
乳癌において差次的に発現される遺伝子

【課題】転移するかまたは転移している腫瘍とあまり転移しそうではない腫瘍との間を識別するためのマーカーを提供する。
【解決手段】腫瘍の作用を予測するための方法であり、より詳細には、腫瘍サンプルが特定の遺伝子配列の発現に関して試験され、それによって転移性の広がりに関する傾向を示す方法。特定の配列に示されるようなポリヌクレオチド配列は、癌細胞(特に乳癌細胞)の転移能力と相関する。腫瘍の転移の危険性を決定するための方法が、腫瘍由来の組織サンプルが、特定の配列に示されるポリヌクレオチドによってコードされたポリペプチドまたはmRNAを発現するかどうかを決定する治療方法および組成物。
【解決手段】腫瘍の作用を予測するための方法であり、より詳細には、腫瘍サンプルが特定の遺伝子配列の発現に関して試験され、それによって転移性の広がりに関する傾向を示す方法。特定の配列に示されるようなポリヌクレオチド配列は、癌細胞(特に乳癌細胞)の転移能力と相関する。腫瘍の転移の危険性を決定するための方法が、腫瘍由来の組織サンプルが、特定の配列に示されるポリヌクレオチドによってコードされたポリペプチドまたはmRNAを発現するかどうかを決定する治療方法および組成物。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、腫瘍の作用を予測するための方法に関する。より詳細には、本発明は、腫瘍サンプルが特定の遺伝子配列の発現に関して試験され、それによって転移性の広がりに関する傾向を示す方法に関する。
【背景技術】
【0002】
(発明の背景)
乳癌は、世界中で一年当たり約1,000,000件の新規症例を伴う、最も一般的な悪性疾患のうちの1つである。多数の組織化学的マーカー、遺伝学的マーカー、および免疫学的マーカーの使用にもかかわらず、臨床医たちが、腫瘍が他の器官に転移する時期を推測することはなお困難である。いくらかの患者は、再発および転移を回避するためにアジュバント治療が必要であるが、他の者は必要ではない。しかし、患者のこれらの亜集団間を識別することは、簡単ではなく、治療の経過は、簡単には図表に表せない。当該分野には、転移するかまたは転移している腫瘍とあまり転移しそうではない腫瘍との間を識別するための新規マーカーの必要性がある。
【発明の概要】
【発明が解決しようとする課題】
【0003】
(発明の要旨)
転移するかまたは転移している腫瘍とあまり転移しそうではない腫瘍との間を識別するためのマーカーを提供することが、本発明の目的である。本発明のこれらおよび他の目的は、以下に記載される1つ以上の実施形態によって提供される。
【課題を解決するための手段】
【0004】
本発明の1つの実施形態は、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列に対して少なくとも85%同一であるアミノ酸配列を有する、単離されそして精製されたヒトタンパク質を提供する。
【0005】
本発明の別の実施形態は、ペプチド結合手段によって互いに融合された、第1のタンパク質セグメントおよび第2のタンパク質セグメントを含む融合タンパク質を提供する。第1のタンパク質セグメントは、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列から選択される、少なくとも6連続するアミノ酸からなり、そして第2のタンパク質セグメントは、配列番号1によってコードされるネイティブなタンパク質において第1のセグメントに隣接することが見出されないアミノ酸配列を含む。
【0006】
本発明のなお別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列を有するヒトタンパク質の少なくとも6連続するアミノ酸からなる、単離されそして精製されたポリペプチドを提供する。
【0007】
本発明のさらに別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列を含むヒトタンパク質に特異的に結合する抗体の調製物を提供する。
【0008】
本発明のなお別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖に対して少なくとも95%同一であるヌクレオチド配列の少なくとも11連続するヌクレオチドを含む、単離されそして精製されたサブゲノムのポリヌクレオチドを提供する。
【0009】
本発明の別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖を含むコード配列を含む、単離されそして精製されたポリヌクレオチドを提供する。
【0010】
本発明のなお別の実施形態は、組織サンプル中の転移を決定するための方法を提供する。配列番号1のコード配列を含む遺伝子の発現産物は、組織サンプル中で測定される。コントロールサンプルよりも高いレベルでこの産物を発現する組織サンプルは、より大きな転移の可能性を有するとして分類される。
【0011】
さらに本発明のさらなる実施形態は、配列番号2をコードするヒト遺伝子を検出するための方法を提供し、この方法は、コンピュータで読み取り可能な形式の配列番号1を得る工程、この配列を、ヒトゲノムのポリヌクレオチド配列と比較する工程、およびパラメータとして12のギャップオープンペナルティー(gap open penalty)および1のギャップエクステンションペナルティー(gap extension penalty)を用いたアフィンギャップサーチ(affine gap serch)を使用するSmith−Watermanアルゴリズムによって決定される場合に、配列番号1に対して少なくとも95%の配列同一性を有する1つ以上のヒトゲノム配列を同定する工程を包含する。
従って、本発明は、転移マーカーとして使用され得る多数のポリヌクレオチドおよびポリペプチドを当該分野に提供する。これらは、乳癌患者の治療経過をより合理的に指示するのに有用である。
例えば、本発明は以下の項目を提供する。
(項目1) 単離された核酸分子であって、以下:
(a)配列番号2の約1〜約273のアミノ酸をコードするポリヌクレオチド;(b)配列番号2の約2〜約273のアミノ酸をコードするポリヌクレオチド;(c)配列番号2の約26〜約273のアミノ酸をコードするポリヌクレオチド;
(d)(a)、(b)、または(c)のポリヌクレオチドのポリヌクレオチド相補体;および
(e)(a)、(b)、(c)、または(d)のポリヌクレオチドと少なくとも90%同一であるポリヌクレオチド、
からなる群より選択されたポリヌクレオチドを含む、単離された核酸分子。
(項目2) 配列番号1のコード領域由来の24〜738の連続したヌクレオチドを含む、単離された核酸分子。
(項目3) 配列番号1のコード領域由来の50〜500の連続したヌクレオチドを含む、項目2に記載の単離された核酸分子。
(項目4) 配列番号1のコード領域由来の75〜250の連続したヌクレオチドを含む、項目3に記載の単離された核酸分子。
(項目5) ポリペプチドをコードするポリヌクレオチドを含む単離された核酸分子であって、ここで、少なくとも1つの保存的アミノ酸置換を除いて、該ポリペプチドが、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;および
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸配列を有する、単離された核酸分子。
(項目6) DNAである、項目1に記載の単離された核酸分子。
(項目7) 組換えベクターを作製する方法であって、項目1に記載の核酸分子をプロモーターと作動可能に連結させた状態でベクター中に挿入する工程を包含する、方法。
(項目8) 項目7に記載の方法によって生成される、組換えベクター。
(項目9) 組換え宿主細胞を作製する方法であって、項目8に記載の組換えベクターを宿主細胞中へ導入する工程を包含する、方法。
(項目10) 項目9に記載の方法によって生成される、組換え宿主細胞。
(項目11) ポリペプチドを生成する組換え方法であって、項目10に記載の組換え宿主細胞を、該ポリペプチドが発現されるような条件下で培養する工程、および該ポリペプチドを回収する工程を包含する、方法。
(項目12) 単離されたポリペプチドであって、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;および
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸と少なくとも95%同一であるアミノ酸を含む、単離されたポリペプチド。
(項目13) 単離されたポリペプチドであって、ここで、少なくとも1つの保存的アミノ酸置換を除いて、該ポリペプチドが、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸配列を有する、単離されたポリペプチド。
(項目14) 単離されたポリペプチドであって、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸を含む、単離されたポリペプチド。
(項目15) 配列番号2のポリペプチドのエピトープ保有部分。
(項目16) 配列番号2の8〜25の連続したアミノ酸を含む、項目15に記載のエピトープ保有部分。
(項目17) 配列番号2の10の連続したアミノ酸を含む、項目15に記載のエピトープ保有部分。
(項目18) 項目12に記載のポリペプチドに特異的に結合する、単離された抗体。
(項目19) 項目13に記載のポリペプチドに特異的に結合する、単離された抗体。
(項目20) 項目14に記載のポリペプチドに特異的に結合する、単離された抗体。
(項目21) 配列番号2をコードするヒト遺伝子を検出するための方法であって、該方法は、コンピュータで読み取り可能なフォーマットで配列番号1を得る工程、該配列とヒトゲノムのポリヌクレオチド配列とを比較する工程、および配列番号1と少なくとも95%同一の配列を有する1以上のヒトゲノム配列を同定する工程を包含し、パラメータとして12のギャップオープンペナルティーおよび1のギャップエクステンションペナルティーを用いるアフィンギャップサーチを使用する、Smith−Waternamアルゴリズムによって測定される、方法。
(項目22) 天然に存在しない融合タンパク質であって、該タンパク質は、ペプチド結合によって互いに融合される、第1タンパク質セグメントおよび第2タンパク質セグメントを含み、ここで、該第1タンパク質セグメントは、配列番号1のヌクレオチド配列またはその相補体によってコードされたアミノ酸配列から選択される、少なくとも6の連続したアミノ酸を含む、天然に存在しない融合タンパク質。
(項目23) 前記第1タンパク質セグメントが、配列番号2の少なくとも6の連続したアミノ酸を含む、項目22に記載の融合タンパク質。
(項目24) 前記第1タンパク質セグメントが、配列番号2の少なくとも12の連続したアミノ酸を含む、項目23に記載の融合タンパク質。
(項目25) 前記第1タンパク質セグメントが、配列番号2のアミノ酸20〜30を含む、項目22に記載の融合タンパク質。
(項目26) 前記第1タンパク質セグメントが、配列番号2の少なくとも50の連続するアミノ酸を含む、項目24に記載の融合タンパク質。
(項目27) 前記第1タンパク質セグメントが、配列番号2の少なくとも100の連続するアミノ酸を含む、項目26に記載の融合タンパク質。
(項目28) 前記第1タンパク質セグメントが、配列番号2のアミノ酸26〜287を含む、項目22に記載の融合タンパク質。
(項目29) 第1組織サンプルおよび第2組織サンプル中の腫瘍細胞の転移能力を比較するための方法であって、該方法は、配列番号1のポリペプチドコード領域を含む遺伝子の発現産物を該組織サンプル中で測定する工程を包含し、ここで、該第1組織サンプル中の該産物の少なくとも2倍大きな発現が、該第2組織サンプルと比較してより大きな転移能力を示す、方法。
(項目30) 前記発現産物がタンパク質である、項目29に記載の方法。
(項目31) 前記タンパク質が該タンパク質に特異的に結合する抗体を使用して測定される、項目30に記載の方法。
(項目32) 前記発現産物がmRNAである、項目29に記載の方法。
(項目33) 項目32に記載の方法であって、前記mRNAがポリペプチドプローブを使用して測定され、該ポリペプチドプローブは、配列番号1のヌクレオチド365〜1173の少なくとも20の連続したヌクレオチドを含む、方法。
(項目34) 哺乳動物の癌細胞によってタンパク質の発現を阻害するための組成物であって、該組成物は、配列番号4のポリヌクレオチドを含む、組成物。
(項目35) 哺乳動物の癌細胞によってタンパク質の発現を阻害するための方法であって、該方法は、配列番号4のポリヌクレオチドを含む組成物と該細胞を接触させる工程を包含する、方法。
【図面の簡単な説明】
【0012】
【図1】図1は、ヒトOut at Firstのポリヌクレオチド配列を示す(配列番号1)。
【図2】図2は、配列番号1によってコードされるアミノ酸配列を示す(配列番号2)。
【図3】図3は、推定シグナルペプチドを示す(配列番号3)。
【図4A】図4Aは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図4B】図4Bは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図4C】図4Cは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図4D】図4Dは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図5】図5は、腫瘍細胞株およびこれらの細胞から作製されたSCIDマウス由来の腫瘍組織における、β−アクチンに対するhsOAFの発現を示す。「PT」は、原発性腫瘍をいう。
【図6】図6は、未処理細胞(WT)に対する、アンチセンスオリゴ(配列番号4)(66−2as)または逆方向コントロール(配列番号5)(66−2rc)を用いた処理の後の、軟寒天におけるMDA−MB−435細胞によるコロニーの増殖を示す。
【図7】図7は、ヒトOAFアミノ酸配列とDrosophila OAFアミノ酸配列との整列である。
【図8】図8Aは、COS−7細胞株およびMCF−7細胞株中のhsOAFタンパク質の発現を示す。図8Bは、乳腺癌(mammory carcinoma)細胞株におけるhsOAFタンパク質の発現を示す。
【図9】図9は、正常ヒト組織中のhsOAFの発現を示す。
【図10A】図10Aは、アンチセンスオリゴ(配列番号4)を用いた処理後にMDA−MB−435細胞中で見出される形態学的変化を示す。AS=アンチセンス;RC=逆方向コントロール(配列番号5);M=馴化培地。
【図10B】図10Bは、AS、RCおよびRC+Mを用いたMDA−MB−435細胞の処置後の、細胞の侵入を示す。
【図11】図11は、ヒトOAFの推定シグナル配列を示す(二重下線)。
【図12】図12Aおよび図12Bは、アンチセンスオリゴ(配列番号4)または逆方向コントロールオリゴ(配列番号5)を用いて処理されたMDA−MB−435細胞による、hsOAFの分泌を示す。
【発明を実施するための形態】
【0013】
(発明の詳細な説明)
乳癌を有する患者の最も致命的な原因は、乳癌の転移および遠方の座位(主に、肺および骨)でのその増殖である。転移は、腫瘍細胞が原発性腫瘍から移住し、血管およびリンパ管を通じて広まり、それから特定の標的器官(ここに、この腫瘍細胞が、再びコロニー形成する)に沈着される、多工程プロセスである。Schirrmacher,V.,Adv.Cancer Res.43:1−73,1985およびLiotta,L.A.ら、Cell 64(2):327−36(1991)。このプロセスの間、腫瘍細胞の侵襲性は、重要である。なぜなら、これらは、多数の基底膜に遭遇してそしてここを通過しなければならないからである。Liotta,L.A.,Am.J.Pathol.117(3):339−48(1984)およびFidler,I.J.,Cancer Res.38(9):2651−60(1978)。従って、腫瘍細胞の侵襲および転移の分子的な原因の解明は、乳癌患者の効果的な処置手順の開発に必須である。乳癌転移において差次的に発現される遺伝子は、転移の間に重要な役割を果たす潜在的な標的である。このような遺伝子およびその生物学的機能の同定は、乳癌の治療および診断の開発に有意に寄与する。
【0014】
乳癌転移に関与するいくつかの重要な遺伝子が、発見されている。エストロゲンレセプターの損失およびビメンチンの存在は、ヒト乳癌の侵襲性および不十分な予後と関連し、そしてまた、ヒト乳癌細胞株の侵襲性および転移可能性と相互に関連する。Aamdal S.ら,Cancer 53(11):2525−9(1984);Clark,G.M.ら,Semin Oncol.,2 補遺 1:20−5(1988);Raymond,W.A.ら,J.Pathol.157(4):299−306(1989);Raymond,W.A.ら,J.Pathol.158(2):107−14(1989);およびThompson,E.W.ら、J.Cell Physiol.150(3):534−44(1992)。E−カドヘリンの発現低下が、乳腺癌の侵襲性に関連している。Vleminckx,K.ら,Cell 66(1):107−19(1991)およびOka,H.ら,Cancer Res.53(7):1696−701(1993)。マプシン(maspin)(正常乳房上皮細胞において発現されるが、ほとんどの乳癌細胞株において発現されないプロテアーゼインヒビター)は、マウスにおいて腫瘍および転移を誘導し、インビトロにおいて基底膜を侵襲するMDA−MB−435細胞の能力を抑制し得た。マプシン発現の損失は、進行癌において最も頻繁に生じた。Zou,Z.ら,Science 263(5146):526−9(1994)およびSeftor,R.E.ら,Cancer Res.58(24):5681−5(1998)。
【0015】
トランスフェクションによるMDA−MB−435細胞中におけるTIMP−4(メタロプロテイナーゼ−4の組織インヒビター)またはCLCA2(Ca2+活性化塩化物チャネル−2)の過剰発現は、細胞の腫瘍形成性、侵襲性および転移性の能力を阻害した。Wang,M.ら,Oncogene 14(23):2767−74(1997)およびGruber,A.D.ら,Cancer
Res.59(21):5488−91(1999)。増殖因子レセプターIGF−IRおよびp185ErbB−2の過剰発現は、乳癌転移に関与することが見出された。Surmacz,E.ら,Breast Cancer Res.Treat 47(3):255−67(1998);Dunn,S.E.ら,Cancer Res.58(15):3353−61(1998);Tan,M.ら,Cancer Res.57(6):1199−205(1997);Dhingra,K.ら,Semin Oncol.23(4):436−45(1996);およびRevillion,F.ら,Eur.J.Cancer 34(6):791−808(1998)。
【0016】
アスパラチルプロテアーゼのカテプシンDは、乳癌患者の不十分な予後のマーカーであると報告され、そして原発性乳癌の細胞質ゾル中の高いカテプシンD濃度と転移の発生との間には有意な相関があるが、カテプシンD分泌と乳癌細胞株の侵襲能力との間の相関は見出されていない。Rochefort,H.,Breast Cancer Res Treat 16(1):3−13(1990);Johnson,M.D.ら,Cancer Res.53(4):873−7(1993);およびRochefort,H.ら,Clin Chim
Acta.291(2):157−70(2000)。オステオポンチン(骨再吸収および骨形成に関与すると考えられる、分泌インテグリン結合糖タンパク質)は、乳腺癌細胞の移動および侵襲を誘導し得る。オステオポンチンレベル(腫瘍細胞または血漿のレベル)は、増強された乳癌の悪性と関連する。Denhardt,D.T.ら,FASEB J 7(15):1475−82(1993);Denhardt,D.T.ら,J.Cell Biochem Suppl.,30−31:92−102(1998);Tuck,A.B.ら,J Cell Biochem.78(3):465−75(2000);Tuck,A.B.ら,Oncogene 18(29):4237−46(1999);およびSinghal,H.ら,Clin Cancer Res.3(4):605−11(1997)。
【0017】
本発明は、非常に転移性のヒト乳癌細胞株において過剰発現された新規遺伝子のクローニングに関連する。これは、分泌タンパク質をコードし、そしてそのタンパク質分泌は、低い転移性の細胞株/非転移性の細胞株よりも、非常に転移性のヒト乳癌細胞株においてより多いことが、確認されている。アンチセンスオリゴ技術による侵襲性(aggressive)MDA−MB−435細胞株のこのタンパク質の分泌ノックアウトは、この細胞の減少した侵襲性および増殖速度を伴って、有意な形態学的変化を引き起こす。この遺伝子は、Drosophila遺伝子OAF(out at first)との相同性に基づいて、hsOAFと称される。Bergstrom,D.E.ら,Genetics 139(3):1331−46(1995)およびMerli,C.ら,Genes Dev.10(10):1260−70(1996)。
【0018】
ポリヌクレオチドが、高い転移性の乳癌細胞と非転移性の癌細胞または低い転移性の癌細胞との間で、差次的に発現されることが、本発明の発見である。従って、このポリヌクレオチドは、転移性マーカー遺伝子に関連する。この情報は、差次的に発現される遺伝子の発現産物に特異的な診断試薬を作製するために利用され得る。これはまた、癌、特に乳癌の適切な処置レジメンを計画する際に臨床医を補助する、診断方法および予後方法にも使用され得る。
【0019】
このポリヌクレオチドは、図1に示され(配列番号1)、そしてこの推定オープンリーディングフレーム(ORF)は、図2に示されるポリペプチド(配列番号2)をコードする。最初の30アミノ酸残基(配列番号3)は、推定シグナルペプチドを含み、そして推定プロテアーゼ切断部位は、「*」によって示される:APLLG*TGAPA(配列番号3の25位と26位との間のアミノ酸)。
【0020】
本発明のポリヌクレオチド配列は、「Out at First」(oaf)として公知のDrosophila遺伝子といくらかの相同性を共有する。oafの転写は、3つのクラスの選択的にポリアデニル化されたRNAを生じ、これらの発現は、発生的に調節される。全てのoaf転写物は、単一のUAG終止コドンによって隔てられた2つの隣接ORFを含む。翻訳の間のUGAコドンの抑制は、同じRNA分子から異なるタンパク質の生成を導き得る。卵形成の間、oaf RNAは、全ての期の栄養細胞で発現され、そして母系的に卵子に寄与する。
【0021】
初期発達の間、oaf遺伝子の接合体転写は、胚帯伸長時にほとんどかまたは全てのセグメントにおいて細胞の小クラスターで起こり、そしてその後は発達中の中枢神経系においてセグメントが反復されるパターンで起こる。oaf遺伝子はまた、両方の性の胎児、幼態および成人の生殖腺で発現される(Bergstrom,D.E.ら,Genetics 139:1331−1346,1995)。
【0022】
