
二重特異性抗体およびその作製方法
本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびにd) 二重特異性抗体を得る段階。本発明はさらに、本発明方法によって得られ得る、二重特異性抗体に関する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、二重特異性抗体の新規作製方法、およびこれらの方法によって得られ得る、二重特異性抗体に関する。
【背景技術】
【0002】
発明の背景
ヒト免疫グロブリンG(IgG)抗体は、異なる構造特性および機能特性を有する4つのサブクラスとして存在する。IgGは2つの重鎖-軽鎖対(半分子)から構成されており、これらはヒンジ領域に位置する重鎖間ジスルフィド結合によって結合している。ヒトIgG4分子は、ヒンジ領域に位置する重鎖間ジスルフィド結合の有無によって異なる種々の分子形態として存在する。IgG4分子は、重鎖間ジスルフィド結合が両方とも形成されているか、またはいずれも形成されていない形態として存在する(6、7)。しかしながら、ヒトIgG4は、CH3ドメイン間、およびCH1ドメインとCH2ドメインとの間の比較的強力な非共有結合性相互作用のために(4)、これらの鎖間ジスルフィド結合の有無に関係なく(6、8)、溶液中では、免疫グロブリンG分子に一般的なIg重鎖2本および軽鎖2本からなる四量体として存在する。SDS-PAGEなどのサイズ決定分析によって実証されるように、非共有結合的に会合した2つの半分子は、非還元条件下で変性させた場合のみ解離する(6、9)。
【0003】
ヒトIgG4抗体が、他のIgGサブクラスとは異なり、抗原との相互作用において一価の分子として挙動することがこの数年間の間にわかってきた。血清由来のヒトIgG4は、架橋できないために、精製抗原を沈降させることができないことが見出された。このような血清由来のIgG4は機能的に一価であるが(1、2)、対照的に、組換えで作製されたIgG4は抗原との相互作用において二価として挙動する(3)。これらの知見に基づき、血清中のIgG4分子は半分子(すなわち、重鎖1本および軽鎖1本からなる分子)を交換して、同一の抗原に架橋することができない二重特異性分子を生じ得ることが提唱された(3〜5)。この半分子の交換の過程を、本明細書では「Fabアーム交換」とも称する。
【0004】
二重特異性抗体は、例えば疾患関連部位にエフェクター機構を再標的化するためのメディエーターとして使用することができるため、治療薬として興味深い可能性を有する。しかしながら、二重特異性抗体を開発する上での大きな障害の1つは、ハイブリッドハイブリドーマ法および化学的結合法などの伝統的な技術によって、十分な質および量の物質を作製することが困難であることであった(10)。
【0005】
WO 2005/062916(特許文献1)は、IgG4に基づいて、マウスにおいてインビボで多量体分子を形成する方法を記載している。さらにWO 2005/062916(特許文献1)は、異なる抗原結合特異性を有する2種類のIgG4抗体を、インビトロで、生理食塩水緩衝液中で同時にインキュベートすると、両抗原と反応し得る産物が形成されることを記載している。しかしながら、WO 2005/062916(特許文献1)では、これらの産物が凝集体であるのか、または二重特異性抗体であるのかは実証されておらず、また使用した条件下において反応の収率は低かった。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】WO 2005/062916
【発明の概要】
【0007】
驚くべきことに、還元条件下において、異なる抗原結合特異性を有する2種類のIgG4抗体またはIgG4様抗体が、高い効率で半分子の交換を行い、したがって凝集体の形成を伴わずに二重特異性抗体を形成し得ることが、今回見出された。
【0008】
したがって第1の主要な局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0009】
いかなる特定の理論によっても縛られることはないが、抗体の2つの領域が、半分子の交換を起こすその能力に重要な影響を及ぼすと考えられる。
【0010】
第一に、IgG4のようなコアヒンジ領域内にCPSC配列を有する抗体が、IgG1のようなCPPCコアヒンジ配列を有する抗体よりもより容易に交換を行うことから、半分子の交換の能力は、分子のコアヒンジ領域内の配列の差によって影響を受ける可能性がある。いかなる理論によっても縛られることはないが、CPSC配列がより可動性の高いコアヒンジをもたらし、鎖内ジスルフィド結合を形成する可能性を引き起こすと仮定される。注目すべきことにコアヒンジの構造は、タンパク質ジスルフィドイソメラーゼ(PDI)の活性ドメイン、CXXCに類似している。PDIの異なるアイソフォームのこれらのCXXCモチーフは、タンパク質におけるジスルフィド結合の形成、還元、および再構成を触媒する。したがって、いかなる特定の理論によっても縛られることはないが、IgG4様コアヒンジ配列を有する抗体は、ジスルフィド結合の再構成に関する内因性の活性を有する可能性があると考えられ、この活性を、本発明の方法において使用する条件によって促進する。
【0011】
第二に、この場合も同様に、いかなる理論によっても縛られることはないが、交換反応が起こるようにするためには、CH3領域の配列はIgG4様であるべきである、すなわち強力な半分子間の相互作用を形成しないようにすべきであることが結果から示される。
【0012】
別の主要な局面において、本発明は、本発明の方法によって得られるか、または得られ得る、単離された二重特異性抗体、およびそのような抗体を含む薬学的組成物に関する。
【0013】
さらなる局面において、本発明は、2つのIgG4様CH3領域を含む、単離された二重特異性抗体、およびそのような抗体を含む薬学的組成物に関する。
【0014】
なおさらなる局面において、本発明は、以下の段階を含む、所望の特性を有する二重特異性抗体を選択する方法に関する:
a) 一組の抗体を提供する段階であって、各抗体が異なる標的特異性を有し、かつ各抗体がIgG4様CH3領域を含む段階、
b) 還元条件下で、該一組の抗体の各抗体を、該一組の別の抗体と共にインキュベートし、それによって一組の抗体混合物を作製する段階であって、各混合物が異なる二重特異性抗体を含む段階、
c) 結果として得られた一組の抗体混合物を所与の所望の特性についてアッセイする段階、および
d) 所望の特性を有する二重特異性抗体混合物を選択する段階。
【図面の簡単な説明】
【0015】
【図1】精製組換えIgG1およびIgG4のSDS-Page解析を示す。精製後、Betv1およびFeld1のIgG1抗体およびIgG4抗体を、非還元SDS-PAGEで解析した。
【図2】様々な時点での、nu/nu Balb/cマウスにおける二重特異性IgGレベルを示す。異種架橋アッセイ法で決定した二重特異性IgGの量を、Bet v 1結合試験で決定したBet v 1特異的IgGの量に対してプロットした。IgG1およびIgG4含有血漿試料によるデータを、それぞれ白記号および黒記号で表す。点線は、IgGの半分子の交換が無作為でかつ完全である場合の、二重特異性IgGの計算された量を表す。
【図3】二重特異性ヒトIgG4分子がインビボで生成されることを示す。(A) SCIDマウスの群(n=5)にキメラ抗体混合物:IgG1-Betv1 100μg/IgG1-Feld1 100μg(四角)、IgG4-Betv1 100μg/IgG4-Feld1 100μg(丸)、または3) IgG4-Betv1 100μg/IgG4-Feld1 100μg+非関連組換えIgG4(IgG4-EGFR) 2,000μg(三角)を注射した。血漿中のBet v 1およびFel d 1に対する二重特異性活性を評価することによって、二重特異性抗体の生成を時間を追って追跡した。全IgG-Bet v 1濃度に対する二重特異性IgGの割合をパーセントとして表した。星印の付いた矢印は、過剰な非関連IgG4の存在下で、IgG4-Betv1/IgG4-Feld1を投与したマウスにおいて予測される二重特異性反応性レベル(4%)を示し、星印のない矢印は、IgG4-Betv1/IgG4-Feld1混合物を投与したマウスにおいて予測される二重特異性反応性レベル(50%)を示す。エラーバーはSEMを表す。(B) マウス血漿における、放射標識Fel d 1の、Fel d 1結合セファロースに対する架橋を評価することによって、単一特異性架橋活性を試験した。単一特異的反応性は、IgGのクリアランスを補正するために、架橋によって結合した放射標識Fel d 1の量と全IgG-Feld1との比として表した。エラーバーはSEMを表す。
【図4】マウス血漿中の二重特異性活性のSEC解析を示す。IgG4混合物を投与したマウスからt=24時間の時点で採取した血漿(10μl)を、Superdex200カラムで分画した。マウスに、Bet v 1結合IgG4 300μgおよびFel d 1結合IgG4 300μgを含む混合物を投与した。その画分中で、Fel d 1特異的IgG(■)の濃度を抗原結合試験で測定し、二重特異性IgG Bet v 1-Fel d 1(●)の濃度をBet v 1-Fel d 1架橋アッセイ法で決定した。IVIgを用いたこのカラムの較正から、単量体、二量体、および凝集型IgGはそれぞれ12.9 ml、11.0 ml、および8.4 mlに溶出されることが明らかになった(データは示さず)。
【図5】全血成分中のIgGの交換を示す。全血、血液細胞、血漿、および血清中で、キメラIgG混合物を37℃で24時間インキュベートし、その後異種架橋アッセイ法(Fel d 1-Bet v 1)で二重特異性活性を測定することによって、IgG4およびIgG1の交換を評価した。血液は、ドナー2名から得た:A(黒色バー)およびB(灰色バー)。キメラIgG4(パネルA)、キメラIgG1(パネルB)を補充した、またはIgGを添加しない(パネルC)混合物において、二重特異性活性を決定した。表示したデータはすべて、37℃で24時間インキュベートした後に測定した。
【図6】ヒト血液細胞によるIgGの交換を示す。キメラIgG混合物を単核細胞(MNC)、血小板(Thr)、および赤血球(Ery)と共に37℃で48時間インキュベートし、その後異種架橋アッセイ法(Fel d 1-Bet v 1)で二重特異性活性を測定することによって、IgG4(黒色バー)およびIgG1(灰色バー)の交換を評価した。対照として、抗体混合物を無血清培地(SFC)中でもインキュベートした。二重特異性活性は、添加した量に対する、結合した125I-Bet v 1の割合(%)として表した。
【図7】HEKおよびマウス細胞株によるIgG4の交換を示す。キメラIgG4混合物をHEK細胞、マウスB細胞(J558)、またはハイブリドーマ細胞と共に37℃でインキュベートすることによって、IgG4半分子の交換を評価した。異種架橋アッセイ法(Fel d 1-Bet v 1)での二重特異性活性を、t=0時間(灰色バー)およびt=24時間(黒色バー)の時点で採取した試料1μl中で測定した。二重特異性活性は、添加した量に対する結合した125I-Bet v 1の割合(%)として表した。
【図8】IgG4の赤血球媒介性の交換を示す。IgG4-Betv1/IgG4-Feld1混合物を、新たに精製した赤血球(ery、黒記号)と共にインキュベートすると、二重特異性抗体が生成されるのに対して、IgG1アイソタイプの混合物では二重特異性は認められなかった。対照として、抗体混合物を赤血球なしでPBS中でインキュベートした(白記号)。矢印は、二重特異性IgGの、最大の予測割合(50%)を示す。エラーバーは、2つ組の測定値の範囲を示す。
【図9】PBS中でのIgG4の交換を示す。二重特異性活性(パネルA)、二価性、および抗原結合を測定することによって、IgG1(白色バー)、IgG4(灰色バー)、および過剰な非関連IgG4の存在下での、IgG4(黒色バー)のPBS中での交換を評価した。パネルAにおけるIgG半分子の交換は、二重特異性IgGの濃度(異種架橋アッセイ法で決定)、およびIgG半分子の交換が無作為でかつ完全である場合の、二重特異性IgGの最大予測濃度から算出した。交換は、100%である最大交換から起算した割合(%)として表した。パネルBに、同種架橋アッセイ法で測定した、時間に伴うFel d 1二価性を示す。二価性IgGの濃度は、t=0時の二価性IgGの濃度を100%と設定することによって正規化した。
【図10】赤血球溶解物によるIgG4の交換を示す。赤血球の溶解物中でキメラIgG4混合物を37℃でインキュベートすることにより、IgG4半分子の交換を評価した。溶解物の希釈を高めながら、IgG4をインキュベートした。異種架橋アッセイ法(Bet v 1-Fel d 1)における二重特異性活性を、表示の時点で採取した試料において測定した。二重特異性は、添加した量に対する結合した125I-Bet v 1の割合(%)として表した。
【図11】赤血球溶解物によって誘導される二重特異性活性のSEC解析を示す。 IgG4を新たに調製した赤血球溶解物と共に37℃で24時間インキュベートし、続いてAKTA HPLCユニット(Amersham Biosciences、スウェーデン、ウプサラ)においてSuperdex200カラムに0.5 ml/分で流して分画した。その画分中で、Bet v 1特異的IgG(■)の濃度を抗原結合試験で測定し、二重特異性IgG Fel d 1-Bet v 1(●)の濃度をBet v 1-Fel d 1架橋アッセイ法で決定した。このカラムの較正から、単量体、二量体、および凝集型IgGはそれぞれ12.1 ml、10.3 ml、および8.3 mlに溶出されることが明らかになった(データは示さず)。
【図12】IgG4のGSH媒介性の交換を示す。PBS/アジド中の、漸増濃度のGSHの存在下でIgG4をインキュベートすることにより、IgG4半分子のGSH媒介性の交換を評価した。表示の時点で試料を採取し、その試料中の抗原結合および二重特異性活性を測定した。IgG4半分子の交換は、二重特異性IgGの濃度(異種架橋アッセイ法で決定)、およびIgG4半分子の交換が無作為でかつ完全である場合の二重特異性IgG4の最大予測濃度から算出した。交換は、100%に設定した最大交換から起算した割合(%)として表した。
【図13】IgG4半分子のGSH媒介性の交換のSECを示す。IgG4をGSH(0.5 mM)と共にインキュベートし、続いてAKTA HPLCユニット(Amersham Biosciences、スウェーデン、ウプサラ)においてSuperdex200カラムに0.5 ml/分で流して分画した。その画分中で、Bet v 1特異的IgG(■)の濃度を抗原結合試験で測定し、二重特異性IgG Fel d 1-Bet v 1(●)の濃度をBet v 1-Fel d 1架橋アッセイ法で決定した。このカラムの較正から、単量体、二量体、および凝集型IgGはそれぞれ12.1 ml、10.3 ml、および8.3 mlに溶出されることが明らかになった(データは示さず)。
【図14】IgG4のGSH媒介性の交換の温度依存性を示す。IgG4-Betv1およびIgG4-Feld1混合物を、PBS中、表示の温度でGSHと共にインキュベートした。t=0時間(灰色バー)およびt=24時間(黒色バー)の時点で、二重特異性IgG4の濃度を評価した。これらのデータから、IgG4 Betv1濃度に対する二重特異性IgGの割合を算出し、割合(%)として表した。エラーバーは、2つ組の測定値の範囲を表す。
【図15】一連の還元剤によって媒介されるIgG4交換を示す。PBS中のIgG4-Betv1およびIgG4-Feld1を、様々な作用物質(GSSG以外はすべて還元性)の存在下で、37℃で24時間インキュベートした。Bet v 1特異的IgGの濃度をBet v 1結合アッセイ法で測定し、二重特異性IgGの濃度を異種架橋アッセイ法(Fel d 1-Bet v 1)で測定した。IgG-Betv1濃度に対する二重特異性IgGの割合(%)を算出した。標準誤差のバーは、3回の測定値から算出されたSEMを表す。
【図16】GSHを用いた完全ヒトIgG4抗体の交換を示す。(A) IgG4-CD20/IgG4-EGFr混合物またはIgG1-CD20/IgG1-EGFr混合物を、0.5 mM GSHと共に、またはこれを伴わずに37℃でインキュベートした。表示の時点で試料を採取した。二重特異性抗体の形成を、サンドイッチELISA法で測定した。Y軸は、二重特異性CD20/EGFR抗体の形成の測定値としての、405 nmの光学密度を示す。(B) IgG4のGSH用量依存的な交換。IgG4-CD20とIgG4-EGFrの混合物を、表示の濃度のGSHと共に、37℃で24時間インキュベートした。二重特異性抗体の形成を、サンドイッチELISA法で測定した。405 nmの光学密度を、二重特異性CD20/EGFR抗体の形成の測定値として、Y軸にプロットする。(C) IgG4半分子のGSH媒介性の交換は、反応に使用した成分によって影響を受け、培地(Freestyle 293)中では、より低いGSH濃度において起こる。(D) IgG4半分子のGSH媒介性の交換は、5 mM GSHよりも0.5 mM GSHにおいて、より高い。(E/F) ESI-TOF質量分析による、IgG4-EGFRとIgG4-CD20との間の、Fabアーム交換の検出。IgG4混合物を、0.5 mM GSHの非存在下(E)または存在下(F)で24時間インキュベートした後、抗体をPNGase Fで脱グリコシル化し、結果として得られた抗体の分子量をESI-TOF質量分析によって決定した。デコンボリューション処理したESI-TOFスペクトルを示す。データは、2回の実験の代表的なものである。
【図17】アカゲザルIVIgが組換えヒトIgG4抗体のFabアーム交換に関与することを示す。(A) 2種類の組換えヒトIgG4抗体(IgG4-CD20およびIgG4-EGFr)の混合物を、精製アカゲザル免疫グロブリンまたはヒトIVIgの存在下または非存在下において、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。(B) 2種類の組換えヒトIgG4抗体(IgG4-CD20およびIgG4-EGFr)の混合物を、数頭の動物(供給源を括弧内に表示)に由来する、過剰な(括弧内に表示)精製アカゲザル免疫グロブリンまたはヒトIVIgの存在下または非存在下で、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。(C) 2種類の組換えヒトIgG4抗体(IgG4-CD20およびIgG4-EGFr)の混合物を、過剰な(括弧内に表示)精製チンパンジー、ヒヒ、カニクイザル、ウマ、およびブタ免疫グロブリン(供給源も括弧内に表示)またはヒトIVIgの存在下または非存在下で、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。
【図18】定常領域の配列を示す。下線の配列はCH3領域を表す。
【図19】IgG1変異体のGSH媒介性の半分子の交換を示す。(A) 0 mM、0.1 mM、1 mM、および10 mM GSHを用いて、様々なIgG1変異体の半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図中ではIgG4 wtと表示);‐IgG1 a-feld1 wtとIgG4 a-betv1 wt(IgG1 wtと表示);‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(IgG1-CPSCと表示);‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(IgG1-CH3(IgG4)と表示);‐IgG1 a-feld1 CPSC-CH3(IgG4)とa-betv1 IgG1 CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示)。(B) 0.5 mMおよび5 mM GSHを用いて、様々なIgG1変異体のIgG4 wt分子との半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG4 a-betv1 wt(IgG1と表示);‐IgG1 a-feld1 CPSCとIgG4 a-betv1 wt(IgG1-CPSCと表示);‐IgG1 a-feld1 CH3(IgG4)とIgG4 a-betv1 wt(IgG1-CH3(IgG4)と表示);‐IgG1 a-feld1 CPSC-CH3(IgG4)とIgG4 a-betv1 wt(IgG1-CPSC-CH3(G4)と表示);‐IgG1 a-feld1 R238QとIgG4 a-betv1 wt(IgG1-R238Qと表示);‐IgG1 a-feld1 K292RとIgG4 a-betv1 wt(IgG1-K292Rと表示);‐IgG1 a-feld1 Q302EとIgG4 a-betv1 wt(IgG1-Q302Eと表示);‐IgG1 a-feld1 P328LとIgG4 a-betv1 wt(IgG1-P328Lと表示);‐IgG1 a-feld1 CPSC-K292RとIgG4 a-betv1 wt(IgG1-CPSC-K292Rと表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示)。(C) 0.5 mMおよび5 mM GSHを用いて、様々なIgG1変異体の半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(IgG1と表示);‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(IgG1-CPSCと表示);‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(IgG1-CH3(IgG4)と表示);‐IgG1 a-feld1 CPSC-CH3(IgG4)とIgG1 a-betv1 CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示);‐IgG1 a-feld1 R238QとIgG1 a-betv1 R238Q(IgG1-R238Qと表示);‐IgG1 a-feld1 K292RとIgG1 a-betv1 K292R(IgG1-K292Rと表示);‐IgG1 a-feld1 Q302EとIgG1 a-betv1 Q302E(IgG1-Q302Eと表示);‐IgG1 a-feld1 P328LとIgG1 a-betv1 P328L(IgG1-P328Lと表示);‐IgG1 a-feld1 CPSC-K292RとIgG1 a-betv1 CPSC-K292R(IgG1-CPSC-K292Rと表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示)。
【図20】0.5 mM GSHにおいて、野生型(IgG4)コアヒンジを有するIgG4分子は、組換えヒトIgG4抗体のFabアーム交換に関与するが、IgG1コアヒンジを有する分子は関与しないことを示す。(A) 2種類の組換えヒトIgG4抗体(上記のIgG4-CD20およびIgG4-EGFr)の混合物を、過剰なTysabri(50および100マイクログラム/ml)の存在下または非存在下で、0.5 mM GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。(B) 2種類の組換えヒトIgG4抗体(上記のIgG4-CD20およびIgG4-EGFr)の混合物を、等モル量(10マイクログラム/ml)のTysabriまたはMylotargの存在下または非存在下で、0.5 mM GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。
【図21】292位にさらなる変異を有するIgG1-CPSC構築物の、半分子の交換を示す。0.5 mM GSHを用いて、様々なIgG1変異体による半分子の交換を試験した。以下の混合物を用いて、交換を試験した:‐IgG1-2F8 wtとIgG1-7D8 wt(IgG1と表示);‐IgG1-2F8-CPSCとIgG1-7D8-CPSC(IgG1-CPSCと表示);‐IgG1-2F8-CH3(IgG4)とIgG1-7D8-CH3(IgG4)(IgG1-CH3(IgG4)と表示);‐IgG1-2F8-CPSC-CH3(IgG4)とIgG1-7D8-CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示);‐IgG1-2F8-CPSC-R238QとIgG1-7D8-CPSC-R238Q(IgG1-CPSC-R238Qと表示);‐IgG1-2F8-CPSC-K292RとIgG1-7D8-CPSC-K292R(IgG1-CPSC-K292Rと表示);‐IgG1-2F8-CPSC-K292YとIgG1-7D8-CPSC-K292Y(IgG1-CPSC-K292Yと表示);‐IgG1-2F8-CPSC-K292FとIgG1-7D8-CPSC-K292F(IgG1-CPSC-K292Fと表示);‐IgG1-2F8-CPSC-K292WとIgG1-7D8-CPSC-K292W(IgG1-CPSC-K292Wと表示);‐IgG1-2F8-CPSC-Q302EとIgG1-7D8-CPSC-Q302E(IgG1-CPSC-Q302Eと表示);‐IgG1-2F8-CPSC-P328LとIgG1-7D8-CPSC-P328L(IgG1-CPSC-P328Lと表示);‐IgG4-2F8 wtとIgG4-7D8 wt(IgG4と表示)。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。
【図22】コアヒンジの安定化が、インビボでのFabアーム交換からIgG4抗体治療薬を保護することを示す。(A) ESI-TOF質量分析による、IgG4-EGFR-CPPCとIgG4-CD20との間のFabアーム交換の検出。IgG4-EGFR-CPPC/IgG4-CD20混合物を、5 mM GSHの存在下(F)で24時間インキュベートした後、抗体をPNGase Fで脱グリコシル化し、結果として得られた抗体の分子量をESI-TOF質量分析によって決定した。デコンボリューション処理したESI-TOFスペクトルを示す。二重特異性EGFR/CD20抗体は、5 mM GSHを使用した場合に検出することができた(GSHなしでの、または0.5 mM GSHの存在下でのインキュベーションでは、二重特異性抗体は得られなかった(データは示さず))。(B) SCIDマウスの群(n=4)に、IgG4-CD20/IgG4-EGFR(白丸)、IgG4-CD20/IgG1-EGFR、およびIgG4-CD20/IgG4-EGFR-CPPCの抗体混合物(各300μg)を注射した。二重特異性抗体の生成を時間と共に追跡し、ELISA法により定量した。インビトロ交換された抗体混合物を参照として用いて、二重特異性抗体を定量した。データ点は、別々の実験で少なくとも2回測定した、マウス4匹の平均値±SEM値を表す。IgG4-CD20/IgG1-EGFR混合物およびIgG4-CD20/IgG4-EGFR-CPPC混合物では、二重特異性抗体は検出することができなかった。アッセイの検出限界を示すが(点線)、これは2000 ng/mlの血清レベルを表す。
【図23】時間に伴うCXXC変異体のFabアーム交換を示す。CXXC変異体抗体の混合物を、0.5 mM GSHと共に37℃でインキュベートした。表示の時点で試料を採取した。二重特異性抗体の形成を測定した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(IgG1と表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示);‐IgG4 a-feld1 CGHCとIgG4 a-betv1 CGHC(CGHCと表示);‐IgG4 a-feld1 CGCとIgG4 a-betv1 CGC(CGCと表示);‐IgG4 a-feld1 CPRCとIgG4 a-betv1 CPRC(CPRCと表示);‐IgG4 a-feld1 CPHCとIgG4 a-betv1 CPHC(CPHCと表示)。
【図24】CXXC変異体のGSH媒介性Fabアーム交換を示す。1〜20,000μM GSHを用いて、CXXC変異体によるFabアーム交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(IgG1と表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示);‐IgG4 a-feld1 CGHCとIgG4 a-betv1 CGHC(CGHCと表示);‐IgG4 a-feld1 CGCとIgG4 a-betv1 CGC(CGCと表示);‐IgG4 a-feld1 CPRCとIgG4 a-betv1 CPRC(CPRCと表示);‐IgG4 a-feld1 CPHCとIgG4 a-betv1 CPHC(CPHCと表示)。
【発明を実施するための形態】
【0016】
発明の詳細な説明
定義
「免疫グロブリン」という用語は、2対のポリペプチド鎖、すなわち1対の低分子量軽(L)鎖および1対の重(H)鎖からなる構造的に関連した糖タンパク質のクラスを指し、これら4つはすべてジスルフィド結合によって相互に結合している。免疫グロブリンの構造は十分に特徴付けられている。例えば、Fundamental Immunology Ch.7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989))(11)を参照されたい。簡潔に説明すると、各重鎖は典型的に、重鎖可変領域(本明細書ではVHまたはVHと略す)および重鎖定常領域からなる。重鎖定常領域は典型的に、3つのドメイン、CH1、CH2、およびCH3からなる。各軽鎖は典型的に、軽鎖可変領域(本明細書ではVLまたはVLと略す)および軽鎖定常領域からなる。軽鎖定常領域は典型的に、1つのドメイン、CLからなる。VH領域およびVL領域は、フレームワーク領域(FR)と称されるより保存された領域が介在する相補性決定領域(CDR)とも称される超可変性の領域(または配列および/もしくは構造的に規定されたループの形態において超可変性であり得る超可変領域)に、さらに細分され得る。各VHおよびVLは典型的に、アミノ末端からカルボキシ末端へ以下の順序:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4で配置された、3つのCDRおよび4つのFRから構成される((12)も参照されたい)。典型的に、この領域におけるアミノ酸残基の番号付けは、Kabat(13)に記載されている方法によって行われる。この番号付けシステムを用いると、ペプチドの実際の線状アミノ酸配列は、可変ドメインのFRまたはCDRの短縮またはその中への挿入に対応する、より少数のまたは付加的なアミノ酸を含み得る。例えば、重鎖可変ドメインは、VH CDR2の残基52の後に単一のアミノ酸の挿入(Kabatによると残基52a)、および重鎖FRの残基82の後に挿入された残基(例えば、Kabatによると残基82a、82b、および82c等)を含み得る。残基のKabat番号付けは、所与の抗体について、この抗体の配列と「標準」Kabat番号付け配列との相同性の領域における整列によって決定することができる。
【0017】
本発明との関連における「抗体」(Ab)という用語は、少なくとも約30分、少なくとも約45分、少なくとも約1時間、少なくとも約2時間、少なくとも約4時間、少なくとも約8時間、少なくとも約12時間、約24時間もしくはそれ以上、約48時間もしくはそれ以上、約3、4、5、6、7日もしくはそれ以上等といったかなりの期間の、または任意の他の関連する、機能的に規定された期間(抗原に対する抗体結合に付随する生理反応を誘導、促進、増強、および/もしくは調節するのに十分な時間、ならびに/または抗体がFc媒介性エフェクター活性を動員するのに十分な時間など)の半減期を有して、典型的な生理的条件下で抗原に特異的に結合する能力を有する、免疫グロブリン分子、免疫グロブリン分子の断片、またはそれらいずれかの誘導体を指す。免疫グロブリン分子の重鎖および軽鎖の可変領域は、抗原と相互作用する結合ドメインを含む。抗体(Ab)の定常領域は、免疫系の様々な細胞(エフェクター細胞など)、および補体活性化の古典的経路の第1成分であるC1qなどの補体系の成分を含む、宿主の組織または因子に対する免疫グロブリンの結合を媒介し得る。上記の通り、本明細書における抗体という用語は、特に明記しない限り、または文脈上明らかに矛盾しない限り、変異型または野生型のコアヒンジ領域を含み、抗原に特異的に結合する能力を保持する抗体の断片を含む。
【0018】
抗体の抗原結合機能は、全長抗体の断片によって行われ得ることが示されている。「抗体」という用語の中に包含される結合断片の例として、例えば、ヒンジ領域においてジスルフィド架橋によって連結された2つのFab断片を含む、二価の断片であるF(ab')2断片が挙げられる。このような断片は一般的に抗体の意味に含まれるが、それらは集合的にかつ各々独立して本発明の独特の特徴であり、異なる生物学的特性および有用性を示す。抗体という用語は、特に明記しない限り、ポリクローナル抗体、モノクローナル抗体(mAb)、キメラ抗体およびヒト化抗体などの抗体様ポリペプチド、ならびに酵素的切断、ペプチド合成、および組換え技法などの任意の公知の技法によって提供される、抗原に特異的に結合する能力を保持する抗体断片(抗原結合断片)も含むこともまた、理解されるべきである。このようにして作製された抗体は、任意のアイソタイプを有し得る。
【0019】
本明細書で使用する「ヒト抗体」という用語は、ヒト生殖系列免疫グロブリン配列に由来する可変領域および定常領域を有する抗体を含むことが意図される。本発明のヒト抗体は、ヒト生殖系列免疫グロブリン配列によってコードされないアミノ酸残基(例えば、インビトロでのランダムな、もしくは部位特異的な突然変異誘発によって、またはインビボでの体細胞突然変異によって導入された変異)を含み得る。しかしながら、本明細書で使用する「ヒト抗体」という用語は、マウスのような別の哺乳動物種の生殖系列に由来するCDR配列がヒトフレームワーク配列上に移植された抗体を含まないことが意図される。
【0020】
本明細書で使用する「単離された抗体」は、異なる抗原特異性を有する他の抗体を実質的に含まない抗体を指すことが意図される。しかしながら、特定のヒト標的抗原のエピトープ、アイソフォーム、または変種に特異的に結合する単離された抗体は、例えば他の種由来の他の関連抗原(種相同体など)に対する交差反応性を有してよい。さらに、単離された抗体は、他の細胞物質および/または化学物質を実質的に含まない可能性がある。本発明の1つの態様では、異なる特異性を有する「単離された」モノクローナル抗体の組み合わせを、明確な組成物中で組み合わせる。
【0021】
本明細書で使用する「モノクローナル抗体」または「モノクローナル抗体組成物」という用語は、単一分子の組成物の、抗体分子の調製物を指す。モノクローナル抗体組成物は、特定のエピトープに対する単一の結合特異性および親和性を示す。したがって、「ヒトモノクローナル抗体」という用語は、ヒト生殖系列免疫グロブリン配列に由来する可変領域および定常領域を有する、単一の結合特異性を示す抗体を指す。ヒトモノクローナル抗体は、不死化細胞と融合した、ヒト重鎖導入遺伝子および軽鎖導入遺伝子を含むゲノムを有する、遺伝子導入マウスなどの、遺伝子導入または染色体導入非ヒト動物から得られたB細胞を含むハイブリドーマによって作製することができる。
【0022】
所定の抗原に対する抗体の結合との関連における、本明細書で使用する「結合」という用語は典型的に、例えば、抗原をリガンドとしておよび抗体を分析物として用いて、BIAcore 3000装置において表面プラズモン共鳴(SPR)技術によって決定した場合の、約10-7 Mもしくはそれ未満、例えば約10-8 Mもしくはそれ未満、例えば約10-9 Mもしくはそれ未満、約10-10 Mもしくはそれ未満、または約10-11 Mもしくはさらにそれ未満のKDに対応する親和性での結合であり、所定の抗原または近縁の抗原以外の非特異的抗原(例えば、BSA、カゼイン)に対する結合の親和性よりも、少なくとも10倍低い、例えば少なくとも100倍低い、例えば少なくとも1000倍低い、例えば少なくとも10,000倍低い、例えば少なくとも100,000倍低いKDに対応する親和性で、所定の抗原に結合する。親和性がより低い量は抗体のKDに依存し、それゆえ抗体のKDが非常に低い(すなわち、抗体が高度に特異的である)場合、抗原に対する親和性が非特異的抗原に対する親和性よりも低い量は少なくとも10,000倍であり得る。
【0023】
本明細書で使用する「kd」(sec-1)という用語は、特定の抗体-抗原相互作用の解離速度定数を指す。この値は、koff値とも称される。
【0024】
本明細書で使用する「ka」(M-1×sec-1)という用語は、特定の抗体-抗原相互作用の会合速度定数を指す。
【0025】
本明細書で使用する「KD」(M)という用語は、特定の抗体-抗原相互作用の解離平衡定数を指す。
【0026】
本明細書で使用する「KA」(M-1)という用語は、特定の抗体-抗原相互作用の会合平衡定数を指し、kaをkdで割ることによって得られる。
【0027】
本明細書で使用する「アイソタイプ」とは、重鎖定常領域遺伝子によってコードされる免疫グロブリンクラス(例えば、IgG1、IgG2、IgG3、IgG4、IgD、IgA、IgE、またはIgM)を指す。
【0028】
「エピトープ」という用語は、抗体に特異的に結合し得るタンパク質決定基を意味する。エピトープは、通常はアミノ酸または糖側鎖などの分子の表面基からなり、通常は特異的な三次元構造特性および特異的な電荷特性を有する。高次構造的エピトープと非高次構造的エピトープは、前者に対する結合は変性溶媒の存在下で失われるが、後者に対する結合は失われないという点で識別される。エピトープは、結合に直接関与するアミノ酸残基(エピトープの免疫優性成分とも称される)、および特異的抗原結合ペプチドによって効果的に阻止されるアミノ酸残基のような、結合に直接関与しない他のアミノ酸残基(すなわち、このアミノ酸残基は特異的抗原結合ペプチドのフットプリント内にある)を含み得る。
【0029】
本明細書で使用する場合のヒト抗体は、例えばヒト免疫グロブリン遺伝子を保有する遺伝子導入マウスを免疫することによって、またはヒト免疫グロブリン遺伝子ライブラリーをスクリーニングすることによって、ヒト免疫グロブリン配列を用いる系から抗体が得られる場合に、特定の生殖系列配列「に由来し」、選択されたヒト抗体は、生殖系列免疫グロブリン遺伝子によりコードされるアミノ酸配列と、アミノ酸配列が少なくとも90%、例えば少なくとも95%、例えば少なくとも96%、例えば少なくとも97%、例えば少なくとも98%、または例えば少なくとも99%同一である。典型的に、重鎖CDR3の外部では、特定のヒト生殖系列配列に由来するヒト抗体は、生殖系列免疫グロブリン遺伝子によってコードされるアミノ酸配列と20アミノ酸以下の差、例えば10アミノ酸以下の差、例えば5以下、例えば4、3、2または1アミノ酸以下の差を示す。
【0030】
「二重特異性抗体」という用語は、2つの異なる結合特異性を有する任意の抗体、すなわち同じ標的抗原上に位置し得るか、またはより一般的には、異なる標的抗原上に位置し得る2つの異なるエピトープに結合する抗体を含むことが意図される。
【0031】
本明細書で使用する「エフェクター細胞」という用語は、免疫応答の認識期および活性化期とは対照的に、免疫応答のエフェクター期に関与する免疫細胞を指す。例示的な免疫細胞には、骨髄性またはリンパ系起源の細胞、例えばリンパ球(B細胞、および細胞傷害性T細胞(CTL)を含むT細胞など)、キラー細胞、ナチュラルキラー細胞、マクロファージ、単球、好酸球、好中球などの多形核細胞、顆粒球、肥満細胞、および好塩基球が含まれる。一部のエフェクター細胞は特定のFc受容体を発現し、特定の免疫機能を行う。いくつかの態様において、エフェクター細胞は、抗体依存性細胞性細胞傷害(ADCC)を誘導し得るナチュラルキラー細胞のように、ADCCを誘導することができる。例えば、FcRを発現する単球、マクロファージは、標的細胞の特異的殺傷、および免疫系の他の成分に対する抗原提示、または抗原を提示する細胞に対する結合に関与する。いくつかの態様において、エフェクター細胞は、標的抗原または標的細胞を貪食することができる。エフェクター細胞における特定のFcRの発現は、サイトカインなどの体液性因子によって調節され得る。例えば、FcγRIの発現はインターフェロンγ(IFN-γ)および/またはG-CSFによって上方制御されることが判明している。この発現増強によって、標的に対するFcγRI保有細胞の細胞傷害活性が高まる。エフェクター細胞は、標的抗原または標的細胞を貪食または溶解することができる。
【0032】
「治療」とは、症状または疾患状態を緩和する、改善する、停止させる、または根絶する(治癒する)目的で、本発明の治療上活性な化合物の有効量を投与することを指す。
【0033】
「有効量」とは、必要な投与量で必要な期間の間の、所望の治療結果を達成するために有効な量を指す。抗体の治療上有効な量は、個体の疾患状態、年齢、性別、および体重、ならびに個体において所望の応答を誘発する抗体の能力などの要因に応じて異なり得る。治療上有効な量とは、治療上有益な効果が、抗体または抗体部分の、任意の毒性または有害効果を上回る量でもある。
【0034】
「IgG4様コアヒンジ領域」という用語は、システイン残基が、抗体分子内の他のシステイン/ジスルフィド架橋よりもさらに著しく還元および/またはジスルフィド結合の異性化を受けやすいコアヒンジ領域を指す。したがって、IgG4様コアヒンジ領域を有する抗体について、抗体内の他のジスルフィド架橋および全般的な抗体構造はそのままに保ちながら、コア領域内のシステイン残基/ジスルフィド架橋が還元され、続いて別の半分子内のコアヒンジシステインとジスルフィド架橋を形成し得る還元状態を見出すことができる。例えば、IgG4様コアヒンジ領域は、IgG4コアヒンジ領域であってよく、またはCPPCからCPSCへの変異のように、コア領域内のCPPC配列のプロリンの1つが、例えばセリンに変異した別のアイソタイプの抗体のコアヒンジ配列であってよい。
【0035】
本出願との関連における「IgG4様CH3領域」という用語は、IgG4、例えばヒトIgG4のCH3と同一であるCH3領域、またはIgG4 CH3領域と機能的に同等であるCH3領域を指す。この状況における機能的に同等とは、CH3領域が、IgG4のCH3領域と同様に、安定した半分子間相互作用を形成しないことを意味する。所与のCH3領域による安定した半分子間の形成は、例えばIgG4のCH3をその所与のCH3領域と置換し、実施例31または32に示す条件下で交換について試験することによって、調べることができる。交換が認められた場合には、安定した半分子間相互作用は形成されていない。例えば、IgG4様CH3領域は、半分子の交換を可能にするという点でIgG4由来のCH3領域と同等に効率的であるCH3領域であってよい。したがって、IgG4様CH3領域は、例えばIgG4のCH3領域の配列と75%を超えて、例えば90%を超えて同一であるなど、IgG4のCH3領域と構造的に類似していてよい。しかしながら、本状況におけるIgG4様CH3領域は、加えてまたはあるいは、IgG4のCH3領域と構造的に近似していないが、ジスルフィド結合または塩の中継など共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するアミノ酸残基を全く含まず、同時に他のペプチドはCH3領域の同一のアミノ酸配列を含むことから同様の機能的特性を有するCH3領域であってもよい。例えば、IgG4様CH3領域は、半分子CH3-CH3間相互作用に関与する1つまたは複数のアミノ酸残基が変化しているかまたは欠失している変異型IgG1 CH3領域であってよい。
【0036】
「還元条件」または「還元環境」という用語は、基質、ここでいう抗体のコア領域内のシステイン残基が、酸化されるより還元された状態になる可能性が高い条件または環境を指す。
【0037】
「還元剤」という用語は、その環境において分子を還元する化合物、すなわち、その環境において分子をより還元した状態に、およびより還元しつつある状態に変える化合物を指す。還元剤は、電子を供与することにより作用し、それによって、基質の還元後にそれ自体は酸化された状態になる。したがって、還元剤とは電子を供与する作用物質である。還元剤の例には、ジチオスレイトール(DTT)、メルカプトエタノール、システイン、チオグリコール酸、システアミン、グルタチオン、および水素化ホウ素ナトリウムが含まれる。1つの態様において、還元剤は酵素を含まない。
【0038】
「ジスルフィド結合形成」とは、1つまたは2つのポリペプチド中に存在する2つのシステイン間で共有結合を形成する過程を指し、この結合は「-S--S-」として図式化される。
【0039】
「ジスルフィド結合の還元」とは、ジスルフィド結合の切断によって、2つのチオール基(-SH基)が生じる過程を指す。
【0040】
「ジスルフィド結合の異性化」という用語は、異なるシステイン間のジスルフィド結合の交換、すなわちジスルフィド結合の再編成を指す。
【0041】
「タンパク質ジスルフィド結合イソメラーゼ」とは、タンパク質におけるジスルフィド結合の異性化を触媒するタンパク質を指す。
【0042】
ジスルフィド結合の還元に関連して使用する場合の「有意でない還元」とは、溶液中の特定のジスルフィド架橋の、一般的には10%未満、例えば5%未満、例えば2%未満または1%未満が還元を起こすことを意味する。
【0043】
発明の局面および態様
上記の通り、第1の主要な局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0044】
好ましい態様において、本発明の方法において使用する第1の抗体および第2の抗体はモノクローナル抗体である。モノクローナル抗体は、例えば、Kohler et al.(14)によって最初に記載されたハイブリドーマ法によって作製することもできるし、または組換えDNA法によって作製することもできる。モノクローナル抗体は、例えばClackson et al.(15)およびMarks et al.(16)に記載されている技法を用いて、ファージ抗体ライブラリーから単離することもできる。モノクローナル抗体は、任意の適切な供給源から得ることができる。したがって、例えば、モノクローナル抗体は、例えば表面上に抗原を発現する細胞の形態をした関心対象の抗原、または関心対象の抗原をコードする核酸で免疫したマウスから得られたマウス脾臓B細胞から調製したハイブリドーマから得ることができる。モノクローナル抗体はまた、免疫したヒト、またはラット、イヌ、霊長動物等のような非ヒト哺乳動物の抗体発現細胞に由来するハイブリドーマから得ることもできる。
【0045】
1つの態様において、本発明の抗体はヒト抗体である。指向化されたヒトモノクローナル抗体は、マウス系ではなくヒト免疫系の一部を保有する遺伝子導入または染色体導入マウスを用いて作製することができる。このような遺伝子導入および染色体導入マウスには、本明細書においてそれぞれHuMAbマウスおよびKMマウスと称するマウスが含まれ、本明細書ではまとめて「遺伝子導入マウス」と称する。
【0046】
HuMAbマウスは、内在性μ鎖およびκ鎖遺伝子座を不活化する標的変異と共に、再編成されていないヒト重鎖(μおよびγ)およびκ軽鎖免疫グロブリン配列をコードするヒト免疫グロブリン遺伝子のミニ遺伝子座を含む(17)。したがって、マウスはマウスIgMまたはκの発現低下を示し、免疫化に応答して、導入されたヒト重鎖および軽鎖導入遺伝子はクラススイッチおよび体細胞突然変異を起こして、高親和性ヒトIgG、κモノクローナル抗体を生成する(17〜20)。HuMAbマウスの調製は、参考文献21〜25に詳述されている。US 5,545,806、US 5,569,825、US 5,625,126、US 5,633,425、US 5,789,650、US 5,877,397、US 5,661,016、US 5,814,318、US 5,874,299、US 5,770,429、US 5,545,807、WO 98/24884、WO 94/25585、WO 93/1227、WO 92/22645、WO 92/03918、およびWO 01/09187もまた参照されたい。
【0047】
HCo7マウスは、(Chen et al.(26)に記載されているような)その内在性軽鎖(κ)遺伝子におけるJKD破壊、(WO 01/14424の実施例1に記載されているような)その内在性重鎖遺伝子におけるCMD破壊、(Fishwild et al.(25))に記載されているような)KCo5ヒトκ軽鎖導入遺伝子、および(US 5,770,429に記載されているような)HCo7ヒト重鎖導入遺伝子を有する。
【0048】
