
不斉酸化反応を有する蛋白質複合体およびその製造方法

【課題】緩和(mild)な条件下で、効果的な酵素類似(enzyme-mimetic)反応を可能とする、有用な材料を提供する。
【解決手段】動植物粗蛋白質を濃縮する第1の工程と、該粗蛋白質をアルギン酸カルシウムゲルに包括して空気酸化させた後、目的の蛋白質複合体を作成し抽出する第2の工程と、得られる蛋白質複合体溶液を、結晶化沈殿させる第3の工程と、沈殿した結晶化蛋白質複合体を架橋固定化する第4の工程により、蛋白質複合体を製造する。また、第5の工程により、該蛋白質複合体を凍結乾燥(FD)処理して水分を取り除き、粉砕する。好適に常温保存に富む不斉酸化触媒を作ることができる。
【解決手段】動植物粗蛋白質を濃縮する第1の工程と、該粗蛋白質をアルギン酸カルシウムゲルに包括して空気酸化させた後、目的の蛋白質複合体を作成し抽出する第2の工程と、得られる蛋白質複合体溶液を、結晶化沈殿させる第3の工程と、沈殿した結晶化蛋白質複合体を架橋固定化する第4の工程により、蛋白質複合体を製造する。また、第5の工程により、該蛋白質複合体を凍結乾燥(FD)処理して水分を取り除き、粉砕する。好適に常温保存に富む不斉酸化触媒を作ることができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、蛋白質およびカルシウムを少なくとも含み、且つ不斉酸化反応を触媒する活性を有する蛋白質複合体、およびその製造方法に関する。この蛋白質複合体は、例えば、動植物組織から平易に分離できる水溶性粗蛋白質をゲル(アルギン酸カルシウムゲル等)に包括して該蛋白質を空気酸化させた後、分離することにより得ることができる。
【背景技術】
【0002】
医薬、食品分野を始めとする種々の分野において、光学異性体を分離し、有用な一方の光学異性体を分取する技術の必要性が高まっている。例えば、光学異性体の一方を選択的に酸化還元又はデラセミ化する機能を有する固定化酵素を用いて、医薬品や香料を生成させる方法について研究されている。
【0003】
生体内の化学変化は、通常は酵素で触媒される。このような酵素の作用・機構を有用物質生産や分析に利用するシステムの一つとして、バイオリアクターが挙げられる。
【0004】
バイオリアクターの中心となるのは、不溶性の担体に結合させた固定化酵素である。これによって,触媒となる酵素と生産物を容易に分離することができる。固定化酵素は,研究,医療,分析,産業に広く利用されている。化学反応を行わせるバイオリアクターの中枢部分は反応素子であり,原料を製品に変換したり化学変化を利用して分析に利用したりする目的で,精製した酵素,オルガネラあるいは細胞そのものが用いられている。反応素子はリアクター中に留まり、反復使用が可能なものでなければならないので,反応素子は種々の方法で固定化されている(このような酵素の活用の詳細に関しては、例えば、「化学増刊119 ハイブリッドプロセスによる有用物質生産」 化学同人 「バイオリアクター」福井三郎編、バイオテクノロジーシリーズ、講談社 「固定化酵素」千畑一郎編、講談社サイエンティフィック等を参照することができる)。
【0005】
しかしながら,酵素は精製するのにコストがかかり,精製された酵素は不安定なことが多い為,安定化する為の経費負担に改善の余地がある。この理由から,精製酵素の代りに目的の酵素を含む微生物をそのまま固定化して用いた例がある。例えば、アスパルターゼを含む微生物を固定化してL−アスパラギン酸を製造する例や、次いで,このプラントでつくられるアスパラギン酸を原料として,連続的にL−アラニンを製造した例などである(「化学増刊119 ハイブリッドプロセスによる有用物質生産」 化学同人; 「バイオリアクター」福井三郎編、バイオテクノロジーシリーズ、講談社; 「固定化酵素」千畑一郎編、講談社サイエンティフィック等を参照)。
【0006】
“化学と工業”Vol.62−1,January 2009、pp44−45(生体触媒開発を通じて)のように、工業的にみる酸化還元触媒は微生物や酵母、植物培養細胞など、細胞内に内在する脱水素酵素と補酵素の機能をそのまま活用する方法がコスト的に優位であり、生物固体から酸化還元酵素や補酵素の単離・精製を行う余分な経費負担は酵素の安定化や、ケトン→アルコール反応と補酵素NADH→NAD+反応、又はその逆反応を共役させる操作コストにおいて見合わないとされている。
【0007】
また、最近の酵素に変わる触媒設計の研究では、「粗蛋白質」内部に金属錯体を導入して酵素様活性ドメインを作成した例が見られる。
【0008】
例えば、「蛋白質 核酸 酵素」2004年,11月号,Vol.49,No.14 −ヘム酵素を中心とした金属酵素の分子設計−」(名古屋大学大学院理学研究科 渡辺芳人)に公知のように、クロロペルオキシダーゼ(CPO)の活性ドメイン内のヘム(鉄)を他の金属錯体に置き換えても酸化活性が起きることが示され、配置する金属近傍の機能性アミノ酸残基の適当な配置を設計することが活性ドメイン構築に重要であることが示されている。
【0009】
また、文献“Tetrahedron Letters”No.44,pp4281−4284(1978)(AsymMetric Reduction of aryl trifluoromethyl ketones with an model compound in a chiral hydrophobic binding site of sodium cholate micelle, β-cyclodextrin and bovine serum albumin)「馬場直道ら;京都大学化学研究所」によれば、酵素以外の成分:界面活性剤様タンジル酸(NAC)、β−シクロデキストリン(β−CD)、牛血清アルブミン(BSA)を触媒として、1−プロピル−1,4−ジハイドロニコチンアミド(NAH)又は水素化ホウ素ナトリウム(NaBH4)の存在下、基質トリフルオロメチル−アセトフェノンを作用させて不斉還元反応している。以上の結果、牛血清アルブミン(BSA)にも活性ドメインが存在(立体配置R、光学純度46.6%ee)することが示されている。
【0010】
100%ee近くの光学純度で、動植物由来粗蛋白から光学活性アルコールを分割した例が特許第3294860号公報(光学活性アルコールの製造方法)に示されており、穀類又は豆類から水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムゲルにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程とを組み合わせて、光学活性アルコール(100%ee、収率50%)を合成している。特許第3683129号公報(光学活性アルコールの製造方法)では、卵白及び卵白から分離されたオボアルブミンから選択される水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程とを組み合わせて、光学活性アルコール(100%ee)を合成している。
【0011】
一般的に、固定化酵素を作製する方法は、下記のような方法が代表的である。
1)該抽出・精製酵素を水不溶性の担体、例えば、セルロース、デキストラン、アガロース等の多糖類の誘導体、及びポリアクリルアミドゲル等に結合させる担体結合法、2)該抽出・精製酵素を2個もしくはそれ以上の官能基を有する試薬を用いて該抽出・精製酵素間に架橋結合を形成させて固定する架橋法、3)該抽出・精製酵素を、ゲル、例えば、アルギン酸塩、デンプン、コンニャク、ポリアクリルアミドゲル及びポリビニルアルコール等のゲルの細かい格子の中に取り入れる(格子型)か、半透明膜の皮膜によって被覆する(マイクロカプセル型)包括法がある。
【0012】
また、1990年代に登場したCLEC(Cross-Linked Enzyme Crystal)法は、該抽出・精製酵素を硫酸アンモニウムやポリエチレングリコール(PEG)などで結晶化し、グルタルアルデヒド(GA)などの多価修飾試薬を用い架橋させる方法で、産業用の固定化酵素の手法として最も活用されている。該CLEC法としては、例えば有機合成用酵素としてAltus株式会社より、ChiroCLEC(キラル化合物合成酵素)が製品化されている。ChiroCLEC−BLはBacillus licheniformis由来のSubtilisinを、ChiroCLEC−CRはCandida rugosa由来のLipaseを固定化し、架橋固相化してある。
【0013】
これらは有機溶媒中でも安定に活性を示すと同時に、熱安定性にも優れている。光学活性を有しながら、カルボン酸、アルコール、アミノ酸、エステル等の加水分解又はアシル化を行うことができる特徴がある。架橋酵素結晶(CLEC)は、1)酵素を架橋固定化する操作条件下で酵素の働きを至適化でき、2)加水分解酵素、酸化還元酵素、脱離酵素など、市販されている様々な酵素の中で開発でき、3)特定のニーズを満たすように手直しされた遺伝子導入微生物産生の酵素などから開発可能である。
【0014】
また、特公平4−68914(酵素の固定化方法)のように多孔性水不溶性担体のアミノ化シリカゲルに酵素を吸着させて、その後、多官能性架橋剤(グルタルアルデヒド)による共有結合反応により酵素の固定化を行った例がある。この場合、担体と酵素の結合力が弱く担体から酵素が脱離しやすい難点を、多官能性架橋剤に架橋することで解決している。多孔質アミノ化シリカゲルや多孔質アミノ化ゼオライトなどの多孔性担体の酵素吸着性に加えて、更に、多官能性架橋剤(グルタルアルデヒド)を利用することで固定化担体の活性半減期の著しい向上を実現している。
【0015】
このような架橋酵素結晶(CLEC)の知られている利点をまとめると、次の様になる。尚、酵素の結晶化とは、酵素分子の周囲に覆われた水分子が硫安やポリエチレングリコール添加により外れてゆき、酵素分子どうしが重合し始めることであり、最終的に分子が大きくなり沈殿することを意味する。有機化合物の凝固点まで冷やして結晶化する意味合いとは異なっている。
【0016】
(架橋酵素結晶の利点)
・高純度の酵素は必要としない(部分的に精製された調製品に適用可能)
・簡単な操作と広い応用
・室温で長期間(1年以上)安定
・実質100%活性なタンパク質(高い容積測定(volumetric)と触媒生産性)
【0017】
・容易な回収と再使用
・単独酵素と比較して高い温度安定性と有機溶媒への耐性
・高い活性と選択性(単独酵素よりも高い場合もある)
・水媒体中で酵素を濾す必要はない
・迅速な至適化(HTEを使用して)は開発時間を短くする
・触媒カスケードプロセス用に1つ以上の酵素を含むコンビCLEA
【0018】
更に架橋酵素結晶(CLEC)の市場と応用の可能性では、CLEC合成(薬剤、香料及び味覚物質、農薬、機能性食品、ファインケミカル、バルクのモノマー)、食品および飲料、パルプおよび紙、化粧品、オイルおよび脂質、織物、排出処理、界面活性剤、バイオセンサー、診断薬、タンパク質輸送などがある。
【0019】
Dehydrogenases については、現在、微生物由来のAlcohol dehydrogenase(ADH)from Rhodococcus erythropolis 、Formate dehydrogenase(FDH)from Candida boidinii がCLEA調整用の酵素類として知られている。
【先行技術文献】
【特許文献】
【0020】
【特許文献1】特許第3294860号公報
【特許文献2】特許第3683129号公報
【特許文献3】特公平4−68914号公報
【非特許文献】
【0021】
【非特許文献1】「化学増刊119 ハイブリッドプロセスによる有用物質生産」化学同人 「バイオリアクター」福井三郎編、
【非特許文献2】バイオテクノロジーシリーズ、講談社「固定化酵素」千畑一郎編、講談社サイエンティフィック
【非特許文献3】“化学と工業”Vol.62−1,January 2009、pp44−45
【0022】
【非特許文献4】「蛋白質 核酸 酵素」2004年、11月号、Vol.49、No14 −ヘム酵素を中心とした金属酵素の分子設計−(名古屋大学大学院理学研究科 渡辺芳人)
【非特許文献5】“Tetrahedron Letters” N0.44,pp4281−4284(1978)(AsymMetric Reduction of aryl trifluoromethyl ketones with an model compound in a chiral hydrophobic binding site of sodium cholate micelle, β-cyclodextrin and bovine serum albumin)「馬場直道ら;京都大学化学研究所」
【非特許文献6】“食品と技術” 12月号,pp1−9(2008)(グルテンタンパク質のネットワーク形成における食塩の役割)「裏出令子;京都大学大学院農学研究科」
【0023】
【非特許文献7】Crystal stracture of the DsbB-DsbA complex reveals a disulfide bond severation mechanism−Cell 127 789−801(2006)(稲葉謙次ら; 九州大学生体防御医学研究所)
【0024】
【非特許文献8】STRUCTURE-FUNCTION STUDIES OF GLUTAREDOXINS AND RELATED OXIDOREDUCTASRE−」(Tobias H. ElGAn; From THE DEPARTMENT OF BIOSCIENCES AND NUTRITION Karolinska Institutet, Stockholm, Sweden)
【発明の概要】
【発明が解決しようとする課題】
【0025】
本発明の目的は、上述した従来技術の欠点を解消し、緩和(mild)な条件下で、効率的な酵素類似(enzyme-mimetic)反応を可能とする、有用な材料を提供することにある。
【0026】
本発明の他の目的は、環境的側面および安全性に優れる、「酵素類似」反応活性を有する材料を提供することにある。
【0027】
本発明の他の目的は、このような「酵素類似」反応活性を有する材料の好適な製造方法を提供することにある。
【課題を解決するための手段】
【0028】
本発明者は鋭意研究の結果、不斉酸化反応を触媒する活性を有する蛋白質複合体を発見した。
【0029】
本発明の蛋白質複合体は、上記発見に基づくものであり、より詳しくは、蛋白質およびカルシウムを少なくとも含む蛋白質複合体であって;且つ、不斉酸化反応を触媒する活性を有することを特徴とするものである。
【0030】
本発明者は、上記知見に基づき更に研究を進めた結果、上記した有用な性質を有する蛋白質複合体が効率的に得られる方法を見出した。本発明の製造方法は上記発見に基づくものであり、より詳しくは、動植物由来水溶性部分から粗蛋白質を濃縮する第1の工程と;前記蛋白質をゲルに包括して空気酸化させた後、該ゲルから蛋白質複合体を抽出する第2の工程と;抽出された蛋白質複合体を水溶液中にて結晶化沈殿させる第3の工程と;沈殿した蛋白質複合体を架橋する第4の工程を含むことを特徴としている。
【0031】
上記製造方法の一態様によれば、動植物の水溶性部分から粗蛋白質を濃縮する第1の工程と;該粗蛋白質をアルギン酸カルシウムゲルに包括して、該ゲルを空気に数時間曝した後(空気酸化)、温水振とうして蛋白質複合体を抽出する第2の工程と;該蛋白質複合体を飽和30%硫安沈殿させる第3の工程と;沈殿した該結晶化蛋白質複合体を架橋する第4の工程を含むことができる。
【0032】
本発明の他の態様によれば、安価な動植物由来粗蛋白質をアルギン酸カルシウムゲルに包括して、該ゲルビーズ(酸素と溶存カルシウム存在下)を空気酸化させて(i)分子間ジスルフィド結合と(ii)分子間凝集(<6.4Å)と、(iii)水溶性変化を誘導させ(蛋白質複合体形成)、引き続き、温水中で振ることにより該複合体を好適に抽出することができる。
【0033】
本発明の一態様によれば、更に、蛋白質複合体又は架橋結晶化された蛋白質複合体を用いてでさえも、基質ラセミ体アルコールの一方の鏡像体を不斉酸化して、100%e.e.近い光学活性アルコールが合成することができることを示しており、また、その具体的な製造法が提供される。
【0034】
本発明の一態様によれば、更に、動植物由来祖蛋白質の蛋白質複合体が、ベンゼン骨格やナフタレン骨格を有する二級アルコール基質に対する一方の鏡像体に選択的な不斉酸化する反応に加えて(収率50%)、それらの骨格を持たないマツタケオールのようなアルキル鎖系二級アルコール類のデラセミ化反応活性(収率>85%)を有することも提供される。
【0035】
本発明においては、上述の精製された微生物由来酵素、遺伝子組み換え微生物由来酵素、又は微生物や遺伝子組み換え微生物、更には動物組織(肝臓、すい臓など)由来の固定化酵素、固定化微生物の製造を対象としているのではなく、高コストの酵素の単離と精製の工程を必要とせず、より安価な動植物資源由来粗蛋白質をその対象としていることが一つの特徴である。このような「植物資源」としては、例えば、蕎麦、アマランサス、米、小麦、大麦、トウモロコシ、エンバク、ライ麦、粟、ヒエ、キビ、ハト麦、モロコシ等の穀類;小豆、インゲン豆、豌豆、リョクトウ、大豆等の豆類、更には一般的な草や雑草をも含めた、これらの種子、葉、茎、根、花、実の各植物組織も挙げられる。
【0036】
更に、「動物資源」としては、卵白又はオボアルブミンとして、鶏卵由来のものが挙げられるが、鶏類、両生類、魚類の卵アルブミンも同様に用いることができ、鶏卵由来のものに限定するものではないし、由来の蛋白質を卵に限定するものでもない。
【0037】
本発明の一態様においては、後述するように、例えば、穀類、豆類、そして草や雑草をも含めたこれらの種子、葉、茎、根、花、実から水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムゲルビーズに包括化して、空気酸化後、10時間以上40℃の温水中で振とうさせて、蛋白質複合体水溶液画分をビーズと分離する第2の工程と、得られる蛋白質複合体水溶性画分を飽和30%硫安にして沈殿させる第3の工程と、得られた結晶化蛋白質複合体をグルタルアルデヒドにて架橋する第4の工程、更に、得られた架橋結晶化蛋白質複合体を乾燥粉体化する第5の工程を組み合わせた、環境に優しく低コストで且つ保存性の高い蛋白質複合体を製造することも可能となる。
【発明の効果】
【0038】
上述したように本発明によれば、実用的な不斉酸化活性を賦与した植物由来の蛋白質複合体、及び/又は動物由来の蛋白質複合体が提供される。本発明によれば、更に、このような動植物由来の架橋結晶化蛋白質に好適な製造方法であって、環境に優しく低コスト生産を実現する蛋白質複合体の製造方法が提供される。
【図面の簡単な説明】
【0039】
【図1】図1は、本発明製造方法の第2の工程において、豌豆蛋白・アルギン酸カルシウムゲルビーズに蒸留水を加えて、振とう開始後所定の時間後に、基質R−2−ナフチルエタノールを加えた時の基質の残量%を示したグラフである。
【図2a】図2aは、本発明製造方法の第2の工程において、空振とう10時間後、種々の基質を添加した際の反応時間に伴う基質と生成物の%比率をモニターリングしたグラフである。
【図2b】図2bは、本発明製造方法の第2の工程において、空振とう10時間後の水溶液等に、基質R−2−ナフチルエタノールを加えて、生成ケトン%をモニターリングしたグラフである。
【0040】
【図2c】図2cは、本発明製造方法の第2の工程において、異なる塩化カルシウム濃度に滴下することで調整した豌豆・アルギン酸カルシウムゲルビーズを空振とう10時間後、基質Rー2−ナフチルエタノールに作用させて蛋白質複合体活性を調べたグラフである。
【図2d】図2dは、本発明製造方法の第2の工程において、所定の豌豆蛋白質量で豌豆・アルギン酸カルシウムゲルビーズを調製したものを、空振盪10時間後にそれぞれ基質R−2−ナフチルエタノール(50mg)を加えて、蛋白質複合体活性を調べたグラフである。
【0041】
【図3】図3は豌豆−Caアルギン酸ゲルを10時間以上温水中にて振とう後、溶出される蛋白質複合体を硫安30%にて結晶化沈殿された、水溶性蛋白成分の分子量決定(SDS−page)である。 ここに、溶出溶液は、10時間以上温水中に振盪後の溶液画分である。硫安沈殿は、溶出溶液を30%硫安溶液にした時の沈殿物である。上清は、硫安沈殿物を水に再溶解したものを遠心分離して得られる上清画分である。
【0042】
【図4】図4は、「食品と技術」2008年12月号総括,裏出令子記述(グルテンタンパク質のネットワーク形成における食塩の役割)に見られる食塩添加あるいは無添加小麦グルテン生地のβ-メルカプトエタノール入りSDS溶液で可溶な成分のSDS−pageである。
【0043】
【図5】図5は、フーリエ変換赤外分光分析(FT−IR)を用いるサンプル(i)〜(Vi)の定性分析結果である。
【図6a】図6aは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【図6b】図6bは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【図6c】図6cは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【図6d】図6dは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【0044】
【図7】図7は、本発明製造方法の第2の工程において、豌豆・アルギン酸カルシウムゲルを空気中で所定時間静置後、蛋白質複合体を抽出して架橋結晶化後、凍結乾燥させて得られる蛋白質複合体の収量と6時間後の不斉酸化活性(右軸)とモニタリングしたグラフである。
【0045】
【図8】図8は第2の工程において、蛋白質複合体に基質ラセミ−1−オクテン−3−オールを作用させて、香料のR−1−オクテン−3−オール(マツタケオール;98.2%ee)を合成したGCクロマトである。
【0046】
【図9a】図9aは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9b】図9bは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0047】
【図9c】図9cは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9d】図9dは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0048】
【図9e】図9eは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9f】図9fは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0049】
【図9g】図9gは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9h】図9hは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0050】
【図10】図10は、蛋白質複合体の全工程製造フローと各工程の簡単な説明を示している。
【0051】
【図11a】図11aは、原核生物(大腸菌)におけるタンパク質ジスルフィド結合形成のための細胞システム(米国化学会誌「Crystal structure of the DsbB-DsbA complex reveals a disulfide bond severation mechanism」Cell, volume 127, Issue 4,789−801,17 Novemver 2006: 稲葉謙次ら(九州大学生体防御医学研究所)の図を引用)。
【図11b】図11bは、真核生物(酵母小胞体)におけるタンパク質ジスルフィド結合形成のための細胞システム(米国化学会誌「Crystal structure of the DsbB-DsbA complex reveals a disulfide bond severation mechanism」Cell, volume 127, Issue 4,789−801,17 Novemver 2006: 稲葉謙次ら(九州大学生体防御医学研究所)の図を引用)。
【図12】細胞内のシステイン酸化によって得られるスルフィド系酵素の種類とその酵素機能(反応性)を示したブロック図である。
【図13】実施例20において得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。
【図14】実施例20において得られた蛋白質複合体の凍結真空乾燥(FD)粉末(50mg、75mg、100mg、200mg、300mg)にラセミ−2−ナフチルエタノール濃度10ppmの50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、4時間置きにGCにて、不斉酸化されるR−2−ナフチルエタノールの残留比(%)について追跡を行った結果を示すグラフである。
【図15】実施例20において得られた蛋白質複合体の凍結真空乾燥(FD)粉末(300mg)にラセミ−2−ナフチルエタノール濃度(10-140ppm)の50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、不斉酸化されるR−2−ナフチルエタノールの残留比(%)について4時間置きにGC追跡を行った結果を示示すグラフである。
【発明を実施するための形態】
【0052】
以下、必要に応じて図面を参照しつつ本発明を更に具体的に説明する。以下の記載において量比を表す「部」および「%」は、特に断らない限り質量基準とする。
【0053】
(蛋白質複合体)
本発明の蛋白質複合体は、蛋白質を少なくとも含み、且つ、FT−IRにおいて、(1085±50cm−1)の領域、および(1411±50cm−1)の領域に、それぞれ少なくとも1個のピークを有することを特徴とする。ここに、FT−IR測定は、後述するように、例えば、Thermo Fisher Scientific製フーリエ変換赤外分光分析装置Magna-750および赤外顕微鏡Nic−Planを用い、顕微ATR法(Geクリスタル)により、例えば波数分解能8cm−1、積算回数32回の条件で測定することができる。(このようなFT−IR測定の詳細に関しては、例えば、文献”Carbohydrate Polymers”66,191−197,(2005)(Enzymatically produced nano-ordered short elements containing cellulose Iβ crystalline domains)「Noriko Hayashiら;Bioconversion Laboratory, Forestry and Forest Products Research Institute (FFPRI)」に実施例があり、吸収波長の評価に関しては、本「ACADEMIC PRESS; New York and London 1971; A Subsidiory of Harcourt Brace Jovanovich, Publishers」”INFRARED SPECTRA OF INORGANIC COMPOUNDS (3800−45cm−1)”「Richard A. Nyquist and Renald O. Kagel; Chemistry Physics Research Laboratory, The Dow Chemical Company Midland MichiGAn」を参照できる。
【0054】
(IRピークの強度)
本発明の蛋白質複合体において、上記FT−IR測定におけるピーク強度は、以下の通りであることが好ましい。
(1)(1085±50cm−1)の領域におけるピーク(以下、「ピーク1」という)は、測定されたIRスペクトル中の最も強度が大きいピークであるか、または最も強度が大きいピークの強度をI0とした際に、(1/10)×I0以上のピーク強度を有することが好ましい。このピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
(2)(1411±50cm−1)の領域におけるピーク(以下、「ピーク2」という)は、測定されたIRスペクトル中の最も強度が大きいピークであるか、または最も強度が大きいピークの強度をI0とした際に、(1/10)×I0以上のピーク強度を有することが好ましい。このピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
【0055】
(他のIRピークの強度)
本発明の蛋白質複合体において、上記した「ピーク1」および「ピーク2」以外のIRピークは特に制限されない。本発明の蛋白質複合体は、例えば、下記の領域にピークを有していても良い。
【0056】
また、本発明の蛋白質複合体において、上記FT−IR測定ピーク強度の評価は、以下の通りであることが更に好ましい。
