
一次代謝産物の量産方法、一次代謝産物の量産菌株及びその製造方法
産業的に有用で環境に優しい生物化学物質であるエタノールなどのアルコール、乳酸及びコハク酸などの一次代謝産物の生産のための最適の菌株と最適の条件を提供し、これを利用した一次代謝産物の量産方法を提供する。
本発明は、微生物の代謝経路中に特定代謝経路を遮断して他の一次代謝産物の生産を増加させる方法、前記特定代謝経路に関与する物質のコーディング遺伝子を変更して他の一次代謝産物の量産が可能な形質転換体及び、このような形質転換体の製造方法に関するものである。前記一次代謝産物は環境に優しい生物化学物質で、産業的効用性の高いエタノールなどのアルコール、乳酸またはコハク酸などであり得る。

本発明は、微生物の代謝経路中に特定代謝経路を遮断して他の一次代謝産物の生産を増加させる方法、前記特定代謝経路に関与する物質のコーディング遺伝子を変更して他の一次代謝産物の量産が可能な形質転換体及び、このような形質転換体の製造方法に関するものである。前記一次代謝産物は環境に優しい生物化学物質で、産業的効用性の高いエタノールなどのアルコール、乳酸またはコハク酸などであり得る。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は2006年2月16日付で出願された韓国特許出願第10−2006−0015116号及び2007年2月6日付で出願された韓国特許出願第10−2007−0011953号を優先権主張し、これら出願明細書は本明細書に言及されたことのように参照として含まれる。
【0002】
本発明は微生物の代謝経路中の特定代謝経路を遮断して他の一次代謝産物の生産を増加させる方法、前記特定代謝経路に関与する物質のコーディング遺伝子が変更されて他の一次代謝産物の量産が可能な形質転換体及び、このような形質転換体の製造方法に関するものである。前記一次代謝産物は環境に優しい生物化学物質で、産業的効用性の高い乳酸、コハク酸またはエタノールなどのアルコールなどであり得る。
【背景技術】
【0003】
産業革命以降、人類は石油化学工業の発達に基づいて著しい発展をなしてきたが、これに伴う副作用を見過ごした無分別な開発と濫用に伴う環境破壊という大きな問題を残し、これは必ず解決しなければならない問題となっている。
【0004】
オゾン層の破壊など、環境破壊で現れる異常気候などに対する認識の変化によって、全世界はこれに対する自救策としてこれ以上の地球環境破壊を防止するために環境保護対策である気候変化協約及び京都議定書(Kyoto Protocol)の採択及び発効などに努力している。しかし、このような一連の環境保護対策は全世界的にエネルギーを多く使用する石油化学工業の広範囲な発達と石油依存度の高い国家に対する経済的及び社会的波紋が大きいことと予想される。
【0005】
現在再生資源から生産できる代替化学製品に対する研究が進められており、この中でも乳酸とコハク酸が有用な生物化学製品としてその可能性を認められている。乳酸の場合、既に生分解性プラスチックとして開発が完了して生産に突入した状態であり、今後市場を形成することが予測されている。先進国では既に政府主導下の研究が活発に進められており、米国のカーギル(Cargill)社とダウ(Dow)社が合作して乳酸重合体(PLA;polylactic acid)生産技術が乳酸の発酵生産研究と共に開発され、デュポン社とDenocor社が合作してPTT(polytrimethylene terephthalate)の原料となるPDO(1,3−propanediol)の生産技術が開発された。PLAは従来に開発された繊維と比較して湿気回復率、弾性回復率及び紫外線吸収面で優れた性能を有すると調査され、生分解性で環境に優しい高分子としての可能性を確保した。従来に開発された繊維として代表的なナイロンとポリエステル及び環境に優しい高分子であるPLAの物性を下記の表1に示した。
【0006】
【表1】
【0007】
前記表1に示されたように、PLAはナイロンとポリエステルなどの従来の合繊と比較して対等であるか優れた物性を有することが分かり、これはPLAが化学的合繊製品を代替する代替材であることを意味する。
【0008】
また、他の1つの有用な生物化学製品であるコハク酸ポリマーはPLAより高い柔軟性を有することが知られ、2004年米国エネルギー局(Department of Energy;DOE)は未来のバイオマス由来の高付加価値化合物のうちの1つとして選定した(NREL、2004)。
【0009】
コハク酸は4個の炭素からなるTCA回路の中間生成物であるジカルボン酸で、低濃度で存在するが、すべての植物細胞及び動物細胞で発見される化学物質である。コハク酸及びそれらの誘導体はプラスチック、食品、医薬品及び化粧品産業などで広範囲に利用されている。
【0010】
コハク酸は石油化学工業の発達と共に生産が増加している合成高分子の弱点である難分解性を克服する生分解性高分子のモノマーとしての利用価値が増加している。現在使用されているプラスチックの1/3程度が比較的短期間に使用される一回包装用途で使用されているので、これら廃棄物による環境汚染問題が深刻に台頭しており、これらプラスチックの相当部分が生分解性で代替されなければならないという環境規制のために、生分解性プラスチック事業は各国の最大の関心事であった。最近、次世代生分解性高分子として思われる生分解性脂肪族ポリエステルであるポリブチレンサクシネートに対する研究が活発に進められており、その主原料がコハク酸である(Kirk−other、1979)。
【0011】
しかし、現在生産されているコハク酸の販売単価は産業体が要求する単価と比較して高く、また生産及び精製が効率的でないのが実情である。このような要因によってコハク酸は現在大部分が化学的合成法によって生産されている。つまり、マレイン酸無水和物(maleican hydride)を水素化してコハク酸無水和物を生産した後、再びこれを水和化してコハク酸を生産する。しかし、既に言及したように、急変する環境規制強化に応じる工程環境の変化によって、前記のような化学的合成方法でない生物学的方法の開発が必要となり、微生物培養技術と遺伝子工学的技術方法の発達によって発酵法によるコハク酸の生産に対する研究が関心を集めている。特に、発酵法によるコハク酸生産法は使用原料が安い再生資源を利用するという点で経済的利点があり、環境に優しい清浄技術であるという長所がある。
【0012】
発酵法によるコハク酸生産のためには高効率の菌株開発が要求される。大部分のコハク酸発酵微生物は絶対嫌気性または通性嫌気性菌株であることが知られている。このような嫌気性微生物は呼気性微生物より外部条件の変化に応じて細胞成長に非常に大きな影響を受けるだけでなく、代謝産物の生成も影響を受けるために、コハク酸生産微生物の生理学的及び環境的研究が重要である。また、このような生理学的及び環境的な研究資料に基づいてコハク酸生産代謝回路の分析を通じてコハク酸が過剰生成されるように最適の発酵条件を確立することが要求される(Cynthia et al.,1996)。
【0013】
一方、2004年米国再生アルコール協会(RFA、Renewable Fuel Association)の調査によると、米国内約80個余りのアルコール生産企業から約3,500,000,000ガロン(gallon)のアルコールが生産され、原料資源の豊富なブラジルでもやはり4,000,000,000ガロンに至るアルコールが生産された。米国内アルコール需要のほとんどは燃料分野に使用され、その規模は約3,000,000,000ガロンに至った。さらに、これら生産量の大部分はとうもろこしなどの原料物質を使用して生産された。とうもろこしなどの原料(feedstock)を利用したアルコール生産が有する最も大きな長所は環境に優しい工程であることである。このような天然物質をアルコール生産原料として使用することによってアルコール生産の時に少ないエネルギーを利用して少ない二酸化炭素を発生させる。また、再生エネルギーを利用するために廃棄物処理などで発生する別途の費用負担とエネルギー消耗が少ないという利点もある。これは韓国のような高い石油依存度を有する国家であるほど必ず解決しなければならない問題として台頭している。
【0014】
代表的なアルコール種類であるエタノールは酒類、産業用または実験室用溶媒、変性アルコール製造、医薬、化粧品製造、有機合成用基質などの多様な用途として使用することが可能であり、これに伴う需要も非常に高い。最近、エタノールはガソリン燃料のノッキング制御を改善させて排煙での一酸化炭素低減効果を有するガソリン添加剤または代替エネルギー源としてその利用性が拡大している。飲み物用アルコールを除いた大部分のエタノールは主に化学合成によって生産されたが、原油価格の上昇で製造原価が上昇して微生物による発酵生産で代替しようとする努力が必要であるのが実情である。
【発明の開示】
【発明が解決しようとする課題】
【0015】
前記の要求に応じるために本発明は、産業的に有用で環境に優しい生物化学物質であるエタノールなどのアルコール、乳酸及びコハク酸などの一次代謝産物の生産のための最適な菌株と最適な条件を提供し、これを利用した一次代謝産物の量産方法を提供することを目的とする。
【発明の効果】
【0016】
本発明は、微生物の代謝経路中において特定経路を遮断することによって、環境に優しく産業的に有用な多様な有機酸を含む一次代謝産物を量産する技術を提供し、本発明によって量産された有機酸は従来の化学合成物質の代替として多様な分野に適用されて、費用節減効果と環境保護効果を得ることができる。
【発明を実施するための最良の形態】
【0017】
後述する発明の詳細な説明によって、本発明がより詳細に説明され、本発明の完全な理解と付随的な利点がより明確になる。
【0018】
本発明は微生物の代謝経路中において特定代謝産物の代謝経路を遮断して他の一次代謝産物の生産を増加させる方法;前記特定代謝産物の代謝経路に関与する物質のコーディング遺伝子を変更して他の一次代謝産物の量産が可能な形質転換体及び、このような形質転換体の製造方法に関するものである。前記一次代謝産物は環境に優しい生物化学物質として産業的効用性の高いアルコール、乳酸またはコハク酸であり得る。
【0019】
本発明では一次代謝産物の量産のための菌株としてザイモモナス・モビリス(Zymomonas mobilis)を使用することができる。ザイモモナス・モビリスはアルコール発酵菌として細胞増殖に比べて産物転換率に優れた菌株として知られている。ザイモモナス・モビリスは理論上の産物生産収率が約98%以上であり、エタノール生産性が5g/g/lで、10g/g/h以上のブドウ糖代謝速度で1モルのブドウ糖から2モルのエタノールを生産する。
【0020】
ザイモモナス・モビリスの物質代謝経路を見ると、当該過程によって生成されたピルビン酸がアセトアルデヒドに転換され、再びアルコール脱水素酵素によって最終的にエタノールを生産する。このような高効率エタノール生産に関与する主な酵素はピルビン酸デカルボキシラーゼ(Pyruvate Decarboxylase)で、ピルビン酸からアセトアルデヒドへの転換を媒介する。したがって、このようなピルビン酸デカルボキシラーゼの生産が阻害されると、ピルビン酸からアセトアルデヒドへの転換が遮断されてアルコールが生産されず、宿主細胞はエネルギー生成のためにアルコール生産以外の他の経路を利用してアルコール以外の他の一次代謝産物を生産する。
【0021】
前記一次代謝産物としては、C2代謝産物であるエタノール、C3代謝産物である乳酸とピルビン酸、C6代謝産物であるクエン酸、C5代謝産物であるグルタミン酸、C4代謝産物であるコハク酸、フマル酸及びマレイン酸などがある。したがって、前記のようにエタノール代謝経路が遮断されると、この他の代謝産物である乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸などの生産が増加し、特に乳酸とコハク酸の生産量が顕著に増加することを観察することができる。この時、乳酸生成経路において、ピルビン酸から乳酸への転換を媒介する乳酸脱水素酵素(Lactate Dehydrogenase)の生成を阻害して乳酸生成を遮断することによって、コハク酸の生産をより増加させることができる。
【0022】
また、ザイモモナス・モビリスは従来に知られていない部分的な呼吸回路を有して乳酸を電子供与体に利用することが明らかになっており、乳酸生産を遮断することによって、より嫌気性発酵を誘導することで細胞増殖を促進し、エタノール生産速度を向上させることができ、さらにピルビン酸デカルボキシラーゼの基質特異性を変化させてブタンジオール(butanediol)などを生産することができる。
【0023】
このように、微生物の代謝経路中において特定代謝経路を遮断させることによって、前記特定代謝経路で生成する一次代謝産物以外の他の一次代謝産物の生産を増加させることができる。
【0024】
このような点に基づいて、本発明はザイモモナス・モビリスでのピルビン酸デカルボキシラーゼ生成及び/または乳酸脱水素酵素の生成を阻害してアルコール生成及び/または乳酸の生産を抑制することによって、他の一次代謝産物、特に、エタノールなどのアルコール、コハク酸及び乳酸の生産を増加させる技術を提供する。
【0025】
より具体的に、本発明はピルビン酸デカルボキシラーゼのコーディング遺伝子であるpdc(pyruvate decarboxylase)遺伝子(SEQ ID NO:1)及び/または乳酸脱水素酵素コーディング遺伝子であるldhA(lactated e hydrogenase)遺伝子(SEQ ID NO:2)を除去してピルビン酸デカルボキシラーゼ及び/または乳酸脱水素酵素の生成を阻害することによって、アルコール以外の一次代謝産物を量産する技術を提供する。
【0026】
アルコール発酵によってエネルギーを得るザイモモナスにおいて、pdc遺伝子は生存に必須の遺伝子として知られており、これを除去するとザイモモナス・モビリス菌株が生存できないことが予測されてきた。しかし、本発明ではpdc遺伝子を除去した場合、正常菌株と比較して約2倍程度の増殖遅延はあるが生存可能であり、アルコール以外の一次代謝産物、例えば、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸の生産が増加することを確認した。
