
二重特異性抗ErbB−2/抗c−Met抗体
本発明は、ヒトErbB−2及びヒトc−Metに対する二重特異性抗体、それらの製造方法、当該抗体を含んでいる医薬組成物、並びにその使用に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒトErbB−2及びヒトc−Metに対する二重特異性抗体、それらの製造方法、前述の抗体を含む医薬組成物、並びにその使用に関する。
【背景技術】
【0002】
ErbBファミリー・タンパク質
ErbBタンパク質ファミリーは、4種類のメンバーであるErbB−1(別名、上皮生長因子受容体(EGFR))、ErbB−2(ヒトではHER2、齧歯動物ではneu)、ErbB−3(別名、HER3)及びErbB−4(別名、HER4)から成る。ErbBファミリー・タンパク質は、受容体チロシンキナーゼであって、細胞増殖、分化、及び生存の重要な伝達物質となっている。
【0003】
ErbB−2と抗ErbB−2抗体
ErbBタンパク質ファミリーの第2のメンバーであるErbB−2(ERBB2、HER2;CD340、HER−2/neu、c−erb B2/neuタンパク質、神経芽細胞腫/神経膠芽腫由来癌遺伝子相同体|v−erb−b2 鳥類赤血球性白血病ウイルス癌遺伝子相同体2;配列番号14としても知られている)は、それ自身のリガンド結合ドメインを持たないタンパク質であるため、成長因子に結合することができない。しかしながら、他のリガンド結合EGF受容体ファミリーメンバーに強く結合してヘテロ二量体を形成し、リガンド結合を安定させ、そして下流のシグナル伝達経路のキナーゼ媒介型活性化を促進する、例えばそれらはマイトジェン活性化プロテインキナーゼやホスファチジルイノシトール−3キナーゼに関与している。本明細書中に示した最も頻度の高い対立遺伝子Ile654/Ile655を含めて、第654及び655位アミノ酸における対立遺伝子変異のアイソフォームa(第624及び625位のアイソフォームb)が報告されている。この遺伝子の増幅、及び/又は過剰発現は、乳房及び卵巣の腫瘍を含めた多数の癌で報告されている。選択的スプライシングはいくつかのさらなる転写物変異体をもたらすが、一部のものは異なるアイソフォームをコードし、他のものは完全に特徴づけされていない。ErB−2は、元々、形質変換遺伝子の産物として化学物質で処理されたラットの神経芽細胞腫から同定された。neu癌原遺伝子の活性型は、コードされたタンパク質の膜貫通領域における(バリンからグルタミン酸への)点突然変異から得られた(Semba, K., et al., PNAS 82 (1985) 6497-501;Coussens, L., et al., Science 230 (1985) 1132-9;Bargmann, C.I., et al., Nature 319 (1986) 226-30;Yamamoto, T., et al., Nature 319 (1986) 230-4)。
【0004】
neuのヒト相同体の増幅は、乳癌及び卵巣癌で観察され、且つ、予後不良と相関する(Slamon, D.J, et al. Science 235 (1987) 177-182;Slamon, D.J., et al., Science 244 (1989) 707-712;及びUS 4,968,603)。これまで、neu癌原遺伝子に対する点突然変異相同体は、ヒトの腫瘍については報告されていない。(一様でないが高い頻度で遺伝子増幅による)HER2の過剰発現は、胃、子宮内膜、唾液腺、肺、腎臓、結腸、甲状腺、膵臓及び膀胱の癌腫を含めた他の癌腫においても観察されている。特に、King, C.R., et al., Science 229 (1985) 974-976;Yokota, J, et al., Lancet 1 (1986) 765-767;Fukushige, S., et al., Mol Cell Biol. 6 (1986) 955-958;Guerin, M., et al., Oncogene Res. 3 (1988) 21-31;Cohen, J.A., et al., Oncogene, 4 (1989) 81-88;Yonemura, Y., et al., Cancer Res. 51 (1991) 1034-1038;Borst, M.P., et al., Gynecol. Oncol. 38 (1990) 364-366;Weiner, D.B., et al., Cancer Res. 50 (1990) 421-425;Kern, J.A., et al., Cancer Res. 50 (1990) 5184-5187;Park, J.B., et al, Cancer Res. 49 (1989) 6605-6609;Zhau, H.E., et al., Mol. Carcinog. 3 (1990) 254-257;Aasland, R., et al., Br. J. Cancer 57 (1988) 358-363;Williams, T.M, et al, Pathobiology 59 (1991) 46-52;及びMcCann, A, et al. Cancer 65 (1990) 88-92を参照のこと。HER2は、前立腺癌において過剰発現されることもある(Gu, K, et al., Cancer Lett. 99 (1996) 185-189;Ross, J.S, et al. Hum. Pathol. 28 (1997) 827-833;Ross, J.S, et al. Cancer 79 (1997) 2162-2170;及びSadasivan, R, et al, J. Urol. 150 (1993) 126-131)。
【0005】
ヒトHER2タンパク質産物に対する抗体が、例えばHudziak, R.M, et al, Mol. Cell. Biol. 9 (1989) 1165-1172によって作出された(前記文献は、ヒト乳房腫瘍細胞株SK−BR−3を使用して特徴づけされた抗HER2抗体のパネルの作出について記載している)。この抗HER2抗体のパネルには、とりわけ、(HER2の細胞外ドメインの異なるエピトープに向けられている)2C4(パーツズマブ)及び4D5(トラスツズマブ、Herceptin(商標))抗体が含まれる。前記抗体への暴露後のSK−BR−3細胞の相対細胞増殖は、72時間後の単分子層のクリスタルバイオレット染色によって測定された。このアッセイを使用すると、細胞増殖を56%阻害した4D5(トラスツズマブ、Herceptin(商標))と呼ばれる抗体で、最大の阻害が得られた。前記パネル中の他の抗体は、このアッセイにおいて細胞増殖をある程度減少させた。抗体4D5については、TNF−αの細胞傷害効果に対するHER2過剰発現乳房腫瘍細胞株の感受性を高めることがさらに見出された(US5,677,171)。Hudziak, R.M.らが議論しているHER2抗体は、例えばFendly, B.M, et al. Cancer Research 50 (1990) 1550-1558においてさらに特徴づけされている。
【0006】
c−Metと抗c−Met抗体
MET(間葉−上皮移行因子)は、タンパク質METをコードする癌原遺伝子であり、(c−Met、肝細胞増殖受容体HGFR;HGF受容体;細胞分散因子受容体;SF受容体;配列番号15としても知られている)(Dean, M., et al., Nature 318 (1985) 385-8;Chan, A.M., et al., Oncogene 1 (1987) 229-33;Bottaro, D.P., et al., Science 251 (1991) 802-4;Naldini, L., et al.5 EMBO J. 10 (1991) 2867-78;Maulik, Gautam., et al., Cytokine Growth Factor Rev. 13 (2002) 41-59)。METは、胚発生と創傷治癒に不可欠な膜受容体である。肝細胞増殖因子(HGF)は、MET受容体の唯一の既知のリガンドである。METは通常、上皮起源の細胞によって発現されるが、HGFの発現は間葉起源の細胞に限られている。HGF刺激により、METは浸潤性増殖として知られているプログラムをまとめて引き起こす数個の生物反応を誘発する。癌における異常なMET活性化は、異常な活性のMETが腫瘍増殖や、腫瘍に栄養を供給する新生血管の形成(血管新生)や、他の臓器への癌の分散(転移癌)を引き起こす、予後不良と関連がある。METは、腎臓、肝臓、胃、乳房、及び脳の癌を含めた多くのタイプのヒト悪性腫瘍において無秩序である。本来なら、幹細胞と始原細胞だけがMETを発現しており、それが胎児において新しい組織を作り出したり、成人において損傷した組織を再生するために、これらの細胞が侵襲的に増殖することを可能にしている。しかしながら、癌幹細胞は、METを発現する正常幹細胞の能力を乗っ取り、それによって癌持続性と体内の他の部位への分散の原因となっている。
【0007】
癌原遺伝子MET産物は、肝細胞増殖因子受容体であり、チロシンキナーゼ活性をコードしている。初期一本鎖前駆タンパク質は、翻訳後に分割されて、α及びβサブユニットを生じ、それがジスルフィド結合して成熟受容体を形成する。METの遺伝子内の様々な突然変異は乳頭状腎癌に関連している。
【0008】
抗c−Met抗体は、例えば、US5,686,292、US7,476,724、WO2004/072117、WO2004/108766、WO2005/016382、WO2005/063816、WO2006/015371、WO2006/104911、WO2007/126799、又はWO2009/007427により知られている。
【0009】
c−Met結合ペプチドは、例えば、Matzke, A., et al., Cancer Res 65 (14) (2005) 6105-10及びTam, Eric, M., et al., J. Mol. Biol. 385 (2009)79-90により知られている。
【0010】
多特異性抗体
広範な種類の組換え抗体フォーマットが最近開発されてきている、例えば、IgG抗体フォーマットと一本鎖ドメインの融合による、例えば、四価二重特異性抗体が開発された(例えば、Coloma M. J. et al., Nature Biotech. 15(1997)159-163;WO2001077342;及びMorrison S. L. Nature Biotech. 25(2007)1233-1234を参照のこと)。
【0011】
また、抗体コア構造(IgA、IgD、IgE、IgG又はIgM)がもはや保持されていないいくつかの新しいフォーマット、例えば、2つ以上の抗原に結合できる、ジ抗体、トリ抗体又はテトラ抗体、ミニ抗体、いくつかの一本鎖フォーマット(scFv、ビス−scFv)が開発された(Holliger P. et al., Nature Biotech. 23(2005)1126-1136;Fischer N., Leger O. Pathobiology 74(2007)3-14;Shen J. et al., J. Immunol. Methods 318(2007)65-74;Wu C. et al., Nature Biotech. 25(2007)1290-1297)。
【0012】
すべてのこのようなフォーマットはリンカーを使用して抗体コア(IgA、IgD、IgE、IgG又はIgM)を更なる結合性タンパク質(例えば、scFv)に融合するか、あるいは、例えば、2つのFabフラグメント又はscFvを融合する(Fischer N., Leger O. Pathobiology 74(2007)3-14)。天然に存在するものに対して高度の類似性を維持することによって、Fc受容体結合を通して介在される、エフェクター機能、例えば、補体依存性細胞毒性(CDC)又は抗体依存性細胞毒性(ADCC)を保持することが望まれることもあり得ることに留意すべきである。
【0013】
WO2007/024715では、改変多価及び多特異性結合タンパク質として二重可変ドメイン免疫グロブリンが報告されている。生物学的に活性な抗体二量体の調製方法は、US6,897,044に報告されている。ペプチド・リンカーを介して互いに連結されている少なくとも4つの可変ドメインを持つ多価性Fv抗体構築物が、US7,129,330で報告されている。二量体及び多量体の抗原結合構築物は、US2005/0079170で報告されている。(天然の免疫グロブリンでない)結合構造によって互いに共有結合した3つ又は4つのFabフラグメントを含んでなる三又は四価の単一特異性抗原結合性タンパク質が、US6,511,663で報告されている。WO2006/020258では、原核生物及び真核生物細胞で効率的に発現され、治療薬や診断法に有用である四価の二重特異性抗体が報告されている。2つのタイプのポリペプチド二量体を含んでなる混合物からの少なくとも1つの鎖間ジスルフィド結合によって連結されず二量体からの少なくとも1つの鎖間ジスルフィド結合によって連結される二量体を分離するか、又は優先的には合成する方法が、US2005/0163782で報告されている。二重特異性の四価の受容体が、US5,959,083で報告されている。三重機能性以上の抗原結合部位を持つ改変抗体が、WO2001/077342で報告されている。
【0014】
多特異性、且つ、多価の抗原結合性ポリペプチドが、WO1997/001580で報告されている。WO1992/004053では、通常、同じ抗原決定基に結合するIgGクラスのモノクローナル抗体から調製されたホモ接合体が合成架橋によって共有結合で連結されていることが報告されている。2つ以上の免疫グロブリンモノマーが共に四価又は六価のIgG分子を形成するのに関係している、(通常IgGクラスのオリゴマーが分泌されることよって)抗原に対して高い親和性を有するオリゴマーモノクローナル抗体が、WO1991/06305で報告されている。ヒツジ由来抗体及び改変抗体構築物が、US6,350,860で報告されていて、それは、インターフェロンγ活性が病因である疾患を治療するのに使用される。US2005/0100543では、二重特異性抗体の多価担体である標的を設定可能な、すなわち、標的を設定可能な構築物の各分子が2以上の二重特異性抗体の担体として機能することができる、構築物が報告されている。遺伝的に改変された二重特異性四価抗体は、WO1995/009917で報告されている。安定しているWO2007/109254には、安定化scFvから成るか、又はそれを含んでなる安定化結合分子が報告されている。US2007/0274985は、一本鎖Fab(scFab)フラグメントを含んでなる抗体フォーマットに関する。
【0015】
WO2008/140493は、抗EGFRファミリーメンバー抗体及び1若しくは複数の抗EGFRファミリーメンバー抗体を含んでなる二重特異性抗体に関する。US2004/0071696は、EGFRタンパク質ファミリーのメンバーに結合する二重特異性抗体分子に関する。
【0016】
WO2009111707(A1)は、Met及びHER拮抗薬を用いた併用療法に関する。WO2009111691(A2A3)は、Met及びEGFR拮抗薬を用いた併用療法に関する。
【0017】
WO2004072117は、c−Metの下方制御/内部移行を誘発するc−Met抗体、及び特に第2抗原としてErbB−2を用いた二重特異性抗体におけるそれらの使用可能性に関する。
【発明の概要】
【0018】
この発明の第一の態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、前記二重特異性抗体は、フローサイトメトリーアッセイにおいて1時間後に計測した場合に、OVCAR−8細胞上での、抗体の不存在下でのc−Metの内部移行と比較して15%未満のc−Metの内部移行を示すことを特徴とする。
【0019】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する1又は2つの抗原結合部位、及びヒトc−Metに特異的に結合する1つの抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二価又は三価の二重特異性抗体である。
【0020】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する2つの抗原結合部位、及びヒトc−Metに特異的に結合する3つ目の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する三価の二重特異性抗体である。
【0021】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する1つの抗原結合部位、及びヒトc−Metに特異的に結合する1つの抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二価の二重特異性抗体である。
【0022】
本発明の一態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位、及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、以下の:
前記第1の抗原結合部位が、重鎖可変ドメイン内に配列番号15のCDR3H領域、配列番号16のCDR2H領域、及び配列番号17のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号18のCDR3L領域、配列番号19のCDR2L領域、及び配列番号20のCDR1L領域を含んでなり;そして
前記第2の抗原結合部位が、重鎖可変ドメイン内に配列番号21のCDR3H領域、配列番号22のCDR2H領域、及び配列番号23のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号24のCDR3L領域、配列番号25のCDR2L領域、配列番号26のCDR1L領域を含んでなる、
を特徴とする前記二重特異性抗体である。
【0023】
前記二重特異性抗体は、好ましくは、以下の:
ErbB−2に特異的に結合する前記第1の抗原結合部位が、重鎖可変ドメインとして配列番号1の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号2の配列を含んでなり;そして
c−Metに特異的に結合する前記第2の抗原結合部位が、重鎖可変ドメインとして配列番号3の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号4の配列を含んでなる、
を特徴とする。
【0024】
本発明の更なる態様は、IgG1又はIgG3サブクラスの定常領域を含んでなることを特徴とする本発明による二重特異性抗体である。
【0025】
一実施形態では、本発明による前記二重特異性抗体は、前記抗体がAsn297にて糖鎖でグリコシル化されていることで、前記糖鎖内のフコース量が65%以下であることを特徴とする。
【0026】
本発明の更なる態様は、前記二重特異性抗体の鎖をコードする核酸分子である。
【0027】
本発明のより更なる態様は、前記二重特異性抗体を含んでなる医薬組成物、癌治療のための前記組成物、癌治療薬の製造のための前記二重特異性抗体の使用、そのような処置を必要としている患者に前記二重特異性抗体を投与することを含めた癌に罹患している患者の治療法である。
【0028】
乳房の腫瘍は高レベルのErbB2を発現していることが多く、そしてErbB2陽性腫瘍のうちのかなりの割合がさらにc−Met陽性でもある。乳房の腫瘍におけるc−Met発現が予後不良と相関があることは、これまで多くの研究で示されてきた(Kang, L, Y., et al., Cancer Res. 63 (2003) 1101-1105;Lengyel, E., et al., Int. J. Cancer 113 (2005) 678-82)。そのため、本発明による二重特異性<ErbB−2−c−Met>抗体は、抗腫瘍効果や癌細胞阻害のような価値のある特性を有する。
【0029】
本発明による抗体は、例えば、とりわけErbB2とc−Metの両受容体が発現されている癌細胞の増殖阻害や癌に罹患している患者に有益な抗腫瘍効果といった、非常に価値のある特性を示す。本発明による二重特異性<ErbB2−c−Met>抗体は、ErbB2とc−Metの両受容体を発現している癌細胞に対してそれらの親である単一特異性の二価<c−Met>抗体と比較すると、低いc−Met受容体の内部移行を示す。
【0030】
発明の詳細な説明
この発明の第一の態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、前記二重特異性抗体は、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、前記二重特異性抗体の不存在下でのc−Metの内部移行と比較して、OVCAR−8細胞に対する15%未満のc−Metの内部移行を示すことを特徴とする。
【0031】
よって、本発明は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体に関するものであり、ここで、前記二重特異性抗体は、フローサイトメトリーアッセイにより、1時間のOVCAR−8細胞−抗体インキュベーション後に測定した場合、前記二重特異性抗体の不存在下でのOVCAR−8細胞上のc−Metの内部移行と比較して15%未満の、OVCAR−8細胞上のc−Metの内部移行を増加させる。
【0032】
一実施形態では、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体は、前記二重特異性抗体が、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して、10%未満のc−Metの内部移行を示すことを特徴とする。
【0033】
一実施形態では、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体は、前記二重特異性抗体が、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して、7%未満のc−Metの内部移行を示すことを特徴とする。
【0034】
一実施形態では、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体は、前記二重特異性抗体が、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して、5%未満のc−Metの内部移行を示すことを特徴とする。
【0035】
「c−Metの内部移行」という用語は、抗体の不存在下でのc−Metの内部移行と比較した場合の、OVCAR−8細胞(NCI Cell Line designation;NCI(国立癌研究所)から購入したOVCAR-8-NCI;Schilder, RJ. et al., Int. J. Cancer 45 (1990) 416-422;Dcediobi, O.N. et al., Mol. Cancer. Ther. 5 (2006) 2606- 2012;Lorenzi, P.L., et al., Mol. Cancer Ther. 8 (2009) 713-724)上での、抗体誘導型のc−Met受容体の内部移行を指す。c−Met受容体のそのような内部移行は、本発明による二重特異性抗体によって引き起こされ、そして実施例11に記載のとおりフローサイトメトリーアッセイ(FACS)によって1時間後に計測される。本発明による二重特異性抗体は、抗体の不存在下でのc−Metの内部移行と比較した場合に、抗体への暴露の1時間後にOVCAR−8細胞における15%未満のc−Metの内部移行を示す。一実施形態では、前記抗体は10%未満のc−Metの内部移行を示す。一実施形態では、前記抗体は7%未満のc−Metの内部移行を示す。一実施形態では、前記抗体は5%未満のc−Metの内部移行を示す。
【0036】
この発明の別の態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体であって、フローサイトメトリーアッセイによって1時間後にOVCAR−8細胞において計測したときに、(対応する)単一特異性の二価の親c−Met抗体によって誘発されたc−Metの内部移行と比較して、前記二重特異性抗体が、50%以上(一実施形態では60%以上;他の実施形態では70%以上、一実施形態では80%以上)c−Metの内部移行を減少させることを特徴とする。c−Metの内部移行の減少は、以下のとおり計算される(OVCAR−8細胞におけるフローサイトメトリーアッセイにより1時間後に計測される内部移行率(%)の値を使用しているが、0より低い内部移行率(%)の値は0%内部移行と設定する、例えばBsAB02に関して(−7%内部移行を0%の内部移行と設定した)):100×(単一特異性の二価の親c−Met抗体によって誘発されたc−Metの内部移行率(%)−二重特異性ErbB−2/cMet抗体によって誘発されたc−Metの内部移行率(%))/単一特異性の二価の親c−Met抗体によって誘発されたc−Metの内部移行率(%)。例えば:二重特異性ErbB2/cMet抗体BsAB02が(0%と設定される)−7%のc−Metの内部移行を示し、且つ、単一特異性の二価の親c−Met抗体Mab 5D5が37%のc−Metの内部移行を示す。よって、前記二重特異性ErbB−2/cMet抗体BsAB02は、100×(40−0)/40%=100%のc−Metの内部移行の減少を示す(実施例11でのOVCAR−8細胞におけるフローサイトメトリーアッセイにより1時間後に計測される内部移行値を参照のこと)。
【0037】
本明細書中に使用される場合、「抗体」は、抗原結合部位を含んでなる結合タンパク質を指す。「結合部位」又は「抗原結合部位」という用語は、本明細書中では、リガンドが実際に結合している抗体分子の(単数若しくは複数の)領域を意味し、抗体に由来する。「抗原結合部位」という用語には、抗体重鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)又はVH/VLの対が含まれ、抗体全体又は抗体フラグメント、例えば一本鎖Fv、VHドメイン及び/又はVLドメイン、Fab、又は(Fab)2などに由来することもある。この発明の一実施形態では、それぞれの抗体結合部位が、抗体重鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)を含んでなり、好ましくは、抗体軽鎖可変ドメイン(VL)と抗体重鎖可変ドメイン(VH)から成る1対によって形成される。
【0038】
抗体由来の抗原結合部位に加えて、例えばMatzke, A., et al., Cancer Res. 65 (14) (2005) 6105-10. July 15, 2005に記載された結合ペプチドもまた、抗原(例えばc−Met)に特異的に結合し得る。よって、この発明の更なる態様は、ヒトErbB−2に特異的に結合する抗原結合部位及びヒトc−Metに特異的に結合する結合ペプチドを含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性結合性分子である。よってこの発明の更なる態様は、ヒトc−Metに特異的に結合する抗原結合部位及びヒトErbB−2に特異的に結合する結合ペプチドを含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性結合性分子である。
【0039】
ErbB−2(ERBB2、HER2;CD340、HER−2/neu、c−erb B2/neuタンパク質、神経芽細胞腫/神経膠芽腫由来癌遺伝子相同体|v−erb−b2 鳥類赤血球性白血病ウイルス癌遺伝子相同体2;配列番号14としても知られている)は、それ自身のリガンド結合ドメインを持たないタンパク質であるため、成長因子に結合することができない。しかしながら、他のリガンド結合EGF受容体ファミリーメンバーに強く結合してヘテロ二量体を形成し、リガンド結合を安定させ、そして下流のシグナル伝達経路のキナーゼ媒介型活性化を促進する、例えばそれらはマイトジェン活性化プロテインキナーゼやホスファチジルイノシトール−3キナーゼに関与している。本明細書中に示した最も頻度の高い対立遺伝子Ile654/Ile655を含めて、第654及び655位アミノ酸における対立遺伝子変異のアイソフォームa(第624及び625位のアイソフォームb)が報告されている。この遺伝子の増幅、及び/又は過剰発現は、乳房及び卵巣の腫瘍を含めた多数の癌で報告されている。選択的スプライシングはいくつかのさらなる転写物変異体をもたらすが、一部のものは異なるアイソフォームをコードし、他のものは完全に特徴づけされていない。ErB−2は、元々、形質変換遺伝子の産物として化学物質で処理されたラットの神経芽細胞腫から同定された。neu癌原遺伝子の活性型は、コードされたタンパク質の膜貫通領域における(バリンからグルタミン酸への)点突然変異から得られた(Semba, K., et al., PNAS 82 (1985) 6497-501;Coussens, L., et al., Science 230 (1985) 1132-9;Bargmann, C.I., et al., Nature 319 (1986) 226-30;Yamamoto, T., et al., Nature 319 (1986) 230-4)。
【0040】
ヒトErbB−2に特異的に結合する抗原結合部位、特に重質鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)は、a)例えば2C4(パーツズマブ;パーツズマブは、マウス抗HER2抗体2C4の遺伝子組み換えヒト化バージョンである、そしてそれぞれの調製方法と共にWO01/00245及びWO2006/007398に記載されている)及び4D5(トラスツズマブ(マウス抗HER2抗体4D5の遺伝子組み換えヒト化バージョン、Herceptin(商標);トラスツズマブ及びその調製方法はUS5,821,337に記載されている))抗体(Hudziak, R., M, et al, Mol. Cell. Biol. 9 (1989) 1165-1172;Fendly, B., M., et al., Cancer Research 50 (1990) 1550-1558)のような既知の抗ErbB−2抗体、あるいは、b)とりわけヒトErbB−2タンパク質又は核酸若しくはその断片のいずれかを使用したデノボ免疫法によって、又はファージ提示法によって得られた新しい抗ErbB−2抗体、に由来することができる。
【0041】
MET(間葉−上皮移行因子)は、タンパク質METをコードする癌原遺伝子であり、(c−Met、肝細胞増殖受容体HGFR;HGF受容体;細胞分散因子受容体;SF受容体;配列番号13としても知られている)(Dean, M., et al., Nature 318 (1985) 385-8;Chan, A.M., et al., Oncogene 1 (1987) 229-33;Bottaro, D.P., et al., Science 251 (1991) 802-4;Naldini, L., et al.5 EMBO J. 10 (1991) 2867-78;Maulik, G., et al., Cytokine Growth Factor Rev. 13 (2002) 41-59)。METは、胚発生と創傷治癒に不可欠な膜受容体である。肝細胞増殖因子(HGF)は、MET受容体の唯一の既知のリガンドである。METは通常、上皮起源の細胞によって発現されるが、HGFの発現は間葉起源の細胞に限られている。HGF刺激により、METは浸潤性増殖として知られているプログラムをまとめて引き起こす数個の生物反応を誘発する。癌における異常なMET活性化は、異常な活性のMETが腫瘍増殖や、腫瘍に栄養を供給する新生血管の形成(血管新生)や、他の臓器への癌の分散(転移癌)を引き起こす、予後不良と関連がある。METは、腎臓、肝臓、胃、乳房、及び脳の癌を含めた多くのタイプのヒト悪性腫瘍において無秩序である。本来なら、幹細胞と始原細胞だけがMETを発現しており、それが胎児において新しい組織を作り出したり、成人において損傷した組織を再生するために、これらの細胞が侵襲的に増殖することを可能にしている。しかしながら、癌幹細胞は、METを発現する正常幹細胞の能力を乗っ取り、それによって癌持続性と体内の他の部位への分散の原因となっている。
【0042】
ヒトc−Metに特異的に結合する抗原結合部位、特に重質鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)は、a)例えばUS5,686,292、US7,476,724、WO2004/072117、WO2004/108766、WO2005/016382、WO2005/063816、WO2006/015371、WO2006/104911、WO2007/126799、又はWO2009/007427に記載の既知の抗c−Met抗体、あるいは、b)例えば、とりわけヒト抗−Metタンパク質又は核酸若しくはその断片のいずれかを使用したデノボ免疫法によって、又はファージ提示法によって得られた新しい抗c−Met抗体、に由来することができる。
【0043】
本発明の更なる態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、以下の:
ErbB−2に特異的に結合する前記第1の抗原結合部位が、重鎖可変ドメインとして配列番号1の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号2の配列を含んでなり;そして
c−Metに特異的に結合する前記第2の抗原結合部位が、重鎖可変ドメインとして配列番号3の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号4の配列を含んでなる、
を特徴とする前記二重特異性抗体である。
【0044】
抗体の特異性は、抗原の特定のエピトープに対する抗体の選択的認識を指す。例えば、天然抗体は単一特異性である。本発明による「二重特異性抗体」は、2つの異なった抗原結合特異性を有する抗体である。抗体が2以上の特異性を有する場合には、認識されたエピトープは単一の抗原又は2以上の抗原と結合することもできる。本発明の抗体は、2つの異なった抗原、すなわち第1の抗原としてのErbB−2及び第2の抗原としてのc−Metに特異的である。
【0045】
用語「単一特異性」抗体は、本明細書中で使用される場合、そのそれぞれが同じ抗原の同じエピトープに結合する1以上の結合部位を持つ抗体を意味する。
【0046】
用語「価」は、本明細書中で使用される場合、抗体分子内に存在している一定数の結合部位を意味する。同様に、用語「二価」、「四価」、及び「六価」は、抗体分子内にそれぞれ2つの結合部位、4つの結合部位、及び6つの結合部位の存在を意味する。本発明による二重特異性抗体は、少なくとも「二価」であり、そして「三価」又は「多価」(例えば「四価」若しくは「六価」)でもあり得る。
【0047】
本発明の抗体の抗原結合部位は、抗原に対する結合部位の親和性に多かれ少なかれ貢献する6つの相補性決定領域(CDRs)を含むことができる。3つの重鎖可変ドメインCDRs(CDRH1、CDRH2及びCDRH3)と3つの軽鎖可変ドメインCDRs(CDRL1、CDRL2及びCDRL3)がある。CDRとフレームワーク領域(FR)の程度は、配列内での可変性によってそれらの領域が規定されたコンパイルされたアミノ酸配列のデータベースとの比較によって決定される。いくつかのCDRを含んでなる(すなわち、結合特異性が3、4又は5つのCDRによって決定される)機能性抗原結合部位もまた、本発明の範囲内に含まれる。例えば、6つのCDRの完全セット以下でも結合に十分であり得る。場合によっては、VH又はVLドメインで十分である。
【0048】
好ましい実施形態では、本発明の抗体は、ヒト起源の1若しくは複数の免疫グロブリンクラスの免疫グロブリン定常領域を更に含む。免疫グロブリンクラスには、IgG、IgM、IgA、IgD、及びIgEアイソタイプ、そしてIgGとIgAの場合には、それらのサブタイプが含まれる。好ましい実施形態では、本発明の抗体は、IgGタイプ抗体の定常ドメイン構造を有するが、4つの抗原結合部位を有する。これは、例えば、c−Metに特異的に結合する1つ(若しくは2つ)の完全抗原結合部位(例えば一本鎖Fabフラグメント若しくは一本鎖Fv)を、ErbB−2に特異的に結合する完全な抗体の重鎖又は軽鎖のN又はC末端のいずれかに連結して、三価の二重特異性抗体(又は四価の二重特異性抗体)を得ることによって達成される。あるいは、例えばEP出願番号第07024867.9号、EP出願番号第07024864.6号、EP出願番号第07024865.3号又はRidgway, J.B., Protein Eng. 9 (1996) 617-621;WO96/027011;Merchant, A.M., et al., Nature Biotech 16 (1998) 677-681;Atwell, S., et al., J. Mol. Biol. 270 (1997) 26-35及びEP1870459ALに記載のとおり、免疫グロブリン定常領域を含んでなる、ヒトErbB−2及びヒトc−Metに対する、IgGのような二重特異性の二価抗体を使用できる。
【0049】
用語「モノクローナル抗体」又は「モノクローナル抗体組成物」とは、本明細書において使用される場合、単一のアミノ酸組成物の抗体分子の調製物を意味する。
【0050】
用語「キメラ抗体」とは、通常組換えDNA技術により調製される、可変領域、すなわち1つの源又は種からの結合領域、及び異なった源又は種に由来する定常領域の少なくとも一部を含んでなる抗体を意味する。ネズミ可変領域及びヒト定常領域を含んでなるキメラ抗体が好ましい。本発明により包含される他の好ましい形の「キメラ抗体」は、定常領域が、特にC1q結合及び/又はFc受容体(FcR)結合に関して、本発明の性質を生じるように、修飾されているか、又は元の抗体のその領域から変更されているそれらの抗体である。そのようなキメラ抗体はまた、「クラス−スイッチ抗体」とも呼ばれる。キメラ抗体は、免疫グロブリン可変領域をコードするDNAセグメント、及び免疫グロブリン定常領域をコードするDNAセグメントを含んでなる免疫グロブリン遺伝子が発現された生成物である。キメラ抗体を生成するための方法は、当業界において現在良く知られている、従来の組換えDNA及び遺伝子トランスフェクション技術を包含する。例えば、Morrison, S.L., et al., Proc. Natl. Acad Sci. USA 81(1984)6851-6855;US 5,202,238及びUS 5,204,244を参照のこと。
【0051】
用語「ヒト化抗体」とは、そのフレームワーク又は「相補性決定領域」(CDR)が、親イムノグロブリンのCDRに比較して、異なった特異性の免疫グロブリンのCDRを含んでなるよう修飾されている抗体を意味する。好ましい実施形態では、ネズミCDRは、「ヒト化抗体」を調製するために、ヒト抗体のフレームワーク領域中に移植される。例えば、Riechmann, L., et al., Nature 332(1988)323-327;及びNeuberger, M.S., et al., Nature 314(1985)268-270を参照のこと。特に好ましいCDRは、キメラ抗体に関して上記に示される抗原を認識する配列を表すものに対応する。本発明により包含される他の形の「ヒト化抗体」とは、定常領域が、特にC1q結合及び/又はFc受容体(FcR)結合に関して、本発明の性質を生じるように、更に修飾されているか、又は元の抗体のその領域から変更されている抗体である。
【0052】
用語「ヒト抗体」とは、本明細書において使用される場合、ヒト生殖系免疫グロブリン配列に由来する可変及び定常領域を有する抗体を包含することを意図する。ヒト抗体は技術的に良く知られている(van Dijk, M.A., and van de Winkel, J. G., Curr. Opin. Chem. Biol. 5(2001)368-374)。ヒト抗体はまた、免疫化に基づいて、内因性免疫グロブリン生成の不在下でヒト抗体の全レパートリー又は選択物を生成できるトランスジェニック動物(例えば、マウス)においても生成され得る。そのような生殖系変異マウス内へのヒト生殖系免疫グロブリン遺伝子アレイのトランスファーは、抗原攻撃に基づくヒト抗体の生成をもたらすであろう(例えば、Jakobovits, A., et al., Proc. Natl. Acad. Sci. USA 90(1993)2551-2555;Jakobovits, A., et al., Nature 362(1993)255-258;Bruggemann, M., et al., Year Immunol. 7(1993)33-40を参照のこと)。ヒト抗体はまた、ファージ提示ライブラリーにおいても生成され得る(Hoogenboom, H. R., and Winter, G., J. Mol. Biol. 227(1992)381-388;Marks, J.D., et al., J. Mol. Biol. 222(1991)581-597)。Cole, S.P.C.ら及びBoerner, Pらの技術はまた、ヒトモノクローナル抗体の調製のためにも利用できる(Cole, S.P.C., et al., Monoclonal Antibodies and Cancer Therapy, Liss, A. L.(1985) 77-96;及びBoerner, P., et al., J. Immunol. 147(1991)86-95)。本発明のキメラ及びヒト化抗体についてすでに言及されたように、用語「ヒト抗体」とは、本明細書において使用される場合、また、特にC1q結合及び/又はFcR結合に関して、例えばFc部分の「クラススイッチ」、すなわち変更又は突然変異誘発(IgG1からIgG4への及び/又はIgG1/IgG4突然変異)により、本発明の性質を生じるように定常領域において修飾されているそのような抗体も含んでなる。
【0053】
用語「組換えヒト抗体」とは、本明細書において使用される場合、組換え手段により調製され、発現され、作出され、又は単離されるすべてのヒト抗体、例えば宿主細胞、例えばNS0又はCHO細胞から又はヒト免疫グロブリン遺伝子に関してトランスジェニックである動物(例えば、マウス)から単離された抗体、又は宿主細胞中にトランスフェクトされた組換え発現ベクターを用いて発現された抗体を意味する。そのような組換えヒト抗体は、再配置された形で、可変及び定常領域を有する。本発明の組換えヒト抗体は、生体内における体細胞超突然変異にゆだねられている。従って、組換え抗体のVH及びVL領域のアミノ酸配列は、ヒト生殖系VH及びVL領域に由来し、そしてその配列に関連しているが、天然においては、ヒト抗体生殖系レパートリー内で生体内には存在することができない配列である。
【0054】
「可変ドメイン」(軽鎖の可変領域(VL)、重鎖の可変領域(VH))は、本明細書において使用される場合、抗原への抗体の結合において直接関与する軽鎖及び重鎖の対の各々を示す。ヒト可変軽鎖及び重鎖のドメインは、同じ一般構造を有し、そして各々のドメインは4個のフレームワーク(FR)領域を含んで成り、それらの配列は広く保存され、3個の「超可変領域」(又は相補性決定領域、CDR)により結合される。フレームワーク領域は、β−シートコンホメーションを採用し、そしてCDRはβ−シート構造体を結合するループを形成することができる。各々の鎖におけるCDRは、フレームワーク領域によりそれらの立体構造を保持され、そして他の鎖からのCDRと共に抗原結合部位を形成する。抗体重鎖及び軽鎖CDR3領域は、本発明の抗体の結合特異性/親和性において特に重要な役割を演じ、従って、本発明の更なる目的を提供する。
【0055】
用語「超可変領域」又は「抗体の抗原−結合部分若しくは抗原結合部位」とは、本明細書において使用される場合、抗原結合を担当できる抗体のアミノ酸残基を指す。超可変領域は「相補性決定領域」又は「CDR」からのアミノ酸残基を含んでなる。「フレームワーク」又は「FR」領域は、本明細書において定義されるように超可変領域残基以外のそれらの可変ドメイン領域である。従って、抗体の軽鎖及び重鎖は、N末端からC末端に向かって、ドメインFR1、CDR1、FR2、CDR2、FR3、CDR3及びFR4を含んでなる。各々の鎖上のCDRはそのようなフレームワークアミノ酸により分離される。特に、重鎖のCDR3は、ほとんど抗原結合に寄与する領域である。CDR及びFR領域は、Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD(1991)の標準的な定義に従って決定される。
【0056】
本明細書において使用される場合、用語「結合」又は「特異的に結合」とは、試験管内におけるアッセイ、好ましくは精製した野生型抗原を用いたプラズモン共鳴アッセイ(BIAcore、GE-Healthcare Uppsala, Sweden)における抗体の抗原(ヒトErbB−2又はヒトc−Metのいずれか)のエピトープへの結合を指す。結合の親和性は、用語ka(抗体/抗原複合体からの抗体結合に関する速度定数)、kD(解離定数)、及びKD(kD/ka)によって規定される。結合又は特異的に結合は、10-8mol/l以下、好ましくは10-9M〜10-13mol/lの結合親和性(KD)を意味する。よって、本発明による二重特異性<ErbB−2−c−Met>抗体は、各抗原に対して10-8mol/l、好ましくは10-9M〜10-13mol/lの結合親和性(KD)を有する特異性をもって特異的に結合する。