(1)(1085±50cm−1)の領域におけるピーク(以下、「ピーク1」という)は、測定されたIRスペクトル中の最も強度が大きいピークであり、最も強度が大きいピークの強度をI0とした際に、2番目に強度が大きいピークが(1411±50cm−1)にあり、このピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
(2)(1411±50cm−1)の領域におけるピーク(以下、「ピーク2」という)は、測定されたIRスペクトル中の2番目に最も強度が大きいピークであり、2番目に強度が大きいピークの強度をI0とした際に、3番目に強度が大きいピークが(1649±50cm−1)にありこのピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
【0057】
(他のIRピークの強度)
上記の場合にも、本発明の蛋白質複合体において、上記した「ピーク1」および「ピーク2」以外のIRピークは特に制限されない。本発明の蛋白質複合体は、例えば、下記の領域にピークを有していても良い。
【0058】
(好ましい蛋白質複合体)
本発明の蛋白質複合体は、蛋白質およびカルシウムを少なくとも含むことが好ましい。また、蛋白質複合体は、不斉酸化反応を触媒する活性を有することが好ましい。このような「蛋白質およびカルシウムを少なくとも含む」こと、および「不斉酸化反応を触媒する活性を有する」は、後述する方法によって好適に確認することができる。
【0059】
(不斉酸化反応の好ましい触媒)
本発明の蛋白質複合体の不斉酸化反応の好ましい態様は、例えば、後述する実施例12の条件下で、基質ラセミ−1−オクテン−3−オール(マツタケオール)を作用させた場合に得られるR−1−オクテン−3−オールの光学純度(%ee)で好適に表すことができる。本発明の蛋白質複合体は、この実施例12の条件で、85%以上の化学収率で、R−1−オクテン−3−オールを与えることが好ましい。更には、この光学純度は、95%ee以上(特に98%ee以上)であることが好ましい。
【0060】
(好適な特製)
本発明の蛋白質複合体は、下記の好適な特製のうち、1以上を有することが好ましい。
(1) 上記蛋白質複合体が、更に糖を含むこと。
(2) 前記蛋白質が、架橋結晶化蛋白質であること。
(3) 前記不斉酸化反応が、基質ラセミ体アルコールの一方の鏡像体を選択的に与える酸化反応であること。
(4) 前記不斉酸化により得られるケトンが、更に不斉酸化されるデラセミ化反応を与える活性を、前記蛋白質複合体が有すること。
【0061】
(本発明の推定メカニズム)
本発明者の知見によれば、本発明において、好適な動植物由来の蛋白質複合体が提供されるのは以下の理由によるものと推定される。
【0062】
安価な材料の動植物由来の粗蛋白質は、カルシウム塩と酸素存在を実現できるアルギン酸カルシウムゲルに包括後、空気中で好適に静置(空気酸化)させることによって、(i)タンパク質内システイン残基(−SH)どうしの分子間ジスルフィド結合(−S−S−)、(ii)タンパク質内分子間凝集(分子間距離の短縮:<6.4Å)、(iii)水溶性への性質変化が生じ、活性ドメイン(チオレドキシンフォールド:Cys−X−Y−Cys配列)の形成のみならず、温水中で振ることにより好適に蛋白質複合体が溶出できる性質になる。
【0063】
温水中に溶出される溶存Ca塩と酸素の存在は、ジスルフィド結合したタンパク質ポリマー(蛋白質複合体)を水溶液に溶け易くする効果があり、温水中で振ることにより容易に抽出される。タンパク質ポリマー(蛋白質複合体)の触媒活性向上と収量向上と収量向上のポイントは、空気中で好適に該ゲルビーズを静置(空気酸化)させる点と、温水中に溶け出す程度の低Caイオン濃度存在下で「滲み出し」易くさせる点にある。
【0064】
本発明の製造方法においては、動植物蛋白質の有する空気酸化に伴うこれらの「蛋白質複合体形成」と「滲み出し」の特製を生かして、アルギン酸カルシウムゲルによる固定ではなくて、好適なジスルフィド結合への誘導といった「逆転の発想」により、酵素様に作用する蛋白質複合体を低コストで、環境に優しく、効果的に作成することに特徴がある。
【0065】
(推定メカニズムの関連文献)
上記した本発明の「推定メカニズム」のサポートに関しては、例えば、以下の文献を挙げることができる。
【0066】
文献“食品と技術” 12月号,pp1−9(2008)(グルテンタンパク質のネットワーク形成における食塩の役割)「裏出令子;京都大学大学院農学研究科」では、食塩や塩化カルシウムには、グルテンタンパク質の相互作用を変化させてグリアジンやグルテニンを水に溶けるような性質に変化させる効果(水溶性効果)と、グルテン構成タンパク質間の相互作用を強めて分子間距離を短縮させる効果(凝集効果)が公知となっている。
【0067】
分子間距離を短縮させる効果(凝集効果)において、グルテン分子間距離は塩無添加では7.7Åであるが、塩添加の場合、分子間距離では6.4Å以内に短縮されることが示されている。ミオグロビンミュータントにみる活性ドメイン内のヒツチジン−鉄間の分子間距離は5.7Åであることから、塩添加による凝集効果は活性ドメインの機能性アミノ酸残基の適当な配置を設計する効果も期待できる。「蛋白質 核酸 酵素」2004年,11月号,Vol.49,No.14 −ヘム酵素を中心とした金属酵素の分子設計−」(名古屋大学大学院理学研究科 渡辺芳人)
【0068】
更に、小麦粉生地の蛋白成分(グルテニンとグリアジン)は、食塩存在下で作製した場合には、この蛋白質分子間の相互作用を変化させ、純水では水溶性となり、食塩水では不溶性を示す性質に変化することにも報告がある。
【0069】
小麦グルテン内グルテニンポリマーの網状構造の形成とは、小麦粉の水の添加により始まるが、空気中の酸素を使って、グルテニンのC末端とN末端システイン残基どうしが自発的に分子間ジスルフィド結合(R1−S−S−R2)する空気酸化の働きが知られている。しかしながら、最近の報告(非特許文献7)「−Crystal structure of the DsbB−DsbA complex reveals a disulfide bond severation mechanism−Cell 127 789-801 (2006)」(稲葉謙次ら; 九州大学生体防御医学研究所)には、大腸菌では、ジスルフィド結合を導入する為の酵素が存在しており、基質蛋白質の2つのシステインを酸化しジスルフィド結合を作る酵素(DsbA)と、使われたDsbAを酸化し酸化能力を回復させる酵素(DsbB)が、細胞内で活性エネルギーを作り出すことに関与する物質であるユビキノンと連動して働くとされている。DsbBが受け取った電子は呼吸鎖成分であるユビキノン(UQ)に受け渡され、最終的にはチトクロム酸化酵素を介して「酸素が電子受容体」となることが分かっている(図11aおよび図11bを参照)。
【0070】
更に、原核生物(大腸菌)の蛋白質ジスルフィド形成システムと真核細胞(酵母小胞体など)のそれとは類似しており、小胞体にはDsbBの機能ホモログとして小胞体膜近傍にEro1pが存在する。大腸菌システムでのジスルフィド結合を創り出すための酸化力は、「ユビキノン」としているのに対し、小胞体システムではFAD(Flavin Adenine Dinucleotide)となる。大腸菌におけるDsbA−DsbB−ユビキノン酸化システムと真核生物のPDI−Ero1p−FADは互いに機能的ホモログの関係にあるとある(図11aおよび図11b参照)。
【0071】
その他、ジスルフィド結合は、多くの細胞表層にある蛋白質にとって、立体構造を正しく形成し、維持する為に必須な化学結合であり、また、ジスルフィド結合の形成と解離は、細胞の機能に重要な蛋白質の活性や細胞内での存在場所に影響を与へたり、細胞の酸化還元環境の変化に起因するストレスに対応している。
【0072】
−STRUCTURE-FUNCTION STUDIES OF GLUTAREDOXINS AND RELATED OXIDOREDUCTASRE−」(Tobias H. ElGAn; From THE DEPARTMENT OF BIOSCIENCES AND NUTRITION Karolinska Institutet, Stockholm, Sweden)では、酵素活性ドメインには2つのシステインからなる共通配列「チオレドキシンフォールド(Cys−X−Y−Cys)」が存在し、この部位がジスルフィド結合の形成や異性化を触媒して酸化還元活性を持つとされている。Cys−X−Y−CysのXとYには疎水性のアミノ酸がくることが多く、還元型ではシステイン残基は2つの自由なチオール基となり、酸化型ではそれらはジスルフィド結合でつながれる(後述する図11aおよび図11bを参照)。
【0073】
小麦タンパク質の約70〜80%を占める成分を分類すると、アルブミン、グロブリン及び小麦プロラミンであるグリアジン(25kda〜50kdaの間にあるバンド)、そして高分子量グルテニン(HMW:75kda〜150kdaの間にある3バンド)と低分子量グルテニン(LMW:25kda〜50kdaの間にあるバンド)からなるグルテニンに分類される。図4にそのSDS−pageを示している。
【0074】
(標的となる触媒活性)
図12は、文献“Biochemical and Biophysical Research ComMunications”300,pp1−4(2003)(Multiple roles of cysteine in biocatalysis)「Niroshini M, Gilesら;School of Chemistry, University of Exeter」で公知のように、細胞内システイン酸化により形成された様々な酵素の機能に関してなど示されている。1)Peptidase、2)glyceraldehyde 3-phosphate dehydrogenase (GAPDH)、3)alcohol dehydrogenase (ADH)、4)bacterial nitrile hydratase (NHase)、5)ribonucleotide reductase (RNRase)、6)pyruvate formate lyase (PFL)、7)benzylsuccinate synthase (BSS)、8)reduction of glutathione disulfide (GSSG)、9)thioredoxin (Trx)、10)glutathion reductase (GR)、11)thioredoxin reductase (TR)、12)NADH oxidase (Nox), 13)NADH peroxidase (Npx)。
【0075】
また、特許第3294860号公報(光学活性アルコールの製造方法)で公知のように、穀類又は豆類から水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムゲルにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程が示されている。また、特許第3683129号公報(光学活性アルコールの製造方法)で公知のように、卵白及び卵白から分離されたオボアルブミンから選択される水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程が示されている。
【0076】
本発明との違いは、上記知見に加えて、1)空気酸化により構築されるタンパク質分子内の活性ドメイン形成メカニズムへの言及と、2)架橋結晶化後に水分を除去して常温保存が可能な粉末触媒の製造法に発展している点にある。
以下、本発明の製造方法を構成する各工程について説明する。
【0077】
(第1の工程)
本発明の第1の工程における水溶性蛋白質の抽出においては、穀類又は豆類を砕き粒の大きい部分と殻部を取り除き、このようにして得られた穀類及び豆類粉砕粉を約20〜60℃、好ましくは約40℃で、pH約6〜8、好ましくは約pH7.0において、穀類または豆類の約7〜15重量倍の水で、30分以上抽出する。約45分で抽出するのが最も効果的であって、これ以上長く抽出しても抽出物の量は変わらない。pHの調製が必要なときは、H2SO4、HCl、H3PO4などの食品級酸、又はNaOHなどの食品級アルカリを用いて上記適性範囲を調製できる。上記水溶性蛋白質抽出液、又はこの抽出液から、デカンター、遠心分離などにより食物繊維部を分離した、タンパクカードをそのまま第2工程に移すか、必要に応じて噴霧乾燥、凍結乾燥、真空乾燥などにより粉末としてから、再溶解して第2工程に移しても良い。種子(外皮(ふすま、ぬか)、胚芽(スプラウト))に関しては、上記の穀類及び豆類粉砕粉の工程にて得ることができる。外皮(ふすま、ぬか)、胚芽(スプラウト)、葉(若葉、新芽)、茎、根、花を凍結乾燥(FD)、熱風乾燥(AD)、真空乾燥などで水分を除去して乾燥後、ボールミルなどを利用して5μm以下まで細かく粉砕する。
【0078】
この粉砕粉をそのまま第2工程に移すことができる。等電点沈殿のpHの選定は沈殿量の多い画分の選定が目的であり、例えば大豆や豌豆や蕎麦蛋白質の場合はpH4.5及びpH9.5付近であり、このカードに5〜10重量倍の水を加え、ミキサー、攪拌機などにより解砕して、蛋白質スラリーを調製し、中和(pH6〜8)し中和スラリーとする。このスラリーを、前記と同様の噴霧乾燥、凍結真空乾燥(FD)、真空乾燥などにより粉末としてから、再溶解して第2工程に移す。
【0079】
しかしながら、多量の蛋白質の処理が必要なときには、上記タンパクガードをH2SO4、HCl、H3PO4などの食品級酸、又はNaOHなどの食品級アルカリを用いて等電点処理し、次いでデカンター、遠心分離機などによりホエイを分離してタンパクカードをえる。この等電点沈殿等は、水溶性蛋白質の濃縮を目的としたものであって、処理後においても水溶性蛋白質抽出液をそのまま噴霧乾燥などにより粉末化した場合と同様な効果を奏するものである。
【0080】
(第2の工程)
第2の工程において、該抽出蛋白質を包括化する方法は、例えば、該抽出蛋白質を、ゲル、例えば、アルギン酸塩、デンプン、コンニャク、ポリアクリルアミドゲル及びポリビニルアルコール等のゲルの細かい格子の中に取り入れる(格子型)か、半透膜性の皮膜によって被覆する(マイクロカプセル型)包括法があるが、蛋白質複合体を作成する上では、海草より抽出するアルギン酸のカルシウムを用いた包括化法が安価で環境に優しく、かつ、包括操作が平易な点で最も好ましい。この包括化されたビーズは、蛋白質複合体を水溶液中に溶出させる為に、温水温度30℃〜45℃で、且つ、溶存酸素濃度がゼロにならないように酸素供給が必要で、且つ、ビーズが振れる回転数で振とうさせることが望ましい。
【0081】
(第3の工程)
第3の工程において、蛋白質を結晶化沈殿するためには、硫酸アンモニウム、ポリエチレングリコール(PEG)、ポリエチレングリコール/塩化リチウム,2−メチル−2,4−ペンタジオール,塩化ナトリウム,マロン酸ナトリウム等があり、いずれの結晶化沈殿剤として本願発明において用いることができる。しかしながら、最も安価な硫酸アンモニウムを用いるのがトータルコストダウンの観点から有益である。蛋白質の結晶化は、通常濃度が10〜50mg/mLのタンパク質溶液で用いて行うことが望ましく、サンプルをセントリコンやアミコンウルトラなどで濃縮する手法ある。
【0082】
(第4の工程)
第4の工程において、結晶化沈殿された蛋白質を架橋するためにはグルタルアルデヒド(以下、「GA」と略称する)、フォルムアルデヒド(以下、「FA」と略称する)等があり、いずれも架橋剤として本願発明において用いることができる。グルタルアルデヒドは、示性式OHC(CH2)3CHO として表される有機化合物で、反応は、タンパク質のリジン残基のε−アミノ基との間で起こるが、α−アミノ基やSH基との間でも起こり、分子間架橋を形成することができる。ひとつのグルタルアルデヒド分子が単独で架橋形成を起こせるとは考えられていない。(参考文献;重中義信/監修『原生動物の観察と実験法』(共立出版株式会社、1998年)ISBN 4320053532)GAは2つのアルデヒド基、FAは1つアルデヒド基を持っており、架橋させることで、1)室温で長期間(1年以上)安定、2)実質100%活性なタンパク質(高い容積測定(volumetric)と触媒生産性)、3)容易な回収と再使用、4)単独酵素と比較して高い温度安定性と有機溶媒への耐性、5)単独酵素よりも高い活性と選択性になるなど、産業用酵素試薬として実用レベルで広く活用できるようになる。
【0083】
GAは2つのアルデヒド基を持っているためか、FAに比べて非常に強い固定力を持っており、固定力の強いGAであるが、難点は浸透速度がとても遅い。FAは固定力が弱いものの浸透速度はGAの数倍あるので、GA固定液にFAをまぜることによって、双方の弱点を補いあうことができる。GA/FA混合固定液はカルノフスキー(Karnovsky,1965)が考案したので(5%GA+4%FA)、「希釈カルノフスキーを使用」などと今でもカルノフスキーの名称が通っている。GAとFAの他に固定液に種々の添加物を入れる場合がある。最も多いのは浸透圧を調整(上げる)ためにショ糖、ブドウ糖、食塩などを入れることができる。架橋による固定時間は、バルク使用の場合(厚さ1mM以下)、おおよそ2時間というのが最小時間としておく。固定液の温度については、室温で固定する場合、4℃(氷水)で固定する場合など様々で可能ですが、再現性を重視して、常温20℃〜40℃で固定することが好ましい。一定温度と言うことであれば、氷を浮かべた水の中に「固定液と試料を入れた試料瓶をいれておけば、ほぼ4℃を保つことができるが、作業トータルコスト等を考慮して、常温20℃〜40℃で固定することが好ましい。固定後は残留GAとFAを、0.1Mリン酸緩衝液(220 mOsm位)やその緩衝液に0.1M〜0.2Mのショ糖やグルコースを加えて洗浄することができる。
【0084】
(第5の工程)
第5の工程において、乾燥粉体化するためには、1)熱風乾燥(AD)、2)凍結真空乾燥(FD)、3)加圧乾燥、4)減圧乾燥、5)自然乾燥など本願発明において用いることができる。しかしながら、凍結真空乾燥(FD)は水分を完全に除去でき、引き続く粉砕の工程に有利であり、尚且つ保存性も高くなる。
【0085】
(各データの説明)
以下、図面に示した各データを説明する。
【0086】
図1は第2の工程において、豌豆蛋白・アルギン酸カルシウムゲルビーズに蒸留水を加えて、空振盪開始後0時間後、5時間後、10時間後、15時間後、20時間後、25時間後、35時間後、45時間後、55時間後に基質(ラセミ)−2−ナフチルエタノールを加えた時の基質の残量%を示したものである。
【0087】
図2aは第2の工程において、空振盪10時間後、a)基質ラセミ−2−ナフチルエタノール、R−2−ナフチルエタノール、2−アセトナフトンをそれぞれ50mg添加したときの反応時間に伴う基質と生成物の%比率をモニターリングしたものである。
【0088】
図2bは第2の工程において、空振盪10時間後の水溶液(120ml、240ml)、空振盪10時間後のビーズ+蒸留水(DW:120ml)、新しいビーズ+空振盪10時間後の水溶液(120ml)に、それぞれ基質R−2−ナフチルエタノール(50mg、75mg、又は100mg)を加えて、生成ケトン%をモニターリングしたものである。
【0089】
図2cは第2の工程において、異なる塩化カルシウム濃度(5g/L、7.5g/L、10g/L、15g/L、20g/L、30g/L)に滴下することで調製した豌豆・アルギン酸カルシウムゲルビーズを空振盪10時間後、基質R−2−ナフチルエタノールに作用させて蛋白質複合体活性を調べたものである。
【0090】
図2dは第2の工程において、豌豆蛋白量2g、3g、4g、5gで豌豆・アルギン酸カルシウムゲルビーズを調整したものを、空振盪10時間後にそれぞれ基質R−2−ナフチルエタノール(50mg)を加えて、蛋白質複合体活性を調べたものである。
【0091】
図3は豌豆−Caアルギン酸ゲルを10時間以上温水中にて振盪して得られる蛋白質複合体を硫安30%にて結晶化沈殿させて、更に、遠心分離(15分、10000rpm)して得られる沈殿物をSDS−pageした図である。
【0092】
SDS−PAGE
サンプル10 μLを,2×サンプルバッファー(0.1 M Tris/HCl pH 6.8,3 % SDS,10 %グリセリン,10% β‐メルカプトエタノール,0.1% BPB)10 μLと混合し,100℃で5 min加熱した。これらについて,SDS−PAGE mini(4−20 %グラジエントゲル,TEFCO社)を用いて分子量マーカー(SDS−PAGEスタンダード Broad,Bio−Rad)と共に泳動(18 mA定電流,泳動バッファー:25 mM Tris/HCl,0.19 M Glycine,0.1 % SDS,pH 8.3)を行った後,CBB染色(PHastGel Blue R,Amersham Biosciences社)を行った。
【0093】
図4は、文献“食品と技術” 12月号、pp1−9(2008)(グルテンタンパク質のネットワーク形成における食塩の役割)「裏出令子;京都大学大学院農学研究科」に記載された「図5 DSTによる架橋効率に食塩添加が及ぼす影響」の抜粋である。(以下説明文)食塩添加あるいは無添加生地をDSTで処理した後、βメルカプトエタノール入りSDS溶液でタンパク質を可溶化した。可溶化液を超遠心し、可溶性画分(S)と不溶性画分(P)に分けた。不溶性画分はメタ過ヨウ素酸ナトリウムで処理した。それぞれの試料は、SDSポリアクリルアミドゲル電気泳動で分離しタンパク質を染色した。
【0094】
図5は、エンドウ蛋白質粉末(i)とアルギン酸ナトリウム粉末(ii)と、定法に従いエンドウ蛋白質・アルギン酸カルシウムゲルを作成し、第2の工程において(iii)恒温振盪培養器による振盪、又は(iV)ジャーファメンタ温水中で攪拌して調整した蛋白質複合体粉末((iii)と(iV))と、第2の包括工程を省き、エンドウ蛋白質(i)を2%塩化カルシウム/50mMトリス塩酸バッファー((V)pH6.0又は(Vi)pH8.0)中で恒温振とう(55rpm、40℃、24時間))して作成した架橋結晶化粉末((V)と(Vi))をFT−IRにより分析した結果を示している。
【0095】
赤外分光分析(FT−IR)は、Thermo Fisher Scientific製フーリエ変換赤外分光分析装置Magna−750および赤外顕微鏡Nic−Planを用い、顕微ATR法(Geクリスタル)により、波数分解能8cm−1、積算回数32回で測定した。
【0096】
図5において、記号A〜Eは、下記の意味を有している。
A:1649cm−1付近・・・ペプチド結合(−C(=O)−N−)に特徴となるピーク。
B:1528cm−1付近・・・ペプチド結合(−C(=O)−N−)に特徴となるピーク。
C:1411cm−1付近・・・カルボキシラート(−C(=O)−O−Na)のケトン基に特徴となるピーク。
D:1122cm−1付近(1085cm−1、1032cm−1)・・・糖エーテル(−C−O−C−)特徴となるピーク。
E:1085cm−1付近・・・硫安((NH4)2SO4)に特徴のピーク。
【0097】
図6は、図5と同サンプルであるが、恒温振とう培養作成した蛋白質複合体粉末(i)と、豌豆蛋白質に2%塩化カルシウム/50mMトリス塩酸バッファー溶液((ii)pH6.0又は(iii)pH8.0)から得られる架橋結晶化粉末((ii)と(iii))、及びエンドウ蛋白質粉末(iV)をX線マイクロアナライザー定性分析を行った結果である。X線マイクロアナライザー分析は、島津製作所製EPMA−1600を用い、試料表面に金コートして測定した。
【0098】
図6において、各サンプルの意味は、以下の通りである。
サンプル(i)・・・振とう培養器製架橋結晶化蛋白質複合体
サンプル(ii)・・・20mM塩化Ca/50mMトリス塩酸バッファー(pH6.0)
サンプル(iii)・・・20mM塩化Ca/50mMトリス塩酸バッファー(pH8.0)
サンプル(iv)・・・エンドウ蛋白質粉末
【0099】
図7は第2の工程において、アルギン酸カルシウムゲルに包括したビーズを空気中で0時間、0.5時間、1時間、3時間、5時間、7時間静置後、温水中で振とうして蛋白質複合体を抽出し、更に架橋結晶化後に凍結乾燥させて粉末化したサンプルの6時間後の不斉酸化活性強度(右軸)と得られた蛋白質複合体の収量(左軸)を示す図である。
【0100】
図8は第2の工程において、図1と同一条件で豌豆・アルギン酸カルシウムゲルビーズに包括化して空振とう10時間後に溶出される蛋白質複合体水溶液に基質ラセミ−1−オクテン−3−オール(マツタケオール)を作用させて、98.2%eeの光学純度で香料のR−1−オクテン−3−オールを合成したGCクロマトグラムである。
【0101】
図9a〜図9h(および表1)は、100Lジャーファメンタを用いたエンドウ蛋白粉末(1kg)・アルギン酸カルシウムゲル(計20L)と加水(20L)における酸素供給有無に関わる、DO(溶存酸素濃度)とpHのモニタリング図、及び得られる蛋白質複合体の収量、活性をまとめた図および表である。
【0102】
図9aおよび図9b:1回目の実験の結果
図9cおよび図9d:2回目の実験の結果
図9eおよび図9f:3回目の実験の結果
図9gおよび図9h:4回目の実験の結果
【0103】
【表1】

(説明)
Ca濃度(FD)・・・FD乾燥後サンプルの酸分解法による原子吸光光度計によるCa濃度(mg/g)測定結果。
×・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間に殆ど不斉酸化されない場合(生成ケトン10%以下)。
△・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間までに不斉酸化が途中で止まる場合(生成ケトン30%以下)。
○・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間で不斉酸化される場合(生成ケトン40%以上)。
【0104】
図10は、蛋白質複合体の製造フローの工程図である。
【0105】
図11aは、原核生物(大腸菌)における、ジスルフィド結合形成の細胞内システムを示す。図11bは、真核生物(酵母小胞体)における、ジスルフィド結合形成の細胞内システムを示す。
【0106】
図12は、細胞内のシステイン酸化によって得られるスルフィド系酵素の種類とその酵素機能(反応性)を示した図である。
【0107】
(不斉酸化反応を有する蛋白質複合体)
本発明の蛋白質複合体は、動植物由来粗蛋白質の空気酸化形成物であって、且つ、不斉酸化反応を示すなど酵素様活性を有することを特徴とする。
【0108】
(不斉酸化活性の測定方法)
本発明の不斉酸化反応を有する蛋白質複合体の活性は、例えば、後述(実施例15「抽出作業の条件」内6.不斉酸化活性)に記載する方法により好適に測定可能である。
【0109】
(蛋白質複合体の精製法)
本発明において、蛋白質複合体の精製方法は特に限定されない。蛋白質複合体の収量を増やす目的から、ろ過の必要はないが、アルギン酸カルシウムゲルビーズとの分離の意味でガラスビーズなどを用いることができる。この場合、口径サイズが40−200μmで可能であるが、口径200μmであることが好ましい。
【0110】
次に、本発明を実施例に基づき具体的に説明するが、これは説明のためのものであって、これにより限定して解すべきではない。
【0111】
実施例1
(架橋結晶化した豌豆由来の蛋白質複合体の一般的な製造方法)
先ず第1の工程として、豌豆を粉砕して殻を取り除き、pH7.0付近の蒸留水(約40℃)9重量倍にて、約45分間に溶解される豌豆蛋白質成分をNaOH水溶液を用いてpH7.0にして沈殿成分の食物繊維を除去し、水溶性蛋白部分をアルカリ条件(pH9.5付近)又は酸性条件(pH4.5付近)にして蛋白質を等電点沈殿させ、蛋白沈殿部をpH7.0の蒸留水にて再溶解して得られる豌豆蛋白水溶液(試料濃度5.0%)を噴霧乾燥処理を行い粉体の豌豆蛋白を調製する。また、3%アルギン酸ナトリウム水溶液はオートクレーブの条件、温度121℃、時間20分で、アルギン酸ナトリウムを水溶液中に溶解して調整する。
【0112】
次に、第2の工程において、豌豆蛋白粉20gに10倍等量の蒸留水200mlを加えて、3%のアルギン酸ナトリウム水溶液を1.0倍等量の200mlを加えて均一になるまで攪拌し、得られた豌豆・アルギン酸ナトリウム混合液を4%の塩化カルシウム水溶液中に滴下して、固定化状態の豌豆蛋白含有・アルギン酸カルシウムゲルビーズを作製する。ビーズは5時間、空気中で静置した。豌豆蛋白含有・アルギン酸カルシウムゲルビーズを蒸留水にて洗浄し、塩化カルシウム水溶液を除去した後に用いた豌豆蛋白粉の20倍等量の蒸留水(400ml)を抽出溶液として添加し、恒温振とう培養(40℃、55rpm)を2日間して水溶性画分を分離回収後、ビーズは更に1日間振とうさせて同様に目的溶液を回収した。更にもう1日間振とう後、目的溶液を分離回収できる。回収された反応溶液に含まれる蛋白質複合体は、引き続き第3の工程で結晶化沈殿させた。
