
上皮膜タンパク質−2(EMP2)および増殖性硝子体網膜症(PVR)
EMP2の活性または機能を拮抗する作用剤を剥離のリスクを有する対象に投与することによる、増殖性硝子体網膜症に関連する網膜剥離の予防方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願に対する相互参照
[0001] 本願は、2009年11月20日に出願された米国仮特許出願第61/263,228号の利益を主張するものであり、当該出願の内容は、参照することによってその全体が本明細書に組み込まれる。本願はまた、2007年10月8日に出願された米国特許出願第11/868,788号および2008年10月9日に出願された米国特許出願第11/884,806号に関する主題を含み、前記各出願の内容は、参照することによってその全体が本明細書に組み込まれる。
【0002】
連邦政府支援研究開発のもとで創出された発明の権利に関する陳述
[0002] 本研究は、米国復員軍人援護局により支援されたものであり、連邦政府は、本発明においてある程度の権利を有する。
【0003】
コンパクトディスクで提出された添付の「配列表」、表、またはコンピュータプログラム表への参照
[0003] 不適用
【背景技術】
【0004】
[0004] 網膜剥離または開放性眼外傷の治療の不成功の最も一般的な理由は、増殖性硝子体網膜症(PVR)(Pastor, J. C. Surv Ophthalmol 43:3-18(1998))と呼ばれる、硝子体腔で形成する細胞膜の増殖および収縮である。PVRは、裂孔原性網膜剥離の最大5〜20%のケースの合併症として生じると推定されている(Pastor, J. C. Surv Ophthalmol 43:3-18(1998); Pastor, J. C.ら, Prog Retin Eye Res 21:127-44(2002))。眼の外傷において、PVRのリスクは非常に高い。2003年〜2005年の間にイラクで眼球内異物による眼の外傷を有していた退役軍人を検査したところ、PVRが患者の21%において視力低下の原因であることが明らかになった(Colyer M.H.ら, Ophthalmology. 2007 Aug;114(8):1439-47)。
【0005】
[0005] 臨床研究によって、硝子体出血、眼球内ガス、大きなまたは長期にわたる網膜剥離、および手術の失敗(特に、それらが凍結療法または光凝固術の過剰な使用に関連する場合)を包含する、PVR発症の複数のリスク因子が同定された(Pastor, J. C. Surv Ophthalmol 43:3-18(1998); Pastor, J. C.ら, Prog Retin Eye Res 21: 127-44(2002))。眼球/網膜の損傷の程度が、PVRを増大させるサイトカイン生成または他のシグナル伝達を増大させる可能性がある。炎症性サイトカイン(例えば、IL-8およびMCP-1)、ならびにIL−1、IL−6、TNF−α、およびIFN−γは、PVRにおいて増大することが明らかにされているが、サイトカインレベルは、PVRの重症度と直接的には相関しない(Asaria, R. H. Compr Ophthalmol Update 7:179-85(2006))。
【0006】
[0006] 網膜色素上皮(RPE)は、PVRに関与する重要な細胞型の1つであると考えられている。PVRの根底にある発症機序は複雑であるが、外傷または網膜剥離の後、RPE細胞が硝子体内に放出されるか、または網膜下の位置から移動するように刺激される可能性が高い。これらの細胞は次に移動し、増殖し、脱分化し、上皮間葉転換(EMT)を受けて、PVRの網膜前膜の生成を促進する(Asaria, R. H. Compr Ophthalmol Update 7:179-85(2006))。RPE細胞は、再発性網膜剥離およびさらなる失明をもたらすことが多い牽引力を生じさせる膜収縮をもたらす可能性が高い(Pastor, J. C.ら, Prog Retin Eye Res 21:127-44(2002); Kroll, P.ら, Ophthalmologica 221:78-94(2007))。
【0007】
[0007] EMP2はRPE細胞において高度に発現することが明らかにされているが、その機能は理解され始めているにすぎない(Wadehra, M.ら, Exp Mol Pathol 74:106-12(2003))。テトラスパニンファミリー(CD9、CD53、CD81、CD82)のメンバーとしては、インテグリンおよび他の細胞表面受容体の調節が一般的である。EMP2は、以前に、FAK.Srcシグナル伝達複合体の活性化の促進を介する細胞の収縮能力の調節において重要であることが示されている(Morales, S. A.ら, Exp Eye Res 85:790-8(2007); Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。さらに、FAK/Src複合体の活性化は、EMP2とFAK/Src複合体との間の物理的関連を介することが証明されている(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。これらのインビトロでの研究によって、EMP2およびFAK/Srcシグナル伝達複合体がRPE介在性のゲル収縮における重要な構成要素であることが明らかにされた。
【0008】
[0008] 本発明者らは、本明細書において、RPE細胞がインビボでの研究およびインビトロでの研究の両方で一致していると報告するが、これまでに報告された観察結果と本研究との間には多くの潜在的な差が存在する。まず、インビトロでの研究は、コラーゲンをRPE細胞の足場として用いるが、インビボでの硝子体におけるRPE細胞は、ヒアルロン酸ならびにII、V/XI、およびIXを包含する多くのコラーゲン型に曝露される。さらに、炎症細胞、ミュラー細胞、線維芽細胞などを包含する、インビボでPVR膜に寄与する多くの細胞型が存在する(Pastor, J. C. Surv Ophthalmol 43:3-18(1998); Pastor, J. C.ら, Prog Retin Eye Res 21:127-44(2002); Asaria, R. H. Compr Ophthalmol Update 7:179-85(2006); Kim, I. K.ら. Ophthalmol Clin North Am 15:81-6(2002))。インビボでは、PVR膜の形成の間に、細胞が多くのサイトカインに曝露されることが知られており、網膜前膜は、血管内皮増殖因子(VEGF)、(PDGF)、IL−6、IL−8、TNF−α、TGF−βなどを包含するサイトカインを含有することが明らかにされている(Harada, C.ら, Prog Retin Eye Res 25:149-64(2006))。これらの細胞およびこれらの細胞によって生じるサイトカインの環境は、細胞および膜の挙動に確実に影響する。
【発明の概要】
【発明が解決しようとする課題】
【0009】
[0009] 今日、手術の進歩にもかかわらず、患者は、PVRおよび牽引性網膜前膜に続発して視力を失うことが非常に多い。本発明は、PVRに関連する網膜剥離の治療および/または予防の必要性を満たすものである。
【課題を解決するための手段】
【0010】
[0010] 1つの態様において、本発明は、対象における増殖性硝子体網膜症または網膜剥離を治療する方法を提供する。いくつかの実施形態において、本方法は、眼におけるEMP2の活性を低減させる抗EMP2剤を対象に投与することを含む。
【0011】
[0011] いくつかの実施形態において、対象はヒトである。いくつかの実施形態において、抗EMP2剤は局所的に投与される。いくつかの実施形態において、抗EMP2剤は、全身投与される。いくつかの実施形態において、抗EMP2剤は、眼に直接投与される。いくつかの実施形態において、抗EMP2剤は、眼内に投与される。いくつかの実施形態において、抗EMP2剤は、眼の硝子体液内に投与される。
【0012】
[0012] いくつかの実施形態において、抗EMP2剤の投与は、網膜剥離のリスクを低減させる。いくつかの実施形態において、抗EMP2剤の投与は、網膜剥離の程度を低減させる。
【0013】
[0013] いくつかの実施形態において、抗EMP2剤は、眼への注入用に製剤化される。いくつかの実施形態において、抗EMP2剤は、眼の外での局所投与用に製剤化される。
【0014】
[0014] いくつかの実施形態において、硝子体網膜症は、裂孔原性の硝子体網膜症である。いくつかの実施形態において、抗EMP2剤を投与する前に、眼は外傷を有するか、または網膜剥離のリスクの原因となる手術を受けている。
【0015】
[0015] 別の態様において、本発明は、生体試料における網膜色素上皮細胞の収縮能力を低下させる方法を提供する。いくつかの実施形態において、本方法は、生体試料と、網膜色素上皮細胞におけるEMP2の活性を低減させる抗EMP2剤とを接触させることを含む。
【0016】
[0016] いくつかの実施形態において、生体試料は組織試料である。いくつかの実施形態において、組織試料は眼の組織である。
【0017】
[0017] いくつかの実施形態において、抗EMP2剤は、配列番号1、配列番号2、または配列番号30に対して少なくとも70%のアミノ酸配列同一性を有するEMP2ポリペプチドに特異的に結合する抗体またはそのフラグメントである。いくつかの実施形態において、抗体は、配列番号4、配列番号6、配列番号8、または配列番号10に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む重鎖を含む。いくつかの実施形態において、抗体は、配列番号5、配列番号7、配列番号9、または配列番号11に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗体は、(1)配列番号4、配列番号6、配列番号8、および配列番号10からなる群から選択される重鎖、ならびに(2)配列番号5、配列番号7、配列番号9、および配列番号11からなる群から選択される軽鎖を含む。いくつかの実施形態において、抗体は、配列番号27、配列番号28、または配列番号29に対して少なくとも70%のアミノ酸配列同一性を有する。いくつかの実施形態において、抗体は、配列番号27、配列番号28、または配列番号29のアミノ酸配列を有する。
【0018】
[0018] いくつかの実施形態において、抗体は、モノクローナル抗体である。いくつかの実施形態において、抗体は、ヒト化モノクローナル抗体である。いくつかの実施形態において、抗体は、ポリクローナル抗体である。いくつかの実施形態において、抗体フラグメントは、ダイアボディ(diabody)またはミニボディとして提供される。いくつかの実施形態において、抗体フラグメントは、Fvフラグメント、F(ab’)フラグメント、またはF(ab’)2フラグメントである。いくつかの実施形態において、抗体は、マウスの抗原結合部位とエフェクター機能を調節するヒト化領域とを有するキメラ抗体である。いくつかの実施形態において、抗体またはそのフラグメントは、細胞傷害剤に結合している。
【0019】
[0019] いくつかの実施形態において、抗EMP2剤は、配列番号1のポリペプチドに対して少なくとも70%のアミノ酸配列同一性を有するEMP2タンパク質をコードする核酸の発現を低減できるsiRNAである。いくつかの実施形態において、siRNAは、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドに対して少なくとも70%のヌクレオチド配列同一性を有する核酸配列を含む。いくつかの実施形態において、siRNAは、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドを含む。
【0020】
[0020] いくつかの実施形態において、抗EMP2剤は、配列番号3の核酸配列に対して少なくとも70%のヌクレオチド配列同一性を有するEMP2ポリヌクレオチド配列を切断できるハンマーヘッドリボザイムである。
【0021】
[0021] いくつかの実施形態において、抗EMP2剤は、プロゲステロン受容体アンタゴニストである。いくつかの実施形態において、抗EMP2剤は、ミフェプリストンである。
【図面の簡単な説明】
【0022】
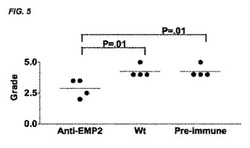
【図1】[0022]表1に示されるようにFastenburgシステムに従って等級付けされた、ウサギPVRモデルの図である。各群3頭のウサギであり、対照は、ベクターのみでトランスフェクトされたARPE19細胞を投与したウサギであり、Riboは、EMP2の発現がより低い、リボザイムでトランスフェクトされた細胞を投与したウサギであり、EMP2は、EMP2を過剰発現するようにトランスフェクトされた細胞を投与したウサギであった。EMP2を過剰発現する細胞を投与したウサギは、PVR形成の速度が増大し、一方、リボザイムノックダウンによってEMP2発現のレベルがより低い細胞を投与した動物(Ribo)は、PVR形成の程度が減少した。示される結果は、細胞注入の5週間後に得たものである。比較した群の上に、スチューデントT検定のp値を示す。
【図2】[0023]図1で示される実験から単離された眼球の肉眼的な病変断面を示す図である。PVR膜形成の程度を、これらの画像で見ることができる。各群の1つの眼球が示されている。図1におけるように、対照は、ベクターのみでトランスフェクトされたARPE19細胞を投与したウサギであり、Riboは、EMP2の発現がより低い、リボザイムでトランスフェクトされた細胞を投与したウサギであり、EMP2は、EMP2を過剰発現するようにトランスフェクトされた細胞を投与したウサギであった。EMP2を過剰発現する細胞を投与したウサギは、PVR膜形成の速度が増大し、一方、リボザイムノックダウンによってEMP2発現のレベルがより低い細胞を投与した動物(Ribo)は、PVR膜形成の程度が減少した。表される結果は、細胞注入の5週間後に得たものである。
【図3】[0024]図1および2で示される実験の網膜前膜を示す組織学的断面を示す図である。図1および2におけるように、対照は、ベクターのみでトランスフェクトされたARPE19細胞を投与したウサギであり(A、B)、Riboは、EMP2の発現がより低い、リボザイムでトランスフェクトされた細胞を投与したウサギであり(C、D)、EMP2は、EMP2を過剰発現するようにトランスフェクトされた細胞を投与したウサギであった(E、F)。矢印は網膜前膜を指し、これは、Ribo群(E、F)におけるよりも対照群(A、B)およびEMP2群(C、D)で顕著であった。
【図4】[0025]網膜前膜において見られるEMP2の発現を示す図である。網膜前PVR膜は、対照ウサギ群およびEMP2ウサギ群で最も顕著であった(図1〜3)。この図は、対照ウサギ(A)およびEMP2を過剰発現するウサギ(B)におけるEMP2の発現(紫)を示す。
【図5】[0026]PVRのウサギモデルは、細胞を抗EMP2ポリクローナル抗体と共にプレインキュベートした場合のPVR膜形成の阻害を示す。各群3頭のウサギであり、抗EMP2は、抗EMP2ポリクローナル抗体と共にプレインキュベートされたARPE19細胞を投与したウサギであり、野生型(Wild-type)(Wt)は、野生型ARPE19細胞を投与したウサギであり、免疫前(Pre-immune)は、ポリクローナル免疫前(対照)抗体と共にプレインキュベートされたARPE19細胞を投与したウサギであった。表される結果は、細胞注入の5週間後に得たものである。比較した群の上に、スチューデントT検定のp値を示す。
【図6】[0027]網膜前膜において見られるEMP2の発現を示す図である。図5において示されるように、網膜前PVR膜は、抗EMP2抗体で前処理されたウサギ群におけるよりも、野生型ウサギ群および免疫前ウサギ群で最も顕著であった。この図は、3つのウサギ群全て、すなわち、Wt(A)、免疫前(B)、および抗EMP2抗体(C)の網膜前PVR膜におけるEMP2の発現(紫)を示す。
【図7】[0028]A.FAKのリン酸化に対するEMP2の影響を示す図である。全FAK、p−FAK(Y397、Y407、Y861、およびY925)、およびβ−アクチンのレベルを、細胞系ARPE−19およびARPE−19/EMP2においてウェスタンブロットによって測定し、デンシトメトリーによって定量した。B. p−Src(Tyr 416およびTyr 527)およびβ−アクチンを、細胞系ARPE−19およびARPE−19/EMP2においてウェスタンブロットによって測定し、デンシトメトリーによって定量した。各部位のリン酸化レベル(平均±標準誤差)を全FAK(A)またはβ−アクチン(B)に対して正規化し、少なくとも3つの独立した実験から計算した。代表的なウェスタンブロットも示す。
【図8】[0029]FAKとEMP2との関連。(A)ARPE−19およびARPE−19/EMP2(107個細胞)の溶解物を、FAKに対する抗体、EMP2に対する抗体、または無関係なウサギ抗体(−対照)を用いて免疫沈降した。SDS−PAGEを、免疫沈降物のアリコートまたは元の溶解物のアリコートを用いて行い(それぞれ、106個および105個細胞当量)、ウェスタンブロットをFAKについて分析した。FAKおよびEMP2の同族(cognate)免疫沈降(データは示していない)が、比較的効率的であった(50〜60%)。EMP2は、ARPE−19およびARPE−19/EMP2において、それぞれ、全FAKの25%および30%を共免疫沈降した。データは、3回以上の実験の代表である。(B)ARPE−19細胞およびARPE−19/EMP2細胞を、EMP2に対する抗体で染色した後に、FITCに結合したロバ抗ウサギ二次IgG(緑)で染色し、かつ、FAKに対する抗体で染色した後に、テキサスレッドに結合したロバ抗ヤギ二次IgG(赤)で染色した。これらの2つのチャネルの併合が示されており(中央右)、同様に、共局在化したピクセルが強調して示されている併合が示されている(右)。少なくとも6個のフィールドが、各試料の分析のためにランダムに選択され、関連のパーセンテージ(percent association)は、複数のフィールドから生じた平均値である。フィールド当たり複数のスライスが取得され、示されているものは、スライドと接触し接着斑を含有する、細胞の底表面の単一のスライスである。ARPE−19およびARPE−19/EMP2において、それぞれ全FAKの68%および96%がEMP2と共局在化した。(C)EMP2とリン酸化型FAKとの間の関連を評価するために、ARPE−19細胞およびARPE−19/EMP2細胞を、EMP2に対する抗体およびp576/577−FAKに対する抗体で染色した。EMP2に対する結合抗体は、FITCに結合した二次抗体を用いて同定され、一方、p−FAK(Y576/577)に対する結合抗体の染色は、テキサスレッドに結合した二次抗体で示される。これらの2つのチャネルの併合が示されており(白)、共局在化したピクセルが観察および定量されている。少なくとも6個のフィールドが、各試料の分析のためにランダムに選択され、関連のパーセンテージは、複数のフィールドから生じた平均値である。フィールド当たり複数のスライスが取得され、表されているものは、スライドと接触し接着斑を含有する、細胞の底表面の単一のスライスである。ARPE−19細胞において、リン酸化型FAKの31%がEMP2と共局在化している。ARPE−19/EMP2細胞において、リン酸化型FAKの97%がEMP2と共局在化している。EMP2レベルが増大すると、リン酸化型FAKとEMP2との関連が増大する。
【図9】[0030]EMP2発現の調節が、FAKのリン酸化および局在化に影響することを示す図である。ARPE−19細胞およびARPE−19/EMP2細胞をp576/577−FAK(A)一次抗体で染色した。細胞をテキサスレッドに結合したロバ抗ヤギ抗体と共にインキュベートし、蛍光顕微鏡法下で観察した。少なくとも4つの個別の試料のリン酸化型FAK(B)をピクセル強度を計算することによって定量し(方法を参照されたい)、データをスチューデントt検定(独立、一方向)によって評価した。
【図10】[0031]EMP2およびリン酸化型FAKの発現の増大によって、F−アクチンの分布が変化することを示す図である。ARPE−19細胞(AおよびC)およびARPE−19/EMP2細胞(BおよびD)をガラスカバースリップ上で一晩平板培養した。細胞をパラホルムアルデヒドで固定し、サポニンで透過処理し、テキサスレッド−Xファロイジンで染色した。代表的な写真をカラー(AおよびB)および白黒(CおよびD)で示す。
【図11】[0032]EMP2発現の増大によって、α−平滑筋アクチンの発現が増大した。(A)ARPE−19細胞およびARPE−19/EMP2細胞におけるα−平滑筋アクチン(α-sma)の定常状態のタンパク質レベルをウェスタンブロット分析によって測定した。β−アクチンをロード対照として用いた。(B)α−smaの量を、β−アクチンと比較して各試料について計算した。少なくとも3つの独立した実験を行い、結果を数値的に表す。発現レベルの差をスチューデントt検定(独立、一方向)を用いて評価した。
【図12】[0033]EMP2レベルの増大によって、コラーゲンへの接着が増大することを示す図である。ARPE−19細胞およびARPE−19/EMP2細胞を、フィブロネクチン、ビトロネクチン、コラーゲンI、およびコラーゲンIVで被覆したプレート上で平板培養した。結合細胞をクリスタルバイオレットの取り込みについて分析し、吸光度の定量的評価をマイクロプレートリーダー(590nm)によって測定した。EMP2の発現が増大すると、コラーゲンI(P<0.001)およびコラーゲンIV(P<0.001)への接着が増大した。フィブロネクチンへの接着は、ARPE−19/EMP2細胞と比較して、ARPE−19細胞において増大した(P=0.02)。EMP2レベルの変化は、ビトロネクチンへの接着に影響しなかった。結果は、少なくとも8回反復した1つの実験から表され、少なくとも3つの独立した実験を行い、同等の結果を得た。スチューデントt検定(独立、一方向)を統計分析に用いた。
【図13】[0034]組換えによるLMP2発現変化を示す図である。A.EMP2の定常状態のタンパク質レベルをARPE−19細胞(対照細胞)、ARPE−19/EMP2細胞(EMP2の増大)、およびARPE−19/Ribo細胞(EMP2の減少)においてウェスタンブロット分析によって測定した。EMP2発現の増大を測定するために、様々な希釈(1:10、1:25、1:50、および1:100)のARPE−19/EMP2細胞の溶解物を評価した。B.ARPE−19細胞をEMP2に特異的なsiRNA(ARPE-19/EMP2 siRNA)または対照のスクランブルsiRNA(ARPE-19/対照siRNA)で一時的にトランスフェクトした。実験は独立して少なくとも3回行い、同様の結果を得た。
【図14】[0035]EMP2の修飾が、コラーゲンゲルの収縮に影響したことを示す図である。コラーゲンゲル収縮アッセイをARPE−19/EMP2細胞、ARPE−19/Ribo細胞、およびARPE−19細胞で行った。EMP2の発現が上昇した細胞系および低下した細胞系では、結果として、それぞれ、ゲル収縮が57%増大および55%減少した(P=0.002)。実験は独立して少なくとも3回行い、同様の結果を得た。
【図15】[0036]コラーゲン結合インテグリンを示す図である。細胞表面の発現を、α1インテグリン、α2インテグリン、またはα3インテグリンに対するモノクローナル抗体を用いるフローサイトメトリーによって測定した。A.代表的な実験のヒストグラムを表す。アイソタイプ対照の染色が白抜きの波形として示され、特異的な染色パターンには影が付けられている。蛍光強度の平均の数値が、各パネルの右上隅に表されている。B.これらの各インテグリンの表面発現を、3つの独立した実験で評価し、結果を表にして平均を表した。2つの細胞系における各インテグリンの発現の統計的比較をスチューデントt検定(独立、一方向)で行った。
【図16】[0037]EMPレベルの増大は増殖および移動を減少させたが、侵入またはコラーゲンの生成に対する影響は有していなかった。A.ARPE−19細胞およびARPE−19/EMP2細胞を正常培地またはコラーゲンI25mg/mLで処理し、48時間にわたってインキュベートし、増殖をBrdUの組み込みによって評価した。B.細胞の移動に対するEMP2発現の影響。ARPE−19細胞およびARPE−19/EMP2細胞を24ウェルプレート内で平板培養し、コンフルエントになったら擦り傷を作り、移動を様々な時点で測定した。ARPE−19細胞およびARPE−19/EMP2細胞は、未処理のままであるか、またはPDGF50ng/mLで処理し、移動を24時間後に測定した。C.ARPE−19細胞およびARPE−19/EMP2細胞を、重合コラーゲンの薄層で被覆した8μmの孔サイズのポリカーボネート膜を有する侵入チャンバインサート上に播種した。10%PBSまたはPDGF50ng/mlを化学誘引物質として用いた。侵襲性細胞は、重合コラーゲン層を通って移動して、ポリカーボネート膜の底に張り付き、染色、抽出、およびマイクロプレートリーダー(560nm)による測定によって検出された。D.ARPE−19細胞およびARPE−19/EMP2細胞を、TGF−β10ng/mLの存在下または非存在下の無血清培地で72時間にわたって成長させた。培地を回収し、コラーゲンの生成をELISAによって測定した。TGF−βの存在下または非存在下でのコラーゲンの生成に統計的に有意な変化はなかった。しかし、TGF−βは、未処理条件と比較して、各細胞系でコラーゲンの生成を増大させた。全ての研究は、試料当たり6ウェルで、個別に少なくとも3回行った。結果は、スチューデントt検定(独立、一方向)を用いて、統計的有意性について評価した。P<0.05のレベルを、統計的に有意であると考えた。
【図17】[0038]EMP2の過剰発現によって、FAKの活性化が増大した。細胞抽出物(タンパク質10μg)を還元条件において4%〜20%のSDS−PAGE勾配ゲルによって分画し、ウェスタン免疫ブロットをFAK、pFAK、およびβ−アクチンに対する抗体で探査した。A:代表的な免疫ブロット、B:実験は独立して少なくとも3回行い、同様の結果を得た。β−アクチンロード対照に対して正規化したバンドの密度を定量した。実験は独立して少なくとも3回行い、同様の結果を得た。
【図18】[0039]EMP2の過剰発現によって、PP2に対する耐性が増大した。細胞を、様々な濃度の低分子阻害剤PP2(FAK/Src阻害剤)で1時間にわたり前処理し、ゲルの収縮を評価した。実験は、試料当たり6回の反復で、個別に少なくとも3回行い、代表的な実験が表されている(A〜C)。統計分析を、媒体のみの対照と比較して、各阻害剤濃度について、スチューデントt検定で行った。A.EMP2の発現が減少した、ARPE−19/EMP2 siRNA細胞。B.ARPE−19細胞。C.EMP2を過剰発現するARPE−19/EMP2細胞。D.各点は、媒体対照に曝露された同一の細胞系の総収縮に対して正規化された、各細胞系についての各濃度での収縮の阻害の平均を表す。実験は独立して少なくとも3回行い、同様の結果を得た。
【図19】[0040]EMP2の過剰発現によって、SU6656による阻害に対する耐性が増大した。細胞を様々な濃度の低分子阻害剤SU6656(FAK/Src阻害剤)で1時間にわたり前処理し、ゲルの収縮を評価した。実験は、試料当たり6回の反復で個別に少なくとも3回行い、代表的な実験が表されている。媒体のみの対照と比較して、各阻害剤濃度について、スチューデントt検定で統計分析を行った。A.ARPE−19/EMP2細胞。B.EMP2を過剰発現するARPE−19/EMP2細胞。C.各点は、媒体対照に曝露された同一の細胞系の総収縮に対して正規化された、各細胞系についての各濃度での収縮の阻害の平均を表す。実験は独立して少なくとも3回行い、同様の結果を得た。
【図20】[0041]抗体KS49、KS83、KS41、およびKS89の配列。A. KS49重鎖(配列番号4)およびKS49軽鎖(配列番号5)のアミノ酸配列。CDR−H1(配列番号12)、CDR−H2(配列番号13)、CDR−H3(配列番号14)、CDR−L1(配列番号15)、CDR−L2(配列番号16)、およびCDR−L3(配列番号17)に下線が引かれている。B. KS83重鎖(配列番号6)およびKS49軽鎖(配列番号7)のアミノ酸配列。CDR−H1(配列番号12)、CDR−H2(配列番号13)、CDR−H3(配列番号18)、CDR−L1(配列番号19)、CDR−L2(配列番号20)、およびCDR−L3(配列番号21)に下線が引かれている。C. KS41重鎖(配列番号8)およびKS41軽鎖(配列番号9)のアミノ酸配列。CDR−H1(配列番号22)、CDR−H2(配列番号23)、CDR−H3(配列番号24)、CDR−L1(配列番号25)、CDR−L2(配列番号26)、およびCDR−L3(配列番号27)に下線が引かれている。D. KS89重鎖(配列番号10)およびKS89軽鎖(配列番号11)のアミノ酸配列。CDR−H1(配列番号22)、CDR−H2(配列番号23)、CDR−H3(配列番号24)、CDR−L1(配列番号25)、CDR−L2(配列番号26)、およびCDR−L3(配列番号27)に下線が引かれている。
【発明を実施するための形態】
【0023】
I.定義
[0042] ここで、本明細書および添付の特許請求の範囲において用いられる場合、単数形「1つの(a)」、「1つの(an)」、および「その(the)」が、別段の指示が内容から明らかでない限り、複数形の言及を包含することに留意する。
【0024】
[0043] 「上皮膜タンパク質2」または「EMP2」は、(1)例えばGenBank受託番号NM_001424(EMP2のmRNA)およびP54851(EMP2タンパク質)で示される、本明細書において記載される個々に言及される核酸配列またはアミノ酸配列によってコードされるポリペプチドに対して、少なくとも約25、50、100、200、500、1000、またはそれを超えるアミノ酸の領域にわたり、約60%を超えるアミノ酸配列同一性、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、またはそれを超えるアミノ酸配列同一性を有するアミノ酸配列を有する、(2)GenBank受託番号P54851(EMP2タンパク質)で示される言及されるアミノ酸配列を含む免疫原に対する抗体、例えばポリクローナル抗体、そのそれぞれの免疫原性フラグメント、およびそのそれぞれの保存的に修飾された変異体に特異的に結合する、(3)GenBank受託番号P54851(EMP2タンパク質)で示される言及されるアミノ酸配列をコードする核酸、およびそのそれぞれの保存的に修飾された変異体に、ストリンジェントなハイブリダイゼーション条件下で特異的にハイブリダイズする、(4)GenBank受託番号NM_001424(EMP2のmRNA)で示される参照核酸配列に対して、好ましくは、少なくとも約15、20、25、50、100、150、200、250、500、1000、またはそれを超えるヌクレオチドの領域にわたり、約95%を超える、好ましくは約96%、97%、98%、99%を超える、またはそれより高いヌクレオチド配列同一性を有する核酸配列を有する、核酸、例えば、遺伝子、mRNA前駆体、mRNA、ならびにポリペプチド、多型変異体、対立遺伝子、突然変異体、および種間ホモログを言う。ポリヌクレオチド配列またはポリペプチド配列は、典型的には、限定はしないが、ヒトなどの霊長類、ラット、マウス、ハムスターなどの齧歯動物、ウシ、ブタ、ウマ、ヒツジ、または任意の哺乳動物を包含する、哺乳動物のものである。本発明の核酸およびタンパク質は、天然の分子または組換え分子の両方を包含する。
【0025】
[0044] 「抗EMP2剤」または「EMP2阻害剤」は、網膜剥離に関与する拘縮を仲介する細胞性EMP2(例えば、配列番号1のアミノ酸配列を有するEMP2)の機能、活性、または組織レベルに干渉する作用剤である。EMP2阻害剤は、EMP2ポリペプチド;抗EMP2抗体(例えば、組換え抗体、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、ヒトモノクローナル抗体、ヒト化もしくは霊長類化(primatized)モノクローナル抗体、または抗体フラグメント);EMP2siRNA分子;EMP2−リボザイム;EMP2への結合またはEMP2の結合と競合する化合物、または宿主細胞におけるEMP2核酸の発現、転写、もしくは翻訳を阻害する作用物質もしくは化合物であり得る。いくつかの実施形態において、EMP2阻害剤は、無菌担体および/または生理学的に許容可能な担体も含む組成物で提供される。
【0026】
[0045] 「調節剤」は、言及される活性を増大または減少させ得る作用物質である。調節剤は、阻害剤と、言及される活性または実体に関して阻害剤と逆の効果を有する(例えば、増大させる、刺激する、増加させる、増強する、加速する)「活性化剤」との両方を包含する。
【0027】
[0046] 用語「ポリペプチド」、「ペプチド」、および「タンパク質」は、本明細書において、アミノ酸残基のポリマーを言うためにほぼ同じ意味で用いられる。この用語は、1つまたは複数のアミノ酸残基が、天然アミノ酸に対応する人工の化学的模倣剤である、アミノ酸ポリマー、ならびに、天然アミノ酸ポリマーおよび非天然アミノ酸ポリマーに適用される。ポリペプチドを得る(例えば、生成する、単離する、精製する、合成する、および組換えによって製造する)ための方法は、当業者に周知である。
【0028】
[0047] 用語「アミノ酸」は、天然アミノ酸および合成アミノ酸、ならびに天然アミノ酸に類似した様式で機能するアミノ酸類似体およびアミノ酸模倣体を言う。天然アミノ酸は、遺伝子コードによってコードされるもの、および、後に修飾されるアミノ酸、例えば、ヒドロキシプロリン、γ−カルボキシグルタミン酸、およびO−ホスホセリンである。アミノ酸類似体は、天然アミノ酸と同一の基本的な化学構造、すなわち水素、カルボキシル基、アミノ基、およびR基に結合しているα炭素を有する化合物、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムを言う。このような類似体は、修飾されたR基(例えば、ノルロイシン)または修飾されたペプチド骨格を有するが、天然アミノ酸と同一の基本的な化学構造を保持している。アミノ酸模倣体は、アミノ酸の一般的な化学構造と異なる構造を有するが天然アミノ酸に類似した様式で機能する化合物を言う。
【0029】
[0048] アミノ酸は、本明細書において、IUPAC−IUB Biochemical Nomenclature Commissionによって推奨されている、一般に知られているその三文字記号または一文字記号によって言及され得る。ヌクレオチドは、同様に、その一般に認められている一文字コードによって言及され得る。
【0030】
[0049] アミノ酸配列の「保存的に修飾された変異体」に関して、当業者には、コードされる配列における単一のアミノ酸または低いパーセンテージのアミノ酸を改変する、付加する、または欠失する、核酸配列、ペプチド配列、ポリペプチド配列、またはタンパク質配列への個別の置換、欠失、または付加が、改変の結果、化学的に類似のアミノ酸でアミノ酸が置換される「保存的に修飾された変異体」であることが認識されよう。機能的に類似のアミノ酸を提供する保存的置換の表は、当技術分野において周知である。このような保存的に修飾された変異体は、本発明の多型変異体、種間ホモログ、および対立遺伝子に追加されるものであり、これらを排除しない。
【0031】
[0050] 以下の8つの群は、それぞれ、互いの保存的置換であるアミノ酸を含有する:1)アラニン(A)、グリシン(G)、2)アスパラギン酸(D)、グルタミン酸(E)、3)アスパラギン(N)、グルタミン(Q)、4)アルギニン(R)、リジン(K)、5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V)、6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)、7)セリン(S)、スレオニン(T)、および8)システイン(C)、メチオニン(M)(例えば、Creighton, Proteins(1984)を参照されたい)。
【0032】
[0051] 「抗体」は、抗原を特異的に結合および認識する免疫グロブリン遺伝子またはそのフラグメントのフレームワーク領域を含むポリペプチドを言う。認知されている免疫グロブリン遺伝子には、κ、λ、α、γ、δ、ε、およびμ定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子が包含される。軽鎖は、κまたはλとして分類される。重鎖は、γ、μ、α、δ、またはεとして分類され、これは、免疫グロブリンクラスIgG、IgM、IgA、IgD、およびIgEをそれぞれ規定する。典型的には、抗体の抗原結合領域は、結合の特異性および親和性において最も重要である。
【0033】
[0052] 典型的な免疫グロブリン(抗体)の構造単位は、四量体を含む。各四量体は、2つの同一のポリペプチド鎖対からなり、各対は、1つの「軽」鎖(約25kD)および1つの「重」鎖(約50〜70kD)を有する。各鎖のN末端は、抗原認識に最も関与する約100〜110、またはそれを超えるアミノ酸からなる可変領域を規定する。可変軽鎖(VL)および可変重鎖(VH)という用語は、それぞれこれらの軽鎖および重鎖を言う。
【0034】
[0053] 用語「相補性決定領域」または「CDR」は、VLおよびVHの超可変領域を言う。CDRは、このような標的タンパク質に対する特異性を有する抗体鎖の標的タンパク質結合部位である。ヒトのVLまたはVHのそれぞれに3つのCDR(N末端から連続的に番号付けされたCDR1〜3)があり、可変ドメインの約15〜20%を構成する。CDRは、標的タンパク質のエピトープに対して構造的に相補性であり、したがって、結合特異性に直接的に関与する。VLまたはVHの残りのストレッチ、いわゆるフレームワーク領域は、アミノ酸配列にあまり変化を有さない(Kuby, Immunology,第4版, 第4章. W.H Freeman & Co., New York, 2000)。
【0035】
[0054] CDRおよびフレームワーク領域の位置は、当技術分野における様々な周知の定義、例えば、Kabat,Chothia,international ImMunoGeneTicsデータベース(IMGT)(ワールドワイドウェブのimgt.cines.fr/にある)およびAbM(例えば、Johnsonら, Nucleic Acids Res., 29:205-206(2001)、ChothiaおよびLesk, J. Mol. Biol., 196:901 -917(1987)、Chothiaら, Nature, 342:877-883(1989)、Chothiaら, J. Mol. Biol., 227:799-817(1992)、Al-Lazikaniら, J.Mol.Biol., 273:927-748(1997)を参照されたい)を用いて決定される。抗原結合部位の定義はまた、Ruizら、Nucleic Acids Res.、28:219〜221(2000);およびLefranc,M.P.、Nucleic Acids Res.、29:207〜209(2001);MacCallumら、J.Mol.Biol.、262:732〜745(1996);およびMartinら、Proc、Natl.Acad.Sci.USA、86:9268〜9272(1989);Martinら、Methods Enzymol.、203:121〜153(1991);およびReesら、In Sternberg M.J.E.(編)、Protein Structure Prediction、Oxford University Press、Oxford、141〜172(1996)において記載されている。
【0036】
[0055] 抗体は、例えば、無傷の免疫グロブリンとして、または様々なペプチダーゼでの消化によって生じる多くの良く特徴付けされたフラグメントとして存在する。したがって、例えば、ペプシンは、ヒンジ領域におけるジスルフィド連結の下で抗体を消化して、それ自体ジスルフィド結合によってVH−CH1に結合した軽鎖であるFabの二量体であるF(ab)’2を生成する。F(ab)’2は、ヒンジ領域におけるジスルフィド連結を破壊して、それによって、F(ab)’2二量体をFab’単量体に変換するための、穏やかな条件下で還元され得る。Fab’単量体は、原則的に、ヒンジ領域の一部を有するFabである(Fundamental Immunology(Paul編, 第3版, 1993)を参照されたい。様々な抗体フラグメントが無傷抗体の消化に関して規定されているが、当業者には、このようなフラグメントが、化学的にまたは組換えDNA法を用いて、デノボ合成され得ることが理解されよう。したがって、本明細書において用いられる抗体という用語はまた、抗体全体の修飾によって生じる抗体フラグメント、または組換えDNA法を用いてデノボ合成される抗体フラグメント(例えば、一本鎖Fv)、またはファージディスプレイライブラリーを用いて同定される抗体フラグメント(例えば、McCaffertyら, Nature 348:552-554(1990)を参照されたい)を包含する。
【0037】
[0056] したがって、抗体という用語はまた、ミニボディ、ダイアボディ、トリアボディ(tirabody)などを包含する。ダイアボディは、高い結合力および特異性を有する、小さな二価の生体特異的抗体フラグメントである。それらの高いシグナル対ノイズ比は、典型的には、特異的抗原の診断的標的化および治療的標的化についてのそれらの潜在能力を増大させる良好な特異性および速い血液クリアランスのため、より良好である(Sundaresanら, J Nucl Med 44:1962-9(2003)。さらに、これらの抗体は、必要に応じて、小さな一本鎖Fvから様々なアイソフォームを有する無傷のIgGまでにわたる、異なるタイプの抗体フラグメントとして操作され得るため、有利である(Wu & Senter, Nat.Biotechnol. 23:1137-1146(2005))。いくつかの実施形態において、抗体フラグメントは、ダイアボディの一部である。いくつかの実施形態において、本発明は、本発明に従って用いるための高い結合力の抗体を提供する。
【0038】
[0057] Hollingerら、PNAS(USA)90(14):6444〜6448(1993)によって最初に記載されたダイアボディは、本明細書において開示される重鎖および軽鎖を用いて、および本明細書において開示される個別のCDR領域を用いて構築され得る。典型的には、ダイアボディフラグメントは、同一鎖上の2つのドメインの間の対合を可能にするには短すぎるリンカーによって軽鎖可変ドメイン(VL)に連結している、重鎖可変ドメイン(VH)を含む。したがって、1つのフラグメントVHドメインおよびVLドメインは、別のフラグメントの相補的なVHドメインおよびVLドメインと対合させられ、それによって、2つの抗原結合部位を形成する。トリアボディは、同様に、3つの抗原結合部位で構築され得る。Fvフラグメントは、非共有結合による相互作用によって結びついているVLドメインおよびVHドメインを含む、完全な抗原結合部位を含有する。本発明によって包含されるFvフラグメントはまた、VHドメインおよびVLドメインがグルタルアルデヒド、分子間ジスルフィド、または他のリンカーを介して架橋されている構築物を包含する。重鎖および軽鎖の可変ドメインは、共に融合されて、親免疫グロブリンの元の特異性を維持している一本鎖可変フラグメント(scFv)を形成し得る。Gruberら、J.Immunol.152(12):5368〜74(1994)によって最初に記載された一本鎖Fv(scFv)二量体は、本明細書において開示される重鎖および軽鎖を用いて、および本明細書において開示される個別のCDR領域を用いて構築され得る。当技術分野において知られている多くの技術を、本発明の特異的結合構築物を調製するために用いることができる(全ての目的で、特にミニボディおよびダイアボディの設計に関する目的で、参照することによってその全体がそれぞれ本明細書に組み込まれる、米国特許出願公開第20070196274号および米国特許出願公開第20050163782号を参照されたい)。
【0039】
[0058] 二重特異性抗体は、化学的架橋またはハイブリッドハイブリドーマ技術によって生成することができる。あるいは、二重特異性抗体分子は、組換え技術によって生成することができる(二重特異性抗体を参照されたい)。二量体化は、VHドメインおよびVLドメインを結合するリンカーの長さを、scFvフラグメントを生成するために通常用いられる約15アミノ酸から約5アミノ酸まで低減することによって促進され得る。これらのリンカーは、VHドメインおよびVLドメインの鎖内組み立てに有利に働く。好適な短いリンカーはSGGGSであるが、他のリンカーを用いることができる。したがって、2つのフラグメントは、二量体分子に組み立てられる。リンカーの長さを0〜2アミノ酸にさらに低減させると、三量体分子(トリアボディ)または四量体分子(テトラボディ)が生成し得る。
【0040】
[0059] 抗体、例えば、組換え抗体、モノクローナル抗体または組換えポリクローナル抗体の調製には、当技術分野において知られている多くの技術を用いることができる(例えば、Kohler & Milstein, Nature 256:495-497(1975); Kozborら, Immunology Today 4:72(1983); Coleら, in Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., 77-96頁(1985); Coligan, Current Protocols in Immunology(1991); Harlow & Lane, Antibodies, A Laboratory Manual(1988);およびGoding, Monoclonal Antibodies: Principles and Practice(第2版, 1986)を参照されたい)。目的の抗体の重鎖および軽鎖をコードする遺伝子は細胞からクローニングすることができ、例えば、モノクローナル抗体をコードする遺伝子はハイブリドーマからクローニングすることができ、組換えモノクローナル抗体を生成するために用いることができる。モノクローナル抗体の重鎖および軽鎖をコードする遺伝子ライブラリーも、ハイブリドーマまたは形質細胞から作製することができる。重鎖および軽鎖の遺伝子産物のランダムな組み合わせは、異なる抗原特異性を有する大きな抗体プールをもたらす(例えば、Kuby, Immunology(第3版, 1997)を参照されたい)。一本鎖抗体または組換え抗体の生成のための技術(米国特許第4,946,778号、米国特許第4,816,567号)を、本発明のポリペプチドに対する抗体を生成するために適用することができる。また、トランスジェニックマウスまたは他の生物、例えば他の哺乳動物を、ヒト化抗体またはヒト抗体を発現させるために用いることができる(例えば、米国特許第5,545,807号、米国特許第5,545,806号、米国特許第5,569,825号、米国特許第5,625,126号、米国特許第5,633,425号、米国特許第5,661,016号、Marksら, Bio/Technology 10:779-783(1992); Lonbergら, Nature 368:856-859(1994); Morrison, Nature 368:812-13(1994); Fishwildら, Nature Biotechnology 14:845-51(1996);Neuberger, Nature Biotechnology 14:826(1996);およびLonberg & Huszar, Intern. Rev. Immunol. 13:65-93(1995)を参照されたい)。あるいは、ファージディスプレイ技術を用いて、選択された抗原に特異的に結合する抗体およびヘテロマーFabフラグメントを同定することができる(例えば、McCaffertyら, Nature 348:552-554(1990); Marksら, Biotechnology 10:779-783(1992)を参照されたい)。抗体はまた、二重特異性にすることができ、すなわち2つの異なる抗原を認識できるようにすることができる(例えば、WO93/08829、Trauneckerら, EMBO J. 10:3655-3659(1991);およびSureshら, Methods in Enzymology 121:210(1986)を参照されたい)。抗体はまた、ヘテロコンジュゲート、例えば共有結合によって結合した2つの抗体、または免疫毒素であり得る(例えば、米国特許第4,676,980号、WO91/00360、WO92/200373、およびEP03089を参照されたい)。
【0041】
[0060] 非ヒト抗体をヒト化または霊長類化するための方法は、当技術分野において周知である。通常、ヒト化抗体は、その中に非ヒトである由来源から導入された1つまたは複数のアミノ酸残基を有する。これらの非ヒトアミノ酸残基は移入(import)残基と呼ばれることが多く、この移入残基は、典型的には、移入可変ドメインから得られる。ヒト化は、原則的には、齧歯動物のCDRまたはCDR配列をヒト抗体の対応する配列で置換することによって、Winter and co−workersの方法に従って行うことができる(例えば、Jonesら, Nature 321:522-525(1986); Riechmannら, Nature 332:323-327(1988); Verhoeyenら, Science 239:1534-1536(1988)、およびPresta, Curr. Op. Struct. Biol. 2:593-596(1992)を参照されたい)。したがって、このようなヒト化抗体は、無傷のヒト可変ドメインほどには実質的には非ヒト種の対応する配列によって置換されていないキメラ抗体である(米国特許第4,816,567号)。実際には、ヒト化抗体は、典型的には、いくつかのCDR残基および場合によってはいくつかのFR残基が、齧歯動物抗体における類似の部位の残基によって置換されている、ヒト抗体である。
【0042】
[0061] 「キメラ抗体」は、(a)抗原結合部位(可変領域)が異なるまたは改変されたクラス、エフェクター機能、および/または種の定常領域に連結するかあるいは酵素、毒素、ホルモン、成長因子、薬剤などのキメラ抗体に新たな特徴を与える完全に異なる分子に連結するように、定常領域もしくはその一部が改変、置換、または交換されている、または(b)可変領域またはその一部が、異なるまたは改変された抗原特異性を有する可変領域で改変、置換、または交換されている、抗体分子である。
【0043】
[0062] 抗体に「特異的に(または選択的に)結合する」という表現、または「特異的に(または選択的に)免疫反応性である」という表現は、タンパク質またはペプチドについて言う場合、多くはタンパク質および他の生物製剤の異種集団における、タンパク質の存在の決定要因である結合反応を言う。したがって、指定した免疫アッセイ条件下において、特定の抗体は、バックグラウンドの少なくとも2倍、さらに典型的には、バックグラウンドの10〜100倍超、特定のタンパク質に結合する。このような条件下での抗体への特異的な結合には、特定のタンパク質への特異性について選択された抗体が必要である。例えば、ポリクローナル抗体は、選択された抗原と特異的に免疫反応性であるが他のタンパク質とはそうではないポリクローナル抗体のみを得るように選択することができる。この選択は、他の分子と交差反応する抗体を取り去ることによって達成され得る。様々な免疫アッセイ形式を用いて、特定のタンパク質と特異的に免疫反応性である抗体を選択することができる。例えば、固相ELISA免疫アッセイが、タンパク質と特異的に免疫反応性である抗体を選択するために通常用いられる(例えば、特異的免疫反応性を決定するために用いられ得る免疫アッセイの形式および条件の説明については、Harlow & Lane, Using Antibodies, A Laboratory Manual(1998)を参照されたい)。
【0044】
