
中胚葉および最終段階内胚葉細胞集団
本発明は、内中胚葉および中胚葉細胞が豊富な細胞集団、および内胚葉が豊富な細胞集団を提供する。本発明の細胞集団は、細胞補充療法用の細胞を作製するのに有用である。本発明はさらに、肝細胞を作製する方法、肝細胞が豊富な細胞集団、および肝細胞補充療法の方法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
連邦政府により支援された研究に関する記載
本発明は、国立衛生研究所によって与えられたGrant Nos.2RO1HL48834-09および2RO1HL65169-02で政府の支援によってなされた。米国政府は、本発明において確かな権利を有することができる。
【背景技術】
【0002】
胚発生中、身体の組織は3つの主な細胞集団:外胚葉、中胚葉および最終段階内胚葉(definitive endoderm)から形成される。一次胚葉としても知られるこれらの細胞集団は、原腸形成として知られているプロセスによって形成される。原腸形成の後、それぞれの一次胚葉は、特異的な一組の細胞集団と組織を生成する。中胚葉は血液細胞、内皮細胞、心臓および骨格筋、および脂肪細胞を生じる。最終段階内胚葉は、肝臓、膵臓および肺を生成する。外胚葉は、神経系、皮膚および副腎組織を生じる。
【0003】
これらの胚細胞層からの組織発生のプロセスは多数の分化段階を含み、複雑な分子の変化を表す。中胚葉およびその誘導体に関しては、3つの異なる段階が同定されてきている。第一段階は、エピブラストとして知られる構造内での細胞からの中胚葉の誘導である。発生期中胚葉としても知られる新たに形成される中胚葉は、初期胚における将来の組織発生の部位であろう異なる位置に移動する。パターン形成として知られるこのプロセスは、特定組織への分化の初期段階を表す可能性がある幾つかの分子の変化を伴う。特化として知られる最終段階は、パターン形成した中胚葉亜集団からの異なる組織の生成を含む。近年の研究は、中胚葉は連続的に誘導され明確な発生能を有する亜集団であることを示唆する証拠を与えている。最初に形成される中胚葉は胚外領域に移動し、造血および内皮細胞を生じ、一方次の集団は発生中の胚では前に移動し、心臓および頭蓋間充組織となる。これらの系統の関係は組織学的分析によって初期に定義されており、細胞を追跡する研究によって大部分は確認されている。この発生運命の分離は発生生物学の分野では充分容認されているが、今日まで、これらの系統に拘束される前に、中胚葉および内胚葉を単離する利用可能な方法は存在していない。
【特許文献1】米国特許第5,843,780号
【特許文献2】米国特許第6,159,462号
【特許文献3】米国特許第6,844,422号
【特許文献4】米国特許第6,110,739号
【非特許文献1】Papaioannouら(1998) Bioessays 20: 9〜19
【非特許文献2】Wilsonら(1995) Development 121: 877〜86
【非特許文献3】Wilkinsonら(1990) Nature 343: 657〜9
【非特許文献4】Herrmannら(1991) Development 113: 913〜7
【非特許文献5】Smith (1997) Current Opinion in Genetics & Development 7: 474〜480
【非特許文献6】Evansら(1981) Nature 292: 154〜156
【非特許文献7】Thomsonら(1995) Proc.Nat'l.Acad.Sci.USA 92;7844
【非特許文献8】Reubinoffら(2000) Nature Biotech.18: 399
【非特許文献9】Glinkaら(1998) Nature 391: 357〜362
【非特許文献10】Odoricoら、(2001) Stem Cells 19: 193〜204
【非特許文献11】Grompeら(1993) Genes & Dev.7: 2298
【非特許文献12】Oversturfら(1996) Nature Genet.12: 266〜273
【非特許文献13】Kennedyら(1997) Nature 386: 488〜93
【非特許文献14】Bradyら(1990) Meth.in Mol.and Cell Bio.2: 17〜25
【非特許文献15】Smith(2001) Annu.Rev.Cell Dev. Biol.17:435〜62
【非特許文献16】Hooperら(1987) Nature 326: 292
【非特許文献17】Rogersら(1991) Development 113: 815〜24
【非特許文献18】Shalabyら(1995) Nature 376: 62〜6
【非特許文献19】Palisら(1999) Development 126: 5073〜84
【非特許文献20】Faloonら(2000) Development 127: 1931〜41
【非特許文献21】Shivdasaniら(1995) Nature 373: 432〜4
【非特許文献22】Wangら(1996) Proc.Natl.Acad. Sci.93:3444〜9
【非特許文献23】Kaestnerら(1994) Genomics 20: 177〜85
【非特許文献24】Weinsteinら(1994) Cell 78: 575〜588
【非特許文献25】Beddington and Robertson (1999) Cell 96: 195〜209
【非特許文献26】Robb and Tam (2004) Sem.Cell & Dev.Biol.15: 543〜554
【非特許文献27】Liuら(1999) Nat.Genet.22: 361〜4
【非特許文献28】Inman et.al.(2002) Mol.Pharmacol.62: 65〜74
【非特許文献29】Zaret(2001) Curr Opin Genet Dev 11: 568〜574
【非特許文献30】Blockら、(1996) J Cell Biol 132: 1133〜1149
【発明の開示】
【課題を解決するための手段】
【0004】
本発明は、中胚葉および最終段階内胚葉細胞集団を単離するための方法を提供する。これらの細胞集団は、細胞の増殖および分化に影響を与える作用物質を同定し、組織発生と関係がある遺伝子を同定し、かつ細胞補充療法用の分化細胞および組織を作製するのに有用である。
【0005】
本発明は、内中胚葉(mesendoderm)および中胚葉細胞が豊富な細胞集団を提供する。内中胚葉細胞は、brachuryを発現し(brach+)、分化誘導条件の存在下で、中胚葉、ならびに、心臓および骨格筋、血管平滑筋、内皮および造血細胞を含めた中胚葉誘導体を生成することができ、内胚葉、ならびに、肝臓細胞および膵臓細胞を含めた内胚葉誘導体も生成することができる細胞として、本明細書では定義する。中胚葉細胞は、brach+であり分化誘導条件の存在下で心臓および骨格筋、血管平滑筋、内皮および造血細胞を生成することができ、内胚葉および内胚葉誘導体を生成することができない細胞として、本明細書では定義する。
【0006】
本発明はさらに、内胚葉細胞が豊富な細胞集団を提供する。内胚葉細胞は、brachuryを発現せず(brach-)、分化誘導条件の存在下で、肺細胞、肝臓細胞および膵臓細胞を生成することができる細胞として、本明細書では定義する。
【0007】
本発明はさらに、内中胚葉および中胚葉細胞が豊富な細胞集団、および内胚葉細胞が豊富な細胞集団を単離する方法を提供する。
【0008】
他の実施形態では本発明は、肝細胞が豊富な細胞集団、およびこのような集団を作製する方法を提供する。
【0009】
他の実施形態では本発明は、肝細胞補充療法の方法を提供する。
【0010】
他の実施形態では本発明は、本発明の細胞集団の増殖、分化または生存に影響を与える作用物質を同定する方法を提供する。特定系統および組織の細胞分化および発生と関係がある遺伝子を同定する方法も提供する。
【0011】
brach+細胞を特異的に認識する抗体も提供する。これらの抗体は、例えば内中胚葉および中胚葉細胞集団を単離するのに有用である。
【0012】
他の実施形態では本発明は、in vitroで細胞を生成するための方法を提供する。このような細胞は、例えば細胞補充療法に有用である。
【0013】
本発明は、1つのbrachury対立遺伝子が不活性状態でありbrachury遺伝子座が転写される細胞中で選択可能なマーカーが発現されるように、選択可能なマーカーをコードするDNAがbrachury遺伝子座に存在するゲノムを有する、トランスジェニック非ヒト哺乳動物も提供する。
【発明を実施するための最良の形態】
【0014】
胚形成中、中胚葉の形成は、体の形成のプランの確立および血液、内皮、心臓および骨格筋などの多臓器系の発生における重要なステップである。中胚葉形成を制御する分子機構は、しかしながらほとんど定義されていない。培養中の胚幹(ES)細胞の分化に基づくモデル系を使用して、造血、内皮、心臓および骨格筋、および脂肪細胞系統を含めた中胚葉由来集団が研究されてきている。in vitroモデルは中胚葉の誘導および特化を支持するが、これらの分化事象はES細胞から生じる胚様体(EB)として知られる複雑なコロニーにおいて起こる。中胚葉形成および組織発生をさらによく理解するために、それらが形成されるときにEBから中胚葉細胞集団を単離することが有利であるはずである。しかしながら、発生期中胚葉細胞集団に特異的な抗体は明確に定義されていないので、抗体を使用する細胞選別によって、これらの集団を単離することはできていない。
【0015】
(Tとしても知られる)brachuryはT-ボックス遺伝子として知られる転写因子のファミリーの基本メンバーであり、マウスにおいて元来存在する変異として最初に同定された。Papaioannouら(1998) Bioessays 20: 9〜19。ヘテロ接合性のマウスは生存可能であるが、野生型動物より短い尾を有する。ホモ接合性のマウスは、day 10 p.c.で死に至り、脊索を欠いており、後部中胚葉組織の発生において欠陥を示す。キメラ動物の分析によって、brachuryは中胚葉細胞の移動性に影響を与えることが示されている。Wilsonら(1995) Development 121: 877〜86。発現の分析によって、brachuryに関する特有かつ興味深いパターンが明らかになった。それは原条を通じて進入する全細胞において、および発生期および初期移動性中胚葉において一過的に発現される。Wilkinsonら(1990) Nature 343: 657〜9; Herrmannら(1991) Development 113: 913〜7。発現は沿軸、側板、および胚外中胚葉において直ちに下方制御され、原条の退行の後は終末芽および脊索に限られる。このパターンを考慮すると、brachuryは、初期中胚葉の最適なマーカーの1つであると考えられ、この系統の発生を追跡するのに使用されている。brachuryは分析した全ての種において同定されており、中胚葉発生におけるその役割は系統発生中保たれることが示唆される。Papaioannouら(1998)。
【0016】
本発明によれば、選択可能なマーカー遺伝子は組換えによってbrachury遺伝子座に標的化されている。ES細胞分化の開始後、brachuryの発現を表すパターンで選択可能なマーカーが発現されることが発見されてきている。選択可能なマーカーは、EBからのbrachury陽性(Brach+)細胞の選別、およびしたがって内中胚葉および中胚葉細胞が豊富な細胞集団の単離および特徴付けを可能にしている。
【0017】
本発明によって例示される選択可能なマーカーは、強化型緑色蛍光タンパク質(EGFPまたはGFP)である。細胞選別を容易にするはずである他の選択可能なマーカーは当業者に知られており、本発明中で使用することができる。GFPをコードするcDNAは当技術分野で知られており(かつ例えばClontech、Palo Alto、CAからプラスミドpEGFP.C1として市販されており)、当技術分野で知られている方法によって標的ベクター(GFP-Bry)を構築することにより、brachury遺伝子座に標的化することができる。ベクターを設計して、brachury遺伝子の第一エクソンの約3分の2とGFP発現カセットを交換することが好ましい。
【0018】
ヒトおよびマウスを含めた多種由来のbrachury遺伝子は当技術分野で知られており、例えばSmith (1997) Current Opinion in Genetics & Development 7: 474〜480によって総説されている。GFP発現カセットは、GFP cDNA、および、下流のbrachuryエクソンの翻訳を妨げるための、1つまたは複数の翻訳停止コドンを含むことが好ましい。brachury遺伝子の下流領域の転写を妨げるために、カセットはSV40ポリアデニル化シグナル配列をコードするエクソンをさらに含むことができる。
【0019】
当技術分野で知られている方法によってES細胞中にベクターを導入して、相同的組換えによってGFP-Bry構築体を組み込む。当技術分野で知られており例えばEvansら(1981) Nature 292: 154〜156: Thomsonら(1995) Proc.Nat'l.Acad.Sci.USA 92;7844;米国特許第5,843,780号;およびReubinoffら(2000) Nature Biotech.18: 399によって開示された方法によって、胚盤胞からES細胞を単離することができる。好ましい実施形態では、ES細胞はマウスまたはヒトES細胞である。標的化成功後、brachury開始コドンはGFPの開始コドンになり、標的brachury対立遺伝子の破壊をもたらす。生成する細胞をGFP-BryES細胞と呼ぶ。GFP-BryES細胞は本明細書では、1つのbrachury対立遺伝子が不活性状態でありGFPがbrachury制御要素の制御下で発現されるES細胞として定義する。
【0020】
1つのbrachury対立遺伝子が不活性状態であるGFP-BryES細胞は生命力があり、正常に発生および分化することが、本発明によって発見されている。さらに、GFP発現は内因性のbrachury発現を反映することが発見されている。したがって、GFPを発現する細胞を選択することによってbrach+細胞を単離することができる。GFPを発現する細胞はフローサイトメトリーによって、例えば蛍光活性化細胞選別(FACS)によって都合良く単離することができる。蛍光性に基づいて細胞を選別するための方法は、当業者によく知られている。
【0021】
本発明は、組換えによってHNF3β遺伝子座に標的化されている選択可能なマーカー遺伝子を有する、GFP-BryES細胞も提供する。HNF3βは大部分の内胚葉細胞型において発現されることが知られている。HNF3βはFoxa2としても知られ、かつこれらの用語は本明細書では交換して使用可能である。このマーカーはHNF3β+細胞の選別を可能にする。本発明によって例示されるマーカーは、細胞内ドメインの大部分が欠けている切断型ヒトCD4である。この細胞表面分子は、シグナルを伝達することができず、抗体がフローサイトメトリーに利用可能であるので好ましい。細胞選別を容易にするはずである他のマーカーは当業者に知られており、本発明中で使用することができる。ヒトCD4をコードするcDNAは当技術分野で知られており、当技術分野で知られている方法によってHNF3β遺伝子座に標的化することができる。HNF3βと相同的な2アーム間にクローニングした切断型ヒトCD4をコードするcDNAを有する標的ベクターを、GFP-BryES細胞中にエレクトロポレーションにより導入して、CD4-HNF/GFP-BryES細胞を得ることができる。これらの細胞中でのヒトCD4の発現はHNF3βの内因性の発現を反映する。
【0022】
本明細書で前に定義した内中胚葉および中胚葉細胞が豊富な細胞集団は、GFP+細胞を得るのに充分な時間、例えばマウス細胞に関して約1〜約4日間、血清の存在下でGFP-BryES細胞を培養し、例えばフローサイトメトリーによってGFP+細胞を選別および単離することによって得ることができる。単離される細胞集団は、少なくとも約50%、および好ましくは少なくとも約75%、およびより好ましくは少なくとも約90%、および最も好ましくは少なくとも約95%、または少なくとも約99%の内中胚葉および中胚葉細胞を含む。内中胚葉および中胚葉の相対量は血清中での培養時間の長さを調節することによって変えることができ、短い培養時間は造血および内皮系統にパターン形成される内中胚葉および中胚葉の存在に好ましく、長い培養時間は心臓および骨格筋系統にパターン形成される中胚葉の存在に好ましい。例えば、中胚葉が豊富な細胞集団は、約2.5〜4.5日間血清中で培養し、次にGFP+細胞を選別および単離することによって得ることができる。血清の存在下での培養は、動物血清、例えばウシ胎児血清(FCS)を補った培地中での培養として本明細書では定義する。好ましい実施形態では、約5%〜約25%の血清を培地に補う。最適濃度は血清バッチ依存性である可能性があり、当業者によって決定することができる。
【0023】
内中胚葉および中胚葉細胞が豊富な細胞集団は、ヒトおよびマウス細胞に関するin vitroでの分化の時間の違いを考慮に入れて血清中での培養時間の長さを延長した類似の方法によって、ヒトES細胞から作製したGFP-BryES細胞から得ることができる。したがって、ヒトES細胞から作製したGFP-BryES細胞は、GFP+細胞を選別および単離する前に、GFP+細胞を得るのに充分な時間、例えば約2〜約18日間血清中で培養する。
【0024】
マウスとヒト細胞集団の両方に関して、例えばチロシンキナーゼ受容体、ヒトKDRまたはマウスFlk-1の存在をアッセイすることによって、単離した細胞が中胚葉より先、例えば血管芽細胞に分化しているかどうか、容易に決定することができる。KDRおよびFlk-1は内中胚葉および発生期中胚葉中では発現されないが、これらの細胞は血管芽細胞/前赤血球集団に分化すると、KDRまたはFlk-1発現は検出可能である。KDR+およびflk-1+細胞は、KDRまたはFlk-1に対する抗体を使用してフローサイトメトリーによって同定することができる。このような抗体は当技術分野で知られており、抗体生成の標準的な方法を使用して作製することもできる。内中胚葉および中胚葉が豊富な細胞集団は、細胞選別によりKDR+またはFlk-1+細胞を除去することによって、さらに豊富な状態にすることができる。
【0025】
図17中に示すように内中胚葉は、内胚葉と中胚葉の両方およびその対応する系統を生じる、以前は同定されていなかった細胞集団であることが、本発明によって発見されている。in vitro培養において血清の存在または不在を使用して、どの系統が内中胚葉から生じるかを決定することができることが、さらに発見されている。特に、内胚葉細胞が豊富な細胞集団は、約2〜4日間血清の存在下でマウスES細胞から作製したGFP-BryES細胞を培養し、例えばフローサイトメトリーによってGFP+細胞を選別および単離し、次に約1〜約10日間血清の不在下でGFPを培養することによって得ることができる。本明細書で前に定義したように、単離される細胞集団は、少なくとも50%、および好ましくは少なくとも約75%、およびより好ましくは少なくとも約90%、および最も好ましくは少なくとも約95%、または少なくとも約99%の内胚葉細胞を含む。
【0026】
内胚葉細胞が豊富な細胞集団は、約2〜10日間血清の存在下でGFP-BryES細胞を培養し、および次いでGFP+細胞を選別および単離し、次に約1〜約15日間血清の不在下でGFP+細胞を培養することによって、ヒトES細胞から作製したGFP-BryES細胞から得ることができる。
【0027】
内胚葉細胞が豊富な集団は、前に記載したようにKDR+またはFlk-1+細胞を同定および選別することによって、さらに豊富な状態にすることができる。
【0028】
内胚葉が豊富な細胞集団は、約2〜約10日間血清の不在下および増殖因子アクチビンまたはノーダルの存在下でGFP-Bry胚幹細胞を培養し、およびbrachuryを発現する細胞を単離することによって得ることができることが、本発明によってさらに発見されている。アクチビンまたはノーダルの量は、胚幹細胞から内胚葉への分化を誘導するのに充分な量である。このような分化は例えばHNF3β、Mixl-1、Sox17、Hex-1またはpdx-1を含めた内胚葉の発生と関係がある遺伝子の発現をアッセイすることによって測定することができる。好ましい実施形態では、アクチビンの濃度は少なくとも約30ng/mlである。他の好ましい実施形態では、アクチビンの濃度は約100ng/mlである。他の実施形態では、約2〜約10日間血清の不在下およびアクチビンまたはノーダルおよびWntシグナル伝達の阻害剤の存在下で、GFP-Bry胚幹細胞を培養し、brachuryを発現する細胞を単離する。好ましい実施形態では、阻害剤はDKK-1である。他の好ましい実施形態では、DKK-1の濃度は少なくとも約30ng/ml、または約100ng/mlである。
【0029】
中胚葉が豊富な細胞集団は、約2〜約10日間血清の不在下およびアクチビンまたはノーダルの存在下でGFP-Bry胚幹細胞を培養し、およびbrachuryを発現する細胞を単離することによって得ることができる。アクチビンまたはノーダルの量は、胚幹細胞から中胚葉への分化を誘導するのに充分であるが、内胚葉への分化を誘導するには不充分な量である。中胚葉への分化は、例えばGATA-1を含めた中胚葉の発生と関係がある遺伝子の発現、および内胚葉の発生と関係がある遺伝子の発現の不在をアッセイすることによって測定することができる。好ましい実施形態では、アクチビンの濃度は30ng/ml未満である。他の好ましい実施形態では、アクチビンの濃度は約3ng/mlである。
【0030】
内胚葉が豊富な細胞集団は、約1〜約6日間血清の不在下およびアクチビンまたは関連タンパク質ノーダルおよびWnt分子の存在下で胚幹細胞を培養し、brach+/HNF3β3+細胞を単離し、および少なくとも約1日間血清の不在下およびWntシグナル伝達の阻害剤の存在下で単離細胞を培養することによって得ることができることが、本発明によってさらに発見されている。brach+/HNF3β3+細胞の単離は、本明細書で前に記載したC4-HNF/GFP-BryES細胞を使用すること、および例えば細胞選別によってCD4およびGFPを選択することによって容易にすることができる。
【0031】
Wntは、当技術分野でよく知られているポリペプチドのファミリーを指す。例えば米国特許第6,159,462号を参照。Wnt増殖因子のファミリーは、マウスにおいて少なくとも10個の遺伝子、およびヒトにおいて少なくとも7個の遺伝子によってコードされるタンパク質を含む。本発明の好ましい実施形態では、Wnt分子は組換えWnt3dである。
【0032】
Wntシグナル伝達の阻害剤も当技術分野でよく知られており、例えば米国特許第6,844,422号中で開示されたWnt-I、およびGlinkaら(1998) Nature 391: 357〜362によって開示されたDickkopf-1(DKK-1)を含む。
【0033】
本発明は、内胚葉細胞が豊富な細胞集団を作製する方法であって、約2〜約4日間血清の不在下でマウス胚幹細胞を、あるいは約2〜約10日間血清の不在下でヒト胚細胞を培養する段階と、brach+/HNF3β3+細胞を単離する段階と、約1日〜約10日間血清の不在下およびアクチビンまたはノーダルおよびWntシグナル伝達の阻害剤の存在下で単離細胞を培養する段階とを含む方法も提供する。brach+/HNF3β3+細胞の単離は、前に記載したC4-HNF/GFP-BryES細胞を使用することによって容易にすることができる。
【0034】
前述の2つの方法のそれぞれにおいて、アクチビンまたはノーダルおよびWnt阻害剤の量は、ES細胞から内胚葉への分化を誘導するのに充分な量である。内胚葉の発生と関係がある遺伝子の発現をアッセイすることによって、分化を測定することができる。好ましい実施形態では、アクチビンの濃度は少なくとも約30ng/mlである。他の好ましい実施形態では、アクチビンの濃度は約100ng/mlである。好ましい実施形態では、Wnt阻害剤はDKK-1であり、DKK-1の濃度は少なくとも約30ng/mlである。他の好ましい実施形態では、DKK-1の濃度は約100ng/mlである。
【0035】
本発明はさらに、肝細胞が豊富な細胞集団を作製する方法であって、約2〜4日間血清の不在下およびアクチビンまたはノーダルの存在下でマウス胚幹細胞を、あるいは約2〜10日間血清の不在下およびアクチビンまたはノーダルの存在下でヒト胚細胞を培養する段階と、brach+/HNF3β3+/cKit+細胞を単離する段階と、肝芽細胞を生成するのに充分な条件下において血清の不在下ならびにBMP-4およびbFGFの存在下で単離細胞を培養する段階と、肝細胞への肝芽細胞の成熟に充分な条件下において肝芽細胞を培養する段階とを含む方法を提供する。アクチビンまたはノーダルの量は、ES細胞から内胚葉への分化を誘導するのに充分な量である。好ましい実施形態では、アクチビンは少なくとも約30ng/mlの濃度で存在し、BMP-4は少なくとも約10ng/mlの濃度で存在し、かつbFGFは少なくとも約1.0ng/mlの濃度で存在する。肝細胞はグリコーゲン貯蔵に関するアッセイによって、あるいは例えば電子顕微鏡によって大きな核/細胞質比、多量のグリコーゲン貯蔵および毛細胆管の存在などの肝細胞の特徴を観察することによって同定することができる。
【0036】
前述の方法は、少なくとも約10%の肝細胞を含む細胞集団を与える。好ましい実施形態では、細胞集団は少なくとも約50%の肝細胞を含む。より好ましい実施形態では、細胞集団は少なくとも約60%、または少なくとも約70%、または少なくとも約80%、または最も好ましくは少なくとも約90%の肝細胞を含む。
【0037】
本発明はさらに、前に記載した細胞集団の増殖、分化または生存に影響を与える作用物質を同定する方法を提供する。この方法は、試験する作用物質の不在下および存在下において本明細書で前に記載した細胞集団の1つ由来の細胞を培養する段階と、作用物質が細胞集団の増殖、分化または生存に対して影響を有するかどうか決定する段階とを含む。試験する作用物質はポリペプチド、多糖、ポリヌクレオチドなど、抗体またはその断片、天然または合成化合物のライブラリー由来の化合物、合理的薬剤設計から得た化合物、あるいは当技術分野で知られているアッセイ、例えば米国特許第6,110,739号中に記載された標準的な増殖および分化アッセイを使用して細胞集団に対するその影響を評価することができる任意の作用物質を含めた、天然または合成、1つの化合物または混合物、小分子またはポリマーであってよい。このような作用物質は、in vivoおよびin vitroで細胞増殖および分化を制御するのに有用である。
【0038】
本発明はさらに、特定系統および組織の細胞分化および発生と関係がある遺伝子を同定する方法を提供する。この方法は、異なる量の培養時間後に本発明のGFP+細胞の集団を単離する段階と、異なる集団中での遺伝子発現プロファイルを比較する段階と、一集団中で特有に発現される遺伝子を同定する段階を含む。好ましい実施形態では、マイクロアレイ分析およびサブトラクティブハイブリダイゼーションを使用して、遺伝子発現プロファイルを比較する。
【0039】
他の実施形態では本発明は、brachury陰性(brach-)細胞ではなくbrachury陽性(brach+)細胞を認識する抗体を作製する方法を提供する。ポリクローナル抗体は、免疫原の形で本発明の細胞を動物に注射することによって作製することができる。さらに、GFP-細胞ではなくGFP+細胞中に存在する細胞表面マーカーを同定する段階、およびマーカーまたはその断片に対する抗体を作製する段階によって、抗体を作製することができる。抗体はモノクローナルまたはポリクローナルであってよく、断片、遺伝子工学処理された抗体、単鎖抗体などであってよい。抗体は当技術分野でよく知られている方法によって作製することができる。このような抗体は、内中胚葉および中胚葉などのbrach+細胞を同定および単離するのに有用である。
【0040】
本発明は、in vitroで哺乳動物細胞を生成するための方法も提供する。一実施形態では、この方法は、心筋、血管平滑筋、内皮または造血細胞への中胚葉の分化に有効な条件下で、内中胚葉および中胚葉細胞が豊富な細胞集団由来の細胞を培養する段階を含む。in vitroでの様々な細胞型への分化に有効な条件は、当技術分野で知られている。他の実施形態では、この方法は、肝臓細胞または膵臓細胞への内胚葉の分化に有効な条件下で、内胚葉細胞が豊富な細胞集団由来の細胞を培養する段階を含む。このような分化に有効な条件は、当技術分野で知られている。インシュリンを生成する膵臓ランゲルハンス島細胞の生成が特に企図される。
【0041】
本発明によって実証されるように、異なる経過日数のEBから単離したbrach+細胞は異なる発生能を有する。約3日齢マウスのEB由来のBrach+/Flk-細胞は造血および内皮系統を効率良く生じ、一方約3日齢〜10日齢のEB由来の細胞は心筋細胞系統の細胞を生じる。したがって、内中胚葉および中胚葉が豊富な細胞集団を得るために使用するES細胞の培養時間を調節することによって、当業者は造血および内皮系統または心筋細胞系統の効率良い生成を選択することができる。
【0042】
このような細胞は、例えば限られた数の細胞型の破壊または機能不全が原因である障害を治療するための細胞補充療法に有用である。これらの障害には、真性糖尿病、肝不全、心不全、心臓血管および他の血管疾患、Duchenne型筋ジストロフィー、骨形成不全症、および骨髄移植によって治療可能な障害、例えば白血病および貧血がある。Odoricoら、(2001) Stem Cells 19: 193〜204を参照。
【0043】
本発明の細胞集団は、細胞補充療法用の分化細胞および組織を生成するのに有用である。細胞補充療法用の本発明の細胞集団の適性は、限られた数の細胞型の破壊または機能不全と関係がある障害の動物モデルに細胞を移植することによって評価することができる。例えば、参照により本明細書に組み込まれるGrompeら(1993) Genes & Dev.7: 2298によって例えば開示されたフマリルアセト酢酸(FAH)欠陥マウスは、肝不全のモデルとなる。FAH欠陥マウスは、NTBC(2-(2-ニトロ-4-トリフルオロメチルベンゾイル)-1,3-シクロヘキセジオン)で治療するか、あるいは正常な肝細胞を移植しない限り、進行性肝不全および尿細管障害に罹患する。したがってこれらのマウスは、EBから生じる未分化肝細胞の特徴を有する細胞の発生能を試験するための理想的なモデルとなる。NTBCを除去したFAH欠陥マウスに肝細胞を移植するための方法は当技術分野で知られており、例えばOversturfら(1996) Nature Genet.12: 266〜273によって開示されている。正常な肝機能はマウスの生存によって示され、血清中アスパラギン酸トランスアミナーゼのレベル、血漿中ビリルビンのレベルを測定すること、および再生肝臓の正常な構造を決定することによって評価することもできる。
【0044】
特定の細胞型の破壊または機能不全が原因である他の障害の動物モデルは、当技術分野で知られている。このようなモデルを同様に使用して、本発明の他の細胞集団を評価することができる。
【0045】
好ましい実施形態では、本発明は、肝細胞補充療法の方法であって、本発明の方法によって生成した肝細胞を含む組成物を、その治療の必要がある対象に投与する段階を含む方法を提供する。好ましい実施形態では、対象はヒトである。例えば注射、移植または注入を含めた肝臓組織への送達をもたらす経路によって組成物を投与することができる。本発明の方法は、肝不全、肝臓系代謝性疾患、および慢性肝臓疾患を治療するのに有用である。
【0046】
本発明は、1つのbrachury対立遺伝子が不活性状態でありbrachury遺伝子座が転写される細胞中で選択可能なマーカーが発現されるように、選択可能なマーカーをコードするDNAがbrachury遺伝子座に存在する、トランスジェニック非ヒト哺乳動物も提供する。好ましい実施形態では、哺乳動物はマウスであり、選択可能なマーカーはGFPである。特に、トランスジェニックマウスは、GFPをコードするDNA配列がbrachury制御要素と動作可能に連結しているトランス遺伝子を含むゲノムを有し、このトランス遺伝子はbrachuryを通常発現する細胞中で発現される。トランスジェニックマウスは、胚盤胞に本明細書で前に記載したGFP-BryES細胞を注射し、次いでそれらを偽妊娠状態のメスに移植することによって得ることができる。トランスジェニック子孫は、brach+/-と関係がある短い尾の表現型によって、および分子分析によって同定する。このようなトランスジェニック動物は、本発明の方法によって使用するための中胚葉を単離する初期胚を得るのに、brachury遺伝子を発現する任意の成体細胞集団を同定、単離および特徴付けするのに有用である。このような細胞は、新規の幹細胞集団となる可能性がある。
【0047】
本明細書に引用する全ての参照文献は、それらの全容を本明細書に組み込む。
【実施例】
【0048】
以下の実施例は、本発明をさらに例示するのに役立つ。
(実施例1)
【0049】
材料および方法