【0113】
第3の工程において、分離回収溶液を弱水溶性成分の沈殿を目的として、飽和30%硫安に調整した後、20時間以上静置させた。蛋白質複合体は、塩化カルシウムの存在下において容易にビーズより溶出されるだけでなく、飽和30%硫安溶液の存在下においては、非水溶性となり凝集沈殿した。
【0114】
第4の工程において、沈殿凝集した結晶化蛋白質複合体は更に、溶液濃度にして0.25%になるように市販25%グルタルアルデヒド溶液を添加し、2時間以上(好ましくは10時間以上)静置した。得られる蛋白質複合体は、冷却遠心機(10,000rpm、15min、10℃)後、デカンテーションにより容易に溶液から分離できた。
【0115】
第5の工程(凍結真空乾燥(FD))は、沈殿物の蛋白質複合体を−50℃にまで冷やして1時間静置後、約0.1mbar以下の真空状態にした後、温度を−50℃から50℃まで昇温させて水分を昇華させて、水分除去を行った。一連のFD所要時間は20時間程度であった。
【0116】
第6の工程において、凍結真空乾燥(FD)により水分除去された蛋白質複合体は、引き続くボールミル粉砕により、常温保存できる粉末状の触媒とできた。
【0117】
実施例2
(第2の工程における豌豆(PP)、大豆(SP)、小麦(WP)蛋白質由来蛋白質複合体の基質特異性の検討)
第2の工程において、豌豆(PP)、大豆(SP)、小麦(WP)由来粗蛋白質は種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し、水溶液画分を噴霧乾燥して得られる粉末20gは定法に従って、10倍等量の蒸留水200mlを加えて、3%のアルギン酸ナトリウム水溶液を1倍等量の200mlを加えて均一になるまで攪拌し、得られた豌豆・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中に滴下して、豌豆蛋白含有・アルギン酸カルシウムゲルビーズを作製する。該作製ビーズに豌豆蛋白粉の20倍等量の蒸留水(400ml)を反応溶液として添加し、恒温振とう培養器の温度設定40℃、振とう数55rpmにて10時間以上の振とう後、基質変換反応を行った。基質としては2−ナフチルエタノール(naphthyl/1)と、メタ、パラ位に塩素(Cl−/2a、2b)、臭素(Br−/3a、3b)、フッ素(F−/4a、4b)、メチル(Me−/5a、5b)、メトキシ(MeO−/6a、6b)、ニトロ(NO2−/7a、7b)を置換させたフェニルエタノールを用いた結果、反応時間、生成物、光学純度、化学収率を以下の表にまとめる。
【0118】
(第2の工程における豌豆(PP)、大豆(SP)、小麦(WP)蛋白質由来蛋白質複合体のメタ位置基質の基質特異性)
【表2】
【0119】
以上の結果から、豌豆(PP)、大豆(SP)、小麦(WP)由来蛋白質由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質であるメタ位置換塩素(Cl−/2a)、臭素(Br−/3a)、フッ素(F−/4a)、メチル(Me−/5a)、メトキシ(MeO−/6a)、ニトロ(NO2−/7a)のラセミ−フェニルエタノール類のR体を選択的酸化し、S体アルコールが化学収率50%程度、光学純度99%e.e.以上で得た。
【0120】
(第2の工程における豌豆(PP)、大豆(SP)、小麦(WP)蛋白質由来蛋白質複合体のパラ位置基質の基質特異性の検討)
【表3】
【0121】
以上の結果から、豌豆(PP)、大豆(SP)、小麦(WP)由来蛋白質由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質パラ位置換塩素(Cl−/2b)、臭素(Br−/3b)、フッ素(F−/4b)、メチル(Me−/5b)、メトキシ(MeO−/6b)、ニトロ(NO2−/7b)のラセミ−フェニルエタノール類のR体を選択的酸化し、S体アルコールが化学収率50%、光学純度99%e.e.以上で得られた。
【0122】
実施例3
(第2の工程におけるその他植物由来組織材料由来蛋白質複合体の基質特異性の検討)
第2の工程において、小麦若葉(YWL)、大麦若葉(YBL)、ヨモギの葉(AVI)、小麦ふすま(WB)、ワカメ(WS)、ニンジン(C)、カボチャ(P)といった草や雑草をも含めた種子、葉、茎、根、花、実からの不斉酸化反応を有する蛋白質複合体の溶出を確認する目的で、温水60℃以上で10分間漬け置き洗浄後、必要により薄くスライスして、凍結真空乾燥(FD)後、ボールミルを用いて細かく粉砕した。これらの植物材料粉末20gは定法に従って、10倍等量の蒸留水200mlを加えて、3%アルギン酸ナトリウム水溶液を1倍等量200ml加えて均一になるまで攪拌し、得られた植物組織含有・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中に滴下して、植物組織含有・アルギン酸カルシウムゲルビーズを作製する。該ビーズに、植物組織含有粉の20倍等量の蒸留水(400ml)を反応溶液として添加し、恒温振とう温度設定40℃、振とう数55rpmの条件にして、10時間以上の振とう後、基質変換反応を行った。基質としてはメタ、パラ位に臭素(Br−/1m、1p)、塩素(Cl−/2m、2p)、フッ素(F−/3m、3p)、メチル(Me−/4m、4p)、メトキシ(MeO−/5m、5p)、2−ナフチルエタノール(naphthyl/6)を置換させたフェニルエタノール類を作用させて、得られた結果については、反応時間、生成物、光学純度、化学収率を以下の表にまとめた。
【0123】
(第2の工程における小麦若葉、大麦若葉、小麦ふすま由来蛋白質複合体のメタ位置換基質の基質特異性の検討)
【表4】
【0124】
以上の結果から、小麦若葉(YWL)、大麦若葉(YBL)、小麦ふすま(WB)由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質であるメタ位置換臭素(Br−/1m)、塩素(Cl−/2m)、フッ素(F−/3m)、メチル(Me−/4m)、メトキシ(MeO−/5m)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のR体選択的に酸化して、S体アルコールが化学収率50%、光学純度99%e.e.以上で得た。
【0125】
(第2の工程における小麦若葉、大麦若葉、小麦ふすま由来蛋白質複合体のパラ位置換基質の基質特異性の検討)
【表5】
【0126】
以上の結果から、小麦若葉(YWL)、大麦若葉(YBL)、小麦ふすま(WB)由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質であるパラ位置換臭素(Br−/1p)、塩素(Cl−/2p)、フッ素(F−/3p)、メチル(Me−/4p)、メトキシ(MeO−/5p)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のR体選択的に酸化して、S体アルコールが化学収率50%程度、光学純度99%e.e.以上で得た。
【0127】
(第2の工程におけるヨモギの葉、ワカメ、ニンジン、カボチャ由来蛋白質複合体の基質特異性の検討)
【表6】
【0128】
以上の結果から、ヨモギの葉(AVI)、ワカメ(WS)、ニンジン(C)、カボチャ(P)由来の蛋白質複合体ゲルより好適に溶出され、同時に、基質であるパラ位置換臭素(Br−/1p)、塩素(Cl−/2p)、フッ素(F−/3p)、メチル(Me−/4p)、メトキシ(MeO−/5p)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のR体選択的な酸化により、S体アルコールが化学収率20〜50%、光学純度99%e.e.以上で得た。
【0129】
実施例4
(第2の工程における卵白アルブミン由来蛋白質複合体のパラ位置置換基質の基質特異性の検討)
第2の工程において、卵白から分離されたオボアルブミンから作成される蛋白質複合体の活性測定を目的として、鶏卵より卵白を分離し、硫安沈殿にてオボアルブミンを分離し、0.1〜0.5%のオボアルブミンの濃度になるように水に溶解後噴霧乾燥処理して、粉末の卵白アルブミン粉末を調整した。これらの卵白アルブミン粉末20gは定法に従って、10倍等量の蒸留水200mlを加えて、3%アルギン酸ナトリウム水溶液を1倍等量200mlを加えて均一にし、得られた卵白アルブミン・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中に滴下して、卵白アルブミン・アルギン酸カルシウムゲルビーズを作製した。該ビーズに、卵白アルブミン粉の20倍等量の蒸留水(400ml)を反応溶液として添加し、恒温振とう培養設定40℃、振とう数55rpmにて、10時間以上振とう後、基質変換反応を行った。基質としてはパラ位に臭素(Br−/1)、塩素(Cl−/2)、メトキシ(MeO−/5)、2−ナフチルエタノール(naphthyl/6)を置換させたラセミ−フェニルエタノール類を作用させて、得られた反応時間、生成物、光学純度、化学収率の結果については以下の表7にまとめた。
【0130】
(第2の工程における卵白アルブミン由来蛋白質複合体のパラ位置換基質の位置特異性の検討)
【表7】
【0131】
以上の結果から、卵白アルブミン由来の蛋白質複合体は、ゲルより好適に溶出され、同時に、基質パラ位置換臭素(Br−/1)、塩素(Cl−/2)、メトキシ(MeO−/5)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のS体選択的に酸化して、R体アルコールが化学収率26%、光学純度85〜95%e.e.で得られた。この結果は、植物資源由来粗蛋白質由来の蛋白質複合体がS体アルコール(99%e.e.)を得るのに対し、卵白アルブミン由来が逆のR体アルコール(85〜95%e.e.)が得られ、動植物の使い分けにより両鏡像体の使い分けが可能となる。
【0132】
実施例5
(第2の工程、恒温振とう器中で10時間の空振とうする理由)
図1は第2の工程において、豌豆蛋白4gを40mlの蒸留水に溶かして、更に3%アルギン酸ナトリウム(40ml)を加えて均一化した溶液を、4.0%の塩化カルシウム水溶液中に滴下して調整された、豌豆蛋白・アルギン酸カルシウムゲルビーズに、120mlの蒸留水を加えて、恒温振とう器設定温度40℃、振とう数55rpmにて振とうさせて、空振とう開始後0時間後、5時間後、10時間後、15時間後、20時間後、25時間後、35時間後、45時間後、そして55時間後に基質R−2−ナフチルエタノールを加えた時の、その豌豆蛋白質由来蛋白質複合体の不斉酸化活性に伴う基質残量%を示している。
【0133】
その結果、豌豆蛋白質由来蛋白質複合体の不斉酸化反応は、空振とう後10時間よりゲルより溶出され、同時に、基質R−2−ナフチルエタノールに好適に作用することが判った。
【0134】
実施例6a
(第2の工程、基質特異性の検討)
図2aは、図1と同一条件で振とう開始後10時間後、a)は基質特異性を調べる目的で、基質ラセミー2−ナフチルエタノール、R−2−ナフチルエタノール、2−アセトナフトンをそれぞれ50mgづつ添加したときの反応時間に伴う基質と生成物の%比率をモニターリングしたものである。その結果、空振とう10時間後に現れる活性は、S体アルコールは酸化せず、基質ラセミー2−ナフチルエタノールのR体のみを立体選択的に不斉酸化することが判った。
【0135】
実施例6b
(第2の工程、反応箇所の検討)
図2bは第2の工程において、図1と同一条件で豌豆・アルギン酸ナトリウムを調製後、空振とう10時間後の水溶液(120ml)、空振とう10時間後の水溶液(240ml)、空振とう10時間後ビーズ+蒸留水(DW:120ml)、新しいビーズ+空振とう10時間後の水溶液(120ml)でそれぞれ基質R−2−ナフチルエタノール(50mg、75mg、又は100mg)を加え、不斉酸化活性をモニタリングした図である。
その結果、空振とう10時間後に現れる蛋白質複合体は、ビーズ中ではなく、水溶液中に反応の場があり、好適に2−ナフチルエタノールのR体のみに作用することが判った。
【0136】
実施例6c
(第2の工程、包括ゲル化塩化カルシウム濃度の活性に及ぼす効果)
図2cは第2の工程において、図1と同一条件で豌豆・アルギン酸ナトリウム混液を調整後、異なる塩化カルシウム濃度(5g/L、7.5g/L、10g/L、15g/L、20g/L、30g/L)中に滴下してゲル化して、同じ条件にて10時間空振とう後、基質R−2−ナフチルエタノールを加えて、その塩化カルシウム濃度に及ぼす蛋白質複合体の活性強度の差をまとめた図である。
【0137】
その結果、塩化カルシウム濃度が低いほど、蛋白質複合体の作成と溶出が効率的であって、不斉酸化活性は高まることが判った。逆に、塩化カルシウム濃度が高いほど、蛋白質複合体が溶出し難い傾向を確認できた。一方、塩化カルシウム濃度10g/L、15g/L、20g/Lでは殆ど活性に差は見られなかった。
【0138】
実施例6d
(第2の工程、豌豆蛋白量の最適化)
図2dは第2の工程における、豌豆アルギン酸ゲルを調製時の豌豆蛋白量(2g、3g、4g、5g)の活性に及ぼす影響について調べた。この場合、塩化カルシウム濃度10g/Lで、豌豆・アルギン酸カルシウムゲルを調製し、空振とう10時間後に得られる蛋白質複合体が及ぼす、基質R−2−ナフチルエタノール(50mg)への影響を示した図である。
【0139】
その結果、好適な蛋白質複合体作成と抽出の条件は、豌豆蛋白量が4gであるが、2g〜5gでは何れも活性に与える蛋白質複合体に差は殆ど見られなかった。
【0140】
実施例7
(第2の工程、香料であるR−1−オクテン−3−オールの合成検討)
図8は第2の工程において、図1と同一条件で豌豆・アルギン酸カルシウムゲルに包括化し、空振とう10時間後に得られる蛋白質複合体の、基質ラセミ−1−オクテン−3−オールの分割に関わる効果を調べた図である。
【0141】
この結果から、蛋白質複合体の触媒作用は、生成ケトンのクロマトピークが2%以下であることと、化学収率が85%以上であることから、図8に示すR−1−オクテン−3−オール(98.2%ee)のGCクロマトは、デラセミ化により得られることが判った(後述する実施例12と表11では連続再利用について検討している)。
【0142】
実施例8
(第3の工程の蛋白質複合体の抽出、好適な硫安濃度の検討)
実施例1の条件で得られる蛋白質複合体溶液を、50mLプラスチック遠心管中に40ml移して、1)各8.8g硫安を加え飽和30%として20時間以上静置後、2)遠心分離(10.000rpm、15min)して、硫安溶液をデカンテーション除去後、3)50mMTris塩酸緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール3ppmを10ml加えて不斉酸化活性を測定した。4)デカンテーション後の溶液画分は更に、3.1g硫安を加え飽和40%にして20時間以上静置後、遠心分離して得られる沈殿物は、その有する不斉酸化活性を同様な方法で検査した。デカンテーション後の溶液画分は更に、3.15g硫安を加え、飽和50%にして静置後、遠心沈殿して同様にその酸化活性を検査した。デカンテーション後の溶液画分は更に、3.3g硫安を加え、飽和60%にして遠心分離して静置後、同様に酸化活性を検査した。デカンテーション後の溶液画分は更に、3.45g硫安を加え、飽和70%にして遠心分離して静置後、同様に酸化活性を調べた。その結果を以下に示す。
【0143】
表8(反応30時間後の結果)
【表8】
【0144】
以上の結果から第3の行程の蛋白質複合体の結晶化の最適な硫安濃度は飽和30%であると判定した。得られる蛋白質複合体が非水溶性グルテン様蛋白質であり、カルシウム存在下で水溶性に変化している性質なので、飽和30%の弱水溶性条件にて沈殿できると推察できた。
【0145】
実施例9
(第3の工程の結晶化蛋白質複合体が処理する基質濃度)
実施例1の条件で得られる結晶化蛋白質複合体水溶液を、50mLプラスチック遠心管に移して、1)各8.8g硫安を加え飽和30%として静置し、2)遠心分離(10.000rpm、15min)してデカンテーション後、沈殿して得られる結晶化蛋白質複合体に、3)50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−2−ナフチルエタノール濃度溶液(1ppm、2ppm、3ppm、4ppm、5ppm)分を各10mL加えて調べた。
【0146】
表9(反応30時間後の結果)
【表9】
【0147】
以上の結果から、第3の工程で得られる水分を含む結晶化蛋白質複合体約50mg/50ml遠心管が酸化できる基質ラセミ−2−ナフチルエタノール濃度は1ppm〜5ppm/10mlで顕著な差は無く不斉酸化できることを確認した。
【0148】
実施例10
(第4の工程の結晶化蛋白質複合体の抽出、及びグルタルアルデヒド架橋濃度0.1%と1.0%の検討)
実施例1の条件で得られる蛋白質複合体溶液を、50mLプラスチック遠心管中に40ml移し取り、1)各8.8g硫安を加え飽和30%として20時間以上静置後、2)25%グルタルアルデヒド溶液をそれぞれ0.1%(0.32ml/40ml)、1.0%(1.6ml/40ml)になるように加えて10時間以上静置後、3)遠心分離(10.000rpm、15min)して、上清の硫安・グルタルアルデヒド溶液をデカンテーション除去した。沈殿の蛋白質複合体は、50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−2−ナフチルエタノール濃度3ppm分を10ml加えて不斉酸化の活性を確認した。
【0149】
表10(反応30時間後と60時間後の結果)
【表10】
【0150】
以上の結果から第4の工程の結晶化蛋白質複合体の架橋のGA濃度は0.1%と1.0%で何れも酸化活性を示すが、1.0%に比べて0.1%のほうがR-アルコール不斉酸化活性がより好適に進んだ。更に、0.1%に近い濃度ほど、デラセミ化反応が起こり易い傾向が確認できた。
【0151】
実施例11
(第4の工程の蛋白質複合体粉末に関わる連続再利用の検討)
実施例1の条件で得られる蛋白質複合体溶液40mlを、50mLプラスチック遠心管4本分にそれぞれ移し、1)各8.8g硫安をそれぞれ加えて飽和30%とし、一昼夜静置後、2)25%グルタルアルデヒド溶液をそれぞれ0.1%になるように0.32ml/40ml加え、2〜4時間静置した。上清の硫安・グルタルアルデヒド溶液はそれぞれ、3)遠心分離(10.000rpm、15min)してデカンテーション除去して、目的の沈殿物蛋白質複合体は更に、凍結真空乾燥(FD)後に粉砕した。蛋白質複合体粉4)358mg分を3ppmラセミ−2−ナフチルエタノール・50mMTrisHCl緩衝液(pH8.0、40mL)に溶かし、活性を測定した。連続再利用は、9時間後と15時間後に行い、遠心分離後に沈殿物を回収して、新たに3ppmラセミ−2−ナフチルエタノール・50mMTrisHCl緩衝液(pH8.0、40mL)を加え、この繰り返し作業にて検討した。その結果は次の通りである。
【0152】
表11(蛋白質複合体の連続再利用の結果)
【表11】
【0153】
以上の結果から第4の工程の蛋白質複合体粉末に関わる連続再利用の可能性について確認した結果、最低16回(8日間)は、不斉酸化活性を維持できて再利用できることが判った。
【0154】
実施例12
(第4の工程の蛋白質複合体粉末の連続再利用に関わる香料(R-1−オクテンー3−オール)の合成)
実施例11で得られる蛋白質複合体の凍結真空乾燥(FD)粉末5gに、ラセミ-1-オクテン-3-オール濃度10ppmの50mMTrisHCl(pH8.0)緩衝液150mlを加えて、500ml三角フラスコ中で不斉酸化反応を試みた。30時間後又は60時間後に5mlの反応溶液を遠心分離(10000rpm、15min)により上清のみを回収し、ジエチルエーテルで基質を抽出後、GC測定を行った。尚、連続再利用の操作は、実施例11と同様な方法にて光学純度と収率を導いた。反応は60時間で止め、遠心分離した上清部をジエチルエーテルにて抽出後、飽和塩水にて洗浄、シリカゲルクロマトグラフィーにて単離・精製を行って化学収率と光学純度を求めた。
【0155】
表12(香料(R−1−オクテンー3−オール)の連続再利用による合成)
【表12】
【0156】
以上の結果から、基質ラセミ−1−オクテン−3−オールの連続的合成に関しては、最低4回は再利用でき、生成ケトンは何れの場合も4%未満で且つ、化学収率が85%以上であることから、デラセミ化反応が生じていると判定できた。得られた光学純度は4回とも98%ee以上の高光学純度でえられ、図5は得られたR−1−オクテン−3−オールのGC/FIDスペクトルの結果を示している。
これまでの結果から、下記の反応式に示すように、ベンゼンやナフタレン骨格を有するアリール系基質に関しては50%の生成ケトンは還元され難いが、1−オクテン−3−オールなどのアルキル鎖系基質に関しては生成ケトンが更に不斉還元されるデラセミ化が生じることが判った。
【0157】
【化1】
【0158】
実施例13
(蛋白質複合体の分子量決定(SDS−page))
図3は豌豆−Caアルギン酸ゲルを10時間以上温水中にて振とう後、溶出される蛋白質複合体を硫安30%にて結晶化沈殿して得られる沈殿物のSDS−pageである。図に示す「酵素溶液」レーンは、10時間以上温水振とう後、溶出される温水溶液そのものであり、「硫安沈殿」レーンは、該温水溶液を硫安30%飽和になるように結晶化沈殿し、該沈殿物に水を加えて再溶解させたものであり、「上清」レーンは、該硫安沈殿物を遠心分離して得られる上清に示すSDS−page結果である。
【0159】
図4は「食品と技術」2008年12月号総説 裏出令子記述(グルテンタンパク質のネットワーク形成における食塩の役割)に掲載済みの「図5(DSTによる架橋効率に食塩添加が及ぼす影響)」の抜粋である。図は、小麦粉に食塩添加又は無添加の水溶液を加えて作成した生地を、βメルカプトエタノール入りSDS溶液で可溶化して得られる液を超遠心し、可溶性画分(S)と不溶性画分(P)についてSDS−pageを行った結果を表す。不溶性画分はメタ過ヨウ素酸ナトリウムで処理し、SDSポリアクリルアミドゲル電気泳動で分離しタンパク質を染色してある。
【0160】
以上、図4について、図3のSDSにより切られたS-S結合蛋白成分のフラグメントのバンド配置の持つ分子量が図3に示すSDS-pageと一致していることから、蛋白質複合体は、酵素ではなく、高分子・低分子グルテニンとグリアジンに似た成分より成ると判定できた。グルテニンは空気酸化(及び/又は内因性酵素)の働きにより、高分子・低分子グルテニン分子間にジスルフィド結合を生じてポリマー化するが、このチオール−ジチオール部位が酸化還元反応の活性ドメインとなり、基質ラセミアルコールの不斉酸化を発現するようになると推察できる。溶存カルシウムは、分子間ジスルフィド結合形成を誘導するだけでなく、高分子・低分子グルテニン分子間凝集(6.4Å以下)、更には、水溶性への性質変化にも関与していると考えられる。
【0161】
実施例14
図5は、フーリエ変換赤外分光分析(FT−IR)を用いるサンプル(i)〜(vi)の定性分析結果である。各サンプル間の分子構造上の違いを比較する目的で行った。尚、サンプル(i)〜(vi)のサンプル情報は以下の通りである。
(1)エンドウ蛋白質粉末・・・前述の実施例1に記載の方法にて得られる。
【0162】
(2)アルギン酸ナトリウム粉末・・・市販品より得られる。
(3)振とう培養器製の架橋結晶化蛋白質複合体粉末・・・後述の実施例15に記載の方法にて得られる。
【0163】
(4)ジャーファメンタ製の架橋結晶化蛋白質複合体粉末(iv)・・・後述の実施例16に記載の方法にて得られる。
【0164】
(5)20mM塩化Ca/50mMトリス塩酸バッファー(pH6.0)及び(pH8.0)・・・実施例17に記載の方法にて得られる。
【0165】
図5の結果、不斉酸化反応を有する「サンプル(iii)(iV)」と不斉酸化反応が起こらない「サンプル(v)(vi)」のフーリエ変換赤外分光分析(FT−IR)上での違いは、1)1411cm−1にあるカルボキシレート(R−C(=O)−O−Ca)の水酸基(−O−Ca)の吸収と、2)1082cm−1の糖エーテル(−C−O−C−)又は硫安(−O−S(=O)2−O−)の吸収の2点にある。不斉酸化反応を有する「サンプル(iii)(iV)」は供にこの2点に強い吸収が見られることから、1)サンプル内にアルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り込んだのか、2)分子間ジスルフィド結合(−S−S−)の際に硫黄酸化物を生じたとすれば説明できる。また、この2点が「不斉酸化反応を有する」か否かの重要なポイントとなる部分であると考えられ、アルギン酸Caに粗蛋白質を包括する意義が存在している。
【0166】
更に、図6は、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。尚、サンプル(i)〜(iV)のサンプル情報は既に説明済みである。
【0167】
図6のX線マイクロアナライザーの結果から、不斉酸化反応を有する「サンプル(i)」には、ナトリウム(Na)が検出されているが、不斉酸化反応を有しない「サンプル(ii)(iii)」には見られていない。従って、アルギン酸ナトリウムを4%塩化カルシウム溶液中に滴下して、アルギン酸カルシウムに包括する第2の工程において、エンドウ蛋白質粉末(iv)由来のナトリウム(Na)が作用を受けずに残ったか、或いはナトリウム(Na)原子がカルシウム(Ca)と置換されなかったカルボキシレート(R−C(=O)−O−Na)のアルギン酸Na分子の残骸/足跡とも推測できる。また、(i)〜(iv)の硫黄(S)の量が(i)の場合に顕著に多いことから、分子間ジスルフィド結合などの形成を経て得られた硫黄酸化物(−O−S(=O)−O−)が相対的に不斉酸化反応の要因になっている可能性が示されている。
【0168】
従って、後の実施例19に記載する酸素(O2)の吸収が、エンドウ蛋白質内システイン(R1−S−H H−S−R2)通しの分子間ジスルフィド結合(R1−S−S−R2)の形成に関与するものであったと推察できた。また、(iv)のナトリウム(Na)原子がアルギン酸Na分子の残骸/足跡である場合、1)サンプル内にアルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り組んでいる可能性も示唆された。
【0169】
実施例15
(図5(iii)及び図6(i)に記載の振とう培養器製による蛋白質複合体の製造法)
第2の工程において、種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し水溶性画分を噴霧乾燥して得られる豌豆(PP)由来蛋白粉末30gに、10倍等量の蒸留水300mlを加えて十分に攪拌・溶解後、3%のアルギン酸ナトリウム水溶液を1倍等量の300mlを加えて均一になるまで攪拌し、得られる豌豆・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中(pH9.16)に滴下してゲルビーズを作製した。該作製ビーズに豌豆蛋白粉の20倍等量の蒸留水(500ml)を反応溶液として添加し、恒温振とう培養器の温度設定40℃、振とう数55rpmにて10時間以上の振とうした。
使用装置
(1)反応槽:恒温振とう培養器3器中にそれぞれ5000ml三角フラスコを設置。
振とう数:55rpm
温度設定:40℃
【0170】
攪拌(培養)作業の条件
1.豌豆蛋白質(30g)・アルギン酸カルシウムゲル(約600mL)
2.蒸留水:500mL×3(器)×3(日分)
3.抽出回数:3回/3日(1日に1回づつ回収)
【0171】
抽出作業の条件
1.一日分の培養水溶液500mL×3(器分)をビーズと分離、回収した。
2.硫安沈殿・・・30%飽和硫安になるように工業用硫安を添加、結晶化沈殿の為に1日置き。
3.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加し、2時間以上置き。
4.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
5.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
6.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール濃度10ppmを40mL加えて酸化活性を検討した。
7.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA−300)にて測定した。
【0172】