本発明のポリヌクレオチドは、7つの高転移性乳癌細胞株対非転移性または低転移性乳癌細胞株の組み合わせにおいて、差次的に発現された。用いられる細胞株は、MDA−MB−361(ヒト乳房腺癌由来)、MDA−MB−231(骨および/または肺への転移性を有するヒト乳癌細胞由来);MDA−MB−468(ヒトエストロゲンレセプター陰性乳癌細胞由来);MCF−7(非転移性ヒト乳癌細胞);ZR−75−1(エストロゲンレセプター陽性のヒト乳癌由来、Engleら,Cancer Res.38:3352−64(1978);およびMDA−MB−435(エストロゲンレセプター陰性のヒト乳癌細胞由来、Rishiら,Cancer Res.56:5246−5252(1996))である。
【0023】
発現プロフィールは、以下のようなものである:
【0024】
【表1】

mRNA発現のアップレギュレーションを、この細胞株由来の総RNAを用いたノーザンブロット分析により確認した(図5)。
【0025】
本発明のポリヌクレオチドの発現を比較した細胞株は、種々の転移能を有するヒト乳癌を表わす。細胞株ZR−75−1培養物は、乳癌患者の悪性の腹水の浸出液に由来した。インビトロで増殖された細胞株は、本来の細胞を有するドナーからの生検または細胞調製物において見られる形態に酷似して再集合した。ZR−75−1細胞は、エストロゲンにより特異的に刺激される。そしてこの細胞は、エストロゲン応答を研究するためのモデル系として用いられてきた(Engel,L.W.ら,Cancer Res.38:3352−3364,1978)。
【0026】
細胞株MDA−MB−435は、ヒト乳癌のモデルとして研究されてきた(例えば、レチノイン酸存在下での増殖阻害の作用機構を研究するための)エストロゲンレセプター陰性の細胞株である(Rishi,A.K.ら,Cancer Res.56:5246−5252,1996)。レチノイドによる増殖阻害はまた、MCF−7細胞およびMDA MB 468細胞においても研究されてきた(Tin−U,C.K.ら,Am.Soc.Clin.Onc.Proceedings,第17巻,2125,1998)。
【0027】
細胞株MDA−MB−361は、ヒト乳房腺癌から、特に悪性の部位から得られた(ATCC番号HTB−27)。ヒトWnt遺伝子の差次的な発現が、この細胞株において研究されてきた(Huguet,E.L.ら,Cancer Res.54:2615−2621,1994)。
【0028】
一旦転移が起こると、乳房の原発腫瘍細胞は、基底膜に侵入し、そして身体の他の器官に広がり、そして乳癌を有する患者の生存見込みをごくわずかにする。臨床的な診断および治療のために、乳癌の侵入および転移に関与する遺伝子を同定することは重要なことである。このような遺伝子は、治療用薬物開発のための診断または候補標的の潜在的な指標である。例えば、ヒト乳房腫瘍におけるビメンチンの存在は、わずかな予後の指標であるエストロゲンレセプターの欠損および腫瘍の浸潤に関連する(Raymond,W.A.ら,J.Pathol.157(4):299−306(1989);Raymond,W.A.ら,J.Pathol.158(2):107−14(1989);およびThompson,E.W.ら,J.Cell Physiol.150(3):534−44(1994))。増大した基質メタロプロテイナーゼの活性は、癌腫、特に乳癌の転移性の表現型に関連する(Basset,P.ら,Nature 348(6303):699−704(1990)およびBasset,P.ら,Cancer 74(3補完):1045−9(1994))。オステオポンチン、分泌型インテグリン結合糖タンパク質、は、ヒト乳房の上皮細胞の浸潤性の増大を誘導し得、そして乳癌における増加した悪性に関連する(Tuck,A.B.ら,J.Cell Biochem.78(3):465−75(2000);Tuck,A.B.ら,Oncogene 18(29):4237−46(1999);およびSinghal,H.ら,Clin Cancer Res.3(4):605−11(1997))。
【0029】
本発明は、乳癌転移に関連する新規の分泌タンパク質(hsOAF)の同定に関する。このタンパク質の役割を解明するために使用されるヒト乳癌細胞株は、これらの転移能に基づいて3つの群に分けられる:高転移性、低転移性、および非転移性。これらの細胞株群の間の異なる転移能を利用することにより、そして発達したマイクロアレイ技術を用いることにより、高転移性のヒト乳癌細胞株と低転移性/非転移性株との間で差次的に発現される遺伝子を同定した。これらの遺伝子は、乳癌転移性の分子機構のより深い理解を導き得る発現の確認および機能の研究のよい候補である。hsOAF遺伝子は、本発明の中心であり、新規の分泌タンパク質をコードする。hsOAF mRNAの発現は、種々のヒト正常組織においてほぼ共通である。しかし、hsOAFタンパク質の分泌は、低転移性/非転移性のヒト乳癌細胞株よりも高転移性株のほうが、ずっと強い。そしてMDA−MB−435は、最も強いhsOAF分泌性を有する。hsOAF遺伝子は、Drosophila OAF(out at first)遺伝子との相同性に基づき命名された。しかし、Drosophila OAFタンパク質は、分泌タンパク質ではあり得ない。というのも、このタンパク質は、N末端に代表的なシグナルペプチド配列を有しないからである(図7)。
【0030】
乳癌転移における分泌hsOAFタンパク質の重要性に取り組むために、アンチセンスオリゴ技術が、特異的なノックアウトhsOAF発現に用いられた。アンチセンスオリゴ技術は、遺伝子機能の研究のための劇的な遺伝子発現の減少のための、効果的で迅速な方法である(Stein,C.A.ら,Science
261(5124):1004−12(1993);Defacque,H.ら,J.Cell Physiol.178(1):109−19(1999))。高転移性のMDA−MB−435細胞のhsOAFタンパク質分泌のノックアウトは、細胞形態の変化、細胞浸潤性の減少、および細胞増殖速度の低下を引き起こす。馴化培地(正常なMDA−MB−435細胞の培養培地)での細胞の処理は、hsOAFタンパク質分泌のノックアウトにより引き起こされた全ての表現型変化をある程度回復させ得た。本発明者らは、特定の機構に束縛されないが、分泌hsOAFタンパク質は、MDA−MB−435細胞の浸潤および増殖に必須であると考えられている。しかし、アンチセンスオリゴ技術による別の高転移性細胞株MDA−MB−231のhsOAFタンパク質分泌のノックアウトは、有意な細胞の変化を全く引き起こさなかった。MDA−MB−435およびMDA−MB−231は、全く異なる転移性の細胞株であり、そしてMDA−MB−435は、MDA−MB−213よりもずっと強いhsOAFタンパク質の分泌性を示す。
【0031】
hsOAF遺伝子は、ヒト乳房腫瘍において異種接合性の喪失が頻繁に発生する染色体11q23領域に配置されている(Negrini,M.ら,Cancer Res 55(14):3003−7(1995)およびTomlinson,I.P.ら,J.Clin.Pathol.48(5):424−8(1995))。原発性ヒト乳房腫瘍における11q23での異種接合性の喪失は、転移後の乏しい生存性に関連することが報告されてきた(Winqvist,R.ら,Cancer Res.55(12):2660−4(1995))。11q23はまた、ATM(Ataxia−telangiectasia,mutated)、およびMLL(これは、急性白血病における染色体の再配置により頻繁に破壊される)のような遺伝子座を含む(Rasio,D.ら,Cancer Res.55(24):6053−7(1995)およびRubnitz,J.E.ら,Leukemia 10(1):74−82(1996))。乳癌転移における染色体11q23での変異とhsOAF遺伝子発現との間の関係は、不明確なままである。
【0032】
分泌hsOAFタンパク質は、乳癌に対する薬物開発のための適切な標的であり得、そして悪性の乳房腫瘍に対する良い診断指標であり得る。したがって、配列番号1およびこの配列を含むポリヌクレオチドは、転移性の指標として有用である。転移性指標ヌクレオチドまたはアミノ酸配列との関連は、非転移性または低転移性細胞と比較して高い転移性の同様の発現パターンを有する改変体を含む。転移性指標ポリペプチドは、長さの点で全長転移性指標タンパク質と相違し得、そして転移性指標タンパク質の少なくとも6、8、10、12、15、18、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、120、140、160、180、200、220、240、260、265、270、または271以上の連続するアミノ酸を含む。例示的なポリヌクレオチドとして、配列番号2の約1〜約273;1〜273;約2〜約273;2〜273;約26〜約273;および26〜273のアミノ酸をコードするポリヌクレオチドが挙げられる。
【0033】
指標タンパク質およびポリペプチドの改変体もまた見出され得る。転移性指標タンパク質またはポリペプチド改変体は、天然に存在することも存在しないこともあり得る。天然に存在する転移性指標タンパク質またはポリペプチド改変体は、ヒトまたは他の種において見出され、そして配列番号1に示されるヌクレオチド配列またはその相補体に対応する遺伝子によりコードされるタンパク質と実質的に同一であるアミノ酸配列を含む。低転移性または非転移性乳癌細胞と比較して、天然に存在する転移性指標タンパク質またはポリペプチド改変体と実質的に同じ高い転移性の発現パターンを保持する天然に存在しない転移性指標タンパク質またはポリペプチド改変体もまた、本発明に含まれる。好ましくは、天然に存在するかまたは天然に存在しない転移性指標タンパク質またはポリペプチド改変体は、配列番号1に示されるヌクレオチド配列によりコードされるアミノ酸配列と、少なくとも85%、90%、91%、92%、93%、94%、または95%同一であるアミノ酸配列を有する。より好ましくは、この分子は、少なくとも96%、97%、98%、または99%同一である。野生型タンパク質またはポリペプチドと改変体との間の配列同一性のパーセントは、野生型タンパク質またはポリペプチドと改変体を整列させて、当業者に公知である、野生型と改変体との間のアミノ酸一致数の計数、および野生型配列の合計アミノ酸残基数による合計一致数の除算によって、最大アミノ酸一致数を獲得することにより決定される。
【0034】
好ましくは、転移性指標タンパク質またはポリペプチド改変体におけるアミノ酸の変化は、保存的アミノ酸変化、すなわち類似の電荷を有するかまたは非電荷のアミノ酸での置換である。保存的アミノ酸変化は、それらの側鎖に関して関連するアミノ酸ファミリーの1つの置換を含む。天然に存在するアミノ酸は、遺伝的に4つのファミリー:酸性(アスパルテート、グルタメート)、塩基性(リジン、アルギニン、ヒスチジン)、非極性(アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、および非荷電極性(グリシン、アスパラギン、グルタミン、シスチン、セリン、スレオニン、チロシン)アミノ酸に分けられる。フェニルアラニン、トリプトファン、およびチロシンは、しばしば芳香族アミノ酸として一緒に分類される。
【0035】
ロイシンのイソロイシンまたはバリンでの孤立性(isolated)置換、アスパラギン酸のグルタミン酸での孤立性置換、トレオニンのセリンでの孤立性置換、またはあるアミノ酸の構造的に関連するアミノ酸での類似の置換が、得られた転移マーカータンパク質または転移マーカーポリペプチドの改変体の生物学的特性に対して大きな影響を有さないことを予測することは妥当である。転移マーカータンパク質または転移マーカーポリペプチドの改変体の特性および機能は、配列番号1に示されるヌクレオチド配列によってコードされるアミノ酸配列を含む転移マーカータンパク質または転移マーカーポリペプチドと同じ型の特性および機能であるが、その改変体の特性および機能は、ある程度異なり得る。アミノ酸変化が、適切な差次的発現パターンを有する転移マーカータンパク質または転移マーカーポリペプチドの改変体を生じるか否かは、容易に決定され得る。例えば、ヌクレオチドプローブは、本明細書中に開示されるマーカー遺伝子配列から選択され得、そして当該分野で公知のように、ノーザンブロットまたは組織切片において、マーカー遺伝子mRNAを検出するために使用され得る。あるいは、転移マーカー遺伝子のタンパク質産物に特異的に結合する抗体が、転移マーカータンパク質またはその改変体の発現を検出するために使用され得る。
【0036】
転移マーカータンパク質改変体としては、グリコシル化形態、他の分子との凝集結合体、および無関係な化学部分との共有結合体が挙げられる。転移マーカー改変体としてはまた、対立遺伝子改変体、種改変体、およびムテインが挙げられる。転移マーカー遺伝子の差次的発現に影響を与えない領域の切断型または欠失型もまた、転移マーカー改変体である。共有結合性の改変体は、当該分野で公知のように、アミノ酸側鎖あるいはN末端残基またはC末端残基に見出される基に官能基を連結させることによって調製され得る。
【0037】
本発明のポリペプチドのいくらかのアミノ酸配列が、そのタンパク質の構造または機能に有意な影響を及ぼすことなく改変され得ることが、当該分野で認識される。このような配列中の差異を意図する場合、そのタンパク質上に活性を決定する重要な領域が存在することは覚えておくべきである。一般に、類似の機能を果たす残基が使用される限り、三次構造を形成する残基を置換することが可能である。他の場合において、その変更がタンパク質の重要でない領域に生じる場合は、残基の型は、全く重要でなくてもよい。アミノ酸の置換はまた、細胞表面レセプターへの結合の選択性を変化させ得る。Ostadeら、Nature 361:266−268(1993)は、TNF−αの2つの公知のTNFレセプターの型のうちの1つの型のみへの、選択的結合を生じる特定の変異を記載する。従って、本発明のポリペプチドは、天然の変異または人為的操作のいずれか由来の、1以上のアミノ酸の置換、欠失または付加を含み得る。
【0038】
本発明はさらに、比較可能な発現パターンを示すか、または抗原性領域を含む、開示されたポリペプチドの改変体を含む。このような変異としては、欠失、挿入、反転、反復、および型置換が挙げられる。どのアミノ酸変化が表現型的にサイレントであるようであるかに関するガイダンスは、Bowie,J.U.ら、「Deciphering the Message in Protein Sequences:Tolerance to Amino Acid Substitutions」、Science 247:1306−1310(1990)に見出され得る。
【0039】
荷電アミノ酸の、別の荷電アミノ酸での置換および中性または負に荷電したアミノ酸での置換は、特に関心深い。後者は、正電荷が減少したタンパク質を生じ、開示されたタンパク質の特性を改善する。凝集の防止が、非常に望ましい。タンパク質の凝集は、活性の損失を生じるだけでなく、それらが免疫原性であり得るので、薬学的処方物を調製する際に問題であり得る(Pinckardら、Clin.Exp.Immunol.2:331−340(1967);Robbinsら、Diabetes 36:838−845(1987);Clelandら、Crit.Rev.Therapeutic Drug Carrier Systems 10:307−377(1993))。
【0040】
機能に必須である本発明のポリペプチド中のアミノ酸は、当該分野で公知の方法(例えば、部位特異的変異誘発またはアラニンスキャニング変異誘発(CunninghamおよびWells,Science 244:1081−1085(1989)))によって同定され得る。後者の手順は、その分子の全ての残基で単一のアラニン変異を導入する。次いで、得られた変異体分子を、レセプター結合またはインビトロ増殖活性のような、生物学的活性について試験する。リガンド−レセプター結合に重要な部位はまた、構造分析(例えば、結晶化、核磁気共鳴または光親和性標識(Smithら、J.Mol.Biol.224:899−904(1992)およびde Vosら、Science 255:306−312(1992))によって決定され得る。
【0041】
示されるように、変化は、好ましくは、わずかな性質の変化である(例えば、タンパク質のフォールディングまたは活性に有意には影響を与えない保存的アミノ酸置換)。もちろん、当業者が作製するアミノ酸置換の数は、上記の因子を含む、多くの因子に依存する。一般的に言えば、任意の所定のポリペプチドについての置換の数は、50、40、30、25、20、15、10、5または3より多くない。
【0042】
全長転移マーカータンパク質は、標準的な生化学的方法を使用して、転移マーカータンパク質を産生するヒト細胞(例えば、転移性乳癌細胞)から抽出され得る。単離および精製された転移マーカータンパク質または転移マーカーポリペプチドは、細胞中で転移マーカータンパク質または転移マーカーポリペプチドが通常会合している他の化合物(例えば、特定のタンパク質、炭水化物、脂質、または細胞内小器官)から分離される。単離および精製された転移マーカータンパク質または転移マーカーポリペプチドの調製物は、少なくとも80%純粋であり;好ましくは、その調製物は、90%、95%、または99%純粋である。
【0043】
配列番号2をコードするヒト遺伝子を、当該分野において公知の方法を使用して同定および単離し得る。1つの方法によれば、配列番号1は、コンピュータで読み取り可能な形式で調製される。この配列が、ヒトゲノムのポリヌクレオチド配列と比較され、そして配列番号1に対して少なくとも95%の配列同一性を有する1つ以上のヒトゲノム配列が、例えば、パラメータとして12のギャップオープンペナルティーおよび1のギャップエクステンションペナルティーを用いるアフィンギャップサーチを使用して、Smith−Watermanアルゴリズムを使用することによって、同定される。当該分野において公知の方法を使用して、配列番号1とヒトゲノム配列との間の相同の領域に基づくプローブが調製され、そしてヒトゲノムDNAからポリヌクレオチドを単離するために使用される。出願日現在、配列番号1の全ポリヌクレオチドに対応するヒトポリヌクレオチドは、公共のデータベースにおいて確認されなかった。従って、本発明は、配列番号1のコード領域を含むヒトゲノムDNA、および配列番号1と相同性を共有せず相同領域と連続する任意の未翻訳領域を包含する。このようなゲノムDNAとしては、イントロン、プロモーター、および配列番号2をコードする領域を有するヒト遺伝子に機能的に関連する他の調節領域が挙げられるが、これらに限定されない。
【0044】
転移マーカータンパク質および転移マーカーポリペプチドはまた、組換えDNA方法または合成化学的方法によって、産生され得る。組換え転移マーカータンパク質または転移マーカーポリペプチドの産生のために、配列番号1に示すヌクレオチド配列、または転移マーカータンパク質をコードする配列の改変体から選択されるコード配列が、公知の原核生物または真核生物の発現系において、発現され得る(以下を参照のこと)。細菌、酵母、昆虫、または哺乳動物の発現系が、当該分野において公知であるように、使用され得る。
【0045】
あるいは、合成化学方法(例えば、固相ペプチド合成)が、転移マーカータンパク質または転移マーカーポリペプチドを合成するために使用され得る。ペプチド、アナログ、または誘導体の産生のための一般的手段は、CHEMISTRY
AND BIOCHEMISTRY OF AMINO ACIDS,PEPTIDES, AND PROTEINS−−A SURVEY OF RECENT DEVELOPMENTS、Weinstein,B.編、Marcell Dekker,Inc.出版、New York(1983)に概説されている。さらに、通常のL−立体異性体に代えてのD−アミノ酸での置換が、その分子の半減期を増加するために実行され得る。転移マーカー改変体は、同様に産生され得る。
【0046】
少なくとも6、8、10、12、15、18、20、25、30、35、40、50、55、60、65、70、75、80、85、90、95、100、120、140、160、180、200、220、240、250、260、265、270もしくは271またはそれより多くの連続した転移マーカーアミノ酸を含む、天然には存在しない融合タンパク質もまた、構築され得る。ヒト転移マーカー融合タンパク質は、転移マーカーアミノ酸配列に対する抗体を生成するため、および種々のアッセイ系において使用するために、有用である。例えば、転移マーカー有効タンパク質を使用して、転移マーカータンパク質と相互作用し、そしてこれらの機能に影響を与えるタンパク質を、同定し得る。タンパク質アフィニティークロマトグラフィーのような物理的方法、または酵母ツーハイブリッドもしくはファージディスプレイ系のようなタンパク質−タンパク質相互作用に関するライブラリーに基づくアッセイもまた、この目的で使用され得る。このような方法は、当該分野において周知であり、そして薬物スクリーニングとしてもまた使用され得る。
【0047】
転移マーカー融合タンパク質は、ペプチド結合によって一緒に融合した2つのタンパク質セグメントを含む。第1のタンパク質セグメントは、転移マーカータンパク質の少なくとも6、8、10、12、15、18、20、25、30、35、40、50、55、60、65、70、75、80、85、90、95、100、120、140、160、180、200、220、240、250、260、265、270もしくは271またはそれより多くの連続したアミノ酸を含む。これらのアミノ酸は、配列番号1に示されるヌクレオチド配列によってコードされるアミノ酸配列から、または上記のもののようなこの配列の改変体から、選択され得る。第1のタンパク質セグメントはまた、全長転移マーカータンパク質を含み得る。
【0048】
1つの好ましい実施形態において、第1のタンパク質セグメントは、配列番号2に示されるポリペプチドを含む。この実施形態の改変において、この第1のタンパク質セグメントは、配列番号2のアミノ酸31〜287からなる。この融合タンパク質は、配列番号2のシグナルペプチドを欠き、そして細胞の内側での発現された融合タンパク質の保持のために適切である。
【0049】
第2のタンパク質セグメントは、配列番号1によってコードされるネイティブタンパク質において第1のタンパク質セグメントに隣接して見出されなかった、全長タンパク質またはタンパク質フラグメントあるいはポリペプチドであり得る。この融合タンパク質は、当該分野において公知であるように、検出可能なマーカー(例えば、放射性マーカー、蛍光マーカー、化学発光マーカー、またはビオチン化マーカー)で標識され得る。第2のタンパク質セグメントは、β−ガラクトシダーゼのような検出可能な産物を生成する酵素であり得る。第1のタンパク質セグメントは、好都合であるように、N末端またはC末端であり得る。
【0050】
2つのタンパク質セグメントを組換え的にかまたは共有結合させることによってかのいずれかで、融合タンパク質を作製するための技術もまた、周知である。組換えDNA法は、以下に記載のように、例えば、第2のタンパク質セグメントをコードしかつ宿主細胞においてDNA構築物を発現するヌクレオチドで、適切なリーディングフレームにおいて配列番号1のコード配列を含むDNA構築物を作製することによって、転移マーカー融合タンパク質を調製するために用いられ得る。配列番号1のオープンリーディングフレームを、図4に示す。
【0051】