[0063] 例として、EMP2に対するポリクローナル抗体は、グルタチオン−S−トランスフェラーゼ−EMP2融合タンパク質を用いて得ることができる。該抗体は、グルタチオン−S−トランスフェラーゼ(GST)−EMP2融合タンパク質として構築された遺伝子の第1の細胞外領域(アミノ酸16〜64)に対して生成することができる。EMP2ペプチドは、以下のプライマー:CGCGGATCCTCTACCATTGACAATGCCTGG(フォワード:下線が引かれたBamHI)、CCGGAATTCTTACGCCTGCATCACAGAATAACC(リバース、下線が引かれたEcoRI)を用いてPCRによってクローニングすることができる。PCR産物は、GST遺伝子を含有するpGEX−4T−1ベクター(Pharmacia)のBamHI部位およびEcoRI部位内に直接的にクローニングすることができる。EMP2フラグメントは、GSTとインフレームでクローニングされて、融合タンパク質を生じさせる。インサートは、配列決定によって確認することができる。GST融合タンパク質は、従前に記載されているように生成することができる(Smith DBら, Gene 67:31-40(1988)を参照されたい)。log相の細菌(OD6000.6〜0.9)をイソプロピル−1−チオ−β−D−ガラクトピラノシド1mMと共に、37℃で2.5〜3時間にわたり誘導することができる。細菌を溶解し、可溶性画分をグルタチオン−セファロースカラム(Pierce, Rockford, IL)にロードする。カラムをリン酸緩衝生理食塩水(PBS)/EDTA10ベッド容量で洗浄する。融合タンパク質を、Tris−Cl50mM(pH8.0)内の還元型グルタチオン(Sigma, St Louis, MO)20mMを用いてカラムから溶出する。抗体の調製のために、ウサギをGST−EMP2融合タンパク質で2回免疫化し、最後の免疫化の2週間後から血清を回収する(Research Genetics, Huntsville, AL)。
【0045】
[0064] EMP2に対する完全にヒトのモノクローナル抗体は、組換えファージディスプレイ技術を用いてヒト抗体ファージディスプレイ遺伝子ライブラリーから生成することができる。ヒトEMP2に対するこのようなモノクローナル抗体は、診断目的で、およびPVRにおけるような網膜剥離の予防または治療のために用いることができる。
【0046】
[0065] 「核酸」は、一本鎖型または二本鎖型のデオキシリボヌクレオチドまたはリボヌクレオチドおよびそのポリマー、ならびにそれらの相補鎖(complement)を言う。この用語は、合成され、天然、および非天然であり、参照核酸と類似の結合特性を有し、かつ参照ヌクレオチドと類似の様式で代謝される、既知のヌクレオチド類似体または修飾された骨格残基もしくは連結を含有する核酸を包含する。このような類似体の例には、限定はしないが、ホスホロチオエート、ホスホルアミデート、メチルホスホネート、キラル−メチルホスホネート、2−O−メチルリボヌクレオチド、ペプチド−核酸(PNA)が包含される。
【0047】
[0066] 別段の指示がない限り、特定の核酸配列はまた、明確に示された配列と同様に、その保存的に修飾された変異体(例えば、縮重コドンの置換)および相補的な配列を暗黙的に包含する。具体的には、縮重コドンの置換は、1つまたは複数の選択された(または全ての)コドンの第3の位置が混合塩基および/またはデオキシイノシン残基で置換されている配列を生成することによって達成することができる(Batzerら, Nucleic Acid Res. 19:5081(1991); Ohtsukaら, J. Biol. Chem. 260:2605-2608(1985); Rossoliniら, Mol. Cell. Probes 8:91-98(1994))。核酸という用語は、遺伝子、cDNA、mRNA、オリゴヌクレオチド、およびポリヌクレオチドとほぼ同じ意味で用いられる。
【0048】
[0067] 特定の核酸配列はまた、「スプライス変異体」を暗黙的に包含する。同様に、核酸によってコードされる特定のタンパク質は、その核酸のスプライス変異体によってコードされる任意のタンパク質を暗黙的に包含する。「スプライス変異体」は、その名前が示唆するように、遺伝子の選択的スプライシングの産物である。転写の後、最初の核酸転写産物はスプライシングされ、その結果、異なる(選択的な)核酸スプライス産物は異なるポリペプチドをコードし得る。スプライス変異体を生成するためのメカニズムは変化するが、エキソンの選択的スプライシングを包含する。読み過し転写(read-through transcription)によって同一の核酸から誘導される別のポリペプチドもこの定義に包含される。スプライス産物の組換え型を包含する任意のスプライシング反応産物が、この定義に包含される。カリウムチャネルスプライス変異体の例が、Leicherら、J.Biol.Chem.273(52):35095〜35101(1998)において論じられている。
【0049】
[0068] 「siRNA」または「RNAi」は、二本鎖RNAを形成する核酸を言い、この二本鎖RNAは、siRNAが遺伝子または標的遺伝子と同一の細胞において発現している場合に、その遺伝子または標的遺伝子の発現を低減させるかまたは阻害する能力を有する。したがって、「siRNA」または「RNAi」は、相補鎖によって形成される二本鎖RNAを言う。ハイブリダイズして二本鎖分子を形成するsiRNAの相補的部分は、典型的には、実質的なまたは完全な同一性を有する。1つの実施形態において、siRNAは、標的遺伝子に対する実質的なまたは完全な同一性を有し、かつ二本鎖siRNAを形成する核酸を言う。典型的には、siRNAは、少なくとも約15〜50ヌクレオチド長である(例えば、二本鎖siRNAの各相補的配列は15〜50ヌクレオチド長であり、二本鎖siRNAは約15〜50塩基対長であり、好ましくは約20〜30塩基のヌクレオチド、好ましくは約20〜25または約24〜29ヌクレオチド長、例えば、20、21、22、23、24、25、26、27、28、29、または30ヌクレオチド長である。
【0050】
[0069] siRNA分子およびベクターの設計および作製は、当業者に周知である。例えば、好適なsiRNAを設計するための効率的なプロセスは、mRNA転写産物のAUG開始コドンで開始して(例えば、図5を参照されたい)、AAジヌクレオチド配列についてスキャンすることである(Elbashirら, EMBO J 20:6877-6888(2001)を参照されたい)。AAおよび3’隣接ヌクレオチドはそれぞれ、潜在的なsiRNA標的部位である。隣接部位配列の長さは、siRNAの長さを決定する。例えば、19の隣接部位は、21ヌクレオチド長のsiRNAをもたらす。3’オーバーハング(overhanging)UUジヌクレオチドを有するsiRNAは、最も効果的であることが多い。このアプローチはまた、ヘアピンsiRNAを転写するためのRNA pol IIIの使用に適合可能である。RNA pol IIIは、4〜6ヌクレオチドのポリ(T)配列で転写を終結させて、短いポリ(U)尾部を有するRNA分子を生じさせる。しかし、他の3’末端のジヌクレオチドオーバーハングを有するsiRNAもまた、RNAiを効果的に誘導し得、配列は経験的に選択され得る。選択性のために、他のコード配列に対して相同性を有する16〜17を超える連続した塩基対を有する標的配列が、BLASTサーチ(www.ncbi.nlm.nih.gov/BLASTを参照されたい)を行うことによって避けられ得る。
【0051】
[0070] siRNAを直接投与するか、またはsiRNA発現ベクターを用いてRNAiを誘導することができる。ベクターは、短いスペーサー配列によって分離されており、かつ転写を終結させることを助けるTストリングで終わる、挿入された2つの逆反復を有し得る。発現したRNA転写産物は、短いヘアピンsiRNAにフォールディングすると予測される。siRNAの標的配列、推定ヘアピンの柄部をコードする逆反復の長さ、逆反復の順序、ヘアピンのループをコードするスペーサー配列の長さおよび組成、ならびに5’−オーバーハングの存在または非存在の選択は、変化し得る。siRNA発現カセットの好ましい順序は、センス鎖、短いスペーサー、およびアンチセンス鎖である。これらの様々な柄部長さ(例えば、15〜30)を有するヘアピンsiRNAが適している。ヘアピンsiRNAのセンス鎖とアンチセンス鎖とを連結するループの長さは、様々な長さであり得る(例えば、3〜9ヌクレオチド、またはそれ以上の長さ)。ベクターは、プロモーターおよび発現エンハンサー、またはsiRNAをコードするヌクレオチド配列に作動可能に連結している他の調節エレメントを含有し得る。
【0052】
[0071] 「制御配列」という表現は、特定の宿主生物における作動可能に連結しているコード配列の発現に必要なDNA配列を言う。原核生物に好適な制御配列には、例えば、プロモーター、任意選択によりオペレーター配列、およびリボソーム結合部位が包含される。真核細胞は、プロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが知られている。これらの制御エレメントは、調節エレメントがそれに対して応答性である外部因子を付加および制御することで臨床医が遺伝子の発現をオフまたはオンにすることができるように設計され得る。
【0053】
[0072] 核酸は、別の核酸配列と機能的関係に置かれているとき、「作動可能に連結している」。例えば、プレ配列もしくは分泌リーダーのDNAは、ポリペプチドの分泌に関与するプレタンパク質として発現している場合、ポリペプチドのDNAに作動可能に連結しており、プロモーターもしくはエンハンサーは、配列の転写に影響する場合、コード配列に作動可能に連結し、あるいはリボソーム結合部位は、翻訳を促進するように位置している場合、コード配列に作動可能に連結している。通常、「作動可能に連結している」は、連結しているDNA配列が互いに近くにあることを意味し、分泌リーダーの場合では、連続しかつリーディング相にあることを意味する。しかし、エンハンサーは連続している必要はない。連結は、好都合な制限部位でのライゲーションによって達成される。このような部位が存在しない場合は、合成オリゴヌクレオチドアダプターまたは合成オリゴヌクレオチドリンカーが、従来の慣例に従って用いられる。
【0054】
[0073] 「保存的に修飾された変異体」は、アミノ酸および核酸配列の両方に適用される。特定の核酸配列に関して、保存的に修飾された変異体は、同一もしくは実質的に同一のアミノ酸配列をコードする核酸を言うか、核酸がアミノ酸配列をコードしていない場合、本質的に同一な配列を言う。遺伝子コードの縮重によって、多数の機能的に同一の核酸は任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUは全て、アミノ酸アラニンをコードする。したがって、アラニンがコドンによって特定されている全ての位置において、コドンは、コードされるポリペプチドを改変することなく、記載された対応するコドンのいずれかに改変され得る。このような核酸の変異は、一種の保存的に修飾された変異である「サイレント変異」である。ポリペプチドをコードする本明細書における全ての核酸配列は、核酸の全ての考えられるサイレント変異も表す。当業者には、核酸における各コドン(通常はメチオニンの唯一のコドンであるAUGと、通常はトリプトファンの唯一のコドンであるTGGとを除く)が修飾されて、機能的に同一の分子がもたらされ得ることが認識されよう。したがって、ポリペプチドをコードする核酸の各サイレント変異は、それぞれの記載されている配列において、発現産物に関しては黙示的であるが、実際のプローブ配列に関しては黙示的ではない。
【0055】
[0074] EMP2ポリヌクレオチドおよびEMP2ポリペプチドを包含する2つ以上の核酸配列またはポリペプチド配列の状況における、用語「同一」またはパーセント「同一性」は、以下に記載されるデフォルトパラメータを有するBLASTまたはBLAST2.0配列比較アルゴリズムを用いて測定され、あるいは手動のアラインメントおよび目視検査によって測定されるときに(例えば、NCBIウェブサイトhttp://www.ncbi.nlm.nih.gov/BLAST/などを参照されたい)、同一であるか、あるいは特定のパーセンテージの同一のアミノ酸残基もしくはヌクレオチド(すなわち、比較窓または指定領域にわたり最大の対応となるよう比較およびアラインしたときに、特定の領域にわたって約60%の同一性、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、またはそれを超える同一性)を有する、2つ以上の配列または部分配列を言う。そして、このような配列は、「実質的に同一である」と言われる。この定義はまた、試験配列の相補鎖を言うか、試験配列の相補鎖に適用することができる。この定義には、欠失および/または付加を有する配列、ならびに置換を有する配列も包含される。以下に記載されるように、好ましいアルゴリズムは、ギャップなどを明らかにし得る。好ましくは、同一性は、少なくとも約25アミノ酸長もしくはヌクレオチド長の領域にわたって存在し、より好ましくは50〜100アミノ酸長もしくはヌクレオチド長の領域にわたって存在する。
【0056】
[0075] 配列比較では、典型的には、ある配列を参照配列とし、これに対して試験配列が比較される。配列比較アルゴリズムを用いる場合、試験配列および参照配列は、コンピュータに入れられ、必要に応じて部分配列の座標が指定され、配列アルゴリズムプログラムのパラメータが指定される。好ましくは、デフォルトのプログラムパラメータを用いることができ、または別のパラメータを指定することができる。次に、配列比較アルゴリズムは、プログラムパラメータに基づいて参照配列に対して試験配列の配列同一性パーセントを計算する。
【0057】
[0076] 本明細書において用いられる「比較窓(comparison window)」には、参照配列の20から完全長までからなる群から選択される連続位置数のいずれか1つ、通常は約25〜100、または50〜約150、さらに通常は約100〜約150を有するセグメントであって、ある配列と同一の連続位置数を有する参照配列とを当該2つの配列が最適にアラインされた後に比較し得るセグメントへの言及が包含される。比較のための配列アラインメントの方法は、当技術分野において周知である。比較のための配列の最適なアラインメントは、例えば、Smith & Waterman、Adv.Appl.Math.2:482(1981)の局所的相同性アルゴリズムによって、Needleman & Wunsch、J.Mol.Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、Pearson & Lipman、Proc.Nat’l.Acad.Sci.USA 85:2444(1988)の類似法のためのサーチによって、これらのアルゴリズムのコンピュータ化された実行(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI におけるGAP、BESTFIT、FASTA、およびTFASTA)によって、あるいは手動のアラインメントおよび目視検査(例えば、Current Protocols in Molecular Biology(Ausubelら編、1995補完)を参照されたい)によって行うことができる。
【0058】
[0077] 配列同一性パーセントおよび配列類似性の決定に好適なアルゴリズムの好ましい例は、BLASTアルゴリズムおよびBLAST2.0アルゴリズムであり、これは、Altschulら、Nuc.Acids Res.25:3389〜3402(1977)およびAltschulら、J.Mol.Biol.215:403〜410(1990)にそれぞれ記載されている。BLASTおよびBLAST2.0は、本明細書において記載されるパラメータと共に用いられて、本発明の核酸およびタンパク質の配列同一性パーセントを決定する。BLAST分析を行うためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に利用可能である。このアルゴリズムは、データベース配列における同一の長さの文字とアラインされた場合にいくつかの正値閾値スコアTに適合するかまたはこれを満たす、検索配列における長さWの短い文字を同定することによって、高スコア配列対(HSP)を最初に同定することを伴う。Tは、隣接文字スコア閾値(上記のAltschulら)と呼ばれる。これらの最初の隣接文字ヒットは、それを含むさらに長いHSPを発見するためのサーチを開始するきっかけとして作用する。文字ヒットは、累積アラインメントスコアが増大し得る限り、各配列に沿って両方向に伸長する。累積スコアは、ヌクレオチド配列では、パラメータM(適合(matching)残基対についてのリワードスコア、常に>0)およびN(ミスマッチ残基についてのペナルティースコア、常に<0)を用いて計算される。アミノ酸配列では、スコアリングマトリクスを用いて累積スコアを計算する。各方向での文字ヒットの伸長は、累積アラインメントスコアが量Xによってその最大達成値から減少する場合、1つまたは複数の負のスコアリングの残基のアラインメントの蓄積に起因して累積スコアがゼロ以下に向かう場合、またはいずれかの配列の末端に達する場合、停止する。BLASTアルゴリズムのパラメータW、T、およびXは、アラインメントの感度および速さを決定する。BLASTNプログラム(ヌクレオチド配列のための)は、デフォルトとして11の文字長(W)、10の期待値(E)、M=5、N=−4、および両鎖の比較を用いる。アミノ酸配列では、BLASTPプログラムは、デフォルトとして、3の文字長および10の期待値(E)を用い、BLOSUM62スコアリングマトリクス(Henikoff & Henikoff, Proc. Natl. Acad. Sci. USA 89:10915(1989)を参照されたい)は、50のアラインメント(B)、10の期待値(E)、M=5、N=−4、および両鎖の比較を用いる。
【0059】
[0078] 用語「非相同」は、核酸の一部に関して用いられる場合、核酸が、天然では互いの同一の関連が見られない2つ以上の部分配列を含むことを示す。例えば、新たな機能的核酸を作製するように配置された、関連しない遺伝子に由来する2つ以上の配列を有する核酸、例えば、1つの由来源に由来するプロモーターおよび別の由来源に由来するコード領域を有する核酸は、典型的には、組換えによって生成される。同様に、非相同タンパク質は、タンパク質が、天然では互いの同一の関連が見られない2つ以上の部分配列を含むことを示す(例えば、融合タンパク質)。
【0060】
[0079] 表現「ストリンジェントなハイブリダイゼーション条件」は、プローブが典型的には核酸の複雑な混合物においてその標的部分配列にハイブリダイズするが、他の配列にはハイブリダイズしない条件を言う。ストリンジェントな条件は、配列依存性であり、異なる状況で異なる。より長い配列は、より高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションに対するさらなる指針は、Tijssen、Techniques in Biochemistry and Molecular Biology − Hybridization with Nucleic Probes、「Overview of principles of hybridization and the strategy of nucleic acid assays」(1993)において見られる。通常、ストリンジェントな条件は、規定されたイオン強度pHでの特異的配列の熱融点(Tm)よりも約5〜10℃低くなるように選択される。Tmは、(規定されたイオン強度、pH、および核酸濃度下で)平衡状態で、標的に相補的なプローブの50%が標的配列にハイブリダイズする(標的配列は過剰に存在するため、Tmでは、平衡状態でプローブの50%が塞がれている)温度である。ストリンジェントな条件はまた、ホルムアミドなどの不安定化剤を添加して達成され得る。選択的または特異的なハイブリダイゼーションでは、正のシグナルは、バックグラウンドの少なくとも2倍であり、好ましくはバックグラウンドハイブリダイゼーションの10倍である。典型的なストリンジェントなハイブリダイゼーション条件は、50%ホルムアミド、5×SSC、および1%SDS、42℃でのインキュベーション、あるいは、5×SSC、1%SDS、65℃でのインキュベーションと、65℃での0.2×SSCおよび0.1%SDSでの洗浄とであり得る。
【0061】
[0080] ストリンジェントな条件下で互いにハイブリダイズしない核酸は、これらの核酸がコードするポリペプチドが実質的に同一であれば、依然として実質的に同一である。これは、例えば、核酸のコピーが遺伝子コードによって認められている最大のコドン縮重を用いて生成する場合に生じる。このようなケースでは、核酸は、典型的には、中度にストリンジェントなハイブリダイゼーション条件下でハイブリダイズする。典型的な「中度にストリンジェントなハイブリダイゼーション条件」には、37℃での、40%ホルムアミド、NaCl1M、1%SDSからなる緩衝液でのハイブリダイゼーション、および45℃での1×SSCにおける洗浄が包含される。正のハイブリダイゼーションは、バックグラウンドの少なくとも2倍である。当業者は、別のハイブリダイゼーション条件および洗浄条件を用いて類似のストリンジェンシーの条件を与え得ることを容易に認識されよう。ハイブリダイゼーションパラメータを決定するためのさらなるガイドラインは、多くの参考文献、例えば、Current Protocols in Molecular Biology、Ausubelら編、John Wiley & Sonsにおいて提供されている。
【0062】
[0081] PCRにおいて、低いストリンジェンシーでの増幅には約36℃の温度が典型的であるが、アニーリング温度は、プライマーの長さに応じて約32℃〜48℃の間で変化し得る。高いストリンジェンシーのPCR増幅では約62℃の温度が典型的であるが、高いストリンジェンシーでのアニーリングの温度は、プライマーの長さおよび特異性に応じて約50℃〜約65℃の範囲であり得る。高いストリンジェンシーでの増幅および低いストリンジェンシーでの増幅の両方にとって典型的なサイクル条件には、90℃〜95℃で30秒間〜2分間にわたる変性相、30秒間〜2分間続くアニーリング相、および約72℃で1〜2分間にわたる伸長相が包含される。低いストリンジェンシーでの増幅反応および高いストリンジェンシーでの増幅反応のためのプロトコルおよびガイドラインは、例えば、Innisら(1990)PCR Protocols、A Guide to Methods and Applications、Academic Press,Inc.N.Y.)において提供されている。
【0063】
[0082] 本発明に従ったEMP2抗体またはEMP2ポリペプチドには、標識または検出可能な部分が結合していてよい。「標識」または「検出可能な部分」は、分光学的手段、光化学的手段、生化学的手段、免疫化学的手段、化学的手段、または他の物理的手段によって検出可能な組成物である。例えば、有用な標識には、32P、蛍光染料、電子密度試薬、酵素(例えば、ELISAで一般に用いられるもの)、ビオチン、ジゴキシゲニン、または例えば放射性標識をペプチドに組み込むことによって検出可能であるか、あるいはペプチドと特異的に反応する抗体を検出するために用いることができるハプテンおよびタンパク質が包含される。
【0064】
[0083] 用語「組換え」は、例えば細胞、または核酸、タンパク質、もしくはベクターを参照して用いられる場合、細胞、核酸、タンパク質、またはベクターが、非相同な核酸もしくはタンパク質の導入またはネイティブな核酸もしくはタンパク質の改変によって修飾されていることを示すか、あるいは細胞がそのように修飾された細胞から誘導されていることを示す。したがって、例えば、組換え細胞は、ネイティブ(非組換え)型の細胞では見られない遺伝子を発現するか、あるいは異常に発現する、低発現である、または全く発現されないネイティブな遺伝子を発現する。
【0065】
[0084] 「生体試料」には、生検試料および剖検試料などの組織の切片、ならびに組織学的目的で採取された凍結切片が包含される。このような試料には、産物の血液および血液画分(例えば、血清、血漿、血小板、赤血球など)、痰、組織、培養細胞、例えば、一次培養物、外植片、および形質転換細胞、便、尿などが包含される。生体試料は、典型的には、真核生物から、最も好ましくは哺乳動物、例えば霊長類、例えばチンパンジーもしくはヒト、ウシ、イヌ、ネコ、齧歯動物、例えば、モルモット、ラット、マウス、ウサギ、トリ、爬虫類、または魚から得られる。
【0066】
[0085] 用語「治療する」または「治療」には、
(1)疾患を予防すること、すなわち、生物に曝露され得るがまだその疾患の症候を経験していないか示していない哺乳動物において、疾患の臨床症候が発症しないようにすること、
(2)疾患を阻害すること、すなわち、疾患またはその臨床症候の発症を止めるか低減させること(これには、観察された剥離の程度または対象の数もしくは対象が剥離を有するリスクを低減させることが包含される)、および/または
(3)疾患を軽減すること、すなわち、疾患またはその臨床症候を退行させること
が包含される。
【0067】
[0086] 本明細書における「治療上有効な用量または量」は、投与の目的である効果をもたらす用量を意味する。正確な用量および製剤は治療の目的に応じ、当業者は既知の技術を用いて認識することができる(例えば、Lieberman, Pharmaceutical Dosage Forms(第1-3巻, 1992); Lloyd, The Art, Science and Technology of Pharmaceutical Compounding(1999); Remington: The Science and Practice of Pharmacy, 第20版、Gennaro, Editor(2003),およびPickar, Dosage Calculations(1999)を参照されたい)。
【0068】
[0087] 「患者」または「対象」は、ヒトおよび非ヒト動物、例えば哺乳動物(例えば、霊長類、ウシ、ウマ、ネコ、イヌ、ウサギ、マウス、およびラット)を言う。したがって、本発明の方法は、ヒトの治療法および獣医学上の適用の両方に適用可能である。
【0069】
[0088] 「薬学的に許容可能な塩」は、投与経路に従って、比較的無毒の酸または塩基と共に調製される活性化合物の塩を包含することを意味する。本発明の阻害剤が比較的酸性の官能基を含有する場合、塩基付加塩は、中性形態のこのような化合物をニートのまたは好適な不活性溶媒中で十分な量の所望の塩基と接触させることによって得ることができる。薬学的に許容可能な塩基付加塩の例には、ナトリウム塩、カリウム塩、カルシウム塩、アンモニウム塩、有機アミノ塩、またはマグネシウム塩、または類似の塩が包含される。本発明の化合物が比較的塩基性の官能基を含有する場合、酸付加塩は、中性形態のこのような化合物をニートのまたは好適な不活性溶媒中で十分な量の所望の酸と接触させることによって得ることができる。薬学的に許容可能な酸付加塩の例には、塩酸、臭化水素酸、硝酸、炭酸、一水素炭酸、リン酸、一水素リン酸、二水素リン酸、硫酸、一水素硫酸、ヨウ化水素酸、または亜リン酸などのような、無機酸から誘導される塩、および、酢酸、プロピオン酸、イソ酪酸、マレイン酸、マロン酸、安息香酸、コハク酸、スベリン酸、フマル酸、乳酸、マンデル酸、フタル酸、ベンゼンスルホン酸、p−トリルスルホン酸、クエン酸、酒石酸、メタンスルホン酸などのような、比較的無毒の有機酸から誘導される塩が包含される。また、アルギン酸塩などのアミノ酸の塩、およびグルクロン酸またはガラクツロン酸などのような有機酸の塩も包含される(例えば、Bergeら, Journal of Pharmaceutical Science 66:1-19(1977)を参照されたい)。特定の具体的な本発明の化合物は、塩基性官能基および酸性官能基の両方を含有し、それにより、化合物が塩基付加塩または酸付加塩に変換されることを可能にする。
【0070】
[0089] 中性形態の化合物は、従来の様式で塩と塩基または酸とを接触させ、親化合物を単離することによって、再生成することができる。親形態の化合物は、極性溶媒における可溶性などの特定の物理的特性において様々な塩形態と異なるが、その他の点では、塩は、本発明の目的では親形態の化合物に等しい。
【0071】
[0090] 塩形態に加えて、本発明は、プロドラッグ形態の化合物を提供する。本明細書において記載される化合物のプロドラッグは、生理学的条件下で化学的変化を容易に受けて本発明の化合物をもたらす化合物である。さらに、プロドラッグは、エクスビボ環境で、化学的または生化学的方法によって、本発明の化合物に変換され得る。例えば、プロドラッグは、好適な酵素試薬または化学試薬を有する経皮パッチ容器内にある場合、本発明の化合物に穏やかに変換され得る。
【0072】
[0091] 本発明の特定の化合物は、非溶媒和形態、および水和形態を包含する溶媒和形態で存在し得る。通常、溶媒和形態は非溶媒和形態と同等であり、本発明の範囲内に包含されることが意図される。本発明の特定の化合物は、複数の結晶形態または非晶質形態で存在し得る。通常、全ての物理的形態は、本発明によって考慮される使用にとって同等であり、本発明の範囲内にあることが意図される。
【0073】
[0092] 核酸およびポリペプチドなどのバイオポリマーとは別に、本発明の特定の化合物は、非対称の炭素原子(光学中心)または二重結合を有し、ラセミ体、ジアステレオマー、幾何異性体、および個別の異性体は全て、本発明の範囲内に包含されることが意図される。化合物がアミノ酸または核酸を含む好ましい実施形態において、アミノ酸および核酸はそれぞれ、主な天然の生物学的鏡像異性体である。
【0074】
[0093] 本明細書において用いられる用語「担体」は、典型的には、インビボまたはインビトロの生物学的系に適用される活性剤のための希釈剤または媒体として用いられる不活性物質を言う(例えば、治療剤などの薬剤)。この用語はまた、組成物に凝集性を付与する、典型的には不活性物質を包含する。
【0075】
II.抗EMP2組成物
[0094] 本発明は、EMP2ポリペプチドの活性を阻害するための組成物を提供し、前記組成物は、対象の眼における増殖性硝子体網膜症または網膜剥離を治療する方法において用いられる。特定の理論に縛られることはないが、EMP2は、網膜色素上皮細胞において網膜剥離および失明をもたらし得る細胞の収縮を正に調節すると考えられている。したがって、本発明は、抗EMP2剤を提供することによって細胞の収縮および網膜剥離を予防および/または治療する方法を提供する。
【0076】
[0095] いくつかの実施形態において、抗EMP2剤は、抗EMP2抗体またはそのフラグメントを含む。いくつかの実施形態において、抗EMP2剤は、EMP2阻害性オリゴヌクレオチド、例えば、抗EMP2siRNAまたは抗EMP2リボザイムを含む。
【0077】
A.抗EMP2抗体
[0096] 本発明は、ヒトEMP2ポリペプチド(KS49, KS83)およびマウスEMP2ポリペプチド(KS83)に特異的な高結合力抗体をコードする抗体配列を同定する。同定された配列は、画像化、診断方法、またはインビボ療法のための、組換え抗EMP2抗体(例えば、モノクローナル抗体、ポリクローナル抗体、ミニボディ、ダイアボディ、およびトリアボディ)またはそのフラグメントの生成に有用である
【0078】
[0097] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、配列番号1、配列番号2、または配列番号30に実質的に同一な(すなわち、少なくとも約15、20、25、50、100、200、500、1000、またはそれを超えるアミノ酸の領域にわたり、約60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%を超える、またはそれを超えるアミノ酸配列同一性を有する)EMP2ポリペプチドに特異的に結合し得るものである。いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、配列番号1、配列番号2、または配列番号30のアミノ酸配列を有するEMP2ポリペプチドに特異的に結合し得る。
【0079】
[0098] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS49重鎖(配列番号4)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS49軽鎖(配列番号5)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号4のアミノ酸配列を有する重鎖および/または配列番号5のアミノ酸配列を有する軽鎖を含む。
【0080】
[0099] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS83重鎖(配列番号6)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS83軽鎖(配列番号7)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号6のアミノ酸配列を有する重鎖および/または配列番号7のアミノ酸配列を有する軽鎖を含む。
【0081】
[0100] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS41重鎖(配列番号8)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS41軽鎖(配列番号9)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号8のアミノ酸配列を有する重鎖および/または配列番号9のアミノ酸配列を有する軽鎖を含む。
【0082】
[0101] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS89重鎖(配列番号10)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS89軽鎖(配列番号11)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号10のアミノ酸配列を有する重鎖および/または配列番号11のアミノ酸配列を有する軽鎖を含む。
【0083】
[0102] いくつかの実施形態において、本発明は、KS49、KS83、KS41、およびKS89から選択される抗体のCDR領域を含む抗EMP2配列を提供する。本発明によって提供されるCDR領域は、限定はしないが抗体、scFv、トリアボディ、ダイアボディ、ミニボディなどを包含する、抗EMP−2抗体またはそのフラグメントを構築するために用いることができる。いくつかの実施形態において、本発明の抗EMP−2結合タンパク質は、KS49、KS83、KS41、およびKS89から選択される抗体の少なくとも1つのCDR領域を含む。抗EMP−2結合タンパク質は、本明細書において提供される抗体の、例えばCDR−H1、CDR−H2、CDR−H3、CDR−L1、CDR−L2、CDR−L3、またはその組み合わせを含み得る。本発明のいくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、本明細書において提供される抗体の3つ全てのCDR−H配列、本明細書において提供される抗体の3つ全てのCDR−L配列、またはその両方を含み得る。抗EMP2CDR配列は、抗体骨格またはそのフラグメント上で用いることができ、同様に、ヒト化抗体、またはヒト化配列を含有する抗体を含み得る。これらの抗体は、例えば、EMP2を検出するため、インビボでEMP−2を発現する細胞を検出するため、またはEMP2機能を妨げるために用いることができる。いくつかの実施形態において、CDR領域は、Kabatの定義、Chothiaの定義、AbMの定義、接触の定義、または任意の他の好適なCDR番号付け系を用いて規定され得る。
【0084】
[0103] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、SYAMH(配列番号12)およびEYPMH(配列番号22)からなる群から選択されるアミノ酸配列を有するCDR−H1、VISYDGSNKYYADSVKG(配列番号13)およびVISYDGEYQKYADSVKG(配列番号23)からなる群から選択されるアミノ酸配列を有するCDR−H2、ならびにDRRGRKSAGIDY(配列番号14)、TVGATGAFDI(配列番号18)、およびTINNGMDV(配列番号24)からなる群から選択されるアミノ酸を有するCDR−H3を含む重鎖領域を含む。
【0085】
[0104] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、QASQDISNYLN(配列番号15)、RASQSIGKWLA(配列番号19)、およびRASQGIRNDLG(配列番号25)からなる群から選択されるアミノ酸配列を有するCDR−L1、AASSLQS(配列番号16)、KASSLEG(配列番号20)、およびGASSLQS(配列番号26)からなる群から選択されるアミノ酸配列を有するCDR−L2、ならびにLQDYNGWT(配列番号17)およびQQSHNFPPT(配列番号21)からなる群から選択されるアミノ酸配列を有するCDR−L3を含む軽鎖領域を含む。
【0086】
[0105] いくつかの実施形態において、EMP2ポリペプチドに特異的に結合し得る抗EMP2抗体またはそのフラグメントは、配列番号27〜29の抗体に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%のアミノ酸配列同一性を有するアミノ酸配列を含む。
【0087】
B.抗EMP2オリゴヌクレオチド
[0106] いくつかの実施形態において、EMP2の活性は、抗EMP2オリゴヌクレオチドを投与することによって阻害される。いくつかの実施形態において、抗EMP2オリゴヌクレオチドはEMP2siRNAであり、該EMP2siRNAは、EMP2ポリペプチドをコードするポリヌクレオチドの発現を低減できる。いくつかの実施形態において、抗EMP2オリゴヌクレオチドはEMP2リボザイムであり、EMP2リボザイムは、細胞におけるEMP2の発現を阻害し得る。
【0088】
[0107] いくつかの実施形態において、EMP2siRNAは、配列番号3の核酸配列またはその相補鎖に対する実質的な同一性を有する(すなわち、約60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%を超える、またはそれを超えるヌクレオチド配列同一性を有する)ポリヌクレオチドに特異的にハイブリダイズするオリゴヌクレオチドを含む。いくつかの実施形態において、EMP2siRNAは、配列番号3の核酸配列またはその相補鎖の少なくとも15、18、20、25、30、35、40、45、50、またはそれを超える連続するヌクレオチドに実質的に同一な核酸配列を含む。
【0089】
[0108] いくつかの実施形態において、EMP2阻害剤は、EMP2リボザイムである。リボザイムは、RNAの特異的切断を触媒し得る酵素的RNA分子である。リボザイムの作用のメカニズムは、相補的な標的RNAへのリボザイム分子の配列特異的ハイブリダイゼーションと、その後のエンドヌクレアーゼ切断とを伴う。配列番号3の核酸配列に対して実質的な同一性を有する(すなわち、約60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%を超える、またはそれを超えるヌクレオチド配列同一性を有する)mRNAを包含するEMP2mRNAのエンドヌクレアーゼ切断を特異的および効率的に触媒する工学操作されたハンマーヘッドモチーフリボザイム分子は、本発明の範囲内である。
【0090】
[0109] 任意の潜在的なRNA標的内の特異的リボザイム切断部位は、配列GUA、GUU、およびGUCを包むリボザイム切断部位の標的分子をスキャンすることによって最初に同定される。同定されると、切断部位を含有する標的遺伝子の領域に対応する15〜20リボヌクレオチドの間の短いRNA配列は、オリゴヌクレオチド配列を不適切にし得る二次構造などの予測される構造的特徴について評価し得る。
【0091】
[0110] 本発明のアンチセンスRNA分子およびアンチセンスDNA分子の両方ならびにリボザイムは、RNA分子を合成するための当技術分野において知られている任意の方法によって調製することができる。これらには、例えば固相ホスホラミダイト化学合成などの当技術分野において周知のオリゴデオキシリボヌクレオチドを化学的に合成するための技術が包含される。あるいは、RNA分子は、アンチセンスRNA分子をコードするDNA配列をインビトロおよびインビボで転写することによって生成することができる。このようなDNA配列は、T7ポリメラーゼプロモーターまたはSP6ポリメラーゼプロモーターなどの好適なRNAポリメラーゼプロモーターを組み込む様々なベクター内に組み込むことができる。リボザイムを作製する方法は、当技術分野において周知である(例えば、米国特許出願公開第20060062785号を参照されたい)。
【0092】
[0111] 所望のEMP2siRNA配列またはEMP2リボザイム配列および制御配列を含有するEMP2siRNAまたはEMP2リボザイムに好適なベクターの構築は、当技術分野において良く理解されている標準的なライゲーション技術および制限技術を採用する(Maniatisら, in Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York(1982)を参照されたい)。単離されたプラスミド、DNA配列、または合成されたオリゴヌクレオチドは、所望の形態に切断、調整、およびライゲーションされる。
【0093】
C.製剤
[0112] 本明細書において記載される治療方法に従って投与するための組成物は、一般に、薬学的に許容可能な担体、好ましくは水性担体に溶解した本明細書において記載される抗EMP2剤を含む。例えば緩衝生理食塩水などの、様々な水性担体を用いることができる。これらの溶液は無菌であり、望ましくない物質を全体として含まない。これらの組成物は、従来の周知の滅菌技術によって滅菌することができる。組成物は、好適な生理学的条件に応じて、薬学的に許容可能な補助物質、例えば、pH調整剤およびpH緩衝剤、毒性調整剤など、例えば、酢酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、乳酸ナトリウムなどを含有し得る。これらの製剤における活性剤の濃度は広く変化し得、選択された特定の投与様式および患者の要求に応じて、流体の容量、粘度、体重などに主に基づいて選択される。
【0094】
[0113] 本発明において用いるための好適な製剤は、参照することによって本明細書に組み込まれる、Remington:The Science and Practice of Pharmacy、第20版、Gennaro,Editor(2003)において見られる。さらに、薬剤送達のための方法の簡単な概説については、参照することによって本明細書に組み込まれる、Langer、Science 249:1527〜1533(1990)を参照されたい。本明細書において記載される医薬組成物は、当業者に知られている様式で、すなわち、従来の混合プロセス、溶解プロセス、粒子化プロセス、糖衣形成プロセス、粉末化プロセス、乳化プロセス、カプセル化プロセス、封入プロセス、または凍結乾燥プロセスによって製造することができる。以下の方法および賦形剤は典型的なものにすぎず、限定するものでは全くない。
【0095】
[0114] 注入では、本発明の化合物は、水溶液内で、好ましくは、ハンクス液、リンゲル液、または緩衝生理食塩水などの生理学的に適合可能な緩衝液内で製剤化することができる。経粘膜投与では、浸透されるバリアに好適な浸透剤が製剤において用いられる。このような浸透剤は、一般に、当技術分野において知られている。
【0096】
[0115] 経口投与では、本発明に従って用いるための阻害剤は、当技術分野において周知の薬学的に許容可能な担体と組み合わせることによって容易に製剤化することができる。このような担体は、化合物を、治療する患者による経口摂取のための錠剤、ピル、糖衣、カプセル、エマルジョン、親油性懸濁液および親水性懸濁液、液体、ゲル、シロップ、スラリー、懸濁液などとして製剤化することを可能にする。経口使用のための医薬製剤は、化合物を固体賦形剤と混合し、得られた混合物を任意選択により粉砕し、必要に応じて、好適な補助剤を添加した後に顆粒の混合物を加工して錠剤または糖衣の核を得ることによって得ることができる。好適な賦形剤は、特に、充填剤、例えば、乳糖、ショ糖、マンニトール、またはソルビトールを包含する糖;セルロース製剤、例えばトウモロコシデンプン、コムギデンプン、コメデンプン、ジャガイモデンプン、ゼラチン、トラガカントゴム、メチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースナトリウム、および/またはポリビニルピロリドン(PVP)などである。必要であれば、架橋型ポリビニルピロリドン、寒天、またはアルギン酸もしくはその塩、例えばアルギン酸ナトリウムなどの崩壊剤を添加することができる。
【0097】
[0116] 糖衣核は、好適な被覆を有する。この目的では、濃縮された糖溶液を用いることができ、これは、アラビアゴム、タルク、ポリビニルピロリドン、カルボポールゲル、ポリエチレングリコール、および/または二酸化チタン、ラッカー溶液、ならびに好適な有機溶媒または溶媒混合物を任意選択により含有し得る。活性化合物用量の異なる組み合わせを同定または特徴付けするために、染料または顔料を錠剤または糖衣の被覆に添加することができる。
【0098】
[0117] 経口で用いることができる医薬製剤には、ゼラチンで作製された押し込み型(push-fit)カプセル、ならびにゼラチンおよびグリセロールまたはソルビトールなどの可塑剤で作製された密封型ソフトカプセルが包含される。押し込み型カプセルは、乳糖などの充填剤、デンプンなどの結合剤、および/またはタルクもしくはステアリン酸マグネシウムなどの潤滑剤、および任意選択により安定剤と混合された活性成分を含有し得る。ソフトカプセルでは、活性化合物は、脂肪油、液体パラフィン、または液体ポリエチレングリコールなどの好適な液体に溶解または懸濁することができる。さらに、安定剤を添加することができる。経口投与のための全ての製剤は、このような投与に好適な用量であるべきである。
【0099】
[0118] いくつかの実施形態において、静脈内投与のための医薬組成物は、1日当たり患者当たり約0.1〜100mgで提供され得る。1日当たり患者当たり0.1から最大約100mgまでの用量を用いることができる。実質的にさらに高い用量が、局所投与で可能である。非経口投与可能な組成物を調製するための実際の方法は、当業者に知られているかまたは明らかであり、Remington:The Science and Practice of Pharmacy、第21版、2005、Lippincott Williams & Wilkins,Publishersなどの刊行物においてさらに詳細に記載されている。
【0100】
[0119] 医薬組成物は、投与方法に応じて、様々な剤形および用量で投与することができる。例えば、経口投与に好適な単位剤形は、限定はしないが粉末、錠剤、ピル、カプセル、およびトローチを包含する。抗体は、経口投与された場合に消化から保護されるべきであると認識される。これは、典型的には、分子を組成物と複合して、分子を酸および酵素による加水分解に対して耐性にすることによって、または適切に耐性である抗体内、例えばリポソームまたはタンパク質バリア内に分子をパッケージングすることによって、達成する。作用剤を消化から保護する手段は、当技術分野において周知である。
【0101】
[0120] 医薬製剤、特に、本発明に従ったポリペプチドEMP2阻害剤および核酸EMP2阻害剤からなる医薬製剤は、所望の純度を有する抗体またはオリゴヌクレオチドを最適な薬学的に許容可能な担体、賦形剤、または安定剤と混合することによって調製することができる。このような製剤は、凍結乾燥製剤または水溶液であり得る。許容可能な担体、賦形剤、または安定剤は、用いられる用量および濃度でレシピエントに対して無毒である。許容可能な担体、賦形剤、または安定剤は、酢酸塩、リン酸塩、クエン酸塩、および他の有機酸;抗酸化剤(例えば、アスコルビン酸);防腐剤;低分子量ポリペプチド;タンパク質、例えば、血清アルブミンもしくはゼラチンまたはポリビニルピロリドンなどの親水性ポリマー;ならびにアミノ酸、単糖、二糖、およびグルコース、マンノース、またはデキストリンを包含する他の炭水化物;キレート化剤;ならびにイオン性界面活性剤および非イオン性界面活性剤(例えば、ポリソルベート);塩形成対イオン、例えばナトリウム;金属錯体(例えば、Zn−タンパク質錯体);および/または非イオン性界面活性剤であり得る。抗体は、0.5〜200mg/mlの間または10〜50mg/mlの間の濃度で製剤化することができる。
【0102】
[0121] いくつかの実施形態において、本発明は、EMP2阻害剤と、細胞レベルまたは生物レベルで生理学的に許容可能な担体とを含む組成物を提供する。典型的には、生理学的に許容可能な担体は、液体、固体、または半固体の形態で存在する。液体担体の例には、生理食塩水、リン酸緩衝液、通常の緩衝生理食塩水(NaCl135〜150mM)、水、緩衝水、0.4%生理食塩水、0.3%グリシン、安定性を増強するための糖タンパク質(例えば、アルブミン、リポタンパク質、グロブリンなど)などが包含される。固体または半固体の担体の例には、マンニトール、ソルビトール、キシリトール、マルトデキストリン、乳糖、デキストロース、ショ糖、グルコース、イノシトール、粉末糖、糖蜜、デンプン、セルロース、微晶質セルロース、ポリビニルピロリドン、アカシアゴム、グアーゴム、トラガカントゴム、アルギン酸塩、アイリッシュモスの抽出物、パンワールゴム、ガッチゴム、イサポールハスクの粘液、Veegum(登録商標)、ラーチアラボガラクタン、ゼラチン、メチルセルロース、エチルセルロース、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ポリアクリル酸(例えば、カルボポール)、ケイ酸カルシウム、リン酸カルシウム、第二リン酸カルシウム、硫酸カルシウム、カオリン、塩化ナトリウム、ポリエチレングリコール、およびその組み合わせが包含される。生理学的に許容可能な担体は一部には、投与される特定の組成物によって、および組成物を投与するために用いられる特定の方法によって決定されるため、本発明の医薬組成物の様々な好適な製剤が存在する(例えば、Remington 's Pharmaceutical Sciences, 第17版, 1989を参照されたい)。担体および組成物は、好ましくは無菌である。
【0103】
[0122] 本発明の組成物は、従来の周知の滅菌技術によって滅菌してもよいし、あるいは無菌条件下で生成してもよい。水溶液は、使用のためにパッケージされ得るか、または無菌条件下で濾過されて凍結乾燥され、凍結乾燥された製剤は、投与の前に無菌水溶液と組み合わされる。組成物は、好適な生理学的条件に応じて、薬学的にまたは生理学的に許容可能な補助剤、例えば、pH調整剤およびpH緩衝剤、張度調節剤、湿潤剤など、例えば、酢酸ナトリウム、乳酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、ソルビタンラウリン酸モノエステル、およびオレイン酸トリエタノールアミンを含有し得る。
【0104】
[0123] 経口投与に好適な製剤は、(a)液体溶液、例えば、希釈剤内、例えば、水、生理食塩水、またはPEG400内に懸濁された、有効量のパッケージングされたプラチナ基剤の薬剤、(b)液体、固体、顆粒、またはゼラチンとして所定量のプラチナ基剤の薬剤をそれぞれ含有する、カプセル、包剤、または錠剤、(c)好適な液体内の懸濁液、ならびに(d)好適なエマルジョンを含み得る。錠剤形態は、乳糖、ショ糖、マンニトール、ソルビトール、リン酸カルシウム、トウモロコシデンプン、ジャガイモデンプン、微晶質セルロース、ゼラチン、コロイド状二酸化ケイ素、タルク、ステアリン酸マグネシウム、ステアリン酸、および他の賦形剤、着色剤、充填剤、結合剤、希釈剤、緩衝剤、湿潤剤、防腐剤、香料、染料、崩壊剤、ならびに薬学的に適合可能な担体の1つまたは複数を包含し得る。
【0105】
[0124] 本発明は、本発明に従ったEMP2阻害剤を含む局所的医薬組成物を提供する。いくつかの実施形態において、阻害剤は、低分子有機化合物、EMP2ポリペプチド、または抗EMP2抗体である。阻害剤は、単位剤形当たり目的の症状に有効な上記のEMP2阻害剤を含む、単位剤形であり得る。
【0106】
[0125] EMP2阻害剤の局所的製剤は、薬学的に許容可能な担体と組み合わせて製剤化することができる。本発明の化合物を局所投与するための剤形には、粉末、スプレー、泡、ゼリー、軟膏、ペースト、クリーム、ローション、ゲル、溶液、パッチ、坐剤、およびリポソーム製剤が包含される。剤形は、泌尿生殖器領域で活性成分を徐放性するための粘膜接着性ポリマーを用いて製剤化することができる。活性化合物は、無菌条件下で、薬学的に許容可能な担体、および必要であり得る任意の防腐剤、緩衝液、または推進剤と混合することができる。局所的製剤は、阻害剤を局所的な乾燥製剤、液体製剤、クリーム製剤、およびエアロゾル製剤で一般に用いられる従来の薬学的希釈剤および薬学的担体と組み合わせることによって調製することができる。軟膏およびクリームは、例えば、好適な増粘剤および/またはゲル化剤を添加した水性基剤または油性基剤を用いて製剤化することができる。このような基剤には、水および/または油、例えば液体パラフィンまたは植物油、例えばピーナッツ油またはヒマシ油が含まれ得る。基剤の性質に従って用いられ得る増粘剤には、ソフトパラフィン、ステアリン酸アルミニウム、セトステアリルアルコール、プロピレングリコール、ポリエチレングリコール、羊毛脂、水添ラノリン、蜜ろうなどが包含される。ローションは、水性基剤または油性基剤を用いて製剤化することができ、通常は、安定化剤、乳化剤、分散剤、懸濁剤、増粘剤、着色剤、香料などの1つまたは複数も包含する。粉末は、任意の好適な粉末基剤、例えば、タルク、乳糖、デンプンなどを用いて製剤化することができる。ドロップは、1つまたは複数の分散剤、懸濁剤、可溶化剤なども含む水性基剤または非水性基剤を用いて製剤化することができる。