以上の結果、表12のように、恒温振とう培養器による架橋結晶化蛋白質複合体の製造法の一例をまとめた表であるが、恒温振とう培養器にそれぞれ、アルミフォイルなどで簡易的に封をした5000ml三角フラスコであるので、振とう数55rpmで十分に培養液が波打つので酸素補給が十分に行われ、グルテニン−グルテニンシステイン残基(R1−S−H・・・O・・・H−S−R2)脱水縮合反応(→H2O)を介するジスルフィド結合((−S−S−))によりポリマー化を誘導し、アルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り込んだ。この蛋白質複合体に形成された活性ドメインが、酵素様にRアルコールを選択的に不斉酸化して、100%e.e.のS−アルコールを得ることができたと考えられる。
【0173】
表13(図5(iii)及び図6(i)に記載の振とう培養器製による蛋白質複合体の活性強度)
【表13】
【0174】
注)・抽出液中Ca濃度(mg/mL)・・・振とう溶液のCa濃度を原子吸光高度計にて測定した。
・Ca濃度(mg/g)・・・FD後の粉末サンプルCa量を酸分解処理後、原子吸光光度計にて測定した。
・R体酸化(%)・・・図9記載のスケールに反応させて7hr後、20hr後のGC測定による生成ケトン(%)を記載した。
(基質ラセミアルコールのR体50%が選択的に酸化されてケトンになる為、ケトン%が50%に近いかそれ以上の場合、もう一方のS体アルコールの光学純度は100%近い。)
・光学純度(%e.e.)・・・GCの「S−アルコール(面積%)−R−アルコール(面積%)」より求める。
【0175】
実施例16(図5(iv)に記載の5Lジャーファメンタ製による架橋結晶化蛋白質複合体の製造法)
第2の工程において、種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し水溶性画分を噴霧乾燥して得られる豌豆(PP)由来蛋白粉末50gに、10倍等量の蒸留水500mlを加えて十分に攪拌・溶解後、3%のアルギン酸ナトリウム水溶液を1倍等量の500mlを加えて均一になるまで攪拌し、得られる豌豆・アルギン酸ナトリウム混合液(1000mL分)を4.0%の塩化カルシウム水溶液中(pH9.16)に滴下してゲルビーズを作製した。該作製ビーズに豌豆蛋白粉の20倍等量の蒸留水(1000mL分)を反応溶液として添加し、高崎科学製5L用ジャーファメンタの温度設定40℃、振とう数800rpmにて振とうした。
【0176】
使用装置
(1)反応槽:高崎科学製5L用ジャーファメンタ
羽根:2枚簡易式棒状パドル 800rpm
温度:40℃
酸素供給:0.5mg/L
攪拌(培養)作業の条件
4.豌豆蛋白質(30g)・アルギン酸カルシウムゲル(約1000mL)
5.蒸留水:1000mL×3(日分)
6.抽出回数:3回/3日(1日に1回づつ回収)
【0177】
抽出作業の条件・・・一日分の培養水溶液1000mLをビーズと分離、回収した。
1.硫安沈殿・・・30%飽和硫安になるように工業用硫安を添加、結晶化沈殿の為に1日置き。
2.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加し、2時間以上置き。
3.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
4.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
5.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール濃度10ppmを40mL加えて酸化活性を検討した。
6.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA−300)にて測定した。
【0178】
以上の結果、表13のように、5L用ジャーファメンタにおいても、恒温振とう培養機を用いた場合と同様、目的のアルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り込み、エンドウ蛋白質内に活性ドメインを作成し、蛋白質複合体が、酵素様にRアルコールを選択的に不斉酸化して、100%e.e.のS−アルコールを得ることができたと考えられた。
【0179】
表14(図5(iv)に記載の5L用ジャーファメンタによる蛋白質複合体の活性強度)
【表14】
【0180】
注)・抽出液中Ca濃度(mg/mL)・・・振とう溶液のCa濃度を原子吸光高度計にて測定した。
・Ca濃度(mg/g)・・・FD後の粉末サンプルCa量を酸分解処理後、原子吸光光度計にて測定した。
・R体酸化(%)・・・図9記載のスケールに反応させて7hr後、20hr後のGC測定による生成ケトン(%)を記載した。
(基質ラセミアルコールのR体50%が選択的に酸化されてケトンになる為、ケトン%が50%に近いかそれ以上の場合、もう一方のS体アルコールの光学純度は100%近い。)
・光学純度(%e.e.)・・・GCの「S−アルコール(面積%)− R−アルコール(面積%)」より求める。
【0181】
実施例17(図5(v)(vi)及び図6(ii)(iii)に記載の「架橋結晶化蛋白質(pH6.0又はpH8.0)」の製造法)
アルギン酸カルシウムゲルに包括する第2の工程の必要性を確認する目的で、第2の工程(アルギン酸カルシウムゲルへ包括)を省き、エンドウ粗蛋白質(4g)を2%塩化カルシウム/50mMトリス塩酸バッファー溶液/120ml((v)pH6.0又は(vi)pH8.0)で恒温振とう(55rpm、40℃、24時間)した。振とう後、エンドウ蛋白質カルシウム溶液は、30%飽和になるように硫安を加えて24時間以上静置後(第3の工程)、結晶化沈殿物を0.25%グルタルアルデヒドにて架橋し(第4の工程)、得られる架橋結晶化したエンドウ粗蛋白質を凍結乾燥(FD)処理して粉末とした。
【0182】
(2)反応槽:恒温振とう培養器にそれぞれ300ml三角フラスコを設置。
振とう数:55rpm
温度設定:40℃
【0183】
攪拌(培養)作業の条件
7.豌豆蛋白質(4g)
8.2%塩化カルシウム/50mMトリス塩酸バッファー溶液(120m)
9.抽出回数:1回/1日
【0184】
抽出作業の条件
1.硫安沈殿・・・恒温振とう(55rpm、40℃、24時間)後、30%飽和になるように硫安を加えた。
2.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加して2時間以上置き。
3.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
4.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
5.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール濃度10ppmを40mL加えて酸化活性を検討した。
6.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA-300)にて測定した。
【0185】
以上、表15に示すように、蛋白質複合体の作成には、グルテニン−グルテニンジスルフィド結合((−S−S−))を誘導する溶存Ca濃度と溶存酸素に加えて、「アルギン酸カルシウムゲル」に包括する第2の工程が不可欠でありことが確認できた。
【0186】
表15.(図5(v)(vi)及び図6(ii)(iii)に記載の「蛋白質複合体(pH6.0又はpH8.0)」の活性強度)
【表15】
【0187】
注)・Ca濃度(mg/g)・・・FD後の粉末サンプルCa量を酸分解処理後、原子吸光光度計にて測定した。
・R体酸化(%)・・・7hr後、20hr後のGC測定による生成ケトン(%)を記載した。
・不斉酸化活性
×・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間に殆ど不斉酸化されない場合(生成ケトン10%以下)。
△・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間までに不斉酸化が途中で止まる場合(生成ケトン30%以下)。
○・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間で不斉酸化される場合(生成ケトン40%以上)。
【0188】
実施例18
(第2の工程のアルギン酸カルシウム包括ビーズの空気酸化の時間に及ぼす6時間後の不斉酸化活性(右軸)と蛋白質複合体収量への影響)
図7は第2の工程において、定法に従い球状のエンドウ蛋白・アルギン酸カルシウムビーズを作製後、該ビーズを空気中に0時間、0.5時間、1時間、3時間、5時間、7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−2−ナフチルエタノール溶液/50mMTrisHCl緩衝液(pH8.0、40mL)を加えて、恒温振とう培養(40℃、55rpm)6時間後に酸化活性を示す。
【0189】
以上の結果から、空気酸化時間を長く取るに従い、架橋結晶化蛋白質複合体の不斉酸化活性は強くなった。一方、得られる蛋白質複合体の収量は一時間後と三時間後に高かった。従って、蛋白質複合体は、空気酸化(及び/又は内因性酵素)の影響により、ゲルビーズ内カルシウム塩と酸素環境下、安価な材料の動植物由来粗蛋白質に(i)タンパク質内分子間ジスルフィド結合(−S−S−)、(ii)蛋白質内分子間凝集(分子間距離の短縮:<6.4Å)、(iii)水溶性への性質変化などを誘導して活性ドメイン(チオレドキシンフォールド:Cys−X−Y−Cys配列)を形成し、好適に不斉酸化反応を有する蛋白質複合体を形成することが確認できた。
【0190】
実施例19
(第2の工程の蛋白質複合体の100Lジャー生産におけるpHとDO(溶存酸素濃度)の推移)
図9は第2の工程において、種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し水溶性画分を噴霧乾燥して得られる豌豆蛋白粉末1kgは定法に従って、10倍等量の蒸留水10Lを加えて攪拌混合後、3%のアルギン酸ナトリウム水溶液を1倍等量の10Lを加えて、均一になるまで攪拌して得られた豌豆・アルギン酸ナトリウム混合液(約20L)を、ゲル調整装置を用いて4.0%の塩化カルシウム水溶液中に滴下して、直径1〜2mm程度の球状のビーズを作製し、架橋結晶化蛋白質複合体を作成する際に加える蒸留水(20L)のpHとDO(溶存酸素)をモニターリングした図である。
【0191】
使用装置
(2)反応槽:400φ×700H(TL−TL)竪型円筒槽
羽根:4枚傾斜パドル 200φ 35rpm
温度:40℃
(3)晶析槽
420φ×460H 竪型円筒槽
羽根:6枚羽根タービン 120φ
A.攪拌(培養)作業の条件
1.豌豆蛋白質(1kg)・アルギン酸カルシウムゲル(約20L)
2.蒸留水: 1回目−20Lで3日間攪拌(酸素供給なし)後第3.4工程
2回目−20Lで2日間攪拌(酸素供給なし)後第3.4工程
3回目−20Lで1日間攪拌(酸素供給あり)後第3.4工程
4回目−20Lで1日間攪拌(酸素供給あり)後第3.4工程
【0192】
B.蛋白質複合体抽出作業の条件
1.各攪拌後の培養水溶液20L分をビーズと分離、回収した。
2.硫安沈殿・・・30%飽和硫安になるように工業用硫安を添加、結晶化沈殿の為に1日置き。
3.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加し、2時間以上置き。
4.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
5.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
6.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−2−ナフチルエタノール濃度10ppmを40mL加えて酸化させて、R体が酸化して得られるケトン%を表している。
7.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA-300)にて測定した。
【0193】
図9のように、4.0%塩化カルシウム水溶液(pH9.16)に滴下して作成したビーズは、酸素供給を行わず蒸留水中で攪拌した場合、pHが5.72(1回目)、6.20(2回目)と弱酸性の中で推移し、溶存酸素(DO)は0.7時間後(1回目)、1時間後(2回目)にゼロとなった。また、酸素供給(0.5mg/L制御)を行ない攪拌すると、pHは弱酸性から7.20(3回目)と弱アルカリ条件側に転じ、4回目は常に弱アルカリのpH7.45付近を推移し、DOは3・4回目ともに、0〜1mg/L付近を推移するようになる。
【0194】
以上の結果、酸素供給のない1・2回目は、高い溶存Caイオン濃度が存在している一方で、溶存酸素濃度がゼロ(酸欠)の状態における蛋白質複合体の形成、即ち、グルテニン−グルテニン分子間ジスルフィド結合と分子間距離短縮の効果に伴う活性ドメイン形成の成否を検討した結果であるが活性は認められなかった。更に、引き続いて3・4回目は、新しい水20Lの切り替え3・4回目であり、低い溶存Caイオン濃度の存在下、空気供給による一定の溶存酸素濃度を維持させた場合における蛋白質複合体の形成、即ち、グルテニン−グルテニン分子間ジスルフィド結合と分子間距離短縮の効果に伴う活性ドメイン形成の成否を検討した結果であるが、この場合も弱い酸化活性は認められる一方で、生成アルコールの光学純度は50%eeを超えるに至らなかった。従って、溶存Caイオン濃度と一定の溶存酸素濃度の維持が、グルテニン−グルテニン分子間ジスルフィド結合と分子間距離短縮の効果により生じる活性ドメインの形成において、重要なポイントとなることが確認できた。
【0195】
(第2の工程の3%アルギン酸カルシウム包括ビーズの空気酸化の時間に及ぼす6時間後の不斉酸化活性(右軸)と蛋白質複合体収量(左軸)への影響)
図7は第2の工程において、定法に従い球状のエンドウ蛋白・アルギン酸カルシウムビーズを作成後、該ビーズを空気中に0時間、0.5時間、1時間、3時間、5時間、7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−ナフチルエタノール溶液/50mMtris HCl緩衝液(pH8.0、40ml)を加えて、恒温振とう培養(40℃、55rpm)6時間後に示す酸化活性である。
【0196】
実施例18a
(第2の工程の3%アルギン酸カルシウム包括ビーズの空気酸化の時間に及ぼす6時間後の不斉酸化活性(右軸)と蛋白質複合体収量(左軸)への影響)
図7は第2の工程において、定法に従い球状のエンドウ蛋白・アルギン酸カルシウムビーズを作成後、該ビーズを空気中に0時間、0.5時間、1時間、3時間、5時間、7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−2−ナフチルエタノール溶液/50mMtris HCl緩衝液(pH8.0、40ml)を加えて、恒温振とう培養(40℃、55rpm)6時間後に示す酸化活性である。
【0197】
以上の結果から、空気酸化時間を長く取るに従い、架橋結晶化蛋白質複合体の不斉酸化活性は強くなった。一方、得られる蛋白質複合体の収量は1時間以降一定して11%〜15%程度の間を推移するが、12%〜13%の収率に集中した。従って、蛋白質複合体は、空気酸化(及び/又は内因性酵素)の影響によりゲルビーズ内カルシウム塩と酸素環境下、安価な材料の動植物由来粗蛋白質に(i)タンパク質内分子間ジスルフィド結合(−S−S−)、(ii)蛋白質内分子間凝集(<6.4Å)、(iii)水溶性への性質変化などを誘導して活性ドメイン(チオレドキシンフォールド:Cys−X−Y−Cys配列)を形成し、好適に不斉酸化反応を有する蛋白質複合体を形成することが確認できた。
【0198】
実施例20
(第2の工程のアルギン酸Na濃度(0.5%、1%、1.5%、2%、3%)に及ぼすアルギン酸カルシウム包括ビーズ空気酸化7時間後の不斉酸化活性6時間後(右軸)と蛋白質複合体収量への影響)
図13は第2の工程において、定法に従い球状又は棒状のエンドウ蛋白・アルギン酸カルシウムビーズ(アルギン酸Na濃度(0.5%、1%、1.5%、2%、3%))を作成後、該ビーズを空気中に7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−2−ナフチルエタノール溶液/50mMtris HCl緩衝液(pH8.0、40ml)を加えて、恒温振とう培養(40℃、55rpm)6時間後に示す酸化活性である。
【0199】
以上の結果から、得られる蛋白質複合体の収量は、アルギン酸Na濃度が1.5%の時が最も高く、それより濃度を濃くしても薄くしても収量は徐々に減少した(0.5%<1%<1.5%>2%>3%)。一方、蛋白質複合体の持つ酸化活性は、アルギン酸Na濃度が高くなるに連れ若干上昇した(0.5%<1%<1.5%<2%<3%)。従って、アルギン酸Na濃度を1.5%付近に設定して作成した蛋白質・アルギン酸カルシウムゲルを5〜7時間又はそれ以上時間空気酸化させた後、定法に従い架橋結晶化した蛋白質複合体(CLPCs)を得ることで収量の向上が見込めることを明らかにした。
【0200】
実施例21
(基質濃度(10ppm)に対する蛋白質複合体(50mg、75mg、100mg、200mg、300mg)の不斉酸化速度の違い)
図14は、実施例20で得られる蛋白質複合体の凍結真空乾燥(FD)粉末(50mg、75mg、100mg、200mg、300mg)にラセミ−2−ナフチルエタノール濃度10ppmの50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、4時間置きにGCにて、不斉酸化されるR体−2−ナフチルエタノールの残留比(%)について追跡を行った結果を示している。
【0201】
以上の結果、基質濃度(10ppm)に対する不斉酸化反応に対し、触媒である蛋白質複合体の添加量を増やせば増やすほど反応速度は上昇している結果を得た(50mg>75mg>100mg>200mg>300mg)。また、この蛋白質複合体反応は従来のパン酵母や植物培養細胞、更には微生物菌体の持つ補酵素NAD(P)に依存する脱水素反応とは異なり、補酵素NAD(P)に非依存型の脱水素反応であるため、原料コストの低減化だけでなく、反応コストの低減化においても期待できる触媒であると考えられる。
【0202】
実施例21a
(蛋白質複合体(300mg)に対する基質濃度(10−30ppm、30−90ppm、90−110ppm、110−140ppm)の不斉酸化速度の違い)
図15は、実施例20で得られる蛋白質複合体の凍結真空乾燥(FD)粉末(300mg)にラセミ−2−ナフチルエタノール濃度(10−140ppm)の50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、不斉酸化されるR体−2−ナフチルエタノールの残留比(%)について4時間置きにGC追跡を行った結果を示している。
【0203】
以上の結果、基質濃度(10〜140ppm)の、触媒の蛋白質複合体(300mg)に対する不斉酸化反応は、図15に示す通り、10−30ppm、30−90ppm、90−110ppm、110−140ppmの、4つのパターンに分類された。即ち、基質濃度10−30ppmについては反応時間20時間、30−90ppmについては40時間、90−110ppmについては60時間、110−140ppmについては140時間を要すことが判った。従って、基質濃度は低いほど反応時間が早く、110ppm以上の濃度では、反応時間が極端に遅くなることが判った。
【産業上の利用可能性】
【0204】
動植物資源、例えば穀類や豆類としては、蕎麦、アマランサス、米、小麦、大麦、トウモロコシ、エンバク、ライ麦、粟、ヒエ、キビ、ハト麦、モロコシ等の穀類、又は小豆、インゲン豆、豌豆、リョクトウ、大豆等の豆類由来、更には草や雑草をも含めた、これら穀類や豆類の種(外皮(ふすま、ぬか)、胚芽(スプラウト))、葉(若葉、新芽)、茎、根、花などの各植物組織、更には、動物由来の卵白や筋肉のような各動物組織を凍結真空乾燥(FD)させて細かく粉砕するか、必要によりその温水に溶解して殻を取り除き、水溶性部分を噴霧乾燥して水溶性蛋白質を濃縮する第1の工程と、該粗蛋白質をアルギン酸カルシウムゲルに包括して空気酸化させた後、温水中にて空振とうしてゲルから目的の蛋白質複合体を抽出する第2の工程と、蛋白質複合体を30%飽和硫安にて結晶化し沈殿させる第3の工程と、蛋白質複合体を架橋する第4の工程と、必要により得られた蛋白質複合体を凍結真空乾燥(FD)して粉末にする第5の工程を組み合わせ、医薬品や香料や食品等のファインケミカル分野における合成する光学活性アルコールなどの光学異性体を製造安価で、環境に優しく、平易に生産することができる。
【技術分野】
【0001】
本発明は、蛋白質およびカルシウムを少なくとも含み、且つ不斉酸化反応を触媒する活性を有する蛋白質複合体、およびその製造方法に関する。この蛋白質複合体は、例えば、動植物組織から平易に分離できる水溶性粗蛋白質をゲル(アルギン酸カルシウムゲル等)に包括して該蛋白質を空気酸化させた後、分離することにより得ることができる。
【背景技術】
【0002】
医薬、食品分野を始めとする種々の分野において、光学異性体を分離し、有用な一方の光学異性体を分取する技術の必要性が高まっている。例えば、光学異性体の一方を選択的に酸化還元又はデラセミ化する機能を有する固定化酵素を用いて、医薬品や香料を生成させる方法について研究されている。
【0003】
生体内の化学変化は、通常は酵素で触媒される。このような酵素の作用・機構を有用物質生産や分析に利用するシステムの一つとして、バイオリアクターが挙げられる。
【0004】
バイオリアクターの中心となるのは、不溶性の担体に結合させた固定化酵素である。これによって,触媒となる酵素と生産物を容易に分離することができる。固定化酵素は,研究,医療,分析,産業に広く利用されている。化学反応を行わせるバイオリアクターの中枢部分は反応素子であり,原料を製品に変換したり化学変化を利用して分析に利用したりする目的で,精製した酵素,オルガネラあるいは細胞そのものが用いられている。反応素子はリアクター中に留まり、反復使用が可能なものでなければならないので,反応素子は種々の方法で固定化されている(このような酵素の活用の詳細に関しては、例えば、「化学増刊119 ハイブリッドプロセスによる有用物質生産」 化学同人 「バイオリアクター」福井三郎編、バイオテクノロジーシリーズ、講談社 「固定化酵素」千畑一郎編、講談社サイエンティフィック等を参照することができる)。
【0005】
しかしながら,酵素は精製するのにコストがかかり,精製された酵素は不安定なことが多い為,安定化する為の経費負担に改善の余地がある。この理由から,精製酵素の代りに目的の酵素を含む微生物をそのまま固定化して用いた例がある。例えば、アスパルターゼを含む微生物を固定化してL−アスパラギン酸を製造する例や、次いで,このプラントでつくられるアスパラギン酸を原料として,連続的にL−アラニンを製造した例などである(「化学増刊119 ハイブリッドプロセスによる有用物質生産」 化学同人; 「バイオリアクター」福井三郎編、バイオテクノロジーシリーズ、講談社; 「固定化酵素」千畑一郎編、講談社サイエンティフィック等を参照)。
【0006】
“化学と工業”Vol.62−1,January 2009、pp44−45(生体触媒開発を通じて)のように、工業的にみる酸化還元触媒は微生物や酵母、植物培養細胞など、細胞内に内在する脱水素酵素と補酵素の機能をそのまま活用する方法がコスト的に優位であり、生物固体から酸化還元酵素や補酵素の単離・精製を行う余分な経費負担は酵素の安定化や、ケトン→アルコール反応と補酵素NADH→NAD+反応、又はその逆反応を共役させる操作コストにおいて見合わないとされている。
【0007】
また、最近の酵素に変わる触媒設計の研究では、「粗蛋白質」内部に金属錯体を導入して酵素様活性ドメインを作成した例が見られる。
【0008】
例えば、「蛋白質 核酸 酵素」2004年,11月号,Vol.49,No.14 −ヘム酵素を中心とした金属酵素の分子設計−」(名古屋大学大学院理学研究科 渡辺芳人)に公知のように、クロロペルオキシダーゼ(CPO)の活性ドメイン内のヘム(鉄)を他の金属錯体に置き換えても酸化活性が起きることが示され、配置する金属近傍の機能性アミノ酸残基の適当な配置を設計することが活性ドメイン構築に重要であることが示されている。
【0009】
また、文献“Tetrahedron Letters”No.44,pp4281−4284(1978)(AsymMetric Reduction of aryl trifluoromethyl ketones with an model compound in a chiral hydrophobic binding site of sodium cholate micelle, β-cyclodextrin and bovine serum albumin)「馬場直道ら;京都大学化学研究所」によれば、酵素以外の成分:界面活性剤様タンジル酸(NAC)、β−シクロデキストリン(β−CD)、牛血清アルブミン(BSA)を触媒として、1−プロピル−1,4−ジハイドロニコチンアミド(NAH)又は水素化ホウ素ナトリウム(NaBH4)の存在下、基質トリフルオロメチル−アセトフェノンを作用させて不斉還元反応している。以上の結果、牛血清アルブミン(BSA)にも活性ドメインが存在(立体配置R、光学純度46.6%ee)することが示されている。
【0010】
100%ee近くの光学純度で、動植物由来粗蛋白から光学活性アルコールを分割した例が特許第3294860号公報(光学活性アルコールの製造方法)に示されており、穀類又は豆類から水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムゲルにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程とを組み合わせて、光学活性アルコール(100%ee、収率50%)を合成している。特許第3683129号公報(光学活性アルコールの製造方法)では、卵白及び卵白から分離されたオボアルブミンから選択される水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程とを組み合わせて、光学活性アルコール(100%ee)を合成している。
【0011】
一般的に、固定化酵素を作製する方法は、下記のような方法が代表的である。
1)該抽出・精製酵素を水不溶性の担体、例えば、セルロース、デキストラン、アガロース等の多糖類の誘導体、及びポリアクリルアミドゲル等に結合させる担体結合法、2)該抽出・精製酵素を2個もしくはそれ以上の官能基を有する試薬を用いて該抽出・精製酵素間に架橋結合を形成させて固定する架橋法、3)該抽出・精製酵素を、ゲル、例えば、アルギン酸塩、デンプン、コンニャク、ポリアクリルアミドゲル及びポリビニルアルコール等のゲルの細かい格子の中に取り入れる(格子型)か、半透明膜の皮膜によって被覆する(マイクロカプセル型)包括法がある。
【0012】
また、1990年代に登場したCLEC(Cross-Linked Enzyme Crystal)法は、該抽出・精製酵素を硫酸アンモニウムやポリエチレングリコール(PEG)などで結晶化し、グルタルアルデヒド(GA)などの多価修飾試薬を用い架橋させる方法で、産業用の固定化酵素の手法として最も活用されている。該CLEC法としては、例えば有機合成用酵素としてAltus株式会社より、ChiroCLEC(キラル化合物合成酵素)が製品化されている。ChiroCLEC−BLはBacillus licheniformis由来のSubtilisinを、ChiroCLEC−CRはCandida rugosa由来のLipaseを固定化し、架橋固相化してある。
【0013】
これらは有機溶媒中でも安定に活性を示すと同時に、熱安定性にも優れている。光学活性を有しながら、カルボン酸、アルコール、アミノ酸、エステル等の加水分解又はアシル化を行うことができる特徴がある。架橋酵素結晶(CLEC)は、1)酵素を架橋固定化する操作条件下で酵素の働きを至適化でき、2)加水分解酵素、酸化還元酵素、脱離酵素など、市販されている様々な酵素の中で開発でき、3)特定のニーズを満たすように手直しされた遺伝子導入微生物産生の酵素などから開発可能である。
【0014】
また、特公平4−68914(酵素の固定化方法)のように多孔性水不溶性担体のアミノ化シリカゲルに酵素を吸着させて、その後、多官能性架橋剤(グルタルアルデヒド)による共有結合反応により酵素の固定化を行った例がある。この場合、担体と酵素の結合力が弱く担体から酵素が脱離しやすい難点を、多官能性架橋剤に架橋することで解決している。多孔質アミノ化シリカゲルや多孔質アミノ化ゼオライトなどの多孔性担体の酵素吸着性に加えて、更に、多官能性架橋剤(グルタルアルデヒド)を利用することで固定化担体の活性半減期の著しい向上を実現している。
【0015】
