
人工多能性幹細胞の作製方法
【課題】人工多能性幹細胞の新規作製方法を提供する。
【解決手段】この発明は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子を、哺乳動物由来の体細胞に導入し、これによって該体細胞から人工多能性幹細胞を誘導することを含み、該初期化因子はSox2及びSox2をコードする核酸を含まない、哺乳動物人工多能性幹細胞の作製方法に関する。
【解決手段】この発明は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子を、哺乳動物由来の体細胞に導入し、これによって該体細胞から人工多能性幹細胞を誘導することを含み、該初期化因子はSox2及びSox2をコードする核酸を含まない、哺乳動物人工多能性幹細胞の作製方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、人工多能性幹細胞の新規作製方法に関する。具体的には、本発明は、Oct3/4及びNanog、或いはそれらをコードする核酸、を主要な初期化因子として使用することを特徴とする体細胞から人工多能性幹細胞を作製する方法に関する。
以下において、「人工多能性幹細胞」は、単に「iPS細胞」と称することもある。
【背景技術】
【0002】
近年、マウス及びヒトのiPS細胞が相次いで樹立された。Takahashi及びYamanaka(非特許文献1)は、Fbx15遺伝子座にネオマイシン耐性遺伝子をノックインしたレポーターマウス由来線維芽細胞に、Oct3/4, Sox2, Klf4及びc-Myc遺伝子を導入し強制発現させることによって、iPS細胞を樹立した。Okitaら(非特許文献2)は、Fbx15よりも多能性細胞に発現が限局しているNanogの遺伝子座に緑色蛍光タンパク質(GFP)及びピューロマイシン耐性遺伝子を組み込んだトランスジェニックマウスを作製し、該マウス由来の線維芽細胞で上記4遺伝子を強制発現させ、ピューロマイシン耐性かつGFP陽性の細胞を選別することにより、遺伝子発現やエピジェネティック修飾が胚性幹(ES)細胞とほぼ同等のiPS細胞(Nanog iPS細胞)を樹立することに成功した。同様の結果が他のグループによっても再現された(非特許文献3、非特許文献4)。その後、c-Myc遺伝子を除いた3因子によってもiPS細胞を作製できることが明らかとなった(非特許文献5)。
【0003】
さらに、Takahashiら(非特許文献6)は、ヒトの皮膚由来線維芽細胞にマウスと同様の4遺伝子を導入することにより、iPS細胞を樹立することに成功した。一方、Yuら(非特許文献7)は、Oct3/4, Sox2, Nanog及びLin28遺伝子を使用してヒトiPS細胞を作製した。このように、体細胞に特定の因子を導入することにより、ヒト及びマウスで、分化多能性においてES細胞と遜色のないiPS細胞を作製できることが示された。
【0004】
前述のYuら (非特許文献7)においては、4種の遺伝子のうちNanogまたはLin28遺伝子を除いた3種の遺伝子導入によってiPS細胞コロニーが得られたが、Oct3/4またはSox2遺伝子を除いた3種の遺伝子導入ではiPS細胞コロニーは得られなかったことから、iPS細胞樹立におけるOct3/4及びSox2遺伝子の重要性が示されている((特許文献1)も参照)。またParkら(非特許文献8)においても、iPS細胞の樹立にはOct3/4とSox2が必須であると述べられている。
【0005】
さらにまた、(特許文献2)においては、Oct3/4, Sox2, Klf4及びc-Mycの各遺伝子を導入し、マウスiPS細胞を作製しているが、その際にDox-inducible systemでiPS細胞を誘導するという手法を開示している。また体細胞として成熟B細胞を用いてiPS細胞をうまく誘導できたことが記載されている。
【0006】
また、体細胞からiPS細胞を誘導することに関する特許文献として、Yamanaka(特許文献3)及びThomsonら(特許文献1)が公開されている。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO2008/118820
【特許文献2】WO2008/124133
【特許文献3】WO2007/069666
【非特許文献】
【0008】
【非特許文献1】Takahashi, K. and Yamanaka, S., Cell, 126: 663-676 (2006)
【非特許文献2】Okita, K. et al., Nature, 448: 313-317 (2007)
【非特許文献3】Wernig, M. et al., Nature, 448: 318-324 (2007)
【非特許文献4】Maherali, N. et al., Cell Stem Cell, 1: 55-70 (2007)
【非特許文献5】Nakagawa, M. et al., Nat. Biotethnol., 26: 101-106 (2008)
【非特許文献6】Takahashi, K. et al., Cell, 131: 861-872 (2007)
【非特許文献7】Yu, J. et al., Science, 318: 1917-1920 (2007)
【非特許文献8】Park I.H et al., Nature, 451: 141-146 (2008)
【発明の概要】
【発明が解決しようとする課題】
【0009】
上記のように、Yamanakaらの発見(Takahashi, K. and Yamanaka, S. Cell 126:663-676 (2006))がブレークスルーとなって、体細胞から人工多能性幹細胞を誘導することを可能にする初期化因子として、いくつかの組み合わせが提案されており、そのいずれも体細胞から人工多能性幹細胞を誘導する。提案された初期化因子の組み合わせのなかで共通する因子は、Oct3/4のみであるが、最近、Oct3/4だけで初期化が起こるとする報告がなされ、これによると、使用された体細胞が神経幹細胞であり、この細胞はSox2遺伝子が内因的に発現されている細胞であるので特殊な例であるといえる(Kim, J.B. et al., Cell 136:411-419 (2009))。また、Oct3/4と他の初期化因子との組み合わせのなかで、Oct3/4とSox2(WO2008/118820)、Oct3/4とKlf4(Kim, J.B. et al., Nature 454:646-650 (2008)、Shi, Y. et al., Cell Stem Cell, 2:525-528(2008))、またはOct3/4とc-Myc(Kim, J.B. et al., Nature 454:646-650 (2008))からなる2因子の組み合わせが報告されている。
【0010】
しかしながら、報告された初期化因子の組み合わせがなぜ体細胞を人工多能性幹細胞に誘導することを可能にするのか明らかでないし、その作用機序についても明らかにされていない。
【0011】
このような情況下において、本発明の目的は、これまでに報告されていない、体細胞の初期化を可能にする初期化因子の新規の組み合わせを見出し、この組み合わせ因子を用いて体細胞から人工多能性幹細胞を作製する方法を提供することである。
【課題を解決するための手段】
【0012】
国際公開WO2008/118820(2008年10月2日公開)には、Oct3/4及びSox2を必須の初期化因子とし、これにNanog及びLin28のうち少なくとも1つの初期化因子を追加しうる、体細胞を人工多能性幹細胞に初期化するための初期化因子の組み合わせが記載されている。これによれば、初期化因子の組み合わせの例は、Oct3/4及びSox2;Oct3/4、Sox2及びNanog;Oct3/4、Sox2及びLin28;並びに、Oct3/4、Sox2、Nanog及びLin28であり、これに対し、Oct3/4、Nanog及びLin28の組み合わせは初期化を起こさないことが示されている。また Huangfu D.et al., nature biotech.,Vol.26, No.11: 1269-1275 (2008)(オンライン公表日:2008年10月12日)においては、(Oct3/4, Sox2及びKlf4のうちの)いずれか2因子をVPAと共にヒト線維芽細胞に導入し、人工多能性幹細胞の樹立を試みているが、Oct3/4及びSox2の組み合わせ以外の組み合わせを使用しても、人工多能性幹細胞は樹立できなかったことが記載されている。
【0013】
本発明者らは、今回、意外にも、Oct3/4及びNanogからなる組み合わせだけで、体細胞を人工多能性幹細胞に初期化することが可能であることを見出した。これまで報告された2つの初期化因子の組み合わせは、Oct3/4及びSox2(国際公開WO2008/118820)、Oct3/4及びKlf4(Kim, J.B. et al., Nature 454:646-650 (2008)、Shi, Y. et al., Cell Stem Cell, 2:525-528(2008))、及びOct3/4及びc-Myc(Kim, J.B. et al., Nature 454:646-650 (2008))のみである。しかし、後者の2つの組み合わせ(Nature 454:646-650 (2008)及び Cell Stem Cell, 2:525-528(2008))については、体細胞がマウス神経幹細胞であり、もともとSox2遺伝子の発現が見られる細胞であり、敢えてSox2遺伝子を初期化因子として加える必要がない。それゆえ、Oct3/4及びSox2が必須であるとするこれまでの知見からすれば、本発明におけるOct3/4及びNanogからなる2つの初期化因子の組み合わせは、当業者にはまったく予想されていなかった意外な組み合わせである。なお、本発明者らが実験に用いた成人皮膚由来線維芽細胞(HDF)はSox2を発現していない細胞であり(Cell,131, 861-872(2007))、この観点からも、本発明におけるOct3/4及びNanogからなる2つの初期化因子の組み合わせは、当業者にはまったく予想されていなかった意外な組み合わせである。
【0014】
したがって、本発明は、以下の特徴を含む。
本発明は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子を、哺乳動物由来の体細胞に導入し、これによって該体細胞から人工多能性幹細胞を誘導することを含み、該初期化因子はSox2及びSox2をコードする核酸を含まない、哺乳動物人工多能性幹細胞の作製方法を提供する。
【0015】
その実施形態において、上記初期化因子は、Lin28、又はLin28をコードする核酸、をさらに含むことができる。
【0016】
別の実施形態において、上記初期化因子は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、からなる。
【0017】
或いは、別の実施形態において、上記初期化因子は、Oct3/4、Nanog及びLin28、或いはOct3/4、Nanog及びLin28をコードする核酸、からなる。
【0018】
別の実施形態において、上記初期化因子及び体細胞が由来する哺乳動物は、互いに同じか又は異なる哺乳動物である。
【0019】
別の実施形態において、上記哺乳動物は、霊長類又はげっ歯類である。
別の実施形態において、上記霊長類はヒトである。
【0020】
別の実施形態において、上記げっ歯類はマウスである。
さらに別の実施形態において、上記核酸はベクターに含まれている。
【0021】
別の実施形態において、上記核酸は、上記体細胞のゲノムに組み込まれるか、或いは、ゲノムに組み込まれない状態で細胞内に存在する。
【0022】
さらに別の実施形態において、上記人工多能性幹細胞の誘導時に、該細胞の樹立効率改善物質をさらに存在させる。
【0023】
別の実施形態において、上記樹立効率改善物質が、ヒストンデアセチラーゼ(HDAC)阻害剤、ヒストンメチルトランスフェラーゼ(G9a)阻害剤又はDNAメチル化酵素(Dnmt)阻害剤である。
【図面の簡単な説明】
【0024】
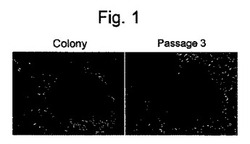
【図1】図1は、樹立したiPS細胞クローンT4F-1の形態を示す写真である。左図はコロニー形成時、右図は3継代目のコロニー像を示す。
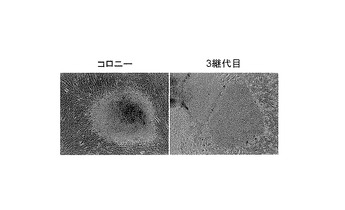
【図2】図2は、樹立したヒトiPS細胞クローンについてGenomic-PCR解析を行った結果を示す写真である。図中「T4F-1」はOct3/4, Nanogの2遺伝子のみがゲノムに挿入されたクローンであり、「T4F-2」はOct3/4, Sox2, Nanog, Lin28の4遺伝子がゲノムに挿入されたクローンである。また図中、「Y4F」はOct3/4, Sox2, Klf4, c-Mycの4遺伝子導入で樹立したクローンを、「Y3F」はOct3/4, Sox2, Klf4の3遺伝子導入で樹立したクローンをそれぞれ示す。「HDF」は導入に用いた成人皮膚由来線維芽細胞を、「H2O」は陰性コントロールとしての溶媒(水)を示す。
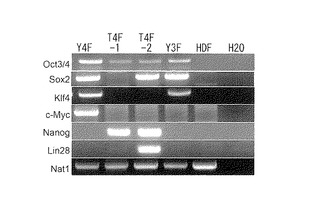
【図3】図3は、樹立したヒトiPS細胞クローンが、ES細胞特異的なマーカー遺伝子(Oct3/4, Nanog, Lin28, PH34, Dnmt3b及びNodal)を発現していることを示すRT-PCRの写真である。図中、「T4F-1」、「T4F-2」は樹立した各クローンを、また「201B6 P19」、「201B7 P37」は、いずれもTakahashi, K.ら, Cell, 131:861-872 (2007)に記載されたOct3/4,Sox2,Klf4,Nanogを用いて樹立されたiPS細胞クローンを示す(P19又はP37は継代数である。)。「ES」はES細胞を、「AHDF」は導入に用いた成人皮膚由来線維芽細胞を、「H2O」は陰性コントロールを示す。
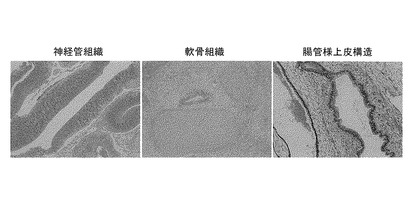
【図4】図4は、2遺伝子(Oct3/4, Nanog)のゲノム中への挿入により樹立したヒトiPS細胞クローン(T4F-1)をScidマウスの精巣内に注射して得られたテラトーマの組織染色像(ヘマトトキシン・エオシン染色)である。左から、神経管組織、軟骨組織、腸管様上皮構造の組織像を示す。
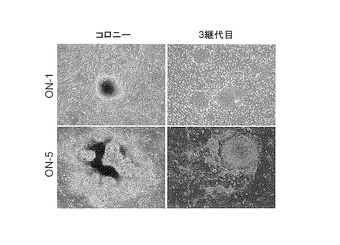
【図5】図5は、2遺伝子(Oct3/4, Nanog)の導入により樹立したiPS細胞クローンON-1及びON-5の形態を示す写真である。左図はコロニー形成時、右図は3継代目のコロニー像を示す。
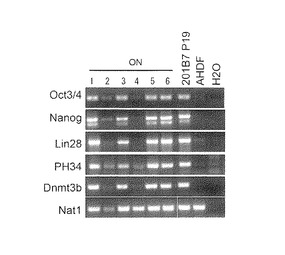
【図6】図6は、樹立したヒトiPS細胞クローンが、ES細胞特異的なマーカー遺伝子(Oct3/4, Nanog, Lin28, PH34及びDnmt3b)を発現していることを示すRT-PCRの写真である。図中、「ON-1〜ON-6」は2遺伝子導入で樹立した各クローンを、また「201B7 P19」は、Takahashi, K.ら, Cell, 131:861-872 (2007)に記載されたOct3/4,Sox2,Klf4,Nanogを用いて樹立されたiPS細胞クローンを示す(P19は継代数である。)。「AHDF」は導入に用いた成人皮膚由来線維芽細胞を、「H2O」は陰性コントロールを示す。
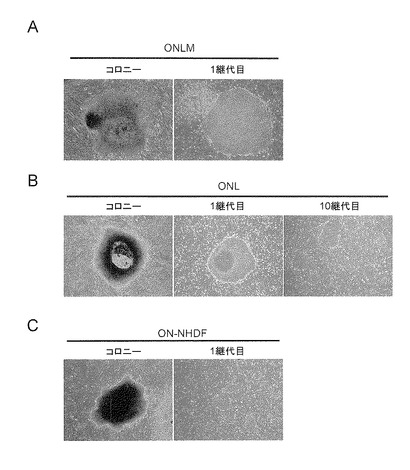
【図7】図7は、ヒト成人皮膚由来線維芽細胞への、4遺伝子(Oct3/4, Nanog, Lin28, c-Myc)、3遺伝子(Oct3/4, Nanog, Lin28)の導入により樹立されたiPS細胞クローンONLM(図7A)及びONL(図7B)、ヒト新生児包皮由来線維芽細胞への、2遺伝子(Oct3/4, Nanog)の導入により樹立されたiPS細胞クローンON(図7C)の各形態を示す写真である。左図はいずれもコロニー形成時、右図及び中央図は1又は10継代目のコロニー像を示す。
【図8】図8は、ヒト歯髄幹細胞DP31への2遺伝子(Oct3/4, Nanog)の導入により樹立されたiPS細胞クローンの形態を示す写真である。左図はコロニー形成時、右図は2継代目のコロニー像を示す。
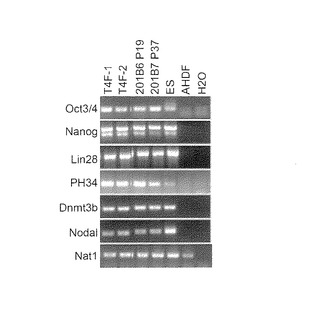
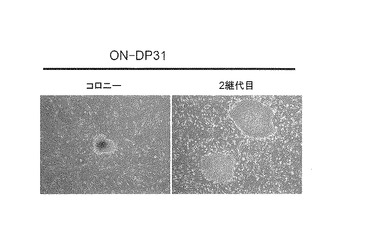
【図9】図9は、樹立したヒトiPS細胞クローンについてGenomic-PCR解析を行った結果を示す写真である。レーン左から以下の細胞の結果を示す:・ONL:Oct3/4, Nanog, Lin28の3遺伝子を成人皮膚由来線維芽細胞に導入して樹立したiPS細胞クローン、・AHDF-ON: Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したiPS細胞クローン、・NHDF-ON: Oct3/4, Nanogの2遺伝子をヒト新生児包皮由来線維芽細胞に導入して樹立したiPS細胞クローン、・201B6:Takahashi, K.ら, Cell, 131:861-872 (2007)に記載されたOct3/4,Sox2,Klf4,Nanogを用いて樹立されたiPS細胞クローン、・AHDF: ヒト成人皮膚由来線維芽細胞、・NHDF: ヒト新生児包皮由来線維芽細胞。
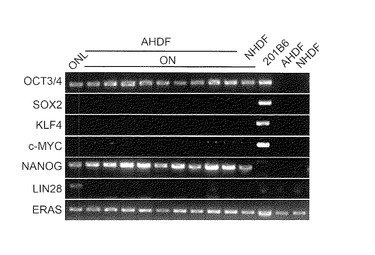
【図10】図10は、樹立したヒトiPS細胞クローンについてGenomic-PCR解析を行った結果を示す写真である。レーン左から以下の細胞の結果を示す:・OSKMNL: Oct3/4, Sox2, Klf4, c-Myc, Nanog, Lin28の6遺伝子を成人皮膚由来線維芽細胞に導入して樹立したiPS細胞クローン、・Dp31 ON:Oct3/4, Nanogの2遺伝子をヒト歯髄幹細胞Dp31に導入して樹立したiPS細胞クローン、・Dp31:ヒト歯髄幹細胞、・H2O:陰性コントロール(水)。
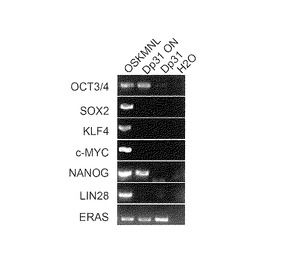
【図11】図11は、Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したヒトiPS細胞クローンをScidマウスの精巣内に注射して得られたテラトーマの組織染色像(ヘマトトキシン・エオシン染色)である。左から、神経組織、腺上皮組織、軟骨組織の組織像を示す。
【図12】図12は、Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したヒトiPS細胞クローンが、幹細胞マーカーを発現することを示した免疫染色の写真である。
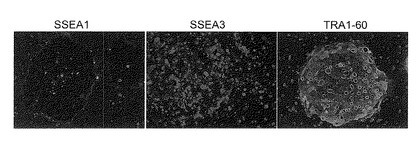
【図13】図13は、Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したヒトiPS細胞クローンが三胚葉系への分化能を有することを、βIII-チューブリン(tubulin)(図中Tubrin)、α-フェトプロテイン(fetoprotein)(図中AFP)、ビメンチン(Vimentin)、平滑筋アクチン(smooth muscle actin)(図中SMA)およびデスミン(Desmin)に対する各抗体を用いた染色により確認した結果を示す写真である。Cy3-Negaおよび488-Negaは陰性コントロールである。
【発明を実施するための形態】
【0025】
本発明をさらに詳細に説明する。
1. 定義
本明細書中で使用される用語は、以下の意味を包含する。その他の用語は、当業界で一般的に使用される意味を含むことが意図されている。
【0026】
