
上皮細胞におけるヒトパピローマウイルスゲノムの特異的なメチル化に関連する化合物および方法
本発明は、HPV関連疾患の検出及び治療に有用な化合物及び方法に関する。本発明は、HPVの複製が自然感染した組織及び細胞において生じるメカニズムを解明したことに基づいている。さらに、本発明は、ウイルス感染細胞における、この病的細胞の前癌及び癌細胞への成長の促進を可能にするウイルスゲノムの異なる後成的変化を確認したことに基づくものである。従って、本発明は、新形成及びその前駆病変を診断する方法並びに悪性腫瘍の発症を予防し、又は腫瘍増殖を抑制する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、HPV関連疾患の検出及び治療に有用な化合物及び方法に関する。本発明は、HPVの複製が自然感染した組織及び細胞において生じるメカニズムを解明したことに基づいている。さらに、本発明は、ウイルス感染細胞における、この病的細胞の前癌及び癌細胞への成長の促進を可能にするウイルスゲノムの異なる後成的変化を確認したことに基づくものである。従って、本発明は、新形成及びその前駆病変を診断する方法並びに悪性腫瘍の発症を予防し、又は腫瘍増殖を抑制する方法を提供する。
【背景技術】
【0002】
人体の単一の正常細胞の表現型は、特定の時点で発現される遺伝子のセットによって規定される。こうした遺伝子発現のパターンは、全ての生細胞及び複雑度の増したレベルで全ての器官に対して構造及び機能を与える蛋白質の組合せ及び個々の濃度を規定している。全ての器官が適切に機能するのに不可欠である分化過程も個々の遺伝子のレベル及び組合せの変更によって調節されている。識別的遺伝子発現パターンは、例えば、大幅に変化した遺伝子発現パターンを有する、完全な分化細胞への分化を経ることができる子孫を生じる幹細胞としての細胞の状態を決定する。こうした分化細胞は、最終的に、さらにその遺伝子発現パターンを変化させ、それにより老化してついには死にゆく細胞の特性を取り入れることによってその寿命を決定する。こうした過程の分子的調節については解明されているとはとても言えないが、いくつかの証拠は、この調節の本質的な部分がヒトゲノム内の特定のGpC島の明確なメチル化によって媒介されていることを示唆している。
【0003】
概念的には、細胞のゲノム内のこの特定のヌクレオチド群へメチル(C(H)3)−基を付加すると、このゲノムのメチル化伸展の凝縮及び強化構造を生じ、その結果として転写の阻害がもたらされる。即ち、ゲノムの高度にメチル化された部分は、通常mRNAに転写されない(発現抑制(silenced))。このメカニズムは、遺伝子の調節及び刷り込みの重要な部分であり、生細胞におけるこうした重要なステップを決定する主要な生物学的特徴の1つである(Collasほか、2007年)。分化過程におけるメチル化の詳細な影響については、これまでごく皮相的に研究されているに過ぎない(Kiefer、2007年)。しかしながら、正常細胞の本格的な新生癌細胞への形質転換に関与する形質転換過程に対してより詳細な洞察が集中されている(Esteller、2007年及びFeinberg、2004年)。特に、重要な腫瘍抑制活性を有する遺伝子群の場合、それらのプロモータ領域群のメチル化が新生癌細胞のこうした各遺伝子の発現低下を招いたことが観察されている。従って、メチル化は、欠失及び/又は突然変異に加えて、腫瘍抑制遺伝子の不活化における主要な分子段階の1つであり、多くの腫瘍抑制遺伝子、例えば、サイクリン依存性キナーゼ抑制因子p16INK4a、DNAミスマッチ修復遺伝子MSH2、MHL1などに関して明白に立証されている(Esteller、2007年及びFeinberg、2004年)。
【0004】
クロマチン構造に対する影響に加えて、異なるヌクレオチドに対してより精密にメチル化を行うことにより特定の転写因子の結合性に影響を与えることもできる(Klose及びBird、2006年)。従って、これによって正又は負に作用する転写因子の親和性に影響を与えることができ、それ故、この場合も上記各遺伝子の発現の活性化又は低下をもたらす、こうした転写因子の作用及び活性が修飾される。
【0005】
ヒト・パピローマウイルスは、皮膚又は粘膜面の上皮の増殖を引き起こす重要な上皮親和性ウイルスである。こうしたウイルスは、特定の他と区別できる型のもの(ハイリスクパピローマウイルス、HR−HPV:high risk papillomavirus)が、特に生殖器官及び最も重要なことだが子宮頸部の粘膜面の腫瘍性病変と関連付けられることが明らかとなった(zur Hausen、2002年)とき、特に科学的な関心を集めた。今日では、HR−HPV型(特に、HPV16及び18)の持続感染が子宮頚癌発症の必須の前提条件であることは一般に受け入れられている。E6及びE7と称する特定のウイルス遺伝子は、子宮頚部その他のHPV関連癌細胞では継続的に発現されていることが分かった。さらに、こうした遺伝子は、インビトロ培養一次ヒト上皮細胞において悪性形質転換を誘導することが明らかにされ、重要なことには、その発現は、子宮頚癌細胞又はその高悪性度前駆細胞の腫瘍性増殖の特徴を誘導し、維持するのに基本的に必要とされる。
【0006】
これまでのところ、約13種のハイリスクHPV型の特性が明らかにされている(Munozほか、2003年)。これらのウイルスは、臨床的に検知可能な病変を発症している(まだ発症していない)女性にもよく見られる。こうしたウイルス感染の罹患率のピークは年齢15乃至30歳の若い女性に見られるのに対して、より上の年齢の女性では低下する(Schiffmanほか、2007年)。このことは、若い男性にも同程度の頻度でみられるHR−HPV感染症が性行為感染の特徴を有することを示している。しかしながら、性別による主な相違は、男性ではHR−HPV感染がその宿主によって自覚されることがまれであり、浸潤性の癌へと進行することがない不顕性感染をもたらすのが通常であるのに対して、女性では明らかに長期の持続感染がかなりより高頻度で生じるため、最終的に浸潤性の癌へと進行する可能性のある子宮移行部の上皮細胞の悪性形質転換がもたらされることがある。興味深いことに、この現象が概して子宮頸部の移行部に存在する数少ない上皮細胞に限定されるように思われるのに対して、同程度の率で感染する上皮部位は、子宮移行部の上皮細胞と比較して悪性形質転換を受ける傾向が実質的に少ない。
【0007】
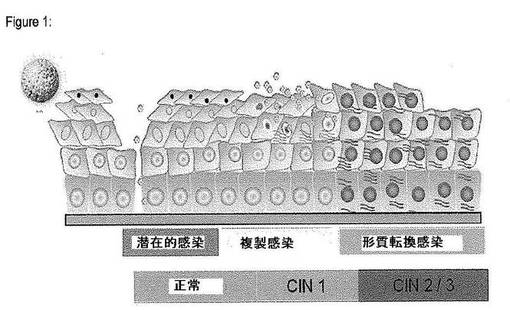
通常の生活環の過程でHPVは先ず上皮の基底及び傍基底細胞に感染する。いくつかの論拠から、このウイルスは先ず潜伏感染の状態に入ることが示唆されている(Longworth及びLaimins、2004年)。この過程では、宿主細胞が一旦分裂すると、ウイルスゲノムは単一の感染細胞当たりでは極めて低いコピー数で複製されるが、この段階ではウイルスゲノムの複製も、場合によっては溶菌感染も開始されない。技術的な限界のために、この種の潜伏感染の存在に関する正確な証拠はまだ得られていない。むしろ、これは、新規感染の証拠がなくても免疫抑制時にウイルス複製活性が再活性化されうることを示す疫学研究から予想される。こうした潜伏感染は、明らかに活動性複製感染に切り替わり得る。この場合、基底及び傍基底細胞ではウイルス遺伝子の発現活性は全くほとんど維持されないが、これらの感染上皮細胞が一旦分化し始め、この上皮の中間細胞層に達すると、こうした分化細胞層においてウイルスゲノムの複製を引き起こす遺伝子発現の増加が観察される。興味深いことに、ウイルスゲノムの複製及びウイルスの初期遺伝子の発現は、増殖能、及び最終分化上皮細胞を予めプログラムされた分化経路に押し込むこうした細胞の活性化遺伝子発現特性を不可逆的に失っていて、最終的に上皮の外表面に扁平上皮細胞残屑として崩壊する最終分化細胞に概して限定されると考えられる(Longworth及びLaimins、2004年)。中間細胞においてウイルスゲノムが複製し、上皮細胞が一旦正常な分化過程で表面細胞層に達すると、ウイルスゲノムがさらに再プログラム化され、全ての初期遺伝子の発現が止められる一方で、各外被蛋白質の翻訳をもたらす後期遺伝子L1及びL2の本格的な発現がここで観察される。これらの蛋白質は自然に集まって、複製されたウイルスゲノムが含まれるウイルスの外被を形成し、最終的に、この新たに合成された成熟ウイルス外被は、上皮の外面の崩壊しつつある扁平上皮細胞の残屑から放出され、更なる感染サイクルを再開することができる(図1)。これらの知見から、ウイルスの複製サイクルは正常な上皮の分化過程と厳格に連関していることが示唆される。しかしながら、上皮細胞の分化の誘因となる分子事象以後に、この複雑な複製過程を制御するウイルス遺伝子発現パターンの調節に関与する分子構造についてはこれまでのところほとんど又は全く知られていない。
【0008】
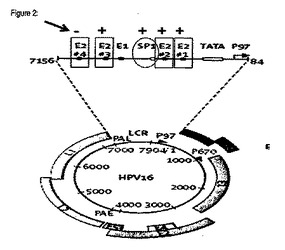
ウイルス遺伝子発現の調節及びウイルスゲノムの複製に重要な意味を持つ構造は、長鎖制御領域(LCR:long control region)又は上流調節領域もしくは(URR:upstream regulatory region)と呼ばれる配列エレメント内に保持されている。このエレメントには、重要な正及び負の転写因子の結合部位及び複製起点が存在する(図2)。
【0009】
初期に発現されるいくつかのパピローマウイルス遺伝子の一部であるパピローマウイルスE2蛋白質は、ウイルスのプロモータ自体に結合することによりウイルス遺伝子の発現に対して実質的な抑制機能を有する(Wellsほか、2000年)。従って、これは転写レプレッサーの働きをし、HPV URRエレメントの制御下の遺伝子の転写を阻止する。この概念は、インビトロで増殖させた非分化上皮細胞、場合によっては基底又は傍基底細胞の類似非分化表現型を保持する癌細胞株も用いて組織培養モデルで実施されている種々の分子研究に基づいたものである。E2蛋白質が発現されると、ウイルスプロモータの下方制御及び隣接遺伝子の発現停止がもたらされることが分かっている(Bouvardほか、1994年)。
【0010】
子宮頚癌細胞では、パピローマウイルスゲノムは、通常、宿主細胞染色体中に組み込まれることが分かっている(Pert及びColeman、2007年b)。ウイルスゲノムの宿主細胞ゲノム中への組込み部位は不規則に選ばれるように思われるが、組み込まれたウイルスDNA断片のウイルスゲノムは、通常、極めて特異な仕方で保存される。まず第一に、URRを含む断片並びに全てこれから分析されるE6及びE7遺伝子は、組み込まれたウイルスゲノムコピーからもE6及びE7遺伝子の発現を可能にする立体配置で保たれるのに対して、下流に位置する遺伝子E2及び後期遺伝子カセットは、通常、URR内の調節エレメントから切り離され、従って、機能的に、又は構造的にも不活化される(zur Hausen、2002年)。興味深いことに、この点に関して調べられた全てのHPV関連癌細胞で、ウイルスE6及びE7の発現を確認することができた。前述のように、これらの遺伝子は、細胞周期の調節、分化過程及び死のカスケードに少なくともある程度関与しているいくつかの宿主細胞蛋白質との複雑な相互作用を通じて複製している細胞に重要な増殖調節特性を付与する。これらの遺伝子の発現は、上皮細胞の悪性形質転換を誘導し、維持するのに重要で十分な必要条件である。これらの遺伝子の発現が形質転換細胞で遮断されれば、この細胞は細胞周期停止に陥り、増殖を止め、最終的には死滅する(von Knebelほか、1988年)。
【0011】
この観察と一致して、HR−HPV型のE2遺伝子をコードしているオープンリーディングフレームはHPV関連癌では通常、破壊されている。従って、HPV形質転換細胞中に機能的に活性なE2蛋白質を再導入すると、悪性表現型、細胞周期停止及び最終的には細胞死の抑制がもたらされる(Wellsほか、2000年)。
【0012】
従って、宿主細胞染色体に組み込まれることによるE2 ORFの破壊がHPV感染上皮細胞の癌化を最終的にもたらす分子カスケードにおいて絶対必要ではないにしても重要なステップであることは、この分野の科学者の間で一般に受け入れられている(Pett及びColeman、2007年a、Pettほか、2006年、Pettほか、2004年並びにAlazawiほか、2002年)。
【0013】
上皮分化の特徴とHPV複製サイクルとが密接に関連していることは、ウイルスの遺伝子発現及び複製の制御に宿主細胞の環境が重要な役割を果たしていることを示唆している(図1参照)。いくつかの証拠から、ウイルスゲノムの遺伝子発現及び複製調節においてメチル化その他の後成的制御の関与が示唆されている。こうしたデータの多くは、人工的な組織培養モデルに基づいたものである。これまでのところいずれのモデルによってもウイルスの遺伝子発現及び複製の分化依存性の調節の詳細を説明することはできなかった。
【0014】
パピローマウイルスDNAのメチル化については既に約20年前に報告されている(Burnett及びSleeman、1984年並びにWettstein及びStevens、1983年)。しかしながら、パピローマウイルス遺伝子発現制御及びこれに関連した発癌効果の調節におけるその生物学的意義については依然として極めて不明のところが多い(Listほか、1994年、Roslほか、1993年並びにThainほか1996年)。子宮頚癌細胞株において、HPV−16及びHPV−18ゲノムにおけるCpGメチル化はLCR領域及び一部のL1 ORF領域ではこのウイルスゲノムのどの他の部分よりも高い頻度で生じるように思われる(Badalほか、2004年及びBadalほか、2003年)。さらに、特にL1遺伝子のメチル化は、発癌時における宿主細胞ゲノム中への外来ウイルスDNAの組込みに関連があるように思われ、このため、HPV感染細胞の腫瘍性変化の生体指標となる可能性があることが示唆された(Kalantariほか、2004年、Turanほか、2006年)。別のデータからは、E2BSにおけるCpGメチル化が、インビトロでのE2蛋白質結合を妨げ(Thainほか、1996年)、転写活性化の際の細胞におけるE2蛋白質機能を調節することも示唆されている(Bhattacharjee及びSengupta、2006年並びにKimほか、2003年)。しかしながら、癌におけるDNAメチル化はHPV DNAに限られるものではない。概略前述したように、これは極めて重要な特徴とみなされるが、どちらかと言えば、宿主ゲノム全体にわたってよく生じる事象である(Esteller、2007年)。多くの細胞遺伝子の過剰メチル化の頻度は、新形成の重篤度が増大するにつれて著しく増加することが分かっている(Bannoほか、2007年、Dongほか、2001年、Fengほか、2005年、Jeongほか、2006年、Kangほか、2006年、Laiほか、2007年、Leaほか、2004年、Reesink−Petersほか、2004年、Sengほか、2007年、Steenbergenほか、2004年、Virmaniほか、2001年、Widschwendterほか、2004年)。
