
人工ペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質
本発明は、少なくとも1つの酵素活性ドメインおよび少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性および細菌溶解活性を有する組換えポリペプチドに関する。本発明はさらに、少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性を有する組換えポリペプチドに関する。さらに、本発明は、組換えポリペプチドをコードするヌクレオチド配列を含む核酸分子、ベクター、および宿主細胞に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、少なくとも1つの酵素活性ドメインおよび少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性および細菌溶解活性を有する組換えポリペプチドに関する。本発明はさらに、少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性を有する組換えポリペプチドに関する。さらに、本発明は、組換えポリペプチドをコードするヌクレオチド配列を含む核酸分子、ベクター、および宿主細胞に関する。
【背景技術】
【0002】
近年、バクテリオファージエンドリシンなどのペプチドグリカン分解酵素が、抗菌剤としてますます注目を受けている。古典的な抗生物質に対する病原性細菌の抵抗性が出現しかつ蔓延することを考慮して、これらの生物を制御する代替的な方法への要求が大きくなっている。特にグラム陽性菌の場合、いわゆるエンザイバイオティクス(enzybiotics)のようなファージエンドリシンの適用が将来有望なアプローチである。グラム陽性においては外膜の欠如のために、これらの酵素がエキソリシンとしても働き、すなわち感受性細胞の溶解を外部から引き起こすことができる。この特性は、リステリア(Listeria)に感染するファージ由来のエンドリシンについて示されたように、例えば、分子生物学において細菌細胞からの核酸およびタンパク質の効率的な回収のために活用することができる。食物由来の病原体であるリステリア・モノサイトゲネス(Listeria monocytogenes)の制御のための適用を目指し、これらのリシンをコードする遺伝子を、チーズ生産においてスターター生物として使用されるラクトコッカス・ラクティス(Lactococcus lactis)およびいくつかの乳酸桿菌を含む多数の乳酸菌中に導入した。これらの細菌は、エンドリシンを過剰発現および分泌し、その結果L.モノサイトゲネス細胞に対して溶解活性を示した。これまでに報告されたファージにコードされるペプチドグリカンヒドロラーゼの医学的適用は、バチルス・アンスラシス(Bacillus anthracis)の検出および殺傷、ならびにインビトロおよびマウスモデルにおけるストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)の制御を含む。
【0003】
エンドリシンは、dsDNAファージの後期遺伝子領域にコードされ、および溶解性増殖サイクルの終わりに産生される細胞壁溶解酵素である。同様の酵素はまた、細菌ゲノム中に組み込まれたプロファージゲノム内でも見出される。それらの機能は内側からの細菌ペプチドグリカンの分解であり、宿主細胞の溶解およびファージ子孫の放出をもたらす。ペプチドグリカン内の異なる標的結合に従い、エンドリシンは5つの異なるクラスに分類することができる:両方ともグリコシダーゼであり、かつグリカン鎖の2つのβ-1,4-グリコシド結合の1つをそれぞれ切断する(i)N-アセチル-β-D-ムラミダーゼ(リゾチームとしても公知である)および(ii)N-アセチル-β-D-グルコサミニダーゼ;(iii)ムラミダーゼと同様の結合を切断するが、異なる機構による溶解性トランスグリコシラーゼ;(iv)グリカンとペプチド部分との間を切るN-アセチルムラモイル-L-アラニンアミダーゼ;および(v)ペプチド部分内部を切断するエンドペプチダーゼ。溶解性トランスグリコシラーゼを除くすべてのエンドリシンはヒドロラーゼである。同様の酵素活性はまた、それ自身の細胞壁または密接に関連した細菌の細胞壁を溶解する細菌酵素、いわゆるオートリシン、およびバクテリオシンなどの他の細菌細胞壁溶解ポリペプチドにおいても見出される。細菌オートリシンは、細胞壁リモデリング、細胞分裂、形質転換において、または病原性因子として重要な役割を果たす細胞壁溶解酵素である。それらは一緒にペプチドグリカン溶解酵素としてまとめることができる。ペプチドグリカン溶解酵素の作用によるペプチドグリカンの酵素分解は、細胞壁の完全性の損失をもたらし、かつ最終的に高い内圧により引き起こされる細胞破裂をもたらす。
【0004】
細菌細胞を殺傷する特徴によって、ペプチドグリカン溶解酵素は、ヒトおよび動物における細菌感染に対する予防剤または治療剤として、抗菌薬またはエンザイバイオティック(enzybiotic)としての使用のための、医学的環境、公的環境、もしくは私的環境における消毒剤としての使用のための、食品産業、動物飼料産業、もしくは化粧品産業における細菌汚染の除染剤として、または細菌で汚染された表面に対する一般の界面活性剤としての使用のための興味深い候補となる。ペプチドグリカン溶解酵素は、別個の細菌群、属、種、株、または血清型由来の細菌細胞に特異的に結合しかつ溶解するため、細菌の診断において同様に有用である。特異的な細胞溶解はしばしば、核酸ベースの方法または免疫学的方法などの検出される細菌細胞の細胞内容物に依拠する追加的な検出方法と併用される。
【0005】
グラム陽性バックグラウンド由来のエンドリシン、オートリシン、および他のペプチドグリカン溶解酵素は、触媒活性と基質認識とが分離され、かつ少なくとも2つの別個の機能ドメインである酵素活性ドメイン(EAD)と細胞結合ドメイン(CBD)とに局限されるモジュラー構成を示す。ファージが感染しているグラム陰性宿主由来の大抵のエンドリシンは、単一ドメインの球状タンパク質である。しかしながら、最近2つの機能ドメインからなるグラム陰性バックグラウンド由来の2つのリシンが報告された:N末端の細胞結合ドメインおよびC末端の触媒ドメインからなるシュードモナス・エルギノーサ(Pseudomonas aeruginosa)ファージΦKZおよびELのエンドリシン(Briers et al. Molecular Microbiology, Volume 65, Number 5, September 2007, pp. 1334-1344(11)(非特許文献1))。対照的に、ファージが感染しているグラム陽性細菌由来のエンドリシンの大部分は、N末端のEADおよびC末端のCBDを有して逆方向のドメインを特徴とする。
【0006】
結合ドメインにより認識される細菌細胞壁におけるリガンドが公知であるのは、非常にわずかなエンドリシンのみである。肺炎球菌ファージCpl-1リゾチームは、ストレプトコッカス・ニューモニエの細胞壁におけるテイコ酸を含有するコリンを特異的に認識し、コリン結合タンパク質(CBP)のファミリーに位置する。これらのタンパク質のコリン結合モジュール(CBM)は、4〜18個に渡る多数のタンデムコピーにおいて見出される約20アミノ酸残基のリピートにより形成される。Cpl-1は、通常短いリンカー領域を介して連結される2つの分離したドメイン(EADおよびCBD)を有するこれらの酵素のモジュラー設計を例証する。モジュラーファージエンドリシンの大部分は1つの触媒ドメインおよび1つの基質結合ドメインからなるが、例えば、ストレプトコッカス・アガラクティエ(Streptococcus agalactiae)ファージB30のエンドリシン(ムラミダーゼおよびエンドペプチダーゼ)、スタフィロコッカス・アウレウス(Staphylococcus aureus)ファージΦ11のエンドリシン(エンドペプチダーゼおよびアミダーゼ)、ストレプトコッカス・アガラクティエファージNCTC 11261のエンドリシン(エンドペプチダーゼおよびムラミダーゼ)、ならびにスタフィロコッカス・ワーネリ(Staphylococcus warneri)MファージΦWMYのエンドリシン(エンドペプチダーゼおよびアミダーゼ)のように2つの異なる酵素活性を有する多数のタンパク質が存在する。
【0007】
ファージおよび細菌起源のペプチドグリカン溶解酵素、エンドリシン、ならびにオートリシンまたはバクテリオリシンが同様のモジュラー構築を示すという事実、ならびに細菌およびファージ由来溶解タンパク質の別個のドメインの間に高い相同性が見出されるという事実は、機能ドメインの相互交換によるこれらのタンパク質の共通の祖先および共進化を示唆する(Garcia et al., 1990, Gene 86, 81-88(非特許文献2))。Diazら(1990, Proc. Natl. Acad. Sci., 87, 8125-8129(非特許文献3))は、組み合わされた生化学特性を示すファージおよび細菌性肺炎球菌酵素のキメラを作製した。ヌクレオチド相同性を欠損する遺伝子由来の組換えキメラがDiazら(1991, J. Biol. Chem., 266, 5464-6571(非特許文献4))において構築され、基質認識におけるCBDの機能も確認された。Crouxら(1993, Mol. Microbiol., 9, 1019-1025(非特許文献5))は、他の細菌ファミリー由来の細胞に対するエンドリシンの酵素活性において切り換えを導く、肺炎球菌およびクロストリジウム細胞壁溶解酵素に基づくキメラまでも作製した。Sanzら(1996, Eur. J. Biochem., 235, 601-605(非特許文献6))は、2つのEADおよび1つのCBDを含むモジュール組み立てにより多機能肺炎球菌ムレインヒドロラーゼを構築した。最近、リゾスタフィン、スタフィロコッカス・シミュランス(Staphylococcus simulans)由来のペプチドグリカンヒドロラーゼ、およびストレプトコッカス・アガラクティエファージB30エンドリシン、ならびにそのC末端が切断されたバーションからなる融合タンパク質が報告された(Donovan et al. 2006, Appl. Environ. Microbiol., 72, 2988-2996(非特許文献7))。この場合にもまた、人工構築物は、スタフィロコッカス細胞およびストレプトコッカス細胞の両方を溶解する、両方の酵素の特性を併せ持った。Loessnerら(2002, Mol. Microbiol., 44, 335-349(非特許文献8))は、リステリア・モノサイトゲネスを役割モデルとして使用し、細菌細胞壁炭水化物への特異的認識および高親和性結合を決定するCBDの概念を記述した。米国特許第2004/0197833号(特許文献1)は、標的細胞の濃縮のための方法における固定化された単離CBDの使用を教示する。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第2004/0197833号
【非特許文献】
【0009】
【非特許文献1】Briers et al. Molecular Microbiology, Volume 65, Number 5, September 2007, pp. 1334-1344(11)
【非特許文献2】Garcia et al., 1990, Gene 86, 81-88
【非特許文献3】Diaz et al., 1990, Proc. Natl. Acad. Sci., 87, 8125-8129
【非特許文献4】Diaz et al., 1991, J. Biol. Chem., 266, 5464-6571
【非特許文献5】Croux et al., 1993, Mol. Microbiol., 9, 1019-1025
【非特許文献6】Sanz et al., 1996, Eur. J. Biochem., 235, 601-605
【非特許文献7】Donovan et al., 2006, Appl. Environ. Microbiol., 72, 2988-2996
【非特許文献8】Loessner et al., 2002, Mol. Microbiol., 44, 335-349
【発明の概要】
【0010】
本発明の目的は、細菌細胞の信頼のおける検出および濃縮、および/または溶解を可能にする、改善されたおよび有利なタンパク質を提供することである。
【0011】
前記目的は、添付の特許請求の範囲において規定されるような内容により解決される。
【図面の簡単な説明】
【0012】
以下の図面は本発明を例証する。
【図1】リステリア細胞に対するGFP-ダブルCBD融合タンパク質、および参照として働くGFP-シングルCBD構築物の模式図。GFP=緑色蛍光タンパク質;CBD500、CBD118、CBDP35=それぞれリステリアファージエンドリシンPly500、Ply118、およびPlyP35の細胞壁結合ドメイン;L=PlyPSAエンドリシンのリンカー領域。
【図2】細胞壁からの解離が減少したためにより高い親和性をもたらす、重複したCBDを有するペプチドグリカン結合タンパク質。(A)ダブルCBD500融合タンパク質、および参照として役立つそれぞれのシングルCBD500構築物の模式図。GFP=緑色蛍光タンパク質;EAD500=リステリアファージエンドリシンPly500の酵素活性ドメイン;CBD500=Ply500の細胞壁結合ドメイン。(B)50 nMの濃度で測定したHGFP_CBD500(黒色)およびHGFP_CBD500-500(灰色)のSPRセンサグラムのオーバーレイ。結合および解離相を示す。RU=相対応答単位。
【図3】様々なNaCl濃度で測光溶解アッセイを用いて測定した、N末端Hisタグを有する野性型ply500(白丸)およびH_EAD_CBD500-500(黒四角)のリステリア・モノサイトゲネスWSLC1042の細胞に対する相対溶解活性。200 mM NaClでの野生型ply500の最適活性が1.0に相当する。すべてのアッセイは3つ組で行った。
【図4】腸球菌に対するペプチドグリカン溶解酵素の最小殺菌濃度(MBC)の測定。エンテロコッカス・フェカリス(Enterococcus faecalis)株17の生存細胞の細菌濃度を、細胞溶解アッセイに存在するFab25VL(四角)またはEADFab25_CBD25_CBD20(丸)のタンパク質濃度に依存して示す。
【発明を実施するための形態】
【0013】
本明細書において使用される「ペプチドグリカン溶解酵素」という用語は、細菌細胞壁を溶解するのに適当である酵素を指す。酵素は、ペプチドグリカン溶解酵素の「酵素活性ドメイン」(EAD)が構成される以下の活性の少なくとも1つを含む:エンドペプチダーゼ、N-アセチル-ムラモイル-L-アラニン-アミダーゼ(アミダーゼ)、N-アセチル-ムラミダーゼ(リゾチームもしくは溶解性トランスグリコシラーゼ)、またはN-アセチル-グルコサミニダーゼ。酵素は、ファージもしくはプロファージにコードされる、いわゆる「エンドリシン」であるか、または、細菌によりコードされる関連した細胞壁溶解酵素、いわゆる「オートリシン」、もしくはバクテリオシンなどの他の細菌ペプチドグリカン溶解酵素、病原性因子もしくは他の抗菌ポリペプチド(例えば、リゾスタフィン、ALE-1リシン、ムタノリシン、エンテロリシン)に由来するかのいずれかである。加えて、ペプチドグリカン溶解酵素はまた、酵素的に不活性であり、および宿主細菌の細胞壁に結合する領域、いわゆるCBD(細胞壁結合ドメイン)も含有する。
【0014】
本明細書において使用される「ペプチドグリカン結合タンパク質」という用語は、ペプチドグリカン溶解酵素について記述される酵素活性のいずれも有さない人工的に構築された細菌細胞結合タンパク質を指す。ペプチドグリカン結合タンパク質は、CBDに由来する1つより多いCBDを含む。ペプチドグリカン結合タンパク質は、天然に存在するCBDのシャッフリングにより、および/または天然に存在するCBDの増倍により構築される。
【0015】
本明細書において使用される「ドメイン」という用語は、特定の機能を与えられ、および構造的または進化的に保存されたドメインともまた一致し得る、ペプチドグリカン溶解酵素のサブユニットを指す。ドメインに関連する特定の機能は、例えば細菌ペプチドグリカン溶解または細菌細胞結合である。機能ドメインは「モジュール」と呼ばれることもある。
【0016】
本明細書において使用される「CBD」という用語は、しばしばタンパク質のC末端に見出されるペプチドグリカン溶解酵素の細胞壁結合ドメインを指す。CBDドメインは、細胞壁を加水分解する点では酵素活性を有さないが、細菌細胞壁へのペプチドグリカン溶解酵素の結合を媒介する。本明細書において使用されるCBDという用語は、天然に存在するペプチドグリカン溶解酵素に由来するポリペプチド鎖内のセグメントを説明する。
【0017】
本明細書において使用される「EAD」という用語は、細菌ペプチドグリカンの加水分解を担うペプチドグリカン溶解酵素の酵素活性ドメインを指す。これは、ペプチドグリカン溶解酵素について記述される酵素活性の少なくとも1つを含有する。本明細書において使用されるEADという用語は、天然に存在するペプチドグリカン溶解酵素に由来するポリペプチド鎖内のセグメントを説明する。
【0018】
「CHAP」ドメイン(システイン、ヒスチジン依存性アミドヒドロラーゼ/ペプチダーゼ(cysteine, histidine-dependent amidohydrolases/peptidases))は、細菌、バクテリオファージ、古細菌、およびトリパノソーマ(Trypanosomidae)科の真核生物由来のタンパク質において見出される約110〜約140アミノ酸残基の領域である。タンパク質はペプチドグリカン加水分解において主に機能する可能性がある。CHAPドメインは一般に、細菌タイプのSH3ドメインおよびアミダーゼドメインのいくつかのファミリーと結合する。CHAPドメインを含有するタンパク質は、求核攻撃機構において触媒性システイン残基を利用する可能性がある。CHAPドメインは2つの不変アミノ酸残基、システインおよびヒスチジン残基を含有する。これらの残基は、CHAPドメインを含有するタンパク質の推定上の活性部位の一部を形成する。
【0019】
本明細書において使用される「ami」という用語は、アミダーゼ活性を示す酵素的に定義されたドメインを説明し、すなわち、ペプチドグリカン骨格におけるN-アセチルムラミンと、ペプチドリンカーにおいて通常L-alaである隣接するアミノ酸残基との間のアミド結合を加水分解する。アミダーゼはしばしば、活性について金属イオン依存性である。
【0020】
本明細書において使用される、Src相同性3ドメインと呼ばれることもある「SH3」ドメインという用語は、他の結合パートナーと相互作用するタンパク質について特徴的である約60アミノ酸残基の小さな非触媒性タンパク質ドメインを説明する。それはプロリンに富むコンセンサスモチーフにより同定される。SH3ドメインはCBD内に位置する。ペプチドグリカン溶解酵素において見出されるSH3ドメインは、しばしばSH3bまたはSH3_5タイプのものである。
【0021】
「野生型」という用語は、配列に関してタンパク質または核酸の天然に存在する形態を指す。
【0022】
本明細書において使用される「シャッフリング」という用語は、新たなキメラポリペプチド構築物中への、異なる野生型酵素由来のポリペプチドの異なる断片の組み合わせを指す。本文脈において、酵素は好ましくはペプチドグリカン溶解酵素であり、および断片は好ましくはEADおよびCBDである。通常、断片は、分子生物学的方法により核酸レベルで組み合わされる。追加的なリンカー配列が、構造上またはクローニングの理由のために断片の間に導入されてもよい。
【0023】
本発明の1つの対象は、少なくとも1つのEADおよび少なくとも2つのCBDから構成されるペプチドグリカン溶解酵素を指す。本発明に係る人工的に作製されたペプチドグリカン溶解酵素は、天然に存在するタンパク質と比較して拡大したかもしくは変化した結合範囲、または細菌細胞壁への増加した結合親和性、または増加したかもしくは変化した溶解活性、またはそれらの組み合わせなどの新たな特性を示す。
【0024】
本発明の別の対象は、少なくとも2つのCBDから構成されるペプチドグリカン結合タンパク質を指す。本発明に係る人工的に作製されたペプチドグリカン結合タンパク質は、天然に存在するタンパク質と比較して拡大したかもしくは変化した結合範囲、または細菌細胞壁への増加した結合親和性、または両方などの新たな特性を示す。
【0025】
本発明に係るペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質において、少なくとも2つのCBDは、2つの異なるペプチドグリカン溶解酵素に由来してもよいし(ドメインシャッフリング)、または、エンドリシンにおいて天然に存在する1つのCBDの増倍によってもよい。1つより多いEADが本発明に係るペプチドグリカン溶解酵素に存在する場合、EADは2つの異なるペプチドグリカン溶解酵素に由来してもよい。
【0026】
一方、グラム陽性およびグラム陰性菌の様々な属、種、または株に対する多数のペプチドグリカン溶解タンパク質が当技術分野において記述されている。ペプチドグリカン溶解タンパク質のモジュラー性質、およびEADとCBDとの間の区別は周知である。ペプチドグリカン溶解タンパク質に存在する多数の保存されたドメインは、機能的に特徴決定されており、および、ポリペプチドまたはヌクレオチド配列内のそれらの存在は、それぞれのタンパク質または核酸データベース、例えば、CDD(Marchler-Bauer et al., 2005; Nucleic Acids Research, 33, D192-D196);Pfam(Finn et al., 2006, Nucleic Acids Research 34, D247-D251)またはSMART(Schultz et al., 1998, Proc. Natl. Acad. Sci. USA 95, 5857-5864、Letunic et al., 2006, Nucleic Acids Res 34, D257-D260)を使用する適切なコンピュータープログラムにより、または欠失変異体を用いた結合アッセイ(Loessner et al., 2002, Mol. Microbiol., 44, 335-349)により予測することができる。本発明に係る人工ペプチドグリカン溶解酵素は、Sambrookら(Molecular cloning. A laboratory manual; 2nd ed. Cold Spring Harbor Laboratory Press 1989)において記述されているような組換えタンパク質のクローニングおよび産生のための標準的な技術を用いて、EADに由来する所望の酵素活性を細胞結合活性のための少なくとも2つのCBDと組み合わせることにより構築される。本発明に係る人工ペプチドグリカン結合タンパク質は、Sambrookら(Molecular cloning. A laboratory manual; 2nd ed. Cold Spring Harbor Laboratory Press 1989)において記述されているような組換えタンパク質のクローニングおよび産生のための標準的な技術を用いて、細胞結合活性のための少なくとも2つのCBDを組み合わせることにより構築される。少なくとも2つのCBDは、シャッフルされたキメラ酵素をもたらす異なるペプチドグリカン溶解酵素に由来していてもよく、または1つの天然に存在する酵素由来のCBDを増倍してもよく、または両方の組み合わせであってもよい。主として、すべての天然に存在するペプチドグリカン溶解酵素が、EADおよびCBDドメインの供給のための潜在的候補である。
【0027】
ペプチドグリカン溶解酵素は好ましくは、アミダーゼ_5(バクテリオファージペプチドグリカンヒドロラーゼ、pfam05382)、アミダーゼ_2(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01510)、アミダーゼ_3(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01520)、Transgly(トランスグリコシラーゼ、pfam00912)、ペプチダーゼ_M23(ペプチダーゼファミリーM23、pfam01551)、エンドリシン_オートリシン(CD00737)、ヒドロラーゼ_2(細胞壁ヒドロラーゼ、pfam07486)、CHAP(アミダーゼ、pfam05257)、トランスグリコシラーゼ(トランスグリコシラーゼ様ドメイン、pfam06737)、MtlB(膜結合型溶解性ムレイントランスグリコシラーゼB、COG2951)、MtlA(膜結合型溶解性ムレイントランスグリコシラーゼA、COG2821)、MtlE(膜結合型溶解性ムレイントランスグリコシラーゼE、COG0741)、バクテリオファージ_λ_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、CD00736)、ペプチダーゼ_M74(ペニシリン非感受性ムレインエンドペプチダーゼ、pfam03411)、SLT(トランスグリコシラーゼSLT、pfam01464)、Lys(C型リゾチーム/α-ラクトアルブミンファミリー、pfam00062)、COG5632(N-アセチルムラモイル-L-アラニンアミダーゼ、COG5632)、MepA(ムレインエンドペプチダーゼ、COG3770)、COG1215(グリコシルトランスフェラーゼ、COG1215)、AmiC(N-アセチルムラモイル-L-アラニンアミダーゼ、COG0860)、Spr(細胞壁結合型ヒドロラーゼ、COG0791)、バクテリオファージ_T4様_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、cd00735)、LT_GEWL(溶解性トランスグリコシラーゼ(LT)およびガチョウ卵白リゾチーム(GEWL)ドメイン、cd00254)、ペプチダーゼ_S66(LD-カルボキシペプチダーゼ、pfam02016)、グリコ_ヒドロ_70(グリコシルヒドロラーゼファミリー70、pfam02324)、グリコ_ヒドロ_25(グリコシルヒドロラーゼファミリー25)、VanY(D-アラニル-D-アラニンカルボキシペプチダーゼ、pfam02557)、およびLYZ2(リゾチームサブファミリー2、smart 00047)から構成される群より選択される少なくとも1つのEADを含む。