【0027】
つまり、本発明において、ザイモモナス・モビリスのゲノムでpdc遺伝子が除去されると、エタノール生産能生が除去されて速い速度で大量蓄積されたピルビン酸を有用産物の量産に利用できる可能性を有し、エタノールでない多様な有用産物を生産できる“Cell Factory Z.mobilis”として開発及び活用することができる。このようにザイモモナス・モビリスが量産できる有用物質としてはピルビン酸、グリセロール及びアセチル−coAから得られる乳酸、ヒドロキシプロピオン酸(3−hydroxypropionic acid)、ヒドロキシブタン酸(3−hydroxybutanoic acid)、プロパンジオール(1,3−propanediol)、グルタミン酸、ポリグルタミン酸、アスパラギン酸、リンゴ酸、フマル酸、コハク酸、クエン酸、アジピン酸、ピルビン酸、グリセロール、キシリトール、ソルビトール、アラビニトル(arabinitol)などがある。また、コエンザイムQ10、ポリプレニルジホスフェート(polyprenyl diphosphates)及びポリテルペン(polyterpene)、ジテルペン(diterpene)、モノテルペン(monoterpene)、トリテルペン(triterpene)、セスキテルペン(sesquiterpene)などのイソプレノイド化合物(isoprenoid compounds)等の量産も可能であり、これらは化粧品、添加剤、保護剤、医薬品の前駆体などとして非常に有用である。
【0028】
この中でもコハク酸の場合には約100%以上の生産向上を示すことを確認した。従来のC2、C3、C5及びC6代謝産物と異なって、C4代謝産物の場合には量産菌株がほとんど開発されていないという点と、前記のようにコハク酸がプラスチック及び樹脂分野、医薬分野、食品分野、化粧品分野、農業分野、洗剤/乳化剤分野、織物分野、写真分野、触媒分野、防食分野及びメッキ分野など、多様な適用分野への適用が可能な産業上有用な物質であるという点を考慮する時、本発明におけるC4代謝産物のコハク酸の生産量向上は非常に意味があると言える。
【0029】
ザイモモナス・モビリスの代謝経路は次の反応式1のようである:
【0030】
【化1】
【0031】
本発明の1つの側面は、ザイモモナス・モビリスのゲノム(Accession No.:AE008692)からpdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子を除去してザイモモナス・モビリスでの一次代謝産物の生産を増加させる方法に関するものである。前記一次代謝産物はエタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであり得る。
【0032】
より具体的に、本発明はザイモモナス・モビリスのゲノムからpdc遺伝子(SEQ ID NO:1)を除去してアルコールの生成経路を遮断することによって、アルコール以外の一次代謝産物の生産を向上させる方法を提供する。前記一次代謝産物は乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであってもよく、より好ましくは乳酸及びコハク酸からなる群の中で選択された1つ以上のものであってもよい。
【0033】
前記pdc遺伝子が除去されたザイモモナス・モビリスでの代謝経路は次の反応式2のようである:
【0034】
【化2】
【0035】
また、本発明はザイモモナス・モビリスのゲノムからldhA遺伝子(SEQ ID NO:2)を除去して乳酸生成経路を遮断することによって、乳酸以外の一次代謝産物の生産を向上させる方法を提供する。前記一次代謝産物はエタノール、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであってもよく、より好ましくはエタノール及び/またはコハク酸であってもよい。
【0036】
前記ldhA遺伝子が除去されたザイモモナス・モビリスでの代謝経路は次の反応式3のようである:
【0037】
【化3】
【0038】
また、本発明はザイモモナス・モビリスのゲノムから、pdc遺伝子(SEQ ID NO:1)とldhA遺伝子(SEQ ID NO:2)を同時に除去してアルコール及び乳酸生成経路を全て遮断することによって、アルコール及び乳酸以外の一次代謝産物の生産を向上させる方法を提供する。前記一次代謝産物はピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであってもよく、より好ましくはコハク酸であってもよい。
【0039】
前記pdc遺伝子及びldhA遺伝子が全て除去されたザイモモナス・モビリスでの代謝経路は次の反応式4のようである:
【0040】
【化4】
【0041】
本発明のまた他の側面は、pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子が除去されたことを特徴とするザイモモナス・モビリス形質転換体に関するものである。
【0042】
より具体的に、本発明はpdc遺伝子(SEQ ID NO:1)が除去されたザイモモナス・モビリスの形質転換体を提供する。前記形質転換体は乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の量産が可能なものであってもよく、より好ましくは乳酸及びコハク酸からなる群の中で選択された1つ以上の量産が可能なものであってもよい。本発明の具体例において、前記pdc遺伝子(SEQ ID NO:1)が除去された形質転換体はKCTC 11012BPであり得る。
【0043】
また、本発明はldhA遺伝子(SEQ ID NO:2)が除去されたザイモモナス・モビリスの形質転換体を提供する。前記形質転換体はエタノール、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の量産が可能なものであってもよく、より好ましくはエタノール、及び/またはコハク酸の量産が可能なものであってもよい。本発明の具体例において、前記ldhA遺伝子(SEQ ID NO:2)が除去された形質転換体はKCTC 11013BPであり得る。
【0044】
また、本発明はpdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)が全て除去されたザイモモナス・モビリスの形質転換体を提供する。前記形質転換体はピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の量産が可能なものであってもよく、より好ましくはコハク酸の量産が可能なものであってもよい。本発明の具体例において、前記pdc遺伝子(SEQ ID NO:1)とldhA遺伝子(SEQ ID NO:2)が除去された形質転換体はKCTC 10908BPであり得る。
【0045】
本発明のまた他の側面は、ザイモモナス・モビリスのゲノムからpdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子を除去する段階を含む、前記ザイモモナス・モビリス形質転換体の製造方法を提供する。
【0046】
より具体的に、本発明のpdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法は、
−ザイモモナス・モビリスのpdc遺伝子(SEQ ID NO:1)を有する遺伝子断片を適切なプラスミドにクローニングし;
−前記プラスミドからpdc遺伝子を除去し;
−前記pdc遺伝子が除去されたプラスミドを利用してザイモモナス・モビリス菌株を形質転換させる段階を含む。
【0047】
前記pdc遺伝子クローニング段階において、前記pdc遺伝子を有する遺伝子断片は、ザイモモナス・モビリスゲノムのpdc遺伝子と、その両側に位置する相同組み換え(homologous recombination)のための相同部位を有するものであり、前記形質転換段階が、前記相同組み換えのための相同部位による相同組み換えによって、前記ザイモモナス・モビリス菌株のpdc遺伝子含有部位を、プラスミドのpdc遺伝子が除去された部位で、代替させるものであり得る。前記相同組み換えのための相同部位は、ザイモモナス・モビリスゲノムのpdc遺伝子の両端に位置する1500乃至5000bpのポリヌクレオチドであってもよく、本発明の具体例では、pdc遺伝子の5'末端から上流(upstream)方向のSacI部位までの2933bpのポリヌクレオチドと、pdc遺伝子の3'末端から下流(downstream)方向のXbaI部位までの2873bpのポリヌクレオチドであってもよい。
【0048】
この時、pdcが除去されたザイモモナス・モビリス形質転換体の選別を容易にするために、前記pdc遺伝子除去段階において、pdc遺伝子を除去し、その位置に適切な選別マーカを代替させることができる。前記選別マーカとしてクロラムフェニコール抵抗遺伝子(cmR)、テトラサイクリン抵抗遺伝子(tetR)、アンピシリン抵抗遺伝子(ampR)またはカナマイシン抵抗遺伝子(kmR)を使用することができる。
【0049】
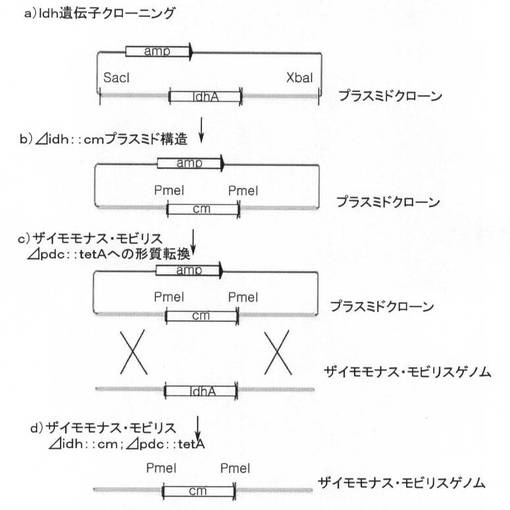
本発明の1つの具体例によるpdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法を図1に模式的に示した。
【0050】
また、本発明のldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法は
−ザイモモナス・モビリスのldhA遺伝子(SEQ ID NO:2)を有する遺伝子断片を適切なプラスミドにクローニングし;
−前記プラスミドからldhA遺伝子を除去し;
−前記ldhA遺伝子が除去されたプラスミドを利用してザイモモナス・モビリス菌株を形質転換させる段階を含む。
【0051】
前記ldhA遺伝子クローニング段階において、前記ldhA遺伝子を有する遺伝子断片は、ザイモモナス・モビリスゲノムのldhA遺伝子とその両側に位置する相同組み換えのための相同部位を有するものであり、前記形質転換段階が、前記相同組み換えのための相同部位による相同組み換えによって、前記ザイモモナス・モビリス菌株のldhA遺伝子含有部位を、プラスミドのldhA遺伝子が除去された部位で、代替させるものであり得る。前記相同組み換えのための相同部位は、ザイモモナス・モビリスゲノムのldhA遺伝子の両端に位置する1500乃至5000bpのポリヌクレオチドであってもよく、本発明の具体例では、ldhA遺伝子の5'末端から上流方向のSacI部位までの4879bpのポリヌクレオチドと、ldhA遺伝子の3'末端から下流方向のXbaI部位までの4984bpのポリヌクレオチドであってもよい。
【0052】
この時、ldhAが除去されたザイモモナス・モビリス形質転換体の選別を容易にするために、前記ldhA遺伝子除去段階において、ldhA遺伝子を除去し、その位置に適切な選別マーカを代替させることができる。前記選別マーカとしてクロラムフェニコール抵抗遺伝子、テトラサイクリン抵抗遺伝子、アンピシリン抵抗遺伝子またはカナマイシン抵抗遺伝子を使用することができる。
【0053】
本発明の1つの具体例によるldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法を図4に模式的に示した。
【0054】
また、本発明は前記pdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法とldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法を連続行って、pdc遺伝子及びldhA遺伝子が全て除去されたザイモモナス・モビリス形質転換体を製造する方法を提供する。
【0055】
また、本発明は前記pdc遺伝子及び/またはldhA遺伝子が除去されたザイモモナス・モビリス形質転換体を培養してエタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の一次代謝産物を量産する方法を提供する。この時、培養温度は30乃至34℃にし、培養時間は10乃至14時間程度にするのが好ましい。
【0056】
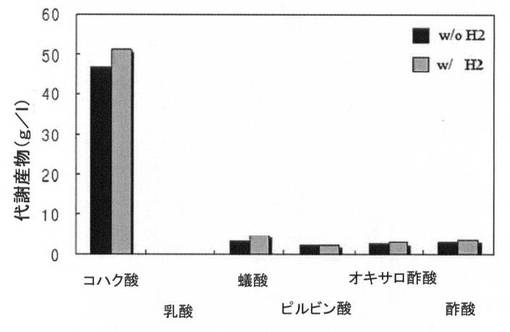
本発明の量産方法は、前記ザイモモナス・モビリス形質転換体を培養する時、培養培地にCO2供給源を追加的に添加して、前記一次代謝産物への転換時に炭素供給源として作用して一次代謝産物の生産量を増加させることができる。例えば、Z.mobilisのコハク酸生産は主にリンゴ酸酵素(malic enzyme)によって行われるが、ピルビン酸(C3)からマレイン酸(C4)が生成されるためにはカルボキシル化されなければならないので、炭素を供給することによってコハク酸生産効率増加効果を得ることができる。
【0057】
前記炭素供給源としてCO2気体または炭酸塩などを使用することができる。前記炭酸塩としては通常のすべての炭酸塩を使用することができ、NaHCO3、Na2CO3及びCaCO3からなる群の中で選択されたものであってもよい。代謝経路における、前記一次代謝産物とのトランスフェラーゼの効果的な作用を考慮する時、CO2気体の場合には0.2乃至1vvm(aeration volume/medium volume/minute)、炭酸塩の場合には1乃至50mM、好ましくは5乃至20mMの量で添加することができる。
【0058】
