
乳酸耐性に優れた生物および乳酸耐性に優れた生物の作製方法
【課題】
乳酸に対し耐性のない微生物に対し、乳酸耐性遺伝子を解明或いは遺伝子導入する事なく、あるいは、突然変異を導入し乳酸耐性微生物を作製する。
【解決手段】
所望の形質を生物に迅速に付与する方法およびシステムを用いて変異を導入し、所望の乳酸存在下で培養することによって、乳酸耐性を有する生物であって、該生物は、該生物の天然型が有する耐性よりも高い耐性を有する、生物、好ましくは、乳酸耐性微生物を作製することができる。
乳酸に対し耐性のない微生物に対し、乳酸耐性遺伝子を解明或いは遺伝子導入する事なく、あるいは、突然変異を導入し乳酸耐性微生物を作製する。
【解決手段】
所望の形質を生物に迅速に付与する方法およびシステムを用いて変異を導入し、所望の乳酸存在下で培養することによって、乳酸耐性を有する生物であって、該生物は、該生物の天然型が有する耐性よりも高い耐性を有する、生物、好ましくは、乳酸耐性微生物を作製することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は乳酸耐性に優れた微生物の作製方法、及び該方法により作製した乳酸耐性に優れた微生物に関する。
【背景技術】
【0002】
乳酸発酵で生産された乳酸が蓄積すると、培地pHと細胞内pHが共に低下するが、微生物は細胞内のpHを一定にするためATPを消費して、細胞外へプロトンを汲み出そうとする。例えば、S.cerevisiaeの場合、細胞外pH3.0の時、細胞内pHは5.5〜5.75であり(非特許文献1=Imai,T. et al.Appl.Environ.Microbiol.61,3604−3608,1995)、細胞外pH6.0〜10.0のとき、細胞内pHは5.9〜6.75に維持されている(非特許文献2=Imai,T. et al.J.Biotechnol.38,165−172,1995)。微生物が細胞内のpH低下を維持できなかった場合、解糖系や糖新生経路のキー・エンザイムであるcAMPプロテインキナーゼの抑制、細胞内cAMP濃度低下、ヒートショックなどの他の生物反応のトリガーなどになり、最終的には増殖が抑制されると考えられている。
【0003】
Lactobacillusに代表される乳酸菌を用いて未利用バイオマスからの乳酸生産は各種試みがなされているが、その乳酸発酵の行われるpHは5.0〜5.5と高く、乳酸のpKa(〜3.8)以下での発酵が望まれている。その理由は、pKa(〜3.8)以下の乳酸は未解離の状態で存在し、発酵培養液から乳酸抽出で直接抽出することが可能だが、高いpHの培地中からの乳酸精製には乳酸を用いた直接抽出が出来ず、抽出精製コストが高くなることにある。乳酸発酵でより高濃度の乳酸を生産することは工業生産上非常に重要である。
【0004】
Lactococcus lactisの低pH下での細胞増殖には、挿入変異解析法を用いて18遺伝子座の関連が示唆され、多数の低pHに対する耐性機構が関与していると予想される(非特許文献3=Rallu,F.et al.Mol.Microbiolo.35,517−528,2000)。また、Lactobacillusの酸への適応のプロテオミクス解析の結果からは、63種のタンパク質の協調的発現誘導が起こることが報告されている(非特許文献4=DeAngelis,M.et al.Microbiology147,1863−1873,2001)。また、Lactobacillus acidophilus NCFMの全ゲノム解析の結果から、酸耐性に関連する遺伝子として2成分から構成される9種の調節システムが推定されている(非特許文献5=Altermann,E.et al.Proc.Natl.Acad.Sci.USA.102(11),3906−3912,2005)。
【0005】
Lactobacillus sanfranciscensisの酸耐性の解析では、ヒートショックタンパク質4種の中でGrpEのみが選択的に発現上昇していた(非特許文献6=Maria DeAngelis et.al.Microbiol.147,1863−1873,2001)。以上の例にあるように、乳酸耐性機構は遺伝子レベルで検討の余地が多分に残されている。
現在、乳酸耐性を付与する、あるいは、乳酸産生能を維持する方法は以下のような方法が用いられている。第一の方法は、新規に乳酸耐性度の高い乳酸耐性菌株を分離する方法である。得られた乳酸耐性菌を宿主として高濃度の乳酸を生産するための遺伝仕組換えを実施するためにはベクター系の開発が必要で実用化までの道のりが遠い。第二の方法は、乳酸菌を化学突然変異誘発剤処理し突然変異を導入し、乳酸耐性変異体を作製する方法だが、大幅な乳酸耐性の付与には成功していない。第三の方法は、異なる性質の乳酸菌の細胞融合で乳酸耐性変異体を作製する方法である。具体例としてpH4.0にて野生株に比べ乳酸を3倍生産する乳酸耐性株を得ている(非特許文献7=Patnaik,R.et al.Nature Biotechnology 20.707−712,2002)が、更に乳酸に対する耐性度の高い株の作製が可能かどうかは不明である。第四の方法は、乳酸耐性菌から単離された乳酸耐性関連遺伝子を遺伝子組み換えで宿主菌に導入し、乳酸耐性を付与するものである。具体例としては、ワイン酵母Oenococcus oeniから単離された多剤耐性遺伝子ファミリーに属するorfC、LysRファミリーの推定上の転写調節因子orfB、低分子ヒートショックタンパク質Lo18の3遺伝子を大腸菌に導入し、酸耐性を付与した例がある(非特許文献8=Lett. Appl. Microbiol.33(2),126−130,2001)。この方法は、乳酸耐性を付与したい生物種に適用可能かどうかが不明である。また、仮に適用が可能な場合でも、乳酸耐性度を更に増強したい場合、乳酸耐性関連遺伝子の発現を増加させる以外に合理的手段がない。第五の方法は、ケモスタットを用いて酸に馴化させる方法である。具体例としては、菌の致死量以下の低pH(pH5.0、60分間)で前処理後、致死pH(pH3.0)処理し、耐性化遺伝子の発現を誘導し、乳酸耐性に馴化させる方法がある(非特許文献9=Lorca GL et.al.J.Mol.Microbiol.Biotechnol,4(6),525−532,2002)。この方法では、馴化して獲得させた形質が安定に保持されるのか、どの生物種に適用可能か、また、馴化可能な乳酸濃度がいくらなのか全く不明である。
その他に、乳酸発酵能を長期間維持する方法としては、培養後期における乳酸菌の乳酸発酵能の低下を防ぐために超伝導磁石で形成された磁場空間内で、静磁場を印加し乳酸菌を活性化する方法が原理的には考えられている(特許文献1=三浦靖、特開2005−34040)。しかし、工業生産する場面で、宿主菌を超伝導の磁場に置くことは、現実的には不可能である。
【非特許文献1】Imai,T. et al.Appl.Environ.Microbiol.61,3604−3608,1995
【非特許文献2】Imai,T. et al.J.Biotechnol.38,165−172,1995
【非特許文献3】Rallu,F.et al.Mol.Microbiolo.35,517−528,2000
【非特許文献4】DeAngelis,M.et al.Microbiology147,1863−1873,2001
【非特許文献5】Altermann,E.et al.Proc.Natl.Acad.Sci.USA.102(11),3906−3912,2005
【非特許文献6】Maria DeAngelis et.al.Microbiol.147,1863−1873,2001
【非特許文献7】Patnaik,R.et al.Nature Biotechnology 20.707−712,2002
【非特許文献8】Lett. Appl. Microbiol.33(2),126−130,2001
【非特許文献9】Lorca GL et.al.J.Mol.Microbiol.Biotechnol,4(6),525−532,2002
【特許文献1】特開2005−34040
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、乳酸耐性の低い実験微生物に対して、低コスト、短時間で、効率よく、さらに高濃度の乳酸に対して耐性に優れた形質を付与する方法を提供することである。
【0007】
また、本発明の別の目的は、乳酸耐性に優れた形質を付与した微生物を提供することである。
【課題を解決するための手段】
【0008】
本発明の上記目的は、鋭意検討の結果、遺伝子の複製において正確さにかける生物を用いて、通常では生育しない乳酸濃度存在下で培養し、生育する生物を単離することを特徴とする乳酸耐性微生物の作製方法によって達成された。
【発明の効果】
【0009】
各種の乳酸に対して、耐性に優れた微生物作製が可能となる。また、本発明は、乳酸耐性機構を解明するためのコストが不要で、これまでに解明された実験微生物のゲノム情報や開発されたベクター系が有効活用されることになる。本発明で作製した乳酸耐性酵母を宿主として乳酸を工業的に生産する場合、乳酸耐性度が増加した分の乳酸発酵が可能となり、コスト削減につながる。
【発明を実施するための最良の形態】
【0010】
本明細書の全体にわたり、単数形の表現は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。また、本明細書において使用される用語は、特に言及しない限り、当該分野で通常用いられる意味で用いられることが理解されるべきである。
【0011】
(用語)
以下に本明細書において特に使用される用語の定義を列挙する。
【0012】
本明細書において「生物」とは、当該分野における最も広義に用いられ、生命現象を営むものをいい、代表的には、細胞構造、増殖(自己再生産)、成長、調節性、物質代謝、修復能力など種々の特性を有し、通常、核酸のつかさどる遺伝と、タンパク質のつかさどる代謝の関与する増殖を基本的な属性として有する。生物には、ウイルス、原核生物、真核生物(酵母のような単細胞生物、植物、動物のような多細胞生物など)などが包含される。本発明の方法は、グラム陽性細菌、および真核生物などの高等生物を含む、どのような生物であっても適用され得ることが理解される。本明細書では、サイズの小さな生物を「微生物」とも呼ぶが、乳酸耐性という点では同一であるので、本明細書では、生物と微生物とは交換可能に使用され得ることが理解される。
【0013】
本明細書において「真核生物」とは、当該分野において通常用いられる意味と同様に用いられ、核膜のある、明確な核構造を持つ細胞からなる生物をいう。真核生物としては、例えば、酵母のような単細胞生物、イネ、コムギ、トウモロコシ、ダイズのような植物、マウス、ラット、ウシ、ウマ、ブタ、サルのような動物、ハエ、カイコなどの昆虫が挙げられるがそれらに限定されない。本明細書では、酵母、線虫、ショウジョウバエ、カイコ、イネ、コムギ、ダイズ、トウモロコシ、シロイヌナズナ、ヒト、マウス、ラット、ウシ、ウマ、ブタ、カエル、魚類(例えば、ゼブラフィッシュ)などがモデルとして使用され得るがそれらに限定されない。
【0014】
本明細書において「原核生物」とは、当該分野において通常用いられる意味と同様に用いられ、明確な核構造を持たない細胞からなる生物をいう。原核生物としては、例えば、大腸菌、サルモネラ菌のようなグラム陰性細菌、枯草菌、放線菌、ブドウ球菌のようなグラム陽性細菌、藍藻類、水素細菌などが挙げられるがそれらに限定されない。本明細書では、代表的に、大腸菌以外に、グラム陽性細菌、として使用され得るがそれに限定されない。
【0015】
本明細書において「動物」は、当該分野において最も広義で用いられ、脊椎動物および無脊椎動物(例えば、節足動物)を含む。動物としては、哺乳綱、鳥綱、爬虫綱、両生綱、魚綱、昆虫綱、蠕虫綱などが挙げられるがそれらに限定されない。動物は、好ましくは、脊椎動物(例えば、メクラウナギ類、ヤツメウナギ類、軟骨魚類、硬骨魚類、両生類、爬虫類、鳥類、哺乳動物など)であり得るが、それらに限定されない。ある一つの実施形態では、動物は、哺乳動物(例えば、単孔類、有袋類、貧歯類、皮翼類、翼手類、食肉類、食虫類、長鼻類、奇蹄類、偶蹄類、管歯類、有鱗類、海牛類、クジラ目、霊長類、齧歯類、ウサギ目など)であり得るがそれらに限定されない。さらに好ましくは、霊長類(たとえば、チンパンジー、ニホンザル、ヒト)または他のモデル動物となり得る種(例えば、奇蹄類、偶蹄類、マウスなどの齧歯類、ウサギ目など)であり得るがそれらに限定されない。本発明の方法は、生物であればどのようなものであっても適用できることが本発明において初めて解明されたことから、どのような生物であっても、対象とされ得ることが理解されるべきである。
【0016】
本明細書において用いられる「植物」とは、植物界に属する生物の総称であり、クロロフィル、かたい細胞壁、豊富な永続性の胚的組織の存在,および運動する能力がない生物により特徴付けられる。代表的には、植物は、細胞壁の形成・クロロフィルによる同化作用をもつ顕花植物をいう。「植物」は、単子葉植物および双子葉植物のいずれも含む。好ましい植物としては、有用植物、例えば、コムギ、トウモロコシ、イネ、オオムギ、ソルガムなどのイネ科に属する単子葉植物が挙げられるがそれらに限定されない。好ましい植物のほかの例としては、タバコ、ピーマン、ナス、メロン、トマト、サツマイモ、キャベツ、ネギ、ブロッコリー、ニンジン、キウリ、柑橘類、白菜、レタス、モモ、ジャガイモおよびリンゴが挙げられる。好ましい植物は作物に限られず、花、樹木、芝生、雑草なども含まれる。特に他で示さない限り、植物は、植物体、植物器官、植物組織、植物細胞、および種子のいずれをも意味する。植物器官の例としては、根、葉、茎、および花などが挙げられる。植物細胞の例としては、カルスおよび懸濁培養細胞が挙げられる。本発明の方法は、生物であればどのようなものであっても適用できることが本発明において初めて解明されたことから、どのような生物であっても、対象とされ得ることが理解されるべきである。
【0017】
本明細書において、「遺伝子」とは、細胞中に存在する核酸の一定の長さの配列をいう。本発明において遺伝子は、遺伝形質を規定するものであっても規定しないものであってもよい。本明細書において、遺伝子は、通常ゲノムに存在するものをさすが、それに限定されず、染色体外の配列、ミトコンドリアの配列なども包含することが理解される。多くの遺伝子は、通常染色体上に一定の順序に配列している。タンパク質の一次構造を規定するものを構造遺伝子といい、その発現を左右するものを調節遺伝子(たとえば、プロモーター)という。本明細書では、遺伝子は、特に言及しない限り、構造遺伝子および調節遺伝子を包含する。したがって、例えば、DNAポリメラーゼ遺伝子というときは、通常、DNAポリメラーゼの構造遺伝子ならびにDNAポリメラーゼのプロモーターなどの転写および/または翻訳の調節配列の両方を包含する。本発明では、構造遺伝子のほか、転写および/または翻訳などの調節配列もまた、本発明が対象とする遺伝子として有用であることが理解される。本明細書では、「遺伝子」は、「ポリヌクレオチド」、「オリゴヌクレオチド」、「核酸」および「核酸分子」ならびに/または「タンパク質」、「ポリペプチド」、「オリゴペプチド」および「ペプチド」を指すことがある。本明細書においてはまた、「遺伝子産物」は、遺伝子によって発現された「ポリヌクレオチド」、「オリゴヌクレオチド」、「核酸」および「核酸分子」ならびに/または「タンパク質」「ポリペプチド」、「オリゴペプチド」および「ペプチド」を包含する。当業者であれば、遺伝子産物が何たるかはその状況に応じて理解することができる。
【0018】
本明細書において「遺伝子の複製」とは、遺伝物質であるDNAまたはRNAにおいて,親の核酸鎖を鋳型として,親核酸と同一の構造と機能を有する新しい核酸分子(DNAであればDNA、RNAであればRNA)を生成することをいう。真核細胞では,二本鎖DNA分子上にある多数の複製開始点に複製酵素(DNAポリメラーゼα)を含む複製開始複合体が形成されることによって複製が開始され、複製反応が複製開始点から両方向へ進行する。複製開始は、細胞周期によって制御される。酵母における自立複製配列(ARS=autonomously replicating sequence)は複製開始点とされる。大腸菌などの原核細胞では、ゲノムの二本鎖環状DNA分子に1個の複製開始点(ori)が存在し、oriに複製開始複合体が形成され、反応はoriから両方向に進行する。複製開始複合体は複製酵素(DNAポリメラーゼIII)を含む10種類以上のタンパク質因子を含む複雑な構造を有する。複製反応は、二本鎖DNAのらせん構造の部分的巻戻しに始まり、その後短いDNAプライマーが合成され、3’−OH基から新たなDNA鎖が伸長され、相補鎖鋳型において岡崎フラグメントが合成され、岡崎フラグメントの連結、鋳型と照らし合わせる校正(プルーフリーティング)などの多段階の反応によって複製反応が進行する。
【0019】
生物の遺伝情報であるゲノムDNAの複製機構は、Kornberg A.and Baker T.,“DNA Replication”,New York,Freeman,1992に詳述されている。代表的には、1つの1本鎖DNAを鋳型として相補的な鎖を合成し、その結果、1つの2本鎖DNAを生じる酵素(DNA複製を行う酵素)を、DNAポリメラーゼという。DNA複製には、少なくとも2つのDNAポリメラーゼが必要である。通常、リーディング鎖およびラギング鎖の合成を同時に行わなければならないからである。DNA複製は、DNA上の決まった位置から開始されるが、その位置を複製開始点(ori)という。例えば、細菌では、通常、環状ゲノムDNAに少なくとも1つの両方向性の複製開始点を有する。これらを総合すると、通常は、1つのゲノムDNA複製には、4つのDNAポリメラーゼが同時に作用する必要があることになる。本発明では、好ましくは、リーディング鎖およびラギング鎖の一方のみの複製誤りが調節されることが有利であり得、あるいは、二本の鎖のあいだの複製誤りの頻度が異なることが有利であり得る。
【0020】
本明細書において「複製誤り」とは、遺伝子(DNAなど)の複製の過程で生じるヌクレオチド取り込みの誤りをいう。複製誤りは、通常、生体では、その頻度は108〜1012回に1回程度できわめて低い。複製誤りの頻度が低い理由としては、ヌクレオチドの取り込みが鋳型DNAと取り込まれるヌクレオチドとが相補的な塩基対を形成することによって複製が起こること、DNAポリメラーゼδ、εなどの酵素の校正機能すなわち3’→5’エキソヌクレアーゼが,鋳型に相補性を示さないヌクレオチドが誤って取り込まれたときそれを察知し直ちに切り出す機能が存在することなどが挙げられる。従って、本発明において複製におけるエラープローン頻度の調節は、特異的塩基対形成の障害、校正機能の障害などによって行うことができる。
【0021】
本明細書において「エラーフリー」とは、遺伝子(DNAなど)の複製にいて誤りがほとんどない、好ましくは実質的に全くない性質をいう。エラーフリーは、主に、校正機能を有する酵素(例えば、DNAポリメラーゼδ、εなど)の校正機能の精度によって影響を受ける。
【0022】
本明細書において「エラープローン」とは、遺伝子(DNAなど)の複製における誤り易い(すなわち、複製誤りの)性質をいう。エラープローンは、主に、校正機能を有する酵素(例えば、DNAポリメラーゼδ、εなど)の校正機能の精度によって影響を受ける。
【0023】
本明細書においてエラープローンとエラーフリーとは、絶対的に(すなわち、エラープローン頻度のレベルなどで決定する)分類することができ、あるいは相対的(2種類以上の遺伝子の複製を担う因子(例えば、DNAポリメラーゼなど)におけるエラープローン頻度について多いほうをエラープローンとし、少ないほうをエラーフリーとする)に分類することができる。
【0024】