本明細書で使用される「人工多能性幹細胞」又は「iPS細胞」という用語は、人工的に作製された分化多能性を有する細胞であり、いわゆる胚性幹(ES)細胞ではないがES細胞に類似した特性を有する細胞である。この細胞は、Takahashi及びYamanaka(Cell 126:663-676 (2006))によってマウス体細胞からはじめて樹立され、その後、ヒト体細胞からも同様に樹立可能であることが示されている(国際公開WO2007/069666;Takahashi, K.ら, Cell 131:861-872 (2007):Yu, J.ら,Science 318:1917-1920 (2007);Nakagawa, M.ら,Nat. Biotechnol. 26:101-106 (2008))。iPS細胞は、動物の体を構成する種々の細胞に分化する能力(すなわち、分化多能性)、核型を保持したまま半永久的に増殖し続ける能力、ES細胞が通常発現する遺伝子群が同様に発現されている、などの特徴を有するために、体細胞の初期化(すなわち、再プログラム化(reprogramming))によって人工的に誘導された細胞であり、分化した体細胞の特性と明らかな相違を示す。
【0027】
本明細書で使用する「初期化」という用語は、「核初期化(nuclear reprogramming)」とも称され、分化した細胞が、未分化細胞、特に分化多能性細胞に誘導、変換される過程又は手段を指す。初期化という現象はもともと、脱核した未受精卵に体細胞の核を注入する、ES細胞を体細胞と融合させる、ES細胞抽出液を体細胞中に浸透化させる、などの方法によって観察されてきたが、本発明における初期化では、卵子、卵母細胞などの生殖系列細胞及びES細胞に依存することなく、体細胞からiPS細胞への初期化を起こすことができる。
【0028】
このような初期化を可能にする因子が、初期化因子である。本明細書で使用される初期化因子は、蛋白質、核酸或いはそれらの組み合わせを含む。これまで報告された初期化因子は、Oct3/4、Sox2及びKlf4;Oct3/4、Klf4及びc-Myc;Oct3/4、Sox2、Klf4及びc-Myc;Oct3/4及びSox2;Oct3/4、Sox2及びNanog;Oct3/4、Sox2及びLin28; Oct3/4及びKlf4などであるが、本発明で使用可能な初期化因子は、少なくともOct3/4及びNanogを含む初期化因子からなり、ただしSox2を含まない。
【0029】
なお、本明細書では、初期化因子の遺伝子名は、哺乳動物の種類を問わず同じ遺伝子名、例えばOct3/4、Nanog、Sox2、Klf4、c-Myc、Lin28などを使用する。すなわち、これらの遺伝子名は、通常マウスの遺伝子名として使用されるが、本明細書ではマウスのみならず、ヒトやその他の哺乳動物の場合にも上記遺伝子名を使用することにする。
【0030】
また、Oct3/4は、Oct3 、Oct4又はPOU5F1と称されることがあり、いずれも同一の転写因子を指す。本明細書では、これらを総称してOct3/4と呼ぶことにする。
【0031】
本明細書で使用する「核酸」という用語は、ゲノムDNA、cDNA、mRNAなどのデオキシリボ核酸(DNA)又はリボ核酸(RNA)を指す。
【0032】
本明細書で使用する「体細胞」とは、卵子、卵母細胞などの生殖系列細胞又は分化全能性細胞を除くあらゆる哺乳動物細胞をいう。
【0033】
本明細書で使用する「哺乳動物」は、すべての哺乳動物を含むことを意図している。特に好ましい哺乳動物には、霊長類、げっ歯類、有蹄類などが含まれる。
【0034】
2. 初期化因子
本発明により体細胞からiPS細胞を誘導するための初期化因子は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子であり、Sox2及びSox2をコードする核酸を除く他の初期化因子又はサイトカインを適宜含むことができる。
【0035】
初期化因子の組み合わせの好ましい例は、Oct3/4及びNanogの組み合わせ、或いは、Oct3/4、Nanog及びLin28の組み合わせである。各初期化因子が由来する哺乳動物の種類は限定されないものとし、任意の哺乳動物である。好ましい哺乳動物は、例えば霊長類(例えばヒト、サル、チンパンジーなど)、げっ歯類(例えばマウス、ラット、モルモット、ハムスターなど)、有蹄類(例えばウシ、ウマ、ヒツジ、ヤギ、ブタなど)、ペット動物(例えばイヌ、ネコなど)などである。通常、ある特定の哺乳動物由来の体細胞を初期化するときには、それと同一の哺乳動物由来の初期化因子を使用することが好ましい。例えばヒト由来の体細胞からiPS細胞を誘導するときには、ヒト由来の初期化因子を使用する。
上記Oct3/4、Nanog、Lin28の各アミノ酸及びヌクレオチド配列は、GenBank(米国NCBI)にアクセスすることによって入手することが可能である。
【0036】
Oct3/4について、例えばヒトOct3/4、マウスOct3/4、ラットOct3/4の配列は、それぞれ、NM_203289又はNM_002701、NM_013633、NM_001009178として登録されている。
【0037】
Nanogについて、例えばヒトNanog、マウスNanog、ラットNanogの配列は、それぞれ、NM_024865、NM_028016、NM_001100781として登録されている。
【0038】
Lin28について、例えばヒトLin28、マウスLin28、ラットLin28の配列は、それぞれ、NM_024674、NM_145833、NM_001109269として登録されている。
【0039】
Lin28に類似の因子として、同じLinファミリーに属するLin28bが知られている。よってLin28に加えて、またはLin28に代えて、Lin28bを使用することができる。Lin28bについて、たとえばヒトLin28b、マウスLin28bの配列は、それぞれ、NM_001004317、NM_001031772として登録されている。
【0040】
上記以外の他の初期化因子の非限定的な例は、ECAT1、ECAT2(ESG1とも称する)、ECAT3(Fbx15とも称する)、ECAT5(Erasとも称する)、ECAT7、ECAT8、ECAT9(Gdf3とも称する)、ECAT10(Sox15とも称する)、ECAT15-1(Dppa4とも称する)、ECAT15-2(Dppa2とも称する)、Fthl17、Sall4、Rex1(Zfp42とも称する)、Utf1、Tcl1、Stella(Dppa3とも称する)、β-catenin(Ctnnb1とも称する)、Stat3、Grb2、c-Myc、Sox1、Sox3、N-Myc、L-Myc、Klf1、Klf2、Klf4、Klf5(以上、国際公開WO2007/069666)、並びに、FoxD3、ZNF206、Mybl2、Otx2(以上、国際公開WO2008/118820)などから選択される1又は複数の初期化因子(群)である。これらの初期化因子の配列のGenBank登録番号(ヒト及びマウス)は以下のとおりである。
【0041】
ECAT1について、ヒトECAT1、マウスECAT1の配列は、それぞれ、AB211062、AB211060として登録されている。
【0042】
ECAT2について、ヒトECAT2、マウスECAT2の配列は、それぞれ、NM_001025290、NM_025274として登録されている。
【0043】
ECAT3について、ヒトECAT3、マウスECAT3の配列は、それぞれ、NM_152676、NM_015798として登録されている。
【0044】
ECAT5について、ヒトECAT5、マウスECAT5の配列は、それぞれ、NM_181532、NM_181548として登録されている。
【0045】
ECAT7について、ヒトECAT7、マウスECAT7の配列は、それぞれ、NM_013369、NM_019448として登録されている。
【0046】
ECAT8について、ヒトECAT8、マウスECAT8の配列は、それぞれ、AB211063、AB211061として登録されている。
【0047】
ECAT9について、ヒトECAT9、マウスECAT9の配列は、それぞれ、NM_020634、NM_008108として登録されている。
【0048】
ECAT10について、ヒトECAT10、マウスECAT10の配列は、それぞれ、NM_006942、NM_009235として登録されている。
【0049】
ECAT15-1について、ヒトECAT15-1、マウスECAT15-1の配列は、それぞれ、NM_018189、NM_028610として登録されている。
【0050】
ECAT15-2について、ヒトECAT15-2、マウスECAT15-2の配列は、それぞれ、NM_138815、NM_028615として登録されている。
【0051】
Fthl17について、ヒトFthl17、マウスFthl17の配列は、NM_031894、NM_031261として登録されている。
【0052】
Sal14について、ヒトSal14、マウスSal14の配列は、それぞれ、NM_020436、NM_175303として登録されている。
【0053】
Rex1について、ヒトRex1、マウスRex1の配列は、それぞれ、NM_174900、NM_009556として登録されている。
【0054】
Utf1について、ヒトUtf1、マウスUtf1の配列は、それぞれ、NM_003577、NM_009482として登録されている。
【0055】
Tcl1について、ヒトTcl1、マウスTcl1の配列は、それぞれ、NM_021966、NM_009337として登録されている。
【0056】
Stellaについて、ヒトStella、マウスStellaの配列は、それぞれ、NM_199286、NM_139218として登録されている。
【0057】
β-cateninについて、ヒトβ-catenin、マウスβ-cateninの配列は、それぞれ、NM_001904、NM_007614として登録されている。
【0058】
Stat3について、ヒトStat3、マウスStat3の配列は、それぞれ、NM_139276、NM_213659として登録されている。
【0059】
Grb2について、ヒトGrb2、マウスGrb2の配列は、それぞれ、NM_002086、NM_008163として登録されている。
【0060】
FoxD3について、ヒトFoxD3、マウスFoxD3の配列は、それぞれ、NM_012183、NM_010425として登録されている。
【0061】
ZNF206について、ヒトZNF206、マウスZNF206の配列は、それぞれ、NM_032805、NM_001033425として登録されている。
【0062】
Mybl2について、ヒトMybl2、マウスMybl2の配列は、それぞれ、NM_002466、NM_008652として登録されている。
【0063】
Otx2について、ヒトOtx2、マウスOtx2の配列は、それぞれ、NM_172337、NM_144841として登録されている。
【0064】
c-Mycについて、ヒトc-Myc、マウスc-Mycの配列は、それぞれ、NM_002467、NM_010849として登録されている。
【0065】
N-Mycについて、ヒトN-Myc、マウスN-Mycの配列は、それぞれ、NM_005378、NM_008709として登録されている。
【0066】
L-Mycについて、ヒトL-Myc、マウスL-Mycの配列は、それぞれ、NM_001033081、NM_008506として登録されている。
【0067】
Sox1について、ヒトSox1、マウスSox1の配列は、それぞれ、NM_005986、NM_009233として登録されている。
【0068】
Sox3について、ヒトSox3、マウスSox3の配列は、それぞれ、NM_005634、NM_009237として登録されている。
【0069】
Klf1について、ヒトKlf1、マウスKlf1の配列は、それぞれNM_006563、NM_010635として登録されている。
【0070】
Klf2について、ヒトKlf2、マウスKlf2の配列は、それぞれ、NM_016270、NM_008452として登録されている。
【0071】
Klf4について、ヒトKlf4、マウスKlf4の配列は、それぞれ、NM_004235、NM_010637として登録されている。Klf5について、ヒトKlf5、マウスKlf5の配列は、それぞれ、NM_001730、NM_009769として登録されている。
【0072】
本発明の方法におけるiPS細胞の誘導系には、上記初期化因子と組み合わせて、以下に例示されるようなヒトを含む哺乳動物由来のサイトカインを少なくとも1種含むことができる。サイトカインの例は、bFGF(basic Fibroblast Growth Factor)、SCF(Stem Cell Factor)、TERT(teromerase reverse transcriptase)、SV40 Large T、HPV16 (human papillomavirus type 16) E6又はE7、Bmi1(ポリコーム遺伝子産物)などを含む。
【0073】
本発明の方法では、初期化因子は、蛋白質(若しくはポリペプチド)又は核酸(若しくはポリヌクレオチド)のいずれかの形態で使用することができる。このような蛋白質又は核酸は、PCR(polymerase chain reaction)技術、遺伝子組換え技術などの慣用技術を使用することによって作製することができる。
【0074】
一般に、核酸は、それを含む哺乳動物細胞由来のゲノムライブラリー又はcDNAライブラリーからPCR技術を利用して増幅されうる。そのために、該核酸のポリヌクレオチド配列に基づいてセンス及びアンチセンスプライマーを作製する。この場合、プライマーは、初期化因子としての核酸のORF(open reading frame)配列の両末端にアニーリングするように設計される。一般に、プライマーのサイズは、約17〜30塩基長、好ましくは約20〜26塩基長である。PCRは、変性段階、アニーリング段階及び伸長段階を1サイクルとして約20〜40サイクルを実施することからなり、変性段階は、二本鎖DNAを変性して各一本鎖DNAに解離する段階(例えば約94〜96℃、約30秒〜約5分)であり、アニーリング段階は、鋳型としての各一本鎖DNAにセンスプライマー又はアンチセンスプライマーをアニーリング(すなわち、結合)する段階(例えば約50〜65℃、約30秒〜1分)であり、伸長段階は、一本鎖DNAを鋳型としてセンス及びアンチセンスプライマーを伸長しDNA鎖を合成する段階(例えば約72℃、約30秒〜10分)である。上記サイクルの実施前に約94〜96℃、約2〜7分程度の前処理を行ってもよいし、また、上記サイクルの完了後に約72℃、約5〜10分程度の後処理を行ってもよい。PCRに使用するバッファーは、PCRバッファー(MgCl2含有)を使用し、反応に際して、該バッファーにプライマー、耐熱性DNAポリメラーゼ酵素(例えばTaq DNAポリメラーゼなど)、4種のデオキシヌクレオチド(dNTP(N=A,T,C,G))を添加する。通常、反応液は約10〜100μl程度である。PCR技術は、例えばSaiki, R.K.ら,Science 230:1350-1354 (1985)、Erlich, H.A.ら,Science 252:1643-1651 (1991);Hughes, S.及びMoody, A., PCR (Methods Express), Scion Publishing (2007)などに記載されている。
【0075】
哺乳動物細胞由来のcDNAライブラリーは、組織又は細胞(体細胞、幹細胞、胚性幹細胞を含む)から全mRNAを抽出し、オリゴdTプライマーを使用する逆転写(RT)-PCR法によって全mRNAから第1鎖DNAを合成し、DNA合成酵素I、DNAリガーゼ、RNaseHにより第2鎖DNAを合成し、T4 DNA合成酵素により末端平滑化を行い、EcoRIアダプターを連結したのち、cDNAをリン酸化し、ファージベクター(例えばλgt11)に該cDNAを連結することによって作製されうる。
【0076】
哺乳動物細胞由来のゲノムライブラリーは、組織又は細胞からゲノムDNAを抽出し、適する制限酵素(例えばSau3A)によって部分分解し、スクロース密度勾配遠心により約40kb前後のDNA断片を回収し、例えばコスミドなどのベクターに該DNA断片を連結することによって作製されうる。
【0077】
上記のようにPCR増幅によって生成された初期化因子としての核酸に対して遺伝子組換え技術を適用し、該核酸を含むベクター、或いは、該核酸によってコードされる蛋白質を作製することができる。核酸(特に、cDNA)は、二本鎖の形態でベクターに挿入され、外来核酸が発現可能なように適当な制御配列によって調節されうる。ベクターは、例えば、プラスミド、ファージ、コスミド、ウイルス(例えばレトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、バキュロウイルスなど)、人工染色体(例えばHAC、BAC、PAC、YACなど)などのベクターを含むことができる。
【0078】
制御配列には、プロモーター、エンハンサー、ターミネーター、ポリアデニル化サイト、リボソーム結合サイト、複製開始点などが適宜含まれる。特に、プロモーターの例は、非限定的に、Oct3/4プロモーター、ヒトサイトメガロウイルス(CMV)プロモーター、アデノウイルス後期プロモーター、ワクシニアウイルス7.5Kプロモーター、SV40プロモーター、ポリヘドリンプロモーター、メタロチオネインプロモーター、カリフラワーモザイクウイルスプロモーター、タバコモザイクウイルスプロモーター、解糖系酵素プロモーターなどを含み、またプロモーターは、組織特異的プロモーター、誘導性プロモーター又は構成性プロモーターでありうる。
【0079】
さらにまた、場合により、ベクターはさらに選択マーカーを含んでもよく、選択マーカーとして、例えば、薬剤耐性遺伝子(例えばG418、ネオマイシン耐性遺伝子、ピューロマイシン耐性遺伝子など)、レポーター遺伝子(例えばGFP(green fluorescence protein)、GUS(β-gluclonidase)、FLAGなど)、ネガティブ選択マーカー(例えばチミジンキナーゼ(TK)遺伝子、ジフテリアトキシン(DT)遺伝子など)などが挙げられる。また、ベクターは、外来核酸の挿入を容易にするための複数の制限酵素認識サイトを有するマルチクローニングサイト、IRES(internal ribosomal entry site)などの配列をさらに含んでいてもよい。
【0080】
本発明の方法で使用される初期化因子としての蛋白質類は、該蛋白質(シグナル配列を含む前駆体蛋白質、又は成熟蛋白質)をコードするDNAを含む発現ベクターを、宿主細胞としての原核細胞又は真核細胞、例えば細菌類(大腸菌、枯草菌、シュードモナス属など)、酵母(サッカロミセス属、カンジダ属、ピキア属など)、昆虫細胞(Sf系細胞など)、哺乳動物細胞(CHO、NIH3T3、HEK293、COS、BHKなど)、植物細胞などに導入し、形質転換又は形質導入された宿主細胞を適当な培地中で培養し、該細胞又は培地から目的の蛋白質を回収することを含む方法によって得ることができる。蛋白質の回収は、クロマトグラフィー(例えばゲルろ過クロマトグラフィー、イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、HPLC、FPLCなど)、電気泳動、等電点電気泳動、限外ろ過、硫安沈殿、有機溶媒沈殿などの慣用手段を適宜組み合わせて行うことができる。
【0081】