ES細胞の増殖および分化。15%のウシ胎児血清(FCS)、ペニシリン、ストレプトマイシン、LIF(1%条件付き培地)および1.5×10-4Mのモノチオグリセロール(MTG;Sigma)を補ったダルベッコ改変イーグル培地(DMEM)中の照射した胚栄養細胞上にES細胞を保った。分化の開始の2日前に、同じ培地中のゼラチンプレートに細胞を移した。EBを生成するために、ES細胞をトリプシン処理し、分化培養物中に様々な密度で平板培養した。EBの分化は60mmのペトリグレード皿中、15%のFCS、2mMのL-グルタミン(Gibco/BRL)、トランスフェリン(200ug/ml)、0.5mMのアスコルビン酸(Sigma)、および4.5×10-4MのMTGを補ったIMDM中で実施した。37℃において5%CO2/空気混合物中の加湿チャンバー内に培養物を保った。
【0050】
無血清培地。2つの異なる無血清培地を以下の実施例の異なる態様において使用した:Knockout SR(Gibco BRL)およびStemPro34(Gibco BRL)を補ったIIMD。
【0051】
メチルセルロースコロニーアッセイ。A)芽細胞コロニー:芽細胞コロニーを生成するために(BL-CFCアッセイ)、EB由来細胞を0.5×〜1.5×105個の細胞/mlで10%のFCS(Hyclone)、血管内皮増殖因子(VEGF;5ng/ml)、c-kitリガンド(KL;1%条件付き培地)、IL-6(5ng/ml)および25%のD4T内皮細胞の条件付き培地を補った1%のメチルセルロースにおいて(Kennedyら(1997) Nature 386: 488〜93)平板培養した。移行性コロニーがVEGFの不在下で生成した。コロニーは培養の4日後に記録した。B)造血コロニー:初期および最終段階造血コロニーを増殖させるために、細胞を10%の血漿由来血清(PDS;Antech)、5%無タンパク質ハイブリドーマ培地(PFHM-II;Gibco-BRL)および以下のサイトカイン:c-kitリガンド(KL;1%条件付き培地)、エリスロポイエチン(2U/ml)、IL-11(25ng/ml)、IL-3(1%条件付き培地)、GM-CSF(3ng/ml)、G-CSF(30ng/ml)、M-CSF(5ng/ml)、IL-6(5ng/ml)およびトロンボポイエチン(TPO;5ng/ml)を含む1%のメチルセルロース中に平板培養した。培養物は37℃、5%CO2で保った。初期赤血球コロニーは培養の第5〜6日で記録し、一方最終段階赤血球(BFU-E)、マクロファージ、および多系統コロニーは培養の第7〜10日で計数した。c-kitリガンドは、KL発現ベクター(Genetics Instituteによって親切にも与えられた)でトランスフェクトしたCHO細胞によって条件付けした培地に由来した。IL-3は、IL-3を発現するベクターでトランスフェクトしたX63AG8-653ミエローマ細胞によって条件付けした培地から得た。VEGF、GM-CSF、M-CSF、IL-6、IL-11、アクチビンBMP2、BMP4、bFGF、FGF8、および1hhはR&D systemsから購入した。
【0052】
再凝集培養物。15%のFCS(またはKnockout SR)、2mMのL-グルタミン(Gibco/BRL)、0.5mMのアスコルビン酸(Sigma)、および4.5×10-4MのMTGを補ったIMDM1ml当たり2×105個で、24ウエルのペトリグレードプレート中で細胞を培養した。これらを使用して、細胞とウエルの底部の接着を妨げた。
【0053】
心筋アッセイ。15%血清物を補ったIMDM中でGFP+細胞を再凝集させた。20時間後、凝集体を24または96ウエルプレートのいずれか中、10%血清物を補ったIMDM中(無血清)で培養した。ウエルはゼラチンで予め処理した。培養物は拍動細胞の出現の発生に関して毎日調べた。拍動細胞は培養第2日と第6日の間に通常検出された。
【0054】
細胞表面マーカーの染色およびFACS分析。標準的な条件を使用して細胞を染色した。染色細胞懸濁液はFACScan(Becton Dickinson、CA)で分析した。
【0055】
遺伝子発現の分析。ポリA+RT-PCR分析用に、Bradyら((1990) Meth.in Mol.and Cell Bio.2: 17〜25)の方法を使用した。逆転写、ポリAテーリングおよびPCR手順を記載されたのと同様に実施した、ただしX-dTオリゴヌクレオチドは5'-GTTAACTCGAGAATTC(T)24-3'に短縮した。PCR反応からの増幅産物はアガロースゲル上で分離し、Zeta-プローブGT膜(Biorad)に移し、あるいはスロットブロット装置(Schleicher & Schuell)を有する膜に移した。生成したブロットは、遺伝子(β-H1以外全て)の3'領域に対応する32Pでランダムにプライマー処理したcDNA断片(Ready-to-Go Labelling、Pharmacia)とハイブリダイズさせた。β-H1特異的プローブは、その3'末端に8塩基の相同性を共有する2つのオリゴヌクレオチド(5'-TGGAGTCAAAGAGGGCATCATAGACACATGGG-3'、5'-CAGTACACTGGCAATCCCATGTG-3')をアニーリングさせることによって調製した。このβ-H1特異的オリゴヌクレオチドは、クレノウ充填反応を使用して32Pで標識した。遺伝子特異的PCR用に、全てのRNAはRNeasyミニキットを用いてそれぞれのサンプルから抽出し、RNase free DNase(Qiagen)で処理した。2マイクログラムの全てのRNAは、Omniscript RTキット(Qiagen)を使用しランダムヘキサマーを用いてcDNAに逆転写した。PCRは適切なオリゴヌクレオチドを使用して実施した。PCR反応は、2.5UのTaqポリメラーゼ(Promega)、PCRバッファー、2.5mMのMgCl2、 0.2uMのそれぞれのプライマーおよび0.2mMのdNTPを用いて実施した。循環条件は以下の通りであった;94℃で5分間、次に35サイクルの増幅(94℃で1分間の変性、60℃で1分間のアニーリング、72℃で1分間の伸長)および72℃で7分間の最終インキュベーション。
(実施例2)
【0056】
標的ES細胞の生成
培養中の適切な条件下で、胚幹(ES)細胞は分化し、広範囲の系統由来の発生段階の細胞集団を含む胚様体(EB)として知られる三次元のコロニーを形成する。Smith(2001) Annu.Rev.Cell Dev. Biol.17: 435〜62。これらのEB由来集団の中では、造血、内皮、心筋および骨格筋系統の誘導体を含めた中胚葉誘導体を検出することができる。
【0057】
EBにおける中胚葉形成の開始を追跡するため、およびこの集団を表す細胞を単離するために、緑色蛍光タンパク質(GFP)をbrachury遺伝子座に標的化した。標的構築体はGFPのcDNA、および人工イントロン、第一エクソン中でneoカセットに隣接するSV40ポリ(A)配列およびloxPを含んでおり、図1中に示す。ランダムな組み込みに対する選択のために、チミジンキナーゼ(TK)遺伝子を標的構築体の3'末端に含めた。標的ベクターは以下のように構築した。
【0058】
完全なマウスbrachury(Bry)遺伝子を有するBACクローンを、プライマー5'-AAGGAGCTAACTAACGAGATGAT-3'および5'-TACCTTCAGCACCGGGAACAT3'を用いて、129/Ola系統ゲノムライブラリー(Genome Systems)のPCRスクリーニングによって単離した。これらのプライマーはそれぞれ第一および第二Bryエクソン内でアニーリングし、〜600bpの診断用バンドを増幅させる。Bry遺伝子の1エクソンおよび2kbを超える5'隣接領域を有する約3kb長のPstI制限断片を同定し、BACからプラスミドpBSK(Strategene)にサブクローニングし、構築体pBSK.Bry-5'を得た。開始コドンの直ぐ上流の約2kbの領域をシークエンスして、ベクターを構築するのに適したプライマーアニーリング部位を同定した。
【0059】
オリゴ5'-GCTAGCTAATGGATCCA-3'/5'-GATCTGGATCCATTAGCTAGCTGCA-3'および5'-GATCTTAATGAACGGCAGGTGGGTGCGCGTCCGGAG-3'/5'TCGACTCCGGACGCGCACCCACCTGCCGTTCATTAA-3'をプラスミドpBSKのPstI/SalI部位中に挿入して、2つの連続した翻訳停止コドンおよび人工3'スプライシング部位を有する新たな、より適切なポリリンカー(構築体pBry-AA)を作製した。プラスミドpEGFP.C1(Clontech)はNheI/BglIIで2回消化し、生成した停止コドンを含まないEGFPをコードする〜760bpのDNA断片はpBRY-AAのNheI/BglII部位にクローニングし、構築体pBry-ABを生成した。loxPに隣接するネオマイシン耐性遺伝子を有するプラスミドpL2-Neo2のXhoI/SalI断片をpBry-ABのSalI部位に挿入して、プラスミドpBry-ACを生成した(EGFPとNeoの転写は同じ方向)。
【0060】
共通のスプライシングドナー部位、人工イントロン、スプライシングアクセプター部位およびSV40ポリアデニル化配列を含む短いエクソンを有する556bpのXmaI/MluI断片を、市販の発現ベクターpBK-CMV(Stratagene)から切除した。この断片は以下の方法でプラスミドpBry-AC中に挿入した:XmaI端を最終EGFPコドンの後のBspEI部位と連結させ、一方Mlu端はリンカーとしてのオリゴ5'-CGCGTTACTAGTAAGACGTCT-3'/5'-CCGGAGACGTCTTACTAGTAA-3'と共にloxP-neo-loxPカセットの直ぐ上流に位置するBspEI部位に挿入した。生成した構築体:pBry-AE。HSVチミジンキナーゼ遺伝子をコードする〜1.9kbのXhoI/SalI断片をpBry-AEのXhoI部位に挿入して、ランダムな組み込みに対する選択を可能にした(構築体pBry-AH)。相同性の「短いアーム」をコードするNotI/Eco47III断片をpBry-AFから切除し、pBry-AHのNotI/Eco47III部位にクローニングして、プラスミドpBry-AIを生成した。相同性の「長いアーム」はSalIを用いてpBry-AKから切除し、正しい方向でpBry-AIのSail部位に挿入して、最終標的ベクターBを生成した。
【0061】
胚幹細胞(E14.1、129/Ola Hooperら(1987) Nature 326: 292)に、NotI線状標的ベクターpBry-AMをエレクトロポレーションにより導入した。トランスフェクト細胞を含む4枚の皿にはG418単独選択を施し、他の4枚の皿にはG418+ガンシクロビル(Ganc)二重選択を施した。相同的組換え事象を経験したクローンは、「相同性の短いアーム」の直ぐ上流のBry遺伝子の5'領域中のゲノム配列とアニーリングする1つのプライマー(5'-CAGGTAGAACCCACAACTCCGAG-3')、EGFPの5'部分(診断用バンド:約1.3kb)に対する他のプライマー(5'-CCGGACACGCTGAACTTGTGGC-3')を用いたPCRによって同定した。正しく標的化されたクローンは、サザンブロット分析によって確認した:候補クローンのゲノムDNAはHincIIで消化し、標的構築体の外側に位置するプローブとハイブリダイズさせた。オリゴヌクレオチド対5'-ACAGGATCCCTAAGCCTCAAAAGAGTCGCT-3'/5'-TCTTGGATCCTCCTATCCTATCCCGAAGCTCCT-3'を使用するPCRにより、プローブはBry5'隣接領域(BryATG開始コドンに関して-2018〜-1249)に由来した。384のG418単独および80 G418+のGanc二重選択クローンをスクリーニングし、そのそれぞれ4個のうちの3個が陽性であることが分かり、1.04%および3.75%の標的化効率に相当した。2個の陽性クローンを改変型Creリコンビナーゼ発現ベクターで一過的にトランスフェクトして、neo遺伝子を除去した。これらの標的クローンは、本明細書では以後GFP-BryES細胞と呼ぶ。
【0062】
brachuryは発生段階のEBにおいて一過的に発現され、その開始は造血および内皮系統の樹立を明確にする遺伝子の発現に先行する。GFP-BryES細胞におけるGFPの発現がEB発生中のbrachury遺伝子の発現を正確に反映するかどうかを決定するために、GFPの発現を評価した。
【0063】
6日間のEB分化期中の典型的な発現パターンは、図2A中に示す。この実験では、低レベルのメッセージを分化の24時間以内に検出した。発現は次の48時間上方制御され、4日間持続し、次いで分化の第6日までに検出不能なレベルまで急激に低下した。FACS分析によって定義したGFPの発現は、同様の経時的パターンをたどった。低レベルのGFP+細胞(〜5%)を分化の第2日で早くも検出した。半分を超える(65%の)EB由来細胞が第3日でGFPを発現し、ほぼ全ての細胞が分化の第4日で陽性であった。PCRによって観察したように、発現はこの時点以後急激に低下し、第6日までは、存在した場合でも少数のGFP+細胞が存在した。GFPの発現のこの急激な低下は、それは長期間細胞内で持続しなかったことを示した。分化の第3日および第4日でのGFP+細胞の高い割合は、これらの条件下でのEB内の中胚葉の発生が広範囲に及ぶことを示唆する。合わせて考えると、これらの発見は、GFPの発現はEB発生中のbrachury遺伝子の発現を正確に反映することを示す。
【0064】
1つのbrachury対立遺伝子の不活性化が、ES細胞のin vitroでの発生能に対して有害な影響を有し得る可能性を評価した。示したように、ヘテロ接合性のマウスは軽度の表現型を示す。GFP-BryES細胞が造血発生中に何らかの検出可能な欠陥を示すかどうかを測定するために、それらから生成したEBを、造血前駆体および芽コロニー形成細胞(BL-CFC)含量、および遺伝子発現パターンに関して分析した。図3Aおよび3B中のデータは、野生型細胞と比較してGFP-BryES細胞が、匹敵する数の初期(Eryp)および最終段階(Eryd、Mac、Mac/Ery、およびMix)造血前駆体およびBL-CFCを生成することを示す。遺伝子発現パターン(図3C)は前駆体の分析を支持し、GFP-Bryから生成したEBと野生型ES細胞の間ではほとんど違いを示さなかったことを確認した。両組のEBが、分化の最初の3日間でRex-1の発現の低下を示した。Rex-1は転写因子であり、ES細胞中で発現され、分化を経ると下方制御される。Rogersら(1991) Development 113: 815〜24。Rex-1の低下は、brachury発現の典型的な一過性の波に続き、それは造血および内皮系統の発生と関係がある遺伝子の発現開始の直前に先行する。これらの系統の樹立に必要不可欠なFlk-1、受容体チロシンキナーゼ(Shalabyら(1995) Nature 376: 62〜6)は、分化の第3日と第6日の間に発現される。GATA-1、造血転写因子、ならびに胚および成体グロビン遺伝子、βH1およびβmajorは、分化の第4日に低レベルで検出した。3つ全ての遺伝子が次の24時間上方制御され、この発生段階での初期赤血球系統の広がりおよび成熟が反映された。Palisら(1999) Development 126: 5073〜84。この実施例で観察した前駆体の数および遺伝子発現パターンは、以前の研究において見られたものと一致し、造血系の樹立をもたらす分子プログラムは標的GFP-BryES細胞において完全な状態であることを示す。
(実施例3)
【0065】
brachury+細胞の単離
GFPの発現に基づいてbrachury+細胞を単離することができるかどうか決定するために、第3.5日EB由来のGFP+集団を適切な遺伝子の発現に関して選別し分析した。図4AはGFP陽性(2)および陰性(1)集団の単離に使用したゲートを示す。RT-PCR分析によって、brachuryの発現はGFP+画分に限られていたことが明らかになり、GFPの発現に基づく細胞選別を使用してbrachury+細胞を単離することができることが示された。Flk-1、造血および内皮発生の初期マーカーの1つは、GFP-画分中よりGFP+中において高レベルで存在し、Flk-1は少なくともbrachury+細胞の亜集団中で発現されたことが示された。brachuryおよびFlk-1とは対照的に、Pax-6、初期神経発生と関係がある遺伝子[79,80]は、brachury陰性であるこの系統の前駆体と一致してGFP-画分中において高レベルで発現した。これらの細胞選別試験は、brachury遺伝子座の制御下でのGFPの発現は、EB由来のbrachury+細胞の単離、特徴付けおよび操作に関する新規のマーカーをもたらすことを示す。
【0066】
この実施例は、GFP+細胞は細胞選別によって第3.5日EBから単離することができることを実証する。GFP+およびGFP-画分の遺伝子発現分析は、brachuryの発現は陽性画分に主に分離すること、GFPに基づく画分化はbrachury陽性細胞を単離するための方法をもたらすことを明らかに実証する発見を示す。brachury以外に、初期造血および内皮発生と関係がある受容体チロシンFlk-1も、陰性画分中より陽性画分中において高レベルで発現される。対照的に、Rex-1およびPax-6、初期外胚葉および神経外胚葉のマーカーは、GFP-画分中で発現される。これらの発見は、brachuryの状況でのGFPの発現を使用して、外胚葉から中胚葉を分離することができることを実証する。
(実施例4)
【0067】
Flk-1の発現に基づく亜集団へのbrachury陽性細胞の分離
Flk-1は、初期胚における造血および内皮系統の樹立に必要不可欠であり、BL-CFCを含めたこれらの系統の初期前駆体において発現されることが示されてきている[Faloonら(2000) Development 127: 1931〜41]。血液および血管発生におけるこの重要な役割を考慮すると、GFP+集団内のその発現は、これらの系統への特化を経た中胚葉の亜集団を明確にすると仮定した。この可能性をさらに調べるために、GFPおよびF1k-1陽性細胞の存在に関して幾つかの発生段階でEBを分析した。図5A中に示した実験では、4.8%の3.0日EB集団がF1k-1ではなくGFPを発現し、一方1.2%の細胞が両方のマーカーを発現した。両画分の大きさは次の12時間で劇的に増大し、GFP+/Flk-1-およびGFP+/Flk-1+細胞はそれぞれ全EB集団の52%と26%を表した。GFPおよびFlk-1発現によって定義される3集団、GFP-/Flk-1-(画分2)、GFP+/Flk-1-(画分3)およびGFP+/Flk-1+(画分4)の発生能を評価するために、細胞を両時点において単離し、BL-CFCおよび2°EB含量に関して、遺伝子発現パターンに関して分析した。これらの画分の発生能を、予め選別した集団(画分1)のそれと比較した。大部分のBL-CFCは、分化の第3.0日と第3.5日の両方でGFP+/Flk-1+画分中において見られた(図5B)。全てのBL-CFCがFlk-1を発現することを以前の試験が示していることを考慮すると、これは驚くことではない。2°EBはGFP-/Flk-1-画分に限られており、これは2°EBが初期EBの残留未分化ES細胞に由来する事実と一致する発見である。GFP+/Flk-1-画分は、これらの培養で使用した条件下でほとんどコロニーを生成しなかった。遺伝子発現の分析によって、2つの時点で単離した集団間の幾つかの興味深い違いが明らかになった(図5C)。Rex-1、ES細胞のマーカーは、GFP-画分よりGFP+において低いレベルで発現され、これらの集団は分化を経ていることが示された。brachuryはGFP+画分において両方の時点で発現された。そのレベルは第3.5日EBから単離したGFP+/Flk-1+中よりGFP+/Flk-1-中で高いようであり、これらの細胞が造血および内皮系統に成熟すると、その発現は下方制御される可能性があることが示唆される。予想したように、Flk-1は両方の時点でGFP+/Flk-1+細胞中において主に発現された。初期と最終段階の両方の造血発生に必要不可欠である(Shivdasaniら(1995) Nature 373: 432〜4)Scl、ヘリックス-ループ-ヘリックス転写因子は、GFP+/Flk-1+画分に限られるようである。同様に、最終段階造血系の樹立に必要とされる転写因子Runx1(Wangら(1996) Proc. Natl. Acad. Sci. 93: 3444〜9)は、GFP+/Flk-1+画分中で最も容易に検出した。第3.0日EBから単離したGFP+/Flk-1-画分中では、ある程度のRunx1の発現があった。ノーダルは分化の第3日で3つ全ての画分中で発現される。第3.5日では、GFP+/Flk-1+画分中での発現のレベルは、他の画分中より有意に低いようである。Wnt3aおよびWnt8aは著しく限られたパターンの発現を示し、それは両方の時点においてGFP+/Flk-1-画分中のみで見られ、系統限定的なマーカーの発現前の初期中胚葉の機能と一致した。BMP2は両方のGFP+画分中で発現され、一方BMP4はGFP+/Flk-1+細胞中で主に見られ、これらの分子はこの系では発生の異なる段階で役割を果たすことが示された。GFP+/Flk-1+細胞のBL-CFC発生能および遺伝子発現パターンは、それらの細胞がマウス胚において見られる胚外中胚葉を表すことを示す。
【0068】
この実施例は、第3日および第3.5日のEBのbrachury画分は、Flk-1発現に基づいて2つの画分:brachury+/Flk-1-(GFP+/Flk-1-)およびbrachury+/Flk-1+(GFP+/Flk-1+)に分けることができることを実証する(図5A)。機能性試験によって、前駆体(BL-CFC)は(GFP+/Flk-1+)画分に分離する造血細胞と内皮細胞の両方を生成することができることを実証し、Flk-1の上方制御はこれらの系統への拘束を示すことが示唆された(図5B)。遺伝子発現試験によって、GFP+/Flk-1-集団とGFP+/Flk-1+集団の間の明確な違いを実証した(図5C)。
(実施例5)
【0069】
GFP/Flk画分間の発生関係
図5中で観察した発現パターンは、3つの画分は、GFP-/Flk-1-細胞はGFP+/Flk-1-細胞に分化し、次いでそれがGFP+/Flk-1+細胞に分化する発生の連続性を示すという解釈と一致する。これらの画分が共通の発生経路内で特異的段階を示すかどうかを測定するために、各画分を第3日EBから単離し、20時間培養し、次いでGFPおよびFlk-1の発現に関して再度分析した。BL-CFCおよびEryp-CFCの発生能を、培養の前後に各集団に関して測定した。単離した細胞は、ペトリグレードの24ウエルプレート中、EB分化用に使用した同じ培地中に、1×105個の細胞以上の密度で培養した。これらの条件下では、細胞は直ぐに再凝集し、EB様構造を形成し、これは細胞数の増大または損失ほとんどなしで正常な発生プログラムに従うようである。20時間の再凝集培養後、GFP-/Flk-1-細胞は相当数のGFP+/Flk-1-細胞集団、および少数のGFP+/Flk-1+に分化した。GFP+/Flk-1-細胞は、同じ培養時間中に相当数のGFP+/Flk-1+細胞集団を生成した。GFP+/Flk-1+集団は、再凝集培養後に幾らかGFPおよびFlk-1の発現を失うようであった。結果は図6中に示す。前駆体の発生能の変化は、表面マーカーの変化と一致していた。3つの中で最も未熟であるGFP-/Flk-1-画分は、培養の前後で検出不能な数のBL-CFCおよびEryp-CFCを含んでいた。GFP+/Flk-1-画分も、培養前にわずかなBL-CFCおよびEryp-CFCを含んでいた。しかしながら、培養後、BL-CFCの発生能は細胞105個当たり74〜1564に劇的に増大し、Flk-1の発現の増大と一致した。Eryp-CFCの頻度は培養期間中変わらなかった。GFP+/Flk-1+画分は、培養前にBL-CFCを含んでいたが、Eryp-CFCはほとんど含んでいなかった。培養後にBL-CFCは検出されなかったが、この集団はここでEryp-CFCを含んでいた。表面マーカーの分析と一緒にすると、これらの前駆体のデータは、前中胚葉(GFP-/Flk-1-)から中胚葉/前血管芽細胞(BL-CFC)集団(GFP+/Flk-1-)、血管芽細胞/前赤血球集団(GFP+/Flk-1+)、後血管芽細胞/赤血球集団(おそらくGFPlo/Flk-1lo)への発生の進行を支持する。最初の表現型を有する細胞がGFP-/Flk-1-およびGFP+/Flk-1-培養物中に残存したので、所与の集団中の全ての細胞が20時間の培養期間後に分化したわけではないようである。
【0070】
この実施例は、単離し20〜24時間再培養すると、第3日EBから単離した3つの集団のそれぞれは分化し続けることを示し、あるパターンでは、これらの集団は発生の連続性を示すことを示す。例えば、GFP-/Flk-1-はGFP+/Flk-1-細胞に分化し、次いでそれはGFP+/Flk-1+に分化した。細胞表面特性のこれらの変化は、予想した発生能の変化と関係があった。GFP+/Flk-1-画分は、培養前にわずかな造血/内皮前駆体(BL-CFC)を含んでいた。培養後、これらの前駆体を検出し、第3日EB由来のGFP+/Flk-1-画分は、Flk-1+細胞に分化する発生能ならびに造血および内皮発生能を含むことが明らかに実証された。
(実施例6)
【0071】
GFP/Flk-1-細胞の発生能
前述の実施例は、第3日EBから単離したGFP+/Flk-1-細胞は、一晩培養の後GFP+/Flk-1+細胞およびBL-CFCを効率良く生成したことを実証した。このプレBL-CFCの発生能が発生のこの段階で単離したGFP+/Flk-1-画分に特異的であったかどうかを決定するために、異なる経過日数のEB由来のGFP+/Flk-1-細胞を、BL-CFCに分化するそれらの能力に関してアッセイした。図7A中に示すように、BL-CFCを生成する能力は第3日のGFP+/Flk-1-細胞において最も顕著であった。この発生能は分化の第3.5日までに劇的に低下し、第4.0日にはほとんど存在しなかった。これらと同じEBから新たに単離したGFP+/Flk-1+画分のBL-CFC含有量はこの期間中に増大し、分化は通常の形式で進行していたことが示された。異なる段階のEB細胞から再凝集した培養物におけるFlk-1発現のパターンは、BL-CFCのデータと一致した。再凝集した第3日のGFP+/Flk-1-細胞由来の培養物は、集団全体の40%より多くを表した独特のFlk-1画分を含んでいた(図7B)。第3.5日および第4.0日培養物におけるFlk-1の発現は有意に低く、独特のピークではなく集団全体のピークからなっていた。
(実施例7)
【0072】
GFPおよびFlk-1亜集団の心筋細胞発生能
造血および内皮中胚葉の発生が心臓および頭蓋中胚葉の発生に続くマウス胚における事象の順序を考慮して、単離集団の心筋細胞の発生能を測定した。この分析のために、異なる集団の培養物からの凝集体をマイクロタイターまたは24ウエルプレート、無血清培地中に移し、心筋細胞を示す拍動細胞塊の発生を調べた。これらの条件は、再凝集細胞からの心筋細胞の効率良い発生を助長することが知られている。これらの塊中の心筋性の細胞の別個の確認として、代表的な群を顕微鏡スライドに移し、固定しトロポニンTの心臓特異的イソ型の存在に関して染色した。分析した全ての拍動細胞塊はトロポニンT陽性細胞を含んでいたことが分かり、それらが心筋細胞であったことが示された。このアッセイを使用して、再凝集した異なる段階のEB由来のGFP+/Flk-1-およびGFP+/Flk-1+画分の心筋細胞の発生能を測定した。比較用に、新たに単離したGFP+/Flk-1+細胞および培養したGFP+/Flk-1-細胞のBL-CFC発生能を分析した。
【0073】
【表1】
【0074】
表1中に示したように、異なる画分のBL-CFC発生能は、前の実験で観察したそれと同等であった。3つの異なるEB集団から単離したGFP+/Flk-1+細胞がBL-CFCを生成し、最高数は第3.5日および第4.0日で見られた。GFP+/Flk-1-細胞におけるプレBL-CFC発生能は第2.75日で最大であり、第3.5日および第4.0日で著しく低下した。画分の心筋細胞発生能は逆のパターンを示した。第3.5日および第4.0日GFP+/Flk-1-細胞由来の相当な割合(>80%)の移動凝集体が、拍動心筋細胞を生成した。初期(第2.75日)GFP+/Flk-1-細胞から生成した凝集体では拍動細胞が観察されなかった。発生の三段階のいずれかでEBから単離したGFP+/Flk-1+細胞から生成した凝集体では、拍動細胞を検出しなかった。この分析からの発見は、異なる段階で単離したGFP+/Flk-1-細胞は異なる発生能を有するという概念と一致する。初期に発生する細胞は血管芽細胞の運命を有しているようであり、一方後期に発生する細胞は、心臓系統およびおそらく他の集団を生成する。GFP+/Flk-1+集団は心筋細胞発生能を失っているようであり、血管芽細胞系統に限られる可能性がある。これらの発見を考慮に入れて、初期発生(第2.75日〜第3.0日)GFP+/Flk-1-細胞は前血管芽細胞中胚葉と呼び、一方第3.5日と第4.0日の間に発生する集団は、前心臓中胚葉と呼ぶ。第3.0日〜第3.5日GFP+/Flk-1+集団はBL-CFCを生成し、一方後期段階のEB(第4.0日)から単離した集団は初期赤血球の原種を含み、造血への拘束の開始が示される。この発生能を考慮して、GFP+/Flk-1+集団は血管芽細胞中胚葉と呼ぶ。
【0075】
実施例5および6は、異なる経過日数のEBから単離したGFP+細胞は異なる発生能を有することを実証する。前の実施例で示したように、第3日EB由来のGFP+/Flk-1-細胞は、造血系統と内皮系統の両方を効率良く生成する。拍動細胞塊の欠如によって実証されたように、これらの細胞は心筋細胞(心臓組織)に分化しなかった。対照的に、第4日EB由来のGFP+細胞は、培養後わずかなFlk-1+細胞およびBL-CFCに分化した。しかしながらこの集団は、心筋細胞系統の細胞を生成した。これらの発見は、異なる経過日数のEBから単離したGFP+(brachury+)画分は、異なる発生運命を有する異なる集団にパターン形成していることを実証する。前述の実施例で使用した条件および観察した発生能以外に、条件および添加剤を変えることによって他の発生能を観察することができる。
(実施例8)
【0076】
血清由来因子の役割
brachury+細胞の発生における血清の役割を評価するために、EBを血清の不在下で分化させた。EBはこれらの条件下で発生したが、それらは通常の条件において見られるEBより幾分小さかった。血清の不在下では、これらのEB内でGFP+細胞は検出されず、これらの条件下では中胚葉は誘導されなかったことが示された(図8)。培養物に血清を加えると、相当数のGFP+/Flk-1-およびGFP+/Flk-1+細胞が発生した。これらの発見は、血清内に見られる成分は、brachury+細胞の発生および分化を誘導することができることを明らかに実証する。このプロセス中で役割を果たす可能性がある因子を同定する際の最初のステップとして、BMP4(20ng/ml)を、無血清培養物内の発生中のEBに加えた。この濃度で、BMP4は分化3日以内に相当なbrachury+細胞集団を誘導した。しかしながら、血清とは対照的に、BMP4はこの期間中GFP+/Flk-1+集団の発生を助長しなかった。血清の存在下で誘導したGFP+/Flk-1-細胞からGFP+/Flk-1+をBMP4が誘導することができるかどうかを測定するために、GFP+/Flk-1-細胞を血清の存在下で3日間分化させたEBから単離した。これらの細胞は培地単独、血清を含む培地中、またはBMP4を含む培地中で再凝集させた。図8の下列中に示すように、血清の不在下で再凝集させたとき、GFP+/Flk-1-細胞は実質的に分化しなかった。予想通り、培養物に血清を加えると、同じ集団は大きなGFP+/Flk-1+集団を生成した。初期分化培養物における発見と一致して、培養したGFP+/Flk-1-細胞からの相当数のGFP+/Flk-1+細胞の発生を、BMP4は誘導することができなかった。
【0077】
図9は、前述の実施例に基づいて中胚葉発生中の段階を要約する。ステップ#1は、中胚葉誘導および前造血および内皮(前血管芽細胞)運命へのパターン形成を表す。ステップ#2は、造血および内皮系統への特化を表す。ステップ#3および#4は、前心臓運命へのパターン形成を表す。
(実施例9)
【0078】
細胞集団の単離および内胚葉発生能の特徴付け