本明細書において「エラープローン頻度」とは、エラープローンの性質のレベルをいう。エラープローン頻度は、例えば、遺伝子配列における変異の絶対数(変異の数そのもの)または相対数(全長における変異の数の比率)で表現することができる。あるいは、ある生物または酵素について言及するとき、エラープローン頻度は、ある生物の生殖または分裂1回あたりの遺伝子配列における変異の絶対数または相対数で表現してもよい。特に言及しない場合、遺伝子配列における複製過程1回あたりの誤差の数で表される。エラープローン頻度は、逆の尺度として本明細書において「精度」ということがある。エラープローン頻度が均一であるとは、複数の遺伝子の複製を担う因子(ポリメラーゼなど)に言及するとき、互いのエラープローン頻度が実質的に等しいことをいう。他方、エラープローン頻度が不均一であるとは、有意な差異が複数の遺伝子の複製を担う因子(ポリメラーゼなど)に存在する場合をいう。
【0025】
本明細書において「エラープローン頻度の調節」とは、エラープローン頻度を変化させることをいう。そのようなエラープローン頻度の調節には、エラープローン頻度の上昇および低減が含まれる。エラープローン頻度の調節のための手法としては、例えば、校正機能を有するDNAポリメラーゼの改変、複製中に重合反応または伸長反応を阻害または抑制するような因子の挿入これらの反応を促進するような因子の阻害、抑制、単数または複数の塩基の欠損、DNA修復酵素の欠損、異常塩基の除去修復因子機能を有する酵素の改変、ミスマッチ塩基対修復因子の改変、複製自体の精度の低減などが挙げられるがそれらに限定されない。エラープローン頻度の調節は、DNAの二本鎖の両方に対して行われてもよいし、片方のみに対して行われてもよい。好ましくは、片方のみに対して行われることが有利であり得る。有害な変異誘発が低減されるからである。
【0026】
本明細書において「DNAポリメラーゼ」またはPolとは、4種類のデオキシリボヌクレオシド5’−三リン酸からピロリン酸を遊離してDNAを重合する働きを有する酵素をいう。DNAポリメラーゼ反応には、鋳型となるDNA、プライマー分子、Mg2+などが必要とされる。プライマーの3’−OH末端に鋳型に相補的なヌクレオチドを順次付加し分子鎖を伸長する。
【0027】
大腸菌にはDNAポリメラーゼI、II、IIIの少なくとも3種類の酵素が知られている。DNAポリメラーゼIはDNA傷害の修復,遺伝的組換えおよびDNA複製に関与する。DNAポリメラーゼIIおよびIIIは補助的機能を有するといわれる。この酵素は数種のタンパク質からなるサブユニット構造をとり,その構成からコア酵素およびホロ酵素の2つに分けられる。コア酵素は,α、εおよびθから構成され、ホロ酵素には、α、εおよびθサブユニットのほかに、τ、γ、δおよびβの成分がある。真核生物細胞も複数のDNAポリメラーゼをもつことが知られており、高等生物ではDNAポリメラーゼα、β、γ、δおよびεなど多種類存在する。動物では、DNAポリメラーゼα(核DNAの複製に関与、細胞増殖期のDNA複製に作用)、DNAポリメラーゼβ(核でのDNAの修復に関与、増殖期、停止期のDNA傷害の修復などの作用を有する)、DNAポリメラーゼγ(ミトコンドリアDNAの複製と修復に関与。エキソヌクレアーゼ活性をもつ)、DNAポリメラーゼδ(DNAの伸長に関与。エキソヌクレアーゼ活性をもつ)、DNAポリメラーゼε(ラギング鎖のすき間の複製に関与。エキソヌクレアーゼをもつ)などが知られている。
【0028】
校正機能を担うDNAポリメラーゼ(グラム陽性細菌、グラム陰性細菌、真核生物など)では、ExoI用のアミノ酸配列が3’→5’エキソヌクレアーゼ活性中心を担うとされており、この部位が校正機能の精度に影響すると考えられている。
【0029】
配列番号1:Pol III:317−IMSFDIECAGRI−328(Saccharomyces cerevisiae);
配列番号2:Pol II:286−VMAFDIETTKPP−297(Saccharomyces cerevisiae);
変異型pol−3遺伝子発現株(AMY128−1、以下pol−3変異株と略す)は、この配列を有しているものと推定される。AMY128−1の代表例は、独立行政法人製品評価技術基盤機構 特許生物寄託センター(〒292−0818 千葉県木更津市かずさ鎌足2−5−8)に受託番号NITE P−130(変異型pol−3遺伝子発現株(1倍体株)(AMY128−1))(寄託日平成17年8月19日)として寄託済みである。また、変異型pol−3遺伝子発現株(2倍体株)(BYD5)は、受託番号 NITE P−131(寄託日平成17年8月19日)として寄託済みである。野生型pol−3遺伝子発現株(1倍体株)(AMY52−3D)は、受託番号 NITE P−129(寄託日平成17年8月19日)として寄託済みである。
【0030】
ここで明らかなように、校正機能を有するDNAポリメラーゼは、アスパラギン酸およびグルタミン酸がよく保存されている。本明細書において、このようなアスパラギン酸およびグルタミン酸を含む領域は、本明細書において校正機能活性部位ということがある。
【0031】
1つの好ましい実施形態では、このような3’→5’エキソヌクレアーゼ活性が破壊されるような変異を、DNAポリメラーゼをコードする遺伝子(DNAポリメラーゼ遺伝子)に導入することにより、校正機能が低下した(すなわち、エラープローン頻度が増加した)DNAポリメラーゼをコードする核酸分子およびポリペプチドを産生することができる。なお、校正機能の3’→5’エキソヌクレアーゼ活性は、単一のDNAポリメラーゼ遺伝子(PolC,POL2,CDC2等)において、DNA重合活性を担う分子中に含まれる場合(例えば、真核生物、グラム陽性細菌等)と、DNA重合活性をコードする遺伝子(例えば、dnaE)とは異なる遺伝子(例えば、dnaQ)にコードされる場合(例えば、グラム陰性細菌等)があることが知られており(Kornberg A.and Baker T.,“DNA Replication ”,New York,Freeman,1992)、当業者は、それらの特性を理解したうえで、本発明におけるエラープローン頻度の調節を適宜行うことができる。例えば、真核生物などでは、校正機能は変化するが、DNA重合活性はほとんど変化させない変異をDNAポリメラーゼに導入することが好ましい。この場合、上述のような校正機能に関与する2つの酸性アミノ酸(Derbyshire et.al.,EMBO J.10,pp.17−24,Jan.1991; Fijalkowska and Schaaper,“Mutants in the Exo I motif of Escherichia coli dnaQ:Defective proofreading and inviability due to error catastrophe”,Proc.Natl.Acad.Sci.U.S.A.Vol.93,pp.2856−2861,Apr.1996)を改変すること(好ましくは、保存されない置換(例えば、アラニン、バリンなどへの置換))が挙げられるがそれらに限定されない。
【0032】
本明細書において「校正機能」とは、細胞が受けたDNA損傷および/または誤りを検知し補修する機能をいう。そのような機能は、脱プリン、脱ピリミジンがある場合はそのまま塩基が挿入されることによるか、あるいは、A−Pエンドヌクレアーゼ(apurinic−apyrimidinic endonuclease)で一本鎖切断が入れられたのち5’→3’エキソヌクレアーゼで除去されることによって達成され得る。除去部分は、DNAポリメラーゼでDNA合成され補充され、正常DNAとの連結はDNAリガーゼが行う。このような反応は、除去修復といわれる。アルキル化剤による化学修飾,異常塩基,放射線,紫外線などによるDNA塩基傷害などは傷害部分をDNAグリコシダーゼで取り除かれた上で上記の反応で修復される(不定期DNA合成)。そのような校正機能を有するDNAポリメラーゼとしては、例えば、真核生物におけるDNAポリメラーゼδ、DNAポリメラーゼεなどが挙げられるがそれらに限定されない。本明細書では、校正機能の程度を表すために、忠実度という用語もまた用いられ得る。この忠実度との用語は、DNA複製の正確さを意味する。正常なDNAポリメラーゼは、通常、忠実度が高いDNAポリメラーゼであり、改変により校正機能が低下したDNAポリメラーゼは、忠実度が低いDNAポリメラーゼであり得る。
【0033】
このようなDNAポリメラーゼの校正機能については、例えば、Kunkel,T.A.:J Biol.Chem.,260,12866−12874(1985);Kunkel,T.A.,Sabotino,R.D.& Bambara,R.A.:Proc.Natl.Acad.Sci.USA,84,4865−4869(1987);Wu,C.I.& Maeda,N.:Nature,327,167−170(1987);Roberts,J.D.& Kunkel,T.Al:Proc.Natl.Acad.Sci.USA,85,7064−7068(1988);Thomas,D.C.,Fitzgerald,M.P.& Kunkel,T.A.:Basic Life Sciences,52,287−297(1990);Trinh,T.Q.& Siden,R.R.Nature,352,544−547(1991);Weston−Hafer,K.,& Berg,D.E.Genetics,127,649−655(1991);Veaute,X.& Fuchs,R.p.p.:Science,261,598−600(1993);Roberts,J.D.,Izuta,S.,Thomas,D.C.& Kunkel,T.A.:J Biol Chem.,269,1711−1717(1994);Roche,W.A.,Trinh,T.Q.& Siden,R.R.J.Bacteriol.,177,4385−4391(1995);Kang,S.,Jaworski,A.,Ohshirna,K.& Wells,Nat.Genet.,10,213−218(1995);Fijalkowska,I.J.,Jonczyk,P.,Maliszewska−Tkaczyk,M.,Bialoskorska,M.& Schaaper,R.M.Proc.NatJ.Acad.Sci.USA.95,10020−10025(1998);Maliszewska−Tkaczyk,M.,Jonezyk,P.,Bialoskorska,M.,Schaaper,M.& Fijalkowska,I.:Proc.Natl.Acad.Sci.USA,97,12678− 12683(2000);Gwel,D.,Jonezyk,P.,Bialoskorska,M.,Schaaper,R.M.& Fijalkowska,I.J.:Mutation Research,501,129−136(2002).Negative;Roberts,J.D.,Thomas,D.C.& Kunkel,T.A.:Proc Nat].Acad Sci.USA,88,3465−3469(1991);Roberts,J.D.,Nguyen,D.& Kunkel,T.A.:Biochemistry;32,4083−4089(1993);Francino,M.P.,Chac,L.,Riley,M.A.& Ochman,H.:Science,272,107−109(1996);A Boulet,M.Simon,G.Faye,GA Bauer & PM Burgers.EMBO J,8,1849−1854,(1989);Morrison A,Araki H,Clark AB,Hamatake RK,& Sugino A.Cell,62(6),1143−1151,(1990)などを参照のこと。
【0034】
本明細書において「DNAポリメラーゼδ」とは、真核生物のものをさす場合、DNAの伸長に関与する酵素であって、エキソヌクレアーゼ活性をもち、これに起因して校正機能を有するといわれる。このDNAポリメラーゼδの校正機能の調節は、上記の校正機能に関連するアミノ酸配列に改変を導入することによって達成することができる。代表的なDNAポリメラーゼδは、核酸配列およびアミノ酸配列として、polδ:X61920 gi/171411/gb/M61710.1/YSCDPB2[171411]に記載されている配列を有する。DNAポリメラーゼδは、Simon,M.et al.、EMBO J.10,2163−2170,1991に記載されており、その内容は本明細書において参考として援用される。
【0035】
本明細書において「DNAポリメラーゼε」とは、真核生物のものをさす場合、ラギング鎖のすき間の複製に関与する酵素であって、エキソヌクレアーゼ活性をもち、これに起因して校正機能を有するといわれる。代表的なDNAポリメラーゼεは、核酸配列およびアミノ酸配列として、polε:M60416 gi/171408/gb/M60416.1/YSCDNAPOL[171408]に記載されている配列を有する。
【0036】
DNAポリメラーゼδおよびεの分類は、HUGO分類によると別名はデルタがPOLD1/POL3,イプシロンがPOLE/POL2となっており、本明細書では、どの命名法をも用い得る。
【0037】
その他DNAポリメラーゼの説明については、例えば、Lawrence C.W.et al.,J.Mol.Biol.122,1−21,1978、Lawrence C.W.et al.,Genetics 92,397−408、Lawrence C.W.et al.MGG,195,487−490,1984、Lawrence C.W.et al.MGG.200,86−91,1985(DNAポリメラーゼβおよびDNAポリメラーゼζ);Maher V.M.et al.Nature 261,593−595,1976、McGregor,W.G.et al.,Mol.Cell.Biol.19,147−154、1999(DNAポリメラーゼη);Strand M.et al.,Nature 365,275−276,1993、Prolla T.A.,et al.,Mol.Cell.Biol.15,407−415,1994、Kat A.,et al.,Proc.Natl.Acad.Sci.USA 90,6424−6428、Bhattacharyya N.P.,et al.,Proc.Natl.Acad.Sci.USA 91,6319−6323,1994、Faber F.A.,et al.,Hum.Mol.Genet.3,253−256,1994、Eshleman,J.R.,et al.,Oncogene 10,33−37,1995、Morrison A.,et al.,Proc.Natl.Acad.Sci.USA 88,9473−9477,1991、Morrison A.,et al.,EMBO J.12,1467−1473,1993、Foury F.,et al.,EMBO J.11,2717−2726,1992(DNAポリメラーゼλ、DNAポリメラーゼμなど)などに記載されており、その内容は本明細書において参考として援用される。
【0038】
本明細書においてDNAポリメラーゼなどの遺伝子および酵母などの生物の「野生型」は、もっとも広汎な定義では、天然に存在するDNAポリメラーゼなどの遺伝子および酵母などの生物を含み、通常、天然に存在するDNAポリメラーゼなどの遺伝子および酵母などの生物のうち、由来となる生物種においてもっとも広汎に存在するものをいう。従って、通常、ある種において最初に同定されるDNAポリメラーゼなどの遺伝子および酵母などの生物の種は野生型といえる。野生型はまた、「天然標準型」ともいう。生物であれば、野生型は、酵素活性が正常であり得、形質が正常であり得、行動が正常であり得、生理が正常であり得、繁殖が正常であり得、ゲノムが正常であり得る。
【0039】
本明細書において校正機能が「野生型のものよりも低い」とは、ある校正機能を有する酵素などについて言及するとき、その酵素の野生型よりも校正機能が低いこと(すなわち、その酵素での校正処理の後に残留する変異の数が野生型による校正処理の後に残留する変異の数よりも多いこと)をいう。そのような野生型との比較は、相対的または絶対的な表示によって行うことができる。そのような比較はまた、エラープローン頻度などによって行うことができる。
【0040】
本明細書において「変異」とは、遺伝子について言及するとき、その遺伝子の配列の変化を生じることまたはその変化によって生じた遺伝子の(核酸またはアミノ酸)配列の状態をいう。本明細書では、例えば、変異は、校正機能について生じる遺伝子配列の変化について用いられる。本明細書では、特に言及しない場合は、変異は、改変と同義で用いられる。
【0041】
有用な変異体を作製するためには、生物において変異誘発を行うことがもっとも一般的である。変異とは、通常、遺伝子をコードする塩基配列の変化をいい、DNA配列の変化が包含される。変異は、それが発生した個体に与える影響により、大きく次の3種類に分けられる:A)中立変異(neutral mutation):この変異は、ほとんどの変異が該当し、生物の成育および代謝にほとんど影響がない。B)有害変異(deleterious mutation):この変異は、中立変異よりは頻度は少ない。生物の成長または代謝を阻害する。有害変異には、生育に必須な遺伝子を破壊するような致死変異(lethal mutation)も含まれる。微生物の場合、種によっても異なるが、通常全変異に占める有害変異の割合は、約1/10〜1/100とされている。C)有益変異(beneficial mutation):この変異は、生物の育種に有益な変異である。その発生頻度は中立変異と比較して極めて低い。したがって、有益変異が導入された生物個体を得るためには、大きな生物集団と、長い時間が必要となる。また、生物の育種の十分な効果は、単一の変異だけで現れることはまれであり、複数の有益変異の蓄積が必要であることが多い。従って、本発明は、乳酸耐性のみを付与する技術としては初めて提供されたことになる。
【0042】
本明細書において「成長(または生長)」とは、ある生物について言及するとき、その生物の個体としての量的増大をいう。成長は、具体的には体長(身長)、体重などの計測値の増加で個体の量的増大を認識することができる。個体の量的増大は細胞の増大および細胞数の増加に依存する。
【0043】
本明細書において「実質的に同じ成長(または生長)」とは、生物について言及するとき、その生物が、比較対象となる生物(例えば、遺伝形質の変換前の生物)と比較して、成長速度がほとんど変化しないことをいう。そのような成長速度がほとんど変化しない範囲には、例えば、通常の成長の統計分布における1偏差分以内に入ることなどが挙げられるがそれに限定されない。また、本発明の生物では、例えば、(1)子供の数が変わらない;(2)形態は変化するが、通常の人為的突然変異と違って、障害的ではなく、変異率が極めて高いにも拘わらず、見た目が“美しい”と予想される。(成長とは直接関係しないが、本発明の方法により創出された変異体の特徴だと考えられる);(3)一度獲得した形質あるいは遺伝型あるいは表現型は後もどりしないなどの効果が得られる。
【0044】
本明細書において生物の「生産」とは、ある生物について言及するとき、その生物の個体を作り出すことをいう。
【0045】
本明細書において生物の「再生産」とは、ある生物について言及するとき、親個体から次の世代の新たな個体を作り出すことをいう。再生産には、生殖、繁殖など天然現象によるもの、クローン(核移植)技術などの人工技術などによるものが包含されるがそれらに限定されない。再生産に用いられる技術としては、例えば、植物の場合、1培養細胞から個体ができる;接木、挿し木などが挙げられるがそれらに限定されない。再生産によって生産された生物は、通常、親に由来する遺伝形質を有する。有性生殖によって再生産される生物では、再生産された生物は、通常2つの性にそれぞれ由来する遺伝形質を有する。通常、そのような由来遺伝形質は、2つの性に由来するものをほぼ等しい割合で有する。無性生殖によって再生される生物では、通常再生産された生物は、親に由来する遺伝形質を有する。
【0046】
本明細書において使用される「細胞」は、当該分野において用いられる最も広義の意味と同様に定義され、多細胞生物の組織の構成単位であって、外界を隔離する膜構造に包まれ、内部に自己再生能を備え、遺伝情報およびその発現機構を有する生命体をいう。本明細書において使用される細胞は、天然に存在する細胞であっても、人工的に改変された細胞(例えば、融合細胞、遺伝子改変細胞)であってもよい。細胞の供給源としては、例えば、単一の細胞培養物であり得、あるいは、正常に成長したトランスジェニック動物の胚、血液、または体組織、または正常に成長した細胞株由来の細胞のような細胞混合物が挙げられるがそれらに限定されない。