また、炭素源(CO2)供給と共に水素供給もまたコハク酸などの一次代謝産物の生産に重要な要素である。水素供給は細胞内電子伝達を促進させてフマル酸還元酵素(fumarate reductase)によるコハク酸などの一次代謝産物の生産効率を増加させる役割を果たす。例えば、ザイモモナス・モビリスは嫌気性細菌であり、細胞内NADHを利用してATPを生成することができないので、細胞内NADH(NADH+H+)は大部分NADH脱水素酵素によってNADに酸化され、この時に生成されたプロトン(H+)はΔpH維持に使用され、電子はキノン(quinone)、シトクローム(cytochrome)などの電子伝達経路を経てフマル酸に伝達され、フマル酸還元酵素によってコハク酸が生成される。外部から供給された水素(H2)は細胞膜に存在するキノンを通って細胞内に流入するが、キノンは細胞膜でキノン回路を通じて水素をプロトンと電子に変えながら電子伝達中継の役割を担当して、水素からプロトンを細胞内に供給し、電子をシトクロームに伝達する。したがって、培養培地に水素を供給することは、NADHがNADに酸化しながら生成されるプロトン(H+)の供給と同一な効果を得ることができるので、電子伝達促進によるコハク酸などの一次代謝産物の生産効率を増進させる効果を得ることができる。この時、水素はガス状態で添加することができ、0.2乃至1vvm(aeration volume/medium volume/minute)の量で添加するのが好ましい。
【0059】
本発明の具体例において、前記ザイモモナス・モビリス形質転換体をRM培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)にNaHCO310mMを添加したり、CO2気体1vvmを添加して、30℃で14時間培養して、より増加されたコハク酸生産効果を得ることができる。このような場合、コハク酸の生産効率が最大5g/g/hまで向上する。
【0060】
以下、本発明をさらに詳しく説明する。但し、これは例として提示されるものに過ぎず、これによって本発明が制限されるわけではない。
【実施例】
【0061】
(実施例1:pdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造)
図1に示された方法によって、pdc遺伝子が除去されたザイモモナス・モビリス形質転換体を製造した。以下、図1を参照して説明する。
【0062】
[1−1.pdc遺伝子クローニング]
ザイモモナス・モビリス(Z.mobilis)のゲノム遺伝子(AE008692)からpdc遺伝子を含んで7513bpに該当する遺伝子断片をポリメラーゼ連鎖反応(polymerase chain reaction;PCR)方法を利用して得た。これに使用されたプライマーは次の通りである。
【0063】
正方向プライマー(pdcF):
5−CCTGAATAGCTGGATCTAGAGCCCGTCAAAGC−3(SEQ ID NO:7)
逆方向プライマー(pdcR):
5−CTGATCAAGGAGAGCTCGGCCTCCAAGC−3(SEQ ID NO:8)
【0064】
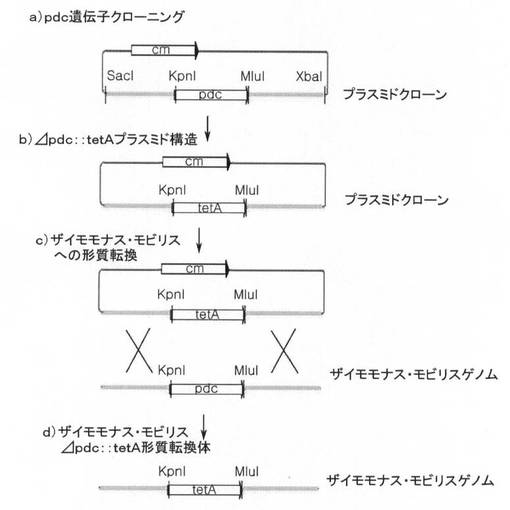
SacI制限酵素(NEB、ニューイングランドバイオラボ社)とXbaI制限酵素(NEB、ニューイングランドバイオラボ社)を用いて、PCRによって得られた遺伝子断片を切断し、その後、同一の制限酵素で処理されたpHSG398ベクター(コアバイオシステム、TAKARA)にリガーゼ(NEB、 ニューイングランドバイオラボ社)を使用して挿入した。図1の段階a)から分かるように、前記遺伝子断片は、pdc遺伝子(1707bp)、前記pdc遺伝子の5'末端側に5'末端から上流方向のSacI部位までのポリヌクレオチド(上流相同部位、2933bp、SEQ ID NO:3)及び前記pdc遺伝子の3'末端側に3'末端から下流方向のXbaI部位までのポリヌクレオチド(下流相同部位、2873bp、SEQ ID NO:4)を有する。前記5'相同部位と3'相同部位は、Z.mobilis菌株の形質転換時に、Z.mobilisのゲノムとの相同組み換えに使用される。
【0065】
[1−2.pdc遺伝子がtetR遺伝子に置換された(Δpdc::tetR)プラスミド製作]
前記段階1−1で得られたプラスミドに、KpnI制限酵素(NEB、ニューイングランドバイオラボ社)とMluI制限酵素(NEB、ニューイングランドバイオラボ社)を用いて処理した後、pBR322ベクター(コアバイオシステム、TAKARA)からポリメラーゼ連鎖反応で増幅されたtetR遺伝子(J01749)を、リガーゼ(NEB、ニューイングランドバイオラボ社)を使用して挿入して、pdc遺伝子がtetR遺伝子に置換されたプラスミドを製作した。
【0066】
[1−3.Z.mobilisの形質転換]
前記段階1−2で得られたプラスミドを使用して、Z.mobilisZM4(ATCC31821)を 電気穿孔法(エレクトロぽレーション)で形質転換した。Z.mobilisZM4をRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)で10時間培養した後、新たなRM液体培地に移して600nm可視光線で0.3〜0.4の吸光度を有するように4時間培養する。培養液を20分間氷に放置し、遠心分離(5000rpm、5分)して上澄液を除去して10%のグリセロールで洗浄した。3回の洗浄過程を経た後、100μlの体積で濃縮されたZ.mobilisZM4を前記プラスミドに形質転換した。遺伝子導入システム(Bio−Rad)を使用し、使用された電気穿孔法条件は3.0kV、25μF、そして400Ωであり、時間定数は8.8〜9.9であった。
【0067】
形質転換時に、前記プラスミドに含まれた5'相同部位及び3'相同部位と、これに対するZ.mobilisZM4ゲノムのそれぞれの相同部位間の相同組み換えによって、Z.mobilisZM4ゲノムのpdc遺伝子が除去され、その代わりに前記プラスミドに存在するtetR遺伝子が挿入されて、pdc遺伝子がtetR遺伝子で置換されたZ.mobilisΔpdc::tetR形質転換体が得られた。このように得られたZ.mobilisΔpdc::tetR形質転換体を2006年10月26日付で大韓民国大田市儒城区魚隠洞に所在する韓国生命工学研究院内生物資源センターに寄託して、寄託番号KCTC 11012BPを受けた。
【0068】
[1−4.Z.mobilisΔpdc::tetR形質転換体の選別及び確認]
前記段階1−3で得られた形質転換体を、テトラサイクリンとエタノールを含むRM固体培地(エタノール、20g/l;グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)で30℃で5日間培養した後、生存細胞を収集した。
【0069】
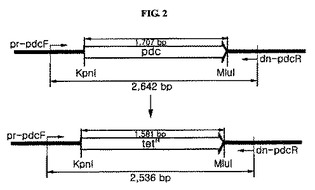
前記で収集された生存細胞がZ.mobilisΔpdc::tetR形質転換体であることを確認するために、図2に示された方法を利用した。図2から分かるように、pdc遺伝子を含む野生型Z.mobilisゲノムの場合、pdc遺伝子の上流側に位置するプライマー(pr−pdcF)部位とpdc遺伝子の下流側に位置するプライマー(dn−pdcR)部位間の塩基の長さが、2536bpである一方で、前記pdc遺伝子がtetR遺伝子に置換されたZ.mobilis形質転換体の場合には、前記プライマー間の塩基の長さが2642bpとなる。したがって、収集された生存細胞のゲノムを前記プライマーを利用してポリメラーゼ連鎖反応で増幅された長さを確認すると、Z.mobilisΔpdc::tetR形質転換体の有無を確認することができる。
【0070】
これを具体的に説明すると、前記収集された生存細胞からDNA Easy Tissue Kit(LRS Labs,QIAGEN)を利用して製造社の方法によってゲノムDNAを分離した。生存細胞のゲノムDNAを鋳型として次のようなプライマーを使用してPCR反応を行った。
【0071】
正方向プライマー(pr−pdcF):
5'−GAGGGAAAGGCTTTGTCAGTGTTGCG−3'(SEQ ID NO:9)
逆方向プライマー(dn−pdcR):
5'−TGACGCGGTTACCGTTAATTTCAGCGC−3'(SEQ ID NO:10)
【0072】
対照群として、野生型Z.mobilisを同一に処理した。前記結果を図3に示した。
【0073】
図3において、WTは対照群の野生型Z.mobilisを示し、M1及びM2は本発明のZ.mobilisΔpdc::tetR形質転換体を示す。図3から分かるように、本発明の場合に2536bpヌクレオチド断片が得られ、これを通じてpdc遺伝子が除去されたことを確認することができた。
【0074】
(実施例2:ldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造)
図4に示された方法によって、ldhA遺伝子が除去されたザイモモナス・モビリス形質転換体を製造した。以下、図4を参照して説明する。
【0075】
[2−1.ldhA遺伝子クローニング]
ザイモモナス・モビリス(Z.mobilis)のゲノム遺伝子(AE008692)からldh遺伝子を含んで10859bpに該当する遺伝子断片をポリメラーゼ連鎖反応(polymerase chain reaction)方法を利用して得た。使用されたプライマーは次の通りである。
【0076】
正方向プライマー(ldhAF):
5−TGGCAGTCCTCCATCTAGATCGAAGGTGC−3(SEQ ID NO:11)
逆方向プライマー(ldhAR):
5−GTGATCTGACGGTGAGCTCAGCATGCAGG−3(SEQ ID NO:12)
【0077】
SacI制限酵素(NEB、ニューイングランドバイオラボ社)とXbaI制限酵素(NEB、ニューイングランドバイオラボ社)を用いて、PCRによって得られた遺伝子断片を切断し、その後、同一な制限酵素で処理されたpGEM−T Easyベクター(Seoulin Bio、Promega)にリガーゼ(NEB、ニューイングランドバイオラボ社)を使用して挿入した。図1の段階a)から分かるように、前記遺伝子断片は、ldhA遺伝子(996bp)、前記ldhA遺伝子の5'末端側に5'末端から上流方向のSacI部位までのポリヌクレオチド(上流相同部位、4879bp、SEQ ID NO:5)及び前記ldhA遺伝子の3'末端側に3'末端から下流方向のXbaI部位までのポリヌクレオチド(下流相同部位、4984bp、SEQ ID NO:6)を有する。前記5'相同部位と3'相同部位は、Z.mobilis菌株の形質転換時に、Z.mobilisのゲノムとの相同組み換えに使用される。
【0078】
[2−2.ldhA遺伝子がcmR遺伝子に置換された(ΔldhA::cmR)プラスミド製作]
前記段階2−1で得られたプラスミドを鋳型としてldhA上流及び下流側のみを同時に増幅するプライマーを製作してポリメラーゼ連鎖反応で遺伝子断片を得た。使用されたプライマーは次の通りである。
【0079】
正方向プライマー(ldhA−PmeI−2F):
5−AACTAGTTTAAACAAGAGCGAAGAATAGCAAAGAAT−3(SEQ ID NO:13)
逆方向プライマー(ldhA−PmeI−2R):
5−CTCTTGTTTAAACTAGTTATGGCATAGGCTATTACG−3(SEQ ID NO:14)
【0080】
前記遺伝子断片をPmeI制限酵素(NEB、ニューイングランドバイオラボ社)処理した後、pHSG398ベクター(コアバイオシステム、TAKARA)からポリメラーゼ連鎖反応で増幅されたcmR遺伝子(U08461)を、リガーゼ(NEB、ニューイングランドバイオラボ社)を使用して挿入して、ldhA遺伝子がcmR遺伝子に置換されたプラスミドを製作した。
【0081】
[2−3.Z.mobilisの形質転換]
前記段階2−2で得られたプラスミドを使用して、Z.mobilisZM4(ATCC31821)を形質転換した。Z.mobilisZM4をRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)で10時間培養した後、新たなRM液体培地に移して600nmの可視光線で0.3‘0.4の吸光度を有するように4時間培養する。培養液を20分間氷に放置し、遠心分離(5000rpm、5分)して上澄液を除去し、10%のグリセロールで洗浄した。3回の洗浄過程を経た後、100μlの体積で濃縮されたZ.mobilisZM4を前記プラスミドに形質転換した。
【0082】
形質転換する時、前記プラスミドに含まれた5'相同部位及び3'相同部位と、これに対するZ.mobilisZM4ゲノムのそれぞれの相同部位間の相同組み換えによってZ.mobilisZM4ゲノムのldhA遺伝子が除去され、その代わりに前記プラスミドに存在するcmR遺伝子が挿入されてldhA遺伝子がcmR遺伝子に置換されたZ.mobilisΔldhA::cmR形質転換体が得られた。このように得られたZ.mobilisΔldhA::cmR形質転換体を2006年10月26日付で大韓民国、大田市、儒城区魚隠洞に所在する韓国生命工学研究院内生物資源センターに寄託して寄託番号KCTC 11013BPを受けた。
【0083】
[2−4.Z.mobilisΔldhA::cmR形質転換体の選別及び確認]
前記段階1〜3で得られた形質転換体を、クロラムフェニコールを含むRM固体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;クロラムフェニコール、75μg/ml;pH5.2)で30℃で5日間培養し、クロラムフェニコールに抵抗性を示す生存細胞を収集した。
【0084】
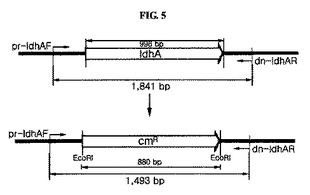
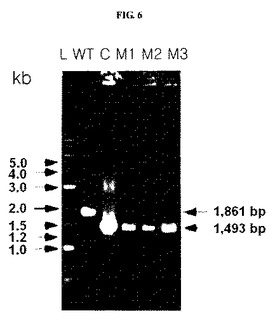
前記で収集された生存細胞がZ.mobilisΔldhA::cmR形質転換体であることを確認するために、図5に示された方法を利用した。クロラムフェニコールに抵抗性を示す生存細胞をRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;クロラムフェニコール、75μg/ml;pH5.2)に30℃で16時間培養し、遠心分離(10、000×g、5分)して上澄液を除去した後、細胞を収集した。図5から分かるように、ldhA遺伝子を含む野生型Z.mobilisゲノムの場合、ldhA遺伝子の上流側に位置するプライマー(pr−ldhAF)部位とldhA遺伝子の下流側に位置するプライマー(dn−ldhAR)部位間の塩基の長さが、1861bpである一方で、前記ldhA遺伝子がcmR遺伝子に置換されたZ.mobilis形質転換体の場合には、前記プライマー間の塩基の長さが1493bpとなる。したがって、収集された生存細胞のゲノムを前記プライマーを利用してポリメラーゼ連鎖反応で増幅された長さを確認すると、Z.mobilisΔldhA::cmR形質転換体の有無が確認できる。
