
代替軽鎖の発現
本発明は、非相同性シグナル配列を有する代替軽鎖配列を含む代替軽鎖(SURROBODY(商標))構造体に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、非相同性シグナル配列を有する代替軽鎖配列を含む代替軽鎖(SURROBODY)(商標)構造体に関する。
【背景技術】
【0002】
Bリンパ球により産生される抗体(Ig)分子は、重(H)鎖および軽(L)鎖で構築されている。HおよびL鎖のアミノ末端ドメインのアミノ酸配列は、特に抗原結合部位を形成する3つの高度可変領域(CDR1、CDR2、CDR3)に可変性(VHおよびVL)がある。HおよびL鎖のアッセンブリは、L鎖の定常領域(CL)と、重鎖の第一の定常領域(CH1)の間のジスルフィド結合により、そしてVHドメインとVLドメインの間の非共有結合的相互作用により安定化されている。
【0003】
ヒトおよび多くの動物、例えばマウスにおいて、抗体のHおよびL鎖をコードする遺伝子は、V領域の遺伝子フラグメントコード化部分の段階的な体細胞再配列により組立てられる。Bリンパ球発達の様々な段階が、Ig遺伝子lociの再配列状態により特徴づけられる(例えば、Melchers,F.&Rolink,A.,B−Lymphocyte Development and Biology,Paul,W.E.ed.,1999,Lippincott,Philadephia参照)。
【0004】
B細胞の前駆体(プレB細胞)は、完全に発達した軽鎖の代わりにVpreB(1〜3)およびλ5と呼ばれる遺伝子組が生成されること、ならびにμ重鎖が同時発現されることにより、骨髄において同定された。
【0005】
ヒトVpreB1(CAG30495)の主なアイソフォームは、145aa長ポリペプチド(配列番号1)である。それは、IgVドメイン様構造を有するが、典型的なVドメインの最後のβ鎖(β7)が欠如しており、任意の別のタンパク質に対して配列相同性を示さないカルボキシル末端を有する。VpreB2は、142アミノ酸のマウスVpreB2ポリペプチド(P13373;配列番号2)およびマウスVpreB2配列の171アミノ酸長スプライスバリアント(CAA019641;配列番号3)をはじめとする複数のアイソフォームを有する。VpreB1およびVpreB2配列は、EP0269127および米国特許第5,182,205号;Collins et al.,Genome Biol.5(10):R84(2004);およびHollins et al.,Proc.Natl.Acad.Sci.USA 86(14):5552−5556(1989)に開示されている。ヒトVpreB3(配列番号4)の主なアイソフォームは、Collins et al.,Genome Biol.5(10):R84(2004)に開示された123aa長タンパク質(CAG30496)である。
【0006】
VpreB(1〜3)は、もう一方タンパク質λ5とは非共有結合的に会合されている。ヒトλ5は、209アミノ酸ポリペプチド(CAA01962:配列番号5)であり、抗体軽鎖と強力な相同性を有するIgCドメイン様構造と、そのアミノ酸末端に向かって、2つの機能的に異なる領域とを含み、その領域の一方は、Vλドメインのβ7鎖に対して強力な相同性を示す。ヒトλ5様タンパク質は、213アミノ酸(NP_064455;配列番号6)を有し、抗体λ軽鎖定常領域に対して約84%の配列同一性を示す。
【0007】
更なる詳細については、以下のレビュー雑誌を参照されたい:Karasuyama et al.,Adv.Immunol.63:1−41(1996);Melchers et al.,Immunology Today 14:60−68(1993);およびMelchers,Proc.Natl.Acad.Sci.USA 96:2571−2573(1999)。
【0008】
VpreBとλ5のポリペプチドは、一緒になって、代替軽鎖または偽軽鎖と呼ばれる非共有結合的に会合されたIg軽鎖様構造を形成している。初期のプレB細胞の表面では、代替軽鎖が、シグナルトランスデューサーCD79a/CD79bヘテロ二量体と会合された膜結合Igμ重鎖にジスルフィド結合して、B細胞受容体様構造、いわゆるプレB細胞受容体(プレBCR)を形成している。
【0009】
Surrobodyは、抗体レパートリーの正常な発達の間に生成されるプレB細胞受容体(プレBCR)に基づいている。抗体とは異なり、プレBCRは、2つの代替軽鎖成分、VpreBおよびλ5と対になった抗体重鎖で構成された三量体である。VpreBおよびλ5は両者とも、遺伝子再配列を受けず、V(D)J組換えの開始前に初期プレB細胞内で発現される遺伝子によりコードされている。プレBCRは、重鎖と、2つの非共有結合的に会合されたタンパク質:VpreBおよびλ5とで構成されている成熟免疫グロブリンと構造的に異なっており、即ちそれらは、抗体内の2つとは対照的に3つの成分を有する。更に、VpreBは、Vλ Igドメインと相同性があり、λ5は、抗体のCλドメインと相同性があるが、それぞれは、非標準のペプチド伸長を有しており、つまりVpreB1は、C末端に追加の21残基を有し、λ5は、N末端に50アミノ酸の伸長を有する。
【0010】
κ様代替軽鎖(κ様SLC)を用いて、κ様β細胞受容体(κ様BCR)が同定された(Frances et al.,EMBO J 13:5937−43(1994);Thompson et al.,Immunogenetics 48:305−11(1998);Rangel et al.,J Biol Chem 280:17807−14(2005))。
【0011】
Rangel et al.,J Biol Chem 280(18):17807−17814(2005)には、Thompson et al.,Immunogenetics 48:308−311(1998)により過去に報告されたcDNA配列と同一であることが判明された、再配列されていないVκ遺伝子の生成物であるVκ様タンパク質の同定または分子特徴づけが報告された。ところが、Fnances et al.,EMBO J 13:5937−43(1994)には、B細胞前駆体の表面でμ重鎖と会合し、それによりB細胞発達のためのλ5経路の代替法を提供する能力を有する、再配列された生殖細胞系JCkの同定および特徴づけが報告されている。
【0012】
κ様およびλ様のプレBCRが協調的に作用して軽鎖再配列を促進し、B細胞始原を確実に成熟させることが提案された。レビューについては、McKeller and Martinez−Valdez Seminars in Immunology 18:4043(2006)を参照されたい。
【0013】
Surrobodyの設計および生成の更なる詳細は、全体の開示が本明細書に参照により明確に組み込まれる、Xu et al.,Proc.Natl.Acad.Sci.USA 2008,105(31):10756−61、2008年10月2日公開のPCT公開第WO2008/118970号、2008年7月11日出願の米国特許仮出願第61/134,929号、およびXu et al.,J.Mol.Biol.2010,397,352−360に提供されている。
【0014】
代替軽鎖は、プレB細胞上でのタンパク質産生および細胞外提示を可能にするリーダー配列を有する。しかし、典型的には、設計された代替軽鎖構造体の組換え発現は、同一の重鎖を用いた抗体よりも低いことが見出されている。それ故、代替軽鎖構造体の組換え発現の効率を改善することが求められている。
【発明の概要】
【発明が解決しようとする課題】
【0015】
本発明は、少なくとも一部は、非相同性リーダー配列を用いることにより、代替軽鎖構造体の組換え発現の効率を顕著に改善しうるという実験的発見に基づく。
【課題を解決するための手段】
【0016】
一態様において、本発明は、代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、SLCポリペプチドまたはSLCポリペプチドを含有するSLC構造体をコードする単離された核酸分子を提供する。一実施形態において、SLCポリペプチドとしては、VpreBポリペプチド、λ5ポリペプチド、またはそれらのフラグメントもしくはバリアントが挙げられる。別の実施形態において、VpreBポリペプチドは、天然型VpreB1配列、天然型VpreB2配列、天然型VpreB3配列、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。幾つかの実施形態において、天然型VpreB配列は、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3、配列番号5のヒトVpreB様ポリペプチド、配列番号6のヒトVpreB dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の一実施形態において、λ5ポリペプチドは、配列番号7のヒトλ5様、配列番号8のヒトλ5ポリペプチド、配列番号9のヒトλ5 dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の実施形態において、SLCポリペプチドとしては、Vκ様ポリペプチド、JCκポリペプチド、またはそれらのフラグメントもしくはバリアントが挙げられる。別の一実施形態において、Vκ様ポリペプチド配列は、配列番号12〜24、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。幾つかの実施形態において、JCκポリペプチド配列は、配列番号26〜39、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。
【0017】
別の態様において、本発明は、代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、SLCポリペプチドをコードした単離された核酸分子を提供し、SLCポリペプチドとしては、SLCポリペプチド融合体、またはそれらのフラグメントもしくはバリアントが挙げられる。一実施形態において、SLC融合体としては、VpreB−λ5ポリペプチド融合体、またはそれらのフラグメントもしくはバリアントが挙げられる。別の実施形態において、VpreBポリペプチド配列およびλ5ポリペプチド配列の融合は、それぞれVpreB配列およびλ5配列のCDR3類似領域で、またはその周辺で起こる。別の一実施形態において、VpreBポリペプチド配列は、カルボキシ末端で、λ5ポリペプチド配列のアミノ末端に連結されている。一実施形態において、SLC融合体としては、Vκ様−JCκポリペプチド融合体、またはそれらのフラグメントもしくはバリアントが挙げられる。別の実施形態において、Vκ様ポリペプチド配列およびJCκポリペプチド配列の融合は、それぞれVκ様配列およびJCκ配列のCDR3類似領域で、またはその周辺で起こる。別の一実施形態において、Vκ様ポリペプチド配列は、カルボキシ末端で、JCκポリペプチド配列のアミノ末端に融合されている。
【0018】
別の一態様において、本発明は、非SLC分子を含有するSLC融合体を提供する。一実施形態において、SLC融合体は、非SLC分子と、VpreB、λ5、Vκ様、およびJCκ配列の少なくとも1つとを含有する。別の実施形態において、非SLC分子は、非SLCポリペプチドであってもよい。一実施形態において、融合体は、非SLCポリペプチドに融合されたλ5配列またはVpreB配列を含む。別の一実施形態において、融合は、VpreB配列またはλ5配列のCDR3類似領域で、またはその周辺で起こる。幾つかの実施形態において、λ5配列のN末端が、非SLCポリペプチドのC末端に融合されているか、またはVpreB配列のV末端が、非SLCポリペプチドのN末端に融合されている。別の一実施形態において、融合体は、非SLCポリペプチドに融合されたVκ様またはJCκ配列を含む。別の一実施形態において、融合は、Vκ様配列またはJCκ配列のCDR3類似領域で、またはその周辺で起こる。幾つかの実施形態において、JCκ配列のN末端が、非SLCポリペプチドのC末端に融合されているか、またはVκ様配列のC末端が、非SLCポリペプチドのN末端に融合されている。一実施形態において、本発明は、SLCポリペプチドが非SLC分子を含有するSLC融合ポリペプチドを含む、SLCポリペプチドをコードする単離された核酸分子を提供する。
【0019】
全ての実施形態において、非相同性分泌リーダー配列は、抗体、サイトカイン、リンフォカイン、モノカイン、ケモカイン、ポリペプチドホルモン、消化酵素、および細胞外マトリックスの成分からなる群より選択される分泌されたポリペプチドのリーダー配列であってもよい。一実施形態において、サイトカインは、成長ホルモン、例えば、ヒト成長ホルモン、N−メチオニルヒト成長ホルモン、およびウシ成長ホルモン;副甲状腺ホルモン;チロキシン;インスリン;プロインスリン;リラキシン;プロリラキシン;糖タンパク質ホルモン、例えば、卵胞刺激ホルモン(FSH)、甲状腺刺激ホルモン(TSH)、および黄体形成ホルモン(LH);肝細胞増殖因子;線維芽細胞増殖因子;プロラクチン;胎盤性ラクトゲン;腫瘍壊死因子−αおよび−β(TNF−αおよび−β);ミュラー管抑制物質;マウスゴナドトロピン関連ペプチド;インヒビン;アクチビン;血管内皮増殖因子;インテグリン;トロンボポエチン(TPO);神経成長因子、例えば、NGF−β;血小板成長因子;トランスフォーミング増殖因子(TGF)、例えば、TGF−αおよびTGF−β;インスリン様成長因子−Iおよび−II;エリスロポエチン(EPO);骨誘導因子;インターフェロン、例えば、インターフェロン−α、−βおよび−γ;コロニー刺激因子(CSF)、例えば、マクロファージ−CSF(M−CSF)、顆粒球−マクロファージ−CSF(GM−CSF)、および顆粒球−CSF(G−CSF);インターロイキン(IL)、例えば、IL−1、IL−1a、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12;腫瘍壊死因子、例えば、TNF−αまたはTNF−β;MIP−1α;MIP−1β;ならびにLIFおよびkitリガンド(KL)をはじめとする他のポリペプチド因子からなる群より選択される分泌された他のポリペプチドからなる群より選択されてもよい。
【0020】
全ての実施形態において、分泌リーダー配列は、ヒトおよび非ヒトホ乳類アルブミン、トランスフェリン、CD36、成長ホルモン、組織プラスミノーゲン活性化因子(t−PA)、エリスロポエチン(EPO)、ならびにニューブラスチンのリーダー配列からなる群より選択されてもよい。
【0021】
全ての実施形態において、分泌リーダー配列は、合成配列であってもよい。
【0022】
