
二量体または多量体のマイクロタンパク質
標的タンパク質に対する特異的な結合活性を有するアミノ酸配列を好ましくは含む、少なくとも2個のマイクロタンパク質を含むポリペプチドを開示する。さらに、このようなポリペプチドをコードするポリヌクレオチド、ならびに薬学的組成物、および該ポリペプチドまたは該ポリヌクレオチドを含むキットも開示する。本明細書では、開示されたポリペプチドまたはポリヌクレオチドを投与する治療法、および第2の医学的使用も開示する。加えて本出願の開示は、開示されたポリペプチドの作製に使用可能なマイクロタンパク質内に共有結合を形成させる方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、標的タンパク質に対する特異的な結合活性を有するアミノ酸配列を好ましくは含む、少なくとも2個のマイクロタンパク質(microprotein)を含むポリペプチドに関する。本発明はさらに、このようなポリペプチドをコードするポリヌクレオチド、ならびに薬学的組成物、および該ポリペプチドまたは該ポリヌクレオチドを含むキットにも関する。本発明は、本発明のポリペプチドまたはポリヌクレオチドを投与する治療法、および第2の医学的使用にも関する。本発明は加えて、本発明のポリペプチドを作製するために使用可能な、マイクロタンパク質内に共有結合を形成させる方法に関する。
【背景技術】
【0002】
本発明は、治療に使用されるペプチドおよびペプチド模倣物の分野に関する。先行技術では、治療に応用されるペプチドおよびポリペプチドならびにこれらの類似体の探索は、確立された方法である。このような方法は、ヒトまたは動物の体内で自然に生じるタンパク質-タンパク質相互作用を修飾する試みに基づく主な局面である。特にリガンド-受容体の相互作用は、アゴニストまたはアンタゴニストである人工リガンド分子によって調節可能であることがわかっている。例えば、リガンド分子の二量体形成時には、二量体の膜結合型受容体は、該リガンドの結合によって刺激可能となる場合がある。この方法は、多くがシグナル伝達カスケードを引き起こすことでアゴニストとして作用するサイトカインおよびホルモンに関する研究に基づく。シグナル伝達過程は、個々のアゴニストが2つの受容体分子に結合することで開始されることがしばしばあることが観察されている。多くの場合、このようなアゴニストは、個々の単量体が単量体型の受容体分子と結合することでシグナル伝達過程を活性化する二量体であるか、または空間的に離れた2つの結合部位を含む単体の単量体である(Frank 2002, Grotzinger 2002, Mellado et al., 2001)。
【0003】
細胞表面受容体の活性化が、受容体リガンドによって2つの受容体分子を空間的に連結されることで生じる周知の例がTpo/TpoR系である(Dower et al., 1998;Geddis et al., 2002)。トロンボポエチン(TPO)は、血小板が骨髄巨核球から産生される過程である巨核球形成の調節に重要な役割を果たす、332アミノ酸のグリコシル化されたポリペプチドである(Kuter et al., 1994, Kaushansky et al., 1994;Wendling et al., 1994, Sauvage et al., 1994)。したがってTPOは、さまざまな血液疾患、例えば、血小板異常または血小板減少性状態に主に起因する疾患の診断および治療の両方における潜在的に有用な用途を有する。数年前には、TPOペプチド模倣物の開発が報告されている(WO 96/40750、WO 98/25965、Cwirla et al., 1997)。このようなペプチドは、TpoRに結合して活性化するが、天然のTPOに対する配列相同性は無いものが設計された。このような分子は、TPO受容体(TPOR)と高親和性で結合し、化学的架橋によって天然のサイトカインであるTPOと同程度の活性を有する二量体を形成することで、さらに最適化可能なTPORアゴニストとして作用することが報告されている。活性が高いことがわかっているペプチド配列はIEGPTLRQWLAARAである(Cwirla et al., 1997)。この配列を含むペプチドは、単量体として弱いTPORアゴニストとして、および2つの単量体が架橋されてホモ二量体を形成すると強いアゴニストとして作用する。しかしながら、このペプチドコンストラクトには、治療に使用する際には、以下のいくつかの短所がある:
I)このようなペプチドは、立体構造的に高度に柔軟なので、それ自体がタンパク質分解を受けやすく、このため血漿半減期は短い。加えて、胃内および腸内における、ならびに腸管刷子縁膜の表面上におけるプロテアーゼ(ペプシン、トリプシン、エラスターゼ、キモトリプシンなど)によるタンパク質分解性の攻撃に対する感受性があるために、例えば血小板減少症の治療目的の経口投与は行われていない。
II)Cwirla et al. (1997)に記載されているようなペプチドを組換え的に作製することは困難である。小型のペプチドは、細胞プロテアーゼによってしばしば速やかに分解されることから、微生物発現系において高収量で過剰発現させることは極めて困難である。したがって、望ましくない副産物の発生などの、その全ての既知の短所(例えば、合成の成熟前の終結に起因する切断;または立体異性のアミノ酸の取り込み)のある、化学的なペプチド合成が選択される方法である。
【0004】
他のTPOアンタゴニストペプチドはWO 03/031589に報告されている。しかしながら、それらのペプチドにも前述の短所はある。
【0005】
上記の概要は、個々の生物学的標的系において、アゴニスト作用またはアンタゴニスト作用を有するエフェクター分子として二量体または多量体(ポリ)ペプチドの作製を試みる際に遭遇する状況の例示的なものである。したがって先行技術では、組換え的に容易に作製可能で、および/または長い半減期、高い安定性、および生物学的利用能などの有利なインビボ特性を有する二量体もしくは多量体のペプチドのエフェクター分子の応用を可能とする手段および方法が必要なことは明らかである。
【0006】
マイクロタンパク質は、多様な小型のタンパク質、典型的には、シスチンノットおよび短い3本鎖のβ-シートからなる共通の構造モチーフを有する、最長50残基の長さのタンパク質であると先行技術で記載されている。このようなタンパク質は、小型タンパク質の阻害剤シスチンノット(ICK)ファミリー(Le-Nguyen et al., 1990)の成員としても知られている。これらは共通の構造を有するが、多様な生物学的活性、および無視できるアミノ酸配列の同一性を有する。例には、(i)電位依存性Ca2+チャネルとの高親和性結合によって神経毒として作用する、イモガイ(Conus magus)の毒液に含まれる26残基のポリペプチドであるω-コノトキシンMVIIa (Kohno et al., 1995);(ii)39アミノ酸のペプチドである、ジャガイモのカルボキシペプチダーゼ阻害因子(PCI)(Rees and Lipscomb, 1982);ならびに(iii)テッポウウリ(Ecballium elaterium)由来のEETI-II(Le-Nguyen et al., 1990)などがある。
【0007】
既知のICKファミリーの全てのマイクロタンパク質が、マイクロタンパク質と個々の標的分子の結合による活性の阻害に基づく阻害性の機能すなわちアンタゴニスト機能を有する。加えて、既知の全てのマイクロタンパク質は、開鎖状か環状にかかわらず、単量体のタンパク質として存在する。
【0008】
前述したように、先行技術では、特にアゴニストペプチドまたはアンタゴニストペプチドのエフェクター分子の作製に特に適した方法が求められている。先行技術は、この目的に原理的には有用な可能性のあるペプチド片の環化または連結の過程を提供している。しかしながら、このような過程は一般に不利に作用する。というのは、ペプチド出発材料が化学合成によって、すなわち組換え的ではなく提供されることが求められるからである。Gaertnerらは、組換え的に作製されたペプチドに応用可能なペプチド片を連結させる過程を提案している(J. Biol. Chem. 269 (1994), 7224-7230)。しかし、マイクロタンパク質に応用可能であることは報告されていない。また先行技術の知識に鑑みて、マイクロタンパク質に応用可能か否かは不明であると言わざるを得ない。Gaertnerらが報告した手順は、カルボキシ末端におけるヒドラジドの存在を必要とし、かつ、先行技術に基づくと、組換え的に作製されたマイクロタンパク質から開始した場合にどのようにしてこれが生じるかは明らかではない。さらに、Gaertnerらの報告した反応スキームは、第1の段階に酸化反応を、そしてペプチド連結後に還元を含む。このような反応条件が、望ましくない再シャッフリング、すなわちマイクロタンパク質内に存在する3つのジスルフィドの開裂を生じる可能性があるか否かは現時点で不明である。
【0009】
以上を総合すると、本発明の基礎となる技術上の問題は、長い半減期、安定性、および/または生物学的利用能などのインビボ特性が改善された二量体または多量体のペプチドもしくはポリペプチドの使用を可能とする、作製が容易なペプチド構造の提供、および対応する作製法の提供である。
【0010】
この技術上の問題は、請求項に記載された態様の提供によって解決される。
【0011】
したがって本発明は、少なくとも2個のマイクロタンパク質を含むポリペプチドに関する。
【発明の開示】
【0012】
本発明のポリペプチドによって、機能特性の改善、特に、先行技術の二量体/多量体ペプチド構造と比較して長い半減期、安定性、および/または生物学的利用能などのインビボ特性の改善をもたらすやり方で二量体または多量体の形状のペプチドの機能性の提供を可能とするフレームワーク構造が提供される。加えて、1つもしくは複数のエフェクターペプチドが付加された本発明のポリペプチドは、ペプチドの立体構造の固定化の上昇、およびこれに伴うコンホメーションエントロピーの低下に起因する親和性および選択性の改善を、先行技術のペプチドの二量体/多量体と比較して示すことが想定される。本発明のポリペプチドはさらに、複数のマイクロタンパク質ループの置換による複数のペプチドの導入によって多価性またはヘテロ機能性を導入する可能性を拓くという利点を有する。本発明のポリペプチドの他の有利な特性は、両端におけるタンパク質分解による切断からの保護が、環状のマイクロタンパク質を使用することで容易に達成可能であるという事実である。このような特性の改善は、極めて密なトポロジー、組み入れられたペプチド配列に加わる強固なマイクロタンパク質の3次元構造によって、およびマイクロタンパク質に内在する有利な特性によって説明可能である。
【0013】
驚くべきことに、先行技術に記載されたペプチド二量体に関して知られている機能性、すなわち機能性ペプチド部分が3次元構造に関して高度の自由度を有することが、対応するペプチドが移植されたマイクロタンパク質の二量体に観察可能なことが報告されている。TPOアゴニストペプチドとハイブリッドのTPO受容体の相互作用をモデルとして使用することで、驚くべきことに、このようなペプチドが、二量体のマイクロタンパク質の足場に移植された場合にTPO受容体を効率的に活性化可能であることが明らかになる(添付の実施例1を参照)。特に、TPOR結合配列(IEGPTLRQWLAARA;SEQ ID NO: 7)がマイクロタンパク質の足場に、TPO受容体に対する親和性を失うことなく移植可能であり、および同受容体結合配列を含むマイクロタンパク質の二量体形成に伴って、巨核球の形成をTPO様に誘導する、完全な活性を有するアゴニストが得られるという意外な知見が得られている。図3からわかるように、マイクロタンパク質の2つの足場が、前述のペプチドの組み入れに使用された。1つは、ヒトアグーチ関連タンパク質(McNulty et al., 2001)であるAGRP (AGRP')のメラノコルチン受容体結合ドメインに由来する、シスチンに富む領域の足場に基づくものであり、もう1つは、テッポウウリ(Ecballium elaterium)のトリプシン阻害因子であるマイクロタンパク質(Christmann et al., 1999)に由来する。AGRP'では、メラノコルチン受容体との結合に関与する結合ループ(-CYCRFFNAFCYC-、SEQ ID NO: 20、Joseph et al., 2003)がTpoR結合配列と置換されている。受容体と結合した場合、同ペプチドの構造情報は利用できないため、さまざまなコンストラクトが、ペプチドを、多様なループ構造に強制的に導入することを目的として作製されている。これは、ループ内に追加の残基を導入することで(実施例1の表1に示すSE-AG-TP-040)、またはジスルフィド結合を形成可能な2つのシステイン残基を導入することで(実施例1の表1のSE-AG-TP-050)達成された。実際に特定のループ構造が必要なことは図3に示されている。というのは、必ずしも全てのコンストラクトがアゴニスト活性を示すわけではないからである。
【0014】
他の利点として、本発明では、二量体ペプチドまたは多量体ペプチドのエフェクター分子を高収率で組換え的に作製することが可能となる。1つの態様では、本発明のポリペプチドは、それが少なくとも2個のマイクロタンパク質を含む融合タンパク質である場合には、組換え的発現を応用することで直接的に作製できる。別の態様では、前駆体、例えばマイクロタンパク質の単量体を組換え的に作製した後に、該前駆体を、先行技術の手法か、または以下に示す方法で二量体化または多量体化することができる。マイクロタンパク質の組換え的発現については例えば、EP 04 02 2455.2、Molina et al. (1992)、またはSchmoldt et al. (2004)に記載されている。
【0015】
本発明と関連して、「少なくとも2個のマイクロタンパク質を含む」という表現は、少なくとも2個のマイクロタンパク質が、ポリペプチドが使用される条件、例えば生理学的条件では結合が破壊されないように相互に結合している状態を意味する。本出願では、本発明のポリペプチドは、同意表現として、「マイクロタンパク質の二量体または多量体」とも表現される。「多量体」という表現は、3個またはこれ以上を意味すると理解される。「オリゴマー」または「オリゴマーの」という表現は、原則的には同じ意味であるが、文脈によっては二量体も含む場合がある。この少なくとも2個のマイクロタンパク質の結合は、共有結合および/または非共有結合による場合がある。本発明のポリペプチド内では、マイクロタンパク質は、意図された応用に依存して、考え得るあらゆる配置で存在する場合がある。特に、マイクロタンパク質は、直線状、環状、もしくは分枝状の配置を取り得るか、またはこれらの混合物である場合がある。直線状の構造では、マイクロタンパク質は、いずれかのタンデム配置に、すなわち一方のマイクロタンパク質のC末端が別のマイクロタンパク質のN末端と連結されるように連結され得る。または、マイクロタンパク質は、「頭部と頭部(head-to-head)」または「尾部と尾部(tail-to-tail)」の配置、すなわち一方のマイクロタンパク質のN末端が別のマイクロタンパク質のN末端と連結されるか、および/または一方のマイクロタンパク質のC末端が別のマイクロタンパク質のC末端と連結される配置を取り得る。
【0016】
好ましい態様では、本発明のポリペプチドは環状である。
【0017】
「環状」という表現は、当業者であれば環状のペプチドまたはポリペプチドの構造を推定可能な任意の意味に準じる。特に、「環状」という表現は、それ自体が環状である、例えば環状のペプチドバックボーン構造とシスチンノットを形成する、本発明のポリペプチドに含まれる1つもしくは複数のマイクロタンパク質の単量体を意味する場合がある。同様に、「環状」という表現は、複数のマイクロタンパク質を含む本発明のポリペプチド内の高次の環状構造を意味する場合がある。このような高次の環状構造の好ましい例は、少なくとも2個のマイクロタンパク質のペプチドバックボーンが連結されて1つの環を形成する「大環状分子」である。このようにマイクロタンパク質は、タンデムすなわち「頭部と頭部」もしくは「尾部と尾部」のいずれかの配置を取り得る。
【0018】
特に好ましい態様では、本発明のポリペプチドは、該ポリペプチドのマイクロタンパク質が、一方のマイクロタンパク質のC末端が別のマイクロタンパク質のN末端に共有結合で結合されるように配置される1つの大環状分子を形成する。
【0019】
本発明のポリペプチド内では、マイクロタンパク質は直接的に、または好ましくはリンカー分子を介する様式のいずれかで連結可能である。好ましくは、リンカー分子は二機能性またはオリゴ機能性(oligofunctional)のリンカー分子である。「二機能性」という表現は、2個のマイクロタンパク質が、化学的手段または酵素的手段によって1つのリンカー分子に共有結合的にカップリング可能であることを意味する。「オリゴ機能性」は、2個以上のマイクロタンパク質が、化学的手段または酵素的手段によって1つのリンカー分子に共有結合的にカップリング可能であることを意味する。2個もしくは2個以上のペプチド部分またはポリペプチド部分の連結に適したリンカー分子は、Baumert, Methods Enzymol. 172 (1989), 584-609;Yoshitake Eur. J. Biochem. 101 (1979), 395-399;Pierce Chemicals (1999), Double agents cross-linking reagents selection guide, Pierce Chemicals, Rockford, IL, USA;Peters, Annu. Rev. Biochem. 46 (1977), 523-551;およびFasold, Angew. Chem. Int. Ed. Engl. 10 (1971), 795-801などの先行技術に広く記載されている。したがって当業者であれば、特にその提供ならびにその用途に関して、適切なリンカー分子を承知している。典型的には、リンカー分子は例えば、2つのタンパク質ドメイン間、この場合はマイクロタンパク質間に空間を提供するよう作用し、それにより、例えばその機能性の3次元構造に本ドメインが正しく折りたたまれることを可能にする。さらにリンカー分子は2つのドメイン間に、2つのドメインの、それらの標的構造、例えば二量体のタンパク質受容体への有効な適合に必要な距離を設けることができる。
【0020】
好ましくは、直接的な連結、またはリンカー分子を介した連結はペプチド結合を介して実現される。リンカー分子は、このような場合は、好ましくはペプチドである場合がある。この態様に従って、マイクロタンパク質の二量体/多量体を組換え的に、すなわち対応する発現コンストラクトにコードされた融合タンパク質として作製することが可能である。ペプチドリンカーは好ましくは、比較的短く、例えば最長10アミノ酸、好ましくは最長5アミノ酸からなる。
【0021】
ある応用では、本発明のポリペプチド内の2個のマイクロタンパク質間の結合を破壊することが必要な場合がある。このような状況では、マイクロタンパク質は、切断可能なリンカーを介して連結されることが好ましい。当業者にとっては、対応する切断可能なリンカーは文献から周知である。例えば、切断可能なリンカーは、例えばトロンビン切断部位などの酵素切断部位を含むペプチドリンカーの場合がある。別の方法では、切断可能なリンカー、好ましくはペプチドリンカーを、酵素的切断以外の切断機構に、例えば化学的手段または物理的手段に感受性をもつように設計することもできる。
【0022】
さらに、本発明のポリペプチド内で2個のマイクロタンパク質を連結するリンカー分子を、リンカーが柔軟なヒンジの場合と同様に、特定の動きが可能なように設計することができる。この場合、当業者であれば、対応するペプチドリンカーまたは非ペプチドリンカーの構造について記載された先行技術の文献を参照することができる。
【0023】
本発明のポリペプチドでは、少なくとも2個のマイクロタンパク質が非ペプチドカップリングによって連結されることが好ましい。非ペプチドカップリングは、二機能性またはオリゴ機能性のリンカー分子を含むことが好ましい。特に好ましいのは、アジピン酸ヒドラジド、ビス-スクシンイミジル-スベラート(DSS)、およびEDTA-ヒドラジドから選択されるリンカー分子である。
【0024】
「マイクロタンパク質」という表現は、先行技術の文献で十分に確立した意味をもち、ならびに一般に、最長50アミノ酸の比較的小さいサイズのポリペプチド、および分子内ジスルフィド結合に基づく定まった構造を意味する。マイクロタンパク質は典型的には、極めて安定であり、かつ熱、pH、およびタンパク質分解による破壊に耐性を有する。マイクロタンパク質に関する現在の知見、特に構造および存在に関する知見は例えば、Craik (2001);Pallaghy (1994);およびCraik (J. Mol. Biol. 294 (1999), 1327-1336)にまとめられている。
【0025】
好ましい態様では、本発明のポリペプチド内の個々のマイクロタンパク質は、少なくとも6個のシステイン残基を含み、このうち6個のシステイン残基が1つのシスチンノットを形成するようにジスルフィド結合を介して連結されている。
【0026】
マイクロタンパク質は、阻害因子シスチンノット(ICK)ポリペプチドとしても知られており、および以下の説明のようにも呼ばれる。
【0027】
「シスチンノット」という表現は、2つのジスルフィド結合によって形成された埋め込まれた環と、これらを連結し、第3のジスルフィド結合がくぐるバックボーンセグメントを含む3つのジスルフィド結合フレームワークによって安定化される、バックボーンセグメントの連結の小さな3つのβ-シートを特徴とする、ICKポリペプチドによって形成される3次元構造を意味する。好ましくは、シスチンノットは、6個の保存されたシステイン残基、およびこれを連結するバックボーンセグメントによって形成され、第1のジスルフィド結合は、第1と第4のシステイン残基の間に位置し、第2のジスルフィド結合は、第2と第5のシステイン残基の間に位置し、ならびに第3のジスルフィド結合は、第3と第6のシステイン残基の間にあり、第3のジスルフィド結合は、他の2つのジスルフィド結合、およびこれらを連結するバックボーンセグメントによって形成された環をくぐる。仮に、適切であれば、ジスルフィド結合は、シスチンノットの全体的なトポロジーを同様に確実に形成する、その化学的等価物と置換され得る。あるマイクロタンパク質が正しいシスチンノットを形成するか否かの検討に際しては、当業者であれば、どのシスチン残基が相互に連結されているかを見極めることができる。これは例えば、Gorasson (J. Biol. Chem. 278 (2003), 48188-48196)、およびHorn (J. Biol. Chem. 279 (2004), 35867-35878)に記載された手法で実施可能である。シスチンノットを有するマイクロタンパク質は例えば、Craik (2001);Pallaghy (1994);およびCraik (J. Mol. Biol. 294 (1999), 1327-1336)に記載されている。
【0028】
本発明と関連して使用されるマイクロタンパク質は、開いた状態または環状の構造を取り得るペプチドバックボーンを有する場合がある。開いた構造は好ましくは、N末端にアミノ基を、およびC末端にカルボキシ基を有するマイクロタンパク質を意味する。しかしながら、当業者であればペプチド化学における最新知見を元に推定する、末端の任意の修飾も想定される。閉じた構造では、マイクロタンパク質のペプチドバックボーンの末端は、好ましくは共有結合を介して、より好ましくはアミド(すなわちペプチド)結合を介して連結される。シスチンノットのトポロジーを有する、閉じた構造のマイクロタンパク質は先行技術で「シクロチド(cyclotide)」として、およびそれらの結び目は「環状シスチンノット(CCK)」として知られる。シクロチドは例えば、WO 01/27147およびCraik (Curr. Opinion in Drug Discovery & Development 5 (2002), 251-260)に記載されている。
【0029】
本発明のポリペプチド内のマイクロタンパク質が、アミノ酸モチーフCX3-CX4-CX4-7-CX1-CX4-5-CX5-7(SEQ ID NO: 8)を含むことがさらに好ましい。上式でXは独立に、相互に任意のアミノ酸残基に由来することを意味する。Cは、標準的な命名法に準じてシステインを意味する。好ましくは、アミノ酸Xはシステインではない。配列内のシステイン残基Cは、前述したようなシスチンノットを形成することがさらに好ましい。
【0030】
本発明のさらに好ましい態様では、マイクロタンパク質は28〜40アミノ酸の長さを有する。
【0031】
特定の応用では、本発明のポリペプチド内の1つもしくは複数のマイクロタンパク質が、特定の最大のサイズを超えないことが有利な場合がある。これは例えば、マイクロタンパク質の単量体が、酵素の触媒部位などの、標的タンパク質のポケットに適合しなければならない場合について言える。したがって、本発明との関連で使用されるマイクロタンパク質は、最長35アミノ酸、より好ましくは最長32アミノ酸、および最も好ましくは最長30アミノ酸の長さを有することが特に好ましい。好ましくは、前述の好ましいサイズの範囲は、本発明のポリペプチド内に1個のマイクロタンパク質であり、より好ましくは2個のマイクロタンパク質であり、可能ならば、さらにより好ましくは少なくとも3個のマイクロタンパク質である。最も好ましくは、本発明のポリペプチド内の全てのマイクロタンパク質は、前述のサイズ制限の任意の1つを示す。
【0032】
本発明のポリペプチドに含まれるマイクロタンパク質は、野生型のマイクロタンパク質か、または修飾型のマイクロタンパク質のいずれかの場合がある。修飾は、欠失、付加、置換による修飾、またはペプチド化学の当業者に既知である、翻訳後修飾による修飾などの他の修飾の場合がある。
【0033】
好ましい態様では、マイクロタンパク質は、機能性のペプチド配列がマイクロタンパク質内に移植される本発明のポリペプチドに使用される。「移植する(grafting)」という表現は、本発明との関連で、マイクロタンパク質配列の一部と、望ましい機能を持たせることが意図される別のペプチド配列との置換を意味する。この機能は、例えば受容体、リガンド、抗体、抗原、酵素、または他の結合タンパク質のような標的タンパク質などの標的分子の結合の場合がある。標的分子にペプチド配列が結合すると、標的分子に阻害性または活性性の作用を及ぼす場合がある。有利には、マイクロタンパク質の一部が、マイクロタンパク質の3次元構造の形成に必須ではない置換に選択される。移植型マイクロタンパク質を作製するための数多くの方法が、単量体のマイクロタンパク質に関する先行技術に記載されている。これについては例えば、Craik(Curr. Op. Drug Discovery Design 5 (2002), 251-260), WO 01/27147, Barry, Structure 12 (2004), 85-94;Craik, Toxicon 39 (2001), 1809-1813;Chiche, Current Protein and Peptide Science 5 (2004), 341-349;Rosengren, J. Biol. Chem. 278 (2003), 8606-8616;およびChristmann, Protein Eng. 9 (1999), 797-806を参照されたい。これらの引用文献は以降、参照により本明細書に組み入れられる。典型的には、移植時には、マイクロタンパク質の3次元構造の形成に重要な要素を含まないマイクロタンパク質のループが所望のアミノ酸配列と置換されるか、または部分的に置換される。
【0034】
移植は、文献に記載された方法で実施できる。一般に、所望のアミノ酸配列の組み入れは、マイクロタンパク質のコード配列に、例えばPCR増幅を含む手法などの一般的なインサイチュー突然変異誘発法で突然変異を導入することによって行われる。移植型のマイクロタンパク質は次に、突然変異が導入されたコード配列を適切な宿主で発現されることで提供され得る。この目的で、Sambrook et al. (2001)に記載された方法などの標準的な組換えDNA操作法および発現法を適用することができる。
【0035】
または、移植型マイクロタンパク質は、組換え発現以外の方法で、例えばマイクロタンパク質の翻訳後の化学的操作によって作製することもできる。好ましい方法では、移植型マイクロタンパク質は、後述するヒドラゾン結合の形成を含む本発明の方法で作製される。
【0036】
本発明との関連で使用されるマイクロタンパク質は、アミノ酸、好ましくは天然のアミノ酸のみからなる場合がある。しかしながら、ペプチド化学およびポリペプチド化学の当業者になじみのある手法で誘導体化されるマイクロタンパク質も含まれる。このような誘導体は例えば、1つもしくは複数のアミノ酸の、化学的に修飾されたアミノ酸などの類似体との置換、N末端およびC末端における環化、または例えばマイクロタンパク質の治療効果を改善する可能性のある機能性部分との結合を含む場合がある。誘導体化部分を含めることで例えば、ポリペプチドの安定性、溶解性、生物学的半減期、または吸収が改善され得る。このような部分は、マイクロタンパク質の任意の望ましくない副作用を減ずるか、または除去可能である。適切な部分の概要は例えば、E. W. MartinによるRemington's Pharmaceutical Sciences (18th ed., Mack Publishing Co., Easton, PA (1990))に記載されている。ポリエチレングリコール(PEG)は、治療的タンパク質の作製に使用可能な、このような化学的部分の一例である。PEGをタンパク質に結合させると、タンパク質分解から保護されることが報告されている(Sada et al., J. Fermentation Bioengineering 71 (1991), 137-139)。タンパク質への特定のPEG部分の結合には、さまざまな方法がある(総説としてAbuchowski et al.、「Enzymes as Drugs」;Holcerberg and Roberts, eds. (1981), 367-383を参照)。一般にPEG分子は、タンパク質内に存在する反応基を介してタンパク質に連結される。例えば、タンパク質のリシン上のアミノ基、またはアミノ末端は特に、この結合に都合がよい。治療に有用なマイクロタンパク質の作製に使用可能な他の化学的修飾は、グルタルアルデヒドなどの架橋試薬の追加、グリコールやエタノールなどのアルコールの追加、またはスルヒドロキシド(sulhydroxide)-ブロッキング試薬、もしくは側鎖残基のリン酸化、アセチル化、酸化、グルコシル化、リボシル化などの修飾試薬の追加、重金属元素および/または最大10個のN末端もしくはC末端における追加的なアミノ酸残基の結合を含む。好ましくは、後者の残基はヒスチジンであるか、または、より好ましくは残基RGS-(His)6である。
【0037】
他の適切な誘導体化は、1つもしくは複数の追加のアミノ酸配列との融合の場合がある。このような融合タンパク質では、追加のアミノ酸配列は、マイクロタンパク質の配列に、共有結合または非共有結合、好ましくはペプチド結合によって連結可能である。連結は、当技術分野で周知の方法による遺伝子融合に基づく場合があるほか、例えばWO 94/04686に記載された手順による化学的な架橋によって実施することができる。追加されるアミノ酸配列は好ましくは、柔軟なリンカー、有利には、追加の配列によって形成される3次構造のC末端と、マイクロタンパク質のN末端の間、またはこの逆の間の距離を確保するのに十分な長さを有する複数の親水性のペプチド結合されたアミノ酸を含む場合のあるポリペプチドリンカーによって連結可能である。融合タンパク質は、切断可能なリンカー、またはタンパク質分解酵素用の切断部位(例えばCNBr切断部位またはトロンビン切断部位;図2参照)を含む場合がある。
【0038】
さらに、追加されるアミノ酸配列は典型的には、所定の特異性または機能、例えば核局在化シグナル、トランス活性化ドメイン、DNA結合ドメイン、ホルモン結合ドメイン、タンパク質タグ(GST、GFP、h-mycペプチド、FLAG、HAペプチド)を有する。
【0039】
好ましい態様では、マイクロタンパク質はバルナーゼ(barnase)と、好ましくは不活性なバルナーゼと融合される。好ましくは、バルナーゼとの融合は、マイクロタンパク質の組換え的な作製を容易にするために用いられ、およびバルナーゼ部分は、本発明のポリペプチドへのマイクロタンパク質の組込みに先だって除去される。
【0040】
「バルナーゼ」は、バチルス・アミロリクエファシエンス(Bacillus amyloliquefaciens)に由来する細胞外リボヌクレアーゼである(Fersht, 1993;Paddon, 1987)。マイクロタンパク質とバルナーゼの融合が、いくつかの利点をもたらし得ることが既に報告されている(Schmoldt et al., 2004)。特に、マイクロタンパク質が大腸菌などの宿主細胞における発現によって組換え的に作製される場合には、融合したバルナーゼ部分は可溶化作用を有する。これは、発現されたマイクロタンパク質を封入体から単離する必要性、および続いてこれを酸化して活性ジスルフィド結合構造を得る必要性を大きく減じるか、または完全に除く可能性がある。他の利点には、バルスター(barstar)-バルナーゼの親和性を利用して、発現されたマイクロタンパク質を粗抽出物から精製可能な点(EP 04 02 2455,2)、ならびに融合タンパク質を結晶化させることで促進型構造モデリング用の入力としての既知のバルナーゼ構造を使用する3次元構造解析が容易になる点(EP 04 02 2455,2)がある。
【0041】
バルナーゼ融合体が、活性型のバルナーゼを使用して構築されるのであれば、バルナーゼ阻害因子であるバルスターを十分な量で同時に発現させることが必要な場合がある。というのは、これを行わないとバルナーゼが宿主細胞に致死的な作用を及ぼすからである(Martsev, 2004)。これに関連して、His-102がAlaと置換されたバルナーゼなどの不活性な突然変異型バルナーゼを使用することが好ましい場合がある(図2、および対応する図のレジェンド参照)。この結果、バルナーゼ融合体に関連する利点が維持されて、バルスターを追加的に同時発現させる必要は無くなる。
【0042】
本発明との関連で使用されるマイクロタンパク質は例えば、天然の状態で純粋な産物か、または化学合成手順の産物、または組換え手法によって原核生物もしくは真核生物の宿主から(例えば、培養した細菌、酵母、高等植物、昆虫、および哺乳類の細胞により)産生される産物の場合がある。
【0043】
組換え発現を介するマイクロタンパク質の提供について、さまざまな発現系に関する総説には例えば、Methods in Enzymology 153 (1987), 385-516, Bitter et al. (Methods in Enzymology 153 (1987), 516-544)、およびSawers et al. (Applied Microbiology and Biotechnology 46 (1996), 1-9), Billman-Jacobe (Current Opinion in Biotechnology 7 (1996), 500-4), Hockney (Trends in Biotechnology 12 (1994), 456-463), Griffiths et al., (Methods in Molecular Biology 75 (1997), 427-440)などがある。酵母の発現系に関する総説には例えば、Hensingら(Antonie van Leuwenhoek 67 (1995), 261-279)、Bussineauら(Developments in Biological Standardization 83 (1994), 13-19)、Gellissenら(Antonie van Leuwenhoek 62 (1992), 79-93、Fleer (Current Opinion in Biotechnology 3 (1992), 486-496)、Vedvick (Current Opinion in Biotechnology 2 (1991), 742-745)、およびBuckholz (Bio/Technology 9 (1991), 1067-1072)などがある。