【0085】
これを具体的に説明すると、前記収集された生存細胞からDNA Easy Tissue Kit(LRS Labs、QIAGEN)を利用して製造会社の方法によってゲノムDNAを分離した。生存細胞のゲノムDNAを鋳型として次のようなプライマーを使用してPCR反応を行った。
【0086】
正方向プライマー(npr−ldhAF):
5'−CAGCAAGTTCGATCTGTCTGGCGATCG−3'(SEQ ID NO:15)
逆方向プライマー(dn−ldhAR):
5'−GATTAAATAATGCGGCGATGGCTAAGCAAGG−3'(SEQ ID NO:16)
【0087】
対照群として野生型Z.mobilisを同一に処理した。
【0088】
前記結果を図6に示した。図6において、WTは対照群の野生型Z.mobilisを示し、M1、M2、そしてM3は本発明のZ.mobilisΔldhA::cmR形質転換体を示す。図6から分かるように、本発明の場合に1493bpヌクレオチド断片が得られ、これを通じてldhA遺伝子が除去されたことを確認することができた。
【0089】
(実施例3:pdc遺伝子及びldhA遺伝子が全て除去されたZ.mobilis形質転換体の製造)
前記実施例1と実施例2の過程を連続的に行って、pdc遺伝子とldhA遺伝子が全て除去されたZ.mobilisΔpdc::tetR/ΔldhA::cmR形質転換体を製造した。このように得られたZ.mobilisΔpdc::tetR/ΔldhA::cmR形質転換体を2006年2月15日付で大韓民国、大田市、儒城区魚隠洞に所在する韓国生命工学研究院内生物資源センターに寄託して寄託番号KCTC 10908BPを受けた。
【0090】
(実施例4:一次代謝産物の生産性試験)
前記実施例1乃至3で製造されたZ.mobilis形質転換体を利用して、これらの一次代謝産物生産能力を試験した。対照群として野生型Z.mobilisZM4を使用した。
【0091】
野生型Z.mobilisZM4(ATCC 31821)、実施例1乃至3で製造されたZ.mobilisΔpdc::tetR形質転換体、Z.mobilisΔldhA::cmR形質転換体及びZ.mobilisΔpdc::tetR/ΔldhA::cmR形質転換体を、NaHCO310mMが添加されたRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;テトラサイクリン、15μg/ml;pH5.2)で30℃で16時間培養した。前記培養された培養液から細胞を除去し、得られた培養上澄液をHPLC(high performance liquid chromatography)を利用して代謝産物を測定した。前記測定において、日立HPLCシステム(モデルD−7000)が使用され、AminexHPX−87Hコラムで代謝産物を分離した。有機酸はUV検出器(日立D−4200)で、糖とエタノールはRI(refractive index)検出器(D−3300)で、確認及び定量した。0.0025Nの硫酸を移動相(溶媒)として使用し、カラム温度は60℃であり、流速は0.6ml/minであった。
【0092】
前記方法を3回行って得られた結果値の平均を、下記の表2と図7A乃至10Bに示した。
【0093】
【表2】
【0094】
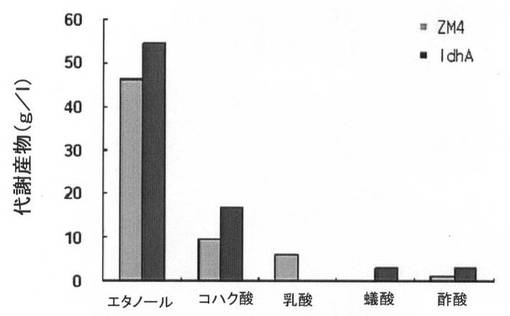
前記表2から分かるように、本発明の実施例1乃至3で製作された形質転換体は、野生型と比較して増加したコハク酸生産能力を示した。
【0095】
また、ΔldhA::cmR形質転換体は、非常に優れたエタノール生産能力を示し、Δpdc::tetR形質転換体は、非常に優れたコハク酸及び乳酸生産能力を示すことが明らかになった。
【0096】
(実施例5:細胞増殖率及び一次代謝産物の生産性試験)
【0097】
前記実施例4と同様な方法で培養して、時間に応じた菌体増殖及び産物生産を示す尺度として活用するために動力学分析を行い、指数増殖期、つまり、最大菌体増殖及び産物生産区間で測定された数値から次のような方法で得た。
【0098】
【数1】
【0099】
【数2】
【0100】
【数3】
【0101】
【数4】
【0102】
【数5】
【0103】
【数6】
【0104】
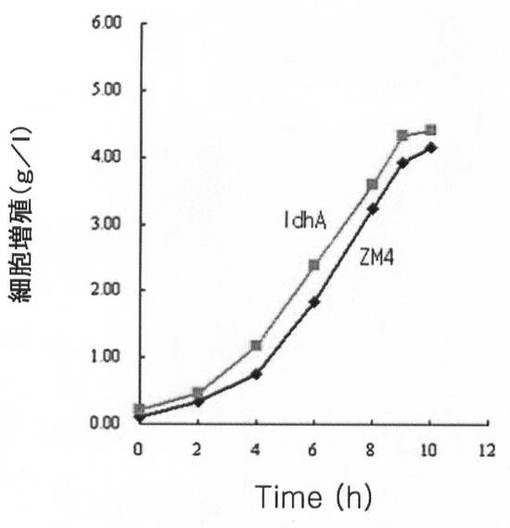
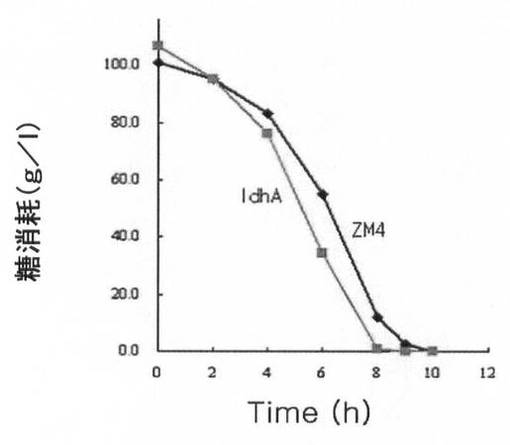
このように得られた野生型(ZM4)、Δpdc形質転換体及びΔpdc/ΔldhA形質転換体の動力学分析(kineti canalysis)結果を下記の表3に示し、細胞増殖、糖消耗量及び代謝産物生産量を、各図11A乃至図11Cに示した。
【0105】
【表3】
【図面の簡単な説明】
【0106】
【図1】本発明の実施例1及び3におけるザイモモナス・モビリスZM4でのpdc遺伝子の除去過程を示す模式図である。
【図2】本発明の実施例1で製作されたZM4形質転換体でのpdc遺伝子除去を確認するためのプライマー設計を示す模式図である。
【図3】本発明の実施例1で製作されたZM4形質転換体と野生型ZM4菌株に対する電気泳動結果を示す図面である。
【図4】本発明の実施例2及び3におけるザイモモナス・モビリスZM4でのldhA遺伝子の除去過程を示す模式図である。
【図5】本発明の実施例2で製作されたZM4形質転換体でのldhA遺伝子除去を確認するためのプライマー設計を示す模式図である。
【図6】本発明の実施例2で製作されたZM4形質転換体と野生型ZM4菌株に対する電気泳動結果を示す図面である。
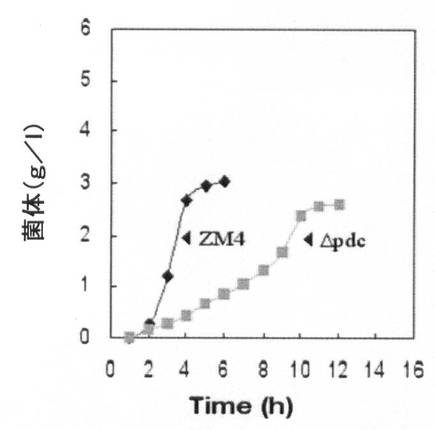
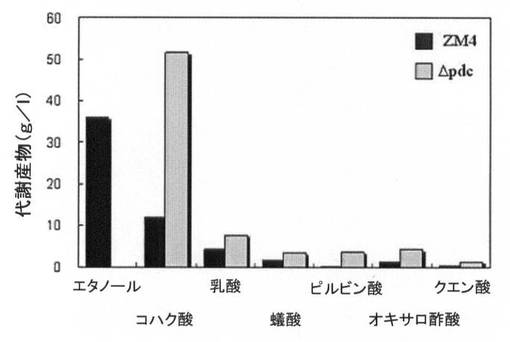
【図7A】水素供給なく培養した場合の、pdc遺伝子が除去された形質転換体(Δpdc)の増殖率(菌体量:g/L)と一次代謝産物生成能力を、野生型ZM4菌株と比較して示すグラフで、図7Aは増殖率を、図7Bは一次代謝産物生成能力を比較すると示す。
【図7B】図7A参照。
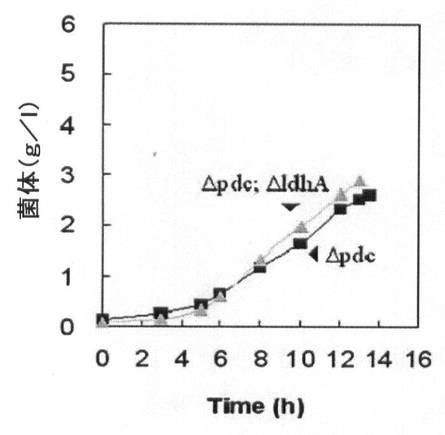
【図8A】水素供給なく培養した場合のpdc遺伝子が除去された形質転換体の増殖率(菌体量:g/L)と一次代謝産物生成能力を、pdc遺伝子とldhA遺伝子が全て除去された形質転換体と比較して示すグラフで、図8Aは増殖率を、図8Bは一次代謝産物生成能力を比較して示す。
【図8B】図8A参照。
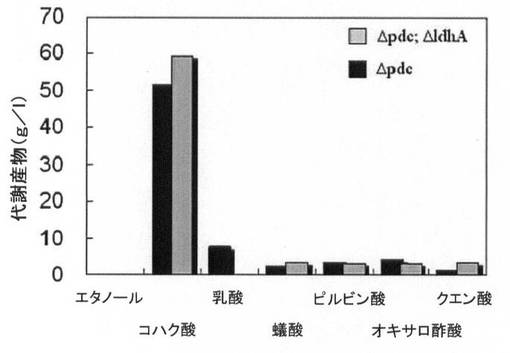
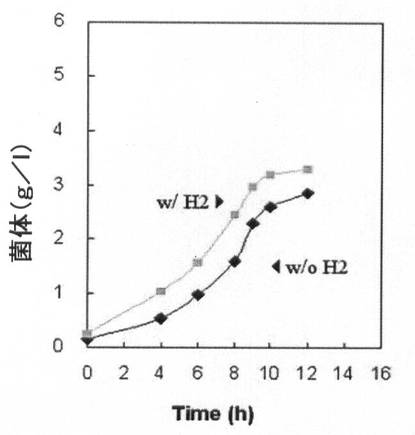
【図9A】水素供給して培養した場合と水素供給のない場合の、pdc遺伝子が除去された形質転換体(KCTC 11012BP)の増殖率(菌体量:g/L)と一次代謝産物生成能力を比較して示すグラフである。
【図9B】図9A参照。
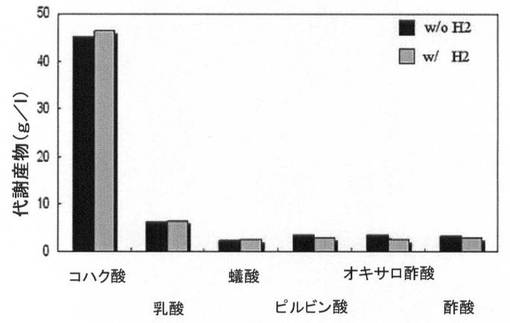
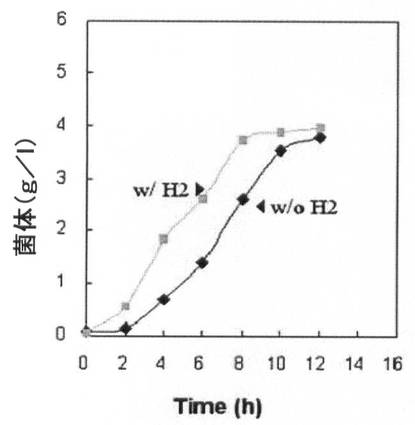
【図10A】水素供給して培養した場合と水素供給のない場合の、pdc遺伝子とldhA遺伝子が除去された形質転換体(KCTC 10908BP)の増殖率(菌体量:g/L)と一次代謝産物生成能力を比較して示すグラフである。
【図10B】図10A参照。
【図11A】ldhA遺伝子が除去された形質転換体の細胞増殖、糖消耗及び代謝産物生産能力を、ザイモモナス・モビリスZM4と比較して示すグラフで、図11Aは細胞増殖を、図11Bは糖消耗量を、図11Cは代謝産物生産能力を示す。
【図11B】図11A参照。
【図11C】図11B参照。
【技術分野】
【0001】
本出願は2006年2月16日付で出願された韓国特許出願第10−2006−0015116号及び2007年2月6日付で出願された韓国特許出願第10−2007−0011953号を優先権主張し、これら出願明細書は本明細書に言及されたことのように参照として含まれる。
【0002】
本発明は微生物の代謝経路中の特定代謝経路を遮断して他の一次代謝産物の生産を増加させる方法、前記特定代謝経路に関与する物質のコーディング遺伝子が変更されて他の一次代謝産物の量産が可能な形質転換体及び、このような形質転換体の製造方法に関するものである。前記一次代謝産物は環境に優しい生物化学物質で、産業的効用性の高い乳酸、コハク酸またはエタノールなどのアルコールなどであり得る。
【背景技術】
【0003】
産業革命以降、人類は石油化学工業の発達に基づいて著しい発展をなしてきたが、これに伴う副作用を見過ごした無分別な開発と濫用に伴う環境破壊という大きな問題を残し、これは必ず解決しなければならない問題となっている。
【0004】
オゾン層の破壊など、環境破壊で現れる異常気候などに対する認識の変化によって、全世界はこれに対する自救策としてこれ以上の地球環境破壊を防止するために環境保護対策である気候変化協約及び京都議定書(Kyoto Protocol)の採択及び発効などに努力している。しかし、このような一連の環境保護対策は全世界的にエネルギーを多く使用する石油化学工業の広範囲な発達と石油依存度の高い国家に対する経済的及び社会的波紋が大きいことと予想される。
【0005】
現在再生資源から生産できる代替化学製品に対する研究が進められており、この中でも乳酸とコハク酸が有用な生物化学製品としてその可能性を認められている。乳酸の場合、既に生分解性プラスチックとして開発が完了して生産に突入した状態であり、今後市場を形成することが予測されている。先進国では既に政府主導下の研究が活発に進められており、米国のカーギル(Cargill)社とダウ(Dow)社が合作して乳酸重合体(PLA;polylactic acid)生産技術が乳酸の発酵生産研究と共に開発され、デュポン社とDenocor社が合作してPTT(polytrimethylene terephthalate)の原料となるPDO(1,3−propanediol)の生産技術が開発された。PLAは従来に開発された繊維と比較して湿気回復率、弾性回復率及び紫外線吸収面で優れた性能を有すると調査され、生分解性で環境に優しい高分子としての可能性を確保した。従来に開発された繊維として代表的なナイロンとポリエステル及び環境に優しい高分子であるPLAの物性を下記の表1に示した。
【0006】
【表1】
【0007】
前記表1に示されたように、PLAはナイロンとポリエステルなどの従来の合繊と比較して対等であるか優れた物性を有することが分かり、これはPLAが化学的合繊製品を代替する代替材であることを意味する。
【0008】
また、他の1つの有用な生物化学製品であるコハク酸ポリマーはPLAより高い柔軟性を有することが知られ、2004年米国エネルギー局(Department of Energy;DOE)は未来のバイオマス由来の高付加価値化合物のうちの1つとして選定した(NREL、2004)。
【0009】
コハク酸は4個の炭素からなるTCA回路の中間生成物であるジカルボン酸で、低濃度で存在するが、すべての植物細胞及び動物細胞で発見される化学物質である。コハク酸及びそれらの誘導体はプラスチック、食品、医薬品及び化粧品産業などで広範囲に利用されている。
【0010】
コハク酸は石油化学工業の発達と共に生産が増加している合成高分子の弱点である難分解性を克服する生分解性高分子のモノマーとしての利用価値が増加している。現在使用されているプラスチックの1/3程度が比較的短期間に使用される一回包装用途で使用されているので、これら廃棄物による環境汚染問題が深刻に台頭しており、これらプラスチックの相当部分が生分解性で代替されなければならないという環境規制のために、生分解性プラスチック事業は各国の最大の関心事であった。最近、次世代生分解性高分子として思われる生分解性脂肪族ポリエステルであるポリブチレンサクシネートに対する研究が活発に進められており、その主原料がコハク酸である(Kirk−other、1979)。
【0011】
しかし、現在生産されているコハク酸の販売単価は産業体が要求する単価と比較して高く、また生産及び精製が効率的でないのが実情である。このような要因によってコハク酸は現在大部分が化学的合成法によって生産されている。つまり、マレイン酸無水和物(maleican hydride)を水素化してコハク酸無水和物を生産した後、再びこれを水和化してコハク酸を生産する。しかし、既に言及したように、急変する環境規制強化に応じる工程環境の変化によって、前記のような化学的合成方法でない生物学的方法の開発が必要となり、微生物培養技術と遺伝子工学的技術方法の発達によって発酵法によるコハク酸の生産に対する研究が関心を集めている。特に、発酵法によるコハク酸生産法は使用原料が安い再生資源を利用するという点で経済的利点があり、環境に優しい清浄技術であるという長所がある。
【0012】
発酵法によるコハク酸生産のためには高効率の菌株開発が要求される。大部分のコハク酸発酵微生物は絶対嫌気性または通性嫌気性菌株であることが知られている。このような嫌気性微生物は呼気性微生物より外部条件の変化に応じて細胞成長に非常に大きな影響を受けるだけでなく、代謝産物の生成も影響を受けるために、コハク酸生産微生物の生理学的及び環境的研究が重要である。また、このような生理学的及び環境的な研究資料に基づいてコハク酸生産代謝回路の分析を通じてコハク酸が過剰生成されるように最適の発酵条件を確立することが要求される(Cynthia et al.,1996)。
【0013】
