
並行オリゴヌクレオチド伸長によるDNA配列決定
【課題】現在利用可能なDNA配列決定技術に対して、オリゴヌクレオチドブロックの連続的な連結により1つ以上のプライマーを段階的に伸長することによってテンプレートにおけるヌクレオチドを同定する方法を提供する。
【解決手段】ポリヌクレオチド中のヌクレオチド配列を同定する方法であって、以下の工程:(a)開始オリゴヌクレオチドを、オリゴヌクレオチドプローブと該開始オリゴヌクレオチドとを連結することによってポリヌクレオチドに沿って伸長させ、伸長した二重鎖を形成させる工程;(b)該ポリヌクレオチドの1つ以上のヌクレオチドを同定する工程;および(c)ヌクレオチドの配列が決定されるまで工程(a)および(b)を繰り返す工程、を包含する、方法。
【解決手段】ポリヌクレオチド中のヌクレオチド配列を同定する方法であって、以下の工程:(a)開始オリゴヌクレオチドを、オリゴヌクレオチドプローブと該開始オリゴヌクレオチドとを連結することによってポリヌクレオチドに沿って伸長させ、伸長した二重鎖を形成させる工程;(b)該ポリヌクレオチドの1つ以上のヌクレオチドを同定する工程;および(c)ヌクレオチドの配列が決定されるまで工程(a)および(b)を繰り返す工程、を包含する、方法。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、一般にポリヌクレオチドのヌクレオチド配列を決定する方法に関し、さらに詳しくは、オリゴヌクレオチドブロックの連続的な連結により1つ以上のプライマーを段階的に伸長することによってテンプレートにおけるヌクレオチドを同定する方法に関する。
【背景技術】
【0002】
背景技術
現在利用可能な技術でのポリヌクレオチドの分析は、試験ポリヌクレオチドが標準または単離されたフラグメントと同一であるかまたは異なるかの確認から、試験ポリヌクレオチドの各ヌクレオチドの明白な同定および順序付けまでの範囲の情報を提供する。このような技術は、遺伝子の機能および制御を理解するのに、および分子生物学の基本的技術の多くを適用するのに非常に重要であるだけでなく、これらはまた、ゲノム解析および非常に多くの非研究的適用(例えば、遺伝的同定、法医学分析、遺伝学カウンセリング、医学診断など)における手段としてますます重要となった。これらの後者の適用において、部分的な配列情報を提供する技術(例えば、フィンガープリント法および配列比較)および完全な配列決定を提供する技術の両方が共に使用されてきた。例えば、Gibbsら,Proc.Natl.Acad.Sci.,86:1919−1923(1989);Gyllenstenら,Proc.Natl.Acad.Sci.,85:7652−7656(1988);Carranoら,Genomics,4:129−136(1989);Caetano−Anollesら,Mol.Gen.Genet.,235:157−165(1992);BrennerおよびLivak,Proc,Natl.Acad.Sci.,86:8902−8906(1989);Greenら,PCR Methods and Applications,1:77−90(1991);およびVersalovicら,Nucleic Acids Research,19:6823−6831(1991)。
【0003】
天然のDNAは2つの線状ポリマー、またはヌクレオチドのストランドよりなる。各ストランドは、ホスホジエステル結合によって連結されたヌクレオシドの鎖である。2つのストランドは、2つのストランドのヌクレオチドの相補的塩基の間の水素結合によって逆平行向きにまとまっている:デオキシアデノシン(A)はチミジン(T)と対になり、そしてデオキシグアノシン(G)は、デオキシシチジン(C)と対になる。
【0004】
現在、DNA配列決定に対する以下の2つの基本的アプローチがある:ジデキオシチェーンターミネーション法(例えば、Sangerら,Proc.Natl.Acad.Sci.,74:5463−5467(1977));および化学的分解法(例えば、Maxamら,Proc.Natl.Acad.Sci.,74:560−564(1977))。チェーンターミネーション法はいくつかの方法で改良されており、そして全ての現在利用可能な自動DNA配列決定機のための基礎として供される。例えば、Sangerら,J.Mol.Biol.,143:161−178(1980);Schreierら,J.Mol.Biol.,129:169−172(1979);Smithら,Nucleic Acids Research,13:2399−2412(1985);Smithら,Nature,321:674−679(1987);Proberら,Science,238:336−341(1987);Section II,Meth.Enzymol.,155:51−334(1987);Churchら,Science,240:185−188(1988);Hunkapillerら,Science,254:59−67(1991);Bevanら,PCR Methods and Applications,1:222−228(1992)。
【0005】
チェーンターミネーション法および化学的分解法は、両方とも、1組以上の標識DNAフラグメントの生成を必要とし、そして各々は共通の起源を有し、各々は既知の塩基で終結する。次いで、1組または複数組のフラグメントは、配列情報を得るために、サイズにより分離されなければならない。両方法において、DNAフラグメントは高分解能ゲル電気泳動によって分離され、この電気泳動は、わずか1ヌクレオチドだけサイズが異なる非常に大きなフラグメントを区別する能力を有しなければならない。残念ながら、この工程は一度に配列決定できるDNA鎖のサイズを厳しく制限する。これらの技術を用いる配列決定は、約400〜450ヌクレオチドまでのDNA鎖に信頼性よく適応できる。Bankierら,Meth.Enzymol.,155:51−93(1987);およびHawkinsら,Electrophoresis,13:552−559(1992)。
【0006】
いくつかの著しい技術的問題が、例えば、500〜600ヌクレオチドを超える長い標的ポリヌクレオチドの配列決定、または高容量の多くの標的ポリヌクレオチドの配列決定へのこのような技術の適用をひどく妨げてきた。このような問題としては、i)大きな労働力を要するゲル電気泳動分離工程は、自動化が困難であり、そしてデータの解析に過剰の変動性(例えば、温度の影響によるバンドの広がり、DNA配列決定フラグメントにおける二次構造による圧縮、分離ゲルにおける不均一性など)を導入する;ii)その特性(例えば、進化性、忠実度、重合速度、チェーンターミネーターの取り込み速度など)がしばしば配列依存性である核酸ポリメラーゼ;iii)ゲル中で空間的に重なるバンドに典型的にはfmol量で存在するDNA配列決定フラグメントの検出および分析;iv)標識部分が単一の均一相に濃縮されずに数百の空間的に分離されたバンドに分布することによる低シグナル;およびv)単一レーン蛍光検出の場合、適切な発光特性および吸収特性、量子収率、およびスペクトル分解性を持つ色素の利用可能性が挙げられる。例えば、Trainor,Anal.Biochem.,62:418−426(1990);Connellら,Biotechniques,5:342−348(1987);Kargerら,Nucleic Acids Research,19:4955−4962(1991);Fungら,米国特許第4,855,225号;およびNishikawaら,Electrophoresis,12:623−631(1991)。
【0007】
もう1つの問題が、診断的配列決定の領域における現在の技術には存在する。常に拡大し続けている多数の障害、障害に対する感受性、疾患状態の予後などは、1つ以上の遺伝子座における、特定のDNA配列の存在、またはDNA配列における変化(または変異)の程度に関連付けられてきた。このような現象の例は、ヒト白血球抗原(HLA)タイピング、膵嚢胞性線維症、腫瘍の進行および不均一性、p53プロトオンコジーン変異、rasプロトオンコジーン変異などを含む。例えば、Gyllenstenら,PCR Methods and Applications,1:91−98(1991);Santamariaら,国際出願第PCT/US92/01675号;Tsuiら,国際出願第PCT/CA90/00267号など。診断情報または予後情報を得るためのこのような病的状態に関連するDNA配列の決定における困難は、複数の亜集団のDNA(例えば、対立遺伝子変異体、多重突然変異体形態など)が頻繁に存在することにある。現在の配列決定技術を用いて複数配列の存在および同一性を区別することは、異なる種類のDNAを単離し、そして恐らくはそれをクローン化するさらなる労力なくしては、実質的に不可能である。
【0008】
DNA配列決定に、DNAフラグメントの高分解能電気泳動分離を要しないで、より分析しやすいシグナルを生じ、そしてヘテロ接合遺伝子座由来のDNAを容易に分析するための手段を提供する別のアプローチが利用可能になれば、配列決定技術において主要な進歩がなされ得る。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明の目的は、現在利用可能なDNA配列決定技術に対してこのような別のアプローチを提供することである。
【課題を解決するための手段】
【0010】
本発明によって、以下が提供される:
1)ポリヌクレオチド中のヌクレオチド配列を同定する方法であって、以下の工程:
(a)開始オリゴヌクレオチドを、オリゴヌクレオチドプローブと該開始オリゴヌクレオチドとを連結することによってポリヌクレオチドに沿って伸長させ、伸長した二重鎖を形成させる工程;
(b)該ポリヌクレオチドの1つ以上のヌクレオチドを同定する工程;および
(c)ヌクレオチドの配列が決定されるまで工程(a)および(b)を繰り返す工程、
を包含する、方法。
(2)前記オリゴヌクレオチドプローブが前記開始オリゴヌクレオチドに対して遠位の末端に鎖終結部分を有する、請求項1に記載の方法。
(3)前記同定の工程が、前記鎖終結部分を除去する工程、および1つ以上の標識した鎖終結ヌクレオシド三リン酸の存在下で核酸ポリメラーゼを用いて該オリゴヌクレオチドプローブを伸長させる工程を包含する、請求項2に記載の方法。
(4)前記伸長した二重鎖上に伸長可能な末端を再生する工程をさらに包含する、請求項3に記載の方法。
(5)前記オリゴヌクレオチドプローブが4つのリボヌクレオチドのサブ配列を包含し、そして前記再生の工程が該オリゴヌクレオチドプローブをRNase Hを用いて切断する工程を包含する、請求項4に記載の方法。
(6)前記鎖終結部分が3’ホスフェートである、請求項5に記載の方法。
(7)伸長した二重鎖または前記開始オリゴヌクレオチドが前記オリゴヌクレオチドプローブに連結できない場合は常に、該伸長した二重鎖または該開始オリゴヌクレオチドにキャップ形成させる工程をさらに包含する、請求項2に記載の方法。
(8)前記伸長した二重鎖上に伸長可能な末端を再生する工程をさらに包含する、請求項2に記載の方法。
(9)前記再生の工程が、前記伸長した二重鎖において化学的に切断可能なヌクレオシド間結合を切断する工程を包含する、請求項8に記載の方法。
(10)前記化学的に切断可能なヌクレオシド間結合がホスホルアミデートである、請求項9に記載の方法。
(11)前記再生の工程が、前記伸長した二重鎖においてヌクレオシド間結合を酵素的に切断する工程を包含する、請求項8に記載の方法。
(12)前記オリゴヌクレオチドプローブが4種のリボヌクレオチドのサブ配列を包含し、そして前記再生の工程が該オリゴヌクレオチドプローブをRNase Hを用いて切断する工程を包含する、請求項11に記載の方法。
(13)ポリヌクレオチドのヌクレオチド配列を決定する方法であって、以下の工程:
(a)該ポリヌクレオチドを含むテンプレートを提供する工程;
(b)該ポリヌクレオチドに近接する該テンプレートと二重鎖を形成する開始オリゴヌクレオチドを提供する工程;
(c)該開始オリゴヌクレオチドに近接する該テンプレートにオリゴヌクレオチドプローブをアニールする工程;
(d)該オリゴヌクレオチドプローブを該開始オリゴヌクレオチドに連結させて、伸長した二重鎖を形成させる工程;
(e)該連結されたオリゴヌクレオチドプローブ上の標識によって該ポリヌクレオチドの1つ以上のヌクレオチドを同定する工程;および
(f)該ポリヌクレオチドのヌクレオチド配列が決定されるまで工程(c)〜(e)を繰り返す工程、
を包含する、方法。
(14)前記オリゴヌクレオチドプローブが前記開始オリゴヌクレオチドに対して遠位の末端に鎖終結部分を有し、そして前記方法が該オリゴヌクレオチドプローブ上に伸長可能な末端を再生する工程をさらに包含する、請求項13に記載の方法。
(15)前記オリゴヌクレオチドプローブに連結できない前記伸長した二重鎖または前記開始オリゴヌクレオチドにキャップ形成する工程をさらに包含する、請求項14に記載の方法。
(16)前記同定の工程が前記ポリヌクレオチドの1つのヌクレオチドを同定することよりなる、請求項14に記載の方法。
(17)前記同定の工程が前記鎖終結部分を除去する工程、および1つ以上の標識された鎖終結ヌクレオシド三リン酸の存在下で核酸ポリメラーゼを用いて前記オリゴヌクレオチドプローブを伸長させる工程を包含する、請求項16に記載の方法。
(18)オリゴヌクレオチドプローブであって、以下の式:
HO−(3’)(B)j(5’)−OP(=O)(O−)NH−(B)k−Bt*
を有し、ここで:
Bはヌクレオシドまたはそのアナログであり;
jは1〜12の範囲であり;
kはjとkとの合計が12以下であるように、0〜12の範囲であり;
Bt*は標識された鎖終結部分である、
オリゴヌクレオチドプローブ。
(19)以下からなる群より選択されるオリゴヌクレオチドプローブであって:
OP(=O)(O−)O−(5’)(B)sRRRR(B)wBt*、
HO−(3’)(B)sRRRR(B)wBt*、および
OP(=O)(O−)O−(5’)(B)sRRRR(B)w(3’)OP(=O)(O−)O
ここで:
Bはデオキシリボヌクレオチドまたはそのアナログであり;
Rはリボヌクレオチドであり;
sは1〜8の範囲であり;
wはjとkとの合計が8以下であるように0〜8の範囲であり;
Bt*は標識された鎖終結部分である、
オリゴヌクレオチドプローブ。
【0011】
発明の要旨
本発明は、一本鎖テンプレートに沿った二重鎖伸長の繰り返しサイクルに基づく核酸配列分析方法を提供する。好ましくは、このような伸長は、開始オリゴヌクレオチドとテンプレートとの間に形成される二重鎖から出発する。開始オリゴヌクレオチドを、最初の伸長サイクルにおいて、オリゴヌクレオチドプローブをその末端に連結することにより伸長し、伸長した二重鎖を形成させる。次いで、伸長した二重鎖を、その後の連結サイクルによって繰り返し伸長させる。各サイクルの間に、首尾よく連結されたオリゴヌクレオチドプローブ上、またはオリゴヌクレオチドプローブと会合した標識によって、テンプレート中の1つ以上ヌクレオチドの同一性を決定する。好ましくは、オリゴヌクレオチドプローブは、1回のサイクルで伸長した二重鎖の伸長が1回だけ起こるように、末端の位置にブロッキング部分(例えば、鎖終結ヌクレオチド)を有する。二重鎖を、次のサイクルでブロッキング部分を除去し、そして伸長可能な末端を再生することによってさらに伸長させる。
【0012】
本発明の1つの局面において、複数の異なる開始オリゴヌクレオチドをテンプレートの別々の試料に提供する。各開始オリゴヌクレオチドは、伸長を受けつつある末端が、複数の他のそれぞれの開始オリゴヌクレオチドと、1つ以上のヌクレオチドと合致しない(out of register)かまたは一致しないように、テンプレートと二重鎖を形成する。言い換えれば、伸長のための出発ヌクレオチドは、異なる開始オリゴヌクレオチドの各々につき、1つ以上のヌクレオチドにより異なる。このようにして、同一の長さのオリゴヌクレオチドプローブを用いる各伸長サイクルの後、同一の相対的相が異なるテンプレート上の開始オリゴヌクレオチドの末端間に存在する。従って、例えば、i)開始オリゴヌクレオチドが1つのヌクレオチドにより一致しない、ii)9マーのオリゴヌクレオチドプローブを伸長工程で用いる、およびiii)9つの異なる開始オリゴヌクレオチドを使用する好ましい実施態様においては、9つのテンプレートヌクレオチドが各伸長サイクルにおいて同時に同定される。
【0013】
定義
ポリヌクレオチドに関して本明細書中で使用する「配列決定」、「ヌクレオチド配列の決定」、「配列決定」および同様の用語は、ポリヌクレオチドの部分配列および全長配列の情報の決定を含む。すなわち、この用語は、配列比較、フィンガープリント法、標識ポリヌクレオチドについての同様のレベルの情報、および試験ポリヌクレオチドの各ヌクレオシドの明確な同定および順序付けを含む。
【0014】
プローブおよび標的ポリヌクレオチドの突出ストランドに関して「完全にマッチした二重鎖」とは、この突出ストランドが、二本鎖構造における各ヌクレオチドが反対のストランド上のヌクレオチドとワトソン−クリック塩基対合をするように、他のものと二本鎖構造を形成することを意味する。この用語はまた、プローブの縮重を減少させるために使用され得る、ヌクレオシドアナログ(デオキシイノシン、2−アミノプリン塩基を有するヌクレオシドなど)の対合を包含する。
【0015】
本明細書中で用いる「オリゴヌクレオチド」という用語は、デオキシリボヌクレオシド、リボヌクレオシドなどを含む、ヌクレオシドまたはそのアナログの線状オリゴマーを含む。通常、オリゴヌクレオチドは、数個のモノマー単位(例えば3〜4)から数百のモノマー単位までの範囲のサイズである。オリゴヌクレオチドを「ATGCCTG」のような文字の配列によって表す場合は、常に、このヌクレオチドは左から右に5’→3’の順序であり、そして特記しない限り、「A」はデオキシアデノシンを示し、「C」はデオキシシチジンを示し、「G」はデオキシグアノシンを示し、そして「T」はチミジンを示すことが理解される。
【0016】
本明細書中で使用する「ヌクレオシド」とは、例えば、KornbergおよびBaker,DNA Replication,第2版(Freeman,San Francisco,1992)に記載されるように、2’−デオキシ形態および2’−ヒドロキシル形態を含む、天然のヌクレオシドを含む。ヌクレオシドに関して「アナログ」とは、例えば、Scheit,Nucleotide Analogs(John Wiley,New York,1980)によって一般的に記載されているように、修飾された塩基部分および/または修飾された糖部分を有する合成ヌクレオシドを含む。このようなアナログは、結合特性の増強、縮重の減少、特異性の増大などのために設計された合成ヌクレオシドを含む。
【0017】
本明細書で使用される「連結」とは、テンプレート駆動反応において、2つ以上の核酸(例えば、オリゴヌクレオチドおよび/またはポリヌクレオチド)の末端間で共有結合または結合(linkage)を形成することを意味する。結合(bond)または結合(linkage)の性質は広範囲に変化し得るし、そして連結は酵素的または化学的に行われ得る。
【図面の簡単な説明】
【0018】
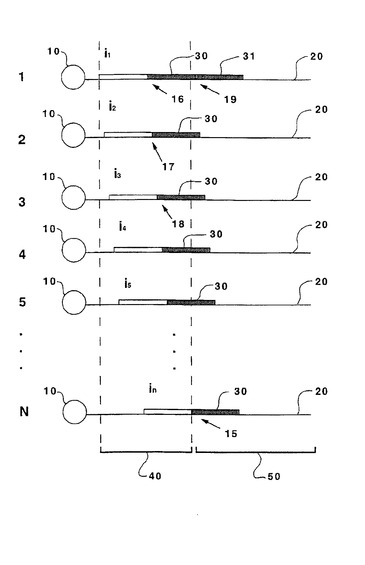
