソン(フィブロインH鎖の1位(メチオニン残基)〜14位(グルタミン残基)からなるタンパク質をコードする塩基配列)であり、1176〜2144bpはフィブロインH鎖遺伝子の第1イントロンであり、2145〜2561bpはFib−H NTDの第2エクソン(フィブロインH鎖の15位(チロシン残基)〜153位(アラニン残基)からなるタンパク質をコードする塩基配列)であり、2562〜2567bpはBamHIサイトであり、2568〜2936bpはVH(抗WASP−scFvのVHをコードする塩基配列)であり、2937〜2981bpはlinker(「フレキシブルなリンカー配列(アミノ酸配列:GGGGS×3)」をコードする塩基配列)であり、2982〜3329bpはVL(抗WASP−scFvのVLをコードする塩基配列)であり、3336〜3365bpはmyc−tag(Myc−tagをコードする塩基配列)であり、3366〜3371bpはSalIサイトであり、3372〜3551bpはFib−H CTD(C−terminal domain(C末端領域))(フィブロインH鎖の5205位(セリン残基)〜5263位(システイン残基)からなるタンパク質をコードする塩基配列)であり、3552〜3851bpはFib−Hp(A)(フィブロインH鎖遺伝子の3’UTR及びポリAシグナル)であり、3852〜3859bpはFseIサイトである。
【0070】
(調製例5)
<遺伝子組換えカイコの作出>
次に、構築したプラスミドpBac[3XP3−DsRed2afm]−HC−anti−WASP−scFv−Mycと、piggyBacヘルパープラスミドpHA3PIGのDNA溶液(DNAの濃度が0.2μg/μlになるよう、0.5mM リン酸緩衝液(pH7.0),5mM KClにて調製)を産卵後4〜10時間のカイコ(品種:w1−pnd)の受精卵396個にマイクロインジェクションした。これを25℃でインキュベーションして幼虫を孵化させた。孵化した126頭の幼虫を人工飼料(日本農産工業)で飼育し、成虫に育てた。得られた成虫を兄妹交配して、85組の受精卵を得た。これらを産卵6〜10日目に蛍光顕微鏡下で観察し、胚の神経系統で赤色蛍光タンパク質の発現が認められる個体すなわち目的の組換えカイコを識別した。得られた遺伝子組換えカイコ個体を、S02系統又はS02と呼称し、飼育を続けた。
【0071】
(実施例2)
<未精練シルクパウダーの調製方法>
実施例1に記載の方法(フィブロインL鎖とscFvとの融合タンパク質において抗体活性が保持されていたパウダー化の方法)に従って、未精練のS02由来の繭からシルクパウダーを調製した。
【0072】
<組換えカイコ産生繭におけるフィブロインH鎖−抗WASP−scFvの発現>
実施例1及び2にて得られた未精練シルクパウダー10mgに対して9M臭化リチウム(LiBr)溶液500μLを加え、ローテーターで室温にて3時間撹拌して完全に該シルクパウダーを溶解させた。次いで、10,000xg、室温にて20分間遠心して上清を回収し、水で2倍希釈した後、等量の2xSDSサンプルバッファーを加えて、65℃で30分間インキュベートした。このように調製したサンプルをSDS−PAGEにて分離し、PVDFメンブレンにトランスファーした後、抗フィブロインH鎖(FibH)抗体(本発明者小島作成)、抗Myc−tag抗体(MBL社製)、抗フィブロインL鎖(FibL)抗体又は抗GFP抗体と室温にて1時間反応させた後、HPR−標識抗ウサギ抗体等の前記一次抗体に対応する各二次抗体と室温にて1時間反応させ、ECL試薬(GEヘルスケア社製)を用いて検出した。得られた結果を図10に示す。
【0073】
図10に示した結果から明らかなように、抗Myc抗体及び抗フィブロインH鎖抗体を用いたウェスタンブロットにおいて、約70kDaの大きさに目的とするFibH−anti−WASP−scFv−Myc融合タンパク質がS02由来の未精錬シルクパウダーには発現していることが確認された。なお、内在性のフィブロインH鎖タンパク質は350kDaの巨大なタンパク質なので、本ウェスタンブロット解析では検出されなかった。また、FibH−anti−WASP−scFv−Myc融合タンパク質を構成していないフィブロインL鎖やGFPに対する抗体を用いたウェスタンブロットにおいて、S02由来の未精錬シルクパウダーには特異的なバンドは検出されなかった。
【0074】
<シルクパウダーの抗原特異的な結合解析>
実施例1及び2にて得られた未精練シルクパウダー(W1由来、S01由来、S02由来)を10mgずつ測り取り、1.5mLチューブに入れ、20%エタノール1mLを加えよく懸濁し、分散させた後、遠心して上清のエタノール溶液を捨て、続けて、非特異的な結合を抑えるために、ブロッキング液(製品名:ブロッキングワン、ナカライテスク社製)1mLを加えて室温にて90分間ローテーターを用いて撹拌しながらインキュベートした。ブロッキング後、大腸菌で発現させた組換えタンパク質GST及びGST−WASP−NT(scFvの抗原)をプローブタンパク質(各々20μg/mLになるようブロッキング液にて調製)として、シルクパウダーと室温にて60分間ローテーターを用いて撹拌しながらインキュベートした。次いで、TBST溶液(10mM Tri−HCl、pH8.0、0.15M NaCl、0.05% Tween−20)にて5回洗浄した後、2xSDSサンプルバッファーを加えて100℃にて10分間ボイルした。このように調製したサンプルをSDS−PAGEにて分離した後、PVDFメンブレンにトランスファーして、抗GST抗体、抗Myc抗体を用いてウェスタンブロットを行った。得られた結果を図11に示す。