【0044】
発現ベクターは、文献に詳細に説明されている。一般に発現ベクターは、選択マーカー遺伝子、および選択された宿主における複製を確実なものとするための複製起点を含むだけでなく、細菌またはウイルスのプロモーター、ならびに多くの場合は転写終結シグナルも含む。プロモーターと終結シグナルの間には一般に、コードDNA配列の挿入を可能とする少なくとも1か所の制限酵素切断部位すなわちポリリンカーが存在する。
【0045】
遺伝子の構成的な発現を確実なものとするためのプロモーター、および対象遺伝子の発現の細かな制御を可能とする誘導型のプロモーターを使用することが可能である。このような特性を有する細菌およびウイルスのプロモーター配列は文献に詳述されている。微生物(例えば大腸菌や出芽酵母(S. cerevisiae))における発現の調節配列は文献に詳しく記載されている。下流に位置する配列の特に強い発現を可能とするプロモーターは例えばT7プロモーター(Studier et al., Methods in Enzymology 185 (1990), 60-89)、lacUV5、trp、trp-lacUV5 (DeBoer et al., in Rodriguez and Chamberlin (Eds), Promoters, Structure and Function;Praeger, New York, (1982), 462-481 ;DeBoer et al., Proc. Natl. Acad. Sci. USA (1983), 21-25)、Ip1、rac (Boros et al., Gene 42 (1986), 97-100)である。誘導型プロモーターは好ましくは、タンパク質の合成に使用される。このようなプロモーターは、構成的プロモーターの場合より多いタンパク質収量にしばしばつながる。最適な量のタンパク質を得るためには、2段階の工程がしばしば使用される。第1段階では、宿主細胞が、比較的高い細胞密度となるまで最適条件で培養される。第2段階では、使用されるプロモーターの型に応じた転写が誘導される。この点に関しては、乳糖またはIPTG(=イソプロピル-β-D-チオガラクトピラノシド)によって誘導可能なtacプロモーターが特に適している(deBoer et al., Proc. Natl. Acad. Sci. USA 80(1983), 21-25)。転写の終結シグナルも文献に記載されている。
【0046】
適切な宿主細胞の形質転換またはトランスフェクションは、前述の方法の1つで実施可能である。宿主細胞を、使用される特定の宿主細胞の要件に、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素などに関して適合する栄養培地で培養する。マイクロタンパク質を組換え細胞培養物から、硫酸アンモニウムまたはエタノールによる沈殿法、酸による抽出、陰イオン交換クロマトグラフィーもしくは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティクロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーを含む方法で回収して精製することができる。タンパク質の立体構造を完成させるためには、タンパク質の再折りたたみ段階を必要であれば使用することができる。また、高処理能液体クロマトグラフィー(HPLC)を最終的な精製段階に行うことができる。
【0047】
組換え的な作製手順に使用される宿主に依存して、発現されるポリペプチドをグリコシル化することが可能なほか、グリコシル化しないことが可能である。ポリペプチドは、先頭にメチオニンアミノ酸残基を含む場合もある。
【0048】
好ましくは、マイクロタンパク質は最初に、融合タンパク質として組換え的に、有利にはバルナーゼによって作製後に、融合連結部における切断と、これに続く分離によって融合パートナーから放出される。
【0049】
同様に、マイクロタンパク質は、例えば実施例1(後述)で使用される手順などの、当技術分野で報告されている任意の適切な標準的なペプチド合成手順で作製することができる(例えばMerrifield, Methods Enzymol. 289 (1997), 3-13;Hancock, Mol. Biotechnol. 4 (1995), 73-86;and Merrifield, Adv. Enzymol. Relat. Areas Mol. Biol. 32 (1969), 221-296を参照)。
【0050】
本発明のポリペプチドは、当業者になじみのある任意の適切な方法、および文献に記載されている方法で作製することができる。したがって、マイクロタンパク質の設計および作製に関する上記の説明は、本発明のポリペプチドに、これが適切な限りにおいて応用することができる。ポリペプチドが、マイクロタンパク質、および1つもしくは複数の任意で追加されるリンカーに加えて他の構造エレメントを含む場合があることに注目すべきである。これは、N末端もしくはC末端に、および/または内部に存在する可能性のある追加のアミノ酸配列を意味する場合がある。このようなアミノ酸配列は例えば、バルナーゼ、好ましくは不活性型のバルナーゼなどの、ポリペプチドもしくはこの一部の発現および/または精製を容易にする融合パートナーである場合がある(図2参照)。追加のアミノ酸配列は、結合、安定化、検出などの他の機能を果たす場合もある。ペプチド工学およびポリペプチド工学に知悉している当業者であれば、個々の目的に叶う適切なアミノ酸配列を理解するであろう。さらに本発明のポリペプチドは、例えばグリコシル化、リン酸化、リボシル化、PEG化(PEGの結合)、またはフルオロフォアや放射性核種などの検出可能または治療的に有効な部分の付加によって生じる他の構造エレメントを含む場合がある。この場合も、ペプチド工学およびポリペプチド工学に知悉している当業者であれば、適切な手段および方法を理解するであろう。
【0051】
特に好ましい態様では、本発明のポリペプチドは、標的タンパク質に対する特異的な結合活性を有するアミノ酸配列を含む、少なくとも2個のマイクロタンパク質を含む。
【0052】
「特異的な結合活性」という表現は、標的タンパク質以外のタンパク質に対する親和性と比較して高い親和性を意味する。好ましくは、標的タンパク質に対する親和性は、他のタンパク質に対するより少なくとも100倍高く、より好ましくは少なくとも1000倍高い。好ましくは、「特異的な」という表現は、アミノ酸配列が、指定の標的タンパク質以外のタンパク質に有意な程度で結合しないことを意味する。結合は従来の方法で、例えば、標的タンパク質を透析バッグ内でリガンドとともに配置し、平衡に達した後に、透析バッグの内外におけるリガンドの分布を測定する平衡透析で測定可能である。または、標的を固体表面に固定化して、リガンドの結合を濃度依存的に、例えば表面プラズモン共鳴(BiaCore)によって、質量の増分を測定することで測定可能である。
【0053】
「標的タンパク質」という表現は、本発明のポリペプチドとの結合が望ましい作用を有する可能性のある、任意の想定可能な1つもしくは複数のタンパク質の場合がある。
【0054】
特に、「標的タンパク質」という表現は、本発明のポリペプチドの結合に起因する、近傍に位置する2個もしくは2個以上のタンパク質の結合が望ましい作用をもたらすタンパク質を意味する。このようなタンパク質は、この場合、ホモ二量体もしくはヘテロ二量体、または多量体の場合があるが、ホモ二量体または多量体であることが好ましく、および、さらにより好ましくはホモ二量体である。主な局面では、本発明のポリペプチドと標的タンパク質の結合に及ぼす作用は、標的タンパク質の少なくとも2個が近傍に位置するように、本発明のポリペプチド内の少なくとも2個のマイクロタンパク質のそれぞれが1つの標的タンパク質に結合するという事実に基づくことが理解されている。標的タンパク質の局所的な結合が、活性作用または阻害作用のいずれかの作用を標的タンパク質に及ぼす可能性があることもさらに理解される。例えば、ホモ二量体の受容体タンパク質の2つのサブユニットの局所的な結合は、例えばシグナル伝達カスケードを引き起こす、その活性化に至る場合がある。一方、多量体型酵素の基質結合部位に対する本発明のポリペプチドの結合は、酵素に阻害作用をもたらす場合がある。好ましい態様では、該ポリペプチドの結合は、標的タンパク質の天然の結合分子(例えばリガンド)と実質的に同じ作用を有する。この場合、本発明のポリペプチドは、天然の結合分子の活性の少なくとも10%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも100%、およびさらにより好ましくは少なくとも150%または200%を有することが好ましい(該分子の活性を決定する標準的な方法で計算時)。
【0055】
本発明のポリペプチドの特に好ましい態様では、標的タンパク質は、膜結合型受容体であり、有利には、受容体の分子が相互に近傍に位置する際にシグナル伝達カスケードの下流の他のタンパク質を活性化する受容体である。
【0056】
「膜結合型受容体」という表現は、細胞の形質膜に、特に膜貫通型タンパク質として位置し、かつリガンドと結合可能な受容体タンパク質を意味する。好ましくは、リガンドはペプチドまたはポリペプチドである。さらに受容体が、ヘテロ二量体もしくはホモ二量体、または多量体の形状、好ましくはホモ二量体または多量体、より好ましくはホモ二量体の形状を有し、ならびにこのような形状の受容体が、単量体の形状で存在する受容体の活性とは異なる活性を有することが好ましい。こうした活性の変化は、本発明のポリペプチドと受容体の結合によって、本発明によって促進されると想定される。有利には受容体は、二量体または多量体の形状で存在する場合に活性である。
【0057】
先行技術の文献には、上記の定義に一致する多くの膜結合型受容体について記載されているので、それらの活性は、本発明のポリペプチドとの結合によって調節可能である。例には、EPO受容体、表皮成長因子受容体、ヒト成長ホルモン受容体、TGF-β受容体、FGF受容体、受容体チロシンキナーゼファミリーの成員、および少なくとも一部のG-タンパク質共役受容体などがある。膜結合型受容体は例えば、Schlessinger, Cell 110 (2002), 669-672;Spivak-Kroizman, J. Biol. Chem. 267 (1992), 8056-8063;Plotnikov, Cell 98 (1999), 641-650;Remy, Science 83 (1999), 990-993;およびAngers, Annual Review of Pharmacology and Toxicology 42, (2002), 409-435に記載されている。
【0058】
当業者であれば、本発明に従って、膜結合型受容体などの任意の標的タンパク質に特異的に結合可能なポリペプチドを作製することができる。好ましくは、標的タンパク質に結合するリガンドの構造は明らかであり、および関連するリガンド構造を、先行技術に記載された方法(前述)でマイクロタンパク質に移植することができる。
【0059】
またはマイクロタンパク質そのものが、望ましい結合活性を有する。例えば、四量体の酵素トリプターゼ(EP 04 02 2455.2)に対して特異的な結合活性を有するマイクロタンパク質が存在することがわかっている。したがって、野生型マイクロタンパク質、または望ましい結合活性を有するこの変異型の二量体化もしくは多量体化が、本発明のポリペプチドの提供につながる可能性もある。
【0060】
さらに、最新の手法を用いることで、任意の標的タンパク質に対して意図された結合活性を有するペプチドまたはポリペプチドの配列を、同族のリガンドの構造の詳細が未だ不明であっても提供することが可能である。この点に関しては、適切なスクリーニング法、3次元構造の決定、およびコンピューターによるモデリング法について以下に言及する。スクリーニングは例えば、ファージディスプレイされたペプチドライブラリーを出発材料として使用することで、従来の方法で実施できる。さらに、酵母ツーハイブリッドスクリーニング、またはこれに由来する手法を摘要できる。標的タンパク質との結合に重要な役割を果たすペプチド構造が明らかになれば、そのペプチドを前述の対応する方法によって、適切なマイクロタンパク質の足場内に移植することができる。
【0061】
特に好ましい態様では、膜結合型受容体は、トロンボポエチン(TPO)受容体である。
【0062】
TPO受容体に対して特異的な結合活性を有する本発明のポリペプチドは、TPOアゴニストとして作用することが想定される。これは、TPO受容体のホモ二量体化が、後の第2のメッセンジャー経路を誘導することが知られているためである。このようなアゴニストは、例えば血小板減少症のような、血小板産生の増加を必要とする病理学的状態の治療に有用な場合がある。
【0063】
したがって、この態様に従って、本発明のポリペプチドはTPO受容体を刺激することが可能である。「TPO受容体」という表現は、天然のTPO受容体(その機能的な対立遺伝子異型およびイソ型の全てを含む)、好ましくは、単離されたか、もしくは組換え的に作製されたヒトTPO受容体、または同じ生物学的活性を実質的に有する天然のTPO受容体の変異型を意味する。この表現は例えば、TPOと結合可能なTPO受容体の細胞外部分を含む融合コンストラクト、およびTPO結合による融合コンストラクトの二量体形成時に、検出可能な細胞シグナルを放出可能な融合パートナーを意味する場合がある。このような変異型TPO受容体は例えば、実施例1(後述)に記載されたアッセイ法に使用されるハイブリッドのTPO受容体である。
【0064】
「TPO受容体を刺激可能な」という表現は、TPO受容体に結合して2つのTPO受容体分子の二量体形成を誘導することでTPO受容体の活性化を引き起こす化合物の活性を意味する。この活性は、Cwirla et al. (1997)に記載されているような、先行技術で既知の方法で測定することができる。好ましくは活性は、実施例1(後述)に記載されたTPOアゴニストアッセイ法で、すなわち結果を図4および図5に示すルシフェラーゼアッセイ法、または結果を図6に示す骨髄細胞増殖アッセイ法、または結果を図7に示すインビボ血小板増殖試験で測定される。
【0065】
TPO受容体結合に関する態様の好ましい形状では、本発明のポリペプチドは、アミノ酸配列IEGPTLRQWLAARA(SEQ ID NO: 7)を含む。このアミノ酸配列は、TPO拮抗性のマイクロタンパク質二量体を作製する際の基礎として良好に使用される(後述する実施例1を参照)。
【0066】
TPO受容体に特異的に結合する能力を有する他のペプチドは、WO 03/031589などの先行技術に記載されている。このようなペプチド配列は、本発明のポリペプチドに含まれるマイクロタンパク質部分内に移植するために同様に使用可能である。
【0067】
特に好ましい態様では、本発明のTPO受容体結合ポリペプチドは、以下からなる群より選択されるアミノ酸配列を含む少なくとも2個のマイクロタンパク質を含む:
(a)SEQ ID NO: 1〜6の任意の1つで表されるアミノ酸配列、
(b)該ポリペプチド内に存在時にTPO受容体を刺激可能な、(a)のアミノ酸配列の断片、ならびに
(c)該ポリペプチド中に存在時にTPO受容体を刺激可能な、(a)もしくは(b)のアミノ酸配列または断片の少なくとも1残基が置換されている、付加されている、および/または欠失されている機能的等価物。
【0068】
好ましくは、本発明のポリペプチドに含まれる少なくとも2個の、およびより好ましくは全てのマイクロタンパク質は、上記の(a)〜(c)で定義されたアミノ酸配列を有する。さらに、上記の(a)〜(c)で定義されるアミノ酸配列を有するマイクロタンパク質の二量体が好ましく、二量体形成は有利には、ビス-スクシンイミジル-スベラート(DSS)リンカーを使用することで実現される。上記の(a)で定義されるマイクロタンパク質の二量体が特に好ましい。というのはTPO受容体を活性化可能であることが報告されているからである(実施例1、後述)。
【0069】
この態様に従って、上記の(a)で定義されるマイクロタンパク質の配列の断片も、本発明のポリペプチド内に存在する場合に該断片がTPO受容体を刺激可能な場合に使用することができる。「本発明のポリペプチド内に存在する場合」という表現は好ましくは、「上記の説明に従って、二量体の状態で存在する場合」を意味することを意図する。「断片」という表現は当業者に明瞭な意味を有し、および断片が定義されるアミノ酸配列内のアミノ酸残基の部分的に連続した配列を意味する。したがって、標準のアミノ酸配列と比較して、このような断片は、少なくとも1つのアミノ酸残基をN末端に、C末端に、または両端において欠く。環状の標準配列の場合は、断片は少なくとも1つのアミノ酸残基を、該配列の1か所に欠くことで、断片は環状または線状となり得る。好ましくは断片は、6個の保存されたシステイン残基を保持し、およびこれらの存在により、シスチンノットのトポロジーを形成可能となる。断片が、(a)の対応する配列に含まれるTPO受容体の結合アミノ酸配列の全体を保持することがさらに好ましい。
【0070】
「機能的等価物」という表現は、ポリペプチド内に存在する場合TPO受容体を刺激可能な、(a)もしくは(b)のアミノ酸配列または断片の少なくとも1つの残基が、置換されているか、付加されているか、および/または欠失されている、(a)もしくは(b)で定義されるマイクロタンパク質の変異型を意味する。好ましくは機能的等価物は、シスチンノットを形成するようにジスルフィド結合を介して連結される6個のシステイン残基を含むアミノ酸配列を有する。
【0071】
本発明で使用される機能的等価物は例えば、本発明のポリペプチド内に存在する場合にTPO受容体を刺激する活性を有する、相補鎖が(a)または(b)で定義されるマイクロタンパク質をコードするヌクレオチド配列とハイブリダイズするポリヌクレオチドにコードされたマイクロタンパク質の場合がある。
【0072】
この文脈において、「ハイブリダイゼーション」という表現は、例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載されているような、従来のハイブリダイゼーション条件、好ましくはストリンジェントな条件におけるハイブリダイゼーションを意味する。特に好ましい態様では、「ハイブリダイゼーション」という表現は、以下の条件においてハイブリダイゼーションが起きることを意味する:
ハイブリダイゼーション用緩衝液:2×SSC;10×デンハート溶液(Fikoll 400 + PEG + BSA;比1:1:1);0.1% SDS;5 mM EDTA;50 mM Na2HPO4;250μg/mlのニシン精子DNA;50μg/mlのtRNA;または0.25 Mリン酸ナトリウム緩衝液、pH 7.2;1 mM EDTA、7% SDS
ハイブリダイゼーション温度T = 60℃
洗浄用緩衝液:2×SSC;0.1% SDS
洗浄温度T = 60℃
【0073】
(a)もしくは(b)で定義されるマイクロタンパク質または断片をコードするヌクレオチド配列とハイブリダイズする機能的等価物をコードするポリヌクレオチドは原理的には、そのようなタンパク質を発現する任意の生物に由来する場合があるか、またはその修飾型バージョンをコードする場合がある。このようなハイブリッド形成性のポリヌクレオチドは例えば、細菌、真菌、植物、または動物のゲノムライブラリーもしくはcDNAライブラリーから単離することができる。このようなハイブリッド形成性のポリヌクレオチドは、本明細書に記載されたマイクロタンパク質をコードするポリヌクレオチド、またはこの一部もしくは逆相補物を使用して、例えば標準的な方法によるハイブリダイゼーションによって同定および単離することができる(例えばSambrook and Russell (2001)、Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAを参照)。
【0074】
このようなハイブリッド形成性のポリヌクレオチドは、ポリヌクレオチドが本発明のポリペプチド内に存在する場合にTPO受容体を刺激可能なポリペプチドをコードする限りにおいて、(a)または(b)で定義されるマイクロタンパク質をコードするポリヌクレオチドの内の1つの断片、誘導体、および対立遺伝子変異型も含む。この文脈における「誘導体」という表現は、このようなポリヌクレオチドの配列が、1か所もしくは複数の位置に関して上記のように定義されるマイクロタンパク質をコードするポリヌクレオチドの1つの配列と異なり、ならびに、これらの配列に対して、好ましくはタンパク質の機能に不可欠な配列範囲内の配列に対して高度の相同性を示すことを意味する。誘導体は、シスチンノットを形成するようにジスルフィド結合を介して連結される6個のシステイン残基を含むアミノ酸配列をコードすることが特に好ましい。
【0075】
ヌクレオチド配列とハイブリダイズするポリヌクレオチドの性質は同様に、ポリヌクレオチドが、上記の(a)または(b)で定義されるマイクロタンパク質のアミノ酸配列に対して、少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも60%、および特に好ましくは少なくとも70%、特に好ましくは少なくとも80%、ならびにさらにより好ましくは少なくとも90%の相同性すなわち配列同一性を有するポリペプチドをコードすることを意味する。さらに、ヌクレオチド配列とハイブリダイズするポリヌクレオチドの性質は、ポリヌクレオチドが、上記の(a)または(b)で定義されるマイクロタンパク質をコードするヌクレオチド配列と比較時に、少なくとも40%、好ましくは少なくとも50%、より好ましくは少なくとも60%、さらにより好ましくは65%以上、特に少なくとも70%、特に好ましくは少なくとも80%、特に少なくとも90%、およびさらにより好ましくは少なくとも95%の相同性すなわち配列同一性を有することを意味する場合がある。
【0076】
好ましくは、相同性の程度は、個々の配列を、SEQ ID NO: 1〜6の任意の1つのアミノ酸配列と比較することで決定される。比較される配列が同じ長さを有しない場合は、相同性の程度は好ましくは、長い方の配列内の個々の残基と同一な、短い方の配列内のアミノ酸残基またはヌクレオチド残基のパーセンテージを意味する。相同性の程度は、ClustalW解析を併用するDNAstarプログラムなどの既知のコンピュータープログラムを使用して、従来の手順で決定することができる。このプログラムは、DNASTAR, Inc., 1228 South Park Street, Madison, WI 53715から、またはDNASTAR, Ltd., Abacus House, West Ealing, London W13 OAS UK (support@dnastar.com)から入手可能であり、およびEMBLの支部に設置されたサーバーからアクセス可能である。
【0077】
Clustal解析法を使用して、特定の配列が、参照配列に対して例えば80%同一であるか否かを判定する場合、好ましくはアミノ酸配列の比較について、以下の設定を用いる:マトリックス:blosum 30;オープンギャップペナルティ(open gap penalty):10.0;ギャップ伸長ペナルティ(extend gap penalty):0.05;ディレイ・ディバージェント(delay divergent):40;ギャップセパレーション距離(gap separation distance):8。ヌクレオチド配列の比較時は、ギャップ伸長ペナルティは好ましくは5.0に設定される。
【0078】
好ましくは、ハイブリダイズするポリヌクレオチドの相同性の程度は、コード配列の全長に対して計算される。さらに、このようなハイブリッド形成性のポリヌクレオチド、および特にこれに含まれるコード配列は、少なくとも75ヌクレオチド、および好ましくは少なくとも100ヌクレオチドの長さを有することが好ましい。
【0079】
好ましくは、本発明と関連して使用されるマイクロタンパク質をコードするポリヌクレオチドとハイブリダイズする配列は、具体的に開示されたマイクロタンパク質をコードするポリヌクレオチドに対して少なくとも90%、好ましくは少なくとも93%、より好ましくは少なくとも95%、さらにより好ましくは少なくとも98%、および特に好ましくは少なくとも99%の同一性を有する領域を含み、この相同性を有する領域は、少なくとも75ヌクレオチド、ならびに好ましくは少なくとも100ヌクレオチドの長さを有する。
【0080】
相同性はさらに、比較対象のポリヌクレオチド、またはこれにコードされるポリペプチドの間の機能的および/または構造的な等価性の存在を意味する。上記の分子に相同であり、ならびに、これらの分子の誘導体であるポリヌクレオチドは通常、同じ生物学的機能を有する、これらの分子の変異体である。これらは、天然の変異、好ましくは上記の(a)もしくは(b)で定義されるマイクロタンパク質をコードするポリヌクレオチドのオーソログ、例えば他の対立遺伝子、品種、種などに由来する配列か、もしくは突然変異を含む場合があり、該突然変異は天然で形成される可能性があるか、または意図的な突然変異誘発によって作製可能である。異型、例えば対立遺伝子変異型は、天然の変異型か、または化学合成によって作製される変異型、もしくは組換えDNA手法によって作製される変異型、もしくはこれらの組み合わせの場合がある。上記の特定のマイクロタンパク質をコードするポリヌクレオチドからの逸脱は例えば、欠失、置換、挿入、および/または組換えによって、例えば2種類もしくは2種類以上の異なるマイクロタンパク質の融合によって作製可能である。DNAまたはRNAのいずれかに対して実施可能な核酸の修飾は、当業者に既知の標準的な手法で実施できる(例えば、SambrookおよびRussell、「Molecular Cloning, A Laboratory Manual」;CSH Press, Cold Spring Harbor, 2001、またはHigginsおよびHames(編)、「Protein expression. A Practical Approach」、Practical Approach Series No. 202. Oxford University Press, 1999)。好ましくはDNAの増幅はポリメラーゼ連鎖反応(PCR)で達成され、および修飾は、例えばテンプレート配列に関する突然変異を含むプライマーオリゴヌクレオチドの適切な選択によって使用される(例えばLandt, Gene 96 (1990), 125-128参照)。
【0081】
本明細書に開示された具体的なマイクロタンパク質の変異型であるポリペプチドは、該マイクロタンパク質と共通して有する特徴を有する。これらは例えば、生物学的活性、分子量、免疫学的反応性、構造など、および例えばゲル電気泳動における移動の挙動、クロマトグラフィー時の挙動、沈降係数、溶解性、分光学特性、安定性、最適pH、最適温度などの物理的特性を含む。
【0082】
さらに好ましい態様では、本発明は、標的タンパク質が二量体またはオリゴマーの酵素であり、およびポリペプチドの少なくともマイクロタンパク質が該酵素の活性部位に特異的に結合するアミノ酸配列を含む、本発明のポリペプチドに関する。
【0083】
この態様は特に、二量体または多量体の構造を有する、およびこれに対応して2か所もしくは多数の活性部位を有する酵素に関する。したがって、本発明のポリペプチドは、該ポリペプチドによって露出された結合ペプチドが酵素の活性部位に適合可能なように直接的に設計可能であることが想定される。ポリペプチドと酵素の結合は主に、基質の活性部位への接近を妨げることで酵素に阻害作用を有すると想定される。二量体もしくはオリゴマーの酵素の例には、代表的なものにトリプターゼ、プロテアソーム、カテプシンC、またはヒトグランザイムAなどがある。このような酵素は、Loidl, Biochemistry 96 (1999), 5418-5422;Turk, Biol. Chem. 378 (1997), 141-150;およびBell, Nat. Struct. Biol. 10 (2003), 527-534などの先行技術の文献に記載されている。
【0084】
特に好ましい態様では、二量体またはオリゴマーの酵素はトリプターゼである。マイクロタンパク質がトリプターゼ(EP 04 02 2455.2)を阻害するのに使用可能であることが最近報告されている。トリプターゼは活性状態では、個々の単量体が活性部位を含む四量体酵素なので、マイクロタンパク質の二量体または多量体がトリプターゼ阻害剤として特に適していると想定される。
【0085】
「トリプターゼ」という表現は、90%〜98%の配列同一性を有するα-トリプターゼ、I-トリプターゼ、II/β-トリプターゼ、およびIII-トリプターゼの、これまで知られている4種類の近縁の酵素を含む(Miller, 1998;Vanderslice, 1990)。トリプターゼは、ヒトのマスト細胞の主要な分泌型プロテアーゼであり、ならびに神経ペプチドのプロセシングおよび組織炎症に関与すると考えられている。成熟型のヒトトリプターゼは、四量体のグリコシル化分子であり、ヘパリン結合性を有し、および不均一な、触媒活性を有するサブユニットを含む(例えばVanderslice et al., 1990;Miller et al., 1989, Sommerhoff et al., 1999を参照)。トリプターゼは、マスト細胞の分泌顆粒中に貯蔵されている。マスト細胞の活性化後にヒトトリプターゼは、さまざまな生体液中に見出される。本発明との関連で、マイクロタンパク質の好ましい標的は、マスト細胞のトリプターゼ、より好ましくはβ-トリプターゼまたはα-トリプターゼである。好ましくは、このようなトリプターゼはヒトトリプターゼである。
【0086】
阻害性のトリプターゼの活性は、先行技術に記載され、および以下に概説する方法で検討することができる。
【0087】
トリプターゼ阻害活性の適切なアッセイ法は、EP 04 02 2455.2で説明されている。したがって、阻害性の活性阻害剤の濃度は、トリプシンによる滴定で決定可能である。この目的で、ウシ膵臓トリプシンが、p-ニトロフェニルp'-グアニジノ安息香酸(Chase & Shaw, 1970)を使用する活性部位の滴定によって標準化可能であり、活性阻害剤の濃度は、阻害剤とトリプシンの相互作用が1:1と仮定して計算される。阻害剤とトリプシンおよびトリプターゼとの複合体の見かけ上の平衡解離定数(Kiapp)は基本的に、Bieth (Bull. Eur. Physiopathol. Respir. 16 (Suppl.)(1980), 183-197)に記載された手順で決定できる。簡単に説明すると、阻害剤の濃度を上昇させながら一定濃度の酵素とインキュベートする。次に基質を添加し、残存する酵素の活性を測定する。Kiapp値は、定常状態の速度を、緊密な結合を示す阻害剤に関する方程式(Morrison, 1969)に、非線形回帰解析で適合させることで計算される。任意のマイクロタンパク質のトリプターゼの阻害活性の指標となる見かけのKi値(Kiappとも表記される)の計算は、Morrison (1969)の手順に従って実施することができる。典型的には、本発明の好ましい態様のポリペプチドは、Kiが最大で1 mM、好ましくは最大で0.5 mM、より好ましくは最大で0.2 mM、さらにより好ましくは最大で0.1 mM、さらに好ましくは最大で0.05 mM、特に好ましくは最大で0.02 mM、特に好ましくは最大で0.005 mMであるトリプターゼ阻害活性を有する。最も好ましいKiは、最大0.002 mMである。活性アッセイ法で決定される値は、用いられる特定のアッセイ法の系に典型的な誤差範囲内を変動し得ると理解すべきであり、好ましくは±20%以内、さらに好ましくは±10%以内、および特に好ましくは5%以内の範囲である。
【0088】
トリプターゼの阻害に使用される本発明のポリペプチドがトリプシンに阻害活性を追加的に示すことがさらに好ましい。トリプシン阻害に関する試験は、正しい折りたたみトポロジーの形成の指標となる場合がある。適切なトリプシン阻害アッセイ法は、Van Nostrand (1990)およびSinha (1991)に記載された方法に基づくEP 04 02 2455.2に記載されている。好ましくはポリペプチドは、最大1 nM、および好ましくは最大0.5 nMの範囲の対トリプシンKiを示す。有利には、治療への応用に望ましい可能性のあるトリプターゼに対する高い選択性を考慮して、ポリペプチドは、トリプシンや血液凝固因子などの他のプロテアーゼに関して比較的低い阻害活性を示すことが好ましい。
【0089】
本発明はさらに、上記の態様に従って本発明のポリペプチドをコードするヌクレオチド配列を含む核酸分子に関する。
【0090】
この態様は、含まれるコード配列が、本発明のポリペプチドの二量体または多量体の構造を反映する核酸分子を意味することが意図される。これは好ましくは核酸分子が、マイクロタンパク質の単量体が直接的に、またはペプチドリンカーを介して連結される融合タンパク質をコードすることを意味する。しかしながら核酸分子が、本発明のポリペプチドの前駆体として2個か、またはこれ以上のマイクロタンパク質をコードし、適切なプロセシング段階後に初めて、例えば化学的な修飾によって初めて、本発明のタンパク質を生じることも意図される。
【0091】
本発明の核酸分子は、任意のタイプのポリヌクレオチド、例えばDNA分子もしくはRNA分子、またはこれらの組み合わせの場合がある。これらのポリヌクレオチドは、当技術分野で周知の任意の適切な手法で得られ、例えば合成的に、またはPCRなどのインビボもしくはインビトロにおける組換え手法で作製することができる。このようなポリヌクレオチドは、従来技術で知られている、任意の修飾を含む場合がある(修飾の例として例えばUS 5525711、US 4711955、US 5792608、またはEP 302175を参照)。このようなポリヌクレオチドは、1本鎖または2本鎖、直鎖状または環状の場合があり、サイズに制限は何らない。好ましくは、核酸分子はDNAまたはmRNAである。
【0092】
本発明の融合タンパク質をコードする核酸分子は一般に、組換え型の核酸分子である。「組換え型の核酸分子」という表現は、事前には、結果として生じる組換え核酸分子のように連結されていない、核酸分子またはこの一部分を人工的に混合するのに有用な手法で作製された任意の核酸分子を意味する。適切な手法は例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y. およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989)に代表される先行技術から利用できる。
【0093】
好ましい態様では、組換え核酸分子に含まれる核酸分子は、原核細胞または真核細胞における発現を可能とする発現制御配列に使用可能に連結される。適切な発現制御配列は、標的宿主生物に応用可能なプロモーターを含む。このようなプロモーターは、原核生物界および真核生物界の多様な宿主に関して当業者に周知であり、ならびに文献に記載されている。例えば、このようなプロモーターは、天然の遺伝子から単離することが可能なほか、合成プロモーターまたはキメラプロモーターの場合がある。同様に、プロモーターは標的ゲノム上に既に存在する場合があり、および技術分野で周知の適切な手法、例えば相同的組換えなどでコード配列に連結可能である。