【0057】
FcγRIIIへの抗体の結合は、BIAcoreアッセイ(GE-Healthcare Uppsala, Sweden)により調査され得る。結合の親和性は、用語ka(抗体/抗原複合体からの抗体結合に関する速度定数)、kD(解離定数)、及びKD(kD/ka)により規定される。
【0058】
用語「エピトープ」とは、抗体に対して特異的に結合できるいずれかのポリペプチド決定基を包含する。ある実施形態においては、エピトープ決定基は、分子、例えばアミノ酸、糖側鎖、ホスホリン又はスルホニルなどの化学的活性表面群を包含し、そしてある実施形態においては、特定の立体構造特性及び/又は比電荷特性を有することができる。エピトープは、抗体により結合される抗原の領域である。
【0059】
ある実施形態においては、抗体は、それがタンパク質及び/又は高分子の複合混合物中でその標的抗原を選択的に認識する場合、抗原を特異的に結合すると言われる。
【0060】
用語「定常領域」は、本明細書中で使用される場合、可変領域以外の抗体のドメインの集合を意味する。定常領域は、抗原の結合に直接的にかかわらないが、様々なエフェクター機能を示す。それらの重鎖の定常領域のアミノ酸配列によって、抗体は以下のクラス:IgA、IgD、IgE、IgG及びIgMに分類され、そしてそのいくつか、例えばIgG1、IgG2、IgG3、及びIgG4、IgA1及びIgA2などは、サブクラスに更に分類されることもできる。異なった抗体クラスに該当する重鎖定常領域は、それぞれα、δ、ε、γ、及びμと呼ばれる。全部で5つの抗体クラスで見ることができる軽鎖定常領域は、κ(カッパ)及びλ(ラムダ)と呼ばれる。定常領域は、好ましくはヒト起源に由来する。
【0061】
用語「ヒト起源由来の定常領域」は、本願で使用される場合、ヒト抗体のサブクラスIgG1、IgG2、IgG3、若しくはIgG4の重鎖定常領域、及び/又は定常軽鎖カッパ又はラムダ領域を規定する。そのような定常領域は、当技術分野の現状において周知であり、例えば、Kabat, E.A.によって説明されている(例えばJohnson, G., and Wu, T.T., Nucleic Acids Res. 28(2000)214-218;Kabat, E.A., et al., Proc, Natl. Acad. Sci. USA 72(1975)2785-2788を参照のこと)。
【0062】
一実施形態では、本発明による二重特異性抗体は、IgG1又はIgG3サブクラス(好ましくはIgG1サブクラス)の定常領域を含んでなり、それは好ましくはヒト起源に由来する。一実施形態では、本発明による二重特異性抗体は、IgG1又はIgG3サブクラス(好ましくはIgG1サブクラス)のFc部分を含んでなり、それは好ましくはヒト起源に由来する。
【0063】
IgG4サブクラスの抗体がFc受容体(FcyRIIIa)結合の低減を示している一方で、他のIgGサブクラスの抗体は強い結合を示している。しかしながら、Pro238、Asp265、Asp270、Asn297(Fc炭水化物の欠如)、Pro329、Leu234、Leu235、Gly236、Gly237、Ile253、Ser254、Lys288、Thr307、Gln311、Asn434、及びHis435は、変更された場合には、同様に低減されたFc受容体結合性を示す残基である(Shields, R.L., et al., J. Biol. Chem. 276(2001)6591-6604;Lund, J., et al., FASEB J. 9(1995)115-119;Morgan, A., et al., Immunology 86(1995)319-324;EP 0 307 434)。
【0064】
1つの実施形態では、本発明による抗体は、IgG1抗体と比較してFcR結合が低減されているので、全長親抗体は、FcR結合に関して、IgG4サブクラスのもの、あるいはS228、L234、L235及び/又はD265に突然変異を伴う、及び/又はPVA236突然変異を含むIgG1又はIgG2サブクラスのものである。1つの実施形態では、全長親抗体内の突然変異は、S228P、L234A、L235A、L235E、及び/又は、PVA236である。他の実施形態では、全長親抗体内の突然変異は、IgG4ではS228PそしてとIgG1ではL234AとL235Aにある。
【0065】
抗体の定常領域は、ADCC(抗体依存性細胞毒性)及びCDC(補体依存性細胞毒性)に直接関与する。補体活性化(CDC)は、大部分のIgG抗体サブクラスの定常領域への補体C1q因子の結合により開始する。抗体へのC1qの結合は、いわゆる結合部位で所定のタンパク質−タンパク質相互作用により起きる。そのような定常領域の結合部位は、当技術分野の現状において公知であり、例えばLukas, T.J., et al., J. Immunol. 127(1981)2555-2560;Brunhouse, R. and Cebra, J.J., Mol. Immunol. 16(1979)907-917;Burton, D.R., et al., Nature 288(1980)338-344;Thommesen, J.E., et al., Mol. Immunol. 37(2000)995-1004;Idusogie, E.E., et al., J. Immunol. 164(2000)4178-4184;Hezareh, M., et al., J. Virol. 75(2001)12161-12168;Morgan, A., et al., Immunology 86(1995)319-324;及びEP 0 307 434により記載されている。そのような定常領域の結合部位は、例えばアミノ酸L234、L235、D270、N297、E318、K320、K322、P331、及びP329(付番は常にKabatのEUインデックスに準じる)を特徴とする。
【0066】
用語「抗体依存性細胞毒性(ADCC)」は、エフェクター細胞の存在下での、本発明よる抗体によるヒト標的細胞の溶解を指す。ADCCは、好ましくは、新たに単離されたPBMC、又は単球若しくはナチュラルキラー(NK)細胞又は恒久的に増殖しているNK細胞株のような精製された軟膜由来エフェクター細胞等のエフェクター細胞の存在下で、本発明よる抗体を用いて、ErB−1及びc−Met発現細胞の調製物を処理することにより測定される。
【0067】
用語「補体依存性細胞毒性(CDC)」は、大部分のIgG抗体サブクラスのFc部への補体C1q因子の結合により開始する過程と規定される。抗体へのC1qの結合は、いわゆる結合部位で所定のタンパク質−タンパク質相互作用により起きる。そのようなFc部の結合部位は、当技術分野の現状において公知である(前記を参照のこと)。そのようなFc部の結合部位は、例えばアミノ酸L234、L235、D270、N297、E318、K320、K322、P331、及びP329(付番は常にKabatのEUインデックスに準じる)を特徴とする。サブクラスIgG1、IgG2、及びIgG3の抗体は、通常はC1q及びC3の結合を含めた補体活性化を示し、一方でIgG4は補体系を活性化せず、C1q及びC3と結合しない。
【0068】
モノクローナル抗体の細胞媒介型エフェクター機能は、Umana, P., et al., Nature Biotechnol. 17(1999)176-180及びUS 6,602,684に記載されているように、それらのオリゴ糖構成要素を操作することによって増強することができる。癌免疫療法において最もよく使用される抗体であるIgG1型抗体は、保存されたN結合型グリコシル化部位を各CH2ドメイン中のAsn297において有する糖タンパク質である。Asn297に結合した2つの複合型二分岐オリゴ糖は、CH2ドメインの間に埋もれて、ポリペプチド骨格との広範囲の接触面を形成し、それらの存在は、抗体依存性細胞毒性(ADCC)のようなエフェクター機能を抗体が媒介するために不可欠である(Lifely, M.R., et al., Glycobiology 5(1995)813-822;Jefferis, R., et al., Immunol. Rev. 163(1998)59-76;Wright, A. and Morrison, S.L., Trends Biotechnol. 15(1997)26-32)。Umana, P., et al., Nature Biotechnol. 17(1999)176-180及びWO 99/54342は、2つに分岐したオリゴ糖の形成を触媒するグリコシルトランスフェラーゼであるβ(1,4)−N−アセチルグルコサミニルトランスフェラーゼIII(「GnTIII」)がチャイニーズハムスター卵巣(CHO)細胞において過剰発現されると、抗体の生体内でのADCC活性が有意に増大することを示した。また、Asn297炭水化物の組成の改変又はその除去も、FcγR及びC1qへの結合に影響を及ぼす(Umana, P., et al., Nature Biotechnol. 17(1999)176-180;Davies, J., et al., Biotechnol. Bioeng. 74(2001)288-294;Mimura, Y., et al., J. Biol. Chem. 276(2001)45539-45547;Radaev, S., et al., J. Biol. Chem. 276(2001)16478-16483;Shields, R.L., et al., J. Biol. Chem. 276(2001)6591-6604;Shields, R.L., et al., J. Biol. Chem. 277(2002)26733-26740;Simmons, L.C., et al., J. Immunol. Methods 263(2002)133-147)。
【0069】
フコースの量を減少させることによってモノクローナル抗体の細胞媒介型エフェクター機能を高める方法は、例えば、WO2005/018572、WO2006/116260、WO2006/114700、WO2004/065540、WO2005/011735、WO2005/027966、WO1997/028267、US2006/0134709、US2005/0054048、US2005/0152894、WO2003/035835、WO2000/061739、Niwa, R., et al., J. Immunol. Methods 306 (2005) 151-160;Shinkawa, T., et al, J Biol Chem, 278 (2003) 3466-3473;WO03/055993又はUS2005/0249722に記載されている。
【0070】
本発明の一実施形態では、本発明による二重特異性抗体は、(IgG1若しくはIgG3サブクラス)Asn297にて糖鎖を用いてグリコシル化され、それによって、前記糖鎖内のフコースの量が65%以下になる(付番はKabatに準じる)。他の実施形態では、前記糖鎖内のフコースの量が、5%〜65%、好ましくは20%〜40%である。本発明による「Asn297」は、Fc領域内の約297位に見られるアミノ酸であるアスパラギンを意味する。抗体のわずかな配列変化により、Asn297はまた、297位の数アミノ酸(通常、±3アミノ酸以下)上流又は下流、すなわち、294位から300位の間でも見られることがある。
【0071】
ヒトIgG1又はIgG3のグリコシル化は、最大2つのGal残基で終わるコアフコース化二本鎖複合型オリゴ糖グリコシル化としてAsn297で起こる。IgG1又はIgG3サブクラスのヒト重鎖定常領域は、Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD.(1991)、及びBruggemann, M., et al., J. Exp. Med. 166(1987)1351-1361;Love, T.W., et al., Methods Enzymol. 178(1989)515-527によって詳細に説明されている。これらの構造は、終末のGal残基の量に依存して、G0、G1(α−1,6−若しくはα−1,3−)又はG2グリカン残基として指定される(Raju, T.S., Bioprocess Int. 1(2003)44-53)。抗体Fc部分のCHOタイプのグリコシル化は、例えば、Routier, F.H., Glycoconjugate J. 14(1997)201-207によって説明されている。非糖修飾CHO宿主細胞において組換え発現される抗体は、通常少なくとも85%の量がAsn297においてフコース化されている。全長親抗体の修飾オリゴ糖は、ハイブリッドであっても複合体であってもよい。好ましくは、分枝、還元/非フコース化オリゴ糖がハイブリッドされる。他の実施形態において、分枝、還元又は非フコース化オリゴ糖が複合体化される。
【0072】
本発明による「フコースの量」は、MALDI−TOF質量分析法によって測定し、平均値として算出した、Asn297に結合したすべての糖残基(例えば、複合体構造物、ハイブリッド構造物、及び高マンノース構造物)の合計に対するAsn297における糖鎖内のフコースの量を意味する。フコースの相対量は、MALDI−TOFによって測定した、N-グリコシダーゼFで処理したサンプルにおいて同定された全糖構造物(例えば、それぞれ複合体構造物、ハイブリッド構造物、並びにオリゴマンノース構造物及び高マンノース構造物)に対するフコース含有構造物のパーセンテージである(例えばWO2008/077546(A1)を参照のこと)。
【0073】
一実施形態は、WO2005/044859、WO2004/065540、WO2007/031875、Umana, P., et al., Nature Biotechnol. 17 (1999) 176-180、WO99/154342、WO2005/018572、WO2006/116260、WO2006/114700、WO2005/011735、WO2005/027966、WO97/028267、US2006/0134709、US2005/0054048、US2005/0152894、WO2003/035835又はWO2000/061739に記載の手順を用いた、Asn297にて糖鎖を用いて(糖鎖が)グリコシル化され、それによって、前記糖鎖内のフコースの量が65%以下になる、IgG1又はIgG3サブクラスの二重特異性抗体の調製法である。
【0074】
一実施形態は、Niwa, R., et al., J. Immunol. Methods 306 (2005) 151-160;Shinkawa, T. et al, J Biol Chem, 278 (2003) 3466-3473;WO03/055993又はUS2005/0249722に記載の手順を用いた、Asn297にて糖鎖を用いて(糖鎖が)グリコシル化され、それによって、前記糖鎖内のフコースの量が65%以下になる、IgG1又はIgG3サブクラスの二重特異性抗体の調製法である。
【0075】
二重特異性抗体フォーマット
本発明の抗体は、2個以上の結合部位を持つので、多特異性であり、好ましくは二重特異性である。すなわち、抗体は、2以上の結合部位がある場合でさえ、二重特異性であり得る(すなわち、抗体は三価又は多価性である)。本発明の二重特異性抗体には、例えば多価性一本鎖抗体、二重特異性抗体、及び三重特異性抗体、並びに1若しくは複数のペプチド・リンカーを介して連結された更なる抗原結合部位(例えば一本鎖Fv、VHドメイン及び/又はVLドメイン、Fab、又は(Fab)2)を持つ全長抗体の定常ドメイン構造を有する抗体が含まれる。抗体は、単一種からの全長であることも、又はキメラ化若しくは又はヒト化されることもできる。3つ以上の抗原結合部位を有する抗体に関しては、タンパク質に2つの異なった抗原のための結合部位がある限り、いくつかの結合部位が同一のであることもあり得る。すなわち、第1の結合部位がErbB−2に特異的である一方で、第2の結合部位はc−Metに特異的であり、そして逆もまた同様である。
【0076】
好ましい実施形態では、本発明によるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体は、(好ましくはIgG1又はIgG3サブクラスの)抗体のFc領域を含んでなる。
【0077】
二価の二重特異性フォーマット
免疫グロブリン定常領域を含んでなるヒトErbB−2及びヒトc−Metに対する二重特異性の二価抗体は、例えば、WO2009/080251、WO2009/080252、WO2009/080253若しくはRidgway, J.B., Protein Eng. 9 (1996) 617-621;WO96/027011;Merchant, A.M., et al. Nature Biotech 16 (1998) 677-681;Atwell, S., et al., J. Mol. Biol. 270 (1997) 26-35及びEP1870459A1に記載のように使用できる。
【0078】
よって、本発明の一実施形態では、本発明による二重特異性<ErbB−2−c−Met>抗体は、以下の:
a)ErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びに
b)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖;
を含んでなり、ここで、定常ドメインCLとCH1、及び/又は可変ドメインVLとVHが互いによって置き換えられている二価の二重特異性抗体である。
【0079】
本発明の他の実施形態において、本発明による二重特異性<ErbB−2−c−Met>抗体は、以下の:
a)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖;並びに
b)ErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;
を含んでなり、ここで、定常ドメインCLとCH1、及び/又は可変ドメインVLとVHが互いによって置き換えられている二価の二重特異性抗体である。
【0080】
以下で記載する「ホール−into−ノブ」技術を用いた代表的な図式的構造については、図2a〜cを参照のこと。
【0081】
そのようなヘテロ二量体の二価の二重特異性抗ErbB−2/抗c−Met抗体の収率を改善するために、前記全長抗体のCH3ドメインは、例えばWO96/027011、Ridgway, J., B., et al., Protein Eng 9 (1996) 617-621;及びMerchant, A., M., et al., Nat Biotechnol 16 (1998) 677-681中にいくつかの実施例を用いて詳細に説明されている「ホール−into−ノブ」技術によって変更され得る。この方法では、2種のCH3ドメインの相互作用表面が、それらの2種のCH3ドメインを含む両重鎖のヘテロダイマー化を高めるために変更される。(2種の重鎖の)2種のCH3ドメインの個々が「ノブ」であり、そして他が「ホール」である。ジスルフィド架橋の導入は、ヘテロダイマーを安定化し(Merchant, A..M., et al, Nature Biotech 16(1998)677-681;Atwell, S., et al., J. Mol. Biol. 270(1997)26-35)、そして収率を高める。
【0082】
従って、本発明の1つの態様では、前記二価の二重特異性抗体は、1つの重鎖のCH3ドメイン及び他の重鎖のCH3ドメインが、抗体CH3ドメイン間に本来の界面を含んでなる界面で各々接触し;
ここで、前記界面は二価の二重特異性抗体の形成を促進するよう変更され、ここで、前記変更が:
a)二価の二重特異性抗体内のもう片方の重鎖のCH3ドメインの本来の界面と接触する、一方の重鎖のCH3ドメインの本来の界面内で、アミノ酸残基がより大きな側鎖体積を有するアミノ残基により置換され、それにより、もう片方の重鎖のCH3ドメインの界面内の腔に位置決定できる、一方の重鎖CH3ドメインの界面内に隆起部を生じるように、一方の重鎖のCH3ドメインが変更され、そして
b)二価の二重特異性抗体内の第1CH3ドメインの本来の界面と接触する、第2CH3ドメインの本来の界面内で、アミノ酸残基がより小さな側鎖体積を有するアミノ酸残基により置換され、それにより、第1CH3ドメインの界面内の隆起部が位置決定できる第2CH3ドメインの界面内に腔を生じるように、もう片方の重鎖のCH3ドメインが変更されることにより更に特徴づけられる。
【0083】
好ましくは、より大きな側鎖体積を有する前記アミノ酸残基が、アルギニン(R)、フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)から成る群から選択される。
【0084】
好ましくは、より小さな側鎖体積を有する前記アミノ酸残基が、アラニン(A)、セリン(S)、トレオニン(T)、バリン(V)から成る群から選択される。
【0085】
本発明の1つの態様においては、両CH3ドメインは更に、それらの両CH3ドメイン間にジスルフィド架橋が形成され得るよう、各々のCH3ドメインのその対応する位置のアミノ酸としてのシステイン(C)の導入により変更される。
【0086】
好ましい実施形態では、前記二価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にT366W突然変異、及び「ホール鎖」のCH3ドメイン内にT366S、L368A、Y407V突然変異を含んでなる。例えば、「ノブ鎖」のCH3ドメイン内にY349C突然変異、及び「ホール鎖」のCH3ドメイン内にE356C突然変異又はS354C突然変異を導入することによって、CH3ドメイン間の追加的な鎖間ジスルフィド架橋もまた使用できる(Merchant, A.M., et al., Nature Biotech 16(1998)677-681)。よって、別の好ましい実施形態では、前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にE356C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなる(鎖間ジスルフィド架橋を形成する、一方のCH3ドメイン内の追加的なY349C突然変異ともう片方のCH3ドメイン内の追加的なE356C又はS354C突然変異)(付番は常にKabatのEUインデックスに準じる)。しかし、EP1870459A1によって記載される他のノブ−into−ホール技術もまた代替的に又は追加的に使用できる。前記二価の二重特異性抗体の好ましい例は、「ノブ鎖」のCH3ドメイン内のR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内のD399K;E357K突然変異である(付番は常にKabatのEUインデックスに準じる)。
【0087】
別の好ましい実施形態では、前記二価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内のT366W突然変異及び「ホール鎖」のCH3ドメイン内のT366S、L368A、Y407V突然変異、そして追加的に「ノブ鎖」のCH3ドメイン内のR409D;K370突然変異及び「ホール鎖」のCH3ドメイン内のD399K;E357KE突然変異を含んでなる。
【0088】
別の好ましい実施形態では、前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなり、そして追加的に「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0089】
三価の二重特異性フォーマット
この発明の別の好ましい態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖及び2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fabフラグメント
を含んでなり、ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、前記全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合される、三価の二重特異性抗体である。
【0090】
以下で記載する「ホール−into−ノブ」技術を用いた代表的な図式的構造は、図5aを参照のこと。
【0091】
この発明の別の好ましい態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖及び2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fvフラグメント
を含んでなり、ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、前記全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合される、三価の二重特異性抗体である。
【0092】
以下で記載する「ホール−into−ノブ」技術を用いた代表的な図式的構造は、図5bを参照のこと。
【0093】
1つの好ましい実施形態では、ヒトc−Metに結合する前記一本鎖Fab又はFvフラグメントは、前記の全長抗体に、前記全長抗体の重鎖のC末端のペプチドコネクタを介して融合される。
【0094】
この発明の別の好ましい態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖及び2つの抗体軽鎖から成る全長抗体;
b)以下の:
ba)抗体重鎖可変ドメイン(VH);又は
bb)抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、から成るポリペプチド、
ここで、前記ポリペプチドは、VHドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちの一方のC末端に融合されていて、
【0095】
c)以下の:
ca)抗体軽鎖可変ドメイン(VL);又は
cb)抗体軽鎖可変ドメイン(VL)と抗体軽鎖定常ドメイン(CL)、
から成るポリペプチド、
ここで、前記ポリペプチドはVLドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちのもう片方のC末端に融合されている、
を含んでなり;そして
ここで、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)が一緒になって、ヒトc−Metに特異的に結合する抗原結合部位を形成する、三価の二重特異性抗体である。
【0096】
好ましくは、前記b)及びc)のペプチドコネクタは、同一であり、且つ、少なくとも25個のアミノ酸、好ましくは30〜50個のアミノ酸から成るペプチドである。
【0097】
代表的な図式的構造については、図3a〜cを参照のこと。
【0098】
任意には、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)は、連結され、そして下記の位置の間へのジスルフィド結合の導入によって鎖間ジスルフィド架橋によって安定化させる:
i)重鎖可変ドメインの第44位〜軽鎖可変ドメインの第100位
ii)重鎖可変ドメインの第105位〜軽鎖可変ドメインの第43位、又は、
iii)重鎖可変ドメインの第101位〜軽鎖可変ドメインの第100位(付番は常にKabatのEUインデックスに準じる)。
【0099】
安定化のために非天然ジスルフィド架橋を導入する技術は、例えば、WO94/029350、Rajagopal, et al., Prot Engin. (1997) 1453-59;Kobayashi, H., et al., Nuclear Medicine & Biology, 25, (1998) 387-393;又はSchmidt, M., et al., Oncogene 18 (1999) 1711-1721に記載されている。1つの実施形態では、b)とc)のポリペプチドの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第44位と軽鎖可変ドメインの第100位の間にある。1つの実施形態では、b)とc)のポリペプチドの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第105位と軽鎖可変ドメインの第43位の間にある(付番は常にKabatのEUインデックスに準じる)。1つの実施形態では、一本鎖Fabフラグメントの可変ドメインVHとVLの間の前記任意のジスルフィドによる安定化のない、三価の二重特異性抗体が好まれる。
【0100】
一方の重鎖への一本鎖Fab、Fvフラグメントの融合によって(図5a又は5b)又は全長抗体の両方の重鎖への別個のポリペプチドの融合によって(図3a〜c)、ヘテロ二量体の三価の二重特異性抗体がもたらされる。そのようなヘテロ二量体の三価の二重特異性抗ErbB−2/Met抗体の収率を高めるために、前記全長抗体のCH3ドメインは、例えばWO96/027011、Ridgway, J., B., et al., Protein Eng 9 (1996) 617-621;及びMerchant, A., M., et al., Nat Biotechnol 16 (1998) 677-681中にいくつかの実施例を用いて詳細に説明されている「ホール−into−ノブ」技術によって変更され得る。この方法では、2種のCH3ドメインの相互作用表面が、それらの2種のCH3ドメインを含む両重鎖のヘテロダイマー化を高めるために変更される。(2種の重鎖の)2種のCH3ドメインの個々が「ノブ」であり、そして他が「ホール」である。ジスルフィド架橋の導入は、ヘテロダイマーを安定化し(Merchant, A..M., et al, Nature Biotech 16(1998)677-681;Atwell, S., et al., J. Mol. Biol. 270(1997)26-35)、そして収率を高める。
【0101】
従って、本発明の1つの態様では、前記三価の二重特異性抗体は、全長抗体の1つの重鎖のCH3ドメイン及び全長抗体の他の重鎖のCH3ドメインが、抗体CH3ドメイン間に本来の界面を含んでなる界面で各々接触し;
ここで、前記界面は二価の二重特異性抗体の形成を促進するよう変更され、ここで、前記変更が:
a)二価の二重特異性抗体内のもう片方の重鎖のCH3ドメインの本来の界面と接触する、一方の重鎖のCH3ドメインの本来の界面内で、アミノ酸残基がより大きな側鎖体積を有するアミノ残基により置換され、それにより、もう片方の重鎖のCH3ドメインの界面内の腔に位置決定できる、一方の重鎖CH3ドメインの界面内に隆起部を生じるように、一方の重鎖のCH3ドメインが変更され、そして
b)三価の二重特異性抗体内の第1CH3ドメインの本来の界面と接触する、第2CH3ドメインの本来の界面内で、アミノ酸残基がより小さな側鎖体積を有するアミノ酸残基により置換され、それにより、第1CH3ドメインの界面内の隆起部が位置決定できる第2CH3ドメインの界面内に腔を生じるように、もう片方の重鎖のCH3ドメインが変更されることにより更に特徴づけられる。
【0102】
好ましくは、より大きな側鎖体積を有する前記アミノ酸残基が、アルギニン(R)、フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)から成る群から選択される。
【0103】
好ましくは、より小さな側鎖体積を有する前記アミノ酸残基が、アラニン(A)、セリン(S)、トレオニン(T)、バリン(V)から成る群から選択される。
【0104】
本発明の1つの態様においては、両CH3ドメインは更に、それらの両CH3ドメイン間にジスルフィド架橋が形成され得るよう、各々のCH3ドメインのその対応する位置のアミノ酸としてのシステイン(C)の導入により変更される。
【0105】
好ましい実施形態では、前記三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にT366W突然変異、及び「ホール鎖」のCH3ドメイン内にT366S、L368A、Y407V突然変異を含んでなる。例えば、「ノブ鎖」のCH3ドメイン内にY349C突然変異、及び「ホール鎖」のCH3ドメイン内にE356C突然変異又はS354C突然変異を導入することによって、CH3ドメイン間の追加的な鎖間ジスルフィド架橋もまた使用できる(Merchant, A.M., et al., Nature Biotech 16(1998)677-681)。よって、別の好ましい実施形態では、前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にE356C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなる(鎖間ジスルフィド架橋を形成する、一方のCH3ドメイン内の追加的なY349C突然変異ともう片方のCH3ドメイン内の追加的なE356C又はS354C突然変異)(付番は常にKabatのEUインデックスに準じる)。しかし、EP1870459A1によって記載される他のノブ−into−ホール技術もまた代替的に又は追加的に使用できる。前記三価の二重特異性抗体の好ましい例は、「ノブ鎖」のCH3ドメイン内のR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内のD399K;E357K突然変異である(付番は常にKabatのEUインデックスに準じる)。
【0106】
別の好ましい実施形態では、前記三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内のT366W突然変異及び「ホール鎖」のCH3ドメイン内のT366S、L368A、Y407V突然変異、そして追加的に「ノブ鎖」のCH3ドメイン内のR409D;K370突然変異及び「ホール鎖」のCH3ドメイン内のD399K;E357KE突然変異を含んでなる。
【0107】
別の好ましい実施形態では、前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなり、そして追加的に「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0108】
この発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fabフラグメント、
ここで、前記一本鎖Fabフラグメントが、抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、抗体軽鎖可変ドメイン(VL)、抗体軽鎖定常ドメイン(CL)及びリンカーから成り、且つ、ここで、前記抗体ドメインと前記リンカーが、N末端からC末端に向かって、以下の順序:
ba)VH−CH1−リンカー−VL−CL、又は
bb)VL−CL−リンカー−VH−CH1、
のうちの一方を有し、
ここで、前記リンカーが少なくとも30個のアミノ酸、好ましくは32〜50個のアミノ酸から成るペプチドである、
を含んでなり、そして
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖ののC又はN末端の(好ましくは重鎖のC末端の)ペプチドコネクタを介して融合されている;
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸、好ましくは10〜50個のアミノ酸から成るペプチドである、三価の二重特異性抗体である。
【0109】
この実施形態の中で、好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にT366W突然変異、そして2つのCH3ドメインのうちのもう片方にT366S、L368A、Y407V突然変異を含んでなり、より好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのなかのもう片方にS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる。任意に、前記実施形態では、三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異を含んでなり、且つ、「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0110】
この発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fvフラグメント、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖ののC又はN末端の(好ましくは重鎖のC末端の)ペプチドコネクタを介して融合され;そして
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸、好ましくは10〜50個のアミノ酸から成るペプチドである、三価の二重特異性抗体である。
【0111】
この実施形態の中で、好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にT366W突然変異、そして2つのCH3ドメインのうちのもう片方にT366S、L368A、Y407V突然変異を含んでなり、より好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのなかのもう片方にS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる。任意に、前記実施形態では、三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異を含んでなり、且つ、「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0112】
よって好ましい実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fvフラグメント、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖のC末端のペプチドコネクタを介して融合され(2つの抗体重鎖−一本鎖Fv融合ペプチドをもたらし);そして
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸である、三価の二重特異性抗体である。
【0113】
本発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖;
から成る全長抗体;並びに
b)以下の:
ba)抗体重鎖可変ドメイン(VH);又は
bb)抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、から成るポリペプチド、
ここで、前記ポリペプチドは、VHドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちの一方のC末端に融合されて(抗体重鎖−VH融合ペプチドをもたらし)、
ここで、前記ペプチドコネクタが5つのアミノ酸、好ましくは25〜50個のアミノ酸から成るペプチドである、
【0114】
c)以下の:
ca)抗体軽鎖可変ドメイン(VL);又は
cb)抗体軽鎖可変ドメイン(VL)と抗体軽鎖定常ドメイン(CL)、
から成るポリペプチド、
ここで、前記ポリペプチドはVLドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちのもう片方のC末端に融合されて(抗体重鎖−VL融合ペプチドをもたらし)、
ここで、前記ペプチドコネクタがb)のペプチドコネクタと同一である、
を含んでなり;そして
ここで、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)が一緒になって、ヒトc−Metに特異的に結合する抗原結合部位を形成する、三価の二重特異性抗体である。
【0115】
この実施形態の中で、好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にT366W突然変異、そして2つのCH3ドメインのうちのもう片方にT366S、L368A、Y407V突然変異を含んでなり、より好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのなかのもう片方にS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる。任意に、前記実施形態では、三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異を含んでなり、且つ、「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0116】
本発明の別の態様では、本発明による三価の二重特異性抗体は、以下の:
a)ヒトErbB−2に結合し、2つの抗体重鎖、VH−CH1−HR−CH2−CH3及び2つの抗体軽鎖VL−CLから成る全長抗体;
(ここで、好ましくは2つのCH3ドメインのうちの一方がY349C、T366W突然変異を含んでなり、そして2つのCH3ドメインのうちのもう片方がS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる);
b)以下の:
ba)抗体重鎖可変ドメイン(VH);又は
bb)抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、
から成るポリペプチド;
ここで、前記ポリペプチドはVHドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちの一方のC末端に融合されている;
【0117】
c)以下の:
ca)抗体軽鎖可変ドメイン(VL)、又は
cb)抗体軽鎖可変ドメイン(VL)と抗体軽鎖定常ドメイン(CL)、から成るポリペプチド、ここで、前記ポリペプチドはVLドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちのもう片方のC末端に融合されている;
を含んでなり、そして
ここで、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)が一緒になって、ヒトc−Metに特異的に結合する抗原結合部位を形成する。
【0118】
四価の二重特異性フォーマット
一実施形態では、本発明による多特異性抗体は四価であり、ここで、ヒトc−Metに特異的に結合する(単数若しくは複数の)抗原結合部位は、(例えばWO2009/007427に記載のように)c−Metの二量化を阻害する。
【0119】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する2つの抗原結合部位及びヒトc−Metに特異的に結合する2つの抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する四価の二重特異性抗体であって、ここで、ここで、ヒトc−Metに特異的に結合する前記抗原結合部位は、(例えばWO2009/007427に記載のように)c−Metの二量化を阻害する。
【0120】
そのため、この発明の別の態様は、以下の:
a)ヒトc−Metに特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ErbB−2に特異的に結合する2つの同一の一本鎖Fabフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0121】
そのため、この発明の別の態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する2つの同一の一本鎖Fabフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0122】
代表的な図式的構造は、図6aを参照のこと。
【0123】
そのため、この発明の別の態様は、以下の:
a)ErbB−2に特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する2つの同一の一本鎖Fvフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0124】
そのため、この発明の別の態様は、以下の:
a)ヒトc−Metに特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ErbB−2に特異的に結合する2つの同一の一本鎖Fvフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0125】
代表的な図式的構造は、図6bを参照のこと。
【0126】
好ましい一実施形態では、ヒトc−Met又はヒトErbB−2に結合する一本鎖Fab又はFvフラグメントが、前記全長抗体に、その全長抗体の重鎖のC末端のペプチドコネクタを介して融合されている。