単離および精製された転移マーカータンパク質、ポリペプチド、改変体、または融合タンパク質は、転移マーカータンパク質に特異的に結合する抗体の調製物を得るために、免疫原として使用され得る。この抗体は、とりわけ、ヒト組織およびその画分において、野生型の転移マーカータンパク質を検出するために使用され得る。この抗体はまた、転移マーカータンパク質の過小発現もしくは過剰発現を生じるかまたはサイズもしくは電気泳動移動度が変化した転移マーカータンパク質の発現を生じる、転移マーカー遺伝子の変異の存在を検出するために使用され得る。
【0052】
ポリクローナル抗体およびモノクローナル抗体の調製は、標準的方法を使用してなされ得る。単鎖抗体もまた、調製され得る。好ましい免疫原は、配列番号2を含むポリペプチドである。転移マーカータンパク質、ポリペプチド、改変体、または融合タンパク質に特異的に結合する単鎖抗体は、当該分野で公知であるように、例えば、単鎖免疫グロブリンディスプレイライブラリーから単離される。そのライブラリーは、配列番号2の転移マーカータンパク質のアミノ酸配列に対して「パニング」され、そして転移マーカータンパク質の異なるエピトープに高い親和性で結合する多数の単鎖抗体が、単離され得る。Hayashiら、1995、Gene 160:129〜30。単鎖抗体はまた、DNA増幅法(例えば、ハイブリドーマcDNAをテンプレートとして使用する、ポリメラーゼ連鎖反応(PCR))を使用して、構築され得る。Thirionら、1996、Eur.J.Cancer Prev.5;507〜11。
【0053】
転移マーカー特異的抗体は、配列番号1に示すヌクレオチド配列によってコードされるアミノ酸配列を有する、全長転移マーカータンパク質に存在するエピトープ、転移マーカーポリペプチド、または転移マーカー改変体に、単独でかまたは融合タンパク質の一部として、特異的に結合する。好ましくは、転移マーカーエピトープは、他のヒトタンパク質には存在しない。代表的には、少なくとも6、8、10、または12の連続したアミノ酸が、エピトープを形成するために必要である。しかし、非連続のアミノ酸を含むエピトープは、より多くのアミノ酸、例えば、少なくとも15、25または50のアミノ酸を必要とし得る。
【0054】
転移マーカータンパク質、ポリペプチド、融合タンパク質、または改変体に特異的に結合する抗体は、ウエスタンブロットまたは他の免疫化学的アッセイにおいて使用される場合に、他のタンパク質を用いて提供される検出シグナルより少なくとも5倍、10倍または20倍高い検出シグナルを提供する。好ましくは、転移マーカーエピトープに特異的に結合する抗体は、免疫化学的アッセイにおいて他のタンパク質を検出せず、そして転移マーカータンパク質、ポリペプチド、融合タンパク質、または変異体を、溶液から免疫沈降させ得る。
【0055】
サブゲノムポリヌクレオチドは、全体染色体未満を含む。好ましくは、ポリヌクレオチドは、イントロンを含まない。好ましい実施形態において、ポリヌクレオチド分子は、配列番号1の10、15、20、25、30、40、50、75、100、150、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700、1800、1900、2000、2100、2200、2300または2350のヌクレオチドの連続した配列、あるいは、その相補体を含む。配列番号1に示されるヌクレオチド配列の相補体は、配列番号1に示される連続したヌクレオチド配列と、ワトソン−クリック塩基対を形成する連続するヌクレオチド配列である。
【0056】
転移マーカータンパク質または改変体のアミノ酸配列をコードする縮重ヌクレオチド配列、ならびに配列番号1に示されるヌクレオチド配列のコード領域に少なくとも85%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%同一であるポリヌクレオチドを含む相同的ヌクレオチド配列もまた、転移マーカーサブゲノムポリヌクレオチドである。代表的には、相同的転移マーカーサブゲノムポリヌクレオチド配列は、当該分野において公知のように、ストリンジェントな条件下でハイブリダイゼーションを行うことによって確認され得る。野生型と相同な改変体との間の配列同一性%は、当該分野で公知なように、野生型ポリヌクレオドをその改変体と整列させて最も多い数のヌクレオチドの一致を得、野生型と改変体との間のヌクレオチドの一致の数を計数し、そして一致の合計数を野生型配列のヌクレオチドの合計数で割ることによって決定される。同一性%を計算するための好ましいアルゴリズムは、Smith−Waterman相同性サーチアルゴリズム(以下のサーチパラメーター:12のギャップオープンペナルティーおよび1のギャップエクステンションペナルティーとともにアフィンギャップサーチを用いてMPSRCHプログラム(Oxford Molecular)を実行する)である。
【0057】
転移マーカータンパク質コード配列を含む転移マーカーサブゲノムポリヌクレオチドは、発現構築物において使用される。好ましくは、転移マーカーサブゲノムポリヌクレオチドは、発現プラスミド(例えば、Ecdyson系、pIND、In Vitoro Gene)に挿入される。転移マーカーサブゲノムポリヌクレオチドは、当該分野で周知の技術を使用してベクターおよび細胞株において増殖され得る。転移マーカーサブゲノムポリヌクレオチドは、直鎖状分子または環状分子上であり得る。これらは、自律的に複製する分子上にあり得るかまたは、複製配列を有さない分子上にあり得る。これらは、それら自身によって、または当該分野で公知の他の調節配列によって調節され得る。
【0058】
次いで、転移マーカー発現構築物を含む宿主細胞を使用して、転移マーカータンパク質の全てまたは一部を発現し得る。転移マーカー発現構築物を含む宿主細胞は、原核生物または真核生物であり得る。種々の宿主細胞は、細菌、酵母、昆虫、およびヒトの発現系における使用に利用可能であり、転移マーカー発現構築物を発現するかまたは増殖するために使用され得る(以下を参照こと)。発現構築物は、当該分野で公知な任意に技術を使用して宿主細胞に導入され得る。これらの技術には、トランスフェリン−ポリカチオン媒介DNA移入、裸の核酸またはカプセル化核酸を用いたトランスフェクション、リポソーム媒介細胞融合、DNAコートラテックスビーズの細胞内輸送、プロトプラスト融合、ウイルス感染、エレクトロポレーション、およびリン酸カルシウム媒介トランスフェクションが挙げられる。
【0059】
転移マーカー発現構築物は、選択された宿主細胞において機能的であるプロモーターを含む。当業者は、当該分野で公知であり使用される多くの数の細胞型特異的プロモーターから適切なプロモーターを容易に選択し得る。この発現構築物はまた、宿主細胞中で機能的である転写ターミネーターを含み得る。この発現構築物は、転移マーカータンパク質の全てまたは一部分、改変体、融合タンパク質、抗体、またはリボザイムをコードするポリヌクレオチドセグメントを含む。このポリヌクレオチドセグメントは、プロモーターから下流に位置する。ポリヌクレオチドセグメントの転写は、プロモーターで開始する。この発現構築物は、直鎖状または環状であり得、そして所望される場合、自律複製配列を含み得る。
【0060】
転移マーカー発現構築物を発現するための細菌系としては、Changら、Nature(1978)275:615、Goeddelら、Nature(1979)281:544、Goeddelら、Nucleic Acids Res.(1980)8:4057、EP36,776、米国特許第4,551,433号、deBoerら、Proc.Nat’l.Acad Sci.USA(1983)80:21−25、およびSiebenlistら、Cell(1980)20:269に記載される発現系が挙げられる。
【0061】
酵母における発現系としては、Hinnenら、Proc.Nat’l.Acad.Sci.USA(1978)75:1929;Itoら、J.Bacteriol.(1983)153:163;Kurtzら、Mol.Cell.Biol.(1986)6:142;Kunzeら、J.Basic Microbiol.(1985)25:141;Gleesonら、J.Gen.Microbiol.(1986)132:3459、Roggenkampら、Mol.Gen.Genet.(1986)202:302)Dasら、J.Bacteriol.(1984)158:1165;De Louvencourtら、J.Bacteriol.(1983)154:737、Van den Bergら、Bio/Technology(1990)8:135;Kunzeら、J.Basic Microbiol.(1985)25:141;Creggら、Mol.Cell.Biol.(1985)5:3376、米国特許第4,837,148号、米国特許第4,929,555号;BeachおよびNurse、Nature(1981)300:706;Davidowら、Curr.Genet.(1985)10:380、Gaillardinら、Curr.Genet.(1985)10:49、Ballanceら、Biochem.Biophys.Res.Commun.(1983)112:284−289;Tilburnら、Gene(1983)26:205−221、Yeltonら、Proc.Nat’l.Acad.Sci.USA(1984)81:1470−1474、KellyおよびHynes、EMBO J.(1985)4:475479;EP244,234およびWO 91/00357に記載される発現系が挙げられる。
【0062】
昆虫における転移マーカー発現構築物の発現は、米国特許第4,745,051号、Friesenら(1986)「バキュロウイルス遺伝子発現の調節」:THE MOLECULAR BIOLOGY OF BACULOVIRUSES(W.Doerfler編)、EP127,839、EP155,476、およびVlakら、J.Gen.Virol.(1988)69:765−776、Millerら、Ann.Rev.Microbiol.(1988)42:177、Carbonellら、Gene(1988)73:409、Maedaら、Nature(1985)315:592−594、Lebacq−Verheydenら、Mol.Call.Biol.(1988)8:3129;Smithら、Proc.Nat’l.Acad.Sci.USA(1985)82:8404、Miyajimaら、Gene(1987)58:273;およびMartinら、DNA(1988)7:99に記載されるように達成され得る。多くのバキュロウイルス株および改変体および対応する宿主由来の許容昆虫宿主細胞は、Luckowら、Bio/Technology(1988)6:47−55、Millerら、GENETIC ENGINEERING(Setlow,J.K.ら編)、第8巻(Plenum Publishing、1986)、277〜279頁、ならびにMaedaら、Nature,(1985)315:592−594に記載される。
【0063】
転移マーカー発現構築物の哺乳動物発現は、Dijkemaら、EMBO J.(1985)4:761、Gormanら、Proc.Nat’l.Acad.Sci.USA(1982b)79:6777、Boshartら、Cell(1985)41:521および米国特許第4,399,216号に記載されるように達成され得る。転移マーカー発現構築物の哺乳動物発現の他の特徴は、HamおよびWallace,Meth.Enz.(1979)58:44、BarnesおよびSato、Anal.Biochem.(1980)102:255、米国特許第4,767,704号、米国特許第4,657,866号、米国特許第4,927,762号、米国特許第4,560,655号、WO 90/103430、WO 87/00195号、ならびに米国特許再発行30,985に記載されるように容易にされ得る。
【0064】
本発明のサブゲノムポリヌクレオチドはまた、mRNAまたはオリゴヌクレオチド(天然の転移マーカーmRNAまたはその相補体のいずれかの配列を有する)、全長転移マーカータンパク質、転移マーカー融合タンパク質、転移マーカーポリペプチド、または転移マーカー特異的リボザイム、または単鎖抗体を細胞(好ましくは真核生物細胞)に送達する目的のために、遺伝子送達ビヒクルにおいて使用され得る。本発明に従って、遺伝子送達ビヒクルは、例えば、裸のプラスミドDNA、転移マーカーサブゲノムポリヌクレオチドを含むウイルス発現ベクター、またはリポソームもしくは縮合試薬と連結した転移マーカーサブゲノムポリヌクレオチドであり得る。
【0065】
本発明は、生物学的サンプルにおいて転移マーカー遺伝子発現を検出する方法を提供する。転移マーカー遺伝子発現の検出は、例えば、転移を同定するため、または、組織サンプル(好ましくは、腫瘍)における転移の可能性を決定するために有用である。次いで、適切な処置レジメンが、身体の他の器官における転移性の癌を発達する危険がある患者について設計され得る。
【0066】
身体サンプルは、例えば、固体組織または流体サンプルであり得る。配列番号1によってコードされるネイティブなポリヌクレオチドは、推定の分泌タンパク質であり、血液およびリンパ流体(特に、身体の腫瘍部位からの排出するもの)を含む体液において検出されるようである。タンパク質または核酸の発現産物は、身体サンプル中で検出され得る。1つの実施形態において、この身体サンプルは、転移マーカータンパク質の存在に関してアッセイされる。この転移マーカータンパク質は、配列番号1に示されるヌクレオチド配列によりコードされる配列またはその相補体を含み、かつ本発明のマーカータンパク質特異抗体を用いて検出され得る。この抗体は、例えば、放射性、蛍光、ビオチン化、または酵素学的タグで標識化され得、そして直接的に検出され得るか、または(標識化した二次抗体を使用する)間接的な免疫化学的方法を用いて検出され得る。この転移マーカータンパク質の存在は、(例えば、免疫細胞化学によって組織切片において、または溶解物中において、当該分野において公知であるようなウェスタンブロットを用いて)アッセイされ得る。マーカータンパク質の存在は、組織サンプルが転移性であることを示す。
【0067】
別の実施形態において、身体サンプルは、マーカータンパク質mRNAの存在についてアッセイされる。サンプルは、選択されたポリペプチドに対応するmRNAとハイブリダイズし得る核酸ハイブリダイゼーションプローブと接触され得る。なおさらに、サンプルは、ポリペプチドの発現を示すmRNAを検出するためにノーザンブロット技術に供され得る。mRNAが検出されるそれらの技術について、サンプルは、核酸増幅プロセスに供され得、それによって、mRNA分子またはその選択された部分は、適切なヌクレオチドプライマーを使用して、増幅される。他のRNA検出技術がまた使用され得、これには、インサイチュウハイブリダイゼーションが挙げられるが、これに限定されない。
【0068】
マーカータンパク質特異的プローブは、配列番号1に開示されるcDNA配列を使用して作製され得る。プローブは、好ましくは、少なくとも15〜50ヌクレオチドの長さであるが、これらは、少なくとも8、10、11、12、20、25、30、35、40、45、60、75、または100以上のヌクレオチドの長さであり得る。プローブを選択するための好ましい領域は、配列番号1の446位〜1173位のヌクレオチド内にある。プローブは、化学的に合成され得るか、または制限酵素を使用してより長いポリヌクレオチドから作製され得る。プローブは、例えば、放射性タグ、ビオチン化タグ、または蛍光タグで標識され得る。
【0069】
必要に応じて、身体サンプル内の特定の転移マーカー発現産物のレベルが定量され得る。定量は、例えば、身体サンプル内で検出される発現産物のレベルを、検量線に存在する生成物の量と比較することによって達成され得る。コンピュータの援助ありでまたはなしで、視覚的にまたは密度測定のような技術を使用して比較がなされ得る。コントロールとしての使用のために、身体サンプルは、他のヒト、試験される患者の他の非癌性器官、または試験される患者からの非転移性乳癌から単離され得る。本明細書中の結果によって示されるように、低い転移性または非転移性の乳癌細胞における配列番号1の発現は、高度に転移性の乳癌細胞における発現レベルの3%と44%との間である。試験サンプルにおける発現が適切なコントロールサンプルの少なくとも二倍である場合、これは、転移性細胞であることを示す。
【0070】
本発明の転移性マーカー特異的試薬(例えば、抗体およびヌクレオチドプローブ)をコードするポリヌクレオチドは、生物学的サンプルにおけるマーカー遺伝子発現産物を検出するためのキットに供給され得る。このキットはまた、緩衝液または標識成分、ならびに生物学的サンプルにおいてマーカー発現産物を検出するために試薬を使用するための説明書を含み得る。
【0071】
転移マーカー遺伝子の発現は、アンチセンスオリゴヌクレオチド配列を使用して変更され得る。アンチセンス配列は、配列番号1に示されるヌクレオチド配列を有する転移マーカー遺伝子のコード配列(ヌクレオチド365〜1173)の少なくとも一部に相補的である。好ましくは、このアンチセンスオリゴヌクレオチド配列は、少なくとも6ヌクレオチドの長さであるが、約8、12、15、20、25、30、35、40、45、または50ヌクレオチドの長さであり得る。より長い配列もまた、用いられ得る。アンチセンスオリゴヌクレオチド分子は、DNA構築物中に提供され得、そして細胞分裂が減少されるべきである細胞中へと導入され得る。このような細胞としては、高度に転移性の乳癌細胞が挙げられる。
【0072】
アンチセンスオリゴヌクレオチドは、デオキシリボヌクレオチド、リボヌクレオチド、または両方の組み合わせで構成され得る。オリゴヌクレオチドは、手動または自動合成機によって、1つのヌクレオチドの5’末端を、アルキルホスホネート、ホスホロチオエート、ホスホロジチオエート、アルキルホスホノチオエート、アルキルホスホネート、ホスホラミデート(phosphoramidates)、リン酸エステル、カルバメート、アセトアミデート(acetamidate)、カルボキシメチルエステル、カーボネート、およびリン酸トリエステルのような、非リン酸ジエステルのヌクレオチド間結合を有する別のヌクレオチドの3’末端と共有結合することにより合成され得る。Brown,1994,Meth.Mol.Biol.20:1−8;Sonveaux,1994,Meth.Mol.Biol.26:1−72;Uhlmannら、1990,Chem.Rev.90:543−583を参照のこと。
【0073】
転移マーカータンパク質に特異的に結合する本発明の抗体はまた、転移マーカー遺伝子発現を変更するために使用され得る。抗体とは、タンパク質に対する特異的な結合を保持する単鎖抗体のような抗体およびその一部または誘導体を意味する。特異的抗体は、転移性マーカータンパク質に結合し、そしてこのタンパク質を細胞中で機能しないようにする。上記のように、本発明の特異的抗体をコードするポリヌクレオチドは、細胞中に導入され得る。
【0074】
本発明のマーカータンパク質は、治療的な抗転移性効果を有する薬物をスクリーニングするために用いられ得る。転移性マーカータンパク質合成に対する試験化合物の効果はまた、転移を調節する試験化合物を同定するために用いられ得る。スクリーニングされ得る試験化合物としては、被験体に投与され得る天然産物または合成のいずれかの任意の物質が挙げられる。化合物のライブラリーまたは混合物が試験され得る。この化合物または物質は、薬学的な効果が以前に公知の物質であっても未知の物質であってもよい。
【0075】
転移性マーカータンパク質の合成は、当該分野で公知の、タンパク質合成を測定する任意の手段(例えば、標識アミノ酸のタンパク質への組み込みおよびポリアクリルアミドゲル中での標識した転移性マーカータンパク質の検出)によって測定され得る。転移性マーカータンパク質の量は、例えば、ウエスタンブロットにおいて、本発明の転移性マーカータンパク質特異的抗体を用いて、検出され得る。試験化合物の存在または非存在下で合成された転移性マーカータンパク質の量は、当該分野で公知の任意の手段(例えば、標準曲線で示される転移性マーカータンパク質の量と、合成した転移性マーカータンパク質の量の比較)によって決定され得る。
【0076】
転移性マーカータンパク質合成に対する試験化合物の効果はまた、当該分野で公知であるように、本発明の転移性マーカータンパク質特異的ヌクレオチドプローブを用いる試験化合物に応答する転移性マーカータンパク質mRNA発現の量を測定することによって、ノーザンブロット分析によって、測定され得る。
【0077】
代表的には、生物学的サンプルが、ある範囲の濃度(例えば、1.0nM、5.0nM、10nM、50nM、100nM、500nM、1mM、10mM、50mM、および100mM)の試験化合物と接触される。好ましくは、試験化合物は、転移性マーカータンパク質の発現を、60%、75%、または80%まで増大または減少する。より詳細には、85%、90%、95%または98%の増大または減少が達成される。
【0078】
本発明は、転移性マーカータンパク質の発現を増大または減少するための組成物を提供する。これらの組成物は、転移性マーカータンパク質遺伝子発現産物の全てまたは少なくとも一部をコードするポリヌクレオチドを含む。好ましくは、治療用組成物は、プロモーターおよび転移性マーカータンパク質の少なくとも一部をコードするポリヌクレオチドセグメントを含む発現構築物を含み、転移能力(力価)を減少させるのに効果的である。細胞の転移能力を減少させるのに有効である転移性マーカー遺伝子またはタンパク質の部分は、当該分野で公知のように、例えば、転移性マーカー遺伝子またはポリペプチドの部分を転移性細胞株(例えば、MDA−MB−231、MDA−MB−435、Km12C、またはKm12L4)に導入すること、およびインビボで移植された場合、細胞の分裂速度または細胞が転移を形成する能力をアッセイすることによって決定され得る。非転移性細胞株(例えば、MCF−7)は、転移性マーカータンパク質の一部が、転移性マーカー遺伝子の発現を増大する能力をアッセイするために用いられ得る。
【0079】
代表的には、治療用転移性マーカー組成物が、注射可能に、流体溶液または懸濁液のいずれかとして、調製され得る;しかし、注射前の流体ビヒクル中の溶液に、または流体ビヒクル中の懸濁液に適切な固体形態がまた調製され得る。転移性マーカー組成物がまた、米国特許第4,853,230号、欧州特許225,189号、AU9,224,296号およびAU9,230,801号に記載のように、当該分野で公知の方法に従って、腸溶コーティング錠剤またはゲルカプセル中に処方され得る。
【0080】
本発明の転移性マーカー治療剤の投与は、局所(local)投与または全身投与(これには、注射、経口投与、粒子銃、またはカテーテル投与を含む)、および局所(topical)投与を含み得る。種々の方法を用いて、身体内の特定の部位に治療用転移性マーカー組成物を直接投与し得る。
【0081】
腫瘍(転移性病変を含む)の処置のために、例えば、治療用転移性マーカー組成物が、腫瘍の本体内のいくつかの異なる位置に数回注射され得る。あるいは、腫瘍を助ける動脈が、同定され得、そして治療用組成物が、この組成物を腫瘍に直接送達するために、このような動脈に注射される。
【0082】
壊死性中心を有する腫瘍は、吸引され得、そしてこの組成物がこの腫瘍のいまや空になった中心に直接注射される。治療用転移性マーカー組成物は、例えば、この組成物の局所適用によって、腫瘍の表面に直接投与され得る。X線画像を用いて、上記の特定の送達方法が補助され得る。転移性マーカータンパク質もしくはポリペプチド、または転移性マーカー小ゲノムポリヌクレオチドおよび他の治療用因子を含む、併用療法の薬剤が、同時にまたは引き続いて投与され得る。