【0028】
ペプチドグリカン溶解酵素は好ましくは、SH3_5(細菌SH3ドメイン、pfam08460)、SH3_4(細菌SH3ドメイン、pfam06347)、SH3_3(細菌SH3ドメイン、pfam08239)、SH3b(細菌SH3ドメインホモログ、smart00287)、LysM(細胞壁分解に関与する種々の酵素において見出されるLysMドメイン、pfam01476およびcd00118)、PG_結合_1(推定上のペプチドグリカン結合ドメイン、pfam01471)、PG_結合_2(推定上のペプチドグリカン結合ドメイン、pfam08823)、MtlA(ムレイン分解トランスグリコシラーゼ由来のペプチドグリカン結合ドメイン、pfam03462)、Cpl-7(Cpl-7リゾチームのC末端ドメイン、pfam08230)、CW_結合_1(推定上の細胞壁結合リピート、pfam01473)、LytB(推定上の細胞壁結合ドメイン、COG2247)、およびLytE(LysMリピート、COG1388)から構成される群より選択される少なくとも1つのCBDを含む。
【0029】
好ましくは、上述のドメインは、約15〜約250アミノ酸残基の範囲のアミノ酸残基長を有し、約20〜約200アミノ酸残基の範囲の長さが好ましい。例として、約15〜約40アミノ酸残基長のドメインが、LysMドメインまたはコリン結合を担うCW_結合_1モチーフなどのペプチドグリカン結合ドメインにおいて見出される。これらの小さなドメインは、野生型細胞壁溶解酵素においても天然の反復モチーフとしてしばしば見出される。これらのドメインは、シャッフルされたキメラの人工ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質を作製するために、他の細胞壁溶解酵素由来の追加的なCBDと組み合わせることができる。
【0030】
通常、ペプチドグリカン溶解酵素の完全なEADまたはCBDドメインは、上述の保存されたドメインより大きい。優先的に、EADまたはCBDは、約50残基〜約400残基長の範囲である。各EADおよびCBDは、ペプチドグリカン溶解または細菌細胞結合の機能を示すために少なくとも1つの機能ドメインを含有するが、1つより多い機能ドメインおよび未知の機能を有する追加的な配列セグメントもまた含むことができる。ペプチドグリカン結合酵素のEADおよびCBDドメインは、上述の保存されたドメインによって常に規定されるわけではない。上述の保存されたドメインのいずれも見出されないが細菌細胞に結合しかつ細菌細胞を溶解する公知のペプチドグリカン結合酵素(例えばPly118)もまた存在する。潜在的ドメインがEADまたはCBDとして機能するか否かは、適切な機能アッセイ(例えば、ペプチドグリカン溶解(EAD)については測光溶解アッセイ、プレート溶解アッセイ、または最小殺菌濃度(MBC)の測定、および細胞結合(CBD)については細胞結合アッセイ、蛍光顕微鏡法、または結合親和性の測定)を用いて試験することができる。EADおよびCBDのドメインの境界は、配列の間の局所的類似性の領域を見出す局所的アラインメント検索手段(例えば、NCBIのBLAST、Altschul et al., 1997, Nucleic Acids Res. 17, 3389-3402)により規定することができる。加えて、多数のペプチドグリカン溶解酵素が、それらのEADおよびCBDドメインに関して既に記述されている。
【0031】
好ましくは、本発明のペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質は、Ply500、Ply511、Ply118、Ply100、PlyP40、Ply3626、phiLM4エンドリシン、PlyCD119、PlyPSAa、Ply21、PlyBA、Ply12、PlyP35、PlyPH、PlyL、PlyB、phi11エンドリシン、phi MR11エンドリシン、phi12エンドリシン、S.アウレウスファージPVLアミダーゼ、plypitti26、ΦSA2usaエンドリシン、スタフィロコッカス・ワーネリMファージΦWMY PlyGBSのエンドリシン、B30エンドリシン、Cpl-1、Cpl-7、Cpl-9、PlyG、PlyC、palアミダーゼ、Fab25、Fab20、エンテロコッカス・フェカリスV583プロファージ由来のエンドリシン、リゾスタフィン、ファージPL-1アミダーゼ、S.カピティス(S. capitis)ALE-1エンドペプチダーゼ、ムタノリシン(ストレプトマイセス・グロビスポラス(Streptomyces globisporus)ATCC 21553のN-アセチルムラミダーゼ)、エンテロリシンA(エンテロコッカス・フェカリスLMG 2333由来の細胞壁分解バクテリオシン)、LysK、LytM、L.モノサイトゲネス由来のAmiオートリシン、シュードモナス・エルギノーサファージΦKZおよびELのエンドリシン、T4リゾチーム、gp61ムラミダーゼ、およびSTM0016ムラミダーゼからなる群より選択される野生型ペプチドグリカン溶解酵素に由来するEADおよびCBDから構成される。
【0032】
野生型ペプチドグリカン溶解酵素PlyP40は、その野生型形態において344アミノ酸残基の長さを有する。それは、他の公知のエンドリシンと最小の相同性のみを有する2つの機能ドメインを所有する。1〜200位のN末端アミノ酸残基は、SEQ ID NO: 103に示される酵素活性ドメイン(EAD)に相当する。PlyP40の細胞結合ドメイン(CBD)は、SEQ ID NO: 104に示されるC末端に位置する227〜344のアミノ酸残基を含む。従って、野生型ペプチドグリカン溶解酵素PlyP40に由来するEADは、好ましくはSEQ ID NO: 103記載のアミノ酸配列を含み、一方野生型ペプチドグリカン溶解酵素PlyP40に由来するCBDは、好ましくはSEQ ID NO: 104記載のアミノ酸配列を含む。
【0033】
本発明に係る酵素およびタンパク質を構築するための天然に存在するペプチドグリカン溶解酵素由来の断片は、上述のような保存された機能ドメインの予測より決定された単なる配列セグメントを併せ持たなくてもよいが、好ましくは異なる機能モジュールを連結する適切なリンカー配列を付加する。リンカー配列は、規定された機能ドメインの近隣の野生型配列に由来してもよく、または当技術分野より公知である外部の適切なリンカー配列であってもよい。適切なリンカーは例えば、x線構造より定義されたリステリアエンドリシンPlyPSA(Korndorfer et al., 2006, J. Mol. Biol., 364, 678-689)由来の配列AAKNPNまたはTGKTVAAKNPNRHS(SEQ ID No: 61および11)を有する短いドメインリンカーである。ポリグリシンリンカーもまた、柔軟なドメインリンカーとして働くことが当技術分野において公知である。好ましいリンカーはまた、グリシンおよびアラニンに富むリンカーである。グリシンおよびアラニンに富むリンカーについての特定の配列は、SEQ ID NO:63、64、および65として示される。例えばエンテロリシンAにおいて、天然のリンカーとして生じるプロリンおよびスレオニンに富む配列もまた好ましい(SEQ ID NO:66)。プロリンおよびスレオニンに富むリンカー配列は、コンセンサスモチーフ(PT)xPまたは(PT)xT(式中、xは1〜10の範囲の整数を意味する)により記述することができる。別のリンカーの可能性は、Crouxら(1993, Molec. Microbiol., 9, 1019-1025)において記述されているEADとCBDとの間のいわゆる「ジャンクションゾーン」である。当業者は、野生型酵素から取り出される機能ドメインについて適切な境界を予測するためのいくつかの方法、例えば、2次構造予測、ドメインリンカーの予測、タンパク質の高解像度X線およびNMR構造におけるタンパク質の3Dモデルの検査、またはドメインリンカーおよび境界の検査を承知している。適切な方法は、例えば、Garnier et al., 1996, Methods in Enzymology 266, 540-553;Miyazaki et al., 2002, J. Struct. Funct. Genomics, 15, 37-51;George und Heringa, 2003, Protein Eng. 15, 871-879;Bae et al., 2005, Bioinformatics, 21, 2264-2270、Altschul et al., 1997, Nucleic Acids Res. 17, 3389-3402;Schwede et al., 2003, Nucleic Acids Research 31, 3381-3385. Lund et al, CPHmodels 2.0: X3M a Computer Program to Extract 3D Models. Abstract at the CASP5 conferenceA102, 2002において記述されている。EADとCBDドメインとの間、またはCBDとCBDドメインとの間のポリペプチドリンカーの長さは、約5〜約150アミノ酸残基、優先的には約6〜約60アミノ酸残基の範囲である。
【0034】
好ましくは、本発明に係るペプチドグリカン溶解酵素におけるEADおよびCBDの組み合わせのための順序は、N末端からC末端へEAD-CBD1-CBD2(-CBDN、N=3またはそれ以上)である。少なくとも2番目のEADがN末端またはC末端においてEADの隣に付加されるバリアントもまた好ましい。少なくとも2つのCBDがN末端に、または中間に位置するEADと共にN末端およびC末端に位置するバリアントもまた好ましい。加えて、両方ともN末端、C末端、または中間に位置し得るが、特にN末端で好ましいマーカー配列またはタグが含まれ得る。
【0035】
本発明に係るペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質は、それらが由来する野生型酵素と比較して新たな特性を示す。
【0036】
ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の結合範囲は、認識される細菌宿主範囲を決定する。天然に存在するペプチドグリカン酵素の大部分は、比較的狭い宿主範囲を示す。ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の技術的適用のためには、それぞれの適用に依存して増加した数の細菌株または細菌種が殺傷、捕捉、または検出され得るように、タンパク質の宿主範囲を拡大することがしばしば有利である。1つのタンパク質内に含まれる拡大した宿主範囲は、同様の適用のための2つまたはそれ以上のタンパク質の使用を回避し、タンパク質産生についての削減された費用、異なるタンパク質について条件を最適化するための削減された努力、より単純な医学的認可手続き、および削減された免疫原性の利点を有する。例えばブドウ球菌(Staphylococci)および腸球菌(Enterococci)を組み合わせる拡大した宿主範囲は、両方の属の多耐性株が増加する問題である院内感染の治療または予防において有用である。拡大した宿主範囲はまた、細菌検出、または食物から有害な細菌を除去する方法においても有用である。例えば、病原性株はリステリアのすべての血清型内で見出される。しかしながら、天然に存在するいずれのリステリアエンドリシンも、すべての血清型由来の細胞を溶解することはできない。本発明に係る1つより多いCBDを併せ持つペプチドグリカン溶解酵素は、すべての血清型を溶解することができる。天然に存在するタンパク質と比較して拡大していないが、何らかの形で変化した宿主範囲は、細菌細胞の与えられたセットを溶解、捕捉、または検出するためのテーラードタンパク質を必要とする適用に有用である可能性がある。ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の結合範囲は、当技術分野で公知のアッセイを用いて、または実施例において説明されるプレート溶解アッセイ、測光溶解アッセイ、結合アッセイ、もしくは蛍光顕微鏡法を用いて測定することができる。
【0037】
ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の野生型タンパク質と比較して増加した結合親和性は、細胞溶解、細胞捕捉、および検出などの細菌細胞の結合に依拠する任意の技術的適用のために必要とされるタンパク質の量を減少させることを助長する。これは費用を削減し、ならびに治療的適用における免疫反応および潜在的な副作用を最小化する。細菌細胞の捕捉に依拠する適用において、アッセイはバックグラウンドシグナルを減少させる洗浄段階についてより感受性が低く、インキュベーション時間を削減することができ、および検出アッセイはより感度が高い。増加した結合親和性は、当技術分野で公知のアッセイを用いて、または実施例において説明される表面プラズモン共鳴解析もしくは最小殺菌濃度の測定のためのアッセイを用いて測定することができる。
【0038】
ペプチドグリカン溶解酵素の野生型酵素と比較して増加した溶解活性は、感染の防護および治療、衛生、細菌検出における最初の段階としての細胞溶解、または食物、飼料、化粧料などからの病原性細菌の除去などの細菌細胞の溶解に依拠するすべての適用において有用である。それぞれの適用のために必要とされるタンパク質の量は、野生型タンパク質と比較して削減され、費用を削減し、ならびに治療的適用における免疫反応および潜在的な副作用を最小化する。野生型と比較して変化した溶解活性は、例えば人工酵素の異なる最適pH、または他の緩衝液組成物でのより高い溶解活性(例えば、高いイオン強度、有機溶媒の存在下における活性、特定のイオンの存在下における活性)であり得る。これはまた、血液、ヒト血清、または他の医学的試料などの特定の試料におけるより高い活性も含む。例えば酪農業において、食物製品または食物加工における中間産物はしばしば低いpH値を有するため、より低いpHに移行した人工酵素の最適pHは、例えば食品産業における人工酵素の適用にとって興味深い。高い塩濃度の下での酵素機能もまた、食品産業、例えばチーズ生産において重要である。増加したかまたは変化した溶解活性は、当技術分野で公知のアッセイを用いて、または実施例において説明されるプレート溶解アッセイ、測光溶解アッセイ、もしくは最小殺菌濃度の測定のためのアッセイを用いて測定することができる。
【0039】
1つの局面において、本発明は、リステリア細菌を溶解、捕捉、および/または検出するために使用することができる人工ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質に関する。本発明者らは、野生型酵素と比較して新たな特性を示す人工ペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質を作製するために、上述の方法を用いてリステリアエンドリシンply500(SEQ ID NO:1)、ply118(SEQ ID NO:3)、およびplyP35(SEQ ID NO:5)のドメインを組み合わせた。ply500は、EADとして、保存されたD-アラニル-D-アラニンカルボキシペプチダーゼ(VanY;pfam02557)ドメインを含む。ply500のCBDは約H133〜Q150の範囲のアミノ酸残基で始まり、かつK289で終わる。Ply118については、アミノ酸配列内に、保存されたドメインは見出されなかった。しかしながら、相同なペプチドグリカン溶解酵素との配列アラインメントより、ply118のCBDは約D90〜K180の範囲のアミノ酸残基で始まり、かつアミノ酸残基K289で終わることが導き出された。CBD118についての好ましいN末端開始アミノ酸残基は、D90、K100、G127、S151、N161、またはK180である。plyP35もまた、EADとして、保存されたD-アラニル-D-アラニンカルボキシペプチダーゼ(VanY;pfam02557)を含む。plyP35のCBDは約P130〜N156の範囲のアミノ酸残基で始まり、かつY281〜K291の範囲のアミノ酸残基で終わる。CBDP35についての好ましいN末端開始アミノ酸残基は、P130、A134、K143、およびN156である。CBDP35についての好ましいC末端アミノ酸残基は、Y281、L286、およびK291である。
【0040】
本発明に係る好ましいペプチドグリカン結合タンパク質は、追加的なリンカー配列を用いずに連結されたN末端位置のply500のCBD(アミノ酸残基H133〜K289)およびC末端位置のply118のCBD(アミノ酸残基D90〜I281)を含むCBD500-118(SEQ ID NO:7)、ならびに、ply500のCBD(アミノ酸残基Q150〜K289)およびply118のCBD(アミノ酸残基K100〜I281)を、2つのドメインを連結するリンカー(L)と共に含むCBD500L118(SEQ ID NO:9)である。この場合のドメインリンカーは、リステリアエンドリシンPlyPSAのアミノ酸残基173〜186に対応するアミノ酸配列TGKTVAAKNPNRHS(SEQ ID NO:11)を有するplyPSAリンカー領域である(Korndorfer et al., 2006, J. Mol. Biol., 364, 678-689)。
【0041】
本発明に係るさらに好ましい態様は、追加的なリンカー配列を用いずに連結されたN末端位置のply500のCBD(アミノ酸残基H133〜K289)およびC末端位置のply118のCBD(アミノ酸残基D90〜I281)を含む人工ペプチドグリカン結合タンパク質CBD118-500(SEQ ID NO:13)、ならびに、ply118のCBD(アミノ酸残基K100〜I281)およびply500のCBD(アミノ酸残基Q150〜K289)を2つのドメインを連結するリンカー(L)と共に含むCBD118L500(SEQ ID NO:15)である。この場合のドメインリンカーはplyPSAリンカー領域(SEQ ID NO:11)である。
【0042】
ペプチドグリカン結合タンパク質CBD500-118、CBD500L118、CBD118-500、およびCBD118L500はすべて、CBDドメインが由来した野生型酵素ply500およびply118と比較して、宿主範囲および結合活性に関して変化した細胞結合活性を示す。構築物CBD500L118およびCBD118L500の細胞結合活性は、2つのドメインの間のリンカーが、野生型酵素の結合特異性を併せ持つ拡大した宿主範囲を達成するのを助長することを表す。
【0043】
本発明に係るさらに好ましいペプチドグリカン結合タンパク質は、N末端位置のply500のCBD(アミノ酸残基Q150〜K289)およびC末端位置のplyP35のCBD(アミノ酸残基P130〜K291)を含むCBD500-P35(SEQ ID NO:17)、ならびに、N末端にplyP35のCBD(アミノ酸残基P130〜K291)およびC末端にply500のCBD(アミノ酸残基Q150〜K289)を含む、逆方向のCBDを有するタンパク質CBDP35-500(SEQ ID NO:19)である。この場合、plyP35のCBDについての断片がplyP35の内部ドメインリンカーを含むため、CBDは外部リンカー配列により連結されない。
【0044】
人工ペプチドグリカン結合タンパク質CBD500-P35およびCBDP35-500は両方とも、CBDドメインが由来した野生型酵素ply500およびplyP35と比較して拡大した宿主範囲を示す。両方のキメラタンパク質は、1つのタンパク質内に2つの野生型酵素の異なる結合特異性を併せ持つ。CBDの方向はこの場合相違を生じない。CBDは両方ともN末端およびC末端に位置することができる。
【0045】
本発明に係るさらに好ましいペプチドグリカン結合タンパク質は、ply500のCBD(アミノ酸残基Q150〜K289)の重複を示すCBD500-500(SEQ ID NO:21)、およびply500において天然に存在するCBDの重複を示す人工ペプチドグリカン溶解酵素EAD-CBD500-500(SEQ ID NO:23)である。
【0046】
本発明に係る両方のタンパク質は、野生型と比較してリステリア細胞に対してより高い結合親和性を示し、およびEAD-CBD500-500は加えて、ply500と比較して高い塩条件の下で増加した溶解活性を示す。
【0047】
別の局面において、本発明は、腸球菌細菌を溶解、捕捉、または検出するために使用することができる人工ペプチドグリカン溶解酵素に関する。本発明者らは、野生型酵素と比較して新たな特性を示す人工ペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質を作製するために、上述の方法を用いて腸球菌エンドリシンFab25VL(SEQ ID NO:25)およびFab20VL(SEQ ID NO:27)のドメインを組み合わせた。
【0048】
Fab25VLは、E.フェシウム(E. faecium)種由来の細菌に優先的に結合しかつ溶解するが、E.フェカリス種由来のいくつかの株にもまた結合しかつ溶解する317アミノ酸残基長のエンドリシンである。Fab25VLのN末端に位置するEAD(アミノ酸残基1〜167)は、N-アセチルムラモイル-L-アラニン-アミダーゼとして機能する保存されたアミダーゼ_2ドメインを示す。CBDはアミノ酸残基200〜317を含む。2つのドメインの間に、アミノ酸残基168〜199を含むリンカー領域が観察される。Fab20VLは、E.フェカリス種由来の細菌に優先的に結合しかつ溶解する365アミノ酸残基長のエンドリシンである。Fab20VLのN末端に位置するEAD(アミノ酸残基40〜194)もまた、N-アセチルムラモイル-L-アラニン-アミダーゼとして機能する保存されたアミダーゼ_2ドメインを示す。Fab20VLのCBD(アミノ酸残基215〜365)は、ブドウ球菌および連鎖球菌ファージ由来のペプチドグリカン溶解酵素と相同性を示す細菌SH3ドメインをC末端部分に含む。アミノ酸残基1〜19の欠失を有する、Fab20VLのN末端が切断されたバリアントであるFab20K(SEQ ID NO:29)を構築した。これはFab20VLと比較して大腸菌(E.coli)においてより良好な発現を示した。
【0049】
本発明に係る好ましいペプチドグリカン溶解酵素は、Fab25のEADおよびCBD(アミノ酸残基1〜317)を、Fab20VLのCBD(アミノ酸残基215〜365)およびFab20VLに由来する短いリンカーセグメント(アミノ酸残基200〜214)と共に組み合わせるSEQ ID NO:31である。構築物はEADFab25_CBD25_CBD20と表される。EADFab25_CBD25_CBD20は、野生型酵素と比較した新たな特徴として、拡大した宿主範囲、生細胞についての増加した溶解活性、および増加した結合活性を示す。
【0050】
本発明に係るさらに好ましいペプチドグリカン結合酵素は、ブドウ球菌細菌を検出することおよび結合することができる少なくとも2つのEADおよび少なくとも2つのCBDから構成される。
【0051】
好ましくは本発明に係るペプチドグリカン溶解タンパク質は、Hisタグ(Nieba et al., 1997, Anal. Biochem., 252, 217-228)、Strepタグ(Voss & Skerra, 1997, Protein Eng., 10, 975-982)、Aviタグ(米国特許第5,723,584号;米国特許第5,874239号)、Mycタグ(Evan et al., Mol&Cell Biol, 5, 3610-3616)、GSTタグ(Peng et al. 1993, Protein Expr. Purif., 412, 95-100)、JSタグ(国際公開公報第2008/077397号)、システインタグ(欧州特許第1399551号、SEQ ID NO:6および7)、HAタグ(アミノ酸配列EQKLISEEDL)、FLAGタグ(Hopp et al., Bio/Technology. 1988;6:1204-1210)、または当技術分野において公知である他のタグなどのタグを含む。好ましくはタグは、本発明に係るペプチドグリカン溶解タンパク質のC末端またはN末端、最も好ましくはN末端に結合される。タグは、ペプチドグリカン溶解タンパク質の発現および/または精製を促進するために、本発明に係るペプチドグリカン溶解タンパク質を表面に固定化するために、または、例えば様々なELISAアッセイフォーマットにおいて抗体結合による、ペプチドグリカン溶解タンパク質の検出用のマーカーとして働くために有用であり得る。