【0015】
最近の研究では、正常上皮細胞及び子宮頚癌細胞においてHPV16のE2結合部位E2BS2乃至4内のCpGのメチル化の有無が調べられた(Bhattacharjee及びSengupta、2006年)。この論文の著者らは、メチル化が、形質転換組織におけるP97プロモータの近位にあるE2BS2乃至4に位置する明確なCpG島に見出されたと報告している。
【0016】
しかしながら、この研究では、特定部位のメチル化パターンの明確な変化で異なる生物学的特徴を説明することができる明瞭で一貫したパターンを示すことができなかった。
【0017】
Kimら(Kimほか、2003年)は、W12細胞を用いたインビトロ組織培養モデルの高度分化細胞集団において低メチル化が関連付けられると報告した。これに対して、低分化基底細胞様細胞からのHPV16 LCRは多数のメチル化シトシンを含み、多くの場合、E2BSがメチル化されていた。
【0018】
さらに、ヒトの癌は特にCpG島においてDNAメチル化のパターン変化を示すことが多いことが先行技術に開示されている。島内のメチル化は連鎖遺伝子の転写抑制を伴うことが分かっている。腫瘍の発症及び進行の全ての局面に関与する遺伝子は、メチル化され、後成的に発現抑制されるようになる可能性がある。このような発現抑制された遺伝子を再発現させると、腫瘍増殖の抑制又は抗癌療法への感受性増大をもたらすことができる。DNAメチル化を抑制することができる後成的薬剤については、現在、前臨床評価及び癌患者での臨床試験が行われている。ヌクレオシド阻害剤5−アザシチジン及びデシタビンは、多くの種類の癌に対して数多くの第1相及び第2相試験で試験されている。しかしながら、両剤の用量規定毒性は骨髄抑制であり、最もよく報告されている非血液学的有害作用は悪心及び嘔吐であった。従って、脱メチル化剤による癌の全身療法は、通常重篤な副作用を引き起こすことになる。
【0019】
美容整形症状及び皮膚症状を処置するのに特異的DNAメチル化阻害剤を用いることができる。5−アザシチジンは正常ヒト線維芽細胞による基礎レベル及びTGFβ誘導コラーゲン生合成を共に阻害することが分かっている。コラーゲンは瘢痕化に対して直接的な影響を示す。従って、ヒトの皮膚においてコラーゲンの過剰産生を抑制すると、瘢痕形成が抑制されることになる。さらに、癌性及び前癌性皮膚病変の処置にUV処置と脱メチル化を併用することができる。
【0020】
本発明は、パピローマウイルスゲノム内の異なる特定の配列エレメントのメチル化がウイルスの生活環、複製及び遺伝子発現パターンを制御しているという知見に関する。さらに、本発明は、このメチル化パターンの特定の変化が一部のパピローマウイルスにより誘発される悪性形質転換の誘導に関与しているという知見に関する。このことは、例えば子宮頚部、膣、陰門、肛門、口腔咽頭管などの体腔の粘膜上皮の病変の場合に特に重要である。脱メチル化剤による全身療法に伴う望ましくない、ある程度耐えられない副作用を克服するために、本発明者らは、特に体腔の粘膜上皮のHPV関連病変及び癌の処置において、脱メチル化剤による局所処置が患者に望ましくない副作用をもたらすことなく癌を治療するのに極めて効果的な方法となりうることを見出した。
【発明の概要】
【課題を解決するための手段】
【0021】
本発明は、例えば体腔の粘膜上皮などの病変の処置及び/又は治療のための脱メチル化剤の局所適用に関する。この点に関して、本発明はさらに、HPV関連病変の処置及び治療の方法に関する。HPV関連病変は、例えば子宮頚癌及びその前駆症状段階並びに皮膚の疣贅を含む。
【0022】
さらに、本発明は、ウイルス複製及び分子の形質転換感染への移行を阻止することによりHPV関連病変及び癌を処置するために体腔の粘膜上皮へ局所適用するための脱メチル化剤含有医薬組成物に関する。この点に関して、本発明はさらに、HPV関連新形成の処置及び予防用医薬組成物に関する。この医薬組成物は、医薬用として許容可能な担体並びにE2BS1、NFIBS、TEF−1BS及び/又はSP1BSにおける特定のメチル化パターンを阻害する後成的阻害剤又は脱メチル化剤を含む。
【0023】
また、本発明は、HPV E2BS1、NFIBS及び/又はTEF−1BSのメチル化パターンを測定することによって潜伏感染した細胞とHPV増殖感染細胞とを識別する方法に関する(図5参照)。
【0024】
配列番号1乃至8の配列を含む単離核酸であって、この核酸は下記配列中、大文字で示した位置でメチル化されているものとする核酸。
【0025】
【化1】

上記配列の全てのヌクレオチドの位置はNCBI受託番号NC_001526のHPV16全ゲノム配列に従って引用した。大文字のCはメチルシトシンに対応する。大文字のGは逆相補鎖にメチルシトシンが位置する部位を示す。
【0026】
下記の配列番号9乃至15の単離核酸
【0027】
【化2】
【図面の簡単な説明】
【0028】
【図1】ウイルスの生活環及び形質転換感染への進行の模式図である。
【図2】HPV16ゲノム及び長鎖制御領域(LCR)又は上流調節領域(URR)の拡大構造の模式図である。ウイルスE2蛋白質及びSP1転写因子の結合部位の相対的位置が示されている。
【図3】正常と腫瘍性上皮との間でE2BS4のメチル化パターンを比較した(Bhattacharjee及びSengupta、2006年)により報告されたメチル化パターンの分析結果並びにE2BS1における非形質転換上皮と形質転換(p16INK4a陽性)上皮との主要な相違を明らかにした本発明者らの結果の模式図である。
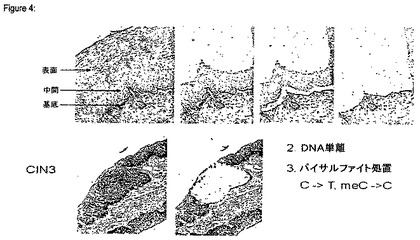
【図4】微小細胞を切り出した子宮頚部上皮の例を示す図である。上部パネル:初期HPV感染(出現しつつあるCIN1の証拠)であるが、形質転換されていないため、p16INK4aが染色されていない上皮。下部パネル:無秩序なHPV E6/E7癌遺伝子発現を示す高悪性度扁平上皮内病変及び高悪性度扁平上皮内病変(HSIL:high grade squamous intraepithelial lesion)の証拠。
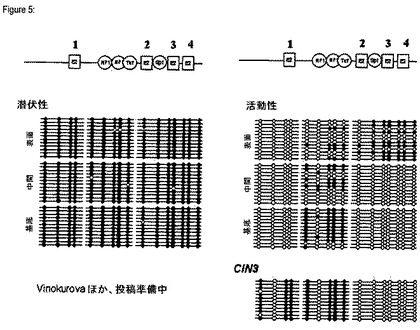
【図5】HPV16 URR内の重要な転写因子結合部位のメチル化パターンを示す模式図である。
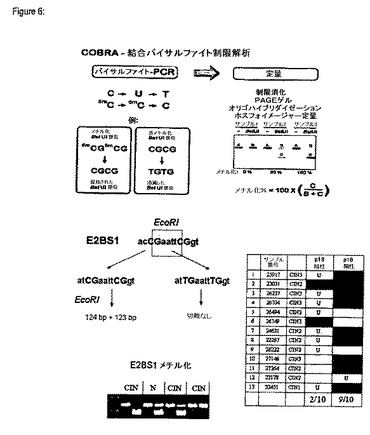
【図6】COBRAアッセイの原理及びHPV16ゲノムのE2BS1のいくつかの臨床サンプルの分析結果を示す図である。
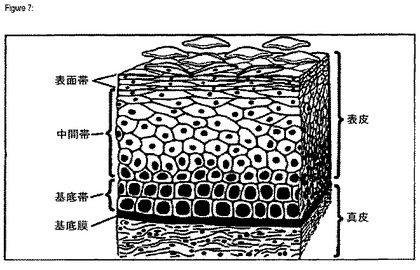
【図7】正常扁平上皮の構造を示す図[KossLGほか、1999年]である。
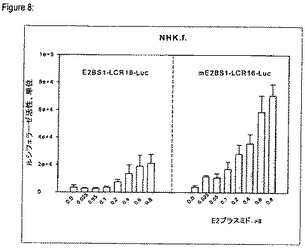
【図8】p97初期プロモータ活性に対するE2BS−1メチル化の影響を示す図である。
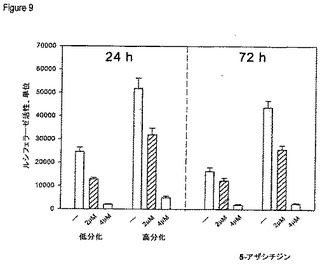
【図9】p97初期HPV16プロモータ活性に対する5−アザシチジンの影響を示す図である。
【発明を実施するための形態】
【0029】
本発明者らは、ヒトパピローマウイルスゲノムのメチル化パターン、特にウイルスの生活環の3つの想定相、即ち、a.潜伏感染、b.複製感染及びc.形質転換感染におけるURR(その同族E2BSを含む)のメチル化パターンについて調べた。分化段階の異なる上皮細胞内のHR−HPVゲノムの詳細なメチル化状態を明らかにするために、組織から微小細胞を切り出し(図4参照)、異なる細胞層からのHPV配列を増幅させて、バイサルファイトシーケンシングPCR(BSP:bisulfite sequencing PCR)を用いたHPVゲノムの異なるメチル化パターンの正確な評価を行った(図5参照)。
【0030】
ここでいうHPV関連病変とは、急性又は持続性HPV感染により誘発された任意の上皮の全ての変化を含む、HPV感染により誘発された病変のことをいう。これらの病変は、臨床的影響のない小さな疣贅から、もっと正確に言えば、皮膚、生殖器官、粘膜表面などの外方増殖パピローマ、内反性パピローマを初めとするコンジローム並びに腫瘍形成性パピローマウイルス遺伝子タイプにより誘発される前癌及び浸潤性病変(それに由来する転移を含む)に及ぶものとすることができる。HPV誘発性前癌又は癌病変の多くは、種々の上皮組織の移行部、特に、例えば、子宮頚部移行部、肛門上皮と直腸粘膜の接合部又は扁桃領域の口腔咽頭部内のような、単層腺上皮が多層扁平上皮と交わる所に生じる。
【0031】
本発明の発明者らは、HPVの細胞変性効果の影響を未だ受けていない正常な上皮では、ウイルスゲノムが基底細胞内に存続することができることを見出し、基底及び傍基底細胞に潜伏期のウイルス感染が存在することが明らかとなった(データは図に示されていない)。この潜伏期のウイルスゲノムは分析することができたすべてのCpGで完全にメチル化されており、ウイルスゲノムの全体的なメチル化によって感染の潜伏が媒介され、ウイルス遺伝子の発現が阻止されることにより感染上皮の細胞病理学的又は組織病理学的変化も防止されることが示唆される。すなわち、本発明者らは、HPVゲノムのメチル化状態と感染の潜伏様式との間に関連性があることを見出した。この時期のメチル化がE2BS1にも見出されることに留意することは重要である。
【0032】
ウイルスゲノムを初期異形成病変(CIN1)の切り出し微小細胞から単離した場合には、宿主細胞の分化状態及びウイルスゲノム内の各CpGに応じてメチル化パターンの異なる変化が観察された(図5参照)。基底細胞層では、URRの異なる部分にメチル化されていないE2BS(1乃至4)が認められた。興味深いことに、E2BS1がメチル化されなかったことによって、恐らくウイルスのエンハンサエレメントの結合及び抑制が可能となる。ウイルスDNAが中間細胞層から単離された場合には、ゲノムの諸部分は脱メチル化されるようになる。これによってNF1、TEF−1などのような正の転写因子(TF:transcription factor)の接近が可能となり、ウイルス遺伝子の発現が刺激される。ウイルスの遺伝子発現の増強は分化細胞内の脱メチル化が増加することに一致して生じ、従って、ウイルスゲノムの複製が開始される(図5)。
【0033】
表面細胞層では、E2(E2BS2、3及び4)並びにSP1結合部位を含む初期プロモータ内の全てのCpGがメチル化され、初期プロモータ活性の不活化が示唆された。
【0034】
すでに高悪性度扁平上皮内病変(HSIL:high grade squamous intraepithelial lesion)へ進行し、従ってp16INK4aマーカ(Klaesほか、2001年)の過剰発現により明らかとなるウイルスの癌遺伝子E6及びE7の大量発現を示す(図4参照)病変では、E2BS1の一貫したメチル化が認められた(図5)。このことから、メチル化は、この特定の結合部位へのE2蛋白質の接近を阻止することによって負の転写調節を無効にし、コントロール不良の高レベルのウイルス遺伝子発現がHPV16 URR(LCR)により媒介されることを可能にすることが示唆される。
【0035】
結合バイサルファイト制限解析(COBRA:Combined Bisulfite Restriction Analysis)(Xiong及びLaird、1997年)を用いてこのメチル化パターンを、無秩序なE6及びE7癌遺伝子の発現を示す病変のp16INK4a陽性部分とHPVE6及びE7遺伝子発現障害の制限を明らかに依然として維持する病変のHPVに感染するもp16INK4a陰性の部分との間で比較した。図6に示したデータは、E2BS1のメチル化はウイルス癌遺伝子の無秩序な発現と同時に起きるという概念を裏付けるものである。
【0036】
E2結合部位は以下の通り定義される:
【0037】
【化3】
NCBIアセッション番号NC_001526のHPV16全ゲノム配列によるBSの位置。
【0038】
正常上皮は、図7に示したように記述することができる。皮膚及び粘膜の最外層は、3つの主な帯域に分類することができる、形態の進行性の変化により区別される多上皮細胞層からなる(図20)[Koss LGほか、Introduction to Gynecologic Cytopathology with Histologic and Clinical Correlations、1999年:p.23−30]。
【0039】
基底帯は、球状であり、比較的大きな核を有する低分化基底細胞の1又は2層(基底層)からなる。正常な扁平上皮では、基底幹細胞は分裂することができる唯一つの細胞である。この基底細胞は真皮から表皮を隔てる基底膜に付着している。
【0040】
上皮の中間帯は上皮の厚さの大部分に相当し、表面に向かって大きさが増大する数層の細胞(傍基底層又は有棘層及び中間扁平層又は顆粒層)からなる。一旦基底膜に付着しなくなると、こうした細胞は細胞周期を出て、分裂を止め、分化し始める。
【0041】
表面帯は、小さな縮合核及びグリコーゲンで満たされている細胞質を有するより大きく扁平な細胞の列(成熟扁平層又は角質層)を含む。表面帯の外層は最終分化細胞からなる。
【0042】
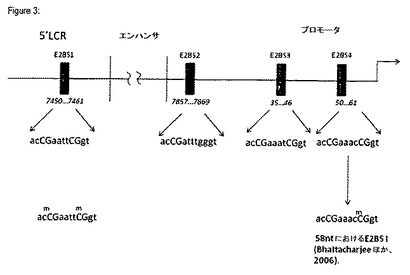
ゲノム位置7,198番目と161番目との間の分析領域には16個のCpGが含まれる(図5)。この分析領域は、5’LCR、エンハンサ及びプロモータと呼ばれる3つの機能的に異なるセグメントに分けることができる。この5’LCRは、E2BS1(7,450−nnnnn−7,461)内のCpG(nt7452、nt7458)を含む4個のCpGを含む。エンハンサは、NFIBS(7541−cttgccatgCGtgccaaatc−7560及び7663−aatcactatgCGccaaC−6379)並びにTEF−1BS(7684−tacatacCGct−7694)内の4個のCpG(nt7550、7673、7679及び7691)を含む5個のCpGを含む。プロモータ内の最後の7個のCpG、これらのうちの6個(nt7859、31、37、43、52、58)は、Sp1(25−taagggCG−32)並びに3つのE2結合部位(E2BS2 7857−acCGatttgggt−7869、E2BS3 35−acCGaaatCGgt−46及びE2BS4 50−acCGaaacCGgt−61)と重複する。