【0047】
本発明において使用される細胞は、どの生物由来の細胞(たとえば、任意の種類の単細胞生物(例えば、細菌、酵母)または多細胞生物(例えば、動物(たとえば、脊椎動物、無脊椎動物)、植物(たとえば、単子葉植物、双子葉植物など)など))でもよい。例えば、脊椎動物(たとえば、メクラウナギ類、ヤツメウナギ類、軟骨魚類、硬骨魚類、両生類、爬虫類、鳥類、哺乳動物など)由来の細胞が用いられ、より詳細には、哺乳動物(例えば、単孔類、有袋類、貧歯類、皮翼類、翼手類、食肉類、食虫類、長鼻類、奇蹄類、偶蹄類、管歯類、有鱗類、海牛類、クジラ目、霊長類、齧歯類、ウサギ目など)由来の細胞が用いられる。1つの実施形態では、霊長類(たとえば、チンパンジー、ニホンザル、ヒト)由来の細胞、特にヒト由来の細胞が用いられるがそれに限定されない。本発明において用いられる細胞は、上記細胞は、幹細胞であってもよく体細胞であってもよい。そのような細胞は、移植目的に使用されるものであってもよい。あるいは、植物細胞としては、好ましくは、顕花植物(単子葉または双子葉)由来の細胞が用いられ、より好ましくは双子葉植物細胞が用いられ、より好ましくはイネ科、ナス科、ウリ科、アブラナ科、セリ科、バラ科、マメ科、ムラサキ科の植物由来の細胞が用いられる。さらに好ましくは、コムギ、トウモロコシ、イネ、オオムギ、ソルガム、タバコ、ピーマン、ナス、メロン、トマト、イチゴ、サツマイモ、アブラナ、キャベツ、ネギ、ブロッコリー、ダイズ、アルファルファ、アマ、ニンジン、キウリ、柑橘類、ハクサイ、レタス、モモ、ジャガイモ、ムラサキ、オウレン、ポプラおよびリンゴ由来の細胞が用いられる。植物細胞は、植物体の一部、器官、組織、培養細胞などであり得る。細胞、組織、器官または個体の形質転換法は、当該分野で周知である。そのような技術は、本発明において引用した文献などに十分記載されている。核酸分子の生物細胞への導入は、一過的であっても恒常的であってもよい。一過性または恒常性の遺伝子導入の技術はそれぞれ当該分野において周知である。本発明において用いられる細胞を分化させて形質転換植物を作出する技術もまた当該分野において周知であり、そのような技術は、本発明において引用した文献などに十分記載されていることが理解される。形質転換植物から種子を得る技術もまた、当該分野において周知であり、そのような技術は、本発明において引用した文献などに記載されている。
【0048】
本明細書において「単離された」とは、通常の環境において天然に付随する物質が少なくとも低減されていること、好ましくは実質的に含まないをいう。従って、単離された細胞とは、天然の環境において付随する他の物質(たとえば、他の細胞、タンパク質、核酸など)を実質的に含まない細胞をいう。核酸またはポリペプチドについていう場合、「単離された」とは、たとえば、組換えDNA技術により作製された場合には細胞物質または培養培地を実質的に含まず、化学合成された場合には前駆体化学物質またはその他の化学物質を実質的に含まない、核酸またはポリペプチドを指す。単離された核酸は、好ましくは、その核酸が由来する生物において天然に該核酸に隣接している(flanking)配列(即ち、該核酸の5’末端および3’末端に位置する配列)を含まない。
【0049】
本明細書において、「樹立された」または「確立された」細胞とは、特定の性質(例えば、多分化能)を維持し、かつ、細胞が培養条件下で安定に増殖し続けるようになった状態をいう。
【0050】
本明細書において「乳酸」とは、α-ヒドロキシプロピオン酸のことであり、L型は、解糖の最終生成物であり,乳酸脱水素酵素の作用によりピルビン酸の還元によって生成される。
【0051】
本明細書において「乳酸耐性」とは、乳酸に対して耐性を有することをいう。そのような耐性とは、微量の乳酸の存在に対して、死滅しないことを少なくとも意味するが、より好ましくは、乳酸の存在下で成長することを意味し得る。
【0052】
本明細書において使用され得る、乳酸としては、L型、D型、ラセミ体など、自然界に存在し得る任意の形態を挙げることができるが、好ましくはL型である。これらはそれぞれ単独で用いてもよく、2種以上を併用してもよい。
【0053】
乳酸耐性は以下のようにして決定する。乳酸耐性度を評価したい微生物を、乳酸を含まない培地で培養し、一晩培養液を準備する。各種濃度の乳酸を含む培地を準備し、一晩培養液を0.1〜10%(v/v)添加し、一晩培養する。加える菌の体積は目的や菌の種類によって決める。一晩培養後の菌濃度を吸光光度計をもいる方法、寒天プレート法、トリパンブルー染色による生存率測定などで決定する。中性付近の培地を用いた場合と同等の増殖性が認められた乳酸濃度の数値を、その濃度の乳酸耐性を持つと評価する。
【0054】
本発明の方法において、微生物に乳酸を加えて選択培養する工程は、寒天培地上に変異処理した乳酸感受性微生物を塗布した上に乳酸を目的の濃度比率で重層しても良く又は液体培養において変異処理した乳酸感受性菌及び目的乳酸を一緒に入れて培養しても良い。また、pH、温度及び通気の割合(通気する場合)等の条件も当業者によって容易に設定され得るが、好ましくは、その微生物が成育する条件が良いことは言うまでもない。
【0055】
(一般生化学・分子生物学)
(一般技術)
本明細書において用いられる分子生物学的手法、生化学的手法、微生物学的手法は、当該分野において周知であり慣用されるものであり、例えば、Sambrook J.et al.(1989).Molecular Cloning:A Laboratory Manual,Cold Spring Harborおよびその3rd Ed.(2001);Ausubel,F.M.(1987).Current Protocols in Molecular Biology,Greene Pub.Associates and Wiley−Interscience;Ausubel,F.M.(1989).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associat ES and Wiley−Interscience;Innis,M.A.(1990).PCR Protocols:A Guide to Methods and Applications,Academic Press;Ausubel,F.M.(1992).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates;Ausubel,F.M.(1995).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates;Innis,M.A.et al.(1995).PCR Strategies,Academic Press;Ausubel,F.M.(1999).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Wiley,and annual updates;Sninsky,J.J.et al.(1999).PCR Applications:Protocols for Functional Genomics,Academic Press、別冊実験医学「遺伝子導入&発現解析実験法」羊土社、1997などに記載されており、これらは本明細書において関連する部分(全部であり得る)が参考として援用される。
【0056】
人工的に合成した遺伝子を作製するためのDNA合成技術および核酸化学については、例えば、Gait,M.J.(1985).Oligonucleotide Synthesis:A Practical Approach,IRLPress;Gait,M.J.(1990).Oligonucleotide Synthesis:A Practical Approach,IRL Press;Eckstein,F.(1991).Oligonucleotides and Analogues:A Practical Approach,IRL Press;Adams,R.L.etal.(1992).The Biochemistry of the Nucleic Acids,Chapman&Hall;Shabarova,Z.et al.(1994).Advanced Organic Chemistry of Nucleic Acids,Weinheim;Blackburn,G.M.et al.(1996).Nucleic Acids in Chemistry and Biology,Oxford University Press;Hermanson,G.T.(I996).Bioconjugate Techniques,Academic Pressなどに記載されており、これらは本明細書において関連する部分が参考として援用される。
【0057】
以下に好ましい実施形態の説明を記載するが、この実施形態は本発明の例示であり、本発明の範囲はそのような好ましい実施形態に限定されないことが理解されるべきである。
【0058】
1つの局面において、本発明は、乳酸耐性を有する生物であって、該生物は、該生物の天然型が有する耐性よりも高い耐性を有する、生物を提供する。
【0059】
1つの実施形態において、本発明において対象となる乳酸は、L型乳酸、D型乳酸、L型・D型乳酸のラセミ体からなる群より選択されるタイプであり得る。
【0060】
1つの実施形態において、本発明は、乳酸に対して少なくとも1%の濃度の耐性を有する生物を提供する。これまで、乳酸に対して多くとも1%未満の濃度までしか耐性がある生物がほとんど存在していなかった。特に乳酸菌以外では、1%未満程度の耐性しか報告されておらず、特に2%以上の耐性を持つ生物を生産することは不可能と考えられていた。また、交叉によっても乳酸耐性が得られたとの報告は皆無である。従って、本発明は、従来存在しなかった生物を提供するという顕著な効果を奏する。好ましい実施形態では、本発明は、乳酸に対して少なくとも2%の、好ましくは、少なくとも3%の、より好ましくは少なくとも4%の、より好ましくは少なくとも5%の、さらに好ましくは少なくとも6%の、さらに好ましくは少なくとも7%の、さらに好ましくは少なくとも8%の、さらに好ましくは少なくとも9%の、さらにより好ましくは少なくとも10%の、さらに好ましくは少なくとも15%の、最も好ましくは少なくとも20%の濃度の耐性を有する生物を提供する。ここで、乳酸に対する耐性は、その耐性とすべき環境中に少なくとも18時間、通常、少なくとも1日間、好ましくは2日間、好ましくは3日間、好ましくは4日間、好ましくは4日間、さらに好ましくは6日間生存している(少なくとも生存率が10%以上であり、好ましくは少なくとも生存率が20%である。本発明は、生存率は、概して80%以上であり、優れた生存率を示す。)ことを意味する。
【0061】
別の実施形態において、本発明の生物は、任意の微生物であり、さらに好ましくは、真核生物であり、さらにより好ましくは酵母である。特に、Saccharomyces俗の生物が好ましく、さらに好ましくはSaccharomyces cervisiaeが選択される。別の実施形態では、本発明の生物は、原核生物であり得る。酵母については、乳酸1%でも耐性が獲得できていなかった。
【0062】
本発明で得られた株の代表例は、〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに受領番号NITE P−122号(寄託日平成17年7月29日;7%乳酸を含むYPD培地中で増殖可能)として寄託済みである。
【0063】
別の局面において、本発明は、乳酸に対する耐性が改善された生物を作製するための方法を提供する。この方法は:A)目的の生物を、該乳酸について、正常状態での最大許容濃度で、好ましくは1日程度培養する工程;B)該目的の生物を、該乳酸について、改善について所望の濃度で培養する工程;およびC)生存する生物を選択し再生産する工程を包含する。このような再生産の方法には、当該分野において公知の技術を応用できることが理解される。一度最大許容濃度よりも低い濃度で培養しておき、その後、所網の最大許容濃度より高い濃度で培養することによって、より効率所望の濃度で耐性を有する生物を効率よく獲得することができたことに特徴がある。より具体的にいうと、本発明の1つの実施形態では、本発明の方法は、A)正常状態での最大許容濃度で1日間培養する工程、B)Aより1%高い乳酸濃度で1日間培養する工程、C)Bより1%高い乳酸濃度で1日間培養する工程、B)〜C)を繰り返して、最終的に耐性を付与したい乳酸濃度で1日間培養することによって、より効率所望の濃度で耐性を有する生物を効率よく獲得することができたことに特徴がある。表3、図4に示されているように、5%乳酸耐性株は同じ乳酸濃度で継代を繰り返すと増殖速度が早くなっていく。培養工程A)と培養工程B)との間にA’)最大許容濃度より1%高い濃度の乳酸中で培養する工程、および必要に応じてA’’)該培養工程A’)における濃度より1%高い濃度の乳酸中で培養する工程を行い、必要に応じてA’’)において1%ずつ高い濃度の乳酸中での培養工程を繰り返して行って所望の濃度での培養を達成することを包含する
好ましい実施形態では、本発明の方法は、D)生物の遺伝子の複製におけるエラープローン頻度を調節する工程をさらに包含する。
【0064】
別の実施形態において、本発明において使用されるエラープローン頻度を調節する工程は、生物のDNAポリメラーゼのエラープローン頻度を調節することを包含することが好ましい。本発明において使用されるDNAポリメラーゼは、真核細胞におけるDNAポリメラーゼα、DNAポリメラーゼβ、DNAポリメラーゼγ、DNAポリメラーゼδおよびDNAポリメラーゼεならびにそれらに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを含む。
【0065】
好ましい実施形態において、本発明において使用されるDNAポリメラーゼは、真核細胞におけるDNAポリメラーゼδおよびDNAポリメラーゼεならびにそれに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを包含する。
【0066】
好ましい実施形態では、本発明において用いられる生物は、真核生物(例えば、動物、植物、真菌または酵母の細胞)であり、より好ましくは酵母である。酵母は、種々の環境因子に対して耐性を獲得しにくいという特徴があり、本発明では、これを乳酸に対する耐性獲得という形で従来不可能であった酵母の生産に結び付けたという点で顕著である。
【0067】
別の実施形態では、本発明において用いられる生物は、分離した細胞の状態で存在する。
【0068】
好ましい実施形態では、本発明において得られる生物は、所望の形質の変換後も、野生型と実質的に同じ成長を示す。古典的な変異誘発方法では、野生型と実施的に同じ成長を示すものはほとんど取れなかった。従って、本発明は、乳酸に対する耐性を持ちながら、かつ、野生型と実施的に同じ成長を呈する(正常型とほぼ同じ)性質を有する生物を提供するという点で従来技術では提供し得なかった生物を提供するという効果を奏する。
【0069】
別の実施形態では、本発明の方法において用いられる所望の濃度は、最大許容濃度よりも高い濃度であることが好ましい。最大許容濃度は、本発明の方法を実施する直前に実際に測定してもよく、あるいは、文献などから既知の数値を入手して使用してもよい。
【0070】
別の局面では、本発明は、本発明の方法によって生産された生物に関する。このような生物は、乳酸に対する耐性が従来技術にも増して増大しており、従来技術では入手できなかった乳酸耐性を有する生物であり得る。
【0071】
別の局面において、本発明は、生物の乳酸耐性能を評価する方法を提供する。この方法は、A)該生物を、該生物が通常生育する条件下で生育させる工程;B)該乳酸存在下または不存在下で該生物を生育させる工程;およびC)該生物が死滅する該乳酸の濃度を決定する工程、を包含する。
【0072】
1つの実施形態において、生物が死滅しているか否かの決定は、その細胞が形を維持している細胞かどうかを判定基準として用いることができる。形を維持しているだけの細胞は、生存しているかどうかが不明であることから、トリパンブルー染色などにより従来はさらなる測定をする必要があったが、本発明において、乳酸中では、形を維持すること自体が生存の重要な指標であることが明らかになった。従って、この形の維持を指標とする評価方法は、従来技術に比して顕著に簡便な判定方法を提供する。
【0073】
ここで、形を維持しているかどうかの判定は、形を維持している(細胞が生存している)と判断する基準は、きれいな球形で細胞の輪郭が明確な場合である。形を維持できていないと判断する基準は、きれいな球形でなく輪郭が薄くしわがよって、細胞内が薄く透けている場合である。
【0074】
別の局面において、本発明は、生物の乳酸耐性能を評価する方法を提供する。この方法は、A)該生物を、該生物が通常生育する条件下で生育させる工程;B)該乳酸存在下または不存在下で該生物を生育させる工程;およびC)該生物が死滅する該乳酸の濃度を決定する工程、を包含する。
【0075】
1つの実施形態において、上記生物が死滅しているか否かの決定は、その細胞が形を維持している細胞かどうかを判定基準として用いてもよい。
【0076】
別の局面では、本発明はまた、菌学的性質は、乳酸耐性以外は野生型と同じである生物を提供する。このような生物は、古典的な変異方法では、入手不可能であった。
【0077】
本明細書において引用された、科学文献、特許、特許出願などの参考文献は、その全体が、各々具体的に記載されたのと同じ程度に本明細書において参考として援用される。
【0078】
以上、本発明を、理解の容易のために好ましい実施形態を示して説明してきた。以下に、実施例に基づいて本発明を説明するが、上述の説明および以下の実施例は、例示の目的のみに提供され、本発明を限定する目的で提供したのではない。従って、本発明の範囲は、本明細書に具体的に記載された実施形態にも実施例にも限定されず、特許請求の範囲によってのみ限定される。
【実施例】
【0079】
以下に実施例を示して本発明をさらに詳しく説明するが、この発明は以下の例に限定されるものではない。以下の実施例において用いられる試薬類、例外を除き、Sigma(St.Louis,USA、和光純薬(大阪、日本)などから市販されるものを用いた。以下において使用した動物は、日本の大学において規定される飼育規準を遵守して飼育および実験した。以下、実施例により本発明をさらに具体的に説明する。
【0080】
(実施例1:乳酸添加による培地pHの変化と細胞増殖への影響の検討)
(酵母株と試薬)
大阪市立大学の下田らから供与された変異型pol−3遺伝子発現株(BYD5、以下pol−3変異株と略す。BYD5は、受託番号 NITE P−131(寄託日平成17年8月19日)として〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに寄託済みである)と野生型pol−3遺伝子発現株(AMY52−3D、以下、野生株と略す。AMY52−3Dは、受託番号 NITE P−129(寄託日平成17年8月19日)として〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに寄託済みである。)を用いた。BYD5の遺伝子型は、〔pol3−01/pol3−01、MATa/MATα、lys1−1/lys1−1〕であり、AMY52−3Dの遺伝子型は、〔MATα、ura3−52、leu2−1、lys1−1、ade2−1、his1−7、hom3−10、trp1−289、canR〕である。
【0081】
試験に用いた液体培地のYPD medium(cat.#630409)および寒天培地のYPD agar medium(cat.#630410)はBD Biosciences Clontechから購入し、オートクレーブ(120℃、15分)により滅菌した。添加したL−乳酸(以下、乳酸と略す。cat.# 129−02666)は和光純薬から購入した。
【0082】
(方法)
液体培地に乳酸濃度が0%〜20%(v/v)となるように添加し、それぞれのpHを測定した。
