
人参から新規な糖リポタンパク質ジントニンを製造する方法、及び前記方法で製造された新規な糖リポタンパク質ジントニン
人参から分離同定した新規な糖リポタンパク質としてのジントニンを製造する方法、前記方法で人参から分離同定した新規な糖リポタンパク質ジントニン、及び前記新規な糖脂質タンパク質ジントニンの用途を提供する。
本発明の新規な糖リポタンパク質ジントニンは、一時的に細胞質内遊離Ca2+の増加を誘発し、細胞内カルシウム濃度の一時的な増加によりアフリカツメガエル卵母細胞(Xenopus oocytes)で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)を活性化して細胞内カルシウム濃度を増加させるので、カルシウム濃度の低下に起因する疾患の予防及び治療に有用に使用できるうえ、強壮作用、免疫力増加、性機能強化、神経系保護及び活性、血管新生、抗糖尿作用などの細胞内カルシウム依存性の様々な生理活性などにおいても優れた効果を示す。

本発明の新規な糖リポタンパク質ジントニンは、一時的に細胞質内遊離Ca2+の増加を誘発し、細胞内カルシウム濃度の一時的な増加によりアフリカツメガエル卵母細胞(Xenopus oocytes)で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)を活性化して細胞内カルシウム濃度を増加させるので、カルシウム濃度の低下に起因する疾患の予防及び治療に有用に使用できるうえ、強壮作用、免疫力増加、性機能強化、神経系保護及び活性、血管新生、抗糖尿作用などの細胞内カルシウム依存性の様々な生理活性などにおいても優れた効果を示す。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、人参に存在する新規な糖リポタンパク質(glycolipoprotein)に係り、さらに詳しくは、人参から分離同定して新規な糖リポタンパク質を製造する方法、前記方法で人参から分離同定した新規な糖リポタンパク質、および前記新規な糖リポタンパク質の用途に関する。
【背景技術】
【0002】
人参は、一般に、適応促進薬(adaptogen)又は生命延長のための強壮剤として用いられており、ストレス、疲労、疾病、癌及び糖尿病に対抗して生体機能を向上させる。このような人参は、韓国だけでなく、中国及び日本でも数百年前から使われてきた。最近、人参は世界で薬草として最も有名なものの一つである(非特許文献1)。
【0003】
ジンセノサイド(Ginsenoside)は、1960年代初めに分離されて特性化されたから、生理学的、薬理学的研究で代表的な成分として多用されるものである(非特許文献2、非特許文献3)。この他にも、最近の研究では、人参にはよく知られていない他の成分、例えば多糖類(polysaccharides)、ポリアセチレン類(polyaccetylenes)、タンパク質(protein)などが含有されていることが明らかになった(非特許文献4)。
【0004】
人参のジンセノサイド成分は、量が少なく、分離過程が複雑であり、純粋なジンセノサイドの場合には価格が高いため、人参の根からブタノール抽出方法で得た粗総サポニン分画(crude ginseng total saponin fraction、CGSF)を使用してきた(非特許文献5、非特許文献6、非特許文献7、非特許文献8、非特許文献9、非特許文献10、非特許文献11、非特許文献12、非特許文献13及び非特許文献14)。
【0005】
粗総サポニン分画(CGSF)は細胞膜シグナル経路(signal pathway)を介してその活性が現れるものと知られているが、例えば、Choi等は、アフリカツメガエル卵母細胞(Xenopus oocytes)に粗総サポニン分画を処理したとき、PLCβ−IP3に連結されているPTX−insentive Gαq/11タンパク質を介してカルシウム依存性塩素イオンチャネル(Ca2+activated Chloride Channel、CaCC)が活性化されることを解明した(非特許文献7)。また、Lee等は、アフリカツメガエル卵母細胞に粗総サポニン分画を持続的に処理したとき、粗総サポニン分画によって活性化されたカルシウム依存性塩素イオンチャネル電流(CaCC currents)が自発的に減少することを報告した(非特許文献9)。
【0006】
カルモジュリン(Calmodulin)をアフリカツメガエル卵母細胞に直接注入したとき、或いは細胞内のカルシウム貯蔵庫を枯渇させたときに、粗総サポニン分画によるカルシウム依存性塩素イオンチャネルの活性がなくなる(非特許文献12)。しかも、アフリカツメガエル卵母細胞に粗総サポニンを処理したときにSOCE(stored-operated Ca2+ entry)が誘発され(非特許文献10)、これにより細胞外或いは細胞内のカルシウム貯蔵庫でカルシウム濃度の増加が引き起こされてアフリカツメガエル卵母細胞内のCaCCが活性化されるものと知られている(非特許文献15)。
【0007】
本発明者らは、ジンセノサイドがCaCCを活性化させることを確認するために、粗総サポニン分画物(CGSF)から純粋なジンセノサイドを分離する過程中に、CGSFよりジンセノサイドに富む分画物の場合、CaCCの活性に対するその効果が急激に消えるか無くなることを見出した。すなわち、CGSFから純粋に分離されたジンセノサイドの場合、アフリカツメガエル卵母細胞内におけるCaCC活性効果がないことを確認した。これはジンセノサイドだけでなく、未知のある物質がCGSFに存在し、この物質がアフリカツメガエル卵母細胞内のCaCCの活性に影響を及ぼすことが分かった。
【0008】
そこで、CGSF内に内因性CaCCの活性を起こすある成分の分離のために鋭意努力した結果、新規な生理活性成分を分離同定し、本発明を完成するに至った。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Tyler,J.Pharm.Technol.11,214-220,1995
【非特許文献2】Shibata,et al.Tetraheadron Letters 1962,1239-1245,1963
【非特許文献3】Wagner-Jauregg and Roth,Pharm Acta Helv 37,352-357,1962
【非特許文献4】Nah,Kor.J.Ginseng Sci.21,1-12,1997
【非特許文献5】Kanzaki,et al.Br J Pharmacol.125(2),255-262,1998
【非特許文献6】Choi,et al.Br J.Pharmacol.132,641-648,2001
【非特許文献7】Choi,et al.,J.Biol.Chem.276,48797-48802,2001
【非特許文献8】Choi,et al.Eur J Pharmacol 468,83-92,2003
【非特許文献9】Lee,et al,J Biol Chem.279,9912-9921,2004
【非特許文献10】Jeong,et al,Br J Pharmacol.142,585-593,2004
【非特許文献11】Reay,et al,J Psychopharmacol.19,357-365,2005
【非特許文献12】Lee,et al,Arch Pharm Res 28,413-420,2005
【非特許文献13】Wei,et al J Ethnopharmacol.111,613-618,2007
【非特許文献14】Eriksson,et al J Ethnopharmacol.119,17-23,2008
【非特許文献15】Dascal,CRC Crit Rev Biochem 22,317-387,1987
【発明の概要】
【発明が解決しようとする課題】
【0010】
したがって、本発明の主な目的は、人参から強壮活性の役目を果たす新規な生理活性物質を分離同定する方法、前記方法で分離同定された生理活性物質、及び前記生理活性物質の用途を提供することにある。
【課題を解決するための手段】
【0011】
上記目的を達成するために、本発明は、人参に存在する新規な糖リポタンパク質(glycolipoprotein)を人参から分離同定して製造する方法を提供する。
【0012】
具体的に、本発明に係る人参から新規な糖リポタンパク質としてのジントニン(以下「ジントニン(gintonin)という」を製造する方法は、(1)人参からメタノール抽出物を製造する段階と、(2)前記メタノール抽出物を水とn−ブタノールの混合溶媒に分画する段階と、(3)前記水分画物とn−ブタノール分画物のうち、n−ブタノール分画物に対して、溶出溶媒としてクロロメタン:メタノール:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離する段階と、(4)前記8個の分画物のうち、CaCC(Ca2+activated Chloride Channel)に対する活性が最も高い第7の分画物に対して、溶出溶媒としてエタノール:酢酸エチル:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、2つの分画に分離する段階と、(5)前記2つの分画物のうち、CaCC活性が高い分画物IIを透析し、最終生成物を得る段階とを含んでなる。
【0013】
また、本発明は、これに加えて、(6)最終分画物をNaClが含有されたpH7.2のリン酸塩緩衝食塩水(phosphate buffer saline、PBS)溶液に溶解し、陰イオン交換クロマトグラフィー及びゲル濾過クロマトグラフィーを行い、2つの分画に分離する段階と、(7)前記2つの分画物に対してそれぞれ、NaClが含有されたpH8.2のTris−HCl及びNaClが含有されたpH7.2のリン酸塩緩衝食塩水(PBS)で陰イオン交換クロマトグラフィーを行い、個別ジントニンを分離する段階とを含んでもよい。
【0014】
また、本発明は、前述したような方法で人参から分離同定した新規な糖リポタンパク質(ジントニン)を提供する。
【0015】
本発明において、前記糖リポタンパク質(ジントニン)の分子量が約67kDaであり、見掛け分子量は約13kDaの五量体であることを特徴とし、アミノ酸組成成分として、システインとシスチン(cysteine and cystine)、アスパラギンとアスパラギン酸(asparagine and aspartic acid)、グルタミンとグルタミン酸(glutamine and glutamic acid)、セリン(serine)、グリシン(glycine)、アルギニン(arginine)、トレオニン(threonine)、アラニン(alanine)、プロリン(proline)、バリン(valine)、イソロイシン(isoleucine)、ロイシン(leucine)、フェニルアラニン(phenylalanine)、トリプトファン(tryptophan)及びリシン(lysine)を含み、炭水化物組成成分として、ラムノース(rhamnose)、アラビノース(arabinose)、グルコース(glucose)、マンノース(mannose)、キシロース(xylose)及びグルコサミン(glucosamine)を含み、脂質組成成分としては、リノール酸(linoleic acid)、パルミチン酸(palmitic acid)、オレイン酸(oleic acid)及びステアリン酸(stearic acid)などを含む。
【0016】
人参は、ウコキ科薬用植物であって、例えば、加工方法に従って加工していない水参(生参)、水参を乾燥させた白参、水参を蒸して乾燥させた紅参などがあり、栽培方法によっては人参の畑で人為的に栽培した栽培参、人参の種を山中に播いて自然状態で栽培した長脳参、自然状態で自生した山参などがあり、本発明において、用語「人参」は前記全ての人参の種類を含む意味で使用される。また、本発明における人参は高麗人参(オタネニンジン、Panax ginseng C.A.Meyer)、西洋参(アメリカニンジン)及び中国人参(田七人参)などを含む。
【0017】
以下、本発明を詳細に説明する。
【0018】
本発明の人参に存在する新規な糖リポタンパク質としてのジントニンは、下記のとおり収得できる。
【0019】
まず、乾燥した人参粉末を、試料重量の約1〜20倍、好ましくは約1〜10倍に達する体積の水、メタノール、エタノール、ブタノールなどのC1〜C4の低級アルコールの極性溶媒又はこれらの約1:0.1〜1:10の混合比を有する混合溶媒で、好ましくは1:0.2〜1:5の混合比(v/v)を有する水とメタノールとの混合溶媒で、70〜120℃で約0.1〜48時間、好ましくは5〜12時間攪拌抽出、熱水抽出、冷浸抽出、加温抽出、還流冷却抽出又は超音波抽出などの抽出方法、好ましくは還流冷却抽出によって行った後、濾過して上層液を回収し、前述の過程を数回、好ましくは2〜5回繰り返し行った後、上層液を集めて減圧濃縮したものを、水とn−ブタノールが同量で混合された混合溶媒で溶媒分画し、水とn−ブタノールの分画物を得る。
【0020】
前記分画物のうち、粗総サポニン分画(CGSF)に該当するn−ブタノール分画物に対して、溶出溶媒としてクロロメタノール:メタノール:水(CHCl3L:MeOH:H2O)の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離した後、分離された分画物に対してアフリカツメガエル卵母細胞におけるカルシウム依存性塩素イオンチャネル(CaCC)の活性確認実験を行って分画物の活性を検索した後、最も活性の高い分画に対してさらにエタノール:酢酸エタノール:水(EtOH:EtOAc:H2O)の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行って2個の分画に分画し、二重活性の高い分画を過量の蒸留水と透析膜(dialysis membrane)を用いて透析し、本発明の粗ジントニンを得ることができる。
【0021】
こうして得られた粗ジントニンは、電気泳動(SDS−PAGE)による分子量分析、タンパク質及びアミノ酸組成分析、炭水化物組成分析、GC−MSによる脂質組成分析などを介して確認することができる。
【0022】
また、本発明の個別ジントニンは、上述したような工程で得られた粗ジントニンをNaClの含有されたリン酸塩緩衝食塩水(phosphate buffer saline、PBS)に溶解し、陰イオン交換クロマトグラフィー及びゲル濾過クロマトグラフィーを行って2個のジントニン分画物を収得する。それぞれのジントニン分画物は、NaClが含有されたTris−HCl(pH8.2)及びNaClが含有されたPBS(pH7.2)を用いて陰イオン交換クロマトグラフィーを行って得ることができる。
【0023】
本発明において、ジントニンの製造のために、山参、長脳参、栽培参、西洋参及び中国人参などの人参を使用することができる。前記人参は、水参、白参、紅参などの形として利用でき、好ましくは4〜6年根の高麗人参(Panax ginseng C.A.Meyer)から製造した紅参を使用することが良いが、これに限定されるものではない。
【0024】
また、本発明は、糖リポタンパク質(ジントニン)をカルシウム濃度の低下に起因する疾患の予防または治療剤としての用途で提供する。
【0025】
具体的に、本発明は、前述の方法で製造された糖リポタンパク質としてのジントニンを有効成分として含む、カルシウム濃度の低下に起因する疾患の予防及び治療のための薬学組成物を提供する。
【0026】
本発明において、前記ジントニンは、一時的に細胞質内遊離Ca2+(Free Ca2+)の増加を誘発し、細胞内カルシウム濃度の一時的な増加により、アフリカツメガエル卵母細胞で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)を活性化して細胞内カルシウム濃度を増加させ、その結果、カルシウム濃度の低下に起因する疾患の予防及び治療に優れた効果を示し、特にカルシウム濃度の低下により引き起こされる神経系疾患の異常を改善することにより、治療及び予防が可能である。
【0027】
カルシウム濃度の低下による神経系疾患は、精神分裂(Schizophrenia)、アルツハイマー病(Alzheimer’s disease)、ハンチントン病(Huntington’s disease)、家族性片麻痺性片頭痛(Familial hemiplegic migraine)、癲癇(epilepsy)、一過性運動失調症(episodic ataxia)、脊髄小脳失調症(spinocerebellar ataxias)などを含むが、これらの疾患に限定されるものではない。
【0028】
また、本発明の組成物は、カルシウム不足による成長阻害などにも効果があり、細胞内カルシウム依存性の様々な生理活性、例えば強壮作用、免疫力増加、性機能強化、神経系保護及び活性、血管新生、抗糖尿作用などにおいても効果を示す。
【0029】
また、人参は長時間生薬として使用されてきたものであり、このような人参から分離した本発明のジントニンも毒性及び副作用などの問題が全くない。
【0030】
本発明のカルシウム濃度の低下による疾患の予防及び治療用薬学組成物は、組成物の全体重量に対し、前記ジントニンは0.0001〜10重量%、好ましくは0.001〜1重量%で含む。
【0031】
また、本発明のジントニンを含む組成物は、薬学的組成物の製造に通常用いる適切な担体、賦形剤及び希釈剤をさらに含んでもよく、本発明のジントニンの薬学的投与形態は単独で或いは他の薬理的活性化合物との結合又は適当な集合として使用できる。
【0032】