【0052】
好ましくは本発明に係るペプチドグリカン溶解タンパク質は、ビオチン、ストレプトアビジン、GFP(緑色蛍光タンパク質)、YFP(黄色蛍光タンパク質)、シアン蛍光タンパク質、RedStarタンパク質、または他の蛍光マーカー、アルカリホスファターゼ、西洋ワサビペルオキシダーゼ、免疫金標識、スピン標識、または当技術分野において公知である他のマーカーおよび標識などの、マーカーまたは標識部分を含む。マーカーは、それらがポリペプチド性質の場合は組換え法により、またはポリペプチド残基の化学修飾により翻訳後に付着され得る。マーカーまたは標識は、本発明に係るペプチドグリカン溶解タンパク質が診断において使用される際に、それらを検出するために特に有用である。
【0053】
さらに好ましいペプチドグリカン溶解タンパク質は、構築物HGFP-CBD118-500(SEQ ID NO:33)、HGFP-CBD500-118(SEQ ID NO:35)、HGFP-CBD118L500(SEQ ID NO:37)、HGFP-CBD500L118(SEQ ID NO:39)、HGFP-CBD500-P35(SEQ ID NO:41)、HGFP-CBDP35-500(SEQ ID NO:43)、HCBD500-GFP-CBD118(SEQ ID NO:45)、HCBD118-GFP-CBD500(SEQ ID NO:47)、HGFP-CBD500-500(SEQ ID NO:49)、およびHEAD-CBD500-500(SEQ ID NO:51)である。Hは6個のヒスチジンを含むhisタグを表し(SEQ ID NO:53)、およびGFPは蛍光マーカーとして導入された緑色蛍光タンパク質を表す(SEQ ID NO:55)。
【0054】
hisタグおよびGFPマーカーを含む上述の融合構築物のすべては、リステリア細胞結合活性を示し、ならびにペプチドグリカン溶解酵素は追加的に溶解活性を示す。しかしながら、構築物HCBD500-GFP-CBD118およびHCBD118-GFP-CBD500は、細胞結合活性、特にCBD118の細胞結合活性がこれらの構築物において減少するため、2つのCBDドメインの間での位置付けと比較してN末端位置のGFPマーカーが好ましいことを示す。
【0055】
まとめると、本発明の目的は、天然に存在するドメインのシャッフリングまたは増倍による機能ドメインの人工的組み合わせにより、野生型ペプチドグリカン溶解酵素の特性を変化および改善することである。本発明に係るペプチドグリカン溶解酵素は、天然に存在するタンパク質の結合範囲を拡大するために、および/または細菌細胞壁への結合親和性を増加するために、および/または溶解活性を増加または修飾するために、少なくとも1つのEADおよび少なくとも2つのCBDから構成される。本発明に係るペプチドグリカン結合タンパク質は、天然に存在するタンパク質の結合範囲を拡大するために、および/または細菌細胞壁への結合親和性を増加するために、少なくとも2つのCBDから構成される。本発明に係るポリペプチドにおいて、少なくとも2つのCBDは、2つの異なるペプチドグリカン溶解酵素に由来する(ドメインシャッフリング)か、または天然に存在するCBDの増倍による。
【0056】
SEQ ID NO: 7、9、13、15、17、19、21、23、31、33、35、37、39、41、43、45、47、49、51、71、73、75、77、79、81、83、85、87、89、91、93、95、97、99、101に示されるような組換えポリペプチドが好ましい。
【0057】
本発明の別の好ましい態様において、上記に列挙されたような本発明に係る組換えポリペプチドは、アミノ酸配列の修飾および/または変化を含む。そのような変化および/または修飾は、欠失、挿入および付加、置換、もしくはそれらの組み合わせなどの変異、ならびに/またはアミノ酸残基の化学変化、例えば、ビオチン化、アセチル化、ペグ化、アミノ基、SH基、もしくはカルボキシル基の化学変化を含み得る。該修飾したおよび/または変化した組換えポリペプチドは、上記に列挙されたようなそれぞれの組換えポリペプチドの単一ドメインの活性を示す。しかしながら、単一ドメインの該活性は各々、上記に列挙されたようなそれぞれの組換えポリペプチドの単一ドメインの活性より高く、またはより低くあり得る。特に、単一ドメインの該活性は、上記に列挙されたようなそれぞれの組換えポリペプチドの単一ドメインの活性の約10、20、30、40、50、60、70、80、90、100、110、120、130、140、150、160、170、180、190、200%またはそれより高くてもよい。単一ドメインの活性は、本明細書において既に説明されたようなCBDおよびEADの活性を測定するためのアッセイにより測定することができる。
【0058】
さらなる局面において、本発明は、本発明に係るポリペプチドをコードするヌクレオチド配列を含む核酸分子に関する。
【0059】
SEQ ID NO: 8、10、14、16、18、20、22、24、32、34、36、38、40、42、44、46、48、50、52、72、74、76、78、80、82、84、86、88、90、92、94、96、98、100、102に示されるようなヌクレオチド配列を含む核酸分子が好ましい。
【0060】
さらなる局面において、本発明は、本発明の核酸配列を含むベクターに関する。好ましくは、該ベクターは、適切な宿主細胞において本発明の該ポリペプチドの発現を提供する。該宿主細胞は、単なる生物工学的な理由、例えば収率、溶解性、費用などのために選択され得るが、該細胞が対象に投与されるべき場合は、医学的観点からも、例えば非病原性細菌または酵母、ヒト細胞が選択され得る。該ベクターは、本発明に係る前記ポリペプチドの構成性または誘導性の発現を提供し得る。
【0061】
本発明のさらなる局面において、上述のポリペプチドおよび/または細胞は、対象における細菌感染の処置または予防のための、特に、ブドウ球菌(例えば、S.アウレウス、S.アウレウス(MRSA)、S.エピデルミディス(S. epidermidis)、S.ヘモリチカス(S. haemolyticus)、S.シミュランス、S.サプロフィチカス(S. saprophyticus)、S.クロモゲネス(S. chromogenes)、S.ヒイカス(S. hyicus)、S.ワーネリ、および/またはS.キシローサス(S. xylosus))、腸球菌(例えば、エンテロコッカス・フェシウム、E.フェシウム(VRE)、エンテロコッカス・フェカリス)、連鎖球菌(ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)、S.ニューモニエ、S.ミュータンス(S. mutans)、S.ウベリス(S. uberis)、S.アガラクティエ、S.ディスガラクティエ(S. dysgalactiae)、ランスフィールド群A、B、Cの連鎖球菌)、クロストリジウム属(例えば、C.パーフリンジェンス(C. perfringens)、C.ディフィシレ(C. difficile)、C.テタニ(C. tetani)、C.ボツリナム(C. botulinum)、C.チロブチリクム(C. tyrobutyricum))、バチルス属(例えば、バチルス・アンスラシス、B.セレウス(B. cereus))、リステリア属(例えば、L.モノサイトゲネス、L.イノキュア(L. innocua))、ヘモフィルス・インフルエンザ(Haemophilus influenza)、コリネバクテリウム・ジフテリエ(Corynebacterium diphteriae)、プロピオニバクテリウム・アクネ(Propionibacterium acne)、マイコバクテリア(例えば、マイコバクテリウム・ツベルクローシス(Mycobacterium tuberculosis)、M.ボビス(M. bovis))などのグラム陽性菌により引き起こされる感染の処置または予防のための方法において使用される。または、本発明に係るポリペプチドおよび/または細胞は、対象における細菌感染の処置または予防のための、特に、腸内細菌科(Enterobacteriaceae)(エシェリキア属(Escherichia)、特に大腸菌(E. coli)、サルモネラ属(Salmonella)、シゲラ属(Shigella)、シトロバクター属(Citrobacter)、エドワードシエラ属(Edwardsiella)、エンテロバクター属(Enterobacter)、ハフニア属(Hafnia)、クレブシエラ属(Klebsiella)、特にK.ニューモニエ(K. pneumoniae)、モルガネラ属(Morganella)、プロテウス属(Proteus)、プロビデンシア属(Providencia)、セラチア属(Serratia)、エルシニア属(Yersinia))、シュードモナス科(Pseudomonadaceae)(シュードモナス属(Pseudomonas)、特にP.エルギノーサ、バークホルデリア属(Burkholderia)、ステノトロフォモナス属(Stenotrophomonas)、シェワネラ属(Shewanella)、スフィンゴモナス属(Sphingomonas)、コマモナス属(Comamonas))、ナイセリア属(Neisseria)、モラクセラ属(Moraxella)、ビブリオ属(Vibrio)、アエロモナス属(Aeromonas)、ブルセラ属(Brucella)、フランシセラ属(Francisella)、ボルデテラ属(Bordetella)、レジオネラ属(Legionella)、バルトネラ属(Bartonella)、コクシエラ属(Coxiella)、ヘモフィルス属(Haemophilus)、パスツレラ属(Pasteurella)、マンヘミア属(Mannheimia)、アクチノバチルス属(Actinobacillus)、ガードネレラ属(Gardnerella)、スピロヘータ科(Spirochaetaceae)(トレポネーマ属(Treponema)およびボレリア属(Borrelia))、レプトスピラ科(Leptospiraceae)、カンピロバクター属(Campylobacter)、ヘリコバクター属(Helicobacter)、スピリルム属(Spirillum)、ストレプトバチルス属(Streptobacillus)、バクテロイデス科(Bacteroidaceae)(バクテロイデス属(Bacteroides)、フソバクテリウム属(Fusobacterium)、プレボテラ属(Prevotella)、ポルフィロモナス属(Porphyromonas))、アシネトバクター属(Acinetobacter)、特にA.バウマニ(A. baumanii)などのヒトまたは動物にとって病原性である株を含む細菌群、科、属、または種のグラム陰性菌により引き起こされる感染の処置または予防のための方法において使用される。
【0062】
前記対象は、ヒト対象または動物、特に畜牛などの畜産および/または酪農業において使用される動物であり得る。前記処置の方法は、感染の部位、または感染に対して予防的に処置される部位へと十分な量の本発明の前記ポリペプチドを適用することを包含する。
【0063】
特に、前記処置の方法は、皮膚、軟部組織、呼吸器系、肺、消化管、眼、耳、歯、鼻咽腔、口、骨、膣、菌血症および/または心内膜炎の創傷の感染の処置または予防のためであり得る。
【0064】
さらに好ましい態様において、本発明に係るポリペプチドは、動物、特に家畜および乳牛におけるブドウ球菌の感染の処置(または予防)の方法において使用される。特に、本出願のポリペプチドは、ウシ乳房炎、特にS.アウレウス、S.エピデルミディス、S.シミュランス、S.クロモゲネス、S.ヒイカス、S.ワーネリ、およびS.キシローサスにより引き起こされるウシ乳房炎の処置(または予防)の方法における使用に適する。
【0065】
さらに、本発明のポリペプチドは、特に外科手術の前もしくは後、または例えば血液透析の間の清浄剤として予防的に使用され得る。同様に、未熟児および免疫不全の人、または補綴装置を必要とする対象が、予防的または急性感染の間のいずれかに本発明のポリペプチドで処置され得る。同様の文脈において、特にスタフィロコッカス・アウレウス(MRSA)、エンテロコッカス・フェシウム(VRE)、シュードモナス・エルギノーサ(FQRP)、または抗生物質耐性クロストリジウム・ディフィシレなどの抗生物質耐性株による院内感染が、予防的または急性期の間に本発明のポリペプチドで処置され得る。本態様において、本発明のポリペプチドは、界面活性剤、テンシド(tensids)、溶剤、抗生物質、ランチビオティクス、またはバクテリオシンなどの殺菌溶液において有用である他の構成要素と組み合わせても殺菌剤として使用され得る。
【0066】
特に好ましい態様において、本発明のポリペプチドは、処置される(または予防される)感染が多耐性細菌株、特に1つまたは複数の以下の抗生物質に対して耐性である株により引き起こされる場合、医学的処置のために使用される:ペニシリン、ストレプトマイシン、テトラサイクリン、メチシリン、セファロチン、ゲンタマイシン、セフォタキシム、セファロスポリン、バンコマイシン、リネゾリド、セフタジジム、イミペネム、またはダプトマイシン。さらに、本発明のポリペプチドは、抗生物質、ランチビオティクス、バクテリオシン、他のエンドリシンなどのような従来の抗菌剤と組み合わせて投与することにより処置の方法において使用され得る。
【0067】
本発明に係る処置(または予防)の方法において使用される投与量および投与の経路は、処置される特定の疾患/感染の部位に依存する。投与の経路は、例えば特定の態様において、経口、局所、鼻咽頭、非経口、静脈内、直腸、または任意の他の投与の経路であり得る。
【0068】
感染の部位(または感染する危険にさらされた部位)への本発明のポリペプチドの適用のために、ペプチドグリカン溶解酵素が感染の部位に到達するまでプロテアーゼ、酸化、免疫応答などのような環境の影響から保護されるような様式で、本発明のポリペプチドが製剤化され得る。
【0069】
従って、本発明のポリペプチドは、カプセル、糖衣丸、丸剤、坐剤、注射可能な溶液、または任意の他の医学的に妥当な生薬製剤として製剤化され得る。いくつかの態様において、これらの生薬製剤は、適切な担体、安定化剤、着香料、緩衝液、または他の適切な試薬を含み得る。
【0070】
例えば、局所適用のために、本発明のポリペプチドはローションまたは硬膏として投与され得る。
【0071】
鼻咽頭適用のために、本発明に係るポリペプチドは、鼻へのスプレーを介して適用されるように生理食塩水において製剤化され得る。
【0072】
腸の処置のために、例えばウシ乳房炎において、坐薬製剤を構想することができる。または、経口投与が考慮され得る。この場合、本発明のポリペプチドは、感染の部位に及ぶまで苛酷な消化環境から保護されなければならない。これは、例えば胃における消化の最初の段階を生き延び、および後に本発明のポリペプチドを腸環境中に分泌する細菌を担体として使用することにより達成することができる。
【0073】
すべての医学的適用は、病原性細菌に遭遇した際に特異的にかつ直ちに病原性細菌を溶解する本発明のポリペプチドの効果に依拠する。これは、病原性細菌および細菌負荷における減少をもたらすことにより処置される対象の健康状態に即時的な影響を有し、ならびに同時的に免疫系を救援する。従って、当業者が直面する主要な課題は、本発明のポリペプチドを、処置されるそれぞれの疾患のために的確に製剤化することである。この目的のために、通常、これらの適用について従来の薬物のために使用されるのと同様の生薬製剤を使用することができる。
【0074】
本発明のさらなる局面において、上述のポリペプチドおよび/または細胞は、担体物質を任意で含む薬学的組成物の成分である。
【0075】
もっとさらなる局面において、ポリペプチドおよび/または細胞は、化粧料組成物の一部である。上述のように、いくつかの細菌種は、皮膚などの患者の体の環境的に曝露された表面上に炎症を引き起こし得る。そのような炎症を予防するために、または前記細菌性病原体の軽症の症状発現を除去するために、既に存在するかまたは新しく定着する病原性細菌を溶解する目的で十分な量の本発明のポリペプチドを含む特別な化粧料調製物が使用され得る。
【0076】
さらなる局面において、本発明は、食材における、食物加工設備上での、食物加工工場における、棚および食物保管領域などの食材と接触する表面上での、ならびに、病原性細菌、通性病原性細菌、または他の望ましくない細菌が潜在的に食物材料に寄生し得るすべての他の状況における、本発明に係る前記ポリペプチドの使用に関する。
【0077】
本発明のさらなる局面は、細菌感染の診断における本発明に係る前記ポリペプチドの使用に関する。本局面において、本発明に係るポリペプチドは、病原性細菌を特異的に溶解する手段として使用される。本発明に係るポリペプチドによる細菌細胞の溶解は、Triton X-100などの界面活性剤、またはポリミキシンBなどの細菌細胞外被を弱くする他の添加物の添加により支持され得る。特異的な細胞溶解は、PCR、核酸ハイブリダイゼーション、もしくはNASBA(核酸配列ベース増幅)などの核酸ベースの方法、IMS、免疫蛍光法、もしくはELISA技術などの免疫学的方法、または、別個の細菌群もしくは種に特異的なタンパク質(例えば、腸内細菌にとってのβ-ガラクトシダーゼ、コアグラーゼ陽性株にとってのコアグラーゼ)を用いる酵素アッセイなどの細菌細胞の細胞内容物に依拠する他の方法を使用するその後の細菌の特異的検出のための最初の段階として必要とされる。
【0078】
本発明の別の局面は、試料由来の病原性細菌またはそうでなければ望ましくない細菌の結合、濃縮、除去、捕捉、および検出のための本発明に係るペプチドグリカン結合タンパク質の使用である。本発明に係る方法に関する試料は、検出、結合、濃縮、除去、または捕捉の標的である細菌を想定しているかまたは含有する任意の材料である。試料は、例えば、食物または飼料材料、表面材料、または、ヒトもしくは獣医学的診断プローブであり得る。細菌の検出は、本発明に係るペプチドグリカン結合タンパク質に付着されたマーカーの検出を介して、または前記タンパク質自体の検出により、例えばELISAなどの免疫学的方法により行われる。本発明に係る方法のために、本発明に係るペプチドグリカン結合タンパク質は、適切な支持構造、例えば、マイクロタイタープレート、試験ストリップ、スライド、ウエハ、フィルター材料、反応管、磁性物質、ガラスもしくはラテックス粒子、ピペットチップ、またはフロースルーセルチャンバー(flow-through cell chamber)上に固定化され得る。支持構造は、例えば、ポリスチレン、ポリプロピレン、ポリカーボネート、PMMA、酢酸セルロース、ニトロセルロース、ガラス、ケイ素ウエハ、ラテックスからなり得る。固定化は、吸着により、共有結合により、またはさらなるタンパク質により達成され得るが、共有結合が好ましい。固定化は機能的なものであり、すなわち、前記ペプチドグリカン結合タンパク質は支持材料に結合するが細菌にとって接触可能な構造を示すことが妥当である。
【実施例】
【0079】
実施例1.DNA技術およびクローニング手順
エンドリシンベースの融合タンパク質をコードするプラスミドの構築のために、Sambrookら(Molecular cloning. A laboratory manual; 2nd ed. Cold Spring Harbor Laboratory Press 1989)によるDNA技術およびクローニング手順を使用した。プラスミドpQE-30(QIAGEN)およびその誘導体pHGFP、pHGFP_CBD118、pHGFP_CBD500(Loessner et al. 2002)、ならびにpHEADPSA(Korndoerfer et al. 2006)を、N末端に6xHisタグが付加された人工融合タンパク質をコードするプラスミドの構築のためのベクター骨格として使用した(HはHisタグを意味する)。プラスミド中への断片の挿入のために必要とされる制限酵素部位は、プライマーを介して導入した。ダブルCBD融合構築物を、2つのCBD断片の別々の増幅およびその後のライゲーションによるか、または代替的にPCRベースのオーバーラップ伸長による遺伝子スプライシング(SOE PCR)法を介して2つの断片を融合することによるかのいずれかで作製した(Horton et al. 1990)。CBD118(SEQ ID NO:57)およびCBD500(SEQ ID NO:58)をコードする断片をEcoRI/MunI部位を介して両方の方向においてライゲーションし、ならびにその後pHGFPのSacI/SalI部位中へ挿入して、pHGFP_CBD118-500およびpHGFP_CBD500-118を産生した。プラスミドpHGFP_CBDP35-500およびpHGFP_CBD500-P35を同様の方法で作製した。pHGFP_CBD118L500およびpHGFP_CBD500L118の場合は、pHGFP中への挿入の前にSOE PCRによりPlyPSAリンカーをコードする断片を2つのCBDの間に導入した。pHCBD500_GFP_118およびpHCBD118_GFP_500のためとしては、5'CBDおよびGFP断片をKpnI部位を介してまたはSOE PCRにより最初に融合し、ならびにその後pHGFP_CBD118およびpHGFP_CBD500のBamHI/SacI部位中へそれぞれライゲーションして、これらのプラスミドの単なるGFP断片と置き換えた。pHGFP_CBD500-500の構築のためには、CBD500断片をpHGFP_CBD500のSacI部位中へクローニングし、CBD500の重複をもたらした。pHEAD_CBD500-500は、完全なply500遺伝子をpHGFP_CBD500のBamHI/SacI部位中へ挿入し、GFP断片と置き換えることにより作製した。すべての構築物について、遺伝子融合を可能にするために3'末端のもの以外のすべての終止コドンを削除した。TAAを一般に3'末端の終止コドンとして導入した。すべての構築物をヌクレオチド配列決定により確認した。
【0080】
実施例2.Hisタグ付き組換えタンパク質の過剰発現および精製
Hisタグ付き(それぞれの構築物において「H」により省略される)融合タンパク質の過剰発現を大腸菌XL1-Blue MRF'(Stratagene)において行った。それぞれの株を、プラスミド選択のために100μg/mlアンピシリンおよび30μg/mlテトラサイクリンを含有する修飾LB培地(15 g/lトリプトース、8 g/l酵母エキス、5 g/l NaCl)において30℃で増殖させ、ひとたび0.5のOD600に到達した後0.1〜1 mMのIPTGを誘導物質として添加した。30℃で4時間のさらなるインキュベーションの後、GFPドメインを含有するタンパク質を産生する培養物を、収集および250 mlの培養物あたり5 mlの緩衝液A(500 mM NaCl、50 mM Na2HPO4、5 mMイミダゾール、0.1% Tween 20、pH 8.0)における再懸濁の前に、一晩4℃で保管した。GFPが存在しなかった場合は、誘導4時間後に細胞をペレットにした。100 MPaでのフレンチプレス20Kセル(SLM Aminco)の2回の通過により細胞を破砕し、ならびに細胞破片を遠心分離および濾過(0.2μM PES膜、Millipore)により除去した。
【0081】
未加工の抽出物における6xHisタグ付き標的タンパク質を、Micro Biospinカラム(BIORAD)を用いてNi-NTA Superflow樹脂(QIAGEN)での固定化金属アフィニティークロマトグラフィー(IMAC)により精製した。緩衝液B(500 mM NaCl、50 mM Na2HPO4、250 mMイミダゾール、0.1% Tween 20、pH 8.0)が溶出緩衝液として働いた。精製したタンパク質を透析緩衝液(100 mM NaCl、50 mM NaH2PO4、0.005%〜0.1% Tween 20、pH 8.0)の2回の交換に対して透析し、濾過(0.2μM PES膜、Millipore)し、および50%(v/v)のグリセロールの添加後−20℃で保管した。各タンパク質について、過剰発現および精製の過程をSDS-PAGEにより解析し、およびタンパク質濃度を分光光度的に測定した(NanoDrop ND-1000 Spectrophotometer)。
【0082】
実施例3.結合アッセイおよび蛍光顕微鏡法
GFP-CBD融合タンパク質の結合特性を、すべての種および血清型のリステリア株の代表的なセット(表1)を用いて結合により検討した。PBST緩衝液(50 mM NaH2PO4、120 mM NaCl、pH 8.0、0.01% Tween 20)における各株の後期対数期細胞を、過剰のGFP-CBDタンパク質と共に5分間室温でインキュベーションした。緩衝液で2回洗浄した後、Axioplan顕微鏡、ならびに励起BP 450〜490 nm、ビームスプリッターFT 510 nm、および発光LP 520 nmのフィルターセット(Carl Zeiss AG)を用いた蛍光顕微鏡法のために細胞を調製した。標識された細胞の写真をLeica DFC320カメラを用いることにより取得した。各アッセイについて、4種類のスコアシステムを用いて目視検査により結合強度を評価した:++、+、(+)、および−はそれぞれ、強い、弱い、非常に弱い、および結合しないを示す。
【0083】