【0107】
[0126] 軟膏、ペースト、クリーム、およびゲルはまた、賦形剤、例えば、動物および植物の脂肪、油、ワックス、パラフィン、デンプン、トラガカント、セルロース誘導体、ポリエチレングリコール、シリコーン、ベントナイト、ケイ酸、タルク、および酸化亜鉛、またはその混合物を含有し得る。粉末およびスプレーはまた、賦形剤、例えば、乳糖、タルク、ケイ酸、水酸化アルミニウム、ケイ酸カルシウム、およびポリアミド粉末、またはこれらの剤の混合物を含有し得る。スプレーはさらに、慣習的な推進剤、例えば、クロロフルオロ炭化水素ならびに揮発性の非置換炭化水素、例えばブタンおよびプロパンを含有し得る。
【0108】
[0127] EMP2阻害剤は、症状の治療に有効な単位剤形の組成物内に存在し得る。EMP2阻害剤の用量は、当業者に周知の多くの因子、例えば、特定の化合物、治療する症状、レシピエント患者の年齢、体重、および臨床症状、ならびに臨床医または治療を管理する専門家の経験および判断に応じる。有効量の化合物は、症候の自覚的な軽減または臨床医もしくは他の資格のある観察者によって認められる客観的に同定可能な改善をもたらすものである。用量範囲は、用いられる化合物、投与経路、および特定の化合物の効力で変化する。
【0109】
[0128] 本発明は、1つまたは複数の上記の薬理学的化合物および薬学的に許容可能な担体を含む、局所的な徐放性医薬組成物および持続放出医薬組成物を提供する。好ましくは、組成物は、このような治療を必要としている対象に単位剤形で投与される。局所的な徐放性組成物および持続放出組成物は、典型的には、1)親水性基剤中の吸収剤、2)疎水性基剤中の吸収剤、および3)好適な媒体内に分散した吸収剤マトリクスを含有する被覆されたビーズを包む種々のものである。
【0110】
III.治療方法
[0129] 本発明はさらに、網膜剥離または増殖性硝子体網膜症を有する対象の眼における網膜剥離を治療、予防、および/または阻害する方法に関する。本方法は、全体として、本明細書において記載される抗EMP2剤(例えば、抗EMP2抗体もしくはそのフラグメント、抗EMP2siRNA、または抗EMP2リボザイム)を、眼におけるEMP2活性を低減させるおよび/または阻害するために十分な条件下および用量で対象に投与することを含む。
【0111】
[0130] 医師または獣医師は、医薬組成物において採用される本発明の抗EMP2剤の投薬を所望の治療効果を達成するために必要なレベルより低いレベルで開始し、所望の効果が達成されるまで徐々に用量を増大させることができる。通常、本発明の組成物の有効量は、治療する具体的な疾患または症状、投与手段、標的部位、患者の生理学的状態、患者がヒトであるか動物であるか、投与される他の医薬品、および治療が予防的であるか治療的であるかを包含する、多くの異なる因子に応じて変化する。治療用量は、安全性および効能を最適化するように滴定される必要がある。組成物の単回投与または複数回投与を、患者によって要求され認容される用量および頻度に応じて投与することができる。
【0112】
[0131] 抗体との投与では、用量は、宿主体重1kg当たり約0.0001〜100mg、より一般的には0.01〜5mgの範囲である。例えば、用量は、体重1kg当たり1mgまたは体重1kg当たり10mgまたは1〜10mg/kgの範囲内であり得る。投薬は、必要に応じて、または要求に応じて、毎日、週に1回、週に2回、月に1回、またはそれを超えるかもしくはそれ未満の頻度であり得る。典型的な治療レジメは、週に1回、2週毎に1回、または月に1回、または3〜6ヶ月に1回の投与を伴う。
【0113】
[0132] 剤が核酸である実施形態において、典型的な用量は、体重1kg当たり約0.1mgから、体重1kg当たり約100mgまで(体重1kg当たり約100mgを含める)の範囲であり得、例えば、体重1kg当たり約1mgから体重1kg当たり約50mgまでの間である。いくつかの実施形態において、体重1kg当たり約1、2、3、4、5、10、15、20、30、40、または50mgである。
【0114】
[0133] 本発明の抗EMP2剤およびその医薬組成物は、任意の投与経路(例えば、静脈内、局所的、腹腔内、非経口、経口、膣内、直腸、眼球、硝子体内、および眼球内)によって投与することができる。これらは、筋肉内経路、腹腔内経路、皮下経路、経口経路、局所的経路、または吸入経路によって、一定期間にわたるボーラス注入または持続注入として投与することができる。抗体の静脈内投与または皮下投与が好ましい。投与は、局所的または全身的であり得る。これらは、対象となる疾患を有している、その疾患の病歴がある、またはその疾患のリスクがあると診断された対象に投与することができる。
【実施例】
【0115】
[0134] 以下の実施例は例示のために提供されるが、特許請求される発明を限定するものではない。
【0116】
(実施例1)
[0135] 増殖性硝子体網膜症(PVR)は、一部には、異常な創傷治癒戦略における、硝子体腔内での細胞の移動、膜の形成、および収縮を伴う、網膜色素上皮(RPE)の脱分化から生じると考えられている。インビトロでのコラーゲン−ゲル収縮アッセイにおいて、網膜色素上皮細胞系(ARPE-19)による接着斑キナーゼ(FAK)の活性化を介して収縮を制御する、4回膜貫通タンパク質である上皮膜タンパク質2(EMP2)を研究した。この研究の目的は、EMP−2発現のレベルがインビボモデル系において臨床PVRを変化させたかどうかを調べることであった。
【0117】
[0136] 方法:ARPE−19細胞系をATCCから得て、EMP2のレベルを、EMP−2を過剰発現する構築物、EMP2リボザイム、またはベクターのみの安定なトランスフェクションを介して調節した。これらの安定にトランスフェクトされた細胞系をPVRのウサギモデルにおいて用いた。PVRの重症度を、二人のマスキングされた観察者が分類した。また、EMP2ブロック抗体を用いて、PVRモデルにおける機能的なEMP2を減少させた。免疫組織化学を用いて、インビボでのEMP2の発現を評価した。
【0118】
[0137] 結果:高レベルのEMP2発現を有するトランスフェクタントは、低レベルのEMP2トランスフェクタントよりも有意に重症度の大きなPVRを誘発した(p=0.05)。さらに、より低レベルのEMP2を有するトランスフェクタントは、野生型細胞によって誘発されるPVRの程度よりも有意に小さなPVRの重症度を有していた(p=0.02)。特異的抗体でEMP2を妨げると、PVR重症度のレベルが有意に減少した(p=0.01)。PVR膜は、EMP2発現について陽性であることが明らかになった。
【0119】
[0138] 結論:これらのインビボでの研究は、EMP2の発現とPVRの重症度との間の直接的な相関を裏付けるものである。これらの結果は、EMP2の発現における変化を介してRPE生物学を制御する可能性を実証し、この疾患の潜在的な治療標的を提供する。
【0120】
[0139] RPEの制御および膜収縮の予防は、深刻なPVR応答の予防における最初の目的である。インビトロでの実験モデルでは、本発明者らの研究室は、インテグリン(α1、α2、およびα3)のライゲーションを介するFAKの活性化をコラーゲンゲル収縮の重大な制御点として同定した(Morales, S. A.ら, Exp Eye Res 85:790-8(2007))。テトラスパンスーパーファミリーは、インテグリンおよび他の受容体複合体と組み立てられた細胞内輸送分子および細胞内シグナル伝達分子のタイプを決定する鍵となるクラスのタンパク質である(Hemler, M. E. T Annu Rev Cell Dev Biol 19:397-422(2003))。特定のテトラスパンファミリーメンバーは、個別の細胞型および分化または活性化の状態において、高度に制御されている。したがって、テトラスパンの組み合わせ発現は、インテグリンの配置およびシグナル伝達の質に対する重要な寄与因子である。上皮膜タンパク質−2(EMP2)は、インテグリンの特異的な配置およびFAKを介するシグナル伝達を制御するように作用する、テトラスパンファミリーメンバーである(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。RPEにおいて高度に発現するEMP2(Wadehra, M.ら, Exp Mol Pathol 74:106-12 (2003))は、異なる生化学的および生理学的役割を有する成長停止特異的遺伝子3/末梢ミエリンタンパク質22(GAS3/PMP22)4−TMタンパク質ファミリーのメンバーである(Wadehra, M.ら, Exp Mol Pathol 74:106-12(2003); Forbes, A.ら, J Biol Chem 282:26542-51(2007); Wadehra, M.ら, Mol Biol Cell 15:2073-83(2004); Wadehra, M.ら, J Biol Chem 277:41094-100(2002); Wadehra, M.ら, Reprod Biol Endocrinol 6:15(2008); Wadehra, M.ら, Clin Immunol 107:129-36(2003); Wang, C. X.ら, Blood 97:3890-5(2001))。
【0121】
[0140] この研究において、本発明者らは、PVRのウサギモデルを用いて、PVRおよび網膜剥離の発症および進行においてEMP2発現が有する役割を研究した。本発明者らは、EMP2の発現の減少または抗体によるEMP2のブロックによって、PVRの形成が減少することを明らかにした。
【0122】
方法および材料
[0141] 細胞系:安定なARPE−19細胞系であるARPE−19は、ATCCから得られる自然発生的なヒト網膜色素上皮(RPE)である(CRL-2302)。ARPE−19細胞を、以下の構築物の1つでトランスフェクトした:pEGFP−N3=ベクター対照、pEGFP−N3−EMP2=EMP2を過剰発現する構築物、pEGFP−N3−Ribol=EMP2の発現が低減した構築物。ARPE−19細胞を、トランスフェクションの前に24時間平板培養した。FuGENE6(Roche Molecular Biochemicals, Indianapolis, Indiana)を用いて、細胞を上記の構築物の1つでトランスフェクトした。安定なクローンをジェネティシン(700mg/ml、Invitrogen Life Technologies)を用いて選択した。これらの安定な細胞系をARPE−19/V、ARPE−19/EMP2、またはARPE−19/Riboと呼ぶ。これらの細胞系によるEMP−2の発現は、以前に報告されていた(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。
【0123】
[0142] 成長培地:ATCC培地#30−2006:1:1で混合した、ダルベッコ変法イーグル培地および重炭酸ナトリウム1.2g/L、L−グルタミン2.5mM、HEPES15mM、およびピルビン酸ナトリウム0.5mMを含有するハムF12培地 90%;ウシ胎児血清#30−2020 10%。
【0124】
[0143] PVRの動物モデル:この研究において用いられたウサギは、UCLA Institutional Review Board(IRB)によって認可された制度的ガイドラインに従って処理した。PVRは、以前に記載されたように、アルビノニュージーランドウサギの右目においてのみ誘発した(Nakagawa, M.ら, Invest Ophthalmol Vis Sci 36:2388-95(1995))。簡潔に述べると、30ゲージの針を用いて、C3F8ガス(100%)0.4mlを、麻酔導入した後に角膜縁の4mm後方の硝子体腔内に注入した。前方穿刺をマイクロブレードで行い、眼球内圧力の上昇を予防した。2日後、成長培地内に再懸濁したARPE−19細胞調製物を、0.1ccの容量で注入した。左目は対照として用いた。ウサギは、週に1回、間接検眼法によって、マスキング様式でPVRの発症および炎症について試験した。5週目に、動物をペントバルビタール(100mg/kg)を用いて犠牲死させた。眼球を10%ホルマリン内に置き、上部毛様体扁平部切開を#11のブレードで行って、眼球内腔に固定剤が届くようにした。眼球を3日間にわたり固定し、二等分し、Fastenbergによって確立されたPVRスケールを用いて肉眼的所見についてマスキング様式でスコア付けした(表1)(Fastenberg, D. M.ら, Am J Ophthalmol 95:663-7(1983); Fastenberg, D. M.ら, Am J Ophthalmology 93:565-72(1982))。眼球をパラフィン内に包埋し、切片化し、ヘマトキシリンおよびエオシンで染色した。切片を顕微鏡によって評価し、マスキング様式でPVRについて等級付けした。
【0125】
[0144] 統計分析:スチューデントt検定(独立、一方向)を用い、P<0.05を統計的に有意であると判断した。
【0126】
【表1】

【0127】
結果
[0145] EMP−2発現の増大を伴うPVR形成の増大:良く特徴付けされたPVRのウサギモデルを用いて、本発明者らは、記載されているARPE−19細胞の硝子体内注入を用いて、PVRを誘発した(Nakagawa, M.ら, Invest Ophthalmol Vis Sci 36:2388-95(1995))。細胞は、野生型、EMP2を過剰発現する修飾型ARPE−19細胞、またはリボザイムトランスフェクションによりEMP2を低発現する修飾型ARPE−19細胞であった(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。眼を、PVR形成の程度について、注入の5週間後に、二人の観察者が等級付けした。最初の3頭のウサギは、以前の報告に類似した、一貫した程度のPVR形成を有することが明らかになった(データは示していない)。野生型RPE細胞を与えたウサギは、EMP2を低発現する細胞を与えたウサギよりも有意に重症なレベルのPVRを有していた(それぞれ2.5対1.3、p=0.02)。(図1)さらに、EMP2を過剰発現する細胞を与えたウサギは、EMP−2を低発現するものよりも有意に多いPVR形成を有していた(それぞれ2.9対1.3、p=0.05)。EMP2発現の増大は、PVRの重症度の増大と相関しており、これと合致して、EMP2レベルの減少は、PVRの重症度の低減と相関していた。図2は、3つの異なるウサギ群から得られた典型的な肉眼的な組織学的標本を示す。
【0128】
[0146] EMP−2の発現の増大を伴う線維症および膜形成の増大:EMP2を過剰発現する細胞を与えた眼の組織学的試験によって、野生型細胞を投与したウサギまたはEMP2を低発現する(リボザイムトランスフェクトされた)細胞を投与したウサギの両方よりも大きなPVR膜形成が確認された(図3)。インビボでのPVR膜は、免疫組織化学によって、5週間目に、EMP2発現について陽性であることが明らかになった(図4)。
【0129】
[0147] EMP−2ブロック抗体で妨げられたPVR膜形成:ARPE−19細胞を、抗ヒトEMP2ポリクローナル抗体、対照抗体(免疫前血清)、または培地のみでプレインキュベートした。眼を細胞注入の5週間後に再び試験した。PVR形成の程度を二人のマスキングされた観察者が等級付けした。抗ヒトEMP2ポリクローナル抗体を用いたARPE−19細胞におけるEMP2のブロックによって、未処理細胞または免疫前対照抗体に曝露された細胞と比較してPVRの重症度が減少した(それぞれ2.6対4.3、p=0.01)(図5)。さらに、組織学的評価によって、線維症および牽引の減少が確認された(図6)。特異的抗体でライゲーションした後のEMP2発現の一時的な減少を記載したマウスモデルにおける本発明者らの以前のインビボでの研究によって予測されたように、EMP2の発現が残りの膜において観察された(Shimazaki, K.ら, Microbes Infect 9:1003-10(2007))。
【0130】
考察
[0148] PVRの発症の間の最初に、RPE細胞は網膜表面に結合し、移動し、増殖し、膜形成すると考えられる(Eibl, K. H.ら, Invest Ophthalmol Vis Sci 47:364-70(2006))。この後に、膜の収縮および牽引性網膜剥離が生じる。
【0131】
[0149] この研究において、本発明者らは、EMP2が膜収縮において重要な調節役割を有するというインビトロでの証拠にまで言及し、ウサギPVRモデルにおけるインビボでのEMP2発現の関連性の証拠を示す。ゲル収縮アッセイは、EMP2発現の増大を伴うコラーゲンゲルの収縮の増大を示した(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。ここで、本発明者らは、PVRを生じさせるRPE細胞におけるEMP2の発現レベルがウサギにおけるPVRの重症度と相関することを明らかにする。PVRにおけるEMP2の関連性についての独立した証拠を、対照抗体とは対照的にPVRの重症度を減少させたEMP2に対するブロック抗体(blocking antibody)を用いて得た。
【0132】
(実施例2)
[0150] 増殖性硝子体網膜症(PVR)は、穿通性損傷または裂孔原性網膜剥離を包含する重症の眼球外傷の合併症として生じる異常な創傷治癒である。上皮膜タンパク質2(EMP2)は、FAK活性化の調節によって、インビトロでのPVRモデルである網膜色素上皮細胞系ARPE−19によるコラーゲンゲルの収縮を調節する。この研究の目的は、ARPE−19細胞によるコラーゲンゲル収縮の阻害におけるEMP2特異的組換えダイアボディの効能を調べることである。
【0133】
方法
[0151] EMP2ダイアボディをファージライブラリーを用いて組換えによって構築し、ヒトEMP2ペプチドに対する反応性について選択した。全ての実験において、ARPE−19細胞を抗EMP2または対照ダイアボディ20μg/mlで2時間にわたり前処理した。毒性、接着、および移動をそれぞれ、Annexin Vを用いるフローサイトメトリー、1型コラーゲンへの結合、および創傷治癒アッセイを介して評価した。コラーゲンゲルの収縮をインビトロでのアッセイを用いて評価した。
【0134】
結果
[0152] 抗EMP2ダイアボディを用いて毒性は観察されなかった。抗EMP2ダイアボディは、接着または移動を有意には改変しなかった。抗EMP2ダイアボディへの曝露によって、4時間目でのEMP2タンパク質レベルが75%低減した。治療前のレベルへのEMP2の回復が16時間後に見られた。それに合致した、活性化型FAKの基底レベルの減少もまた観察された。コラーゲン刺激性のFAK活性化は、抗EMP2ダイアボディで処理したARPE−19細胞において25%低減した。抗EMP2ダイアボディ治療は、ARPE−19細胞によるコラーゲンゲルの収縮を有意に低減させ(p<0.001)、用量依存性であった。
【0135】
結論
[0153] 抗EMP2ダイアボディは、EMP2のダウンレギュレーションを介して収縮能力を減少させて、FAKの活性化を減少させ、コラーゲンゲルの収縮を阻害する。
【0136】
(実施例3)
導入
[0154] 接着斑キナーゼ(FAK)およびリン酸化を介するその活性化は、細胞周期の進行、増殖、侵入、移動、生存、および収縮にとって重要である(Schaller MDら, Proc Natl Acad Sci USA, 89:5192-5196(1992))。FAKのリン酸化は、インテグリン受容体が細胞外マトリクスの構成要素にライゲーションすることによって開始し得る。さらに、受容体チロシンキナーゼシグナル伝達を介するEGFおよびPDGFのライゲーション、ならびにG−タンパク質連結型の受容体を介するリゾホスファチジン酸(LPA)、ボンベシン、およびスフィンゴシルホスホリルコリン(SPC)への受容体のライゲーションを包含する多くの非インテグリンシグナル伝達経路が、FAKの活性化を促進する(Rankin SおよびRozengurt E, J Biol Chem, 269:704-710(1994); Seufferlein TおよびRozengurt E, J Biol Chem, 269:9345-9351(1994); Seufferlein TおよびRozengurt E, J Biol Chem, 270:24334-24342(1995); Sieg DJら, Nat Cell Biol, 2:249-256(2000); Sinnett-Smith Jら, J Biol Chem, 268:14261-14268(1993))。FAKの活性化は、複数の部位でのリン酸化を必要とする複雑なプロセスである。リン酸化は、Tyr−397およびTyr−407(N末端ドメイン)、Tyr−576およびTyr−577(キナーゼドメイン活性化ループ)、ならびにTyr−861およびTyr−925(C末端ドメイン)で生じる。FAKの活性化における最初の工程は、Tyr−397での自己リン酸化であり、これによりSrcチロシンキナーゼのSH2ドメインのための結合部位が生じ、その結果、Srcキナーゼドメインを活性化させる。残りのFAKチロシンリン酸化部位は、Srcによって優先的にリン酸化される(Calalb MBら, Mol Cell Biol, 15:954-963(1995); Schlaepfer DDら, Prog Biophys Mol Biol, 71:435-478(1999); Schlaepfer DDおよびHunter T, Trends Cell Biol, 8:151-157(1998))。キナーゼドメインの活性化ループにおけるFAKのTyr−576およびTyr−577のリン酸化は、最大のFAKキナーゼ活性に必要であり(Calalb MBら, Mol Cell Biol, 15:954-963(1995); Mitra SKおよびSchlaepfer DD, Curr Opin Cell Biol, 18:516-523(2006))、FAKのTyr−925のリン酸化によって、Grb2低分子アダプタータンパク質へのSH2結合部位が生じる(Schlaepfer DDら, Nature, 372:786-791(1994))。FAKへのGrb2の結合は、ERK2/MAPキナーゼカスケードなどの下流標的の活性化をもたらすいくつかのシグナル伝達経路の1つである。Tyr−407およびTyr−861でのリン酸化は、上皮間葉分化転換(EMT)に関与している(Nakatnoto Tら, Mol Cell Biol, 17:3884-3897(1997))。Tyr−861でのFAKのリン酸化は、F−アクチンの構成にも関与している(Lunn JAら, J Biol Chem, 282:10370-10379(2007))。
【0137】
[0155] 上皮膜タンパク質2(EMP2)は、成長停止特異的−3/末梢ミエリンタンパク質−22(GAS3/PMP22)ファミリーに属するテトラスパン(4回膜貫通)タンパク質である。EMP2は、皮膚、肺、子宮、心臓、甲状腺、および眼に局在する(Wang CXら, Blood, 97:3890-3895(2001))。眼では、EMP2は、角膜、毛様体、および網膜色素上皮(RPE)を包含する複数の上皮層に存在する(Wadehra Mら, Exp Mol Pathol, 74:106-112(2003))。EMP2は、インテグリン、グリコシルホスファチジルイノシトールを足場としたタンパク質、およびMHCクラスIタンパク質の輸送を調節することが示されている(Wadehra Mら, Dev Biol, 287:336-345(2005); Wadehra Mら, Mol Biol Cell, 15:2073-2083(2004); Wadehra Mら, J Biol Chem, 277:41094-41100(2002); Wadehra Mら, Clin Immunol, 107:129-136(2003))。EMP2は、胚盤胞移植などの生理学的環境、ならびに細胞の増殖、侵入、接着、および転移のモデル環境において、特定のインテグリンアイソフォームの機能に物理的に関連し、それを調節する(Wadehra Mら, Exp Mol Pathol, 74:106-112(2003); Wadehra Mら, Dev Biol, 287:336-345(2005); Wadehra Mら, Mol Biol Cell, 15:2073-2083(2004); Wadehra Mら, J Biol Chem, 277:41094-41100(2002); Wadehra Mら, Clin Immunol, 107:129-136(2003); Wadehra Mら, Dev Biol, 292:430-441(2006))。
【0138】
[0156] EMP2機能に関与する生化学的メカニズムは、明らかにされていない。しかし、他のテトラスパンタンパク質において、顕著なテーマは、シグナル伝達経路に関与する非相同テトラスパンおよび他の膜タンパク質との多重の関連である(Boucheix Cら, Cell Mol Life Sci, 58:1189-1205(2001); Hemler ME, J Cell Biol, 155:1103-1107(2001))。これは、テトラスパンタンパク質が、膜におけるシグナル伝達複合体の機能的組み立て体を支持する分子アダプターとして作用することを示唆する(Hong IKら, J Biol Chem, 281:24279-24292(2006))。おそらく、PI−4キナーゼを特定の膜位置に動員しホスホイノシチド依存性のシグナル伝達を誘発することが示されている、テトラスパンタンパク質のサブセットが最も良く規定されている(Yauch RLおよびHemler ME, Biochem J, 351 Pt 3:629-637(2000))。
【0139】
[0157] EMP2は、Tyr−576および577でのリン酸化によって測定されるFAKの活性化を介してARPE−19細胞系のコラーゲンゲル収縮を増大させることが知られている(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。この研究の目的は、EMP2とFAK活性化との関連をさらに調査することであった。ここで、本発明者らは、EMP2がFAKと物理的に関連して、複数の部位でFAKのリン酸化を増大させることを示す。このFAKのリン酸化の増大によって、機能的な機械的な細胞の改変が生じ、細胞の収縮能力が増大する。これらの細胞の改変には、接着斑密度の増大、アクチン細胞骨格における立体配座の変化、アクチン組成物の改変、および細胞の接着能力の増大が包含される。これらの所見は、EMP2がFAKの活性化に重要な膜シグナル伝達複合体の機能的組み立てにおけるテトラスパンの新たな役割を表し得ることを示唆する。
【0140】
方法
[0158] 細胞系−RPE特異的マーカーであるCRALBPおよびRPE−65を発現する自然発生的な網膜色素上皮(RPE)細胞系であるARPE−19を、American Type Culture Collectionから得た(CRL-2302, ATCC, Manassas, VA)。EMP2を過剰発現する細胞系であるARPE−19/EMP2を、75倍高いタンパク質レベルのEMP2を発現するEMP2過剰発現のレトロウイルス構築物の安定な感染を介して生成した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。ARPE−19細胞を5%CO2の湿潤チャンバにおいて37℃で、10%ウシ胎児血清(FBS)(ATCC, Manassas, VA)を補ったDMEM−F12培地において培養した。全ての実験において細胞を平板培養し、一晩インキュベートした。全ての細胞は、別段の指示がない限り、各実験の開始時点で約75%コンフルエントであった。
【0141】
[0159] 抗体−ヒトFAKについての全ての抗体は、Santa Cruz Biotechnology(Santa Cruz, CA)から得た:リン酸化型FAK(p-FAK)(クローンTyr 576/577)についてのヤギ抗体、ならびにFAK(クローンC-20)、p−FAK(Tyr 397)、(Tyr 407)、(Tyr 861)、および(Tyr 925)についてのウサギ抗体。ヒトp−Src(Tyr 416およびTyr 527)についてのウサギ抗体は、Cell Signaling Technology(Danvers, MA)から得た。ヒトβ−アクチン(クローン2A2.1)を認識するマウス抗体は、US Biological(Swampscott, MA)から得た。ヒトα−平滑筋アクチン(α-sma)(クローン1A4)に対するマウス抗体は、Sigma(St. Louis, Mo)から得た。F−アクチンを同定するために用いるテキサスレッド−Xファロイジン(T7471)は、Invitrogen(Carlsbad, Ca)から得た。ヒトEMP2に対するウサギ抗血清は、ヒトEMP2の第2の細胞外ループ(EDIHDKNAKFYPVTREGSYG)に結合した多抗原ペプチドで動物を免疫化して生成した(Research Genetics, Huntsville, AL)。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体は、Southern Biotech(Birmingham, Alabama)から得た。ホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体は、BD Biosciences(San Diego, CA)から得た。テキサスレッド結合ロバ抗ヤギ抗体(705-075-147)およびFITC結合ロバ抗ウサギ抗体(711-095-152)は、Jackson ImmunoResearch Laboratories(West Grove, PA)から得た。
【0142】
[0160] ウェスタンブロット分析−ウェスタン分析を、以前記述されているように行った(Morales S Aら, Experimental Eye Research(2007))。簡潔に述べると、細胞タンパク質を、プロテアーゼ阻害剤およびホスファターゼ阻害剤(Upstate, Charlottesville, VA)を含有するRIPA緩衝液を用いて単離し、タンパク質濃度をBCAタンパク質アッセイ(Bio Rad, Hercules, CA)で決定した。EMP2の検出のために、N−連結グリカンをPNgaseを用いて切断した(New England Bioloabs, Beverly, MA)。溶解物を製造者の指示に従って、37℃で2時間にわたり処理した。全部で10μgのタンパク質を各レーンにロードし、タンパク質を還元条件下で4〜20%SDS−PAGE勾配ゲルによって分画した。タンパク質をニトロセルロース膜(Amersham Life Sciences, Buckinghamshire, UK)に移し、転移の妥当性をPonceau Sレッド染色(Sigma Chemical Co., St. Louis, MO)を用いて確認した。
【0143】
[0161] 膜を、TBS Tween(TBST; Upstate, Charlottesville, VA)内の無脂肪乳でブロックした。ブロットを、α−sma、FAK、およびp−FAK(Tyr 576/577)、(Tyr 397)、(Tyr 407)、(Tyr 861)、(Tyr 925)では1:200の希釈、EMP2およびp−Src(Tyr 416)、(Tyr 527)では1:1000の希釈、およびβ−アクチンでは1:5000の希釈の一次抗体と共に、1時間にわたりインキュベートした。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体またはホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体を、1:2000の希釈でブロットに曝露した。ブロットをECL(Pierce, Rockford IL)で発現させ、ブロットのスキャンおよびNIH Image Jソフトウェアでのバンド密度の測定によって定量した。β−アクチンまたは全FAKのバンドの強度を内部対照として用い、全てのバンドを、定量による密度の測定のために線形範囲で分析した。少なくとも3つの独立した実験を行い、スチューデントt検定(独立、一方向)を用いて統計的に評価した。p<0.05を統計的に有意であると考えた。共免疫沈降−ARPE−19細胞およびARPE−19/EMP2細胞を10cmの皿内に平板培養した(Corning, Corning, NY)。細胞をPBSで2回洗浄し、4℃で30分間にわたり溶解し(アプロチニン10μg/ml、ペプスタチン2μg/ml、EDTA0.1mM、HEPES10mM、およびKCl10mMを含有する1%Nonidet P-40、ならびにRoche Applied Sciences, Mannheim, Germanyから入手したComplete Mini Protease Inhibitor Cocktail Tablet)、次に、15秒間にわたり超音波処理した。細胞溶解物をタンパク質Aアガロースビーズ(Santa Cruz Biotechnology, Santa Cruz, CA)と共にインキュベートして事前確認し、次に、アガロースビーズおよび抗FAKポリクローナル抗体(Santa Cruz Biotechnology, Santa Cruz, CA)または抗EMP2ウサギポリクローナル抗体と共に一晩インキュベートした。ビーズを溶解溶液内で4回洗浄し、Tris62.5mM(pH6.8)内で2回洗浄した。免疫複合体を、Laemmli試料緩衝液(Tris-Cl62.5mM(pH6.8)、10%グリセロール、2%SDS、0.01%ブロモフェノールブルー、2%β−メルカプトエタノール)内で5分間にわたり煮沸してビーズから溶出した。試料を、ウェスタンブロット分析を用いて分析した。EMP2の検出のために、N−連結グリカンを、PNgase(New England Bioloabs, Beverly, MA)を用いて切断した。溶出物を製造者の指示に従って、37℃で2時間にわたり処理した。
【0144】
[0162] 共免疫沈降実験におけるFAKおよびEMP2の化学量論を以下の様式で決定した。まず、細胞全体の溶解物および同族免疫沈降物の滴定をデンシトメトリーによって定量した。このデータを用いて、本発明者らは、同族免疫沈降物における各タンパク質のパーセントを計算した(投入した細胞当量当たり)。第2に、共免疫沈降物を同様に分析し、共免疫沈降物内に捕捉された各タンパク質のパーセントを決定した。最後に、本発明者らは、共免疫沈降したタンパク質の化学量論を表すこれらの2つの値(同族のパーセントおよび共免疫沈降物のパーセント)の比率を計算した。
【0145】
[0163] 免疫蛍光−ARPE−19細胞およびARPE−19/EMP2細胞をガラスカバースリップ(Fisher)上で一晩平板培養した。細胞を4%パラホルムアルデヒドで20分間にわたり固定し、0.075%サポニンで15分間にわたり透過処理した。細胞を10%正常ロバ血清で30分間にわたりブロックした。細胞を一次抗体と共に湿潤チャンバにおいて4℃で一晩インキュベートし、次に、PBSに0.01%Triton X−100(PBST)を加えたもので3〜4回洗浄した。細胞を、湿潤チャンバにおいて室温で、フルオレセインイソチオシアネート(FITC)結合ロバ抗ウサギIgGおよびテキサスレッド結合ロバ抗ヤギ抗体と共に1時間にわたりインキュベートした。細胞をPBSTで洗浄し、2回蒸留したH2Oで簡単にすすぎ、VECTASHIELD Mounting Medium(Vector Laboratories, Burlingame, CA)を用いて顕微鏡スライド上に載せた。
【0146】
[0164] Zeiss LSM 510レーザー走査共焦点顕微鏡(Thornwood, NY)を用いて、タンパク質の分布および共局在化を評価した。FITCで標識された細胞およびテキサスレッドで標識された細胞を検出するために、試料をそれぞれ488nmおよび568nmのアルゴンレーザーおよびクリプトンレーザーで励起させた。LSMソフトウェアを、顕微鏡の制御、走査およびレーザーモジュール、画像記録、ならびに画像データの分析に用いた。
【0147】
[0165] LSMソフトウェアを用いて共局在化分析を行い、関連のパーセントを得た。強調されたピクセルを、Image J(NIHソフトウェア)を用いて決定した。少なくとも6つのフィールドを各試料の分析のためにランダムに選択し、関連のパーセントは、複数のフィールドから生じた平均値である。フィールド当たり複数のスライスを評価したが、スライドと接触し接着斑を含有する細胞底表面の単一のスライスのみを接着斑とEMP2との共局在化の定量に用いた。全ての実験において、細胞は60倍の油浸対物レンズを用いて観察した。各実験を少なくとも4回反復した。
【0148】
[0166] 接着アッセイ−ARPE−19細胞およびARPE−19/EMP2細胞を24ウェルコラーゲン被覆プレート(BD Biosciences, San Diego, CA)、またはフィブロネクチン、ビトロネクチン、コラーゲンI、およびコラーゲンIVで事前被覆されたCytoMatrix Screeningプレート(Millipore, Billerica, MA)上で、ウェル当たり2×105個細胞の濃度で平板培養した。細胞を5%CO2の湿潤チャンバにおいて37℃で2時間にわたりインキュベートした。次にプレートをPBSで3回洗浄し、全ての未付着細胞を除去した。結合細胞をクリスタルバイオレットの取り込みについて分析し、可溶化した後、吸光度を、Bio−Radマイクロプレートリーダー550(Hercules, CA)によって595nmで測定した。各実験は少なくとも8回の反復を含み、少なくとも3つの独立した実験を行い、同等の結果を得た。スチューデントt検定(独立、一方向)を用い、P<0.05を統計的に有意であると判断した。
【0149】
結果
[0167] ARPE−19細胞におけるp−FAKとEMP2との関連。Srcの相互作用および相互リン酸化事象を介して生じた、FAKのリン酸化は、多様な細胞応答を推進するシグナル伝達タンパク質およびアダプタータンパク質を動員する(Jacamo ROおよびRozengurt E., Biochem Biophys Res Commun, 334:1299-1304(2005))。本発明者らは、以前に、ARPE−19細胞におけるEMP2の過剰発現がFAK/Src活性の改変をもたらすかどうかを調べた。本発明者らは、全FAKレベルがARPE−19細胞およびARPE−19/EMP2細胞の両方において等しいことを示し(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))、図7において確認したが、EMP2の過剰発現は、対照ARPE−19細胞と比較して、活性化型FAK(Tyr-576/577)のレベルがほぼ2倍増大することを実証した。各活性化型FAKをそれ自身のβ−アクチンロード対照に正規化して行った複数の評価は、EMP2レベルの増大とFAK活性化との間の統計的に有意な相関を示した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。FAKリン酸化の他の部位を、ARPE−19細胞およびARPE−19/EMP2細胞において定量的ウェスタンブロットによって調べ、全FAKに正規化した(図7A)。EMP2の過剰発現は、Tyr−397、Tyr−407、Tyr−861、およびTyr−925でのFAKのリン酸化を2〜3倍増強した。
【0150】
[0168] Srcの酵素活性は、Tyr−416(増加)およびTyr−527(抑制)でのチロシンリン酸化によって相互調節される(Harvey Rら, Mol Cell Biol, 9:3647-3656(1989))。対照細胞と比較して、EMP2の過剰発現は、Tyr−416でのSrcリン酸化の増大と関連していた(図7B)。このSrcキナーゼドメインの活性化がTyr−397でのFAKのリン酸化と関連しているため、この結果は予測された。重要なことには、キナーゼドメインには存在しないSrc Tyr−527では、リン酸化レベルに変化はなかった(図7B)。総合すると、これらの所見は、EMP2の上昇によって、機械的収縮性を増大させることが知られているFAKおよびSrcのリン酸化が変化することを示す。
【0151】
[0169] EMP2はFAKと物理的に関連する。EMP2の発現は、効率的なインテグリン介在性の細胞応答に必要であり、EMP2は、特定のインテグリンアイソフォームと物理的に関連する(Wadehra Mら, Dev Biol, 287:336-345(2005); Wadehra Mら, Mol Biol Cell, 15:2073-2083(2004); Wadehra Mら, J Biol Chem, 277:41094-41100(2002); Wadehra Mら, Clin Immunol, 107:129-136(2003))。FAKは、インテグリンの重要なシグナル伝達パートナーであり、その役割は、インビトロでのβ1インテグリンの細胞質尾部とFAKのN末端ドメインとの物理的関連を包含する(Lyman Sら, J Biol Chem, 272:22538-22547(1997); Schaller MDら, J Cell Biol, 130:1181-1187(1995))。したがって、本発明者らは、FAKのリン酸化に対するEMP2レベルの正の影響がFAKとEMP2との間の物理的関連を反映し得ると仮定した。この問題に取り組むため、本発明者らは、物理的関連の2つの要件、すなわち、免疫沈降を直接的または間接的に介する結合、および共焦点顕微鏡法を介する同一の細胞空間内での局在化を試験した。
【0152】
[0170] 本発明者らは、まず、EMP2およびFAKが物理的に関連しているかを試験することによってこの見解を評価した(図8A)。EMP2に対する抗体を用いる免疫沈降物には、ARPE−19細胞およびARPE−19/EMP2細胞の両方において容易に検出可能なFAKが包含される。関連の化学量論を決定するために、本発明者らは3つの独立した実験を用いて、免疫沈降物におけるEMP2およびFAKの定量されたレベルを表にした。まず、細胞全体の溶解物および同族免疫沈降物の滴定を、デンシトメトリーによって定量した。このデータを用いて、本発明者らは、同族免疫沈降物における各タンパク質のパーセントを計算した(投入した細胞当量当たり)。第2に、共免疫沈降物を同様に分析して、共免疫沈降物内に捕捉された各タンパク質のパーセントを決定した。最後に、本発明者らは、共免疫沈降したタンパク質の化学量論を表すこれらの2つの値(同族のパーセントおよび共免疫沈降物のパーセント)の比率を計算した。FAK(図8A)およびEMP2の同族の免疫沈降(データは示していない)は、比較的効率的であった(50〜60%)。EMP2免疫沈降物におけるFAKのレベルは、ARPE−19およびARPE−19/EMP2においてそれぞれ12%および15%であり、全FAKの24%〜30%がそれぞれ、これらの2つの細胞系においてEMP2と関連した。
【0153】
[0171] EMP2およびFAKが同一の細胞空間内に局在しているかを評価するために、in situでのそれらの共局在化を共焦点顕微鏡法によって試験した。フィールド当たり複数のレベルを評価したが、スライドと接触し接着斑を含有する細胞底表面の単一のスライスのみを定量に用いた。EMP2(緑)およびFAK(赤)を免疫蛍光によって検出し、共焦点顕微鏡法によって可視化した(図8B)。これらの2つのチャネルの併合が示されており、同様に、強調された共局在化したピクセルが白いシグナルで示されている併合が示されている(右)。検証によると、EMP2およびFAKは、両細胞型において顕著に共局在化していた。3つの独立した実験のピクセル定量によると、ガラススライドに接触している細胞表面に存在しているFAKの68%±9%および96%±1%がそれぞれ、ARPE−19細胞およびARPE−19/EMP2細胞においてEMP2と共局在化していた。全FAKレベルが、ARPE−19/EMP2細胞系において、わずかな、しかし有意ではない増大を示した(図7A)。このことは、EMP2レベルが全FAKタンパク質発現に影響しないことを示す、本発明者らが以前に公表したデータを裏付けるものである(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。
【0154】
[0172] EMP2がリン酸化型FAKと関連するかどうかを試験するために、細胞をEMP2およびp576/577−FAK一次抗体で染色した。フィールド当たり複数のスライスを評価したが、スライドと接触し接着斑を含有する細胞底表面の単一のスライスのみを定量に用いた。EMP2をFITCで検出し、リン酸化型FAKをテキサスレッドで検出した(図8C)。ARPE−19では、p−FAK(Y576/577)の31%±8%がEMP2と共局在化している。細胞内には、共局在化を示す中心領域があるが、この細胞内領域の同定にはさらなる研究が必要である。ARPE−19/EMP2では、p−FAK(Y576/577)の97%±0.4%がこのレベルでEMP2と共局在化している。EMP2およびFAK(全FAKおよびp−FAK(Y576/577)はin situで高度に共局在化しており、EMP2が上昇すると共局在化の程度が増大する。しかし、EMP2が過剰発現している程度を考慮すると、これはFAKの特異的な強化を示唆していない可能性がある。
【0155】
[0173] EMP2レベルが増大するとリン酸化型FAKの分布が改変した。ARPE−19細胞およびARPE−19/EMP2細胞(図9A)をリン酸化型FAK(Y576/577)タンパク質について染色した。EMP2レベルが増大すると、リン酸化型FAKの分布が改変した。ARPE−19細胞では、リン酸化型FAKは主に細胞の周辺で見られたが(図9A)、一方、ARPE−19/EMP2細胞では、リン酸化型FAKが細胞全体にわたり分布しているのが見られる(図9A)。少なくとも4つの個別の試料のピクセル強度を、スチューデントt検定(独立、一方向)によって測定し、タンパク質検出にウェスタンブロットを用いた本発明者らの以前の報告(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))と一致するこの評価も、ARPE−19/EMP2細胞におけるリン酸化型FAKの有意な増大を示した(図9B)。
【0156】
[0174] EMP2およびリン酸化型FAKの発現が増大すると、F−アクチンの分布が改変する。EMP2の発現が増大すると、F−アクチン構成に関連しているTyr−861を包含する複数の部位でFAKリン酸化が増大する。したがって、本発明者らは、EMP2発現の増大と関連する観察可能なアクチン構成における付随する変化が存在したかを試験した。ARPE−19細胞(図10A、C)およびARPE−19/EMP2細胞(図10B、D)をテキサスレッド−Xファロイジン一次抗体で染色した。EMP2発現が増大すると、F−アクチンの発現が改変した。F−アクチンは、ARPE−19/EMP細胞において細胞の周辺に沿って発現したが、一方、F−アクチンは、ARPE−19細胞では細胞の全体にわたり発現した。本発明者らは、ゲル収縮アッセイにおいて試験したものに類似した、活発に成長している細胞におけるアクチン線維を試験することを選択した。コンフルエント細胞における配置を検討するためのさらなる研究を、今後行うことができる。
【0157】
[0175] EMP2の発現が増大すると、α−平滑筋アクチンの発現が増大した。EMP2の発現が増大すると、共に細胞EMTにおいて重要な部位として関与しているTyr−861およびTyr−407でのFAKのリン酸化が増大する。EMTのマーカーは、α−smaの発現である。関連するFAKリン酸化を伴うEMP2発現の増大がα−smaの発現に機能的な影響を有するかを試験するために、α−smaのレベルをARPE−19細胞およびARPE−19/EMP2細胞においてウェスタンブロット分析によって測定した(図11A)。次にブロットをECLで発現させて、結合抗体を視覚化し、β−アクチンを内部対照として用いて定量した(図11B)。EMP2の発現が増大すると、α−smaの発現が有意に増大した。
【0158】
[0176] EMP2レベルが増大すると、I型コラーゲンおよびIV型コラーゲンへの細胞接着が増大する。ARPE−19/EMP2細胞は、ARPE−19細胞と比較して、細胞全体にわたりリン酸化型FAKの大きな分布を示した。本発明者らは、リン酸化型FAKの分布の増大は、ARPE−19/EMP2細胞へのさらなる接着能力をもたらす機能的接着斑に相当すると仮定した。この仮説を試験するために、本発明者らは、ARPE−19細胞およびARPE−19/EMP2細胞を、フィブロネクチン、ビトロネクチン、コラーゲンI、およびコラーゲンIVで事前被覆したプレート上で平板培養した。結合細胞をクリスタルバイオレットの取り込みについて、Bio−Radマイクロプレートリーダー550によって分析した。EMP2の発現が増大すると、I型コラーゲンおよびN型コラーゲンへの接着の付着相が特異的に増大し(P<0.001)、フィブロネクチンへの付着が減少し(P<0.02)、ビトロネクチンへの付着には影響はなかった(図12)。
【0159】
考察
[0177] この研究は、EMP2が、他のテトラスパンファミリーメンバーのように(Hong IKら, J Biol Chem, 281:24279-24292(2006))、特定のインテグリンアイソフォームとその関連するシグナル伝達分子との間の分子アダプターであるという仮説によって導かれた。本発明者らの以前の研究は、FAK−Src経路の活性化が、RPE細胞系ARPE−19を用いるPVRのインビトロモデルでのコラーゲンゲルの収縮の生成において重要であることを同定した(Morales SAら, Experimental Eye Research(2007))。さらに、本発明者らの研究は、EMP2が、FAK(y-576/577)の活性化を介してコラーゲンゲル収縮を制御し得ることを実証した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。本発明者らは、以前に、EMP2がインテグリンβ1と関連することを実証し(Wadehra Mら, J Biol Chem, 277:41094-41100(2002))、他のグループは、FAKがインビトロでインテグリンβ1サブユニットに結合することを示している(Schlaepfer DDら, Prog Biophys Mol Biol, 71:435-478(1999); Parsons JTら, Oncogene, 19:5606-5613(2000))。この研究において、本発明者らは、EMP2がFAKの活性化を増強し、FAKと物理的に関連し、その機能的な下流の結果は、接着斑の密度、細胞の接着能力、F−アクチンの立体配座、およびアクチンの組成に影響するという証拠を提供する。したがって、EMP2は、効率的なインテグリン介在性のFAK活性化およびFAKに関連する細胞機能に対するその影響のための分子アダプターとして作用し得る。共免疫沈降および共局在化の研究は、多タンパク質複合体の形成を介する直接的なまたは間接的な関連を裏付けるが、この関連の具体的な詳細を規定するためには、さらなる生化学的研究が必要であろう。
【0160】
[0178] FAKの活性化のための重要なメカニズムは、複数部位での自己リン酸化、Srcの動員、およびFAKのリン酸化をもたらす、インテグリン受容体介在性のFAKのクラスタリングである。活性化型FAK/Src複合体は、その後、Grb2、p130Cas、パキシリン、Talin、CAP、Graf、Src、およびPI3−キナーゼのp85サブユニットなどの、種々のタンパク質のための分子足場として作用し、多くのシグナル伝達経路に関与し得る(Schlaepfer DDら, Prog Biophys Mol Biol, 71:435-478(1999); Parsons JTら, Oncogene, 19:5606-5613(2000); Cox BDら, J Cell Biochem, 99:35-52(2006))。
【0161】