このような架橋酵素結晶(CLEC)の知られている利点をまとめると、次の様になる。尚、酵素の結晶化とは、酵素分子の周囲に覆われた水分子が硫安やポリエチレングリコール添加により外れてゆき、酵素分子どうしが重合し始めることであり、最終的に分子が大きくなり沈殿することを意味する。有機化合物の凝固点まで冷やして結晶化する意味合いとは異なっている。
【0016】
(架橋酵素結晶の利点)
・高純度の酵素は必要としない(部分的に精製された調製品に適用可能)
・簡単な操作と広い応用
・室温で長期間(1年以上)安定
・実質100%活性なタンパク質(高い容積測定(volumetric)と触媒生産性)
【0017】
・容易な回収と再使用
・単独酵素と比較して高い温度安定性と有機溶媒への耐性
・高い活性と選択性(単独酵素よりも高い場合もある)
・水媒体中で酵素を濾す必要はない
・迅速な至適化(HTEを使用して)は開発時間を短くする
・触媒カスケードプロセス用に1つ以上の酵素を含むコンビCLEA
【0018】
更に架橋酵素結晶(CLEC)の市場と応用の可能性では、CLEC合成(薬剤、香料及び味覚物質、農薬、機能性食品、ファインケミカル、バルクのモノマー)、食品および飲料、パルプおよび紙、化粧品、オイルおよび脂質、織物、排出処理、界面活性剤、バイオセンサー、診断薬、タンパク質輸送などがある。
【0019】
Dehydrogenases については、現在、微生物由来のAlcohol dehydrogenase(ADH)from Rhodococcus erythropolis 、Formate dehydrogenase(FDH)from Candida boidinii がCLEA調整用の酵素類として知られている。
【先行技術文献】
【特許文献】
【0020】
【特許文献1】特許第3294860号公報
【特許文献2】特許第3683129号公報
【特許文献3】特公平4−68914号公報
【非特許文献】
【0021】
【非特許文献1】「化学増刊119 ハイブリッドプロセスによる有用物質生産」化学同人 「バイオリアクター」福井三郎編、
【非特許文献2】バイオテクノロジーシリーズ、講談社「固定化酵素」千畑一郎編、講談社サイエンティフィック
【非特許文献3】“化学と工業”Vol.62−1,January 2009、pp44−45
【0022】
【非特許文献4】「蛋白質 核酸 酵素」2004年、11月号、Vol.49、No14 −ヘム酵素を中心とした金属酵素の分子設計−(名古屋大学大学院理学研究科 渡辺芳人)
【非特許文献5】“Tetrahedron Letters” N0.44,pp4281−4284(1978)(AsymMetric Reduction of aryl trifluoromethyl ketones with an model compound in a chiral hydrophobic binding site of sodium cholate micelle, β-cyclodextrin and bovine serum albumin)「馬場直道ら;京都大学化学研究所」
【非特許文献6】“食品と技術” 12月号,pp1−9(2008)(グルテンタンパク質のネットワーク形成における食塩の役割)「裏出令子;京都大学大学院農学研究科」
【0023】
【非特許文献7】Crystal stracture of the DsbB-DsbA complex reveals a disulfide bond severation mechanism−Cell 127 789−801(2006)(稲葉謙次ら; 九州大学生体防御医学研究所)
【0024】
【非特許文献8】STRUCTURE-FUNCTION STUDIES OF GLUTAREDOXINS AND RELATED OXIDOREDUCTASRE−」(Tobias H. ElGAn; From THE DEPARTMENT OF BIOSCIENCES AND NUTRITION Karolinska Institutet, Stockholm, Sweden)
【発明の概要】
【発明が解決しようとする課題】
【0025】
本発明の目的は、上述した従来技術の欠点を解消し、緩和(mild)な条件下で、効率的な酵素類似(enzyme-mimetic)反応を可能とする、有用な材料を提供することにある。
【0026】
本発明の他の目的は、環境的側面および安全性に優れる、「酵素類似」反応活性を有する材料を提供することにある。
【0027】
本発明の他の目的は、このような「酵素類似」反応活性を有する材料の好適な製造方法を提供することにある。
【課題を解決するための手段】
【0028】
本発明者は鋭意研究の結果、不斉酸化反応を触媒する活性を有する蛋白質複合体を発見した。
【0029】
本発明の蛋白質複合体は、上記発見に基づくものであり、より詳しくは、蛋白質およびカルシウムを少なくとも含む蛋白質複合体であって;且つ、不斉酸化反応を触媒する活性を有することを特徴とするものである。
【0030】
本発明者は、上記知見に基づき更に研究を進めた結果、上記した有用な性質を有する蛋白質複合体が効率的に得られる方法を見出した。本発明の製造方法は上記発見に基づくものであり、より詳しくは、動植物由来水溶性部分から粗蛋白質を濃縮する第1の工程と;前記蛋白質をゲルに包括して空気酸化させた後、該ゲルから蛋白質複合体を抽出する第2の工程と;抽出された蛋白質複合体を水溶液中にて結晶化沈殿させる第3の工程と;沈殿した蛋白質複合体を架橋する第4の工程を含むことを特徴としている。
【0031】
上記製造方法の一態様によれば、動植物の水溶性部分から粗蛋白質を濃縮する第1の工程と;該粗蛋白質をアルギン酸カルシウムゲルに包括して、該ゲルを空気に数時間曝した後(空気酸化)、温水振とうして蛋白質複合体を抽出する第2の工程と;該蛋白質複合体を飽和30%硫安沈殿させる第3の工程と;沈殿した該結晶化蛋白質複合体を架橋する第4の工程を含むことができる。
【0032】
本発明の他の態様によれば、安価な動植物由来粗蛋白質をアルギン酸カルシウムゲルに包括して、該ゲルビーズ(酸素と溶存カルシウム存在下)を空気酸化させて(i)分子間ジスルフィド結合と(ii)分子間凝集(<6.4Å)と、(iii)水溶性変化を誘導させ(蛋白質複合体形成)、引き続き、温水中で振ることにより該複合体を好適に抽出することができる。
【0033】
本発明の一態様によれば、更に、蛋白質複合体又は架橋結晶化された蛋白質複合体を用いてでさえも、基質ラセミ体アルコールの一方の鏡像体を不斉酸化して、100%e.e.近い光学活性アルコールが合成することができることを示しており、また、その具体的な製造法が提供される。
【0034】
本発明の一態様によれば、更に、動植物由来祖蛋白質の蛋白質複合体が、ベンゼン骨格やナフタレン骨格を有する二級アルコール基質に対する一方の鏡像体に選択的な不斉酸化する反応に加えて(収率50%)、それらの骨格を持たないマツタケオールのようなアルキル鎖系二級アルコール類のデラセミ化反応活性(収率>85%)を有することも提供される。
【0035】
本発明においては、上述の精製された微生物由来酵素、遺伝子組み換え微生物由来酵素、又は微生物や遺伝子組み換え微生物、更には動物組織(肝臓、すい臓など)由来の固定化酵素、固定化微生物の製造を対象としているのではなく、高コストの酵素の単離と精製の工程を必要とせず、より安価な動植物資源由来粗蛋白質をその対象としていることが一つの特徴である。このような「植物資源」としては、例えば、蕎麦、アマランサス、米、小麦、大麦、トウモロコシ、エンバク、ライ麦、粟、ヒエ、キビ、ハト麦、モロコシ等の穀類;小豆、インゲン豆、豌豆、リョクトウ、大豆等の豆類、更には一般的な草や雑草をも含めた、これらの種子、葉、茎、根、花、実の各植物組織も挙げられる。
【0036】
更に、「動物資源」としては、卵白又はオボアルブミンとして、鶏卵由来のものが挙げられるが、鶏類、両生類、魚類の卵アルブミンも同様に用いることができ、鶏卵由来のものに限定するものではないし、由来の蛋白質を卵に限定するものでもない。
【0037】
本発明の一態様においては、後述するように、例えば、穀類、豆類、そして草や雑草をも含めたこれらの種子、葉、茎、根、花、実から水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムゲルビーズに包括化して、空気酸化後、10時間以上40℃の温水中で振とうさせて、蛋白質複合体水溶液画分をビーズと分離する第2の工程と、得られる蛋白質複合体水溶性画分を飽和30%硫安にして沈殿させる第3の工程と、得られた結晶化蛋白質複合体をグルタルアルデヒドにて架橋する第4の工程、更に、得られた架橋結晶化蛋白質複合体を乾燥粉体化する第5の工程を組み合わせた、環境に優しく低コストで且つ保存性の高い蛋白質複合体を製造することも可能となる。
【発明の効果】
【0038】
上述したように本発明によれば、実用的な不斉酸化活性を賦与した植物由来の蛋白質複合体、及び/又は動物由来の蛋白質複合体が提供される。本発明によれば、更に、このような動植物由来の架橋結晶化蛋白質に好適な製造方法であって、環境に優しく低コスト生産を実現する蛋白質複合体の製造方法が提供される。
【図面の簡単な説明】
【0039】
【図1】図1は、本発明製造方法の第2の工程において、豌豆蛋白・アルギン酸カルシウムゲルビーズに蒸留水を加えて、振とう開始後所定の時間後に、基質R−2−ナフチルエタノールを加えた時の基質の残量%を示したグラフである。
【図2a】図2aは、本発明製造方法の第2の工程において、空振とう10時間後、種々の基質を添加した際の反応時間に伴う基質と生成物の%比率をモニターリングしたグラフである。
【図2b】図2bは、本発明製造方法の第2の工程において、空振とう10時間後の水溶液等に、基質R−2−ナフチルエタノールを加えて、生成ケトン%をモニターリングしたグラフである。
【0040】
【図2c】図2cは、本発明製造方法の第2の工程において、異なる塩化カルシウム濃度に滴下することで調整した豌豆・アルギン酸カルシウムゲルビーズを空振とう10時間後、基質Rー2−ナフチルエタノールに作用させて蛋白質複合体活性を調べたグラフである。
【図2d】図2dは、本発明製造方法の第2の工程において、所定の豌豆蛋白質量で豌豆・アルギン酸カルシウムゲルビーズを調製したものを、空振盪10時間後にそれぞれ基質R−2−ナフチルエタノール(50mg)を加えて、蛋白質複合体活性を調べたグラフである。
【0041】
【図3】図3は豌豆−Caアルギン酸ゲルを10時間以上温水中にて振とう後、溶出される蛋白質複合体を硫安30%にて結晶化沈殿された、水溶性蛋白成分の分子量決定(SDS−page)である。 ここに、溶出溶液は、10時間以上温水中に振盪後の溶液画分である。硫安沈殿は、溶出溶液を30%硫安溶液にした時の沈殿物である。上清は、硫安沈殿物を水に再溶解したものを遠心分離して得られる上清画分である。
【0042】
【図4】図4は、「食品と技術」2008年12月号総括,裏出令子記述(グルテンタンパク質のネットワーク形成における食塩の役割)に見られる食塩添加あるいは無添加小麦グルテン生地のβ-メルカプトエタノール入りSDS溶液で可溶な成分のSDS−pageである。
【0043】
【図5】図5は、フーリエ変換赤外分光分析(FT−IR)を用いるサンプル(i)〜(Vi)の定性分析結果である。
【図6a】図6aは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【図6b】図6bは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【図6c】図6cは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【図6d】図6dは、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。
【0044】
【図7】図7は、本発明製造方法の第2の工程において、豌豆・アルギン酸カルシウムゲルを空気中で所定時間静置後、蛋白質複合体を抽出して架橋結晶化後、凍結乾燥させて得られる蛋白質複合体の収量と6時間後の不斉酸化活性(右軸)とモニタリングしたグラフである。
【0045】
【図8】図8は第2の工程において、蛋白質複合体に基質ラセミ−1−オクテン−3−オールを作用させて、香料のR−1−オクテン−3−オール(マツタケオール;98.2%ee)を合成したGCクロマトである。
【0046】
【図9a】図9aは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9b】図9bは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0047】
【図9c】図9cは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9d】図9dは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0048】
【図9e】図9eは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9f】図9fは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0049】
【図9g】図9gは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるDO(溶存酸素)の推移を示している。
【図9h】図9hは、本発明製造方法の第2の工程において、蛋白質複合体の100Lジャー生産におけるpHの推移を示している。
【0050】
【図10】図10は、蛋白質複合体の全工程製造フローと各工程の簡単な説明を示している。
【0051】
【図11a】図11aは、原核生物(大腸菌)におけるタンパク質ジスルフィド結合形成のための細胞システム(米国化学会誌「Crystal structure of the DsbB-DsbA complex reveals a disulfide bond severation mechanism」Cell, volume 127, Issue 4,789−801,17 Novemver 2006: 稲葉謙次ら(九州大学生体防御医学研究所)の図を引用)。
【図11b】図11bは、真核生物(酵母小胞体)におけるタンパク質ジスルフィド結合形成のための細胞システム(米国化学会誌「Crystal structure of the DsbB-DsbA complex reveals a disulfide bond severation mechanism」Cell, volume 127, Issue 4,789−801,17 Novemver 2006: 稲葉謙次ら(九州大学生体防御医学研究所)の図を引用)。
【図12】細胞内のシステイン酸化によって得られるスルフィド系酵素の種類とその酵素機能(反応性)を示したブロック図である。
【図13】実施例20において得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。
【図14】実施例20において得られた蛋白質複合体の凍結真空乾燥(FD)粉末(50mg、75mg、100mg、200mg、300mg)にラセミ−2−ナフチルエタノール濃度10ppmの50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、4時間置きにGCにて、不斉酸化されるR−2−ナフチルエタノールの残留比(%)について追跡を行った結果を示すグラフである。
【図15】実施例20において得られた蛋白質複合体の凍結真空乾燥(FD)粉末(300mg)にラセミ−2−ナフチルエタノール濃度(10-140ppm)の50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、不斉酸化されるR−2−ナフチルエタノールの残留比(%)について4時間置きにGC追跡を行った結果を示示すグラフである。
【発明を実施するための形態】
【0052】
以下、必要に応じて図面を参照しつつ本発明を更に具体的に説明する。以下の記載において量比を表す「部」および「%」は、特に断らない限り質量基準とする。
【0053】
(蛋白質複合体)
本発明の蛋白質複合体は、蛋白質を少なくとも含み、且つ、FT−IRにおいて、(1085±50cm−1)の領域、および(1411±50cm−1)の領域に、それぞれ少なくとも1個のピークを有することを特徴とする。ここに、FT−IR測定は、後述するように、例えば、Thermo Fisher Scientific製フーリエ変換赤外分光分析装置Magna-750および赤外顕微鏡Nic−Planを用い、顕微ATR法(Geクリスタル)により、例えば波数分解能8cm−1、積算回数32回の条件で測定することができる。(このようなFT−IR測定の詳細に関しては、例えば、文献”Carbohydrate Polymers”66,191−197,(2005)(Enzymatically produced nano-ordered short elements containing cellulose Iβ crystalline domains)「Noriko Hayashiら;Bioconversion Laboratory, Forestry and Forest Products Research Institute (FFPRI)」に実施例があり、吸収波長の評価に関しては、本「ACADEMIC PRESS; New York and London 1971; A Subsidiory of Harcourt Brace Jovanovich, Publishers」”INFRARED SPECTRA OF INORGANIC COMPOUNDS (3800−45cm−1)”「Richard A. Nyquist and Renald O. Kagel; Chemistry Physics Research Laboratory, The Dow Chemical Company Midland MichiGAn」を参照できる。
【0054】
(IRピークの強度)
本発明の蛋白質複合体において、上記FT−IR測定におけるピーク強度は、以下の通りであることが好ましい。
(1)(1085±50cm−1)の領域におけるピーク(以下、「ピーク1」という)は、測定されたIRスペクトル中の最も強度が大きいピークであるか、または最も強度が大きいピークの強度をI0とした際に、(1/10)×I0以上のピーク強度を有することが好ましい。このピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
(2)(1411±50cm−1)の領域におけるピーク(以下、「ピーク2」という)は、測定されたIRスペクトル中の最も強度が大きいピークであるか、または最も強度が大きいピークの強度をI0とした際に、(1/10)×I0以上のピーク強度を有することが好ましい。このピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
【0055】
(他のIRピークの強度)
本発明の蛋白質複合体において、上記した「ピーク1」および「ピーク2」以外のIRピークは特に制限されない。本発明の蛋白質複合体は、例えば、下記の領域にピークを有していても良い。
【0056】
また、本発明の蛋白質複合体において、上記FT−IR測定ピーク強度の評価は、以下の通りであることが更に好ましい。
(1)(1085±50cm−1)の領域におけるピーク(以下、「ピーク1」という)は、測定されたIRスペクトル中の最も強度が大きいピークであり、最も強度が大きいピークの強度をI0とした際に、2番目に強度が大きいピークが(1411±50cm−1)にあり、このピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
(2)(1411±50cm−1)の領域におけるピーク(以下、「ピーク2」という)は、測定されたIRスペクトル中の2番目に最も強度が大きいピークであり、2番目に強度が大きいピークの強度をI0とした際に、3番目に強度が大きいピークが(1649±50cm−1)にありこのピーク強度は、更には(1/5)×I0以上(特に(1/3)×I0以上)であることが好ましい。
【0057】
(他のIRピークの強度)
上記の場合にも、本発明の蛋白質複合体において、上記した「ピーク1」および「ピーク2」以外のIRピークは特に制限されない。本発明の蛋白質複合体は、例えば、下記の領域にピークを有していても良い。
【0058】
(好ましい蛋白質複合体)
本発明の蛋白質複合体は、蛋白質およびカルシウムを少なくとも含むことが好ましい。また、蛋白質複合体は、不斉酸化反応を触媒する活性を有することが好ましい。このような「蛋白質およびカルシウムを少なくとも含む」こと、および「不斉酸化反応を触媒する活性を有する」は、後述する方法によって好適に確認することができる。
【0059】
(不斉酸化反応の好ましい触媒)
本発明の蛋白質複合体の不斉酸化反応の好ましい態様は、例えば、後述する実施例12の条件下で、基質ラセミ−1−オクテン−3−オール(マツタケオール)を作用させた場合に得られるR−1−オクテン−3−オールの光学純度(%ee)で好適に表すことができる。本発明の蛋白質複合体は、この実施例12の条件で、85%以上の化学収率で、R−1−オクテン−3−オールを与えることが好ましい。更には、この光学純度は、95%ee以上(特に98%ee以上)であることが好ましい。
【0060】
(好適な特製)
本発明の蛋白質複合体は、下記の好適な特製のうち、1以上を有することが好ましい。
(1) 上記蛋白質複合体が、更に糖を含むこと。
(2) 前記蛋白質が、架橋結晶化蛋白質であること。
(3) 前記不斉酸化反応が、基質ラセミ体アルコールの一方の鏡像体を選択的に与える酸化反応であること。
(4) 前記不斉酸化により得られるケトンが、更に不斉酸化されるデラセミ化反応を与える活性を、前記蛋白質複合体が有すること。
【0061】
(本発明の推定メカニズム)
本発明者の知見によれば、本発明において、好適な動植物由来の蛋白質複合体が提供されるのは以下の理由によるものと推定される。
【0062】
安価な材料の動植物由来の粗蛋白質は、カルシウム塩と酸素存在を実現できるアルギン酸カルシウムゲルに包括後、空気中で好適に静置(空気酸化)させることによって、(i)タンパク質内システイン残基(−SH)どうしの分子間ジスルフィド結合(−S−S−)、(ii)タンパク質内分子間凝集(分子間距離の短縮:<6.4Å)、(iii)水溶性への性質変化が生じ、活性ドメイン(チオレドキシンフォールド:Cys−X−Y−Cys配列)の形成のみならず、温水中で振ることにより好適に蛋白質複合体が溶出できる性質になる。
【0063】
温水中に溶出される溶存Ca塩と酸素の存在は、ジスルフィド結合したタンパク質ポリマー(蛋白質複合体)を水溶液に溶け易くする効果があり、温水中で振ることにより容易に抽出される。タンパク質ポリマー(蛋白質複合体)の触媒活性向上と収量向上と収量向上のポイントは、空気中で好適に該ゲルビーズを静置(空気酸化)させる点と、温水中に溶け出す程度の低Caイオン濃度存在下で「滲み出し」易くさせる点にある。
【0064】
本発明の製造方法においては、動植物蛋白質の有する空気酸化に伴うこれらの「蛋白質複合体形成」と「滲み出し」の特製を生かして、アルギン酸カルシウムゲルによる固定ではなくて、好適なジスルフィド結合への誘導といった「逆転の発想」により、酵素様に作用する蛋白質複合体を低コストで、環境に優しく、効果的に作成することに特徴がある。
【0065】
(推定メカニズムの関連文献)
上記した本発明の「推定メカニズム」のサポートに関しては、例えば、以下の文献を挙げることができる。
【0066】
文献“食品と技術” 12月号,pp1−9(2008)(グルテンタンパク質のネットワーク形成における食塩の役割)「裏出令子;京都大学大学院農学研究科」では、食塩や塩化カルシウムには、グルテンタンパク質の相互作用を変化させてグリアジンやグルテニンを水に溶けるような性質に変化させる効果(水溶性効果)と、グルテン構成タンパク質間の相互作用を強めて分子間距離を短縮させる効果(凝集効果)が公知となっている。
【0067】
分子間距離を短縮させる効果(凝集効果)において、グルテン分子間距離は塩無添加では7.7Åであるが、塩添加の場合、分子間距離では6.4Å以内に短縮されることが示されている。ミオグロビンミュータントにみる活性ドメイン内のヒツチジン−鉄間の分子間距離は5.7Åであることから、塩添加による凝集効果は活性ドメインの機能性アミノ酸残基の適当な配置を設計する効果も期待できる。「蛋白質 核酸 酵素」2004年,11月号,Vol.49,No.14 −ヘム酵素を中心とした金属酵素の分子設計−」(名古屋大学大学院理学研究科 渡辺芳人)
【0068】
更に、小麦粉生地の蛋白成分(グルテニンとグリアジン)は、食塩存在下で作製した場合には、この蛋白質分子間の相互作用を変化させ、純水では水溶性となり、食塩水では不溶性を示す性質に変化することにも報告がある。
【0069】
小麦グルテン内グルテニンポリマーの網状構造の形成とは、小麦粉の水の添加により始まるが、空気中の酸素を使って、グルテニンのC末端とN末端システイン残基どうしが自発的に分子間ジスルフィド結合(R1−S−S−R2)する空気酸化の働きが知られている。しかしながら、最近の報告(非特許文献7)「−Crystal structure of the DsbB−DsbA complex reveals a disulfide bond severation mechanism−Cell 127 789-801 (2006)」(稲葉謙次ら; 九州大学生体防御医学研究所)には、大腸菌では、ジスルフィド結合を導入する為の酵素が存在しており、基質蛋白質の2つのシステインを酸化しジスルフィド結合を作る酵素(DsbA)と、使われたDsbAを酸化し酸化能力を回復させる酵素(DsbB)が、細胞内で活性エネルギーを作り出すことに関与する物質であるユビキノンと連動して働くとされている。DsbBが受け取った電子は呼吸鎖成分であるユビキノン(UQ)に受け渡され、最終的にはチトクロム酸化酵素を介して「酸素が電子受容体」となることが分かっている(図11aおよび図11bを参照)。
【0070】
更に、原核生物(大腸菌)の蛋白質ジスルフィド形成システムと真核細胞(酵母小胞体など)のそれとは類似しており、小胞体にはDsbBの機能ホモログとして小胞体膜近傍にEro1pが存在する。大腸菌システムでのジスルフィド結合を創り出すための酸化力は、「ユビキノン」としているのに対し、小胞体システムではFAD(Flavin Adenine Dinucleotide)となる。大腸菌におけるDsbA−DsbB−ユビキノン酸化システムと真核生物のPDI−Ero1p−FADは互いに機能的ホモログの関係にあるとある(図11aおよび図11b参照)。
【0071】
その他、ジスルフィド結合は、多くの細胞表層にある蛋白質にとって、立体構造を正しく形成し、維持する為に必須な化学結合であり、また、ジスルフィド結合の形成と解離は、細胞の機能に重要な蛋白質の活性や細胞内での存在場所に影響を与へたり、細胞の酸化還元環境の変化に起因するストレスに対応している。
【0072】
−STRUCTURE-FUNCTION STUDIES OF GLUTAREDOXINS AND RELATED OXIDOREDUCTASRE−」(Tobias H. ElGAn; From THE DEPARTMENT OF BIOSCIENCES AND NUTRITION Karolinska Institutet, Stockholm, Sweden)では、酵素活性ドメインには2つのシステインからなる共通配列「チオレドキシンフォールド(Cys−X−Y−Cys)」が存在し、この部位がジスルフィド結合の形成や異性化を触媒して酸化還元活性を持つとされている。Cys−X−Y−CysのXとYには疎水性のアミノ酸がくることが多く、還元型ではシステイン残基は2つの自由なチオール基となり、酸化型ではそれらはジスルフィド結合でつながれる(後述する図11aおよび図11bを参照)。
【0073】
小麦タンパク質の約70〜80%を占める成分を分類すると、アルブミン、グロブリン及び小麦プロラミンであるグリアジン(25kda〜50kdaの間にあるバンド)、そして高分子量グルテニン(HMW:75kda〜150kdaの間にある3バンド)と低分子量グルテニン(LMW:25kda〜50kdaの間にあるバンド)からなるグルテニンに分類される。図4にそのSDS−pageを示している。
【0074】
(標的となる触媒活性)