【図1】図1は、本発明による複数のテンプレートの並行伸長を模式的に示す。
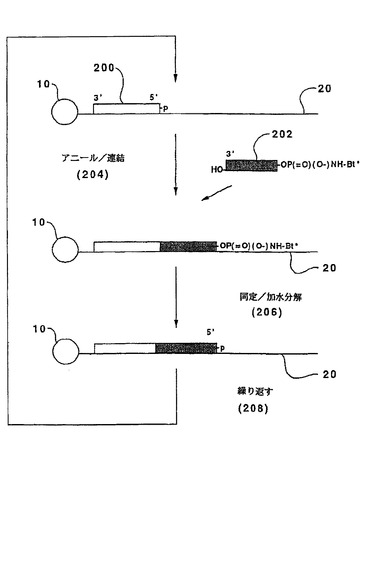
【図2】図2は、酸不安定結合を使用する本発明の1つの実施態様を模式的に示す。
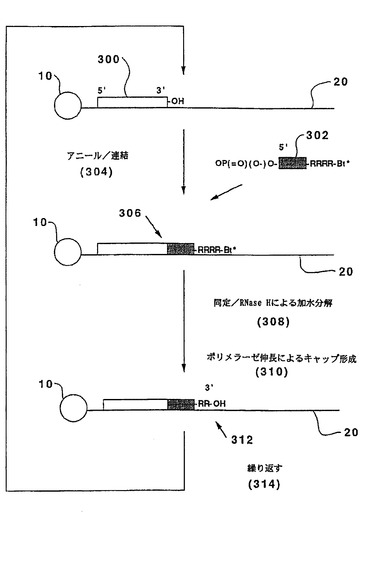
【図3A】図3Aは、3’→5’伸長でRNase H不安定オリゴヌクレオチドを使用する本発明の1つの実施態様を模式的に示す。
【図3B】図3Bは、5’→3’伸長でRNase H不安定オリゴヌクレオチドを使用する本発明の1つの実施態様を模式的に示す。
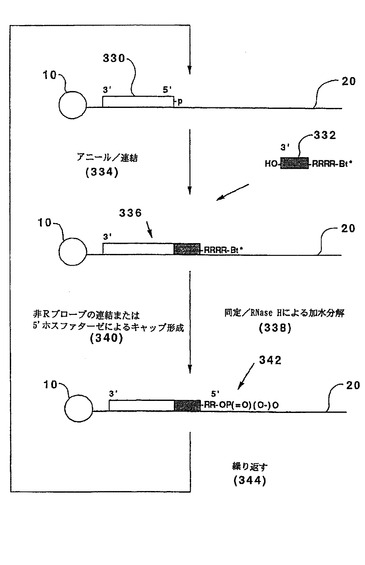
【図4】図4は、連結、その後のポリメラーゼ伸長および切断を使用する本発明の1つの実施態様を模式的に示す。
【発明を実施するための形態】
【0019】
発明の詳細な説明
本発明は、同様のサイズのDNAフラグメントの電気泳動による分離を不要とし、そしてゲルまたは同様の媒体中のDNAフラグメントの空間的に重なるバンドの検出および分析に関する困難をなくす核酸の配列決定方法を提供する。本発明はまた、DNAポリメラーゼを用いて長い一本鎖テンプレートからDNAフラグメントを作製する必要もない。
【0020】
本発明の1つの局面の一般的なスキームを図1に模式的に示す。以下により十分に記載するように、本発明は、この実施態様の特定の特徴によって限定されることを意図しない。配列未知のポリヌクレオチド(50)および結合領域(40)を含むテンプレート(20)を固相支持体(10)に付着させる。好ましくは、Nマーのプローブを使用する実施態様では、テンプレートをN個のアリコートに分け、そして各アリコートに、他の開始オリゴヌクレオチドの位置とは異なる結合領域(40)中のある位置で完全にマッチした二重鎖を形成する異なる開始オリゴヌクレオチドikを提供する。すなわち、開始オリゴヌクレオチドi1〜iNは、未知配列に対して近い側の二重鎖の末端が未知配列のはじまりから0〜N−1ヌクレオチドであるように、結合領域(40)においてテンプレートと二重鎖の組を形成する。従って、Nマーのプローブを用いる連結の第1サイクルにおいて、図1中のi1に連結したプローブ(30)の末端ヌクレオチド(16)は、結合領域(40)のN−1ヌクレオチドに対して相補的である。同様に、図1中のi2に連結したプローブ(30)の末端ヌクレオチド(17)は、結合領域(40)のN−2ヌクレオチドに対して相補的であり;図1中のi3に連結したプローブ(30)の末端ヌクレオチド(18)は、結合領域(40)のN−3ヌクレオチドに対して相補的である。以下同様である。最後に、inに連結したプローブ(30)の末端ヌクレオチド(15)は、未知配列(50)の最初のヌクレオチドに対して相補的である。連結の第2サイクルにおいて、プローブ(31)の末端ヌクレオチド(19)は、開始オリゴヌクレオチドi1で出発する二重鎖における未知配列(50)の2番目のヌクレオチド(19)に対して相補的である。同様に、開始オリゴヌクレオチドi2、i3、i4などで出発する二重鎖に連結したプローブの末端ヌクレオチドは、未知配列(50)の3番目、4番目、および5番目のヌクレオチドに対して相補的である。
【0021】
上記の実施態様において、オリゴヌクレオチドプローブは、伸長した二重鎖に隣接するヌクレオチドの同一性が標識から決定できるように標識される。
【0022】
結合領域(40)は既知配列を有するが、長さおよび組成は大いに変化し得る。結合領域は開始オリゴヌクレオチドのハイブリダイゼーションを適応させるために十分に長くなければならない。異なる結合領域を同一かまたは異なるかのいずれかの開始オリゴヌクレオチドとともに使用できるが、調製の便宜のためには、同一の結合領域および異なる開始オリゴヌクレオチドを提供するのが好ましい。従って、全てのテンプレートを同一に調製し、次いで、異なる開始オリゴヌクレオチドでの使用のためにアリコートに分ける。好ましくは、結合領域は、異なる開始オリゴヌクレオチドの組を適応させるのに十分な長さであるべきであって、各々はテンプレートにハイブリダイズしてその後の連結のための異なる出発点を生じる。最も好ましくは、結合領域は、約20〜50ヌクレオチドの間の長さである。
【0023】
開始オリゴヌクレオチドは、伸長サイクルのいずれの洗浄工程の間にも無傷のままでいる結合領域との高度に安定な二重鎖を形成するように選択される。これは、都合よく、開始オリゴヌクレオチドの長さが、オリゴヌクレオチドプローブの長さよりもかなり長いように選択することによって、および/またはそれらがGCリッチとなるように選択することによって達成される。開始オリゴヌクレオチドはまた、種々の技術(例えば、Summertonら,米国特許第4,123,610号)によってテンプレートストランドに架橋することもでき;または、それらは、その天然の対応物(例えば、ペプチド核酸)よりも安定性の大きな二重鎖を形成するヌクレオチドアナログよりなることもできる。Science,254:1497−1500(1991);Hanveyら,Science,258:1481−1485(1992);およびPCT出願第PCT/EP92/01219号および同第PCT/EP92/01220号。
【0024】
好ましくは、開始オリゴヌクレオチドの長さは約20〜30ヌクレオチドであり、そしてその組成は、使用されるオリゴヌクレオチドプローブの融解温度を約10〜50℃だけ超える二重鎖融解温度を提供するために十分なパーセントのGおよびCを含む。より好ましくは、開始オリゴヌクレオチドの二重鎖融解温度は、オリゴヌクレオチドプローブの融解温度を約20〜50℃だけ超える。配列決定操作で使用される異なる開始オリゴヌクレオチドの数Nは、各サイクルで1つのヌクレオチドが同定される場合の1から、実際的に使用できるオリゴヌクレオチドプローブのサイズによってのみサイズが制限される多数まで変化し得る。オリゴヌクレオチドプローブのサイズを制限する因子は、妥当な速度でハイブリダイゼーション反応を駆動するために十分に高濃度の個々のプローブを有する混合物を調製する際の困難、二次構造の形成に対するより長いプローブの感受性、1塩基ミスマッチに対する感受性の低下などを含む。好ましくは、Nは1〜16の範囲であり;より好ましくは、Nは1〜12の範囲であり;そして最も好ましくは、Nは1〜8の範囲である。
【0025】
広範囲の種々のオリゴヌクレオチドプローブを本発明で使用することができる。一般に、オリゴヌクレオチドプローブは、開始オリゴヌクレオチドまたは伸長した二重鎖に連結して、次の伸長サイクルの伸長した二重鎖を生じることができるべきである;連結は、プローブが連結の前にテンプレートと二重鎖を形成するべきであるのでテンプレートに駆動されるべきである;プローブは、1回の伸長サイクルにおいて同一のテンプレートに複数のプローブが連結するのを防ぐためにブロッキング部分を有すべきであり、プローブは連結後に伸長可能な末端を再生するために処理または修飾され得るべきであり、そしてプローブは成功した連結後にテンプレートに関連する配列情報の獲得を可能とするシグナリング部分を有すべきである。以下でより十分に記載するように、実施態様に応じて、伸長した二重鎖または開始オリゴヌクレオチドは、オリゴヌクレオチドプローブによって、5’→3’方向または3’→5’方向のいずれかに伸長され得る。一般に、オリゴヌクレオチドプローブはテンプレートと完全にマッチした二重鎖を形成する必要はないが、このような結合が通常は好ましい。テンプレート中の1つのヌクレオチドが各伸長サイクルで同定される好ましい実施態様において、完全な塩基対合は、その特定のヌクレオチドを同定するために必要とされるに過ぎない。例えば、オリゴヌクレオチドプローブを伸長した二重鎖に酵素的に連結する実施態様において、完全な塩基対合(すなわち、適切なワトソン−クリック塩基対合)が、連結されるプローブの末端ヌクレオチドとテンプレート中のその相補物との間で必要とされる。一般に、このような実施態様において、プローブの残りのヌクレオチドは、次の連結がテンプレートに沿って所定の部位または塩基数で起こることを確実とする「スペーサー」として働く。すなわち、それらの対合、またはその欠落は、さらなる配列情報を提供しない。同様に、塩基同定についてポリメラーゼ伸長に頼る実施態様において、プローブは主としてスペーサーとして働き、従って、テンプレートに対する特異的ハイブリダイゼーションは、望ましいとはいえ、重要ではない。
【0026】
好ましくは、オリゴヌクレオチドプローブは、所定の長さの全ての可能な配列のオリゴヌクレオチドを含む混合物としてテンプレートに適用される。このような混合物の複雑さは、例えば、Kong Thoo Linら,Nucleic Acids Research,20:5149−5152;米国特許第5,002,867号;Nicholsら,Nature,369:492−493(1994)によって教示されるデオキシイノシンなどのようないわゆる縮重低下アナログを用いることを含む多数の方法によって;またはオリゴヌクレオチドプローブの多数の混合物(例えば、一緒にすると所定の長さの全ての可能な配列を含むオリゴヌクレオチド配列の4つのばらばらのサブセットを含む4つの混合物)を別々に適用することによって低下させることができる。
【0027】
本発明の開始オリゴヌクレオチドおよびオリゴヌクレオチドプローブは、都合よく、自動DNA合成機(例えば、Applied Biosystems,Inc.(Foster City,California)392型または394型のDNA/RNA合成機)で、例えば、以下の文献に開示されるホスホルアミダイト化学のような標準的な化学を用いて合成される:BeaucageおよびIyer,Tetrahedron,48:2223−2311(1992);Molkoら,米国特許第4,980,460号;Kosterら,米国特許第4,725,677号;Caruthersら,米国特許第4,415,732号;同第4,458,066号;および同第4,973,679号など。例えば、ホスホロチオエート、ホスホルアミデートなどのような非天然の骨格基が得られる別の化学を使用することもできる。但し、得られたオリゴヌクレオチドは特定の実施態様の連結および他の試薬に適合するものとする。オリゴヌクレオチドプローブの混合物は、例えば、Teleniusら,Genomics,13:718−725(1992);Welshら,Nucleic Acids Research,19:5275−5279(1991);Grothuesら,Nucleic Acids Research,21:1321−1322(1993);Hartley,欧州特許出願第90304496.4号などに開示されるような周知の技術を用いて容易に合成される。一般に、これらの技術は、縮重を導入することが望まれるカップリング工程の間の成長するオリゴヌクレオチドへの活性化モノマーの混合物の適用を必要とするだけである。
【0028】
従来のリガーゼを本発明で使用する場合、以下でより十分に記載するように、いくつかの実施態様ではプローブの5’末端をリン酸化し得る。5’モノホスフェートを化学的または酵素的のいずれかでキナーゼを用いてオリゴヌクレオチドに結合させることができる。例えば、Sambrookら,Molecular Cloning;A Laboratory Manual,第2版(Cold Spring Harbor Laboratory,New York,1989)。化学的リン酸化は、HornおよびUrdea,Tetrahedron Lett.,27:4705(1986)によって記載されており、そして開示されたプロトコルを実施するための試薬は、例えば、Clontech Laboratories(Palo Alto,California)から5’Phosphate−ONTMとして市販されている。好ましくは、必要な場合、オリゴヌクレオチドプローブは化学的にリン酸化される。
【0029】
本発明のプローブは、蛍光部分、発色部分などの直接的または間接的な結合を含む種々の方法で標識され得る。DNAを標識し、そしてDNAプローブを構築するための方法論の多くの総合的な概説が、本発明のプローブを構築するために適用可能な手引きを提供する。このような概説は、Matthewsら,Anal.Biochem.,第169巻,1−25頁(1988);Haugland,Handbook of Fluorescent Probes and Research Chemicals(Molecular Probes,Inc.,Eugene,1992);KellerおよびManak,DNA Probes,第2版(Stockton Press,New York,1993);およびEckstein編,Oligonucleotides and Analogues:A Practical Approach(IRL Press,Oxford,1991)などを含む。本発明に適用できる多くのより詳細な方法は、以下の文献の見本に開示される:Fungら,米国特許第4,757,141号;Hobbs,Jr.ら,米国特許第5,151,507号;Cruickshank,米国特許第5,091,519号;(レポーター基の結合のための官能化されたオリゴヌクレオチドの合成);Jablonskiら,Nucleic Acids Research,14:6115−6128(1986)(酵素−オリゴヌクレオチドコンジュゲート);およびUrdeaら,米国特許第5,124,246号(分岐DNA)。
【0030】
好ましくは、プローブは、例えば、Menchenら,米国特許第5,188,934号;Begotら,PCT出願第PCT/US90/05565号によって開示されるような1つ以上の蛍光色素で標識される。
【0031】
テンプレートに対するオリゴヌクレオチドプローブの適用のためのハイブリダイゼーション条件の選択における手引きは、多数の文献(例えば、Wetmur,Critical Reviews in Biochemistry and Molecular Biology,26:227−259(1991);DoveおよびDavidson,J.Mol.Biol.5:467−478(1962);Hutton,Nucleic Acids Research,10:3537−3555(1977);Breslauerら,Proc.Natl.Acad.Sci.83:3746−3750(1986);Innisら編,PCR Protocols(Academic Press,New York,1990)など)に見い出すことができる。
【0032】
一般に、オリゴヌクレオチドプローブを、伸長した二重鎖の末端に並列してテンプレートにアニールする場合、二重鎖とプローブとを連結する、すなわち、相互の共有結合を生じさせる。連結は酵素的または化学的のいずれかで達成され得る。化学的連結方法は当該分野で周知である。例えば、Ferrisら,Nucleosides&Nucleotides,8:407−414(1989);Shabarovaら,Nucleic Acids Research,19:4247−4251(1991)など。好ましくは、酵素的連結は標準的プロトコルでリガーゼを用いて実施される。多くのリガーゼが知られており、そして本発明で使用するのに適切である。例えば、Lehman,Science,186:790−797(1974);Englerら,DNA Ligases,3−30頁,Boyer編,The Enzymes,第15B巻(Academic Press,New York,1982)など。好ましいリガーゼは、T4 DNAリガーゼ、T7 DNAリガーゼ、E.coli DNAリガーゼ、Taqリガーゼ、Pfuリガーゼ、およびTthリガーゼを含む。それらの使用についてのプロトコルは周知である。例えば、Sambrookら(上記);Barany,PCR Methods and Applications,1:5−16(1991);Marshら,Strategies,5:73−76(1992)など。一般に、リガーゼは、隣接するストランドの3'ヒドロキシルへの連結のために5’リン酸基が存在することを必要とする。
【0033】
標的ポリヌクレオチドの調製
好ましくは、標的ポリヌクレオチドを結合領域に連結してテンプレートを形成し、そしてテンプレートを複雑かつ時間を消費する精製工程を伴わない試薬の連続的適用を可能とする固相支持体(例えば、磁性粒子、ポリマーマイクロスフィア、フィルター物質など)に付着させる。標識ポリヌクレオチドの長さは広範囲に変化し得る;しかし、調製の便宜のためには、従来の配列決定で使用される長さが好ましい。例えば、数百塩基対(200〜300)から1〜2キロ塩基対までの範囲の長さが好ましい。
【0034】
標的ポリヌクレオチドは、種々の常法によって調製され得る。例えば、標的ポリヌクレオチドは、従来のDNA配列決定で使用されるものを含む、従来の任意のクローニングベクターのインサートとして調製され得る。適切なクローニングベクターの選択および使用のための広範な手引きがSambrookら,Molecular Cloning:A Laboratory Manual,第2版(Cold Spring Harbor Laboratory,New York,1989)、および同様の文献で見い出される。SambrookらおよびInnisら編,PCR Protocols(Academic Press,New York,1990)もまた、標的ポリヌクレオチドを調製するためのポリメラーゼ連鎖反応の使用について手引きを提供する。好ましくは、この方法で使用する他の試薬から標的ポリヌクレオチドを分離するのを容易にするために、磁性ビーズまたは他の固体支持体への付着を可能とするクローン化標的ポリヌクレオチドまたはPCR増幅標的ポリヌクレオチドを調製する。このような調製技術についてのプロトコルは、Wahlbergら,Electrophoresis,13:547−551(1992);Tongら,Anal. Chem.,64;2672−2677(1992);Hultmanら,Nucleic Acids Research,17:4937−4946(1989);Hultmanら,Biotechniques,10:84−93(1991);Syvanenら,Nucleic Acids Research,16:11327−11338(1988);Dattaguptaら,米国特許第4,734,363号;Uhlen,PCT出願第PCT/GB89/00304号;および同様の文献に十分に記載される。キットもまた、このような方法を実施するために、例えば、Dynal AS.(Oslo,Norway)からDynabeadsTMテンプレート調製キットとして市販される。
【0035】
一般に、本発明の方法で使用する微粒子またはビーズのサイズおよび形状は重要ではない;しかし、直径数m(例えば、1〜2m)から直径数百m(例えば、200〜1000m)の範囲のサイズの微粒子が好ましい。なぜなら、それらは、例えば蛍光標識プローブからの容易に検出できるシグナルの生成を可能としつつ、試薬および試料の使用量を最小にするからである。
【0036】
伸長可能な末端の連結、キャップ形成、および再生についてのスキーム