【0127】
この発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの同一の抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの同一の抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する2つの一本鎖Fabフラグメント、
ここで、前記一本鎖Fabフラグメントが、抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、抗体軽鎖可変ドメイン(VL)、抗体軽鎖定常ドメイン(CL)及びリンカーから成り、且つ、ここで、前記抗体ドメインと前記リンカーが、N末端からC末端に向かって、以下の順序:
ba)VH−CH1−リンカー−VL−CL、又は
bb)VL−CL−リンカー−VH−CH1、
のうちの一方を有し、
ここで、前記リンカーが少なくとも30個のアミノ酸、好ましくは32〜50個のアミノ酸から成るペプチドである、
を含んでなり;そして
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖ののC又はN末端のペプチドコネクタを介して融合されている;
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸、好ましくは10〜50個のアミノ酸から成るペプチドである、四価の二重特異性抗体である。
【0128】
三価又は四価フォーマットのいずれかで使用される場合に、用語「全長抗体」は、2つの「全長抗体重鎖」と2つの「全長抗体軽鎖」から成る抗体を意味する(図1を参照のこと)。「全長抗体重鎖」は、N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成り、VH−CH1−HR−CH2−CH3と略記される、ポリペプチドであり、更に任意に、IgEサブクラスの抗体の場合には、抗体重鎖定常ドメイン4(CH4)から成るポリペプチドである。好ましくは「全長抗体重鎖」は、N末端からC末端に向かって、VH、CH1、HR、CH2及びCH3から成るポリペプチドである。「全長抗体軽鎖」は、N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成り、VL−CLと略記される、ポリペプチドである。抗体軽鎖定常ドメイン(CL)は、κ(カッパ)又はλ(ラムダ)であり得る。2つの全長抗体鎖は、CLドメインとCH1ドメインの間、及び全長抗体重鎖のヒンジ領域の間のポリペプチド間ジスルフィド結合を介して一緒につながっている。典型的な全長抗体の例は、IgG(例えば、IgG1及びIgG2)、IgM、IgA、IgD、並びにIgEのような天然抗体である。本発明による全長抗体は、単一種、例えばヒトからのものであることができ、又はそれらは、キメラ化若しくはヒト化抗体であることもできる。本発明による全長抗体は、一対のVHとVLによってそれぞれ形成された抗原結合部位を2つ含んでなるが、それらは、ともに同じ抗原に特異的に結合する。前記全長抗体の重鎖又は軽鎖のC末端は、前記重鎖又は軽鎖のC末端における最後のアミノ酸を意味する。前記全長抗体の重鎖又は軽鎖のN末端は、前記重鎖又は軽鎖のN末端における最後のアミノ酸を意味する。
【0129】
本発明の中で使用される場合、「ペプチドコネクタ」という用語は、アミノ酸配列を有するペプチドを意味し、それは、好ましくは合成起源のものである。本発明によるこれらのペプチドコネクタは、一本鎖Fabフラグメントを、全長抗体のC又はN末端に融合して、本発明による多特異性抗体を形成するのに使用される。好ましくは、前記b)のペプチドコネクタは、少なくとも5個のアミノ酸の長さ、好ましくは5〜100個、より好ましくは10〜50個のアミノ酸の長さを有するアミノ酸配列を持つペプチドである。一実施形態では、前記ペプチドコネクタは、(GxS)n又は(GxS)nGmであって、G=グリシン、S=セリン、及び(x=3、n=3、4、5若しくは6、及びm=0、1、2若しくは3)又は(x=4、n=2、3、4若しくは5及びm=0、1、2若しくは3)、好ましくはx=4及びn=2又は3、より好ましくはx=4、n=2を示す。好ましくは、三価の二重特異性抗体(ここで、VH若しくはVH−CH1ポリペプチドとVL若しくはVL−CLポリペプチド(図7a〜c)が全長抗体のC末端に2つの同一のペプチドコネクタを介して融合されている)において、前記ペプチドコネクタは、少なくとも25個のアミノ酸のペプチドであり、好ましくは30〜50個のアミノ酸のペプチド、そしてより好ましくは、前記ペプチドコネクタは、(GxS)n又は(GxS)nGmであって、G=グリシン、S=セリン、及び(x=3、n=6、7若しくは8、及びm=0、1、2若しくは3)又は(x=4、n=5、6、若しくは7及びm=0、1、2、若しくは3)、好ましくはx=4及びn=5、6、7を示す。
【0130】
「一本鎖Fabフラグメント」(図2aを参照のこと)は、抗体重鎖可変ドメイン(VH)、抗体定常ドメイン1(CH1)、抗体軽鎖可変ドメイン(VL)、抗体軽鎖定常ドメイン(CL)及びリンカーから成るポリペプチドであって、ここで、前記抗体ドメイン及び前記リンカーは、N末端からC末端に向かって、以下の順序:a)VH−CH1−リンカー−VL−CL、b)VL−CL−リンカー−VH−CH1、c)VH−CL−リンカー−VL−CH1又はd)VL−CH1−リンカー−VH−CLのうちのいずれか1つであり;且つ、ここで、前記リンカーは、少なくとも30個のアミノ酸、好ましくは32〜50個のアミノ酸から成るポリペプチドである。前記一本鎖Fabフラグメントa)VH−CH1−リンカー−VL−CL、b)VL−CL−リンカー−VH−CH1、c)VH−CL−リンカー−VL−CH1及びd)VL−CH1−リンカー−VH−CLは、CLドメインとCH1ドメインの間の天然のジスルフィド結合によって安定化される。「N末端」という用語は、N末端の最後のアミノ酸を意味し、「C末端」という用語は、C末端の最後のアミノ酸を意味する。
【0131】
「リンカー」という用語は、本発明の中で一本鎖Fabフラグメントに関して使用され、そして(好ましくは合成起源のものである)アミノ酸配列を持つペプチドを意味する。本発明によるこれらのペプチドは、a)VH−CH1をVL−CLと、b)VL−CLをVH−CH1と、c)VH−CLをVL−CH1と又はd)VL−CH1をVH−CLと連結して、以下の本発明による一本鎖Fabフラグメントa)VH−CH1−リンカー−VL−CL、b)VL−CL−リンカー−VH−CH1、c)VH−CL−リンカー−VL−CH1又はd)VL−CH1−リンカー−VH−CLを形成するのに使用される。一本鎖Fabフラグメントの中の前記リンカーは、少なくとも30個のアミノ酸の長さ、好ましくは32〜50個のアミノ酸の長さを有するアミノ酸配列を持つペプチドである。一実施形態では、前記リンカーは、(GxS)nであって、G=グリシン、S=セリン、(x=3、n=8、9若しくは10及びm=0、1、2若しくは3)又は(x=4、n=6、7若しくは8及びm=0、1、2若しくは3)を示し、好ましくはx=4、n=6若しくは7及びm=0、1、2若しくは3を示し、より好ましくはx=4、n=7及びm=2を示す。一実施形態では、前記リンカーは(G4S)6G2である。
【0132】
好ましい実施形態では、前記一本鎖Fabフラグメントの中の前記抗体ドメイン及び前記リンカーは、N末端からにC末端に向かって、以下の順序:a)VH−CH1−リンカー−VL−CL、又はb)VL−CL−リンカー−VH−CH1のいずれか一方であり、より好ましくはVL−CL−リンカー−VH−CH1である。
【0133】
別の好ましい実施形態では、前記一本鎖Fabフラグメントの中の前記抗体ドメイン及び前記リンカーは、N末端からC末端に向かって、以下の順序:a)VH−CL−リンカー−VL−CH1又はb)VL−CH1−リンカー−VH−CLのいずれか一方である。
【0134】
任意には、前記一本鎖Fabフラグメントにおいて、CLドメインとCH1ドメインの間の天然のジスルフィド結合に加えて、抗体重鎖可変ドメイン(VH)と抗体軽鎖可変ドメイン(VL)にも、以下の位置間のジスルフィド結合の導入によってジスルフィドにより安定化される:
i)重鎖可変ドメインの第44位〜軽鎖可変ドメインの第100位
ii)重鎖可変ドメインの第105位〜軽鎖可変ドメインの第43位、又は、
iii)重鎖可変ドメインの第101位〜軽鎖可変ドメインの第100位(付番は常にKabatのEUインデックスに準じる)。
【0135】
一本鎖Fabフラグメントのそういった更なるジスルフィドによる安定化は、その一本鎖Fabフラグメントの可変ドメインVHとVLの間のジスルフィド結合の導入によって達成される。一本鎖Fabフラグメントの安定化のために非天然ジスルフィド架橋を導入する技術は、例えば、WO94/029350、Rajagopal, et al., Prot Engin. (1997) 1453-59;Kobayashi, H., et al., Nuclear Medicine & Biology, 25, (1998) 387-393;又はSchmidt, M., et al., Oncogene 18 (1999) 1711-1721に記載されている。1つの実施形態では、本発明による抗体内に含まれている一本鎖Fabフラグメントの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第44位と軽鎖可変ドメインの第100位の間にある。1つの実施形態では、本発明による抗体内に含まれている一本鎖Fabフラグメントの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第105位と軽鎖可変ドメインの第43位の間にある(付番は常にKabatのEUインデックスに準じる)。
【0136】
一実施形態では、一本鎖Fabフラグメントの可変ドメインVHとVLの間の前記任意のジスルフィドによる安定化のない一本鎖Fabフラグメントが好まれる。
【0137】
「一本鎖Fvフラグメント」(図2bを参照のこと)は、抗体重鎖可変ドメイン(VH)、抗体軽鎖可変ドメイン(VL)、及び一本鎖Fvリンカーから成るポリペプチドであって、ここで、前記抗体ドメイン及び前記一本鎖Fvリンカーは、N末端からC末端に向かって、以下の順序:a)VH−一本鎖Fvリンカー−VL、b)VL−一本鎖Fvリンカー−VHであり;好ましくは、a)VH−一本鎖Fvリンカー−VLであり;且つ、ここで、前記一本鎖Fvリンカーは、少なくとも15個のアミノ酸の長さを有し、一実施形態では、少なくとも20個のアミノ酸を有するアミノ酸配列を持つポリペプチドである。「N末端」という用語は、N末端の最後のアミノ酸を意味し、「C末端」という用語は、C末端の最後のアミノ酸を意味する。
【0138】
一本鎖Fvフラグメントの中で使用される「一本鎖Fvリンカー」という用語は、(好ましくは合成起源のものである)アミノ酸配列を持つペプチドを意味する。前記一本鎖Fvリンカーは、少なくとも15個のアミノ酸の長さを有し、一実施形態では、少なくとも20個のアミノ酸の長さ、好ましくは15〜30個のアミノ酸の長さを有するアミノ酸配列を持つペプチドである。一実施形態では、前記一本鎖リンカーは、(GxS)nであって、G=グリシン、S=セリン、(x=3及びn=4、5若しくは6)又は(x=4及びn=3、4、5若しくは6)を示し、好ましくはx=4、n=3、4若しくは5を示し、より好ましくはx=4、n=3若しくは4を示す。一実施形態では、前記一本鎖Fvリンカーは(G4S)3又は(G4S)4である。
【0139】
更に、前記一本鎖Fvフラグメントは、好ましくは、ジスルフィドにより安定化されている。一本鎖抗体のこうした更なるジスルフィドによる安定化は、その一本鎖抗体の可変ドメインの間のジスルフィド結合の導入によって達成され、そしてそれは、例えば、WO 94/029350、Rajagopal, V., et al., Prot. Engin. 10 (1997) 1453-59;Kobayashi, R, et al., Nuclear Medicine & Biology 25 (1998) 387-393;又はSchmidt, M., et al., Oncogene 18 (1999) 1711-1721に記載されている。
【0140】
ジスルフィドにより安定化した一本鎖Fvフラグメントの一実施形態では、本発明による抗体に含まれている一本鎖Fvフラグメントの可変ドメインの間のジスルフィド結合は、各一本鎖Fvフラグメントごとに独立に、以下のものから選択される:
i)重鎖可変ドメインの第44位〜軽鎖可変ドメインの第100位
ii)重鎖可変ドメインの第105位〜軽鎖可変ドメインの第43位、又は、
iii)重鎖可変ドメインの第101位〜軽鎖可変ドメインの第100位。
【0141】
一実施形態では、本発明による抗体に含まれている一本鎖Fvフラグメントの可変ドメインの間のジスルフィド結合は、重鎖可変ドメインの第44位と軽鎖可変ドメインの第100位の間にある。
【0142】
本発明に記載の抗体は、リコンビナント手段により産生される。よって、本発明の1つ態様は、本発明による抗体をコードする核酸であり、そして更なる態様は、本発明による抗体をコードする前記核酸を含んでなる細胞である。リコンビナント産生のための方法は、当技術分野の現状において広く公知であり、抗体ポリペプチドのその後の単離及び通常は医薬として許容され得る純度への精製を伴う、原核細胞及び真核細胞におけるタンパク質の発現を含む。宿主細胞における前述の抗体の発現のために、個別に改変した軽鎖及び重鎖をコードする核酸が、標準的な方法により発現ベクターに挿入される。発現は、CHO細胞、NS0細胞、SP2/0細胞、HEK293細胞、COS細胞、PER.C6細胞、酵母、又はE.コリ(E. coli)細胞などの適切な原核又は真核宿主細胞において行われ、そして抗体は、その細胞から回収される(上清又は細胞溶解後に)。抗体のリコンビナント産生のための一般的な方法は、当技術分野の現状において周知であり、例えばMakrides, S.C., Protein Expr. Purif. 17(1999)183-202;Geisse, S., et al., Protein Expr. Purif. 8(1996)271-282;Kaufman, R.J., Mol. Biotechnol. 16(2000)151-160;Werner, R.G., Drug Res. 48(1998)870-880の総説に記載されている。
【0143】
二重特異性抗体は、例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、又はアフィニティークロマトグラフィーなどの通常の免疫グロブリン精製手順により培養液から適切に分離される。モノクローナル抗体をコードするDNA及びRNAは、通常の手順を使用して容易に単離及び配列決定される。ハイブリドーマ細胞は、そのようなDNA及びRNAの起源として役立つことができる。いったん単離されれば、DNAを発現ベクターに挿入し、次いでそれをHEK293細胞、CHO細胞、又は骨髄腫細胞などの宿主細胞にトランスフェクトして、この宿主細胞におけるリコンビナントモノクローナル抗体の合成を達成することができ、この宿主細胞は、さもなければ免疫グロブリンタンパク質を産生しない。
【0144】
二重特異性抗体のアミノ酸配列変異体(突然変異体)は、抗体のDNA内に適切なヌクレオチドの変更を導入することによって、又はヌクレオチド合成によって調製される。しかし、そのような改変は、例えば上記のように非常に限られた範囲内でのみ行うことができる。例えば、改変は、IgGアイソタイプ及びエピトープの結合などの前述の抗体の特徴を変化させるのではなく、リコンビナント産生の収率又はタンパク質の安定性を改善することがあり、精製を容易にすることもある。
【0145】
用語「宿主細胞」は、本願で使用される場合、本発明による抗体を産生するために設計され得るあらゆる種類の細胞系を意味する。1つの実施形態では、HEK293細胞及びCHO細胞が宿主細胞として使用される。本明細書において使用される場合、用語「細胞」、「細胞株」及び「細胞培地」とは、互換的に使用され、そしてすべてのそのような名称は子孫も包含する。従って、単語「形質転換体」及び「形質転換細胞」とは、トランスファーの数に関係なく、それらに由来する初代対象細胞及び培養物を包含する。すべての子孫は、故意の又は偶然な突然変異のために、DNA含有において正確には同一でないこともまた理解されている。元の形質転換された細胞についてスクリーンされる場合、同じ機能的又は生物学的活性を有する変異体子孫が包含される。
【0146】
NS0細胞における発現は、例えばBarnes, L.M., et al., Cytotechnology 32(2000)109-123;Barnes, L.M., et al., Biotech. Bioeng. 73(2001)261-270により記載されている。一過性発現が、例えばDurocher, Y., et al., Nucl. Acids. Res. 30(2002)E9により記載される。可変ドメインのクローニングは、Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86(1989)3833-3837;Carter, P., et al., Proc. Natl. Acad. Sci. USA 89(1992)4285-4289;及びNorderhaug, L., et al., J. Immunol. Methods 204(1997)77-87により記載されている。好ましい一過性発現システム(HEK293)は、Schlaeger, E.-J., and Christensen, K., in Cytotechnology 30(1999)71-83 及びSchlaeger, E.-J., in J. Immunol. Methods 194(1996)191-199により記載される。
【0147】
原核生物のために適切である制御配列には、例えばプロモーター、任意にはオペレーター配列及びリボソームに結合部位が含まれる。真核細胞は、プロモーター、エンハンサー及びポリアデニル化シグナルを使用することが知られている。
【0148】
核酸は、それが他の核酸配列と機能的な関連性をもって配置されるとき、「操作可能な形で連結され」ている。例えば、プレ配列又は分泌リーダーのDNAは、それがポリペプチドの分泌に関与するプレタンパク質として発現されるとき、ポリペプチドに関してDNAと操作可能な形で連結している;プロモーター又はエンハンサーは、それがその配列の転写に影響するとき、コード配列と操作可能な形で連結されている;又は、リボソーム結合部位は、それが転写を促進するように配置されているとき、コード配列に操作可能な形で連結されている。一般的に、「操作可能な形で連結された」は、連結されたDNA配列が隣接し、そして、分泌リーダーの場合は、隣接し、且つ、読み枠内にあることを意味する。しかしながら、エンハンサーは隣接している必要は無い。連結は、都合の良い制限部位でライゲーションを行うことにより達成される。そのような部位が存在しなければ、確立された手段に従い、合成オリゴヌクレオチドアダプター又はリンカーが使用される。
【0149】
抗体の精製は、アルカリ/SDS処理、CsClバンディング、カラムクロマトグラフィー、アガロースゲル電気泳動、及び当該技術分野において周知な他の方法を含む標準的な技術により、他の細胞成分又は他の汚染物、例えば、他の細胞核酸又はタンパク質を除去するために実行される。Ausubel, F., et al. (ed.), Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York(1987)を参照のこと。別の方法、例えば、微生物タンパク質を用いた親和性クロマトグラフィー(例えば、プロテインA又はプロテインG親和性クロマトグラフィー)、イオン交換クロマトグラフィー(例えば、カチオン交換(カルボキシメチル樹脂)、陰イオン交換性(アミノエチル樹脂)及び混合モード交換)、thiophilic吸着(例えば、β−メルカプトエタノールと他のSHリガンド類を用いて)、疎水性相互作用又は芳香族吸着クロマトグラフィー(例えば、フェニル−セファロース、アザ−アレノフィリック樹脂、又はm−アミノフェニルボロン酸を用いて)、金属キレート親和性クロマトグラフィー(例えば、ニッケル(II)及びCu(II)親和性材料を用いて)、サイズ排除クロマトグラフィー、及び電気泳動法(ゲル電気泳動、毛細管電気泳動など)などが、確立されており、そしてタンパク質精製のために幅広く使用されている(Vijayalakshmi, M.A., Appl. Biochem. Biotech. 75 (1998) 93-102)。
【0150】
本明細書において使用される場合、用語「細胞」、「細胞株」及び「細胞培地」とは、互換的に使用され、そしてすべてのそのような名称は子孫も包含する。従って、単語「形質転換体」及び「形質転換細胞」とは、トランスファーの数に関係なく、それらに由来する初代対象細胞及び培養物を包含する。すべての子孫は、故意の又は偶然な突然変異のために、DNA含有において正確には同一でないこともまた理解されている。元の形質転換された細胞についてスクリーンされる場合、同じ機能的又は生物学的活性を有する変異体子孫が包含される。異なった命名法が意図される場合は、その内容から明白であろう。
【0151】
用語「形質転換」とは、本明細書において使用される場合、宿主細胞中へのベクター/核酸のトランスファー工程を指す。強い細胞壁バリアを有さない細胞が宿主細胞として使用される場合、トランスフェクションが、例えばGraham, F.L., and van der Ed, A.J. Virology 52(1978)456-467により記載されるようなリン酸カルシウム沈殿方法により実施される。しかしながら、核注入又はプロトプラスト融合による細胞中へのDNAの導入のための他の方法がまた使用され得る。原核細胞、又は実質的な細胞壁構造を含む細胞が使用される場合、1つのトランスフェクション方法は、Cohen, S.N, et al, PNAS 69(1972)2110-2114により記載されるような、塩化カルシウムを用いるカルシウム処理法である。
【0152】
本明細書において使用される場合、「発現」とは、核酸がmRNAに転換される方法、及び/又は転写されたmRNA(転写体とも呼ばれる)がペプチド、ポリペプチド又はタンパク質に連続的に翻訳される過程を指す。転写体及びコードされるポリペプチドは、集合的には、遺伝子生成物とも呼ばれる。ポリヌクレオチドがゲノムDNAに由来する場合、真核細胞における発現は、mRNAのスプライシングを含む。
【0153】
「ベクター」とは、挿入される核酸分子を、宿主細胞中に及び/又は宿主細胞間にトランスファーする、核酸分子、特に自己複製する核酸分子である。この用語は、DNAの細胞中への挿入(例えば、染色体組込み)のために主に機能するベクター、DNA又はRNAの複製のために主に機能するベクターの複製、及びDNA又はRNAの転写及び/又は翻訳のために機能する発現ベクターを包含する。記載されるような機能のうちの2つ以上を提供するベクターもまた包含される。
【0154】
「発現ベクター」とは、適切な宿主細胞中に導入される場合、転写され、そしてポリペプチドに翻訳され得るポリヌクレオチドである。「発現システム」とは、所望する発現生成物を生成するために機能することができる発現ベクターを含んでなる適切な宿主細胞を通常指す。
【0155】
医薬組成物
本発明の1つの態様は、本発明による抗体を含んでなる医薬組成物である。本発明の別の態様は、医薬組成物の製造のための本発明による抗体の使用である。本発明の更なる態様は、本発明による抗体を含んでなる医薬組成物の製造のための方法である。別の態様では、本発明は、医薬担体と一緒に処方される本発明による抗体を含んでなる組成物、例えば医薬組成物を提供する。
【0156】
本発明の1つの態様は、癌治療のための本発明による二重特異性抗体である。
【0157】
本発明の別の態様は、癌治療のための前記医薬組成物である。
【0158】
本発明の別の態様は、癌処置用の治療薬の製造のための本発明による抗体の使用である。
【0159】
本発明の別の態様は、本発明による抗体をそういった処置を必要としている患者に投与することによる、癌に罹患している患者の処置方法である。
【0160】
本明細書中で使用されるとき、「医薬担体」には、生理学的に適合する任意の及びすべての溶媒、分散媒、被覆剤、抗細菌剤及び抗真菌剤、等張剤及び吸収遅延剤などが含まれる。好ましくは、前記担体は、(例えば注射又は点滴による)静脈内投与、筋肉内投与、皮下投与、非経口投与、脊髄投与又は上皮投与に適切である。
【0161】
本発明の組成物は、当該技術分野において知られる様々な方法により投与され得る。当業者に認識されるように、投与の経路及び/又は様式は、所望される結果に依存して異なる。ある投与径路により本発明の組成物を投与するためには、その組成物を、その不活性化を防止する材料で被覆し、又はその組成物と共に同時に投与する必要がある。例えば、前記組成物は、例えばリポソーム、又は希釈剤などの適切な担体に加えられて対象に投与され得る。医薬として許容される希釈剤には、生理食塩水及び水性緩衝溶液が含まれる。医薬として許容される担体には、滅菌水性溶液又は分散剤、滅菌済みの注射可能な溶液又は分散剤を即時調製するための滅菌粉末が含まれる。そのような医薬活性物質のための媒体及び薬剤は、当該技術分野において知られている。
【0162】
「非経口投与」及び「非経口的に投与される」という語句は、本明細書中で使用される場合、腸内投与及び局所投与以外の投与様式を意味し、通常は注射により、これだけに限定されるものではないが、静脈内、筋肉内、動脈内、鞘内、嚢内、眼窩内、心臓内、皮内、腹腔内、経気管、皮下、表皮下、関節内、皮下、くも膜下、髄腔内、硬膜外及び胸骨内注射及び点滴を含む。
【0163】
用語「癌」は、本明細書中で使用される場合、例えばリンパ腫、リンパ球性白血病、肺癌、非小細胞肺(NSCL)癌、細気管支肺胞細胞性肺癌、骨癌、膵臓癌、皮膚癌、頭頚部の癌、皮膚又は眼内の黒色腫、子宮癌、卵巣癌、直腸癌、肛門部の癌、胃癌、胃せん癌、結腸癌、乳癌、子宮癌、卵管の癌腫、子宮内膜の癌腫、子宮頚部の癌腫、膣癌、外陰部癌、ホジキン病、食道癌、小腸癌、内分泌系の癌、甲状腺癌、副甲状腺癌、副腎癌、軟組織の肉腫、尿道の癌、陰茎の癌、前立腺癌、膀胱癌、腎臓又は輸尿管の癌、腎細胞癌、腎盂癌、中皮腫、肝細胞性癌、胆管癌、中枢神経系(CNS)の新生物、脊椎軸腫瘍、脳幹膠腫、多形膠芽腫、星状細胞腫、神経鞘腫、脳室上衣腫、髄芽腫、髄腹腫、扁平上皮癌、脳下垂体腺腫及びユーイング肉腫、そして前記癌のうちのいずれかの難治性バーション、又は前記癌のうちの1つ若しくは複数の組合せを含めたものであり得る。
【0164】
本発明の別の態様は、血管新生阻害剤としての本発明による二重特異性抗体又は前記医薬組成物である。そのような血管新生阻害剤は、癌、特に固形腫瘍、及び他の血管疾患の治療に使用できる。
【0165】
本発明の1つの態様は、血管疾患の治療のための本発明による二重特異性抗体である。
【0166】
本発明の別の態様は、血管疾患処置用の治療薬の製造のための本発明による抗体の使用である。
【0167】
本発明の別の態様は、本発明による抗体をそういった処置を必要としている患者に投与することによる、血管疾患に罹患している患者の処置方法である。
【0168】
「血管疾患」という用語には、癌、炎症性疾患、アテローム性動脈硬化症、虚血、外傷、敗血症、COPD、喘息、糖尿病、AMD、網膜障害、脳卒中、肥満症、急性肺傷害、出血、血管漏出、例えばサイトカイン誘発性血管漏出、アレルギー、グレーブス病、橋本の自己免疫性甲状腺炎、特発性血小板減少性紫斑病、巨細胞性動脈炎、関節リウマチ、全身性エリテマトーデス(SLE)、ループス腎炎、クローン病、多発性硬化症、潰瘍性大腸炎、特に固形腫瘍、眼内の新生血管症候群、例えば増殖性網膜症又は加齢性黄斑変性症(AMD)など、関節リウマチ、及び乾癬が含まれる(Folkman, J., et al., J. Biol. Chem. 267 (1992) 10931-10934;Klagsbrun, et al., Annu. Rev. Physiol. 53 (1991) 217-239;及びGarner, A., Vascular diseases, In: Pathobiology of ocular disease, A dynamic approach, Garner, A., and Klintworth, G. K., (eds.), 2nd edition, Marcel Dekker, New York (1994) 1625-1710)。
【0169】
これらの組成物は更に、保存料、湿潤剤、乳化剤及び分散剤などの補助剤を含むこともできる。微生物の存在の予防は、前掲の殺菌手段によって、及び様々な抗菌及び抗真菌剤、例えばパラベン、クロロブタノール、フェノール、ソルビン酸などの含有によって確保できる。組成物中に等張剤、例えば糖、塩化ナトリウムなどを含むことが望ましいこともある。加えて、注射用医薬形態の持続的吸収は、モノステアリン酸アルミニウムやゼラチンなどの吸収を遅らせる作用物質の含有によって引き起こすこともできる。
【0170】
選択した投与経路にかかわらず、(好適な水和形態で使用することもできる)本発明の化合物、及び/又は本発明の医薬組成物を、当業者に知られている従来の方法によって医薬的に許容し得る剤形に処方する。
【0171】
本発明の医薬組成物中の有効成分の実際の投与量レベルは、特定の患者、組成物、及び投与様式について、その患者にとって有毒であることなしに、所望の治療応答を達成するのに有効である有効成分量を得るように変更することもできる。選択される投与量レベルは、用いられる本発明の特定の組成物の活性、、投与経路、投与時間、用いられる特定の化合物の排泄速度、処置期間、用いられる特定の組成物と組合せて使用されるその他の薬物、化合物及び/又は材料、処置される患者の年齢、性別、体重、身体状態、健康全般、及び前病歴、並びに医療分野で周知の要因など、を含めた様々な薬物動態学的要因に依存するであろう。
【0172】
組成物は、それがシリンジによって提出できる程度に無菌であり、且つ、液状でなければならない。水に加えて、好ましくは、担体は等張緩衝生理食塩溶液である。
【0173】
適切な流動性は、例えば、レシチンなどのコーティングの使用によって、分散液の場合には所望の粒度の維持によって、そして界面活性剤の使用によって、維持できる。多くの場合、組成物中に等張剤、例えば、糖、マンニトールやソルビトールなどのポリアルコール、及び塩化ナトリウムを含むことが望ましい。
【0174】
ヒトErbB−2及びヒトc−Metに対するこの発明による二重特異性抗体が、価値のある特徴、例えば生物学的又は薬理学的な活性などを有することがここでわかった。
【0175】
以下の実施例、配列表及び図面は、本発明の理解を助けるために提供されるが、本発明の真の範囲は添付の特許請求の範囲に示される。本発明の要旨から逸脱することなしに、示された手順に改変を加えることができると理解されている。
【0176】
アミノ酸配列の説明
配列番号1 重鎖可変ドメイン<ErbB−2>トラスツズマブ
配列番号2 軽鎖可変ドメイン<ErbB−2>トラスツズマブ
配列番号3 重鎖可変ドメイン<c−Met>Mab 5D5
配列番号4 軽鎖可変ドメイン<c−Met>Mab 5D5
配列番号5 重鎖<c−Met>Mab 5D5
配列番号6 軽鎖<c−Met>Mab 5D5
配列番号7 重鎖<c−Met>Fab 5D5
配列番号8 軽鎖<c−Met>Fab 5D5
配列番号9 ヒトIgG1の重鎖定常領域
配列番号10 ヒトIgG3の重鎖定常領域
配列番号11 ヒト軽鎖カッパ定常領域
配列番号12 ヒト軽鎖ラムダ定常領域
配列番号13 ヒトc−Met
配列番号14 ヒトErbB−2
配列番号15 重鎖CDR3H、<ErbB−2>トラスツズマブ
配列番号16 重鎖CDR2H、<ErbB−2>トラスツズマブ
配列番号17 重鎖CDR1H、<ErbB−2>トラスツズマブ
配列番号18 軽鎖CDR3L、<ErbB−2>トラスツズマブ
配列番号19 軽鎖CDR2L、<ErbB−2>トラスツズマブ
配列番号20 軽鎖CDR1L、<ErbB−2>トラスツズマブ
配列番号21 重鎖CDR3H、<c−Met>Mab 5D5
配列番号22 重鎖CDR2H、<c−Met>Mab 5D5
配列番号23 重鎖CDR1H、<c−Met>Mab 5D5
配列番号24 軽鎖CDR3L、<c−Met>Mab 5D5
配列番号25 軽鎖CDR2L、<c−Met>Mab 5D5
配列番号26 軽鎖CDR1L、<c−Met>Mab 5D5
【図面の簡単な説明】
【0177】
【図1】典型的な順序で可変ドメインと定常ドメインを含んでなる2対の重鎖と軽鎖を持ち、第1の抗原1に特異的に結合するCH4ドメインを含まない全長抗体の図式的構造。
【図2a】以下の:a)ヒトErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びにb)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖を含んでなり、ここで、前記可変ドメインVL及びVH、及び/又は前記定常ドメインCL及びCH1が互いによって置き換えられ、それらがノブ−into−ホール技術で修飾されている、二価の二重特異性<ErbB−2/c−Met>抗体の図式的構造。
【図2b】以下の:a)ヒトErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びにb)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖を含んでなり、ここで、前記可変ドメインVL及びVH、及び/又は前記定常ドメインCL及びCH1が互いによって置き換えられ、それらがノブ−into−ホール技術で修飾されている、二価の二重特異性<ErbB−2/c−Met>抗体の図式的構造。
【図2c】以下の:a)ヒトErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びにb)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖を含んでなり、ここで、前記可変ドメインVL及びVH、及び/又は前記定常ドメインCL及びCH1が互いによって置き換えられ、それらがノブ−into−ホール技術で修飾されている、二価の二重特異性<ErbB−2/c−Met>抗体の図式的構造。
【図3a】a)図3a:2つのポリペプチドVHとVLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する); b)図3b:2つのポリペプチドVH−CH1とVL−CLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性<ErbB−2/c−Met>抗体の略図。
【図3b】a)図3a:2つのポリペプチドVHとVLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する); b)図3b:2つのポリペプチドVH−CH1とVL−CLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性<ErbB−2/c−Met>抗体の略図。
【図3c】図3c:「ノブとホール」を備えた2つのポリペプチドVHとVLがそこに融合されている(VHドメインとVLドメインは両方が一緒になってc−Metに特異的に結合する抗原結合部位を形成する)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性抗体の略図。
【図3d】図3d:「ノブとホール」を備えた2つのポリペプチドVHとVLがそれに融合されている(VHドメインとVLドメインは両方が一緒になってc−Metに特異的に結合する抗原結合部位を形成する、ここで、それらのVHドメイン及びVLドメインはVH44位とVL100位の間に鎖間ジスルフィド架橋を含んでなる)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性抗体の略図。
【図4a】4a:4種類の考えられる一本鎖Fabフラグメントの図式的構造。
【図4b】4b:2種類の一本鎖Fvフラグメントの図式的構造。
【図5a】1つの一本鎖Fabフラグメント(図5a)又は1つの一本鎖Fvフラグメント(図5b)と全長抗体を含んでなる三価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−ノブとホールを備えた二重特異性三価の例。
【図5b】1つの一本鎖Fabフラグメント(図5a)又は1つの一本鎖Fvフラグメント(図5b)と全長抗体を含んでなる三価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−ノブとホールを備えた二重特異性三価の例。
【図6a】2つの一本鎖がFabフラグメント(図6a)又は2つの一本鎖Fvフラグメント(図6b)と全長抗体を含んでなる四価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−c−Met結合部位はc−Met二量化阻害抗体に由来する。
【図6b】2つの一本鎖がFabフラグメント(図6a)又は2つの一本鎖Fvフラグメント(図6b)と全長抗体を含んでなる四価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−c−Met結合部位はc−Met二量化阻害抗体に由来する。
【図7a】類上皮腫癌細胞株A431におけるErbB2/2/3及びc−Metの細胞表面発現のフローサイトメトリー分析。
【図7b】卵巣癌細胞株OVCAR−8におけるErbB2/2/3及びc−Metの細胞表面発現のフローサイトメトリー分析。
【図8】0、30、60及び120分(=0、0.5、1、及び2時間)に計測したOVCAR−8癌細胞における内部移行アッセイ。
【図9a】OVCAR−8癌細胞における増殖アッセイ。単一特異性の親<HER2>及び<c−Met>抗体と比較した、本発明による二重特異性<HER2/c−Met>抗体BsAB02(BsAb)の癌細胞増殖の阻害。
【図9b】HGFの存在下での癌細胞株Ovcar−8における増殖アッセイ。単一特異性の親<HER2>及び<c−Met>抗体と比較した、本発明による二重特異性<HER2/c−Met>抗体BsAB02(BsAb)の癌細胞増殖の阻害。
【実施例】
【0178】
実験手順
材料と方法
組換えDNA技術
Sambrook, J. et al., Molecular cloning: A laboratory manual;Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989に記載の標準的な方法を、DNAを操作するために使用した。分子生物学的試薬を製造業者の取扱説明書に従って使用した。
【0179】
DNAとタンパク質の配列分析、及び配列データ・マネジメント
以下でヒト免疫グロブリン軽鎖及び重鎖のヌクレオチド配列に関する概説は:Kabat, E.A. et al.,(1991)Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242にある。抗体鎖のアミノ酸には、EUナンバリングに従って付番した(Edelman, G.M., et al., PNAS 63(1969)78-85;Kabat, E.A., et al.,(1991)Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242)。GCG(Genetics Computer Group、Madison, Wisconsin)のソフトウェアパッケージ バージョン10.2及びInfomaxのVector NTI Advance suiteバージョン8.0を、配列新規作成、マッピング、分析、アノテーション及び図示に使用した。
【0180】
DNA配列決定
DNA配列を、SequiServe(Vaterstetten, Germany)及びGeneart AG(Regensburg, Germany)にて実施された二本鎖配列決定法によって決定した。
【0181】
遺伝子合成
所望の遺伝子セグメントは、自動化遺伝子合成によって合成オリゴヌクレオチドとPCR産物からGeneart AG(Regensburg, Germany)で調製された。1つの制限エンドヌクレアーゼ切断部位に隣接した遺伝子セグメントを、pGA18(ampR)プラスミド内にクローン化した。プラスミドDNAを形質転換細菌から精製し、そしてUV分光法によって濃度決定した。サブクローニングした遺伝子フラグメントのDNA配列を、DNA配列決定法で確認した。同じように、ペプチドコネクタで連結されているC末端<c−Met>5D5 scFab VH領域を持つ/持たないCH3ドメイン内にS354C及びT366Wが突然変異を保有する改変「ノブ−into−ホール」<ErbB−2>抗体重鎖、並びにペプチドコネクタで連結されているC末端<c−Met>5D5 scFab VL領域を持つ/持たない、Y349C、T366S、L368A、及びY407V突然変異を保有する「ノブ−into−ホール」<ErbB−2>抗体重鎖をコードするDNA配列を、隣接するBamHI及びXbaI制限部位を用いて遺伝子合成によって調製した。最後に、<ErbB−2>抗体及び<c−Met>5D5抗体の無改変重鎖及び軽鎖をコードするDNA配列を、隣接するBamHI及びXbaI制限部位によって合成した。すべての構築物をリーダーペプチド(MGWSCIILFLVATATGVHS)をコードする5’末端DNA配列を用いて設計し、そして真核細胞内での分泌のためのタンパク質を目的とした。
【0182】
発現プラスミドの構築
Roche発現ベクターを、すべての重及び軽鎖scFv融合タンパク質をコードする発現プラスミドの構築に使用した。ベクターは、以下の要素:
− 選択マーカーとしてのハイグロマイシン耐性遺伝子、
− エプスタイン−バールウイルス(EBV)の複製開始点、oriP、
− E.コリ内でのこのプラスミドの複製を可能にするベクターpUC18からの複製開始点、
− E.コリ内でアンピシリン耐性を付与するβ−ラクタマーゼ遺伝子、
− ヒト・サイトメガロウイルス(HCMV)からの前初期エンハンサー及びプロモーター
− ヒト1−免疫グロブリン・ポリアデニル化(「ポリA」)シグナル配列、及び
− 固有のBamHI及びXbaI制限部位、
から構成される。
【0183】
記載したように、重鎖又は軽鎖構築物、並びにC末端VH及びVLドメインを持つ「ノブ−into−ホール」構築物を含んでなる免疫グロブリン融合遺伝子を、遺伝子合成によって調製し、そしてpGA18(ampR)プラスミド内にクローン化した。合成DNAセグメントとRoche発現ベクターを保有するpG18(ampR)プラスミドを、BamHI及びXbaI制限酵素(Roche Molecular Biochemicals)によって消化し、そしてアガロースゲル電気泳動法にかけた。次いで、精製した、重鎖又は軽鎖をコードするDNAセグメントを、単離したRoche発現ベクターBamHI/XbaI断片に連結して、最終的な発現ベクターを得た。最終的な発現ベクターをE.コリ細胞内に形質転換し、発現プラスミドDNAを単離し(Miniprep)、そして制限酵素分析とDNA配列決定にかけた。正しいクローンを、150mlのLB−Amp培地中で培養し、再びプラスミドDNAを単離し(Maxiprep)、そして配列完全性をDNA配列決定によって確認した。
【0184】
HEK293細胞における免疫グロブリン変異体の一過性発現
遺伝子組み換え免疫グロブリン変異体を、製造業者の取扱説明書(Invitrogen, USA)に従ってFreeStyle(商標)293発現系を使用したヒト胎児腎臓293−F細胞の一過性トランスフェクションによって発現した。簡単に言えば、懸濁液FreeStyle(商標)293−F細胞を、37℃/8%のCO2にてFreeStyle(商標)293発現培地中で培養し、そしてその細胞をトランスフェクションの日に1〜2×l06生存細胞/mlの密度で新しい培地中に植え付けた。DNA−293fectin(商標)複合体を、250mlの最終的なトランスフェクション体積に対して1:1モル比で、325μlの293fectin(商標)(Invitrogen, Germany)と250μgの重鎖又は軽鎖プラスミドDNAを使用したOpti-MEM(登録商標)I培地(Invitrogen, USA)中で調製した。「ノブ−into−ホール」DNA−293fectin複合体を、250mlの最終的なトランスフェクション体積に対して1:1:2モル比で、325μlの293fectin(商標)(Invitrogen, Germany)と、250μgの「ノブ−into−ホール」重鎖1及び2と、軽鎖プラスミドDNAを使用したOpti-MEM(登録商標)I培地(Invitrogen, USA)中で調製した。抗体を含有している細胞培養上清を、14000gで30分間の遠心分離及び滅菌濾紙(0.22μm)を通して濾過してトランスフェクションの7日後に採取した。上清を−20℃にて精製まで保存した。