【0094】
本発明の核酸分子は、遺伝子工学の領域で従来から使用されている、特にプラスミド、コスミド、ウイルス、およびバクテリオファージなどのベクター上に存在する場合がある。このようなベクターも本発明の一部である。
【0095】
本発明の好ましい態様では、本発明のベクターは、真菌細胞、植物細胞、微生物の細胞、または動物細胞、特に哺乳類細胞の形質転換に適している。好ましくは、このようなベクターは、酵母や細菌、特に大腸菌などの微生物の形質転換に適している。当業者に周知の方法で組換えベクターを構築することができる;これについては例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y.、およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989)に記載された手法を参照されたい。またはベクターは、標的細胞への輸送目的で内部に本発明の組換え核酸分子を再構成可能なリポソームの場合がある。
【0096】
有利には、ベクターに含まれる核酸分子、および本発明のポリペプチドをコードする核酸分子は、宿主細胞における該ポリペプチドの発現を可能とする1つもしくは複数の発現制御エレメントに使用可能に連結される。
【0097】
原核細胞または真核細胞における、例えば大腸菌における本発明の核酸分子の発現には興味深い点がある。というのは、これらの分子にコードされたタンパク質の生物学的活性のより正確な解析が可能となるからである。加えて、改変された生物学的特性を有することになるタンパク質の合成に至る、分子生物学分野で一般的な方法(例えばSambrook and Russell, 2001, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYを参照)で、多様な追加的な突然変異を核酸分子中に挿入することが可能である。この点に関しては、一方では核酸分子が、コードDNA配列の5'端または3'端からの進行的な欠失によって作られ、および該核酸分子が、これに対応して短縮化されたタンパク質の合成に至る欠失突然変異体を作製することが可能である。他方、点突然変異の導入も、アミノ酸配列の修飾が例えばタンパク質の生物学的活性に影響を及ぼす位置において想定される。
【0098】
原核細胞を対象とした遺伝子工学では、本発明の核酸分子、またはこれらの分子の一部を、DNA配列の組換えによる突然変異誘発または配列修飾を可能とするプラスミド中に導入することができる。標準的な方法(Sambrook and Russell, 2001, Molecular Cloning: A laboratory manual, Cold Spring Harbor Laboratory Press, NY, USAを参照)では、塩基を交換したり、天然配列または合成配列を追加したりすることができる。アダプターおよびリンカーを断片に加えることでDNA断片を相互に連結することができる。さらに、適切な制限酵素切断部位を提供するか、または過剰なDNAもしくは制限酵素切断部位を除去することができる工学的手段を用いることができる。挿入、欠失、または置換が可能な、これらの場合、インビトロ突然変異誘発、「プライマー修復」、制限、または連結を使用することができる。一般に、配列解析、制限酵素解析、ならびに生化学および分子生物学の他の方法が解析法として実施される。
【0099】
他の局面では、本発明は、本発明のポリペプチドまたは本発明の核酸分子、ならびに任意で、薬学的に許容される担体を含む薬学的組成物に関する。
【0100】
このような薬学的組成物は、治療的有効量のポリペプチドまたは核酸分子、および任意で、薬学的に許容される担体を含む。薬学的組成物は、生理学的に許容される担体とともに、本明細書に記載された手順で患者に投与することができる。特定の態様では、「薬学的に許容される」という表現は、規制当局または他の一般に認められている、動物、具体的には人間を対象とした使用に関する薬局方によって承認されていることを意味する。「担体」という表現は、治療薬とともに投与される希釈物、アジュバント、賦形剤、または溶媒を意味する。このような薬学的担体は、水および、石油、動物油、植物油、またはピーナッツオイル、ダイズ油、鉱油、ゴマ油などの合成油を含む油などの無菌性の液体の場合がある。薬学的組成物が静脈内に投与される場合は、水が好ましい担体である。生理食塩水溶液および水性デキストロースおよびグリセロール溶液を、特に注射用溶液用の液体担体として使用することもできる。適切な薬学的賦形剤は、デンプン、グルコース、乳糖、ショ糖、ゼラチン、マルト、コメ、コムギ、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリン酸グリセロール、タルク、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピレングリコール、水、エタノールなどを含む。このような組成物は、望ましいならば、微量の湿潤剤もしくは乳化剤、またはpH緩衝剤を含む場合もある。このような組成物は、溶液、懸濁液、乳濁液、錠剤、丸剤、カプセル、粉末、徐放性製剤などの形状を取り得る。このような組成物は、トリグリセリドなどの従来の結合剤および担体を使用して坐剤として製剤化することができる。経口用製剤は、薬学グレードのマンニトール、乳糖、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどの標準的な担体を含む場合がある。適切な薬学的担体の例は、E.W. Martinの「Remington's Pharmaceutical Sciences」(前述)に記載されている。このような組成物は、治療的有効量の前述のマイクロタンパク質を、患者への適切な投与用の形状として提供するために、適量の担体とともに、好ましくは純粋な状態で含む場合がある。剤形は、投与様式に適したものとすべきである。
【0101】
他の好ましい態様では、組成物は、常用の手順に従って、ヒトへの静脈内投与に適合する薬学的組成物として製剤化される。典型的には、静脈内投与用の組成物は、無菌性の等張性の水性緩衝液を溶媒とする溶液である。必要であれば、組成物は可溶化剤、および注射部位における痛みを和らげるためのリグノカインなどの局所麻酔薬も含んでもよい。一般に、このような内容物は個別に、または単位投与剤形として混合状態で、例えば、活性薬剤量が表示されたアンプルやサシェ(sachette)などの密封容器中の凍結乾燥粉末、または水を含まない濃縮物のいずれかによって提供される。組成物が注射で投与される場合は、無菌性の薬学グレードの水または生理食塩水を含む注入ボトルに分注することが可能である。組成物を注射で投与する場合は、内容物を投与前に混合可能なように、注射用の無菌水または生理食塩水を含むアンプルを提供することができる。
【0102】
本発明との関連で使用される薬学的組成物は、中性または塩の状態として製剤化することができる。薬学的に許容される塩は、塩酸、リン酸、酢酸、シュウ酸、酒石酸などに由来する塩などの陰イオンによって形成される塩、およびナトリウム、カリウム、アンモニウム、カルシウム、水酸化第2鉄、イソプロピルアミン、トリエチルアミン、2-エチルアミノエタノール、ヒスチジン、プロカインなどに由来する塩などの陽イオンによって形成される塩を含む。
【0103】
最適投与量範囲の決定の一助とするために、インビトロアッセイ法を任意で利用できる。製剤化に使用される正確な用量は、投与経路、および疾患または障害の重傷度に依存することもあり、ならびに実施者の判断、および個々の患者の置かれた状況に従って決定されるべきである。有効用量は、インビトロモデルまたは動物モデルの試験系から得られた用量反応曲線から推定することができる。好ましくは、薬学的組成物は、直接的に投与されるか、またはアジュバントと組み合わせて投与される。
【0104】
本発明と関連して、「対象」という表現は、本発明のポリペプチドを個体に投与することで関連づけられる、治療が必要とされる個体を意味する。好ましくは、対象は脊椎動物であり、さらにより好ましくは哺乳類であり、特に好ましくはヒトである。
【0105】
「投与される」という表現は、マイクロタンパク質を含む前述の薬学的組成物の治療的に有効な用量の個体への投与を意味する。「治療的有効量」という表現は、投与されることで効果を生じる用量を意味する。正確な用量は、治療の目的に依存し、および当業者によって既知の手法で確認される。当技術分野で周知なように、および上述したように、全身か局所への輸送、年齢、体重、全般的健康、性別、食事、投与時間、薬剤相互作用、および状態の重症度を考慮した調整が必要な場合があり、当業者によって常用の実験法で確認される。
【0106】
このような方法は、ヒトの治療と獣医学的応用の両方に応用される。所望の治療的活性を有する、本明細書に記載された、生理学的に許容される担体に含まれる化合物を患者に、本明細書に記載された手順で投与することができる。導入様式に依存して、化合物を後述するさまざまな方法で製剤化することができる。製剤中の治療的活性化合物の濃度は、約0.1〜100重量%を変動し得る。薬剤は単独で、または他の治療法と組み合わせて投与することができる。
【0107】
薬学的組成物の投与は、経口、皮下、静脈内、動脈内、節内、髄内、髄腔内、脳室内、鼻腔内、気管支内、経皮、節内、直腸内、腹腔内、筋肉内、肺内、膣内、直腸内、または眼内を含むがこれらに限定されない、前述の多様な方法で実施できる。例えば創傷や炎症の治療における状況によっては、薬学的に有効な薬剤は、溶液乾燥スプレーとして直接的に投与される場合がある。
【0108】
担当医および臨床的因子が投与法を決定する。医学分野で周知なように、任意の1人の患者あたりの投与量は、患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、投与時間および投与径路、全般的健康、ならびに同時に投与される他の薬剤を含む数多くの因子に依存する。典型的な用量は例えば、0.001〜1000μgの範囲の範囲を取り得る;しかしながら、この例示的な範囲に満たない用量、または上回る用量が、特に前述の諸因子を考慮して想定される。
【0109】
投与量は、好ましくは週に1回投与されるが、治療の進行中は、投与量は、さらにより長い間隔を設けて投与される場合があり、必要であれば、さらにより短い間隔、例えば毎日投与することができる。好ましい例では、免疫応答が当業者に既知の方法でモニタリングされ、投与量が例えば、時間、量、および/または組成に関して最適化される。進行状況は、定期的な評価によってモニタリング可能である。薬学的組成物は、局所的または全身的に投与することができる。投与は好ましくは、非経口的に、例えば静脈内に行われる。非経口的投与用の調製物は、無菌性の水性または非水性の溶液、懸濁液、および乳濁液を含む。非水性溶媒の例には、プロピレングリコール、ポリエチレングリコール、オリーブオイルなどの植物油、およびオレイン酸エチルなどの注射可能な有機エステルがある。水性担体には、水、アルコール/水溶液、乳濁液、または生理食塩水および緩衝用溶媒を含む懸濁液などがある。非経口的な溶媒には、塩化ナトリウム溶液、リンゲルデキストロース液、デキストロースおよび塩化ナトリウム、乳酸加リンゲル液、または不揮発性油などがある。静脈注射用の溶媒には、溶液、および栄養補充液、電解質補充液(例えばリンゲルデキストロース液を元にした溶液)などがある。保存剤、ならびに例えば抗菌剤、抗酸化剤、キレート化剤、および不活性ガスなどの他の添加剤を含めることもできる。
【0110】
好ましい態様では、薬学的組成物は、吸入用のエアロゾルとして製剤化される。
【0111】
さらに好ましい態様では、薬学的組成物は、経口投与経路用に製剤化される。
【0112】
別の好ましい態様では、薬学的組成物は、経皮投与用に製剤化される。
【0113】
好ましい態様では、本発明の薬学的組成物は、本発明のポリペプチドを発現する遺伝子輸送ベクターの形状で患者に投与可能なように考案される。エクスビボでベクターによって細胞を形質転換して、形質転換細胞を患者に投与することも好ましい。
【0114】
これらの態様では、本発明の薬学的組成物は、前述した本発明のポリペプチドをコードするポリヌクレオチドを含み、および発現を可能とするベクターである。このようなベクターは、発現ベクターおよび/または遺伝子輸送ベクターの場合がある。発現ベクターは、この文脈では、エクスビボにおける遺伝子治療法で使用されることを意味し、すなわち適切な宿主細胞が体外でトランスフェクトされた後に対象に投与される。遺伝子輸送ベクターは、本明細書では、インビボにおける遺伝子治療への応用に適したベクターを意味し、すなわちベクターは、全身的または局所的のいずれかで対象に直接的に投与される。本明細書で言及されるベクターは、核酸のみからなる場合があるか、または例えば、標的細胞への輸送、例えば循環系における標的輸送、安定性、および/または、生物学的利用能を高める追加的な化合物との複合体である場合がある。このような追加的な化合物の例には、脂質物質、ポリカチオン、膜破壊性ペプチド、または他の化合物、抗体もしくはこの断片、または標的細胞を特異的に認識する受容体結合分子などがある。発現ベクターもしくは遺伝子輸送ベクターは好ましくは、レトロウイルス、ワクシニアウイルス、アデノ関連ウイルス、ヘルペスウイルス、またはウシパピローマウイルスなどのウイルスに由来する場合があり、および標的細胞集団に、例えば呼吸器の細胞への輸送に使用される場合がある。当業者に周知の方法で、組換え発現ベクターまたは遺伝子輸送ベクターを構築することができる;これについては例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y.、およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989)に記載された手法を参照されたい。またはベクターを、標的細胞への輸送用に、リポソーム中に再構成することができる。本発明のポリペプチドをコードするポリヌクレオチドを含むベクターは、細胞宿主のタイプによって変動する周知の方法で宿主細胞に輸送することができる。例えば塩化カルシウムトランスフェクションは、原核細胞に広く使用されており、リン酸カルシウム処理またはエレクトロポレーションは他の細胞宿主に使用可能である(Sambrook、前掲を参照)。
【0115】
エクスビボまたはインビボの遺伝子治療に適したベクターおよび方法は文献に記載されており、当業者に既知である;これについては例えば、Giordano, Nature Medicine 2 (1996), 534-539;Schaper, Circ. Res. 79 (1996), 911-919;Anderson, Science 256 (1992), 808-813;Isner, Lancet 348 (1996), 370-374;Muhlhauser, Circ. Res. 77 (1995), 1077-1086;Wang, Nature Medicine 2 (1996), 714-716;WO 94/29469;WO 97/00957、またはSchaper, Current Opinion in Biotechnology 7 (1996), 635-640、および各文献で引用された参考文献を参照されたい。本発明のこの態様で使用されるベクターは、細胞への直接導入用に、またはリポソームもしくはウイルスベクター(例えばアデノウイルスやレトロウイルス)を介する導入用に設計可能である。
【0116】
好ましい遺伝子輸送ベクターには、バキュロウイルス、アデノウイルス、およびワクシニアウイルスをベースとしたベクターなどがある。これらは好ましくは、非複製型である。
【0117】
本発明は、TPO受容体を刺激することで治療または予防が可能な疾患もしくは状態の治療用または予防用の薬学的組成物を作製するための、前述のTPO受容体に結合してこれを刺激可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用にも関する。
【0118】
したがって本発明は同様に、該ポリペプチドまたは該核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、TPO受容体を刺激することで治療可能な疾患もしくは状態に罹患した個体の治療法に関する。
【0119】
このような医学的応用は、TPO受容体に有効に拮抗するマイクロタンパク質の二量体が作製可能であるという知見に基づく(実施例1、後述)。
【0120】
TPOは、骨髄巨核球から血小板が産生される過程である、巨核球形成の調節に重要な役割を果たす(Kuter et al., 1994, Kaushansky et al., 1994;Wendling et al., 1994, Sauvage et al., 1994)。TPOは肝臓で産生されるが、その主な機能を骨髄で発揮し、骨髄で、幹細胞の巨核球前駆細胞への分化、ならびに巨核球の増殖、倍数体化、および究極的には循環性の血小板体への断片化を刺激する。TPOは、血小板減少症を含む状態の主要調節因子でもあり、および、いくつかの研究で、以下の複数の方法で巨核球形成を亢進させることが明らかにされている:(1)巨核球のサイズおよび数の増加を生じる;(2)巨核球中で倍数性のDNA量の増加を生じる;(3)巨核球の核内分裂を高める;(4)巨核球を成熟させる;ならびに(5)骨髄内で少数のアセチルコリンエステラーゼ陽性細胞の状態の前駆細胞のパーセンテージを高める。TPOのこれらの機能を考慮して、当業者であれば、TPO受容体を刺激することで治療または予防可能な疾患を容易に理解するであろう。このような疾患には、以下に挙げるような多様な形状の血小板減少症、再生不良性貧血などの血液学的疾患、さまざまな状態の骨髄不全、骨髄異形成症候群、および肝疾患が含まれる。TPOの刺激は、輸液と関連して、特に、末梢幹細胞の可動化および分離、血小板の分離(アフェレシス)、インビトロにおける巨核球もしくは血小板の産生、または血小板保存の改善との関連で、または外科手術、特に大手術(例えば心臓もしくは全身の手術)との関連でも有用である。
【0121】
血小板は血液の凝固に必要であり、その数が極めて少なくなると、大出血によって患者が死亡するリスクが患者に生じる。したがって、TPOアゴニストである本発明のポリペプチドは、さまざまな血液疾患、例えば主に血小板異常に起因する疾患の診断と治療の両方における潜在的に有用な応用が期待される。同様に、このようなTPOアゴニストには、血小板減少性状態の治療に、特に、癌もしくはリンパ腫の治療としての化学療法、放射線療法、または骨髄移植に由来する状態の治療に潜在的に応用可能である。実際に、癌患者を対象とした進行中の臨床試験から、高用量カルボプラチン化学療法による投与時に、組換え型のヒトTPOが、血小板減少時期の減少、および血小板回復の促進に有効なことが報告されている(Basser et al., 1997)。同様の結果は、PEG化された巨核球分化因子(peg-MGDF、ヒトTPOのPEG化された短縮型のN末端断片)に関する臨床試験でも得られている(Fanucchi et al., 1997)。血小板減少症の患者における血小板レベルの回復が緩やかなことは重大な問題なので、本発明のポリペプチドがTPO模倣物として作用することで血小板減少症の治療に良好に使用可能であることが想定される。
【0122】
前述の第2の医学的使用または治療法の好ましい態様では、疾患もしくは状態は、血小板減少症、再生不良性貧血、骨髄不全、骨髄異形成症候群、および肝疾患からなる群より選択される。
【0123】
「血小板減少症」という表現は、当分野の専門家によって認識される明瞭な意味を有する。本発明の好ましい態様は特に、例えば化学療法(血液学的な悪性腫瘍または固形腫瘍)、放射線療法、骨髄移植(同種もしくは自家)、または末梢幹細胞移植によって引き起こされる血小板減少症の一形態に関する。さらに、先天性の血小板減少症、免疫による血小板減少症(免疫性血小板減少性紫斑病(ITP))、HIVによる血小板減少症、または薬剤による血小板減少症も対象となる。
【0124】
以上のことから本発明は、TPO受容体を刺激するための、前述したTPO受容体と結合してこれを刺激することが可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用にも関する。好ましくは、使用はインビトロである。
【0125】
他の局面では、本発明は、トリプターゼの活性を阻害することで治療または予防が可能な疾患もしくは状態の治療用または予防用の薬学的組成物を作製するための、前述したトリプターゼに結合してこれを阻害可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用に関する。
【0126】
したがって本発明は同様に、該ポリペプチドまたは該核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、トリプターゼの活性を阻害することで治療可能な疾患または状態に罹患した個体の治療法に関する。
【0127】
本発明のこの局面、すなわち本発明のトリプターゼ阻害性ポリペプチドの治療目的における使用を提供することで、低分子量トリプターゼ阻害剤に関して知られている短所は克服される(例えばNewhouse, 2002を参照)。例えば、このような小型分子は、トリプターゼ以外の分子への結合を引き起こす比較的低い結合特異性のために、投与対象の生物体に毒性作用を示す場合がある。このような小型分子と比較して、本発明のポリペプチド内のマイクロタンパク質は、より大きな相互作用表面積を示すので、より選択性の高い結合が予想される。さらに、タンパク質ベースの結合分子は典型的には、低分子量分子より低い解離速度定数を有するので、標的との、より長期間の結合を有し、このため、より有益な結合特性を有する。
【0128】
加えて、低分子量トリプターゼ阻害剤を凌ぐ他の利点は、本発明のポリペプチドが膜障壁を越えることができないと予想されるという事実である。これは、該ポリペプチドの、マスト細胞の生理学的状態に潜在的に負に影響し得るマスト細胞内に保存されたトリプターゼへの結合を妨げる。小型分子は対照的に、しばしば膜を越える場合がある。さらに、特にシスチンノットタンパク質は、酵素による分解または熱による分解に対して極めて安定なことでよく知られている。
【0129】
治療へのトリプターゼ阻害剤の使用に関する先行技術の文献(例えばNewhouse (2002)、および同文献に引用された参考文献を参照)は広範なため、当業者であれば、どのような疾患が、トリプターゼの活性を阻害することで治療可能または予防可能かを理解すると思われる。
【0130】
好ましくは、上記の第2の医学的使用または治療法は、喘息、炎症、乾癬、肺線維症、間質性肺疾患、慢性関節リウマチ、歯肉炎、歯周炎、アレルギー反応、アレルギー性鼻炎、骨関節症、アテローム性動脈硬化症、血管形成、多発性硬化症、および癌からなる群より選択される疾患もしくは状態に関する。
【0131】
上記と一致して、本発明は、トリプターゼの活性を阻害するための、前述のトリプターゼに結合してこれを阻害可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用にも関する。好ましくは、使用はインビトロである。
【0132】
他の局面では、本発明は、本発明のポリペプチド、または本発明の核酸分子を含むキットにも関する。
【0133】
本発明のキットの成分は、任意で緩衝液および/または溶液を含むバイアルなどの容器内に包装され得る。適切であれば、1つもしくは複数の成分は、1つの同じ容器内に包装され得る。加算的または択一的に、1つもしくは複数の成分を例えばニトロセルロースフィルターやナイロン膜などの固相支持体に、またはマイクロタイタープレートのウェルに吸収させることが可能である。
【0134】
他の局面では、本発明は、以下の段階を含む、マイクロタンパク質内に共有結合を形成させる方法に関する:
(a)N末端に反応性カルボニル基を含み、およびC末端にホモセリンラクトン残基を含むマイクロタンパク質基質を提供する段階;ならびに
(b)前記N末端基およびC末端の残基がヒドラゾン結合に変換するように、マイクロタンパク質基質を反応させる段階。
【0135】
この方法は、マイクロタンパク質の環化、またはマイクロタンパク質へのペプチド配列の移植を含む、マイクロタンパク質の処理に特に有用である。さらに、この方法は、本発明の二量体または多量体の作製に特に有用である。
【0136】
この方法の提供によって、マイクロタンパク質への共有結合の導入に関して、先行技術の手法と比較してマイクロタンパク質の扱いは極めて容易になる。例えば、マイクロタンパク質を環化させる現行の方法は、C末端にチオエステルを導入する化学合成に基づく(Davies, 2003を参照)。このような合成戦略には、収量を上げるための最適化時に問題が生じる場合がしばしばあり、および達成時に問題となることがしばしばあるインビトロにおけるジスルフィド結合の形成を必要とする。というのは、誤ったジスルフィドを有する、かなりの量の副反応産物が生じるからである(Price-Carter et al., 1996)。インテインによる環化にも、この点で問題がある。というのは、インテイン-マイクロタンパク質の融合タンパク質は、還元性条件がジスルフィド結合の形成を除去してしまう、細菌の細胞質に存在するからである。
【0137】
バックボーンの環化が自然に生じる仕組みは未だ不明であるが、複数の手法が、合成環状タンパク質を作製させるために開発されている(Evans et al., 1999, Scott et al., 1999 Tam and Lu, 1998)。一般に、N末端とC末端の連結を達成するには2つの主な戦略がある:第1の方法では、ペプチド結合をC末端およびN末端においてそれぞれ切断するインテインと呼ばれる、修飾型タンパク質のスプライシングエレメントを利用する(Evans et al., 1999;Williams et al., 2002)。仮に、同じ読み枠で標的タンパク質のN末端およびC末端にクローニングされた場合、N末端とC末端が連結されたペプチド結合の形成に自然につながる、N末端のシステインおよびC末端のチオエステルが生じる。
【0138】
第2の戦略は、N末端にシステイン残基を、およびC末端にα-チオエステルを有する標的タンパク質の固相合成に拠っている。仮に両端が相互に近傍に位置すれば、自発的な反応が最初にチオエステル結合型中間体を生じ、これが分子内再編成を受けて、最終的に天然のペプチド結合が生じる(Tam and Lu, 1998, Muir 2003)。この戦略は、標的タンパク質の固相合成と、これに続く環化-酸化、またはこの逆である酸化-環化に拠っている。第1の方法では、直鎖状のBoc合成ペプチドおよび完全脱保護状態のペプチドが最初に大環状分子中に「閉じられ(zipped)」、次に酸化される(Tam and Lu, 1998)。別の方法では、完全に脱保護されたペプチドから、「頭部-頭部」の環化に適切な立体構造を得るための、折りたたまれた開鎖状態の前駆体を作製する。いずれの方法にも要件および限界がある。特に、これらの方法では、方法が化学的に合成されたマイクロタンパク質の使用に制限される、マイクロタンパク質への追加的な機能の化学的な導入が必要とされる。したがって、これらは、組換え的に作製されたマイクロタンパク質では作用しない。というのは、このようなタンパク質は天然のアミノ酸しか含まないからである。しかしながら、コストを考えれば、および望ましくない副産物を避けることを考えれば、組換え的な作製が一般に有利である。後者は通常、切断に至る合成エラーのために、および立体異性体(ラセミ化合物)の形成のために、マイクロタンパク質の化学合成に伴って生じる。本発明の方法では、これらの制限が迂回され、および、適切な発現宿主で正しいジスルフィド結合の結合性を伴って産生される、組換え的に合成されたマイクロタンパク質の環化が合成後に可能となる。
【0139】
マイクロタンパク質の組換え的な作製は、先行技術に記載された任意の適切な方法で、および好ましくは本明細書に記載された手順で実施可能である。本発明の好ましい発現宿主は、大腸菌およびピキア・パストリス(Pichia pastoris)である。
【0140】
本発明の方法は、マイクロタンパク質基質内のN末端とC末端の間のヒドラゾン結合の形成に基づいている。「マイクロタンパク質基質」という表現は、その一次構造、二次構造、および/または三次構造に基づいて、マイクロタンパク質として、多数のマイクロタンパク質として、またはこの一部として認識され得るペプチドもしくはポリペプチドを意味する。好ましくは、この表現は、共有結合の閉鎖に伴ってマイクロタンパク質となる前駆体を意味する。
【0141】
N末端の反応性カルボニル基およびC末端のホモセリンラクトン残基の提供は、当技術分野で周知の適切な方法で可能である。本発明に即して、反応性カルボニル基は、C末端のホモセリンラクトン残基とヒドラゾン結合を形成可能でなければならない。好ましくは、N末端の反応性カルボニル基は、グリオキシリル基またはケト基である。ケト基は、例えばDixon (Methods Enzymol. 25 (1972), 409-419)に記載されているように、N末端のアミノ酸残基の、金属を触媒とする適切なアミノ基転移によって形成され得る。
【0142】
好ましい態様では、グリオキシル基は、使用されるマイクロタンパク質基質内に存在するN末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される。
【0143】
好ましい態様では、マイクロタンパク質基質のN末端のセリン残基またはスレオニン残基は、該マイクロタンパク質基質を含む前駆体ポリペプチドの、メチオニンと後続のセリン残基もしくはスレオニン残基との間のペプチド結合の臭化シアンの使用による切断によって提供される。
【0144】
マイクロタンパク質基質のC末端のホモセリンラクトン残基が、メチオニンと後続のアミノ酸残基間におけるペプチド結合の臭化シアンによる切断によって提供されることも好ましい。この態様、および前述の好ましい態様では、臭化シアンが切断用薬剤として使用されるので、いずれの切断反応も同時に進行可能である。したがって、これらの段階が、例えば、出発材料として個々のマイクロタンパク質を含む1つの担体タンパク質を使用して同時に実施されることが可能であり、および特に好ましい。この過程を図8に示す。
【0145】
段階(b)のヒドラゾン形成は、当技術分野で既知の任意の適切な方法で実施可能である。
【0146】
好ましい態様では、段階(b)は、以下の段階を含む:
(i)C末端のホモセリンラクトン残基をホモセリンヒドラジドと反応させる段階;
(ii)ホモセリンヒドラジドとN末端の反応性カルボニル基を反応させてヒドラゾンを生じさせる段階;ならびに
(iii)任意でヒドラゾンを還元させる段階。
【0147】
これらの方法の段階を、図9および図10に示したマイクロタンパク質の環化の例で説明する。
【0148】
マイクロタンパク質は担体タンパク質から、臭化シアン(CNBr)を使用する化学的切断によって切り離すことができる(図10a)。CNBrはタンパク質のペプチド結合をメチオニンのC末端で切断する。この反応によって、メチオニン残基がホモセリンラクトンに変換される。融合タンパク質が臭化シアンによって切断されると、担体タンパク質が放出され、およびアミンと容易に反応することが知られている(Majerle et al., 2000)ホモセリンラクトンをカルボキシ末端に含む、アミノ末端にセリンを有するマイクロタンパク質が放出される(図10b)。本発明の主な局面は、このラクトンとヒドラジンの反応によるC末端のヒドラジドの生成に関する(図10c)。ヒドラジド形成の他の方法は、活性化エステルの化学合成か、またはヒドラジンの存在下における制限的なタンパク質分解による切断のいずれかに拠る(Gaertner et al., 1992、Rose et al., 1996)。本発明に従って、ヒドラジド形成は好ましくは、臭化シアンによる切断と、これに続くヒドラジン分解によって容易に達成され得る。アミノ末端のセリンは、Geoghegan and Stroh (1992)に記載された手順で、過ヨウ素酸ナトリウムを使用する酸化によって選択的に活性化され得る。この結果、C末端のヒドラジドと容易に反応してヒドラゾンを形成するグリオキシリル基が形成される(図10d)。ヒドラゾンは、シアノホウ化水素ナトリウムによって還元されて、安定な環状のマイクロタンパク質分子が得られる(図10e)。N末端のセリン残基の代わりに、例えばスレオニン残基も、本質的に同じ段階および反応条件で使用可能であることが理解されている。
【0149】
以上のことから、本発明の方法の好ましい態様は、マイクロタンパク質基質が1つの単体のマイクロタンパク質であること、および共有結合の形成が、マイクロタンパク質のペプチドバックボーンの環化のためになされることである。
【0150】
別の好ましい態様では、マイクロタンパク質基質は、1つの単体のマイクロタンパク質であり、および共有結合の形成は、グラフトペプチド(graft peptide)をマイクロタンパク質に連結するためになされる。この応用を、図11に図示する。
【0151】
したがって、特に好ましい態様では、方法は、以下の段階を含む:
(A)グラフトペプチドと置換されるマイクロタンパク質のループアミノ酸の配列を、メチオニンと後続のアミノ酸残基の間の適切なペプチド結合において臭化シアンで切断する段階(ループアミノ酸配列のC末端における後続のアミノ酸残基はセリン残基またはスレオニン残基である);
(B)セリン残基またはスレオニン残基を、グリオキシリル基を形成するように穏やかな酸化によって反応させる段階;
(C)N末端の反応性カルボニル基およびC末端のホモセリンラクトンヒドラジドを含むグラフトペプチド配列を、マイクロタンパク質のグリオキシリル基と反応させてヒドラゾンを生じさせる段階;
(D)段階(A)の切断によって生じたC末端のホモセリンラクトン残基を、ホモセリンヒドラジドと反応させる段階;ならびに
(E)(D)のホモセリンヒドラジドを、グラフトペプチドのN末端の反応性カルボニル基と反応させてヒドラゾンを生じさせる段階。
【0152】
グラフトペプチドのN末端の反応性カルボニル基は好ましくはグリオキシリル基またはケト基であり、グリオキシル基は好ましくは、N末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される。
【0153】
本発明の方法のさらに好ましい態様では、マイクロタンパク質基質は、少なくとも2個のマイクロタンパク質を含み、および共有結合の形成が、マイクロタンパク質の二量体またはオリゴマーの形成のためになされる。
【0154】
同様に、マイクロタンパク質基質は、少なくとも2個のマイクロタンパク質を含み、および共有結合の形成が、大環状分子の二量体またはオリゴマーのマイクロタンパク質の形成のためになされることが好ましい。
【0155】
これらの態様は特に、上述の本発明のポリペプチドの作製に適している。特に、臭化シアンによる切断を介するC末端の活性化と、これに続くヒドラジン分解、およびアミノ末端のアルデヒドによるヒドラゾン形成を含む方法を、ホモ二量体またはヘテロ二量体のマイクロタンパク質の形成に応用することもできる。このために、マイクロタンパク質は例えば、セリン残基またはスレオニン残基をそのアミノ末端に含み、およびメチオニン残基をそのカルボキシ末端に含むように、組換え的に作製することができる。臭化シアンによる切断によって、活性化されたホモセリンラクトンが形成される。マイクロタンパク質調製物は、次に2つの等量部分に分けることができる。一方の半分にはヒドラジン水和物を添加し、これがカルボキシ末端におけるホモセリンラクトンの形成につながる。もう一方の半分については、過ヨウ素酸ナトリウムでアミノ末端を酸化して、アミノ末端にグリオキサル部分を得る。両調製物を混合する。ヒドラゾン形成時には、シアノホウ化水素ナトリウムによって還元可能な直鎖状の二量体が形成される。ホモセリンラクトンのヒドラジン分解およびセリンの酸化の手順は繰り返すことができる。この結果、二量体のアミノ末端におけるグリオキサル形成、およびカルボキシ末端におけるヒドラジド形成に至る。分子内におけるヒドラゾン形成によって大環状分子が形成される。