全ての実施形態において、分泌リーダー配列は、天然型分泌リーダー配列のコンセンサス配列であってもよい。
【0023】
全ての実施形態において、非相同性シグナル配列は、配列番号36(METDTLLLWVLLLWVPGSTG)であってもよい。
【0024】
全ての実施形態において、本発明は、代替軽鎖(SLC)構造体をコードする単離された核酸分子を提供する。
【0025】
一態様において、本発明は、ベクターおよび組換え宿主細胞を提供する。全ての実施形態において、ベクターは、本明細書に記載された核酸分子を含有していてもよい。全ての実施形態において、組換え宿主細胞は、本明細書に記載された核酸で形質転換されていてもよい。
【0026】
別の態様において、本発明は、組換え宿主細胞内での代替軽鎖(SLC)ポリペプチドまたはSLC構造体の発現方法を提供する。一実施形態において、その方法は、SLCポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、組換え宿主細胞を、SLCポリペプチドまたはSLC構造体をコードする核酸分子で形質転換するステップを含む。別の一実施形態において、組換え宿主細胞は、真核細胞である。別の一実施形態において、組換え宿主細胞は、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞である。幾つかの実施形態において、SLCポリペプチドまたはSLC構造体は、VpreBポリペプチド、λ5ポリペプチド、VpreB−λ5ポリペプチド融合体、Vκ様ポリペプチド、JCκポリペプチド、およびVκ様−JCκポリペプチド融合体の1種以上を含むSLCポリペプチドからなる群より選択される。
【図面の簡単な説明】
【0027】
【図1】天然型リーダー配列を有する配列番号1のヒトVpreB1アミノ酸配列;配列番号2および3のマウスVpreB2配列;配列番号4のヒトVpreB3様配列;図11で「VpreB dTail」(配列番号5)と称された「三量体」内のトランケート型VpreB1の配列;ならびにマウスIgκリーダー配列を有する配列番号6のヒトVpreB1アミノ酸配列を示す。下線は、VpreBアミノ酸配列内のリーダー配列を示す。
【図2】配列番号7のヒトλ5様配列;配列番号8のヒトλ5配列;図11で「λ5 dTail」(配列番号9)と称された「三量体」内のトランケート型λ5配列;ならびにマウスIgκリーダー配列を有する配列番号10のヒトλ5 dTail配列を示す。下線は、λ5アミノ酸配列内のリーダー配列を示す。
【図3】配列番号35としてのヒトVpreB1−λ5キメラアミノ酸配列を示す(マウスIgκリーダー配列に下線)。
【図4A】および
【図4B】(A)配列番号11のヒトVκ様ヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(AJ004956;配列番号12)(天然型リーダー配列に下線)、ならびに(B)それぞれがAJ004956Vκ様プロトタイプ配列(配列番号12)に直線状に並んだ、異なる長さの伸長(配列番号13〜24)を含む、Vκファミリー全てからの可能なVκ様タンパク質の予測される成熟アミノ酸配列を示す。
【図5A】(A)配列番号25のヒトJCκヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(配列番号26)(予測される成熟JCkタンパク質と比較したユニーク配列には2本の下線を、潜在的リーダー開裂配列には1本の下線を付す)、(B)残りのκJ定常領域再配列(J1〜J5Cκ)からの予測されるJCκ様アミノ酸配列(配列番号27〜31)、ならびに(C)追加されたマウスIgκリーダー配列に下線を付したJCκ(配列番号32)、追加されたマウスIgκリーダー配列のみに下線を付した組換えJCκ(配列番号33)、および追加されたマウスIgκリーダー配列に下線を付した予測されるプロセッシングJCκ(配列番号34)、を含むJCkを設計された分泌最適化バリアント示す。
【図5B】(A)配列番号25のヒトJCκヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(配列番号26)(予測される成熟JCkタンパク質と比較したユニーク配列には2本の下線を、潜在的リーダー開裂配列には1本の下線を付す)、(B)残りのκJ定常領域再配列(J1〜J5Cκ)からの予測されるJCκ様アミノ酸配列(配列番号27〜31)、ならびに(C)追加されたマウスIgκリーダー配列に下線を付したJCκ(配列番号32)、追加されたマウスIgκリーダー配列のみに下線を付した組換えJCκ(配列番号33)、および追加されたマウスIgκリーダー配列に下線を付した予測されるプロセッシングJCκ(配列番号34)、を含むJCkを設計された分泌最適化バリアント示す。
【図5C】(A)配列番号25のヒトJCκヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(配列番号26)(予測される成熟JCkタンパク質と比較したユニーク配列には2本の下線を、潜在的リーダー開裂配列には1本の下線を付す)、(B)残りのκJ定常領域再配列(J1〜J5Cκ)からの予測されるJCκ様アミノ酸配列(配列番号27〜31)、ならびに(C)追加されたマウスIgκリーダー配列に下線を付したJCκ(配列番号32)、追加されたマウスIgκリーダー配列のみに下線を付した組換えJCκ(配列番号33)、および追加されたマウスIgκリーダー配列に下線を付した予測されるプロセッシングJCκ(配列番号34)、を含むJCkを設計された分泌最適化バリアント示す。
【図6】VpreBおよびλ5配列により形成された代替軽鎖の略図、代替軽鎖配列を含む例示的な融合ポリペプチド、ならびにV−J結合から得られた抗体軽鎖構造である。
【図7】様々な代替軽鎖欠失および一本鎖構造体の略図である。
【図8】代替軽鎖構造体への組み合わせ的機能多様性(combinatorial functional diversity)の組込みを概略的に図示している。
【図9】様々な例示的代替軽鎖構造体の遺伝子およびタンパク質構造を示す。
【図10】代替軽鎖(SLC)成分に機能性を加える様々な代表的方法を示す。
【図11】様々な三量体および二量体代替軽鎖(SLC)構造体を示す。
【図12】様々なヘテロ二量体代替κ軽鎖欠失バリアントの略図である。「完全長」構造体において、Vκ様およびJCκ配列は両者とも、それぞれC−末端およびN−末端の伸長(テール)を保持している。dJバリアントでは、JCκのN−末端伸長が欠失されている。dVκテールバリアントでは、Vκ様配列のC−末端伸長が、除去されているが、JCκのN−末端伸長は保持されている。「短κ」バリアントでは、Vκ様配列のC末端テールおよびJCκ配列のN−末端伸長は両者とも、保持されている。
【図13】個別に、または別のタンパク質、例えば、抗体重鎖またはそのフラグメントと一緒に用いることができる、κ様軽鎖欠失および一本鎖構造体。
【図14】κ様代替軽鎖構造体への組み合わせ的機能多様性の組込み。赤線は、追加された多様性、例えば、ペプチドライブラリーを示す。
【図15】軽鎖は、遺伝子再配列およびRNAプロセッシングの生成物である。
【図16A】Vκ様タンパク質が再配列されていないVκIV−遺伝子転写および翻訳から得られることを示す。VκIVは、71のVL生殖細胞系遺伝子のうちの1つである。Vκ様タンパク質を作製することが可能なVL生殖細胞系遺伝子は更に70存在するため、κV遺伝子は更に39、そしてλV遺伝子は更に31存在する。
【図16B】JCκが、再配列されていないJおよびC生殖細胞系からのプロセッシングRNAの生成物であることを示す。JCκは、45のJC生殖細胞系の組み合わせのうちの1つである。JCκ様タンパク質を作製することが可能なVL生殖細胞系遺伝子が更に44存在し、つまりCκを組み合わせるJκ遺伝子が更に4、そして10のCλ遺伝子と組み合わせるJλ遺伝子が更に4存在する(合計40)。
【図17】κ様代替軽鎖成分に機能性を加えた略図を示す。二機能性および三機能性構造を示す。A:scFvを制約された融合体;B:Vκ様−scFv融合体;C:JCκ−scFv融合体;SLC二重融合。
【図18】代替軽鎖の機能的テール伸長のタイプを示す。
【図19】κ様およびλ様代替軽鎖機能的キメラを示す。
【図20】(A)Surrobody構成、(B)二機能性および二重特異性Surrobody構成、ならびに(C)(A)および(B)に表わされた分子のクローニング方策を示す。
【発明を実施するための形態】
【0028】
発明の詳細な説明
A.定義
他に断りがなければ、本明細書で用いられる技術的および科学的用語は、当業者により一般に理解されるものと同じ意味を有する。Singleton et al.,Dictionary of Microbiology and Molecular Biology 2nd ed.,J.Wiley&Sons(New York,NY 1994)は、本願で用いられる多くの用語の概説を当業者に提供する。
【0029】
当業者には、本発明の実践で用いられうる、本明細書に記載されたものと類似または同一の多くの方法および材料が認識されよう。事実、本発明は、記載された方法および材料に限定されない。本発明の目的では、次の用語を以下の通り定義する。
【0030】
本願全体を通して、単数形の使用は、他に明記されない限り、複数形を包含する。
【0031】
本願において、「または」の使用は、他に明記されない限り、「および/または」を包含する。
【0032】
更に用語「包含する」、「はじめとする」、および「包含された」は、限定されていない。
【0033】
本発明の文脈において、用語「抗体(Ab)」は、V(D)J遺伝子組換えから得られた伝統的組換え重鎖およびVJ遺伝子組換えから得られた伝統的組換え軽鎖、またはそれらのフラグメントからの天然型抗体を指すために用いられる。
【0034】
「天然型抗体」は、2つの同一軽(L)鎖および2つの同一重(H)鎖で構成された約150,000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は、ジスルフィド共有結合により重鎖に連結されているが、ジスルフィド結合の数は、異なる免疫グロブリンアイソタイプの重鎖間で変動する。各重鎖および軽鎖は、規則的な間隔の鎖内ジスルフィド架橋も有する。各重鎖は、一方の末端に、可変ドメイン(VH)と、それに続く多数の定常ドメインを有する。各軽鎖は、一方の末端の可変ドメイン(VL)および他方の末端の定常ドメインを有し、軽鎖の定常ドメインは、重鎖の第一の定常ドメインと直線状に並べられ、軽鎖の可変ドメインは、重鎖の可変ドメインと直線状に並べられている。特定のアミノ酸残基が、軽鎖可変ドメインと重鎖可変ドメインの間に境界を形成すると考えられている(Chothia et al.,J.Mol.Biol.186:651(1985);Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0035】
抗体鎖に関連する用語「可変」は、抗体の間で配列が広範囲にわたって異なり、特定抗原に対する各特定抗体の結合および特異性に関与する、抗体鎖の部分を指すために用いられる。そのような可変性は、軽鎖可変ドメインと重鎖可変ドメインの両方の高度可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存された部分は、フレームワーク領域(FR)と呼ばれる。天然型重鎖および軽鎖の可変領域はそれぞれ、4つのFR(それぞれFR1、FR2、FR3およびFR4)を含み、3つの高度可変領域により接続されたβシート配置を大きく採用し、それがループ接続を形成し、幾つかの例ではβシート構造の一部を形成している。各鎖の高度可変領域は、FRによって接近してまとめられていて、別の鎖の高度可変領域と共に抗体の抗原結合部位の形成に寄与する(Kabat et al.,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.(1991),p647−669参照)。定常ドメインは、抗原への抗体の結合に直接関与せず、様々なエフェクター機能、例えば、抗体依存性細胞毒性における抗体の関与などを示す。
【0036】
本明細書で用いられる用語「高度可変領域」は、抗原結合を担う抗体のアミノ酸残基を指す。高度可変領域は、「相補性決定領域」または「CDR」からのアミノ酸残基(即ち、軽鎖可変ドメイン内の30〜36(L1)、46〜55(L2)および86〜96(L3)の残基、ならびに重鎖可変ドメイン内の30〜35(HI)、47〜58(H2)、および93〜101(H3)の残基;MacCallum et al.,J Mol Biol.262(5):732−45(1996))を含む。
【0037】
用語「フレームワーク領域」は、より異なるCDR領域の間に存在する、当該技術分野で認識された抗体可変領域の部分を指す。そのようなフレームワーク領域は、典型的には、フレームワーク1〜4(FR1、FR2、FR3、およびFR4)と呼ばれ、重鎖または軽鎖抗体可変領域内に見出される3つのCDRを、三次元空間に保持するための足場を提供することで、CDRが抗原結合表面を形成することができる。
【0038】
それらの重鎖の定常ドメインのアミノ酸配列に応じて、抗体を異なる分類に割り付けることができる。抗体には5つの主要な分類、IgA、IgD、IgE、IgG、およびIgMが存在し、これらの複数は、亜分類(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgAおよびIgA2に更に分別される場合がある。好ましい実施形態において、本発明のイムノアドヘシンの構築に用いられる免疫グロブリン配列は、IgG免疫グロブリン重鎖ドメインのものである。ヒトイムノアドヘシンでは、ヒトIgG1およびIgG3免疫グロブリン配列の使用が好ましい。IgG1を用いることの主な利点は、IgG1イムノアドヘシンが固定化プロテインA上で効率的に精製されうることである。しかし、特定のイムノアドヘシン構築用にIg融合パートナーを選択する場合には、別の構造的および機能的性質を考慮すべきである。例えば、IgG3ヒンジは、より長く、より可撓性があるため、より大きな「アドヘシン」ドメインを収容することができ、IgG1に融合されると適切にフォールディングまたは機能しない可能性がある。別の考慮すべき事柄は、結合価かもしれず、IgGイムノアドヘシンは、二価ホモ二量体であるが、IgAおよびIgMのようなIgサブタイプは、基本的なIgホモ二量体単位の、それぞれ二量体または五量体構造をもたらす可能性がある。インビボ適用のために設計されたVEGF受容体Ig様ドメイン/免疫グロブリンキメラでは、Fc領域に特有の薬物動態性およびエフェクター機能も重要となる。IgG1、IgG2およびIgG4は全て、インビボ半減期21日を有するが、補体系を活性化させる際のそれらの相対的能力は異なる。その上、様々な免疫グロブリンが、様々な数のアロタイピックアイソタイプ(allotypic isotypes)を有する。