一方、2004年米国再生アルコール協会(RFA、Renewable Fuel Association)の調査によると、米国内約80個余りのアルコール生産企業から約3,500,000,000ガロン(gallon)のアルコールが生産され、原料資源の豊富なブラジルでもやはり4,000,000,000ガロンに至るアルコールが生産された。米国内アルコール需要のほとんどは燃料分野に使用され、その規模は約3,000,000,000ガロンに至った。さらに、これら生産量の大部分はとうもろこしなどの原料物質を使用して生産された。とうもろこしなどの原料(feedstock)を利用したアルコール生産が有する最も大きな長所は環境に優しい工程であることである。このような天然物質をアルコール生産原料として使用することによってアルコール生産の時に少ないエネルギーを利用して少ない二酸化炭素を発生させる。また、再生エネルギーを利用するために廃棄物処理などで発生する別途の費用負担とエネルギー消耗が少ないという利点もある。これは韓国のような高い石油依存度を有する国家であるほど必ず解決しなければならない問題として台頭している。
【0014】
代表的なアルコール種類であるエタノールは酒類、産業用または実験室用溶媒、変性アルコール製造、医薬、化粧品製造、有機合成用基質などの多様な用途として使用することが可能であり、これに伴う需要も非常に高い。最近、エタノールはガソリン燃料のノッキング制御を改善させて排煙での一酸化炭素低減効果を有するガソリン添加剤または代替エネルギー源としてその利用性が拡大している。飲み物用アルコールを除いた大部分のエタノールは主に化学合成によって生産されたが、原油価格の上昇で製造原価が上昇して微生物による発酵生産で代替しようとする努力が必要であるのが実情である。
【発明の開示】
【発明が解決しようとする課題】
【0015】
前記の要求に応じるために本発明は、産業的に有用で環境に優しい生物化学物質であるエタノールなどのアルコール、乳酸及びコハク酸などの一次代謝産物の生産のための最適な菌株と最適な条件を提供し、これを利用した一次代謝産物の量産方法を提供することを目的とする。
【発明の効果】
【0016】
本発明は、微生物の代謝経路中において特定経路を遮断することによって、環境に優しく産業的に有用な多様な有機酸を含む一次代謝産物を量産する技術を提供し、本発明によって量産された有機酸は従来の化学合成物質の代替として多様な分野に適用されて、費用節減効果と環境保護効果を得ることができる。
【発明を実施するための最良の形態】
【0017】
後述する発明の詳細な説明によって、本発明がより詳細に説明され、本発明の完全な理解と付随的な利点がより明確になる。
【0018】
本発明は微生物の代謝経路中において特定代謝産物の代謝経路を遮断して他の一次代謝産物の生産を増加させる方法;前記特定代謝産物の代謝経路に関与する物質のコーディング遺伝子を変更して他の一次代謝産物の量産が可能な形質転換体及び、このような形質転換体の製造方法に関するものである。前記一次代謝産物は環境に優しい生物化学物質として産業的効用性の高いアルコール、乳酸またはコハク酸であり得る。
【0019】
本発明では一次代謝産物の量産のための菌株としてザイモモナス・モビリス(Zymomonas mobilis)を使用することができる。ザイモモナス・モビリスはアルコール発酵菌として細胞増殖に比べて産物転換率に優れた菌株として知られている。ザイモモナス・モビリスは理論上の産物生産収率が約98%以上であり、エタノール生産性が5g/g/lで、10g/g/h以上のブドウ糖代謝速度で1モルのブドウ糖から2モルのエタノールを生産する。
【0020】
ザイモモナス・モビリスの物質代謝経路を見ると、当該過程によって生成されたピルビン酸がアセトアルデヒドに転換され、再びアルコール脱水素酵素によって最終的にエタノールを生産する。このような高効率エタノール生産に関与する主な酵素はピルビン酸デカルボキシラーゼ(Pyruvate Decarboxylase)で、ピルビン酸からアセトアルデヒドへの転換を媒介する。したがって、このようなピルビン酸デカルボキシラーゼの生産が阻害されると、ピルビン酸からアセトアルデヒドへの転換が遮断されてアルコールが生産されず、宿主細胞はエネルギー生成のためにアルコール生産以外の他の経路を利用してアルコール以外の他の一次代謝産物を生産する。
【0021】
前記一次代謝産物としては、C2代謝産物であるエタノール、C3代謝産物である乳酸とピルビン酸、C6代謝産物であるクエン酸、C5代謝産物であるグルタミン酸、C4代謝産物であるコハク酸、フマル酸及びマレイン酸などがある。したがって、前記のようにエタノール代謝経路が遮断されると、この他の代謝産物である乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸などの生産が増加し、特に乳酸とコハク酸の生産量が顕著に増加することを観察することができる。この時、乳酸生成経路において、ピルビン酸から乳酸への転換を媒介する乳酸脱水素酵素(Lactate Dehydrogenase)の生成を阻害して乳酸生成を遮断することによって、コハク酸の生産をより増加させることができる。
【0022】
また、ザイモモナス・モビリスは従来に知られていない部分的な呼吸回路を有して乳酸を電子供与体に利用することが明らかになっており、乳酸生産を遮断することによって、より嫌気性発酵を誘導することで細胞増殖を促進し、エタノール生産速度を向上させることができ、さらにピルビン酸デカルボキシラーゼの基質特異性を変化させてブタンジオール(butanediol)などを生産することができる。
【0023】
このように、微生物の代謝経路中において特定代謝経路を遮断させることによって、前記特定代謝経路で生成する一次代謝産物以外の他の一次代謝産物の生産を増加させることができる。
【0024】
このような点に基づいて、本発明はザイモモナス・モビリスでのピルビン酸デカルボキシラーゼ生成及び/または乳酸脱水素酵素の生成を阻害してアルコール生成及び/または乳酸の生産を抑制することによって、他の一次代謝産物、特に、エタノールなどのアルコール、コハク酸及び乳酸の生産を増加させる技術を提供する。
【0025】
より具体的に、本発明はピルビン酸デカルボキシラーゼのコーディング遺伝子であるpdc(pyruvate decarboxylase)遺伝子(SEQ ID NO:1)及び/または乳酸脱水素酵素コーディング遺伝子であるldhA(lactated e hydrogenase)遺伝子(SEQ ID NO:2)を除去してピルビン酸デカルボキシラーゼ及び/または乳酸脱水素酵素の生成を阻害することによって、アルコール以外の一次代謝産物を量産する技術を提供する。
【0026】
アルコール発酵によってエネルギーを得るザイモモナスにおいて、pdc遺伝子は生存に必須の遺伝子として知られており、これを除去するとザイモモナス・モビリス菌株が生存できないことが予測されてきた。しかし、本発明ではpdc遺伝子を除去した場合、正常菌株と比較して約2倍程度の増殖遅延はあるが生存可能であり、アルコール以外の一次代謝産物、例えば、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸の生産が増加することを確認した。
【0027】
つまり、本発明において、ザイモモナス・モビリスのゲノムでpdc遺伝子が除去されると、エタノール生産能生が除去されて速い速度で大量蓄積されたピルビン酸を有用産物の量産に利用できる可能性を有し、エタノールでない多様な有用産物を生産できる“Cell Factory Z.mobilis”として開発及び活用することができる。このようにザイモモナス・モビリスが量産できる有用物質としてはピルビン酸、グリセロール及びアセチル−coAから得られる乳酸、ヒドロキシプロピオン酸(3−hydroxypropionic acid)、ヒドロキシブタン酸(3−hydroxybutanoic acid)、プロパンジオール(1,3−propanediol)、グルタミン酸、ポリグルタミン酸、アスパラギン酸、リンゴ酸、フマル酸、コハク酸、クエン酸、アジピン酸、ピルビン酸、グリセロール、キシリトール、ソルビトール、アラビニトル(arabinitol)などがある。また、コエンザイムQ10、ポリプレニルジホスフェート(polyprenyl diphosphates)及びポリテルペン(polyterpene)、ジテルペン(diterpene)、モノテルペン(monoterpene)、トリテルペン(triterpene)、セスキテルペン(sesquiterpene)などのイソプレノイド化合物(isoprenoid compounds)等の量産も可能であり、これらは化粧品、添加剤、保護剤、医薬品の前駆体などとして非常に有用である。
【0028】
この中でもコハク酸の場合には約100%以上の生産向上を示すことを確認した。従来のC2、C3、C5及びC6代謝産物と異なって、C4代謝産物の場合には量産菌株がほとんど開発されていないという点と、前記のようにコハク酸がプラスチック及び樹脂分野、医薬分野、食品分野、化粧品分野、農業分野、洗剤/乳化剤分野、織物分野、写真分野、触媒分野、防食分野及びメッキ分野など、多様な適用分野への適用が可能な産業上有用な物質であるという点を考慮する時、本発明におけるC4代謝産物のコハク酸の生産量向上は非常に意味があると言える。
【0029】
ザイモモナス・モビリスの代謝経路は次の反応式1のようである:
【0030】
【化1】
【0031】
本発明の1つの側面は、ザイモモナス・モビリスのゲノム(Accession No.:AE008692)からpdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子を除去してザイモモナス・モビリスでの一次代謝産物の生産を増加させる方法に関するものである。前記一次代謝産物はエタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであり得る。
【0032】
より具体的に、本発明はザイモモナス・モビリスのゲノムからpdc遺伝子(SEQ ID NO:1)を除去してアルコールの生成経路を遮断することによって、アルコール以外の一次代謝産物の生産を向上させる方法を提供する。前記一次代謝産物は乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであってもよく、より好ましくは乳酸及びコハク酸からなる群の中で選択された1つ以上のものであってもよい。
【0033】
前記pdc遺伝子が除去されたザイモモナス・モビリスでの代謝経路は次の反応式2のようである:
【0034】
【化2】
【0035】
また、本発明はザイモモナス・モビリスのゲノムからldhA遺伝子(SEQ ID NO:2)を除去して乳酸生成経路を遮断することによって、乳酸以外の一次代謝産物の生産を向上させる方法を提供する。前記一次代謝産物はエタノール、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであってもよく、より好ましくはエタノール及び/またはコハク酸であってもよい。
【0036】
前記ldhA遺伝子が除去されたザイモモナス・モビリスでの代謝経路は次の反応式3のようである:
【0037】
【化3】
【0038】
また、本発明はザイモモナス・モビリスのゲノムから、pdc遺伝子(SEQ ID NO:1)とldhA遺伝子(SEQ ID NO:2)を同時に除去してアルコール及び乳酸生成経路を全て遮断することによって、アルコール及び乳酸以外の一次代謝産物の生産を向上させる方法を提供する。前記一次代謝産物はピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上のものであってもよく、より好ましくはコハク酸であってもよい。
【0039】
前記pdc遺伝子及びldhA遺伝子が全て除去されたザイモモナス・モビリスでの代謝経路は次の反応式4のようである:
【0040】
【化4】
【0041】
本発明のまた他の側面は、pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子が除去されたことを特徴とするザイモモナス・モビリス形質転換体に関するものである。
【0042】
より具体的に、本発明はpdc遺伝子(SEQ ID NO:1)が除去されたザイモモナス・モビリスの形質転換体を提供する。前記形質転換体は乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の量産が可能なものであってもよく、より好ましくは乳酸及びコハク酸からなる群の中で選択された1つ以上の量産が可能なものであってもよい。本発明の具体例において、前記pdc遺伝子(SEQ ID NO:1)が除去された形質転換体はKCTC 11012BPであり得る。
【0043】
また、本発明はldhA遺伝子(SEQ ID NO:2)が除去されたザイモモナス・モビリスの形質転換体を提供する。前記形質転換体はエタノール、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の量産が可能なものであってもよく、より好ましくはエタノール、及び/またはコハク酸の量産が可能なものであってもよい。本発明の具体例において、前記ldhA遺伝子(SEQ ID NO:2)が除去された形質転換体はKCTC 11013BPであり得る。
【0044】
また、本発明はpdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)が全て除去されたザイモモナス・モビリスの形質転換体を提供する。前記形質転換体はピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の量産が可能なものであってもよく、より好ましくはコハク酸の量産が可能なものであってもよい。本発明の具体例において、前記pdc遺伝子(SEQ ID NO:1)とldhA遺伝子(SEQ ID NO:2)が除去された形質転換体はKCTC 10908BPであり得る。
【0045】
本発明のまた他の側面は、ザイモモナス・モビリスのゲノムからpdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子を除去する段階を含む、前記ザイモモナス・モビリス形質転換体の製造方法を提供する。
【0046】
より具体的に、本発明のpdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法は、