【0185】
二重特異性抗体と対照抗体の精製
三価の二重特異性抗体及び対照抗体を、Protein A-Sepharose(商標)(GE Healthcare、Sweden)を使用した親和性クロマトグラフィーとSuperdex200サイズ排除クロマトグラフィーによって細胞培養上清から精製した。簡単に言えば、滅菌濾過した細胞培養上清を、PBSバッファー(10mMのNa2HPO4、1mMのKH2PO4、137mMのNaCl及び2.7mMのKCl、pH、7.4)で平衡化したHiTrap ProteinA HP(5ml)カラムにかけた。非結合タンパク質を平衡化バッファーで洗い落とした。抗体と抗体変異体を、0.1Mのクエン酸バッファー、pH2.8で溶出し、そしてタンパク質含有画分を0.1mlの1M Tris(pH8.5)で中和した。次いで、溶出したタンパク質画分を貯留し、Amiconの超遠心濾過機デバイス(MWCO:30K、Millipore)を用いて3mlの体積まで濃縮し、20mMのヒスチジン、140mMのNaCl、pH6.0で平衡化したSuperdex200 HiLoad 120ml 16/60ゲル濾過カラム(GE Healthcare、Sweden)に投入した。高分子量集合体が5%未満しかない、精製された二重特異性抗体と対照抗体を含有する画分を貯留し、そして1.0mg/mlのアリコートとして−80℃にて保存した。Fabフラグメントを、精製した5D5モノクローナル抗体のパパイン消化と、それに続くプロテインAクロマトグラフィーによる夾雑Fcドメインの除去によって作り出した。非結合Fabフラグメントを、20mMのヒスチジン、140mMのNaCl、pH6.0で平衡化したSuperdex200 HiLoad 120ml 16/60ゲル濾過カラム(GE Healthcare、Sweden)で更に精製し、貯留し、そして1.0mg/mlのアリコートとして−80℃にて保存した。
【0186】
精製タンパク質の分析
精製タンパク質サンプルのタンパク質濃度を、アミノ酸配列に基づいて算出したモル吸光係数を使用し、280nmにて吸光度(OD)を計測することによって測定した。二重特異性抗体及び対照抗体の純度と分子量を、還元剤(5mMの1,4−ジチオスレイトール)の存在下と不存在下でのSDS−PAGE分析、そしてクーマシーブリリアントブルーでの染色によって分析した。NuPAGE(登録商標)Pre-Castゲルシステム(Invitrogen, USA)を、製造業者の取扱説明書に従って使用した(4−20%のトリスグリシン・ゲル)。二重特異性抗体及び対照抗体サンプルの凝集体量を、200mMのKH2PO4、250mMのKCl、pH7.0の泳動バッファー中、25℃にてSuperdex200分析用サイズ排除カラム(GE Healthcare、Sweden)を使用した高性能SECによって分析した。25μgのタンパク質を、0.5ml/分の流量にてカラム上に注入し、そして50分間かけて定組成で溶出した。安定性分析のために、1mg/mlの濃度の精製タンパク質を、4℃と40℃にて7日間インキュベートし、その後、高性能SECによって評価した。還元した二重特異性抗体軽鎖及び重鎖のアミノ酸骨格の完全性を、ペプチド−N−グリコシダーゼF(Roche Molecular Biochemicals)での酵素的処理によるN−グリカンの除去後のNanoElectrospray Q-TOF質量分析によって確認した。
【0187】
c−Metリン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のA549細胞を、0.5%のFCS(ウシ胎仔血清)を含むRPMI中、HGF刺激の前日に植え付けた。翌日に、成長培地を、0.2%のBSA(ウシ血清アルブミン)を含むRPMIで一時間、置き換えた。次いで、5μg/mLの二重特異性抗体を培地に加え、そして細胞を、HGFを加えるまで10分間インキュベートし、50ng/mlの終濃度で更に10分間インキュベートした。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを、100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解した。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進めた。タンパク質濃度を、BCA方法(Pierce)を使用して測定した。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移した。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Y1230、1234、1235(44-888、Biosource)に対するリン酸化部位特異的c−Met抗体を用いて現像した。免疫ブロットを、非リン酸化c−Metに結合する抗体(AF276、R&D)で再検出した。
【0188】
ErbB2/Her2リン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のSk−Br3細胞を、10%のFCS(ウシ胎仔血清)を含むRPMI中、抗体追加の前日に植え付ける。翌日に、5μg/mLの対照又は二重特異性抗体を培地に加え、そして細胞を、更に1時間インキュベートする。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解する。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進める。タンパク質濃度を、BCA方法(Pierce)を使用して測定する。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移す。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Y1221/22(Cell Signaling、2243)に対するリン酸化部位特異的Her2抗体を用いて現像する。免疫ブロットを、非リン酸化Her2に結合する抗体(Cell Signaling、2165)で再検出する。
【0189】
AKTリン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のA431細胞を、10%のFCS(ウシ胎仔血清)を含むRPMI中、抗体追加の前日に植え付ける。翌日に、5μg/mLの対照又は二重特異性抗体を培地に加え、そして細胞を、更に1時間インキュベートする。次いで、一部の細胞を、25ng/mLのHGF(R&D、294-HGN)で更に15分間刺激する。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解する。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進める。タンパク質濃度を、BCA方法(Pierce)を使用して測定する。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移す。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Thr308(Cell signaling、9275)に対するリン酸化部位特異的AKT抗体を用いて現像する。免疫ブロットを、アクチンに結合する抗体(Abcam、ab20272)で再検出する。
【0190】
ERK1/2リン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のA431細胞を、10%のFCS(ウシ胎仔血清)を含むRPMI中、抗体追加の前日に植え付ける。翌日に、5μg/mLの対照又は二重特異性抗体を培地に加え、そして細胞を、更に1時間インキュベートする。次いで、一部の細胞を、25ng/mLのHGF(R&D、294-HGN)で更に15分間刺激する。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解する。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進める。タンパク質濃度を、BCA方法(Pierce)を使用して測定する。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移す。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Thr202/Tyr204(CellSignaling、Nr.9106)に対するリン酸化部位特異的Erk1/2抗体を用いて現像する。免疫ブロットを、アクチンに結合する抗体(Abcam、ab20272)で再検出する。
【0191】
細胞−細胞播種(dissemination)(分散アッセイ)
化合物処置の前日に、A549(1ウェルあたり4000個の細胞)又はA431(1ウェルあたり8000個の細胞)を、0.5%のFCSを含むRPMI中、96ウェルのE-Plate(Roche、05232368001)内の200μLの全容量の中に植え付けた。接着と細胞成長を、電気抵抗を観察する15分毎のスイープによりReal Time Cell Analyzer機器で一晩観察した。翌日、細胞を、5分毎にスイープしながらPBS中のそれぞれの抗体希釈物5μLと共にプレインキュベートした。30分後に、20ng/mLの終濃度をもたらす2.5μLのHGF溶液を加え、そして実験を更に72時間続けた。急激な変化を、180分間の1分毎のスイープで観察し、続いて残りの期間、15分毎のスイープで観察した。
【0192】
HUVEC増殖検定
HUVEC細胞(Promocell、C-12200)を、0.5% FCS含有EBM−2培地(Promocell、C-22211)中でコラーゲンコートした96ウェル内に植え付ける。翌日、対照又は二重特異性抗体の希釈系列をその細胞に加える。30分のインキュベーションの後に、25ng/mLのHGF(R&D、294-HGN)を加え、そして細胞をさらに72時間インキュベートした後に、cell titer glowアッセイ(Promega、G7571/2/3)を用いて、ATP量の形で細胞増殖を製造業者の助言に従って測定する。
【0193】
Sk−Br3増殖アッセイ
a)増殖研究のために、96ウェル細胞培養プレートの1ウェルあたり10000個の細胞を、血清削減培地(RPMI 1640+4%のPCS)中で植え付けた。翌日、親Her2又はc−Met抗体、並びに二重特異性抗体を加え、そして細胞をさらに48時間培養した後に、細胞増殖の指標としてATPを、cell titer glowアッセイ(Promega)を用いて測定する。
【0194】
b)HGF存在下での増殖研究のために、96ウェル細胞培養プレートの1ウェルあたり10000個の細胞を、血清削減培地(RPMI 1640+4%のPCS)中で植え付けた。翌日、親Her2又はc−Met抗体、並びに二重特異性抗体のほかに、25ng/mLのHGF(R&D、294-HGN)も加え、そして細胞をさらに48時間培養した後に、細胞増殖の指標としてATPを、cell titer glowアッセイ(Promega)を用いて測定する。
【0195】
フローサイトメトリーアッセイ(FACS)
a)結合アッセイ
c−Met及びErbB−2発現性細胞を剥離し、そしてカウントした。96ウェルのコニカルプレートの1ウェルあたり1.5×10e5個の細胞を植え付けた。細胞を、遠沈し(1500rpm、4℃、5分間)、そして2%のFCS(ウシ胎仔血清)を含むPBS中のそれぞれの二重特異性抗体の希釈系列50μL中で、氷上において30分間インキュベートした。細胞を、再度遠沈し、そして2%のFCSを含有するPBS200μLで洗浄し、続いて2%のFCS(Jackson Immunoresearch、109116098)を含有するPBSに希釈した、ヒトFcに対するフィコエリトリン結合抗体と一緒に30分間の2回目のインキュベーションを行った。細胞を、2%のFCSを含有するPBS200μLで2回、遠沈洗浄し、BD CellFix溶液(BD Biosciences)で再懸濁し、そして氷上で少なくとも10分間インキュベートした。細胞の平均蛍光強度(mfi)を、フローサイトメトリー(FACS Canto、BD)によって測定した。Mfiを、少なくとも2つの独立した染色の二重反復試験で決定した。フローサイトメトリースペクトルを、FlowJoソフトウェア(TreeStar)を使用して更に加工した。最大半減結合を、XLFit4.0(IDBS)及び用量応答ワンサイトモデル205を使用して決定した。
【0196】
b)内部移行アッセイ
細胞は、剥離し、そしてカウントした。5×10e5個の細胞を、エッペンドルフチューブ内の50μLの完全培地中に入れ、5μg/mLのそれぞれの二重特異性抗体と一緒に37℃にてインキュベートした。表示した時点で細胞を氷上に保存し、タイムコースが完了するまで保存した。その後、細胞を、FACSチューブに移し、遠沈し(1500rpm、4℃、5分間)、PBS+2%のFCSで洗浄し、そして2%のFCS(Jackson Immunoresearch、109116098)を含有したPBS中に希釈した、ヒトFcに対するフィコエリトリン結合二次抗体50μLと一緒に30分間インキュベートした。細胞を、再度遠沈し、PBS+2%のFCSで洗浄し、そして蛍光強度をフローサイトメトリー(FACS Canto、BD)によって測定した。
【0197】
Cell Titer Glowアッセイ
細胞生存率と増殖をcell titer glowアッセイ(Promega)を使用することで定量化した。製造業者の取扱説明書に従ってアッセイを実施した。簡単に言えば、細胞を、96ウェルプレート内の100μLの全容量中で所望の期間、培養した。増殖アッセイのために、細胞を、インキュベーターから取り出し、そして室温にて30分間、静置した。100μLのcell titer glow試薬を加え、そしてマルチウェルプレートを2分間、オービタル振盪機にかけた。発光をマイクロプレートリーダー(Tecan)により15分後に定量化した。
【0198】
Wst−1アッセイ
Wst−1生存率と細胞増殖アッセイをエンドポイント分析として実施し、代謝的に活性な細胞の数を検出した。簡単に言えば、20μLのWst−1試薬(Roche、1164480700)を200μLの培地に加えた。色素の完全な現像まで、96ウェルプレートを更に30分間〜1時間インキュベートした。染色強度を、450nmの波長にてマイクロプレートリーダー(Tecan)により定量化した。
【0199】
二重特異性<ErbB2−c−Met>抗体の設計
以下に発現され、そして精製された二重特異性<ErbB2−c−Met>抗体はすべて、以下に示すように最終的に修飾されたIgG1サブクラスの定常領域又は少なくともFcの一部(配列番号9のヒトIgG1定常領域)を含んでなる。
【0200】
表1において:表1に示したそれぞれの特徴を有する、全長ErbB−2抗体(トラスツズマブ)とc−Met抗体由来の1つの一本鎖Fabフラグメント(cMet 5D5)(基本的な構造スキームについては図5aを参照のこと)に基づく三価の二重特異性<ErbB2−c−Met>抗体は、先に記載した一般的な方法に従って発現及び精製されるか、又は発現及び精製できる。トラスツズマブ及びcMet 5D5の対応するVH及びVLを、配列表に示す。
【0201】
【表1】
【0202】
実施例1:
ErbB−2及びc−Metへの二重特異性抗体の結合(表面プラズモン共鳴法)
結合親和力を、25℃にて標準的な結合アッセイ、例えば表面プラズモン共鳴法などによって測定した(BIAcore(登録商標)、GE-Healthcare、Uppsala, Sweden)。親和性測定のために、30μg/mlの抗Fcγ抗体(ヤギ由来、Jackson Immuno Research)を、標準的なアミン・カップリングによってCM−5センサーチップの表面に結合させ、そしてSPR器具(Biacore T100)により化学的にブロッキングした。結合後に、単一又は二重特異性ErbB2/cMet抗体を、25℃、5μL/分の流速にて注入し、続いて30μL/分にてヒトErbB2又はc−Met ECDの希釈系列(0nM〜1000nM)を注入した。結合実験のためのランニングバッファーとしてPBS/0.1% BSAを使用した。次いで、前記チップを、10mMのグリシンHCl、pH2.0溶液の60秒パルスを用いて再生した。
【0203】
【表2】
【0204】
実施例2:
二重特異性Her2/c−Met抗体フォーマットによるHGF誘発c−Met受容体リン酸化の阻害
二重特異性抗体のc−Met部分の機能性を確認するために、c−Metリン酸化アッセイを実施する。この実験では、A549肺癌細胞又はHT29大腸癌細胞を、二重特異性抗体又は対照抗体で処理した後にHGFに晒した。次いで、細胞を溶解し、そしてc−Met受容体のリン酸化を調べる。両細胞株は、免疫ブロットにおけるリン酸化c−Metに特異的なバンドの出現によって観察できるように、HGFで刺激され得る。親又は二重特異性抗体の結合が、受容体リン酸化の阻害につながる。あるいは、当業者は、オートクリンHGFループを有する細胞、例えばU87MGを使用して、親又は二重特異性抗体の不存在下又は存在下でのc−Met受容体リン酸化を評価できる。
【0205】
実施例3:
Her2/cMet二重特異性抗体での処置後のHer2受容体リン酸化の分析
二重特異性Her2/cMet抗体Sk−Br3のHer2結合部分の機能性を確認するために、A431を、親EGFR抗体又は二重特異性Her2/cMet抗体のいずれかと一緒にインキュベートする。親又は二重特異性抗体が結合して、関係のないIgG対照抗体が結合しないことで、受容体リン酸化の阻害につながる。あるいは、当業者は更に、親又は二重特異性抗体の存在又は不存在下、NRGで刺激されてErbB2/Her2受容体リン酸化を引き起こす細胞も使用できる。
【0206】
実施例4:
Her2/cMet二重特異性抗体での処置後のPI3Kシグナル伝達の分析
Her2並びにc−Met受容体は、細胞増殖シグナルを伝えるPI3K経路を介してシグナル伝達できる。AKTのHer2及びc−Met受容体リン酸化の同時標的化を実証するために、PI3K経路の下流標的を観察することもできる。このために、非刺激細胞、NRG若しくはHGFで処理した細胞又は両サイトカインで処理した細胞を、非特異的、親対照又は二重特異性抗体と一緒に並行してインキュベートした。あるいは、当業者は更に、ErbB2/Her2を過剰発現する細胞、及び/又はc−Metシグナル伝達を活性化するオートクリンHGFループを有する細胞を評価することもできる。AKTはPI3K経路の主要な下流シグナル伝達成分であり、そしてこのタンパク質のリン酸化はこの経路を介したシグナル伝達の重要な指標である。
【0207】
実施例5:
Her2/cMet二重特異性抗体での処置後のMAPKシグナル伝達の分析
c−Met受容体は、MAPK経路を介してシグナル伝達できる。c−Met受容体の標的化を実証するために、MAPK経路の主要な下流標的であるERK1/2のリン酸化を観察することができる。このために、非刺激細胞又はHGFで処理した細胞を、非特異的、親対照又は二重特異性抗体と一緒に並行してインキュベートした。あるいは、当業者は更に、c−Metシグナル伝達を活性化するオートクリンHGFループを有する細胞を評価することもできる。
【0208】
実施例6:
二重特異性Her2/c−Met抗体フォーマットによるHGF誘発HUVEC増殖の阻害
HUVEC増殖検定は、HGFの血管新生及び細胞分裂効果を実証するために実施することができる。HUVECに対するHGFの添加は、c−Met結合抗体によって用量依存様式で阻害され得る細胞増殖の増加をもたらす。
【0209】
実施例7:
二重特異性Her2/c−Met抗体によるSk−Br3増殖の阻害
a)フローサイトメトリーにより独立に確認されているように、Sk−Br3細胞は、高いHer2の細胞表面レベル及び中程度に高いc−Metの細胞表面発現を示している。親Her2結合抗体又は二重特異性Her2/c−Met抗体の添加が増殖の減少につながったのに対して、c−Met結合抗体は増殖に対してわずかな効果しか持っていなかった。
【0210】
b)活性なHer2−c−Met受容体シグナル伝達ネットワークが増殖を起こす状況をシミュレートするために、アッセイを、HGF条件培地の存在下ではあるが、記載のとおり行う。この状況において、いずれか一方の親抗体の添加がcell titer glow分析によって測定されるように、細胞増殖に対してわずかな効果しか持っていない一方で、二重特異性抗体又は親抗体の組合せ物の添加は細胞増殖の減少につながる。
【0211】
実施例8:
二重特異性Her2/c−Met抗体フォーマットによる癌細胞株DU14SにおけるHGF誘発細胞−細胞播種(分散)阻害の分析
細胞のHGF誘発分散は細胞の形態学的変化を誘導し、そして細胞の円形化、糸状仮足様の突起、紡錘状の構造、及び細胞の特定の運動性をもたらす。二重特異性Her2/cMet抗体は、HGF誘発細胞−細胞播種を抑制した。
【0212】
実施例9:
二重特異性HER2/c−Met抗体フォーマットによるHGF誘発HUVEC増殖の阻害
HUVEC増殖検定は、HGFの細胞分裂効果を実証するために実施することができる。HUVECに対するHGFの添加は、増殖の二倍の増加につながる。二重特異性抗体と同じ濃度領域でのヒトIgG対照抗体の添加が細胞増殖に対して影響を持たなかった一方で、5D5 FabフラグメントはHGF誘発増殖を阻害する。
【0213】
実施例10:
二重特異性HER2/c−Met抗体による癌細胞株A431におけるHGF誘発細胞−細胞播種(分散)阻害の分析
HGF誘導分散には細胞の形態学的変化が含まれ、細胞の円形化、糸状仮足様の突起、紡錘状の構造、及び細胞の特定の運動性をもたらす。Real Time Cell Analyzer(Roche)は、所定の細胞培養ウェルのインピーダンスを計測し、それにより細胞の形態と増殖の変化を間接的に観察することができる。A431及びA549細胞に対するHGFの添加は、時間の関数として観察されるインピーダンスの変化をもたらす。
【0214】
実施例11:
ErbB−2及びc−Mct発現癌細胞株における抗体媒介型受容体内部移行の分析
細胞の、Her2又はc−Metに特異的に結合する抗体とのインキュベーションが、受容体の内部移行を引き起こすことを示した。二重特異性抗体の内部移行能力を評価するために、抗体誘発受容体内部移行を検討するために実験構成を設計する。このために、OVCAR−8細胞((NCI Cell Line designation;NCI(国立癌研究所)から購入したOVCAR-8-NCI;Schilder RJ, et al Int J Cancer, 1990 Mar 15;45(3):416-22;Ikediobi ON, et al, Mol Cancer Ther. 2006;5;2606-12;Lorenzi, P.L., et al., Mol Cancer Ther. 2009;8(4):713-24))(フローサイトメトリーで確認されたとおり、この細胞はHer2並びにcMetを発現する−図7bを参照のこと)を、37℃にてそれぞれの一次抗体と一緒に異なった期間(例えば0、30、60、120分=0、0.5、1、2時間(h))インキュベートした。細胞プロセスは、細胞を4℃に急速に冷やすことによって停止する。一次抗体のFcに特異的に結合する蛍光体を結合した二次抗体を、細胞表面に結合した抗体を検出するのに使用した。抗体−受容体複合体の内部移行は、細胞表面の抗体−受容体複合体を激減させるので、平均蛍光強度の減少を結果的にもたらす。内部移行はOvcar−8細胞において検討した。結果を以下の表及び図8に示す。それぞれの受容体の内部移行率(%)を、それぞれの抗体の内部移行を介して計測する(図8において、二重特異性<ErbB2−cMet>抗体BsAB02はcMet/HER2と示されており、親単一特異性の二価抗体は<HER2>及び<cMet>と示されている)。
【0215】
【表3】
【0216】
実施例12
糖鎖操作されたバージョンの二重特異性Her2/c−Met抗体の調製
二重特異性Her2/c−Met抗体のDNA配列を、各ベクターがEBV OriP配列を担持している、MPSVプロモーター及び合成ポリA部位の上流の制御下にある哺乳動物発現ベクター内にサブクローニングする。
【0217】
リン酸カルシウムトランスフェクション・アプローチを使用した、哺乳動物二重特異性抗体発現ベクターでのHEK293−EBNA細胞の同時トランスフェクションによって、二重特異性抗体を作り出す。対数増殖期のHEK293−EBNA細胞をリン酸カルシウム法によってトランスフェクトする。糖鎖操作された抗体の製造のために、前記細胞を、1つが融合GnTIIIポリペプチド発現のためのもの(GnT−III発現ベクター)であり、1つがマンノシダーゼII発現のためのもの(ゴルジマンノシダーゼII発現ベクター)である、2つの追加のプラスミドを用いて、それぞれ1:1:4:4の比で同時トランスフェクトした。細胞を、10%のPCSを補ったDMEM培地を使用してTフラスコ内で接着性単層培養として培養し、そしてそれらが50〜80%集密になるとトランスフェクトする。T150フラスコのトランスフェクションのために、トランスフェクションの24時間前に、1500万個の細胞を、(最終的に10%V/Vに)FCSを補った25mlのDMEM培地中に植え付け、そして細胞を37℃、5%のCO2雰囲気のインキュベーター内に一晩置く。トランスフェクトされるそれぞれのT150フラスコについて、DNAの溶液、CaCl2、及び水を、軽鎖及び重鎖発現ベクターの間で等しく分配される94μgの総プラスミドベクタDNA、469μlの終量までの水、及び469μlの1M CaCl2溶液を混合することによって調製する。この溶液に、938μlの50mM HEPES、280mMのNaCl、1.5mM Na2HPO4溶液、pH7.05を加え、すぐに10秒間混合し、そして室温にて20秒間静置する。その懸濁液を、2%のPCSを補った10mlのDMEMで希釈し、そして既存の培地に代えてT150に加える。更に13mlのトランスフェクション培地を加える。細胞を、37℃、5%のCO2にて約17〜20時間インキュベートし、次いで、培地を25mlのDMEM、10%のFCSで置き換える。条件培養培地を、210×gにて15分間の遠心分離によってトランスフェクトの7日後に回収し、その溶液を濾過滅菌(0.22μmフィルター)し、0.01%w/vの終濃度にアジ化ナトリウムを加え、そして4℃に保った。
【0218】
分泌された二重特異性の非フコシル(afocusylated)糖鎖操作された抗体を、プロテインA親和性クロマトグラフィーとそれに続く、陽イオン交換クロマトグラフィー、そしてSuperdex 200カラム(Amersham Pharmacia)による最終的なサイズ排除クロマトグラフィーステップによって精製し、バッファーを25mMのリン酸カリウム、125mMの塩化ナトリウム、100mMのグリシン溶液、pH6.7に交換し、且つ、純粋な単量体のIgG1抗体を回収した。抗体濃度を、280nmの吸光度から分光光度計を使用して推定した。
【0219】
抗体のFc領域に結合されたオリゴ糖を、記載したようにMALDI/TOF−MSによって分析する。PVDF膜に固定されるか又は溶液中の抗体を用いて、PNGaseF消化によって、抗体からオリゴ糖を酵素的に放出させる。放出されたオリゴ糖を含む得られた消化産物溶液を、最高MALDI/TOF−MS分析法のためにそのまま調製するか、又はMALDI/TOF−MS分析法のためのサンプル調製前にEndoHグリコシダーゼで更に消化する。
【0220】
実施例13:
二重特異性Her2/c−Met抗体の糖構造の分析
フコース及び非フコース(a−フコース)含有オリゴ糖構造の相対比の測定のために、精製した抗体材料の放出されたグリカンをMALDI−Tof−質量分析法によって分析する。今回、タンパク質骨格からオリゴ糖を放出するために、抗体サンプル(約50μg)を、0.1Mのリン酸ナトリウム緩衝液、pH6.0中の5mU N−グリコシダーゼF(Prozyme# GKE-5010B)と一緒に37℃にて一晩インキュベートする。それに続いて、放出されたグリカン構造物を、NuTip−Carbonピペットチップ(Glygenから入手:NuTip1〜10μl、Cat.Nr#NTlCAR)を使用して分離し、そして脱塩する。第1ステップとして、NuTip−Carbonピペットチップを、3μlの1M NaOHに続き、20μLの純水(例えばBaker製のHPLCグラジエントグレード、#4218)、3μLの30% 酢酸、そして再び20μLの純水で洗浄することによって、オリゴ糖の結合に向けて準備する。今回、それぞれの溶液を、NuTip−Carbonピペットチップ内のクロマトグラフィー材料の上部に添加し、そして圧力をかけてそれを通過させる。その後、10μgの抗体に相当するグリカン構造物を、先に記載のN−グリコシダーゼF消化産物を4〜5回ほど吸い上げたり吸い出したりすることによって、NuTip−Carbonピペットチップ内の材料に結合させる。NuTip−Carbonピペットチップ内の材料に結合したグリカンを、先に記載したやり方で20μLの純水で洗浄し、そしてそれぞれ0.5μLの10%と2.0μLの20% アセトニトリルを用いて段階的に溶出する。このステップのために、溶出溶液を、0.5mLの反応ベイル(reaction vails)内に充填し、そして各回毎に4〜5回吸い上げたり吸い出したりする。MALDI−Tof質量分析法による分析のために、両方の溶出液を組合せる。この計測において、0.4μLの組合せた溶出液を、MALDI標的上で1.6μLのSDHBマトリックス溶液(5mg/mlで20%のエタノール/5mMのNaCl中に溶解した2.5−ジヒドロキシ安息香酸/2−ヒドロキシ−5−メトキシ安息香酸[Bruker Daltonics#209813])と混合し、そして、適当に調整したBruker Ultraflex TOF/TOF装置で分析した。日常的には、50〜300ショットが、記録され、そして1回の実験にまとめられる。得られたスペクトルを、フレックス分析ソフトウェア(Bruker Daltonics)によって評価し、そして質量を検出したピークのそれぞれについて決定する。それに続いて、計算した質量を、それぞれの構造物(例えばそれぞれフコースを含む及び含まない複合体、ハイブリッド、及びオリゴ又は高マンノース)について理論的に予想した質量を比較することによって、ピークをフコース又はa−フコース(非フコース)含有グリコール構造物に割り当てる。
【0221】
ハイブリッド構造物の比の測定のために、抗体サンプルを、N−グリコシダーゼFとエンド−グリコシダーゼHを併用して消化する。N−グリコシダーゼFは、タンパク質骨格からすべてのN結合型グリカン構造(複合体、ハイブリッド、並びにオリゴ及び高マンノース構造)を放出し、そしてエンド−グリコシダーゼHは、グリカンの還元末端の2つのGlcNAc残基の間で更にすべてのハイブリッドタイプのグリカンを開裂する。この消化産物は、N−グリコシダーゼF消化したサンプルについて先に記載したのと同様のやり方で、続けて処理し、そしてMALDI−Tof質量分析法によって分析する。N−グリコシダーゼF消化産物からのパターンと、併用N−グリコシダーゼF/エンドH消化産物からのパターンを比較することによって、糖構造に特異的なシグナルの減少度が、ハイブリッド構造の相対内容を推測するのに使用される。
【0222】
それぞれの糖構造の相対量は、個々のグリコール構造のピークの高さと、検出したすべての糖構造のピークの高さの比から計算される。フコースの量は、N−グリコシダーゼF処理サンプルにおいて特定した全糖構造(例えば、それぞれ複合体、ハイブリッド、並びにオリゴ及び高マンノース構造)に対するフコース含有構造のパーセンテージである。非フコース化の量は、N−グリコシダーゼF処理サンプルにおいて特定した全糖構造(例えば、それぞれ複合体、ハイブリッド、並びにオリゴ及び高マンノース構造)に対するフコースを欠く構造のパーセンテージである。
【0223】
実施例14:
Her2/cMet二重特異性抗体での処置後の細胞遊走の分析
活性なc−Metシグナル伝達の1つの重要な側面は、遊走と侵襲的なプログラムの誘導である。c−Met阻害抗体の有効性は、HGF誘発細胞遊走の阻害を計測することによって測定できる。このために、HGF誘導性癌細胞株A431を、二重特異性抗体又はIgG対照抗体の存在下又は不存在下、HGFで処理し、そして8μmの細孔を遊走して通過した細胞の数を、インピーダンス読み出しを備えたCIMプレートを使用するAceaリアルタイム細胞分析装置により時間依存様式で計測する。
【0224】
実施例15:
二重特異性Her2/c−Met抗体のインビトロADCC
本発明によるHer2/cMet二重特異性抗体は、両受容体を発現している細胞において少ない内部移行を示す。少ない内部移行は、細胞表面の抗体−受容体複合体の長期暴露がNk細胞により認識されやすいので、これらの抗体を糖鎖操作することの理論的根拠を大いに裏付けている。少ない内部移行と糖鎖操作は、親抗体と比較して高い抗体依存性細胞傷害(ADCC)に置き換えられる。これらの効果を実証するインビトロ実験構成は、細胞表面にHer2とcMetの両方を発現する癌細胞、例えばA431、及びNk細胞株又はPBMCのもののようなエフェクター細胞を使用して設計できる。腫瘍細胞を、親単一特異性抗体又は二重特異性抗体と一緒に最長24hプレインキュベートし、続いてエフェクター細胞株を加える。細胞溶解物を定量し、そして単一特異性抗体と二重特異性抗体の識別を可能にする。
【0225】
標的細胞、例えばPC−3(DSMZ#ACC465、前立腺腺癌、Ham’s F12 Nutrient mixture+2mMのL−アラニル−L−グルタミン+10%のPCS中で培養)を、指数増殖期にトリプシン/EDTA(Gibco#25300-054)を用いて回収する。洗浄ステップ、及び細胞数と生存率の確認ステップの後に、必要とされるアリコートを、細胞インキュベーター内で37℃にて30分間、カルセイン(Invitrogen#C3100MP;5mlの培地中の5Mio細胞に対して、1バイアルを50μlのDMSO中に再懸濁した)で標識する。その後、細胞を、AIM培地で3回洗浄し、そして、細胞数と生存率を確認し、そして細胞数を0.3Mio/mlに調整する。
【0226】
その間に、エフェクター細胞としてのPBMCを、製造業者のプロトコール(洗浄ステップ、1×400gにて、そして2×350gにて、それぞれ10分間)に従って密度勾配遠心分離法(Histopaque-1077、Sigma#H8889)によって調製する。細胞数と生存率を確認し、そして細胞数を15Mio/mlに調整する。
【0227】
100μlのカルセイン染色標的細胞を、丸底96ウェルプレート内に入れ、50μlの希釈した抗体を加え、そして50μlのエフェクター細胞を加える。いくつかの実験では、標的細胞を、10mg/ml Redimuneの濃度にてRedimune(登録商標)NF Liquid(ZLB Behring)と混合する。
【0228】
抗体なしで同時培養される標的及びエフェクター細胞によって測定される自然発生的溶解、及び標的細胞だけを溶解する1% Triton X100によって測定される最大溶解が、対照の役割を担う。プレートを、加湿した細胞インキュベーター内で37℃にて4時間インキュベートする。
【0229】
標的細胞の殺滅を、製造業者の取扱説明書に従って細胞毒性検出キット(LDH Detection Kit、Roche#1 644 793)を使用して、損傷細胞から放出されるLDHを計測することによって評価する。簡単に言えば、各ウェルからの100μlの上清を、透明な平底96ウェルプレート内で、キットからの100μlの基質と混合する。基質の呈色反応のVmax値を、少なくとも10分間、490nmにてELISAリーダーにより測定する。特有の抗体媒介型殺滅のパーセンテージを、以下のとおり計算する:((A−SR)/(MR−SR)×100、式中、Aは特定の抗体濃度におけるVmaxの平均であり、SRは自然発生的放出のVmaxの平均であり、そしてMRは最大放出のVmaxの平均である。
【0230】
実施例16:
パラクリンHGFループを有する皮下異種移植モデルにおける二重特異性Her2/cMet抗体のインビボ有効性
Mrc−5細胞と同時注入される皮下のKPL4モデルは、c−Metのパラクリン活性化ループによく似ている。KPL4は、細胞表面にc−Met並びにHer2をある程度発現する。KPL4及びMrc−5細胞は、対数増殖期において標準的な細胞培養条件下で維持される。KPL4とMrc−5細胞は、1000万個のKPL4細胞と100万個のMrc−5による10:1の比で注射される。細胞を、SCIDベージュマウスに移植する。腫瘍が定着し、そして100〜150mm3のサイズに達した後、処置を始める。マウスを、20mg/kgの抗体/マウスの負荷量で処置し、その後毎週一回、10mg/kgの抗体/マウスで処置する。腫瘍体積を1週間に二度計測し、そして動物の体重を平行して観察する。単一抗体の単独処置と組合せを、二重特異性抗体での治療法と比較する。
【0231】
実施例17:
二重特異性Her2/cMet抗体によるOVCAR−8増殖の阻害
a)フローサイトメトリーにより独立に確認されているように、OVCAR−8細胞(NCI Cell Line designation;NCI(国立癌研究所)から購入したOVCAR-8-NCI;Schilder, RJ. et al., Int. J. Cancer 1990 Mar 15;45(3): 416-422;Ikediobi, O.N., et al., Mol. Cancer. Ther. 2006;5;2606- 2012;Lorenzi, P.L., et al., Mol. Cancer Ther. 2009; 8(4): 713-724)は、顕著なHer2及びc−Metの細胞表面レベルを示している(図7bを参照のこと)。二重特異性Her2/c−Met抗体によるOVCAR−8細胞増殖の阻害は、48時間後にCell Titer Glow(商標)アッセイにより計測した。結果を図9aに示す。対照はPBSバッファー(リン酸緩衝生理食塩水)であった。
【0232】
計測では、(0%の阻害に設定したバッファー対照と比較して)6%の阻害というHER2抗体トラスツズマブの阻害が明らかになった。二重特異性Her2/c−Met BsAB02(BsAb)抗体は、癌細胞増殖のより顕著な阻害を導いた(11%の阻害)。一価のc−Met抗体である片腕5D5(OA5D5)は、増殖に対して効果がないことが明らかになった。HER2抗体トラスツズマブと一価のc−Met抗体の片腕5D5(OA5D5)の組合せは、それほど顕著ではない減少しか導かなかった(6%の阻害)。
【0233】
b)OVCAR−8細胞はHER2シグナル伝達に依存している。活性なHER−c−Met受容体シグナル伝達ネットワークが更なる増殖を起こす状況をシミュレートするために、アッセイをa)(48時間後のCell Titer Glow(商標)アッセイ)で記載したように、しかしHGF条件培地の存在下で行った。結果を図9bに示す。
【0234】
計測では、0%の阻害に設定したHGF処理細胞と比較すると、Her2抗体トラスツズマブ(2%の阻害)と一価のc−Met抗体の片腕5D5(OA5D5)(3%の阻害)の阻害効果はほとんどないことが明らかになった。二重特異性Her2/c−Met抗体BsAB02(BsAb)は、Ovcar−8細胞の癌細胞増殖に対して顕著な阻害を示した(17%の阻害)。Her2抗体のトラスツズマブと一価のc−Met抗体の片腕5D5(OA5D5)の組合せは、細胞増殖のそれほど顕著ではない減少につながった(10%の阻害)。
【技術分野】
【0001】
本発明は、ヒトErbB−2及びヒトc−Metに対する二重特異性抗体、それらの製造方法、前述の抗体を含む医薬組成物、並びにその使用に関する。
【背景技術】
【0002】
ErbBファミリー・タンパク質
ErbBタンパク質ファミリーは、4種類のメンバーであるErbB−1(別名、上皮生長因子受容体(EGFR))、ErbB−2(ヒトではHER2、齧歯動物ではneu)、ErbB−3(別名、HER3)及びErbB−4(別名、HER4)から成る。ErbBファミリー・タンパク質は、受容体チロシンキナーゼであって、細胞増殖、分化、及び生存の重要な伝達物質となっている。
【0003】
ErbB−2と抗ErbB−2抗体
ErbBタンパク質ファミリーの第2のメンバーであるErbB−2(ERBB2、HER2;CD340、HER−2/neu、c−erb B2/neuタンパク質、神経芽細胞腫/神経膠芽腫由来癌遺伝子相同体|v−erb−b2 鳥類赤血球性白血病ウイルス癌遺伝子相同体2;配列番号14としても知られている)は、それ自身のリガンド結合ドメインを持たないタンパク質であるため、成長因子に結合することができない。しかしながら、他のリガンド結合EGF受容体ファミリーメンバーに強く結合してヘテロ二量体を形成し、リガンド結合を安定させ、そして下流のシグナル伝達経路のキナーゼ媒介型活性化を促進する、例えばそれらはマイトジェン活性化プロテインキナーゼやホスファチジルイノシトール−3キナーゼに関与している。本明細書中に示した最も頻度の高い対立遺伝子Ile654/Ile655を含めて、第654及び655位アミノ酸における対立遺伝子変異のアイソフォームa(第624及び625位のアイソフォームb)が報告されている。この遺伝子の増幅、及び/又は過剰発現は、乳房及び卵巣の腫瘍を含めた多数の癌で報告されている。選択的スプライシングはいくつかのさらなる転写物変異体をもたらすが、一部のものは異なるアイソフォームをコードし、他のものは完全に特徴づけされていない。ErB−2は、元々、形質変換遺伝子の産物として化学物質で処理されたラットの神経芽細胞腫から同定された。neu癌原遺伝子の活性型は、コードされたタンパク質の膜貫通領域における(バリンからグルタミン酸への)点突然変異から得られた(Semba, K., et al., PNAS 82 (1985) 6497-501;Coussens, L., et al., Science 230 (1985) 1132-9;Bargmann, C.I., et al., Nature 319 (1986) 226-30;Yamamoto, T., et al., Nature 319 (1986) 230-4)。
【0004】
neuのヒト相同体の増幅は、乳癌及び卵巣癌で観察され、且つ、予後不良と相関する(Slamon, D.J, et al. Science 235 (1987) 177-182;Slamon, D.J., et al., Science 244 (1989) 707-712;及びUS 4,968,603)。これまで、neu癌原遺伝子に対する点突然変異相同体は、ヒトの腫瘍については報告されていない。(一様でないが高い頻度で遺伝子増幅による)HER2の過剰発現は、胃、子宮内膜、唾液腺、肺、腎臓、結腸、甲状腺、膵臓及び膀胱の癌腫を含めた他の癌腫においても観察されている。特に、King, C.R., et al., Science 229 (1985) 974-976;Yokota, J, et al., Lancet 1 (1986) 765-767;Fukushige, S., et al., Mol Cell Biol. 6 (1986) 955-958;Guerin, M., et al., Oncogene Res. 3 (1988) 21-31;Cohen, J.A., et al., Oncogene, 4 (1989) 81-88;Yonemura, Y., et al., Cancer Res. 51 (1991) 1034-1038;Borst, M.P., et al., Gynecol. Oncol. 38 (1990) 364-366;Weiner, D.B., et al., Cancer Res. 50 (1990) 421-425;Kern, J.A., et al., Cancer Res. 50 (1990) 5184-5187;Park, J.B., et al, Cancer Res. 49 (1989) 6605-6609;Zhau, H.E., et al., Mol. Carcinog. 3 (1990) 254-257;Aasland, R., et al., Br. J. Cancer 57 (1988) 358-363;Williams, T.M, et al, Pathobiology 59 (1991) 46-52;及びMcCann, A, et al. Cancer 65 (1990) 88-92を参照のこと。HER2は、前立腺癌において過剰発現されることもある(Gu, K, et al., Cancer Lett. 99 (1996) 185-189;Ross, J.S, et al. Hum. Pathol. 28 (1997) 827-833;Ross, J.S, et al. Cancer 79 (1997) 2162-2170;及びSadasivan, R, et al, J. Urol. 150 (1993) 126-131)。
【0005】