本発明に係るジントニンを含む組成物は、それぞれ通常の方法によって散剤、顆粒剤、錠剤、カプセル剤、懸濁液、エマルジョン、シロップ、エアロゾールなどの経口剤形、外用剤、坐剤及び滅菌注射溶液の形に剤形化して使用できる。前記組成物に含まれる担体、賦形剤及び希釈剤としては、ラクトース、デキストロース、スクロース、ソルビトール、マンニトール、キシリトール、エリトリトール、マルチトール、澱粉、アカシアゴム、アルギン酸塩、ゼラチン、リン酸カルシウム、珪酸カルシウム、セルロース、メチルセルロース、微晶質セルロース、ポリビニルピロリドン、水、ヒドロキシ安息香酸メチル、ヒドロキシ安息香酸プロピル、タルク、ステアリン酸マグネシウム、及びミネラル類を挙げることができる。製剤化する場合には、通常使用する充填剤、増量剤、結合剤、湿潤剤、崩壊剤、界面活性剤などの希釈剤又は賦形剤を用いて調製される。経口投与のための固形製剤には、錠剤、丸薬、散剤、顆粒剤、カプセル剤などが含まれる。このような固形製剤は、前記ジントニン又は分画物に少なくとも一つの賦形剤、例えば澱粉、カルボン酸カルシウム、スクロース又はラクトース、ゼラチンなどを混ぜて調製される。また、単純な賦形剤以外に、ステアリン酸マグネシウム、タルクなどの潤滑剤も使用される。経口のための液相製剤としては、懸濁剤、内溶液剤、乳剤、シロップ剤などが該当するが、よく使用される単純希釈剤としての水、流動パラフィン以外に、様々な賦形剤、例えば湿潤剤、甘味剤、芳香剤、保存剤などが含まれてもよい。非経口投与のための製剤には、滅菌した水溶液、非水性溶剤、懸濁剤、乳剤、凍結乾燥製剤、坐剤が含まれる。非水性溶剤、懸濁剤としてはプロピレングリコール、ポリエチレングリコール、オリーブ油などの植物性油、オレイン酸エチルなどの注射可能なエステルなどが使用でき、坐剤の基剤としてはウイテプソル(witepsol)、マクロゴール(macrogol)、ツイン(tween)61、カカオ脂、ラウリン脂、グリセロール、ゼラチンなどが使用できる。
【0033】
本発明の組成物の好ましい投与量は、患者の状態及び体重、病気の重症程度、薬物形態、投与経路及び期間によって異なるが、当業者によって適切に選択できる。ところが、望ましい効果のために、本発明の組成物は1日0.0001〜100mg/kgで投与することがよい。投与は1日に1回投与してもよく、数回に分けて投与してもよい。前記投与量はいずれの面においても本発明の範囲を限定するものではない。
【0034】
本発明の薬学組成物は、ラット、マウス、家畜、ヒトなどの哺乳動物に様々な経路で投与できる。投与の全ての方式は予想できるが、例えば、経口、直腸、静脈、筋肉、皮下、子宮内膜または脳室内(intracerebroventricular)注射によって投与できる。
【0035】
本発明のジントニンを含む組成物は、前述したような剤形で、カルシウム濃度の低下に起因する疾患の予防及び治療のための薬剤、食品及び飲料などに様々に利用できる。前記ジントニンを添加することが可能な食品としては、例えば、各種食品類、飲料、チューインガム、茶、ビタミン複合剤、健康機能性食品類などがある。
【0036】
本発明のジントニンは、毒性及び副作用が殆どないので、予防の目的で長期間服用する場合にも安心して使用することができる。
【0037】
本発明のジントニンは、カルシウム濃度の低下に起因する疾患の予防を目的として食品又は飲料に添加できるが、この際、食品又は飲料中に前記ジントニンは全体重量に対して0.01〜15重量%で加えることができ、健康飲料組成物は100mLを基準として0.02〜5g、好ましくは0.3〜1gの割合で加えることができる。
【0038】
本発明の健康機能性飲料組成物は、指示された比率で必須成分として前記ジントニンを含有する以外は他の成分には特別な制限がなく、通常の飲料と同様に、様々な香味剤又は天然炭水化物などをさらに含んでもよい。この際、前記天然炭水化物としては、例えば、ブドウ糖、果糖などの単糖類、マルトース、スクロースなどの二糖類、及びデキストリン、シクロデキストリンなどの多糖類のような通常の糖、並びにキシリトール、ソルビトール、エリトリトールなどの糖アルコールなどが挙げられる。また、香味剤としては天然香味剤(タウマチン、ステビア抽出物、例えばレバウジオシドA、グリシルリジンなど)及び合成香味剤(サッカリン、アスパルテームなど)を有利に使用することができる。前記天然炭水化物の比率は本発明の組成物100mLあたり一般に約1〜20g、好ましくは約5〜12gである。
【0039】
この他に、本発明の抽出物は、様々な栄養剤、ビタミン、ミネラル(電解質)、合成矯味剤、及び天然矯味剤などの矯味剤、着色剤及び増音剤(チーズ、チョコレートなど)、ペクチン酸及びその塩、アルギン酸及びその塩、有機酸、保護性コロイド増粘剤、pH調節剤、安定化剤、防腐剤、グリセリン、アルコール、炭酸飲料に使用される炭酸化剤などを含有することができる。その他に、本発明の抽出物は天然果物ジュース飲料、果物ジュース飲料及び野菜医療の製造のための果肉を含有することができ、このような成分を独立に或いは組み合わせて使用することができる。また、前記添加剤の比率は、あまり重要ではないが、本発明の抽出物100重量部当たり0〜約20重量部の範囲で選択されることが一般的である。
【発明の効果】
【0040】
本発明は、人参に存在する新規な糖リポタンパク質としてのジントニンを分離同定して製造する方法、前記方法で製造された新規な糖リポタンパク質ジントニン、及び前記新規な糖リポタンパク質ジントニンのカルシウム濃度の低下に起因する疾患の予防及び治療剤としての用途を提供するという効果がある。
【0041】
本発明のジントニンは、一時的に細胞質内遊離Ca2+の増加を誘発し、細胞内カルシウム濃度の一時的な増加により、アフリカツメガエル卵母細胞で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)を活性化して細胞内カルシウム濃度を増加させるので、カルシウム濃度の低下に起因する疾患の予防及び治療に有用に使用できる。
【0042】
また、本発明のジントニンは、細胞内カルシウム依存性の様々な生理活性、例えば強壮作用、免疫力増加、性機能強化、神経系保護及び活性、血管新生、抗糖尿作用などにおいても優れた効果を示す。
【図面の簡単な説明】
【0043】
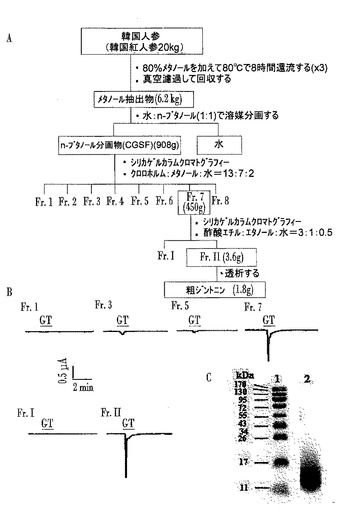
【図1】Aは人参から新規な糖リポタンパク質としてのジントニンを分離同定する方法を図式化した図であり、Bはジントニン分画物のアフリカツメガエル卵母細胞におけるCaCCの活性を確認したクロマトグラムであり、Cは粗ジントニンのSDS−PAGE結果である。
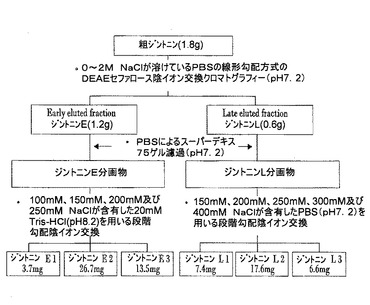
【図2】粗ジントニンから個別ジントニンを分離同定する方法を図式化した図である。
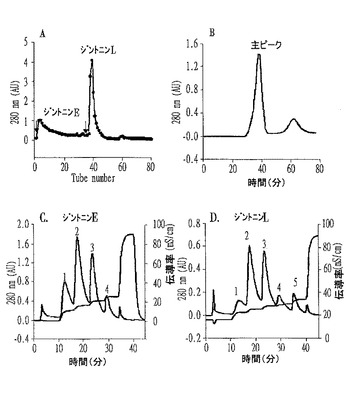
【図3】個別ジントニンの陰イオン交換及びゲルクロマトグラムであって、Aは粗ジントニンから分離したジントニンE分画とジントニンL分画の陰イオン交換クロマトグラムであり、Bは前記Aから得たジントニンL分画のゲル濾過クロマトグラムであり、CはジントニンE分画の陰イオン交換クロマトグラムであり、DはジントニンL分画の陰イオン交換クロマトグラムである。
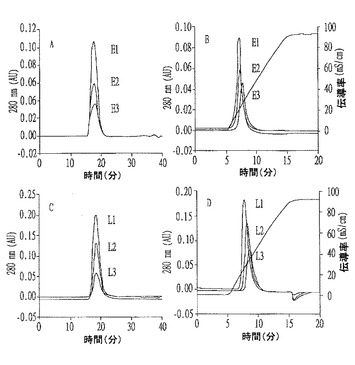
【図4】ゲル又は連続陰イオン交換クロマトグラフィーによる個別ジントニンの溶出パターンを示すもので、AとCは精製されたジントニンE1〜E3及びジントニンL1〜L3のゲル濾過クロマトグラムであり、BとDは精製されたジントニンE1〜E3及びジントニンL1〜L3の陰イオン交換クロマトグラムである。
【図5】ゲル濾過クロマトグラフィーを用いたジントニンの分子量決定較正曲線であって、各点は使用された標準タンパク質であり(IgG:160kD、BSA:67kDa、β−ラクトグロブリン:35kDa、シトクロムC:12kDa、アプロチニン:6.5kDa)、矢印はジントニンである。
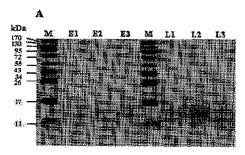
【図6】Aは個別ジントニンのSDS−PAGEであり、Bは前記個別ジントニンの炭水化物成分のゲル染色SDS−PAGE結果である。
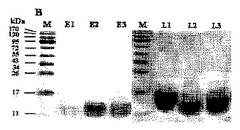
【図7】HPAED−PADクロマトグラムによる個別ジントニンの炭水化物成分分析結果を示すもので、Aは中性糖(Neutral sugar)の成分分析であり(標準物質:1.L−果糖、2.L−ラムノース、3.D−アラビアノース、4.D−ガラクトース、5.D−グルコース、6.D−マンノース、7.D−キシロース)、Bはアミノ糖の成分分析である(標準物質:1.D−ガラクトサミン、2.D−グルコサミン)。
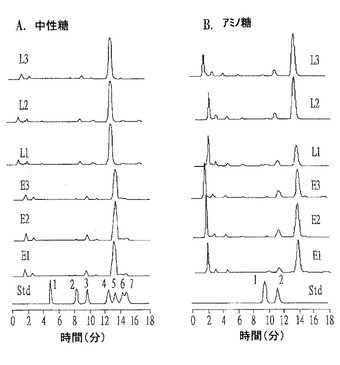
【図8】個別ジントニンE1〜E3及びL1〜L3の脂質成分をGC−MSスペクトル分析結果である。
【図9】ジントニン処理の際にマウスEAT細胞における内因性、内向性CaCCの電流流れを示すもので、Aは−80mVの保持電位におけるジントニンとジンセノサイドの電流流れを示すグラフ、BとCはそれぞれ粗ジントニンと個別ジントニンの濃度別電流流れを示すグラフである。
【図10】ジントニン処理の際にマウスEAT細胞における細胞内カルシウムの増加を確認したグラフであって、AはCa2+緩衝液に培養したFura−2に混ぜたEAT細胞における時間による細胞内カルシウムの増加を確認したものであり、BはCa2+遊離緩衝液に培養したFura−2に混ぜたEAT細胞におけるジントニン濃度による細胞内カルシウムの増加を確認したものである。
【図11】ジントニンによるCaCCの活性及びマウスEAT細胞における細胞内カルシウムの増加に対するPLC抑制剤、IP3受容体拮抗剤及びCa2+キレート剤の効果を示すもので、AはPLC抑制剤(U−73343/U−73122)によって、BとCはそれぞれIP3受容体拮抗剤(2−APB)及びCa2+キレート剤(BAPTA−AM)によってCaCCの活性化が減少することを確認した電流流れの棒グラフである。
【図12】マウスEAT細胞でジントニンによって増加する細胞内カルシウム濃度に対する活性型又は非活性型PLC抑制剤の効果を示すもので、AはCa2+緩衝液に培養したFura−2に混ぜたEAT細胞における細胞内カルシウム増加の抑制を確認したものであり、BはCa2+遊離緩衝液に培養したFura−2に混ぜたEAT細胞におけるジントニン濃度による細胞内カルシウム増加の抑制を確認したグラフである。
【発明を実施するための形態】
【0044】
以下、実施例によって本発明をさらに詳細に説明する。これらの実施例が本発明を例示するためのものに過ぎず、本発明の範囲を限定するものではないのは、当業者にとっては自明なことであろう。
【0045】
実施例1:人参からの粗ジントニン(crude gintonin)の分離
韓国人参公社(大田、韓国)から購入した6年根紅参(Panax ginseng C.A.Meyer)20kgを細かく粉砕(>3mm)し、80%メタノールを30L加えて80℃で8時間還流冷却抽出した後、真空濾過して上層液を回収した。この過程を3回繰り返し行って上層液を集めた後、減圧濃縮し(メタノール抽出物の総6.2kgを収得)、水とn−ブタノールの混合溶媒(1:1)で溶媒分画して水とn−ブタノールの分画物を収得した(n−ブタノール分画物を総908g収得)。
【0046】
前記n−ブタノール分画物(粗総サポニン分画、CGSF)は濃縮してクロロホルム:メタノール:水(CHCl3:MeOH:H2O=13:7:2)の混合溶媒で溶離してシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離した。その後、それぞれの分画物に対して、アフリカツメガエル卵母細胞で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)に対する活性を実験し、CaCCの活性が最も高く示された第7の分画(図1のB参照)に対して、さらに酢酸エチル:エタノール:水(EtOAc:EtOH:H2O=1:3:0.5)の混合溶媒で溶離してシリカゲルカラムクロマトグラフィーを行い、2個の分画物を得た。
【0047】
前記2個の分画物のうち、EAT(Ehrlich Ascites Tumor)細胞における細胞内カルシウム放出実験の評価とアフリカツメガエル卵母細胞で強いCaCC活性を示した分画物IIを4℃で8時間過量の蒸留水(distilled water、DW)とSpectra/Por透析膜(molecular weight cut of 6,000〜8,000)(Spectrum Laboratories、Inc.,Califoinia、USA)を用いて透析し、ジンセノサド及び他の低分子物質を除去した後、最終的に分画物を得た。
【0048】
前記得た分画物に対して電気泳動(SDS−PAGE)を行った結果、クマシーブリリアントブルー(Coomassie Brilliant Blue)で染色され、見掛け分子量が約13kDaであることを確認した。これは、EAT細胞における細胞内カルシウム放出実験の評価とアフリカツメガエル卵母細胞でCaCC活性を起こす新規物質が、ジンセノサイドではなく、タンパク質の1種類であることを示唆する。前記タンパク質をジントニンと命名し、これから得た最終分画物は粗ジントニン(crude gintonin)と命名した。
【0049】
ところが、人参のタンパク質または多糖類を含んでいる水分画物、および脂質(lipid)成分を含有している石油エーテル分画物は、EAT細胞の細胞内カルシウム放出実験とアフリカツメガエル卵母細胞におけるCaCC活性効果が全くなかった(図式しない)。
【0050】
実施例2:個別ジントニンの分離
粗ジントニン(crude gintonin)から個別ジントニン(gintonins)を分離するために、前記実施例1で得た粗ジントニンをリン酸塩緩衝食塩水(phosphate buffer saline、PBS)(pH7.2)に溶解し、DEAEセファロースCL−6Bカラムが装着された陰イオン交換樹脂クロマトグラフィー(GE Healthcare、Uppsala、Sweden)を行い、2個の主なピーク(major peak)を得た。この際、溶媒は、0〜2M NaClがPBSに溶けているものを使用した。1mL/分の流速にて280nmでモニタリングしながら線形勾配(linear gradient)方式で分離して得た2個の主なピークはそれぞれジントニンE(early eluted gintonin)、ジントニンL(late eluted gintonin)と命名した。
【0051】
前記ジントニンEの分画物と前記ジントニンLの分画物はCentriVap
DNA真空濃縮器(Labconco、Missouri、USA)で濃縮した後、PBS(pH7.2)を溶離液とし、スーパーデキス(Superdex)75ゲル濾過カラムを用いて0.5mL/分の流速にて280nmでモニタリングしながらクロマトグラフィーを行い、分子量の大きい主ピークと副ピークを得た(図3のB参照)。
【0052】
こうして得たジントニンEとジントニンLは、アフリカツメガエル卵母細胞でCaCC活性を示すことを確認した後、ジントニンE分画物は再び100、150、200、250mM NaClが含有された20mM Tris−HCl(pH8.2)を用いて段階勾配(step gradient)DEAE陰イオン交換カラム(anion exchange column:HiTrapTMDEAE FF、1mL)に1mL/分の流速にてロードして4個のピークを得た(図3のC参照)。これらのうち、100、150及び200mM NaCl溶離液の分画物を大量透析(extensive dialysis)及び濃縮した後、さらに0〜1M NaClが含有された20mM Tris−HCl(pH8.2)を用いて段階勾配DEAE陰イオン交換カラムを行い、しかる後に、最終的にスーパーデキス75ゲル濾過クロマトグラフィーを行って最終分画物をそれぞれ3.7、26.7及び13.5mgずつ得た。これらはそれぞれジントニンE1、E2及びE3と命名した。
【0053】
一方、ジントニンL分画物は150、200、250、300、400mM NaClが含有されたPBS(pH7.2)を用いて前記と同様の方法で得た5個のピーク(図3のD参照)のうち、150、200及び250mM NaCl溶離液の分画物に対して最終分画物を7.4、17.6及び6.6mgずつ得た。これらはそれぞれジントニンL1、L2及びL3と命名した。
【0054】
また、粗総サポニン分画(CGSF)908gからそれぞれ、ジントニンE1の場合は0.0004%(3.7mg)、ジントニンE2の場合は0.0029%(26.7mg)、ジントニンE3の場合は0.0015%(13.5mg)、ジントニンL1の場合は0.0008%(7.4mg)、ジントニンL2の場合は0.0019%(17.6mg)、ジントニンL3の場合は0.007%(6.6mg)の収率で得られた。
【0055】
実施例3:ジントニンの分子量決定