【0083】
あるいは、転移性マーカー治療組成物が、エクソビボでヒト細胞に導入され得、次いで、この細胞はヒトの中に入れられる。細胞は、種々の場所から(例えば、以下を含む:選択された腫瘍からまたは罹患した器官から)取り出され得る。さらに、治療組成物は、罹患していない細胞、例えば、真皮線維芽細胞または末梢血リンパ球中に挿入され得る。所望の場合、T細胞サブセットまたは幹細胞のような細胞の特定の画分がまた、血液から特異的に取り除かれ得る(例えば、PCT WO 91/16116を参照のこと)。次いで、除去された細胞を、上記の任意の技術を用い、続いてヒトに(好ましくは、処置されるべき腫瘍または他の部位の近傍に、またはその中に)この細胞を戻し、転移性マーカー治療組成物と接触させ得る。上記の方法は、さらに、線維芽細胞または他の混入のない腫瘍細胞を枯渇し、続いてヒトから腫瘍細胞を取り出す工程、および/または例えば、照射によって細胞を不活性化する工程を包含し得る。
【0084】
転移性マーカー組成物の投与の用量および投与の手段の両方は、治療用組成物の特定の質、患者の状態、年齢、および体重、疾患の進行、ならびに他の関連要因に基づいて決定され得る。好ましくは、本発明の治療用組成物は、転移性マーカー遺伝子の発現を50%、60%、70%、または80%まで減少する。より好ましくは、転移性マーカー遺伝子の発現は、90%、95%、99%または100%まで減少される。転移性マーカー遺伝子の発現を変更するために選択される機構の有効性は、当該分野で周知の方法(例えば、転移性マーカー遺伝子のmRNAに対するヌクレオチドプローブのハイブリダイゼーション、定量的RT−PCR、または本発明の特定の抗体を用いる転移性マーカータンパク質の検出)を用いて評価され得る。
【0085】
この組成物が、転移性マーカータンパク質、ポリペプチド、または抗体を含む場合、有効な用量の組成物が、患者の体重あたり約5μg〜約50μg/kg、患者の体重あたり約50μg〜約5mg/kg、約100μg〜約500μg/kg、そして約200〜約250μg/kgの範囲である。
【0086】
転移性マーカーサブゲノムポリヌクレオチドを含む治療用組成物は、局所投与について約100ng〜約200mgのDNAの範囲で投与され得る。約500ng〜約50mg、約1μg〜約2mg、約5μg〜約500μg、そして約20μg〜約100μgのDNAの濃度範囲がまた、遺伝子治療プロトコールの間に用いられ得る。形質転換および発現の作用および有効性の方法のような要因が、転移性マーカーサブゲノムポリヌクレオチドの最終的な有効性に必要である投薬量に影響することが考慮される。より大きい発現が組織のより大きい面積にわたって所望される場合、例えば、腫瘍部位の異なる隣接部位または近接する組織部位への、より大量の転移性マーカーサブゲノムポリヌクレオチドもしくは同じ量(投与の引き続くプロトコールで再投与される)、または、数回の投与が、正の治療結果をもたらすために必要であり得る。全ての場合において、臨床試験における慣用的実験が、至適の治療効果についての特定の範囲を決定する。
【0087】
内因性転移性マーカー遺伝子の細胞内での発現がまた、相同組み換えによって、内因性転移性マーカー遺伝子とインフレームでDNA構築物(転移性マーカータンパク質標的化配列、調節性配列、エキソン、および不対スプライスドナー部位を含む)を導入することによって変更され得、これによって、DNA構築物を含む相同組み換え細胞が形成される。新しい転写単位が、所望の場合、転移性マーカー遺伝子をターンオンまたはターンオフするために用いられ得る。内因性遺伝子発現に影響する方法は、本明細書において参考として援用される、米国特許第5,641,670号に教示されている。
【0088】
転移性マーカーサブゲノムポリヌクレオチドはまた、細胞に対する転移性マーカーサブゲノム性ポリヌクレオチドの移入を増強するか、または細胞内の転移性マーカーサブゲノム性ポリヌクレオチドの引き続く生物学的効果を増強するのに有用である試験化合物をスクリーニングする目的で被験体に送達され得る。このような生物学的効果としては、相補的転移性マーカーmRNAに対するハイブリダイゼーション、およびその翻訳の阻害、転移性マーカーmRNAおよび/または転移性マーカータンパク質を形成するための転移性マーカーサブゲノムポリヌクレオチドの発現、ならびに転移性マーカーサブゲノムポリヌクレオチドの複製および組み込みが挙げられる。被験体は、細胞培養物または動物、好ましくは哺乳動物、より好ましくはヒトであり得る。
【0089】
上記の開示は、本発明を一般的に記載する。より完全な理解が、以下の特定の実施例を参照して得られ得る。以下の実施例は、本明細書において例示のみの目的で提供されるものであり、本発明の範囲を限定することを意図しない。
【0090】
(実施例)
(材料および方法)
細胞培養。MDA−MB−435、MDA−MB−231、ALAB、MDA−MB−468、MDA−MB−361、ZR−75−1、MCF−7、MDA−MB−453およびSK−BR−3ヒト乳癌細胞株(Chiron Master Culture Collection、Chiron Corporationから入手)を、37℃で5%CO2で、以下の中で増殖した:DMEM+HAM’S F−12(1:1)(Bio*Whittaker,Walkersville,MD)(2mM Lグルタミン、1mM Sodium Pyruvate、100U/mlペニシリンおよび100μg/ml ストレプトマイシン(Bio*Whittaker,Walkersville,MD)、1×ビタミン溶液、1×非必須アミノ酸(Irvine Scientific,Santa Ana,CA)および10%熱不活化ウシ血清(Life Technologies,Rockville,MD)を含有)。COS−7細胞をATCCから入手し、そして10%熱不活化ウシ胎仔血清(Life Technologies)を含有するDMEM中で37℃、5%CO2中において増殖した。
【0091】
Opti−MEM1上清の濃度。Centricon YM−10および/またはMicron YM−10カラム(Millipore Corporation,Bedford,MA)を通して、Opti−MEM1(Life Technologies)培養培地を濃縮した。次いで、SDS−PAGEサンプルローディング緩衝液を添加し、そしてサンプルを煮沸した。
【0092】
ノーザンブロットハイブリダイゼーション。培養した乳癌細胞株、およびRNeasy Maxi Kit(Qiagen,Valencia,CA)を用いて、乳癌細胞株を移植したSCIDマウスの腫瘍組織から、総RNAを調製した。1レーンあたり、約20μgの総RNAを、電気泳動のためにホルムアルデヒド/アガロースゲル上にロードし、次いで、Hybond−N+ナイロンメンブレン(Amersham Life Science,Little Chalfont,England)に移した。このブロットをUV照射で固定した。5mg/mlの変性一本鎖精子DNAとともに、Rapid−Hyb緩衝液(Amersham Life Science)を65℃に予熱し、そして65℃で30分間振盪しながらこの緩衝液中でブロットをプレハイブリダイズした。[α−32P]dCTP(3000Ci/mmol,Amersham Pharmcia Biotech Inc.,Piscataway,NJ)(Prime−It RmT Kit,Stratagene,La Jolla,CA)で標識し、そしてProbeQuantTMG−50 Micro Column(Amersham Pharmacia Biotech Inc.)で精製したプローブとして、hsOAF cDNAフラグメントまたはβアクチンcDNAフラグメントを添加し、そしてこのブロットにハイブリダイズさせて、65℃で一晩振盪した。このブロットを2×SSC、0.1%(w/v)SDSを用いて、室温で20分間、1×SSC、0.1%(w/v)SDSで、65℃15分間2回洗浄し、次いでHyperfilms(Amersham Life
Science)に曝露した。
【0093】
イムノブロッティング。タンパク質サンプルを10〜20% SDS−PAGEゲル上の電気泳動に供し、次いで、25mM Tris,192mMグリシン、20%(v/v)メタノール、pH8.3における電気ブロッティングによってPVDF膜(0.2μm)に転写した。膜を10%脱脂乳を含有するTBST(pH7.5)中でブロックし、次いで、1%BSAを含有するPBS(pH7.4)中でウサギ抗hs−OAF血清(1:1000)とともに、ブロッティングし、次いで、二次抗体アルカリホスファターゼ結合体化ヤギ抗ウサギIgG(1:2000)(Santa Cruz Biotechnology,Inc.,Santa Cruz,CAを用いてプローブした。次いで、タンパク質バンドをNBT/BCIP試薬(Boehringer Mannheim,Germany)によって可視化した。
【0094】
一過性トランスフェクション。hsOAF cDNAのコード領域(356〜1174)を、改変発現ベクターpRetro−On(Clontech,Palo Alto,CA)中にクローニングした。hsOAFを保有するpRetro−OnベクターまたはコントロールのpRetro−Onベクター(GFPを有する)を、製造業者によって提供されたプロトコール中に指示されたように、EffecteneTMTransfection Reagent Kit(Qiagen)を用いて100mm培養プレート上のCOS−7細胞中にトランスフェクトした。細胞を10%FBSを含有するDMEM中で一晩回収し、次いでOpti−MEM1に切り替えた。さらに2日後、この上清を収集し、ウエスタンブロット分析用に濃縮した。
【0095】
アンチセンスオリゴトランスフェクション。MDA−MB−435細胞をトランスフェクションの1日前に6−ウェルの培養プレート上に播種し、トランスフェクションで90%の密度を得た。100μMアンチセンスまたは逆のコントロールオリゴを、トランスフェクション用に、Opti−MEM1中で2μMに希釈した。0.5mM滅菌リピトイド1(lipitoid1)を、同量のOpti−MEM1中で、1.5nmolリピトイド1:1μgオリゴの比に希釈した。希釈したオリゴおよび希釈したリピトイド1を混合して、直ちに、培養培地中の細胞に加え、最終濃度100、200、または300nMオリゴとした。6時間後、トランスフェクション混合物を正常培養培地で置換し、そして細胞を一晩、回収のためにインキュベートした。アンチセンスオリゴの配列は、AGCTGCGGATGCCACACTTGTAGG(配列番号4)であり、そして逆のコントロールオリゴの配列は、GGATGTTCACACCGTAGGCGTCGA(配列番号5)である。
【0096】
Matrigel侵襲アッセイ。 細胞をトリプシン処理し、洗浄し、そして、計測のために培地に再懸濁した。4×104細胞を洗浄し、氷上の100μlの培地中で再懸濁した。200μlのMatrigel(Callaborative Biomedical Products,Bedford,MA)を氷上で細胞に添加した。Matrigelおよび細胞を注意深く混合し、次いで、24ウェル培養プレートのウェル中に分配し、そして、37℃で30分間凝固した。Matrigel−細胞混合物を0.5mlの培地で被い、そして、6日間5%CO2中で37℃で、インキュベートした。培地を2日毎に補充した。
【0097】
増殖アッセイ。 細胞をトリプシン処理し、洗浄し、そして、計測のために培地中で再懸濁した。次いで、細胞をインキュベーションのために96ウェルプレート(5000細胞/ウェル)に移した。細胞数をQuantosTM細胞増殖アッセイキット(Stratagene,La Jolla,CA)を用いて毎日計測した。
【0098】
(実施例1)
(ヒトcDNA配列の同定)
ショウジョウバエ Out at First(oaf)遺伝子の推定ヒトホモログをコードするDNAを配列番号1に示す。hsOAFおよびショウジョウバエOAFのアライメントを図7に示す。ポリヌクレオチドは、2366塩基対を含み、そして、オープンリーディングフレームが同定されている。ORFの翻訳、273アミノ酸のポリペプチドを、配列番号2に示す。図4は、DNAおよびアミノ酸配列を提供し、ORFの位置を示す。シグナルペプチド由来の最初の30アミノ酸は、このタンパク質が分泌され得ることを示す。シグナルペプチドのアミノ酸配列は以下のようである:MRLPGVPLARPALLLLLPLLAPLLG#TGAPA(配列番号3)。「#」は、推定プロテアーゼ切断部位の位置を示す。
【0099】
(実施例2)
(乳癌細胞株における配列番号1の差次的発現)
以下のヒト乳癌細胞株における配列番号1の発現を比較した:
ヒト乳腺癌に由来するMDA−MB−361;
骨および/または肺に転移性のヒト乳癌に由来するMDA−MB−231;
エストロゲンレセプターネガティブなヒト乳癌細胞に由来するMDA−MB−468;
エストロゲンレセプターネガティブなヒト乳癌細胞に由来するMDA−MB−435;
転移性でないヒト乳癌細胞に由来するMCF−7;および
エストロゲンレセプターポジティブなヒト乳癌細胞に由来するZR−75−1。
【0100】
配列番号1の発現を、高度に転移性の乳癌細胞株MDA−MB231およびMDA−MB−435において測定し、そして、低い転移性または非転移性の乳癌細胞株と比較した。MDA−MB−361における発現は、MDA−MB−231のレベルの11%であり;MDA−MD−468における発現は、MDA−MB−231におけるレベルの44%であり;MCF−7における発現は、MDA−MB−231におけるレベルの17%であり;そしてZR−75−1における発現は、MDA−MB−231におけるレベルの12%であった。
【0101】
MDA−MB−361における発現は、MDA−MB−435におけるレベルの6%であり;MDA−MB−468における発現は、MDA−MB−435におけるレベルの36%であり;そしてMCF−7における発現は、MDA−MB−435におけるレベルの3%であった。従って、表2に示されるように、高い転移可能性を有するヒト腫瘍に由来する乳癌細胞株における配列番号1の増加した発現の明らかな傾向が存在する。
【0102】
【表2】
この遺伝子の同様の発現パターンは、腫瘍化乳癌細胞株で移植したSCIDマウス由来の腫瘍組織サンプル中で、残ったままである(図6)。
【0103】
(実施例3)
(分泌タンパク質をコードするhsOAFおよびhsOAFタンパク質分泌レベルは、乳癌細胞株のhsOAF mRNA発現レベルと一貫している)
推定シグナルペプチド配列は、hsOAF遺伝子の推定アミノ酸配列のN末端に位置する(図3)。hsOAFタンパク質の分泌を検証するために、COS−7細胞およびMCF−7細胞の一過性のトランスフェクションを、hsOAF cDNAを保有するベクターpRetro−Onを用いて行なった。一方、GFPを保有するベクターpRetoro−Onをコントロールとして使用した。hsOAFウサギ抗血清を使用して、分泌されたhsOAFタンパク質を、免疫ブロッティングによるhsOAFでのトランスフェクション後、両方の細胞株のOpti−MEM1培地中で検出した(図8A)。分泌されたhsOAFタンパク質は、おそらくグリコシル化されている。なぜなら、より大きいみかけの分子量を有する複数のバンドが見出されたためである(分泌されたhsOAFタンパク質の推定MWは、28Kdaである)。同じhsOAF抗血清を使用して、種々の乳癌細胞株によるhsOAFタンパク質の分泌を検出する。hsOAFタンパク質の分泌レベルは、これらの細胞株全体の中でhsOAF mRNA発現レベルと一貫していた;高度に転移性の細胞株は、低い転移性/非転移性細胞株よりも、はるかに強力なhsOAF分泌を示した(図8B)。MDA−MB−435は、最も強力なhsOAFタンパク質分泌を有した。
【0104】
(実施例4)
(アンチセンスオリゴによるMDA−MB−435細胞におけるhsOAF発現のノックアウトは、形態学的変化、減少した細胞侵襲性およびより緩徐な増殖速度を引き起こした)
高レベルのhsOAF遺伝子発現が、ヒト乳癌細胞の転移可能性について必須であるか否かを決定するために、アンチセンスオリゴ技術を使用して、hsOAF発現をノックアウトし、次いで、結果として生じた効果を観察した。MDA−MB−435の高度に転移性の細胞株は、調査された乳癌細胞株のすべての中で、最も強力なhsOAFタンパク質分泌を示したので、MDA−MB−435を選択した。hsOAFアンチセンス(AS)およびリバースコントロール(RC)オリゴのいくつかの対を選択して、mRNAレベルにおけるhsOAF遺伝子発現を中断するそれらの能力について試験した。Lightcycler(Roche Diagnostics,Indianapolis,IN)においてリアルタイム定量的RT−PCR分析を行ない、細胞中のhsOAF mRNAレベルを測定する。Kang,Sら、Cancer Research 60(18):5296−5302(2000)。次いで、オリゴの作用する濃度の力価について、最高の対を選択した。低いオリゴ濃度は、細胞に対する潜在的なオリゴ毒性を減少するために好ましい。結果は、100nMのアンチセンスオリゴを用いる処理は、MDA−MB−435細胞のhsOAFタンパク質の分泌を有意に減少するのに十分であることを示した(図12)。100nMの作用する濃度でのオリゴ(配列番号4(AS)および5(RC))の対を以下のすべての実験について使用した。
【0105】
hsOAFアンチセンスオリゴでのMDA−MB−435細胞の処理後、細胞の劇的な形態学的変化が、減少したhsOAFタンパク質の分泌と共に観察された(図10A)。細胞は、より球状になり、そして、それらの広がっていく突出(spreading protrusion)を失う。一方、リバースコントロールオリゴで処理した細胞は、正常な細胞培養されたMDA−MB−435細胞に類似したままである。さらに、アンチセンスオリゴで処理された細胞に添加された馴化培地として、高レベルのhsOAFタンパク質を含む、正常なMDA−MB−435細胞の培地は、完全ではないが、この形態学的変化を防止し得る。細胞形態の変化は、細胞の減少した侵襲能力の指標であり得る。
【0106】
次いで、Mrtrigel侵襲アッセイを行ない、細胞の侵襲性を評価した。matrigelに包埋された乳癌細胞の星状の侵襲性形態学は、それらの転移可能性と相関することが報告されている(Thompson,E.W.ら、J.Cell Physiol.150(3)534−44(1992);Sugiura,T.,ら、J.Cell Biol,146(6):1375−89(1999);Abini,A.,ら、Cancer Res.47(12):3239−45(1987);およびKramer,R.H.,ら、Cancer
Res.46(4 Pt 2):1980−89(1986))、そして、これは、matrigel中で増殖された種々の癌細胞株で確認された。細胞をトリプシン処理し、計測し、そして、matrigelと混合した。次いで、培地を細胞−matrigel混合物上に被せた。インキュベーションの6日後、細胞侵襲を調査した(図10B)。結果は、hsOAFリバースコントロールオリゴで処理した細胞が、正常なMDA−MB−435細胞のように、貫通性、侵襲性、ネットワーク様三次元構造を形成することを示した;他方、hsOAFアンチセンスオリゴで処理した細胞は、平らな球状のコロニーのみを生じた。再び、貫通性コロニーがまた、馴化培地においてインキュベートしたhsOAFアンチセンスオリゴ処理細胞において観察された。これらのデータは、分泌されたhsタンパク質が、MDA−MB−435細胞の侵襲性および転移可能性について必要であることを実証する。
【0107】
さらなる実験を行ない、分泌されたhsOAFタンパク質が、MDA−MB−435細胞増殖に関与するか否かを調査した。細胞増殖アッセイ結果は、hsOAFタンパク質分泌のノックアウトが、変化は穏やかであるが、確かに、MDA−MB−435細胞の増殖速度を落とした。
【0108】
(実施例5)
(ヒト乳癌細胞およびヒト組織におけるRNA発現のノーザンブロット分析)
図5に示されるように、mRNA発現を転移性細胞株MDA−MB−231およびMDA−MB−435においてアップレギュレートした。総RNAを、QuiagenからのRNeasy Kitを使用して調製した。ノーザンブロット分析を、原発性腫瘍または肺における転移由来の細胞からのグアニジニウムチオシアネート/フェノールクロロホルム抽出によって単離した20〜30μgの総RNAを使用して行なった。原発性腫瘍および肺転移は、当業者に周知の方法に従って、scidマウスに注射した細胞株から発症した。pCR2.0−TAベクター(In vitorogen)にクローン化されたhOAFの部分的cDNAクローンを含むプラスミドを、放射線標識し、そして、Expresshyb(Clontech)中で65℃で、ハイブリダイズした。調査された全ての組織の中で、肝臓、膵臓、脾臓、卵巣および小腸は、有意なhsOAF発現を示した。HsOAF mRNA発現はまた、心臓、骨格筋、腎臓、前立腺、結腸および骨髄において検出された(図9)。
【0109】
表3は、種々の腫瘍および正常組織におけるhsOAFポジティブの割合を示す。
【0110】
【表3】
(実施例6)
(軟寒天アッセイ)
軟寒天アッセイ:底層は、細胞上の層化(layering)の数時間以内に新たにプレートされた培地中の2mlの0.6%寒天からなる。細胞の層について、MDA−MB−435細胞を、上記のように、0.05%トリプシン中のプレートから取り出し、そして培地中で2回洗浄した。細胞を犂先(coulter)カウンターで計測し、そして、培地中で1mlあたり106に再懸濁した。10mlのアリコートを、96ウェルプレート中の培地と共に配置する(WST1で計測をチェックするために)か、または、軟寒天アッセイのためにさらに希釈した。2000個の細胞を、0.6%寒天の底層上に、二連のウェルで、800mlの0.4%寒天中に希釈した。
【0111】
培地層:細胞層寒天を凝固した後、2mlの培地を最上層に流し、そして、アンチセンスオリゴまたはリバースコントロールオリゴを、送達ビヒクルを用いずに添加した。新鮮な培地およびオリゴを3〜4日毎に添加した。
【0112】
10日から3週間で、コロニーを計測した。コロニーの視野を眼で計測した。Wst−1代謝値を使用して、出発細胞数における小さな差異を補完した。より大きい視野は、差異の視覚的な記録についてスキャンし得る。結果を図6に示し、ここで、コントロールオリゴヌクレオチドに曝露された細胞と比較して、アンチセンスで処理したMDA−MB−435細胞は、より少ないコロニーを形成した。
【0113】
当業者は、慣例的な実験以上のものは使用せず、本明細書中に記載される本発明の特定の実施形態に対する多数の等価物を認識するか、または確認し得る。このような特定の実施形態および等価物は、上記の特許請求の範囲によって含まれることが意図される。
【0114】
全ての特許、公開された特許出願、および本明細書中で援用された刊行物は、本明細書中で十分に開示されるように、参考として援用される。
【技術分野】
【0001】
(発明の分野)
本発明は、腫瘍の作用を予測するための方法に関する。より詳細には、本発明は、腫瘍サンプルが特定の遺伝子配列の発現に関して試験され、それによって転移性の広がりに関する傾向を示す方法に関する。
【背景技術】
【0002】
(発明の背景)