アフリカツメガエルおよびゼブラフィッシュなどのモデル系を使用した試験は、中胚葉および内胚葉は、内中胚葉として知られる共通の前駆体集団から発生することを示唆している。発生の内中胚葉段階がEBにおいて存在するかしないか判定するために、内胚葉系統の発生に関する条件を樹立した。この系統の発生に関する培養条件を樹立する際の最初のステップとして、分化培養物中の血清の量を変えた。EBを血清(SP34、次いでSRを添加したIMDM)の存在下および不在下で生成させ、外胚葉、内胚葉および中胚葉発生と関係がある遺伝子の発現に関して異なる段階でアッセイした。外胚葉系統に関しては、神経系統の発生を、PAX6、Wntl、neruoDおよびニューロフィラメント(NFL)の発現を分析して評価した。これらの遺伝子は、神経発生の異なる段階で発現されることが知られている。内胚葉発生の初期段階はHNF3βの発現によって調べた。内胚葉発生および特化の後期段階を評価するために、肝臓の発生と関係がある遺伝子を分析した。これらはHex、α-フェトプロテイン(AFP)、HNF4、アルブミン(Alb)、α-1-アンチトリプシン(AAT)およびチロシンアミノトランスフェラーゼ(TAT)を含んでいた。中胚葉発生は、brachuryおよびGATA-1の発現によって調べた。遺伝子の発現パターン以外に、EBからの神経突起の発達を調べることによって神経発生をアッセイした。これらの神経突起の神経性はβIII-チューブリンの発現によって実証した。中胚葉発生は、EBにおける造血細胞の原種を列挙することによっても評価した。図10は、10日間の分化期間のEBの発生能に対する血清の影響を示す。血清の存在下(血清)では、この系統の発生と関係がある遺伝子の発現の欠如によって実証されるように、神経外胚葉の分化はほとんどない。HNF3βは分化の初期段階(第2〜3日)で発現され、次いで下方制御される。予想通り、GATA-1はこれらの条件下で生成したEBにおいて発現される。これらの遺伝子の発現パターンは、血清の不在下(血清-)で増殖させたEBでは基本的に逆であった。これらのEBは神経外胚葉と関係がある全ての遺伝子を発現したが、中胚葉/造血マーカーGATA-1は発現しなかった。HNF3βはこれらの条件下で増殖させた後期段階のEB(第10日)において発現した。GFPの発現によって調べたbrachury発現のパターンは、これらの発見と一致した。血清の存在下で生成したEBは、分化の第2日と第5日の間に存在した相当数のbrachury+集団を生成した(図11、-B-ライン)。血清の不在下で増殖させたEBではbrachuryは検出されなかった(図11、-H-ライン)。造血細胞前駆体のアッセイによってこれらの発見を確認した。血清中で生成したEBは前駆体を含んでおり(図12、まだら線)、一方血清なしで増殖させたEBは含んでいなかった(図12、色塗り線、示さず)。最後に、EBの神経突起の発生能の評価は、これらの様々な分析と一致した。血清中で増殖させたEBは神経突起を生成しなかった。対照的に、血清の不在下で生成したEBの85%は、この活性を示した(図13)。一緒に考えると、これらの発見は、特定の系統を生成するための培養条件(血清)の重要性を実証する。これらの発見はさらに、血清完全条件も無血清条件も内胚葉発生には最適ではなかったことを実証する。
【0079】
血清中で生成した初期段階のEBにおけるHNF3βの強い上方制御は、血清は内胚葉系統の成熟ではなく、樹立に重要である可能性があることを示唆した。この可能性を試験するために、EBは血清の存在下で2日間分化開始させ、次いで無血清状態(SR)に移した。図14中に示すように、これらの条件(血清+/-)下で生成したEBは、分化の第3日と第5日の間でHNF3βを発現した。AFPは分化の第5日で上方制御された。GATA-1の発現レベルは、血清刺激したEBにおいて見られたレベルと比較して低下した。次に、血清+/-条件下で生成した第6日EBを、増殖因子bFGFの存在下で組織培養グレード皿に平板培養し(それらを接着させ)た。これらの皿をゼラチンまたはマトリゲルのいずれかでコーティングして、さらなる内胚葉分化に対して基質が何らかの影響を有していたかどうかを測定した。5日後、培地を交換し、他の因子をこれらの培養物に加えて、肝臓系統の発生を促進した。実験の概要およびデータは図15中に示す。この実験では、第6日のEB段階でAFPは発現されなかった。ゼラチンまたはマトリゲルのいずれか上でbFGFの存在下において培養すると、その発現レベルは上方制御された。低レベルのALBもこの段階で検出した。試験した全ての条件において追加的な培養期間後、ALBの発現は増大した。最終培養ステップ後にAATおよびTATも発現された。EB由来細胞をbFGFおよびDexの存在下で培養したとき、最高レベルのTATが見られた。これらの発見は、内胚葉系統の細胞を生成することは可能であること、および適切な条件下において、それらの細胞は発生中の肝臓と関係がある遺伝子を発現する細胞に分化することを明らかに示す。
【0080】
これらの内胚葉細胞がbrachury+またはbrachury-細胞から発生したかどうかを、さらに測定した。この問題に取り組むため、GFP (Bry)+およびGFP(Bry)-細胞を、細胞選別によって第2.5日EBから単離した。これらの集団を再凝集させ、第6日まで集合体として培養した。第6日に、それらを組織培養グレード皿へ、bFGFを含む培地中に4日間移した(合計10日間)。遺伝子発現の分析によって、HNF3βを発現する細胞はGFP+画分(d2.5)に分離したことを示した(図16)。培養時間と共に、この遺伝子はGFP-画分から生成した細胞中で発現された。これはおそらく、発現の後期段階でHNF3βが非内胚葉集団中で発現される事実を反映する。AFP、HEX、ALBおよびHNF4はいずれもGFP+画分の誘導体中で発現されたが、GFP-細胞から生成した細胞集団中では発現されなかった。対照的に、PAX6およびneruoD、神経外胚葉のマーカーは、GFP-画分から生成した細胞中で主に見られた。これらの発見は、内胚葉系統はbrachury+集団から樹立され、中胚葉系統にさらに分化し、これらの系統は共通の前駆体、内中胚葉に由来することを示す。
【0081】
brachury発現細胞の肝臓発生能をさらに評価するために、bry+画分とbry-画分の両方由来の細胞集団を、α-フェトプロテイン(AFP)、アルブミン(ALB)およびトランスチレチン(TTR) などの初期肝細胞発生を表す遺伝子、およびα1-アンチトリプシン(AAT)、チロシンアミノトランスフェラーゼ(TAT)およびカルバモイルリン酸シンテターゼI(CPase)を含めたこの系統の成熟を示す遺伝子の発現に関して分析した。β-アクチンの発現は対照として使用した。選別前に細胞を分析し、後にbry+集団とbry-集団に選別した。胎児の肝臓および成体の肝臓対照も分析した。全てのこれらの遺伝子の発現はbry+集団由来の細胞に限られており、肝細胞の特徴を有する細胞はbrachury発現細胞から発生することが示された。
(実施例10)
【0082】
EBにおける中胚葉および内胚葉発生の動態
実施例6中に記載した事前動態分析によって、明確な発生運命を有する中胚葉の亜集団は明確で一時的な形式で生成したことを実証した。より詳細な動態分析によって、分化の第2.5日と第4.0日の間にGFP+Flk1-(これ以後はGFP+集団と呼ぶ)集団の動的発生を示した(図18)。単離し再凝集させると、第2.5日および第3.0日GFP+画分は、造血および内皮の拘束の初期段階(図19)を表す芽細胞コロニーを生成した(図19中に血管芽細胞の発生能として示す)。このGFP+細胞集団は、心筋細胞(心臓)発生能をほとんど示さなかった。初期のGFP+細胞とは対照的に、第3.5日EBから単離した細胞は著しく低下したBL-CFC発生能を示したが、心筋細胞への分化においては効率が良かった。第4日EBから単離したGFP+細胞は心筋細胞には分化せず、BL-CFCを生成する能力をほとんど有しておらず、それらは何か他の中胚葉系統になるよう運命付けられている可能性があることが示唆された。遺伝子発現の分析はこれらの機能アッセイを支持し、4つのGFP+画分間の分子差異を実証した(図20)。幾つかの遺伝子は全集団中で発現されたが、他の遺伝子は興味深い分化パターンを示した。造血発生および心臓分化の阻害に重要であると考えられる遺伝子Wnt3aは、第2.5日および第3.0日集団において発現され、第3.5日と第4.0日細胞において下方制御された。このパターンは、造血/血管から心筋へのこれらの集団の発生能の変化と一致する。当該の第二のパターンは遺伝子Mixl-1のパターンである。中胚葉および内胚葉の発生において役割を果たすこの遺伝子は、第2.5日および第3.0日GFP集団において発現されたが、第3.5日と第4.0日画分では発現されなかった。一緒に考えると、この実施例の発見は、異なる発生能を有する中胚葉集団はEB内では明確で一時的な形式で生成することを、明らかに実証する。さらに、中胚葉/内胚葉遺伝子Mixl-1の発現分析によって、内胚葉発生能を有する細胞も特定の時期、すなわち分化の第2.5日と第3.0日の間に生成することを示す。
【0083】
内胚葉発生の動態をさらに調べるために、第3.0日および第4.0日EB由来のGFP+細胞を単離し、肝細胞様細胞への分化を促進する条件下で培養した。図21中に示すように、第4.0日由来のEBではなく第3.0日EBから単離したGFP+細胞(+/-)が、HNF3βの発現によって定義されるように内胚葉発生能を示した。これらの発見は、内胚葉が特定の時期、分化の第4日の前にGFP+集団内で生成することを示す。
(実施例11)
【0084】
in vivoでのGFP+集団の発生能
GFP+集団の内胚葉発生能をさらに評価するために、GFP+およびGFP-第2.5日EB細胞を肝細胞の分化を促進することが知られている条件下で14日間培養し、レシピエントSCID-ベージュマウスの腎被膜下に次いで移植した。数匹のマウスを移植直後に殺傷し、移植片を有する腎臓を切片化し、切片はHep1およびAFPに対する抗体で染色した。Hep1は肝細胞の特異的マーカーであり、一方AFPは肝細胞系統の最終段階内胚葉および未熟細胞において発現される。この切片内の幾つかの細胞はHep1に関して陽性染色され、一方Hep1+細胞の近辺の他の細胞はAFPを発現することが分かった。Hep1+またはAFP+細胞は、GFP-陰性細胞の移植片内には見られなかった。Hep1の発現によって定義されるように、これらの知見はPCRデータを支持し、その培養条件が未熟肝細胞の特徴を有する細胞の発生を支持することを実証する。
【0085】
これらの移植実験は移植直後の移植片中の肝細胞様細胞の存在を実証するが、移植した組織が、奇形腫として知られる腫瘍様の塊を生成したので、経時的にこれらの集団の成熟を調べることは困難であった。奇形腫はおそらく汚染した未分化ES細胞から、あるいは中胚葉起源でありbrachuryを発現することが知られているGEP+原始生殖細胞から発生する。
(実施例12)
【0086】
アクチビンによる中胚葉および内胚葉の誘導
内胚葉発生能を有する細胞をさらに豊富にするために、他のモデル系においてこの細胞集団を誘導することが知られている増殖因子の影響を試験した。アフリカツメガエルにおける試験によって、アクチビンが培養中に外胚葉から中胚葉と内胚葉の両方を誘導することが示されてきている。特に興味深かったのは、それが使用した異なる濃度で異なる細胞型を誘導した点で、アクチビンはこの系においてモルホゲンとして働いたという観察結果であった。アクチビンがES/EB系において同様の発生能を示したかどうかを測定するために、以下のプロトコルを使用してアクチビンをEB培養物に加えた。ES細胞は、血清を含まないStem Pro 34培地中で2日間分化した。この段階で、発生中のEBを採取し、血清物(無血清)および100ng/mlの濃度でアクチビンを補ったIMDM中で再度培養した。EBは異なる日数で採取し、GFP+の発現および外胚葉、中胚葉および内胚葉発生を示す遺伝子の発現に関してアッセイした。図22A、B中に示すように、GFPにより測定して、この量のアクチビンはbrachuryを誘導した。GFP誘導の動態は血清中で分化したEBと比較して遅れたが、この濃度のアクチビンは、分化の第6日までに相当数のbrachury陽性細胞(60%)を誘導した。分子分析によって、アクチビン誘導型細胞はHNF3β、Mixl-1、Sox17、Hex-1、およびpdx-1を含めた内胚葉発生と関係がある広範囲の遺伝子を発現したことを示した(図23C)。この遺伝子は膵臓発生に必要不可欠であるので、pdx-1の誘導は興味深い。GATA-1などの造血発生と関係がある遺伝子、およびPAX6などの神経外胚葉分化を示す遺伝子は、アクチビンによって誘導されなかった。
【0087】
アクチビンがES分化モデルにおいてモルホゲンの性質を示したかどうかを測定するために、異なる濃度の因子をEB培養物に加えた。わずか1ng/mlのアクチビンが、培養の7日までにGFP+の発現(集団全体の10%)を誘導した(図23A)。GFP+細胞の頻度は3ng/mlで刺激した培養物中では40%まで増大し、30ng/mlで50%を超える安定レベルに達した。これらの集団の遺伝子発現分析によって、異なる濃度のアクチビンは異なる発生プログラムを誘導したことを示した。1または3ng/mlのアクチビンの存在下で分化したEBは、内胚葉発生を示す遺伝子の、存在した場合でも弱い発現を示した(図23A)。HNF3β、Sox17、Hex-1はいずれも、10、30、または100ng/mlのアクチビンで刺激した培養物において誘導された。Pdx-1の発現は最高量のアクチビンを必要とし、100ng/mlで刺激したEBにおいて最も誘導された。いずれの濃度のアクチビンにおいてもGATA1およびc-fmsは発現されなかった。PAX6発現のパターンは内胚葉遺伝子のそれとは逆であり、アクチビンの増大する濃度と共に下方制御された。
【0088】
アクチビン誘導型EBは、造血への拘束と関係がある遺伝子を発現せず、この発生プログラムは誘導されなかったことが示された。これらのEBの造血発生能をさらに評価するために、原種の発生能に関してこれらを分析した。分子分析から予想したように、これらのEBは測定可能な数の初期(Ep)または最終段階(mac/mix)造血細胞原種を含んでいなかった(図24A)。これらのアクチビン誘導型EBを2.5日間血清でさらに刺激すると、しかしながら、それらは幾らかの造血細胞原種を生成し、それらは中胚葉発生能を含むことが示された(図24B)。
【0089】
PCR分析によって、アクチビンはHNF3β、および内胚葉分化と関係あることが知られている他の遺伝子の発現を誘導したことを実証した。アクチビン誘導型EBにおける内胚葉原種の割合をさらに推定するために、100ng/mlのアクチビンで刺激した培養物由来の細胞を、HNF3βおよびHex1に対する抗体で染色した。非誘導型培養物由来のEBは対照として使用した(アクチビン-)。相当な割合のアクチビン誘導型集団(推定で全体の50〜60%)は、HNF3βとHex1の両方を発現した。非誘導型EBにおける細胞は、これらのタンパク質を発現しなかった。これらの発見は、これらのEB内の相当数の細胞が内胚葉系統であることを明らかに実証する。
【0090】
これらのアクチビン誘導型集団の発生能をさらに調べるために、GFP+細胞を3ngまたは100ngで刺激したEBから単離し、肝細胞誘導状態でさらに培養した(合計14日間)。図25中に示すように、100ng培養物由来の細胞のみがアルブミンを発現した細胞に分化し、肝臓の分化と一致した。一緒に考えると、これらの試験からの発見は、ES/EB系におけるモルホゲンとしてアクチビンが機能すること、および内胚葉誘導には高濃度が必要とされることを示す。
【0091】
血清誘導型brachury+細胞によって生成される奇形腫が、原始生殖細胞の存在に原因がある場合、生殖細胞のプログラムは、これらの条件下では誘導されない可能性があるのでアクチビン誘導型細胞が移植元の良い供給源となり得ることが考えられる。この仮説を試験するために、GFP+およびGFP-細胞を100ng/mlのアクチビンで誘導したEBから単離し、14日間培養して肝細胞様細胞の分化を促進した。この培養期間後、細胞を採取しレシピエント動物の腎被膜下に移植した。移植後3週間で、マウスを殺傷し腎臓を分析した。結果は図26A〜C中に示す。GFP-細胞を移植した全てのマウスが、3つ全ての生殖層由来の細胞からなる大きな多系統の奇形腫を発生した。対照的に、GFP+細胞を移植した動物中では奇形腫を検出しなかった。これらの細胞は、腸上皮、骨および骨格筋を含めた内胚葉および中胚葉由来組織からなる分化した細胞塊を生成する。幾つかの場合、GFP+細胞からの移植片中に皮膚も観察され、この系統がbry+細胞から発生する可能性もあることが示唆された。これらの発見は、移植後に奇形腫を形成せずに分化組織を生成するGFP+集団を作製することができることを示す。
(実施例13)
【0092】
bry+/c-kit-およびbry+/c-kit+細胞の発生能
前述の実施例は、中胚葉と内胚葉の両方の発生能を有するbry+細胞集団から肝細胞系統が発生することを明らかに示す。bry+画分の分析によって、これらの細胞の亜集団が受容体チロシンキナーゼc-kitを発現したこと(図27A)、およびこの集団がFlk-1を発現した細胞とは異なっていたことが明らかになった。c-kitの発現が内胚葉発生能を有する細胞の分離の有用なマーカーであるかどうか決定するために、第3日の血清刺激したEB由来のbry+/c-kit-(+/-)およびbry+/c-kit+(+/+)細胞を肝細胞発生能に関してアッセイした。図27B中に示すように、HNF3β、AFPおよびALB発現細胞は、いずれもbry+/c-kit+集団に由来した。この画分の内胚葉発生能を推定するために、選別した細胞をカバーガラスに平板培養し、HNF3βに対する抗体で染色した。80%を超えるbry+/c-kit+細胞がHNF3βタンパク質を発現し、一方10%未満のbry+/c-kit-が陽性であった。これらの発見は、内胚葉原種はbrachuryとc-kitの両方を発現すること、およびこの集団は内胚葉発生能を有する細胞が非常に豊富であることを示す。brachuryおよびc-kitの発現に基づく細胞の単離は、内胚葉原種を単離するための新規な戦略となる。
(実施例14)
【0093】
アクチビン誘導型細胞の発生能
図23中に表したPCR分析は、異なる濃度のアクチビンは異なる発生プログラムを誘導すること、および内胚葉発生能を有する細胞は最高レベルのこの因子によって誘導されることを示す。アクチビンの分化応答を定量化するために、異なる濃度のアクチビンで刺激した細胞をカバーガラスに接着させ、抗HNF3β抗体で染色した。100ng/mlのアクチビンで刺激したEB集団全体の50%より多くがHNF3βを発現し、一方3ng/mlで刺激した細胞のわずか10%が陽性であった。バックグラウンドレベルの染色のみを、非刺激集団において観察した。これらの発見は、高濃度のアクチビンは、EB集団全体の相当な割合が示す確かな内胚葉プログラムを刺激し得ることを実証する。
【0094】
アクチビン処理細胞の発生能の他の評価として、異なる濃度のこの因子の存在下で分化させた第6日EBを4日間血清物培地に移し、次いで肝細胞状態でさらに4日間再度平板培養した。培養の第14日に、各群由来の細胞を採取しPCRによる発現分析に供した。Myf5および骨格筋アクチンの発現を調べて、他の中胚葉由来系統を表す骨格筋の発生を評価した。サーファクタントプロテインC(SP-C)、肺特異的遺伝子は、AFPおよびALBに加えて内胚葉分化のマーカーとして含めた。図28中に示すように、Myf5および骨格筋アクチンはわずか1ng/mlのアクチビンで刺激した培養物中で発現し、この発現は広範囲の因子濃度で検出した。両遺伝子の発現は、しかしながら最高濃度(100ng/ml)のアクチビンで下方制御された。少量のアクチビンで刺激した培養物は、骨格筋の形態を有する細胞群を含んでいた。免疫染色によって、これらの細胞が骨格筋ミオシンとα-アクチニンの両方を発現したことを実証し、これらが骨格筋系統であることを示した。図28中に示すように、3および10ng/mlで刺激したEBは最も確かな骨格筋の発生を示したので、骨格筋の発達をもたらした再度平板培養したEBの割合の評価は遺伝子発現の分析と一致した。3つの内胚葉遺伝子の発現パターンは、骨格筋遺伝子に関して観察した発現パターンとは異なっていた。いずれも低いアクチビン濃度では発現されず、最高濃度の因子で刺激した培養物においていずれも容易に検出した。PAX6の発現は、未処理培養物および低濃度の因子で刺激した培養物に限られた。この分析からの発見によって、異なる濃度のアクチビンが異なる発生プログラムを誘導し、低濃度は中胚葉の運命に好ましく、高濃度は内胚葉の運命に好ましいことを実証した際の、本明細書中の前の実施例12からの発見が確認され広がる。さらに、これらの結果は、アクチビンによって誘導された内胚葉細胞は、肝細胞および肺の特徴を有する細胞に分化しそれらを生成することができることを示す。
【0095】
低濃度および高濃度の因子で刺激したEBから単離したbrachury陽性および陰性集団も、骨格筋および内胚葉遺伝子の発現に関して分析した。図29中に示すように、myf5および骨格筋アクチンの発現は、3ng/mlのアクチビンで刺激したEBから単離したbry+集団から生成した集団に限られた。同様に、内胚葉遺伝子は、100ng/mlの因子の存在下で生成したEBから単離したbrachury+由来細胞中で発現された。これらの発見は、中胚葉と内胚葉の両方がbrachury+細胞から発生することをさらに実証する。
(実施例15)
【0096】
HNF-CD4標的ES細胞の生成
EB分化中に内胚葉および中胚葉集団を追跡し単離するために、brachury遺伝子座に標的化したGFPを既に有するES細胞を、ヒトCD4(hCD4)タンパク質と共にHNF3β遺伝子座に標的化した。HNF3βは大部分の内胚葉細胞型において発現され、胚発生中の非常に初期に始まることが知られている。Kaestnerら(1994) Genomics 20: 177〜85。標的構築体は、アミノ酸番号424で切断され細胞内ドメインの大部分を欠くヒトCD4のcDNAを含む。シグナルを伝達することができず、hCD4を認識し内因性のマウスCD4と交差反応しないはずである、フローサイトメトリー用の容易に入手可能な抗体が存在するので、この細胞表面分子を選択した。図30A中に示すように、このベクターは、マウスHNF3β遺伝子座と相同性がある2つの領域、ウシ成長ホルモンポリ(A)配列、およびloxpに隣接するピューロマイシンカセットも含む。ジフテリアトキシンA(DTA)遺伝子を標的ベクターの5'末端に含めて、ランダムな組み込みを選択した。標的ベクターは以下のように構築した。
【0097】
相同性のある短いアームと長いアームを、公開済みのHNF3β標的ベクターから得た。Weinsteinら(1994) Cell78: 575〜588。同じ長さのアームを使用し、一方1.8kb断片の短いアームを使用してベクター中にhCD4ノックを作製した。短いアームは、HNF3βプロモーターの一部分、エクソン1、イントロン1、およびエクソン2の一部分、内因性ATG開始部位の直前に末端を含む。長いアームは、最終エクソンの末端の3kb下流で始まる7kbの配列からなる。標準的な分子生物学の技法を使用して、コザック配列次にウシ成長ホルモンポリ(A)配列およびピューロマイシンカセットを有するhCD4を、図30A中に示すように相同性のある2つのアーム間にクローニングした。
【0098】
実施例2中に記載したGFP-BryES細胞に、HNF3β標的ベクターをエレクトロポレーションにより導入した。次いでピューロマイシンを用いて細胞を選択し、クローンを採取し増大させた。ゲノムDNAを単離し、制限酵素SacIで切断し、図30B中に示すように内因性ATGの上流のプローブを用いてサザンブロット分析を実施した。野生型対立遺伝子は3.3kbのバンドを与え、一方標的対立遺伝子は2.8kbの大きさのバンドを与える。スクリーニングした48のクローンのうち、2つの標的クローンが生成し、4.2%の標的化効率に相当した。両方の標的クローンは、改変型Creリコンビナーゼ発現ベクターを用いて一過的にトランスフェクトしてpuro遺伝子を除去した。この標的クローンは、本明細書では以後CD4-HNF/GFP-T ES細胞と呼ぶ。
【0099】
HNF3βは、血清誘導分化中にEBにおいて一過的に発現される。CD4-HNF/GFP-T ES細胞中でのhCD4の発現が、EB発生中のHNF3β遺伝子の発現を正確に反映するかどうか測定するために、CD4対GFPの発現を評価した。
【0100】
5日間血清誘導EBの分化中のGFP対hCD4の典型的な発現パターンを図31A中に示す。ES細胞はhCD4発現が低いから陰性である。分化後、hCD4の発現は特に第3日のGFP+細胞において急激に上方制御される。発現は第3.5日までに急速に低下し、第5日までにはほぼ検出不能である。対照的に、GFPの発現は第4日まで持続し、第5日までに低下する。GFPおよびhCD4発現の平均蛍光強度は、図31B中に示す。hCD4のピークは第3日であり直ぐに消失し、一方GFPのピークは第3.5日〜第4日であることが分かる。HNF3βおよびbrachury発現に関する半定量RT PCRも、これらのサンプルにおいて実施した(図31C)。内因性遺伝子に関するPCRは、GFPおよびhCD4に関するフローサイトメトリーを使用して見られたものに従う。HNF3βとbrachuryはいずれも第3日までに発現され、一方HNF3βの発現は第3.5日までに低下し、第5日までには検出不能である。brachuryの発現は第4日まで高いレベルを保ち続ける。これらのデータによって、hCD4は内因性HNF3βの発現を反映していることが確認される。さらに、hCD4の発現はHNF3βのRNAレベルと共に低下することが分かり、hCD4タンパク質の半減期は過度に長いわけではなく、それを使用してHNF3βを発現する細胞を正確に選択することができることが示唆される。全てのGFP+細胞が最初はhCD4+であるが、次いでhCD4の発現が急速に消失することも分かる。このデータは内胚葉と中胚葉の両方に対する内中胚葉前駆体を示唆し、実施例9中のデータを支持する。この集団はc.elegansなどの低級生物において実証されてきているが、哺乳動物においては樹立されていない。前に使用された条件を、確かな中胚葉誘導のために選択した。内中胚葉が形成されるが、急速に中胚葉に転換すると考えられる。
(実施例16)
【0101】
原条形成のモデルとしてのEB分化
胚発生中、中胚葉系統と内胚葉系統の両方が、原条と呼ばれる構造から誘導される。Beddington and Robertson (1999) Cell 96: 195〜209。原条全体で発現されるbrachuryなどの特定の遺伝子の発現によって、原条は特徴付けられる。HNF3βも原条中で発現されるが、それは前部でのみ検出されている。hCD4によって実証されたHNF3βの発現は、Bry-GFP+集団中では異種であることが分かった。血清誘導は原条様細胞の群の発生を可能にし、GFP+HNF3βlo細胞は原条後部になり、一方GFP+HNF3βhi細胞は原条前部に対応すると考えられる。
【0102】
GFPおよびhCD4の発現を使用して、CD4-HNF/GFP-T ES細胞から後部および前部原条様細胞を分離することができるかどうか測定するために、3.25日間分化したEBを3つの集団:GFP+hCD4lo、GFP+hCD4med、およびGFP+hCD4hiに選別した(図32A)。原条中で非対称的に発現されることが認められている、多くの遺伝子が存在する。原条後部または原条前部のいずれかにおいて発現されることが知られている遺伝子に関して、選別集団においてRT PCRを実施した(図32B)。原条後部特異的遺伝子Fgf8、Evx1、HoxB1、およびTbx6はいずれも、hCD4hi細胞と比較してhCD4loおよびhCD4med集団中において高レベルで発現された。対照的に、原条前部の遺伝子Chordin、Cerl、およびGscはいずれも、hCD4hi細胞において高レベルで発現され、hCD4med細胞においては発現が低いから陰性であり、hCD4lo細胞においては全く発現されなかった。HNF3βの発現も調べて、hCD4選別によって異なるレベルのHNF3β遺伝子を発現する細胞を単離したことを確認した。HNF3βの最高レベルはhCD4hi集団中で見られ、予想通りhCD4medおよびhCD4lo集団中では量が減少した。様々なhCD4集団中の発現パターンが、原条中の遺伝子発現を正確に表すものであることを確認するために、7.25日齢マウス胚の原条を切開し、遺伝子発現に関して分析した。GFP-BryES細胞由来のマウス胚を使用した。前部、中部、および原条後部組織を次いで蛍光下の切開範囲で切開して、得られた組織が胚のGFP+(原条)部分由来であったことを確実にした。図32Cは、切開した原条組織から実施したRT PCRの結果を示す。これらのデータは、選別したhCD4集団が、調べた原条前部および原条後部遺伝子に関して同様の相対的遺伝子発現プロファイルを示すことを示す。これらのデータは、CD4-HNF/GFP-T ES細胞におけるhCD4およびGFPの発現を使用して、原条前部および原条後部を示す遺伝子発現パターンを有する細胞を単離することができることを示す。
(実施例17)
【0103】
hCD4発現細胞の異なる集団の機能的発生能
系統追跡実験によって、原条前部と原条後部は異なる系統に発生することが実証されている。原条後部は卵黄嚢および造血細胞に分化し、一方原条前部は内胚葉および心臓組織などの前部中胚葉を形成する。Robb and Tam (2004) Sem.Cell & Dev.Biol.15: 543〜554。EB由来の原条様細胞が内因性原条と同様に機能的に働くかどうかを測定するために、CD4-HNF/GFP-T ES細胞を3.25日間分化させ、GFP+hCD4loおよびGFP+hCD4hi集団に選別し、造血、心臓、および内胚葉発生能に関してアッセイした。全てのアッセイ用に、選別した集団を無血清状態で48時間再凝集させた。図33Aは、再凝集後にマトリゲル上に平板培養した選別細胞の心臓発生能を示す。接着後24〜48時間の拍動細胞の存在に関して個々のEBを記録した。拍動EBはこの時点ではhCD4hi集団からのみ見られた。図33Bは、選別した集団の造血発生能を実証する。細胞は再凝集後にトリプシン処理し、造血増殖因子を含むメチルセルロース中に平板培養した。造血コロニーの存在を5〜7日後に計数した。造血コロニーの大部分はhCD4lo集団中で見られた。図33Cは、再凝集後に無血清状態でマトリゲル上に平板培養した選別集団の、内胚葉特異的遺伝子に関するRT PCRを示す。hCD4hi細胞のみが、マトリゲル上で5日後に内胚葉遺伝子HNF4、Pdx1、AAT、ALB、およびHNF3βを発現した。これらのデータは、hCD4およびGFPの発現を使用して、異なる発生能を有する集団を分離することができることを実証する。これらの発生能は、それぞれ原条後部および原条前部と同等であるhCD4loおよびhCD4hi細胞と一致する。
(実施例18)
【0104】
アクチビンシグナル伝達とWntシグナル伝達の両方が、ES由来原条様細胞に必要とされる
マウスにおける試験によって、原腸形成および内胚葉/中胚葉誘導における、Wntシグナル伝達およびノーダル/アクチビンシグナル伝達の重要性が実証されてきている。Liuら(1999) Nat.Genet.22: 361〜4。本明細書における前の実施例12は、アクチビンは無血清培養においてES細胞から、中胚葉集団と内胚葉集団の両方を誘導することができることを実証する。アクチビンおよびWntシグナル伝達のES由来原条様細胞の分化に対する影響を測定するために、組換えWnt3aまたはアクチビンAのいずれかを用いて、無血清培養においてCD4-HNF/GFP-T細胞を分化させた。アクチビンとWntの両方が最初にGFP+CD4+細胞を誘導することができるが、Wntがより速い動態を示した(図34A)。アクチビン刺激は、第6日のGFP-Bry発現の消失後持続する高レベルのCD4発現をもたらす。対照的に、Wntで誘導したGFP+細胞は低レベルのCD4を有しており、第5日までに発現が急速に消失する。アクチビンはGscおよびCer1などの他の原条前部マーカーの確かな発現をさらに誘導するが、一方Wntは誘導しない(図34B)。しかしながらWntは原条後部マーカーHoxB1およびTbx6の発現を刺激し、一方アクチビンはTbx6のみを誘導する。これらのデータは、アクチビンはES原条前部細胞を誘導し、WntはES原条後部細胞を誘導することを示唆する。第6日に、アクチビンまたはWnt3aのいずれかで刺激したEBを、マトリゲル上無血清培地中に平板培養して、さらに発生させた。さらに6日後、これらの培養物を採取し、内胚葉遺伝子の発現をRT-PCRによって測定した(図34C)。アクチビンによる誘導によって、内胚葉遺伝子Alb、Aat、HNF4、およびPdx1を発現する細胞を発生させることができ、実施例12における発見を確認した。しかしながら、Wnt3aで誘導した培養物はこれらのマーカーのいずれも発現せず、アクチビンシグナル伝達が内胚葉発生に必要とされることが示唆された。
【0105】
Wntによる刺激がノーダルを誘導し、アクチビン処理がWnt3を誘導することを考慮すると(図34B)、アクチビン/ノーダルシグナル伝達またはWntシグナル伝達単独で原条形成を誘導することができるかどうか、あるいは両方の因子が共に必要とされるかどうかは分からない。CD4-HNF/GFP-T ES細胞は、これらの経路の特異的阻害剤と共にアクチビンまたはWntのいずれかで誘導した。DKK1を使用してWntシグナル伝達を阻害し、一方で小分子阻害剤SB-431542(SB)を使用してノーダル/TGFβシグナル伝達を阻害した。DKK1はWntの共受容体、LRP5およびLRP6と結合することによって機能し、これらの受容体はβカテニンシグナル伝達経路の活性化に必要不可欠である。SBは受容体ALK4、5、および7の特異的阻害剤であり、したがってTGFβ、アクチビン、およびノーダルシグナル伝達を阻害することができる。しかしながらSBは、BMPシグナル伝達を阻害しない。Inman et.al. (2002) Mol.Pharmacol.62: 65〜74。両方の阻害剤共に、いずれかの誘導物質を用いたGFP+CD4+細胞の発生を完全に阻害した(図34A)。原条マーカーGsc、Cer1、HoxB1、およびTbx6も不在であったか、あるいは大幅に減少した(図34B)。これは、ES由来原条様細胞を誘導するために、Wntおよびアクチビン/ノーダルシグナル伝達が共に必要とされることを示す。したがって、中胚葉および内胚葉生殖層およびそれらの誘導組織を誘導するためには、アクチビン/ノーダルとWntシグナルの両方が最初に必要とされる。