【0043】
これまでの報告に所載されていることとは対照的に、本発明者らは、HPVの細胞変性効果の影響を未だ受けていない正常上皮では、ウイルスゲノムが基底細胞内に存続することができることを見出し、基底及び傍基底細胞に潜伏期のウイルス感染が存在することが明らかに示された(データは図に示されていない)。この潜伏期のウイルスゲノムは、分析することができた16個のCpGの全てにおいて完全にメチル化されており(図5)、このことから、ウイルスゲノムの全体的なメチル化によって感染の潜伏が媒介されてウイルス遺伝子の発現が阻止され、これによって感染上皮の細胞病理学的又は組織病理学的変化も防止されることが示唆される。この知見は、これまでHPVゲノムのメチル化状態と潜伏様式の感染とを関連づけるデータは報告されていないので、新規なものである。この時期において、メチル化がE2BS1においても見られることに留意することは重要である。
【0044】
ウイルスゲノムを初期異形成病変(CIN1)の切り出し微小細胞から単離した場合には、宿主細胞の分化状態及びウイルスゲノム内の各CpGに応じてメチル化パターンの異なる変化が観察された(図5参照)。
【0045】
プロモータ領域では、分析した12のアンプリマークローンの、基底細胞及び中間層からの細胞画分から単離したDNAは全て、メチル化されていないCpGを含んでいた。表面細胞層の分化のより進んだ細胞から単離したHPV16DNAでは著しく異なった結果が観察された。分化細胞に富んだこの表面層から単離したHPV16DNAから作製した12種のアンプリマークローンのうちの8種において、E2BS(2、3及び4)及びSP1部位を含むプロモータ内のCpGがメチル化された。
【0046】
エンハンサ領域では、2個のNFIBS及び1個のTEF−1BS内の4個のCpGが基底細胞内で高度にメチル化されていたが、分化のより進んだ細胞内ではメチル化の程度はより低かった。これによってNF1、TEF−1などのような正の転写因子(TF)の接近が可能となり、ウイルス遺伝子の発現が刺激される。ウイルスの遺伝子発現の増強は分化細胞内の脱メチル化が増加することに一致して生じる(図5)。
【0047】
5’LCRセグメントでは、4個のCpGの全てが分化段階と関係なくメチル化されていなかった。興味深いことに、E2BS1がメチル化されなかったことによって、恐らくウイルスのエンハンサエレメントの結合及び抑制が可能となる。
【0048】
すでに高悪性度扁平上皮内病変(HSIL)へ進行し、従ってp16INK4aマーカ(Klaesほか、2001年)の過剰発現により明らかとなるウイルスの癌遺伝子E6及びE7の大量発現を示す(図4参照)病変では、特有のメチル化パターンが認められた(図5)。プロモータ及びエンハンサ領域では、NFIBS及びTEF−1BS内の4個のCpGがメチル化された。5’LCRでは、E2BS1内の2個のCpGを含む3個のCpGがメチル化された。このことから、メチル化は、この特定の結合部位へのE2蛋白質の接近を阻止することによって負の転写調節を無効にし、コントロール不良の高レベルのウイルス遺伝子発現がHPV16 URR(LCR)により媒介されることを可能にすることが示唆される。
【0049】
これによって、本発明者らは、こうした宿主細胞組織内での各ウイルスゲノムの生物学的特性を用いてウイルスゲノムの異なるメチル化パターンを示すことができた。この研究の重要な知見は以下のとおりである。
【0050】
ウイルス活性を示さない潜伏感染細胞では、HPVゲノムは、全てではないにしても殆ど全てのCpG島において高度にメチル化され、それによってウイルス遺伝子の任意の生物学的に意義のある発現が阻止される。例えば、基底帯、中間帯及び表面帯の細胞においてCpG島の80%超、好ましくは90%、好ましくは95%、より好ましくは98%以上がメチル化される。これによって数少ない上皮幹細胞における潜伏HPV感染の完全な隠蔽が媒介され、その後、それは不確定の期間持続することができる(図5参照)。
【0051】
まだ特徴付けられていない条件下では、個々の細胞におけるHPV感染の潜伏状態は複製感染様式に切り替わり得る。基底細胞では、(E2BS2、3及び4並びにSP1結合部位並びに(E2BS1を含む)5’長鎖制御領域内のCpGを含む)初期HPVプロモータはメチル化されていないCpGを含んでいた。従って、エンハンサ領域では、NF1及びTEF−1部位内の4個のCpGは高度にメチル化されていた。このメチル化のパターンは、p97プロモータの活性が低いことを示唆するものである。(図5)
中間細胞では、HPVエンハンサのメチル化の程度は、基底細胞に比し、分化のより進んだ細胞ではより低く、これによって、核因子1(NF1:nuclear factor I)、転写エンハンサ因子TEF−1(transcriptional enhancer factor−1)などの転写活性化因子の結合によるp97プロモータの活性増大がもたらされる。しかしながら、初期プロモータの活性は、E2の負の制御(E2BS1脱メチル化)のために一定のレベルに維持される。
【0052】
最終分化表面細胞では、E2(E2BS2、3及び4)並びにSP1結合部位を含む初期プロモータ内の全てのCpGがメチル化され、初期プロモータ活性の不活化が示唆された。
【0053】
ウイルスゲノムのメチル化状態が実質的に宿主細胞の上皮分化の程度によって変化するという事実は、DNAのメチル化がウイルスの生活環及び恐らくまた上皮分化において重要な役割を有することを示唆している。さらに、機能的に異なる高リスク及び低リスクウイルス型の両者は、増殖性生活環を調節するための共通のメカニズムを共有していると考えられる。
【0054】
複製感染が長期持続すると、基底及び傍基底細胞のうちの感染細胞は、基底及び傍基底細胞のメチル化プロフィルを主にEBS1、nt7459のメチル化を特徴とするHPV感染の形質転換様式に切り替えることができる。このCpG島のメチル化は、E2BS1へのE2結合の低下及び興味深いことにURRに対するその抑制機能の低下をもたらす。その結果、これによって各HPVゲノムは細胞内監視制御を緩め、基底細胞コンパートメント内でウイルス癌遺伝子E6及びE7の十分なレベルの発現を開始する。
【0055】
バイサルフェートゲノム配列決定(BGS:bisulfate genomic sequencing)法によるHPV16のURRにおけるメチル化CpGのマッピング。
【0056】
レーザーにより切り出した分化程度の異なる(基底、中間及び表面層)微小細胞から単離したDNAについて、バイサルファイトゲノム配列決定(BGS)法によりHPV16のURRのメチル化状態を分析した。
【0057】
EZDNAメチル化キット(Zymo Research社、Orange、CA)を用い、メーカーの推奨する方法に従ってDNAのバイサルファイト修飾を行った。対照としてCaski及びSiHa細胞株からのDNAを用い、サンプルと同時に処置して完全なバイサルファイト処置を保証した。
【0058】
HPV16のURR(nt7198からnt161まで;NC_001526)に架かるように設計したプライマを用いて入れ子BSM−PCR系を開発し、実施した(表1)。
【0059】
【表1】
PCR反応混液は、10×PCR緩衝液、5μlの50mM MgCl2、0.5μlの2mMデオキシリボヌクレオチド三リン酸、0.5mMの各PCRプライマ(1.5μlのプライマ(25pmol/μl))、2.0UのPlatinum Taq(Invitrogen社)及び2μlのバイサルファイト修飾DNAを含む計50μlとして作製した。各分析ではDNAを含まない陰性対照を置いた。増幅条件は以下の通りとした:最初の94℃2分間の変性後、94℃30秒間の入れ子PCR、50℃30秒間のアニーリング、72℃40秒間及び最後に72℃4分間の伸長の40サイクル及び30サイクル。PCR産物は、電気泳動により分離し、臭化エチジウムで染色した2%アガロースゲルから単離した。次いで、QIAquick Gel Extraction Kit(Qiagen社、Hilden)によりメーカーの使用説明書に従って単離PCR産物を精製した。精製PCR断片は、TA Cloning System(Invitrogen社)を用いてクローニングし、12の個々のクローンについて配列決定を行い、HPV16 DNA内のメチル化CpGの有無及びパターンを明らかにした。バイサルファイト修飾サンプルDNAの配列決定は、BigDyeターミネータ配列決定キット(Applied Biosystems社、Foster City、CA)を用い、メーカーの推奨する方法に従って実施した。配列決定PCR産物はABI Prism 310 Genetic Analyzerで解析した。
【0060】
HPV16 E2BS1のメチル化状態の測定:結合バイサルファイト制限解析(COBRA)
HPV16のP97プロモータに対して遠位にあるE2BS−1[7450−acCGaattCGgt−7461]のメチル化状態を測定するために、結合バイサルファイト制限解析(COBRA)(Xiong及びLaird、1997年)を用いた。
【0061】
p16陽性及びp16陰性病変部から単離したDNAを、EZ DNAメチル化キット(Zymo Research社、Orange、CA)を用い、メーカーの推奨する方法に従ってバイサルファイト処理した。
【0062】
処理後、5μlのアリコートを、1×緩衝液、1.25mMデオキシヌクレオチド三リン酸混合物、2.5pmolの各プライマ及び1.5単位のTaq DNAポリメラーゼ(Life Technologies社)を含有する溶液50μl中で増幅させた。PCRは以下の通り行った。ホットスタート後のサイクルパラメータは、45サイクルでは94℃30秒間、50℃30秒間及び72℃60秒間とし、最終伸長では72℃4分間とした。COBRAに用いたプライマは以下の通りとした:mHPV16_E2BS1順方向 5’AATTGTGTTGTGGTTATTTATTG3’及びmHPV16_E2BS1逆方向 5−CAAATTTAAACCATAATTACTAAC3’。
【0063】
増幅後、制限酵素EcoRI(New England Biolabs社)でPCR産物を消化した。EcoRIは、メチル化及びバイサルファイト変換DNAに特有のE2BS1配列を認識する(図6)。DNAは2%アガロースゲル中で電気泳動した。ゲルは臭化エチジウムで染色した。
【0064】
本発明者らは組織特異性を説明づけることができる。何故なら、HPVはウイルスのサイクルを支えるための特定のメチル化機構を必要とするからである。
【0065】
これらのデータから、ウイルスゲノムのメチル化状態は実質的に宿主細胞の上皮分化の程度によって変化することが示唆される。さらに、我々の結果は、E2結合部位1のメチル化は子宮頚癌進行の初期段階において重要な役割を果たしているとも考えられることを示している。
【0066】
従って、正常なウイルスのサイクルを妨害することができる脱メチル化剤を用いることは、結局のところ比較的理にかなっていると考えられる。本明細書でいう脱メチル化剤は、核酸のメチル化状態に悪影響を及ぼす任意の薬剤とすることができ、例えば、5−アザシチジン(Vidaza)、5−アザ−20−デオキシシチジン(Decitabine、dacogen)、アラビノシル−5−アザシチジン(Fazarabine)、5−6−ジヒドロ−5−アザシチジン(DHAC)、5−フルオロ−20−デオキシシチジン(Gemcitabine)、エピガロカテキン−3−没食子酸塩(EGCG)、ヒドララジン、プロカインアミド、プロカイン、ゼブラリン又はこれらの組合せとすることができる。別のクラスの脱メチル化剤としては、特定のオリゴヌクレオチド類(例えば、EGX30P)、特定のRNAi又はDNMT1アンチセンス(MG98)がある。好ましい実施態様では、5−アザシチジン、5−アザ−2−デオキシシチジン又はこれらの組合せが用いられる。
【0067】
本発明による局所投与用医薬組成物は、エアロゾル剤、クリーム剤、ゲル剤、液剤、軟膏剤、ペースト剤、パッチ剤、タンポン剤、キャップ剤及び任意の他のデバイス及び/又は脱メチル化剤の放出制御用製剤のような形式で提供することができる。
【0068】
本発明の医薬組成物は活性成分をさらに含むことができる。本発明の活性薬剤を用いて、本発明の1種以上の化合物を活性成分として含有し、この活性成分の放出を制御及び調節することができる放出制御医薬製剤を形成することができる。放出制御組成物は、例えば、ポリ(ジメチルシロキサン)、エチレン酢酸ビニルコポリマー類、ポリカロフィル(polycarophil)、ヒドロキシプロピルメチルセルロース及びポリアクリル酸類などの適切なポリマー担体を選ぶことによって得ることができる。また、薬物放出速度及び作用持続時間は、ヒドロゲルなどのポリマー物質及びその他上記のポリマーの粒子(例えば、マイクロカプセル)に活性成分を取り込ませることによって制御することもできる。放出制御はリポソーム、ミクロスフェア、マイクロエマルションなどのようなコロイド状薬物送達系を含む方法によって達成することができる。
【0069】
脱メチル化剤は、例えば、5−アザシチジン又は塩酸5,6−ジヒドロ−5−アザシチジンとすることができる。また、親水性クリーム又は親水性ゲルを基剤として5−アザシチジンの化学的に安定で可溶性のアナログを用いることもできる。例えば、1%クリーム剤は、ステアリン酸、Aquaphor(登録商標)、パルミチン酸イソプロピル、ステアリン酸ポリオキシル40、プロピレングリコール、ソルビン酸カリウム、ソルビン酸、ラウリル硫酸トリエタノールアミン及び精製水を含む親水性クリーム基剤中に1%w/wの5−アザシチジン又は1%w/wの塩酸5,6−ジヒドロ−5−アザシチジンを含有する局所製剤である。
【0070】
添加剤としては、(燐酸ナトリウムなどの)緩衝剤、(グリセリン又はソルビトールなどの)湿潤剤及び当業者に周知の他の賦形剤を挙げることができる。1%水性ゲルは、ベータデクス、エデト酸二ナトリウム、ヒドロキシエチルセルロース、メチルパラベン、ナイアシンアミド、フェノキシエタノール、プロピレングリコール、プロピルパラベン及び精製水の基剤中に1%w/wの5−アザシチジン又は1%w/w又は塩酸5,6−ジヒドロ−5−アザシチジンを含有する。5−アザシチジン(1%)親水性ゲルは、蒸留水にヒドロキシエチルセルロース(Natrosol1)を1%溶かした溶液に重量で1%の5−アザシチジンを添合することによって調製することができる。