【0156】
以上の態様および他の態様が開示され、ならびに本発明の記述および実施例に含まれる。本発明に用いられる方法、用途、および化合物の任意の1つに関する他の文献は公共図書館から、例えば電子機器を使用して入手可能である。例えば、インターネット上で利用可能な公的データベース「Medline」を、例えばhttp://www.ncbi.nlm.nih.gov/PubMed/medline.htmlのアドレスから利用することができる。http://www.ncbi.nlm.nih.gov/、http://www.infobiogen.fr/、http://www.fmi.ch/biology/research_tools.html、http://www.tigr.org/などの他のデータベースおよびアドレスは当業者には既知であり、ならびに例えばhttp://www.google.de経由で入手できる。生物工学における特許情報の概要、ならびに遡及的な検索および現状認識に有用な特許情報の提供先の調査については、Berks, TIBTECH 12 (1994), 352-364に記載されている。
【0157】
さらに、「および/または」という表現は、本明細書で使用される場合は、「および」、「または」、および「この表現によって関連づけられる要素の全て、または任意の他の組み合わせ」の意味を含む。
【0158】
本発明を、以下の非制限的な図および実施例を参照することで、さらに説明する。
【0159】
本発明を、以下の実施例で説明する。
【0160】
実施例1:
TPOアゴニストおよびアンタゴニストの産生および測定
材料および方法
分子生物学的手法
実施例において特に言及されない限り、全ての組換えDNA手法は、Sambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USA、またはAusubel et al. (1994), Current Protocols in Molecular Biology, Current Protocolsの第1巻および第2巻に記載されたプロトコルに従って実施される。
【0161】
DNAコンストラクト
バチルス・アミロリクエファシエンス(B. amyloliquefaciens)のRNAseの酵素的に不活性な異型であるバルナーゼ'、および個々のマイクロタンパク質のコード配列からなる融合遺伝子の構築は、基本的に文献に記載された方法で行った。マイクロタンパク質の遺伝子を、標準的なクローニング法(Sambrook et al., 2001)によるポリメラーゼ連鎖反応によって発現ベクターpBar100中に集合させた(図2、EP 04 02 2455.2)。ET-TP-020をコードする遺伝子を、プラスミドpBar100-EETI-II M7I(Schmoldt et al., 2004)をテンプレートとして、ならびにオリゴヌクレオチド対のbarmitte-up

およびETTP21-SOE1lo
、ならびにETTP21-SOE2up-new
、およびcathindmitte-lo
を使用して、Taq DNAポリメラーゼ(NEB)を使用する2段階のSOE PCRで集合させた。結果として得られた産物をゲル電気泳動と、これに続くQIAquick Gel抽出キット(Quiagen)を使用したゲル抽出で精製した。次にこれらを、隣接するオリゴヌクレオチドbarmitte-upおよびcathindmitte-loによる第2のPCR反応のテンプレートとして使用した。得られた生成物をフェノール/クロロフォルム抽出法およびエタノール沈殿法で精製し、NcoIおよびHindIIIで切断し、同様に消化されたpBar100-EETI-II M7I中に連結した。
【0162】
タンパク質の発現および二量体形成
文献に記載された手順で融合タンパク質を発現させて精製した。個々のマイクロタンパク質を、融合パートナーであるバルナーゼ'から、臭化シアン処理によって放出させた。マイクロタンパク質を逆相HPLCによって、文献(Schmoldt et al., 2004、およびEP 04 02 2455.2)に記載された手順で精製した。
【0163】
二量体の変異型を得るために、2等量の個々のマイクロタンパク質をDMF/DMSO(1:1)に溶解した。トリエチルアミンを最終濃度が1%となるように、二機能性クロスリンカーである、1等量のDSS(ビス-スクシンイミジル-スベラート、Pierce)とともに添加した。一晩のインキュベーション後に、結果として得られた二量体の架橋型マイクロタンパク質をRP-HPLCで精製した。
【0164】
Ba/F3レポーター細胞アッセイ法
TPOR/4Rαハイブリッド受容体用の発現ベクターであるpcDNATPOR/4Rαを以下の手順で構築した。ヒトTPO受容体のコード配列を含むcDNAクローンを対象に、オリゴヌクレオチドTpoR-Xho-lo
およびTpoR-Xho-up
を使用してPCR反応を行った。結果として得られたPCR産物をXhoIで消化し、同様に消化されたベクターpASKcDNA-NHに連結してpASKcDNA-NH-TPORを得た。pcDNA/4RαのNheI/HindIII断片(Krause et al., 2004)をpASK21TETIsendc1/2 (Christmann et al., 1999)にサブクローニングすることでpASKcDNA-NHを得た。ベクターpASKcDNA-NH-TPORから、TPORのコード配列を含むNheI/HindIII断片を、同様に消化されたpcDNA/4Rαに連結してpcDNATPOR/4Rαを得た。STAT6レポーター遺伝子コンストラクトpIεTATALucを、ヒトIεプロモーター(Ezernieks et al., 1996)のIL-4応答領域に由来するプロモーター/エンハンサー配列の全体を含む合成HindIII/BamHI断片をルシフェラーゼ発現プラスミドpTATALuc+ (Altschmied et al., 1997)に挿入して作製した。
【0165】
細胞培養、トランスフェクション、およびレポーター遺伝子アッセイ法
マウスのプレB細胞系列Ba/F3を文献(Lischke et al., 1995)に記載された手順で培養した。細胞を、細胞系列Nucleofector(商標)キットV(Amaxa, Germany)を使用してトランスフェクトした。簡単に説明すると、8×106個の細胞をRPMI 1640/10% FCS中で2時間、飢餓状態におき、遠心分離し、4μgの発現ベクターpcDNATPOR/4Rαおよび1μgのレポーター遺伝子コンストラクトpIεTATALucを添加した100μlのトランスフェクション試薬Vに再懸濁した。トランスフェクションは、プログラムT16を使用するNucleofector(商標)装置で実施した。個々のトランスフェクションバッチを3.5 mlのRPMI 1640/10% FCS中に回収し、1ウェルあたり2×105個の細胞/100μlとなるように96ウェル細胞培養プレートに添加した。細胞を何ら処理せずに1時間、静置した後に、さまざまな濃度のhTPO (Immunotools, Germany)、および個々のマイクロタンパク質候補をそれぞれ総容量が200μlとなるように添加した。TPO競合アッセイ法に関しては、細胞のアリコートを個々のマイクロタンパク質試料と1時間プレインキュベートした後にhTPOで刺激した。37℃、5% CO2で12時間のインキュベーション後に、細胞溶解物を調製し、ルシフェラーゼ活性を文献(Krause et al., 2004)に記載された手順で測定した。
【0166】
結果
TpoR結合配列(小文字で示す)が移植されたマイクロタンパク質のアミノ酸配列を表1に示す。マイクロタンパク質の足場として、マイクロタンパク質AGRP'、ヒトアグーチ関連タンパク質(McNulty et al., 2001)のメラノコルチン受容体結合ドメイン、およびマイクロタンパク質EETI-II(Christmann et al., 1999)をそれぞれ使用した。下線を付したアミノ酸をスペーサー配列として、またはSE-AG-TP-050の場合にはTPOR結合ペプチドの配列から外れるように導入した。アンタゴニスト活性の決定に際しては、マイクロタンパク質をバルナーゼ'との融合タンパク質として使用した(図2およびEP 04 02 2455.2)。アゴニスト活性の決定に関しては、精製済みのマイクロタンパク質を二量体として使用した。
表1:トロンボポエチン受容体(TPOR)結合配列によるループ置換を有するマイクロタンパク質のアミノ酸配列。タンパク質を、酵素的に不活性なバルナーゼ(バルナーゼ')との融合タンパク質として産生させ、hTPO受容体の活性化の阻害能力または誘導能力(拮抗作用)を検討した。アゴニスト活性の検討対象のマイクロタンパク質は化学的に二量体化されている。適用した試験はTPORの細胞外ドメインの二量体形成を誘導する能力の判定を含んだ。n.d.:決定せず。
【0167】
図3は、マウスの造血前駆細胞系列Ba/F3を使用したレポーター遺伝子アッセイ法の結果を示す。これらの細胞は、ヒトIL-4受容体由来のhTPOR細胞外ドメインおよび細胞内4Rαドメインの融合体をコードする遺伝子でトランスフェクトされ、ならびにレポーター遺伝子コンストラクトpIεTATALuc (Krause et al., 2004)と同時にトランスフェクトされた。TPO受容体の活性化の阻害を、5 nMのTPOおよび表1に示した500 nMの各化合物を使用して測定した。結果として、直鎖状のペプチド(SN-LN-TP-020)、ならびにマイクロタンパク質SE-AG-TP-030、SE-AG-TP-040、SE-ET-TP-020、およびSE-ET-TP-030は、TPOを介する効率的な受容体活性化に関してTPOのアンタゴニストとして作用可能であった。同様の結果は、いくつかのアッセイ法でSE-AG-TP-040に関して得られた。一方、アゴニスト活性は、検討された全てのマイクロタンパク質に関して認められた(表1)。
【0168】
図4は、SE-ET-TP-020のアゴニスト作用を示す。SE-ET-TP-020マイクロタンパク質は、二機能性の化学リンカーDSSの使用により二量体化させた。Ba/F3を対象としたTPO受容体活性化アッセイ法では、組換え型のヒトTPOと比較するために、精製済みの二量体を使用した。図4は、二量体であるSE-ET-TP-020による活性化と比較して、ヒトTPOによってTPORが活性化することを示す。この結果から、二量体のSE-ET-TP-020がTPOと同じ用量範囲で活性を示すことがわかる。
【0169】
同様に、二量体のSE-AG-TP-040およびSE-AG-TP-050を提供し、TPOのアゴニスト活性を検討した。これらは図5に示す。この結果から、これらのマイクロタンパク質二量体がハイブリッドTPO受容体を効率的に活性化したことがわかる。
【0170】
二量体のSE-ET-TP-020 (「SE-ET-TP-020d」とも呼ばれる)が骨髄細胞の増殖に及ぼす作用を評価するために、初代ヒト骨髄細胞を、50 ng/mlもしくは10 ng/mlのrhuTPO、または30 nM、10 nM、もしくは3 nMの二量体のSE-ET-TP-020dのいずれかを含む、またはいずれも含まない半固体培地で培養した。培養開始から10〜12日後に、大きな細胞(巨核球と推定される)のコロニーを数えた。結果を図6に示す。これらの結果から、二量体マイクロタンパク質SE-ET-TP-020dが、Mk前駆細胞集団の増殖を刺激する能力に関してTPOと似ていることがわかる。
【0171】
SE-ET-TP-020のインビボにおける機能を評価するために、Balb/cマウス(n=5)に1日2回、それぞれ10μg/kgおよび1000μg/kgのSE-ET-TP-020を皮下注射で投与した。5日後に血小板数を、対応するプラセボ実験と比較して測定した。図7からわかるように、SE-ET-TP-020が投与されたマウスでは血小板数が倍加した。
【0172】
実施例2:
マイクロタンパク質の合成後の環化
実験手順
省略形
Boc:tert-ブトキシカルボニル;DTT:ジチオトレイトール;ESI-MS:エレクトロスプレーイオン化質量分析;Fmoc:9-フルオレニルメチルオキシカルボニル;HATU:2-(1H-9-アザベンゾトリアゾール-1-イル)-1,3,3,3-テトラメチルウロニウムヘキサフルオロリン酸;HBTU:2-(1H-ベンゾトリアゾール-1-イル)-1,3,3,3-テトラメチルウロニウムヘキサフルオロリン酸;HOBt:1-ヒドロキシ-1H-ベンゾトリアゾール;HPLC:高圧液体クロマトグラフィー;SPPS:固相ペプチド合成;TFA:トリフルオロ酢酸。
【0173】
材料
試薬および溶媒は最高品質の市販品とし、さらなる精製を行わずに使用した。シアノホウ化水素ナトリウムおよびm-過ヨウ素酸ナトリウムはSIGMA-Aldrichから購入し、ヒドラジン一水和物および臭化シアンはFluka (Taufkirchen, Germany)から購入した。ESI質量スペクトルは、TSQ 700 Finnegan分光器で測定した。HPLCは、YMC J'sphere ODS H-80、RP C-18カラムを調製実験(250×4.6 mm、4μm、80Å)、および解析用試料(250×4.6μm、80Å)に使用して、Pharmacia Actaをベースとするシステムで実施した。
【0174】
発現ベクターpBar100-cyclo-McoEeTIの構築
発現ベクターpBar100-cyclo-McoEeTIの作製では、McoEeTIのコード配列を、Taqポリメラーゼ(Eppendorf)を使用する2段階のSOE-PCRで増幅した。初回のPCRは、プラスミドpBar100-McoEeTI(図2;EP 04 02 2455.2;Schmoldt et al., 2004)をテンプレートとして、ならびにオリゴヌクレオチドBspHI-McoTI-MSDGG-up
およびMCoTI-MSDGGhinten-SOE-lo
、ならびにMCoTI-MSDGGhinten-SOE-up
およびcat-hind-Mitte-lo
をそれぞれ使用して行い、2つの重複する断片を得た。これらの断片を、隣接するオリゴヌクレオチドBspHI-McoTI-MSDGG-upおよびcat-hind-Mitte-loを使用する第2のPCRテンプレートとして使用した。得られた産物をPagIおよびHindIII (MBI Fermentas)で消化し、事前にNcoIおよびHindIIIで切断されたpBar100-EETI-II M7I (Schmoldt et al., 2004)に連結して、発現ベクターpBar100-cyclo-McoEeTIを得た。このプラスミドは、tacプロモーターの制御下に、精製ハンドルとして使用される、バチルス・アミロリクエファシエンス(Bacillus amyloliquefaciens)のRNaseバルナーゼの不活性化突然変異体であるバルナーゼ'、phoA周辺質リーダー配列、およびcyclo-McoEeTI遺伝子の遺伝子融合体を有する。結果として得られるバルナーゼ'-cyclo-McoEeTI融合タンパク質は、2つのメチオニン残基を、バルナーゼ'とcyclo-McoEeTIの境界、およびcyclo-McoEeTIのC末端に有する。これが臭化シアンによって切断されると、融合パートナーであるバルナーゼ'が除去され、ならびにN末端にセリンを、およびC末端にホモセリンラクトンを有するcyclo-McoEeTIペプチドが得られる(例えば図10のa、b参照)。
【0175】
バルナーゼ'-cyclo-McoEeTI融合体の産生および精製
lacI遺伝子を含むヘルパープラスミドpRep4 (Qiagen)を含む大腸菌株71-18[F' lacIq lacZΔM15 proA+B+ Δlac-proAB supE thi1](入手先はB. Muller-Hill)をエレクトロポレーションによってpBar100-cyclo-McoEeTIで形質転換し、25μg/mlのクロラムフェニコールおよび37.5μg/mlのカナマイシンを含む50 mlのリッチ培地中で37℃で一晩、成長させた。産生は、5 Lの培養槽(Bioengineering)中で、文献(Schmoldt et al., 2004)に記載された手順で実施し、精製は、Schmoldt et al.に記載された手順に若干の変更を施して、バルナーゼ'を精製ハンドルとして使用して実施した。簡単に説明すると、バルナーゼ'-cyclo-McoEeTI融合タンパク質を、細胞培養液1リットルあたり55 mlの氷酢酸による酸性化後に培養液から精製した。この培養液を濾過し、H2Oで1:5の割合で希釈し、600 mlのSP-Sepharose XL (Amersham Biosciences)を含む、直径100 mmのガラス製カラムにアプライした。バルナーゼ'-融合タンパク質を100〜1000 mMのNaClの段階的な勾配で溶出し、および融合タンパク質を含むフラクションを、130 mlのAmberchrom CG-300M(Tosoh Bioscience)を含む26 mmのガラス製カラムに直接アプライした。H2O/0.1%(v/v)酢酸で洗浄後に、融合タンパク質をカラムから、0%(v/v)〜90%イソプロパノール/0.1%(v/v)酢酸の勾配を用いて溶出した。約25〜40%(v/v)のイソプロパノールの範囲の融合タンパク質含有フラクションを混合して凍結乾燥した。8 Mの尿素に再溶解し、および50 mMの酢酸アンモニウムに対して透析した後に、別の陽イオン交換クロマトグラフィーを、Vision BioCadワークステーション(PerSeptive Biosystems)に取り付けられたSP-Sepharose XL (Amersham Biosciences)を含むXK26カラム(2.6 x 20 cm、ベッドボリューム100 ml)を用いて流速8 mL/分で行った。溶出は、0〜0.5 M NaClの勾配を設けて行った。フラクションを含む融合タンパク質を混合し、および1/10容量の37% HClを添加してタンパク質を沈殿させ、ならびにHaereus Omnifuge 3L-R中で4000 rpmで10分間、遠心分離した。
【0176】
バルナーゼ'-cyclo-McoEeTIのCNBrによる切断
沈殿した融合タンパク質を、タンパク質1 mgあたり20 mlの0.2 M HCl/8 M尿素に可溶化し、1μlの5 M臭化シアン溶液(Fluka)を添加した(Kaiser and Metzka, 1999)。一晩のインキュベーション後に、試料を、Amberchrom CG-300M(Tosoh Bioscience、ベッドボリューム100 ml)を含むXK26カラム(Amersham Biosciences)に直接アプライした。H2O/0.1%(v/v) TFAで洗浄後に、切断されたcyclo-McoEeTIペプチドをバルナーゼ'から、5%(v/v)〜90%アセトニトリル/0.1%(v/v) TFAの勾配を利用して分離した。約20〜30%(v/v)アセトニトリルの範囲のcyclo-McoEeTIを含むフラクションを混合して凍結乾燥した。追加の精製を、Pharmacia Actaをベースとしたシステムで、YMC J'sphere ODS H-80、RP C-18調製用カラムを使用して行い、5 mgの純粋なMcoEeTI-セリンラクトンを得た。
【0177】
McoEeTIのヒドラジドの形成
ヒドラジン水和物(7μL、140μmol)を、水に溶解したMcoEeTIホモセリンラクトン(2,4.7 mg、1.4μmol)溶液(2 mL)に添加した。この混合物を室温で1時間、攪拌した。反応は、解析HPLCで制御した。開始時の試料に含まれるラクトンのピークが消失した後に、反応混合物を凍結乾燥して過剰なヒドラジンを除去した。凍結乾燥後の乾燥残渣を水-アセトニトリル混合液に再溶解し、および調製用HPLCで精製した。純粋な収率:2.1 mg(44.3%)。HPLC:tR=ESI MS(メタノール)の結果は以下の通りであった:m/z 856.0 ([M+4H]4+, 100)、1152.6 ([M+3H]3+, 33)、692.2 ([M+5H]5+, 27)、1728.6 ([M+2H]2+, 5)。
【0178】
過ヨウ素酸による酸化および環化
McoeETIヒドラジド(1.6 mg、0.46μmol)をリン酸緩衝液(1 mL、0.01 mmol、pH 7)に溶解した。NaIO4(1 mg、4.6μmol)を、リン酸緩衝液を溶媒とする溶液(1 mL)として室温で添加した。5分後にHPLCに注入することで反応を終了させた。モニタリングは、215 nmおよび280 nm(大環状分子の吸光度)で行った。純度は1 mg(63.7%)であった。
【0179】
参考文献
【図面の簡単な説明】
【0180】
【図1】本発明に従って連結された環状の二量体または多量体のマイクロタンパク質の例。(A)C-N結合型の二量体マイクロタンパク質;(B)二量体マイクロタンパク質の大環状分子;および(C)架橋型のマイクロタンパク質オリゴマー。
【図2】バルナーゼ'(barnase')-ICKペプチド融合コンストラクトの設計。(A)酵素的に不活性なH102A変異型の発現に至るバルナーゼ'遺伝子を含むプラスミドpBar100の略図;f1、複製起点;cat、クロラムフェニコール耐性マーカー;tetR、テトラサイクリンリプレッサーのコード遺伝子;colE1、colE1複製起点;tac、tacプロモーター配列;phoAs、アルカリホスファターゼの周辺質シグナル配列;ICK、ICKペプチドのコード配列。(B)バルナーゼ'-ICKペプチド融合体のDNAおよびタンパク質の配列。バルナーゼの#102位において置換されたアミノ酸(H102A)を太字で示す。pBar100シリーズの発現ベクターでは、臭化シアンによる融合タンパク質の化学的切断に使用可能な1つのメチオニンコドンが、バルナーゼ'の配列とICKペプチドのコード配列の境界に存在する。pBar100Thromベクターは、トロンビン認識部位(LVPRGS;SEQ ID NO: 9)を追加的にコードする。
【図3】さまざまなマイクロタンパク質が、TPOを介する受容体の活性化に及ぼす作用。このアッセイ法では、ルシフェラーゼレポーター遺伝子アッセイ法によってTPOR細胞外ドメインの二量体形成が検出される。灰色:500 nMの各マイクロタンパク質の添加後の発光の測定値。緑色:組換え型のヒトTPO(5 nM)を各化合物とともに添加した際の発光。
【図4】組換え型ヒトTPOによる活性化(A)と比較時の、二量体SE-ET-TP-020(B)の添加による、ハイブリッドTPO受容体の用量依存性の活性化。
【図5】二量体のSE-AG-TP-050および二量体SE-AG-TP-020(図5A)および二量体SE-AG-TP-040(図5B)の添加による、ハイブリッドTPO受容体の用量依存性の活性化。
【図6】組換え型ヒトTPOまたは二量体SE-ET-TP-020の添加による骨髄細胞増殖の誘導。大きな細胞のコロニー数を示す。
【図7】プラセボ投与を受けた対照マウスと比較時の、二量体SE-ET-TP-020が投与されたBalb/cマウスにおける血小板力価の約2倍の増加。
【図8】担体タンパク質、および両端でメチオニン残基と隣接するマイクロタンパク質を含む融合タンパク質の略図。
【図9】カルボキシ末端と隣接するメチオニンにおける臭化シアンによる切断によるマイクロタンパク質の環化と、これに続くヒドラジド形成、アミノ末端のセリンの酸化、分子内環化、および結果として生じるヒドラゾンの還元のスキームの概略。
【図10】カルボキシ末端と隣接するメチオニンにおける臭化シアンによる切断によるマイクロタンパク質の環化と、これに続くヒドラジド形成、アミノ末端のセリンの酸化、分子内環化、およびヒドラゾン結合の還元を示す略図。
【図11】ヒドラゾン形成に基づく化学的連結によるマイクロタンパク質のループの置換。Z:ホモセリンラクトンヒドラジド;B:ホモセリンラクトン;X:セリンアルデヒド;およびJ:ホモセリン。組み入れられたペプチドは化学的に合成され、およびC末端にヒドラジド(Z)を有する点に留意されたい。
【図12】cyclo-McoEeTIホモセリンラクトン(A)のcyclo-McoEeTIヒドラジド(B)への変換のRP-HPLC解析。RPカラムからの個々の溶出時間をクロマトグラムに示す。
【図13】10分間のインキュベーション(A)後のcyclo-McoEeTIヒドラジドの過ヨウ素酸ナトリウムによる反応のRP-HPLC解析、および17分で溶出される環状産物のRP-HPLC解析(B)。
【技術分野】
【0001】
本発明は、標的タンパク質に対する特異的な結合活性を有するアミノ酸配列を好ましくは含む、少なくとも2個のマイクロタンパク質(microprotein)を含むポリペプチドに関する。本発明はさらに、このようなポリペプチドをコードするポリヌクレオチド、ならびに薬学的組成物、および該ポリペプチドまたは該ポリヌクレオチドを含むキットにも関する。本発明は、本発明のポリペプチドまたはポリヌクレオチドを投与する治療法、および第2の医学的使用にも関する。本発明は加えて、本発明のポリペプチドを作製するために使用可能な、マイクロタンパク質内に共有結合を形成させる方法に関する。
【背景技術】
【0002】
本発明は、治療に使用されるペプチドおよびペプチド模倣物の分野に関する。先行技術では、治療に応用されるペプチドおよびポリペプチドならびにこれらの類似体の探索は、確立された方法である。このような方法は、ヒトまたは動物の体内で自然に生じるタンパク質-タンパク質相互作用を修飾する試みに基づく主な局面である。特にリガンド-受容体の相互作用は、アゴニストまたはアンタゴニストである人工リガンド分子によって調節可能であることがわかっている。例えば、リガンド分子の二量体形成時には、二量体の膜結合型受容体は、該リガンドの結合によって刺激可能となる場合がある。この方法は、多くがシグナル伝達カスケードを引き起こすことでアゴニストとして作用するサイトカインおよびホルモンに関する研究に基づく。シグナル伝達過程は、個々のアゴニストが2つの受容体分子に結合することで開始されることがしばしばあることが観察されている。多くの場合、このようなアゴニストは、個々の単量体が単量体型の受容体分子と結合することでシグナル伝達過程を活性化する二量体であるか、または空間的に離れた2つの結合部位を含む単体の単量体である(Frank 2002, Grotzinger 2002, Mellado et al., 2001)。
【0003】
細胞表面受容体の活性化が、受容体リガンドによって2つの受容体分子を空間的に連結されることで生じる周知の例がTpo/TpoR系である(Dower et al., 1998;Geddis et al., 2002)。トロンボポエチン(TPO)は、血小板が骨髄巨核球から産生される過程である巨核球形成の調節に重要な役割を果たす、332アミノ酸のグリコシル化されたポリペプチドである(Kuter et al., 1994, Kaushansky et al., 1994;Wendling et al., 1994, Sauvage et al., 1994)。したがってTPOは、さまざまな血液疾患、例えば、血小板異常または血小板減少性状態に主に起因する疾患の診断および治療の両方における潜在的に有用な用途を有する。数年前には、TPOペプチド模倣物の開発が報告されている(WO 96/40750、WO 98/25965、Cwirla et al., 1997)。このようなペプチドは、TpoRに結合して活性化するが、天然のTPOに対する配列相同性は無いものが設計された。このような分子は、TPO受容体(TPOR)と高親和性で結合し、化学的架橋によって天然のサイトカインであるTPOと同程度の活性を有する二量体を形成することで、さらに最適化可能なTPORアゴニストとして作用することが報告されている。活性が高いことがわかっているペプチド配列はIEGPTLRQWLAARAである(Cwirla et al., 1997)。この配列を含むペプチドは、単量体として弱いTPORアゴニストとして、および2つの単量体が架橋されてホモ二量体を形成すると強いアゴニストとして作用する。しかしながら、このペプチドコンストラクトには、治療に使用する際には、以下のいくつかの短所がある:
I)このようなペプチドは、立体構造的に高度に柔軟なので、それ自体がタンパク質分解を受けやすく、このため血漿半減期は短い。加えて、胃内および腸内における、ならびに腸管刷子縁膜の表面上におけるプロテアーゼ(ペプシン、トリプシン、エラスターゼ、キモトリプシンなど)によるタンパク質分解性の攻撃に対する感受性があるために、例えば血小板減少症の治療目的の経口投与は行われていない。
II)Cwirla et al. (1997)に記載されているようなペプチドを組換え的に作製することは困難である。小型のペプチドは、細胞プロテアーゼによってしばしば速やかに分解されることから、微生物発現系において高収量で過剰発現させることは極めて困難である。したがって、望ましくない副産物の発生などの、その全ての既知の短所(例えば、合成の成熟前の終結に起因する切断;または立体異性のアミノ酸の取り込み)のある、化学的なペプチド合成が選択される方法である。
【0004】
他のTPOアンタゴニストペプチドはWO 03/031589に報告されている。しかしながら、それらのペプチドにも前述の短所はある。
【0005】
上記の概要は、個々の生物学的標的系において、アゴニスト作用またはアンタゴニスト作用を有するエフェクター分子として二量体または多量体(ポリ)ペプチドの作製を試みる際に遭遇する状況の例示的なものである。したがって先行技術では、組換え的に容易に作製可能で、および/または長い半減期、高い安定性、および生物学的利用能などの有利なインビボ特性を有する二量体もしくは多量体のペプチドのエフェクター分子の応用を可能とする手段および方法が必要なことは明らかである。
【0006】
マイクロタンパク質は、多様な小型のタンパク質、典型的には、シスチンノットおよび短い3本鎖のβ-シートからなる共通の構造モチーフを有する、最長50残基の長さのタンパク質であると先行技術で記載されている。このようなタンパク質は、小型タンパク質の阻害剤シスチンノット(ICK)ファミリー(Le-Nguyen et al., 1990)の成員としても知られている。これらは共通の構造を有するが、多様な生物学的活性、および無視できるアミノ酸配列の同一性を有する。例には、(i)電位依存性Ca2+チャネルとの高親和性結合によって神経毒として作用する、イモガイ(Conus magus)の毒液に含まれる26残基のポリペプチドであるω-コノトキシンMVIIa (Kohno et al., 1995);(ii)39アミノ酸のペプチドである、ジャガイモのカルボキシペプチダーゼ阻害因子(PCI)(Rees and Lipscomb, 1982);ならびに(iii)テッポウウリ(Ecballium elaterium)由来のEETI-II(Le-Nguyen et al., 1990)などがある。
【0007】
既知のICKファミリーの全てのマイクロタンパク質が、マイクロタンパク質と個々の標的分子の結合による活性の阻害に基づく阻害性の機能すなわちアンタゴニスト機能を有する。加えて、既知の全てのマイクロタンパク質は、開鎖状か環状にかかわらず、単量体のタンパク質として存在する。
【0008】
前述したように、先行技術では、特にアゴニストペプチドまたはアンタゴニストペプチドのエフェクター分子の作製に特に適した方法が求められている。先行技術は、この目的に原理的には有用な可能性のあるペプチド片の環化または連結の過程を提供している。しかしながら、このような過程は一般に不利に作用する。というのは、ペプチド出発材料が化学合成によって、すなわち組換え的ではなく提供されることが求められるからである。Gaertnerらは、組換え的に作製されたペプチドに応用可能なペプチド片を連結させる過程を提案している(J. Biol. Chem. 269 (1994), 7224-7230)。しかし、マイクロタンパク質に応用可能であることは報告されていない。また先行技術の知識に鑑みて、マイクロタンパク質に応用可能か否かは不明であると言わざるを得ない。Gaertnerらが報告した手順は、カルボキシ末端におけるヒドラジドの存在を必要とし、かつ、先行技術に基づくと、組換え的に作製されたマイクロタンパク質から開始した場合にどのようにしてこれが生じるかは明らかではない。さらに、Gaertnerらの報告した反応スキームは、第1の段階に酸化反応を、そしてペプチド連結後に還元を含む。このような反応条件が、望ましくない再シャッフリング、すなわちマイクロタンパク質内に存在する3つのジスルフィドの開裂を生じる可能性があるか否かは現時点で不明である。
【0009】
以上を総合すると、本発明の基礎となる技術上の問題は、長い半減期、安定性、および/または生物学的利用能などのインビボ特性が改善された二量体または多量体のペプチドもしくはポリペプチドの使用を可能とする、作製が容易なペプチド構造の提供、および対応する作製法の提供である。
【0010】
この技術上の問題は、請求項に記載された態様の提供によって解決される。
【0011】
したがって本発明は、少なくとも2個のマイクロタンパク質を含むポリペプチドに関する。
【発明の開示】
【0012】
本発明のポリペプチドによって、機能特性の改善、特に、先行技術の二量体/多量体ペプチド構造と比較して長い半減期、安定性、および/または生物学的利用能などのインビボ特性の改善をもたらすやり方で二量体または多量体の形状のペプチドの機能性の提供を可能とするフレームワーク構造が提供される。加えて、1つもしくは複数のエフェクターペプチドが付加された本発明のポリペプチドは、ペプチドの立体構造の固定化の上昇、およびこれに伴うコンホメーションエントロピーの低下に起因する親和性および選択性の改善を、先行技術のペプチドの二量体/多量体と比較して示すことが想定される。本発明のポリペプチドはさらに、複数のマイクロタンパク質ループの置換による複数のペプチドの導入によって多価性またはヘテロ機能性を導入する可能性を拓くという利点を有する。本発明のポリペプチドの他の有利な特性は、両端におけるタンパク質分解による切断からの保護が、環状のマイクロタンパク質を使用することで容易に達成可能であるという事実である。このような特性の改善は、極めて密なトポロジー、組み入れられたペプチド配列に加わる強固なマイクロタンパク質の3次元構造によって、およびマイクロタンパク質に内在する有利な特性によって説明可能である。
【0013】