両方の細胞壁結合ドメインが両方の方向において互いに直接融合し、かつN末端のGFPドメインに付着している構築物HGFP_CBD500-118およびHGFP_CBD118-500は、両方とも試験された血清型4、5、および6株のすべての株に弱い結合を示した。これは単なるCBD500の結合パターンに相当するため、これらの結果は、CBD118がこれらの構築物において機能的でないことを示唆した。他方で、CBD500が機能性を保持するためにタンパク質のC末端に位置する必要はないことが示された。融合構築物における両方の結合ドメインの増強された柔軟性がCBD118を機能的にさせる可能性があると仮定して、本発明者らはCBDを分離するPlyPSAのリンカーペプチドを含むタンパク質HGFP_CBD500L118およびHGFP_CBD118L500を作製した。再び、両方の構築物は血清型4、5、および6株に属するすべての株を標識したが、追加的に7種類の血清型1/2株のうち4種類もまた試験された。蛍光顕微鏡法より、後者は、HGFP_CBD118について観察されたように主に極および隔壁で標識されることが観察された。対照的に、両方のタンパク質は、血清型4、5、および6の株をHGFP_CBD500のような細胞表面に渡る一様な分布において装飾(decorate)した。従って、これらのダブルCBD構築物は、血清型1/2、3、および「7」内の結合範囲はHGFP_CBD118のものよりも狭いが、両方のCBDの特性を併せ持った。短いリンカーの導入は、CBD118がリガンドに接触することを可能にしただけでなく、C末端位置におけるCBD500の結合も増強した。融合タンパク質HGFP_CBD118L500は、HGFP_CBD500のように、血清型4、5、および6細胞の大部分について同等に強い装飾を示した。加えて、GFPが中央の位置に配置され、一方CBD500およびCBD118がN末端またはC末端のいずれかに位置する2つの融合タンパク質を生成した。HCBD500_GFP_CBD118においては、CBD118をGFPのC末端に直接付着させ、HGFP_CBD118の場合と同様の環境に置いた。このタンパク質は、装飾が非常に弱かったが、試験したすべての血清型1/2株を標識することができた。CBDが逆向きである構築物HCBD118_GFP_CBD500は、血清型4、5、および6株の大部分に強く結合したが、1つの血清型1/2株を弱く標識したのみであった。再び、CBD500はC末端に位置した際により強い結合を示した。しかしながら、N末端位置においてもまた機能的であることが示された(HCBD500_GFP_CBD118)。
【0084】
(表1)様々な種および血清型のリステリア細胞への様々なリステリアエンドリシン由来のGFPタグ付きCBDおよびダブルCBD融合タンパク質の結合
500G118および118G500は、HCBD500_GFP_118およびHCBD118_GFP_500をそれぞれ意味する。「L」はシャッフリングされたCBDの間に導入されたリンカーを意味する。++強い、+弱い、(+)非常に弱い、−結合しない;WLSC:Weihenstephan Listeria Strain Collection;SV:リステリア血清型
【0085】
さらなるアプローチにおいて、ダブルCBD融合構築物におけるCBD118をCBDP35により置き換えた(SEQ ID NO:59)。ファージP35のエンドリシンのCBDは、完全な細胞表面に渡る一様な分布で結合し、血清型1/2および3の大部分の株ならびに血清型4、5、および6のいくつかの株を強く標識した。新たに構築されたタンパク質HGFP_CBD500-P35およびHGFP_CBDP35-500の結合パターンは、Ply500およびPlyP35のシングルCBDの結合パターンのほぼ正確な組み合わせを表した:それらは、融合物内のシングルCBDの位置にかかわらず、CBD500またはCBDP35または両方のいずれかが結合するすべての株に強い結合を示した。これらの結果は、異なるペプチドグリカン溶解酵素由来の2つの細胞壁結合ドメインの組み合わせは、それらがC末端位置でない時でさえ人工融合タンパク質において完全に機能的であり得ることを証明した。
【0086】
実施例4.表面プラズモン共鳴解析(SPR)による結合親和性の測定
HGFP_CBD500(SEQ ID NO:60)およびHGFP_CBD500-500のL.モノサイトゲネスWSLC 1042の細胞壁への親和性を、BIAcore X機器およびC1センサーチップ(BIAcore, Uppsala, Sweden)を用いて表面プラズモン共鳴解析により測定した。チップ表面をアミンカップリング法で活性化し、および両方のフローセルにおいてHGFP-CBD500分子を用いてコーティングした(5μl/minの流速で、70μlの10 mM酢酸ナトリウム緩衝液、pH 5における0.5 mg/mlタンパク質)。HBS緩衝液(10 mM HEPES、150 mM NaCl、3.4 mM EDTA、0.005% Tween 20、pH 7.8)中の熱不活性化WSLC 1042細胞をその後、フローセルFc2において固定化したCBDに結合させた(1 mlあたり3.0×1010細胞;3μl/minの流速で15μl)。最終的に、HBS緩衝液において固定化した細胞と3種類の異なる濃度のHGFP_CBD500(50 nM、100 nM、200 nM)およびHGFP_CBD500-500(12.5 nM、25 nM、50 nM)両方との間の相互作用を測定し(10μl/minで30μl)、Fc1を対照セルとした。結合相は3分間測定し、解離相は12分間測定した。すべての段階を25℃で行った。動態学的データの評価は、「物質移動を伴う1:1結合」モデルを使用してBIAevaluation software,version 4.1(BIAcore)で行った。各タンパク質について測定した3種類の濃度に対して取得された平衡結合定数を表2に示す。
【0087】
(表2)リステリア・モノサイトゲネスWSLC 1042の細胞壁へのHGFP_CBD500およびHGFP_CBD500-500の結合の平衡親和性定数(KA)
【0088】
人工ダブルCBDを含む構築物HGFP_CBD500-500は、ファージA500 - ply500のエンドリシンの天然のCBDを含むHGFP_CBD500と比較して約50倍高い親和性で固定化リステリア細胞に結合することが示された。シングルおよびダブルCBDタンパク質構築物の両方のセンサグラムを比較すると、両方の構築物が主に解離相において異なることが明らかであった。ひとたび細胞表面に結合すると、HGFP_CBD500はHGFP_CBD500-500よりずっと急速に脱離し、ダブルCBD構築物はより高い全体的な親和性をもたらした。
【0089】
実施例5.測光溶解アッセイ
野生型およびキメラペプチドグリカン溶解酵素の溶解活性を測光溶解アッセイにより測定した。リステリア・モノサイトゲネス株WSLC 1001(血清型1/2 c)およびWSLC 1042(血清型4 b)の基質細胞を、細菌をTB培地において後期対数期まで増殖させ、およびPBS緩衝液(50 mM NaH2PO4、120 mM NaCl、pH 8.0)において50倍濃度で凍結することにより調製した。アッセイは、PBSにおいて約1.0の初期OD600に希釈した細胞を用いて、1 mlの総体積中で行った。比較するすべての精製した天然のエンドリシンおよびキメラタンパク質を、20μlの体積において等モル量で細胞に添加し、および600 nmでのODを15秒の間隔で最大10分間測定した。使用した酵素濃度は30〜152 pmol/mlに渡った。陰性対照のために、20μl緩衝液を細胞に添加した。すべてのアッセイは3つ組で行った。Loessnerら(2002, Mol. Microbiol. 44, 335-349)は、細胞壁におけるリガンドへのCBDの結合についての分子基盤としてイオン性相互作用を示唆した。エンドリシンply500のCBDは約100 mMのNaCl濃度で最適結合を示し、および増加する塩濃度と共に減少する結合能力を示した。それに基づいて、hisタグ付き野生型ply500(HPL500)およびply500の天然に存在するCBD500の重複を用いた本発明に係る構築物H_EAD_CBD500-500の溶解活性を、高い塩条件の下で比較した。NaCl濃度(1Mと2Mの間)を用いる以外は、アッセイは上述のように行った。光度曲線を標準化し、および対照アッセイのデータにより補正した(補正値=値+(1−対照値))。結果として生じた曲線をソフトウェアSigmaPlot 9.0(Systat Software, Inc.)を用いて以下のシグモイド関数と適合させた:f=y0+a / (1+exp(-(x−x0) / b))∧c。相対酵素活性に相当する関数の最も急な傾きを測定した。
【0090】
驚くべきことに、1 M NaClまたはより高い塩濃度で、本発明に係るペプチドグリカン溶解酵素H_EAD_CBD500-500は、天然に存在する酵素ply500よりも高い溶解活性を示した。
【0091】
実施例6.腸球菌エンドリシンのタンパク質発現および精製
腸球菌エンドリシンFab25VL、Fab20VL、Fab20K、および本発明に係るペプチドグリカン溶解酵素EADFab25_CBD25_CBD20を大腸菌HMS174 DE3において発現させ、およびそれから単離した。タンパク質発現は、1 mM IPTGでの誘導後、3時間37℃で行った。細菌細胞ペレットを遠心分離(5000 rpm、15分、4℃)により収集し、25 ml緩衝液A(25mM トリス、pH 8.0、500 mM NaCl、20 mMイミダゾール、0.1% Tween 20、10%グリセロール)に再懸濁し、および細胞をマイクロフルイダイザー(microfluidizer)において破砕した。細菌細胞破片を遠心分離(12000 rpm、5分、4℃)により除去した。上清を30%飽和の硫酸アンモニウム沈澱に供した。沈殿物を遠心分離(12000 rpm、5分、4℃)により回収した。エンドリシンを含む上清を5 mlフェニルセファロースカラム(High Sub FF, Amersham)を用いた疎水性クロマトグラフィーに適用した。カラムを10容量の緩衝液B(25mMトリス、pH 7.0、500 mM NaCl、30%硫酸アンモニウム、10%グリセロール)で洗浄した。エンドリシンを10カラム容量の緩衝液C(25mMトリス、pH 7.0、500 mM NaCl、10%グリセロール)で溶出した。タンパク質を含有する画分をクマシー染色SDSゲル上でエンドリシンについて解析した。エンドリシンを含有する画分をプールし、および実施例7記載のプレート溶解アッセイにおいて溶解活性について解析した。
【0092】
実施例7.腸球菌細菌に対するペプチドグリカン溶解タンパク質の溶解活性および宿主範囲を試験するためのプレート溶解アッセイ
医学的に関連する種であるエンテロコッカス・フェシウムおよびエンテロコッカス・フェカリス由来の種々の腸球菌細菌を、3 ml BHI培地の前培養において一晩37℃で増殖させた。各株について、2 mlの前培養物を25 mlの新鮮培地中に播種し、および約1のOD600 nmまでインキュベーションした。細菌細胞を4500 rpm、15分間、4℃での遠心分離により収集した。細胞ペレットを500μl BHI培地に再懸濁した。熱不活性化細胞に対する溶解活性の試験(表3)のために、細胞を85℃で45分間インキュベーションし、および1400 rpmでの遠心分離により収集した。細胞ペレットを10 ml LB上層寒天に再懸濁し、および上層寒天をLBプレート上に注いだ。生細胞に対する溶解活性の試験(表4)のためには、腸球菌前培養物を1 ml BHI培地にて一晩増殖させ、前培養物を10 ml BHI上層寒天と混合し、LBプレート上へ注ぎ、および2時間30℃でインキュベーションした。腸球菌エンドリシンFab25VL、Fab20K、エンドリシンFab25VLおよびFab20Kの等モルの組み合わせ、ならびに本発明に係る人工酵素EADFab25_CBD25_CBD20を使用した。5μlのペプチドグリカン溶解タンパク質溶液各々を、上層寒天中に浸漬した菌叢の上にピペットでスポットした。スポットしたタンパク質溶液の周囲の溶解ゾーンの出現を、4種類のスコアシステムを用いて30℃でのプレートのインキュベーションの18時間後に解析した:+++、++、+、ならびに−は、それぞれ強い、中程度、弱い、弱くおよび溶解しないを示す。
【0093】
(表3)熱不活性化腸球菌細胞に対してペプチドグリカン溶解酵素を用いたプレート溶解アッセイ
【0094】
(表4)生存腸球菌細胞に対してペプチドグリカン溶解酵素を用いたプレート溶解アッセイ
【0095】
熱不活性化腸球菌細胞を用いた場合(表3)、エンドリシンFab20KはすべてのE.フェカリス株を高効率で溶解するが、E.フェシウムのいずれの株も溶解しないことが判明した。Fab20エンドリシンはE.フェカリス株に特異的であるように見えた。Fab25VLはE.フェシウムのすべての株を高効率で溶解したが、E.フェカリスの2種類の株のみしか溶解しなかった。他のE.フェカリス株は中程度の効率で溶解されるか、または全く溶解されなかった。従って、天然に存在するエンドリシンFab25VLは、E.フェシウムに厳密に特異的ではなかったが、この種由来の株をE.フェカリス種由来の株よりも確実に溶解した。2種類のエンドリシンFab25VLおよびFab20Kの1:1混合物は試験したすべての株を高効率で溶解したが、本発明に係る1種類の酵素のみ、すなわち、Fab25VLおよびFab20KのCBDを併せ持つがFab25VLのEADのみを有するEADFab25_CBD25_CBD20を用いて、同様のことが達成された。2種類の代わりに1種類の酵素のみを使用することは、酵素産生を容易にし、費用を削減し、および治療的適用における免疫学的反応を最小化する。生細胞を用いるアッセイを行った場合(表4)、本発明に係る酵素であるEADFab25_CBD25_CBD20を用いる利点がより明白であった。これらのアッセイ条件の下でFab25VLはE.フェシウム細胞を主に溶解し、およびいくつかのE.フェカリス細胞のみを低効率で溶解したが、エンドリシンFab20Kは全く作用しなかった。生細胞のいずれも全く溶解されず、恐らくエンドリシン結合のための細胞表面受容体またはペプチドグリカン溶解のための基質分子が、生細胞において接触可能でないことを示唆した。2種類の酵素の組み合わせは状況を改善しなかったが、EADFab25_CBD25_CBD20を用いた細胞溶解はずっとより良好な結果を示した。試験したすべての株は少なくとも低効率で溶解されたが、株の大部分は高効率で溶解された。この予想外の結果は、たとえ最初の特徴決定が細菌宿主範囲に関して使用される1つより多いCBDの純粋に付加的な効果を示唆するとしても、本発明に係る人工ペプチドグリカン溶解酵素が特定の使用において好都合の効果を有し得ることを示す。
【0096】
実施例8.腸球菌に対するペプチドグリカン溶解酵素の最小細菌濃度(MBC)の測定
エンテロコッカス・フェカリス株17を一晩37℃でBHI培地において増殖させた。前培養物を25 ml新鮮培地中に1:10で希釈し、および37℃で約1のOD600 nmまでインキュベーションした。細菌細胞を4500 rpmで5分間、4℃での遠心分離により収集し、および細胞ペレットを105 cfu/mlの濃度になるように溶解緩衝液(2 mM CaCl2、10 mM BSAを含むPBS(2.25 mM NaH2PO3、7.75 mM Na2HPO3、150 mM NaCl))に再懸濁した。Fab25VLおよびEADFab25_CBD25_CBD20のタンパク質溶液(1 mg/ml)を溶解緩衝液において50μg/ml、5μg/ml、0.5μg/ml、0.05μg/ml、0.005μg/ml、0.0005μg/ml、および0.00005μg/mlへ連続希釈した。450μlの細胞懸濁液および50μlの各濃度のタンパク質溶液を混合し、および1時間37℃でインキュベーションした。対照として、添加されたタンパク質を含まない溶解緩衝液をインキュベーションした。100μlの1:10および1:100希釈の溶解試料をLB寒天プレートにプレーティングし、1日37℃でインキュベーションし、およびペプチドグリカン溶解酵素による溶解を生き延びた細胞を計数した。例えば、MBC99.9%は、最初の細菌細胞濃度が1000分の一に減少する最も低い酵素濃度として定義される。この場合、生存細胞のcfu/mlは102より低くなければならない。
【0097】
エンテロコッカス・フェカリス株17に対するMBCは、天然に存在する酵素Fab25 VLよりも本発明に係るペプチドグリカン溶解酵素EADFab25_CBD25_CBD20を用いた場合により低いことが観察された。MBC99.9%はEADFab25_CBD25_CBD20で0.05μg/mlであり、一方Fab25VLを用いて5μg/mlであった。この結果は、EADFab25_CBD25_CBD20の、細菌細胞へのより高い結合親和性および/または増加した溶解活性を示唆する。本発明に係るポリペプチドを用いる場合、病原性細菌を殺傷するためには100分の一に減らしたタンパク質を使用する必要がある。
【技術分野】
【0001】
本発明は、少なくとも1つの酵素活性ドメインおよび少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性および細菌溶解活性を有する組換えポリペプチドに関する。本発明はさらに、少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性を有する組換えポリペプチドに関する。さらに、本発明は、組換えポリペプチドをコードするヌクレオチド配列を含む核酸分子、ベクター、および宿主細胞に関する。
【背景技術】
【0002】
近年、バクテリオファージエンドリシンなどのペプチドグリカン分解酵素が、抗菌剤としてますます注目を受けている。古典的な抗生物質に対する病原性細菌の抵抗性が出現しかつ蔓延することを考慮して、これらの生物を制御する代替的な方法への要求が大きくなっている。特にグラム陽性菌の場合、いわゆるエンザイバイオティクス(enzybiotics)のようなファージエンドリシンの適用が将来有望なアプローチである。グラム陽性においては外膜の欠如のために、これらの酵素がエキソリシンとしても働き、すなわち感受性細胞の溶解を外部から引き起こすことができる。この特性は、リステリア(Listeria)に感染するファージ由来のエンドリシンについて示されたように、例えば、分子生物学において細菌細胞からの核酸およびタンパク質の効率的な回収のために活用することができる。食物由来の病原体であるリステリア・モノサイトゲネス(Listeria monocytogenes)の制御のための適用を目指し、これらのリシンをコードする遺伝子を、チーズ生産においてスターター生物として使用されるラクトコッカス・ラクティス(Lactococcus lactis)およびいくつかの乳酸桿菌を含む多数の乳酸菌中に導入した。これらの細菌は、エンドリシンを過剰発現および分泌し、その結果L.モノサイトゲネス細胞に対して溶解活性を示した。これまでに報告されたファージにコードされるペプチドグリカンヒドロラーゼの医学的適用は、バチルス・アンスラシス(Bacillus anthracis)の検出および殺傷、ならびにインビトロおよびマウスモデルにおけるストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)の制御を含む。
【0003】
エンドリシンは、dsDNAファージの後期遺伝子領域にコードされ、および溶解性増殖サイクルの終わりに産生される細胞壁溶解酵素である。同様の酵素はまた、細菌ゲノム中に組み込まれたプロファージゲノム内でも見出される。それらの機能は内側からの細菌ペプチドグリカンの分解であり、宿主細胞の溶解およびファージ子孫の放出をもたらす。ペプチドグリカン内の異なる標的結合に従い、エンドリシンは5つの異なるクラスに分類することができる:両方ともグリコシダーゼであり、かつグリカン鎖の2つのβ-1,4-グリコシド結合の1つをそれぞれ切断する(i)N-アセチル-β-D-ムラミダーゼ(リゾチームとしても公知である)および(ii)N-アセチル-β-D-グルコサミニダーゼ;(iii)ムラミダーゼと同様の結合を切断するが、異なる機構による溶解性トランスグリコシラーゼ;(iv)グリカンとペプチド部分との間を切るN-アセチルムラモイル-L-アラニンアミダーゼ;および(v)ペプチド部分内部を切断するエンドペプチダーゼ。溶解性トランスグリコシラーゼを除くすべてのエンドリシンはヒドロラーゼである。同様の酵素活性はまた、それ自身の細胞壁または密接に関連した細菌の細胞壁を溶解する細菌酵素、いわゆるオートリシン、およびバクテリオシンなどの他の細菌細胞壁溶解ポリペプチドにおいても見出される。細菌オートリシンは、細胞壁リモデリング、細胞分裂、形質転換において、または病原性因子として重要な役割を果たす細胞壁溶解酵素である。それらは一緒にペプチドグリカン溶解酵素としてまとめることができる。ペプチドグリカン溶解酵素の作用によるペプチドグリカンの酵素分解は、細胞壁の完全性の損失をもたらし、かつ最終的に高い内圧により引き起こされる細胞破裂をもたらす。
【0004】
細菌細胞を殺傷する特徴によって、ペプチドグリカン溶解酵素は、ヒトおよび動物における細菌感染に対する予防剤または治療剤として、抗菌薬またはエンザイバイオティック(enzybiotic)としての使用のための、医学的環境、公的環境、もしくは私的環境における消毒剤としての使用のための、食品産業、動物飼料産業、もしくは化粧品産業における細菌汚染の除染剤として、または細菌で汚染された表面に対する一般の界面活性剤としての使用のための興味深い候補となる。ペプチドグリカン溶解酵素は、別個の細菌群、属、種、株、または血清型由来の細菌細胞に特異的に結合しかつ溶解するため、細菌の診断において同様に有用である。特異的な細胞溶解はしばしば、核酸ベースの方法または免疫学的方法などの検出される細菌細胞の細胞内容物に依拠する追加的な検出方法と併用される。
【0005】
グラム陽性バックグラウンド由来のエンドリシン、オートリシン、および他のペプチドグリカン溶解酵素は、触媒活性と基質認識とが分離され、かつ少なくとも2つの別個の機能ドメインである酵素活性ドメイン(EAD)と細胞結合ドメイン(CBD)とに局限されるモジュラー構成を示す。ファージが感染しているグラム陰性宿主由来の大抵のエンドリシンは、単一ドメインの球状タンパク質である。しかしながら、最近2つの機能ドメインからなるグラム陰性バックグラウンド由来の2つのリシンが報告された:N末端の細胞結合ドメインおよびC末端の触媒ドメインからなるシュードモナス・エルギノーサ(Pseudomonas aeruginosa)ファージΦKZおよびELのエンドリシン(Briers et al. Molecular Microbiology, Volume 65, Number 5, September 2007, pp. 1334-1344(11)(非特許文献1))。対照的に、ファージが感染しているグラム陽性細菌由来のエンドリシンの大部分は、N末端のEADおよびC末端のCBDを有して逆方向のドメインを特徴とする。
【0006】
結合ドメインにより認識される細菌細胞壁におけるリガンドが公知であるのは、非常にわずかなエンドリシンのみである。肺炎球菌ファージCpl-1リゾチームは、ストレプトコッカス・ニューモニエの細胞壁におけるテイコ酸を含有するコリンを特異的に認識し、コリン結合タンパク質(CBP)のファミリーに位置する。これらのタンパク質のコリン結合モジュール(CBM)は、4〜18個に渡る多数のタンデムコピーにおいて見出される約20アミノ酸残基のリピートにより形成される。Cpl-1は、通常短いリンカー領域を介して連結される2つの分離したドメイン(EADおよびCBD)を有するこれらの酵素のモジュラー設計を例証する。モジュラーファージエンドリシンの大部分は1つの触媒ドメインおよび1つの基質結合ドメインからなるが、例えば、ストレプトコッカス・アガラクティエ(Streptococcus agalactiae)ファージB30のエンドリシン(ムラミダーゼおよびエンドペプチダーゼ)、スタフィロコッカス・アウレウス(Staphylococcus aureus)ファージΦ11のエンドリシン(エンドペプチダーゼおよびアミダーゼ)、ストレプトコッカス・アガラクティエファージNCTC 11261のエンドリシン(エンドペプチダーゼおよびムラミダーゼ)、ならびにスタフィロコッカス・ワーネリ(Staphylococcus warneri)MファージΦWMYのエンドリシン(エンドペプチダーゼおよびアミダーゼ)のように2つの異なる酵素活性を有する多数のタンパク質が存在する。
【0007】