ヒトHER2タンパク質産物に対する抗体が、例えばHudziak, R.M, et al, Mol. Cell. Biol. 9 (1989) 1165-1172によって作出された(前記文献は、ヒト乳房腫瘍細胞株SK−BR−3を使用して特徴づけされた抗HER2抗体のパネルの作出について記載している)。この抗HER2抗体のパネルには、とりわけ、(HER2の細胞外ドメインの異なるエピトープに向けられている)2C4(パーツズマブ)及び4D5(トラスツズマブ、Herceptin(商標))抗体が含まれる。前記抗体への暴露後のSK−BR−3細胞の相対細胞増殖は、72時間後の単分子層のクリスタルバイオレット染色によって測定された。このアッセイを使用すると、細胞増殖を56%阻害した4D5(トラスツズマブ、Herceptin(商標))と呼ばれる抗体で、最大の阻害が得られた。前記パネル中の他の抗体は、このアッセイにおいて細胞増殖をある程度減少させた。抗体4D5については、TNF−αの細胞傷害効果に対するHER2過剰発現乳房腫瘍細胞株の感受性を高めることがさらに見出された(US5,677,171)。Hudziak, R.M.らが議論しているHER2抗体は、例えばFendly, B.M, et al. Cancer Research 50 (1990) 1550-1558においてさらに特徴づけされている。
【0006】
c−Metと抗c−Met抗体
MET(間葉−上皮移行因子)は、タンパク質METをコードする癌原遺伝子であり、(c−Met、肝細胞増殖受容体HGFR;HGF受容体;細胞分散因子受容体;SF受容体;配列番号15としても知られている)(Dean, M., et al., Nature 318 (1985) 385-8;Chan, A.M., et al., Oncogene 1 (1987) 229-33;Bottaro, D.P., et al., Science 251 (1991) 802-4;Naldini, L., et al.5 EMBO J. 10 (1991) 2867-78;Maulik, Gautam., et al., Cytokine Growth Factor Rev. 13 (2002) 41-59)。METは、胚発生と創傷治癒に不可欠な膜受容体である。肝細胞増殖因子(HGF)は、MET受容体の唯一の既知のリガンドである。METは通常、上皮起源の細胞によって発現されるが、HGFの発現は間葉起源の細胞に限られている。HGF刺激により、METは浸潤性増殖として知られているプログラムをまとめて引き起こす数個の生物反応を誘発する。癌における異常なMET活性化は、異常な活性のMETが腫瘍増殖や、腫瘍に栄養を供給する新生血管の形成(血管新生)や、他の臓器への癌の分散(転移癌)を引き起こす、予後不良と関連がある。METは、腎臓、肝臓、胃、乳房、及び脳の癌を含めた多くのタイプのヒト悪性腫瘍において無秩序である。本来なら、幹細胞と始原細胞だけがMETを発現しており、それが胎児において新しい組織を作り出したり、成人において損傷した組織を再生するために、これらの細胞が侵襲的に増殖することを可能にしている。しかしながら、癌幹細胞は、METを発現する正常幹細胞の能力を乗っ取り、それによって癌持続性と体内の他の部位への分散の原因となっている。
【0007】
癌原遺伝子MET産物は、肝細胞増殖因子受容体であり、チロシンキナーゼ活性をコードしている。初期一本鎖前駆タンパク質は、翻訳後に分割されて、α及びβサブユニットを生じ、それがジスルフィド結合して成熟受容体を形成する。METの遺伝子内の様々な突然変異は乳頭状腎癌に関連している。
【0008】
抗c−Met抗体は、例えば、US5,686,292、US7,476,724、WO2004/072117、WO2004/108766、WO2005/016382、WO2005/063816、WO2006/015371、WO2006/104911、WO2007/126799、又はWO2009/007427により知られている。
【0009】
c−Met結合ペプチドは、例えば、Matzke, A., et al., Cancer Res 65 (14) (2005) 6105-10及びTam, Eric, M., et al., J. Mol. Biol. 385 (2009)79-90により知られている。
【0010】
多特異性抗体
広範な種類の組換え抗体フォーマットが最近開発されてきている、例えば、IgG抗体フォーマットと一本鎖ドメインの融合による、例えば、四価二重特異性抗体が開発された(例えば、Coloma M. J. et al., Nature Biotech. 15(1997)159-163;WO2001077342;及びMorrison S. L. Nature Biotech. 25(2007)1233-1234を参照のこと)。
【0011】
また、抗体コア構造(IgA、IgD、IgE、IgG又はIgM)がもはや保持されていないいくつかの新しいフォーマット、例えば、2つ以上の抗原に結合できる、ジ抗体、トリ抗体又はテトラ抗体、ミニ抗体、いくつかの一本鎖フォーマット(scFv、ビス−scFv)が開発された(Holliger P. et al., Nature Biotech. 23(2005)1126-1136;Fischer N., Leger O. Pathobiology 74(2007)3-14;Shen J. et al., J. Immunol. Methods 318(2007)65-74;Wu C. et al., Nature Biotech. 25(2007)1290-1297)。
【0012】
すべてのこのようなフォーマットはリンカーを使用して抗体コア(IgA、IgD、IgE、IgG又はIgM)を更なる結合性タンパク質(例えば、scFv)に融合するか、あるいは、例えば、2つのFabフラグメント又はscFvを融合する(Fischer N., Leger O. Pathobiology 74(2007)3-14)。天然に存在するものに対して高度の類似性を維持することによって、Fc受容体結合を通して介在される、エフェクター機能、例えば、補体依存性細胞毒性(CDC)又は抗体依存性細胞毒性(ADCC)を保持することが望まれることもあり得ることに留意すべきである。
【0013】
WO2007/024715では、改変多価及び多特異性結合タンパク質として二重可変ドメイン免疫グロブリンが報告されている。生物学的に活性な抗体二量体の調製方法は、US6,897,044に報告されている。ペプチド・リンカーを介して互いに連結されている少なくとも4つの可変ドメインを持つ多価性Fv抗体構築物が、US7,129,330で報告されている。二量体及び多量体の抗原結合構築物は、US2005/0079170で報告されている。(天然の免疫グロブリンでない)結合構造によって互いに共有結合した3つ又は4つのFabフラグメントを含んでなる三又は四価の単一特異性抗原結合性タンパク質が、US6,511,663で報告されている。WO2006/020258では、原核生物及び真核生物細胞で効率的に発現され、治療薬や診断法に有用である四価の二重特異性抗体が報告されている。2つのタイプのポリペプチド二量体を含んでなる混合物からの少なくとも1つの鎖間ジスルフィド結合によって連結されず二量体からの少なくとも1つの鎖間ジスルフィド結合によって連結される二量体を分離するか、又は優先的には合成する方法が、US2005/0163782で報告されている。二重特異性の四価の受容体が、US5,959,083で報告されている。三重機能性以上の抗原結合部位を持つ改変抗体が、WO2001/077342で報告されている。
【0014】
多特異性、且つ、多価の抗原結合性ポリペプチドが、WO1997/001580で報告されている。WO1992/004053では、通常、同じ抗原決定基に結合するIgGクラスのモノクローナル抗体から調製されたホモ接合体が合成架橋によって共有結合で連結されていることが報告されている。2つ以上の免疫グロブリンモノマーが共に四価又は六価のIgG分子を形成するのに関係している、(通常IgGクラスのオリゴマーが分泌されることよって)抗原に対して高い親和性を有するオリゴマーモノクローナル抗体が、WO1991/06305で報告されている。ヒツジ由来抗体及び改変抗体構築物が、US6,350,860で報告されていて、それは、インターフェロンγ活性が病因である疾患を治療するのに使用される。US2005/0100543では、二重特異性抗体の多価担体である標的を設定可能な、すなわち、標的を設定可能な構築物の各分子が2以上の二重特異性抗体の担体として機能することができる、構築物が報告されている。遺伝的に改変された二重特異性四価抗体は、WO1995/009917で報告されている。安定しているWO2007/109254には、安定化scFvから成るか、又はそれを含んでなる安定化結合分子が報告されている。US2007/0274985は、一本鎖Fab(scFab)フラグメントを含んでなる抗体フォーマットに関する。
【0015】
WO2008/140493は、抗EGFRファミリーメンバー抗体及び1若しくは複数の抗EGFRファミリーメンバー抗体を含んでなる二重特異性抗体に関する。US2004/0071696は、EGFRタンパク質ファミリーのメンバーに結合する二重特異性抗体分子に関する。
【0016】
WO2009111707(A1)は、Met及びHER拮抗薬を用いた併用療法に関する。WO2009111691(A2A3)は、Met及びEGFR拮抗薬を用いた併用療法に関する。
【0017】
WO2004072117は、c−Metの下方制御/内部移行を誘発するc−Met抗体、及び特に第2抗原としてErbB−2を用いた二重特異性抗体におけるそれらの使用可能性に関する。
【発明の概要】
【0018】
この発明の第一の態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、前記二重特異性抗体は、フローサイトメトリーアッセイにおいて1時間後に計測した場合に、OVCAR−8細胞上での、抗体の不存在下でのc−Metの内部移行と比較して15%未満のc−Metの内部移行を示すことを特徴とする。
【0019】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する1又は2つの抗原結合部位、及びヒトc−Metに特異的に結合する1つの抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二価又は三価の二重特異性抗体である。
【0020】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する2つの抗原結合部位、及びヒトc−Metに特異的に結合する3つ目の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する三価の二重特異性抗体である。
【0021】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する1つの抗原結合部位、及びヒトc−Metに特異的に結合する1つの抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二価の二重特異性抗体である。
【0022】
本発明の一態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位、及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、以下の:
前記第1の抗原結合部位が、重鎖可変ドメイン内に配列番号15のCDR3H領域、配列番号16のCDR2H領域、及び配列番号17のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号18のCDR3L領域、配列番号19のCDR2L領域、及び配列番号20のCDR1L領域を含んでなり;そして
前記第2の抗原結合部位が、重鎖可変ドメイン内に配列番号21のCDR3H領域、配列番号22のCDR2H領域、及び配列番号23のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号24のCDR3L領域、配列番号25のCDR2L領域、配列番号26のCDR1L領域を含んでなる、
を特徴とする前記二重特異性抗体である。
【0023】
前記二重特異性抗体は、好ましくは、以下の:
ErbB−2に特異的に結合する前記第1の抗原結合部位が、重鎖可変ドメインとして配列番号1の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号2の配列を含んでなり;そして
c−Metに特異的に結合する前記第2の抗原結合部位が、重鎖可変ドメインとして配列番号3の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号4の配列を含んでなる、
を特徴とする。
【0024】
本発明の更なる態様は、IgG1又はIgG3サブクラスの定常領域を含んでなることを特徴とする本発明による二重特異性抗体である。
【0025】
一実施形態では、本発明による前記二重特異性抗体は、前記抗体がAsn297にて糖鎖でグリコシル化されていることで、前記糖鎖内のフコース量が65%以下であることを特徴とする。
【0026】
本発明の更なる態様は、前記二重特異性抗体の鎖をコードする核酸分子である。
【0027】
本発明のより更なる態様は、前記二重特異性抗体を含んでなる医薬組成物、癌治療のための前記組成物、癌治療薬の製造のための前記二重特異性抗体の使用、そのような処置を必要としている患者に前記二重特異性抗体を投与することを含めた癌に罹患している患者の治療法である。
【0028】
乳房の腫瘍は高レベルのErbB2を発現していることが多く、そしてErbB2陽性腫瘍のうちのかなりの割合がさらにc−Met陽性でもある。乳房の腫瘍におけるc−Met発現が予後不良と相関があることは、これまで多くの研究で示されてきた(Kang, L, Y., et al., Cancer Res. 63 (2003) 1101-1105;Lengyel, E., et al., Int. J. Cancer 113 (2005) 678-82)。そのため、本発明による二重特異性<ErbB−2−c−Met>抗体は、抗腫瘍効果や癌細胞阻害のような価値のある特性を有する。
【0029】
本発明による抗体は、例えば、とりわけErbB2とc−Metの両受容体が発現されている癌細胞の増殖阻害や癌に罹患している患者に有益な抗腫瘍効果といった、非常に価値のある特性を示す。本発明による二重特異性<ErbB2−c−Met>抗体は、ErbB2とc−Metの両受容体を発現している癌細胞に対してそれらの親である単一特異性の二価<c−Met>抗体と比較すると、低いc−Met受容体の内部移行を示す。
【0030】
発明の詳細な説明
この発明の第一の態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、前記二重特異性抗体は、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、前記二重特異性抗体の不存在下でのc−Metの内部移行と比較して、OVCAR−8細胞に対する15%未満のc−Metの内部移行を示すことを特徴とする。
【0031】
よって、本発明は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体に関するものであり、ここで、前記二重特異性抗体は、フローサイトメトリーアッセイにより、1時間のOVCAR−8細胞−抗体インキュベーション後に測定した場合、前記二重特異性抗体の不存在下でのOVCAR−8細胞上のc−Metの内部移行と比較して15%未満の、OVCAR−8細胞上のc−Metの内部移行を増加させる。
【0032】
一実施形態では、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体は、前記二重特異性抗体が、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して、10%未満のc−Metの内部移行を示すことを特徴とする。
【0033】
一実施形態では、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体は、前記二重特異性抗体が、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して、7%未満のc−Metの内部移行を示すことを特徴とする。
【0034】
一実施形態では、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体は、前記二重特異性抗体が、フローサイトメトリーアッセイにより1時間後に計測した場合、OVCAR−8細胞上での、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して、5%未満のc−Metの内部移行を示すことを特徴とする。
【0035】
「c−Metの内部移行」という用語は、抗体の不存在下でのc−Metの内部移行と比較した場合の、OVCAR−8細胞(NCI Cell Line designation;NCI(国立癌研究所)から購入したOVCAR-8-NCI;Schilder, RJ. et al., Int. J. Cancer 45 (1990) 416-422;Dcediobi, O.N. et al., Mol. Cancer. Ther. 5 (2006) 2606- 2012;Lorenzi, P.L., et al., Mol. Cancer Ther. 8 (2009) 713-724)上での、抗体誘導型のc−Met受容体の内部移行を指す。c−Met受容体のそのような内部移行は、本発明による二重特異性抗体によって引き起こされ、そして実施例11に記載のとおりフローサイトメトリーアッセイ(FACS)によって1時間後に計測される。本発明による二重特異性抗体は、抗体の不存在下でのc−Metの内部移行と比較した場合に、抗体への暴露の1時間後にOVCAR−8細胞における15%未満のc−Metの内部移行を示す。一実施形態では、前記抗体は10%未満のc−Metの内部移行を示す。一実施形態では、前記抗体は7%未満のc−Metの内部移行を示す。一実施形態では、前記抗体は5%未満のc−Metの内部移行を示す。
【0036】
この発明の別の態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位とヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなるヒトErbB−2及びヒトc−Metに特異的に結合する前記二重特異性抗体であって、フローサイトメトリーアッセイによって1時間後にOVCAR−8細胞において計測したときに、(対応する)単一特異性の二価の親c−Met抗体によって誘発されたc−Metの内部移行と比較して、前記二重特異性抗体が、50%以上(一実施形態では60%以上;他の実施形態では70%以上、一実施形態では80%以上)c−Metの内部移行を減少させることを特徴とする。c−Metの内部移行の減少は、以下のとおり計算される(OVCAR−8細胞におけるフローサイトメトリーアッセイにより1時間後に計測される内部移行率(%)の値を使用しているが、0より低い内部移行率(%)の値は0%内部移行と設定する、例えばBsAB02に関して(−7%内部移行を0%の内部移行と設定した)):100×(単一特異性の二価の親c−Met抗体によって誘発されたc−Metの内部移行率(%)−二重特異性ErbB−2/cMet抗体によって誘発されたc−Metの内部移行率(%))/単一特異性の二価の親c−Met抗体によって誘発されたc−Metの内部移行率(%)。例えば:二重特異性ErbB2/cMet抗体BsAB02が(0%と設定される)−7%のc−Metの内部移行を示し、且つ、単一特異性の二価の親c−Met抗体Mab 5D5が37%のc−Metの内部移行を示す。よって、前記二重特異性ErbB−2/cMet抗体BsAB02は、100×(40−0)/40%=100%のc−Metの内部移行の減少を示す(実施例11でのOVCAR−8細胞におけるフローサイトメトリーアッセイにより1時間後に計測される内部移行値を参照のこと)。
【0037】
本明細書中に使用される場合、「抗体」は、抗原結合部位を含んでなる結合タンパク質を指す。「結合部位」又は「抗原結合部位」という用語は、本明細書中では、リガンドが実際に結合している抗体分子の(単数若しくは複数の)領域を意味し、抗体に由来する。「抗原結合部位」という用語には、抗体重鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)又はVH/VLの対が含まれ、抗体全体又は抗体フラグメント、例えば一本鎖Fv、VHドメイン及び/又はVLドメイン、Fab、又は(Fab)2などに由来することもある。この発明の一実施形態では、それぞれの抗体結合部位が、抗体重鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)を含んでなり、好ましくは、抗体軽鎖可変ドメイン(VL)と抗体重鎖可変ドメイン(VH)から成る1対によって形成される。
【0038】
抗体由来の抗原結合部位に加えて、例えばMatzke, A., et al., Cancer Res. 65 (14) (2005) 6105-10. July 15, 2005に記載された結合ペプチドもまた、抗原(例えばc−Met)に特異的に結合し得る。よって、この発明の更なる態様は、ヒトErbB−2に特異的に結合する抗原結合部位及びヒトc−Metに特異的に結合する結合ペプチドを含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性結合性分子である。よってこの発明の更なる態様は、ヒトc−Metに特異的に結合する抗原結合部位及びヒトErbB−2に特異的に結合する結合ペプチドを含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性結合性分子である。
【0039】
ErbB−2(ERBB2、HER2;CD340、HER−2/neu、c−erb B2/neuタンパク質、神経芽細胞腫/神経膠芽腫由来癌遺伝子相同体|v−erb−b2 鳥類赤血球性白血病ウイルス癌遺伝子相同体2;配列番号14としても知られている)は、それ自身のリガンド結合ドメインを持たないタンパク質であるため、成長因子に結合することができない。しかしながら、他のリガンド結合EGF受容体ファミリーメンバーに強く結合してヘテロ二量体を形成し、リガンド結合を安定させ、そして下流のシグナル伝達経路のキナーゼ媒介型活性化を促進する、例えばそれらはマイトジェン活性化プロテインキナーゼやホスファチジルイノシトール−3キナーゼに関与している。本明細書中に示した最も頻度の高い対立遺伝子Ile654/Ile655を含めて、第654及び655位アミノ酸における対立遺伝子変異のアイソフォームa(第624及び625位のアイソフォームb)が報告されている。この遺伝子の増幅、及び/又は過剰発現は、乳房及び卵巣の腫瘍を含めた多数の癌で報告されている。選択的スプライシングはいくつかのさらなる転写物変異体をもたらすが、一部のものは異なるアイソフォームをコードし、他のものは完全に特徴づけされていない。ErB−2は、元々、形質変換遺伝子の産物として化学物質で処理されたラットの神経芽細胞腫から同定された。neu癌原遺伝子の活性型は、コードされたタンパク質の膜貫通領域における(バリンからグルタミン酸への)点突然変異から得られた(Semba, K., et al., PNAS 82 (1985) 6497-501;Coussens, L., et al., Science 230 (1985) 1132-9;Bargmann, C.I., et al., Nature 319 (1986) 226-30;Yamamoto, T., et al., Nature 319 (1986) 230-4)。
【0040】
ヒトErbB−2に特異的に結合する抗原結合部位、特に重質鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)は、a)例えば2C4(パーツズマブ;パーツズマブは、マウス抗HER2抗体2C4の遺伝子組み換えヒト化バージョンである、そしてそれぞれの調製方法と共にWO01/00245及びWO2006/007398に記載されている)及び4D5(トラスツズマブ(マウス抗HER2抗体4D5の遺伝子組み換えヒト化バージョン、Herceptin(商標);トラスツズマブ及びその調製方法はUS5,821,337に記載されている))抗体(Hudziak, R., M, et al, Mol. Cell. Biol. 9 (1989) 1165-1172;Fendly, B., M., et al., Cancer Research 50 (1990) 1550-1558)のような既知の抗ErbB−2抗体、あるいは、b)とりわけヒトErbB−2タンパク質又は核酸若しくはその断片のいずれかを使用したデノボ免疫法によって、又はファージ提示法によって得られた新しい抗ErbB−2抗体、に由来することができる。
【0041】
MET(間葉−上皮移行因子)は、タンパク質METをコードする癌原遺伝子であり、(c−Met、肝細胞増殖受容体HGFR;HGF受容体;細胞分散因子受容体;SF受容体;配列番号13としても知られている)(Dean, M., et al., Nature 318 (1985) 385-8;Chan, A.M., et al., Oncogene 1 (1987) 229-33;Bottaro, D.P., et al., Science 251 (1991) 802-4;Naldini, L., et al.5 EMBO J. 10 (1991) 2867-78;Maulik, G., et al., Cytokine Growth Factor Rev. 13 (2002) 41-59)。METは、胚発生と創傷治癒に不可欠な膜受容体である。肝細胞増殖因子(HGF)は、MET受容体の唯一の既知のリガンドである。METは通常、上皮起源の細胞によって発現されるが、HGFの発現は間葉起源の細胞に限られている。HGF刺激により、METは浸潤性増殖として知られているプログラムをまとめて引き起こす数個の生物反応を誘発する。癌における異常なMET活性化は、異常な活性のMETが腫瘍増殖や、腫瘍に栄養を供給する新生血管の形成(血管新生)や、他の臓器への癌の分散(転移癌)を引き起こす、予後不良と関連がある。METは、腎臓、肝臓、胃、乳房、及び脳の癌を含めた多くのタイプのヒト悪性腫瘍において無秩序である。本来なら、幹細胞と始原細胞だけがMETを発現しており、それが胎児において新しい組織を作り出したり、成人において損傷した組織を再生するために、これらの細胞が侵襲的に増殖することを可能にしている。しかしながら、癌幹細胞は、METを発現する正常幹細胞の能力を乗っ取り、それによって癌持続性と体内の他の部位への分散の原因となっている。
【0042】
ヒトc−Metに特異的に結合する抗原結合部位、特に重質鎖可変ドメイン(VH)及び/又は抗体軽鎖可変ドメイン(VL)は、a)例えばUS5,686,292、US7,476,724、WO2004/072117、WO2004/108766、WO2005/016382、WO2005/063816、WO2006/015371、WO2006/104911、WO2007/126799、又はWO2009/007427に記載の既知の抗c−Met抗体、あるいは、b)例えば、とりわけヒト抗−Metタンパク質又は核酸若しくはその断片のいずれかを使用したデノボ免疫法によって、又はファージ提示法によって得られた新しい抗c−Met抗体、に由来することができる。
【0043】
本発明の更なる態様は、ヒトErbB−2に特異的に結合する第1の抗原結合部位及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、以下の:
ErbB−2に特異的に結合する前記第1の抗原結合部位が、重鎖可変ドメインとして配列番号1の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号2の配列を含んでなり;そして
c−Metに特異的に結合する前記第2の抗原結合部位が、重鎖可変ドメインとして配列番号3の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号4の配列を含んでなる、
を特徴とする前記二重特異性抗体である。
【0044】
抗体の特異性は、抗原の特定のエピトープに対する抗体の選択的認識を指す。例えば、天然抗体は単一特異性である。本発明による「二重特異性抗体」は、2つの異なった抗原結合特異性を有する抗体である。抗体が2以上の特異性を有する場合には、認識されたエピトープは単一の抗原又は2以上の抗原と結合することもできる。本発明の抗体は、2つの異なった抗原、すなわち第1の抗原としてのErbB−2及び第2の抗原としてのc−Metに特異的である。
【0045】
用語「単一特異性」抗体は、本明細書中で使用される場合、そのそれぞれが同じ抗原の同じエピトープに結合する1以上の結合部位を持つ抗体を意味する。
【0046】
用語「価」は、本明細書中で使用される場合、抗体分子内に存在している一定数の結合部位を意味する。同様に、用語「二価」、「四価」、及び「六価」は、抗体分子内にそれぞれ2つの結合部位、4つの結合部位、及び6つの結合部位の存在を意味する。本発明による二重特異性抗体は、少なくとも「二価」であり、そして「三価」又は「多価」(例えば「四価」若しくは「六価」)でもあり得る。
【0047】
本発明の抗体の抗原結合部位は、抗原に対する結合部位の親和性に多かれ少なかれ貢献する6つの相補性決定領域(CDRs)を含むことができる。3つの重鎖可変ドメインCDRs(CDRH1、CDRH2及びCDRH3)と3つの軽鎖可変ドメインCDRs(CDRL1、CDRL2及びCDRL3)がある。CDRとフレームワーク領域(FR)の程度は、配列内での可変性によってそれらの領域が規定されたコンパイルされたアミノ酸配列のデータベースとの比較によって決定される。いくつかのCDRを含んでなる(すなわち、結合特異性が3、4又は5つのCDRによって決定される)機能性抗原結合部位もまた、本発明の範囲内に含まれる。例えば、6つのCDRの完全セット以下でも結合に十分であり得る。場合によっては、VH又はVLドメインで十分である。
【0048】
好ましい実施形態では、本発明の抗体は、ヒト起源の1若しくは複数の免疫グロブリンクラスの免疫グロブリン定常領域を更に含む。免疫グロブリンクラスには、IgG、IgM、IgA、IgD、及びIgEアイソタイプ、そしてIgGとIgAの場合には、それらのサブタイプが含まれる。好ましい実施形態では、本発明の抗体は、IgGタイプ抗体の定常ドメイン構造を有するが、4つの抗原結合部位を有する。これは、例えば、c−Metに特異的に結合する1つ(若しくは2つ)の完全抗原結合部位(例えば一本鎖Fabフラグメント若しくは一本鎖Fv)を、ErbB−2に特異的に結合する完全な抗体の重鎖又は軽鎖のN又はC末端のいずれかに連結して、三価の二重特異性抗体(又は四価の二重特異性抗体)を得ることによって達成される。あるいは、例えばEP出願番号第07024867.9号、EP出願番号第07024864.6号、EP出願番号第07024865.3号又はRidgway, J.B., Protein Eng. 9 (1996) 617-621;WO96/027011;Merchant, A.M., et al., Nature Biotech 16 (1998) 677-681;Atwell, S., et al., J. Mol. Biol. 270 (1997) 26-35及びEP1870459ALに記載のとおり、免疫グロブリン定常領域を含んでなる、ヒトErbB−2及びヒトc−Metに対する、IgGのような二重特異性の二価抗体を使用できる。
【0049】
用語「モノクローナル抗体」又は「モノクローナル抗体組成物」とは、本明細書において使用される場合、単一のアミノ酸組成物の抗体分子の調製物を意味する。
【0050】
用語「キメラ抗体」とは、通常組換えDNA技術により調製される、可変領域、すなわち1つの源又は種からの結合領域、及び異なった源又は種に由来する定常領域の少なくとも一部を含んでなる抗体を意味する。ネズミ可変領域及びヒト定常領域を含んでなるキメラ抗体が好ましい。本発明により包含される他の好ましい形の「キメラ抗体」は、定常領域が、特にC1q結合及び/又はFc受容体(FcR)結合に関して、本発明の性質を生じるように、修飾されているか、又は元の抗体のその領域から変更されているそれらの抗体である。そのようなキメラ抗体はまた、「クラス−スイッチ抗体」とも呼ばれる。キメラ抗体は、免疫グロブリン可変領域をコードするDNAセグメント、及び免疫グロブリン定常領域をコードするDNAセグメントを含んでなる免疫グロブリン遺伝子が発現された生成物である。キメラ抗体を生成するための方法は、当業界において現在良く知られている、従来の組換えDNA及び遺伝子トランスフェクション技術を包含する。例えば、Morrison, S.L., et al., Proc. Natl. Acad Sci. USA 81(1984)6851-6855;US 5,202,238及びUS 5,204,244を参照のこと。
【0051】
用語「ヒト化抗体」とは、そのフレームワーク又は「相補性決定領域」(CDR)が、親イムノグロブリンのCDRに比較して、異なった特異性の免疫グロブリンのCDRを含んでなるよう修飾されている抗体を意味する。好ましい実施形態では、ネズミCDRは、「ヒト化抗体」を調製するために、ヒト抗体のフレームワーク領域中に移植される。例えば、Riechmann, L., et al., Nature 332(1988)323-327;及びNeuberger, M.S., et al., Nature 314(1985)268-270を参照のこと。特に好ましいCDRは、キメラ抗体に関して上記に示される抗原を認識する配列を表すものに対応する。本発明により包含される他の形の「ヒト化抗体」とは、定常領域が、特にC1q結合及び/又はFc受容体(FcR)結合に関して、本発明の性質を生じるように、更に修飾されているか、又は元の抗体のその領域から変更されている抗体である。
【0052】
用語「ヒト抗体」とは、本明細書において使用される場合、ヒト生殖系免疫グロブリン配列に由来する可変及び定常領域を有する抗体を包含することを意図する。ヒト抗体は技術的に良く知られている(van Dijk, M.A., and van de Winkel, J. G., Curr. Opin. Chem. Biol. 5(2001)368-374)。ヒト抗体はまた、免疫化に基づいて、内因性免疫グロブリン生成の不在下でヒト抗体の全レパートリー又は選択物を生成できるトランスジェニック動物(例えば、マウス)においても生成され得る。そのような生殖系変異マウス内へのヒト生殖系免疫グロブリン遺伝子アレイのトランスファーは、抗原攻撃に基づくヒト抗体の生成をもたらすであろう(例えば、Jakobovits, A., et al., Proc. Natl. Acad. Sci. USA 90(1993)2551-2555;Jakobovits, A., et al., Nature 362(1993)255-258;Bruggemann, M., et al., Year Immunol. 7(1993)33-40を参照のこと)。ヒト抗体はまた、ファージ提示ライブラリーにおいても生成され得る(Hoogenboom, H. R., and Winter, G., J. Mol. Biol. 227(1992)381-388;Marks, J.D., et al., J. Mol. Biol. 222(1991)581-597)。Cole, S.P.C.ら及びBoerner, Pらの技術はまた、ヒトモノクローナル抗体の調製のためにも利用できる(Cole, S.P.C., et al., Monoclonal Antibodies and Cancer Therapy, Liss, A. L.(1985) 77-96;及びBoerner, P., et al., J. Immunol. 147(1991)86-95)。本発明のキメラ及びヒト化抗体についてすでに言及されたように、用語「ヒト抗体」とは、本明細書において使用される場合、また、特にC1q結合及び/又はFcR結合に関して、例えばFc部分の「クラススイッチ」、すなわち変更又は突然変異誘発(IgG1からIgG4への及び/又はIgG1/IgG4突然変異)により、本発明の性質を生じるように定常領域において修飾されているそのような抗体も含んでなる。
【0053】
用語「組換えヒト抗体」とは、本明細書において使用される場合、組換え手段により調製され、発現され、作出され、又は単離されるすべてのヒト抗体、例えば宿主細胞、例えばNS0又はCHO細胞から又はヒト免疫グロブリン遺伝子に関してトランスジェニックである動物(例えば、マウス)から単離された抗体、又は宿主細胞中にトランスフェクトされた組換え発現ベクターを用いて発現された抗体を意味する。そのような組換えヒト抗体は、再配置された形で、可変及び定常領域を有する。本発明の組換えヒト抗体は、生体内における体細胞超突然変異にゆだねられている。従って、組換え抗体のVH及びVL領域のアミノ酸配列は、ヒト生殖系VH及びVL領域に由来し、そしてその配列に関連しているが、天然においては、ヒト抗体生殖系レパートリー内で生体内には存在することができない配列である。
【0054】
「可変ドメイン」(軽鎖の可変領域(VL)、重鎖の可変領域(VH))は、本明細書において使用される場合、抗原への抗体の結合において直接関与する軽鎖及び重鎖の対の各々を示す。ヒト可変軽鎖及び重鎖のドメインは、同じ一般構造を有し、そして各々のドメインは4個のフレームワーク(FR)領域を含んで成り、それらの配列は広く保存され、3個の「超可変領域」(又は相補性決定領域、CDR)により結合される。フレームワーク領域は、β−シートコンホメーションを採用し、そしてCDRはβ−シート構造体を結合するループを形成することができる。各々の鎖におけるCDRは、フレームワーク領域によりそれらの立体構造を保持され、そして他の鎖からのCDRと共に抗原結合部位を形成する。抗体重鎖及び軽鎖CDR3領域は、本発明の抗体の結合特異性/親和性において特に重要な役割を演じ、従って、本発明の更なる目的を提供する。
【0055】
用語「超可変領域」又は「抗体の抗原−結合部分若しくは抗原結合部位」とは、本明細書において使用される場合、抗原結合を担当できる抗体のアミノ酸残基を指す。超可変領域は「相補性決定領域」又は「CDR」からのアミノ酸残基を含んでなる。「フレームワーク」又は「FR」領域は、本明細書において定義されるように超可変領域残基以外のそれらの可変ドメイン領域である。従って、抗体の軽鎖及び重鎖は、N末端からC末端に向かって、ドメインFR1、CDR1、FR2、CDR2、FR3、CDR3及びFR4を含んでなる。各々の鎖上のCDRはそのようなフレームワークアミノ酸により分離される。特に、重鎖のCDR3は、ほとんど抗原結合に寄与する領域である。CDR及びFR領域は、Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD(1991)の標準的な定義に従って決定される。
【0056】
本明細書において使用される場合、用語「結合」又は「特異的に結合」とは、試験管内におけるアッセイ、好ましくは精製した野生型抗原を用いたプラズモン共鳴アッセイ(BIAcore、GE-Healthcare Uppsala, Sweden)における抗体の抗原(ヒトErbB−2又はヒトc−Metのいずれか)のエピトープへの結合を指す。結合の親和性は、用語ka(抗体/抗原複合体からの抗体結合に関する速度定数)、kD(解離定数)、及びKD(kD/ka)によって規定される。結合又は特異的に結合は、10-8mol/l以下、好ましくは10-9M〜10-13mol/lの結合親和性(KD)を意味する。よって、本発明による二重特異性<ErbB−2−c−Met>抗体は、各抗原に対して10-8mol/l、好ましくは10-9M〜10-13mol/lの結合親和性(KD)を有する特異性をもって特異的に結合する。
【0057】
FcγRIIIへの抗体の結合は、BIAcoreアッセイ(GE-Healthcare Uppsala, Sweden)により調査され得る。結合の親和性は、用語ka(抗体/抗原複合体からの抗体結合に関する速度定数)、kD(解離定数)、及びKD(kD/ka)により規定される。
【0058】
用語「エピトープ」とは、抗体に対して特異的に結合できるいずれかのポリペプチド決定基を包含する。ある実施形態においては、エピトープ決定基は、分子、例えばアミノ酸、糖側鎖、ホスホリン又はスルホニルなどの化学的活性表面群を包含し、そしてある実施形態においては、特定の立体構造特性及び/又は比電荷特性を有することができる。エピトープは、抗体により結合される抗原の領域である。
【0059】
ある実施形態においては、抗体は、それがタンパク質及び/又は高分子の複合混合物中でその標的抗原を選択的に認識する場合、抗原を特異的に結合すると言われる。
【0060】
用語「定常領域」は、本明細書中で使用される場合、可変領域以外の抗体のドメインの集合を意味する。定常領域は、抗原の結合に直接的にかかわらないが、様々なエフェクター機能を示す。それらの重鎖の定常領域のアミノ酸配列によって、抗体は以下のクラス:IgA、IgD、IgE、IgG及びIgMに分類され、そしてそのいくつか、例えばIgG1、IgG2、IgG3、及びIgG4、IgA1及びIgA2などは、サブクラスに更に分類されることもできる。異なった抗体クラスに該当する重鎖定常領域は、それぞれα、δ、ε、γ、及びμと呼ばれる。全部で5つの抗体クラスで見ることができる軽鎖定常領域は、κ(カッパ)及びλ(ラムダ)と呼ばれる。定常領域は、好ましくはヒト起源に由来する。
【0061】
用語「ヒト起源由来の定常領域」は、本願で使用される場合、ヒト抗体のサブクラスIgG1、IgG2、IgG3、若しくはIgG4の重鎖定常領域、及び/又は定常軽鎖カッパ又はラムダ領域を規定する。そのような定常領域は、当技術分野の現状において周知であり、例えば、Kabat, E.A.によって説明されている(例えばJohnson, G., and Wu, T.T., Nucleic Acids Res. 28(2000)214-218;Kabat, E.A., et al., Proc, Natl. Acad. Sci. USA 72(1975)2785-2788を参照のこと)。
【0062】
一実施形態では、本発明による二重特異性抗体は、IgG1又はIgG3サブクラス(好ましくはIgG1サブクラス)の定常領域を含んでなり、それは好ましくはヒト起源に由来する。一実施形態では、本発明による二重特異性抗体は、IgG1又はIgG3サブクラス(好ましくはIgG1サブクラス)のFc部分を含んでなり、それは好ましくはヒト起源に由来する。
【0063】
IgG4サブクラスの抗体がFc受容体(FcyRIIIa)結合の低減を示している一方で、他のIgGサブクラスの抗体は強い結合を示している。しかしながら、Pro238、Asp265、Asp270、Asn297(Fc炭水化物の欠如)、Pro329、Leu234、Leu235、Gly236、Gly237、Ile253、Ser254、Lys288、Thr307、Gln311、Asn434、及びHis435は、変更された場合には、同様に低減されたFc受容体結合性を示す残基である(Shields, R.L., et al., J. Biol. Chem. 276(2001)6591-6604;Lund, J., et al., FASEB J. 9(1995)115-119;Morgan, A., et al., Immunology 86(1995)319-324;EP 0 307 434)。
【0064】
1つの実施形態では、本発明による抗体は、IgG1抗体と比較してFcR結合が低減されているので、全長親抗体は、FcR結合に関して、IgG4サブクラスのもの、あるいはS228、L234、L235及び/又はD265に突然変異を伴う、及び/又はPVA236突然変異を含むIgG1又はIgG2サブクラスのものである。1つの実施形態では、全長親抗体内の突然変異は、S228P、L234A、L235A、L235E、及び/又は、PVA236である。他の実施形態では、全長親抗体内の突然変異は、IgG4ではS228PそしてとIgG1ではL234AとL235Aにある。
【0065】