(実施例19)
【0106】
アクチビンシグナル伝達およびWnt阻害は、内胚葉分化を誘導する可能性がある
実施例12は、高レベルのアクチビンは、完全に無血清の状態で分化したEBからの内胚葉遺伝子の発現を誘導する可能性があることを実証した。in vivoでの中胚葉/内胚葉誘導と関係がある重要な分子であるノーダルは、アクチビンとして同じ受容体を介して伝わる。ノーダルおよびWntシグナル伝達の阻害剤は、原条前部中あるいはその周辺で優先的に発現されることも知られている。内胚葉が原条前部から形成されることを考慮すると、Wntシグナルの阻害と組み合わせたノーダル/アクチビンシグナル伝達は、内胚葉形成を誘導すると考えられる。この仮説を試験するために、CD4-HNF/GFP-T ES細胞を、血清中で3.25日間分化させ、GFP+hCD4med細胞を選別し、因子を含まず、DKK1(Wnt阻害剤)、アクチビン、あるいはアクチビンとDKK1を含む無血清培地中で再凝集させた。hCD4-HNF3βおよびGFP-Bryレベルに対するこれらの培養条件の影響は、フローサイトメトリーによって分析した(図35A)。無血清培地単独では、hCD4の発現は再凝集1日後に消失した。DKK1を加えることによるWntシグナル伝達の阻害によって、幾つかの細胞は低レベルのhCD4を保持することができた。アクチビンの添加は、新たに選別した細胞と同等なHNF3βレベルの保持を可能にした。最後に、アクチビンとDKK1は相乗的に働いて、選別後のレベルよりhCD4のレベルをほぼ1ログ増大させた。GFPレベルは試験した全ての状態で消失し、全ての培養物において分化が進行していたが、より多くのhCD4を発現する培養物が内胚葉系統に分化していたことが示唆された。この概念をさらに試験するために、前述の培養条件で2日間再凝集させた選別細胞を、因子を含まない無血清培地で洗浄し、マトリゲル上無血清培地中に平板培養した。5日後、培養物を採取し、遺伝子の発現をRT PCRによって測定した。図35Bは膵臓、肝臓、および腸に特異的な幾つかの内胚葉遺伝子の発現を示す。培地単独またはDKK1処理培養物のいずれにおいても、これらの遺伝子は検出されなかった。非常に低レベルの2つの遺伝子、Pdx1およびALBのみを、アクチビンのみで処理した培養物において検出した。アクチビンとDKK1の両方を用いた処理は、Pdx1、AAT、HNF4、ALB、Cdx2、およびIFABPの高発現をもたらした。これらのデータは、アクチビン受容体を介したシグナル伝達とDKK1処理によるWntシグナルの阻害の組合せは、培地単独で培養すると内胚葉を形成しないはずである細胞集団からの内胚葉形成を誘導することを実証する。これらの培養条件には、膵臓および肝臓などの内胚葉由来組織の大規模な生成を可能にする可能性がある。これらの実験は、定量的に内胚葉発生を評価するためのツールとしての、CD4-HNF/GFP-T ES細胞の有用性をさらに実証する。
(実施例20)
【0107】
内胚葉由来マウス胚幹細胞からの肝細胞の作製
本明細書における前の実施例15中に記載したGFP-Bry、hCD4-Foxa2ES細胞系を使用することによって、内胚葉細胞をフローサイトメトリーによって、ES細胞分化時のそれらのGFPおよびhCD4発現によって追跡した(用語HNF3βとFoxa2は同義的に使用される)。
【0108】
GFP-Bry、hCD4-Foxa2ES細胞は血清の不在下に保ち、フィーダー細胞はNeurobasal、DMEM/F12、N2サプリメント、B27およびレチノイン酸、10%BSA、MTG(1.5×10-4M)、グルタミン、LIF(1000U/ml)およびBMP-4(10ng/ml)からなる培地中に保った。これらの条件において、細胞は依然未分化状態である堅いコロニーを形成する。内胚葉のプログラムを誘導するために、ES細胞を採取し、アクチビン-A(50ng/ml)の存在下、無血清培地(IMDM、F12培地、B27およびレチノイン酸、N2サプリメント、BSA、MTG、グルタミン、アスコルビン酸)中、低密度(35,000個細胞/5ml)、超低接着性の皿中で培養して、胚様体(EB)形成を可能にした。ES細胞が分化するとき、GFP-Bry、hCD4-Foxa2および幹細胞リガンドc-Kitの受容体の発現パターンを定義した。大きな細胞集団GFP-Bry+、hCD4-Foxa2highおよびcKithigh(名称+++)が第4日に出現し、それらの相当集団GFP-Bry+、HCD4lowおよびc-Kit-(名称+L-)に対して、その内胚葉発生能およびしたがって肝細胞運命に関してアッセイした。この問題に取り組むため、胚中の内胚葉細胞を肝臓に特化させることが知られている2つのサイトカイン、BMP-4(30ng/ml)およびbFGF(10ng/ml)の存在下において、前に示した無血清培地中で、両集団を2日間再凝集させた。Zaret(2001) Curr Opin Genet Dev 11: 568〜574。この2つのサイトカインに加えて、アクチビン-A(50ng/ml)を加えて、中胚葉運命と対抗する細胞の内胚葉発生能を保った。肝細胞原種(肝芽細胞)の成熟を可能にするために、次いで凝集体を、肝芽細胞/肝細胞の成熟および増殖に適した培地中のゼラチンコーティング皿上に平板培養した。この培地は、VEGF(10ng/ml)、デキサメタソン、および肝芽細胞/肝細胞の増殖を促進することが知られているブロック培地中に存在する他のサイトカインを補った、前述の分化用基礎培地からなる。これらはヒト組換え体HGF(20ng/ml)、EGF(10ng/ml)、bFGF(10ng/ml)およびTGFα(20ng/ml)を含む。Blockら、(1996) J Cell Biol 132: 1133〜1149。6日後、+++凝集体は皿に接着した無数の「肝細胞コロニー」、および接着しなかった多数の浮遊する凝集体に分化した。全ての+L-凝集体が皿に接着し非常に少数の「肝細胞コロニー」を形成したが、培養物中に浮遊する凝集体は見られなかった。これらの結果は、第4日の+++集団は内胚葉細胞が非常に豊富であることを示唆し、BMP-4、bFGFとアクチビン-Aの組合せは、第4日+++細胞の肝細胞系統への特化を強く促進することを実証する。
【0109】
ゼラチンコーティング皿上での平板培養後1日で、+++凝集体の2/3が培養物中に依然として浮遊する状態であり、一方他の1/3は接着し、他の細胞によって時折囲まれた肝細胞コロニーを形成した。この初期の時点では、免疫組織化学法によりアッセイして、コロニー内の多くの細胞がアルブミンおよびAFPを発現した。周囲の細胞はいずれもCD31陽性細胞であり、それらが内皮細胞であることが示唆された。
【0110】
第12日以後、肝細胞コロニーはさらに大きく成長した。この後期の時点では、コロニー内の大部分の細胞はAFPおよびアルブミン陽性であり、コロニー周囲の大部分の細胞は予定内皮CD31陽性細胞である。hCD4に関する免疫組織化学法によって、肝細胞コロニー中に含まれていた全ての細胞がfoxa2を発現したこと、およびこの発現がこれらのコロニーに限られていたことを示した。これらの結果によって、内胚葉起源の肝細胞コロニーを確認した。
【0111】
同様に、第12日に浮遊する+++凝集体はさらに大きく成長し、AFPおよびアルブミンに関する免疫組織化学法用に処理した。大部分の凝集体は中空状態であり、凝集体を構成する細胞はAFPおよびアルブミンを発現し、一方IgG対照部分は陰性であった。さらに、凝集体内のAFPおよびアルブミンも染色され、この2つのタンパク質の分泌が示唆された。
【0112】
第12日の+++培養物をさらに特徴付けるために、フローサイトメトリー分析を接着した培養物と浮遊する凝集体の両方に実施して、内胚葉由来集団(hCD4-Foxa2+)、内皮細胞集団(CD31+またはPECAM-1)、肝芽細胞/肝細胞集団(hCD4++alb+、またはhCD4+/AFP+)を定量化した(図36A、36B)。接着した培養物中に含まれていた細胞の30%〜40%は内胚葉マーカーhCD4-Foxa2に陽性であり、一方50%はCD31+内皮細胞であった。浮遊する凝集体は、大きな(50%〜60%)hCD4-Foxa2+細胞、およびCD31+内皮細胞の小さな細胞集団(30%)からなっていた(図36A)。大部分のhCD4-Foxa2+細胞はさらに、細胞質間染色によりアッセイしてアルブミン+およびAFP+であった(図36B)。RT-PCRによる遺伝子発現の分析によって、第12日の+++培養物由来の接着した培養物(a)および浮遊する凝集体(f)はfoxa2、AFPおよびアルブミンを発現したことを確認した(図36C)。
【0113】
機能アッセイを実施して、肝細胞の同一性を証明した。Acid period Schiffアッセイは、成熟肝細胞に特徴的な細胞質中のグリコーゲン貯蔵の存在を示す。第12日の+++の接着した培養物は皿中で直接染色し、グリコーゲンに関する典型的な赤い細胞質染色を含むコロニー内に多数の細胞を示した。さらに、第12日+++の浮遊する凝集体に電子顕微鏡による分析を実施した。浮遊する凝集体を構成した大部分の細胞は、大きな核/細胞質比、多量のグリコーゲン貯蔵および毛細胆管の存在を含めた肝細胞の特徴を示した。
【0114】
全体としてこれらのデータによって、アクチビン-A誘導によるES細胞から内胚葉段階への発生進行、BMP-4、アクチビンAおよびbFGFの組合せによる肝臓特化ステップ、および最後に肝芽細胞/肝細胞集団の増殖/成熟をin vitroで模倣することにより、マウスES細胞から肝細胞が効率良く生成することを示す。
【図面の簡単な説明】
【0115】
【図1】緑色蛍光タンパク質(GFP)のbrachury遺伝子座への標的化のために使用するベクターのスキームおよび戦略を示す図である。
【図2A】発生段階の胚様体(EB)におけるGFPおよびbrachuryの発現を示す図である。逆転写酵素ポリメラーゼ連鎖反応(RT-PCR)によって測定したbrachuryの発現の動態を示す図である。
【図2B】発生段階の胚様体(EB)におけるGFPおよびbrachuryの発現を示す図である。蛍光活性化細胞選別(FACS)分析によって測定したGFP発現の動態を示す図である。前の図2A中の図および図2B中のヒストグラムにおける数字はEB分化の日数を表す。
【図3A】野生型およびGFP-BryES細胞の発生能を示す図である。第6日のEBの発生能を示すヒストグラムである(Mac/Ery:マクロファージおよび最終段階赤血球細胞のコロニー;Mac:純粋なマクロファージコロニー;Eryd:最終段階赤血球細胞のコロニー; Mix:多系統コロニー;Eryp:初期赤血球コロニー)。
【図3B】野生型およびGFP-BryES細胞の発生能を示す図である。EBの胚芽コロニー形成細胞(BL-CFC)の発生能を示すヒストグラムである。
【図3C】野生型およびGFP-BryES細胞の発生能を示す図である。野生型およびGFP-Bry細胞に関するEB発生中の遺伝子発現パターンを示す図である。レーンの上の数字はEB分化の日数を表す。
【図4A】GFPに基づいて単離したEB画分の遺伝子発現プロファイルを示す図である。第3.5日EBにおけるGFP発現のプロファイルを示す図である。1および2は、GFP-画分とGFP+画分を単離するために使用したゲートを表す。
【図4B】GFPに基づいて単離したEB画分の遺伝子発現プロファイルを示す図である。単離画分のRT-PCRによる発現分析を示す図である。
【図5A】GFPおよびFlk-1集団の単離および特徴付けを示す図である。第3.0日および第3.5日EBからGFP-/Flk-1-、GFP+/Flk-1-およびGFP+/Flk-1+画分を単離するために使用した、プロファイルおよびゲートを示す図である。ゲートの隣の数字は3つの異なる集団を表す。
【図5B】GFPおよびFlk-1集団の単離および特徴付けを示す図である。異なる画分の胚芽細胞コロニー(Blast)および二次EB(2°)発生能を示す図である。
【図5C】GFPおよびFlk-1集団の単離および特徴付けを示す図である。単離画分の発現分析を示す図である。上図に示す発現は、Bradyら(1990) Meth. In Mol. And Cell Bio.2: 17〜25によって記載されたポリA+全体増幅PCR法を使用して評価した。下図中のデータは、遺伝子特異的オリゴヌクレオチドを使用するRT-PCR分析によって得た。各列の上の数字は、図5A中に表した細胞集団を示す。
【図6】単離した第3日EB由来の画分におけるGFPおよびFlk-1の発現を示す図である。上列は、培養前(pre)の3つの画分の発現プロファイルを示す。下列は、培養20時間後(post)の同じ細胞集団のプロファイルを示す。各プロファイル下の数字は、各集団のBL-CFCおよび初期赤血球(Eryp-CFC)の発生能(平板培養した1×105個の細胞当たりの前駆体数)を示す。
【図7A】培養前後の単離した細胞集団のBL-CFC発生能およびFlk-1発現を示す図である。底部の数字は細胞集団を指す:1は予め選別した画分であり、3はGFP+/Flk-1-画分であり、4はGFP+/Flk-1+画分である。細胞は20時間培養し、次いで凝集体を解離させ、BL-CFCに関して分析した。データは第3、3.5および4.0日EBから単離した細胞に関して示す。
【図7B】培養前後の単離した細胞集団のBL-CFC発生能およびFlk-1発現を示す図である。上列は、培養前(pre)の第3、3.5および4.0日EBから単離したGFP+/Flk-1-細胞を表す。下列は、培養後(post)の同じ画分のFlk-1の発現パターンを示す。バーの上の数字はFlk-1+細胞の割合を表す。
【図8A】各ヒストグラムの上に示した条件下での、第3.0日EB由来細胞におけるbrachuryおよびFlk-1+の発生に対するBMP-4およびウシ胎児血清(FCS)の影響を示す図である。
【図8B】各ヒストグラムの上に示した条件下での、第3.0日EB由来細胞におけるbrachuryおよびFlk-1+の発生に対するBMP-4およびウシ胎児血清(FCS)の影響を示す図である。示した条件下で20時間培養したGFP+//Flk-1-細胞から作製した細胞集団における、brachuryおよびFlk-1の発現を示す図である。
【図9】EBにおける中胚葉形成および特化のスキームモデルである。
【図10】血清の存在および不在下でのEBにおける遺伝子の発現を示す図である。
【図11】異なる条件下で作製したEBにおけるbrachuryの発現を示すグラフである。
【図12】血清中で生成したEBは前駆体を含んでいる(まだら線)ことを示す図である。
【図13】血清の存在および不在下での神経分化を示すスキーム図である。
【図14】血清中2日間で始め次いで無血清状態に移したEBにおける、遺伝子の発現を示す図である。
【図15】bFGFの存在下で培養したEBにおける遺伝子発現を示す図である。
【図16】bFGFの存在下で培養したBry+およびBry-細胞における、遺伝子発現パターンを示す図である。
【図17】本発明の中胚葉および内胚葉細胞集団、および誘導体細胞型へのこれらの集団の分化の図である。
【図18】2.5、3.0、3.5および4.0日間分化したEBにおけるGFP(brachury)およびFlk-1の発現の動態を示す図である。矢印は、後の試験中の分析に使用した単離GFP+集団を示す。
【図19】EB分化の4段階から単離したGFP+集団の、血管芽細胞および心臓細胞発生能を示す図である。各段階からの細胞を細胞選別によって単離し、24時間再凝集させ、造血および心臓細胞発生能に関して分析した。再凝集培養物から回収した1×105個の細胞当たりの芽細胞コロニー(血管芽細胞)として、あるいは心筋分化を示す拍動細胞塊に分化した凝集体の割合として、データを示す。
【図20】4つのGFP+EB由来細胞集団における示した遺伝子の、RT-PCRによる発現分析の図である。数字はEB分化の日数を示す。
【図21】第3.0日および4.0日EBから単離したGFP+集団におけるHNF3βの発現を示す図である。Preは選別前の細胞を表し、-/-はGFPまたはFlk-1を発現しない細胞であり、かつ+/-はGFP+Flk-1-集団を表す。
【図22A】無血清培養物中のEBの発生に対するアクチビンの影響を示す図である。100ng/mlのアクチビンの存在下で分化した第6日EBにおけるGFPの発現を示すFACSプロファイル。
【図22B】無血清培養中のEBの発生に対するアクチビンの影響を示す図である。100ng/mlのアクチビンを含む培養物中のGFP誘導の動態。白丸はアクチビンの存在下で分化したEBであり、黒い四角形はアクチビンの不在下で分化したEBである。
【図22C】無血清培養中のEBの発生に対するアクチビンの影響を示す図である。アクチビンの存在下(+アクチビン)または不在下(-アクチビン)で増殖させた第6日EBにおける示した遺伝子の、RT-PCRによる発現分析。数字はEB分化の日数を示す。
【図23A】EBの発生能に対する異なる濃度のアクチビンの影響を示す図である。異なる濃度のアクチビンを用いて誘導した第7日EBにおけるGFPの発現。
【図23B】EBの発生能に対する異なる濃度のアクチビンの影響を示す図である。異なる濃度のアクチビンを用いて誘導した第7日EBのRT-PCRによる発現分析。
【図24A】異なる濃度のアクチビンの存在下で分化したEBの、造血細胞の原種の含有量を示す図である。第7日EBの原種の発生能。Epは初期赤血球の原種であり、mac/mixは最終段階造血細胞の原種を表す。
【図24B】異なる濃度のアクチビンの存在下で分化したEBの、造血細胞の原種の含有量を示す図である。血清に2.5日曝した後の、第7日アクチビン誘導型EBの原種の発生能。
【図25】3ng/mlまたは100ng/mlのいずれかのアクチビンを用いて誘導したGFP+細胞由来のアルブミン発現細胞の発生を示す図である。GFP+およびGFP-細胞を分化の第6日で単離し、前に記載した条件でさらに8日間培養して肝細胞の分化を助長した。
【図26A】bry+細胞集団の3週齢の腎臓移植片を示す図である。
【図26B】bry-細胞集団の3週齢の腎臓移植片を示す図である。
【図26C】bry+およびbry-集団の移植片の切片を示す図である。
【図27A】第3日に血清刺激したEBから単離したbry+/c-kit+(+/+)およびbry+/c-kit-(+/-)画分を示す、FACSプロファイルの図である。数字はそれぞれの画分中の細胞の割合を表す。
【図27B】それぞれの画分の発現の分析を示す図である。第3日は選別の直後に分析した細胞を表す。第15日は肝細胞状態で15日間培養した細胞集団を表す。
【図28】異なる濃度のアクチビンを用いて誘導したEB由来の細胞集団の発現の分析を示す図である。図面の上部の数字は、アクチビン濃度を示す。図面の底部の数字は、骨格筋が発達したEBの割合の推定値を表す。
【図29】アクチビン誘導集団から単離したbrachury画分の発現の分析を示す図である。図面の上部の数字は、アクチビン濃度を示す。
【図30A】ヒトCD4(hCD4)のHNF3β遺伝子座への標的化のために使用する方法を示す図である。CD4の標的化のために使用するベクターのスキームおよび戦略を示す図である。
【図30B】ヒトCD4(hCD4)のHNF3β遺伝子座への標的化のために使用する方法を示す図である。標的ベクターをエレクトロポレーションにより導入したES細胞クローンのサザンブロットを示す図である。
【図31A】血清で誘導した発生中のEBにおけるGFP、hCD4、brachury、およびHNF3βの発現を示す図である。フローサイトメトリー分析によって測定したGFPのhCD4に対する発現のドットプロットを示す図である。
【図31B】血清で誘導した発生中のEBにおけるGFP、hCD4、brachury、およびHNF3βの発現を示す図である。図31A中のプロットのGFP発現とhCD4発現の両方の平均蛍光強度(MFI)を示す図である。MFIは、各マーカーに関して経時変化中に検出した最大率として示す。
【図31C】血清で誘導した発生中のEBにおけるGFP、hCD4、brachury、およびHNF3βの発現を示す図である。図31A中のフローサイトメトリーに使用したのと同じサンプル中の、RT PCRによるbrachuryおよびHNF3βの発現を示す図である。
【図32A】異なるGFP hCD4発現集団およびマウス胚の原条から切開した組織における、遺伝子発現を示す図である。EBは血清中で3.25日間分化させ、GFPおよびhCD4の発現によって選別した。これらの集団に関する細胞選別の前後のGFP対hCD4発現のドットプロットを示す図である。GFP+hCD4lo、GFP+hCD4med、およびGFP+hCD4hi集団を示した純度に選別した。
【図32B】異なるGFP hCD4発現集団およびマウス胚の原条から切開した組織における、遺伝子発現を示す図である。EBは血清中で3.25日間分化させ、GFPおよびhCD4の発現によって選別した。図32Aにおいて選別したサンプル中のRT PCRによる遺伝子の発現を示す図である。
【図32C】異なるGFP hCD4発現集団およびマウス胚の原条から切開した組織における、遺伝子発現を示す図である。EBは血清中で3.25日間分化させ、GFPおよびhCD4の発現によって選別した。第7.25日胚から切開した原条から切開した組織中の、RT PCRによる遺伝子の発現を示す図である。使用した略称:Pre、選別前、;ant、原条前部組織;mid、原条中部組織、pos、原条後部組織。
【図33A】異なるGFP hCD4発現集団の発生能を示す図である。図32中と同様にEBを分化させ細胞選別した。2集団、GFP+hCD4loおよびGFP+hCD4hi細胞のみを選別した。選別後、2日間血清物を含む培地中で細胞を再凝集させた。さらに24〜48時間マトリゲル上に平板培養した後の、選別した集団の心臓発生能を示す図である。拍動細胞を含むEBの割合を計算した。
【図33B】異なるGFP hCD4発現集団の発生能を示す図である。図32中と同様にEBを分化させ細胞選別した。2集団、GFP+hCD4loおよびGFP+hCD4hi細胞のみを選別した。選別後、2日間血清物を含む培地中で細胞を再凝集させた。造血サイトカインを含むメチルセルロース中に平板培養することによって測定した、選別した集団の造血発生能を示す図である。
【図33C】異なるGFP hCD4発現集団の発生能を示す図である。図32中と同様にEBを分化させ細胞選別した。2集団、GFP+hCD4loおよびGFP+hCD4hi細胞のみを選別した。選別後、2日間血清物を含む培地中で細胞を再凝集させた。無血清培地中でさらに5日間マトリゲル上に平板培養した後の、選別した集団のRT PCRによる内胚葉遺伝子の発現を示す図である。
【図34A】アクチビンA Wnt3aおよび/またはこれらの経路の阻害剤を含む無血清状態中で分化させたES細胞を示す図である。ES細胞は無血清培地中で2日間培養し、次いでトリプシン処理して単細胞集団を得た。次いでこれらの細胞を示した条件で再凝集させた。第6日で、EBをマトリゲルコーティング皿上、無血清培地に平板培養した。培地単独、アクチビンA 25ng/ml、Wnt3a 100ng/ml、あるいはこれらの刺激およびDKK1 100ng/mlまたはSB-431542 6uMのいずれかで再凝集させたEBを用いて第3日〜第6日のGFP対hCD4発現のドットプロットを示す図である。
【図34B】アクチビンA Wnt3aおよび/またはこれらの経路の阻害剤を含む無血清状態中で分化させたES細胞を示す図である。ES細胞は無血清培地中で2日間培養し、次いでトリプシン処理して単細胞集団を得た。次いでこれらの細胞を示した条件で再凝集させた。第6日で、EBをマトリゲルコーティング皿上、無血清培地に平板培養した。図34A中に示した培養物から得たRNAからの、RT PCRによる遺伝子発現データを示す図である。
【図34C】アクチビンA Wnt3aおよび/またはこれらの経路の阻害剤を含む無血清状態中で分化させたES細胞を示す図である。ES細胞は無血清培地中で2日間培養し、次いでトリプシン処理して単細胞集団を得た。次いでこれらの細胞を示した条件で再凝集させた。第6日で、EBをマトリゲルコーティング皿上、無血清培地に平板培養した。前に示したように培地単独、アクチビンまたはWnt3aで最初に刺激しマトリゲル上でさらに6日間平板培養したEBから得たRNAからの、RT PCRによる遺伝子発現データを示す図である。
【図35A】EBの分化に対するDKK1およびアクチビンの影響を示す図である。EBは図32中のGFP+hCD4medと同様に分化させ選別し、血清物のみを含む培地、あるいはヒトDKK1 100ng/ml、アクチビン100ng/ml、または両方の因子を一緒に加えた培地中で再凝集させた。再凝集1日後の選別した細胞のhCD4およびGFPの発現に関するヒストグラムを示す図である。
【図35B】EBの分化に対するDKK1およびアクチビンの影響を示す図である。EBは図32中のGFP+hCD4medと同様に分化させ選別し、血清物のみを含む培地、あるいはヒトDKK1 100ng/ml、アクチビン100ng/ml、または両方の因子を一緒に加えた培地中で再凝集させた。RT PCRによる内胚葉遺伝子の発現を示す図である。選別した集団は前に示したように2日間処理し、次いでマトリゲル上、因子を加えていない無血清培地中に平板培養した。遺伝子の発現は、さらに5日間平板培養後に分析した。
【図36A】フローサイトメトリー分析を示す図である。
【図36B】フローサイトメトリー分析を示す図である。
【図36C】肝細胞コロニーの遺伝子発現の分析を示す図である。
【技術分野】
【0001】
連邦政府により支援された研究に関する記載
本発明は、国立衛生研究所によって与えられたGrant Nos.2RO1HL48834-09および2RO1HL65169-02で政府の支援によってなされた。米国政府は、本発明において確かな権利を有することができる。
【背景技術】
【0002】
胚発生中、身体の組織は3つの主な細胞集団:外胚葉、中胚葉および最終段階内胚葉(definitive endoderm)から形成される。一次胚葉としても知られるこれらの細胞集団は、原腸形成として知られているプロセスによって形成される。原腸形成の後、それぞれの一次胚葉は、特異的な一組の細胞集団と組織を生成する。中胚葉は血液細胞、内皮細胞、心臓および骨格筋、および脂肪細胞を生じる。最終段階内胚葉は、肝臓、膵臓および肺を生成する。外胚葉は、神経系、皮膚および副腎組織を生じる。
【0003】
これらの胚細胞層からの組織発生のプロセスは多数の分化段階を含み、複雑な分子の変化を表す。中胚葉およびその誘導体に関しては、3つの異なる段階が同定されてきている。第一段階は、エピブラストとして知られる構造内での細胞からの中胚葉の誘導である。発生期中胚葉としても知られる新たに形成される中胚葉は、初期胚における将来の組織発生の部位であろう異なる位置に移動する。パターン形成として知られるこのプロセスは、特定組織への分化の初期段階を表す可能性がある幾つかの分子の変化を伴う。特化として知られる最終段階は、パターン形成した中胚葉亜集団からの異なる組織の生成を含む。近年の研究は、中胚葉は連続的に誘導され明確な発生能を有する亜集団であることを示唆する証拠を与えている。最初に形成される中胚葉は胚外領域に移動し、造血および内皮細胞を生じ、一方次の集団は発生中の胚では前に移動し、心臓および頭蓋間充組織となる。これらの系統の関係は組織学的分析によって初期に定義されており、細胞を追跡する研究によって大部分は確認されている。この発生運命の分離は発生生物学の分野では充分容認されているが、今日まで、これらの系統に拘束される前に、中胚葉および内胚葉を単離する利用可能な方法は存在していない。
【特許文献1】米国特許第5,843,780号
【特許文献2】米国特許第6,159,462号
【特許文献3】米国特許第6,844,422号
【特許文献4】米国特許第6,110,739号
【非特許文献1】Papaioannouら(1998) Bioessays 20: 9〜19
【非特許文献2】Wilsonら(1995) Development 121: 877〜86
【非特許文献3】Wilkinsonら(1990) Nature 343: 657〜9
【非特許文献4】Herrmannら(1991) Development 113: 913〜7
【非特許文献5】Smith (1997) Current Opinion in Genetics & Development 7: 474〜480
【非特許文献6】Evansら(1981) Nature 292: 154〜156
【非特許文献7】Thomsonら(1995) Proc.Nat'l.Acad.Sci.USA 92;7844
【非特許文献8】Reubinoffら(2000) Nature Biotech.18: 399
【非特許文献9】Glinkaら(1998) Nature 391: 357〜362
【非特許文献10】Odoricoら、(2001) Stem Cells 19: 193〜204
【非特許文献11】Grompeら(1993) Genes & Dev.7: 2298
【非特許文献12】Oversturfら(1996) Nature Genet.12: 266〜273
【非特許文献13】Kennedyら(1997) Nature 386: 488〜93
【非特許文献14】Bradyら(1990) Meth.in Mol.and Cell Bio.2: 17〜25
【非特許文献15】Smith(2001) Annu.Rev.Cell Dev. Biol.17:435〜62
【非特許文献16】Hooperら(1987) Nature 326: 292
【非特許文献17】Rogersら(1991) Development 113: 815〜24
【非特許文献18】Shalabyら(1995) Nature 376: 62〜6
【非特許文献19】Palisら(1999) Development 126: 5073〜84
【非特許文献20】Faloonら(2000) Development 127: 1931〜41
【非特許文献21】Shivdasaniら(1995) Nature 373: 432〜4
【非特許文献22】Wangら(1996) Proc.Natl.Acad. Sci.93:3444〜9
【非特許文献23】Kaestnerら(1994) Genomics 20: 177〜85
【非特許文献24】Weinsteinら(1994) Cell 78: 575〜588
【非特許文献25】Beddington and Robertson (1999) Cell 96: 195〜209
【非特許文献26】Robb and Tam (2004) Sem.Cell & Dev.Biol.15: 543〜554
【非特許文献27】Liuら(1999) Nat.Genet.22: 361〜4
【非特許文献28】Inman et.al.(2002) Mol.Pharmacol.62: 65〜74
【非特許文献29】Zaret(2001) Curr Opin Genet Dev 11: 568〜574
【非特許文献30】Blockら、(1996) J Cell Biol 132: 1133〜1149
【発明の開示】
【課題を解決するための手段】
【0004】
本発明は、中胚葉および最終段階内胚葉細胞集団を単離するための方法を提供する。これらの細胞集団は、細胞の増殖および分化に影響を与える作用物質を同定し、組織発生と関係がある遺伝子を同定し、かつ細胞補充療法用の分化細胞および組織を作製するのに有用である。
【0005】
本発明は、内中胚葉(mesendoderm)および中胚葉細胞が豊富な細胞集団を提供する。内中胚葉細胞は、brachuryを発現し(brach+)、分化誘導条件の存在下で、中胚葉、ならびに、心臓および骨格筋、血管平滑筋、内皮および造血細胞を含めた中胚葉誘導体を生成することができ、内胚葉、ならびに、肝臓細胞および膵臓細胞を含めた内胚葉誘導体も生成することができる細胞として、本明細書では定義する。中胚葉細胞は、brach+であり分化誘導条件の存在下で心臓および骨格筋、血管平滑筋、内皮および造血細胞を生成することができ、内胚葉および内胚葉誘導体を生成することができない細胞として、本明細書では定義する。
【0006】
本発明はさらに、内胚葉細胞が豊富な細胞集団を提供する。内胚葉細胞は、brachuryを発現せず(brach-)、分化誘導条件の存在下で、肺細胞、肝臓細胞および膵臓細胞を生成することができる細胞として、本明細書では定義する。
【0007】
本発明はさらに、内中胚葉および中胚葉細胞が豊富な細胞集団、および内胚葉細胞が豊富な細胞集団を単離する方法を提供する。
【0008】
他の実施形態では本発明は、肝細胞が豊富な細胞集団、およびこのような集団を作製する方法を提供する。
【0009】
他の実施形態では本発明は、肝細胞補充療法の方法を提供する。
【0010】
他の実施形態では本発明は、本発明の細胞集団の増殖、分化または生存に影響を与える作用物質を同定する方法を提供する。特定系統および組織の細胞分化および発生と関係がある遺伝子を同定する方法も提供する。
【0011】
brach+細胞を特異的に認識する抗体も提供する。これらの抗体は、例えば内中胚葉および中胚葉細胞集団を単離するのに有用である。
【0012】
他の実施形態では本発明は、in vitroで細胞を生成するための方法を提供する。このような細胞は、例えば細胞補充療法に有用である。
【0013】
本発明は、1つのbrachury対立遺伝子が不活性状態でありbrachury遺伝子座が転写される細胞中で選択可能なマーカーが発現されるように、選択可能なマーカーをコードするDNAがbrachury遺伝子座に存在するゲノムを有する、トランスジェニック非ヒト哺乳動物も提供する。