[0179] この報告において、本発明者らは、EMP2がFAKと関連して、接着斑の形成を増強することによってFAKの活性化を増大させることを実証し、EMP2がインテグリンのライゲーションとFAKの活性化との間の分子アダプターとして作用し得ると仮定する。本発明者らは、以前、増殖性硝子体網膜症(PVR)のインビトロでの相関要因であるコラーゲンゲル収縮におけるEMP2の役割を試験した。PVRは、患者の最大10%において、裂孔原性網膜剥離の外科的修復に続発する合併症である(Charteris DGら, Eye, 16:369-374(2002); Kirchhof B., Graefes Arch Clin Exp Ophthalmol, 242:699-703(2004); Lewis GPら, Eye, 16:375-387(2002))。PVRは、RPEを包含する複数の細胞型の、硝子体腔内への複雑な移動の最終結果に相当する(Baudouin Cら, Ophthalmologica, 203:38-46(1991); Casaroli-Marano RPら, Invest Ophthalmol Vis Sci, 40:2062-2072(1999); Charteris DGら, Br J Ophthalmol, 79:953-960(1995); Vinores SAら, Invest Ophthalmol Vis Sci, 31:2529-2545(1990))。牽引性網膜剥離をもたらす収縮性の細胞の力に関連する移動、膜形成、および異常な創傷治癒戦略をもたらすRPE細胞の上皮間葉移行(EMT)の証拠が存在する。ARPE−19細胞によって行われるコラーゲンゲル収縮を用いて、本発明者らは、FAKを介するインテグリン受容体の活性化およびシグナル伝達をARPE−19介在性の収縮に必要な非常に重要なプロセスであると同定した(Morales SAら, Experimental Eye Research(2007))。本発明者らは、最近、EMP2の発現レベルがコラーゲンゲルの収縮と正に相関すること、およびこの表現型がFAK活性化の促進を介して達成されることを観察した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。
【0162】
[0180] PVRの発症機序の状況において、EMTは、鍵となる構成要素である。Tyr−407およびTyr−861でのFAKのリン酸化は、EMTの進行において重要であると同定される(Nakamura Kら, Oncogene, 20:2626-2635(2001))。EMP2の過剰発現は、Tyr−407およびTyr−861でのリン酸化を増大させ、したがって、EMTを進行させると予測される。EMTの1つのマーカーはα−smaのアップレギュレーションであり、これは、EMP2を過剰発現する細胞系であるARP−19/EMP2細胞において観察された。EMP2の増大およびFAK活性化の増大に関連するこれらの変化は、さらなる収縮性の表現型を促進すると予測される。F−アクチンの構成はまた、コラーゲンゲルに接触する細胞の能力に影響し得る。ARPE−19細胞では、F−アクチンの発現は細胞全体にわたり観察され、これはおそらく、より堅固な細胞構造をもたらし、コラーゲンゲルを収縮させる細胞の能力を低減させる。ARPE−19/EMP2細胞では、F−アクチンは、皮質内分布している細胞の縁に沿って見られるのみであり、これは、さらなる収縮能力を可能にするさらに柔軟な立体配座をもたらし得る。
【0163】
[0181] テトラスパンタンパク質は、様々な複合体の形成に関与して「テトラスパン網」を形成することが示されており(Levy SおよびShoham T, Physiology(Bethesda)20:218-224(2005); Levy SおよびShoham T, Nat Rev Immunol, 5:136-148(2005))、これは、シグナル伝達および分類プロセスを調節する足場および膜ドメインの生成である(Levy SおよびShoham T, Physiology(Bethesda)20:218-224(2005); Hemler ME, Nat Rev Mol Cell Biol, 6:801-811(2005))。これらの複合体は、それらの膜タンパク質成分のシグナル伝達、輸送、および構造的特徴を調節し得る。テトラスパンがインテグリン分子と複合体を形成する場合、テトラスパンは細胞の接着および移動性を調節し得る(Caplan MIら, Curr Opin Nephrol Hypertens, 16:353-358(2007))。本発明者らの報告はテトラスパンのパズルにピースを加えるものであり、EMP2が、さらに大きな細胞収縮能力を有するEMT表現型に向かわせる物理的関連を介するFAK活性化を調節することを実証する。EMP2−FAKの関連は、PVRのインビトロモデルの状況における有意な機能的細胞応答を実証する、これまでに報告されていない新規なタンパク質−タンパク質相互作用に相当する。
【0164】
(実施例4)
導入
[0182] 4回膜貫通(テトラスパン)タンパク質であるEMP2は、眼、肺、心臓、甲状腺、および子宮における異なる位置で発現する(Wang CXら, Blood, 97(12):3890-3895(2001))。眼では、EMP2は、角膜、毛様体、および網膜色素上皮(RPE)を包含する複数の上皮層に局在化している(Wadehra Mら, Exp Mol Pathol, 74(2):106-112(2003))。複数の非眼球細胞型では、EMP2は、選択的な受容体輸送において重要な役割を有し、増殖、侵入、接着、および移動において重要な分子に影響する(Wadehra Mら, Exp Mol Pathol, 74(2):106-112(2003); Wadehra Mら, Dev Biol, 287(2):336-345(2005); Wadehra Mら, Mol Biol Cell, 15(5):2073-2083(2004); Wadehra Mら, J Biol. Chem, 277(43):41094-41100(2002); Wadehra Mら, Cancer 107(1):90-98(2006); Wadehra Mら, Clin Immunol, 107(2):129-136(2003))。これらの研究は、眼球由来の細胞における異なる重要な細胞プロセスの協調的な制御におけるEMP2の潜在的な中心的役割を示唆する。
【0165】
[0183] PVRは、患者の最大10%において裂孔原性網膜剥離の後に観察され、失明をもたらす可能性がある合併症である(Charteris DGら, Eye, 16(4):369-374(2002); Kirchhof B., Graefes Arch Clin Exp Ophthalmol, 242(8):699-703(2004); Lewis GPら, Eye, 16(4):375-387(2002))。PVRの根幹にある病態生理学は複雑であり、RPEを包含する複数の細胞型が、この疾患における重要な役割を有すると考えられている(Baudouin Cら, Ophthalmologica, 203(1):38-46(1991); Casaroli-Marano RPら, Invest Ophthalmol Vis Sci, 40(9):2062-2072(1999); Charteris DG., Br J Ophthalmol, 79(10):953-960(1995); Vinores SAら, Invest Ophthalmol Vis Sci, 31(12):2529-2545(1990))。牽引性網膜剥離をもたらし得る収縮性の細胞の力に関連する、細胞の移動、膜形成、および異常な創傷治癒プロセスをもたらす、上皮間葉移行の証拠が存在する。PVRの予防のための新たな戦略には、複雑な病態生理学の理解を進めることが必要である。PVRの1つのインビトロでの相関要因は、コラーゲンゲルの収縮である。
【0166】
[0184] 一次ヒト細胞(Hunt RCら, Invest Ophthalmol Vis Sci, 35(3):955-963(1994))、ヒトARPE−19細胞(Bando Hら, Exp Eye Res, 82(3):529-537(2006))、ウシ(Ando Aら, Br J Ophthalmol, 84(11):1306- 1311(2000))、およびウサギ(Zheng Yら, Invest Ophthalmol Vis Sci, 45(2):668-674(2004))を包含する種々の種のRPEが、コラーゲンゲル収縮の研究において用いられている。この研究は、いかにEMP2が、ARPE−19細胞系におけるEMP2の発現を組換えによって改変することを介して、コラーゲンゲル収縮を制御するかを試験するために設計された。本発明者らの以前の研究(Morales SAら, Exp Eye Res, 85(6):790-798(2007))および他者による研究(Bando Hら, Exp Eye Res, 82(3):529-537(2006); Carver Wら, J Cell Physiol, 165(2):425-437(1995); Cooke MEら, J Cell Sci, 113:2375-2383(2000); Kieffer JDら, Biochem Biophys Res Commun, 217(2):466-474(1995); Langholz Oら, J Cell Biol , 131:1903-1915(1995); Moulin VおよびPlamondon M., Br J Dermatol, 147(5):886-892(2002); Robbins SGら, Invest Ophthalmol Vis Sci, 35(9):3475-3485(1994); Zhang ZGら, J Cell Sci, 119:1886-1895(2006))は、特異的なインテグリンアイソフォームを同定し、コラーゲンマトリクスがコラーゲンゲル収縮において非常に重要であることが確実であることを見出した。本発明者らは、以前、FAK/Src経路の活性が、外因性の炎症性刺激の存在下または非存在下でのコラーゲンゲル収縮の観点からARPE−19細胞系において必須であると同定した。本研究では、EMP2の発現レベルはコラーゲンゲルの収縮を制御し、EMP2の増大は、ARPE−19細胞系におけるFAK活性化の増強に関連していた。
【0167】
方法
[0185] EMP2構築物: 以前に報告されているように、ハンマーヘッドリボザイムを生成して、ヒトEMP2転写産物を切断した(Wadehra Mら, Dev Biol, 287(2):336-345(2005))。EMP2の発現を低減させるためのトランスフェクションにおいて良好に機能することが実証されているhRZ2 EMP2ハンマーヘッドリボザイムを、この研究において用いた。簡潔に述べると、pEGFPにおけるhRZ2構築物(BD-Clontech, Palo Alto, CA)をARPE−19内にトランスフェクトし、安定なクローンを選択した(Wadehra Mら, Dev Biol, 287(2):336-345(2005))。
【0168】
[0186] ヒトEMP2の完全長cDNAをレトロウイルスベクターpMSCV−IRES−GFP内のEcoRI部位でクローニングした(Hawley RGら, Gene Ther, 1(2):136-138(1994))。EMP2の発現は、5’の長い末端反復(LTR)によって推進した。このベクターはまた、内部リボソーム侵入部位を介する緑色蛍光タンパク質(GFP)の発現を可能にする。高力価のヘルパーフリーレトロウイルスストックを、以前に記載されているように、293T細胞の一時的なコトランスフェクションによって調製した(Pear W., In: Ausubel FM, Brent R, Lingston RE,ら編, Current Protocols in Molecular Biology, 第2巻. New York: John Wiley & Sons; 9.11-9.15(1996))。
【0169】
[0187] 細胞系: RPE特異的マーカーであるCRALBPおよびRPE−65を発現する自然発生的な網膜色素上皮(RPE)細胞系であるARPE−19を、American Type Culture Collectionから得た(CRL-2302; ATCC, Manassas, VA)。ARPE−19細胞を、5%CO2の湿潤チャンバにおいて37℃で、10%ウシ胎児血清(FBS; ATCC)を補ったDMEM−F12培地において培養した。培養培地を週に2回交換した。コンフルエントになった後、培養物を0.05%(重量/容量)トリプシン内で解離することによって継代した。EMP2のレベルは、EMP−2を過剰発現するレトロウイルス構築物の安定な感染を介してAFPE−19細胞において増大し、GFP陽性細胞の流動選別によって選択された。EMP2の発現が増大するように安定に感染したこれらの細胞は、ARPE−19/EMP2と呼ばれる。EMP2のレベルは、pEGFP−hRZ2リボザイム構築物3.4μgおよびトランスフェクション試薬(FuGENE 6; Roche, Basel, Switzerland)でのARPE−19細胞の安定なトランスフェクションにより減少した。ARPE−19/Ribo細胞を、GFP陽性細胞について流動選別した。とりわけ、トランスフェクション試薬は、トリパンブルー排除によって決定したところ、ARPE−19細胞に対する検出可能な毒性を生じさせなかった(データは示していない)。
【0170】
[0188] EMP2のレベルは、ARPE−19細胞をEMP2siRNA75ピコモル(L-016226-00; Dharmacon, Lafayette, CO)および親油性トランスフェクション試薬(Lipofectamine 2000; Invitrogen, Carlsbad, CA)で一時的にトランスフェクトすることによって減少し、48時間後に分析された。陰性対照として、細胞をスクランブル対照siRNA75ピコモル(D-001206-13-05; Dharmacon)でトランスフェクトした。EMP2siRNAおよび対照siRNAはそれぞれ、EMP2を標的化する4つのsiRNAのプールまたは4つの非標的化siRNAのプールである。EMP2発現のレベルを、ウェスタンブロットによって定量した。
【0171】
[0189] 抗体: ヒトα1(クローンSR84)、α2(クローンAK-7)、およびα3(クローンC3II.1)インテグリンアイソフォームに特異的なモノクローナル抗体(mAb)は、BD Biosciences(San Diego, CA)から得た。ヒトEMP2に対するウサギ抗血清は、ヒトEMP2の第2の細胞外ループ(EDIHDKNAKFYPVTREGSYG)に結合した多抗原ペプチドで動物を免疫化した後に生成した(Research Genetics, Huntsville, AL)(Wadehra Mら, Exp Mol Pathol, 74(2):106-112(2003))。ヒトFAK(クローンC-20)およびpFAK(クローンTyr 576/577)に特異的なウサギ抗体を、Santa Cruz Biotechnology(Santa Cruz, CA)から得た。ヒトβ−アクチン(クローン2A2.1)に特異的なマウス抗体を、US Biological(Swampscott, MA)から得た。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体を、Southern Biotech(Birmingham, AL)から購入した。ホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体、およびマウスIgGに特異的なR−PE結合抗体を、BD Biosciencesから得た。
【0172】
[0190] フローサイトメトリー: α1、α2、およびα3インテグリンサブユニットの膜発現を、フローサイトメトリーによって評価した。細胞を、PBS内の2%パラホルムアルデヒド(重量/容量)を用いて20分間にわたり氷上で固定したが、透過処理はせず、次に、一次抗体と共に30分間にわたり氷上で、PBS+2%FCS内でインキュベートした。それらを2回洗浄し、R−PE結合抗マウスIgG抗体と共に30分間にわたり氷上でインキュベートした(BD Biosciences)。R−PEを100万個細胞当たり0.25μgで用いた。陰性対照として、細胞をアイソタイプ対照抗体のみと共にインキュベートした。2回連続して洗浄した後、細胞をPBS内に再懸濁し、フローサイトメトリーで分析した(FACScan; BD Biosciences)。細胞集団における発現の反映である、平均蛍光強度(MFI)として計算されたインテグリンの発現レベルを、複数の独立した実験において決定した。
【0173】
[0191] コラーゲンゲルの収縮:コラーゲンゲル収縮アッセイを以前報告されているように行った(Morales SAら, Exp Eye Res, 85(6):790-798(2007))。簡潔に述べると、コラーゲンゲルを、I型コラーゲン(BD Biosciences)、10×DMEM、およびDMEM/F12を組み合わせることによって調製した。I型コラーゲン混合物の最終濃度は、2.5mg/mLであった。コラーゲン溶液(500μL)を24ウェルプレートの各ウェルに添加し、5%C02で、37℃で1時間にわたりインキュベートした。EMP2レベルが変化した、培養したARPE−19細胞を採取し、無血清DMEM/F12内に5×105個/mLの最終濃度で再懸濁した。EMP2レベルが変化したARPE−19細胞を、ウェル当たり2.5×105個細胞の濃度でコラーゲンゲル上に播種し、収縮のパーセンテージを24時間目に測定した。各ゲルの領域を画像キャプチャ(Gel Doc 2000; Bio-Rad, Hercules, CA)を用いてゲルの写真を撮ることによって得、NIH Image J(Wayne Rasband, National Institutes of Health, Bethesda, MDにより開発; http://rsb.info.nih.gov/ij/index.htmlで入手可能)で定量した。ゲルの領域を測定するために、本発明者らは、各ゲルの輪郭を描くための楕円形測定ツールを用いた。時点0でのゲルの領域を24時間後のゲルの領域と比較して、各試料についての収縮のパーセンテージを生成した。各実験は少なくとも6回の反復を含み、少なくとも3つの独立した実験を行い、同等の結果を得た。スチューデントt検定(独立、一方向)を用いた。P<0.05を、統計的に有意であると判断した。
【0174】
[0192] 増殖アッセイ: 細胞を96ウェルプレート上に播種し、一晩インキュベートした。培地を除去し、次に、正常培地またはコラーゲンI25mg/mLを含有する培地で置き換えた。次に細胞を48時間にわたりインキュベートし、非同位体比色免疫アッセイであるCalbiochem(San Diego、CA)のBrdU細胞増殖アッセイによって測定される、BrdUの組み込みによって増殖を評価した。反応生成物をマイクロプレートリーダー(モデル550、Bio-Rad)により595nmの波長で定量した。
【0175】
[0193] 移動アッセイ:ARPE−19細胞およびARPE−19/EMP2細胞を24ウェルプレート上に播種し、細胞がコンフルエントに達するまで3日間にわたりインキュベートした。細胞をPBSで洗浄し、無血清培地を添加し、細胞を一晩インキュベートした。ピペットチップ10μLを用いて単層内に擦り傷を作り、培地を除去して、無血清培地またはPDGF50ng/mLを含有する無血清培地で置き換えた。創傷の写真を様々な時点で撮り、擦り傷の閉鎖のパーセンテージをNIH Image Jソフトウェアで定量した。擦り傷の領域を創傷が生じた直後に測定した。時間が経つにつれて細胞はクリアな領域に移動したが、ギャップは依然として24時間後にも視認可能であった。ギャップのサイズを測定し、元の擦り傷のサイズで割り、この値を閉鎖のパーセンテージとして表した。
【0176】
[0194] 侵入アッセイ:細胞侵入アッセイ(QCM Collagen Cell Invasion Assay; Chemicon, Temecula, CA)を用いて、全ての侵入アッセイを行った。ARPE−19細胞およびARPE−19/EMP2細胞を、重合コラーゲンの薄層で被覆された8μmの孔サイズのポリカーボネート膜を含有する侵入チャンバインサート上に播種した。重合コラーゲン層を介して侵入および移動する細胞は、ポリカーボネート膜の底に張り付く。インサート膜の底にある侵入細胞を比色分析によって同定し、マイクロプレートリーダー(モデル550; Bio-Rad)を用いた560nmでの検出を用いて定量的に分析した。
【0177】
[0195] コラーゲンの生成:ARPE−19細胞およびARPE−19/EMP2細胞を、TCP−β10ng/mLの存在下または非存在下の無血清培地内で、6ウェルプレート上に7×105個細胞/mLの濃度で播種した。培地を72時間後に回収した。分泌されたコラーゲンを測定した(Sircol Collagen Assay; Accurate Chemical & Scientific, Westbury, NY)。コラーゲンアッセイは、比色分析手順である。反応生成物をマイクロプレートリーダーで540nmの波長で定量した。各実験は少なくとも6回の反復を含み、少なくとも3つの独立した実験を行った。スチューデントt検定(独立、一方向)を用いた。P<0.05を統計的に有意であると判断した。
【0178】
[0196] FAK/Srcの阻害:コラーゲンゲルをDMEM/F12内のI型コラーゲン(BD Biosciences)で、2.5mg/mLの最終濃度で調製した。新たに調製したコラーゲン溶液を、24ウェルプレートの各ウェルに添加し、5%CO2で、37℃で1時間にわたりインキュベートした。培養したARPE−19細胞、ARPE−19/EMP2細胞、およびARPE−19/EMP2siRNA細胞を採取し、無血清DMEM/F12内に5×105個/mLの最終濃度で再懸濁した。細胞を様々な濃度の低分子阻害剤で1時間にわたり前処理した。阻害剤PP2(FAK/Src阻害剤)およびSU6656(Src阻害剤)は、DMSO中に希釈して用いた(Calbiochem)。細胞を、媒体対照としてDMSOのみで処理した。ARPE−19細胞、ARPE−19/EMP2細胞、およびARPE−19/EMP2siRNA細胞を、コラーゲンゲル上にウェル当たり2.5×105個細胞の濃度で播種し、収縮のパーセンテージを特定の時間間隔で測定した。少なくとも3つの独立した実験を行い、示されているところでは、結果を、スチューデントt検定(独立、一方向)で統計的有意性について評価した。P<0.05のレベルを、統計的に有意であると考えた。
【0179】
[0197] ウェスタンブロット分析:ウェスタンブロット分析を、以前記載されているように行った(Wadehra Mら. Dev Biol, 292(2):430-441(2006))。簡潔に述べると、細胞タンパク質を、プロテアーゼ阻害剤およびホスファターゼ阻害剤を含有するRIPA緩衝液(Upstate, Charlottesville, VA)を用いて単離し、タンパク質濃度をタンパク質アッセイ(BCA; Bio-Rad)で決定した。全部でタンパク質10μgを各レーンにロードし、タンパク質を、還元条件下で、4%〜20%SDS−PAGE勾配ゲルによって分画した。タンパク質をニトロセルロース膜(GE Healthcare, Buckinghamshire, UK)に移し、転移の妥当性を、Ponceau Sレッド染色(Sigma-Aldrich)によって確認した。膜を、次に、TBS Tween(TBST; Upstate)内の無脂肪乳でブロックした。ブロットを、FAKおよびp−FAK(Tyr 576/577)では1:200の希釈、EMP2では1:1000の希釈、β−アクチンでは1:5000の希釈の一次抗体と共に、1時間にわたりインキュベートした。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体またはホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体を、1:2000の希釈でブロットに曝露した。ブロットを、次に、化学発光で発現させて、結合抗体を視覚化し(ECL; Pierce, Rockford, IL)、内部対照としてβ−アクチンで定量した。ウェスタンブロット分析を、NIH Image Jで定量した。ブロットをフラットベッドスキャナでデジタル化し、バンドの密度を、Image Jを用いて測定した。ロードのばらつきを明らかにするために、β−アクチンを用いて各試料を正規化した。少なくとも3つの独立した実験を行い、示されているところでは、結果を、スチューデントt検定(独立、一方向)で統計的有意性について評価した。P<0.05のレベルを統計的に有意であると考えた。
【0180】
結果
[0198] ARPE−19細胞におけるEMP2発現の組換えによる変化: EMP2は、網膜色素上皮細胞において高度に発現し、これは、膜の形成およびコラーゲンゲルの収縮において重要であると考えられている。コラーゲンゲルの収縮に対するEMP2の影響を研究するために、EMP2レベルをARPE−19細胞系において変化させて、2つのさらなるARPE−19系、すなわちARPE−19/EMP2と呼ばれる過剰発現系およびARPE−19/Riboと呼ばれる安定なノックダウンを生じた。EMP2の定常状態のタンパク質レベルを、ARPE−19細胞、ARPE−19/EMP2細胞、およびARPE−19/Ribo細胞においてウェスタンブロット分析によって測定した。実験は独立して少なくとも3回行い、同様の結果を得、1つの代表的な実験を図13Aに示す。ARPE−19/EMP2細胞は、連続希釈を用いて比較すると、野生型細胞と比較して、EMP2の発現において75〜100倍の増大を示した。ARPE−19/Ribo細胞は、ARPE−19細胞と比較して、EMP2発現において5倍の減少を示した。
【0181】
[0199] EMP2発現を減少させるための別の方法では、EMP2に特異的なsiRNA(ARPE-19/EMP2siRNA)または対照スクランブルsiRNA(ARPE-19/対照siRNA)で一時的にトランスフェクトしたARPE−19細胞を用いた。特異的siRNAを用いるこの技術は、対照ARPE−19における発現のおよそ40%まで、EMP2の発現を特異的に減少させた(図13B)。スクランブルsiRNAは、EMP2の発現においていかなる変化ももたらさなかった。
【0182】
[0200] コラーゲンゲルの収縮のEMP2による修飾:ARPE−19/EMP2細胞、ARPE19/Ribo細胞、およびARPE−19細胞のコラーゲンゲル収縮アッセイを、試料当たり6回の反復で少なくとも3回の個別の回数行った。EMP2レベルの改変は、コラーゲンゲルを収縮させる細胞の能力に有意に影響した。EMP2発現の増大は、未処理の野生型細胞と比較して、収縮を有意に増大させた(図14)。これと合致して、ゲル収縮の減少が、ARPE−19/Ribo細胞において観察された。EMP2構築物を有さない、レトロウイルスに感染した細胞系である、さらなる対照ARPE−19細胞系、およびリボザイムを有さないトランスフェクトされたEGFP−N3ベクターは、野生型対照ARPE−19細胞系と比較して収縮を改変しなかった(データは示していない)。
【0183】
[0201] インテグリンα1、α2、またはα3における変化を介する収縮に対するEMP2の影響:以前の結果は、EMP2の発現が増大するとコラーゲンゲルの収縮が増大することを示した。EMP2は、マウスNIH3T3およびヒト子宮内膜癌系HEC1Aを包含する様々な細胞型における特異的なインテグリンアイソフォームの細胞内輸送および細胞表面発現を調節することが知られている(Wadehra Mら, Dev Biol, 287(2):336-345(2005); Wadehra Mら, J Biol. Chem, 277(43):41094-41100(2002))。本発明者らおよび他者は、α1、α2、およびα3インテグリンの係合の、ARPE−19細胞系におけるコラーゲンゲル収縮における重要な工程としての重要性を実証し(Bando Hら, Exp Eye Res, 82(3):529-537(2006); Morales SAら, Exp Eye Res, 85(6):790-798(2007); Carver Wら, J Cell Physiol, 165(2):425-437(1995); Cooke MEら, J Cell Sci, 113:2375-2383(2000); Kieffer JDら, Biochem Biophys Res Commun, 217(2):466-474(1995); Langholz Oら, J Cell Biol , 131:1903-1915(1995); Moulin VおよびPlamondon M., Br J Dermatol, 147(5):886-892(2002); Robbins SGら, Invest Ophthalmol Vis Sci, 35(9):3475-3485(1994); Zhang ZGら, J Cell Sci, 119:1886-1895(2006))、それによって、EMP2が、コラーゲンを結合するこれらのインテグリンの1つの細胞表面発現を増大し得ると仮定した。
【0184】
[0202] EMP2の発現が改変した細胞系を、これらのインテグリンアイソフォームの表面発現についてフローサイトメトリーによって評価した(図15)。予測した結果とは対照的に、EMP2を過剰発現するように操作された細胞は実際に、インテグリンα1(P=0.003)およびα2(P=0.002)の細胞表面発現のおよそ65%の減少を示した。ARPE−19/EMP2細胞におけるインテグリンα3の発現では、わずかな、しかし統計的に有意ではない減少があった(図15B)。この結果によって、本発明者らは、EMP2がこの細胞系におけるコラーゲンゲル収縮を調節し得る他のメカニズムを調べた。
【0185】
[0203] 増殖、移動、侵入、およびコラーゲン生成における変化と収縮に対するEMP2の影響との関連:EMP2がコラーゲンの収縮を調節し得る潜在的なメカニズムは、収縮の増大をもたらす細胞増殖の増大によるものである。BrdUの組み込みによってアッセイされた増殖を、コラーゲン刺激の非存在下または存在下で、ARPF−19細胞およびARPE−19/EMP2細胞において評価した(図16A)。コラーゲンの非存在下では、EMP2の過剰発現によって、増殖が穏やかに減少した(20%)。コラーゲン刺激の後、両細胞系の増殖に統計的に有意な差はなく、したがって、増殖の増大によってARPE−19/EMP2細胞によるコラーゲンゲルの収縮が増強され得るという仮説を反証した。
【0186】
[0204] 移動に対する、EMP2発現の増大の考えられる影響を調べるために、本発明者らは、創傷治癒アッセイを行った。ARPE−19細胞およびARPE−19/EMP2細胞を、PDGF50ng/mLで処理しないままにするかまたは処理し、閉鎖のパーセンテージを、創傷閉鎖の代用的評価として、6、12、および24時間目に測定した。6時間および12時間の時点で、PDGFの非存在下または存在下で、2つの細胞系の間に差はなかった(データは示していない)。24時間の時点では、PDGF刺激の非存在下で、ARPE−19/EMP2細胞は、ARPE−19細胞と比較して、ギャップの閉鎖において23%の減少を示した。PDGF処理はこの影響を無効にし、その結果、2つの細胞系において同等の創傷治癒が生じた(図16B)。増殖および移動の両方が、創傷の閉鎖に関与する。ARPE−19/EMP2細胞のより遅い増殖速度は創傷閉鎖の減少に関与し得るが、EMP2を過剰発現する細胞があまり運動性を示さない可能性もある。運動性の表現型を促進することが知られているPDGF処理は、ARPE−19細胞の創傷閉鎖を刺激し、さらに、ARPE−19/EMP2細胞の創傷閉鎖の減少を克服した。ARPE−19/EMP2細胞によるコラーゲンゲル収縮の増大は、EMP2の発現の改変に起因する移動の増大によって説明することはできない。
【0187】
[0205] 本発明者らは、別のメカニズム、すなわち、EMP2の発現がコラーゲンマトリクスに侵入する細胞の能力に影響し得るという可能性を試験した。ARFE−19細胞およびARPE−19/EMP2細胞をポリカーボネート膜上に播種し、10%FESまたはPDGF50ngを化学誘引物質として用いた。侵入を24および48時間目に測定した。いずれの条件下でも、ARPE−19細胞とARPE−19/EMP2細胞との間で侵入に差はなかった(図16C)。コラーゲンゲル収縮のEMP2介在性の増強は、改変した侵入能力によるものではなかった。
【0188】
[0206] 本発明者らが調べたさらなるメカニズムは、EMP2レベルがコラーゲンの生成に影響するかどうかであった。ARPE−19細胞およびARPE−19/EMP2細胞を、24ウェルプレートにおいて72時間にわたり成長させた。培地を回収し、コラーゲン生成を分析した。2つの細胞系の間でコラーゲンの生成に統計的な差はなかった(図16D)。ARPE−19/EMP2細胞による細胞収縮能力の増大は、コラーゲン生成の改変によって説明することはできない。
【0189】
[0207] ARPE−19細胞におけるFAK活性化およびコラーゲンゲル収縮のEMP2による制御:インテグリンの発現、増殖、移動、または侵入における変化は、いかにしてEMP2発現の増大がゲルコラーゲンの収縮を増大させたかを説明するものではない。以前に、本発明者らは、インテグリンのライゲーションおよびFAK/Src複合体の活性化がARPE−19細胞によるコラーゲンゲルの収縮に必要であることを観察した(Morales SAら, Exp Eye Res, 85(6):790-798(2007))。本発明者らは、ARPE−19細胞におけるEMP2の過剰発現によってFAK/Srcの活性が改変するかを調べた。全FAKレベルはARPE−19細胞およびARPE−19/EMP2細胞の両方において等しかったが、過剰発現するARP−19/EMP2細胞は、対照ARPE−19細胞と比較して、活性化型FAKのレベルがほぼ2倍増大することを示した(図17A)。各活性化型FAKをそれ自身のβ−アクチンロード対照に正規化して行った複数の評価は、EMP2レベルの増大とFAK活性化との間の統計的に有意な相関を示した(図17B)。
【0190】
[0208] FAK/Src阻害剤によるコラーゲンゲル収縮の阻害に対する耐性に対するEMP2過剰発現の影響:EMP2の過剰発現がFAK活性化の増大に関連するという観察を受けて、FAK/Src経路の阻害剤を用いて、ゲル収縮の状況におけるこの観察の機能的有意性を試験するためのさらなる研究を行った。もしEMP2の影響がFAKの活性化に続発するものであれば、ARPE−19/EMP細胞における観察されたコラーゲンゲルの収縮は、対照ARPE−19細胞と比較して、FAK/Srcの阻害に対する耐性の増大を示すはずである。この予測を調べるために、本発明者らは、様々な濃度の、ARPE−19細胞におけるコラーゲンゲルの収縮を予防することが実証されているFAK/Src低分子阻害剤であるPP2およびSU6656を用いた(Morales SAら, Exp Eye Res, 85(6):790-798(2007))(図18、19)。EMP2特異的siRNAでのEMP2発現の減少は、対照スクランブルsiRNAと比較して、コラーゲンゲル収縮の減少を示した。この結果は、ARPE−19/Ribo細胞系において観察されるコラーゲンゲル収縮の減少と合致した(図18A)。FAK阻害剤PP2に対するコラーゲンゲル収縮アッセイの感受性はEMP2のレベルと逆相関し、EMP2siRNAで処理した細胞においては感受性が増大し、EMP2を過剰発現する細胞においては耐性が増大した(図18A〜C)。媒体対照に正規化された収縮の阻害のパーセンテージを異なる濃度の阻害剤で各細胞系について決定した。ゲル収縮の50%の阻害を達成するために必要なPP2の濃度は、ARPE−19細胞では2.5μMであり、ARPE−19/EMP2siRNA細胞では1μM未満であり、過剰発現するARPE−19/EMP2細胞では5μMであり、これらの細胞系において観察されるFAK活性化の変化と一致した(図18D)。FAK/Src経路の第2の阻害剤であるSU6656は、ARPE−19細胞およびARPE−19/EMP2細胞で類似の結果を示し、したがって、コラーゲンゲル収縮のEMP2による制御についてのメカニズムがFAKの活性化の変化を介することの独立した裏付けを提供した(図19)。
【0191】
考察
[0209] PVRについてのインビトロでの相関要因であるコラーゲンゲルの収縮は、FAK介在性のインテグリンシグナル伝達に依存する細胞プロセスである(Morales SAら, Exp Eye Res, 85(6):790-798(2007))。この記事において、本発明者らは、EMP2がコラーゲンゲルの収縮をFAK活性化の増強に依存するプロセスで調節することを実証した。FAK活性化のEMP2による調節は新たな観察であるが、様々なインテグリンシグナル伝達メカニズムのテトラスパニン関連性の調節についての以前の観察は、このファミリーの膜関連性タンパク質に共通する潜在的な機能を示唆する。
【0192】
[0210] テトラスパニンがFAK依存性経路または他のシグナル伝達経路の活性化を調節するメカニズムは、いまだ明らかにされていない。特定のテトラスパニン(CD9、CD53、CD81、CD82、およびCD151)は互いに関連するか、またはさらなる膜タンパク質とのヘテロ複合体で関連し、結果として、チロシンのリン酸化を増大させる(Boucheix Cら, Cell Mol Life Sci, 58(9):1189-1205(2001); Hemler ME., J Cell Biol, 155(7):1103-1107(2001); Hong IKら, J Biol Chem, 281(34):24279-24292(2006); Lagaudriere-Gesbert Cら, Cell Immunol, 182(2):105-112(1997); Todd SCら, J Exp Med, 184(5):2055-2060(1996))。これは、4回膜貫通タンパク質が、膜におけるシグナル伝達複合体の機能的組み立て体を支持する分子アダプターとして作用することを示唆する(Hong IKら., J Biol Chem, 281(34):24279-24292(2006))。
【0193】
[0211] インテグリンおよび他の細胞表面受容体の機能的調節は、テトラスパンファミリーの複数のタンパク質に共通の、再発的に観察される特徴である。いくつかのテトラスパニン(CD9、CD53、CD81、およびCD82)は、インテグリン(α3β1、α43β1、およびα63β1)とのタンパク質−タンパク質相互作用に関与し、接着および細胞活性化を改変させる(Lagaudriere-Gesbert Cら, Cell Immunol, 182(2):105-112(1997); Todd SCら, J Exp Med, 184(5):2055-2060(1996))。CD9、CD63、CD81、CD151、およびA15/TALLA1を包含する複数のテトラスパニンは、PI−4キナーゼを特異的な膜位置に動員し、ホスホイノシチド依存性のシグナル伝達を誘発する(Yauch RLおよびHemler ME., Biochem J, 3:629-637(2000))。さらに、CD81と抗CD81抗体との架橋は、TCR/CD3複合体を介するシグナル伝達にとって共刺激性である(Todd SCら, J Exp Med, 184(5):2055-2060(1996))。発現レベルの変化またはテトラスパニンの架橋との間の他の関連は、JNKの活性化を誘発するCD53の発現またはライゲーションのアップレギュレーションを包含するシグナル変換の活性化(Yunta Mら, Eur J Biochem, 269(3):1012-1021(2002))、線維肉腫細胞におけるFAKのチロシンリン酸化を誘発するCD9による接着の制御(Berditchevski FおよびOdintsova E, J Cell Biol, 146(2):477-492(1999))、ならびにヒト黒色腫細胞におけるFAK、Src、およびc−Junキナーゼの接着依存性の活性化を誘発する同族親和性のCD151相互作用(Hong IKら, J Biol Chem, 281(34):24279-24292(2006))に関連している。さらに、CD151の同族親和性のタンパク質−タンパク質相互作用は、FAK−SrcおよびMAPキナーゼを伴う経路を介するc−Junへのインテグリン依存性のシグナル伝達を調節する(Hong IKら, J Biol Chem, 281 (34):24279-24292 (2006))。
【0194】
[0212] RPE介在性のコラーゲンゲル収縮は、いくつかの受容体およびシグナル伝達経路の相互作用を伴う(Bando Hら, Exp Eye Res, 82(3):529-537(2006); Carver Wら, J Cell Physiol, 165(2):425-437(1995); Cooke MEら, J Cell Sci, 113:2375-2383(2000); Kieffer JDら, Biochem Biophys Res Commun, 217(2):466-474(1995); Langholz Oら, J Cell Biol. , 131:1903-1915(1995); Moulin VおよびPlamondon M., Br J Dermatol, 147(5):886-892(2002); Robbins SGら, Invest Ophthalmol Vis Sci, 35(9):3475-3485(1994); Zhang ZGら, J Cell Sci, 119:1886-1895(2006); Xia Hら, J Biol Chem, 279(31):33024-33034(2004))。この研究の結果は、コラーゲン収縮をもたらす、FAK/Src複合体の活性化の促進におけるEMP2の役割を裏付けるものである。
【0195】
[0213] 本明細書において引用される各刊行物、特許出願、特許、および他の参考文献は、それが本開示と矛盾しない限り、参照することによってその全体が組み込まれる。特に、本明細書において引用される全ての刊行物は、本発明に関連して用いられ得る、その刊行物において報告されている方法、試薬、およびツールを記載および開示することを目的として、参照することによってその全体が本明細書に組み込まれる。本明細書における何物も、本発明が先行発明に基づいてこのような開示に先行する資格を有さないということを承認するものとしては解釈されない。
【0196】
[0214] 前述の発明は、理解を明確にすることを目的とした実例および実施例によってある程度詳細に記載されているが、当業者には、本発明の教示に照らして、添付の特許請求の範囲の趣旨または範囲から逸脱することなく、ある程度の変化および変更を本発明に行い得ることが容易に明らかとなろう。
【技術分野】
【0001】
関連出願に対する相互参照
[0001] 本願は、2009年11月20日に出願された米国仮特許出願第61/263,228号の利益を主張するものであり、当該出願の内容は、参照することによってその全体が本明細書に組み込まれる。本願はまた、2007年10月8日に出願された米国特許出願第11/868,788号および2008年10月9日に出願された米国特許出願第11/884,806号に関する主題を含み、前記各出願の内容は、参照することによってその全体が本明細書に組み込まれる。
【0002】
連邦政府支援研究開発のもとで創出された発明の権利に関する陳述
[0002] 本研究は、米国復員軍人援護局により支援されたものであり、連邦政府は、本発明においてある程度の権利を有する。
【0003】
コンパクトディスクで提出された添付の「配列表」、表、またはコンピュータプログラム表への参照
[0003] 不適用
【背景技術】
【0004】
[0004] 網膜剥離または開放性眼外傷の治療の不成功の最も一般的な理由は、増殖性硝子体網膜症(PVR)(Pastor, J. C. Surv Ophthalmol 43:3-18(1998))と呼ばれる、硝子体腔で形成する細胞膜の増殖および収縮である。PVRは、裂孔原性網膜剥離の最大5〜20%のケースの合併症として生じると推定されている(Pastor, J. C. Surv Ophthalmol 43:3-18(1998); Pastor, J. C.ら, Prog Retin Eye Res 21:127-44(2002))。眼の外傷において、PVRのリスクは非常に高い。2003年〜2005年の間にイラクで眼球内異物による眼の外傷を有していた退役軍人を検査したところ、PVRが患者の21%において視力低下の原因であることが明らかになった(Colyer M.H.ら, Ophthalmology. 2007 Aug;114(8):1439-47)。
【0005】
[0005] 臨床研究によって、硝子体出血、眼球内ガス、大きなまたは長期にわたる網膜剥離、および手術の失敗(特に、それらが凍結療法または光凝固術の過剰な使用に関連する場合)を包含する、PVR発症の複数のリスク因子が同定された(Pastor, J. C. Surv Ophthalmol 43:3-18(1998); Pastor, J. C.ら, Prog Retin Eye Res 21: 127-44(2002))。眼球/網膜の損傷の程度が、PVRを増大させるサイトカイン生成または他のシグナル伝達を増大させる可能性がある。炎症性サイトカイン(例えば、IL-8およびMCP-1)、ならびにIL−1、IL−6、TNF−α、およびIFN−γは、PVRにおいて増大することが明らかにされているが、サイトカインレベルは、PVRの重症度と直接的には相関しない(Asaria, R. H. Compr Ophthalmol Update 7:179-85(2006))。
【0006】
[0006] 網膜色素上皮(RPE)は、PVRに関与する重要な細胞型の1つであると考えられている。PVRの根底にある発症機序は複雑であるが、外傷または網膜剥離の後、RPE細胞が硝子体内に放出されるか、または網膜下の位置から移動するように刺激される可能性が高い。これらの細胞は次に移動し、増殖し、脱分化し、上皮間葉転換(EMT)を受けて、PVRの網膜前膜の生成を促進する(Asaria, R. H. Compr Ophthalmol Update 7:179-85(2006))。RPE細胞は、再発性網膜剥離およびさらなる失明をもたらすことが多い牽引力を生じさせる膜収縮をもたらす可能性が高い(Pastor, J. C.ら, Prog Retin Eye Res 21:127-44(2002); Kroll, P.ら, Ophthalmologica 221:78-94(2007))。
【0007】
[0007] EMP2はRPE細胞において高度に発現することが明らかにされているが、その機能は理解され始めているにすぎない(Wadehra, M.ら, Exp Mol Pathol 74:106-12(2003))。テトラスパニンファミリー(CD9、CD53、CD81、CD82)のメンバーとしては、インテグリンおよび他の細胞表面受容体の調節が一般的である。EMP2は、以前に、FAK.Srcシグナル伝達複合体の活性化の促進を介する細胞の収縮能力の調節において重要であることが示されている(Morales, S. A.ら, Exp Eye Res 85:790-8(2007); Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。さらに、FAK/Src複合体の活性化は、EMP2とFAK/Src複合体との間の物理的関連を介することが証明されている(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。これらのインビトロでの研究によって、EMP2およびFAK/Srcシグナル伝達複合体がRPE介在性のゲル収縮における重要な構成要素であることが明らかにされた。
【0008】
[0008] 本発明者らは、本明細書において、RPE細胞がインビボでの研究およびインビトロでの研究の両方で一致していると報告するが、これまでに報告された観察結果と本研究との間には多くの潜在的な差が存在する。まず、インビトロでの研究は、コラーゲンをRPE細胞の足場として用いるが、インビボでの硝子体におけるRPE細胞は、ヒアルロン酸ならびにII、V/XI、およびIXを包含する多くのコラーゲン型に曝露される。さらに、炎症細胞、ミュラー細胞、線維芽細胞などを包含する、インビボでPVR膜に寄与する多くの細胞型が存在する(Pastor, J. C. Surv Ophthalmol 43:3-18(1998); Pastor, J. C.ら, Prog Retin Eye Res 21:127-44(2002); Asaria, R. H. Compr Ophthalmol Update 7:179-85(2006); Kim, I. K.ら. Ophthalmol Clin North Am 15:81-6(2002))。インビボでは、PVR膜の形成の間に、細胞が多くのサイトカインに曝露されることが知られており、網膜前膜は、血管内皮増殖因子(VEGF)、(PDGF)、IL−6、IL−8、TNF−α、TGF−βなどを包含するサイトカインを含有することが明らかにされている(Harada, C.ら, Prog Retin Eye Res 25:149-64(2006))。これらの細胞およびこれらの細胞によって生じるサイトカインの環境は、細胞および膜の挙動に確実に影響する。
【発明の概要】
【発明が解決しようとする課題】
【0009】
[0009] 今日、手術の進歩にもかかわらず、患者は、PVRおよび牽引性網膜前膜に続発して視力を失うことが非常に多い。本発明は、PVRに関連する網膜剥離の治療および/または予防の必要性を満たすものである。
【課題を解決するための手段】
【0010】
[0010] 1つの態様において、本発明は、対象における増殖性硝子体網膜症または網膜剥離を治療する方法を提供する。いくつかの実施形態において、本方法は、眼におけるEMP2の活性を低減させる抗EMP2剤を対象に投与することを含む。
【0011】
[0011] いくつかの実施形態において、対象はヒトである。いくつかの実施形態において、抗EMP2剤は局所的に投与される。いくつかの実施形態において、抗EMP2剤は、全身投与される。いくつかの実施形態において、抗EMP2剤は、眼に直接投与される。いくつかの実施形態において、抗EMP2剤は、眼内に投与される。いくつかの実施形態において、抗EMP2剤は、眼の硝子体液内に投与される。
【0012】
[0012] いくつかの実施形態において、抗EMP2剤の投与は、網膜剥離のリスクを低減させる。いくつかの実施形態において、抗EMP2剤の投与は、網膜剥離の程度を低減させる。
【0013】
[0013] いくつかの実施形態において、抗EMP2剤は、眼への注入用に製剤化される。いくつかの実施形態において、抗EMP2剤は、眼の外での局所投与用に製剤化される。
【0014】
[0014] いくつかの実施形態において、硝子体網膜症は、裂孔原性の硝子体網膜症である。いくつかの実施形態において、抗EMP2剤を投与する前に、眼は外傷を有するか、または網膜剥離のリスクの原因となる手術を受けている。
【0015】
[0015] 別の態様において、本発明は、生体試料における網膜色素上皮細胞の収縮能力を低下させる方法を提供する。いくつかの実施形態において、本方法は、生体試料と、網膜色素上皮細胞におけるEMP2の活性を低減させる抗EMP2剤とを接触させることを含む。
【0016】
[0016] いくつかの実施形態において、生体試料は組織試料である。いくつかの実施形態において、組織試料は眼の組織である。
【0017】
[0017] いくつかの実施形態において、抗EMP2剤は、配列番号1、配列番号2、または配列番号30に対して少なくとも70%のアミノ酸配列同一性を有するEMP2ポリペプチドに特異的に結合する抗体またはそのフラグメントである。いくつかの実施形態において、抗体は、配列番号4、配列番号6、配列番号8、または配列番号10に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む重鎖を含む。いくつかの実施形態において、抗体は、配列番号5、配列番号7、配列番号9、または配列番号11に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗体は、(1)配列番号4、配列番号6、配列番号8、および配列番号10からなる群から選択される重鎖、ならびに(2)配列番号5、配列番号7、配列番号9、および配列番号11からなる群から選択される軽鎖を含む。いくつかの実施形態において、抗体は、配列番号27、配列番号28、または配列番号29に対して少なくとも70%のアミノ酸配列同一性を有する。いくつかの実施形態において、抗体は、配列番号27、配列番号28、または配列番号29のアミノ酸配列を有する。
【0018】