前述したように得た6個のそれぞれ異なるジントニンは、ヤマモト等の方法(Yamamoto,Y.,Nunome,T.,Yamauchi,R.,Kato,K.,and Sone,Y.(1995) Carbohydr Res.275,319-332)に準じてゲル濾過クロマトグラフィーを行って分子量を決定するが、PBS(pH7.2)で平衡を維持したスーパーデキス75カラム(10×300mm)を装着したバイオロジックデュオフロークロマトグラフィーシステム(BioLogic DuoFlowTMChromatograpgy、BIO−RAD、California、USA)を用いて280nmでモニタリングしながら流速0.5mL/分で分画物を集めた後、免疫グロブリンG(IgG;160,000Da)、牛血清アルブミン(BSA;67,000Da)、β−ラクトグロブリン(35,000Da)、シトクロムC(12,327Da)、及びアプロチニン(6,512Da)などの標準タンパク質からなる較正曲線(calibration curve)を用いて分子量を決定した。
【0056】
その結果、ジントニンの帯びる電荷が異なってNaCl勾配溶出によって溶出(elution)パターンが異なったが、ゲル濾過クロマトグラフィーではほぼ同時に溶出して分子量がほぼ同一であった(図4のB及びD参照)。
【0057】
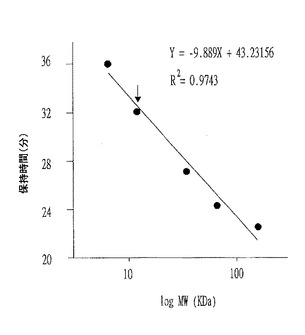
また、標準タンパク質を用いて決定したジントニンL2のネイティブ(native)分子量も約67kDaであった(図5参照)。他のジントニンの分子量も約67kDaであると確認された(図式せず)。
【0058】
また、SDS−PAGEの結果、6個のジントニンがブロード(broad)であるが、単一な主バンド(single major band)を示し、その見掛け分子量(apparent molecular weight)は約13kDaであると確認された。
【0059】
前述したような結果は、本発明のジントニンが五量体(pentamer)であることを示唆する(図6のA参照)。
【0060】
実施例4:ジントニンの純度決定
前記粗ジントニン及び個別ジントニンの純度を決定するために、12.0%分離ゲル(separating gel)を用いて電気泳動(SDS−PAGE)を行った(Laemmli,U.K.(1970) Nature 227,680-685)。
【0061】
具体的に、粗ジントニンと個別ジントニンE1〜E3、L1〜L3を100μgずつ取って各レイン(lane)にロードして電気泳動した後、ジントニンバンドをクマシーブリリアントブルーR−250染色で視覚化した。また、糖タンパク質(glycoprotein)検出のために、ゲルはPAS染色(periodic acid-schiff based staining)技法を用いて染色し、40%エタノール−7%酢酸水溶液で30分間培養した後、1%過ヨウ素酸−3%酢酸で1時間培養し、しかる後に、過ヨウ素酸溶液を除去し、Schiff試薬を添加して1時間培養した。染色された糖タンパク質のバンドは桃色を呈し、PAS染色されたゲルは7.5%酢酸で浸漬(soaking)した後、保管した。
【0062】
実施例5:ジントニンのアミノ酸組成分析
一般アミノ酸(general amino acid)分析のために、ジントニン30μgを真空内(in vacuo)で6N HClを用いて110℃で24時間加水分解した。また、システインの分析のためには、ジントニンを過酸化(peroxidation)させた後、6N HClを用いて110℃で24時間加水分解し、蟻酸:過酸化水素(10:1)溶液で処理した。トリプトファンの分析のためには、試料を4Mメタンスルホン酸(methanesulfonic acid)で加水分解し、4M KOHを加えた。
【0063】
また、フェニルイソチオシアネート(Phenylisothiocyanate、PITOC)誘導体に隠されたアミノ酸は、大田所在の韓国基礎科学支援研究所に依頼し、Waters Nova−Pak C18カラム(3.9×300mm)を装着したHPLC(Hewlett Packard 1100 series)で分析した。
【0064】
タンパク質成分は、標準として牛血清アルブミン(BSA)を用いたBradford法(Bradford,M.M.(1976).Anal.Biochem.72,248-254)で決定した。
【0065】
実施例6:ジントニンの部分アミノ酸配列
ジントニンのN末端アミノ酸配列は、自動エドマン分解アプライドバイオシステムモデル477Aガス相タンパク質配列(automated Edman degration on an Applied Biosystem model 477A gas-phase protein sequence)及びミリゲン6600固体相配列(MilliGen 6600 solid-phase sequence)によって行われた。
【0066】
ジントニン100μgに対して12%分離ゲルを用いてSDS−PAGEを行い、タンパク質バンドはクマシーブリリアントブルーR−250染色で視覚化した。前記バンドを切断してトリプシンで消化(digesting)し、前記消化産物(digestion product)は脱塩及び濃縮した。
【0067】
こうして誘導された全てのペプチドのMS/MSは、韓国基礎科学支援研究所で、ナノ−ESIを装着した質量分析計(Mass spectrometer)(QTOF II、Micromass、UK)を用いて行った。この際、生成されたイオン(product ion)は、反射体(reflector)、マイクロチャネル検出器(microchannel plate detector)及び時間対デジタル変換器(time-to-digital converter)が固定された直角TOF分析器(orthogonal TOF analyzer)を用いて分析した。データはウィンドウNT環境でMass Lynx softwareが駆動されるPCを用いて処理した。配列同族体はBLAST(www.ncbi.nln.nih.gov/BLAST.cgi)を用いて検索した。
【0068】
表1はジントニンのde novoアミノ酸配列と組成を示すものであり、純粋分離されたジントニンは自動エドマン分解によるN末端アミノ酸配列からなるものと確認された。ところが、ジントニンのN末端がブロッキング(blocking)されて全体配列を得ることができなかったため、MALDI−TOF−MS/MSを用いてトリプシン処理したジントニンから18個の内部ペプチド配列(internal peptide sequence)を推定した結果、6個の個別ジントニンは一般に幾つかのペプチドを含有し、アミノ酸配列はヒトグルコキナーゼ(human glucokinase)及びグルコキナーゼイソ型2(glucokinase isoform 2)と部分的相同性を示した(図式せず)。
【0069】
また、Bradford方法によるジントニンの総タンパク質の含量は、ジントニンE1、E2、E3、L1、L2及びL3でそれぞれ9.4、24.1、20.5、35.8、37.1及び39.6%であった。ジントニンL1〜L3のタンパク質含量はジントニンE1のそれより約3倍高い含量である。
【0070】
また、ジントニンにおける疎水性アミノ酸(hydrophobic amino acid)は、親水性アミノ酸(hydrophilic amino acid)よりさらに多かったが、ヒスチジン(His)、チロシン(tyr)及びメチオニン(Met)は検出されなかった。これらの中でも、メチオニンはジントニンのN末端がブロッキングされて検出されなかったものと思われる。
【0071】
一方、ジントニンE1〜E3及びL1〜L3では、共通にフェニルアラニン(Phe)が最も高く含有されるものと確認された。また、ジントニンL1〜L3におけるリシン(Lys)はE1〜E3のそれより高く示された(表1参照)。
【0072】
【表1】
実施例7:ジントニンの炭水化物組成分析
図6のAに示すように、SDS−PAGEにおけるジントニンのバンドは単一(single)であるがブロード(broad)であり、クマシーブリリアントブルー染色で強く染色されなかったため、炭水化物を含んでいる可能性がある。
【0073】
したがって、ジントニンの炭水化物組成を確認するために、ジントニンを2Mトリフルオロ酢酸(trifluoroacetic acid)を用いて100℃で4時間加水分解した後、中性糖(neutral sugar)を得た。また、ジントニンをガラスアンプルで6N HClを用いて100℃で4時間加水分解によってアミノ糖(amino sugar)と酸性糖(acid sugar)を得た。
【0074】
ジントニンの炭水化物組成は、ソウル所在の世宗大学校炭水化物素材研究所に依頼し、PAS染色(periodic acid-schiff based staining)技法を用いて染色した後、CarboPacTMPAlカラムを装着したHPAEC−PADシステム(high performance anion exchange chromatography-plused amphermetric dectection system、Dionex、California、USA)を用いて分析した。単糖類のモル重量(molar weight)はピーク面積(peak area)から計算した。炭水化物の含量は、中性糖の場合にはフェノール−スルホン酸方法(Hounsell,E.F.,Davies,M.J.,and Smith,K.D.(1997) Protein protocol handbood,Humanna press,Totawa,803-804)で決定し、酸性糖の場合にはAnthrone方法で決定した(Scott,T.A.,and Melvin,E.H.(1953)Anal.Biochem.25,1656-1660)。
【0075】
その結果、ジントニンは、ラムノース(rhamnose)、アラビノース(arabinose)、グルコース(glucose)、マンノース(mannose)、キシロース(xylose)などの5個の異なる種類の中性糖、及びグルコサミンなどの1個のアミノ糖から構成されるものと確認された。6個のジントニンは互いにほぼ類似した炭水化物構成を有することが分かった(表2及び図7参照)。ところが、ジントニンE1〜E3からマンノースは検出されなかった。
【0076】
一方、1−N−アセチル−ノイラミン酸(1-N-acetyl-neuraminic acid)、2−ガラクツロン酸(2-galacturonic acid)、3−グルクロン酸(3-glucuronic acid)などの酸性糖は検出されなかった(図式せず)。
【0077】
最後に、ジントニンの総炭水化物含量は、ジントニンE1、E2、E3、L1、L2及びL3でそれぞれ約34.3、45.3、38.1、36.0、29.8及び45.2%であった。その結果より、本発明のジントニンは糖タンパク質であることを再確認することができた。
【0078】
【表2】
実施例8:ジントニンの脂質成分分析
本発明のジントニンは、n−ブタノール抽出によってジンセノサイドと同様の分画にあったため、ジントニンも脂質部分(lipid moiety)を含有するものと予想することができる。
【0079】
これを確認するために、ジントニンを6N HClを用いて4時間100℃で加水分解し或いはリポタンパク質リパーゼ(lipoprotein lipase)に消化させて脂質(lipid)及び疎水性部分(hydrophobic moiety)を確認した。
【0080】
具体的に、粗総サポニン分画(CGFS)を酸加水分解又は消化させたものを濃縮し、さらに水とn−ヘキサンにさらに分画した後、n−ヘキサン分画物に対して、DB5−MS毛細管カラム(capillary column)(30cm×250μm×0.25μm)を装着したAgilent 6890N GC−MSシステム(California、USA)を用いて、韓国基礎科学支援研究所で脂質(lipid)及び疎水性部分を分析した。この際、GC(6890N)は、火イオン化検出器(flame ionization detector)と分離注入システム(split injection system)が装着されており、supelco SPB−1毛細管カラム(内径:15m×0.32mm、厚さ:0.25m)を固定した。
【0081】
その結果、表3及び図8に示すように、ジントニンは、エステル型(ester form)又は自由型(free form)のパルミチン酸(plamitic acid)、ステアリン酸(stearic acid)、オレイン酸(oleic acid)、リノール酸(linoleic acid)などを含有するものと確認された。
【0082】
また、ジントニンE1又はL1における多価不飽和脂肪酸としてのリノール酸の比率は飽和脂肪酸としてのパルミチン酸とステアリン酸又はオレイン酸より高かった。パルミチン酸はジントニンE2、E3、L1、L2で多量含有されていた。
【0083】
また、GCによって測定したジントニンに含有された総脂質含量は、ジントニンE1、E2、E3、L1、L2及びL3がそれぞれ34.3、45.3、38.1、36.0、29.8及び45.2%であった。これらの中でも、ジントニンE1の脂質含量が最も高かった。
【0084】
前述したような結果は、本発明のジントニンが新規な糖リポタンパク質であることを示唆する。
【0085】
【表3】
測定例1:アフリカツメガエル卵母細胞(Xenopus laevis oocytes)からのCaCC活性の測定
1−(1).卵母細胞(Oocyte)の準備
Xenopus I(Ann Arbor、MI、USA)から得たアフリカツメガエル(Xenopus laevis)は、最上の規格指針に従って保管及び処理し、卵子(oocytes)を分離するために、カエルを3−アミノ安息香酸エチルエステル(3-amino benzoic acid ethyl ester)の通気溶液(aerated solution)で麻酔させて手術した後、コラゲナーゼで処理し、しかる後に、82.5mM NaCl、2mM KCl、1mM MgCl2、5mM HEPES、2.5mMピルビン酸ナトリウム(sodium pyruvate)、100units/mLフェニリシン及び100μg/mLストレプトマイシンが含有されたCa2+遊離培地で2時間攪拌して分離した。
【0086】
V−VI段階の卵子を収集し、ゲンタマイシン(gentamicin)50μg/mLを追加したND96(96mM NaCl、2mM KCl、1mM MgCl2、1.8mM CaCl2、及び5mM HEPES、pH7.4)で保管した。前記卵子を含む溶液は、連続的に軽く振とうしながら18℃に維持し、毎日交換した。
【0087】
1−(2).CaCCの測定
2−電極電圧固定(two-electrode voltage clamp)記録は、小型プレキシガラス(plexiglass)ネットチャンバー(0.5mL)に置かれた個別的な卵子から得た。電気生理学実験(electrophysiological experiments)は、3M KClで充填したマイクロ電極(抵抗:0.2〜0.7MΩ)と卵子固定増幅器(Oocyte Clamp amplifier、OC−725C、Warner Instrument、CT、USA)を用いて室温で行った後、CaCCを−80mV保持電位(holding potential)で記録した。
【0088】
ジントニンは、バス灌流(bath perfusion)によって卵子に適用した(Lee,J.H.,Jeong,S.M.,Lee,B.H.,Noh,H.S.,Kim,B.K.,Kim,J.I.,Rhim,H.,Kim,H.C.,Kim,K.M.,and Nah,S.Y.(2004)J Biol Chem 279,9912-9921)。
【0089】
測定例2:マウスEAT細胞からの細胞内遊離カルシウム([Ca2+]i)の測定
2−(1).EAT(Ehrlich Ascites Tumor)細胞の準備
EAT細胞は、Jorgensen等の方法(Jorgensen,N.K.,Pedersen,S.F.,Hoffmann,E.K.(1999)Am J Physiol 276.C26-C37)によってICRマウスの腹腔内移植(interaperitoneal transplantation)から毎週収集した。
【0090】
具体的に、腹水(ascites fluid)は、マウス腹膜腔(peritoneal cavity)から2.5U/mLのヘパリンが含有されたHank’s溶液に収集した。RBCからEAT細胞を分離するためには、EAT細胞を10及び20%フィコール溶液(Ficoll solution)の2層を含む遠心分離管にロードして1,800rpmで20分間遠心分離し、EAT細胞のみを得た。前記細胞は、所望の細胞数となるように浮遊させた後、実験を開始する前に約30分間培養した。
【0091】
2−(2).Fura 2とEAT細胞との混合
Fura 2−負荷マウスEAT細胞におけるG−タンパク質結合受容体作用剤(G-protein coupled receptor agonist)の処理は、細胞内遊離カルシウム([Ca2+]i)の一時的な増加を誘導することが明らかになった(Scott and Melvin.Anal.Biochem.,25,1656-1660,1953)。
【0092】
本発明では、ジントニン又はジンセノサイドRa、Rb1、Rb2、Rc、Rg1がFura2−負荷マウスEAT細胞における[Ca2+]iの一時的増加に効果を示すかを確認するために、Fura−2に混ぜたEAT細胞(2〜4×106/mL)は1.5mM Ca2+緩衝液またはCa2+遊離緩衝液で10分間培養した後、ジントニンとジンセノサイドを添加し、EAT細胞における細胞内カルシウム濃度に及ぼす効果を確認した。但し、利用可能な純粋分離された個別ジントニンの量の限界のために、代表的なジントニンとしてジントニンE2を用いてマウスEAT細胞(Riken BRC Cell Bank、Tsukuba、Japan)における[Ca2+]iの増加について実験した。