【0039】
異なる分類の免疫グロブリンに対応する重鎖定常ドメインは、それぞれα、δ、ε、γ、およびμと呼ばれる。
【0040】
任意の脊椎動物種からの抗体の「軽鎖」は、定常ドメインのアミノ酸配列に基づいて、カッパ(κ)およびラムダ(λ)と呼ばれる2種の明確に異なる型の一方に割り付けることができる。本明細書における抗体軽鎖の任意の参照は、κおよびλの両方の軽鎖を包含する。
【0041】
「抗体フラグメント」は、一般に抗原が結合した、完全長抗体の一部、またはその可変ドメインを含む。抗体フラグメントの例としては、非限定的に、Fab、Fab’、F(ab’)2、scFv、および(scFv)2フラグメントが挙げられる。
【0042】
本明細書で用いられる用語「抗体結合領域」は、抗原に結合しうる免疫グロブリンまたは抗体可変領域の1部分以上を指す。典型的には、抗体結合領域は、例えば、抗体軽鎖(VL)(またはその可変領域)、抗体重鎖(VH)(またはその可変領域)、重鎖Fd領域、抗体軽鎖と重鎖の混合体(またはその可変領域)、例えば、Fab、F(ab’)2、単一ドメイン、もしくは一本鎖抗体(scFv)、または完全長抗体、例えば、IgG(例えば、IgG1、IgG2、IgG3、またはIgG4サブタイプ)、IgA1、IgA2、IgD、IgE、またはIgM抗体である。
【0043】
本明細書で用いられる用語「エピトープ」は、アミノ酸が少なくとも約3〜5、好ましくは少なくとも約5〜10、または少なくとも約5〜15、典型的には約500以下または約1000以下の配列を指し、単独で、またはより大きな配列の一部として、そのような配列に応答して生成された抗体に結合している配列を定義する。エピトープは、由来する親タンパク質の部分と同一の配列を有するポリペプチドに限定されない。事実、ウイルスゲノムは、不断に変化する状態にあり、単離物の間に比較的高度の変動性が示される。したがって用語「エピトープ」は、天然型配列と同一の配列に加え、天然型配列への修飾、例えば欠失、置換および/または挿入を包含する。一般にそのような修飾は、本質的に保存的であるが、非保存的修飾も企図される。その用語は、具体的には「ミモトープ」、即ち、連続した直線状天然型配列を画定しない配列、または天然型タンパク質中には必ずしも生じないが、天然型タンパク質上のエピトープを機能的に模倣している配列を包含する。用語「エピトープ」は、具体的には線形および立体配座エピトープを包含する。
【0044】
用語「代替軽鎖ポリペプチド」または「SLCポリペプチド」は、本明細書では、VpreBポリペプチド、λ5ポリペプチド、Vκ様ポリペプチド、JCκポリペプチド、またはそれらのバリアントを指すために用いられる。
【0045】
用語「非代替軽鎖分子」または「非SLC分子」は、本明細書では、SLCポリペプチドでない分子を指すために用いられる。非SLC分子は、ポリペプチド、例えば、サイトカインまたは抗体フラグメントであってもよい。
【0046】
用語「VpreB」は、本明細書では最も広い意味で用いられ、詳細には、非限定的に、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3様配列、配列番号5のヒトVpreB dTをはじめとする任意の天然型配列またはバリアントVpreBポリペプチド、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、そのような天然型配列ポリペプチドのバリアントをはじめとするアイソフォームを指す。
【0047】
用語「λ5」は、本明細書では最も広い意味で用いられ、詳細には、非限定的に、配列番号6のヒトλ5、配列番号7のヒトλ5様タンパク質、配列番号9として示されるヒトλ5 dT、配列番号10のヒトVpreB1アミノ酸配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、そのような天然型配列λ5ポリペプチドのバリアントをはじめとするアイソフォームを指す。
【0048】
用語「バリアントVpreBポリペプチド」および「VpreBポリペプチドのバリアント」は、互換的に用いられ、本明細書ではアミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列VpreBポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントVpreBポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントVpreBポリペプチド」は、好ましくは天然型配列VpreBポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントVpreBポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントVpreBポリペプチドとしては、非限定的に、VpreB配列のC末端の非Ig様ユニークテールが部分的または完全に除去されているVpreBポリペプチドが挙げられる。
【0049】
用語「バリアントλ5ポリペプチド」および「λ5ポリペプチドのバリアント」は、互換的に用いられ、本明細書では、アミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列λ5ポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントλ5ポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントλ5ポリペプチド」は、好ましくは天然型配列λ5ポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントλ5ポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントλ5ポリペプチドとしては、非限定的に、λ5配列のN末端のユニークテールが部分的または完全に除去されているλ5ポリペプチドが挙げられる。
【0050】
用語「バリアントVκ様ポリペプチド」および「Vκ様ポリペプチドのバリアント」は、互換的に用いられ、本明細書では、アミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列Vκ様ポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントVκ様ポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントVκ様ポリペプチド」は、好ましくは天然型配列Vκ様ポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントVκ様ポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントVκ様ポリペプチドとしては、非限定的に、Vκ様配列のC末端の非Ig様ユニークテールが部分的または完全に除去されているVκ様ポリペプチドが挙げられる。
【0051】
用語「バリアントJCκポリペプチド」および「JCκポリペプチドのバリアント」は、互換的に用いられ、本明細書では、アミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列JCκポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントJCκポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントJCκポリペプチド」は、好ましくは天然型配列JCκポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントJCκポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントJCκポリペプチドとしては、非限定的に、JCκ配列のN末端のユニークテールが部分的または完全に除去されているJCκポリペプチドが挙げられる。
【0052】
アミノ酸配列同一性の百分率は、配列比較プログラムNCBI−BLAST2を用いて決定してもよい(Altschul et al.,Nucleic Acids Res.25:3389−3402(1997))。NCBI−BLAST2配列比較プログラムは、からダウンロードしてもよく、さもなければメリーランド州ベセスダのアメリカ国立衛生研究所から得てもよい。NCBI−BLAST2は、複数の研究パラメータを利用しており、それらの研究パラメータの全てが、例えば、unmask=yes、strand=all、expected occurrences=10、minimum low complexity length=15/5、multi−pass e−value=0.01、constant for multi−pass=25、dropoff for final gapped alignment=25、scoring matrix=BLOSUM62、をはじめとするデフォルト値に設定される。
【0053】
用語「VpreB配列」は、本明細書では、本明細書の先に定義された「VpreB」またはそのフラグメントの配列を指すために用いられる。
【0054】
用語「λ5配列」は、本明細書では、本明細書の先に定義された「λ5」またはそのフラグメントの配列を指すために用いられる。
【0055】
用語「Vκ様配列」は、本明細書では、本明細書の先に定義された「Vκ様」またはそのフラグメントの配列を指すために用いられる。
【0056】
用語「JCκ配列」は、本明細書では、本明細書の先に定義された「JCκ」またはそのフラグメントの配列を指すために用いられる。
【0057】
本明細書で用いられる用語「λ様代替軽鎖」は、VpreBとλ5タンパク質との非共有結合的会合により形成される二量体を指す。
【0058】
本明細書で用いられる用語「κ様代替軽鎖」は、Vκ様とJCκタンパク質との非共有結合的会合により形成される二量体を指す。
【0059】
本明細書に定義される用語「λ様代替軽鎖」は、本明細書の先に定義された「VpreB配列」および/または「λ5配列」を含む任意のポリペプチド配列を意味する。具体的には、本明細書に定義される「λ様代替軽鎖配列」としては、非限定的に、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3配列、配列番号5のヒトVpreB dT;および配列番号6のヒトVpreB1アミノ酸配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、それらのフラグメントおよびバリアントをはじめとする様々なアイソフォームが挙げられる。加えて用語「λ様代替軽鎖配列」としては、非限定的に、配列番号7のヒトλ5配列、配列番号8のヒトλ5様配列、配列番号9として示されるヒトλ5 dTail、配列番号10のヒトλ5 dTail、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、それらのフラグメントおよびバリアントをはじめとするアイソフォームが挙げられる。加えて用語「λ様代替軽鎖配列」としては、本明細書の先に定義されたVpreBおよびλ5の両方の配列を含む配列が挙げられる。
【0060】
本明細書に定義される用語「κ様代替軽鎖配列」は、本明細書の先に定義された「Vκ様配列」および/または「JCκ」を含む任意のポリペプチド配列を意味する。具体的には、本明細書に定義される「κ様代替軽鎖配列」としては、非限定的に、配列番号12〜24のいずれかのヒトVκ様配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、それらのフラグメントおよびバリアントをはじめとする様々なアイソフォームが挙げられる。加えて「κ様代替軽鎖配列」としては、非限定的に、配列番号12〜24のいずれかのヒトVκ様配列、配列番号25〜35のいずれかのヒトJCκ配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、そのフラグメントおよびバリアントをはじめとするアイソフォームが挙げられる。加えて用語「κ様代替軽鎖配列」としては、本明細書の先に定義されたVκ様およびJCκの両方の配列を含む配列が挙げられる。
【0061】
用語「代替軽鎖構造体」は、最も広い意味で用いられ、異種アミノ酸配列、核酸、および代替軽鎖配列にコンジュゲートされた別の分子をはじめとする任意の、そして全ての追加の異種成分を包含し、「コンジュゲーション」は、以下に定義する。
【0062】
「代替軽鎖構造体」は、本明細書では、「Surrobody(商標)」または「Surrobody」とも呼ばれ、その2つの用語は、互換的に用いられる。特定のSurrobody(商標)λ様代替軽鎖構造体は、Xu et al.,Proc.Natl.Acad.Sci.USA 2008,105(31):10756−61および2008年10月2日公開のPCT公開第WO2008/118970号に開示されている。同じく企図されるのは、米国特許公開第2010−0062950号、およびXu et al.,J.Mol.Biol.2010,397,352−360に記載されたκ様代替軽鎖構造体であり、それらの開示の全体は、本明細書に参照により明確に組み込まれる。
【0063】
本発明のポリペプチドの文脈において、第一のアミノ酸配列に関する用語「異種アミノ酸配列」は、第一のアミノ酸配列に自然に会合されておらず、少なくとも第一のアミノ酸配列が本明細書の代替軽鎖構造体内に存在する形態ではないアミノ酸配列を指すために用いられる。したがって、VpreB、λ5、Vκ様、またはJCκに関する「異種アミノ酸配列」は、その天然の環境において、天然型VpreB、λ5、Vκ様、またはJCκと会合されていない任意のアミノ酸配列である。これらには、非限定的に、i)VpreBと共に発達中のB細胞上に代替軽鎖を形成するそれらのλ5配列とは異なるλ5配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたλ5配列;ii)λ5と共に発達中のB細胞上に代替軽鎖を形成するそれらのVpreB配列とは異なるVpreB配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたVpreB配列;iii)JCκ様と共に発達中のB細胞上にκ様代替軽鎖を形成するそれらのVκ様配列とは異なるVκ様配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたVκ様配列;ならびにiv)Vκ様と共に発達中のB細胞上にκ様代替軽鎖を形成するそれらのJCκ配列とは異なるJCκ配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたJCκ配列が挙げられる。