【発明を実施するための最良の形態】
【0014】
胚形成中、中胚葉の形成は、体の形成のプランの確立および血液、内皮、心臓および骨格筋などの多臓器系の発生における重要なステップである。中胚葉形成を制御する分子機構は、しかしながらほとんど定義されていない。培養中の胚幹(ES)細胞の分化に基づくモデル系を使用して、造血、内皮、心臓および骨格筋、および脂肪細胞系統を含めた中胚葉由来集団が研究されてきている。in vitroモデルは中胚葉の誘導および特化を支持するが、これらの分化事象はES細胞から生じる胚様体(EB)として知られる複雑なコロニーにおいて起こる。中胚葉形成および組織発生をさらによく理解するために、それらが形成されるときにEBから中胚葉細胞集団を単離することが有利であるはずである。しかしながら、発生期中胚葉細胞集団に特異的な抗体は明確に定義されていないので、抗体を使用する細胞選別によって、これらの集団を単離することはできていない。
【0015】
(Tとしても知られる)brachuryはT-ボックス遺伝子として知られる転写因子のファミリーの基本メンバーであり、マウスにおいて元来存在する変異として最初に同定された。Papaioannouら(1998) Bioessays 20: 9〜19。ヘテロ接合性のマウスは生存可能であるが、野生型動物より短い尾を有する。ホモ接合性のマウスは、day 10 p.c.で死に至り、脊索を欠いており、後部中胚葉組織の発生において欠陥を示す。キメラ動物の分析によって、brachuryは中胚葉細胞の移動性に影響を与えることが示されている。Wilsonら(1995) Development 121: 877〜86。発現の分析によって、brachuryに関する特有かつ興味深いパターンが明らかになった。それは原条を通じて進入する全細胞において、および発生期および初期移動性中胚葉において一過的に発現される。Wilkinsonら(1990) Nature 343: 657〜9; Herrmannら(1991) Development 113: 913〜7。発現は沿軸、側板、および胚外中胚葉において直ちに下方制御され、原条の退行の後は終末芽および脊索に限られる。このパターンを考慮すると、brachuryは、初期中胚葉の最適なマーカーの1つであると考えられ、この系統の発生を追跡するのに使用されている。brachuryは分析した全ての種において同定されており、中胚葉発生におけるその役割は系統発生中保たれることが示唆される。Papaioannouら(1998)。
【0016】
本発明によれば、選択可能なマーカー遺伝子は組換えによってbrachury遺伝子座に標的化されている。ES細胞分化の開始後、brachuryの発現を表すパターンで選択可能なマーカーが発現されることが発見されてきている。選択可能なマーカーは、EBからのbrachury陽性(Brach+)細胞の選別、およびしたがって内中胚葉および中胚葉細胞が豊富な細胞集団の単離および特徴付けを可能にしている。
【0017】
本発明によって例示される選択可能なマーカーは、強化型緑色蛍光タンパク質(EGFPまたはGFP)である。細胞選別を容易にするはずである他の選択可能なマーカーは当業者に知られており、本発明中で使用することができる。GFPをコードするcDNAは当技術分野で知られており(かつ例えばClontech、Palo Alto、CAからプラスミドpEGFP.C1として市販されており)、当技術分野で知られている方法によって標的ベクター(GFP-Bry)を構築することにより、brachury遺伝子座に標的化することができる。ベクターを設計して、brachury遺伝子の第一エクソンの約3分の2とGFP発現カセットを交換することが好ましい。
【0018】
ヒトおよびマウスを含めた多種由来のbrachury遺伝子は当技術分野で知られており、例えばSmith (1997) Current Opinion in Genetics & Development 7: 474〜480によって総説されている。GFP発現カセットは、GFP cDNA、および、下流のbrachuryエクソンの翻訳を妨げるための、1つまたは複数の翻訳停止コドンを含むことが好ましい。brachury遺伝子の下流領域の転写を妨げるために、カセットはSV40ポリアデニル化シグナル配列をコードするエクソンをさらに含むことができる。
【0019】
当技術分野で知られている方法によってES細胞中にベクターを導入して、相同的組換えによってGFP-Bry構築体を組み込む。当技術分野で知られており例えばEvansら(1981) Nature 292: 154〜156: Thomsonら(1995) Proc.Nat'l.Acad.Sci.USA 92;7844;米国特許第5,843,780号;およびReubinoffら(2000) Nature Biotech.18: 399によって開示された方法によって、胚盤胞からES細胞を単離することができる。好ましい実施形態では、ES細胞はマウスまたはヒトES細胞である。標的化成功後、brachury開始コドンはGFPの開始コドンになり、標的brachury対立遺伝子の破壊をもたらす。生成する細胞をGFP-BryES細胞と呼ぶ。GFP-BryES細胞は本明細書では、1つのbrachury対立遺伝子が不活性状態でありGFPがbrachury制御要素の制御下で発現されるES細胞として定義する。
【0020】
1つのbrachury対立遺伝子が不活性状態であるGFP-BryES細胞は生命力があり、正常に発生および分化することが、本発明によって発見されている。さらに、GFP発現は内因性のbrachury発現を反映することが発見されている。したがって、GFPを発現する細胞を選択することによってbrach+細胞を単離することができる。GFPを発現する細胞はフローサイトメトリーによって、例えば蛍光活性化細胞選別(FACS)によって都合良く単離することができる。蛍光性に基づいて細胞を選別するための方法は、当業者によく知られている。
【0021】
本発明は、組換えによってHNF3β遺伝子座に標的化されている選択可能なマーカー遺伝子を有する、GFP-BryES細胞も提供する。HNF3βは大部分の内胚葉細胞型において発現されることが知られている。HNF3βはFoxa2としても知られ、かつこれらの用語は本明細書では交換して使用可能である。このマーカーはHNF3β+細胞の選別を可能にする。本発明によって例示されるマーカーは、細胞内ドメインの大部分が欠けている切断型ヒトCD4である。この細胞表面分子は、シグナルを伝達することができず、抗体がフローサイトメトリーに利用可能であるので好ましい。細胞選別を容易にするはずである他のマーカーは当業者に知られており、本発明中で使用することができる。ヒトCD4をコードするcDNAは当技術分野で知られており、当技術分野で知られている方法によってHNF3β遺伝子座に標的化することができる。HNF3βと相同的な2アーム間にクローニングした切断型ヒトCD4をコードするcDNAを有する標的ベクターを、GFP-BryES細胞中にエレクトロポレーションにより導入して、CD4-HNF/GFP-BryES細胞を得ることができる。これらの細胞中でのヒトCD4の発現はHNF3βの内因性の発現を反映する。
【0022】
本明細書で前に定義した内中胚葉および中胚葉細胞が豊富な細胞集団は、GFP+細胞を得るのに充分な時間、例えばマウス細胞に関して約1〜約4日間、血清の存在下でGFP-BryES細胞を培養し、例えばフローサイトメトリーによってGFP+細胞を選別および単離することによって得ることができる。単離される細胞集団は、少なくとも約50%、および好ましくは少なくとも約75%、およびより好ましくは少なくとも約90%、および最も好ましくは少なくとも約95%、または少なくとも約99%の内中胚葉および中胚葉細胞を含む。内中胚葉および中胚葉の相対量は血清中での培養時間の長さを調節することによって変えることができ、短い培養時間は造血および内皮系統にパターン形成される内中胚葉および中胚葉の存在に好ましく、長い培養時間は心臓および骨格筋系統にパターン形成される中胚葉の存在に好ましい。例えば、中胚葉が豊富な細胞集団は、約2.5〜4.5日間血清中で培養し、次にGFP+細胞を選別および単離することによって得ることができる。血清の存在下での培養は、動物血清、例えばウシ胎児血清(FCS)を補った培地中での培養として本明細書では定義する。好ましい実施形態では、約5%〜約25%の血清を培地に補う。最適濃度は血清バッチ依存性である可能性があり、当業者によって決定することができる。
【0023】
内中胚葉および中胚葉細胞が豊富な細胞集団は、ヒトおよびマウス細胞に関するin vitroでの分化の時間の違いを考慮に入れて血清中での培養時間の長さを延長した類似の方法によって、ヒトES細胞から作製したGFP-BryES細胞から得ることができる。したがって、ヒトES細胞から作製したGFP-BryES細胞は、GFP+細胞を選別および単離する前に、GFP+細胞を得るのに充分な時間、例えば約2〜約18日間血清中で培養する。
【0024】
マウスとヒト細胞集団の両方に関して、例えばチロシンキナーゼ受容体、ヒトKDRまたはマウスFlk-1の存在をアッセイすることによって、単離した細胞が中胚葉より先、例えば血管芽細胞に分化しているかどうか、容易に決定することができる。KDRおよびFlk-1は内中胚葉および発生期中胚葉中では発現されないが、これらの細胞は血管芽細胞/前赤血球集団に分化すると、KDRまたはFlk-1発現は検出可能である。KDR+およびflk-1+細胞は、KDRまたはFlk-1に対する抗体を使用してフローサイトメトリーによって同定することができる。このような抗体は当技術分野で知られており、抗体生成の標準的な方法を使用して作製することもできる。内中胚葉および中胚葉が豊富な細胞集団は、細胞選別によりKDR+またはFlk-1+細胞を除去することによって、さらに豊富な状態にすることができる。
【0025】
図17中に示すように内中胚葉は、内胚葉と中胚葉の両方およびその対応する系統を生じる、以前は同定されていなかった細胞集団であることが、本発明によって発見されている。in vitro培養において血清の存在または不在を使用して、どの系統が内中胚葉から生じるかを決定することができることが、さらに発見されている。特に、内胚葉細胞が豊富な細胞集団は、約2〜4日間血清の存在下でマウスES細胞から作製したGFP-BryES細胞を培養し、例えばフローサイトメトリーによってGFP+細胞を選別および単離し、次に約1〜約10日間血清の不在下でGFPを培養することによって得ることができる。本明細書で前に定義したように、単離される細胞集団は、少なくとも50%、および好ましくは少なくとも約75%、およびより好ましくは少なくとも約90%、および最も好ましくは少なくとも約95%、または少なくとも約99%の内胚葉細胞を含む。
【0026】
内胚葉細胞が豊富な細胞集団は、約2〜10日間血清の存在下でGFP-BryES細胞を培養し、および次いでGFP+細胞を選別および単離し、次に約1〜約15日間血清の不在下でGFP+細胞を培養することによって、ヒトES細胞から作製したGFP-BryES細胞から得ることができる。
【0027】
内胚葉細胞が豊富な集団は、前に記載したようにKDR+またはFlk-1+細胞を同定および選別することによって、さらに豊富な状態にすることができる。
【0028】
内胚葉が豊富な細胞集団は、約2〜約10日間血清の不在下および増殖因子アクチビンまたはノーダルの存在下でGFP-Bry胚幹細胞を培養し、およびbrachuryを発現する細胞を単離することによって得ることができることが、本発明によってさらに発見されている。アクチビンまたはノーダルの量は、胚幹細胞から内胚葉への分化を誘導するのに充分な量である。このような分化は例えばHNF3β、Mixl-1、Sox17、Hex-1またはpdx-1を含めた内胚葉の発生と関係がある遺伝子の発現をアッセイすることによって測定することができる。好ましい実施形態では、アクチビンの濃度は少なくとも約30ng/mlである。他の好ましい実施形態では、アクチビンの濃度は約100ng/mlである。他の実施形態では、約2〜約10日間血清の不在下およびアクチビンまたはノーダルおよびWntシグナル伝達の阻害剤の存在下で、GFP-Bry胚幹細胞を培養し、brachuryを発現する細胞を単離する。好ましい実施形態では、阻害剤はDKK-1である。他の好ましい実施形態では、DKK-1の濃度は少なくとも約30ng/ml、または約100ng/mlである。
【0029】
中胚葉が豊富な細胞集団は、約2〜約10日間血清の不在下およびアクチビンまたはノーダルの存在下でGFP-Bry胚幹細胞を培養し、およびbrachuryを発現する細胞を単離することによって得ることができる。アクチビンまたはノーダルの量は、胚幹細胞から中胚葉への分化を誘導するのに充分であるが、内胚葉への分化を誘導するには不充分な量である。中胚葉への分化は、例えばGATA-1を含めた中胚葉の発生と関係がある遺伝子の発現、および内胚葉の発生と関係がある遺伝子の発現の不在をアッセイすることによって測定することができる。好ましい実施形態では、アクチビンの濃度は30ng/ml未満である。他の好ましい実施形態では、アクチビンの濃度は約3ng/mlである。
【0030】
内胚葉が豊富な細胞集団は、約1〜約6日間血清の不在下およびアクチビンまたは関連タンパク質ノーダルおよびWnt分子の存在下で胚幹細胞を培養し、brach+/HNF3β3+細胞を単離し、および少なくとも約1日間血清の不在下およびWntシグナル伝達の阻害剤の存在下で単離細胞を培養することによって得ることができることが、本発明によってさらに発見されている。brach+/HNF3β3+細胞の単離は、本明細書で前に記載したC4-HNF/GFP-BryES細胞を使用すること、および例えば細胞選別によってCD4およびGFPを選択することによって容易にすることができる。
【0031】
Wntは、当技術分野でよく知られているポリペプチドのファミリーを指す。例えば米国特許第6,159,462号を参照。Wnt増殖因子のファミリーは、マウスにおいて少なくとも10個の遺伝子、およびヒトにおいて少なくとも7個の遺伝子によってコードされるタンパク質を含む。本発明の好ましい実施形態では、Wnt分子は組換えWnt3dである。
【0032】
Wntシグナル伝達の阻害剤も当技術分野でよく知られており、例えば米国特許第6,844,422号中で開示されたWnt-I、およびGlinkaら(1998) Nature 391: 357〜362によって開示されたDickkopf-1(DKK-1)を含む。
【0033】
本発明は、内胚葉細胞が豊富な細胞集団を作製する方法であって、約2〜約4日間血清の不在下でマウス胚幹細胞を、あるいは約2〜約10日間血清の不在下でヒト胚細胞を培養する段階と、brach+/HNF3β3+細胞を単離する段階と、約1日〜約10日間血清の不在下およびアクチビンまたはノーダルおよびWntシグナル伝達の阻害剤の存在下で単離細胞を培養する段階とを含む方法も提供する。brach+/HNF3β3+細胞の単離は、前に記載したC4-HNF/GFP-BryES細胞を使用することによって容易にすることができる。
【0034】
前述の2つの方法のそれぞれにおいて、アクチビンまたはノーダルおよびWnt阻害剤の量は、ES細胞から内胚葉への分化を誘導するのに充分な量である。内胚葉の発生と関係がある遺伝子の発現をアッセイすることによって、分化を測定することができる。好ましい実施形態では、アクチビンの濃度は少なくとも約30ng/mlである。他の好ましい実施形態では、アクチビンの濃度は約100ng/mlである。好ましい実施形態では、Wnt阻害剤はDKK-1であり、DKK-1の濃度は少なくとも約30ng/mlである。他の好ましい実施形態では、DKK-1の濃度は約100ng/mlである。
【0035】
本発明はさらに、肝細胞が豊富な細胞集団を作製する方法であって、約2〜4日間血清の不在下およびアクチビンまたはノーダルの存在下でマウス胚幹細胞を、あるいは約2〜10日間血清の不在下およびアクチビンまたはノーダルの存在下でヒト胚細胞を培養する段階と、brach+/HNF3β3+/cKit+細胞を単離する段階と、肝芽細胞を生成するのに充分な条件下において血清の不在下ならびにBMP-4およびbFGFの存在下で単離細胞を培養する段階と、肝細胞への肝芽細胞の成熟に充分な条件下において肝芽細胞を培養する段階とを含む方法を提供する。アクチビンまたはノーダルの量は、ES細胞から内胚葉への分化を誘導するのに充分な量である。好ましい実施形態では、アクチビンは少なくとも約30ng/mlの濃度で存在し、BMP-4は少なくとも約10ng/mlの濃度で存在し、かつbFGFは少なくとも約1.0ng/mlの濃度で存在する。肝細胞はグリコーゲン貯蔵に関するアッセイによって、あるいは例えば電子顕微鏡によって大きな核/細胞質比、多量のグリコーゲン貯蔵および毛細胆管の存在などの肝細胞の特徴を観察することによって同定することができる。
【0036】
前述の方法は、少なくとも約10%の肝細胞を含む細胞集団を与える。好ましい実施形態では、細胞集団は少なくとも約50%の肝細胞を含む。より好ましい実施形態では、細胞集団は少なくとも約60%、または少なくとも約70%、または少なくとも約80%、または最も好ましくは少なくとも約90%の肝細胞を含む。
【0037】
本発明はさらに、前に記載した細胞集団の増殖、分化または生存に影響を与える作用物質を同定する方法を提供する。この方法は、試験する作用物質の不在下および存在下において本明細書で前に記載した細胞集団の1つ由来の細胞を培養する段階と、作用物質が細胞集団の増殖、分化または生存に対して影響を有するかどうか決定する段階とを含む。試験する作用物質はポリペプチド、多糖、ポリヌクレオチドなど、抗体またはその断片、天然または合成化合物のライブラリー由来の化合物、合理的薬剤設計から得た化合物、あるいは当技術分野で知られているアッセイ、例えば米国特許第6,110,739号中に記載された標準的な増殖および分化アッセイを使用して細胞集団に対するその影響を評価することができる任意の作用物質を含めた、天然または合成、1つの化合物または混合物、小分子またはポリマーであってよい。このような作用物質は、in vivoおよびin vitroで細胞増殖および分化を制御するのに有用である。
【0038】
本発明はさらに、特定系統および組織の細胞分化および発生と関係がある遺伝子を同定する方法を提供する。この方法は、異なる量の培養時間後に本発明のGFP+細胞の集団を単離する段階と、異なる集団中での遺伝子発現プロファイルを比較する段階と、一集団中で特有に発現される遺伝子を同定する段階を含む。好ましい実施形態では、マイクロアレイ分析およびサブトラクティブハイブリダイゼーションを使用して、遺伝子発現プロファイルを比較する。
【0039】
他の実施形態では本発明は、brachury陰性(brach-)細胞ではなくbrachury陽性(brach+)細胞を認識する抗体を作製する方法を提供する。ポリクローナル抗体は、免疫原の形で本発明の細胞を動物に注射することによって作製することができる。さらに、GFP-細胞ではなくGFP+細胞中に存在する細胞表面マーカーを同定する段階、およびマーカーまたはその断片に対する抗体を作製する段階によって、抗体を作製することができる。抗体はモノクローナルまたはポリクローナルであってよく、断片、遺伝子工学処理された抗体、単鎖抗体などであってよい。抗体は当技術分野でよく知られている方法によって作製することができる。このような抗体は、内中胚葉および中胚葉などのbrach+細胞を同定および単離するのに有用である。
【0040】
本発明は、in vitroで哺乳動物細胞を生成するための方法も提供する。一実施形態では、この方法は、心筋、血管平滑筋、内皮または造血細胞への中胚葉の分化に有効な条件下で、内中胚葉および中胚葉細胞が豊富な細胞集団由来の細胞を培養する段階を含む。in vitroでの様々な細胞型への分化に有効な条件は、当技術分野で知られている。他の実施形態では、この方法は、肝臓細胞または膵臓細胞への内胚葉の分化に有効な条件下で、内胚葉細胞が豊富な細胞集団由来の細胞を培養する段階を含む。このような分化に有効な条件は、当技術分野で知られている。インシュリンを生成する膵臓ランゲルハンス島細胞の生成が特に企図される。
【0041】
本発明によって実証されるように、異なる経過日数のEBから単離したbrach+細胞は異なる発生能を有する。約3日齢マウスのEB由来のBrach+/Flk-細胞は造血および内皮系統を効率良く生じ、一方約3日齢〜10日齢のEB由来の細胞は心筋細胞系統の細胞を生じる。したがって、内中胚葉および中胚葉が豊富な細胞集団を得るために使用するES細胞の培養時間を調節することによって、当業者は造血および内皮系統または心筋細胞系統の効率良い生成を選択することができる。
【0042】
このような細胞は、例えば限られた数の細胞型の破壊または機能不全が原因である障害を治療するための細胞補充療法に有用である。これらの障害には、真性糖尿病、肝不全、心不全、心臓血管および他の血管疾患、Duchenne型筋ジストロフィー、骨形成不全症、および骨髄移植によって治療可能な障害、例えば白血病および貧血がある。Odoricoら、(2001) Stem Cells 19: 193〜204を参照。
【0043】
本発明の細胞集団は、細胞補充療法用の分化細胞および組織を生成するのに有用である。細胞補充療法用の本発明の細胞集団の適性は、限られた数の細胞型の破壊または機能不全と関係がある障害の動物モデルに細胞を移植することによって評価することができる。例えば、参照により本明細書に組み込まれるGrompeら(1993) Genes & Dev.7: 2298によって例えば開示されたフマリルアセト酢酸(FAH)欠陥マウスは、肝不全のモデルとなる。FAH欠陥マウスは、NTBC(2-(2-ニトロ-4-トリフルオロメチルベンゾイル)-1,3-シクロヘキセジオン)で治療するか、あるいは正常な肝細胞を移植しない限り、進行性肝不全および尿細管障害に罹患する。したがってこれらのマウスは、EBから生じる未分化肝細胞の特徴を有する細胞の発生能を試験するための理想的なモデルとなる。NTBCを除去したFAH欠陥マウスに肝細胞を移植するための方法は当技術分野で知られており、例えばOversturfら(1996) Nature Genet.12: 266〜273によって開示されている。正常な肝機能はマウスの生存によって示され、血清中アスパラギン酸トランスアミナーゼのレベル、血漿中ビリルビンのレベルを測定すること、および再生肝臓の正常な構造を決定することによって評価することもできる。
【0044】
特定の細胞型の破壊または機能不全が原因である他の障害の動物モデルは、当技術分野で知られている。このようなモデルを同様に使用して、本発明の他の細胞集団を評価することができる。
【0045】
好ましい実施形態では、本発明は、肝細胞補充療法の方法であって、本発明の方法によって生成した肝細胞を含む組成物を、その治療の必要がある対象に投与する段階を含む方法を提供する。好ましい実施形態では、対象はヒトである。例えば注射、移植または注入を含めた肝臓組織への送達をもたらす経路によって組成物を投与することができる。本発明の方法は、肝不全、肝臓系代謝性疾患、および慢性肝臓疾患を治療するのに有用である。
【0046】
本発明は、1つのbrachury対立遺伝子が不活性状態でありbrachury遺伝子座が転写される細胞中で選択可能なマーカーが発現されるように、選択可能なマーカーをコードするDNAがbrachury遺伝子座に存在する、トランスジェニック非ヒト哺乳動物も提供する。好ましい実施形態では、哺乳動物はマウスであり、選択可能なマーカーはGFPである。特に、トランスジェニックマウスは、GFPをコードするDNA配列がbrachury制御要素と動作可能に連結しているトランス遺伝子を含むゲノムを有し、このトランス遺伝子はbrachuryを通常発現する細胞中で発現される。トランスジェニックマウスは、胚盤胞に本明細書で前に記載したGFP-BryES細胞を注射し、次いでそれらを偽妊娠状態のメスに移植することによって得ることができる。トランスジェニック子孫は、brach+/-と関係がある短い尾の表現型によって、および分子分析によって同定する。このようなトランスジェニック動物は、本発明の方法によって使用するための中胚葉を単離する初期胚を得るのに、brachury遺伝子を発現する任意の成体細胞集団を同定、単離および特徴付けするのに有用である。このような細胞は、新規の幹細胞集団となる可能性がある。
【0047】
本明細書に引用する全ての参照文献は、それらの全容を本明細書に組み込む。
【実施例】
【0048】
以下の実施例は、本発明をさらに例示するのに役立つ。
(実施例1)
【0049】
材料および方法
ES細胞の増殖および分化。15%のウシ胎児血清(FCS)、ペニシリン、ストレプトマイシン、LIF(1%条件付き培地)および1.5×10-4Mのモノチオグリセロール(MTG;Sigma)を補ったダルベッコ改変イーグル培地(DMEM)中の照射した胚栄養細胞上にES細胞を保った。分化の開始の2日前に、同じ培地中のゼラチンプレートに細胞を移した。EBを生成するために、ES細胞をトリプシン処理し、分化培養物中に様々な密度で平板培養した。EBの分化は60mmのペトリグレード皿中、15%のFCS、2mMのL-グルタミン(Gibco/BRL)、トランスフェリン(200ug/ml)、0.5mMのアスコルビン酸(Sigma)、および4.5×10-4MのMTGを補ったIMDM中で実施した。37℃において5%CO2/空気混合物中の加湿チャンバー内に培養物を保った。
【0050】
無血清培地。2つの異なる無血清培地を以下の実施例の異なる態様において使用した:Knockout SR(Gibco BRL)およびStemPro34(Gibco BRL)を補ったIIMD。
【0051】
メチルセルロースコロニーアッセイ。A)芽細胞コロニー:芽細胞コロニーを生成するために(BL-CFCアッセイ)、EB由来細胞を0.5×〜1.5×105個の細胞/mlで10%のFCS(Hyclone)、血管内皮増殖因子(VEGF;5ng/ml)、c-kitリガンド(KL;1%条件付き培地)、IL-6(5ng/ml)および25%のD4T内皮細胞の条件付き培地を補った1%のメチルセルロースにおいて(Kennedyら(1997) Nature 386: 488〜93)平板培養した。移行性コロニーがVEGFの不在下で生成した。コロニーは培養の4日後に記録した。B)造血コロニー:初期および最終段階造血コロニーを増殖させるために、細胞を10%の血漿由来血清(PDS;Antech)、5%無タンパク質ハイブリドーマ培地(PFHM-II;Gibco-BRL)および以下のサイトカイン:c-kitリガンド(KL;1%条件付き培地)、エリスロポイエチン(2U/ml)、IL-11(25ng/ml)、IL-3(1%条件付き培地)、GM-CSF(3ng/ml)、G-CSF(30ng/ml)、M-CSF(5ng/ml)、IL-6(5ng/ml)およびトロンボポイエチン(TPO;5ng/ml)を含む1%のメチルセルロース中に平板培養した。培養物は37℃、5%CO2で保った。初期赤血球コロニーは培養の第5〜6日で記録し、一方最終段階赤血球(BFU-E)、マクロファージ、および多系統コロニーは培養の第7〜10日で計数した。c-kitリガンドは、KL発現ベクター(Genetics Instituteによって親切にも与えられた)でトランスフェクトしたCHO細胞によって条件付けした培地に由来した。IL-3は、IL-3を発現するベクターでトランスフェクトしたX63AG8-653ミエローマ細胞によって条件付けした培地から得た。VEGF、GM-CSF、M-CSF、IL-6、IL-11、アクチビンBMP2、BMP4、bFGF、FGF8、および1hhはR&D systemsから購入した。
【0052】
再凝集培養物。15%のFCS(またはKnockout SR)、2mMのL-グルタミン(Gibco/BRL)、0.5mMのアスコルビン酸(Sigma)、および4.5×10-4MのMTGを補ったIMDM1ml当たり2×105個で、24ウエルのペトリグレードプレート中で細胞を培養した。これらを使用して、細胞とウエルの底部の接着を妨げた。
【0053】
心筋アッセイ。15%血清物を補ったIMDM中でGFP+細胞を再凝集させた。20時間後、凝集体を24または96ウエルプレートのいずれか中、10%血清物を補ったIMDM中(無血清)で培養した。ウエルはゼラチンで予め処理した。培養物は拍動細胞の出現の発生に関して毎日調べた。拍動細胞は培養第2日と第6日の間に通常検出された。
【0054】
細胞表面マーカーの染色およびFACS分析。標準的な条件を使用して細胞を染色した。染色細胞懸濁液はFACScan(Becton Dickinson、CA)で分析した。
【0055】
遺伝子発現の分析。ポリA+RT-PCR分析用に、Bradyら((1990) Meth.in Mol.and Cell Bio.2: 17〜25)の方法を使用した。逆転写、ポリAテーリングおよびPCR手順を記載されたのと同様に実施した、ただしX-dTオリゴヌクレオチドは5'-GTTAACTCGAGAATTC(T)24-3'に短縮した。PCR反応からの増幅産物はアガロースゲル上で分離し、Zeta-プローブGT膜(Biorad)に移し、あるいはスロットブロット装置(Schleicher & Schuell)を有する膜に移した。生成したブロットは、遺伝子(β-H1以外全て)の3'領域に対応する32Pでランダムにプライマー処理したcDNA断片(Ready-to-Go Labelling、Pharmacia)とハイブリダイズさせた。β-H1特異的プローブは、その3'末端に8塩基の相同性を共有する2つのオリゴヌクレオチド(5'-TGGAGTCAAAGAGGGCATCATAGACACATGGG-3'、5'-CAGTACACTGGCAATCCCATGTG-3')をアニーリングさせることによって調製した。このβ-H1特異的オリゴヌクレオチドは、クレノウ充填反応を使用して32Pで標識した。遺伝子特異的PCR用に、全てのRNAはRNeasyミニキットを用いてそれぞれのサンプルから抽出し、RNase free DNase(Qiagen)で処理した。2マイクログラムの全てのRNAは、Omniscript RTキット(Qiagen)を使用しランダムヘキサマーを用いてcDNAに逆転写した。PCRは適切なオリゴヌクレオチドを使用して実施した。PCR反応は、2.5UのTaqポリメラーゼ(Promega)、PCRバッファー、2.5mMのMgCl2、 0.2uMのそれぞれのプライマーおよび0.2mMのdNTPを用いて実施した。循環条件は以下の通りであった;94℃で5分間、次に35サイクルの増幅(94℃で1分間の変性、60℃で1分間のアニーリング、72℃で1分間の伸長)および72℃で7分間の最終インキュベーション。
(実施例2)
【0056】
標的ES細胞の生成
培養中の適切な条件下で、胚幹(ES)細胞は分化し、広範囲の系統由来の発生段階の細胞集団を含む胚様体(EB)として知られる三次元のコロニーを形成する。Smith(2001) Annu.Rev.Cell Dev. Biol.17: 435〜62。これらのEB由来集団の中では、造血、内皮、心筋および骨格筋系統の誘導体を含めた中胚葉誘導体を検出することができる。
【0057】
EBにおける中胚葉形成の開始を追跡するため、およびこの集団を表す細胞を単離するために、緑色蛍光タンパク質(GFP)をbrachury遺伝子座に標的化した。標的構築体はGFPのcDNA、および人工イントロン、第一エクソン中でneoカセットに隣接するSV40ポリ(A)配列およびloxPを含んでおり、図1中に示す。ランダムな組み込みに対する選択のために、チミジンキナーゼ(TK)遺伝子を標的構築体の3'末端に含めた。標的ベクターは以下のように構築した。
【0058】
完全なマウスbrachury(Bry)遺伝子を有するBACクローンを、プライマー5'-AAGGAGCTAACTAACGAGATGAT-3'および5'-TACCTTCAGCACCGGGAACAT3'を用いて、129/Ola系統ゲノムライブラリー(Genome Systems)のPCRスクリーニングによって単離した。これらのプライマーはそれぞれ第一および第二Bryエクソン内でアニーリングし、〜600bpの診断用バンドを増幅させる。Bry遺伝子の1エクソンおよび2kbを超える5'隣接領域を有する約3kb長のPstI制限断片を同定し、BACからプラスミドpBSK(Strategene)にサブクローニングし、構築体pBSK.Bry-5'を得た。開始コドンの直ぐ上流の約2kbの領域をシークエンスして、ベクターを構築するのに適したプライマーアニーリング部位を同定した。
【0059】
オリゴ5'-GCTAGCTAATGGATCCA-3'/5'-GATCTGGATCCATTAGCTAGCTGCA-3'および5'-GATCTTAATGAACGGCAGGTGGGTGCGCGTCCGGAG-3'/5'TCGACTCCGGACGCGCACCCACCTGCCGTTCATTAA-3'をプラスミドpBSKのPstI/SalI部位中に挿入して、2つの連続した翻訳停止コドンおよび人工3'スプライシング部位を有する新たな、より適切なポリリンカー(構築体pBry-AA)を作製した。プラスミドpEGFP.C1(Clontech)はNheI/BglIIで2回消化し、生成した停止コドンを含まないEGFPをコードする〜760bpのDNA断片はpBRY-AAのNheI/BglII部位にクローニングし、構築体pBry-ABを生成した。loxPに隣接するネオマイシン耐性遺伝子を有するプラスミドpL2-Neo2のXhoI/SalI断片をpBry-ABのSalI部位に挿入して、プラスミドpBry-ACを生成した(EGFPとNeoの転写は同じ方向)。