驚くべきことに、先行技術に記載されたペプチド二量体に関して知られている機能性、すなわち機能性ペプチド部分が3次元構造に関して高度の自由度を有することが、対応するペプチドが移植されたマイクロタンパク質の二量体に観察可能なことが報告されている。TPOアゴニストペプチドとハイブリッドのTPO受容体の相互作用をモデルとして使用することで、驚くべきことに、このようなペプチドが、二量体のマイクロタンパク質の足場に移植された場合にTPO受容体を効率的に活性化可能であることが明らかになる(添付の実施例1を参照)。特に、TPOR結合配列(IEGPTLRQWLAARA;SEQ ID NO: 7)がマイクロタンパク質の足場に、TPO受容体に対する親和性を失うことなく移植可能であり、および同受容体結合配列を含むマイクロタンパク質の二量体形成に伴って、巨核球の形成をTPO様に誘導する、完全な活性を有するアゴニストが得られるという意外な知見が得られている。図3からわかるように、マイクロタンパク質の2つの足場が、前述のペプチドの組み入れに使用された。1つは、ヒトアグーチ関連タンパク質(McNulty et al., 2001)であるAGRP (AGRP')のメラノコルチン受容体結合ドメインに由来する、シスチンに富む領域の足場に基づくものであり、もう1つは、テッポウウリ(Ecballium elaterium)のトリプシン阻害因子であるマイクロタンパク質(Christmann et al., 1999)に由来する。AGRP'では、メラノコルチン受容体との結合に関与する結合ループ(-CYCRFFNAFCYC-、SEQ ID NO: 20、Joseph et al., 2003)がTpoR結合配列と置換されている。受容体と結合した場合、同ペプチドの構造情報は利用できないため、さまざまなコンストラクトが、ペプチドを、多様なループ構造に強制的に導入することを目的として作製されている。これは、ループ内に追加の残基を導入することで(実施例1の表1に示すSE-AG-TP-040)、またはジスルフィド結合を形成可能な2つのシステイン残基を導入することで(実施例1の表1のSE-AG-TP-050)達成された。実際に特定のループ構造が必要なことは図3に示されている。というのは、必ずしも全てのコンストラクトがアゴニスト活性を示すわけではないからである。
【0014】
他の利点として、本発明では、二量体ペプチドまたは多量体ペプチドのエフェクター分子を高収率で組換え的に作製することが可能となる。1つの態様では、本発明のポリペプチドは、それが少なくとも2個のマイクロタンパク質を含む融合タンパク質である場合には、組換え的発現を応用することで直接的に作製できる。別の態様では、前駆体、例えばマイクロタンパク質の単量体を組換え的に作製した後に、該前駆体を、先行技術の手法か、または以下に示す方法で二量体化または多量体化することができる。マイクロタンパク質の組換え的発現については例えば、EP 04 02 2455.2、Molina et al. (1992)、またはSchmoldt et al. (2004)に記載されている。
【0015】
本発明と関連して、「少なくとも2個のマイクロタンパク質を含む」という表現は、少なくとも2個のマイクロタンパク質が、ポリペプチドが使用される条件、例えば生理学的条件では結合が破壊されないように相互に結合している状態を意味する。本出願では、本発明のポリペプチドは、同意表現として、「マイクロタンパク質の二量体または多量体」とも表現される。「多量体」という表現は、3個またはこれ以上を意味すると理解される。「オリゴマー」または「オリゴマーの」という表現は、原則的には同じ意味であるが、文脈によっては二量体も含む場合がある。この少なくとも2個のマイクロタンパク質の結合は、共有結合および/または非共有結合による場合がある。本発明のポリペプチド内では、マイクロタンパク質は、意図された応用に依存して、考え得るあらゆる配置で存在する場合がある。特に、マイクロタンパク質は、直線状、環状、もしくは分枝状の配置を取り得るか、またはこれらの混合物である場合がある。直線状の構造では、マイクロタンパク質は、いずれかのタンデム配置に、すなわち一方のマイクロタンパク質のC末端が別のマイクロタンパク質のN末端と連結されるように連結され得る。または、マイクロタンパク質は、「頭部と頭部(head-to-head)」または「尾部と尾部(tail-to-tail)」の配置、すなわち一方のマイクロタンパク質のN末端が別のマイクロタンパク質のN末端と連結されるか、および/または一方のマイクロタンパク質のC末端が別のマイクロタンパク質のC末端と連結される配置を取り得る。
【0016】
好ましい態様では、本発明のポリペプチドは環状である。
【0017】
「環状」という表現は、当業者であれば環状のペプチドまたはポリペプチドの構造を推定可能な任意の意味に準じる。特に、「環状」という表現は、それ自体が環状である、例えば環状のペプチドバックボーン構造とシスチンノットを形成する、本発明のポリペプチドに含まれる1つもしくは複数のマイクロタンパク質の単量体を意味する場合がある。同様に、「環状」という表現は、複数のマイクロタンパク質を含む本発明のポリペプチド内の高次の環状構造を意味する場合がある。このような高次の環状構造の好ましい例は、少なくとも2個のマイクロタンパク質のペプチドバックボーンが連結されて1つの環を形成する「大環状分子」である。このようにマイクロタンパク質は、タンデムすなわち「頭部と頭部」もしくは「尾部と尾部」のいずれかの配置を取り得る。
【0018】
特に好ましい態様では、本発明のポリペプチドは、該ポリペプチドのマイクロタンパク質が、一方のマイクロタンパク質のC末端が別のマイクロタンパク質のN末端に共有結合で結合されるように配置される1つの大環状分子を形成する。
【0019】
本発明のポリペプチド内では、マイクロタンパク質は直接的に、または好ましくはリンカー分子を介する様式のいずれかで連結可能である。好ましくは、リンカー分子は二機能性またはオリゴ機能性(oligofunctional)のリンカー分子である。「二機能性」という表現は、2個のマイクロタンパク質が、化学的手段または酵素的手段によって1つのリンカー分子に共有結合的にカップリング可能であることを意味する。「オリゴ機能性」は、2個以上のマイクロタンパク質が、化学的手段または酵素的手段によって1つのリンカー分子に共有結合的にカップリング可能であることを意味する。2個もしくは2個以上のペプチド部分またはポリペプチド部分の連結に適したリンカー分子は、Baumert, Methods Enzymol. 172 (1989), 584-609;Yoshitake Eur. J. Biochem. 101 (1979), 395-399;Pierce Chemicals (1999), Double agents cross-linking reagents selection guide, Pierce Chemicals, Rockford, IL, USA;Peters, Annu. Rev. Biochem. 46 (1977), 523-551;およびFasold, Angew. Chem. Int. Ed. Engl. 10 (1971), 795-801などの先行技術に広く記載されている。したがって当業者であれば、特にその提供ならびにその用途に関して、適切なリンカー分子を承知している。典型的には、リンカー分子は例えば、2つのタンパク質ドメイン間、この場合はマイクロタンパク質間に空間を提供するよう作用し、それにより、例えばその機能性の3次元構造に本ドメインが正しく折りたたまれることを可能にする。さらにリンカー分子は2つのドメイン間に、2つのドメインの、それらの標的構造、例えば二量体のタンパク質受容体への有効な適合に必要な距離を設けることができる。
【0020】
好ましくは、直接的な連結、またはリンカー分子を介した連結はペプチド結合を介して実現される。リンカー分子は、このような場合は、好ましくはペプチドである場合がある。この態様に従って、マイクロタンパク質の二量体/多量体を組換え的に、すなわち対応する発現コンストラクトにコードされた融合タンパク質として作製することが可能である。ペプチドリンカーは好ましくは、比較的短く、例えば最長10アミノ酸、好ましくは最長5アミノ酸からなる。
【0021】
ある応用では、本発明のポリペプチド内の2個のマイクロタンパク質間の結合を破壊することが必要な場合がある。このような状況では、マイクロタンパク質は、切断可能なリンカーを介して連結されることが好ましい。当業者にとっては、対応する切断可能なリンカーは文献から周知である。例えば、切断可能なリンカーは、例えばトロンビン切断部位などの酵素切断部位を含むペプチドリンカーの場合がある。別の方法では、切断可能なリンカー、好ましくはペプチドリンカーを、酵素的切断以外の切断機構に、例えば化学的手段または物理的手段に感受性をもつように設計することもできる。
【0022】
さらに、本発明のポリペプチド内で2個のマイクロタンパク質を連結するリンカー分子を、リンカーが柔軟なヒンジの場合と同様に、特定の動きが可能なように設計することができる。この場合、当業者であれば、対応するペプチドリンカーまたは非ペプチドリンカーの構造について記載された先行技術の文献を参照することができる。
【0023】
本発明のポリペプチドでは、少なくとも2個のマイクロタンパク質が非ペプチドカップリングによって連結されることが好ましい。非ペプチドカップリングは、二機能性またはオリゴ機能性のリンカー分子を含むことが好ましい。特に好ましいのは、アジピン酸ヒドラジド、ビス-スクシンイミジル-スベラート(DSS)、およびEDTA-ヒドラジドから選択されるリンカー分子である。
【0024】
「マイクロタンパク質」という表現は、先行技術の文献で十分に確立した意味をもち、ならびに一般に、最長50アミノ酸の比較的小さいサイズのポリペプチド、および分子内ジスルフィド結合に基づく定まった構造を意味する。マイクロタンパク質は典型的には、極めて安定であり、かつ熱、pH、およびタンパク質分解による破壊に耐性を有する。マイクロタンパク質に関する現在の知見、特に構造および存在に関する知見は例えば、Craik (2001);Pallaghy (1994);およびCraik (J. Mol. Biol. 294 (1999), 1327-1336)にまとめられている。
【0025】
好ましい態様では、本発明のポリペプチド内の個々のマイクロタンパク質は、少なくとも6個のシステイン残基を含み、このうち6個のシステイン残基が1つのシスチンノットを形成するようにジスルフィド結合を介して連結されている。
【0026】
マイクロタンパク質は、阻害因子シスチンノット(ICK)ポリペプチドとしても知られており、および以下の説明のようにも呼ばれる。
【0027】
「シスチンノット」という表現は、2つのジスルフィド結合によって形成された埋め込まれた環と、これらを連結し、第3のジスルフィド結合がくぐるバックボーンセグメントを含む3つのジスルフィド結合フレームワークによって安定化される、バックボーンセグメントの連結の小さな3つのβ-シートを特徴とする、ICKポリペプチドによって形成される3次元構造を意味する。好ましくは、シスチンノットは、6個の保存されたシステイン残基、およびこれを連結するバックボーンセグメントによって形成され、第1のジスルフィド結合は、第1と第4のシステイン残基の間に位置し、第2のジスルフィド結合は、第2と第5のシステイン残基の間に位置し、ならびに第3のジスルフィド結合は、第3と第6のシステイン残基の間にあり、第3のジスルフィド結合は、他の2つのジスルフィド結合、およびこれらを連結するバックボーンセグメントによって形成された環をくぐる。仮に、適切であれば、ジスルフィド結合は、シスチンノットの全体的なトポロジーを同様に確実に形成する、その化学的等価物と置換され得る。あるマイクロタンパク質が正しいシスチンノットを形成するか否かの検討に際しては、当業者であれば、どのシスチン残基が相互に連結されているかを見極めることができる。これは例えば、Gorasson (J. Biol. Chem. 278 (2003), 48188-48196)、およびHorn (J. Biol. Chem. 279 (2004), 35867-35878)に記載された手法で実施可能である。シスチンノットを有するマイクロタンパク質は例えば、Craik (2001);Pallaghy (1994);およびCraik (J. Mol. Biol. 294 (1999), 1327-1336)に記載されている。
【0028】
本発明と関連して使用されるマイクロタンパク質は、開いた状態または環状の構造を取り得るペプチドバックボーンを有する場合がある。開いた構造は好ましくは、N末端にアミノ基を、およびC末端にカルボキシ基を有するマイクロタンパク質を意味する。しかしながら、当業者であればペプチド化学における最新知見を元に推定する、末端の任意の修飾も想定される。閉じた構造では、マイクロタンパク質のペプチドバックボーンの末端は、好ましくは共有結合を介して、より好ましくはアミド(すなわちペプチド)結合を介して連結される。シスチンノットのトポロジーを有する、閉じた構造のマイクロタンパク質は先行技術で「シクロチド(cyclotide)」として、およびそれらの結び目は「環状シスチンノット(CCK)」として知られる。シクロチドは例えば、WO 01/27147およびCraik (Curr. Opinion in Drug Discovery & Development 5 (2002), 251-260)に記載されている。
【0029】
本発明のポリペプチド内のマイクロタンパク質が、アミノ酸モチーフCX3-CX4-CX4-7-CX1-CX4-5-CX5-7(SEQ ID NO: 8)を含むことがさらに好ましい。上式でXは独立に、相互に任意のアミノ酸残基に由来することを意味する。Cは、標準的な命名法に準じてシステインを意味する。好ましくは、アミノ酸Xはシステインではない。配列内のシステイン残基Cは、前述したようなシスチンノットを形成することがさらに好ましい。
【0030】
本発明のさらに好ましい態様では、マイクロタンパク質は28〜40アミノ酸の長さを有する。
【0031】
特定の応用では、本発明のポリペプチド内の1つもしくは複数のマイクロタンパク質が、特定の最大のサイズを超えないことが有利な場合がある。これは例えば、マイクロタンパク質の単量体が、酵素の触媒部位などの、標的タンパク質のポケットに適合しなければならない場合について言える。したがって、本発明との関連で使用されるマイクロタンパク質は、最長35アミノ酸、より好ましくは最長32アミノ酸、および最も好ましくは最長30アミノ酸の長さを有することが特に好ましい。好ましくは、前述の好ましいサイズの範囲は、本発明のポリペプチド内に1個のマイクロタンパク質であり、より好ましくは2個のマイクロタンパク質であり、可能ならば、さらにより好ましくは少なくとも3個のマイクロタンパク質である。最も好ましくは、本発明のポリペプチド内の全てのマイクロタンパク質は、前述のサイズ制限の任意の1つを示す。
【0032】
本発明のポリペプチドに含まれるマイクロタンパク質は、野生型のマイクロタンパク質か、または修飾型のマイクロタンパク質のいずれかの場合がある。修飾は、欠失、付加、置換による修飾、またはペプチド化学の当業者に既知である、翻訳後修飾による修飾などの他の修飾の場合がある。
【0033】
好ましい態様では、マイクロタンパク質は、機能性のペプチド配列がマイクロタンパク質内に移植される本発明のポリペプチドに使用される。「移植する(grafting)」という表現は、本発明との関連で、マイクロタンパク質配列の一部と、望ましい機能を持たせることが意図される別のペプチド配列との置換を意味する。この機能は、例えば受容体、リガンド、抗体、抗原、酵素、または他の結合タンパク質のような標的タンパク質などの標的分子の結合の場合がある。標的分子にペプチド配列が結合すると、標的分子に阻害性または活性性の作用を及ぼす場合がある。有利には、マイクロタンパク質の一部が、マイクロタンパク質の3次元構造の形成に必須ではない置換に選択される。移植型マイクロタンパク質を作製するための数多くの方法が、単量体のマイクロタンパク質に関する先行技術に記載されている。これについては例えば、Craik(Curr. Op. Drug Discovery Design 5 (2002), 251-260), WO 01/27147, Barry, Structure 12 (2004), 85-94;Craik, Toxicon 39 (2001), 1809-1813;Chiche, Current Protein and Peptide Science 5 (2004), 341-349;Rosengren, J. Biol. Chem. 278 (2003), 8606-8616;およびChristmann, Protein Eng. 9 (1999), 797-806を参照されたい。これらの引用文献は以降、参照により本明細書に組み入れられる。典型的には、移植時には、マイクロタンパク質の3次元構造の形成に重要な要素を含まないマイクロタンパク質のループが所望のアミノ酸配列と置換されるか、または部分的に置換される。
【0034】
移植は、文献に記載された方法で実施できる。一般に、所望のアミノ酸配列の組み入れは、マイクロタンパク質のコード配列に、例えばPCR増幅を含む手法などの一般的なインサイチュー突然変異誘発法で突然変異を導入することによって行われる。移植型のマイクロタンパク質は次に、突然変異が導入されたコード配列を適切な宿主で発現されることで提供され得る。この目的で、Sambrook et al. (2001)に記載された方法などの標準的な組換えDNA操作法および発現法を適用することができる。
【0035】
または、移植型マイクロタンパク質は、組換え発現以外の方法で、例えばマイクロタンパク質の翻訳後の化学的操作によって作製することもできる。好ましい方法では、移植型マイクロタンパク質は、後述するヒドラゾン結合の形成を含む本発明の方法で作製される。
【0036】
本発明との関連で使用されるマイクロタンパク質は、アミノ酸、好ましくは天然のアミノ酸のみからなる場合がある。しかしながら、ペプチド化学およびポリペプチド化学の当業者になじみのある手法で誘導体化されるマイクロタンパク質も含まれる。このような誘導体は例えば、1つもしくは複数のアミノ酸の、化学的に修飾されたアミノ酸などの類似体との置換、N末端およびC末端における環化、または例えばマイクロタンパク質の治療効果を改善する可能性のある機能性部分との結合を含む場合がある。誘導体化部分を含めることで例えば、ポリペプチドの安定性、溶解性、生物学的半減期、または吸収が改善され得る。このような部分は、マイクロタンパク質の任意の望ましくない副作用を減ずるか、または除去可能である。適切な部分の概要は例えば、E. W. MartinによるRemington's Pharmaceutical Sciences (18th ed., Mack Publishing Co., Easton, PA (1990))に記載されている。ポリエチレングリコール(PEG)は、治療的タンパク質の作製に使用可能な、このような化学的部分の一例である。PEGをタンパク質に結合させると、タンパク質分解から保護されることが報告されている(Sada et al., J. Fermentation Bioengineering 71 (1991), 137-139)。タンパク質への特定のPEG部分の結合には、さまざまな方法がある(総説としてAbuchowski et al.、「Enzymes as Drugs」;Holcerberg and Roberts, eds. (1981), 367-383を参照)。一般にPEG分子は、タンパク質内に存在する反応基を介してタンパク質に連結される。例えば、タンパク質のリシン上のアミノ基、またはアミノ末端は特に、この結合に都合がよい。治療に有用なマイクロタンパク質の作製に使用可能な他の化学的修飾は、グルタルアルデヒドなどの架橋試薬の追加、グリコールやエタノールなどのアルコールの追加、またはスルヒドロキシド(sulhydroxide)-ブロッキング試薬、もしくは側鎖残基のリン酸化、アセチル化、酸化、グルコシル化、リボシル化などの修飾試薬の追加、重金属元素および/または最大10個のN末端もしくはC末端における追加的なアミノ酸残基の結合を含む。好ましくは、後者の残基はヒスチジンであるか、または、より好ましくは残基RGS-(His)6である。
【0037】
他の適切な誘導体化は、1つもしくは複数の追加のアミノ酸配列との融合の場合がある。このような融合タンパク質では、追加のアミノ酸配列は、マイクロタンパク質の配列に、共有結合または非共有結合、好ましくはペプチド結合によって連結可能である。連結は、当技術分野で周知の方法による遺伝子融合に基づく場合があるほか、例えばWO 94/04686に記載された手順による化学的な架橋によって実施することができる。追加されるアミノ酸配列は好ましくは、柔軟なリンカー、有利には、追加の配列によって形成される3次構造のC末端と、マイクロタンパク質のN末端の間、またはこの逆の間の距離を確保するのに十分な長さを有する複数の親水性のペプチド結合されたアミノ酸を含む場合のあるポリペプチドリンカーによって連結可能である。融合タンパク質は、切断可能なリンカー、またはタンパク質分解酵素用の切断部位(例えばCNBr切断部位またはトロンビン切断部位;図2参照)を含む場合がある。
【0038】
さらに、追加されるアミノ酸配列は典型的には、所定の特異性または機能、例えば核局在化シグナル、トランス活性化ドメイン、DNA結合ドメイン、ホルモン結合ドメイン、タンパク質タグ(GST、GFP、h-mycペプチド、FLAG、HAペプチド)を有する。
【0039】
好ましい態様では、マイクロタンパク質はバルナーゼ(barnase)と、好ましくは不活性なバルナーゼと融合される。好ましくは、バルナーゼとの融合は、マイクロタンパク質の組換え的な作製を容易にするために用いられ、およびバルナーゼ部分は、本発明のポリペプチドへのマイクロタンパク質の組込みに先だって除去される。
【0040】
「バルナーゼ」は、バチルス・アミロリクエファシエンス(Bacillus amyloliquefaciens)に由来する細胞外リボヌクレアーゼである(Fersht, 1993;Paddon, 1987)。マイクロタンパク質とバルナーゼの融合が、いくつかの利点をもたらし得ることが既に報告されている(Schmoldt et al., 2004)。特に、マイクロタンパク質が大腸菌などの宿主細胞における発現によって組換え的に作製される場合には、融合したバルナーゼ部分は可溶化作用を有する。これは、発現されたマイクロタンパク質を封入体から単離する必要性、および続いてこれを酸化して活性ジスルフィド結合構造を得る必要性を大きく減じるか、または完全に除く可能性がある。他の利点には、バルスター(barstar)-バルナーゼの親和性を利用して、発現されたマイクロタンパク質を粗抽出物から精製可能な点(EP 04 02 2455,2)、ならびに融合タンパク質を結晶化させることで促進型構造モデリング用の入力としての既知のバルナーゼ構造を使用する3次元構造解析が容易になる点(EP 04 02 2455,2)がある。
【0041】
バルナーゼ融合体が、活性型のバルナーゼを使用して構築されるのであれば、バルナーゼ阻害因子であるバルスターを十分な量で同時に発現させることが必要な場合がある。というのは、これを行わないとバルナーゼが宿主細胞に致死的な作用を及ぼすからである(Martsev, 2004)。これに関連して、His-102がAlaと置換されたバルナーゼなどの不活性な突然変異型バルナーゼを使用することが好ましい場合がある(図2、および対応する図のレジェンド参照)。この結果、バルナーゼ融合体に関連する利点が維持されて、バルスターを追加的に同時発現させる必要は無くなる。
【0042】
本発明との関連で使用されるマイクロタンパク質は例えば、天然の状態で純粋な産物か、または化学合成手順の産物、または組換え手法によって原核生物もしくは真核生物の宿主から(例えば、培養した細菌、酵母、高等植物、昆虫、および哺乳類の細胞により)産生される産物の場合がある。
【0043】
組換え発現を介するマイクロタンパク質の提供について、さまざまな発現系に関する総説には例えば、Methods in Enzymology 153 (1987), 385-516, Bitter et al. (Methods in Enzymology 153 (1987), 516-544)、およびSawers et al. (Applied Microbiology and Biotechnology 46 (1996), 1-9), Billman-Jacobe (Current Opinion in Biotechnology 7 (1996), 500-4), Hockney (Trends in Biotechnology 12 (1994), 456-463), Griffiths et al., (Methods in Molecular Biology 75 (1997), 427-440)などがある。酵母の発現系に関する総説には例えば、Hensingら(Antonie van Leuwenhoek 67 (1995), 261-279)、Bussineauら(Developments in Biological Standardization 83 (1994), 13-19)、Gellissenら(Antonie van Leuwenhoek 62 (1992), 79-93、Fleer (Current Opinion in Biotechnology 3 (1992), 486-496)、Vedvick (Current Opinion in Biotechnology 2 (1991), 742-745)、およびBuckholz (Bio/Technology 9 (1991), 1067-1072)などがある。
【0044】
発現ベクターは、文献に詳細に説明されている。一般に発現ベクターは、選択マーカー遺伝子、および選択された宿主における複製を確実なものとするための複製起点を含むだけでなく、細菌またはウイルスのプロモーター、ならびに多くの場合は転写終結シグナルも含む。プロモーターと終結シグナルの間には一般に、コードDNA配列の挿入を可能とする少なくとも1か所の制限酵素切断部位すなわちポリリンカーが存在する。
【0045】
遺伝子の構成的な発現を確実なものとするためのプロモーター、および対象遺伝子の発現の細かな制御を可能とする誘導型のプロモーターを使用することが可能である。このような特性を有する細菌およびウイルスのプロモーター配列は文献に詳述されている。微生物(例えば大腸菌や出芽酵母(S. cerevisiae))における発現の調節配列は文献に詳しく記載されている。下流に位置する配列の特に強い発現を可能とするプロモーターは例えばT7プロモーター(Studier et al., Methods in Enzymology 185 (1990), 60-89)、lacUV5、trp、trp-lacUV5 (DeBoer et al., in Rodriguez and Chamberlin (Eds), Promoters, Structure and Function;Praeger, New York, (1982), 462-481 ;DeBoer et al., Proc. Natl. Acad. Sci. USA (1983), 21-25)、Ip1、rac (Boros et al., Gene 42 (1986), 97-100)である。誘導型プロモーターは好ましくは、タンパク質の合成に使用される。このようなプロモーターは、構成的プロモーターの場合より多いタンパク質収量にしばしばつながる。最適な量のタンパク質を得るためには、2段階の工程がしばしば使用される。第1段階では、宿主細胞が、比較的高い細胞密度となるまで最適条件で培養される。第2段階では、使用されるプロモーターの型に応じた転写が誘導される。この点に関しては、乳糖またはIPTG(=イソプロピル-β-D-チオガラクトピラノシド)によって誘導可能なtacプロモーターが特に適している(deBoer et al., Proc. Natl. Acad. Sci. USA 80(1983), 21-25)。転写の終結シグナルも文献に記載されている。
【0046】
適切な宿主細胞の形質転換またはトランスフェクションは、前述の方法の1つで実施可能である。宿主細胞を、使用される特定の宿主細胞の要件に、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素などに関して適合する栄養培地で培養する。マイクロタンパク質を組換え細胞培養物から、硫酸アンモニウムまたはエタノールによる沈殿法、酸による抽出、陰イオン交換クロマトグラフィーもしくは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティクロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーを含む方法で回収して精製することができる。タンパク質の立体構造を完成させるためには、タンパク質の再折りたたみ段階を必要であれば使用することができる。また、高処理能液体クロマトグラフィー(HPLC)を最終的な精製段階に行うことができる。
【0047】
組換え的な作製手順に使用される宿主に依存して、発現されるポリペプチドをグリコシル化することが可能なほか、グリコシル化しないことが可能である。ポリペプチドは、先頭にメチオニンアミノ酸残基を含む場合もある。
【0048】
好ましくは、マイクロタンパク質は最初に、融合タンパク質として組換え的に、有利にはバルナーゼによって作製後に、融合連結部における切断と、これに続く分離によって融合パートナーから放出される。
【0049】
同様に、マイクロタンパク質は、例えば実施例1(後述)で使用される手順などの、当技術分野で報告されている任意の適切な標準的なペプチド合成手順で作製することができる(例えばMerrifield, Methods Enzymol. 289 (1997), 3-13;Hancock, Mol. Biotechnol. 4 (1995), 73-86;and Merrifield, Adv. Enzymol. Relat. Areas Mol. Biol. 32 (1969), 221-296を参照)。
【0050】
本発明のポリペプチドは、当業者になじみのある任意の適切な方法、および文献に記載されている方法で作製することができる。したがって、マイクロタンパク質の設計および作製に関する上記の説明は、本発明のポリペプチドに、これが適切な限りにおいて応用することができる。ポリペプチドが、マイクロタンパク質、および1つもしくは複数の任意で追加されるリンカーに加えて他の構造エレメントを含む場合があることに注目すべきである。これは、N末端もしくはC末端に、および/または内部に存在する可能性のある追加のアミノ酸配列を意味する場合がある。このようなアミノ酸配列は例えば、バルナーゼ、好ましくは不活性型のバルナーゼなどの、ポリペプチドもしくはこの一部の発現および/または精製を容易にする融合パートナーである場合がある(図2参照)。追加のアミノ酸配列は、結合、安定化、検出などの他の機能を果たす場合もある。ペプチド工学およびポリペプチド工学に知悉している当業者であれば、個々の目的に叶う適切なアミノ酸配列を理解するであろう。さらに本発明のポリペプチドは、例えばグリコシル化、リン酸化、リボシル化、PEG化(PEGの結合)、またはフルオロフォアや放射性核種などの検出可能または治療的に有効な部分の付加によって生じる他の構造エレメントを含む場合がある。この場合も、ペプチド工学およびポリペプチド工学に知悉している当業者であれば、適切な手段および方法を理解するであろう。
【0051】
特に好ましい態様では、本発明のポリペプチドは、標的タンパク質に対する特異的な結合活性を有するアミノ酸配列を含む、少なくとも2個のマイクロタンパク質を含む。
【0052】
「特異的な結合活性」という表現は、標的タンパク質以外のタンパク質に対する親和性と比較して高い親和性を意味する。好ましくは、標的タンパク質に対する親和性は、他のタンパク質に対するより少なくとも100倍高く、より好ましくは少なくとも1000倍高い。好ましくは、「特異的な」という表現は、アミノ酸配列が、指定の標的タンパク質以外のタンパク質に有意な程度で結合しないことを意味する。結合は従来の方法で、例えば、標的タンパク質を透析バッグ内でリガンドとともに配置し、平衡に達した後に、透析バッグの内外におけるリガンドの分布を測定する平衡透析で測定可能である。または、標的を固体表面に固定化して、リガンドの結合を濃度依存的に、例えば表面プラズモン共鳴(BiaCore)によって、質量の増分を測定することで測定可能である。
【0053】
「標的タンパク質」という表現は、本発明のポリペプチドとの結合が望ましい作用を有する可能性のある、任意の想定可能な1つもしくは複数のタンパク質の場合がある。
【0054】
特に、「標的タンパク質」という表現は、本発明のポリペプチドの結合に起因する、近傍に位置する2個もしくは2個以上のタンパク質の結合が望ましい作用をもたらすタンパク質を意味する。このようなタンパク質は、この場合、ホモ二量体もしくはヘテロ二量体、または多量体の場合があるが、ホモ二量体または多量体であることが好ましく、および、さらにより好ましくはホモ二量体である。主な局面では、本発明のポリペプチドと標的タンパク質の結合に及ぼす作用は、標的タンパク質の少なくとも2個が近傍に位置するように、本発明のポリペプチド内の少なくとも2個のマイクロタンパク質のそれぞれが1つの標的タンパク質に結合するという事実に基づくことが理解されている。標的タンパク質の局所的な結合が、活性作用または阻害作用のいずれかの作用を標的タンパク質に及ぼす可能性があることもさらに理解される。例えば、ホモ二量体の受容体タンパク質の2つのサブユニットの局所的な結合は、例えばシグナル伝達カスケードを引き起こす、その活性化に至る場合がある。一方、多量体型酵素の基質結合部位に対する本発明のポリペプチドの結合は、酵素に阻害作用をもたらす場合がある。好ましい態様では、該ポリペプチドの結合は、標的タンパク質の天然の結合分子(例えばリガンド)と実質的に同じ作用を有する。この場合、本発明のポリペプチドは、天然の結合分子の活性の少なくとも10%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも100%、およびさらにより好ましくは少なくとも150%または200%を有することが好ましい(該分子の活性を決定する標準的な方法で計算時)。
【0055】
本発明のポリペプチドの特に好ましい態様では、標的タンパク質は、膜結合型受容体であり、有利には、受容体の分子が相互に近傍に位置する際にシグナル伝達カスケードの下流の他のタンパク質を活性化する受容体である。
【0056】
「膜結合型受容体」という表現は、細胞の形質膜に、特に膜貫通型タンパク質として位置し、かつリガンドと結合可能な受容体タンパク質を意味する。好ましくは、リガンドはペプチドまたはポリペプチドである。さらに受容体が、ヘテロ二量体もしくはホモ二量体、または多量体の形状、好ましくはホモ二量体または多量体、より好ましくはホモ二量体の形状を有し、ならびにこのような形状の受容体が、単量体の形状で存在する受容体の活性とは異なる活性を有することが好ましい。こうした活性の変化は、本発明のポリペプチドと受容体の結合によって、本発明によって促進されると想定される。有利には受容体は、二量体または多量体の形状で存在する場合に活性である。
【0057】
先行技術の文献には、上記の定義に一致する多くの膜結合型受容体について記載されているので、それらの活性は、本発明のポリペプチドとの結合によって調節可能である。例には、EPO受容体、表皮成長因子受容体、ヒト成長ホルモン受容体、TGF-β受容体、FGF受容体、受容体チロシンキナーゼファミリーの成員、および少なくとも一部のG-タンパク質共役受容体などがある。膜結合型受容体は例えば、Schlessinger, Cell 110 (2002), 669-672;Spivak-Kroizman, J. Biol. Chem. 267 (1992), 8056-8063;Plotnikov, Cell 98 (1999), 641-650;Remy, Science 83 (1999), 990-993;およびAngers, Annual Review of Pharmacology and Toxicology 42, (2002), 409-435に記載されている。
【0058】