【0083】
寒天培地プレートに展開したpol−3変異株および野生株のコロニーを白金耳で掻きとり、YPD培地4mLを入れた試験管(直径18mm)に植菌した。これらを30℃、180 rpmで一晩(16時間以上)振とう培養した。
【0084】
それぞれの培養液の一部をとり、血球計算盤を用いて細胞密度を測定し、両株の細胞密度が等しくなるように液体培地で希釈した。
【0085】
培養開始時の細胞密度が約2.0×106 cells/mLとなるように各培養液をマイクロチューブにとり、5000xgで3分間遠心して細胞を回収した。
【0086】
乳酸濃度1%、3%、5%、6%、7%、8%、9%、10%、11%、13%、15%、17%、19%、20%(v/v)の液体培地4mLに回収した細胞を再懸濁し、30℃、180 rpmで18時間振とう培養した。なお、培養は直径18mmの試験管を使用した。
【0087】
培養後、血球計算盤を用いて細胞密度を測定し、乳酸添加により細胞増殖に与える影響を比較した。
【0088】
(結果)
乳酸添加による培地pHの検討
乳酸を液体培地に添加したときのpHの変化(表1・図2)および培地の変化(図1)を以下に記す。
【0089】
乳酸を添加することによりYPD培地のpHが著しく低下することが明らかになった(表1)。
【0090】
表1 乳酸添加によるYPD培地のpH変化
【0091】
【表1】

。
【0092】
乳酸を添加することにより培地成分の一部が析出し、培地が濁ることが分かった(図1)。
【0093】
乳酸添加による細胞増殖に与える影響の検討
野生株およびpol−3変異株を乳酸含有培地で18時間培養したときの細胞密度を測定した結果を以下に記す(表2および図2)。
【0094】
表2 乳酸添加による野生株およびpol−3変異株の細胞密度の比較(18時間培養)
【0095】
【表2】
。
【0096】
野生株のおよびpol−3変異株の細胞密度は乳酸添加によって低下した。特に、乳酸の添加量が5%を超えるとほとんど増殖できないことが明らかになった。
【0097】
乳酸添加による培地pHの変化および両株の細胞増殖に相関が認められることから、培地のpH低下により両株の細胞増殖が阻害されていることが示された。
【0098】
(実施例2:乳酸耐性酵母の作製方法)
本実施例では、pol−3変異株に乳酸耐性を付与した方法を以下に記す。
【0099】
(使用した酵母株)
実施例1に同じく、pol−3変異株(BYD5)を使用した。
【0100】
(方法)
(1)pol−3変異株のコロニーをかきとり、液体培地4mL を入れた試験管(直径18mm)に植菌し、30℃、180 rpmで一晩培養した。
【0101】
(2)培養液を15mL容の遠沈管(cat.#430791/コーニング)にとり、1000xgで5分間遠心して全ての細胞を回収した。
【0102】
(3)液体培地100mLを入れた300mL容三角フラスコに回収した全ての細胞を再懸濁し、30℃、180 rpmで一晩培養した。
【0103】
(4)培養液を50mL容遠沈管(cat.#430829/コーニング)にとり、1000xgで5分間遠心して全ての細胞を回収した。
【0104】
(5)3%(v/v)乳酸培地150mLを入れた300mL容三角フラスコに回収した全ての細胞を懸濁し、30℃、180 rpmで一晩培養した。
【0105】
(6)3%乳酸培養液の一部をとり、等量の0.1%トリパンブルー溶液(cat.# 345−07421/DOJINDO)と混合して細胞を染色した。青色に染まった細胞は死滅細胞、染まっていない細胞は生存細胞であるため、生存細胞の割合を観察した。
【0106】
(7)生存細胞の存在を確認した後、3%乳酸培養液の1/3量(50mL)を50mL容遠沈管にとり、1000xgで5分間遠心して細胞を回収した。
【0107】
(8)4%(v/v)乳酸培地150mLを入れた300mL容三角フラスコに回収した細胞を懸濁し、30℃、180 rpmで一晩培養した。
【0108】
(9)4%乳酸培養液の一部をとり、トリパンブルー染色により細胞を観察した。生存細胞を確認した後、1/3量(50mL)の培養液を50mL容遠沈管にとり、1000xgで5分間遠心して細胞を回収した。
【0109】
(10)5%(v/v)乳酸培地150mLを入れた300mL容三角フラスコに回収した細胞を懸濁し、30℃、180 rpmで一晩培養した。
【0110】
(11)5%乳酸培養液の一部をとり、トリパンブルー染色により生存細胞の確認を行った。
【0111】
(12)5%乳酸培養液1/10量(15mL)をとり、1000xgで5分間遠心して細胞を回収した。5%乳酸培地70mLを入れた200mL容振とうフラスコ(cat.# 4070FK 200/IWAKI)に回収した細胞を懸濁した後、細胞密度を測定した。測定後、これらの培養液は30℃、180 rpmで振とう培養した。
【0112】
(13)24時間ごとに培養液の細胞密度を測定し、培養開始直後の細胞密度に対して約5〜10倍になるまで培養を行った。
【0113】
(14)5%乳酸培地で増殖が確認された培養液から1/10量(7mL)をとり、1000xgで5分間遠心して細胞を回収した。これらを再び5%乳酸培地に懸濁し、30℃、180 rpmで振とう培養した。
【0114】
(11)〜(14)の操作を繰り返すことで継代培養を行った。なお、継代培養の基準として、1.トリパンブルー染色により、生存細胞を確認すること 2.培養開始直後の細胞密度に対して約5〜10倍増殖していること、以上の2点を設定した。
【0115】
(結果)
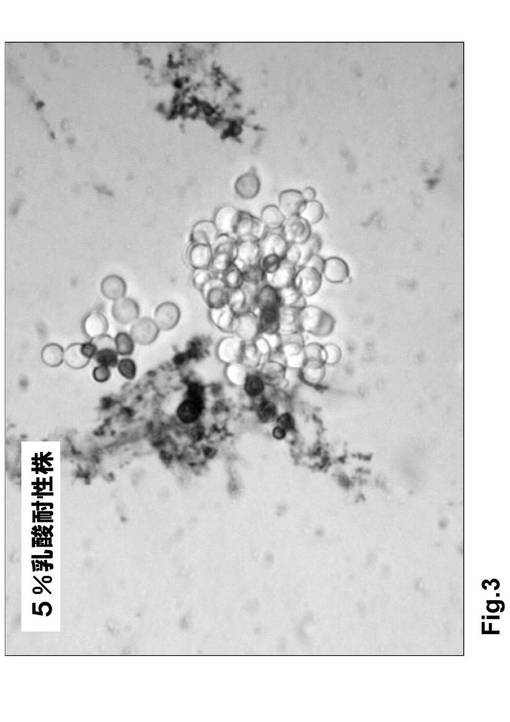
培地の乳酸濃度を徐々に上げ、最終的に5%(v/v)乳酸培地で継代培養を行ったときの細胞密度を以下に記す(表3・図4)。
【0116】
(1)pol−3変異株を3%乳酸培地で培養した結果、細胞密度が増大し、また、生存細胞も多く認められた。
【0117】
(2)3%乳酸培養で増殖した株を4%乳酸培地へ移植した結果、細胞密度の増大および生存細胞が認められた。
【0118】
(3)4%乳酸培養で増殖した株を5%乳酸培地へ移植した結果、培養開始から4日目に細胞密度の増大が認められた。
【0119】
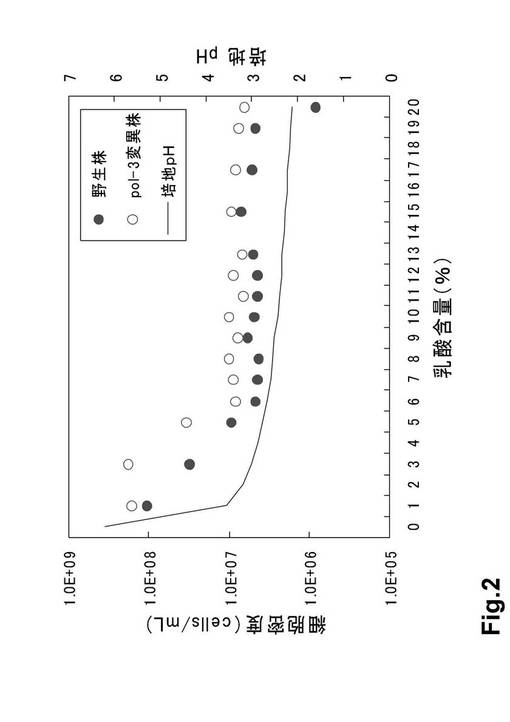
(4)トリパンブルー染色の結果、5%乳酸培養において多くの生存細胞が確認された(図3)。
【0120】
(5)5%乳酸培養で増殖した株を再び5%乳酸培地へ移植し、同培地で継代培養を繰り返した。この結果、継代回数が増すごとに細胞増殖の速度が高まる傾向が認められた(表3・図4)。
以上の結果、本方法によりpol−3変異株に5%乳酸耐性を付与することができることが示された。
表3 5%乳酸培養におけるpol−3変異株の細胞増殖
【0121】
【表3】
(実施例3:5%乳酸耐性株の培養特性)
5%乳酸に対して耐性を獲得したpol−3変異株(5%乳酸耐性株と記す)の培養特性を検討した方法を以下に記す。
【0122】
(使用した酵母株)
実施例1に同じく、野生株および、実施例2によって獲得した5%乳酸耐性株を使用した。
【0123】
(方法)
実施例2によって獲得した5%乳酸耐性株は、5%乳酸培地70mLを入れた200mL容バッフル付き三角フラスコで継代培養を行っている。培養は30℃、200 rpmで行い、前培養液の1/10程度の細胞を継代している。
【0124】
(1)野生株のコロニーをかきとり、液体培地4mLを入れた試験管(φ18mm)に植菌し、30℃、180 rpmで一晩培養した。
【0125】
(2)野生株の培養液を5000xg、3分間遠心して細胞を回収し、液体培地70mLを入れた200mL容バッフル付き三角フラスコに懸濁した。これを30℃、200 rpmで回転振とう培養した。
【0126】
(3)5%乳酸培地で継代している5%乳酸耐性株、および前培養した野生株を50mL容遠沈管にとり、1000xgで5分間遠心して細胞を回収した。
【0127】
(4)回収した細胞を数mLの乳酸無添加の液体培地に再懸濁した後、血球計算盤で細胞密度を測定した。
【0128】
(5)測定した細胞密度をもとに、野生株および5%乳酸耐性株の細胞密度が等しくなるように乳酸無添加の液体培地で希釈した。
【0129】
(6)最終細胞密度が約2.0×107cells/mLとなるように細胞希釈液を15mL容の遠沈管にとり、1000xgで5分間遠心して細胞を回収した。5%、6%および7%の乳酸培地70mLを入れた200mL容バッフル付き三角フラスコに回収した細胞を懸濁し、30℃、200 rpmで回転振とう培養した。なお、野生株については5%のみ培養を行った。また、これらの乳酸培地は0.45μmの滅菌フィルター(ADVANTEC/cat.# 25CS045AS)で濾過して使用した。
【0130】
(7)懸濁直後を0日目とし、24時間ごとに3日間の細胞密度を測定した。なお、血球計算盤による測定は、1測定につき3回行い、平均値および標準偏差を算出した。
【0131】
(結果)
野生株および5%乳酸耐性株を各乳酸培地で培養したときの細胞密度の変化を以下に記す(図5および表4・図6)。
【0132】
(1)5%乳酸培養の結果、5%乳酸耐性株は野生株に比べて顕著な増殖が認められ、また、野生株はほとんど増殖していないことが分かった(図5)。
【0133】
(2)5%乳酸耐性株は、5%および6%乳酸を含む培養条件において速やかに増殖し、培養3日目では約10倍に増殖した(表4、図6)。
【0134】
(3)5%乳酸耐性株を7%乳酸培地で3日間培養したが顕著な増殖は認められなかった(表4、図6)。
【0135】
以上の結果、pol−3変異株を乳酸に馴化させることによって取得した5%乳酸耐性株は、5%および6%乳酸に対して耐性を獲得していることが分かった。
表4 野生株および5%乳酸耐性株の乳酸添加による細胞増殖への影響
【0136】
【表4】
(実施例4:7%乳酸耐性株の作製)
実施例2および3に記した5%乳酸耐性株に7%乳酸への耐性を付与した方法と結果を以下に記す。
【0137】
(方法)
実施例2および3によって取得した5%乳酸耐性株(pol−3変異株)は、5%(v/v)乳酸培地70mLを入れた200mL容バッフル付き三角フラスコを用いて30℃、200 rpmで継代培養を行っている。新しい培地への継代は2〜3日ごとに行い、全培養液に対して1/10量の細胞を回収し、移植している。
【0138】
(1)5%乳酸耐性株の継代培養から全培養液の1/5量をとり、1000xgで5分間遠心して細胞を回収した。7%(v/v)乳酸培地70mLを入れた200mL容バッフル付き三角フラスコに回収した細胞を懸濁し、30℃、200 rpmで一晩培養した。(継代1回目)
(2)全培養液の1/5量をとり、1000xgで5分間遠心して細胞を回収した。新鮮な7%乳酸培地70mLに回収した細胞を懸濁し、30℃、200 rpmで5日間培養した。(継代2回目)
(3)全培養液の1/10量をとり、1000xgで5分間遠心して細胞を回収した。新鮮な7%乳酸培地70mLに回収した細胞を懸濁し、30℃、200 rpmで5日間培養した。(継代3回目)
(4)全培養液の1/10量をとり、1000xgで5分間遠心して細胞を回収した。新鮮な7%乳酸培地70mLに回収した細胞を懸濁し、30℃、200 rpmで4日間培養した。(継代4回目)
(5)4回目の継代培養株および液体培地で一晩前培養した野生株の一部を7%乳酸培地に移植し、30℃、200 rpmで一晩回転振とう培養した。これらの細胞についてトリパンブルー染色を行い、生存細胞を顕微鏡観察することによって7%乳酸への耐性を確認した。
【0139】
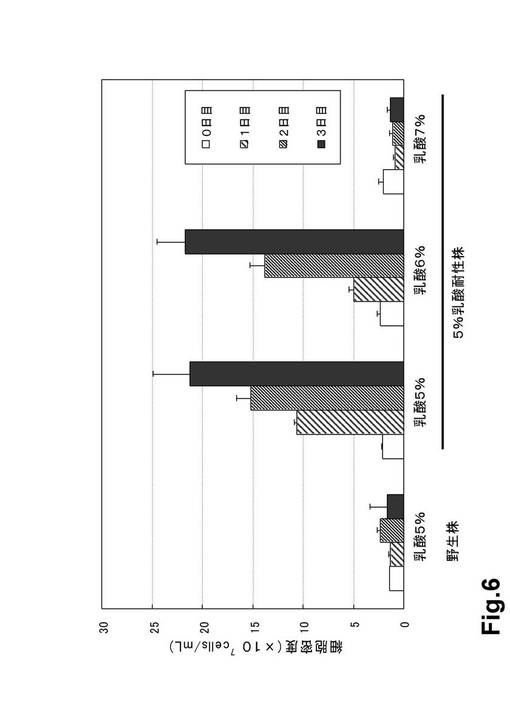
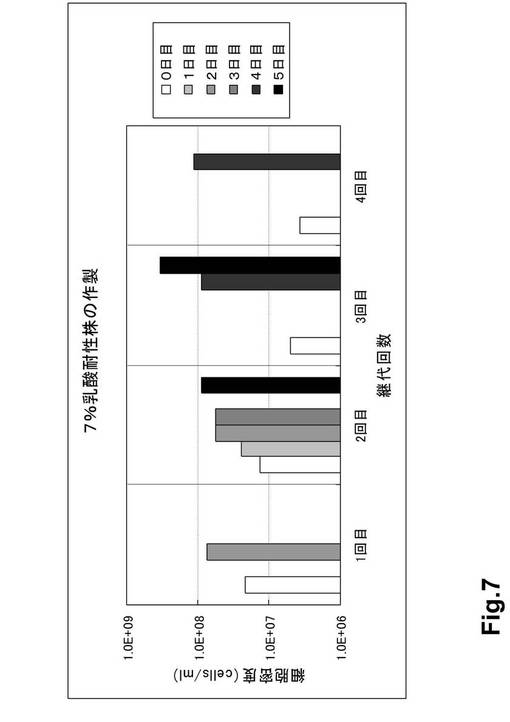
(結果)
5%乳酸耐性株を7%乳酸培地で継代培養したときの細胞密度の変化を以下に記す(表5・図7)。
【0140】
(1)継代培養2回目において5日間培養したところ、継代直後に比べて約6.7倍の増殖が認められた(表5・図7)。
【0141】
(2)継代培養3回目において5日間培養したところ、継代直後に比べて約66.9倍の増殖が認められた(表5・図7)。
【0142】
(3)継代培養4回目において4日間培養したところ、継代直後に比べて約310倍の増殖が認められた(表5・図7)。
【0143】
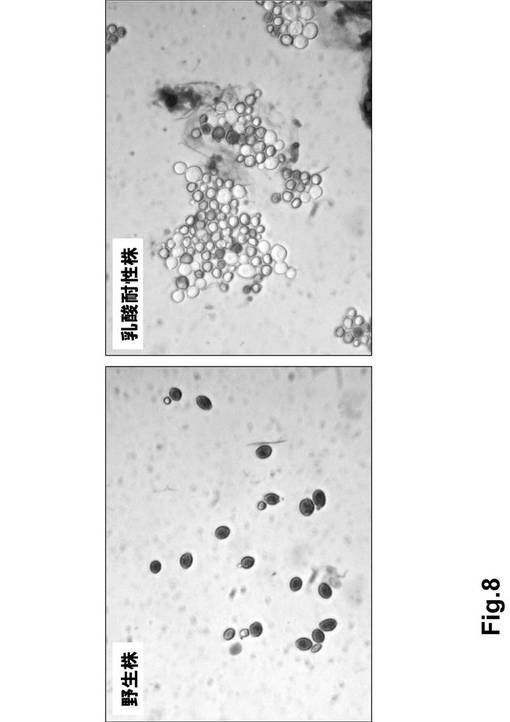
(4)トリパンブルー染色の結果、野生株はほぼ全ての細胞が死滅したにも関わらず、乳酸耐性株では多くの生存細胞が観察された(図8)。
【0144】
上記の結果、pol−3変異株から取得した5%乳酸耐性株は7%乳酸培養に馴化させることにより、速やかに増殖することが可能となった。すなわち、本株は7%乳酸に対して耐性を獲得した。
表5 5%乳酸耐性株を7%乳酸培地で継代培養したときの細胞密度の変化
【0145】
【表5】
本実施例で得られた株の代表例は、〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに受領番号NITE P−122(乳酸耐性株(BYD5-LA7)受託番号 7%乳酸を含むYPD培地中で増殖可能)として寄託済み(平成17年7月29日)である。
【0146】
以上のように、本発明の好ましい実施形態を用いて本発明を例示してきたが、本発明は、特許請求の範囲によってのみその範囲が解釈されるべきであることが理解される。当業者は、本発明の具体的な好ましい実施形態の記載から、本発明の記載および技術常識に基づいて等価な範囲を実施することができることが理解される。本明細書において引用した特許、特許出願および文献は、その内容自体が具体的に本明細書に記載されているのと同様にその内容が本明細書に対する参考として援用されるべきであることが理解される。
【産業上の利用可能性】
【0147】
乳酸耐性酵母は高濃度の乳酸を産生させるための宿主として利用する。乳酸や乳酸ナトリウム、乳酸カルシウムは、すでに輸液等の医薬品原料・食品添加物・化粧品原料として使用されている。また、乳酸の重合体であるポリ乳酸は、生分解性を有するポリマーとして注目され、フィルム・樹脂製品・医療材料その他多くの分野で用途開発が進められている。
【0148】
また、微生物の乳酸耐性を増強する本発明手法を乳酸菌に適用し、経口摂取後、胃酸で死滅せず小腸もしくは結腸に生きたまま到達する酸耐性を増強させる。その結果、腸管病原菌感染に対する防御機能と免疫応答に対する効果のプロバイオテイック特性(欧州、No.92810516−2)を高めた機能性食品として利用できる。その他に、乳酸菌は潰瘍性大腸炎予防の可能性、糖尿病予防の可能性、感染症の低減効果、インフルエンザ抗体価上昇の増強作用、ピロリ菌の抑制効果などの医学的応用が期待されている。
【図面の簡単な説明】
【0149】
【図1】図1は、乳酸添加による培地の変化(左:無添加のYPD培地 右:3%乳酸を加えたYPD培地)である。
【図2】図2¥は、 乳酸添加による野生株およびpol−3変異株の細胞増殖の比較(緑丸:野生株の細胞密度、赤丸:pol−3変異株の細胞密度、青線:培地pH)である。
【図3】図3は、5%乳酸耐性株のトリパンブルー染色による観察(5%乳酸培養の2日目、青色細胞:死滅細胞 無色細胞:生存細胞)である。
【図4】図4は、5%乳酸培養におけるpol−3変異株の細胞増殖(表3より)である。
【図5】図5は、5%乳酸培養における細胞の増殖比較(1日目 左:野生株、右:5%乳酸耐性株)である。
【図6】図6は、野生株および5%乳酸耐性株の乳酸添加による細胞増殖への影響(表3より)である。
【図7】図7は、5%乳酸耐性株を7%乳酸培地で継代培養したときの細胞密度の変化(表5より)である。
【図8】図8は、7%乳酸培地で培養した野生株および乳酸耐性株のトリパンブルー染色写真(左:野生株 右:7%乳酸耐性株 ※7%乳酸培養1日目、白色が生存細胞で青色が死滅細胞を示している。)である。
【技術分野】
【0001】
本発明は乳酸耐性に優れた微生物の作製方法、及び該方法により作製した乳酸耐性に優れた微生物に関する。
【背景技術】
【0002】
乳酸発酵で生産された乳酸が蓄積すると、培地pHと細胞内pHが共に低下するが、微生物は細胞内のpHを一定にするためATPを消費して、細胞外へプロトンを汲み出そうとする。例えば、S.cerevisiaeの場合、細胞外pH3.0の時、細胞内pHは5.5〜5.75であり(非特許文献1=Imai,T. et al.Appl.Environ.Microbiol.61,3604−3608,1995)、細胞外pH6.0〜10.0のとき、細胞内pHは5.9〜6.75に維持されている(非特許文献2=Imai,T. et al.J.Biotechnol.38,165−172,1995)。微生物が細胞内のpH低下を維持できなかった場合、解糖系や糖新生経路のキー・エンザイムであるcAMPプロテインキナーゼの抑制、細胞内cAMP濃度低下、ヒートショックなどの他の生物反応のトリガーなどになり、最終的には増殖が抑制されると考えられている。
【0003】
Lactobacillusに代表される乳酸菌を用いて未利用バイオマスからの乳酸生産は各種試みがなされているが、その乳酸発酵の行われるpHは5.0〜5.5と高く、乳酸のpKa(〜3.8)以下での発酵が望まれている。その理由は、pKa(〜3.8)以下の乳酸は未解離の状態で存在し、発酵培養液から乳酸抽出で直接抽出することが可能だが、高いpHの培地中からの乳酸精製には乳酸を用いた直接抽出が出来ず、抽出精製コストが高くなることにある。乳酸発酵でより高濃度の乳酸を生産することは工業生産上非常に重要である。
【0004】
Lactococcus lactisの低pH下での細胞増殖には、挿入変異解析法を用いて18遺伝子座の関連が示唆され、多数の低pHに対する耐性機構が関与していると予想される(非特許文献3=Rallu,F.et al.Mol.Microbiolo.35,517−528,2000)。また、Lactobacillusの酸への適応のプロテオミクス解析の結果からは、63種のタンパク質の協調的発現誘導が起こることが報告されている(非特許文献4=DeAngelis,M.et al.Microbiology147,1863−1873,2001)。また、Lactobacillus acidophilus NCFMの全ゲノム解析の結果から、酸耐性に関連する遺伝子として2成分から構成される9種の調節システムが推定されている(非特許文献5=Altermann,E.et al.Proc.Natl.Acad.Sci.USA.102(11),3906−3912,2005)。
【0005】
Lactobacillus sanfranciscensisの酸耐性の解析では、ヒートショックタンパク質4種の中でGrpEのみが選択的に発現上昇していた(非特許文献6=Maria DeAngelis et.al.Microbiol.147,1863−1873,2001)。以上の例にあるように、乳酸耐性機構は遺伝子レベルで検討の余地が多分に残されている。
現在、乳酸耐性を付与する、あるいは、乳酸産生能を維持する方法は以下のような方法が用いられている。第一の方法は、新規に乳酸耐性度の高い乳酸耐性菌株を分離する方法である。得られた乳酸耐性菌を宿主として高濃度の乳酸を生産するための遺伝仕組換えを実施するためにはベクター系の開発が必要で実用化までの道のりが遠い。第二の方法は、乳酸菌を化学突然変異誘発剤処理し突然変異を導入し、乳酸耐性変異体を作製する方法だが、大幅な乳酸耐性の付与には成功していない。第三の方法は、異なる性質の乳酸菌の細胞融合で乳酸耐性変異体を作製する方法である。具体例としてpH4.0にて野生株に比べ乳酸を3倍生産する乳酸耐性株を得ている(非特許文献7=Patnaik,R.et al.Nature Biotechnology 20.707−712,2002)が、更に乳酸に対する耐性度の高い株の作製が可能かどうかは不明である。第四の方法は、乳酸耐性菌から単離された乳酸耐性関連遺伝子を遺伝子組み換えで宿主菌に導入し、乳酸耐性を付与するものである。具体例としては、ワイン酵母Oenococcus oeniから単離された多剤耐性遺伝子ファミリーに属するorfC、LysRファミリーの推定上の転写調節因子orfB、低分子ヒートショックタンパク質Lo18の3遺伝子を大腸菌に導入し、酸耐性を付与した例がある(非特許文献8=Lett. Appl. Microbiol.33(2),126−130,2001)。この方法は、乳酸耐性を付与したい生物種に適用可能かどうかが不明である。また、仮に適用が可能な場合でも、乳酸耐性度を更に増強したい場合、乳酸耐性関連遺伝子の発現を増加させる以外に合理的手段がない。第五の方法は、ケモスタットを用いて酸に馴化させる方法である。具体例としては、菌の致死量以下の低pH(pH5.0、60分間)で前処理後、致死pH(pH3.0)処理し、耐性化遺伝子の発現を誘導し、乳酸耐性に馴化させる方法がある(非特許文献9=Lorca GL et.al.J.Mol.Microbiol.Biotechnol,4(6),525−532,2002)。この方法では、馴化して獲得させた形質が安定に保持されるのか、どの生物種に適用可能か、また、馴化可能な乳酸濃度がいくらなのか全く不明である。