ファージおよび細菌起源のペプチドグリカン溶解酵素、エンドリシン、ならびにオートリシンまたはバクテリオリシンが同様のモジュラー構築を示すという事実、ならびに細菌およびファージ由来溶解タンパク質の別個のドメインの間に高い相同性が見出されるという事実は、機能ドメインの相互交換によるこれらのタンパク質の共通の祖先および共進化を示唆する(Garcia et al., 1990, Gene 86, 81-88(非特許文献2))。Diazら(1990, Proc. Natl. Acad. Sci., 87, 8125-8129(非特許文献3))は、組み合わされた生化学特性を示すファージおよび細菌性肺炎球菌酵素のキメラを作製した。ヌクレオチド相同性を欠損する遺伝子由来の組換えキメラがDiazら(1991, J. Biol. Chem., 266, 5464-6571(非特許文献4))において構築され、基質認識におけるCBDの機能も確認された。Crouxら(1993, Mol. Microbiol., 9, 1019-1025(非特許文献5))は、他の細菌ファミリー由来の細胞に対するエンドリシンの酵素活性において切り換えを導く、肺炎球菌およびクロストリジウム細胞壁溶解酵素に基づくキメラまでも作製した。Sanzら(1996, Eur. J. Biochem., 235, 601-605(非特許文献6))は、2つのEADおよび1つのCBDを含むモジュール組み立てにより多機能肺炎球菌ムレインヒドロラーゼを構築した。最近、リゾスタフィン、スタフィロコッカス・シミュランス(Staphylococcus simulans)由来のペプチドグリカンヒドロラーゼ、およびストレプトコッカス・アガラクティエファージB30エンドリシン、ならびにそのC末端が切断されたバーションからなる融合タンパク質が報告された(Donovan et al. 2006, Appl. Environ. Microbiol., 72, 2988-2996(非特許文献7))。この場合にもまた、人工構築物は、スタフィロコッカス細胞およびストレプトコッカス細胞の両方を溶解する、両方の酵素の特性を併せ持った。Loessnerら(2002, Mol. Microbiol., 44, 335-349(非特許文献8))は、リステリア・モノサイトゲネスを役割モデルとして使用し、細菌細胞壁炭水化物への特異的認識および高親和性結合を決定するCBDの概念を記述した。米国特許第2004/0197833号(特許文献1)は、標的細胞の濃縮のための方法における固定化された単離CBDの使用を教示する。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第2004/0197833号
【非特許文献】
【0009】
【非特許文献1】Briers et al. Molecular Microbiology, Volume 65, Number 5, September 2007, pp. 1334-1344(11)
【非特許文献2】Garcia et al., 1990, Gene 86, 81-88
【非特許文献3】Diaz et al., 1990, Proc. Natl. Acad. Sci., 87, 8125-8129
【非特許文献4】Diaz et al., 1991, J. Biol. Chem., 266, 5464-6571
【非特許文献5】Croux et al., 1993, Mol. Microbiol., 9, 1019-1025
【非特許文献6】Sanz et al., 1996, Eur. J. Biochem., 235, 601-605
【非特許文献7】Donovan et al., 2006, Appl. Environ. Microbiol., 72, 2988-2996
【非特許文献8】Loessner et al., 2002, Mol. Microbiol., 44, 335-349
【発明の概要】
【0010】
本発明の目的は、細菌細胞の信頼のおける検出および濃縮、および/または溶解を可能にする、改善されたおよび有利なタンパク質を提供することである。
【0011】
前記目的は、添付の特許請求の範囲において規定されるような内容により解決される。
【図面の簡単な説明】
【0012】
以下の図面は本発明を例証する。
【図1】リステリア細胞に対するGFP-ダブルCBD融合タンパク質、および参照として働くGFP-シングルCBD構築物の模式図。GFP=緑色蛍光タンパク質;CBD500、CBD118、CBDP35=それぞれリステリアファージエンドリシンPly500、Ply118、およびPlyP35の細胞壁結合ドメイン;L=PlyPSAエンドリシンのリンカー領域。
【図2】細胞壁からの解離が減少したためにより高い親和性をもたらす、重複したCBDを有するペプチドグリカン結合タンパク質。(A)ダブルCBD500融合タンパク質、および参照として役立つそれぞれのシングルCBD500構築物の模式図。GFP=緑色蛍光タンパク質;EAD500=リステリアファージエンドリシンPly500の酵素活性ドメイン;CBD500=Ply500の細胞壁結合ドメイン。(B)50 nMの濃度で測定したHGFP_CBD500(黒色)およびHGFP_CBD500-500(灰色)のSPRセンサグラムのオーバーレイ。結合および解離相を示す。RU=相対応答単位。
【図3】様々なNaCl濃度で測光溶解アッセイを用いて測定した、N末端Hisタグを有する野性型ply500(白丸)およびH_EAD_CBD500-500(黒四角)のリステリア・モノサイトゲネスWSLC1042の細胞に対する相対溶解活性。200 mM NaClでの野生型ply500の最適活性が1.0に相当する。すべてのアッセイは3つ組で行った。
【図4】腸球菌に対するペプチドグリカン溶解酵素の最小殺菌濃度(MBC)の測定。エンテロコッカス・フェカリス(Enterococcus faecalis)株17の生存細胞の細菌濃度を、細胞溶解アッセイに存在するFab25VL(四角)またはEADFab25_CBD25_CBD20(丸)のタンパク質濃度に依存して示す。
【発明を実施するための形態】
【0013】
本明細書において使用される「ペプチドグリカン溶解酵素」という用語は、細菌細胞壁を溶解するのに適当である酵素を指す。酵素は、ペプチドグリカン溶解酵素の「酵素活性ドメイン」(EAD)が構成される以下の活性の少なくとも1つを含む:エンドペプチダーゼ、N-アセチル-ムラモイル-L-アラニン-アミダーゼ(アミダーゼ)、N-アセチル-ムラミダーゼ(リゾチームもしくは溶解性トランスグリコシラーゼ)、またはN-アセチル-グルコサミニダーゼ。酵素は、ファージもしくはプロファージにコードされる、いわゆる「エンドリシン」であるか、または、細菌によりコードされる関連した細胞壁溶解酵素、いわゆる「オートリシン」、もしくはバクテリオシンなどの他の細菌ペプチドグリカン溶解酵素、病原性因子もしくは他の抗菌ポリペプチド(例えば、リゾスタフィン、ALE-1リシン、ムタノリシン、エンテロリシン)に由来するかのいずれかである。加えて、ペプチドグリカン溶解酵素はまた、酵素的に不活性であり、および宿主細菌の細胞壁に結合する領域、いわゆるCBD(細胞壁結合ドメイン)も含有する。
【0014】
本明細書において使用される「ペプチドグリカン結合タンパク質」という用語は、ペプチドグリカン溶解酵素について記述される酵素活性のいずれも有さない人工的に構築された細菌細胞結合タンパク質を指す。ペプチドグリカン結合タンパク質は、CBDに由来する1つより多いCBDを含む。ペプチドグリカン結合タンパク質は、天然に存在するCBDのシャッフリングにより、および/または天然に存在するCBDの増倍により構築される。
【0015】
本明細書において使用される「ドメイン」という用語は、特定の機能を与えられ、および構造的または進化的に保存されたドメインともまた一致し得る、ペプチドグリカン溶解酵素のサブユニットを指す。ドメインに関連する特定の機能は、例えば細菌ペプチドグリカン溶解または細菌細胞結合である。機能ドメインは「モジュール」と呼ばれることもある。
【0016】
本明細書において使用される「CBD」という用語は、しばしばタンパク質のC末端に見出されるペプチドグリカン溶解酵素の細胞壁結合ドメインを指す。CBDドメインは、細胞壁を加水分解する点では酵素活性を有さないが、細菌細胞壁へのペプチドグリカン溶解酵素の結合を媒介する。本明細書において使用されるCBDという用語は、天然に存在するペプチドグリカン溶解酵素に由来するポリペプチド鎖内のセグメントを説明する。
【0017】
本明細書において使用される「EAD」という用語は、細菌ペプチドグリカンの加水分解を担うペプチドグリカン溶解酵素の酵素活性ドメインを指す。これは、ペプチドグリカン溶解酵素について記述される酵素活性の少なくとも1つを含有する。本明細書において使用されるEADという用語は、天然に存在するペプチドグリカン溶解酵素に由来するポリペプチド鎖内のセグメントを説明する。
【0018】
「CHAP」ドメイン(システイン、ヒスチジン依存性アミドヒドロラーゼ/ペプチダーゼ(cysteine, histidine-dependent amidohydrolases/peptidases))は、細菌、バクテリオファージ、古細菌、およびトリパノソーマ(Trypanosomidae)科の真核生物由来のタンパク質において見出される約110〜約140アミノ酸残基の領域である。タンパク質はペプチドグリカン加水分解において主に機能する可能性がある。CHAPドメインは一般に、細菌タイプのSH3ドメインおよびアミダーゼドメインのいくつかのファミリーと結合する。CHAPドメインを含有するタンパク質は、求核攻撃機構において触媒性システイン残基を利用する可能性がある。CHAPドメインは2つの不変アミノ酸残基、システインおよびヒスチジン残基を含有する。これらの残基は、CHAPドメインを含有するタンパク質の推定上の活性部位の一部を形成する。
【0019】
本明細書において使用される「ami」という用語は、アミダーゼ活性を示す酵素的に定義されたドメインを説明し、すなわち、ペプチドグリカン骨格におけるN-アセチルムラミンと、ペプチドリンカーにおいて通常L-alaである隣接するアミノ酸残基との間のアミド結合を加水分解する。アミダーゼはしばしば、活性について金属イオン依存性である。
【0020】
本明細書において使用される、Src相同性3ドメインと呼ばれることもある「SH3」ドメインという用語は、他の結合パートナーと相互作用するタンパク質について特徴的である約60アミノ酸残基の小さな非触媒性タンパク質ドメインを説明する。それはプロリンに富むコンセンサスモチーフにより同定される。SH3ドメインはCBD内に位置する。ペプチドグリカン溶解酵素において見出されるSH3ドメインは、しばしばSH3bまたはSH3_5タイプのものである。
【0021】
「野生型」という用語は、配列に関してタンパク質または核酸の天然に存在する形態を指す。
【0022】
本明細書において使用される「シャッフリング」という用語は、新たなキメラポリペプチド構築物中への、異なる野生型酵素由来のポリペプチドの異なる断片の組み合わせを指す。本文脈において、酵素は好ましくはペプチドグリカン溶解酵素であり、および断片は好ましくはEADおよびCBDである。通常、断片は、分子生物学的方法により核酸レベルで組み合わされる。追加的なリンカー配列が、構造上またはクローニングの理由のために断片の間に導入されてもよい。
【0023】
本発明の1つの対象は、少なくとも1つのEADおよび少なくとも2つのCBDから構成されるペプチドグリカン溶解酵素を指す。本発明に係る人工的に作製されたペプチドグリカン溶解酵素は、天然に存在するタンパク質と比較して拡大したかもしくは変化した結合範囲、または細菌細胞壁への増加した結合親和性、または増加したかもしくは変化した溶解活性、またはそれらの組み合わせなどの新たな特性を示す。
【0024】
本発明の別の対象は、少なくとも2つのCBDから構成されるペプチドグリカン結合タンパク質を指す。本発明に係る人工的に作製されたペプチドグリカン結合タンパク質は、天然に存在するタンパク質と比較して拡大したかもしくは変化した結合範囲、または細菌細胞壁への増加した結合親和性、または両方などの新たな特性を示す。
【0025】
本発明に係るペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質において、少なくとも2つのCBDは、2つの異なるペプチドグリカン溶解酵素に由来してもよいし(ドメインシャッフリング)、または、エンドリシンにおいて天然に存在する1つのCBDの増倍によってもよい。1つより多いEADが本発明に係るペプチドグリカン溶解酵素に存在する場合、EADは2つの異なるペプチドグリカン溶解酵素に由来してもよい。
【0026】
一方、グラム陽性およびグラム陰性菌の様々な属、種、または株に対する多数のペプチドグリカン溶解タンパク質が当技術分野において記述されている。ペプチドグリカン溶解タンパク質のモジュラー性質、およびEADとCBDとの間の区別は周知である。ペプチドグリカン溶解タンパク質に存在する多数の保存されたドメインは、機能的に特徴決定されており、および、ポリペプチドまたはヌクレオチド配列内のそれらの存在は、それぞれのタンパク質または核酸データベース、例えば、CDD(Marchler-Bauer et al., 2005; Nucleic Acids Research, 33, D192-D196);Pfam(Finn et al., 2006, Nucleic Acids Research 34, D247-D251)またはSMART(Schultz et al., 1998, Proc. Natl. Acad. Sci. USA 95, 5857-5864、Letunic et al., 2006, Nucleic Acids Res 34, D257-D260)を使用する適切なコンピュータープログラムにより、または欠失変異体を用いた結合アッセイ(Loessner et al., 2002, Mol. Microbiol., 44, 335-349)により予測することができる。本発明に係る人工ペプチドグリカン溶解酵素は、Sambrookら(Molecular cloning. A laboratory manual; 2nd ed. Cold Spring Harbor Laboratory Press 1989)において記述されているような組換えタンパク質のクローニングおよび産生のための標準的な技術を用いて、EADに由来する所望の酵素活性を細胞結合活性のための少なくとも2つのCBDと組み合わせることにより構築される。本発明に係る人工ペプチドグリカン結合タンパク質は、Sambrookら(Molecular cloning. A laboratory manual; 2nd ed. Cold Spring Harbor Laboratory Press 1989)において記述されているような組換えタンパク質のクローニングおよび産生のための標準的な技術を用いて、細胞結合活性のための少なくとも2つのCBDを組み合わせることにより構築される。少なくとも2つのCBDは、シャッフルされたキメラ酵素をもたらす異なるペプチドグリカン溶解酵素に由来していてもよく、または1つの天然に存在する酵素由来のCBDを増倍してもよく、または両方の組み合わせであってもよい。主として、すべての天然に存在するペプチドグリカン溶解酵素が、EADおよびCBDドメインの供給のための潜在的候補である。
【0027】
ペプチドグリカン溶解酵素は好ましくは、アミダーゼ_5(バクテリオファージペプチドグリカンヒドロラーゼ、pfam05382)、アミダーゼ_2(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01510)、アミダーゼ_3(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01520)、Transgly(トランスグリコシラーゼ、pfam00912)、ペプチダーゼ_M23(ペプチダーゼファミリーM23、pfam01551)、エンドリシン_オートリシン(CD00737)、ヒドロラーゼ_2(細胞壁ヒドロラーゼ、pfam07486)、CHAP(アミダーゼ、pfam05257)、トランスグリコシラーゼ(トランスグリコシラーゼ様ドメイン、pfam06737)、MtlB(膜結合型溶解性ムレイントランスグリコシラーゼB、COG2951)、MtlA(膜結合型溶解性ムレイントランスグリコシラーゼA、COG2821)、MtlE(膜結合型溶解性ムレイントランスグリコシラーゼE、COG0741)、バクテリオファージ_λ_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、CD00736)、ペプチダーゼ_M74(ペニシリン非感受性ムレインエンドペプチダーゼ、pfam03411)、SLT(トランスグリコシラーゼSLT、pfam01464)、Lys(C型リゾチーム/α-ラクトアルブミンファミリー、pfam00062)、COG5632(N-アセチルムラモイル-L-アラニンアミダーゼ、COG5632)、MepA(ムレインエンドペプチダーゼ、COG3770)、COG1215(グリコシルトランスフェラーゼ、COG1215)、AmiC(N-アセチルムラモイル-L-アラニンアミダーゼ、COG0860)、Spr(細胞壁結合型ヒドロラーゼ、COG0791)、バクテリオファージ_T4様_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、cd00735)、LT_GEWL(溶解性トランスグリコシラーゼ(LT)およびガチョウ卵白リゾチーム(GEWL)ドメイン、cd00254)、ペプチダーゼ_S66(LD-カルボキシペプチダーゼ、pfam02016)、グリコ_ヒドロ_70(グリコシルヒドロラーゼファミリー70、pfam02324)、グリコ_ヒドロ_25(グリコシルヒドロラーゼファミリー25)、VanY(D-アラニル-D-アラニンカルボキシペプチダーゼ、pfam02557)、およびLYZ2(リゾチームサブファミリー2、smart 00047)から構成される群より選択される少なくとも1つのEADを含む。
【0028】
ペプチドグリカン溶解酵素は好ましくは、SH3_5(細菌SH3ドメイン、pfam08460)、SH3_4(細菌SH3ドメイン、pfam06347)、SH3_3(細菌SH3ドメイン、pfam08239)、SH3b(細菌SH3ドメインホモログ、smart00287)、LysM(細胞壁分解に関与する種々の酵素において見出されるLysMドメイン、pfam01476およびcd00118)、PG_結合_1(推定上のペプチドグリカン結合ドメイン、pfam01471)、PG_結合_2(推定上のペプチドグリカン結合ドメイン、pfam08823)、MtlA(ムレイン分解トランスグリコシラーゼ由来のペプチドグリカン結合ドメイン、pfam03462)、Cpl-7(Cpl-7リゾチームのC末端ドメイン、pfam08230)、CW_結合_1(推定上の細胞壁結合リピート、pfam01473)、LytB(推定上の細胞壁結合ドメイン、COG2247)、およびLytE(LysMリピート、COG1388)から構成される群より選択される少なくとも1つのCBDを含む。
【0029】
好ましくは、上述のドメインは、約15〜約250アミノ酸残基の範囲のアミノ酸残基長を有し、約20〜約200アミノ酸残基の範囲の長さが好ましい。例として、約15〜約40アミノ酸残基長のドメインが、LysMドメインまたはコリン結合を担うCW_結合_1モチーフなどのペプチドグリカン結合ドメインにおいて見出される。これらの小さなドメインは、野生型細胞壁溶解酵素においても天然の反復モチーフとしてしばしば見出される。これらのドメインは、シャッフルされたキメラの人工ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質を作製するために、他の細胞壁溶解酵素由来の追加的なCBDと組み合わせることができる。
【0030】
通常、ペプチドグリカン溶解酵素の完全なEADまたはCBDドメインは、上述の保存されたドメインより大きい。優先的に、EADまたはCBDは、約50残基〜約400残基長の範囲である。各EADおよびCBDは、ペプチドグリカン溶解または細菌細胞結合の機能を示すために少なくとも1つの機能ドメインを含有するが、1つより多い機能ドメインおよび未知の機能を有する追加的な配列セグメントもまた含むことができる。ペプチドグリカン結合酵素のEADおよびCBDドメインは、上述の保存されたドメインによって常に規定されるわけではない。上述の保存されたドメインのいずれも見出されないが細菌細胞に結合しかつ細菌細胞を溶解する公知のペプチドグリカン結合酵素(例えばPly118)もまた存在する。潜在的ドメインがEADまたはCBDとして機能するか否かは、適切な機能アッセイ(例えば、ペプチドグリカン溶解(EAD)については測光溶解アッセイ、プレート溶解アッセイ、または最小殺菌濃度(MBC)の測定、および細胞結合(CBD)については細胞結合アッセイ、蛍光顕微鏡法、または結合親和性の測定)を用いて試験することができる。EADおよびCBDのドメインの境界は、配列の間の局所的類似性の領域を見出す局所的アラインメント検索手段(例えば、NCBIのBLAST、Altschul et al., 1997, Nucleic Acids Res. 17, 3389-3402)により規定することができる。加えて、多数のペプチドグリカン溶解酵素が、それらのEADおよびCBDドメインに関して既に記述されている。
【0031】
好ましくは、本発明のペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質は、Ply500、Ply511、Ply118、Ply100、PlyP40、Ply3626、phiLM4エンドリシン、PlyCD119、PlyPSAa、Ply21、PlyBA、Ply12、PlyP35、PlyPH、PlyL、PlyB、phi11エンドリシン、phi MR11エンドリシン、phi12エンドリシン、S.アウレウスファージPVLアミダーゼ、plypitti26、ΦSA2usaエンドリシン、スタフィロコッカス・ワーネリMファージΦWMY PlyGBSのエンドリシン、B30エンドリシン、Cpl-1、Cpl-7、Cpl-9、PlyG、PlyC、palアミダーゼ、Fab25、Fab20、エンテロコッカス・フェカリスV583プロファージ由来のエンドリシン、リゾスタフィン、ファージPL-1アミダーゼ、S.カピティス(S. capitis)ALE-1エンドペプチダーゼ、ムタノリシン(ストレプトマイセス・グロビスポラス(Streptomyces globisporus)ATCC 21553のN-アセチルムラミダーゼ)、エンテロリシンA(エンテロコッカス・フェカリスLMG 2333由来の細胞壁分解バクテリオシン)、LysK、LytM、L.モノサイトゲネス由来のAmiオートリシン、シュードモナス・エルギノーサファージΦKZおよびELのエンドリシン、T4リゾチーム、gp61ムラミダーゼ、およびSTM0016ムラミダーゼからなる群より選択される野生型ペプチドグリカン溶解酵素に由来するEADおよびCBDから構成される。
【0032】
野生型ペプチドグリカン溶解酵素PlyP40は、その野生型形態において344アミノ酸残基の長さを有する。それは、他の公知のエンドリシンと最小の相同性のみを有する2つの機能ドメインを所有する。1〜200位のN末端アミノ酸残基は、SEQ ID NO: 103に示される酵素活性ドメイン(EAD)に相当する。PlyP40の細胞結合ドメイン(CBD)は、SEQ ID NO: 104に示されるC末端に位置する227〜344のアミノ酸残基を含む。従って、野生型ペプチドグリカン溶解酵素PlyP40に由来するEADは、好ましくはSEQ ID NO: 103記載のアミノ酸配列を含み、一方野生型ペプチドグリカン溶解酵素PlyP40に由来するCBDは、好ましくはSEQ ID NO: 104記載のアミノ酸配列を含む。
【0033】
本発明に係る酵素およびタンパク質を構築するための天然に存在するペプチドグリカン溶解酵素由来の断片は、上述のような保存された機能ドメインの予測より決定された単なる配列セグメントを併せ持たなくてもよいが、好ましくは異なる機能モジュールを連結する適切なリンカー配列を付加する。リンカー配列は、規定された機能ドメインの近隣の野生型配列に由来してもよく、または当技術分野より公知である外部の適切なリンカー配列であってもよい。適切なリンカーは例えば、x線構造より定義されたリステリアエンドリシンPlyPSA(Korndorfer et al., 2006, J. Mol. Biol., 364, 678-689)由来の配列AAKNPNまたはTGKTVAAKNPNRHS(SEQ ID No: 61および11)を有する短いドメインリンカーである。ポリグリシンリンカーもまた、柔軟なドメインリンカーとして働くことが当技術分野において公知である。好ましいリンカーはまた、グリシンおよびアラニンに富むリンカーである。グリシンおよびアラニンに富むリンカーについての特定の配列は、SEQ ID NO:63、64、および65として示される。例えばエンテロリシンAにおいて、天然のリンカーとして生じるプロリンおよびスレオニンに富む配列もまた好ましい(SEQ ID NO:66)。プロリンおよびスレオニンに富むリンカー配列は、コンセンサスモチーフ(PT)xPまたは(PT)xT(式中、xは1〜10の範囲の整数を意味する)により記述することができる。別のリンカーの可能性は、Crouxら(1993, Molec. Microbiol., 9, 1019-1025)において記述されているEADとCBDとの間のいわゆる「ジャンクションゾーン」である。当業者は、野生型酵素から取り出される機能ドメインについて適切な境界を予測するためのいくつかの方法、例えば、2次構造予測、ドメインリンカーの予測、タンパク質の高解像度X線およびNMR構造におけるタンパク質の3Dモデルの検査、またはドメインリンカーおよび境界の検査を承知している。適切な方法は、例えば、Garnier et al., 1996, Methods in Enzymology 266, 540-553;Miyazaki et al., 2002, J. Struct. Funct. Genomics, 15, 37-51;George und Heringa, 2003, Protein Eng. 15, 871-879;Bae et al., 2005, Bioinformatics, 21, 2264-2270、Altschul et al., 1997, Nucleic Acids Res. 17, 3389-3402;Schwede et al., 2003, Nucleic Acids Research 31, 3381-3385. Lund et al, CPHmodels 2.0: X3M a Computer Program to Extract 3D Models. Abstract at the CASP5 conferenceA102, 2002において記述されている。EADとCBDドメインとの間、またはCBDとCBDドメインとの間のポリペプチドリンカーの長さは、約5〜約150アミノ酸残基、優先的には約6〜約60アミノ酸残基の範囲である。