1つの局面において、本発明は、オリゴヌクレオチドプローブの連結および同定の繰り返し工程を必要とする。しかしながら、同一工程における同一の伸長した二重鎖に対する複数プローブの連結は、通常は同定の問題を誘引するであろうから、多重伸長を防止しそして伸長可能な末端を再生するのに有用である。さらに、もし連結工程が100%効果的でなければ、それらがいずれのさらなる連結工程にも参画しないように、連結を受けない伸長二重鎖にキャップ形成するのが望ましいであろう。すなわち、キャップ形成工程は、好ましくは、ポリヌクレオチド合成のような他の合成化学プロセスから類推して、連結工程の後に起こる(例えば、Andrusら、米国特許第4,816,571号)。これは、その後の同定工程で生じるシグナルから潜在的に有意なノイズの源を除去するであろう。
【0037】
以下、本発明の連結、キャップ形成、再生および同定工程を実施するためのいくつかの例示的スキームを記載する。それらは、手引きの目的で提示するものであって、限定することを意図しない。
【0038】
開始オリゴヌクレオチドまたは3’→5’方向に伸長した二重鎖を伸長するためのスキームは図2に示す。テンプレート(20)をその5’末端により、固相支持体(10)に付着させる。これは、常法技術を用い、ビオチンまたは同様の連結部分を介して、丁度都合よく達成され得る。5’リン酸基を有する開始オリゴヌクレオチド(200)を、連結および同定の最初のサイクル前に、前記のようにテンプレート(20)にアニールする。以下の形態のオリゴヌクレオチドプローブ(202)を使用する:
HO−(3’)BBB...BBB(5’)−OP(=O)(O−)NH−Bt*
ここで、BBB...BBBはオリゴヌクレオチドプローブ(202)のヌクレオチドの配列を表し、Bt*はホスホルアミデート基、または光切断結合のような、他の不安定結合を介してオリゴヌクレオチドの5’炭素に連結した標識された鎖終結部分である。Bt*の性質は広く変化し得る。それは連続的連結を防止する限り、標識されたヌクレオシド(例えば、5’P3’Nホスホルアミデートを介してカップリングしたもの)または他の部分であり得る。それは、AgrawalおよびTang、国際出願第PCT/US91/08347号に記載されるように、単にリンカーによって結合された標識であり得る。オリゴヌクレオチドプローブの重要な特徴は、アニーリングおよび連結(204)の後に、標識を除去でき、例えば、Letsingerら、J.Am.Chem.Soc.,94:292−293(1971);Letsingerら、Biochem.,15:2810−2816(1976);Gryaznovら,Nucleic Acid Research,20:3403−3409(1992);および同様の文献によって教示されるように、ホスホルアミデート結合を酸で処理することによって、伸長可能な末端を再生できることである。例えば、ホスホルアミデートの加水分解は、室温における40分間のジクロロメタン中の0.8%トリフルオロ酢酸での処理によって達成され得る。このようにして、Bt*上の標識を介して連結したプローブをアニーリングし、連結し、そして同定した後、酸加水分解(206)により鎖終結部分を切断し、それによりリン結合を破壊し、連結したオリゴヌクレオチド上に5’モノホスフェートを残しておく。この工程は連続的サイクルで繰り返され得る(208)。この実施態様の1つの局面において、単一の開始オリゴヌクレオチドを、1つのヌクレオチドのみが各配列決定サイクルで同定されるように使用し得る。このような実施態様では、このプローブは好ましくは以下の形態を有する:
HO−(3’)B(5’)−OP(=O)(O−)NHBB...BBB−Bt*。
【0039】
このようにして、各連結および酸切断工程の後、二重鎖は1ヌクレオチドだけ伸長される。
【0040】
加水分解の前に、キャップ形成工程が導入され得る。例えば、プローブ(202)は以下の形態を有する:
HO−(3’)BB...Bp^B...BB(5’)−OP(=O)(O−)NH−Bt*
ここで、「p^」はホスホロチオエート、メチルホスホネートなどのようなエキソヌクレアーゼ耐性結合である。このような実施態様において、キャップ形成は、未連結伸長二重鎖を切断してエキソヌクレアーゼ耐性結合に戻すλエキソヌクレアーゼのようなエキソヌクレアーゼで伸長二重鎖を処理することによって達成され得る。次いで、伸長二重鎖の5’末端におけるこの結合の存在は、その後の連結にそれが関与することから防止する。明らかに、多くの他のキャップ形成方法、例えばアシル化、不活性オリゴヌクレオチドの連結などを使用し得る。遊離3’ヒドロキシルが関与する場合、キャップ形成は、鎖終結ヌクレオシド三リン酸、例えばジデオキシヌクレオシド三リン酸などの存在下で、DNAポリメラーゼで二重鎖を伸長させることによって達成され得る。
【0041】
前記したホスホルアミデート結合は、本明細書中では「化学的に切断可能なヌクレオシド間結合」と呼ばれるヌクレオシド間結合の一般的クラスの例である。これらは、酸化性環境、還元性環境、特徴的波長の光(光不安定性結合用)などのような特徴的な化学的または物理的条件でそれらを処理することによって切断され得るヌクレオシド間結合である。本発明に従い使用され得る化学的に切断可能なヌクレオシド間結合の他の例は、Urdea第5,380,833号;Gryaznovら、Nucleic Acids Research,21:1403−1408(1993)(ジスルフィド);Gryaznovら、Nucleic Acids Research,22:2366−2369(1994)(ブロモアセチル);Urdeaら、国際出願PCT/US91/05287(光不安定性);および同様の文献に記載されている。
【0042】
本発明で使用され得るさらなる化学的に切断可能な結合は、伸長可能なヌクレオシドに化学的に変換され得る鎖終結ヌクレオチドを含む。このような化合物の例は以下の文献に記載されている:Canardら、国際出願PCT/FR94/00345;Ansorge,独国特許出願第DE 4141178 A1号;Metzkerら、Nucleic Acids Research,22:4259−4267(1994);Cheeseman、米国特許第5,302,509号;Rossら,国際出願第PCT/US90/06178号など。
【0043】
開始オリゴヌクレオチドまたは伸長した二重鎖を5’→3’方向に伸長させるためのスキームが図3Aに示される。テンプレート(20)は、その3’末端によって固相支持体(10)に結合させる。前記のように、これは、常法技術を用い、ビオチンまたは同様の連結部分を介して、丁度都合よく達成され得る。3’ヒドロキシル基を有する開始オリゴヌクレオチド(300)は、連結および同定の最初のサイクル前に、前記のようにテンプレート(20)にアニールされる。以下の形態のオリゴヌクレオチドプローブ(302)を使用する:
OP(=O)(O−)O−(5’)BBB...BBBRRRRBt*。
【0044】
ここで、BBB...BBBRRRRはオリゴヌクレオチドプローブ(302)の2’−デオキシヌクレオチドの配列を表し、「RRRR」はプローブ(302)の4つのリボヌクレオチドの配列を表し、そしてBt*は前記のように標識された鎖終結部分である。このような混合されたRNA−DNAオリゴヌクレオチドは通常の自動DNA合成機を用いて容易に合成される(例えば、Duckら、米国特許第5,011,769号)。RNase Hは、4つのリボヌクレオチドセグメントの中心において特異的にプローブを切断し(Hogrefeら、J.Biol.Chem.,265:5561−5566(1990))、伸長した二重鎖上に3’ヒドロキシル(312)を残し、これはその後の連結工程に参加し得る。このようにして、本実施態様におけるサイクルは、テンプレート(20)にプローブ(302)をアニールされそして、連結(304)させて、伸長した二重鎖(306)を形成することにより進行する。Bt*を介する同定の後、伸長した二重鎖は、標識を切断し、伸長可能な末端を再生するために、RNase Hで処理される。次いで、サイクルが繰り返される(314)。キャップ形成(310)は、RNase H処理の前に、4つのジデオキシヌクレオシド三リン酸、ddATP、ddCTP、ddGTPおよびddTTPの存在下、DNAポリメラーゼで未連結末端を伸長させることによって実施され得る。
【0045】
図3Bで説明したように、同様のスキームが3’5’伸長のために使用され得る。このような実施例において、開始オリゴヌクレオチドまたは伸長した二重鎖(330)は、5’一リン酸を有し、オリゴヌクレオチドプローブ(332)は以下の形態を有する:
HO−(3’)BBB...BBBRRRRB..BBt*。
【0046】
前記のように、アニーリング、連結(334)、および同定(338)の後、伸長した二重鎖(336)は、この場合は伸長した二重鎖の末端で5’一リン酸(342)を残すRNase Hにより、切断される。再生した伸長可能な末端を用いて、サイクルは繰り返され得る(344)。キャップ形成工程は、RNase H加水分解前に、未標識非RNA含有プローブを連結することにより、またはホスファターゼでの処理による任意の残存する5’モノホスフェートを除去することのいずれかによって含まれ得る。
【0047】
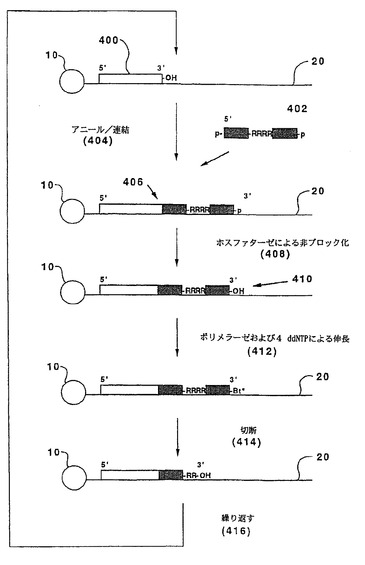
ヌクレオチドの同定は、連結後のポリメラーゼ伸長により達成され得る。図4に例示するように、この実施態様については、テンプレート(20)を前記のように固相支持体(10)に結合させ、3’ヒドロキシルを有する開始オリゴヌクレオチド(400)を最初のサイクル前にテンプレートにアニールさせる。以下の形態を有するオリゴヌクレオチドプローブ(402):
OP(=O)(O−)O−(5’)BBB...BBBRRRRB...B(3’)OP(=O)(O−)O
がテンプレート(20)にアニールされ、そして連結(404)され、伸長した二重鎖(406)が形成される。同一サイクルにおいてプローブの連続的連結を防ぐ3’一リン酸がホスファターゼ(408)で除去され、遊離の3’ヒドロキシル(410)が露出される。明らかに、別のブロッキングアプローチも使用され得る。伸長した二重鎖(406)は、標識したジデオキシヌクレオシド三リン酸(412)の存在下で、核酸ポリメラーゼによってさらに伸長し、それにより、取り込まれたジデオキシヌクレオシドの標識によってテンプレート(20)のヌクレオチドの同定が可能となる。次いで、標識されたジデオキシヌクレオチドおよびプローブ(402)の一部は、伸長した二重鎖(406)上に伸長可能な末端を再生するために、例えば、RNase H処理によって切断される(414)。次いで、サイクルが繰り返される(416)。
【0048】
実施しなければならない別々のアニーリング反応の数を減少させるために、オリゴヌクレオチドプローブは、その相補的配列と完全にマッチした二重鎖が、同様の安定性または結合の自由エネルギーを有するプローブの混合物またはサブセットにグループ分けされ得る。同様の二重鎖安定性を有するオリゴヌクレオチドプローブのこのようなサブセットは、本明細書中ではオリゴヌクレオチドプローブの「ストリンジェンシークラス」といわれる。次いで、オリゴヌクレオチドプローブの混合物またはストリンジェンシークラスは、実質的に標的ポリヌクレオチドに相補的なオリゴヌクレオチドプローブのみが二重鎖を形成するような条件下で、標的ポリヌクレオチドと別々に混ぜ合わされる。すなわち、ハイブリダイゼーション反応のストリンジェンシーは、実質的に完全に相補的なオリゴヌクレオチドプローブのみが二重鎖を形成するように選択される。次いでこれらの完全にマッチした二重鎖が、連結され、伸長した二重鎖が形成される。所与のオリゴヌクレオチドプローブ長さにつき、各ストリンジェンシークラス内のオリゴヌクレオチドプローブの数は、広く変化し得る。オリゴヌクレオチドプローブの長さおよびストリンジェンシークラスサイズの選択は、標識配列の長さ、およびそれがどのように調製されるか、ハイブリダイゼーション反応が自動化され得る程度、ハイブリダイゼーション反応のストリンジェンシーが制御され得る程度、相補的配列を有するオリゴヌクレオチドプローブの存在または不存在などのようないくつかの因子に依存する。特定の実施態様のためのストリンジェンシークラスの適切なサイズを選択する手引きは、核酸ハイブリダイゼーションおよびポリメラーゼ連鎖反応法についての一般的文献中に見い出され得る(例えば、Gotoh,Adv.Biophys.16:1−52(1983);Wetmer,Critical Reviews in Biochemistry and Molecular Biology 26:227−259(1991);Breslauerら,Proc.Natl.Acad.Sci. 83:3746−3750(1986);Wolfら、Nucleic Acids Research,15:2911−2926(1987);Innisら編,PCR Protocols(Academic Press,New York,1990);McGrawら、Biotechniques,8:674−678(1990)など)。ストリンジェンシーは、温度、塩濃度、ホルムアミドのような特定の有機溶媒の濃度などを含むいくつかの変化するパラメーターによって制御され得る。好ましくは、用いられる種々のポリメラーゼまたはリガーゼの活性が、塩濃度または有機溶媒濃度がオリゴヌクレオチドプローブの特異的アニーリングを保証するために変えられ得る程度を制限するために、温度がストリンジェンシークラスを規定するために用いられる。
【0049】
一般に、ストリンジェンシークラスが大きければ大きいほど、ハイブリダイズする混合物の複雑性が大きくなり、そして混合物におけるいずれかの特定のオリゴヌクレオチドプローブの濃度が低下する。標的ポリヌクレオチド上に相補的部位を有する、より低濃度のオリゴヌクレオチドプローブはハイブリダイズし連結されるオリゴヌクレオチドプローブの相対的公算(relative likelihood)を低下させる。これは、今度は、感受性の低下を導く。また、ストリンジェンシークラスが大きければ大きいほど、オリゴヌクレオチドプローブと相補的配列との間に形成される二重鎖の安定性に大きな変動を有する。他方、ストリンジェンシークラスが小さければ小さいほど、1セットの全てのオリゴヌクレオチドプローブが標的ポリヌクレオチドにハイブリダイズすることを保証するために、より多数のハイブリダイゼーション反応を必要とする。
【0050】
例えば、8マーのオリゴヌクレオチドプローブが使用される場合、ストリンジェンシークラスは、各々約50から約500の間のオリゴヌクレオチドプローブを含み得る。このようにして、数百〜数千のハイブリダイゼーション/連結反応が必要とされる。より大きなサイズのオリゴヌクレオチドプローブについては、より大きなストリンジェンシークラスが、ハイブリダイゼーション/伸長反応の数を実際的なもの(例えば、104〜105またはそれ以上)とするために必要とされる。
【0051】
同一ストリンジェンシークラスのオリゴヌクレオチドプローブは、十分にランダムなオリゴヌクレオチドプローブが合成されるのと類似した様式で(例えば、Teleniusら、Genomics,13:718−725(1992);Welshら、Nucleic Acids Research,19:5275−5279(1991);Grothuesら、Nucleic Acuds Research,21:1321−1322(1993);Hartley,欧州特許出願90304496.4などに開示されているように)、同時に合成され得る。差異は、各サイクルにおいて、モノマーの異なる混合物を、増殖するオリゴヌクレオチドプローブ鎖に適用することであり、ここで、混合物中の各モノマーの割合は、ストリンジェンシークラスにおけるオリゴヌクレオチドプローブの位置における各ヌクレオシドの割合によって指示される。ストリンジェンシークラスは、利用可能なアルゴリズム(例えば、Breslauerら、Proc.Natl.Acad.Sci.,83:3746−3750(1986);Loweら、Nucleic Acids Research,18:1757−1761(1990)など)により二重鎖形成の自由エネルギーを計算することによって容易に形成される。オリゴヌクレオチドプローブは、標準的な反応条件下(例えば、標準的なバブルソート、Base,Computer Algorithms(Addison−Wesley,Menlo Park,1978))でのそれらの相補体への結合の自由エネルギーにより順序付けられる。例えば、以下のものは標準的なハイブリダイゼーション条件下での二重鎖形成の自由エネルギーによる最大安定性(頂部から底部にかけて)を有する、および二重鎖形成の自由エネルギーの最低安定性を持つ10の6マーのリストである(自由エネルギーはBreslauerら(前出)により計算される)。
【0052】
オリゴヌクレオチドプローブ
ランキング配列(5’→3’)
1 GCGCGC
2 CGCGCG
3 CCCGCG
4 CGCCCG
5 CGCGCC
6 CGCGGC
7 CGGCGC
8 GCCGCG
9 GCGCCG
10 GCGCGG
・ ・
・ ・
・ ・
4087 TCATAT
4088 TGATAT
4089 CATATA
4090 TATATG
4091 ATCATG
4092 ATGATG
4093 CATCAT
4094 CATGAT
4095 CATATG
4096 ATATAT
このように、もしストリンジェンシークラスが最初の10個の6マーからなるのであれば、最初の(最も3’側の)位置についての混合物モノマーは、0:4:6:0(A:C:G:T)となるであろうし、第2の位置については、それは0:6:4:0となるであろう(以下、同様)。もしストリンジェンシークラスが最後の10個の6マーからなるならば、最初の位置についてのモノマーの混合物は1:0:4:5となるであろうし、第2の位置については、それは5:0:0:5であろう(以下、同様)。次いで、得られた混合物は、加熱溶出によって所望のストリンジェンシークラスの配列につきさらに富化され得る(例えば、Miyazawaら、J.Mol.Biol.,11:223−237(1965))。
【0053】
より便宜的には、数百〜数千のオリゴヌクレオチドを含むストリンジェンシークラスは、種々の並行合成アプローチによって直接的に合成され得る(例えば、Frankら、米国特許第4,689,405号;Matsonら、Anal.Biochem.,224;110−116(1995);Fodorら、国際出願第PCT/US93/04145号;Peaseら、Proc.Natl.Acad.Sci.、91:5022−5026(1994);Southernら、J.Biotechnology,35:217−227(1994),Brennan、国際出願第PCT/US94/05896号など)。
【0054】
いくつかの場合において、オリゴヌクレオチドプローブ−ダイマーを形成するのに感受性のサブセットまたはオリゴヌクレオチドプローブにおいて、他のオリゴヌクレオチドプローブに対する相補的配列を有する別々のサブセットのオリゴヌクレオチドプローブに配置することによって、オリゴヌクレオチドプローブのさらなるストリンジェンシークラスを形成することが所望され得る。
【0055】
明らかに、当業者は、先に明示的に記載されていないが、本発明のなおさらなる実施態様を設計するために、前記の実施態様の特徴を組み合わせることができる。
【0056】