その他に、乳酸発酵能を長期間維持する方法としては、培養後期における乳酸菌の乳酸発酵能の低下を防ぐために超伝導磁石で形成された磁場空間内で、静磁場を印加し乳酸菌を活性化する方法が原理的には考えられている(特許文献1=三浦靖、特開2005−34040)。しかし、工業生産する場面で、宿主菌を超伝導の磁場に置くことは、現実的には不可能である。
【非特許文献1】Imai,T. et al.Appl.Environ.Microbiol.61,3604−3608,1995
【非特許文献2】Imai,T. et al.J.Biotechnol.38,165−172,1995
【非特許文献3】Rallu,F.et al.Mol.Microbiolo.35,517−528,2000
【非特許文献4】DeAngelis,M.et al.Microbiology147,1863−1873,2001
【非特許文献5】Altermann,E.et al.Proc.Natl.Acad.Sci.USA.102(11),3906−3912,2005
【非特許文献6】Maria DeAngelis et.al.Microbiol.147,1863−1873,2001
【非特許文献7】Patnaik,R.et al.Nature Biotechnology 20.707−712,2002
【非特許文献8】Lett. Appl. Microbiol.33(2),126−130,2001
【非特許文献9】Lorca GL et.al.J.Mol.Microbiol.Biotechnol,4(6),525−532,2002
【特許文献1】特開2005−34040
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、乳酸耐性の低い実験微生物に対して、低コスト、短時間で、効率よく、さらに高濃度の乳酸に対して耐性に優れた形質を付与する方法を提供することである。
【0007】
また、本発明の別の目的は、乳酸耐性に優れた形質を付与した微生物を提供することである。
【課題を解決するための手段】
【0008】
本発明の上記目的は、鋭意検討の結果、遺伝子の複製において正確さにかける生物を用いて、通常では生育しない乳酸濃度存在下で培養し、生育する生物を単離することを特徴とする乳酸耐性微生物の作製方法によって達成された。
【発明の効果】
【0009】
各種の乳酸に対して、耐性に優れた微生物作製が可能となる。また、本発明は、乳酸耐性機構を解明するためのコストが不要で、これまでに解明された実験微生物のゲノム情報や開発されたベクター系が有効活用されることになる。本発明で作製した乳酸耐性酵母を宿主として乳酸を工業的に生産する場合、乳酸耐性度が増加した分の乳酸発酵が可能となり、コスト削減につながる。
【発明を実施するための最良の形態】
【0010】
本明細書の全体にわたり、単数形の表現は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。また、本明細書において使用される用語は、特に言及しない限り、当該分野で通常用いられる意味で用いられることが理解されるべきである。
【0011】
(用語)
以下に本明細書において特に使用される用語の定義を列挙する。
【0012】
本明細書において「生物」とは、当該分野における最も広義に用いられ、生命現象を営むものをいい、代表的には、細胞構造、増殖(自己再生産)、成長、調節性、物質代謝、修復能力など種々の特性を有し、通常、核酸のつかさどる遺伝と、タンパク質のつかさどる代謝の関与する増殖を基本的な属性として有する。生物には、ウイルス、原核生物、真核生物(酵母のような単細胞生物、植物、動物のような多細胞生物など)などが包含される。本発明の方法は、グラム陽性細菌、および真核生物などの高等生物を含む、どのような生物であっても適用され得ることが理解される。本明細書では、サイズの小さな生物を「微生物」とも呼ぶが、乳酸耐性という点では同一であるので、本明細書では、生物と微生物とは交換可能に使用され得ることが理解される。
【0013】
本明細書において「真核生物」とは、当該分野において通常用いられる意味と同様に用いられ、核膜のある、明確な核構造を持つ細胞からなる生物をいう。真核生物としては、例えば、酵母のような単細胞生物、イネ、コムギ、トウモロコシ、ダイズのような植物、マウス、ラット、ウシ、ウマ、ブタ、サルのような動物、ハエ、カイコなどの昆虫が挙げられるがそれらに限定されない。本明細書では、酵母、線虫、ショウジョウバエ、カイコ、イネ、コムギ、ダイズ、トウモロコシ、シロイヌナズナ、ヒト、マウス、ラット、ウシ、ウマ、ブタ、カエル、魚類(例えば、ゼブラフィッシュ)などがモデルとして使用され得るがそれらに限定されない。
【0014】
本明細書において「原核生物」とは、当該分野において通常用いられる意味と同様に用いられ、明確な核構造を持たない細胞からなる生物をいう。原核生物としては、例えば、大腸菌、サルモネラ菌のようなグラム陰性細菌、枯草菌、放線菌、ブドウ球菌のようなグラム陽性細菌、藍藻類、水素細菌などが挙げられるがそれらに限定されない。本明細書では、代表的に、大腸菌以外に、グラム陽性細菌、として使用され得るがそれに限定されない。
【0015】
本明細書において「動物」は、当該分野において最も広義で用いられ、脊椎動物および無脊椎動物(例えば、節足動物)を含む。動物としては、哺乳綱、鳥綱、爬虫綱、両生綱、魚綱、昆虫綱、蠕虫綱などが挙げられるがそれらに限定されない。動物は、好ましくは、脊椎動物(例えば、メクラウナギ類、ヤツメウナギ類、軟骨魚類、硬骨魚類、両生類、爬虫類、鳥類、哺乳動物など)であり得るが、それらに限定されない。ある一つの実施形態では、動物は、哺乳動物(例えば、単孔類、有袋類、貧歯類、皮翼類、翼手類、食肉類、食虫類、長鼻類、奇蹄類、偶蹄類、管歯類、有鱗類、海牛類、クジラ目、霊長類、齧歯類、ウサギ目など)であり得るがそれらに限定されない。さらに好ましくは、霊長類(たとえば、チンパンジー、ニホンザル、ヒト)または他のモデル動物となり得る種(例えば、奇蹄類、偶蹄類、マウスなどの齧歯類、ウサギ目など)であり得るがそれらに限定されない。本発明の方法は、生物であればどのようなものであっても適用できることが本発明において初めて解明されたことから、どのような生物であっても、対象とされ得ることが理解されるべきである。
【0016】
本明細書において用いられる「植物」とは、植物界に属する生物の総称であり、クロロフィル、かたい細胞壁、豊富な永続性の胚的組織の存在,および運動する能力がない生物により特徴付けられる。代表的には、植物は、細胞壁の形成・クロロフィルによる同化作用をもつ顕花植物をいう。「植物」は、単子葉植物および双子葉植物のいずれも含む。好ましい植物としては、有用植物、例えば、コムギ、トウモロコシ、イネ、オオムギ、ソルガムなどのイネ科に属する単子葉植物が挙げられるがそれらに限定されない。好ましい植物のほかの例としては、タバコ、ピーマン、ナス、メロン、トマト、サツマイモ、キャベツ、ネギ、ブロッコリー、ニンジン、キウリ、柑橘類、白菜、レタス、モモ、ジャガイモおよびリンゴが挙げられる。好ましい植物は作物に限られず、花、樹木、芝生、雑草なども含まれる。特に他で示さない限り、植物は、植物体、植物器官、植物組織、植物細胞、および種子のいずれをも意味する。植物器官の例としては、根、葉、茎、および花などが挙げられる。植物細胞の例としては、カルスおよび懸濁培養細胞が挙げられる。本発明の方法は、生物であればどのようなものであっても適用できることが本発明において初めて解明されたことから、どのような生物であっても、対象とされ得ることが理解されるべきである。
【0017】
本明細書において、「遺伝子」とは、細胞中に存在する核酸の一定の長さの配列をいう。本発明において遺伝子は、遺伝形質を規定するものであっても規定しないものであってもよい。本明細書において、遺伝子は、通常ゲノムに存在するものをさすが、それに限定されず、染色体外の配列、ミトコンドリアの配列なども包含することが理解される。多くの遺伝子は、通常染色体上に一定の順序に配列している。タンパク質の一次構造を規定するものを構造遺伝子といい、その発現を左右するものを調節遺伝子(たとえば、プロモーター)という。本明細書では、遺伝子は、特に言及しない限り、構造遺伝子および調節遺伝子を包含する。したがって、例えば、DNAポリメラーゼ遺伝子というときは、通常、DNAポリメラーゼの構造遺伝子ならびにDNAポリメラーゼのプロモーターなどの転写および/または翻訳の調節配列の両方を包含する。本発明では、構造遺伝子のほか、転写および/または翻訳などの調節配列もまた、本発明が対象とする遺伝子として有用であることが理解される。本明細書では、「遺伝子」は、「ポリヌクレオチド」、「オリゴヌクレオチド」、「核酸」および「核酸分子」ならびに/または「タンパク質」、「ポリペプチド」、「オリゴペプチド」および「ペプチド」を指すことがある。本明細書においてはまた、「遺伝子産物」は、遺伝子によって発現された「ポリヌクレオチド」、「オリゴヌクレオチド」、「核酸」および「核酸分子」ならびに/または「タンパク質」「ポリペプチド」、「オリゴペプチド」および「ペプチド」を包含する。当業者であれば、遺伝子産物が何たるかはその状況に応じて理解することができる。
【0018】
本明細書において「遺伝子の複製」とは、遺伝物質であるDNAまたはRNAにおいて,親の核酸鎖を鋳型として,親核酸と同一の構造と機能を有する新しい核酸分子(DNAであればDNA、RNAであればRNA)を生成することをいう。真核細胞では,二本鎖DNA分子上にある多数の複製開始点に複製酵素(DNAポリメラーゼα)を含む複製開始複合体が形成されることによって複製が開始され、複製反応が複製開始点から両方向へ進行する。複製開始は、細胞周期によって制御される。酵母における自立複製配列(ARS=autonomously replicating sequence)は複製開始点とされる。大腸菌などの原核細胞では、ゲノムの二本鎖環状DNA分子に1個の複製開始点(ori)が存在し、oriに複製開始複合体が形成され、反応はoriから両方向に進行する。複製開始複合体は複製酵素(DNAポリメラーゼIII)を含む10種類以上のタンパク質因子を含む複雑な構造を有する。複製反応は、二本鎖DNAのらせん構造の部分的巻戻しに始まり、その後短いDNAプライマーが合成され、3’−OH基から新たなDNA鎖が伸長され、相補鎖鋳型において岡崎フラグメントが合成され、岡崎フラグメントの連結、鋳型と照らし合わせる校正(プルーフリーティング)などの多段階の反応によって複製反応が進行する。
【0019】
生物の遺伝情報であるゲノムDNAの複製機構は、Kornberg A.and Baker T.,“DNA Replication”,New York,Freeman,1992に詳述されている。代表的には、1つの1本鎖DNAを鋳型として相補的な鎖を合成し、その結果、1つの2本鎖DNAを生じる酵素(DNA複製を行う酵素)を、DNAポリメラーゼという。DNA複製には、少なくとも2つのDNAポリメラーゼが必要である。通常、リーディング鎖およびラギング鎖の合成を同時に行わなければならないからである。DNA複製は、DNA上の決まった位置から開始されるが、その位置を複製開始点(ori)という。例えば、細菌では、通常、環状ゲノムDNAに少なくとも1つの両方向性の複製開始点を有する。これらを総合すると、通常は、1つのゲノムDNA複製には、4つのDNAポリメラーゼが同時に作用する必要があることになる。本発明では、好ましくは、リーディング鎖およびラギング鎖の一方のみの複製誤りが調節されることが有利であり得、あるいは、二本の鎖のあいだの複製誤りの頻度が異なることが有利であり得る。
【0020】
本明細書において「複製誤り」とは、遺伝子(DNAなど)の複製の過程で生じるヌクレオチド取り込みの誤りをいう。複製誤りは、通常、生体では、その頻度は108〜1012回に1回程度できわめて低い。複製誤りの頻度が低い理由としては、ヌクレオチドの取り込みが鋳型DNAと取り込まれるヌクレオチドとが相補的な塩基対を形成することによって複製が起こること、DNAポリメラーゼδ、εなどの酵素の校正機能すなわち3’→5’エキソヌクレアーゼが,鋳型に相補性を示さないヌクレオチドが誤って取り込まれたときそれを察知し直ちに切り出す機能が存在することなどが挙げられる。従って、本発明において複製におけるエラープローン頻度の調節は、特異的塩基対形成の障害、校正機能の障害などによって行うことができる。
【0021】
本明細書において「エラーフリー」とは、遺伝子(DNAなど)の複製にいて誤りがほとんどない、好ましくは実質的に全くない性質をいう。エラーフリーは、主に、校正機能を有する酵素(例えば、DNAポリメラーゼδ、εなど)の校正機能の精度によって影響を受ける。
【0022】
本明細書において「エラープローン」とは、遺伝子(DNAなど)の複製における誤り易い(すなわち、複製誤りの)性質をいう。エラープローンは、主に、校正機能を有する酵素(例えば、DNAポリメラーゼδ、εなど)の校正機能の精度によって影響を受ける。
【0023】
本明細書においてエラープローンとエラーフリーとは、絶対的に(すなわち、エラープローン頻度のレベルなどで決定する)分類することができ、あるいは相対的(2種類以上の遺伝子の複製を担う因子(例えば、DNAポリメラーゼなど)におけるエラープローン頻度について多いほうをエラープローンとし、少ないほうをエラーフリーとする)に分類することができる。
【0024】
本明細書において「エラープローン頻度」とは、エラープローンの性質のレベルをいう。エラープローン頻度は、例えば、遺伝子配列における変異の絶対数(変異の数そのもの)または相対数(全長における変異の数の比率)で表現することができる。あるいは、ある生物または酵素について言及するとき、エラープローン頻度は、ある生物の生殖または分裂1回あたりの遺伝子配列における変異の絶対数または相対数で表現してもよい。特に言及しない場合、遺伝子配列における複製過程1回あたりの誤差の数で表される。エラープローン頻度は、逆の尺度として本明細書において「精度」ということがある。エラープローン頻度が均一であるとは、複数の遺伝子の複製を担う因子(ポリメラーゼなど)に言及するとき、互いのエラープローン頻度が実質的に等しいことをいう。他方、エラープローン頻度が不均一であるとは、有意な差異が複数の遺伝子の複製を担う因子(ポリメラーゼなど)に存在する場合をいう。
【0025】
本明細書において「エラープローン頻度の調節」とは、エラープローン頻度を変化させることをいう。そのようなエラープローン頻度の調節には、エラープローン頻度の上昇および低減が含まれる。エラープローン頻度の調節のための手法としては、例えば、校正機能を有するDNAポリメラーゼの改変、複製中に重合反応または伸長反応を阻害または抑制するような因子の挿入これらの反応を促進するような因子の阻害、抑制、単数または複数の塩基の欠損、DNA修復酵素の欠損、異常塩基の除去修復因子機能を有する酵素の改変、ミスマッチ塩基対修復因子の改変、複製自体の精度の低減などが挙げられるがそれらに限定されない。エラープローン頻度の調節は、DNAの二本鎖の両方に対して行われてもよいし、片方のみに対して行われてもよい。好ましくは、片方のみに対して行われることが有利であり得る。有害な変異誘発が低減されるからである。
【0026】
本明細書において「DNAポリメラーゼ」またはPolとは、4種類のデオキシリボヌクレオシド5’−三リン酸からピロリン酸を遊離してDNAを重合する働きを有する酵素をいう。DNAポリメラーゼ反応には、鋳型となるDNA、プライマー分子、Mg2+などが必要とされる。プライマーの3’−OH末端に鋳型に相補的なヌクレオチドを順次付加し分子鎖を伸長する。
【0027】
大腸菌にはDNAポリメラーゼI、II、IIIの少なくとも3種類の酵素が知られている。DNAポリメラーゼIはDNA傷害の修復,遺伝的組換えおよびDNA複製に関与する。DNAポリメラーゼIIおよびIIIは補助的機能を有するといわれる。この酵素は数種のタンパク質からなるサブユニット構造をとり,その構成からコア酵素およびホロ酵素の2つに分けられる。コア酵素は,α、εおよびθから構成され、ホロ酵素には、α、εおよびθサブユニットのほかに、τ、γ、δおよびβの成分がある。真核生物細胞も複数のDNAポリメラーゼをもつことが知られており、高等生物ではDNAポリメラーゼα、β、γ、δおよびεなど多種類存在する。動物では、DNAポリメラーゼα(核DNAの複製に関与、細胞増殖期のDNA複製に作用)、DNAポリメラーゼβ(核でのDNAの修復に関与、増殖期、停止期のDNA傷害の修復などの作用を有する)、DNAポリメラーゼγ(ミトコンドリアDNAの複製と修復に関与。エキソヌクレアーゼ活性をもつ)、DNAポリメラーゼδ(DNAの伸長に関与。エキソヌクレアーゼ活性をもつ)、DNAポリメラーゼε(ラギング鎖のすき間の複製に関与。エキソヌクレアーゼをもつ)などが知られている。
【0028】
校正機能を担うDNAポリメラーゼ(グラム陽性細菌、グラム陰性細菌、真核生物など)では、ExoI用のアミノ酸配列が3’→5’エキソヌクレアーゼ活性中心を担うとされており、この部位が校正機能の精度に影響すると考えられている。
【0029】
配列番号1:Pol III:317−IMSFDIECAGRI−328(Saccharomyces cerevisiae);
配列番号2:Pol II:286−VMAFDIETTKPP−297(Saccharomyces cerevisiae);
変異型pol−3遺伝子発現株(AMY128−1、以下pol−3変異株と略す)は、この配列を有しているものと推定される。AMY128−1の代表例は、独立行政法人製品評価技術基盤機構 特許生物寄託センター(〒292−0818 千葉県木更津市かずさ鎌足2−5−8)に受託番号NITE P−130(変異型pol−3遺伝子発現株(1倍体株)(AMY128−1))(寄託日平成17年8月19日)として寄託済みである。また、変異型pol−3遺伝子発現株(2倍体株)(BYD5)は、受託番号 NITE P−131(寄託日平成17年8月19日)として寄託済みである。野生型pol−3遺伝子発現株(1倍体株)(AMY52−3D)は、受託番号 NITE P−129(寄託日平成17年8月19日)として寄託済みである。
【0030】
ここで明らかなように、校正機能を有するDNAポリメラーゼは、アスパラギン酸およびグルタミン酸がよく保存されている。本明細書において、このようなアスパラギン酸およびグルタミン酸を含む領域は、本明細書において校正機能活性部位ということがある。
【0031】