【0064】
VpreBまたはλ5に関する「異種アミノ酸配列」は、天然の環境では、VpreBまたはλ5配列は互いに共有結合的に会合、例えば融合されていないため、天然型配列VpreBまたはλ5をはじめとする対応するVpreBまたはλ5と共有結合的に会合、例えば融合されている、VpreBまたはλ5配列も包含する。同様に、Vκ様またはJCκに関する「異種アミノ酸配列」は、天然の環境では、Vκ様またはJCκ配列は互いに共有的に会合、例えば融合されていないため、天然型配列Vκ様またはJCκをはじめとする対応するVκ様またはJCκと共有結合的に会合、例えば融合されている、Vκ様またはJCκ配列も包含する。異種アミノ酸配列としては、非限定的に、抗体および重鎖配列をはじめとする抗体配列ならびにそれらのフラグメント、例えば、抗体軽鎖および重鎖可変領域配列、ならびに抗体軽鎖および重鎖定常領域配列も挙げられる。
【0065】
用語「コンジュゲート」、「コンジュゲートされた」および「コンジュゲーション」は、共有結合または非共有結合の任意の、そして全ての形態を指し、非限定的に、直接遺伝子融合または化学的融合、リンカーまたは架橋剤を介したカップリング、そして例えば、ファンデルワールス力を介した、またはロイシンジッパーを用いた非共有結合的会合が挙げられる。
【0066】
用語「可撓性リンカー」は、本明細書では、化学構造に基づけば、意図された文脈および環境において三次元空間に固定されていると予測されない任意のリンカーを指すために用いられる。
【0067】
用語「融合」は、本明細書では、1本のポリペプチド鎖内の異なる起点のアミノ酸配列をコードするヌクレオチド配列をインフレームで組み合わせることによって、それらのアミノ酸配列を組み合わせることを指すのに用いられる。用語「融合」は、その末端の1つと融合することに加えて、内部融合、即ちポリペプチド鎖内の異なる起点の配列を挿入することを明示的に包含する。
【0068】
本明細書で用いられる用語「ペプチド」、「ポリペプチド」および「タンパク質」は全て、共有結合的「ペプチド結合」によって連結されたアミノ酸の一次配列を指す。一般に、ペプチドは、少数のアミノ酸、典型的には約2〜約50のアミノ酸からなり、タンパク質よりも短い。本明細書で定義される用語「ポリペプチド」は、ペプチドおよびタンパク質を包含する。
【0069】
用語「アミノ酸」または「アミノ酸残基」は、典型的には、当該技術分野で認識された定義を有するアミノ酸、例えば、アラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Gln)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ile)、ロイシン(Leu)、リシン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、トレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)およびバリン(Val)からなる群より選択されるアミノ酸を指すが、修飾アミノ酸、合成アミノ酸または希アミノ酸を所望どおり使用してもよい。したがって、具体的には37 CFR 1.822(b)(4)に列挙された修飾および異常アミノ酸は、この定義に含まれ、本明細書に参照により明確に組み込まれる。アミノ酸は、様々な亜群に細分することができる。したがってアミノ酸は、非極性側鎖(例えば、Ala、Cys、Ile、Leu、Met、Phe、Pro、Val)、負電荷の側鎖(例えば、Asp、Glu)、正電荷の側鎖(例えば、Arg、His、Lys)、または非荷電の極性側鎖(例えば、Asn、Cys、Gln、Gly、His、Met、Phe、Ser、Thr、TrpおよびTyr)を有するとして分類することができる。アミノ酸は、小アミノ酸(Gly、Ala)、求核性アミノ酸(Ser、His、Thr、Cys)、疎水性アミノ酸(Val、Leu、Ile、Met、Pro)、芳香族アミノ酸(Phe、Tyr、Trp、Asp、Glu)、アミド(Asp、Glu)および塩基性アミノ酸(Lys、Arg)として分類することもできる。
【0070】
用語「ポリヌクレオチド」は、核酸、例えば、DNA分子およびRNA分子、ならびにそれらの類似体(例えば、ヌクレオチド類似体または核酸化学を利用して生成されるDNAまたはRNA)を指す。ポリヌクレオチドは、所望通り、例えば当該技術分野で認識された核酸化学を利用して、合成的に生成し、または例えばポリメラーゼを用いて、酵素的に生成し、そして所望なら修飾してもよい。典型的な修飾としては、メチル化、ビオチン化、および当該技術分野で公知の別の修飾が挙げられる。加えて核酸分子は、一本鎖または二本鎖であってもよく、所望ならば、検出可能な部分と連結させることができる。
【0071】
参照ポリペプチドに関する用語「バリアント」は、天然型ポリペプチドと比較して、少なくとも1つのアミノ酸変異または修飾(すなわち、改変)を有するポリペプチドを指す。「アミノ酸修飾」により生成されたバリアントは、例えば、天然型アミノ酸配列中の少なくとも1つのアミノ酸を置換、欠失、挿入および/または化学修飾することによって生成することができる。
【0072】
「アミノ酸修飾」は、所定のアミノ酸配列のアミノ酸配列における変化を指す。例示的な修飾としては、アミノ酸置換、挿入および/または欠失が挙げられる
【0073】
特定の位置「でのアミノ酸修飾」は、特定の残基の置換もしくは欠失、または特定の残基に隣接する少なくとも1つのアミノ酸残基の挿入を指す。特定の残基に「隣接する」挿入は、その1つまたは2つの残基の内部の挿入を意味する。挿入は、N末端またはC末端から特定の残基までに存在してもよい。
【0074】
「アミノ酸置換」は、所定のアミノ酸配列内の少なくとも1つの既存のアミノ酸残基を、別の異なる「交換」アミノ酸残基で交換することを指す。交換残基は、「天然アミノ酸残基」(即ち、遺伝子コードによりコードされている)であってもよく、アラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リシン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);トレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)からなる群より選択されてもよい。1つ以上の非天然アミノ酸残基での置換も、本明細書でのアミノ酸置換の定義に包含される。
【0075】
「非天然アミノ酸残基」は、先に列挙された天然アミノ酸残基以外の、ポリペプチド鎖内の隣接アミノ酸残基と共有結合できる残基を指す。非天然アミノ酸残基の例としては、ノルロイシン、オルニチン、ノルバリン、ホモセリンおよびEllman et al.,Meth.Enzym.202:301 336(1991)に記載されるような別のアミノ酸残基類似体が挙げられる。そのような非天然アミノ酸残基を生成するために、Noren et al.Science 224:182(1989)およびEllman et al.の上記文献の手順を用いることができる。簡潔に述べると、これらの手順は、非天然アミノ酸残基によりサプレッサーtRNAを化学的に活性化させて、RNAのインビトロ転写および翻訳を行うことを含む。
【0076】
「アミノ酸挿入」は、少なくとも1つのアミノ酸を所定のアミノ酸配列に組込むことを指す。挿入は通常、1つまたは2つのアミノ酸残基の挿入からなるが、本願は、より大きな「ペプチド挿入」、例えば、約3〜約5または最大約10のアミノ酸残基の挿入を企図する。挿入された残基は、先に開示された通り天然または非天然のものであってもよい。
【0077】
「アミノ酸欠失」は、所定のアミノ酸配列からの少なくとも1つのアミノ酸残基の除去を指す。
【0078】
用語「突然変異誘発」は、他に明記されていなければ、ポリヌクレオチドまたはポリペプチド配列を改変するための、任意の当該技術分野で認識された技術を指す。突然変異誘発の好ましいタイプとしては、エラープローンPCR突然変異誘発、飽和突然変異誘発、または別の部位特異的突然変異誘発が挙げられる。
【0079】
「部位特異的突然変異誘発」は、当該技術分野における技術標準であり、一本鎖ファージDNAに相補的な合成オリゴヌクレオチドプライマーを用いて実施して、限定されたミスマッチ以外の変異を誘発させると、所望の突然変異が現れる。簡潔に述べると、合成オリゴヌクレオチドを、一本鎖ファージDNAに相補的な鎖の合成を導くプライマーとして用い、得られた二本鎖DNAをファージ支持宿主細菌(phage−supporting host bacterium)内に形質変換させる。形質転換された細菌の培養物を上層寒天に塗布し、ファージを含む単細胞からのプラーク形成を可能にする。理論的には、新しいプラークの50%が、突然変異形態を一本鎖として有するファージを含有し、50%が本来の配列を有する。完全一致のハイブリダイゼーションが可能であるが、本来の鎖とのミスマッチがハイブリダイゼーションを防止するのに十分となる温度で、該当するプラークを、キナーゼ処理した合成プライマーとハイブリダイズすることによって選択する。その後、プローブとハイブリダイズするプラークが選択され、配列決定および培養されて、DNAが回収される。
【0080】
用語「ベクター」は、細胞内での自己複製が可能で、DNAセグメント、例えば遺伝子またはポリヌクレオチドが作動可能に連結されることで、結合したセグメントの複製をもたらすことができるrDNA分子を指すために用いられる。1種以上のポリペプチドをコードする遺伝子の発現を導くことが可能なベクターを、本明細書では「発現ベクター」と呼ぶ。用語「制御配列(control sequences)」は、特定の宿主生物中の作動可能に連結されたコード配列の発現に必要なDNA配列を指す。例えば、原核生物に適した制御配列としては、プロモータ、場合によりオペレータ配列、およびリボソーム結合部位が挙げられる。真核細胞は、プロモータ、ポリアデニル化シグナル、およびエンハンサーを用いることが公知である。ベクターは、追加のDNAセグメントにライゲートされている場合がある環状二本鎖DNAループを指す「プラスミド」であってもよい。ベクターは、追加のDNAセグメントがウイルスゲノムにライゲートされている場合があるファージベクターまたはウイルスベクターであってもよい。適切なベクター、例えば、細菌起源を有する細菌ベクターまたは複製およびエピソームホ乳類ベクターは、導入される宿主細胞内で自己複製が可能である。ベクターは、宿主細胞に導入する際、宿主細胞ゲノム、例えば、非エピソームホ乳類ベクターに合体させて、宿主ゲノムと一緒に複製してもよい。
【0081】
核酸は、別の核酸配列と機能的関係にある場合に、「作動可能に連結」されている。例えば、プレ配列または分泌リーダーのDNAは、ポリペプチドの分泌に関与するプレタンパク質として発現される場合、ポリペプチドのDNAに作動可能に連結されており、プロモータもしくはエンハンサーは、コード配列の転写に影響を及ぼす場合、コード配列に作動可能に連結されており、またはリボソーム結合部位は、翻訳を促進するように位置する場合、コード配列に作動可能に連結されている。一般に、「作動可能に連結された」とは、連結されているDNA配列が隣接すること、そして分泌リーダーの場合、隣接し、かつ読み取り期(reading phase)にあることを意味する。しかし、エンハンサーは、隣接している必要はない。連結は、簡便な制限部位でのライゲーションによって果される。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーを従来の実践法に従って使用する。
【0082】
「ファージディスプレイライブラリー」は、クローン化されたタンパク質配列の回収物をファージコートタンパク質との融合体として発現するタンパク質発現ライブラリーである。したがって、語句「ファージディスプレイライブラリー」は、本明細書では、外来(一般には非相同性)タンパク質を発現するファージ(例えば、繊維状ファージ)の回収物を指す。外来タンパク質は、ファージと接触している別の部分と自由に相互作用(結合)することができる。外来タンパク質を提示する各ファージは、ファージディスプレイライブラリーの「メンバー」である。
【0083】
用語「繊維状ファージ」は、異種ポリペプチドを表面に提示することができるウイルス粒子を指し、非限定的に、fl、fd、Pf1、およびM13が挙げられる。繊維状ファージは、選択マーカー、例えば、テトラサイクリン(例えば、「fd−tet」)を含有していてもよい。様々な繊維状ファージディスプレイシステムが、当業者に周知である(例えば、Zacher et al.,Gene 9:127−140(1980),Smith et al.,Science 228:1315−1317(1985);およびParmley and Smith Gene 73:305−318(1988)参照)。
【0084】
用語「パンニング」は、標的に対する親和力および特異性が高い、抗体などの化合物を運搬する化合物を同定および単離する際の複数回のスクリーニング工程を指すために用いられる。
【0085】
「リーダー配列」、「シグナルペプチド」、または「分泌リーダー」は、互換的に用いられ、一部であるポリペプチドの細胞内輸送を導くアミノ酸残基を含む配列を含有する。ポリペプチドは、典型的にはN末端に、分泌リーダー、シグナルペプチド、またはリーダー配列を含有する。これらのポリペプチドは、リーダー配列がシグナルエンドペプチターゼによりポリペプチドの残り部分から開裂されうる開裂部位を含んでいてもよい。そのような開裂により、成熟ポリペプチドが生成する。開裂は、典型的には、分泌の間に、またはインタクトポリペプチドが適切な細胞内コンパートメントに導かれた後に起こる。
【0086】
「宿主細胞」は、本明細書に記載された分子をコードする核酸および/または核酸含有ベクターの形質転換のレシピエントとなりうる、またはレシピエントになった個々の細胞または細胞培養物を包含する。本発明の方法において、宿主細胞は、真核細胞、例えば、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞であってもよい。別の適切な宿主細胞は、当業者に公知である。
【0087】
B.詳細な説明