本発明の方法で使用される初期化因子としての核酸類は、好ましくは、上記蛋白質(シグナル配列を含む前駆体蛋白質、又は成熟蛋白質)をコードするDNAを発現可能に含むベクターの形態である。特に、このようなベクターをヒト体細胞の初期化に使用するときには、初期化の完了後に上記DNAのサイレンシングを起こすか、或いは発現が一過性であることが望ましい。ベクターは、レトロウイルス、レンチウイルスなどのような細胞のゲノムに組み込まれ易いベクター、或いは、アデノウイルス、プラスミド、人工染色体などのような細胞のゲノムに組み込まれ難いベクターのいずれでもよい(Stadtfeld, M.ら, Science, 322, 945-949 (2008)(オンライン公表日:2008年9月25日); Okita, K.ら, Science, 322, 949-953 (2008)( オンライン公表日:2008年10月9日); Takahashi, K.ら, Cell 131:861-872 (2007);Yu, J.ら, Science 318:1917-1920 (2007))。
【0082】
上記の遺伝子組換え技術の一般的な手順は、Sambrook, J.ら, Molecular Cloning: A Laboratory Manual, 2nd edition, Cold Spring Harbor Laboratory Press (1989)、Ausubel, F.M.ら,Short protocols in Molecular Biology: A Compendium Methods from Current Protocols in Molecular Biology, John Wiley & Sons (1999)などに記載されている。
【0083】
3. 人工多能性幹(iPS)細胞の樹立効率改善物質
本発明の方法においては、Oct3/4及びNanog、又はそれらをコードする核酸、からなる初期化因子によって、或いは、これらの初期化因子に加えて、Sox2及びSox2をコードする核酸を除く他の初期化因子やサイトカインによって、哺乳動物体細胞からiPS細胞を樹立することができる。腫瘍遺伝子の一種であるc-Myc及びKlf4が初期化因子として使用される場合には、iPS細胞の樹立効率は高まるが、逆に、これらのいずれか又は両方が不在である場合には、iPS細胞の樹立効率が著しく低下することが知られている。
【0084】
しかし、このような場合、iPS細胞の樹立効率を改善する物質を、iPS細胞誘導系に添加することによって、iPS細胞の樹立を促進することが可能である。このような物質には、例えば塩基性線維芽細胞増殖因子(bFGF)、幹細胞因子(SCF)などのサイトカインが含まれる。また、例えばヒストンデアセチラーゼ(HDAC)阻害剤、例えばバルプロ酸(VPA)(Huangfu, D.ら,Nat. Biotechnol., 26(7):795-797 (2008))、ヒストンメチルトランスフェラーゼ(G9a)阻害剤、例えばBIX-01294 (BIX) (Shi, Y.ら,Cell Stem Cell, 2:525-528 (2008);Kubicek, S.ら,Mol. Cell 25:473-481 (2007);Shi, Y.ら, Cell Stem Cell, 3, 568-574 (2008))、DNAメチル化酵素(Dnmt)阻害剤、例えば5’-アザシチジン(Huangfu, D.ら,上記)などの低分子化合物が含まれる。またp53に対するshRNAやsiRNAなどのp53阻害剤や、UTF1を細胞内に導入してもよい(Yang Zhaoら, Cell Stem Cell, 3, pp475-479, 2008)。他にもシグナル伝達に関して、Wntシグナルの活性化(Marson A.ら, Cell Stem Cell, 3, pp132-135, 2008)、マイトジェン活性化プロテインキナーゼおよびグリコーゲン合成酵素キナーゼ-3シグナル伝達の阻害(Silva J.ら, PloS Biology, 6, pp2237-2247 2008)、ES細胞特異的miRNA (例えば、miR-302-367クラスター (Mol. Cell Biol. Doi:10. 1128/MCB. 00398-08), miR-302 (RNA 14:1-10 (2008)), miR-291-3p, Mir-294及びmiR-295 (Nat. Biotechnol. 27:459-461 (2009))なども、iPS細胞の樹立効率を改善することができる。
【0085】
上記のような樹立効率改善物質は、分化多能性に関わる遺伝子の活性化及び分化細胞で特異的に発現される遺伝子の不活性化の両方に何らかの作用をすることが、推定されている(Huangfu, D.ら,上記)。本発明の方法においても、樹立効率改善物質を使用することによって、体細胞からiPS細胞への初期化(すなわち、再プログラム化)を促進することが可能である。
【0086】
さらに、体細胞の初期化段階で、低酸素条件下で細胞を培養することによってiPS細胞の樹立効率を向上させることができる (Yoshida, Y. et al., Cell Stem Cell 5:237-241 (2009))。本明細書で使用する「低酸素条件」という用語は、細胞培養の際の酸素濃度が空気中の酸素濃度と比べてかなり低いことを意味している。具体的には、このような条件として、5〜10% CO2 / 95〜90%空気の雰囲気中の酸素濃度より低い酸素濃度、例えば18%以下の酸素濃度からなる条件が挙げられる。前記雰囲気中の酸素濃度の好ましい例は、15%以下(例えば14%以下、13%以下、12%以下又は11%以下)、10%以下(例えば、9%以下、8%以下、7%以下又は6%以下)、或いは5%以下(例えば、4%以下、3%以下又は2%以下)である。或いは、前記雰囲気中の好ましい酸素濃度は、0.1%以上(例えば、0.2%以上、0.3%以上又は0.4%以上)、0.5%以上 (例えば、0.6%以上、0.7%以上、0.8%以上又は0.95%以上)、或いは1%以上 (例えば、1.1%以上、1.2%以上、1.3%以上又は1.4%以上)である。
【0087】
細胞環境において前記低酸素状態を作るための手法としては、以下のものに限定されないが、酸素濃度を調節することが可能なCO2インキュベーター中で細胞を培養するという最も簡単な方法でありかつ好ましい例としてのこの方法を挙げることができる。このようなインキュベーターは種々の装置メーカー(例えば、Thermo Scientific, 池本理化工業(Ikemoto Scientific Technology), 十慈フィールド(Juji Field), 和研薬(Wakenyaku)など)から市販されており、上記の目的で使用しうる。
【0088】
4. 体細胞
本発明で使用可能な体細胞は、上記定義のとおり、生殖系列細胞(卵子、卵母細胞、胚性幹(ES)細胞など)又は分化全能性細胞を除く、哺乳動物由来のすべての細胞である。
【0089】
体細胞が由来する哺乳動物は、特に制限されずいかなる種類の動物も包含する。好ましい哺乳動物は、霊長類(例えばヒト、サル、チンパンジー等)、げっ歯類(例えばマウス、ラット、ハムスター、モルモット等)、及び有蹄類(ウシ、ヒツジ、ヤギ、ウマ、ブタ等)、ペット動物(イヌ、ネコ等)から選択され、さらに好ましい哺乳動物はヒト及びマウスである。
【0090】
体細胞には、非限定的に、胎児(仔)期の体細胞、新生児(仔)の体細胞、及び成熟した体細胞のいずれも包含されるし、また、初代培養細胞、継代細胞、及び株化細胞のいずれも包含されるし、さらにまた、組織幹細胞や組織前駆細胞も包含される。
【0091】
具体的には、体細胞は、非限定的に、例えば(1)神経幹細胞、造血幹細胞、間葉系幹細胞、歯髄幹細胞等の組織幹細胞(体性幹細胞)、(2)組織前駆細胞、(3)リンパ球、上皮細胞、内皮細胞、筋肉細胞、線維芽細胞(皮膚細胞等)、毛細胞、肝細胞、胃粘膜細胞、腸細胞、脾細胞、膵細胞(膵外分泌細胞等)、脳細胞、肺細胞、腎細胞、皮膚細胞等の分化した細胞などを包含する。
【0092】
体細胞を得るための起源として適する哺乳動物個体は、非限定的に、好ましくは患者自身、或いは、得られたiPS細胞が再生医療のために使用されたときに拒絶反応が生じないという観点から、同一の又は実質的に同一のHLA型をもつ他人である。本明細書で使用するHLA型に関連した「実質的に同一」という用語は、体細胞由来のiPS細胞の分化を誘導することによって作製された細胞が患者に移植されるときに、HLA型が、ドナーとレシピエントとの間で、主要HLA類(例えば、HLA-A, HLA-B及びHLA-DRの3つの遺伝子の遺伝子座)が同じである場合のように、移植された細胞が生存可能である程度に一致することを意味する。この定義は以下においても同様に使用される。iPS細胞が投与されないか又は移植されないとき、すなわち例えば、iPS細胞を、患者での薬剤の感受性や副作用の有無を評価するスクリーニング用の細胞供給源として使用する場合、体細胞を、患者自身から、或いは薬剤の感受性や副作用と相関する同一の遺伝子多型を有する他人から、得ることが望ましい。
【0093】
iPS細胞の作製に先立ち、ヒトを含む哺乳動物から体細胞を取得し、これを動物細胞培養用培地にて培養し、必要であれば継代培養に付し、これにより初代培養細胞又は継代培養細胞を得る。このようにして得られた培養細胞を、iPS細胞の作製のために使用する。
【0094】
体細胞の培養は、例えばDMEM(Dulbecco’s modified Eagle medium)、MEM (minimum essential medium)、α-MEM(minimum essential medium alpha modification)、Ham’s F12、RPMI1640、それらの混合培地などの基本培地に、血清(10% FBS等)、抗生物質(penicillin、streptomycin等)、ピルビン酸Na、グルタミン、非必須アミノ酸、L-デキストロースなどの物質を適宜選択して含有させた培地上で約37℃の温度、5% CO2存在下にて行うことができる。
【0095】
5. 人工多能性幹(iPS)細胞の作製
本発明によるiPS細胞の作製は、初期化因子が、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子(ただし、Sox2及びSox2をコードする核酸を含まない)であることを除けば、従来公知の手順(WO2007/069666; WO2008/11820; WO2008/124133;Takahashi, K.ら, Cell 131:861-872 (2007))で行うことができる。
【0096】
本発明で使用しうる初期化因子は、上記2節に例示されたような種々の初期化因子の組み合わせ(ただし、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、は必須である。)を含み、それらは蛋白質又は核酸からなる。該核酸の好適な形態はベクターであり、初期化因子は、通常、その発現が可能なように上記制御配列と連結される。この場合、Oct3/4及びNanogを含む2以上の初期化因子は、同じ又は異なるベクターに発現可能なように挿入される。通常、初期化因子の数が約2〜4程度であれば、1つのベクター中に複数の初期化因子をタンデムに連結することができる。各初期化因子の5’側にはプロモーター配列、3’側にはターミネーター配列、ポリA配列などの配列がそれぞれ連結され、初期化因子間にはリンカー配列を配置してもよい。好ましくは、複数の初期化因子が発現可能なようにタンデムに連結されて、それらを含むカセットを作製し、このカセットをベクターに組み込むとよい。2つ又はそれ以上の初期化因子(又遺伝子)の連結のために、ポリシストロニックな発現を可能にするあらゆる配列、例えば口蹄疫ウイルス由来2A配列 (PLoS ONE3, e2532, 2008, Stem Cells 25, 1707, 2007)、IRES配列 (米国特許第4,937,190号)、好ましくは前記2A配列、を使用することができる。
【0097】
本発明で使用可能なベクターは、上で例示されるような、プラスミド、ウイルス、人工染色体などから選択されうる。これまで、iPS細胞の作製に使用されたベクターは、レトロウイルス、レンチウイルス、アデノウイルスなどのウイルスベクター、及びプラスミドであり、これらのベクターは、本発明でも同様に使用できる(WO2007/069666; WO2008/11820; WO2008/124133; Takahashi, K.ら, Cell 131:861-872 (2007); Stadtfeld, M.ら,Science, 322, 945-949 (2008)(オンライン公表日:2008年9月25日); Okita, K.ら, Science, 322, 949-953 (2008)( オンライン公表日:2008年10月9日))。さらに使用可能なベクターとして、エピソーマルベクター (Yu et al., Science, 324, 797-801 (2009))、トランスポゾン (例えば、ピギーバックトランスポゾン(piggyback transposone): Kaji, K. et al., Nature, 458:771-775 (2009); Woltjen et al., Nature, 458:766-770 (2009)、センダイウイルスベクター (J. Biol. Chem., 282:27383-27391 (2007);日本特許第3,602,058号)が挙げられる。
【0098】
他のベクターとして、人工染色体も使用できる。人工染色体の例には、ヒト人工染色体(HAC)、酵母人工染色体(YAC)、細菌人工染色体(BAC、PAC)などが含まれる。このうちHAC及びYACは、セントロメア、2つのテロメア及び染色体断片を含むミニ染色体である。この染色体断片に、上記複数の初期化因子を含むカセットを挿入する。
【0099】
哺乳動物由来の培養体細胞に、核酸をコードする初期化因子を導入する方法には、慣用の手法が含まれ、エレクトロポレーション法、マイクロインジェクション法、リン酸カルシウム法、ウイルス感染法、リポフェクション法などが含まれる。
【0100】
蛋白質の形態で初期化因子を培養哺乳動物体細胞に導入する方法には、従来の方法、例えば、蛋白質デリバリー試薬を使用する方法、蛋白質輸送ドメイン(PTD)又は細胞透過性ペプチド (CPP)との融合蛋白質を使用する方法、マイクロインジェクション法、リポソーム法、リポフェクション法などが含まれる。蛋白質デリバリー剤は、例えば、カチオン性脂質ベースのBioPOTER Protein Delivery Reagent (Gene Therapy Systems)、Pro-JectTM Protein Transfection Reagent (PIERCE)、ProVectin (IMGENEX)、脂質ベースのProfect-1 (Targeting System)、膜透過性ペプチドベースのPenetrain Peptide (Q biogene)、Chariot Kit (Active Motif)、及び(不活性化センダイウイルス由来の)HVJエンベロープを使用するGenomONE (石原産業、日本)として市販されている。細胞内への蛋白質又はペプチドの導入は、それぞれの試薬に添付された使用説明書に従って行うことができ、その一般的な手順は以下のとおりである。すなわち、初期化因子を適当な溶媒 (例えば、PBS又はHEPESなどのバッファー)で希釈し、その後、これにデリバリー試薬を加え、混合物を室温で5〜15分間インキュベーションして、初期化因子とデリバリー試薬との間で複合体を形成する。この複合体を、予め交換された血清不含有培地中の細胞に添加し、その後、該細胞を37℃、1時間〜数時間インキュベーションする。このあと、培地を除去し、血清含有培地と交換する。
【0101】
使用されるPTDとして開示されるものは、Drosophila由来AntP; HIV由来TAT (Frankel, A. et al., Cell 55:1189-1193 (1988)); Green, M.と Loewenstein, P.M. Cell 55:1179-1188 (1988)); Penetratin (Derossei, D. et al., J. Biol. Chem. 269:10444-10450 (1994)); Buforin II (Park, C.B. et al., Proc. Natl. Acad. Sci. USA 97:8245-8250 (2000)); Transportan (Pooga, M. et al., FASEB J. 12:67-77 (1998); MAP (model amphipathic peptide) (Oehlke, J. et al., Biochim. Biophys. Acta. 1414:127-139(1998)); K-FGF (Lin, Z. et al., J. Biol. Chem. 270:14255-14258 (1995)); Ku70 (Sawada, M. et al., Nature Cell Biol. 5:352-357 (2003)); Prion (Lundberg, P. et al., Biochem. Biophys. Res. Commun. 299:85-90 (2002)); pVEC (Elmquist, A. et al., Exp. Cell Res. 269:237-244 (2001)); Pep-1 (Morris, M.C. et al., Nature Biotechnol. 19:1173-1176 (2001)); Pep-7 (Gao, C. et al., Bioorg. Med. Chem. 10:4057-4065 (2002)); SynB1 (Rousselle, C. et al., Mol. Pharmacol. 57:679-686 (2000)); HN-1 (Hong, F.D. & Clayman, G.L., Cancer Res. 60:6551-6556 (2000));及びHSV由来VP22などの蛋白質の細胞通過ドメインである。PTD由来のCPPには、11R (Cell Stem Cell 4:381-384 (2009))及び9R (Cell Stem Cell 4:472-476 (2009))などのポリアルギニンが含まれる。
【0102】
融合蛋白質を発現する、並びに、初期化因子をコードするcDNA及びPTD又はCPP配列が組み込まれたベクターが作製されて、該融合蛋白質をコードするDNAを発現し、その後、該融合蛋白質を回収して細胞に導入する。該導入は、蛋白質デリバリー試薬を添加しないことを除いて上記と同様の方法で行うことができる。
【0103】
マイクロインジェクションは、先端径約1μmを有するガラス針の中に蛋白質溶液を入れ、針を細胞に刺すことによってこの溶液を細胞に導入し、これによって蛋白質を確実に細胞内に導入できる、方法である。
【0104】
蛋白質を導入する操作は、1回又はそれ以上、例えば1〜10回、1〜5回など、好ましくは2回以上繰り返して行うことができる。操作の繰り返しの間の時間間隔は、例えば6〜48時間、好ましくは12〜24時間である。
【0105】
iPS細胞誘導のために、培養体細胞と上記初期化因子を、適する動物細胞培養培地中で接触させて、該初期化因子を体細胞内に導入し、該細胞を形質転換又は形質導入する。培養培地としては、非限定的に、上記4節に記載したような培地、例えば(1) 10〜15%FBSを含有するDMEM、DMEM/F12又はDME培地(これらの培地にはさらに、LIF(leukemia inhibiting factor)、ペニシリン/ストレプトマイシン、ピューロマイシン、L-グルタミン、非必須アミノ酸類、β-メルカプトエタノールなどを適宜含むことができる。)、(2) bFGF又はSCFを含有するES細胞培養用培地、例えばマウスES細胞培養用培地(例えばTX-WESTM、コスモバイオ)又は霊長類ES細胞培養用培地(例えばReproCELLTM、コスモバイオ)、などが含まれる。
【0106】