図12は、文献“Biochemical and Biophysical Research ComMunications”300,pp1−4(2003)(Multiple roles of cysteine in biocatalysis)「Niroshini M, Gilesら;School of Chemistry, University of Exeter」で公知のように、細胞内システイン酸化により形成された様々な酵素の機能に関してなど示されている。1)Peptidase、2)glyceraldehyde 3-phosphate dehydrogenase (GAPDH)、3)alcohol dehydrogenase (ADH)、4)bacterial nitrile hydratase (NHase)、5)ribonucleotide reductase (RNRase)、6)pyruvate formate lyase (PFL)、7)benzylsuccinate synthase (BSS)、8)reduction of glutathione disulfide (GSSG)、9)thioredoxin (Trx)、10)glutathion reductase (GR)、11)thioredoxin reductase (TR)、12)NADH oxidase (Nox), 13)NADH peroxidase (Npx)。
【0075】
また、特許第3294860号公報(光学活性アルコールの製造方法)で公知のように、穀類又は豆類から水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムゲルにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程が示されている。また、特許第3683129号公報(光学活性アルコールの製造方法)で公知のように、卵白及び卵白から分離されたオボアルブミンから選択される水溶性蛋白質を抽出する第1の工程と、前記蛋白質をアルギン酸カルシウムにて包括化する第2の工程と、前記包括化された蛋白質を触媒として基質の不斉酸化変換反応を行う第3の工程が示されている。
【0076】
本発明との違いは、上記知見に加えて、1)空気酸化により構築されるタンパク質分子内の活性ドメイン形成メカニズムへの言及と、2)架橋結晶化後に水分を除去して常温保存が可能な粉末触媒の製造法に発展している点にある。
以下、本発明の製造方法を構成する各工程について説明する。
【0077】
(第1の工程)
本発明の第1の工程における水溶性蛋白質の抽出においては、穀類又は豆類を砕き粒の大きい部分と殻部を取り除き、このようにして得られた穀類及び豆類粉砕粉を約20〜60℃、好ましくは約40℃で、pH約6〜8、好ましくは約pH7.0において、穀類または豆類の約7〜15重量倍の水で、30分以上抽出する。約45分で抽出するのが最も効果的であって、これ以上長く抽出しても抽出物の量は変わらない。pHの調製が必要なときは、H2SO4、HCl、H3PO4などの食品級酸、又はNaOHなどの食品級アルカリを用いて上記適性範囲を調製できる。上記水溶性蛋白質抽出液、又はこの抽出液から、デカンター、遠心分離などにより食物繊維部を分離した、タンパクカードをそのまま第2工程に移すか、必要に応じて噴霧乾燥、凍結乾燥、真空乾燥などにより粉末としてから、再溶解して第2工程に移しても良い。種子(外皮(ふすま、ぬか)、胚芽(スプラウト))に関しては、上記の穀類及び豆類粉砕粉の工程にて得ることができる。外皮(ふすま、ぬか)、胚芽(スプラウト)、葉(若葉、新芽)、茎、根、花を凍結乾燥(FD)、熱風乾燥(AD)、真空乾燥などで水分を除去して乾燥後、ボールミルなどを利用して5μm以下まで細かく粉砕する。
【0078】
この粉砕粉をそのまま第2工程に移すことができる。等電点沈殿のpHの選定は沈殿量の多い画分の選定が目的であり、例えば大豆や豌豆や蕎麦蛋白質の場合はpH4.5及びpH9.5付近であり、このカードに5〜10重量倍の水を加え、ミキサー、攪拌機などにより解砕して、蛋白質スラリーを調製し、中和(pH6〜8)し中和スラリーとする。このスラリーを、前記と同様の噴霧乾燥、凍結真空乾燥(FD)、真空乾燥などにより粉末としてから、再溶解して第2工程に移す。
【0079】
しかしながら、多量の蛋白質の処理が必要なときには、上記タンパクガードをH2SO4、HCl、H3PO4などの食品級酸、又はNaOHなどの食品級アルカリを用いて等電点処理し、次いでデカンター、遠心分離機などによりホエイを分離してタンパクカードをえる。この等電点沈殿等は、水溶性蛋白質の濃縮を目的としたものであって、処理後においても水溶性蛋白質抽出液をそのまま噴霧乾燥などにより粉末化した場合と同様な効果を奏するものである。
【0080】
(第2の工程)
第2の工程において、該抽出蛋白質を包括化する方法は、例えば、該抽出蛋白質を、ゲル、例えば、アルギン酸塩、デンプン、コンニャク、ポリアクリルアミドゲル及びポリビニルアルコール等のゲルの細かい格子の中に取り入れる(格子型)か、半透膜性の皮膜によって被覆する(マイクロカプセル型)包括法があるが、蛋白質複合体を作成する上では、海草より抽出するアルギン酸のカルシウムを用いた包括化法が安価で環境に優しく、かつ、包括操作が平易な点で最も好ましい。この包括化されたビーズは、蛋白質複合体を水溶液中に溶出させる為に、温水温度30℃〜45℃で、且つ、溶存酸素濃度がゼロにならないように酸素供給が必要で、且つ、ビーズが振れる回転数で振とうさせることが望ましい。
【0081】
(第3の工程)
第3の工程において、蛋白質を結晶化沈殿するためには、硫酸アンモニウム、ポリエチレングリコール(PEG)、ポリエチレングリコール/塩化リチウム,2−メチル−2,4−ペンタジオール,塩化ナトリウム,マロン酸ナトリウム等があり、いずれの結晶化沈殿剤として本願発明において用いることができる。しかしながら、最も安価な硫酸アンモニウムを用いるのがトータルコストダウンの観点から有益である。蛋白質の結晶化は、通常濃度が10〜50mg/mLのタンパク質溶液で用いて行うことが望ましく、サンプルをセントリコンやアミコンウルトラなどで濃縮する手法ある。
【0082】
(第4の工程)
第4の工程において、結晶化沈殿された蛋白質を架橋するためにはグルタルアルデヒド(以下、「GA」と略称する)、フォルムアルデヒド(以下、「FA」と略称する)等があり、いずれも架橋剤として本願発明において用いることができる。グルタルアルデヒドは、示性式OHC(CH2)3CHO として表される有機化合物で、反応は、タンパク質のリジン残基のε−アミノ基との間で起こるが、α−アミノ基やSH基との間でも起こり、分子間架橋を形成することができる。ひとつのグルタルアルデヒド分子が単独で架橋形成を起こせるとは考えられていない。(参考文献;重中義信/監修『原生動物の観察と実験法』(共立出版株式会社、1998年)ISBN 4320053532)GAは2つのアルデヒド基、FAは1つアルデヒド基を持っており、架橋させることで、1)室温で長期間(1年以上)安定、2)実質100%活性なタンパク質(高い容積測定(volumetric)と触媒生産性)、3)容易な回収と再使用、4)単独酵素と比較して高い温度安定性と有機溶媒への耐性、5)単独酵素よりも高い活性と選択性になるなど、産業用酵素試薬として実用レベルで広く活用できるようになる。
【0083】
GAは2つのアルデヒド基を持っているためか、FAに比べて非常に強い固定力を持っており、固定力の強いGAであるが、難点は浸透速度がとても遅い。FAは固定力が弱いものの浸透速度はGAの数倍あるので、GA固定液にFAをまぜることによって、双方の弱点を補いあうことができる。GA/FA混合固定液はカルノフスキー(Karnovsky,1965)が考案したので(5%GA+4%FA)、「希釈カルノフスキーを使用」などと今でもカルノフスキーの名称が通っている。GAとFAの他に固定液に種々の添加物を入れる場合がある。最も多いのは浸透圧を調整(上げる)ためにショ糖、ブドウ糖、食塩などを入れることができる。架橋による固定時間は、バルク使用の場合(厚さ1mM以下)、おおよそ2時間というのが最小時間としておく。固定液の温度については、室温で固定する場合、4℃(氷水)で固定する場合など様々で可能ですが、再現性を重視して、常温20℃〜40℃で固定することが好ましい。一定温度と言うことであれば、氷を浮かべた水の中に「固定液と試料を入れた試料瓶をいれておけば、ほぼ4℃を保つことができるが、作業トータルコスト等を考慮して、常温20℃〜40℃で固定することが好ましい。固定後は残留GAとFAを、0.1Mリン酸緩衝液(220 mOsm位)やその緩衝液に0.1M〜0.2Mのショ糖やグルコースを加えて洗浄することができる。
【0084】
(第5の工程)
第5の工程において、乾燥粉体化するためには、1)熱風乾燥(AD)、2)凍結真空乾燥(FD)、3)加圧乾燥、4)減圧乾燥、5)自然乾燥など本願発明において用いることができる。しかしながら、凍結真空乾燥(FD)は水分を完全に除去でき、引き続く粉砕の工程に有利であり、尚且つ保存性も高くなる。
【0085】
(各データの説明)
以下、図面に示した各データを説明する。
【0086】
図1は第2の工程において、豌豆蛋白・アルギン酸カルシウムゲルビーズに蒸留水を加えて、空振盪開始後0時間後、5時間後、10時間後、15時間後、20時間後、25時間後、35時間後、45時間後、55時間後に基質(ラセミ)−2−ナフチルエタノールを加えた時の基質の残量%を示したものである。
【0087】
図2aは第2の工程において、空振盪10時間後、a)基質ラセミ−2−ナフチルエタノール、R−2−ナフチルエタノール、2−アセトナフトンをそれぞれ50mg添加したときの反応時間に伴う基質と生成物の%比率をモニターリングしたものである。
【0088】
図2bは第2の工程において、空振盪10時間後の水溶液(120ml、240ml)、空振盪10時間後のビーズ+蒸留水(DW:120ml)、新しいビーズ+空振盪10時間後の水溶液(120ml)に、それぞれ基質R−2−ナフチルエタノール(50mg、75mg、又は100mg)を加えて、生成ケトン%をモニターリングしたものである。
【0089】
図2cは第2の工程において、異なる塩化カルシウム濃度(5g/L、7.5g/L、10g/L、15g/L、20g/L、30g/L)に滴下することで調製した豌豆・アルギン酸カルシウムゲルビーズを空振盪10時間後、基質R−2−ナフチルエタノールに作用させて蛋白質複合体活性を調べたものである。
【0090】
図2dは第2の工程において、豌豆蛋白量2g、3g、4g、5gで豌豆・アルギン酸カルシウムゲルビーズを調整したものを、空振盪10時間後にそれぞれ基質R−2−ナフチルエタノール(50mg)を加えて、蛋白質複合体活性を調べたものである。
【0091】
図3は豌豆−Caアルギン酸ゲルを10時間以上温水中にて振盪して得られる蛋白質複合体を硫安30%にて結晶化沈殿させて、更に、遠心分離(15分、10000rpm)して得られる沈殿物をSDS−pageした図である。
【0092】
SDS−PAGE
サンプル10 μLを,2×サンプルバッファー(0.1 M Tris/HCl pH 6.8,3 % SDS,10 %グリセリン,10% β‐メルカプトエタノール,0.1% BPB)10 μLと混合し,100℃で5 min加熱した。これらについて,SDS−PAGE mini(4−20 %グラジエントゲル,TEFCO社)を用いて分子量マーカー(SDS−PAGEスタンダード Broad,Bio−Rad)と共に泳動(18 mA定電流,泳動バッファー:25 mM Tris/HCl,0.19 M Glycine,0.1 % SDS,pH 8.3)を行った後,CBB染色(PHastGel Blue R,Amersham Biosciences社)を行った。
【0093】
図4は、文献“食品と技術” 12月号、pp1−9(2008)(グルテンタンパク質のネットワーク形成における食塩の役割)「裏出令子;京都大学大学院農学研究科」に記載された「図5 DSTによる架橋効率に食塩添加が及ぼす影響」の抜粋である。(以下説明文)食塩添加あるいは無添加生地をDSTで処理した後、βメルカプトエタノール入りSDS溶液でタンパク質を可溶化した。可溶化液を超遠心し、可溶性画分(S)と不溶性画分(P)に分けた。不溶性画分はメタ過ヨウ素酸ナトリウムで処理した。それぞれの試料は、SDSポリアクリルアミドゲル電気泳動で分離しタンパク質を染色した。
【0094】
図5は、エンドウ蛋白質粉末(i)とアルギン酸ナトリウム粉末(ii)と、定法に従いエンドウ蛋白質・アルギン酸カルシウムゲルを作成し、第2の工程において(iii)恒温振盪培養器による振盪、又は(iV)ジャーファメンタ温水中で攪拌して調整した蛋白質複合体粉末((iii)と(iV))と、第2の包括工程を省き、エンドウ蛋白質(i)を2%塩化カルシウム/50mMトリス塩酸バッファー((V)pH6.0又は(Vi)pH8.0)中で恒温振とう(55rpm、40℃、24時間))して作成した架橋結晶化粉末((V)と(Vi))をFT−IRにより分析した結果を示している。
【0095】
赤外分光分析(FT−IR)は、Thermo Fisher Scientific製フーリエ変換赤外分光分析装置Magna−750および赤外顕微鏡Nic−Planを用い、顕微ATR法(Geクリスタル)により、波数分解能8cm−1、積算回数32回で測定した。
【0096】
図5において、記号A〜Eは、下記の意味を有している。
A:1649cm−1付近・・・ペプチド結合(−C(=O)−N−)に特徴となるピーク。
B:1528cm−1付近・・・ペプチド結合(−C(=O)−N−)に特徴となるピーク。
C:1411cm−1付近・・・カルボキシラート(−C(=O)−O−Na)のケトン基に特徴となるピーク。
D:1122cm−1付近(1085cm−1、1032cm−1)・・・糖エーテル(−C−O−C−)特徴となるピーク。
E:1085cm−1付近・・・硫安((NH4)2SO4)に特徴のピーク。
【0097】
図6は、図5と同サンプルであるが、恒温振とう培養作成した蛋白質複合体粉末(i)と、豌豆蛋白質に2%塩化カルシウム/50mMトリス塩酸バッファー溶液((ii)pH6.0又は(iii)pH8.0)から得られる架橋結晶化粉末((ii)と(iii))、及びエンドウ蛋白質粉末(iV)をX線マイクロアナライザー定性分析を行った結果である。X線マイクロアナライザー分析は、島津製作所製EPMA−1600を用い、試料表面に金コートして測定した。
【0098】
図6において、各サンプルの意味は、以下の通りである。
サンプル(i)・・・振とう培養器製架橋結晶化蛋白質複合体
サンプル(ii)・・・20mM塩化Ca/50mMトリス塩酸バッファー(pH6.0)
サンプル(iii)・・・20mM塩化Ca/50mMトリス塩酸バッファー(pH8.0)
サンプル(iv)・・・エンドウ蛋白質粉末
【0099】
図7は第2の工程において、アルギン酸カルシウムゲルに包括したビーズを空気中で0時間、0.5時間、1時間、3時間、5時間、7時間静置後、温水中で振とうして蛋白質複合体を抽出し、更に架橋結晶化後に凍結乾燥させて粉末化したサンプルの6時間後の不斉酸化活性強度(右軸)と得られた蛋白質複合体の収量(左軸)を示す図である。
【0100】
図8は第2の工程において、図1と同一条件で豌豆・アルギン酸カルシウムゲルビーズに包括化して空振とう10時間後に溶出される蛋白質複合体水溶液に基質ラセミ−1−オクテン−3−オール(マツタケオール)を作用させて、98.2%eeの光学純度で香料のR−1−オクテン−3−オールを合成したGCクロマトグラムである。
【0101】
図9a〜図9h(および表1)は、100Lジャーファメンタを用いたエンドウ蛋白粉末(1kg)・アルギン酸カルシウムゲル(計20L)と加水(20L)における酸素供給有無に関わる、DO(溶存酸素濃度)とpHのモニタリング図、及び得られる蛋白質複合体の収量、活性をまとめた図および表である。
【0102】
図9aおよび図9b:1回目の実験の結果
図9cおよび図9d:2回目の実験の結果
図9eおよび図9f:3回目の実験の結果
図9gおよび図9h:4回目の実験の結果
【0103】
【表1】
(説明)
Ca濃度(FD)・・・FD乾燥後サンプルの酸分解法による原子吸光光度計によるCa濃度(mg/g)測定結果。
×・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間に殆ど不斉酸化されない場合(生成ケトン10%以下)。
△・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間までに不斉酸化が途中で止まる場合(生成ケトン30%以下)。
○・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間で不斉酸化される場合(生成ケトン40%以上)。
【0104】
図10は、蛋白質複合体の製造フローの工程図である。
【0105】
図11aは、原核生物(大腸菌)における、ジスルフィド結合形成の細胞内システムを示す。図11bは、真核生物(酵母小胞体)における、ジスルフィド結合形成の細胞内システムを示す。
【0106】
図12は、細胞内のシステイン酸化によって得られるスルフィド系酵素の種類とその酵素機能(反応性)を示した図である。
【0107】
(不斉酸化反応を有する蛋白質複合体)
本発明の蛋白質複合体は、動植物由来粗蛋白質の空気酸化形成物であって、且つ、不斉酸化反応を示すなど酵素様活性を有することを特徴とする。
【0108】
(不斉酸化活性の測定方法)
本発明の不斉酸化反応を有する蛋白質複合体の活性は、例えば、後述(実施例15「抽出作業の条件」内6.不斉酸化活性)に記載する方法により好適に測定可能である。
【0109】
(蛋白質複合体の精製法)
本発明において、蛋白質複合体の精製方法は特に限定されない。蛋白質複合体の収量を増やす目的から、ろ過の必要はないが、アルギン酸カルシウムゲルビーズとの分離の意味でガラスビーズなどを用いることができる。この場合、口径サイズが40−200μmで可能であるが、口径200μmであることが好ましい。
【0110】
次に、本発明を実施例に基づき具体的に説明するが、これは説明のためのものであって、これにより限定して解すべきではない。
【0111】
実施例1
(架橋結晶化した豌豆由来の蛋白質複合体の一般的な製造方法)
先ず第1の工程として、豌豆を粉砕して殻を取り除き、pH7.0付近の蒸留水(約40℃)9重量倍にて、約45分間に溶解される豌豆蛋白質成分をNaOH水溶液を用いてpH7.0にして沈殿成分の食物繊維を除去し、水溶性蛋白部分をアルカリ条件(pH9.5付近)又は酸性条件(pH4.5付近)にして蛋白質を等電点沈殿させ、蛋白沈殿部をpH7.0の蒸留水にて再溶解して得られる豌豆蛋白水溶液(試料濃度5.0%)を噴霧乾燥処理を行い粉体の豌豆蛋白を調製する。また、3%アルギン酸ナトリウム水溶液はオートクレーブの条件、温度121℃、時間20分で、アルギン酸ナトリウムを水溶液中に溶解して調整する。
【0112】
次に、第2の工程において、豌豆蛋白粉20gに10倍等量の蒸留水200mlを加えて、3%のアルギン酸ナトリウム水溶液を1.0倍等量の200mlを加えて均一になるまで攪拌し、得られた豌豆・アルギン酸ナトリウム混合液を4%の塩化カルシウム水溶液中に滴下して、固定化状態の豌豆蛋白含有・アルギン酸カルシウムゲルビーズを作製する。ビーズは5時間、空気中で静置した。豌豆蛋白含有・アルギン酸カルシウムゲルビーズを蒸留水にて洗浄し、塩化カルシウム水溶液を除去した後に用いた豌豆蛋白粉の20倍等量の蒸留水(400ml)を抽出溶液として添加し、恒温振とう培養(40℃、55rpm)を2日間して水溶性画分を分離回収後、ビーズは更に1日間振とうさせて同様に目的溶液を回収した。更にもう1日間振とう後、目的溶液を分離回収できる。回収された反応溶液に含まれる蛋白質複合体は、引き続き第3の工程で結晶化沈殿させた。
【0113】
第3の工程において、分離回収溶液を弱水溶性成分の沈殿を目的として、飽和30%硫安に調整した後、20時間以上静置させた。蛋白質複合体は、塩化カルシウムの存在下において容易にビーズより溶出されるだけでなく、飽和30%硫安溶液の存在下においては、非水溶性となり凝集沈殿した。
【0114】
第4の工程において、沈殿凝集した結晶化蛋白質複合体は更に、溶液濃度にして0.25%になるように市販25%グルタルアルデヒド溶液を添加し、2時間以上(好ましくは10時間以上)静置した。得られる蛋白質複合体は、冷却遠心機(10,000rpm、15min、10℃)後、デカンテーションにより容易に溶液から分離できた。
【0115】
第5の工程(凍結真空乾燥(FD))は、沈殿物の蛋白質複合体を−50℃にまで冷やして1時間静置後、約0.1mbar以下の真空状態にした後、温度を−50℃から50℃まで昇温させて水分を昇華させて、水分除去を行った。一連のFD所要時間は20時間程度であった。
【0116】
第6の工程において、凍結真空乾燥(FD)により水分除去された蛋白質複合体は、引き続くボールミル粉砕により、常温保存できる粉末状の触媒とできた。
【0117】
実施例2
(第2の工程における豌豆(PP)、大豆(SP)、小麦(WP)蛋白質由来蛋白質複合体の基質特異性の検討)
第2の工程において、豌豆(PP)、大豆(SP)、小麦(WP)由来粗蛋白質は種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し、水溶液画分を噴霧乾燥して得られる粉末20gは定法に従って、10倍等量の蒸留水200mlを加えて、3%のアルギン酸ナトリウム水溶液を1倍等量の200mlを加えて均一になるまで攪拌し、得られた豌豆・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中に滴下して、豌豆蛋白含有・アルギン酸カルシウムゲルビーズを作製する。該作製ビーズに豌豆蛋白粉の20倍等量の蒸留水(400ml)を反応溶液として添加し、恒温振とう培養器の温度設定40℃、振とう数55rpmにて10時間以上の振とう後、基質変換反応を行った。基質としては2−ナフチルエタノール(naphthyl/1)と、メタ、パラ位に塩素(Cl−/2a、2b)、臭素(Br−/3a、3b)、フッ素(F−/4a、4b)、メチル(Me−/5a、5b)、メトキシ(MeO−/6a、6b)、ニトロ(NO2−/7a、7b)を置換させたフェニルエタノールを用いた結果、反応時間、生成物、光学純度、化学収率を以下の表にまとめる。
【0118】
(第2の工程における豌豆(PP)、大豆(SP)、小麦(WP)蛋白質由来蛋白質複合体のメタ位置基質の基質特異性)
【表2】
【0119】
以上の結果から、豌豆(PP)、大豆(SP)、小麦(WP)由来蛋白質由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質であるメタ位置換塩素(Cl−/2a)、臭素(Br−/3a)、フッ素(F−/4a)、メチル(Me−/5a)、メトキシ(MeO−/6a)、ニトロ(NO2−/7a)のラセミ−フェニルエタノール類のR体を選択的酸化し、S体アルコールが化学収率50%程度、光学純度99%e.e.以上で得た。
【0120】
(第2の工程における豌豆(PP)、大豆(SP)、小麦(WP)蛋白質由来蛋白質複合体のパラ位置基質の基質特異性の検討)
【表3】
【0121】
以上の結果から、豌豆(PP)、大豆(SP)、小麦(WP)由来蛋白質由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質パラ位置換塩素(Cl−/2b)、臭素(Br−/3b)、フッ素(F−/4b)、メチル(Me−/5b)、メトキシ(MeO−/6b)、ニトロ(NO2−/7b)のラセミ−フェニルエタノール類のR体を選択的酸化し、S体アルコールが化学収率50%、光学純度99%e.e.以上で得られた。
【0122】
実施例3
(第2の工程におけるその他植物由来組織材料由来蛋白質複合体の基質特異性の検討)
第2の工程において、小麦若葉(YWL)、大麦若葉(YBL)、ヨモギの葉(AVI)、小麦ふすま(WB)、ワカメ(WS)、ニンジン(C)、カボチャ(P)といった草や雑草をも含めた種子、葉、茎、根、花、実からの不斉酸化反応を有する蛋白質複合体の溶出を確認する目的で、温水60℃以上で10分間漬け置き洗浄後、必要により薄くスライスして、凍結真空乾燥(FD)後、ボールミルを用いて細かく粉砕した。これらの植物材料粉末20gは定法に従って、10倍等量の蒸留水200mlを加えて、3%アルギン酸ナトリウム水溶液を1倍等量200ml加えて均一になるまで攪拌し、得られた植物組織含有・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中に滴下して、植物組織含有・アルギン酸カルシウムゲルビーズを作製する。該ビーズに、植物組織含有粉の20倍等量の蒸留水(400ml)を反応溶液として添加し、恒温振とう温度設定40℃、振とう数55rpmの条件にして、10時間以上の振とう後、基質変換反応を行った。基質としてはメタ、パラ位に臭素(Br−/1m、1p)、塩素(Cl−/2m、2p)、フッ素(F−/3m、3p)、メチル(Me−/4m、4p)、メトキシ(MeO−/5m、5p)、2−ナフチルエタノール(naphthyl/6)を置換させたフェニルエタノール類を作用させて、得られた結果については、反応時間、生成物、光学純度、化学収率を以下の表にまとめた。
【0123】
(第2の工程における小麦若葉、大麦若葉、小麦ふすま由来蛋白質複合体のメタ位置換基質の基質特異性の検討)
【表4】
【0124】
以上の結果から、小麦若葉(YWL)、大麦若葉(YBL)、小麦ふすま(WB)由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質であるメタ位置換臭素(Br−/1m)、塩素(Cl−/2m)、フッ素(F−/3m)、メチル(Me−/4m)、メトキシ(MeO−/5m)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のR体選択的に酸化して、S体アルコールが化学収率50%、光学純度99%e.e.以上で得た。
【0125】
(第2の工程における小麦若葉、大麦若葉、小麦ふすま由来蛋白質複合体のパラ位置換基質の基質特異性の検討)
【表5】
【0126】
以上の結果から、小麦若葉(YWL)、大麦若葉(YBL)、小麦ふすま(WB)由来の蛋白質複合体はゲルより好適に溶出され、同時に、基質であるパラ位置換臭素(Br−/1p)、塩素(Cl−/2p)、フッ素(F−/3p)、メチル(Me−/4p)、メトキシ(MeO−/5p)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のR体選択的に酸化して、S体アルコールが化学収率50%程度、光学純度99%e.e.以上で得た。
【0127】
(第2の工程におけるヨモギの葉、ワカメ、ニンジン、カボチャ由来蛋白質複合体の基質特異性の検討)
【表6】
【0128】
以上の結果から、ヨモギの葉(AVI)、ワカメ(WS)、ニンジン(C)、カボチャ(P)由来の蛋白質複合体ゲルより好適に溶出され、同時に、基質であるパラ位置換臭素(Br−/1p)、塩素(Cl−/2p)、フッ素(F−/3p)、メチル(Me−/4p)、メトキシ(MeO−/5p)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のR体選択的な酸化により、S体アルコールが化学収率20〜50%、光学純度99%e.e.以上で得た。
【0129】
実施例4
(第2の工程における卵白アルブミン由来蛋白質複合体のパラ位置置換基質の基質特異性の検討)
第2の工程において、卵白から分離されたオボアルブミンから作成される蛋白質複合体の活性測定を目的として、鶏卵より卵白を分離し、硫安沈殿にてオボアルブミンを分離し、0.1〜0.5%のオボアルブミンの濃度になるように水に溶解後噴霧乾燥処理して、粉末の卵白アルブミン粉末を調整した。これらの卵白アルブミン粉末20gは定法に従って、10倍等量の蒸留水200mlを加えて、3%アルギン酸ナトリウム水溶液を1倍等量200mlを加えて均一にし、得られた卵白アルブミン・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中に滴下して、卵白アルブミン・アルギン酸カルシウムゲルビーズを作製した。該ビーズに、卵白アルブミン粉の20倍等量の蒸留水(400ml)を反応溶液として添加し、恒温振とう培養設定40℃、振とう数55rpmにて、10時間以上振とう後、基質変換反応を行った。基質としてはパラ位に臭素(Br−/1)、塩素(Cl−/2)、メトキシ(MeO−/5)、2−ナフチルエタノール(naphthyl/6)を置換させたラセミ−フェニルエタノール類を作用させて、得られた反応時間、生成物、光学純度、化学収率の結果については以下の表7にまとめた。
【0130】
(第2の工程における卵白アルブミン由来蛋白質複合体のパラ位置換基質の位置特異性の検討)
【表7】
【0131】
以上の結果から、卵白アルブミン由来の蛋白質複合体は、ゲルより好適に溶出され、同時に、基質パラ位置換臭素(Br−/1)、塩素(Cl−/2)、メトキシ(MeO−/5)、2−ナフチルエタノール(naphthyl/6)のラセミ−フェニルエタノール類のS体選択的に酸化して、R体アルコールが化学収率26%、光学純度85〜95%e.e.で得られた。この結果は、植物資源由来粗蛋白質由来の蛋白質複合体がS体アルコール(99%e.e.)を得るのに対し、卵白アルブミン由来が逆のR体アルコール(85〜95%e.e.)が得られ、動植物の使い分けにより両鏡像体の使い分けが可能となる。