−ザイモモナス・モビリスのpdc遺伝子(SEQ ID NO:1)を有する遺伝子断片を適切なプラスミドにクローニングし;
−前記プラスミドからpdc遺伝子を除去し;
−前記pdc遺伝子が除去されたプラスミドを利用してザイモモナス・モビリス菌株を形質転換させる段階を含む。
【0047】
前記pdc遺伝子クローニング段階において、前記pdc遺伝子を有する遺伝子断片は、ザイモモナス・モビリスゲノムのpdc遺伝子と、その両側に位置する相同組み換え(homologous recombination)のための相同部位を有するものであり、前記形質転換段階が、前記相同組み換えのための相同部位による相同組み換えによって、前記ザイモモナス・モビリス菌株のpdc遺伝子含有部位を、プラスミドのpdc遺伝子が除去された部位で、代替させるものであり得る。前記相同組み換えのための相同部位は、ザイモモナス・モビリスゲノムのpdc遺伝子の両端に位置する1500乃至5000bpのポリヌクレオチドであってもよく、本発明の具体例では、pdc遺伝子の5'末端から上流(upstream)方向のSacI部位までの2933bpのポリヌクレオチドと、pdc遺伝子の3'末端から下流(downstream)方向のXbaI部位までの2873bpのポリヌクレオチドであってもよい。
【0048】
この時、pdcが除去されたザイモモナス・モビリス形質転換体の選別を容易にするために、前記pdc遺伝子除去段階において、pdc遺伝子を除去し、その位置に適切な選別マーカを代替させることができる。前記選別マーカとしてクロラムフェニコール抵抗遺伝子(cmR)、テトラサイクリン抵抗遺伝子(tetR)、アンピシリン抵抗遺伝子(ampR)またはカナマイシン抵抗遺伝子(kmR)を使用することができる。
【0049】
本発明の1つの具体例によるpdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法を図1に模式的に示した。
【0050】
また、本発明のldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法は
−ザイモモナス・モビリスのldhA遺伝子(SEQ ID NO:2)を有する遺伝子断片を適切なプラスミドにクローニングし;
−前記プラスミドからldhA遺伝子を除去し;
−前記ldhA遺伝子が除去されたプラスミドを利用してザイモモナス・モビリス菌株を形質転換させる段階を含む。
【0051】
前記ldhA遺伝子クローニング段階において、前記ldhA遺伝子を有する遺伝子断片は、ザイモモナス・モビリスゲノムのldhA遺伝子とその両側に位置する相同組み換えのための相同部位を有するものであり、前記形質転換段階が、前記相同組み換えのための相同部位による相同組み換えによって、前記ザイモモナス・モビリス菌株のldhA遺伝子含有部位を、プラスミドのldhA遺伝子が除去された部位で、代替させるものであり得る。前記相同組み換えのための相同部位は、ザイモモナス・モビリスゲノムのldhA遺伝子の両端に位置する1500乃至5000bpのポリヌクレオチドであってもよく、本発明の具体例では、ldhA遺伝子の5'末端から上流方向のSacI部位までの4879bpのポリヌクレオチドと、ldhA遺伝子の3'末端から下流方向のXbaI部位までの4984bpのポリヌクレオチドであってもよい。
【0052】
この時、ldhAが除去されたザイモモナス・モビリス形質転換体の選別を容易にするために、前記ldhA遺伝子除去段階において、ldhA遺伝子を除去し、その位置に適切な選別マーカを代替させることができる。前記選別マーカとしてクロラムフェニコール抵抗遺伝子、テトラサイクリン抵抗遺伝子、アンピシリン抵抗遺伝子またはカナマイシン抵抗遺伝子を使用することができる。
【0053】
本発明の1つの具体例によるldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法を図4に模式的に示した。
【0054】
また、本発明は前記pdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法とldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造方法を連続行って、pdc遺伝子及びldhA遺伝子が全て除去されたザイモモナス・モビリス形質転換体を製造する方法を提供する。
【0055】
また、本発明は前記pdc遺伝子及び/またはldhA遺伝子が除去されたザイモモナス・モビリス形質転換体を培養してエタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1つ以上の一次代謝産物を量産する方法を提供する。この時、培養温度は30乃至34℃にし、培養時間は10乃至14時間程度にするのが好ましい。
【0056】
本発明の量産方法は、前記ザイモモナス・モビリス形質転換体を培養する時、培養培地にCO2供給源を追加的に添加して、前記一次代謝産物への転換時に炭素供給源として作用して一次代謝産物の生産量を増加させることができる。例えば、Z.mobilisのコハク酸生産は主にリンゴ酸酵素(malic enzyme)によって行われるが、ピルビン酸(C3)からマレイン酸(C4)が生成されるためにはカルボキシル化されなければならないので、炭素を供給することによってコハク酸生産効率増加効果を得ることができる。
【0057】
前記炭素供給源としてCO2気体または炭酸塩などを使用することができる。前記炭酸塩としては通常のすべての炭酸塩を使用することができ、NaHCO3、Na2CO3及びCaCO3からなる群の中で選択されたものであってもよい。代謝経路における、前記一次代謝産物とのトランスフェラーゼの効果的な作用を考慮する時、CO2気体の場合には0.2乃至1vvm(aeration volume/medium volume/minute)、炭酸塩の場合には1乃至50mM、好ましくは5乃至20mMの量で添加することができる。
【0058】
また、炭素源(CO2)供給と共に水素供給もまたコハク酸などの一次代謝産物の生産に重要な要素である。水素供給は細胞内電子伝達を促進させてフマル酸還元酵素(fumarate reductase)によるコハク酸などの一次代謝産物の生産効率を増加させる役割を果たす。例えば、ザイモモナス・モビリスは嫌気性細菌であり、細胞内NADHを利用してATPを生成することができないので、細胞内NADH(NADH+H+)は大部分NADH脱水素酵素によってNADに酸化され、この時に生成されたプロトン(H+)はΔpH維持に使用され、電子はキノン(quinone)、シトクローム(cytochrome)などの電子伝達経路を経てフマル酸に伝達され、フマル酸還元酵素によってコハク酸が生成される。外部から供給された水素(H2)は細胞膜に存在するキノンを通って細胞内に流入するが、キノンは細胞膜でキノン回路を通じて水素をプロトンと電子に変えながら電子伝達中継の役割を担当して、水素からプロトンを細胞内に供給し、電子をシトクロームに伝達する。したがって、培養培地に水素を供給することは、NADHがNADに酸化しながら生成されるプロトン(H+)の供給と同一な効果を得ることができるので、電子伝達促進によるコハク酸などの一次代謝産物の生産効率を増進させる効果を得ることができる。この時、水素はガス状態で添加することができ、0.2乃至1vvm(aeration volume/medium volume/minute)の量で添加するのが好ましい。
【0059】
本発明の具体例において、前記ザイモモナス・モビリス形質転換体をRM培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)にNaHCO310mMを添加したり、CO2気体1vvmを添加して、30℃で14時間培養して、より増加されたコハク酸生産効果を得ることができる。このような場合、コハク酸の生産効率が最大5g/g/hまで向上する。
【0060】
以下、本発明をさらに詳しく説明する。但し、これは例として提示されるものに過ぎず、これによって本発明が制限されるわけではない。
【実施例】
【0061】
(実施例1:pdc遺伝子が除去されたザイモモナス・モビリス形質転換体の製造)
図1に示された方法によって、pdc遺伝子が除去されたザイモモナス・モビリス形質転換体を製造した。以下、図1を参照して説明する。
【0062】
[1−1.pdc遺伝子クローニング]
ザイモモナス・モビリス(Z.mobilis)のゲノム遺伝子(AE008692)からpdc遺伝子を含んで7513bpに該当する遺伝子断片をポリメラーゼ連鎖反応(polymerase chain reaction;PCR)方法を利用して得た。これに使用されたプライマーは次の通りである。
【0063】
正方向プライマー(pdcF):
5−CCTGAATAGCTGGATCTAGAGCCCGTCAAAGC−3(SEQ ID NO:7)
逆方向プライマー(pdcR):
5−CTGATCAAGGAGAGCTCGGCCTCCAAGC−3(SEQ ID NO:8)
【0064】
SacI制限酵素(NEB、ニューイングランドバイオラボ社)とXbaI制限酵素(NEB、ニューイングランドバイオラボ社)を用いて、PCRによって得られた遺伝子断片を切断し、その後、同一の制限酵素で処理されたpHSG398ベクター(コアバイオシステム、TAKARA)にリガーゼ(NEB、 ニューイングランドバイオラボ社)を使用して挿入した。図1の段階a)から分かるように、前記遺伝子断片は、pdc遺伝子(1707bp)、前記pdc遺伝子の5'末端側に5'末端から上流方向のSacI部位までのポリヌクレオチド(上流相同部位、2933bp、SEQ ID NO:3)及び前記pdc遺伝子の3'末端側に3'末端から下流方向のXbaI部位までのポリヌクレオチド(下流相同部位、2873bp、SEQ ID NO:4)を有する。前記5'相同部位と3'相同部位は、Z.mobilis菌株の形質転換時に、Z.mobilisのゲノムとの相同組み換えに使用される。
【0065】
[1−2.pdc遺伝子がtetR遺伝子に置換された(Δpdc::tetR)プラスミド製作]
前記段階1−1で得られたプラスミドに、KpnI制限酵素(NEB、ニューイングランドバイオラボ社)とMluI制限酵素(NEB、ニューイングランドバイオラボ社)を用いて処理した後、pBR322ベクター(コアバイオシステム、TAKARA)からポリメラーゼ連鎖反応で増幅されたtetR遺伝子(J01749)を、リガーゼ(NEB、ニューイングランドバイオラボ社)を使用して挿入して、pdc遺伝子がtetR遺伝子に置換されたプラスミドを製作した。
【0066】
[1−3.Z.mobilisの形質転換]
前記段階1−2で得られたプラスミドを使用して、Z.mobilisZM4(ATCC31821)を 電気穿孔法(エレクトロぽレーション)で形質転換した。Z.mobilisZM4をRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)で10時間培養した後、新たなRM液体培地に移して600nm可視光線で0.3〜0.4の吸光度を有するように4時間培養する。培養液を20分間氷に放置し、遠心分離(5000rpm、5分)して上澄液を除去して10%のグリセロールで洗浄した。3回の洗浄過程を経た後、100μlの体積で濃縮されたZ.mobilisZM4を前記プラスミドに形質転換した。遺伝子導入システム(Bio−Rad)を使用し、使用された電気穿孔法条件は3.0kV、25μF、そして400Ωであり、時間定数は8.8〜9.9であった。
【0067】
形質転換時に、前記プラスミドに含まれた5'相同部位及び3'相同部位と、これに対するZ.mobilisZM4ゲノムのそれぞれの相同部位間の相同組み換えによって、Z.mobilisZM4ゲノムのpdc遺伝子が除去され、その代わりに前記プラスミドに存在するtetR遺伝子が挿入されて、pdc遺伝子がtetR遺伝子で置換されたZ.mobilisΔpdc::tetR形質転換体が得られた。このように得られたZ.mobilisΔpdc::tetR形質転換体を2006年10月26日付で大韓民国大田市儒城区魚隠洞に所在する韓国生命工学研究院内生物資源センターに寄託して、寄託番号KCTC 11012BPを受けた。
【0068】
[1−4.Z.mobilisΔpdc::tetR形質転換体の選別及び確認]
前記段階1−3で得られた形質転換体を、テトラサイクリンとエタノールを含むRM固体培地(エタノール、20g/l;グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)で30℃で5日間培養した後、生存細胞を収集した。
【0069】
前記で収集された生存細胞がZ.mobilisΔpdc::tetR形質転換体であることを確認するために、図2に示された方法を利用した。図2から分かるように、pdc遺伝子を含む野生型Z.mobilisゲノムの場合、pdc遺伝子の上流側に位置するプライマー(pr−pdcF)部位とpdc遺伝子の下流側に位置するプライマー(dn−pdcR)部位間の塩基の長さが、2536bpである一方で、前記pdc遺伝子がtetR遺伝子に置換されたZ.mobilis形質転換体の場合には、前記プライマー間の塩基の長さが2642bpとなる。したがって、収集された生存細胞のゲノムを前記プライマーを利用してポリメラーゼ連鎖反応で増幅された長さを確認すると、Z.mobilisΔpdc::tetR形質転換体の有無を確認することができる。
【0070】
これを具体的に説明すると、前記収集された生存細胞からDNA Easy Tissue Kit(LRS Labs,QIAGEN)を利用して製造社の方法によってゲノムDNAを分離した。生存細胞のゲノムDNAを鋳型として次のようなプライマーを使用してPCR反応を行った。
【0071】
正方向プライマー(pr−pdcF):
5'−GAGGGAAAGGCTTTGTCAGTGTTGCG−3'(SEQ ID NO:9)
逆方向プライマー(dn−pdcR):
5'−TGACGCGGTTACCGTTAATTTCAGCGC−3'(SEQ ID NO:10)
【0072】