【0093】
具体的に、Jorgensen等の方法によって、NaCl 120mM、KCl 5mM、MgCl21mM、CaCl21.5mM、グルコース10mM、及びHEPES 25mMからなるCa2+緩衝液(pH7.2)と、NaCl 120mM、KCl 5mM、MgCl21mM、EGTA 0.2mM、グルコース10mM及びHEPES 25mMよりなるCa2+遊離緩衝液(pH7.4)を用いて、37℃の水浴で30分間振とうしながら2μN Fura 2−AMにEAT細胞(2〜4×106/mL)を混合し、過量のfura−2は細胞をCa2+緩衝液又はCa2+遊離緩衝液で3回洗浄して除去した。
【0094】
2−(3).細胞浮遊物内[Ca2+]iの蛍光測定
[Ca2+]iは、RF−5300PC細胞内イオン測定システム(Intracellular Ion Measurement System)(Shimadzu Corporation、Japan)を用いてFura 2−負荷セルから推定した。具体的に、Fura 2−負荷セルは、最終的に2〜4×106/mLまで実験培地で希釈した後、ポリスチレンキュベット(Elkay Ultra−UV)に移した。細胞はテフロン(登録商標)でコートされた磁石を用いて攪拌した。キュベットハウジングは37℃に調節した。励起波長(excitation wavelengths)は、コンピュータ制御の下で340nmと380nmを交互にし、放出(emission)は510nmで検出した。この際、励起及び放出の隙間幅は5nmとし、背景補正はJorgensen等の方法に準じて行うが、ジギトニン(digitonin)及びEGTAはfura−2が完全にCa2+と化合し、Ca2+から解離される状態を作るための濃縮反応試薬(concentration adjustment reagent)として使用した。
【0095】
2−(4).Fura 2測定値からの[Ca2+]iの推定
測定された340nm対380nmの比率値は、Hounsell等の式(Hounsell,E.F.,Davies,M.J.,and Smith,K.D.(1997)Protein protocol handbood,Humanna press,Totawa,803-804)を用いて[Ca2+]iの値に変換した。
【0096】
[Ca2+]=Kd[(R−Rmin)/(Rmax−R)](Sf380/Sb380)
但し、Kdは解離定数(224nM)であり、Rは測定された340nm対380nmの蛍光比(fluorescence ratio)であり、Rmax及びRminは50μg/mLのジギトニンを添加した飽和濃度でのR値及び20mM EGTAを添加した遊離培地でのR値である。また、Sf380とSb380はジギトニンとEGTAを添加したときの380nmでの蛍光強度であり、比率はこれらの値が表示されるときの最大及び最小値である(Grynkiewicz,G.,M.Poenie,and R.Y.Tsien.(1985) J Biol Chem 260:3440-3450)。
【0097】
2−(5).データ分析
InsP6を含む濃度反応曲線を得るために、観察されたピークの振幅を標準化及び図表化した後、Origin software(Northampton、MA)を用いて下記のHill式に対応させた。
【0098】
y/ymax=[A]n/([A]n+[EC50]n)
式中、yはジントニンの与えられた濃度における%活性であり、ymaxは最大ピーク電流(maximum peak current)であり、[EC50]は最大反応の50%を示すことが可能なジントニンの濃度であり、[A]はジントニンの濃度である。また、nは相互作用係数(interaction coefficient)であり、全ての値は平均±標準誤差で示し、対照群のデータとジントニン処理群のデータ間の平均値はunpaired Student’s t−testを用いて分析した。統計学的有意性はP<0.05で検定した。
【0099】
実験例1.アフリカツメガエル卵母細胞(Xenopus laevis oocytes)で内因性に存在するCaCCの活性に対する粗ジントニン及び個別ジントニンの効果確認
アフリカツメガエル卵母細胞(Xenopus laevis oocytes)で内因性に存在するCaCCの活性に対する粗ジントニン及び個別ジントニンの効果を確認するために、前記測定例1の夫々の方法を用いて、CaCCの活性増加として記録される保持電位を測定した。
【0100】
その結果、粗ジントニンを処理した群では−80mV保持電位で内向性(inward)のCl−電流を誘導したが、ジンセノサイドは誘導効果が全く確認されなかった。ED50は4.4±0.5μg/mLであった(図9のA及びB参照)。
【0101】
また、個別ジントニンに対しても、アフリカツメガエル卵母細胞におけるCaCC活性効果を実験した結果、ジントニンE1〜E3及びL1はほぼ同一程度のCaCC活性を示したが、ジントニンL2とL3は相対的に弱いCaCC活性を示した(図9のC参照)。この際、ED50は、それぞれジントニンE1〜E3が1.5±0.1、2.3±0.1、1.5±0.1であり、ジントニンL1〜L3が1.0±0.1、3.2±0.2及び4.3±0.2μg/mLであった。
【0102】
実験例2:マウスEAT細胞における細胞内遊離カルシウム([Ca2+]i)に対する効果の確認
前記測定例2のそれぞれの方法に従い、マウスEAT細胞における細胞内[Ca2+]iを測定した結果、浮遊させたEAT細胞にジンセノサイドを単独でそれぞれ100μMずつ処理したとき、実験したあるジンセノサイドもEAT細胞に反応を起こさなかった反面、これと対照的に、100ng/mLのジントニンE2で処理した実験群では[Ca2+]iを一時的に増加させることが確認された(図10のA参照)。
【0103】
図10のBはジントニンE2によって誘導された[Ca2+]iの増加が処理濃度依存的であることを示す。興味深く、ジントニンE2によって誘導された[Ca2+]iの一時的な増加は外部のCa2+(0.2mM EGTAを含むCa2+遊離緩衝液)を除去することにより大きく減少した(P<0.001、細胞外Ca2+の存在対比)。これは細胞外Ca2+もジントニン処理の際に細胞内に入ることを意味する。ところが、ジントニンE2の処理は依然として濃度依存的に[Ca2+]iを増加させた。これより、ジントニンE2によって誘導された[Ca2+]iの増加は細胞内貯蔵Ca2+の放出と細胞外におけるCa2+の流入によるものであることが分かった。
【0104】
この際、EC50値は、細胞外Ca2+の存在と不在によってそれぞれ33.6±8.2ng/mLと43.6±14.2ng/mL(平均±標準誤差、n=5〜6)であった。このような結果は、哺乳動物細胞における[Ca2+]iの増加を起こすものがジンセノサイドではなくジントニンであることを示す。
【0105】
実験例3:粗ジントニンによるCaCC活性とEAT細胞における[Ca2+]i上昇効果に対するPLC抑制剤、IP3受容体拮抗剤又はCa2+キレート剤効果の確認
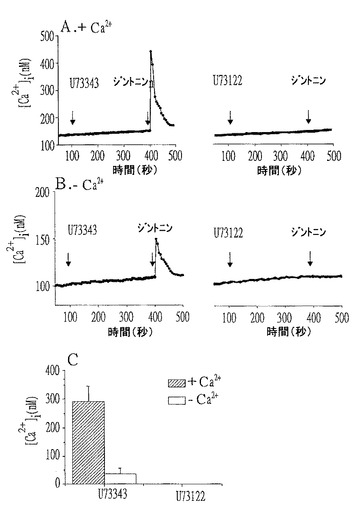
細胞質の[Ca2+]iとCaCC活性化を増加させるジントニン誘導シグナル伝達経路におけるリン脂質分解酵素C(phospholipase C、PLC)の可能な役目を説明するために、まず、活性PLC抑制剤としてのU−73122と非活性類似体としてのU−73343の効果を、アフリカツメガエル卵母細胞(Xenopus oocytes)とマウスEAT細胞を用いて確認した。
【0106】
アフリカツメガエル卵母細胞において、粗ジントニン誘導CaCC電流はU−73122で対照群が3.9±0.3であったが、U−73343では対照群の97.2±0.2%であった(図11のA参照)。このような結果は、ジントニンによって誘導された[Ca2+]iの上昇とジントニンによって媒介されたCaCC活性化がPLC活性化を介して媒介されることを意味する。
【0107】
次の段階として、IP3受容体を介しての細胞内Ca2+放出がジントニン誘導CaCC活性化を起こすか否かを検査した(Grynkiewicz,et al.,J.Biol.Chem.260:3440-3450,1985;Parekh,A.B.Pflugers Arch.-Eur.J.Physiol.,430,954-963,1995;Broad,et al.,J.Biol.Chem.276,15945-15952,2001)。
【0108】
このために、IP3受容体拮抗剤としての膜透過性2−APB(2-aminoethoxydiphenylborate)のジントニン誘導CaCC活性に対する効果を検査した。
【0109】
図11のBに示すように、2−APBの添加はジントニンによって誘導された内向性C1−電流(1.3±0.2〜0.03±0.01μAの対照電流、P<0.01)をほぼ完全に消滅させた。また、膜透過性BAPTA(1,2-bis(o-aminophenoxy)ethane-N,N,N’,N’-tetraacetic acid)−AM(10μM最終)処理で細胞内遊離Ca2+を除去することもジントニン媒介CaCC活性化を殆ど消滅させた(図11のC参照)。
【0110】
マウスEAT細胞の場合でも、U−73122の処理は細胞外Ca2+の存否を問わずジントニンの作用を殆ど消滅させるものと確認されたが、U−73343はそうではなかった(図12参照)。
【0111】
このような結果より、アフリカツメガエル卵母細胞とマウスEAT細胞でPLC−IP3−Ca2+経路を介して起るCaCC活性を起こす粗総サポニン分画(CGSF)の主要活性物質がジントニンであることを再び確認することができた。
【0112】
結論的に、本発明のジントニンは、細胞内カルシウム濃度を増加させる作用だけでなく、細胞外からカルシウムの流入増加を誘発して細胞内カルシウム濃度を増加させるシグナル伝達経路を介してその効能を示すことが分かった。
【0113】
本発明のジントニンを含む薬学組成物の製剤例を下記のとおり例示する。ところが、これらの例は本発明を限定するためのものではなく、具体的に説明するためのものである。
【0114】
製剤例1:錠剤の製造
ジントニン 100mg
乳糖 100mg
澱粉 100mg
ステアリン酸マグネシウム 2mg
これらの成分を混合し、通常の錠剤の製造方法に従って打錠して錠剤を製造した。
【0115】
製剤例2:散剤の製造
ジントニン 100mg
乳糖 100mg
タルク 10mg
これらの成分を混合し、気密布に充填して散剤を製造する。
【0116】
製剤例3:カプセル剤の製造
ジントニン 100mg
結晶性セルロース 3mg
ラクトース 14.8mg
ステアリン酸マグネシウム 0.2mg
通常のカプセル剤の製造方法に従って、これらの成分を混合し、ゼラチンカプセルに充填してカプセル剤を製造する。
【0117】
製剤例4:注射剤の製造
ジントニン 20mg
注射用滅菌蒸留水 適量
pH調節剤 適量
通常の注射剤の製造方法に従って、1アンプル当たり(2mL)前記の成分含量で注射剤を製造する。
【0118】
製剤例5:液剤の製造
ジントニン 1000mg
異性化糖 10g
マンニトール 5g
精製水 適量
通常の液剤の製造方法に従って、精製水にそれぞれの成分を加えて溶解させ、レモン香を適量加えた後、前述の成分を混合し、しかる後に、精製水を加えて全体100mLに調節した後、褐色瓶に充填して滅菌させて液剤を製造する。
【0119】
製剤例6:健康食品の製造
ジントニン 100mg
ビタミン混合物 適量
ビタミンAアセテート 70μL
ビタミンE 1.0mg
ビタミンB1 0.13mg
ビタミンB2 0.15mg
ビタミンB6 0.5mg
ビタミンB12 0.2μg
ビタミンC 10mg
ビオチン 10μg
ニコチン酸アミド 1.7mg
葉酸 50μg
パントテン酸カルシウム 0.5mg
無機質混合物 適量
硫酸第1鉄 1.75mg
酸化亜鉛 0.82mg
炭酸マグネシウム 25.3mg
第1リン酸カリウム 15mg
第2リン酸カルシウム 55mg
クエン酸カリウム 90mg
炭酸カルシウム 100mg
塩化マグネシウム 24.8mg
前記ビタミン及びミネラル混合物の組成比は、比較的健康食品に適した成分を好適な実施例で混合組成したが、その配合比を任意に変形実施しても構わず、通常の健康食品製造方法に従って前述の成分を混合した後、顆粒を製造し、通常の方法によって健康食品組成物の製造に使用することができる。
【0120】
製剤例7:健康飲料の製造
ジントニン 100g
ビタミンC 15g
ビタミンE(粉末) 100g
乳酸鉄 19.75g
酸化亜鉛 3.5g
ニコチン酸アミド 3.5g
ビタミンA 1.0g
ビタミンB1 0.13g
ビタミンB2 0.15g
通常の健康飲料の製造方法に従って前述の成分を混合した後、約1時間85℃で攪拌加熱し、しかる後に、作られた溶液を濾過し、滅菌した2Lの容器に取得し、密封滅菌させた後、冷蔵保管して本発明の健康飲料組成物の製造に使用する。
【0121】
前記組成比は、比較的嗜好飲料に適した成分を好適な実施例として混合組成したが、需要階層や需要国家、使用用途などの地域的、民族的嗜好度に応じてその配合比を任意に変形して実施しても構わない。
以上、本発明の内容の特定部分について詳細に記述したが、このような具体的記述は好適な実施様態に過ぎないものであり、これらの実施様態に本発明の範囲が限定されないのは当該分野における通常の知識を有する者には明白なことであろう。よって、本発明の実質的な範囲は添付された請求の範囲とそれらの等価物によって定められるべきであるといえる。
【技術分野】
【0001】
本発明は、人参に存在する新規な糖リポタンパク質(glycolipoprotein)に係り、さらに詳しくは、人参から分離同定して新規な糖リポタンパク質を製造する方法、前記方法で人参から分離同定した新規な糖リポタンパク質、および前記新規な糖リポタンパク質の用途に関する。
【背景技術】
【0002】
人参は、一般に、適応促進薬(adaptogen)又は生命延長のための強壮剤として用いられており、ストレス、疲労、疾病、癌及び糖尿病に対抗して生体機能を向上させる。このような人参は、韓国だけでなく、中国及び日本でも数百年前から使われてきた。最近、人参は世界で薬草として最も有名なものの一つである(非特許文献1)。
【0003】
ジンセノサイド(Ginsenoside)は、1960年代初めに分離されて特性化されたから、生理学的、薬理学的研究で代表的な成分として多用されるものである(非特許文献2、非特許文献3)。この他にも、最近の研究では、人参にはよく知られていない他の成分、例えば多糖類(polysaccharides)、ポリアセチレン類(polyaccetylenes)、タンパク質(protein)などが含有されていることが明らかになった(非特許文献4)。
【0004】
人参のジンセノサイド成分は、量が少なく、分離過程が複雑であり、純粋なジンセノサイドの場合には価格が高いため、人参の根からブタノール抽出方法で得た粗総サポニン分画(crude ginseng total saponin fraction、CGSF)を使用してきた(非特許文献5、非特許文献6、非特許文献7、非特許文献8、非特許文献9、非特許文献10、非特許文献11、非特許文献12、非特許文献13及び非特許文献14)。
【0005】
粗総サポニン分画(CGSF)は細胞膜シグナル経路(signal pathway)を介してその活性が現れるものと知られているが、例えば、Choi等は、アフリカツメガエル卵母細胞(Xenopus oocytes)に粗総サポニン分画を処理したとき、PLCβ−IP3に連結されているPTX−insentive Gαq/11タンパク質を介してカルシウム依存性塩素イオンチャネル(Ca2+activated Chloride Channel、CaCC)が活性化されることを解明した(非特許文献7)。また、Lee等は、アフリカツメガエル卵母細胞に粗総サポニン分画を持続的に処理したとき、粗総サポニン分画によって活性化されたカルシウム依存性塩素イオンチャネル電流(CaCC currents)が自発的に減少することを報告した(非特許文献9)。
【0006】
カルモジュリン(Calmodulin)をアフリカツメガエル卵母細胞に直接注入したとき、或いは細胞内のカルシウム貯蔵庫を枯渇させたときに、粗総サポニン分画によるカルシウム依存性塩素イオンチャネルの活性がなくなる(非特許文献12)。しかも、アフリカツメガエル卵母細胞に粗総サポニンを処理したときにSOCE(stored-operated Ca2+ entry)が誘発され(非特許文献10)、これにより細胞外或いは細胞内のカルシウム貯蔵庫でカルシウム濃度の増加が引き起こされてアフリカツメガエル卵母細胞内のCaCCが活性化されるものと知られている(非特許文献15)。
【0007】
本発明者らは、ジンセノサイドがCaCCを活性化させることを確認するために、粗総サポニン分画物(CGSF)から純粋なジンセノサイドを分離する過程中に、CGSFよりジンセノサイドに富む分画物の場合、CaCCの活性に対するその効果が急激に消えるか無くなることを見出した。すなわち、CGSFから純粋に分離されたジンセノサイドの場合、アフリカツメガエル卵母細胞内におけるCaCC活性効果がないことを確認した。これはジンセノサイドだけでなく、未知のある物質がCGSFに存在し、この物質がアフリカツメガエル卵母細胞内のCaCCの活性に影響を及ぼすことが分かった。
【0008】