培養手順の例は次のとおりである。37℃、5%CO2存在下にて、10%FBS含有DMEM又はDMEM/F12培地上で体細胞と初期化因子を接触させ約6〜7日間培養し、その後、細胞をフィーダー細胞(マイトマイシンC処理STO細胞、SNL細胞等)上に蒔きなおし、体細胞と初期化因子の接触から約10日後からbFGF含有霊長類ES細胞培養用培地で培養し、該接触から約30〜約45日又はそれ以上ののちにiPS様コロニーを生じさせることができる。この培養手順は、ヒトiPS細胞などの霊長類iPS細胞の誘導に適している。
【0107】
或いは、その代替例として、37℃、5% CO2存在下にて、フィーダー細胞(マイトマイシンC処理STO細胞、SNL細胞等)上で10%FBS含有DMEM培地(これにはさらに、LIF、ペニシリン/ストレプトマイシン、ピューロマイシン、L-グルタミン、非必須アミノ酸類、β-メルカプトエタノールなどを適宜含むことができる。)で培養し、約25〜約30日又はそれ以上ののちにiPS様コロニーを生じさせることができる。この培養手順は、マウスiPS細胞などのげっ歯類iPS細胞の誘導に使用される。
【0108】
上記培養の間には、培養開始2日目以降から毎日1回新鮮な培地と培地交換を行う。また、初期化に使用する体細胞の細胞数は、限定されないが、培養ディッシュ100cm2あたり約5×103〜約5×106細胞の範囲である。
【0109】
ディッシュ上の上記iPS様コロニーを、トリプシン及びコラゲナーゼIVを含む溶液(CTK溶液)を用いて処理し、残ったコロニーを上記フィーダー細胞上に蒔いてES細胞培養用培地で同様に培養することにより、iPS細胞又はiPS細胞コロニーを得ることができる。さらに同様の培養条件でiPS細胞を継代することができる。
【0110】
iPS細胞の同定は、ES細胞等の分化多能性細胞の特性に基づく試験によって行うことができる。具体的には、ES細胞特異的マーカー遺伝子の発現、半永久的細胞増殖能、分化多能性(三胚葉の形成)などの特性について細胞を試験し、細胞がこれらの特性をもつ場合に、該細胞がiPS細胞であると同定する(Takahashi, K.ら, Cell 131:861-872 (2007))。
【0111】
ES細胞特異的マーカー遺伝子としては、例えばOct3/4, Nanog, Lin28, PH34, Dnmt3b, Noda1, SSEA3, SSEA4, Tra-1-60, Tra-1-81, Tra2-49/6F(アルカリホスファターゼ)などを挙げることができる。これらの遺伝子の増幅に特異的なプライマーを使用するRT-PCR法によって、細胞内の上記マーカー遺伝子の発現を検出することができる。後述の実施例(ヒトiPS細胞の樹立)では、Oct3/4, Nanog, Lin28, PH34, Dnmt3b, Nodalの遺伝子発現が確認された。
【0112】
半永久的細胞増殖能については、約4ヶ月〜6ヶ月以上にわたる細胞の培養試験において、該細胞が指数的増殖をすることを確認する。ヒトiPS細胞のコロニー倍加時間(又は集団倍加時間(population doubling time))は、例えば46.9±12.4時間、47.8±6.6時間又は43.2±11.5時間程度であることが知られているので、この値を増殖能の指標とすることができる(Takahashi, K.ら, Cell 131:861-872 (2007))。また、iPS細胞は、高いテロメラーゼ活性をもつため、例えばTRAP(telomeric repeat amplification protocol)法によって該活性を検出してもよい。後述の実施例(ヒトiPS細胞の樹立)では、作製された細胞は指数的増殖を示した。
【0113】
分化多能性(3胚葉の形成)は、例えばテラトーマの形成及びテラトーマ組織内の3胚葉(内胚葉、中胚葉及び外胚葉)系の各組織(もしくは細胞)の同定によって確認することができる。具体的には、マウスiPS細胞の場合はヌードマウスの皮下に、またヒトiPS細胞の場合はScidマウスの精巣内に細胞を注射して腫瘍の形成を確認し、さらに腫瘍組織が、例えば軟骨組織(もしくは細胞)、神経管組織(もしくは細胞)、筋肉組織(もしくは細胞)、脂肪組織(もしくは細胞)、腸管様組織(もしくは細胞)などから構成されていることを確認する。後述の実施例(ヒトiPS細胞の樹立)では、形成されたテラトーマが、神経管組織(外胚葉)、軟骨組織(中胚葉)、腸管様組織(内胚葉)などを含むことが、組織染色法によって確認された。
上記の各試験に基づいて、細胞がiPS細胞であることを確認し、iPS細胞コロニーを選択することができる。
【0114】
6. 人工多能性幹(iPS)細胞の応用
本発明の方法によって作製されたiPS細胞は、分化多能性を有し、ES細胞と非常に類似した特性を有している。このため、iPS細胞は、ES細胞の代替細胞として利用することが可能である。
【0115】
iPS細胞は、それが分化多能性をもつことから、種々の分化細胞、前駆細胞及び組織に誘導することができる。具体的には、iPS細胞から、例えばactivin A/BMP4(bone morphogenetic protein 4)、VEGF(vascular endothelial growth factor)などの因子の存在下で神経細胞、心筋細胞などの種々の分化細胞を誘導することができる。このようにして得られた分化細胞は、患者に、例えば患者の欠陥組織に、移植することによって患者の治療(いわゆる、再生医療)のために使用できる。
【0116】
また、哺乳動物(ヒトを除く)由来の胚の胚盤胞にiPS細胞を導入し、その胚を、同じ動物種の仮親の子宮に移植することによって、iPS細胞の遺伝子型及び形質の一部を受け継いだキメラ動物を作製することができる (WO2007/069666)。iPS細胞における特定の遺伝子の改変、ノックアウト(KO)又はノックイン(KI)による遺伝子機能の解明、疾患モデルの作製、物質(蛋白質等)生産などが可能になる。このとき、予め遺伝子改変した体細胞から誘導されたiPS細胞も使用できる。
【0117】
したがって、本発明は、例えば、上記の方法で作製されたiPS細胞及びiPS細胞集団、遺伝子改変されたiPS細胞、並びに、そのようなiPS細胞を使用するキメラ動物の作製法及びそれによって得られたキメラ動物や子孫動物なども包含する。
【実施例】
【0118】
以下の実施例によって本発明をさらに具体的に説明する。しかし、本発明の範囲は、その実施例によって制限されないものとする。
【0119】
[実施例1]
Oct3/4及びNanogによるヒトiPS細胞の樹立(1)
36歳の成人皮膚由来線維芽細胞(HDF)に対して、Takahashi, K.ら, Cell, 131:861-872 (2007)に記載の方法に従い、レンチウイルス(pLenti6/UbC-Slc7a1)を用いて、マウスエコトロピックウイルスレセプターSlc7a1遺伝子を発現させた。この細胞(1×105個/6ウエルプレートの1ウエル)に対して、Takahashi, K.ら, Cell, 131:861-872 (2007) に記載の方法に従いヒト由来の4 遺伝子(Oct3/4,Sox2,Nanog,Lin28)をレトロウイルスで導入した。ウイルス感染から7日後に細胞を回収し、フィーダー細胞上への蒔き直しを行った(5×105個/100 mmディッシュ)。フィーダー細胞にはマイトマイシンCで処理して、細胞分裂を止めたSNL細胞(McMahon, A. P. & Bradley, A. Cell 62, 1073-1085 (1990))を用いた。感染10日後から霊長類ES細胞培養用培地 (ReproCELL、日本) に4 ng/mlの組換えヒトbFGF(WAKO、日本)を加えた培地で培養を行った。感染35日後にヒトiPS細胞様のコロニーが2個出現した。iPS細胞のコロニーから樹立したクローンはヒトES細胞様の形態を示し、フィーダー細胞上で増殖を続けることができた(図1)。
【0120】
これらの樹立したヒトiPS細胞のクローンについて、Cell, 131, 861-872(2007)の記載に準じてGenomic-PCR解析を行い、使用した4遺伝子がゲノム上に挿入されているか検討した。樹立したiPSクローンのうち1クローン(T4F-2)は4因子共にゲノムに挿入されていたが、他の1クローン(T4F-1)ではOct3/4,Nanogのみがゲノムに挿入されていた(図2)。レトロウイルスベクターはゲノムに挿入されなければ安定的に発現しないので、このクローンT4F-1はOct3/4,Nanogのみの発現で樹立されたと考えられた。
【0121】
樹立したクローンについてRever Tra Ace kit(Takara、日本)を使用してRT-PCR解析を行った結果、これらのクローン(T4F-1、T4F-2)はヒトES細胞特異的なマーカー遺伝子であるOct3/4, Nanog, Lin28, PH34, Dnmt3b及びNodalを発現しており、その発現量はヒトES細胞や既存のiPS細胞(201B6及び201B7:Takahashi, K.ら, Cell, 131:861-872 (2007))と同等であることが示された(図3)。
【0122】
さらにこのクローンT4F-1をScidマウスの精巣に挿入し分化多能性を調べた。具体的にはまず、ヒトiPS細胞クローンT4F-1を、組換えヒトbFGF(4ng/ml)及びRhoキナーゼ阻害剤Y-27632(10μM)を含有する霊長類ES細胞培養用培地 (ReproCELL、コスモバイオ、ともに日本)中で培養した。1時間後、collagen IVで処理して細胞を採取後、遠心して回収し、Y-27632(10μM)を含有するDMEM/F12中に浮遊させた。コンフルエントになった細胞(100mmディッシュ)の1/4量をScidマウスの精巣内に注射した。2〜3ヶ月後、腫瘍を切り刻んで4%フォルムアルデヒドを含有するPBSバッファーで固定した。パラフィン包埋組織をスライスし、ヘマトキシリン・エオシンで染色した。結果を図4に示す。組織学的に見ると腫瘍は複数の種類の細胞から構成されており、神経管組織、軟骨組織、腸管様上皮構造など3胚葉系に分化していたことから、iPS細胞の多能性が証明された (図4)。
以上の結果より、Oct3/4, Nanogの発現のみで樹立されたクローンは、iPS細胞であることが確認された。
【0123】
[実施例2]
Oct3/4及びNanogによるヒトiPS細胞の樹立(2)
Oct3/4,Nanogのみの導入でiPS細胞が樹立できるか検討した。上記同様、Slc7a1を発現させた36歳の成人皮膚由来線維芽細胞にレトロウイルスを用いてOct3/4,Nanogのみを導入し、感染7日後フィーダー細胞上へ蒔き直し、感染10日後から霊長類ES細胞培養用培地(ReproCELL(日本))に4 ng/mlの組換えヒトbFGF(WAKO、日本)を加えた培地で培養を行った。感染45日後にヒトiPS様のコロニーが1つ出現した(ON-1)。さらに、非ヒトiPS様コロニーを4個(ON-2、ON-4、ON-5、ON-6)ひろい、これを培養していくと4クローン中2クローン(ON-5、ON-6)がヒトES様の形態を示すように変化した。他の2クローンは非ES様の形態であった(ON-2、ON-4)。さらに47日目に、iPS誘導を行っている培養皿をCTK溶液(2.5%トリプシン 5 ml, 1 mg/mlコラゲナーゼIV 5 ml, 0.1 M CaCl2 0.5 ml, KSR 10 ml を30 mlの滅菌水に溶解したもの)を用いて処理し、その後PBSを用いて培養皿を洗ったところ、培養皿上に数個のコロニーが残った。これらをセルスクレイパーでかき集めフィーダー細胞上にまいて培養すると、ヒトES様のコロニーが得られた(ON-3)。これら樹立した4クローン(ON-1, ON-3, ON-5, ON-6)はフィーダー細胞上で増殖を続けることができた。クローンON-1とON-5のES様コロニー像を図5に示す。
【0124】
これらの樹立したヒトiPS細胞のクローンについて、Rever Tra Ace kit(Takara)を使用してRT-PCR解析を行った。その結果、これらのクローン(ON-1, ON-3, ON-5, ON-6)は、いずれもヒトES細胞特異的なマーカー遺伝子であるOct3/4, Nanog, Lin28, PH34及びDnmt3bを発現しており、その発現量はヒトES細胞や201B7と同等であった(図6)。
以上の結果から、iPS細胞をOct3/4,Nanogのみで樹立できることが明らかとなった。
【0125】
[実施例3]
ヒトiPS細胞の樹立
初期化因子を(a) Oct3/4, Nanog, Lin28及びc-Myc、又は(b) Oct3/4, Nanog及びLin28に代えたことを除いて実施例2と同様の手順で、36歳の成人皮膚由来線維芽細胞からヒトiPS細胞(ONLM及びONL)を樹立した(図7A及び図7B)。
【0126】
さらにまた、体細胞を成人皮膚由来線維芽細胞からヒト新生児包皮由来線維芽細胞に代えたことを除いて実施例2と同様の手順で、初期化因子Oct3/4及びNanogのみにより、該細胞からヒトiPS細胞を樹立した(図7C)。また体細胞を成人皮膚由来線維芽細胞から歯髄幹細胞(J.Dent.Res., 87(7): 676-681 (2008))に代えたことを除いて実施例2と同様の手順で、初期化因子Oct3/4及びNanogのみにより、該細胞からヒトiPS細胞を樹立した(図8)。
[実施例4]
Oct3/4及びNanogのみで樹立したヒトiPS細胞の解析
(1)Genomic-PCR解析
実施例2および3においてOct3/4及びNanogのみで樹立した成人皮膚線維芽細胞由来のiPS細胞、ヒト新生児包皮線維芽細胞由来のiPS細胞、および歯髄幹細胞由来のiPS細胞を用いて、Cell, 131, 861-872(2007)の記載に準じてGenomic-PCR解析を行った。結果を図9および図10に示す。いずれのiPS細胞においても導入したOct3/4遺伝子およびNanog遺伝子がゲノムに挿入されていた。また、導入に用いなかったSox2、Klf4、c-MycおよびLin28の各遺伝子がゲノムに挿入されていないことも確認した(図9、図10)。
【0127】
(2)テラトーマ形成能
成人皮膚由来の線維芽細胞からOct3/4及びNanogのみで樹立したiPS細胞を用いて、実施例1と同様にして分化多能性を調べた。結果を図11に示す。組織学的に見ると腫瘍は複数の種類の細胞から構成されており、神経組織、腺上皮組織、軟骨組織など三胚葉系に分化していたことから、iPS細胞の分化多能性が確認された (図11)。
【0128】
(3)免疫染色により観察された未分化マーカーの発現
成人皮膚由来の線維芽細胞からOct3/4及びNanogのみで樹立したiPS細胞を、マイトマイシンCで処理したSNLフィーダー上に播いた。5日間培養した後、細胞を4% パラフォルムアルデヒドで固定し、5% normal goat serum、1% BSA および0.2% TritonX-100を含むPBSバッファーで処理した。一次抗体として、抗ヒトSSEA1抗体 (DHSB, 1:100)、抗ヒトSSEA3抗体 (Peter W. Andrews博士より供与, 1:10), 抗ヒトTRA-1-60抗体(Peter W. Andrews博士より供与, 1:50)を用いた。また二次抗体としてAlexa488またはCy3(cyanine 3)でラベルされた抗体を用いた。核はHoechst 33342で染色した。結果を図12に示す。樹立したiPS細胞はSSEA3およびTRA1-60を発現していた。一方、抗SSEA1抗体ではコロニーの縁部しか染まらなかった。ヒトES細胞および過去に樹立済みのiPS細胞で同様な発現パターンが報告されていることから、2因子で樹立したiPS細胞は幹細胞マーカーの発現においてES細胞と同等であることが示された。
【0129】
(4)in vitro分化誘導
成人皮膚線維芽細胞からOct3/4及びNanogのみで樹立したiPS細胞をlow-binding dishに播き、Cell, 131, 861-872 (2007)に記載の方法でpoly-hydroxyethyl methacrylate (HEMA)でコートしたディッシュ上で8日間浮遊培養し、ゼラチンコートされたプレート上で胚様体(embryoid body:EB)を形成させた(100mmディッシュ)。8日間培養後、細胞を4% パラフォルムアルデヒドで固定し、5% normal goat serum、1% BSA および0.2% TritonX-100を含むPBSで処理した。内胚葉系細胞の分化マーカーであるAFP (α-fetoprotein)(R&D systems製)、中胚葉系細胞の分化マーカーであるSMA (smooth muscle actin)(DAKO製)、Desmin (NeoMarkers製)、Vimentin(Santa Cruz製)、外胚葉系の分化マーカーであるβIII-tubulin (Chemicon製)の各抗体を用いた染色を行った。二次抗体としてAlexa488またはCy3(cyanine 3)でラベルされた抗体を用いた。核はHoechst 33342で染色した。結果を図13に示す。
【0130】
染色によりこれらのマーカーの発現が確認され、樹立されたヒトiPS細胞はin vitroで三胚葉系への分化能を有することが確認された。
【0131】
(5) 核型の分析
樹立された3種のiPS細胞クローンについて、核型分析を日本遺伝子研究所(Nihon Gene Research Laboratories Inc.,日本)で行った。その結果、すべての核型が正常範囲内であった。
【0132】
以上の実施例1〜4で行ったGenomic-PCRおよびRT-PCRのプライマーのリストを表1、表2および配列表(配列番号:1〜20)に示した。
【0133】
【表1】

【0134】
【表2】
【技術分野】
【0001】
本発明は、人工多能性幹細胞の新規作製方法に関する。具体的には、本発明は、Oct3/4及びNanog、或いはそれらをコードする核酸、を主要な初期化因子として使用することを特徴とする体細胞から人工多能性幹細胞を作製する方法に関する。
以下において、「人工多能性幹細胞」は、単に「iPS細胞」と称することもある。
【背景技術】
【0002】
近年、マウス及びヒトのiPS細胞が相次いで樹立された。Takahashi及びYamanaka(非特許文献1)は、Fbx15遺伝子座にネオマイシン耐性遺伝子をノックインしたレポーターマウス由来線維芽細胞に、Oct3/4, Sox2, Klf4及びc-Myc遺伝子を導入し強制発現させることによって、iPS細胞を樹立した。Okitaら(非特許文献2)は、Fbx15よりも多能性細胞に発現が限局しているNanogの遺伝子座に緑色蛍光タンパク質(GFP)及びピューロマイシン耐性遺伝子を組み込んだトランスジェニックマウスを作製し、該マウス由来の線維芽細胞で上記4遺伝子を強制発現させ、ピューロマイシン耐性かつGFP陽性の細胞を選別することにより、遺伝子発現やエピジェネティック修飾が胚性幹(ES)細胞とほぼ同等のiPS細胞(Nanog iPS細胞)を樹立することに成功した。同様の結果が他のグループによっても再現された(非特許文献3、非特許文献4)。その後、c-Myc遺伝子を除いた3因子によってもiPS細胞を作製できることが明らかとなった(非特許文献5)。
【0003】
さらに、Takahashiら(非特許文献6)は、ヒトの皮膚由来線維芽細胞にマウスと同様の4遺伝子を導入することにより、iPS細胞を樹立することに成功した。一方、Yuら(非特許文献7)は、Oct3/4, Sox2, Nanog及びLin28遺伝子を使用してヒトiPS細胞を作製した。このように、体細胞に特定の因子を導入することにより、ヒト及びマウスで、分化多能性においてES細胞と遜色のないiPS細胞を作製できることが示された。
【0004】