当業者であれば、本発明に従って、膜結合型受容体などの任意の標的タンパク質に特異的に結合可能なポリペプチドを作製することができる。好ましくは、標的タンパク質に結合するリガンドの構造は明らかであり、および関連するリガンド構造を、先行技術に記載された方法(前述)でマイクロタンパク質に移植することができる。
【0059】
またはマイクロタンパク質そのものが、望ましい結合活性を有する。例えば、四量体の酵素トリプターゼ(EP 04 02 2455.2)に対して特異的な結合活性を有するマイクロタンパク質が存在することがわかっている。したがって、野生型マイクロタンパク質、または望ましい結合活性を有するこの変異型の二量体化もしくは多量体化が、本発明のポリペプチドの提供につながる可能性もある。
【0060】
さらに、最新の手法を用いることで、任意の標的タンパク質に対して意図された結合活性を有するペプチドまたはポリペプチドの配列を、同族のリガンドの構造の詳細が未だ不明であっても提供することが可能である。この点に関しては、適切なスクリーニング法、3次元構造の決定、およびコンピューターによるモデリング法について以下に言及する。スクリーニングは例えば、ファージディスプレイされたペプチドライブラリーを出発材料として使用することで、従来の方法で実施できる。さらに、酵母ツーハイブリッドスクリーニング、またはこれに由来する手法を摘要できる。標的タンパク質との結合に重要な役割を果たすペプチド構造が明らかになれば、そのペプチドを前述の対応する方法によって、適切なマイクロタンパク質の足場内に移植することができる。
【0061】
特に好ましい態様では、膜結合型受容体は、トロンボポエチン(TPO)受容体である。
【0062】
TPO受容体に対して特異的な結合活性を有する本発明のポリペプチドは、TPOアゴニストとして作用することが想定される。これは、TPO受容体のホモ二量体化が、後の第2のメッセンジャー経路を誘導することが知られているためである。このようなアゴニストは、例えば血小板減少症のような、血小板産生の増加を必要とする病理学的状態の治療に有用な場合がある。
【0063】
したがって、この態様に従って、本発明のポリペプチドはTPO受容体を刺激することが可能である。「TPO受容体」という表現は、天然のTPO受容体(その機能的な対立遺伝子異型およびイソ型の全てを含む)、好ましくは、単離されたか、もしくは組換え的に作製されたヒトTPO受容体、または同じ生物学的活性を実質的に有する天然のTPO受容体の変異型を意味する。この表現は例えば、TPOと結合可能なTPO受容体の細胞外部分を含む融合コンストラクト、およびTPO結合による融合コンストラクトの二量体形成時に、検出可能な細胞シグナルを放出可能な融合パートナーを意味する場合がある。このような変異型TPO受容体は例えば、実施例1(後述)に記載されたアッセイ法に使用されるハイブリッドのTPO受容体である。
【0064】
「TPO受容体を刺激可能な」という表現は、TPO受容体に結合して2つのTPO受容体分子の二量体形成を誘導することでTPO受容体の活性化を引き起こす化合物の活性を意味する。この活性は、Cwirla et al. (1997)に記載されているような、先行技術で既知の方法で測定することができる。好ましくは活性は、実施例1(後述)に記載されたTPOアゴニストアッセイ法で、すなわち結果を図4および図5に示すルシフェラーゼアッセイ法、または結果を図6に示す骨髄細胞増殖アッセイ法、または結果を図7に示すインビボ血小板増殖試験で測定される。
【0065】
TPO受容体結合に関する態様の好ましい形状では、本発明のポリペプチドは、アミノ酸配列IEGPTLRQWLAARA(SEQ ID NO: 7)を含む。このアミノ酸配列は、TPO拮抗性のマイクロタンパク質二量体を作製する際の基礎として良好に使用される(後述する実施例1を参照)。
【0066】
TPO受容体に特異的に結合する能力を有する他のペプチドは、WO 03/031589などの先行技術に記載されている。このようなペプチド配列は、本発明のポリペプチドに含まれるマイクロタンパク質部分内に移植するために同様に使用可能である。
【0067】
特に好ましい態様では、本発明のTPO受容体結合ポリペプチドは、以下からなる群より選択されるアミノ酸配列を含む少なくとも2個のマイクロタンパク質を含む:
(a)SEQ ID NO: 1〜6の任意の1つで表されるアミノ酸配列、
(b)該ポリペプチド内に存在時にTPO受容体を刺激可能な、(a)のアミノ酸配列の断片、ならびに
(c)該ポリペプチド中に存在時にTPO受容体を刺激可能な、(a)もしくは(b)のアミノ酸配列または断片の少なくとも1残基が置換されている、付加されている、および/または欠失されている機能的等価物。
【0068】
好ましくは、本発明のポリペプチドに含まれる少なくとも2個の、およびより好ましくは全てのマイクロタンパク質は、上記の(a)〜(c)で定義されたアミノ酸配列を有する。さらに、上記の(a)〜(c)で定義されるアミノ酸配列を有するマイクロタンパク質の二量体が好ましく、二量体形成は有利には、ビス-スクシンイミジル-スベラート(DSS)リンカーを使用することで実現される。上記の(a)で定義されるマイクロタンパク質の二量体が特に好ましい。というのはTPO受容体を活性化可能であることが報告されているからである(実施例1、後述)。
【0069】
この態様に従って、上記の(a)で定義されるマイクロタンパク質の配列の断片も、本発明のポリペプチド内に存在する場合に該断片がTPO受容体を刺激可能な場合に使用することができる。「本発明のポリペプチド内に存在する場合」という表現は好ましくは、「上記の説明に従って、二量体の状態で存在する場合」を意味することを意図する。「断片」という表現は当業者に明瞭な意味を有し、および断片が定義されるアミノ酸配列内のアミノ酸残基の部分的に連続した配列を意味する。したがって、標準のアミノ酸配列と比較して、このような断片は、少なくとも1つのアミノ酸残基をN末端に、C末端に、または両端において欠く。環状の標準配列の場合は、断片は少なくとも1つのアミノ酸残基を、該配列の1か所に欠くことで、断片は環状または線状となり得る。好ましくは断片は、6個の保存されたシステイン残基を保持し、およびこれらの存在により、シスチンノットのトポロジーを形成可能となる。断片が、(a)の対応する配列に含まれるTPO受容体の結合アミノ酸配列の全体を保持することがさらに好ましい。
【0070】
「機能的等価物」という表現は、ポリペプチド内に存在する場合TPO受容体を刺激可能な、(a)もしくは(b)のアミノ酸配列または断片の少なくとも1つの残基が、置換されているか、付加されているか、および/または欠失されている、(a)もしくは(b)で定義されるマイクロタンパク質の変異型を意味する。好ましくは機能的等価物は、シスチンノットを形成するようにジスルフィド結合を介して連結される6個のシステイン残基を含むアミノ酸配列を有する。
【0071】
本発明で使用される機能的等価物は例えば、本発明のポリペプチド内に存在する場合にTPO受容体を刺激する活性を有する、相補鎖が(a)または(b)で定義されるマイクロタンパク質をコードするヌクレオチド配列とハイブリダイズするポリヌクレオチドにコードされたマイクロタンパク質の場合がある。
【0072】
この文脈において、「ハイブリダイゼーション」という表現は、例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載されているような、従来のハイブリダイゼーション条件、好ましくはストリンジェントな条件におけるハイブリダイゼーションを意味する。特に好ましい態様では、「ハイブリダイゼーション」という表現は、以下の条件においてハイブリダイゼーションが起きることを意味する:
ハイブリダイゼーション用緩衝液:2×SSC;10×デンハート溶液(Fikoll 400 + PEG + BSA;比1:1:1);0.1% SDS;5 mM EDTA;50 mM Na2HPO4;250μg/mlのニシン精子DNA;50μg/mlのtRNA;または0.25 Mリン酸ナトリウム緩衝液、pH 7.2;1 mM EDTA、7% SDS
ハイブリダイゼーション温度T = 60℃
洗浄用緩衝液:2×SSC;0.1% SDS
洗浄温度T = 60℃
【0073】
(a)もしくは(b)で定義されるマイクロタンパク質または断片をコードするヌクレオチド配列とハイブリダイズする機能的等価物をコードするポリヌクレオチドは原理的には、そのようなタンパク質を発現する任意の生物に由来する場合があるか、またはその修飾型バージョンをコードする場合がある。このようなハイブリッド形成性のポリヌクレオチドは例えば、細菌、真菌、植物、または動物のゲノムライブラリーもしくはcDNAライブラリーから単離することができる。このようなハイブリッド形成性のポリヌクレオチドは、本明細書に記載されたマイクロタンパク質をコードするポリヌクレオチド、またはこの一部もしくは逆相補物を使用して、例えば標準的な方法によるハイブリダイゼーションによって同定および単離することができる(例えばSambrook and Russell (2001)、Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAを参照)。
【0074】
このようなハイブリッド形成性のポリヌクレオチドは、ポリヌクレオチドが本発明のポリペプチド内に存在する場合にTPO受容体を刺激可能なポリペプチドをコードする限りにおいて、(a)または(b)で定義されるマイクロタンパク質をコードするポリヌクレオチドの内の1つの断片、誘導体、および対立遺伝子変異型も含む。この文脈における「誘導体」という表現は、このようなポリヌクレオチドの配列が、1か所もしくは複数の位置に関して上記のように定義されるマイクロタンパク質をコードするポリヌクレオチドの1つの配列と異なり、ならびに、これらの配列に対して、好ましくはタンパク質の機能に不可欠な配列範囲内の配列に対して高度の相同性を示すことを意味する。誘導体は、シスチンノットを形成するようにジスルフィド結合を介して連結される6個のシステイン残基を含むアミノ酸配列をコードすることが特に好ましい。
【0075】
ヌクレオチド配列とハイブリダイズするポリヌクレオチドの性質は同様に、ポリヌクレオチドが、上記の(a)または(b)で定義されるマイクロタンパク質のアミノ酸配列に対して、少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも60%、および特に好ましくは少なくとも70%、特に好ましくは少なくとも80%、ならびにさらにより好ましくは少なくとも90%の相同性すなわち配列同一性を有するポリペプチドをコードすることを意味する。さらに、ヌクレオチド配列とハイブリダイズするポリヌクレオチドの性質は、ポリヌクレオチドが、上記の(a)または(b)で定義されるマイクロタンパク質をコードするヌクレオチド配列と比較時に、少なくとも40%、好ましくは少なくとも50%、より好ましくは少なくとも60%、さらにより好ましくは65%以上、特に少なくとも70%、特に好ましくは少なくとも80%、特に少なくとも90%、およびさらにより好ましくは少なくとも95%の相同性すなわち配列同一性を有することを意味する場合がある。
【0076】
好ましくは、相同性の程度は、個々の配列を、SEQ ID NO: 1〜6の任意の1つのアミノ酸配列と比較することで決定される。比較される配列が同じ長さを有しない場合は、相同性の程度は好ましくは、長い方の配列内の個々の残基と同一な、短い方の配列内のアミノ酸残基またはヌクレオチド残基のパーセンテージを意味する。相同性の程度は、ClustalW解析を併用するDNAstarプログラムなどの既知のコンピュータープログラムを使用して、従来の手順で決定することができる。このプログラムは、DNASTAR, Inc., 1228 South Park Street, Madison, WI 53715から、またはDNASTAR, Ltd., Abacus House, West Ealing, London W13 OAS UK (support@dnastar.com)から入手可能であり、およびEMBLの支部に設置されたサーバーからアクセス可能である。
【0077】
Clustal解析法を使用して、特定の配列が、参照配列に対して例えば80%同一であるか否かを判定する場合、好ましくはアミノ酸配列の比較について、以下の設定を用いる:マトリックス:blosum 30;オープンギャップペナルティ(open gap penalty):10.0;ギャップ伸長ペナルティ(extend gap penalty):0.05;ディレイ・ディバージェント(delay divergent):40;ギャップセパレーション距離(gap separation distance):8。ヌクレオチド配列の比較時は、ギャップ伸長ペナルティは好ましくは5.0に設定される。
【0078】
好ましくは、ハイブリダイズするポリヌクレオチドの相同性の程度は、コード配列の全長に対して計算される。さらに、このようなハイブリッド形成性のポリヌクレオチド、および特にこれに含まれるコード配列は、少なくとも75ヌクレオチド、および好ましくは少なくとも100ヌクレオチドの長さを有することが好ましい。
【0079】
好ましくは、本発明と関連して使用されるマイクロタンパク質をコードするポリヌクレオチドとハイブリダイズする配列は、具体的に開示されたマイクロタンパク質をコードするポリヌクレオチドに対して少なくとも90%、好ましくは少なくとも93%、より好ましくは少なくとも95%、さらにより好ましくは少なくとも98%、および特に好ましくは少なくとも99%の同一性を有する領域を含み、この相同性を有する領域は、少なくとも75ヌクレオチド、ならびに好ましくは少なくとも100ヌクレオチドの長さを有する。
【0080】
相同性はさらに、比較対象のポリヌクレオチド、またはこれにコードされるポリペプチドの間の機能的および/または構造的な等価性の存在を意味する。上記の分子に相同であり、ならびに、これらの分子の誘導体であるポリヌクレオチドは通常、同じ生物学的機能を有する、これらの分子の変異体である。これらは、天然の変異、好ましくは上記の(a)もしくは(b)で定義されるマイクロタンパク質をコードするポリヌクレオチドのオーソログ、例えば他の対立遺伝子、品種、種などに由来する配列か、もしくは突然変異を含む場合があり、該突然変異は天然で形成される可能性があるか、または意図的な突然変異誘発によって作製可能である。異型、例えば対立遺伝子変異型は、天然の変異型か、または化学合成によって作製される変異型、もしくは組換えDNA手法によって作製される変異型、もしくはこれらの組み合わせの場合がある。上記の特定のマイクロタンパク質をコードするポリヌクレオチドからの逸脱は例えば、欠失、置換、挿入、および/または組換えによって、例えば2種類もしくは2種類以上の異なるマイクロタンパク質の融合によって作製可能である。DNAまたはRNAのいずれかに対して実施可能な核酸の修飾は、当業者に既知の標準的な手法で実施できる(例えば、SambrookおよびRussell、「Molecular Cloning, A Laboratory Manual」;CSH Press, Cold Spring Harbor, 2001、またはHigginsおよびHames(編)、「Protein expression. A Practical Approach」、Practical Approach Series No. 202. Oxford University Press, 1999)。好ましくはDNAの増幅はポリメラーゼ連鎖反応(PCR)で達成され、および修飾は、例えばテンプレート配列に関する突然変異を含むプライマーオリゴヌクレオチドの適切な選択によって使用される(例えばLandt, Gene 96 (1990), 125-128参照)。
【0081】
本明細書に開示された具体的なマイクロタンパク質の変異型であるポリペプチドは、該マイクロタンパク質と共通して有する特徴を有する。これらは例えば、生物学的活性、分子量、免疫学的反応性、構造など、および例えばゲル電気泳動における移動の挙動、クロマトグラフィー時の挙動、沈降係数、溶解性、分光学特性、安定性、最適pH、最適温度などの物理的特性を含む。
【0082】
さらに好ましい態様では、本発明は、標的タンパク質が二量体またはオリゴマーの酵素であり、およびポリペプチドの少なくともマイクロタンパク質が該酵素の活性部位に特異的に結合するアミノ酸配列を含む、本発明のポリペプチドに関する。
【0083】
この態様は特に、二量体または多量体の構造を有する、およびこれに対応して2か所もしくは多数の活性部位を有する酵素に関する。したがって、本発明のポリペプチドは、該ポリペプチドによって露出された結合ペプチドが酵素の活性部位に適合可能なように直接的に設計可能であることが想定される。ポリペプチドと酵素の結合は主に、基質の活性部位への接近を妨げることで酵素に阻害作用を有すると想定される。二量体もしくはオリゴマーの酵素の例には、代表的なものにトリプターゼ、プロテアソーム、カテプシンC、またはヒトグランザイムAなどがある。このような酵素は、Loidl, Biochemistry 96 (1999), 5418-5422;Turk, Biol. Chem. 378 (1997), 141-150;およびBell, Nat. Struct. Biol. 10 (2003), 527-534などの先行技術の文献に記載されている。
【0084】
特に好ましい態様では、二量体またはオリゴマーの酵素はトリプターゼである。マイクロタンパク質がトリプターゼ(EP 04 02 2455.2)を阻害するのに使用可能であることが最近報告されている。トリプターゼは活性状態では、個々の単量体が活性部位を含む四量体酵素なので、マイクロタンパク質の二量体または多量体がトリプターゼ阻害剤として特に適していると想定される。
【0085】
「トリプターゼ」という表現は、90%〜98%の配列同一性を有するα-トリプターゼ、I-トリプターゼ、II/β-トリプターゼ、およびIII-トリプターゼの、これまで知られている4種類の近縁の酵素を含む(Miller, 1998;Vanderslice, 1990)。トリプターゼは、ヒトのマスト細胞の主要な分泌型プロテアーゼであり、ならびに神経ペプチドのプロセシングおよび組織炎症に関与すると考えられている。成熟型のヒトトリプターゼは、四量体のグリコシル化分子であり、ヘパリン結合性を有し、および不均一な、触媒活性を有するサブユニットを含む(例えばVanderslice et al., 1990;Miller et al., 1989, Sommerhoff et al., 1999を参照)。トリプターゼは、マスト細胞の分泌顆粒中に貯蔵されている。マスト細胞の活性化後にヒトトリプターゼは、さまざまな生体液中に見出される。本発明との関連で、マイクロタンパク質の好ましい標的は、マスト細胞のトリプターゼ、より好ましくはβ-トリプターゼまたはα-トリプターゼである。好ましくは、このようなトリプターゼはヒトトリプターゼである。
【0086】
阻害性のトリプターゼの活性は、先行技術に記載され、および以下に概説する方法で検討することができる。
【0087】
トリプターゼ阻害活性の適切なアッセイ法は、EP 04 02 2455.2で説明されている。したがって、阻害性の活性阻害剤の濃度は、トリプシンによる滴定で決定可能である。この目的で、ウシ膵臓トリプシンが、p-ニトロフェニルp'-グアニジノ安息香酸(Chase & Shaw, 1970)を使用する活性部位の滴定によって標準化可能であり、活性阻害剤の濃度は、阻害剤とトリプシンの相互作用が1:1と仮定して計算される。阻害剤とトリプシンおよびトリプターゼとの複合体の見かけ上の平衡解離定数(Kiapp)は基本的に、Bieth (Bull. Eur. Physiopathol. Respir. 16 (Suppl.)(1980), 183-197)に記載された手順で決定できる。簡単に説明すると、阻害剤の濃度を上昇させながら一定濃度の酵素とインキュベートする。次に基質を添加し、残存する酵素の活性を測定する。Kiapp値は、定常状態の速度を、緊密な結合を示す阻害剤に関する方程式(Morrison, 1969)に、非線形回帰解析で適合させることで計算される。任意のマイクロタンパク質のトリプターゼの阻害活性の指標となる見かけのKi値(Kiappとも表記される)の計算は、Morrison (1969)の手順に従って実施することができる。典型的には、本発明の好ましい態様のポリペプチドは、Kiが最大で1 mM、好ましくは最大で0.5 mM、より好ましくは最大で0.2 mM、さらにより好ましくは最大で0.1 mM、さらに好ましくは最大で0.05 mM、特に好ましくは最大で0.02 mM、特に好ましくは最大で0.005 mMであるトリプターゼ阻害活性を有する。最も好ましいKiは、最大0.002 mMである。活性アッセイ法で決定される値は、用いられる特定のアッセイ法の系に典型的な誤差範囲内を変動し得ると理解すべきであり、好ましくは±20%以内、さらに好ましくは±10%以内、および特に好ましくは5%以内の範囲である。
【0088】
トリプターゼの阻害に使用される本発明のポリペプチドがトリプシンに阻害活性を追加的に示すことがさらに好ましい。トリプシン阻害に関する試験は、正しい折りたたみトポロジーの形成の指標となる場合がある。適切なトリプシン阻害アッセイ法は、Van Nostrand (1990)およびSinha (1991)に記載された方法に基づくEP 04 02 2455.2に記載されている。好ましくはポリペプチドは、最大1 nM、および好ましくは最大0.5 nMの範囲の対トリプシンKiを示す。有利には、治療への応用に望ましい可能性のあるトリプターゼに対する高い選択性を考慮して、ポリペプチドは、トリプシンや血液凝固因子などの他のプロテアーゼに関して比較的低い阻害活性を示すことが好ましい。
【0089】
本発明はさらに、上記の態様に従って本発明のポリペプチドをコードするヌクレオチド配列を含む核酸分子に関する。
【0090】
この態様は、含まれるコード配列が、本発明のポリペプチドの二量体または多量体の構造を反映する核酸分子を意味することが意図される。これは好ましくは核酸分子が、マイクロタンパク質の単量体が直接的に、またはペプチドリンカーを介して連結される融合タンパク質をコードすることを意味する。しかしながら核酸分子が、本発明のポリペプチドの前駆体として2個か、またはこれ以上のマイクロタンパク質をコードし、適切なプロセシング段階後に初めて、例えば化学的な修飾によって初めて、本発明のタンパク質を生じることも意図される。
【0091】
本発明の核酸分子は、任意のタイプのポリヌクレオチド、例えばDNA分子もしくはRNA分子、またはこれらの組み合わせの場合がある。これらのポリヌクレオチドは、当技術分野で周知の任意の適切な手法で得られ、例えば合成的に、またはPCRなどのインビボもしくはインビトロにおける組換え手法で作製することができる。このようなポリヌクレオチドは、従来技術で知られている、任意の修飾を含む場合がある(修飾の例として例えばUS 5525711、US 4711955、US 5792608、またはEP 302175を参照)。このようなポリヌクレオチドは、1本鎖または2本鎖、直鎖状または環状の場合があり、サイズに制限は何らない。好ましくは、核酸分子はDNAまたはmRNAである。
【0092】
本発明の融合タンパク質をコードする核酸分子は一般に、組換え型の核酸分子である。「組換え型の核酸分子」という表現は、事前には、結果として生じる組換え核酸分子のように連結されていない、核酸分子またはこの一部分を人工的に混合するのに有用な手法で作製された任意の核酸分子を意味する。適切な手法は例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y. およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989)に代表される先行技術から利用できる。
【0093】
好ましい態様では、組換え核酸分子に含まれる核酸分子は、原核細胞または真核細胞における発現を可能とする発現制御配列に使用可能に連結される。適切な発現制御配列は、標的宿主生物に応用可能なプロモーターを含む。このようなプロモーターは、原核生物界および真核生物界の多様な宿主に関して当業者に周知であり、ならびに文献に記載されている。例えば、このようなプロモーターは、天然の遺伝子から単離することが可能なほか、合成プロモーターまたはキメラプロモーターの場合がある。同様に、プロモーターは標的ゲノム上に既に存在する場合があり、および技術分野で周知の適切な手法、例えば相同的組換えなどでコード配列に連結可能である。
【0094】
本発明の核酸分子は、遺伝子工学の領域で従来から使用されている、特にプラスミド、コスミド、ウイルス、およびバクテリオファージなどのベクター上に存在する場合がある。このようなベクターも本発明の一部である。
【0095】
本発明の好ましい態様では、本発明のベクターは、真菌細胞、植物細胞、微生物の細胞、または動物細胞、特に哺乳類細胞の形質転換に適している。好ましくは、このようなベクターは、酵母や細菌、特に大腸菌などの微生物の形質転換に適している。当業者に周知の方法で組換えベクターを構築することができる;これについては例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y.、およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989)に記載された手法を参照されたい。またはベクターは、標的細胞への輸送目的で内部に本発明の組換え核酸分子を再構成可能なリポソームの場合がある。
【0096】
有利には、ベクターに含まれる核酸分子、および本発明のポリペプチドをコードする核酸分子は、宿主細胞における該ポリペプチドの発現を可能とする1つもしくは複数の発現制御エレメントに使用可能に連結される。
【0097】
原核細胞または真核細胞における、例えば大腸菌における本発明の核酸分子の発現には興味深い点がある。というのは、これらの分子にコードされたタンパク質の生物学的活性のより正確な解析が可能となるからである。加えて、改変された生物学的特性を有することになるタンパク質の合成に至る、分子生物学分野で一般的な方法(例えばSambrook and Russell, 2001, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYを参照)で、多様な追加的な突然変異を核酸分子中に挿入することが可能である。この点に関しては、一方では核酸分子が、コードDNA配列の5'端または3'端からの進行的な欠失によって作られ、および該核酸分子が、これに対応して短縮化されたタンパク質の合成に至る欠失突然変異体を作製することが可能である。他方、点突然変異の導入も、アミノ酸配列の修飾が例えばタンパク質の生物学的活性に影響を及ぼす位置において想定される。
【0098】
原核細胞を対象とした遺伝子工学では、本発明の核酸分子、またはこれらの分子の一部を、DNA配列の組換えによる突然変異誘発または配列修飾を可能とするプラスミド中に導入することができる。標準的な方法(Sambrook and Russell, 2001, Molecular Cloning: A laboratory manual, Cold Spring Harbor Laboratory Press, NY, USAを参照)では、塩基を交換したり、天然配列または合成配列を追加したりすることができる。アダプターおよびリンカーを断片に加えることでDNA断片を相互に連結することができる。さらに、適切な制限酵素切断部位を提供するか、または過剰なDNAもしくは制限酵素切断部位を除去することができる工学的手段を用いることができる。挿入、欠失、または置換が可能な、これらの場合、インビトロ突然変異誘発、「プライマー修復」、制限、または連結を使用することができる。一般に、配列解析、制限酵素解析、ならびに生化学および分子生物学の他の方法が解析法として実施される。
【0099】
他の局面では、本発明は、本発明のポリペプチドまたは本発明の核酸分子、ならびに任意で、薬学的に許容される担体を含む薬学的組成物に関する。
【0100】
このような薬学的組成物は、治療的有効量のポリペプチドまたは核酸分子、および任意で、薬学的に許容される担体を含む。薬学的組成物は、生理学的に許容される担体とともに、本明細書に記載された手順で患者に投与することができる。特定の態様では、「薬学的に許容される」という表現は、規制当局または他の一般に認められている、動物、具体的には人間を対象とした使用に関する薬局方によって承認されていることを意味する。「担体」という表現は、治療薬とともに投与される希釈物、アジュバント、賦形剤、または溶媒を意味する。このような薬学的担体は、水および、石油、動物油、植物油、またはピーナッツオイル、ダイズ油、鉱油、ゴマ油などの合成油を含む油などの無菌性の液体の場合がある。薬学的組成物が静脈内に投与される場合は、水が好ましい担体である。生理食塩水溶液および水性デキストロースおよびグリセロール溶液を、特に注射用溶液用の液体担体として使用することもできる。適切な薬学的賦形剤は、デンプン、グルコース、乳糖、ショ糖、ゼラチン、マルト、コメ、コムギ、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリン酸グリセロール、タルク、塩化ナトリウム、乾燥スキムミルク、グリセロール、プロピレングリコール、水、エタノールなどを含む。このような組成物は、望ましいならば、微量の湿潤剤もしくは乳化剤、またはpH緩衝剤を含む場合もある。このような組成物は、溶液、懸濁液、乳濁液、錠剤、丸剤、カプセル、粉末、徐放性製剤などの形状を取り得る。このような組成物は、トリグリセリドなどの従来の結合剤および担体を使用して坐剤として製剤化することができる。経口用製剤は、薬学グレードのマンニトール、乳糖、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどの標準的な担体を含む場合がある。適切な薬学的担体の例は、E.W. Martinの「Remington's Pharmaceutical Sciences」(前述)に記載されている。このような組成物は、治療的有効量の前述のマイクロタンパク質を、患者への適切な投与用の形状として提供するために、適量の担体とともに、好ましくは純粋な状態で含む場合がある。剤形は、投与様式に適したものとすべきである。
【0101】
他の好ましい態様では、組成物は、常用の手順に従って、ヒトへの静脈内投与に適合する薬学的組成物として製剤化される。典型的には、静脈内投与用の組成物は、無菌性の等張性の水性緩衝液を溶媒とする溶液である。必要であれば、組成物は可溶化剤、および注射部位における痛みを和らげるためのリグノカインなどの局所麻酔薬も含んでもよい。一般に、このような内容物は個別に、または単位投与剤形として混合状態で、例えば、活性薬剤量が表示されたアンプルやサシェ(sachette)などの密封容器中の凍結乾燥粉末、または水を含まない濃縮物のいずれかによって提供される。組成物が注射で投与される場合は、無菌性の薬学グレードの水または生理食塩水を含む注入ボトルに分注することが可能である。組成物を注射で投与する場合は、内容物を投与前に混合可能なように、注射用の無菌水または生理食塩水を含むアンプルを提供することができる。
【0102】
本発明との関連で使用される薬学的組成物は、中性または塩の状態として製剤化することができる。薬学的に許容される塩は、塩酸、リン酸、酢酸、シュウ酸、酒石酸などに由来する塩などの陰イオンによって形成される塩、およびナトリウム、カリウム、アンモニウム、カルシウム、水酸化第2鉄、イソプロピルアミン、トリエチルアミン、2-エチルアミノエタノール、ヒスチジン、プロカインなどに由来する塩などの陽イオンによって形成される塩を含む。
【0103】
最適投与量範囲の決定の一助とするために、インビトロアッセイ法を任意で利用できる。製剤化に使用される正確な用量は、投与経路、および疾患または障害の重傷度に依存することもあり、ならびに実施者の判断、および個々の患者の置かれた状況に従って決定されるべきである。有効用量は、インビトロモデルまたは動物モデルの試験系から得られた用量反応曲線から推定することができる。好ましくは、薬学的組成物は、直接的に投与されるか、またはアジュバントと組み合わせて投与される。
【0104】
本発明と関連して、「対象」という表現は、本発明のポリペプチドを個体に投与することで関連づけられる、治療が必要とされる個体を意味する。好ましくは、対象は脊椎動物であり、さらにより好ましくは哺乳類であり、特に好ましくはヒトである。
【0105】
「投与される」という表現は、マイクロタンパク質を含む前述の薬学的組成物の治療的に有効な用量の個体への投与を意味する。「治療的有効量」という表現は、投与されることで効果を生じる用量を意味する。正確な用量は、治療の目的に依存し、および当業者によって既知の手法で確認される。当技術分野で周知なように、および上述したように、全身か局所への輸送、年齢、体重、全般的健康、性別、食事、投与時間、薬剤相互作用、および状態の重症度を考慮した調整が必要な場合があり、当業者によって常用の実験法で確認される。
【0106】
このような方法は、ヒトの治療と獣医学的応用の両方に応用される。所望の治療的活性を有する、本明細書に記載された、生理学的に許容される担体に含まれる化合物を患者に、本明細書に記載された手順で投与することができる。導入様式に依存して、化合物を後述するさまざまな方法で製剤化することができる。製剤中の治療的活性化合物の濃度は、約0.1〜100重量%を変動し得る。薬剤は単独で、または他の治療法と組み合わせて投与することができる。
【0107】
薬学的組成物の投与は、経口、皮下、静脈内、動脈内、節内、髄内、髄腔内、脳室内、鼻腔内、気管支内、経皮、節内、直腸内、腹腔内、筋肉内、肺内、膣内、直腸内、または眼内を含むがこれらに限定されない、前述の多様な方法で実施できる。例えば創傷や炎症の治療における状況によっては、薬学的に有効な薬剤は、溶液乾燥スプレーとして直接的に投与される場合がある。
【0108】
担当医および臨床的因子が投与法を決定する。医学分野で周知なように、任意の1人の患者あたりの投与量は、患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、投与時間および投与径路、全般的健康、ならびに同時に投与される他の薬剤を含む数多くの因子に依存する。典型的な用量は例えば、0.001〜1000μgの範囲の範囲を取り得る;しかしながら、この例示的な範囲に満たない用量、または上回る用量が、特に前述の諸因子を考慮して想定される。
【0109】
投与量は、好ましくは週に1回投与されるが、治療の進行中は、投与量は、さらにより長い間隔を設けて投与される場合があり、必要であれば、さらにより短い間隔、例えば毎日投与することができる。好ましい例では、免疫応答が当業者に既知の方法でモニタリングされ、投与量が例えば、時間、量、および/または組成に関して最適化される。進行状況は、定期的な評価によってモニタリング可能である。薬学的組成物は、局所的または全身的に投与することができる。投与は好ましくは、非経口的に、例えば静脈内に行われる。非経口的投与用の調製物は、無菌性の水性または非水性の溶液、懸濁液、および乳濁液を含む。非水性溶媒の例には、プロピレングリコール、ポリエチレングリコール、オリーブオイルなどの植物油、およびオレイン酸エチルなどの注射可能な有機エステルがある。水性担体には、水、アルコール/水溶液、乳濁液、または生理食塩水および緩衝用溶媒を含む懸濁液などがある。非経口的な溶媒には、塩化ナトリウム溶液、リンゲルデキストロース液、デキストロースおよび塩化ナトリウム、乳酸加リンゲル液、または不揮発性油などがある。静脈注射用の溶媒には、溶液、および栄養補充液、電解質補充液(例えばリンゲルデキストロース液を元にした溶液)などがある。保存剤、ならびに例えば抗菌剤、抗酸化剤、キレート化剤、および不活性ガスなどの他の添加剤を含めることもできる。