1つの好ましい実施形態では、このような3’→5’エキソヌクレアーゼ活性が破壊されるような変異を、DNAポリメラーゼをコードする遺伝子(DNAポリメラーゼ遺伝子)に導入することにより、校正機能が低下した(すなわち、エラープローン頻度が増加した)DNAポリメラーゼをコードする核酸分子およびポリペプチドを産生することができる。なお、校正機能の3’→5’エキソヌクレアーゼ活性は、単一のDNAポリメラーゼ遺伝子(PolC,POL2,CDC2等)において、DNA重合活性を担う分子中に含まれる場合(例えば、真核生物、グラム陽性細菌等)と、DNA重合活性をコードする遺伝子(例えば、dnaE)とは異なる遺伝子(例えば、dnaQ)にコードされる場合(例えば、グラム陰性細菌等)があることが知られており(Kornberg A.and Baker T.,“DNA Replication ”,New York,Freeman,1992)、当業者は、それらの特性を理解したうえで、本発明におけるエラープローン頻度の調節を適宜行うことができる。例えば、真核生物などでは、校正機能は変化するが、DNA重合活性はほとんど変化させない変異をDNAポリメラーゼに導入することが好ましい。この場合、上述のような校正機能に関与する2つの酸性アミノ酸(Derbyshire et.al.,EMBO J.10,pp.17−24,Jan.1991; Fijalkowska and Schaaper,“Mutants in the Exo I motif of Escherichia coli dnaQ:Defective proofreading and inviability due to error catastrophe”,Proc.Natl.Acad.Sci.U.S.A.Vol.93,pp.2856−2861,Apr.1996)を改変すること(好ましくは、保存されない置換(例えば、アラニン、バリンなどへの置換))が挙げられるがそれらに限定されない。
【0032】
本明細書において「校正機能」とは、細胞が受けたDNA損傷および/または誤りを検知し補修する機能をいう。そのような機能は、脱プリン、脱ピリミジンがある場合はそのまま塩基が挿入されることによるか、あるいは、A−Pエンドヌクレアーゼ(apurinic−apyrimidinic endonuclease)で一本鎖切断が入れられたのち5’→3’エキソヌクレアーゼで除去されることによって達成され得る。除去部分は、DNAポリメラーゼでDNA合成され補充され、正常DNAとの連結はDNAリガーゼが行う。このような反応は、除去修復といわれる。アルキル化剤による化学修飾,異常塩基,放射線,紫外線などによるDNA塩基傷害などは傷害部分をDNAグリコシダーゼで取り除かれた上で上記の反応で修復される(不定期DNA合成)。そのような校正機能を有するDNAポリメラーゼとしては、例えば、真核生物におけるDNAポリメラーゼδ、DNAポリメラーゼεなどが挙げられるがそれらに限定されない。本明細書では、校正機能の程度を表すために、忠実度という用語もまた用いられ得る。この忠実度との用語は、DNA複製の正確さを意味する。正常なDNAポリメラーゼは、通常、忠実度が高いDNAポリメラーゼであり、改変により校正機能が低下したDNAポリメラーゼは、忠実度が低いDNAポリメラーゼであり得る。
【0033】
このようなDNAポリメラーゼの校正機能については、例えば、Kunkel,T.A.:J Biol.Chem.,260,12866−12874(1985);Kunkel,T.A.,Sabotino,R.D.& Bambara,R.A.:Proc.Natl.Acad.Sci.USA,84,4865−4869(1987);Wu,C.I.& Maeda,N.:Nature,327,167−170(1987);Roberts,J.D.& Kunkel,T.Al:Proc.Natl.Acad.Sci.USA,85,7064−7068(1988);Thomas,D.C.,Fitzgerald,M.P.& Kunkel,T.A.:Basic Life Sciences,52,287−297(1990);Trinh,T.Q.& Siden,R.R.Nature,352,544−547(1991);Weston−Hafer,K.,& Berg,D.E.Genetics,127,649−655(1991);Veaute,X.& Fuchs,R.p.p.:Science,261,598−600(1993);Roberts,J.D.,Izuta,S.,Thomas,D.C.& Kunkel,T.A.:J Biol Chem.,269,1711−1717(1994);Roche,W.A.,Trinh,T.Q.& Siden,R.R.J.Bacteriol.,177,4385−4391(1995);Kang,S.,Jaworski,A.,Ohshirna,K.& Wells,Nat.Genet.,10,213−218(1995);Fijalkowska,I.J.,Jonczyk,P.,Maliszewska−Tkaczyk,M.,Bialoskorska,M.& Schaaper,R.M.Proc.NatJ.Acad.Sci.USA.95,10020−10025(1998);Maliszewska−Tkaczyk,M.,Jonezyk,P.,Bialoskorska,M.,Schaaper,M.& Fijalkowska,I.:Proc.Natl.Acad.Sci.USA,97,12678− 12683(2000);Gwel,D.,Jonezyk,P.,Bialoskorska,M.,Schaaper,R.M.& Fijalkowska,I.J.:Mutation Research,501,129−136(2002).Negative;Roberts,J.D.,Thomas,D.C.& Kunkel,T.A.:Proc Nat].Acad Sci.USA,88,3465−3469(1991);Roberts,J.D.,Nguyen,D.& Kunkel,T.A.:Biochemistry;32,4083−4089(1993);Francino,M.P.,Chac,L.,Riley,M.A.& Ochman,H.:Science,272,107−109(1996);A Boulet,M.Simon,G.Faye,GA Bauer & PM Burgers.EMBO J,8,1849−1854,(1989);Morrison A,Araki H,Clark AB,Hamatake RK,& Sugino A.Cell,62(6),1143−1151,(1990)などを参照のこと。
【0034】
本明細書において「DNAポリメラーゼδ」とは、真核生物のものをさす場合、DNAの伸長に関与する酵素であって、エキソヌクレアーゼ活性をもち、これに起因して校正機能を有するといわれる。このDNAポリメラーゼδの校正機能の調節は、上記の校正機能に関連するアミノ酸配列に改変を導入することによって達成することができる。代表的なDNAポリメラーゼδは、核酸配列およびアミノ酸配列として、polδ:X61920 gi/171411/gb/M61710.1/YSCDPB2[171411]に記載されている配列を有する。DNAポリメラーゼδは、Simon,M.et al.、EMBO J.10,2163−2170,1991に記載されており、その内容は本明細書において参考として援用される。
【0035】
本明細書において「DNAポリメラーゼε」とは、真核生物のものをさす場合、ラギング鎖のすき間の複製に関与する酵素であって、エキソヌクレアーゼ活性をもち、これに起因して校正機能を有するといわれる。代表的なDNAポリメラーゼεは、核酸配列およびアミノ酸配列として、polε:M60416 gi/171408/gb/M60416.1/YSCDNAPOL[171408]に記載されている配列を有する。
【0036】
DNAポリメラーゼδおよびεの分類は、HUGO分類によると別名はデルタがPOLD1/POL3,イプシロンがPOLE/POL2となっており、本明細書では、どの命名法をも用い得る。
【0037】
その他DNAポリメラーゼの説明については、例えば、Lawrence C.W.et al.,J.Mol.Biol.122,1−21,1978、Lawrence C.W.et al.,Genetics 92,397−408、Lawrence C.W.et al.MGG,195,487−490,1984、Lawrence C.W.et al.MGG.200,86−91,1985(DNAポリメラーゼβおよびDNAポリメラーゼζ);Maher V.M.et al.Nature 261,593−595,1976、McGregor,W.G.et al.,Mol.Cell.Biol.19,147−154、1999(DNAポリメラーゼη);Strand M.et al.,Nature 365,275−276,1993、Prolla T.A.,et al.,Mol.Cell.Biol.15,407−415,1994、Kat A.,et al.,Proc.Natl.Acad.Sci.USA 90,6424−6428、Bhattacharyya N.P.,et al.,Proc.Natl.Acad.Sci.USA 91,6319−6323,1994、Faber F.A.,et al.,Hum.Mol.Genet.3,253−256,1994、Eshleman,J.R.,et al.,Oncogene 10,33−37,1995、Morrison A.,et al.,Proc.Natl.Acad.Sci.USA 88,9473−9477,1991、Morrison A.,et al.,EMBO J.12,1467−1473,1993、Foury F.,et al.,EMBO J.11,2717−2726,1992(DNAポリメラーゼλ、DNAポリメラーゼμなど)などに記載されており、その内容は本明細書において参考として援用される。
【0038】
本明細書においてDNAポリメラーゼなどの遺伝子および酵母などの生物の「野生型」は、もっとも広汎な定義では、天然に存在するDNAポリメラーゼなどの遺伝子および酵母などの生物を含み、通常、天然に存在するDNAポリメラーゼなどの遺伝子および酵母などの生物のうち、由来となる生物種においてもっとも広汎に存在するものをいう。従って、通常、ある種において最初に同定されるDNAポリメラーゼなどの遺伝子および酵母などの生物の種は野生型といえる。野生型はまた、「天然標準型」ともいう。生物であれば、野生型は、酵素活性が正常であり得、形質が正常であり得、行動が正常であり得、生理が正常であり得、繁殖が正常であり得、ゲノムが正常であり得る。
【0039】
本明細書において校正機能が「野生型のものよりも低い」とは、ある校正機能を有する酵素などについて言及するとき、その酵素の野生型よりも校正機能が低いこと(すなわち、その酵素での校正処理の後に残留する変異の数が野生型による校正処理の後に残留する変異の数よりも多いこと)をいう。そのような野生型との比較は、相対的または絶対的な表示によって行うことができる。そのような比較はまた、エラープローン頻度などによって行うことができる。
【0040】
本明細書において「変異」とは、遺伝子について言及するとき、その遺伝子の配列の変化を生じることまたはその変化によって生じた遺伝子の(核酸またはアミノ酸)配列の状態をいう。本明細書では、例えば、変異は、校正機能について生じる遺伝子配列の変化について用いられる。本明細書では、特に言及しない場合は、変異は、改変と同義で用いられる。
【0041】
有用な変異体を作製するためには、生物において変異誘発を行うことがもっとも一般的である。変異とは、通常、遺伝子をコードする塩基配列の変化をいい、DNA配列の変化が包含される。変異は、それが発生した個体に与える影響により、大きく次の3種類に分けられる:A)中立変異(neutral mutation):この変異は、ほとんどの変異が該当し、生物の成育および代謝にほとんど影響がない。B)有害変異(deleterious mutation):この変異は、中立変異よりは頻度は少ない。生物の成長または代謝を阻害する。有害変異には、生育に必須な遺伝子を破壊するような致死変異(lethal mutation)も含まれる。微生物の場合、種によっても異なるが、通常全変異に占める有害変異の割合は、約1/10〜1/100とされている。C)有益変異(beneficial mutation):この変異は、生物の育種に有益な変異である。その発生頻度は中立変異と比較して極めて低い。したがって、有益変異が導入された生物個体を得るためには、大きな生物集団と、長い時間が必要となる。また、生物の育種の十分な効果は、単一の変異だけで現れることはまれであり、複数の有益変異の蓄積が必要であることが多い。従って、本発明は、乳酸耐性のみを付与する技術としては初めて提供されたことになる。
【0042】
本明細書において「成長(または生長)」とは、ある生物について言及するとき、その生物の個体としての量的増大をいう。成長は、具体的には体長(身長)、体重などの計測値の増加で個体の量的増大を認識することができる。個体の量的増大は細胞の増大および細胞数の増加に依存する。
【0043】
本明細書において「実質的に同じ成長(または生長)」とは、生物について言及するとき、その生物が、比較対象となる生物(例えば、遺伝形質の変換前の生物)と比較して、成長速度がほとんど変化しないことをいう。そのような成長速度がほとんど変化しない範囲には、例えば、通常の成長の統計分布における1偏差分以内に入ることなどが挙げられるがそれに限定されない。また、本発明の生物では、例えば、(1)子供の数が変わらない;(2)形態は変化するが、通常の人為的突然変異と違って、障害的ではなく、変異率が極めて高いにも拘わらず、見た目が“美しい”と予想される。(成長とは直接関係しないが、本発明の方法により創出された変異体の特徴だと考えられる);(3)一度獲得した形質あるいは遺伝型あるいは表現型は後もどりしないなどの効果が得られる。
【0044】
本明細書において生物の「生産」とは、ある生物について言及するとき、その生物の個体を作り出すことをいう。
【0045】
本明細書において生物の「再生産」とは、ある生物について言及するとき、親個体から次の世代の新たな個体を作り出すことをいう。再生産には、生殖、繁殖など天然現象によるもの、クローン(核移植)技術などの人工技術などによるものが包含されるがそれらに限定されない。再生産に用いられる技術としては、例えば、植物の場合、1培養細胞から個体ができる;接木、挿し木などが挙げられるがそれらに限定されない。再生産によって生産された生物は、通常、親に由来する遺伝形質を有する。有性生殖によって再生産される生物では、再生産された生物は、通常2つの性にそれぞれ由来する遺伝形質を有する。通常、そのような由来遺伝形質は、2つの性に由来するものをほぼ等しい割合で有する。無性生殖によって再生される生物では、通常再生産された生物は、親に由来する遺伝形質を有する。
【0046】
本明細書において使用される「細胞」は、当該分野において用いられる最も広義の意味と同様に定義され、多細胞生物の組織の構成単位であって、外界を隔離する膜構造に包まれ、内部に自己再生能を備え、遺伝情報およびその発現機構を有する生命体をいう。本明細書において使用される細胞は、天然に存在する細胞であっても、人工的に改変された細胞(例えば、融合細胞、遺伝子改変細胞)であってもよい。細胞の供給源としては、例えば、単一の細胞培養物であり得、あるいは、正常に成長したトランスジェニック動物の胚、血液、または体組織、または正常に成長した細胞株由来の細胞のような細胞混合物が挙げられるがそれらに限定されない。
【0047】
本発明において使用される細胞は、どの生物由来の細胞(たとえば、任意の種類の単細胞生物(例えば、細菌、酵母)または多細胞生物(例えば、動物(たとえば、脊椎動物、無脊椎動物)、植物(たとえば、単子葉植物、双子葉植物など)など))でもよい。例えば、脊椎動物(たとえば、メクラウナギ類、ヤツメウナギ類、軟骨魚類、硬骨魚類、両生類、爬虫類、鳥類、哺乳動物など)由来の細胞が用いられ、より詳細には、哺乳動物(例えば、単孔類、有袋類、貧歯類、皮翼類、翼手類、食肉類、食虫類、長鼻類、奇蹄類、偶蹄類、管歯類、有鱗類、海牛類、クジラ目、霊長類、齧歯類、ウサギ目など)由来の細胞が用いられる。1つの実施形態では、霊長類(たとえば、チンパンジー、ニホンザル、ヒト)由来の細胞、特にヒト由来の細胞が用いられるがそれに限定されない。本発明において用いられる細胞は、上記細胞は、幹細胞であってもよく体細胞であってもよい。そのような細胞は、移植目的に使用されるものであってもよい。あるいは、植物細胞としては、好ましくは、顕花植物(単子葉または双子葉)由来の細胞が用いられ、より好ましくは双子葉植物細胞が用いられ、より好ましくはイネ科、ナス科、ウリ科、アブラナ科、セリ科、バラ科、マメ科、ムラサキ科の植物由来の細胞が用いられる。さらに好ましくは、コムギ、トウモロコシ、イネ、オオムギ、ソルガム、タバコ、ピーマン、ナス、メロン、トマト、イチゴ、サツマイモ、アブラナ、キャベツ、ネギ、ブロッコリー、ダイズ、アルファルファ、アマ、ニンジン、キウリ、柑橘類、ハクサイ、レタス、モモ、ジャガイモ、ムラサキ、オウレン、ポプラおよびリンゴ由来の細胞が用いられる。植物細胞は、植物体の一部、器官、組織、培養細胞などであり得る。細胞、組織、器官または個体の形質転換法は、当該分野で周知である。そのような技術は、本発明において引用した文献などに十分記載されている。核酸分子の生物細胞への導入は、一過的であっても恒常的であってもよい。一過性または恒常性の遺伝子導入の技術はそれぞれ当該分野において周知である。本発明において用いられる細胞を分化させて形質転換植物を作出する技術もまた当該分野において周知であり、そのような技術は、本発明において引用した文献などに十分記載されていることが理解される。形質転換植物から種子を得る技術もまた、当該分野において周知であり、そのような技術は、本発明において引用した文献などに記載されている。
【0048】
本明細書において「単離された」とは、通常の環境において天然に付随する物質が少なくとも低減されていること、好ましくは実質的に含まないをいう。従って、単離された細胞とは、天然の環境において付随する他の物質(たとえば、他の細胞、タンパク質、核酸など)を実質的に含まない細胞をいう。核酸またはポリペプチドについていう場合、「単離された」とは、たとえば、組換えDNA技術により作製された場合には細胞物質または培養培地を実質的に含まず、化学合成された場合には前駆体化学物質またはその他の化学物質を実質的に含まない、核酸またはポリペプチドを指す。単離された核酸は、好ましくは、その核酸が由来する生物において天然に該核酸に隣接している(flanking)配列(即ち、該核酸の5’末端および3’末端に位置する配列)を含まない。
【0049】
本明細書において、「樹立された」または「確立された」細胞とは、特定の性質(例えば、多分化能)を維持し、かつ、細胞が培養条件下で安定に増殖し続けるようになった状態をいう。
【0050】
本明細書において「乳酸」とは、α-ヒドロキシプロピオン酸のことであり、L型は、解糖の最終生成物であり,乳酸脱水素酵素の作用によりピルビン酸の還元によって生成される。
【0051】
本明細書において「乳酸耐性」とは、乳酸に対して耐性を有することをいう。そのような耐性とは、微量の乳酸の存在に対して、死滅しないことを少なくとも意味するが、より好ましくは、乳酸の存在下で成長することを意味し得る。
【0052】
本明細書において使用され得る、乳酸としては、L型、D型、ラセミ体など、自然界に存在し得る任意の形態を挙げることができるが、好ましくはL型である。これらはそれぞれ単独で用いてもよく、2種以上を併用してもよい。
【0053】
乳酸耐性は以下のようにして決定する。乳酸耐性度を評価したい微生物を、乳酸を含まない培地で培養し、一晩培養液を準備する。各種濃度の乳酸を含む培地を準備し、一晩培養液を0.1〜10%(v/v)添加し、一晩培養する。加える菌の体積は目的や菌の種類によって決める。一晩培養後の菌濃度を吸光光度計をもいる方法、寒天プレート法、トリパンブルー染色による生存率測定などで決定する。中性付近の培地を用いた場合と同等の増殖性が認められた乳酸濃度の数値を、その濃度の乳酸耐性を持つと評価する。
【0054】
本発明の方法において、微生物に乳酸を加えて選択培養する工程は、寒天培地上に変異処理した乳酸感受性微生物を塗布した上に乳酸を目的の濃度比率で重層しても良く又は液体培養において変異処理した乳酸感受性菌及び目的乳酸を一緒に入れて培養しても良い。また、pH、温度及び通気の割合(通気する場合)等の条件も当業者によって容易に設定され得るが、好ましくは、その微生物が成育する条件が良いことは言うまでもない。
【0055】
(一般生化学・分子生物学)
(一般技術)