また、本発明は、本発明の方法を自動的に実施するためのシステムおよび装置を含む。このようなシステムおよび装置は、i)標的ポリヌクレオチドをつなぎ止めるために使用される固相支持体の性質、ii)所望の並行操作の程度、iii)使用される検出スキーム;iv)試薬を再使用するか捨てるかなどを含めたいくつかの設計製薬に依存して種々の形態をとり得る。一般に、装置は一連の試薬貯蔵器、好ましくは固相支持体(例えば、磁気ビーズ)に付着させた標的ポリヌクレオチドを含有する1つ以上の反応容器、1つ以上の検出ステーション、および試薬貯蔵器から反応容器へおよび反応容器から検出ステーションへ、予め決定した様式で試薬を移すためのコンピュータ制御手段を含む。コンピュータ温度を制御するためのコンピューター制御手段は、Harrisonら、Biotechniques,14:88−97(1993);Fujitaら、Biotechniques,9:584−591(1990);Wadaら、Rev.Sci.Instrum,54:1569−1572(1983)などに開示されているもののような、種々の一般目的の実験室ロボットによって実行され得る。このような実験室ロボットはまた市販されている(例えば、Applied Biosystemsモデル800Catalyst(Foster City,CA))。
【0057】
本発明の異なる実施態様を実施するために種々のキットが提供され得る。一般に、本発明のキットは、オリゴヌクレオチドプローブ、開始オリゴヌクレオチド、および検出システムを含む。キットはさらに、連結試薬および本発明の特別の実施態様を実施するための指示を含む。タンパク質リガーゼ、RNase H、核酸ポリメラーゼ、または他の酵素を使用する実施態様においては、それらの各緩衝液が含ませられ得る。いくつかの場合においては、これらの緩衝液は同一であり得る。好ましくは、キットはまた、テンプレートをつなぎ止めるための固相支持体(例えば、磁気ビーズ)を含む。1つの好ましいキットにおいて、標的ポリヌクレオチドの異なる末端ヌクレオチドに対応するプローブが、明瞭なスペクトル的に分解できる蛍光色素を保持するように、蛍光的に標識されたオリゴヌクレオチドプローブが提供される。本明細書で用いられる「スペクトル的に分解可能な」とは、その色素が、操作条件下で、それらのスペクトル特性、特に蛍光放射波長に基づいて区別され得ることを意味する。このようにして、1つまたはそれ以上の末端ヌクレオチドの同一性は異なる色彩、またはおそらくは異なる波長における強度の比率に相関する。より好ましくは、4つのこのようなプローブは、標的ポリヌクレオチド上の4つのスペクトル的に分解可能な蛍光色素と4つの可能な末端ヌクレオチドの各々の間の一対一の対応を可能とするように提供される。スペクトル的に分解可能な色素のセットは、米国特許第4,855,225号および第5,188,934号:国際出願第PCT/US90/05565号;およびLeeら、Nucleic Acids Research,20:2471−2483(1992)に開示されている。
【実施例】
【0058】
実施例1
4つの開始オリゴヌクレオチドを用いてpUC19から増幅された
標的ポリヌクレオチドの配列決定
本実施例においては、結合領域およびpUC19プラスミドの一部を含むテンプレートをPCRによって増幅し、そして磁気ビーズに付着させる。4つの開始オリゴヌクレオチドを、下記のように別々の反応で使用する。以下の式に示すように、4つの中央リボヌクレオチドならびに、両5’および3’一リン酸を有する8マーのオリゴヌクレオチドプローブを使用する:
OP(=O)(O−)O−(5’)BBRRRRBB(3’)−OP(=O)(O−)O。
【0059】
アニーリングの後、プローブを開始オリゴヌクレオチドに酵素的に連結し、磁気ビーズ支持体を洗浄する。連結したプローブの3’リン酸をホスファターゼで除去し、その後、4つの標識したジデオキシヌクレオシド三リン酸鎖終結剤の存在下で、プローブをDNAポリメラーゼで伸長させる。伸長したヌクレオチドの洗浄および同定の後、連結されたプローブをRNase Hでリボヌクレオチド部分で切断して標識を除去し、そして伸長可能な末端を再生する。
【0060】
36マーの結合領域を含む以下の二本鎖のフラグメントをSacI/XmaI消化したpUC19に連結する:
CCTCTCCCTTCCCTCTCCTCCCTCTCCCCTCTCCCTC
TCGAGGAGAGGGAAGGGAGAGGAGGGAGAGGGGAGAGGGAGGGCC。
【0061】
単離および増幅の後、改変されたpUC19の402塩基対フラグメントを、テンプレートとして使用するために、PCRによって増幅する。該フラグメントは、41位からポリリンカー領域中のSacI部位に隣接して挿入された結合領域(未改変pUC19の413位)のpUC19の領域にわたる(Yanisch−Perronら、Gene,33:103−119(1985))。配列5’−CCCTCTCCCCTCTCCCTCx−3’および5’−GCAGCTCCCGGAGACGGT−3’(ここで「x」は製造業者のプロトコル付きの市販されている試薬(例えば、3’ビオチン−ON CPG(Clonetech Laboratories,Palo Alto,California))を用い、合成の間に付着される3’ビオチン部分である)を有する2つの18マーのオリゴヌクレオチドプローブを使用する。増幅されたテンプレートを単離し、M280−ストレプトアビジン(Dynal,Inc.,Great Neck,New York)とともに、製造業者のプロトコル(Dynabeads Template Preparation Kit)を用い、ストレプトアビジン被覆磁気ビーズ(Dynabeads)に付着させる。約300gのDynabeads M280−ストレプトアビジンをロードするために十分な量のビオチニル化313塩基対フラグメントを提供する。
【0062】
開始オリゴヌクレオチドとで形成される二重鎖が、二重鎖安定性を増強させるために、約66%GCの組成を有するように、結合領域配列を選択する。また、二次構造形成および結合領域内の1以上の位置への開始オリゴヌクレオチドの偶発的ハイブリダイゼーションを防ぐように配列を選択する。結合領域内の所与の開始オリゴヌクレオチドの位置のあらゆるシフティングも、有意数のミスマッチ塩基を生じる。
【0063】
ローディングの後、テンプレートの非ビオチニル化ストランドを加熱変性によって除去し、その後、磁気ビーズを洗浄し、そして4つのアリコートに分ける。磁気ビーズに付着させたテンプレートは以下の配列を有する:
(磁気ビーズ)−(リンカー)−(3’)−CTCCCTCTCCCCTCTCCCTCCTC−TCCCTTCCTCTCCTCGAGCTTAAGT...CTCGACG−(5’)。
【0064】
以下の4つのオリゴヌクレオチドを、テンプレートの別々のアリコートの各々における開始オリゴヌクレオチドとして使用する:
5’−GAGGAGAGGGAAGGAGAGGAG
5’−GGAGGAGAGGGAAGGAGAGGA
5’−GGGAGGAGAGGGAAGGAGAGG
5’−AGGGAGGAGAGGGAAGGAGAG。
【0065】
以下の反応および洗浄は、一般に、特記しない限り、使用する酵素のための50L容量の製造業者(New England Biolabs)の推奨する緩衝液中で実施する。また、標準的な緩衝液はSambrookら、Molecular Cloning,第2版(Cold Spring Harbor Laboratory Press,1989)に記載されている。
【0066】
4つのアリコートの各々用の8マーのプローブ全てを一緒に含む、96ストリンジェンシークラスの684または682オリゴヌクレオチドプローブ各々(48の異なるアニーリング温度の各々のための2のサブセット)を形成する。96のクラスの各々のプローブを、同一成分を有する反応混合物中の標的オリゴヌクレオチドに別々にアニールするが、37℃未満の温度でSequenaseおよびT4 DNAリガーゼで行った伸長および連結、およびTaq Stoffelフラグメントおよび他の熱安定性リガーゼで行った伸長および連結は例外である。
【0067】
48のストリンジェンシー条件は、同一温度のサブセットの各グループが、次の最高および次の最低ストリンジェンシークラスを含むサブセットグループのそれと1℃だけアニーリング温度が異なるように、22℃〜70℃の範囲のアニーリング温度によって規定される。アニーリング温度の範囲(22〜70℃)は、各々、最も不安定なおよび最も安定な8マーが、標準的なPCR緩衝溶液中で、約50パーセント最大アニーリングを有することが予想される温度より5〜10℃低いの温度によっておおよそ境界を決める。
【0068】
80℃における5〜10分間のインキュベーションの後、反応混合物を20〜30分間にかけて、それらの各アニーリング温度にもってゆく。連結、洗浄、およびホスファターゼでの処理の後、2単位のポリメラーゼおよび標識ジデオキシヌクレオチド三リン酸(0.08mM最終反応濃度、およびTAMRA(テトラメチルローダミン)、FAM(フルオレセイン)、ROX(ローダミンX)、およびJOE(2’,7’−ジメトキシ−4’,5’−ジクロロフルオレセイン)で標識)を添加する。15分後、ビーズをH2Oで洗浄し、そして各反応混合物を標準的な波長(例えば、Users Manual,モデル373DNAシーケンサー(Applied Biosystems,Foster City,CA))を用いて照射することにより伸長したヌクレオチドの同一性を決定する。
【0069】
同定後、反応混合物を製造業者の推奨のプロトコルを用いRNase Hで処理し、そして洗浄する。RNase H処理した伸長二重鎖は、再生された3’ヒドロキシルを有し、次の連結/伸長/切断のサイクルの準備ができている。試験配列の全てのヌクレオチドが同定されるまでサイクルを実施する。
【0070】
実施例2
ある開始オリゴヌクレオチドを用いるpUC19から増幅された標的ポリヌクレオチドの配列決定
この実施例において伸長は5’→3’方向であるので、ビオチン部分を結合領域のCTリッチストランドにハイブリダイズするプライマーの5’末端に付着させる以外は、本実施例では、実施例1に従ってテンプレートを調製する。このようにして、本実施例では、一本鎖テンプレートの結合領域はGAリッチなセグメントである(本質的には、実施例1の結合領域の相補体)。配列5’−xGAGGGAGAGGGGAGAGGG−3’および5’−ACCGTCTCCGGGAGCTGC−3’(ここで「x」は製造業者のプロトコル付きの市販されている試薬(例えば、Aminolinkアミノアルキルホスホルアミダイト連結剤(Applied Biosystems,Foster City,California)およびClontech Laboratories(Palo Alto,California)から入手可能なビオチン−X−NHSエステル)を用い、合成の間に付着される5’ビオチン部分である)を有する2つの18マーのオリゴヌクレオチドプローブを使用する。
【0071】
以下の配列を有する単一の12マーの開始オリゴヌクレオチドを使用する:
5’−OP(=O)(O−)O−CCTCTCCCTTCCCTCTCCTCC−3’。
以下の式に示される、プローブの最も3’側と3’側から2番目のヌクレオシドとの間に酸不安定ホスホルアミデート結合を有する6マーのオリゴヌクレオチドプローブを使用する:
HO−(3’)B(5’)−OP(=O)(O−)NH−(3’)BBBBBt*
ここで、標識は最も3’側の同一性に対応するように(従って、16の異なる標識ジデオキシヌクレオシドをプローブの合成に使用する)、Bt*はJOE−、FAM−、TAMRA−、またはROX−標識ジデオキシヌクレオシドである。
【0072】
前記のように、6マーのプローブを、各々42または43プローブを含む、96ストリンジェンシークラス(48の異なるアニーリング温度の各々につき2つのサブセット)に調製する。ハイブリダイゼーションおよび連結は、前記のように行う。連結および洗浄の後、オリゴヌクレオチドプローブの蛍光シグナルによって標的ポリヌクレオチド中のヌクレオシドを同定する。次いで、室温にてジクロロメタン中の0.8%トリフルオロ酢酸で、伸長した二重鎖を40分間処理することによって、酸切断を行い、伸長した二重鎖上で伸長可能な末端を再生する。標的ポリヌクレオチドの配列が決定されるまで該プロセスを継続する。
【技術分野】
【0001】
発明の分野
本発明は、一般にポリヌクレオチドのヌクレオチド配列を決定する方法に関し、さらに詳しくは、オリゴヌクレオチドブロックの連続的な連結により1つ以上のプライマーを段階的に伸長することによってテンプレートにおけるヌクレオチドを同定する方法に関する。
【背景技術】
【0002】
背景技術
現在利用可能な技術でのポリヌクレオチドの分析は、試験ポリヌクレオチドが標準または単離されたフラグメントと同一であるかまたは異なるかの確認から、試験ポリヌクレオチドの各ヌクレオチドの明白な同定および順序付けまでの範囲の情報を提供する。このような技術は、遺伝子の機能および制御を理解するのに、および分子生物学の基本的技術の多くを適用するのに非常に重要であるだけでなく、これらはまた、ゲノム解析および非常に多くの非研究的適用(例えば、遺伝的同定、法医学分析、遺伝学カウンセリング、医学診断など)における手段としてますます重要となった。これらの後者の適用において、部分的な配列情報を提供する技術(例えば、フィンガープリント法および配列比較)および完全な配列決定を提供する技術の両方が共に使用されてきた。例えば、Gibbsら,Proc.Natl.Acad.Sci.,86:1919−1923(1989);Gyllenstenら,Proc.Natl.Acad.Sci.,85:7652−7656(1988);Carranoら,Genomics,4:129−136(1989);Caetano−Anollesら,Mol.Gen.Genet.,235:157−165(1992);BrennerおよびLivak,Proc,Natl.Acad.Sci.,86:8902−8906(1989);Greenら,PCR Methods and Applications,1:77−90(1991);およびVersalovicら,Nucleic Acids Research,19:6823−6831(1991)。
【0003】
天然のDNAは2つの線状ポリマー、またはヌクレオチドのストランドよりなる。各ストランドは、ホスホジエステル結合によって連結されたヌクレオシドの鎖である。2つのストランドは、2つのストランドのヌクレオチドの相補的塩基の間の水素結合によって逆平行向きにまとまっている:デオキシアデノシン(A)はチミジン(T)と対になり、そしてデオキシグアノシン(G)は、デオキシシチジン(C)と対になる。
【0004】
現在、DNA配列決定に対する以下の2つの基本的アプローチがある:ジデキオシチェーンターミネーション法(例えば、Sangerら,Proc.Natl.Acad.Sci.,74:5463−5467(1977));および化学的分解法(例えば、Maxamら,Proc.Natl.Acad.Sci.,74:560−564(1977))。チェーンターミネーション法はいくつかの方法で改良されており、そして全ての現在利用可能な自動DNA配列決定機のための基礎として供される。例えば、Sangerら,J.Mol.Biol.,143:161−178(1980);Schreierら,J.Mol.Biol.,129:169−172(1979);Smithら,Nucleic Acids Research,13:2399−2412(1985);Smithら,Nature,321:674−679(1987);Proberら,Science,238:336−341(1987);Section II,Meth.Enzymol.,155:51−334(1987);Churchら,Science,240:185−188(1988);Hunkapillerら,Science,254:59−67(1991);Bevanら,PCR Methods and Applications,1:222−228(1992)。
【0005】
チェーンターミネーション法および化学的分解法は、両方とも、1組以上の標識DNAフラグメントの生成を必要とし、そして各々は共通の起源を有し、各々は既知の塩基で終結する。次いで、1組または複数組のフラグメントは、配列情報を得るために、サイズにより分離されなければならない。両方法において、DNAフラグメントは高分解能ゲル電気泳動によって分離され、この電気泳動は、わずか1ヌクレオチドだけサイズが異なる非常に大きなフラグメントを区別する能力を有しなければならない。残念ながら、この工程は一度に配列決定できるDNA鎖のサイズを厳しく制限する。これらの技術を用いる配列決定は、約400〜450ヌクレオチドまでのDNA鎖に信頼性よく適応できる。Bankierら,Meth.Enzymol.,155:51−93(1987);およびHawkinsら,Electrophoresis,13:552−559(1992)。
【0006】
いくつかの著しい技術的問題が、例えば、500〜600ヌクレオチドを超える長い標的ポリヌクレオチドの配列決定、または高容量の多くの標的ポリヌクレオチドの配列決定へのこのような技術の適用をひどく妨げてきた。このような問題としては、i)大きな労働力を要するゲル電気泳動分離工程は、自動化が困難であり、そしてデータの解析に過剰の変動性(例えば、温度の影響によるバンドの広がり、DNA配列決定フラグメントにおける二次構造による圧縮、分離ゲルにおける不均一性など)を導入する;ii)その特性(例えば、進化性、忠実度、重合速度、チェーンターミネーターの取り込み速度など)がしばしば配列依存性である核酸ポリメラーゼ;iii)ゲル中で空間的に重なるバンドに典型的にはfmol量で存在するDNA配列決定フラグメントの検出および分析;iv)標識部分が単一の均一相に濃縮されずに数百の空間的に分離されたバンドに分布することによる低シグナル;およびv)単一レーン蛍光検出の場合、適切な発光特性および吸収特性、量子収率、およびスペクトル分解性を持つ色素の利用可能性が挙げられる。例えば、Trainor,Anal.Biochem.,62:418−426(1990);Connellら,Biotechniques,5:342−348(1987);Kargerら,Nucleic Acids Research,19:4955−4962(1991);Fungら,米国特許第4,855,225号;およびNishikawaら,Electrophoresis,12:623−631(1991)。
【0007】
もう1つの問題が、診断的配列決定の領域における現在の技術には存在する。常に拡大し続けている多数の障害、障害に対する感受性、疾患状態の予後などは、1つ以上の遺伝子座における、特定のDNA配列の存在、またはDNA配列における変化(または変異)の程度に関連付けられてきた。このような現象の例は、ヒト白血球抗原(HLA)タイピング、膵嚢胞性線維症、腫瘍の進行および不均一性、p53プロトオンコジーン変異、rasプロトオンコジーン変異などを含む。例えば、Gyllenstenら,PCR Methods and Applications,1:91−98(1991);Santamariaら,国際出願第PCT/US92/01675号;Tsuiら,国際出願第PCT/CA90/00267号など。診断情報または予後情報を得るためのこのような病的状態に関連するDNA配列の決定における困難は、複数の亜集団のDNA(例えば、対立遺伝子変異体、多重突然変異体形態など)が頻繁に存在することにある。現在の配列決定技術を用いて複数配列の存在および同一性を区別することは、異なる種類のDNAを単離し、そして恐らくはそれをクローン化するさらなる労力なくしては、実質的に不可能である。