[0018] いくつかの実施形態において、抗体は、モノクローナル抗体である。いくつかの実施形態において、抗体は、ヒト化モノクローナル抗体である。いくつかの実施形態において、抗体は、ポリクローナル抗体である。いくつかの実施形態において、抗体フラグメントは、ダイアボディ(diabody)またはミニボディとして提供される。いくつかの実施形態において、抗体フラグメントは、Fvフラグメント、F(ab’)フラグメント、またはF(ab’)2フラグメントである。いくつかの実施形態において、抗体は、マウスの抗原結合部位とエフェクター機能を調節するヒト化領域とを有するキメラ抗体である。いくつかの実施形態において、抗体またはそのフラグメントは、細胞傷害剤に結合している。
【0019】
[0019] いくつかの実施形態において、抗EMP2剤は、配列番号1のポリペプチドに対して少なくとも70%のアミノ酸配列同一性を有するEMP2タンパク質をコードする核酸の発現を低減できるsiRNAである。いくつかの実施形態において、siRNAは、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドに対して少なくとも70%のヌクレオチド配列同一性を有する核酸配列を含む。いくつかの実施形態において、siRNAは、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドを含む。
【0020】
[0020] いくつかの実施形態において、抗EMP2剤は、配列番号3の核酸配列に対して少なくとも70%のヌクレオチド配列同一性を有するEMP2ポリヌクレオチド配列を切断できるハンマーヘッドリボザイムである。
【0021】
[0021] いくつかの実施形態において、抗EMP2剤は、プロゲステロン受容体アンタゴニストである。いくつかの実施形態において、抗EMP2剤は、ミフェプリストンである。
【図面の簡単な説明】
【0022】
【図1】[0022]表1に示されるようにFastenburgシステムに従って等級付けされた、ウサギPVRモデルの図である。各群3頭のウサギであり、対照は、ベクターのみでトランスフェクトされたARPE19細胞を投与したウサギであり、Riboは、EMP2の発現がより低い、リボザイムでトランスフェクトされた細胞を投与したウサギであり、EMP2は、EMP2を過剰発現するようにトランスフェクトされた細胞を投与したウサギであった。EMP2を過剰発現する細胞を投与したウサギは、PVR形成の速度が増大し、一方、リボザイムノックダウンによってEMP2発現のレベルがより低い細胞を投与した動物(Ribo)は、PVR形成の程度が減少した。示される結果は、細胞注入の5週間後に得たものである。比較した群の上に、スチューデントT検定のp値を示す。
【図2】[0023]図1で示される実験から単離された眼球の肉眼的な病変断面を示す図である。PVR膜形成の程度を、これらの画像で見ることができる。各群の1つの眼球が示されている。図1におけるように、対照は、ベクターのみでトランスフェクトされたARPE19細胞を投与したウサギであり、Riboは、EMP2の発現がより低い、リボザイムでトランスフェクトされた細胞を投与したウサギであり、EMP2は、EMP2を過剰発現するようにトランスフェクトされた細胞を投与したウサギであった。EMP2を過剰発現する細胞を投与したウサギは、PVR膜形成の速度が増大し、一方、リボザイムノックダウンによってEMP2発現のレベルがより低い細胞を投与した動物(Ribo)は、PVR膜形成の程度が減少した。表される結果は、細胞注入の5週間後に得たものである。
【図3】[0024]図1および2で示される実験の網膜前膜を示す組織学的断面を示す図である。図1および2におけるように、対照は、ベクターのみでトランスフェクトされたARPE19細胞を投与したウサギであり(A、B)、Riboは、EMP2の発現がより低い、リボザイムでトランスフェクトされた細胞を投与したウサギであり(C、D)、EMP2は、EMP2を過剰発現するようにトランスフェクトされた細胞を投与したウサギであった(E、F)。矢印は網膜前膜を指し、これは、Ribo群(E、F)におけるよりも対照群(A、B)およびEMP2群(C、D)で顕著であった。
【図4】[0025]網膜前膜において見られるEMP2の発現を示す図である。網膜前PVR膜は、対照ウサギ群およびEMP2ウサギ群で最も顕著であった(図1〜3)。この図は、対照ウサギ(A)およびEMP2を過剰発現するウサギ(B)におけるEMP2の発現(紫)を示す。
【図5】[0026]PVRのウサギモデルは、細胞を抗EMP2ポリクローナル抗体と共にプレインキュベートした場合のPVR膜形成の阻害を示す。各群3頭のウサギであり、抗EMP2は、抗EMP2ポリクローナル抗体と共にプレインキュベートされたARPE19細胞を投与したウサギであり、野生型(Wild-type)(Wt)は、野生型ARPE19細胞を投与したウサギであり、免疫前(Pre-immune)は、ポリクローナル免疫前(対照)抗体と共にプレインキュベートされたARPE19細胞を投与したウサギであった。表される結果は、細胞注入の5週間後に得たものである。比較した群の上に、スチューデントT検定のp値を示す。
【図6】[0027]網膜前膜において見られるEMP2の発現を示す図である。図5において示されるように、網膜前PVR膜は、抗EMP2抗体で前処理されたウサギ群におけるよりも、野生型ウサギ群および免疫前ウサギ群で最も顕著であった。この図は、3つのウサギ群全て、すなわち、Wt(A)、免疫前(B)、および抗EMP2抗体(C)の網膜前PVR膜におけるEMP2の発現(紫)を示す。
【図7】[0028]A.FAKのリン酸化に対するEMP2の影響を示す図である。全FAK、p−FAK(Y397、Y407、Y861、およびY925)、およびβ−アクチンのレベルを、細胞系ARPE−19およびARPE−19/EMP2においてウェスタンブロットによって測定し、デンシトメトリーによって定量した。B. p−Src(Tyr 416およびTyr 527)およびβ−アクチンを、細胞系ARPE−19およびARPE−19/EMP2においてウェスタンブロットによって測定し、デンシトメトリーによって定量した。各部位のリン酸化レベル(平均±標準誤差)を全FAK(A)またはβ−アクチン(B)に対して正規化し、少なくとも3つの独立した実験から計算した。代表的なウェスタンブロットも示す。
【図8】[0029]FAKとEMP2との関連。(A)ARPE−19およびARPE−19/EMP2(107個細胞)の溶解物を、FAKに対する抗体、EMP2に対する抗体、または無関係なウサギ抗体(−対照)を用いて免疫沈降した。SDS−PAGEを、免疫沈降物のアリコートまたは元の溶解物のアリコートを用いて行い(それぞれ、106個および105個細胞当量)、ウェスタンブロットをFAKについて分析した。FAKおよびEMP2の同族(cognate)免疫沈降(データは示していない)が、比較的効率的であった(50〜60%)。EMP2は、ARPE−19およびARPE−19/EMP2において、それぞれ、全FAKの25%および30%を共免疫沈降した。データは、3回以上の実験の代表である。(B)ARPE−19細胞およびARPE−19/EMP2細胞を、EMP2に対する抗体で染色した後に、FITCに結合したロバ抗ウサギ二次IgG(緑)で染色し、かつ、FAKに対する抗体で染色した後に、テキサスレッドに結合したロバ抗ヤギ二次IgG(赤)で染色した。これらの2つのチャネルの併合が示されており(中央右)、同様に、共局在化したピクセルが強調して示されている併合が示されている(右)。少なくとも6個のフィールドが、各試料の分析のためにランダムに選択され、関連のパーセンテージ(percent association)は、複数のフィールドから生じた平均値である。フィールド当たり複数のスライスが取得され、示されているものは、スライドと接触し接着斑を含有する、細胞の底表面の単一のスライスである。ARPE−19およびARPE−19/EMP2において、それぞれ全FAKの68%および96%がEMP2と共局在化した。(C)EMP2とリン酸化型FAKとの間の関連を評価するために、ARPE−19細胞およびARPE−19/EMP2細胞を、EMP2に対する抗体およびp576/577−FAKに対する抗体で染色した。EMP2に対する結合抗体は、FITCに結合した二次抗体を用いて同定され、一方、p−FAK(Y576/577)に対する結合抗体の染色は、テキサスレッドに結合した二次抗体で示される。これらの2つのチャネルの併合が示されており(白)、共局在化したピクセルが観察および定量されている。少なくとも6個のフィールドが、各試料の分析のためにランダムに選択され、関連のパーセンテージは、複数のフィールドから生じた平均値である。フィールド当たり複数のスライスが取得され、表されているものは、スライドと接触し接着斑を含有する、細胞の底表面の単一のスライスである。ARPE−19細胞において、リン酸化型FAKの31%がEMP2と共局在化している。ARPE−19/EMP2細胞において、リン酸化型FAKの97%がEMP2と共局在化している。EMP2レベルが増大すると、リン酸化型FAKとEMP2との関連が増大する。
【図9】[0030]EMP2発現の調節が、FAKのリン酸化および局在化に影響することを示す図である。ARPE−19細胞およびARPE−19/EMP2細胞をp576/577−FAK(A)一次抗体で染色した。細胞をテキサスレッドに結合したロバ抗ヤギ抗体と共にインキュベートし、蛍光顕微鏡法下で観察した。少なくとも4つの個別の試料のリン酸化型FAK(B)をピクセル強度を計算することによって定量し(方法を参照されたい)、データをスチューデントt検定(独立、一方向)によって評価した。
【図10】[0031]EMP2およびリン酸化型FAKの発現の増大によって、F−アクチンの分布が変化することを示す図である。ARPE−19細胞(AおよびC)およびARPE−19/EMP2細胞(BおよびD)をガラスカバースリップ上で一晩平板培養した。細胞をパラホルムアルデヒドで固定し、サポニンで透過処理し、テキサスレッド−Xファロイジンで染色した。代表的な写真をカラー(AおよびB)および白黒(CおよびD)で示す。
【図11】[0032]EMP2発現の増大によって、α−平滑筋アクチンの発現が増大した。(A)ARPE−19細胞およびARPE−19/EMP2細胞におけるα−平滑筋アクチン(α-sma)の定常状態のタンパク質レベルをウェスタンブロット分析によって測定した。β−アクチンをロード対照として用いた。(B)α−smaの量を、β−アクチンと比較して各試料について計算した。少なくとも3つの独立した実験を行い、結果を数値的に表す。発現レベルの差をスチューデントt検定(独立、一方向)を用いて評価した。
【図12】[0033]EMP2レベルの増大によって、コラーゲンへの接着が増大することを示す図である。ARPE−19細胞およびARPE−19/EMP2細胞を、フィブロネクチン、ビトロネクチン、コラーゲンI、およびコラーゲンIVで被覆したプレート上で平板培養した。結合細胞をクリスタルバイオレットの取り込みについて分析し、吸光度の定量的評価をマイクロプレートリーダー(590nm)によって測定した。EMP2の発現が増大すると、コラーゲンI(P<0.001)およびコラーゲンIV(P<0.001)への接着が増大した。フィブロネクチンへの接着は、ARPE−19/EMP2細胞と比較して、ARPE−19細胞において増大した(P=0.02)。EMP2レベルの変化は、ビトロネクチンへの接着に影響しなかった。結果は、少なくとも8回反復した1つの実験から表され、少なくとも3つの独立した実験を行い、同等の結果を得た。スチューデントt検定(独立、一方向)を統計分析に用いた。
【図13】[0034]組換えによるLMP2発現変化を示す図である。A.EMP2の定常状態のタンパク質レベルをARPE−19細胞(対照細胞)、ARPE−19/EMP2細胞(EMP2の増大)、およびARPE−19/Ribo細胞(EMP2の減少)においてウェスタンブロット分析によって測定した。EMP2発現の増大を測定するために、様々な希釈(1:10、1:25、1:50、および1:100)のARPE−19/EMP2細胞の溶解物を評価した。B.ARPE−19細胞をEMP2に特異的なsiRNA(ARPE-19/EMP2 siRNA)または対照のスクランブルsiRNA(ARPE-19/対照siRNA)で一時的にトランスフェクトした。実験は独立して少なくとも3回行い、同様の結果を得た。
【図14】[0035]EMP2の修飾が、コラーゲンゲルの収縮に影響したことを示す図である。コラーゲンゲル収縮アッセイをARPE−19/EMP2細胞、ARPE−19/Ribo細胞、およびARPE−19細胞で行った。EMP2の発現が上昇した細胞系および低下した細胞系では、結果として、それぞれ、ゲル収縮が57%増大および55%減少した(P=0.002)。実験は独立して少なくとも3回行い、同様の結果を得た。
【図15】[0036]コラーゲン結合インテグリンを示す図である。細胞表面の発現を、α1インテグリン、α2インテグリン、またはα3インテグリンに対するモノクローナル抗体を用いるフローサイトメトリーによって測定した。A.代表的な実験のヒストグラムを表す。アイソタイプ対照の染色が白抜きの波形として示され、特異的な染色パターンには影が付けられている。蛍光強度の平均の数値が、各パネルの右上隅に表されている。B.これらの各インテグリンの表面発現を、3つの独立した実験で評価し、結果を表にして平均を表した。2つの細胞系における各インテグリンの発現の統計的比較をスチューデントt検定(独立、一方向)で行った。
【図16】[0037]EMPレベルの増大は増殖および移動を減少させたが、侵入またはコラーゲンの生成に対する影響は有していなかった。A.ARPE−19細胞およびARPE−19/EMP2細胞を正常培地またはコラーゲンI25mg/mLで処理し、48時間にわたってインキュベートし、増殖をBrdUの組み込みによって評価した。B.細胞の移動に対するEMP2発現の影響。ARPE−19細胞およびARPE−19/EMP2細胞を24ウェルプレート内で平板培養し、コンフルエントになったら擦り傷を作り、移動を様々な時点で測定した。ARPE−19細胞およびARPE−19/EMP2細胞は、未処理のままであるか、またはPDGF50ng/mLで処理し、移動を24時間後に測定した。C.ARPE−19細胞およびARPE−19/EMP2細胞を、重合コラーゲンの薄層で被覆した8μmの孔サイズのポリカーボネート膜を有する侵入チャンバインサート上に播種した。10%PBSまたはPDGF50ng/mlを化学誘引物質として用いた。侵襲性細胞は、重合コラーゲン層を通って移動して、ポリカーボネート膜の底に張り付き、染色、抽出、およびマイクロプレートリーダー(560nm)による測定によって検出された。D.ARPE−19細胞およびARPE−19/EMP2細胞を、TGF−β10ng/mLの存在下または非存在下の無血清培地で72時間にわたって成長させた。培地を回収し、コラーゲンの生成をELISAによって測定した。TGF−βの存在下または非存在下でのコラーゲンの生成に統計的に有意な変化はなかった。しかし、TGF−βは、未処理条件と比較して、各細胞系でコラーゲンの生成を増大させた。全ての研究は、試料当たり6ウェルで、個別に少なくとも3回行った。結果は、スチューデントt検定(独立、一方向)を用いて、統計的有意性について評価した。P<0.05のレベルを、統計的に有意であると考えた。
【図17】[0038]EMP2の過剰発現によって、FAKの活性化が増大した。細胞抽出物(タンパク質10μg)を還元条件において4%〜20%のSDS−PAGE勾配ゲルによって分画し、ウェスタン免疫ブロットをFAK、pFAK、およびβ−アクチンに対する抗体で探査した。A:代表的な免疫ブロット、B:実験は独立して少なくとも3回行い、同様の結果を得た。β−アクチンロード対照に対して正規化したバンドの密度を定量した。実験は独立して少なくとも3回行い、同様の結果を得た。
【図18】[0039]EMP2の過剰発現によって、PP2に対する耐性が増大した。細胞を、様々な濃度の低分子阻害剤PP2(FAK/Src阻害剤)で1時間にわたり前処理し、ゲルの収縮を評価した。実験は、試料当たり6回の反復で、個別に少なくとも3回行い、代表的な実験が表されている(A〜C)。統計分析を、媒体のみの対照と比較して、各阻害剤濃度について、スチューデントt検定で行った。A.EMP2の発現が減少した、ARPE−19/EMP2 siRNA細胞。B.ARPE−19細胞。C.EMP2を過剰発現するARPE−19/EMP2細胞。D.各点は、媒体対照に曝露された同一の細胞系の総収縮に対して正規化された、各細胞系についての各濃度での収縮の阻害の平均を表す。実験は独立して少なくとも3回行い、同様の結果を得た。
【図19】[0040]EMP2の過剰発現によって、SU6656による阻害に対する耐性が増大した。細胞を様々な濃度の低分子阻害剤SU6656(FAK/Src阻害剤)で1時間にわたり前処理し、ゲルの収縮を評価した。実験は、試料当たり6回の反復で個別に少なくとも3回行い、代表的な実験が表されている。媒体のみの対照と比較して、各阻害剤濃度について、スチューデントt検定で統計分析を行った。A.ARPE−19/EMP2細胞。B.EMP2を過剰発現するARPE−19/EMP2細胞。C.各点は、媒体対照に曝露された同一の細胞系の総収縮に対して正規化された、各細胞系についての各濃度での収縮の阻害の平均を表す。実験は独立して少なくとも3回行い、同様の結果を得た。
【図20】[0041]抗体KS49、KS83、KS41、およびKS89の配列。A. KS49重鎖(配列番号4)およびKS49軽鎖(配列番号5)のアミノ酸配列。CDR−H1(配列番号12)、CDR−H2(配列番号13)、CDR−H3(配列番号14)、CDR−L1(配列番号15)、CDR−L2(配列番号16)、およびCDR−L3(配列番号17)に下線が引かれている。B. KS83重鎖(配列番号6)およびKS49軽鎖(配列番号7)のアミノ酸配列。CDR−H1(配列番号12)、CDR−H2(配列番号13)、CDR−H3(配列番号18)、CDR−L1(配列番号19)、CDR−L2(配列番号20)、およびCDR−L3(配列番号21)に下線が引かれている。C. KS41重鎖(配列番号8)およびKS41軽鎖(配列番号9)のアミノ酸配列。CDR−H1(配列番号22)、CDR−H2(配列番号23)、CDR−H3(配列番号24)、CDR−L1(配列番号25)、CDR−L2(配列番号26)、およびCDR−L3(配列番号27)に下線が引かれている。D. KS89重鎖(配列番号10)およびKS89軽鎖(配列番号11)のアミノ酸配列。CDR−H1(配列番号22)、CDR−H2(配列番号23)、CDR−H3(配列番号24)、CDR−L1(配列番号25)、CDR−L2(配列番号26)、およびCDR−L3(配列番号27)に下線が引かれている。
【発明を実施するための形態】
【0023】
I.定義
[0042] ここで、本明細書および添付の特許請求の範囲において用いられる場合、単数形「1つの(a)」、「1つの(an)」、および「その(the)」が、別段の指示が内容から明らかでない限り、複数形の言及を包含することに留意する。
【0024】
[0043] 「上皮膜タンパク質2」または「EMP2」は、(1)例えばGenBank受託番号NM_001424(EMP2のmRNA)およびP54851(EMP2タンパク質)で示される、本明細書において記載される個々に言及される核酸配列またはアミノ酸配列によってコードされるポリペプチドに対して、少なくとも約25、50、100、200、500、1000、またはそれを超えるアミノ酸の領域にわたり、約60%を超えるアミノ酸配列同一性、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、またはそれを超えるアミノ酸配列同一性を有するアミノ酸配列を有する、(2)GenBank受託番号P54851(EMP2タンパク質)で示される言及されるアミノ酸配列を含む免疫原に対する抗体、例えばポリクローナル抗体、そのそれぞれの免疫原性フラグメント、およびそのそれぞれの保存的に修飾された変異体に特異的に結合する、(3)GenBank受託番号P54851(EMP2タンパク質)で示される言及されるアミノ酸配列をコードする核酸、およびそのそれぞれの保存的に修飾された変異体に、ストリンジェントなハイブリダイゼーション条件下で特異的にハイブリダイズする、(4)GenBank受託番号NM_001424(EMP2のmRNA)で示される参照核酸配列に対して、好ましくは、少なくとも約15、20、25、50、100、150、200、250、500、1000、またはそれを超えるヌクレオチドの領域にわたり、約95%を超える、好ましくは約96%、97%、98%、99%を超える、またはそれより高いヌクレオチド配列同一性を有する核酸配列を有する、核酸、例えば、遺伝子、mRNA前駆体、mRNA、ならびにポリペプチド、多型変異体、対立遺伝子、突然変異体、および種間ホモログを言う。ポリヌクレオチド配列またはポリペプチド配列は、典型的には、限定はしないが、ヒトなどの霊長類、ラット、マウス、ハムスターなどの齧歯動物、ウシ、ブタ、ウマ、ヒツジ、または任意の哺乳動物を包含する、哺乳動物のものである。本発明の核酸およびタンパク質は、天然の分子または組換え分子の両方を包含する。
【0025】
[0044] 「抗EMP2剤」または「EMP2阻害剤」は、網膜剥離に関与する拘縮を仲介する細胞性EMP2(例えば、配列番号1のアミノ酸配列を有するEMP2)の機能、活性、または組織レベルに干渉する作用剤である。EMP2阻害剤は、EMP2ポリペプチド;抗EMP2抗体(例えば、組換え抗体、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、ヒトモノクローナル抗体、ヒト化もしくは霊長類化(primatized)モノクローナル抗体、または抗体フラグメント);EMP2siRNA分子;EMP2−リボザイム;EMP2への結合またはEMP2の結合と競合する化合物、または宿主細胞におけるEMP2核酸の発現、転写、もしくは翻訳を阻害する作用物質もしくは化合物であり得る。いくつかの実施形態において、EMP2阻害剤は、無菌担体および/または生理学的に許容可能な担体も含む組成物で提供される。
【0026】
[0045] 「調節剤」は、言及される活性を増大または減少させ得る作用物質である。調節剤は、阻害剤と、言及される活性または実体に関して阻害剤と逆の効果を有する(例えば、増大させる、刺激する、増加させる、増強する、加速する)「活性化剤」との両方を包含する。
【0027】
[0046] 用語「ポリペプチド」、「ペプチド」、および「タンパク質」は、本明細書において、アミノ酸残基のポリマーを言うためにほぼ同じ意味で用いられる。この用語は、1つまたは複数のアミノ酸残基が、天然アミノ酸に対応する人工の化学的模倣剤である、アミノ酸ポリマー、ならびに、天然アミノ酸ポリマーおよび非天然アミノ酸ポリマーに適用される。ポリペプチドを得る(例えば、生成する、単離する、精製する、合成する、および組換えによって製造する)ための方法は、当業者に周知である。
【0028】
[0047] 用語「アミノ酸」は、天然アミノ酸および合成アミノ酸、ならびに天然アミノ酸に類似した様式で機能するアミノ酸類似体およびアミノ酸模倣体を言う。天然アミノ酸は、遺伝子コードによってコードされるもの、および、後に修飾されるアミノ酸、例えば、ヒドロキシプロリン、γ−カルボキシグルタミン酸、およびO−ホスホセリンである。アミノ酸類似体は、天然アミノ酸と同一の基本的な化学構造、すなわち水素、カルボキシル基、アミノ基、およびR基に結合しているα炭素を有する化合物、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムを言う。このような類似体は、修飾されたR基(例えば、ノルロイシン)または修飾されたペプチド骨格を有するが、天然アミノ酸と同一の基本的な化学構造を保持している。アミノ酸模倣体は、アミノ酸の一般的な化学構造と異なる構造を有するが天然アミノ酸に類似した様式で機能する化合物を言う。
【0029】
[0048] アミノ酸は、本明細書において、IUPAC−IUB Biochemical Nomenclature Commissionによって推奨されている、一般に知られているその三文字記号または一文字記号によって言及され得る。ヌクレオチドは、同様に、その一般に認められている一文字コードによって言及され得る。
【0030】
[0049] アミノ酸配列の「保存的に修飾された変異体」に関して、当業者には、コードされる配列における単一のアミノ酸または低いパーセンテージのアミノ酸を改変する、付加する、または欠失する、核酸配列、ペプチド配列、ポリペプチド配列、またはタンパク質配列への個別の置換、欠失、または付加が、改変の結果、化学的に類似のアミノ酸でアミノ酸が置換される「保存的に修飾された変異体」であることが認識されよう。機能的に類似のアミノ酸を提供する保存的置換の表は、当技術分野において周知である。このような保存的に修飾された変異体は、本発明の多型変異体、種間ホモログ、および対立遺伝子に追加されるものであり、これらを排除しない。
【0031】
[0050] 以下の8つの群は、それぞれ、互いの保存的置換であるアミノ酸を含有する:1)アラニン(A)、グリシン(G)、2)アスパラギン酸(D)、グルタミン酸(E)、3)アスパラギン(N)、グルタミン(Q)、4)アルギニン(R)、リジン(K)、5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V)、6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)、7)セリン(S)、スレオニン(T)、および8)システイン(C)、メチオニン(M)(例えば、Creighton, Proteins(1984)を参照されたい)。
【0032】
[0051] 「抗体」は、抗原を特異的に結合および認識する免疫グロブリン遺伝子またはそのフラグメントのフレームワーク領域を含むポリペプチドを言う。認知されている免疫グロブリン遺伝子には、κ、λ、α、γ、δ、ε、およびμ定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子が包含される。軽鎖は、κまたはλとして分類される。重鎖は、γ、μ、α、δ、またはεとして分類され、これは、免疫グロブリンクラスIgG、IgM、IgA、IgD、およびIgEをそれぞれ規定する。典型的には、抗体の抗原結合領域は、結合の特異性および親和性において最も重要である。
【0033】
[0052] 典型的な免疫グロブリン(抗体)の構造単位は、四量体を含む。各四量体は、2つの同一のポリペプチド鎖対からなり、各対は、1つの「軽」鎖(約25kD)および1つの「重」鎖(約50〜70kD)を有する。各鎖のN末端は、抗原認識に最も関与する約100〜110、またはそれを超えるアミノ酸からなる可変領域を規定する。可変軽鎖(VL)および可変重鎖(VH)という用語は、それぞれこれらの軽鎖および重鎖を言う。
【0034】
[0053] 用語「相補性決定領域」または「CDR」は、VLおよびVHの超可変領域を言う。CDRは、このような標的タンパク質に対する特異性を有する抗体鎖の標的タンパク質結合部位である。ヒトのVLまたはVHのそれぞれに3つのCDR(N末端から連続的に番号付けされたCDR1〜3)があり、可変ドメインの約15〜20%を構成する。CDRは、標的タンパク質のエピトープに対して構造的に相補性であり、したがって、結合特異性に直接的に関与する。VLまたはVHの残りのストレッチ、いわゆるフレームワーク領域は、アミノ酸配列にあまり変化を有さない(Kuby, Immunology,第4版, 第4章. W.H Freeman & Co., New York, 2000)。
【0035】
[0054] CDRおよびフレームワーク領域の位置は、当技術分野における様々な周知の定義、例えば、Kabat,Chothia,international ImMunoGeneTicsデータベース(IMGT)(ワールドワイドウェブのimgt.cines.fr/にある)およびAbM(例えば、Johnsonら, Nucleic Acids Res., 29:205-206(2001)、ChothiaおよびLesk, J. Mol. Biol., 196:901 -917(1987)、Chothiaら, Nature, 342:877-883(1989)、Chothiaら, J. Mol. Biol., 227:799-817(1992)、Al-Lazikaniら, J.Mol.Biol., 273:927-748(1997)を参照されたい)を用いて決定される。抗原結合部位の定義はまた、Ruizら、Nucleic Acids Res.、28:219〜221(2000);およびLefranc,M.P.、Nucleic Acids Res.、29:207〜209(2001);MacCallumら、J.Mol.Biol.、262:732〜745(1996);およびMartinら、Proc、Natl.Acad.Sci.USA、86:9268〜9272(1989);Martinら、Methods Enzymol.、203:121〜153(1991);およびReesら、In Sternberg M.J.E.(編)、Protein Structure Prediction、Oxford University Press、Oxford、141〜172(1996)において記載されている。
【0036】
[0055] 抗体は、例えば、無傷の免疫グロブリンとして、または様々なペプチダーゼでの消化によって生じる多くの良く特徴付けされたフラグメントとして存在する。したがって、例えば、ペプシンは、ヒンジ領域におけるジスルフィド連結の下で抗体を消化して、それ自体ジスルフィド結合によってVH−CH1に結合した軽鎖であるFabの二量体であるF(ab)’2を生成する。F(ab)’2は、ヒンジ領域におけるジスルフィド連結を破壊して、それによって、F(ab)’2二量体をFab’単量体に変換するための、穏やかな条件下で還元され得る。Fab’単量体は、原則的に、ヒンジ領域の一部を有するFabである(Fundamental Immunology(Paul編, 第3版, 1993)を参照されたい。様々な抗体フラグメントが無傷抗体の消化に関して規定されているが、当業者には、このようなフラグメントが、化学的にまたは組換えDNA法を用いて、デノボ合成され得ることが理解されよう。したがって、本明細書において用いられる抗体という用語はまた、抗体全体の修飾によって生じる抗体フラグメント、または組換えDNA法を用いてデノボ合成される抗体フラグメント(例えば、一本鎖Fv)、またはファージディスプレイライブラリーを用いて同定される抗体フラグメント(例えば、McCaffertyら, Nature 348:552-554(1990)を参照されたい)を包含する。
【0037】
[0056] したがって、抗体という用語はまた、ミニボディ、ダイアボディ、トリアボディ(tirabody)などを包含する。ダイアボディは、高い結合力および特異性を有する、小さな二価の生体特異的抗体フラグメントである。それらの高いシグナル対ノイズ比は、典型的には、特異的抗原の診断的標的化および治療的標的化についてのそれらの潜在能力を増大させる良好な特異性および速い血液クリアランスのため、より良好である(Sundaresanら, J Nucl Med 44:1962-9(2003)。さらに、これらの抗体は、必要に応じて、小さな一本鎖Fvから様々なアイソフォームを有する無傷のIgGまでにわたる、異なるタイプの抗体フラグメントとして操作され得るため、有利である(Wu & Senter, Nat.Biotechnol. 23:1137-1146(2005))。いくつかの実施形態において、抗体フラグメントは、ダイアボディの一部である。いくつかの実施形態において、本発明は、本発明に従って用いるための高い結合力の抗体を提供する。
【0038】
[0057] Hollingerら、PNAS(USA)90(14):6444〜6448(1993)によって最初に記載されたダイアボディは、本明細書において開示される重鎖および軽鎖を用いて、および本明細書において開示される個別のCDR領域を用いて構築され得る。典型的には、ダイアボディフラグメントは、同一鎖上の2つのドメインの間の対合を可能にするには短すぎるリンカーによって軽鎖可変ドメイン(VL)に連結している、重鎖可変ドメイン(VH)を含む。したがって、1つのフラグメントVHドメインおよびVLドメインは、別のフラグメントの相補的なVHドメインおよびVLドメインと対合させられ、それによって、2つの抗原結合部位を形成する。トリアボディは、同様に、3つの抗原結合部位で構築され得る。Fvフラグメントは、非共有結合による相互作用によって結びついているVLドメインおよびVHドメインを含む、完全な抗原結合部位を含有する。本発明によって包含されるFvフラグメントはまた、VHドメインおよびVLドメインがグルタルアルデヒド、分子間ジスルフィド、または他のリンカーを介して架橋されている構築物を包含する。重鎖および軽鎖の可変ドメインは、共に融合されて、親免疫グロブリンの元の特異性を維持している一本鎖可変フラグメント(scFv)を形成し得る。Gruberら、J.Immunol.152(12):5368〜74(1994)によって最初に記載された一本鎖Fv(scFv)二量体は、本明細書において開示される重鎖および軽鎖を用いて、および本明細書において開示される個別のCDR領域を用いて構築され得る。当技術分野において知られている多くの技術を、本発明の特異的結合構築物を調製するために用いることができる(全ての目的で、特にミニボディおよびダイアボディの設計に関する目的で、参照することによってその全体がそれぞれ本明細書に組み込まれる、米国特許出願公開第20070196274号および米国特許出願公開第20050163782号を参照されたい)。
【0039】
[0058] 二重特異性抗体は、化学的架橋またはハイブリッドハイブリドーマ技術によって生成することができる。あるいは、二重特異性抗体分子は、組換え技術によって生成することができる(二重特異性抗体を参照されたい)。二量体化は、VHドメインおよびVLドメインを結合するリンカーの長さを、scFvフラグメントを生成するために通常用いられる約15アミノ酸から約5アミノ酸まで低減することによって促進され得る。これらのリンカーは、VHドメインおよびVLドメインの鎖内組み立てに有利に働く。好適な短いリンカーはSGGGSであるが、他のリンカーを用いることができる。したがって、2つのフラグメントは、二量体分子に組み立てられる。リンカーの長さを0〜2アミノ酸にさらに低減させると、三量体分子(トリアボディ)または四量体分子(テトラボディ)が生成し得る。
【0040】
[0059] 抗体、例えば、組換え抗体、モノクローナル抗体または組換えポリクローナル抗体の調製には、当技術分野において知られている多くの技術を用いることができる(例えば、Kohler & Milstein, Nature 256:495-497(1975); Kozborら, Immunology Today 4:72(1983); Coleら, in Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., 77-96頁(1985); Coligan, Current Protocols in Immunology(1991); Harlow & Lane, Antibodies, A Laboratory Manual(1988);およびGoding, Monoclonal Antibodies: Principles and Practice(第2版, 1986)を参照されたい)。目的の抗体の重鎖および軽鎖をコードする遺伝子は細胞からクローニングすることができ、例えば、モノクローナル抗体をコードする遺伝子はハイブリドーマからクローニングすることができ、組換えモノクローナル抗体を生成するために用いることができる。モノクローナル抗体の重鎖および軽鎖をコードする遺伝子ライブラリーも、ハイブリドーマまたは形質細胞から作製することができる。重鎖および軽鎖の遺伝子産物のランダムな組み合わせは、異なる抗原特異性を有する大きな抗体プールをもたらす(例えば、Kuby, Immunology(第3版, 1997)を参照されたい)。一本鎖抗体または組換え抗体の生成のための技術(米国特許第4,946,778号、米国特許第4,816,567号)を、本発明のポリペプチドに対する抗体を生成するために適用することができる。また、トランスジェニックマウスまたは他の生物、例えば他の哺乳動物を、ヒト化抗体またはヒト抗体を発現させるために用いることができる(例えば、米国特許第5,545,807号、米国特許第5,545,806号、米国特許第5,569,825号、米国特許第5,625,126号、米国特許第5,633,425号、米国特許第5,661,016号、Marksら, Bio/Technology 10:779-783(1992); Lonbergら, Nature 368:856-859(1994); Morrison, Nature 368:812-13(1994); Fishwildら, Nature Biotechnology 14:845-51(1996);Neuberger, Nature Biotechnology 14:826(1996);およびLonberg & Huszar, Intern. Rev. Immunol. 13:65-93(1995)を参照されたい)。あるいは、ファージディスプレイ技術を用いて、選択された抗原に特異的に結合する抗体およびヘテロマーFabフラグメントを同定することができる(例えば、McCaffertyら, Nature 348:552-554(1990); Marksら, Biotechnology 10:779-783(1992)を参照されたい)。抗体はまた、二重特異性にすることができ、すなわち2つの異なる抗原を認識できるようにすることができる(例えば、WO93/08829、Trauneckerら, EMBO J. 10:3655-3659(1991);およびSureshら, Methods in Enzymology 121:210(1986)を参照されたい)。抗体はまた、ヘテロコンジュゲート、例えば共有結合によって結合した2つの抗体、または免疫毒素であり得る(例えば、米国特許第4,676,980号、WO91/00360、WO92/200373、およびEP03089を参照されたい)。
【0041】
[0060] 非ヒト抗体をヒト化または霊長類化するための方法は、当技術分野において周知である。通常、ヒト化抗体は、その中に非ヒトである由来源から導入された1つまたは複数のアミノ酸残基を有する。これらの非ヒトアミノ酸残基は移入(import)残基と呼ばれることが多く、この移入残基は、典型的には、移入可変ドメインから得られる。ヒト化は、原則的には、齧歯動物のCDRまたはCDR配列をヒト抗体の対応する配列で置換することによって、Winter and co−workersの方法に従って行うことができる(例えば、Jonesら, Nature 321:522-525(1986); Riechmannら, Nature 332:323-327(1988); Verhoeyenら, Science 239:1534-1536(1988)、およびPresta, Curr. Op. Struct. Biol. 2:593-596(1992)を参照されたい)。したがって、このようなヒト化抗体は、無傷のヒト可変ドメインほどには実質的には非ヒト種の対応する配列によって置換されていないキメラ抗体である(米国特許第4,816,567号)。実際には、ヒト化抗体は、典型的には、いくつかのCDR残基および場合によってはいくつかのFR残基が、齧歯動物抗体における類似の部位の残基によって置換されている、ヒト抗体である。
【0042】
[0061] 「キメラ抗体」は、(a)抗原結合部位(可変領域)が異なるまたは改変されたクラス、エフェクター機能、および/または種の定常領域に連結するかあるいは酵素、毒素、ホルモン、成長因子、薬剤などのキメラ抗体に新たな特徴を与える完全に異なる分子に連結するように、定常領域もしくはその一部が改変、置換、または交換されている、または(b)可変領域またはその一部が、異なるまたは改変された抗原特異性を有する可変領域で改変、置換、または交換されている、抗体分子である。
【0043】
[0062] 抗体に「特異的に(または選択的に)結合する」という表現、または「特異的に(または選択的に)免疫反応性である」という表現は、タンパク質またはペプチドについて言う場合、多くはタンパク質および他の生物製剤の異種集団における、タンパク質の存在の決定要因である結合反応を言う。したがって、指定した免疫アッセイ条件下において、特定の抗体は、バックグラウンドの少なくとも2倍、さらに典型的には、バックグラウンドの10〜100倍超、特定のタンパク質に結合する。このような条件下での抗体への特異的な結合には、特定のタンパク質への特異性について選択された抗体が必要である。例えば、ポリクローナル抗体は、選択された抗原と特異的に免疫反応性であるが他のタンパク質とはそうではないポリクローナル抗体のみを得るように選択することができる。この選択は、他の分子と交差反応する抗体を取り去ることによって達成され得る。様々な免疫アッセイ形式を用いて、特定のタンパク質と特異的に免疫反応性である抗体を選択することができる。例えば、固相ELISA免疫アッセイが、タンパク質と特異的に免疫反応性である抗体を選択するために通常用いられる(例えば、特異的免疫反応性を決定するために用いられ得る免疫アッセイの形式および条件の説明については、Harlow & Lane, Using Antibodies, A Laboratory Manual(1998)を参照されたい)。
【0044】
[0063] 例として、EMP2に対するポリクローナル抗体は、グルタチオン−S−トランスフェラーゼ−EMP2融合タンパク質を用いて得ることができる。該抗体は、グルタチオン−S−トランスフェラーゼ(GST)−EMP2融合タンパク質として構築された遺伝子の第1の細胞外領域(アミノ酸16〜64)に対して生成することができる。EMP2ペプチドは、以下のプライマー:CGCGGATCCTCTACCATTGACAATGCCTGG(フォワード:下線が引かれたBamHI)、CCGGAATTCTTACGCCTGCATCACAGAATAACC(リバース、下線が引かれたEcoRI)を用いてPCRによってクローニングすることができる。PCR産物は、GST遺伝子を含有するpGEX−4T−1ベクター(Pharmacia)のBamHI部位およびEcoRI部位内に直接的にクローニングすることができる。EMP2フラグメントは、GSTとインフレームでクローニングされて、融合タンパク質を生じさせる。インサートは、配列決定によって確認することができる。GST融合タンパク質は、従前に記載されているように生成することができる(Smith DBら, Gene 67:31-40(1988)を参照されたい)。log相の細菌(OD6000.6〜0.9)をイソプロピル−1−チオ−β−D−ガラクトピラノシド1mMと共に、37℃で2.5〜3時間にわたり誘導することができる。細菌を溶解し、可溶性画分をグルタチオン−セファロースカラム(Pierce, Rockford, IL)にロードする。カラムをリン酸緩衝生理食塩水(PBS)/EDTA10ベッド容量で洗浄する。融合タンパク質を、Tris−Cl50mM(pH8.0)内の還元型グルタチオン(Sigma, St Louis, MO)20mMを用いてカラムから溶出する。抗体の調製のために、ウサギをGST−EMP2融合タンパク質で2回免疫化し、最後の免疫化の2週間後から血清を回収する(Research Genetics, Huntsville, AL)。
【0045】
[0064] EMP2に対する完全にヒトのモノクローナル抗体は、組換えファージディスプレイ技術を用いてヒト抗体ファージディスプレイ遺伝子ライブラリーから生成することができる。ヒトEMP2に対するこのようなモノクローナル抗体は、診断目的で、およびPVRにおけるような網膜剥離の予防または治療のために用いることができる。
【0046】
[0065] 「核酸」は、一本鎖型または二本鎖型のデオキシリボヌクレオチドまたはリボヌクレオチドおよびそのポリマー、ならびにそれらの相補鎖(complement)を言う。この用語は、合成され、天然、および非天然であり、参照核酸と類似の結合特性を有し、かつ参照ヌクレオチドと類似の様式で代謝される、既知のヌクレオチド類似体または修飾された骨格残基もしくは連結を含有する核酸を包含する。このような類似体の例には、限定はしないが、ホスホロチオエート、ホスホルアミデート、メチルホスホネート、キラル−メチルホスホネート、2−O−メチルリボヌクレオチド、ペプチド−核酸(PNA)が包含される。
【0047】
[0066] 別段の指示がない限り、特定の核酸配列はまた、明確に示された配列と同様に、その保存的に修飾された変異体(例えば、縮重コドンの置換)および相補的な配列を暗黙的に包含する。具体的には、縮重コドンの置換は、1つまたは複数の選択された(または全ての)コドンの第3の位置が混合塩基および/またはデオキシイノシン残基で置換されている配列を生成することによって達成することができる(Batzerら, Nucleic Acid Res. 19:5081(1991); Ohtsukaら, J. Biol. Chem. 260:2605-2608(1985); Rossoliniら, Mol. Cell. Probes 8:91-98(1994))。核酸という用語は、遺伝子、cDNA、mRNA、オリゴヌクレオチド、およびポリヌクレオチドとほぼ同じ意味で用いられる。
【0048】
[0067] 特定の核酸配列はまた、「スプライス変異体」を暗黙的に包含する。同様に、核酸によってコードされる特定のタンパク質は、その核酸のスプライス変異体によってコードされる任意のタンパク質を暗黙的に包含する。「スプライス変異体」は、その名前が示唆するように、遺伝子の選択的スプライシングの産物である。転写の後、最初の核酸転写産物はスプライシングされ、その結果、異なる(選択的な)核酸スプライス産物は異なるポリペプチドをコードし得る。スプライス変異体を生成するためのメカニズムは変化するが、エキソンの選択的スプライシングを包含する。読み過し転写(read-through transcription)によって同一の核酸から誘導される別のポリペプチドもこの定義に包含される。スプライス産物の組換え型を包含する任意のスプライシング反応産物が、この定義に包含される。カリウムチャネルスプライス変異体の例が、Leicherら、J.Biol.Chem.273(52):35095〜35101(1998)において論じられている。
【0049】
[0068] 「siRNA」または「RNAi」は、二本鎖RNAを形成する核酸を言い、この二本鎖RNAは、siRNAが遺伝子または標的遺伝子と同一の細胞において発現している場合に、その遺伝子または標的遺伝子の発現を低減させるかまたは阻害する能力を有する。したがって、「siRNA」または「RNAi」は、相補鎖によって形成される二本鎖RNAを言う。ハイブリダイズして二本鎖分子を形成するsiRNAの相補的部分は、典型的には、実質的なまたは完全な同一性を有する。1つの実施形態において、siRNAは、標的遺伝子に対する実質的なまたは完全な同一性を有し、かつ二本鎖siRNAを形成する核酸を言う。典型的には、siRNAは、少なくとも約15〜50ヌクレオチド長である(例えば、二本鎖siRNAの各相補的配列は15〜50ヌクレオチド長であり、二本鎖siRNAは約15〜50塩基対長であり、好ましくは約20〜30塩基のヌクレオチド、好ましくは約20〜25または約24〜29ヌクレオチド長、例えば、20、21、22、23、24、25、26、27、28、29、または30ヌクレオチド長である。
【0050】
[0069] siRNA分子およびベクターの設計および作製は、当業者に周知である。例えば、好適なsiRNAを設計するための効率的なプロセスは、mRNA転写産物のAUG開始コドンで開始して(例えば、図5を参照されたい)、AAジヌクレオチド配列についてスキャンすることである(Elbashirら, EMBO J 20:6877-6888(2001)を参照されたい)。AAおよび3’隣接ヌクレオチドはそれぞれ、潜在的なsiRNA標的部位である。隣接部位配列の長さは、siRNAの長さを決定する。例えば、19の隣接部位は、21ヌクレオチド長のsiRNAをもたらす。3’オーバーハング(overhanging)UUジヌクレオチドを有するsiRNAは、最も効果的であることが多い。このアプローチはまた、ヘアピンsiRNAを転写するためのRNA pol IIIの使用に適合可能である。RNA pol IIIは、4〜6ヌクレオチドのポリ(T)配列で転写を終結させて、短いポリ(U)尾部を有するRNA分子を生じさせる。しかし、他の3’末端のジヌクレオチドオーバーハングを有するsiRNAもまた、RNAiを効果的に誘導し得、配列は経験的に選択され得る。選択性のために、他のコード配列に対して相同性を有する16〜17を超える連続した塩基対を有する標的配列が、BLASTサーチ(www.ncbi.nlm.nih.gov/BLASTを参照されたい)を行うことによって避けられ得る。
【0051】
[0070] siRNAを直接投与するか、またはsiRNA発現ベクターを用いてRNAiを誘導することができる。ベクターは、短いスペーサー配列によって分離されており、かつ転写を終結させることを助けるTストリングで終わる、挿入された2つの逆反復を有し得る。発現したRNA転写産物は、短いヘアピンsiRNAにフォールディングすると予測される。siRNAの標的配列、推定ヘアピンの柄部をコードする逆反復の長さ、逆反復の順序、ヘアピンのループをコードするスペーサー配列の長さおよび組成、ならびに5’−オーバーハングの存在または非存在の選択は、変化し得る。siRNA発現カセットの好ましい順序は、センス鎖、短いスペーサー、およびアンチセンス鎖である。これらの様々な柄部長さ(例えば、15〜30)を有するヘアピンsiRNAが適している。ヘアピンsiRNAのセンス鎖とアンチセンス鎖とを連結するループの長さは、様々な長さであり得る(例えば、3〜9ヌクレオチド、またはそれ以上の長さ)。ベクターは、プロモーターおよび発現エンハンサー、またはsiRNAをコードするヌクレオチド配列に作動可能に連結している他の調節エレメントを含有し得る。
【0052】
[0071] 「制御配列」という表現は、特定の宿主生物における作動可能に連結しているコード配列の発現に必要なDNA配列を言う。原核生物に好適な制御配列には、例えば、プロモーター、任意選択によりオペレーター配列、およびリボソーム結合部位が包含される。真核細胞は、プロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが知られている。これらの制御エレメントは、調節エレメントがそれに対して応答性である外部因子を付加および制御することで臨床医が遺伝子の発現をオフまたはオンにすることができるように設計され得る。
【0053】
[0072] 核酸は、別の核酸配列と機能的関係に置かれているとき、「作動可能に連結している」。例えば、プレ配列もしくは分泌リーダーのDNAは、ポリペプチドの分泌に関与するプレタンパク質として発現している場合、ポリペプチドのDNAに作動可能に連結しており、プロモーターもしくはエンハンサーは、配列の転写に影響する場合、コード配列に作動可能に連結し、あるいはリボソーム結合部位は、翻訳を促進するように位置している場合、コード配列に作動可能に連結している。通常、「作動可能に連結している」は、連結しているDNA配列が互いに近くにあることを意味し、分泌リーダーの場合では、連続しかつリーディング相にあることを意味する。しかし、エンハンサーは連続している必要はない。連結は、好都合な制限部位でのライゲーションによって達成される。このような部位が存在しない場合は、合成オリゴヌクレオチドアダプターまたは合成オリゴヌクレオチドリンカーが、従来の慣例に従って用いられる。
【0054】
[0073] 「保存的に修飾された変異体」は、アミノ酸および核酸配列の両方に適用される。特定の核酸配列に関して、保存的に修飾された変異体は、同一もしくは実質的に同一のアミノ酸配列をコードする核酸を言うか、核酸がアミノ酸配列をコードしていない場合、本質的に同一な配列を言う。遺伝子コードの縮重によって、多数の機能的に同一の核酸は任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUは全て、アミノ酸アラニンをコードする。したがって、アラニンがコドンによって特定されている全ての位置において、コドンは、コードされるポリペプチドを改変することなく、記載された対応するコドンのいずれかに改変され得る。このような核酸の変異は、一種の保存的に修飾された変異である「サイレント変異」である。ポリペプチドをコードする本明細書における全ての核酸配列は、核酸の全ての考えられるサイレント変異も表す。当業者には、核酸における各コドン(通常はメチオニンの唯一のコドンであるAUGと、通常はトリプトファンの唯一のコドンであるTGGとを除く)が修飾されて、機能的に同一の分子がもたらされ得ることが認識されよう。したがって、ポリペプチドをコードする核酸の各サイレント変異は、それぞれの記載されている配列において、発現産物に関しては黙示的であるが、実際のプローブ配列に関しては黙示的ではない。