本明細書において用いられる分子生物学的手法、生化学的手法、微生物学的手法は、当該分野において周知であり慣用されるものであり、例えば、Sambrook J.et al.(1989).Molecular Cloning:A Laboratory Manual,Cold Spring Harborおよびその3rd Ed.(2001);Ausubel,F.M.(1987).Current Protocols in Molecular Biology,Greene Pub.Associates and Wiley−Interscience;Ausubel,F.M.(1989).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associat ES and Wiley−Interscience;Innis,M.A.(1990).PCR Protocols:A Guide to Methods and Applications,Academic Press;Ausubel,F.M.(1992).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates;Ausubel,F.M.(1995).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates;Innis,M.A.et al.(1995).PCR Strategies,Academic Press;Ausubel,F.M.(1999).Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Wiley,and annual updates;Sninsky,J.J.et al.(1999).PCR Applications:Protocols for Functional Genomics,Academic Press、別冊実験医学「遺伝子導入&発現解析実験法」羊土社、1997などに記載されており、これらは本明細書において関連する部分(全部であり得る)が参考として援用される。
【0056】
人工的に合成した遺伝子を作製するためのDNA合成技術および核酸化学については、例えば、Gait,M.J.(1985).Oligonucleotide Synthesis:A Practical Approach,IRLPress;Gait,M.J.(1990).Oligonucleotide Synthesis:A Practical Approach,IRL Press;Eckstein,F.(1991).Oligonucleotides and Analogues:A Practical Approach,IRL Press;Adams,R.L.etal.(1992).The Biochemistry of the Nucleic Acids,Chapman&Hall;Shabarova,Z.et al.(1994).Advanced Organic Chemistry of Nucleic Acids,Weinheim;Blackburn,G.M.et al.(1996).Nucleic Acids in Chemistry and Biology,Oxford University Press;Hermanson,G.T.(I996).Bioconjugate Techniques,Academic Pressなどに記載されており、これらは本明細書において関連する部分が参考として援用される。
【0057】
以下に好ましい実施形態の説明を記載するが、この実施形態は本発明の例示であり、本発明の範囲はそのような好ましい実施形態に限定されないことが理解されるべきである。
【0058】
1つの局面において、本発明は、乳酸耐性を有する生物であって、該生物は、該生物の天然型が有する耐性よりも高い耐性を有する、生物を提供する。
【0059】
1つの実施形態において、本発明において対象となる乳酸は、L型乳酸、D型乳酸、L型・D型乳酸のラセミ体からなる群より選択されるタイプであり得る。
【0060】
1つの実施形態において、本発明は、乳酸に対して少なくとも1%の濃度の耐性を有する生物を提供する。これまで、乳酸に対して多くとも1%未満の濃度までしか耐性がある生物がほとんど存在していなかった。特に乳酸菌以外では、1%未満程度の耐性しか報告されておらず、特に2%以上の耐性を持つ生物を生産することは不可能と考えられていた。また、交叉によっても乳酸耐性が得られたとの報告は皆無である。従って、本発明は、従来存在しなかった生物を提供するという顕著な効果を奏する。好ましい実施形態では、本発明は、乳酸に対して少なくとも2%の、好ましくは、少なくとも3%の、より好ましくは少なくとも4%の、より好ましくは少なくとも5%の、さらに好ましくは少なくとも6%の、さらに好ましくは少なくとも7%の、さらに好ましくは少なくとも8%の、さらに好ましくは少なくとも9%の、さらにより好ましくは少なくとも10%の、さらに好ましくは少なくとも15%の、最も好ましくは少なくとも20%の濃度の耐性を有する生物を提供する。ここで、乳酸に対する耐性は、その耐性とすべき環境中に少なくとも18時間、通常、少なくとも1日間、好ましくは2日間、好ましくは3日間、好ましくは4日間、好ましくは4日間、さらに好ましくは6日間生存している(少なくとも生存率が10%以上であり、好ましくは少なくとも生存率が20%である。本発明は、生存率は、概して80%以上であり、優れた生存率を示す。)ことを意味する。
【0061】
別の実施形態において、本発明の生物は、任意の微生物であり、さらに好ましくは、真核生物であり、さらにより好ましくは酵母である。特に、Saccharomyces俗の生物が好ましく、さらに好ましくはSaccharomyces cervisiaeが選択される。別の実施形態では、本発明の生物は、原核生物であり得る。酵母については、乳酸1%でも耐性が獲得できていなかった。
【0062】
本発明で得られた株の代表例は、〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに受領番号NITE P−122号(寄託日平成17年7月29日;7%乳酸を含むYPD培地中で増殖可能)として寄託済みである。
【0063】
別の局面において、本発明は、乳酸に対する耐性が改善された生物を作製するための方法を提供する。この方法は:A)目的の生物を、該乳酸について、正常状態での最大許容濃度で、好ましくは1日程度培養する工程;B)該目的の生物を、該乳酸について、改善について所望の濃度で培養する工程;およびC)生存する生物を選択し再生産する工程を包含する。このような再生産の方法には、当該分野において公知の技術を応用できることが理解される。一度最大許容濃度よりも低い濃度で培養しておき、その後、所網の最大許容濃度より高い濃度で培養することによって、より効率所望の濃度で耐性を有する生物を効率よく獲得することができたことに特徴がある。より具体的にいうと、本発明の1つの実施形態では、本発明の方法は、A)正常状態での最大許容濃度で1日間培養する工程、B)Aより1%高い乳酸濃度で1日間培養する工程、C)Bより1%高い乳酸濃度で1日間培養する工程、B)〜C)を繰り返して、最終的に耐性を付与したい乳酸濃度で1日間培養することによって、より効率所望の濃度で耐性を有する生物を効率よく獲得することができたことに特徴がある。表3、図4に示されているように、5%乳酸耐性株は同じ乳酸濃度で継代を繰り返すと増殖速度が早くなっていく。培養工程A)と培養工程B)との間にA’)最大許容濃度より1%高い濃度の乳酸中で培養する工程、および必要に応じてA’’)該培養工程A’)における濃度より1%高い濃度の乳酸中で培養する工程を行い、必要に応じてA’’)において1%ずつ高い濃度の乳酸中での培養工程を繰り返して行って所望の濃度での培養を達成することを包含する
好ましい実施形態では、本発明の方法は、D)生物の遺伝子の複製におけるエラープローン頻度を調節する工程をさらに包含する。
【0064】
別の実施形態において、本発明において使用されるエラープローン頻度を調節する工程は、生物のDNAポリメラーゼのエラープローン頻度を調節することを包含することが好ましい。本発明において使用されるDNAポリメラーゼは、真核細胞におけるDNAポリメラーゼα、DNAポリメラーゼβ、DNAポリメラーゼγ、DNAポリメラーゼδおよびDNAポリメラーゼεならびにそれらに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを含む。
【0065】
好ましい実施形態において、本発明において使用されるDNAポリメラーゼは、真核細胞におけるDNAポリメラーゼδおよびDNAポリメラーゼεならびにそれに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを包含する。
【0066】
好ましい実施形態では、本発明において用いられる生物は、真核生物(例えば、動物、植物、真菌または酵母の細胞)であり、より好ましくは酵母である。酵母は、種々の環境因子に対して耐性を獲得しにくいという特徴があり、本発明では、これを乳酸に対する耐性獲得という形で従来不可能であった酵母の生産に結び付けたという点で顕著である。
【0067】
別の実施形態では、本発明において用いられる生物は、分離した細胞の状態で存在する。
【0068】
好ましい実施形態では、本発明において得られる生物は、所望の形質の変換後も、野生型と実質的に同じ成長を示す。古典的な変異誘発方法では、野生型と実施的に同じ成長を示すものはほとんど取れなかった。従って、本発明は、乳酸に対する耐性を持ちながら、かつ、野生型と実施的に同じ成長を呈する(正常型とほぼ同じ)性質を有する生物を提供するという点で従来技術では提供し得なかった生物を提供するという効果を奏する。
【0069】
別の実施形態では、本発明の方法において用いられる所望の濃度は、最大許容濃度よりも高い濃度であることが好ましい。最大許容濃度は、本発明の方法を実施する直前に実際に測定してもよく、あるいは、文献などから既知の数値を入手して使用してもよい。
【0070】
別の局面では、本発明は、本発明の方法によって生産された生物に関する。このような生物は、乳酸に対する耐性が従来技術にも増して増大しており、従来技術では入手できなかった乳酸耐性を有する生物であり得る。
【0071】
別の局面において、本発明は、生物の乳酸耐性能を評価する方法を提供する。この方法は、A)該生物を、該生物が通常生育する条件下で生育させる工程;B)該乳酸存在下または不存在下で該生物を生育させる工程;およびC)該生物が死滅する該乳酸の濃度を決定する工程、を包含する。
【0072】
1つの実施形態において、生物が死滅しているか否かの決定は、その細胞が形を維持している細胞かどうかを判定基準として用いることができる。形を維持しているだけの細胞は、生存しているかどうかが不明であることから、トリパンブルー染色などにより従来はさらなる測定をする必要があったが、本発明において、乳酸中では、形を維持すること自体が生存の重要な指標であることが明らかになった。従って、この形の維持を指標とする評価方法は、従来技術に比して顕著に簡便な判定方法を提供する。
【0073】
ここで、形を維持しているかどうかの判定は、形を維持している(細胞が生存している)と判断する基準は、きれいな球形で細胞の輪郭が明確な場合である。形を維持できていないと判断する基準は、きれいな球形でなく輪郭が薄くしわがよって、細胞内が薄く透けている場合である。
【0074】
別の局面において、本発明は、生物の乳酸耐性能を評価する方法を提供する。この方法は、A)該生物を、該生物が通常生育する条件下で生育させる工程;B)該乳酸存在下または不存在下で該生物を生育させる工程;およびC)該生物が死滅する該乳酸の濃度を決定する工程、を包含する。
【0075】
1つの実施形態において、上記生物が死滅しているか否かの決定は、その細胞が形を維持している細胞かどうかを判定基準として用いてもよい。
【0076】
別の局面では、本発明はまた、菌学的性質は、乳酸耐性以外は野生型と同じである生物を提供する。このような生物は、古典的な変異方法では、入手不可能であった。
【0077】
本明細書において引用された、科学文献、特許、特許出願などの参考文献は、その全体が、各々具体的に記載されたのと同じ程度に本明細書において参考として援用される。
【0078】
以上、本発明を、理解の容易のために好ましい実施形態を示して説明してきた。以下に、実施例に基づいて本発明を説明するが、上述の説明および以下の実施例は、例示の目的のみに提供され、本発明を限定する目的で提供したのではない。従って、本発明の範囲は、本明細書に具体的に記載された実施形態にも実施例にも限定されず、特許請求の範囲によってのみ限定される。
【実施例】
【0079】
以下に実施例を示して本発明をさらに詳しく説明するが、この発明は以下の例に限定されるものではない。以下の実施例において用いられる試薬類、例外を除き、Sigma(St.Louis,USA、和光純薬(大阪、日本)などから市販されるものを用いた。以下において使用した動物は、日本の大学において規定される飼育規準を遵守して飼育および実験した。以下、実施例により本発明をさらに具体的に説明する。
【0080】
(実施例1:乳酸添加による培地pHの変化と細胞増殖への影響の検討)
(酵母株と試薬)
大阪市立大学の下田らから供与された変異型pol−3遺伝子発現株(BYD5、以下pol−3変異株と略す。BYD5は、受託番号 NITE P−131(寄託日平成17年8月19日)として〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに寄託済みである)と野生型pol−3遺伝子発現株(AMY52−3D、以下、野生株と略す。AMY52−3Dは、受託番号 NITE P−129(寄託日平成17年8月19日)として〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに寄託済みである。)を用いた。BYD5の遺伝子型は、〔pol3−01/pol3−01、MATa/MATα、lys1−1/lys1−1〕であり、AMY52−3Dの遺伝子型は、〔MATα、ura3−52、leu2−1、lys1−1、ade2−1、his1−7、hom3−10、trp1−289、canR〕である。
【0081】
試験に用いた液体培地のYPD medium(cat.#630409)および寒天培地のYPD agar medium(cat.#630410)はBD Biosciences Clontechから購入し、オートクレーブ(120℃、15分)により滅菌した。添加したL−乳酸(以下、乳酸と略す。cat.# 129−02666)は和光純薬から購入した。
【0082】
(方法)
液体培地に乳酸濃度が0%〜20%(v/v)となるように添加し、それぞれのpHを測定した。
【0083】
寒天培地プレートに展開したpol−3変異株および野生株のコロニーを白金耳で掻きとり、YPD培地4mLを入れた試験管(直径18mm)に植菌した。これらを30℃、180 rpmで一晩(16時間以上)振とう培養した。
【0084】
それぞれの培養液の一部をとり、血球計算盤を用いて細胞密度を測定し、両株の細胞密度が等しくなるように液体培地で希釈した。
【0085】
培養開始時の細胞密度が約2.0×106 cells/mLとなるように各培養液をマイクロチューブにとり、5000xgで3分間遠心して細胞を回収した。
【0086】
乳酸濃度1%、3%、5%、6%、7%、8%、9%、10%、11%、13%、15%、17%、19%、20%(v/v)の液体培地4mLに回収した細胞を再懸濁し、30℃、180 rpmで18時間振とう培養した。なお、培養は直径18mmの試験管を使用した。
【0087】
培養後、血球計算盤を用いて細胞密度を測定し、乳酸添加により細胞増殖に与える影響を比較した。
【0088】
(結果)
乳酸添加による培地pHの検討
乳酸を液体培地に添加したときのpHの変化(表1・図2)および培地の変化(図1)を以下に記す。
【0089】
乳酸を添加することによりYPD培地のpHが著しく低下することが明らかになった(表1)。
【0090】
表1 乳酸添加によるYPD培地のpH変化
【0091】
【表1】
。
【0092】
乳酸を添加することにより培地成分の一部が析出し、培地が濁ることが分かった(図1)。
【0093】
乳酸添加による細胞増殖に与える影響の検討
野生株およびpol−3変異株を乳酸含有培地で18時間培養したときの細胞密度を測定した結果を以下に記す(表2および図2)。
【0094】
表2 乳酸添加による野生株およびpol−3変異株の細胞密度の比較(18時間培養)
【0095】
【表2】
。
【0096】
野生株のおよびpol−3変異株の細胞密度は乳酸添加によって低下した。特に、乳酸の添加量が5%を超えるとほとんど増殖できないことが明らかになった。
【0097】
乳酸添加による培地pHの変化および両株の細胞増殖に相関が認められることから、培地のpH低下により両株の細胞増殖が阻害されていることが示された。
【0098】
(実施例2:乳酸耐性酵母の作製方法)
本実施例では、pol−3変異株に乳酸耐性を付与した方法を以下に記す。
【0099】
(使用した酵母株)
実施例1に同じく、pol−3変異株(BYD5)を使用した。
【0100】
(方法)
(1)pol−3変異株のコロニーをかきとり、液体培地4mL を入れた試験管(直径18mm)に植菌し、30℃、180 rpmで一晩培養した。
【0101】
(2)培養液を15mL容の遠沈管(cat.#430791/コーニング)にとり、1000xgで5分間遠心して全ての細胞を回収した。
【0102】
(3)液体培地100mLを入れた300mL容三角フラスコに回収した全ての細胞を再懸濁し、30℃、180 rpmで一晩培養した。
【0103】
(4)培養液を50mL容遠沈管(cat.#430829/コーニング)にとり、1000xgで5分間遠心して全ての細胞を回収した。
【0104】
(5)3%(v/v)乳酸培地150mLを入れた300mL容三角フラスコに回収した全ての細胞を懸濁し、30℃、180 rpmで一晩培養した。
【0105】
(6)3%乳酸培養液の一部をとり、等量の0.1%トリパンブルー溶液(cat.# 345−07421/DOJINDO)と混合して細胞を染色した。青色に染まった細胞は死滅細胞、染まっていない細胞は生存細胞であるため、生存細胞の割合を観察した。
【0106】
(7)生存細胞の存在を確認した後、3%乳酸培養液の1/3量(50mL)を50mL容遠沈管にとり、1000xgで5分間遠心して細胞を回収した。
【0107】
(8)4%(v/v)乳酸培地150mLを入れた300mL容三角フラスコに回収した細胞を懸濁し、30℃、180 rpmで一晩培養した。
【0108】
(9)4%乳酸培養液の一部をとり、トリパンブルー染色により細胞を観察した。生存細胞を確認した後、1/3量(50mL)の培養液を50mL容遠沈管にとり、1000xgで5分間遠心して細胞を回収した。
【0109】
(10)5%(v/v)乳酸培地150mLを入れた300mL容三角フラスコに回収した細胞を懸濁し、30℃、180 rpmで一晩培養した。
【0110】
(11)5%乳酸培養液の一部をとり、トリパンブルー染色により生存細胞の確認を行った。
【0111】
(12)5%乳酸培養液1/10量(15mL)をとり、1000xgで5分間遠心して細胞を回収した。5%乳酸培地70mLを入れた200mL容振とうフラスコ(cat.# 4070FK 200/IWAKI)に回収した細胞を懸濁した後、細胞密度を測定した。測定後、これらの培養液は30℃、180 rpmで振とう培養した。
【0112】
(13)24時間ごとに培養液の細胞密度を測定し、培養開始直後の細胞密度に対して約5〜10倍になるまで培養を行った。
【0113】
(14)5%乳酸培地で増殖が確認された培養液から1/10量(7mL)をとり、1000xgで5分間遠心して細胞を回収した。これらを再び5%乳酸培地に懸濁し、30℃、180 rpmで振とう培養した。
【0114】
(11)〜(14)の操作を繰り返すことで継代培養を行った。なお、継代培養の基準として、1.トリパンブルー染色により、生存細胞を確認すること 2.培養開始直後の細胞密度に対して約5〜10倍増殖していること、以上の2点を設定した。