【0060】
共通のスプライシングドナー部位、人工イントロン、スプライシングアクセプター部位およびSV40ポリアデニル化配列を含む短いエクソンを有する556bpのXmaI/MluI断片を、市販の発現ベクターpBK-CMV(Stratagene)から切除した。この断片は以下の方法でプラスミドpBry-AC中に挿入した:XmaI端を最終EGFPコドンの後のBspEI部位と連結させ、一方Mlu端はリンカーとしてのオリゴ5'-CGCGTTACTAGTAAGACGTCT-3'/5'-CCGGAGACGTCTTACTAGTAA-3'と共にloxP-neo-loxPカセットの直ぐ上流に位置するBspEI部位に挿入した。生成した構築体:pBry-AE。HSVチミジンキナーゼ遺伝子をコードする〜1.9kbのXhoI/SalI断片をpBry-AEのXhoI部位に挿入して、ランダムな組み込みに対する選択を可能にした(構築体pBry-AH)。相同性の「短いアーム」をコードするNotI/Eco47III断片をpBry-AFから切除し、pBry-AHのNotI/Eco47III部位にクローニングして、プラスミドpBry-AIを生成した。相同性の「長いアーム」はSalIを用いてpBry-AKから切除し、正しい方向でpBry-AIのSail部位に挿入して、最終標的ベクターBを生成した。
【0061】
胚幹細胞(E14.1、129/Ola Hooperら(1987) Nature 326: 292)に、NotI線状標的ベクターpBry-AMをエレクトロポレーションにより導入した。トランスフェクト細胞を含む4枚の皿にはG418単独選択を施し、他の4枚の皿にはG418+ガンシクロビル(Ganc)二重選択を施した。相同的組換え事象を経験したクローンは、「相同性の短いアーム」の直ぐ上流のBry遺伝子の5'領域中のゲノム配列とアニーリングする1つのプライマー(5'-CAGGTAGAACCCACAACTCCGAG-3')、EGFPの5'部分(診断用バンド:約1.3kb)に対する他のプライマー(5'-CCGGACACGCTGAACTTGTGGC-3')を用いたPCRによって同定した。正しく標的化されたクローンは、サザンブロット分析によって確認した:候補クローンのゲノムDNAはHincIIで消化し、標的構築体の外側に位置するプローブとハイブリダイズさせた。オリゴヌクレオチド対5'-ACAGGATCCCTAAGCCTCAAAAGAGTCGCT-3'/5'-TCTTGGATCCTCCTATCCTATCCCGAAGCTCCT-3'を使用するPCRにより、プローブはBry5'隣接領域(BryATG開始コドンに関して-2018〜-1249)に由来した。384のG418単独および80 G418+のGanc二重選択クローンをスクリーニングし、そのそれぞれ4個のうちの3個が陽性であることが分かり、1.04%および3.75%の標的化効率に相当した。2個の陽性クローンを改変型Creリコンビナーゼ発現ベクターで一過的にトランスフェクトして、neo遺伝子を除去した。これらの標的クローンは、本明細書では以後GFP-BryES細胞と呼ぶ。
【0062】
brachuryは発生段階のEBにおいて一過的に発現され、その開始は造血および内皮系統の樹立を明確にする遺伝子の発現に先行する。GFP-BryES細胞におけるGFPの発現がEB発生中のbrachury遺伝子の発現を正確に反映するかどうかを決定するために、GFPの発現を評価した。
【0063】
6日間のEB分化期中の典型的な発現パターンは、図2A中に示す。この実験では、低レベルのメッセージを分化の24時間以内に検出した。発現は次の48時間上方制御され、4日間持続し、次いで分化の第6日までに検出不能なレベルまで急激に低下した。FACS分析によって定義したGFPの発現は、同様の経時的パターンをたどった。低レベルのGFP+細胞(〜5%)を分化の第2日で早くも検出した。半分を超える(65%の)EB由来細胞が第3日でGFPを発現し、ほぼ全ての細胞が分化の第4日で陽性であった。PCRによって観察したように、発現はこの時点以後急激に低下し、第6日までは、存在した場合でも少数のGFP+細胞が存在した。GFPの発現のこの急激な低下は、それは長期間細胞内で持続しなかったことを示した。分化の第3日および第4日でのGFP+細胞の高い割合は、これらの条件下でのEB内の中胚葉の発生が広範囲に及ぶことを示唆する。合わせて考えると、これらの発見は、GFPの発現はEB発生中のbrachury遺伝子の発現を正確に反映することを示す。
【0064】
1つのbrachury対立遺伝子の不活性化が、ES細胞のin vitroでの発生能に対して有害な影響を有し得る可能性を評価した。示したように、ヘテロ接合性のマウスは軽度の表現型を示す。GFP-BryES細胞が造血発生中に何らかの検出可能な欠陥を示すかどうかを測定するために、それらから生成したEBを、造血前駆体および芽コロニー形成細胞(BL-CFC)含量、および遺伝子発現パターンに関して分析した。図3Aおよび3B中のデータは、野生型細胞と比較してGFP-BryES細胞が、匹敵する数の初期(Eryp)および最終段階(Eryd、Mac、Mac/Ery、およびMix)造血前駆体およびBL-CFCを生成することを示す。遺伝子発現パターン(図3C)は前駆体の分析を支持し、GFP-Bryから生成したEBと野生型ES細胞の間ではほとんど違いを示さなかったことを確認した。両組のEBが、分化の最初の3日間でRex-1の発現の低下を示した。Rex-1は転写因子であり、ES細胞中で発現され、分化を経ると下方制御される。Rogersら(1991) Development 113: 815〜24。Rex-1の低下は、brachury発現の典型的な一過性の波に続き、それは造血および内皮系統の発生と関係がある遺伝子の発現開始の直前に先行する。これらの系統の樹立に必要不可欠なFlk-1、受容体チロシンキナーゼ(Shalabyら(1995) Nature 376: 62〜6)は、分化の第3日と第6日の間に発現される。GATA-1、造血転写因子、ならびに胚および成体グロビン遺伝子、βH1およびβmajorは、分化の第4日に低レベルで検出した。3つ全ての遺伝子が次の24時間上方制御され、この発生段階での初期赤血球系統の広がりおよび成熟が反映された。Palisら(1999) Development 126: 5073〜84。この実施例で観察した前駆体の数および遺伝子発現パターンは、以前の研究において見られたものと一致し、造血系の樹立をもたらす分子プログラムは標的GFP-BryES細胞において完全な状態であることを示す。
(実施例3)
【0065】
brachury+細胞の単離
GFPの発現に基づいてbrachury+細胞を単離することができるかどうか決定するために、第3.5日EB由来のGFP+集団を適切な遺伝子の発現に関して選別し分析した。図4AはGFP陽性(2)および陰性(1)集団の単離に使用したゲートを示す。RT-PCR分析によって、brachuryの発現はGFP+画分に限られていたことが明らかになり、GFPの発現に基づく細胞選別を使用してbrachury+細胞を単離することができることが示された。Flk-1、造血および内皮発生の初期マーカーの1つは、GFP-画分中よりGFP+中において高レベルで存在し、Flk-1は少なくともbrachury+細胞の亜集団中で発現されたことが示された。brachuryおよびFlk-1とは対照的に、Pax-6、初期神経発生と関係がある遺伝子[79,80]は、brachury陰性であるこの系統の前駆体と一致してGFP-画分中において高レベルで発現した。これらの細胞選別試験は、brachury遺伝子座の制御下でのGFPの発現は、EB由来のbrachury+細胞の単離、特徴付けおよび操作に関する新規のマーカーをもたらすことを示す。
【0066】
この実施例は、GFP+細胞は細胞選別によって第3.5日EBから単離することができることを実証する。GFP+およびGFP-画分の遺伝子発現分析は、brachuryの発現は陽性画分に主に分離すること、GFPに基づく画分化はbrachury陽性細胞を単離するための方法をもたらすことを明らかに実証する発見を示す。brachury以外に、初期造血および内皮発生と関係がある受容体チロシンFlk-1も、陰性画分中より陽性画分中において高レベルで発現される。対照的に、Rex-1およびPax-6、初期外胚葉および神経外胚葉のマーカーは、GFP-画分中で発現される。これらの発見は、brachuryの状況でのGFPの発現を使用して、外胚葉から中胚葉を分離することができることを実証する。
(実施例4)
【0067】
Flk-1の発現に基づく亜集団へのbrachury陽性細胞の分離
Flk-1は、初期胚における造血および内皮系統の樹立に必要不可欠であり、BL-CFCを含めたこれらの系統の初期前駆体において発現されることが示されてきている[Faloonら(2000) Development 127: 1931〜41]。血液および血管発生におけるこの重要な役割を考慮すると、GFP+集団内のその発現は、これらの系統への特化を経た中胚葉の亜集団を明確にすると仮定した。この可能性をさらに調べるために、GFPおよびF1k-1陽性細胞の存在に関して幾つかの発生段階でEBを分析した。図5A中に示した実験では、4.8%の3.0日EB集団がF1k-1ではなくGFPを発現し、一方1.2%の細胞が両方のマーカーを発現した。両画分の大きさは次の12時間で劇的に増大し、GFP+/Flk-1-およびGFP+/Flk-1+細胞はそれぞれ全EB集団の52%と26%を表した。GFPおよびFlk-1発現によって定義される3集団、GFP-/Flk-1-(画分2)、GFP+/Flk-1-(画分3)およびGFP+/Flk-1+(画分4)の発生能を評価するために、細胞を両時点において単離し、BL-CFCおよび2°EB含量に関して、遺伝子発現パターンに関して分析した。これらの画分の発生能を、予め選別した集団(画分1)のそれと比較した。大部分のBL-CFCは、分化の第3.0日と第3.5日の両方でGFP+/Flk-1+画分中において見られた(図5B)。全てのBL-CFCがFlk-1を発現することを以前の試験が示していることを考慮すると、これは驚くことではない。2°EBはGFP-/Flk-1-画分に限られており、これは2°EBが初期EBの残留未分化ES細胞に由来する事実と一致する発見である。GFP+/Flk-1-画分は、これらの培養で使用した条件下でほとんどコロニーを生成しなかった。遺伝子発現の分析によって、2つの時点で単離した集団間の幾つかの興味深い違いが明らかになった(図5C)。Rex-1、ES細胞のマーカーは、GFP-画分よりGFP+において低いレベルで発現され、これらの集団は分化を経ていることが示された。brachuryはGFP+画分において両方の時点で発現された。そのレベルは第3.5日EBから単離したGFP+/Flk-1+中よりGFP+/Flk-1-中で高いようであり、これらの細胞が造血および内皮系統に成熟すると、その発現は下方制御される可能性があることが示唆される。予想したように、Flk-1は両方の時点でGFP+/Flk-1+細胞中において主に発現された。初期と最終段階の両方の造血発生に必要不可欠である(Shivdasaniら(1995) Nature 373: 432〜4)Scl、ヘリックス-ループ-ヘリックス転写因子は、GFP+/Flk-1+画分に限られるようである。同様に、最終段階造血系の樹立に必要とされる転写因子Runx1(Wangら(1996) Proc. Natl. Acad. Sci. 93: 3444〜9)は、GFP+/Flk-1+画分中で最も容易に検出した。第3.0日EBから単離したGFP+/Flk-1-画分中では、ある程度のRunx1の発現があった。ノーダルは分化の第3日で3つ全ての画分中で発現される。第3.5日では、GFP+/Flk-1+画分中での発現のレベルは、他の画分中より有意に低いようである。Wnt3aおよびWnt8aは著しく限られたパターンの発現を示し、それは両方の時点においてGFP+/Flk-1-画分中のみで見られ、系統限定的なマーカーの発現前の初期中胚葉の機能と一致した。BMP2は両方のGFP+画分中で発現され、一方BMP4はGFP+/Flk-1+細胞中で主に見られ、これらの分子はこの系では発生の異なる段階で役割を果たすことが示された。GFP+/Flk-1+細胞のBL-CFC発生能および遺伝子発現パターンは、それらの細胞がマウス胚において見られる胚外中胚葉を表すことを示す。
【0068】
この実施例は、第3日および第3.5日のEBのbrachury画分は、Flk-1発現に基づいて2つの画分:brachury+/Flk-1-(GFP+/Flk-1-)およびbrachury+/Flk-1+(GFP+/Flk-1+)に分けることができることを実証する(図5A)。機能性試験によって、前駆体(BL-CFC)は(GFP+/Flk-1+)画分に分離する造血細胞と内皮細胞の両方を生成することができることを実証し、Flk-1の上方制御はこれらの系統への拘束を示すことが示唆された(図5B)。遺伝子発現試験によって、GFP+/Flk-1-集団とGFP+/Flk-1+集団の間の明確な違いを実証した(図5C)。
(実施例5)
【0069】
GFP/Flk画分間の発生関係
図5中で観察した発現パターンは、3つの画分は、GFP-/Flk-1-細胞はGFP+/Flk-1-細胞に分化し、次いでそれがGFP+/Flk-1+細胞に分化する発生の連続性を示すという解釈と一致する。これらの画分が共通の発生経路内で特異的段階を示すかどうかを測定するために、各画分を第3日EBから単離し、20時間培養し、次いでGFPおよびFlk-1の発現に関して再度分析した。BL-CFCおよびEryp-CFCの発生能を、培養の前後に各集団に関して測定した。単離した細胞は、ペトリグレードの24ウエルプレート中、EB分化用に使用した同じ培地中に、1×105個の細胞以上の密度で培養した。これらの条件下では、細胞は直ぐに再凝集し、EB様構造を形成し、これは細胞数の増大または損失ほとんどなしで正常な発生プログラムに従うようである。20時間の再凝集培養後、GFP-/Flk-1-細胞は相当数のGFP+/Flk-1-細胞集団、および少数のGFP+/Flk-1+に分化した。GFP+/Flk-1-細胞は、同じ培養時間中に相当数のGFP+/Flk-1+細胞集団を生成した。GFP+/Flk-1+集団は、再凝集培養後に幾らかGFPおよびFlk-1の発現を失うようであった。結果は図6中に示す。前駆体の発生能の変化は、表面マーカーの変化と一致していた。3つの中で最も未熟であるGFP-/Flk-1-画分は、培養の前後で検出不能な数のBL-CFCおよびEryp-CFCを含んでいた。GFP+/Flk-1-画分も、培養前にわずかなBL-CFCおよびEryp-CFCを含んでいた。しかしながら、培養後、BL-CFCの発生能は細胞105個当たり74〜1564に劇的に増大し、Flk-1の発現の増大と一致した。Eryp-CFCの頻度は培養期間中変わらなかった。GFP+/Flk-1+画分は、培養前にBL-CFCを含んでいたが、Eryp-CFCはほとんど含んでいなかった。培養後にBL-CFCは検出されなかったが、この集団はここでEryp-CFCを含んでいた。表面マーカーの分析と一緒にすると、これらの前駆体のデータは、前中胚葉(GFP-/Flk-1-)から中胚葉/前血管芽細胞(BL-CFC)集団(GFP+/Flk-1-)、血管芽細胞/前赤血球集団(GFP+/Flk-1+)、後血管芽細胞/赤血球集団(おそらくGFPlo/Flk-1lo)への発生の進行を支持する。最初の表現型を有する細胞がGFP-/Flk-1-およびGFP+/Flk-1-培養物中に残存したので、所与の集団中の全ての細胞が20時間の培養期間後に分化したわけではないようである。
【0070】
この実施例は、単離し20〜24時間再培養すると、第3日EBから単離した3つの集団のそれぞれは分化し続けることを示し、あるパターンでは、これらの集団は発生の連続性を示すことを示す。例えば、GFP-/Flk-1-はGFP+/Flk-1-細胞に分化し、次いでそれはGFP+/Flk-1+に分化した。細胞表面特性のこれらの変化は、予想した発生能の変化と関係があった。GFP+/Flk-1-画分は、培養前にわずかな造血/内皮前駆体(BL-CFC)を含んでいた。培養後、これらの前駆体を検出し、第3日EB由来のGFP+/Flk-1-画分は、Flk-1+細胞に分化する発生能ならびに造血および内皮発生能を含むことが明らかに実証された。
(実施例6)
【0071】
GFP/Flk-1-細胞の発生能
前述の実施例は、第3日EBから単離したGFP+/Flk-1-細胞は、一晩培養の後GFP+/Flk-1+細胞およびBL-CFCを効率良く生成したことを実証した。このプレBL-CFCの発生能が発生のこの段階で単離したGFP+/Flk-1-画分に特異的であったかどうかを決定するために、異なる経過日数のEB由来のGFP+/Flk-1-細胞を、BL-CFCに分化するそれらの能力に関してアッセイした。図7A中に示すように、BL-CFCを生成する能力は第3日のGFP+/Flk-1-細胞において最も顕著であった。この発生能は分化の第3.5日までに劇的に低下し、第4.0日にはほとんど存在しなかった。これらと同じEBから新たに単離したGFP+/Flk-1+画分のBL-CFC含有量はこの期間中に増大し、分化は通常の形式で進行していたことが示された。異なる段階のEB細胞から再凝集した培養物におけるFlk-1発現のパターンは、BL-CFCのデータと一致した。再凝集した第3日のGFP+/Flk-1-細胞由来の培養物は、集団全体の40%より多くを表した独特のFlk-1画分を含んでいた(図7B)。第3.5日および第4.0日培養物におけるFlk-1の発現は有意に低く、独特のピークではなく集団全体のピークからなっていた。
(実施例7)
【0072】
GFPおよびFlk-1亜集団の心筋細胞発生能
造血および内皮中胚葉の発生が心臓および頭蓋中胚葉の発生に続くマウス胚における事象の順序を考慮して、単離集団の心筋細胞の発生能を測定した。この分析のために、異なる集団の培養物からの凝集体をマイクロタイターまたは24ウエルプレート、無血清培地中に移し、心筋細胞を示す拍動細胞塊の発生を調べた。これらの条件は、再凝集細胞からの心筋細胞の効率良い発生を助長することが知られている。これらの塊中の心筋性の細胞の別個の確認として、代表的な群を顕微鏡スライドに移し、固定しトロポニンTの心臓特異的イソ型の存在に関して染色した。分析した全ての拍動細胞塊はトロポニンT陽性細胞を含んでいたことが分かり、それらが心筋細胞であったことが示された。このアッセイを使用して、再凝集した異なる段階のEB由来のGFP+/Flk-1-およびGFP+/Flk-1+画分の心筋細胞の発生能を測定した。比較用に、新たに単離したGFP+/Flk-1+細胞および培養したGFP+/Flk-1-細胞のBL-CFC発生能を分析した。
【0073】
【表1】
【0074】
表1中に示したように、異なる画分のBL-CFC発生能は、前の実験で観察したそれと同等であった。3つの異なるEB集団から単離したGFP+/Flk-1+細胞がBL-CFCを生成し、最高数は第3.5日および第4.0日で見られた。GFP+/Flk-1-細胞におけるプレBL-CFC発生能は第2.75日で最大であり、第3.5日および第4.0日で著しく低下した。画分の心筋細胞発生能は逆のパターンを示した。第3.5日および第4.0日GFP+/Flk-1-細胞由来の相当な割合(>80%)の移動凝集体が、拍動心筋細胞を生成した。初期(第2.75日)GFP+/Flk-1-細胞から生成した凝集体では拍動細胞が観察されなかった。発生の三段階のいずれかでEBから単離したGFP+/Flk-1+細胞から生成した凝集体では、拍動細胞を検出しなかった。この分析からの発見は、異なる段階で単離したGFP+/Flk-1-細胞は異なる発生能を有するという概念と一致する。初期に発生する細胞は血管芽細胞の運命を有しているようであり、一方後期に発生する細胞は、心臓系統およびおそらく他の集団を生成する。GFP+/Flk-1+集団は心筋細胞発生能を失っているようであり、血管芽細胞系統に限られる可能性がある。これらの発見を考慮に入れて、初期発生(第2.75日〜第3.0日)GFP+/Flk-1-細胞は前血管芽細胞中胚葉と呼び、一方第3.5日と第4.0日の間に発生する集団は、前心臓中胚葉と呼ぶ。第3.0日〜第3.5日GFP+/Flk-1+集団はBL-CFCを生成し、一方後期段階のEB(第4.0日)から単離した集団は初期赤血球の原種を含み、造血への拘束の開始が示される。この発生能を考慮して、GFP+/Flk-1+集団は血管芽細胞中胚葉と呼ぶ。
【0075】
実施例5および6は、異なる経過日数のEBから単離したGFP+細胞は異なる発生能を有することを実証する。前の実施例で示したように、第3日EB由来のGFP+/Flk-1-細胞は、造血系統と内皮系統の両方を効率良く生成する。拍動細胞塊の欠如によって実証されたように、これらの細胞は心筋細胞(心臓組織)に分化しなかった。対照的に、第4日EB由来のGFP+細胞は、培養後わずかなFlk-1+細胞およびBL-CFCに分化した。しかしながらこの集団は、心筋細胞系統の細胞を生成した。これらの発見は、異なる経過日数のEBから単離したGFP+(brachury+)画分は、異なる発生運命を有する異なる集団にパターン形成していることを実証する。前述の実施例で使用した条件および観察した発生能以外に、条件および添加剤を変えることによって他の発生能を観察することができる。
(実施例8)
【0076】
血清由来因子の役割
brachury+細胞の発生における血清の役割を評価するために、EBを血清の不在下で分化させた。EBはこれらの条件下で発生したが、それらは通常の条件において見られるEBより幾分小さかった。血清の不在下では、これらのEB内でGFP+細胞は検出されず、これらの条件下では中胚葉は誘導されなかったことが示された(図8)。培養物に血清を加えると、相当数のGFP+/Flk-1-およびGFP+/Flk-1+細胞が発生した。これらの発見は、血清内に見られる成分は、brachury+細胞の発生および分化を誘導することができることを明らかに実証する。このプロセス中で役割を果たす可能性がある因子を同定する際の最初のステップとして、BMP4(20ng/ml)を、無血清培養物内の発生中のEBに加えた。この濃度で、BMP4は分化3日以内に相当なbrachury+細胞集団を誘導した。しかしながら、血清とは対照的に、BMP4はこの期間中GFP+/Flk-1+集団の発生を助長しなかった。血清の存在下で誘導したGFP+/Flk-1-細胞からGFP+/Flk-1+をBMP4が誘導することができるかどうかを測定するために、GFP+/Flk-1-細胞を血清の存在下で3日間分化させたEBから単離した。これらの細胞は培地単独、血清を含む培地中、またはBMP4を含む培地中で再凝集させた。図8の下列中に示すように、血清の不在下で再凝集させたとき、GFP+/Flk-1-細胞は実質的に分化しなかった。予想通り、培養物に血清を加えると、同じ集団は大きなGFP+/Flk-1+集団を生成した。初期分化培養物における発見と一致して、培養したGFP+/Flk-1-細胞からの相当数のGFP+/Flk-1+細胞の発生を、BMP4は誘導することができなかった。
【0077】
図9は、前述の実施例に基づいて中胚葉発生中の段階を要約する。ステップ#1は、中胚葉誘導および前造血および内皮(前血管芽細胞)運命へのパターン形成を表す。ステップ#2は、造血および内皮系統への特化を表す。ステップ#3および#4は、前心臓運命へのパターン形成を表す。
(実施例9)
【0078】
細胞集団の単離および内胚葉発生能の特徴付け
アフリカツメガエルおよびゼブラフィッシュなどのモデル系を使用した試験は、中胚葉および内胚葉は、内中胚葉として知られる共通の前駆体集団から発生することを示唆している。発生の内中胚葉段階がEBにおいて存在するかしないか判定するために、内胚葉系統の発生に関する条件を樹立した。この系統の発生に関する培養条件を樹立する際の最初のステップとして、分化培養物中の血清の量を変えた。EBを血清(SP34、次いでSRを添加したIMDM)の存在下および不在下で生成させ、外胚葉、内胚葉および中胚葉発生と関係がある遺伝子の発現に関して異なる段階でアッセイした。外胚葉系統に関しては、神経系統の発生を、PAX6、Wntl、neruoDおよびニューロフィラメント(NFL)の発現を分析して評価した。これらの遺伝子は、神経発生の異なる段階で発現されることが知られている。内胚葉発生の初期段階はHNF3βの発現によって調べた。内胚葉発生および特化の後期段階を評価するために、肝臓の発生と関係がある遺伝子を分析した。これらはHex、α-フェトプロテイン(AFP)、HNF4、アルブミン(Alb)、α-1-アンチトリプシン(AAT)およびチロシンアミノトランスフェラーゼ(TAT)を含んでいた。中胚葉発生は、brachuryおよびGATA-1の発現によって調べた。遺伝子の発現パターン以外に、EBからの神経突起の発達を調べることによって神経発生をアッセイした。これらの神経突起の神経性はβIII-チューブリンの発現によって実証した。中胚葉発生は、EBにおける造血細胞の原種を列挙することによっても評価した。図10は、10日間の分化期間のEBの発生能に対する血清の影響を示す。血清の存在下(血清)では、この系統の発生と関係がある遺伝子の発現の欠如によって実証されるように、神経外胚葉の分化はほとんどない。HNF3βは分化の初期段階(第2〜3日)で発現され、次いで下方制御される。予想通り、GATA-1はこれらの条件下で生成したEBにおいて発現される。これらの遺伝子の発現パターンは、血清の不在下(血清-)で増殖させたEBでは基本的に逆であった。これらのEBは神経外胚葉と関係がある全ての遺伝子を発現したが、中胚葉/造血マーカーGATA-1は発現しなかった。HNF3βはこれらの条件下で増殖させた後期段階のEB(第10日)において発現した。GFPの発現によって調べたbrachury発現のパターンは、これらの発見と一致した。血清の存在下で生成したEBは、分化の第2日と第5日の間に存在した相当数のbrachury+集団を生成した(図11、-B-ライン)。血清の不在下で増殖させたEBではbrachuryは検出されなかった(図11、-H-ライン)。造血細胞前駆体のアッセイによってこれらの発見を確認した。血清中で生成したEBは前駆体を含んでおり(図12、まだら線)、一方血清なしで増殖させたEBは含んでいなかった(図12、色塗り線、示さず)。最後に、EBの神経突起の発生能の評価は、これらの様々な分析と一致した。血清中で増殖させたEBは神経突起を生成しなかった。対照的に、血清の不在下で生成したEBの85%は、この活性を示した(図13)。一緒に考えると、これらの発見は、特定の系統を生成するための培養条件(血清)の重要性を実証する。これらの発見はさらに、血清完全条件も無血清条件も内胚葉発生には最適ではなかったことを実証する。
【0079】
血清中で生成した初期段階のEBにおけるHNF3βの強い上方制御は、血清は内胚葉系統の成熟ではなく、樹立に重要である可能性があることを示唆した。この可能性を試験するために、EBは血清の存在下で2日間分化開始させ、次いで無血清状態(SR)に移した。図14中に示すように、これらの条件(血清+/-)下で生成したEBは、分化の第3日と第5日の間でHNF3βを発現した。AFPは分化の第5日で上方制御された。GATA-1の発現レベルは、血清刺激したEBにおいて見られたレベルと比較して低下した。次に、血清+/-条件下で生成した第6日EBを、増殖因子bFGFの存在下で組織培養グレード皿に平板培養し(それらを接着させ)た。これらの皿をゼラチンまたはマトリゲルのいずれかでコーティングして、さらなる内胚葉分化に対して基質が何らかの影響を有していたかどうかを測定した。5日後、培地を交換し、他の因子をこれらの培養物に加えて、肝臓系統の発生を促進した。実験の概要およびデータは図15中に示す。この実験では、第6日のEB段階でAFPは発現されなかった。ゼラチンまたはマトリゲルのいずれか上でbFGFの存在下において培養すると、その発現レベルは上方制御された。低レベルのALBもこの段階で検出した。試験した全ての条件において追加的な培養期間後、ALBの発現は増大した。最終培養ステップ後にAATおよびTATも発現された。EB由来細胞をbFGFおよびDexの存在下で培養したとき、最高レベルのTATが見られた。これらの発見は、内胚葉系統の細胞を生成することは可能であること、および適切な条件下において、それらの細胞は発生中の肝臓と関係がある遺伝子を発現する細胞に分化することを明らかに示す。
【0080】
これらの内胚葉細胞がbrachury+またはbrachury-細胞から発生したかどうかを、さらに測定した。この問題に取り組むため、GFP (Bry)+およびGFP(Bry)-細胞を、細胞選別によって第2.5日EBから単離した。これらの集団を再凝集させ、第6日まで集合体として培養した。第6日に、それらを組織培養グレード皿へ、bFGFを含む培地中に4日間移した(合計10日間)。遺伝子発現の分析によって、HNF3βを発現する細胞はGFP+画分(d2.5)に分離したことを示した(図16)。培養時間と共に、この遺伝子はGFP-画分から生成した細胞中で発現された。これはおそらく、発現の後期段階でHNF3βが非内胚葉集団中で発現される事実を反映する。AFP、HEX、ALBおよびHNF4はいずれもGFP+画分の誘導体中で発現されたが、GFP-細胞から生成した細胞集団中では発現されなかった。対照的に、PAX6およびneruoD、神経外胚葉のマーカーは、GFP-画分から生成した細胞中で主に見られた。これらの発見は、内胚葉系統はbrachury+集団から樹立され、中胚葉系統にさらに分化し、これらの系統は共通の前駆体、内中胚葉に由来することを示す。
【0081】
brachury発現細胞の肝臓発生能をさらに評価するために、bry+画分とbry-画分の両方由来の細胞集団を、α-フェトプロテイン(AFP)、アルブミン(ALB)およびトランスチレチン(TTR) などの初期肝細胞発生を表す遺伝子、およびα1-アンチトリプシン(AAT)、チロシンアミノトランスフェラーゼ(TAT)およびカルバモイルリン酸シンテターゼI(CPase)を含めたこの系統の成熟を示す遺伝子の発現に関して分析した。β-アクチンの発現は対照として使用した。選別前に細胞を分析し、後にbry+集団とbry-集団に選別した。胎児の肝臓および成体の肝臓対照も分析した。全てのこれらの遺伝子の発現はbry+集団由来の細胞に限られており、肝細胞の特徴を有する細胞はbrachury発現細胞から発生することが示された。
(実施例10)
【0082】
EBにおける中胚葉および内胚葉発生の動態
実施例6中に記載した事前動態分析によって、明確な発生運命を有する中胚葉の亜集団は明確で一時的な形式で生成したことを実証した。より詳細な動態分析によって、分化の第2.5日と第4.0日の間にGFP+Flk1-(これ以後はGFP+集団と呼ぶ)集団の動的発生を示した(図18)。単離し再凝集させると、第2.5日および第3.0日GFP+画分は、造血および内皮の拘束の初期段階(図19)を表す芽細胞コロニーを生成した(図19中に血管芽細胞の発生能として示す)。このGFP+細胞集団は、心筋細胞(心臓)発生能をほとんど示さなかった。初期のGFP+細胞とは対照的に、第3.5日EBから単離した細胞は著しく低下したBL-CFC発生能を示したが、心筋細胞への分化においては効率が良かった。第4日EBから単離したGFP+細胞は心筋細胞には分化せず、BL-CFCを生成する能力をほとんど有しておらず、それらは何か他の中胚葉系統になるよう運命付けられている可能性があることが示唆された。遺伝子発現の分析はこれらの機能アッセイを支持し、4つのGFP+画分間の分子差異を実証した(図20)。幾つかの遺伝子は全集団中で発現されたが、他の遺伝子は興味深い分化パターンを示した。造血発生および心臓分化の阻害に重要であると考えられる遺伝子Wnt3aは、第2.5日および第3.0日集団において発現され、第3.5日と第4.0日細胞において下方制御された。このパターンは、造血/血管から心筋へのこれらの集団の発生能の変化と一致する。当該の第二のパターンは遺伝子Mixl-1のパターンである。中胚葉および内胚葉の発生において役割を果たすこの遺伝子は、第2.5日および第3.0日GFP集団において発現されたが、第3.5日と第4.0日画分では発現されなかった。一緒に考えると、この実施例の発見は、異なる発生能を有する中胚葉集団はEB内では明確で一時的な形式で生成することを、明らかに実証する。さらに、中胚葉/内胚葉遺伝子Mixl-1の発現分析によって、内胚葉発生能を有する細胞も特定の時期、すなわち分化の第2.5日と第3.0日の間に生成することを示す。
【0083】
内胚葉発生の動態をさらに調べるために、第3.0日および第4.0日EB由来のGFP+細胞を単離し、肝細胞様細胞への分化を促進する条件下で培養した。図21中に示すように、第4.0日由来のEBではなく第3.0日EBから単離したGFP+細胞(+/-)が、HNF3βの発現によって定義されるように内胚葉発生能を示した。これらの発見は、内胚葉が特定の時期、分化の第4日の前にGFP+集団内で生成することを示す。
(実施例11)
【0084】
in vivoでのGFP+集団の発生能