乳癌は、世界中で一年当たり約1,000,000件の新規症例を伴う、最も一般的な悪性疾患のうちの1つである。多数の組織化学的マーカー、遺伝学的マーカー、および免疫学的マーカーの使用にもかかわらず、臨床医たちが、腫瘍が他の器官に転移する時期を推測することはなお困難である。いくらかの患者は、再発および転移を回避するためにアジュバント治療が必要であるが、他の者は必要ではない。しかし、患者のこれらの亜集団間を識別することは、簡単ではなく、治療の経過は、簡単には図表に表せない。当該分野には、転移するかまたは転移している腫瘍とあまり転移しそうではない腫瘍との間を識別するための新規マーカーの必要性がある。
【発明の概要】
【発明が解決しようとする課題】
【0003】
(発明の要旨)
転移するかまたは転移している腫瘍とあまり転移しそうではない腫瘍との間を識別するためのマーカーを提供することが、本発明の目的である。本発明のこれらおよび他の目的は、以下に記載される1つ以上の実施形態によって提供される。
【課題を解決するための手段】
【0004】
本発明の1つの実施形態は、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列に対して少なくとも85%同一であるアミノ酸配列を有する、単離されそして精製されたヒトタンパク質を提供する。
【0005】
本発明の別の実施形態は、ペプチド結合手段によって互いに融合された、第1のタンパク質セグメントおよび第2のタンパク質セグメントを含む融合タンパク質を提供する。第1のタンパク質セグメントは、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列から選択される、少なくとも6連続するアミノ酸からなり、そして第2のタンパク質セグメントは、配列番号1によってコードされるネイティブなタンパク質において第1のセグメントに隣接することが見出されないアミノ酸配列を含む。
【0006】
本発明のなお別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列を有するヒトタンパク質の少なくとも6連続するアミノ酸からなる、単離されそして精製されたポリペプチドを提供する。
【0007】
本発明のさらに別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖によってコードされるアミノ酸配列を含むヒトタンパク質に特異的に結合する抗体の調製物を提供する。
【0008】
本発明のなお別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖に対して少なくとも95%同一であるヌクレオチド配列の少なくとも11連続するヌクレオチドを含む、単離されそして精製されたサブゲノムのポリヌクレオチドを提供する。
【0009】
本発明の別の実施形態は、配列番号1のヌクレオチド配列またはその相補鎖を含むコード配列を含む、単離されそして精製されたポリヌクレオチドを提供する。
【0010】
本発明のなお別の実施形態は、組織サンプル中の転移を決定するための方法を提供する。配列番号1のコード配列を含む遺伝子の発現産物は、組織サンプル中で測定される。コントロールサンプルよりも高いレベルでこの産物を発現する組織サンプルは、より大きな転移の可能性を有するとして分類される。
【0011】
さらに本発明のさらなる実施形態は、配列番号2をコードするヒト遺伝子を検出するための方法を提供し、この方法は、コンピュータで読み取り可能な形式の配列番号1を得る工程、この配列を、ヒトゲノムのポリヌクレオチド配列と比較する工程、およびパラメータとして12のギャップオープンペナルティー(gap open penalty)および1のギャップエクステンションペナルティー(gap extension penalty)を用いたアフィンギャップサーチ(affine gap serch)を使用するSmith−Watermanアルゴリズムによって決定される場合に、配列番号1に対して少なくとも95%の配列同一性を有する1つ以上のヒトゲノム配列を同定する工程を包含する。
従って、本発明は、転移マーカーとして使用され得る多数のポリヌクレオチドおよびポリペプチドを当該分野に提供する。これらは、乳癌患者の治療経過をより合理的に指示するのに有用である。
例えば、本発明は以下の項目を提供する。
(項目1) 単離された核酸分子であって、以下:
(a)配列番号2の約1〜約273のアミノ酸をコードするポリヌクレオチド;(b)配列番号2の約2〜約273のアミノ酸をコードするポリヌクレオチド;(c)配列番号2の約26〜約273のアミノ酸をコードするポリヌクレオチド;
(d)(a)、(b)、または(c)のポリヌクレオチドのポリヌクレオチド相補体;および
(e)(a)、(b)、(c)、または(d)のポリヌクレオチドと少なくとも90%同一であるポリヌクレオチド、
からなる群より選択されたポリヌクレオチドを含む、単離された核酸分子。
(項目2) 配列番号1のコード領域由来の24〜738の連続したヌクレオチドを含む、単離された核酸分子。
(項目3) 配列番号1のコード領域由来の50〜500の連続したヌクレオチドを含む、項目2に記載の単離された核酸分子。
(項目4) 配列番号1のコード領域由来の75〜250の連続したヌクレオチドを含む、項目3に記載の単離された核酸分子。
(項目5) ポリペプチドをコードするポリヌクレオチドを含む単離された核酸分子であって、ここで、少なくとも1つの保存的アミノ酸置換を除いて、該ポリペプチドが、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;および
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸配列を有する、単離された核酸分子。
(項目6) DNAである、項目1に記載の単離された核酸分子。
(項目7) 組換えベクターを作製する方法であって、項目1に記載の核酸分子をプロモーターと作動可能に連結させた状態でベクター中に挿入する工程を包含する、方法。
(項目8) 項目7に記載の方法によって生成される、組換えベクター。
(項目9) 組換え宿主細胞を作製する方法であって、項目8に記載の組換えベクターを宿主細胞中へ導入する工程を包含する、方法。
(項目10) 項目9に記載の方法によって生成される、組換え宿主細胞。
(項目11) ポリペプチドを生成する組換え方法であって、項目10に記載の組換え宿主細胞を、該ポリペプチドが発現されるような条件下で培養する工程、および該ポリペプチドを回収する工程を包含する、方法。
(項目12) 単離されたポリペプチドであって、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;および
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸と少なくとも95%同一であるアミノ酸を含む、単離されたポリペプチド。
(項目13) 単離されたポリペプチドであって、ここで、少なくとも1つの保存的アミノ酸置換を除いて、該ポリペプチドが、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸配列を有する、単離されたポリペプチド。
(項目14) 単離されたポリペプチドであって、以下:
(a)配列番号2の約1〜約273のアミノ酸;
(b)配列番号2の約2〜約273のアミノ酸;
(c)配列番号2の約26〜約273のアミノ酸、
からなる群より選択されたアミノ酸を含む、単離されたポリペプチド。
(項目15) 配列番号2のポリペプチドのエピトープ保有部分。
(項目16) 配列番号2の8〜25の連続したアミノ酸を含む、項目15に記載のエピトープ保有部分。
(項目17) 配列番号2の10の連続したアミノ酸を含む、項目15に記載のエピトープ保有部分。
(項目18) 項目12に記載のポリペプチドに特異的に結合する、単離された抗体。
(項目19) 項目13に記載のポリペプチドに特異的に結合する、単離された抗体。
(項目20) 項目14に記載のポリペプチドに特異的に結合する、単離された抗体。
(項目21) 配列番号2をコードするヒト遺伝子を検出するための方法であって、該方法は、コンピュータで読み取り可能なフォーマットで配列番号1を得る工程、該配列とヒトゲノムのポリヌクレオチド配列とを比較する工程、および配列番号1と少なくとも95%同一の配列を有する1以上のヒトゲノム配列を同定する工程を包含し、パラメータとして12のギャップオープンペナルティーおよび1のギャップエクステンションペナルティーを用いるアフィンギャップサーチを使用する、Smith−Waternamアルゴリズムによって測定される、方法。
(項目22) 天然に存在しない融合タンパク質であって、該タンパク質は、ペプチド結合によって互いに融合される、第1タンパク質セグメントおよび第2タンパク質セグメントを含み、ここで、該第1タンパク質セグメントは、配列番号1のヌクレオチド配列またはその相補体によってコードされたアミノ酸配列から選択される、少なくとも6の連続したアミノ酸を含む、天然に存在しない融合タンパク質。
(項目23) 前記第1タンパク質セグメントが、配列番号2の少なくとも6の連続したアミノ酸を含む、項目22に記載の融合タンパク質。
(項目24) 前記第1タンパク質セグメントが、配列番号2の少なくとも12の連続したアミノ酸を含む、項目23に記載の融合タンパク質。
(項目25) 前記第1タンパク質セグメントが、配列番号2のアミノ酸20〜30を含む、項目22に記載の融合タンパク質。
(項目26) 前記第1タンパク質セグメントが、配列番号2の少なくとも50の連続するアミノ酸を含む、項目24に記載の融合タンパク質。
(項目27) 前記第1タンパク質セグメントが、配列番号2の少なくとも100の連続するアミノ酸を含む、項目26に記載の融合タンパク質。
(項目28) 前記第1タンパク質セグメントが、配列番号2のアミノ酸26〜287を含む、項目22に記載の融合タンパク質。
(項目29) 第1組織サンプルおよび第2組織サンプル中の腫瘍細胞の転移能力を比較するための方法であって、該方法は、配列番号1のポリペプチドコード領域を含む遺伝子の発現産物を該組織サンプル中で測定する工程を包含し、ここで、該第1組織サンプル中の該産物の少なくとも2倍大きな発現が、該第2組織サンプルと比較してより大きな転移能力を示す、方法。
(項目30) 前記発現産物がタンパク質である、項目29に記載の方法。
(項目31) 前記タンパク質が該タンパク質に特異的に結合する抗体を使用して測定される、項目30に記載の方法。
(項目32) 前記発現産物がmRNAである、項目29に記載の方法。
(項目33) 項目32に記載の方法であって、前記mRNAがポリペプチドプローブを使用して測定され、該ポリペプチドプローブは、配列番号1のヌクレオチド365〜1173の少なくとも20の連続したヌクレオチドを含む、方法。
(項目34) 哺乳動物の癌細胞によってタンパク質の発現を阻害するための組成物であって、該組成物は、配列番号4のポリヌクレオチドを含む、組成物。
(項目35) 哺乳動物の癌細胞によってタンパク質の発現を阻害するための方法であって、該方法は、配列番号4のポリヌクレオチドを含む組成物と該細胞を接触させる工程を包含する、方法。
【図面の簡単な説明】
【0012】
【図1】図1は、ヒトOut at Firstのポリヌクレオチド配列を示す(配列番号1)。
【図2】図2は、配列番号1によってコードされるアミノ酸配列を示す(配列番号2)。
【図3】図3は、推定シグナルペプチドを示す(配列番号3)。
【図4A】図4Aは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図4B】図4Bは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図4C】図4Cは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図4D】図4Dは、配列番号1の翻訳を示す(配列番号1、ポリヌクレオチド;配列番号2、アミノ酸配列)。
【図5】図5は、腫瘍細胞株およびこれらの細胞から作製されたSCIDマウス由来の腫瘍組織における、β−アクチンに対するhsOAFの発現を示す。「PT」は、原発性腫瘍をいう。
【図6】図6は、未処理細胞(WT)に対する、アンチセンスオリゴ(配列番号4)(66−2as)または逆方向コントロール(配列番号5)(66−2rc)を用いた処理の後の、軟寒天におけるMDA−MB−435細胞によるコロニーの増殖を示す。
【図7】図7は、ヒトOAFアミノ酸配列とDrosophila OAFアミノ酸配列との整列である。
【図8】図8Aは、COS−7細胞株およびMCF−7細胞株中のhsOAFタンパク質の発現を示す。図8Bは、乳腺癌(mammory carcinoma)細胞株におけるhsOAFタンパク質の発現を示す。
【図9】図9は、正常ヒト組織中のhsOAFの発現を示す。
【図10A】図10Aは、アンチセンスオリゴ(配列番号4)を用いた処理後にMDA−MB−435細胞中で見出される形態学的変化を示す。AS=アンチセンス;RC=逆方向コントロール(配列番号5);M=馴化培地。
【図10B】図10Bは、AS、RCおよびRC+Mを用いたMDA−MB−435細胞の処置後の、細胞の侵入を示す。
【図11】図11は、ヒトOAFの推定シグナル配列を示す(二重下線)。
【図12】図12Aおよび図12Bは、アンチセンスオリゴ(配列番号4)または逆方向コントロールオリゴ(配列番号5)を用いて処理されたMDA−MB−435細胞による、hsOAFの分泌を示す。
【発明を実施するための形態】
【0013】
(発明の詳細な説明)
乳癌を有する患者の最も致命的な原因は、乳癌の転移および遠方の座位(主に、肺および骨)でのその増殖である。転移は、腫瘍細胞が原発性腫瘍から移住し、血管およびリンパ管を通じて広まり、それから特定の標的器官(ここに、この腫瘍細胞が、再びコロニー形成する)に沈着される、多工程プロセスである。Schirrmacher,V.,Adv.Cancer Res.43:1−73,1985およびLiotta,L.A.ら、Cell 64(2):327−36(1991)。このプロセスの間、腫瘍細胞の侵襲性は、重要である。なぜなら、これらは、多数の基底膜に遭遇してそしてここを通過しなければならないからである。Liotta,L.A.,Am.J.Pathol.117(3):339−48(1984)およびFidler,I.J.,Cancer Res.38(9):2651−60(1978)。従って、腫瘍細胞の侵襲および転移の分子的な原因の解明は、乳癌患者の効果的な処置手順の開発に必須である。乳癌転移において差次的に発現される遺伝子は、転移の間に重要な役割を果たす潜在的な標的である。このような遺伝子およびその生物学的機能の同定は、乳癌の治療および診断の開発に有意に寄与する。
【0014】
乳癌転移に関与するいくつかの重要な遺伝子が、発見されている。エストロゲンレセプターの損失およびビメンチンの存在は、ヒト乳癌の侵襲性および不十分な予後と関連し、そしてまた、ヒト乳癌細胞株の侵襲性および転移可能性と相互に関連する。Aamdal S.ら,Cancer 53(11):2525−9(1984);Clark,G.M.ら,Semin Oncol.,2 補遺 1:20−5(1988);Raymond,W.A.ら,J.Pathol.157(4):299−306(1989);Raymond,W.A.ら,J.Pathol.158(2):107−14(1989);およびThompson,E.W.ら、J.Cell Physiol.150(3):534−44(1992)。E−カドヘリンの発現低下が、乳腺癌の侵襲性に関連している。Vleminckx,K.ら,Cell 66(1):107−19(1991)およびOka,H.ら,Cancer Res.53(7):1696−701(1993)。マプシン(maspin)(正常乳房上皮細胞において発現されるが、ほとんどの乳癌細胞株において発現されないプロテアーゼインヒビター)は、マウスにおいて腫瘍および転移を誘導し、インビトロにおいて基底膜を侵襲するMDA−MB−435細胞の能力を抑制し得た。マプシン発現の損失は、進行癌において最も頻繁に生じた。Zou,Z.ら,Science 263(5146):526−9(1994)およびSeftor,R.E.ら,Cancer Res.58(24):5681−5(1998)。
【0015】
トランスフェクションによるMDA−MB−435細胞中におけるTIMP−4(メタロプロテイナーゼ−4の組織インヒビター)またはCLCA2(Ca2+活性化塩化物チャネル−2)の過剰発現は、細胞の腫瘍形成性、侵襲性および転移性の能力を阻害した。Wang,M.ら,Oncogene 14(23):2767−74(1997)およびGruber,A.D.ら,Cancer
Res.59(21):5488−91(1999)。増殖因子レセプターIGF−IRおよびp185ErbB−2の過剰発現は、乳癌転移に関与することが見出された。Surmacz,E.ら,Breast Cancer Res.Treat 47(3):255−67(1998);Dunn,S.E.ら,Cancer Res.58(15):3353−61(1998);Tan,M.ら,Cancer Res.57(6):1199−205(1997);Dhingra,K.ら,Semin Oncol.23(4):436−45(1996);およびRevillion,F.ら,Eur.J.Cancer 34(6):791−808(1998)。
【0016】
アスパラチルプロテアーゼのカテプシンDは、乳癌患者の不十分な予後のマーカーであると報告され、そして原発性乳癌の細胞質ゾル中の高いカテプシンD濃度と転移の発生との間には有意な相関があるが、カテプシンD分泌と乳癌細胞株の侵襲能力との間の相関は見出されていない。Rochefort,H.,Breast Cancer Res Treat 16(1):3−13(1990);Johnson,M.D.ら,Cancer Res.53(4):873−7(1993);およびRochefort,H.ら,Clin Chim
Acta.291(2):157−70(2000)。オステオポンチン(骨再吸収および骨形成に関与すると考えられる、分泌インテグリン結合糖タンパク質)は、乳腺癌細胞の移動および侵襲を誘導し得る。オステオポンチンレベル(腫瘍細胞または血漿のレベル)は、増強された乳癌の悪性と関連する。Denhardt,D.T.ら,FASEB J 7(15):1475−82(1993);Denhardt,D.T.ら,J.Cell Biochem Suppl.,30−31:92−102(1998);Tuck,A.B.ら,J Cell Biochem.78(3):465−75(2000);Tuck,A.B.ら,Oncogene 18(29):4237−46(1999);およびSinghal,H.ら,Clin Cancer Res.3(4):605−11(1997)。
【0017】
本発明は、非常に転移性のヒト乳癌細胞株において過剰発現された新規遺伝子のクローニングに関連する。これは、分泌タンパク質をコードし、そしてそのタンパク質分泌は、低い転移性の細胞株/非転移性の細胞株よりも、非常に転移性のヒト乳癌細胞株においてより多いことが、確認されている。アンチセンスオリゴ技術による侵襲性(aggressive)MDA−MB−435細胞株のこのタンパク質の分泌ノックアウトは、この細胞の減少した侵襲性および増殖速度を伴って、有意な形態学的変化を引き起こす。この遺伝子は、Drosophila遺伝子OAF(out at first)との相同性に基づいて、hsOAFと称される。Bergstrom,D.E.ら,Genetics 139(3):1331−46(1995)およびMerli,C.ら,Genes Dev.10(10):1260−70(1996)。
【0018】
ポリヌクレオチドが、高い転移性の乳癌細胞と非転移性の癌細胞または低い転移性の癌細胞との間で、差次的に発現されることが、本発明の発見である。従って、このポリヌクレオチドは、転移性マーカー遺伝子に関連する。この情報は、差次的に発現される遺伝子の発現産物に特異的な診断試薬を作製するために利用され得る。これはまた、癌、特に乳癌の適切な処置レジメンを計画する際に臨床医を補助する、診断方法および予後方法にも使用され得る。
【0019】
このポリヌクレオチドは、図1に示され(配列番号1)、そしてこの推定オープンリーディングフレーム(ORF)は、図2に示されるポリペプチド(配列番号2)をコードする。最初の30アミノ酸残基(配列番号3)は、推定シグナルペプチドを含み、そして推定プロテアーゼ切断部位は、「*」によって示される:APLLG*TGAPA(配列番号3の25位と26位との間のアミノ酸)。
【0020】
本発明のポリヌクレオチド配列は、「Out at First」(oaf)として公知のDrosophila遺伝子といくらかの相同性を共有する。oafの転写は、3つのクラスの選択的にポリアデニル化されたRNAを生じ、これらの発現は、発生的に調節される。全てのoaf転写物は、単一のUAG終止コドンによって隔てられた2つの隣接ORFを含む。翻訳の間のUGAコドンの抑制は、同じRNA分子から異なるタンパク質の生成を導き得る。卵形成の間、oaf RNAは、全ての期の栄養細胞で発現され、そして母系的に卵子に寄与する。
【0021】
初期発達の間、oaf遺伝子の接合体転写は、胚帯伸長時にほとんどかまたは全てのセグメントにおいて細胞の小クラスターで起こり、そしてその後は発達中の中枢神経系においてセグメントが反復されるパターンで起こる。oaf遺伝子はまた、両方の性の胎児、幼態および成人の生殖腺で発現される(Bergstrom,D.E.ら,Genetics 139:1331−1346,1995)。
【0022】
本発明のポリヌクレオチドは、7つの高転移性乳癌細胞株対非転移性または低転移性乳癌細胞株の組み合わせにおいて、差次的に発現された。用いられる細胞株は、MDA−MB−361(ヒト乳房腺癌由来)、MDA−MB−231(骨および/または肺への転移性を有するヒト乳癌細胞由来);MDA−MB−468(ヒトエストロゲンレセプター陰性乳癌細胞由来);MCF−7(非転移性ヒト乳癌細胞);ZR−75−1(エストロゲンレセプター陽性のヒト乳癌由来、Engleら,Cancer Res.38:3352−64(1978);およびMDA−MB−435(エストロゲンレセプター陰性のヒト乳癌細胞由来、Rishiら,Cancer Res.56:5246−5252(1996))である。
【0023】
発現プロフィールは、以下のようなものである:
【0024】
【表1】
mRNA発現のアップレギュレーションを、この細胞株由来の総RNAを用いたノーザンブロット分析により確認した(図5)。
【0025】
本発明のポリヌクレオチドの発現を比較した細胞株は、種々の転移能を有するヒト乳癌を表わす。細胞株ZR−75−1培養物は、乳癌患者の悪性の腹水の浸出液に由来した。インビトロで増殖された細胞株は、本来の細胞を有するドナーからの生検または細胞調製物において見られる形態に酷似して再集合した。ZR−75−1細胞は、エストロゲンにより特異的に刺激される。そしてこの細胞は、エストロゲン応答を研究するためのモデル系として用いられてきた(Engel,L.W.ら,Cancer Res.38:3352−3364,1978)。
【0026】