HCo12マウスは、(Chen et al.(26)に記載されているような)その内在性軽鎖(κ)遺伝子におけるJKD破壊、(WO 01/14424の実施例1に記載されているような)その内在性重鎖遺伝子におけるCMD破壊、(Fishwild et al.(25)に記載されているような)KCo5ヒトκ軽鎖導入遺伝子、および(WO 01/14424の実施例2に記載されているような)HCo12ヒト重鎖導入遺伝子を有する。
【0049】
KMマウス系統において、内在性マウスκ軽鎖遺伝子は、Chen et al.(26)に記載されている通りにホモ接合性に破壊されており、かつ内在性マウス重鎖遺伝子も、WO 01/09187の実施例1に記載されている通りにホモ接合性に破壊されている。このマウス系統は、Fishwild et al.(25)に記載されている通りに、ヒトκ軽鎖導入遺伝子、KCo5を保有する。このマウス系統はまた、WO 02/43478に記載されている通りに、第14染色体断片hCF(SC20)からなるヒト重鎖導入染色体を保有する。
【0050】
これらの遺伝子導入マウス由来の脾臓細胞を用いて、周知の技法に従って、ヒトモノクローナル抗体を分泌するハイブリドーマを作製することができる。このような遺伝子導入非ヒト動物、本発明において使用する抗体の発現をコードする操作可能な核酸配列を含む非ヒト動物、1つまたは複数の標的コード核酸配列を安定にトランスフェクトした非ヒト動物等は、本発明のさらなる特色である。
【0051】
本発明において使用しようとするヒトモノクローナルもしくはポリクローナル抗体、またはその他の種が起源である本発明において使用する抗体はまた、関心対象の免疫グロブリン重鎖および軽鎖配列についてトランスジェニックである別の非ヒト哺乳動物または植物の作製、ならびにそれらから回収可能な形態での抗体の生産を通じて、トランスジェニックで作製することも可能である。哺乳動物におけるトランスジェニック生産に関連して、抗体をヤギ、ウシ、またはその他の哺乳動物の乳において生産し、そこから回収することができる。例えば、US 5,827,690、US 5,756,687、US 5,750,172、US 5,741,957を参照されたい。
【0052】
さらに、本発明において使用しようとするヒトまたはその他の抗体は、当技術分野で周知の技法を用いて、ファージディスプレイ、レトロウイルスディスプレイ、リボソームディスプレイ、およびその他の技法を含むがこれらに限定されないディスプレイ型の技術を通じて作製することができ、結果として得られた分子は、そのような技法が当技術分野で周知であるので、親和性成熟などのさらなる成熟に供することができる(例えば、参考文献27、28、および30(ファージディスプレイ)、29(リボソームディスプレイ)、31〜35、ならびにUS 5,733,743を参照されたい)。ディスプレイ技術を利用してヒトではない抗体を生産する場合には、このような抗体をヒト化することができる。
【0053】
上記の通りに、いくつかの態様において、本発明の方法において使用する第1および/または第2の抗体はIgG4抗体である。しかしながら、本発明において使用する抗体は、原則的には、CH3領域の配列が半分子の交換を可能にするという条件で任意のアイソタイプのものであってよい。例えば、本発明の方法において使用するまたは得られる抗体は、(任意の特定の変異位置以外では)SEQ ID NO:19〜22に示す定常領域のいずれかを含み得る。
【0054】
したがって、本発明の方法の1つの態様において、第1および/または第2の抗体はコアヒンジ領域内にCPPC配列を含む。別の態様において、第1および/または第2の抗体はIgG4様コアヒンジ領域を含む。例えば、いくつかの態様において、第1および/または第2の抗体は、コアヒンジ領域内に、式中、X1およびX2の両方がプロリンではないという条件で、X1およびX2は任意のアミノ酸であってよい、CX1X2C配列を含む抗体である。別の態様において、第1および/または第2の抗体は、コアヒンジ領域内に、式中、X3はプロリンを除く任意のアミノ酸であってよい、CX3PCまたはCPX3C配列を含む抗体である。さらなる態様において、第1および/または第2の抗体は、コアヒンジ領域内にCSPC、CPSC、CRPC、CPRC、CGHC、またはCPHC配列を含む抗体である。上記の変異は、例えば当技術分野で周知の部位特異的突然変異誘発によって導入することができる。
【0055】
アイソタイプの選択は典型的に、CDC誘導またはADCCにおける活性などの所望のエフェクター機能によって導かれる。例示的なアイソタイプは、IgG1、IgG2、IgG3、およびIgG4である(例えば、SEQ ID NO:19〜22を参照されたい)。ヒト軽鎖定常領域であるκまたはλのいずれかを使用することができる。必要に応じて、本発明において使用するための抗体のクラスを、公知の方法によってスイッチすることができる。例えば、本来IgM、IgG1、またはIgG2であった本発明において使用しようとする抗体を、本発明のIgG4抗体にクラススイッチすることができる。したがって、様々な治療用途のために、本発明の抗体のエフェクター機能を、例えばIgG1、IgG2、IgG3、IgG4、IgD、IgA、IgE、またはIgM抗体へのアイソタイプスイッチによって変更することができる。
【0056】
1つの態様において、本発明において使用する第1および/または第2の抗体は全長抗体である。別の態様において、使用する本発明の第1および/または第2の抗体は抗体断片である。
【0057】
本発明の方法の1つの態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO:22)を有するIgG4 CH3領域などの、IgG4 CH3領域を含む。
【0058】
しかしながら、本発明の方法の別の態様において、第1および/または第2の抗体は非IgG4アイソタイプのCH3領域を含み、そのCH3配列は、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている。
【0059】
例えば、本発明の1つのさらなる態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 19)を有するCH3領域を含み、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:238位のArg(R)がGln(Q)によって置換されている;239位のAsp(D)がGlu(E)によって置換されている;292位のLys(K)がArg(R)によって置換されている;302位のGln(Q)がGlu(E)によって置換されている;および328位のPro(P)がLeu(L)によって置換されている。
【0060】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 19)を有するCH3領域を含み、292位のLys(K)がArg(R)によって置換されている。
【0061】
別の態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 19)を有するCH3領域を含むが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている。
【0062】
別のさらなる態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 20)を有するCH3領域を含み、このCH3領域は、以下のアミノ酸置換のうちの1つもしくは複数または5つすべてがなされるように改変されている:234位のArg(R)がGln(Q)によって置換されている;276位のMet(M)がVal(V)によって置換されている;288位のLys(K)がArg(R)によって置換されている;298位のGln(Q)がGlu(E)によって置換されている;および324位のPro(P)がLeu(L)によって置換されている。
【0063】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 20)を有するCH3領域を含み、234位のArg(R)がGln(Q)によって置換されている。
【0064】
さらなる好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 20)を有するCH3領域を含み、234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている。
【0065】
別のさらなる態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 21)を有するCH3領域を含み、このCH3領域は、以下のアミノ酸置換のうちの1つもしくは複数または10個すべてがなされるように改変されている:285位のArg(R)がGln(Q)によって置換されている;314位のSer(S)がAsn(N)によって置換されている;322位のAsn(N)がLys(K)によって置換されている;327位のMet(M)がVal(V)によって置換されている;339位のLys(K)がArg(R)によって置換されている;349位のGln(Q)がGlu(E)によって置換されている;352位のIle(I)がVal(V)によって置換されている;365位のArg(R)がHis(H)によって置換されている;366位のPhe(F)がTyr(Y)によって置換されている;および375位のPro(P)がLeu(L)によって置換されている。
【0066】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 21)を有するCH3領域を含み、285位のArg(R)がGln(Q)によって置換されている。
【0067】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 21)を有するCH3領域を含み、285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている。
【0068】
本発明の方法のさらなる態様において、第1の抗体はコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含み、第2の抗体もコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含む。
【0069】
上記の通りに、主要な局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0070】
本発明の方法の1つの態様において、段階c)における条件は、コアヒンジ領域外のジスルフィド架橋の有意な還元または異性化が起こらないように選択する。
【0071】
別の態様において、段階c)における還元条件は、ジスルフィド結合交換を行うコアヒンジ領域の内因性の活性を促進する条件である。
【0072】
本発明のさらなる態様において、段階c)は還元剤の添加を含む。さらなる態様において、段階c)は、グルタチオン、L-システイン、ジチオスレイトール、β-メルカプト-エタノール、およびシステアミンからなる群より選択される作用物質の添加を含む。
【0073】
本発明の方法の1つの態様において、還元剤の濃度は、段階c)で生じる溶液の酸化還元電位が、実施例31に記載する条件下で、1マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、実施例31に記載する条件下で、例えば10マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば50マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば0.1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性であるといった濃度である。
【0074】
さらなる態様において、還元剤の濃度は、段階c)で生じる溶液の酸化還元電位が、
‐実施例31に記載する条件下で、1マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、実施例31に記載する条件下で、例えば10マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば50マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば0.1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、かつ
‐実施例31に記載する条件下で、1 Mのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、例えば100 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、15 mMのグルタチオンによって生じる酸化還元電位と等しいかもしくはより還元性が低い
濃度である。
【0075】
第1の抗体がコアヒンジ領域内にCPPC配列を有する、および/または第2の抗体がコアヒンジ領域内にCPPC配列を有する場合の1つの態様において、段階c)で生じる溶液の酸化還元電位は、実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性であることが好ましい。
【0076】
さらなる態様において、還元剤の濃度は、段階c)で生じる溶液の酸化還元電位が、
‐実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性であり、かつ
‐1 Mのグルタチオンによって生じる酸化還元電位と等しいかまたはそれよりも還元性が低い、例えば100 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはそれよりも還元性が低い、15 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはそれよりも還元性が低い
濃度である。
【0077】
本発明の方法の1つの態様において、段階c)は、還元型グルタチオンの存在下で、20℃またはそれ以上、例えば37℃の温度における、少なくとも1時間、例えば少なくとも2時間、例えば少なくとも5時間、例えば少なくとも10時間の抗体のインキュベーションを含む。
【0078】
本発明の方法のさらなる態様において、段階c)において選択する条件は、本明細書に記載するサイズ排除クロマトグラフィーにより決定されるように(ここで出発材料の抗体よりも早く溶出するピークは、凝集体の形成を示す)、結果として得られた組成物中の抗体分子の10%未満、例えば5%未満、例えば2%未満、例えば1%未満が凝集状態である条件である。
【0079】
本発明のエクスビボ方法の1つの態様において、本方法は、PDIなどのタンパク質ジスルフィドイソメラーゼ活性を有するタンパク質の添加を含む。本発明のエクスビボ方法の別の態様において、本方法は、PDIなどのタンパク質ジスルフィドイソメラーゼ活性を有するタンパク質の添加を含まない。
【0080】
本発明のエクスビボ方法の1つの態様において、本方法は生細胞または細胞抽出物の添加を含まない。
【0081】
上記の通り、本発明の方法において使用する第1の抗体および第2の抗体は、結合特異性が異なる、すなわち異なるエピトープに結合する。原則として、本発明の方法では、出発材料として特異性の任意の組み合わせを使用することができる。本発明の方法はまた、2種類の異なる抗体のみを出発材料とすることに限定されない。したがって、本発明の方法はまた、出発材料として3つまたはそれ以上の抗体を用いて行うこともできる。このような態様において、本発明の方法の段階d)で得られた組成物は、複数の二重特異性抗体を含むことになる。
【0082】
本発明の方法の1つの態様において、第1の抗体は、腫瘍細胞に対する、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、CD38、もしくはCXCR5などの腫瘍細胞タンパク質に対する、またはB細胞受容体のシグナル伝達成分である、CD79aもしくはCD79bに対する結合特異性を有する。別の態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は、erbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、またはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有する。
【0083】
さらなる態様において、第1の抗体はerbB1に対する結合特異性を有し、かつ第2の抗体はerbB2に対する結合特異性を有する。
【0084】
別の態様において、第1の抗体はCD19に対する結合特異性を有し、かつ第2の抗体はCD20に対する結合特異性を有する。
【0085】
さらなる態様において、第1の抗体はCD38に対する結合特異性を有し、かつ第2の抗体はCD34に対する結合特異性を有する。
【0086】
なおさらなる態様において、第1の抗体はCD4に対する結合特異性を有し、かつ第2の抗体はCXCR5に対する結合特異性を有する。
【0087】
本発明の方法の別の態様において、第1の抗体は病原微生物に対する結合特異性を有する。さらなる態様において、第1の抗体は病原微生物に対する結合特異性を有し、かつ第2の抗体は、CD3、CD25、CD28、CD16、CD89、CD32、またはCD1などのエフェクター細胞タンパク質に対する結合特異性を有する。
【0088】
二重特異性抗体を用いて、化学療法薬が作用すべき細胞に対し、この薬剤をより特異的に標的化することもできる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は化学療法薬に対する結合特異性を有する。
【0089】
さらに、二重特異性抗体において血清タンパク質に対する結合特異性を含ませることによって、抗体の血清半減期を変更することができる。例えば、二重特異性抗体において血清アルブミンに対する結合特異性を含ませることによって、血清半減期を延長することができる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は血清アルブミンなどの血液タンパク質に対する結合特異性を有する。
【0090】
第2の結合特異性を用いて、抗体を脳または肝臓などの特定の組織に標的化することもできる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体はトランスフェリンなどの脳タンパク質または肝臓タンパク質に対する結合特異性を有する。
【0091】
さらに、第2の結合特異性を用いて、血液凝固因子を特定の所望の作用部位に標的化することもできる。例えば、腫瘍細胞に対する第1の結合特異性および血液凝固因子に対する第2の結合特異性を有する二重特異性抗体は、腫瘍に血液凝固を導き、したがって腫瘍増殖を停止させることができる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は組織因子などの血液凝固に関与するタンパク質に対する結合特異性を有する。
【0092】
本発明のさらなる態様では、第1および/または第2の抗体を、細胞毒性薬;放射性同位体;プロドラッグまたは薬物、例えばタキサン;サイトカイン;ケモカイン、および補体、例えばC1qからなる群より選択される化合物に連結する。このような化合物は、例えば癌治療において、標的細胞をより効果的に殺傷することができる。または化合物は、結果として得られた二重特異性抗体に、すなわち半分子の交換が起こった後に、結合させることも可能である。
【0093】
本発明の方法のさらなる態様において、本方法は、さらなる半分子の交換を停止させるために、段階c)において得られた組成物を非還元条件または還元性のより低い条件に導くさらなる段階を含む。これは、例えば小分子還元剤を除去するための、結果として得られた組成物の、透析またはサイズベースのクロマトグラフィーなど、当技術分野で公知の種々の方法によって行うことができる。
【0094】
本発明の方法のなおさらなる態様においては、2つの半分子の化学的架橋を行い、二重特異性抗体がその後、そうしなければこの抗体が半分子の交換を起こし得るインビボ条件などの条件下で用いられる場合でさえもさらなる交換を防ぐことによって、結果として得られた二重特異性抗体を安定化する。したがって、1つの態様において、本発明の方法は以下のさらなる段階を含む:
a) 例えばビス-マレイミドヘキサンなどのマレイミドを含む化合物を用いて、ヒンジ領域内のシステインを化学的に架橋する段階、
b) 例えば過ヨウ素酸酸化、およびその後のアジピンジヒドラジドなどの適切な架橋剤によるアルデヒド基の反応を介して、半分子上の炭水化物側鎖を化学的に架橋する段階、または
c) 例えば以下の組み合わせのうちの1つまたは複数を用いて(SEQ ID NO:19を参照されたい)、例えばMerchant et al.(36)(参照により本明細書に組み入れられる)に記載されている通りに、CH3領域内に非対称的に導入されたシステインを架橋する段階:
‐第1の抗体中のD282Cと第2の抗体中のK275C、
‐第1の抗体中のD282Sと第2の抗体中のK275S、
‐第1の抗体中のY232Cと第2の抗体中のS237C、
‐第1の抗体中のY232Cと第2の抗体中のD239C、
‐第1の抗体中のY232Cと第2の抗体中のE240C、
‐第1の抗体中のL234Cと第2の抗体中のS237C、
‐第1の抗体中のT277Cと第2の抗体中のV280C、
‐第1の抗体中のV280Cと第2の抗体中のK275C。
【0095】
さらなる局面において、本発明は、架橋法によって、例えば上記の3つの架橋法のいずれかによって得られるか、または得られ得る、安定した二重特異性抗体に関する。
【0096】
結果として得られた二重特異性抗体が架橋によって安定されたかどうかにかかわらず、本発明の方法はいくつかの態様において、二重特異性抗体を精製するさらなる段階を含み得る。(これらに限定されないが)標準的なプロテインAクロマトグラフィー、プロテインG、プロテインL、陽イオン/陰イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、疎水性相互作用クロマトグラフィー、チオフィリッククロマトグラフィーなどの標準的なクロマトグラフィー技法を用いて、またはIgG分子を捕獲するように設計されたリガンド(プロテインA模倣体、ラマVHHリガンド等)を用いて、二重特異性抗体を含む混合物を精製することができる。または、塩が誘導する沈殿(硫酸アンモニウム)、有機溶媒(DMSO、エタノール)の添加、pHの変更、または非イオン性ポリマー(ポリエチレングリコール)などの標準的な技法を用いて、IgG混合物を沈殿させることができる。別の設定においては、混合物を、IgG分子の濃縮を可能にする膜を用いる濾過技法に供することができる。ある種の混合物は二重特異性抗体の他に親IgG分子をなお含んでいる場合があるため、二重特異性抗体を完全な均一性になるまで精製するためには、これらすべての技法の組み合わせが必要である場合がある。親の単一特異性IgG分子から二重特異性抗体を分離するには、さらなる精製段階が次に必要となる場合がある。これは、例えば、第1の結合特異性に対するアフィニティーカラムを用いて結合および溶出し、次に第2の結合特異性に対するアフィニティーカラムを用いて結合および溶出することによる精製によって行うことができる。特に化学的架橋を行わなかった場合の好ましい態様では、非還元条件など、さらなる半分子の交換を妨げる条件下で精製を行う。
【0097】
(精製された)二重特異性抗体の量、質、および純度は、吸光度測定、HP-SEC、SDS-PAGE、未変性PAGE、およびRP-HPLCなどの日常的な生化学的技法を用いて解析することができる。特に関心対象となるのは、親IgG分子から二重特異性抗体を識別できる技法である。このような技法の例は、(これらに限定されないが)、電荷および/または質量に基づいて、分子の高度に正確な分離および検出を可能にするIEF、cIEF、CIEX、および質量分析(ESI、MALDI)である。二重特異性抗体の二重結合特異性は、例えばELISA、RIA、表面プラズモン共鳴(SPR)、バイオレイヤー干渉法、DELFIA、FRET、ECL、Gyros、およびAlfaScreenを使用する種々の異なる結合アッセイ形式を用いて評価することができる。
【0098】
1つの態様において、半分子の交換は、関心対象の2種類の抗原の一方に対する二重特異性抗体の形成を支持する条件下で行うことができる。例えば、抗原XおよびYに対する抗体を考えてみる。5倍過剰または10倍過剰などの、抗原Xに対する過剰な抗体を用いて交換を行う場合、抗体Yに対する、ほとんどまたはすべての抗体は二重特異性(すなわち、抗原XおよびYを認識する)となる。
【0099】
この工程の後に、マトリックス固定化抗原Yおよびアフィニティーカラムクロマトグラフィーにおいて、二重特異性抗体を精製することができる。結合した抗体は、所望の二重特異性抗体について高度に濃縮されている。抗原Xに対する非結合抗体は、上記のサイクルの反復に使用することができる。
【0100】
インビボにおける交換を防ぐために安定化が必要である場合には、上記の通りに二重特異性抗体を架橋することができる。化学的架橋後に、抗原Zに対する過剰な抗体とのさらなる交換反応、およびその後のマトリックス固定化抗原Zに対する抗Z含有抗体の吸収(アフィニティーカラムクロマトグラフィーなど)を行うことによって、非安定化抗体を安定化抗体から精製することができる。そこで、非結合画分は所望の安定化二重結合抗体を含んでいる。
【0101】
本発明の方法のなおさらなる態様において、本方法は、治療用途のために、結果として得られた二重特異性抗体を製剤化するさらなる段階を含む。これは、ヒト使用に適している、特に静脈内投与などの非経口投与に適している水溶液中での、治療上有効な量の二重特異性抗体の製剤化を含む。
【0102】
さらなる局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1抗体を提供する段階であって、該第1の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0103】
好ましくは、段階c)において還元剤を添加するが、この剤の濃度は、段階c)で生じる溶液の酸化還元電位が、実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である。
【0104】
さらなる局面において、本発明は、本明細書に記載する本発明の方法のいずれかによって得られるか、または得られ得る、二重特異性抗体を含む組成物に関する。
【0105】
さらなる主要な局面において、本発明は、2つのIgG4様CH3領域を含む単離された二重特異性抗体に関する。
【0106】
1つの態様において、該抗体はコアヒンジ領域内に1つまたは2つのCPPC配列を含む。
【0107】
別の態様において、該抗体は、コアヒンジ領域内に、式中、X1およびX2は両方がプロリンではないという条件で、X1およびX2は任意のアミノ酸であってよい、1つまたは2つのCX1X2C配列を含む。
【0108】
さらなる態様において、該抗体はコアヒンジ領域内に、式中、X3はプロリンを除く任意のアミノ酸であってよい、1つまたは2つのCX3PCまたはCPX3C配列を含む。
【0109】
なおさらなる態様において、該抗体は、コアヒンジ領域内に1つまたは2つのCSPC、CPSC、CRPC、またはCPRC配列を含む。
【0110】
単離された二重特異性抗体のいくつかの態様において、第1および/または第2のCH3領域は非IgG4アイソタイプのものであって、このCH3配列は、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている。
【0111】
その1つのさらなる態様において、第1および/または第2のCH3領域は、図18に示す配列(SEQ ID NO: 19)を有し、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:238位のArg(R)がGln(Q)によって置換されている;239位のAsp(D)がGlu(E)によって置換されている;292位のLys(K)がArg(R)によって置換されている;302位のGln(Q)がGlu(E)によって置換されている;および328位のPro(P)がLeu(L)によって置換されている。
【0112】
別のさらなる態様において、第1および/または第2のCH3領域は、図18に示す配列(SEQ ID NO: 20)を有し、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:234位のArg(R)がGln(Q)によって置換されている;276位のMet(M)がVal(V)によって置換されている;288位のLys(K)がArg(R)によって置換されている;298位のGln(Q)がGlu(E)によって置換されている;および324位のPro(P)がLeu(L)によって置換されている。
【0113】
なおさらなる態様において、第1および/または第2のCH3領域は、図18に示す配列(SEQ ID NO: 21)を有し、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:285位のArg(R)がGln(Q)によって置換されている;314位のSer(S)がAsn(N)によって置換されている;322位のAsn(N)がLys(K)によって置換されている;327位のMet(M)がVal(V)によって置換されている;339位のLys(K)がArg(R)によって置換されている;349位のGln(Q)がGlu(E)によって置換されている;352位のIle(I)がVal(V)によって置換されている;365位のArg(R)がHis(H)によって置換されている;366位のPhe(F)がTyr(Y)によって置換されている;および375位のPro(P)がLeu(L)によって置換されている。
【0114】
なおさらなる態様において、本発明の抗体の第1および/または第2のCH3領域はIgG4 CH3領域である。
【0115】
なおさらなる局面において、本発明は、薬として使用するための、例えば癌または感染症の治療用の薬として使用するためなどの、本発明の二重特異性抗体、または本明細書に記載する本発明の方法のいずれかによって得られるか、もしくは得られ得る、二重特異性抗体を含む薬学的組成物などの組成物に関する。
【0116】
なおさらなる局面において、本発明は、癌または感染症の治療用の薬を調製するための、本発明の二重特異性抗体、または本発明の方法のいずれかによって得られるか、もしくは得られ得る、二重特異性抗体を含む組成物の使用に関する。
【0117】
本発明の方法を用いて、標的結合特異性の、特に興味深いまたは効果的な組み合わせを選択することもできる。例えば、本発明の方法を用いて、異なる結合特異性を有する一組の抗体から、異なる二重特異性抗体の一組または「マトリックス」を作製することができる。次に、結果として得られた二重特異性抗体の一組またはマトリックスを所望の生物学的特性について試験して、最良の組み合わせを選択することができる。
【0118】
したがって、なおさらなる局面において、本発明は、以下の段階を含む、所望の特性を有する二重特異性抗体を選択する方法に関する:
a) 一組の抗体を提供する段階であって、各抗体が異なる標的特異性を有し、かつ各抗体がIgG4様CH3領域を含む段階、
b) 還元条件下で、該一組の抗体の各抗体を該一組の別の抗体と共にインキュベートし、それによって一組の抗体混合物を作製する段階であって、各混合物が異なる二重特異性抗体を含む段階、
c) 結果として得られた一組の抗体混合物を所与の所望の特性についてアッセイする段階、および
d) 所望の特性を有する二重特異性抗体混合物を選択する段階。
【0119】
上記の方法における段階b)は、段階c)について先に上で記した通りに行うことができる。
【0120】
1つの態様において、試験すべき所望の特性は腫瘍細胞殺傷である。
【0121】
参考文献

【0122】
本発明を以下の実施例によってさらに例証するが、実施例はさらなる限定と解釈されるべきではない。
【0123】
実施例
実施例1:オリゴヌクレオチドプライマーおよびPCR増幅
オリゴヌクレオチドプライマーは、Isogen Bioscience(オランダ、マールセン)によって合成および定量された。プライマーを100 pmol/μlとなるようにH2Oに溶解し、-20℃で保存した。すべてのPCRプライマーおよび配列決定プライマーの概要を以下に示す。PCRについては、PfuTurbo(登録商標) Hotstart DNAポリメラーゼ(Stratagene、オランダ、アムステルダム)を製造業者の説明書に従って使用した。各反応混合物は、全量20μlのPCR反応緩衝液(ポリメラーゼと共に供給)中に、200μM混合dNTP(Roche Diagnostics、オランダ、アルメレ)、フォワードプライマーおよびリバースプライマーいずれも6.7 pmol、ゲノムDNA 100 ngまたはプラスミドDNA 1 ng、およびPfuTurbo(登録商標) Hotstart DNAポリメラーゼ1単位を含んだ。32サイクルプログラム:95℃で2分の変性;95℃で30秒、60〜70℃勾配(または別の特定のアニーリング温度)で30秒、および72℃で3分の30サイクル;72℃で10分の最終伸長を用いて、TGradient Thermocycler 96(Whatman Biometra、ドイツ、ゲッティンゲン)でPCR反応を行った。必要に応じて、さらなる解析または処理時までPCR混合物を4℃で保存した。
【0124】
実施例2:アガロースゲル電気泳動
1×Tris酢酸EDTA緩衝剤中で50 mlのゲルを用いて、Sambrook(37)に従ってアガロースゲル電気泳動を行った。ゲル中にエチジウムブロマイドを含め、UV光下で観察することによって、DNAを可視化した。CCDカメラおよび画像解析システム(GeneGnome;Syngene、Westburg B.V.、オランダ、ルースデン経由)によりゲル画像を記録した。
【0125】
実施例3:PCR産物の解析および精製ならびに酵素消化
製造業者の説明書に従ってMinElute PCR精製キット(Qiagen、Westburg、オランダ、ルースデン経由;製品# 28006)を用いて、所望のPCR断片の精製を行った。単離したDNAをUV分光法によって定量し、アガロースゲル電気泳動によってその質を評価した。
【0126】
または、1% Tris酢酸EDTAアガロースゲルを用いてアガロース電気泳動によって(例えば、複数の断片が存在する場合)、PCR産物または消化産物を分離した。所望の断片をゲルから切り出し、製造業者の説明書に従ってQIAEX IIゲル抽出キット(Qiagen;製品# 20051)を用いて回収した。
【0127】
実施例4:UV分光法によるDNAの定量
製造業者の説明書に従ってNanoDrop ND-1000分光光度計(Isogen Life Science、オランダ、マールセン)を用いて、核酸の光学密度を決定した。260 nmにおける光学密度(OD)の解析により(1 OD260 nm単位=50μg/ml)、DNA濃度を測定した。すべての試料について、核酸を溶解した緩衝液を参照として使用した。
【0128】
実施例5:制限酵素消化
制限酵素および補充物は、New England Biolabs(米国、マサチューセッツ州、ベバリー)またはFermetas(リトアニア、ヴィルニアス)から入手し、製造業者の説明書に従って使用した。
【0129】
DNA(100 ng)を、最終量10μlの適切な緩衝液中で酵素5単位で消化した(必要に応じて、反応量を増やした)。消化物を、推奨される温度で最低60分間インキュベートした。適合しない緩衝液または温度の要求を伴う制限酵素による二重消化を必要とする断片については、消化を連続して行った。必要に応じて、消化産物をアガロースゲル電気泳動およびゲル抽出によって精製した。
【0130】
実施例6:DNA断片のライゲーション
製造業者の説明書に従って、迅速ライゲーションキット(New England Biolabs)によってDNA断片のライゲーションを行った。各ライゲーションについて、ベクターDNAをおよそ3倍モル濃度の過剰の挿入物DNAと混合した
【0131】
実施例7:大腸菌(E. coli)の形質転換
製造業者の説明書に従って熱ショック法を用いて、プラスミドDNA(DNA溶液1〜5μl、典型的にはDNAライゲーション混合物2μl)をOne Shot DH5α-T1RまたはMACH-1 T1Rコンピテント大腸菌細胞(Invitrogen、オランダ、ブレダ;製品# 12297-016)に形質転換した。次に、50μg/mlアンピシリンを含むルリア-ベルターニ(LB)寒天プレートに細胞をプレーティングした。細菌コロニーが明白になるまで、プレートを37℃で16〜18時間インキュベートした。
【0132】
実施例8:PCRによる細菌コロニーのスクリーニング
HotStarTaqマスターミックスキット(Qiagen;製品# 203445)ならびに適切なフォワードプライマーおよびリバースプライマー(付属書1)を用いるコロニーPCRにより、所望の配列を含むベクターの存在について細菌コロニーをスクリーニングした。選択したコロニーを20μlピペットチップで軽く触れ、小規模培養用の2 ml LB中に短時間触れさせ、次いでPCR混合物中に再懸濁した。35サイクルプログラム:95℃で15分の変性;94℃で30秒、55℃で30秒、および72℃で2分の35サイクル;その後の72℃で10分の最終伸長段階を用いて、TGradient Thermocycler 96でPCRを行った。必要に応じて、アガロースゲル電気泳動による解析時まで、PCR混合物を4℃で保存した。
【0133】
実施例9:大腸菌培養物からのプラスミドDNA単離
製造業者の説明書に従ってQiagen(Westburg、オランダ、ルースデン経由)による以下のキットを用いて、プラスミドDNAを大腸菌培養物から単離した。大量プラスミド調製(50〜150 ml培養物)については、HiSpeedプラスミドMaxiキット(製品# 12663)またはHiSpeedプラスミドMidiキット(製品# 12643)のいずれかを使用した。小規模プラスミド調製(±2 ml培養物)については、Qiaprepスピンミニプレップキット(製品# 27106)を使用し、DNAを溶出緩衝液(キットにより供給) 50μlで溶出した。
【0134】
実施例10:DNA配列決定
当技術分野で公知の標準的な手順を用いて、プラスミドDNAの配列を決定した。配列は、Vector NTIソフトウェア(Informax、英国、オックスフォード)を用いて解析した。
【0135】
実施例11:HEK-293F細胞における一過性発現
Freestyle(商標) 293-F(懸濁増殖および既知組成Freestyle培地に適応したHEK-293サブクローン、例えばHEK-293F)細胞をInvtrogenから入手し、293fectin(Invitrogen)を用いて製造業者のプロトコールに従ってトランスフェクトした。
【0136】
実施例12:pTomG4;可変重鎖領域とヒトIgG4の定常領域の発現用ベクターの構築
ゲノムDNAをボランティアの血液試料から単離し、プライマーIGG4gene2fおよびIGG4gene2r(以下の表を参照されたい)を用いるPCRにおいて、鋳型として使用し、IgG4の重鎖の完全なゲノム定常領域を増幅し、哺乳動物発現ベクターpEE6.4(Lonza Biologics)にクローニングするための適切な制限部位を導入した。PCR断片を精製し、pEE6.4にクローニングした。このために、PCR産物をHindIIIおよびEcoRIで消化し、続いて制限酵素を加熱不活化した。pEE6.4ベクターもHindIIIおよびEcoRIで消化し、続いて制限酵素を加熱不活化し、エビアルカリ性ホスファターゼでベクター断片を脱リン酸化し、次いでホスファターゼを加熱不活化した。IgG4断片およびpEE6.4HindIII/EcoRI脱リン酸化ベクターを連結し、コンピテントMACH1-T1R細胞(Invitrogen)に形質転換した。3つのクローンをLB中で培養し、プラスミドDNAを少量の培養物(1.5 mL)から単離した。制限消化から、pEE6.4ベクター中にIgG4断片がクローニングされたことと合致するパターンが明らかになった。2つのクローンによるプラスミドDNAをDH5α-T1R大腸菌に形質転換し、プラスミドDNAを単離し、構築物を挿入物の配列解析によって調べ、1つのクローンが、イントロン中のいくつかの軽微な違いを除いて、GenbankデータベースによるゲノムIgG4クローンと同一であることが判明した。これらの違いは、おそらくは多型またはGenbank配列における配列の誤りのいずれかである。このプラスミドをpTomG4と命名した。
【0137】
(表1)プライマー配列
【0138】
実施例13:マウス抗Betv1抗体および抗Feld1抗体の可変領域のクローニング
製造業者のプロトコールに従ってRNeasyキット(Qiagen、Westburg、オランダ、ルースデン)を用いて、0.3×105個(Betv1)または0.9×105個(Feld1)のマウスハイブリドーマ細胞(Betv1については:参考文献38によるクローン2H8、およびFeld1については:参考文献39によるクローン4F7)から全RNAを調製した。
【0139】
製造業者のプロトコールに従ってSMART RACE cDNA増幅キット(BD Biosciences Clontech、米国、カリフォルニア州、マウンテンビュー)を用いて、全RNA約100 ngからRNAの5'-RACE-相補的DNA(cDNA)を調製した。
【0140】
Betv1抗体およびFeld1抗体のVL領域およびVH領域をPCRによって増幅した。このために、製造業者の説明書に従ってPfuTurbo(登録商標) Hotstart DNAポリメラーゼ(Stratagene)を使用した。各反応混合物は、全量30μlのPCR反応緩衝液(ポリメラーゼと共に供給)中に、200μM混合dNTP(Roche Diagnostics)、リバースプライマー(VH領域用のRACEG1mm1およびVL領域用のRACEKmm1) 12 pmol、UPM-ミックス(UPM-ミックス:2μM ShortUPMH3オリゴヌクレオチドおよび0.4μM LongUPMH3オリゴヌクレオチド) 7.2pmol、上記の5'RACE cDNA鋳型0.6μl、およびPfuTurbo(登録商標) Hotstart DNAポリメラーゼ1.5単位を含んだ。
【0141】
35サイクルプログラム:95℃で2分の変性;95℃で30秒、55℃で30秒、および72℃で1.5分の35サイクル; 72℃で10分の最終伸長を用いて、TGradient Thermocycler 96(Whatman Biometra)でPCR反応を行った。反応産物を1% TAEアガロースゲルでのアガロースゲル電気泳動によって分離し、エチジウムブロマイドで染色した。正確な大きさのバンドをゲルから切り出し、QiaexIIゲル抽出キット(Qiagen)を用いてDNAをアガロースから単離した。
【0142】
200μM dATPおよびAmplitaq(Perkin Elmer) 2.5単位と共に72℃で10分間インキュベートすることによって、ゲルから単離したPCR断片にA尾部を付加し、minieluteカラム(Qiagen)を用いて精製した。製造業者のプロトコールに従ってpGEMT easyベクター系IIキット(Promega)を用いて、A尾部を付加したPCR断片をpGEMTeasyベクター(Promega)にクローニングした。ライゲーション混合物2μlをOneShot DH5αT1Rコンピテント大腸菌(Invitrogen)に形質転換し、LB/Amp/IPTG/Xgalプレートにプレーティングした。VH配列およびVL配列それぞれについて挿入物を含む4つの白色コロニーを拾い、挿入物の配列を決定した。Betv1のVHおよびVLの推定アミノ酸配列をSEQ ID NO:15および16に、ならびにFeld1の推定アミノ酸配列をSEQ ID NO:17および18に示す。
【0143】
実施例14:pConG1fBetV1:Betv1-IgG1の重鎖の産生用ベクターの構築
マウス抗BetV1抗体のVHコード領域を、プライマーVHexbetv1forおよびVHexbetv1revを用いてこの領域を含むプラスミド(実施例13)からPCRにより増幅し、pConG1f0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。VH断片をゲル精製し、pConG1f0.4にクローニングした。このために、PCR産物およびpConKappa0.4ベクターをHindIIIおよびApaIで消化し、精製した。VH断片およびpConG1f0.4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、正確な配列を確認した。このプラスミドをpConG1fBetv1と命名した。
【0144】
実施例15:pConKBetv1:Betv1の軽鎖の産生用ベクターの構築
VLコード領域マウス抗BetV1抗体を、プライマーVLexbetv1forおよびVLexbetv1revを用いてこの領域を含むプラスミド(実施例13)から増幅し、pConK0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。PCR産物およびpConKappa0.4ベクターをHindIIIおよびBsiWIで消化し、精製した。VL断片およびpConKappa0.4HindIII-BsiWI消化ベクターを連結し、コンピテントDH5α T1R大腸菌に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpConKBetv1と命名した。
【0145】
実施例16:pTomG4Betv1:Betv1-IgG4の重鎖の産生用ベクターの構築
Betv1-IgG4発現用ベクターを構築するため、BetV1のVH領域をpTomG4にクローニングした。このために、pTomG4およびpConG1fBetv1をHindIIIおよびApaIで消化し、関連する断片を単離した。Betv1 VH断片およびpTomG4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpTomG4Betv1と命名した。
【0146】
実施例17:pConG1fFeld1:Feld1-IgG1の重鎖の産生用ベクターの構築
マウス抗Feld1抗体のVHコード領域を、プライマーVHexfeld1forおよびVHexfeld1revを用いてこの領域を含むプラスミド(実施例13)からPCRにより増幅し、pConG1f0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。VH断片をゲル精製し、pConG1f0.4にクローニングした。このために、PCR産物およびpConKappa0.4ベクターをHindIIIおよびApaIで消化し、精製した。VH断片およびpConG1f0.4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、正確な配列を確認した。このプラスミドをpConG1fFeld1と命名した。
【0147】
実施例18:pConKFeld1:Feld1の軽鎖の産生用ベクターの構築
VLコード領域マウス抗Feld1抗体を、プライマーVLexfeld1forおよびVLexfeld1revを用いてこの領域を含むプラスミド(実施例13)から増幅し、pConK0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。PCR産物およびpConKappa0.4ベクターをHindIIIおよびBsiWIで消化し、精製した。VL断片およびpConKappa0.4HindIII-BsiWI消化ベクターを連結し、コンピテントDH5α T1R大腸菌に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpConKFeld1と命名した。
【0148】
実施例19:pTomG4Feld1:Feld1-IgG4の重鎖の産生用ベクターの構築
Feld1-IgG4発現用ベクターを構築するため、Feld1のVH領域をpTomG4にクローニングした。このために、pTomG4およびpConG1f Feld1をHindIIIおよびApaIで消化し、関連する断片を単離した。Feld1 VH断片およびpTomG4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpTomG4Feld1と命名した。
【0149】
実施例20:2F8-IgG4および7D8-IgG4の発現用の抗体発現ベクターの構築