【0071】
このプリンシパルは、本発明の種々の活性薬剤間の組合せ又は本発明の種々の活性薬剤と抗HPV活性を示す他の薬物との組合せに適用することができる。
【0072】
脱メチル化剤は単独で投与することができるが、医薬製剤として与えることが好ましい。本発明の医薬組成物はa)脱メチル化剤及びb)医薬用として許容可能な担体を含む。
【0073】
別の実施態様では、この医薬組成物はa)脱メチル化剤、b)ウイルス感染に対して相乗効果をもたらす抗ウイルス剤及びc)医薬用として許容可能な担体を含む。
【0074】
5−アザシチジン、5−アザ−2−デオキシシチジン又はこれらの組合せは、現在知られているか将来開発されるかもしれない種々の医薬製剤に調製することができる。
【0075】
特定の実施態様では、5’−アザシチジンは0.001%w/w乃至50%w/w、好ましくは0.005%w/w乃至20%w/w、より好ましくは0.005%w/w乃至10%w/wの濃度で局所医薬組成物として適用される。この脱メチル化剤は約0.001%乃至10%の濃度で脱メチル化剤を含むクリーム剤である。
【0076】
【表2】
各種形態の適用スケジュール:ブラシ剤−8時間毎、タンポン剤−毎日、子宮頚管キャップ剤−2日毎。
【0077】
5’−アザシチジンは、0.1ng乃至100μgの投与量、別の実施態様では0.5ng乃至50μgの各単回適用投与量、さらに別の実施態様では1ng乃至10μgの投与量、別の実施態様では500ng乃至2μgの投与量で使用する。本発明の特定の実施態様では、1日当たり1投与量、1日当たり2投与量、1日当たり3投与量、1日当たり4投与量又は1日当たり5以上もの投与量が適用される。本発明の特定の実施態様では、適用は、脱メチル化剤を連続的に放出する媒体を用いて連続的となるようにデザインされる。別の実施態様では、こうした投与量は毎週、週2回、週3回、隔週又は毎月与えられる。このような投与量は、例えば、計1週間、計2週間、計3週間、計4週間、計5週間、計6週間、計2ヶ月間、計3ヶ月間又はそれ以上もの間適用することができる。特定の実施態様では、適用期間は半年、1年又はそれ以上もの期間とすることができる。
【0078】
子宮頚部への薬物送達に用いることができるデバイスの1例は綿棒又はブラシである。
【0079】
薬物送達に用いることができるデバイスの他の例は子宮頚管キャップである。例えば、キャビティーリムキャップ(cavity rim cap)は子宮頚部に付着する。このキャビティーリムキャップはPrentif(登録商標)Capsである。このPrentif(登録商標)Cervical Capは患者の子宮頚部を覆い、避妊の障壁方法として用いられる。同様に、TodayR(登録商標)避妊用スポンジは、患者の子宮頚部にフィットするように形成されたスポンジ状デバイスであった。
【0080】
患者の子宮頚部に関連して用いられる他のタイプのデバイスは、膣タンポン、膣リング、膣ストリップ、膣カプセル、生体付着性フィルム又はスポンジである。デバイスには、約1乃至約10mgの後成的薬剤、約10%のヒドロキシプロピルメチルセルロース、約70%の親水性薬物用飽和脂肪酸トリグリセリド又は脂溶性薬物用PEG6000/PEG400及び約15%のエトキシジグリコールを含む組成物を含浸させる。
【実施例】
【0081】
(実施例1)
メチル化パターンの検出
HPV16 URRのメチル化が宿主上皮細胞の分化状態によって影響されるかどうかを調べるために、レーザーにより切り出した分化程度の異なる正常上皮微小細胞から単離したHPV16のDNAをバイサルファイトゲノム配列決定(BGS)法によって分析する。
【0082】
正常上皮由来の基底細胞及び中間層からの細胞画分中のp97HPV16プロモータはメチル化されていないCpGを含有していた。これに対して、分化細胞に富んだ表面層におけるHPV16プロモータは、E2(E2BS2、3及び4)並びにSP1結合部位内のCpGを含め、メチル化されていた。エンハンサ領域では、全ての分化段階において1つのCpG部位(7454)のメチル化の低下が見られた。従って、その他の4個のCpGは、基底細胞では高度にメチル化されたが、より分化の進んだ細胞ではメチル化の程度は低い。5’LCR領域は全ての分化段階で低メチル化状態であった。
【0083】
さらに、結合バイサルファイト制限解析(COBRA)を用い、HPV形質転換上皮細胞の生体指標としてのサイクリン依存性キナーゼ抑制因子p16INK4aの過剰発現によって、HSIL病変部中のHPV16 URRのメチル化パターンと対応する正常上皮とを比較する。
【0084】
これらのデータから、E2BS1のメチル化は複製HPV感染から形質転換HPV感染への移行過程で特異的に生じることが分かる。
【0085】
(実施例2)
診断の評価
HPV16に感染した、細胞学的、組織学的に確認された低悪性度(LSIL、n=53)及び高悪性度(HSIL、n=48)病変部から得られた101個のサンプル(塗抹及びパンチ生検標本)について、結合バイサルファイト制限解析(COBRA)法によりE2BS1メチル化の有無を調べた。
【0086】
特異的なLCR HPV16メチル化の検出は、癌への進行の高いリスクを有する前癌病変を検出するための分子マーカとなる。
【0087】
【表3】
E2BS1のメチル化は高悪性度病変部の75%に存在したが、低悪性度上皮内病変部では22.6%に過ぎなかった。従って、特異的なLCR HPV16メチル化の検出は、癌への進行のリスクが高い前癌病変を検出するための分子マーカとなり得る。
【0088】
E2BS1へのE2の結合がp97プロモータの活性化に関与しているかどうかを確認するために、一過性導入実験においてP97の活性に対するE2BS1メチル化の影響を分析した。野生型HPV16 LCR中にE2BS1内の2個のCpGジヌクレオチドの選択的メチル化を生じさせた。正常なヒト陰茎包皮の角化細胞に、HPV16 E2のための漸増量の発現ベクターと、ルシフェラーゼ遺伝子の直前にHPV16 LCR全体(E2BS1−LCR16−Luc)を含むか、E2BS1内にメチル化を有するLCR(methE2BS1−LCR16−Luc)を含むレポータプラスミド約50ngとを同時導入した。E2BS1のメチル化により初期p97HPV16プロモータは4乃至6倍活性化された(図8)。このデータから、この特定結合部位へのE2蛋白質の結合を調節することで、メチル化はこのp97プロモータを活性化し、コントロール不良の高レベルのウイルス遺伝子発現を可能にすることが示唆される。
【0089】
さらに、こうしたデータから、E2BS1の特異的なメチル化の検出を行うことにより急性HR−HPV感染から形質転換HR−HPV感染へ既に移行している病変部を特定できることが分かる。従って、E2BS1のメチル化はHSILの生体指標となりうる。
【0090】
(実施例3)
細胞培養における5’−アザシチジンの治療的適用
p97プロモータの活性に対する後成的阻害剤(脱メチル化剤)による処置の効果をルシフェラーゼアッセイを用いて評価する。ルシフェラーゼ遺伝子の直前にクローニングしたHPV16の完全なLCR断片を含有するレポータプラスミドLCR16_Lucを導入した正常ヒト角化細胞を約50乃至60%の集密となるように増殖させた。2μM及び4μMの5−アザシチジンで細胞を24及び72時間処置した。細胞に5−アザシチジンを添加すると、初期p97プロモータの活性が用量依存性に減少した。
【0091】
図9に示したように、未分化又は分化細胞を2μMの5−アザシチジンで処置すると、p97プロモータの活性は40乃至60%まで低下した)。4μMの5’−アザシチジンで処置した両細胞とも、プロモータ活性が基底レベルまで著しく低下した。
【0092】
これらのデータから、脱メチル化剤の1例としての5−アザシチジンの適用によってHPV16 URRの活性の大幅な抑制がもたらされることが分かる。従って、これは、ウイルスの生活環を遮断し、急性HPV感染から形質転換HPV感染への切り替えを阻止する大いなる可能性を有している。こうしたデータに基づき、このような薬剤を用いてウイルスへの複製及びそのより高悪性度の異形成さらには癌への進行を阻止することができる。
【0093】
(実施例4)
5アザシチジンによる低悪性度子宮頚部病変の処置
低悪性度病変(CIN1)HPV16陽性病変を有する患者及び低リスクHPV関連病変(コンジローム、疣贅)を有する患者を5’−アザシチジンで局所処置した。
【0094】
重量で1%の5アザシチジンを(蒸留水にヒドロキシエチルセルロース(Natrosol1)を1%溶かした溶液)に添合して5’−アザシチジン(1%)親水性ゲルを調製するとともに、5アザシチジンを含有しない対応するプラセボゲルを作製した。そのまま使用できるゲル(実薬及びプラセボ)を、使用直前に子宮頚部キャップの内面に付着させた活性のないスポンジによって子宮頚部キャップの内面に塗布した。調製してすぐに、医療関係者がその子宮頚部キャップを子宮頚部に設置する。従って、子宮頚部キャップの内面に付着した治療剤を子宮頚部へ直接拡散させて処置する。子宮頚部キャップは4週間2日毎に子宮頚部に設置した。薬物に接触させた最後の日の1週後に、病変並びに薬物関連副作用の兆候と症状の有無について患者を評価した。治癒は、病変が完全に消失し、コルポスコピーにより感染の臨床的兆候が存在しないことと定義した。退縮率を未処置対照群と比較する。
【0095】
5’−アザシチジン(1%)ゲルは、プラセボ(病変の治癒率:22.2%(2/9))よりも有意に多くの病変(85.7%(6/7))を治癒させた。16名の患者のうち、10名では薬物関連有害症状の訴えがなかった。主に実薬ゲル群の6名の患者が軽度の紅斑及び浮腫をきたしたが、多くの場合、そのような症状の発現は48時間以内に無くなった。
【0096】
(実施例5)
脱メチル化剤による皮膚疣贅の処置
脱メチル化剤を含む種々の医薬組成物を皮膚疣贅に適用した。これらの組成物は以下の通りとした:親水性クリーム基剤中1%5’−アザシチジン、10%5,6−ジヒドロ−5−アザシチジン及び10%エピガロカテキン3−没食子酸塩(EGCG)。こうした医薬組成物を皮膚疣贅に局所適用した。5名の患者に1%5’−アザシチジン、4名の患者に10%5,6−ジヒドロ−5−アザシチジン及び7名の患者に10%エピガロカテキン3−没食子酸塩(EGCG)並びに10名の患者にプラセボを処置した。このクリーム剤は、4週間閉鎖により毎日適用した。全ての患者を1名の医師が調べた。臨床評価は処置開始後1、2及び4週目に実施された。
【0097】
【表4】
退縮率は、未処置対照群に比し、処置患者でより高かった。この処置の開始2週後には、疣贅は炎症、発赤の軽微な兆候を示し、実質的により平坦になった。処置開始4週後には疣贅は治癒した。
【0098】
塩酸5,6−ジヒドロ−5−アザシチジン及びエピガロカテキン3−没食子酸塩(EGCG)は皮膚疣贅に対して同様な活性を示したが、それらの濃度は5’−アザシチジンの10倍であった。
【0099】
(実施例6)
5アザシチジンによる高悪性度子宮頚部病変の処置
実施例4に記載した5’−アザシチジン(1%)親水性ゲルを子宮頚部のCIN2病変に適用した。
【0100】
この治療剤を12週間2日毎に子宮頚部キャップ及びスポンジで子宮頚部に送達してこの頚部に設置した。
【0101】
全ての患者に対して、薬物に接触させた最後の日の1週後に反復コルポスコープ評価を行った。以前に組織像でCIN2と判定されたのと同じ部位から頚部生検又は塗抹標本が採取された。注目される転帰は、対象者が処置を必要とする疾患、即ち、CIN2以上の悪性度の病変を依然として有するかどうかであった。
【0102】
13週目で無疾患又はCIN1への退縮は、5’−アザシチジンで66.7%(4/6)、プラセボで14.2%(1/7)であった。
【技術分野】
【0001】
本発明は、HPV関連疾患の検出及び治療に有用な化合物及び方法に関する。本発明は、HPVの複製が自然感染した組織及び細胞において生じるメカニズムを解明したことに基づいている。さらに、本発明は、ウイルス感染細胞における、この病的細胞の前癌及び癌細胞への成長の促進を可能にするウイルスゲノムの異なる後成的変化を確認したことに基づくものである。従って、本発明は、新形成及びその前駆病変を診断する方法並びに悪性腫瘍の発症を予防し、又は腫瘍増殖を抑制する方法を提供する。
【背景技術】
【0002】
人体の単一の正常細胞の表現型は、特定の時点で発現される遺伝子のセットによって規定される。こうした遺伝子発現のパターンは、全ての生細胞及び複雑度の増したレベルで全ての器官に対して構造及び機能を与える蛋白質の組合せ及び個々の濃度を規定している。全ての器官が適切に機能するのに不可欠である分化過程も個々の遺伝子のレベル及び組合せの変更によって調節されている。識別的遺伝子発現パターンは、例えば、大幅に変化した遺伝子発現パターンを有する、完全な分化細胞への分化を経ることができる子孫を生じる幹細胞としての細胞の状態を決定する。こうした分化細胞は、最終的に、さらにその遺伝子発現パターンを変化させ、それにより老化してついには死にゆく細胞の特性を取り入れることによってその寿命を決定する。こうした過程の分子的調節については解明されているとはとても言えないが、いくつかの証拠は、この調節の本質的な部分がヒトゲノム内の特定のGpC島の明確なメチル化によって媒介されていることを示唆している。
【0003】
概念的には、細胞のゲノム内のこの特定のヌクレオチド群へメチル(C(H)3)−基を付加すると、このゲノムのメチル化伸展の凝縮及び強化構造を生じ、その結果として転写の阻害がもたらされる。即ち、ゲノムの高度にメチル化された部分は、通常mRNAに転写されない(発現抑制(silenced))。このメカニズムは、遺伝子の調節及び刷り込みの重要な部分であり、生細胞におけるこうした重要なステップを決定する主要な生物学的特徴の1つである(Collasほか、2007年)。分化過程におけるメチル化の詳細な影響については、これまでごく皮相的に研究されているに過ぎない(Kiefer、2007年)。しかしながら、正常細胞の本格的な新生癌細胞への形質転換に関与する形質転換過程に対してより詳細な洞察が集中されている(Esteller、2007年及びFeinberg、2004年)。特に、重要な腫瘍抑制活性を有する遺伝子群の場合、それらのプロモータ領域群のメチル化が新生癌細胞のこうした各遺伝子の発現低下を招いたことが観察されている。従って、メチル化は、欠失及び/又は突然変異に加えて、腫瘍抑制遺伝子の不活化における主要な分子段階の1つであり、多くの腫瘍抑制遺伝子、例えば、サイクリン依存性キナーゼ抑制因子p16INK4a、DNAミスマッチ修復遺伝子MSH2、MHL1などに関して明白に立証されている(Esteller、2007年及びFeinberg、2004年)。