対照群として、野生型Z.mobilisを同一に処理した。前記結果を図3に示した。
【0073】
図3において、WTは対照群の野生型Z.mobilisを示し、M1及びM2は本発明のZ.mobilisΔpdc::tetR形質転換体を示す。図3から分かるように、本発明の場合に2536bpヌクレオチド断片が得られ、これを通じてpdc遺伝子が除去されたことを確認することができた。
【0074】
(実施例2:ldhA遺伝子が除去されたザイモモナス・モビリス形質転換体の製造)
図4に示された方法によって、ldhA遺伝子が除去されたザイモモナス・モビリス形質転換体を製造した。以下、図4を参照して説明する。
【0075】
[2−1.ldhA遺伝子クローニング]
ザイモモナス・モビリス(Z.mobilis)のゲノム遺伝子(AE008692)からldh遺伝子を含んで10859bpに該当する遺伝子断片をポリメラーゼ連鎖反応(polymerase chain reaction)方法を利用して得た。使用されたプライマーは次の通りである。
【0076】
正方向プライマー(ldhAF):
5−TGGCAGTCCTCCATCTAGATCGAAGGTGC−3(SEQ ID NO:11)
逆方向プライマー(ldhAR):
5−GTGATCTGACGGTGAGCTCAGCATGCAGG−3(SEQ ID NO:12)
【0077】
SacI制限酵素(NEB、ニューイングランドバイオラボ社)とXbaI制限酵素(NEB、ニューイングランドバイオラボ社)を用いて、PCRによって得られた遺伝子断片を切断し、その後、同一な制限酵素で処理されたpGEM−T Easyベクター(Seoulin Bio、Promega)にリガーゼ(NEB、ニューイングランドバイオラボ社)を使用して挿入した。図1の段階a)から分かるように、前記遺伝子断片は、ldhA遺伝子(996bp)、前記ldhA遺伝子の5'末端側に5'末端から上流方向のSacI部位までのポリヌクレオチド(上流相同部位、4879bp、SEQ ID NO:5)及び前記ldhA遺伝子の3'末端側に3'末端から下流方向のXbaI部位までのポリヌクレオチド(下流相同部位、4984bp、SEQ ID NO:6)を有する。前記5'相同部位と3'相同部位は、Z.mobilis菌株の形質転換時に、Z.mobilisのゲノムとの相同組み換えに使用される。
【0078】
[2−2.ldhA遺伝子がcmR遺伝子に置換された(ΔldhA::cmR)プラスミド製作]
前記段階2−1で得られたプラスミドを鋳型としてldhA上流及び下流側のみを同時に増幅するプライマーを製作してポリメラーゼ連鎖反応で遺伝子断片を得た。使用されたプライマーは次の通りである。
【0079】
正方向プライマー(ldhA−PmeI−2F):
5−AACTAGTTTAAACAAGAGCGAAGAATAGCAAAGAAT−3(SEQ ID NO:13)
逆方向プライマー(ldhA−PmeI−2R):
5−CTCTTGTTTAAACTAGTTATGGCATAGGCTATTACG−3(SEQ ID NO:14)
【0080】
前記遺伝子断片をPmeI制限酵素(NEB、ニューイングランドバイオラボ社)処理した後、pHSG398ベクター(コアバイオシステム、TAKARA)からポリメラーゼ連鎖反応で増幅されたcmR遺伝子(U08461)を、リガーゼ(NEB、ニューイングランドバイオラボ社)を使用して挿入して、ldhA遺伝子がcmR遺伝子に置換されたプラスミドを製作した。
【0081】
[2−3.Z.mobilisの形質転換]
前記段階2−2で得られたプラスミドを使用して、Z.mobilisZM4(ATCC31821)を形質転換した。Z.mobilisZM4をRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;pH5.2)で10時間培養した後、新たなRM液体培地に移して600nmの可視光線で0.3‘0.4の吸光度を有するように4時間培養する。培養液を20分間氷に放置し、遠心分離(5000rpm、5分)して上澄液を除去し、10%のグリセロールで洗浄した。3回の洗浄過程を経た後、100μlの体積で濃縮されたZ.mobilisZM4を前記プラスミドに形質転換した。
【0082】
形質転換する時、前記プラスミドに含まれた5'相同部位及び3'相同部位と、これに対するZ.mobilisZM4ゲノムのそれぞれの相同部位間の相同組み換えによってZ.mobilisZM4ゲノムのldhA遺伝子が除去され、その代わりに前記プラスミドに存在するcmR遺伝子が挿入されてldhA遺伝子がcmR遺伝子に置換されたZ.mobilisΔldhA::cmR形質転換体が得られた。このように得られたZ.mobilisΔldhA::cmR形質転換体を2006年10月26日付で大韓民国、大田市、儒城区魚隠洞に所在する韓国生命工学研究院内生物資源センターに寄託して寄託番号KCTC 11013BPを受けた。
【0083】
[2−4.Z.mobilisΔldhA::cmR形質転換体の選別及び確認]
前記段階1〜3で得られた形質転換体を、クロラムフェニコールを含むRM固体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;クロラムフェニコール、75μg/ml;pH5.2)で30℃で5日間培養し、クロラムフェニコールに抵抗性を示す生存細胞を収集した。
【0084】
前記で収集された生存細胞がZ.mobilisΔldhA::cmR形質転換体であることを確認するために、図5に示された方法を利用した。クロラムフェニコールに抵抗性を示す生存細胞をRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;クロラムフェニコール、75μg/ml;pH5.2)に30℃で16時間培養し、遠心分離(10、000×g、5分)して上澄液を除去した後、細胞を収集した。図5から分かるように、ldhA遺伝子を含む野生型Z.mobilisゲノムの場合、ldhA遺伝子の上流側に位置するプライマー(pr−ldhAF)部位とldhA遺伝子の下流側に位置するプライマー(dn−ldhAR)部位間の塩基の長さが、1861bpである一方で、前記ldhA遺伝子がcmR遺伝子に置換されたZ.mobilis形質転換体の場合には、前記プライマー間の塩基の長さが1493bpとなる。したがって、収集された生存細胞のゲノムを前記プライマーを利用してポリメラーゼ連鎖反応で増幅された長さを確認すると、Z.mobilisΔldhA::cmR形質転換体の有無が確認できる。
【0085】
これを具体的に説明すると、前記収集された生存細胞からDNA Easy Tissue Kit(LRS Labs、QIAGEN)を利用して製造会社の方法によってゲノムDNAを分離した。生存細胞のゲノムDNAを鋳型として次のようなプライマーを使用してPCR反応を行った。
【0086】
正方向プライマー(npr−ldhAF):
5'−CAGCAAGTTCGATCTGTCTGGCGATCG−3'(SEQ ID NO:15)
逆方向プライマー(dn−ldhAR):
5'−GATTAAATAATGCGGCGATGGCTAAGCAAGG−3'(SEQ ID NO:16)
【0087】
対照群として野生型Z.mobilisを同一に処理した。
【0088】
前記結果を図6に示した。図6において、WTは対照群の野生型Z.mobilisを示し、M1、M2、そしてM3は本発明のZ.mobilisΔldhA::cmR形質転換体を示す。図6から分かるように、本発明の場合に1493bpヌクレオチド断片が得られ、これを通じてldhA遺伝子が除去されたことを確認することができた。
【0089】
(実施例3:pdc遺伝子及びldhA遺伝子が全て除去されたZ.mobilis形質転換体の製造)
前記実施例1と実施例2の過程を連続的に行って、pdc遺伝子とldhA遺伝子が全て除去されたZ.mobilisΔpdc::tetR/ΔldhA::cmR形質転換体を製造した。このように得られたZ.mobilisΔpdc::tetR/ΔldhA::cmR形質転換体を2006年2月15日付で大韓民国、大田市、儒城区魚隠洞に所在する韓国生命工学研究院内生物資源センターに寄託して寄託番号KCTC 10908BPを受けた。
【0090】
(実施例4:一次代謝産物の生産性試験)
前記実施例1乃至3で製造されたZ.mobilis形質転換体を利用して、これらの一次代謝産物生産能力を試験した。対照群として野生型Z.mobilisZM4を使用した。
【0091】
野生型Z.mobilisZM4(ATCC 31821)、実施例1乃至3で製造されたZ.mobilisΔpdc::tetR形質転換体、Z.mobilisΔldhA::cmR形質転換体及びZ.mobilisΔpdc::tetR/ΔldhA::cmR形質転換体を、NaHCO310mMが添加されたRM液体培地(グルコース、50g/l;酵母抽出物、10g/l;MgSO4、1g/l;(NH4)2SO4、1g/l;KH2PO4、2g/l;テトラサイクリン、15μg/ml;pH5.2)で30℃で16時間培養した。前記培養された培養液から細胞を除去し、得られた培養上澄液をHPLC(high performance liquid chromatography)を利用して代謝産物を測定した。前記測定において、日立HPLCシステム(モデルD−7000)が使用され、AminexHPX−87Hコラムで代謝産物を分離した。有機酸はUV検出器(日立D−4200)で、糖とエタノールはRI(refractive index)検出器(D−3300)で、確認及び定量した。0.0025Nの硫酸を移動相(溶媒)として使用し、カラム温度は60℃であり、流速は0.6ml/minであった。
【0092】
前記方法を3回行って得られた結果値の平均を、下記の表2と図7A乃至10Bに示した。
【0093】
【表2】
【0094】
前記表2から分かるように、本発明の実施例1乃至3で製作された形質転換体は、野生型と比較して増加したコハク酸生産能力を示した。
【0095】
また、ΔldhA::cmR形質転換体は、非常に優れたエタノール生産能力を示し、Δpdc::tetR形質転換体は、非常に優れたコハク酸及び乳酸生産能力を示すことが明らかになった。
【0096】
(実施例5:細胞増殖率及び一次代謝産物の生産性試験)
【0097】
前記実施例4と同様な方法で培養して、時間に応じた菌体増殖及び産物生産を示す尺度として活用するために動力学分析を行い、指数増殖期、つまり、最大菌体増殖及び産物生産区間で測定された数値から次のような方法で得た。
【0098】
【数1】
【0099】
【数2】
【0100】
【数3】
【0101】
【数4】
【0102】
【数5】
【0103】
【数6】
【0104】
このように得られた野生型(ZM4)、Δpdc形質転換体及びΔpdc/ΔldhA形質転換体の動力学分析(kineti canalysis)結果を下記の表3に示し、細胞増殖、糖消耗量及び代謝産物生産量を、各図11A乃至図11Cに示した。
【0105】
【表3】
【図面の簡単な説明】
【0106】
【図1】本発明の実施例1及び3におけるザイモモナス・モビリスZM4でのpdc遺伝子の除去過程を示す模式図である。
【図2】本発明の実施例1で製作されたZM4形質転換体でのpdc遺伝子除去を確認するためのプライマー設計を示す模式図である。
【図3】本発明の実施例1で製作されたZM4形質転換体と野生型ZM4菌株に対する電気泳動結果を示す図面である。
【図4】本発明の実施例2及び3におけるザイモモナス・モビリスZM4でのldhA遺伝子の除去過程を示す模式図である。
【図5】本発明の実施例2で製作されたZM4形質転換体でのldhA遺伝子除去を確認するためのプライマー設計を示す模式図である。
【図6】本発明の実施例2で製作されたZM4形質転換体と野生型ZM4菌株に対する電気泳動結果を示す図面である。
【図7A】水素供給なく培養した場合の、pdc遺伝子が除去された形質転換体(Δpdc)の増殖率(菌体量:g/L)と一次代謝産物生成能力を、野生型ZM4菌株と比較して示すグラフで、図7Aは増殖率を、図7Bは一次代謝産物生成能力を比較すると示す。
【図7B】図7A参照。
【図8A】水素供給なく培養した場合のpdc遺伝子が除去された形質転換体の増殖率(菌体量:g/L)と一次代謝産物生成能力を、pdc遺伝子とldhA遺伝子が全て除去された形質転換体と比較して示すグラフで、図8Aは増殖率を、図8Bは一次代謝産物生成能力を比較して示す。
【図8B】図8A参照。
【図9A】水素供給して培養した場合と水素供給のない場合の、pdc遺伝子が除去された形質転換体(KCTC 11012BP)の増殖率(菌体量:g/L)と一次代謝産物生成能力を比較して示すグラフである。
【図9B】図9A参照。
【図10A】水素供給して培養した場合と水素供給のない場合の、pdc遺伝子とldhA遺伝子が除去された形質転換体(KCTC 10908BP)の増殖率(菌体量:g/L)と一次代謝産物生成能力を比較して示すグラフである。
【図10B】図10A参照。
【図11A】ldhA遺伝子が除去された形質転換体の細胞増殖、糖消耗及び代謝産物生産能力を、ザイモモナス・モビリスZM4と比較して示すグラフで、図11Aは細胞増殖を、図11Bは糖消耗量を、図11Cは代謝産物生産能力を示す。
【図11B】図11A参照。
【図11C】図11B参照。
【特許請求の範囲】
【請求項1】
ザイモモナス・モビリス(Zymomonas mobilis)のゲノムから、ピルビン酸デカルボキシラーゼをコーディングするpdc遺伝子(SEQ ID NO:1)及び乳酸脱水素酵素をコーディングするldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子を除去することによって、ザイモモナス・モビリスによるエタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1種以上の一次代謝産物の生産を増加させる方法。
【請求項2】
pdc遺伝子(SEQ ID NO:1)を除去してコハク酸及び乳酸からなる群の中で選択された1種以上の一次代謝産物の生産を増加させることを特徴とする、請求項1に記載の方法。
【請求項3】