HuMab 2F8(IgG1-EGFR)およびHuMab 7D8(IgG1-CD20)の発現用の発現ベクターを構築した。HuMab 2F8(WO 02/100348)およびHuMab 7D8(WO 04/035607)のVHおよびVLコード領域を、IgG1重鎖の産生用の発現ベクターpConG1f(Lonza Biologics)およびκ軽鎖の産生用のpConKappaにクローニングし、ベクターpConG1f2F8、pConG1f7D8、pConKappa2F8、およびpConKappa7D8を得た。pConG1f2F8およびpConG1f7D8のVH領域をHindIII/ApaI消化によってこれらのベクターから除去し、HidIII/ApaI消化したpTomG4ベクターに挿入し、それぞれpTomG42F8およびpTomG47D8を得た。
【0150】
実施例21:HEK-293F細胞における一過的発現によるBetv1-IgG1、Betv1-IgG4、Feld1-IgG1、およびFeld1-IgG4の産生
製造業者の説明書に従って293fectinを用いて、関連する重鎖ベクターおよび軽鎖ベクターをHEK-293F細胞に同時トランスフェクトすることによって、すべての構築物から抗体を産生させた。Betv1-IgG1については、pConG1Betv1およびpConKBetv1を同時発現させた。Betv1-IgG4については、pTomG4Betv1およびpConKBetv1を同時発現させた。Feld1-IgG1については、pConG1Feld1およびpConKFeld1を同時発現させた。Feld1-IgG4については、pTomG4Feld1およびpConKFeld1を同時発現させた。IgG1-EGFrについては、pConG1f2F8およびpConKappa2F8を同時発現させた。IgG4-EGFrについては、pTomG42F8およびpConKappa2F8を同時発現させた。IgG1-CD20については、pConG1f7D8およびpConKappa7D8を同時発現させた。IgG4-CD20については、pTomG47D8およびpConKappa7D8を同時発現させた。
【0151】
実施例22:IgG1抗体およびIgG4抗体の精製
IgG1抗体およびIgG4抗体を、プロテインAアフィニティークロマトグラフィーで精製した。細胞培養上清を0.20μMデッドエンドフィルターを通して濾過し、次に5 mlプロテインAカラム(rProtein A FF、GE Healthvcare)に負荷し、0.1 Mクエン酸-NaOH、pH3でIgGを溶出した。溶出液を直ちに2 M Tris-HCl、pH9で中和し、12.6 mMリン酸ナトリウム、140 mM NaCl、pH 7.4(B. Braun、オランダ、オッス)に対して一晩透析した。透析後、試料を0.20μMデッドエンドフィルターを通して滅菌濾過した。比濁法および280 nmの吸光度によって、精製IgGの濃度を決定した。SDS-PAGE、IEF、質量分析、および糖解析によって、精製タンパク質を解析した。
【0152】
実施例23:精製IgGのSDS-PAGE解析
精製後、Betv1およびFeld1のIgG1抗体およびIgG4抗体を非還元SDS-PAGEで解析した。使用したBis-Tris電気泳動法はLaemmli法(Laemmli 1970 Nature 227(5259): 680-5)の改良法であり、試料を中性pHで泳動した。SDS-PAGEゲルをクマシーで染色し、GeneGenius(Synoptics、英国、ケンブリッジ)を用いてデジタル画像化した。
【0153】
図1に示され得るように、Betv1およびFeld1のIgG1は、全長四量体(2本の重鎖および2本の軽鎖)のFeld1およびBetv1のIgG1分子を表す1本の主要なバンドを示した。Betv1およびFeld1のIgG4は、四量体IgG4分子を表す主要なバンドに加えて、相当量の半分子(すなわち、1本の重鎖および1本の軽鎖)を有することが示された。
【0154】
実施例24:マウスにおけるIgG4半分子の交換の評価
6〜8週齢のnu/nu Balb/cマウス5匹を用いて、IgG4半分子の交換を追跡した。マウスをCentral Laboratory Animal Facility(オランダ、ユトレヒト)の隔離施設に収容し、水および食物を自由に与えながらフィルタートップケージで飼育した。実験はすべて、ユトレヒト大学動物倫理委員会によって承認された。
【0155】
キメラ抗体を腹腔内投与した。投与の4.25時間、24時間、48時間、および72時間後に血液試料(75〜100μl)を採取した。血液をヘパリン含有バイアルに回収し、10,000 gで5分間遠心分離して、細胞から血漿を分離した。抗原特異的抗体および二重特異性抗体のレベルの決定用に、血漿を-20℃で保存した。
【0156】
本実験では、キメラIgG4半分子の交換(n=2)をIgG1半分子の交換(n=3)と比較した。200μl中600μg(各抗原特異的抗体300μg)/マウスの用量で、Bet v 1特異的抗体とFel d 1特異的抗体(IgG1またはIgG4)の混合物をマウスに投与した。
【0157】
Bet v 1結合抗体またはFel d 1結合抗体の血漿濃度を抗原結合試験で測定した。この目的のために、血漿試料を、125I標識Bet v 1または125I標識Fel d 1の存在下で、PBS/IAT(1μg/ml IVIg、0.3%ウシ血清アルブミン、0.1% Tween-20、0.05%(w/v) NaN3を補充したPBS) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体またはFel d 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGまたはFel d 1特異的IgGの濃度を算出した。二重特異性IgGの濃度は、異種架橋アッセイ法の2つの変法において測定した。第1のアッセイ法では、血漿を全量300μlのPBS-IAT中、セファロース結合Bet v 1(0.5 mg)と共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Fel d 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Bet v 1-Fel d 1)の濃度を、精製Fel d 1結合rIgGから得られたFel d 1結合試験の検量線を用いて算出した。第2のアッセイ法では、Fel d 1-Bet v 1架橋活性を、セファロース結合rFel d 1(0.5 mg)および125I標識Bet v 1を用いて同様の手順で測定した。二重特異性IgG(Fel d 1-Bet v 1)の濃度を、標準物質として精製Bet v 1特異的rIgGを用いて(Bet v 1結合試験における検量線と同じ検量線)算出した。
【0158】
図2において、様々な時点における二重特異性IgG(Fel d 1-Bet v 1)の濃度をBet v 1結合IgGの濃度に対してプロットする。二重特異性IgGは、IgG4を投与したマウスとは対照的に、IgG1混合物を投与したマウスでは認められなかった。24時間後に二重特異性IgG4の生成は最大であり、これは100%の交換に相当した。
【0159】
図3Aでは、二重特異性ヒトIgG4の形成を時間を追って追跡する。IgG4の混合物を注射したマウスの血漿中には二重特異性抗体が時間と共に出現したが、IgG1を注射したマウスでは出現せず、二重特異性反応性は1〜2日間のインキュベーション後に最大でほぼ50%に達した(注記:等量のIgG4-Betv1とIgG4-Feld1が交換された場合、半抗体の無作為でかつ完全な交換後に、IgG4-Betv1半抗体の最大50%が二重特異性画分に取り込まれる)。等量のIgG4-Betv1とIgG4-Feld1間の無作為的なFabアーム交換は、IgG4分子の約半分が二重特異性を獲得することと一致する。対照として、非関連抗原に対する20倍過剰のさらなるIgG4(抗EGFr抗体2F8から作製したIgG4)を、IgG4-Betv1およびIgG4-Feld1と共にマウスに注射した。過剰な非関連IgG4は、Betv1-Feld1二重特異性IgG4の生成と競合した。
【0160】
別の実験(図3B)において、同じマウス血漿試料を、放射標識可溶性Fel d 1をセファロース固定化Fel d 1に架橋する能力について試験した。単一特異性架橋活性は、IgG4の同等の混合物を投与したマウスでは減少したが、IgG1の同等の混合物を投与したマウスでは減少しないことが判明し、単一特異性架橋活性の喪失が示された。約1日後に、約50%の最大減少に達した。さらなる過剰な非関連IgG4を投与したマウスでは、単一特異性架橋活性は同様の動態学でほぼ完全に消失した。
【0161】
サイズ排除クロマトグラフィーを行って、IgG4を投与したマウスにおいて認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した(図4を参照されたい)。この目的のために、血漿試料(t=24時間の時点で採取)をSuperdex200カラムで分画し、その後その画分中でFel d 1結合IgGおよびBet v 1-Fel d 1架橋IgGを測定した。Fel d 1結合抗体は、保持容量約12.9 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Bet v 1-Fel d 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された。rIgG1含有血漿中には、分画前にBet v 1-Fel d 1架橋活性は存在しなかった。同様に溶出画分においても、異種架橋活性は測定されなかった(データは示さず)。
【0162】
実施例25:全血(成分)における交換活性の評価
キメラ抗体を混合し、続いて全血、血液細胞、血漿、または血清と共にインキュベートして、全血(成分)の交換活性を調べた。
【0163】
本実験では、IgG4半分子の交換を、2名の健常な血液ドナーであるAおよびB由来の全血において評価し、このドナーのIgG4の内因性血漿レベルは比濁法によって決定した(それぞれ、346μg/mlおよび554μg/mlであった)。最終濃度40μg/mlのTFPI(Chiron Corporation、カリフォルニア州、エメリービルによる組織因子経路阻害剤)を補充したバキュテイナ中に、全血を採取した。全血の遠心分離により、血液細胞および血漿を取得した。細胞画分をOptimem(Invitrogen、オランダ、ブレダ)で3回洗浄し、続いてOptimemに再懸濁した。血清は、全血をガラスバキュテイナ中で凝固活性化剤と共に37℃で30分間インキュベートし、その後凝固した血液を遠心沈殿させることによって取得した。IgG4半分子の交換を評価し、IgG1半分子の交換と比較した。対照として、血液試料をキメラ抗体の非存在下で同様にインキュベートした。以下の抗体混合物をPBS中で調製した:
1. Bet v 1特異的IgG4(10μg)およびFel d 1特異的IgG4(10μg)
2. Bet v 1特異的IgG1(10μg)およびFel d 1特異的IgG1(10μg)。
【0164】
これらの抗体混合物を、水平軌道振盪機(125 rpm)上で、全量100μl中で血液、血液細胞、血漿、または血清と共に37℃でインキュベートした(各抗体の最終濃度は0.1μg/mlであった)。全血および血液細胞を伴うインキュベーション混合物中の最終ヘマトクリットは、約40%であった。24時間後、インキュベーション混合物をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料10μlをPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS) 500μl中に採取した。試料は必要に応じて4℃で保存した。
【0165】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合組換えFel d 1 0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0166】
図5において、二重特異性活性を、異種架橋アッセイ法で決定された、結合した125I標識Bet v 1の割合(%)として表す。二重特異性活性はIgG4半分子の交換の尺度であり、これは主に全血および全血の細胞画分において認められた(図5A)。細胞画分における二重特異性レベルは、全血よりもさらに高かった。これは、細胞画分内に、添加したキメラIgG4抗体と同様に交換され得る内因性IgG4がもはや存在しないという事実によって説明される可能性が最も高い。いくらかの二重特異性活性は血漿および血清においても認められたが、この活性は全血において認められた活性よりもはるかに低く、またOptimem中でIgG4混合物をインキュベートすることによって得られた1.7%というバックグラウンドレベルよりもわずかに高いにすぎなかった。二重特異性活性は、IgG1を含むインキュベーションのいずれにおいても認められなかった(図5B)。同様にキメラ抗体を含まない対照インキュベーションにおいても、二重特異性活性は認められなかった(図5C)。サイズ排除クロマトグラフィーを行って、IgG4混合物において認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した。この目的のために、試料(t=24時間の時点で採取)をSuperdex200カラムで分画し、その後その画分中でFel d 1結合IgGおよびBet v 1-Fel d 1架橋IgGを測定した。Fel d 1結合抗体は、保持容量約12.9 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Bet v 1-Fel d 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された(データは示さず)。
【0167】
実施例26:血液細胞媒介性IgG4交換活性の評価
キメラ抗体を混合し、続いて3種の異なる型のヒト血液細胞(すなわち、単核細胞(MNC)、赤血球、および血小板)と共にインキュベートして、IgG4交換活性を調べた。
【0168】
匿名ドナー由来の全血をヘパリン含有バキュテイナ中に採取し、続いてPercoll(Pharmacia Fine Chemicals、スウェーデン、ウプサラ)中で遠心分離して、MNCを単離した。使用前に、単離したMNCをOptimem無血清培地(Invitrogen、オランダ、ブレダ)に再懸濁した。新たに精製された赤血球および血小板(SanquinのBlood Cell Research Departmentにより提供)は、2名の異なる匿名ドナーから得られた。これらの細胞も、3回洗浄した後にOptimemに再懸濁した。加えて、血小板には10 mMグルコースを補充した。
【0169】
IgG4半分子の交換を評価し、IgG1半分子の交換と比較した。以下の抗体混合物をPBS中で調製した:
‐Bet v 1特異的IgG4(10μg)およびFel d 1特異的IgG4(10μg)
‐Bet v 1特異的IgG1(10μg)およびFel d 1特異的IgG1(10μg)。
【0170】
これらの抗体混合物を、水平軌道振盪機(125 rpm)上で、全量100μl中で1.8×104個MNC、4.0×108個赤血球、または3.5×104個血小板と共に37℃でインキュベートした(各抗体の最終濃度は0.1μg/mlであった)。48時間後、インキュベーション混合物をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料10μlをPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS) 500μl中に採取した。試料は必要に応じて4℃で保存した。
【0171】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合組換えFel d 1 0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0172】
図6において、二重特異性活性を、異種架橋アッセイ法で決定された、結合した125I標識Bet v 1の割合(%)として表す。3種の細胞型はすべて、二重特異性活性を誘導することができた。いくらかの二重特異性活性がOptimem無血清培地においても認められたが、この活性は、血液細胞の存在下で認められた活性よりもはるかに低かった。試験した細胞のいずれも、IgG1半分子を交換することはできなかった。
【0173】
実施例27:ヒト細胞株およびマウス細胞株によるIgG4交換の評価
キメラIgG4抗体を混合し、続いて3種の異なる細胞株(すなわち、ヒト胚腎(HEK)細胞、マウスB細胞、またはハイブリドーマ)と共にインキュベートして、IgG4交換活性を調べた。
【0174】
マウスB細胞の供給源として、細胞株J558(SanquinのAntigen Presentation Research Groupにより提供)を選択した。抗C1エステラーゼ阻害因子を産生するハイブリドーマは、SanquinのAutoimmune Research Groupより入手した。懸濁HEK(293F)細胞は、Invitrogen、オランダ、ブレダより入手した。細胞はすべてPBSで3回洗浄した後、PBSに再懸濁した。
【0175】
Bet v 1特異的IgG4(2μg)およびFel d 1特異的IgG4(2μg)からなるIgG4抗体混合物を上記の細胞と共にインキュベートすることによって、IgG4半分子の交換を評価した。抗体混合物を、水平軌道振盪機(125 rpm)上で、全量50μl中で24×105個HEK細胞、25×105個マウスB細胞、または21×105個ハイブリドーマと共に37℃でインキュベートした(各抗体の最終濃度は80μg/mlであった)。0時間および24時間後、インキュベーション混合物をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料をPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に採取した。試料は必要に応じて4℃で保存した。
【0176】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料希釈物を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合組換えFel d 1 0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0177】
図7において、二重特異性活性を、異種架橋アッセイ法で決定された、結合した125I標識Bet v 1の割合(%)として表す。3種の細胞型はすべて、IgG4半分子を交換することができた。
【0178】
実施例28:赤血球によるIgG4半分子の交換の評価
キメラ抗体を混合し、続いてヒト赤血球と共にインキュベートして、IgG4半分子の交換を調べた。赤血球はドナー1名から精製し、SAGM(生理食塩水アデニングルコースマンニトール)緩衝液中で4℃で保存した。使用前に、細胞をPBSで3回洗浄した。
【0179】
本実験では、IgG4半分子の交換をIgG1半分子の交換と比較した。また、過剰な非関連IgG4の存在下におけるIgG4の交換も評価した。以下の抗体混合物をPBS中で調製した:
‐Bet v 1特異的IgG4(4μg)およびFel d 1特異的IgG4(4μg)
‐Bet v 1特異的IgG1(4μg)およびFel d 1特異的IgG1(4μg)
‐Bet v 1特異的IgG4(4μg)、Fel d 1特異的IgG4(4μg)、および抗原Xに特異的な非関連IgG4(80μg)。
【0180】
これらの混合物を全量100μlの、0.05%(w/v) NaN3を補充したPBS中で赤血球と共にインキュベートし(最終ヘマトクリットは約40%であった)、続いて水平軌道振盪機(125 rpm)上で37℃でインキュベートした。表示の時点で、赤血球をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料10μlをPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS) 500μl中に採取した。試料は、二重特異性活性、二価性、および抗原結合を測定する前に4℃で保存した。対照として、赤血球を含まないPBS中で同じ混合物を同様にインキュベートした。
【0181】
Bet v 1結合抗体のレベルを抗原結合試験で測定した。この目的のために、試料を、125I標識Bet v 1の存在下で、PBS-IAT(1μg/ml IVIgを補充したPBS-AT) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGの濃度を算出した。Fel d 1特異的抗体およびBet v 1特異的抗体を用いる実験における二重特異性活性は、Feld1-Betv1架橋アッセイ法で測定した。このアッセイ法では、IgG含有試料を全量300μlのPBS-AT中、セファロース結合ネコ抽出物(0.5 mg)と共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Feld1-Betv1)の濃度を、標準物質として精製IgG1-Betv1を用いて(Prot Gセファロースを用いるBet v 1結合試験で得られる)算出した。
【0182】
図8に、赤血球媒介性の交換から得られたデータを示す。赤血球の存在下でIgG1半分子の交換が認められなかったのに対して、IgG4半分子のほぼ最大の交換が72時間後に認められた(パネルA)(注記:等量のIgG4-Betv1とIgG4-Feld1が交換された場合、半抗体の無作為でかつ完全な交換後に、IgG4-Betv1半抗体の最大50%が二重特異性画分に取り込まれる)。過剰な非関連IgG4の存在下では、IgG4半分子の交換はほとんど測定されず、これはBet v 1特異的IgG4およびFel d 1特異的IgG4と非関連IgG4との予測される交換と一致する。サイズ排除クロマトグラフィーを行って、IgG4混合物において認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した。この目的のために、試料(t=72時間の時点で採取)をSuperdex200カラムで分画し、その後その画分中でFel d 1結合IgGおよびBet v 1-Fel d 1架橋IgGを測定した。Fel d 1結合抗体は、保持容量約12.9 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Bet v 1-Fel d 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された(データは示さず)。
【0183】
理論的には、IgG4半分子の交換は二価性の減少も伴う。これを試験するために、インキュベーション混合物中の二価性を測定した。IgG1混合物ではFel d 1二価性の減少は認められなかったのに対して、IgG4混合物では約50%の減少が認められた。この減少は、1対1比で混合した2種類の異なるIgG4分子の最大交換と一致する。予想通り、過剰な非関連IgG4を添加したIgG4混合物における二価性の減少はより高く(約80%)、これは過剰な非関連IgG4半分子の存在下で2つの相同的な半分子(Bet v 1またはFel d1特異的)の再ハイブリダイゼーションの確率が低くなるためである。72時間のインキュベーション後に抗原結合はわずかに(約10%)減少するにすぎなかったため(データは示さず)、二価性の強い減少は、インキュベーション中に抗原結合が失われたためではなかった。
【0184】
PBS(0.05%(w/v) NaN3を補充)中でのIgGの交換も評価して、IgG4半分子が自発的に交換され得るかどうかを調べた。本実験の設定は、赤血球を添加しなかったこと以外は、赤血球の存在下での交換と同様であった。図9Aに示されるように、PBS中での37℃でのインキュベーション中に、IgG1半分子またはIgG4半分子の自発的な交換は認められなかった。しかしながら、IgG4混合物においていくらかのバックグラウンドが認められ、これは赤血球とのインキュベーション中にも存在した。PBS中でのインキュベーション中に、二価性の減少は認められなかった(図9B)。
【0185】
実施例29:赤血球溶解物によるIgG4交換の評価
キメラIgG4抗体を混合し、続いて赤血球溶解物の漸増希釈物と共にインキュベートした。赤血球は健常ドナーから単離し、ヘマトクリット60.7%の状態でSAGM(生理食塩水アデニングルコースマンニトール)緩衝液中で4℃で保存した。溶解物を得るために、細胞をPBS-アジド(0.05%(w/v) NaN3を補充したPBS)で3回洗浄し、保存緩衝液の量の2倍量の水に再懸濁した。結果として、非希釈赤血球溶解物はヘマトクリット30%に相当した。
【0186】
Bet v 1特異的IgG4(1μg)およびFel d 1特異的IgG4(1μg)からなるIgG4抗体混合物を、新たに調製した溶解物50μl(全量100μlとなるようにPBS/アジドを補充)と共に37℃でインキュベートすることによって、IgG4半分子の交換を評価した。各抗体の最終濃度は10μg/mlであった。表示の時点で、インキュベーション混合物からPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に試料を採取して、二重特異性活性を測定した。試料は必要に応じて4℃で保存した。
【0187】
二重特異性活性(すなわち、Bet v 1-Fel d 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料希釈物を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合カバノキ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Fel d 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Bet v 1-Fel d 1)の濃度を、精製Fel d 1結合rIgGから得られたFel d 1結合試験の検量線を用いて算出した。
【0188】
図10において、時間に伴う二重特異性活性の生成を、異種架橋アッセイ法で決定された、結合した125I標識Fel d 1の割合(%)として示す。これらのデータから、赤血球の溶解物が交換活性を含むことが明白である。最も高い交換率は非希釈溶解物で認められ、希釈が高いほど交換率は低下した。実際に、PBS中での対照インキュベーションでは二重特異性活性は認められなかった。
【0189】
サイズ排除クロマトグラフィーを行って、赤血球溶解物によって誘導された二重特異性活性がIgG凝集の結果であるという可能性を排除した(図11)。この目的のために、Bet v 1結合IgG4 10μg、Fel d 1結合IgG4 10μg、および赤血球溶解物50μlからなるインキュベーション混合物を調製し、最終量100μlとなるようにPBS/アジドを補充した。この混合物を37℃で24時間インキュベートし、その後70μlをSuperdex200カラムで分画した。その画分中で、Bet v 1結合IgGおよびFel d 1-Bet v 1架橋IgGを測定した。Bet v 1結合抗体のレベルは、抗原結合試験で測定した。試料を、125I標識Bet v 1の存在下で、PBS-IAT(1μg/ml IVIg、0.3%ウシ血清アルブミン、0.1% Tween-20、0.05%(w/v) NaN3を補充したPBS) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGの濃度を算出した。二重特異性IgGの濃度(すなわち、Fel d 1-Bet v 1架橋活性)は、異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT中、Fel d 1抗原が存在するセファロース結合ネコ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Fel d 1-Bet v 1)の濃度を、精製Bet v 1結合rIgGから得られた、Bet v 1結合試験で使用したのと同じ検量線を用いて算出した。
【0190】
Bet v 1結合抗体は、保持容量約12.6 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した(図11)。異種Fel d 1-Bet v 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された。
【0191】
実施例30:透析赤血球溶解物におけるIgG4交換活性の評価
赤血球を健常ドナーから単離し、ヘマトクリット60.7%の状態でSAGM(生理食塩水アデニングルコースマンニトール)緩衝液中で4℃で保存した。溶解物を得るために、細胞をPBS-アジド(0.05%(w/v) NaN3を補充したPBS)で3回洗浄し、保存緩衝液の量の2倍量の水に再懸濁した。したがって、非希釈赤血球溶解物はヘマトクリット30%に相当した。Pierceによる透析膜カセット(3.5 kDカットオフ)を用いて、溶解物の一部をPBS-アジドに対して透析した。非透析溶解物をAmiconフィルター(3.5 kDカットオフ)中で遠心分離することにより、限外濾過液を得た。
【0192】
IgG4抗体混合物(Bet v 1特異的IgG4(0.5μg)およびFel d 1特異的IgG4(0.5μg))を新たに調製した赤血球溶解物(25μl)または透析溶解物(25μl)と共に37℃でインキュベートすることによって、IgG4半分子の交換を評価した。各インキュベーションの全量は50μlであり、各抗体の最終濃度は10μg/mlであった。以下の補充物を使用した:Sigmaによる還元型グルタチオン(GSH)、グルコース-6-リン酸(G-6-P)、およびNADPH(いずれもRocheによる)。これらの化合物は、使用前に水に溶解した。24時間のインキュベーション後、インキュベーション混合物からPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に試料を採取して、二重特異性活性を測定した。試料は必要に応じて4℃で保存した。
【0193】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料希釈物を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合ネコ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0194】
交換レベルを、新たに調製した溶解物によって生じた二重特異性活性と比較した(表2)。
【0195】
(表2)透析した赤血球溶解物において二重特異性活性を回復する因子の概要
透析した赤血球溶解物の交換活性を、新たに調製した溶解物と比較した。透析した溶解物に限外濾過液5□lを補充した。G-6-P、NADPH、およびGSHの濃度は、それぞれ5 mM、0.1 mM、および0.5 mMであった。
【0196】
これらのデータから、赤血球溶解物の活性が透析後に失われたことが明白である。限外濾過液を添加することによって、交換の大部分が回復された。この結果から、透析中に成分(<3.5 kD)が失われ、これが交換反応に必須であることが示唆された。IgG4半分子の交換にジスルフィド架橋の還元および酸化が必要であることから、このような成分は酸化還元サイクルに関与している可能性が高い。したがって、酸化還元サイクルの3つの「補助因子」(G-6-P、NADPH、およびGSH)を透析した溶解物に添加して、これらの化合物が交換活性を回復し得るかどうかを調べた。G-6-P、NADPH、およびGSHを共に補充した場合に、交換活性は回復され得た。個々の因子の存在下で透析溶解物をインキュベートすることにより、交換活性は、G-6-PでもNADPHでもなく、GSHによって回復されることが明らかになった。
【0197】
実施例31:還元型グルタチオンによるIgG4半分子の交換の評価
キメラ抗体を混合し、続いて還元型グルタチオン(GSH)と共にインキュベートして、IgG4半分子の交換を調べた。GSH(Sigma-Aldrich、ミズーリ州、セントルイス)は、使用前に水に溶解した。
【0198】
本実験では、Bet v 1特異的IgG4(1μg)およびFel d 1特異的IgG4(1μg)からなるIgG4抗体混合物を、GSHを含むPBS/アジド中で37℃でインキュベートすることによって、IgG4半分子の交換を評価した。全インキュベーション量は100μlであり、各抗体の最終濃度は10μg/mlであった。表示の時点で、インキュベーション混合物からPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に試料を採取した。抗原結合および二重特異性活性の測定用に、試料を4℃で保存した。
【0199】
Bet v 1結合抗体のレベルを抗原結合試験で測定した。試料を、125I標識Bet v 1の存在下で、PBS-IAT(1μg/ml IVIgを補充したPBS-AT) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGの濃度を算出した。二重特異性IgGの濃度(すなわち、Fel d 1-Bet v 1架橋活性)は、異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT中、Fel d 1抗原が存在するセファロース結合ネコ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Fel d 1-Bet v 1)の濃度を、精製Bet v 1結合IgGから得られた、Bet v 1結合試験で使用したのと同じ検量線を用いて算出した。
【0200】
図12に、IgG4半分子のGSH媒介性の交換の時間経過を示す。これらのデータから、IgG4半分子がGSHの存在下で交換されることが明らかである。本実験では、最適な交換は0.1 mM GSHと1 mM GSHとの間で認められ、0.5 mM GSHを用いて24時間後に最も高い交換(約90%)に達した。
【0201】
サイズ排除クロマトグラフィーを行って、IgG4のGSH媒介性の交換後に認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した(図13)。この目的のために、Bet v 1結合IgG4およびFel d 1結合IgG4(各抗体10μg)の混合物を、PBS/アジド中で0.5 mM GSHと共にインキュベートした。この混合物(最終量100μl)を37℃で24時間インキュベートし、その後70μlをSuperdex200カラムで分画した。その画分中で、Bet v 1結合IgGおよびFel d 1-Bet v 1架橋IgGを測定した。Bet v 1結合抗体は保持容量約12.6 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Fel d 1-Bet v 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された。交換が4℃よりも37℃でより効率的に起こったことから、GSHの存在下での二重特異性IgG4分子の生成は、温度依存的であることが判明した(図14)。
【0202】
実施例32. 他の作用物質の存在下における二重特異性IgGの生成
IgG1-Betv1およびIgG1-Feld1またはIgG4-Betv1およびIgG4-Feld1を抗体の最終濃度10μg/mlで混合し、全量50μl中で還元剤と共にインキュベートした。GSHとは別に、以下の作用物質を試験した(インキュベーション混合物中の最終濃度):L-システインはSigmaによる(100μM)、ジチオスレイトール(DTT)はBioradによる(50μM)、β-メルカプト-エタノール(BME)はBioradによる(100μM)、および酸化型グルタチオン(GSSG、一連の作用物質のうち、他のすべてが還元性であるのに対して、この作用物質は還元性ではないことに留意されたい)はSigmaによる(100μM)。混合物を37℃で24時間インキュベートし、試料をPBS/AT中に採取し、そこで(二重)特異性IgG濃度を測定した。図15から、精製IgG4-Betv1およびIgG4-Feld1の混合物にGSHまたは他の還元剤(GSSG以外)を添加することは、Fabアーム交換および二重特異性IgG4の生成を誘導するのに十分であることが示される。対照的に、対照IgG1混合物では二重特異性反応性は誘導されなかった。
【0203】
実施例33. GSHを用いる完全ヒトIgG4抗体の交換
IgG1-CD20、IgG4-CD20、IgG1-EGFr、およびIgG4-EGFrを混合し、全量1 ml中でGSHと共にインキュベートした。各抗体の最終濃度は50μg/mlであった;GSHの最終濃度は0.5 mMであった。混合物を37℃で24時間インキュベートし、試料をPBS-AT中に採取し、そこで(二重)特異性IgG濃度を測定した。
【0204】
サンドイッチELISA法を用いて二重特異性活性を決定した。このアッセイのために、ELISAプレート(Greiner bio-one、ドイツ、フリッケンハウゼン)を、PBS中のEGFRの組換え細胞外ドメイン1μg/ml(100μl/ウェル)で4℃でコーティングした。プレートをPBS/0.05% Tween 20(PBT)で3回洗浄した。試料をPBT/0.2% BSA(PBTB)で希釈し、ELISAプレートに移した(100μl/ウェル)。プレート振盪機(300 rpm)上で室温(RT)で90分間インキュベートした後、試料を廃棄し、プレートをPBTで3回洗浄した。次に、PBTB中の2μg/mlのマウス抗イディオタイプモノクローナル抗体2F2 SAB1.1(抗CD20抗体7D8に対する;Genmab) 100μlを添加し、プレート振盪機(300 rpm)にてRTで90分間インキュベートした。抗イディオタイプ抗体を廃棄し、プレートをPBTで3回洗浄し、続いてPBTBで1000×希釈したHRP結合ヤギ抗マウスIgG(Jackson ImmunoResearch Laboratories、米国、ペンシルバニア州、ウェストグローブ) 100μl/ウェルを添加し、プレート振盪機(300 rpm)にてRTで90分間インキュベートした。検出抗体を廃棄し、プレートをPBTで3回洗浄した。50 mg ABTS錠剤(Roche Diagnostics GmbH、ドイツ、マンハイム)をABTS緩衝液(Roche)に溶解し、ELISAプレートに添加した(100μl/ウェル)。ELISAプレートをアルミホイルで覆って、プレート振盪機(300 rpm)上でRTで30分間(または必要に応じてそれよりも長く)インキュベートし、シュウ酸(Riedel de Haen Seelze、ドイツ) 100μl/ウェルで反応を停止させた。ELISAプレートをRTで10分間置いた後、ELISAプレートリーダーで405 nmの吸光度を読み取った。
【0205】
図16Aは、GSHの存在下でIgG4-EGFrおよびIgG4-CD20の混合物をインキュベートした場合には、二重特異性抗EGFR/CD20抗体が時間と共に形成されたが、GSHの非存在下では形成されなかったことを示す。Fabアーム交換は、IgG1抗体の混合物では、GSHの存在下でも非存在下でも起こらなかった。
【0206】
IgG4半分子のGSH媒介性の交換のダイナミックレンジを調べるため、GSHの完全濃度曲線(0.5〜1,000μM)を用いて交換を解析した。IgG4-CD20およびIgG4-EGFrを混合し、全量1 ml中でGSHと共にインキュベートした。各抗体の最終濃度は50μg/mlであった;GSHの最終濃度は図16Bに示す通りであった。混合物を37℃で24時間インキュベートし、試料をPBS-AT中に採取し、そこで(二重)特異性IgG濃度を測定した。
【0207】
図16Bは、IgG4半分子の交換の明白なGSH用量依存性を示す。どのように反応成分がGSH媒介性IgG4半分子の交換に影響するのかを調べるために、PBS、ならびに血清およびタンパク質を含まない既知組成培地(FreeStyle 293発現培地、GIBCO/Invitrogen Corporation)中で交換を試験した。この組織培地中では、GSH媒介性の交換がより低いGSH濃度で起こることが判明した(図16C)。5 mM GSHと共にインキュベートすると、0.5 mMの場合よりも低い交換が起こったことから、GSH媒介性IgG4半分子の交換には最適条件が存在することも判明した(図16D)。
【0208】
IgG4-EGFrおよびIgG4-CD20の混合物をGSHの非存在下または存在下で24時間インキュベートし、質量分析(ESI-TOF MS)によって評価した。各抗体200μg/mlを含む試料50μlを、N-グリコシダーゼF(Roche Diagnostics NL BV、オランダ、アルメレ) 1μlで一晩かけて脱グリコシル化した。BEH C8、1.7μm、2.1×50 mmカラムを用いてAcquity UPLC(商標)(Waters、米国、ミルフォード)において、試料を60℃で脱塩した。5μlを注入し、5%〜95%溶離液Bの勾配で溶出した。溶離液AはMilliQ水(Millipore Synthesis A10装置)であり、溶離液BはLC-MS等級アセトニトリル(Biosolve、オランダ、ファルケンスワールト)であった。いずれの溶離液も、有機変性剤として0.05%ギ酸(Fluka Riedel-de Han、ドイツ、ブーフ)を含んだ。micrOTOF(商標)質量分析計(Bruker、ドイツ、ブレーメン)を陽イオンモードで操作して、飛行時間型エレクトロスプレーイオン化質量スペクトルをオンラインで記録した。各解析において、500〜5000 m/zスケールをES調整混合物(Agilent Technologies、米国、サンタクララ)で内部較正した。DataAnalysis(商標)ソフトウェアv.3.3(Bruker)によって提供される最大エントロピーアルゴリズムを用いることにより、質量スペクトルをデコンボリューション処理した。
【0209】
図16Eは、GSHの非存在下で、IgG4-CD20(145.5 kD)およびIgG4-EGFR(145.9 kD)の分子量が変化しないままであったことを示す。しかしながらGSHの存在下では(図16F)、Fabアーム交換分子に対応する質量を有する新たなピークが出現した(145.7 kD)。新たな質量は、二重特異性抗EGFR/CD20抗体の予測質量に相当した。さらに、MSスペクトルのピーク高から、二重特異性抗体が混合物中の全抗体量の50%を示すことが推定され得、無作為的交換が24時間以内に平衡に達したことが示された。
【0210】
実施例34. 組換えヒトIgG4抗体のFabアーム交換に関与するアカゲザル(およびその他の種)由来のポリクローナル免疫グロブリン
2種類の組換えヒトIgG4抗体(上記と同様のIgG4-CD20およびIgG4-EGFr)の混合物を、アカゲザル(6×)、チンパンジー(2×)、カニクイザル、ヒヒ、ウマ、およびブタ由来の精製免疫グロブリンまたはヒトIVIgの存在下または非存在下で、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、上記のサンドイッチELISA法で測定した。加えて、ヤギ、ウサギ、およびヒツジ免疫グロブリンもこのアッセイ法で試験した。
【0211】
図17Aは、アカゲザルポリクローナル免疫グロブリンが、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換を阻害する能力において、ヒトポリクローナル免疫グロブリン(IVIg)に匹敵することを示す。これは、アカゲザル免疫グロブリンの成分がFabアーム交換に関与することを意味する。アカゲザル免疫グロブリン、おそらくはアカゲザルIgG4は、組換えヒトIgG4とFabアームを交換することができる。
【0212】
図17Bは、数頭のその他のアカゲザルに由来するポリクローナル免疫グロブリンが、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換を異なる効力で阻害することを示す。これは、Fabアーム交換に関与するアカゲザル免疫グロブリンの成分が異なる濃度で存在すること、またはすべてのアカゲザル中にその成分が存在するわけではないことを意味する。
【0213】
図17Cは、数頭のその他のサル種(ヒヒ、チンパンジー、カニクイザル)由来のポリクローナル免疫グロブリン、ならびにウマおよびブタ由来の免疫グロブリンが、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換を異なる効力で阻害することを示す。これは、Fabアーム交換に関与する成分が、これらの種において異なる濃度で存在することを意味する。ヤギ、ウサギ、およびヒツジ免疫グロブリンは、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換に影響を及ぼさなかった(データは示さず)。
【0214】
実施例35. ヒンジ領域変異体またはCH3ドメイン変異体の半分子の交換
3種類のIgG1変異体を作製した:IgG4コアヒンジを有するIgG1(IgG1-CPSC)、および2種類のCH3ドメイン交換変異体(IgG1-CH3(IgG4)およびIgG1-CPSC-CH3(IgG4))。
【0215】
部位特異的突然変異誘発を用いて、pEE-G1-wt a Bet v 1を鋳型として使用して、IgG1のヒンジ内にP228S変異を導入した(228は抗体アミノ酸残基のEU番号を参照する。同位置はKabat番号付けでは241番を有し、SEQ ID NO:19では111番(CPPCコアヒンジ配列における3位)を有する)。Vector NTI Advance 10により、突然変異誘発プライマー、フォワードおよびリバースを設計した:
P228S Mutプライマー-F:SEQ ID NO:22
P228S Mutプライマー-R:SEQ ID NO:23。
【0216】
Quickchange部位特異的突然変異誘発キット(Stratagene)を用いて、pEE-G1-CPSC変異体を作製した。ポリメラーゼ連鎖反応(PCR)混合物は、pEE-G1 a Betv1 DNA鋳型5μl(約35 ng)、突然変異誘発プライマー-フォワード1.5μl(約150 ng)、突然変異誘発プライマー-リバース1.5μl(約150 ng)、dNTP混合物1μl、反応緩衝液(10×) 5μl、H2O 36μl、および最後にPfu Turbo DNAポリメラーゼ1μlからなった。次に混合物をPCR:95℃で30秒、95℃で30秒(変性)、55℃で1分(アニーリング)、および68℃で17分(伸長)に供した。このサイクルを20回繰り返した。
【0217】
DNAの消化およびライゲーションを用いて、CH3ドメイン交換変異体構築物IgG1-CH3(IgG4)およびIgG1-CPSC-CH3(IgG4)を作製した。CH3ドメインおよびCH3ドメインを含まないベクターを得るための消化反応は、以下の通りであった:DNA(pEE-G1-betv1、pEE-G1-CPSC、およびpEE-G4-betv1) 約1500 ng、BSA 2μl、Neb3緩衝液2μl、SalI 1μl、およびH20を20μl量になるまで添加した。37℃で30分間インキュベートした。DNAを精製し、H20 30μlで溶出してから、SanDI 1μlおよびユニバーサル緩衝液3μlを添加し、37℃で30分間インキュベートした。断片を、エチジウムブロマイドを含む1%アガロースゲルでのゲル電気泳動に供した。紫外線下で断片をゲルから切り出し、DNA精製キット(Amersham)を用いて溶解した。pEE-G4-wt SalI/SanDI(IgG4 CH3ドメインを含む)断片を、以下の手順を用いてpEE-G1-wtおよびpEE-G1-CPSCに連結した:全量20μl中の鋳型DNA(SalI/SanDI消化したpEE-G1-wtおよびpEE-G1-CPSC) 1μl、SalI/SanDI挿入物5μl、Ligate-it緩衝液4μl、H20 9μl、およびリガーゼ1μl。5分後にライゲーションを停止させた。
【0218】
DNAの消化(ApaIおよびHindIIIを使用)およびライゲーションを用いて、上記と同様の手順に従って、bet v 1変異体抗体のVHドメインをpEE-G4-a-feld1 wtのVHドメインと置換した。
【0219】