【0004】
クロマチン構造に対する影響に加えて、異なるヌクレオチドに対してより精密にメチル化を行うことにより特定の転写因子の結合性に影響を与えることもできる(Klose及びBird、2006年)。従って、これによって正又は負に作用する転写因子の親和性に影響を与えることができ、それ故、この場合も上記各遺伝子の発現の活性化又は低下をもたらす、こうした転写因子の作用及び活性が修飾される。
【0005】
ヒト・パピローマウイルスは、皮膚又は粘膜面の上皮の増殖を引き起こす重要な上皮親和性ウイルスである。こうしたウイルスは、特定の他と区別できる型のもの(ハイリスクパピローマウイルス、HR−HPV:high risk papillomavirus)が、特に生殖器官及び最も重要なことだが子宮頸部の粘膜面の腫瘍性病変と関連付けられることが明らかとなった(zur Hausen、2002年)とき、特に科学的な関心を集めた。今日では、HR−HPV型(特に、HPV16及び18)の持続感染が子宮頚癌発症の必須の前提条件であることは一般に受け入れられている。E6及びE7と称する特定のウイルス遺伝子は、子宮頚部その他のHPV関連癌細胞では継続的に発現されていることが分かった。さらに、こうした遺伝子は、インビトロ培養一次ヒト上皮細胞において悪性形質転換を誘導することが明らかにされ、重要なことには、その発現は、子宮頚癌細胞又はその高悪性度前駆細胞の腫瘍性増殖の特徴を誘導し、維持するのに基本的に必要とされる。
【0006】
これまでのところ、約13種のハイリスクHPV型の特性が明らかにされている(Munozほか、2003年)。これらのウイルスは、臨床的に検知可能な病変を発症している(まだ発症していない)女性にもよく見られる。こうしたウイルス感染の罹患率のピークは年齢15乃至30歳の若い女性に見られるのに対して、より上の年齢の女性では低下する(Schiffmanほか、2007年)。このことは、若い男性にも同程度の頻度でみられるHR−HPV感染症が性行為感染の特徴を有することを示している。しかしながら、性別による主な相違は、男性ではHR−HPV感染がその宿主によって自覚されることがまれであり、浸潤性の癌へと進行することがない不顕性感染をもたらすのが通常であるのに対して、女性では明らかに長期の持続感染がかなりより高頻度で生じるため、最終的に浸潤性の癌へと進行する可能性のある子宮移行部の上皮細胞の悪性形質転換がもたらされることがある。興味深いことに、この現象が概して子宮頸部の移行部に存在する数少ない上皮細胞に限定されるように思われるのに対して、同程度の率で感染する上皮部位は、子宮移行部の上皮細胞と比較して悪性形質転換を受ける傾向が実質的に少ない。
【0007】
通常の生活環の過程でHPVは先ず上皮の基底及び傍基底細胞に感染する。いくつかの論拠から、このウイルスは先ず潜伏感染の状態に入ることが示唆されている(Longworth及びLaimins、2004年)。この過程では、宿主細胞が一旦分裂すると、ウイルスゲノムは単一の感染細胞当たりでは極めて低いコピー数で複製されるが、この段階ではウイルスゲノムの複製も、場合によっては溶菌感染も開始されない。技術的な限界のために、この種の潜伏感染の存在に関する正確な証拠はまだ得られていない。むしろ、これは、新規感染の証拠がなくても免疫抑制時にウイルス複製活性が再活性化されうることを示す疫学研究から予想される。こうした潜伏感染は、明らかに活動性複製感染に切り替わり得る。この場合、基底及び傍基底細胞ではウイルス遺伝子の発現活性は全くほとんど維持されないが、これらの感染上皮細胞が一旦分化し始め、この上皮の中間細胞層に達すると、こうした分化細胞層においてウイルスゲノムの複製を引き起こす遺伝子発現の増加が観察される。興味深いことに、ウイルスゲノムの複製及びウイルスの初期遺伝子の発現は、増殖能、及び最終分化上皮細胞を予めプログラムされた分化経路に押し込むこうした細胞の活性化遺伝子発現特性を不可逆的に失っていて、最終的に上皮の外表面に扁平上皮細胞残屑として崩壊する最終分化細胞に概して限定されると考えられる(Longworth及びLaimins、2004年)。中間細胞においてウイルスゲノムが複製し、上皮細胞が一旦正常な分化過程で表面細胞層に達すると、ウイルスゲノムがさらに再プログラム化され、全ての初期遺伝子の発現が止められる一方で、各外被蛋白質の翻訳をもたらす後期遺伝子L1及びL2の本格的な発現がここで観察される。これらの蛋白質は自然に集まって、複製されたウイルスゲノムが含まれるウイルスの外被を形成し、最終的に、この新たに合成された成熟ウイルス外被は、上皮の外面の崩壊しつつある扁平上皮細胞の残屑から放出され、更なる感染サイクルを再開することができる(図1)。これらの知見から、ウイルスの複製サイクルは正常な上皮の分化過程と厳格に連関していることが示唆される。しかしながら、上皮細胞の分化の誘因となる分子事象以後に、この複雑な複製過程を制御するウイルス遺伝子発現パターンの調節に関与する分子構造についてはこれまでのところほとんど又は全く知られていない。
【0008】
ウイルス遺伝子発現の調節及びウイルスゲノムの複製に重要な意味を持つ構造は、長鎖制御領域(LCR:long control region)又は上流調節領域もしくは(URR:upstream regulatory region)と呼ばれる配列エレメント内に保持されている。このエレメントには、重要な正及び負の転写因子の結合部位及び複製起点が存在する(図2)。
【0009】
初期に発現されるいくつかのパピローマウイルス遺伝子の一部であるパピローマウイルスE2蛋白質は、ウイルスのプロモータ自体に結合することによりウイルス遺伝子の発現に対して実質的な抑制機能を有する(Wellsほか、2000年)。従って、これは転写レプレッサーの働きをし、HPV URRエレメントの制御下の遺伝子の転写を阻止する。この概念は、インビトロで増殖させた非分化上皮細胞、場合によっては基底又は傍基底細胞の類似非分化表現型を保持する癌細胞株も用いて組織培養モデルで実施されている種々の分子研究に基づいたものである。E2蛋白質が発現されると、ウイルスプロモータの下方制御及び隣接遺伝子の発現停止がもたらされることが分かっている(Bouvardほか、1994年)。
【0010】
子宮頚癌細胞では、パピローマウイルスゲノムは、通常、宿主細胞染色体中に組み込まれることが分かっている(Pert及びColeman、2007年b)。ウイルスゲノムの宿主細胞ゲノム中への組込み部位は不規則に選ばれるように思われるが、組み込まれたウイルスDNA断片のウイルスゲノムは、通常、極めて特異な仕方で保存される。まず第一に、URRを含む断片並びに全てこれから分析されるE6及びE7遺伝子は、組み込まれたウイルスゲノムコピーからもE6及びE7遺伝子の発現を可能にする立体配置で保たれるのに対して、下流に位置する遺伝子E2及び後期遺伝子カセットは、通常、URR内の調節エレメントから切り離され、従って、機能的に、又は構造的にも不活化される(zur Hausen、2002年)。興味深いことに、この点に関して調べられた全てのHPV関連癌細胞で、ウイルスE6及びE7の発現を確認することができた。前述のように、これらの遺伝子は、細胞周期の調節、分化過程及び死のカスケードに少なくともある程度関与しているいくつかの宿主細胞蛋白質との複雑な相互作用を通じて複製している細胞に重要な増殖調節特性を付与する。これらの遺伝子の発現は、上皮細胞の悪性形質転換を誘導し、維持するのに重要で十分な必要条件である。これらの遺伝子の発現が形質転換細胞で遮断されれば、この細胞は細胞周期停止に陥り、増殖を止め、最終的には死滅する(von Knebelほか、1988年)。
【0011】
この観察と一致して、HR−HPV型のE2遺伝子をコードしているオープンリーディングフレームはHPV関連癌では通常、破壊されている。従って、HPV形質転換細胞中に機能的に活性なE2蛋白質を再導入すると、悪性表現型、細胞周期停止及び最終的には細胞死の抑制がもたらされる(Wellsほか、2000年)。
【0012】
従って、宿主細胞染色体に組み込まれることによるE2 ORFの破壊がHPV感染上皮細胞の癌化を最終的にもたらす分子カスケードにおいて絶対必要ではないにしても重要なステップであることは、この分野の科学者の間で一般に受け入れられている(Pett及びColeman、2007年a、Pettほか、2006年、Pettほか、2004年並びにAlazawiほか、2002年)。
【0013】
上皮分化の特徴とHPV複製サイクルとが密接に関連していることは、ウイルスの遺伝子発現及び複製の制御に宿主細胞の環境が重要な役割を果たしていることを示唆している(図1参照)。いくつかの証拠から、ウイルスゲノムの遺伝子発現及び複製調節においてメチル化その他の後成的制御の関与が示唆されている。こうしたデータの多くは、人工的な組織培養モデルに基づいたものである。これまでのところいずれのモデルによってもウイルスの遺伝子発現及び複製の分化依存性の調節の詳細を説明することはできなかった。
【0014】
パピローマウイルスDNAのメチル化については既に約20年前に報告されている(Burnett及びSleeman、1984年並びにWettstein及びStevens、1983年)。しかしながら、パピローマウイルス遺伝子発現制御及びこれに関連した発癌効果の調節におけるその生物学的意義については依然として極めて不明のところが多い(Listほか、1994年、Roslほか、1993年並びにThainほか1996年)。子宮頚癌細胞株において、HPV−16及びHPV−18ゲノムにおけるCpGメチル化はLCR領域及び一部のL1 ORF領域ではこのウイルスゲノムのどの他の部分よりも高い頻度で生じるように思われる(Badalほか、2004年及びBadalほか、2003年)。さらに、特にL1遺伝子のメチル化は、発癌時における宿主細胞ゲノム中への外来ウイルスDNAの組込みに関連があるように思われ、このため、HPV感染細胞の腫瘍性変化の生体指標となる可能性があることが示唆された(Kalantariほか、2004年、Turanほか、2006年)。別のデータからは、E2BSにおけるCpGメチル化が、インビトロでのE2蛋白質結合を妨げ(Thainほか、1996年)、転写活性化の際の細胞におけるE2蛋白質機能を調節することも示唆されている(Bhattacharjee及びSengupta、2006年並びにKimほか、2003年)。しかしながら、癌におけるDNAメチル化はHPV DNAに限られるものではない。概略前述したように、これは極めて重要な特徴とみなされるが、どちらかと言えば、宿主ゲノム全体にわたってよく生じる事象である(Esteller、2007年)。多くの細胞遺伝子の過剰メチル化の頻度は、新形成の重篤度が増大するにつれて著しく増加することが分かっている(Bannoほか、2007年、Dongほか、2001年、Fengほか、2005年、Jeongほか、2006年、Kangほか、2006年、Laiほか、2007年、Leaほか、2004年、Reesink−Petersほか、2004年、Sengほか、2007年、Steenbergenほか、2004年、Virmaniほか、2001年、Widschwendterほか、2004年)。
【0015】
最近の研究では、正常上皮細胞及び子宮頚癌細胞においてHPV16のE2結合部位E2BS2乃至4内のCpGのメチル化の有無が調べられた(Bhattacharjee及びSengupta、2006年)。この論文の著者らは、メチル化が、形質転換組織におけるP97プロモータの近位にあるE2BS2乃至4に位置する明確なCpG島に見出されたと報告している。
【0016】
しかしながら、この研究では、特定部位のメチル化パターンの明確な変化で異なる生物学的特徴を説明することができる明瞭で一貫したパターンを示すことができなかった。
【0017】
Kimら(Kimほか、2003年)は、W12細胞を用いたインビトロ組織培養モデルの高度分化細胞集団において低メチル化が関連付けられると報告した。これに対して、低分化基底細胞様細胞からのHPV16 LCRは多数のメチル化シトシンを含み、多くの場合、E2BSがメチル化されていた。
【0018】
さらに、ヒトの癌は特にCpG島においてDNAメチル化のパターン変化を示すことが多いことが先行技術に開示されている。島内のメチル化は連鎖遺伝子の転写抑制を伴うことが分かっている。腫瘍の発症及び進行の全ての局面に関与する遺伝子は、メチル化され、後成的に発現抑制されるようになる可能性がある。このような発現抑制された遺伝子を再発現させると、腫瘍増殖の抑制又は抗癌療法への感受性増大をもたらすことができる。DNAメチル化を抑制することができる後成的薬剤については、現在、前臨床評価及び癌患者での臨床試験が行われている。ヌクレオシド阻害剤5−アザシチジン及びデシタビンは、多くの種類の癌に対して数多くの第1相及び第2相試験で試験されている。しかしながら、両剤の用量規定毒性は骨髄抑制であり、最もよく報告されている非血液学的有害作用は悪心及び嘔吐であった。従って、脱メチル化剤による癌の全身療法は、通常重篤な副作用を引き起こすことになる。
【0019】
美容整形症状及び皮膚症状を処置するのに特異的DNAメチル化阻害剤を用いることができる。5−アザシチジンは正常ヒト線維芽細胞による基礎レベル及びTGFβ誘導コラーゲン生合成を共に阻害することが分かっている。コラーゲンは瘢痕化に対して直接的な影響を示す。従って、ヒトの皮膚においてコラーゲンの過剰産生を抑制すると、瘢痕形成が抑制されることになる。さらに、癌性及び前癌性皮膚病変の処置にUV処置と脱メチル化を併用することができる。
【0020】
本発明は、パピローマウイルスゲノム内の異なる特定の配列エレメントのメチル化がウイルスの生活環、複製及び遺伝子発現パターンを制御しているという知見に関する。さらに、本発明は、このメチル化パターンの特定の変化が一部のパピローマウイルスにより誘発される悪性形質転換の誘導に関与しているという知見に関する。このことは、例えば子宮頚部、膣、陰門、肛門、口腔咽頭管などの体腔の粘膜上皮の病変の場合に特に重要である。脱メチル化剤による全身療法に伴う望ましくない、ある程度耐えられない副作用を克服するために、本発明者らは、特に体腔の粘膜上皮のHPV関連病変及び癌の処置において、脱メチル化剤による局所処置が患者に望ましくない副作用をもたらすことなく癌を治療するのに極めて効果的な方法となりうることを見出した。