細胞株MDA−MB−435は、ヒト乳癌のモデルとして研究されてきた(例えば、レチノイン酸存在下での増殖阻害の作用機構を研究するための)エストロゲンレセプター陰性の細胞株である(Rishi,A.K.ら,Cancer Res.56:5246−5252,1996)。レチノイドによる増殖阻害はまた、MCF−7細胞およびMDA MB 468細胞においても研究されてきた(Tin−U,C.K.ら,Am.Soc.Clin.Onc.Proceedings,第17巻,2125,1998)。
【0027】
細胞株MDA−MB−361は、ヒト乳房腺癌から、特に悪性の部位から得られた(ATCC番号HTB−27)。ヒトWnt遺伝子の差次的な発現が、この細胞株において研究されてきた(Huguet,E.L.ら,Cancer Res.54:2615−2621,1994)。
【0028】
一旦転移が起こると、乳房の原発腫瘍細胞は、基底膜に侵入し、そして身体の他の器官に広がり、そして乳癌を有する患者の生存見込みをごくわずかにする。臨床的な診断および治療のために、乳癌の侵入および転移に関与する遺伝子を同定することは重要なことである。このような遺伝子は、治療用薬物開発のための診断または候補標的の潜在的な指標である。例えば、ヒト乳房腫瘍におけるビメンチンの存在は、わずかな予後の指標であるエストロゲンレセプターの欠損および腫瘍の浸潤に関連する(Raymond,W.A.ら,J.Pathol.157(4):299−306(1989);Raymond,W.A.ら,J.Pathol.158(2):107−14(1989);およびThompson,E.W.ら,J.Cell Physiol.150(3):534−44(1994))。増大した基質メタロプロテイナーゼの活性は、癌腫、特に乳癌の転移性の表現型に関連する(Basset,P.ら,Nature 348(6303):699−704(1990)およびBasset,P.ら,Cancer 74(3補完):1045−9(1994))。オステオポンチン、分泌型インテグリン結合糖タンパク質、は、ヒト乳房の上皮細胞の浸潤性の増大を誘導し得、そして乳癌における増加した悪性に関連する(Tuck,A.B.ら,J.Cell Biochem.78(3):465−75(2000);Tuck,A.B.ら,Oncogene 18(29):4237−46(1999);およびSinghal,H.ら,Clin Cancer Res.3(4):605−11(1997))。
【0029】
本発明は、乳癌転移に関連する新規の分泌タンパク質(hsOAF)の同定に関する。このタンパク質の役割を解明するために使用されるヒト乳癌細胞株は、これらの転移能に基づいて3つの群に分けられる:高転移性、低転移性、および非転移性。これらの細胞株群の間の異なる転移能を利用することにより、そして発達したマイクロアレイ技術を用いることにより、高転移性のヒト乳癌細胞株と低転移性/非転移性株との間で差次的に発現される遺伝子を同定した。これらの遺伝子は、乳癌転移性の分子機構のより深い理解を導き得る発現の確認および機能の研究のよい候補である。hsOAF遺伝子は、本発明の中心であり、新規の分泌タンパク質をコードする。hsOAF mRNAの発現は、種々のヒト正常組織においてほぼ共通である。しかし、hsOAFタンパク質の分泌は、低転移性/非転移性のヒト乳癌細胞株よりも高転移性株のほうが、ずっと強い。そしてMDA−MB−435は、最も強いhsOAF分泌性を有する。hsOAF遺伝子は、Drosophila OAF(out at first)遺伝子との相同性に基づき命名された。しかし、Drosophila OAFタンパク質は、分泌タンパク質ではあり得ない。というのも、このタンパク質は、N末端に代表的なシグナルペプチド配列を有しないからである(図7)。
【0030】
乳癌転移における分泌hsOAFタンパク質の重要性に取り組むために、アンチセンスオリゴ技術が、特異的なノックアウトhsOAF発現に用いられた。アンチセンスオリゴ技術は、遺伝子機能の研究のための劇的な遺伝子発現の減少のための、効果的で迅速な方法である(Stein,C.A.ら,Science
261(5124):1004−12(1993);Defacque,H.ら,J.Cell Physiol.178(1):109−19(1999))。高転移性のMDA−MB−435細胞のhsOAFタンパク質分泌のノックアウトは、細胞形態の変化、細胞浸潤性の減少、および細胞増殖速度の低下を引き起こす。馴化培地(正常なMDA−MB−435細胞の培養培地)での細胞の処理は、hsOAFタンパク質分泌のノックアウトにより引き起こされた全ての表現型変化をある程度回復させ得た。本発明者らは、特定の機構に束縛されないが、分泌hsOAFタンパク質は、MDA−MB−435細胞の浸潤および増殖に必須であると考えられている。しかし、アンチセンスオリゴ技術による別の高転移性細胞株MDA−MB−231のhsOAFタンパク質分泌のノックアウトは、有意な細胞の変化を全く引き起こさなかった。MDA−MB−435およびMDA−MB−231は、全く異なる転移性の細胞株であり、そしてMDA−MB−435は、MDA−MB−213よりもずっと強いhsOAFタンパク質の分泌性を示す。
【0031】
hsOAF遺伝子は、ヒト乳房腫瘍において異種接合性の喪失が頻繁に発生する染色体11q23領域に配置されている(Negrini,M.ら,Cancer Res 55(14):3003−7(1995)およびTomlinson,I.P.ら,J.Clin.Pathol.48(5):424−8(1995))。原発性ヒト乳房腫瘍における11q23での異種接合性の喪失は、転移後の乏しい生存性に関連することが報告されてきた(Winqvist,R.ら,Cancer Res.55(12):2660−4(1995))。11q23はまた、ATM(Ataxia−telangiectasia,mutated)、およびMLL(これは、急性白血病における染色体の再配置により頻繁に破壊される)のような遺伝子座を含む(Rasio,D.ら,Cancer Res.55(24):6053−7(1995)およびRubnitz,J.E.ら,Leukemia 10(1):74−82(1996))。乳癌転移における染色体11q23での変異とhsOAF遺伝子発現との間の関係は、不明確なままである。
【0032】
分泌hsOAFタンパク質は、乳癌に対する薬物開発のための適切な標的であり得、そして悪性の乳房腫瘍に対する良い診断指標であり得る。したがって、配列番号1およびこの配列を含むポリヌクレオチドは、転移性の指標として有用である。転移性指標ヌクレオチドまたはアミノ酸配列との関連は、非転移性または低転移性細胞と比較して高い転移性の同様の発現パターンを有する改変体を含む。転移性指標ポリペプチドは、長さの点で全長転移性指標タンパク質と相違し得、そして転移性指標タンパク質の少なくとも6、8、10、12、15、18、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、120、140、160、180、200、220、240、260、265、270、または271以上の連続するアミノ酸を含む。例示的なポリヌクレオチドとして、配列番号2の約1〜約273;1〜273;約2〜約273;2〜273;約26〜約273;および26〜273のアミノ酸をコードするポリヌクレオチドが挙げられる。
【0033】
指標タンパク質およびポリペプチドの改変体もまた見出され得る。転移性指標タンパク質またはポリペプチド改変体は、天然に存在することも存在しないこともあり得る。天然に存在する転移性指標タンパク質またはポリペプチド改変体は、ヒトまたは他の種において見出され、そして配列番号1に示されるヌクレオチド配列またはその相補体に対応する遺伝子によりコードされるタンパク質と実質的に同一であるアミノ酸配列を含む。低転移性または非転移性乳癌細胞と比較して、天然に存在する転移性指標タンパク質またはポリペプチド改変体と実質的に同じ高い転移性の発現パターンを保持する天然に存在しない転移性指標タンパク質またはポリペプチド改変体もまた、本発明に含まれる。好ましくは、天然に存在するかまたは天然に存在しない転移性指標タンパク質またはポリペプチド改変体は、配列番号1に示されるヌクレオチド配列によりコードされるアミノ酸配列と、少なくとも85%、90%、91%、92%、93%、94%、または95%同一であるアミノ酸配列を有する。より好ましくは、この分子は、少なくとも96%、97%、98%、または99%同一である。野生型タンパク質またはポリペプチドと改変体との間の配列同一性のパーセントは、野生型タンパク質またはポリペプチドと改変体を整列させて、当業者に公知である、野生型と改変体との間のアミノ酸一致数の計数、および野生型配列の合計アミノ酸残基数による合計一致数の除算によって、最大アミノ酸一致数を獲得することにより決定される。
【0034】
好ましくは、転移性指標タンパク質またはポリペプチド改変体におけるアミノ酸の変化は、保存的アミノ酸変化、すなわち類似の電荷を有するかまたは非電荷のアミノ酸での置換である。保存的アミノ酸変化は、それらの側鎖に関して関連するアミノ酸ファミリーの1つの置換を含む。天然に存在するアミノ酸は、遺伝的に4つのファミリー:酸性(アスパルテート、グルタメート)、塩基性(リジン、アルギニン、ヒスチジン)、非極性(アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、および非荷電極性(グリシン、アスパラギン、グルタミン、シスチン、セリン、スレオニン、チロシン)アミノ酸に分けられる。フェニルアラニン、トリプトファン、およびチロシンは、しばしば芳香族アミノ酸として一緒に分類される。
【0035】
ロイシンのイソロイシンまたはバリンでの孤立性(isolated)置換、アスパラギン酸のグルタミン酸での孤立性置換、トレオニンのセリンでの孤立性置換、またはあるアミノ酸の構造的に関連するアミノ酸での類似の置換が、得られた転移マーカータンパク質または転移マーカーポリペプチドの改変体の生物学的特性に対して大きな影響を有さないことを予測することは妥当である。転移マーカータンパク質または転移マーカーポリペプチドの改変体の特性および機能は、配列番号1に示されるヌクレオチド配列によってコードされるアミノ酸配列を含む転移マーカータンパク質または転移マーカーポリペプチドと同じ型の特性および機能であるが、その改変体の特性および機能は、ある程度異なり得る。アミノ酸変化が、適切な差次的発現パターンを有する転移マーカータンパク質または転移マーカーポリペプチドの改変体を生じるか否かは、容易に決定され得る。例えば、ヌクレオチドプローブは、本明細書中に開示されるマーカー遺伝子配列から選択され得、そして当該分野で公知のように、ノーザンブロットまたは組織切片において、マーカー遺伝子mRNAを検出するために使用され得る。あるいは、転移マーカー遺伝子のタンパク質産物に特異的に結合する抗体が、転移マーカータンパク質またはその改変体の発現を検出するために使用され得る。
【0036】
転移マーカータンパク質改変体としては、グリコシル化形態、他の分子との凝集結合体、および無関係な化学部分との共有結合体が挙げられる。転移マーカー改変体としてはまた、対立遺伝子改変体、種改変体、およびムテインが挙げられる。転移マーカー遺伝子の差次的発現に影響を与えない領域の切断型または欠失型もまた、転移マーカー改変体である。共有結合性の改変体は、当該分野で公知のように、アミノ酸側鎖あるいはN末端残基またはC末端残基に見出される基に官能基を連結させることによって調製され得る。
【0037】
本発明のポリペプチドのいくらかのアミノ酸配列が、そのタンパク質の構造または機能に有意な影響を及ぼすことなく改変され得ることが、当該分野で認識される。このような配列中の差異を意図する場合、そのタンパク質上に活性を決定する重要な領域が存在することは覚えておくべきである。一般に、類似の機能を果たす残基が使用される限り、三次構造を形成する残基を置換することが可能である。他の場合において、その変更がタンパク質の重要でない領域に生じる場合は、残基の型は、全く重要でなくてもよい。アミノ酸の置換はまた、細胞表面レセプターへの結合の選択性を変化させ得る。Ostadeら、Nature 361:266−268(1993)は、TNF−αの2つの公知のTNFレセプターの型のうちの1つの型のみへの、選択的結合を生じる特定の変異を記載する。従って、本発明のポリペプチドは、天然の変異または人為的操作のいずれか由来の、1以上のアミノ酸の置換、欠失または付加を含み得る。
【0038】
本発明はさらに、比較可能な発現パターンを示すか、または抗原性領域を含む、開示されたポリペプチドの改変体を含む。このような変異としては、欠失、挿入、反転、反復、および型置換が挙げられる。どのアミノ酸変化が表現型的にサイレントであるようであるかに関するガイダンスは、Bowie,J.U.ら、「Deciphering the Message in Protein Sequences:Tolerance to Amino Acid Substitutions」、Science 247:1306−1310(1990)に見出され得る。
【0039】
荷電アミノ酸の、別の荷電アミノ酸での置換および中性または負に荷電したアミノ酸での置換は、特に関心深い。後者は、正電荷が減少したタンパク質を生じ、開示されたタンパク質の特性を改善する。凝集の防止が、非常に望ましい。タンパク質の凝集は、活性の損失を生じるだけでなく、それらが免疫原性であり得るので、薬学的処方物を調製する際に問題であり得る(Pinckardら、Clin.Exp.Immunol.2:331−340(1967);Robbinsら、Diabetes 36:838−845(1987);Clelandら、Crit.Rev.Therapeutic Drug Carrier Systems 10:307−377(1993))。
【0040】
機能に必須である本発明のポリペプチド中のアミノ酸は、当該分野で公知の方法(例えば、部位特異的変異誘発またはアラニンスキャニング変異誘発(CunninghamおよびWells,Science 244:1081−1085(1989)))によって同定され得る。後者の手順は、その分子の全ての残基で単一のアラニン変異を導入する。次いで、得られた変異体分子を、レセプター結合またはインビトロ増殖活性のような、生物学的活性について試験する。リガンド−レセプター結合に重要な部位はまた、構造分析(例えば、結晶化、核磁気共鳴または光親和性標識(Smithら、J.Mol.Biol.224:899−904(1992)およびde Vosら、Science 255:306−312(1992))によって決定され得る。
【0041】
示されるように、変化は、好ましくは、わずかな性質の変化である(例えば、タンパク質のフォールディングまたは活性に有意には影響を与えない保存的アミノ酸置換)。もちろん、当業者が作製するアミノ酸置換の数は、上記の因子を含む、多くの因子に依存する。一般的に言えば、任意の所定のポリペプチドについての置換の数は、50、40、30、25、20、15、10、5または3より多くない。
【0042】
全長転移マーカータンパク質は、標準的な生化学的方法を使用して、転移マーカータンパク質を産生するヒト細胞(例えば、転移性乳癌細胞)から抽出され得る。単離および精製された転移マーカータンパク質または転移マーカーポリペプチドは、細胞中で転移マーカータンパク質または転移マーカーポリペプチドが通常会合している他の化合物(例えば、特定のタンパク質、炭水化物、脂質、または細胞内小器官)から分離される。単離および精製された転移マーカータンパク質または転移マーカーポリペプチドの調製物は、少なくとも80%純粋であり;好ましくは、その調製物は、90%、95%、または99%純粋である。
【0043】
配列番号2をコードするヒト遺伝子を、当該分野において公知の方法を使用して同定および単離し得る。1つの方法によれば、配列番号1は、コンピュータで読み取り可能な形式で調製される。この配列が、ヒトゲノムのポリヌクレオチド配列と比較され、そして配列番号1に対して少なくとも95%の配列同一性を有する1つ以上のヒトゲノム配列が、例えば、パラメータとして12のギャップオープンペナルティーおよび1のギャップエクステンションペナルティーを用いるアフィンギャップサーチを使用して、Smith−Watermanアルゴリズムを使用することによって、同定される。当該分野において公知の方法を使用して、配列番号1とヒトゲノム配列との間の相同の領域に基づくプローブが調製され、そしてヒトゲノムDNAからポリヌクレオチドを単離するために使用される。出願日現在、配列番号1の全ポリヌクレオチドに対応するヒトポリヌクレオチドは、公共のデータベースにおいて確認されなかった。従って、本発明は、配列番号1のコード領域を含むヒトゲノムDNA、および配列番号1と相同性を共有せず相同領域と連続する任意の未翻訳領域を包含する。このようなゲノムDNAとしては、イントロン、プロモーター、および配列番号2をコードする領域を有するヒト遺伝子に機能的に関連する他の調節領域が挙げられるが、これらに限定されない。
【0044】
転移マーカータンパク質および転移マーカーポリペプチドはまた、組換えDNA方法または合成化学的方法によって、産生され得る。組換え転移マーカータンパク質または転移マーカーポリペプチドの産生のために、配列番号1に示すヌクレオチド配列、または転移マーカータンパク質をコードする配列の改変体から選択されるコード配列が、公知の原核生物または真核生物の発現系において、発現され得る(以下を参照のこと)。細菌、酵母、昆虫、または哺乳動物の発現系が、当該分野において公知であるように、使用され得る。
【0045】
あるいは、合成化学方法(例えば、固相ペプチド合成)が、転移マーカータンパク質または転移マーカーポリペプチドを合成するために使用され得る。ペプチド、アナログ、または誘導体の産生のための一般的手段は、CHEMISTRY
AND BIOCHEMISTRY OF AMINO ACIDS,PEPTIDES, AND PROTEINS−−A SURVEY OF RECENT DEVELOPMENTS、Weinstein,B.編、Marcell Dekker,Inc.出版、New York(1983)に概説されている。さらに、通常のL−立体異性体に代えてのD−アミノ酸での置換が、その分子の半減期を増加するために実行され得る。転移マーカー改変体は、同様に産生され得る。
【0046】
少なくとも6、8、10、12、15、18、20、25、30、35、40、50、55、60、65、70、75、80、85、90、95、100、120、140、160、180、200、220、240、250、260、265、270もしくは271またはそれより多くの連続した転移マーカーアミノ酸を含む、天然には存在しない融合タンパク質もまた、構築され得る。ヒト転移マーカー融合タンパク質は、転移マーカーアミノ酸配列に対する抗体を生成するため、および種々のアッセイ系において使用するために、有用である。例えば、転移マーカー有効タンパク質を使用して、転移マーカータンパク質と相互作用し、そしてこれらの機能に影響を与えるタンパク質を、同定し得る。タンパク質アフィニティークロマトグラフィーのような物理的方法、または酵母ツーハイブリッドもしくはファージディスプレイ系のようなタンパク質−タンパク質相互作用に関するライブラリーに基づくアッセイもまた、この目的で使用され得る。このような方法は、当該分野において周知であり、そして薬物スクリーニングとしてもまた使用され得る。
【0047】
転移マーカー融合タンパク質は、ペプチド結合によって一緒に融合した2つのタンパク質セグメントを含む。第1のタンパク質セグメントは、転移マーカータンパク質の少なくとも6、8、10、12、15、18、20、25、30、35、40、50、55、60、65、70、75、80、85、90、95、100、120、140、160、180、200、220、240、250、260、265、270もしくは271またはそれより多くの連続したアミノ酸を含む。これらのアミノ酸は、配列番号1に示されるヌクレオチド配列によってコードされるアミノ酸配列から、または上記のもののようなこの配列の改変体から、選択され得る。第1のタンパク質セグメントはまた、全長転移マーカータンパク質を含み得る。
【0048】
1つの好ましい実施形態において、第1のタンパク質セグメントは、配列番号2に示されるポリペプチドを含む。この実施形態の改変において、この第1のタンパク質セグメントは、配列番号2のアミノ酸31〜287からなる。この融合タンパク質は、配列番号2のシグナルペプチドを欠き、そして細胞の内側での発現された融合タンパク質の保持のために適切である。
【0049】
第2のタンパク質セグメントは、配列番号1によってコードされるネイティブタンパク質において第1のタンパク質セグメントに隣接して見出されなかった、全長タンパク質またはタンパク質フラグメントあるいはポリペプチドであり得る。この融合タンパク質は、当該分野において公知であるように、検出可能なマーカー(例えば、放射性マーカー、蛍光マーカー、化学発光マーカー、またはビオチン化マーカー)で標識され得る。第2のタンパク質セグメントは、β−ガラクトシダーゼのような検出可能な産物を生成する酵素であり得る。第1のタンパク質セグメントは、好都合であるように、N末端またはC末端であり得る。
【0050】
2つのタンパク質セグメントを組換え的にかまたは共有結合させることによってかのいずれかで、融合タンパク質を作製するための技術もまた、周知である。組換えDNA法は、以下に記載のように、例えば、第2のタンパク質セグメントをコードしかつ宿主細胞においてDNA構築物を発現するヌクレオチドで、適切なリーディングフレームにおいて配列番号1のコード配列を含むDNA構築物を作製することによって、転移マーカー融合タンパク質を調製するために用いられ得る。配列番号1のオープンリーディングフレームを、図4に示す。
【0051】
単離および精製された転移マーカータンパク質、ポリペプチド、改変体、または融合タンパク質は、転移マーカータンパク質に特異的に結合する抗体の調製物を得るために、免疫原として使用され得る。この抗体は、とりわけ、ヒト組織およびその画分において、野生型の転移マーカータンパク質を検出するために使用され得る。この抗体はまた、転移マーカータンパク質の過小発現もしくは過剰発現を生じるかまたはサイズもしくは電気泳動移動度が変化した転移マーカータンパク質の発現を生じる、転移マーカー遺伝子の変異の存在を検出するために使用され得る。
【0052】
ポリクローナル抗体およびモノクローナル抗体の調製は、標準的方法を使用してなされ得る。単鎖抗体もまた、調製され得る。好ましい免疫原は、配列番号2を含むポリペプチドである。転移マーカータンパク質、ポリペプチド、改変体、または融合タンパク質に特異的に結合する単鎖抗体は、当該分野で公知であるように、例えば、単鎖免疫グロブリンディスプレイライブラリーから単離される。そのライブラリーは、配列番号2の転移マーカータンパク質のアミノ酸配列に対して「パニング」され、そして転移マーカータンパク質の異なるエピトープに高い親和性で結合する多数の単鎖抗体が、単離され得る。Hayashiら、1995、Gene 160:129〜30。単鎖抗体はまた、DNA増幅法(例えば、ハイブリドーマcDNAをテンプレートとして使用する、ポリメラーゼ連鎖反応(PCR))を使用して、構築され得る。Thirionら、1996、Eur.J.Cancer Prev.5;507〜11。
【0053】