【0132】
実施例5
(第2の工程、恒温振とう器中で10時間の空振とうする理由)
図1は第2の工程において、豌豆蛋白4gを40mlの蒸留水に溶かして、更に3%アルギン酸ナトリウム(40ml)を加えて均一化した溶液を、4.0%の塩化カルシウム水溶液中に滴下して調整された、豌豆蛋白・アルギン酸カルシウムゲルビーズに、120mlの蒸留水を加えて、恒温振とう器設定温度40℃、振とう数55rpmにて振とうさせて、空振とう開始後0時間後、5時間後、10時間後、15時間後、20時間後、25時間後、35時間後、45時間後、そして55時間後に基質R−2−ナフチルエタノールを加えた時の、その豌豆蛋白質由来蛋白質複合体の不斉酸化活性に伴う基質残量%を示している。
【0133】
その結果、豌豆蛋白質由来蛋白質複合体の不斉酸化反応は、空振とう後10時間よりゲルより溶出され、同時に、基質R−2−ナフチルエタノールに好適に作用することが判った。
【0134】
実施例6a
(第2の工程、基質特異性の検討)
図2aは、図1と同一条件で振とう開始後10時間後、a)は基質特異性を調べる目的で、基質ラセミー2−ナフチルエタノール、R−2−ナフチルエタノール、2−アセトナフトンをそれぞれ50mgづつ添加したときの反応時間に伴う基質と生成物の%比率をモニターリングしたものである。その結果、空振とう10時間後に現れる活性は、S体アルコールは酸化せず、基質ラセミー2−ナフチルエタノールのR体のみを立体選択的に不斉酸化することが判った。
【0135】
実施例6b
(第2の工程、反応箇所の検討)
図2bは第2の工程において、図1と同一条件で豌豆・アルギン酸ナトリウムを調製後、空振とう10時間後の水溶液(120ml)、空振とう10時間後の水溶液(240ml)、空振とう10時間後ビーズ+蒸留水(DW:120ml)、新しいビーズ+空振とう10時間後の水溶液(120ml)でそれぞれ基質R−2−ナフチルエタノール(50mg、75mg、又は100mg)を加え、不斉酸化活性をモニタリングした図である。
その結果、空振とう10時間後に現れる蛋白質複合体は、ビーズ中ではなく、水溶液中に反応の場があり、好適に2−ナフチルエタノールのR体のみに作用することが判った。
【0136】
実施例6c
(第2の工程、包括ゲル化塩化カルシウム濃度の活性に及ぼす効果)
図2cは第2の工程において、図1と同一条件で豌豆・アルギン酸ナトリウム混液を調整後、異なる塩化カルシウム濃度(5g/L、7.5g/L、10g/L、15g/L、20g/L、30g/L)中に滴下してゲル化して、同じ条件にて10時間空振とう後、基質R−2−ナフチルエタノールを加えて、その塩化カルシウム濃度に及ぼす蛋白質複合体の活性強度の差をまとめた図である。
【0137】
その結果、塩化カルシウム濃度が低いほど、蛋白質複合体の作成と溶出が効率的であって、不斉酸化活性は高まることが判った。逆に、塩化カルシウム濃度が高いほど、蛋白質複合体が溶出し難い傾向を確認できた。一方、塩化カルシウム濃度10g/L、15g/L、20g/Lでは殆ど活性に差は見られなかった。
【0138】
実施例6d
(第2の工程、豌豆蛋白量の最適化)
図2dは第2の工程における、豌豆アルギン酸ゲルを調製時の豌豆蛋白量(2g、3g、4g、5g)の活性に及ぼす影響について調べた。この場合、塩化カルシウム濃度10g/Lで、豌豆・アルギン酸カルシウムゲルを調製し、空振とう10時間後に得られる蛋白質複合体が及ぼす、基質R−2−ナフチルエタノール(50mg)への影響を示した図である。
【0139】
その結果、好適な蛋白質複合体作成と抽出の条件は、豌豆蛋白量が4gであるが、2g〜5gでは何れも活性に与える蛋白質複合体に差は殆ど見られなかった。
【0140】
実施例7
(第2の工程、香料であるR−1−オクテン−3−オールの合成検討)
図8は第2の工程において、図1と同一条件で豌豆・アルギン酸カルシウムゲルに包括化し、空振とう10時間後に得られる蛋白質複合体の、基質ラセミ−1−オクテン−3−オールの分割に関わる効果を調べた図である。
【0141】
この結果から、蛋白質複合体の触媒作用は、生成ケトンのクロマトピークが2%以下であることと、化学収率が85%以上であることから、図8に示すR−1−オクテン−3−オール(98.2%ee)のGCクロマトは、デラセミ化により得られることが判った(後述する実施例12と表11では連続再利用について検討している)。
【0142】
実施例8
(第3の工程の蛋白質複合体の抽出、好適な硫安濃度の検討)
実施例1の条件で得られる蛋白質複合体溶液を、50mLプラスチック遠心管中に40ml移して、1)各8.8g硫安を加え飽和30%として20時間以上静置後、2)遠心分離(10.000rpm、15min)して、硫安溶液をデカンテーション除去後、3)50mMTris塩酸緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール3ppmを10ml加えて不斉酸化活性を測定した。4)デカンテーション後の溶液画分は更に、3.1g硫安を加え飽和40%にして20時間以上静置後、遠心分離して得られる沈殿物は、その有する不斉酸化活性を同様な方法で検査した。デカンテーション後の溶液画分は更に、3.15g硫安を加え、飽和50%にして静置後、遠心沈殿して同様にその酸化活性を検査した。デカンテーション後の溶液画分は更に、3.3g硫安を加え、飽和60%にして遠心分離して静置後、同様に酸化活性を検査した。デカンテーション後の溶液画分は更に、3.45g硫安を加え、飽和70%にして遠心分離して静置後、同様に酸化活性を調べた。その結果を以下に示す。
【0143】
表8(反応30時間後の結果)
【表8】
【0144】
以上の結果から第3の行程の蛋白質複合体の結晶化の最適な硫安濃度は飽和30%であると判定した。得られる蛋白質複合体が非水溶性グルテン様蛋白質であり、カルシウム存在下で水溶性に変化している性質なので、飽和30%の弱水溶性条件にて沈殿できると推察できた。
【0145】
実施例9
(第3の工程の結晶化蛋白質複合体が処理する基質濃度)
実施例1の条件で得られる結晶化蛋白質複合体水溶液を、50mLプラスチック遠心管に移して、1)各8.8g硫安を加え飽和30%として静置し、2)遠心分離(10.000rpm、15min)してデカンテーション後、沈殿して得られる結晶化蛋白質複合体に、3)50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−2−ナフチルエタノール濃度溶液(1ppm、2ppm、3ppm、4ppm、5ppm)分を各10mL加えて調べた。
【0146】
表9(反応30時間後の結果)
【表9】
【0147】
以上の結果から、第3の工程で得られる水分を含む結晶化蛋白質複合体約50mg/50ml遠心管が酸化できる基質ラセミ−2−ナフチルエタノール濃度は1ppm〜5ppm/10mlで顕著な差は無く不斉酸化できることを確認した。
【0148】
実施例10
(第4の工程の結晶化蛋白質複合体の抽出、及びグルタルアルデヒド架橋濃度0.1%と1.0%の検討)
実施例1の条件で得られる蛋白質複合体溶液を、50mLプラスチック遠心管中に40ml移し取り、1)各8.8g硫安を加え飽和30%として20時間以上静置後、2)25%グルタルアルデヒド溶液をそれぞれ0.1%(0.32ml/40ml)、1.0%(1.6ml/40ml)になるように加えて10時間以上静置後、3)遠心分離(10.000rpm、15min)して、上清の硫安・グルタルアルデヒド溶液をデカンテーション除去した。沈殿の蛋白質複合体は、50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−2−ナフチルエタノール濃度3ppm分を10ml加えて不斉酸化の活性を確認した。
【0149】
表10(反応30時間後と60時間後の結果)
【表10】
【0150】
以上の結果から第4の工程の結晶化蛋白質複合体の架橋のGA濃度は0.1%と1.0%で何れも酸化活性を示すが、1.0%に比べて0.1%のほうがR-アルコール不斉酸化活性がより好適に進んだ。更に、0.1%に近い濃度ほど、デラセミ化反応が起こり易い傾向が確認できた。
【0151】
実施例11
(第4の工程の蛋白質複合体粉末に関わる連続再利用の検討)
実施例1の条件で得られる蛋白質複合体溶液40mlを、50mLプラスチック遠心管4本分にそれぞれ移し、1)各8.8g硫安をそれぞれ加えて飽和30%とし、一昼夜静置後、2)25%グルタルアルデヒド溶液をそれぞれ0.1%になるように0.32ml/40ml加え、2〜4時間静置した。上清の硫安・グルタルアルデヒド溶液はそれぞれ、3)遠心分離(10.000rpm、15min)してデカンテーション除去して、目的の沈殿物蛋白質複合体は更に、凍結真空乾燥(FD)後に粉砕した。蛋白質複合体粉4)358mg分を3ppmラセミ−2−ナフチルエタノール・50mMTrisHCl緩衝液(pH8.0、40mL)に溶かし、活性を測定した。連続再利用は、9時間後と15時間後に行い、遠心分離後に沈殿物を回収して、新たに3ppmラセミ−2−ナフチルエタノール・50mMTrisHCl緩衝液(pH8.0、40mL)を加え、この繰り返し作業にて検討した。その結果は次の通りである。
【0152】
表11(蛋白質複合体の連続再利用の結果)
【表11】
【0153】
以上の結果から第4の工程の蛋白質複合体粉末に関わる連続再利用の可能性について確認した結果、最低16回(8日間)は、不斉酸化活性を維持できて再利用できることが判った。
【0154】
実施例12
(第4の工程の蛋白質複合体粉末の連続再利用に関わる香料(R-1−オクテンー3−オール)の合成)
実施例11で得られる蛋白質複合体の凍結真空乾燥(FD)粉末5gに、ラセミ-1-オクテン-3-オール濃度10ppmの50mMTrisHCl(pH8.0)緩衝液150mlを加えて、500ml三角フラスコ中で不斉酸化反応を試みた。30時間後又は60時間後に5mlの反応溶液を遠心分離(10000rpm、15min)により上清のみを回収し、ジエチルエーテルで基質を抽出後、GC測定を行った。尚、連続再利用の操作は、実施例11と同様な方法にて光学純度と収率を導いた。反応は60時間で止め、遠心分離した上清部をジエチルエーテルにて抽出後、飽和塩水にて洗浄、シリカゲルクロマトグラフィーにて単離・精製を行って化学収率と光学純度を求めた。
【0155】
表12(香料(R−1−オクテンー3−オール)の連続再利用による合成)
【表12】
【0156】
以上の結果から、基質ラセミ−1−オクテン−3−オールの連続的合成に関しては、最低4回は再利用でき、生成ケトンは何れの場合も4%未満で且つ、化学収率が85%以上であることから、デラセミ化反応が生じていると判定できた。得られた光学純度は4回とも98%ee以上の高光学純度でえられ、図5は得られたR−1−オクテン−3−オールのGC/FIDスペクトルの結果を示している。
これまでの結果から、下記の反応式に示すように、ベンゼンやナフタレン骨格を有するアリール系基質に関しては50%の生成ケトンは還元され難いが、1−オクテン−3−オールなどのアルキル鎖系基質に関しては生成ケトンが更に不斉還元されるデラセミ化が生じることが判った。
【0157】
【化1】
【0158】
実施例13
(蛋白質複合体の分子量決定(SDS−page))
図3は豌豆−Caアルギン酸ゲルを10時間以上温水中にて振とう後、溶出される蛋白質複合体を硫安30%にて結晶化沈殿して得られる沈殿物のSDS−pageである。図に示す「酵素溶液」レーンは、10時間以上温水振とう後、溶出される温水溶液そのものであり、「硫安沈殿」レーンは、該温水溶液を硫安30%飽和になるように結晶化沈殿し、該沈殿物に水を加えて再溶解させたものであり、「上清」レーンは、該硫安沈殿物を遠心分離して得られる上清に示すSDS−page結果である。
【0159】
図4は「食品と技術」2008年12月号総説 裏出令子記述(グルテンタンパク質のネットワーク形成における食塩の役割)に掲載済みの「図5(DSTによる架橋効率に食塩添加が及ぼす影響)」の抜粋である。図は、小麦粉に食塩添加又は無添加の水溶液を加えて作成した生地を、βメルカプトエタノール入りSDS溶液で可溶化して得られる液を超遠心し、可溶性画分(S)と不溶性画分(P)についてSDS−pageを行った結果を表す。不溶性画分はメタ過ヨウ素酸ナトリウムで処理し、SDSポリアクリルアミドゲル電気泳動で分離しタンパク質を染色してある。
【0160】
以上、図4について、図3のSDSにより切られたS-S結合蛋白成分のフラグメントのバンド配置の持つ分子量が図3に示すSDS-pageと一致していることから、蛋白質複合体は、酵素ではなく、高分子・低分子グルテニンとグリアジンに似た成分より成ると判定できた。グルテニンは空気酸化(及び/又は内因性酵素)の働きにより、高分子・低分子グルテニン分子間にジスルフィド結合を生じてポリマー化するが、このチオール−ジチオール部位が酸化還元反応の活性ドメインとなり、基質ラセミアルコールの不斉酸化を発現するようになると推察できる。溶存カルシウムは、分子間ジスルフィド結合形成を誘導するだけでなく、高分子・低分子グルテニン分子間凝集(6.4Å以下)、更には、水溶性への性質変化にも関与していると考えられる。
【0161】
実施例14
図5は、フーリエ変換赤外分光分析(FT−IR)を用いるサンプル(i)〜(vi)の定性分析結果である。各サンプル間の分子構造上の違いを比較する目的で行った。尚、サンプル(i)〜(vi)のサンプル情報は以下の通りである。
(1)エンドウ蛋白質粉末・・・前述の実施例1に記載の方法にて得られる。
【0162】
(2)アルギン酸ナトリウム粉末・・・市販品より得られる。
(3)振とう培養器製の架橋結晶化蛋白質複合体粉末・・・後述の実施例15に記載の方法にて得られる。
【0163】
(4)ジャーファメンタ製の架橋結晶化蛋白質複合体粉末(iv)・・・後述の実施例16に記載の方法にて得られる。
【0164】
(5)20mM塩化Ca/50mMトリス塩酸バッファー(pH6.0)及び(pH8.0)・・・実施例17に記載の方法にて得られる。
【0165】
図5の結果、不斉酸化反応を有する「サンプル(iii)(iV)」と不斉酸化反応が起こらない「サンプル(v)(vi)」のフーリエ変換赤外分光分析(FT−IR)上での違いは、1)1411cm−1にあるカルボキシレート(R−C(=O)−O−Ca)の水酸基(−O−Ca)の吸収と、2)1082cm−1の糖エーテル(−C−O−C−)又は硫安(−O−S(=O)2−O−)の吸収の2点にある。不斉酸化反応を有する「サンプル(iii)(iV)」は供にこの2点に強い吸収が見られることから、1)サンプル内にアルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り込んだのか、2)分子間ジスルフィド結合(−S−S−)の際に硫黄酸化物を生じたとすれば説明できる。また、この2点が「不斉酸化反応を有する」か否かの重要なポイントとなる部分であると考えられ、アルギン酸Caに粗蛋白質を包括する意義が存在している。
【0166】
更に、図6は、X線マイクロアナライザーEPMA−1600を用いるサンプル(i)〜(iV)の定性分析結果である。尚、サンプル(i)〜(iV)のサンプル情報は既に説明済みである。
【0167】
図6のX線マイクロアナライザーの結果から、不斉酸化反応を有する「サンプル(i)」には、ナトリウム(Na)が検出されているが、不斉酸化反応を有しない「サンプル(ii)(iii)」には見られていない。従って、アルギン酸ナトリウムを4%塩化カルシウム溶液中に滴下して、アルギン酸カルシウムに包括する第2の工程において、エンドウ蛋白質粉末(iv)由来のナトリウム(Na)が作用を受けずに残ったか、或いはナトリウム(Na)原子がカルシウム(Ca)と置換されなかったカルボキシレート(R−C(=O)−O−Na)のアルギン酸Na分子の残骸/足跡とも推測できる。また、(i)〜(iv)の硫黄(S)の量が(i)の場合に顕著に多いことから、分子間ジスルフィド結合などの形成を経て得られた硫黄酸化物(−O−S(=O)−O−)が相対的に不斉酸化反応の要因になっている可能性が示されている。
【0168】
従って、後の実施例19に記載する酸素(O2)の吸収が、エンドウ蛋白質内システイン(R1−S−H H−S−R2)通しの分子間ジスルフィド結合(R1−S−S−R2)の形成に関与するものであったと推察できた。また、(iv)のナトリウム(Na)原子がアルギン酸Na分子の残骸/足跡である場合、1)サンプル内にアルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り組んでいる可能性も示唆された。
【0169】
実施例15
(図5(iii)及び図6(i)に記載の振とう培養器製による蛋白質複合体の製造法)
第2の工程において、種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し水溶性画分を噴霧乾燥して得られる豌豆(PP)由来蛋白粉末30gに、10倍等量の蒸留水300mlを加えて十分に攪拌・溶解後、3%のアルギン酸ナトリウム水溶液を1倍等量の300mlを加えて均一になるまで攪拌し、得られる豌豆・アルギン酸ナトリウム混合液を4.0%の塩化カルシウム水溶液中(pH9.16)に滴下してゲルビーズを作製した。該作製ビーズに豌豆蛋白粉の20倍等量の蒸留水(500ml)を反応溶液として添加し、恒温振とう培養器の温度設定40℃、振とう数55rpmにて10時間以上の振とうした。
使用装置
(1)反応槽:恒温振とう培養器3器中にそれぞれ5000ml三角フラスコを設置。
振とう数:55rpm
温度設定:40℃
【0170】
攪拌(培養)作業の条件
1.豌豆蛋白質(30g)・アルギン酸カルシウムゲル(約600mL)
2.蒸留水:500mL×3(器)×3(日分)
3.抽出回数:3回/3日(1日に1回づつ回収)
【0171】
抽出作業の条件
1.一日分の培養水溶液500mL×3(器分)をビーズと分離、回収した。
2.硫安沈殿・・・30%飽和硫安になるように工業用硫安を添加、結晶化沈殿の為に1日置き。
3.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加し、2時間以上置き。
4.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
5.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
6.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール濃度10ppmを40mL加えて酸化活性を検討した。
7.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA−300)にて測定した。
【0172】
以上の結果、表12のように、恒温振とう培養器による架橋結晶化蛋白質複合体の製造法の一例をまとめた表であるが、恒温振とう培養器にそれぞれ、アルミフォイルなどで簡易的に封をした5000ml三角フラスコであるので、振とう数55rpmで十分に培養液が波打つので酸素補給が十分に行われ、グルテニン−グルテニンシステイン残基(R1−S−H・・・O・・・H−S−R2)脱水縮合反応(→H2O)を介するジスルフィド結合((−S−S−))によりポリマー化を誘導し、アルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り込んだ。この蛋白質複合体に形成された活性ドメインが、酵素様にRアルコールを選択的に不斉酸化して、100%e.e.のS−アルコールを得ることができたと考えられる。
【0173】
表13(図5(iii)及び図6(i)に記載の振とう培養器製による蛋白質複合体の活性強度)
【表13】
【0174】
注)・抽出液中Ca濃度(mg/mL)・・・振とう溶液のCa濃度を原子吸光高度計にて測定した。
・Ca濃度(mg/g)・・・FD後の粉末サンプルCa量を酸分解処理後、原子吸光光度計にて測定した。
・R体酸化(%)・・・図9記載のスケールに反応させて7hr後、20hr後のGC測定による生成ケトン(%)を記載した。
(基質ラセミアルコールのR体50%が選択的に酸化されてケトンになる為、ケトン%が50%に近いかそれ以上の場合、もう一方のS体アルコールの光学純度は100%近い。)
・光学純度(%e.e.)・・・GCの「S−アルコール(面積%)−R−アルコール(面積%)」より求める。
【0175】
実施例16(図5(iv)に記載の5Lジャーファメンタ製による架橋結晶化蛋白質複合体の製造法)
第2の工程において、種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し水溶性画分を噴霧乾燥して得られる豌豆(PP)由来蛋白粉末50gに、10倍等量の蒸留水500mlを加えて十分に攪拌・溶解後、3%のアルギン酸ナトリウム水溶液を1倍等量の500mlを加えて均一になるまで攪拌し、得られる豌豆・アルギン酸ナトリウム混合液(1000mL分)を4.0%の塩化カルシウム水溶液中(pH9.16)に滴下してゲルビーズを作製した。該作製ビーズに豌豆蛋白粉の20倍等量の蒸留水(1000mL分)を反応溶液として添加し、高崎科学製5L用ジャーファメンタの温度設定40℃、振とう数800rpmにて振とうした。
【0176】
使用装置
(1)反応槽:高崎科学製5L用ジャーファメンタ
羽根:2枚簡易式棒状パドル 800rpm
温度:40℃
酸素供給:0.5mg/L
攪拌(培養)作業の条件
4.豌豆蛋白質(30g)・アルギン酸カルシウムゲル(約1000mL)
5.蒸留水:1000mL×3(日分)
6.抽出回数:3回/3日(1日に1回づつ回収)
【0177】
抽出作業の条件・・・一日分の培養水溶液1000mLをビーズと分離、回収した。
1.硫安沈殿・・・30%飽和硫安になるように工業用硫安を添加、結晶化沈殿の為に1日置き。
2.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加し、2時間以上置き。
3.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
4.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
5.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール濃度10ppmを40mL加えて酸化活性を検討した。
6.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA−300)にて測定した。
【0178】
以上の結果、表13のように、5L用ジャーファメンタにおいても、恒温振とう培養機を用いた場合と同様、目的のアルギン酸Ca由来カルボキシレート(R−C(=O)−O−Ca)と糖エーテル(−C−O−C−)を包括して取り込み、エンドウ蛋白質内に活性ドメインを作成し、蛋白質複合体が、酵素様にRアルコールを選択的に不斉酸化して、100%e.e.のS−アルコールを得ることができたと考えられた。
【0179】
表14(図5(iv)に記載の5L用ジャーファメンタによる蛋白質複合体の活性強度)
【表14】
【0180】
注)・抽出液中Ca濃度(mg/mL)・・・振とう溶液のCa濃度を原子吸光高度計にて測定した。
・Ca濃度(mg/g)・・・FD後の粉末サンプルCa量を酸分解処理後、原子吸光光度計にて測定した。
・R体酸化(%)・・・図9記載のスケールに反応させて7hr後、20hr後のGC測定による生成ケトン(%)を記載した。
(基質ラセミアルコールのR体50%が選択的に酸化されてケトンになる為、ケトン%が50%に近いかそれ以上の場合、もう一方のS体アルコールの光学純度は100%近い。)
・光学純度(%e.e.)・・・GCの「S−アルコール(面積%)− R−アルコール(面積%)」より求める。
【0181】
実施例17(図5(v)(vi)及び図6(ii)(iii)に記載の「架橋結晶化蛋白質(pH6.0又はpH8.0)」の製造法)
アルギン酸カルシウムゲルに包括する第2の工程の必要性を確認する目的で、第2の工程(アルギン酸カルシウムゲルへ包括)を省き、エンドウ粗蛋白質(4g)を2%塩化カルシウム/50mMトリス塩酸バッファー溶液/120ml((v)pH6.0又は(vi)pH8.0)で恒温振とう(55rpm、40℃、24時間)した。振とう後、エンドウ蛋白質カルシウム溶液は、30%飽和になるように硫安を加えて24時間以上静置後(第3の工程)、結晶化沈殿物を0.25%グルタルアルデヒドにて架橋し(第4の工程)、得られる架橋結晶化したエンドウ粗蛋白質を凍結乾燥(FD)処理して粉末とした。
【0182】
(2)反応槽:恒温振とう培養器にそれぞれ300ml三角フラスコを設置。
振とう数:55rpm
温度設定:40℃
【0183】
攪拌(培養)作業の条件
7.豌豆蛋白質(4g)
8.2%塩化カルシウム/50mMトリス塩酸バッファー溶液(120m)
9.抽出回数:1回/1日
【0184】
抽出作業の条件
1.硫安沈殿・・・恒温振とう(55rpm、40℃、24時間)後、30%飽和になるように硫安を加えた。
2.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加して2時間以上置き。
3.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
4.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
5.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−ナフチルエタノール濃度10ppmを40mL加えて酸化活性を検討した。
6.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA-300)にて測定した。
【0185】
以上、表15に示すように、蛋白質複合体の作成には、グルテニン−グルテニンジスルフィド結合((−S−S−))を誘導する溶存Ca濃度と溶存酸素に加えて、「アルギン酸カルシウムゲル」に包括する第2の工程が不可欠でありことが確認できた。
【0186】
表15.(図5(v)(vi)及び図6(ii)(iii)に記載の「蛋白質複合体(pH6.0又はpH8.0)」の活性強度)
【表15】
【0187】
注)・Ca濃度(mg/g)・・・FD後の粉末サンプルCa量を酸分解処理後、原子吸光光度計にて測定した。
・R体酸化(%)・・・7hr後、20hr後のGC測定による生成ケトン(%)を記載した。
・不斉酸化活性
×・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間に殆ど不斉酸化されない場合(生成ケトン10%以下)。
△・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間までに不斉酸化が途中で止まる場合(生成ケトン30%以下)。
○・・・「蛋白質複合体」(350mg)によって、ラセミ−2−ナフチルエタノール(10ppm、40ml)溶液のR体のみが20時間で不斉酸化される場合(生成ケトン40%以上)。
【0188】
実施例18
(第2の工程のアルギン酸カルシウム包括ビーズの空気酸化の時間に及ぼす6時間後の不斉酸化活性(右軸)と蛋白質複合体収量への影響)
図7は第2の工程において、定法に従い球状のエンドウ蛋白・アルギン酸カルシウムビーズを作製後、該ビーズを空気中に0時間、0.5時間、1時間、3時間、5時間、7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−2−ナフチルエタノール溶液/50mMTrisHCl緩衝液(pH8.0、40mL)を加えて、恒温振とう培養(40℃、55rpm)6時間後に酸化活性を示す。
【0189】
以上の結果から、空気酸化時間を長く取るに従い、架橋結晶化蛋白質複合体の不斉酸化活性は強くなった。一方、得られる蛋白質複合体の収量は一時間後と三時間後に高かった。従って、蛋白質複合体は、空気酸化(及び/又は内因性酵素)の影響により、ゲルビーズ内カルシウム塩と酸素環境下、安価な材料の動植物由来粗蛋白質に(i)タンパク質内分子間ジスルフィド結合(−S−S−)、(ii)蛋白質内分子間凝集(分子間距離の短縮:<6.4Å)、(iii)水溶性への性質変化などを誘導して活性ドメイン(チオレドキシンフォールド:Cys−X−Y−Cys配列)を形成し、好適に不斉酸化反応を有する蛋白質複合体を形成することが確認できた。
【0190】
実施例19
(第2の工程の蛋白質複合体の100Lジャー生産におけるpHとDO(溶存酸素濃度)の推移)