本発明の方法を実施する技術は、当該技術分野で周知であり、例えば、Ausubel et al.,Current Protocols of Molecular Biology,John Wiley and Sons(1997);Molecular Cloning:A Laboratory Manual,Third Edition,J.Sambrook and D.W.Russell,eds,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;O’Brian et al.,Analytical Chemistry of Bacillus Thuringiensis,Hickle and Fitch,eds,Am.Chem.Soc.,1990;Bacillus thuringiensis:biology,ecology and safety,T.R.Glare and M.O’Callaghan,eds,John Wiley,2000;Antibody Phage Display,Methods and Protocols,Humana Press,2001;およびAntibodies,G.Subramanian,ed,Kluwer Academic,2004をはじめとする標準的な実験教書に記載されている。突然変異誘発は、例えば、部位特異的突然変異誘発(Kunkel et al.,Proc.Natl.Acad.Sci.USA 82:488−492(1985))を利用して実施することができる。PCR増幅法は、米国特許第4,683,192号、同第4,683,202号、同第4,800,159号、および同第4,965,188号に、ならびに「PCR Technology:Principles and Applications for DNA Amplification」,H.Erlich,ed,Stockton Press,New York(1989);およびPCR Protocols:A Guide to Methods and Applications,Innis et al.,eds,Academic Press,San Diego,Calif.(1990)をはじめとする複数の教書に記載されている。
【0088】
非相同性リーダー配列
ヒトVpreB1(CAG30495)の主なアイソフォームは、19アミノ酸のリーダー配列を含む145アミノ酸長のポリペプチド(図1の配列番号1)である。類似のリーダー配列が、別のVpreBポリペプチド内に存在する。ヒトトランケート型VpreB1配列(天然型VpreB1のC末端にある特徴的「テール」が欠如)は、「VpreB1 dTail配列」とも呼ばれ、配列番号5として示される。
【0089】
ヒトλ5(CAA10962)の主なアイソフォームは、30アミノ酸のリーダー配列を含む209アミノ酸長のポリペプチド(配列番号7)である。類似のリーダー配列が、別のλ5ポリペプチド内に存在する。ヒトトランケート型λ5配列(天然型λ5のN末端にある特徴的「テール」が欠如)は、「λ5 dTail配列」とも呼ばれ、配列番号9として示される。
【0090】
具体的には、天然型ヒトVκ様ポリペプチド配列としては、非限定的に、20アミノ酸のリーダー配列を含む配列番号11として示されるAJ004956のポリヌクレオチドによりコードされた、ヒトκ様ポリペプチド(配列番号12)が挙げられる。類似のリーダー配列が、別のVκ様ポリペプチド内に存在する。
【0091】
天然型配列JCκ様ポリペプチドとしては、非限定的に、潜在的な22アミノ酸のリーダー配列を含み、そのうちの15のアミノ酸が伝統的に組換えられたJCκ配列に特有に付加されている、プロトタイプリーダー配列(配列番号26)が欠如した、図5Aに示されたAAB32987 ヒトJCκポリペプチドが挙げられる。類似の組換えリーダー配列が、別のJCκポリペプチド内に存在する。
【0092】
本発明は、代替軽鎖構造体が、内在性リーダーVpreBリーダー配列および/もしくはλ5リーダー配列、または内在性Vκ様リーダー配列および/もしくはJCκリーダー配列を含む配列から生成された場合よりも高い収率で、代替軽鎖構造体を生成するための核酸およびポリペプチド構造体を提供する。本発明は、代替軽鎖構造体が、VpreB配列および/もしくはλ5の内在性リーダー、またはVκ様および/もしくはJCκの内在性リーダーのコード配列を含むか、あるいは内在性リーダー配列を含まずにDNA配列から生成された場合よりも高い収率で、代替軽鎖構造体を生成するためのベクター、宿主細胞および方法も提供する。より高い収率は、少なくとも1種の内在性分泌リーダー配列を、本発明の非相同性リーダー配列と交換することにより実現される。したがって本発明は、非相同性リーダー配列を含む、代替軽鎖および代替軽鎖構造体を提供する。
【0093】
好ましくは、非相同性リーダーペプチドにより実現される発現レベルは、発現が本質的に同じ条件下で実施される場合に、相同性リーダー配列を用いて実現される発現レベルよりも、少なくとも約5%高い、少なくとも約10%高い、少なくとも約20%高い、少なくとも約30%高い、少なくとも約40%高い、または少なくとも約50%高い。
【0094】
本発明において、非相同性リーダー配列は、天然型VpreBリーダー配列および/もしくは天然型λ5リーダー配列の代わりに代替軽鎖ポリペプチドのアミノ末端に融合されるか、または天然型Vκ様リーダー配列および/もしくは天然型JCκリーダー配列の代わりにκ様代替軽鎖ポリペプチドのアミノ末端に融合される。本発明人は、代替軽鎖配列(一緒に融合されるか、または非共有結合的に会合されている、VpreB/λ5配列またはVκ様/JCκ配列)および抗体重鎖配列を含む代替軽鎖配列構造体の生成時に、代替軽鎖の天然型リーダー配列とは対照的に、特定の非相同性リーダー配列が意外にも良好に機能することを発見した。
【0095】
本発明によれば、非相同性リーダー配列は、抗体軽鎖ならびにヒトおよび非ヒトホ乳類分泌タンパク質のリーダー配列をはじめとする、高度翻訳タンパク質からの任意のリーダー配列であってもよい。分泌タンパク質が含まれており、それらの配列は、一般のデータベース、例えばSwiss−Prot、UniProt、TrEMBL、RefSeq、EnsemblおよびCBI−Geneから入手できる。加えて、SPD、つまりウェブに基づく分泌タンパク質データベース、http://spd.cbi.pku.edu.cn.で入手できる、そのような配列の供給源である(Chen et al.,Nucleic Acids Res.,2005,33:D169−D173)。そのような分泌タンパク質としては、非限定的に、抗体、サイトカイン、リンフォカイン、モノカイン、ケモカイン、ポリペプチドホルモン、消化酵素、および細胞外マトリックスの成分が挙げられる。
【0096】
サイトカインのうちで含まれるのは、成長ホルモン、例えば、ヒト成長ホルモン、N−メチオニルヒト成長ホルモン、およびウシ成長ホルモン;副甲状腺ホルモン;チロキシン;インスリン;プロインスリン;リラキシン;プロリラキシン;糖タンパク質ホルモン、例えば、卵胞刺激ホルモン(FSH)、甲状腺刺激ホルモン(TSH)、および黄体形成ホルモン(LH);肝細胞増殖因子;線維芽細胞増殖因子;プロラクチン;胎盤性ラクトゲン;腫瘍壊死因子−αおよび−β(TNF−αおよび−β);ミュラー管抑制物質;マウスゴナドトロピン関連ペプチド;インヒビン;アクチビン;血管内皮増殖因子;インテグリン;トロンボポエチン(TPO);神経成長因子、例えば、NGF−β;血小板成長因子;トランスフォーミング増殖因子(TGF)、例えば、TGF−αおよびTGF−β;インスリン様成長因子−Iおよび−II;エリスロポエチン(EPO);骨誘導因子;インターフェロン、例えば、インターフェロン−α、−βおよび−γ;コロニー刺激因子(CSF)、例えば、マクロファージ−CSF(M−CSF);顆粒球−マクロファージ−CSF(GM−CSF);ならびに顆粒球−CSF(G−CSF);インターロイキン(IL)、例えば、IL−1、IL−1a、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12;腫瘍壊死因子、例えば、TNF−αまたはTNF−β;MIP−1α;MIP−1β;ならびにLIFおよびkitリガンド(KL)をはじめとする他のポリペプチド因子である。
【0097】
本発明の構造体での使用に適した追加のリーダー配列は、一般に入手可能なシグナルペプチドデータベース、例えば、http://proline.bis.nus.edu.sq/spdbでアクセス可能なSPdbシグナルペプチドデータベースに含まれる(Choo et al.,BMC Bioinformatics 2005,6:249参照)。
【0098】
適切な非相同性リーダー配列の具体的な例としては、非限定的に、ヒトおよび非ヒトホ乳類アルブミン、トランスフェリン、CD36、成長ホルモン、組織プラスミノーゲン活性化因子(t−PA)、エリスロポエチン(EPO)、ニューブラスチンのリーダー配列、ならびに別の分泌ヒトおよび非ヒトタンパク質からのリーダーペプチドが挙げられる。
【0099】
マウスIgκリーダー配列は、非相同性リーダー配列として用いられてもよい(METDTLLLWVLLLWVPGSTG−配列番号36)。
【0100】
非相同性リーダー配列が、i)VpreBおよびλ5の両方の代替軽鎖構造体、またはii)Vκ様およびJCκの両方の代替軽鎖構造体、に存在する場合、i)またはii)内の各非相同性リーダー配列は、他方と同一であってもよく、または他方と異なっていてもよい。
【0101】
天然型タンパク質からのシグナルペプチドに加えて、本発明の非相同性リーダー配列としては、消化されて、本質的に生じるリーダー配列の性能を更に改善することができ、本発明の代替軽鎖構造体の発現で用いられる宿主生物中での最良の性能のために特別に適合させることができる、合成およびコンセンサスリーダー配列が挙げられる。
【0102】
代替軽鎖構造体
本明細書の代替軽鎖(SLC)構造体は、抗体レパートリーの通常の発達時に生成されるプレB細胞受容体(プレBCR)に基づいている。抗体とは異なり、プレBCRは、2つの代替軽鎖成分、VpreBおよびλ5と対になった抗体重鎖で構成された三量体である。VpreBおよびλ5は両者とも、遺伝子再配列を受けない遺伝子によりコードされ、V(D)J組換えの開始前に初期のプロB細胞内で発現される。プレBCRは、1つの重鎖と、2つの非共有結合的に会合されたタンパク質:VpreBおよびλ5とで構成された成熟免疫グロブリンとは、構造的に異なっており、即ちそれらは、抗体内の2つの成分とは対照的に3つの成分を有する。更に、VpreBはVλIgドメインと相同性があり、λ5は抗体のCλドメインと相同性があるが、それぞれは、非標準のペプチド伸長を有し、VpreB1は、C末端に追加の21の残基を有し;λ5は、N末端に50アミノ酸の伸長を有する。Surrobodyの設計および生成の更なる詳細については、Xu et al.,Proc.Natl.Acad.Sci.USA 2008,105(31):10756−61、2008年10月2日公開のPCT公開第WO2008/118970号に提供されている。
【0103】
同様に、本明細書に記載されたκ様代替軽鎖構造体は、プレB細胞受容体(プレBCR)に基づいている。κ様軽鎖は、JCκ融合遺伝子と組になった生殖細胞系VκIV遺伝子である。これらの遺伝子のそれぞれにおいて、ペプチド伸長は、CDR3と類似した部位の付近に存在する。これらの2種のタンパク質は、ゲノムレベルで組換えられていないようであるため、重鎖への会合が相互に排他的で、λ様代替軽鎖に記載された会合と類似していると思われる。κ様代替軽鎖構造体の設計および生成の更なる詳細については、全体の開示が本明細書に参照により明確に組み込まれる、米国特許公開第2010−0062950号、およびXu et al.,J.Mol.Biol.2010,397,352−360に見出すことができる。
【0104】
本発明は、代替軽鎖(SLC)ポリペプチドと、非相同性シグナル配列を含む代替軽鎖配列を有するSLCポリペプチドを含有するSLC構造体とを企図する。一実施形態において、SLC構造体は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、λ5配列にコンジュゲートされたVpreB配列を含んでいてもよい。別の実施形態において、VpreB配列は、天然型VpreB1配列、天然型VpreB2配列、天然型VpreB3配列ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の一実施形態において、天然型VpreB配列は、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3、配列番号5のヒトVpreB様ポリペプチド、配列番号6のヒトVpreB dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の一実施形態において、λ5配列は、配列番号7のヒトλ5様、配列番号8のヒトλ5ポリペプチド、または配列番号9のヒトλ5 dTailポリペプチドの全てまたは一部を含む。
【0105】
本発明は、λ5配列およびVpreB配列が共有結合リンカーにより結合されている、SLC構造体も企図する。一実施形態において、本発明は、λ5配列がVpreB配列と非共有結合的に会合されている、SLC構造体を提供する。別の一実施形態において、本発明は、前記VpreB配列とλ5配列とのコンジュゲートが抗体重鎖配列と非共有結合的に会合されている、SLC構造体を企図する。
【0106】
本明細書に記載された通り、本発明は、SLCポリペプチドをコードする単離された核酸分子、およびSLCポリペプチドを含むSLC構造体に関する。一実施形態において、本発明は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、λ5配列に融合されたVpreB配列を含む代替軽鎖をコードする核酸を提供する。別の実施形態において、本発明は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、ペプチドまたはペプチドリンカーによりλ5配列に結合されたVpreB配列を含む代替軽鎖をコードする核酸を提供する。別の一実施形態において、本発明は、核酸を含むベクターを提供する。別の実施形態において、本発明は、核酸で形質転換された組換え宿主細胞を提供する。
【0107】
別の態様において、本発明は、代替軽鎖構造体のライブラリーを提供する。別の実施形態において、ライブラリーは、SLCをコードする核酸を含む。別の一実施形態において、ライブラリーは、ディスプレイの形態であってもよい。
【0108】
別の一態様において、本発明は、κ様代替軽鎖ポリペプチドと、κ様SLCポリペプチドを含むSLC構造体とを企図する。一実施形態において、本発明は、前記Vκ配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列にコンジュゲートされたVκ様配列を含むκ様SLC構造体に関する。別の実施形態において、Vκ様配列は、配列番号12〜24、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。他の一実施形態において、JCκ配列は、配列番号26〜39、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。