【0034】
好ましくは、本発明に係るペプチドグリカン溶解酵素におけるEADおよびCBDの組み合わせのための順序は、N末端からC末端へEAD-CBD1-CBD2(-CBDN、N=3またはそれ以上)である。少なくとも2番目のEADがN末端またはC末端においてEADの隣に付加されるバリアントもまた好ましい。少なくとも2つのCBDがN末端に、または中間に位置するEADと共にN末端およびC末端に位置するバリアントもまた好ましい。加えて、両方ともN末端、C末端、または中間に位置し得るが、特にN末端で好ましいマーカー配列またはタグが含まれ得る。
【0035】
本発明に係るペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質は、それらが由来する野生型酵素と比較して新たな特性を示す。
【0036】
ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の結合範囲は、認識される細菌宿主範囲を決定する。天然に存在するペプチドグリカン酵素の大部分は、比較的狭い宿主範囲を示す。ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の技術的適用のためには、それぞれの適用に依存して増加した数の細菌株または細菌種が殺傷、捕捉、または検出され得るように、タンパク質の宿主範囲を拡大することがしばしば有利である。1つのタンパク質内に含まれる拡大した宿主範囲は、同様の適用のための2つまたはそれ以上のタンパク質の使用を回避し、タンパク質産生についての削減された費用、異なるタンパク質について条件を最適化するための削減された努力、より単純な医学的認可手続き、および削減された免疫原性の利点を有する。例えばブドウ球菌(Staphylococci)および腸球菌(Enterococci)を組み合わせる拡大した宿主範囲は、両方の属の多耐性株が増加する問題である院内感染の治療または予防において有用である。拡大した宿主範囲はまた、細菌検出、または食物から有害な細菌を除去する方法においても有用である。例えば、病原性株はリステリアのすべての血清型内で見出される。しかしながら、天然に存在するいずれのリステリアエンドリシンも、すべての血清型由来の細胞を溶解することはできない。本発明に係る1つより多いCBDを併せ持つペプチドグリカン溶解酵素は、すべての血清型を溶解することができる。天然に存在するタンパク質と比較して拡大していないが、何らかの形で変化した宿主範囲は、細菌細胞の与えられたセットを溶解、捕捉、または検出するためのテーラードタンパク質を必要とする適用に有用である可能性がある。ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の結合範囲は、当技術分野で公知のアッセイを用いて、または実施例において説明されるプレート溶解アッセイ、測光溶解アッセイ、結合アッセイ、もしくは蛍光顕微鏡法を用いて測定することができる。
【0037】
ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質の野生型タンパク質と比較して増加した結合親和性は、細胞溶解、細胞捕捉、および検出などの細菌細胞の結合に依拠する任意の技術的適用のために必要とされるタンパク質の量を減少させることを助長する。これは費用を削減し、ならびに治療的適用における免疫反応および潜在的な副作用を最小化する。細菌細胞の捕捉に依拠する適用において、アッセイはバックグラウンドシグナルを減少させる洗浄段階についてより感受性が低く、インキュベーション時間を削減することができ、および検出アッセイはより感度が高い。増加した結合親和性は、当技術分野で公知のアッセイを用いて、または実施例において説明される表面プラズモン共鳴解析もしくは最小殺菌濃度の測定のためのアッセイを用いて測定することができる。
【0038】
ペプチドグリカン溶解酵素の野生型酵素と比較して増加した溶解活性は、感染の防護および治療、衛生、細菌検出における最初の段階としての細胞溶解、または食物、飼料、化粧料などからの病原性細菌の除去などの細菌細胞の溶解に依拠するすべての適用において有用である。それぞれの適用のために必要とされるタンパク質の量は、野生型タンパク質と比較して削減され、費用を削減し、ならびに治療的適用における免疫反応および潜在的な副作用を最小化する。野生型と比較して変化した溶解活性は、例えば人工酵素の異なる最適pH、または他の緩衝液組成物でのより高い溶解活性(例えば、高いイオン強度、有機溶媒の存在下における活性、特定のイオンの存在下における活性)であり得る。これはまた、血液、ヒト血清、または他の医学的試料などの特定の試料におけるより高い活性も含む。例えば酪農業において、食物製品または食物加工における中間産物はしばしば低いpH値を有するため、より低いpHに移行した人工酵素の最適pHは、例えば食品産業における人工酵素の適用にとって興味深い。高い塩濃度の下での酵素機能もまた、食品産業、例えばチーズ生産において重要である。増加したかまたは変化した溶解活性は、当技術分野で公知のアッセイを用いて、または実施例において説明されるプレート溶解アッセイ、測光溶解アッセイ、もしくは最小殺菌濃度の測定のためのアッセイを用いて測定することができる。
【0039】
1つの局面において、本発明は、リステリア細菌を溶解、捕捉、および/または検出するために使用することができる人工ペプチドグリカン溶解酵素またはペプチドグリカン結合タンパク質に関する。本発明者らは、野生型酵素と比較して新たな特性を示す人工ペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質を作製するために、上述の方法を用いてリステリアエンドリシンply500(SEQ ID NO:1)、ply118(SEQ ID NO:3)、およびplyP35(SEQ ID NO:5)のドメインを組み合わせた。ply500は、EADとして、保存されたD-アラニル-D-アラニンカルボキシペプチダーゼ(VanY;pfam02557)ドメインを含む。ply500のCBDは約H133〜Q150の範囲のアミノ酸残基で始まり、かつK289で終わる。Ply118については、アミノ酸配列内に、保存されたドメインは見出されなかった。しかしながら、相同なペプチドグリカン溶解酵素との配列アラインメントより、ply118のCBDは約D90〜K180の範囲のアミノ酸残基で始まり、かつアミノ酸残基K289で終わることが導き出された。CBD118についての好ましいN末端開始アミノ酸残基は、D90、K100、G127、S151、N161、またはK180である。plyP35もまた、EADとして、保存されたD-アラニル-D-アラニンカルボキシペプチダーゼ(VanY;pfam02557)を含む。plyP35のCBDは約P130〜N156の範囲のアミノ酸残基で始まり、かつY281〜K291の範囲のアミノ酸残基で終わる。CBDP35についての好ましいN末端開始アミノ酸残基は、P130、A134、K143、およびN156である。CBDP35についての好ましいC末端アミノ酸残基は、Y281、L286、およびK291である。
【0040】
本発明に係る好ましいペプチドグリカン結合タンパク質は、追加的なリンカー配列を用いずに連結されたN末端位置のply500のCBD(アミノ酸残基H133〜K289)およびC末端位置のply118のCBD(アミノ酸残基D90〜I281)を含むCBD500-118(SEQ ID NO:7)、ならびに、ply500のCBD(アミノ酸残基Q150〜K289)およびply118のCBD(アミノ酸残基K100〜I281)を、2つのドメインを連結するリンカー(L)と共に含むCBD500L118(SEQ ID NO:9)である。この場合のドメインリンカーは、リステリアエンドリシンPlyPSAのアミノ酸残基173〜186に対応するアミノ酸配列TGKTVAAKNPNRHS(SEQ ID NO:11)を有するplyPSAリンカー領域である(Korndorfer et al., 2006, J. Mol. Biol., 364, 678-689)。
【0041】
本発明に係るさらに好ましい態様は、追加的なリンカー配列を用いずに連結されたN末端位置のply500のCBD(アミノ酸残基H133〜K289)およびC末端位置のply118のCBD(アミノ酸残基D90〜I281)を含む人工ペプチドグリカン結合タンパク質CBD118-500(SEQ ID NO:13)、ならびに、ply118のCBD(アミノ酸残基K100〜I281)およびply500のCBD(アミノ酸残基Q150〜K289)を2つのドメインを連結するリンカー(L)と共に含むCBD118L500(SEQ ID NO:15)である。この場合のドメインリンカーはplyPSAリンカー領域(SEQ ID NO:11)である。
【0042】
ペプチドグリカン結合タンパク質CBD500-118、CBD500L118、CBD118-500、およびCBD118L500はすべて、CBDドメインが由来した野生型酵素ply500およびply118と比較して、宿主範囲および結合活性に関して変化した細胞結合活性を示す。構築物CBD500L118およびCBD118L500の細胞結合活性は、2つのドメインの間のリンカーが、野生型酵素の結合特異性を併せ持つ拡大した宿主範囲を達成するのを助長することを表す。
【0043】
本発明に係るさらに好ましいペプチドグリカン結合タンパク質は、N末端位置のply500のCBD(アミノ酸残基Q150〜K289)およびC末端位置のplyP35のCBD(アミノ酸残基P130〜K291)を含むCBD500-P35(SEQ ID NO:17)、ならびに、N末端にplyP35のCBD(アミノ酸残基P130〜K291)およびC末端にply500のCBD(アミノ酸残基Q150〜K289)を含む、逆方向のCBDを有するタンパク質CBDP35-500(SEQ ID NO:19)である。この場合、plyP35のCBDについての断片がplyP35の内部ドメインリンカーを含むため、CBDは外部リンカー配列により連結されない。
【0044】
人工ペプチドグリカン結合タンパク質CBD500-P35およびCBDP35-500は両方とも、CBDドメインが由来した野生型酵素ply500およびplyP35と比較して拡大した宿主範囲を示す。両方のキメラタンパク質は、1つのタンパク質内に2つの野生型酵素の異なる結合特異性を併せ持つ。CBDの方向はこの場合相違を生じない。CBDは両方ともN末端およびC末端に位置することができる。
【0045】
本発明に係るさらに好ましいペプチドグリカン結合タンパク質は、ply500のCBD(アミノ酸残基Q150〜K289)の重複を示すCBD500-500(SEQ ID NO:21)、およびply500において天然に存在するCBDの重複を示す人工ペプチドグリカン溶解酵素EAD-CBD500-500(SEQ ID NO:23)である。
【0046】
本発明に係る両方のタンパク質は、野生型と比較してリステリア細胞に対してより高い結合親和性を示し、およびEAD-CBD500-500は加えて、ply500と比較して高い塩条件の下で増加した溶解活性を示す。
【0047】
別の局面において、本発明は、腸球菌細菌を溶解、捕捉、または検出するために使用することができる人工ペプチドグリカン溶解酵素に関する。本発明者らは、野生型酵素と比較して新たな特性を示す人工ペプチドグリカン溶解酵素およびペプチドグリカン結合タンパク質を作製するために、上述の方法を用いて腸球菌エンドリシンFab25VL(SEQ ID NO:25)およびFab20VL(SEQ ID NO:27)のドメインを組み合わせた。
【0048】
Fab25VLは、E.フェシウム(E. faecium)種由来の細菌に優先的に結合しかつ溶解するが、E.フェカリス種由来のいくつかの株にもまた結合しかつ溶解する317アミノ酸残基長のエンドリシンである。Fab25VLのN末端に位置するEAD(アミノ酸残基1〜167)は、N-アセチルムラモイル-L-アラニン-アミダーゼとして機能する保存されたアミダーゼ_2ドメインを示す。CBDはアミノ酸残基200〜317を含む。2つのドメインの間に、アミノ酸残基168〜199を含むリンカー領域が観察される。Fab20VLは、E.フェカリス種由来の細菌に優先的に結合しかつ溶解する365アミノ酸残基長のエンドリシンである。Fab20VLのN末端に位置するEAD(アミノ酸残基40〜194)もまた、N-アセチルムラモイル-L-アラニン-アミダーゼとして機能する保存されたアミダーゼ_2ドメインを示す。Fab20VLのCBD(アミノ酸残基215〜365)は、ブドウ球菌および連鎖球菌ファージ由来のペプチドグリカン溶解酵素と相同性を示す細菌SH3ドメインをC末端部分に含む。アミノ酸残基1〜19の欠失を有する、Fab20VLのN末端が切断されたバリアントであるFab20K(SEQ ID NO:29)を構築した。これはFab20VLと比較して大腸菌(E.coli)においてより良好な発現を示した。
【0049】
本発明に係る好ましいペプチドグリカン溶解酵素は、Fab25のEADおよびCBD(アミノ酸残基1〜317)を、Fab20VLのCBD(アミノ酸残基215〜365)およびFab20VLに由来する短いリンカーセグメント(アミノ酸残基200〜214)と共に組み合わせるSEQ ID NO:31である。構築物はEADFab25_CBD25_CBD20と表される。EADFab25_CBD25_CBD20は、野生型酵素と比較した新たな特徴として、拡大した宿主範囲、生細胞についての増加した溶解活性、および増加した結合活性を示す。
【0050】
本発明に係るさらに好ましいペプチドグリカン結合酵素は、ブドウ球菌細菌を検出することおよび結合することができる少なくとも2つのEADおよび少なくとも2つのCBDから構成される。
【0051】
好ましくは本発明に係るペプチドグリカン溶解タンパク質は、Hisタグ(Nieba et al., 1997, Anal. Biochem., 252, 217-228)、Strepタグ(Voss & Skerra, 1997, Protein Eng., 10, 975-982)、Aviタグ(米国特許第5,723,584号;米国特許第5,874239号)、Mycタグ(Evan et al., Mol&Cell Biol, 5, 3610-3616)、GSTタグ(Peng et al. 1993, Protein Expr. Purif., 412, 95-100)、JSタグ(国際公開公報第2008/077397号)、システインタグ(欧州特許第1399551号、SEQ ID NO:6および7)、HAタグ(アミノ酸配列EQKLISEEDL)、FLAGタグ(Hopp et al., Bio/Technology. 1988;6:1204-1210)、または当技術分野において公知である他のタグなどのタグを含む。好ましくはタグは、本発明に係るペプチドグリカン溶解タンパク質のC末端またはN末端、最も好ましくはN末端に結合される。タグは、ペプチドグリカン溶解タンパク質の発現および/または精製を促進するために、本発明に係るペプチドグリカン溶解タンパク質を表面に固定化するために、または、例えば様々なELISAアッセイフォーマットにおいて抗体結合による、ペプチドグリカン溶解タンパク質の検出用のマーカーとして働くために有用であり得る。
【0052】
好ましくは本発明に係るペプチドグリカン溶解タンパク質は、ビオチン、ストレプトアビジン、GFP(緑色蛍光タンパク質)、YFP(黄色蛍光タンパク質)、シアン蛍光タンパク質、RedStarタンパク質、または他の蛍光マーカー、アルカリホスファターゼ、西洋ワサビペルオキシダーゼ、免疫金標識、スピン標識、または当技術分野において公知である他のマーカーおよび標識などの、マーカーまたは標識部分を含む。マーカーは、それらがポリペプチド性質の場合は組換え法により、またはポリペプチド残基の化学修飾により翻訳後に付着され得る。マーカーまたは標識は、本発明に係るペプチドグリカン溶解タンパク質が診断において使用される際に、それらを検出するために特に有用である。
【0053】
さらに好ましいペプチドグリカン溶解タンパク質は、構築物HGFP-CBD118-500(SEQ ID NO:33)、HGFP-CBD500-118(SEQ ID NO:35)、HGFP-CBD118L500(SEQ ID NO:37)、HGFP-CBD500L118(SEQ ID NO:39)、HGFP-CBD500-P35(SEQ ID NO:41)、HGFP-CBDP35-500(SEQ ID NO:43)、HCBD500-GFP-CBD118(SEQ ID NO:45)、HCBD118-GFP-CBD500(SEQ ID NO:47)、HGFP-CBD500-500(SEQ ID NO:49)、およびHEAD-CBD500-500(SEQ ID NO:51)である。Hは6個のヒスチジンを含むhisタグを表し(SEQ ID NO:53)、およびGFPは蛍光マーカーとして導入された緑色蛍光タンパク質を表す(SEQ ID NO:55)。
【0054】
hisタグおよびGFPマーカーを含む上述の融合構築物のすべては、リステリア細胞結合活性を示し、ならびにペプチドグリカン溶解酵素は追加的に溶解活性を示す。しかしながら、構築物HCBD500-GFP-CBD118およびHCBD118-GFP-CBD500は、細胞結合活性、特にCBD118の細胞結合活性がこれらの構築物において減少するため、2つのCBDドメインの間での位置付けと比較してN末端位置のGFPマーカーが好ましいことを示す。
【0055】
まとめると、本発明の目的は、天然に存在するドメインのシャッフリングまたは増倍による機能ドメインの人工的組み合わせにより、野生型ペプチドグリカン溶解酵素の特性を変化および改善することである。本発明に係るペプチドグリカン溶解酵素は、天然に存在するタンパク質の結合範囲を拡大するために、および/または細菌細胞壁への結合親和性を増加するために、および/または溶解活性を増加または修飾するために、少なくとも1つのEADおよび少なくとも2つのCBDから構成される。本発明に係るペプチドグリカン結合タンパク質は、天然に存在するタンパク質の結合範囲を拡大するために、および/または細菌細胞壁への結合親和性を増加するために、少なくとも2つのCBDから構成される。本発明に係るポリペプチドにおいて、少なくとも2つのCBDは、2つの異なるペプチドグリカン溶解酵素に由来する(ドメインシャッフリング)か、または天然に存在するCBDの増倍による。
【0056】
SEQ ID NO: 7、9、13、15、17、19、21、23、31、33、35、37、39、41、43、45、47、49、51、71、73、75、77、79、81、83、85、87、89、91、93、95、97、99、101に示されるような組換えポリペプチドが好ましい。
【0057】
本発明の別の好ましい態様において、上記に列挙されたような本発明に係る組換えポリペプチドは、アミノ酸配列の修飾および/または変化を含む。そのような変化および/または修飾は、欠失、挿入および付加、置換、もしくはそれらの組み合わせなどの変異、ならびに/またはアミノ酸残基の化学変化、例えば、ビオチン化、アセチル化、ペグ化、アミノ基、SH基、もしくはカルボキシル基の化学変化を含み得る。該修飾したおよび/または変化した組換えポリペプチドは、上記に列挙されたようなそれぞれの組換えポリペプチドの単一ドメインの活性を示す。しかしながら、単一ドメインの該活性は各々、上記に列挙されたようなそれぞれの組換えポリペプチドの単一ドメインの活性より高く、またはより低くあり得る。特に、単一ドメインの該活性は、上記に列挙されたようなそれぞれの組換えポリペプチドの単一ドメインの活性の約10、20、30、40、50、60、70、80、90、100、110、120、130、140、150、160、170、180、190、200%またはそれより高くてもよい。単一ドメインの活性は、本明細書において既に説明されたようなCBDおよびEADの活性を測定するためのアッセイにより測定することができる。
【0058】
さらなる局面において、本発明は、本発明に係るポリペプチドをコードするヌクレオチド配列を含む核酸分子に関する。
【0059】
SEQ ID NO: 8、10、14、16、18、20、22、24、32、34、36、38、40、42、44、46、48、50、52、72、74、76、78、80、82、84、86、88、90、92、94、96、98、100、102に示されるようなヌクレオチド配列を含む核酸分子が好ましい。
【0060】
さらなる局面において、本発明は、本発明の核酸配列を含むベクターに関する。好ましくは、該ベクターは、適切な宿主細胞において本発明の該ポリペプチドの発現を提供する。該宿主細胞は、単なる生物工学的な理由、例えば収率、溶解性、費用などのために選択され得るが、該細胞が対象に投与されるべき場合は、医学的観点からも、例えば非病原性細菌または酵母、ヒト細胞が選択され得る。該ベクターは、本発明に係る前記ポリペプチドの構成性または誘導性の発現を提供し得る。
【0061】
本発明のさらなる局面において、上述のポリペプチドおよび/または細胞は、対象における細菌感染の処置または予防のための、特に、ブドウ球菌(例えば、S.アウレウス、S.アウレウス(MRSA)、S.エピデルミディス(S. epidermidis)、S.ヘモリチカス(S. haemolyticus)、S.シミュランス、S.サプロフィチカス(S. saprophyticus)、S.クロモゲネス(S. chromogenes)、S.ヒイカス(S. hyicus)、S.ワーネリ、および/またはS.キシローサス(S. xylosus))、腸球菌(例えば、エンテロコッカス・フェシウム、E.フェシウム(VRE)、エンテロコッカス・フェカリス)、連鎖球菌(ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)、S.ニューモニエ、S.ミュータンス(S. mutans)、S.ウベリス(S. uberis)、S.アガラクティエ、S.ディスガラクティエ(S. dysgalactiae)、ランスフィールド群A、B、Cの連鎖球菌)、クロストリジウム属(例えば、C.パーフリンジェンス(C. perfringens)、C.ディフィシレ(C. difficile)、C.テタニ(C. tetani)、C.ボツリナム(C. botulinum)、C.チロブチリクム(C. tyrobutyricum))、バチルス属(例えば、バチルス・アンスラシス、B.セレウス(B. cereus))、リステリア属(例えば、L.モノサイトゲネス、L.イノキュア(L. innocua))、ヘモフィルス・インフルエンザ(Haemophilus influenza)、コリネバクテリウム・ジフテリエ(Corynebacterium diphteriae)、プロピオニバクテリウム・アクネ(Propionibacterium acne)、マイコバクテリア(例えば、マイコバクテリウム・ツベルクローシス(Mycobacterium tuberculosis)、M.ボビス(M. bovis))などのグラム陽性菌により引き起こされる感染の処置または予防のための方法において使用される。または、本発明に係るポリペプチドおよび/または細胞は、対象における細菌感染の処置または予防のための、特に、腸内細菌科(Enterobacteriaceae)(エシェリキア属(Escherichia)、特に大腸菌(E. coli)、サルモネラ属(Salmonella)、シゲラ属(Shigella)、シトロバクター属(Citrobacter)、エドワードシエラ属(Edwardsiella)、エンテロバクター属(Enterobacter)、ハフニア属(Hafnia)、クレブシエラ属(Klebsiella)、特にK.ニューモニエ(K. pneumoniae)、モルガネラ属(Morganella)、プロテウス属(Proteus)、プロビデンシア属(Providencia)、セラチア属(Serratia)、エルシニア属(Yersinia))、シュードモナス科(Pseudomonadaceae)(シュードモナス属(Pseudomonas)、特にP.エルギノーサ、バークホルデリア属(Burkholderia)、ステノトロフォモナス属(Stenotrophomonas)、シェワネラ属(Shewanella)、スフィンゴモナス属(Sphingomonas)、コマモナス属(Comamonas))、ナイセリア属(Neisseria)、モラクセラ属(Moraxella)、ビブリオ属(Vibrio)、アエロモナス属(Aeromonas)、ブルセラ属(Brucella)、フランシセラ属(Francisella)、ボルデテラ属(Bordetella)、レジオネラ属(Legionella)、バルトネラ属(Bartonella)、コクシエラ属(Coxiella)、ヘモフィルス属(Haemophilus)、パスツレラ属(Pasteurella)、マンヘミア属(Mannheimia)、アクチノバチルス属(Actinobacillus)、ガードネレラ属(Gardnerella)、スピロヘータ科(Spirochaetaceae)(トレポネーマ属(Treponema)およびボレリア属(Borrelia))、レプトスピラ科(Leptospiraceae)、カンピロバクター属(Campylobacter)、ヘリコバクター属(Helicobacter)、スピリルム属(Spirillum)、ストレプトバチルス属(Streptobacillus)、バクテロイデス科(Bacteroidaceae)(バクテロイデス属(Bacteroides)、フソバクテリウム属(Fusobacterium)、プレボテラ属(Prevotella)、ポルフィロモナス属(Porphyromonas))、アシネトバクター属(Acinetobacter)、特にA.バウマニ(A. baumanii)などのヒトまたは動物にとって病原性である株を含む細菌群、科、属、または種のグラム陰性菌により引き起こされる感染の処置または予防のための方法において使用される。