【0008】
DNA配列決定に、DNAフラグメントの高分解能電気泳動分離を要しないで、より分析しやすいシグナルを生じ、そしてヘテロ接合遺伝子座由来のDNAを容易に分析するための手段を提供する別のアプローチが利用可能になれば、配列決定技術において主要な進歩がなされ得る。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明の目的は、現在利用可能なDNA配列決定技術に対してこのような別のアプローチを提供することである。
【課題を解決するための手段】
【0010】
本発明によって、以下が提供される:
1)ポリヌクレオチド中のヌクレオチド配列を同定する方法であって、以下の工程:
(a)開始オリゴヌクレオチドを、オリゴヌクレオチドプローブと該開始オリゴヌクレオチドとを連結することによってポリヌクレオチドに沿って伸長させ、伸長した二重鎖を形成させる工程;
(b)該ポリヌクレオチドの1つ以上のヌクレオチドを同定する工程;および
(c)ヌクレオチドの配列が決定されるまで工程(a)および(b)を繰り返す工程、
を包含する、方法。
(2)前記オリゴヌクレオチドプローブが前記開始オリゴヌクレオチドに対して遠位の末端に鎖終結部分を有する、請求項1に記載の方法。
(3)前記同定の工程が、前記鎖終結部分を除去する工程、および1つ以上の標識した鎖終結ヌクレオシド三リン酸の存在下で核酸ポリメラーゼを用いて該オリゴヌクレオチドプローブを伸長させる工程を包含する、請求項2に記載の方法。
(4)前記伸長した二重鎖上に伸長可能な末端を再生する工程をさらに包含する、請求項3に記載の方法。
(5)前記オリゴヌクレオチドプローブが4つのリボヌクレオチドのサブ配列を包含し、そして前記再生の工程が該オリゴヌクレオチドプローブをRNase Hを用いて切断する工程を包含する、請求項4に記載の方法。
(6)前記鎖終結部分が3’ホスフェートである、請求項5に記載の方法。
(7)伸長した二重鎖または前記開始オリゴヌクレオチドが前記オリゴヌクレオチドプローブに連結できない場合は常に、該伸長した二重鎖または該開始オリゴヌクレオチドにキャップ形成させる工程をさらに包含する、請求項2に記載の方法。
(8)前記伸長した二重鎖上に伸長可能な末端を再生する工程をさらに包含する、請求項2に記載の方法。
(9)前記再生の工程が、前記伸長した二重鎖において化学的に切断可能なヌクレオシド間結合を切断する工程を包含する、請求項8に記載の方法。
(10)前記化学的に切断可能なヌクレオシド間結合がホスホルアミデートである、請求項9に記載の方法。
(11)前記再生の工程が、前記伸長した二重鎖においてヌクレオシド間結合を酵素的に切断する工程を包含する、請求項8に記載の方法。
(12)前記オリゴヌクレオチドプローブが4種のリボヌクレオチドのサブ配列を包含し、そして前記再生の工程が該オリゴヌクレオチドプローブをRNase Hを用いて切断する工程を包含する、請求項11に記載の方法。
(13)ポリヌクレオチドのヌクレオチド配列を決定する方法であって、以下の工程:
(a)該ポリヌクレオチドを含むテンプレートを提供する工程;
(b)該ポリヌクレオチドに近接する該テンプレートと二重鎖を形成する開始オリゴヌクレオチドを提供する工程;
(c)該開始オリゴヌクレオチドに近接する該テンプレートにオリゴヌクレオチドプローブをアニールする工程;
(d)該オリゴヌクレオチドプローブを該開始オリゴヌクレオチドに連結させて、伸長した二重鎖を形成させる工程;
(e)該連結されたオリゴヌクレオチドプローブ上の標識によって該ポリヌクレオチドの1つ以上のヌクレオチドを同定する工程;および
(f)該ポリヌクレオチドのヌクレオチド配列が決定されるまで工程(c)〜(e)を繰り返す工程、
を包含する、方法。
(14)前記オリゴヌクレオチドプローブが前記開始オリゴヌクレオチドに対して遠位の末端に鎖終結部分を有し、そして前記方法が該オリゴヌクレオチドプローブ上に伸長可能な末端を再生する工程をさらに包含する、請求項13に記載の方法。
(15)前記オリゴヌクレオチドプローブに連結できない前記伸長した二重鎖または前記開始オリゴヌクレオチドにキャップ形成する工程をさらに包含する、請求項14に記載の方法。
(16)前記同定の工程が前記ポリヌクレオチドの1つのヌクレオチドを同定することよりなる、請求項14に記載の方法。
(17)前記同定の工程が前記鎖終結部分を除去する工程、および1つ以上の標識された鎖終結ヌクレオシド三リン酸の存在下で核酸ポリメラーゼを用いて前記オリゴヌクレオチドプローブを伸長させる工程を包含する、請求項16に記載の方法。
(18)オリゴヌクレオチドプローブであって、以下の式:
HO−(3’)(B)j(5’)−OP(=O)(O−)NH−(B)k−Bt*
を有し、ここで:
Bはヌクレオシドまたはそのアナログであり;
jは1〜12の範囲であり;
kはjとkとの合計が12以下であるように、0〜12の範囲であり;
Bt*は標識された鎖終結部分である、
オリゴヌクレオチドプローブ。
(19)以下からなる群より選択されるオリゴヌクレオチドプローブであって:
OP(=O)(O−)O−(5’)(B)sRRRR(B)wBt*、
HO−(3’)(B)sRRRR(B)wBt*、および
OP(=O)(O−)O−(5’)(B)sRRRR(B)w(3’)OP(=O)(O−)O
ここで:
Bはデオキシリボヌクレオチドまたはそのアナログであり;
Rはリボヌクレオチドであり;
sは1〜8の範囲であり;
wはjとkとの合計が8以下であるように0〜8の範囲であり;
Bt*は標識された鎖終結部分である、
オリゴヌクレオチドプローブ。
【0011】
発明の要旨
本発明は、一本鎖テンプレートに沿った二重鎖伸長の繰り返しサイクルに基づく核酸配列分析方法を提供する。好ましくは、このような伸長は、開始オリゴヌクレオチドとテンプレートとの間に形成される二重鎖から出発する。開始オリゴヌクレオチドを、最初の伸長サイクルにおいて、オリゴヌクレオチドプローブをその末端に連結することにより伸長し、伸長した二重鎖を形成させる。次いで、伸長した二重鎖を、その後の連結サイクルによって繰り返し伸長させる。各サイクルの間に、首尾よく連結されたオリゴヌクレオチドプローブ上、またはオリゴヌクレオチドプローブと会合した標識によって、テンプレート中の1つ以上ヌクレオチドの同一性を決定する。好ましくは、オリゴヌクレオチドプローブは、1回のサイクルで伸長した二重鎖の伸長が1回だけ起こるように、末端の位置にブロッキング部分(例えば、鎖終結ヌクレオチド)を有する。二重鎖を、次のサイクルでブロッキング部分を除去し、そして伸長可能な末端を再生することによってさらに伸長させる。
【0012】
本発明の1つの局面において、複数の異なる開始オリゴヌクレオチドをテンプレートの別々の試料に提供する。各開始オリゴヌクレオチドは、伸長を受けつつある末端が、複数の他のそれぞれの開始オリゴヌクレオチドと、1つ以上のヌクレオチドと合致しない(out of register)かまたは一致しないように、テンプレートと二重鎖を形成する。言い換えれば、伸長のための出発ヌクレオチドは、異なる開始オリゴヌクレオチドの各々につき、1つ以上のヌクレオチドにより異なる。このようにして、同一の長さのオリゴヌクレオチドプローブを用いる各伸長サイクルの後、同一の相対的相が異なるテンプレート上の開始オリゴヌクレオチドの末端間に存在する。従って、例えば、i)開始オリゴヌクレオチドが1つのヌクレオチドにより一致しない、ii)9マーのオリゴヌクレオチドプローブを伸長工程で用いる、およびiii)9つの異なる開始オリゴヌクレオチドを使用する好ましい実施態様においては、9つのテンプレートヌクレオチドが各伸長サイクルにおいて同時に同定される。
【0013】
定義
ポリヌクレオチドに関して本明細書中で使用する「配列決定」、「ヌクレオチド配列の決定」、「配列決定」および同様の用語は、ポリヌクレオチドの部分配列および全長配列の情報の決定を含む。すなわち、この用語は、配列比較、フィンガープリント法、標識ポリヌクレオチドについての同様のレベルの情報、および試験ポリヌクレオチドの各ヌクレオシドの明確な同定および順序付けを含む。
【0014】
プローブおよび標的ポリヌクレオチドの突出ストランドに関して「完全にマッチした二重鎖」とは、この突出ストランドが、二本鎖構造における各ヌクレオチドが反対のストランド上のヌクレオチドとワトソン−クリック塩基対合をするように、他のものと二本鎖構造を形成することを意味する。この用語はまた、プローブの縮重を減少させるために使用され得る、ヌクレオシドアナログ(デオキシイノシン、2−アミノプリン塩基を有するヌクレオシドなど)の対合を包含する。
【0015】
本明細書中で用いる「オリゴヌクレオチド」という用語は、デオキシリボヌクレオシド、リボヌクレオシドなどを含む、ヌクレオシドまたはそのアナログの線状オリゴマーを含む。通常、オリゴヌクレオチドは、数個のモノマー単位(例えば3〜4)から数百のモノマー単位までの範囲のサイズである。オリゴヌクレオチドを「ATGCCTG」のような文字の配列によって表す場合は、常に、このヌクレオチドは左から右に5’→3’の順序であり、そして特記しない限り、「A」はデオキシアデノシンを示し、「C」はデオキシシチジンを示し、「G」はデオキシグアノシンを示し、そして「T」はチミジンを示すことが理解される。
【0016】
本明細書中で使用する「ヌクレオシド」とは、例えば、KornbergおよびBaker,DNA Replication,第2版(Freeman,San Francisco,1992)に記載されるように、2’−デオキシ形態および2’−ヒドロキシル形態を含む、天然のヌクレオシドを含む。ヌクレオシドに関して「アナログ」とは、例えば、Scheit,Nucleotide Analogs(John Wiley,New York,1980)によって一般的に記載されているように、修飾された塩基部分および/または修飾された糖部分を有する合成ヌクレオシドを含む。このようなアナログは、結合特性の増強、縮重の減少、特異性の増大などのために設計された合成ヌクレオシドを含む。
【0017】
本明細書で使用される「連結」とは、テンプレート駆動反応において、2つ以上の核酸(例えば、オリゴヌクレオチドおよび/またはポリヌクレオチド)の末端間で共有結合または結合(linkage)を形成することを意味する。結合(bond)または結合(linkage)の性質は広範囲に変化し得るし、そして連結は酵素的または化学的に行われ得る。
【図面の簡単な説明】
【0018】
【図1】図1は、本発明による複数のテンプレートの並行伸長を模式的に示す。
【図2】図2は、酸不安定結合を使用する本発明の1つの実施態様を模式的に示す。
【図3A】図3Aは、3’→5’伸長でRNase H不安定オリゴヌクレオチドを使用する本発明の1つの実施態様を模式的に示す。
【図3B】図3Bは、5’→3’伸長でRNase H不安定オリゴヌクレオチドを使用する本発明の1つの実施態様を模式的に示す。
【図4】図4は、連結、その後のポリメラーゼ伸長および切断を使用する本発明の1つの実施態様を模式的に示す。
【発明を実施するための形態】
【0019】
発明の詳細な説明
本発明は、同様のサイズのDNAフラグメントの電気泳動による分離を不要とし、そしてゲルまたは同様の媒体中のDNAフラグメントの空間的に重なるバンドの検出および分析に関する困難をなくす核酸の配列決定方法を提供する。本発明はまた、DNAポリメラーゼを用いて長い一本鎖テンプレートからDNAフラグメントを作製する必要もない。
【0020】
本発明の1つの局面の一般的なスキームを図1に模式的に示す。以下により十分に記載するように、本発明は、この実施態様の特定の特徴によって限定されることを意図しない。配列未知のポリヌクレオチド(50)および結合領域(40)を含むテンプレート(20)を固相支持体(10)に付着させる。好ましくは、Nマーのプローブを使用する実施態様では、テンプレートをN個のアリコートに分け、そして各アリコートに、他の開始オリゴヌクレオチドの位置とは異なる結合領域(40)中のある位置で完全にマッチした二重鎖を形成する異なる開始オリゴヌクレオチドikを提供する。すなわち、開始オリゴヌクレオチドi1〜iNは、未知配列に対して近い側の二重鎖の末端が未知配列のはじまりから0〜N−1ヌクレオチドであるように、結合領域(40)においてテンプレートと二重鎖の組を形成する。従って、Nマーのプローブを用いる連結の第1サイクルにおいて、図1中のi1に連結したプローブ(30)の末端ヌクレオチド(16)は、結合領域(40)のN−1ヌクレオチドに対して相補的である。同様に、図1中のi2に連結したプローブ(30)の末端ヌクレオチド(17)は、結合領域(40)のN−2ヌクレオチドに対して相補的であり;図1中のi3に連結したプローブ(30)の末端ヌクレオチド(18)は、結合領域(40)のN−3ヌクレオチドに対して相補的である。以下同様である。最後に、inに連結したプローブ(30)の末端ヌクレオチド(15)は、未知配列(50)の最初のヌクレオチドに対して相補的である。連結の第2サイクルにおいて、プローブ(31)の末端ヌクレオチド(19)は、開始オリゴヌクレオチドi1で出発する二重鎖における未知配列(50)の2番目のヌクレオチド(19)に対して相補的である。同様に、開始オリゴヌクレオチドi2、i3、i4などで出発する二重鎖に連結したプローブの末端ヌクレオチドは、未知配列(50)の3番目、4番目、および5番目のヌクレオチドに対して相補的である。
【0021】
上記の実施態様において、オリゴヌクレオチドプローブは、伸長した二重鎖に隣接するヌクレオチドの同一性が標識から決定できるように標識される。
【0022】
結合領域(40)は既知配列を有するが、長さおよび組成は大いに変化し得る。結合領域は開始オリゴヌクレオチドのハイブリダイゼーションを適応させるために十分に長くなければならない。異なる結合領域を同一かまたは異なるかのいずれかの開始オリゴヌクレオチドとともに使用できるが、調製の便宜のためには、同一の結合領域および異なる開始オリゴヌクレオチドを提供するのが好ましい。従って、全てのテンプレートを同一に調製し、次いで、異なる開始オリゴヌクレオチドでの使用のためにアリコートに分ける。好ましくは、結合領域は、異なる開始オリゴヌクレオチドの組を適応させるのに十分な長さであるべきであって、各々はテンプレートにハイブリダイズしてその後の連結のための異なる出発点を生じる。最も好ましくは、結合領域は、約20〜50ヌクレオチドの間の長さである。
【0023】
開始オリゴヌクレオチドは、伸長サイクルのいずれの洗浄工程の間にも無傷のままでいる結合領域との高度に安定な二重鎖を形成するように選択される。これは、都合よく、開始オリゴヌクレオチドの長さが、オリゴヌクレオチドプローブの長さよりもかなり長いように選択することによって、および/またはそれらがGCリッチとなるように選択することによって達成される。開始オリゴヌクレオチドはまた、種々の技術(例えば、Summertonら,米国特許第4,123,610号)によってテンプレートストランドに架橋することもでき;または、それらは、その天然の対応物(例えば、ペプチド核酸)よりも安定性の大きな二重鎖を形成するヌクレオチドアナログよりなることもできる。Science,254:1497−1500(1991);Hanveyら,Science,258:1481−1485(1992);およびPCT出願第PCT/EP92/01219号および同第PCT/EP92/01220号。
【0024】
好ましくは、開始オリゴヌクレオチドの長さは約20〜30ヌクレオチドであり、そしてその組成は、使用されるオリゴヌクレオチドプローブの融解温度を約10〜50℃だけ超える二重鎖融解温度を提供するために十分なパーセントのGおよびCを含む。より好ましくは、開始オリゴヌクレオチドの二重鎖融解温度は、オリゴヌクレオチドプローブの融解温度を約20〜50℃だけ超える。配列決定操作で使用される異なる開始オリゴヌクレオチドの数Nは、各サイクルで1つのヌクレオチドが同定される場合の1から、実際的に使用できるオリゴヌクレオチドプローブのサイズによってのみサイズが制限される多数まで変化し得る。オリゴヌクレオチドプローブのサイズを制限する因子は、妥当な速度でハイブリダイゼーション反応を駆動するために十分に高濃度の個々のプローブを有する混合物を調製する際の困難、二次構造の形成に対するより長いプローブの感受性、1塩基ミスマッチに対する感受性の低下などを含む。好ましくは、Nは1〜16の範囲であり;より好ましくは、Nは1〜12の範囲であり;そして最も好ましくは、Nは1〜8の範囲である。
【0025】
広範囲の種々のオリゴヌクレオチドプローブを本発明で使用することができる。一般に、オリゴヌクレオチドプローブは、開始オリゴヌクレオチドまたは伸長した二重鎖に連結して、次の伸長サイクルの伸長した二重鎖を生じることができるべきである;連結は、プローブが連結の前にテンプレートと二重鎖を形成するべきであるのでテンプレートに駆動されるべきである;プローブは、1回の伸長サイクルにおいて同一のテンプレートに複数のプローブが連結するのを防ぐためにブロッキング部分を有すべきであり、プローブは連結後に伸長可能な末端を再生するために処理または修飾され得るべきであり、そしてプローブは成功した連結後にテンプレートに関連する配列情報の獲得を可能とするシグナリング部分を有すべきである。以下でより十分に記載するように、実施態様に応じて、伸長した二重鎖または開始オリゴヌクレオチドは、オリゴヌクレオチドプローブによって、5’→3’方向または3’→5’方向のいずれかに伸長され得る。一般に、オリゴヌクレオチドプローブはテンプレートと完全にマッチした二重鎖を形成する必要はないが、このような結合が通常は好ましい。テンプレート中の1つのヌクレオチドが各伸長サイクルで同定される好ましい実施態様において、完全な塩基対合は、その特定のヌクレオチドを同定するために必要とされるに過ぎない。例えば、オリゴヌクレオチドプローブを伸長した二重鎖に酵素的に連結する実施態様において、完全な塩基対合(すなわち、適切なワトソン−クリック塩基対合)が、連結されるプローブの末端ヌクレオチドとテンプレート中のその相補物との間で必要とされる。一般に、このような実施態様において、プローブの残りのヌクレオチドは、次の連結がテンプレートに沿って所定の部位または塩基数で起こることを確実とする「スペーサー」として働く。すなわち、それらの対合、またはその欠落は、さらなる配列情報を提供しない。同様に、塩基同定についてポリメラーゼ伸長に頼る実施態様において、プローブは主としてスペーサーとして働き、従って、テンプレートに対する特異的ハイブリダイゼーションは、望ましいとはいえ、重要ではない。
【0026】