【0109】
一実施形態において、本発明は、Vκ様配列が前記JCκ配列に融合されている、κ様SLC構造体を企図する。別の実施形態において、融合は、それぞれ前記Vκ様配列および前記JCκ配列のCDR3類似領域で、またはその周辺で起こる。一実施形態において、本発明は、前記Vκ様配列および前記JCκ配列が共有結合リンカーにより結合されている、κ様SLC構造体を企図する。
【0110】
一実施形態において、本発明は、前記Vκ様配列が前記JCκと非共有結合的に会合されている、κ様SLC構造体を提供する。一実施形態において、本発明は、前記Vκ様配列と前記JCκ配列とのコンジュゲートが抗体重鎖配列と非共有結合的に会合されている、κ様SLC構造体を提供する。
【0111】
一実施形態において、本発明は、前記分泌リーダー配列が合成配列であってもよい、κ様SLC構造体を提供する。一実施形態において、本発明は、前記分泌リーダー配列が天然型分泌リーダー配列のコンセンサス配列であってもよい、κ様SLC構造体を提供する。
【0112】
別の態様において、本発明は、κ様SLC構造体をコードする、単離された核酸を提供する。一実施形態において、本発明は、前記Vκ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列に融合されたVκ様配列を含むκ様代替軽鎖をコードする核酸を提供する。別の実施形態において、本発明は、前記Vκ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、ペプチドまたはペプチドリンカーによりJCκ配列に結合されたVκ様配列を含むκ様代替軽鎖をコードする核酸を提供する。別の一実施形態において、本発明は、核酸を含むベクターを提供する。別の実施形態において、本発明は、核酸で形質転換された組換え宿主細胞を提供する。
【0113】
一実施形態において、本発明は、κ様代替軽鎖構造体のライブラリーを提供する。別の実施形態において、ライブラリーは、κ様SLCをコードする核酸を含む。別の一実施形態において、ライブラリーは、ディスプレイの形態であってもよい。
【0114】
別の一態様において、本発明は、κ様SLCの発現方法を提供する。一実施形態において、本発明は、前記Vκ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列に共有結合されたVκ様配列を含むキメラ分子をコードする核酸で前記組換え宿主細胞を形質転換することを含む、組換え宿主細胞におけるκ様代替軽鎖の発現方法を提供する。一実施形態において、Vκ様配列は、JCκ配列に融合されている。別の実施形態において、Vκ様配列は、ペプチドまたはペプチドリンカーを介してJCκ配列に結合されている。別の実施形態において、組換え宿主細胞は、真核細胞である。一実施形態において、組換え宿主細胞は、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞である。
【0115】
別の一実施形態において、本発明は、配列番号6として示されたVpreB配列を含むSLC構造体を提供する。別の実施形態において、本発明は、配列番号10として示されたλ5配列を含むSLC構造体を提供する。一実施形態において、本発明は、配列番号35として示されたポリペプチドを含むSLC構造体を提供する。
【0116】
λ様Surrobodyの具体的な例としては、天然型配列のフラグメントおよびバリアントを含むVpreB配列、例えば、VpreB1、VpreB2、またはVpreB3配列が、天然型配列のフラグメントおよびバリアントを含むλ5配列にコンジュゲートされている、ポリペプチドが挙げられる。このタイプの代表的融合は、全体の開示が本明細書に参照により明確に組み込まれる、2008年10月2日公開のPCT公開第WO2008/118970号に提供されている。非相同性リーダー配列での融合の例は、図3(配列番号35)に示されている。
【0117】
直接の融合において、典型的にはVpreB配列のC末端(例えば、VpreB1、VpreB2、またはVpreB3配列)は、λ5配列のN末端に融合されている。完全長の天然型VpreB配列を完全長λ5配列に融合することは可能であるが(例えば、図7の最初の図解を参照)、典型的には融合は、2つのポリペプチドそれぞれのCDR3類似部位で、またはその周辺で起こる。VpreB1およびλ5の類似CDR3部位に基づく代表的な融合構造体を、図6に示す。この実施形態において、融合は、CDR3類似領域の片側の約10のアミノ酸残基の内部で、またはその内部のある位置で起こってもよい。好ましい実施形態において、融合は、天然型ヒトVpreB1配列(配列番号1)の約116アミノ酸残基から126アミノ酸残基の間、および天然型ヒトλ5配列(配列番号7)の約82アミノ酸残基から93アミノ酸残基の間で起こる。
【0118】
VpreB配列またはλ5配列を、それぞれ抗体λ軽鎖のCDR3領域または抗体軽鎖の可変領域に融合することもできる。VpreBおよびλ5の一方だけがトランケートされている追加の構造体も、図7に示す。類似の構造体を、抗体κ軽鎖配列を用いて調製することができる。κ様代替軽鎖構造体の例示を、図12〜19に見出すことができる。
【0119】
更なる直接融合構造を、図11の右側に示す。「SLC融合1」と称される構造は、トランケート型VpreB1配列(天然型VpreB1配列のC末端にある特徴的「テール」が欠如)の、同様にトランケートされたλ5配列への融合体が、抗体重鎖と非共有結合的に会合されている、2つの二量体で構成された四量体である。「SLC融合2」と称される構造は、トランケート型VpreB1配列(天然型VpreB1配列のC末端にある特徴的「テール」が欠如)の、抗体軽鎖構造体領域への融合体が、抗体重鎖と非共有結合的に会合されている、2つの二量体で構成された四量体である。「SLC融合3」と称される構造は、抗体軽鎖可変領域のトランケート型λ5配列(天然型λ5のN末端にある特徴的「テール」が欠如)への融合体が、抗体重鎖と非共有結合的に会合されている、2つの二量体で構成された四量体である。
【0120】
先に注記された通り、本発明のポリペプチド構造体は、直接融合に加えて、VpreB配列(天然型配列のフラグメントまたはバリアントを含む)と、異種配列、例えばλ5配列(天然型配列のフラグメントまたはバリアントを含む)および/または抗体配列との非共有結合的会合を包含する。したがって、例えば完全長VpreB配列が、トランケート型λ5配列と非共有結合的に会合されていてもよい。あるいはトランケート型VpreB配列が、完全長λ5配列と非共有結合的に会合されていてもよい。
【0121】
抗体重鎖との非共有結合的会合において、非共有結合的に会合されたVpreB1およびλ5配列を含む代替軽鎖構造体を、図11の左側に示す。様々な例示を示したが、構造が、例えば完全長VpreB1およびλ5配列、トランケート型λ5配列と会合された完全長VpreB1配列(「λ5 dT」)、完全長λ5配列と会合されたトランケート型VpreB1配列(「VpreB dT」)およびトランケート型λ5配列と会合されたトランケート型VpreB1配列(「短」)を包含してもよい。
【0122】
図11は、特定の具体的構造体を示しているが、様々な他の構造体を類似の様式で製造および使用しうることは、当業者には理解されよう。例えば構造は、図11に示された構造とは対照的に、非対称で、各アームに異なる代替軽鎖配列を含んでいても、そして/または三量体もしくは五量体構造を有していてもよい。
【0123】
本明細書の代替軽鎖構造体(Surrobody)の全てが、抗体配列と会合されていてもよい。例えば図9に示す通り、VpreB−λ5融合体は、ペプチドリンカーにより、抗体重鎖の可変領域配列に連結することができる。別の実施形態において、VpreB−λ5融合体は、抗体重鎖、または可変領域配列を含むそのフラグメントと非共有結合的に会合されて、二量体複合体を形成している。更に別の実施形態において、VpreBおよびλ5配列は、互いに、そして抗体重鎖または可変領域配列を含むそのフラグメントに、非共有結合的に会合されており、それにより三量体複合体を形成している。抗体重鎖を含む実例の構造体を、図11に示す。
【0124】
κ様Surrobodyの具体的例としては、天然型配列のフラグメントまたはバリアントを含むVκ様配列が、天然型配列のフラグメントまたはバリアントを含むJCκ配列にコンジュゲートされているポリペプチドが挙げられる。この型の代表的な融合体は、全体の開示が本明細書に参照により明確に組み込まれる、米国特許公開第2001−0062950号、およびXu et al.,J.Mol.Biol.2010,397,352−360に示されている。
【0125】
本明細書のポリペプチド構造体の具体的例としては、Vκ様および/またはJCκ配列が抗体重鎖、またはそのフラグメントに会合されているポリペプチドが挙げられる。Vκ様配列およびJCκ配列の両方を含む具体的なヘテロ二量体構造体を、図12に示す。図12に示す通り、本発明のκ様代替軽鎖構造体では、Vκ様ポリペプチドおよび/またはJCκポリペプチドは、類似の抗体配列内に存在しない、それぞれC−およびN−末端伸長を含有していてもよい。あるいは伸長の一部または全体が、本明細書のκ様代替軽鎖構造体から除去されていてもよい。
【0126】
個別に用いられうる、または追加の異種配列、例えば抗体重鎖配列、例えば、完全長抗体重鎖またはそのフラグメントにより、更に誘導体化および/または会合されうる他のκ様代替軽鎖構造体。
【0127】
Vκ様ポリペプチドおよび/またはJCκポリペプチドのC−およびN−末端伸長は、本発明の構造体に存在する必要はないが、図14に示す通り、ループライブラリーをスクリーニングした結果、それらは直線伸長、または例えば多様性に制約のある形態のいずれかにより、組み合わせ的機能多様性を創り出すための特有の好機を提供するため、少なくとも1種のそのような付加物の少なくとも一部を保持することが有益となる。加えて、Vκ様ポリペプチドおよび/またはJCκポリペプチドの「テール」部分が、他のペプチドおよび/またはポリペプチドに融合されて、様々な所望の特性、例えば、結合の強化、追加の結合特異性、pKの上昇、半減期の改善、半減期の短縮、細胞表面への固定、細胞内トランスロケーションの増加、ドミナントネガティブ活性などを提供することができる。具体的な機能的テール伸長を、図18に列挙する。
【0128】
所望なら、本発明の構造体を、例えば、公知の治療抗体をはじめとする抗体のCDR1、CDR2および/またはCDR3領域の公知配列または配列モチーフを、κ様代替軽鎖配列のCDR1、CDR2および/またはCDR3類似領域に組込む、または付加することにより、設計することができる。これにより、抗体ではないが、公知の治療抗体と同様の、またはそれを超えた結合特異性または親和力を示す分子を作製することができる。
【0129】
Vκ様およびJCκ遺伝子は、独立したタンパク質として、そして代替軽鎖として機能することができるポリペプチドをコードしているため、代替様の軽鎖を、真の軽鎖から設計することができ、そして設計された真の代替軽鎖のために提案されたこれまでの適用例に用いることができる。これは、VpreBまたはVκ様遺伝子のいずれかに類似したペプチド伸長を含む可変軽領域を発現することにより、果たすことができる。同様に、定常領域を、λ5またはJCκ遺伝子のいずれかおよびそれらのペプチド伸長と似せるように設計することができる。更に任意のキメラまたはヘテロ二量体の組み合わせは、本明細書の範囲内である。
【0130】
幾つかの実施形態において、SLC構造体は、異種アミノ酸配列または非SLCポリペプチドを含む。特定の実施形態において、異種アミノ酸配列は、1種以上の追加の機能性を、本発明の構造体に付加する。SLC構造体は、非SLCポリペプチドを含むように設計されてもよい。一実施形態において、非SLCポリペプチドは、第一のSLC成分および/または第二のSLC成分に融合されている。非SLCポリペプチドのアミノ末端が、第一のSLC成分のカルボキシ末端に融合されていてもよく、そして/または非SLCポリペプチドのカルボキシ末端が、第二のSLC成分のアミノ末端に融合されていてもよい。別の実施形態において、第一のSLC成分は、VpreBポリペプチドまたはVκ様ポリペプチドである。別の一実施形態において、第二のSLC成分は、λ5ポリペプチドまたはJCκポリペプチドである。
【0131】
所望の結合特異性を有する抗体可変領域配列を含む、追加の機能性を有するλ様SLC構造体を、図10に示す。詳細には図10は、VpreBおよびλ5に連結する融合タンパク質を作製するための抗VEGF一本鎖Fv(scFv)の挿入を示している(図10A)。こうして得られた、設計されたSLC制約scFvは、抗TNF−α抗体の重鎖と対をなしている。図10Bは、VpreBのC末端への抗VEGF scFvの融合を表わしている。図10Cは、λ5のアミノ末端への抗オボアルブミンscFvの融合を表わしている。TNF−αおよびオボアルブミンの両方に結合する能力を有する3部のタンパク質複合体を形成することができる。図10Dは、三特異性分子を作製するための、2つの融合構造体(VpreB−抗VEGF scFvおよびλ5−抗オボアルブミン)と抗TNF−α抗体の重鎖との組み合わせを表わしている。VpreBおよびλ5ポリペプチド配列を含む様々な二機能性および三機能性構造体を、そのような方策を用いて構築してもよい。加えて図8に表わされる通り、組み合わせ的機能多様性を、λ様SLC構造体に組込んでもよい。
【0132】
所望の結合特異性を有する抗体可変領域配列を含む追加の機能性を有するκ様SLC構造体を、図17に示す。詳細には図17は、本明細書の先に記載された、Vκ様およびJCκポリペプチド配列を含む様々な二機能性および三機能性構造体を示している。
【0133】
本発明の代替軽鎖(SLC)構造体は、二量体または2部構成で提供されてもよい。この構成の例を図9に提供し、それは、2部構成に対応するVpreB−λ5融合体および抗体重鎖(右側、下から二番目の図)のタンパク質構造を示している。SLC構造体は、三量体または3部構成で提供されてもよい。図9は、3部構成に対応するVpreB、λ5、および抗体重鎖(右側、一番下の図)のタンパク質構造を示している。
【0134】