そこで、CGSF内に内因性CaCCの活性を起こすある成分の分離のために鋭意努力した結果、新規な生理活性成分を分離同定し、本発明を完成するに至った。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Tyler,J.Pharm.Technol.11,214-220,1995
【非特許文献2】Shibata,et al.Tetraheadron Letters 1962,1239-1245,1963
【非特許文献3】Wagner-Jauregg and Roth,Pharm Acta Helv 37,352-357,1962
【非特許文献4】Nah,Kor.J.Ginseng Sci.21,1-12,1997
【非特許文献5】Kanzaki,et al.Br J Pharmacol.125(2),255-262,1998
【非特許文献6】Choi,et al.Br J.Pharmacol.132,641-648,2001
【非特許文献7】Choi,et al.,J.Biol.Chem.276,48797-48802,2001
【非特許文献8】Choi,et al.Eur J Pharmacol 468,83-92,2003
【非特許文献9】Lee,et al,J Biol Chem.279,9912-9921,2004
【非特許文献10】Jeong,et al,Br J Pharmacol.142,585-593,2004
【非特許文献11】Reay,et al,J Psychopharmacol.19,357-365,2005
【非特許文献12】Lee,et al,Arch Pharm Res 28,413-420,2005
【非特許文献13】Wei,et al J Ethnopharmacol.111,613-618,2007
【非特許文献14】Eriksson,et al J Ethnopharmacol.119,17-23,2008
【非特許文献15】Dascal,CRC Crit Rev Biochem 22,317-387,1987
【発明の概要】
【発明が解決しようとする課題】
【0010】
したがって、本発明の主な目的は、人参から強壮活性の役目を果たす新規な生理活性物質を分離同定する方法、前記方法で分離同定された生理活性物質、及び前記生理活性物質の用途を提供することにある。
【課題を解決するための手段】
【0011】
上記目的を達成するために、本発明は、人参に存在する新規な糖リポタンパク質(glycolipoprotein)を人参から分離同定して製造する方法を提供する。
【0012】
具体的に、本発明に係る人参から新規な糖リポタンパク質としてのジントニン(以下「ジントニン(gintonin)という」を製造する方法は、(1)人参からメタノール抽出物を製造する段階と、(2)前記メタノール抽出物を水とn−ブタノールの混合溶媒に分画する段階と、(3)前記水分画物とn−ブタノール分画物のうち、n−ブタノール分画物に対して、溶出溶媒としてクロロメタン:メタノール:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離する段階と、(4)前記8個の分画物のうち、CaCC(Ca2+activated Chloride Channel)に対する活性が最も高い第7の分画物に対して、溶出溶媒としてエタノール:酢酸エチル:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、2つの分画に分離する段階と、(5)前記2つの分画物のうち、CaCC活性が高い分画物IIを透析し、最終生成物を得る段階とを含んでなる。
【0013】
また、本発明は、これに加えて、(6)最終分画物をNaClが含有されたpH7.2のリン酸塩緩衝食塩水(phosphate buffer saline、PBS)溶液に溶解し、陰イオン交換クロマトグラフィー及びゲル濾過クロマトグラフィーを行い、2つの分画に分離する段階と、(7)前記2つの分画物に対してそれぞれ、NaClが含有されたpH8.2のTris−HCl及びNaClが含有されたpH7.2のリン酸塩緩衝食塩水(PBS)で陰イオン交換クロマトグラフィーを行い、個別ジントニンを分離する段階とを含んでもよい。
【0014】
また、本発明は、前述したような方法で人参から分離同定した新規な糖リポタンパク質(ジントニン)を提供する。
【0015】
本発明において、前記糖リポタンパク質(ジントニン)の分子量が約67kDaであり、見掛け分子量は約13kDaの五量体であることを特徴とし、アミノ酸組成成分として、システインとシスチン(cysteine and cystine)、アスパラギンとアスパラギン酸(asparagine and aspartic acid)、グルタミンとグルタミン酸(glutamine and glutamic acid)、セリン(serine)、グリシン(glycine)、アルギニン(arginine)、トレオニン(threonine)、アラニン(alanine)、プロリン(proline)、バリン(valine)、イソロイシン(isoleucine)、ロイシン(leucine)、フェニルアラニン(phenylalanine)、トリプトファン(tryptophan)及びリシン(lysine)を含み、炭水化物組成成分として、ラムノース(rhamnose)、アラビノース(arabinose)、グルコース(glucose)、マンノース(mannose)、キシロース(xylose)及びグルコサミン(glucosamine)を含み、脂質組成成分としては、リノール酸(linoleic acid)、パルミチン酸(palmitic acid)、オレイン酸(oleic acid)及びステアリン酸(stearic acid)などを含む。
【0016】
人参は、ウコキ科薬用植物であって、例えば、加工方法に従って加工していない水参(生参)、水参を乾燥させた白参、水参を蒸して乾燥させた紅参などがあり、栽培方法によっては人参の畑で人為的に栽培した栽培参、人参の種を山中に播いて自然状態で栽培した長脳参、自然状態で自生した山参などがあり、本発明において、用語「人参」は前記全ての人参の種類を含む意味で使用される。また、本発明における人参は高麗人参(オタネニンジン、Panax ginseng C.A.Meyer)、西洋参(アメリカニンジン)及び中国人参(田七人参)などを含む。
【0017】
以下、本発明を詳細に説明する。
【0018】
本発明の人参に存在する新規な糖リポタンパク質としてのジントニンは、下記のとおり収得できる。
【0019】
まず、乾燥した人参粉末を、試料重量の約1〜20倍、好ましくは約1〜10倍に達する体積の水、メタノール、エタノール、ブタノールなどのC1〜C4の低級アルコールの極性溶媒又はこれらの約1:0.1〜1:10の混合比を有する混合溶媒で、好ましくは1:0.2〜1:5の混合比(v/v)を有する水とメタノールとの混合溶媒で、70〜120℃で約0.1〜48時間、好ましくは5〜12時間攪拌抽出、熱水抽出、冷浸抽出、加温抽出、還流冷却抽出又は超音波抽出などの抽出方法、好ましくは還流冷却抽出によって行った後、濾過して上層液を回収し、前述の過程を数回、好ましくは2〜5回繰り返し行った後、上層液を集めて減圧濃縮したものを、水とn−ブタノールが同量で混合された混合溶媒で溶媒分画し、水とn−ブタノールの分画物を得る。
【0020】
前記分画物のうち、粗総サポニン分画(CGSF)に該当するn−ブタノール分画物に対して、溶出溶媒としてクロロメタノール:メタノール:水(CHCl3L:MeOH:H2O)の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離した後、分離された分画物に対してアフリカツメガエル卵母細胞におけるカルシウム依存性塩素イオンチャネル(CaCC)の活性確認実験を行って分画物の活性を検索した後、最も活性の高い分画に対してさらにエタノール:酢酸エタノール:水(EtOH:EtOAc:H2O)の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行って2個の分画に分画し、二重活性の高い分画を過量の蒸留水と透析膜(dialysis membrane)を用いて透析し、本発明の粗ジントニンを得ることができる。
【0021】
こうして得られた粗ジントニンは、電気泳動(SDS−PAGE)による分子量分析、タンパク質及びアミノ酸組成分析、炭水化物組成分析、GC−MSによる脂質組成分析などを介して確認することができる。
【0022】
また、本発明の個別ジントニンは、上述したような工程で得られた粗ジントニンをNaClの含有されたリン酸塩緩衝食塩水(phosphate buffer saline、PBS)に溶解し、陰イオン交換クロマトグラフィー及びゲル濾過クロマトグラフィーを行って2個のジントニン分画物を収得する。それぞれのジントニン分画物は、NaClが含有されたTris−HCl(pH8.2)及びNaClが含有されたPBS(pH7.2)を用いて陰イオン交換クロマトグラフィーを行って得ることができる。
【0023】
本発明において、ジントニンの製造のために、山参、長脳参、栽培参、西洋参及び中国人参などの人参を使用することができる。前記人参は、水参、白参、紅参などの形として利用でき、好ましくは4〜6年根の高麗人参(Panax ginseng C.A.Meyer)から製造した紅参を使用することが良いが、これに限定されるものではない。
【0024】
また、本発明は、糖リポタンパク質(ジントニン)をカルシウム濃度の低下に起因する疾患の予防または治療剤としての用途で提供する。
【0025】
具体的に、本発明は、前述の方法で製造された糖リポタンパク質としてのジントニンを有効成分として含む、カルシウム濃度の低下に起因する疾患の予防及び治療のための薬学組成物を提供する。
【0026】
本発明において、前記ジントニンは、一時的に細胞質内遊離Ca2+(Free Ca2+)の増加を誘発し、細胞内カルシウム濃度の一時的な増加により、アフリカツメガエル卵母細胞で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)を活性化して細胞内カルシウム濃度を増加させ、その結果、カルシウム濃度の低下に起因する疾患の予防及び治療に優れた効果を示し、特にカルシウム濃度の低下により引き起こされる神経系疾患の異常を改善することにより、治療及び予防が可能である。
【0027】
カルシウム濃度の低下による神経系疾患は、精神分裂(Schizophrenia)、アルツハイマー病(Alzheimer’s disease)、ハンチントン病(Huntington’s disease)、家族性片麻痺性片頭痛(Familial hemiplegic migraine)、癲癇(epilepsy)、一過性運動失調症(episodic ataxia)、脊髄小脳失調症(spinocerebellar ataxias)などを含むが、これらの疾患に限定されるものではない。
【0028】
また、本発明の組成物は、カルシウム不足による成長阻害などにも効果があり、細胞内カルシウム依存性の様々な生理活性、例えば強壮作用、免疫力増加、性機能強化、神経系保護及び活性、血管新生、抗糖尿作用などにおいても効果を示す。
【0029】
また、人参は長時間生薬として使用されてきたものであり、このような人参から分離した本発明のジントニンも毒性及び副作用などの問題が全くない。
【0030】
本発明のカルシウム濃度の低下による疾患の予防及び治療用薬学組成物は、組成物の全体重量に対し、前記ジントニンは0.0001〜10重量%、好ましくは0.001〜1重量%で含む。
【0031】
また、本発明のジントニンを含む組成物は、薬学的組成物の製造に通常用いる適切な担体、賦形剤及び希釈剤をさらに含んでもよく、本発明のジントニンの薬学的投与形態は単独で或いは他の薬理的活性化合物との結合又は適当な集合として使用できる。
【0032】
本発明に係るジントニンを含む組成物は、それぞれ通常の方法によって散剤、顆粒剤、錠剤、カプセル剤、懸濁液、エマルジョン、シロップ、エアロゾールなどの経口剤形、外用剤、坐剤及び滅菌注射溶液の形に剤形化して使用できる。前記組成物に含まれる担体、賦形剤及び希釈剤としては、ラクトース、デキストロース、スクロース、ソルビトール、マンニトール、キシリトール、エリトリトール、マルチトール、澱粉、アカシアゴム、アルギン酸塩、ゼラチン、リン酸カルシウム、珪酸カルシウム、セルロース、メチルセルロース、微晶質セルロース、ポリビニルピロリドン、水、ヒドロキシ安息香酸メチル、ヒドロキシ安息香酸プロピル、タルク、ステアリン酸マグネシウム、及びミネラル類を挙げることができる。製剤化する場合には、通常使用する充填剤、増量剤、結合剤、湿潤剤、崩壊剤、界面活性剤などの希釈剤又は賦形剤を用いて調製される。経口投与のための固形製剤には、錠剤、丸薬、散剤、顆粒剤、カプセル剤などが含まれる。このような固形製剤は、前記ジントニン又は分画物に少なくとも一つの賦形剤、例えば澱粉、カルボン酸カルシウム、スクロース又はラクトース、ゼラチンなどを混ぜて調製される。また、単純な賦形剤以外に、ステアリン酸マグネシウム、タルクなどの潤滑剤も使用される。経口のための液相製剤としては、懸濁剤、内溶液剤、乳剤、シロップ剤などが該当するが、よく使用される単純希釈剤としての水、流動パラフィン以外に、様々な賦形剤、例えば湿潤剤、甘味剤、芳香剤、保存剤などが含まれてもよい。非経口投与のための製剤には、滅菌した水溶液、非水性溶剤、懸濁剤、乳剤、凍結乾燥製剤、坐剤が含まれる。非水性溶剤、懸濁剤としてはプロピレングリコール、ポリエチレングリコール、オリーブ油などの植物性油、オレイン酸エチルなどの注射可能なエステルなどが使用でき、坐剤の基剤としてはウイテプソル(witepsol)、マクロゴール(macrogol)、ツイン(tween)61、カカオ脂、ラウリン脂、グリセロール、ゼラチンなどが使用できる。
【0033】
本発明の組成物の好ましい投与量は、患者の状態及び体重、病気の重症程度、薬物形態、投与経路及び期間によって異なるが、当業者によって適切に選択できる。ところが、望ましい効果のために、本発明の組成物は1日0.0001〜100mg/kgで投与することがよい。投与は1日に1回投与してもよく、数回に分けて投与してもよい。前記投与量はいずれの面においても本発明の範囲を限定するものではない。
【0034】
本発明の薬学組成物は、ラット、マウス、家畜、ヒトなどの哺乳動物に様々な経路で投与できる。投与の全ての方式は予想できるが、例えば、経口、直腸、静脈、筋肉、皮下、子宮内膜または脳室内(intracerebroventricular)注射によって投与できる。
【0035】
本発明のジントニンを含む組成物は、前述したような剤形で、カルシウム濃度の低下に起因する疾患の予防及び治療のための薬剤、食品及び飲料などに様々に利用できる。前記ジントニンを添加することが可能な食品としては、例えば、各種食品類、飲料、チューインガム、茶、ビタミン複合剤、健康機能性食品類などがある。
【0036】
本発明のジントニンは、毒性及び副作用が殆どないので、予防の目的で長期間服用する場合にも安心して使用することができる。
【0037】
本発明のジントニンは、カルシウム濃度の低下に起因する疾患の予防を目的として食品又は飲料に添加できるが、この際、食品又は飲料中に前記ジントニンは全体重量に対して0.01〜15重量%で加えることができ、健康飲料組成物は100mLを基準として0.02〜5g、好ましくは0.3〜1gの割合で加えることができる。
【0038】
本発明の健康機能性飲料組成物は、指示された比率で必須成分として前記ジントニンを含有する以外は他の成分には特別な制限がなく、通常の飲料と同様に、様々な香味剤又は天然炭水化物などをさらに含んでもよい。この際、前記天然炭水化物としては、例えば、ブドウ糖、果糖などの単糖類、マルトース、スクロースなどの二糖類、及びデキストリン、シクロデキストリンなどの多糖類のような通常の糖、並びにキシリトール、ソルビトール、エリトリトールなどの糖アルコールなどが挙げられる。また、香味剤としては天然香味剤(タウマチン、ステビア抽出物、例えばレバウジオシドA、グリシルリジンなど)及び合成香味剤(サッカリン、アスパルテームなど)を有利に使用することができる。前記天然炭水化物の比率は本発明の組成物100mLあたり一般に約1〜20g、好ましくは約5〜12gである。
【0039】
この他に、本発明の抽出物は、様々な栄養剤、ビタミン、ミネラル(電解質)、合成矯味剤、及び天然矯味剤などの矯味剤、着色剤及び増音剤(チーズ、チョコレートなど)、ペクチン酸及びその塩、アルギン酸及びその塩、有機酸、保護性コロイド増粘剤、pH調節剤、安定化剤、防腐剤、グリセリン、アルコール、炭酸飲料に使用される炭酸化剤などを含有することができる。その他に、本発明の抽出物は天然果物ジュース飲料、果物ジュース飲料及び野菜医療の製造のための果肉を含有することができ、このような成分を独立に或いは組み合わせて使用することができる。また、前記添加剤の比率は、あまり重要ではないが、本発明の抽出物100重量部当たり0〜約20重量部の範囲で選択されることが一般的である。
【発明の効果】
【0040】
本発明は、人参に存在する新規な糖リポタンパク質としてのジントニンを分離同定して製造する方法、前記方法で製造された新規な糖リポタンパク質ジントニン、及び前記新規な糖リポタンパク質ジントニンのカルシウム濃度の低下に起因する疾患の予防及び治療剤としての用途を提供するという効果がある。
【0041】