【0075】
図11に示した結果から明らかなように、S01由来の未精錬シルクパウダーのみならず、S02由来の未精錬シルクパウダーにおいても抗原特異的な結合活性が確認された(図11上部パネル WB:抗GST抗体 参照)。また、かかる結合活性の強さは、各々のパウダーにおける一本鎖抗体(scFv)の発現量に比例していることが示唆された(図11下部パネル WB:抗Myc抗体 参照)。
【0076】
以上の結果から、本発明の方法によって調製されたシルクパウダー上の一本鎖抗体は、フィブロインタンパク質のL鎖及びH鎖のいずれに融合させた形で発現させても、優れた抗原特異性を維持して、抗体分子として機能していることが確かめられた。
【産業上の利用可能性】
【0077】
以上説明したように、本発明によれば、抗体をその結合性や抗原特異性を維持させたまま、非常に簡便に絹糸から調製することが可能となる。しかも、本発明の製造方法によって得られる一本鎖抗体は、該一本鎖抗体の元になったモノクローナル抗体(親抗体)と同程度の極めて高い結合性及び抗原特異性を有しており、さらには該一本鎖抗体に融合しているフィブロインタンパク質等から構成されるシルクパウダーを担体として利用することもできる。したがって、抗体生産・精製・担体への固定化のプロセスが簡略化され、従来の方法と比較して安価な抗体活性を利用したアフィニティー担体を提供することが可能となり、様々な病原体の検出・同定や疾病診断を目的とした新しいバイオマテリアルとしての有効である。
【特許請求の範囲】
【請求項1】
一本鎖抗体(scFV)とフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解する工程と、
該工程で得られた前記融合タンパク質の溶解液に再構成処理を施す工程と、
を含むことを特徴とする、scFVの製造方法。
【請求項2】
分子内又は分子間の水素結合を切断する性質を有する溶液で絹糸を溶解する、請求項1に記載の製造方法。
【請求項3】
前記再構成処理を施した融合タンパク質の溶解液を凍結乾燥して粉砕する工程をさらに含むことを特徴とする、請求項1又は2に記載の製造方法。
【請求項4】
前記フィブロインタンパク質が、フィブロインL鎖及びフィブロインH鎖からなる群から選択される少なくとも一のタンパク質であることを特徴とする、請求項1〜3のうちのいずれか一項に記載の製造方法。
【請求項5】
前記再構成処理が透析処理であることを特徴とする、請求項1〜4のうちのいずれか一項に記載の製造方法。
【請求項1】
一本鎖抗体(scFV)とフィブロインタンパク質との融合タンパク質を発現させた絹糸を精錬せずに溶解する工程と、
該工程で得られた前記融合タンパク質の溶解液に再構成処理を施す工程と、
を含むことを特徴とする、scFVの製造方法。
【請求項2】
分子内又は分子間の水素結合を切断する性質を有する溶液で絹糸を溶解する、請求項1に記載の製造方法。
【請求項3】
前記再構成処理を施した融合タンパク質の溶解液を凍結乾燥して粉砕する工程をさらに含むことを特徴とする、請求項1又は2に記載の製造方法。
【請求項4】
前記フィブロインタンパク質が、フィブロインL鎖及びフィブロインH鎖からなる群から選択される少なくとも一のタンパク質であることを特徴とする、請求項1〜3のうちのいずれか一項に記載の製造方法。
【請求項5】
前記再構成処理が透析処理であることを特徴とする、請求項1〜4のうちのいずれか一項に記載の製造方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】

【図8】
【図9】
【図10】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−239436(P2012−239436A)
【公開日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願番号】特願2011−113813(P2011−113813)
【出願日】平成23年5月20日(2011.5.20)
【国等の委託研究の成果に係る記載事項】(出願人による申告)農業・食品産業技術総合研究機構生物系特定産業技術研究支援センター平成21年度基礎的試験研究委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
【公開日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願日】平成23年5月20日(2011.5.20)
【国等の委託研究の成果に係る記載事項】(出願人による申告)農業・食品産業技術総合研究機構生物系特定産業技術研究支援センター平成21年度基礎的試験研究委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
[ Back to top ]