【0055】
[0074] EMP2ポリヌクレオチドおよびEMP2ポリペプチドを包含する2つ以上の核酸配列またはポリペプチド配列の状況における、用語「同一」またはパーセント「同一性」は、以下に記載されるデフォルトパラメータを有するBLASTまたはBLAST2.0配列比較アルゴリズムを用いて測定され、あるいは手動のアラインメントおよび目視検査によって測定されるときに(例えば、NCBIウェブサイトhttp://www.ncbi.nlm.nih.gov/BLAST/などを参照されたい)、同一であるか、あるいは特定のパーセンテージの同一のアミノ酸残基もしくはヌクレオチド(すなわち、比較窓または指定領域にわたり最大の対応となるよう比較およびアラインしたときに、特定の領域にわたって約60%の同一性、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、またはそれを超える同一性)を有する、2つ以上の配列または部分配列を言う。そして、このような配列は、「実質的に同一である」と言われる。この定義はまた、試験配列の相補鎖を言うか、試験配列の相補鎖に適用することができる。この定義には、欠失および/または付加を有する配列、ならびに置換を有する配列も包含される。以下に記載されるように、好ましいアルゴリズムは、ギャップなどを明らかにし得る。好ましくは、同一性は、少なくとも約25アミノ酸長もしくはヌクレオチド長の領域にわたって存在し、より好ましくは50〜100アミノ酸長もしくはヌクレオチド長の領域にわたって存在する。
【0056】
[0075] 配列比較では、典型的には、ある配列を参照配列とし、これに対して試験配列が比較される。配列比較アルゴリズムを用いる場合、試験配列および参照配列は、コンピュータに入れられ、必要に応じて部分配列の座標が指定され、配列アルゴリズムプログラムのパラメータが指定される。好ましくは、デフォルトのプログラムパラメータを用いることができ、または別のパラメータを指定することができる。次に、配列比較アルゴリズムは、プログラムパラメータに基づいて参照配列に対して試験配列の配列同一性パーセントを計算する。
【0057】
[0076] 本明細書において用いられる「比較窓(comparison window)」には、参照配列の20から完全長までからなる群から選択される連続位置数のいずれか1つ、通常は約25〜100、または50〜約150、さらに通常は約100〜約150を有するセグメントであって、ある配列と同一の連続位置数を有する参照配列とを当該2つの配列が最適にアラインされた後に比較し得るセグメントへの言及が包含される。比較のための配列アラインメントの方法は、当技術分野において周知である。比較のための配列の最適なアラインメントは、例えば、Smith & Waterman、Adv.Appl.Math.2:482(1981)の局所的相同性アルゴリズムによって、Needleman & Wunsch、J.Mol.Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、Pearson & Lipman、Proc.Nat’l.Acad.Sci.USA 85:2444(1988)の類似法のためのサーチによって、これらのアルゴリズムのコンピュータ化された実行(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI におけるGAP、BESTFIT、FASTA、およびTFASTA)によって、あるいは手動のアラインメントおよび目視検査(例えば、Current Protocols in Molecular Biology(Ausubelら編、1995補完)を参照されたい)によって行うことができる。
【0058】
[0077] 配列同一性パーセントおよび配列類似性の決定に好適なアルゴリズムの好ましい例は、BLASTアルゴリズムおよびBLAST2.0アルゴリズムであり、これは、Altschulら、Nuc.Acids Res.25:3389〜3402(1977)およびAltschulら、J.Mol.Biol.215:403〜410(1990)にそれぞれ記載されている。BLASTおよびBLAST2.0は、本明細書において記載されるパラメータと共に用いられて、本発明の核酸およびタンパク質の配列同一性パーセントを決定する。BLAST分析を行うためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に利用可能である。このアルゴリズムは、データベース配列における同一の長さの文字とアラインされた場合にいくつかの正値閾値スコアTに適合するかまたはこれを満たす、検索配列における長さWの短い文字を同定することによって、高スコア配列対(HSP)を最初に同定することを伴う。Tは、隣接文字スコア閾値(上記のAltschulら)と呼ばれる。これらの最初の隣接文字ヒットは、それを含むさらに長いHSPを発見するためのサーチを開始するきっかけとして作用する。文字ヒットは、累積アラインメントスコアが増大し得る限り、各配列に沿って両方向に伸長する。累積スコアは、ヌクレオチド配列では、パラメータM(適合(matching)残基対についてのリワードスコア、常に>0)およびN(ミスマッチ残基についてのペナルティースコア、常に<0)を用いて計算される。アミノ酸配列では、スコアリングマトリクスを用いて累積スコアを計算する。各方向での文字ヒットの伸長は、累積アラインメントスコアが量Xによってその最大達成値から減少する場合、1つまたは複数の負のスコアリングの残基のアラインメントの蓄積に起因して累積スコアがゼロ以下に向かう場合、またはいずれかの配列の末端に達する場合、停止する。BLASTアルゴリズムのパラメータW、T、およびXは、アラインメントの感度および速さを決定する。BLASTNプログラム(ヌクレオチド配列のための)は、デフォルトとして11の文字長(W)、10の期待値(E)、M=5、N=−4、および両鎖の比較を用いる。アミノ酸配列では、BLASTPプログラムは、デフォルトとして、3の文字長および10の期待値(E)を用い、BLOSUM62スコアリングマトリクス(Henikoff & Henikoff, Proc. Natl. Acad. Sci. USA 89:10915(1989)を参照されたい)は、50のアラインメント(B)、10の期待値(E)、M=5、N=−4、および両鎖の比較を用いる。
【0059】
[0078] 用語「非相同」は、核酸の一部に関して用いられる場合、核酸が、天然では互いの同一の関連が見られない2つ以上の部分配列を含むことを示す。例えば、新たな機能的核酸を作製するように配置された、関連しない遺伝子に由来する2つ以上の配列を有する核酸、例えば、1つの由来源に由来するプロモーターおよび別の由来源に由来するコード領域を有する核酸は、典型的には、組換えによって生成される。同様に、非相同タンパク質は、タンパク質が、天然では互いの同一の関連が見られない2つ以上の部分配列を含むことを示す(例えば、融合タンパク質)。
【0060】
[0079] 表現「ストリンジェントなハイブリダイゼーション条件」は、プローブが典型的には核酸の複雑な混合物においてその標的部分配列にハイブリダイズするが、他の配列にはハイブリダイズしない条件を言う。ストリンジェントな条件は、配列依存性であり、異なる状況で異なる。より長い配列は、より高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションに対するさらなる指針は、Tijssen、Techniques in Biochemistry and Molecular Biology − Hybridization with Nucleic Probes、「Overview of principles of hybridization and the strategy of nucleic acid assays」(1993)において見られる。通常、ストリンジェントな条件は、規定されたイオン強度pHでの特異的配列の熱融点(Tm)よりも約5〜10℃低くなるように選択される。Tmは、(規定されたイオン強度、pH、および核酸濃度下で)平衡状態で、標的に相補的なプローブの50%が標的配列にハイブリダイズする(標的配列は過剰に存在するため、Tmでは、平衡状態でプローブの50%が塞がれている)温度である。ストリンジェントな条件はまた、ホルムアミドなどの不安定化剤を添加して達成され得る。選択的または特異的なハイブリダイゼーションでは、正のシグナルは、バックグラウンドの少なくとも2倍であり、好ましくはバックグラウンドハイブリダイゼーションの10倍である。典型的なストリンジェントなハイブリダイゼーション条件は、50%ホルムアミド、5×SSC、および1%SDS、42℃でのインキュベーション、あるいは、5×SSC、1%SDS、65℃でのインキュベーションと、65℃での0.2×SSCおよび0.1%SDSでの洗浄とであり得る。
【0061】
[0080] ストリンジェントな条件下で互いにハイブリダイズしない核酸は、これらの核酸がコードするポリペプチドが実質的に同一であれば、依然として実質的に同一である。これは、例えば、核酸のコピーが遺伝子コードによって認められている最大のコドン縮重を用いて生成する場合に生じる。このようなケースでは、核酸は、典型的には、中度にストリンジェントなハイブリダイゼーション条件下でハイブリダイズする。典型的な「中度にストリンジェントなハイブリダイゼーション条件」には、37℃での、40%ホルムアミド、NaCl1M、1%SDSからなる緩衝液でのハイブリダイゼーション、および45℃での1×SSCにおける洗浄が包含される。正のハイブリダイゼーションは、バックグラウンドの少なくとも2倍である。当業者は、別のハイブリダイゼーション条件および洗浄条件を用いて類似のストリンジェンシーの条件を与え得ることを容易に認識されよう。ハイブリダイゼーションパラメータを決定するためのさらなるガイドラインは、多くの参考文献、例えば、Current Protocols in Molecular Biology、Ausubelら編、John Wiley & Sonsにおいて提供されている。
【0062】
[0081] PCRにおいて、低いストリンジェンシーでの増幅には約36℃の温度が典型的であるが、アニーリング温度は、プライマーの長さに応じて約32℃〜48℃の間で変化し得る。高いストリンジェンシーのPCR増幅では約62℃の温度が典型的であるが、高いストリンジェンシーでのアニーリングの温度は、プライマーの長さおよび特異性に応じて約50℃〜約65℃の範囲であり得る。高いストリンジェンシーでの増幅および低いストリンジェンシーでの増幅の両方にとって典型的なサイクル条件には、90℃〜95℃で30秒間〜2分間にわたる変性相、30秒間〜2分間続くアニーリング相、および約72℃で1〜2分間にわたる伸長相が包含される。低いストリンジェンシーでの増幅反応および高いストリンジェンシーでの増幅反応のためのプロトコルおよびガイドラインは、例えば、Innisら(1990)PCR Protocols、A Guide to Methods and Applications、Academic Press,Inc.N.Y.)において提供されている。
【0063】
[0082] 本発明に従ったEMP2抗体またはEMP2ポリペプチドには、標識または検出可能な部分が結合していてよい。「標識」または「検出可能な部分」は、分光学的手段、光化学的手段、生化学的手段、免疫化学的手段、化学的手段、または他の物理的手段によって検出可能な組成物である。例えば、有用な標識には、32P、蛍光染料、電子密度試薬、酵素(例えば、ELISAで一般に用いられるもの)、ビオチン、ジゴキシゲニン、または例えば放射性標識をペプチドに組み込むことによって検出可能であるか、あるいはペプチドと特異的に反応する抗体を検出するために用いることができるハプテンおよびタンパク質が包含される。
【0064】
[0083] 用語「組換え」は、例えば細胞、または核酸、タンパク質、もしくはベクターを参照して用いられる場合、細胞、核酸、タンパク質、またはベクターが、非相同な核酸もしくはタンパク質の導入またはネイティブな核酸もしくはタンパク質の改変によって修飾されていることを示すか、あるいは細胞がそのように修飾された細胞から誘導されていることを示す。したがって、例えば、組換え細胞は、ネイティブ(非組換え)型の細胞では見られない遺伝子を発現するか、あるいは異常に発現する、低発現である、または全く発現されないネイティブな遺伝子を発現する。
【0065】
[0084] 「生体試料」には、生検試料および剖検試料などの組織の切片、ならびに組織学的目的で採取された凍結切片が包含される。このような試料には、産物の血液および血液画分(例えば、血清、血漿、血小板、赤血球など)、痰、組織、培養細胞、例えば、一次培養物、外植片、および形質転換細胞、便、尿などが包含される。生体試料は、典型的には、真核生物から、最も好ましくは哺乳動物、例えば霊長類、例えばチンパンジーもしくはヒト、ウシ、イヌ、ネコ、齧歯動物、例えば、モルモット、ラット、マウス、ウサギ、トリ、爬虫類、または魚から得られる。
【0066】
[0085] 用語「治療する」または「治療」には、
(1)疾患を予防すること、すなわち、生物に曝露され得るがまだその疾患の症候を経験していないか示していない哺乳動物において、疾患の臨床症候が発症しないようにすること、
(2)疾患を阻害すること、すなわち、疾患またはその臨床症候の発症を止めるか低減させること(これには、観察された剥離の程度または対象の数もしくは対象が剥離を有するリスクを低減させることが包含される)、および/または
(3)疾患を軽減すること、すなわち、疾患またはその臨床症候を退行させること
が包含される。
【0067】
[0086] 本明細書における「治療上有効な用量または量」は、投与の目的である効果をもたらす用量を意味する。正確な用量および製剤は治療の目的に応じ、当業者は既知の技術を用いて認識することができる(例えば、Lieberman, Pharmaceutical Dosage Forms(第1-3巻, 1992); Lloyd, The Art, Science and Technology of Pharmaceutical Compounding(1999); Remington: The Science and Practice of Pharmacy, 第20版、Gennaro, Editor(2003),およびPickar, Dosage Calculations(1999)を参照されたい)。
【0068】
[0087] 「患者」または「対象」は、ヒトおよび非ヒト動物、例えば哺乳動物(例えば、霊長類、ウシ、ウマ、ネコ、イヌ、ウサギ、マウス、およびラット)を言う。したがって、本発明の方法は、ヒトの治療法および獣医学上の適用の両方に適用可能である。
【0069】
[0088] 「薬学的に許容可能な塩」は、投与経路に従って、比較的無毒の酸または塩基と共に調製される活性化合物の塩を包含することを意味する。本発明の阻害剤が比較的酸性の官能基を含有する場合、塩基付加塩は、中性形態のこのような化合物をニートのまたは好適な不活性溶媒中で十分な量の所望の塩基と接触させることによって得ることができる。薬学的に許容可能な塩基付加塩の例には、ナトリウム塩、カリウム塩、カルシウム塩、アンモニウム塩、有機アミノ塩、またはマグネシウム塩、または類似の塩が包含される。本発明の化合物が比較的塩基性の官能基を含有する場合、酸付加塩は、中性形態のこのような化合物をニートのまたは好適な不活性溶媒中で十分な量の所望の酸と接触させることによって得ることができる。薬学的に許容可能な酸付加塩の例には、塩酸、臭化水素酸、硝酸、炭酸、一水素炭酸、リン酸、一水素リン酸、二水素リン酸、硫酸、一水素硫酸、ヨウ化水素酸、または亜リン酸などのような、無機酸から誘導される塩、および、酢酸、プロピオン酸、イソ酪酸、マレイン酸、マロン酸、安息香酸、コハク酸、スベリン酸、フマル酸、乳酸、マンデル酸、フタル酸、ベンゼンスルホン酸、p−トリルスルホン酸、クエン酸、酒石酸、メタンスルホン酸などのような、比較的無毒の有機酸から誘導される塩が包含される。また、アルギン酸塩などのアミノ酸の塩、およびグルクロン酸またはガラクツロン酸などのような有機酸の塩も包含される(例えば、Bergeら, Journal of Pharmaceutical Science 66:1-19(1977)を参照されたい)。特定の具体的な本発明の化合物は、塩基性官能基および酸性官能基の両方を含有し、それにより、化合物が塩基付加塩または酸付加塩に変換されることを可能にする。
【0070】
[0089] 中性形態の化合物は、従来の様式で塩と塩基または酸とを接触させ、親化合物を単離することによって、再生成することができる。親形態の化合物は、極性溶媒における可溶性などの特定の物理的特性において様々な塩形態と異なるが、その他の点では、塩は、本発明の目的では親形態の化合物に等しい。
【0071】
[0090] 塩形態に加えて、本発明は、プロドラッグ形態の化合物を提供する。本明細書において記載される化合物のプロドラッグは、生理学的条件下で化学的変化を容易に受けて本発明の化合物をもたらす化合物である。さらに、プロドラッグは、エクスビボ環境で、化学的または生化学的方法によって、本発明の化合物に変換され得る。例えば、プロドラッグは、好適な酵素試薬または化学試薬を有する経皮パッチ容器内にある場合、本発明の化合物に穏やかに変換され得る。
【0072】
[0091] 本発明の特定の化合物は、非溶媒和形態、および水和形態を包含する溶媒和形態で存在し得る。通常、溶媒和形態は非溶媒和形態と同等であり、本発明の範囲内に包含されることが意図される。本発明の特定の化合物は、複数の結晶形態または非晶質形態で存在し得る。通常、全ての物理的形態は、本発明によって考慮される使用にとって同等であり、本発明の範囲内にあることが意図される。
【0073】
[0092] 核酸およびポリペプチドなどのバイオポリマーとは別に、本発明の特定の化合物は、非対称の炭素原子(光学中心)または二重結合を有し、ラセミ体、ジアステレオマー、幾何異性体、および個別の異性体は全て、本発明の範囲内に包含されることが意図される。化合物がアミノ酸または核酸を含む好ましい実施形態において、アミノ酸および核酸はそれぞれ、主な天然の生物学的鏡像異性体である。
【0074】
[0093] 本明細書において用いられる用語「担体」は、典型的には、インビボまたはインビトロの生物学的系に適用される活性剤のための希釈剤または媒体として用いられる不活性物質を言う(例えば、治療剤などの薬剤)。この用語はまた、組成物に凝集性を付与する、典型的には不活性物質を包含する。
【0075】
II.抗EMP2組成物
[0094] 本発明は、EMP2ポリペプチドの活性を阻害するための組成物を提供し、前記組成物は、対象の眼における増殖性硝子体網膜症または網膜剥離を治療する方法において用いられる。特定の理論に縛られることはないが、EMP2は、網膜色素上皮細胞において網膜剥離および失明をもたらし得る細胞の収縮を正に調節すると考えられている。したがって、本発明は、抗EMP2剤を提供することによって細胞の収縮および網膜剥離を予防および/または治療する方法を提供する。
【0076】
[0095] いくつかの実施形態において、抗EMP2剤は、抗EMP2抗体またはそのフラグメントを含む。いくつかの実施形態において、抗EMP2剤は、EMP2阻害性オリゴヌクレオチド、例えば、抗EMP2siRNAまたは抗EMP2リボザイムを含む。
【0077】
A.抗EMP2抗体
[0096] 本発明は、ヒトEMP2ポリペプチド(KS49, KS83)およびマウスEMP2ポリペプチド(KS83)に特異的な高結合力抗体をコードする抗体配列を同定する。同定された配列は、画像化、診断方法、またはインビボ療法のための、組換え抗EMP2抗体(例えば、モノクローナル抗体、ポリクローナル抗体、ミニボディ、ダイアボディ、およびトリアボディ)またはそのフラグメントの生成に有用である
【0078】
[0097] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、配列番号1、配列番号2、または配列番号30に実質的に同一な(すなわち、少なくとも約15、20、25、50、100、200、500、1000、またはそれを超えるアミノ酸の領域にわたり、約60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%を超える、またはそれを超えるアミノ酸配列同一性を有する)EMP2ポリペプチドに特異的に結合し得るものである。いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、配列番号1、配列番号2、または配列番号30のアミノ酸配列を有するEMP2ポリペプチドに特異的に結合し得る。
【0079】
[0098] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS49重鎖(配列番号4)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS49軽鎖(配列番号5)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号4のアミノ酸配列を有する重鎖および/または配列番号5のアミノ酸配列を有する軽鎖を含む。
【0080】
[0099] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS83重鎖(配列番号6)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS83軽鎖(配列番号7)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号6のアミノ酸配列を有する重鎖および/または配列番号7のアミノ酸配列を有する軽鎖を含む。
【0081】
[0100] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS41重鎖(配列番号8)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS41軽鎖(配列番号9)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号8のアミノ酸配列を有する重鎖および/または配列番号9のアミノ酸配列を有する軽鎖を含む。
【0082】
[0101] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、KS89重鎖(配列番号10)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む重鎖、および/またはKS89軽鎖(配列番号11)に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、もしくは99%の配列同一性を有するアミノ酸配列を含む軽鎖を含む。いくつかの実施形態において、抗EMP2抗体は、配列番号10のアミノ酸配列を有する重鎖および/または配列番号11のアミノ酸配列を有する軽鎖を含む。
【0083】
[0102] いくつかの実施形態において、本発明は、KS49、KS83、KS41、およびKS89から選択される抗体のCDR領域を含む抗EMP2配列を提供する。本発明によって提供されるCDR領域は、限定はしないが抗体、scFv、トリアボディ、ダイアボディ、ミニボディなどを包含する、抗EMP−2抗体またはそのフラグメントを構築するために用いることができる。いくつかの実施形態において、本発明の抗EMP−2結合タンパク質は、KS49、KS83、KS41、およびKS89から選択される抗体の少なくとも1つのCDR領域を含む。抗EMP−2結合タンパク質は、本明細書において提供される抗体の、例えばCDR−H1、CDR−H2、CDR−H3、CDR−L1、CDR−L2、CDR−L3、またはその組み合わせを含み得る。本発明のいくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、本明細書において提供される抗体の3つ全てのCDR−H配列、本明細書において提供される抗体の3つ全てのCDR−L配列、またはその両方を含み得る。抗EMP2CDR配列は、抗体骨格またはそのフラグメント上で用いることができ、同様に、ヒト化抗体、またはヒト化配列を含有する抗体を含み得る。これらの抗体は、例えば、EMP2を検出するため、インビボでEMP−2を発現する細胞を検出するため、またはEMP2機能を妨げるために用いることができる。いくつかの実施形態において、CDR領域は、Kabatの定義、Chothiaの定義、AbMの定義、接触の定義、または任意の他の好適なCDR番号付け系を用いて規定され得る。
【0084】
[0103] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、SYAMH(配列番号12)およびEYPMH(配列番号22)からなる群から選択されるアミノ酸配列を有するCDR−H1、VISYDGSNKYYADSVKG(配列番号13)およびVISYDGEYQKYADSVKG(配列番号23)からなる群から選択されるアミノ酸配列を有するCDR−H2、ならびにDRRGRKSAGIDY(配列番号14)、TVGATGAFDI(配列番号18)、およびTINNGMDV(配列番号24)からなる群から選択されるアミノ酸を有するCDR−H3を含む重鎖領域を含む。
【0085】
[0104] いくつかの実施形態において、抗EMP2抗体またはそのフラグメントは、QASQDISNYLN(配列番号15)、RASQSIGKWLA(配列番号19)、およびRASQGIRNDLG(配列番号25)からなる群から選択されるアミノ酸配列を有するCDR−L1、AASSLQS(配列番号16)、KASSLEG(配列番号20)、およびGASSLQS(配列番号26)からなる群から選択されるアミノ酸配列を有するCDR−L2、ならびにLQDYNGWT(配列番号17)およびQQSHNFPPT(配列番号21)からなる群から選択されるアミノ酸配列を有するCDR−L3を含む軽鎖領域を含む。
【0086】
[0105] いくつかの実施形態において、EMP2ポリペプチドに特異的に結合し得る抗EMP2抗体またはそのフラグメントは、配列番号27〜29の抗体に対して少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%のアミノ酸配列同一性を有するアミノ酸配列を含む。
【0087】
B.抗EMP2オリゴヌクレオチド
[0106] いくつかの実施形態において、EMP2の活性は、抗EMP2オリゴヌクレオチドを投与することによって阻害される。いくつかの実施形態において、抗EMP2オリゴヌクレオチドはEMP2siRNAであり、該EMP2siRNAは、EMP2ポリペプチドをコードするポリヌクレオチドの発現を低減できる。いくつかの実施形態において、抗EMP2オリゴヌクレオチドはEMP2リボザイムであり、EMP2リボザイムは、細胞におけるEMP2の発現を阻害し得る。
【0088】
[0107] いくつかの実施形態において、EMP2siRNAは、配列番号3の核酸配列またはその相補鎖に対する実質的な同一性を有する(すなわち、約60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%を超える、またはそれを超えるヌクレオチド配列同一性を有する)ポリヌクレオチドに特異的にハイブリダイズするオリゴヌクレオチドを含む。いくつかの実施形態において、EMP2siRNAは、配列番号3の核酸配列またはその相補鎖の少なくとも15、18、20、25、30、35、40、45、50、またはそれを超える連続するヌクレオチドに実質的に同一な核酸配列を含む。
【0089】
[0108] いくつかの実施形態において、EMP2阻害剤は、EMP2リボザイムである。リボザイムは、RNAの特異的切断を触媒し得る酵素的RNA分子である。リボザイムの作用のメカニズムは、相補的な標的RNAへのリボザイム分子の配列特異的ハイブリダイゼーションと、その後のエンドヌクレアーゼ切断とを伴う。配列番号3の核酸配列に対して実質的な同一性を有する(すなわち、約60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%を超える、またはそれを超えるヌクレオチド配列同一性を有する)mRNAを包含するEMP2mRNAのエンドヌクレアーゼ切断を特異的および効率的に触媒する工学操作されたハンマーヘッドモチーフリボザイム分子は、本発明の範囲内である。
【0090】
[0109] 任意の潜在的なRNA標的内の特異的リボザイム切断部位は、配列GUA、GUU、およびGUCを包むリボザイム切断部位の標的分子をスキャンすることによって最初に同定される。同定されると、切断部位を含有する標的遺伝子の領域に対応する15〜20リボヌクレオチドの間の短いRNA配列は、オリゴヌクレオチド配列を不適切にし得る二次構造などの予測される構造的特徴について評価し得る。
【0091】
[0110] 本発明のアンチセンスRNA分子およびアンチセンスDNA分子の両方ならびにリボザイムは、RNA分子を合成するための当技術分野において知られている任意の方法によって調製することができる。これらには、例えば固相ホスホラミダイト化学合成などの当技術分野において周知のオリゴデオキシリボヌクレオチドを化学的に合成するための技術が包含される。あるいは、RNA分子は、アンチセンスRNA分子をコードするDNA配列をインビトロおよびインビボで転写することによって生成することができる。このようなDNA配列は、T7ポリメラーゼプロモーターまたはSP6ポリメラーゼプロモーターなどの好適なRNAポリメラーゼプロモーターを組み込む様々なベクター内に組み込むことができる。リボザイムを作製する方法は、当技術分野において周知である(例えば、米国特許出願公開第20060062785号を参照されたい)。
【0092】
[0111] 所望のEMP2siRNA配列またはEMP2リボザイム配列および制御配列を含有するEMP2siRNAまたはEMP2リボザイムに好適なベクターの構築は、当技術分野において良く理解されている標準的なライゲーション技術および制限技術を採用する(Maniatisら, in Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York(1982)を参照されたい)。単離されたプラスミド、DNA配列、または合成されたオリゴヌクレオチドは、所望の形態に切断、調整、およびライゲーションされる。
【0093】
C.製剤
[0112] 本明細書において記載される治療方法に従って投与するための組成物は、一般に、薬学的に許容可能な担体、好ましくは水性担体に溶解した本明細書において記載される抗EMP2剤を含む。例えば緩衝生理食塩水などの、様々な水性担体を用いることができる。これらの溶液は無菌であり、望ましくない物質を全体として含まない。これらの組成物は、従来の周知の滅菌技術によって滅菌することができる。組成物は、好適な生理学的条件に応じて、薬学的に許容可能な補助物質、例えば、pH調整剤およびpH緩衝剤、毒性調整剤など、例えば、酢酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、乳酸ナトリウムなどを含有し得る。これらの製剤における活性剤の濃度は広く変化し得、選択された特定の投与様式および患者の要求に応じて、流体の容量、粘度、体重などに主に基づいて選択される。
【0094】
[0113] 本発明において用いるための好適な製剤は、参照することによって本明細書に組み込まれる、Remington:The Science and Practice of Pharmacy、第20版、Gennaro,Editor(2003)において見られる。さらに、薬剤送達のための方法の簡単な概説については、参照することによって本明細書に組み込まれる、Langer、Science 249:1527〜1533(1990)を参照されたい。本明細書において記載される医薬組成物は、当業者に知られている様式で、すなわち、従来の混合プロセス、溶解プロセス、粒子化プロセス、糖衣形成プロセス、粉末化プロセス、乳化プロセス、カプセル化プロセス、封入プロセス、または凍結乾燥プロセスによって製造することができる。以下の方法および賦形剤は典型的なものにすぎず、限定するものでは全くない。
【0095】
[0114] 注入では、本発明の化合物は、水溶液内で、好ましくは、ハンクス液、リンゲル液、または緩衝生理食塩水などの生理学的に適合可能な緩衝液内で製剤化することができる。経粘膜投与では、浸透されるバリアに好適な浸透剤が製剤において用いられる。このような浸透剤は、一般に、当技術分野において知られている。
【0096】
[0115] 経口投与では、本発明に従って用いるための阻害剤は、当技術分野において周知の薬学的に許容可能な担体と組み合わせることによって容易に製剤化することができる。このような担体は、化合物を、治療する患者による経口摂取のための錠剤、ピル、糖衣、カプセル、エマルジョン、親油性懸濁液および親水性懸濁液、液体、ゲル、シロップ、スラリー、懸濁液などとして製剤化することを可能にする。経口使用のための医薬製剤は、化合物を固体賦形剤と混合し、得られた混合物を任意選択により粉砕し、必要に応じて、好適な補助剤を添加した後に顆粒の混合物を加工して錠剤または糖衣の核を得ることによって得ることができる。好適な賦形剤は、特に、充填剤、例えば、乳糖、ショ糖、マンニトール、またはソルビトールを包含する糖;セルロース製剤、例えばトウモロコシデンプン、コムギデンプン、コメデンプン、ジャガイモデンプン、ゼラチン、トラガカントゴム、メチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースナトリウム、および/またはポリビニルピロリドン(PVP)などである。必要であれば、架橋型ポリビニルピロリドン、寒天、またはアルギン酸もしくはその塩、例えばアルギン酸ナトリウムなどの崩壊剤を添加することができる。
【0097】
[0116] 糖衣核は、好適な被覆を有する。この目的では、濃縮された糖溶液を用いることができ、これは、アラビアゴム、タルク、ポリビニルピロリドン、カルボポールゲル、ポリエチレングリコール、および/または二酸化チタン、ラッカー溶液、ならびに好適な有機溶媒または溶媒混合物を任意選択により含有し得る。活性化合物用量の異なる組み合わせを同定または特徴付けするために、染料または顔料を錠剤または糖衣の被覆に添加することができる。
【0098】
[0117] 経口で用いることができる医薬製剤には、ゼラチンで作製された押し込み型(push-fit)カプセル、ならびにゼラチンおよびグリセロールまたはソルビトールなどの可塑剤で作製された密封型ソフトカプセルが包含される。押し込み型カプセルは、乳糖などの充填剤、デンプンなどの結合剤、および/またはタルクもしくはステアリン酸マグネシウムなどの潤滑剤、および任意選択により安定剤と混合された活性成分を含有し得る。ソフトカプセルでは、活性化合物は、脂肪油、液体パラフィン、または液体ポリエチレングリコールなどの好適な液体に溶解または懸濁することができる。さらに、安定剤を添加することができる。経口投与のための全ての製剤は、このような投与に好適な用量であるべきである。
【0099】
[0118] いくつかの実施形態において、静脈内投与のための医薬組成物は、1日当たり患者当たり約0.1〜100mgで提供され得る。1日当たり患者当たり0.1から最大約100mgまでの用量を用いることができる。実質的にさらに高い用量が、局所投与で可能である。非経口投与可能な組成物を調製するための実際の方法は、当業者に知られているかまたは明らかであり、Remington:The Science and Practice of Pharmacy、第21版、2005、Lippincott Williams & Wilkins,Publishersなどの刊行物においてさらに詳細に記載されている。
【0100】
[0119] 医薬組成物は、投与方法に応じて、様々な剤形および用量で投与することができる。例えば、経口投与に好適な単位剤形は、限定はしないが粉末、錠剤、ピル、カプセル、およびトローチを包含する。抗体は、経口投与された場合に消化から保護されるべきであると認識される。これは、典型的には、分子を組成物と複合して、分子を酸および酵素による加水分解に対して耐性にすることによって、または適切に耐性である抗体内、例えばリポソームまたはタンパク質バリア内に分子をパッケージングすることによって、達成する。作用剤を消化から保護する手段は、当技術分野において周知である。
【0101】
[0120] 医薬製剤、特に、本発明に従ったポリペプチドEMP2阻害剤および核酸EMP2阻害剤からなる医薬製剤は、所望の純度を有する抗体またはオリゴヌクレオチドを最適な薬学的に許容可能な担体、賦形剤、または安定剤と混合することによって調製することができる。このような製剤は、凍結乾燥製剤または水溶液であり得る。許容可能な担体、賦形剤、または安定剤は、用いられる用量および濃度でレシピエントに対して無毒である。許容可能な担体、賦形剤、または安定剤は、酢酸塩、リン酸塩、クエン酸塩、および他の有機酸;抗酸化剤(例えば、アスコルビン酸);防腐剤;低分子量ポリペプチド;タンパク質、例えば、血清アルブミンもしくはゼラチンまたはポリビニルピロリドンなどの親水性ポリマー;ならびにアミノ酸、単糖、二糖、およびグルコース、マンノース、またはデキストリンを包含する他の炭水化物;キレート化剤;ならびにイオン性界面活性剤および非イオン性界面活性剤(例えば、ポリソルベート);塩形成対イオン、例えばナトリウム;金属錯体(例えば、Zn−タンパク質錯体);および/または非イオン性界面活性剤であり得る。抗体は、0.5〜200mg/mlの間または10〜50mg/mlの間の濃度で製剤化することができる。
【0102】
[0121] いくつかの実施形態において、本発明は、EMP2阻害剤と、細胞レベルまたは生物レベルで生理学的に許容可能な担体とを含む組成物を提供する。典型的には、生理学的に許容可能な担体は、液体、固体、または半固体の形態で存在する。液体担体の例には、生理食塩水、リン酸緩衝液、通常の緩衝生理食塩水(NaCl135〜150mM)、水、緩衝水、0.4%生理食塩水、0.3%グリシン、安定性を増強するための糖タンパク質(例えば、アルブミン、リポタンパク質、グロブリンなど)などが包含される。固体または半固体の担体の例には、マンニトール、ソルビトール、キシリトール、マルトデキストリン、乳糖、デキストロース、ショ糖、グルコース、イノシトール、粉末糖、糖蜜、デンプン、セルロース、微晶質セルロース、ポリビニルピロリドン、アカシアゴム、グアーゴム、トラガカントゴム、アルギン酸塩、アイリッシュモスの抽出物、パンワールゴム、ガッチゴム、イサポールハスクの粘液、Veegum(登録商標)、ラーチアラボガラクタン、ゼラチン、メチルセルロース、エチルセルロース、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース、ポリアクリル酸(例えば、カルボポール)、ケイ酸カルシウム、リン酸カルシウム、第二リン酸カルシウム、硫酸カルシウム、カオリン、塩化ナトリウム、ポリエチレングリコール、およびその組み合わせが包含される。生理学的に許容可能な担体は一部には、投与される特定の組成物によって、および組成物を投与するために用いられる特定の方法によって決定されるため、本発明の医薬組成物の様々な好適な製剤が存在する(例えば、Remington 's Pharmaceutical Sciences, 第17版, 1989を参照されたい)。担体および組成物は、好ましくは無菌である。
【0103】
[0122] 本発明の組成物は、従来の周知の滅菌技術によって滅菌してもよいし、あるいは無菌条件下で生成してもよい。水溶液は、使用のためにパッケージされ得るか、または無菌条件下で濾過されて凍結乾燥され、凍結乾燥された製剤は、投与の前に無菌水溶液と組み合わされる。組成物は、好適な生理学的条件に応じて、薬学的にまたは生理学的に許容可能な補助剤、例えば、pH調整剤およびpH緩衝剤、張度調節剤、湿潤剤など、例えば、酢酸ナトリウム、乳酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、ソルビタンラウリン酸モノエステル、およびオレイン酸トリエタノールアミンを含有し得る。
【0104】
[0123] 経口投与に好適な製剤は、(a)液体溶液、例えば、希釈剤内、例えば、水、生理食塩水、またはPEG400内に懸濁された、有効量のパッケージングされたプラチナ基剤の薬剤、(b)液体、固体、顆粒、またはゼラチンとして所定量のプラチナ基剤の薬剤をそれぞれ含有する、カプセル、包剤、または錠剤、(c)好適な液体内の懸濁液、ならびに(d)好適なエマルジョンを含み得る。錠剤形態は、乳糖、ショ糖、マンニトール、ソルビトール、リン酸カルシウム、トウモロコシデンプン、ジャガイモデンプン、微晶質セルロース、ゼラチン、コロイド状二酸化ケイ素、タルク、ステアリン酸マグネシウム、ステアリン酸、および他の賦形剤、着色剤、充填剤、結合剤、希釈剤、緩衝剤、湿潤剤、防腐剤、香料、染料、崩壊剤、ならびに薬学的に適合可能な担体の1つまたは複数を包含し得る。
【0105】
[0124] 本発明は、本発明に従ったEMP2阻害剤を含む局所的医薬組成物を提供する。いくつかの実施形態において、阻害剤は、低分子有機化合物、EMP2ポリペプチド、または抗EMP2抗体である。阻害剤は、単位剤形当たり目的の症状に有効な上記のEMP2阻害剤を含む、単位剤形であり得る。
【0106】
[0125] EMP2阻害剤の局所的製剤は、薬学的に許容可能な担体と組み合わせて製剤化することができる。本発明の化合物を局所投与するための剤形には、粉末、スプレー、泡、ゼリー、軟膏、ペースト、クリーム、ローション、ゲル、溶液、パッチ、坐剤、およびリポソーム製剤が包含される。剤形は、泌尿生殖器領域で活性成分を徐放性するための粘膜接着性ポリマーを用いて製剤化することができる。活性化合物は、無菌条件下で、薬学的に許容可能な担体、および必要であり得る任意の防腐剤、緩衝液、または推進剤と混合することができる。局所的製剤は、阻害剤を局所的な乾燥製剤、液体製剤、クリーム製剤、およびエアロゾル製剤で一般に用いられる従来の薬学的希釈剤および薬学的担体と組み合わせることによって調製することができる。軟膏およびクリームは、例えば、好適な増粘剤および/またはゲル化剤を添加した水性基剤または油性基剤を用いて製剤化することができる。このような基剤には、水および/または油、例えば液体パラフィンまたは植物油、例えばピーナッツ油またはヒマシ油が含まれ得る。基剤の性質に従って用いられ得る増粘剤には、ソフトパラフィン、ステアリン酸アルミニウム、セトステアリルアルコール、プロピレングリコール、ポリエチレングリコール、羊毛脂、水添ラノリン、蜜ろうなどが包含される。ローションは、水性基剤または油性基剤を用いて製剤化することができ、通常は、安定化剤、乳化剤、分散剤、懸濁剤、増粘剤、着色剤、香料などの1つまたは複数も包含する。粉末は、任意の好適な粉末基剤、例えば、タルク、乳糖、デンプンなどを用いて製剤化することができる。ドロップは、1つまたは複数の分散剤、懸濁剤、可溶化剤なども含む水性基剤または非水性基剤を用いて製剤化することができる。
【0107】
[0126] 軟膏、ペースト、クリーム、およびゲルはまた、賦形剤、例えば、動物および植物の脂肪、油、ワックス、パラフィン、デンプン、トラガカント、セルロース誘導体、ポリエチレングリコール、シリコーン、ベントナイト、ケイ酸、タルク、および酸化亜鉛、またはその混合物を含有し得る。粉末およびスプレーはまた、賦形剤、例えば、乳糖、タルク、ケイ酸、水酸化アルミニウム、ケイ酸カルシウム、およびポリアミド粉末、またはこれらの剤の混合物を含有し得る。スプレーはさらに、慣習的な推進剤、例えば、クロロフルオロ炭化水素ならびに揮発性の非置換炭化水素、例えばブタンおよびプロパンを含有し得る。
【0108】
[0127] EMP2阻害剤は、症状の治療に有効な単位剤形の組成物内に存在し得る。EMP2阻害剤の用量は、当業者に周知の多くの因子、例えば、特定の化合物、治療する症状、レシピエント患者の年齢、体重、および臨床症状、ならびに臨床医または治療を管理する専門家の経験および判断に応じる。有効量の化合物は、症候の自覚的な軽減または臨床医もしくは他の資格のある観察者によって認められる客観的に同定可能な改善をもたらすものである。用量範囲は、用いられる化合物、投与経路、および特定の化合物の効力で変化する。
【0109】
[0128] 本発明は、1つまたは複数の上記の薬理学的化合物および薬学的に許容可能な担体を含む、局所的な徐放性医薬組成物および持続放出医薬組成物を提供する。好ましくは、組成物は、このような治療を必要としている対象に単位剤形で投与される。局所的な徐放性組成物および持続放出組成物は、典型的には、1)親水性基剤中の吸収剤、2)疎水性基剤中の吸収剤、および3)好適な媒体内に分散した吸収剤マトリクスを含有する被覆されたビーズを包む種々のものである。
【0110】
III.治療方法
[0129] 本発明はさらに、網膜剥離または増殖性硝子体網膜症を有する対象の眼における網膜剥離を治療、予防、および/または阻害する方法に関する。本方法は、全体として、本明細書において記載される抗EMP2剤(例えば、抗EMP2抗体もしくはそのフラグメント、抗EMP2siRNA、または抗EMP2リボザイム)を、眼におけるEMP2活性を低減させるおよび/または阻害するために十分な条件下および用量で対象に投与することを含む。
【0111】
[0130] 医師または獣医師は、医薬組成物において採用される本発明の抗EMP2剤の投薬を所望の治療効果を達成するために必要なレベルより低いレベルで開始し、所望の効果が達成されるまで徐々に用量を増大させることができる。通常、本発明の組成物の有効量は、治療する具体的な疾患または症状、投与手段、標的部位、患者の生理学的状態、患者がヒトであるか動物であるか、投与される他の医薬品、および治療が予防的であるか治療的であるかを包含する、多くの異なる因子に応じて変化する。治療用量は、安全性および効能を最適化するように滴定される必要がある。組成物の単回投与または複数回投与を、患者によって要求され認容される用量および頻度に応じて投与することができる。
【0112】
[0131] 抗体との投与では、用量は、宿主体重1kg当たり約0.0001〜100mg、より一般的には0.01〜5mgの範囲である。例えば、用量は、体重1kg当たり1mgまたは体重1kg当たり10mgまたは1〜10mg/kgの範囲内であり得る。投薬は、必要に応じて、または要求に応じて、毎日、週に1回、週に2回、月に1回、またはそれを超えるかもしくはそれ未満の頻度であり得る。典型的な治療レジメは、週に1回、2週毎に1回、または月に1回、または3〜6ヶ月に1回の投与を伴う。
【0113】
[0132] 剤が核酸である実施形態において、典型的な用量は、体重1kg当たり約0.1mgから、体重1kg当たり約100mgまで(体重1kg当たり約100mgを含める)の範囲であり得、例えば、体重1kg当たり約1mgから体重1kg当たり約50mgまでの間である。いくつかの実施形態において、体重1kg当たり約1、2、3、4、5、10、15、20、30、40、または50mgである。
【0114】
[0133] 本発明の抗EMP2剤およびその医薬組成物は、任意の投与経路(例えば、静脈内、局所的、腹腔内、非経口、経口、膣内、直腸、眼球、硝子体内、および眼球内)によって投与することができる。これらは、筋肉内経路、腹腔内経路、皮下経路、経口経路、局所的経路、または吸入経路によって、一定期間にわたるボーラス注入または持続注入として投与することができる。抗体の静脈内投与または皮下投与が好ましい。投与は、局所的または全身的であり得る。これらは、対象となる疾患を有している、その疾患の病歴がある、またはその疾患のリスクがあると診断された対象に投与することができる。
【実施例】
【0115】
[0134] 以下の実施例は例示のために提供されるが、特許請求される発明を限定するものではない。
【0116】
(実施例1)
[0135] 増殖性硝子体網膜症(PVR)は、一部には、異常な創傷治癒戦略における、硝子体腔内での細胞の移動、膜の形成、および収縮を伴う、網膜色素上皮(RPE)の脱分化から生じると考えられている。インビトロでのコラーゲン−ゲル収縮アッセイにおいて、網膜色素上皮細胞系(ARPE-19)による接着斑キナーゼ(FAK)の活性化を介して収縮を制御する、4回膜貫通タンパク質である上皮膜タンパク質2(EMP2)を研究した。この研究の目的は、EMP−2発現のレベルがインビボモデル系において臨床PVRを変化させたかどうかを調べることであった。
【0117】
[0136] 方法:ARPE−19細胞系をATCCから得て、EMP2のレベルを、EMP−2を過剰発現する構築物、EMP2リボザイム、またはベクターのみの安定なトランスフェクションを介して調節した。これらの安定にトランスフェクトされた細胞系をPVRのウサギモデルにおいて用いた。PVRの重症度を、二人のマスキングされた観察者が分類した。また、EMP2ブロック抗体を用いて、PVRモデルにおける機能的なEMP2を減少させた。免疫組織化学を用いて、インビボでのEMP2の発現を評価した。
【0118】
[0137] 結果:高レベルのEMP2発現を有するトランスフェクタントは、低レベルのEMP2トランスフェクタントよりも有意に重症度の大きなPVRを誘発した(p=0.05)。さらに、より低レベルのEMP2を有するトランスフェクタントは、野生型細胞によって誘発されるPVRの程度よりも有意に小さなPVRの重症度を有していた(p=0.02)。特異的抗体でEMP2を妨げると、PVR重症度のレベルが有意に減少した(p=0.01)。PVR膜は、EMP2発現について陽性であることが明らかになった。
【0119】
[0138] 結論:これらのインビボでの研究は、EMP2の発現とPVRの重症度との間の直接的な相関を裏付けるものである。これらの結果は、EMP2の発現における変化を介してRPE生物学を制御する可能性を実証し、この疾患の潜在的な治療標的を提供する。
【0120】
[0139] RPEの制御および膜収縮の予防は、深刻なPVR応答の予防における最初の目的である。インビトロでの実験モデルでは、本発明者らの研究室は、インテグリン(α1、α2、およびα3)のライゲーションを介するFAKの活性化をコラーゲンゲル収縮の重大な制御点として同定した(Morales, S. A.ら, Exp Eye Res 85:790-8(2007))。テトラスパンスーパーファミリーは、インテグリンおよび他の受容体複合体と組み立てられた細胞内輸送分子および細胞内シグナル伝達分子のタイプを決定する鍵となるクラスのタンパク質である(Hemler, M. E. T Annu Rev Cell Dev Biol 19:397-422(2003))。特定のテトラスパンファミリーメンバーは、個別の細胞型および分化または活性化の状態において、高度に制御されている。したがって、テトラスパンの組み合わせ発現は、インテグリンの配置およびシグナル伝達の質に対する重要な寄与因子である。上皮膜タンパク質−2(EMP2)は、インテグリンの特異的な配置およびFAKを介するシグナル伝達を制御するように作用する、テトラスパンファミリーメンバーである(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。RPEにおいて高度に発現するEMP2(Wadehra, M.ら, Exp Mol Pathol 74:106-12 (2003))は、異なる生化学的および生理学的役割を有する成長停止特異的遺伝子3/末梢ミエリンタンパク質22(GAS3/PMP22)4−TMタンパク質ファミリーのメンバーである(Wadehra, M.ら, Exp Mol Pathol 74:106-12(2003); Forbes, A.ら, J Biol Chem 282:26542-51(2007); Wadehra, M.ら, Mol Biol Cell 15:2073-83(2004); Wadehra, M.ら, J Biol Chem 277:41094-100(2002); Wadehra, M.ら, Reprod Biol Endocrinol 6:15(2008); Wadehra, M.ら, Clin Immunol 107:129-36(2003); Wang, C. X.ら, Blood 97:3890-5(2001))。
【0121】
[0140] この研究において、本発明者らは、PVRのウサギモデルを用いて、PVRおよび網膜剥離の発症および進行においてEMP2発現が有する役割を研究した。本発明者らは、EMP2の発現の減少または抗体によるEMP2のブロックによって、PVRの形成が減少することを明らかにした。
【0122】
方法および材料