本発明のジントニンは、一時的に細胞質内遊離Ca2+の増加を誘発し、細胞内カルシウム濃度の一時的な増加により、アフリカツメガエル卵母細胞で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)を活性化して細胞内カルシウム濃度を増加させるので、カルシウム濃度の低下に起因する疾患の予防及び治療に有用に使用できる。
【0042】
また、本発明のジントニンは、細胞内カルシウム依存性の様々な生理活性、例えば強壮作用、免疫力増加、性機能強化、神経系保護及び活性、血管新生、抗糖尿作用などにおいても優れた効果を示す。
【図面の簡単な説明】
【0043】
【図1】Aは人参から新規な糖リポタンパク質としてのジントニンを分離同定する方法を図式化した図であり、Bはジントニン分画物のアフリカツメガエル卵母細胞におけるCaCCの活性を確認したクロマトグラムであり、Cは粗ジントニンのSDS−PAGE結果である。
【図2】粗ジントニンから個別ジントニンを分離同定する方法を図式化した図である。
【図3】個別ジントニンの陰イオン交換及びゲルクロマトグラムであって、Aは粗ジントニンから分離したジントニンE分画とジントニンL分画の陰イオン交換クロマトグラムであり、Bは前記Aから得たジントニンL分画のゲル濾過クロマトグラムであり、CはジントニンE分画の陰イオン交換クロマトグラムであり、DはジントニンL分画の陰イオン交換クロマトグラムである。
【図4】ゲル又は連続陰イオン交換クロマトグラフィーによる個別ジントニンの溶出パターンを示すもので、AとCは精製されたジントニンE1〜E3及びジントニンL1〜L3のゲル濾過クロマトグラムであり、BとDは精製されたジントニンE1〜E3及びジントニンL1〜L3の陰イオン交換クロマトグラムである。
【図5】ゲル濾過クロマトグラフィーを用いたジントニンの分子量決定較正曲線であって、各点は使用された標準タンパク質であり(IgG:160kD、BSA:67kDa、β−ラクトグロブリン:35kDa、シトクロムC:12kDa、アプロチニン:6.5kDa)、矢印はジントニンである。
【図6】Aは個別ジントニンのSDS−PAGEであり、Bは前記個別ジントニンの炭水化物成分のゲル染色SDS−PAGE結果である。
【図7】HPAED−PADクロマトグラムによる個別ジントニンの炭水化物成分分析結果を示すもので、Aは中性糖(Neutral sugar)の成分分析であり(標準物質:1.L−果糖、2.L−ラムノース、3.D−アラビアノース、4.D−ガラクトース、5.D−グルコース、6.D−マンノース、7.D−キシロース)、Bはアミノ糖の成分分析である(標準物質:1.D−ガラクトサミン、2.D−グルコサミン)。
【図8】個別ジントニンE1〜E3及びL1〜L3の脂質成分をGC−MSスペクトル分析結果である。
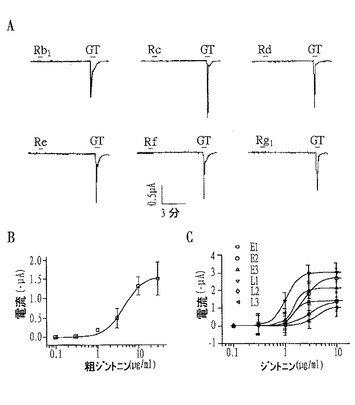
【図9】ジントニン処理の際にマウスEAT細胞における内因性、内向性CaCCの電流流れを示すもので、Aは−80mVの保持電位におけるジントニンとジンセノサイドの電流流れを示すグラフ、BとCはそれぞれ粗ジントニンと個別ジントニンの濃度別電流流れを示すグラフである。
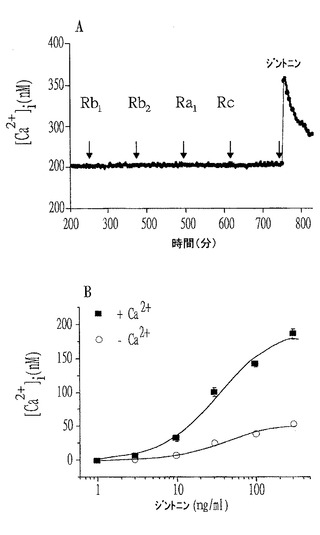
【図10】ジントニン処理の際にマウスEAT細胞における細胞内カルシウムの増加を確認したグラフであって、AはCa2+緩衝液に培養したFura−2に混ぜたEAT細胞における時間による細胞内カルシウムの増加を確認したものであり、BはCa2+遊離緩衝液に培養したFura−2に混ぜたEAT細胞におけるジントニン濃度による細胞内カルシウムの増加を確認したものである。
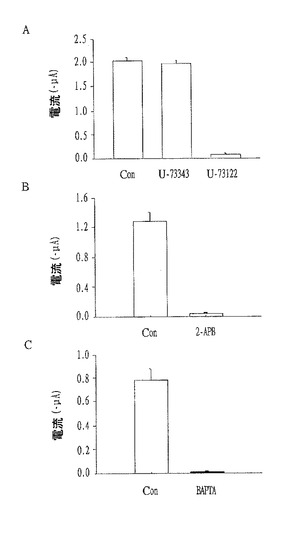
【図11】ジントニンによるCaCCの活性及びマウスEAT細胞における細胞内カルシウムの増加に対するPLC抑制剤、IP3受容体拮抗剤及びCa2+キレート剤の効果を示すもので、AはPLC抑制剤(U−73343/U−73122)によって、BとCはそれぞれIP3受容体拮抗剤(2−APB)及びCa2+キレート剤(BAPTA−AM)によってCaCCの活性化が減少することを確認した電流流れの棒グラフである。
【図12】マウスEAT細胞でジントニンによって増加する細胞内カルシウム濃度に対する活性型又は非活性型PLC抑制剤の効果を示すもので、AはCa2+緩衝液に培養したFura−2に混ぜたEAT細胞における細胞内カルシウム増加の抑制を確認したものであり、BはCa2+遊離緩衝液に培養したFura−2に混ぜたEAT細胞におけるジントニン濃度による細胞内カルシウム増加の抑制を確認したグラフである。
【発明を実施するための形態】
【0044】
以下、実施例によって本発明をさらに詳細に説明する。これらの実施例が本発明を例示するためのものに過ぎず、本発明の範囲を限定するものではないのは、当業者にとっては自明なことであろう。
【0045】
実施例1:人参からの粗ジントニン(crude gintonin)の分離
韓国人参公社(大田、韓国)から購入した6年根紅参(Panax ginseng C.A.Meyer)20kgを細かく粉砕(>3mm)し、80%メタノールを30L加えて80℃で8時間還流冷却抽出した後、真空濾過して上層液を回収した。この過程を3回繰り返し行って上層液を集めた後、減圧濃縮し(メタノール抽出物の総6.2kgを収得)、水とn−ブタノールの混合溶媒(1:1)で溶媒分画して水とn−ブタノールの分画物を収得した(n−ブタノール分画物を総908g収得)。
【0046】
前記n−ブタノール分画物(粗総サポニン分画、CGSF)は濃縮してクロロホルム:メタノール:水(CHCl3:MeOH:H2O=13:7:2)の混合溶媒で溶離してシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離した。その後、それぞれの分画物に対して、アフリカツメガエル卵母細胞で内因性に存在するカルシウム依存性塩素イオンチャネル(CaCC)に対する活性を実験し、CaCCの活性が最も高く示された第7の分画(図1のB参照)に対して、さらに酢酸エチル:エタノール:水(EtOAc:EtOH:H2O=1:3:0.5)の混合溶媒で溶離してシリカゲルカラムクロマトグラフィーを行い、2個の分画物を得た。
【0047】
前記2個の分画物のうち、EAT(Ehrlich Ascites Tumor)細胞における細胞内カルシウム放出実験の評価とアフリカツメガエル卵母細胞で強いCaCC活性を示した分画物IIを4℃で8時間過量の蒸留水(distilled water、DW)とSpectra/Por透析膜(molecular weight cut of 6,000〜8,000)(Spectrum Laboratories、Inc.,Califoinia、USA)を用いて透析し、ジンセノサド及び他の低分子物質を除去した後、最終的に分画物を得た。
【0048】
前記得た分画物に対して電気泳動(SDS−PAGE)を行った結果、クマシーブリリアントブルー(Coomassie Brilliant Blue)で染色され、見掛け分子量が約13kDaであることを確認した。これは、EAT細胞における細胞内カルシウム放出実験の評価とアフリカツメガエル卵母細胞でCaCC活性を起こす新規物質が、ジンセノサイドではなく、タンパク質の1種類であることを示唆する。前記タンパク質をジントニンと命名し、これから得た最終分画物は粗ジントニン(crude gintonin)と命名した。
【0049】
ところが、人参のタンパク質または多糖類を含んでいる水分画物、および脂質(lipid)成分を含有している石油エーテル分画物は、EAT細胞の細胞内カルシウム放出実験とアフリカツメガエル卵母細胞におけるCaCC活性効果が全くなかった(図式しない)。
【0050】
実施例2:個別ジントニンの分離
粗ジントニン(crude gintonin)から個別ジントニン(gintonins)を分離するために、前記実施例1で得た粗ジントニンをリン酸塩緩衝食塩水(phosphate buffer saline、PBS)(pH7.2)に溶解し、DEAEセファロースCL−6Bカラムが装着された陰イオン交換樹脂クロマトグラフィー(GE Healthcare、Uppsala、Sweden)を行い、2個の主なピーク(major peak)を得た。この際、溶媒は、0〜2M NaClがPBSに溶けているものを使用した。1mL/分の流速にて280nmでモニタリングしながら線形勾配(linear gradient)方式で分離して得た2個の主なピークはそれぞれジントニンE(early eluted gintonin)、ジントニンL(late eluted gintonin)と命名した。
【0051】
前記ジントニンEの分画物と前記ジントニンLの分画物はCentriVap
DNA真空濃縮器(Labconco、Missouri、USA)で濃縮した後、PBS(pH7.2)を溶離液とし、スーパーデキス(Superdex)75ゲル濾過カラムを用いて0.5mL/分の流速にて280nmでモニタリングしながらクロマトグラフィーを行い、分子量の大きい主ピークと副ピークを得た(図3のB参照)。
【0052】
こうして得たジントニンEとジントニンLは、アフリカツメガエル卵母細胞でCaCC活性を示すことを確認した後、ジントニンE分画物は再び100、150、200、250mM NaClが含有された20mM Tris−HCl(pH8.2)を用いて段階勾配(step gradient)DEAE陰イオン交換カラム(anion exchange column:HiTrapTMDEAE FF、1mL)に1mL/分の流速にてロードして4個のピークを得た(図3のC参照)。これらのうち、100、150及び200mM NaCl溶離液の分画物を大量透析(extensive dialysis)及び濃縮した後、さらに0〜1M NaClが含有された20mM Tris−HCl(pH8.2)を用いて段階勾配DEAE陰イオン交換カラムを行い、しかる後に、最終的にスーパーデキス75ゲル濾過クロマトグラフィーを行って最終分画物をそれぞれ3.7、26.7及び13.5mgずつ得た。これらはそれぞれジントニンE1、E2及びE3と命名した。
【0053】
一方、ジントニンL分画物は150、200、250、300、400mM NaClが含有されたPBS(pH7.2)を用いて前記と同様の方法で得た5個のピーク(図3のD参照)のうち、150、200及び250mM NaCl溶離液の分画物に対して最終分画物を7.4、17.6及び6.6mgずつ得た。これらはそれぞれジントニンL1、L2及びL3と命名した。
【0054】
また、粗総サポニン分画(CGSF)908gからそれぞれ、ジントニンE1の場合は0.0004%(3.7mg)、ジントニンE2の場合は0.0029%(26.7mg)、ジントニンE3の場合は0.0015%(13.5mg)、ジントニンL1の場合は0.0008%(7.4mg)、ジントニンL2の場合は0.0019%(17.6mg)、ジントニンL3の場合は0.007%(6.6mg)の収率で得られた。
【0055】
実施例3:ジントニンの分子量決定
前述したように得た6個のそれぞれ異なるジントニンは、ヤマモト等の方法(Yamamoto,Y.,Nunome,T.,Yamauchi,R.,Kato,K.,and Sone,Y.(1995) Carbohydr Res.275,319-332)に準じてゲル濾過クロマトグラフィーを行って分子量を決定するが、PBS(pH7.2)で平衡を維持したスーパーデキス75カラム(10×300mm)を装着したバイオロジックデュオフロークロマトグラフィーシステム(BioLogic DuoFlowTMChromatograpgy、BIO−RAD、California、USA)を用いて280nmでモニタリングしながら流速0.5mL/分で分画物を集めた後、免疫グロブリンG(IgG;160,000Da)、牛血清アルブミン(BSA;67,000Da)、β−ラクトグロブリン(35,000Da)、シトクロムC(12,327Da)、及びアプロチニン(6,512Da)などの標準タンパク質からなる較正曲線(calibration curve)を用いて分子量を決定した。
【0056】
その結果、ジントニンの帯びる電荷が異なってNaCl勾配溶出によって溶出(elution)パターンが異なったが、ゲル濾過クロマトグラフィーではほぼ同時に溶出して分子量がほぼ同一であった(図4のB及びD参照)。
【0057】
また、標準タンパク質を用いて決定したジントニンL2のネイティブ(native)分子量も約67kDaであった(図5参照)。他のジントニンの分子量も約67kDaであると確認された(図式せず)。
【0058】
また、SDS−PAGEの結果、6個のジントニンがブロード(broad)であるが、単一な主バンド(single major band)を示し、その見掛け分子量(apparent molecular weight)は約13kDaであると確認された。
【0059】
前述したような結果は、本発明のジントニンが五量体(pentamer)であることを示唆する(図6のA参照)。
【0060】
実施例4:ジントニンの純度決定
前記粗ジントニン及び個別ジントニンの純度を決定するために、12.0%分離ゲル(separating gel)を用いて電気泳動(SDS−PAGE)を行った(Laemmli,U.K.(1970) Nature 227,680-685)。
【0061】
具体的に、粗ジントニンと個別ジントニンE1〜E3、L1〜L3を100μgずつ取って各レイン(lane)にロードして電気泳動した後、ジントニンバンドをクマシーブリリアントブルーR−250染色で視覚化した。また、糖タンパク質(glycoprotein)検出のために、ゲルはPAS染色(periodic acid-schiff based staining)技法を用いて染色し、40%エタノール−7%酢酸水溶液で30分間培養した後、1%過ヨウ素酸−3%酢酸で1時間培養し、しかる後に、過ヨウ素酸溶液を除去し、Schiff試薬を添加して1時間培養した。染色された糖タンパク質のバンドは桃色を呈し、PAS染色されたゲルは7.5%酢酸で浸漬(soaking)した後、保管した。
【0062】
実施例5:ジントニンのアミノ酸組成分析
一般アミノ酸(general amino acid)分析のために、ジントニン30μgを真空内(in vacuo)で6N HClを用いて110℃で24時間加水分解した。また、システインの分析のためには、ジントニンを過酸化(peroxidation)させた後、6N HClを用いて110℃で24時間加水分解し、蟻酸:過酸化水素(10:1)溶液で処理した。トリプトファンの分析のためには、試料を4Mメタンスルホン酸(methanesulfonic acid)で加水分解し、4M KOHを加えた。
【0063】
また、フェニルイソチオシアネート(Phenylisothiocyanate、PITOC)誘導体に隠されたアミノ酸は、大田所在の韓国基礎科学支援研究所に依頼し、Waters Nova−Pak C18カラム(3.9×300mm)を装着したHPLC(Hewlett Packard 1100 series)で分析した。
【0064】
タンパク質成分は、標準として牛血清アルブミン(BSA)を用いたBradford法(Bradford,M.M.(1976).Anal.Biochem.72,248-254)で決定した。
【0065】
実施例6:ジントニンの部分アミノ酸配列
ジントニンのN末端アミノ酸配列は、自動エドマン分解アプライドバイオシステムモデル477Aガス相タンパク質配列(automated Edman degration on an Applied Biosystem model 477A gas-phase protein sequence)及びミリゲン6600固体相配列(MilliGen 6600 solid-phase sequence)によって行われた。
【0066】
ジントニン100μgに対して12%分離ゲルを用いてSDS−PAGEを行い、タンパク質バンドはクマシーブリリアントブルーR−250染色で視覚化した。前記バンドを切断してトリプシンで消化(digesting)し、前記消化産物(digestion product)は脱塩及び濃縮した。
【0067】
こうして誘導された全てのペプチドのMS/MSは、韓国基礎科学支援研究所で、ナノ−ESIを装着した質量分析計(Mass spectrometer)(QTOF II、Micromass、UK)を用いて行った。この際、生成されたイオン(product ion)は、反射体(reflector)、マイクロチャネル検出器(microchannel plate detector)及び時間対デジタル変換器(time-to-digital converter)が固定された直角TOF分析器(orthogonal TOF analyzer)を用いて分析した。データはウィンドウNT環境でMass Lynx softwareが駆動されるPCを用いて処理した。配列同族体はBLAST(www.ncbi.nln.nih.gov/BLAST.cgi)を用いて検索した。
【0068】
表1はジントニンのde novoアミノ酸配列と組成を示すものであり、純粋分離されたジントニンは自動エドマン分解によるN末端アミノ酸配列からなるものと確認された。ところが、ジントニンのN末端がブロッキング(blocking)されて全体配列を得ることができなかったため、MALDI−TOF−MS/MSを用いてトリプシン処理したジントニンから18個の内部ペプチド配列(internal peptide sequence)を推定した結果、6個の個別ジントニンは一般に幾つかのペプチドを含有し、アミノ酸配列はヒトグルコキナーゼ(human glucokinase)及びグルコキナーゼイソ型2(glucokinase isoform 2)と部分的相同性を示した(図式せず)。