GFP+集団の内胚葉発生能をさらに評価するために、GFP+およびGFP-第2.5日EB細胞を肝細胞の分化を促進することが知られている条件下で14日間培養し、レシピエントSCID-ベージュマウスの腎被膜下に次いで移植した。数匹のマウスを移植直後に殺傷し、移植片を有する腎臓を切片化し、切片はHep1およびAFPに対する抗体で染色した。Hep1は肝細胞の特異的マーカーであり、一方AFPは肝細胞系統の最終段階内胚葉および未熟細胞において発現される。この切片内の幾つかの細胞はHep1に関して陽性染色され、一方Hep1+細胞の近辺の他の細胞はAFPを発現することが分かった。Hep1+またはAFP+細胞は、GFP-陰性細胞の移植片内には見られなかった。Hep1の発現によって定義されるように、これらの知見はPCRデータを支持し、その培養条件が未熟肝細胞の特徴を有する細胞の発生を支持することを実証する。
【0085】
これらの移植実験は移植直後の移植片中の肝細胞様細胞の存在を実証するが、移植した組織が、奇形腫として知られる腫瘍様の塊を生成したので、経時的にこれらの集団の成熟を調べることは困難であった。奇形腫はおそらく汚染した未分化ES細胞から、あるいは中胚葉起源でありbrachuryを発現することが知られているGEP+原始生殖細胞から発生する。
(実施例12)
【0086】
アクチビンによる中胚葉および内胚葉の誘導
内胚葉発生能を有する細胞をさらに豊富にするために、他のモデル系においてこの細胞集団を誘導することが知られている増殖因子の影響を試験した。アフリカツメガエルにおける試験によって、アクチビンが培養中に外胚葉から中胚葉と内胚葉の両方を誘導することが示されてきている。特に興味深かったのは、それが使用した異なる濃度で異なる細胞型を誘導した点で、アクチビンはこの系においてモルホゲンとして働いたという観察結果であった。アクチビンがES/EB系において同様の発生能を示したかどうかを測定するために、以下のプロトコルを使用してアクチビンをEB培養物に加えた。ES細胞は、血清を含まないStem Pro 34培地中で2日間分化した。この段階で、発生中のEBを採取し、血清物(無血清)および100ng/mlの濃度でアクチビンを補ったIMDM中で再度培養した。EBは異なる日数で採取し、GFP+の発現および外胚葉、中胚葉および内胚葉発生を示す遺伝子の発現に関してアッセイした。図22A、B中に示すように、GFPにより測定して、この量のアクチビンはbrachuryを誘導した。GFP誘導の動態は血清中で分化したEBと比較して遅れたが、この濃度のアクチビンは、分化の第6日までに相当数のbrachury陽性細胞(60%)を誘導した。分子分析によって、アクチビン誘導型細胞はHNF3β、Mixl-1、Sox17、Hex-1、およびpdx-1を含めた内胚葉発生と関係がある広範囲の遺伝子を発現したことを示した(図23C)。この遺伝子は膵臓発生に必要不可欠であるので、pdx-1の誘導は興味深い。GATA-1などの造血発生と関係がある遺伝子、およびPAX6などの神経外胚葉分化を示す遺伝子は、アクチビンによって誘導されなかった。
【0087】
アクチビンがES分化モデルにおいてモルホゲンの性質を示したかどうかを測定するために、異なる濃度の因子をEB培養物に加えた。わずか1ng/mlのアクチビンが、培養の7日までにGFP+の発現(集団全体の10%)を誘導した(図23A)。GFP+細胞の頻度は3ng/mlで刺激した培養物中では40%まで増大し、30ng/mlで50%を超える安定レベルに達した。これらの集団の遺伝子発現分析によって、異なる濃度のアクチビンは異なる発生プログラムを誘導したことを示した。1または3ng/mlのアクチビンの存在下で分化したEBは、内胚葉発生を示す遺伝子の、存在した場合でも弱い発現を示した(図23A)。HNF3β、Sox17、Hex-1はいずれも、10、30、または100ng/mlのアクチビンで刺激した培養物において誘導された。Pdx-1の発現は最高量のアクチビンを必要とし、100ng/mlで刺激したEBにおいて最も誘導された。いずれの濃度のアクチビンにおいてもGATA1およびc-fmsは発現されなかった。PAX6発現のパターンは内胚葉遺伝子のそれとは逆であり、アクチビンの増大する濃度と共に下方制御された。
【0088】
アクチビン誘導型EBは、造血への拘束と関係がある遺伝子を発現せず、この発生プログラムは誘導されなかったことが示された。これらのEBの造血発生能をさらに評価するために、原種の発生能に関してこれらを分析した。分子分析から予想したように、これらのEBは測定可能な数の初期(Ep)または最終段階(mac/mix)造血細胞原種を含んでいなかった(図24A)。これらのアクチビン誘導型EBを2.5日間血清でさらに刺激すると、しかしながら、それらは幾らかの造血細胞原種を生成し、それらは中胚葉発生能を含むことが示された(図24B)。
【0089】
PCR分析によって、アクチビンはHNF3β、および内胚葉分化と関係あることが知られている他の遺伝子の発現を誘導したことを実証した。アクチビン誘導型EBにおける内胚葉原種の割合をさらに推定するために、100ng/mlのアクチビンで刺激した培養物由来の細胞を、HNF3βおよびHex1に対する抗体で染色した。非誘導型培養物由来のEBは対照として使用した(アクチビン-)。相当な割合のアクチビン誘導型集団(推定で全体の50〜60%)は、HNF3βとHex1の両方を発現した。非誘導型EBにおける細胞は、これらのタンパク質を発現しなかった。これらの発見は、これらのEB内の相当数の細胞が内胚葉系統であることを明らかに実証する。
【0090】
これらのアクチビン誘導型集団の発生能をさらに調べるために、GFP+細胞を3ngまたは100ngで刺激したEBから単離し、肝細胞誘導状態でさらに培養した(合計14日間)。図25中に示すように、100ng培養物由来の細胞のみがアルブミンを発現した細胞に分化し、肝臓の分化と一致した。一緒に考えると、これらの試験からの発見は、ES/EB系におけるモルホゲンとしてアクチビンが機能すること、および内胚葉誘導には高濃度が必要とされることを示す。
【0091】
血清誘導型brachury+細胞によって生成される奇形腫が、原始生殖細胞の存在に原因がある場合、生殖細胞のプログラムは、これらの条件下では誘導されない可能性があるのでアクチビン誘導型細胞が移植元の良い供給源となり得ることが考えられる。この仮説を試験するために、GFP+およびGFP-細胞を100ng/mlのアクチビンで誘導したEBから単離し、14日間培養して肝細胞様細胞の分化を促進した。この培養期間後、細胞を採取しレシピエント動物の腎被膜下に移植した。移植後3週間で、マウスを殺傷し腎臓を分析した。結果は図26A〜C中に示す。GFP-細胞を移植した全てのマウスが、3つ全ての生殖層由来の細胞からなる大きな多系統の奇形腫を発生した。対照的に、GFP+細胞を移植した動物中では奇形腫を検出しなかった。これらの細胞は、腸上皮、骨および骨格筋を含めた内胚葉および中胚葉由来組織からなる分化した細胞塊を生成する。幾つかの場合、GFP+細胞からの移植片中に皮膚も観察され、この系統がbry+細胞から発生する可能性もあることが示唆された。これらの発見は、移植後に奇形腫を形成せずに分化組織を生成するGFP+集団を作製することができることを示す。
(実施例13)
【0092】
bry+/c-kit-およびbry+/c-kit+細胞の発生能
前述の実施例は、中胚葉と内胚葉の両方の発生能を有するbry+細胞集団から肝細胞系統が発生することを明らかに示す。bry+画分の分析によって、これらの細胞の亜集団が受容体チロシンキナーゼc-kitを発現したこと(図27A)、およびこの集団がFlk-1を発現した細胞とは異なっていたことが明らかになった。c-kitの発現が内胚葉発生能を有する細胞の分離の有用なマーカーであるかどうか決定するために、第3日の血清刺激したEB由来のbry+/c-kit-(+/-)およびbry+/c-kit+(+/+)細胞を肝細胞発生能に関してアッセイした。図27B中に示すように、HNF3β、AFPおよびALB発現細胞は、いずれもbry+/c-kit+集団に由来した。この画分の内胚葉発生能を推定するために、選別した細胞をカバーガラスに平板培養し、HNF3βに対する抗体で染色した。80%を超えるbry+/c-kit+細胞がHNF3βタンパク質を発現し、一方10%未満のbry+/c-kit-が陽性であった。これらの発見は、内胚葉原種はbrachuryとc-kitの両方を発現すること、およびこの集団は内胚葉発生能を有する細胞が非常に豊富であることを示す。brachuryおよびc-kitの発現に基づく細胞の単離は、内胚葉原種を単離するための新規な戦略となる。
(実施例14)
【0093】
アクチビン誘導型細胞の発生能
図23中に表したPCR分析は、異なる濃度のアクチビンは異なる発生プログラムを誘導すること、および内胚葉発生能を有する細胞は最高レベルのこの因子によって誘導されることを示す。アクチビンの分化応答を定量化するために、異なる濃度のアクチビンで刺激した細胞をカバーガラスに接着させ、抗HNF3β抗体で染色した。100ng/mlのアクチビンで刺激したEB集団全体の50%より多くがHNF3βを発現し、一方3ng/mlで刺激した細胞のわずか10%が陽性であった。バックグラウンドレベルの染色のみを、非刺激集団において観察した。これらの発見は、高濃度のアクチビンは、EB集団全体の相当な割合が示す確かな内胚葉プログラムを刺激し得ることを実証する。
【0094】
アクチビン処理細胞の発生能の他の評価として、異なる濃度のこの因子の存在下で分化させた第6日EBを4日間血清物培地に移し、次いで肝細胞状態でさらに4日間再度平板培養した。培養の第14日に、各群由来の細胞を採取しPCRによる発現分析に供した。Myf5および骨格筋アクチンの発現を調べて、他の中胚葉由来系統を表す骨格筋の発生を評価した。サーファクタントプロテインC(SP-C)、肺特異的遺伝子は、AFPおよびALBに加えて内胚葉分化のマーカーとして含めた。図28中に示すように、Myf5および骨格筋アクチンはわずか1ng/mlのアクチビンで刺激した培養物中で発現し、この発現は広範囲の因子濃度で検出した。両遺伝子の発現は、しかしながら最高濃度(100ng/ml)のアクチビンで下方制御された。少量のアクチビンで刺激した培養物は、骨格筋の形態を有する細胞群を含んでいた。免疫染色によって、これらの細胞が骨格筋ミオシンとα-アクチニンの両方を発現したことを実証し、これらが骨格筋系統であることを示した。図28中に示すように、3および10ng/mlで刺激したEBは最も確かな骨格筋の発生を示したので、骨格筋の発達をもたらした再度平板培養したEBの割合の評価は遺伝子発現の分析と一致した。3つの内胚葉遺伝子の発現パターンは、骨格筋遺伝子に関して観察した発現パターンとは異なっていた。いずれも低いアクチビン濃度では発現されず、最高濃度の因子で刺激した培養物においていずれも容易に検出した。PAX6の発現は、未処理培養物および低濃度の因子で刺激した培養物に限られた。この分析からの発見によって、異なる濃度のアクチビンが異なる発生プログラムを誘導し、低濃度は中胚葉の運命に好ましく、高濃度は内胚葉の運命に好ましいことを実証した際の、本明細書中の前の実施例12からの発見が確認され広がる。さらに、これらの結果は、アクチビンによって誘導された内胚葉細胞は、肝細胞および肺の特徴を有する細胞に分化しそれらを生成することができることを示す。
【0095】
低濃度および高濃度の因子で刺激したEBから単離したbrachury陽性および陰性集団も、骨格筋および内胚葉遺伝子の発現に関して分析した。図29中に示すように、myf5および骨格筋アクチンの発現は、3ng/mlのアクチビンで刺激したEBから単離したbry+集団から生成した集団に限られた。同様に、内胚葉遺伝子は、100ng/mlの因子の存在下で生成したEBから単離したbrachury+由来細胞中で発現された。これらの発見は、中胚葉と内胚葉の両方がbrachury+細胞から発生することをさらに実証する。
(実施例15)
【0096】
HNF-CD4標的ES細胞の生成
EB分化中に内胚葉および中胚葉集団を追跡し単離するために、brachury遺伝子座に標的化したGFPを既に有するES細胞を、ヒトCD4(hCD4)タンパク質と共にHNF3β遺伝子座に標的化した。HNF3βは大部分の内胚葉細胞型において発現され、胚発生中の非常に初期に始まることが知られている。Kaestnerら(1994) Genomics 20: 177〜85。標的構築体は、アミノ酸番号424で切断され細胞内ドメインの大部分を欠くヒトCD4のcDNAを含む。シグナルを伝達することができず、hCD4を認識し内因性のマウスCD4と交差反応しないはずである、フローサイトメトリー用の容易に入手可能な抗体が存在するので、この細胞表面分子を選択した。図30A中に示すように、このベクターは、マウスHNF3β遺伝子座と相同性がある2つの領域、ウシ成長ホルモンポリ(A)配列、およびloxpに隣接するピューロマイシンカセットも含む。ジフテリアトキシンA(DTA)遺伝子を標的ベクターの5'末端に含めて、ランダムな組み込みを選択した。標的ベクターは以下のように構築した。
【0097】
相同性のある短いアームと長いアームを、公開済みのHNF3β標的ベクターから得た。Weinsteinら(1994) Cell78: 575〜588。同じ長さのアームを使用し、一方1.8kb断片の短いアームを使用してベクター中にhCD4ノックを作製した。短いアームは、HNF3βプロモーターの一部分、エクソン1、イントロン1、およびエクソン2の一部分、内因性ATG開始部位の直前に末端を含む。長いアームは、最終エクソンの末端の3kb下流で始まる7kbの配列からなる。標準的な分子生物学の技法を使用して、コザック配列次にウシ成長ホルモンポリ(A)配列およびピューロマイシンカセットを有するhCD4を、図30A中に示すように相同性のある2つのアーム間にクローニングした。
【0098】
実施例2中に記載したGFP-BryES細胞に、HNF3β標的ベクターをエレクトロポレーションにより導入した。次いでピューロマイシンを用いて細胞を選択し、クローンを採取し増大させた。ゲノムDNAを単離し、制限酵素SacIで切断し、図30B中に示すように内因性ATGの上流のプローブを用いてサザンブロット分析を実施した。野生型対立遺伝子は3.3kbのバンドを与え、一方標的対立遺伝子は2.8kbの大きさのバンドを与える。スクリーニングした48のクローンのうち、2つの標的クローンが生成し、4.2%の標的化効率に相当した。両方の標的クローンは、改変型Creリコンビナーゼ発現ベクターを用いて一過的にトランスフェクトしてpuro遺伝子を除去した。この標的クローンは、本明細書では以後CD4-HNF/GFP-T ES細胞と呼ぶ。
【0099】
HNF3βは、血清誘導分化中にEBにおいて一過的に発現される。CD4-HNF/GFP-T ES細胞中でのhCD4の発現が、EB発生中のHNF3β遺伝子の発現を正確に反映するかどうか測定するために、CD4対GFPの発現を評価した。
【0100】
5日間血清誘導EBの分化中のGFP対hCD4の典型的な発現パターンを図31A中に示す。ES細胞はhCD4発現が低いから陰性である。分化後、hCD4の発現は特に第3日のGFP+細胞において急激に上方制御される。発現は第3.5日までに急速に低下し、第5日までにはほぼ検出不能である。対照的に、GFPの発現は第4日まで持続し、第5日までに低下する。GFPおよびhCD4発現の平均蛍光強度は、図31B中に示す。hCD4のピークは第3日であり直ぐに消失し、一方GFPのピークは第3.5日〜第4日であることが分かる。HNF3βおよびbrachury発現に関する半定量RT PCRも、これらのサンプルにおいて実施した(図31C)。内因性遺伝子に関するPCRは、GFPおよびhCD4に関するフローサイトメトリーを使用して見られたものに従う。HNF3βとbrachuryはいずれも第3日までに発現され、一方HNF3βの発現は第3.5日までに低下し、第5日までには検出不能である。brachuryの発現は第4日まで高いレベルを保ち続ける。これらのデータによって、hCD4は内因性HNF3βの発現を反映していることが確認される。さらに、hCD4の発現はHNF3βのRNAレベルと共に低下することが分かり、hCD4タンパク質の半減期は過度に長いわけではなく、それを使用してHNF3βを発現する細胞を正確に選択することができることが示唆される。全てのGFP+細胞が最初はhCD4+であるが、次いでhCD4の発現が急速に消失することも分かる。このデータは内胚葉と中胚葉の両方に対する内中胚葉前駆体を示唆し、実施例9中のデータを支持する。この集団はc.elegansなどの低級生物において実証されてきているが、哺乳動物においては樹立されていない。前に使用された条件を、確かな中胚葉誘導のために選択した。内中胚葉が形成されるが、急速に中胚葉に転換すると考えられる。
(実施例16)
【0101】
原条形成のモデルとしてのEB分化
胚発生中、中胚葉系統と内胚葉系統の両方が、原条と呼ばれる構造から誘導される。Beddington and Robertson (1999) Cell 96: 195〜209。原条全体で発現されるbrachuryなどの特定の遺伝子の発現によって、原条は特徴付けられる。HNF3βも原条中で発現されるが、それは前部でのみ検出されている。hCD4によって実証されたHNF3βの発現は、Bry-GFP+集団中では異種であることが分かった。血清誘導は原条様細胞の群の発生を可能にし、GFP+HNF3βlo細胞は原条後部になり、一方GFP+HNF3βhi細胞は原条前部に対応すると考えられる。
【0102】
GFPおよびhCD4の発現を使用して、CD4-HNF/GFP-T ES細胞から後部および前部原条様細胞を分離することができるかどうか測定するために、3.25日間分化したEBを3つの集団:GFP+hCD4lo、GFP+hCD4med、およびGFP+hCD4hiに選別した(図32A)。原条中で非対称的に発現されることが認められている、多くの遺伝子が存在する。原条後部または原条前部のいずれかにおいて発現されることが知られている遺伝子に関して、選別集団においてRT PCRを実施した(図32B)。原条後部特異的遺伝子Fgf8、Evx1、HoxB1、およびTbx6はいずれも、hCD4hi細胞と比較してhCD4loおよびhCD4med集団中において高レベルで発現された。対照的に、原条前部の遺伝子Chordin、Cerl、およびGscはいずれも、hCD4hi細胞において高レベルで発現され、hCD4med細胞においては発現が低いから陰性であり、hCD4lo細胞においては全く発現されなかった。HNF3βの発現も調べて、hCD4選別によって異なるレベルのHNF3β遺伝子を発現する細胞を単離したことを確認した。HNF3βの最高レベルはhCD4hi集団中で見られ、予想通りhCD4medおよびhCD4lo集団中では量が減少した。様々なhCD4集団中の発現パターンが、原条中の遺伝子発現を正確に表すものであることを確認するために、7.25日齢マウス胚の原条を切開し、遺伝子発現に関して分析した。GFP-BryES細胞由来のマウス胚を使用した。前部、中部、および原条後部組織を次いで蛍光下の切開範囲で切開して、得られた組織が胚のGFP+(原条)部分由来であったことを確実にした。図32Cは、切開した原条組織から実施したRT PCRの結果を示す。これらのデータは、選別したhCD4集団が、調べた原条前部および原条後部遺伝子に関して同様の相対的遺伝子発現プロファイルを示すことを示す。これらのデータは、CD4-HNF/GFP-T ES細胞におけるhCD4およびGFPの発現を使用して、原条前部および原条後部を示す遺伝子発現パターンを有する細胞を単離することができることを示す。
(実施例17)
【0103】
hCD4発現細胞の異なる集団の機能的発生能
系統追跡実験によって、原条前部と原条後部は異なる系統に発生することが実証されている。原条後部は卵黄嚢および造血細胞に分化し、一方原条前部は内胚葉および心臓組織などの前部中胚葉を形成する。Robb and Tam (2004) Sem.Cell & Dev.Biol.15: 543〜554。EB由来の原条様細胞が内因性原条と同様に機能的に働くかどうかを測定するために、CD4-HNF/GFP-T ES細胞を3.25日間分化させ、GFP+hCD4loおよびGFP+hCD4hi集団に選別し、造血、心臓、および内胚葉発生能に関してアッセイした。全てのアッセイ用に、選別した集団を無血清状態で48時間再凝集させた。図33Aは、再凝集後にマトリゲル上に平板培養した選別細胞の心臓発生能を示す。接着後24〜48時間の拍動細胞の存在に関して個々のEBを記録した。拍動EBはこの時点ではhCD4hi集団からのみ見られた。図33Bは、選別した集団の造血発生能を実証する。細胞は再凝集後にトリプシン処理し、造血増殖因子を含むメチルセルロース中に平板培養した。造血コロニーの存在を5〜7日後に計数した。造血コロニーの大部分はhCD4lo集団中で見られた。図33Cは、再凝集後に無血清状態でマトリゲル上に平板培養した選別集団の、内胚葉特異的遺伝子に関するRT PCRを示す。hCD4hi細胞のみが、マトリゲル上で5日後に内胚葉遺伝子HNF4、Pdx1、AAT、ALB、およびHNF3βを発現した。これらのデータは、hCD4およびGFPの発現を使用して、異なる発生能を有する集団を分離することができることを実証する。これらの発生能は、それぞれ原条後部および原条前部と同等であるhCD4loおよびhCD4hi細胞と一致する。
(実施例18)
【0104】
アクチビンシグナル伝達とWntシグナル伝達の両方が、ES由来原条様細胞に必要とされる
マウスにおける試験によって、原腸形成および内胚葉/中胚葉誘導における、Wntシグナル伝達およびノーダル/アクチビンシグナル伝達の重要性が実証されてきている。Liuら(1999) Nat.Genet.22: 361〜4。本明細書における前の実施例12は、アクチビンは無血清培養においてES細胞から、中胚葉集団と内胚葉集団の両方を誘導することができることを実証する。アクチビンおよびWntシグナル伝達のES由来原条様細胞の分化に対する影響を測定するために、組換えWnt3aまたはアクチビンAのいずれかを用いて、無血清培養においてCD4-HNF/GFP-T細胞を分化させた。アクチビンとWntの両方が最初にGFP+CD4+細胞を誘導することができるが、Wntがより速い動態を示した(図34A)。アクチビン刺激は、第6日のGFP-Bry発現の消失後持続する高レベルのCD4発現をもたらす。対照的に、Wntで誘導したGFP+細胞は低レベルのCD4を有しており、第5日までに発現が急速に消失する。アクチビンはGscおよびCer1などの他の原条前部マーカーの確かな発現をさらに誘導するが、一方Wntは誘導しない(図34B)。しかしながらWntは原条後部マーカーHoxB1およびTbx6の発現を刺激し、一方アクチビンはTbx6のみを誘導する。これらのデータは、アクチビンはES原条前部細胞を誘導し、WntはES原条後部細胞を誘導することを示唆する。第6日に、アクチビンまたはWnt3aのいずれかで刺激したEBを、マトリゲル上無血清培地中に平板培養して、さらに発生させた。さらに6日後、これらの培養物を採取し、内胚葉遺伝子の発現をRT-PCRによって測定した(図34C)。アクチビンによる誘導によって、内胚葉遺伝子Alb、Aat、HNF4、およびPdx1を発現する細胞を発生させることができ、実施例12における発見を確認した。しかしながら、Wnt3aで誘導した培養物はこれらのマーカーのいずれも発現せず、アクチビンシグナル伝達が内胚葉発生に必要とされることが示唆された。
【0105】
Wntによる刺激がノーダルを誘導し、アクチビン処理がWnt3を誘導することを考慮すると(図34B)、アクチビン/ノーダルシグナル伝達またはWntシグナル伝達単独で原条形成を誘導することができるかどうか、あるいは両方の因子が共に必要とされるかどうかは分からない。CD4-HNF/GFP-T ES細胞は、これらの経路の特異的阻害剤と共にアクチビンまたはWntのいずれかで誘導した。DKK1を使用してWntシグナル伝達を阻害し、一方で小分子阻害剤SB-431542(SB)を使用してノーダル/TGFβシグナル伝達を阻害した。DKK1はWntの共受容体、LRP5およびLRP6と結合することによって機能し、これらの受容体はβカテニンシグナル伝達経路の活性化に必要不可欠である。SBは受容体ALK4、5、および7の特異的阻害剤であり、したがってTGFβ、アクチビン、およびノーダルシグナル伝達を阻害することができる。しかしながらSBは、BMPシグナル伝達を阻害しない。Inman et.al. (2002) Mol.Pharmacol.62: 65〜74。両方の阻害剤共に、いずれかの誘導物質を用いたGFP+CD4+細胞の発生を完全に阻害した(図34A)。原条マーカーGsc、Cer1、HoxB1、およびTbx6も不在であったか、あるいは大幅に減少した(図34B)。これは、ES由来原条様細胞を誘導するために、Wntおよびアクチビン/ノーダルシグナル伝達が共に必要とされることを示す。したがって、中胚葉および内胚葉生殖層およびそれらの誘導組織を誘導するためには、アクチビン/ノーダルとWntシグナルの両方が最初に必要とされる。
(実施例19)
【0106】
アクチビンシグナル伝達およびWnt阻害は、内胚葉分化を誘導する可能性がある
実施例12は、高レベルのアクチビンは、完全に無血清の状態で分化したEBからの内胚葉遺伝子の発現を誘導する可能性があることを実証した。in vivoでの中胚葉/内胚葉誘導と関係がある重要な分子であるノーダルは、アクチビンとして同じ受容体を介して伝わる。ノーダルおよびWntシグナル伝達の阻害剤は、原条前部中あるいはその周辺で優先的に発現されることも知られている。内胚葉が原条前部から形成されることを考慮すると、Wntシグナルの阻害と組み合わせたノーダル/アクチビンシグナル伝達は、内胚葉形成を誘導すると考えられる。この仮説を試験するために、CD4-HNF/GFP-T ES細胞を、血清中で3.25日間分化させ、GFP+hCD4med細胞を選別し、因子を含まず、DKK1(Wnt阻害剤)、アクチビン、あるいはアクチビンとDKK1を含む無血清培地中で再凝集させた。hCD4-HNF3βおよびGFP-Bryレベルに対するこれらの培養条件の影響は、フローサイトメトリーによって分析した(図35A)。無血清培地単独では、hCD4の発現は再凝集1日後に消失した。DKK1を加えることによるWntシグナル伝達の阻害によって、幾つかの細胞は低レベルのhCD4を保持することができた。アクチビンの添加は、新たに選別した細胞と同等なHNF3βレベルの保持を可能にした。最後に、アクチビンとDKK1は相乗的に働いて、選別後のレベルよりhCD4のレベルをほぼ1ログ増大させた。GFPレベルは試験した全ての状態で消失し、全ての培養物において分化が進行していたが、より多くのhCD4を発現する培養物が内胚葉系統に分化していたことが示唆された。この概念をさらに試験するために、前述の培養条件で2日間再凝集させた選別細胞を、因子を含まない無血清培地で洗浄し、マトリゲル上無血清培地中に平板培養した。5日後、培養物を採取し、遺伝子の発現をRT PCRによって測定した。図35Bは膵臓、肝臓、および腸に特異的な幾つかの内胚葉遺伝子の発現を示す。培地単独またはDKK1処理培養物のいずれにおいても、これらの遺伝子は検出されなかった。非常に低レベルの2つの遺伝子、Pdx1およびALBのみを、アクチビンのみで処理した培養物において検出した。アクチビンとDKK1の両方を用いた処理は、Pdx1、AAT、HNF4、ALB、Cdx2、およびIFABPの高発現をもたらした。これらのデータは、アクチビン受容体を介したシグナル伝達とDKK1処理によるWntシグナルの阻害の組合せは、培地単独で培養すると内胚葉を形成しないはずである細胞集団からの内胚葉形成を誘導することを実証する。これらの培養条件には、膵臓および肝臓などの内胚葉由来組織の大規模な生成を可能にする可能性がある。これらの実験は、定量的に内胚葉発生を評価するためのツールとしての、CD4-HNF/GFP-T ES細胞の有用性をさらに実証する。
(実施例20)
【0107】
内胚葉由来マウス胚幹細胞からの肝細胞の作製
本明細書における前の実施例15中に記載したGFP-Bry、hCD4-Foxa2ES細胞系を使用することによって、内胚葉細胞をフローサイトメトリーによって、ES細胞分化時のそれらのGFPおよびhCD4発現によって追跡した(用語HNF3βとFoxa2は同義的に使用される)。
【0108】
GFP-Bry、hCD4-Foxa2ES細胞は血清の不在下に保ち、フィーダー細胞はNeurobasal、DMEM/F12、N2サプリメント、B27およびレチノイン酸、10%BSA、MTG(1.5×10-4M)、グルタミン、LIF(1000U/ml)およびBMP-4(10ng/ml)からなる培地中に保った。これらの条件において、細胞は依然未分化状態である堅いコロニーを形成する。内胚葉のプログラムを誘導するために、ES細胞を採取し、アクチビン-A(50ng/ml)の存在下、無血清培地(IMDM、F12培地、B27およびレチノイン酸、N2サプリメント、BSA、MTG、グルタミン、アスコルビン酸)中、低密度(35,000個細胞/5ml)、超低接着性の皿中で培養して、胚様体(EB)形成を可能にした。ES細胞が分化するとき、GFP-Bry、hCD4-Foxa2および幹細胞リガンドc-Kitの受容体の発現パターンを定義した。大きな細胞集団GFP-Bry+、hCD4-Foxa2highおよびcKithigh(名称+++)が第4日に出現し、それらの相当集団GFP-Bry+、HCD4lowおよびc-Kit-(名称+L-)に対して、その内胚葉発生能およびしたがって肝細胞運命に関してアッセイした。この問題に取り組むため、胚中の内胚葉細胞を肝臓に特化させることが知られている2つのサイトカイン、BMP-4(30ng/ml)およびbFGF(10ng/ml)の存在下において、前に示した無血清培地中で、両集団を2日間再凝集させた。Zaret(2001) Curr Opin Genet Dev 11: 568〜574。この2つのサイトカインに加えて、アクチビン-A(50ng/ml)を加えて、中胚葉運命と対抗する細胞の内胚葉発生能を保った。肝細胞原種(肝芽細胞)の成熟を可能にするために、次いで凝集体を、肝芽細胞/肝細胞の成熟および増殖に適した培地中のゼラチンコーティング皿上に平板培養した。この培地は、VEGF(10ng/ml)、デキサメタソン、および肝芽細胞/肝細胞の増殖を促進することが知られているブロック培地中に存在する他のサイトカインを補った、前述の分化用基礎培地からなる。これらはヒト組換え体HGF(20ng/ml)、EGF(10ng/ml)、bFGF(10ng/ml)およびTGFα(20ng/ml)を含む。Blockら、(1996) J Cell Biol 132: 1133〜1149。6日後、+++凝集体は皿に接着した無数の「肝細胞コロニー」、および接着しなかった多数の浮遊する凝集体に分化した。全ての+L-凝集体が皿に接着し非常に少数の「肝細胞コロニー」を形成したが、培養物中に浮遊する凝集体は見られなかった。これらの結果は、第4日の+++集団は内胚葉細胞が非常に豊富であることを示唆し、BMP-4、bFGFとアクチビン-Aの組合せは、第4日+++細胞の肝細胞系統への特化を強く促進することを実証する。
【0109】
ゼラチンコーティング皿上での平板培養後1日で、+++凝集体の2/3が培養物中に依然として浮遊する状態であり、一方他の1/3は接着し、他の細胞によって時折囲まれた肝細胞コロニーを形成した。この初期の時点では、免疫組織化学法によりアッセイして、コロニー内の多くの細胞がアルブミンおよびAFPを発現した。周囲の細胞はいずれもCD31陽性細胞であり、それらが内皮細胞であることが示唆された。
【0110】
第12日以後、肝細胞コロニーはさらに大きく成長した。この後期の時点では、コロニー内の大部分の細胞はAFPおよびアルブミン陽性であり、コロニー周囲の大部分の細胞は予定内皮CD31陽性細胞である。hCD4に関する免疫組織化学法によって、肝細胞コロニー中に含まれていた全ての細胞がfoxa2を発現したこと、およびこの発現がこれらのコロニーに限られていたことを示した。これらの結果によって、内胚葉起源の肝細胞コロニーを確認した。
【0111】
同様に、第12日に浮遊する+++凝集体はさらに大きく成長し、AFPおよびアルブミンに関する免疫組織化学法用に処理した。大部分の凝集体は中空状態であり、凝集体を構成する細胞はAFPおよびアルブミンを発現し、一方IgG対照部分は陰性であった。さらに、凝集体内のAFPおよびアルブミンも染色され、この2つのタンパク質の分泌が示唆された。
【0112】
第12日の+++培養物をさらに特徴付けるために、フローサイトメトリー分析を接着した培養物と浮遊する凝集体の両方に実施して、内胚葉由来集団(hCD4-Foxa2+)、内皮細胞集団(CD31+またはPECAM-1)、肝芽細胞/肝細胞集団(hCD4++alb+、またはhCD4+/AFP+)を定量化した(図36A、36B)。接着した培養物中に含まれていた細胞の30%〜40%は内胚葉マーカーhCD4-Foxa2に陽性であり、一方50%はCD31+内皮細胞であった。浮遊する凝集体は、大きな(50%〜60%)hCD4-Foxa2+細胞、およびCD31+内皮細胞の小さな細胞集団(30%)からなっていた(図36A)。大部分のhCD4-Foxa2+細胞はさらに、細胞質間染色によりアッセイしてアルブミン+およびAFP+であった(図36B)。RT-PCRによる遺伝子発現の分析によって、第12日の+++培養物由来の接着した培養物(a)および浮遊する凝集体(f)はfoxa2、AFPおよびアルブミンを発現したことを確認した(図36C)。