前述のYuら (非特許文献7)においては、4種の遺伝子のうちNanogまたはLin28遺伝子を除いた3種の遺伝子導入によってiPS細胞コロニーが得られたが、Oct3/4またはSox2遺伝子を除いた3種の遺伝子導入ではiPS細胞コロニーは得られなかったことから、iPS細胞樹立におけるOct3/4及びSox2遺伝子の重要性が示されている((特許文献1)も参照)。またParkら(非特許文献8)においても、iPS細胞の樹立にはOct3/4とSox2が必須であると述べられている。
【0005】
さらにまた、(特許文献2)においては、Oct3/4, Sox2, Klf4及びc-Mycの各遺伝子を導入し、マウスiPS細胞を作製しているが、その際にDox-inducible systemでiPS細胞を誘導するという手法を開示している。また体細胞として成熟B細胞を用いてiPS細胞をうまく誘導できたことが記載されている。
【0006】
また、体細胞からiPS細胞を誘導することに関する特許文献として、Yamanaka(特許文献3)及びThomsonら(特許文献1)が公開されている。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO2008/118820
【特許文献2】WO2008/124133
【特許文献3】WO2007/069666
【非特許文献】
【0008】
【非特許文献1】Takahashi, K. and Yamanaka, S., Cell, 126: 663-676 (2006)
【非特許文献2】Okita, K. et al., Nature, 448: 313-317 (2007)
【非特許文献3】Wernig, M. et al., Nature, 448: 318-324 (2007)
【非特許文献4】Maherali, N. et al., Cell Stem Cell, 1: 55-70 (2007)
【非特許文献5】Nakagawa, M. et al., Nat. Biotethnol., 26: 101-106 (2008)
【非特許文献6】Takahashi, K. et al., Cell, 131: 861-872 (2007)
【非特許文献7】Yu, J. et al., Science, 318: 1917-1920 (2007)
【非特許文献8】Park I.H et al., Nature, 451: 141-146 (2008)
【発明の概要】
【発明が解決しようとする課題】
【0009】
上記のように、Yamanakaらの発見(Takahashi, K. and Yamanaka, S. Cell 126:663-676 (2006))がブレークスルーとなって、体細胞から人工多能性幹細胞を誘導することを可能にする初期化因子として、いくつかの組み合わせが提案されており、そのいずれも体細胞から人工多能性幹細胞を誘導する。提案された初期化因子の組み合わせのなかで共通する因子は、Oct3/4のみであるが、最近、Oct3/4だけで初期化が起こるとする報告がなされ、これによると、使用された体細胞が神経幹細胞であり、この細胞はSox2遺伝子が内因的に発現されている細胞であるので特殊な例であるといえる(Kim, J.B. et al., Cell 136:411-419 (2009))。また、Oct3/4と他の初期化因子との組み合わせのなかで、Oct3/4とSox2(WO2008/118820)、Oct3/4とKlf4(Kim, J.B. et al., Nature 454:646-650 (2008)、Shi, Y. et al., Cell Stem Cell, 2:525-528(2008))、またはOct3/4とc-Myc(Kim, J.B. et al., Nature 454:646-650 (2008))からなる2因子の組み合わせが報告されている。
【0010】
しかしながら、報告された初期化因子の組み合わせがなぜ体細胞を人工多能性幹細胞に誘導することを可能にするのか明らかでないし、その作用機序についても明らかにされていない。
【0011】
このような情況下において、本発明の目的は、これまでに報告されていない、体細胞の初期化を可能にする初期化因子の新規の組み合わせを見出し、この組み合わせ因子を用いて体細胞から人工多能性幹細胞を作製する方法を提供することである。
【課題を解決するための手段】
【0012】
国際公開WO2008/118820(2008年10月2日公開)には、Oct3/4及びSox2を必須の初期化因子とし、これにNanog及びLin28のうち少なくとも1つの初期化因子を追加しうる、体細胞を人工多能性幹細胞に初期化するための初期化因子の組み合わせが記載されている。これによれば、初期化因子の組み合わせの例は、Oct3/4及びSox2;Oct3/4、Sox2及びNanog;Oct3/4、Sox2及びLin28;並びに、Oct3/4、Sox2、Nanog及びLin28であり、これに対し、Oct3/4、Nanog及びLin28の組み合わせは初期化を起こさないことが示されている。また Huangfu D.et al., nature biotech.,Vol.26, No.11: 1269-1275 (2008)(オンライン公表日:2008年10月12日)においては、(Oct3/4, Sox2及びKlf4のうちの)いずれか2因子をVPAと共にヒト線維芽細胞に導入し、人工多能性幹細胞の樹立を試みているが、Oct3/4及びSox2の組み合わせ以外の組み合わせを使用しても、人工多能性幹細胞は樹立できなかったことが記載されている。
【0013】
本発明者らは、今回、意外にも、Oct3/4及びNanogからなる組み合わせだけで、体細胞を人工多能性幹細胞に初期化することが可能であることを見出した。これまで報告された2つの初期化因子の組み合わせは、Oct3/4及びSox2(国際公開WO2008/118820)、Oct3/4及びKlf4(Kim, J.B. et al., Nature 454:646-650 (2008)、Shi, Y. et al., Cell Stem Cell, 2:525-528(2008))、及びOct3/4及びc-Myc(Kim, J.B. et al., Nature 454:646-650 (2008))のみである。しかし、後者の2つの組み合わせ(Nature 454:646-650 (2008)及び Cell Stem Cell, 2:525-528(2008))については、体細胞がマウス神経幹細胞であり、もともとSox2遺伝子の発現が見られる細胞であり、敢えてSox2遺伝子を初期化因子として加える必要がない。それゆえ、Oct3/4及びSox2が必須であるとするこれまでの知見からすれば、本発明におけるOct3/4及びNanogからなる2つの初期化因子の組み合わせは、当業者にはまったく予想されていなかった意外な組み合わせである。なお、本発明者らが実験に用いた成人皮膚由来線維芽細胞(HDF)はSox2を発現していない細胞であり(Cell,131, 861-872(2007))、この観点からも、本発明におけるOct3/4及びNanogからなる2つの初期化因子の組み合わせは、当業者にはまったく予想されていなかった意外な組み合わせである。
【0014】
したがって、本発明は、以下の特徴を含む。
本発明は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子を、哺乳動物由来の体細胞に導入し、これによって該体細胞から人工多能性幹細胞を誘導することを含み、該初期化因子はSox2及びSox2をコードする核酸を含まない、哺乳動物人工多能性幹細胞の作製方法を提供する。
【0015】
その実施形態において、上記初期化因子は、Lin28、又はLin28をコードする核酸、をさらに含むことができる。
【0016】
別の実施形態において、上記初期化因子は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、からなる。
【0017】
或いは、別の実施形態において、上記初期化因子は、Oct3/4、Nanog及びLin28、或いはOct3/4、Nanog及びLin28をコードする核酸、からなる。
【0018】
別の実施形態において、上記初期化因子及び体細胞が由来する哺乳動物は、互いに同じか又は異なる哺乳動物である。
【0019】
別の実施形態において、上記哺乳動物は、霊長類又はげっ歯類である。
別の実施形態において、上記霊長類はヒトである。
【0020】
別の実施形態において、上記げっ歯類はマウスである。
さらに別の実施形態において、上記核酸はベクターに含まれている。
【0021】
別の実施形態において、上記核酸は、上記体細胞のゲノムに組み込まれるか、或いは、ゲノムに組み込まれない状態で細胞内に存在する。
【0022】
さらに別の実施形態において、上記人工多能性幹細胞の誘導時に、該細胞の樹立効率改善物質をさらに存在させる。
【0023】
別の実施形態において、上記樹立効率改善物質が、ヒストンデアセチラーゼ(HDAC)阻害剤、ヒストンメチルトランスフェラーゼ(G9a)阻害剤又はDNAメチル化酵素(Dnmt)阻害剤である。
【図面の簡単な説明】
【0024】
【図1】図1は、樹立したiPS細胞クローンT4F-1の形態を示す写真である。左図はコロニー形成時、右図は3継代目のコロニー像を示す。
【図2】図2は、樹立したヒトiPS細胞クローンについてGenomic-PCR解析を行った結果を示す写真である。図中「T4F-1」はOct3/4, Nanogの2遺伝子のみがゲノムに挿入されたクローンであり、「T4F-2」はOct3/4, Sox2, Nanog, Lin28の4遺伝子がゲノムに挿入されたクローンである。また図中、「Y4F」はOct3/4, Sox2, Klf4, c-Mycの4遺伝子導入で樹立したクローンを、「Y3F」はOct3/4, Sox2, Klf4の3遺伝子導入で樹立したクローンをそれぞれ示す。「HDF」は導入に用いた成人皮膚由来線維芽細胞を、「H2O」は陰性コントロールとしての溶媒(水)を示す。
【図3】図3は、樹立したヒトiPS細胞クローンが、ES細胞特異的なマーカー遺伝子(Oct3/4, Nanog, Lin28, PH34, Dnmt3b及びNodal)を発現していることを示すRT-PCRの写真である。図中、「T4F-1」、「T4F-2」は樹立した各クローンを、また「201B6 P19」、「201B7 P37」は、いずれもTakahashi, K.ら, Cell, 131:861-872 (2007)に記載されたOct3/4,Sox2,Klf4,Nanogを用いて樹立されたiPS細胞クローンを示す(P19又はP37は継代数である。)。「ES」はES細胞を、「AHDF」は導入に用いた成人皮膚由来線維芽細胞を、「H2O」は陰性コントロールを示す。
【図4】図4は、2遺伝子(Oct3/4, Nanog)のゲノム中への挿入により樹立したヒトiPS細胞クローン(T4F-1)をScidマウスの精巣内に注射して得られたテラトーマの組織染色像(ヘマトトキシン・エオシン染色)である。左から、神経管組織、軟骨組織、腸管様上皮構造の組織像を示す。
【図5】図5は、2遺伝子(Oct3/4, Nanog)の導入により樹立したiPS細胞クローンON-1及びON-5の形態を示す写真である。左図はコロニー形成時、右図は3継代目のコロニー像を示す。
【図6】図6は、樹立したヒトiPS細胞クローンが、ES細胞特異的なマーカー遺伝子(Oct3/4, Nanog, Lin28, PH34及びDnmt3b)を発現していることを示すRT-PCRの写真である。図中、「ON-1〜ON-6」は2遺伝子導入で樹立した各クローンを、また「201B7 P19」は、Takahashi, K.ら, Cell, 131:861-872 (2007)に記載されたOct3/4,Sox2,Klf4,Nanogを用いて樹立されたiPS細胞クローンを示す(P19は継代数である。)。「AHDF」は導入に用いた成人皮膚由来線維芽細胞を、「H2O」は陰性コントロールを示す。
【図7】図7は、ヒト成人皮膚由来線維芽細胞への、4遺伝子(Oct3/4, Nanog, Lin28, c-Myc)、3遺伝子(Oct3/4, Nanog, Lin28)の導入により樹立されたiPS細胞クローンONLM(図7A)及びONL(図7B)、ヒト新生児包皮由来線維芽細胞への、2遺伝子(Oct3/4, Nanog)の導入により樹立されたiPS細胞クローンON(図7C)の各形態を示す写真である。左図はいずれもコロニー形成時、右図及び中央図は1又は10継代目のコロニー像を示す。
【図8】図8は、ヒト歯髄幹細胞DP31への2遺伝子(Oct3/4, Nanog)の導入により樹立されたiPS細胞クローンの形態を示す写真である。左図はコロニー形成時、右図は2継代目のコロニー像を示す。
【図9】図9は、樹立したヒトiPS細胞クローンについてGenomic-PCR解析を行った結果を示す写真である。レーン左から以下の細胞の結果を示す:・ONL:Oct3/4, Nanog, Lin28の3遺伝子を成人皮膚由来線維芽細胞に導入して樹立したiPS細胞クローン、・AHDF-ON: Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したiPS細胞クローン、・NHDF-ON: Oct3/4, Nanogの2遺伝子をヒト新生児包皮由来線維芽細胞に導入して樹立したiPS細胞クローン、・201B6:Takahashi, K.ら, Cell, 131:861-872 (2007)に記載されたOct3/4,Sox2,Klf4,Nanogを用いて樹立されたiPS細胞クローン、・AHDF: ヒト成人皮膚由来線維芽細胞、・NHDF: ヒト新生児包皮由来線維芽細胞。
【図10】図10は、樹立したヒトiPS細胞クローンについてGenomic-PCR解析を行った結果を示す写真である。レーン左から以下の細胞の結果を示す:・OSKMNL: Oct3/4, Sox2, Klf4, c-Myc, Nanog, Lin28の6遺伝子を成人皮膚由来線維芽細胞に導入して樹立したiPS細胞クローン、・Dp31 ON:Oct3/4, Nanogの2遺伝子をヒト歯髄幹細胞Dp31に導入して樹立したiPS細胞クローン、・Dp31:ヒト歯髄幹細胞、・H2O:陰性コントロール(水)。
【図11】図11は、Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したヒトiPS細胞クローンをScidマウスの精巣内に注射して得られたテラトーマの組織染色像(ヘマトトキシン・エオシン染色)である。左から、神経組織、腺上皮組織、軟骨組織の組織像を示す。
【図12】図12は、Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したヒトiPS細胞クローンが、幹細胞マーカーを発現することを示した免疫染色の写真である。
【図13】図13は、Oct3/4, Nanogの2遺伝子を成人皮膚由来線維芽細胞に導入して樹立したヒトiPS細胞クローンが三胚葉系への分化能を有することを、βIII-チューブリン(tubulin)(図中Tubrin)、α-フェトプロテイン(fetoprotein)(図中AFP)、ビメンチン(Vimentin)、平滑筋アクチン(smooth muscle actin)(図中SMA)およびデスミン(Desmin)に対する各抗体を用いた染色により確認した結果を示す写真である。Cy3-Negaおよび488-Negaは陰性コントロールである。
【発明を実施するための形態】
【0025】
本発明をさらに詳細に説明する。
1. 定義
本明細書中で使用される用語は、以下の意味を包含する。その他の用語は、当業界で一般的に使用される意味を含むことが意図されている。
【0026】
本明細書で使用される「人工多能性幹細胞」又は「iPS細胞」という用語は、人工的に作製された分化多能性を有する細胞であり、いわゆる胚性幹(ES)細胞ではないがES細胞に類似した特性を有する細胞である。この細胞は、Takahashi及びYamanaka(Cell 126:663-676 (2006))によってマウス体細胞からはじめて樹立され、その後、ヒト体細胞からも同様に樹立可能であることが示されている(国際公開WO2007/069666;Takahashi, K.ら, Cell 131:861-872 (2007):Yu, J.ら,Science 318:1917-1920 (2007);Nakagawa, M.ら,Nat. Biotechnol. 26:101-106 (2008))。iPS細胞は、動物の体を構成する種々の細胞に分化する能力(すなわち、分化多能性)、核型を保持したまま半永久的に増殖し続ける能力、ES細胞が通常発現する遺伝子群が同様に発現されている、などの特徴を有するために、体細胞の初期化(すなわち、再プログラム化(reprogramming))によって人工的に誘導された細胞であり、分化した体細胞の特性と明らかな相違を示す。
【0027】
本明細書で使用する「初期化」という用語は、「核初期化(nuclear reprogramming)」とも称され、分化した細胞が、未分化細胞、特に分化多能性細胞に誘導、変換される過程又は手段を指す。初期化という現象はもともと、脱核した未受精卵に体細胞の核を注入する、ES細胞を体細胞と融合させる、ES細胞抽出液を体細胞中に浸透化させる、などの方法によって観察されてきたが、本発明における初期化では、卵子、卵母細胞などの生殖系列細胞及びES細胞に依存することなく、体細胞からiPS細胞への初期化を起こすことができる。
【0028】
このような初期化を可能にする因子が、初期化因子である。本明細書で使用される初期化因子は、蛋白質、核酸或いはそれらの組み合わせを含む。これまで報告された初期化因子は、Oct3/4、Sox2及びKlf4;Oct3/4、Klf4及びc-Myc;Oct3/4、Sox2、Klf4及びc-Myc;Oct3/4及びSox2;Oct3/4、Sox2及びNanog;Oct3/4、Sox2及びLin28; Oct3/4及びKlf4などであるが、本発明で使用可能な初期化因子は、少なくともOct3/4及びNanogを含む初期化因子からなり、ただしSox2を含まない。
【0029】
なお、本明細書では、初期化因子の遺伝子名は、哺乳動物の種類を問わず同じ遺伝子名、例えばOct3/4、Nanog、Sox2、Klf4、c-Myc、Lin28などを使用する。すなわち、これらの遺伝子名は、通常マウスの遺伝子名として使用されるが、本明細書ではマウスのみならず、ヒトやその他の哺乳動物の場合にも上記遺伝子名を使用することにする。
【0030】
また、Oct3/4は、Oct3 、Oct4又はPOU5F1と称されることがあり、いずれも同一の転写因子を指す。本明細書では、これらを総称してOct3/4と呼ぶことにする。
【0031】
本明細書で使用する「核酸」という用語は、ゲノムDNA、cDNA、mRNAなどのデオキシリボ核酸(DNA)又はリボ核酸(RNA)を指す。
【0032】
本明細書で使用する「体細胞」とは、卵子、卵母細胞などの生殖系列細胞又は分化全能性細胞を除くあらゆる哺乳動物細胞をいう。
【0033】
本明細書で使用する「哺乳動物」は、すべての哺乳動物を含むことを意図している。特に好ましい哺乳動物には、霊長類、げっ歯類、有蹄類などが含まれる。
【0034】
2. 初期化因子
本発明により体細胞からiPS細胞を誘導するための初期化因子は、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子であり、Sox2及びSox2をコードする核酸を除く他の初期化因子又はサイトカインを適宜含むことができる。
【0035】