【0069】
また、Bradford方法によるジントニンの総タンパク質の含量は、ジントニンE1、E2、E3、L1、L2及びL3でそれぞれ9.4、24.1、20.5、35.8、37.1及び39.6%であった。ジントニンL1〜L3のタンパク質含量はジントニンE1のそれより約3倍高い含量である。
【0070】
また、ジントニンにおける疎水性アミノ酸(hydrophobic amino acid)は、親水性アミノ酸(hydrophilic amino acid)よりさらに多かったが、ヒスチジン(His)、チロシン(tyr)及びメチオニン(Met)は検出されなかった。これらの中でも、メチオニンはジントニンのN末端がブロッキングされて検出されなかったものと思われる。
【0071】
一方、ジントニンE1〜E3及びL1〜L3では、共通にフェニルアラニン(Phe)が最も高く含有されるものと確認された。また、ジントニンL1〜L3におけるリシン(Lys)はE1〜E3のそれより高く示された(表1参照)。
【0072】
【表1】
実施例7:ジントニンの炭水化物組成分析
図6のAに示すように、SDS−PAGEにおけるジントニンのバンドは単一(single)であるがブロード(broad)であり、クマシーブリリアントブルー染色で強く染色されなかったため、炭水化物を含んでいる可能性がある。
【0073】
したがって、ジントニンの炭水化物組成を確認するために、ジントニンを2Mトリフルオロ酢酸(trifluoroacetic acid)を用いて100℃で4時間加水分解した後、中性糖(neutral sugar)を得た。また、ジントニンをガラスアンプルで6N HClを用いて100℃で4時間加水分解によってアミノ糖(amino sugar)と酸性糖(acid sugar)を得た。
【0074】
ジントニンの炭水化物組成は、ソウル所在の世宗大学校炭水化物素材研究所に依頼し、PAS染色(periodic acid-schiff based staining)技法を用いて染色した後、CarboPacTMPAlカラムを装着したHPAEC−PADシステム(high performance anion exchange chromatography-plused amphermetric dectection system、Dionex、California、USA)を用いて分析した。単糖類のモル重量(molar weight)はピーク面積(peak area)から計算した。炭水化物の含量は、中性糖の場合にはフェノール−スルホン酸方法(Hounsell,E.F.,Davies,M.J.,and Smith,K.D.(1997) Protein protocol handbood,Humanna press,Totawa,803-804)で決定し、酸性糖の場合にはAnthrone方法で決定した(Scott,T.A.,and Melvin,E.H.(1953)Anal.Biochem.25,1656-1660)。
【0075】
その結果、ジントニンは、ラムノース(rhamnose)、アラビノース(arabinose)、グルコース(glucose)、マンノース(mannose)、キシロース(xylose)などの5個の異なる種類の中性糖、及びグルコサミンなどの1個のアミノ糖から構成されるものと確認された。6個のジントニンは互いにほぼ類似した炭水化物構成を有することが分かった(表2及び図7参照)。ところが、ジントニンE1〜E3からマンノースは検出されなかった。
【0076】
一方、1−N−アセチル−ノイラミン酸(1-N-acetyl-neuraminic acid)、2−ガラクツロン酸(2-galacturonic acid)、3−グルクロン酸(3-glucuronic acid)などの酸性糖は検出されなかった(図式せず)。
【0077】
最後に、ジントニンの総炭水化物含量は、ジントニンE1、E2、E3、L1、L2及びL3でそれぞれ約34.3、45.3、38.1、36.0、29.8及び45.2%であった。その結果より、本発明のジントニンは糖タンパク質であることを再確認することができた。
【0078】
【表2】
実施例8:ジントニンの脂質成分分析
本発明のジントニンは、n−ブタノール抽出によってジンセノサイドと同様の分画にあったため、ジントニンも脂質部分(lipid moiety)を含有するものと予想することができる。
【0079】
これを確認するために、ジントニンを6N HClを用いて4時間100℃で加水分解し或いはリポタンパク質リパーゼ(lipoprotein lipase)に消化させて脂質(lipid)及び疎水性部分(hydrophobic moiety)を確認した。
【0080】
具体的に、粗総サポニン分画(CGFS)を酸加水分解又は消化させたものを濃縮し、さらに水とn−ヘキサンにさらに分画した後、n−ヘキサン分画物に対して、DB5−MS毛細管カラム(capillary column)(30cm×250μm×0.25μm)を装着したAgilent 6890N GC−MSシステム(California、USA)を用いて、韓国基礎科学支援研究所で脂質(lipid)及び疎水性部分を分析した。この際、GC(6890N)は、火イオン化検出器(flame ionization detector)と分離注入システム(split injection system)が装着されており、supelco SPB−1毛細管カラム(内径:15m×0.32mm、厚さ:0.25m)を固定した。
【0081】
その結果、表3及び図8に示すように、ジントニンは、エステル型(ester form)又は自由型(free form)のパルミチン酸(plamitic acid)、ステアリン酸(stearic acid)、オレイン酸(oleic acid)、リノール酸(linoleic acid)などを含有するものと確認された。
【0082】
また、ジントニンE1又はL1における多価不飽和脂肪酸としてのリノール酸の比率は飽和脂肪酸としてのパルミチン酸とステアリン酸又はオレイン酸より高かった。パルミチン酸はジントニンE2、E3、L1、L2で多量含有されていた。
【0083】
また、GCによって測定したジントニンに含有された総脂質含量は、ジントニンE1、E2、E3、L1、L2及びL3がそれぞれ34.3、45.3、38.1、36.0、29.8及び45.2%であった。これらの中でも、ジントニンE1の脂質含量が最も高かった。
【0084】
前述したような結果は、本発明のジントニンが新規な糖リポタンパク質であることを示唆する。
【0085】
【表3】
測定例1:アフリカツメガエル卵母細胞(Xenopus laevis oocytes)からのCaCC活性の測定
1−(1).卵母細胞(Oocyte)の準備
Xenopus I(Ann Arbor、MI、USA)から得たアフリカツメガエル(Xenopus laevis)は、最上の規格指針に従って保管及び処理し、卵子(oocytes)を分離するために、カエルを3−アミノ安息香酸エチルエステル(3-amino benzoic acid ethyl ester)の通気溶液(aerated solution)で麻酔させて手術した後、コラゲナーゼで処理し、しかる後に、82.5mM NaCl、2mM KCl、1mM MgCl2、5mM HEPES、2.5mMピルビン酸ナトリウム(sodium pyruvate)、100units/mLフェニリシン及び100μg/mLストレプトマイシンが含有されたCa2+遊離培地で2時間攪拌して分離した。
【0086】
V−VI段階の卵子を収集し、ゲンタマイシン(gentamicin)50μg/mLを追加したND96(96mM NaCl、2mM KCl、1mM MgCl2、1.8mM CaCl2、及び5mM HEPES、pH7.4)で保管した。前記卵子を含む溶液は、連続的に軽く振とうしながら18℃に維持し、毎日交換した。
【0087】
1−(2).CaCCの測定
2−電極電圧固定(two-electrode voltage clamp)記録は、小型プレキシガラス(plexiglass)ネットチャンバー(0.5mL)に置かれた個別的な卵子から得た。電気生理学実験(electrophysiological experiments)は、3M KClで充填したマイクロ電極(抵抗:0.2〜0.7MΩ)と卵子固定増幅器(Oocyte Clamp amplifier、OC−725C、Warner Instrument、CT、USA)を用いて室温で行った後、CaCCを−80mV保持電位(holding potential)で記録した。
【0088】
ジントニンは、バス灌流(bath perfusion)によって卵子に適用した(Lee,J.H.,Jeong,S.M.,Lee,B.H.,Noh,H.S.,Kim,B.K.,Kim,J.I.,Rhim,H.,Kim,H.C.,Kim,K.M.,and Nah,S.Y.(2004)J Biol Chem 279,9912-9921)。
【0089】
測定例2:マウスEAT細胞からの細胞内遊離カルシウム([Ca2+]i)の測定
2−(1).EAT(Ehrlich Ascites Tumor)細胞の準備
EAT細胞は、Jorgensen等の方法(Jorgensen,N.K.,Pedersen,S.F.,Hoffmann,E.K.(1999)Am J Physiol 276.C26-C37)によってICRマウスの腹腔内移植(interaperitoneal transplantation)から毎週収集した。
【0090】
具体的に、腹水(ascites fluid)は、マウス腹膜腔(peritoneal cavity)から2.5U/mLのヘパリンが含有されたHank’s溶液に収集した。RBCからEAT細胞を分離するためには、EAT細胞を10及び20%フィコール溶液(Ficoll solution)の2層を含む遠心分離管にロードして1,800rpmで20分間遠心分離し、EAT細胞のみを得た。前記細胞は、所望の細胞数となるように浮遊させた後、実験を開始する前に約30分間培養した。
【0091】
2−(2).Fura 2とEAT細胞との混合
Fura 2−負荷マウスEAT細胞におけるG−タンパク質結合受容体作用剤(G-protein coupled receptor agonist)の処理は、細胞内遊離カルシウム([Ca2+]i)の一時的な増加を誘導することが明らかになった(Scott and Melvin.Anal.Biochem.,25,1656-1660,1953)。
【0092】
本発明では、ジントニン又はジンセノサイドRa、Rb1、Rb2、Rc、Rg1がFura2−負荷マウスEAT細胞における[Ca2+]iの一時的増加に効果を示すかを確認するために、Fura−2に混ぜたEAT細胞(2〜4×106/mL)は1.5mM Ca2+緩衝液またはCa2+遊離緩衝液で10分間培養した後、ジントニンとジンセノサイドを添加し、EAT細胞における細胞内カルシウム濃度に及ぼす効果を確認した。但し、利用可能な純粋分離された個別ジントニンの量の限界のために、代表的なジントニンとしてジントニンE2を用いてマウスEAT細胞(Riken BRC Cell Bank、Tsukuba、Japan)における[Ca2+]iの増加について実験した。
【0093】
具体的に、Jorgensen等の方法によって、NaCl 120mM、KCl 5mM、MgCl21mM、CaCl21.5mM、グルコース10mM、及びHEPES 25mMからなるCa2+緩衝液(pH7.2)と、NaCl 120mM、KCl 5mM、MgCl21mM、EGTA 0.2mM、グルコース10mM及びHEPES 25mMよりなるCa2+遊離緩衝液(pH7.4)を用いて、37℃の水浴で30分間振とうしながら2μN Fura 2−AMにEAT細胞(2〜4×106/mL)を混合し、過量のfura−2は細胞をCa2+緩衝液又はCa2+遊離緩衝液で3回洗浄して除去した。
【0094】
2−(3).細胞浮遊物内[Ca2+]iの蛍光測定
[Ca2+]iは、RF−5300PC細胞内イオン測定システム(Intracellular Ion Measurement System)(Shimadzu Corporation、Japan)を用いてFura 2−負荷セルから推定した。具体的に、Fura 2−負荷セルは、最終的に2〜4×106/mLまで実験培地で希釈した後、ポリスチレンキュベット(Elkay Ultra−UV)に移した。細胞はテフロン(登録商標)でコートされた磁石を用いて攪拌した。キュベットハウジングは37℃に調節した。励起波長(excitation wavelengths)は、コンピュータ制御の下で340nmと380nmを交互にし、放出(emission)は510nmで検出した。この際、励起及び放出の隙間幅は5nmとし、背景補正はJorgensen等の方法に準じて行うが、ジギトニン(digitonin)及びEGTAはfura−2が完全にCa2+と化合し、Ca2+から解離される状態を作るための濃縮反応試薬(concentration adjustment reagent)として使用した。
【0095】
2−(4).Fura 2測定値からの[Ca2+]iの推定
測定された340nm対380nmの比率値は、Hounsell等の式(Hounsell,E.F.,Davies,M.J.,and Smith,K.D.(1997)Protein protocol handbood,Humanna press,Totawa,803-804)を用いて[Ca2+]iの値に変換した。
【0096】
[Ca2+]=Kd[(R−Rmin)/(Rmax−R)](Sf380/Sb380)
但し、Kdは解離定数(224nM)であり、Rは測定された340nm対380nmの蛍光比(fluorescence ratio)であり、Rmax及びRminは50μg/mLのジギトニンを添加した飽和濃度でのR値及び20mM EGTAを添加した遊離培地でのR値である。また、Sf380とSb380はジギトニンとEGTAを添加したときの380nmでの蛍光強度であり、比率はこれらの値が表示されるときの最大及び最小値である(Grynkiewicz,G.,M.Poenie,and R.Y.Tsien.(1985) J Biol Chem 260:3440-3450)。
【0097】
2−(5).データ分析
InsP6を含む濃度反応曲線を得るために、観察されたピークの振幅を標準化及び図表化した後、Origin software(Northampton、MA)を用いて下記のHill式に対応させた。
【0098】
y/ymax=[A]n/([A]n+[EC50]n)
式中、yはジントニンの与えられた濃度における%活性であり、ymaxは最大ピーク電流(maximum peak current)であり、[EC50]は最大反応の50%を示すことが可能なジントニンの濃度であり、[A]はジントニンの濃度である。また、nは相互作用係数(interaction coefficient)であり、全ての値は平均±標準誤差で示し、対照群のデータとジントニン処理群のデータ間の平均値はunpaired Student’s t−testを用いて分析した。統計学的有意性はP<0.05で検定した。
【0099】
実験例1.アフリカツメガエル卵母細胞(Xenopus laevis oocytes)で内因性に存在するCaCCの活性に対する粗ジントニン及び個別ジントニンの効果確認
アフリカツメガエル卵母細胞(Xenopus laevis oocytes)で内因性に存在するCaCCの活性に対する粗ジントニン及び個別ジントニンの効果を確認するために、前記測定例1の夫々の方法を用いて、CaCCの活性増加として記録される保持電位を測定した。
【0100】
その結果、粗ジントニンを処理した群では−80mV保持電位で内向性(inward)のCl−電流を誘導したが、ジンセノサイドは誘導効果が全く確認されなかった。ED50は4.4±0.5μg/mLであった(図9のA及びB参照)。
【0101】
また、個別ジントニンに対しても、アフリカツメガエル卵母細胞におけるCaCC活性効果を実験した結果、ジントニンE1〜E3及びL1はほぼ同一程度のCaCC活性を示したが、ジントニンL2とL3は相対的に弱いCaCC活性を示した(図9のC参照)。この際、ED50は、それぞれジントニンE1〜E3が1.5±0.1、2.3±0.1、1.5±0.1であり、ジントニンL1〜L3が1.0±0.1、3.2±0.2及び4.3±0.2μg/mLであった。
【0102】
実験例2:マウスEAT細胞における細胞内遊離カルシウム([Ca2+]i)に対する効果の確認
前記測定例2のそれぞれの方法に従い、マウスEAT細胞における細胞内[Ca2+]iを測定した結果、浮遊させたEAT細胞にジンセノサイドを単独でそれぞれ100μMずつ処理したとき、実験したあるジンセノサイドもEAT細胞に反応を起こさなかった反面、これと対照的に、100ng/mLのジントニンE2で処理した実験群では[Ca2+]iを一時的に増加させることが確認された(図10のA参照)。
【0103】