また、1つのIgG4変異体を作製した:IgG4-S228Pnew。この変異体では、228位のセリン(SEQ ID NO:19における111位)をプロリン(IgG1コアヒンジ)に置換することによって、ヒンジが安定化される。製造業者の説明書に従ってQuickChange II XL部位特異的突然変異誘発キット(Stratagene、オランダ、アムステルダム)を用いて、部位特異的突然変異誘発を行った。この方法は、突然変異誘発に成功したものをスクリーニングするために、サイレントな余分なXmaI部位の導入を含んだ。簡潔に説明すると、10×反応緩衝液5μl、オリゴヌクレオチドS228Pfcorrect(100 pmol/μl) 1μl、オリゴヌクレオチドS228Prcorrect(100 pmol/μl) 1μl、dNTP混合物1μl、Quicksolution 3μl、プラスミドpTomG42F8HG(50 ng/μl)(2006年11月28に出願された「Recombinant monovalent antibodies and methods for production thereof」という表題のPCT出願(RO/DK (Genmab)に記載されている) 1μl、およびPfuUltra HF DNAポリメラーゼ1μlを全量50μl中で混合し、18サイクルプログラム:95℃で1分の変性;95℃で50秒、60℃で50秒、および68℃で10分間の18サイクルを用いて、TGradient Thermocycler 96(Whatman Biometra、ドイツ、ゲッティンゲン;製品# 050-801)で増幅した:PCR混合物は、さらなる処理時まで4℃で保存した。次に、PCR混合物をDpnI 1μlと共に37℃で60分間インキュベートして、pTomG42F8HGベクターを消化し、さらなる処理時まで4℃で保存した。反応混合物を3 M NaAc 5μlおよびエタノール125μlで沈殿させ、-20℃で20分間インキュベートし、14000×gで4℃で20分間遠心沈殿させた。DNAペレットを70%エタノールで洗浄し、乾燥させ、水4μlに溶解した。全部で4μlの反応量を、製造業者の説明書(Invitrogen)に従ってOne Shot DNH5α T1Rコンピテント大腸菌細胞(Invitrogen、オランダ、ブレダ)に形質転換した。次に、50μg/mlアンピシリンを含むルリア-ベルターニ(LB)寒天プレートに細胞をプレーティングした。細菌コロニーが明白になるまで、プレートを37℃で16〜18時間インキュベートした。
【0220】
コロニーPCRおよびXmaI(突然変異誘発によってXmaI部位が失われる)消化によるスクリーニング後に、プラスミドを細菌から単離し、DNA配列決定によって変異を確認した。望ましくない余分な変異が導入されたかどうかを調べるために、全HCコード領域の配列を決定したが、さらなる変異は全く含んでいなかった。最終的な構築物をpTomG42F8S228PNewと命名した。
【0221】
トランスフェクション試薬として293 Fectin(Invitrogen)を用いて、これらの構築物による組換え抗体を、3 ml、6ウェルプレート(NUNC)または125 ml三角フラスコ(Corning)においてHEK 293細胞で一過性に発現させた。
【0222】
非精製抗体(FreeStyle 293発現培地、GIBCO/Invitrogen Corporation)の以下の混合物を0.1 mM GSHと共に37℃で24時間インキュベートし、試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(以下、IgG1 CPSC - IgG1 CPSCと表示)
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CH3(IgG4)(IgG1 CPSC - IgG1 CH3(IgG4))
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC/CH3(IgG4)(IgG1 CPSC - IgG1 CPSC/CH3(IgG4))
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(IgG1 CH3(IgG4) - IgG1 CH3(IgG4))
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CPSC/CH3(IgG4)(IgG1 CH3(IgG4) - IgG1 CPSC/CH3(IgG4))
‐IgG1 a-feld1 CPSC/CH3(IgG4)とa-betv1 IgG1 CPSC/CH3(IgG4)(IgG1 CPSC/CH3(IgG4) - IgG1 CPSC/CH3(IgG4))
‐IgG1 a-feld1 CPSC/CH3(IgG4)とIgG4 a-betv1 wt(IgG1 CPSC/CH3(IgG4) - IgG4 wt
‐IgG4 a-bet1 S228PnewとIgG4 wt。
【0223】
結果から、これらのインビトロ条件(0.1 mM GSH)において、抗体の一方がCPSCヒンジを含み、かつ両抗体がIgG4様CH3を含む場合に、半分子の交換が起こることが示された。また、半分子の交換は、IgG1ヒンジを含むIgG4分子とIgG4 wt分子の間でも起こる。
-=交換なし
+=交換が起こる
±=限られた交換(約5%)
空白欄=未検
【0224】
0 mM、0.1 mM、1 mM、および10 mM GSHを用いて、様々な変異体による半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)
‐IgG1 a-feld1 CPSC/CH3(IgG4)とa-betv1 IgG1 CPSC/CH3(IgG4))。
【0225】
1 mMまでのGSH濃度については、結果(図19A)から上記のことが確認された。10 mM GSHでは、IgG1 a-feld1 CH3(IgG4)およびIgG1 a-betv1 CH3(IgG4)を含む反応においても半分子の交換が認められた。
【0226】
先の実施例に記載された通りに、サイズ排除クロマトグラフィーを行って、適切なIgG1変異体のGSH媒介性の交換後に認められた二重特異性活性が、IgG凝集の結果であるという可能性を排除した。異種Fel d 1-Bet v 1架橋活性は、単量体IgGの保持容量に対応する画分中に検出された。
【0227】
半分子を交換する能力に関与するCH3ドメイン内のアミノ酸残基を同定するために、IgG1とIgG4との間で異なる位置でIgG4様残基をIgG1のCH3に導入した。したがって、基本的に上記の通りに、鋳型としてpEE-G1-wt a Bet v 1またはpEE-G1-wt a Fel d 1を用いて、R238Q、K292R、Q302E、またはP328L変異(番号はSEQ ID NO:19を参照する)をIgG1のCH3ドメインに導入した。さらに、鋳型としてpEE-G1-CPSC betv1またはpEE-G1-CPSC feld1を用いて、K292R変異をIgG1 CPSCのCH3ドメインにも導入した。手短に説明すると、Vector NTI Advance 10により、突然変異誘発プライマー、フォワードおよびリバースを設計した。Quickchange部位特異的突然変異誘発キット(Stratagene)を用いて、構築物を作製した。トランスフェクション試薬として293 Fectin(Invitrogen)を用いて、これらの構築物による組換え抗体を、3 ml、6ウェルプレート(NUNC)または125 ml三角フラスコ(Corning)においてHEK 293細胞で一過性に発現させた。非精製抗体(FreeStyle 293発現培地、GIBCO/Invitrogen Corporation)の以下の混合物を0.5 mMまたは5 mM GSHと共に37℃で24時間インキュベートし、試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG1 a-feld1 wtとIgG4 a-betv1 wt(図19BではIgG1と表示)
‐IgG1 a-feld1 CPSCとIgG4 a-betv1 wt(図19BではIgG1-CPSCと表示)
‐IgG1 a-feld1 CH3(IgG4)とIgG4 a-betv1 wt(図19BではIgG1-CH3(G4)と表示)
‐IgG1 a-feld1 CPSC/CH3(IgG4)とIgG4 a-betv1 wt(図19BではIgG1-CPSC/CH3(G4)と表示)
‐IgG1 a-feld1 R238QとIgG4 a-betv1 wt(図19BではIgG1-R238Qと表示)
‐IgG1 a-feld1 K292RとIgG4 a-betv1 wt(図19BではIgG1-K292Rと表示)
‐IgG1 a-feld1 Q302EとIgG4 a-betv1 wt(図19BではIgG1-Q302Eと表示)
‐IgG1 a-feld1 P328LとIgG4 a-betv1 wt(図19BではIgG1-P328Lと表示)
‐IgG1 a-feld1 CPSC/K292RとIgG4 a-betv1 wt(図19BではIgG1-CPSC/K292Rと表示)
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図19BではIgG4と表示)
‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(図19CではIgG1と表示)
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(図19CではIgG1-CPSCと表示)
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(図19CではIgG1-CH3(G4)と表示)
‐IgG1 a-feld1 CPSC/CH3(IgG4)とIgG1 a-betv1 CPSC/CH3(IgG4)(図19CではIgG1-CPSC/CH3(G4)と表示)
‐IgG1 a-feld1 R238QとIgG1 a-betv1 R238Q(図19CではIgG1-R238Qと表示)
‐IgG1 a-feld1 K292RとIgG1 a-betv1 K292R(図19CではIgG1-K292Rと表示)
‐IgG1 a-feld1 Q302EとIgG1 a-betv1 Q302E(図19CではIgG1-Q302Eと表示)
‐IgG1 a-feld1 P328LとIgG1 a-betv1 P328L(図19CではIgG1-P328Lと表示)
‐IgG1 a-feld1 CPSC/K292RとIgG1 a-betv1 CPSC/K292R(図19CではIgG1-CPSC/K292Rと表示)
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図19CではIgG4と表示)。
【0228】
結果から、試験したインビトロ条件(0.5 mMおよび5 mM GSH)において、両抗体が292位にRを含む場合に半分子の交換が起こることが示された(図19B+C)。238位のRまたはQ、302位のQまたはE、および328位のPまたはLは、この実験設定においてIgG1が半分子を交換できないことに影響しない。
【0229】
実施例36. 0.5 mM GSHにおいて、組換えヒトIgG4抗体のFabアーム交換反応に関与しない安定化IgG1様コアヒンジを有するIgG4分子
2種類の組換えヒトIgG4抗体(上記のIgG4-CD20およびIgG4-EGFr)を、過剰(10、50、および100マイクログラム/ml)のTysabriまたは過剰(10マイクログラム/ml)のMylotargの存在下または非存在下において、0.5 mM GSHと共に37℃で24時間インキュベートした。Tysabriは野生型IgG4コアヒンジを含む市販のヒト化IgG4抗体であり、Mylotargは安定化IgG1様コアヒンジを含む市販のヒト化IgG4抗体である。Fabアーム交換を介した二重特異性抗体の形成を、上記の通りにサンドイッチELISA法で測定した。
【0230】
図20Aは、過剰なTysabriの存在下において、組換えCD20抗体およびEGFr抗体のFabアームの交換が阻害されたことを示す。
【0231】
図20Bは、過剰なTysabriの存在下では組換えCD20抗体およびEGFr抗体のFabアームの交換が阻害されたが、過剰なMylotargの存在下ではそのように阻害されなかったことを示す。
【0232】
このことは、TysabriはFabアーム交換反応に関与するがMylotargは関与しないこと、および安定化IgG1様コアヒンジは0.5 mM GSHを使用するインビトロ条件下でFabアーム交換に関与しないことを示す。
【0233】
実施例37. 292位にさらなる変異を有するIgG1-CPSC構築物の半分子の交換
実施例35と同様に、pConG1f2F8(EGFRに特異的)およびpConG1f7D8(CD20に特異的)において3種類のIgG1変異体を作製した:IgG4コアヒンジを有するIgG1(IgG1-CPSC)、および2種類のCH3ドメイン交換変異体(IgG1-CH3(IgG4)およびIgG1-CPSC-CH3(IgG4))(すなわち、IgG1のCH3領域をIgG4のCH3領域によって置換した構築物)。これにより、以下の構築物pG1f-2F8CPSC、pG1f-7D8CPSC、pG1f-2F8-CH3(G4)、pG1f-7D8-CH3(G4)、pG1f-2F8CPSC-CH3(G4)、およびpG1f-7D8CPSC-CH3(G4)を得た。
【0234】
続いて、基本的に上記の通りに、R238Q、K292R、K292Y、K292F、K292W、Q302E、またはP328L変異(SEQ ID NO: 19を参照されたい)を、pG1f-2F8CPSC構築物およびpG1f-7D8CPSC構築物の両方のCH3ドメインに導入した。手短に説明すると、Vector NTI Advance 10により、突然変異誘発プライマー、フォワードおよびリバースを設計した。Quickchange部位特異的突然変異誘発キット(Stratagene)を用いて、構築物を作製した。
【0235】
トランスフェクション試薬として293 Fectin(Invitrogen)を用いて、これらの構築物による組換え抗体を、3 ml、6ウェルプレート(NUNC)または125 ml三角フラスコ(Corning)においてHEK 293細胞で一過性に発現させた。続いて培養上清をPBSに対して透析し、比濁法により濃度を測定した(上記を参照されたい)。非精製の緩衝液交換抗体の以下の混合物を0.5 GSHと共に37℃で24時間インキュベートし、試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG1-2F8 wtとIgG1-7D8 wt(IgG1と表示)
‐IgG1-2F8-CPSCとIgG1-7D8-CPSC(IgG1-CPSCと表示)
‐IgG1-2F8-CH3(IgG4)とIgG1-7D8-CH3(IgG4)(IgG1-CH3(IgG4)と表示)
‐IgG1-2F8-CPSC-CH3(IgG4)とIgG1-7D8-CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示)
‐IgG1-2F8-CPSC-R238QとIgG1-7D8-CPSC-R238Q(IgG1-CPSC-R238Qと表示)
‐IgG1-2F8-CPSC-K292RとIgG1-7D8-CPSC-K292R(IgG1-CPSC-K292Rと表示)
‐IgG1-2F8-CPSC-K292YとIgG1-7D8-CPSC-K292Y(IgG1-CPSC-K292Yと表示)
‐IgG1-2F8-CPSC-K292FとIgG1-7D8-CPSC-K292F(IgG1-CPSC-K292Fと表示)
‐IgG1-2F8-CPSC-K292WとIgG1-7D8-CPSC-K292W(IgG1-CPSC-K292Wと表示)
‐IgG1-2F8-CPSC-Q302EとIgG1-7D8-CPSC-Q302E(IgG1-CPSC-Q302Eと表示)
‐IgG1-2F8-CPSC-P328LとIgG1-7D8-CPSC-P328L(IgG1-CPSC-P328Lと表示)
IgG4-2F8 wtとIgG4-7D8 wt(IgG4と表示)。
【0236】
図21は、試験したインビトロ条件(0.5 mM)において、CPSCヒンジが存在し、かつ292位がRである場合に、半分子の交換が起こることを示す。加えて、結果から、292位のYまたはFもまた、程度はより低いものの半分子の交換を促進するが、Wは促進しないことが示される。238位のRまたはQ、302位のQまたはE、および328位のPまたはLは、IgG1-CPSCが半分子を交換できないことに影響しない。
【0237】
実施例38 (5 mM GSHと共に)インビトロではFabアーム交換ができるが、インビボではできない、安定化CPPCヒンジを有するIgG4分子
IgG4-EGFR-CPPCおよびIgG4-CD20の混合物を、5 mM GSHの存在下で24時間インキュベートし、質量分析(ESI-TOF MS)によって評価した。各抗体200μg/mlを含む試料50μlを、N-グリコシダーゼF(Roche Diagnostics NL BV、オランダ、アルメレ) 1μlで一晩かけて脱グリコシル化した。BEH C8、1.7μm、2.1×50 mmカラムを用いてAcquity UPLC(商標)(Waters、米国、ミルフォード)において、試料を60℃で脱塩した。5μlを注入し、5%〜95%溶離液Bの勾配で溶出した。溶離液AはMilliQ水(Millipore Synthesis A10装置)であり、溶離液BはLC-MS等級アセトニトリル(Biosolve、オランダ、ファルケンスワールト)であった。いずれの溶離液も、有機変性剤として0.05%ギ酸(Fluka Riedel-de Han、ドイツ、ブーフ)を含んだ。micrOTOF(商標)質量分析計(Bruker、ドイツ、ブレーメン)を陽イオンモードで操作して、飛行時間型エレクトロスプレーイオン化質量スペクトルをオンラインで記録した。各解析において、500〜5000 m/zスケールをES調整混合物(Agilent Technologies、米国、サンタクララ)で内部較正した。DataAnalysis(商標)ソフトウェアv.3.3(Bruker)によって提供される最大エントロピーアルゴリズムを用いることにより、質量スペクトルをデコンボリューション処理した。
【0238】
図22Aは、5 mM GSHの存在下において、Fabアーム交換分子に対応する中間質量を有する新たなピークが出現した(145.7 kDa)ことを示す。新たな質量は、二重特異性抗EGFR/CD20抗体の予測質量に相当した。GSHを使用しなかった場合または0.5 mM GSHを使用した場合には、二重特異性抗体のピークは出現しなかった(データは示さず)。これは、IgG1様のCPPCヒンジおよびIgG4様CH3領域を含む変異体が、(実施例35、36、37においても示される通り)より高いGSH濃度においてインビトロで半分子を交換できることを示す。
【0239】
IgG1様のCPPCヒンジおよびIgG4様CH3を含む安定化ヒンジ変異体のFabアーム交換がインビボで起こるかどうかを調べるために、本発明者らは、IgG4-CD20とIgG1-EGFR、IgG4-EGFR、IgG4-EGFR-CPPCの同等の混合物を免疫不全マウスに注射した。様々な時点で血液試料を採取し、インビトロ交換された混合物(IgG4-EGFR/IgG4-CD20)を参照標準物質として用いて、二重特異性抗体を(上記の通りに)ELISA法で定量した。
【0240】
図22Bは、野生型IgG4分子(IgG4-EGFR)を含む混合物を注射したマウスの血液中に、二重特異性抗体が出現したことを示す。ヒンジ安定化IgG4(IgG4-EGFR-CPPC)またはIgG1分子(IgG1-EGFR)を含む混合物では、二重特異性抗体は検出不能であった((図中に記号は示されていない))。これは、コアヒンジ安定化がインビボでのIgG4 Fabアーム交換を妨げ、インビボで半分子を交換できないことを示す(しかし本発明者らは、ヒンジ安定化IgG4の検出レベル(72時間で<8%)未満の低レベル交換が起こるということを排除できない)。
【0241】
これにより、安定化CPPCヒンジを含む二重特異性抗体を、インビトロでのFabアーム交換によって得ることができることが示唆される。これらの二重特異性抗体をその後特異的に精製した後、これらの抗体は、インビボでの注射に際して安定したままである(すなわち、Fabアーム交換しない)。
【0242】
実施例39. CXXC変異体のFabアーム交換
コアヒンジ内に様々なCXXCモチーフを含む抗体のFabアームを交換する能力を試験した。上記の通りに部位特異的突然変異誘発技法を用いて、以下のCXXCモチーフをIgG4 bet v 1およびIgG4 feld 1に導入した:
‐CGHC(タンパク質ジスルフィドイソメラーゼ、PDIについて記載されている活性部位配列)
‐CGC(ジスルフィド還元電位を有すると記載されているペプチド)
‐CPRC(ゴリラIgG4のコアヒンジ配列)
‐CPHC(ヒトチオレドキシンについて記載されている活性部位配列)。
【0243】
精製抗体の以下の混合物を0.5 mM GSHと共に37℃でインキュベートし、0〜24時間の様々な時点で試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(図23および24においてIgG1と表示)
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図23および24においてIgG4と表示)
‐IgG4 a-feld1 CGHCとIgG4 a-betv1 CGHC(図23および24においてCGHCと表示)
‐IgG4 a-feld1 CGCとIgG4 a-betv1 CGC(図23および24においてCGCと表示)
‐IgG4 a-feld1 CPRCとIgG4 a-betv1 CPRC(図23および24においてCPRCと表示)
‐IgG4 a-feld1 CPHCとIgG4 a-betv1 CPHC(図23および24においてCPHCと表示)。
【0244】
結果(図23)から、時間が経過しても、CGCモチーフまたはIgG1コアヒンジを含む抗体のFabアーム交換は起こらないことが示された。CGHCモチーフを含む抗体のFabアーム交換は、IgG4 wt抗体と同程度に効果的であった。いくらかゆっくりではあるものの、CPRCモチーフを含む抗体のFabアーム交換も起こり、程度はより低いがCPHCモチーフを含む抗体のFabアーム交換も起こった。
【0245】
また、これらの混合物が37℃での24時間のインキュベーション後にFabアーム交換を行う能力に及ぼすGSH濃度(1〜20,000μM)の影響を試験した。CPHC、CPRC、およびCGHCモチーフ含有抗体ならびにIgG4 wt抗体のFabアーム交換は、GSH濃度に依存し(図24)、最適値は100μM〜1,000μM GSHであることが判明した。
【技術分野】
【0001】
発明の分野
本発明は、二重特異性抗体の新規作製方法、およびこれらの方法によって得られ得る、二重特異性抗体に関する。
【背景技術】
【0002】
発明の背景
ヒト免疫グロブリンG(IgG)抗体は、異なる構造特性および機能特性を有する4つのサブクラスとして存在する。IgGは2つの重鎖-軽鎖対(半分子)から構成されており、これらはヒンジ領域に位置する重鎖間ジスルフィド結合によって結合している。ヒトIgG4分子は、ヒンジ領域に位置する重鎖間ジスルフィド結合の有無によって異なる種々の分子形態として存在する。IgG4分子は、重鎖間ジスルフィド結合が両方とも形成されているか、またはいずれも形成されていない形態として存在する(6、7)。しかしながら、ヒトIgG4は、CH3ドメイン間、およびCH1ドメインとCH2ドメインとの間の比較的強力な非共有結合性相互作用のために(4)、これらの鎖間ジスルフィド結合の有無に関係なく(6、8)、溶液中では、免疫グロブリンG分子に一般的なIg重鎖2本および軽鎖2本からなる四量体として存在する。SDS-PAGEなどのサイズ決定分析によって実証されるように、非共有結合的に会合した2つの半分子は、非還元条件下で変性させた場合のみ解離する(6、9)。
【0003】
ヒトIgG4抗体が、他のIgGサブクラスとは異なり、抗原との相互作用において一価の分子として挙動することがこの数年間の間にわかってきた。血清由来のヒトIgG4は、架橋できないために、精製抗原を沈降させることができないことが見出された。このような血清由来のIgG4は機能的に一価であるが(1、2)、対照的に、組換えで作製されたIgG4は抗原との相互作用において二価として挙動する(3)。これらの知見に基づき、血清中のIgG4分子は半分子(すなわち、重鎖1本および軽鎖1本からなる分子)を交換して、同一の抗原に架橋することができない二重特異性分子を生じ得ることが提唱された(3〜5)。この半分子の交換の過程を、本明細書では「Fabアーム交換」とも称する。
【0004】
二重特異性抗体は、例えば疾患関連部位にエフェクター機構を再標的化するためのメディエーターとして使用することができるため、治療薬として興味深い可能性を有する。しかしながら、二重特異性抗体を開発する上での大きな障害の1つは、ハイブリッドハイブリドーマ法および化学的結合法などの伝統的な技術によって、十分な質および量の物質を作製することが困難であることであった(10)。
【0005】
WO 2005/062916(特許文献1)は、IgG4に基づいて、マウスにおいてインビボで多量体分子を形成する方法を記載している。さらにWO 2005/062916(特許文献1)は、異なる抗原結合特異性を有する2種類のIgG4抗体を、インビトロで、生理食塩水緩衝液中で同時にインキュベートすると、両抗原と反応し得る産物が形成されることを記載している。しかしながら、WO 2005/062916(特許文献1)では、これらの産物が凝集体であるのか、または二重特異性抗体であるのかは実証されておらず、また使用した条件下において反応の収率は低かった。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】WO 2005/062916
【発明の概要】
【0007】
驚くべきことに、還元条件下において、異なる抗原結合特異性を有する2種類のIgG4抗体またはIgG4様抗体が、高い効率で半分子の交換を行い、したがって凝集体の形成を伴わずに二重特異性抗体を形成し得ることが、今回見出された。
【0008】
したがって第1の主要な局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0009】
いかなる特定の理論によっても縛られることはないが、抗体の2つの領域が、半分子の交換を起こすその能力に重要な影響を及ぼすと考えられる。
【0010】
第一に、IgG4のようなコアヒンジ領域内にCPSC配列を有する抗体が、IgG1のようなCPPCコアヒンジ配列を有する抗体よりもより容易に交換を行うことから、半分子の交換の能力は、分子のコアヒンジ領域内の配列の差によって影響を受ける可能性がある。いかなる理論によっても縛られることはないが、CPSC配列がより可動性の高いコアヒンジをもたらし、鎖内ジスルフィド結合を形成する可能性を引き起こすと仮定される。注目すべきことにコアヒンジの構造は、タンパク質ジスルフィドイソメラーゼ(PDI)の活性ドメイン、CXXCに類似している。PDIの異なるアイソフォームのこれらのCXXCモチーフは、タンパク質におけるジスルフィド結合の形成、還元、および再構成を触媒する。したがって、いかなる特定の理論によっても縛られることはないが、IgG4様コアヒンジ配列を有する抗体は、ジスルフィド結合の再構成に関する内因性の活性を有する可能性があると考えられ、この活性を、本発明の方法において使用する条件によって促進する。
【0011】
第二に、この場合も同様に、いかなる理論によっても縛られることはないが、交換反応が起こるようにするためには、CH3領域の配列はIgG4様であるべきである、すなわち強力な半分子間の相互作用を形成しないようにすべきであることが結果から示される。
【0012】
別の主要な局面において、本発明は、本発明の方法によって得られるか、または得られ得る、単離された二重特異性抗体、およびそのような抗体を含む薬学的組成物に関する。
【0013】
さらなる局面において、本発明は、2つのIgG4様CH3領域を含む、単離された二重特異性抗体、およびそのような抗体を含む薬学的組成物に関する。
【0014】
なおさらなる局面において、本発明は、以下の段階を含む、所望の特性を有する二重特異性抗体を選択する方法に関する:
a) 一組の抗体を提供する段階であって、各抗体が異なる標的特異性を有し、かつ各抗体がIgG4様CH3領域を含む段階、
b) 還元条件下で、該一組の抗体の各抗体を、該一組の別の抗体と共にインキュベートし、それによって一組の抗体混合物を作製する段階であって、各混合物が異なる二重特異性抗体を含む段階、
c) 結果として得られた一組の抗体混合物を所与の所望の特性についてアッセイする段階、および
d) 所望の特性を有する二重特異性抗体混合物を選択する段階。
【図面の簡単な説明】
【0015】
【図1】精製組換えIgG1およびIgG4のSDS-Page解析を示す。精製後、Betv1およびFeld1のIgG1抗体およびIgG4抗体を、非還元SDS-PAGEで解析した。
【図2】様々な時点での、nu/nu Balb/cマウスにおける二重特異性IgGレベルを示す。異種架橋アッセイ法で決定した二重特異性IgGの量を、Bet v 1結合試験で決定したBet v 1特異的IgGの量に対してプロットした。IgG1およびIgG4含有血漿試料によるデータを、それぞれ白記号および黒記号で表す。点線は、IgGの半分子の交換が無作為でかつ完全である場合の、二重特異性IgGの計算された量を表す。
【図3】二重特異性ヒトIgG4分子がインビボで生成されることを示す。(A) SCIDマウスの群(n=5)にキメラ抗体混合物:IgG1-Betv1 100μg/IgG1-Feld1 100μg(四角)、IgG4-Betv1 100μg/IgG4-Feld1 100μg(丸)、または3) IgG4-Betv1 100μg/IgG4-Feld1 100μg+非関連組換えIgG4(IgG4-EGFR) 2,000μg(三角)を注射した。血漿中のBet v 1およびFel d 1に対する二重特異性活性を評価することによって、二重特異性抗体の生成を時間を追って追跡した。全IgG-Bet v 1濃度に対する二重特異性IgGの割合をパーセントとして表した。星印の付いた矢印は、過剰な非関連IgG4の存在下で、IgG4-Betv1/IgG4-Feld1を投与したマウスにおいて予測される二重特異性反応性レベル(4%)を示し、星印のない矢印は、IgG4-Betv1/IgG4-Feld1混合物を投与したマウスにおいて予測される二重特異性反応性レベル(50%)を示す。エラーバーはSEMを表す。(B) マウス血漿における、放射標識Fel d 1の、Fel d 1結合セファロースに対する架橋を評価することによって、単一特異性架橋活性を試験した。単一特異的反応性は、IgGのクリアランスを補正するために、架橋によって結合した放射標識Fel d 1の量と全IgG-Feld1との比として表した。エラーバーはSEMを表す。
【図4】マウス血漿中の二重特異性活性のSEC解析を示す。IgG4混合物を投与したマウスからt=24時間の時点で採取した血漿(10μl)を、Superdex200カラムで分画した。マウスに、Bet v 1結合IgG4 300μgおよびFel d 1結合IgG4 300μgを含む混合物を投与した。その画分中で、Fel d 1特異的IgG(■)の濃度を抗原結合試験で測定し、二重特異性IgG Bet v 1-Fel d 1(●)の濃度をBet v 1-Fel d 1架橋アッセイ法で決定した。IVIgを用いたこのカラムの較正から、単量体、二量体、および凝集型IgGはそれぞれ12.9 ml、11.0 ml、および8.4 mlに溶出されることが明らかになった(データは示さず)。
【図5】全血成分中のIgGの交換を示す。全血、血液細胞、血漿、および血清中で、キメラIgG混合物を37℃で24時間インキュベートし、その後異種架橋アッセイ法(Fel d 1-Bet v 1)で二重特異性活性を測定することによって、IgG4およびIgG1の交換を評価した。血液は、ドナー2名から得た:A(黒色バー)およびB(灰色バー)。キメラIgG4(パネルA)、キメラIgG1(パネルB)を補充した、またはIgGを添加しない(パネルC)混合物において、二重特異性活性を決定した。表示したデータはすべて、37℃で24時間インキュベートした後に測定した。
【図6】ヒト血液細胞によるIgGの交換を示す。キメラIgG混合物を単核細胞(MNC)、血小板(Thr)、および赤血球(Ery)と共に37℃で48時間インキュベートし、その後異種架橋アッセイ法(Fel d 1-Bet v 1)で二重特異性活性を測定することによって、IgG4(黒色バー)およびIgG1(灰色バー)の交換を評価した。対照として、抗体混合物を無血清培地(SFC)中でもインキュベートした。二重特異性活性は、添加した量に対する、結合した125I-Bet v 1の割合(%)として表した。
【図7】HEKおよびマウス細胞株によるIgG4の交換を示す。キメラIgG4混合物をHEK細胞、マウスB細胞(J558)、またはハイブリドーマ細胞と共に37℃でインキュベートすることによって、IgG4半分子の交換を評価した。異種架橋アッセイ法(Fel d 1-Bet v 1)での二重特異性活性を、t=0時間(灰色バー)およびt=24時間(黒色バー)の時点で採取した試料1μl中で測定した。二重特異性活性は、添加した量に対する結合した125I-Bet v 1の割合(%)として表した。
【図8】IgG4の赤血球媒介性の交換を示す。IgG4-Betv1/IgG4-Feld1混合物を、新たに精製した赤血球(ery、黒記号)と共にインキュベートすると、二重特異性抗体が生成されるのに対して、IgG1アイソタイプの混合物では二重特異性は認められなかった。対照として、抗体混合物を赤血球なしでPBS中でインキュベートした(白記号)。矢印は、二重特異性IgGの、最大の予測割合(50%)を示す。エラーバーは、2つ組の測定値の範囲を示す。
【図9】PBS中でのIgG4の交換を示す。二重特異性活性(パネルA)、二価性、および抗原結合を測定することによって、IgG1(白色バー)、IgG4(灰色バー)、および過剰な非関連IgG4の存在下での、IgG4(黒色バー)のPBS中での交換を評価した。パネルAにおけるIgG半分子の交換は、二重特異性IgGの濃度(異種架橋アッセイ法で決定)、およびIgG半分子の交換が無作為でかつ完全である場合の、二重特異性IgGの最大予測濃度から算出した。交換は、100%である最大交換から起算した割合(%)として表した。パネルBに、同種架橋アッセイ法で測定した、時間に伴うFel d 1二価性を示す。二価性IgGの濃度は、t=0時の二価性IgGの濃度を100%と設定することによって正規化した。
【図10】赤血球溶解物によるIgG4の交換を示す。赤血球の溶解物中でキメラIgG4混合物を37℃でインキュベートすることにより、IgG4半分子の交換を評価した。溶解物の希釈を高めながら、IgG4をインキュベートした。異種架橋アッセイ法(Bet v 1-Fel d 1)における二重特異性活性を、表示の時点で採取した試料において測定した。二重特異性は、添加した量に対する結合した125I-Bet v 1の割合(%)として表した。
【図11】赤血球溶解物によって誘導される二重特異性活性のSEC解析を示す。 IgG4を新たに調製した赤血球溶解物と共に37℃で24時間インキュベートし、続いてAKTA HPLCユニット(Amersham Biosciences、スウェーデン、ウプサラ)においてSuperdex200カラムに0.5 ml/分で流して分画した。その画分中で、Bet v 1特異的IgG(■)の濃度を抗原結合試験で測定し、二重特異性IgG Fel d 1-Bet v 1(●)の濃度をBet v 1-Fel d 1架橋アッセイ法で決定した。このカラムの較正から、単量体、二量体、および凝集型IgGはそれぞれ12.1 ml、10.3 ml、および8.3 mlに溶出されることが明らかになった(データは示さず)。
【図12】IgG4のGSH媒介性の交換を示す。PBS/アジド中の、漸増濃度のGSHの存在下でIgG4をインキュベートすることにより、IgG4半分子のGSH媒介性の交換を評価した。表示の時点で試料を採取し、その試料中の抗原結合および二重特異性活性を測定した。IgG4半分子の交換は、二重特異性IgGの濃度(異種架橋アッセイ法で決定)、およびIgG4半分子の交換が無作為でかつ完全である場合の二重特異性IgG4の最大予測濃度から算出した。交換は、100%に設定した最大交換から起算した割合(%)として表した。
【図13】IgG4半分子のGSH媒介性の交換のSECを示す。IgG4をGSH(0.5 mM)と共にインキュベートし、続いてAKTA HPLCユニット(Amersham Biosciences、スウェーデン、ウプサラ)においてSuperdex200カラムに0.5 ml/分で流して分画した。その画分中で、Bet v 1特異的IgG(■)の濃度を抗原結合試験で測定し、二重特異性IgG Fel d 1-Bet v 1(●)の濃度をBet v 1-Fel d 1架橋アッセイ法で決定した。このカラムの較正から、単量体、二量体、および凝集型IgGはそれぞれ12.1 ml、10.3 ml、および8.3 mlに溶出されることが明らかになった(データは示さず)。
【図14】IgG4のGSH媒介性の交換の温度依存性を示す。IgG4-Betv1およびIgG4-Feld1混合物を、PBS中、表示の温度でGSHと共にインキュベートした。t=0時間(灰色バー)およびt=24時間(黒色バー)の時点で、二重特異性IgG4の濃度を評価した。これらのデータから、IgG4 Betv1濃度に対する二重特異性IgGの割合を算出し、割合(%)として表した。エラーバーは、2つ組の測定値の範囲を表す。
【図15】一連の還元剤によって媒介されるIgG4交換を示す。PBS中のIgG4-Betv1およびIgG4-Feld1を、様々な作用物質(GSSG以外はすべて還元性)の存在下で、37℃で24時間インキュベートした。Bet v 1特異的IgGの濃度をBet v 1結合アッセイ法で測定し、二重特異性IgGの濃度を異種架橋アッセイ法(Fel d 1-Bet v 1)で測定した。IgG-Betv1濃度に対する二重特異性IgGの割合(%)を算出した。標準誤差のバーは、3回の測定値から算出されたSEMを表す。
【図16】GSHを用いた完全ヒトIgG4抗体の交換を示す。(A) IgG4-CD20/IgG4-EGFr混合物またはIgG1-CD20/IgG1-EGFr混合物を、0.5 mM GSHと共に、またはこれを伴わずに37℃でインキュベートした。表示の時点で試料を採取した。二重特異性抗体の形成を、サンドイッチELISA法で測定した。Y軸は、二重特異性CD20/EGFR抗体の形成の測定値としての、405 nmの光学密度を示す。(B) IgG4のGSH用量依存的な交換。IgG4-CD20とIgG4-EGFrの混合物を、表示の濃度のGSHと共に、37℃で24時間インキュベートした。二重特異性抗体の形成を、サンドイッチELISA法で測定した。405 nmの光学密度を、二重特異性CD20/EGFR抗体の形成の測定値として、Y軸にプロットする。(C) IgG4半分子のGSH媒介性の交換は、反応に使用した成分によって影響を受け、培地(Freestyle 293)中では、より低いGSH濃度において起こる。(D) IgG4半分子のGSH媒介性の交換は、5 mM GSHよりも0.5 mM GSHにおいて、より高い。(E/F) ESI-TOF質量分析による、IgG4-EGFRとIgG4-CD20との間の、Fabアーム交換の検出。IgG4混合物を、0.5 mM GSHの非存在下(E)または存在下(F)で24時間インキュベートした後、抗体をPNGase Fで脱グリコシル化し、結果として得られた抗体の分子量をESI-TOF質量分析によって決定した。デコンボリューション処理したESI-TOFスペクトルを示す。データは、2回の実験の代表的なものである。
【図17】アカゲザルIVIgが組換えヒトIgG4抗体のFabアーム交換に関与することを示す。(A) 2種類の組換えヒトIgG4抗体(IgG4-CD20およびIgG4-EGFr)の混合物を、精製アカゲザル免疫グロブリンまたはヒトIVIgの存在下または非存在下において、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。(B) 2種類の組換えヒトIgG4抗体(IgG4-CD20およびIgG4-EGFr)の混合物を、数頭の動物(供給源を括弧内に表示)に由来する、過剰な(括弧内に表示)精製アカゲザル免疫グロブリンまたはヒトIVIgの存在下または非存在下で、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。(C) 2種類の組換えヒトIgG4抗体(IgG4-CD20およびIgG4-EGFr)の混合物を、過剰な(括弧内に表示)精製チンパンジー、ヒヒ、カニクイザル、ウマ、およびブタ免疫グロブリン(供給源も括弧内に表示)またはヒトIVIgの存在下または非存在下で、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。
【図18】定常領域の配列を示す。下線の配列はCH3領域を表す。
【図19】IgG1変異体のGSH媒介性の半分子の交換を示す。(A) 0 mM、0.1 mM、1 mM、および10 mM GSHを用いて、様々なIgG1変異体の半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図中ではIgG4 wtと表示);‐IgG1 a-feld1 wtとIgG4 a-betv1 wt(IgG1 wtと表示);‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(IgG1-CPSCと表示);‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(IgG1-CH3(IgG4)と表示);‐IgG1 a-feld1 CPSC-CH3(IgG4)とa-betv1 IgG1 CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示)。(B) 0.5 mMおよび5 mM GSHを用いて、様々なIgG1変異体のIgG4 wt分子との半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG4 a-betv1 wt(IgG1と表示);‐IgG1 a-feld1 CPSCとIgG4 a-betv1 wt(IgG1-CPSCと表示);‐IgG1 a-feld1 CH3(IgG4)とIgG4 a-betv1 wt(IgG1-CH3(IgG4)と表示);‐IgG1 a-feld1 CPSC-CH3(IgG4)とIgG4 a-betv1 wt(IgG1-CPSC-CH3(G4)と表示);‐IgG1 a-feld1 R238QとIgG4 a-betv1 wt(IgG1-R238Qと表示);‐IgG1 a-feld1 K292RとIgG4 a-betv1 wt(IgG1-K292Rと表示);‐IgG1 a-feld1 Q302EとIgG4 a-betv1 wt(IgG1-Q302Eと表示);‐IgG1 a-feld1 P328LとIgG4 a-betv1 wt(IgG1-P328Lと表示);‐IgG1 a-feld1 CPSC-K292RとIgG4 a-betv1 wt(IgG1-CPSC-K292Rと表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示)。(C) 0.5 mMおよび5 mM GSHを用いて、様々なIgG1変異体の半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(IgG1と表示);‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(IgG1-CPSCと表示);‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(IgG1-CH3(IgG4)と表示);‐IgG1 a-feld1 CPSC-CH3(IgG4)とIgG1 a-betv1 CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示);‐IgG1 a-feld1 R238QとIgG1 a-betv1 R238Q(IgG1-R238Qと表示);‐IgG1 a-feld1 K292RとIgG1 a-betv1 K292R(IgG1-K292Rと表示);‐IgG1 a-feld1 Q302EとIgG1 a-betv1 Q302E(IgG1-Q302Eと表示);‐IgG1 a-feld1 P328LとIgG1 a-betv1 P328L(IgG1-P328Lと表示);‐IgG1 a-feld1 CPSC-K292RとIgG1 a-betv1 CPSC-K292R(IgG1-CPSC-K292Rと表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示)。
【図20】0.5 mM GSHにおいて、野生型(IgG4)コアヒンジを有するIgG4分子は、組換えヒトIgG4抗体のFabアーム交換に関与するが、IgG1コアヒンジを有する分子は関与しないことを示す。(A) 2種類の組換えヒトIgG4抗体(上記のIgG4-CD20およびIgG4-EGFr)の混合物を、過剰なTysabri(50および100マイクログラム/ml)の存在下または非存在下で、0.5 mM GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。(B) 2種類の組換えヒトIgG4抗体(上記のIgG4-CD20およびIgG4-EGFr)の混合物を、等モル量(10マイクログラム/ml)のTysabriまたはMylotargの存在下または非存在下で、0.5 mM GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。
【図21】292位にさらなる変異を有するIgG1-CPSC構築物の、半分子の交換を示す。0.5 mM GSHを用いて、様々なIgG1変異体による半分子の交換を試験した。以下の混合物を用いて、交換を試験した:‐IgG1-2F8 wtとIgG1-7D8 wt(IgG1と表示);‐IgG1-2F8-CPSCとIgG1-7D8-CPSC(IgG1-CPSCと表示);‐IgG1-2F8-CH3(IgG4)とIgG1-7D8-CH3(IgG4)(IgG1-CH3(IgG4)と表示);‐IgG1-2F8-CPSC-CH3(IgG4)とIgG1-7D8-CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示);‐IgG1-2F8-CPSC-R238QとIgG1-7D8-CPSC-R238Q(IgG1-CPSC-R238Qと表示);‐IgG1-2F8-CPSC-K292RとIgG1-7D8-CPSC-K292R(IgG1-CPSC-K292Rと表示);‐IgG1-2F8-CPSC-K292YとIgG1-7D8-CPSC-K292Y(IgG1-CPSC-K292Yと表示);‐IgG1-2F8-CPSC-K292FとIgG1-7D8-CPSC-K292F(IgG1-CPSC-K292Fと表示);‐IgG1-2F8-CPSC-K292WとIgG1-7D8-CPSC-K292W(IgG1-CPSC-K292Wと表示);‐IgG1-2F8-CPSC-Q302EとIgG1-7D8-CPSC-Q302E(IgG1-CPSC-Q302Eと表示);‐IgG1-2F8-CPSC-P328LとIgG1-7D8-CPSC-P328L(IgG1-CPSC-P328Lと表示);‐IgG4-2F8 wtとIgG4-7D8 wt(IgG4と表示)。Fabアーム交換を介した二重特異性抗体の形成を、サンドイッチELISA法で測定した。
【図22】コアヒンジの安定化が、インビボでのFabアーム交換からIgG4抗体治療薬を保護することを示す。(A) ESI-TOF質量分析による、IgG4-EGFR-CPPCとIgG4-CD20との間のFabアーム交換の検出。IgG4-EGFR-CPPC/IgG4-CD20混合物を、5 mM GSHの存在下(F)で24時間インキュベートした後、抗体をPNGase Fで脱グリコシル化し、結果として得られた抗体の分子量をESI-TOF質量分析によって決定した。デコンボリューション処理したESI-TOFスペクトルを示す。二重特異性EGFR/CD20抗体は、5 mM GSHを使用した場合に検出することができた(GSHなしでの、または0.5 mM GSHの存在下でのインキュベーションでは、二重特異性抗体は得られなかった(データは示さず))。(B) SCIDマウスの群(n=4)に、IgG4-CD20/IgG4-EGFR(白丸)、IgG4-CD20/IgG1-EGFR、およびIgG4-CD20/IgG4-EGFR-CPPCの抗体混合物(各300μg)を注射した。二重特異性抗体の生成を時間と共に追跡し、ELISA法により定量した。インビトロ交換された抗体混合物を参照として用いて、二重特異性抗体を定量した。データ点は、別々の実験で少なくとも2回測定した、マウス4匹の平均値±SEM値を表す。IgG4-CD20/IgG1-EGFR混合物およびIgG4-CD20/IgG4-EGFR-CPPC混合物では、二重特異性抗体は検出することができなかった。アッセイの検出限界を示すが(点線)、これは2000 ng/mlの血清レベルを表す。