ldhA遺伝子(SEQ ID NO:2)を除去してエタノール及びコハク酸からなる群の中で選択された1種以上の一次代謝産物の生産を増加させることを特徴とする、請求項1に記載の方法。
【請求項4】
pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)を全て除去してコハク酸の生産を増加させることを特徴とする、請求項1に記載の方法。
【請求項5】
ゲノムから、pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子が除去されて、エタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1種以上の一次代謝産物の量産が可能なザイモモナス・モビリス形質転換体。
【請求項6】
pdc遺伝子(SEQ ID NO:1)が除去されて、コハク酸及び乳酸からなる群の中で選択された1種以上の一次代謝産物の量産が可能であることを特徴とする、請求項5に記載の形質転換体。
【請求項7】
ldhA遺伝子(SEQ ID NO:2)が除去されて、エタノール及びコハク酸からなる群の中で選択された1種以上の一次代謝産物の量産が可能であることを特徴とする、請求項5に記載の形質転換体。
【請求項8】
pdc遺伝子(SEQ ID NO:1)とldhA遺伝子(SEQ ID NO:2)が全て除去されて、コハク酸の量産が可能であることを特徴とする、請求項5に記載の形質転換体。
【請求項9】
KCTC11012BP、KCTC11013BP及びKCTC10908BPからなる群の中で選択された株であることを特徴とする、請求項5に記載の形質転換体。
【請求項10】
ザイモモナス・モビリスのゲノムから、pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1種以上の遺伝子を除去する段階
を含む、請求項5に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項11】
ザイモモナス・モビリスのpdc遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記pdc遺伝子がクローニングされたプラスミドからpdc遺伝子を除去し;
前記pdc遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階
を含むことを特徴とする、請求項10に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項12】
前記pdc遺伝子を有する遺伝子断片は、ザイモモナス・モビリスのpdc遺伝子と、その5’及び3’末端の両方に位置する相同組み換えのための1500乃至5000bpの相同部位を有することを特徴とする、請求項11に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項13】
ザイモモナス・モビリスのldhA遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記ldhA遺伝子がクローニングされたプラスミドからldhA遺伝子を除去し;
前記ldhA遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階を含むことを特徴とする、請求項10に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項14】
前記ldhA遺伝子を有する遺伝子断片は、ザイモモナス・モビリスのldhA遺伝子と、その5’及び3’末端の両方に位置する相同組み換えのための1500乃至5000bpの相同部位を有することを特徴とする、請求項13に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項15】
ザイモモナス・モビリスのpdc遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記pdc遺伝子がクローニングされたプラスミドからpdc遺伝子を除去し;
前記pdc遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階;
ザイモモナス・モビリスのldhA遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記ldhA遺伝子がクローニングされたプラスミドからldhA遺伝子を除去し;
前記ldhA遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階
を連続して行うことを特徴とする、請求項10に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項16】
pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子が除去されたザイモモナス・モビリス形質転換体を製作する段階;及び
前記ザイモモナス・モビリス形質転換体を30乃至34℃で10乃至14時間培養する段階を含む、
エタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1種以上の一次代謝産物の生産方法。
【請求項17】
前記ザイモモナス・モビリス形質転換体の培養時に、CO2気体を0.2乃至1vvmの量で添加したり、培養培地に炭酸塩を1乃至50mMの濃度で追加的に添加して培養することを特徴とする、請求項16に記載の一次代謝産物の生産方法。
【請求項18】
前記炭酸塩がNAHCO3、NA2CO3及びCaCO3からなる群の中で選択されたことを特徴とする、請求項17に記載の一次代謝産物の生産方法。
【請求項19】
前記ザイモモナス・モビリス形質転換体の培養時に、水素気体を0.2乃至1vvmの量で追加的に添加して培養することを特徴とする、請求項16乃至18のいずれかに記載の一次代謝産物の生産方法。
【請求項1】
ザイモモナス・モビリス(Zymomonas mobilis)のゲノムから、ピルビン酸デカルボキシラーゼをコーディングするpdc遺伝子(SEQ ID NO:1)及び乳酸脱水素酵素をコーディングするldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子を除去することによって、ザイモモナス・モビリスによるエタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1種以上の一次代謝産物の生産を増加させる方法。
【請求項2】
pdc遺伝子(SEQ ID NO:1)を除去してコハク酸及び乳酸からなる群の中で選択された1種以上の一次代謝産物の生産を増加させることを特徴とする、請求項1に記載の方法。
【請求項3】
ldhA遺伝子(SEQ ID NO:2)を除去してエタノール及びコハク酸からなる群の中で選択された1種以上の一次代謝産物の生産を増加させることを特徴とする、請求項1に記載の方法。
【請求項4】
pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)を全て除去してコハク酸の生産を増加させることを特徴とする、請求項1に記載の方法。
【請求項5】
ゲノムから、pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子が除去されて、エタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1種以上の一次代謝産物の量産が可能なザイモモナス・モビリス形質転換体。
【請求項6】
pdc遺伝子(SEQ ID NO:1)が除去されて、コハク酸及び乳酸からなる群の中で選択された1種以上の一次代謝産物の量産が可能であることを特徴とする、請求項5に記載の形質転換体。
【請求項7】
ldhA遺伝子(SEQ ID NO:2)が除去されて、エタノール及びコハク酸からなる群の中で選択された1種以上の一次代謝産物の量産が可能であることを特徴とする、請求項5に記載の形質転換体。
【請求項8】
pdc遺伝子(SEQ ID NO:1)とldhA遺伝子(SEQ ID NO:2)が全て除去されて、コハク酸の量産が可能であることを特徴とする、請求項5に記載の形質転換体。
【請求項9】
KCTC11012BP、KCTC11013BP及びKCTC10908BPからなる群の中で選択された株であることを特徴とする、請求項5に記載の形質転換体。
【請求項10】
ザイモモナス・モビリスのゲノムから、pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1種以上の遺伝子を除去する段階
を含む、請求項5に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項11】
ザイモモナス・モビリスのpdc遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記pdc遺伝子がクローニングされたプラスミドからpdc遺伝子を除去し;
前記pdc遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階
を含むことを特徴とする、請求項10に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項12】
前記pdc遺伝子を有する遺伝子断片は、ザイモモナス・モビリスのpdc遺伝子と、その5’及び3’末端の両方に位置する相同組み換えのための1500乃至5000bpの相同部位を有することを特徴とする、請求項11に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項13】
ザイモモナス・モビリスのldhA遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記ldhA遺伝子がクローニングされたプラスミドからldhA遺伝子を除去し;
前記ldhA遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階を含むことを特徴とする、請求項10に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項14】
前記ldhA遺伝子を有する遺伝子断片は、ザイモモナス・モビリスのldhA遺伝子と、その5’及び3’末端の両方に位置する相同組み換えのための1500乃至5000bpの相同部位を有することを特徴とする、請求項13に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項15】
ザイモモナス・モビリスのpdc遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記pdc遺伝子がクローニングされたプラスミドからpdc遺伝子を除去し;
前記pdc遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階;
ザイモモナス・モビリスのldhA遺伝子(SEQ ID NO:1)を有する遺伝子断片をプラスミドにクローニングし;
前記ldhA遺伝子がクローニングされたプラスミドからldhA遺伝子を除去し;
前記ldhA遺伝子が除去されたプラスミドを利用してザイモモナス・モビリスを形質転換させる段階
を連続して行うことを特徴とする、請求項10に記載のザイモモナス・モビリス形質転換体の製造方法。
【請求項16】
pdc遺伝子(SEQ ID NO:1)及びldhA遺伝子(SEQ ID NO:2)からなる群の中で選択された1つ以上の遺伝子が除去されたザイモモナス・モビリス形質転換体を製作する段階;及び
前記ザイモモナス・モビリス形質転換体を30乃至34℃で10乃至14時間培養する段階を含む、
エタノール、乳酸、ピルビン酸、クエン酸、グルタミン酸、コハク酸、フマル酸及びマレイン酸からなる群の中で選択された1種以上の一次代謝産物の生産方法。
【請求項17】
前記ザイモモナス・モビリス形質転換体の培養時に、CO2気体を0.2乃至1vvmの量で添加したり、培養培地に炭酸塩を1乃至50mMの濃度で追加的に添加して培養することを特徴とする、請求項16に記載の一次代謝産物の生産方法。
【請求項18】
前記炭酸塩がNAHCO3、NA2CO3及びCaCO3からなる群の中で選択されたことを特徴とする、請求項17に記載の一次代謝産物の生産方法。
【請求項19】
前記ザイモモナス・モビリス形質転換体の培養時に、水素気体を0.2乃至1vvmの量で追加的に添加して培養することを特徴とする、請求項16乃至18のいずれかに記載の一次代謝産物の生産方法。
【図2】

【図3】

【図5】
【図6】
【図1】
【図4】
【図7A】
【図7B】
【図8A】
【図8B】
【図9A】
【図9B】
【図10A】
【図10B】
【図11A】
【図11B】
【図11C】
【図3】
【図5】
【図6】
【図1】
【図4】
【図7A】
【図7B】
【図8A】
【図8B】
【図9A】
【図9B】
【図10A】
【図10B】
【図11A】
【図11B】
【図11C】
【公表番号】特表2009−526547(P2009−526547A)
【公表日】平成21年7月23日(2009.7.23)
【国際特許分類】
【出願番号】特願2008−555167(P2008−555167)
【出願日】平成19年2月16日(2007.2.16)
【国際出願番号】PCT/KR2007/000860
【国際公開番号】WO2007/094646
【国際公開日】平成19年8月23日(2007.8.23)
【出願人】(503137827)マクロジェン・インコーポレーテッド (2)
【Fターム(参考)】
【公表日】平成21年7月23日(2009.7.23)
【国際特許分類】
【出願日】平成19年2月16日(2007.2.16)
【国際出願番号】PCT/KR2007/000860
【国際公開番号】WO2007/094646
【国際公開日】平成19年8月23日(2007.8.23)
【出願人】(503137827)マクロジェン・インコーポレーテッド (2)
【Fターム(参考)】
[ Back to top ]