初期化因子の組み合わせの好ましい例は、Oct3/4及びNanogの組み合わせ、或いは、Oct3/4、Nanog及びLin28の組み合わせである。各初期化因子が由来する哺乳動物の種類は限定されないものとし、任意の哺乳動物である。好ましい哺乳動物は、例えば霊長類(例えばヒト、サル、チンパンジーなど)、げっ歯類(例えばマウス、ラット、モルモット、ハムスターなど)、有蹄類(例えばウシ、ウマ、ヒツジ、ヤギ、ブタなど)、ペット動物(例えばイヌ、ネコなど)などである。通常、ある特定の哺乳動物由来の体細胞を初期化するときには、それと同一の哺乳動物由来の初期化因子を使用することが好ましい。例えばヒト由来の体細胞からiPS細胞を誘導するときには、ヒト由来の初期化因子を使用する。
上記Oct3/4、Nanog、Lin28の各アミノ酸及びヌクレオチド配列は、GenBank(米国NCBI)にアクセスすることによって入手することが可能である。
【0036】
Oct3/4について、例えばヒトOct3/4、マウスOct3/4、ラットOct3/4の配列は、それぞれ、NM_203289又はNM_002701、NM_013633、NM_001009178として登録されている。
【0037】
Nanogについて、例えばヒトNanog、マウスNanog、ラットNanogの配列は、それぞれ、NM_024865、NM_028016、NM_001100781として登録されている。
【0038】
Lin28について、例えばヒトLin28、マウスLin28、ラットLin28の配列は、それぞれ、NM_024674、NM_145833、NM_001109269として登録されている。
【0039】
Lin28に類似の因子として、同じLinファミリーに属するLin28bが知られている。よってLin28に加えて、またはLin28に代えて、Lin28bを使用することができる。Lin28bについて、たとえばヒトLin28b、マウスLin28bの配列は、それぞれ、NM_001004317、NM_001031772として登録されている。
【0040】
上記以外の他の初期化因子の非限定的な例は、ECAT1、ECAT2(ESG1とも称する)、ECAT3(Fbx15とも称する)、ECAT5(Erasとも称する)、ECAT7、ECAT8、ECAT9(Gdf3とも称する)、ECAT10(Sox15とも称する)、ECAT15-1(Dppa4とも称する)、ECAT15-2(Dppa2とも称する)、Fthl17、Sall4、Rex1(Zfp42とも称する)、Utf1、Tcl1、Stella(Dppa3とも称する)、β-catenin(Ctnnb1とも称する)、Stat3、Grb2、c-Myc、Sox1、Sox3、N-Myc、L-Myc、Klf1、Klf2、Klf4、Klf5(以上、国際公開WO2007/069666)、並びに、FoxD3、ZNF206、Mybl2、Otx2(以上、国際公開WO2008/118820)などから選択される1又は複数の初期化因子(群)である。これらの初期化因子の配列のGenBank登録番号(ヒト及びマウス)は以下のとおりである。
【0041】
ECAT1について、ヒトECAT1、マウスECAT1の配列は、それぞれ、AB211062、AB211060として登録されている。
【0042】
ECAT2について、ヒトECAT2、マウスECAT2の配列は、それぞれ、NM_001025290、NM_025274として登録されている。
【0043】
ECAT3について、ヒトECAT3、マウスECAT3の配列は、それぞれ、NM_152676、NM_015798として登録されている。
【0044】
ECAT5について、ヒトECAT5、マウスECAT5の配列は、それぞれ、NM_181532、NM_181548として登録されている。
【0045】
ECAT7について、ヒトECAT7、マウスECAT7の配列は、それぞれ、NM_013369、NM_019448として登録されている。
【0046】
ECAT8について、ヒトECAT8、マウスECAT8の配列は、それぞれ、AB211063、AB211061として登録されている。
【0047】
ECAT9について、ヒトECAT9、マウスECAT9の配列は、それぞれ、NM_020634、NM_008108として登録されている。
【0048】
ECAT10について、ヒトECAT10、マウスECAT10の配列は、それぞれ、NM_006942、NM_009235として登録されている。
【0049】
ECAT15-1について、ヒトECAT15-1、マウスECAT15-1の配列は、それぞれ、NM_018189、NM_028610として登録されている。
【0050】
ECAT15-2について、ヒトECAT15-2、マウスECAT15-2の配列は、それぞれ、NM_138815、NM_028615として登録されている。
【0051】
Fthl17について、ヒトFthl17、マウスFthl17の配列は、NM_031894、NM_031261として登録されている。
【0052】
Sal14について、ヒトSal14、マウスSal14の配列は、それぞれ、NM_020436、NM_175303として登録されている。
【0053】
Rex1について、ヒトRex1、マウスRex1の配列は、それぞれ、NM_174900、NM_009556として登録されている。
【0054】
Utf1について、ヒトUtf1、マウスUtf1の配列は、それぞれ、NM_003577、NM_009482として登録されている。
【0055】
Tcl1について、ヒトTcl1、マウスTcl1の配列は、それぞれ、NM_021966、NM_009337として登録されている。
【0056】
Stellaについて、ヒトStella、マウスStellaの配列は、それぞれ、NM_199286、NM_139218として登録されている。
【0057】
β-cateninについて、ヒトβ-catenin、マウスβ-cateninの配列は、それぞれ、NM_001904、NM_007614として登録されている。
【0058】
Stat3について、ヒトStat3、マウスStat3の配列は、それぞれ、NM_139276、NM_213659として登録されている。
【0059】
Grb2について、ヒトGrb2、マウスGrb2の配列は、それぞれ、NM_002086、NM_008163として登録されている。
【0060】
FoxD3について、ヒトFoxD3、マウスFoxD3の配列は、それぞれ、NM_012183、NM_010425として登録されている。
【0061】
ZNF206について、ヒトZNF206、マウスZNF206の配列は、それぞれ、NM_032805、NM_001033425として登録されている。
【0062】
Mybl2について、ヒトMybl2、マウスMybl2の配列は、それぞれ、NM_002466、NM_008652として登録されている。
【0063】
Otx2について、ヒトOtx2、マウスOtx2の配列は、それぞれ、NM_172337、NM_144841として登録されている。
【0064】
c-Mycについて、ヒトc-Myc、マウスc-Mycの配列は、それぞれ、NM_002467、NM_010849として登録されている。
【0065】
N-Mycについて、ヒトN-Myc、マウスN-Mycの配列は、それぞれ、NM_005378、NM_008709として登録されている。
【0066】
L-Mycについて、ヒトL-Myc、マウスL-Mycの配列は、それぞれ、NM_001033081、NM_008506として登録されている。
【0067】
Sox1について、ヒトSox1、マウスSox1の配列は、それぞれ、NM_005986、NM_009233として登録されている。
【0068】
Sox3について、ヒトSox3、マウスSox3の配列は、それぞれ、NM_005634、NM_009237として登録されている。
【0069】
Klf1について、ヒトKlf1、マウスKlf1の配列は、それぞれNM_006563、NM_010635として登録されている。
【0070】
Klf2について、ヒトKlf2、マウスKlf2の配列は、それぞれ、NM_016270、NM_008452として登録されている。
【0071】
Klf4について、ヒトKlf4、マウスKlf4の配列は、それぞれ、NM_004235、NM_010637として登録されている。Klf5について、ヒトKlf5、マウスKlf5の配列は、それぞれ、NM_001730、NM_009769として登録されている。
【0072】
本発明の方法におけるiPS細胞の誘導系には、上記初期化因子と組み合わせて、以下に例示されるようなヒトを含む哺乳動物由来のサイトカインを少なくとも1種含むことができる。サイトカインの例は、bFGF(basic Fibroblast Growth Factor)、SCF(Stem Cell Factor)、TERT(teromerase reverse transcriptase)、SV40 Large T、HPV16 (human papillomavirus type 16) E6又はE7、Bmi1(ポリコーム遺伝子産物)などを含む。
【0073】
本発明の方法では、初期化因子は、蛋白質(若しくはポリペプチド)又は核酸(若しくはポリヌクレオチド)のいずれかの形態で使用することができる。このような蛋白質又は核酸は、PCR(polymerase chain reaction)技術、遺伝子組換え技術などの慣用技術を使用することによって作製することができる。
【0074】
一般に、核酸は、それを含む哺乳動物細胞由来のゲノムライブラリー又はcDNAライブラリーからPCR技術を利用して増幅されうる。そのために、該核酸のポリヌクレオチド配列に基づいてセンス及びアンチセンスプライマーを作製する。この場合、プライマーは、初期化因子としての核酸のORF(open reading frame)配列の両末端にアニーリングするように設計される。一般に、プライマーのサイズは、約17〜30塩基長、好ましくは約20〜26塩基長である。PCRは、変性段階、アニーリング段階及び伸長段階を1サイクルとして約20〜40サイクルを実施することからなり、変性段階は、二本鎖DNAを変性して各一本鎖DNAに解離する段階(例えば約94〜96℃、約30秒〜約5分)であり、アニーリング段階は、鋳型としての各一本鎖DNAにセンスプライマー又はアンチセンスプライマーをアニーリング(すなわち、結合)する段階(例えば約50〜65℃、約30秒〜1分)であり、伸長段階は、一本鎖DNAを鋳型としてセンス及びアンチセンスプライマーを伸長しDNA鎖を合成する段階(例えば約72℃、約30秒〜10分)である。上記サイクルの実施前に約94〜96℃、約2〜7分程度の前処理を行ってもよいし、また、上記サイクルの完了後に約72℃、約5〜10分程度の後処理を行ってもよい。PCRに使用するバッファーは、PCRバッファー(MgCl2含有)を使用し、反応に際して、該バッファーにプライマー、耐熱性DNAポリメラーゼ酵素(例えばTaq DNAポリメラーゼなど)、4種のデオキシヌクレオチド(dNTP(N=A,T,C,G))を添加する。通常、反応液は約10〜100μl程度である。PCR技術は、例えばSaiki, R.K.ら,Science 230:1350-1354 (1985)、Erlich, H.A.ら,Science 252:1643-1651 (1991);Hughes, S.及びMoody, A., PCR (Methods Express), Scion Publishing (2007)などに記載されている。
【0075】
哺乳動物細胞由来のcDNAライブラリーは、組織又は細胞(体細胞、幹細胞、胚性幹細胞を含む)から全mRNAを抽出し、オリゴdTプライマーを使用する逆転写(RT)-PCR法によって全mRNAから第1鎖DNAを合成し、DNA合成酵素I、DNAリガーゼ、RNaseHにより第2鎖DNAを合成し、T4 DNA合成酵素により末端平滑化を行い、EcoRIアダプターを連結したのち、cDNAをリン酸化し、ファージベクター(例えばλgt11)に該cDNAを連結することによって作製されうる。
【0076】
哺乳動物細胞由来のゲノムライブラリーは、組織又は細胞からゲノムDNAを抽出し、適する制限酵素(例えばSau3A)によって部分分解し、スクロース密度勾配遠心により約40kb前後のDNA断片を回収し、例えばコスミドなどのベクターに該DNA断片を連結することによって作製されうる。
【0077】
上記のようにPCR増幅によって生成された初期化因子としての核酸に対して遺伝子組換え技術を適用し、該核酸を含むベクター、或いは、該核酸によってコードされる蛋白質を作製することができる。核酸(特に、cDNA)は、二本鎖の形態でベクターに挿入され、外来核酸が発現可能なように適当な制御配列によって調節されうる。ベクターは、例えば、プラスミド、ファージ、コスミド、ウイルス(例えばレトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、バキュロウイルスなど)、人工染色体(例えばHAC、BAC、PAC、YACなど)などのベクターを含むことができる。
【0078】
制御配列には、プロモーター、エンハンサー、ターミネーター、ポリアデニル化サイト、リボソーム結合サイト、複製開始点などが適宜含まれる。特に、プロモーターの例は、非限定的に、Oct3/4プロモーター、ヒトサイトメガロウイルス(CMV)プロモーター、アデノウイルス後期プロモーター、ワクシニアウイルス7.5Kプロモーター、SV40プロモーター、ポリヘドリンプロモーター、メタロチオネインプロモーター、カリフラワーモザイクウイルスプロモーター、タバコモザイクウイルスプロモーター、解糖系酵素プロモーターなどを含み、またプロモーターは、組織特異的プロモーター、誘導性プロモーター又は構成性プロモーターでありうる。
【0079】
さらにまた、場合により、ベクターはさらに選択マーカーを含んでもよく、選択マーカーとして、例えば、薬剤耐性遺伝子(例えばG418、ネオマイシン耐性遺伝子、ピューロマイシン耐性遺伝子など)、レポーター遺伝子(例えばGFP(green fluorescence protein)、GUS(β-gluclonidase)、FLAGなど)、ネガティブ選択マーカー(例えばチミジンキナーゼ(TK)遺伝子、ジフテリアトキシン(DT)遺伝子など)などが挙げられる。また、ベクターは、外来核酸の挿入を容易にするための複数の制限酵素認識サイトを有するマルチクローニングサイト、IRES(internal ribosomal entry site)などの配列をさらに含んでいてもよい。
【0080】
本発明の方法で使用される初期化因子としての蛋白質類は、該蛋白質(シグナル配列を含む前駆体蛋白質、又は成熟蛋白質)をコードするDNAを含む発現ベクターを、宿主細胞としての原核細胞又は真核細胞、例えば細菌類(大腸菌、枯草菌、シュードモナス属など)、酵母(サッカロミセス属、カンジダ属、ピキア属など)、昆虫細胞(Sf系細胞など)、哺乳動物細胞(CHO、NIH3T3、HEK293、COS、BHKなど)、植物細胞などに導入し、形質転換又は形質導入された宿主細胞を適当な培地中で培養し、該細胞又は培地から目的の蛋白質を回収することを含む方法によって得ることができる。蛋白質の回収は、クロマトグラフィー(例えばゲルろ過クロマトグラフィー、イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、HPLC、FPLCなど)、電気泳動、等電点電気泳動、限外ろ過、硫安沈殿、有機溶媒沈殿などの慣用手段を適宜組み合わせて行うことができる。
【0081】
本発明の方法で使用される初期化因子としての核酸類は、好ましくは、上記蛋白質(シグナル配列を含む前駆体蛋白質、又は成熟蛋白質)をコードするDNAを発現可能に含むベクターの形態である。特に、このようなベクターをヒト体細胞の初期化に使用するときには、初期化の完了後に上記DNAのサイレンシングを起こすか、或いは発現が一過性であることが望ましい。ベクターは、レトロウイルス、レンチウイルスなどのような細胞のゲノムに組み込まれ易いベクター、或いは、アデノウイルス、プラスミド、人工染色体などのような細胞のゲノムに組み込まれ難いベクターのいずれでもよい(Stadtfeld, M.ら, Science, 322, 945-949 (2008)(オンライン公表日:2008年9月25日); Okita, K.ら, Science, 322, 949-953 (2008)( オンライン公表日:2008年10月9日); Takahashi, K.ら, Cell 131:861-872 (2007);Yu, J.ら, Science 318:1917-1920 (2007))。
【0082】
上記の遺伝子組換え技術の一般的な手順は、Sambrook, J.ら, Molecular Cloning: A Laboratory Manual, 2nd edition, Cold Spring Harbor Laboratory Press (1989)、Ausubel, F.M.ら,Short protocols in Molecular Biology: A Compendium Methods from Current Protocols in Molecular Biology, John Wiley & Sons (1999)などに記載されている。
【0083】
3. 人工多能性幹(iPS)細胞の樹立効率改善物質
本発明の方法においては、Oct3/4及びNanog、又はそれらをコードする核酸、からなる初期化因子によって、或いは、これらの初期化因子に加えて、Sox2及びSox2をコードする核酸を除く他の初期化因子やサイトカインによって、哺乳動物体細胞からiPS細胞を樹立することができる。腫瘍遺伝子の一種であるc-Myc及びKlf4が初期化因子として使用される場合には、iPS細胞の樹立効率は高まるが、逆に、これらのいずれか又は両方が不在である場合には、iPS細胞の樹立効率が著しく低下することが知られている。
【0084】
しかし、このような場合、iPS細胞の樹立効率を改善する物質を、iPS細胞誘導系に添加することによって、iPS細胞の樹立を促進することが可能である。このような物質には、例えば塩基性線維芽細胞増殖因子(bFGF)、幹細胞因子(SCF)などのサイトカインが含まれる。また、例えばヒストンデアセチラーゼ(HDAC)阻害剤、例えばバルプロ酸(VPA)(Huangfu, D.ら,Nat. Biotechnol., 26(7):795-797 (2008))、ヒストンメチルトランスフェラーゼ(G9a)阻害剤、例えばBIX-01294 (BIX) (Shi, Y.ら,Cell Stem Cell, 2:525-528 (2008);Kubicek, S.ら,Mol. Cell 25:473-481 (2007);Shi, Y.ら, Cell Stem Cell, 3, 568-574 (2008))、DNAメチル化酵素(Dnmt)阻害剤、例えば5’-アザシチジン(Huangfu, D.ら,上記)などの低分子化合物が含まれる。またp53に対するshRNAやsiRNAなどのp53阻害剤や、UTF1を細胞内に導入してもよい(Yang Zhaoら, Cell Stem Cell, 3, pp475-479, 2008)。他にもシグナル伝達に関して、Wntシグナルの活性化(Marson A.ら, Cell Stem Cell, 3, pp132-135, 2008)、マイトジェン活性化プロテインキナーゼおよびグリコーゲン合成酵素キナーゼ-3シグナル伝達の阻害(Silva J.ら, PloS Biology, 6, pp2237-2247 2008)、ES細胞特異的miRNA (例えば、miR-302-367クラスター (Mol. Cell Biol. Doi:10. 1128/MCB. 00398-08), miR-302 (RNA 14:1-10 (2008)), miR-291-3p, Mir-294及びmiR-295 (Nat. Biotechnol. 27:459-461 (2009))なども、iPS細胞の樹立効率を改善することができる。