好ましくは、オリゴヌクレオチドプローブは、所定の長さの全ての可能な配列のオリゴヌクレオチドを含む混合物としてテンプレートに適用される。このような混合物の複雑さは、例えば、Kong Thoo Linら,Nucleic Acids Research,20:5149−5152;米国特許第5,002,867号;Nicholsら,Nature,369:492−493(1994)によって教示されるデオキシイノシンなどのようないわゆる縮重低下アナログを用いることを含む多数の方法によって;またはオリゴヌクレオチドプローブの多数の混合物(例えば、一緒にすると所定の長さの全ての可能な配列を含むオリゴヌクレオチド配列の4つのばらばらのサブセットを含む4つの混合物)を別々に適用することによって低下させることができる。
【0027】
本発明の開始オリゴヌクレオチドおよびオリゴヌクレオチドプローブは、都合よく、自動DNA合成機(例えば、Applied Biosystems,Inc.(Foster City,California)392型または394型のDNA/RNA合成機)で、例えば、以下の文献に開示されるホスホルアミダイト化学のような標準的な化学を用いて合成される:BeaucageおよびIyer,Tetrahedron,48:2223−2311(1992);Molkoら,米国特許第4,980,460号;Kosterら,米国特許第4,725,677号;Caruthersら,米国特許第4,415,732号;同第4,458,066号;および同第4,973,679号など。例えば、ホスホロチオエート、ホスホルアミデートなどのような非天然の骨格基が得られる別の化学を使用することもできる。但し、得られたオリゴヌクレオチドは特定の実施態様の連結および他の試薬に適合するものとする。オリゴヌクレオチドプローブの混合物は、例えば、Teleniusら,Genomics,13:718−725(1992);Welshら,Nucleic Acids Research,19:5275−5279(1991);Grothuesら,Nucleic Acids Research,21:1321−1322(1993);Hartley,欧州特許出願第90304496.4号などに開示されるような周知の技術を用いて容易に合成される。一般に、これらの技術は、縮重を導入することが望まれるカップリング工程の間の成長するオリゴヌクレオチドへの活性化モノマーの混合物の適用を必要とするだけである。
【0028】
従来のリガーゼを本発明で使用する場合、以下でより十分に記載するように、いくつかの実施態様ではプローブの5’末端をリン酸化し得る。5’モノホスフェートを化学的または酵素的のいずれかでキナーゼを用いてオリゴヌクレオチドに結合させることができる。例えば、Sambrookら,Molecular Cloning;A Laboratory Manual,第2版(Cold Spring Harbor Laboratory,New York,1989)。化学的リン酸化は、HornおよびUrdea,Tetrahedron Lett.,27:4705(1986)によって記載されており、そして開示されたプロトコルを実施するための試薬は、例えば、Clontech Laboratories(Palo Alto,California)から5’Phosphate−ONTMとして市販されている。好ましくは、必要な場合、オリゴヌクレオチドプローブは化学的にリン酸化される。
【0029】
本発明のプローブは、蛍光部分、発色部分などの直接的または間接的な結合を含む種々の方法で標識され得る。DNAを標識し、そしてDNAプローブを構築するための方法論の多くの総合的な概説が、本発明のプローブを構築するために適用可能な手引きを提供する。このような概説は、Matthewsら,Anal.Biochem.,第169巻,1−25頁(1988);Haugland,Handbook of Fluorescent Probes and Research Chemicals(Molecular Probes,Inc.,Eugene,1992);KellerおよびManak,DNA Probes,第2版(Stockton Press,New York,1993);およびEckstein編,Oligonucleotides and Analogues:A Practical Approach(IRL Press,Oxford,1991)などを含む。本発明に適用できる多くのより詳細な方法は、以下の文献の見本に開示される:Fungら,米国特許第4,757,141号;Hobbs,Jr.ら,米国特許第5,151,507号;Cruickshank,米国特許第5,091,519号;(レポーター基の結合のための官能化されたオリゴヌクレオチドの合成);Jablonskiら,Nucleic Acids Research,14:6115−6128(1986)(酵素−オリゴヌクレオチドコンジュゲート);およびUrdeaら,米国特許第5,124,246号(分岐DNA)。
【0030】
好ましくは、プローブは、例えば、Menchenら,米国特許第5,188,934号;Begotら,PCT出願第PCT/US90/05565号によって開示されるような1つ以上の蛍光色素で標識される。
【0031】
テンプレートに対するオリゴヌクレオチドプローブの適用のためのハイブリダイゼーション条件の選択における手引きは、多数の文献(例えば、Wetmur,Critical Reviews in Biochemistry and Molecular Biology,26:227−259(1991);DoveおよびDavidson,J.Mol.Biol.5:467−478(1962);Hutton,Nucleic Acids Research,10:3537−3555(1977);Breslauerら,Proc.Natl.Acad.Sci.83:3746−3750(1986);Innisら編,PCR Protocols(Academic Press,New York,1990)など)に見い出すことができる。
【0032】
一般に、オリゴヌクレオチドプローブを、伸長した二重鎖の末端に並列してテンプレートにアニールする場合、二重鎖とプローブとを連結する、すなわち、相互の共有結合を生じさせる。連結は酵素的または化学的のいずれかで達成され得る。化学的連結方法は当該分野で周知である。例えば、Ferrisら,Nucleosides&Nucleotides,8:407−414(1989);Shabarovaら,Nucleic Acids Research,19:4247−4251(1991)など。好ましくは、酵素的連結は標準的プロトコルでリガーゼを用いて実施される。多くのリガーゼが知られており、そして本発明で使用するのに適切である。例えば、Lehman,Science,186:790−797(1974);Englerら,DNA Ligases,3−30頁,Boyer編,The Enzymes,第15B巻(Academic Press,New York,1982)など。好ましいリガーゼは、T4 DNAリガーゼ、T7 DNAリガーゼ、E.coli DNAリガーゼ、Taqリガーゼ、Pfuリガーゼ、およびTthリガーゼを含む。それらの使用についてのプロトコルは周知である。例えば、Sambrookら(上記);Barany,PCR Methods and Applications,1:5−16(1991);Marshら,Strategies,5:73−76(1992)など。一般に、リガーゼは、隣接するストランドの3'ヒドロキシルへの連結のために5’リン酸基が存在することを必要とする。
【0033】
標的ポリヌクレオチドの調製
好ましくは、標的ポリヌクレオチドを結合領域に連結してテンプレートを形成し、そしてテンプレートを複雑かつ時間を消費する精製工程を伴わない試薬の連続的適用を可能とする固相支持体(例えば、磁性粒子、ポリマーマイクロスフィア、フィルター物質など)に付着させる。標識ポリヌクレオチドの長さは広範囲に変化し得る;しかし、調製の便宜のためには、従来の配列決定で使用される長さが好ましい。例えば、数百塩基対(200〜300)から1〜2キロ塩基対までの範囲の長さが好ましい。
【0034】
標的ポリヌクレオチドは、種々の常法によって調製され得る。例えば、標的ポリヌクレオチドは、従来のDNA配列決定で使用されるものを含む、従来の任意のクローニングベクターのインサートとして調製され得る。適切なクローニングベクターの選択および使用のための広範な手引きがSambrookら,Molecular Cloning:A Laboratory Manual,第2版(Cold Spring Harbor Laboratory,New York,1989)、および同様の文献で見い出される。SambrookらおよびInnisら編,PCR Protocols(Academic Press,New York,1990)もまた、標的ポリヌクレオチドを調製するためのポリメラーゼ連鎖反応の使用について手引きを提供する。好ましくは、この方法で使用する他の試薬から標的ポリヌクレオチドを分離するのを容易にするために、磁性ビーズまたは他の固体支持体への付着を可能とするクローン化標的ポリヌクレオチドまたはPCR増幅標的ポリヌクレオチドを調製する。このような調製技術についてのプロトコルは、Wahlbergら,Electrophoresis,13:547−551(1992);Tongら,Anal. Chem.,64;2672−2677(1992);Hultmanら,Nucleic Acids Research,17:4937−4946(1989);Hultmanら,Biotechniques,10:84−93(1991);Syvanenら,Nucleic Acids Research,16:11327−11338(1988);Dattaguptaら,米国特許第4,734,363号;Uhlen,PCT出願第PCT/GB89/00304号;および同様の文献に十分に記載される。キットもまた、このような方法を実施するために、例えば、Dynal AS.(Oslo,Norway)からDynabeadsTMテンプレート調製キットとして市販される。
【0035】
一般に、本発明の方法で使用する微粒子またはビーズのサイズおよび形状は重要ではない;しかし、直径数m(例えば、1〜2m)から直径数百m(例えば、200〜1000m)の範囲のサイズの微粒子が好ましい。なぜなら、それらは、例えば蛍光標識プローブからの容易に検出できるシグナルの生成を可能としつつ、試薬および試料の使用量を最小にするからである。
【0036】
伸長可能な末端の連結、キャップ形成、および再生についてのスキーム
1つの局面において、本発明は、オリゴヌクレオチドプローブの連結および同定の繰り返し工程を必要とする。しかしながら、同一工程における同一の伸長した二重鎖に対する複数プローブの連結は、通常は同定の問題を誘引するであろうから、多重伸長を防止しそして伸長可能な末端を再生するのに有用である。さらに、もし連結工程が100%効果的でなければ、それらがいずれのさらなる連結工程にも参画しないように、連結を受けない伸長二重鎖にキャップ形成するのが望ましいであろう。すなわち、キャップ形成工程は、好ましくは、ポリヌクレオチド合成のような他の合成化学プロセスから類推して、連結工程の後に起こる(例えば、Andrusら、米国特許第4,816,571号)。これは、その後の同定工程で生じるシグナルから潜在的に有意なノイズの源を除去するであろう。
【0037】
以下、本発明の連結、キャップ形成、再生および同定工程を実施するためのいくつかの例示的スキームを記載する。それらは、手引きの目的で提示するものであって、限定することを意図しない。
【0038】
開始オリゴヌクレオチドまたは3’→5’方向に伸長した二重鎖を伸長するためのスキームは図2に示す。テンプレート(20)をその5’末端により、固相支持体(10)に付着させる。これは、常法技術を用い、ビオチンまたは同様の連結部分を介して、丁度都合よく達成され得る。5’リン酸基を有する開始オリゴヌクレオチド(200)を、連結および同定の最初のサイクル前に、前記のようにテンプレート(20)にアニールする。以下の形態のオリゴヌクレオチドプローブ(202)を使用する:
HO−(3’)BBB...BBB(5’)−OP(=O)(O−)NH−Bt*
ここで、BBB...BBBはオリゴヌクレオチドプローブ(202)のヌクレオチドの配列を表し、Bt*はホスホルアミデート基、または光切断結合のような、他の不安定結合を介してオリゴヌクレオチドの5’炭素に連結した標識された鎖終結部分である。Bt*の性質は広く変化し得る。それは連続的連結を防止する限り、標識されたヌクレオシド(例えば、5’P3’Nホスホルアミデートを介してカップリングしたもの)または他の部分であり得る。それは、AgrawalおよびTang、国際出願第PCT/US91/08347号に記載されるように、単にリンカーによって結合された標識であり得る。オリゴヌクレオチドプローブの重要な特徴は、アニーリングおよび連結(204)の後に、標識を除去でき、例えば、Letsingerら、J.Am.Chem.Soc.,94:292−293(1971);Letsingerら、Biochem.,15:2810−2816(1976);Gryaznovら,Nucleic Acid Research,20:3403−3409(1992);および同様の文献によって教示されるように、ホスホルアミデート結合を酸で処理することによって、伸長可能な末端を再生できることである。例えば、ホスホルアミデートの加水分解は、室温における40分間のジクロロメタン中の0.8%トリフルオロ酢酸での処理によって達成され得る。このようにして、Bt*上の標識を介して連結したプローブをアニーリングし、連結し、そして同定した後、酸加水分解(206)により鎖終結部分を切断し、それによりリン結合を破壊し、連結したオリゴヌクレオチド上に5’モノホスフェートを残しておく。この工程は連続的サイクルで繰り返され得る(208)。この実施態様の1つの局面において、単一の開始オリゴヌクレオチドを、1つのヌクレオチドのみが各配列決定サイクルで同定されるように使用し得る。このような実施態様では、このプローブは好ましくは以下の形態を有する:
HO−(3’)B(5’)−OP(=O)(O−)NHBB...BBB−Bt*。
【0039】
このようにして、各連結および酸切断工程の後、二重鎖は1ヌクレオチドだけ伸長される。
【0040】
加水分解の前に、キャップ形成工程が導入され得る。例えば、プローブ(202)は以下の形態を有する:
HO−(3’)BB...Bp^B...BB(5’)−OP(=O)(O−)NH−Bt*
ここで、「p^」はホスホロチオエート、メチルホスホネートなどのようなエキソヌクレアーゼ耐性結合である。このような実施態様において、キャップ形成は、未連結伸長二重鎖を切断してエキソヌクレアーゼ耐性結合に戻すλエキソヌクレアーゼのようなエキソヌクレアーゼで伸長二重鎖を処理することによって達成され得る。次いで、伸長二重鎖の5’末端におけるこの結合の存在は、その後の連結にそれが関与することから防止する。明らかに、多くの他のキャップ形成方法、例えばアシル化、不活性オリゴヌクレオチドの連結などを使用し得る。遊離3’ヒドロキシルが関与する場合、キャップ形成は、鎖終結ヌクレオシド三リン酸、例えばジデオキシヌクレオシド三リン酸などの存在下で、DNAポリメラーゼで二重鎖を伸長させることによって達成され得る。
【0041】
前記したホスホルアミデート結合は、本明細書中では「化学的に切断可能なヌクレオシド間結合」と呼ばれるヌクレオシド間結合の一般的クラスの例である。これらは、酸化性環境、還元性環境、特徴的波長の光(光不安定性結合用)などのような特徴的な化学的または物理的条件でそれらを処理することによって切断され得るヌクレオシド間結合である。本発明に従い使用され得る化学的に切断可能なヌクレオシド間結合の他の例は、Urdea第5,380,833号;Gryaznovら、Nucleic Acids Research,21:1403−1408(1993)(ジスルフィド);Gryaznovら、Nucleic Acids Research,22:2366−2369(1994)(ブロモアセチル);Urdeaら、国際出願PCT/US91/05287(光不安定性);および同様の文献に記載されている。
【0042】
本発明で使用され得るさらなる化学的に切断可能な結合は、伸長可能なヌクレオシドに化学的に変換され得る鎖終結ヌクレオチドを含む。このような化合物の例は以下の文献に記載されている:Canardら、国際出願PCT/FR94/00345;Ansorge,独国特許出願第DE 4141178 A1号;Metzkerら、Nucleic Acids Research,22:4259−4267(1994);Cheeseman、米国特許第5,302,509号;Rossら,国際出願第PCT/US90/06178号など。
【0043】
開始オリゴヌクレオチドまたは伸長した二重鎖を5’→3’方向に伸長させるためのスキームが図3Aに示される。テンプレート(20)は、その3’末端によって固相支持体(10)に結合させる。前記のように、これは、常法技術を用い、ビオチンまたは同様の連結部分を介して、丁度都合よく達成され得る。3’ヒドロキシル基を有する開始オリゴヌクレオチド(300)は、連結および同定の最初のサイクル前に、前記のようにテンプレート(20)にアニールされる。以下の形態のオリゴヌクレオチドプローブ(302)を使用する:
OP(=O)(O−)O−(5’)BBB...BBBRRRRBt*。
【0044】
ここで、BBB...BBBRRRRはオリゴヌクレオチドプローブ(302)の2’−デオキシヌクレオチドの配列を表し、「RRRR」はプローブ(302)の4つのリボヌクレオチドの配列を表し、そしてBt*は前記のように標識された鎖終結部分である。このような混合されたRNA−DNAオリゴヌクレオチドは通常の自動DNA合成機を用いて容易に合成される(例えば、Duckら、米国特許第5,011,769号)。RNase Hは、4つのリボヌクレオチドセグメントの中心において特異的にプローブを切断し(Hogrefeら、J.Biol.Chem.,265:5561−5566(1990))、伸長した二重鎖上に3’ヒドロキシル(312)を残し、これはその後の連結工程に参加し得る。このようにして、本実施態様におけるサイクルは、テンプレート(20)にプローブ(302)をアニールされそして、連結(304)させて、伸長した二重鎖(306)を形成することにより進行する。Bt*を介する同定の後、伸長した二重鎖は、標識を切断し、伸長可能な末端を再生するために、RNase Hで処理される。次いで、サイクルが繰り返される(314)。キャップ形成(310)は、RNase H処理の前に、4つのジデオキシヌクレオシド三リン酸、ddATP、ddCTP、ddGTPおよびddTTPの存在下、DNAポリメラーゼで未連結末端を伸長させることによって実施され得る。
【0045】
図3Bで説明したように、同様のスキームが3’5’伸長のために使用され得る。このような実施例において、開始オリゴヌクレオチドまたは伸長した二重鎖(330)は、5’一リン酸を有し、オリゴヌクレオチドプローブ(332)は以下の形態を有する:
HO−(3’)BBB...BBBRRRRB..BBt*。
【0046】
前記のように、アニーリング、連結(334)、および同定(338)の後、伸長した二重鎖(336)は、この場合は伸長した二重鎖の末端で5’一リン酸(342)を残すRNase Hにより、切断される。再生した伸長可能な末端を用いて、サイクルは繰り返され得る(344)。キャップ形成工程は、RNase H加水分解前に、未標識非RNA含有プローブを連結することにより、またはホスファターゼでの処理による任意の残存する5’モノホスフェートを除去することのいずれかによって含まれ得る。