【発明の概要】
【課題を解決するための手段】
【0021】
本発明は、例えば体腔の粘膜上皮などの病変の処置及び/又は治療のための脱メチル化剤の局所適用に関する。この点に関して、本発明はさらに、HPV関連病変の処置及び治療の方法に関する。HPV関連病変は、例えば子宮頚癌及びその前駆症状段階並びに皮膚の疣贅を含む。
【0022】
さらに、本発明は、ウイルス複製及び分子の形質転換感染への移行を阻止することによりHPV関連病変及び癌を処置するために体腔の粘膜上皮へ局所適用するための脱メチル化剤含有医薬組成物に関する。この点に関して、本発明はさらに、HPV関連新形成の処置及び予防用医薬組成物に関する。この医薬組成物は、医薬用として許容可能な担体並びにE2BS1、NFIBS、TEF−1BS及び/又はSP1BSにおける特定のメチル化パターンを阻害する後成的阻害剤又は脱メチル化剤を含む。
【0023】
また、本発明は、HPV E2BS1、NFIBS及び/又はTEF−1BSのメチル化パターンを測定することによって潜伏感染した細胞とHPV増殖感染細胞とを識別する方法に関する(図5参照)。
【0024】
配列番号1乃至8の配列を含む単離核酸であって、この核酸は下記配列中、大文字で示した位置でメチル化されているものとする核酸。
【0025】
【化1】
上記配列の全てのヌクレオチドの位置はNCBI受託番号NC_001526のHPV16全ゲノム配列に従って引用した。大文字のCはメチルシトシンに対応する。大文字のGは逆相補鎖にメチルシトシンが位置する部位を示す。
【0026】
下記の配列番号9乃至15の単離核酸
【0027】
【化2】
【図面の簡単な説明】
【0028】
【図1】ウイルスの生活環及び形質転換感染への進行の模式図である。
【図2】HPV16ゲノム及び長鎖制御領域(LCR)又は上流調節領域(URR)の拡大構造の模式図である。ウイルスE2蛋白質及びSP1転写因子の結合部位の相対的位置が示されている。
【図3】正常と腫瘍性上皮との間でE2BS4のメチル化パターンを比較した(Bhattacharjee及びSengupta、2006年)により報告されたメチル化パターンの分析結果並びにE2BS1における非形質転換上皮と形質転換(p16INK4a陽性)上皮との主要な相違を明らかにした本発明者らの結果の模式図である。
【図4】微小細胞を切り出した子宮頚部上皮の例を示す図である。上部パネル:初期HPV感染(出現しつつあるCIN1の証拠)であるが、形質転換されていないため、p16INK4aが染色されていない上皮。下部パネル:無秩序なHPV E6/E7癌遺伝子発現を示す高悪性度扁平上皮内病変及び高悪性度扁平上皮内病変(HSIL:high grade squamous intraepithelial lesion)の証拠。
【図5】HPV16 URR内の重要な転写因子結合部位のメチル化パターンを示す模式図である。
【図6】COBRAアッセイの原理及びHPV16ゲノムのE2BS1のいくつかの臨床サンプルの分析結果を示す図である。
【図7】正常扁平上皮の構造を示す図[KossLGほか、1999年]である。
【図8】p97初期プロモータ活性に対するE2BS−1メチル化の影響を示す図である。
【図9】p97初期HPV16プロモータ活性に対する5−アザシチジンの影響を示す図である。
【発明を実施するための形態】
【0029】
本発明者らは、ヒトパピローマウイルスゲノムのメチル化パターン、特にウイルスの生活環の3つの想定相、即ち、a.潜伏感染、b.複製感染及びc.形質転換感染におけるURR(その同族E2BSを含む)のメチル化パターンについて調べた。分化段階の異なる上皮細胞内のHR−HPVゲノムの詳細なメチル化状態を明らかにするために、組織から微小細胞を切り出し(図4参照)、異なる細胞層からのHPV配列を増幅させて、バイサルファイトシーケンシングPCR(BSP:bisulfite sequencing PCR)を用いたHPVゲノムの異なるメチル化パターンの正確な評価を行った(図5参照)。
【0030】
ここでいうHPV関連病変とは、急性又は持続性HPV感染により誘発された任意の上皮の全ての変化を含む、HPV感染により誘発された病変のことをいう。これらの病変は、臨床的影響のない小さな疣贅から、もっと正確に言えば、皮膚、生殖器官、粘膜表面などの外方増殖パピローマ、内反性パピローマを初めとするコンジローム並びに腫瘍形成性パピローマウイルス遺伝子タイプにより誘発される前癌及び浸潤性病変(それに由来する転移を含む)に及ぶものとすることができる。HPV誘発性前癌又は癌病変の多くは、種々の上皮組織の移行部、特に、例えば、子宮頚部移行部、肛門上皮と直腸粘膜の接合部又は扁桃領域の口腔咽頭部内のような、単層腺上皮が多層扁平上皮と交わる所に生じる。
【0031】
本発明の発明者らは、HPVの細胞変性効果の影響を未だ受けていない正常な上皮では、ウイルスゲノムが基底細胞内に存続することができることを見出し、基底及び傍基底細胞に潜伏期のウイルス感染が存在することが明らかとなった(データは図に示されていない)。この潜伏期のウイルスゲノムは分析することができたすべてのCpGで完全にメチル化されており、ウイルスゲノムの全体的なメチル化によって感染の潜伏が媒介され、ウイルス遺伝子の発現が阻止されることにより感染上皮の細胞病理学的又は組織病理学的変化も防止されることが示唆される。すなわち、本発明者らは、HPVゲノムのメチル化状態と感染の潜伏様式との間に関連性があることを見出した。この時期のメチル化がE2BS1にも見出されることに留意することは重要である。
【0032】
ウイルスゲノムを初期異形成病変(CIN1)の切り出し微小細胞から単離した場合には、宿主細胞の分化状態及びウイルスゲノム内の各CpGに応じてメチル化パターンの異なる変化が観察された(図5参照)。基底細胞層では、URRの異なる部分にメチル化されていないE2BS(1乃至4)が認められた。興味深いことに、E2BS1がメチル化されなかったことによって、恐らくウイルスのエンハンサエレメントの結合及び抑制が可能となる。ウイルスDNAが中間細胞層から単離された場合には、ゲノムの諸部分は脱メチル化されるようになる。これによってNF1、TEF−1などのような正の転写因子(TF:transcription factor)の接近が可能となり、ウイルス遺伝子の発現が刺激される。ウイルスの遺伝子発現の増強は分化細胞内の脱メチル化が増加することに一致して生じ、従って、ウイルスゲノムの複製が開始される(図5)。
【0033】
表面細胞層では、E2(E2BS2、3及び4)並びにSP1結合部位を含む初期プロモータ内の全てのCpGがメチル化され、初期プロモータ活性の不活化が示唆された。
【0034】
すでに高悪性度扁平上皮内病変(HSIL:high grade squamous intraepithelial lesion)へ進行し、従ってp16INK4aマーカ(Klaesほか、2001年)の過剰発現により明らかとなるウイルスの癌遺伝子E6及びE7の大量発現を示す(図4参照)病変では、E2BS1の一貫したメチル化が認められた(図5)。このことから、メチル化は、この特定の結合部位へのE2蛋白質の接近を阻止することによって負の転写調節を無効にし、コントロール不良の高レベルのウイルス遺伝子発現がHPV16 URR(LCR)により媒介されることを可能にすることが示唆される。
【0035】
結合バイサルファイト制限解析(COBRA:Combined Bisulfite Restriction Analysis)(Xiong及びLaird、1997年)を用いてこのメチル化パターンを、無秩序なE6及びE7癌遺伝子の発現を示す病変のp16INK4a陽性部分とHPVE6及びE7遺伝子発現障害の制限を明らかに依然として維持する病変のHPVに感染するもp16INK4a陰性の部分との間で比較した。図6に示したデータは、E2BS1のメチル化はウイルス癌遺伝子の無秩序な発現と同時に起きるという概念を裏付けるものである。
【0036】
E2結合部位は以下の通り定義される:
【0037】
【化3】
NCBIアセッション番号NC_001526のHPV16全ゲノム配列によるBSの位置。
【0038】
正常上皮は、図7に示したように記述することができる。皮膚及び粘膜の最外層は、3つの主な帯域に分類することができる、形態の進行性の変化により区別される多上皮細胞層からなる(図20)[Koss LGほか、Introduction to Gynecologic Cytopathology with Histologic and Clinical Correlations、1999年:p.23−30]。
【0039】
基底帯は、球状であり、比較的大きな核を有する低分化基底細胞の1又は2層(基底層)からなる。正常な扁平上皮では、基底幹細胞は分裂することができる唯一つの細胞である。この基底細胞は真皮から表皮を隔てる基底膜に付着している。
【0040】
上皮の中間帯は上皮の厚さの大部分に相当し、表面に向かって大きさが増大する数層の細胞(傍基底層又は有棘層及び中間扁平層又は顆粒層)からなる。一旦基底膜に付着しなくなると、こうした細胞は細胞周期を出て、分裂を止め、分化し始める。
【0041】
表面帯は、小さな縮合核及びグリコーゲンで満たされている細胞質を有するより大きく扁平な細胞の列(成熟扁平層又は角質層)を含む。表面帯の外層は最終分化細胞からなる。
【0042】
ゲノム位置7,198番目と161番目との間の分析領域には16個のCpGが含まれる(図5)。この分析領域は、5’LCR、エンハンサ及びプロモータと呼ばれる3つの機能的に異なるセグメントに分けることができる。この5’LCRは、E2BS1(7,450−nnnnn−7,461)内のCpG(nt7452、nt7458)を含む4個のCpGを含む。エンハンサは、NFIBS(7541−cttgccatgCGtgccaaatc−7560及び7663−aatcactatgCGccaaC−6379)並びにTEF−1BS(7684−tacatacCGct−7694)内の4個のCpG(nt7550、7673、7679及び7691)を含む5個のCpGを含む。プロモータ内の最後の7個のCpG、これらのうちの6個(nt7859、31、37、43、52、58)は、Sp1(25−taagggCG−32)並びに3つのE2結合部位(E2BS2 7857−acCGatttgggt−7869、E2BS3 35−acCGaaatCGgt−46及びE2BS4 50−acCGaaacCGgt−61)と重複する。
【0043】
これまでの報告に所載されていることとは対照的に、本発明者らは、HPVの細胞変性効果の影響を未だ受けていない正常上皮では、ウイルスゲノムが基底細胞内に存続することができることを見出し、基底及び傍基底細胞に潜伏期のウイルス感染が存在することが明らかに示された(データは図に示されていない)。この潜伏期のウイルスゲノムは、分析することができた16個のCpGの全てにおいて完全にメチル化されており(図5)、このことから、ウイルスゲノムの全体的なメチル化によって感染の潜伏が媒介されてウイルス遺伝子の発現が阻止され、これによって感染上皮の細胞病理学的又は組織病理学的変化も防止されることが示唆される。この知見は、これまでHPVゲノムのメチル化状態と潜伏様式の感染とを関連づけるデータは報告されていないので、新規なものである。この時期において、メチル化がE2BS1においても見られることに留意することは重要である。
【0044】
ウイルスゲノムを初期異形成病変(CIN1)の切り出し微小細胞から単離した場合には、宿主細胞の分化状態及びウイルスゲノム内の各CpGに応じてメチル化パターンの異なる変化が観察された(図5参照)。
【0045】
プロモータ領域では、分析した12のアンプリマークローンの、基底細胞及び中間層からの細胞画分から単離したDNAは全て、メチル化されていないCpGを含んでいた。表面細胞層の分化のより進んだ細胞から単離したHPV16DNAでは著しく異なった結果が観察された。分化細胞に富んだこの表面層から単離したHPV16DNAから作製した12種のアンプリマークローンのうちの8種において、E2BS(2、3及び4)及びSP1部位を含むプロモータ内のCpGがメチル化された。
【0046】
エンハンサ領域では、2個のNFIBS及び1個のTEF−1BS内の4個のCpGが基底細胞内で高度にメチル化されていたが、分化のより進んだ細胞内ではメチル化の程度はより低かった。これによってNF1、TEF−1などのような正の転写因子(TF)の接近が可能となり、ウイルス遺伝子の発現が刺激される。ウイルスの遺伝子発現の増強は分化細胞内の脱メチル化が増加することに一致して生じる(図5)。
【0047】
5’LCRセグメントでは、4個のCpGの全てが分化段階と関係なくメチル化されていなかった。興味深いことに、E2BS1がメチル化されなかったことによって、恐らくウイルスのエンハンサエレメントの結合及び抑制が可能となる。
【0048】
すでに高悪性度扁平上皮内病変(HSIL)へ進行し、従ってp16INK4aマーカ(Klaesほか、2001年)の過剰発現により明らかとなるウイルスの癌遺伝子E6及びE7の大量発現を示す(図4参照)病変では、特有のメチル化パターンが認められた(図5)。プロモータ及びエンハンサ領域では、NFIBS及びTEF−1BS内の4個のCpGがメチル化された。5’LCRでは、E2BS1内の2個のCpGを含む3個のCpGがメチル化された。このことから、メチル化は、この特定の結合部位へのE2蛋白質の接近を阻止することによって負の転写調節を無効にし、コントロール不良の高レベルのウイルス遺伝子発現がHPV16 URR(LCR)により媒介されることを可能にすることが示唆される。
【0049】
これによって、本発明者らは、こうした宿主細胞組織内での各ウイルスゲノムの生物学的特性を用いてウイルスゲノムの異なるメチル化パターンを示すことができた。この研究の重要な知見は以下のとおりである。
【0050】
ウイルス活性を示さない潜伏感染細胞では、HPVゲノムは、全てではないにしても殆ど全てのCpG島において高度にメチル化され、それによってウイルス遺伝子の任意の生物学的に意義のある発現が阻止される。例えば、基底帯、中間帯及び表面帯の細胞においてCpG島の80%超、好ましくは90%、好ましくは95%、より好ましくは98%以上がメチル化される。これによって数少ない上皮幹細胞における潜伏HPV感染の完全な隠蔽が媒介され、その後、それは不確定の期間持続することができる(図5参照)。