【0085】
上記のような樹立効率改善物質は、分化多能性に関わる遺伝子の活性化及び分化細胞で特異的に発現される遺伝子の不活性化の両方に何らかの作用をすることが、推定されている(Huangfu, D.ら,上記)。本発明の方法においても、樹立効率改善物質を使用することによって、体細胞からiPS細胞への初期化(すなわち、再プログラム化)を促進することが可能である。
【0086】
さらに、体細胞の初期化段階で、低酸素条件下で細胞を培養することによってiPS細胞の樹立効率を向上させることができる (Yoshida, Y. et al., Cell Stem Cell 5:237-241 (2009))。本明細書で使用する「低酸素条件」という用語は、細胞培養の際の酸素濃度が空気中の酸素濃度と比べてかなり低いことを意味している。具体的には、このような条件として、5〜10% CO2 / 95〜90%空気の雰囲気中の酸素濃度より低い酸素濃度、例えば18%以下の酸素濃度からなる条件が挙げられる。前記雰囲気中の酸素濃度の好ましい例は、15%以下(例えば14%以下、13%以下、12%以下又は11%以下)、10%以下(例えば、9%以下、8%以下、7%以下又は6%以下)、或いは5%以下(例えば、4%以下、3%以下又は2%以下)である。或いは、前記雰囲気中の好ましい酸素濃度は、0.1%以上(例えば、0.2%以上、0.3%以上又は0.4%以上)、0.5%以上 (例えば、0.6%以上、0.7%以上、0.8%以上又は0.95%以上)、或いは1%以上 (例えば、1.1%以上、1.2%以上、1.3%以上又は1.4%以上)である。
【0087】
細胞環境において前記低酸素状態を作るための手法としては、以下のものに限定されないが、酸素濃度を調節することが可能なCO2インキュベーター中で細胞を培養するという最も簡単な方法でありかつ好ましい例としてのこの方法を挙げることができる。このようなインキュベーターは種々の装置メーカー(例えば、Thermo Scientific, 池本理化工業(Ikemoto Scientific Technology), 十慈フィールド(Juji Field), 和研薬(Wakenyaku)など)から市販されており、上記の目的で使用しうる。
【0088】
4. 体細胞
本発明で使用可能な体細胞は、上記定義のとおり、生殖系列細胞(卵子、卵母細胞、胚性幹(ES)細胞など)又は分化全能性細胞を除く、哺乳動物由来のすべての細胞である。
【0089】
体細胞が由来する哺乳動物は、特に制限されずいかなる種類の動物も包含する。好ましい哺乳動物は、霊長類(例えばヒト、サル、チンパンジー等)、げっ歯類(例えばマウス、ラット、ハムスター、モルモット等)、及び有蹄類(ウシ、ヒツジ、ヤギ、ウマ、ブタ等)、ペット動物(イヌ、ネコ等)から選択され、さらに好ましい哺乳動物はヒト及びマウスである。
【0090】
体細胞には、非限定的に、胎児(仔)期の体細胞、新生児(仔)の体細胞、及び成熟した体細胞のいずれも包含されるし、また、初代培養細胞、継代細胞、及び株化細胞のいずれも包含されるし、さらにまた、組織幹細胞や組織前駆細胞も包含される。
【0091】
具体的には、体細胞は、非限定的に、例えば(1)神経幹細胞、造血幹細胞、間葉系幹細胞、歯髄幹細胞等の組織幹細胞(体性幹細胞)、(2)組織前駆細胞、(3)リンパ球、上皮細胞、内皮細胞、筋肉細胞、線維芽細胞(皮膚細胞等)、毛細胞、肝細胞、胃粘膜細胞、腸細胞、脾細胞、膵細胞(膵外分泌細胞等)、脳細胞、肺細胞、腎細胞、皮膚細胞等の分化した細胞などを包含する。
【0092】
体細胞を得るための起源として適する哺乳動物個体は、非限定的に、好ましくは患者自身、或いは、得られたiPS細胞が再生医療のために使用されたときに拒絶反応が生じないという観点から、同一の又は実質的に同一のHLA型をもつ他人である。本明細書で使用するHLA型に関連した「実質的に同一」という用語は、体細胞由来のiPS細胞の分化を誘導することによって作製された細胞が患者に移植されるときに、HLA型が、ドナーとレシピエントとの間で、主要HLA類(例えば、HLA-A, HLA-B及びHLA-DRの3つの遺伝子の遺伝子座)が同じである場合のように、移植された細胞が生存可能である程度に一致することを意味する。この定義は以下においても同様に使用される。iPS細胞が投与されないか又は移植されないとき、すなわち例えば、iPS細胞を、患者での薬剤の感受性や副作用の有無を評価するスクリーニング用の細胞供給源として使用する場合、体細胞を、患者自身から、或いは薬剤の感受性や副作用と相関する同一の遺伝子多型を有する他人から、得ることが望ましい。
【0093】
iPS細胞の作製に先立ち、ヒトを含む哺乳動物から体細胞を取得し、これを動物細胞培養用培地にて培養し、必要であれば継代培養に付し、これにより初代培養細胞又は継代培養細胞を得る。このようにして得られた培養細胞を、iPS細胞の作製のために使用する。
【0094】
体細胞の培養は、例えばDMEM(Dulbecco’s modified Eagle medium)、MEM (minimum essential medium)、α-MEM(minimum essential medium alpha modification)、Ham’s F12、RPMI1640、それらの混合培地などの基本培地に、血清(10% FBS等)、抗生物質(penicillin、streptomycin等)、ピルビン酸Na、グルタミン、非必須アミノ酸、L-デキストロースなどの物質を適宜選択して含有させた培地上で約37℃の温度、5% CO2存在下にて行うことができる。
【0095】
5. 人工多能性幹(iPS)細胞の作製
本発明によるiPS細胞の作製は、初期化因子が、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子(ただし、Sox2及びSox2をコードする核酸を含まない)であることを除けば、従来公知の手順(WO2007/069666; WO2008/11820; WO2008/124133;Takahashi, K.ら, Cell 131:861-872 (2007))で行うことができる。
【0096】
本発明で使用しうる初期化因子は、上記2節に例示されたような種々の初期化因子の組み合わせ(ただし、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、は必須である。)を含み、それらは蛋白質又は核酸からなる。該核酸の好適な形態はベクターであり、初期化因子は、通常、その発現が可能なように上記制御配列と連結される。この場合、Oct3/4及びNanogを含む2以上の初期化因子は、同じ又は異なるベクターに発現可能なように挿入される。通常、初期化因子の数が約2〜4程度であれば、1つのベクター中に複数の初期化因子をタンデムに連結することができる。各初期化因子の5’側にはプロモーター配列、3’側にはターミネーター配列、ポリA配列などの配列がそれぞれ連結され、初期化因子間にはリンカー配列を配置してもよい。好ましくは、複数の初期化因子が発現可能なようにタンデムに連結されて、それらを含むカセットを作製し、このカセットをベクターに組み込むとよい。2つ又はそれ以上の初期化因子(又遺伝子)の連結のために、ポリシストロニックな発現を可能にするあらゆる配列、例えば口蹄疫ウイルス由来2A配列 (PLoS ONE3, e2532, 2008, Stem Cells 25, 1707, 2007)、IRES配列 (米国特許第4,937,190号)、好ましくは前記2A配列、を使用することができる。
【0097】
本発明で使用可能なベクターは、上で例示されるような、プラスミド、ウイルス、人工染色体などから選択されうる。これまで、iPS細胞の作製に使用されたベクターは、レトロウイルス、レンチウイルス、アデノウイルスなどのウイルスベクター、及びプラスミドであり、これらのベクターは、本発明でも同様に使用できる(WO2007/069666; WO2008/11820; WO2008/124133; Takahashi, K.ら, Cell 131:861-872 (2007); Stadtfeld, M.ら,Science, 322, 945-949 (2008)(オンライン公表日:2008年9月25日); Okita, K.ら, Science, 322, 949-953 (2008)( オンライン公表日:2008年10月9日))。さらに使用可能なベクターとして、エピソーマルベクター (Yu et al., Science, 324, 797-801 (2009))、トランスポゾン (例えば、ピギーバックトランスポゾン(piggyback transposone): Kaji, K. et al., Nature, 458:771-775 (2009); Woltjen et al., Nature, 458:766-770 (2009)、センダイウイルスベクター (J. Biol. Chem., 282:27383-27391 (2007);日本特許第3,602,058号)が挙げられる。
【0098】
他のベクターとして、人工染色体も使用できる。人工染色体の例には、ヒト人工染色体(HAC)、酵母人工染色体(YAC)、細菌人工染色体(BAC、PAC)などが含まれる。このうちHAC及びYACは、セントロメア、2つのテロメア及び染色体断片を含むミニ染色体である。この染色体断片に、上記複数の初期化因子を含むカセットを挿入する。
【0099】
哺乳動物由来の培養体細胞に、核酸をコードする初期化因子を導入する方法には、慣用の手法が含まれ、エレクトロポレーション法、マイクロインジェクション法、リン酸カルシウム法、ウイルス感染法、リポフェクション法などが含まれる。
【0100】
蛋白質の形態で初期化因子を培養哺乳動物体細胞に導入する方法には、従来の方法、例えば、蛋白質デリバリー試薬を使用する方法、蛋白質輸送ドメイン(PTD)又は細胞透過性ペプチド (CPP)との融合蛋白質を使用する方法、マイクロインジェクション法、リポソーム法、リポフェクション法などが含まれる。蛋白質デリバリー剤は、例えば、カチオン性脂質ベースのBioPOTER Protein Delivery Reagent (Gene Therapy Systems)、Pro-JectTM Protein Transfection Reagent (PIERCE)、ProVectin (IMGENEX)、脂質ベースのProfect-1 (Targeting System)、膜透過性ペプチドベースのPenetrain Peptide (Q biogene)、Chariot Kit (Active Motif)、及び(不活性化センダイウイルス由来の)HVJエンベロープを使用するGenomONE (石原産業、日本)として市販されている。細胞内への蛋白質又はペプチドの導入は、それぞれの試薬に添付された使用説明書に従って行うことができ、その一般的な手順は以下のとおりである。すなわち、初期化因子を適当な溶媒 (例えば、PBS又はHEPESなどのバッファー)で希釈し、その後、これにデリバリー試薬を加え、混合物を室温で5〜15分間インキュベーションして、初期化因子とデリバリー試薬との間で複合体を形成する。この複合体を、予め交換された血清不含有培地中の細胞に添加し、その後、該細胞を37℃、1時間〜数時間インキュベーションする。このあと、培地を除去し、血清含有培地と交換する。
【0101】
使用されるPTDとして開示されるものは、Drosophila由来AntP; HIV由来TAT (Frankel, A. et al., Cell 55:1189-1193 (1988)); Green, M.と Loewenstein, P.M. Cell 55:1179-1188 (1988)); Penetratin (Derossei, D. et al., J. Biol. Chem. 269:10444-10450 (1994)); Buforin II (Park, C.B. et al., Proc. Natl. Acad. Sci. USA 97:8245-8250 (2000)); Transportan (Pooga, M. et al., FASEB J. 12:67-77 (1998); MAP (model amphipathic peptide) (Oehlke, J. et al., Biochim. Biophys. Acta. 1414:127-139(1998)); K-FGF (Lin, Z. et al., J. Biol. Chem. 270:14255-14258 (1995)); Ku70 (Sawada, M. et al., Nature Cell Biol. 5:352-357 (2003)); Prion (Lundberg, P. et al., Biochem. Biophys. Res. Commun. 299:85-90 (2002)); pVEC (Elmquist, A. et al., Exp. Cell Res. 269:237-244 (2001)); Pep-1 (Morris, M.C. et al., Nature Biotechnol. 19:1173-1176 (2001)); Pep-7 (Gao, C. et al., Bioorg. Med. Chem. 10:4057-4065 (2002)); SynB1 (Rousselle, C. et al., Mol. Pharmacol. 57:679-686 (2000)); HN-1 (Hong, F.D. & Clayman, G.L., Cancer Res. 60:6551-6556 (2000));及びHSV由来VP22などの蛋白質の細胞通過ドメインである。PTD由来のCPPには、11R (Cell Stem Cell 4:381-384 (2009))及び9R (Cell Stem Cell 4:472-476 (2009))などのポリアルギニンが含まれる。
【0102】
融合蛋白質を発現する、並びに、初期化因子をコードするcDNA及びPTD又はCPP配列が組み込まれたベクターが作製されて、該融合蛋白質をコードするDNAを発現し、その後、該融合蛋白質を回収して細胞に導入する。該導入は、蛋白質デリバリー試薬を添加しないことを除いて上記と同様の方法で行うことができる。
【0103】
マイクロインジェクションは、先端径約1μmを有するガラス針の中に蛋白質溶液を入れ、針を細胞に刺すことによってこの溶液を細胞に導入し、これによって蛋白質を確実に細胞内に導入できる、方法である。
【0104】
蛋白質を導入する操作は、1回又はそれ以上、例えば1〜10回、1〜5回など、好ましくは2回以上繰り返して行うことができる。操作の繰り返しの間の時間間隔は、例えば6〜48時間、好ましくは12〜24時間である。
【0105】
iPS細胞誘導のために、培養体細胞と上記初期化因子を、適する動物細胞培養培地中で接触させて、該初期化因子を体細胞内に導入し、該細胞を形質転換又は形質導入する。培養培地としては、非限定的に、上記4節に記載したような培地、例えば(1) 10〜15%FBSを含有するDMEM、DMEM/F12又はDME培地(これらの培地にはさらに、LIF(leukemia inhibiting factor)、ペニシリン/ストレプトマイシン、ピューロマイシン、L-グルタミン、非必須アミノ酸類、β-メルカプトエタノールなどを適宜含むことができる。)、(2) bFGF又はSCFを含有するES細胞培養用培地、例えばマウスES細胞培養用培地(例えばTX-WESTM、コスモバイオ)又は霊長類ES細胞培養用培地(例えばReproCELLTM、コスモバイオ)、などが含まれる。
【0106】
培養手順の例は次のとおりである。37℃、5%CO2存在下にて、10%FBS含有DMEM又はDMEM/F12培地上で体細胞と初期化因子を接触させ約6〜7日間培養し、その後、細胞をフィーダー細胞(マイトマイシンC処理STO細胞、SNL細胞等)上に蒔きなおし、体細胞と初期化因子の接触から約10日後からbFGF含有霊長類ES細胞培養用培地で培養し、該接触から約30〜約45日又はそれ以上ののちにiPS様コロニーを生じさせることができる。この培養手順は、ヒトiPS細胞などの霊長類iPS細胞の誘導に適している。
【0107】
或いは、その代替例として、37℃、5% CO2存在下にて、フィーダー細胞(マイトマイシンC処理STO細胞、SNL細胞等)上で10%FBS含有DMEM培地(これにはさらに、LIF、ペニシリン/ストレプトマイシン、ピューロマイシン、L-グルタミン、非必須アミノ酸類、β-メルカプトエタノールなどを適宜含むことができる。)で培養し、約25〜約30日又はそれ以上ののちにiPS様コロニーを生じさせることができる。この培養手順は、マウスiPS細胞などのげっ歯類iPS細胞の誘導に使用される。
【0108】
上記培養の間には、培養開始2日目以降から毎日1回新鮮な培地と培地交換を行う。また、初期化に使用する体細胞の細胞数は、限定されないが、培養ディッシュ100cm2あたり約5×103〜約5×106細胞の範囲である。
【0109】
ディッシュ上の上記iPS様コロニーを、トリプシン及びコラゲナーゼIVを含む溶液(CTK溶液)を用いて処理し、残ったコロニーを上記フィーダー細胞上に蒔いてES細胞培養用培地で同様に培養することにより、iPS細胞又はiPS細胞コロニーを得ることができる。さらに同様の培養条件でiPS細胞を継代することができる。
【0110】
iPS細胞の同定は、ES細胞等の分化多能性細胞の特性に基づく試験によって行うことができる。具体的には、ES細胞特異的マーカー遺伝子の発現、半永久的細胞増殖能、分化多能性(三胚葉の形成)などの特性について細胞を試験し、細胞がこれらの特性をもつ場合に、該細胞がiPS細胞であると同定する(Takahashi, K.ら, Cell 131:861-872 (2007))。
【0111】
ES細胞特異的マーカー遺伝子としては、例えばOct3/4, Nanog, Lin28, PH34, Dnmt3b, Noda1, SSEA3, SSEA4, Tra-1-60, Tra-1-81, Tra2-49/6F(アルカリホスファターゼ)などを挙げることができる。これらの遺伝子の増幅に特異的なプライマーを使用するRT-PCR法によって、細胞内の上記マーカー遺伝子の発現を検出することができる。後述の実施例(ヒトiPS細胞の樹立)では、Oct3/4, Nanog, Lin28, PH34, Dnmt3b, Nodalの遺伝子発現が確認された。
【0112】
半永久的細胞増殖能については、約4ヶ月〜6ヶ月以上にわたる細胞の培養試験において、該細胞が指数的増殖をすることを確認する。ヒトiPS細胞のコロニー倍加時間(又は集団倍加時間(population doubling time))は、例えば46.9±12.4時間、47.8±6.6時間又は43.2±11.5時間程度であることが知られているので、この値を増殖能の指標とすることができる(Takahashi, K.ら, Cell 131:861-872 (2007))。また、iPS細胞は、高いテロメラーゼ活性をもつため、例えばTRAP(telomeric repeat amplification protocol)法によって該活性を検出してもよい。後述の実施例(ヒトiPS細胞の樹立)では、作製された細胞は指数的増殖を示した。
【0113】
分化多能性(3胚葉の形成)は、例えばテラトーマの形成及びテラトーマ組織内の3胚葉(内胚葉、中胚葉及び外胚葉)系の各組織(もしくは細胞)の同定によって確認することができる。具体的には、マウスiPS細胞の場合はヌードマウスの皮下に、またヒトiPS細胞の場合はScidマウスの精巣内に細胞を注射して腫瘍の形成を確認し、さらに腫瘍組織が、例えば軟骨組織(もしくは細胞)、神経管組織(もしくは細胞)、筋肉組織(もしくは細胞)、脂肪組織(もしくは細胞)、腸管様組織(もしくは細胞)などから構成されていることを確認する。後述の実施例(ヒトiPS細胞の樹立)では、形成されたテラトーマが、神経管組織(外胚葉)、軟骨組織(中胚葉)、腸管様組織(内胚葉)などを含むことが、組織染色法によって確認された。