[0141] 細胞系:安定なARPE−19細胞系であるARPE−19は、ATCCから得られる自然発生的なヒト網膜色素上皮(RPE)である(CRL-2302)。ARPE−19細胞を、以下の構築物の1つでトランスフェクトした:pEGFP−N3=ベクター対照、pEGFP−N3−EMP2=EMP2を過剰発現する構築物、pEGFP−N3−Ribol=EMP2の発現が低減した構築物。ARPE−19細胞を、トランスフェクションの前に24時間平板培養した。FuGENE6(Roche Molecular Biochemicals, Indianapolis, Indiana)を用いて、細胞を上記の構築物の1つでトランスフェクトした。安定なクローンをジェネティシン(700mg/ml、Invitrogen Life Technologies)を用いて選択した。これらの安定な細胞系をARPE−19/V、ARPE−19/EMP2、またはARPE−19/Riboと呼ぶ。これらの細胞系によるEMP−2の発現は、以前に報告されていた(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。
【0123】
[0142] 成長培地:ATCC培地#30−2006:1:1で混合した、ダルベッコ変法イーグル培地および重炭酸ナトリウム1.2g/L、L−グルタミン2.5mM、HEPES15mM、およびピルビン酸ナトリウム0.5mMを含有するハムF12培地 90%;ウシ胎児血清#30−2020 10%。
【0124】
[0143] PVRの動物モデル:この研究において用いられたウサギは、UCLA Institutional Review Board(IRB)によって認可された制度的ガイドラインに従って処理した。PVRは、以前に記載されたように、アルビノニュージーランドウサギの右目においてのみ誘発した(Nakagawa, M.ら, Invest Ophthalmol Vis Sci 36:2388-95(1995))。簡潔に述べると、30ゲージの針を用いて、C3F8ガス(100%)0.4mlを、麻酔導入した後に角膜縁の4mm後方の硝子体腔内に注入した。前方穿刺をマイクロブレードで行い、眼球内圧力の上昇を予防した。2日後、成長培地内に再懸濁したARPE−19細胞調製物を、0.1ccの容量で注入した。左目は対照として用いた。ウサギは、週に1回、間接検眼法によって、マスキング様式でPVRの発症および炎症について試験した。5週目に、動物をペントバルビタール(100mg/kg)を用いて犠牲死させた。眼球を10%ホルマリン内に置き、上部毛様体扁平部切開を#11のブレードで行って、眼球内腔に固定剤が届くようにした。眼球を3日間にわたり固定し、二等分し、Fastenbergによって確立されたPVRスケールを用いて肉眼的所見についてマスキング様式でスコア付けした(表1)(Fastenberg, D. M.ら, Am J Ophthalmol 95:663-7(1983); Fastenberg, D. M.ら, Am J Ophthalmology 93:565-72(1982))。眼球をパラフィン内に包埋し、切片化し、ヘマトキシリンおよびエオシンで染色した。切片を顕微鏡によって評価し、マスキング様式でPVRについて等級付けした。
【0125】
[0144] 統計分析:スチューデントt検定(独立、一方向)を用い、P<0.05を統計的に有意であると判断した。
【0126】
【表1】
【0127】
結果
[0145] EMP−2発現の増大を伴うPVR形成の増大:良く特徴付けされたPVRのウサギモデルを用いて、本発明者らは、記載されているARPE−19細胞の硝子体内注入を用いて、PVRを誘発した(Nakagawa, M.ら, Invest Ophthalmol Vis Sci 36:2388-95(1995))。細胞は、野生型、EMP2を過剰発現する修飾型ARPE−19細胞、またはリボザイムトランスフェクションによりEMP2を低発現する修飾型ARPE−19細胞であった(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。眼を、PVR形成の程度について、注入の5週間後に、二人の観察者が等級付けした。最初の3頭のウサギは、以前の報告に類似した、一貫した程度のPVR形成を有することが明らかになった(データは示していない)。野生型RPE細胞を与えたウサギは、EMP2を低発現する細胞を与えたウサギよりも有意に重症なレベルのPVRを有していた(それぞれ2.5対1.3、p=0.02)。(図1)さらに、EMP2を過剰発現する細胞を与えたウサギは、EMP−2を低発現するものよりも有意に多いPVR形成を有していた(それぞれ2.9対1.3、p=0.05)。EMP2発現の増大は、PVRの重症度の増大と相関しており、これと合致して、EMP2レベルの減少は、PVRの重症度の低減と相関していた。図2は、3つの異なるウサギ群から得られた典型的な肉眼的な組織学的標本を示す。
【0128】
[0146] EMP−2の発現の増大を伴う線維症および膜形成の増大:EMP2を過剰発現する細胞を与えた眼の組織学的試験によって、野生型細胞を投与したウサギまたはEMP2を低発現する(リボザイムトランスフェクトされた)細胞を投与したウサギの両方よりも大きなPVR膜形成が確認された(図3)。インビボでのPVR膜は、免疫組織化学によって、5週間目に、EMP2発現について陽性であることが明らかになった(図4)。
【0129】
[0147] EMP−2ブロック抗体で妨げられたPVR膜形成:ARPE−19細胞を、抗ヒトEMP2ポリクローナル抗体、対照抗体(免疫前血清)、または培地のみでプレインキュベートした。眼を細胞注入の5週間後に再び試験した。PVR形成の程度を二人のマスキングされた観察者が等級付けした。抗ヒトEMP2ポリクローナル抗体を用いたARPE−19細胞におけるEMP2のブロックによって、未処理細胞または免疫前対照抗体に曝露された細胞と比較してPVRの重症度が減少した(それぞれ2.6対4.3、p=0.01)(図5)。さらに、組織学的評価によって、線維症および牽引の減少が確認された(図6)。特異的抗体でライゲーションした後のEMP2発現の一時的な減少を記載したマウスモデルにおける本発明者らの以前のインビボでの研究によって予測されたように、EMP2の発現が残りの膜において観察された(Shimazaki, K.ら, Microbes Infect 9:1003-10(2007))。
【0130】
考察
[0148] PVRの発症の間の最初に、RPE細胞は網膜表面に結合し、移動し、増殖し、膜形成すると考えられる(Eibl, K. H.ら, Invest Ophthalmol Vis Sci 47:364-70(2006))。この後に、膜の収縮および牽引性網膜剥離が生じる。
【0131】
[0149] この研究において、本発明者らは、EMP2が膜収縮において重要な調節役割を有するというインビトロでの証拠にまで言及し、ウサギPVRモデルにおけるインビボでのEMP2発現の関連性の証拠を示す。ゲル収縮アッセイは、EMP2発現の増大を伴うコラーゲンゲルの収縮の増大を示した(Morales, S. A.ら, Invest Ophthalmol Vis Sci 50:462-9(2009))。ここで、本発明者らは、PVRを生じさせるRPE細胞におけるEMP2の発現レベルがウサギにおけるPVRの重症度と相関することを明らかにする。PVRにおけるEMP2の関連性についての独立した証拠を、対照抗体とは対照的にPVRの重症度を減少させたEMP2に対するブロック抗体(blocking antibody)を用いて得た。
【0132】
(実施例2)
[0150] 増殖性硝子体網膜症(PVR)は、穿通性損傷または裂孔原性網膜剥離を包含する重症の眼球外傷の合併症として生じる異常な創傷治癒である。上皮膜タンパク質2(EMP2)は、FAK活性化の調節によって、インビトロでのPVRモデルである網膜色素上皮細胞系ARPE−19によるコラーゲンゲルの収縮を調節する。この研究の目的は、ARPE−19細胞によるコラーゲンゲル収縮の阻害におけるEMP2特異的組換えダイアボディの効能を調べることである。
【0133】
方法
[0151] EMP2ダイアボディをファージライブラリーを用いて組換えによって構築し、ヒトEMP2ペプチドに対する反応性について選択した。全ての実験において、ARPE−19細胞を抗EMP2または対照ダイアボディ20μg/mlで2時間にわたり前処理した。毒性、接着、および移動をそれぞれ、Annexin Vを用いるフローサイトメトリー、1型コラーゲンへの結合、および創傷治癒アッセイを介して評価した。コラーゲンゲルの収縮をインビトロでのアッセイを用いて評価した。
【0134】
結果
[0152] 抗EMP2ダイアボディを用いて毒性は観察されなかった。抗EMP2ダイアボディは、接着または移動を有意には改変しなかった。抗EMP2ダイアボディへの曝露によって、4時間目でのEMP2タンパク質レベルが75%低減した。治療前のレベルへのEMP2の回復が16時間後に見られた。それに合致した、活性化型FAKの基底レベルの減少もまた観察された。コラーゲン刺激性のFAK活性化は、抗EMP2ダイアボディで処理したARPE−19細胞において25%低減した。抗EMP2ダイアボディ治療は、ARPE−19細胞によるコラーゲンゲルの収縮を有意に低減させ(p<0.001)、用量依存性であった。
【0135】
結論
[0153] 抗EMP2ダイアボディは、EMP2のダウンレギュレーションを介して収縮能力を減少させて、FAKの活性化を減少させ、コラーゲンゲルの収縮を阻害する。
【0136】
(実施例3)
導入
[0154] 接着斑キナーゼ(FAK)およびリン酸化を介するその活性化は、細胞周期の進行、増殖、侵入、移動、生存、および収縮にとって重要である(Schaller MDら, Proc Natl Acad Sci USA, 89:5192-5196(1992))。FAKのリン酸化は、インテグリン受容体が細胞外マトリクスの構成要素にライゲーションすることによって開始し得る。さらに、受容体チロシンキナーゼシグナル伝達を介するEGFおよびPDGFのライゲーション、ならびにG−タンパク質連結型の受容体を介するリゾホスファチジン酸(LPA)、ボンベシン、およびスフィンゴシルホスホリルコリン(SPC)への受容体のライゲーションを包含する多くの非インテグリンシグナル伝達経路が、FAKの活性化を促進する(Rankin SおよびRozengurt E, J Biol Chem, 269:704-710(1994); Seufferlein TおよびRozengurt E, J Biol Chem, 269:9345-9351(1994); Seufferlein TおよびRozengurt E, J Biol Chem, 270:24334-24342(1995); Sieg DJら, Nat Cell Biol, 2:249-256(2000); Sinnett-Smith Jら, J Biol Chem, 268:14261-14268(1993))。FAKの活性化は、複数の部位でのリン酸化を必要とする複雑なプロセスである。リン酸化は、Tyr−397およびTyr−407(N末端ドメイン)、Tyr−576およびTyr−577(キナーゼドメイン活性化ループ)、ならびにTyr−861およびTyr−925(C末端ドメイン)で生じる。FAKの活性化における最初の工程は、Tyr−397での自己リン酸化であり、これによりSrcチロシンキナーゼのSH2ドメインのための結合部位が生じ、その結果、Srcキナーゼドメインを活性化させる。残りのFAKチロシンリン酸化部位は、Srcによって優先的にリン酸化される(Calalb MBら, Mol Cell Biol, 15:954-963(1995); Schlaepfer DDら, Prog Biophys Mol Biol, 71:435-478(1999); Schlaepfer DDおよびHunter T, Trends Cell Biol, 8:151-157(1998))。キナーゼドメインの活性化ループにおけるFAKのTyr−576およびTyr−577のリン酸化は、最大のFAKキナーゼ活性に必要であり(Calalb MBら, Mol Cell Biol, 15:954-963(1995); Mitra SKおよびSchlaepfer DD, Curr Opin Cell Biol, 18:516-523(2006))、FAKのTyr−925のリン酸化によって、Grb2低分子アダプタータンパク質へのSH2結合部位が生じる(Schlaepfer DDら, Nature, 372:786-791(1994))。FAKへのGrb2の結合は、ERK2/MAPキナーゼカスケードなどの下流標的の活性化をもたらすいくつかのシグナル伝達経路の1つである。Tyr−407およびTyr−861でのリン酸化は、上皮間葉分化転換(EMT)に関与している(Nakatnoto Tら, Mol Cell Biol, 17:3884-3897(1997))。Tyr−861でのFAKのリン酸化は、F−アクチンの構成にも関与している(Lunn JAら, J Biol Chem, 282:10370-10379(2007))。
【0137】
[0155] 上皮膜タンパク質2(EMP2)は、成長停止特異的−3/末梢ミエリンタンパク質−22(GAS3/PMP22)ファミリーに属するテトラスパン(4回膜貫通)タンパク質である。EMP2は、皮膚、肺、子宮、心臓、甲状腺、および眼に局在する(Wang CXら, Blood, 97:3890-3895(2001))。眼では、EMP2は、角膜、毛様体、および網膜色素上皮(RPE)を包含する複数の上皮層に存在する(Wadehra Mら, Exp Mol Pathol, 74:106-112(2003))。EMP2は、インテグリン、グリコシルホスファチジルイノシトールを足場としたタンパク質、およびMHCクラスIタンパク質の輸送を調節することが示されている(Wadehra Mら, Dev Biol, 287:336-345(2005); Wadehra Mら, Mol Biol Cell, 15:2073-2083(2004); Wadehra Mら, J Biol Chem, 277:41094-41100(2002); Wadehra Mら, Clin Immunol, 107:129-136(2003))。EMP2は、胚盤胞移植などの生理学的環境、ならびに細胞の増殖、侵入、接着、および転移のモデル環境において、特定のインテグリンアイソフォームの機能に物理的に関連し、それを調節する(Wadehra Mら, Exp Mol Pathol, 74:106-112(2003); Wadehra Mら, Dev Biol, 287:336-345(2005); Wadehra Mら, Mol Biol Cell, 15:2073-2083(2004); Wadehra Mら, J Biol Chem, 277:41094-41100(2002); Wadehra Mら, Clin Immunol, 107:129-136(2003); Wadehra Mら, Dev Biol, 292:430-441(2006))。
【0138】
[0156] EMP2機能に関与する生化学的メカニズムは、明らかにされていない。しかし、他のテトラスパンタンパク質において、顕著なテーマは、シグナル伝達経路に関与する非相同テトラスパンおよび他の膜タンパク質との多重の関連である(Boucheix Cら, Cell Mol Life Sci, 58:1189-1205(2001); Hemler ME, J Cell Biol, 155:1103-1107(2001))。これは、テトラスパンタンパク質が、膜におけるシグナル伝達複合体の機能的組み立て体を支持する分子アダプターとして作用することを示唆する(Hong IKら, J Biol Chem, 281:24279-24292(2006))。おそらく、PI−4キナーゼを特定の膜位置に動員しホスホイノシチド依存性のシグナル伝達を誘発することが示されている、テトラスパンタンパク質のサブセットが最も良く規定されている(Yauch RLおよびHemler ME, Biochem J, 351 Pt 3:629-637(2000))。
【0139】
[0157] EMP2は、Tyr−576および577でのリン酸化によって測定されるFAKの活性化を介してARPE−19細胞系のコラーゲンゲル収縮を増大させることが知られている(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。この研究の目的は、EMP2とFAK活性化との関連をさらに調査することであった。ここで、本発明者らは、EMP2がFAKと物理的に関連して、複数の部位でFAKのリン酸化を増大させることを示す。このFAKのリン酸化の増大によって、機能的な機械的な細胞の改変が生じ、細胞の収縮能力が増大する。これらの細胞の改変には、接着斑密度の増大、アクチン細胞骨格における立体配座の変化、アクチン組成物の改変、および細胞の接着能力の増大が包含される。これらの所見は、EMP2がFAKの活性化に重要な膜シグナル伝達複合体の機能的組み立てにおけるテトラスパンの新たな役割を表し得ることを示唆する。
【0140】
方法
[0158] 細胞系−RPE特異的マーカーであるCRALBPおよびRPE−65を発現する自然発生的な網膜色素上皮(RPE)細胞系であるARPE−19を、American Type Culture Collectionから得た(CRL-2302, ATCC, Manassas, VA)。EMP2を過剰発現する細胞系であるARPE−19/EMP2を、75倍高いタンパク質レベルのEMP2を発現するEMP2過剰発現のレトロウイルス構築物の安定な感染を介して生成した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。ARPE−19細胞を5%CO2の湿潤チャンバにおいて37℃で、10%ウシ胎児血清(FBS)(ATCC, Manassas, VA)を補ったDMEM−F12培地において培養した。全ての実験において細胞を平板培養し、一晩インキュベートした。全ての細胞は、別段の指示がない限り、各実験の開始時点で約75%コンフルエントであった。
【0141】
[0159] 抗体−ヒトFAKについての全ての抗体は、Santa Cruz Biotechnology(Santa Cruz, CA)から得た:リン酸化型FAK(p-FAK)(クローンTyr 576/577)についてのヤギ抗体、ならびにFAK(クローンC-20)、p−FAK(Tyr 397)、(Tyr 407)、(Tyr 861)、および(Tyr 925)についてのウサギ抗体。ヒトp−Src(Tyr 416およびTyr 527)についてのウサギ抗体は、Cell Signaling Technology(Danvers, MA)から得た。ヒトβ−アクチン(クローン2A2.1)を認識するマウス抗体は、US Biological(Swampscott, MA)から得た。ヒトα−平滑筋アクチン(α-sma)(クローン1A4)に対するマウス抗体は、Sigma(St. Louis, Mo)から得た。F−アクチンを同定するために用いるテキサスレッド−Xファロイジン(T7471)は、Invitrogen(Carlsbad, Ca)から得た。ヒトEMP2に対するウサギ抗血清は、ヒトEMP2の第2の細胞外ループ(EDIHDKNAKFYPVTREGSYG)に結合した多抗原ペプチドで動物を免疫化して生成した(Research Genetics, Huntsville, AL)。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体は、Southern Biotech(Birmingham, Alabama)から得た。ホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体は、BD Biosciences(San Diego, CA)から得た。テキサスレッド結合ロバ抗ヤギ抗体(705-075-147)およびFITC結合ロバ抗ウサギ抗体(711-095-152)は、Jackson ImmunoResearch Laboratories(West Grove, PA)から得た。
【0142】
[0160] ウェスタンブロット分析−ウェスタン分析を、以前記述されているように行った(Morales S Aら, Experimental Eye Research(2007))。簡潔に述べると、細胞タンパク質を、プロテアーゼ阻害剤およびホスファターゼ阻害剤(Upstate, Charlottesville, VA)を含有するRIPA緩衝液を用いて単離し、タンパク質濃度をBCAタンパク質アッセイ(Bio Rad, Hercules, CA)で決定した。EMP2の検出のために、N−連結グリカンをPNgaseを用いて切断した(New England Bioloabs, Beverly, MA)。溶解物を製造者の指示に従って、37℃で2時間にわたり処理した。全部で10μgのタンパク質を各レーンにロードし、タンパク質を還元条件下で4〜20%SDS−PAGE勾配ゲルによって分画した。タンパク質をニトロセルロース膜(Amersham Life Sciences, Buckinghamshire, UK)に移し、転移の妥当性をPonceau Sレッド染色(Sigma Chemical Co., St. Louis, MO)を用いて確認した。
【0143】
[0161] 膜を、TBS Tween(TBST; Upstate, Charlottesville, VA)内の無脂肪乳でブロックした。ブロットを、α−sma、FAK、およびp−FAK(Tyr 576/577)、(Tyr 397)、(Tyr 407)、(Tyr 861)、(Tyr 925)では1:200の希釈、EMP2およびp−Src(Tyr 416)、(Tyr 527)では1:1000の希釈、およびβ−アクチンでは1:5000の希釈の一次抗体と共に、1時間にわたりインキュベートした。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体またはホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体を、1:2000の希釈でブロットに曝露した。ブロットをECL(Pierce, Rockford IL)で発現させ、ブロットのスキャンおよびNIH Image Jソフトウェアでのバンド密度の測定によって定量した。β−アクチンまたは全FAKのバンドの強度を内部対照として用い、全てのバンドを、定量による密度の測定のために線形範囲で分析した。少なくとも3つの独立した実験を行い、スチューデントt検定(独立、一方向)を用いて統計的に評価した。p<0.05を統計的に有意であると考えた。共免疫沈降−ARPE−19細胞およびARPE−19/EMP2細胞を10cmの皿内に平板培養した(Corning, Corning, NY)。細胞をPBSで2回洗浄し、4℃で30分間にわたり溶解し(アプロチニン10μg/ml、ペプスタチン2μg/ml、EDTA0.1mM、HEPES10mM、およびKCl10mMを含有する1%Nonidet P-40、ならびにRoche Applied Sciences, Mannheim, Germanyから入手したComplete Mini Protease Inhibitor Cocktail Tablet)、次に、15秒間にわたり超音波処理した。細胞溶解物をタンパク質Aアガロースビーズ(Santa Cruz Biotechnology, Santa Cruz, CA)と共にインキュベートして事前確認し、次に、アガロースビーズおよび抗FAKポリクローナル抗体(Santa Cruz Biotechnology, Santa Cruz, CA)または抗EMP2ウサギポリクローナル抗体と共に一晩インキュベートした。ビーズを溶解溶液内で4回洗浄し、Tris62.5mM(pH6.8)内で2回洗浄した。免疫複合体を、Laemmli試料緩衝液(Tris-Cl62.5mM(pH6.8)、10%グリセロール、2%SDS、0.01%ブロモフェノールブルー、2%β−メルカプトエタノール)内で5分間にわたり煮沸してビーズから溶出した。試料を、ウェスタンブロット分析を用いて分析した。EMP2の検出のために、N−連結グリカンを、PNgase(New England Bioloabs, Beverly, MA)を用いて切断した。溶出物を製造者の指示に従って、37℃で2時間にわたり処理した。
【0144】
[0162] 共免疫沈降実験におけるFAKおよびEMP2の化学量論を以下の様式で決定した。まず、細胞全体の溶解物および同族免疫沈降物の滴定をデンシトメトリーによって定量した。このデータを用いて、本発明者らは、同族免疫沈降物における各タンパク質のパーセントを計算した(投入した細胞当量当たり)。第2に、共免疫沈降物を同様に分析し、共免疫沈降物内に捕捉された各タンパク質のパーセントを決定した。最後に、本発明者らは、共免疫沈降したタンパク質の化学量論を表すこれらの2つの値(同族のパーセントおよび共免疫沈降物のパーセント)の比率を計算した。
【0145】
[0163] 免疫蛍光−ARPE−19細胞およびARPE−19/EMP2細胞をガラスカバースリップ(Fisher)上で一晩平板培養した。細胞を4%パラホルムアルデヒドで20分間にわたり固定し、0.075%サポニンで15分間にわたり透過処理した。細胞を10%正常ロバ血清で30分間にわたりブロックした。細胞を一次抗体と共に湿潤チャンバにおいて4℃で一晩インキュベートし、次に、PBSに0.01%Triton X−100(PBST)を加えたもので3〜4回洗浄した。細胞を、湿潤チャンバにおいて室温で、フルオレセインイソチオシアネート(FITC)結合ロバ抗ウサギIgGおよびテキサスレッド結合ロバ抗ヤギ抗体と共に1時間にわたりインキュベートした。細胞をPBSTで洗浄し、2回蒸留したH2Oで簡単にすすぎ、VECTASHIELD Mounting Medium(Vector Laboratories, Burlingame, CA)を用いて顕微鏡スライド上に載せた。
【0146】
[0164] Zeiss LSM 510レーザー走査共焦点顕微鏡(Thornwood, NY)を用いて、タンパク質の分布および共局在化を評価した。FITCで標識された細胞およびテキサスレッドで標識された細胞を検出するために、試料をそれぞれ488nmおよび568nmのアルゴンレーザーおよびクリプトンレーザーで励起させた。LSMソフトウェアを、顕微鏡の制御、走査およびレーザーモジュール、画像記録、ならびに画像データの分析に用いた。
【0147】
[0165] LSMソフトウェアを用いて共局在化分析を行い、関連のパーセントを得た。強調されたピクセルを、Image J(NIHソフトウェア)を用いて決定した。少なくとも6つのフィールドを各試料の分析のためにランダムに選択し、関連のパーセントは、複数のフィールドから生じた平均値である。フィールド当たり複数のスライスを評価したが、スライドと接触し接着斑を含有する細胞底表面の単一のスライスのみを接着斑とEMP2との共局在化の定量に用いた。全ての実験において、細胞は60倍の油浸対物レンズを用いて観察した。各実験を少なくとも4回反復した。
【0148】
[0166] 接着アッセイ−ARPE−19細胞およびARPE−19/EMP2細胞を24ウェルコラーゲン被覆プレート(BD Biosciences, San Diego, CA)、またはフィブロネクチン、ビトロネクチン、コラーゲンI、およびコラーゲンIVで事前被覆されたCytoMatrix Screeningプレート(Millipore, Billerica, MA)上で、ウェル当たり2×105個細胞の濃度で平板培養した。細胞を5%CO2の湿潤チャンバにおいて37℃で2時間にわたりインキュベートした。次にプレートをPBSで3回洗浄し、全ての未付着細胞を除去した。結合細胞をクリスタルバイオレットの取り込みについて分析し、可溶化した後、吸光度を、Bio−Radマイクロプレートリーダー550(Hercules, CA)によって595nmで測定した。各実験は少なくとも8回の反復を含み、少なくとも3つの独立した実験を行い、同等の結果を得た。スチューデントt検定(独立、一方向)を用い、P<0.05を統計的に有意であると判断した。
【0149】
結果
[0167] ARPE−19細胞におけるp−FAKとEMP2との関連。Srcの相互作用および相互リン酸化事象を介して生じた、FAKのリン酸化は、多様な細胞応答を推進するシグナル伝達タンパク質およびアダプタータンパク質を動員する(Jacamo ROおよびRozengurt E., Biochem Biophys Res Commun, 334:1299-1304(2005))。本発明者らは、以前に、ARPE−19細胞におけるEMP2の過剰発現がFAK/Src活性の改変をもたらすかどうかを調べた。本発明者らは、全FAKレベルがARPE−19細胞およびARPE−19/EMP2細胞の両方において等しいことを示し(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))、図7において確認したが、EMP2の過剰発現は、対照ARPE−19細胞と比較して、活性化型FAK(Tyr-576/577)のレベルがほぼ2倍増大することを実証した。各活性化型FAKをそれ自身のβ−アクチンロード対照に正規化して行った複数の評価は、EMP2レベルの増大とFAK活性化との間の統計的に有意な相関を示した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。FAKリン酸化の他の部位を、ARPE−19細胞およびARPE−19/EMP2細胞において定量的ウェスタンブロットによって調べ、全FAKに正規化した(図7A)。EMP2の過剰発現は、Tyr−397、Tyr−407、Tyr−861、およびTyr−925でのFAKのリン酸化を2〜3倍増強した。
【0150】
[0168] Srcの酵素活性は、Tyr−416(増加)およびTyr−527(抑制)でのチロシンリン酸化によって相互調節される(Harvey Rら, Mol Cell Biol, 9:3647-3656(1989))。対照細胞と比較して、EMP2の過剰発現は、Tyr−416でのSrcリン酸化の増大と関連していた(図7B)。このSrcキナーゼドメインの活性化がTyr−397でのFAKのリン酸化と関連しているため、この結果は予測された。重要なことには、キナーゼドメインには存在しないSrc Tyr−527では、リン酸化レベルに変化はなかった(図7B)。総合すると、これらの所見は、EMP2の上昇によって、機械的収縮性を増大させることが知られているFAKおよびSrcのリン酸化が変化することを示す。
【0151】
[0169] EMP2はFAKと物理的に関連する。EMP2の発現は、効率的なインテグリン介在性の細胞応答に必要であり、EMP2は、特定のインテグリンアイソフォームと物理的に関連する(Wadehra Mら, Dev Biol, 287:336-345(2005); Wadehra Mら, Mol Biol Cell, 15:2073-2083(2004); Wadehra Mら, J Biol Chem, 277:41094-41100(2002); Wadehra Mら, Clin Immunol, 107:129-136(2003))。FAKは、インテグリンの重要なシグナル伝達パートナーであり、その役割は、インビトロでのβ1インテグリンの細胞質尾部とFAKのN末端ドメインとの物理的関連を包含する(Lyman Sら, J Biol Chem, 272:22538-22547(1997); Schaller MDら, J Cell Biol, 130:1181-1187(1995))。したがって、本発明者らは、FAKのリン酸化に対するEMP2レベルの正の影響がFAKとEMP2との間の物理的関連を反映し得ると仮定した。この問題に取り組むため、本発明者らは、物理的関連の2つの要件、すなわち、免疫沈降を直接的または間接的に介する結合、および共焦点顕微鏡法を介する同一の細胞空間内での局在化を試験した。
【0152】
[0170] 本発明者らは、まず、EMP2およびFAKが物理的に関連しているかを試験することによってこの見解を評価した(図8A)。EMP2に対する抗体を用いる免疫沈降物には、ARPE−19細胞およびARPE−19/EMP2細胞の両方において容易に検出可能なFAKが包含される。関連の化学量論を決定するために、本発明者らは3つの独立した実験を用いて、免疫沈降物におけるEMP2およびFAKの定量されたレベルを表にした。まず、細胞全体の溶解物および同族免疫沈降物の滴定を、デンシトメトリーによって定量した。このデータを用いて、本発明者らは、同族免疫沈降物における各タンパク質のパーセントを計算した(投入した細胞当量当たり)。第2に、共免疫沈降物を同様に分析して、共免疫沈降物内に捕捉された各タンパク質のパーセントを決定した。最後に、本発明者らは、共免疫沈降したタンパク質の化学量論を表すこれらの2つの値(同族のパーセントおよび共免疫沈降物のパーセント)の比率を計算した。FAK(図8A)およびEMP2の同族の免疫沈降(データは示していない)は、比較的効率的であった(50〜60%)。EMP2免疫沈降物におけるFAKのレベルは、ARPE−19およびARPE−19/EMP2においてそれぞれ12%および15%であり、全FAKの24%〜30%がそれぞれ、これらの2つの細胞系においてEMP2と関連した。
【0153】
[0171] EMP2およびFAKが同一の細胞空間内に局在しているかを評価するために、in situでのそれらの共局在化を共焦点顕微鏡法によって試験した。フィールド当たり複数のレベルを評価したが、スライドと接触し接着斑を含有する細胞底表面の単一のスライスのみを定量に用いた。EMP2(緑)およびFAK(赤)を免疫蛍光によって検出し、共焦点顕微鏡法によって可視化した(図8B)。これらの2つのチャネルの併合が示されており、同様に、強調された共局在化したピクセルが白いシグナルで示されている併合が示されている(右)。検証によると、EMP2およびFAKは、両細胞型において顕著に共局在化していた。3つの独立した実験のピクセル定量によると、ガラススライドに接触している細胞表面に存在しているFAKの68%±9%および96%±1%がそれぞれ、ARPE−19細胞およびARPE−19/EMP2細胞においてEMP2と共局在化していた。全FAKレベルが、ARPE−19/EMP2細胞系において、わずかな、しかし有意ではない増大を示した(図7A)。このことは、EMP2レベルが全FAKタンパク質発現に影響しないことを示す、本発明者らが以前に公表したデータを裏付けるものである(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。
【0154】
[0172] EMP2がリン酸化型FAKと関連するかどうかを試験するために、細胞をEMP2およびp576/577−FAK一次抗体で染色した。フィールド当たり複数のスライスを評価したが、スライドと接触し接着斑を含有する細胞底表面の単一のスライスのみを定量に用いた。EMP2をFITCで検出し、リン酸化型FAKをテキサスレッドで検出した(図8C)。ARPE−19では、p−FAK(Y576/577)の31%±8%がEMP2と共局在化している。細胞内には、共局在化を示す中心領域があるが、この細胞内領域の同定にはさらなる研究が必要である。ARPE−19/EMP2では、p−FAK(Y576/577)の97%±0.4%がこのレベルでEMP2と共局在化している。EMP2およびFAK(全FAKおよびp−FAK(Y576/577)はin situで高度に共局在化しており、EMP2が上昇すると共局在化の程度が増大する。しかし、EMP2が過剰発現している程度を考慮すると、これはFAKの特異的な強化を示唆していない可能性がある。
【0155】
[0173] EMP2レベルが増大するとリン酸化型FAKの分布が改変した。ARPE−19細胞およびARPE−19/EMP2細胞(図9A)をリン酸化型FAK(Y576/577)タンパク質について染色した。EMP2レベルが増大すると、リン酸化型FAKの分布が改変した。ARPE−19細胞では、リン酸化型FAKは主に細胞の周辺で見られたが(図9A)、一方、ARPE−19/EMP2細胞では、リン酸化型FAKが細胞全体にわたり分布しているのが見られる(図9A)。少なくとも4つの個別の試料のピクセル強度を、スチューデントt検定(独立、一方向)によって測定し、タンパク質検出にウェスタンブロットを用いた本発明者らの以前の報告(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))と一致するこの評価も、ARPE−19/EMP2細胞におけるリン酸化型FAKの有意な増大を示した(図9B)。
【0156】
[0174] EMP2およびリン酸化型FAKの発現が増大すると、F−アクチンの分布が改変する。EMP2の発現が増大すると、F−アクチン構成に関連しているTyr−861を包含する複数の部位でFAKリン酸化が増大する。したがって、本発明者らは、EMP2発現の増大と関連する観察可能なアクチン構成における付随する変化が存在したかを試験した。ARPE−19細胞(図10A、C)およびARPE−19/EMP2細胞(図10B、D)をテキサスレッド−Xファロイジン一次抗体で染色した。EMP2発現が増大すると、F−アクチンの発現が改変した。F−アクチンは、ARPE−19/EMP細胞において細胞の周辺に沿って発現したが、一方、F−アクチンは、ARPE−19細胞では細胞の全体にわたり発現した。本発明者らは、ゲル収縮アッセイにおいて試験したものに類似した、活発に成長している細胞におけるアクチン線維を試験することを選択した。コンフルエント細胞における配置を検討するためのさらなる研究を、今後行うことができる。
【0157】
[0175] EMP2の発現が増大すると、α−平滑筋アクチンの発現が増大した。EMP2の発現が増大すると、共に細胞EMTにおいて重要な部位として関与しているTyr−861およびTyr−407でのFAKのリン酸化が増大する。EMTのマーカーは、α−smaの発現である。関連するFAKリン酸化を伴うEMP2発現の増大がα−smaの発現に機能的な影響を有するかを試験するために、α−smaのレベルをARPE−19細胞およびARPE−19/EMP2細胞においてウェスタンブロット分析によって測定した(図11A)。次にブロットをECLで発現させて、結合抗体を視覚化し、β−アクチンを内部対照として用いて定量した(図11B)。EMP2の発現が増大すると、α−smaの発現が有意に増大した。
【0158】
[0176] EMP2レベルが増大すると、I型コラーゲンおよびIV型コラーゲンへの細胞接着が増大する。ARPE−19/EMP2細胞は、ARPE−19細胞と比較して、細胞全体にわたりリン酸化型FAKの大きな分布を示した。本発明者らは、リン酸化型FAKの分布の増大は、ARPE−19/EMP2細胞へのさらなる接着能力をもたらす機能的接着斑に相当すると仮定した。この仮説を試験するために、本発明者らは、ARPE−19細胞およびARPE−19/EMP2細胞を、フィブロネクチン、ビトロネクチン、コラーゲンI、およびコラーゲンIVで事前被覆したプレート上で平板培養した。結合細胞をクリスタルバイオレットの取り込みについて、Bio−Radマイクロプレートリーダー550によって分析した。EMP2の発現が増大すると、I型コラーゲンおよびN型コラーゲンへの接着の付着相が特異的に増大し(P<0.001)、フィブロネクチンへの付着が減少し(P<0.02)、ビトロネクチンへの付着には影響はなかった(図12)。
【0159】
考察
[0177] この研究は、EMP2が、他のテトラスパンファミリーメンバーのように(Hong IKら, J Biol Chem, 281:24279-24292(2006))、特定のインテグリンアイソフォームとその関連するシグナル伝達分子との間の分子アダプターであるという仮説によって導かれた。本発明者らの以前の研究は、FAK−Src経路の活性化が、RPE細胞系ARPE−19を用いるPVRのインビトロモデルでのコラーゲンゲルの収縮の生成において重要であることを同定した(Morales SAら, Experimental Eye Research(2007))。さらに、本発明者らの研究は、EMP2が、FAK(y-576/577)の活性化を介してコラーゲンゲル収縮を制御し得ることを実証した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。本発明者らは、以前に、EMP2がインテグリンβ1と関連することを実証し(Wadehra Mら, J Biol Chem, 277:41094-41100(2002))、他のグループは、FAKがインビトロでインテグリンβ1サブユニットに結合することを示している(Schlaepfer DDら, Prog Biophys Mol Biol, 71:435-478(1999); Parsons JTら, Oncogene, 19:5606-5613(2000))。この研究において、本発明者らは、EMP2がFAKの活性化を増強し、FAKと物理的に関連し、その機能的な下流の結果は、接着斑の密度、細胞の接着能力、F−アクチンの立体配座、およびアクチンの組成に影響するという証拠を提供する。したがって、EMP2は、効率的なインテグリン介在性のFAK活性化およびFAKに関連する細胞機能に対するその影響のための分子アダプターとして作用し得る。共免疫沈降および共局在化の研究は、多タンパク質複合体の形成を介する直接的なまたは間接的な関連を裏付けるが、この関連の具体的な詳細を規定するためには、さらなる生化学的研究が必要であろう。
【0160】
[0178] FAKの活性化のための重要なメカニズムは、複数部位での自己リン酸化、Srcの動員、およびFAKのリン酸化をもたらす、インテグリン受容体介在性のFAKのクラスタリングである。活性化型FAK/Src複合体は、その後、Grb2、p130Cas、パキシリン、Talin、CAP、Graf、Src、およびPI3−キナーゼのp85サブユニットなどの、種々のタンパク質のための分子足場として作用し、多くのシグナル伝達経路に関与し得る(Schlaepfer DDら, Prog Biophys Mol Biol, 71:435-478(1999); Parsons JTら, Oncogene, 19:5606-5613(2000); Cox BDら, J Cell Biochem, 99:35-52(2006))。
【0161】
[0179] この報告において、本発明者らは、EMP2がFAKと関連して、接着斑の形成を増強することによってFAKの活性化を増大させることを実証し、EMP2がインテグリンのライゲーションとFAKの活性化との間の分子アダプターとして作用し得ると仮定する。本発明者らは、以前、増殖性硝子体網膜症(PVR)のインビトロでの相関要因であるコラーゲンゲル収縮におけるEMP2の役割を試験した。PVRは、患者の最大10%において、裂孔原性網膜剥離の外科的修復に続発する合併症である(Charteris DGら, Eye, 16:369-374(2002); Kirchhof B., Graefes Arch Clin Exp Ophthalmol, 242:699-703(2004); Lewis GPら, Eye, 16:375-387(2002))。PVRは、RPEを包含する複数の細胞型の、硝子体腔内への複雑な移動の最終結果に相当する(Baudouin Cら, Ophthalmologica, 203:38-46(1991); Casaroli-Marano RPら, Invest Ophthalmol Vis Sci, 40:2062-2072(1999); Charteris DGら, Br J Ophthalmol, 79:953-960(1995); Vinores SAら, Invest Ophthalmol Vis Sci, 31:2529-2545(1990))。牽引性網膜剥離をもたらす収縮性の細胞の力に関連する移動、膜形成、および異常な創傷治癒戦略をもたらすRPE細胞の上皮間葉移行(EMT)の証拠が存在する。ARPE−19細胞によって行われるコラーゲンゲル収縮を用いて、本発明者らは、FAKを介するインテグリン受容体の活性化およびシグナル伝達をARPE−19介在性の収縮に必要な非常に重要なプロセスであると同定した(Morales SAら, Experimental Eye Research(2007))。本発明者らは、最近、EMP2の発現レベルがコラーゲンゲルの収縮と正に相関すること、およびこの表現型がFAK活性化の促進を介して達成されることを観察した(Morales SAら, Invest Ophthalmol Vis Sci, 9:9(2008))。
【0162】
[0180] PVRの発症機序の状況において、EMTは、鍵となる構成要素である。Tyr−407およびTyr−861でのFAKのリン酸化は、EMTの進行において重要であると同定される(Nakamura Kら, Oncogene, 20:2626-2635(2001))。EMP2の過剰発現は、Tyr−407およびTyr−861でのリン酸化を増大させ、したがって、EMTを進行させると予測される。EMTの1つのマーカーはα−smaのアップレギュレーションであり、これは、EMP2を過剰発現する細胞系であるARP−19/EMP2細胞において観察された。EMP2の増大およびFAK活性化の増大に関連するこれらの変化は、さらなる収縮性の表現型を促進すると予測される。F−アクチンの構成はまた、コラーゲンゲルに接触する細胞の能力に影響し得る。ARPE−19細胞では、F−アクチンの発現は細胞全体にわたり観察され、これはおそらく、より堅固な細胞構造をもたらし、コラーゲンゲルを収縮させる細胞の能力を低減させる。ARPE−19/EMP2細胞では、F−アクチンは、皮質内分布している細胞の縁に沿って見られるのみであり、これは、さらなる収縮能力を可能にするさらに柔軟な立体配座をもたらし得る。
【0163】
[0181] テトラスパンタンパク質は、様々な複合体の形成に関与して「テトラスパン網」を形成することが示されており(Levy SおよびShoham T, Physiology(Bethesda)20:218-224(2005); Levy SおよびShoham T, Nat Rev Immunol, 5:136-148(2005))、これは、シグナル伝達および分類プロセスを調節する足場および膜ドメインの生成である(Levy SおよびShoham T, Physiology(Bethesda)20:218-224(2005); Hemler ME, Nat Rev Mol Cell Biol, 6:801-811(2005))。これらの複合体は、それらの膜タンパク質成分のシグナル伝達、輸送、および構造的特徴を調節し得る。テトラスパンがインテグリン分子と複合体を形成する場合、テトラスパンは細胞の接着および移動性を調節し得る(Caplan MIら, Curr Opin Nephrol Hypertens, 16:353-358(2007))。本発明者らの報告はテトラスパンのパズルにピースを加えるものであり、EMP2が、さらに大きな細胞収縮能力を有するEMT表現型に向かわせる物理的関連を介するFAK活性化を調節することを実証する。EMP2−FAKの関連は、PVRのインビトロモデルの状況における有意な機能的細胞応答を実証する、これまでに報告されていない新規なタンパク質−タンパク質相互作用に相当する。
【0164】
(実施例4)
導入
[0182] 4回膜貫通(テトラスパン)タンパク質であるEMP2は、眼、肺、心臓、甲状腺、および子宮における異なる位置で発現する(Wang CXら, Blood, 97(12):3890-3895(2001))。眼では、EMP2は、角膜、毛様体、および網膜色素上皮(RPE)を包含する複数の上皮層に局在化している(Wadehra Mら, Exp Mol Pathol, 74(2):106-112(2003))。複数の非眼球細胞型では、EMP2は、選択的な受容体輸送において重要な役割を有し、増殖、侵入、接着、および移動において重要な分子に影響する(Wadehra Mら, Exp Mol Pathol, 74(2):106-112(2003); Wadehra Mら, Dev Biol, 287(2):336-345(2005); Wadehra Mら, Mol Biol Cell, 15(5):2073-2083(2004); Wadehra Mら, J Biol. Chem, 277(43):41094-41100(2002); Wadehra Mら, Cancer 107(1):90-98(2006); Wadehra Mら, Clin Immunol, 107(2):129-136(2003))。これらの研究は、眼球由来の細胞における異なる重要な細胞プロセスの協調的な制御におけるEMP2の潜在的な中心的役割を示唆する。
【0165】
[0183] PVRは、患者の最大10%において裂孔原性網膜剥離の後に観察され、失明をもたらす可能性がある合併症である(Charteris DGら, Eye, 16(4):369-374(2002); Kirchhof B., Graefes Arch Clin Exp Ophthalmol, 242(8):699-703(2004); Lewis GPら, Eye, 16(4):375-387(2002))。PVRの根幹にある病態生理学は複雑であり、RPEを包含する複数の細胞型が、この疾患における重要な役割を有すると考えられている(Baudouin Cら, Ophthalmologica, 203(1):38-46(1991); Casaroli-Marano RPら, Invest Ophthalmol Vis Sci, 40(9):2062-2072(1999); Charteris DG., Br J Ophthalmol, 79(10):953-960(1995); Vinores SAら, Invest Ophthalmol Vis Sci, 31(12):2529-2545(1990))。牽引性網膜剥離をもたらし得る収縮性の細胞の力に関連する、細胞の移動、膜形成、および異常な創傷治癒プロセスをもたらす、上皮間葉移行の証拠が存在する。PVRの予防のための新たな戦略には、複雑な病態生理学の理解を進めることが必要である。PVRの1つのインビトロでの相関要因は、コラーゲンゲルの収縮である。
【0166】
[0184] 一次ヒト細胞(Hunt RCら, Invest Ophthalmol Vis Sci, 35(3):955-963(1994))、ヒトARPE−19細胞(Bando Hら, Exp Eye Res, 82(3):529-537(2006))、ウシ(Ando Aら, Br J Ophthalmol, 84(11):1306- 1311(2000))、およびウサギ(Zheng Yら, Invest Ophthalmol Vis Sci, 45(2):668-674(2004))を包含する種々の種のRPEが、コラーゲンゲル収縮の研究において用いられている。この研究は、いかにEMP2が、ARPE−19細胞系におけるEMP2の発現を組換えによって改変することを介して、コラーゲンゲル収縮を制御するかを試験するために設計された。本発明者らの以前の研究(Morales SAら, Exp Eye Res, 85(6):790-798(2007))および他者による研究(Bando Hら, Exp Eye Res, 82(3):529-537(2006); Carver Wら, J Cell Physiol, 165(2):425-437(1995); Cooke MEら, J Cell Sci, 113:2375-2383(2000); Kieffer JDら, Biochem Biophys Res Commun, 217(2):466-474(1995); Langholz Oら, J Cell Biol , 131:1903-1915(1995); Moulin VおよびPlamondon M., Br J Dermatol, 147(5):886-892(2002); Robbins SGら, Invest Ophthalmol Vis Sci, 35(9):3475-3485(1994); Zhang ZGら, J Cell Sci, 119:1886-1895(2006))は、特異的なインテグリンアイソフォームを同定し、コラーゲンマトリクスがコラーゲンゲル収縮において非常に重要であることが確実であることを見出した。本発明者らは、以前、FAK/Src経路の活性が、外因性の炎症性刺激の存在下または非存在下でのコラーゲンゲル収縮の観点からARPE−19細胞系において必須であると同定した。本研究では、EMP2の発現レベルはコラーゲンゲルの収縮を制御し、EMP2の増大は、ARPE−19細胞系におけるFAK活性化の増強に関連していた。
【0167】
方法
[0185] EMP2構築物: 以前に報告されているように、ハンマーヘッドリボザイムを生成して、ヒトEMP2転写産物を切断した(Wadehra Mら, Dev Biol, 287(2):336-345(2005))。EMP2の発現を低減させるためのトランスフェクションにおいて良好に機能することが実証されているhRZ2 EMP2ハンマーヘッドリボザイムを、この研究において用いた。簡潔に述べると、pEGFPにおけるhRZ2構築物(BD-Clontech, Palo Alto, CA)をARPE−19内にトランスフェクトし、安定なクローンを選択した(Wadehra Mら, Dev Biol, 287(2):336-345(2005))。