抗体の定常領域は、ADCC(抗体依存性細胞毒性)及びCDC(補体依存性細胞毒性)に直接関与する。補体活性化(CDC)は、大部分のIgG抗体サブクラスの定常領域への補体C1q因子の結合により開始する。抗体へのC1qの結合は、いわゆる結合部位で所定のタンパク質−タンパク質相互作用により起きる。そのような定常領域の結合部位は、当技術分野の現状において公知であり、例えばLukas, T.J., et al., J. Immunol. 127(1981)2555-2560;Brunhouse, R. and Cebra, J.J., Mol. Immunol. 16(1979)907-917;Burton, D.R., et al., Nature 288(1980)338-344;Thommesen, J.E., et al., Mol. Immunol. 37(2000)995-1004;Idusogie, E.E., et al., J. Immunol. 164(2000)4178-4184;Hezareh, M., et al., J. Virol. 75(2001)12161-12168;Morgan, A., et al., Immunology 86(1995)319-324;及びEP 0 307 434により記載されている。そのような定常領域の結合部位は、例えばアミノ酸L234、L235、D270、N297、E318、K320、K322、P331、及びP329(付番は常にKabatのEUインデックスに準じる)を特徴とする。
【0066】
用語「抗体依存性細胞毒性(ADCC)」は、エフェクター細胞の存在下での、本発明よる抗体によるヒト標的細胞の溶解を指す。ADCCは、好ましくは、新たに単離されたPBMC、又は単球若しくはナチュラルキラー(NK)細胞又は恒久的に増殖しているNK細胞株のような精製された軟膜由来エフェクター細胞等のエフェクター細胞の存在下で、本発明よる抗体を用いて、ErB−1及びc−Met発現細胞の調製物を処理することにより測定される。
【0067】
用語「補体依存性細胞毒性(CDC)」は、大部分のIgG抗体サブクラスのFc部への補体C1q因子の結合により開始する過程と規定される。抗体へのC1qの結合は、いわゆる結合部位で所定のタンパク質−タンパク質相互作用により起きる。そのようなFc部の結合部位は、当技術分野の現状において公知である(前記を参照のこと)。そのようなFc部の結合部位は、例えばアミノ酸L234、L235、D270、N297、E318、K320、K322、P331、及びP329(付番は常にKabatのEUインデックスに準じる)を特徴とする。サブクラスIgG1、IgG2、及びIgG3の抗体は、通常はC1q及びC3の結合を含めた補体活性化を示し、一方でIgG4は補体系を活性化せず、C1q及びC3と結合しない。
【0068】
モノクローナル抗体の細胞媒介型エフェクター機能は、Umana, P., et al., Nature Biotechnol. 17(1999)176-180及びUS 6,602,684に記載されているように、それらのオリゴ糖構成要素を操作することによって増強することができる。癌免疫療法において最もよく使用される抗体であるIgG1型抗体は、保存されたN結合型グリコシル化部位を各CH2ドメイン中のAsn297において有する糖タンパク質である。Asn297に結合した2つの複合型二分岐オリゴ糖は、CH2ドメインの間に埋もれて、ポリペプチド骨格との広範囲の接触面を形成し、それらの存在は、抗体依存性細胞毒性(ADCC)のようなエフェクター機能を抗体が媒介するために不可欠である(Lifely, M.R., et al., Glycobiology 5(1995)813-822;Jefferis, R., et al., Immunol. Rev. 163(1998)59-76;Wright, A. and Morrison, S.L., Trends Biotechnol. 15(1997)26-32)。Umana, P., et al., Nature Biotechnol. 17(1999)176-180及びWO 99/54342は、2つに分岐したオリゴ糖の形成を触媒するグリコシルトランスフェラーゼであるβ(1,4)−N−アセチルグルコサミニルトランスフェラーゼIII(「GnTIII」)がチャイニーズハムスター卵巣(CHO)細胞において過剰発現されると、抗体の生体内でのADCC活性が有意に増大することを示した。また、Asn297炭水化物の組成の改変又はその除去も、FcγR及びC1qへの結合に影響を及ぼす(Umana, P., et al., Nature Biotechnol. 17(1999)176-180;Davies, J., et al., Biotechnol. Bioeng. 74(2001)288-294;Mimura, Y., et al., J. Biol. Chem. 276(2001)45539-45547;Radaev, S., et al., J. Biol. Chem. 276(2001)16478-16483;Shields, R.L., et al., J. Biol. Chem. 276(2001)6591-6604;Shields, R.L., et al., J. Biol. Chem. 277(2002)26733-26740;Simmons, L.C., et al., J. Immunol. Methods 263(2002)133-147)。
【0069】
フコースの量を減少させることによってモノクローナル抗体の細胞媒介型エフェクター機能を高める方法は、例えば、WO2005/018572、WO2006/116260、WO2006/114700、WO2004/065540、WO2005/011735、WO2005/027966、WO1997/028267、US2006/0134709、US2005/0054048、US2005/0152894、WO2003/035835、WO2000/061739、Niwa, R., et al., J. Immunol. Methods 306 (2005) 151-160;Shinkawa, T., et al, J Biol Chem, 278 (2003) 3466-3473;WO03/055993又はUS2005/0249722に記載されている。
【0070】
本発明の一実施形態では、本発明による二重特異性抗体は、(IgG1若しくはIgG3サブクラス)Asn297にて糖鎖を用いてグリコシル化され、それによって、前記糖鎖内のフコースの量が65%以下になる(付番はKabatに準じる)。他の実施形態では、前記糖鎖内のフコースの量が、5%〜65%、好ましくは20%〜40%である。本発明による「Asn297」は、Fc領域内の約297位に見られるアミノ酸であるアスパラギンを意味する。抗体のわずかな配列変化により、Asn297はまた、297位の数アミノ酸(通常、±3アミノ酸以下)上流又は下流、すなわち、294位から300位の間でも見られることがある。
【0071】
ヒトIgG1又はIgG3のグリコシル化は、最大2つのGal残基で終わるコアフコース化二本鎖複合型オリゴ糖グリコシル化としてAsn297で起こる。IgG1又はIgG3サブクラスのヒト重鎖定常領域は、Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD.(1991)、及びBruggemann, M., et al., J. Exp. Med. 166(1987)1351-1361;Love, T.W., et al., Methods Enzymol. 178(1989)515-527によって詳細に説明されている。これらの構造は、終末のGal残基の量に依存して、G0、G1(α−1,6−若しくはα−1,3−)又はG2グリカン残基として指定される(Raju, T.S., Bioprocess Int. 1(2003)44-53)。抗体Fc部分のCHOタイプのグリコシル化は、例えば、Routier, F.H., Glycoconjugate J. 14(1997)201-207によって説明されている。非糖修飾CHO宿主細胞において組換え発現される抗体は、通常少なくとも85%の量がAsn297においてフコース化されている。全長親抗体の修飾オリゴ糖は、ハイブリッドであっても複合体であってもよい。好ましくは、分枝、還元/非フコース化オリゴ糖がハイブリッドされる。他の実施形態において、分枝、還元又は非フコース化オリゴ糖が複合体化される。
【0072】
本発明による「フコースの量」は、MALDI−TOF質量分析法によって測定し、平均値として算出した、Asn297に結合したすべての糖残基(例えば、複合体構造物、ハイブリッド構造物、及び高マンノース構造物)の合計に対するAsn297における糖鎖内のフコースの量を意味する。フコースの相対量は、MALDI−TOFによって測定した、N-グリコシダーゼFで処理したサンプルにおいて同定された全糖構造物(例えば、それぞれ複合体構造物、ハイブリッド構造物、並びにオリゴマンノース構造物及び高マンノース構造物)に対するフコース含有構造物のパーセンテージである(例えばWO2008/077546(A1)を参照のこと)。
【0073】
一実施形態は、WO2005/044859、WO2004/065540、WO2007/031875、Umana, P., et al., Nature Biotechnol. 17 (1999) 176-180、WO99/154342、WO2005/018572、WO2006/116260、WO2006/114700、WO2005/011735、WO2005/027966、WO97/028267、US2006/0134709、US2005/0054048、US2005/0152894、WO2003/035835又はWO2000/061739に記載の手順を用いた、Asn297にて糖鎖を用いて(糖鎖が)グリコシル化され、それによって、前記糖鎖内のフコースの量が65%以下になる、IgG1又はIgG3サブクラスの二重特異性抗体の調製法である。
【0074】
一実施形態は、Niwa, R., et al., J. Immunol. Methods 306 (2005) 151-160;Shinkawa, T. et al, J Biol Chem, 278 (2003) 3466-3473;WO03/055993又はUS2005/0249722に記載の手順を用いた、Asn297にて糖鎖を用いて(糖鎖が)グリコシル化され、それによって、前記糖鎖内のフコースの量が65%以下になる、IgG1又はIgG3サブクラスの二重特異性抗体の調製法である。
【0075】
二重特異性抗体フォーマット
本発明の抗体は、2個以上の結合部位を持つので、多特異性であり、好ましくは二重特異性である。すなわち、抗体は、2以上の結合部位がある場合でさえ、二重特異性であり得る(すなわち、抗体は三価又は多価性である)。本発明の二重特異性抗体には、例えば多価性一本鎖抗体、二重特異性抗体、及び三重特異性抗体、並びに1若しくは複数のペプチド・リンカーを介して連結された更なる抗原結合部位(例えば一本鎖Fv、VHドメイン及び/又はVLドメイン、Fab、又は(Fab)2)を持つ全長抗体の定常ドメイン構造を有する抗体が含まれる。抗体は、単一種からの全長であることも、又はキメラ化若しくは又はヒト化されることもできる。3つ以上の抗原結合部位を有する抗体に関しては、タンパク質に2つの異なった抗原のための結合部位がある限り、いくつかの結合部位が同一のであることもあり得る。すなわち、第1の結合部位がErbB−2に特異的である一方で、第2の結合部位はc−Metに特異的であり、そして逆もまた同様である。
【0076】
好ましい実施形態では、本発明によるヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体は、(好ましくはIgG1又はIgG3サブクラスの)抗体のFc領域を含んでなる。
【0077】
二価の二重特異性フォーマット
免疫グロブリン定常領域を含んでなるヒトErbB−2及びヒトc−Metに対する二重特異性の二価抗体は、例えば、WO2009/080251、WO2009/080252、WO2009/080253若しくはRidgway, J.B., Protein Eng. 9 (1996) 617-621;WO96/027011;Merchant, A.M., et al. Nature Biotech 16 (1998) 677-681;Atwell, S., et al., J. Mol. Biol. 270 (1997) 26-35及びEP1870459A1に記載のように使用できる。
【0078】
よって、本発明の一実施形態では、本発明による二重特異性<ErbB−2−c−Met>抗体は、以下の:
a)ErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びに
b)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖;
を含んでなり、ここで、定常ドメインCLとCH1、及び/又は可変ドメインVLとVHが互いによって置き換えられている二価の二重特異性抗体である。
【0079】
本発明の他の実施形態において、本発明による二重特異性<ErbB−2−c−Met>抗体は、以下の:
a)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖;並びに
b)ErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;
を含んでなり、ここで、定常ドメインCLとCH1、及び/又は可変ドメインVLとVHが互いによって置き換えられている二価の二重特異性抗体である。
【0080】
以下で記載する「ホール−into−ノブ」技術を用いた代表的な図式的構造については、図2a〜cを参照のこと。
【0081】
そのようなヘテロ二量体の二価の二重特異性抗ErbB−2/抗c−Met抗体の収率を改善するために、前記全長抗体のCH3ドメインは、例えばWO96/027011、Ridgway, J., B., et al., Protein Eng 9 (1996) 617-621;及びMerchant, A., M., et al., Nat Biotechnol 16 (1998) 677-681中にいくつかの実施例を用いて詳細に説明されている「ホール−into−ノブ」技術によって変更され得る。この方法では、2種のCH3ドメインの相互作用表面が、それらの2種のCH3ドメインを含む両重鎖のヘテロダイマー化を高めるために変更される。(2種の重鎖の)2種のCH3ドメインの個々が「ノブ」であり、そして他が「ホール」である。ジスルフィド架橋の導入は、ヘテロダイマーを安定化し(Merchant, A..M., et al, Nature Biotech 16(1998)677-681;Atwell, S., et al., J. Mol. Biol. 270(1997)26-35)、そして収率を高める。
【0082】
従って、本発明の1つの態様では、前記二価の二重特異性抗体は、1つの重鎖のCH3ドメイン及び他の重鎖のCH3ドメインが、抗体CH3ドメイン間に本来の界面を含んでなる界面で各々接触し;
ここで、前記界面は二価の二重特異性抗体の形成を促進するよう変更され、ここで、前記変更が:
a)二価の二重特異性抗体内のもう片方の重鎖のCH3ドメインの本来の界面と接触する、一方の重鎖のCH3ドメインの本来の界面内で、アミノ酸残基がより大きな側鎖体積を有するアミノ残基により置換され、それにより、もう片方の重鎖のCH3ドメインの界面内の腔に位置決定できる、一方の重鎖CH3ドメインの界面内に隆起部を生じるように、一方の重鎖のCH3ドメインが変更され、そして
b)二価の二重特異性抗体内の第1CH3ドメインの本来の界面と接触する、第2CH3ドメインの本来の界面内で、アミノ酸残基がより小さな側鎖体積を有するアミノ酸残基により置換され、それにより、第1CH3ドメインの界面内の隆起部が位置決定できる第2CH3ドメインの界面内に腔を生じるように、もう片方の重鎖のCH3ドメインが変更されることにより更に特徴づけられる。
【0083】
好ましくは、より大きな側鎖体積を有する前記アミノ酸残基が、アルギニン(R)、フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)から成る群から選択される。
【0084】
好ましくは、より小さな側鎖体積を有する前記アミノ酸残基が、アラニン(A)、セリン(S)、トレオニン(T)、バリン(V)から成る群から選択される。
【0085】
本発明の1つの態様においては、両CH3ドメインは更に、それらの両CH3ドメイン間にジスルフィド架橋が形成され得るよう、各々のCH3ドメインのその対応する位置のアミノ酸としてのシステイン(C)の導入により変更される。
【0086】
好ましい実施形態では、前記二価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にT366W突然変異、及び「ホール鎖」のCH3ドメイン内にT366S、L368A、Y407V突然変異を含んでなる。例えば、「ノブ鎖」のCH3ドメイン内にY349C突然変異、及び「ホール鎖」のCH3ドメイン内にE356C突然変異又はS354C突然変異を導入することによって、CH3ドメイン間の追加的な鎖間ジスルフィド架橋もまた使用できる(Merchant, A.M., et al., Nature Biotech 16(1998)677-681)。よって、別の好ましい実施形態では、前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にE356C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなる(鎖間ジスルフィド架橋を形成する、一方のCH3ドメイン内の追加的なY349C突然変異ともう片方のCH3ドメイン内の追加的なE356C又はS354C突然変異)(付番は常にKabatのEUインデックスに準じる)。しかし、EP1870459A1によって記載される他のノブ−into−ホール技術もまた代替的に又は追加的に使用できる。前記二価の二重特異性抗体の好ましい例は、「ノブ鎖」のCH3ドメイン内のR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内のD399K;E357K突然変異である(付番は常にKabatのEUインデックスに準じる)。
【0087】
別の好ましい実施形態では、前記二価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内のT366W突然変異及び「ホール鎖」のCH3ドメイン内のT366S、L368A、Y407V突然変異、そして追加的に「ノブ鎖」のCH3ドメイン内のR409D;K370突然変異及び「ホール鎖」のCH3ドメイン内のD399K;E357KE突然変異を含んでなる。
【0088】
別の好ましい実施形態では、前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記二価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなり、そして追加的に「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0089】
三価の二重特異性フォーマット
この発明の別の好ましい態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖及び2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fabフラグメント
を含んでなり、ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、前記全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合される、三価の二重特異性抗体である。
【0090】
以下で記載する「ホール−into−ノブ」技術を用いた代表的な図式的構造は、図5aを参照のこと。
【0091】
この発明の別の好ましい態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖及び2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fvフラグメント
を含んでなり、ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、前記全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合される、三価の二重特異性抗体である。
【0092】
以下で記載する「ホール−into−ノブ」技術を用いた代表的な図式的構造は、図5bを参照のこと。
【0093】
1つの好ましい実施形態では、ヒトc−Metに結合する前記一本鎖Fab又はFvフラグメントは、前記の全長抗体に、前記全長抗体の重鎖のC末端のペプチドコネクタを介して融合される。
【0094】
この発明の別の好ましい態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖及び2つの抗体軽鎖から成る全長抗体;
b)以下の:
ba)抗体重鎖可変ドメイン(VH);又は
bb)抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、から成るポリペプチド、
ここで、前記ポリペプチドは、VHドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちの一方のC末端に融合されていて、
【0095】
c)以下の:
ca)抗体軽鎖可変ドメイン(VL);又は
cb)抗体軽鎖可変ドメイン(VL)と抗体軽鎖定常ドメイン(CL)、
から成るポリペプチド、
ここで、前記ポリペプチドはVLドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちのもう片方のC末端に融合されている、
を含んでなり;そして
ここで、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)が一緒になって、ヒトc−Metに特異的に結合する抗原結合部位を形成する、三価の二重特異性抗体である。
【0096】
好ましくは、前記b)及びc)のペプチドコネクタは、同一であり、且つ、少なくとも25個のアミノ酸、好ましくは30〜50個のアミノ酸から成るペプチドである。
【0097】
代表的な図式的構造については、図3a〜cを参照のこと。
【0098】
任意には、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)は、連結され、そして下記の位置の間へのジスルフィド結合の導入によって鎖間ジスルフィド架橋によって安定化させる:
i)重鎖可変ドメインの第44位〜軽鎖可変ドメインの第100位
ii)重鎖可変ドメインの第105位〜軽鎖可変ドメインの第43位、又は、
iii)重鎖可変ドメインの第101位〜軽鎖可変ドメインの第100位(付番は常にKabatのEUインデックスに準じる)。
【0099】
安定化のために非天然ジスルフィド架橋を導入する技術は、例えば、WO94/029350、Rajagopal, et al., Prot Engin. (1997) 1453-59;Kobayashi, H., et al., Nuclear Medicine & Biology, 25, (1998) 387-393;又はSchmidt, M., et al., Oncogene 18 (1999) 1711-1721に記載されている。1つの実施形態では、b)とc)のポリペプチドの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第44位と軽鎖可変ドメインの第100位の間にある。1つの実施形態では、b)とc)のポリペプチドの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第105位と軽鎖可変ドメインの第43位の間にある(付番は常にKabatのEUインデックスに準じる)。1つの実施形態では、一本鎖Fabフラグメントの可変ドメインVHとVLの間の前記任意のジスルフィドによる安定化のない、三価の二重特異性抗体が好まれる。
【0100】
一方の重鎖への一本鎖Fab、Fvフラグメントの融合によって(図5a又は5b)又は全長抗体の両方の重鎖への別個のポリペプチドの融合によって(図3a〜c)、ヘテロ二量体の三価の二重特異性抗体がもたらされる。そのようなヘテロ二量体の三価の二重特異性抗ErbB−2/Met抗体の収率を高めるために、前記全長抗体のCH3ドメインは、例えばWO96/027011、Ridgway, J., B., et al., Protein Eng 9 (1996) 617-621;及びMerchant, A., M., et al., Nat Biotechnol 16 (1998) 677-681中にいくつかの実施例を用いて詳細に説明されている「ホール−into−ノブ」技術によって変更され得る。この方法では、2種のCH3ドメインの相互作用表面が、それらの2種のCH3ドメインを含む両重鎖のヘテロダイマー化を高めるために変更される。(2種の重鎖の)2種のCH3ドメインの個々が「ノブ」であり、そして他が「ホール」である。ジスルフィド架橋の導入は、ヘテロダイマーを安定化し(Merchant, A..M., et al, Nature Biotech 16(1998)677-681;Atwell, S., et al., J. Mol. Biol. 270(1997)26-35)、そして収率を高める。
【0101】
従って、本発明の1つの態様では、前記三価の二重特異性抗体は、全長抗体の1つの重鎖のCH3ドメイン及び全長抗体の他の重鎖のCH3ドメインが、抗体CH3ドメイン間に本来の界面を含んでなる界面で各々接触し;
ここで、前記界面は二価の二重特異性抗体の形成を促進するよう変更され、ここで、前記変更が:
a)二価の二重特異性抗体内のもう片方の重鎖のCH3ドメインの本来の界面と接触する、一方の重鎖のCH3ドメインの本来の界面内で、アミノ酸残基がより大きな側鎖体積を有するアミノ残基により置換され、それにより、もう片方の重鎖のCH3ドメインの界面内の腔に位置決定できる、一方の重鎖CH3ドメインの界面内に隆起部を生じるように、一方の重鎖のCH3ドメインが変更され、そして
b)三価の二重特異性抗体内の第1CH3ドメインの本来の界面と接触する、第2CH3ドメインの本来の界面内で、アミノ酸残基がより小さな側鎖体積を有するアミノ酸残基により置換され、それにより、第1CH3ドメインの界面内の隆起部が位置決定できる第2CH3ドメインの界面内に腔を生じるように、もう片方の重鎖のCH3ドメインが変更されることにより更に特徴づけられる。
【0102】
好ましくは、より大きな側鎖体積を有する前記アミノ酸残基が、アルギニン(R)、フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)から成る群から選択される。
【0103】
好ましくは、より小さな側鎖体積を有する前記アミノ酸残基が、アラニン(A)、セリン(S)、トレオニン(T)、バリン(V)から成る群から選択される。
【0104】
本発明の1つの態様においては、両CH3ドメインは更に、それらの両CH3ドメイン間にジスルフィド架橋が形成され得るよう、各々のCH3ドメインのその対応する位置のアミノ酸としてのシステイン(C)の導入により変更される。
【0105】
好ましい実施形態では、前記三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にT366W突然変異、及び「ホール鎖」のCH3ドメイン内にT366S、L368A、Y407V突然変異を含んでなる。例えば、「ノブ鎖」のCH3ドメイン内にY349C突然変異、及び「ホール鎖」のCH3ドメイン内にE356C突然変異又はS354C突然変異を導入することによって、CH3ドメイン間の追加的な鎖間ジスルフィド架橋もまた使用できる(Merchant, A.M., et al., Nature Biotech 16(1998)677-681)。よって、別の好ましい実施形態では、前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にE356C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなる(鎖間ジスルフィド架橋を形成する、一方のCH3ドメイン内の追加的なY349C突然変異ともう片方のCH3ドメイン内の追加的なE356C又はS354C突然変異)(付番は常にKabatのEUインデックスに準じる)。しかし、EP1870459A1によって記載される他のノブ−into−ホール技術もまた代替的に又は追加的に使用できる。前記三価の二重特異性抗体の好ましい例は、「ノブ鎖」のCH3ドメイン内のR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内のD399K;E357K突然変異である(付番は常にKabatのEUインデックスに準じる)。
【0106】
別の好ましい実施形態では、前記三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内のT366W突然変異及び「ホール鎖」のCH3ドメイン内のT366S、L368A、Y407V突然変異、そして追加的に「ノブ鎖」のCH3ドメイン内のR409D;K370突然変異及び「ホール鎖」のCH3ドメイン内のD399K;E357KE突然変異を含んでなる。
【0107】
別の好ましい実施形態では、前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなるか、又は前記三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのうちのもう片方にS354C、T366S、L368A、Y407V突然変異を含んでなり、そして追加的に「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異、及び「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0108】
この発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fabフラグメント、
ここで、前記一本鎖Fabフラグメントが、抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、抗体軽鎖可変ドメイン(VL)、抗体軽鎖定常ドメイン(CL)及びリンカーから成り、且つ、ここで、前記抗体ドメインと前記リンカーが、N末端からC末端に向かって、以下の順序:
ba)VH−CH1−リンカー−VL−CL、又は
bb)VL−CL−リンカー−VH−CH1、
のうちの一方を有し、
ここで、前記リンカーが少なくとも30個のアミノ酸、好ましくは32〜50個のアミノ酸から成るペプチドである、
を含んでなり、そして
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖ののC又はN末端の(好ましくは重鎖のC末端の)ペプチドコネクタを介して融合されている;
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸、好ましくは10〜50個のアミノ酸から成るペプチドである、三価の二重特異性抗体である。
【0109】
この実施形態の中で、好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にT366W突然変異、そして2つのCH3ドメインのうちのもう片方にT366S、L368A、Y407V突然変異を含んでなり、より好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのなかのもう片方にS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる。任意に、前記実施形態では、三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異を含んでなり、且つ、「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0110】
この発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fvフラグメント、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖ののC又はN末端の(好ましくは重鎖のC末端の)ペプチドコネクタを介して融合され;そして
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸、好ましくは10〜50個のアミノ酸から成るペプチドである、三価の二重特異性抗体である。
【0111】
この実施形態の中で、好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にT366W突然変異、そして2つのCH3ドメインのうちのもう片方にT366S、L368A、Y407V突然変異を含んでなり、より好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのなかのもう片方にS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる。任意に、前記実施形態では、三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異を含んでなり、且つ、「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0112】
よって好ましい実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fvフラグメント、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖のC末端のペプチドコネクタを介して融合され(2つの抗体重鎖−一本鎖Fv融合ペプチドをもたらし);そして
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸である、三価の二重特異性抗体である。
【0113】
本発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの抗体軽鎖;
から成る全長抗体;並びに
b)以下の:
ba)抗体重鎖可変ドメイン(VH);又は
bb)抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、から成るポリペプチド、
ここで、前記ポリペプチドは、VHドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちの一方のC末端に融合されて(抗体重鎖−VH融合ペプチドをもたらし)、
ここで、前記ペプチドコネクタが5つのアミノ酸、好ましくは25〜50個のアミノ酸から成るペプチドである、
【0114】
c)以下の:
ca)抗体軽鎖可変ドメイン(VL);又は
cb)抗体軽鎖可変ドメイン(VL)と抗体軽鎖定常ドメイン(CL)、
から成るポリペプチド、
ここで、前記ポリペプチドはVLドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちのもう片方のC末端に融合されて(抗体重鎖−VL融合ペプチドをもたらし)、
ここで、前記ペプチドコネクタがb)のペプチドコネクタと同一である、
を含んでなり;そして
ここで、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)が一緒になって、ヒトc−Metに特異的に結合する抗原結合部位を形成する、三価の二重特異性抗体である。
【0115】
この実施形態の中で、好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にT366W突然変異、そして2つのCH3ドメインのうちのもう片方にT366S、L368A、Y407V突然変異を含んでなり、より好ましくは三価の二重特異性抗体は、2つのCH3ドメインのうちの一方にY349C、T366W突然変異、そして2つのCH3ドメインのなかのもう片方にS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる。任意に、前記実施形態では、三価の二重特異性抗体は、「ノブ鎖」のCH3ドメイン内にR409D;K370E突然変異を含んでなり、且つ、「ホール鎖」のCH3ドメイン内にD399K;E357K突然変異を含んでなる。
【0116】
本発明の別の態様では、本発明による三価の二重特異性抗体は、以下の:
a)ヒトErbB−2に結合し、2つの抗体重鎖、VH−CH1−HR−CH2−CH3及び2つの抗体軽鎖VL−CLから成る全長抗体;
(ここで、好ましくは2つのCH3ドメインのうちの一方がY349C、T366W突然変異を含んでなり、そして2つのCH3ドメインのうちのもう片方がS354C(又はE356C)、T366S、L368A、Y407V突然変異を含んでなる);
b)以下の:
ba)抗体重鎖可変ドメイン(VH);又は
bb)抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、
から成るポリペプチド;
ここで、前記ポリペプチドはVHドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちの一方のC末端に融合されている;
【0117】
c)以下の:
ca)抗体軽鎖可変ドメイン(VL)、又は
cb)抗体軽鎖可変ドメイン(VL)と抗体軽鎖定常ドメイン(CL)、から成るポリペプチド、ここで、前記ポリペプチドはVLドメインのN末端で、ペプチドコネクタを介して前記全長抗体の2つの重鎖のうちのもう片方のC末端に融合されている;
を含んでなり、そして
ここで、b)のポリペプチドの抗体重鎖可変ドメイン(VH)とc)のポリペプチドの抗体軽鎖可変ドメイン(VL)が一緒になって、ヒトc−Metに特異的に結合する抗原結合部位を形成する。
【0118】
四価の二重特異性フォーマット
一実施形態では、本発明による多特異性抗体は四価であり、ここで、ヒトc−Metに特異的に結合する(単数若しくは複数の)抗原結合部位は、(例えばWO2009/007427に記載のように)c−Metの二量化を阻害する。
【0119】
本発明の一実施形態では、前記抗体は、ヒトErbB−2に特異的に結合する2つの抗原結合部位及びヒトc−Metに特異的に結合する2つの抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する四価の二重特異性抗体であって、ここで、ここで、ヒトc−Metに特異的に結合する前記抗原結合部位は、(例えばWO2009/007427に記載のように)c−Metの二量化を阻害する。
【0120】
そのため、この発明の別の態様は、以下の:
a)ヒトc−Metに特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ErbB−2に特異的に結合する2つの同一の一本鎖Fabフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0121】
そのため、この発明の別の態様は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する2つの同一の一本鎖Fabフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0122】
代表的な図式的構造は、図6aを参照のこと。
【0123】
そのため、この発明の別の態様は、以下の:
a)ErbB−2に特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する2つの同一の一本鎖Fvフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0124】
そのため、この発明の別の態様は、以下の:
a)ヒトc−Metに特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ErbB−2に特異的に結合する2つの同一の一本鎖Fvフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fvフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されている、四価の二重特異性抗体である。
【0125】
代表的な図式的構造は、図6bを参照のこと。
【0126】
好ましい一実施形態では、ヒトc−Met又はヒトErbB−2に結合する一本鎖Fab又はFvフラグメントが、前記全長抗体に、その全長抗体の重鎖のC末端のペプチドコネクタを介して融合されている。
【0127】
この発明の他の実施形態は、以下の:
a)ヒトErbB−2に特異的に結合し、且つ、以下の:
aa)N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成る2つの同一の抗体重鎖;及び
ab)N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成る2つの同一の抗体軽鎖(VL−CL);
から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する2つの一本鎖Fabフラグメント、
ここで、前記一本鎖Fabフラグメントが、抗体重鎖可変ドメイン(VH)と抗体定常ドメイン1(CH1)、抗体軽鎖可変ドメイン(VL)、抗体軽鎖定常ドメイン(CL)及びリンカーから成り、且つ、ここで、前記抗体ドメインと前記リンカーが、N末端からC末端に向かって、以下の順序:
ba)VH−CH1−リンカー−VL−CL、又は
bb)VL−CL−リンカー−VH−CH1、
のうちの一方を有し、
ここで、前記リンカーが少なくとも30個のアミノ酸、好ましくは32〜50個のアミノ酸から成るペプチドである、
を含んでなり;そして
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、その全長抗体の重鎖又は軽鎖ののC又はN末端のペプチドコネクタを介して融合されている;
ここで、前記ペプチドコネクタは、少なくとも5個のアミノ酸、好ましくは10〜50個のアミノ酸から成るペプチドである、四価の二重特異性抗体である。
【0128】
三価又は四価フォーマットのいずれかで使用される場合に、用語「全長抗体」は、2つの「全長抗体重鎖」と2つの「全長抗体軽鎖」から成る抗体を意味する(図1を参照のこと)。「全長抗体重鎖」は、N末端からC末端に向かって、抗体重鎖可変ドメイン(VH)、抗体定常重鎖ドメイン1(CH1)、抗体ヒンジ領域(HR)、抗体重鎖定常ドメイン2(CH2)、及び抗体重鎖定常ドメイン3(CH3)から成り、VH−CH1−HR−CH2−CH3と略記される、ポリペプチドであり、更に任意に、IgEサブクラスの抗体の場合には、抗体重鎖定常ドメイン4(CH4)から成るポリペプチドである。好ましくは「全長抗体重鎖」は、N末端からC末端に向かって、VH、CH1、HR、CH2及びCH3から成るポリペプチドである。「全長抗体軽鎖」は、N末端からC末端に向かって、抗体軽鎖可変ドメイン(VL)、及び抗体軽鎖定常ドメイン(CL)から成り、VL−CLと略記される、ポリペプチドである。抗体軽鎖定常ドメイン(CL)は、κ(カッパ)又はλ(ラムダ)であり得る。2つの全長抗体鎖は、CLドメインとCH1ドメインの間、及び全長抗体重鎖のヒンジ領域の間のポリペプチド間ジスルフィド結合を介して一緒につながっている。典型的な全長抗体の例は、IgG(例えば、IgG1及びIgG2)、IgM、IgA、IgD、並びにIgEのような天然抗体である。本発明による全長抗体は、単一種、例えばヒトからのものであることができ、又はそれらは、キメラ化若しくはヒト化抗体であることもできる。本発明による全長抗体は、一対のVHとVLによってそれぞれ形成された抗原結合部位を2つ含んでなるが、それらは、ともに同じ抗原に特異的に結合する。前記全長抗体の重鎖又は軽鎖のC末端は、前記重鎖又は軽鎖のC末端における最後のアミノ酸を意味する。前記全長抗体の重鎖又は軽鎖のN末端は、前記重鎖又は軽鎖のN末端における最後のアミノ酸を意味する。
【0129】
本発明の中で使用される場合、「ペプチドコネクタ」という用語は、アミノ酸配列を有するペプチドを意味し、それは、好ましくは合成起源のものである。本発明によるこれらのペプチドコネクタは、一本鎖Fabフラグメントを、全長抗体のC又はN末端に融合して、本発明による多特異性抗体を形成するのに使用される。好ましくは、前記b)のペプチドコネクタは、少なくとも5個のアミノ酸の長さ、好ましくは5〜100個、より好ましくは10〜50個のアミノ酸の長さを有するアミノ酸配列を持つペプチドである。一実施形態では、前記ペプチドコネクタは、(GxS)n又は(GxS)nGmであって、G=グリシン、S=セリン、及び(x=3、n=3、4、5若しくは6、及びm=0、1、2若しくは3)又は(x=4、n=2、3、4若しくは5及びm=0、1、2若しくは3)、好ましくはx=4及びn=2又は3、より好ましくはx=4、n=2を示す。好ましくは、三価の二重特異性抗体(ここで、VH若しくはVH−CH1ポリペプチドとVL若しくはVL−CLポリペプチド(図7a〜c)が全長抗体のC末端に2つの同一のペプチドコネクタを介して融合されている)において、前記ペプチドコネクタは、少なくとも25個のアミノ酸のペプチドであり、好ましくは30〜50個のアミノ酸のペプチド、そしてより好ましくは、前記ペプチドコネクタは、(GxS)n又は(GxS)nGmであって、G=グリシン、S=セリン、及び(x=3、n=6、7若しくは8、及びm=0、1、2若しくは3)又は(x=4、n=5、6、若しくは7及びm=0、1、2、若しくは3)、好ましくはx=4及びn=5、6、7を示す。