上記の各試験に基づいて、細胞がiPS細胞であることを確認し、iPS細胞コロニーを選択することができる。
【0114】
6. 人工多能性幹(iPS)細胞の応用
本発明の方法によって作製されたiPS細胞は、分化多能性を有し、ES細胞と非常に類似した特性を有している。このため、iPS細胞は、ES細胞の代替細胞として利用することが可能である。
【0115】
iPS細胞は、それが分化多能性をもつことから、種々の分化細胞、前駆細胞及び組織に誘導することができる。具体的には、iPS細胞から、例えばactivin A/BMP4(bone morphogenetic protein 4)、VEGF(vascular endothelial growth factor)などの因子の存在下で神経細胞、心筋細胞などの種々の分化細胞を誘導することができる。このようにして得られた分化細胞は、患者に、例えば患者の欠陥組織に、移植することによって患者の治療(いわゆる、再生医療)のために使用できる。
【0116】
また、哺乳動物(ヒトを除く)由来の胚の胚盤胞にiPS細胞を導入し、その胚を、同じ動物種の仮親の子宮に移植することによって、iPS細胞の遺伝子型及び形質の一部を受け継いだキメラ動物を作製することができる (WO2007/069666)。iPS細胞における特定の遺伝子の改変、ノックアウト(KO)又はノックイン(KI)による遺伝子機能の解明、疾患モデルの作製、物質(蛋白質等)生産などが可能になる。このとき、予め遺伝子改変した体細胞から誘導されたiPS細胞も使用できる。
【0117】
したがって、本発明は、例えば、上記の方法で作製されたiPS細胞及びiPS細胞集団、遺伝子改変されたiPS細胞、並びに、そのようなiPS細胞を使用するキメラ動物の作製法及びそれによって得られたキメラ動物や子孫動物なども包含する。
【実施例】
【0118】
以下の実施例によって本発明をさらに具体的に説明する。しかし、本発明の範囲は、その実施例によって制限されないものとする。
【0119】
[実施例1]
Oct3/4及びNanogによるヒトiPS細胞の樹立(1)
36歳の成人皮膚由来線維芽細胞(HDF)に対して、Takahashi, K.ら, Cell, 131:861-872 (2007)に記載の方法に従い、レンチウイルス(pLenti6/UbC-Slc7a1)を用いて、マウスエコトロピックウイルスレセプターSlc7a1遺伝子を発現させた。この細胞(1×105個/6ウエルプレートの1ウエル)に対して、Takahashi, K.ら, Cell, 131:861-872 (2007) に記載の方法に従いヒト由来の4 遺伝子(Oct3/4,Sox2,Nanog,Lin28)をレトロウイルスで導入した。ウイルス感染から7日後に細胞を回収し、フィーダー細胞上への蒔き直しを行った(5×105個/100 mmディッシュ)。フィーダー細胞にはマイトマイシンCで処理して、細胞分裂を止めたSNL細胞(McMahon, A. P. & Bradley, A. Cell 62, 1073-1085 (1990))を用いた。感染10日後から霊長類ES細胞培養用培地 (ReproCELL、日本) に4 ng/mlの組換えヒトbFGF(WAKO、日本)を加えた培地で培養を行った。感染35日後にヒトiPS細胞様のコロニーが2個出現した。iPS細胞のコロニーから樹立したクローンはヒトES細胞様の形態を示し、フィーダー細胞上で増殖を続けることができた(図1)。
【0120】
これらの樹立したヒトiPS細胞のクローンについて、Cell, 131, 861-872(2007)の記載に準じてGenomic-PCR解析を行い、使用した4遺伝子がゲノム上に挿入されているか検討した。樹立したiPSクローンのうち1クローン(T4F-2)は4因子共にゲノムに挿入されていたが、他の1クローン(T4F-1)ではOct3/4,Nanogのみがゲノムに挿入されていた(図2)。レトロウイルスベクターはゲノムに挿入されなければ安定的に発現しないので、このクローンT4F-1はOct3/4,Nanogのみの発現で樹立されたと考えられた。
【0121】
樹立したクローンについてRever Tra Ace kit(Takara、日本)を使用してRT-PCR解析を行った結果、これらのクローン(T4F-1、T4F-2)はヒトES細胞特異的なマーカー遺伝子であるOct3/4, Nanog, Lin28, PH34, Dnmt3b及びNodalを発現しており、その発現量はヒトES細胞や既存のiPS細胞(201B6及び201B7:Takahashi, K.ら, Cell, 131:861-872 (2007))と同等であることが示された(図3)。
【0122】
さらにこのクローンT4F-1をScidマウスの精巣に挿入し分化多能性を調べた。具体的にはまず、ヒトiPS細胞クローンT4F-1を、組換えヒトbFGF(4ng/ml)及びRhoキナーゼ阻害剤Y-27632(10μM)を含有する霊長類ES細胞培養用培地 (ReproCELL、コスモバイオ、ともに日本)中で培養した。1時間後、collagen IVで処理して細胞を採取後、遠心して回収し、Y-27632(10μM)を含有するDMEM/F12中に浮遊させた。コンフルエントになった細胞(100mmディッシュ)の1/4量をScidマウスの精巣内に注射した。2〜3ヶ月後、腫瘍を切り刻んで4%フォルムアルデヒドを含有するPBSバッファーで固定した。パラフィン包埋組織をスライスし、ヘマトキシリン・エオシンで染色した。結果を図4に示す。組織学的に見ると腫瘍は複数の種類の細胞から構成されており、神経管組織、軟骨組織、腸管様上皮構造など3胚葉系に分化していたことから、iPS細胞の多能性が証明された (図4)。
以上の結果より、Oct3/4, Nanogの発現のみで樹立されたクローンは、iPS細胞であることが確認された。
【0123】
[実施例2]
Oct3/4及びNanogによるヒトiPS細胞の樹立(2)
Oct3/4,Nanogのみの導入でiPS細胞が樹立できるか検討した。上記同様、Slc7a1を発現させた36歳の成人皮膚由来線維芽細胞にレトロウイルスを用いてOct3/4,Nanogのみを導入し、感染7日後フィーダー細胞上へ蒔き直し、感染10日後から霊長類ES細胞培養用培地(ReproCELL(日本))に4 ng/mlの組換えヒトbFGF(WAKO、日本)を加えた培地で培養を行った。感染45日後にヒトiPS様のコロニーが1つ出現した(ON-1)。さらに、非ヒトiPS様コロニーを4個(ON-2、ON-4、ON-5、ON-6)ひろい、これを培養していくと4クローン中2クローン(ON-5、ON-6)がヒトES様の形態を示すように変化した。他の2クローンは非ES様の形態であった(ON-2、ON-4)。さらに47日目に、iPS誘導を行っている培養皿をCTK溶液(2.5%トリプシン 5 ml, 1 mg/mlコラゲナーゼIV 5 ml, 0.1 M CaCl2 0.5 ml, KSR 10 ml を30 mlの滅菌水に溶解したもの)を用いて処理し、その後PBSを用いて培養皿を洗ったところ、培養皿上に数個のコロニーが残った。これらをセルスクレイパーでかき集めフィーダー細胞上にまいて培養すると、ヒトES様のコロニーが得られた(ON-3)。これら樹立した4クローン(ON-1, ON-3, ON-5, ON-6)はフィーダー細胞上で増殖を続けることができた。クローンON-1とON-5のES様コロニー像を図5に示す。
【0124】
これらの樹立したヒトiPS細胞のクローンについて、Rever Tra Ace kit(Takara)を使用してRT-PCR解析を行った。その結果、これらのクローン(ON-1, ON-3, ON-5, ON-6)は、いずれもヒトES細胞特異的なマーカー遺伝子であるOct3/4, Nanog, Lin28, PH34及びDnmt3bを発現しており、その発現量はヒトES細胞や201B7と同等であった(図6)。
以上の結果から、iPS細胞をOct3/4,Nanogのみで樹立できることが明らかとなった。
【0125】
[実施例3]
ヒトiPS細胞の樹立
初期化因子を(a) Oct3/4, Nanog, Lin28及びc-Myc、又は(b) Oct3/4, Nanog及びLin28に代えたことを除いて実施例2と同様の手順で、36歳の成人皮膚由来線維芽細胞からヒトiPS細胞(ONLM及びONL)を樹立した(図7A及び図7B)。
【0126】
さらにまた、体細胞を成人皮膚由来線維芽細胞からヒト新生児包皮由来線維芽細胞に代えたことを除いて実施例2と同様の手順で、初期化因子Oct3/4及びNanogのみにより、該細胞からヒトiPS細胞を樹立した(図7C)。また体細胞を成人皮膚由来線維芽細胞から歯髄幹細胞(J.Dent.Res., 87(7): 676-681 (2008))に代えたことを除いて実施例2と同様の手順で、初期化因子Oct3/4及びNanogのみにより、該細胞からヒトiPS細胞を樹立した(図8)。
[実施例4]
Oct3/4及びNanogのみで樹立したヒトiPS細胞の解析
(1)Genomic-PCR解析
実施例2および3においてOct3/4及びNanogのみで樹立した成人皮膚線維芽細胞由来のiPS細胞、ヒト新生児包皮線維芽細胞由来のiPS細胞、および歯髄幹細胞由来のiPS細胞を用いて、Cell, 131, 861-872(2007)の記載に準じてGenomic-PCR解析を行った。結果を図9および図10に示す。いずれのiPS細胞においても導入したOct3/4遺伝子およびNanog遺伝子がゲノムに挿入されていた。また、導入に用いなかったSox2、Klf4、c-MycおよびLin28の各遺伝子がゲノムに挿入されていないことも確認した(図9、図10)。
【0127】
(2)テラトーマ形成能
成人皮膚由来の線維芽細胞からOct3/4及びNanogのみで樹立したiPS細胞を用いて、実施例1と同様にして分化多能性を調べた。結果を図11に示す。組織学的に見ると腫瘍は複数の種類の細胞から構成されており、神経組織、腺上皮組織、軟骨組織など三胚葉系に分化していたことから、iPS細胞の分化多能性が確認された (図11)。
【0128】
(3)免疫染色により観察された未分化マーカーの発現
成人皮膚由来の線維芽細胞からOct3/4及びNanogのみで樹立したiPS細胞を、マイトマイシンCで処理したSNLフィーダー上に播いた。5日間培養した後、細胞を4% パラフォルムアルデヒドで固定し、5% normal goat serum、1% BSA および0.2% TritonX-100を含むPBSバッファーで処理した。一次抗体として、抗ヒトSSEA1抗体 (DHSB, 1:100)、抗ヒトSSEA3抗体 (Peter W. Andrews博士より供与, 1:10), 抗ヒトTRA-1-60抗体(Peter W. Andrews博士より供与, 1:50)を用いた。また二次抗体としてAlexa488またはCy3(cyanine 3)でラベルされた抗体を用いた。核はHoechst 33342で染色した。結果を図12に示す。樹立したiPS細胞はSSEA3およびTRA1-60を発現していた。一方、抗SSEA1抗体ではコロニーの縁部しか染まらなかった。ヒトES細胞および過去に樹立済みのiPS細胞で同様な発現パターンが報告されていることから、2因子で樹立したiPS細胞は幹細胞マーカーの発現においてES細胞と同等であることが示された。
【0129】
(4)in vitro分化誘導
成人皮膚線維芽細胞からOct3/4及びNanogのみで樹立したiPS細胞をlow-binding dishに播き、Cell, 131, 861-872 (2007)に記載の方法でpoly-hydroxyethyl methacrylate (HEMA)でコートしたディッシュ上で8日間浮遊培養し、ゼラチンコートされたプレート上で胚様体(embryoid body:EB)を形成させた(100mmディッシュ)。8日間培養後、細胞を4% パラフォルムアルデヒドで固定し、5% normal goat serum、1% BSA および0.2% TritonX-100を含むPBSで処理した。内胚葉系細胞の分化マーカーであるAFP (α-fetoprotein)(R&D systems製)、中胚葉系細胞の分化マーカーであるSMA (smooth muscle actin)(DAKO製)、Desmin (NeoMarkers製)、Vimentin(Santa Cruz製)、外胚葉系の分化マーカーであるβIII-tubulin (Chemicon製)の各抗体を用いた染色を行った。二次抗体としてAlexa488またはCy3(cyanine 3)でラベルされた抗体を用いた。核はHoechst 33342で染色した。結果を図13に示す。
【0130】
染色によりこれらのマーカーの発現が確認され、樹立されたヒトiPS細胞はin vitroで三胚葉系への分化能を有することが確認された。
【0131】
(5) 核型の分析
樹立された3種のiPS細胞クローンについて、核型分析を日本遺伝子研究所(Nihon Gene Research Laboratories Inc.,日本)で行った。その結果、すべての核型が正常範囲内であった。
【0132】
以上の実施例1〜4で行ったGenomic-PCRおよびRT-PCRのプライマーのリストを表1、表2および配列表(配列番号:1〜20)に示した。
【0133】
【表1】
【0134】
【表2】
【特許請求の範囲】
【請求項1】
Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子を、哺乳動物由来の体細胞に導入し、これによって該体細胞から人工多能性幹細胞を誘導することを含み、該初期化因子はSox2及びSox2をコードする核酸を含まない、哺乳動物人工多能性幹細胞の作製方法。
【請求項2】
前記初期化因子が、Lin28、又はLin28をコードする核酸、をさらに含む、請求項1に記載の方法。
【請求項3】
前記初期化因子が、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、からなる、請求項1に記載の方法。
【請求項4】
前記初期化因子が、Oct3/4、Nanog及びLin28、或いはOct3/4、Nanog及びLin28をコードする核酸、からなる、請求項1に記載の方法。
【請求項5】
前記初期化因子及び体細胞が由来する哺乳動物は、互いに同じか又は異なる哺乳動物である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記哺乳動物が、霊長類又はげっ歯類である、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記霊長類がヒトである、請求項6に記載の方法。
【請求項8】
前記げっ歯類がマウスである、請求項6に記載の方法。
【請求項9】
前記核酸が1つ又は複数のベクターに含まれる、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記核酸が、前記体細胞のゲノムに組み込まれるか、或いは、ゲノムに組み込まれない状態で細胞内に存在する、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記人工多能性幹細胞の誘導時に、該細胞の樹立効率改善物質をさらに存在させる、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記樹立効率改善物質が、ヒストンデアセチラーゼ(HDAC)阻害剤、ヒストンメチルトランスフェラーゼ(G9a)阻害剤又はDNAメチル化酵素(Dnmt)阻害剤である、請求項11に記載の方法。
【請求項1】
Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、を含む哺乳動物由来の初期化因子を、哺乳動物由来の体細胞に導入し、これによって該体細胞から人工多能性幹細胞を誘導することを含み、該初期化因子はSox2及びSox2をコードする核酸を含まない、哺乳動物人工多能性幹細胞の作製方法。
【請求項2】
前記初期化因子が、Lin28、又はLin28をコードする核酸、をさらに含む、請求項1に記載の方法。
【請求項3】
前記初期化因子が、Oct3/4及びNanog、或いはOct3/4及びNanogをコードする核酸、からなる、請求項1に記載の方法。
【請求項4】
前記初期化因子が、Oct3/4、Nanog及びLin28、或いはOct3/4、Nanog及びLin28をコードする核酸、からなる、請求項1に記載の方法。
【請求項5】
前記初期化因子及び体細胞が由来する哺乳動物は、互いに同じか又は異なる哺乳動物である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記哺乳動物が、霊長類又はげっ歯類である、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記霊長類がヒトである、請求項6に記載の方法。
【請求項8】
前記げっ歯類がマウスである、請求項6に記載の方法。
【請求項9】
前記核酸が1つ又は複数のベクターに含まれる、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記核酸が、前記体細胞のゲノムに組み込まれるか、或いは、ゲノムに組み込まれない状態で細胞内に存在する、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記人工多能性幹細胞の誘導時に、該細胞の樹立効率改善物質をさらに存在させる、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記樹立効率改善物質が、ヒストンデアセチラーゼ(HDAC)阻害剤、ヒストンメチルトランスフェラーゼ(G9a)阻害剤又はDNAメチル化酵素(Dnmt)阻害剤である、請求項11に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2012−507258(P2012−507258A)
【公表日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願番号】特願2011−508739(P2011−508739)
【出願日】平成21年10月30日(2009.10.30)
【国際出願番号】PCT/JP2009/069015
【国際公開番号】WO2010/050626
【国際公開日】平成22年5月6日(2010.5.6)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度、文部科学省、「科学技術試験研究」委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504132272)国立大学法人京都大学 (1,269)
【Fターム(参考)】
【公表日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願日】平成21年10月30日(2009.10.30)
【国際出願番号】PCT/JP2009/069015
【国際公開番号】WO2010/050626
【国際公開日】平成22年5月6日(2010.5.6)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度、文部科学省、「科学技術試験研究」委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504132272)国立大学法人京都大学 (1,269)
【Fターム(参考)】
[ Back to top ]