【0047】
ヌクレオチドの同定は、連結後のポリメラーゼ伸長により達成され得る。図4に例示するように、この実施態様については、テンプレート(20)を前記のように固相支持体(10)に結合させ、3’ヒドロキシルを有する開始オリゴヌクレオチド(400)を最初のサイクル前にテンプレートにアニールさせる。以下の形態を有するオリゴヌクレオチドプローブ(402):
OP(=O)(O−)O−(5’)BBB...BBBRRRRB...B(3’)OP(=O)(O−)O
がテンプレート(20)にアニールされ、そして連結(404)され、伸長した二重鎖(406)が形成される。同一サイクルにおいてプローブの連続的連結を防ぐ3’一リン酸がホスファターゼ(408)で除去され、遊離の3’ヒドロキシル(410)が露出される。明らかに、別のブロッキングアプローチも使用され得る。伸長した二重鎖(406)は、標識したジデオキシヌクレオシド三リン酸(412)の存在下で、核酸ポリメラーゼによってさらに伸長し、それにより、取り込まれたジデオキシヌクレオシドの標識によってテンプレート(20)のヌクレオチドの同定が可能となる。次いで、標識されたジデオキシヌクレオチドおよびプローブ(402)の一部は、伸長した二重鎖(406)上に伸長可能な末端を再生するために、例えば、RNase H処理によって切断される(414)。次いで、サイクルが繰り返される(416)。
【0048】
実施しなければならない別々のアニーリング反応の数を減少させるために、オリゴヌクレオチドプローブは、その相補的配列と完全にマッチした二重鎖が、同様の安定性または結合の自由エネルギーを有するプローブの混合物またはサブセットにグループ分けされ得る。同様の二重鎖安定性を有するオリゴヌクレオチドプローブのこのようなサブセットは、本明細書中ではオリゴヌクレオチドプローブの「ストリンジェンシークラス」といわれる。次いで、オリゴヌクレオチドプローブの混合物またはストリンジェンシークラスは、実質的に標的ポリヌクレオチドに相補的なオリゴヌクレオチドプローブのみが二重鎖を形成するような条件下で、標的ポリヌクレオチドと別々に混ぜ合わされる。すなわち、ハイブリダイゼーション反応のストリンジェンシーは、実質的に完全に相補的なオリゴヌクレオチドプローブのみが二重鎖を形成するように選択される。次いでこれらの完全にマッチした二重鎖が、連結され、伸長した二重鎖が形成される。所与のオリゴヌクレオチドプローブ長さにつき、各ストリンジェンシークラス内のオリゴヌクレオチドプローブの数は、広く変化し得る。オリゴヌクレオチドプローブの長さおよびストリンジェンシークラスサイズの選択は、標識配列の長さ、およびそれがどのように調製されるか、ハイブリダイゼーション反応が自動化され得る程度、ハイブリダイゼーション反応のストリンジェンシーが制御され得る程度、相補的配列を有するオリゴヌクレオチドプローブの存在または不存在などのようないくつかの因子に依存する。特定の実施態様のためのストリンジェンシークラスの適切なサイズを選択する手引きは、核酸ハイブリダイゼーションおよびポリメラーゼ連鎖反応法についての一般的文献中に見い出され得る(例えば、Gotoh,Adv.Biophys.16:1−52(1983);Wetmer,Critical Reviews in Biochemistry and Molecular Biology 26:227−259(1991);Breslauerら,Proc.Natl.Acad.Sci. 83:3746−3750(1986);Wolfら、Nucleic Acids Research,15:2911−2926(1987);Innisら編,PCR Protocols(Academic Press,New York,1990);McGrawら、Biotechniques,8:674−678(1990)など)。ストリンジェンシーは、温度、塩濃度、ホルムアミドのような特定の有機溶媒の濃度などを含むいくつかの変化するパラメーターによって制御され得る。好ましくは、用いられる種々のポリメラーゼまたはリガーゼの活性が、塩濃度または有機溶媒濃度がオリゴヌクレオチドプローブの特異的アニーリングを保証するために変えられ得る程度を制限するために、温度がストリンジェンシークラスを規定するために用いられる。
【0049】
一般に、ストリンジェンシークラスが大きければ大きいほど、ハイブリダイズする混合物の複雑性が大きくなり、そして混合物におけるいずれかの特定のオリゴヌクレオチドプローブの濃度が低下する。標的ポリヌクレオチド上に相補的部位を有する、より低濃度のオリゴヌクレオチドプローブはハイブリダイズし連結されるオリゴヌクレオチドプローブの相対的公算(relative likelihood)を低下させる。これは、今度は、感受性の低下を導く。また、ストリンジェンシークラスが大きければ大きいほど、オリゴヌクレオチドプローブと相補的配列との間に形成される二重鎖の安定性に大きな変動を有する。他方、ストリンジェンシークラスが小さければ小さいほど、1セットの全てのオリゴヌクレオチドプローブが標的ポリヌクレオチドにハイブリダイズすることを保証するために、より多数のハイブリダイゼーション反応を必要とする。
【0050】
例えば、8マーのオリゴヌクレオチドプローブが使用される場合、ストリンジェンシークラスは、各々約50から約500の間のオリゴヌクレオチドプローブを含み得る。このようにして、数百〜数千のハイブリダイゼーション/連結反応が必要とされる。より大きなサイズのオリゴヌクレオチドプローブについては、より大きなストリンジェンシークラスが、ハイブリダイゼーション/伸長反応の数を実際的なもの(例えば、104〜105またはそれ以上)とするために必要とされる。
【0051】
同一ストリンジェンシークラスのオリゴヌクレオチドプローブは、十分にランダムなオリゴヌクレオチドプローブが合成されるのと類似した様式で(例えば、Teleniusら、Genomics,13:718−725(1992);Welshら、Nucleic Acids Research,19:5275−5279(1991);Grothuesら、Nucleic Acuds Research,21:1321−1322(1993);Hartley,欧州特許出願90304496.4などに開示されているように)、同時に合成され得る。差異は、各サイクルにおいて、モノマーの異なる混合物を、増殖するオリゴヌクレオチドプローブ鎖に適用することであり、ここで、混合物中の各モノマーの割合は、ストリンジェンシークラスにおけるオリゴヌクレオチドプローブの位置における各ヌクレオシドの割合によって指示される。ストリンジェンシークラスは、利用可能なアルゴリズム(例えば、Breslauerら、Proc.Natl.Acad.Sci.,83:3746−3750(1986);Loweら、Nucleic Acids Research,18:1757−1761(1990)など)により二重鎖形成の自由エネルギーを計算することによって容易に形成される。オリゴヌクレオチドプローブは、標準的な反応条件下(例えば、標準的なバブルソート、Base,Computer Algorithms(Addison−Wesley,Menlo Park,1978))でのそれらの相補体への結合の自由エネルギーにより順序付けられる。例えば、以下のものは標準的なハイブリダイゼーション条件下での二重鎖形成の自由エネルギーによる最大安定性(頂部から底部にかけて)を有する、および二重鎖形成の自由エネルギーの最低安定性を持つ10の6マーのリストである(自由エネルギーはBreslauerら(前出)により計算される)。
【0052】
オリゴヌクレオチドプローブ
ランキング配列(5’→3’)
1 GCGCGC
2 CGCGCG
3 CCCGCG
4 CGCCCG
5 CGCGCC
6 CGCGGC
7 CGGCGC
8 GCCGCG
9 GCGCCG
10 GCGCGG
・ ・
・ ・
・ ・
4087 TCATAT
4088 TGATAT
4089 CATATA
4090 TATATG
4091 ATCATG
4092 ATGATG
4093 CATCAT
4094 CATGAT
4095 CATATG
4096 ATATAT
このように、もしストリンジェンシークラスが最初の10個の6マーからなるのであれば、最初の(最も3’側の)位置についての混合物モノマーは、0:4:6:0(A:C:G:T)となるであろうし、第2の位置については、それは0:6:4:0となるであろう(以下、同様)。もしストリンジェンシークラスが最後の10個の6マーからなるならば、最初の位置についてのモノマーの混合物は1:0:4:5となるであろうし、第2の位置については、それは5:0:0:5であろう(以下、同様)。次いで、得られた混合物は、加熱溶出によって所望のストリンジェンシークラスの配列につきさらに富化され得る(例えば、Miyazawaら、J.Mol.Biol.,11:223−237(1965))。
【0053】
より便宜的には、数百〜数千のオリゴヌクレオチドを含むストリンジェンシークラスは、種々の並行合成アプローチによって直接的に合成され得る(例えば、Frankら、米国特許第4,689,405号;Matsonら、Anal.Biochem.,224;110−116(1995);Fodorら、国際出願第PCT/US93/04145号;Peaseら、Proc.Natl.Acad.Sci.、91:5022−5026(1994);Southernら、J.Biotechnology,35:217−227(1994),Brennan、国際出願第PCT/US94/05896号など)。
【0054】
いくつかの場合において、オリゴヌクレオチドプローブ−ダイマーを形成するのに感受性のサブセットまたはオリゴヌクレオチドプローブにおいて、他のオリゴヌクレオチドプローブに対する相補的配列を有する別々のサブセットのオリゴヌクレオチドプローブに配置することによって、オリゴヌクレオチドプローブのさらなるストリンジェンシークラスを形成することが所望され得る。
【0055】
明らかに、当業者は、先に明示的に記載されていないが、本発明のなおさらなる実施態様を設計するために、前記の実施態様の特徴を組み合わせることができる。
【0056】
また、本発明は、本発明の方法を自動的に実施するためのシステムおよび装置を含む。このようなシステムおよび装置は、i)標的ポリヌクレオチドをつなぎ止めるために使用される固相支持体の性質、ii)所望の並行操作の程度、iii)使用される検出スキーム;iv)試薬を再使用するか捨てるかなどを含めたいくつかの設計製薬に依存して種々の形態をとり得る。一般に、装置は一連の試薬貯蔵器、好ましくは固相支持体(例えば、磁気ビーズ)に付着させた標的ポリヌクレオチドを含有する1つ以上の反応容器、1つ以上の検出ステーション、および試薬貯蔵器から反応容器へおよび反応容器から検出ステーションへ、予め決定した様式で試薬を移すためのコンピュータ制御手段を含む。コンピュータ温度を制御するためのコンピューター制御手段は、Harrisonら、Biotechniques,14:88−97(1993);Fujitaら、Biotechniques,9:584−591(1990);Wadaら、Rev.Sci.Instrum,54:1569−1572(1983)などに開示されているもののような、種々の一般目的の実験室ロボットによって実行され得る。このような実験室ロボットはまた市販されている(例えば、Applied Biosystemsモデル800Catalyst(Foster City,CA))。
【0057】
本発明の異なる実施態様を実施するために種々のキットが提供され得る。一般に、本発明のキットは、オリゴヌクレオチドプローブ、開始オリゴヌクレオチド、および検出システムを含む。キットはさらに、連結試薬および本発明の特別の実施態様を実施するための指示を含む。タンパク質リガーゼ、RNase H、核酸ポリメラーゼ、または他の酵素を使用する実施態様においては、それらの各緩衝液が含ませられ得る。いくつかの場合においては、これらの緩衝液は同一であり得る。好ましくは、キットはまた、テンプレートをつなぎ止めるための固相支持体(例えば、磁気ビーズ)を含む。1つの好ましいキットにおいて、標的ポリヌクレオチドの異なる末端ヌクレオチドに対応するプローブが、明瞭なスペクトル的に分解できる蛍光色素を保持するように、蛍光的に標識されたオリゴヌクレオチドプローブが提供される。本明細書で用いられる「スペクトル的に分解可能な」とは、その色素が、操作条件下で、それらのスペクトル特性、特に蛍光放射波長に基づいて区別され得ることを意味する。このようにして、1つまたはそれ以上の末端ヌクレオチドの同一性は異なる色彩、またはおそらくは異なる波長における強度の比率に相関する。より好ましくは、4つのこのようなプローブは、標的ポリヌクレオチド上の4つのスペクトル的に分解可能な蛍光色素と4つの可能な末端ヌクレオチドの各々の間の一対一の対応を可能とするように提供される。スペクトル的に分解可能な色素のセットは、米国特許第4,855,225号および第5,188,934号:国際出願第PCT/US90/05565号;およびLeeら、Nucleic Acids Research,20:2471−2483(1992)に開示されている。
【実施例】
【0058】
実施例1
4つの開始オリゴヌクレオチドを用いてpUC19から増幅された
標的ポリヌクレオチドの配列決定
本実施例においては、結合領域およびpUC19プラスミドの一部を含むテンプレートをPCRによって増幅し、そして磁気ビーズに付着させる。4つの開始オリゴヌクレオチドを、下記のように別々の反応で使用する。以下の式に示すように、4つの中央リボヌクレオチドならびに、両5’および3’一リン酸を有する8マーのオリゴヌクレオチドプローブを使用する:
OP(=O)(O−)O−(5’)BBRRRRBB(3’)−OP(=O)(O−)O。
【0059】
アニーリングの後、プローブを開始オリゴヌクレオチドに酵素的に連結し、磁気ビーズ支持体を洗浄する。連結したプローブの3’リン酸をホスファターゼで除去し、その後、4つの標識したジデオキシヌクレオシド三リン酸鎖終結剤の存在下で、プローブをDNAポリメラーゼで伸長させる。伸長したヌクレオチドの洗浄および同定の後、連結されたプローブをRNase Hでリボヌクレオチド部分で切断して標識を除去し、そして伸長可能な末端を再生する。
【0060】
36マーの結合領域を含む以下の二本鎖のフラグメントをSacI/XmaI消化したpUC19に連結する:
CCTCTCCCTTCCCTCTCCTCCCTCTCCCCTCTCCCTC
TCGAGGAGAGGGAAGGGAGAGGAGGGAGAGGGGAGAGGGAGGGCC。
【0061】
単離および増幅の後、改変されたpUC19の402塩基対フラグメントを、テンプレートとして使用するために、PCRによって増幅する。該フラグメントは、41位からポリリンカー領域中のSacI部位に隣接して挿入された結合領域(未改変pUC19の413位)のpUC19の領域にわたる(Yanisch−Perronら、Gene,33:103−119(1985))。配列5’−CCCTCTCCCCTCTCCCTCx−3’および5’−GCAGCTCCCGGAGACGGT−3’(ここで「x」は製造業者のプロトコル付きの市販されている試薬(例えば、3’ビオチン−ON CPG(Clonetech Laboratories,Palo Alto,California))を用い、合成の間に付着される3’ビオチン部分である)を有する2つの18マーのオリゴヌクレオチドプローブを使用する。増幅されたテンプレートを単離し、M280−ストレプトアビジン(Dynal,Inc.,Great Neck,New York)とともに、製造業者のプロトコル(Dynabeads Template Preparation Kit)を用い、ストレプトアビジン被覆磁気ビーズ(Dynabeads)に付着させる。約300gのDynabeads M280−ストレプトアビジンをロードするために十分な量のビオチニル化313塩基対フラグメントを提供する。
【0062】
開始オリゴヌクレオチドとで形成される二重鎖が、二重鎖安定性を増強させるために、約66%GCの組成を有するように、結合領域配列を選択する。また、二次構造形成および結合領域内の1以上の位置への開始オリゴヌクレオチドの偶発的ハイブリダイゼーションを防ぐように配列を選択する。結合領域内の所与の開始オリゴヌクレオチドの位置のあらゆるシフティングも、有意数のミスマッチ塩基を生じる。
【0063】
ローディングの後、テンプレートの非ビオチニル化ストランドを加熱変性によって除去し、その後、磁気ビーズを洗浄し、そして4つのアリコートに分ける。磁気ビーズに付着させたテンプレートは以下の配列を有する:
(磁気ビーズ)−(リンカー)−(3’)−CTCCCTCTCCCCTCTCCCTCCTC−TCCCTTCCTCTCCTCGAGCTTAAGT...CTCGACG−(5’)。
【0064】
以下の4つのオリゴヌクレオチドを、テンプレートの別々のアリコートの各々における開始オリゴヌクレオチドとして使用する:
5’−GAGGAGAGGGAAGGAGAGGAG
5’−GGAGGAGAGGGAAGGAGAGGA
5’−GGGAGGAGAGGGAAGGAGAGG
5’−AGGGAGGAGAGGGAAGGAGAG。
【0065】
以下の反応および洗浄は、一般に、特記しない限り、使用する酵素のための50L容量の製造業者(New England Biolabs)の推奨する緩衝液中で実施する。また、標準的な緩衝液はSambrookら、Molecular Cloning,第2版(Cold Spring Harbor Laboratory Press,1989)に記載されている。
【0066】
4つのアリコートの各々用の8マーのプローブ全てを一緒に含む、96ストリンジェンシークラスの684または682オリゴヌクレオチドプローブ各々(48の異なるアニーリング温度の各々のための2のサブセット)を形成する。96のクラスの各々のプローブを、同一成分を有する反応混合物中の標的オリゴヌクレオチドに別々にアニールするが、37℃未満の温度でSequenaseおよびT4 DNAリガーゼで行った伸長および連結、およびTaq Stoffelフラグメントおよび他の熱安定性リガーゼで行った伸長および連結は例外である。
【0067】
48のストリンジェンシー条件は、同一温度のサブセットの各グループが、次の最高および次の最低ストリンジェンシークラスを含むサブセットグループのそれと1℃だけアニーリング温度が異なるように、22℃〜70℃の範囲のアニーリング温度によって規定される。アニーリング温度の範囲(22〜70℃)は、各々、最も不安定なおよび最も安定な8マーが、標準的なPCR緩衝溶液中で、約50パーセント最大アニーリングを有することが予想される温度より5〜10℃低いの温度によっておおよそ境界を決める。
【0068】