【図23】時間に伴うCXXC変異体のFabアーム交換を示す。CXXC変異体抗体の混合物を、0.5 mM GSHと共に37℃でインキュベートした。表示の時点で試料を採取した。二重特異性抗体の形成を測定した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(IgG1と表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示);‐IgG4 a-feld1 CGHCとIgG4 a-betv1 CGHC(CGHCと表示);‐IgG4 a-feld1 CGCとIgG4 a-betv1 CGC(CGCと表示);‐IgG4 a-feld1 CPRCとIgG4 a-betv1 CPRC(CPRCと表示);‐IgG4 a-feld1 CPHCとIgG4 a-betv1 CPHC(CPHCと表示)。
【図24】CXXC変異体のGSH媒介性Fabアーム交換を示す。1〜20,000μM GSHを用いて、CXXC変異体によるFabアーム交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(IgG1と表示);‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(IgG4と表示);‐IgG4 a-feld1 CGHCとIgG4 a-betv1 CGHC(CGHCと表示);‐IgG4 a-feld1 CGCとIgG4 a-betv1 CGC(CGCと表示);‐IgG4 a-feld1 CPRCとIgG4 a-betv1 CPRC(CPRCと表示);‐IgG4 a-feld1 CPHCとIgG4 a-betv1 CPHC(CPHCと表示)。
【発明を実施するための形態】
【0016】
発明の詳細な説明
定義
「免疫グロブリン」という用語は、2対のポリペプチド鎖、すなわち1対の低分子量軽(L)鎖および1対の重(H)鎖からなる構造的に関連した糖タンパク質のクラスを指し、これら4つはすべてジスルフィド結合によって相互に結合している。免疫グロブリンの構造は十分に特徴付けられている。例えば、Fundamental Immunology Ch.7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989))(11)を参照されたい。簡潔に説明すると、各重鎖は典型的に、重鎖可変領域(本明細書ではVHまたはVHと略す)および重鎖定常領域からなる。重鎖定常領域は典型的に、3つのドメイン、CH1、CH2、およびCH3からなる。各軽鎖は典型的に、軽鎖可変領域(本明細書ではVLまたはVLと略す)および軽鎖定常領域からなる。軽鎖定常領域は典型的に、1つのドメイン、CLからなる。VH領域およびVL領域は、フレームワーク領域(FR)と称されるより保存された領域が介在する相補性決定領域(CDR)とも称される超可変性の領域(または配列および/もしくは構造的に規定されたループの形態において超可変性であり得る超可変領域)に、さらに細分され得る。各VHおよびVLは典型的に、アミノ末端からカルボキシ末端へ以下の順序:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4で配置された、3つのCDRおよび4つのFRから構成される((12)も参照されたい)。典型的に、この領域におけるアミノ酸残基の番号付けは、Kabat(13)に記載されている方法によって行われる。この番号付けシステムを用いると、ペプチドの実際の線状アミノ酸配列は、可変ドメインのFRまたはCDRの短縮またはその中への挿入に対応する、より少数のまたは付加的なアミノ酸を含み得る。例えば、重鎖可変ドメインは、VH CDR2の残基52の後に単一のアミノ酸の挿入(Kabatによると残基52a)、および重鎖FRの残基82の後に挿入された残基(例えば、Kabatによると残基82a、82b、および82c等)を含み得る。残基のKabat番号付けは、所与の抗体について、この抗体の配列と「標準」Kabat番号付け配列との相同性の領域における整列によって決定することができる。
【0017】
本発明との関連における「抗体」(Ab)という用語は、少なくとも約30分、少なくとも約45分、少なくとも約1時間、少なくとも約2時間、少なくとも約4時間、少なくとも約8時間、少なくとも約12時間、約24時間もしくはそれ以上、約48時間もしくはそれ以上、約3、4、5、6、7日もしくはそれ以上等といったかなりの期間の、または任意の他の関連する、機能的に規定された期間(抗原に対する抗体結合に付随する生理反応を誘導、促進、増強、および/もしくは調節するのに十分な時間、ならびに/または抗体がFc媒介性エフェクター活性を動員するのに十分な時間など)の半減期を有して、典型的な生理的条件下で抗原に特異的に結合する能力を有する、免疫グロブリン分子、免疫グロブリン分子の断片、またはそれらいずれかの誘導体を指す。免疫グロブリン分子の重鎖および軽鎖の可変領域は、抗原と相互作用する結合ドメインを含む。抗体(Ab)の定常領域は、免疫系の様々な細胞(エフェクター細胞など)、および補体活性化の古典的経路の第1成分であるC1qなどの補体系の成分を含む、宿主の組織または因子に対する免疫グロブリンの結合を媒介し得る。上記の通り、本明細書における抗体という用語は、特に明記しない限り、または文脈上明らかに矛盾しない限り、変異型または野生型のコアヒンジ領域を含み、抗原に特異的に結合する能力を保持する抗体の断片を含む。
【0018】
抗体の抗原結合機能は、全長抗体の断片によって行われ得ることが示されている。「抗体」という用語の中に包含される結合断片の例として、例えば、ヒンジ領域においてジスルフィド架橋によって連結された2つのFab断片を含む、二価の断片であるF(ab')2断片が挙げられる。このような断片は一般的に抗体の意味に含まれるが、それらは集合的にかつ各々独立して本発明の独特の特徴であり、異なる生物学的特性および有用性を示す。抗体という用語は、特に明記しない限り、ポリクローナル抗体、モノクローナル抗体(mAb)、キメラ抗体およびヒト化抗体などの抗体様ポリペプチド、ならびに酵素的切断、ペプチド合成、および組換え技法などの任意の公知の技法によって提供される、抗原に特異的に結合する能力を保持する抗体断片(抗原結合断片)も含むこともまた、理解されるべきである。このようにして作製された抗体は、任意のアイソタイプを有し得る。
【0019】
本明細書で使用する「ヒト抗体」という用語は、ヒト生殖系列免疫グロブリン配列に由来する可変領域および定常領域を有する抗体を含むことが意図される。本発明のヒト抗体は、ヒト生殖系列免疫グロブリン配列によってコードされないアミノ酸残基(例えば、インビトロでのランダムな、もしくは部位特異的な突然変異誘発によって、またはインビボでの体細胞突然変異によって導入された変異)を含み得る。しかしながら、本明細書で使用する「ヒト抗体」という用語は、マウスのような別の哺乳動物種の生殖系列に由来するCDR配列がヒトフレームワーク配列上に移植された抗体を含まないことが意図される。
【0020】
本明細書で使用する「単離された抗体」は、異なる抗原特異性を有する他の抗体を実質的に含まない抗体を指すことが意図される。しかしながら、特定のヒト標的抗原のエピトープ、アイソフォーム、または変種に特異的に結合する単離された抗体は、例えば他の種由来の他の関連抗原(種相同体など)に対する交差反応性を有してよい。さらに、単離された抗体は、他の細胞物質および/または化学物質を実質的に含まない可能性がある。本発明の1つの態様では、異なる特異性を有する「単離された」モノクローナル抗体の組み合わせを、明確な組成物中で組み合わせる。
【0021】
本明細書で使用する「モノクローナル抗体」または「モノクローナル抗体組成物」という用語は、単一分子の組成物の、抗体分子の調製物を指す。モノクローナル抗体組成物は、特定のエピトープに対する単一の結合特異性および親和性を示す。したがって、「ヒトモノクローナル抗体」という用語は、ヒト生殖系列免疫グロブリン配列に由来する可変領域および定常領域を有する、単一の結合特異性を示す抗体を指す。ヒトモノクローナル抗体は、不死化細胞と融合した、ヒト重鎖導入遺伝子および軽鎖導入遺伝子を含むゲノムを有する、遺伝子導入マウスなどの、遺伝子導入または染色体導入非ヒト動物から得られたB細胞を含むハイブリドーマによって作製することができる。
【0022】
所定の抗原に対する抗体の結合との関連における、本明細書で使用する「結合」という用語は典型的に、例えば、抗原をリガンドとしておよび抗体を分析物として用いて、BIAcore 3000装置において表面プラズモン共鳴(SPR)技術によって決定した場合の、約10-7 Mもしくはそれ未満、例えば約10-8 Mもしくはそれ未満、例えば約10-9 Mもしくはそれ未満、約10-10 Mもしくはそれ未満、または約10-11 Mもしくはさらにそれ未満のKDに対応する親和性での結合であり、所定の抗原または近縁の抗原以外の非特異的抗原(例えば、BSA、カゼイン)に対する結合の親和性よりも、少なくとも10倍低い、例えば少なくとも100倍低い、例えば少なくとも1000倍低い、例えば少なくとも10,000倍低い、例えば少なくとも100,000倍低いKDに対応する親和性で、所定の抗原に結合する。親和性がより低い量は抗体のKDに依存し、それゆえ抗体のKDが非常に低い(すなわち、抗体が高度に特異的である)場合、抗原に対する親和性が非特異的抗原に対する親和性よりも低い量は少なくとも10,000倍であり得る。
【0023】
本明細書で使用する「kd」(sec-1)という用語は、特定の抗体-抗原相互作用の解離速度定数を指す。この値は、koff値とも称される。
【0024】
本明細書で使用する「ka」(M-1×sec-1)という用語は、特定の抗体-抗原相互作用の会合速度定数を指す。
【0025】
本明細書で使用する「KD」(M)という用語は、特定の抗体-抗原相互作用の解離平衡定数を指す。
【0026】
本明細書で使用する「KA」(M-1)という用語は、特定の抗体-抗原相互作用の会合平衡定数を指し、kaをkdで割ることによって得られる。
【0027】
本明細書で使用する「アイソタイプ」とは、重鎖定常領域遺伝子によってコードされる免疫グロブリンクラス(例えば、IgG1、IgG2、IgG3、IgG4、IgD、IgA、IgE、またはIgM)を指す。
【0028】
「エピトープ」という用語は、抗体に特異的に結合し得るタンパク質決定基を意味する。エピトープは、通常はアミノ酸または糖側鎖などの分子の表面基からなり、通常は特異的な三次元構造特性および特異的な電荷特性を有する。高次構造的エピトープと非高次構造的エピトープは、前者に対する結合は変性溶媒の存在下で失われるが、後者に対する結合は失われないという点で識別される。エピトープは、結合に直接関与するアミノ酸残基(エピトープの免疫優性成分とも称される)、および特異的抗原結合ペプチドによって効果的に阻止されるアミノ酸残基のような、結合に直接関与しない他のアミノ酸残基(すなわち、このアミノ酸残基は特異的抗原結合ペプチドのフットプリント内にある)を含み得る。
【0029】
本明細書で使用する場合のヒト抗体は、例えばヒト免疫グロブリン遺伝子を保有する遺伝子導入マウスを免疫することによって、またはヒト免疫グロブリン遺伝子ライブラリーをスクリーニングすることによって、ヒト免疫グロブリン配列を用いる系から抗体が得られる場合に、特定の生殖系列配列「に由来し」、選択されたヒト抗体は、生殖系列免疫グロブリン遺伝子によりコードされるアミノ酸配列と、アミノ酸配列が少なくとも90%、例えば少なくとも95%、例えば少なくとも96%、例えば少なくとも97%、例えば少なくとも98%、または例えば少なくとも99%同一である。典型的に、重鎖CDR3の外部では、特定のヒト生殖系列配列に由来するヒト抗体は、生殖系列免疫グロブリン遺伝子によってコードされるアミノ酸配列と20アミノ酸以下の差、例えば10アミノ酸以下の差、例えば5以下、例えば4、3、2または1アミノ酸以下の差を示す。
【0030】
「二重特異性抗体」という用語は、2つの異なる結合特異性を有する任意の抗体、すなわち同じ標的抗原上に位置し得るか、またはより一般的には、異なる標的抗原上に位置し得る2つの異なるエピトープに結合する抗体を含むことが意図される。
【0031】
本明細書で使用する「エフェクター細胞」という用語は、免疫応答の認識期および活性化期とは対照的に、免疫応答のエフェクター期に関与する免疫細胞を指す。例示的な免疫細胞には、骨髄性またはリンパ系起源の細胞、例えばリンパ球(B細胞、および細胞傷害性T細胞(CTL)を含むT細胞など)、キラー細胞、ナチュラルキラー細胞、マクロファージ、単球、好酸球、好中球などの多形核細胞、顆粒球、肥満細胞、および好塩基球が含まれる。一部のエフェクター細胞は特定のFc受容体を発現し、特定の免疫機能を行う。いくつかの態様において、エフェクター細胞は、抗体依存性細胞性細胞傷害(ADCC)を誘導し得るナチュラルキラー細胞のように、ADCCを誘導することができる。例えば、FcRを発現する単球、マクロファージは、標的細胞の特異的殺傷、および免疫系の他の成分に対する抗原提示、または抗原を提示する細胞に対する結合に関与する。いくつかの態様において、エフェクター細胞は、標的抗原または標的細胞を貪食することができる。エフェクター細胞における特定のFcRの発現は、サイトカインなどの体液性因子によって調節され得る。例えば、FcγRIの発現はインターフェロンγ(IFN-γ)および/またはG-CSFによって上方制御されることが判明している。この発現増強によって、標的に対するFcγRI保有細胞の細胞傷害活性が高まる。エフェクター細胞は、標的抗原または標的細胞を貪食または溶解することができる。
【0032】
「治療」とは、症状または疾患状態を緩和する、改善する、停止させる、または根絶する(治癒する)目的で、本発明の治療上活性な化合物の有効量を投与することを指す。
【0033】
「有効量」とは、必要な投与量で必要な期間の間の、所望の治療結果を達成するために有効な量を指す。抗体の治療上有効な量は、個体の疾患状態、年齢、性別、および体重、ならびに個体において所望の応答を誘発する抗体の能力などの要因に応じて異なり得る。治療上有効な量とは、治療上有益な効果が、抗体または抗体部分の、任意の毒性または有害効果を上回る量でもある。
【0034】
「IgG4様コアヒンジ領域」という用語は、システイン残基が、抗体分子内の他のシステイン/ジスルフィド架橋よりもさらに著しく還元および/またはジスルフィド結合の異性化を受けやすいコアヒンジ領域を指す。したがって、IgG4様コアヒンジ領域を有する抗体について、抗体内の他のジスルフィド架橋および全般的な抗体構造はそのままに保ちながら、コア領域内のシステイン残基/ジスルフィド架橋が還元され、続いて別の半分子内のコアヒンジシステインとジスルフィド架橋を形成し得る還元状態を見出すことができる。例えば、IgG4様コアヒンジ領域は、IgG4コアヒンジ領域であってよく、またはCPPCからCPSCへの変異のように、コア領域内のCPPC配列のプロリンの1つが、例えばセリンに変異した別のアイソタイプの抗体のコアヒンジ配列であってよい。
【0035】
本出願との関連における「IgG4様CH3領域」という用語は、IgG4、例えばヒトIgG4のCH3と同一であるCH3領域、またはIgG4 CH3領域と機能的に同等であるCH3領域を指す。この状況における機能的に同等とは、CH3領域が、IgG4のCH3領域と同様に、安定した半分子間相互作用を形成しないことを意味する。所与のCH3領域による安定した半分子間の形成は、例えばIgG4のCH3をその所与のCH3領域と置換し、実施例31または32に示す条件下で交換について試験することによって、調べることができる。交換が認められた場合には、安定した半分子間相互作用は形成されていない。例えば、IgG4様CH3領域は、半分子の交換を可能にするという点でIgG4由来のCH3領域と同等に効率的であるCH3領域であってよい。したがって、IgG4様CH3領域は、例えばIgG4のCH3領域の配列と75%を超えて、例えば90%を超えて同一であるなど、IgG4のCH3領域と構造的に類似していてよい。しかしながら、本状況におけるIgG4様CH3領域は、加えてまたはあるいは、IgG4のCH3領域と構造的に近似していないが、ジスルフィド結合または塩の中継など共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するアミノ酸残基を全く含まず、同時に他のペプチドはCH3領域の同一のアミノ酸配列を含むことから同様の機能的特性を有するCH3領域であってもよい。例えば、IgG4様CH3領域は、半分子CH3-CH3間相互作用に関与する1つまたは複数のアミノ酸残基が変化しているかまたは欠失している変異型IgG1 CH3領域であってよい。
【0036】
「還元条件」または「還元環境」という用語は、基質、ここでいう抗体のコア領域内のシステイン残基が、酸化されるより還元された状態になる可能性が高い条件または環境を指す。
【0037】
「還元剤」という用語は、その環境において分子を還元する化合物、すなわち、その環境において分子をより還元した状態に、およびより還元しつつある状態に変える化合物を指す。還元剤は、電子を供与することにより作用し、それによって、基質の還元後にそれ自体は酸化された状態になる。したがって、還元剤とは電子を供与する作用物質である。還元剤の例には、ジチオスレイトール(DTT)、メルカプトエタノール、システイン、チオグリコール酸、システアミン、グルタチオン、および水素化ホウ素ナトリウムが含まれる。1つの態様において、還元剤は酵素を含まない。
【0038】
「ジスルフィド結合形成」とは、1つまたは2つのポリペプチド中に存在する2つのシステイン間で共有結合を形成する過程を指し、この結合は「-S--S-」として図式化される。
【0039】
「ジスルフィド結合の還元」とは、ジスルフィド結合の切断によって、2つのチオール基(-SH基)が生じる過程を指す。
【0040】
「ジスルフィド結合の異性化」という用語は、異なるシステイン間のジスルフィド結合の交換、すなわちジスルフィド結合の再編成を指す。
【0041】
「タンパク質ジスルフィド結合イソメラーゼ」とは、タンパク質におけるジスルフィド結合の異性化を触媒するタンパク質を指す。
【0042】
ジスルフィド結合の還元に関連して使用する場合の「有意でない還元」とは、溶液中の特定のジスルフィド架橋の、一般的には10%未満、例えば5%未満、例えば2%未満または1%未満が還元を起こすことを意味する。
【0043】
発明の局面および態様
上記の通り、第1の主要な局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0044】
好ましい態様において、本発明の方法において使用する第1の抗体および第2の抗体はモノクローナル抗体である。モノクローナル抗体は、例えば、Kohler et al.(14)によって最初に記載されたハイブリドーマ法によって作製することもできるし、または組換えDNA法によって作製することもできる。モノクローナル抗体は、例えばClackson et al.(15)およびMarks et al.(16)に記載されている技法を用いて、ファージ抗体ライブラリーから単離することもできる。モノクローナル抗体は、任意の適切な供給源から得ることができる。したがって、例えば、モノクローナル抗体は、例えば表面上に抗原を発現する細胞の形態をした関心対象の抗原、または関心対象の抗原をコードする核酸で免疫したマウスから得られたマウス脾臓B細胞から調製したハイブリドーマから得ることができる。モノクローナル抗体はまた、免疫したヒト、またはラット、イヌ、霊長動物等のような非ヒト哺乳動物の抗体発現細胞に由来するハイブリドーマから得ることもできる。
【0045】
1つの態様において、本発明の抗体はヒト抗体である。指向化されたヒトモノクローナル抗体は、マウス系ではなくヒト免疫系の一部を保有する遺伝子導入または染色体導入マウスを用いて作製することができる。このような遺伝子導入および染色体導入マウスには、本明細書においてそれぞれHuMAbマウスおよびKMマウスと称するマウスが含まれ、本明細書ではまとめて「遺伝子導入マウス」と称する。
【0046】
HuMAbマウスは、内在性μ鎖およびκ鎖遺伝子座を不活化する標的変異と共に、再編成されていないヒト重鎖(μおよびγ)およびκ軽鎖免疫グロブリン配列をコードするヒト免疫グロブリン遺伝子のミニ遺伝子座を含む(17)。したがって、マウスはマウスIgMまたはκの発現低下を示し、免疫化に応答して、導入されたヒト重鎖および軽鎖導入遺伝子はクラススイッチおよび体細胞突然変異を起こして、高親和性ヒトIgG、κモノクローナル抗体を生成する(17〜20)。HuMAbマウスの調製は、参考文献21〜25に詳述されている。US 5,545,806、US 5,569,825、US 5,625,126、US 5,633,425、US 5,789,650、US 5,877,397、US 5,661,016、US 5,814,318、US 5,874,299、US 5,770,429、US 5,545,807、WO 98/24884、WO 94/25585、WO 93/1227、WO 92/22645、WO 92/03918、およびWO 01/09187もまた参照されたい。
【0047】
HCo7マウスは、(Chen et al.(26)に記載されているような)その内在性軽鎖(κ)遺伝子におけるJKD破壊、(WO 01/14424の実施例1に記載されているような)その内在性重鎖遺伝子におけるCMD破壊、(Fishwild et al.(25))に記載されているような)KCo5ヒトκ軽鎖導入遺伝子、および(US 5,770,429に記載されているような)HCo7ヒト重鎖導入遺伝子を有する。
【0048】
HCo12マウスは、(Chen et al.(26)に記載されているような)その内在性軽鎖(κ)遺伝子におけるJKD破壊、(WO 01/14424の実施例1に記載されているような)その内在性重鎖遺伝子におけるCMD破壊、(Fishwild et al.(25)に記載されているような)KCo5ヒトκ軽鎖導入遺伝子、および(WO 01/14424の実施例2に記載されているような)HCo12ヒト重鎖導入遺伝子を有する。
【0049】
KMマウス系統において、内在性マウスκ軽鎖遺伝子は、Chen et al.(26)に記載されている通りにホモ接合性に破壊されており、かつ内在性マウス重鎖遺伝子も、WO 01/09187の実施例1に記載されている通りにホモ接合性に破壊されている。このマウス系統は、Fishwild et al.(25)に記載されている通りに、ヒトκ軽鎖導入遺伝子、KCo5を保有する。このマウス系統はまた、WO 02/43478に記載されている通りに、第14染色体断片hCF(SC20)からなるヒト重鎖導入染色体を保有する。
【0050】
これらの遺伝子導入マウス由来の脾臓細胞を用いて、周知の技法に従って、ヒトモノクローナル抗体を分泌するハイブリドーマを作製することができる。このような遺伝子導入非ヒト動物、本発明において使用する抗体の発現をコードする操作可能な核酸配列を含む非ヒト動物、1つまたは複数の標的コード核酸配列を安定にトランスフェクトした非ヒト動物等は、本発明のさらなる特色である。
【0051】
本発明において使用しようとするヒトモノクローナルもしくはポリクローナル抗体、またはその他の種が起源である本発明において使用する抗体はまた、関心対象の免疫グロブリン重鎖および軽鎖配列についてトランスジェニックである別の非ヒト哺乳動物または植物の作製、ならびにそれらから回収可能な形態での抗体の生産を通じて、トランスジェニックで作製することも可能である。哺乳動物におけるトランスジェニック生産に関連して、抗体をヤギ、ウシ、またはその他の哺乳動物の乳において生産し、そこから回収することができる。例えば、US 5,827,690、US 5,756,687、US 5,750,172、US 5,741,957を参照されたい。
【0052】
さらに、本発明において使用しようとするヒトまたはその他の抗体は、当技術分野で周知の技法を用いて、ファージディスプレイ、レトロウイルスディスプレイ、リボソームディスプレイ、およびその他の技法を含むがこれらに限定されないディスプレイ型の技術を通じて作製することができ、結果として得られた分子は、そのような技法が当技術分野で周知であるので、親和性成熟などのさらなる成熟に供することができる(例えば、参考文献27、28、および30(ファージディスプレイ)、29(リボソームディスプレイ)、31〜35、ならびにUS 5,733,743を参照されたい)。ディスプレイ技術を利用してヒトではない抗体を生産する場合には、このような抗体をヒト化することができる。
【0053】
上記の通りに、いくつかの態様において、本発明の方法において使用する第1および/または第2の抗体はIgG4抗体である。しかしながら、本発明において使用する抗体は、原則的には、CH3領域の配列が半分子の交換を可能にするという条件で任意のアイソタイプのものであってよい。例えば、本発明の方法において使用するまたは得られる抗体は、(任意の特定の変異位置以外では)SEQ ID NO:19〜22に示す定常領域のいずれかを含み得る。
【0054】
したがって、本発明の方法の1つの態様において、第1および/または第2の抗体はコアヒンジ領域内にCPPC配列を含む。別の態様において、第1および/または第2の抗体はIgG4様コアヒンジ領域を含む。例えば、いくつかの態様において、第1および/または第2の抗体は、コアヒンジ領域内に、式中、X1およびX2の両方がプロリンではないという条件で、X1およびX2は任意のアミノ酸であってよい、CX1X2C配列を含む抗体である。別の態様において、第1および/または第2の抗体は、コアヒンジ領域内に、式中、X3はプロリンを除く任意のアミノ酸であってよい、CX3PCまたはCPX3C配列を含む抗体である。さらなる態様において、第1および/または第2の抗体は、コアヒンジ領域内にCSPC、CPSC、CRPC、CPRC、CGHC、またはCPHC配列を含む抗体である。上記の変異は、例えば当技術分野で周知の部位特異的突然変異誘発によって導入することができる。
【0055】
アイソタイプの選択は典型的に、CDC誘導またはADCCにおける活性などの所望のエフェクター機能によって導かれる。例示的なアイソタイプは、IgG1、IgG2、IgG3、およびIgG4である(例えば、SEQ ID NO:19〜22を参照されたい)。ヒト軽鎖定常領域であるκまたはλのいずれかを使用することができる。必要に応じて、本発明において使用するための抗体のクラスを、公知の方法によってスイッチすることができる。例えば、本来IgM、IgG1、またはIgG2であった本発明において使用しようとする抗体を、本発明のIgG4抗体にクラススイッチすることができる。したがって、様々な治療用途のために、本発明の抗体のエフェクター機能を、例えばIgG1、IgG2、IgG3、IgG4、IgD、IgA、IgE、またはIgM抗体へのアイソタイプスイッチによって変更することができる。
【0056】
1つの態様において、本発明において使用する第1および/または第2の抗体は全長抗体である。別の態様において、使用する本発明の第1および/または第2の抗体は抗体断片である。
【0057】
本発明の方法の1つの態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO:22)を有するIgG4 CH3領域などの、IgG4 CH3領域を含む。
【0058】
しかしながら、本発明の方法の別の態様において、第1および/または第2の抗体は非IgG4アイソタイプのCH3領域を含み、そのCH3配列は、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている。
【0059】
例えば、本発明の1つのさらなる態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 19)を有するCH3領域を含み、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:238位のArg(R)がGln(Q)によって置換されている;239位のAsp(D)がGlu(E)によって置換されている;292位のLys(K)がArg(R)によって置換されている;302位のGln(Q)がGlu(E)によって置換されている;および328位のPro(P)がLeu(L)によって置換されている。
【0060】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 19)を有するCH3領域を含み、292位のLys(K)がArg(R)によって置換されている。
【0061】
別の態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 19)を有するCH3領域を含むが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている。
【0062】
別のさらなる態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 20)を有するCH3領域を含み、このCH3領域は、以下のアミノ酸置換のうちの1つもしくは複数または5つすべてがなされるように改変されている:234位のArg(R)がGln(Q)によって置換されている;276位のMet(M)がVal(V)によって置換されている;288位のLys(K)がArg(R)によって置換されている;298位のGln(Q)がGlu(E)によって置換されている;および324位のPro(P)がLeu(L)によって置換されている。
【0063】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 20)を有するCH3領域を含み、234位のArg(R)がGln(Q)によって置換されている。
【0064】
さらなる好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 20)を有するCH3領域を含み、234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている。
【0065】
別のさらなる態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 21)を有するCH3領域を含み、このCH3領域は、以下のアミノ酸置換のうちの1つもしくは複数または10個すべてがなされるように改変されている:285位のArg(R)がGln(Q)によって置換されている;314位のSer(S)がAsn(N)によって置換されている;322位のAsn(N)がLys(K)によって置換されている;327位のMet(M)がVal(V)によって置換されている;339位のLys(K)がArg(R)によって置換されている;349位のGln(Q)がGlu(E)によって置換されている;352位のIle(I)がVal(V)によって置換されている;365位のArg(R)がHis(H)によって置換されている;366位のPhe(F)がTyr(Y)によって置換されている;および375位のPro(P)がLeu(L)によって置換されている。
【0066】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 21)を有するCH3領域を含み、285位のArg(R)がGln(Q)によって置換されている。
【0067】
好ましい態様において、第1および/または第2の抗体は、図18に示す配列(SEQ ID NO: 21)を有するCH3領域を含み、285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている。
【0068】
本発明の方法のさらなる態様において、第1の抗体はコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含み、第2の抗体もコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含む。
【0069】
上記の通りに、主要な局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0070】
本発明の方法の1つの態様において、段階c)における条件は、コアヒンジ領域外のジスルフィド架橋の有意な還元または異性化が起こらないように選択する。
【0071】
別の態様において、段階c)における還元条件は、ジスルフィド結合交換を行うコアヒンジ領域の内因性の活性を促進する条件である。
【0072】
本発明のさらなる態様において、段階c)は還元剤の添加を含む。さらなる態様において、段階c)は、グルタチオン、L-システイン、ジチオスレイトール、β-メルカプト-エタノール、およびシステアミンからなる群より選択される作用物質の添加を含む。
【0073】
本発明の方法の1つの態様において、還元剤の濃度は、段階c)で生じる溶液の酸化還元電位が、実施例31に記載する条件下で、1マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、実施例31に記載する条件下で、例えば10マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば50マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば0.1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性であるといった濃度である。
【0074】
さらなる態様において、還元剤の濃度は、段階c)で生じる溶液の酸化還元電位が、
‐実施例31に記載する条件下で、1マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、実施例31に記載する条件下で、例えば10マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば50マイクロMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば0.1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、かつ
‐実施例31に記載する条件下で、1 Mのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、例えば100 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、15 mMのグルタチオンによって生じる酸化還元電位と等しいかもしくはより還元性が低い
濃度である。
【0075】
第1の抗体がコアヒンジ領域内にCPPC配列を有する、および/または第2の抗体がコアヒンジ領域内にCPPC配列を有する場合の1つの態様において、段階c)で生じる溶液の酸化還元電位は、実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性であることが好ましい。
【0076】
さらなる態様において、還元剤の濃度は、段階c)で生じる溶液の酸化還元電位が、
‐実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性であり、かつ
‐1 Mのグルタチオンによって生じる酸化還元電位と等しいかまたはそれよりも還元性が低い、例えば100 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはそれよりも還元性が低い、15 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはそれよりも還元性が低い
濃度である。
【0077】
本発明の方法の1つの態様において、段階c)は、還元型グルタチオンの存在下で、20℃またはそれ以上、例えば37℃の温度における、少なくとも1時間、例えば少なくとも2時間、例えば少なくとも5時間、例えば少なくとも10時間の抗体のインキュベーションを含む。
【0078】
本発明の方法のさらなる態様において、段階c)において選択する条件は、本明細書に記載するサイズ排除クロマトグラフィーにより決定されるように(ここで出発材料の抗体よりも早く溶出するピークは、凝集体の形成を示す)、結果として得られた組成物中の抗体分子の10%未満、例えば5%未満、例えば2%未満、例えば1%未満が凝集状態である条件である。
【0079】
本発明のエクスビボ方法の1つの態様において、本方法は、PDIなどのタンパク質ジスルフィドイソメラーゼ活性を有するタンパク質の添加を含む。本発明のエクスビボ方法の別の態様において、本方法は、PDIなどのタンパク質ジスルフィドイソメラーゼ活性を有するタンパク質の添加を含まない。
【0080】
本発明のエクスビボ方法の1つの態様において、本方法は生細胞または細胞抽出物の添加を含まない。
【0081】
上記の通り、本発明の方法において使用する第1の抗体および第2の抗体は、結合特異性が異なる、すなわち異なるエピトープに結合する。原則として、本発明の方法では、出発材料として特異性の任意の組み合わせを使用することができる。本発明の方法はまた、2種類の異なる抗体のみを出発材料とすることに限定されない。したがって、本発明の方法はまた、出発材料として3つまたはそれ以上の抗体を用いて行うこともできる。このような態様において、本発明の方法の段階d)で得られた組成物は、複数の二重特異性抗体を含むことになる。
【0082】
本発明の方法の1つの態様において、第1の抗体は、腫瘍細胞に対する、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、CD38、もしくはCXCR5などの腫瘍細胞タンパク質に対する、またはB細胞受容体のシグナル伝達成分である、CD79aもしくはCD79bに対する結合特異性を有する。別の態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は、erbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、またはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有する。
【0083】
さらなる態様において、第1の抗体はerbB1に対する結合特異性を有し、かつ第2の抗体はerbB2に対する結合特異性を有する。
【0084】
別の態様において、第1の抗体はCD19に対する結合特異性を有し、かつ第2の抗体はCD20に対する結合特異性を有する。
【0085】
さらなる態様において、第1の抗体はCD38に対する結合特異性を有し、かつ第2の抗体はCD34に対する結合特異性を有する。
【0086】
なおさらなる態様において、第1の抗体はCD4に対する結合特異性を有し、かつ第2の抗体はCXCR5に対する結合特異性を有する。
【0087】
本発明の方法の別の態様において、第1の抗体は病原微生物に対する結合特異性を有する。さらなる態様において、第1の抗体は病原微生物に対する結合特異性を有し、かつ第2の抗体は、CD3、CD25、CD28、CD16、CD89、CD32、またはCD1などのエフェクター細胞タンパク質に対する結合特異性を有する。
【0088】
二重特異性抗体を用いて、化学療法薬が作用すべき細胞に対し、この薬剤をより特異的に標的化することもできる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は化学療法薬に対する結合特異性を有する。
【0089】
さらに、二重特異性抗体において血清タンパク質に対する結合特異性を含ませることによって、抗体の血清半減期を変更することができる。例えば、二重特異性抗体において血清アルブミンに対する結合特異性を含ませることによって、血清半減期を延長することができる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は血清アルブミンなどの血液タンパク質に対する結合特異性を有する。
【0090】
第2の結合特異性を用いて、抗体を脳または肝臓などの特定の組織に標的化することもできる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体はトランスフェリンなどの脳タンパク質または肝臓タンパク質に対する結合特異性を有する。
【0091】
さらに、第2の結合特異性を用いて、血液凝固因子を特定の所望の作用部位に標的化することもできる。例えば、腫瘍細胞に対する第1の結合特異性および血液凝固因子に対する第2の結合特異性を有する二重特異性抗体は、腫瘍に血液凝固を導き、したがって腫瘍増殖を停止させることができる。したがって、本発明の方法のさらなる態様において、第1の抗体は、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有し、かつ第2の抗体は組織因子などの血液凝固に関与するタンパク質に対する結合特異性を有する。
【0092】
本発明のさらなる態様では、第1および/または第2の抗体を、細胞毒性薬;放射性同位体;プロドラッグまたは薬物、例えばタキサン;サイトカイン;ケモカイン、および補体、例えばC1qからなる群より選択される化合物に連結する。このような化合物は、例えば癌治療において、標的細胞をより効果的に殺傷することができる。または化合物は、結果として得られた二重特異性抗体に、すなわち半分子の交換が起こった後に、結合させることも可能である。
【0093】
本発明の方法のさらなる態様において、本方法は、さらなる半分子の交換を停止させるために、段階c)において得られた組成物を非還元条件または還元性のより低い条件に導くさらなる段階を含む。これは、例えば小分子還元剤を除去するための、結果として得られた組成物の、透析またはサイズベースのクロマトグラフィーなど、当技術分野で公知の種々の方法によって行うことができる。
【0094】
本発明の方法のなおさらなる態様においては、2つの半分子の化学的架橋を行い、二重特異性抗体がその後、そうしなければこの抗体が半分子の交換を起こし得るインビボ条件などの条件下で用いられる場合でさえもさらなる交換を防ぐことによって、結果として得られた二重特異性抗体を安定化する。したがって、1つの態様において、本発明の方法は以下のさらなる段階を含む:
a) 例えばビス-マレイミドヘキサンなどのマレイミドを含む化合物を用いて、ヒンジ領域内のシステインを化学的に架橋する段階、
b) 例えば過ヨウ素酸酸化、およびその後のアジピンジヒドラジドなどの適切な架橋剤によるアルデヒド基の反応を介して、半分子上の炭水化物側鎖を化学的に架橋する段階、または
c) 例えば以下の組み合わせのうちの1つまたは複数を用いて(SEQ ID NO:19を参照されたい)、例えばMerchant et al.(36)(参照により本明細書に組み入れられる)に記載されている通りに、CH3領域内に非対称的に導入されたシステインを架橋する段階:
‐第1の抗体中のD282Cと第2の抗体中のK275C、
‐第1の抗体中のD282Sと第2の抗体中のK275S、
‐第1の抗体中のY232Cと第2の抗体中のS237C、
‐第1の抗体中のY232Cと第2の抗体中のD239C、
‐第1の抗体中のY232Cと第2の抗体中のE240C、
‐第1の抗体中のL234Cと第2の抗体中のS237C、
‐第1の抗体中のT277Cと第2の抗体中のV280C、
‐第1の抗体中のV280Cと第2の抗体中のK275C。
【0095】
さらなる局面において、本発明は、架橋法によって、例えば上記の3つの架橋法のいずれかによって得られるか、または得られ得る、安定した二重特異性抗体に関する。
【0096】
結果として得られた二重特異性抗体が架橋によって安定されたかどうかにかかわらず、本発明の方法はいくつかの態様において、二重特異性抗体を精製するさらなる段階を含み得る。(これらに限定されないが)標準的なプロテインAクロマトグラフィー、プロテインG、プロテインL、陽イオン/陰イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、疎水性相互作用クロマトグラフィー、チオフィリッククロマトグラフィーなどの標準的なクロマトグラフィー技法を用いて、またはIgG分子を捕獲するように設計されたリガンド(プロテインA模倣体、ラマVHHリガンド等)を用いて、二重特異性抗体を含む混合物を精製することができる。または、塩が誘導する沈殿(硫酸アンモニウム)、有機溶媒(DMSO、エタノール)の添加、pHの変更、または非イオン性ポリマー(ポリエチレングリコール)などの標準的な技法を用いて、IgG混合物を沈殿させることができる。別の設定においては、混合物を、IgG分子の濃縮を可能にする膜を用いる濾過技法に供することができる。ある種の混合物は二重特異性抗体の他に親IgG分子をなお含んでいる場合があるため、二重特異性抗体を完全な均一性になるまで精製するためには、これらすべての技法の組み合わせが必要である場合がある。親の単一特異性IgG分子から二重特異性抗体を分離するには、さらなる精製段階が次に必要となる場合がある。これは、例えば、第1の結合特異性に対するアフィニティーカラムを用いて結合および溶出し、次に第2の結合特異性に対するアフィニティーカラムを用いて結合および溶出することによる精製によって行うことができる。特に化学的架橋を行わなかった場合の好ましい態様では、非還元条件など、さらなる半分子の交換を妨げる条件下で精製を行う。
【0097】
(精製された)二重特異性抗体の量、質、および純度は、吸光度測定、HP-SEC、SDS-PAGE、未変性PAGE、およびRP-HPLCなどの日常的な生化学的技法を用いて解析することができる。特に関心対象となるのは、親IgG分子から二重特異性抗体を識別できる技法である。このような技法の例は、(これらに限定されないが)、電荷および/または質量に基づいて、分子の高度に正確な分離および検出を可能にするIEF、cIEF、CIEX、および質量分析(ESI、MALDI)である。二重特異性抗体の二重結合特異性は、例えばELISA、RIA、表面プラズモン共鳴(SPR)、バイオレイヤー干渉法、DELFIA、FRET、ECL、Gyros、およびAlfaScreenを使用する種々の異なる結合アッセイ形式を用いて評価することができる。
【0098】
1つの態様において、半分子の交換は、関心対象の2種類の抗原の一方に対する二重特異性抗体の形成を支持する条件下で行うことができる。例えば、抗原XおよびYに対する抗体を考えてみる。5倍過剰または10倍過剰などの、抗原Xに対する過剰な抗体を用いて交換を行う場合、抗体Yに対する、ほとんどまたはすべての抗体は二重特異性(すなわち、抗原XおよびYを認識する)となる。
【0099】
この工程の後に、マトリックス固定化抗原Yおよびアフィニティーカラムクロマトグラフィーにおいて、二重特異性抗体を精製することができる。結合した抗体は、所望の二重特異性抗体について高度に濃縮されている。抗原Xに対する非結合抗体は、上記のサイクルの反復に使用することができる。
【0100】
インビボにおける交換を防ぐために安定化が必要である場合には、上記の通りに二重特異性抗体を架橋することができる。化学的架橋後に、抗原Zに対する過剰な抗体とのさらなる交換反応、およびその後のマトリックス固定化抗原Zに対する抗Z含有抗体の吸収(アフィニティーカラムクロマトグラフィーなど)を行うことによって、非安定化抗体を安定化抗体から精製することができる。そこで、非結合画分は所望の安定化二重結合抗体を含んでいる。
【0101】
本発明の方法のなおさらなる態様において、本方法は、治療用途のために、結果として得られた二重特異性抗体を製剤化するさらなる段階を含む。これは、ヒト使用に適している、特に静脈内投与などの非経口投与に適している水溶液中での、治療上有効な量の二重特異性抗体の製剤化を含む。
【0102】
さらなる局面において、本発明は、以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法に関する:
a) 第1の結合特異性を有する第1抗体を提供する段階であって、該第1の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【0103】
好ましくは、段階c)において還元剤を添加するが、この剤の濃度は、段階c)で生じる溶液の酸化還元電位が、実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である。
【0104】
さらなる局面において、本発明は、本明細書に記載する本発明の方法のいずれかによって得られるか、または得られ得る、二重特異性抗体を含む組成物に関する。
【0105】