【0062】
前記対象は、ヒト対象または動物、特に畜牛などの畜産および/または酪農業において使用される動物であり得る。前記処置の方法は、感染の部位、または感染に対して予防的に処置される部位へと十分な量の本発明の前記ポリペプチドを適用することを包含する。
【0063】
特に、前記処置の方法は、皮膚、軟部組織、呼吸器系、肺、消化管、眼、耳、歯、鼻咽腔、口、骨、膣、菌血症および/または心内膜炎の創傷の感染の処置または予防のためであり得る。
【0064】
さらに好ましい態様において、本発明に係るポリペプチドは、動物、特に家畜および乳牛におけるブドウ球菌の感染の処置(または予防)の方法において使用される。特に、本出願のポリペプチドは、ウシ乳房炎、特にS.アウレウス、S.エピデルミディス、S.シミュランス、S.クロモゲネス、S.ヒイカス、S.ワーネリ、およびS.キシローサスにより引き起こされるウシ乳房炎の処置(または予防)の方法における使用に適する。
【0065】
さらに、本発明のポリペプチドは、特に外科手術の前もしくは後、または例えば血液透析の間の清浄剤として予防的に使用され得る。同様に、未熟児および免疫不全の人、または補綴装置を必要とする対象が、予防的または急性感染の間のいずれかに本発明のポリペプチドで処置され得る。同様の文脈において、特にスタフィロコッカス・アウレウス(MRSA)、エンテロコッカス・フェシウム(VRE)、シュードモナス・エルギノーサ(FQRP)、または抗生物質耐性クロストリジウム・ディフィシレなどの抗生物質耐性株による院内感染が、予防的または急性期の間に本発明のポリペプチドで処置され得る。本態様において、本発明のポリペプチドは、界面活性剤、テンシド(tensids)、溶剤、抗生物質、ランチビオティクス、またはバクテリオシンなどの殺菌溶液において有用である他の構成要素と組み合わせても殺菌剤として使用され得る。
【0066】
特に好ましい態様において、本発明のポリペプチドは、処置される(または予防される)感染が多耐性細菌株、特に1つまたは複数の以下の抗生物質に対して耐性である株により引き起こされる場合、医学的処置のために使用される:ペニシリン、ストレプトマイシン、テトラサイクリン、メチシリン、セファロチン、ゲンタマイシン、セフォタキシム、セファロスポリン、バンコマイシン、リネゾリド、セフタジジム、イミペネム、またはダプトマイシン。さらに、本発明のポリペプチドは、抗生物質、ランチビオティクス、バクテリオシン、他のエンドリシンなどのような従来の抗菌剤と組み合わせて投与することにより処置の方法において使用され得る。
【0067】
本発明に係る処置(または予防)の方法において使用される投与量および投与の経路は、処置される特定の疾患/感染の部位に依存する。投与の経路は、例えば特定の態様において、経口、局所、鼻咽頭、非経口、静脈内、直腸、または任意の他の投与の経路であり得る。
【0068】
感染の部位(または感染する危険にさらされた部位)への本発明のポリペプチドの適用のために、ペプチドグリカン溶解酵素が感染の部位に到達するまでプロテアーゼ、酸化、免疫応答などのような環境の影響から保護されるような様式で、本発明のポリペプチドが製剤化され得る。
【0069】
従って、本発明のポリペプチドは、カプセル、糖衣丸、丸剤、坐剤、注射可能な溶液、または任意の他の医学的に妥当な生薬製剤として製剤化され得る。いくつかの態様において、これらの生薬製剤は、適切な担体、安定化剤、着香料、緩衝液、または他の適切な試薬を含み得る。
【0070】
例えば、局所適用のために、本発明のポリペプチドはローションまたは硬膏として投与され得る。
【0071】
鼻咽頭適用のために、本発明に係るポリペプチドは、鼻へのスプレーを介して適用されるように生理食塩水において製剤化され得る。
【0072】
腸の処置のために、例えばウシ乳房炎において、坐薬製剤を構想することができる。または、経口投与が考慮され得る。この場合、本発明のポリペプチドは、感染の部位に及ぶまで苛酷な消化環境から保護されなければならない。これは、例えば胃における消化の最初の段階を生き延び、および後に本発明のポリペプチドを腸環境中に分泌する細菌を担体として使用することにより達成することができる。
【0073】
すべての医学的適用は、病原性細菌に遭遇した際に特異的にかつ直ちに病原性細菌を溶解する本発明のポリペプチドの効果に依拠する。これは、病原性細菌および細菌負荷における減少をもたらすことにより処置される対象の健康状態に即時的な影響を有し、ならびに同時的に免疫系を救援する。従って、当業者が直面する主要な課題は、本発明のポリペプチドを、処置されるそれぞれの疾患のために的確に製剤化することである。この目的のために、通常、これらの適用について従来の薬物のために使用されるのと同様の生薬製剤を使用することができる。
【0074】
本発明のさらなる局面において、上述のポリペプチドおよび/または細胞は、担体物質を任意で含む薬学的組成物の成分である。
【0075】
もっとさらなる局面において、ポリペプチドおよび/または細胞は、化粧料組成物の一部である。上述のように、いくつかの細菌種は、皮膚などの患者の体の環境的に曝露された表面上に炎症を引き起こし得る。そのような炎症を予防するために、または前記細菌性病原体の軽症の症状発現を除去するために、既に存在するかまたは新しく定着する病原性細菌を溶解する目的で十分な量の本発明のポリペプチドを含む特別な化粧料調製物が使用され得る。
【0076】
さらなる局面において、本発明は、食材における、食物加工設備上での、食物加工工場における、棚および食物保管領域などの食材と接触する表面上での、ならびに、病原性細菌、通性病原性細菌、または他の望ましくない細菌が潜在的に食物材料に寄生し得るすべての他の状況における、本発明に係る前記ポリペプチドの使用に関する。
【0077】
本発明のさらなる局面は、細菌感染の診断における本発明に係る前記ポリペプチドの使用に関する。本局面において、本発明に係るポリペプチドは、病原性細菌を特異的に溶解する手段として使用される。本発明に係るポリペプチドによる細菌細胞の溶解は、Triton X-100などの界面活性剤、またはポリミキシンBなどの細菌細胞外被を弱くする他の添加物の添加により支持され得る。特異的な細胞溶解は、PCR、核酸ハイブリダイゼーション、もしくはNASBA(核酸配列ベース増幅)などの核酸ベースの方法、IMS、免疫蛍光法、もしくはELISA技術などの免疫学的方法、または、別個の細菌群もしくは種に特異的なタンパク質(例えば、腸内細菌にとってのβ-ガラクトシダーゼ、コアグラーゼ陽性株にとってのコアグラーゼ)を用いる酵素アッセイなどの細菌細胞の細胞内容物に依拠する他の方法を使用するその後の細菌の特異的検出のための最初の段階として必要とされる。
【0078】
本発明の別の局面は、試料由来の病原性細菌またはそうでなければ望ましくない細菌の結合、濃縮、除去、捕捉、および検出のための本発明に係るペプチドグリカン結合タンパク質の使用である。本発明に係る方法に関する試料は、検出、結合、濃縮、除去、または捕捉の標的である細菌を想定しているかまたは含有する任意の材料である。試料は、例えば、食物または飼料材料、表面材料、または、ヒトもしくは獣医学的診断プローブであり得る。細菌の検出は、本発明に係るペプチドグリカン結合タンパク質に付着されたマーカーの検出を介して、または前記タンパク質自体の検出により、例えばELISAなどの免疫学的方法により行われる。本発明に係る方法のために、本発明に係るペプチドグリカン結合タンパク質は、適切な支持構造、例えば、マイクロタイタープレート、試験ストリップ、スライド、ウエハ、フィルター材料、反応管、磁性物質、ガラスもしくはラテックス粒子、ピペットチップ、またはフロースルーセルチャンバー(flow-through cell chamber)上に固定化され得る。支持構造は、例えば、ポリスチレン、ポリプロピレン、ポリカーボネート、PMMA、酢酸セルロース、ニトロセルロース、ガラス、ケイ素ウエハ、ラテックスからなり得る。固定化は、吸着により、共有結合により、またはさらなるタンパク質により達成され得るが、共有結合が好ましい。固定化は機能的なものであり、すなわち、前記ペプチドグリカン結合タンパク質は支持材料に結合するが細菌にとって接触可能な構造を示すことが妥当である。
【実施例】
【0079】
実施例1.DNA技術およびクローニング手順
エンドリシンベースの融合タンパク質をコードするプラスミドの構築のために、Sambrookら(Molecular cloning. A laboratory manual; 2nd ed. Cold Spring Harbor Laboratory Press 1989)によるDNA技術およびクローニング手順を使用した。プラスミドpQE-30(QIAGEN)およびその誘導体pHGFP、pHGFP_CBD118、pHGFP_CBD500(Loessner et al. 2002)、ならびにpHEADPSA(Korndoerfer et al. 2006)を、N末端に6xHisタグが付加された人工融合タンパク質をコードするプラスミドの構築のためのベクター骨格として使用した(HはHisタグを意味する)。プラスミド中への断片の挿入のために必要とされる制限酵素部位は、プライマーを介して導入した。ダブルCBD融合構築物を、2つのCBD断片の別々の増幅およびその後のライゲーションによるか、または代替的にPCRベースのオーバーラップ伸長による遺伝子スプライシング(SOE PCR)法を介して2つの断片を融合することによるかのいずれかで作製した(Horton et al. 1990)。CBD118(SEQ ID NO:57)およびCBD500(SEQ ID NO:58)をコードする断片をEcoRI/MunI部位を介して両方の方向においてライゲーションし、ならびにその後pHGFPのSacI/SalI部位中へ挿入して、pHGFP_CBD118-500およびpHGFP_CBD500-118を産生した。プラスミドpHGFP_CBDP35-500およびpHGFP_CBD500-P35を同様の方法で作製した。pHGFP_CBD118L500およびpHGFP_CBD500L118の場合は、pHGFP中への挿入の前にSOE PCRによりPlyPSAリンカーをコードする断片を2つのCBDの間に導入した。pHCBD500_GFP_118およびpHCBD118_GFP_500のためとしては、5'CBDおよびGFP断片をKpnI部位を介してまたはSOE PCRにより最初に融合し、ならびにその後pHGFP_CBD118およびpHGFP_CBD500のBamHI/SacI部位中へそれぞれライゲーションして、これらのプラスミドの単なるGFP断片と置き換えた。pHGFP_CBD500-500の構築のためには、CBD500断片をpHGFP_CBD500のSacI部位中へクローニングし、CBD500の重複をもたらした。pHEAD_CBD500-500は、完全なply500遺伝子をpHGFP_CBD500のBamHI/SacI部位中へ挿入し、GFP断片と置き換えることにより作製した。すべての構築物について、遺伝子融合を可能にするために3'末端のもの以外のすべての終止コドンを削除した。TAAを一般に3'末端の終止コドンとして導入した。すべての構築物をヌクレオチド配列決定により確認した。
【0080】
実施例2.Hisタグ付き組換えタンパク質の過剰発現および精製
Hisタグ付き(それぞれの構築物において「H」により省略される)融合タンパク質の過剰発現を大腸菌XL1-Blue MRF'(Stratagene)において行った。それぞれの株を、プラスミド選択のために100μg/mlアンピシリンおよび30μg/mlテトラサイクリンを含有する修飾LB培地(15 g/lトリプトース、8 g/l酵母エキス、5 g/l NaCl)において30℃で増殖させ、ひとたび0.5のOD600に到達した後0.1〜1 mMのIPTGを誘導物質として添加した。30℃で4時間のさらなるインキュベーションの後、GFPドメインを含有するタンパク質を産生する培養物を、収集および250 mlの培養物あたり5 mlの緩衝液A(500 mM NaCl、50 mM Na2HPO4、5 mMイミダゾール、0.1% Tween 20、pH 8.0)における再懸濁の前に、一晩4℃で保管した。GFPが存在しなかった場合は、誘導4時間後に細胞をペレットにした。100 MPaでのフレンチプレス20Kセル(SLM Aminco)の2回の通過により細胞を破砕し、ならびに細胞破片を遠心分離および濾過(0.2μM PES膜、Millipore)により除去した。
【0081】
未加工の抽出物における6xHisタグ付き標的タンパク質を、Micro Biospinカラム(BIORAD)を用いてNi-NTA Superflow樹脂(QIAGEN)での固定化金属アフィニティークロマトグラフィー(IMAC)により精製した。緩衝液B(500 mM NaCl、50 mM Na2HPO4、250 mMイミダゾール、0.1% Tween 20、pH 8.0)が溶出緩衝液として働いた。精製したタンパク質を透析緩衝液(100 mM NaCl、50 mM NaH2PO4、0.005%〜0.1% Tween 20、pH 8.0)の2回の交換に対して透析し、濾過(0.2μM PES膜、Millipore)し、および50%(v/v)のグリセロールの添加後−20℃で保管した。各タンパク質について、過剰発現および精製の過程をSDS-PAGEにより解析し、およびタンパク質濃度を分光光度的に測定した(NanoDrop ND-1000 Spectrophotometer)。
【0082】
実施例3.結合アッセイおよび蛍光顕微鏡法
GFP-CBD融合タンパク質の結合特性を、すべての種および血清型のリステリア株の代表的なセット(表1)を用いて結合により検討した。PBST緩衝液(50 mM NaH2PO4、120 mM NaCl、pH 8.0、0.01% Tween 20)における各株の後期対数期細胞を、過剰のGFP-CBDタンパク質と共に5分間室温でインキュベーションした。緩衝液で2回洗浄した後、Axioplan顕微鏡、ならびに励起BP 450〜490 nm、ビームスプリッターFT 510 nm、および発光LP 520 nmのフィルターセット(Carl Zeiss AG)を用いた蛍光顕微鏡法のために細胞を調製した。標識された細胞の写真をLeica DFC320カメラを用いることにより取得した。各アッセイについて、4種類のスコアシステムを用いて目視検査により結合強度を評価した:++、+、(+)、および−はそれぞれ、強い、弱い、非常に弱い、および結合しないを示す。
【0083】
両方の細胞壁結合ドメインが両方の方向において互いに直接融合し、かつN末端のGFPドメインに付着している構築物HGFP_CBD500-118およびHGFP_CBD118-500は、両方とも試験された血清型4、5、および6株のすべての株に弱い結合を示した。これは単なるCBD500の結合パターンに相当するため、これらの結果は、CBD118がこれらの構築物において機能的でないことを示唆した。他方で、CBD500が機能性を保持するためにタンパク質のC末端に位置する必要はないことが示された。融合構築物における両方の結合ドメインの増強された柔軟性がCBD118を機能的にさせる可能性があると仮定して、本発明者らはCBDを分離するPlyPSAのリンカーペプチドを含むタンパク質HGFP_CBD500L118およびHGFP_CBD118L500を作製した。再び、両方の構築物は血清型4、5、および6株に属するすべての株を標識したが、追加的に7種類の血清型1/2株のうち4種類もまた試験された。蛍光顕微鏡法より、後者は、HGFP_CBD118について観察されたように主に極および隔壁で標識されることが観察された。対照的に、両方のタンパク質は、血清型4、5、および6の株をHGFP_CBD500のような細胞表面に渡る一様な分布において装飾(decorate)した。従って、これらのダブルCBD構築物は、血清型1/2、3、および「7」内の結合範囲はHGFP_CBD118のものよりも狭いが、両方のCBDの特性を併せ持った。短いリンカーの導入は、CBD118がリガンドに接触することを可能にしただけでなく、C末端位置におけるCBD500の結合も増強した。融合タンパク質HGFP_CBD118L500は、HGFP_CBD500のように、血清型4、5、および6細胞の大部分について同等に強い装飾を示した。加えて、GFPが中央の位置に配置され、一方CBD500およびCBD118がN末端またはC末端のいずれかに位置する2つの融合タンパク質を生成した。HCBD500_GFP_CBD118においては、CBD118をGFPのC末端に直接付着させ、HGFP_CBD118の場合と同様の環境に置いた。このタンパク質は、装飾が非常に弱かったが、試験したすべての血清型1/2株を標識することができた。CBDが逆向きである構築物HCBD118_GFP_CBD500は、血清型4、5、および6株の大部分に強く結合したが、1つの血清型1/2株を弱く標識したのみであった。再び、CBD500はC末端に位置した際により強い結合を示した。しかしながら、N末端位置においてもまた機能的であることが示された(HCBD500_GFP_CBD118)。
【0084】
(表1)様々な種および血清型のリステリア細胞への様々なリステリアエンドリシン由来のGFPタグ付きCBDおよびダブルCBD融合タンパク質の結合
500G118および118G500は、HCBD500_GFP_118およびHCBD118_GFP_500をそれぞれ意味する。「L」はシャッフリングされたCBDの間に導入されたリンカーを意味する。++強い、+弱い、(+)非常に弱い、−結合しない;WLSC:Weihenstephan Listeria Strain Collection;SV:リステリア血清型
【0085】
さらなるアプローチにおいて、ダブルCBD融合構築物におけるCBD118をCBDP35により置き換えた(SEQ ID NO:59)。ファージP35のエンドリシンのCBDは、完全な細胞表面に渡る一様な分布で結合し、血清型1/2および3の大部分の株ならびに血清型4、5、および6のいくつかの株を強く標識した。新たに構築されたタンパク質HGFP_CBD500-P35およびHGFP_CBDP35-500の結合パターンは、Ply500およびPlyP35のシングルCBDの結合パターンのほぼ正確な組み合わせを表した:それらは、融合物内のシングルCBDの位置にかかわらず、CBD500またはCBDP35または両方のいずれかが結合するすべての株に強い結合を示した。これらの結果は、異なるペプチドグリカン溶解酵素由来の2つの細胞壁結合ドメインの組み合わせは、それらがC末端位置でない時でさえ人工融合タンパク質において完全に機能的であり得ることを証明した。
【0086】
実施例4.表面プラズモン共鳴解析(SPR)による結合親和性の測定
HGFP_CBD500(SEQ ID NO:60)およびHGFP_CBD500-500のL.モノサイトゲネスWSLC 1042の細胞壁への親和性を、BIAcore X機器およびC1センサーチップ(BIAcore, Uppsala, Sweden)を用いて表面プラズモン共鳴解析により測定した。チップ表面をアミンカップリング法で活性化し、および両方のフローセルにおいてHGFP-CBD500分子を用いてコーティングした(5μl/minの流速で、70μlの10 mM酢酸ナトリウム緩衝液、pH 5における0.5 mg/mlタンパク質)。HBS緩衝液(10 mM HEPES、150 mM NaCl、3.4 mM EDTA、0.005% Tween 20、pH 7.8)中の熱不活性化WSLC 1042細胞をその後、フローセルFc2において固定化したCBDに結合させた(1 mlあたり3.0×1010細胞;3μl/minの流速で15μl)。最終的に、HBS緩衝液において固定化した細胞と3種類の異なる濃度のHGFP_CBD500(50 nM、100 nM、200 nM)およびHGFP_CBD500-500(12.5 nM、25 nM、50 nM)両方との間の相互作用を測定し(10μl/minで30μl)、Fc1を対照セルとした。結合相は3分間測定し、解離相は12分間測定した。すべての段階を25℃で行った。動態学的データの評価は、「物質移動を伴う1:1結合」モデルを使用してBIAevaluation software,version 4.1(BIAcore)で行った。各タンパク質について測定した3種類の濃度に対して取得された平衡結合定数を表2に示す。
【0087】
(表2)リステリア・モノサイトゲネスWSLC 1042の細胞壁へのHGFP_CBD500およびHGFP_CBD500-500の結合の平衡親和性定数(KA)
【0088】
人工ダブルCBDを含む構築物HGFP_CBD500-500は、ファージA500 - ply500のエンドリシンの天然のCBDを含むHGFP_CBD500と比較して約50倍高い親和性で固定化リステリア細胞に結合することが示された。シングルおよびダブルCBDタンパク質構築物の両方のセンサグラムを比較すると、両方の構築物が主に解離相において異なることが明らかであった。ひとたび細胞表面に結合すると、HGFP_CBD500はHGFP_CBD500-500よりずっと急速に脱離し、ダブルCBD構築物はより高い全体的な親和性をもたらした。
【0089】
実施例5.測光溶解アッセイ
野生型およびキメラペプチドグリカン溶解酵素の溶解活性を測光溶解アッセイにより測定した。リステリア・モノサイトゲネス株WSLC 1001(血清型1/2 c)およびWSLC 1042(血清型4 b)の基質細胞を、細菌をTB培地において後期対数期まで増殖させ、およびPBS緩衝液(50 mM NaH2PO4、120 mM NaCl、pH 8.0)において50倍濃度で凍結することにより調製した。アッセイは、PBSにおいて約1.0の初期OD600に希釈した細胞を用いて、1 mlの総体積中で行った。比較するすべての精製した天然のエンドリシンおよびキメラタンパク質を、20μlの体積において等モル量で細胞に添加し、および600 nmでのODを15秒の間隔で最大10分間測定した。使用した酵素濃度は30〜152 pmol/mlに渡った。陰性対照のために、20μl緩衝液を細胞に添加した。すべてのアッセイは3つ組で行った。Loessnerら(2002, Mol. Microbiol. 44, 335-349)は、細胞壁におけるリガンドへのCBDの結合についての分子基盤としてイオン性相互作用を示唆した。エンドリシンply500のCBDは約100 mMのNaCl濃度で最適結合を示し、および増加する塩濃度と共に減少する結合能力を示した。それに基づいて、hisタグ付き野生型ply500(HPL500)およびply500の天然に存在するCBD500の重複を用いた本発明に係る構築物H_EAD_CBD500-500の溶解活性を、高い塩条件の下で比較した。NaCl濃度(1Mと2Mの間)を用いる以外は、アッセイは上述のように行った。光度曲線を標準化し、および対照アッセイのデータにより補正した(補正値=値+(1−対照値))。結果として生じた曲線をソフトウェアSigmaPlot 9.0(Systat Software, Inc.)を用いて以下のシグモイド関数と適合させた:f=y0+a / (1+exp(-(x−x0) / b))∧c。相対酵素活性に相当する関数の最も急な傾きを測定した。
【0090】
驚くべきことに、1 M NaClまたはより高い塩濃度で、本発明に係るペプチドグリカン溶解酵素H_EAD_CBD500-500は、天然に存在する酵素ply500よりも高い溶解活性を示した。
【0091】
実施例6.腸球菌エンドリシンのタンパク質発現および精製