【0110】
好ましい態様では、薬学的組成物は、吸入用のエアロゾルとして製剤化される。
【0111】
さらに好ましい態様では、薬学的組成物は、経口投与経路用に製剤化される。
【0112】
別の好ましい態様では、薬学的組成物は、経皮投与用に製剤化される。
【0113】
好ましい態様では、本発明の薬学的組成物は、本発明のポリペプチドを発現する遺伝子輸送ベクターの形状で患者に投与可能なように考案される。エクスビボでベクターによって細胞を形質転換して、形質転換細胞を患者に投与することも好ましい。
【0114】
これらの態様では、本発明の薬学的組成物は、前述した本発明のポリペプチドをコードするポリヌクレオチドを含み、および発現を可能とするベクターである。このようなベクターは、発現ベクターおよび/または遺伝子輸送ベクターの場合がある。発現ベクターは、この文脈では、エクスビボにおける遺伝子治療法で使用されることを意味し、すなわち適切な宿主細胞が体外でトランスフェクトされた後に対象に投与される。遺伝子輸送ベクターは、本明細書では、インビボにおける遺伝子治療への応用に適したベクターを意味し、すなわちベクターは、全身的または局所的のいずれかで対象に直接的に投与される。本明細書で言及されるベクターは、核酸のみからなる場合があるか、または例えば、標的細胞への輸送、例えば循環系における標的輸送、安定性、および/または、生物学的利用能を高める追加的な化合物との複合体である場合がある。このような追加的な化合物の例には、脂質物質、ポリカチオン、膜破壊性ペプチド、または他の化合物、抗体もしくはこの断片、または標的細胞を特異的に認識する受容体結合分子などがある。発現ベクターもしくは遺伝子輸送ベクターは好ましくは、レトロウイルス、ワクシニアウイルス、アデノ関連ウイルス、ヘルペスウイルス、またはウシパピローマウイルスなどのウイルスに由来する場合があり、および標的細胞集団に、例えば呼吸器の細胞への輸送に使用される場合がある。当業者に周知の方法で、組換え発現ベクターまたは遺伝子輸送ベクターを構築することができる;これについては例えば、Sambrook and Russell, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y.、およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989)に記載された手法を参照されたい。またはベクターを、標的細胞への輸送用に、リポソーム中に再構成することができる。本発明のポリペプチドをコードするポリヌクレオチドを含むベクターは、細胞宿主のタイプによって変動する周知の方法で宿主細胞に輸送することができる。例えば塩化カルシウムトランスフェクションは、原核細胞に広く使用されており、リン酸カルシウム処理またはエレクトロポレーションは他の細胞宿主に使用可能である(Sambrook、前掲を参照)。
【0115】
エクスビボまたはインビボの遺伝子治療に適したベクターおよび方法は文献に記載されており、当業者に既知である;これについては例えば、Giordano, Nature Medicine 2 (1996), 534-539;Schaper, Circ. Res. 79 (1996), 911-919;Anderson, Science 256 (1992), 808-813;Isner, Lancet 348 (1996), 370-374;Muhlhauser, Circ. Res. 77 (1995), 1077-1086;Wang, Nature Medicine 2 (1996), 714-716;WO 94/29469;WO 97/00957、またはSchaper, Current Opinion in Biotechnology 7 (1996), 635-640、および各文献で引用された参考文献を参照されたい。本発明のこの態様で使用されるベクターは、細胞への直接導入用に、またはリポソームもしくはウイルスベクター(例えばアデノウイルスやレトロウイルス)を介する導入用に設計可能である。
【0116】
好ましい遺伝子輸送ベクターには、バキュロウイルス、アデノウイルス、およびワクシニアウイルスをベースとしたベクターなどがある。これらは好ましくは、非複製型である。
【0117】
本発明は、TPO受容体を刺激することで治療または予防が可能な疾患もしくは状態の治療用または予防用の薬学的組成物を作製するための、前述のTPO受容体に結合してこれを刺激可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用にも関する。
【0118】
したがって本発明は同様に、該ポリペプチドまたは該核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、TPO受容体を刺激することで治療可能な疾患もしくは状態に罹患した個体の治療法に関する。
【0119】
このような医学的応用は、TPO受容体に有効に拮抗するマイクロタンパク質の二量体が作製可能であるという知見に基づく(実施例1、後述)。
【0120】
TPOは、骨髄巨核球から血小板が産生される過程である、巨核球形成の調節に重要な役割を果たす(Kuter et al., 1994, Kaushansky et al., 1994;Wendling et al., 1994, Sauvage et al., 1994)。TPOは肝臓で産生されるが、その主な機能を骨髄で発揮し、骨髄で、幹細胞の巨核球前駆細胞への分化、ならびに巨核球の増殖、倍数体化、および究極的には循環性の血小板体への断片化を刺激する。TPOは、血小板減少症を含む状態の主要調節因子でもあり、および、いくつかの研究で、以下の複数の方法で巨核球形成を亢進させることが明らかにされている:(1)巨核球のサイズおよび数の増加を生じる;(2)巨核球中で倍数性のDNA量の増加を生じる;(3)巨核球の核内分裂を高める;(4)巨核球を成熟させる;ならびに(5)骨髄内で少数のアセチルコリンエステラーゼ陽性細胞の状態の前駆細胞のパーセンテージを高める。TPOのこれらの機能を考慮して、当業者であれば、TPO受容体を刺激することで治療または予防可能な疾患を容易に理解するであろう。このような疾患には、以下に挙げるような多様な形状の血小板減少症、再生不良性貧血などの血液学的疾患、さまざまな状態の骨髄不全、骨髄異形成症候群、および肝疾患が含まれる。TPOの刺激は、輸液と関連して、特に、末梢幹細胞の可動化および分離、血小板の分離(アフェレシス)、インビトロにおける巨核球もしくは血小板の産生、または血小板保存の改善との関連で、または外科手術、特に大手術(例えば心臓もしくは全身の手術)との関連でも有用である。
【0121】
血小板は血液の凝固に必要であり、その数が極めて少なくなると、大出血によって患者が死亡するリスクが患者に生じる。したがって、TPOアゴニストである本発明のポリペプチドは、さまざまな血液疾患、例えば主に血小板異常に起因する疾患の診断と治療の両方における潜在的に有用な応用が期待される。同様に、このようなTPOアゴニストには、血小板減少性状態の治療に、特に、癌もしくはリンパ腫の治療としての化学療法、放射線療法、または骨髄移植に由来する状態の治療に潜在的に応用可能である。実際に、癌患者を対象とした進行中の臨床試験から、高用量カルボプラチン化学療法による投与時に、組換え型のヒトTPOが、血小板減少時期の減少、および血小板回復の促進に有効なことが報告されている(Basser et al., 1997)。同様の結果は、PEG化された巨核球分化因子(peg-MGDF、ヒトTPOのPEG化された短縮型のN末端断片)に関する臨床試験でも得られている(Fanucchi et al., 1997)。血小板減少症の患者における血小板レベルの回復が緩やかなことは重大な問題なので、本発明のポリペプチドがTPO模倣物として作用することで血小板減少症の治療に良好に使用可能であることが想定される。
【0122】
前述の第2の医学的使用または治療法の好ましい態様では、疾患もしくは状態は、血小板減少症、再生不良性貧血、骨髄不全、骨髄異形成症候群、および肝疾患からなる群より選択される。
【0123】
「血小板減少症」という表現は、当分野の専門家によって認識される明瞭な意味を有する。本発明の好ましい態様は特に、例えば化学療法(血液学的な悪性腫瘍または固形腫瘍)、放射線療法、骨髄移植(同種もしくは自家)、または末梢幹細胞移植によって引き起こされる血小板減少症の一形態に関する。さらに、先天性の血小板減少症、免疫による血小板減少症(免疫性血小板減少性紫斑病(ITP))、HIVによる血小板減少症、または薬剤による血小板減少症も対象となる。
【0124】
以上のことから本発明は、TPO受容体を刺激するための、前述したTPO受容体と結合してこれを刺激することが可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用にも関する。好ましくは、使用はインビトロである。
【0125】
他の局面では、本発明は、トリプターゼの活性を阻害することで治療または予防が可能な疾患もしくは状態の治療用または予防用の薬学的組成物を作製するための、前述したトリプターゼに結合してこれを阻害可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用に関する。
【0126】
したがって本発明は同様に、該ポリペプチドまたは該核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、トリプターゼの活性を阻害することで治療可能な疾患または状態に罹患した個体の治療法に関する。
【0127】
本発明のこの局面、すなわち本発明のトリプターゼ阻害性ポリペプチドの治療目的における使用を提供することで、低分子量トリプターゼ阻害剤に関して知られている短所は克服される(例えばNewhouse, 2002を参照)。例えば、このような小型分子は、トリプターゼ以外の分子への結合を引き起こす比較的低い結合特異性のために、投与対象の生物体に毒性作用を示す場合がある。このような小型分子と比較して、本発明のポリペプチド内のマイクロタンパク質は、より大きな相互作用表面積を示すので、より選択性の高い結合が予想される。さらに、タンパク質ベースの結合分子は典型的には、低分子量分子より低い解離速度定数を有するので、標的との、より長期間の結合を有し、このため、より有益な結合特性を有する。
【0128】
加えて、低分子量トリプターゼ阻害剤を凌ぐ他の利点は、本発明のポリペプチドが膜障壁を越えることができないと予想されるという事実である。これは、該ポリペプチドの、マスト細胞の生理学的状態に潜在的に負に影響し得るマスト細胞内に保存されたトリプターゼへの結合を妨げる。小型分子は対照的に、しばしば膜を越える場合がある。さらに、特にシスチンノットタンパク質は、酵素による分解または熱による分解に対して極めて安定なことでよく知られている。
【0129】
治療へのトリプターゼ阻害剤の使用に関する先行技術の文献(例えばNewhouse (2002)、および同文献に引用された参考文献を参照)は広範なため、当業者であれば、どのような疾患が、トリプターゼの活性を阻害することで治療可能または予防可能かを理解すると思われる。
【0130】
好ましくは、上記の第2の医学的使用または治療法は、喘息、炎症、乾癬、肺線維症、間質性肺疾患、慢性関節リウマチ、歯肉炎、歯周炎、アレルギー反応、アレルギー性鼻炎、骨関節症、アテローム性動脈硬化症、血管形成、多発性硬化症、および癌からなる群より選択される疾患もしくは状態に関する。
【0131】
上記と一致して、本発明は、トリプターゼの活性を阻害するための、前述のトリプターゼに結合してこれを阻害可能な本発明のポリペプチド、または該ポリペプチドをコードする本発明の核酸分子の使用にも関する。好ましくは、使用はインビトロである。
【0132】
他の局面では、本発明は、本発明のポリペプチド、または本発明の核酸分子を含むキットにも関する。
【0133】
本発明のキットの成分は、任意で緩衝液および/または溶液を含むバイアルなどの容器内に包装され得る。適切であれば、1つもしくは複数の成分は、1つの同じ容器内に包装され得る。加算的または択一的に、1つもしくは複数の成分を例えばニトロセルロースフィルターやナイロン膜などの固相支持体に、またはマイクロタイタープレートのウェルに吸収させることが可能である。
【0134】
他の局面では、本発明は、以下の段階を含む、マイクロタンパク質内に共有結合を形成させる方法に関する:
(a)N末端に反応性カルボニル基を含み、およびC末端にホモセリンラクトン残基を含むマイクロタンパク質基質を提供する段階;ならびに
(b)前記N末端基およびC末端の残基がヒドラゾン結合に変換するように、マイクロタンパク質基質を反応させる段階。
【0135】
この方法は、マイクロタンパク質の環化、またはマイクロタンパク質へのペプチド配列の移植を含む、マイクロタンパク質の処理に特に有用である。さらに、この方法は、本発明の二量体または多量体の作製に特に有用である。
【0136】
この方法の提供によって、マイクロタンパク質への共有結合の導入に関して、先行技術の手法と比較してマイクロタンパク質の扱いは極めて容易になる。例えば、マイクロタンパク質を環化させる現行の方法は、C末端にチオエステルを導入する化学合成に基づく(Davies, 2003を参照)。このような合成戦略には、収量を上げるための最適化時に問題が生じる場合がしばしばあり、および達成時に問題となることがしばしばあるインビトロにおけるジスルフィド結合の形成を必要とする。というのは、誤ったジスルフィドを有する、かなりの量の副反応産物が生じるからである(Price-Carter et al., 1996)。インテインによる環化にも、この点で問題がある。というのは、インテイン-マイクロタンパク質の融合タンパク質は、還元性条件がジスルフィド結合の形成を除去してしまう、細菌の細胞質に存在するからである。
【0137】
バックボーンの環化が自然に生じる仕組みは未だ不明であるが、複数の手法が、合成環状タンパク質を作製させるために開発されている(Evans et al., 1999, Scott et al., 1999 Tam and Lu, 1998)。一般に、N末端とC末端の連結を達成するには2つの主な戦略がある:第1の方法では、ペプチド結合をC末端およびN末端においてそれぞれ切断するインテインと呼ばれる、修飾型タンパク質のスプライシングエレメントを利用する(Evans et al., 1999;Williams et al., 2002)。仮に、同じ読み枠で標的タンパク質のN末端およびC末端にクローニングされた場合、N末端とC末端が連結されたペプチド結合の形成に自然につながる、N末端のシステインおよびC末端のチオエステルが生じる。
【0138】
第2の戦略は、N末端にシステイン残基を、およびC末端にα-チオエステルを有する標的タンパク質の固相合成に拠っている。仮に両端が相互に近傍に位置すれば、自発的な反応が最初にチオエステル結合型中間体を生じ、これが分子内再編成を受けて、最終的に天然のペプチド結合が生じる(Tam and Lu, 1998, Muir 2003)。この戦略は、標的タンパク質の固相合成と、これに続く環化-酸化、またはこの逆である酸化-環化に拠っている。第1の方法では、直鎖状のBoc合成ペプチドおよび完全脱保護状態のペプチドが最初に大環状分子中に「閉じられ(zipped)」、次に酸化される(Tam and Lu, 1998)。別の方法では、完全に脱保護されたペプチドから、「頭部-頭部」の環化に適切な立体構造を得るための、折りたたまれた開鎖状態の前駆体を作製する。いずれの方法にも要件および限界がある。特に、これらの方法では、方法が化学的に合成されたマイクロタンパク質の使用に制限される、マイクロタンパク質への追加的な機能の化学的な導入が必要とされる。したがって、これらは、組換え的に作製されたマイクロタンパク質では作用しない。というのは、このようなタンパク質は天然のアミノ酸しか含まないからである。しかしながら、コストを考えれば、および望ましくない副産物を避けることを考えれば、組換え的な作製が一般に有利である。後者は通常、切断に至る合成エラーのために、および立体異性体(ラセミ化合物)の形成のために、マイクロタンパク質の化学合成に伴って生じる。本発明の方法では、これらの制限が迂回され、および、適切な発現宿主で正しいジスルフィド結合の結合性を伴って産生される、組換え的に合成されたマイクロタンパク質の環化が合成後に可能となる。
【0139】
マイクロタンパク質の組換え的な作製は、先行技術に記載された任意の適切な方法で、および好ましくは本明細書に記載された手順で実施可能である。本発明の好ましい発現宿主は、大腸菌およびピキア・パストリス(Pichia pastoris)である。
【0140】
本発明の方法は、マイクロタンパク質基質内のN末端とC末端の間のヒドラゾン結合の形成に基づいている。「マイクロタンパク質基質」という表現は、その一次構造、二次構造、および/または三次構造に基づいて、マイクロタンパク質として、多数のマイクロタンパク質として、またはこの一部として認識され得るペプチドもしくはポリペプチドを意味する。好ましくは、この表現は、共有結合の閉鎖に伴ってマイクロタンパク質となる前駆体を意味する。
【0141】
N末端の反応性カルボニル基およびC末端のホモセリンラクトン残基の提供は、当技術分野で周知の適切な方法で可能である。本発明に即して、反応性カルボニル基は、C末端のホモセリンラクトン残基とヒドラゾン結合を形成可能でなければならない。好ましくは、N末端の反応性カルボニル基は、グリオキシリル基またはケト基である。ケト基は、例えばDixon (Methods Enzymol. 25 (1972), 409-419)に記載されているように、N末端のアミノ酸残基の、金属を触媒とする適切なアミノ基転移によって形成され得る。
【0142】
好ましい態様では、グリオキシル基は、使用されるマイクロタンパク質基質内に存在するN末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される。
【0143】
好ましい態様では、マイクロタンパク質基質のN末端のセリン残基またはスレオニン残基は、該マイクロタンパク質基質を含む前駆体ポリペプチドの、メチオニンと後続のセリン残基もしくはスレオニン残基との間のペプチド結合の臭化シアンの使用による切断によって提供される。
【0144】
マイクロタンパク質基質のC末端のホモセリンラクトン残基が、メチオニンと後続のアミノ酸残基間におけるペプチド結合の臭化シアンによる切断によって提供されることも好ましい。この態様、および前述の好ましい態様では、臭化シアンが切断用薬剤として使用されるので、いずれの切断反応も同時に進行可能である。したがって、これらの段階が、例えば、出発材料として個々のマイクロタンパク質を含む1つの担体タンパク質を使用して同時に実施されることが可能であり、および特に好ましい。この過程を図8に示す。
【0145】
段階(b)のヒドラゾン形成は、当技術分野で既知の任意の適切な方法で実施可能である。
【0146】
好ましい態様では、段階(b)は、以下の段階を含む:
(i)C末端のホモセリンラクトン残基をホモセリンヒドラジドと反応させる段階;
(ii)ホモセリンヒドラジドとN末端の反応性カルボニル基を反応させてヒドラゾンを生じさせる段階;ならびに
(iii)任意でヒドラゾンを還元させる段階。
【0147】
これらの方法の段階を、図9および図10に示したマイクロタンパク質の環化の例で説明する。
【0148】
マイクロタンパク質は担体タンパク質から、臭化シアン(CNBr)を使用する化学的切断によって切り離すことができる(図10a)。CNBrはタンパク質のペプチド結合をメチオニンのC末端で切断する。この反応によって、メチオニン残基がホモセリンラクトンに変換される。融合タンパク質が臭化シアンによって切断されると、担体タンパク質が放出され、およびアミンと容易に反応することが知られている(Majerle et al., 2000)ホモセリンラクトンをカルボキシ末端に含む、アミノ末端にセリンを有するマイクロタンパク質が放出される(図10b)。本発明の主な局面は、このラクトンとヒドラジンの反応によるC末端のヒドラジドの生成に関する(図10c)。ヒドラジド形成の他の方法は、活性化エステルの化学合成か、またはヒドラジンの存在下における制限的なタンパク質分解による切断のいずれかに拠る(Gaertner et al., 1992、Rose et al., 1996)。本発明に従って、ヒドラジド形成は好ましくは、臭化シアンによる切断と、これに続くヒドラジン分解によって容易に達成され得る。アミノ末端のセリンは、Geoghegan and Stroh (1992)に記載された手順で、過ヨウ素酸ナトリウムを使用する酸化によって選択的に活性化され得る。この結果、C末端のヒドラジドと容易に反応してヒドラゾンを形成するグリオキシリル基が形成される(図10d)。ヒドラゾンは、シアノホウ化水素ナトリウムによって還元されて、安定な環状のマイクロタンパク質分子が得られる(図10e)。N末端のセリン残基の代わりに、例えばスレオニン残基も、本質的に同じ段階および反応条件で使用可能であることが理解されている。
【0149】
以上のことから、本発明の方法の好ましい態様は、マイクロタンパク質基質が1つの単体のマイクロタンパク質であること、および共有結合の形成が、マイクロタンパク質のペプチドバックボーンの環化のためになされることである。
【0150】
別の好ましい態様では、マイクロタンパク質基質は、1つの単体のマイクロタンパク質であり、および共有結合の形成は、グラフトペプチド(graft peptide)をマイクロタンパク質に連結するためになされる。この応用を、図11に図示する。
【0151】
したがって、特に好ましい態様では、方法は、以下の段階を含む:
(A)グラフトペプチドと置換されるマイクロタンパク質のループアミノ酸の配列を、メチオニンと後続のアミノ酸残基の間の適切なペプチド結合において臭化シアンで切断する段階(ループアミノ酸配列のC末端における後続のアミノ酸残基はセリン残基またはスレオニン残基である);
(B)セリン残基またはスレオニン残基を、グリオキシリル基を形成するように穏やかな酸化によって反応させる段階;
(C)N末端の反応性カルボニル基およびC末端のホモセリンラクトンヒドラジドを含むグラフトペプチド配列を、マイクロタンパク質のグリオキシリル基と反応させてヒドラゾンを生じさせる段階;
(D)段階(A)の切断によって生じたC末端のホモセリンラクトン残基を、ホモセリンヒドラジドと反応させる段階;ならびに
(E)(D)のホモセリンヒドラジドを、グラフトペプチドのN末端の反応性カルボニル基と反応させてヒドラゾンを生じさせる段階。
【0152】
グラフトペプチドのN末端の反応性カルボニル基は好ましくはグリオキシリル基またはケト基であり、グリオキシル基は好ましくは、N末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される。
【0153】
本発明の方法のさらに好ましい態様では、マイクロタンパク質基質は、少なくとも2個のマイクロタンパク質を含み、および共有結合の形成が、マイクロタンパク質の二量体またはオリゴマーの形成のためになされる。
【0154】
同様に、マイクロタンパク質基質は、少なくとも2個のマイクロタンパク質を含み、および共有結合の形成が、大環状分子の二量体またはオリゴマーのマイクロタンパク質の形成のためになされることが好ましい。
【0155】
これらの態様は特に、上述の本発明のポリペプチドの作製に適している。特に、臭化シアンによる切断を介するC末端の活性化と、これに続くヒドラジン分解、およびアミノ末端のアルデヒドによるヒドラゾン形成を含む方法を、ホモ二量体またはヘテロ二量体のマイクロタンパク質の形成に応用することもできる。このために、マイクロタンパク質は例えば、セリン残基またはスレオニン残基をそのアミノ末端に含み、およびメチオニン残基をそのカルボキシ末端に含むように、組換え的に作製することができる。臭化シアンによる切断によって、活性化されたホモセリンラクトンが形成される。マイクロタンパク質調製物は、次に2つの等量部分に分けることができる。一方の半分にはヒドラジン水和物を添加し、これがカルボキシ末端におけるホモセリンラクトンの形成につながる。もう一方の半分については、過ヨウ素酸ナトリウムでアミノ末端を酸化して、アミノ末端にグリオキサル部分を得る。両調製物を混合する。ヒドラゾン形成時には、シアノホウ化水素ナトリウムによって還元可能な直鎖状の二量体が形成される。ホモセリンラクトンのヒドラジン分解およびセリンの酸化の手順は繰り返すことができる。この結果、二量体のアミノ末端におけるグリオキサル形成、およびカルボキシ末端におけるヒドラジド形成に至る。分子内におけるヒドラゾン形成によって大環状分子が形成される。
【0156】
以上の態様および他の態様が開示され、ならびに本発明の記述および実施例に含まれる。本発明に用いられる方法、用途、および化合物の任意の1つに関する他の文献は公共図書館から、例えば電子機器を使用して入手可能である。例えば、インターネット上で利用可能な公的データベース「Medline」を、例えばhttp://www.ncbi.nlm.nih.gov/PubMed/medline.htmlのアドレスから利用することができる。http://www.ncbi.nlm.nih.gov/、http://www.infobiogen.fr/、http://www.fmi.ch/biology/research_tools.html、http://www.tigr.org/などの他のデータベースおよびアドレスは当業者には既知であり、ならびに例えばhttp://www.google.de経由で入手できる。生物工学における特許情報の概要、ならびに遡及的な検索および現状認識に有用な特許情報の提供先の調査については、Berks, TIBTECH 12 (1994), 352-364に記載されている。
【0157】
さらに、「および/または」という表現は、本明細書で使用される場合は、「および」、「または」、および「この表現によって関連づけられる要素の全て、または任意の他の組み合わせ」の意味を含む。
【0158】
本発明を、以下の非制限的な図および実施例を参照することで、さらに説明する。
【0159】
本発明を、以下の実施例で説明する。
【0160】
実施例1:
TPOアゴニストおよびアンタゴニストの産生および測定
材料および方法
分子生物学的手法
実施例において特に言及されない限り、全ての組換えDNA手法は、Sambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USA、またはAusubel et al. (1994), Current Protocols in Molecular Biology, Current Protocolsの第1巻および第2巻に記載されたプロトコルに従って実施される。
【0161】
DNAコンストラクト
バチルス・アミロリクエファシエンス(B. amyloliquefaciens)のRNAseの酵素的に不活性な異型であるバルナーゼ'、および個々のマイクロタンパク質のコード配列からなる融合遺伝子の構築は、基本的に文献に記載された方法で行った。マイクロタンパク質の遺伝子を、標準的なクローニング法(Sambrook et al., 2001)によるポリメラーゼ連鎖反応によって発現ベクターpBar100中に集合させた(図2、EP 04 02 2455.2)。ET-TP-020をコードする遺伝子を、プラスミドpBar100-EETI-II M7I(Schmoldt et al., 2004)をテンプレートとして、ならびにオリゴヌクレオチド対のbarmitte-up
およびETTP21-SOE1lo
、ならびにETTP21-SOE2up-new
、およびcathindmitte-lo
を使用して、Taq DNAポリメラーゼ(NEB)を使用する2段階のSOE PCRで集合させた。結果として得られた産物をゲル電気泳動と、これに続くQIAquick Gel抽出キット(Quiagen)を使用したゲル抽出で精製した。次にこれらを、隣接するオリゴヌクレオチドbarmitte-upおよびcathindmitte-loによる第2のPCR反応のテンプレートとして使用した。得られた生成物をフェノール/クロロフォルム抽出法およびエタノール沈殿法で精製し、NcoIおよびHindIIIで切断し、同様に消化されたpBar100-EETI-II M7I中に連結した。
【0162】
タンパク質の発現および二量体形成
文献に記載された手順で融合タンパク質を発現させて精製した。個々のマイクロタンパク質を、融合パートナーであるバルナーゼ'から、臭化シアン処理によって放出させた。マイクロタンパク質を逆相HPLCによって、文献(Schmoldt et al., 2004、およびEP 04 02 2455.2)に記載された手順で精製した。
【0163】
二量体の変異型を得るために、2等量の個々のマイクロタンパク質をDMF/DMSO(1:1)に溶解した。トリエチルアミンを最終濃度が1%となるように、二機能性クロスリンカーである、1等量のDSS(ビス-スクシンイミジル-スベラート、Pierce)とともに添加した。一晩のインキュベーション後に、結果として得られた二量体の架橋型マイクロタンパク質をRP-HPLCで精製した。
【0164】
Ba/F3レポーター細胞アッセイ法
TPOR/4Rαハイブリッド受容体用の発現ベクターであるpcDNATPOR/4Rαを以下の手順で構築した。ヒトTPO受容体のコード配列を含むcDNAクローンを対象に、オリゴヌクレオチドTpoR-Xho-lo
およびTpoR-Xho-up
を使用してPCR反応を行った。結果として得られたPCR産物をXhoIで消化し、同様に消化されたベクターpASKcDNA-NHに連結してpASKcDNA-NH-TPORを得た。pcDNA/4RαのNheI/HindIII断片(Krause et al., 2004)をpASK21TETIsendc1/2 (Christmann et al., 1999)にサブクローニングすることでpASKcDNA-NHを得た。ベクターpASKcDNA-NH-TPORから、TPORのコード配列を含むNheI/HindIII断片を、同様に消化されたpcDNA/4Rαに連結してpcDNATPOR/4Rαを得た。STAT6レポーター遺伝子コンストラクトpIεTATALucを、ヒトIεプロモーター(Ezernieks et al., 1996)のIL-4応答領域に由来するプロモーター/エンハンサー配列の全体を含む合成HindIII/BamHI断片をルシフェラーゼ発現プラスミドpTATALuc+ (Altschmied et al., 1997)に挿入して作製した。
【0165】
細胞培養、トランスフェクション、およびレポーター遺伝子アッセイ法
マウスのプレB細胞系列Ba/F3を文献(Lischke et al., 1995)に記載された手順で培養した。細胞を、細胞系列Nucleofector(商標)キットV(Amaxa, Germany)を使用してトランスフェクトした。簡単に説明すると、8×106個の細胞をRPMI 1640/10% FCS中で2時間、飢餓状態におき、遠心分離し、4μgの発現ベクターpcDNATPOR/4Rαおよび1μgのレポーター遺伝子コンストラクトpIεTATALucを添加した100μlのトランスフェクション試薬Vに再懸濁した。トランスフェクションは、プログラムT16を使用するNucleofector(商標)装置で実施した。個々のトランスフェクションバッチを3.5 mlのRPMI 1640/10% FCS中に回収し、1ウェルあたり2×105個の細胞/100μlとなるように96ウェル細胞培養プレートに添加した。細胞を何ら処理せずに1時間、静置した後に、さまざまな濃度のhTPO (Immunotools, Germany)、および個々のマイクロタンパク質候補をそれぞれ総容量が200μlとなるように添加した。TPO競合アッセイ法に関しては、細胞のアリコートを個々のマイクロタンパク質試料と1時間プレインキュベートした後にhTPOで刺激した。37℃、5% CO2で12時間のインキュベーション後に、細胞溶解物を調製し、ルシフェラーゼ活性を文献(Krause et al., 2004)に記載された手順で測定した。
【0166】
結果
TpoR結合配列(小文字で示す)が移植されたマイクロタンパク質のアミノ酸配列を表1に示す。マイクロタンパク質の足場として、マイクロタンパク質AGRP'、ヒトアグーチ関連タンパク質(McNulty et al., 2001)のメラノコルチン受容体結合ドメイン、およびマイクロタンパク質EETI-II(Christmann et al., 1999)をそれぞれ使用した。下線を付したアミノ酸をスペーサー配列として、またはSE-AG-TP-050の場合にはTPOR結合ペプチドの配列から外れるように導入した。アンタゴニスト活性の決定に際しては、マイクロタンパク質をバルナーゼ'との融合タンパク質として使用した(図2およびEP 04 02 2455.2)。アゴニスト活性の決定に関しては、精製済みのマイクロタンパク質を二量体として使用した。
表1:トロンボポエチン受容体(TPOR)結合配列によるループ置換を有するマイクロタンパク質のアミノ酸配列。タンパク質を、酵素的に不活性なバルナーゼ(バルナーゼ')との融合タンパク質として産生させ、hTPO受容体の活性化の阻害能力または誘導能力(拮抗作用)を検討した。アゴニスト活性の検討対象のマイクロタンパク質は化学的に二量体化されている。適用した試験はTPORの細胞外ドメインの二量体形成を誘導する能力の判定を含んだ。n.d.:決定せず。
【0167】
図3は、マウスの造血前駆細胞系列Ba/F3を使用したレポーター遺伝子アッセイ法の結果を示す。これらの細胞は、ヒトIL-4受容体由来のhTPOR細胞外ドメインおよび細胞内4Rαドメインの融合体をコードする遺伝子でトランスフェクトされ、ならびにレポーター遺伝子コンストラクトpIεTATALuc (Krause et al., 2004)と同時にトランスフェクトされた。TPO受容体の活性化の阻害を、5 nMのTPOおよび表1に示した500 nMの各化合物を使用して測定した。結果として、直鎖状のペプチド(SN-LN-TP-020)、ならびにマイクロタンパク質SE-AG-TP-030、SE-AG-TP-040、SE-ET-TP-020、およびSE-ET-TP-030は、TPOを介する効率的な受容体活性化に関してTPOのアンタゴニストとして作用可能であった。同様の結果は、いくつかのアッセイ法でSE-AG-TP-040に関して得られた。一方、アゴニスト活性は、検討された全てのマイクロタンパク質に関して認められた(表1)。
【0168】
図4は、SE-ET-TP-020のアゴニスト作用を示す。SE-ET-TP-020マイクロタンパク質は、二機能性の化学リンカーDSSの使用により二量体化させた。Ba/F3を対象としたTPO受容体活性化アッセイ法では、組換え型のヒトTPOと比較するために、精製済みの二量体を使用した。図4は、二量体であるSE-ET-TP-020による活性化と比較して、ヒトTPOによってTPORが活性化することを示す。この結果から、二量体のSE-ET-TP-020がTPOと同じ用量範囲で活性を示すことがわかる。
【0169】
同様に、二量体のSE-AG-TP-040およびSE-AG-TP-050を提供し、TPOのアゴニスト活性を検討した。これらは図5に示す。この結果から、これらのマイクロタンパク質二量体がハイブリッドTPO受容体を効率的に活性化したことがわかる。
【0170】