転移マーカー特異的抗体は、配列番号1に示すヌクレオチド配列によってコードされるアミノ酸配列を有する、全長転移マーカータンパク質に存在するエピトープ、転移マーカーポリペプチド、または転移マーカー改変体に、単独でかまたは融合タンパク質の一部として、特異的に結合する。好ましくは、転移マーカーエピトープは、他のヒトタンパク質には存在しない。代表的には、少なくとも6、8、10、または12の連続したアミノ酸が、エピトープを形成するために必要である。しかし、非連続のアミノ酸を含むエピトープは、より多くのアミノ酸、例えば、少なくとも15、25または50のアミノ酸を必要とし得る。
【0054】
転移マーカータンパク質、ポリペプチド、融合タンパク質、または改変体に特異的に結合する抗体は、ウエスタンブロットまたは他の免疫化学的アッセイにおいて使用される場合に、他のタンパク質を用いて提供される検出シグナルより少なくとも5倍、10倍または20倍高い検出シグナルを提供する。好ましくは、転移マーカーエピトープに特異的に結合する抗体は、免疫化学的アッセイにおいて他のタンパク質を検出せず、そして転移マーカータンパク質、ポリペプチド、融合タンパク質、または変異体を、溶液から免疫沈降させ得る。
【0055】
サブゲノムポリヌクレオチドは、全体染色体未満を含む。好ましくは、ポリヌクレオチドは、イントロンを含まない。好ましい実施形態において、ポリヌクレオチド分子は、配列番号1の10、15、20、25、30、40、50、75、100、150、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700、1800、1900、2000、2100、2200、2300または2350のヌクレオチドの連続した配列、あるいは、その相補体を含む。配列番号1に示されるヌクレオチド配列の相補体は、配列番号1に示される連続したヌクレオチド配列と、ワトソン−クリック塩基対を形成する連続するヌクレオチド配列である。
【0056】
転移マーカータンパク質または改変体のアミノ酸配列をコードする縮重ヌクレオチド配列、ならびに配列番号1に示されるヌクレオチド配列のコード領域に少なくとも85%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%同一であるポリヌクレオチドを含む相同的ヌクレオチド配列もまた、転移マーカーサブゲノムポリヌクレオチドである。代表的には、相同的転移マーカーサブゲノムポリヌクレオチド配列は、当該分野において公知のように、ストリンジェントな条件下でハイブリダイゼーションを行うことによって確認され得る。野生型と相同な改変体との間の配列同一性%は、当該分野で公知なように、野生型ポリヌクレオドをその改変体と整列させて最も多い数のヌクレオチドの一致を得、野生型と改変体との間のヌクレオチドの一致の数を計数し、そして一致の合計数を野生型配列のヌクレオチドの合計数で割ることによって決定される。同一性%を計算するための好ましいアルゴリズムは、Smith−Waterman相同性サーチアルゴリズム(以下のサーチパラメーター:12のギャップオープンペナルティーおよび1のギャップエクステンションペナルティーとともにアフィンギャップサーチを用いてMPSRCHプログラム(Oxford Molecular)を実行する)である。
【0057】
転移マーカータンパク質コード配列を含む転移マーカーサブゲノムポリヌクレオチドは、発現構築物において使用される。好ましくは、転移マーカーサブゲノムポリヌクレオチドは、発現プラスミド(例えば、Ecdyson系、pIND、In Vitoro Gene)に挿入される。転移マーカーサブゲノムポリヌクレオチドは、当該分野で周知の技術を使用してベクターおよび細胞株において増殖され得る。転移マーカーサブゲノムポリヌクレオチドは、直鎖状分子または環状分子上であり得る。これらは、自律的に複製する分子上にあり得るかまたは、複製配列を有さない分子上にあり得る。これらは、それら自身によって、または当該分野で公知の他の調節配列によって調節され得る。
【0058】
次いで、転移マーカー発現構築物を含む宿主細胞を使用して、転移マーカータンパク質の全てまたは一部を発現し得る。転移マーカー発現構築物を含む宿主細胞は、原核生物または真核生物であり得る。種々の宿主細胞は、細菌、酵母、昆虫、およびヒトの発現系における使用に利用可能であり、転移マーカー発現構築物を発現するかまたは増殖するために使用され得る(以下を参照こと)。発現構築物は、当該分野で公知な任意に技術を使用して宿主細胞に導入され得る。これらの技術には、トランスフェリン−ポリカチオン媒介DNA移入、裸の核酸またはカプセル化核酸を用いたトランスフェクション、リポソーム媒介細胞融合、DNAコートラテックスビーズの細胞内輸送、プロトプラスト融合、ウイルス感染、エレクトロポレーション、およびリン酸カルシウム媒介トランスフェクションが挙げられる。
【0059】
転移マーカー発現構築物は、選択された宿主細胞において機能的であるプロモーターを含む。当業者は、当該分野で公知であり使用される多くの数の細胞型特異的プロモーターから適切なプロモーターを容易に選択し得る。この発現構築物はまた、宿主細胞中で機能的である転写ターミネーターを含み得る。この発現構築物は、転移マーカータンパク質の全てまたは一部分、改変体、融合タンパク質、抗体、またはリボザイムをコードするポリヌクレオチドセグメントを含む。このポリヌクレオチドセグメントは、プロモーターから下流に位置する。ポリヌクレオチドセグメントの転写は、プロモーターで開始する。この発現構築物は、直鎖状または環状であり得、そして所望される場合、自律複製配列を含み得る。
【0060】
転移マーカー発現構築物を発現するための細菌系としては、Changら、Nature(1978)275:615、Goeddelら、Nature(1979)281:544、Goeddelら、Nucleic Acids Res.(1980)8:4057、EP36,776、米国特許第4,551,433号、deBoerら、Proc.Nat’l.Acad Sci.USA(1983)80:21−25、およびSiebenlistら、Cell(1980)20:269に記載される発現系が挙げられる。
【0061】
酵母における発現系としては、Hinnenら、Proc.Nat’l.Acad.Sci.USA(1978)75:1929;Itoら、J.Bacteriol.(1983)153:163;Kurtzら、Mol.Cell.Biol.(1986)6:142;Kunzeら、J.Basic Microbiol.(1985)25:141;Gleesonら、J.Gen.Microbiol.(1986)132:3459、Roggenkampら、Mol.Gen.Genet.(1986)202:302)Dasら、J.Bacteriol.(1984)158:1165;De Louvencourtら、J.Bacteriol.(1983)154:737、Van den Bergら、Bio/Technology(1990)8:135;Kunzeら、J.Basic Microbiol.(1985)25:141;Creggら、Mol.Cell.Biol.(1985)5:3376、米国特許第4,837,148号、米国特許第4,929,555号;BeachおよびNurse、Nature(1981)300:706;Davidowら、Curr.Genet.(1985)10:380、Gaillardinら、Curr.Genet.(1985)10:49、Ballanceら、Biochem.Biophys.Res.Commun.(1983)112:284−289;Tilburnら、Gene(1983)26:205−221、Yeltonら、Proc.Nat’l.Acad.Sci.USA(1984)81:1470−1474、KellyおよびHynes、EMBO J.(1985)4:475479;EP244,234およびWO 91/00357に記載される発現系が挙げられる。
【0062】
昆虫における転移マーカー発現構築物の発現は、米国特許第4,745,051号、Friesenら(1986)「バキュロウイルス遺伝子発現の調節」:THE MOLECULAR BIOLOGY OF BACULOVIRUSES(W.Doerfler編)、EP127,839、EP155,476、およびVlakら、J.Gen.Virol.(1988)69:765−776、Millerら、Ann.Rev.Microbiol.(1988)42:177、Carbonellら、Gene(1988)73:409、Maedaら、Nature(1985)315:592−594、Lebacq−Verheydenら、Mol.Call.Biol.(1988)8:3129;Smithら、Proc.Nat’l.Acad.Sci.USA(1985)82:8404、Miyajimaら、Gene(1987)58:273;およびMartinら、DNA(1988)7:99に記載されるように達成され得る。多くのバキュロウイルス株および改変体および対応する宿主由来の許容昆虫宿主細胞は、Luckowら、Bio/Technology(1988)6:47−55、Millerら、GENETIC ENGINEERING(Setlow,J.K.ら編)、第8巻(Plenum Publishing、1986)、277〜279頁、ならびにMaedaら、Nature,(1985)315:592−594に記載される。
【0063】
転移マーカー発現構築物の哺乳動物発現は、Dijkemaら、EMBO J.(1985)4:761、Gormanら、Proc.Nat’l.Acad.Sci.USA(1982b)79:6777、Boshartら、Cell(1985)41:521および米国特許第4,399,216号に記載されるように達成され得る。転移マーカー発現構築物の哺乳動物発現の他の特徴は、HamおよびWallace,Meth.Enz.(1979)58:44、BarnesおよびSato、Anal.Biochem.(1980)102:255、米国特許第4,767,704号、米国特許第4,657,866号、米国特許第4,927,762号、米国特許第4,560,655号、WO 90/103430、WO 87/00195号、ならびに米国特許再発行30,985に記載されるように容易にされ得る。
【0064】
本発明のサブゲノムポリヌクレオチドはまた、mRNAまたはオリゴヌクレオチド(天然の転移マーカーmRNAまたはその相補体のいずれかの配列を有する)、全長転移マーカータンパク質、転移マーカー融合タンパク質、転移マーカーポリペプチド、または転移マーカー特異的リボザイム、または単鎖抗体を細胞(好ましくは真核生物細胞)に送達する目的のために、遺伝子送達ビヒクルにおいて使用され得る。本発明に従って、遺伝子送達ビヒクルは、例えば、裸のプラスミドDNA、転移マーカーサブゲノムポリヌクレオチドを含むウイルス発現ベクター、またはリポソームもしくは縮合試薬と連結した転移マーカーサブゲノムポリヌクレオチドであり得る。
【0065】
本発明は、生物学的サンプルにおいて転移マーカー遺伝子発現を検出する方法を提供する。転移マーカー遺伝子発現の検出は、例えば、転移を同定するため、または、組織サンプル(好ましくは、腫瘍)における転移の可能性を決定するために有用である。次いで、適切な処置レジメンが、身体の他の器官における転移性の癌を発達する危険がある患者について設計され得る。
【0066】
身体サンプルは、例えば、固体組織または流体サンプルであり得る。配列番号1によってコードされるネイティブなポリヌクレオチドは、推定の分泌タンパク質であり、血液およびリンパ流体(特に、身体の腫瘍部位からの排出するもの)を含む体液において検出されるようである。タンパク質または核酸の発現産物は、身体サンプル中で検出され得る。1つの実施形態において、この身体サンプルは、転移マーカータンパク質の存在に関してアッセイされる。この転移マーカータンパク質は、配列番号1に示されるヌクレオチド配列によりコードされる配列またはその相補体を含み、かつ本発明のマーカータンパク質特異抗体を用いて検出され得る。この抗体は、例えば、放射性、蛍光、ビオチン化、または酵素学的タグで標識化され得、そして直接的に検出され得るか、または(標識化した二次抗体を使用する)間接的な免疫化学的方法を用いて検出され得る。この転移マーカータンパク質の存在は、(例えば、免疫細胞化学によって組織切片において、または溶解物中において、当該分野において公知であるようなウェスタンブロットを用いて)アッセイされ得る。マーカータンパク質の存在は、組織サンプルが転移性であることを示す。
【0067】
別の実施形態において、身体サンプルは、マーカータンパク質mRNAの存在についてアッセイされる。サンプルは、選択されたポリペプチドに対応するmRNAとハイブリダイズし得る核酸ハイブリダイゼーションプローブと接触され得る。なおさらに、サンプルは、ポリペプチドの発現を示すmRNAを検出するためにノーザンブロット技術に供され得る。mRNAが検出されるそれらの技術について、サンプルは、核酸増幅プロセスに供され得、それによって、mRNA分子またはその選択された部分は、適切なヌクレオチドプライマーを使用して、増幅される。他のRNA検出技術がまた使用され得、これには、インサイチュウハイブリダイゼーションが挙げられるが、これに限定されない。
【0068】
マーカータンパク質特異的プローブは、配列番号1に開示されるcDNA配列を使用して作製され得る。プローブは、好ましくは、少なくとも15〜50ヌクレオチドの長さであるが、これらは、少なくとも8、10、11、12、20、25、30、35、40、45、60、75、または100以上のヌクレオチドの長さであり得る。プローブを選択するための好ましい領域は、配列番号1の446位〜1173位のヌクレオチド内にある。プローブは、化学的に合成され得るか、または制限酵素を使用してより長いポリヌクレオチドから作製され得る。プローブは、例えば、放射性タグ、ビオチン化タグ、または蛍光タグで標識され得る。
【0069】
必要に応じて、身体サンプル内の特定の転移マーカー発現産物のレベルが定量され得る。定量は、例えば、身体サンプル内で検出される発現産物のレベルを、検量線に存在する生成物の量と比較することによって達成され得る。コンピュータの援助ありでまたはなしで、視覚的にまたは密度測定のような技術を使用して比較がなされ得る。コントロールとしての使用のために、身体サンプルは、他のヒト、試験される患者の他の非癌性器官、または試験される患者からの非転移性乳癌から単離され得る。本明細書中の結果によって示されるように、低い転移性または非転移性の乳癌細胞における配列番号1の発現は、高度に転移性の乳癌細胞における発現レベルの3%と44%との間である。試験サンプルにおける発現が適切なコントロールサンプルの少なくとも二倍である場合、これは、転移性細胞であることを示す。
【0070】
本発明の転移性マーカー特異的試薬(例えば、抗体およびヌクレオチドプローブ)をコードするポリヌクレオチドは、生物学的サンプルにおけるマーカー遺伝子発現産物を検出するためのキットに供給され得る。このキットはまた、緩衝液または標識成分、ならびに生物学的サンプルにおいてマーカー発現産物を検出するために試薬を使用するための説明書を含み得る。
【0071】
転移マーカー遺伝子の発現は、アンチセンスオリゴヌクレオチド配列を使用して変更され得る。アンチセンス配列は、配列番号1に示されるヌクレオチド配列を有する転移マーカー遺伝子のコード配列(ヌクレオチド365〜1173)の少なくとも一部に相補的である。好ましくは、このアンチセンスオリゴヌクレオチド配列は、少なくとも6ヌクレオチドの長さであるが、約8、12、15、20、25、30、35、40、45、または50ヌクレオチドの長さであり得る。より長い配列もまた、用いられ得る。アンチセンスオリゴヌクレオチド分子は、DNA構築物中に提供され得、そして細胞分裂が減少されるべきである細胞中へと導入され得る。このような細胞としては、高度に転移性の乳癌細胞が挙げられる。
【0072】
アンチセンスオリゴヌクレオチドは、デオキシリボヌクレオチド、リボヌクレオチド、または両方の組み合わせで構成され得る。オリゴヌクレオチドは、手動または自動合成機によって、1つのヌクレオチドの5’末端を、アルキルホスホネート、ホスホロチオエート、ホスホロジチオエート、アルキルホスホノチオエート、アルキルホスホネート、ホスホラミデート(phosphoramidates)、リン酸エステル、カルバメート、アセトアミデート(acetamidate)、カルボキシメチルエステル、カーボネート、およびリン酸トリエステルのような、非リン酸ジエステルのヌクレオチド間結合を有する別のヌクレオチドの3’末端と共有結合することにより合成され得る。Brown,1994,Meth.Mol.Biol.20:1−8;Sonveaux,1994,Meth.Mol.Biol.26:1−72;Uhlmannら、1990,Chem.Rev.90:543−583を参照のこと。
【0073】
転移マーカータンパク質に特異的に結合する本発明の抗体はまた、転移マーカー遺伝子発現を変更するために使用され得る。抗体とは、タンパク質に対する特異的な結合を保持する単鎖抗体のような抗体およびその一部または誘導体を意味する。特異的抗体は、転移性マーカータンパク質に結合し、そしてこのタンパク質を細胞中で機能しないようにする。上記のように、本発明の特異的抗体をコードするポリヌクレオチドは、細胞中に導入され得る。
【0074】
本発明のマーカータンパク質は、治療的な抗転移性効果を有する薬物をスクリーニングするために用いられ得る。転移性マーカータンパク質合成に対する試験化合物の効果はまた、転移を調節する試験化合物を同定するために用いられ得る。スクリーニングされ得る試験化合物としては、被験体に投与され得る天然産物または合成のいずれかの任意の物質が挙げられる。化合物のライブラリーまたは混合物が試験され得る。この化合物または物質は、薬学的な効果が以前に公知の物質であっても未知の物質であってもよい。
【0075】
転移性マーカータンパク質の合成は、当該分野で公知の、タンパク質合成を測定する任意の手段(例えば、標識アミノ酸のタンパク質への組み込みおよびポリアクリルアミドゲル中での標識した転移性マーカータンパク質の検出)によって測定され得る。転移性マーカータンパク質の量は、例えば、ウエスタンブロットにおいて、本発明の転移性マーカータンパク質特異的抗体を用いて、検出され得る。試験化合物の存在または非存在下で合成された転移性マーカータンパク質の量は、当該分野で公知の任意の手段(例えば、標準曲線で示される転移性マーカータンパク質の量と、合成した転移性マーカータンパク質の量の比較)によって決定され得る。
【0076】
転移性マーカータンパク質合成に対する試験化合物の効果はまた、当該分野で公知であるように、本発明の転移性マーカータンパク質特異的ヌクレオチドプローブを用いる試験化合物に応答する転移性マーカータンパク質mRNA発現の量を測定することによって、ノーザンブロット分析によって、測定され得る。
【0077】
代表的には、生物学的サンプルが、ある範囲の濃度(例えば、1.0nM、5.0nM、10nM、50nM、100nM、500nM、1mM、10mM、50mM、および100mM)の試験化合物と接触される。好ましくは、試験化合物は、転移性マーカータンパク質の発現を、60%、75%、または80%まで増大または減少する。より詳細には、85%、90%、95%または98%の増大または減少が達成される。
【0078】
本発明は、転移性マーカータンパク質の発現を増大または減少するための組成物を提供する。これらの組成物は、転移性マーカータンパク質遺伝子発現産物の全てまたは少なくとも一部をコードするポリヌクレオチドを含む。好ましくは、治療用組成物は、プロモーターおよび転移性マーカータンパク質の少なくとも一部をコードするポリヌクレオチドセグメントを含む発現構築物を含み、転移能力(力価)を減少させるのに効果的である。細胞の転移能力を減少させるのに有効である転移性マーカー遺伝子またはタンパク質の部分は、当該分野で公知のように、例えば、転移性マーカー遺伝子またはポリペプチドの部分を転移性細胞株(例えば、MDA−MB−231、MDA−MB−435、Km12C、またはKm12L4)に導入すること、およびインビボで移植された場合、細胞の分裂速度または細胞が転移を形成する能力をアッセイすることによって決定され得る。非転移性細胞株(例えば、MCF−7)は、転移性マーカータンパク質の一部が、転移性マーカー遺伝子の発現を増大する能力をアッセイするために用いられ得る。
【0079】
代表的には、治療用転移性マーカー組成物が、注射可能に、流体溶液または懸濁液のいずれかとして、調製され得る;しかし、注射前の流体ビヒクル中の溶液に、または流体ビヒクル中の懸濁液に適切な固体形態がまた調製され得る。転移性マーカー組成物がまた、米国特許第4,853,230号、欧州特許225,189号、AU9,224,296号およびAU9,230,801号に記載のように、当該分野で公知の方法に従って、腸溶コーティング錠剤またはゲルカプセル中に処方され得る。
【0080】
本発明の転移性マーカー治療剤の投与は、局所(local)投与または全身投与(これには、注射、経口投与、粒子銃、またはカテーテル投与を含む)、および局所(topical)投与を含み得る。種々の方法を用いて、身体内の特定の部位に治療用転移性マーカー組成物を直接投与し得る。
【0081】
腫瘍(転移性病変を含む)の処置のために、例えば、治療用転移性マーカー組成物が、腫瘍の本体内のいくつかの異なる位置に数回注射され得る。あるいは、腫瘍を助ける動脈が、同定され得、そして治療用組成物が、この組成物を腫瘍に直接送達するために、このような動脈に注射される。
【0082】
壊死性中心を有する腫瘍は、吸引され得、そしてこの組成物がこの腫瘍のいまや空になった中心に直接注射される。治療用転移性マーカー組成物は、例えば、この組成物の局所適用によって、腫瘍の表面に直接投与され得る。X線画像を用いて、上記の特定の送達方法が補助され得る。転移性マーカータンパク質もしくはポリペプチド、または転移性マーカー小ゲノムポリヌクレオチドおよび他の治療用因子を含む、併用療法の薬剤が、同時にまたは引き続いて投与され得る。
【0083】
あるいは、転移性マーカー治療組成物が、エクソビボでヒト細胞に導入され得、次いで、この細胞はヒトの中に入れられる。細胞は、種々の場所から(例えば、以下を含む:選択された腫瘍からまたは罹患した器官から)取り出され得る。さらに、治療組成物は、罹患していない細胞、例えば、真皮線維芽細胞または末梢血リンパ球中に挿入され得る。所望の場合、T細胞サブセットまたは幹細胞のような細胞の特定の画分がまた、血液から特異的に取り除かれ得る(例えば、PCT WO 91/16116を参照のこと)。次いで、除去された細胞を、上記の任意の技術を用い、続いてヒトに(好ましくは、処置されるべき腫瘍または他の部位の近傍に、またはその中に)この細胞を戻し、転移性マーカー治療組成物と接触させ得る。上記の方法は、さらに、線維芽細胞または他の混入のない腫瘍細胞を枯渇し、続いてヒトから腫瘍細胞を取り出す工程、および/または例えば、照射によって細胞を不活性化する工程を包含し得る。
【0084】
転移性マーカー組成物の投与の用量および投与の手段の両方は、治療用組成物の特定の質、患者の状態、年齢、および体重、疾患の進行、ならびに他の関連要因に基づいて決定され得る。好ましくは、本発明の治療用組成物は、転移性マーカー遺伝子の発現を50%、60%、70%、または80%まで減少する。より好ましくは、転移性マーカー遺伝子の発現は、90%、95%、99%または100%まで減少される。転移性マーカー遺伝子の発現を変更するために選択される機構の有効性は、当該分野で周知の方法(例えば、転移性マーカー遺伝子のmRNAに対するヌクレオチドプローブのハイブリダイゼーション、定量的RT−PCR、または本発明の特定の抗体を用いる転移性マーカータンパク質の検出)を用いて評価され得る。