図10のBはジントニンE2によって誘導された[Ca2+]iの増加が処理濃度依存的であることを示す。興味深く、ジントニンE2によって誘導された[Ca2+]iの一時的な増加は外部のCa2+(0.2mM EGTAを含むCa2+遊離緩衝液)を除去することにより大きく減少した(P<0.001、細胞外Ca2+の存在対比)。これは細胞外Ca2+もジントニン処理の際に細胞内に入ることを意味する。ところが、ジントニンE2の処理は依然として濃度依存的に[Ca2+]iを増加させた。これより、ジントニンE2によって誘導された[Ca2+]iの増加は細胞内貯蔵Ca2+の放出と細胞外におけるCa2+の流入によるものであることが分かった。
【0104】
この際、EC50値は、細胞外Ca2+の存在と不在によってそれぞれ33.6±8.2ng/mLと43.6±14.2ng/mL(平均±標準誤差、n=5〜6)であった。このような結果は、哺乳動物細胞における[Ca2+]iの増加を起こすものがジンセノサイドではなくジントニンであることを示す。
【0105】
実験例3:粗ジントニンによるCaCC活性とEAT細胞における[Ca2+]i上昇効果に対するPLC抑制剤、IP3受容体拮抗剤又はCa2+キレート剤効果の確認
細胞質の[Ca2+]iとCaCC活性化を増加させるジントニン誘導シグナル伝達経路におけるリン脂質分解酵素C(phospholipase C、PLC)の可能な役目を説明するために、まず、活性PLC抑制剤としてのU−73122と非活性類似体としてのU−73343の効果を、アフリカツメガエル卵母細胞(Xenopus oocytes)とマウスEAT細胞を用いて確認した。
【0106】
アフリカツメガエル卵母細胞において、粗ジントニン誘導CaCC電流はU−73122で対照群が3.9±0.3であったが、U−73343では対照群の97.2±0.2%であった(図11のA参照)。このような結果は、ジントニンによって誘導された[Ca2+]iの上昇とジントニンによって媒介されたCaCC活性化がPLC活性化を介して媒介されることを意味する。
【0107】
次の段階として、IP3受容体を介しての細胞内Ca2+放出がジントニン誘導CaCC活性化を起こすか否かを検査した(Grynkiewicz,et al.,J.Biol.Chem.260:3440-3450,1985;Parekh,A.B.Pflugers Arch.-Eur.J.Physiol.,430,954-963,1995;Broad,et al.,J.Biol.Chem.276,15945-15952,2001)。
【0108】
このために、IP3受容体拮抗剤としての膜透過性2−APB(2-aminoethoxydiphenylborate)のジントニン誘導CaCC活性に対する効果を検査した。
【0109】
図11のBに示すように、2−APBの添加はジントニンによって誘導された内向性C1−電流(1.3±0.2〜0.03±0.01μAの対照電流、P<0.01)をほぼ完全に消滅させた。また、膜透過性BAPTA(1,2-bis(o-aminophenoxy)ethane-N,N,N’,N’-tetraacetic acid)−AM(10μM最終)処理で細胞内遊離Ca2+を除去することもジントニン媒介CaCC活性化を殆ど消滅させた(図11のC参照)。
【0110】
マウスEAT細胞の場合でも、U−73122の処理は細胞外Ca2+の存否を問わずジントニンの作用を殆ど消滅させるものと確認されたが、U−73343はそうではなかった(図12参照)。
【0111】
このような結果より、アフリカツメガエル卵母細胞とマウスEAT細胞でPLC−IP3−Ca2+経路を介して起るCaCC活性を起こす粗総サポニン分画(CGSF)の主要活性物質がジントニンであることを再び確認することができた。
【0112】
結論的に、本発明のジントニンは、細胞内カルシウム濃度を増加させる作用だけでなく、細胞外からカルシウムの流入増加を誘発して細胞内カルシウム濃度を増加させるシグナル伝達経路を介してその効能を示すことが分かった。
【0113】
本発明のジントニンを含む薬学組成物の製剤例を下記のとおり例示する。ところが、これらの例は本発明を限定するためのものではなく、具体的に説明するためのものである。
【0114】
製剤例1:錠剤の製造
ジントニン 100mg
乳糖 100mg
澱粉 100mg
ステアリン酸マグネシウム 2mg
これらの成分を混合し、通常の錠剤の製造方法に従って打錠して錠剤を製造した。
【0115】
製剤例2:散剤の製造
ジントニン 100mg
乳糖 100mg
タルク 10mg
これらの成分を混合し、気密布に充填して散剤を製造する。
【0116】
製剤例3:カプセル剤の製造
ジントニン 100mg
結晶性セルロース 3mg
ラクトース 14.8mg
ステアリン酸マグネシウム 0.2mg
通常のカプセル剤の製造方法に従って、これらの成分を混合し、ゼラチンカプセルに充填してカプセル剤を製造する。
【0117】
製剤例4:注射剤の製造
ジントニン 20mg
注射用滅菌蒸留水 適量
pH調節剤 適量
通常の注射剤の製造方法に従って、1アンプル当たり(2mL)前記の成分含量で注射剤を製造する。
【0118】
製剤例5:液剤の製造
ジントニン 1000mg
異性化糖 10g
マンニトール 5g
精製水 適量
通常の液剤の製造方法に従って、精製水にそれぞれの成分を加えて溶解させ、レモン香を適量加えた後、前述の成分を混合し、しかる後に、精製水を加えて全体100mLに調節した後、褐色瓶に充填して滅菌させて液剤を製造する。
【0119】
製剤例6:健康食品の製造
ジントニン 100mg
ビタミン混合物 適量
ビタミンAアセテート 70μL
ビタミンE 1.0mg
ビタミンB1 0.13mg
ビタミンB2 0.15mg
ビタミンB6 0.5mg
ビタミンB12 0.2μg
ビタミンC 10mg
ビオチン 10μg
ニコチン酸アミド 1.7mg
葉酸 50μg
パントテン酸カルシウム 0.5mg
無機質混合物 適量
硫酸第1鉄 1.75mg
酸化亜鉛 0.82mg
炭酸マグネシウム 25.3mg
第1リン酸カリウム 15mg
第2リン酸カルシウム 55mg
クエン酸カリウム 90mg
炭酸カルシウム 100mg
塩化マグネシウム 24.8mg
前記ビタミン及びミネラル混合物の組成比は、比較的健康食品に適した成分を好適な実施例で混合組成したが、その配合比を任意に変形実施しても構わず、通常の健康食品製造方法に従って前述の成分を混合した後、顆粒を製造し、通常の方法によって健康食品組成物の製造に使用することができる。
【0120】
製剤例7:健康飲料の製造
ジントニン 100g
ビタミンC 15g
ビタミンE(粉末) 100g
乳酸鉄 19.75g
酸化亜鉛 3.5g
ニコチン酸アミド 3.5g
ビタミンA 1.0g
ビタミンB1 0.13g
ビタミンB2 0.15g
通常の健康飲料の製造方法に従って前述の成分を混合した後、約1時間85℃で攪拌加熱し、しかる後に、作られた溶液を濾過し、滅菌した2Lの容器に取得し、密封滅菌させた後、冷蔵保管して本発明の健康飲料組成物の製造に使用する。
【0121】
前記組成比は、比較的嗜好飲料に適した成分を好適な実施例として混合組成したが、需要階層や需要国家、使用用途などの地域的、民族的嗜好度に応じてその配合比を任意に変形して実施しても構わない。
以上、本発明の内容の特定部分について詳細に記述したが、このような具体的記述は好適な実施様態に過ぎないものであり、これらの実施様態に本発明の範囲が限定されないのは当該分野における通常の知識を有する者には明白なことであろう。よって、本発明の実質的な範囲は添付された請求の範囲とそれらの等価物によって定められるべきであるといえる。
【特許請求の範囲】
【請求項1】
(1)人参からメタノール抽出物を製造する段階と、
(2)前記メタノール抽出物を水とn−ブタノールの混合溶媒で分画する段階と、
(3)前記水分画物とn−ブタノール分画物のうち、n−ブタノール分画物に対して、溶出溶媒としてクロロメタン:メタノール:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離する段階と、
(4)前記8個の分画物のうち、CaCC(Ca2+activated Chloride Channel)に対する活性が最も高い第7の分画物に対して、溶出溶媒としてエタノール:酢酸エチル:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、2つの分画に分離する段階と、
(5)前記2つの分画物のうち、CaCC活性が高い分画物IIを透析して最終生成物を得る段階とを含んでなる、新規な糖リポタンパク質ジントニンの製造方法。
【請求項2】
前記(5)段階の後、
(6)最終分画物をNaClが含有されたpH7.2のリン酸塩緩衝食塩水(PBS)溶液に溶解し、陰イオン交換クロマトグラフィー及びゲル濾過クロマトグラフィーを行い、2つの分画に分離する段階と、
(7)前記2つの分画物に対して、それぞれ、NaClが含有されたpH8.2のTris−HClとNaClが含有されたpH7.2のリン酸塩緩衝食塩水(PBS)溶液で陰イオン交換クロマトグラフィーを行い、個別糖リポタンパク質を分離する段階とをさらに含む、請求項1に記載の個別ジントニンE1、E2、E3、L1、L2及びL3の製造方法。
【請求項3】
前記人参は紅参、水参、白参、栽培参、長脳参及び山参よりなる群から選ばれることを特徴とする、請求項1に記載の新規な糖リポタンパク質ジントニンの製造方法。
【請求項4】
請求項2の方法で製造され、分子量67kDa及び見掛け分子量13kDaの五量体であることを特徴とする、人参から分離同定した新規な糖リポタンパク質ジントニンE1、E2、E3、L1、L2及びL3。
【請求項5】
前記糖リポタンパク質は、アミノ酸組成成分として、システインとシスチン(cysteine and cystine)、アスパラギンとアスパラギン酸(asparagine and aspartic acid)、グルタミンとグルタミン酸(glutamine and glutamic acid)、セリン(serine)、グリシン(glycine)、アルギニン(arginine)、トレオニン(threonine)、アラニン(alanine)、プロリン(proline)、バリン(valine)、イソロイシン(isoleucine)、ロイシン(leucine)、フェニルアラニン(phenylalanine)、トリプトファン(tryptophan)及びリシン(lysine)を含むことを特徴とする、請求項4に記載の人参から分離同定した新規な糖リポタンパク質ジントニン。
【請求項6】
前記糖リポタンパク質は、炭水化物組成成分として、ラムノース(rhamnose)、アラビノース(arabinose)、グルコース(glucose)、マンノース(mannose)、キシロース(xylose)及びグルコサミン(glucosamine)を含むことを特徴とする、請求項4に記載の人参から分離同定した新規な糖リポタンパク質ジントニン。
【請求項7】
前記糖リポタンパク質は、脂質組成成分として、リノール酸(linoleic acid)、パルミチン酸(palmitic acid)、オレイン酸(oleic acid)及びステアリン酸(stearic acid)を含むことを特徴とする、請求項4に記載の人参から分離同定した新規な糖リポタンパク質。
【請求項8】
請求項4の糖リポタンパク質を有効成分として含む、精神分裂(Schizophrenia)、アルツハイマー病(Alzheimer’s disease)、ハンチントン病(Huntington’s disease)、家族性片麻痺性片頭痛(Familial hemiplegic migraine)、癲癇(epilepsy)、一過性運動失調症(episodic ataxia)、脊髄小脳失調症(spinocerebellar ataxias)及びカルシウム不足による成長阻害の中から選ばれる、カルシウム濃度の低下に起因する疾患の予防及び治療のための薬学組成物。
【請求項9】
請求項4の糖リポタンパク質及び食品学的に許容される食品補助添加剤を含む、健康機能食品。
【請求項1】
(1)人参からメタノール抽出物を製造する段階と、
(2)前記メタノール抽出物を水とn−ブタノールの混合溶媒で分画する段階と、
(3)前記水分画物とn−ブタノール分画物のうち、n−ブタノール分画物に対して、溶出溶媒としてクロロメタン:メタノール:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、8個の分画に分離する段階と、
(4)前記8個の分画物のうち、CaCC(Ca2+activated Chloride Channel)に対する活性が最も高い第7の分画物に対して、溶出溶媒としてエタノール:酢酸エチル:水の混合溶媒を用いてシリカゲルカラムクロマトグラフィーを行い、2つの分画に分離する段階と、
(5)前記2つの分画物のうち、CaCC活性が高い分画物IIを透析して最終生成物を得る段階とを含んでなる、新規な糖リポタンパク質ジントニンの製造方法。
【請求項2】
前記(5)段階の後、
(6)最終分画物をNaClが含有されたpH7.2のリン酸塩緩衝食塩水(PBS)溶液に溶解し、陰イオン交換クロマトグラフィー及びゲル濾過クロマトグラフィーを行い、2つの分画に分離する段階と、
(7)前記2つの分画物に対して、それぞれ、NaClが含有されたpH8.2のTris−HClとNaClが含有されたpH7.2のリン酸塩緩衝食塩水(PBS)溶液で陰イオン交換クロマトグラフィーを行い、個別糖リポタンパク質を分離する段階とをさらに含む、請求項1に記載の個別ジントニンE1、E2、E3、L1、L2及びL3の製造方法。
【請求項3】
前記人参は紅参、水参、白参、栽培参、長脳参及び山参よりなる群から選ばれることを特徴とする、請求項1に記載の新規な糖リポタンパク質ジントニンの製造方法。
【請求項4】
請求項2の方法で製造され、分子量67kDa及び見掛け分子量13kDaの五量体であることを特徴とする、人参から分離同定した新規な糖リポタンパク質ジントニンE1、E2、E3、L1、L2及びL3。
【請求項5】
前記糖リポタンパク質は、アミノ酸組成成分として、システインとシスチン(cysteine and cystine)、アスパラギンとアスパラギン酸(asparagine and aspartic acid)、グルタミンとグルタミン酸(glutamine and glutamic acid)、セリン(serine)、グリシン(glycine)、アルギニン(arginine)、トレオニン(threonine)、アラニン(alanine)、プロリン(proline)、バリン(valine)、イソロイシン(isoleucine)、ロイシン(leucine)、フェニルアラニン(phenylalanine)、トリプトファン(tryptophan)及びリシン(lysine)を含むことを特徴とする、請求項4に記載の人参から分離同定した新規な糖リポタンパク質ジントニン。
【請求項6】
前記糖リポタンパク質は、炭水化物組成成分として、ラムノース(rhamnose)、アラビノース(arabinose)、グルコース(glucose)、マンノース(mannose)、キシロース(xylose)及びグルコサミン(glucosamine)を含むことを特徴とする、請求項4に記載の人参から分離同定した新規な糖リポタンパク質ジントニン。
【請求項7】
前記糖リポタンパク質は、脂質組成成分として、リノール酸(linoleic acid)、パルミチン酸(palmitic acid)、オレイン酸(oleic acid)及びステアリン酸(stearic acid)を含むことを特徴とする、請求項4に記載の人参から分離同定した新規な糖リポタンパク質。
【請求項8】
請求項4の糖リポタンパク質を有効成分として含む、精神分裂(Schizophrenia)、アルツハイマー病(Alzheimer’s disease)、ハンチントン病(Huntington’s disease)、家族性片麻痺性片頭痛(Familial hemiplegic migraine)、癲癇(epilepsy)、一過性運動失調症(episodic ataxia)、脊髄小脳失調症(spinocerebellar ataxias)及びカルシウム不足による成長阻害の中から選ばれる、カルシウム濃度の低下に起因する疾患の予防及び治療のための薬学組成物。
【請求項9】
請求項4の糖リポタンパク質及び食品学的に許容される食品補助添加剤を含む、健康機能食品。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2013−502409(P2013−502409A)
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−525469(P2012−525469)
【出願日】平成22年8月12日(2010.8.12)
【国際出願番号】PCT/KR2010/005304
【国際公開番号】WO2011/062354
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(509080082)コングック ユニヴァーシティ インダストリアル コーペレーション コーポレーション (4)
【Fターム(参考)】
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願日】平成22年8月12日(2010.8.12)
【国際出願番号】PCT/KR2010/005304
【国際公開番号】WO2011/062354
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(509080082)コングック ユニヴァーシティ インダストリアル コーペレーション コーポレーション (4)
【Fターム(参考)】
[ Back to top ]