【0130】
「一本鎖Fabフラグメント」(図2aを参照のこと)は、抗体重鎖可変ドメイン(VH)、抗体定常ドメイン1(CH1)、抗体軽鎖可変ドメイン(VL)、抗体軽鎖定常ドメイン(CL)及びリンカーから成るポリペプチドであって、ここで、前記抗体ドメイン及び前記リンカーは、N末端からC末端に向かって、以下の順序:a)VH−CH1−リンカー−VL−CL、b)VL−CL−リンカー−VH−CH1、c)VH−CL−リンカー−VL−CH1又はd)VL−CH1−リンカー−VH−CLのうちのいずれか1つであり;且つ、ここで、前記リンカーは、少なくとも30個のアミノ酸、好ましくは32〜50個のアミノ酸から成るポリペプチドである。前記一本鎖Fabフラグメントa)VH−CH1−リンカー−VL−CL、b)VL−CL−リンカー−VH−CH1、c)VH−CL−リンカー−VL−CH1及びd)VL−CH1−リンカー−VH−CLは、CLドメインとCH1ドメインの間の天然のジスルフィド結合によって安定化される。「N末端」という用語は、N末端の最後のアミノ酸を意味し、「C末端」という用語は、C末端の最後のアミノ酸を意味する。
【0131】
「リンカー」という用語は、本発明の中で一本鎖Fabフラグメントに関して使用され、そして(好ましくは合成起源のものである)アミノ酸配列を持つペプチドを意味する。本発明によるこれらのペプチドは、a)VH−CH1をVL−CLと、b)VL−CLをVH−CH1と、c)VH−CLをVL−CH1と又はd)VL−CH1をVH−CLと連結して、以下の本発明による一本鎖Fabフラグメントa)VH−CH1−リンカー−VL−CL、b)VL−CL−リンカー−VH−CH1、c)VH−CL−リンカー−VL−CH1又はd)VL−CH1−リンカー−VH−CLを形成するのに使用される。一本鎖Fabフラグメントの中の前記リンカーは、少なくとも30個のアミノ酸の長さ、好ましくは32〜50個のアミノ酸の長さを有するアミノ酸配列を持つペプチドである。一実施形態では、前記リンカーは、(GxS)nであって、G=グリシン、S=セリン、(x=3、n=8、9若しくは10及びm=0、1、2若しくは3)又は(x=4、n=6、7若しくは8及びm=0、1、2若しくは3)を示し、好ましくはx=4、n=6若しくは7及びm=0、1、2若しくは3を示し、より好ましくはx=4、n=7及びm=2を示す。一実施形態では、前記リンカーは(G4S)6G2である。
【0132】
好ましい実施形態では、前記一本鎖Fabフラグメントの中の前記抗体ドメイン及び前記リンカーは、N末端からにC末端に向かって、以下の順序:a)VH−CH1−リンカー−VL−CL、又はb)VL−CL−リンカー−VH−CH1のいずれか一方であり、より好ましくはVL−CL−リンカー−VH−CH1である。
【0133】
別の好ましい実施形態では、前記一本鎖Fabフラグメントの中の前記抗体ドメイン及び前記リンカーは、N末端からC末端に向かって、以下の順序:a)VH−CL−リンカー−VL−CH1又はb)VL−CH1−リンカー−VH−CLのいずれか一方である。
【0134】
任意には、前記一本鎖Fabフラグメントにおいて、CLドメインとCH1ドメインの間の天然のジスルフィド結合に加えて、抗体重鎖可変ドメイン(VH)と抗体軽鎖可変ドメイン(VL)にも、以下の位置間のジスルフィド結合の導入によってジスルフィドにより安定化される:
i)重鎖可変ドメインの第44位〜軽鎖可変ドメインの第100位
ii)重鎖可変ドメインの第105位〜軽鎖可変ドメインの第43位、又は、
iii)重鎖可変ドメインの第101位〜軽鎖可変ドメインの第100位(付番は常にKabatのEUインデックスに準じる)。
【0135】
一本鎖Fabフラグメントのそういった更なるジスルフィドによる安定化は、その一本鎖Fabフラグメントの可変ドメインVHとVLの間のジスルフィド結合の導入によって達成される。一本鎖Fabフラグメントの安定化のために非天然ジスルフィド架橋を導入する技術は、例えば、WO94/029350、Rajagopal, et al., Prot Engin. (1997) 1453-59;Kobayashi, H., et al., Nuclear Medicine & Biology, 25, (1998) 387-393;又はSchmidt, M., et al., Oncogene 18 (1999) 1711-1721に記載されている。1つの実施形態では、本発明による抗体内に含まれている一本鎖Fabフラグメントの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第44位と軽鎖可変ドメインの第100位の間にある。1つの実施形態では、本発明による抗体内に含まれている一本鎖Fabフラグメントの可変ドメインの間の任意のジスルフィド結合は、重鎖可変ドメインの第105位と軽鎖可変ドメインの第43位の間にある(付番は常にKabatのEUインデックスに準じる)。
【0136】
一実施形態では、一本鎖Fabフラグメントの可変ドメインVHとVLの間の前記任意のジスルフィドによる安定化のない一本鎖Fabフラグメントが好まれる。
【0137】
「一本鎖Fvフラグメント」(図2bを参照のこと)は、抗体重鎖可変ドメイン(VH)、抗体軽鎖可変ドメイン(VL)、及び一本鎖Fvリンカーから成るポリペプチドであって、ここで、前記抗体ドメイン及び前記一本鎖Fvリンカーは、N末端からC末端に向かって、以下の順序:a)VH−一本鎖Fvリンカー−VL、b)VL−一本鎖Fvリンカー−VHであり;好ましくは、a)VH−一本鎖Fvリンカー−VLであり;且つ、ここで、前記一本鎖Fvリンカーは、少なくとも15個のアミノ酸の長さを有し、一実施形態では、少なくとも20個のアミノ酸を有するアミノ酸配列を持つポリペプチドである。「N末端」という用語は、N末端の最後のアミノ酸を意味し、「C末端」という用語は、C末端の最後のアミノ酸を意味する。
【0138】
一本鎖Fvフラグメントの中で使用される「一本鎖Fvリンカー」という用語は、(好ましくは合成起源のものである)アミノ酸配列を持つペプチドを意味する。前記一本鎖Fvリンカーは、少なくとも15個のアミノ酸の長さを有し、一実施形態では、少なくとも20個のアミノ酸の長さ、好ましくは15〜30個のアミノ酸の長さを有するアミノ酸配列を持つペプチドである。一実施形態では、前記一本鎖リンカーは、(GxS)nであって、G=グリシン、S=セリン、(x=3及びn=4、5若しくは6)又は(x=4及びn=3、4、5若しくは6)を示し、好ましくはx=4、n=3、4若しくは5を示し、より好ましくはx=4、n=3若しくは4を示す。一実施形態では、前記一本鎖Fvリンカーは(G4S)3又は(G4S)4である。
【0139】
更に、前記一本鎖Fvフラグメントは、好ましくは、ジスルフィドにより安定化されている。一本鎖抗体のこうした更なるジスルフィドによる安定化は、その一本鎖抗体の可変ドメインの間のジスルフィド結合の導入によって達成され、そしてそれは、例えば、WO 94/029350、Rajagopal, V., et al., Prot. Engin. 10 (1997) 1453-59;Kobayashi, R, et al., Nuclear Medicine & Biology 25 (1998) 387-393;又はSchmidt, M., et al., Oncogene 18 (1999) 1711-1721に記載されている。
【0140】
ジスルフィドにより安定化した一本鎖Fvフラグメントの一実施形態では、本発明による抗体に含まれている一本鎖Fvフラグメントの可変ドメインの間のジスルフィド結合は、各一本鎖Fvフラグメントごとに独立に、以下のものから選択される:
i)重鎖可変ドメインの第44位〜軽鎖可変ドメインの第100位
ii)重鎖可変ドメインの第105位〜軽鎖可変ドメインの第43位、又は、
iii)重鎖可変ドメインの第101位〜軽鎖可変ドメインの第100位。
【0141】
一実施形態では、本発明による抗体に含まれている一本鎖Fvフラグメントの可変ドメインの間のジスルフィド結合は、重鎖可変ドメインの第44位と軽鎖可変ドメインの第100位の間にある。
【0142】
本発明に記載の抗体は、リコンビナント手段により産生される。よって、本発明の1つ態様は、本発明による抗体をコードする核酸であり、そして更なる態様は、本発明による抗体をコードする前記核酸を含んでなる細胞である。リコンビナント産生のための方法は、当技術分野の現状において広く公知であり、抗体ポリペプチドのその後の単離及び通常は医薬として許容され得る純度への精製を伴う、原核細胞及び真核細胞におけるタンパク質の発現を含む。宿主細胞における前述の抗体の発現のために、個別に改変した軽鎖及び重鎖をコードする核酸が、標準的な方法により発現ベクターに挿入される。発現は、CHO細胞、NS0細胞、SP2/0細胞、HEK293細胞、COS細胞、PER.C6細胞、酵母、又はE.コリ(E. coli)細胞などの適切な原核又は真核宿主細胞において行われ、そして抗体は、その細胞から回収される(上清又は細胞溶解後に)。抗体のリコンビナント産生のための一般的な方法は、当技術分野の現状において周知であり、例えばMakrides, S.C., Protein Expr. Purif. 17(1999)183-202;Geisse, S., et al., Protein Expr. Purif. 8(1996)271-282;Kaufman, R.J., Mol. Biotechnol. 16(2000)151-160;Werner, R.G., Drug Res. 48(1998)870-880の総説に記載されている。
【0143】
二重特異性抗体は、例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、又はアフィニティークロマトグラフィーなどの通常の免疫グロブリン精製手順により培養液から適切に分離される。モノクローナル抗体をコードするDNA及びRNAは、通常の手順を使用して容易に単離及び配列決定される。ハイブリドーマ細胞は、そのようなDNA及びRNAの起源として役立つことができる。いったん単離されれば、DNAを発現ベクターに挿入し、次いでそれをHEK293細胞、CHO細胞、又は骨髄腫細胞などの宿主細胞にトランスフェクトして、この宿主細胞におけるリコンビナントモノクローナル抗体の合成を達成することができ、この宿主細胞は、さもなければ免疫グロブリンタンパク質を産生しない。
【0144】
二重特異性抗体のアミノ酸配列変異体(突然変異体)は、抗体のDNA内に適切なヌクレオチドの変更を導入することによって、又はヌクレオチド合成によって調製される。しかし、そのような改変は、例えば上記のように非常に限られた範囲内でのみ行うことができる。例えば、改変は、IgGアイソタイプ及びエピトープの結合などの前述の抗体の特徴を変化させるのではなく、リコンビナント産生の収率又はタンパク質の安定性を改善することがあり、精製を容易にすることもある。
【0145】
用語「宿主細胞」は、本願で使用される場合、本発明による抗体を産生するために設計され得るあらゆる種類の細胞系を意味する。1つの実施形態では、HEK293細胞及びCHO細胞が宿主細胞として使用される。本明細書において使用される場合、用語「細胞」、「細胞株」及び「細胞培地」とは、互換的に使用され、そしてすべてのそのような名称は子孫も包含する。従って、単語「形質転換体」及び「形質転換細胞」とは、トランスファーの数に関係なく、それらに由来する初代対象細胞及び培養物を包含する。すべての子孫は、故意の又は偶然な突然変異のために、DNA含有において正確には同一でないこともまた理解されている。元の形質転換された細胞についてスクリーンされる場合、同じ機能的又は生物学的活性を有する変異体子孫が包含される。
【0146】
NS0細胞における発現は、例えばBarnes, L.M., et al., Cytotechnology 32(2000)109-123;Barnes, L.M., et al., Biotech. Bioeng. 73(2001)261-270により記載されている。一過性発現が、例えばDurocher, Y., et al., Nucl. Acids. Res. 30(2002)E9により記載される。可変ドメインのクローニングは、Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86(1989)3833-3837;Carter, P., et al., Proc. Natl. Acad. Sci. USA 89(1992)4285-4289;及びNorderhaug, L., et al., J. Immunol. Methods 204(1997)77-87により記載されている。好ましい一過性発現システム(HEK293)は、Schlaeger, E.-J., and Christensen, K., in Cytotechnology 30(1999)71-83 及びSchlaeger, E.-J., in J. Immunol. Methods 194(1996)191-199により記載される。
【0147】
原核生物のために適切である制御配列には、例えばプロモーター、任意にはオペレーター配列及びリボソームに結合部位が含まれる。真核細胞は、プロモーター、エンハンサー及びポリアデニル化シグナルを使用することが知られている。
【0148】
核酸は、それが他の核酸配列と機能的な関連性をもって配置されるとき、「操作可能な形で連結され」ている。例えば、プレ配列又は分泌リーダーのDNAは、それがポリペプチドの分泌に関与するプレタンパク質として発現されるとき、ポリペプチドに関してDNAと操作可能な形で連結している;プロモーター又はエンハンサーは、それがその配列の転写に影響するとき、コード配列と操作可能な形で連結されている;又は、リボソーム結合部位は、それが転写を促進するように配置されているとき、コード配列に操作可能な形で連結されている。一般的に、「操作可能な形で連結された」は、連結されたDNA配列が隣接し、そして、分泌リーダーの場合は、隣接し、且つ、読み枠内にあることを意味する。しかしながら、エンハンサーは隣接している必要は無い。連結は、都合の良い制限部位でライゲーションを行うことにより達成される。そのような部位が存在しなければ、確立された手段に従い、合成オリゴヌクレオチドアダプター又はリンカーが使用される。
【0149】
抗体の精製は、アルカリ/SDS処理、CsClバンディング、カラムクロマトグラフィー、アガロースゲル電気泳動、及び当該技術分野において周知な他の方法を含む標準的な技術により、他の細胞成分又は他の汚染物、例えば、他の細胞核酸又はタンパク質を除去するために実行される。Ausubel, F., et al. (ed.), Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York(1987)を参照のこと。別の方法、例えば、微生物タンパク質を用いた親和性クロマトグラフィー(例えば、プロテインA又はプロテインG親和性クロマトグラフィー)、イオン交換クロマトグラフィー(例えば、カチオン交換(カルボキシメチル樹脂)、陰イオン交換性(アミノエチル樹脂)及び混合モード交換)、thiophilic吸着(例えば、β−メルカプトエタノールと他のSHリガンド類を用いて)、疎水性相互作用又は芳香族吸着クロマトグラフィー(例えば、フェニル−セファロース、アザ−アレノフィリック樹脂、又はm−アミノフェニルボロン酸を用いて)、金属キレート親和性クロマトグラフィー(例えば、ニッケル(II)及びCu(II)親和性材料を用いて)、サイズ排除クロマトグラフィー、及び電気泳動法(ゲル電気泳動、毛細管電気泳動など)などが、確立されており、そしてタンパク質精製のために幅広く使用されている(Vijayalakshmi, M.A., Appl. Biochem. Biotech. 75 (1998) 93-102)。
【0150】
本明細書において使用される場合、用語「細胞」、「細胞株」及び「細胞培地」とは、互換的に使用され、そしてすべてのそのような名称は子孫も包含する。従って、単語「形質転換体」及び「形質転換細胞」とは、トランスファーの数に関係なく、それらに由来する初代対象細胞及び培養物を包含する。すべての子孫は、故意の又は偶然な突然変異のために、DNA含有において正確には同一でないこともまた理解されている。元の形質転換された細胞についてスクリーンされる場合、同じ機能的又は生物学的活性を有する変異体子孫が包含される。異なった命名法が意図される場合は、その内容から明白であろう。
【0151】
用語「形質転換」とは、本明細書において使用される場合、宿主細胞中へのベクター/核酸のトランスファー工程を指す。強い細胞壁バリアを有さない細胞が宿主細胞として使用される場合、トランスフェクションが、例えばGraham, F.L., and van der Ed, A.J. Virology 52(1978)456-467により記載されるようなリン酸カルシウム沈殿方法により実施される。しかしながら、核注入又はプロトプラスト融合による細胞中へのDNAの導入のための他の方法がまた使用され得る。原核細胞、又は実質的な細胞壁構造を含む細胞が使用される場合、1つのトランスフェクション方法は、Cohen, S.N, et al, PNAS 69(1972)2110-2114により記載されるような、塩化カルシウムを用いるカルシウム処理法である。
【0152】
本明細書において使用される場合、「発現」とは、核酸がmRNAに転換される方法、及び/又は転写されたmRNA(転写体とも呼ばれる)がペプチド、ポリペプチド又はタンパク質に連続的に翻訳される過程を指す。転写体及びコードされるポリペプチドは、集合的には、遺伝子生成物とも呼ばれる。ポリヌクレオチドがゲノムDNAに由来する場合、真核細胞における発現は、mRNAのスプライシングを含む。
【0153】
「ベクター」とは、挿入される核酸分子を、宿主細胞中に及び/又は宿主細胞間にトランスファーする、核酸分子、特に自己複製する核酸分子である。この用語は、DNAの細胞中への挿入(例えば、染色体組込み)のために主に機能するベクター、DNA又はRNAの複製のために主に機能するベクターの複製、及びDNA又はRNAの転写及び/又は翻訳のために機能する発現ベクターを包含する。記載されるような機能のうちの2つ以上を提供するベクターもまた包含される。
【0154】
「発現ベクター」とは、適切な宿主細胞中に導入される場合、転写され、そしてポリペプチドに翻訳され得るポリヌクレオチドである。「発現システム」とは、所望する発現生成物を生成するために機能することができる発現ベクターを含んでなる適切な宿主細胞を通常指す。
【0155】
医薬組成物
本発明の1つの態様は、本発明による抗体を含んでなる医薬組成物である。本発明の別の態様は、医薬組成物の製造のための本発明による抗体の使用である。本発明の更なる態様は、本発明による抗体を含んでなる医薬組成物の製造のための方法である。別の態様では、本発明は、医薬担体と一緒に処方される本発明による抗体を含んでなる組成物、例えば医薬組成物を提供する。
【0156】
本発明の1つの態様は、癌治療のための本発明による二重特異性抗体である。
【0157】
本発明の別の態様は、癌治療のための前記医薬組成物である。
【0158】
本発明の別の態様は、癌処置用の治療薬の製造のための本発明による抗体の使用である。
【0159】
本発明の別の態様は、本発明による抗体をそういった処置を必要としている患者に投与することによる、癌に罹患している患者の処置方法である。
【0160】
本明細書中で使用されるとき、「医薬担体」には、生理学的に適合する任意の及びすべての溶媒、分散媒、被覆剤、抗細菌剤及び抗真菌剤、等張剤及び吸収遅延剤などが含まれる。好ましくは、前記担体は、(例えば注射又は点滴による)静脈内投与、筋肉内投与、皮下投与、非経口投与、脊髄投与又は上皮投与に適切である。
【0161】
本発明の組成物は、当該技術分野において知られる様々な方法により投与され得る。当業者に認識されるように、投与の経路及び/又は様式は、所望される結果に依存して異なる。ある投与径路により本発明の組成物を投与するためには、その組成物を、その不活性化を防止する材料で被覆し、又はその組成物と共に同時に投与する必要がある。例えば、前記組成物は、例えばリポソーム、又は希釈剤などの適切な担体に加えられて対象に投与され得る。医薬として許容される希釈剤には、生理食塩水及び水性緩衝溶液が含まれる。医薬として許容される担体には、滅菌水性溶液又は分散剤、滅菌済みの注射可能な溶液又は分散剤を即時調製するための滅菌粉末が含まれる。そのような医薬活性物質のための媒体及び薬剤は、当該技術分野において知られている。
【0162】
「非経口投与」及び「非経口的に投与される」という語句は、本明細書中で使用される場合、腸内投与及び局所投与以外の投与様式を意味し、通常は注射により、これだけに限定されるものではないが、静脈内、筋肉内、動脈内、鞘内、嚢内、眼窩内、心臓内、皮内、腹腔内、経気管、皮下、表皮下、関節内、皮下、くも膜下、髄腔内、硬膜外及び胸骨内注射及び点滴を含む。
【0163】
用語「癌」は、本明細書中で使用される場合、例えばリンパ腫、リンパ球性白血病、肺癌、非小細胞肺(NSCL)癌、細気管支肺胞細胞性肺癌、骨癌、膵臓癌、皮膚癌、頭頚部の癌、皮膚又は眼内の黒色腫、子宮癌、卵巣癌、直腸癌、肛門部の癌、胃癌、胃せん癌、結腸癌、乳癌、子宮癌、卵管の癌腫、子宮内膜の癌腫、子宮頚部の癌腫、膣癌、外陰部癌、ホジキン病、食道癌、小腸癌、内分泌系の癌、甲状腺癌、副甲状腺癌、副腎癌、軟組織の肉腫、尿道の癌、陰茎の癌、前立腺癌、膀胱癌、腎臓又は輸尿管の癌、腎細胞癌、腎盂癌、中皮腫、肝細胞性癌、胆管癌、中枢神経系(CNS)の新生物、脊椎軸腫瘍、脳幹膠腫、多形膠芽腫、星状細胞腫、神経鞘腫、脳室上衣腫、髄芽腫、髄腹腫、扁平上皮癌、脳下垂体腺腫及びユーイング肉腫、そして前記癌のうちのいずれかの難治性バーション、又は前記癌のうちの1つ若しくは複数の組合せを含めたものであり得る。
【0164】
本発明の別の態様は、血管新生阻害剤としての本発明による二重特異性抗体又は前記医薬組成物である。そのような血管新生阻害剤は、癌、特に固形腫瘍、及び他の血管疾患の治療に使用できる。
【0165】
本発明の1つの態様は、血管疾患の治療のための本発明による二重特異性抗体である。
【0166】
本発明の別の態様は、血管疾患処置用の治療薬の製造のための本発明による抗体の使用である。
【0167】
本発明の別の態様は、本発明による抗体をそういった処置を必要としている患者に投与することによる、血管疾患に罹患している患者の処置方法である。
【0168】
「血管疾患」という用語には、癌、炎症性疾患、アテローム性動脈硬化症、虚血、外傷、敗血症、COPD、喘息、糖尿病、AMD、網膜障害、脳卒中、肥満症、急性肺傷害、出血、血管漏出、例えばサイトカイン誘発性血管漏出、アレルギー、グレーブス病、橋本の自己免疫性甲状腺炎、特発性血小板減少性紫斑病、巨細胞性動脈炎、関節リウマチ、全身性エリテマトーデス(SLE)、ループス腎炎、クローン病、多発性硬化症、潰瘍性大腸炎、特に固形腫瘍、眼内の新生血管症候群、例えば増殖性網膜症又は加齢性黄斑変性症(AMD)など、関節リウマチ、及び乾癬が含まれる(Folkman, J., et al., J. Biol. Chem. 267 (1992) 10931-10934;Klagsbrun, et al., Annu. Rev. Physiol. 53 (1991) 217-239;及びGarner, A., Vascular diseases, In: Pathobiology of ocular disease, A dynamic approach, Garner, A., and Klintworth, G. K., (eds.), 2nd edition, Marcel Dekker, New York (1994) 1625-1710)。
【0169】
これらの組成物は更に、保存料、湿潤剤、乳化剤及び分散剤などの補助剤を含むこともできる。微生物の存在の予防は、前掲の殺菌手段によって、及び様々な抗菌及び抗真菌剤、例えばパラベン、クロロブタノール、フェノール、ソルビン酸などの含有によって確保できる。組成物中に等張剤、例えば糖、塩化ナトリウムなどを含むことが望ましいこともある。加えて、注射用医薬形態の持続的吸収は、モノステアリン酸アルミニウムやゼラチンなどの吸収を遅らせる作用物質の含有によって引き起こすこともできる。
【0170】
選択した投与経路にかかわらず、(好適な水和形態で使用することもできる)本発明の化合物、及び/又は本発明の医薬組成物を、当業者に知られている従来の方法によって医薬的に許容し得る剤形に処方する。
【0171】
本発明の医薬組成物中の有効成分の実際の投与量レベルは、特定の患者、組成物、及び投与様式について、その患者にとって有毒であることなしに、所望の治療応答を達成するのに有効である有効成分量を得るように変更することもできる。選択される投与量レベルは、用いられる本発明の特定の組成物の活性、、投与経路、投与時間、用いられる特定の化合物の排泄速度、処置期間、用いられる特定の組成物と組合せて使用されるその他の薬物、化合物及び/又は材料、処置される患者の年齢、性別、体重、身体状態、健康全般、及び前病歴、並びに医療分野で周知の要因など、を含めた様々な薬物動態学的要因に依存するであろう。
【0172】
組成物は、それがシリンジによって提出できる程度に無菌であり、且つ、液状でなければならない。水に加えて、好ましくは、担体は等張緩衝生理食塩溶液である。
【0173】
適切な流動性は、例えば、レシチンなどのコーティングの使用によって、分散液の場合には所望の粒度の維持によって、そして界面活性剤の使用によって、維持できる。多くの場合、組成物中に等張剤、例えば、糖、マンニトールやソルビトールなどのポリアルコール、及び塩化ナトリウムを含むことが望ましい。
【0174】
ヒトErbB−2及びヒトc−Metに対するこの発明による二重特異性抗体が、価値のある特徴、例えば生物学的又は薬理学的な活性などを有することがここでわかった。
【0175】
以下の実施例、配列表及び図面は、本発明の理解を助けるために提供されるが、本発明の真の範囲は添付の特許請求の範囲に示される。本発明の要旨から逸脱することなしに、示された手順に改変を加えることができると理解されている。
【0176】
アミノ酸配列の説明
配列番号1 重鎖可変ドメイン<ErbB−2>トラスツズマブ
配列番号2 軽鎖可変ドメイン<ErbB−2>トラスツズマブ
配列番号3 重鎖可変ドメイン<c−Met>Mab 5D5
配列番号4 軽鎖可変ドメイン<c−Met>Mab 5D5
配列番号5 重鎖<c−Met>Mab 5D5
配列番号6 軽鎖<c−Met>Mab 5D5
配列番号7 重鎖<c−Met>Fab 5D5
配列番号8 軽鎖<c−Met>Fab 5D5
配列番号9 ヒトIgG1の重鎖定常領域
配列番号10 ヒトIgG3の重鎖定常領域
配列番号11 ヒト軽鎖カッパ定常領域
配列番号12 ヒト軽鎖ラムダ定常領域
配列番号13 ヒトc−Met
配列番号14 ヒトErbB−2
配列番号15 重鎖CDR3H、<ErbB−2>トラスツズマブ
配列番号16 重鎖CDR2H、<ErbB−2>トラスツズマブ
配列番号17 重鎖CDR1H、<ErbB−2>トラスツズマブ
配列番号18 軽鎖CDR3L、<ErbB−2>トラスツズマブ
配列番号19 軽鎖CDR2L、<ErbB−2>トラスツズマブ
配列番号20 軽鎖CDR1L、<ErbB−2>トラスツズマブ
配列番号21 重鎖CDR3H、<c−Met>Mab 5D5
配列番号22 重鎖CDR2H、<c−Met>Mab 5D5
配列番号23 重鎖CDR1H、<c−Met>Mab 5D5
配列番号24 軽鎖CDR3L、<c−Met>Mab 5D5
配列番号25 軽鎖CDR2L、<c−Met>Mab 5D5
配列番号26 軽鎖CDR1L、<c−Met>Mab 5D5
【図面の簡単な説明】
【0177】
【図1】典型的な順序で可変ドメインと定常ドメインを含んでなる2対の重鎖と軽鎖を持ち、第1の抗原1に特異的に結合するCH4ドメインを含まない全長抗体の図式的構造。
【図2a】以下の:a)ヒトErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びにb)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖を含んでなり、ここで、前記可変ドメインVL及びVH、及び/又は前記定常ドメインCL及びCH1が互いによって置き換えられ、それらがノブ−into−ホール技術で修飾されている、二価の二重特異性<ErbB−2/c−Met>抗体の図式的構造。
【図2b】以下の:a)ヒトErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びにb)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖を含んでなり、ここで、前記可変ドメインVL及びVH、及び/又は前記定常ドメインCL及びCH1が互いによって置き換えられ、それらがノブ−into−ホール技術で修飾されている、二価の二重特異性<ErbB−2/c−Met>抗体の図式的構造。
【図2c】以下の:a)ヒトErbB−2に特異的に結合する全長抗体の軽鎖及び重鎖;並びにb)ヒトc−Metに特異的に結合する全長抗体の軽鎖及び重鎖を含んでなり、ここで、前記可変ドメインVL及びVH、及び/又は前記定常ドメインCL及びCH1が互いによって置き換えられ、それらがノブ−into−ホール技術で修飾されている、二価の二重特異性<ErbB−2/c−Met>抗体の図式的構造。
【図3a】a)図3a:2つのポリペプチドVHとVLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する); b)図3b:2つのポリペプチドVH−CH1とVL−CLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性<ErbB−2/c−Met>抗体の略図。
【図3b】a)図3a:2つのポリペプチドVHとVLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する); b)図3b:2つのポリペプチドVH−CH1とVL−CLがそれに融合されている(VHドメインとVLドメインは両方が一緒になって、c−Metに特異的に結合する抗原結合部位を形成する)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性<ErbB−2/c−Met>抗体の略図。
【図3c】図3c:「ノブとホール」を備えた2つのポリペプチドVHとVLがそこに融合されている(VHドメインとVLドメインは両方が一緒になってc−Metに特異的に結合する抗原結合部位を形成する)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性抗体の略図。
【図3d】図3d:「ノブとホール」を備えた2つのポリペプチドVHとVLがそれに融合されている(VHドメインとVLドメインは両方が一緒になってc−Metに特異的に結合する抗原結合部位を形成する、ここで、それらのVHドメイン及びVLドメインはVH44位とVL100位の間に鎖間ジスルフィド架橋を含んでなる)、ErbB−2に特異的に結合する全長抗体を含んでなる本発明による三価の二重特異性抗体の略図。
【図4a】4a:4種類の考えられる一本鎖Fabフラグメントの図式的構造。
【図4b】4b:2種類の一本鎖Fvフラグメントの図式的構造。
【図5a】1つの一本鎖Fabフラグメント(図5a)又は1つの一本鎖Fvフラグメント(図5b)と全長抗体を含んでなる三価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−ノブとホールを備えた二重特異性三価の例。
【図5b】1つの一本鎖Fabフラグメント(図5a)又は1つの一本鎖Fvフラグメント(図5b)と全長抗体を含んでなる三価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−ノブとホールを備えた二重特異性三価の例。
【図6a】2つの一本鎖がFabフラグメント(図6a)又は2つの一本鎖Fvフラグメント(図6b)と全長抗体を含んでなる四価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−c−Met結合部位はc−Met二量化阻害抗体に由来する。
【図6b】2つの一本鎖がFabフラグメント(図6a)又は2つの一本鎖Fvフラグメント(図6b)と全長抗体を含んでなる四価の二重特異性<ErbB−2/c−Met>抗体の図式的構造−c−Met結合部位はc−Met二量化阻害抗体に由来する。
【図7a】類上皮腫癌細胞株A431におけるErbB2/2/3及びc−Metの細胞表面発現のフローサイトメトリー分析。
【図7b】卵巣癌細胞株OVCAR−8におけるErbB2/2/3及びc−Metの細胞表面発現のフローサイトメトリー分析。
【図8】0、30、60及び120分(=0、0.5、1、及び2時間)に計測したOVCAR−8癌細胞における内部移行アッセイ。
【図9a】OVCAR−8癌細胞における増殖アッセイ。単一特異性の親<HER2>及び<c−Met>抗体と比較した、本発明による二重特異性<HER2/c−Met>抗体BsAB02(BsAb)の癌細胞増殖の阻害。
【図9b】HGFの存在下での癌細胞株Ovcar−8における増殖アッセイ。単一特異性の親<HER2>及び<c−Met>抗体と比較した、本発明による二重特異性<HER2/c−Met>抗体BsAB02(BsAb)の癌細胞増殖の阻害。
【実施例】
【0178】
実験手順
材料と方法
組換えDNA技術
Sambrook, J. et al., Molecular cloning: A laboratory manual;Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989に記載の標準的な方法を、DNAを操作するために使用した。分子生物学的試薬を製造業者の取扱説明書に従って使用した。
【0179】
DNAとタンパク質の配列分析、及び配列データ・マネジメント
以下でヒト免疫グロブリン軽鎖及び重鎖のヌクレオチド配列に関する概説は:Kabat, E.A. et al.,(1991)Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242にある。抗体鎖のアミノ酸には、EUナンバリングに従って付番した(Edelman, G.M., et al., PNAS 63(1969)78-85;Kabat, E.A., et al.,(1991)Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242)。GCG(Genetics Computer Group、Madison, Wisconsin)のソフトウェアパッケージ バージョン10.2及びInfomaxのVector NTI Advance suiteバージョン8.0を、配列新規作成、マッピング、分析、アノテーション及び図示に使用した。
【0180】
DNA配列決定
DNA配列を、SequiServe(Vaterstetten, Germany)及びGeneart AG(Regensburg, Germany)にて実施された二本鎖配列決定法によって決定した。
【0181】
遺伝子合成
所望の遺伝子セグメントは、自動化遺伝子合成によって合成オリゴヌクレオチドとPCR産物からGeneart AG(Regensburg, Germany)で調製された。1つの制限エンドヌクレアーゼ切断部位に隣接した遺伝子セグメントを、pGA18(ampR)プラスミド内にクローン化した。プラスミドDNAを形質転換細菌から精製し、そしてUV分光法によって濃度決定した。サブクローニングした遺伝子フラグメントのDNA配列を、DNA配列決定法で確認した。同じように、ペプチドコネクタで連結されているC末端<c−Met>5D5 scFab VH領域を持つ/持たないCH3ドメイン内にS354C及びT366Wが突然変異を保有する改変「ノブ−into−ホール」<ErbB−2>抗体重鎖、並びにペプチドコネクタで連結されているC末端<c−Met>5D5 scFab VL領域を持つ/持たない、Y349C、T366S、L368A、及びY407V突然変異を保有する「ノブ−into−ホール」<ErbB−2>抗体重鎖をコードするDNA配列を、隣接するBamHI及びXbaI制限部位を用いて遺伝子合成によって調製した。最後に、<ErbB−2>抗体及び<c−Met>5D5抗体の無改変重鎖及び軽鎖をコードするDNA配列を、隣接するBamHI及びXbaI制限部位によって合成した。すべての構築物をリーダーペプチド(MGWSCIILFLVATATGVHS)をコードする5’末端DNA配列を用いて設計し、そして真核細胞内での分泌のためのタンパク質を目的とした。
【0182】
発現プラスミドの構築
Roche発現ベクターを、すべての重及び軽鎖scFv融合タンパク質をコードする発現プラスミドの構築に使用した。ベクターは、以下の要素:
− 選択マーカーとしてのハイグロマイシン耐性遺伝子、
− エプスタイン−バールウイルス(EBV)の複製開始点、oriP、
− E.コリ内でのこのプラスミドの複製を可能にするベクターpUC18からの複製開始点、
− E.コリ内でアンピシリン耐性を付与するβ−ラクタマーゼ遺伝子、
− ヒト・サイトメガロウイルス(HCMV)からの前初期エンハンサー及びプロモーター
− ヒト1−免疫グロブリン・ポリアデニル化(「ポリA」)シグナル配列、及び
− 固有のBamHI及びXbaI制限部位、
から構成される。
【0183】
記載したように、重鎖又は軽鎖構築物、並びにC末端VH及びVLドメインを持つ「ノブ−into−ホール」構築物を含んでなる免疫グロブリン融合遺伝子を、遺伝子合成によって調製し、そしてpGA18(ampR)プラスミド内にクローン化した。合成DNAセグメントとRoche発現ベクターを保有するpG18(ampR)プラスミドを、BamHI及びXbaI制限酵素(Roche Molecular Biochemicals)によって消化し、そしてアガロースゲル電気泳動法にかけた。次いで、精製した、重鎖又は軽鎖をコードするDNAセグメントを、単離したRoche発現ベクターBamHI/XbaI断片に連結して、最終的な発現ベクターを得た。最終的な発現ベクターをE.コリ細胞内に形質転換し、発現プラスミドDNAを単離し(Miniprep)、そして制限酵素分析とDNA配列決定にかけた。正しいクローンを、150mlのLB−Amp培地中で培養し、再びプラスミドDNAを単離し(Maxiprep)、そして配列完全性をDNA配列決定によって確認した。
【0184】
HEK293細胞における免疫グロブリン変異体の一過性発現
遺伝子組み換え免疫グロブリン変異体を、製造業者の取扱説明書(Invitrogen, USA)に従ってFreeStyle(商標)293発現系を使用したヒト胎児腎臓293−F細胞の一過性トランスフェクションによって発現した。簡単に言えば、懸濁液FreeStyle(商標)293−F細胞を、37℃/8%のCO2にてFreeStyle(商標)293発現培地中で培養し、そしてその細胞をトランスフェクションの日に1〜2×l06生存細胞/mlの密度で新しい培地中に植え付けた。DNA−293fectin(商標)複合体を、250mlの最終的なトランスフェクション体積に対して1:1モル比で、325μlの293fectin(商標)(Invitrogen, Germany)と250μgの重鎖又は軽鎖プラスミドDNAを使用したOpti-MEM(登録商標)I培地(Invitrogen, USA)中で調製した。「ノブ−into−ホール」DNA−293fectin複合体を、250mlの最終的なトランスフェクション体積に対して1:1:2モル比で、325μlの293fectin(商標)(Invitrogen, Germany)と、250μgの「ノブ−into−ホール」重鎖1及び2と、軽鎖プラスミドDNAを使用したOpti-MEM(登録商標)I培地(Invitrogen, USA)中で調製した。抗体を含有している細胞培養上清を、14000gで30分間の遠心分離及び滅菌濾紙(0.22μm)を通して濾過してトランスフェクションの7日後に採取した。上清を−20℃にて精製まで保存した。
【0185】
二重特異性抗体と対照抗体の精製
三価の二重特異性抗体及び対照抗体を、Protein A-Sepharose(商標)(GE Healthcare、Sweden)を使用した親和性クロマトグラフィーとSuperdex200サイズ排除クロマトグラフィーによって細胞培養上清から精製した。簡単に言えば、滅菌濾過した細胞培養上清を、PBSバッファー(10mMのNa2HPO4、1mMのKH2PO4、137mMのNaCl及び2.7mMのKCl、pH、7.4)で平衡化したHiTrap ProteinA HP(5ml)カラムにかけた。非結合タンパク質を平衡化バッファーで洗い落とした。抗体と抗体変異体を、0.1Mのクエン酸バッファー、pH2.8で溶出し、そしてタンパク質含有画分を0.1mlの1M Tris(pH8.5)で中和した。次いで、溶出したタンパク質画分を貯留し、Amiconの超遠心濾過機デバイス(MWCO:30K、Millipore)を用いて3mlの体積まで濃縮し、20mMのヒスチジン、140mMのNaCl、pH6.0で平衡化したSuperdex200 HiLoad 120ml 16/60ゲル濾過カラム(GE Healthcare、Sweden)に投入した。高分子量集合体が5%未満しかない、精製された二重特異性抗体と対照抗体を含有する画分を貯留し、そして1.0mg/mlのアリコートとして−80℃にて保存した。Fabフラグメントを、精製した5D5モノクローナル抗体のパパイン消化と、それに続くプロテインAクロマトグラフィーによる夾雑Fcドメインの除去によって作り出した。非結合Fabフラグメントを、20mMのヒスチジン、140mMのNaCl、pH6.0で平衡化したSuperdex200 HiLoad 120ml 16/60ゲル濾過カラム(GE Healthcare、Sweden)で更に精製し、貯留し、そして1.0mg/mlのアリコートとして−80℃にて保存した。
【0186】
精製タンパク質の分析
精製タンパク質サンプルのタンパク質濃度を、アミノ酸配列に基づいて算出したモル吸光係数を使用し、280nmにて吸光度(OD)を計測することによって測定した。二重特異性抗体及び対照抗体の純度と分子量を、還元剤(5mMの1,4−ジチオスレイトール)の存在下と不存在下でのSDS−PAGE分析、そしてクーマシーブリリアントブルーでの染色によって分析した。NuPAGE(登録商標)Pre-Castゲルシステム(Invitrogen, USA)を、製造業者の取扱説明書に従って使用した(4−20%のトリスグリシン・ゲル)。二重特異性抗体及び対照抗体サンプルの凝集体量を、200mMのKH2PO4、250mMのKCl、pH7.0の泳動バッファー中、25℃にてSuperdex200分析用サイズ排除カラム(GE Healthcare、Sweden)を使用した高性能SECによって分析した。25μgのタンパク質を、0.5ml/分の流量にてカラム上に注入し、そして50分間かけて定組成で溶出した。安定性分析のために、1mg/mlの濃度の精製タンパク質を、4℃と40℃にて7日間インキュベートし、その後、高性能SECによって評価した。還元した二重特異性抗体軽鎖及び重鎖のアミノ酸骨格の完全性を、ペプチド−N−グリコシダーゼF(Roche Molecular Biochemicals)での酵素的処理によるN−グリカンの除去後のNanoElectrospray Q-TOF質量分析によって確認した。
【0187】
c−Metリン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のA549細胞を、0.5%のFCS(ウシ胎仔血清)を含むRPMI中、HGF刺激の前日に植え付けた。翌日に、成長培地を、0.2%のBSA(ウシ血清アルブミン)を含むRPMIで一時間、置き換えた。次いで、5μg/mLの二重特異性抗体を培地に加え、そして細胞を、HGFを加えるまで10分間インキュベートし、50ng/mlの終濃度で更に10分間インキュベートした。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを、100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解した。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進めた。タンパク質濃度を、BCA方法(Pierce)を使用して測定した。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移した。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Y1230、1234、1235(44-888、Biosource)に対するリン酸化部位特異的c−Met抗体を用いて現像した。免疫ブロットを、非リン酸化c−Metに結合する抗体(AF276、R&D)で再検出した。