【0113】
機能アッセイを実施して、肝細胞の同一性を証明した。Acid period Schiffアッセイは、成熟肝細胞に特徴的な細胞質中のグリコーゲン貯蔵の存在を示す。第12日の+++の接着した培養物は皿中で直接染色し、グリコーゲンに関する典型的な赤い細胞質染色を含むコロニー内に多数の細胞を示した。さらに、第12日+++の浮遊する凝集体に電子顕微鏡による分析を実施した。浮遊する凝集体を構成した大部分の細胞は、大きな核/細胞質比、多量のグリコーゲン貯蔵および毛細胆管の存在を含めた肝細胞の特徴を示した。
【0114】
全体としてこれらのデータによって、アクチビン-A誘導によるES細胞から内胚葉段階への発生進行、BMP-4、アクチビンAおよびbFGFの組合せによる肝臓特化ステップ、および最後に肝芽細胞/肝細胞集団の増殖/成熟をin vitroで模倣することにより、マウスES細胞から肝細胞が効率良く生成することを示す。
【図面の簡単な説明】
【0115】
【図1】緑色蛍光タンパク質(GFP)のbrachury遺伝子座への標的化のために使用するベクターのスキームおよび戦略を示す図である。
【図2A】発生段階の胚様体(EB)におけるGFPおよびbrachuryの発現を示す図である。逆転写酵素ポリメラーゼ連鎖反応(RT-PCR)によって測定したbrachuryの発現の動態を示す図である。
【図2B】発生段階の胚様体(EB)におけるGFPおよびbrachuryの発現を示す図である。蛍光活性化細胞選別(FACS)分析によって測定したGFP発現の動態を示す図である。前の図2A中の図および図2B中のヒストグラムにおける数字はEB分化の日数を表す。
【図3A】野生型およびGFP-BryES細胞の発生能を示す図である。第6日のEBの発生能を示すヒストグラムである(Mac/Ery:マクロファージおよび最終段階赤血球細胞のコロニー;Mac:純粋なマクロファージコロニー;Eryd:最終段階赤血球細胞のコロニー; Mix:多系統コロニー;Eryp:初期赤血球コロニー)。
【図3B】野生型およびGFP-BryES細胞の発生能を示す図である。EBの胚芽コロニー形成細胞(BL-CFC)の発生能を示すヒストグラムである。
【図3C】野生型およびGFP-BryES細胞の発生能を示す図である。野生型およびGFP-Bry細胞に関するEB発生中の遺伝子発現パターンを示す図である。レーンの上の数字はEB分化の日数を表す。
【図4A】GFPに基づいて単離したEB画分の遺伝子発現プロファイルを示す図である。第3.5日EBにおけるGFP発現のプロファイルを示す図である。1および2は、GFP-画分とGFP+画分を単離するために使用したゲートを表す。
【図4B】GFPに基づいて単離したEB画分の遺伝子発現プロファイルを示す図である。単離画分のRT-PCRによる発現分析を示す図である。
【図5A】GFPおよびFlk-1集団の単離および特徴付けを示す図である。第3.0日および第3.5日EBからGFP-/Flk-1-、GFP+/Flk-1-およびGFP+/Flk-1+画分を単離するために使用した、プロファイルおよびゲートを示す図である。ゲートの隣の数字は3つの異なる集団を表す。
【図5B】GFPおよびFlk-1集団の単離および特徴付けを示す図である。異なる画分の胚芽細胞コロニー(Blast)および二次EB(2°)発生能を示す図である。
【図5C】GFPおよびFlk-1集団の単離および特徴付けを示す図である。単離画分の発現分析を示す図である。上図に示す発現は、Bradyら(1990) Meth. In Mol. And Cell Bio.2: 17〜25によって記載されたポリA+全体増幅PCR法を使用して評価した。下図中のデータは、遺伝子特異的オリゴヌクレオチドを使用するRT-PCR分析によって得た。各列の上の数字は、図5A中に表した細胞集団を示す。
【図6】単離した第3日EB由来の画分におけるGFPおよびFlk-1の発現を示す図である。上列は、培養前(pre)の3つの画分の発現プロファイルを示す。下列は、培養20時間後(post)の同じ細胞集団のプロファイルを示す。各プロファイル下の数字は、各集団のBL-CFCおよび初期赤血球(Eryp-CFC)の発生能(平板培養した1×105個の細胞当たりの前駆体数)を示す。
【図7A】培養前後の単離した細胞集団のBL-CFC発生能およびFlk-1発現を示す図である。底部の数字は細胞集団を指す:1は予め選別した画分であり、3はGFP+/Flk-1-画分であり、4はGFP+/Flk-1+画分である。細胞は20時間培養し、次いで凝集体を解離させ、BL-CFCに関して分析した。データは第3、3.5および4.0日EBから単離した細胞に関して示す。
【図7B】培養前後の単離した細胞集団のBL-CFC発生能およびFlk-1発現を示す図である。上列は、培養前(pre)の第3、3.5および4.0日EBから単離したGFP+/Flk-1-細胞を表す。下列は、培養後(post)の同じ画分のFlk-1の発現パターンを示す。バーの上の数字はFlk-1+細胞の割合を表す。
【図8A】各ヒストグラムの上に示した条件下での、第3.0日EB由来細胞におけるbrachuryおよびFlk-1+の発生に対するBMP-4およびウシ胎児血清(FCS)の影響を示す図である。
【図8B】各ヒストグラムの上に示した条件下での、第3.0日EB由来細胞におけるbrachuryおよびFlk-1+の発生に対するBMP-4およびウシ胎児血清(FCS)の影響を示す図である。示した条件下で20時間培養したGFP+//Flk-1-細胞から作製した細胞集団における、brachuryおよびFlk-1の発現を示す図である。
【図9】EBにおける中胚葉形成および特化のスキームモデルである。
【図10】血清の存在および不在下でのEBにおける遺伝子の発現を示す図である。
【図11】異なる条件下で作製したEBにおけるbrachuryの発現を示すグラフである。
【図12】血清中で生成したEBは前駆体を含んでいる(まだら線)ことを示す図である。
【図13】血清の存在および不在下での神経分化を示すスキーム図である。
【図14】血清中2日間で始め次いで無血清状態に移したEBにおける、遺伝子の発現を示す図である。
【図15】bFGFの存在下で培養したEBにおける遺伝子発現を示す図である。
【図16】bFGFの存在下で培養したBry+およびBry-細胞における、遺伝子発現パターンを示す図である。
【図17】本発明の中胚葉および内胚葉細胞集団、および誘導体細胞型へのこれらの集団の分化の図である。
【図18】2.5、3.0、3.5および4.0日間分化したEBにおけるGFP(brachury)およびFlk-1の発現の動態を示す図である。矢印は、後の試験中の分析に使用した単離GFP+集団を示す。
【図19】EB分化の4段階から単離したGFP+集団の、血管芽細胞および心臓細胞発生能を示す図である。各段階からの細胞を細胞選別によって単離し、24時間再凝集させ、造血および心臓細胞発生能に関して分析した。再凝集培養物から回収した1×105個の細胞当たりの芽細胞コロニー(血管芽細胞)として、あるいは心筋分化を示す拍動細胞塊に分化した凝集体の割合として、データを示す。
【図20】4つのGFP+EB由来細胞集団における示した遺伝子の、RT-PCRによる発現分析の図である。数字はEB分化の日数を示す。
【図21】第3.0日および4.0日EBから単離したGFP+集団におけるHNF3βの発現を示す図である。Preは選別前の細胞を表し、-/-はGFPまたはFlk-1を発現しない細胞であり、かつ+/-はGFP+Flk-1-集団を表す。
【図22A】無血清培養物中のEBの発生に対するアクチビンの影響を示す図である。100ng/mlのアクチビンの存在下で分化した第6日EBにおけるGFPの発現を示すFACSプロファイル。
【図22B】無血清培養中のEBの発生に対するアクチビンの影響を示す図である。100ng/mlのアクチビンを含む培養物中のGFP誘導の動態。白丸はアクチビンの存在下で分化したEBであり、黒い四角形はアクチビンの不在下で分化したEBである。
【図22C】無血清培養中のEBの発生に対するアクチビンの影響を示す図である。アクチビンの存在下(+アクチビン)または不在下(-アクチビン)で増殖させた第6日EBにおける示した遺伝子の、RT-PCRによる発現分析。数字はEB分化の日数を示す。
【図23A】EBの発生能に対する異なる濃度のアクチビンの影響を示す図である。異なる濃度のアクチビンを用いて誘導した第7日EBにおけるGFPの発現。
【図23B】EBの発生能に対する異なる濃度のアクチビンの影響を示す図である。異なる濃度のアクチビンを用いて誘導した第7日EBのRT-PCRによる発現分析。
【図24A】異なる濃度のアクチビンの存在下で分化したEBの、造血細胞の原種の含有量を示す図である。第7日EBの原種の発生能。Epは初期赤血球の原種であり、mac/mixは最終段階造血細胞の原種を表す。
【図24B】異なる濃度のアクチビンの存在下で分化したEBの、造血細胞の原種の含有量を示す図である。血清に2.5日曝した後の、第7日アクチビン誘導型EBの原種の発生能。
【図25】3ng/mlまたは100ng/mlのいずれかのアクチビンを用いて誘導したGFP+細胞由来のアルブミン発現細胞の発生を示す図である。GFP+およびGFP-細胞を分化の第6日で単離し、前に記載した条件でさらに8日間培養して肝細胞の分化を助長した。
【図26A】bry+細胞集団の3週齢の腎臓移植片を示す図である。
【図26B】bry-細胞集団の3週齢の腎臓移植片を示す図である。
【図26C】bry+およびbry-集団の移植片の切片を示す図である。
【図27A】第3日に血清刺激したEBから単離したbry+/c-kit+(+/+)およびbry+/c-kit-(+/-)画分を示す、FACSプロファイルの図である。数字はそれぞれの画分中の細胞の割合を表す。
【図27B】それぞれの画分の発現の分析を示す図である。第3日は選別の直後に分析した細胞を表す。第15日は肝細胞状態で15日間培養した細胞集団を表す。
【図28】異なる濃度のアクチビンを用いて誘導したEB由来の細胞集団の発現の分析を示す図である。図面の上部の数字は、アクチビン濃度を示す。図面の底部の数字は、骨格筋が発達したEBの割合の推定値を表す。
【図29】アクチビン誘導集団から単離したbrachury画分の発現の分析を示す図である。図面の上部の数字は、アクチビン濃度を示す。
【図30A】ヒトCD4(hCD4)のHNF3β遺伝子座への標的化のために使用する方法を示す図である。CD4の標的化のために使用するベクターのスキームおよび戦略を示す図である。
【図30B】ヒトCD4(hCD4)のHNF3β遺伝子座への標的化のために使用する方法を示す図である。標的ベクターをエレクトロポレーションにより導入したES細胞クローンのサザンブロットを示す図である。
【図31A】血清で誘導した発生中のEBにおけるGFP、hCD4、brachury、およびHNF3βの発現を示す図である。フローサイトメトリー分析によって測定したGFPのhCD4に対する発現のドットプロットを示す図である。
【図31B】血清で誘導した発生中のEBにおけるGFP、hCD4、brachury、およびHNF3βの発現を示す図である。図31A中のプロットのGFP発現とhCD4発現の両方の平均蛍光強度(MFI)を示す図である。MFIは、各マーカーに関して経時変化中に検出した最大率として示す。
【図31C】血清で誘導した発生中のEBにおけるGFP、hCD4、brachury、およびHNF3βの発現を示す図である。図31A中のフローサイトメトリーに使用したのと同じサンプル中の、RT PCRによるbrachuryおよびHNF3βの発現を示す図である。
【図32A】異なるGFP hCD4発現集団およびマウス胚の原条から切開した組織における、遺伝子発現を示す図である。EBは血清中で3.25日間分化させ、GFPおよびhCD4の発現によって選別した。これらの集団に関する細胞選別の前後のGFP対hCD4発現のドットプロットを示す図である。GFP+hCD4lo、GFP+hCD4med、およびGFP+hCD4hi集団を示した純度に選別した。
【図32B】異なるGFP hCD4発現集団およびマウス胚の原条から切開した組織における、遺伝子発現を示す図である。EBは血清中で3.25日間分化させ、GFPおよびhCD4の発現によって選別した。図32Aにおいて選別したサンプル中のRT PCRによる遺伝子の発現を示す図である。
【図32C】異なるGFP hCD4発現集団およびマウス胚の原条から切開した組織における、遺伝子発現を示す図である。EBは血清中で3.25日間分化させ、GFPおよびhCD4の発現によって選別した。第7.25日胚から切開した原条から切開した組織中の、RT PCRによる遺伝子の発現を示す図である。使用した略称:Pre、選別前、;ant、原条前部組織;mid、原条中部組織、pos、原条後部組織。
【図33A】異なるGFP hCD4発現集団の発生能を示す図である。図32中と同様にEBを分化させ細胞選別した。2集団、GFP+hCD4loおよびGFP+hCD4hi細胞のみを選別した。選別後、2日間血清物を含む培地中で細胞を再凝集させた。さらに24〜48時間マトリゲル上に平板培養した後の、選別した集団の心臓発生能を示す図である。拍動細胞を含むEBの割合を計算した。
【図33B】異なるGFP hCD4発現集団の発生能を示す図である。図32中と同様にEBを分化させ細胞選別した。2集団、GFP+hCD4loおよびGFP+hCD4hi細胞のみを選別した。選別後、2日間血清物を含む培地中で細胞を再凝集させた。造血サイトカインを含むメチルセルロース中に平板培養することによって測定した、選別した集団の造血発生能を示す図である。
【図33C】異なるGFP hCD4発現集団の発生能を示す図である。図32中と同様にEBを分化させ細胞選別した。2集団、GFP+hCD4loおよびGFP+hCD4hi細胞のみを選別した。選別後、2日間血清物を含む培地中で細胞を再凝集させた。無血清培地中でさらに5日間マトリゲル上に平板培養した後の、選別した集団のRT PCRによる内胚葉遺伝子の発現を示す図である。
【図34A】アクチビンA Wnt3aおよび/またはこれらの経路の阻害剤を含む無血清状態中で分化させたES細胞を示す図である。ES細胞は無血清培地中で2日間培養し、次いでトリプシン処理して単細胞集団を得た。次いでこれらの細胞を示した条件で再凝集させた。第6日で、EBをマトリゲルコーティング皿上、無血清培地に平板培養した。培地単独、アクチビンA 25ng/ml、Wnt3a 100ng/ml、あるいはこれらの刺激およびDKK1 100ng/mlまたはSB-431542 6uMのいずれかで再凝集させたEBを用いて第3日〜第6日のGFP対hCD4発現のドットプロットを示す図である。
【図34B】アクチビンA Wnt3aおよび/またはこれらの経路の阻害剤を含む無血清状態中で分化させたES細胞を示す図である。ES細胞は無血清培地中で2日間培養し、次いでトリプシン処理して単細胞集団を得た。次いでこれらの細胞を示した条件で再凝集させた。第6日で、EBをマトリゲルコーティング皿上、無血清培地に平板培養した。図34A中に示した培養物から得たRNAからの、RT PCRによる遺伝子発現データを示す図である。
【図34C】アクチビンA Wnt3aおよび/またはこれらの経路の阻害剤を含む無血清状態中で分化させたES細胞を示す図である。ES細胞は無血清培地中で2日間培養し、次いでトリプシン処理して単細胞集団を得た。次いでこれらの細胞を示した条件で再凝集させた。第6日で、EBをマトリゲルコーティング皿上、無血清培地に平板培養した。前に示したように培地単独、アクチビンまたはWnt3aで最初に刺激しマトリゲル上でさらに6日間平板培養したEBから得たRNAからの、RT PCRによる遺伝子発現データを示す図である。
【図35A】EBの分化に対するDKK1およびアクチビンの影響を示す図である。EBは図32中のGFP+hCD4medと同様に分化させ選別し、血清物のみを含む培地、あるいはヒトDKK1 100ng/ml、アクチビン100ng/ml、または両方の因子を一緒に加えた培地中で再凝集させた。再凝集1日後の選別した細胞のhCD4およびGFPの発現に関するヒストグラムを示す図である。
【図35B】EBの分化に対するDKK1およびアクチビンの影響を示す図である。EBは図32中のGFP+hCD4medと同様に分化させ選別し、血清物のみを含む培地、あるいはヒトDKK1 100ng/ml、アクチビン100ng/ml、または両方の因子を一緒に加えた培地中で再凝集させた。RT PCRによる内胚葉遺伝子の発現を示す図である。選別した集団は前に示したように2日間処理し、次いでマトリゲル上、因子を加えていない無血清培地中に平板培養した。遺伝子の発現は、さらに5日間平板培養後に分析した。
【図36A】フローサイトメトリー分析を示す図である。
【図36B】フローサイトメトリー分析を示す図である。
【図36C】肝細胞コロニーの遺伝子発現の分析を示す図である。
【特許請求の範囲】
【請求項1】
内胚葉細胞が豊富な細胞集団を作製する方法であって、約1日〜約6日間アクチビンまたはノーダルおよびWntの存在下で胚幹細胞を培養する段階と、brach+/HNF3β+細胞を単離する段階と、少なくとも約1日間アクチビンおよびWntシグナル伝達の阻害剤の存在下で前記単離細胞を培養する段階とを含む方法。
【請求項2】
胚幹細胞を血清の不在下で培養する、請求項1に記載の方法。
【請求項3】
単離細胞を血清の不在下で培養する、請求項1に記載の方法。
【請求項4】
胚幹細胞がヒト胚幹細胞である、請求項1に記載の方法。
【請求項5】
WntがWnt3dである、請求項1に記載の方法。
【請求項6】
Wntシグナル伝達の阻害剤がDKK-1である、請求項1に記載の方法。
【請求項7】
アクチビンが少なくとも約30ng/mlの濃度で存在する、請求項1に記載の方法。
【請求項8】
内胚葉細胞が豊富な細胞集団を作製する方法であって、約2日〜約10日間ヒト胚幹細胞を培養する段階と、brach+/HNF3β+細胞を単離する段階と、少なくとも約1日間アクチビンまたはノーダルおよびWntシグナル伝達の阻害剤の存在下で前記brach+/HNF3β+細胞を培養する段階とを含む方法。
【請求項9】
胚幹細胞を血清の不在下で培養する、請求項8に記載の方法。
【請求項10】
brach+/HNF3β+細胞を血清の不在下で培養する、請求項8に記載の方法。
【請求項11】
アクチビンの濃度が少なくとも約30ng/mlである、請求項8に記載の方法。
【請求項12】
アクチビンの濃度が約100ng/mlである、請求項8に記載の方法。
【請求項13】
Wntシグナル伝達の阻害剤がDKK-1である、請求項8に記載の方法。
【請求項14】
DKK-1の濃度が少なくとも約30ng/mlである、請求項8に記載の方法。
【請求項15】
選択可能なマーカーをコードする核酸がHNF3β遺伝子座に存在する、胚幹細胞。
【請求項16】
選択可能なマーカーがヒトCD4である、請求項15に記載の幹細胞。
【請求項17】
第二の選択可能なマーカーをコードする第二の核酸がbrachury遺伝子座に存在する、請求項15に記載の幹細胞。
【請求項18】
第二の選択可能なマーカーが緑色蛍光タンパク質である、請求項15に記載の幹細胞。
【請求項19】
肝細胞が豊富な細胞集団を作製する方法であって、アクチビンまたはノーダルの存在下で胚幹細胞を培養してbrach+/HNF3β3+/cKit+細胞を得る段階と、brach+/HNF3β3+/cKit+細胞を単離する段階と、肝芽細胞を生成するのに充分な条件下においてBMP-4およびbFGFの存在下で前記細胞を培養する段階と、肝細胞への肝芽細胞の成熟に充分な条件下において肝芽細胞を培養する段階とを含む方法。
【請求項20】
胚幹細胞がヒト胚幹細胞である、請求項19に記載の方法。
【請求項21】
胚幹細胞を約2日〜約10日間アクチビンまたはノーダルの存在下で培養する、請求項19に記載の方法。
【請求項22】
胚幹細胞を血清の不在下で培養する、請求項19に記載の方法。
【請求項23】
brach+/HNF3β3+/cKit+細胞を血清の不在下で培養する、請求項19に記載の方法。
【請求項24】
請求項19に記載の方法によって生成される少なくとも約10%の肝細胞を含む細胞集団。
【請求項25】
少なくとも約50%の肝細胞を含む請求項24に記載の細胞集団。
【請求項26】
少なくとも約70%の肝細胞を含む請求項24に記載の細胞集団。
【請求項27】
肝細胞に対して影響を有する作用物質についてスクリーニングする方法であって、試験する作用物質の存在下および不在下において請求項19に記載の方法によって生成した肝細胞を培養する段階と、作用物質が肝細胞に対して影響を有するかどうか決定する段階とを含む方法。
【請求項28】
肝細胞補充療法の方法であって、請求項19に記載の方法によって生成した肝細胞を含む組成物を、そのような治療の必要がある対象に投与する段階を含む方法。
【請求項29】
組成物を注射、移植または注入によって投与する、請求項28に記載の方法。
【請求項1】
内胚葉細胞が豊富な細胞集団を作製する方法であって、約1日〜約6日間アクチビンまたはノーダルおよびWntの存在下で胚幹細胞を培養する段階と、brach+/HNF3β+細胞を単離する段階と、少なくとも約1日間アクチビンおよびWntシグナル伝達の阻害剤の存在下で前記単離細胞を培養する段階とを含む方法。
【請求項2】
胚幹細胞を血清の不在下で培養する、請求項1に記載の方法。
【請求項3】
単離細胞を血清の不在下で培養する、請求項1に記載の方法。
【請求項4】
胚幹細胞がヒト胚幹細胞である、請求項1に記載の方法。
【請求項5】
WntがWnt3dである、請求項1に記載の方法。
【請求項6】
Wntシグナル伝達の阻害剤がDKK-1である、請求項1に記載の方法。
【請求項7】
アクチビンが少なくとも約30ng/mlの濃度で存在する、請求項1に記載の方法。
【請求項8】
内胚葉細胞が豊富な細胞集団を作製する方法であって、約2日〜約10日間ヒト胚幹細胞を培養する段階と、brach+/HNF3β+細胞を単離する段階と、少なくとも約1日間アクチビンまたはノーダルおよびWntシグナル伝達の阻害剤の存在下で前記brach+/HNF3β+細胞を培養する段階とを含む方法。
【請求項9】
胚幹細胞を血清の不在下で培養する、請求項8に記載の方法。
【請求項10】
brach+/HNF3β+細胞を血清の不在下で培養する、請求項8に記載の方法。
【請求項11】
アクチビンの濃度が少なくとも約30ng/mlである、請求項8に記載の方法。
【請求項12】
アクチビンの濃度が約100ng/mlである、請求項8に記載の方法。
【請求項13】
Wntシグナル伝達の阻害剤がDKK-1である、請求項8に記載の方法。
【請求項14】
DKK-1の濃度が少なくとも約30ng/mlである、請求項8に記載の方法。
【請求項15】
選択可能なマーカーをコードする核酸がHNF3β遺伝子座に存在する、胚幹細胞。
【請求項16】
選択可能なマーカーがヒトCD4である、請求項15に記載の幹細胞。
【請求項17】
第二の選択可能なマーカーをコードする第二の核酸がbrachury遺伝子座に存在する、請求項15に記載の幹細胞。
【請求項18】
第二の選択可能なマーカーが緑色蛍光タンパク質である、請求項15に記載の幹細胞。
【請求項19】
肝細胞が豊富な細胞集団を作製する方法であって、アクチビンまたはノーダルの存在下で胚幹細胞を培養してbrach+/HNF3β3+/cKit+細胞を得る段階と、brach+/HNF3β3+/cKit+細胞を単離する段階と、肝芽細胞を生成するのに充分な条件下においてBMP-4およびbFGFの存在下で前記細胞を培養する段階と、肝細胞への肝芽細胞の成熟に充分な条件下において肝芽細胞を培養する段階とを含む方法。
【請求項20】
胚幹細胞がヒト胚幹細胞である、請求項19に記載の方法。
【請求項21】
胚幹細胞を約2日〜約10日間アクチビンまたはノーダルの存在下で培養する、請求項19に記載の方法。
【請求項22】
胚幹細胞を血清の不在下で培養する、請求項19に記載の方法。
【請求項23】
brach+/HNF3β3+/cKit+細胞を血清の不在下で培養する、請求項19に記載の方法。
【請求項24】
請求項19に記載の方法によって生成される少なくとも約10%の肝細胞を含む細胞集団。
【請求項25】
少なくとも約50%の肝細胞を含む請求項24に記載の細胞集団。
【請求項26】
少なくとも約70%の肝細胞を含む請求項24に記載の細胞集団。
【請求項27】
肝細胞に対して影響を有する作用物質についてスクリーニングする方法であって、試験する作用物質の存在下および不在下において請求項19に記載の方法によって生成した肝細胞を培養する段階と、作用物質が肝細胞に対して影響を有するかどうか決定する段階とを含む方法。
【請求項28】
肝細胞補充療法の方法であって、請求項19に記載の方法によって生成した肝細胞を含む組成物を、そのような治療の必要がある対象に投与する段階を含む方法。
【請求項29】
組成物を注射、移植または注入によって投与する、請求項28に記載の方法。
【図1】

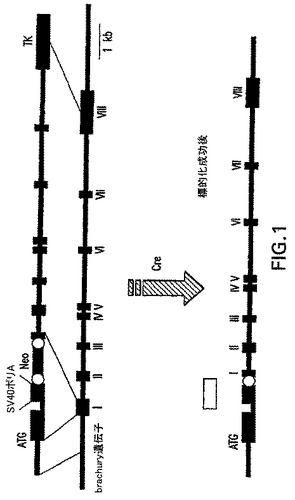
【図2A】

【図2B】
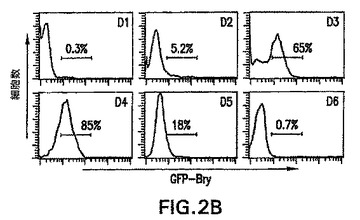
【図3A】
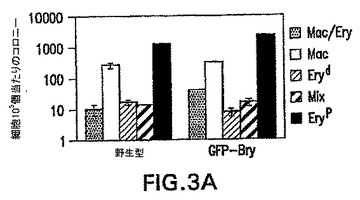
【図3B】
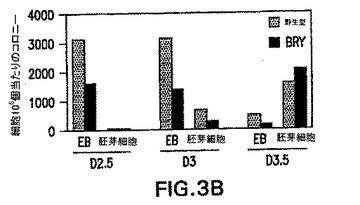
【図3C】
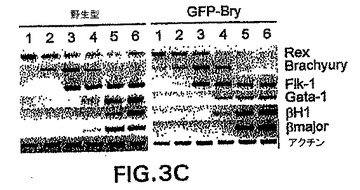
【図4A】
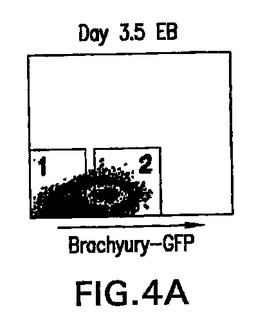
【図4B】
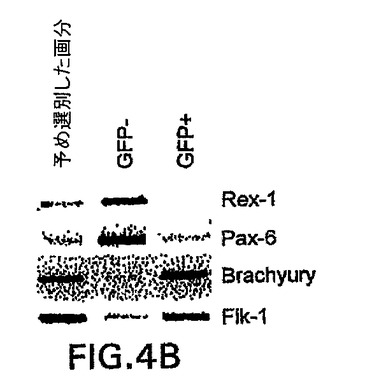
【図5A】
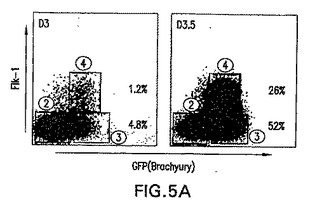
【図5B】
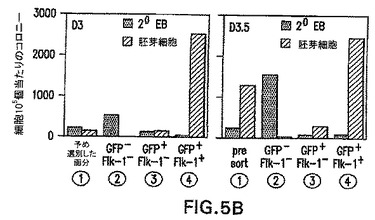
【図5C】
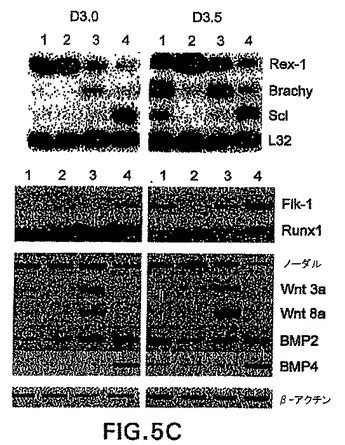
【図6】
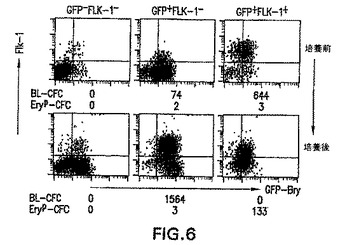
【図7A】
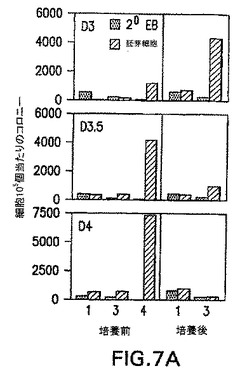
【図7B】
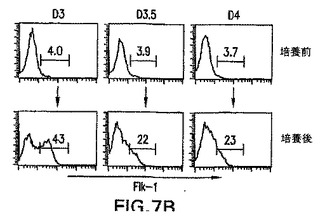
【図8A】
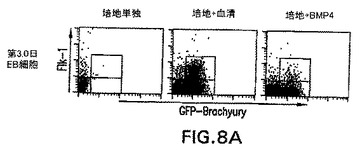
【図8B】
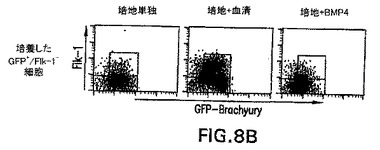
【図9】
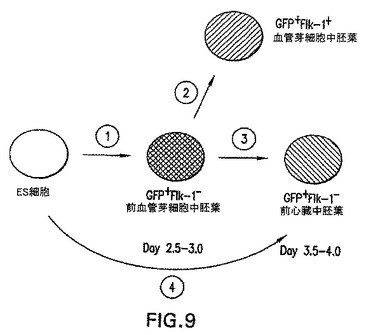
【図10】
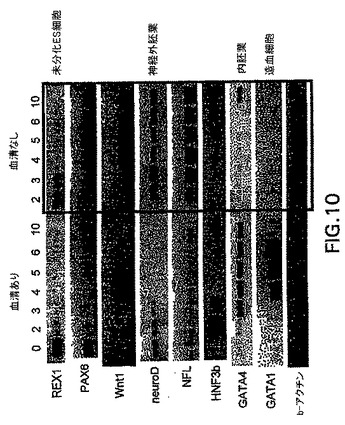
【図11】
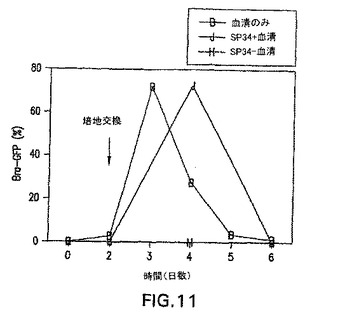
【図12】
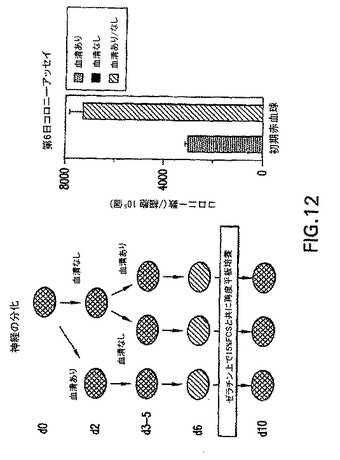
【図13】
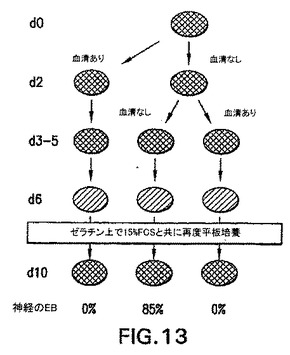
【図14】
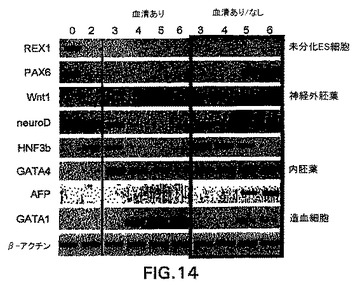
【図15】
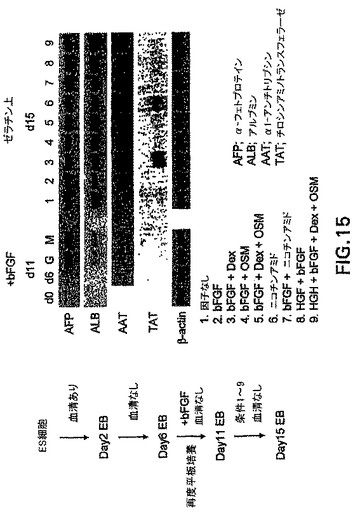
【図16】
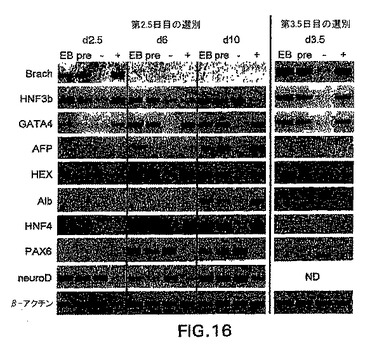
【図17】
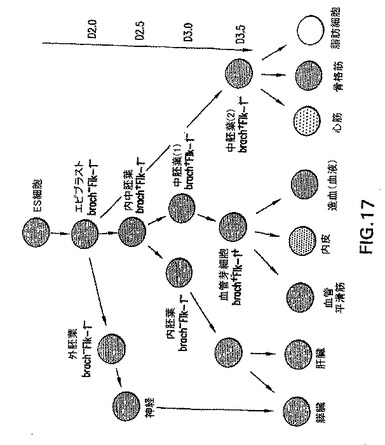
【図18】
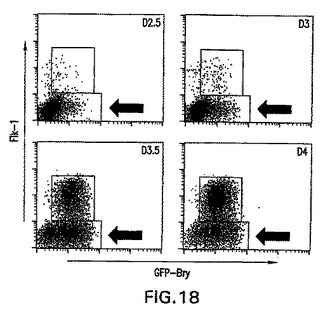
【図19】
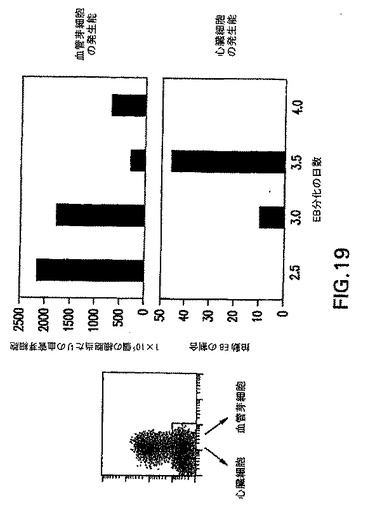
【図20】
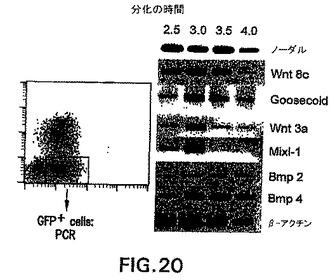
【図21】
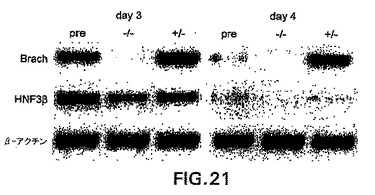
【図22A】
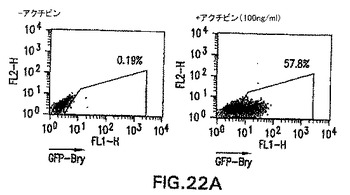
【図22B】
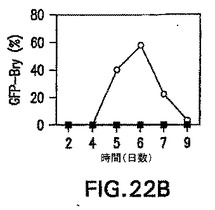
【図22C】
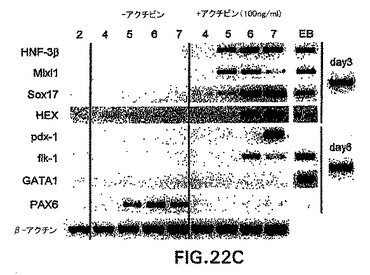
【図23A】
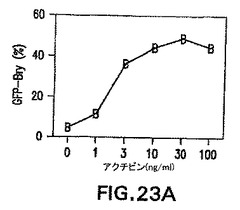
【図23B】
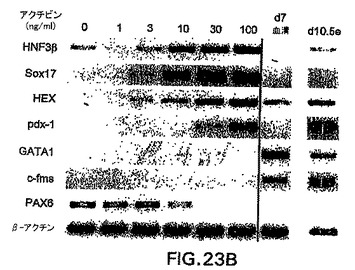
【図24A】
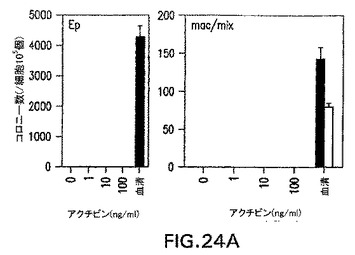
【図24B】
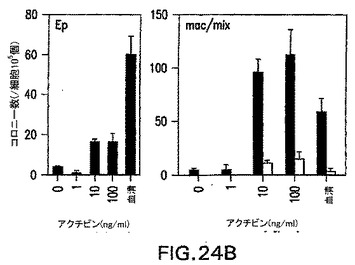
【図25】
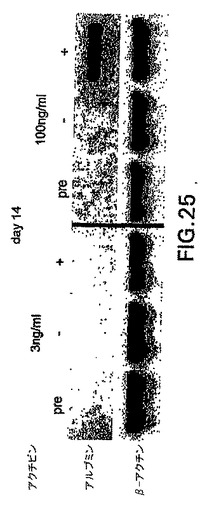
【図26A】
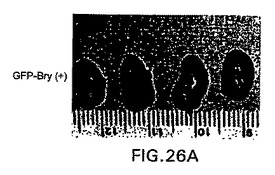
【図26B】

【図26C】
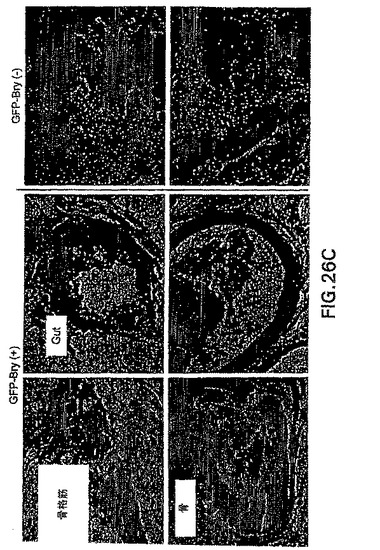
【図27A】
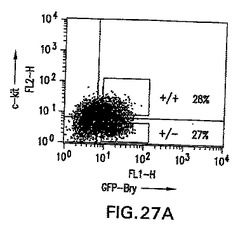
【図27B】
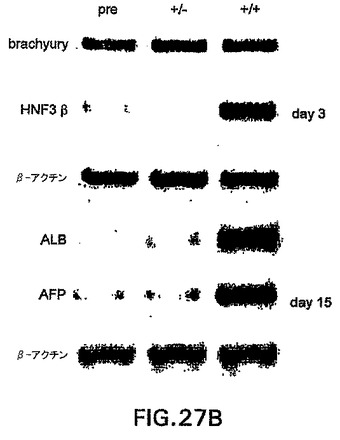
【図28】
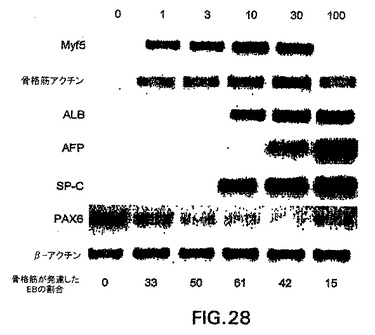
【図29】
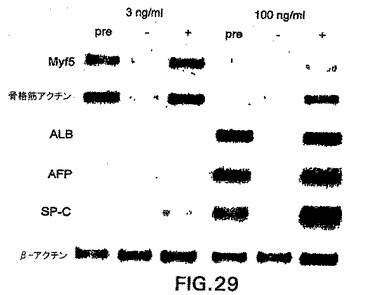
【図30A】
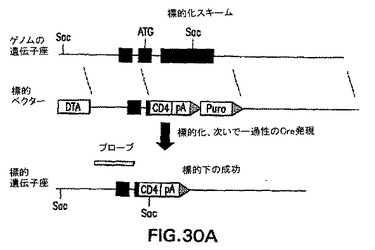
【図30B】

【図31A】
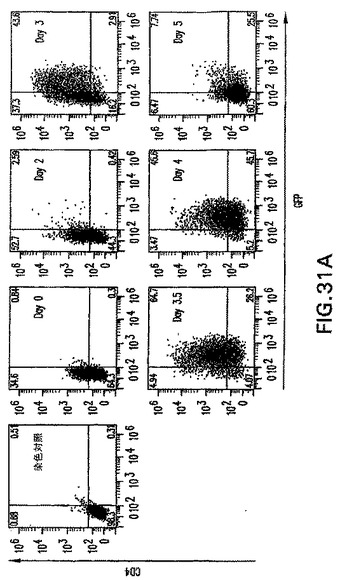
【図31B】
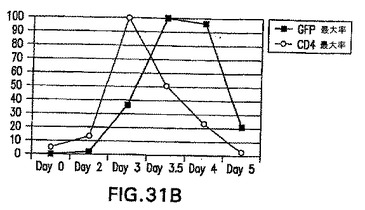
【図31C】

【図32A】
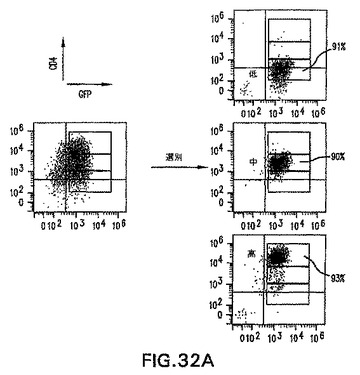
【図32B】
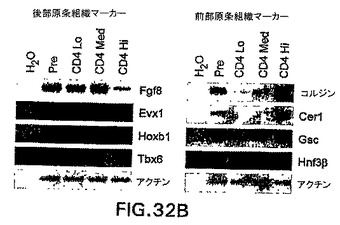
【図32C】
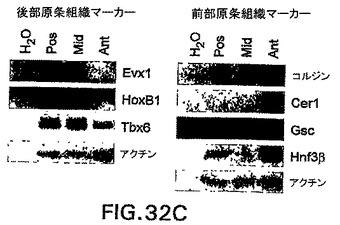
【図33A】
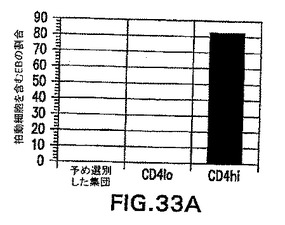
【図33B】
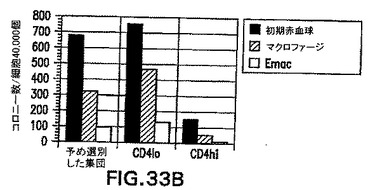
【図33C】
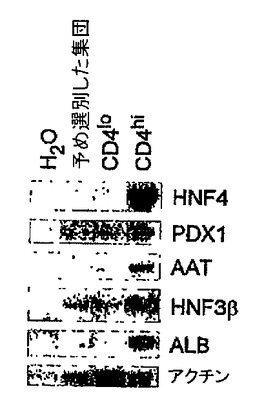
【図34A】
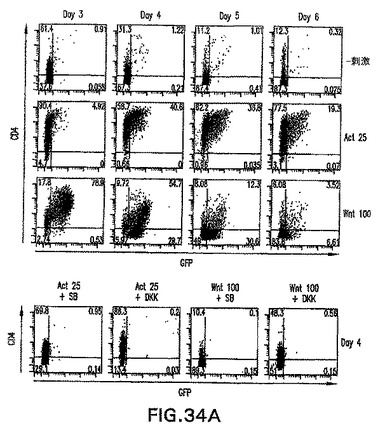
【図34B】
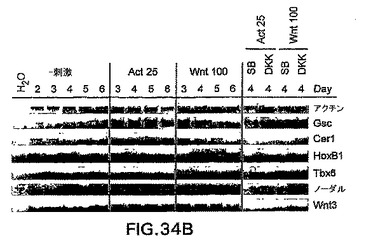
【図34C】
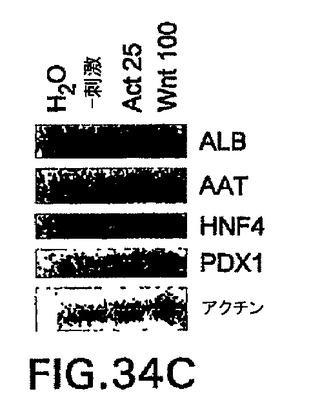
【図35A】
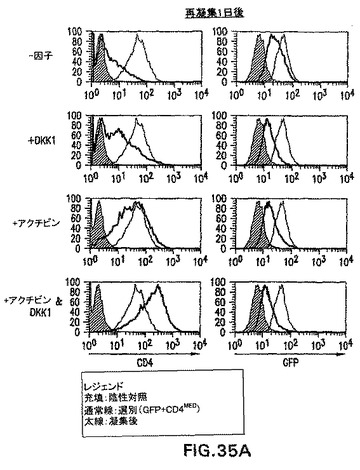
【図35B】
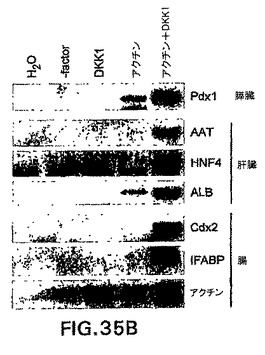
【図36A】
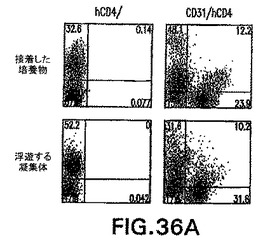
【図36B】
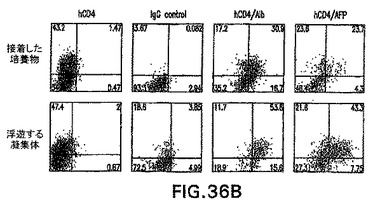
【図36C】
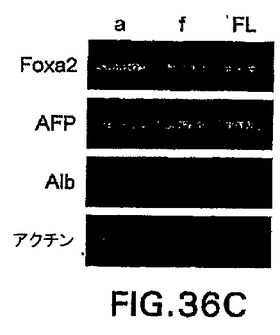
【図2A】
【図2B】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図5A】
【図5B】
【図5C】
【図6】
【図7A】
【図7B】
【図8A】
【図8B】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22A】
【図22B】
【図22C】
【図23A】
【図23B】
【図24A】
【図24B】
【図25】
【図26A】
【図26B】
【図26C】
【図27A】
【図27B】
【図28】
【図29】
【図30A】
【図30B】
【図31A】
【図31B】
【図31C】
【図32A】
【図32B】
【図32C】
【図33A】
【図33B】
【図33C】
【図34A】
【図34B】
【図34C】
【図35A】
【図35B】
【図36A】
【図36B】
【図36C】
【公表番号】特表2008−546414(P2008−546414A)
【公表日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願番号】特願2008−518426(P2008−518426)
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/US2006/024462
【国際公開番号】WO2007/002385
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(502375437)マウント シナイ スクール オブ メディスン オブ ニューヨーク ユニバーシティー (11)
【Fターム(参考)】
【公表日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/US2006/024462
【国際公開番号】WO2007/002385
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(502375437)マウント シナイ スクール オブ メディスン オブ ニューヨーク ユニバーシティー (11)
【Fターム(参考)】
[ Back to top ]