【0051】
まだ特徴付けられていない条件下では、個々の細胞におけるHPV感染の潜伏状態は複製感染様式に切り替わり得る。基底細胞では、(E2BS2、3及び4並びにSP1結合部位並びに(E2BS1を含む)5’長鎖制御領域内のCpGを含む)初期HPVプロモータはメチル化されていないCpGを含んでいた。従って、エンハンサ領域では、NF1及びTEF−1部位内の4個のCpGは高度にメチル化されていた。このメチル化のパターンは、p97プロモータの活性が低いことを示唆するものである。(図5)
中間細胞では、HPVエンハンサのメチル化の程度は、基底細胞に比し、分化のより進んだ細胞ではより低く、これによって、核因子1(NF1:nuclear factor I)、転写エンハンサ因子TEF−1(transcriptional enhancer factor−1)などの転写活性化因子の結合によるp97プロモータの活性増大がもたらされる。しかしながら、初期プロモータの活性は、E2の負の制御(E2BS1脱メチル化)のために一定のレベルに維持される。
【0052】
最終分化表面細胞では、E2(E2BS2、3及び4)並びにSP1結合部位を含む初期プロモータ内の全てのCpGがメチル化され、初期プロモータ活性の不活化が示唆された。
【0053】
ウイルスゲノムのメチル化状態が実質的に宿主細胞の上皮分化の程度によって変化するという事実は、DNAのメチル化がウイルスの生活環及び恐らくまた上皮分化において重要な役割を有することを示唆している。さらに、機能的に異なる高リスク及び低リスクウイルス型の両者は、増殖性生活環を調節するための共通のメカニズムを共有していると考えられる。
【0054】
複製感染が長期持続すると、基底及び傍基底細胞のうちの感染細胞は、基底及び傍基底細胞のメチル化プロフィルを主にEBS1、nt7459のメチル化を特徴とするHPV感染の形質転換様式に切り替えることができる。このCpG島のメチル化は、E2BS1へのE2結合の低下及び興味深いことにURRに対するその抑制機能の低下をもたらす。その結果、これによって各HPVゲノムは細胞内監視制御を緩め、基底細胞コンパートメント内でウイルス癌遺伝子E6及びE7の十分なレベルの発現を開始する。
【0055】
バイサルフェートゲノム配列決定(BGS:bisulfate genomic sequencing)法によるHPV16のURRにおけるメチル化CpGのマッピング。
【0056】
レーザーにより切り出した分化程度の異なる(基底、中間及び表面層)微小細胞から単離したDNAについて、バイサルファイトゲノム配列決定(BGS)法によりHPV16のURRのメチル化状態を分析した。
【0057】
EZDNAメチル化キット(Zymo Research社、Orange、CA)を用い、メーカーの推奨する方法に従ってDNAのバイサルファイト修飾を行った。対照としてCaski及びSiHa細胞株からのDNAを用い、サンプルと同時に処置して完全なバイサルファイト処置を保証した。
【0058】
HPV16のURR(nt7198からnt161まで;NC_001526)に架かるように設計したプライマを用いて入れ子BSM−PCR系を開発し、実施した(表1)。
【0059】
【表1】
PCR反応混液は、10×PCR緩衝液、5μlの50mM MgCl2、0.5μlの2mMデオキシリボヌクレオチド三リン酸、0.5mMの各PCRプライマ(1.5μlのプライマ(25pmol/μl))、2.0UのPlatinum Taq(Invitrogen社)及び2μlのバイサルファイト修飾DNAを含む計50μlとして作製した。各分析ではDNAを含まない陰性対照を置いた。増幅条件は以下の通りとした:最初の94℃2分間の変性後、94℃30秒間の入れ子PCR、50℃30秒間のアニーリング、72℃40秒間及び最後に72℃4分間の伸長の40サイクル及び30サイクル。PCR産物は、電気泳動により分離し、臭化エチジウムで染色した2%アガロースゲルから単離した。次いで、QIAquick Gel Extraction Kit(Qiagen社、Hilden)によりメーカーの使用説明書に従って単離PCR産物を精製した。精製PCR断片は、TA Cloning System(Invitrogen社)を用いてクローニングし、12の個々のクローンについて配列決定を行い、HPV16 DNA内のメチル化CpGの有無及びパターンを明らかにした。バイサルファイト修飾サンプルDNAの配列決定は、BigDyeターミネータ配列決定キット(Applied Biosystems社、Foster City、CA)を用い、メーカーの推奨する方法に従って実施した。配列決定PCR産物はABI Prism 310 Genetic Analyzerで解析した。
【0060】
HPV16 E2BS1のメチル化状態の測定:結合バイサルファイト制限解析(COBRA)
HPV16のP97プロモータに対して遠位にあるE2BS−1[7450−acCGaattCGgt−7461]のメチル化状態を測定するために、結合バイサルファイト制限解析(COBRA)(Xiong及びLaird、1997年)を用いた。
【0061】
p16陽性及びp16陰性病変部から単離したDNAを、EZ DNAメチル化キット(Zymo Research社、Orange、CA)を用い、メーカーの推奨する方法に従ってバイサルファイト処理した。
【0062】
処理後、5μlのアリコートを、1×緩衝液、1.25mMデオキシヌクレオチド三リン酸混合物、2.5pmolの各プライマ及び1.5単位のTaq DNAポリメラーゼ(Life Technologies社)を含有する溶液50μl中で増幅させた。PCRは以下の通り行った。ホットスタート後のサイクルパラメータは、45サイクルでは94℃30秒間、50℃30秒間及び72℃60秒間とし、最終伸長では72℃4分間とした。COBRAに用いたプライマは以下の通りとした:mHPV16_E2BS1順方向 5’AATTGTGTTGTGGTTATTTATTG3’及びmHPV16_E2BS1逆方向 5−CAAATTTAAACCATAATTACTAAC3’。
【0063】
増幅後、制限酵素EcoRI(New England Biolabs社)でPCR産物を消化した。EcoRIは、メチル化及びバイサルファイト変換DNAに特有のE2BS1配列を認識する(図6)。DNAは2%アガロースゲル中で電気泳動した。ゲルは臭化エチジウムで染色した。
【0064】
本発明者らは組織特異性を説明づけることができる。何故なら、HPVはウイルスのサイクルを支えるための特定のメチル化機構を必要とするからである。
【0065】
これらのデータから、ウイルスゲノムのメチル化状態は実質的に宿主細胞の上皮分化の程度によって変化することが示唆される。さらに、我々の結果は、E2結合部位1のメチル化は子宮頚癌進行の初期段階において重要な役割を果たしているとも考えられることを示している。
【0066】
従って、正常なウイルスのサイクルを妨害することができる脱メチル化剤を用いることは、結局のところ比較的理にかなっていると考えられる。本明細書でいう脱メチル化剤は、核酸のメチル化状態に悪影響を及ぼす任意の薬剤とすることができ、例えば、5−アザシチジン(Vidaza)、5−アザ−20−デオキシシチジン(Decitabine、dacogen)、アラビノシル−5−アザシチジン(Fazarabine)、5−6−ジヒドロ−5−アザシチジン(DHAC)、5−フルオロ−20−デオキシシチジン(Gemcitabine)、エピガロカテキン−3−没食子酸塩(EGCG)、ヒドララジン、プロカインアミド、プロカイン、ゼブラリン又はこれらの組合せとすることができる。別のクラスの脱メチル化剤としては、特定のオリゴヌクレオチド類(例えば、EGX30P)、特定のRNAi又はDNMT1アンチセンス(MG98)がある。好ましい実施態様では、5−アザシチジン、5−アザ−2−デオキシシチジン又はこれらの組合せが用いられる。
【0067】
本発明による局所投与用医薬組成物は、エアロゾル剤、クリーム剤、ゲル剤、液剤、軟膏剤、ペースト剤、パッチ剤、タンポン剤、キャップ剤及び任意の他のデバイス及び/又は脱メチル化剤の放出制御用製剤のような形式で提供することができる。
【0068】
本発明の医薬組成物は活性成分をさらに含むことができる。本発明の活性薬剤を用いて、本発明の1種以上の化合物を活性成分として含有し、この活性成分の放出を制御及び調節することができる放出制御医薬製剤を形成することができる。放出制御組成物は、例えば、ポリ(ジメチルシロキサン)、エチレン酢酸ビニルコポリマー類、ポリカロフィル(polycarophil)、ヒドロキシプロピルメチルセルロース及びポリアクリル酸類などの適切なポリマー担体を選ぶことによって得ることができる。また、薬物放出速度及び作用持続時間は、ヒドロゲルなどのポリマー物質及びその他上記のポリマーの粒子(例えば、マイクロカプセル)に活性成分を取り込ませることによって制御することもできる。放出制御はリポソーム、ミクロスフェア、マイクロエマルションなどのようなコロイド状薬物送達系を含む方法によって達成することができる。
【0069】
脱メチル化剤は、例えば、5−アザシチジン又は塩酸5,6−ジヒドロ−5−アザシチジンとすることができる。また、親水性クリーム又は親水性ゲルを基剤として5−アザシチジンの化学的に安定で可溶性のアナログを用いることもできる。例えば、1%クリーム剤は、ステアリン酸、Aquaphor(登録商標)、パルミチン酸イソプロピル、ステアリン酸ポリオキシル40、プロピレングリコール、ソルビン酸カリウム、ソルビン酸、ラウリル硫酸トリエタノールアミン及び精製水を含む親水性クリーム基剤中に1%w/wの5−アザシチジン又は1%w/wの塩酸5,6−ジヒドロ−5−アザシチジンを含有する局所製剤である。
【0070】
添加剤としては、(燐酸ナトリウムなどの)緩衝剤、(グリセリン又はソルビトールなどの)湿潤剤及び当業者に周知の他の賦形剤を挙げることができる。1%水性ゲルは、ベータデクス、エデト酸二ナトリウム、ヒドロキシエチルセルロース、メチルパラベン、ナイアシンアミド、フェノキシエタノール、プロピレングリコール、プロピルパラベン及び精製水の基剤中に1%w/wの5−アザシチジン又は1%w/w又は塩酸5,6−ジヒドロ−5−アザシチジンを含有する。5−アザシチジン(1%)親水性ゲルは、蒸留水にヒドロキシエチルセルロース(Natrosol1)を1%溶かした溶液に重量で1%の5−アザシチジンを添合することによって調製することができる。
【0071】
このプリンシパルは、本発明の種々の活性薬剤間の組合せ又は本発明の種々の活性薬剤と抗HPV活性を示す他の薬物との組合せに適用することができる。
【0072】
脱メチル化剤は単独で投与することができるが、医薬製剤として与えることが好ましい。本発明の医薬組成物はa)脱メチル化剤及びb)医薬用として許容可能な担体を含む。
【0073】
別の実施態様では、この医薬組成物はa)脱メチル化剤、b)ウイルス感染に対して相乗効果をもたらす抗ウイルス剤及びc)医薬用として許容可能な担体を含む。
【0074】
5−アザシチジン、5−アザ−2−デオキシシチジン又はこれらの組合せは、現在知られているか将来開発されるかもしれない種々の医薬製剤に調製することができる。
【0075】
特定の実施態様では、5’−アザシチジンは0.001%w/w乃至50%w/w、好ましくは0.005%w/w乃至20%w/w、より好ましくは0.005%w/w乃至10%w/wの濃度で局所医薬組成物として適用される。この脱メチル化剤は約0.001%乃至10%の濃度で脱メチル化剤を含むクリーム剤である。
【0076】
【表2】
各種形態の適用スケジュール:ブラシ剤−8時間毎、タンポン剤−毎日、子宮頚管キャップ剤−2日毎。
【0077】
5’−アザシチジンは、0.1ng乃至100μgの投与量、別の実施態様では0.5ng乃至50μgの各単回適用投与量、さらに別の実施態様では1ng乃至10μgの投与量、別の実施態様では500ng乃至2μgの投与量で使用する。本発明の特定の実施態様では、1日当たり1投与量、1日当たり2投与量、1日当たり3投与量、1日当たり4投与量又は1日当たり5以上もの投与量が適用される。本発明の特定の実施態様では、適用は、脱メチル化剤を連続的に放出する媒体を用いて連続的となるようにデザインされる。別の実施態様では、こうした投与量は毎週、週2回、週3回、隔週又は毎月与えられる。このような投与量は、例えば、計1週間、計2週間、計3週間、計4週間、計5週間、計6週間、計2ヶ月間、計3ヶ月間又はそれ以上もの間適用することができる。特定の実施態様では、適用期間は半年、1年又はそれ以上もの期間とすることができる。
【0078】
子宮頚部への薬物送達に用いることができるデバイスの1例は綿棒又はブラシである。
【0079】
薬物送達に用いることができるデバイスの他の例は子宮頚管キャップである。例えば、キャビティーリムキャップ(cavity rim cap)は子宮頚部に付着する。このキャビティーリムキャップはPrentif(登録商標)Capsである。このPrentif(登録商標)Cervical Capは患者の子宮頚部を覆い、避妊の障壁方法として用いられる。同様に、TodayR(登録商標)避妊用スポンジは、患者の子宮頚部にフィットするように形成されたスポンジ状デバイスであった。
【0080】
患者の子宮頚部に関連して用いられる他のタイプのデバイスは、膣タンポン、膣リング、膣ストリップ、膣カプセル、生体付着性フィルム又はスポンジである。デバイスには、約1乃至約10mgの後成的薬剤、約10%のヒドロキシプロピルメチルセルロース、約70%の親水性薬物用飽和脂肪酸トリグリセリド又は脂溶性薬物用PEG6000/PEG400及び約15%のエトキシジグリコールを含む組成物を含浸させる。
【実施例】
【0081】
(実施例1)
メチル化パターンの検出
HPV16 URRのメチル化が宿主上皮細胞の分化状態によって影響されるかどうかを調べるために、レーザーにより切り出した分化程度の異なる正常上皮微小細胞から単離したHPV16のDNAをバイサルファイトゲノム配列決定(BGS)法によって分析する。
【0082】
正常上皮由来の基底細胞及び中間層からの細胞画分中のp97HPV16プロモータはメチル化されていないCpGを含有していた。これに対して、分化細胞に富んだ表面層におけるHPV16プロモータは、E2(E2BS2、3及び4)並びにSP1結合部位内のCpGを含め、メチル化されていた。エンハンサ領域では、全ての分化段階において1つのCpG部位(7454)のメチル化の低下が見られた。従って、その他の4個のCpGは、基底細胞では高度にメチル化されたが、より分化の進んだ細胞ではメチル化の程度は低い。5’LCR領域は全ての分化段階で低メチル化状態であった。