【0115】
(結果)
培地の乳酸濃度を徐々に上げ、最終的に5%(v/v)乳酸培地で継代培養を行ったときの細胞密度を以下に記す(表3・図4)。
【0116】
(1)pol−3変異株を3%乳酸培地で培養した結果、細胞密度が増大し、また、生存細胞も多く認められた。
【0117】
(2)3%乳酸培養で増殖した株を4%乳酸培地へ移植した結果、細胞密度の増大および生存細胞が認められた。
【0118】
(3)4%乳酸培養で増殖した株を5%乳酸培地へ移植した結果、培養開始から4日目に細胞密度の増大が認められた。
【0119】
(4)トリパンブルー染色の結果、5%乳酸培養において多くの生存細胞が確認された(図3)。
【0120】
(5)5%乳酸培養で増殖した株を再び5%乳酸培地へ移植し、同培地で継代培養を繰り返した。この結果、継代回数が増すごとに細胞増殖の速度が高まる傾向が認められた(表3・図4)。
以上の結果、本方法によりpol−3変異株に5%乳酸耐性を付与することができることが示された。
表3 5%乳酸培養におけるpol−3変異株の細胞増殖
【0121】
【表3】
(実施例3:5%乳酸耐性株の培養特性)
5%乳酸に対して耐性を獲得したpol−3変異株(5%乳酸耐性株と記す)の培養特性を検討した方法を以下に記す。
【0122】
(使用した酵母株)
実施例1に同じく、野生株および、実施例2によって獲得した5%乳酸耐性株を使用した。
【0123】
(方法)
実施例2によって獲得した5%乳酸耐性株は、5%乳酸培地70mLを入れた200mL容バッフル付き三角フラスコで継代培養を行っている。培養は30℃、200 rpmで行い、前培養液の1/10程度の細胞を継代している。
【0124】
(1)野生株のコロニーをかきとり、液体培地4mLを入れた試験管(φ18mm)に植菌し、30℃、180 rpmで一晩培養した。
【0125】
(2)野生株の培養液を5000xg、3分間遠心して細胞を回収し、液体培地70mLを入れた200mL容バッフル付き三角フラスコに懸濁した。これを30℃、200 rpmで回転振とう培養した。
【0126】
(3)5%乳酸培地で継代している5%乳酸耐性株、および前培養した野生株を50mL容遠沈管にとり、1000xgで5分間遠心して細胞を回収した。
【0127】
(4)回収した細胞を数mLの乳酸無添加の液体培地に再懸濁した後、血球計算盤で細胞密度を測定した。
【0128】
(5)測定した細胞密度をもとに、野生株および5%乳酸耐性株の細胞密度が等しくなるように乳酸無添加の液体培地で希釈した。
【0129】
(6)最終細胞密度が約2.0×107cells/mLとなるように細胞希釈液を15mL容の遠沈管にとり、1000xgで5分間遠心して細胞を回収した。5%、6%および7%の乳酸培地70mLを入れた200mL容バッフル付き三角フラスコに回収した細胞を懸濁し、30℃、200 rpmで回転振とう培養した。なお、野生株については5%のみ培養を行った。また、これらの乳酸培地は0.45μmの滅菌フィルター(ADVANTEC/cat.# 25CS045AS)で濾過して使用した。
【0130】
(7)懸濁直後を0日目とし、24時間ごとに3日間の細胞密度を測定した。なお、血球計算盤による測定は、1測定につき3回行い、平均値および標準偏差を算出した。
【0131】
(結果)
野生株および5%乳酸耐性株を各乳酸培地で培養したときの細胞密度の変化を以下に記す(図5および表4・図6)。
【0132】
(1)5%乳酸培養の結果、5%乳酸耐性株は野生株に比べて顕著な増殖が認められ、また、野生株はほとんど増殖していないことが分かった(図5)。
【0133】
(2)5%乳酸耐性株は、5%および6%乳酸を含む培養条件において速やかに増殖し、培養3日目では約10倍に増殖した(表4、図6)。
【0134】
(3)5%乳酸耐性株を7%乳酸培地で3日間培養したが顕著な増殖は認められなかった(表4、図6)。
【0135】
以上の結果、pol−3変異株を乳酸に馴化させることによって取得した5%乳酸耐性株は、5%および6%乳酸に対して耐性を獲得していることが分かった。
表4 野生株および5%乳酸耐性株の乳酸添加による細胞増殖への影響
【0136】
【表4】
(実施例4:7%乳酸耐性株の作製)
実施例2および3に記した5%乳酸耐性株に7%乳酸への耐性を付与した方法と結果を以下に記す。
【0137】
(方法)
実施例2および3によって取得した5%乳酸耐性株(pol−3変異株)は、5%(v/v)乳酸培地70mLを入れた200mL容バッフル付き三角フラスコを用いて30℃、200 rpmで継代培養を行っている。新しい培地への継代は2〜3日ごとに行い、全培養液に対して1/10量の細胞を回収し、移植している。
【0138】
(1)5%乳酸耐性株の継代培養から全培養液の1/5量をとり、1000xgで5分間遠心して細胞を回収した。7%(v/v)乳酸培地70mLを入れた200mL容バッフル付き三角フラスコに回収した細胞を懸濁し、30℃、200 rpmで一晩培養した。(継代1回目)
(2)全培養液の1/5量をとり、1000xgで5分間遠心して細胞を回収した。新鮮な7%乳酸培地70mLに回収した細胞を懸濁し、30℃、200 rpmで5日間培養した。(継代2回目)
(3)全培養液の1/10量をとり、1000xgで5分間遠心して細胞を回収した。新鮮な7%乳酸培地70mLに回収した細胞を懸濁し、30℃、200 rpmで5日間培養した。(継代3回目)
(4)全培養液の1/10量をとり、1000xgで5分間遠心して細胞を回収した。新鮮な7%乳酸培地70mLに回収した細胞を懸濁し、30℃、200 rpmで4日間培養した。(継代4回目)
(5)4回目の継代培養株および液体培地で一晩前培養した野生株の一部を7%乳酸培地に移植し、30℃、200 rpmで一晩回転振とう培養した。これらの細胞についてトリパンブルー染色を行い、生存細胞を顕微鏡観察することによって7%乳酸への耐性を確認した。
【0139】
(結果)
5%乳酸耐性株を7%乳酸培地で継代培養したときの細胞密度の変化を以下に記す(表5・図7)。
【0140】
(1)継代培養2回目において5日間培養したところ、継代直後に比べて約6.7倍の増殖が認められた(表5・図7)。
【0141】
(2)継代培養3回目において5日間培養したところ、継代直後に比べて約66.9倍の増殖が認められた(表5・図7)。
【0142】
(3)継代培養4回目において4日間培養したところ、継代直後に比べて約310倍の増殖が認められた(表5・図7)。
【0143】
(4)トリパンブルー染色の結果、野生株はほぼ全ての細胞が死滅したにも関わらず、乳酸耐性株では多くの生存細胞が観察された(図8)。
【0144】
上記の結果、pol−3変異株から取得した5%乳酸耐性株は7%乳酸培養に馴化させることにより、速やかに増殖することが可能となった。すなわち、本株は7%乳酸に対して耐性を獲得した。
表5 5%乳酸耐性株を7%乳酸培地で継代培養したときの細胞密度の変化
【0145】
【表5】
本実施例で得られた株の代表例は、〒292−0818 千葉県木更津市かずさ鎌足2−5−8 独立行政法人製品評価技術基盤機構 特許生物寄託センターに受領番号NITE P−122(乳酸耐性株(BYD5-LA7)受託番号 7%乳酸を含むYPD培地中で増殖可能)として寄託済み(平成17年7月29日)である。
【0146】
以上のように、本発明の好ましい実施形態を用いて本発明を例示してきたが、本発明は、特許請求の範囲によってのみその範囲が解釈されるべきであることが理解される。当業者は、本発明の具体的な好ましい実施形態の記載から、本発明の記載および技術常識に基づいて等価な範囲を実施することができることが理解される。本明細書において引用した特許、特許出願および文献は、その内容自体が具体的に本明細書に記載されているのと同様にその内容が本明細書に対する参考として援用されるべきであることが理解される。
【産業上の利用可能性】
【0147】
乳酸耐性酵母は高濃度の乳酸を産生させるための宿主として利用する。乳酸や乳酸ナトリウム、乳酸カルシウムは、すでに輸液等の医薬品原料・食品添加物・化粧品原料として使用されている。また、乳酸の重合体であるポリ乳酸は、生分解性を有するポリマーとして注目され、フィルム・樹脂製品・医療材料その他多くの分野で用途開発が進められている。
【0148】
また、微生物の乳酸耐性を増強する本発明手法を乳酸菌に適用し、経口摂取後、胃酸で死滅せず小腸もしくは結腸に生きたまま到達する酸耐性を増強させる。その結果、腸管病原菌感染に対する防御機能と免疫応答に対する効果のプロバイオテイック特性(欧州、No.92810516−2)を高めた機能性食品として利用できる。その他に、乳酸菌は潰瘍性大腸炎予防の可能性、糖尿病予防の可能性、感染症の低減効果、インフルエンザ抗体価上昇の増強作用、ピロリ菌の抑制効果などの医学的応用が期待されている。
【図面の簡単な説明】
【0149】
【図1】図1は、乳酸添加による培地の変化(左:無添加のYPD培地 右:3%乳酸を加えたYPD培地)である。
【図2】図2¥は、 乳酸添加による野生株およびpol−3変異株の細胞増殖の比較(緑丸:野生株の細胞密度、赤丸:pol−3変異株の細胞密度、青線:培地pH)である。
【図3】図3は、5%乳酸耐性株のトリパンブルー染色による観察(5%乳酸培養の2日目、青色細胞:死滅細胞 無色細胞:生存細胞)である。
【図4】図4は、5%乳酸培養におけるpol−3変異株の細胞増殖(表3より)である。
【図5】図5は、5%乳酸培養における細胞の増殖比較(1日目 左:野生株、右:5%乳酸耐性株)である。
【図6】図6は、野生株および5%乳酸耐性株の乳酸添加による細胞増殖への影響(表3より)である。
【図7】図7は、5%乳酸耐性株を7%乳酸培地で継代培養したときの細胞密度の変化(表5より)である。
【図8】図8は、7%乳酸培地で培養した野生株および乳酸耐性株のトリパンブルー染色写真(左:野生株 右:7%乳酸耐性株 ※7%乳酸培養1日目、白色が生存細胞で青色が死滅細胞を示している。)である。
【特許請求の範囲】
【請求項1】
乳酸耐性を有する生物であって、該生物は、該生物の天然型が有する耐性よりも高い耐性を有する、生物。
【請求項2】
前記乳酸は、D型、L型およびラセミ型からなる群より選択されるタイプである、請求項1に記載の生物
【請求項3】
前記耐性は、乳酸に対して少なくとも1%の濃度での耐性である、請求項1に記載の生物。
【請求項4】
前記耐性は、乳酸に対して少なくとも3%の濃度での耐性である、請求項1に記載の生物。
【請求項5】
前記耐性は、乳酸に対して少なくとも5%の濃度での耐性である、請求項1に記載の生物。
【請求項6】
前記耐性は、乳酸に対して少なくとも7%の濃度での耐性である、請求項1に記載の生物。
【請求項7】
前記耐性は、乳酸に対して少なくとも10%の濃度での耐性である、請求項1に記載の生物。
【請求項8】
前記耐性は、乳酸環境において少なくとも18時間以上生存することを包含する、請求項1に記載の生物。
【請求項9】
前記耐性は、乳酸環境において少なくとも4日以上生存することを包含する、請求項1に記載の生物。
【請求項10】
前記生物は、酵母である、請求項1に記載の生物。
【請求項11】
前記生物は、Saccharomycesに属する生物である、請求項1に記載の生物。
【請求項12】
前記生物は、Saccharomyces cerevisiaeである、請求項1に記載の生物。
【請求項13】
乳酸に対する耐性が改善された生物を作製するための方法であって、該方法は:
A)目的の生物を、該乳酸について、正常状態での最大許容濃度で培養する工程;
B)該目的の生物を、該乳酸について、改善について所望の濃度で培養する工程;および
C)生存する生物を選択し再生産する工程、
を包含する、方法。
【請求項14】
前記培養工程A)は、1日間行われる、請求項13に記載の方法。
【請求項15】
前記培養工程A)と前記培養工程B)との間にA’)前記最大許容濃度より1%高い濃度の乳酸中で培養する工程、および必要に応じてA’’)該培養工程A’)における濃度より1%高い濃度の乳酸中で培養する工程を行い、必要に応じてA’’)において1%ずつ高い濃度の乳酸中での培養工程を繰り返して行って前記所望の濃度での培養を達成することを包含する、請求項13に記載の方法。
【請求項16】
D)前記生物の遺伝子の複製におけるエラープローン頻度を調節する工程をさらに包含する、請求項13に記載の方法。
【請求項17】
前記エラープローン頻度を調節する工程は、前記生物のDNAポリメラーゼのエラープローン頻度を調節することを包含する、請求項16に記載の方法。
【請求項18】
前記DNAポリメラーゼは、真核細胞におけるDNAポリメラーゼα、DNAポリメラーゼβ、DNAポリメラーゼγ、DNAポリメラーゼδおよびDNAポリメラーゼεならびにそれらに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを含む、請求項17に記載の方法。
【請求項19】
前記DNAポリメラーゼは、真核細胞におけるDNAポリメラーゼδおよびDNAポリメラーゼεならびにそれに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを包含する、請求項15に記載の方法。
【請求項20】
前記生物は、真核生物である、請求項13に記載の方法。
【請求項21】
前記生物は、分離した細胞の状態で存在する、請求項13に記載の方法。
【請求項22】
前記生物は、動物、植物、真菌または酵母の細胞である、請求項13に記載の方法。
【請求項23】
前記生物は、酵母である、請求項13に記載の方法。
【請求項24】
前記生物は、Saccharomycesに属する生物である、請求項13に記載の方法。
【請求項25】
前記生物は、Saccharomyces cerevisiaeである、請求項13に記載の方法。
【請求項26】
前記生物は、所望の形質の変換後も、野生型と実質的に同じ成長を示す、請求項13に記載の方法。
【請求項27】
前記所望の濃度は、前記最大許容濃度よりも高い濃度である、請求項13に記載の方法。
【請求項28】
請求項13〜27のいずれか1項に記載の方法によって生産された生物。
【請求項29】
生物の乳酸耐性能を評価する方法であって、
A)該生物を、該生物が通常生育する条件下で生育させる工程;
B)該乳酸存在下または不存在下で該生物を生育させる工程;および
C)該生物が死滅する該乳酸の濃度を決定する工程、
を包含する、方法。
【請求項30】
前記生物が死滅しているか否かの決定は、その細胞が形を維持している細胞かどうかを判定基準として用いることを特徴とする、請求項29に記載の方法。
【請求項31】
菌学的性質は、乳酸耐性以外は野生型と同じである生物。
【請求項1】
乳酸耐性を有する生物であって、該生物は、該生物の天然型が有する耐性よりも高い耐性を有する、生物。
【請求項2】
前記乳酸は、D型、L型およびラセミ型からなる群より選択されるタイプである、請求項1に記載の生物
【請求項3】
前記耐性は、乳酸に対して少なくとも1%の濃度での耐性である、請求項1に記載の生物。
【請求項4】
前記耐性は、乳酸に対して少なくとも3%の濃度での耐性である、請求項1に記載の生物。
【請求項5】
前記耐性は、乳酸に対して少なくとも5%の濃度での耐性である、請求項1に記載の生物。
【請求項6】
前記耐性は、乳酸に対して少なくとも7%の濃度での耐性である、請求項1に記載の生物。
【請求項7】
前記耐性は、乳酸に対して少なくとも10%の濃度での耐性である、請求項1に記載の生物。
【請求項8】
前記耐性は、乳酸環境において少なくとも18時間以上生存することを包含する、請求項1に記載の生物。
【請求項9】
前記耐性は、乳酸環境において少なくとも4日以上生存することを包含する、請求項1に記載の生物。
【請求項10】
前記生物は、酵母である、請求項1に記載の生物。
【請求項11】
前記生物は、Saccharomycesに属する生物である、請求項1に記載の生物。
【請求項12】
前記生物は、Saccharomyces cerevisiaeである、請求項1に記載の生物。
【請求項13】
乳酸に対する耐性が改善された生物を作製するための方法であって、該方法は:
A)目的の生物を、該乳酸について、正常状態での最大許容濃度で培養する工程;
B)該目的の生物を、該乳酸について、改善について所望の濃度で培養する工程;および
C)生存する生物を選択し再生産する工程、
を包含する、方法。
【請求項14】
前記培養工程A)は、1日間行われる、請求項13に記載の方法。
【請求項15】
前記培養工程A)と前記培養工程B)との間にA’)前記最大許容濃度より1%高い濃度の乳酸中で培養する工程、および必要に応じてA’’)該培養工程A’)における濃度より1%高い濃度の乳酸中で培養する工程を行い、必要に応じてA’’)において1%ずつ高い濃度の乳酸中での培養工程を繰り返して行って前記所望の濃度での培養を達成することを包含する、請求項13に記載の方法。
【請求項16】
D)前記生物の遺伝子の複製におけるエラープローン頻度を調節する工程をさらに包含する、請求項13に記載の方法。
【請求項17】
前記エラープローン頻度を調節する工程は、前記生物のDNAポリメラーゼのエラープローン頻度を調節することを包含する、請求項16に記載の方法。
【請求項18】
前記DNAポリメラーゼは、真核細胞におけるDNAポリメラーゼα、DNAポリメラーゼβ、DNAポリメラーゼγ、DNAポリメラーゼδおよびDNAポリメラーゼεならびにそれらに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを含む、請求項17に記載の方法。
【請求項19】
前記DNAポリメラーゼは、真核細胞におけるDNAポリメラーゼδおよびDNAポリメラーゼεならびにそれに対応するDNAポリメラーゼからなる群より選択される少なくとも1つのポリメラーゼを包含する、請求項15に記載の方法。
【請求項20】
前記生物は、真核生物である、請求項13に記載の方法。
【請求項21】
前記生物は、分離した細胞の状態で存在する、請求項13に記載の方法。
【請求項22】
前記生物は、動物、植物、真菌または酵母の細胞である、請求項13に記載の方法。
【請求項23】
前記生物は、酵母である、請求項13に記載の方法。
【請求項24】
前記生物は、Saccharomycesに属する生物である、請求項13に記載の方法。
【請求項25】
前記生物は、Saccharomyces cerevisiaeである、請求項13に記載の方法。
【請求項26】
前記生物は、所望の形質の変換後も、野生型と実質的に同じ成長を示す、請求項13に記載の方法。
【請求項27】
前記所望の濃度は、前記最大許容濃度よりも高い濃度である、請求項13に記載の方法。
【請求項28】
請求項13〜27のいずれか1項に記載の方法によって生産された生物。
【請求項29】
生物の乳酸耐性能を評価する方法であって、
A)該生物を、該生物が通常生育する条件下で生育させる工程;
B)該乳酸存在下または不存在下で該生物を生育させる工程;および
C)該生物が死滅する該乳酸の濃度を決定する工程、
を包含する、方法。
【請求項30】
前記生物が死滅しているか否かの決定は、その細胞が形を維持している細胞かどうかを判定基準として用いることを特徴とする、請求項29に記載の方法。
【請求項31】
菌学的性質は、乳酸耐性以外は野生型と同じである生物。
【図1】


【図2】
【図3】
【図4】
【図5】

【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2007−61024(P2007−61024A)
【公開日】平成19年3月15日(2007.3.15)
【国際特許分類】
【出願番号】特願2005−252843(P2005−252843)
【出願日】平成17年8月31日(2005.8.31)
【出願人】(503116693)株式会社ネオ・モルガン研究所 (10)
【Fターム(参考)】
【公開日】平成19年3月15日(2007.3.15)
【国際特許分類】
【出願日】平成17年8月31日(2005.8.31)
【出願人】(503116693)株式会社ネオ・モルガン研究所 (10)
【Fターム(参考)】
[ Back to top ]