さらなる主要な局面において、本発明は、2つのIgG4様CH3領域を含む単離された二重特異性抗体に関する。
【0106】
1つの態様において、該抗体はコアヒンジ領域内に1つまたは2つのCPPC配列を含む。
【0107】
別の態様において、該抗体は、コアヒンジ領域内に、式中、X1およびX2は両方がプロリンではないという条件で、X1およびX2は任意のアミノ酸であってよい、1つまたは2つのCX1X2C配列を含む。
【0108】
さらなる態様において、該抗体はコアヒンジ領域内に、式中、X3はプロリンを除く任意のアミノ酸であってよい、1つまたは2つのCX3PCまたはCPX3C配列を含む。
【0109】
なおさらなる態様において、該抗体は、コアヒンジ領域内に1つまたは2つのCSPC、CPSC、CRPC、またはCPRC配列を含む。
【0110】
単離された二重特異性抗体のいくつかの態様において、第1および/または第2のCH3領域は非IgG4アイソタイプのものであって、このCH3配列は、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている。
【0111】
その1つのさらなる態様において、第1および/または第2のCH3領域は、図18に示す配列(SEQ ID NO: 19)を有し、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:238位のArg(R)がGln(Q)によって置換されている;239位のAsp(D)がGlu(E)によって置換されている;292位のLys(K)がArg(R)によって置換されている;302位のGln(Q)がGlu(E)によって置換されている;および328位のPro(P)がLeu(L)によって置換されている。
【0112】
別のさらなる態様において、第1および/または第2のCH3領域は、図18に示す配列(SEQ ID NO: 20)を有し、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:234位のArg(R)がGln(Q)によって置換されている;276位のMet(M)がVal(V)によって置換されている;288位のLys(K)がArg(R)によって置換されている;298位のGln(Q)がGlu(E)によって置換されている;および324位のPro(P)がLeu(L)によって置換されている。
【0113】
なおさらなる態様において、第1および/または第2のCH3領域は、図18に示す配列(SEQ ID NO: 21)を有し、このCH3領域は、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている:285位のArg(R)がGln(Q)によって置換されている;314位のSer(S)がAsn(N)によって置換されている;322位のAsn(N)がLys(K)によって置換されている;327位のMet(M)がVal(V)によって置換されている;339位のLys(K)がArg(R)によって置換されている;349位のGln(Q)がGlu(E)によって置換されている;352位のIle(I)がVal(V)によって置換されている;365位のArg(R)がHis(H)によって置換されている;366位のPhe(F)がTyr(Y)によって置換されている;および375位のPro(P)がLeu(L)によって置換されている。
【0114】
なおさらなる態様において、本発明の抗体の第1および/または第2のCH3領域はIgG4 CH3領域である。
【0115】
なおさらなる局面において、本発明は、薬として使用するための、例えば癌または感染症の治療用の薬として使用するためなどの、本発明の二重特異性抗体、または本明細書に記載する本発明の方法のいずれかによって得られるか、もしくは得られ得る、二重特異性抗体を含む薬学的組成物などの組成物に関する。
【0116】
なおさらなる局面において、本発明は、癌または感染症の治療用の薬を調製するための、本発明の二重特異性抗体、または本発明の方法のいずれかによって得られるか、もしくは得られ得る、二重特異性抗体を含む組成物の使用に関する。
【0117】
本発明の方法を用いて、標的結合特異性の、特に興味深いまたは効果的な組み合わせを選択することもできる。例えば、本発明の方法を用いて、異なる結合特異性を有する一組の抗体から、異なる二重特異性抗体の一組または「マトリックス」を作製することができる。次に、結果として得られた二重特異性抗体の一組またはマトリックスを所望の生物学的特性について試験して、最良の組み合わせを選択することができる。
【0118】
したがって、なおさらなる局面において、本発明は、以下の段階を含む、所望の特性を有する二重特異性抗体を選択する方法に関する:
a) 一組の抗体を提供する段階であって、各抗体が異なる標的特異性を有し、かつ各抗体がIgG4様CH3領域を含む段階、
b) 還元条件下で、該一組の抗体の各抗体を該一組の別の抗体と共にインキュベートし、それによって一組の抗体混合物を作製する段階であって、各混合物が異なる二重特異性抗体を含む段階、
c) 結果として得られた一組の抗体混合物を所与の所望の特性についてアッセイする段階、および
d) 所望の特性を有する二重特異性抗体混合物を選択する段階。
【0119】
上記の方法における段階b)は、段階c)について先に上で記した通りに行うことができる。
【0120】
1つの態様において、試験すべき所望の特性は腫瘍細胞殺傷である。
【0121】
参考文献
【0122】
本発明を以下の実施例によってさらに例証するが、実施例はさらなる限定と解釈されるべきではない。
【0123】
実施例
実施例1:オリゴヌクレオチドプライマーおよびPCR増幅
オリゴヌクレオチドプライマーは、Isogen Bioscience(オランダ、マールセン)によって合成および定量された。プライマーを100 pmol/μlとなるようにH2Oに溶解し、-20℃で保存した。すべてのPCRプライマーおよび配列決定プライマーの概要を以下に示す。PCRについては、PfuTurbo(登録商標) Hotstart DNAポリメラーゼ(Stratagene、オランダ、アムステルダム)を製造業者の説明書に従って使用した。各反応混合物は、全量20μlのPCR反応緩衝液(ポリメラーゼと共に供給)中に、200μM混合dNTP(Roche Diagnostics、オランダ、アルメレ)、フォワードプライマーおよびリバースプライマーいずれも6.7 pmol、ゲノムDNA 100 ngまたはプラスミドDNA 1 ng、およびPfuTurbo(登録商標) Hotstart DNAポリメラーゼ1単位を含んだ。32サイクルプログラム:95℃で2分の変性;95℃で30秒、60〜70℃勾配(または別の特定のアニーリング温度)で30秒、および72℃で3分の30サイクル;72℃で10分の最終伸長を用いて、TGradient Thermocycler 96(Whatman Biometra、ドイツ、ゲッティンゲン)でPCR反応を行った。必要に応じて、さらなる解析または処理時までPCR混合物を4℃で保存した。
【0124】
実施例2:アガロースゲル電気泳動
1×Tris酢酸EDTA緩衝剤中で50 mlのゲルを用いて、Sambrook(37)に従ってアガロースゲル電気泳動を行った。ゲル中にエチジウムブロマイドを含め、UV光下で観察することによって、DNAを可視化した。CCDカメラおよび画像解析システム(GeneGnome;Syngene、Westburg B.V.、オランダ、ルースデン経由)によりゲル画像を記録した。
【0125】
実施例3:PCR産物の解析および精製ならびに酵素消化
製造業者の説明書に従ってMinElute PCR精製キット(Qiagen、Westburg、オランダ、ルースデン経由;製品# 28006)を用いて、所望のPCR断片の精製を行った。単離したDNAをUV分光法によって定量し、アガロースゲル電気泳動によってその質を評価した。
【0126】
または、1% Tris酢酸EDTAアガロースゲルを用いてアガロース電気泳動によって(例えば、複数の断片が存在する場合)、PCR産物または消化産物を分離した。所望の断片をゲルから切り出し、製造業者の説明書に従ってQIAEX IIゲル抽出キット(Qiagen;製品# 20051)を用いて回収した。
【0127】
実施例4:UV分光法によるDNAの定量
製造業者の説明書に従ってNanoDrop ND-1000分光光度計(Isogen Life Science、オランダ、マールセン)を用いて、核酸の光学密度を決定した。260 nmにおける光学密度(OD)の解析により(1 OD260 nm単位=50μg/ml)、DNA濃度を測定した。すべての試料について、核酸を溶解した緩衝液を参照として使用した。
【0128】
実施例5:制限酵素消化
制限酵素および補充物は、New England Biolabs(米国、マサチューセッツ州、ベバリー)またはFermetas(リトアニア、ヴィルニアス)から入手し、製造業者の説明書に従って使用した。
【0129】
DNA(100 ng)を、最終量10μlの適切な緩衝液中で酵素5単位で消化した(必要に応じて、反応量を増やした)。消化物を、推奨される温度で最低60分間インキュベートした。適合しない緩衝液または温度の要求を伴う制限酵素による二重消化を必要とする断片については、消化を連続して行った。必要に応じて、消化産物をアガロースゲル電気泳動およびゲル抽出によって精製した。
【0130】
実施例6:DNA断片のライゲーション
製造業者の説明書に従って、迅速ライゲーションキット(New England Biolabs)によってDNA断片のライゲーションを行った。各ライゲーションについて、ベクターDNAをおよそ3倍モル濃度の過剰の挿入物DNAと混合した
【0131】
実施例7:大腸菌(E. coli)の形質転換
製造業者の説明書に従って熱ショック法を用いて、プラスミドDNA(DNA溶液1〜5μl、典型的にはDNAライゲーション混合物2μl)をOne Shot DH5α-T1RまたはMACH-1 T1Rコンピテント大腸菌細胞(Invitrogen、オランダ、ブレダ;製品# 12297-016)に形質転換した。次に、50μg/mlアンピシリンを含むルリア-ベルターニ(LB)寒天プレートに細胞をプレーティングした。細菌コロニーが明白になるまで、プレートを37℃で16〜18時間インキュベートした。
【0132】
実施例8:PCRによる細菌コロニーのスクリーニング
HotStarTaqマスターミックスキット(Qiagen;製品# 203445)ならびに適切なフォワードプライマーおよびリバースプライマー(付属書1)を用いるコロニーPCRにより、所望の配列を含むベクターの存在について細菌コロニーをスクリーニングした。選択したコロニーを20μlピペットチップで軽く触れ、小規模培養用の2 ml LB中に短時間触れさせ、次いでPCR混合物中に再懸濁した。35サイクルプログラム:95℃で15分の変性;94℃で30秒、55℃で30秒、および72℃で2分の35サイクル;その後の72℃で10分の最終伸長段階を用いて、TGradient Thermocycler 96でPCRを行った。必要に応じて、アガロースゲル電気泳動による解析時まで、PCR混合物を4℃で保存した。
【0133】
実施例9:大腸菌培養物からのプラスミドDNA単離
製造業者の説明書に従ってQiagen(Westburg、オランダ、ルースデン経由)による以下のキットを用いて、プラスミドDNAを大腸菌培養物から単離した。大量プラスミド調製(50〜150 ml培養物)については、HiSpeedプラスミドMaxiキット(製品# 12663)またはHiSpeedプラスミドMidiキット(製品# 12643)のいずれかを使用した。小規模プラスミド調製(±2 ml培養物)については、Qiaprepスピンミニプレップキット(製品# 27106)を使用し、DNAを溶出緩衝液(キットにより供給) 50μlで溶出した。
【0134】
実施例10:DNA配列決定
当技術分野で公知の標準的な手順を用いて、プラスミドDNAの配列を決定した。配列は、Vector NTIソフトウェア(Informax、英国、オックスフォード)を用いて解析した。
【0135】
実施例11:HEK-293F細胞における一過性発現
Freestyle(商標) 293-F(懸濁増殖および既知組成Freestyle培地に適応したHEK-293サブクローン、例えばHEK-293F)細胞をInvtrogenから入手し、293fectin(Invitrogen)を用いて製造業者のプロトコールに従ってトランスフェクトした。
【0136】
実施例12:pTomG4;可変重鎖領域とヒトIgG4の定常領域の発現用ベクターの構築
ゲノムDNAをボランティアの血液試料から単離し、プライマーIGG4gene2fおよびIGG4gene2r(以下の表を参照されたい)を用いるPCRにおいて、鋳型として使用し、IgG4の重鎖の完全なゲノム定常領域を増幅し、哺乳動物発現ベクターpEE6.4(Lonza Biologics)にクローニングするための適切な制限部位を導入した。PCR断片を精製し、pEE6.4にクローニングした。このために、PCR産物をHindIIIおよびEcoRIで消化し、続いて制限酵素を加熱不活化した。pEE6.4ベクターもHindIIIおよびEcoRIで消化し、続いて制限酵素を加熱不活化し、エビアルカリ性ホスファターゼでベクター断片を脱リン酸化し、次いでホスファターゼを加熱不活化した。IgG4断片およびpEE6.4HindIII/EcoRI脱リン酸化ベクターを連結し、コンピテントMACH1-T1R細胞(Invitrogen)に形質転換した。3つのクローンをLB中で培養し、プラスミドDNAを少量の培養物(1.5 mL)から単離した。制限消化から、pEE6.4ベクター中にIgG4断片がクローニングされたことと合致するパターンが明らかになった。2つのクローンによるプラスミドDNAをDH5α-T1R大腸菌に形質転換し、プラスミドDNAを単離し、構築物を挿入物の配列解析によって調べ、1つのクローンが、イントロン中のいくつかの軽微な違いを除いて、GenbankデータベースによるゲノムIgG4クローンと同一であることが判明した。これらの違いは、おそらくは多型またはGenbank配列における配列の誤りのいずれかである。このプラスミドをpTomG4と命名した。
【0137】
(表1)プライマー配列
【0138】
実施例13:マウス抗Betv1抗体および抗Feld1抗体の可変領域のクローニング
製造業者のプロトコールに従ってRNeasyキット(Qiagen、Westburg、オランダ、ルースデン)を用いて、0.3×105個(Betv1)または0.9×105個(Feld1)のマウスハイブリドーマ細胞(Betv1については:参考文献38によるクローン2H8、およびFeld1については:参考文献39によるクローン4F7)から全RNAを調製した。
【0139】
製造業者のプロトコールに従ってSMART RACE cDNA増幅キット(BD Biosciences Clontech、米国、カリフォルニア州、マウンテンビュー)を用いて、全RNA約100 ngからRNAの5'-RACE-相補的DNA(cDNA)を調製した。
【0140】
Betv1抗体およびFeld1抗体のVL領域およびVH領域をPCRによって増幅した。このために、製造業者の説明書に従ってPfuTurbo(登録商標) Hotstart DNAポリメラーゼ(Stratagene)を使用した。各反応混合物は、全量30μlのPCR反応緩衝液(ポリメラーゼと共に供給)中に、200μM混合dNTP(Roche Diagnostics)、リバースプライマー(VH領域用のRACEG1mm1およびVL領域用のRACEKmm1) 12 pmol、UPM-ミックス(UPM-ミックス:2μM ShortUPMH3オリゴヌクレオチドおよび0.4μM LongUPMH3オリゴヌクレオチド) 7.2pmol、上記の5'RACE cDNA鋳型0.6μl、およびPfuTurbo(登録商標) Hotstart DNAポリメラーゼ1.5単位を含んだ。
【0141】
35サイクルプログラム:95℃で2分の変性;95℃で30秒、55℃で30秒、および72℃で1.5分の35サイクル; 72℃で10分の最終伸長を用いて、TGradient Thermocycler 96(Whatman Biometra)でPCR反応を行った。反応産物を1% TAEアガロースゲルでのアガロースゲル電気泳動によって分離し、エチジウムブロマイドで染色した。正確な大きさのバンドをゲルから切り出し、QiaexIIゲル抽出キット(Qiagen)を用いてDNAをアガロースから単離した。
【0142】
200μM dATPおよびAmplitaq(Perkin Elmer) 2.5単位と共に72℃で10分間インキュベートすることによって、ゲルから単離したPCR断片にA尾部を付加し、minieluteカラム(Qiagen)を用いて精製した。製造業者のプロトコールに従ってpGEMT easyベクター系IIキット(Promega)を用いて、A尾部を付加したPCR断片をpGEMTeasyベクター(Promega)にクローニングした。ライゲーション混合物2μlをOneShot DH5αT1Rコンピテント大腸菌(Invitrogen)に形質転換し、LB/Amp/IPTG/Xgalプレートにプレーティングした。VH配列およびVL配列それぞれについて挿入物を含む4つの白色コロニーを拾い、挿入物の配列を決定した。Betv1のVHおよびVLの推定アミノ酸配列をSEQ ID NO:15および16に、ならびにFeld1の推定アミノ酸配列をSEQ ID NO:17および18に示す。
【0143】
実施例14:pConG1fBetV1:Betv1-IgG1の重鎖の産生用ベクターの構築
マウス抗BetV1抗体のVHコード領域を、プライマーVHexbetv1forおよびVHexbetv1revを用いてこの領域を含むプラスミド(実施例13)からPCRにより増幅し、pConG1f0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。VH断片をゲル精製し、pConG1f0.4にクローニングした。このために、PCR産物およびpConKappa0.4ベクターをHindIIIおよびApaIで消化し、精製した。VH断片およびpConG1f0.4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、正確な配列を確認した。このプラスミドをpConG1fBetv1と命名した。
【0144】
実施例15:pConKBetv1:Betv1の軽鎖の産生用ベクターの構築
VLコード領域マウス抗BetV1抗体を、プライマーVLexbetv1forおよびVLexbetv1revを用いてこの領域を含むプラスミド(実施例13)から増幅し、pConK0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。PCR産物およびpConKappa0.4ベクターをHindIIIおよびBsiWIで消化し、精製した。VL断片およびpConKappa0.4HindIII-BsiWI消化ベクターを連結し、コンピテントDH5α T1R大腸菌に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpConKBetv1と命名した。
【0145】
実施例16:pTomG4Betv1:Betv1-IgG4の重鎖の産生用ベクターの構築
Betv1-IgG4発現用ベクターを構築するため、BetV1のVH領域をpTomG4にクローニングした。このために、pTomG4およびpConG1fBetv1をHindIIIおよびApaIで消化し、関連する断片を単離した。Betv1 VH断片およびpTomG4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpTomG4Betv1と命名した。
【0146】
実施例17:pConG1fFeld1:Feld1-IgG1の重鎖の産生用ベクターの構築
マウス抗Feld1抗体のVHコード領域を、プライマーVHexfeld1forおよびVHexfeld1revを用いてこの領域を含むプラスミド(実施例13)からPCRにより増幅し、pConG1f0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。VH断片をゲル精製し、pConG1f0.4にクローニングした。このために、PCR産物およびpConKappa0.4ベクターをHindIIIおよびApaIで消化し、精製した。VH断片およびpConG1f0.4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、正確な配列を確認した。このプラスミドをpConG1fFeld1と命名した。
【0147】
実施例18:pConKFeld1:Feld1の軽鎖の産生用ベクターの構築
VLコード領域マウス抗Feld1抗体を、プライマーVLexfeld1forおよびVLexfeld1revを用いてこの領域を含むプラスミド(実施例13)から増幅し、pConK0.4へのクローニングに適した制限部位および理想的なコザック配列を導入した。PCR産物およびpConKappa0.4ベクターをHindIIIおよびBsiWIで消化し、精製した。VL断片およびpConKappa0.4HindIII-BsiWI消化ベクターを連結し、コンピテントDH5α T1R大腸菌に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpConKFeld1と命名した。
【0148】
実施例19:pTomG4Feld1:Feld1-IgG4の重鎖の産生用ベクターの構築
Feld1-IgG4発現用ベクターを構築するため、Feld1のVH領域をpTomG4にクローニングした。このために、pTomG4およびpConG1f Feld1をHindIIIおよびApaIで消化し、関連する断片を単離した。Feld1 VH断片およびpTomG4HindIII-ApaI消化ベクターを連結し、コンピテントDH5α-T1R細胞に形質転換した。正確な挿入物の大きさを含むクローンを選択し、配列を確認した。このプラスミドをpTomG4Feld1と命名した。
【0149】
実施例20:2F8-IgG4および7D8-IgG4の発現用の抗体発現ベクターの構築
HuMab 2F8(IgG1-EGFR)およびHuMab 7D8(IgG1-CD20)の発現用の発現ベクターを構築した。HuMab 2F8(WO 02/100348)およびHuMab 7D8(WO 04/035607)のVHおよびVLコード領域を、IgG1重鎖の産生用の発現ベクターpConG1f(Lonza Biologics)およびκ軽鎖の産生用のpConKappaにクローニングし、ベクターpConG1f2F8、pConG1f7D8、pConKappa2F8、およびpConKappa7D8を得た。pConG1f2F8およびpConG1f7D8のVH領域をHindIII/ApaI消化によってこれらのベクターから除去し、HidIII/ApaI消化したpTomG4ベクターに挿入し、それぞれpTomG42F8およびpTomG47D8を得た。
【0150】
実施例21:HEK-293F細胞における一過的発現によるBetv1-IgG1、Betv1-IgG4、Feld1-IgG1、およびFeld1-IgG4の産生
製造業者の説明書に従って293fectinを用いて、関連する重鎖ベクターおよび軽鎖ベクターをHEK-293F細胞に同時トランスフェクトすることによって、すべての構築物から抗体を産生させた。Betv1-IgG1については、pConG1Betv1およびpConKBetv1を同時発現させた。Betv1-IgG4については、pTomG4Betv1およびpConKBetv1を同時発現させた。Feld1-IgG1については、pConG1Feld1およびpConKFeld1を同時発現させた。Feld1-IgG4については、pTomG4Feld1およびpConKFeld1を同時発現させた。IgG1-EGFrについては、pConG1f2F8およびpConKappa2F8を同時発現させた。IgG4-EGFrについては、pTomG42F8およびpConKappa2F8を同時発現させた。IgG1-CD20については、pConG1f7D8およびpConKappa7D8を同時発現させた。IgG4-CD20については、pTomG47D8およびpConKappa7D8を同時発現させた。
【0151】
実施例22:IgG1抗体およびIgG4抗体の精製
IgG1抗体およびIgG4抗体を、プロテインAアフィニティークロマトグラフィーで精製した。細胞培養上清を0.20μMデッドエンドフィルターを通して濾過し、次に5 mlプロテインAカラム(rProtein A FF、GE Healthvcare)に負荷し、0.1 Mクエン酸-NaOH、pH3でIgGを溶出した。溶出液を直ちに2 M Tris-HCl、pH9で中和し、12.6 mMリン酸ナトリウム、140 mM NaCl、pH 7.4(B. Braun、オランダ、オッス)に対して一晩透析した。透析後、試料を0.20μMデッドエンドフィルターを通して滅菌濾過した。比濁法および280 nmの吸光度によって、精製IgGの濃度を決定した。SDS-PAGE、IEF、質量分析、および糖解析によって、精製タンパク質を解析した。
【0152】
実施例23:精製IgGのSDS-PAGE解析
精製後、Betv1およびFeld1のIgG1抗体およびIgG4抗体を非還元SDS-PAGEで解析した。使用したBis-Tris電気泳動法はLaemmli法(Laemmli 1970 Nature 227(5259): 680-5)の改良法であり、試料を中性pHで泳動した。SDS-PAGEゲルをクマシーで染色し、GeneGenius(Synoptics、英国、ケンブリッジ)を用いてデジタル画像化した。
【0153】
図1に示され得るように、Betv1およびFeld1のIgG1は、全長四量体(2本の重鎖および2本の軽鎖)のFeld1およびBetv1のIgG1分子を表す1本の主要なバンドを示した。Betv1およびFeld1のIgG4は、四量体IgG4分子を表す主要なバンドに加えて、相当量の半分子(すなわち、1本の重鎖および1本の軽鎖)を有することが示された。
【0154】
実施例24:マウスにおけるIgG4半分子の交換の評価
6〜8週齢のnu/nu Balb/cマウス5匹を用いて、IgG4半分子の交換を追跡した。マウスをCentral Laboratory Animal Facility(オランダ、ユトレヒト)の隔離施設に収容し、水および食物を自由に与えながらフィルタートップケージで飼育した。実験はすべて、ユトレヒト大学動物倫理委員会によって承認された。
【0155】
キメラ抗体を腹腔内投与した。投与の4.25時間、24時間、48時間、および72時間後に血液試料(75〜100μl)を採取した。血液をヘパリン含有バイアルに回収し、10,000 gで5分間遠心分離して、細胞から血漿を分離した。抗原特異的抗体および二重特異性抗体のレベルの決定用に、血漿を-20℃で保存した。
【0156】
本実験では、キメラIgG4半分子の交換(n=2)をIgG1半分子の交換(n=3)と比較した。200μl中600μg(各抗原特異的抗体300μg)/マウスの用量で、Bet v 1特異的抗体とFel d 1特異的抗体(IgG1またはIgG4)の混合物をマウスに投与した。
【0157】
Bet v 1結合抗体またはFel d 1結合抗体の血漿濃度を抗原結合試験で測定した。この目的のために、血漿試料を、125I標識Bet v 1または125I標識Fel d 1の存在下で、PBS/IAT(1μg/ml IVIg、0.3%ウシ血清アルブミン、0.1% Tween-20、0.05%(w/v) NaN3を補充したPBS) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体またはFel d 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGまたはFel d 1特異的IgGの濃度を算出した。二重特異性IgGの濃度は、異種架橋アッセイ法の2つの変法において測定した。第1のアッセイ法では、血漿を全量300μlのPBS-IAT中、セファロース結合Bet v 1(0.5 mg)と共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Fel d 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Bet v 1-Fel d 1)の濃度を、精製Fel d 1結合rIgGから得られたFel d 1結合試験の検量線を用いて算出した。第2のアッセイ法では、Fel d 1-Bet v 1架橋活性を、セファロース結合rFel d 1(0.5 mg)および125I標識Bet v 1を用いて同様の手順で測定した。二重特異性IgG(Fel d 1-Bet v 1)の濃度を、標準物質として精製Bet v 1特異的rIgGを用いて(Bet v 1結合試験における検量線と同じ検量線)算出した。
【0158】
図2において、様々な時点における二重特異性IgG(Fel d 1-Bet v 1)の濃度をBet v 1結合IgGの濃度に対してプロットする。二重特異性IgGは、IgG4を投与したマウスとは対照的に、IgG1混合物を投与したマウスでは認められなかった。24時間後に二重特異性IgG4の生成は最大であり、これは100%の交換に相当した。
【0159】
図3Aでは、二重特異性ヒトIgG4の形成を時間を追って追跡する。IgG4の混合物を注射したマウスの血漿中には二重特異性抗体が時間と共に出現したが、IgG1を注射したマウスでは出現せず、二重特異性反応性は1〜2日間のインキュベーション後に最大でほぼ50%に達した(注記:等量のIgG4-Betv1とIgG4-Feld1が交換された場合、半抗体の無作為でかつ完全な交換後に、IgG4-Betv1半抗体の最大50%が二重特異性画分に取り込まれる)。等量のIgG4-Betv1とIgG4-Feld1間の無作為的なFabアーム交換は、IgG4分子の約半分が二重特異性を獲得することと一致する。対照として、非関連抗原に対する20倍過剰のさらなるIgG4(抗EGFr抗体2F8から作製したIgG4)を、IgG4-Betv1およびIgG4-Feld1と共にマウスに注射した。過剰な非関連IgG4は、Betv1-Feld1二重特異性IgG4の生成と競合した。
【0160】
別の実験(図3B)において、同じマウス血漿試料を、放射標識可溶性Fel d 1をセファロース固定化Fel d 1に架橋する能力について試験した。単一特異性架橋活性は、IgG4の同等の混合物を投与したマウスでは減少したが、IgG1の同等の混合物を投与したマウスでは減少しないことが判明し、単一特異性架橋活性の喪失が示された。約1日後に、約50%の最大減少に達した。さらなる過剰な非関連IgG4を投与したマウスでは、単一特異性架橋活性は同様の動態学でほぼ完全に消失した。
【0161】
サイズ排除クロマトグラフィーを行って、IgG4を投与したマウスにおいて認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した(図4を参照されたい)。この目的のために、血漿試料(t=24時間の時点で採取)をSuperdex200カラムで分画し、その後その画分中でFel d 1結合IgGおよびBet v 1-Fel d 1架橋IgGを測定した。Fel d 1結合抗体は、保持容量約12.9 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Bet v 1-Fel d 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された。rIgG1含有血漿中には、分画前にBet v 1-Fel d 1架橋活性は存在しなかった。同様に溶出画分においても、異種架橋活性は測定されなかった(データは示さず)。
【0162】
実施例25:全血(成分)における交換活性の評価
キメラ抗体を混合し、続いて全血、血液細胞、血漿、または血清と共にインキュベートして、全血(成分)の交換活性を調べた。
【0163】
本実験では、IgG4半分子の交換を、2名の健常な血液ドナーであるAおよびB由来の全血において評価し、このドナーのIgG4の内因性血漿レベルは比濁法によって決定した(それぞれ、346μg/mlおよび554μg/mlであった)。最終濃度40μg/mlのTFPI(Chiron Corporation、カリフォルニア州、エメリービルによる組織因子経路阻害剤)を補充したバキュテイナ中に、全血を採取した。全血の遠心分離により、血液細胞および血漿を取得した。細胞画分をOptimem(Invitrogen、オランダ、ブレダ)で3回洗浄し、続いてOptimemに再懸濁した。血清は、全血をガラスバキュテイナ中で凝固活性化剤と共に37℃で30分間インキュベートし、その後凝固した血液を遠心沈殿させることによって取得した。IgG4半分子の交換を評価し、IgG1半分子の交換と比較した。対照として、血液試料をキメラ抗体の非存在下で同様にインキュベートした。以下の抗体混合物をPBS中で調製した:
1. Bet v 1特異的IgG4(10μg)およびFel d 1特異的IgG4(10μg)
2. Bet v 1特異的IgG1(10μg)およびFel d 1特異的IgG1(10μg)。
【0164】
これらの抗体混合物を、水平軌道振盪機(125 rpm)上で、全量100μl中で血液、血液細胞、血漿、または血清と共に37℃でインキュベートした(各抗体の最終濃度は0.1μg/mlであった)。全血および血液細胞を伴うインキュベーション混合物中の最終ヘマトクリットは、約40%であった。24時間後、インキュベーション混合物をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料10μlをPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS) 500μl中に採取した。試料は必要に応じて4℃で保存した。
【0165】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合組換えFel d 1 0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0166】
図5において、二重特異性活性を、異種架橋アッセイ法で決定された、結合した125I標識Bet v 1の割合(%)として表す。二重特異性活性はIgG4半分子の交換の尺度であり、これは主に全血および全血の細胞画分において認められた(図5A)。細胞画分における二重特異性レベルは、全血よりもさらに高かった。これは、細胞画分内に、添加したキメラIgG4抗体と同様に交換され得る内因性IgG4がもはや存在しないという事実によって説明される可能性が最も高い。いくらかの二重特異性活性は血漿および血清においても認められたが、この活性は全血において認められた活性よりもはるかに低く、またOptimem中でIgG4混合物をインキュベートすることによって得られた1.7%というバックグラウンドレベルよりもわずかに高いにすぎなかった。二重特異性活性は、IgG1を含むインキュベーションのいずれにおいても認められなかった(図5B)。同様にキメラ抗体を含まない対照インキュベーションにおいても、二重特異性活性は認められなかった(図5C)。サイズ排除クロマトグラフィーを行って、IgG4混合物において認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した。この目的のために、試料(t=24時間の時点で採取)をSuperdex200カラムで分画し、その後その画分中でFel d 1結合IgGおよびBet v 1-Fel d 1架橋IgGを測定した。Fel d 1結合抗体は、保持容量約12.9 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Bet v 1-Fel d 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された(データは示さず)。
【0167】
実施例26:血液細胞媒介性IgG4交換活性の評価
キメラ抗体を混合し、続いて3種の異なる型のヒト血液細胞(すなわち、単核細胞(MNC)、赤血球、および血小板)と共にインキュベートして、IgG4交換活性を調べた。
【0168】
匿名ドナー由来の全血をヘパリン含有バキュテイナ中に採取し、続いてPercoll(Pharmacia Fine Chemicals、スウェーデン、ウプサラ)中で遠心分離して、MNCを単離した。使用前に、単離したMNCをOptimem無血清培地(Invitrogen、オランダ、ブレダ)に再懸濁した。新たに精製された赤血球および血小板(SanquinのBlood Cell Research Departmentにより提供)は、2名の異なる匿名ドナーから得られた。これらの細胞も、3回洗浄した後にOptimemに再懸濁した。加えて、血小板には10 mMグルコースを補充した。
【0169】
IgG4半分子の交換を評価し、IgG1半分子の交換と比較した。以下の抗体混合物をPBS中で調製した:
‐Bet v 1特異的IgG4(10μg)およびFel d 1特異的IgG4(10μg)
‐Bet v 1特異的IgG1(10μg)およびFel d 1特異的IgG1(10μg)。
【0170】
これらの抗体混合物を、水平軌道振盪機(125 rpm)上で、全量100μl中で1.8×104個MNC、4.0×108個赤血球、または3.5×104個血小板と共に37℃でインキュベートした(各抗体の最終濃度は0.1μg/mlであった)。48時間後、インキュベーション混合物をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料10μlをPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS) 500μl中に採取した。試料は必要に応じて4℃で保存した。
【0171】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合組換えFel d 1 0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0172】
図6において、二重特異性活性を、異種架橋アッセイ法で決定された、結合した125I標識Bet v 1の割合(%)として表す。3種の細胞型はすべて、二重特異性活性を誘導することができた。いくらかの二重特異性活性がOptimem無血清培地においても認められたが、この活性は、血液細胞の存在下で認められた活性よりもはるかに低かった。試験した細胞のいずれも、IgG1半分子を交換することはできなかった。
【0173】
実施例27:ヒト細胞株およびマウス細胞株によるIgG4交換の評価
キメラIgG4抗体を混合し、続いて3種の異なる細胞株(すなわち、ヒト胚腎(HEK)細胞、マウスB細胞、またはハイブリドーマ)と共にインキュベートして、IgG4交換活性を調べた。
【0174】
マウスB細胞の供給源として、細胞株J558(SanquinのAntigen Presentation Research Groupにより提供)を選択した。抗C1エステラーゼ阻害因子を産生するハイブリドーマは、SanquinのAutoimmune Research Groupより入手した。懸濁HEK(293F)細胞は、Invitrogen、オランダ、ブレダより入手した。細胞はすべてPBSで3回洗浄した後、PBSに再懸濁した。
【0175】
Bet v 1特異的IgG4(2μg)およびFel d 1特異的IgG4(2μg)からなるIgG4抗体混合物を上記の細胞と共にインキュベートすることによって、IgG4半分子の交換を評価した。抗体混合物を、水平軌道振盪機(125 rpm)上で、全量50μl中で24×105個HEK細胞、25×105個マウスB細胞、または21×105個ハイブリドーマと共に37℃でインキュベートした(各抗体の最終濃度は80μg/mlであった)。0時間および24時間後、インキュベーション混合物をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料をPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に採取した。試料は必要に応じて4℃で保存した。
【0176】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料希釈物を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合組換えFel d 1 0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0177】
図7において、二重特異性活性を、異種架橋アッセイ法で決定された、結合した125I標識Bet v 1の割合(%)として表す。3種の細胞型はすべて、IgG4半分子を交換することができた。
【0178】
実施例28:赤血球によるIgG4半分子の交換の評価
キメラ抗体を混合し、続いてヒト赤血球と共にインキュベートして、IgG4半分子の交換を調べた。赤血球はドナー1名から精製し、SAGM(生理食塩水アデニングルコースマンニトール)緩衝液中で4℃で保存した。使用前に、細胞をPBSで3回洗浄した。
【0179】
本実験では、IgG4半分子の交換をIgG1半分子の交換と比較した。また、過剰な非関連IgG4の存在下におけるIgG4の交換も評価した。以下の抗体混合物をPBS中で調製した:
‐Bet v 1特異的IgG4(4μg)およびFel d 1特異的IgG4(4μg)
‐Bet v 1特異的IgG1(4μg)およびFel d 1特異的IgG1(4μg)
‐Bet v 1特異的IgG4(4μg)、Fel d 1特異的IgG4(4μg)、および抗原Xに特異的な非関連IgG4(80μg)。
【0180】
これらの混合物を全量100μlの、0.05%(w/v) NaN3を補充したPBS中で赤血球と共にインキュベートし(最終ヘマトクリットは約40%であった)、続いて水平軌道振盪機(125 rpm)上で37℃でインキュベートした。表示の時点で、赤血球をエッペンドルフ遠心機において2800 rpmで1分間遠心分離し、その後試料10μlをPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS) 500μl中に採取した。試料は、二重特異性活性、二価性、および抗原結合を測定する前に4℃で保存した。対照として、赤血球を含まないPBS中で同じ混合物を同様にインキュベートした。
【0181】
Bet v 1結合抗体のレベルを抗原結合試験で測定した。この目的のために、試料を、125I標識Bet v 1の存在下で、PBS-IAT(1μg/ml IVIgを補充したPBS-AT) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGの濃度を算出した。Fel d 1特異的抗体およびBet v 1特異的抗体を用いる実験における二重特異性活性は、Feld1-Betv1架橋アッセイ法で測定した。このアッセイ法では、IgG含有試料を全量300μlのPBS-AT中、セファロース結合ネコ抽出物(0.5 mg)と共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Feld1-Betv1)の濃度を、標準物質として精製IgG1-Betv1を用いて(Prot Gセファロースを用いるBet v 1結合試験で得られる)算出した。
【0182】
図8に、赤血球媒介性の交換から得られたデータを示す。赤血球の存在下でIgG1半分子の交換が認められなかったのに対して、IgG4半分子のほぼ最大の交換が72時間後に認められた(パネルA)(注記:等量のIgG4-Betv1とIgG4-Feld1が交換された場合、半抗体の無作為でかつ完全な交換後に、IgG4-Betv1半抗体の最大50%が二重特異性画分に取り込まれる)。過剰な非関連IgG4の存在下では、IgG4半分子の交換はほとんど測定されず、これはBet v 1特異的IgG4およびFel d 1特異的IgG4と非関連IgG4との予測される交換と一致する。サイズ排除クロマトグラフィーを行って、IgG4混合物において認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した。この目的のために、試料(t=72時間の時点で採取)をSuperdex200カラムで分画し、その後その画分中でFel d 1結合IgGおよびBet v 1-Fel d 1架橋IgGを測定した。Fel d 1結合抗体は、保持容量約12.9 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Bet v 1-Fel d 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された(データは示さず)。
【0183】
理論的には、IgG4半分子の交換は二価性の減少も伴う。これを試験するために、インキュベーション混合物中の二価性を測定した。IgG1混合物ではFel d 1二価性の減少は認められなかったのに対して、IgG4混合物では約50%の減少が認められた。この減少は、1対1比で混合した2種類の異なるIgG4分子の最大交換と一致する。予想通り、過剰な非関連IgG4を添加したIgG4混合物における二価性の減少はより高く(約80%)、これは過剰な非関連IgG4半分子の存在下で2つの相同的な半分子(Bet v 1またはFel d1特異的)の再ハイブリダイゼーションの確率が低くなるためである。72時間のインキュベーション後に抗原結合はわずかに(約10%)減少するにすぎなかったため(データは示さず)、二価性の強い減少は、インキュベーション中に抗原結合が失われたためではなかった。
【0184】
PBS(0.05%(w/v) NaN3を補充)中でのIgGの交換も評価して、IgG4半分子が自発的に交換され得るかどうかを調べた。本実験の設定は、赤血球を添加しなかったこと以外は、赤血球の存在下での交換と同様であった。図9Aに示されるように、PBS中での37℃でのインキュベーション中に、IgG1半分子またはIgG4半分子の自発的な交換は認められなかった。しかしながら、IgG4混合物においていくらかのバックグラウンドが認められ、これは赤血球とのインキュベーション中にも存在した。PBS中でのインキュベーション中に、二価性の減少は認められなかった(図9B)。
【0185】
実施例29:赤血球溶解物によるIgG4交換の評価
キメラIgG4抗体を混合し、続いて赤血球溶解物の漸増希釈物と共にインキュベートした。赤血球は健常ドナーから単離し、ヘマトクリット60.7%の状態でSAGM(生理食塩水アデニングルコースマンニトール)緩衝液中で4℃で保存した。溶解物を得るために、細胞をPBS-アジド(0.05%(w/v) NaN3を補充したPBS)で3回洗浄し、保存緩衝液の量の2倍量の水に再懸濁した。結果として、非希釈赤血球溶解物はヘマトクリット30%に相当した。
【0186】
Bet v 1特異的IgG4(1μg)およびFel d 1特異的IgG4(1μg)からなるIgG4抗体混合物を、新たに調製した溶解物50μl(全量100μlとなるようにPBS/アジドを補充)と共に37℃でインキュベートすることによって、IgG4半分子の交換を評価した。各抗体の最終濃度は10μg/mlであった。表示の時点で、インキュベーション混合物からPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に試料を採取して、二重特異性活性を測定した。試料は必要に応じて4℃で保存した。
【0187】
二重特異性活性(すなわち、Bet v 1-Fel d 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料希釈物を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合カバノキ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Fel d 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Bet v 1-Fel d 1)の濃度を、精製Fel d 1結合rIgGから得られたFel d 1結合試験の検量線を用いて算出した。
【0188】
図10において、時間に伴う二重特異性活性の生成を、異種架橋アッセイ法で決定された、結合した125I標識Fel d 1の割合(%)として示す。これらのデータから、赤血球の溶解物が交換活性を含むことが明白である。最も高い交換率は非希釈溶解物で認められ、希釈が高いほど交換率は低下した。実際に、PBS中での対照インキュベーションでは二重特異性活性は認められなかった。
【0189】