腸球菌エンドリシンFab25VL、Fab20VL、Fab20K、および本発明に係るペプチドグリカン溶解酵素EADFab25_CBD25_CBD20を大腸菌HMS174 DE3において発現させ、およびそれから単離した。タンパク質発現は、1 mM IPTGでの誘導後、3時間37℃で行った。細菌細胞ペレットを遠心分離(5000 rpm、15分、4℃)により収集し、25 ml緩衝液A(25mM トリス、pH 8.0、500 mM NaCl、20 mMイミダゾール、0.1% Tween 20、10%グリセロール)に再懸濁し、および細胞をマイクロフルイダイザー(microfluidizer)において破砕した。細菌細胞破片を遠心分離(12000 rpm、5分、4℃)により除去した。上清を30%飽和の硫酸アンモニウム沈澱に供した。沈殿物を遠心分離(12000 rpm、5分、4℃)により回収した。エンドリシンを含む上清を5 mlフェニルセファロースカラム(High Sub FF, Amersham)を用いた疎水性クロマトグラフィーに適用した。カラムを10容量の緩衝液B(25mMトリス、pH 7.0、500 mM NaCl、30%硫酸アンモニウム、10%グリセロール)で洗浄した。エンドリシンを10カラム容量の緩衝液C(25mMトリス、pH 7.0、500 mM NaCl、10%グリセロール)で溶出した。タンパク質を含有する画分をクマシー染色SDSゲル上でエンドリシンについて解析した。エンドリシンを含有する画分をプールし、および実施例7記載のプレート溶解アッセイにおいて溶解活性について解析した。
【0092】
実施例7.腸球菌細菌に対するペプチドグリカン溶解タンパク質の溶解活性および宿主範囲を試験するためのプレート溶解アッセイ
医学的に関連する種であるエンテロコッカス・フェシウムおよびエンテロコッカス・フェカリス由来の種々の腸球菌細菌を、3 ml BHI培地の前培養において一晩37℃で増殖させた。各株について、2 mlの前培養物を25 mlの新鮮培地中に播種し、および約1のOD600 nmまでインキュベーションした。細菌細胞を4500 rpm、15分間、4℃での遠心分離により収集した。細胞ペレットを500μl BHI培地に再懸濁した。熱不活性化細胞に対する溶解活性の試験(表3)のために、細胞を85℃で45分間インキュベーションし、および1400 rpmでの遠心分離により収集した。細胞ペレットを10 ml LB上層寒天に再懸濁し、および上層寒天をLBプレート上に注いだ。生細胞に対する溶解活性の試験(表4)のためには、腸球菌前培養物を1 ml BHI培地にて一晩増殖させ、前培養物を10 ml BHI上層寒天と混合し、LBプレート上へ注ぎ、および2時間30℃でインキュベーションした。腸球菌エンドリシンFab25VL、Fab20K、エンドリシンFab25VLおよびFab20Kの等モルの組み合わせ、ならびに本発明に係る人工酵素EADFab25_CBD25_CBD20を使用した。5μlのペプチドグリカン溶解タンパク質溶液各々を、上層寒天中に浸漬した菌叢の上にピペットでスポットした。スポットしたタンパク質溶液の周囲の溶解ゾーンの出現を、4種類のスコアシステムを用いて30℃でのプレートのインキュベーションの18時間後に解析した:+++、++、+、ならびに−は、それぞれ強い、中程度、弱い、弱くおよび溶解しないを示す。
【0093】
(表3)熱不活性化腸球菌細胞に対してペプチドグリカン溶解酵素を用いたプレート溶解アッセイ
【0094】
(表4)生存腸球菌細胞に対してペプチドグリカン溶解酵素を用いたプレート溶解アッセイ
【0095】
熱不活性化腸球菌細胞を用いた場合(表3)、エンドリシンFab20KはすべてのE.フェカリス株を高効率で溶解するが、E.フェシウムのいずれの株も溶解しないことが判明した。Fab20エンドリシンはE.フェカリス株に特異的であるように見えた。Fab25VLはE.フェシウムのすべての株を高効率で溶解したが、E.フェカリスの2種類の株のみしか溶解しなかった。他のE.フェカリス株は中程度の効率で溶解されるか、または全く溶解されなかった。従って、天然に存在するエンドリシンFab25VLは、E.フェシウムに厳密に特異的ではなかったが、この種由来の株をE.フェカリス種由来の株よりも確実に溶解した。2種類のエンドリシンFab25VLおよびFab20Kの1:1混合物は試験したすべての株を高効率で溶解したが、本発明に係る1種類の酵素のみ、すなわち、Fab25VLおよびFab20KのCBDを併せ持つがFab25VLのEADのみを有するEADFab25_CBD25_CBD20を用いて、同様のことが達成された。2種類の代わりに1種類の酵素のみを使用することは、酵素産生を容易にし、費用を削減し、および治療的適用における免疫学的反応を最小化する。生細胞を用いるアッセイを行った場合(表4)、本発明に係る酵素であるEADFab25_CBD25_CBD20を用いる利点がより明白であった。これらのアッセイ条件の下でFab25VLはE.フェシウム細胞を主に溶解し、およびいくつかのE.フェカリス細胞のみを低効率で溶解したが、エンドリシンFab20Kは全く作用しなかった。生細胞のいずれも全く溶解されず、恐らくエンドリシン結合のための細胞表面受容体またはペプチドグリカン溶解のための基質分子が、生細胞において接触可能でないことを示唆した。2種類の酵素の組み合わせは状況を改善しなかったが、EADFab25_CBD25_CBD20を用いた細胞溶解はずっとより良好な結果を示した。試験したすべての株は少なくとも低効率で溶解されたが、株の大部分は高効率で溶解された。この予想外の結果は、たとえ最初の特徴決定が細菌宿主範囲に関して使用される1つより多いCBDの純粋に付加的な効果を示唆するとしても、本発明に係る人工ペプチドグリカン溶解酵素が特定の使用において好都合の効果を有し得ることを示す。
【0096】
実施例8.腸球菌に対するペプチドグリカン溶解酵素の最小細菌濃度(MBC)の測定
エンテロコッカス・フェカリス株17を一晩37℃でBHI培地において増殖させた。前培養物を25 ml新鮮培地中に1:10で希釈し、および37℃で約1のOD600 nmまでインキュベーションした。細菌細胞を4500 rpmで5分間、4℃での遠心分離により収集し、および細胞ペレットを105 cfu/mlの濃度になるように溶解緩衝液(2 mM CaCl2、10 mM BSAを含むPBS(2.25 mM NaH2PO3、7.75 mM Na2HPO3、150 mM NaCl))に再懸濁した。Fab25VLおよびEADFab25_CBD25_CBD20のタンパク質溶液(1 mg/ml)を溶解緩衝液において50μg/ml、5μg/ml、0.5μg/ml、0.05μg/ml、0.005μg/ml、0.0005μg/ml、および0.00005μg/mlへ連続希釈した。450μlの細胞懸濁液および50μlの各濃度のタンパク質溶液を混合し、および1時間37℃でインキュベーションした。対照として、添加されたタンパク質を含まない溶解緩衝液をインキュベーションした。100μlの1:10および1:100希釈の溶解試料をLB寒天プレートにプレーティングし、1日37℃でインキュベーションし、およびペプチドグリカン溶解酵素による溶解を生き延びた細胞を計数した。例えば、MBC99.9%は、最初の細菌細胞濃度が1000分の一に減少する最も低い酵素濃度として定義される。この場合、生存細胞のcfu/mlは102より低くなければならない。
【0097】
エンテロコッカス・フェカリス株17に対するMBCは、天然に存在する酵素Fab25 VLよりも本発明に係るペプチドグリカン溶解酵素EADFab25_CBD25_CBD20を用いた場合により低いことが観察された。MBC99.9%はEADFab25_CBD25_CBD20で0.05μg/mlであり、一方Fab25VLを用いて5μg/mlであった。この結果は、EADFab25_CBD25_CBD20の、細菌細胞へのより高い結合親和性および/または増加した溶解活性を示唆する。本発明に係るポリペプチドを用いる場合、病原性細菌を殺傷するためには100分の一に減らしたタンパク質を使用する必要がある。
【特許請求の範囲】
【請求項1】
少なくとも1つの酵素活性ドメインおよび少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性および細菌溶解活性を有する組換えポリペプチド。
【請求項2】
少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性を有する組換えポリペプチド。
【請求項3】
2つの酵素活性ドメインを含む、請求項1記載の組換えポリペプチド。
【請求項4】
酵素活性ドメインおよび/または細菌細胞結合ドメインが、2つの異なるペプチドグリカン溶解酵素に由来する、請求項1、2、または3のいずれか一項記載の組換えポリペプチド。
【請求項5】
酵素活性ドメインが、アミダーゼ_5(バクテリオファージペプチドグリカンヒドロラーゼ、pfam05382)、アミダーゼ_2(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01510)、アミダーゼ_3(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01520)、Transgly(トランスグリコシラーゼ、pfam00912)、ペプチダーゼ_M23(ペプチダーゼファミリーM23、pfam01551)、エンドリシン_オートリシン(CD00737)、ヒドロラーゼ_2(細胞壁ヒドロラーゼ、pfam07486)、CHAP(アミダーゼ、pfam05257)、トランスグリコシラーゼ(トランスグリコシラーゼ様ドメイン、pfam06737)、MtlB(膜結合型溶解性ムレイントランスグリコシラーゼB、COG2951)、MtlA(膜結合型溶解性ムレイントランスグリコシラーゼA、COG2821)、MtlE(膜結合型溶解性ムレイントランスグリコシラーゼE、COG0741)、バクテリオファージ_λ_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、CD00736)、ペプチダーゼ_M74(ペニシリン非感受性ムレインエンドペプチダーゼ、pfam03411)、SLT(トランスグリコシラーゼSLT、pfam01464)、Lys(C型リゾチーム/α-ラクトアルブミンファミリー、pfam00062)、COG5632(N-アセチルムラモイル-L-アラニンアミダーゼ、COG5632)、MepA(ムレインエンドペプチダーゼ、COG3770)、COG1215(グリコシルトランスフェラーゼ、COG1215)、AmiC(N-アセチルムラモイル-L-アラニンアミダーゼ、COG0860)、Spr(細胞壁結合型ヒドロラーゼ、COG0791)、バクテリオファージ_T4様_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、cd00735)、LT_GEWL(溶解性トランスグリコシラーゼ(LT)およびガチョウ卵白リゾチーム(GEWL)ドメイン、cd00254)、ペプチダーゼ_S66(LD-カルボキシペプチダーゼ、pfam02016)、グリコ_ヒドロ_70(グリコシルヒドロラーゼファミリー70、pfam02324)、グリコ_ヒドロ_25(グリコシルヒドロラーゼファミリー25)、VanY(D-アラニル-D-アラニンカルボキシペプチダーゼ、pfam02557)、およびLYZ2(リゾチームサブファミリー2、smart 00047)からなる群より選択される、請求項1〜4のいずれか一項記載の組換えポリペプチド。
【請求項6】
細菌細胞結合ドメインが、SH3_5(細菌SH3ドメイン、pfam08460)、SH3_4(細菌SH3ドメイン、pfam06347)、SH3_3(細菌SH3ドメイン、pfam08239)、SH3b(細菌SH3ドメインホモログ、smart00287)、LysM(細胞壁分解に関与する種々の酵素において見出されるLysMドメイン、pfam01476およびcd00118)、PG_結合_1(推定上のペプチドグリカン結合ドメイン、pfam01471)、PG_結合_2(推定上のペプチドグリカン結合ドメイン、pfam08823)、MtlA(ムレイン分解トランスグリコシラーゼ由来のペプチドグリカン結合ドメイン、pfam03462)、Cpl-7(Cpl-7リゾチームのC末端ドメイン、pfam08230)、CW_結合_1(推定上の細胞壁結合リピート、pfam01473)、LytB(推定上の細胞壁結合ドメイン、COG2247)、およびLytE(LysMリピート、COG1388)からなる群より選択される、請求項1〜4のいずれか一項記載の組換えポリペプチド。
【請求項7】
ドメインが、約15〜約250アミノ酸残基長の範囲、特に約20〜約200アミノ酸残基長の範囲、およびより特に約15〜約40アミノ酸残基長である、請求項1〜6のいずれか一項記載の組換えポリペプチド。
【請求項8】
酵素活性ドメインおよび/または細菌細胞結合ドメインが、Ply500、Ply511、Ply118、Ply100、PlyP40、Ply3626、phiLM4エンドリシン、PlyCD119、PlyPSAa、Ply21、PlyBA、Ply12、PlyP35、PlyPH、PlyL、PlyB、phi11エンドリシン、phi MR11エンドリシン、phi12エンドリシン、S.アウレウス(S. aureus)ファージPVLアミダーゼ、plypitti26、ΦSA2usaエンドリシン、スタフィロコッカス・ワーネリ(Staphylococcus warneri)MファージΦWMY PlyGBSのエンドリシン、B30エンドリシン、Cpl-1、Cpl-7、Cpl-9、PlyG、PlyC、palアミダーゼ、Fab25、Fab20、エンテロコッカス・フェカリス(Enterococcus faecalis)V583プロファージ由来のエンドリシン、リゾスタフィン、ファージPL-1アミダーゼ、S.カピティス(S. capitis)ALE-1エンドペプチダーゼ、ムタノリシン(ストレプトマイセス・グロビスポラス(Streptomyces globisporus)ATCC 21553のN-アセチルムラミダーゼ)、エンテロリシンA(エンテロコッカス・フェカリスLMG 2333由来の細胞壁分解バクテリオシン)、LysK、LytM、L.モノサイトゲネス(L. monocytogenes)由来のAmiオートリシン、シュードモナス・エルギノーサ(Pseudomonas aeruginosa)ファージΦKZおよびELのエンドリシン、T4リゾチーム、gp61ムラミダーゼ、およびSTM0016ムラミダーゼからなる群より選択される野生型ペプチドグリカン溶解酵素に由来する、請求項1〜7のいずれか一項記載の組換えポリペプチド。
【請求項9】
PlyP40由来の酵素活性ドメインがSEQ ID NO: 103記載のアミノ酸配列によりコードされ、および/またはPlyP40由来の細菌細胞結合ドメインがSEQ ID NO: 104記載のアミノ酸配列によりコードされる、請求項8記載の組換えポリペプチド。
【請求項10】
SEQ ID NO: 7、9、13、15、17、19、21、23、31、33、35、37、39、41、43、45、47、49、51、71、73、75、77、79、81、83、85、87、89、91、93、95、97、99、101において示される組換えポリペプチド。
【請求項11】
請求項1〜10のいずれか一項記載の組換えポリペプチドをコードするヌクレオチド配列を含む核酸分子。
【請求項12】
SEQ ID NO: 8、10、14、16、18、20、22、24、32、34、36、38、40、42、44、46、48、50、52、72、74、76、78、80、82、84、86、88、90、92、94、96、98、100、102において示されるヌクレオチド配列を含む、請求項11記載の核酸分子。
【請求項13】
請求項11または12記載の核酸分子を含むベクター。
【請求項14】
薬剤としての使用のための、請求項1〜9のいずれか一項記載の組換えポリペプチド。
【請求項15】
医学的環境、公的環境、もしくは私的環境における消毒剤としての、食品産業、動物飼料産業、もしくは化粧品産業における細菌汚染の除染剤としての、または細菌で汚染された表面に対する界面活性剤としての、請求項1〜9のいずれか一項記載の組換えポリペプチドの使用。
【請求項16】
医薬、食物、もしくは飼料の診断、または環境の診断における診断手段としての、請求項1〜9のいずれか一項記載の組換えポリペプチドの使用。
【請求項1】
少なくとも1つの酵素活性ドメインおよび少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性および細菌溶解活性を有する組換えポリペプチド。
【請求項2】
少なくとも2つの細菌細胞結合ドメインを含む、細菌結合活性を有する組換えポリペプチド。
【請求項3】
2つの酵素活性ドメインを含む、請求項1記載の組換えポリペプチド。
【請求項4】
酵素活性ドメインおよび/または細菌細胞結合ドメインが、2つの異なるペプチドグリカン溶解酵素に由来する、請求項1、2、または3のいずれか一項記載の組換えポリペプチド。
【請求項5】
酵素活性ドメインが、アミダーゼ_5(バクテリオファージペプチドグリカンヒドロラーゼ、pfam05382)、アミダーゼ_2(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01510)、アミダーゼ_3(N-アセチルムラモイル-L-アラニンアミダーゼ、pfam01520)、Transgly(トランスグリコシラーゼ、pfam00912)、ペプチダーゼ_M23(ペプチダーゼファミリーM23、pfam01551)、エンドリシン_オートリシン(CD00737)、ヒドロラーゼ_2(細胞壁ヒドロラーゼ、pfam07486)、CHAP(アミダーゼ、pfam05257)、トランスグリコシラーゼ(トランスグリコシラーゼ様ドメイン、pfam06737)、MtlB(膜結合型溶解性ムレイントランスグリコシラーゼB、COG2951)、MtlA(膜結合型溶解性ムレイントランスグリコシラーゼA、COG2821)、MtlE(膜結合型溶解性ムレイントランスグリコシラーゼE、COG0741)、バクテリオファージ_λ_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、CD00736)、ペプチダーゼ_M74(ペニシリン非感受性ムレインエンドペプチダーゼ、pfam03411)、SLT(トランスグリコシラーゼSLT、pfam01464)、Lys(C型リゾチーム/α-ラクトアルブミンファミリー、pfam00062)、COG5632(N-アセチルムラモイル-L-アラニンアミダーゼ、COG5632)、MepA(ムレインエンドペプチダーゼ、COG3770)、COG1215(グリコシルトランスフェラーゼ、COG1215)、AmiC(N-アセチルムラモイル-L-アラニンアミダーゼ、COG0860)、Spr(細胞壁結合型ヒドロラーゼ、COG0791)、バクテリオファージ_T4様_リゾチーム(N-アセチルムラミン酸とN-アセチルグルコサミンとの間の結合の溶解、cd00735)、LT_GEWL(溶解性トランスグリコシラーゼ(LT)およびガチョウ卵白リゾチーム(GEWL)ドメイン、cd00254)、ペプチダーゼ_S66(LD-カルボキシペプチダーゼ、pfam02016)、グリコ_ヒドロ_70(グリコシルヒドロラーゼファミリー70、pfam02324)、グリコ_ヒドロ_25(グリコシルヒドロラーゼファミリー25)、VanY(D-アラニル-D-アラニンカルボキシペプチダーゼ、pfam02557)、およびLYZ2(リゾチームサブファミリー2、smart 00047)からなる群より選択される、請求項1〜4のいずれか一項記載の組換えポリペプチド。
【請求項6】
細菌細胞結合ドメインが、SH3_5(細菌SH3ドメイン、pfam08460)、SH3_4(細菌SH3ドメイン、pfam06347)、SH3_3(細菌SH3ドメイン、pfam08239)、SH3b(細菌SH3ドメインホモログ、smart00287)、LysM(細胞壁分解に関与する種々の酵素において見出されるLysMドメイン、pfam01476およびcd00118)、PG_結合_1(推定上のペプチドグリカン結合ドメイン、pfam01471)、PG_結合_2(推定上のペプチドグリカン結合ドメイン、pfam08823)、MtlA(ムレイン分解トランスグリコシラーゼ由来のペプチドグリカン結合ドメイン、pfam03462)、Cpl-7(Cpl-7リゾチームのC末端ドメイン、pfam08230)、CW_結合_1(推定上の細胞壁結合リピート、pfam01473)、LytB(推定上の細胞壁結合ドメイン、COG2247)、およびLytE(LysMリピート、COG1388)からなる群より選択される、請求項1〜4のいずれか一項記載の組換えポリペプチド。
【請求項7】
ドメインが、約15〜約250アミノ酸残基長の範囲、特に約20〜約200アミノ酸残基長の範囲、およびより特に約15〜約40アミノ酸残基長である、請求項1〜6のいずれか一項記載の組換えポリペプチド。
【請求項8】
酵素活性ドメインおよび/または細菌細胞結合ドメインが、Ply500、Ply511、Ply118、Ply100、PlyP40、Ply3626、phiLM4エンドリシン、PlyCD119、PlyPSAa、Ply21、PlyBA、Ply12、PlyP35、PlyPH、PlyL、PlyB、phi11エンドリシン、phi MR11エンドリシン、phi12エンドリシン、S.アウレウス(S. aureus)ファージPVLアミダーゼ、plypitti26、ΦSA2usaエンドリシン、スタフィロコッカス・ワーネリ(Staphylococcus warneri)MファージΦWMY PlyGBSのエンドリシン、B30エンドリシン、Cpl-1、Cpl-7、Cpl-9、PlyG、PlyC、palアミダーゼ、Fab25、Fab20、エンテロコッカス・フェカリス(Enterococcus faecalis)V583プロファージ由来のエンドリシン、リゾスタフィン、ファージPL-1アミダーゼ、S.カピティス(S. capitis)ALE-1エンドペプチダーゼ、ムタノリシン(ストレプトマイセス・グロビスポラス(Streptomyces globisporus)ATCC 21553のN-アセチルムラミダーゼ)、エンテロリシンA(エンテロコッカス・フェカリスLMG 2333由来の細胞壁分解バクテリオシン)、LysK、LytM、L.モノサイトゲネス(L. monocytogenes)由来のAmiオートリシン、シュードモナス・エルギノーサ(Pseudomonas aeruginosa)ファージΦKZおよびELのエンドリシン、T4リゾチーム、gp61ムラミダーゼ、およびSTM0016ムラミダーゼからなる群より選択される野生型ペプチドグリカン溶解酵素に由来する、請求項1〜7のいずれか一項記載の組換えポリペプチド。
【請求項9】
PlyP40由来の酵素活性ドメインがSEQ ID NO: 103記載のアミノ酸配列によりコードされ、および/またはPlyP40由来の細菌細胞結合ドメインがSEQ ID NO: 104記載のアミノ酸配列によりコードされる、請求項8記載の組換えポリペプチド。
【請求項10】
SEQ ID NO: 7、9、13、15、17、19、21、23、31、33、35、37、39、41、43、45、47、49、51、71、73、75、77、79、81、83、85、87、89、91、93、95、97、99、101において示される組換えポリペプチド。
【請求項11】
請求項1〜10のいずれか一項記載の組換えポリペプチドをコードするヌクレオチド配列を含む核酸分子。
【請求項12】
SEQ ID NO: 8、10、14、16、18、20、22、24、32、34、36、38、40、42、44、46、48、50、52、72、74、76、78、80、82、84、86、88、90、92、94、96、98、100、102において示されるヌクレオチド配列を含む、請求項11記載の核酸分子。
【請求項13】
請求項11または12記載の核酸分子を含むベクター。
【請求項14】
薬剤としての使用のための、請求項1〜9のいずれか一項記載の組換えポリペプチド。
【請求項15】
医学的環境、公的環境、もしくは私的環境における消毒剤としての、食品産業、動物飼料産業、もしくは化粧品産業における細菌汚染の除染剤としての、または細菌で汚染された表面に対する界面活性剤としての、請求項1〜9のいずれか一項記載の組換えポリペプチドの使用。
【請求項16】
医薬、食物、もしくは飼料の診断、または環境の診断における診断手段としての、請求項1〜9のいずれか一項記載の組換えポリペプチドの使用。
【図1】

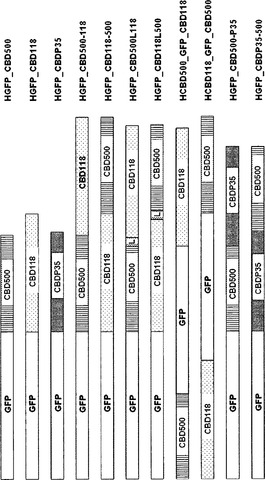
【図2】
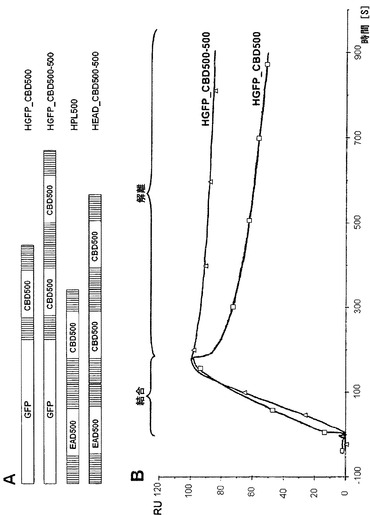
【図3】
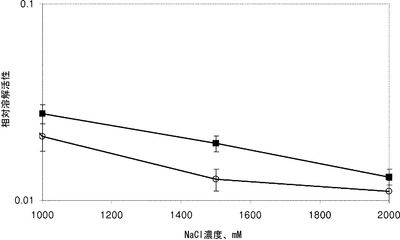
【図4】
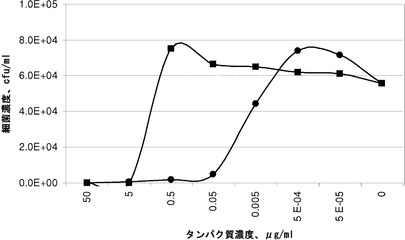
【図2】
【図3】
【図4】
【公表番号】特表2012−500015(P2012−500015A)
【公表日】平成24年1月5日(2012.1.5)
【国際特許分類】
【出願番号】特願2011−523419(P2011−523419)
【出願日】平成21年8月19日(2009.8.19)
【国際出願番号】PCT/EP2009/060716
【国際公開番号】WO2010/020657
【国際公開日】平成22年2月25日(2010.2.25)
【出願人】(309037974)バイオメリュー エス.エー. (4)
【出願人】(510046402)ヒグロス インベスト ゲーエムベーハー (5)
【Fターム(参考)】
【公表日】平成24年1月5日(2012.1.5)
【国際特許分類】
【出願日】平成21年8月19日(2009.8.19)
【国際出願番号】PCT/EP2009/060716
【国際公開番号】WO2010/020657
【国際公開日】平成22年2月25日(2010.2.25)
【出願人】(309037974)バイオメリュー エス.エー. (4)
【出願人】(510046402)ヒグロス インベスト ゲーエムベーハー (5)
【Fターム(参考)】
[ Back to top ]