図9は第2の工程において、種子を粉砕して、水を加えて1時間後に沈殿物のオカラ画分を除去し水溶性画分を噴霧乾燥して得られる豌豆蛋白粉末1kgは定法に従って、10倍等量の蒸留水10Lを加えて攪拌混合後、3%のアルギン酸ナトリウム水溶液を1倍等量の10Lを加えて、均一になるまで攪拌して得られた豌豆・アルギン酸ナトリウム混合液(約20L)を、ゲル調整装置を用いて4.0%の塩化カルシウム水溶液中に滴下して、直径1〜2mm程度の球状のビーズを作製し、架橋結晶化蛋白質複合体を作成する際に加える蒸留水(20L)のpHとDO(溶存酸素)をモニターリングした図である。
【0191】
使用装置
(2)反応槽:400φ×700H(TL−TL)竪型円筒槽
羽根:4枚傾斜パドル 200φ 35rpm
温度:40℃
(3)晶析槽
420φ×460H 竪型円筒槽
羽根:6枚羽根タービン 120φ
A.攪拌(培養)作業の条件
1.豌豆蛋白質(1kg)・アルギン酸カルシウムゲル(約20L)
2.蒸留水: 1回目−20Lで3日間攪拌(酸素供給なし)後第3.4工程
2回目−20Lで2日間攪拌(酸素供給なし)後第3.4工程
3回目−20Lで1日間攪拌(酸素供給あり)後第3.4工程
4回目−20Lで1日間攪拌(酸素供給あり)後第3.4工程
【0192】
B.蛋白質複合体抽出作業の条件
1.各攪拌後の培養水溶液20L分をビーズと分離、回収した。
2.硫安沈殿・・・30%飽和硫安になるように工業用硫安を添加、結晶化沈殿の為に1日置き。
3.架橋・・・結晶化沈殿溶液に25%グルタルアルデヒド(GA)溶液濃度を0.25%に成る様に添加し、2時間以上置き。
4.遠心分離(洗浄)・・・10000rpm・15minにて沈殿物を回収、上清を除去後、蒸留水を加えて2回繰り返して不純物の洗浄。
5.フリーズドライ(粉末化)・・・凍結真空乾燥機(FD)により実施。
6.不斉酸化活性・・・凍結真空乾燥(FD)粉350mgを50mMTrisHCl緩衝液(pH8.0)に溶かしたラセミ−2−ナフチルエタノール濃度10ppmを40mL加えて酸化させて、R体が酸化して得られるケトン%を表している。
7.Ca濃度(mg/g)・・・FD後の粉末サンプル酸分解処理後、Ca量を原子吸光光度計(Contr.AA-300)にて測定した。
【0193】
図9のように、4.0%塩化カルシウム水溶液(pH9.16)に滴下して作成したビーズは、酸素供給を行わず蒸留水中で攪拌した場合、pHが5.72(1回目)、6.20(2回目)と弱酸性の中で推移し、溶存酸素(DO)は0.7時間後(1回目)、1時間後(2回目)にゼロとなった。また、酸素供給(0.5mg/L制御)を行ない攪拌すると、pHは弱酸性から7.20(3回目)と弱アルカリ条件側に転じ、4回目は常に弱アルカリのpH7.45付近を推移し、DOは3・4回目ともに、0〜1mg/L付近を推移するようになる。
【0194】
以上の結果、酸素供給のない1・2回目は、高い溶存Caイオン濃度が存在している一方で、溶存酸素濃度がゼロ(酸欠)の状態における蛋白質複合体の形成、即ち、グルテニン−グルテニン分子間ジスルフィド結合と分子間距離短縮の効果に伴う活性ドメイン形成の成否を検討した結果であるが活性は認められなかった。更に、引き続いて3・4回目は、新しい水20Lの切り替え3・4回目であり、低い溶存Caイオン濃度の存在下、空気供給による一定の溶存酸素濃度を維持させた場合における蛋白質複合体の形成、即ち、グルテニン−グルテニン分子間ジスルフィド結合と分子間距離短縮の効果に伴う活性ドメイン形成の成否を検討した結果であるが、この場合も弱い酸化活性は認められる一方で、生成アルコールの光学純度は50%eeを超えるに至らなかった。従って、溶存Caイオン濃度と一定の溶存酸素濃度の維持が、グルテニン−グルテニン分子間ジスルフィド結合と分子間距離短縮の効果により生じる活性ドメインの形成において、重要なポイントとなることが確認できた。
【0195】
(第2の工程の3%アルギン酸カルシウム包括ビーズの空気酸化の時間に及ぼす6時間後の不斉酸化活性(右軸)と蛋白質複合体収量(左軸)への影響)
図7は第2の工程において、定法に従い球状のエンドウ蛋白・アルギン酸カルシウムビーズを作成後、該ビーズを空気中に0時間、0.5時間、1時間、3時間、5時間、7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−ナフチルエタノール溶液/50mMtris HCl緩衝液(pH8.0、40ml)を加えて、恒温振とう培養(40℃、55rpm)6時間後に示す酸化活性である。
【0196】
実施例18a
(第2の工程の3%アルギン酸カルシウム包括ビーズの空気酸化の時間に及ぼす6時間後の不斉酸化活性(右軸)と蛋白質複合体収量(左軸)への影響)
図7は第2の工程において、定法に従い球状のエンドウ蛋白・アルギン酸カルシウムビーズを作成後、該ビーズを空気中に0時間、0.5時間、1時間、3時間、5時間、7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−2−ナフチルエタノール溶液/50mMtris HCl緩衝液(pH8.0、40ml)を加えて、恒温振とう培養(40℃、55rpm)6時間後に示す酸化活性である。
【0197】
以上の結果から、空気酸化時間を長く取るに従い、架橋結晶化蛋白質複合体の不斉酸化活性は強くなった。一方、得られる蛋白質複合体の収量は1時間以降一定して11%〜15%程度の間を推移するが、12%〜13%の収率に集中した。従って、蛋白質複合体は、空気酸化(及び/又は内因性酵素)の影響によりゲルビーズ内カルシウム塩と酸素環境下、安価な材料の動植物由来粗蛋白質に(i)タンパク質内分子間ジスルフィド結合(−S−S−)、(ii)蛋白質内分子間凝集(<6.4Å)、(iii)水溶性への性質変化などを誘導して活性ドメイン(チオレドキシンフォールド:Cys−X−Y−Cys配列)を形成し、好適に不斉酸化反応を有する蛋白質複合体を形成することが確認できた。
【0198】
実施例20
(第2の工程のアルギン酸Na濃度(0.5%、1%、1.5%、2%、3%)に及ぼすアルギン酸カルシウム包括ビーズ空気酸化7時間後の不斉酸化活性6時間後(右軸)と蛋白質複合体収量への影響)
図13は第2の工程において、定法に従い球状又は棒状のエンドウ蛋白・アルギン酸カルシウムビーズ(アルギン酸Na濃度(0.5%、1%、1.5%、2%、3%))を作成後、該ビーズを空気中に7時間放置後、定法の恒温振とう培養(40℃、55rpm)にて温水振とう抽出(2日間(1回目)、更に1日間(2回目))を行い、温水抽出液を定法に従い飽和30%硫安で一昼夜静置して沈殿させて、更に、0.25%グルタルアルデヒド濃度にて架橋させ、得られた架橋結晶化蛋白質複合体(CLPCs)を凍結乾燥処理した後、ボールミル粉砕したものの収量及び酸化活性をまとめたグラフである。不斉酸化活性は、架橋結晶化蛋白質複合体350mgに10ppmラセミ−2−ナフチルエタノール溶液/50mMtris HCl緩衝液(pH8.0、40ml)を加えて、恒温振とう培養(40℃、55rpm)6時間後に示す酸化活性である。
【0199】
以上の結果から、得られる蛋白質複合体の収量は、アルギン酸Na濃度が1.5%の時が最も高く、それより濃度を濃くしても薄くしても収量は徐々に減少した(0.5%<1%<1.5%>2%>3%)。一方、蛋白質複合体の持つ酸化活性は、アルギン酸Na濃度が高くなるに連れ若干上昇した(0.5%<1%<1.5%<2%<3%)。従って、アルギン酸Na濃度を1.5%付近に設定して作成した蛋白質・アルギン酸カルシウムゲルを5〜7時間又はそれ以上時間空気酸化させた後、定法に従い架橋結晶化した蛋白質複合体(CLPCs)を得ることで収量の向上が見込めることを明らかにした。
【0200】
実施例21
(基質濃度(10ppm)に対する蛋白質複合体(50mg、75mg、100mg、200mg、300mg)の不斉酸化速度の違い)
図14は、実施例20で得られる蛋白質複合体の凍結真空乾燥(FD)粉末(50mg、75mg、100mg、200mg、300mg)にラセミ−2−ナフチルエタノール濃度10ppmの50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、4時間置きにGCにて、不斉酸化されるR体−2−ナフチルエタノールの残留比(%)について追跡を行った結果を示している。
【0201】
以上の結果、基質濃度(10ppm)に対する不斉酸化反応に対し、触媒である蛋白質複合体の添加量を増やせば増やすほど反応速度は上昇している結果を得た(50mg>75mg>100mg>200mg>300mg)。また、この蛋白質複合体反応は従来のパン酵母や植物培養細胞、更には微生物菌体の持つ補酵素NAD(P)に依存する脱水素反応とは異なり、補酵素NAD(P)に非依存型の脱水素反応であるため、原料コストの低減化だけでなく、反応コストの低減化においても期待できる触媒であると考えられる。
【0202】
実施例21a
(蛋白質複合体(300mg)に対する基質濃度(10−30ppm、30−90ppm、90−110ppm、110−140ppm)の不斉酸化速度の違い)
図15は、実施例20で得られる蛋白質複合体の凍結真空乾燥(FD)粉末(300mg)にラセミ−2−ナフチルエタノール濃度(10−140ppm)の50mMTrisHCl(pH8.0)緩衝液40mlを200ml三角フラスコ中に加えて、恒温振とう培養機(40℃、55rpm)にて温水振とうして反応させ、不斉酸化されるR体−2−ナフチルエタノールの残留比(%)について4時間置きにGC追跡を行った結果を示している。
【0203】
以上の結果、基質濃度(10〜140ppm)の、触媒の蛋白質複合体(300mg)に対する不斉酸化反応は、図15に示す通り、10−30ppm、30−90ppm、90−110ppm、110−140ppmの、4つのパターンに分類された。即ち、基質濃度10−30ppmについては反応時間20時間、30−90ppmについては40時間、90−110ppmについては60時間、110−140ppmについては140時間を要すことが判った。従って、基質濃度は低いほど反応時間が早く、110ppm以上の濃度では、反応時間が極端に遅くなることが判った。
【産業上の利用可能性】
【0204】
動植物資源、例えば穀類や豆類としては、蕎麦、アマランサス、米、小麦、大麦、トウモロコシ、エンバク、ライ麦、粟、ヒエ、キビ、ハト麦、モロコシ等の穀類、又は小豆、インゲン豆、豌豆、リョクトウ、大豆等の豆類由来、更には草や雑草をも含めた、これら穀類や豆類の種(外皮(ふすま、ぬか)、胚芽(スプラウト))、葉(若葉、新芽)、茎、根、花などの各植物組織、更には、動物由来の卵白や筋肉のような各動物組織を凍結真空乾燥(FD)させて細かく粉砕するか、必要によりその温水に溶解して殻を取り除き、水溶性部分を噴霧乾燥して水溶性蛋白質を濃縮する第1の工程と、該粗蛋白質をアルギン酸カルシウムゲルに包括して空気酸化させた後、温水中にて空振とうしてゲルから目的の蛋白質複合体を抽出する第2の工程と、蛋白質複合体を30%飽和硫安にて結晶化し沈殿させる第3の工程と、蛋白質複合体を架橋する第4の工程と、必要により得られた蛋白質複合体を凍結真空乾燥(FD)して粉末にする第5の工程を組み合わせ、医薬品や香料や食品等のファインケミカル分野における合成する光学活性アルコールなどの光学異性体を製造安価で、環境に優しく、平易に生産することができる。
【特許請求の範囲】
【請求項1】
動植物由来水溶性部分から粗蛋白質を濃縮する第1の工程と、
前記蛋白質をゲルに包括して空気酸化させた後、該ゲルから蛋白質複合体を抽出する第2の工程と、
抽出された蛋白質複合体を水溶液中にて結晶化沈殿させる第3の工程と、
沈殿した蛋白質複合体を架橋化する第4の工程を含むことを特徴とする架橋結晶化蛋白質複合体の製造方法。
【請求項2】
前記蛋白質複合体が、蛋白質およびカルシウムを少なくとも含む蛋白質複合体である請求項1に記載の製造方法。
【請求項3】
前記蛋白質複合体が、更に糖を含む蛋白質複合体である請求項1または2に記載の製造方法。
【請求項4】
前記蛋白質複合体が、不斉酸化反応を触媒する活性を有する蛋白質複合体である請求項1〜3のいずれか一項に記載の製造方法。
【請求項5】
前記第1の工程において、動植物由来組織を凍結真空乾燥(FD)させて細かく粉砕した後、水にて溶出される粗蛋白質を沈殿して濃縮する請求項1〜4のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項6】
前記第1の工程において、動植物由来組織を凍結真空乾燥(FD)させて細かく粉砕した後、水にて溶出される粗蛋白質を噴霧乾燥して濃縮する請求項1〜5のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項7】
前記第2の工程におけるゲルがアルギン酸カルシウムゲルである請求項1〜6のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項8】
前記第2の工程における空気酸化が、前記ゲルを空気中で静置させることにより行われる請求項1〜7のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項9】
前記第2の工程において、前記ゲルから温水中に蛋白質複合体を抽出する請求項1〜8のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項10】
前記アルギン酸カルシウムゲルを作成するための塩化カルシウム水溶液の濃度が50mM以上である請求項7に記載の蛋白質複合体の製造方法。
【請求項11】
前記第2の工程において、前記アルギン酸カルシウムゲルを温水中で振るときの温水量が、アルギン酸カルシウムゲル量の1.5倍以下である請求項7または10に記載の蛋白質複合体の製造方法。
【請求項12】
前記第2の工程における空気酸化が酸化活性強度に関連している請求項1〜11のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項13】
前記第2の工程において、ゲルを空気中で30分以上静置させて空気酸化する請求項1〜12のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項14】
前記空気酸化が、蛋白質複合体の形成および該複合体の水溶性を高める効果を与える請求項1〜13のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項15】
前記第2の工程において、前記温水中で振る工程が、恒温振とう培養器を用いて行われる請求項11に記載の蛋白質複合体の製造方法。
【請求項16】
前記第2の工程、前記温水中で振る工程が、ジャーファメンターを用いることによる請求項11に記載の蛋白質複合体の製造方法。
【請求項17】
前記第2の工程において、前記蛋白質複合体が高分子グルテニン、低分子グルテニン、及びグリアジンに類似する性質を有する蛋白質複合体である請求項1〜16のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項18】
前記第2の工程において、蛋白質複合体が微量の塩の存在下において凝集し、分子間距離を短縮させる性質を有する請求項1〜17のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項19】
前記微量の塩が、カルシウムイオンを与える塩である請求項18に記載の蛋白質複合体の製造方法。
【請求項20】
前記第3の工程において得られる蛋白質複合体を結晶化させる際に、硫安を用いる請求項1〜19のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項21】
前記硫安の濃度が飽和30%以下である請求項20に記載の蛋白質複合体の製造方法。
【請求項22】
前記第4の工程において、蛋白質複合体を架橋結晶化させる架橋剤が、グルタルアルデヒド(GA)及び/又はフォルムアルデヒド(FA)である請求項1〜21のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項23】
前記第1の工程における植物が、蕎麦、アマランサス、米、小麦、大麦、トウモロコシ、エンバク、ライ麦、粟、ヒエ、キビ、ハト麦、モロコシの穀類;および小豆、インゲン豆、豌豆、リョクトウ、大豆等の豆類から選ばれる請求項1〜22のいずれか一項に記載の架橋結晶化蛋白質複合体の製造方法。
【請求項24】
前記穀類や豆類が、粉砕時の組織として得られる種子外皮、胚芽、葉、茎、根、花の植物資源を含む請求項23に記載の蛋白質複合体の製造方法。
【請求項25】
前記第1の工程における動物が、鶏類、両生類、魚類、鶏卵由来の卵アルブミンや筋肉から選ばれる請求項1〜22のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項26】
基質たるラセミ体アルコールを、蛋白質複合体と反応させて、該ラセミ体アルコールの一方の鏡像体を選択的に得る光学活性アルコールの製造方法。
【請求項27】
蛋白質複合体を用いて、基質ラセミ体アルコールの一方の鏡像体を立体選択的にケトンへ不斉酸化することにより、反応に関わらない他方の鏡像体を高い光学純度で得る光学活性アルコールの製造方法。
【請求項28】
蛋白質を少なくとも含む蛋白質複合体であって;且つ、FT−IRにおいて、(1085±50cm−1)の領域、および(1411±50cm−1)の領域に、それぞれ少なくとも1個のピークを有することを特徴とする蛋白質複合体。
【請求項29】
不斉酸化反応を触媒する活性を有する請求項28に記載の蛋白質複合体。
【請求項30】
更に糖を含む請求項28または29に記載の蛋白質複合体。
【請求項31】
前記蛋白質が、架橋結晶化蛋白質である請求項28〜30のいずれか一項に記載の蛋白質複合体。
【請求項32】
前記不斉酸化反応が、基質ラセミ体アルコールの一方の鏡像体を選択的に与える酸化反応である請求項29に記載の蛋白質複合体。
【請求項33】
前記不斉酸化反応により得られるケトンが、更に不斉還元されるデラセミ化反応を与える活性を有する請求項29または32に記載の蛋白質複合体。
【請求項1】
動植物由来水溶性部分から粗蛋白質を濃縮する第1の工程と、
前記蛋白質をゲルに包括して空気酸化させた後、該ゲルから蛋白質複合体を抽出する第2の工程と、
抽出された蛋白質複合体を水溶液中にて結晶化沈殿させる第3の工程と、
沈殿した蛋白質複合体を架橋化する第4の工程を含むことを特徴とする架橋結晶化蛋白質複合体の製造方法。
【請求項2】
前記蛋白質複合体が、蛋白質およびカルシウムを少なくとも含む蛋白質複合体である請求項1に記載の製造方法。
【請求項3】
前記蛋白質複合体が、更に糖を含む蛋白質複合体である請求項1または2に記載の製造方法。
【請求項4】
前記蛋白質複合体が、不斉酸化反応を触媒する活性を有する蛋白質複合体である請求項1〜3のいずれか一項に記載の製造方法。
【請求項5】
前記第1の工程において、動植物由来組織を凍結真空乾燥(FD)させて細かく粉砕した後、水にて溶出される粗蛋白質を沈殿して濃縮する請求項1〜4のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項6】
前記第1の工程において、動植物由来組織を凍結真空乾燥(FD)させて細かく粉砕した後、水にて溶出される粗蛋白質を噴霧乾燥して濃縮する請求項1〜5のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項7】
前記第2の工程におけるゲルがアルギン酸カルシウムゲルである請求項1〜6のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項8】
前記第2の工程における空気酸化が、前記ゲルを空気中で静置させることにより行われる請求項1〜7のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項9】
前記第2の工程において、前記ゲルから温水中に蛋白質複合体を抽出する請求項1〜8のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項10】
前記アルギン酸カルシウムゲルを作成するための塩化カルシウム水溶液の濃度が50mM以上である請求項7に記載の蛋白質複合体の製造方法。
【請求項11】
前記第2の工程において、前記アルギン酸カルシウムゲルを温水中で振るときの温水量が、アルギン酸カルシウムゲル量の1.5倍以下である請求項7または10に記載の蛋白質複合体の製造方法。
【請求項12】
前記第2の工程における空気酸化が酸化活性強度に関連している請求項1〜11のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項13】
前記第2の工程において、ゲルを空気中で30分以上静置させて空気酸化する請求項1〜12のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項14】
前記空気酸化が、蛋白質複合体の形成および該複合体の水溶性を高める効果を与える請求項1〜13のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項15】
前記第2の工程において、前記温水中で振る工程が、恒温振とう培養器を用いて行われる請求項11に記載の蛋白質複合体の製造方法。
【請求項16】
前記第2の工程、前記温水中で振る工程が、ジャーファメンターを用いることによる請求項11に記載の蛋白質複合体の製造方法。
【請求項17】
前記第2の工程において、前記蛋白質複合体が高分子グルテニン、低分子グルテニン、及びグリアジンに類似する性質を有する蛋白質複合体である請求項1〜16のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項18】
前記第2の工程において、蛋白質複合体が微量の塩の存在下において凝集し、分子間距離を短縮させる性質を有する請求項1〜17のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項19】
前記微量の塩が、カルシウムイオンを与える塩である請求項18に記載の蛋白質複合体の製造方法。
【請求項20】
前記第3の工程において得られる蛋白質複合体を結晶化させる際に、硫安を用いる請求項1〜19のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項21】
前記硫安の濃度が飽和30%以下である請求項20に記載の蛋白質複合体の製造方法。
【請求項22】
前記第4の工程において、蛋白質複合体を架橋結晶化させる架橋剤が、グルタルアルデヒド(GA)及び/又はフォルムアルデヒド(FA)である請求項1〜21のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項23】
前記第1の工程における植物が、蕎麦、アマランサス、米、小麦、大麦、トウモロコシ、エンバク、ライ麦、粟、ヒエ、キビ、ハト麦、モロコシの穀類;および小豆、インゲン豆、豌豆、リョクトウ、大豆等の豆類から選ばれる請求項1〜22のいずれか一項に記載の架橋結晶化蛋白質複合体の製造方法。
【請求項24】
前記穀類や豆類が、粉砕時の組織として得られる種子外皮、胚芽、葉、茎、根、花の植物資源を含む請求項23に記載の蛋白質複合体の製造方法。
【請求項25】
前記第1の工程における動物が、鶏類、両生類、魚類、鶏卵由来の卵アルブミンや筋肉から選ばれる請求項1〜22のいずれか一項に記載の蛋白質複合体の製造方法。
【請求項26】
基質たるラセミ体アルコールを、蛋白質複合体と反応させて、該ラセミ体アルコールの一方の鏡像体を選択的に得る光学活性アルコールの製造方法。
【請求項27】
蛋白質複合体を用いて、基質ラセミ体アルコールの一方の鏡像体を立体選択的にケトンへ不斉酸化することにより、反応に関わらない他方の鏡像体を高い光学純度で得る光学活性アルコールの製造方法。
【請求項28】
蛋白質を少なくとも含む蛋白質複合体であって;且つ、FT−IRにおいて、(1085±50cm−1)の領域、および(1411±50cm−1)の領域に、それぞれ少なくとも1個のピークを有することを特徴とする蛋白質複合体。
【請求項29】
不斉酸化反応を触媒する活性を有する請求項28に記載の蛋白質複合体。
【請求項30】
更に糖を含む請求項28または29に記載の蛋白質複合体。
【請求項31】
前記蛋白質が、架橋結晶化蛋白質である請求項28〜30のいずれか一項に記載の蛋白質複合体。
【請求項32】
前記不斉酸化反応が、基質ラセミ体アルコールの一方の鏡像体を選択的に与える酸化反応である請求項29に記載の蛋白質複合体。
【請求項33】
前記不斉酸化反応により得られるケトンが、更に不斉還元されるデラセミ化反応を与える活性を有する請求項29または32に記載の蛋白質複合体。
【図1】

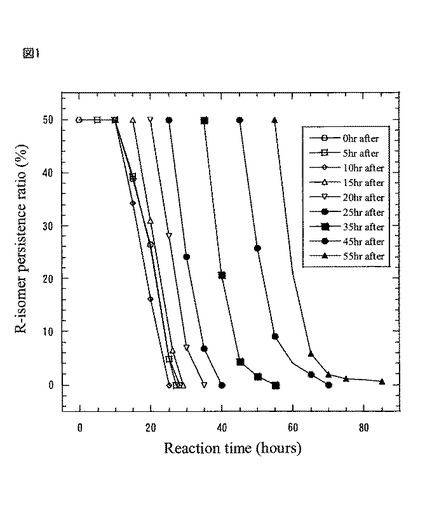
【図2a】
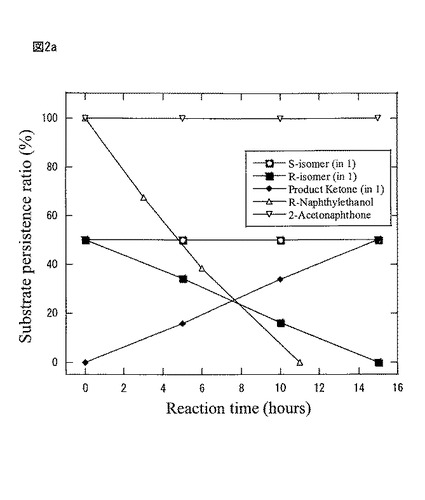
【図2b】
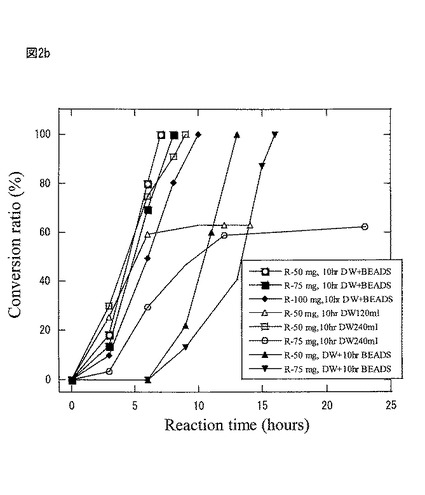
【図2c】
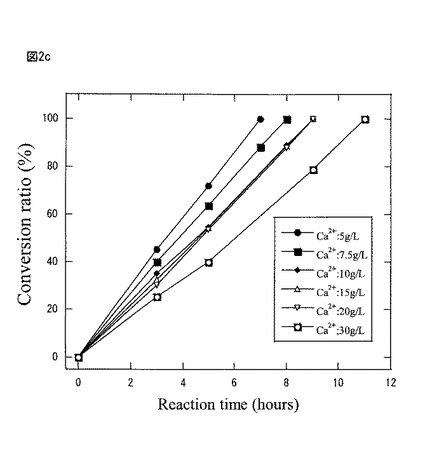
【図2d】
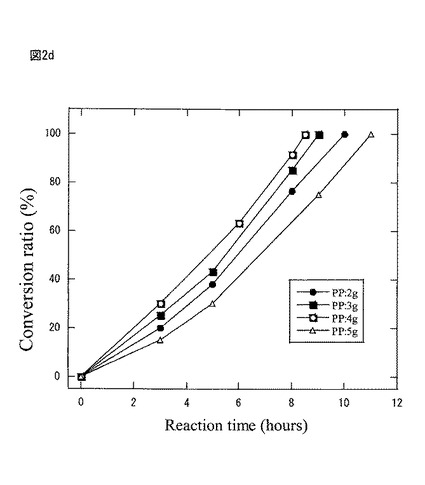
【図5】
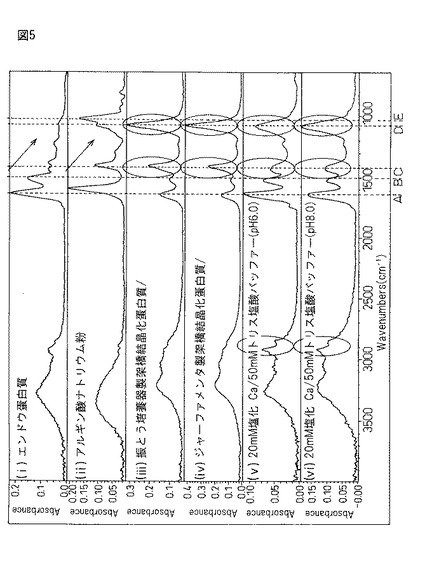
【図6a】
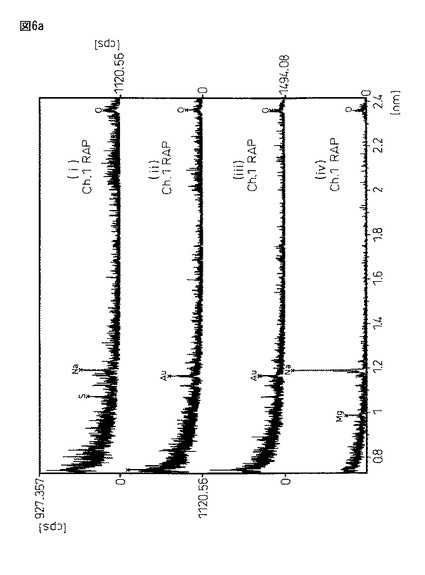
【図6b】
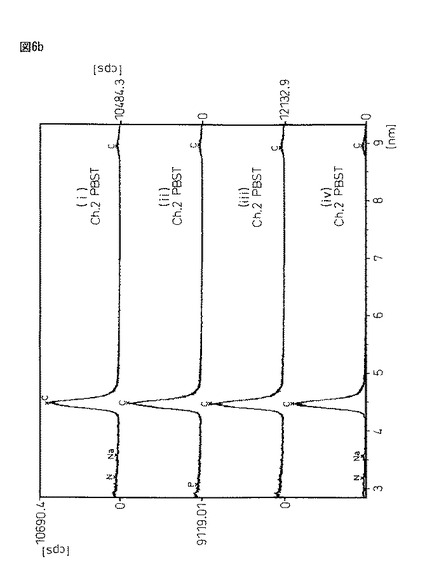
【図6c】
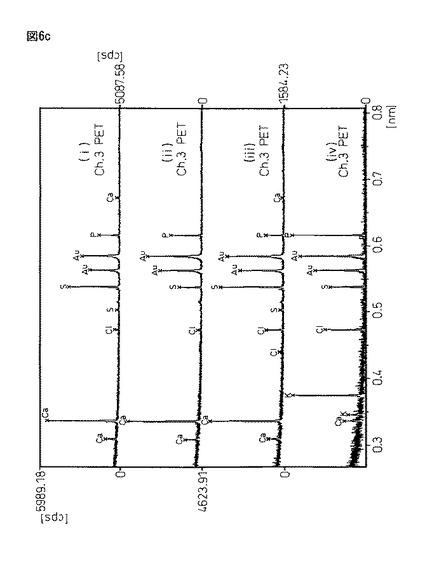
【図6d】
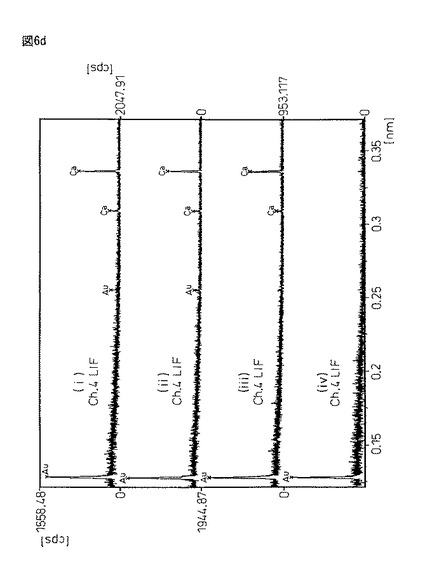
【図7】
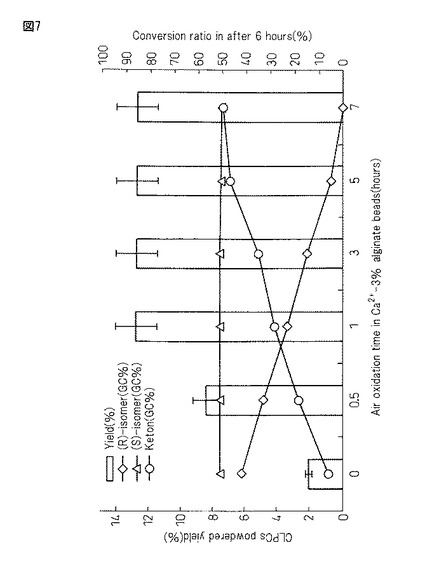
【図8】
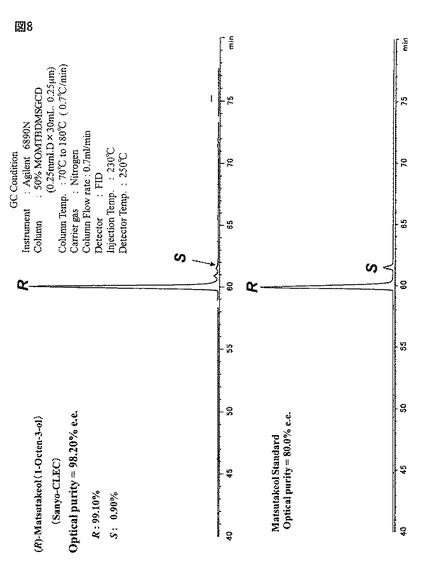
【図9a】
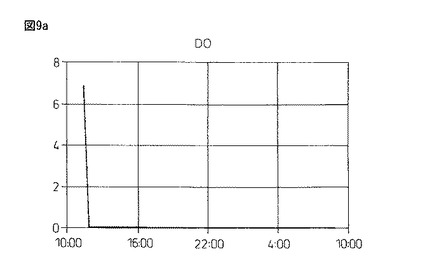
【図9b】
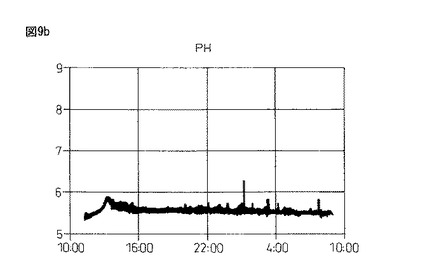
【図9c】
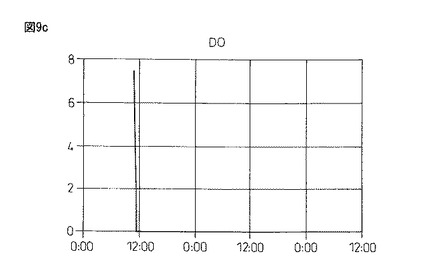
【図9d】
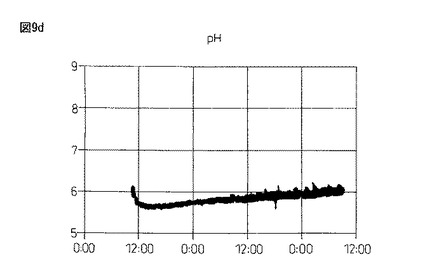
【図9e】
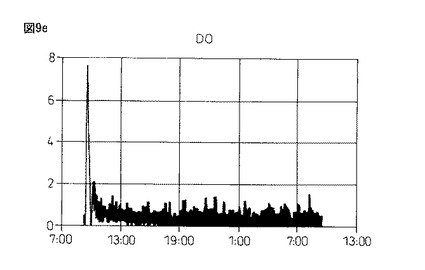
【図9f】
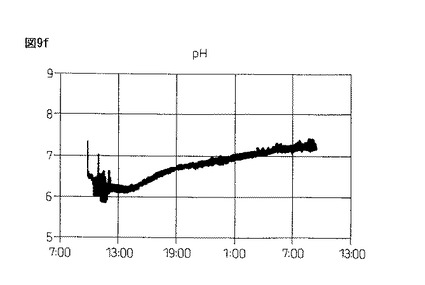
【図9g】
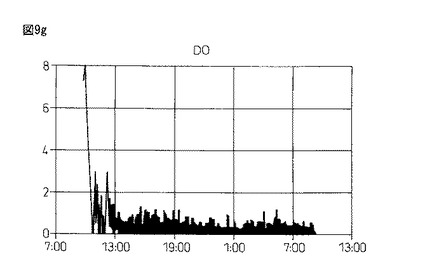
【図9h】
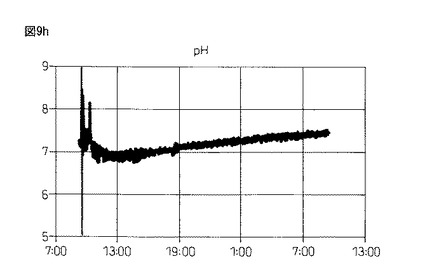
【図10】

【図11a】
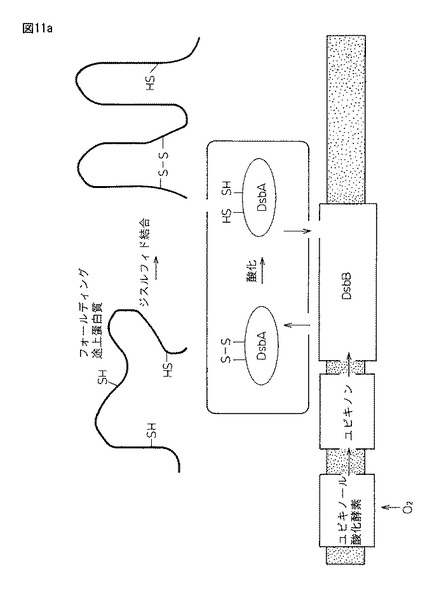
【図11b】
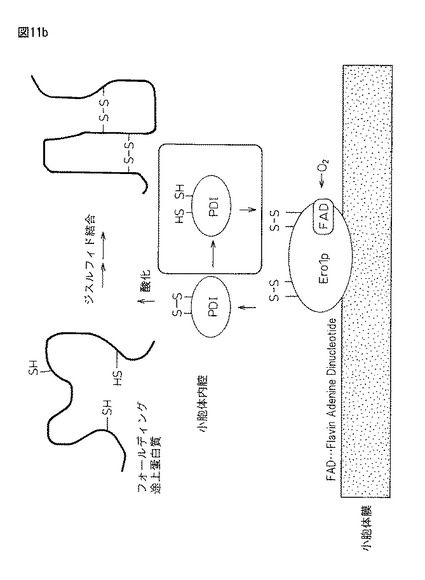
【図12】
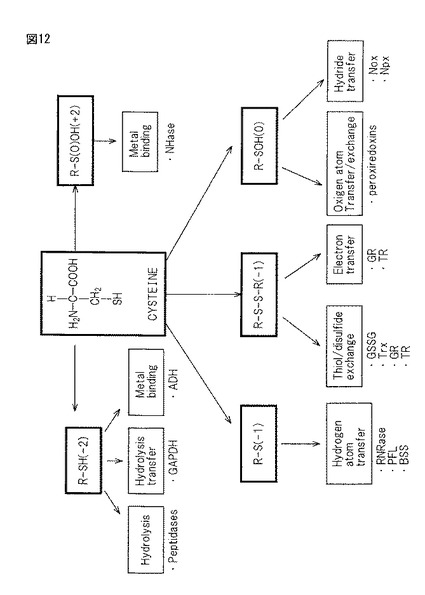
【図13】
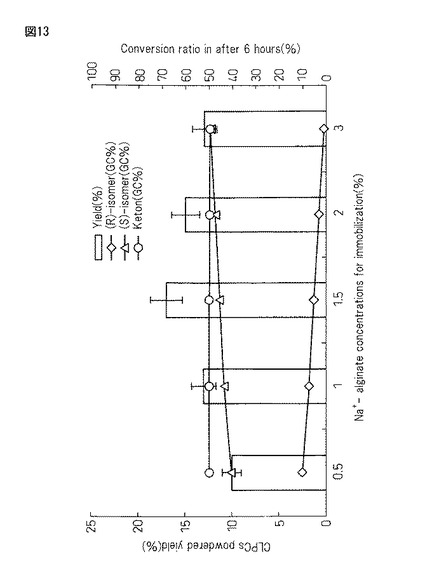
【図14】
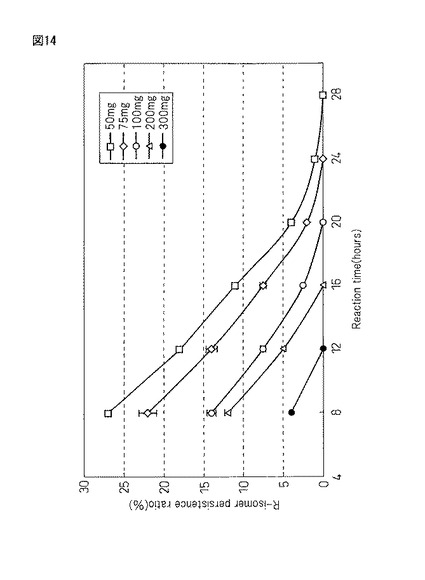
【図15】
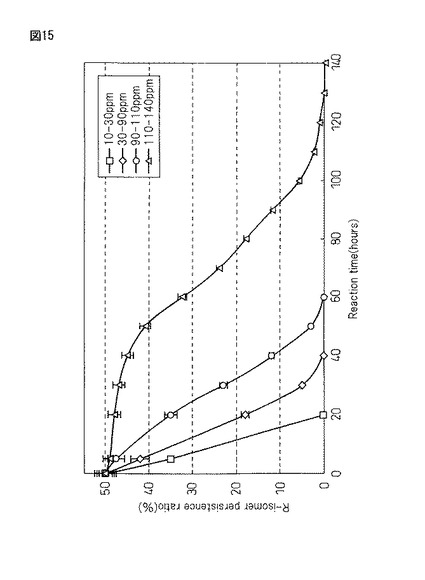
【図3】
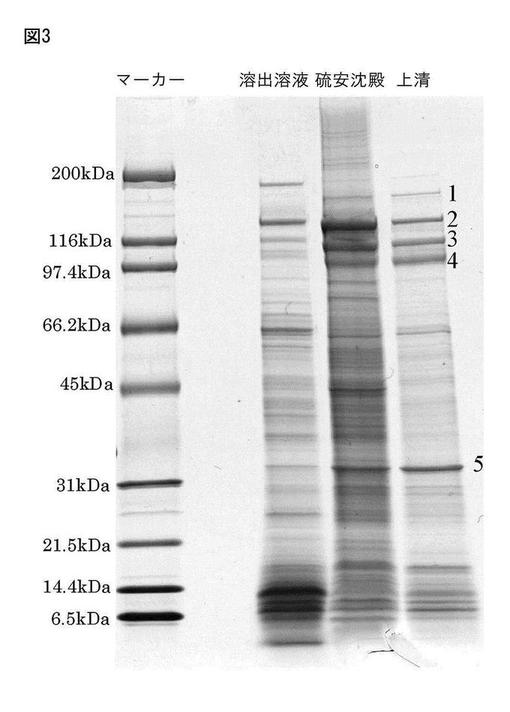
【図4】
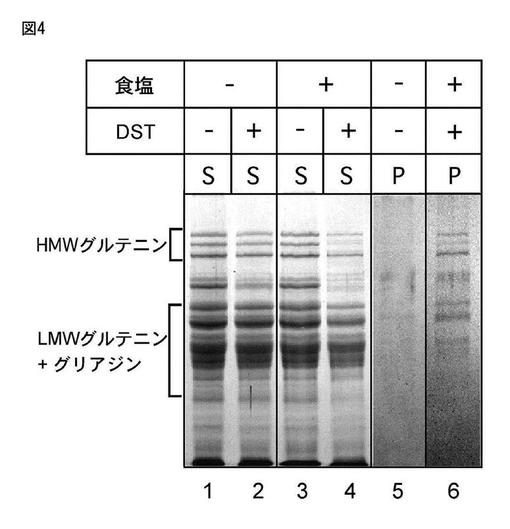
【図2a】
【図2b】
【図2c】
【図2d】
【図5】
【図6a】
【図6b】
【図6c】
【図6d】
【図7】
【図8】
【図9a】
【図9b】
【図9c】
【図9d】
【図9e】
【図9f】
【図9g】
【図9h】
【図10】
【図11a】
【図11b】
【図12】
【図13】
【図14】
【図15】
【図3】
【図4】
【公開番号】特開2013−99353(P2013−99353A)
【公開日】平成25年5月23日(2013.5.23)
【国際特許分類】
【出願番号】特願2013−13467(P2013−13467)
【出願日】平成25年1月28日(2013.1.28)
【分割の表示】特願2011−514475(P2011−514475)の分割
【原出願日】平成22年5月20日(2010.5.20)
【出願人】(000106531)サンヨー食品株式会社 (17)
【Fターム(参考)】
【公開日】平成25年5月23日(2013.5.23)
【国際特許分類】
【出願日】平成25年1月28日(2013.1.28)
【分割の表示】特願2011−514475(P2011−514475)の分割
【原出願日】平成22年5月20日(2010.5.20)
【出願人】(000106531)サンヨー食品株式会社 (17)
【Fターム(参考)】
[ Back to top ]