本発明の代替軽鎖(SLC)構造体は、二機能または二特異性形態で提供されてもよい。図20は、この例を示し、(A)は、Surrobody構成を表しており、(B)は、二機能性および二特異性Surrobody構成を表わしている。図20(B)に示される通り、SLC構造体は、SLCポリペプチド成分(例えば、VpreB、λ5、Vκ様、JCκポリペプチド、またはそれらのフラグメントもしくはバリアント)、および非SLC分子を有するSLC融合ポリペプチドを含んでいてもよい。一実施形態において、非SLC分子は、特定の機能を有する任意のポリペプチドであってもよい。別の実施形態において、ポリペプチドは、追加の機能性を提供しうるサイトカインであってもよい。別の実施形態において、非SLCポリペプチドは、追加の特異性を提供しうる抗体フラグメントであってもよい。図20(C)は、実例のSLC融合クローニング方策およびその各アミノ酸を表わしている。斜線のエリアは、VpreB(アミノ酸120〜145)およびλ5(アミノ酸38〜92)の非免疫グロブリンテール領域を表す。「L」は、内在性リーダー配列を示し、「mL」は、合成Igκリーダー配列を示す。「融合」は、非SLC分子など、該当する遺伝子の融合部位を示す。
【0135】
代替軽鎖構造体の調製
核酸コード代替軽鎖、例えば、VpreBおよびλ5ポリペプチド、またはVκ様もしくはJCκポリペプチドを、天然供給源、例えば、発達中のB細胞から単離することができ、そして/または合成もしくは半合成法により得ることができる。このDNAが同定され単離されるか、さもなければ生成されれば、更なるクローニングまたは発現のために複製可能なベクターにライゲートすることができる。
【0136】
本明細書のポリペプチドのコード配列を発現するために用いうるクローニングおよび発現ベクターは、当該技術分野で周知であり、市販されている。一般にベクター成分は、非限定的に、以下のものの1つ以上が挙げられる:シグナル配列、複製起点、1つ以上のマーカー遺伝子、エンハンサー要素、プロモータ、および転写終結配列。本明細書のベクター内の代替軽鎖構造体をコードするDNAをクローニングまたは発現するのに適した宿主細胞は、原核生物、酵母、またはより高等な真核生物(ホ乳類)細胞であり、ホ乳類細胞が好ましい。
【0137】
適切なホ乳類宿主細胞系の例としては、非限定的に、SV40により形質転換されたサル腎臓CV1系(COS−7、ATCC CRL 1651);懸濁培養での増殖用にサブクローニングされたヒト胚性腎(HEK)細胞系293(HEK293細胞)、(Graham et al.J.Gen Virol.36:59(1977));ベビーハムスター腎臓細胞(BHK、ATCC CCL 10);チャイニーズハムスター卵巣細胞/−DHFR(CHO、Urlaub et al.,Proc.Natl.Acad.Sci.USA 77:4216(1980));マウスセルトリ細胞(TM4、Mather,Biol.Reprod.23:243−251(1980));サル腎臓細胞(CV1 ATCC CCL 70);アフリカミドリザル腎臓細胞(VERO−76、ATCC CRL−1587);ヒト子宮頸癌細胞(HELA、ATCC CCL 2);イヌ腎臓細胞(MDCK、ATCC CCL 34);バッファロー系ラット肝臓細胞(BRL 3A、ATCC CRL 1442);ヒト肺細胞(W138、ATCC CCL 75);ヒト肝臓細胞(Hep G2、HB 8065);マウス乳腺腫(MMT 060562、ATCC CCL 51);TRI細胞(Mather et al.,Annals N.Y.Acad.Sci.383:44−68(1982));MRC5細胞;FS4細胞;およびヒトヘパトーマ系(Hep G2)が挙げられる。
【0138】
ホ乳類細胞内での使用に関しては、発現ベクターでの制御機能が、多くの場合、ウイルス材料により提供される。したがって、一般に用いられるプロモータは、ポリオーマ、アデノウイルス2、レトロウイルス、サイトメガロウイルス、およびシミアンウイルス40(SV40)のゲノムから得ることができる。他のプロモータ、例えば、β−アクチンプロモータは、非相同性供給源から生成させる。適切なプロモータの例としては、非限定的に、SV40ウイルスの初期または後期プロモータ(Fiers et al.,Nature,273:113(1978))、ヒトサイトメガロウイルスの最初期プロモータ(Greenaway et al.,Gene,18;355−360(1982))、ならびに通常は所望の遺伝子配列に関連するプロモータおよび/または制御配列が挙げられるが、但し、そのような制御配列が、宿主細胞系と適合することが前提である。
【0139】
高等な真核生物による、所望の非相同性ポリペプチドをコードするDNAの転写は、エンハンサー配列をベクターに挿入することにより増加する。エンハンサーは、プロモータ上で作用してその転写開始活性を高める、通常は10〜300bpのDNAのシス作用性要素である。エンハンサーは、相対的に配列および位置非依存性であるが、好ましくは発現ベクター内に存在するプロモータ配列の上流に位置する。エンハンサーは、プロモータと同じ供給源、例えば真核細胞ウイルスから、例えば、複製起点の後期側のSV40エンハンサー(bp100〜270)、サイトメガロウイルス初期プロモータエンハンサー、複製起点の後期側のポリオーマエンハンサー、およびアデノウイルスエンハンサーから生成させてもよい。
【0140】
ホ乳類宿主細胞内で用いられる発現ベクターは、ポリアデニル化部位、例えば、SV40(初期および後期)またはHBVなどのウイルス由来のものも含んでいる。
【0141】
複製起点は、外来起源、例えばSV40または他のウイルス(例えば、ポリオーマ、アデノ、VSV、BPV)供給源から得ることができる、または宿主細胞により提供することができる、ベクターの構築により提供されてよい。
【0142】
発現ベクターは、通常、ベクターで形質転換された宿主細胞の生存または発育に必要なタンパク質をコードする選択マーカーを含む。ホ乳類細胞に適した選択マーカーの例としては、ジヒドロ葉酸還元酵素(DHFR)、チミジンキナーゼ(TK)、およびネオマイシンが挙げられる。
【0143】
適切なホ乳類発現ベクターは、当該技術分野で周知であり、市販されている。したがって例えば、本発明の代替軽鎖構造体は、ヒトサイトメガロウイルス(CMV)最初期エンハンサー/プロモータ領域を含むpCI発現ベクター(プロメガ)を用いてホ乳類宿主細胞内で産生させて、DNA挿入部の構成的発現を促進することができる。ベクターは、pTT5発現ベクター(カナダ国立研究機構)であってもよい。ベクターは、ネオマイシンホスホトランスフェラーゼ遺伝子を選択マーカーとして含むことができる。
【0144】
本発明の代替軽鎖構造体は、細菌宿主細胞内で産生することもできる。細菌系内で使用される制御要素としては、場合によりオペレータ−配列を含むプロモータ、およびリボソーム結合部位が挙げられる。適切なプロモータとしては、非限定的に、ガラクトース(gal)、ラクトース(lac)、マルトース、トリプトファン(trp)、β−ラクタマーゼプロモータ、バクテリオファージλ、およびT7プロモータが挙げられる。加えて合成プロモータ、例えばtacプロモータを用いることができる。細菌系内で使用されるプロモータは、一般に、Fab分子をコードするDNAに作動可能に連結されたシャイン・ダルガルノ(SD)配列も含有する。プラスミドpBR322からの複製起点は、ほとんどのグラム陰性菌に適している。
【0145】
抗体代替軽鎖配列を含む多重鎖構造体内の各鎖のコード配列は、真核生物および原核生物の宿主をはじめとする所望の宿主細胞をコトランスフェクトするのに用いられる、別個の調節配列の制御下では、または別個の発現ベクター内では、同じ発現ベクター内に存在することができる。したがって、ノバゲン(Novagen)から市販されるDuet(商標)ベクターを用いて、同義遺伝子を共発現させることができる。
【0146】
形質転換された宿主細胞は、様々な培地で培養させてもよい。ホ乳類宿主細胞を培養するための市販の培地としては、Ham’s F10(シグマ)、最小必須培地(MEM)(シグマ)、RPMI−1640(シグマ)、およびダルベッコ改変イーグル培地((DMEM),(シグマ)が挙げられる。加えてHam et al.,Meth.Enz.58:44(1979)およびBarnes et al.,Anal.Biochem.102:255(1980)に記載された培地のいずれも、宿主細胞用の培地として用いることができる。培養条件、例えば温度、pHなどは、発現用に選択される宿主細胞によって過去に利用されたものであり、製造業者の説明書に含まれるか、さもなければ当業者には自明であろう。
【0147】
ホ乳類、細菌(例えば大腸菌)、または他の宿主細胞を培養するのに更に適した培地は、標準の教書、例えば、Sambrook et al.の上記文献またはAusubel et al.の上記文献にも記載されている。
【0148】
一態様において、本発明は、組換え宿主細胞内での代替軽鎖の発現方法を提供する。一実施形態において、その方法は、SLCポリペプチドまたはSLC融合ポリペプチドをコードする核酸を提供するステップを含む。別の実施形態において、その方法は、組換え宿主細胞を、SLCポリペプチドまたはSLC融合ポリペプチドをコードする核酸で形質転換またはトランスフェクトするステップを含む。一実施形態において、SLC融合ポリペプチドをコードする核酸は、第一のSLC配列および/または第二のSLC配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、第二のSLC配列に共有結合された第一のSLC配列を含むキメラ分子である。第一のSLC配列は、VpreB配列、Vκ様配列、またはそれらの融合ポリペプチドであってもよい。第二のSLC配列は、λ5配列、JCκ配列、またはそれらの融合ポリペプチドであってもよい。
【0149】
一実施形態において、VpreB配列は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、λ5配列に共有結合されている。別の実施形態において、VpreB配列は、λ5配列に融合されている。別の一実施形態において、VpreB配列は、ペプチドまたはペプチドリンカーを介して、λ5配列に結合されている。別の一実施形態において、Vκ様配列は、前記κ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列に共有結合されている。別の医師実施形態において、Vκ様配列は、JCκ配列に融合されている。別の実施形態において、Vκ様配列は、ペプチドまたはペプチドリンカーを介して、JCκ配列に結合されている。
【0150】
全ての実施形態において、発現方法は、代替軽鎖ポリペプチドおよび/または代替軽鎖融合ポリペプチドをはじめとする、代替軽鎖ポリペプチドをコードする1つ以上の核酸で宿主細胞を形質転換またはトランスフェクトするステップを含んでいてもよい。
【0151】
全ての実施形態において、その方法は、抗体重鎖をコードする核酸で宿主細胞を形質転換またはトランスフェクトするステップを更に含んでいてもよい。
【0152】
一態様において、本発明は、改善された収率を有する代替軽鎖ポリペプチドおよび/または代替軽鎖融合ポリペプチドの発現方法を提供する。一実施形態において、天然型リーダー配列の代わりに非相同性リーダー配列を用いる本発明の方法は、天然型リーダー配列を非相同性リーダー配列で交換されていない方法よりも、大きなポリペプチド発現および収率を特徴とする。
【0153】
一実施形態において、組換え宿主細胞は、細菌細胞である。別の実施形態において、宿主細胞は、真核細胞である。一実施形態において、組換え宿主細胞は、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞である。
【0154】
一態様において、本発明は、本明細書に記載された核酸を含有する宿主細胞を提供する。一実施形態において、本発明は、本明細書に記載された少なくとも1種の核酸で形質転換された組換え宿主細胞を提供する。別の一実施形態において、宿主細胞は、非SLC分子を含んでいても、または含んでいなくてもよい、SLC融合をコードする核酸で形質転換されている。
【0155】
全ての実施形態において、宿主細胞は、抗体重鎖をコードする核酸で更に形質転換されている。
【0156】
全ての実施形態において、本発明は、本明細書に記載された核酸を含有するベクターを提供する。全ての実施形態において、宿主細胞は、本明細書に記載された核酸を含有する少なくとも1種のベクターで形質転換されている。
【0157】
精製は、当該技術分野で公知の方法により実施することができる。好ましい実施形態において、代替軽鎖構造体は、Ni−NTA精製システム(インビトロジェン)を用いて6×Hisタグ形態で精製される。
【0158】
κ様SLC分子は、既存の軽鎖V遺伝子および軽鎖定常遺伝子から設計することができる。図15に示す通り、軽鎖は、遺伝子再配列およびRNAプロセッシングの生成物である。κ様SLC分子の成分は、再配列されていない軽鎖V遺伝子および再配列された軽鎖JC遺伝子からの別の機能を提供するため、残りのκおよびλ軽鎖V遺伝子全てから類似の翻訳タンパク質を設計して、Vκ様分子(図16A)、ならびに残りのκJC再配列(4JCκ様)(図16B)と、λJC再配列(4“J”×10“定数”=40JCλ様)(図16B)との全ての組み合わせを作製することが実行可能である。これらの設計分子のそれぞれ1つは、VpreBおよびλ5、ならびにそれらの組み合わせおよびキメラを用いて、Vκ様およびJCκを用いたもの、および2008年10月2日公開のPCT公開WO2008/118970に含まれるものと類似の目的を果たすことができる。
【0159】
本発明の代替軽鎖を用いて、疾患の予防および/または処置のための分子を構築することができる。そのような適用では、代替軽鎖を含有する分子は、通常、医薬組成物の形態で用いられる。技術および配合は、一般に、Remington’s Pharmaceutical Sciences,18th Edition,Mack Publishing Co.(Easton,PA.1990)に見出してもよい。Wang and Hanson「Parenteral Formulations of Proteins and Peptides:Stability and Stabilizers,」Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)も参照されたい。
【0160】