【0085】
この組成物が、転移性マーカータンパク質、ポリペプチド、または抗体を含む場合、有効な用量の組成物が、患者の体重あたり約5μg〜約50μg/kg、患者の体重あたり約50μg〜約5mg/kg、約100μg〜約500μg/kg、そして約200〜約250μg/kgの範囲である。
【0086】
転移性マーカーサブゲノムポリヌクレオチドを含む治療用組成物は、局所投与について約100ng〜約200mgのDNAの範囲で投与され得る。約500ng〜約50mg、約1μg〜約2mg、約5μg〜約500μg、そして約20μg〜約100μgのDNAの濃度範囲がまた、遺伝子治療プロトコールの間に用いられ得る。形質転換および発現の作用および有効性の方法のような要因が、転移性マーカーサブゲノムポリヌクレオチドの最終的な有効性に必要である投薬量に影響することが考慮される。より大きい発現が組織のより大きい面積にわたって所望される場合、例えば、腫瘍部位の異なる隣接部位または近接する組織部位への、より大量の転移性マーカーサブゲノムポリヌクレオチドもしくは同じ量(投与の引き続くプロトコールで再投与される)、または、数回の投与が、正の治療結果をもたらすために必要であり得る。全ての場合において、臨床試験における慣用的実験が、至適の治療効果についての特定の範囲を決定する。
【0087】
内因性転移性マーカー遺伝子の細胞内での発現がまた、相同組み換えによって、内因性転移性マーカー遺伝子とインフレームでDNA構築物(転移性マーカータンパク質標的化配列、調節性配列、エキソン、および不対スプライスドナー部位を含む)を導入することによって変更され得、これによって、DNA構築物を含む相同組み換え細胞が形成される。新しい転写単位が、所望の場合、転移性マーカー遺伝子をターンオンまたはターンオフするために用いられ得る。内因性遺伝子発現に影響する方法は、本明細書において参考として援用される、米国特許第5,641,670号に教示されている。
【0088】
転移性マーカーサブゲノムポリヌクレオチドはまた、細胞に対する転移性マーカーサブゲノム性ポリヌクレオチドの移入を増強するか、または細胞内の転移性マーカーサブゲノム性ポリヌクレオチドの引き続く生物学的効果を増強するのに有用である試験化合物をスクリーニングする目的で被験体に送達され得る。このような生物学的効果としては、相補的転移性マーカーmRNAに対するハイブリダイゼーション、およびその翻訳の阻害、転移性マーカーmRNAおよび/または転移性マーカータンパク質を形成するための転移性マーカーサブゲノムポリヌクレオチドの発現、ならびに転移性マーカーサブゲノムポリヌクレオチドの複製および組み込みが挙げられる。被験体は、細胞培養物または動物、好ましくは哺乳動物、より好ましくはヒトであり得る。
【0089】
上記の開示は、本発明を一般的に記載する。より完全な理解が、以下の特定の実施例を参照して得られ得る。以下の実施例は、本明細書において例示のみの目的で提供されるものであり、本発明の範囲を限定することを意図しない。
【0090】
(実施例)
(材料および方法)
細胞培養。MDA−MB−435、MDA−MB−231、ALAB、MDA−MB−468、MDA−MB−361、ZR−75−1、MCF−7、MDA−MB−453およびSK−BR−3ヒト乳癌細胞株(Chiron Master Culture Collection、Chiron Corporationから入手)を、37℃で5%CO2で、以下の中で増殖した:DMEM+HAM’S F−12(1:1)(Bio*Whittaker,Walkersville,MD)(2mM Lグルタミン、1mM Sodium Pyruvate、100U/mlペニシリンおよび100μg/ml ストレプトマイシン(Bio*Whittaker,Walkersville,MD)、1×ビタミン溶液、1×非必須アミノ酸(Irvine Scientific,Santa Ana,CA)および10%熱不活化ウシ血清(Life Technologies,Rockville,MD)を含有)。COS−7細胞をATCCから入手し、そして10%熱不活化ウシ胎仔血清(Life Technologies)を含有するDMEM中で37℃、5%CO2中において増殖した。
【0091】
Opti−MEM1上清の濃度。Centricon YM−10および/またはMicron YM−10カラム(Millipore Corporation,Bedford,MA)を通して、Opti−MEM1(Life Technologies)培養培地を濃縮した。次いで、SDS−PAGEサンプルローディング緩衝液を添加し、そしてサンプルを煮沸した。
【0092】
ノーザンブロットハイブリダイゼーション。培養した乳癌細胞株、およびRNeasy Maxi Kit(Qiagen,Valencia,CA)を用いて、乳癌細胞株を移植したSCIDマウスの腫瘍組織から、総RNAを調製した。1レーンあたり、約20μgの総RNAを、電気泳動のためにホルムアルデヒド/アガロースゲル上にロードし、次いで、Hybond−N+ナイロンメンブレン(Amersham Life Science,Little Chalfont,England)に移した。このブロットをUV照射で固定した。5mg/mlの変性一本鎖精子DNAとともに、Rapid−Hyb緩衝液(Amersham Life Science)を65℃に予熱し、そして65℃で30分間振盪しながらこの緩衝液中でブロットをプレハイブリダイズした。[α−32P]dCTP(3000Ci/mmol,Amersham Pharmcia Biotech Inc.,Piscataway,NJ)(Prime−It RmT Kit,Stratagene,La Jolla,CA)で標識し、そしてProbeQuantTMG−50 Micro Column(Amersham Pharmacia Biotech Inc.)で精製したプローブとして、hsOAF cDNAフラグメントまたはβアクチンcDNAフラグメントを添加し、そしてこのブロットにハイブリダイズさせて、65℃で一晩振盪した。このブロットを2×SSC、0.1%(w/v)SDSを用いて、室温で20分間、1×SSC、0.1%(w/v)SDSで、65℃15分間2回洗浄し、次いでHyperfilms(Amersham Life
Science)に曝露した。
【0093】
イムノブロッティング。タンパク質サンプルを10〜20% SDS−PAGEゲル上の電気泳動に供し、次いで、25mM Tris,192mMグリシン、20%(v/v)メタノール、pH8.3における電気ブロッティングによってPVDF膜(0.2μm)に転写した。膜を10%脱脂乳を含有するTBST(pH7.5)中でブロックし、次いで、1%BSAを含有するPBS(pH7.4)中でウサギ抗hs−OAF血清(1:1000)とともに、ブロッティングし、次いで、二次抗体アルカリホスファターゼ結合体化ヤギ抗ウサギIgG(1:2000)(Santa Cruz Biotechnology,Inc.,Santa Cruz,CAを用いてプローブした。次いで、タンパク質バンドをNBT/BCIP試薬(Boehringer Mannheim,Germany)によって可視化した。
【0094】
一過性トランスフェクション。hsOAF cDNAのコード領域(356〜1174)を、改変発現ベクターpRetro−On(Clontech,Palo Alto,CA)中にクローニングした。hsOAFを保有するpRetro−OnベクターまたはコントロールのpRetro−Onベクター(GFPを有する)を、製造業者によって提供されたプロトコール中に指示されたように、EffecteneTMTransfection Reagent Kit(Qiagen)を用いて100mm培養プレート上のCOS−7細胞中にトランスフェクトした。細胞を10%FBSを含有するDMEM中で一晩回収し、次いでOpti−MEM1に切り替えた。さらに2日後、この上清を収集し、ウエスタンブロット分析用に濃縮した。
【0095】
アンチセンスオリゴトランスフェクション。MDA−MB−435細胞をトランスフェクションの1日前に6−ウェルの培養プレート上に播種し、トランスフェクションで90%の密度を得た。100μMアンチセンスまたは逆のコントロールオリゴを、トランスフェクション用に、Opti−MEM1中で2μMに希釈した。0.5mM滅菌リピトイド1(lipitoid1)を、同量のOpti−MEM1中で、1.5nmolリピトイド1:1μgオリゴの比に希釈した。希釈したオリゴおよび希釈したリピトイド1を混合して、直ちに、培養培地中の細胞に加え、最終濃度100、200、または300nMオリゴとした。6時間後、トランスフェクション混合物を正常培養培地で置換し、そして細胞を一晩、回収のためにインキュベートした。アンチセンスオリゴの配列は、AGCTGCGGATGCCACACTTGTAGG(配列番号4)であり、そして逆のコントロールオリゴの配列は、GGATGTTCACACCGTAGGCGTCGA(配列番号5)である。
【0096】
Matrigel侵襲アッセイ。 細胞をトリプシン処理し、洗浄し、そして、計測のために培地に再懸濁した。4×104細胞を洗浄し、氷上の100μlの培地中で再懸濁した。200μlのMatrigel(Callaborative Biomedical Products,Bedford,MA)を氷上で細胞に添加した。Matrigelおよび細胞を注意深く混合し、次いで、24ウェル培養プレートのウェル中に分配し、そして、37℃で30分間凝固した。Matrigel−細胞混合物を0.5mlの培地で被い、そして、6日間5%CO2中で37℃で、インキュベートした。培地を2日毎に補充した。
【0097】
増殖アッセイ。 細胞をトリプシン処理し、洗浄し、そして、計測のために培地中で再懸濁した。次いで、細胞をインキュベーションのために96ウェルプレート(5000細胞/ウェル)に移した。細胞数をQuantosTM細胞増殖アッセイキット(Stratagene,La Jolla,CA)を用いて毎日計測した。
【0098】
(実施例1)
(ヒトcDNA配列の同定)
ショウジョウバエ Out at First(oaf)遺伝子の推定ヒトホモログをコードするDNAを配列番号1に示す。hsOAFおよびショウジョウバエOAFのアライメントを図7に示す。ポリヌクレオチドは、2366塩基対を含み、そして、オープンリーディングフレームが同定されている。ORFの翻訳、273アミノ酸のポリペプチドを、配列番号2に示す。図4は、DNAおよびアミノ酸配列を提供し、ORFの位置を示す。シグナルペプチド由来の最初の30アミノ酸は、このタンパク質が分泌され得ることを示す。シグナルペプチドのアミノ酸配列は以下のようである:MRLPGVPLARPALLLLLPLLAPLLG#TGAPA(配列番号3)。「#」は、推定プロテアーゼ切断部位の位置を示す。
【0099】
(実施例2)
(乳癌細胞株における配列番号1の差次的発現)
以下のヒト乳癌細胞株における配列番号1の発現を比較した:
ヒト乳腺癌に由来するMDA−MB−361;
骨および/または肺に転移性のヒト乳癌に由来するMDA−MB−231;
エストロゲンレセプターネガティブなヒト乳癌細胞に由来するMDA−MB−468;
エストロゲンレセプターネガティブなヒト乳癌細胞に由来するMDA−MB−435;
転移性でないヒト乳癌細胞に由来するMCF−7;および
エストロゲンレセプターポジティブなヒト乳癌細胞に由来するZR−75−1。
【0100】
配列番号1の発現を、高度に転移性の乳癌細胞株MDA−MB231およびMDA−MB−435において測定し、そして、低い転移性または非転移性の乳癌細胞株と比較した。MDA−MB−361における発現は、MDA−MB−231のレベルの11%であり;MDA−MD−468における発現は、MDA−MB−231におけるレベルの44%であり;MCF−7における発現は、MDA−MB−231におけるレベルの17%であり;そしてZR−75−1における発現は、MDA−MB−231におけるレベルの12%であった。
【0101】
MDA−MB−361における発現は、MDA−MB−435におけるレベルの6%であり;MDA−MB−468における発現は、MDA−MB−435におけるレベルの36%であり;そしてMCF−7における発現は、MDA−MB−435におけるレベルの3%であった。従って、表2に示されるように、高い転移可能性を有するヒト腫瘍に由来する乳癌細胞株における配列番号1の増加した発現の明らかな傾向が存在する。
【0102】
【表2】
この遺伝子の同様の発現パターンは、腫瘍化乳癌細胞株で移植したSCIDマウス由来の腫瘍組織サンプル中で、残ったままである(図6)。
【0103】
(実施例3)
(分泌タンパク質をコードするhsOAFおよびhsOAFタンパク質分泌レベルは、乳癌細胞株のhsOAF mRNA発現レベルと一貫している)
推定シグナルペプチド配列は、hsOAF遺伝子の推定アミノ酸配列のN末端に位置する(図3)。hsOAFタンパク質の分泌を検証するために、COS−7細胞およびMCF−7細胞の一過性のトランスフェクションを、hsOAF cDNAを保有するベクターpRetro−Onを用いて行なった。一方、GFPを保有するベクターpRetoro−Onをコントロールとして使用した。hsOAFウサギ抗血清を使用して、分泌されたhsOAFタンパク質を、免疫ブロッティングによるhsOAFでのトランスフェクション後、両方の細胞株のOpti−MEM1培地中で検出した(図8A)。分泌されたhsOAFタンパク質は、おそらくグリコシル化されている。なぜなら、より大きいみかけの分子量を有する複数のバンドが見出されたためである(分泌されたhsOAFタンパク質の推定MWは、28Kdaである)。同じhsOAF抗血清を使用して、種々の乳癌細胞株によるhsOAFタンパク質の分泌を検出する。hsOAFタンパク質の分泌レベルは、これらの細胞株全体の中でhsOAF mRNA発現レベルと一貫していた;高度に転移性の細胞株は、低い転移性/非転移性細胞株よりも、はるかに強力なhsOAF分泌を示した(図8B)。MDA−MB−435は、最も強力なhsOAFタンパク質分泌を有した。
【0104】
(実施例4)
(アンチセンスオリゴによるMDA−MB−435細胞におけるhsOAF発現のノックアウトは、形態学的変化、減少した細胞侵襲性およびより緩徐な増殖速度を引き起こした)
高レベルのhsOAF遺伝子発現が、ヒト乳癌細胞の転移可能性について必須であるか否かを決定するために、アンチセンスオリゴ技術を使用して、hsOAF発現をノックアウトし、次いで、結果として生じた効果を観察した。MDA−MB−435の高度に転移性の細胞株は、調査された乳癌細胞株のすべての中で、最も強力なhsOAFタンパク質分泌を示したので、MDA−MB−435を選択した。hsOAFアンチセンス(AS)およびリバースコントロール(RC)オリゴのいくつかの対を選択して、mRNAレベルにおけるhsOAF遺伝子発現を中断するそれらの能力について試験した。Lightcycler(Roche Diagnostics,Indianapolis,IN)においてリアルタイム定量的RT−PCR分析を行ない、細胞中のhsOAF mRNAレベルを測定する。Kang,Sら、Cancer Research 60(18):5296−5302(2000)。次いで、オリゴの作用する濃度の力価について、最高の対を選択した。低いオリゴ濃度は、細胞に対する潜在的なオリゴ毒性を減少するために好ましい。結果は、100nMのアンチセンスオリゴを用いる処理は、MDA−MB−435細胞のhsOAFタンパク質の分泌を有意に減少するのに十分であることを示した(図12)。100nMの作用する濃度でのオリゴ(配列番号4(AS)および5(RC))の対を以下のすべての実験について使用した。
【0105】
hsOAFアンチセンスオリゴでのMDA−MB−435細胞の処理後、細胞の劇的な形態学的変化が、減少したhsOAFタンパク質の分泌と共に観察された(図10A)。細胞は、より球状になり、そして、それらの広がっていく突出(spreading protrusion)を失う。一方、リバースコントロールオリゴで処理した細胞は、正常な細胞培養されたMDA−MB−435細胞に類似したままである。さらに、アンチセンスオリゴで処理された細胞に添加された馴化培地として、高レベルのhsOAFタンパク質を含む、正常なMDA−MB−435細胞の培地は、完全ではないが、この形態学的変化を防止し得る。細胞形態の変化は、細胞の減少した侵襲能力の指標であり得る。
【0106】
次いで、Mrtrigel侵襲アッセイを行ない、細胞の侵襲性を評価した。matrigelに包埋された乳癌細胞の星状の侵襲性形態学は、それらの転移可能性と相関することが報告されている(Thompson,E.W.ら、J.Cell Physiol.150(3)534−44(1992);Sugiura,T.,ら、J.Cell Biol,146(6):1375−89(1999);Abini,A.,ら、Cancer Res.47(12):3239−45(1987);およびKramer,R.H.,ら、Cancer
Res.46(4 Pt 2):1980−89(1986))、そして、これは、matrigel中で増殖された種々の癌細胞株で確認された。細胞をトリプシン処理し、計測し、そして、matrigelと混合した。次いで、培地を細胞−matrigel混合物上に被せた。インキュベーションの6日後、細胞侵襲を調査した(図10B)。結果は、hsOAFリバースコントロールオリゴで処理した細胞が、正常なMDA−MB−435細胞のように、貫通性、侵襲性、ネットワーク様三次元構造を形成することを示した;他方、hsOAFアンチセンスオリゴで処理した細胞は、平らな球状のコロニーのみを生じた。再び、貫通性コロニーがまた、馴化培地においてインキュベートしたhsOAFアンチセンスオリゴ処理細胞において観察された。これらのデータは、分泌されたhsタンパク質が、MDA−MB−435細胞の侵襲性および転移可能性について必要であることを実証する。
【0107】
さらなる実験を行ない、分泌されたhsOAFタンパク質が、MDA−MB−435細胞増殖に関与するか否かを調査した。細胞増殖アッセイ結果は、hsOAFタンパク質分泌のノックアウトが、変化は穏やかであるが、確かに、MDA−MB−435細胞の増殖速度を落とした。
【0108】
(実施例5)
(ヒト乳癌細胞およびヒト組織におけるRNA発現のノーザンブロット分析)
図5に示されるように、mRNA発現を転移性細胞株MDA−MB−231およびMDA−MB−435においてアップレギュレートした。総RNAを、QuiagenからのRNeasy Kitを使用して調製した。ノーザンブロット分析を、原発性腫瘍または肺における転移由来の細胞からのグアニジニウムチオシアネート/フェノールクロロホルム抽出によって単離した20〜30μgの総RNAを使用して行なった。原発性腫瘍および肺転移は、当業者に周知の方法に従って、scidマウスに注射した細胞株から発症した。pCR2.0−TAベクター(In vitorogen)にクローン化されたhOAFの部分的cDNAクローンを含むプラスミドを、放射線標識し、そして、Expresshyb(Clontech)中で65℃で、ハイブリダイズした。調査された全ての組織の中で、肝臓、膵臓、脾臓、卵巣および小腸は、有意なhsOAF発現を示した。HsOAF mRNA発現はまた、心臓、骨格筋、腎臓、前立腺、結腸および骨髄において検出された(図9)。
【0109】
表3は、種々の腫瘍および正常組織におけるhsOAFポジティブの割合を示す。
【0110】
【表3】
(実施例6)
(軟寒天アッセイ)
軟寒天アッセイ:底層は、細胞上の層化(layering)の数時間以内に新たにプレートされた培地中の2mlの0.6%寒天からなる。細胞の層について、MDA−MB−435細胞を、上記のように、0.05%トリプシン中のプレートから取り出し、そして培地中で2回洗浄した。細胞を犂先(coulter)カウンターで計測し、そして、培地中で1mlあたり106に再懸濁した。10mlのアリコートを、96ウェルプレート中の培地と共に配置する(WST1で計測をチェックするために)か、または、軟寒天アッセイのためにさらに希釈した。2000個の細胞を、0.6%寒天の底層上に、二連のウェルで、800mlの0.4%寒天中に希釈した。
【0111】
培地層:細胞層寒天を凝固した後、2mlの培地を最上層に流し、そして、アンチセンスオリゴまたはリバースコントロールオリゴを、送達ビヒクルを用いずに添加した。新鮮な培地およびオリゴを3〜4日毎に添加した。
【0112】
10日から3週間で、コロニーを計測した。コロニーの視野を眼で計測した。Wst−1代謝値を使用して、出発細胞数における小さな差異を補完した。より大きい視野は、差異の視覚的な記録についてスキャンし得る。結果を図6に示し、ここで、コントロールオリゴヌクレオチドに曝露された細胞と比較して、アンチセンスで処理したMDA−MB−435細胞は、より少ないコロニーを形成した。
【0113】
当業者は、慣例的な実験以上のものは使用せず、本明細書中に記載される本発明の特定の実施形態に対する多数の等価物を認識するか、または確認し得る。このような特定の実施形態および等価物は、上記の特許請求の範囲によって含まれることが意図される。
【0114】
全ての特許、公開された特許出願、および本明細書中で援用された刊行物は、本明細書中で十分に開示されるように、参考として援用される。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】


【図2】

【図3】

【図4A】

【図4B】

【図4C】

【図4D】

【図5】
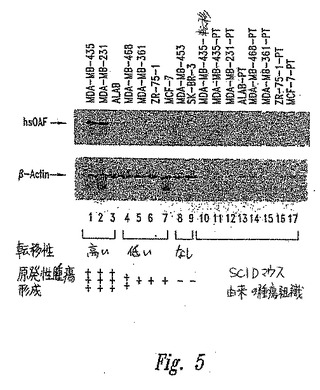
【図6】
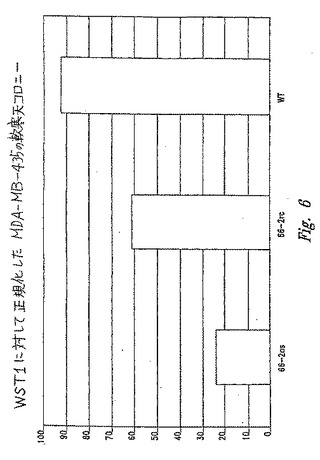
【図7】

【図8】
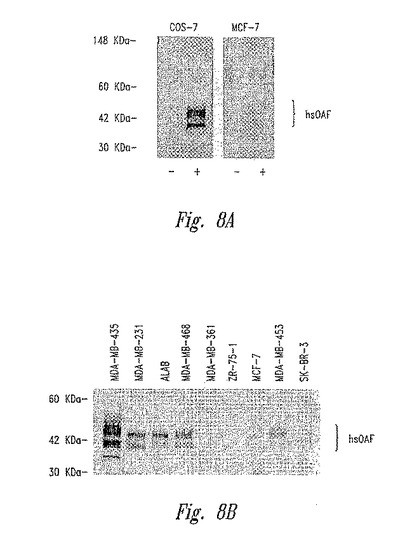
【図9】

【図10A】

【図10B】

【図11】

【図12】
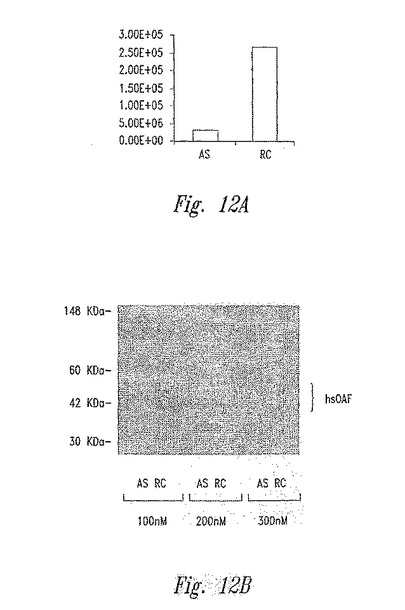
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図11】
【図12】
【公開番号】特開2012−143242(P2012−143242A)
【公開日】平成24年8月2日(2012.8.2)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−71880(P2012−71880)
【出願日】平成24年3月27日(2012.3.27)
【分割の表示】特願2001−551099(P2001−551099)の分割
【原出願日】平成13年1月9日(2001.1.9)
【出願人】(591076811)ノバルティス バクシンズ アンド ダイアグノスティックス,インコーポレーテッド (265)
【Fターム(参考)】
【公開日】平成24年8月2日(2012.8.2)
【国際特許分類】
【出願番号】特願2012−71880(P2012−71880)
【出願日】平成24年3月27日(2012.3.27)
【分割の表示】特願2001−551099(P2001−551099)の分割
【原出願日】平成13年1月9日(2001.1.9)
【出願人】(591076811)ノバルティス バクシンズ アンド ダイアグノスティックス,インコーポレーテッド (265)
【Fターム(参考)】
[ Back to top ]