【0188】
ErbB2/Her2リン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のSk−Br3細胞を、10%のFCS(ウシ胎仔血清)を含むRPMI中、抗体追加の前日に植え付ける。翌日に、5μg/mLの対照又は二重特異性抗体を培地に加え、そして細胞を、更に1時間インキュベートする。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解する。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進める。タンパク質濃度を、BCA方法(Pierce)を使用して測定する。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移す。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Y1221/22(Cell Signaling、2243)に対するリン酸化部位特異的Her2抗体を用いて現像する。免疫ブロットを、非リン酸化Her2に結合する抗体(Cell Signaling、2165)で再検出する。
【0189】
AKTリン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のA431細胞を、10%のFCS(ウシ胎仔血清)を含むRPMI中、抗体追加の前日に植え付ける。翌日に、5μg/mLの対照又は二重特異性抗体を培地に加え、そして細胞を、更に1時間インキュベートする。次いで、一部の細胞を、25ng/mLのHGF(R&D、294-HGN)で更に15分間刺激する。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解する。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進める。タンパク質濃度を、BCA方法(Pierce)を使用して測定する。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移す。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Thr308(Cell signaling、9275)に対するリン酸化部位特異的AKT抗体を用いて現像する。免疫ブロットを、アクチンに結合する抗体(Abcam、ab20272)で再検出する。
【0190】
ERK1/2リン酸化アッセイ
6ウェルプレートの1ウェルあたり5x10e5個のA431細胞を、10%のFCS(ウシ胎仔血清)を含むRPMI中、抗体追加の前日に植え付ける。翌日に、5μg/mLの対照又は二重特異性抗体を培地に加え、そして細胞を、更に1時間インキュベートする。次いで、一部の細胞を、25ng/mLのHGF(R&D、294-HGN)で更に15分間刺激する。それらを氷上に置いてすぐに、細胞を1mMのバナジウム酸ナトリウムを含有する氷冷PBSで一度洗浄し、そしてそれらを100μLの溶解バッファー(50mMのTris−Cl pH7.5、150mMのNaCl、1%のNP40、0.5%のDOC、アプロチニン、0.5mMのPMSF、1mMのバナジウム酸ナトリウム)を用いて細胞培養プレート内で溶解する。細胞溶解物をエッペンドルフチューブに移し、そして溶解を氷上で30分間進める。タンパク質濃度を、BCA方法(Pierce)を使用して測定する。30〜50μgの溶解物を、4−12%のBis-Tris NuPageゲル(Invitrogen)により分離し、そしてゲル上のタンパク質をニトロセルロース膜に移す。膜を、5%のBSAを含有するTBS−Tで一時間ブロッキングし、そして製造業者の取扱説明書に従って、Thr202/Tyr204(CellSignaling、Nr.9106)に対するリン酸化部位特異的Erk1/2抗体を用いて現像する。免疫ブロットを、アクチンに結合する抗体(Abcam、ab20272)で再検出する。
【0191】
細胞−細胞播種(dissemination)(分散アッセイ)
化合物処置の前日に、A549(1ウェルあたり4000個の細胞)又はA431(1ウェルあたり8000個の細胞)を、0.5%のFCSを含むRPMI中、96ウェルのE-Plate(Roche、05232368001)内の200μLの全容量の中に植え付けた。接着と細胞成長を、電気抵抗を観察する15分毎のスイープによりReal Time Cell Analyzer機器で一晩観察した。翌日、細胞を、5分毎にスイープしながらPBS中のそれぞれの抗体希釈物5μLと共にプレインキュベートした。30分後に、20ng/mLの終濃度をもたらす2.5μLのHGF溶液を加え、そして実験を更に72時間続けた。急激な変化を、180分間の1分毎のスイープで観察し、続いて残りの期間、15分毎のスイープで観察した。
【0192】
HUVEC増殖検定
HUVEC細胞(Promocell、C-12200)を、0.5% FCS含有EBM−2培地(Promocell、C-22211)中でコラーゲンコートした96ウェル内に植え付ける。翌日、対照又は二重特異性抗体の希釈系列をその細胞に加える。30分のインキュベーションの後に、25ng/mLのHGF(R&D、294-HGN)を加え、そして細胞をさらに72時間インキュベートした後に、cell titer glowアッセイ(Promega、G7571/2/3)を用いて、ATP量の形で細胞増殖を製造業者の助言に従って測定する。
【0193】
Sk−Br3増殖アッセイ
a)増殖研究のために、96ウェル細胞培養プレートの1ウェルあたり10000個の細胞を、血清削減培地(RPMI 1640+4%のPCS)中で植え付けた。翌日、親Her2又はc−Met抗体、並びに二重特異性抗体を加え、そして細胞をさらに48時間培養した後に、細胞増殖の指標としてATPを、cell titer glowアッセイ(Promega)を用いて測定する。
【0194】
b)HGF存在下での増殖研究のために、96ウェル細胞培養プレートの1ウェルあたり10000個の細胞を、血清削減培地(RPMI 1640+4%のPCS)中で植え付けた。翌日、親Her2又はc−Met抗体、並びに二重特異性抗体のほかに、25ng/mLのHGF(R&D、294-HGN)も加え、そして細胞をさらに48時間培養した後に、細胞増殖の指標としてATPを、cell titer glowアッセイ(Promega)を用いて測定する。
【0195】
フローサイトメトリーアッセイ(FACS)
a)結合アッセイ
c−Met及びErbB−2発現性細胞を剥離し、そしてカウントした。96ウェルのコニカルプレートの1ウェルあたり1.5×10e5個の細胞を植え付けた。細胞を、遠沈し(1500rpm、4℃、5分間)、そして2%のFCS(ウシ胎仔血清)を含むPBS中のそれぞれの二重特異性抗体の希釈系列50μL中で、氷上において30分間インキュベートした。細胞を、再度遠沈し、そして2%のFCSを含有するPBS200μLで洗浄し、続いて2%のFCS(Jackson Immunoresearch、109116098)を含有するPBSに希釈した、ヒトFcに対するフィコエリトリン結合抗体と一緒に30分間の2回目のインキュベーションを行った。細胞を、2%のFCSを含有するPBS200μLで2回、遠沈洗浄し、BD CellFix溶液(BD Biosciences)で再懸濁し、そして氷上で少なくとも10分間インキュベートした。細胞の平均蛍光強度(mfi)を、フローサイトメトリー(FACS Canto、BD)によって測定した。Mfiを、少なくとも2つの独立した染色の二重反復試験で決定した。フローサイトメトリースペクトルを、FlowJoソフトウェア(TreeStar)を使用して更に加工した。最大半減結合を、XLFit4.0(IDBS)及び用量応答ワンサイトモデル205を使用して決定した。
【0196】
b)内部移行アッセイ
細胞は、剥離し、そしてカウントした。5×10e5個の細胞を、エッペンドルフチューブ内の50μLの完全培地中に入れ、5μg/mLのそれぞれの二重特異性抗体と一緒に37℃にてインキュベートした。表示した時点で細胞を氷上に保存し、タイムコースが完了するまで保存した。その後、細胞を、FACSチューブに移し、遠沈し(1500rpm、4℃、5分間)、PBS+2%のFCSで洗浄し、そして2%のFCS(Jackson Immunoresearch、109116098)を含有したPBS中に希釈した、ヒトFcに対するフィコエリトリン結合二次抗体50μLと一緒に30分間インキュベートした。細胞を、再度遠沈し、PBS+2%のFCSで洗浄し、そして蛍光強度をフローサイトメトリー(FACS Canto、BD)によって測定した。
【0197】
Cell Titer Glowアッセイ
細胞生存率と増殖をcell titer glowアッセイ(Promega)を使用することで定量化した。製造業者の取扱説明書に従ってアッセイを実施した。簡単に言えば、細胞を、96ウェルプレート内の100μLの全容量中で所望の期間、培養した。増殖アッセイのために、細胞を、インキュベーターから取り出し、そして室温にて30分間、静置した。100μLのcell titer glow試薬を加え、そしてマルチウェルプレートを2分間、オービタル振盪機にかけた。発光をマイクロプレートリーダー(Tecan)により15分後に定量化した。
【0198】
Wst−1アッセイ
Wst−1生存率と細胞増殖アッセイをエンドポイント分析として実施し、代謝的に活性な細胞の数を検出した。簡単に言えば、20μLのWst−1試薬(Roche、1164480700)を200μLの培地に加えた。色素の完全な現像まで、96ウェルプレートを更に30分間〜1時間インキュベートした。染色強度を、450nmの波長にてマイクロプレートリーダー(Tecan)により定量化した。
【0199】
二重特異性<ErbB2−c−Met>抗体の設計
以下に発現され、そして精製された二重特異性<ErbB2−c−Met>抗体はすべて、以下に示すように最終的に修飾されたIgG1サブクラスの定常領域又は少なくともFcの一部(配列番号9のヒトIgG1定常領域)を含んでなる。
【0200】
表1において:表1に示したそれぞれの特徴を有する、全長ErbB−2抗体(トラスツズマブ)とc−Met抗体由来の1つの一本鎖Fabフラグメント(cMet 5D5)(基本的な構造スキームについては図5aを参照のこと)に基づく三価の二重特異性<ErbB2−c−Met>抗体は、先に記載した一般的な方法に従って発現及び精製されるか、又は発現及び精製できる。トラスツズマブ及びcMet 5D5の対応するVH及びVLを、配列表に示す。
【0201】
【表1】
【0202】
実施例1:
ErbB−2及びc−Metへの二重特異性抗体の結合(表面プラズモン共鳴法)
結合親和力を、25℃にて標準的な結合アッセイ、例えば表面プラズモン共鳴法などによって測定した(BIAcore(登録商標)、GE-Healthcare、Uppsala, Sweden)。親和性測定のために、30μg/mlの抗Fcγ抗体(ヤギ由来、Jackson Immuno Research)を、標準的なアミン・カップリングによってCM−5センサーチップの表面に結合させ、そしてSPR器具(Biacore T100)により化学的にブロッキングした。結合後に、単一又は二重特異性ErbB2/cMet抗体を、25℃、5μL/分の流速にて注入し、続いて30μL/分にてヒトErbB2又はc−Met ECDの希釈系列(0nM〜1000nM)を注入した。結合実験のためのランニングバッファーとしてPBS/0.1% BSAを使用した。次いで、前記チップを、10mMのグリシンHCl、pH2.0溶液の60秒パルスを用いて再生した。
【0203】
【表2】
【0204】
実施例2:
二重特異性Her2/c−Met抗体フォーマットによるHGF誘発c−Met受容体リン酸化の阻害
二重特異性抗体のc−Met部分の機能性を確認するために、c−Metリン酸化アッセイを実施する。この実験では、A549肺癌細胞又はHT29大腸癌細胞を、二重特異性抗体又は対照抗体で処理した後にHGFに晒した。次いで、細胞を溶解し、そしてc−Met受容体のリン酸化を調べる。両細胞株は、免疫ブロットにおけるリン酸化c−Metに特異的なバンドの出現によって観察できるように、HGFで刺激され得る。親又は二重特異性抗体の結合が、受容体リン酸化の阻害につながる。あるいは、当業者は、オートクリンHGFループを有する細胞、例えばU87MGを使用して、親又は二重特異性抗体の不存在下又は存在下でのc−Met受容体リン酸化を評価できる。
【0205】
実施例3:
Her2/cMet二重特異性抗体での処置後のHer2受容体リン酸化の分析
二重特異性Her2/cMet抗体Sk−Br3のHer2結合部分の機能性を確認するために、A431を、親EGFR抗体又は二重特異性Her2/cMet抗体のいずれかと一緒にインキュベートする。親又は二重特異性抗体が結合して、関係のないIgG対照抗体が結合しないことで、受容体リン酸化の阻害につながる。あるいは、当業者は更に、親又は二重特異性抗体の存在又は不存在下、NRGで刺激されてErbB2/Her2受容体リン酸化を引き起こす細胞も使用できる。
【0206】
実施例4:
Her2/cMet二重特異性抗体での処置後のPI3Kシグナル伝達の分析
Her2並びにc−Met受容体は、細胞増殖シグナルを伝えるPI3K経路を介してシグナル伝達できる。AKTのHer2及びc−Met受容体リン酸化の同時標的化を実証するために、PI3K経路の下流標的を観察することもできる。このために、非刺激細胞、NRG若しくはHGFで処理した細胞又は両サイトカインで処理した細胞を、非特異的、親対照又は二重特異性抗体と一緒に並行してインキュベートした。あるいは、当業者は更に、ErbB2/Her2を過剰発現する細胞、及び/又はc−Metシグナル伝達を活性化するオートクリンHGFループを有する細胞を評価することもできる。AKTはPI3K経路の主要な下流シグナル伝達成分であり、そしてこのタンパク質のリン酸化はこの経路を介したシグナル伝達の重要な指標である。
【0207】
実施例5:
Her2/cMet二重特異性抗体での処置後のMAPKシグナル伝達の分析
c−Met受容体は、MAPK経路を介してシグナル伝達できる。c−Met受容体の標的化を実証するために、MAPK経路の主要な下流標的であるERK1/2のリン酸化を観察することができる。このために、非刺激細胞又はHGFで処理した細胞を、非特異的、親対照又は二重特異性抗体と一緒に並行してインキュベートした。あるいは、当業者は更に、c−Metシグナル伝達を活性化するオートクリンHGFループを有する細胞を評価することもできる。
【0208】
実施例6:
二重特異性Her2/c−Met抗体フォーマットによるHGF誘発HUVEC増殖の阻害
HUVEC増殖検定は、HGFの血管新生及び細胞分裂効果を実証するために実施することができる。HUVECに対するHGFの添加は、c−Met結合抗体によって用量依存様式で阻害され得る細胞増殖の増加をもたらす。
【0209】
実施例7:
二重特異性Her2/c−Met抗体によるSk−Br3増殖の阻害
a)フローサイトメトリーにより独立に確認されているように、Sk−Br3細胞は、高いHer2の細胞表面レベル及び中程度に高いc−Metの細胞表面発現を示している。親Her2結合抗体又は二重特異性Her2/c−Met抗体の添加が増殖の減少につながったのに対して、c−Met結合抗体は増殖に対してわずかな効果しか持っていなかった。
【0210】
b)活性なHer2−c−Met受容体シグナル伝達ネットワークが増殖を起こす状況をシミュレートするために、アッセイを、HGF条件培地の存在下ではあるが、記載のとおり行う。この状況において、いずれか一方の親抗体の添加がcell titer glow分析によって測定されるように、細胞増殖に対してわずかな効果しか持っていない一方で、二重特異性抗体又は親抗体の組合せ物の添加は細胞増殖の減少につながる。
【0211】
実施例8:
二重特異性Her2/c−Met抗体フォーマットによる癌細胞株DU14SにおけるHGF誘発細胞−細胞播種(分散)阻害の分析
細胞のHGF誘発分散は細胞の形態学的変化を誘導し、そして細胞の円形化、糸状仮足様の突起、紡錘状の構造、及び細胞の特定の運動性をもたらす。二重特異性Her2/cMet抗体は、HGF誘発細胞−細胞播種を抑制した。
【0212】
実施例9:
二重特異性HER2/c−Met抗体フォーマットによるHGF誘発HUVEC増殖の阻害
HUVEC増殖検定は、HGFの細胞分裂効果を実証するために実施することができる。HUVECに対するHGFの添加は、増殖の二倍の増加につながる。二重特異性抗体と同じ濃度領域でのヒトIgG対照抗体の添加が細胞増殖に対して影響を持たなかった一方で、5D5 FabフラグメントはHGF誘発増殖を阻害する。
【0213】
実施例10:
二重特異性HER2/c−Met抗体による癌細胞株A431におけるHGF誘発細胞−細胞播種(分散)阻害の分析
HGF誘導分散には細胞の形態学的変化が含まれ、細胞の円形化、糸状仮足様の突起、紡錘状の構造、及び細胞の特定の運動性をもたらす。Real Time Cell Analyzer(Roche)は、所定の細胞培養ウェルのインピーダンスを計測し、それにより細胞の形態と増殖の変化を間接的に観察することができる。A431及びA549細胞に対するHGFの添加は、時間の関数として観察されるインピーダンスの変化をもたらす。
【0214】
実施例11:
ErbB−2及びc−Mct発現癌細胞株における抗体媒介型受容体内部移行の分析
細胞の、Her2又はc−Metに特異的に結合する抗体とのインキュベーションが、受容体の内部移行を引き起こすことを示した。二重特異性抗体の内部移行能力を評価するために、抗体誘発受容体内部移行を検討するために実験構成を設計する。このために、OVCAR−8細胞((NCI Cell Line designation;NCI(国立癌研究所)から購入したOVCAR-8-NCI;Schilder RJ, et al Int J Cancer, 1990 Mar 15;45(3):416-22;Ikediobi ON, et al, Mol Cancer Ther. 2006;5;2606-12;Lorenzi, P.L., et al., Mol Cancer Ther. 2009;8(4):713-24))(フローサイトメトリーで確認されたとおり、この細胞はHer2並びにcMetを発現する−図7bを参照のこと)を、37℃にてそれぞれの一次抗体と一緒に異なった期間(例えば0、30、60、120分=0、0.5、1、2時間(h))インキュベートした。細胞プロセスは、細胞を4℃に急速に冷やすことによって停止する。一次抗体のFcに特異的に結合する蛍光体を結合した二次抗体を、細胞表面に結合した抗体を検出するのに使用した。抗体−受容体複合体の内部移行は、細胞表面の抗体−受容体複合体を激減させるので、平均蛍光強度の減少を結果的にもたらす。内部移行はOvcar−8細胞において検討した。結果を以下の表及び図8に示す。それぞれの受容体の内部移行率(%)を、それぞれの抗体の内部移行を介して計測する(図8において、二重特異性<ErbB2−cMet>抗体BsAB02はcMet/HER2と示されており、親単一特異性の二価抗体は<HER2>及び<cMet>と示されている)。
【0215】
【表3】
【0216】
実施例12
糖鎖操作されたバージョンの二重特異性Her2/c−Met抗体の調製
二重特異性Her2/c−Met抗体のDNA配列を、各ベクターがEBV OriP配列を担持している、MPSVプロモーター及び合成ポリA部位の上流の制御下にある哺乳動物発現ベクター内にサブクローニングする。
【0217】
リン酸カルシウムトランスフェクション・アプローチを使用した、哺乳動物二重特異性抗体発現ベクターでのHEK293−EBNA細胞の同時トランスフェクションによって、二重特異性抗体を作り出す。対数増殖期のHEK293−EBNA細胞をリン酸カルシウム法によってトランスフェクトする。糖鎖操作された抗体の製造のために、前記細胞を、1つが融合GnTIIIポリペプチド発現のためのもの(GnT−III発現ベクター)であり、1つがマンノシダーゼII発現のためのもの(ゴルジマンノシダーゼII発現ベクター)である、2つの追加のプラスミドを用いて、それぞれ1:1:4:4の比で同時トランスフェクトした。細胞を、10%のPCSを補ったDMEM培地を使用してTフラスコ内で接着性単層培養として培養し、そしてそれらが50〜80%集密になるとトランスフェクトする。T150フラスコのトランスフェクションのために、トランスフェクションの24時間前に、1500万個の細胞を、(最終的に10%V/Vに)FCSを補った25mlのDMEM培地中に植え付け、そして細胞を37℃、5%のCO2雰囲気のインキュベーター内に一晩置く。トランスフェクトされるそれぞれのT150フラスコについて、DNAの溶液、CaCl2、及び水を、軽鎖及び重鎖発現ベクターの間で等しく分配される94μgの総プラスミドベクタDNA、469μlの終量までの水、及び469μlの1M CaCl2溶液を混合することによって調製する。この溶液に、938μlの50mM HEPES、280mMのNaCl、1.5mM Na2HPO4溶液、pH7.05を加え、すぐに10秒間混合し、そして室温にて20秒間静置する。その懸濁液を、2%のPCSを補った10mlのDMEMで希釈し、そして既存の培地に代えてT150に加える。更に13mlのトランスフェクション培地を加える。細胞を、37℃、5%のCO2にて約17〜20時間インキュベートし、次いで、培地を25mlのDMEM、10%のFCSで置き換える。条件培養培地を、210×gにて15分間の遠心分離によってトランスフェクトの7日後に回収し、その溶液を濾過滅菌(0.22μmフィルター)し、0.01%w/vの終濃度にアジ化ナトリウムを加え、そして4℃に保った。
【0218】
分泌された二重特異性の非フコシル(afocusylated)糖鎖操作された抗体を、プロテインA親和性クロマトグラフィーとそれに続く、陽イオン交換クロマトグラフィー、そしてSuperdex 200カラム(Amersham Pharmacia)による最終的なサイズ排除クロマトグラフィーステップによって精製し、バッファーを25mMのリン酸カリウム、125mMの塩化ナトリウム、100mMのグリシン溶液、pH6.7に交換し、且つ、純粋な単量体のIgG1抗体を回収した。抗体濃度を、280nmの吸光度から分光光度計を使用して推定した。
【0219】
抗体のFc領域に結合されたオリゴ糖を、記載したようにMALDI/TOF−MSによって分析する。PVDF膜に固定されるか又は溶液中の抗体を用いて、PNGaseF消化によって、抗体からオリゴ糖を酵素的に放出させる。放出されたオリゴ糖を含む得られた消化産物溶液を、最高MALDI/TOF−MS分析法のためにそのまま調製するか、又はMALDI/TOF−MS分析法のためのサンプル調製前にEndoHグリコシダーゼで更に消化する。
【0220】
実施例13:
二重特異性Her2/c−Met抗体の糖構造の分析
フコース及び非フコース(a−フコース)含有オリゴ糖構造の相対比の測定のために、精製した抗体材料の放出されたグリカンをMALDI−Tof−質量分析法によって分析する。今回、タンパク質骨格からオリゴ糖を放出するために、抗体サンプル(約50μg)を、0.1Mのリン酸ナトリウム緩衝液、pH6.0中の5mU N−グリコシダーゼF(Prozyme# GKE-5010B)と一緒に37℃にて一晩インキュベートする。それに続いて、放出されたグリカン構造物を、NuTip−Carbonピペットチップ(Glygenから入手:NuTip1〜10μl、Cat.Nr#NTlCAR)を使用して分離し、そして脱塩する。第1ステップとして、NuTip−Carbonピペットチップを、3μlの1M NaOHに続き、20μLの純水(例えばBaker製のHPLCグラジエントグレード、#4218)、3μLの30% 酢酸、そして再び20μLの純水で洗浄することによって、オリゴ糖の結合に向けて準備する。今回、それぞれの溶液を、NuTip−Carbonピペットチップ内のクロマトグラフィー材料の上部に添加し、そして圧力をかけてそれを通過させる。その後、10μgの抗体に相当するグリカン構造物を、先に記載のN−グリコシダーゼF消化産物を4〜5回ほど吸い上げたり吸い出したりすることによって、NuTip−Carbonピペットチップ内の材料に結合させる。NuTip−Carbonピペットチップ内の材料に結合したグリカンを、先に記載したやり方で20μLの純水で洗浄し、そしてそれぞれ0.5μLの10%と2.0μLの20% アセトニトリルを用いて段階的に溶出する。このステップのために、溶出溶液を、0.5mLの反応ベイル(reaction vails)内に充填し、そして各回毎に4〜5回吸い上げたり吸い出したりする。MALDI−Tof質量分析法による分析のために、両方の溶出液を組合せる。この計測において、0.4μLの組合せた溶出液を、MALDI標的上で1.6μLのSDHBマトリックス溶液(5mg/mlで20%のエタノール/5mMのNaCl中に溶解した2.5−ジヒドロキシ安息香酸/2−ヒドロキシ−5−メトキシ安息香酸[Bruker Daltonics#209813])と混合し、そして、適当に調整したBruker Ultraflex TOF/TOF装置で分析した。日常的には、50〜300ショットが、記録され、そして1回の実験にまとめられる。得られたスペクトルを、フレックス分析ソフトウェア(Bruker Daltonics)によって評価し、そして質量を検出したピークのそれぞれについて決定する。それに続いて、計算した質量を、それぞれの構造物(例えばそれぞれフコースを含む及び含まない複合体、ハイブリッド、及びオリゴ又は高マンノース)について理論的に予想した質量を比較することによって、ピークをフコース又はa−フコース(非フコース)含有グリコール構造物に割り当てる。
【0221】
ハイブリッド構造物の比の測定のために、抗体サンプルを、N−グリコシダーゼFとエンド−グリコシダーゼHを併用して消化する。N−グリコシダーゼFは、タンパク質骨格からすべてのN結合型グリカン構造(複合体、ハイブリッド、並びにオリゴ及び高マンノース構造)を放出し、そしてエンド−グリコシダーゼHは、グリカンの還元末端の2つのGlcNAc残基の間で更にすべてのハイブリッドタイプのグリカンを開裂する。この消化産物は、N−グリコシダーゼF消化したサンプルについて先に記載したのと同様のやり方で、続けて処理し、そしてMALDI−Tof質量分析法によって分析する。N−グリコシダーゼF消化産物からのパターンと、併用N−グリコシダーゼF/エンドH消化産物からのパターンを比較することによって、糖構造に特異的なシグナルの減少度が、ハイブリッド構造の相対内容を推測するのに使用される。
【0222】
それぞれの糖構造の相対量は、個々のグリコール構造のピークの高さと、検出したすべての糖構造のピークの高さの比から計算される。フコースの量は、N−グリコシダーゼF処理サンプルにおいて特定した全糖構造(例えば、それぞれ複合体、ハイブリッド、並びにオリゴ及び高マンノース構造)に対するフコース含有構造のパーセンテージである。非フコース化の量は、N−グリコシダーゼF処理サンプルにおいて特定した全糖構造(例えば、それぞれ複合体、ハイブリッド、並びにオリゴ及び高マンノース構造)に対するフコースを欠く構造のパーセンテージである。
【0223】
実施例14:
Her2/cMet二重特異性抗体での処置後の細胞遊走の分析
活性なc−Metシグナル伝達の1つの重要な側面は、遊走と侵襲的なプログラムの誘導である。c−Met阻害抗体の有効性は、HGF誘発細胞遊走の阻害を計測することによって測定できる。このために、HGF誘導性癌細胞株A431を、二重特異性抗体又はIgG対照抗体の存在下又は不存在下、HGFで処理し、そして8μmの細孔を遊走して通過した細胞の数を、インピーダンス読み出しを備えたCIMプレートを使用するAceaリアルタイム細胞分析装置により時間依存様式で計測する。
【0224】
実施例15:
二重特異性Her2/c−Met抗体のインビトロADCC
本発明によるHer2/cMet二重特異性抗体は、両受容体を発現している細胞において少ない内部移行を示す。少ない内部移行は、細胞表面の抗体−受容体複合体の長期暴露がNk細胞により認識されやすいので、これらの抗体を糖鎖操作することの理論的根拠を大いに裏付けている。少ない内部移行と糖鎖操作は、親抗体と比較して高い抗体依存性細胞傷害(ADCC)に置き換えられる。これらの効果を実証するインビトロ実験構成は、細胞表面にHer2とcMetの両方を発現する癌細胞、例えばA431、及びNk細胞株又はPBMCのもののようなエフェクター細胞を使用して設計できる。腫瘍細胞を、親単一特異性抗体又は二重特異性抗体と一緒に最長24hプレインキュベートし、続いてエフェクター細胞株を加える。細胞溶解物を定量し、そして単一特異性抗体と二重特異性抗体の識別を可能にする。
【0225】
標的細胞、例えばPC−3(DSMZ#ACC465、前立腺腺癌、Ham’s F12 Nutrient mixture+2mMのL−アラニル−L−グルタミン+10%のPCS中で培養)を、指数増殖期にトリプシン/EDTA(Gibco#25300-054)を用いて回収する。洗浄ステップ、及び細胞数と生存率の確認ステップの後に、必要とされるアリコートを、細胞インキュベーター内で37℃にて30分間、カルセイン(Invitrogen#C3100MP;5mlの培地中の5Mio細胞に対して、1バイアルを50μlのDMSO中に再懸濁した)で標識する。その後、細胞を、AIM培地で3回洗浄し、そして、細胞数と生存率を確認し、そして細胞数を0.3Mio/mlに調整する。
【0226】
その間に、エフェクター細胞としてのPBMCを、製造業者のプロトコール(洗浄ステップ、1×400gにて、そして2×350gにて、それぞれ10分間)に従って密度勾配遠心分離法(Histopaque-1077、Sigma#H8889)によって調製する。細胞数と生存率を確認し、そして細胞数を15Mio/mlに調整する。
【0227】
100μlのカルセイン染色標的細胞を、丸底96ウェルプレート内に入れ、50μlの希釈した抗体を加え、そして50μlのエフェクター細胞を加える。いくつかの実験では、標的細胞を、10mg/ml Redimuneの濃度にてRedimune(登録商標)NF Liquid(ZLB Behring)と混合する。
【0228】
抗体なしで同時培養される標的及びエフェクター細胞によって測定される自然発生的溶解、及び標的細胞だけを溶解する1% Triton X100によって測定される最大溶解が、対照の役割を担う。プレートを、加湿した細胞インキュベーター内で37℃にて4時間インキュベートする。
【0229】
標的細胞の殺滅を、製造業者の取扱説明書に従って細胞毒性検出キット(LDH Detection Kit、Roche#1 644 793)を使用して、損傷細胞から放出されるLDHを計測することによって評価する。簡単に言えば、各ウェルからの100μlの上清を、透明な平底96ウェルプレート内で、キットからの100μlの基質と混合する。基質の呈色反応のVmax値を、少なくとも10分間、490nmにてELISAリーダーにより測定する。特有の抗体媒介型殺滅のパーセンテージを、以下のとおり計算する:((A−SR)/(MR−SR)×100、式中、Aは特定の抗体濃度におけるVmaxの平均であり、SRは自然発生的放出のVmaxの平均であり、そしてMRは最大放出のVmaxの平均である。
【0230】
実施例16:
パラクリンHGFループを有する皮下異種移植モデルにおける二重特異性Her2/cMet抗体のインビボ有効性
Mrc−5細胞と同時注入される皮下のKPL4モデルは、c−Metのパラクリン活性化ループによく似ている。KPL4は、細胞表面にc−Met並びにHer2をある程度発現する。KPL4及びMrc−5細胞は、対数増殖期において標準的な細胞培養条件下で維持される。KPL4とMrc−5細胞は、1000万個のKPL4細胞と100万個のMrc−5による10:1の比で注射される。細胞を、SCIDベージュマウスに移植する。腫瘍が定着し、そして100〜150mm3のサイズに達した後、処置を始める。マウスを、20mg/kgの抗体/マウスの負荷量で処置し、その後毎週一回、10mg/kgの抗体/マウスで処置する。腫瘍体積を1週間に二度計測し、そして動物の体重を平行して観察する。単一抗体の単独処置と組合せを、二重特異性抗体での治療法と比較する。
【0231】
実施例17:
二重特異性Her2/cMet抗体によるOVCAR−8増殖の阻害
a)フローサイトメトリーにより独立に確認されているように、OVCAR−8細胞(NCI Cell Line designation;NCI(国立癌研究所)から購入したOVCAR-8-NCI;Schilder, RJ. et al., Int. J. Cancer 1990 Mar 15;45(3): 416-422;Ikediobi, O.N., et al., Mol. Cancer. Ther. 2006;5;2606- 2012;Lorenzi, P.L., et al., Mol. Cancer Ther. 2009; 8(4): 713-724)は、顕著なHer2及びc−Metの細胞表面レベルを示している(図7bを参照のこと)。二重特異性Her2/c−Met抗体によるOVCAR−8細胞増殖の阻害は、48時間後にCell Titer Glow(商標)アッセイにより計測した。結果を図9aに示す。対照はPBSバッファー(リン酸緩衝生理食塩水)であった。
【0232】
計測では、(0%の阻害に設定したバッファー対照と比較して)6%の阻害というHER2抗体トラスツズマブの阻害が明らかになった。二重特異性Her2/c−Met BsAB02(BsAb)抗体は、癌細胞増殖のより顕著な阻害を導いた(11%の阻害)。一価のc−Met抗体である片腕5D5(OA5D5)は、増殖に対して効果がないことが明らかになった。HER2抗体トラスツズマブと一価のc−Met抗体の片腕5D5(OA5D5)の組合せは、それほど顕著ではない減少しか導かなかった(6%の阻害)。
【0233】
b)OVCAR−8細胞はHER2シグナル伝達に依存している。活性なHER−c−Met受容体シグナル伝達ネットワークが更なる増殖を起こす状況をシミュレートするために、アッセイをa)(48時間後のCell Titer Glow(商標)アッセイ)で記載したように、しかしHGF条件培地の存在下で行った。結果を図9bに示す。
【0234】
計測では、0%の阻害に設定したHGF処理細胞と比較すると、Her2抗体トラスツズマブ(2%の阻害)と一価のc−Met抗体の片腕5D5(OA5D5)(3%の阻害)の阻害効果はほとんどないことが明らかになった。二重特異性Her2/c−Met抗体BsAB02(BsAb)は、Ovcar−8細胞の癌細胞増殖に対して顕著な阻害を示した(17%の阻害)。Her2抗体のトラスツズマブと一価のc−Met抗体の片腕5D5(OA5D5)の組合せは、細胞増殖のそれほど顕著ではない減少につながった(10%の阻害)。
【特許請求の範囲】
【請求項1】
ヒトErbB−2に特異的に結合する第1の抗原結合部位及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、フローサイトメトリーアッセイにおいて1時間後に計測した場合に、OVCAR−8細胞上で、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して15%未満のc−Metの内部移行を示すことを特徴とする前記二重特異性抗体。
【請求項2】
ヒトErbB−2に特異的に結合する1つ若しくは2つの抗原結合部位及びヒトc−Metに特異的に結合する3つ目の抗原結合部位を含んでなり、二価又は三価であることを特徴とする、請求項1に記載の二重特異性抗体。
【請求項3】
以下の:
a)ErbB−2に特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fabフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、当該全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されていることを特徴とする、請求項2に記載の抗体。
【請求項4】
ヒトErbB−2に特異的に結合する第1の抗原結合部位及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、
前記第1の抗原結合部位が、重鎖可変ドメイン内に配列番号15のCDR3H領域、配列番号16のCDR2H領域、及び配列番号17のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号18のCDR3L領域、配列番号19のCDR2L領域、及び配列番号20のCDR1L領域を含んでなり;そして
前記第2の抗原結合部位が、重鎖可変ドメイン内に配列番号21のCDR3H領域、配列番号22のCDR2H領域、及び配列番号23のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号24のCDR3L領域、配列番号25のCDR2L領域、配列番号26のCDR1L領域を含んでなること、を特徴とする前記二重特異性抗体。
【請求項5】
以下の:
ErbB−2に特異的に結合する前記第1の抗原結合部位が、重鎖可変ドメインとして配列番号1の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号2の配列を含んでなり;そして
c−Metに特異的に結合する前記第2の抗原結合部位が、重鎖可変ドメインとして配列番号3の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号4の配列を含んでなること、を特徴とする、請求項4に記載の二重特異性抗体。
【請求項6】
IgG1又はIgG3サブクラスの定常領域を含んでなることを特徴とする、請求項1〜5のいずれか1項に記載の二重特異性抗体。
【請求項7】
前記抗体がAsn297にて糖鎖でグリコシル化されていることで、当該糖鎖内のフコース量が65%以下であることを特徴とする、請求項1〜6のいずれか1項に記載の二重特異性抗体。
【請求項8】
請求項1〜7のいずれか1項に記載の二重特異性抗体をコードする核酸。
【請求項9】
請求項1〜7のいずれか1項に記載の二重特異性抗体を含んでなる医薬組成物。
【請求項10】
癌治療のための請求項9に記載の医薬組成物。
【請求項11】
癌治療のための、請求項1〜7のいずれか1項に記載の二重特異性抗体。
【請求項12】
癌の治療薬の製造のための、請求項1〜7のいずれか1項に記載の二重特異性抗体の使用。
【請求項13】
癌に罹患している患者の治療方法であって、こういった処置を必要としている患者に、請求項1〜7のいずれか1項に記載の二重特異性抗体を投与することによる前記治療方法。
【請求項1】
ヒトErbB−2に特異的に結合する第1の抗原結合部位及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、フローサイトメトリーアッセイにおいて1時間後に計測した場合に、OVCAR−8細胞上で、当該二重特異性抗体の不存在下でのc−Metの内部移行と比較して15%未満のc−Metの内部移行を示すことを特徴とする前記二重特異性抗体。
【請求項2】
ヒトErbB−2に特異的に結合する1つ若しくは2つの抗原結合部位及びヒトc−Metに特異的に結合する3つ目の抗原結合部位を含んでなり、二価又は三価であることを特徴とする、請求項1に記載の二重特異性抗体。
【請求項3】
以下の:
a)ErbB−2に特異的に結合し、且つ、2つの抗体重鎖と2つの抗体軽鎖から成る全長抗体;並びに
b)ヒトc−Metに特異的に結合する1つの一本鎖Fabフラグメント、
を含んでなり、
ここで、前記のb)の一本鎖Fabフラグメントが、前記のa)の全長抗体に、当該全長抗体の重鎖又は軽鎖のC又はN末端のペプチドコネクタを介して融合されていることを特徴とする、請求項2に記載の抗体。
【請求項4】
ヒトErbB−2に特異的に結合する第1の抗原結合部位及びヒトc−Metに特異的に結合する第2の抗原結合部位を含んでなる、ヒトErbB−2及びヒトc−Metに特異的に結合する二重特異性抗体であって、
前記第1の抗原結合部位が、重鎖可変ドメイン内に配列番号15のCDR3H領域、配列番号16のCDR2H領域、及び配列番号17のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号18のCDR3L領域、配列番号19のCDR2L領域、及び配列番号20のCDR1L領域を含んでなり;そして
前記第2の抗原結合部位が、重鎖可変ドメイン内に配列番号21のCDR3H領域、配列番号22のCDR2H領域、及び配列番号23のCDR1H領域を含んでなり、且つ、軽鎖可変ドメイン内に配列番号24のCDR3L領域、配列番号25のCDR2L領域、配列番号26のCDR1L領域を含んでなること、を特徴とする前記二重特異性抗体。
【請求項5】
以下の:
ErbB−2に特異的に結合する前記第1の抗原結合部位が、重鎖可変ドメインとして配列番号1の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号2の配列を含んでなり;そして
c−Metに特異的に結合する前記第2の抗原結合部位が、重鎖可変ドメインとして配列番号3の配列を含んでなり、且つ、軽鎖可変ドメインとして配列番号4の配列を含んでなること、を特徴とする、請求項4に記載の二重特異性抗体。
【請求項6】
IgG1又はIgG3サブクラスの定常領域を含んでなることを特徴とする、請求項1〜5のいずれか1項に記載の二重特異性抗体。
【請求項7】
前記抗体がAsn297にて糖鎖でグリコシル化されていることで、当該糖鎖内のフコース量が65%以下であることを特徴とする、請求項1〜6のいずれか1項に記載の二重特異性抗体。
【請求項8】
請求項1〜7のいずれか1項に記載の二重特異性抗体をコードする核酸。
【請求項9】
請求項1〜7のいずれか1項に記載の二重特異性抗体を含んでなる医薬組成物。
【請求項10】
癌治療のための請求項9に記載の医薬組成物。
【請求項11】
癌治療のための、請求項1〜7のいずれか1項に記載の二重特異性抗体。
【請求項12】
癌の治療薬の製造のための、請求項1〜7のいずれか1項に記載の二重特異性抗体の使用。
【請求項13】
癌に罹患している患者の治療方法であって、こういった処置を必要としている患者に、請求項1〜7のいずれか1項に記載の二重特異性抗体を投与することによる前記治療方法。
【図1】

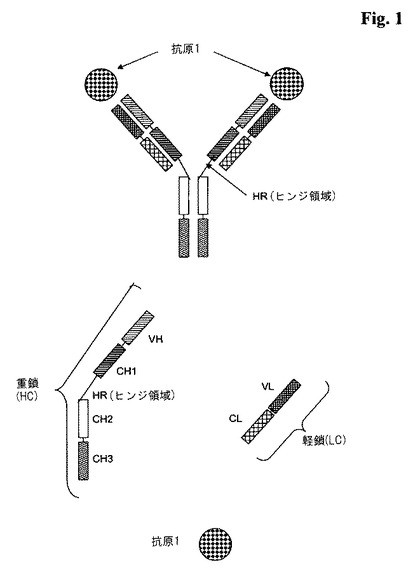
【図2a】

【図2b】

【図2c】
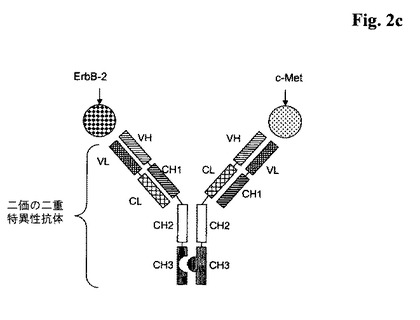
【図3a】
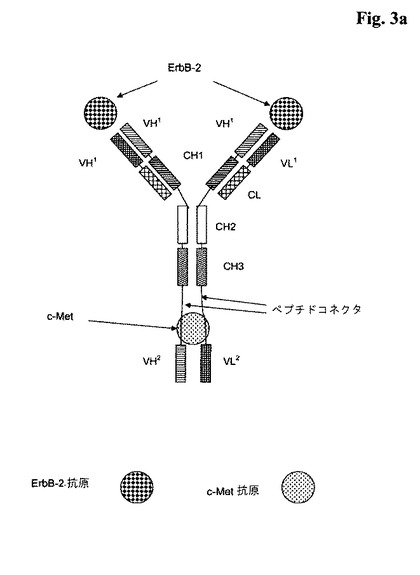
【図3b】
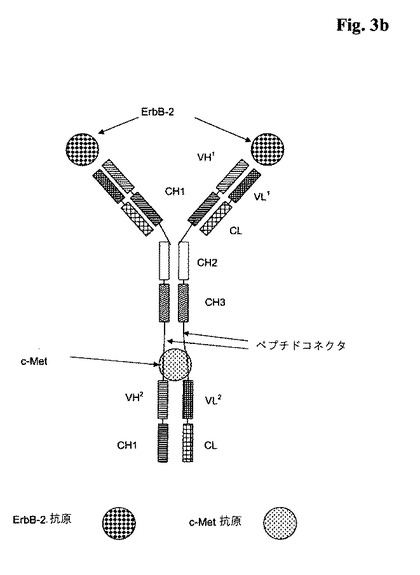
【図3c】
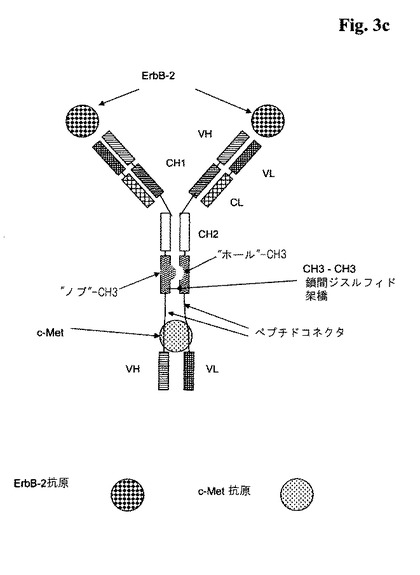
【図3d】
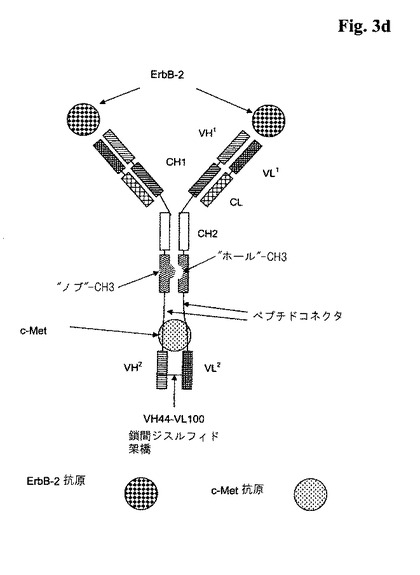
【図4a】

【図4b】
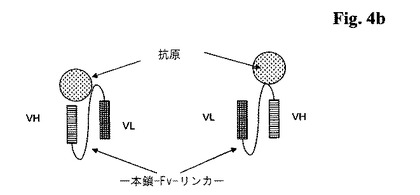
【図5a】

【図5b】
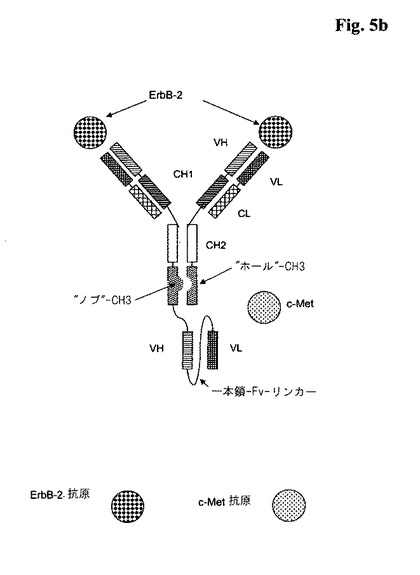
【図6a】
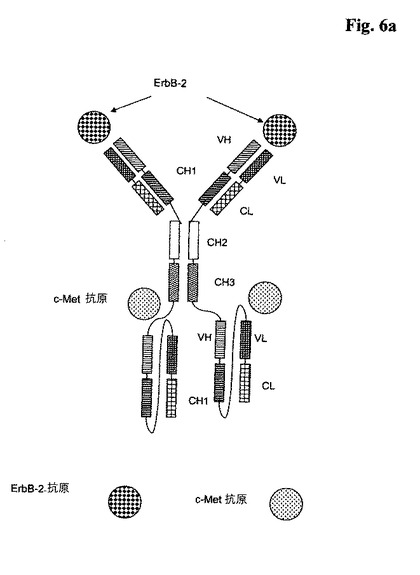
【図6b】

【図7a】
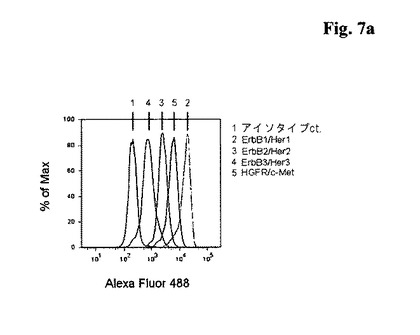
【図7b】

【図8】

【図9a】

【図9b】
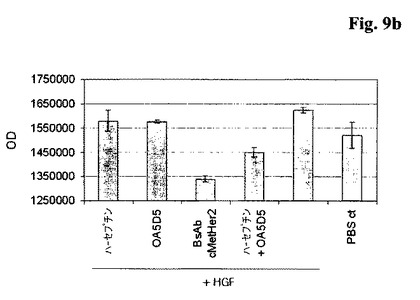
【図2a】
【図2b】
【図2c】
【図3a】
【図3b】
【図3c】
【図3d】
【図4a】
【図4b】
【図5a】
【図5b】
【図6a】
【図6b】
【図7a】
【図7b】
【図8】
【図9a】
【図9b】
【公表番号】特表2012−522525(P2012−522525A)
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2012−503897(P2012−503897)
【出願日】平成22年3月30日(2010.3.30)
【国際出願番号】PCT/EP2010/002006
【国際公開番号】WO2010/115553
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(506153815)ロシュ グリクアート アクチェンゲゼルシャフト (25)
【Fターム(参考)】
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成22年3月30日(2010.3.30)
【国際出願番号】PCT/EP2010/002006
【国際公開番号】WO2010/115553
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(506153815)ロシュ グリクアート アクチェンゲゼルシャフト (25)
【Fターム(参考)】
[ Back to top ]