【0083】
さらに、結合バイサルファイト制限解析(COBRA)を用い、HPV形質転換上皮細胞の生体指標としてのサイクリン依存性キナーゼ抑制因子p16INK4aの過剰発現によって、HSIL病変部中のHPV16 URRのメチル化パターンと対応する正常上皮とを比較する。
【0084】
これらのデータから、E2BS1のメチル化は複製HPV感染から形質転換HPV感染への移行過程で特異的に生じることが分かる。
【0085】
(実施例2)
診断の評価
HPV16に感染した、細胞学的、組織学的に確認された低悪性度(LSIL、n=53)及び高悪性度(HSIL、n=48)病変部から得られた101個のサンプル(塗抹及びパンチ生検標本)について、結合バイサルファイト制限解析(COBRA)法によりE2BS1メチル化の有無を調べた。
【0086】
特異的なLCR HPV16メチル化の検出は、癌への進行の高いリスクを有する前癌病変を検出するための分子マーカとなる。
【0087】
【表3】
E2BS1のメチル化は高悪性度病変部の75%に存在したが、低悪性度上皮内病変部では22.6%に過ぎなかった。従って、特異的なLCR HPV16メチル化の検出は、癌への進行のリスクが高い前癌病変を検出するための分子マーカとなり得る。
【0088】
E2BS1へのE2の結合がp97プロモータの活性化に関与しているかどうかを確認するために、一過性導入実験においてP97の活性に対するE2BS1メチル化の影響を分析した。野生型HPV16 LCR中にE2BS1内の2個のCpGジヌクレオチドの選択的メチル化を生じさせた。正常なヒト陰茎包皮の角化細胞に、HPV16 E2のための漸増量の発現ベクターと、ルシフェラーゼ遺伝子の直前にHPV16 LCR全体(E2BS1−LCR16−Luc)を含むか、E2BS1内にメチル化を有するLCR(methE2BS1−LCR16−Luc)を含むレポータプラスミド約50ngとを同時導入した。E2BS1のメチル化により初期p97HPV16プロモータは4乃至6倍活性化された(図8)。このデータから、この特定結合部位へのE2蛋白質の結合を調節することで、メチル化はこのp97プロモータを活性化し、コントロール不良の高レベルのウイルス遺伝子発現を可能にすることが示唆される。
【0089】
さらに、こうしたデータから、E2BS1の特異的なメチル化の検出を行うことにより急性HR−HPV感染から形質転換HR−HPV感染へ既に移行している病変部を特定できることが分かる。従って、E2BS1のメチル化はHSILの生体指標となりうる。
【0090】
(実施例3)
細胞培養における5’−アザシチジンの治療的適用
p97プロモータの活性に対する後成的阻害剤(脱メチル化剤)による処置の効果をルシフェラーゼアッセイを用いて評価する。ルシフェラーゼ遺伝子の直前にクローニングしたHPV16の完全なLCR断片を含有するレポータプラスミドLCR16_Lucを導入した正常ヒト角化細胞を約50乃至60%の集密となるように増殖させた。2μM及び4μMの5−アザシチジンで細胞を24及び72時間処置した。細胞に5−アザシチジンを添加すると、初期p97プロモータの活性が用量依存性に減少した。
【0091】
図9に示したように、未分化又は分化細胞を2μMの5−アザシチジンで処置すると、p97プロモータの活性は40乃至60%まで低下した)。4μMの5’−アザシチジンで処置した両細胞とも、プロモータ活性が基底レベルまで著しく低下した。
【0092】
これらのデータから、脱メチル化剤の1例としての5−アザシチジンの適用によってHPV16 URRの活性の大幅な抑制がもたらされることが分かる。従って、これは、ウイルスの生活環を遮断し、急性HPV感染から形質転換HPV感染への切り替えを阻止する大いなる可能性を有している。こうしたデータに基づき、このような薬剤を用いてウイルスへの複製及びそのより高悪性度の異形成さらには癌への進行を阻止することができる。
【0093】
(実施例4)
5アザシチジンによる低悪性度子宮頚部病変の処置
低悪性度病変(CIN1)HPV16陽性病変を有する患者及び低リスクHPV関連病変(コンジローム、疣贅)を有する患者を5’−アザシチジンで局所処置した。
【0094】
重量で1%の5アザシチジンを(蒸留水にヒドロキシエチルセルロース(Natrosol1)を1%溶かした溶液)に添合して5’−アザシチジン(1%)親水性ゲルを調製するとともに、5アザシチジンを含有しない対応するプラセボゲルを作製した。そのまま使用できるゲル(実薬及びプラセボ)を、使用直前に子宮頚部キャップの内面に付着させた活性のないスポンジによって子宮頚部キャップの内面に塗布した。調製してすぐに、医療関係者がその子宮頚部キャップを子宮頚部に設置する。従って、子宮頚部キャップの内面に付着した治療剤を子宮頚部へ直接拡散させて処置する。子宮頚部キャップは4週間2日毎に子宮頚部に設置した。薬物に接触させた最後の日の1週後に、病変並びに薬物関連副作用の兆候と症状の有無について患者を評価した。治癒は、病変が完全に消失し、コルポスコピーにより感染の臨床的兆候が存在しないことと定義した。退縮率を未処置対照群と比較する。
【0095】
5’−アザシチジン(1%)ゲルは、プラセボ(病変の治癒率:22.2%(2/9))よりも有意に多くの病変(85.7%(6/7))を治癒させた。16名の患者のうち、10名では薬物関連有害症状の訴えがなかった。主に実薬ゲル群の6名の患者が軽度の紅斑及び浮腫をきたしたが、多くの場合、そのような症状の発現は48時間以内に無くなった。
【0096】
(実施例5)
脱メチル化剤による皮膚疣贅の処置
脱メチル化剤を含む種々の医薬組成物を皮膚疣贅に適用した。これらの組成物は以下の通りとした:親水性クリーム基剤中1%5’−アザシチジン、10%5,6−ジヒドロ−5−アザシチジン及び10%エピガロカテキン3−没食子酸塩(EGCG)。こうした医薬組成物を皮膚疣贅に局所適用した。5名の患者に1%5’−アザシチジン、4名の患者に10%5,6−ジヒドロ−5−アザシチジン及び7名の患者に10%エピガロカテキン3−没食子酸塩(EGCG)並びに10名の患者にプラセボを処置した。このクリーム剤は、4週間閉鎖により毎日適用した。全ての患者を1名の医師が調べた。臨床評価は処置開始後1、2及び4週目に実施された。
【0097】
【表4】
退縮率は、未処置対照群に比し、処置患者でより高かった。この処置の開始2週後には、疣贅は炎症、発赤の軽微な兆候を示し、実質的により平坦になった。処置開始4週後には疣贅は治癒した。
【0098】
塩酸5,6−ジヒドロ−5−アザシチジン及びエピガロカテキン3−没食子酸塩(EGCG)は皮膚疣贅に対して同様な活性を示したが、それらの濃度は5’−アザシチジンの10倍であった。
【0099】
(実施例6)
5アザシチジンによる高悪性度子宮頚部病変の処置
実施例4に記載した5’−アザシチジン(1%)親水性ゲルを子宮頚部のCIN2病変に適用した。
【0100】
この治療剤を12週間2日毎に子宮頚部キャップ及びスポンジで子宮頚部に送達してこの頚部に設置した。
【0101】
全ての患者に対して、薬物に接触させた最後の日の1週後に反復コルポスコープ評価を行った。以前に組織像でCIN2と判定されたのと同じ部位から頚部生検又は塗抹標本が採取された。注目される転帰は、対象者が処置を必要とする疾患、即ち、CIN2以上の悪性度の病変を依然として有するかどうかであった。
【0102】
13週目で無疾患又はCIN1への退縮は、5’−アザシチジンで66.7%(4/6)、プラセボで14.2%(1/7)であった。
【特許請求の範囲】
【請求項1】
配列番号1乃至16からなる群から選ばれる配列を含む単離核酸。
【請求項2】
HPV関連病変の処置のための局所適用用薬剤としての脱メチル化剤。
【請求項3】
5−アザシチジン、5−アザ−20−デオキシシチジン、アラビノシル−5−アザシチジン、5−6−ジヒドロ−5−アザシチジン、5−フルオロ−20−デオキシシチジン、エピガロカテキン3−没食子酸塩、ヒドララジン、プロカインアミド、プロカイン及びゼブラリンからなる群から選ばれる請求項2に記載の脱メチル化剤。
【請求項4】
前記HPV関連病変が疣贅、外方増殖パピローマ、コンジローム、内反性パピローマ、前癌性又は癌性HPV誘発病変、子宮頚部新形成、直腸新形成、肛門上皮の新形成、口腔咽頭の新形成又は扁桃の新形成からなる群から選ばれる、請求項2又は3に記載の脱メチル化剤。
【請求項5】
a)脱メチル化剤及びb)医薬用として許容可能な担体からなる医薬組成物。
【請求項6】
緩衝剤、湿潤剤、賦形剤、保存剤及び抗ウイルス物質からなる群から選ばれる活性成分をさらに含む、請求項5に記載の医薬組成物。
【請求項7】
前記脱メチル化剤が5−アザシチジン、5−アザ−20−デオキシシチジン、アラビノシル−5−アザシチジン、5−6−ジヒドロ−5−アザシチジン、5−フルオロ−20−デオキシシチジン、エピガロカテキン3−没食子酸塩、ヒドララジン、プロカインアミド、プロカイン及びゼブラリンからなる群から選ばれる、請求項5又は6に記載の医薬組成物。
【請求項8】
前記脱メチル化剤が0.001乃至50%の濃度で存在する、請求項5乃至7のいずれか1項に記載の医薬組成物。
【請求項9】
前記医薬用として許容可能な担体がステアリン酸、Aquaphor(登録商標)、パルミチン酸イソプロピル、ステアリン酸ポリオキシル40、プロピレングリコール、ソルビン酸カリウム、ソルビン酸、ラウリル硫酸トリエタノールアミン、DMSO、ベータデクス、エデト酸二ナトリウム、ヒドロキシエチルセルロース、メチルパラベン、ナイアシンアミド、フェノキシエタノール、プロピルパラベン及び精製水から選ばれる少なくとも1成分を含む、請求項5乃至8のいずれか1項に記載の医薬組成物。
【請求項10】
綿、セルロース、デキストラン、ポリエチレン、ポリプロピレン、ポリ(ジメチルシロキサン)、エチレン酢酸ビニルコポリマー類、ポリカロフィル、ヒドロキシプロピルメチルセルロース及びポリアクリル酸類からなるポリマー担体上で提供される、請求項5乃至9のいずれか1項に記載の医薬組成物。
【請求項11】
前記ポリマー担体が子宮頚管キャップ、綿棒、タンポン、膣タンポン、膣リング、膣ストリップ、膣カプセル、生体付着性フィルム、スポンジ又はブラシである、請求項5乃至10のいずれか1項に記載の医薬組成物。
【請求項1】
配列番号1乃至16からなる群から選ばれる配列を含む単離核酸。
【請求項2】
HPV関連病変の処置のための局所適用用薬剤としての脱メチル化剤。
【請求項3】
5−アザシチジン、5−アザ−20−デオキシシチジン、アラビノシル−5−アザシチジン、5−6−ジヒドロ−5−アザシチジン、5−フルオロ−20−デオキシシチジン、エピガロカテキン3−没食子酸塩、ヒドララジン、プロカインアミド、プロカイン及びゼブラリンからなる群から選ばれる請求項2に記載の脱メチル化剤。
【請求項4】
前記HPV関連病変が疣贅、外方増殖パピローマ、コンジローム、内反性パピローマ、前癌性又は癌性HPV誘発病変、子宮頚部新形成、直腸新形成、肛門上皮の新形成、口腔咽頭の新形成又は扁桃の新形成からなる群から選ばれる、請求項2又は3に記載の脱メチル化剤。
【請求項5】
a)脱メチル化剤及びb)医薬用として許容可能な担体からなる医薬組成物。
【請求項6】
緩衝剤、湿潤剤、賦形剤、保存剤及び抗ウイルス物質からなる群から選ばれる活性成分をさらに含む、請求項5に記載の医薬組成物。
【請求項7】
前記脱メチル化剤が5−アザシチジン、5−アザ−20−デオキシシチジン、アラビノシル−5−アザシチジン、5−6−ジヒドロ−5−アザシチジン、5−フルオロ−20−デオキシシチジン、エピガロカテキン3−没食子酸塩、ヒドララジン、プロカインアミド、プロカイン及びゼブラリンからなる群から選ばれる、請求項5又は6に記載の医薬組成物。
【請求項8】
前記脱メチル化剤が0.001乃至50%の濃度で存在する、請求項5乃至7のいずれか1項に記載の医薬組成物。
【請求項9】
前記医薬用として許容可能な担体がステアリン酸、Aquaphor(登録商標)、パルミチン酸イソプロピル、ステアリン酸ポリオキシル40、プロピレングリコール、ソルビン酸カリウム、ソルビン酸、ラウリル硫酸トリエタノールアミン、DMSO、ベータデクス、エデト酸二ナトリウム、ヒドロキシエチルセルロース、メチルパラベン、ナイアシンアミド、フェノキシエタノール、プロピルパラベン及び精製水から選ばれる少なくとも1成分を含む、請求項5乃至8のいずれか1項に記載の医薬組成物。
【請求項10】
綿、セルロース、デキストラン、ポリエチレン、ポリプロピレン、ポリ(ジメチルシロキサン)、エチレン酢酸ビニルコポリマー類、ポリカロフィル、ヒドロキシプロピルメチルセルロース及びポリアクリル酸類からなるポリマー担体上で提供される、請求項5乃至9のいずれか1項に記載の医薬組成物。
【請求項11】
前記ポリマー担体が子宮頚管キャップ、綿棒、タンポン、膣タンポン、膣リング、膣ストリップ、膣カプセル、生体付着性フィルム、スポンジ又はブラシである、請求項5乃至10のいずれか1項に記載の医薬組成物。
【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図1】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図1】
【公表番号】特表2011−502474(P2011−502474A)
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2010−531461(P2010−531461)
【出願日】平成20年10月31日(2008.10.31)
【国際出願番号】PCT/EP2008/009197
【国際公開番号】WO2009/062604
【国際公開日】平成21年5月22日(2009.5.22)
【出願人】(510123242)
【Fターム(参考)】
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願日】平成20年10月31日(2008.10.31)
【国際出願番号】PCT/EP2008/009197
【国際公開番号】WO2009/062604
【国際公開日】平成21年5月22日(2009.5.22)
【出願人】(510123242)
【Fターム(参考)】
[ Back to top ]