二量体のSE-ET-TP-020 (「SE-ET-TP-020d」とも呼ばれる)が骨髄細胞の増殖に及ぼす作用を評価するために、初代ヒト骨髄細胞を、50 ng/mlもしくは10 ng/mlのrhuTPO、または30 nM、10 nM、もしくは3 nMの二量体のSE-ET-TP-020dのいずれかを含む、またはいずれも含まない半固体培地で培養した。培養開始から10〜12日後に、大きな細胞(巨核球と推定される)のコロニーを数えた。結果を図6に示す。これらの結果から、二量体マイクロタンパク質SE-ET-TP-020dが、Mk前駆細胞集団の増殖を刺激する能力に関してTPOと似ていることがわかる。
【0171】
SE-ET-TP-020のインビボにおける機能を評価するために、Balb/cマウス(n=5)に1日2回、それぞれ10μg/kgおよび1000μg/kgのSE-ET-TP-020を皮下注射で投与した。5日後に血小板数を、対応するプラセボ実験と比較して測定した。図7からわかるように、SE-ET-TP-020が投与されたマウスでは血小板数が倍加した。
【0172】
実施例2:
マイクロタンパク質の合成後の環化
実験手順
省略形
Boc:tert-ブトキシカルボニル;DTT:ジチオトレイトール;ESI-MS:エレクトロスプレーイオン化質量分析;Fmoc:9-フルオレニルメチルオキシカルボニル;HATU:2-(1H-9-アザベンゾトリアゾール-1-イル)-1,3,3,3-テトラメチルウロニウムヘキサフルオロリン酸;HBTU:2-(1H-ベンゾトリアゾール-1-イル)-1,3,3,3-テトラメチルウロニウムヘキサフルオロリン酸;HOBt:1-ヒドロキシ-1H-ベンゾトリアゾール;HPLC:高圧液体クロマトグラフィー;SPPS:固相ペプチド合成;TFA:トリフルオロ酢酸。
【0173】
材料
試薬および溶媒は最高品質の市販品とし、さらなる精製を行わずに使用した。シアノホウ化水素ナトリウムおよびm-過ヨウ素酸ナトリウムはSIGMA-Aldrichから購入し、ヒドラジン一水和物および臭化シアンはFluka (Taufkirchen, Germany)から購入した。ESI質量スペクトルは、TSQ 700 Finnegan分光器で測定した。HPLCは、YMC J'sphere ODS H-80、RP C-18カラムを調製実験(250×4.6 mm、4μm、80Å)、および解析用試料(250×4.6μm、80Å)に使用して、Pharmacia Actaをベースとするシステムで実施した。
【0174】
発現ベクターpBar100-cyclo-McoEeTIの構築
発現ベクターpBar100-cyclo-McoEeTIの作製では、McoEeTIのコード配列を、Taqポリメラーゼ(Eppendorf)を使用する2段階のSOE-PCRで増幅した。初回のPCRは、プラスミドpBar100-McoEeTI(図2;EP 04 02 2455.2;Schmoldt et al., 2004)をテンプレートとして、ならびにオリゴヌクレオチドBspHI-McoTI-MSDGG-up
およびMCoTI-MSDGGhinten-SOE-lo
、ならびにMCoTI-MSDGGhinten-SOE-up
およびcat-hind-Mitte-lo
をそれぞれ使用して行い、2つの重複する断片を得た。これらの断片を、隣接するオリゴヌクレオチドBspHI-McoTI-MSDGG-upおよびcat-hind-Mitte-loを使用する第2のPCRテンプレートとして使用した。得られた産物をPagIおよびHindIII (MBI Fermentas)で消化し、事前にNcoIおよびHindIIIで切断されたpBar100-EETI-II M7I (Schmoldt et al., 2004)に連結して、発現ベクターpBar100-cyclo-McoEeTIを得た。このプラスミドは、tacプロモーターの制御下に、精製ハンドルとして使用される、バチルス・アミロリクエファシエンス(Bacillus amyloliquefaciens)のRNaseバルナーゼの不活性化突然変異体であるバルナーゼ'、phoA周辺質リーダー配列、およびcyclo-McoEeTI遺伝子の遺伝子融合体を有する。結果として得られるバルナーゼ'-cyclo-McoEeTI融合タンパク質は、2つのメチオニン残基を、バルナーゼ'とcyclo-McoEeTIの境界、およびcyclo-McoEeTIのC末端に有する。これが臭化シアンによって切断されると、融合パートナーであるバルナーゼ'が除去され、ならびにN末端にセリンを、およびC末端にホモセリンラクトンを有するcyclo-McoEeTIペプチドが得られる(例えば図10のa、b参照)。
【0175】
バルナーゼ'-cyclo-McoEeTI融合体の産生および精製
lacI遺伝子を含むヘルパープラスミドpRep4 (Qiagen)を含む大腸菌株71-18[F' lacIq lacZΔM15 proA+B+ Δlac-proAB supE thi1](入手先はB. Muller-Hill)をエレクトロポレーションによってpBar100-cyclo-McoEeTIで形質転換し、25μg/mlのクロラムフェニコールおよび37.5μg/mlのカナマイシンを含む50 mlのリッチ培地中で37℃で一晩、成長させた。産生は、5 Lの培養槽(Bioengineering)中で、文献(Schmoldt et al., 2004)に記載された手順で実施し、精製は、Schmoldt et al.に記載された手順に若干の変更を施して、バルナーゼ'を精製ハンドルとして使用して実施した。簡単に説明すると、バルナーゼ'-cyclo-McoEeTI融合タンパク質を、細胞培養液1リットルあたり55 mlの氷酢酸による酸性化後に培養液から精製した。この培養液を濾過し、H2Oで1:5の割合で希釈し、600 mlのSP-Sepharose XL (Amersham Biosciences)を含む、直径100 mmのガラス製カラムにアプライした。バルナーゼ'-融合タンパク質を100〜1000 mMのNaClの段階的な勾配で溶出し、および融合タンパク質を含むフラクションを、130 mlのAmberchrom CG-300M(Tosoh Bioscience)を含む26 mmのガラス製カラムに直接アプライした。H2O/0.1%(v/v)酢酸で洗浄後に、融合タンパク質をカラムから、0%(v/v)〜90%イソプロパノール/0.1%(v/v)酢酸の勾配を用いて溶出した。約25〜40%(v/v)のイソプロパノールの範囲の融合タンパク質含有フラクションを混合して凍結乾燥した。8 Mの尿素に再溶解し、および50 mMの酢酸アンモニウムに対して透析した後に、別の陽イオン交換クロマトグラフィーを、Vision BioCadワークステーション(PerSeptive Biosystems)に取り付けられたSP-Sepharose XL (Amersham Biosciences)を含むXK26カラム(2.6 x 20 cm、ベッドボリューム100 ml)を用いて流速8 mL/分で行った。溶出は、0〜0.5 M NaClの勾配を設けて行った。フラクションを含む融合タンパク質を混合し、および1/10容量の37% HClを添加してタンパク質を沈殿させ、ならびにHaereus Omnifuge 3L-R中で4000 rpmで10分間、遠心分離した。
【0176】
バルナーゼ'-cyclo-McoEeTIのCNBrによる切断
沈殿した融合タンパク質を、タンパク質1 mgあたり20 mlの0.2 M HCl/8 M尿素に可溶化し、1μlの5 M臭化シアン溶液(Fluka)を添加した(Kaiser and Metzka, 1999)。一晩のインキュベーション後に、試料を、Amberchrom CG-300M(Tosoh Bioscience、ベッドボリューム100 ml)を含むXK26カラム(Amersham Biosciences)に直接アプライした。H2O/0.1%(v/v) TFAで洗浄後に、切断されたcyclo-McoEeTIペプチドをバルナーゼ'から、5%(v/v)〜90%アセトニトリル/0.1%(v/v) TFAの勾配を利用して分離した。約20〜30%(v/v)アセトニトリルの範囲のcyclo-McoEeTIを含むフラクションを混合して凍結乾燥した。追加の精製を、Pharmacia Actaをベースとしたシステムで、YMC J'sphere ODS H-80、RP C-18調製用カラムを使用して行い、5 mgの純粋なMcoEeTI-セリンラクトンを得た。
【0177】
McoEeTIのヒドラジドの形成
ヒドラジン水和物(7μL、140μmol)を、水に溶解したMcoEeTIホモセリンラクトン(2,4.7 mg、1.4μmol)溶液(2 mL)に添加した。この混合物を室温で1時間、攪拌した。反応は、解析HPLCで制御した。開始時の試料に含まれるラクトンのピークが消失した後に、反応混合物を凍結乾燥して過剰なヒドラジンを除去した。凍結乾燥後の乾燥残渣を水-アセトニトリル混合液に再溶解し、および調製用HPLCで精製した。純粋な収率:2.1 mg(44.3%)。HPLC:tR=ESI MS(メタノール)の結果は以下の通りであった:m/z 856.0 ([M+4H]4+, 100)、1152.6 ([M+3H]3+, 33)、692.2 ([M+5H]5+, 27)、1728.6 ([M+2H]2+, 5)。
【0178】
過ヨウ素酸による酸化および環化
McoeETIヒドラジド(1.6 mg、0.46μmol)をリン酸緩衝液(1 mL、0.01 mmol、pH 7)に溶解した。NaIO4(1 mg、4.6μmol)を、リン酸緩衝液を溶媒とする溶液(1 mL)として室温で添加した。5分後にHPLCに注入することで反応を終了させた。モニタリングは、215 nmおよび280 nm(大環状分子の吸光度)で行った。純度は1 mg(63.7%)であった。
【0179】
参考文献
【図面の簡単な説明】
【0180】
【図1】本発明に従って連結された環状の二量体または多量体のマイクロタンパク質の例。(A)C-N結合型の二量体マイクロタンパク質;(B)二量体マイクロタンパク質の大環状分子;および(C)架橋型のマイクロタンパク質オリゴマー。
【図2】バルナーゼ'(barnase')-ICKペプチド融合コンストラクトの設計。(A)酵素的に不活性なH102A変異型の発現に至るバルナーゼ'遺伝子を含むプラスミドpBar100の略図;f1、複製起点;cat、クロラムフェニコール耐性マーカー;tetR、テトラサイクリンリプレッサーのコード遺伝子;colE1、colE1複製起点;tac、tacプロモーター配列;phoAs、アルカリホスファターゼの周辺質シグナル配列;ICK、ICKペプチドのコード配列。(B)バルナーゼ'-ICKペプチド融合体のDNAおよびタンパク質の配列。バルナーゼの#102位において置換されたアミノ酸(H102A)を太字で示す。pBar100シリーズの発現ベクターでは、臭化シアンによる融合タンパク質の化学的切断に使用可能な1つのメチオニンコドンが、バルナーゼ'の配列とICKペプチドのコード配列の境界に存在する。pBar100Thromベクターは、トロンビン認識部位(LVPRGS;SEQ ID NO: 9)を追加的にコードする。
【図3】さまざまなマイクロタンパク質が、TPOを介する受容体の活性化に及ぼす作用。このアッセイ法では、ルシフェラーゼレポーター遺伝子アッセイ法によってTPOR細胞外ドメインの二量体形成が検出される。灰色:500 nMの各マイクロタンパク質の添加後の発光の測定値。緑色:組換え型のヒトTPO(5 nM)を各化合物とともに添加した際の発光。
【図4】組換え型ヒトTPOによる活性化(A)と比較時の、二量体SE-ET-TP-020(B)の添加による、ハイブリッドTPO受容体の用量依存性の活性化。
【図5】二量体のSE-AG-TP-050および二量体SE-AG-TP-020(図5A)および二量体SE-AG-TP-040(図5B)の添加による、ハイブリッドTPO受容体の用量依存性の活性化。
【図6】組換え型ヒトTPOまたは二量体SE-ET-TP-020の添加による骨髄細胞増殖の誘導。大きな細胞のコロニー数を示す。
【図7】プラセボ投与を受けた対照マウスと比較時の、二量体SE-ET-TP-020が投与されたBalb/cマウスにおける血小板力価の約2倍の増加。
【図8】担体タンパク質、および両端でメチオニン残基と隣接するマイクロタンパク質を含む融合タンパク質の略図。
【図9】カルボキシ末端と隣接するメチオニンにおける臭化シアンによる切断によるマイクロタンパク質の環化と、これに続くヒドラジド形成、アミノ末端のセリンの酸化、分子内環化、および結果として生じるヒドラゾンの還元のスキームの概略。
【図10】カルボキシ末端と隣接するメチオニンにおける臭化シアンによる切断によるマイクロタンパク質の環化と、これに続くヒドラジド形成、アミノ末端のセリンの酸化、分子内環化、およびヒドラゾン結合の還元を示す略図。
【図11】ヒドラゾン形成に基づく化学的連結によるマイクロタンパク質のループの置換。Z:ホモセリンラクトンヒドラジド;B:ホモセリンラクトン;X:セリンアルデヒド;およびJ:ホモセリン。組み入れられたペプチドは化学的に合成され、およびC末端にヒドラジド(Z)を有する点に留意されたい。
【図12】cyclo-McoEeTIホモセリンラクトン(A)のcyclo-McoEeTIヒドラジド(B)への変換のRP-HPLC解析。RPカラムからの個々の溶出時間をクロマトグラムに示す。
【図13】10分間のインキュベーション(A)後のcyclo-McoEeTIヒドラジドの過ヨウ素酸ナトリウムによる反応のRP-HPLC解析、および17分で溶出される環状産物のRP-HPLC解析(B)。
【特許請求の範囲】
【請求項1】
少なくとも2個のマイクロタンパク質(microprotein)を含むポリペプチド。
【請求項2】
環状である、請求項1記載のポリペプチド。
【請求項3】
ポリペプチドのマイクロタンパク質が、1つのマイクロタンパク質のC末端が別のマイクロタンパク質のN末端に共有結合で結合されるように配置されている大環状分子を形成する、請求項2記載のポリペプチド。
【請求項4】
少なくとも2個のマイクロタンパク質が、非ペプチドカップリングによって連結されている、請求項1〜3のいずれか一項記載のポリペプチド。
【請求項5】
非ペプチドカップリングが、二機能性またはオリゴ機能性(oligofunctional)のリンカー分子を含む、請求項4記載のポリペプチド。
【請求項6】
リンカー分子が、アジピン酸ヒドラジド、ビス-スクシンイミジル-スベラート(DSS)、およびEDTA-ヒドラジドから選択される、請求項5記載のポリペプチド。
【請求項7】
個々のマイクロタンパク質が少なくとも6個のシステイン残基を含み、このうち6個のシステイン残基がシスチンノットを形成するように、ジスルフィド結合を介して連結されている、請求項1〜6のいずれか一項記載のポリペプチド。
【請求項8】
マイクロタンパク質の少なくとも1個が、アミノ酸モチーフCX3-CX4-CX4-7-CX1-CX4-5-CX5-7(SEQ ID NO: 8)を含み、Xが相互に独立して任意のアミノ酸残基であることを意味する、請求項1〜7のいずれか一項記載のポリペプチド。
【請求項9】
マイクロタンパク質が28〜40アミノ酸の長さを有する、請求項1〜8のいずれか一項記載のポリペプチド。
【請求項10】
少なくとも2個のマイクロタンパク質が、標的タンパク質に対する特異的な結合活性を有するアミノ酸配列を含む、請求項1〜9のいずれか一項記載のポリペプチド。
【請求項11】
標的タンパク質が膜結合型受容体である、請求項10記載のポリペプチド。
【請求項12】
前記受容体が、該受容体の分子が相互に近接する場合にシグナル伝達カスケードの下流の他のタンパク質を活性化する、請求項11記載のポリペプチド。
【請求項13】
前記受容体がトロンボポエチン(TPO)受容体である、請求項12記載のポリペプチド。
【請求項14】
TPO受容体に対する結合活性を有するアミノ酸配列が、アミノ酸配列IEGPTLRQWLAARA (SEQ ID NO: 7)を含む、請求項13記載のポリペプチド。
【請求項15】
少なくとも2個のマイクロタンパク質が、以下からなる群より選択されるアミノ酸配列を含む、請求項13または14のいずれか一項記載のポリペプチド:
(a)SEQ ID NO: 1〜6の任意の1つで表されるアミノ酸配列、
(b)該ポリペプチド内に存在する場合にTPO受容体を刺激可能な、(a)のアミノ酸配列の断片、ならびに
(c)該ポリペプチド内に存在する場合にTPO受容体を刺激可能な、(a)もしくは(b)のアミノ酸配列または断片の少なくとも1残基が、置換されているか、付加されているか、および/または欠失している機能的等価物。
【請求項16】
標的タンパク質が二量体またはオリゴマーの酵素であり、および特異的な結合活性を有するアミノ酸配列が、該酵素の活性部位に結合する、請求項10記載のポリペプチド。
【請求項17】
前記酵素がトリプターゼである、請求項16記載のポリペプチド。
【請求項18】
請求項1〜17のいずれか一項記載のポリペプチドをコードするヌクレオチド配列を含む核酸分子。
【請求項19】
請求項1〜17のいずれか一項記載のポリペプチド、または請求項18記載の核酸分子、および任意で薬学的に許容される担体を含む薬学的組成物。
【請求項20】
TPO受容体を刺激することで処置または予防が可能な疾患もしくは状態の処置用または予防用の薬学的組成物を作製するための、請求項13〜17のいずれか一項記載のポリペプチドまたは該ポリペプチドをコードする請求項18記載の核酸分子の使用。
【請求項21】
請求項13〜17のいずれか一項記載のポリペプチド、または該タンパク質をコードする請求項18記載の核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、TPO受容体を刺激することで処置可能な疾患もしくは状態に罹患した個体の処置法。
【請求項22】
疾患または状態が、血小板減少症、再生不良性貧血、骨髄不全、骨髄異形成症候群、および肝疾患からなる群より選択される、請求項20の使用または請求項21記載の方法。
【請求項23】
TPO受容体を刺激するための、請求項13〜17のいずれか一項記載のポリペプチド、または該ポリペプチドをコードする、請求項18記載の核酸分子の使用。
【請求項24】
インビトロである、請求項23の使用。
【請求項25】
トリプターゼの活性を阻害することで処置または予防が可能な疾患もしくは状態の処置用または予防用の薬学的組成物を作製するための、請求項17記載のポリペプチド、または該ポリペプチドをコードする、請求項18記載の核酸分子の使用。
【請求項26】
請求項17記載のポリペプチド、または該ポリペプチドをコードする、請求項18記載の核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、トリプターゼの活性を阻害することで処置可能な疾患もしくは状態に罹患した個体の処置法。
【請求項27】
疾患もしくは状態が、喘息、炎症、乾癬、肺線維症、間質性肺疾患、慢性関節リウマチ、歯肉炎、歯周炎、アレルギー反応、アレルギー性鼻炎、骨関節症、アテローム性動脈硬化症、血管形成、多発性硬化症、および癌からなる群より選択される、請求項25の使用または請求項26記載の方法。
【請求項28】
トリプターゼの活性を阻害するための、請求項17記載のポリペプチドまたは該ポリペプチドをコードする請求項18記載の核酸分子の使用。
【請求項29】
インビトロである、請求項28の使用。
【請求項30】
請求項1〜17のいずれか一項記載のポリペプチド、または請求項18記載の核酸分子を含むキット。
【請求項31】
以下の段階を含む、マイクロタンパク質内に共有結合を形成させる方法:
(a)N末端の反応性カルボニル基、およびC末端のホモセリンラクトン残基を含むマイクロタンパク質基質を提供する段階;ならびに
(b)前記N末端基およびC末端の残基をヒドラゾン結合に変換させるようにマイクロタンパク質基質を反応させる段階。
【請求項32】
N末端の反応性カルボニル基がグリオキシリル基またはケト基である、請求項31記載の方法。
【請求項33】
グリオキシリル基が、N末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される、請求項32記載の方法。
【請求項34】
マイクロタンパク質基質のN末端のセリン残基またはスレオニン残基が、メチオニン残基と、これに続くセリン残基またはスレオニン残基との間のペプチド結合において該マイクロタンパク質基質を含む前駆体ポリペプチドを臭化シアンを使用して切断することで提供される、請求項33記載の方法。
【請求項35】
マイクロタンパク質基質のC末端のホモセリンラクトン残基が、メチオニン残基と、これに続くアミノ酸残基との間のペプチド結合を臭化シアンを使用して切断することで提供される、請求項31〜34のいずれか一項記載の方法。
【請求項36】
段階(b)が、以下の段階を含む、請求項31〜35のいずれか一項記載の方法:
(i)C末端のホモセリンラクトン残基をホモセリンヒドラジドに反応させる段階;
(ii)ホモセリンヒドラジドおよびN末端の反応性カルボニル基を反応させてヒドラゾンを生じさせる段階;ならびに
(iii)任意でヒドラゾンを還元する段階。
【請求項37】
マイクロタンパク質基質が1つの単体のマイクロタンパク質であり、かつ共有結合の形成が、該マイクロタンパク質のペプチドバックボーンの環化のためである、請求項31〜36のいずれか一項記載の方法。
【請求項38】
マイクロタンパク質基質が1つの単体のマイクロタンパク質であり、および共有結合の形成が、グラフトペプチド(graft peptide)をマイクロタンパク質内に連結させるためである、請求項31〜36のいずれか一項記載の方法。
【請求項39】
以下の段階を含む、請求項38記載の方法:
(A)グラフトペプチドで置換されるマイクロタンパク質のループ状のアミノ酸配列を、メチオニン残基と後続のアミノ酸残基との間の適切なペプチド結合において臭化シアンを使用して切断する段階であって、ループ状のアミノ酸配列のC末端における該後続のアミノ酸残基がセリン残基またはスレオニン残基である段階;
(B)グリオキシリル基を形成させるために、セリン残基またはスレオニン残基を穏やかな酸化によって反応させる段階;
(C)N末端の反応性カルボニル基、およびC末端のホモセリンラクトンヒドラジドを含むグラフトペプチド配列を、マイクロタンパク質のグリオキシリル基と反応させて、ヒドラゾンを生じさせる段階;
(D)段階(A)の切断によって生じるC末端のホモセリンラクトン残基をホモセリンヒドラジドと反応させる段階;ならびに、
(E)(D)のホモセリンヒドラジドをグラフトペプチドのN末端の反応性カルボニル基と反応させてヒドラゾンを生じさせる段階。
【請求項40】
グラフトペプチドのN末端の反応性カルボニル基がグリオキシリル基またはケト基である、請求項39記載の方法。
【請求項41】
グリオキシリル基が、N末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される、請求項40記載の方法。
【請求項42】
マイクロタンパク質基質が少なくとも2個のマイクロタンパク質を含み、および共有結合の形成が、該マイクロタンパク質の二量体またはオリゴマーを生じさせるためになされる、請求項31〜36のいずれか一項記載の方法。
【請求項43】
マイクロタンパク質基質が少なくとも2個のマイクロタンパク質を含み、ならびに共有結合の形成が、大環状分子の二量体またはオリゴマーのマイクロタンパク質を生じさせるためになされる、請求項31〜36および42のいずれか一項記載の方法。
【請求項44】
請求項31〜43のいずれか一項記載の方法によって得ることができる1個または複数のマイクロタンパク質を含むポリペプチド。
【請求項1】
少なくとも2個のマイクロタンパク質(microprotein)を含むポリペプチド。
【請求項2】
環状である、請求項1記載のポリペプチド。
【請求項3】
ポリペプチドのマイクロタンパク質が、1つのマイクロタンパク質のC末端が別のマイクロタンパク質のN末端に共有結合で結合されるように配置されている大環状分子を形成する、請求項2記載のポリペプチド。
【請求項4】
少なくとも2個のマイクロタンパク質が、非ペプチドカップリングによって連結されている、請求項1〜3のいずれか一項記載のポリペプチド。
【請求項5】
非ペプチドカップリングが、二機能性またはオリゴ機能性(oligofunctional)のリンカー分子を含む、請求項4記載のポリペプチド。
【請求項6】
リンカー分子が、アジピン酸ヒドラジド、ビス-スクシンイミジル-スベラート(DSS)、およびEDTA-ヒドラジドから選択される、請求項5記載のポリペプチド。
【請求項7】
個々のマイクロタンパク質が少なくとも6個のシステイン残基を含み、このうち6個のシステイン残基がシスチンノットを形成するように、ジスルフィド結合を介して連結されている、請求項1〜6のいずれか一項記載のポリペプチド。
【請求項8】
マイクロタンパク質の少なくとも1個が、アミノ酸モチーフCX3-CX4-CX4-7-CX1-CX4-5-CX5-7(SEQ ID NO: 8)を含み、Xが相互に独立して任意のアミノ酸残基であることを意味する、請求項1〜7のいずれか一項記載のポリペプチド。
【請求項9】
マイクロタンパク質が28〜40アミノ酸の長さを有する、請求項1〜8のいずれか一項記載のポリペプチド。
【請求項10】
少なくとも2個のマイクロタンパク質が、標的タンパク質に対する特異的な結合活性を有するアミノ酸配列を含む、請求項1〜9のいずれか一項記載のポリペプチド。
【請求項11】
標的タンパク質が膜結合型受容体である、請求項10記載のポリペプチド。
【請求項12】
前記受容体が、該受容体の分子が相互に近接する場合にシグナル伝達カスケードの下流の他のタンパク質を活性化する、請求項11記載のポリペプチド。
【請求項13】
前記受容体がトロンボポエチン(TPO)受容体である、請求項12記載のポリペプチド。
【請求項14】
TPO受容体に対する結合活性を有するアミノ酸配列が、アミノ酸配列IEGPTLRQWLAARA (SEQ ID NO: 7)を含む、請求項13記載のポリペプチド。
【請求項15】
少なくとも2個のマイクロタンパク質が、以下からなる群より選択されるアミノ酸配列を含む、請求項13または14のいずれか一項記載のポリペプチド:
(a)SEQ ID NO: 1〜6の任意の1つで表されるアミノ酸配列、
(b)該ポリペプチド内に存在する場合にTPO受容体を刺激可能な、(a)のアミノ酸配列の断片、ならびに
(c)該ポリペプチド内に存在する場合にTPO受容体を刺激可能な、(a)もしくは(b)のアミノ酸配列または断片の少なくとも1残基が、置換されているか、付加されているか、および/または欠失している機能的等価物。
【請求項16】
標的タンパク質が二量体またはオリゴマーの酵素であり、および特異的な結合活性を有するアミノ酸配列が、該酵素の活性部位に結合する、請求項10記載のポリペプチド。
【請求項17】
前記酵素がトリプターゼである、請求項16記載のポリペプチド。
【請求項18】
請求項1〜17のいずれか一項記載のポリペプチドをコードするヌクレオチド配列を含む核酸分子。
【請求項19】
請求項1〜17のいずれか一項記載のポリペプチド、または請求項18記載の核酸分子、および任意で薬学的に許容される担体を含む薬学的組成物。
【請求項20】
TPO受容体を刺激することで処置または予防が可能な疾患もしくは状態の処置用または予防用の薬学的組成物を作製するための、請求項13〜17のいずれか一項記載のポリペプチドまたは該ポリペプチドをコードする請求項18記載の核酸分子の使用。
【請求項21】
請求項13〜17のいずれか一項記載のポリペプチド、または該タンパク質をコードする請求項18記載の核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、TPO受容体を刺激することで処置可能な疾患もしくは状態に罹患した個体の処置法。
【請求項22】
疾患または状態が、血小板減少症、再生不良性貧血、骨髄不全、骨髄異形成症候群、および肝疾患からなる群より選択される、請求項20の使用または請求項21記載の方法。
【請求項23】
TPO受容体を刺激するための、請求項13〜17のいずれか一項記載のポリペプチド、または該ポリペプチドをコードする、請求項18記載の核酸分子の使用。
【請求項24】
インビトロである、請求項23の使用。
【請求項25】
トリプターゼの活性を阻害することで処置または予防が可能な疾患もしくは状態の処置用または予防用の薬学的組成物を作製するための、請求項17記載のポリペプチド、または該ポリペプチドをコードする、請求項18記載の核酸分子の使用。
【請求項26】
請求項17記載のポリペプチド、または該ポリペプチドをコードする、請求項18記載の核酸分子、および任意で薬学的に許容される担体を含む有効量の薬学的組成物を個体に投与する段階を含む、トリプターゼの活性を阻害することで処置可能な疾患もしくは状態に罹患した個体の処置法。
【請求項27】
疾患もしくは状態が、喘息、炎症、乾癬、肺線維症、間質性肺疾患、慢性関節リウマチ、歯肉炎、歯周炎、アレルギー反応、アレルギー性鼻炎、骨関節症、アテローム性動脈硬化症、血管形成、多発性硬化症、および癌からなる群より選択される、請求項25の使用または請求項26記載の方法。
【請求項28】
トリプターゼの活性を阻害するための、請求項17記載のポリペプチドまたは該ポリペプチドをコードする請求項18記載の核酸分子の使用。
【請求項29】
インビトロである、請求項28の使用。
【請求項30】
請求項1〜17のいずれか一項記載のポリペプチド、または請求項18記載の核酸分子を含むキット。
【請求項31】
以下の段階を含む、マイクロタンパク質内に共有結合を形成させる方法:
(a)N末端の反応性カルボニル基、およびC末端のホモセリンラクトン残基を含むマイクロタンパク質基質を提供する段階;ならびに
(b)前記N末端基およびC末端の残基をヒドラゾン結合に変換させるようにマイクロタンパク質基質を反応させる段階。
【請求項32】
N末端の反応性カルボニル基がグリオキシリル基またはケト基である、請求項31記載の方法。
【請求項33】
グリオキシリル基が、N末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される、請求項32記載の方法。
【請求項34】
マイクロタンパク質基質のN末端のセリン残基またはスレオニン残基が、メチオニン残基と、これに続くセリン残基またはスレオニン残基との間のペプチド結合において該マイクロタンパク質基質を含む前駆体ポリペプチドを臭化シアンを使用して切断することで提供される、請求項33記載の方法。
【請求項35】
マイクロタンパク質基質のC末端のホモセリンラクトン残基が、メチオニン残基と、これに続くアミノ酸残基との間のペプチド結合を臭化シアンを使用して切断することで提供される、請求項31〜34のいずれか一項記載の方法。
【請求項36】
段階(b)が、以下の段階を含む、請求項31〜35のいずれか一項記載の方法:
(i)C末端のホモセリンラクトン残基をホモセリンヒドラジドに反応させる段階;
(ii)ホモセリンヒドラジドおよびN末端の反応性カルボニル基を反応させてヒドラゾンを生じさせる段階;ならびに
(iii)任意でヒドラゾンを還元する段階。
【請求項37】
マイクロタンパク質基質が1つの単体のマイクロタンパク質であり、かつ共有結合の形成が、該マイクロタンパク質のペプチドバックボーンの環化のためである、請求項31〜36のいずれか一項記載の方法。
【請求項38】
マイクロタンパク質基質が1つの単体のマイクロタンパク質であり、および共有結合の形成が、グラフトペプチド(graft peptide)をマイクロタンパク質内に連結させるためである、請求項31〜36のいずれか一項記載の方法。
【請求項39】
以下の段階を含む、請求項38記載の方法:
(A)グラフトペプチドで置換されるマイクロタンパク質のループ状のアミノ酸配列を、メチオニン残基と後続のアミノ酸残基との間の適切なペプチド結合において臭化シアンを使用して切断する段階であって、ループ状のアミノ酸配列のC末端における該後続のアミノ酸残基がセリン残基またはスレオニン残基である段階;
(B)グリオキシリル基を形成させるために、セリン残基またはスレオニン残基を穏やかな酸化によって反応させる段階;
(C)N末端の反応性カルボニル基、およびC末端のホモセリンラクトンヒドラジドを含むグラフトペプチド配列を、マイクロタンパク質のグリオキシリル基と反応させて、ヒドラゾンを生じさせる段階;
(D)段階(A)の切断によって生じるC末端のホモセリンラクトン残基をホモセリンヒドラジドと反応させる段階;ならびに、
(E)(D)のホモセリンヒドラジドをグラフトペプチドのN末端の反応性カルボニル基と反応させてヒドラゾンを生じさせる段階。
【請求項40】
グラフトペプチドのN末端の反応性カルボニル基がグリオキシリル基またはケト基である、請求項39記載の方法。
【請求項41】
グリオキシリル基が、N末端のセリン残基、スレオニン残基、またはヒドロキシリシン残基の穏やかな酸化によって形成される、請求項40記載の方法。
【請求項42】
マイクロタンパク質基質が少なくとも2個のマイクロタンパク質を含み、および共有結合の形成が、該マイクロタンパク質の二量体またはオリゴマーを生じさせるためになされる、請求項31〜36のいずれか一項記載の方法。
【請求項43】
マイクロタンパク質基質が少なくとも2個のマイクロタンパク質を含み、ならびに共有結合の形成が、大環状分子の二量体またはオリゴマーのマイクロタンパク質を生じさせるためになされる、請求項31〜36および42のいずれか一項記載の方法。
【請求項44】
請求項31〜43のいずれか一項記載の方法によって得ることができる1個または複数のマイクロタンパク質を含むポリペプチド。
【図1】

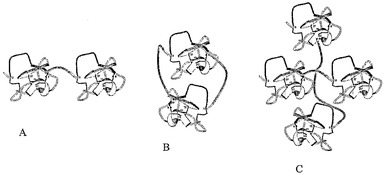
【図2】

【図3】
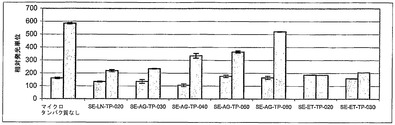
【図4】
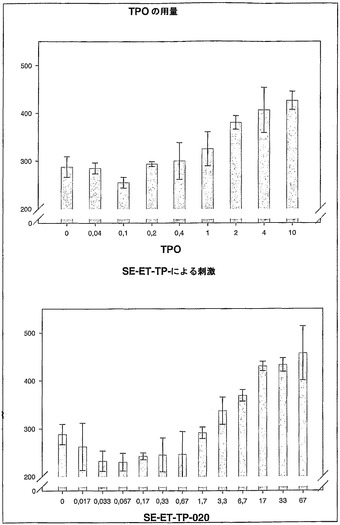
【図5】

【図6】
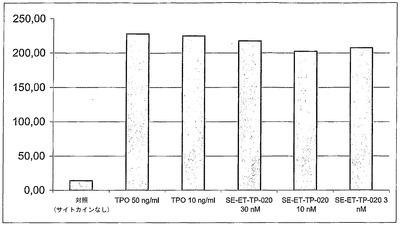
【図7】
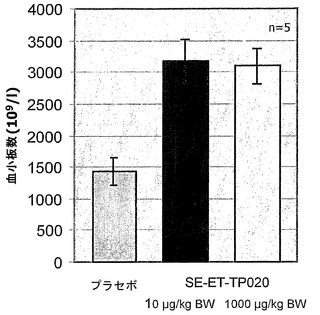
【図8】
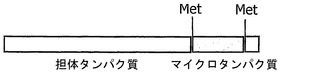
【図9】
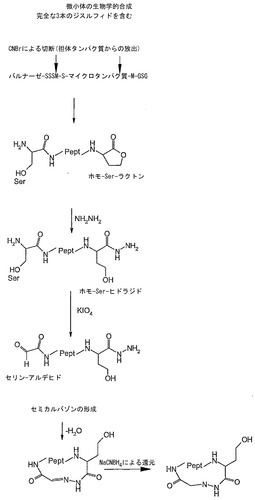
【図10】

【図11】
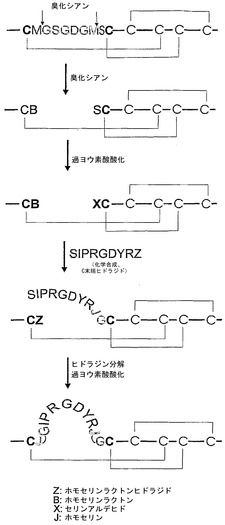
【図12】
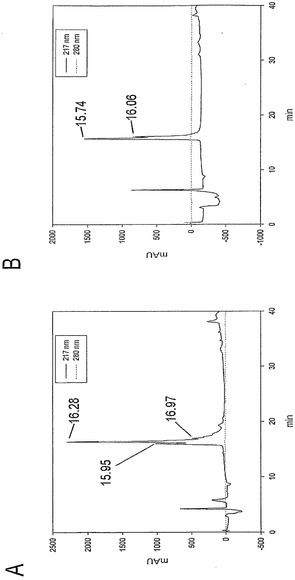
【図13】
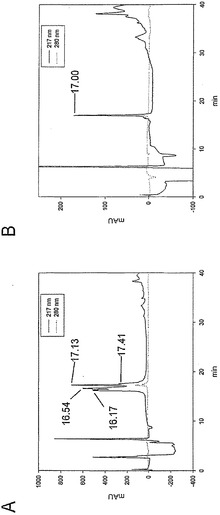
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2008−532505(P2008−532505A)
【公表日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願番号】特願2008−500131(P2008−500131)
【出願日】平成18年3月9日(2006.3.9)
【国際出願番号】PCT/EP2006/002188
【国際公開番号】WO2006/094813
【国際公開日】平成18年9月14日(2006.9.14)
【出願人】(507302210)ナスカセル テクノロジーズ アーゲー. (1)
【Fターム(参考)】
【公表日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願日】平成18年3月9日(2006.3.9)
【国際出願番号】PCT/EP2006/002188
【国際公開番号】WO2006/094813
【国際公開日】平成18年9月14日(2006.9.14)
【出願人】(507302210)ナスカセル テクノロジーズ アーゲー. (1)
【Fターム(参考)】
[ Back to top ]