80℃における5〜10分間のインキュベーションの後、反応混合物を20〜30分間にかけて、それらの各アニーリング温度にもってゆく。連結、洗浄、およびホスファターゼでの処理の後、2単位のポリメラーゼおよび標識ジデオキシヌクレオチド三リン酸(0.08mM最終反応濃度、およびTAMRA(テトラメチルローダミン)、FAM(フルオレセイン)、ROX(ローダミンX)、およびJOE(2’,7’−ジメトキシ−4’,5’−ジクロロフルオレセイン)で標識)を添加する。15分後、ビーズをH2Oで洗浄し、そして各反応混合物を標準的な波長(例えば、Users Manual,モデル373DNAシーケンサー(Applied Biosystems,Foster City,CA))を用いて照射することにより伸長したヌクレオチドの同一性を決定する。
【0069】
同定後、反応混合物を製造業者の推奨のプロトコルを用いRNase Hで処理し、そして洗浄する。RNase H処理した伸長二重鎖は、再生された3’ヒドロキシルを有し、次の連結/伸長/切断のサイクルの準備ができている。試験配列の全てのヌクレオチドが同定されるまでサイクルを実施する。
【0070】
実施例2
ある開始オリゴヌクレオチドを用いるpUC19から増幅された標的ポリヌクレオチドの配列決定
この実施例において伸長は5’→3’方向であるので、ビオチン部分を結合領域のCTリッチストランドにハイブリダイズするプライマーの5’末端に付着させる以外は、本実施例では、実施例1に従ってテンプレートを調製する。このようにして、本実施例では、一本鎖テンプレートの結合領域はGAリッチなセグメントである(本質的には、実施例1の結合領域の相補体)。配列5’−xGAGGGAGAGGGGAGAGGG−3’および5’−ACCGTCTCCGGGAGCTGC−3’(ここで「x」は製造業者のプロトコル付きの市販されている試薬(例えば、Aminolinkアミノアルキルホスホルアミダイト連結剤(Applied Biosystems,Foster City,California)およびClontech Laboratories(Palo Alto,California)から入手可能なビオチン−X−NHSエステル)を用い、合成の間に付着される5’ビオチン部分である)を有する2つの18マーのオリゴヌクレオチドプローブを使用する。
【0071】
以下の配列を有する単一の12マーの開始オリゴヌクレオチドを使用する:
5’−OP(=O)(O−)O−CCTCTCCCTTCCCTCTCCTCC−3’。
以下の式に示される、プローブの最も3’側と3’側から2番目のヌクレオシドとの間に酸不安定ホスホルアミデート結合を有する6マーのオリゴヌクレオチドプローブを使用する:
HO−(3’)B(5’)−OP(=O)(O−)NH−(3’)BBBBBt*
ここで、標識は最も3’側の同一性に対応するように(従って、16の異なる標識ジデオキシヌクレオシドをプローブの合成に使用する)、Bt*はJOE−、FAM−、TAMRA−、またはROX−標識ジデオキシヌクレオシドである。
【0072】
前記のように、6マーのプローブを、各々42または43プローブを含む、96ストリンジェンシークラス(48の異なるアニーリング温度の各々につき2つのサブセット)に調製する。ハイブリダイゼーションおよび連結は、前記のように行う。連結および洗浄の後、オリゴヌクレオチドプローブの蛍光シグナルによって標的ポリヌクレオチド中のヌクレオシドを同定する。次いで、室温にてジクロロメタン中の0.8%トリフルオロ酢酸で、伸長した二重鎖を40分間処理することによって、酸切断を行い、伸長した二重鎖上で伸長可能な末端を再生する。標的ポリヌクレオチドの配列が決定されるまで該プロセスを継続する。
【特許請求の範囲】
【請求項1】
ポリヌクレオチドのヌクレオチド配列を決定する方法であって、以下の工程:
(a)既知配列を有する結合領域及び未知配列を有する標的領域を含む第一のテンプレートポリヌクレオチドを提供する工程;
(b)テンプレートに沿った二重鎖伸長の繰り返しサイクルによって、異なる配列の開始オリゴヌクレオチドを伸長する工程であって、特定の開始オリゴヌクレオチドの伸長方法は、
工程(a)で提供される第一のテンプレートポリヌクレオチドと開始オリゴヌクレオチドの配列を含む第二のポリヌクレオチドとの間に二重鎖を形成する工程であって、開始オリゴヌクレオチドは第一のテンプレートポリヌクレオチドの結合領域の一部とハイブリダイズする工程と
以下の工程を含む二重鎖伸長の繰り返しサイクルを行う工程
(1)異なる配列を有する複数の異なるオリゴヌクレオチドプローブを提供する工程
(2)異なるオリゴヌクレオチドプローブの一つを第二のポリヌクレオチドに連結することにより、第二のポリヌクレオチドを伸長し、より長い第二のポリヌクレオチドを作り出す工程であって、連結されたオリゴヌクレオチドプローブは第一のテンプレートポリヌクレオチドの配列に依存し、連結されたオリゴヌクレオチドプローブは工程(b)(1)で提供されるオリゴヌクレオチドプローブ中の別のオリゴヌクレオチドプローブとはさらに連結される能力を有しない、工程
(3)工程(b)(2)で連結されたオリゴヌクレオチドプローブに会合した標識を検出する工程であって、標識は連結されたオリゴヌクレオチドプローブの塩基に対応するが、連結されたオリゴヌクレオチドプローブの全配列には対応しない、工程および
(4)第二のポリヌクレオチド上に伸長可能な末端を再生して、第二のポリヌクレオチドの伸長の引き続く回においてさらなるオリゴヌクレオチドプローブが第二のポリヌクレオチドに付加できるようになる工程
を含む工程;および
(c)異なる開始オリゴヌクレオチドの一連の伸長の組み合わせに基づいて、未知配列の塩基の連続する配列を決定する工程
を備える、方法。
【請求項2】
工程(b)(4)は工程(b)(2)で連結されたオリゴヌクレオチドプローブを2ヌクレオチド内側で切断する工程を含む、請求項1記載の方法。
【請求項3】
切断は、第二のポリヌクレオチドから標識をも除去する、請求項2記載の方法。
【請求項4】
第二のポリヌクレオチドが工程(b)(2)のオリゴヌクレオチドプローブへ連結できないときに、工程(b)の個々の開始オリゴヌクレオチドの一連の伸長は第二のポリヌクレオチドをキャッピングすることをさらに含む、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
工程(b)(1)で提供されるオリゴヌクレオチドプローブのうち少なくともいくつかは、少なくとも一つのデオキシイノシン塩基を含む、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
工程(b)(1)で提供されるオリゴヌクレオチドプローブは8ヌクレオチド、9ヌクレオチド及び12ヌクレオチドから選択される所定の長さである、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
開始オリゴヌクレオチドは互いに相殺される、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
未知配列の連続する塩基の配列を決定するのに使用する開始オリゴヌクレオチドは、互いに一塩基ずつ相がずれている、請求項7に記載の方法。
【請求項9】
第一のポリヌクレオチドのヌクレオチド配列を決定する方法であって、以下の工程:
(a)オリゴヌクレオチドプローブと開始オリゴヌクレオチドとを連結することによって、開始オリゴヌクレオチドをポリヌクレオチドに沿って伸長させ、さらなるオリゴヌクレオチドプローブとは連結される能力を有しない伸長した二重鎖を形成させる工程;
(b)連結したオリゴヌクレオチドプローブに会合する標識を検出する工程;
(c)連結したオリゴヌクレオチドプローブの2塩基の間の位置で伸長した二重鎖を切断することにより、伸長した二重鎖上の伸長可能な末端を再生する工程;および
(d)ヌクレオチドの配列が決定されるまで工程(a)〜(c)を繰り返す工程;
を備える、方法。
【請求項10】
連結したオリゴヌクレオチドプローブが連結した末端とは反対側の連結したプローブの末端ヌクレオチドに、標識が結合している、請求項1〜9のいずれか一項に記載の方法。
【請求項11】
第一のポリヌクレオチドを増幅して、工程(b)で使用する第一のポリヌクレオチドを作製する工程をさらに含む、請求項1〜10のいずれか一項に記載の方法。
【請求項12】
増幅した第一のポリヌクレオチドは、ビーズに担持された提供された第一のポリヌクレオチドの複数のコピーを有する、請求項11に記載の方法。
【請求項13】
複数の異なる第一のポリヌクレオチドの伸長、同定及び再生工程を並行して複数サイクル行うことに基づいて、異なるポリヌクレオチドの異なる未知配列の塩基の連続する配列を決定する工程をさらに備える、請求項1〜12のいずれか一項に記載の方法。
【請求項14】
複数の異なる第一のポリヌクレオチドは、お互いに共通の結合領域における既知配列を有する、請求項13に記載の方法。
【請求項15】
配列決定されるポリヌクレオチドにおける各ヌクレオシドの順序を決定することをさらに含む、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
開始オリゴヌクレオチドは、少なくとも20塩基の長さであり、かつ、結合領域は少なくとも20塩基の長さであって一連の異なる配列の開始オリゴヌクレオチドの開始点を提供するのに十分な長さである、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
工程(b)は複数の異なるオリゴヌクレオチドプローブを提供し、提供されるオリゴヌクレオチドプローブのひとつが開始オリゴヌクレオチドに連結されて伸長した二重鎖を形成する工程をさらに含む、請求項9に記載の方法。
【請求項18】
提供されるオリゴヌクレオチドプローブの少なくともいくつかは、少なくとも一つのデオキシイノシン塩基を含む、請求項17に記載の方法。
【請求項19】
提供されるプローブは、8ヌクレオチド、9ヌクレオチド及び12ヌクレオチドから選択される所定の長さである、請求項17又は18に記載の方法。
【請求項20】
請求項1〜19のいずれか一項に記載の方法を実施するためのシステムであって、
方法を実施するための試薬を含む、一連の試薬リザーバ;
固相支持体に結合した第一のポリヌクレオチドを含む、少なくとも一つの反応容器;
繰り返しサイクルにおいて、標識プローブ及び連結試薬を反応容器に分注して開始オリゴヌクレオチドを含むポリヌクレオチドに少なくとも一つのオリゴヌクレオチドプローブを連結することを制御し、切断試薬を反応容器に分注し、切断試薬は分注される標識オリゴヌクレオチドプローブの塩基間の結合を切断して伸長可能な末端を再生することを制御するように設定されているコンピュータ制御手段;及び
繰り返しサイクルにおいて、連結したオリゴヌクレオチドプローブと会合した標識を検出するように設定された検出ステーション;
を備えるシステム。
【請求項21】
標識を検出する繰り返しサイクルから塩基の連続する配列を決定するよう設定されたプロセッサをさらに備える、請求項20に記載のシステム。
【請求項1】
ポリヌクレオチドのヌクレオチド配列を決定する方法であって、以下の工程:
(a)既知配列を有する結合領域及び未知配列を有する標的領域を含む第一のテンプレートポリヌクレオチドを提供する工程;
(b)テンプレートに沿った二重鎖伸長の繰り返しサイクルによって、異なる配列の開始オリゴヌクレオチドを伸長する工程であって、特定の開始オリゴヌクレオチドの伸長方法は、
工程(a)で提供される第一のテンプレートポリヌクレオチドと開始オリゴヌクレオチドの配列を含む第二のポリヌクレオチドとの間に二重鎖を形成する工程であって、開始オリゴヌクレオチドは第一のテンプレートポリヌクレオチドの結合領域の一部とハイブリダイズする工程と
以下の工程を含む二重鎖伸長の繰り返しサイクルを行う工程
(1)異なる配列を有する複数の異なるオリゴヌクレオチドプローブを提供する工程
(2)異なるオリゴヌクレオチドプローブの一つを第二のポリヌクレオチドに連結することにより、第二のポリヌクレオチドを伸長し、より長い第二のポリヌクレオチドを作り出す工程であって、連結されたオリゴヌクレオチドプローブは第一のテンプレートポリヌクレオチドの配列に依存し、連結されたオリゴヌクレオチドプローブは工程(b)(1)で提供されるオリゴヌクレオチドプローブ中の別のオリゴヌクレオチドプローブとはさらに連結される能力を有しない、工程
(3)工程(b)(2)で連結されたオリゴヌクレオチドプローブに会合した標識を検出する工程であって、標識は連結されたオリゴヌクレオチドプローブの塩基に対応するが、連結されたオリゴヌクレオチドプローブの全配列には対応しない、工程および
(4)第二のポリヌクレオチド上に伸長可能な末端を再生して、第二のポリヌクレオチドの伸長の引き続く回においてさらなるオリゴヌクレオチドプローブが第二のポリヌクレオチドに付加できるようになる工程
を含む工程;および
(c)異なる開始オリゴヌクレオチドの一連の伸長の組み合わせに基づいて、未知配列の塩基の連続する配列を決定する工程
を備える、方法。
【請求項2】
工程(b)(4)は工程(b)(2)で連結されたオリゴヌクレオチドプローブを2ヌクレオチド内側で切断する工程を含む、請求項1記載の方法。
【請求項3】
切断は、第二のポリヌクレオチドから標識をも除去する、請求項2記載の方法。
【請求項4】
第二のポリヌクレオチドが工程(b)(2)のオリゴヌクレオチドプローブへ連結できないときに、工程(b)の個々の開始オリゴヌクレオチドの一連の伸長は第二のポリヌクレオチドをキャッピングすることをさらに含む、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
工程(b)(1)で提供されるオリゴヌクレオチドプローブのうち少なくともいくつかは、少なくとも一つのデオキシイノシン塩基を含む、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
工程(b)(1)で提供されるオリゴヌクレオチドプローブは8ヌクレオチド、9ヌクレオチド及び12ヌクレオチドから選択される所定の長さである、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
開始オリゴヌクレオチドは互いに相殺される、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
未知配列の連続する塩基の配列を決定するのに使用する開始オリゴヌクレオチドは、互いに一塩基ずつ相がずれている、請求項7に記載の方法。
【請求項9】
第一のポリヌクレオチドのヌクレオチド配列を決定する方法であって、以下の工程:
(a)オリゴヌクレオチドプローブと開始オリゴヌクレオチドとを連結することによって、開始オリゴヌクレオチドをポリヌクレオチドに沿って伸長させ、さらなるオリゴヌクレオチドプローブとは連結される能力を有しない伸長した二重鎖を形成させる工程;
(b)連結したオリゴヌクレオチドプローブに会合する標識を検出する工程;
(c)連結したオリゴヌクレオチドプローブの2塩基の間の位置で伸長した二重鎖を切断することにより、伸長した二重鎖上の伸長可能な末端を再生する工程;および
(d)ヌクレオチドの配列が決定されるまで工程(a)〜(c)を繰り返す工程;
を備える、方法。
【請求項10】
連結したオリゴヌクレオチドプローブが連結した末端とは反対側の連結したプローブの末端ヌクレオチドに、標識が結合している、請求項1〜9のいずれか一項に記載の方法。
【請求項11】
第一のポリヌクレオチドを増幅して、工程(b)で使用する第一のポリヌクレオチドを作製する工程をさらに含む、請求項1〜10のいずれか一項に記載の方法。
【請求項12】
増幅した第一のポリヌクレオチドは、ビーズに担持された提供された第一のポリヌクレオチドの複数のコピーを有する、請求項11に記載の方法。
【請求項13】
複数の異なる第一のポリヌクレオチドの伸長、同定及び再生工程を並行して複数サイクル行うことに基づいて、異なるポリヌクレオチドの異なる未知配列の塩基の連続する配列を決定する工程をさらに備える、請求項1〜12のいずれか一項に記載の方法。
【請求項14】
複数の異なる第一のポリヌクレオチドは、お互いに共通の結合領域における既知配列を有する、請求項13に記載の方法。
【請求項15】
配列決定されるポリヌクレオチドにおける各ヌクレオシドの順序を決定することをさらに含む、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
開始オリゴヌクレオチドは、少なくとも20塩基の長さであり、かつ、結合領域は少なくとも20塩基の長さであって一連の異なる配列の開始オリゴヌクレオチドの開始点を提供するのに十分な長さである、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
工程(b)は複数の異なるオリゴヌクレオチドプローブを提供し、提供されるオリゴヌクレオチドプローブのひとつが開始オリゴヌクレオチドに連結されて伸長した二重鎖を形成する工程をさらに含む、請求項9に記載の方法。
【請求項18】
提供されるオリゴヌクレオチドプローブの少なくともいくつかは、少なくとも一つのデオキシイノシン塩基を含む、請求項17に記載の方法。
【請求項19】
提供されるプローブは、8ヌクレオチド、9ヌクレオチド及び12ヌクレオチドから選択される所定の長さである、請求項17又は18に記載の方法。
【請求項20】
請求項1〜19のいずれか一項に記載の方法を実施するためのシステムであって、
方法を実施するための試薬を含む、一連の試薬リザーバ;
固相支持体に結合した第一のポリヌクレオチドを含む、少なくとも一つの反応容器;
繰り返しサイクルにおいて、標識プローブ及び連結試薬を反応容器に分注して開始オリゴヌクレオチドを含むポリヌクレオチドに少なくとも一つのオリゴヌクレオチドプローブを連結することを制御し、切断試薬を反応容器に分注し、切断試薬は分注される標識オリゴヌクレオチドプローブの塩基間の結合を切断して伸長可能な末端を再生することを制御するように設定されているコンピュータ制御手段;及び
繰り返しサイクルにおいて、連結したオリゴヌクレオチドプローブと会合した標識を検出するように設定された検出ステーション;
を備えるシステム。
【請求項21】
標識を検出する繰り返しサイクルから塩基の連続する配列を決定するよう設定されたプロセッサをさらに備える、請求項20に記載のシステム。
【図1】


【図2】
【図3A】
【図3B】
【図4】
【図2】
【図3A】
【図3B】
【図4】
【公開番号】特開2011−92192(P2011−92192A)
【公開日】平成23年5月12日(2011.5.12)
【国際特許分類】
【出願番号】特願2010−257927(P2010−257927)
【出願日】平成22年11月18日(2010.11.18)
【分割の表示】特願2007−185093(P2007−185093)の分割
【原出願日】平成8年4月16日(1996.4.16)
【出願人】(598114930)ソレクサ・インコーポレイテッド (10)
【氏名又は名称原語表記】Solexa,Inc.
【Fターム(参考)】
【公開日】平成23年5月12日(2011.5.12)
【国際特許分類】
【出願日】平成22年11月18日(2010.11.18)
【分割の表示】特願2007−185093(P2007−185093)の分割
【原出願日】平成8年4月16日(1996.4.16)
【出願人】(598114930)ソレクサ・インコーポレイテッド (10)
【氏名又は名称原語表記】Solexa,Inc.
【Fターム(参考)】
[ Back to top ]