【0168】
[0186] ヒトEMP2の完全長cDNAをレトロウイルスベクターpMSCV−IRES−GFP内のEcoRI部位でクローニングした(Hawley RGら, Gene Ther, 1(2):136-138(1994))。EMP2の発現は、5’の長い末端反復(LTR)によって推進した。このベクターはまた、内部リボソーム侵入部位を介する緑色蛍光タンパク質(GFP)の発現を可能にする。高力価のヘルパーフリーレトロウイルスストックを、以前に記載されているように、293T細胞の一時的なコトランスフェクションによって調製した(Pear W., In: Ausubel FM, Brent R, Lingston RE,ら編, Current Protocols in Molecular Biology, 第2巻. New York: John Wiley & Sons; 9.11-9.15(1996))。
【0169】
[0187] 細胞系: RPE特異的マーカーであるCRALBPおよびRPE−65を発現する自然発生的な網膜色素上皮(RPE)細胞系であるARPE−19を、American Type Culture Collectionから得た(CRL-2302; ATCC, Manassas, VA)。ARPE−19細胞を、5%CO2の湿潤チャンバにおいて37℃で、10%ウシ胎児血清(FBS; ATCC)を補ったDMEM−F12培地において培養した。培養培地を週に2回交換した。コンフルエントになった後、培養物を0.05%(重量/容量)トリプシン内で解離することによって継代した。EMP2のレベルは、EMP−2を過剰発現するレトロウイルス構築物の安定な感染を介してAFPE−19細胞において増大し、GFP陽性細胞の流動選別によって選択された。EMP2の発現が増大するように安定に感染したこれらの細胞は、ARPE−19/EMP2と呼ばれる。EMP2のレベルは、pEGFP−hRZ2リボザイム構築物3.4μgおよびトランスフェクション試薬(FuGENE 6; Roche, Basel, Switzerland)でのARPE−19細胞の安定なトランスフェクションにより減少した。ARPE−19/Ribo細胞を、GFP陽性細胞について流動選別した。とりわけ、トランスフェクション試薬は、トリパンブルー排除によって決定したところ、ARPE−19細胞に対する検出可能な毒性を生じさせなかった(データは示していない)。
【0170】
[0188] EMP2のレベルは、ARPE−19細胞をEMP2siRNA75ピコモル(L-016226-00; Dharmacon, Lafayette, CO)および親油性トランスフェクション試薬(Lipofectamine 2000; Invitrogen, Carlsbad, CA)で一時的にトランスフェクトすることによって減少し、48時間後に分析された。陰性対照として、細胞をスクランブル対照siRNA75ピコモル(D-001206-13-05; Dharmacon)でトランスフェクトした。EMP2siRNAおよび対照siRNAはそれぞれ、EMP2を標的化する4つのsiRNAのプールまたは4つの非標的化siRNAのプールである。EMP2発現のレベルを、ウェスタンブロットによって定量した。
【0171】
[0189] 抗体: ヒトα1(クローンSR84)、α2(クローンAK-7)、およびα3(クローンC3II.1)インテグリンアイソフォームに特異的なモノクローナル抗体(mAb)は、BD Biosciences(San Diego, CA)から得た。ヒトEMP2に対するウサギ抗血清は、ヒトEMP2の第2の細胞外ループ(EDIHDKNAKFYPVTREGSYG)に結合した多抗原ペプチドで動物を免疫化した後に生成した(Research Genetics, Huntsville, AL)(Wadehra Mら, Exp Mol Pathol, 74(2):106-112(2003))。ヒトFAK(クローンC-20)およびpFAK(クローンTyr 576/577)に特異的なウサギ抗体を、Santa Cruz Biotechnology(Santa Cruz, CA)から得た。ヒトβ−アクチン(クローン2A2.1)に特異的なマウス抗体を、US Biological(Swampscott, MA)から得た。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体を、Southern Biotech(Birmingham, AL)から購入した。ホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体、およびマウスIgGに特異的なR−PE結合抗体を、BD Biosciencesから得た。
【0172】
[0190] フローサイトメトリー: α1、α2、およびα3インテグリンサブユニットの膜発現を、フローサイトメトリーによって評価した。細胞を、PBS内の2%パラホルムアルデヒド(重量/容量)を用いて20分間にわたり氷上で固定したが、透過処理はせず、次に、一次抗体と共に30分間にわたり氷上で、PBS+2%FCS内でインキュベートした。それらを2回洗浄し、R−PE結合抗マウスIgG抗体と共に30分間にわたり氷上でインキュベートした(BD Biosciences)。R−PEを100万個細胞当たり0.25μgで用いた。陰性対照として、細胞をアイソタイプ対照抗体のみと共にインキュベートした。2回連続して洗浄した後、細胞をPBS内に再懸濁し、フローサイトメトリーで分析した(FACScan; BD Biosciences)。細胞集団における発現の反映である、平均蛍光強度(MFI)として計算されたインテグリンの発現レベルを、複数の独立した実験において決定した。
【0173】
[0191] コラーゲンゲルの収縮:コラーゲンゲル収縮アッセイを以前報告されているように行った(Morales SAら, Exp Eye Res, 85(6):790-798(2007))。簡潔に述べると、コラーゲンゲルを、I型コラーゲン(BD Biosciences)、10×DMEM、およびDMEM/F12を組み合わせることによって調製した。I型コラーゲン混合物の最終濃度は、2.5mg/mLであった。コラーゲン溶液(500μL)を24ウェルプレートの各ウェルに添加し、5%C02で、37℃で1時間にわたりインキュベートした。EMP2レベルが変化した、培養したARPE−19細胞を採取し、無血清DMEM/F12内に5×105個/mLの最終濃度で再懸濁した。EMP2レベルが変化したARPE−19細胞を、ウェル当たり2.5×105個細胞の濃度でコラーゲンゲル上に播種し、収縮のパーセンテージを24時間目に測定した。各ゲルの領域を画像キャプチャ(Gel Doc 2000; Bio-Rad, Hercules, CA)を用いてゲルの写真を撮ることによって得、NIH Image J(Wayne Rasband, National Institutes of Health, Bethesda, MDにより開発; http://rsb.info.nih.gov/ij/index.htmlで入手可能)で定量した。ゲルの領域を測定するために、本発明者らは、各ゲルの輪郭を描くための楕円形測定ツールを用いた。時点0でのゲルの領域を24時間後のゲルの領域と比較して、各試料についての収縮のパーセンテージを生成した。各実験は少なくとも6回の反復を含み、少なくとも3つの独立した実験を行い、同等の結果を得た。スチューデントt検定(独立、一方向)を用いた。P<0.05を、統計的に有意であると判断した。
【0174】
[0192] 増殖アッセイ: 細胞を96ウェルプレート上に播種し、一晩インキュベートした。培地を除去し、次に、正常培地またはコラーゲンI25mg/mLを含有する培地で置き換えた。次に細胞を48時間にわたりインキュベートし、非同位体比色免疫アッセイであるCalbiochem(San Diego、CA)のBrdU細胞増殖アッセイによって測定される、BrdUの組み込みによって増殖を評価した。反応生成物をマイクロプレートリーダー(モデル550、Bio-Rad)により595nmの波長で定量した。
【0175】
[0193] 移動アッセイ:ARPE−19細胞およびARPE−19/EMP2細胞を24ウェルプレート上に播種し、細胞がコンフルエントに達するまで3日間にわたりインキュベートした。細胞をPBSで洗浄し、無血清培地を添加し、細胞を一晩インキュベートした。ピペットチップ10μLを用いて単層内に擦り傷を作り、培地を除去して、無血清培地またはPDGF50ng/mLを含有する無血清培地で置き換えた。創傷の写真を様々な時点で撮り、擦り傷の閉鎖のパーセンテージをNIH Image Jソフトウェアで定量した。擦り傷の領域を創傷が生じた直後に測定した。時間が経つにつれて細胞はクリアな領域に移動したが、ギャップは依然として24時間後にも視認可能であった。ギャップのサイズを測定し、元の擦り傷のサイズで割り、この値を閉鎖のパーセンテージとして表した。
【0176】
[0194] 侵入アッセイ:細胞侵入アッセイ(QCM Collagen Cell Invasion Assay; Chemicon, Temecula, CA)を用いて、全ての侵入アッセイを行った。ARPE−19細胞およびARPE−19/EMP2細胞を、重合コラーゲンの薄層で被覆された8μmの孔サイズのポリカーボネート膜を含有する侵入チャンバインサート上に播種した。重合コラーゲン層を介して侵入および移動する細胞は、ポリカーボネート膜の底に張り付く。インサート膜の底にある侵入細胞を比色分析によって同定し、マイクロプレートリーダー(モデル550; Bio-Rad)を用いた560nmでの検出を用いて定量的に分析した。
【0177】
[0195] コラーゲンの生成:ARPE−19細胞およびARPE−19/EMP2細胞を、TCP−β10ng/mLの存在下または非存在下の無血清培地内で、6ウェルプレート上に7×105個細胞/mLの濃度で播種した。培地を72時間後に回収した。分泌されたコラーゲンを測定した(Sircol Collagen Assay; Accurate Chemical & Scientific, Westbury, NY)。コラーゲンアッセイは、比色分析手順である。反応生成物をマイクロプレートリーダーで540nmの波長で定量した。各実験は少なくとも6回の反復を含み、少なくとも3つの独立した実験を行った。スチューデントt検定(独立、一方向)を用いた。P<0.05を統計的に有意であると判断した。
【0178】
[0196] FAK/Srcの阻害:コラーゲンゲルをDMEM/F12内のI型コラーゲン(BD Biosciences)で、2.5mg/mLの最終濃度で調製した。新たに調製したコラーゲン溶液を、24ウェルプレートの各ウェルに添加し、5%CO2で、37℃で1時間にわたりインキュベートした。培養したARPE−19細胞、ARPE−19/EMP2細胞、およびARPE−19/EMP2siRNA細胞を採取し、無血清DMEM/F12内に5×105個/mLの最終濃度で再懸濁した。細胞を様々な濃度の低分子阻害剤で1時間にわたり前処理した。阻害剤PP2(FAK/Src阻害剤)およびSU6656(Src阻害剤)は、DMSO中に希釈して用いた(Calbiochem)。細胞を、媒体対照としてDMSOのみで処理した。ARPE−19細胞、ARPE−19/EMP2細胞、およびARPE−19/EMP2siRNA細胞を、コラーゲンゲル上にウェル当たり2.5×105個細胞の濃度で播種し、収縮のパーセンテージを特定の時間間隔で測定した。少なくとも3つの独立した実験を行い、示されているところでは、結果を、スチューデントt検定(独立、一方向)で統計的有意性について評価した。P<0.05のレベルを、統計的に有意であると考えた。
【0179】
[0197] ウェスタンブロット分析:ウェスタンブロット分析を、以前記載されているように行った(Wadehra Mら. Dev Biol, 292(2):430-441(2006))。簡潔に述べると、細胞タンパク質を、プロテアーゼ阻害剤およびホスファターゼ阻害剤を含有するRIPA緩衝液(Upstate, Charlottesville, VA)を用いて単離し、タンパク質濃度をタンパク質アッセイ(BCA; Bio-Rad)で決定した。全部でタンパク質10μgを各レーンにロードし、タンパク質を、還元条件下で、4%〜20%SDS−PAGE勾配ゲルによって分画した。タンパク質をニトロセルロース膜(GE Healthcare, Buckinghamshire, UK)に移し、転移の妥当性を、Ponceau Sレッド染色(Sigma-Aldrich)によって確認した。膜を、次に、TBS Tween(TBST; Upstate)内の無脂肪乳でブロックした。ブロットを、FAKおよびp−FAK(Tyr 576/577)では1:200の希釈、EMP2では1:1000の希釈、β−アクチンでは1:5000の希釈の一次抗体と共に、1時間にわたりインキュベートした。ホースラディッシュペルオキシダーゼ結合ヤギ抗ウサギ抗体またはホースラディッシュペルオキシダーゼ結合ヤギ抗マウス抗体を、1:2000の希釈でブロットに曝露した。ブロットを、次に、化学発光で発現させて、結合抗体を視覚化し(ECL; Pierce, Rockford, IL)、内部対照としてβ−アクチンで定量した。ウェスタンブロット分析を、NIH Image Jで定量した。ブロットをフラットベッドスキャナでデジタル化し、バンドの密度を、Image Jを用いて測定した。ロードのばらつきを明らかにするために、β−アクチンを用いて各試料を正規化した。少なくとも3つの独立した実験を行い、示されているところでは、結果を、スチューデントt検定(独立、一方向)で統計的有意性について評価した。P<0.05のレベルを統計的に有意であると考えた。
【0180】
結果
[0198] ARPE−19細胞におけるEMP2発現の組換えによる変化: EMP2は、網膜色素上皮細胞において高度に発現し、これは、膜の形成およびコラーゲンゲルの収縮において重要であると考えられている。コラーゲンゲルの収縮に対するEMP2の影響を研究するために、EMP2レベルをARPE−19細胞系において変化させて、2つのさらなるARPE−19系、すなわちARPE−19/EMP2と呼ばれる過剰発現系およびARPE−19/Riboと呼ばれる安定なノックダウンを生じた。EMP2の定常状態のタンパク質レベルを、ARPE−19細胞、ARPE−19/EMP2細胞、およびARPE−19/Ribo細胞においてウェスタンブロット分析によって測定した。実験は独立して少なくとも3回行い、同様の結果を得、1つの代表的な実験を図13Aに示す。ARPE−19/EMP2細胞は、連続希釈を用いて比較すると、野生型細胞と比較して、EMP2の発現において75〜100倍の増大を示した。ARPE−19/Ribo細胞は、ARPE−19細胞と比較して、EMP2発現において5倍の減少を示した。
【0181】
[0199] EMP2発現を減少させるための別の方法では、EMP2に特異的なsiRNA(ARPE-19/EMP2siRNA)または対照スクランブルsiRNA(ARPE-19/対照siRNA)で一時的にトランスフェクトしたARPE−19細胞を用いた。特異的siRNAを用いるこの技術は、対照ARPE−19における発現のおよそ40%まで、EMP2の発現を特異的に減少させた(図13B)。スクランブルsiRNAは、EMP2の発現においていかなる変化ももたらさなかった。
【0182】
[0200] コラーゲンゲルの収縮のEMP2による修飾:ARPE−19/EMP2細胞、ARPE19/Ribo細胞、およびARPE−19細胞のコラーゲンゲル収縮アッセイを、試料当たり6回の反復で少なくとも3回の個別の回数行った。EMP2レベルの改変は、コラーゲンゲルを収縮させる細胞の能力に有意に影響した。EMP2発現の増大は、未処理の野生型細胞と比較して、収縮を有意に増大させた(図14)。これと合致して、ゲル収縮の減少が、ARPE−19/Ribo細胞において観察された。EMP2構築物を有さない、レトロウイルスに感染した細胞系である、さらなる対照ARPE−19細胞系、およびリボザイムを有さないトランスフェクトされたEGFP−N3ベクターは、野生型対照ARPE−19細胞系と比較して収縮を改変しなかった(データは示していない)。
【0183】
[0201] インテグリンα1、α2、またはα3における変化を介する収縮に対するEMP2の影響:以前の結果は、EMP2の発現が増大するとコラーゲンゲルの収縮が増大することを示した。EMP2は、マウスNIH3T3およびヒト子宮内膜癌系HEC1Aを包含する様々な細胞型における特異的なインテグリンアイソフォームの細胞内輸送および細胞表面発現を調節することが知られている(Wadehra Mら, Dev Biol, 287(2):336-345(2005); Wadehra Mら, J Biol. Chem, 277(43):41094-41100(2002))。本発明者らおよび他者は、α1、α2、およびα3インテグリンの係合の、ARPE−19細胞系におけるコラーゲンゲル収縮における重要な工程としての重要性を実証し(Bando Hら, Exp Eye Res, 82(3):529-537(2006); Morales SAら, Exp Eye Res, 85(6):790-798(2007); Carver Wら, J Cell Physiol, 165(2):425-437(1995); Cooke MEら, J Cell Sci, 113:2375-2383(2000); Kieffer JDら, Biochem Biophys Res Commun, 217(2):466-474(1995); Langholz Oら, J Cell Biol , 131:1903-1915(1995); Moulin VおよびPlamondon M., Br J Dermatol, 147(5):886-892(2002); Robbins SGら, Invest Ophthalmol Vis Sci, 35(9):3475-3485(1994); Zhang ZGら, J Cell Sci, 119:1886-1895(2006))、それによって、EMP2が、コラーゲンを結合するこれらのインテグリンの1つの細胞表面発現を増大し得ると仮定した。
【0184】
[0202] EMP2の発現が改変した細胞系を、これらのインテグリンアイソフォームの表面発現についてフローサイトメトリーによって評価した(図15)。予測した結果とは対照的に、EMP2を過剰発現するように操作された細胞は実際に、インテグリンα1(P=0.003)およびα2(P=0.002)の細胞表面発現のおよそ65%の減少を示した。ARPE−19/EMP2細胞におけるインテグリンα3の発現では、わずかな、しかし統計的に有意ではない減少があった(図15B)。この結果によって、本発明者らは、EMP2がこの細胞系におけるコラーゲンゲル収縮を調節し得る他のメカニズムを調べた。
【0185】
[0203] 増殖、移動、侵入、およびコラーゲン生成における変化と収縮に対するEMP2の影響との関連:EMP2がコラーゲンの収縮を調節し得る潜在的なメカニズムは、収縮の増大をもたらす細胞増殖の増大によるものである。BrdUの組み込みによってアッセイされた増殖を、コラーゲン刺激の非存在下または存在下で、ARPF−19細胞およびARPE−19/EMP2細胞において評価した(図16A)。コラーゲンの非存在下では、EMP2の過剰発現によって、増殖が穏やかに減少した(20%)。コラーゲン刺激の後、両細胞系の増殖に統計的に有意な差はなく、したがって、増殖の増大によってARPE−19/EMP2細胞によるコラーゲンゲルの収縮が増強され得るという仮説を反証した。
【0186】
[0204] 移動に対する、EMP2発現の増大の考えられる影響を調べるために、本発明者らは、創傷治癒アッセイを行った。ARPE−19細胞およびARPE−19/EMP2細胞を、PDGF50ng/mLで処理しないままにするかまたは処理し、閉鎖のパーセンテージを、創傷閉鎖の代用的評価として、6、12、および24時間目に測定した。6時間および12時間の時点で、PDGFの非存在下または存在下で、2つの細胞系の間に差はなかった(データは示していない)。24時間の時点では、PDGF刺激の非存在下で、ARPE−19/EMP2細胞は、ARPE−19細胞と比較して、ギャップの閉鎖において23%の減少を示した。PDGF処理はこの影響を無効にし、その結果、2つの細胞系において同等の創傷治癒が生じた(図16B)。増殖および移動の両方が、創傷の閉鎖に関与する。ARPE−19/EMP2細胞のより遅い増殖速度は創傷閉鎖の減少に関与し得るが、EMP2を過剰発現する細胞があまり運動性を示さない可能性もある。運動性の表現型を促進することが知られているPDGF処理は、ARPE−19細胞の創傷閉鎖を刺激し、さらに、ARPE−19/EMP2細胞の創傷閉鎖の減少を克服した。ARPE−19/EMP2細胞によるコラーゲンゲル収縮の増大は、EMP2の発現の改変に起因する移動の増大によって説明することはできない。
【0187】
[0205] 本発明者らは、別のメカニズム、すなわち、EMP2の発現がコラーゲンマトリクスに侵入する細胞の能力に影響し得るという可能性を試験した。ARFE−19細胞およびARPE−19/EMP2細胞をポリカーボネート膜上に播種し、10%FESまたはPDGF50ngを化学誘引物質として用いた。侵入を24および48時間目に測定した。いずれの条件下でも、ARPE−19細胞とARPE−19/EMP2細胞との間で侵入に差はなかった(図16C)。コラーゲンゲル収縮のEMP2介在性の増強は、改変した侵入能力によるものではなかった。
【0188】
[0206] 本発明者らが調べたさらなるメカニズムは、EMP2レベルがコラーゲンの生成に影響するかどうかであった。ARPE−19細胞およびARPE−19/EMP2細胞を、24ウェルプレートにおいて72時間にわたり成長させた。培地を回収し、コラーゲン生成を分析した。2つの細胞系の間でコラーゲンの生成に統計的な差はなかった(図16D)。ARPE−19/EMP2細胞による細胞収縮能力の増大は、コラーゲン生成の改変によって説明することはできない。
【0189】
[0207] ARPE−19細胞におけるFAK活性化およびコラーゲンゲル収縮のEMP2による制御:インテグリンの発現、増殖、移動、または侵入における変化は、いかにしてEMP2発現の増大がゲルコラーゲンの収縮を増大させたかを説明するものではない。以前に、本発明者らは、インテグリンのライゲーションおよびFAK/Src複合体の活性化がARPE−19細胞によるコラーゲンゲルの収縮に必要であることを観察した(Morales SAら, Exp Eye Res, 85(6):790-798(2007))。本発明者らは、ARPE−19細胞におけるEMP2の過剰発現によってFAK/Srcの活性が改変するかを調べた。全FAKレベルはARPE−19細胞およびARPE−19/EMP2細胞の両方において等しかったが、過剰発現するARP−19/EMP2細胞は、対照ARPE−19細胞と比較して、活性化型FAKのレベルがほぼ2倍増大することを示した(図17A)。各活性化型FAKをそれ自身のβ−アクチンロード対照に正規化して行った複数の評価は、EMP2レベルの増大とFAK活性化との間の統計的に有意な相関を示した(図17B)。
【0190】
[0208] FAK/Src阻害剤によるコラーゲンゲル収縮の阻害に対する耐性に対するEMP2過剰発現の影響:EMP2の過剰発現がFAK活性化の増大に関連するという観察を受けて、FAK/Src経路の阻害剤を用いて、ゲル収縮の状況におけるこの観察の機能的有意性を試験するためのさらなる研究を行った。もしEMP2の影響がFAKの活性化に続発するものであれば、ARPE−19/EMP細胞における観察されたコラーゲンゲルの収縮は、対照ARPE−19細胞と比較して、FAK/Srcの阻害に対する耐性の増大を示すはずである。この予測を調べるために、本発明者らは、様々な濃度の、ARPE−19細胞におけるコラーゲンゲルの収縮を予防することが実証されているFAK/Src低分子阻害剤であるPP2およびSU6656を用いた(Morales SAら, Exp Eye Res, 85(6):790-798(2007))(図18、19)。EMP2特異的siRNAでのEMP2発現の減少は、対照スクランブルsiRNAと比較して、コラーゲンゲル収縮の減少を示した。この結果は、ARPE−19/Ribo細胞系において観察されるコラーゲンゲル収縮の減少と合致した(図18A)。FAK阻害剤PP2に対するコラーゲンゲル収縮アッセイの感受性はEMP2のレベルと逆相関し、EMP2siRNAで処理した細胞においては感受性が増大し、EMP2を過剰発現する細胞においては耐性が増大した(図18A〜C)。媒体対照に正規化された収縮の阻害のパーセンテージを異なる濃度の阻害剤で各細胞系について決定した。ゲル収縮の50%の阻害を達成するために必要なPP2の濃度は、ARPE−19細胞では2.5μMであり、ARPE−19/EMP2siRNA細胞では1μM未満であり、過剰発現するARPE−19/EMP2細胞では5μMであり、これらの細胞系において観察されるFAK活性化の変化と一致した(図18D)。FAK/Src経路の第2の阻害剤であるSU6656は、ARPE−19細胞およびARPE−19/EMP2細胞で類似の結果を示し、したがって、コラーゲンゲル収縮のEMP2による制御についてのメカニズムがFAKの活性化の変化を介することの独立した裏付けを提供した(図19)。
【0191】
考察
[0209] PVRについてのインビトロでの相関要因であるコラーゲンゲルの収縮は、FAK介在性のインテグリンシグナル伝達に依存する細胞プロセスである(Morales SAら, Exp Eye Res, 85(6):790-798(2007))。この記事において、本発明者らは、EMP2がコラーゲンゲルの収縮をFAK活性化の増強に依存するプロセスで調節することを実証した。FAK活性化のEMP2による調節は新たな観察であるが、様々なインテグリンシグナル伝達メカニズムのテトラスパニン関連性の調節についての以前の観察は、このファミリーの膜関連性タンパク質に共通する潜在的な機能を示唆する。
【0192】
[0210] テトラスパニンがFAK依存性経路または他のシグナル伝達経路の活性化を調節するメカニズムは、いまだ明らかにされていない。特定のテトラスパニン(CD9、CD53、CD81、CD82、およびCD151)は互いに関連するか、またはさらなる膜タンパク質とのヘテロ複合体で関連し、結果として、チロシンのリン酸化を増大させる(Boucheix Cら, Cell Mol Life Sci, 58(9):1189-1205(2001); Hemler ME., J Cell Biol, 155(7):1103-1107(2001); Hong IKら, J Biol Chem, 281(34):24279-24292(2006); Lagaudriere-Gesbert Cら, Cell Immunol, 182(2):105-112(1997); Todd SCら, J Exp Med, 184(5):2055-2060(1996))。これは、4回膜貫通タンパク質が、膜におけるシグナル伝達複合体の機能的組み立て体を支持する分子アダプターとして作用することを示唆する(Hong IKら., J Biol Chem, 281(34):24279-24292(2006))。
【0193】
[0211] インテグリンおよび他の細胞表面受容体の機能的調節は、テトラスパンファミリーの複数のタンパク質に共通の、再発的に観察される特徴である。いくつかのテトラスパニン(CD9、CD53、CD81、およびCD82)は、インテグリン(α3β1、α43β1、およびα63β1)とのタンパク質−タンパク質相互作用に関与し、接着および細胞活性化を改変させる(Lagaudriere-Gesbert Cら, Cell Immunol, 182(2):105-112(1997); Todd SCら, J Exp Med, 184(5):2055-2060(1996))。CD9、CD63、CD81、CD151、およびA15/TALLA1を包含する複数のテトラスパニンは、PI−4キナーゼを特異的な膜位置に動員し、ホスホイノシチド依存性のシグナル伝達を誘発する(Yauch RLおよびHemler ME., Biochem J, 3:629-637(2000))。さらに、CD81と抗CD81抗体との架橋は、TCR/CD3複合体を介するシグナル伝達にとって共刺激性である(Todd SCら, J Exp Med, 184(5):2055-2060(1996))。発現レベルの変化またはテトラスパニンの架橋との間の他の関連は、JNKの活性化を誘発するCD53の発現またはライゲーションのアップレギュレーションを包含するシグナル変換の活性化(Yunta Mら, Eur J Biochem, 269(3):1012-1021(2002))、線維肉腫細胞におけるFAKのチロシンリン酸化を誘発するCD9による接着の制御(Berditchevski FおよびOdintsova E, J Cell Biol, 146(2):477-492(1999))、ならびにヒト黒色腫細胞におけるFAK、Src、およびc−Junキナーゼの接着依存性の活性化を誘発する同族親和性のCD151相互作用(Hong IKら, J Biol Chem, 281(34):24279-24292(2006))に関連している。さらに、CD151の同族親和性のタンパク質−タンパク質相互作用は、FAK−SrcおよびMAPキナーゼを伴う経路を介するc−Junへのインテグリン依存性のシグナル伝達を調節する(Hong IKら, J Biol Chem, 281 (34):24279-24292 (2006))。
【0194】
[0212] RPE介在性のコラーゲンゲル収縮は、いくつかの受容体およびシグナル伝達経路の相互作用を伴う(Bando Hら, Exp Eye Res, 82(3):529-537(2006); Carver Wら, J Cell Physiol, 165(2):425-437(1995); Cooke MEら, J Cell Sci, 113:2375-2383(2000); Kieffer JDら, Biochem Biophys Res Commun, 217(2):466-474(1995); Langholz Oら, J Cell Biol. , 131:1903-1915(1995); Moulin VおよびPlamondon M., Br J Dermatol, 147(5):886-892(2002); Robbins SGら, Invest Ophthalmol Vis Sci, 35(9):3475-3485(1994); Zhang ZGら, J Cell Sci, 119:1886-1895(2006); Xia Hら, J Biol Chem, 279(31):33024-33034(2004))。この研究の結果は、コラーゲン収縮をもたらす、FAK/Src複合体の活性化の促進におけるEMP2の役割を裏付けるものである。
【0195】
[0213] 本明細書において引用される各刊行物、特許出願、特許、および他の参考文献は、それが本開示と矛盾しない限り、参照することによってその全体が組み込まれる。特に、本明細書において引用される全ての刊行物は、本発明に関連して用いられ得る、その刊行物において報告されている方法、試薬、およびツールを記載および開示することを目的として、参照することによってその全体が本明細書に組み込まれる。本明細書における何物も、本発明が先行発明に基づいてこのような開示に先行する資格を有さないということを承認するものとしては解釈されない。
【0196】
[0214] 前述の発明は、理解を明確にすることを目的とした実例および実施例によってある程度詳細に記載されているが、当業者には、本発明の教示に照らして、添付の特許請求の範囲の趣旨または範囲から逸脱することなく、ある程度の変化および変更を本発明に行い得ることが容易に明らかとなろう。
【特許請求の範囲】
【請求項1】
眼におけるEMP2活性を低減させる抗EMP2剤を投与することを含む、対象における増殖性硝子体網膜症または網膜剥離を治療する方法。
【請求項2】
前記対象がヒトである、請求項1に記載の方法。
【請求項3】
前記抗EMP2剤が局所的に投与される、請求項1に記載の方法。
【請求項4】
眼に直接的に投与する、請求項1に記載の方法。
【請求項5】
前記抗EMP2剤が眼球内に投与される、請求項1に記載の方法。
【請求項6】
前記抗EMP2剤が眼の硝子体液に注入される、請求項1に記載の方法。
【請求項7】
前記抗EMP2剤が、配列番号1、配列番号2、または配列番号30に対して少なくとも70%のアミノ酸配列同一性を有するEMP2ポリペプチドに特異的に結合する抗体またはそのフラグメントである、請求項1に記載の方法。
【請求項8】
前記抗EMP2剤が、EMP2の機能を妨げる抗体またはそのフラグメントである、請求項1に記載の方法。
【請求項9】
前記抗EMP2剤が、眼における1つまたは複数の細胞の表面からのEMP2のクリアランスを促進する抗体またはそのフラグメントである、請求項1に記載の方法。
【請求項10】
前記抗体が、配列番号4、配列番号6、配列番号8、または配列番号10に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む重鎖を含む、請求項7〜9のいずれか1項に記載の方法。
【請求項11】
前記抗体が、配列番号5、配列番号7、配列番号9、または配列番号11に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む軽鎖を含む、請求項7〜9のいずれか1項に記載の方法。
【請求項12】
前記抗体が、(1)配列番号4、配列番号6、配列番号8、および配列番号10からなる群から選択される重鎖、ならびに(2)配列番号5、配列番号7、配列番号9、および配列番号11からなる群から選択される軽鎖を含む、請求項7〜9のいずれか1項に記載の方法。
【請求項13】
前記抗体が、配列番号27、配列番号28、または配列番号29に対して少なくとも70%のアミノ酸配列同一性を有する、請求項7〜9のいずれか1項に記載の方法。
【請求項14】
前記抗体が、配列番号27、配列番号28、または配列番号29のアミノ酸配列を有する、請求項13に記載の方法。
【請求項15】
前記抗体が、モノクローナル抗体である、請求項7〜14のいずれか1項に記載の方法。
【請求項16】
前記抗体が、ヒト化モノクローナル抗体である、請求項7〜14のいずれか1項に記載の方法。
【請求項17】
フラグメントが、ダイアボディまたはミニボディとして提供される、請求項7〜14のいずれか1項に記載の方法。
【請求項18】
前記フラグメントが、Fvフラグメント、F(ab’)フラグメント、またはF(ab’)2フラグメントである、請求項7〜14のいずれか1項に記載の方法。
【請求項19】
前記抗体が、マウス抗原結合部位と、エフェクター機能を調節するヒト化領域とを有するキメラ抗体である、請求項7〜14のいずれか1項に記載の方法。
【請求項20】
前記抗体またはそのフラグメントが、細胞傷害性物質に結合している、請求項7〜14のいずれか1項に記載の方法。
【請求項21】
前記抗EMP2剤が、配列番号1のポリペプチドに対して少なくとも70%のアミノ酸配列同一性を有するEMP2タンパク質をコードする核酸の発現を低減できるsiRNAである、請求項1に記載の方法。
【請求項22】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドに対して少なくとも70%のヌクレオチド配列同一性を有する核酸配列を含む、請求項21に記載の方法。
【請求項23】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドを含む、請求項22に記載の方法。
【請求項24】
前記抗EMP2剤が、配列番号3の核酸配列に対して少なくとも70%のヌクレオチド配列同一性を有するEMP2ポリヌクレオチド配列を切断できるハンマーヘッドリボザイムである、請求項1に記載の方法。
【請求項25】
前記抗EMP2剤の投与により、網膜剥離のリスクが低減する、請求項1に記載の方法。
【請求項26】
前記抗EMP2剤が、眼への注入用に製剤化される、請求項1に記載の方法。
【請求項27】
前記抗EMP2剤が、眼の外での局所投与用に製剤化される、請求項1に記載の方法。
【請求項28】
前記硝子体網膜症が、裂孔原性硝子体網膜症である、請求項1に記載の方法。
【請求項29】
前記抗EMP2剤を投与する前に、眼が外傷を有するか、または網膜剥離のリスクの原因となる手術を受けている、請求項1に記載の方法。
【請求項30】
前記抗EMP2剤が、プロゲステロン受容体アンタゴニストである、請求項1に記載の方法。
【請求項31】
生体試料と、網膜色素上皮細胞におけるEMP2の活性を低減させる抗EMP2剤とを接触させることを含む、生体試料における網膜色素上皮細胞の収縮能力を低下させる方法。
【請求項32】
前記抗EMP2剤が、配列番号1、配列番号2、または配列番号30に対して少なくとも70%のアミノ酸配列同一性を有するEMP2ポリペプチドに特異的に結合する抗体またはそのフラグメントである、請求項31に記載の方法。
【請求項33】
前記抗EMP2剤が、EMP2の機能を妨げる抗体またはそのフラグメントである、請求項31に記載の方法。
【請求項34】
前記抗EMP2剤が、網膜色素上皮細胞の表面からのEMP2のクリアランスを促進する抗体またはそのフラグメントである、請求項31に記載の方法。
【請求項35】
前記抗体が、配列番号4、配列番号6、配列番号8、または配列番号10に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む重鎖を含む、請求項32〜34のいずれか1項に記載の方法。
【請求項36】
前記抗体が、配列番号5、配列番号7、配列番号9、または配列番号11に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む軽鎖を含む、請求項32〜34のいずれか1項に記載の方法。
【請求項37】
前記抗体が、(1)配列番号4、配列番号6、配列番号8、および配列番号10からなる群から選択される重鎖、ならびに(2)配列番号5、配列番号7、配列番号9、および配列番号11からなる群から選択される軽鎖を含む、請求項32〜34のいずれか1項に記載の方法。
【請求項38】
前記抗体が、配列番号27、配列番号28、または配列番号29に対して少なくとも70%のアミノ酸配列同一性を有する、請求項32〜34のいずれか1項に記載の方法。
【請求項39】
前記抗体が、配列番号27、配列番号28、または配列番号29のアミノ酸配列を有する、請求項32〜34のいずれか1項に記載の方法。
【請求項40】
前記抗体が、モノクローナル抗体である、請求項32〜39のいずれか1項に記載の方法。
【請求項41】
前記抗体が、ヒト化モノクローナル抗体である、請求項32〜39のいずれか1項に記載の方法。
【請求項42】
前記フラグメントが、ダイアボディまたはミニボディとして提供される、請求項32〜39のいずれか1項に記載の方法。
【請求項43】
前記フラグメントが、Fvフラグメント、F(ab’)フラグメント、またはF(ab’)2フラグメントである、請求項32〜39のいずれか1項に記載の方法。
【請求項44】
前記抗体が、マウス抗原結合部位と、エフェクター機能を調節するヒト化領域とを有するキメラ抗体である、請求項32〜39のいずれか1項に記載の方法。
【請求項45】
前記抗体またはそのフラグメントが、細胞傷害剤に結合している、請求項32〜39のいずれか1項に記載の方法。
【請求項46】
前記抗EMP2剤が、配列番号1のポリペプチドに対して少なくとも70%のアミノ酸配列同一性を有するEMP2タンパク質をコードする核酸の発現を低減できるsiRNAである、請求項31に記載の方法。
【請求項47】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドに対して少なくとも70%のヌクレオチド配列同一性を有する核酸配列を含む、請求項46に記載の方法。
【請求項48】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドを含む、請求項47に記載の方法。
【請求項49】
前記抗EMP2剤が、配列番号3の核酸配列に対して少なくとも70%のヌクレオチド配列同一性を有するEMP2ポリヌクレオチド配列を切断できるハンマーヘッドリボザイムである、請求項31に記載の方法。
【請求項50】
前記生体試料が、組織試料である、請求項31に記載の方法。
【請求項51】
前記組織試料が、眼の組織である、請求項50に記載の方法。
【請求項1】
眼におけるEMP2活性を低減させる抗EMP2剤を投与することを含む、対象における増殖性硝子体網膜症または網膜剥離を治療する方法。
【請求項2】
前記対象がヒトである、請求項1に記載の方法。
【請求項3】
前記抗EMP2剤が局所的に投与される、請求項1に記載の方法。
【請求項4】
眼に直接的に投与する、請求項1に記載の方法。
【請求項5】
前記抗EMP2剤が眼球内に投与される、請求項1に記載の方法。
【請求項6】
前記抗EMP2剤が眼の硝子体液に注入される、請求項1に記載の方法。
【請求項7】
前記抗EMP2剤が、配列番号1、配列番号2、または配列番号30に対して少なくとも70%のアミノ酸配列同一性を有するEMP2ポリペプチドに特異的に結合する抗体またはそのフラグメントである、請求項1に記載の方法。
【請求項8】
前記抗EMP2剤が、EMP2の機能を妨げる抗体またはそのフラグメントである、請求項1に記載の方法。
【請求項9】
前記抗EMP2剤が、眼における1つまたは複数の細胞の表面からのEMP2のクリアランスを促進する抗体またはそのフラグメントである、請求項1に記載の方法。
【請求項10】
前記抗体が、配列番号4、配列番号6、配列番号8、または配列番号10に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む重鎖を含む、請求項7〜9のいずれか1項に記載の方法。
【請求項11】
前記抗体が、配列番号5、配列番号7、配列番号9、または配列番号11に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む軽鎖を含む、請求項7〜9のいずれか1項に記載の方法。
【請求項12】
前記抗体が、(1)配列番号4、配列番号6、配列番号8、および配列番号10からなる群から選択される重鎖、ならびに(2)配列番号5、配列番号7、配列番号9、および配列番号11からなる群から選択される軽鎖を含む、請求項7〜9のいずれか1項に記載の方法。
【請求項13】
前記抗体が、配列番号27、配列番号28、または配列番号29に対して少なくとも70%のアミノ酸配列同一性を有する、請求項7〜9のいずれか1項に記載の方法。
【請求項14】
前記抗体が、配列番号27、配列番号28、または配列番号29のアミノ酸配列を有する、請求項13に記載の方法。
【請求項15】
前記抗体が、モノクローナル抗体である、請求項7〜14のいずれか1項に記載の方法。
【請求項16】
前記抗体が、ヒト化モノクローナル抗体である、請求項7〜14のいずれか1項に記載の方法。
【請求項17】
フラグメントが、ダイアボディまたはミニボディとして提供される、請求項7〜14のいずれか1項に記載の方法。
【請求項18】
前記フラグメントが、Fvフラグメント、F(ab’)フラグメント、またはF(ab’)2フラグメントである、請求項7〜14のいずれか1項に記載の方法。
【請求項19】
前記抗体が、マウス抗原結合部位と、エフェクター機能を調節するヒト化領域とを有するキメラ抗体である、請求項7〜14のいずれか1項に記載の方法。
【請求項20】
前記抗体またはそのフラグメントが、細胞傷害性物質に結合している、請求項7〜14のいずれか1項に記載の方法。
【請求項21】
前記抗EMP2剤が、配列番号1のポリペプチドに対して少なくとも70%のアミノ酸配列同一性を有するEMP2タンパク質をコードする核酸の発現を低減できるsiRNAである、請求項1に記載の方法。
【請求項22】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドに対して少なくとも70%のヌクレオチド配列同一性を有する核酸配列を含む、請求項21に記載の方法。
【請求項23】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドを含む、請求項22に記載の方法。
【請求項24】
前記抗EMP2剤が、配列番号3の核酸配列に対して少なくとも70%のヌクレオチド配列同一性を有するEMP2ポリヌクレオチド配列を切断できるハンマーヘッドリボザイムである、請求項1に記載の方法。
【請求項25】
前記抗EMP2剤の投与により、網膜剥離のリスクが低減する、請求項1に記載の方法。
【請求項26】
前記抗EMP2剤が、眼への注入用に製剤化される、請求項1に記載の方法。
【請求項27】
前記抗EMP2剤が、眼の外での局所投与用に製剤化される、請求項1に記載の方法。
【請求項28】
前記硝子体網膜症が、裂孔原性硝子体網膜症である、請求項1に記載の方法。
【請求項29】
前記抗EMP2剤を投与する前に、眼が外傷を有するか、または網膜剥離のリスクの原因となる手術を受けている、請求項1に記載の方法。
【請求項30】
前記抗EMP2剤が、プロゲステロン受容体アンタゴニストである、請求項1に記載の方法。
【請求項31】
生体試料と、網膜色素上皮細胞におけるEMP2の活性を低減させる抗EMP2剤とを接触させることを含む、生体試料における網膜色素上皮細胞の収縮能力を低下させる方法。
【請求項32】
前記抗EMP2剤が、配列番号1、配列番号2、または配列番号30に対して少なくとも70%のアミノ酸配列同一性を有するEMP2ポリペプチドに特異的に結合する抗体またはそのフラグメントである、請求項31に記載の方法。
【請求項33】
前記抗EMP2剤が、EMP2の機能を妨げる抗体またはそのフラグメントである、請求項31に記載の方法。
【請求項34】
前記抗EMP2剤が、網膜色素上皮細胞の表面からのEMP2のクリアランスを促進する抗体またはそのフラグメントである、請求項31に記載の方法。
【請求項35】
前記抗体が、配列番号4、配列番号6、配列番号8、または配列番号10に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む重鎖を含む、請求項32〜34のいずれか1項に記載の方法。
【請求項36】
前記抗体が、配列番号5、配列番号7、配列番号9、または配列番号11に対して少なくとも70%のアミノ酸配列同一性を有するアミノ酸配列を含む軽鎖を含む、請求項32〜34のいずれか1項に記載の方法。
【請求項37】
前記抗体が、(1)配列番号4、配列番号6、配列番号8、および配列番号10からなる群から選択される重鎖、ならびに(2)配列番号5、配列番号7、配列番号9、および配列番号11からなる群から選択される軽鎖を含む、請求項32〜34のいずれか1項に記載の方法。
【請求項38】
前記抗体が、配列番号27、配列番号28、または配列番号29に対して少なくとも70%のアミノ酸配列同一性を有する、請求項32〜34のいずれか1項に記載の方法。
【請求項39】
前記抗体が、配列番号27、配列番号28、または配列番号29のアミノ酸配列を有する、請求項32〜34のいずれか1項に記載の方法。
【請求項40】
前記抗体が、モノクローナル抗体である、請求項32〜39のいずれか1項に記載の方法。
【請求項41】
前記抗体が、ヒト化モノクローナル抗体である、請求項32〜39のいずれか1項に記載の方法。
【請求項42】
前記フラグメントが、ダイアボディまたはミニボディとして提供される、請求項32〜39のいずれか1項に記載の方法。
【請求項43】
前記フラグメントが、Fvフラグメント、F(ab’)フラグメント、またはF(ab’)2フラグメントである、請求項32〜39のいずれか1項に記載の方法。
【請求項44】
前記抗体が、マウス抗原結合部位と、エフェクター機能を調節するヒト化領域とを有するキメラ抗体である、請求項32〜39のいずれか1項に記載の方法。
【請求項45】
前記抗体またはそのフラグメントが、細胞傷害剤に結合している、請求項32〜39のいずれか1項に記載の方法。
【請求項46】
前記抗EMP2剤が、配列番号1のポリペプチドに対して少なくとも70%のアミノ酸配列同一性を有するEMP2タンパク質をコードする核酸の発現を低減できるsiRNAである、請求項31に記載の方法。
【請求項47】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドに対して少なくとも70%のヌクレオチド配列同一性を有する核酸配列を含む、請求項46に記載の方法。
【請求項48】
前記siRNAが、配列番号3の核酸配列またはその相補鎖の少なくとも15個の連続するヌクレオチドを含む、請求項47に記載の方法。
【請求項49】
前記抗EMP2剤が、配列番号3の核酸配列に対して少なくとも70%のヌクレオチド配列同一性を有するEMP2ポリヌクレオチド配列を切断できるハンマーヘッドリボザイムである、請求項31に記載の方法。
【請求項50】
前記生体試料が、組織試料である、請求項31に記載の方法。
【請求項51】
前記組織試料が、眼の組織である、請求項50に記載の方法。
【図1】

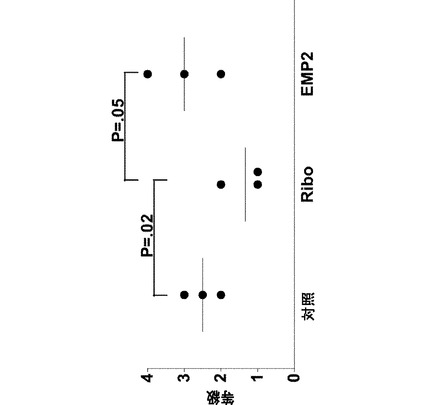
【図2】
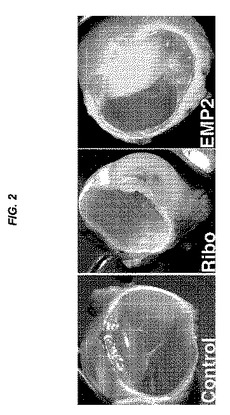
【図3】
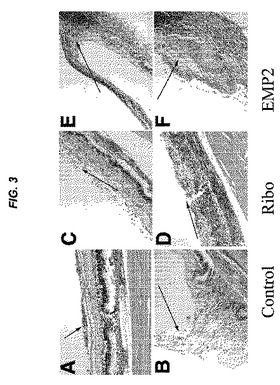
【図4】

【図5】
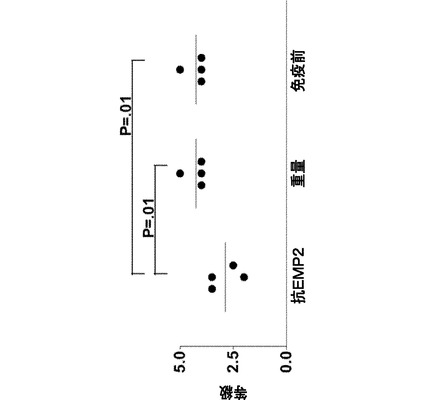
【図6】
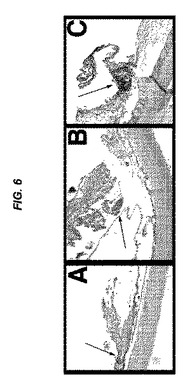
【図7】
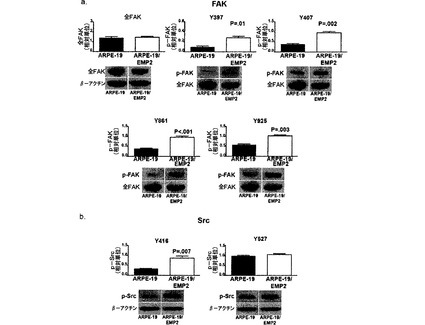
【図8】
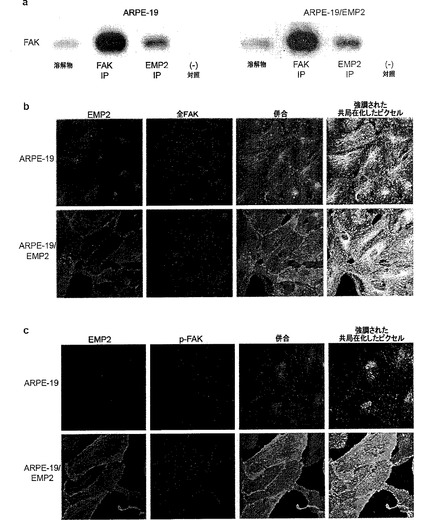
【図9】
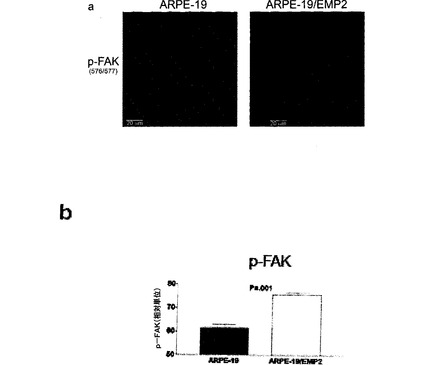
【図10】
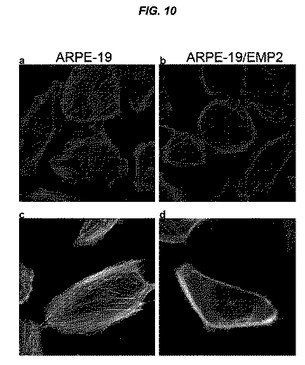
【図11】
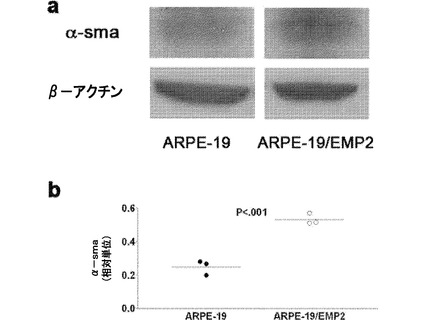
【図12】
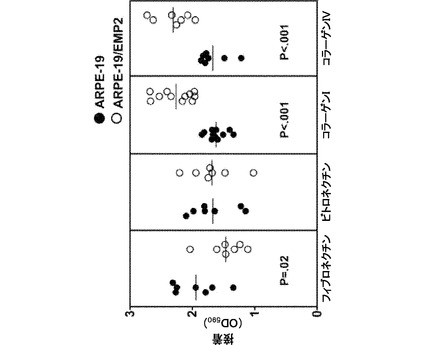
【図13】
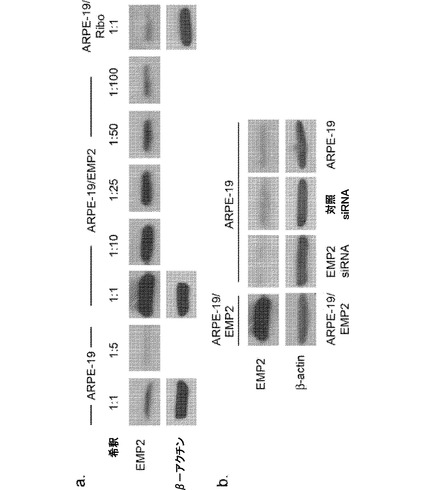
【図14】
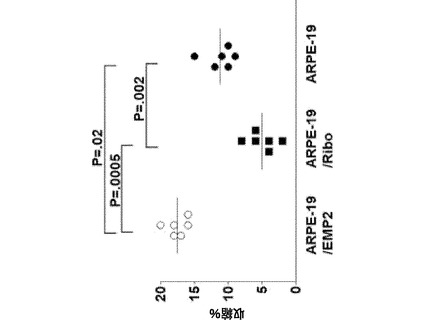
【図15】
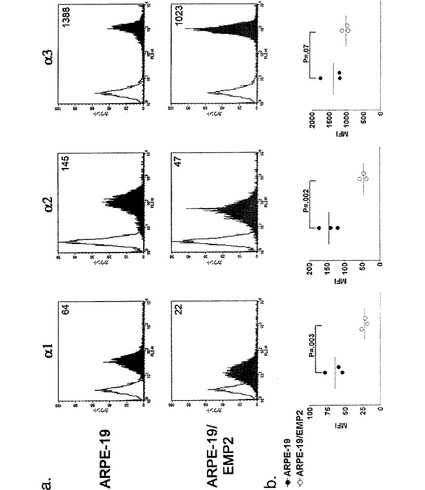
【図16】
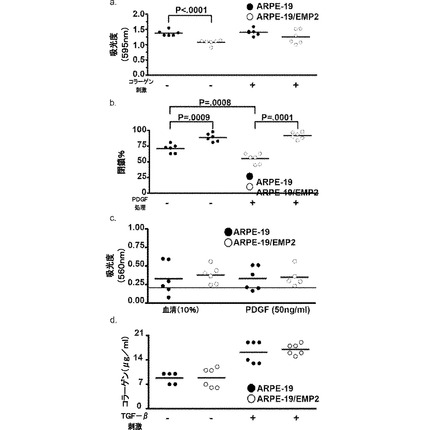
【図17】
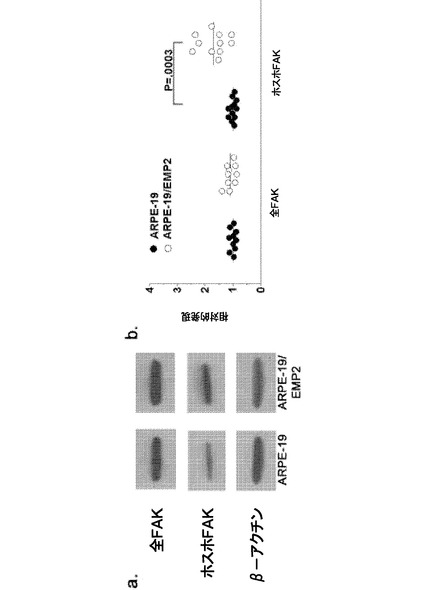
【図18】
【図19】
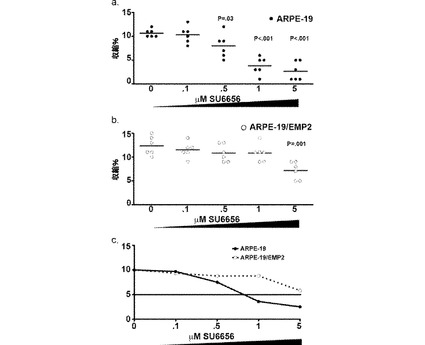
【図20A】

【図20B】

【図20C】
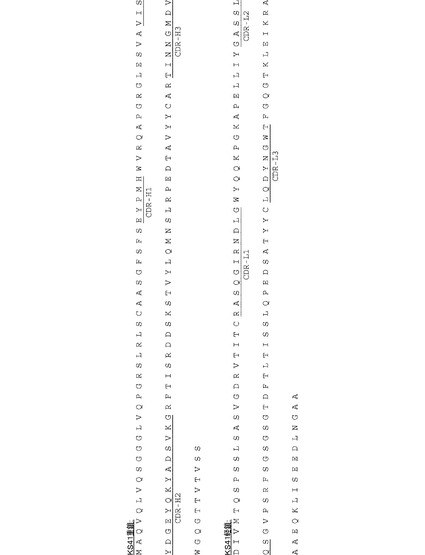
【図20D】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20A】
【図20B】
【図20C】
【図20D】
【公表番号】特表2013−511543(P2013−511543A)
【公表日】平成25年4月4日(2013.4.4)
【国際特許分類】
【出願番号】特願2012−540068(P2012−540068)
【出願日】平成22年11月18日(2010.11.18)
【国際出願番号】PCT/US2010/057296
【国際公開番号】WO2011/063161
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(506115514)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (87)
【出願人】(505133696)ザ・ユナイテッド・ステイツ・ガバメント・アズ・リプレゼンティッド・バイ・ザ・デパートメント・オヴ・ヴェテランズ・アフェアズ (2)
【Fターム(参考)】
【公表日】平成25年4月4日(2013.4.4)
【国際特許分類】
【出願日】平成22年11月18日(2010.11.18)
【国際出願番号】PCT/US2010/057296
【国際公開番号】WO2011/063161
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(506115514)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (87)
【出願人】(505133696)ザ・ユナイテッド・ステイツ・ガバメント・アズ・リプレゼンティッド・バイ・ザ・デパートメント・オヴ・ヴェテランズ・アフェアズ (2)
【Fターム(参考)】
[ Back to top ]