サイズ排除クロマトグラフィーを行って、赤血球溶解物によって誘導された二重特異性活性がIgG凝集の結果であるという可能性を排除した(図11)。この目的のために、Bet v 1結合IgG4 10μg、Fel d 1結合IgG4 10μg、および赤血球溶解物50μlからなるインキュベーション混合物を調製し、最終量100μlとなるようにPBS/アジドを補充した。この混合物を37℃で24時間インキュベートし、その後70μlをSuperdex200カラムで分画した。その画分中で、Bet v 1結合IgGおよびFel d 1-Bet v 1架橋IgGを測定した。Bet v 1結合抗体のレベルは、抗原結合試験で測定した。試料を、125I標識Bet v 1の存在下で、PBS-IAT(1μg/ml IVIg、0.3%ウシ血清アルブミン、0.1% Tween-20、0.05%(w/v) NaN3を補充したPBS) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGの濃度を算出した。二重特異性IgGの濃度(すなわち、Fel d 1-Bet v 1架橋活性)は、異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT中、Fel d 1抗原が存在するセファロース結合ネコ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Fel d 1-Bet v 1)の濃度を、精製Bet v 1結合rIgGから得られた、Bet v 1結合試験で使用したのと同じ検量線を用いて算出した。
【0190】
Bet v 1結合抗体は、保持容量約12.6 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した(図11)。異種Fel d 1-Bet v 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された。
【0191】
実施例30:透析赤血球溶解物におけるIgG4交換活性の評価
赤血球を健常ドナーから単離し、ヘマトクリット60.7%の状態でSAGM(生理食塩水アデニングルコースマンニトール)緩衝液中で4℃で保存した。溶解物を得るために、細胞をPBS-アジド(0.05%(w/v) NaN3を補充したPBS)で3回洗浄し、保存緩衝液の量の2倍量の水に再懸濁した。したがって、非希釈赤血球溶解物はヘマトクリット30%に相当した。Pierceによる透析膜カセット(3.5 kDカットオフ)を用いて、溶解物の一部をPBS-アジドに対して透析した。非透析溶解物をAmiconフィルター(3.5 kDカットオフ)中で遠心分離することにより、限外濾過液を得た。
【0192】
IgG4抗体混合物(Bet v 1特異的IgG4(0.5μg)およびFel d 1特異的IgG4(0.5μg))を新たに調製した赤血球溶解物(25μl)または透析溶解物(25μl)と共に37℃でインキュベートすることによって、IgG4半分子の交換を評価した。各インキュベーションの全量は50μlであり、各抗体の最終濃度は10μg/mlであった。以下の補充物を使用した:Sigmaによる還元型グルタチオン(GSH)、グルコース-6-リン酸(G-6-P)、およびNADPH(いずれもRocheによる)。これらの化合物は、使用前に水に溶解した。24時間のインキュベーション後、インキュベーション混合物からPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に試料を採取して、二重特異性活性を測定した。試料は必要に応じて4℃で保存した。
【0193】
二重特異性活性(すなわち、Fel d 1-Bet v 1架橋活性)を異種架橋アッセイ法で測定した。このアッセイ法では、試料希釈物を全量300μlのPBS-IAT(1μg/ml IVIgを補充したPBS-AT)中、セファロース結合ネコ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。
【0194】
交換レベルを、新たに調製した溶解物によって生じた二重特異性活性と比較した(表2)。
【0195】
(表2)透析した赤血球溶解物において二重特異性活性を回復する因子の概要
透析した赤血球溶解物の交換活性を、新たに調製した溶解物と比較した。透析した溶解物に限外濾過液5□lを補充した。G-6-P、NADPH、およびGSHの濃度は、それぞれ5 mM、0.1 mM、および0.5 mMであった。
【0196】
これらのデータから、赤血球溶解物の活性が透析後に失われたことが明白である。限外濾過液を添加することによって、交換の大部分が回復された。この結果から、透析中に成分(<3.5 kD)が失われ、これが交換反応に必須であることが示唆された。IgG4半分子の交換にジスルフィド架橋の還元および酸化が必要であることから、このような成分は酸化還元サイクルに関与している可能性が高い。したがって、酸化還元サイクルの3つの「補助因子」(G-6-P、NADPH、およびGSH)を透析した溶解物に添加して、これらの化合物が交換活性を回復し得るかどうかを調べた。G-6-P、NADPH、およびGSHを共に補充した場合に、交換活性は回復され得た。個々の因子の存在下で透析溶解物をインキュベートすることにより、交換活性は、G-6-PでもNADPHでもなく、GSHによって回復されることが明らかになった。
【0197】
実施例31:還元型グルタチオンによるIgG4半分子の交換の評価
キメラ抗体を混合し、続いて還元型グルタチオン(GSH)と共にインキュベートして、IgG4半分子の交換を調べた。GSH(Sigma-Aldrich、ミズーリ州、セントルイス)は、使用前に水に溶解した。
【0198】
本実験では、Bet v 1特異的IgG4(1μg)およびFel d 1特異的IgG4(1μg)からなるIgG4抗体混合物を、GSHを含むPBS/アジド中で37℃でインキュベートすることによって、IgG4半分子の交換を評価した。全インキュベーション量は100μlであり、各抗体の最終濃度は10μg/mlであった。表示の時点で、インキュベーション混合物からPBS-AT(0.3%ウシ血清アルブミン、0.1% Tween-20、および0.05%(w/v) NaN3を補充したPBS)中に試料を採取した。抗原結合および二重特異性活性の測定用に、試料を4℃で保存した。
【0199】
Bet v 1結合抗体のレベルを抗原結合試験で測定した。試料を、125I標識Bet v 1の存在下で、PBS-IAT(1μg/ml IVIgを補充したPBS-AT) 750μl中のプロテインGセファロース(Amersham Biosciences、スウェーデン、ウプサラ) 0.75 mgと共に24時間インキュベートした。次に、セファロースをPBS-T(0.1% Tween-20および0.05%(w/v) NaN3を補充したPBS)で洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。精製Bet v 1特異的抗体を標準物質として用いて(比濁計により決定して0〜200 ng/試験の範囲)、Bet v 1特異的IgGの濃度を算出した。二重特異性IgGの濃度(すなわち、Fel d 1-Bet v 1架橋活性)は、異種架橋アッセイ法で測定した。このアッセイ法では、試料を全量300μlのPBS-IAT中、Fel d 1抗原が存在するセファロース結合ネコ抽出物0.5 mgと共に24時間インキュベートした。続いてセファロースをPBS-Tで洗浄し、125I標識Bet v 1と共に24時間インキュベートし、その後セファロースをPBS-Tで洗浄し、添加した放射能の量に対する結合した放射能の量を測定した。二重特異性IgG(Fel d 1-Bet v 1)の濃度を、精製Bet v 1結合IgGから得られた、Bet v 1結合試験で使用したのと同じ検量線を用いて算出した。
【0200】
図12に、IgG4半分子のGSH媒介性の交換の時間経過を示す。これらのデータから、IgG4半分子がGSHの存在下で交換されることが明らかである。本実験では、最適な交換は0.1 mM GSHと1 mM GSHとの間で認められ、0.5 mM GSHを用いて24時間後に最も高い交換(約90%)に達した。
【0201】
サイズ排除クロマトグラフィーを行って、IgG4のGSH媒介性の交換後に認められた二重特異性活性がIgG凝集の結果であるという可能性を排除した(図13)。この目的のために、Bet v 1結合IgG4およびFel d 1結合IgG4(各抗体10μg)の混合物を、PBS/アジド中で0.5 mM GSHと共にインキュベートした。この混合物(最終量100μl)を37℃で24時間インキュベートし、その後70μlをSuperdex200カラムで分画した。その画分中で、Bet v 1結合IgGおよびFel d 1-Bet v 1架橋IgGを測定した。Bet v 1結合抗体は保持容量約12.6 mlの1つのピークとして溶出され、これは単量体IgGの保持容量に相当した。異種Fel d 1-Bet v 1架橋活性も同じ画分中に検出され、二重特異性活性が単量体IgGに付随していることが示された。交換が4℃よりも37℃でより効率的に起こったことから、GSHの存在下での二重特異性IgG4分子の生成は、温度依存的であることが判明した(図14)。
【0202】
実施例32. 他の作用物質の存在下における二重特異性IgGの生成
IgG1-Betv1およびIgG1-Feld1またはIgG4-Betv1およびIgG4-Feld1を抗体の最終濃度10μg/mlで混合し、全量50μl中で還元剤と共にインキュベートした。GSHとは別に、以下の作用物質を試験した(インキュベーション混合物中の最終濃度):L-システインはSigmaによる(100μM)、ジチオスレイトール(DTT)はBioradによる(50μM)、β-メルカプト-エタノール(BME)はBioradによる(100μM)、および酸化型グルタチオン(GSSG、一連の作用物質のうち、他のすべてが還元性であるのに対して、この作用物質は還元性ではないことに留意されたい)はSigmaによる(100μM)。混合物を37℃で24時間インキュベートし、試料をPBS/AT中に採取し、そこで(二重)特異性IgG濃度を測定した。図15から、精製IgG4-Betv1およびIgG4-Feld1の混合物にGSHまたは他の還元剤(GSSG以外)を添加することは、Fabアーム交換および二重特異性IgG4の生成を誘導するのに十分であることが示される。対照的に、対照IgG1混合物では二重特異性反応性は誘導されなかった。
【0203】
実施例33. GSHを用いる完全ヒトIgG4抗体の交換
IgG1-CD20、IgG4-CD20、IgG1-EGFr、およびIgG4-EGFrを混合し、全量1 ml中でGSHと共にインキュベートした。各抗体の最終濃度は50μg/mlであった;GSHの最終濃度は0.5 mMであった。混合物を37℃で24時間インキュベートし、試料をPBS-AT中に採取し、そこで(二重)特異性IgG濃度を測定した。
【0204】
サンドイッチELISA法を用いて二重特異性活性を決定した。このアッセイのために、ELISAプレート(Greiner bio-one、ドイツ、フリッケンハウゼン)を、PBS中のEGFRの組換え細胞外ドメイン1μg/ml(100μl/ウェル)で4℃でコーティングした。プレートをPBS/0.05% Tween 20(PBT)で3回洗浄した。試料をPBT/0.2% BSA(PBTB)で希釈し、ELISAプレートに移した(100μl/ウェル)。プレート振盪機(300 rpm)上で室温(RT)で90分間インキュベートした後、試料を廃棄し、プレートをPBTで3回洗浄した。次に、PBTB中の2μg/mlのマウス抗イディオタイプモノクローナル抗体2F2 SAB1.1(抗CD20抗体7D8に対する;Genmab) 100μlを添加し、プレート振盪機(300 rpm)にてRTで90分間インキュベートした。抗イディオタイプ抗体を廃棄し、プレートをPBTで3回洗浄し、続いてPBTBで1000×希釈したHRP結合ヤギ抗マウスIgG(Jackson ImmunoResearch Laboratories、米国、ペンシルバニア州、ウェストグローブ) 100μl/ウェルを添加し、プレート振盪機(300 rpm)にてRTで90分間インキュベートした。検出抗体を廃棄し、プレートをPBTで3回洗浄した。50 mg ABTS錠剤(Roche Diagnostics GmbH、ドイツ、マンハイム)をABTS緩衝液(Roche)に溶解し、ELISAプレートに添加した(100μl/ウェル)。ELISAプレートをアルミホイルで覆って、プレート振盪機(300 rpm)上でRTで30分間(または必要に応じてそれよりも長く)インキュベートし、シュウ酸(Riedel de Haen Seelze、ドイツ) 100μl/ウェルで反応を停止させた。ELISAプレートをRTで10分間置いた後、ELISAプレートリーダーで405 nmの吸光度を読み取った。
【0205】
図16Aは、GSHの存在下でIgG4-EGFrおよびIgG4-CD20の混合物をインキュベートした場合には、二重特異性抗EGFR/CD20抗体が時間と共に形成されたが、GSHの非存在下では形成されなかったことを示す。Fabアーム交換は、IgG1抗体の混合物では、GSHの存在下でも非存在下でも起こらなかった。
【0206】
IgG4半分子のGSH媒介性の交換のダイナミックレンジを調べるため、GSHの完全濃度曲線(0.5〜1,000μM)を用いて交換を解析した。IgG4-CD20およびIgG4-EGFrを混合し、全量1 ml中でGSHと共にインキュベートした。各抗体の最終濃度は50μg/mlであった;GSHの最終濃度は図16Bに示す通りであった。混合物を37℃で24時間インキュベートし、試料をPBS-AT中に採取し、そこで(二重)特異性IgG濃度を測定した。
【0207】
図16Bは、IgG4半分子の交換の明白なGSH用量依存性を示す。どのように反応成分がGSH媒介性IgG4半分子の交換に影響するのかを調べるために、PBS、ならびに血清およびタンパク質を含まない既知組成培地(FreeStyle 293発現培地、GIBCO/Invitrogen Corporation)中で交換を試験した。この組織培地中では、GSH媒介性の交換がより低いGSH濃度で起こることが判明した(図16C)。5 mM GSHと共にインキュベートすると、0.5 mMの場合よりも低い交換が起こったことから、GSH媒介性IgG4半分子の交換には最適条件が存在することも判明した(図16D)。
【0208】
IgG4-EGFrおよびIgG4-CD20の混合物をGSHの非存在下または存在下で24時間インキュベートし、質量分析(ESI-TOF MS)によって評価した。各抗体200μg/mlを含む試料50μlを、N-グリコシダーゼF(Roche Diagnostics NL BV、オランダ、アルメレ) 1μlで一晩かけて脱グリコシル化した。BEH C8、1.7μm、2.1×50 mmカラムを用いてAcquity UPLC(商標)(Waters、米国、ミルフォード)において、試料を60℃で脱塩した。5μlを注入し、5%〜95%溶離液Bの勾配で溶出した。溶離液AはMilliQ水(Millipore Synthesis A10装置)であり、溶離液BはLC-MS等級アセトニトリル(Biosolve、オランダ、ファルケンスワールト)であった。いずれの溶離液も、有機変性剤として0.05%ギ酸(Fluka Riedel-de Han、ドイツ、ブーフ)を含んだ。micrOTOF(商標)質量分析計(Bruker、ドイツ、ブレーメン)を陽イオンモードで操作して、飛行時間型エレクトロスプレーイオン化質量スペクトルをオンラインで記録した。各解析において、500〜5000 m/zスケールをES調整混合物(Agilent Technologies、米国、サンタクララ)で内部較正した。DataAnalysis(商標)ソフトウェアv.3.3(Bruker)によって提供される最大エントロピーアルゴリズムを用いることにより、質量スペクトルをデコンボリューション処理した。
【0209】
図16Eは、GSHの非存在下で、IgG4-CD20(145.5 kD)およびIgG4-EGFR(145.9 kD)の分子量が変化しないままであったことを示す。しかしながらGSHの存在下では(図16F)、Fabアーム交換分子に対応する質量を有する新たなピークが出現した(145.7 kD)。新たな質量は、二重特異性抗EGFR/CD20抗体の予測質量に相当した。さらに、MSスペクトルのピーク高から、二重特異性抗体が混合物中の全抗体量の50%を示すことが推定され得、無作為的交換が24時間以内に平衡に達したことが示された。
【0210】
実施例34. 組換えヒトIgG4抗体のFabアーム交換に関与するアカゲザル(およびその他の種)由来のポリクローナル免疫グロブリン
2種類の組換えヒトIgG4抗体(上記と同様のIgG4-CD20およびIgG4-EGFr)の混合物を、アカゲザル(6×)、チンパンジー(2×)、カニクイザル、ヒヒ、ウマ、およびブタ由来の精製免疫グロブリンまたはヒトIVIgの存在下または非存在下で、GSHと共に37℃で24時間インキュベートした。Fabアーム交換を介した二重特異性抗体の形成を、上記のサンドイッチELISA法で測定した。加えて、ヤギ、ウサギ、およびヒツジ免疫グロブリンもこのアッセイ法で試験した。
【0211】
図17Aは、アカゲザルポリクローナル免疫グロブリンが、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換を阻害する能力において、ヒトポリクローナル免疫グロブリン(IVIg)に匹敵することを示す。これは、アカゲザル免疫グロブリンの成分がFabアーム交換に関与することを意味する。アカゲザル免疫グロブリン、おそらくはアカゲザルIgG4は、組換えヒトIgG4とFabアームを交換することができる。
【0212】
図17Bは、数頭のその他のアカゲザルに由来するポリクローナル免疫グロブリンが、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換を異なる効力で阻害することを示す。これは、Fabアーム交換に関与するアカゲザル免疫グロブリンの成分が異なる濃度で存在すること、またはすべてのアカゲザル中にその成分が存在するわけではないことを意味する。
【0213】
図17Cは、数頭のその他のサル種(ヒヒ、チンパンジー、カニクイザル)由来のポリクローナル免疫グロブリン、ならびにウマおよびブタ由来の免疫グロブリンが、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換を異なる効力で阻害することを示す。これは、Fabアーム交換に関与する成分が、これらの種において異なる濃度で存在することを意味する。ヤギ、ウサギ、およびヒツジ免疫グロブリンは、還元型グルタチオンの存在下におけるインビトロでの組換え抗体のFabアーム交換に影響を及ぼさなかった(データは示さず)。
【0214】
実施例35. ヒンジ領域変異体またはCH3ドメイン変異体の半分子の交換
3種類のIgG1変異体を作製した:IgG4コアヒンジを有するIgG1(IgG1-CPSC)、および2種類のCH3ドメイン交換変異体(IgG1-CH3(IgG4)およびIgG1-CPSC-CH3(IgG4))。
【0215】
部位特異的突然変異誘発を用いて、pEE-G1-wt a Bet v 1を鋳型として使用して、IgG1のヒンジ内にP228S変異を導入した(228は抗体アミノ酸残基のEU番号を参照する。同位置はKabat番号付けでは241番を有し、SEQ ID NO:19では111番(CPPCコアヒンジ配列における3位)を有する)。Vector NTI Advance 10により、突然変異誘発プライマー、フォワードおよびリバースを設計した:
P228S Mutプライマー-F:SEQ ID NO:22
P228S Mutプライマー-R:SEQ ID NO:23。
【0216】
Quickchange部位特異的突然変異誘発キット(Stratagene)を用いて、pEE-G1-CPSC変異体を作製した。ポリメラーゼ連鎖反応(PCR)混合物は、pEE-G1 a Betv1 DNA鋳型5μl(約35 ng)、突然変異誘発プライマー-フォワード1.5μl(約150 ng)、突然変異誘発プライマー-リバース1.5μl(約150 ng)、dNTP混合物1μl、反応緩衝液(10×) 5μl、H2O 36μl、および最後にPfu Turbo DNAポリメラーゼ1μlからなった。次に混合物をPCR:95℃で30秒、95℃で30秒(変性)、55℃で1分(アニーリング)、および68℃で17分(伸長)に供した。このサイクルを20回繰り返した。
【0217】
DNAの消化およびライゲーションを用いて、CH3ドメイン交換変異体構築物IgG1-CH3(IgG4)およびIgG1-CPSC-CH3(IgG4)を作製した。CH3ドメインおよびCH3ドメインを含まないベクターを得るための消化反応は、以下の通りであった:DNA(pEE-G1-betv1、pEE-G1-CPSC、およびpEE-G4-betv1) 約1500 ng、BSA 2μl、Neb3緩衝液2μl、SalI 1μl、およびH20を20μl量になるまで添加した。37℃で30分間インキュベートした。DNAを精製し、H20 30μlで溶出してから、SanDI 1μlおよびユニバーサル緩衝液3μlを添加し、37℃で30分間インキュベートした。断片を、エチジウムブロマイドを含む1%アガロースゲルでのゲル電気泳動に供した。紫外線下で断片をゲルから切り出し、DNA精製キット(Amersham)を用いて溶解した。pEE-G4-wt SalI/SanDI(IgG4 CH3ドメインを含む)断片を、以下の手順を用いてpEE-G1-wtおよびpEE-G1-CPSCに連結した:全量20μl中の鋳型DNA(SalI/SanDI消化したpEE-G1-wtおよびpEE-G1-CPSC) 1μl、SalI/SanDI挿入物5μl、Ligate-it緩衝液4μl、H20 9μl、およびリガーゼ1μl。5分後にライゲーションを停止させた。
【0218】
DNAの消化(ApaIおよびHindIIIを使用)およびライゲーションを用いて、上記と同様の手順に従って、bet v 1変異体抗体のVHドメインをpEE-G4-a-feld1 wtのVHドメインと置換した。
【0219】
また、1つのIgG4変異体を作製した:IgG4-S228Pnew。この変異体では、228位のセリン(SEQ ID NO:19における111位)をプロリン(IgG1コアヒンジ)に置換することによって、ヒンジが安定化される。製造業者の説明書に従ってQuickChange II XL部位特異的突然変異誘発キット(Stratagene、オランダ、アムステルダム)を用いて、部位特異的突然変異誘発を行った。この方法は、突然変異誘発に成功したものをスクリーニングするために、サイレントな余分なXmaI部位の導入を含んだ。簡潔に説明すると、10×反応緩衝液5μl、オリゴヌクレオチドS228Pfcorrect(100 pmol/μl) 1μl、オリゴヌクレオチドS228Prcorrect(100 pmol/μl) 1μl、dNTP混合物1μl、Quicksolution 3μl、プラスミドpTomG42F8HG(50 ng/μl)(2006年11月28に出願された「Recombinant monovalent antibodies and methods for production thereof」という表題のPCT出願(RO/DK (Genmab)に記載されている) 1μl、およびPfuUltra HF DNAポリメラーゼ1μlを全量50μl中で混合し、18サイクルプログラム:95℃で1分の変性;95℃で50秒、60℃で50秒、および68℃で10分間の18サイクルを用いて、TGradient Thermocycler 96(Whatman Biometra、ドイツ、ゲッティンゲン;製品# 050-801)で増幅した:PCR混合物は、さらなる処理時まで4℃で保存した。次に、PCR混合物をDpnI 1μlと共に37℃で60分間インキュベートして、pTomG42F8HGベクターを消化し、さらなる処理時まで4℃で保存した。反応混合物を3 M NaAc 5μlおよびエタノール125μlで沈殿させ、-20℃で20分間インキュベートし、14000×gで4℃で20分間遠心沈殿させた。DNAペレットを70%エタノールで洗浄し、乾燥させ、水4μlに溶解した。全部で4μlの反応量を、製造業者の説明書(Invitrogen)に従ってOne Shot DNH5α T1Rコンピテント大腸菌細胞(Invitrogen、オランダ、ブレダ)に形質転換した。次に、50μg/mlアンピシリンを含むルリア-ベルターニ(LB)寒天プレートに細胞をプレーティングした。細菌コロニーが明白になるまで、プレートを37℃で16〜18時間インキュベートした。
【0220】
コロニーPCRおよびXmaI(突然変異誘発によってXmaI部位が失われる)消化によるスクリーニング後に、プラスミドを細菌から単離し、DNA配列決定によって変異を確認した。望ましくない余分な変異が導入されたかどうかを調べるために、全HCコード領域の配列を決定したが、さらなる変異は全く含んでいなかった。最終的な構築物をpTomG42F8S228PNewと命名した。
【0221】
トランスフェクション試薬として293 Fectin(Invitrogen)を用いて、これらの構築物による組換え抗体を、3 ml、6ウェルプレート(NUNC)または125 ml三角フラスコ(Corning)においてHEK 293細胞で一過性に発現させた。
【0222】
非精製抗体(FreeStyle 293発現培地、GIBCO/Invitrogen Corporation)の以下の混合物を0.1 mM GSHと共に37℃で24時間インキュベートし、試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(以下、IgG1 CPSC - IgG1 CPSCと表示)
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CH3(IgG4)(IgG1 CPSC - IgG1 CH3(IgG4))
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC/CH3(IgG4)(IgG1 CPSC - IgG1 CPSC/CH3(IgG4))
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(IgG1 CH3(IgG4) - IgG1 CH3(IgG4))
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CPSC/CH3(IgG4)(IgG1 CH3(IgG4) - IgG1 CPSC/CH3(IgG4))
‐IgG1 a-feld1 CPSC/CH3(IgG4)とa-betv1 IgG1 CPSC/CH3(IgG4)(IgG1 CPSC/CH3(IgG4) - IgG1 CPSC/CH3(IgG4))
‐IgG1 a-feld1 CPSC/CH3(IgG4)とIgG4 a-betv1 wt(IgG1 CPSC/CH3(IgG4) - IgG4 wt
‐IgG4 a-bet1 S228PnewとIgG4 wt。
【0223】
結果から、これらのインビトロ条件(0.1 mM GSH)において、抗体の一方がCPSCヒンジを含み、かつ両抗体がIgG4様CH3を含む場合に、半分子の交換が起こることが示された。また、半分子の交換は、IgG1ヒンジを含むIgG4分子とIgG4 wt分子の間でも起こる。
-=交換なし
+=交換が起こる
±=限られた交換(約5%)
空白欄=未検
【0224】
0 mM、0.1 mM、1 mM、および10 mM GSHを用いて、様々な変異体による半分子の交換に及ぼすGSH濃度の影響を試験した。以下の混合物を用いて、交換を試験した:
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 wtとIgG4 a-betv1 wt
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)
‐IgG1 a-feld1 CPSC/CH3(IgG4)とa-betv1 IgG1 CPSC/CH3(IgG4))。
【0225】
1 mMまでのGSH濃度については、結果(図19A)から上記のことが確認された。10 mM GSHでは、IgG1 a-feld1 CH3(IgG4)およびIgG1 a-betv1 CH3(IgG4)を含む反応においても半分子の交換が認められた。
【0226】
先の実施例に記載された通りに、サイズ排除クロマトグラフィーを行って、適切なIgG1変異体のGSH媒介性の交換後に認められた二重特異性活性が、IgG凝集の結果であるという可能性を排除した。異種Fel d 1-Bet v 1架橋活性は、単量体IgGの保持容量に対応する画分中に検出された。
【0227】
半分子を交換する能力に関与するCH3ドメイン内のアミノ酸残基を同定するために、IgG1とIgG4との間で異なる位置でIgG4様残基をIgG1のCH3に導入した。したがって、基本的に上記の通りに、鋳型としてpEE-G1-wt a Bet v 1またはpEE-G1-wt a Fel d 1を用いて、R238Q、K292R、Q302E、またはP328L変異(番号はSEQ ID NO:19を参照する)をIgG1のCH3ドメインに導入した。さらに、鋳型としてpEE-G1-CPSC betv1またはpEE-G1-CPSC feld1を用いて、K292R変異をIgG1 CPSCのCH3ドメインにも導入した。手短に説明すると、Vector NTI Advance 10により、突然変異誘発プライマー、フォワードおよびリバースを設計した。Quickchange部位特異的突然変異誘発キット(Stratagene)を用いて、構築物を作製した。トランスフェクション試薬として293 Fectin(Invitrogen)を用いて、これらの構築物による組換え抗体を、3 ml、6ウェルプレート(NUNC)または125 ml三角フラスコ(Corning)においてHEK 293細胞で一過性に発現させた。非精製抗体(FreeStyle 293発現培地、GIBCO/Invitrogen Corporation)の以下の混合物を0.5 mMまたは5 mM GSHと共に37℃で24時間インキュベートし、試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG1 a-feld1 wtとIgG4 a-betv1 wt(図19BではIgG1と表示)
‐IgG1 a-feld1 CPSCとIgG4 a-betv1 wt(図19BではIgG1-CPSCと表示)
‐IgG1 a-feld1 CH3(IgG4)とIgG4 a-betv1 wt(図19BではIgG1-CH3(G4)と表示)
‐IgG1 a-feld1 CPSC/CH3(IgG4)とIgG4 a-betv1 wt(図19BではIgG1-CPSC/CH3(G4)と表示)
‐IgG1 a-feld1 R238QとIgG4 a-betv1 wt(図19BではIgG1-R238Qと表示)
‐IgG1 a-feld1 K292RとIgG4 a-betv1 wt(図19BではIgG1-K292Rと表示)
‐IgG1 a-feld1 Q302EとIgG4 a-betv1 wt(図19BではIgG1-Q302Eと表示)
‐IgG1 a-feld1 P328LとIgG4 a-betv1 wt(図19BではIgG1-P328Lと表示)
‐IgG1 a-feld1 CPSC/K292RとIgG4 a-betv1 wt(図19BではIgG1-CPSC/K292Rと表示)
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図19BではIgG4と表示)
‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(図19CではIgG1と表示)
‐IgG1 a-feld1 CPSCとIgG1 a-betv1 CPSC(図19CではIgG1-CPSCと表示)
‐IgG1 a-feld1 CH3(IgG4)とIgG1 a-betv1 CH3(IgG4)(図19CではIgG1-CH3(G4)と表示)
‐IgG1 a-feld1 CPSC/CH3(IgG4)とIgG1 a-betv1 CPSC/CH3(IgG4)(図19CではIgG1-CPSC/CH3(G4)と表示)
‐IgG1 a-feld1 R238QとIgG1 a-betv1 R238Q(図19CではIgG1-R238Qと表示)
‐IgG1 a-feld1 K292RとIgG1 a-betv1 K292R(図19CではIgG1-K292Rと表示)
‐IgG1 a-feld1 Q302EとIgG1 a-betv1 Q302E(図19CではIgG1-Q302Eと表示)
‐IgG1 a-feld1 P328LとIgG1 a-betv1 P328L(図19CではIgG1-P328Lと表示)
‐IgG1 a-feld1 CPSC/K292RとIgG1 a-betv1 CPSC/K292R(図19CではIgG1-CPSC/K292Rと表示)
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図19CではIgG4と表示)。
【0228】
結果から、試験したインビトロ条件(0.5 mMおよび5 mM GSH)において、両抗体が292位にRを含む場合に半分子の交換が起こることが示された(図19B+C)。238位のRまたはQ、302位のQまたはE、および328位のPまたはLは、この実験設定においてIgG1が半分子を交換できないことに影響しない。
【0229】
実施例36. 0.5 mM GSHにおいて、組換えヒトIgG4抗体のFabアーム交換反応に関与しない安定化IgG1様コアヒンジを有するIgG4分子
2種類の組換えヒトIgG4抗体(上記のIgG4-CD20およびIgG4-EGFr)を、過剰(10、50、および100マイクログラム/ml)のTysabriまたは過剰(10マイクログラム/ml)のMylotargの存在下または非存在下において、0.5 mM GSHと共に37℃で24時間インキュベートした。Tysabriは野生型IgG4コアヒンジを含む市販のヒト化IgG4抗体であり、Mylotargは安定化IgG1様コアヒンジを含む市販のヒト化IgG4抗体である。Fabアーム交換を介した二重特異性抗体の形成を、上記の通りにサンドイッチELISA法で測定した。
【0230】
図20Aは、過剰なTysabriの存在下において、組換えCD20抗体およびEGFr抗体のFabアームの交換が阻害されたことを示す。
【0231】
図20Bは、過剰なTysabriの存在下では組換えCD20抗体およびEGFr抗体のFabアームの交換が阻害されたが、過剰なMylotargの存在下ではそのように阻害されなかったことを示す。
【0232】
このことは、TysabriはFabアーム交換反応に関与するがMylotargは関与しないこと、および安定化IgG1様コアヒンジは0.5 mM GSHを使用するインビトロ条件下でFabアーム交換に関与しないことを示す。
【0233】
実施例37. 292位にさらなる変異を有するIgG1-CPSC構築物の半分子の交換
実施例35と同様に、pConG1f2F8(EGFRに特異的)およびpConG1f7D8(CD20に特異的)において3種類のIgG1変異体を作製した:IgG4コアヒンジを有するIgG1(IgG1-CPSC)、および2種類のCH3ドメイン交換変異体(IgG1-CH3(IgG4)およびIgG1-CPSC-CH3(IgG4))(すなわち、IgG1のCH3領域をIgG4のCH3領域によって置換した構築物)。これにより、以下の構築物pG1f-2F8CPSC、pG1f-7D8CPSC、pG1f-2F8-CH3(G4)、pG1f-7D8-CH3(G4)、pG1f-2F8CPSC-CH3(G4)、およびpG1f-7D8CPSC-CH3(G4)を得た。
【0234】
続いて、基本的に上記の通りに、R238Q、K292R、K292Y、K292F、K292W、Q302E、またはP328L変異(SEQ ID NO: 19を参照されたい)を、pG1f-2F8CPSC構築物およびpG1f-7D8CPSC構築物の両方のCH3ドメインに導入した。手短に説明すると、Vector NTI Advance 10により、突然変異誘発プライマー、フォワードおよびリバースを設計した。Quickchange部位特異的突然変異誘発キット(Stratagene)を用いて、構築物を作製した。
【0235】
トランスフェクション試薬として293 Fectin(Invitrogen)を用いて、これらの構築物による組換え抗体を、3 ml、6ウェルプレート(NUNC)または125 ml三角フラスコ(Corning)においてHEK 293細胞で一過性に発現させた。続いて培養上清をPBSに対して透析し、比濁法により濃度を測定した(上記を参照されたい)。非精製の緩衝液交換抗体の以下の混合物を0.5 GSHと共に37℃で24時間インキュベートし、試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG1-2F8 wtとIgG1-7D8 wt(IgG1と表示)
‐IgG1-2F8-CPSCとIgG1-7D8-CPSC(IgG1-CPSCと表示)
‐IgG1-2F8-CH3(IgG4)とIgG1-7D8-CH3(IgG4)(IgG1-CH3(IgG4)と表示)
‐IgG1-2F8-CPSC-CH3(IgG4)とIgG1-7D8-CPSC-CH3(IgG4)(IgG1-CPSC-CH3(IgG4)と表示)
‐IgG1-2F8-CPSC-R238QとIgG1-7D8-CPSC-R238Q(IgG1-CPSC-R238Qと表示)
‐IgG1-2F8-CPSC-K292RとIgG1-7D8-CPSC-K292R(IgG1-CPSC-K292Rと表示)
‐IgG1-2F8-CPSC-K292YとIgG1-7D8-CPSC-K292Y(IgG1-CPSC-K292Yと表示)
‐IgG1-2F8-CPSC-K292FとIgG1-7D8-CPSC-K292F(IgG1-CPSC-K292Fと表示)
‐IgG1-2F8-CPSC-K292WとIgG1-7D8-CPSC-K292W(IgG1-CPSC-K292Wと表示)
‐IgG1-2F8-CPSC-Q302EとIgG1-7D8-CPSC-Q302E(IgG1-CPSC-Q302Eと表示)
‐IgG1-2F8-CPSC-P328LとIgG1-7D8-CPSC-P328L(IgG1-CPSC-P328Lと表示)
IgG4-2F8 wtとIgG4-7D8 wt(IgG4と表示)。
【0236】
図21は、試験したインビトロ条件(0.5 mM)において、CPSCヒンジが存在し、かつ292位がRである場合に、半分子の交換が起こることを示す。加えて、結果から、292位のYまたはFもまた、程度はより低いものの半分子の交換を促進するが、Wは促進しないことが示される。238位のRまたはQ、302位のQまたはE、および328位のPまたはLは、IgG1-CPSCが半分子を交換できないことに影響しない。
【0237】
実施例38 (5 mM GSHと共に)インビトロではFabアーム交換ができるが、インビボではできない、安定化CPPCヒンジを有するIgG4分子
IgG4-EGFR-CPPCおよびIgG4-CD20の混合物を、5 mM GSHの存在下で24時間インキュベートし、質量分析(ESI-TOF MS)によって評価した。各抗体200μg/mlを含む試料50μlを、N-グリコシダーゼF(Roche Diagnostics NL BV、オランダ、アルメレ) 1μlで一晩かけて脱グリコシル化した。BEH C8、1.7μm、2.1×50 mmカラムを用いてAcquity UPLC(商標)(Waters、米国、ミルフォード)において、試料を60℃で脱塩した。5μlを注入し、5%〜95%溶離液Bの勾配で溶出した。溶離液AはMilliQ水(Millipore Synthesis A10装置)であり、溶離液BはLC-MS等級アセトニトリル(Biosolve、オランダ、ファルケンスワールト)であった。いずれの溶離液も、有機変性剤として0.05%ギ酸(Fluka Riedel-de Han、ドイツ、ブーフ)を含んだ。micrOTOF(商標)質量分析計(Bruker、ドイツ、ブレーメン)を陽イオンモードで操作して、飛行時間型エレクトロスプレーイオン化質量スペクトルをオンラインで記録した。各解析において、500〜5000 m/zスケールをES調整混合物(Agilent Technologies、米国、サンタクララ)で内部較正した。DataAnalysis(商標)ソフトウェアv.3.3(Bruker)によって提供される最大エントロピーアルゴリズムを用いることにより、質量スペクトルをデコンボリューション処理した。
【0238】
図22Aは、5 mM GSHの存在下において、Fabアーム交換分子に対応する中間質量を有する新たなピークが出現した(145.7 kDa)ことを示す。新たな質量は、二重特異性抗EGFR/CD20抗体の予測質量に相当した。GSHを使用しなかった場合または0.5 mM GSHを使用した場合には、二重特異性抗体のピークは出現しなかった(データは示さず)。これは、IgG1様のCPPCヒンジおよびIgG4様CH3領域を含む変異体が、(実施例35、36、37においても示される通り)より高いGSH濃度においてインビトロで半分子を交換できることを示す。
【0239】
IgG1様のCPPCヒンジおよびIgG4様CH3を含む安定化ヒンジ変異体のFabアーム交換がインビボで起こるかどうかを調べるために、本発明者らは、IgG4-CD20とIgG1-EGFR、IgG4-EGFR、IgG4-EGFR-CPPCの同等の混合物を免疫不全マウスに注射した。様々な時点で血液試料を採取し、インビトロ交換された混合物(IgG4-EGFR/IgG4-CD20)を参照標準物質として用いて、二重特異性抗体を(上記の通りに)ELISA法で定量した。
【0240】
図22Bは、野生型IgG4分子(IgG4-EGFR)を含む混合物を注射したマウスの血液中に、二重特異性抗体が出現したことを示す。ヒンジ安定化IgG4(IgG4-EGFR-CPPC)またはIgG1分子(IgG1-EGFR)を含む混合物では、二重特異性抗体は検出不能であった((図中に記号は示されていない))。これは、コアヒンジ安定化がインビボでのIgG4 Fabアーム交換を妨げ、インビボで半分子を交換できないことを示す(しかし本発明者らは、ヒンジ安定化IgG4の検出レベル(72時間で<8%)未満の低レベル交換が起こるということを排除できない)。
【0241】
これにより、安定化CPPCヒンジを含む二重特異性抗体を、インビトロでのFabアーム交換によって得ることができることが示唆される。これらの二重特異性抗体をその後特異的に精製した後、これらの抗体は、インビボでの注射に際して安定したままである(すなわち、Fabアーム交換しない)。
【0242】
実施例39. CXXC変異体のFabアーム交換
コアヒンジ内に様々なCXXCモチーフを含む抗体のFabアームを交換する能力を試験した。上記の通りに部位特異的突然変異誘発技法を用いて、以下のCXXCモチーフをIgG4 bet v 1およびIgG4 feld 1に導入した:
‐CGHC(タンパク質ジスルフィドイソメラーゼ、PDIについて記載されている活性部位配列)
‐CGC(ジスルフィド還元電位を有すると記載されているペプチド)
‐CPRC(ゴリラIgG4のコアヒンジ配列)
‐CPHC(ヒトチオレドキシンについて記載されている活性部位配列)。
【0243】
精製抗体の以下の混合物を0.5 mM GSHと共に37℃でインキュベートし、0〜24時間の様々な時点で試料をPBS-AT中に採取し、先の実施例に記載した通りにそこで(二重)特異性IgG濃度を測定した:
‐IgG1 a-feld1 wtとIgG1 a-betv1 wt(図23および24においてIgG1と表示)
‐IgG4 a-feld1 wtとIgG4 a-betv1 wt(図23および24においてIgG4と表示)
‐IgG4 a-feld1 CGHCとIgG4 a-betv1 CGHC(図23および24においてCGHCと表示)
‐IgG4 a-feld1 CGCとIgG4 a-betv1 CGC(図23および24においてCGCと表示)
‐IgG4 a-feld1 CPRCとIgG4 a-betv1 CPRC(図23および24においてCPRCと表示)
‐IgG4 a-feld1 CPHCとIgG4 a-betv1 CPHC(図23および24においてCPHCと表示)。
【0244】
結果(図23)から、時間が経過しても、CGCモチーフまたはIgG1コアヒンジを含む抗体のFabアーム交換は起こらないことが示された。CGHCモチーフを含む抗体のFabアーム交換は、IgG4 wt抗体と同程度に効果的であった。いくらかゆっくりではあるものの、CPRCモチーフを含む抗体のFabアーム交換も起こり、程度はより低いがCPHCモチーフを含む抗体のFabアーム交換も起こった。
【0245】
また、これらの混合物が37℃での24時間のインキュベーション後にFabアーム交換を行う能力に及ぼすGSH濃度(1〜20,000μM)の影響を試験した。CPHC、CPRC、およびCGHCモチーフ含有抗体ならびにIgG4 wt抗体のFabアーム交換は、GSH濃度に依存し(図24)、最適値は100μM〜1,000μM GSHであることが判明した。
【特許請求の範囲】
【請求項1】
以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【請求項2】
第1の抗体がコアヒンジ領域内にCPPC配列を含む、請求項1記載のエクスビボ方法。
【請求項3】
第1の抗体がIgG4様コアヒンジ領域を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項4】
第1の抗体が、コアヒンジ領域内に、X1およびX2の両方がプロリンではないという条件で、X1およびX2が任意のアミノ酸であってよい、CX1X2C配列を含む抗体である、請求項1または3のいずれか一項記載のエクスビボ方法。
【請求項5】
第1の抗体が、コアヒンジ領域内に、X3がプロリンを除く任意のアミノ酸であってよい、CX3PCまたはCPX3C配列を含む抗体である、請求項4記載のエクスビボ方法。
【請求項6】
第1の抗体が、コアヒンジ領域内にCSPC、CPSC、CRPC、CPRC、CGHC、またはCPHC配列を含む抗体である、請求項5記載のエクスビボ方法。
【請求項7】
第1の抗体がIgG4 CH3領域を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項8】
第1の抗体が非IgG4アイソタイプのCH3領域を含み、このCH3配列が、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている、請求項1〜6のいずれか一項記載のエクスビボ方法。
【請求項9】
CH3領域がSEQ ID NO: 19に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項8記載のエクスビボ方法:
238位のArg(R)がGln(Q)によって置換;239位のAsp(D)がGlu(E)によって置換;292位のLys(K)がArg(R)によって置換;302位のGln(Q)がGlu(E)によって置換;および328位のPro(P)がLeu(L)によって置換。
【請求項10】
292位のLys(K)がArg(R)によって置換されている、請求項9記載のエクスビボ方法。
【請求項11】
CH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている、請求項1〜6または8のいずれか一項記載のエクスビボ方法。
【請求項12】
CH3領域がSEQ ID NO: 20に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項8記載のエクスビボ方法:
234位のArg(R)がGln(Q)によって置換;276位のMet(M)がVal(V)によって置換;288位のLys(K)がArg(R)によって置換;298位のGln(Q)がGlu(E)によって置換;および324位のPro(P)がLeu(L)によって置換。
【請求項13】
234位のArg(R)がGln(Q)によって置換されている、請求項12記載のエクスビボ方法。
【請求項14】
234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている、請求項12記載のエクスビボ方法。
【請求項15】
5つすべての置換がなされている、請求項12記載のエクスビボ方法。
【請求項16】
CH3領域がSEQ ID NO: 21に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項8記載のエクスビボ方法:
285位のArg(R)がGln(Q)によって置換;314位のSer(S)がAsn(N)によって置換;322位のAsn(N)がLys(K)によって置換;327位のMet(M)がVal(V)によって置換;339位のLys(K)がArg(R)によって置換;349位のGln(Q)がGlu(E)によって置換;352位のIle(I)がVal(V)によって置換;365位のArg(R)がHis(H)によって置換;366位のPhe(F)がTyr(Y)によって置換;および375位のPro(P)がLeu(L)によって置換。
【請求項17】
285位のArg(R)がGln(Q)によって置換されている、請求項16記載のエクスビボ方法。
【請求項18】
285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている、請求項16記載のエクスビボ方法。
【請求項19】
10個すべての置換がなされている、請求項16記載のエクスビボ方法。
【請求項20】
第1の抗体がIgG4抗体である、請求項1、3〜7のいずれか一項記載のエクスビボ方法。
【請求項21】
第2の抗体がコアヒンジ領域内にCPPC配列を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項22】
第2の抗体がIgG4様コアヒンジ領域を含む、請求項1〜20のいずれか一項記載のエクスビボ方法。
【請求項23】
第2の抗体が、コアヒンジ領域内に、X1およびX2の両方がプロリンではないという条件で、X1およびX2が任意のアミノ酸であってよい、CX1X2C配列を含む抗体である、請求項1〜20または22のいずれか一項記載のエクスビボ方法。
【請求項24】
第2の抗体が、コアヒンジ領域内に、X3がプロリンを除く任意のアミノ酸であってよい、CX3PCまたはCPX3C配列を含む抗体である、請求項23記載のエクスビボ方法。
【請求項25】
第2の抗体が、コアヒンジ領域内にCSPC、CPSC、CRPC、またはCPRC配列を含む抗体である、請求項23記載のエクスビボ方法。
【請求項26】
第2の抗体がIgG4 CH3領域を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項27】
第2の抗体が非IgG4アイソタイプのCH3領域を含み、このCH3配列が、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている、請求項1〜25のいずれか一項記載のエクスビボ方法。
【請求項28】
CH3領域がSEQ ID NO: 19に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項27記載のエクスビボ方法:
238位のArg(R)がGln(Q)によって置換;239位のAsp(D)がGlu(E)によって置換;292位のLys(K)がArg(R)によって置換;302位のGln(Q)がGlu(E)によって置換;および328位のPro(P)がLeu(L)によって置換。
【請求項29】
292位のLys(K)がArg(R)によって置換されている、請求項28記載のエクスビボ方法。
【請求項30】
CH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている、請求項21〜27のいずれか一項記載のエクスビボ方法。
【請求項31】
CH3領域がSEQ ID NO: 20に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項27記載のエクスビボ方法:
234位のArg(R)がGln(Q)によって置換;276位のMet(M)がVal(V)によって置換;288位のLys(K)がArg(R)によって置換;298位のGln(Q)がGlu(E)によって置換;および324位のPro(P)がLeu(L)によって置換。
【請求項32】
234位のArg(R)がGln(Q)によって置換されている、請求項31記載のエクスビボ方法。
【請求項33】
234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている、請求項31記載のエクスビボ方法。
【請求項34】
5つすべての置換がなされている、請求項31記載のエクスビボ方法。
【請求項35】
CH3領域がSEQ ID NO: 21に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項27記載のエクスビボ方法:
285位のArg(R)がGln(Q)によって置換;314位のSer(S)がAsn(N)によって置換;322位のAsn(N)がLys(K)によって置換;327位のMet(M)がVal(V)によって置換;339位のLys(K)がArg(R)によって置換;349位のGln(Q)がGlu(E)によって置換;352位のIle(I)がVal(V)によって置換;365位のArg(R)がHis(H)によって置換;366位のPhe(F)がTyr(Y)によって置換;および375位のPro(P)がLeu(L)によって置換。
【請求項36】
285位のArg(R)がGln(Q)によって置換されている、請求項35記載のエクスビボ方法。
【請求項37】
285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている、請求項35記載のエクスビボ方法。
【請求項38】
10個すべての置換がなされている、請求項35記載のエクスビボ方法。
【請求項39】
第2の抗体がIgG4抗体である、請求項1〜20または22〜26のいずれか一項記載のエクスビボ方法。
【請求項40】
第1の抗体がコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含み、および、第2抗体がコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含む、請求項1または2記載のエクスビボ方法。
【請求項41】
第1の抗体および/または第2の抗体がヒト抗体である、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項42】
段階c)における条件が、コアヒンジ領域外のジスルフィド架橋の有意な還元または異性化が起こらないように選択される、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項43】
段階c)が還元剤の添加を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項44】
段階c)が、グルタチオン、L-システイン、ジチオスレイトール、β-メルカプト-エタノール、およびシステアミンからなる群より選択される作用物質の添加を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項45】
作用物質の濃度が、段階c)で生じる溶液の酸化還元電位が実施例31に記載する条件下で、1μMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10μMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば50μMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば0.1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である、請求項43または44記載のエクスビボ方法。
【請求項46】
作用物質の濃度が、段階c)で生じる溶液の酸化還元電位が実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である、請求項43または44記載のエクスビボ方法。
【請求項47】
作用物質の濃度が、段階c)で生じる溶液の酸化還元電位が1 Mのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、例えば100 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、15 mMのグルタチオンによって生じる酸化還元電位と等しいかもしくはより還元性が低い濃度である、請求項45または46記載のエクスビボ方法。
【請求項48】
段階c)が、還元型グルタチオンの存在下での、20℃またはそれ以上、例えば37℃の温度における、少なくとも1時間、例えば少なくとも2時間、例えば少なくとも5時間、例えば少なくとも10時間の、抗体のインキュベーションを含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項49】
結果として得られた組成物中の抗体分子の10%未満、例えば5%未満、例えば2%未満、例えば1%未満が凝集状態である、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項50】
半分子のさらなる交換を停止させるために、段階c)において得られた組成物を非還元条件に導くさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項51】
好ましくは以下から選択される方法を用いて、二重特異性抗体を安定化するさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法:
a) ヒンジ領域内のシステインを化学的に架橋する、
b) 半分子上の炭水化物側鎖を化学的に架橋する、および
c) CH3領域内に非対称的に導入されたシステインを架橋する。
【請求項52】
好ましくは非還元条件下で、二重特異性抗体を精製するさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項53】
治療用途のために、結果として得られた二重特異性抗体を薬学的組成物に製剤化するさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項54】
第1の抗体が、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD38、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する、またはCD79aもしくはCD79bに対する結合特異性を有する、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項55】
第2の抗体が、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項56】
第1の抗体がerbB1に対する結合特異性を有し、かつ第2の抗体がerbB2に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項57】
第1の抗体がCD19に対する結合特異性を有し、かつ第2の抗体がCD20に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項58】
第1の抗体がCD4に対する結合特異性を有し、かつ第2の抗体がCXCR5に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項59】
第1の抗体がCD38に対する結合特異性を有し、かつ第2の抗体がCD34に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項60】
第1の抗体が病原微生物に対する結合特異性を有する、請求項1〜53のいずれか一項記載のエクスビボ方法。
【請求項61】
第2の抗体が、CD3、CD25、CD28、CD16、CD89、CD32、またはCD1などのエフェクター細胞タンパク質に対する結合特異性を有する、請求項54または60記載のエクスビボ方法。
【請求項62】
第2の抗体が化学療法薬に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項63】
第2の抗体が、血清アルブミンなどの血液タンパク質、またはトランスフェリンなどの脳タンパク質、または肝臓タンパク質に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項64】
第2の抗体が組織因子などの血液凝固に関与するタンパク質に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項65】
第1および/または第2の抗体が、細胞毒性剤;放射性同位体;プロドラッグまたは薬物、例えばタキサンなど;サイトカイン;ケモカイン、および補体、例えばC1qなどからなる群より選択される化合物に連結されている、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項66】
以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【請求項67】
段階c)において還元剤が添加され、該剤の濃度が、段階c)で生じる溶液の酸化還元電位が実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である、請求項66記載のエクスビボ方法。
【請求項68】
前記請求項のいずれか一項記載の方法によって得られるか、または得られ得る、単離された二重特異性抗体。
【請求項69】
2つのIgG4様CH3領域を含む、単離された二重特異性抗体。
【請求項70】
コアヒンジ領域内に1つまたは2つのCPPC配列を含む、請求項69記載の二重特異性抗体。
【請求項71】
コアヒンジ領域内に、X1およびX2の両方がプロリンではないという条件で、X1およびX2が任意のアミノ酸であってよい、1つまたは2つのCX1X2C配列を含む、請求項69記載の二重特異性抗体。
【請求項72】
コアヒンジ領域内に、X3がプロリンを除く任意のアミノ酸であってよい、1つまたは2つのCX3PCまたはCPX3C配列を含む、請求項69記載の二重特異性抗体。
【請求項73】
コアヒンジ領域内に1つまたは2つのCSPC、CPSC、CRPC、またはCPRC配列を含む、請求項69記載の二重特異性抗体。
【請求項74】
第1および/または第2のCH3領域が非IgG4アイソタイプのものであって、このCH3配列が、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている、請求項69〜73のいずれか一項記載の二重特異性抗体。
【請求項75】
第1および/または第2のCH3領域がSEQ ID NO: 19に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項74記載の二重特異性抗体:
238位のArg(R)がGln(Q)によって置換;239位のAsp(D)がGlu(E)によって置換;292位のLys(K)がArg(R)によって置換;302位のGln(Q)がGlu(E)によって置換;および328位のPro(P)がLeu(L)によって置換。
【請求項76】
第1および/または第2のCH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がArg(R)によって置換されている、請求項75記載の二重特異性抗体。
【請求項77】
第1および/または第2のCH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている、請求項74記載の二重特異性抗体。
【請求項78】
第1および/または第2のCH3領域がSEQ ID NO: 20に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項74記載の二重特異性抗体:
234位のArg(R)がGln(Q)によって置換;276位のMet(M)がVal(V)によって置換;288位のLys(K)がArg(R)によって置換;298位のGln(Q)がGlu(E)によって置換;および324位のPro(P)がLeu(L)によって置換。
【請求項79】
234位のArg(R)がGln(Q)によって置換されている、請求項74記載の二重特異性抗体。
【請求項80】
234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている、請求項74記載の二重特異性抗体。
【請求項81】
第1および/または第2のCH3領域がSEQ ID NO: 21に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項74記載の二重特異性抗体:
285位のArg(R)がGln(Q)によって置換;314位のSer(S)がAsn(N)によって置換;322位のAsn(N)がLys(K)によって置換;327位のMet(M)がVal(V)によって置換;339位のLys(K)がArg(R)によって置換;349位のGln(Q)がGlu(E)によって置換;352位のIle(I)がVal(V)によって置換;365位のArg(R)がHis(H)によって置換;366位のPhe(F)がTyr(Y)によって置換;および375位のPro(P)がLeu(L)によって置換。
【請求項82】
285位のArg(R)がGln(Q)によって置換されている、請求項81記載の二重特異性抗体。
【請求項83】
285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている、請求項81記載の二重特異性抗体。
【請求項84】
第1および/または第2のCH3領域がIgG4 CH3領域である、請求項69〜73のいずれか一項記載の二重特異性抗体。
【請求項85】
コアヒンジ領域外およびCH3領域外の配列が、IgG1、IgG2、IgG3、およびIgG4からなる群より選択されるアイソタイプのものである、請求項68〜84のいずれか一項記載の二重特異性抗体。
【請求項86】
請求項69〜85のいずれか一項記載の二重特異性抗体を含む薬学的組成物。
【請求項87】
薬として使用するための、請求項86記載の組成物。
【請求項88】
癌の治療用の薬として使用するための、請求項86記載の組成物。
【請求項89】
癌の治療用の薬を調製するための、請求項1〜67のいずれか一項記載の方法によって得られるかもしくは得られ得る二重特異性抗体かまたは請求項68〜85のいずれか一項記載の二重特異性抗体の、使用。
【請求項90】
以下の段階を含む、所望の特性を有する二重特異性抗体を選択する方法:
a) 一組の抗体を提供する段階であって、各抗体が異なる標的特異性を有し、かつ各抗体がIgG4様CH3領域を含む段階、
b) 還元条件下で、該一組の抗体の各抗体を該一組の別の抗体と共にインキュベートし、それによって一組の抗体混合物を作製する段階であって、各混合物が異なる二重特異性抗体を含む段階、
c) 結果として得られた一組の抗体混合物を所与の所望の特性についてアッセイする段階、および
d) 所望の特性を有する二重特異性抗体混合物を選択する段階。
【請求項91】
段階b)が、請求項2〜67における段階c)について規定されるさらなる特性の1つまたは複数を有する、請求項90記載の方法。
【請求項92】
所望の特性が腫瘍細胞殺傷である、請求項90または91記載の方法。
【請求項1】
以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がIgG4様CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がIgG4様CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【請求項2】
第1の抗体がコアヒンジ領域内にCPPC配列を含む、請求項1記載のエクスビボ方法。
【請求項3】
第1の抗体がIgG4様コアヒンジ領域を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項4】
第1の抗体が、コアヒンジ領域内に、X1およびX2の両方がプロリンではないという条件で、X1およびX2が任意のアミノ酸であってよい、CX1X2C配列を含む抗体である、請求項1または3のいずれか一項記載のエクスビボ方法。
【請求項5】
第1の抗体が、コアヒンジ領域内に、X3がプロリンを除く任意のアミノ酸であってよい、CX3PCまたはCPX3C配列を含む抗体である、請求項4記載のエクスビボ方法。
【請求項6】
第1の抗体が、コアヒンジ領域内にCSPC、CPSC、CRPC、CPRC、CGHC、またはCPHC配列を含む抗体である、請求項5記載のエクスビボ方法。
【請求項7】
第1の抗体がIgG4 CH3領域を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項8】
第1の抗体が非IgG4アイソタイプのCH3領域を含み、このCH3配列が、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている、請求項1〜6のいずれか一項記載のエクスビボ方法。
【請求項9】
CH3領域がSEQ ID NO: 19に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項8記載のエクスビボ方法:
238位のArg(R)がGln(Q)によって置換;239位のAsp(D)がGlu(E)によって置換;292位のLys(K)がArg(R)によって置換;302位のGln(Q)がGlu(E)によって置換;および328位のPro(P)がLeu(L)によって置換。
【請求項10】
292位のLys(K)がArg(R)によって置換されている、請求項9記載のエクスビボ方法。
【請求項11】
CH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている、請求項1〜6または8のいずれか一項記載のエクスビボ方法。
【請求項12】
CH3領域がSEQ ID NO: 20に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項8記載のエクスビボ方法:
234位のArg(R)がGln(Q)によって置換;276位のMet(M)がVal(V)によって置換;288位のLys(K)がArg(R)によって置換;298位のGln(Q)がGlu(E)によって置換;および324位のPro(P)がLeu(L)によって置換。
【請求項13】
234位のArg(R)がGln(Q)によって置換されている、請求項12記載のエクスビボ方法。
【請求項14】
234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている、請求項12記載のエクスビボ方法。
【請求項15】
5つすべての置換がなされている、請求項12記載のエクスビボ方法。
【請求項16】
CH3領域がSEQ ID NO: 21に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項8記載のエクスビボ方法:
285位のArg(R)がGln(Q)によって置換;314位のSer(S)がAsn(N)によって置換;322位のAsn(N)がLys(K)によって置換;327位のMet(M)がVal(V)によって置換;339位のLys(K)がArg(R)によって置換;349位のGln(Q)がGlu(E)によって置換;352位のIle(I)がVal(V)によって置換;365位のArg(R)がHis(H)によって置換;366位のPhe(F)がTyr(Y)によって置換;および375位のPro(P)がLeu(L)によって置換。
【請求項17】
285位のArg(R)がGln(Q)によって置換されている、請求項16記載のエクスビボ方法。
【請求項18】
285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている、請求項16記載のエクスビボ方法。
【請求項19】
10個すべての置換がなされている、請求項16記載のエクスビボ方法。
【請求項20】
第1の抗体がIgG4抗体である、請求項1、3〜7のいずれか一項記載のエクスビボ方法。
【請求項21】
第2の抗体がコアヒンジ領域内にCPPC配列を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項22】
第2の抗体がIgG4様コアヒンジ領域を含む、請求項1〜20のいずれか一項記載のエクスビボ方法。
【請求項23】
第2の抗体が、コアヒンジ領域内に、X1およびX2の両方がプロリンではないという条件で、X1およびX2が任意のアミノ酸であってよい、CX1X2C配列を含む抗体である、請求項1〜20または22のいずれか一項記載のエクスビボ方法。
【請求項24】
第2の抗体が、コアヒンジ領域内に、X3がプロリンを除く任意のアミノ酸であってよい、CX3PCまたはCPX3C配列を含む抗体である、請求項23記載のエクスビボ方法。
【請求項25】
第2の抗体が、コアヒンジ領域内にCSPC、CPSC、CRPC、またはCPRC配列を含む抗体である、請求項23記載のエクスビボ方法。
【請求項26】
第2の抗体がIgG4 CH3領域を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項27】
第2の抗体が非IgG4アイソタイプのCH3領域を含み、このCH3配列が、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている、請求項1〜25のいずれか一項記載のエクスビボ方法。
【請求項28】
CH3領域がSEQ ID NO: 19に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項27記載のエクスビボ方法:
238位のArg(R)がGln(Q)によって置換;239位のAsp(D)がGlu(E)によって置換;292位のLys(K)がArg(R)によって置換;302位のGln(Q)がGlu(E)によって置換;および328位のPro(P)がLeu(L)によって置換。
【請求項29】
292位のLys(K)がArg(R)によって置換されている、請求項28記載のエクスビボ方法。
【請求項30】
CH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている、請求項21〜27のいずれか一項記載のエクスビボ方法。
【請求項31】
CH3領域がSEQ ID NO: 20に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項27記載のエクスビボ方法:
234位のArg(R)がGln(Q)によって置換;276位のMet(M)がVal(V)によって置換;288位のLys(K)がArg(R)によって置換;298位のGln(Q)がGlu(E)によって置換;および324位のPro(P)がLeu(L)によって置換。
【請求項32】
234位のArg(R)がGln(Q)によって置換されている、請求項31記載のエクスビボ方法。
【請求項33】
234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている、請求項31記載のエクスビボ方法。
【請求項34】
5つすべての置換がなされている、請求項31記載のエクスビボ方法。
【請求項35】
CH3領域がSEQ ID NO: 21に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項27記載のエクスビボ方法:
285位のArg(R)がGln(Q)によって置換;314位のSer(S)がAsn(N)によって置換;322位のAsn(N)がLys(K)によって置換;327位のMet(M)がVal(V)によって置換;339位のLys(K)がArg(R)によって置換;349位のGln(Q)がGlu(E)によって置換;352位のIle(I)がVal(V)によって置換;365位のArg(R)がHis(H)によって置換;366位のPhe(F)がTyr(Y)によって置換;および375位のPro(P)がLeu(L)によって置換。
【請求項36】
285位のArg(R)がGln(Q)によって置換されている、請求項35記載のエクスビボ方法。
【請求項37】
285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている、請求項35記載のエクスビボ方法。
【請求項38】
10個すべての置換がなされている、請求項35記載のエクスビボ方法。
【請求項39】
第2の抗体がIgG4抗体である、請求項1〜20または22〜26のいずれか一項記載のエクスビボ方法。
【請求項40】
第1の抗体がコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含み、および、第2抗体がコアヒンジ領域内にCPPCを含み、かつIgG4様CH3領域を含む、請求項1または2記載のエクスビボ方法。
【請求項41】
第1の抗体および/または第2の抗体がヒト抗体である、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項42】
段階c)における条件が、コアヒンジ領域外のジスルフィド架橋の有意な還元または異性化が起こらないように選択される、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項43】
段階c)が還元剤の添加を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項44】
段階c)が、グルタチオン、L-システイン、ジチオスレイトール、β-メルカプト-エタノール、およびシステアミンからなる群より選択される作用物質の添加を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項45】
作用物質の濃度が、段階c)で生じる溶液の酸化還元電位が実施例31に記載する条件下で、1μMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10μMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば50μMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば0.1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である、請求項43または44記載のエクスビボ方法。
【請求項46】
作用物質の濃度が、段階c)で生じる溶液の酸化還元電位が実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である、請求項43または44記載のエクスビボ方法。
【請求項47】
作用物質の濃度が、段階c)で生じる溶液の酸化還元電位が1 Mのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、例えば100 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性が低い、15 mMのグルタチオンによって生じる酸化還元電位と等しいかもしくはより還元性が低い濃度である、請求項45または46記載のエクスビボ方法。
【請求項48】
段階c)が、還元型グルタチオンの存在下での、20℃またはそれ以上、例えば37℃の温度における、少なくとも1時間、例えば少なくとも2時間、例えば少なくとも5時間、例えば少なくとも10時間の、抗体のインキュベーションを含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項49】
結果として得られた組成物中の抗体分子の10%未満、例えば5%未満、例えば2%未満、例えば1%未満が凝集状態である、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項50】
半分子のさらなる交換を停止させるために、段階c)において得られた組成物を非還元条件に導くさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項51】
好ましくは以下から選択される方法を用いて、二重特異性抗体を安定化するさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法:
a) ヒンジ領域内のシステインを化学的に架橋する、
b) 半分子上の炭水化物側鎖を化学的に架橋する、および
c) CH3領域内に非対称的に導入されたシステインを架橋する。
【請求項52】
好ましくは非還元条件下で、二重特異性抗体を精製するさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項53】
治療用途のために、結果として得られた二重特異性抗体を薬学的組成物に製剤化するさらなる段階を含む、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項54】
第1の抗体が、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD38、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する、またはCD79aもしくはCD79bに対する結合特異性を有する、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項55】
第2の抗体が、腫瘍細胞、またはerbB1、erbB2、erbB3、erbB4、MUC-1、CD19、CD20、CD4、もしくはCXCR5などの腫瘍細胞タンパク質に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項56】
第1の抗体がerbB1に対する結合特異性を有し、かつ第2の抗体がerbB2に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項57】
第1の抗体がCD19に対する結合特異性を有し、かつ第2の抗体がCD20に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項58】
第1の抗体がCD4に対する結合特異性を有し、かつ第2の抗体がCXCR5に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項59】
第1の抗体がCD38に対する結合特異性を有し、かつ第2の抗体がCD34に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項60】
第1の抗体が病原微生物に対する結合特異性を有する、請求項1〜53のいずれか一項記載のエクスビボ方法。
【請求項61】
第2の抗体が、CD3、CD25、CD28、CD16、CD89、CD32、またはCD1などのエフェクター細胞タンパク質に対する結合特異性を有する、請求項54または60記載のエクスビボ方法。
【請求項62】
第2の抗体が化学療法薬に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項63】
第2の抗体が、血清アルブミンなどの血液タンパク質、またはトランスフェリンなどの脳タンパク質、または肝臓タンパク質に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項64】
第2の抗体が組織因子などの血液凝固に関与するタンパク質に対する結合特異性を有する、請求項54記載のエクスビボ方法。
【請求項65】
第1および/または第2の抗体が、細胞毒性剤;放射性同位体;プロドラッグまたは薬物、例えばタキサンなど;サイトカイン;ケモカイン、および補体、例えばC1qなどからなる群より選択される化合物に連結されている、前記請求項のいずれか一項記載のエクスビボ方法。
【請求項66】
以下の段階を含む、二重特異性抗体を作製するためのエクスビボ方法:
a) 第1の結合特異性を有する第1の抗体を提供する段階であって、該第1の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
b) 該第1の結合特異性と異なる、第2の結合特異性を有する第2の抗体を提供する段階であって、該第2の抗体がコアヒンジ領域内のCPPC配列およびIgG4 CH3領域を含む段階、
c) コアヒンジ領域内のシステインがジスルフィド結合の異性化を起こすことを可能にする還元条件下で、該第1の抗体および該第2の抗体を共にインキュベートする段階、ならびに
d) 二重特異性抗体を得る段階。
【請求項67】
段階c)において還元剤が添加され、該剤の濃度が、段階c)で生じる溶液の酸化還元電位が実施例35に記載する条件下で、1 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば2 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば4 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば6 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば8 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である、例えば10 mMのグルタチオンによって生じる酸化還元電位と等しいかまたはより還元性である濃度である、請求項66記載のエクスビボ方法。
【請求項68】
前記請求項のいずれか一項記載の方法によって得られるか、または得られ得る、単離された二重特異性抗体。
【請求項69】
2つのIgG4様CH3領域を含む、単離された二重特異性抗体。
【請求項70】
コアヒンジ領域内に1つまたは2つのCPPC配列を含む、請求項69記載の二重特異性抗体。
【請求項71】
コアヒンジ領域内に、X1およびX2の両方がプロリンではないという条件で、X1およびX2が任意のアミノ酸であってよい、1つまたは2つのCX1X2C配列を含む、請求項69記載の二重特異性抗体。
【請求項72】
コアヒンジ領域内に、X3がプロリンを除く任意のアミノ酸であってよい、1つまたは2つのCX3PCまたはCPX3C配列を含む、請求項69記載の二重特異性抗体。
【請求項73】
コアヒンジ領域内に1つまたは2つのCSPC、CPSC、CRPC、またはCPRC配列を含む、請求項69記載の二重特異性抗体。
【請求項74】
第1および/または第2のCH3領域が非IgG4アイソタイプのものであって、このCH3配列が、ジスルフィド結合または共有結合性のもしくは安定した非共有結合性の重鎖間結合の形成に関与するいかなるアミノ酸残基も含まないが他のペプチドはCH3領域の同一のアミノ酸配列を含む配列であるか、またはそのように改変されている、請求項69〜73のいずれか一項記載の二重特異性抗体。
【請求項75】
第1および/または第2のCH3領域がSEQ ID NO: 19に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項74記載の二重特異性抗体:
238位のArg(R)がGln(Q)によって置換;239位のAsp(D)がGlu(E)によって置換;292位のLys(K)がArg(R)によって置換;302位のGln(Q)がGlu(E)によって置換;および328位のPro(P)がLeu(L)によって置換。
【請求項76】
第1および/または第2のCH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がArg(R)によって置換されている、請求項75記載の二重特異性抗体。
【請求項77】
第1および/または第2のCH3領域がSEQ ID NO: 19に示す配列を有するが、292位のLys(K)がTyr(W)またはPhe(F)によって置換されている、請求項74記載の二重特異性抗体。
【請求項78】
第1および/または第2のCH3領域がSEQ ID NO: 20に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項74記載の二重特異性抗体:
234位のArg(R)がGln(Q)によって置換;276位のMet(M)がVal(V)によって置換;288位のLys(K)がArg(R)によって置換;298位のGln(Q)がGlu(E)によって置換;および324位のPro(P)がLeu(L)によって置換。
【請求項79】
234位のArg(R)がGln(Q)によって置換されている、請求項74記載の二重特異性抗体。
【請求項80】
234位のArg(R)がGln(Q)によって置換されており;かつ324位のPro(P)がLeu(L)によって置換されている、請求項74記載の二重特異性抗体。
【請求項81】
第1および/または第2のCH3領域がSEQ ID NO: 21に示す配列を有し、このCH3領域が、以下のアミノ酸置換のうちの1つまたは複数がなされるように改変されている、請求項74記載の二重特異性抗体:
285位のArg(R)がGln(Q)によって置換;314位のSer(S)がAsn(N)によって置換;322位のAsn(N)がLys(K)によって置換;327位のMet(M)がVal(V)によって置換;339位のLys(K)がArg(R)によって置換;349位のGln(Q)がGlu(E)によって置換;352位のIle(I)がVal(V)によって置換;365位のArg(R)がHis(H)によって置換;366位のPhe(F)がTyr(Y)によって置換;および375位のPro(P)がLeu(L)によって置換。
【請求項82】
285位のArg(R)がGln(Q)によって置換されている、請求項81記載の二重特異性抗体。
【請求項83】
285位のArg(R)がGln(Q)によって置換されており;かつ375位のPro(P)がLeu(L)によって置換されている、請求項81記載の二重特異性抗体。
【請求項84】
第1および/または第2のCH3領域がIgG4 CH3領域である、請求項69〜73のいずれか一項記載の二重特異性抗体。
【請求項85】
コアヒンジ領域外およびCH3領域外の配列が、IgG1、IgG2、IgG3、およびIgG4からなる群より選択されるアイソタイプのものである、請求項68〜84のいずれか一項記載の二重特異性抗体。
【請求項86】
請求項69〜85のいずれか一項記載の二重特異性抗体を含む薬学的組成物。
【請求項87】
薬として使用するための、請求項86記載の組成物。
【請求項88】
癌の治療用の薬として使用するための、請求項86記載の組成物。
【請求項89】
癌の治療用の薬を調製するための、請求項1〜67のいずれか一項記載の方法によって得られるかもしくは得られ得る二重特異性抗体かまたは請求項68〜85のいずれか一項記載の二重特異性抗体の、使用。
【請求項90】
以下の段階を含む、所望の特性を有する二重特異性抗体を選択する方法:
a) 一組の抗体を提供する段階であって、各抗体が異なる標的特異性を有し、かつ各抗体がIgG4様CH3領域を含む段階、
b) 還元条件下で、該一組の抗体の各抗体を該一組の別の抗体と共にインキュベートし、それによって一組の抗体混合物を作製する段階であって、各混合物が異なる二重特異性抗体を含む段階、
c) 結果として得られた一組の抗体混合物を所与の所望の特性についてアッセイする段階、および
d) 所望の特性を有する二重特異性抗体混合物を選択する段階。
【請求項91】
段階b)が、請求項2〜67における段階c)について規定されるさらなる特性の1つまたは複数を有する、請求項90記載の方法。
【請求項92】
所望の特性が腫瘍細胞殺傷である、請求項90または91記載の方法。
【図1】

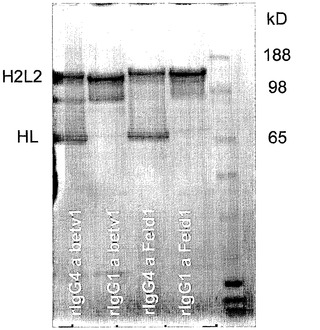
【図2】
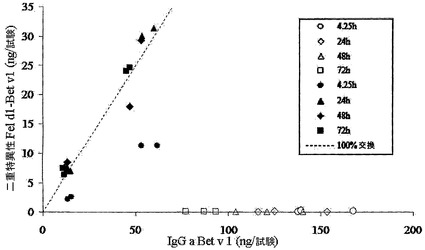
【図3A】
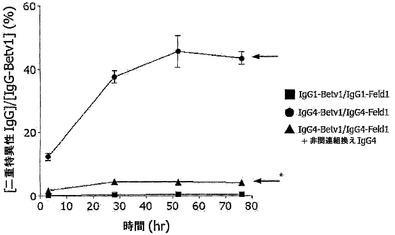
【図3B】
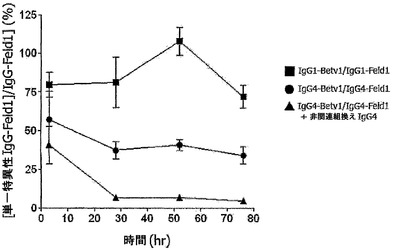
【図4】
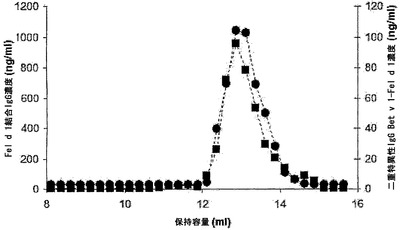
【図5A】
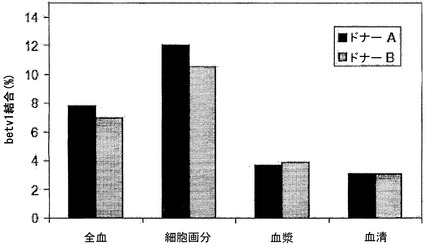
【図5B】
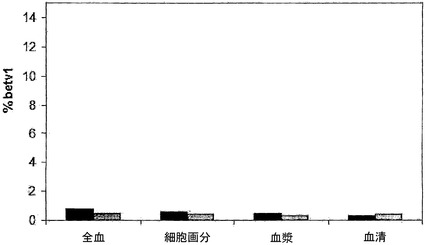
【図5C】
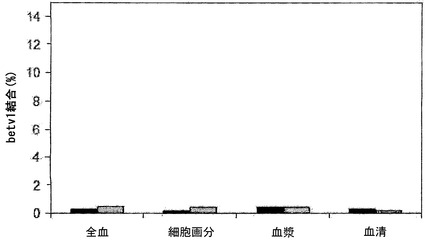
【図6】
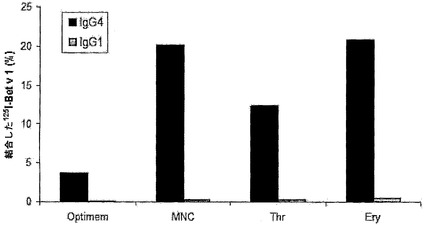
【図7】
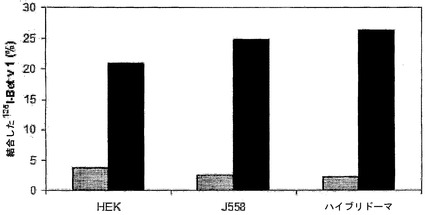
【図8】
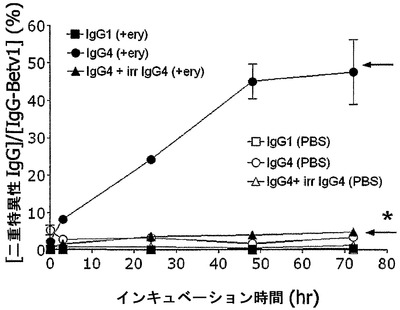
【図9A】
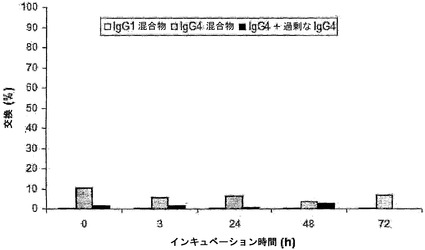
【図9B】
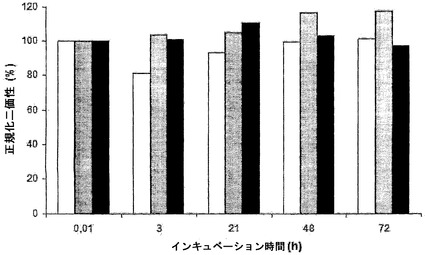
【図10】
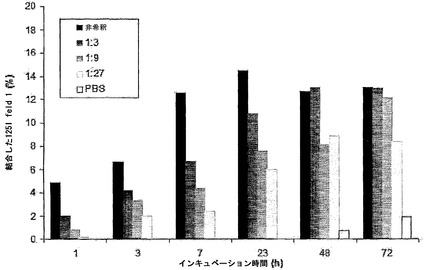
【図11】
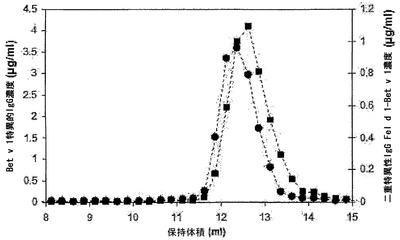
【図12】
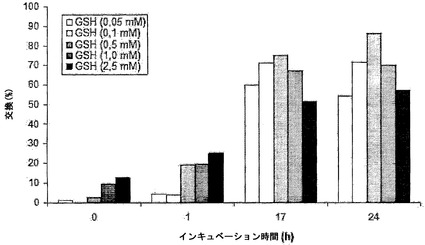
【図13】
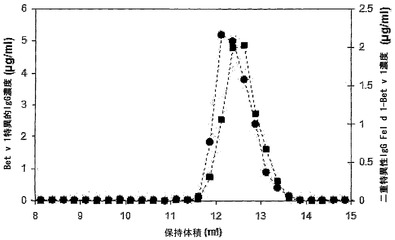
【図14】
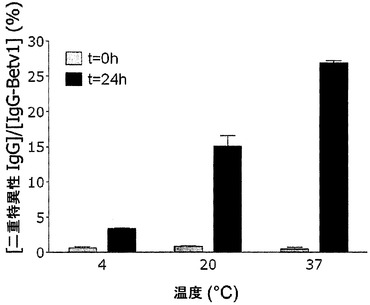
【図15】
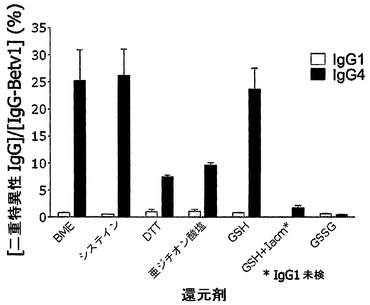
【図16A】
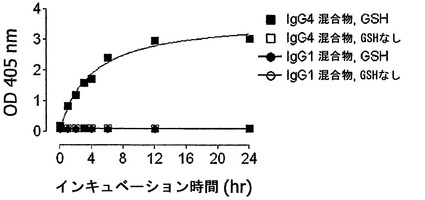
【図16B】
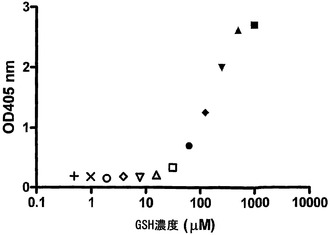
【図16C】
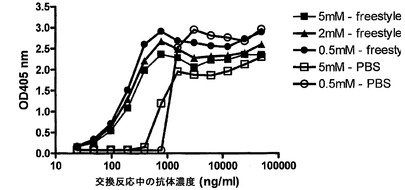
【図16D】

【図16E】
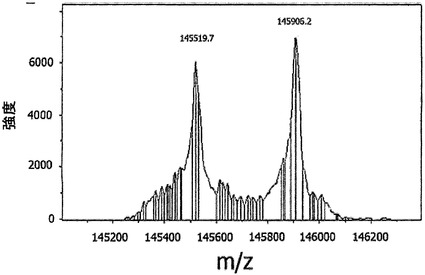
【図16F】
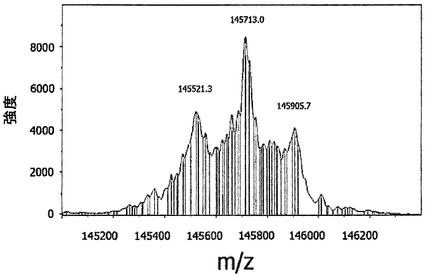
【図17A】
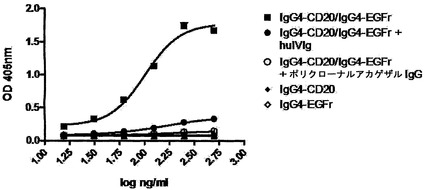
【図17B】
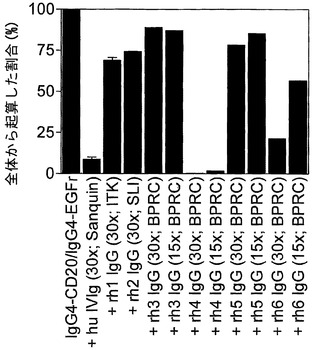
【図17C】
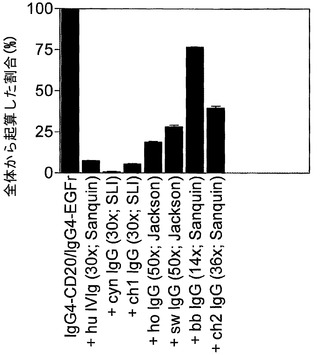
【図18】

【図19A】
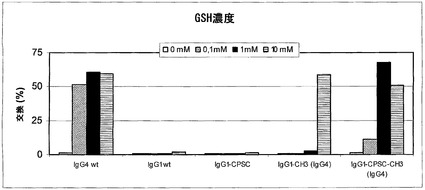
【図19B】
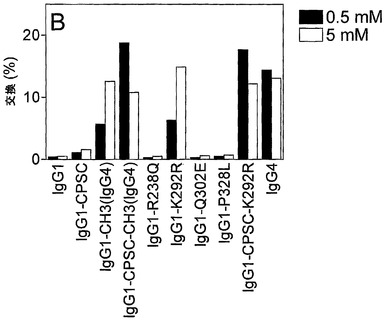
【図19C】
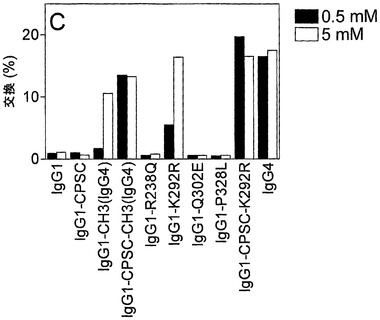
【図20A】
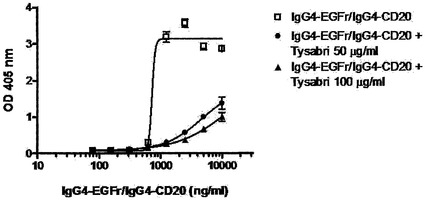
【図20B】
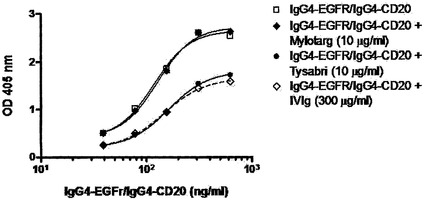
【図21】
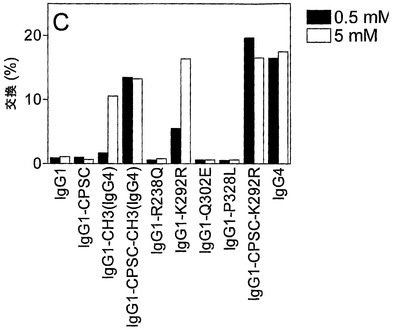
【図22A】
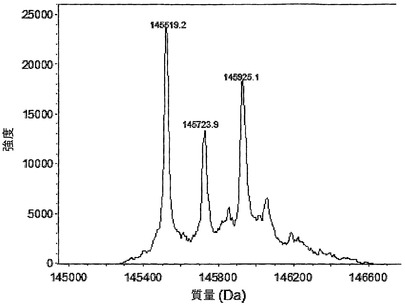
【図22B】
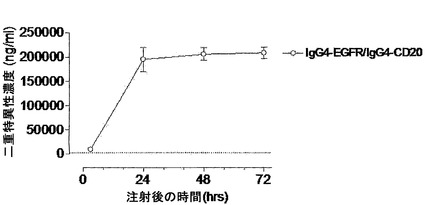
【図23】
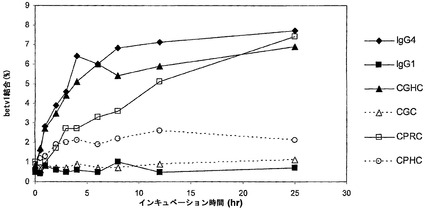
【図24】

【図2】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図5C】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16A】
【図16B】
【図16C】
【図16D】
【図16E】
【図16F】
【図17A】
【図17B】
【図17C】
【図18】
【図19A】
【図19B】
【図19C】
【図20A】
【図20B】
【図21】
【図22A】
【図22B】
【図23】
【図24】
【公表番号】特表2010−522701(P2010−522701A)
【公表日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願番号】特願2010−500070(P2010−500070)
【出願日】平成20年3月28日(2008.3.28)
【国際出願番号】PCT/DK2008/000124
【国際公開番号】WO2008/119353
【国際公開日】平成20年10月9日(2008.10.9)
【出願人】(507316398)ゲンマブ エー/エス (18)
【Fターム(参考)】
【公表日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願日】平成20年3月28日(2008.3.28)
【国際出願番号】PCT/DK2008/000124
【国際公開番号】WO2008/119353
【国際公開日】平成20年10月9日(2008.10.9)
【出願人】(507316398)ゲンマブ エー/エス (18)
【Fターム(参考)】
[ Back to top ]