ポリペプチドを基剤とする医薬組成物は、典型的には、凍結乾燥された配合剤または水性配合剤の形態で配合される。許容されうる担体、賦形剤、または安定化剤は、用いられる投与量および濃度ではレシピエントに対して非毒性であり、緩衝剤、例えば、リン酸塩、クエン酸塩、および他の有機酸;アスコルビン酸およびメチオニンをはじめとする抗酸化剤;防腐剤(例えば、塩化オクタデシルジメチルベンジルアンモニウム;塩化ヘキサメトニウム;塩化ベンザルコニウム;塩化ベンゼトニウム;フェノール、ブチルもしくはベンジルアルコール;アルキルパラベン、例えば、メチルもしくはプロピルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;およびクレゾール);低分子量(約10残基未満)のポリペプチド;タンパク質、例えば、血清アルブミン、ゼラチン、もしくは免疫グロブリン;親水性ポリマー、例えば、ポリビニルピロリドン;アミノ酸、例えば、グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、もしくはリシン;単糖類、二糖類、およびグルコース、マンノース、もしくはデキストリンをはじめとする他の炭水化物;キレート化剤、例えば、EDTA;糖類、例えば、ショ糖、マンニトール、トレハロースもしくはソルビトール;塩形成対イオン、例えば、ナトリウム;金属錯体(例えば、Zn−タンパク質錯体);ならびに/または非イオン性界面活性剤、例えば、TWEEN(商標)、PLURONICS(商標)もしくはポリエチレングリコール(PEG)が挙げられる。
【0161】
その分子は、例えば、コロイド状薬物送達系(例えば、リポソーム、アルブミンマイクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)、またはマクロエマルジョン中で、コアセルベーション技術、または界面重合(例えば、ヒドロキシメチルセリロースまたはゼラチン−マイクロカプセルおよびポリ(メチルメタクリラート)マイクロカプセル)により、マイクロカプセルに封入してもよい。そのような技術は、上記のRemington’s Pharmaceutical Sciencesに開示されている。
【0162】
本明細書に開示された代替軽鎖を含有する分子は、免疫リポソームとして配合されてもよい。分子を含有するリポソームは、Epstein et al.,Proc.Natl.Acad.Sci.USA 82:3688;Hwang et al,Proc.Natl.Acad.Sci.USA 77:4030(1980);米国特許第4,485,045号、同第4,544,545号および1997年10月23日公開のWO97/38731に記載されたものなど、当該技術分野で公知の方法により調製される。循環時間を高めたリポソームは、米国特許第5,013,556号に開示されている。
【0163】
特に有用なリポソームは、ホスファチジルコリン、コレステロールおよびPEG誘導体化ホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を用いる逆相エバポレーション法により生成させることができる。リポソームは、画定された孔径のフィルターから押出されて、所望の口径を有するリポソームが生成される。本発明の分子のフラグメントは、ジスルフィド交換反応を介してリポソームにコンジュゲートすることができる(Martin et al.J.Biol.Chem.257:286−288(1982))。化学療法剤が、場合によりリポソーム中に含有されている。Gabizon et al.J.National Cancer Inst.81(19)1484(1989)を参照されたい。
【0164】
疾患の予防または処置では、分子の適切な投与量は、処置される感染のタイプ、疾患の重症度および経過、ならびに抗体が予防目的または治療目的で投与されるか否かに依存する。分子は、一度に、または一連の処置を通して、適切に患者に投与される。疾患のタイプおよび重症度にもよるが、例えば、1回以上の別々の投与または連続注入のいずれにしろ、抗体約1μg/kg〜約15mg/kgが、患者に投与される典型的な初期候補投与量である。
【0165】
本発明の代替軽鎖を含有する分子は、疾患の処置または予防における使用に適している。一実施形態において、本発明は、医薬として使用される、または疾患の処置に使用される代替軽鎖含有分子を提供する。別の実施形態において、本発明は、疾患を処置する医薬の製造のための、代替軽鎖含有分子の使用を提供する。その分子は、SLCペプチドまたはSLC融合体をコードする核酸であってもよい。
【0166】
一態様において、本発明は、治療上効果的な量の代替軽鎖含有分子をホ乳類に投与するステップを含む、ホ乳類における疾患を処置するのに有用な方法を提供する。治療組成物を、医師の指示通り、短期(急性)もしくは慢性で、または間欠的に投与することができる。
【0167】
本発明は、疾患の処置、予防および/または診断に有用な材料を含有する製造品のキットおよび物品も提供する。キットは、容器と、容器の表面に設置しうる、または容器と結合しうるラベルとを含んでいる。容器は、瓶、バイアル、注射器、または任意の他の適切な容器であってもよく、様々な材料、例えばガラスまたはプラスチックから形成させてもよい。容器は、本明細書に記載された代替軽鎖含有分子を有する組成物を保持しており、滅菌されたアクセスポートを有していてもよい。容器の例としては、静注液バッグ、または皮下注射針により穿孔することができるストッパー付きバイアルが挙げられる。キットは、様々な試薬、例えば希釈剤および緩衝剤を保持する追加の容器を有していてもよい。ラベルは、組成物および目的の使用のための説明書の記載を提供してもよい。分子を含有するキットは、細胞からのポリペプチドの精製または免疫沈降法のための、例えば細胞アッセイ用に、使用が見出される。例えば、タンパク質の単離および精製では、キットは、ビーズ(セファロースビーズ)に結び付いたタンパク質に結合する代替軽鎖含有分子を含有することができる。インビトロで、例えばELISAまたはウェスタンブロットにおいて、タンパク質の検出および定量用の分子を含有するキットを提供することができる。検出に有用なそのような分子は、標識、例えば蛍光または放射線標識と共に提供されてもよい。
【0168】
キットは、本明細書に記載された代替軽鎖を活性剤として含む分子を含む少なくとも1つの容器を有する。ラベルを提供して、組成物が疾患の処置に使用しうることを示してもよい。ラベルは、処置を必要とする対象へ投与するための説明書を提供してもよい。キットは、薬学的に許容しうる緩衝剤、例えば、注射用静菌水(BWFI)、リン酸緩衝生理食塩水、リンガー溶液およびデキストロース溶液を有する追加の容器を更に含んでいてもよい。最後に、キットは、別の緩衝剤、希釈剤、フィルター、針、および注射器をはじめとする任意の別の適切な材料を含有していてもよい。
【0169】
本発明の更なる詳細を、以下の非限定的実施例において提供する。
【0170】
実施例1−HEK293細胞における一過性発現
Surrobodyの上清を、ヒト胚性腎293(HEK293)細胞において一過性に生成させた。pTT5プラスミド(カナダ国立研究機構)を用いて、組換えSurrobody軽鎖構造体を提供した。非相同性リーダー配列METDTLLLWVLLLWVPGSTG(配列番号36−マウスIgκリーダー配列)での天然型リーダー配列の置換を行った、または行わなかったプラスミドの代替軽鎖核酸配列を提供した。以下の核酸配列を含有するpTT5プラスミドを用いた:(a)天然型リーダー配列(配列番号1−図1)もしくは配列番号36の非相同性マウスIgκリーダー配列を含むヒトVpreB1、(b)天然型リーダー配列(配列番号8−図2)もしくは配列番号36の非相同性マウスIgκリーダー配列を含むヒトλ5、または(c)VpreB1およびλ5と、天然型VpreB1リーダー配列もしくは配列番号36の非相同性マウスIgκリーダー配列との融合(配列番号35−図3)。(a)および(b)のプラスミドは、3部Surrobody構成に対応し、(c)のプラスミドは、3部Surrobody構成に対応する。これらのプラスミドを、抗体重鎖を含有するpTT5プラスミドでコトランスフェクトした。
【0171】
Surrobodyを、本質的には過去にXu et al.,(2008).Proc.Natl.Acad.Sci.USA,105:10756−10761;Kashyap et al.,(2008).Proc.Natl.Acad.Sci.USA,105:5986−5991に記載された通り、HEK293のFreestyleに基づく系(インビトロジェン)で一過性に生成させた。HEK293細胞を0.25〜2.0×106細胞/mlの密度で増殖培地中で増殖させ、その後、トランスフェクションの1日前に、増殖培地90mlを含む新しい振とうフラスコに0.75×106細胞/mlの密度で接種した。一晩発育させた後、細胞密度を1〜1.5×106細胞/mlであることを検証した。発現では、pTT5発現ベクターを用いた。次に、DNAトランスフェクション剤混合物を、以下の通り調製した。プラスミドDNA(pTT5−SLC分子)合計0.1mgに対応するDNA溶液を10ml遠沈管内で混合して、増殖培地で最終容量を5mLにした。2部Surrobody構成では、抗体重鎖を含有するプラスミド0.05mgを、VpreB1−λ5キメラプラスミド(Surrobody融合体)0.05mgと混合した。3部Surrobody構成では、抗体重鎖を含有するプラスミド0.033mgを、VpreB1プラスミド0.033mgおよびλ5プラスミド0.033mgと混合した。次に、増殖培地4.8mlをPEI 0.2mlと混和してプラスミドDNA溶液に添加することにより、ポリエチレンイミン(PEI)トランスフェクション溶液を調製した。混合物を力強く1〜2秒間ボルテックス処理した。室温で15分間インキュベートした後、プラスミドDNA−PEI混合物を、10mlピペットで1〜1.5×106細胞/mlの密度のHEK293細胞を含むフラスコに移した。フラスコを直ちに回転させて、振とうインキュベータに移動させた。125rpmのシェーカープラットホームの付いた、37℃、5%CO2の加湿インキュベータで、細胞を6日間発育させた。得られた培養上清におけるタンパク質生成レベルを、定量的反応速度分析(フォルテバイオのOctet:抗Fcセンサー)により決定した。下の表に示す通り、マウスIgκ軽鎖リーダー配列を置換したことで、一過性組換えSurrobody発現レベルが改善している。表1(mg/L)に示す通り、タンパク質レベルが少なくとも20倍改善された。
【0172】
表1

【0173】
3部および2部Surrobodyの複数回のトランスフェクションから得られた精製タンパク質を更に分析すれば、高レベルの収量が裏づけられる。非相同性リーダーを用いた、独立したトランスフェクションのタンパク質収量を、4ヶ月の期間にわたりモニタリングした。プロテインAまたはプロテインGのいずれかのクロマトグラフィー担体および低pHでの溶出を用いた高速タンパク質液体クロマトグラフィー(FPLC)システムで、タンパク質を精製した。表2(mg/L)に示される通り、どちらの構成においても、平均収量は、内在性代替軽鎖リーダー配列を用いて認められたものよりも実質的に高くなった。
【0174】
表2
【0175】
以下の表3は、表2で平均をとった、様々な2部および3部SLC形態で測定された各濃度を提供している。先に記載された通り、一般に2部構成は、Surrobody軽鎖融合体および1つの抗体重鎖を含み、3部構成は、2つのSurrobodyポリペプチドおよび抗体重鎖を含む。1列目の1〜47行目は、2部構成を有する47の異なる構造体に対応し、1列目の49〜63行目は、3部構成を有する15の異なる構造体に対応する。4列目は、検査されたSurrobodyの特徴の幾つかを提供している。「Surrobody」は、2つのプロペプチドおよび1つの重鎖で作製されたSurrobody構造体である。「融合体」は、2つのSLCポリペプチドおよび1つの重鎖の融合体で作製されたSurrobody構造体である。「ペプチドタグを含む融合体」は、エピトープタグが組込まれたSurrobody構造体である。「機能性ペプチド融合体」は、特定の機能を有する少なくとも1つの非SLCポリペプチドが組込まれているSurrobody構造体である。
【0176】
表3
【0177】
表3は、収率の改善が、SLCポリペプチドと、SLC融合ポリペプチドと、非SLC分子を含有するSLC融合ポリペプチドと、を含むSurrobodyをはじめとする複数のSurrobody構成で得ることができる証拠を提供している。
【0178】
Surrobody分子を、チャイニーズハムスター卵巣K1(CHO−K1)細胞中でも一過性に発現させることができる。Surrobodyの上清を、(CHO−K1)細胞中で一過性に産生してもよい。組換えSLCポリペプチドまたは非SLCポリペプチドのプラスミドを、製造業者の説明書に従って、10%ウシ胎仔血清を補充されたダルベッコ改変イーグル培地/F12培地中のLipofectamine−2000(インビトロジェン、カタログ番号11668−027)を用いて、抗体重鎖を含有するプラスミドとコトランスフェクトした。37℃、5%CO2で一晩インキュベートした後、培地をGlutamax−1(インビトロジェン、カタログ番号12362)を含む新しいOpti−MEMI血清使用量低減培地と交換した。トランスフェクトされた上清を72時間後に回収して、0.22μmフィルター装置でろ過した。
【0179】
実施例2−CHO細胞における安定発現
真核生物コドンを最適化させた可溶性分泌遺伝子(DNA2.0)として、ホ乳類で発現された、またはデノボ合成で生成された代替軽鎖構造体を、ホ乳類タンパク質発現のために、pCIプラスミド(プロメガ)にサブクローニングした。製造業者の指導書に従ってチャイニーズハムスター卵巣(CHO−K1)細胞(インビトロジェン)にトランスフェクトする前に、配列を検証した。製造業者の指導書に従って、DNAおよびLipofectamine 2000(インビトロジェン)を合計32μgとし、所望の代替軽鎖を同量用いて、T−75フラスコ中の80%コンフルエントの細胞のトランスフェクションを実施した。トランスフェクションごとにOpti−MEMI 20ml中に、細胞にタンパク質を産生させた。4日後に、ニッケルキレートクロマトグラフィー(Ni−NTAアガロース、キアゲン)を用いて、分泌されたSurrobodyを培養上清から精製した。得られた精製Surrobodyを、遠心式サイズろ過(Centricon Plus−20)を用いて滅菌PBSに緩衝液交換して、A280読み取り、SDSゲル、またはウェスタンブロット分析により、公知の標準物質と比較して、それらのタンパク質濃度を決定した。
【0180】
前述の記載では本発明は、特定の実施形態を参照して示したが、それはそのように限定されるものではない。事実、図示されたものおよび本明細書に記載されたものに加えて、本発明の様々な改変が、前述の説明から当業者に自明となり、それは添付の特許請求の範囲に含まれる。
【0181】
本明細書に引用された個々の発行物、特許および特許出願が、具体的かつ個別に参考として本明細書に組込まれていることを示されたのと同程度に、全ての発行物、特許および特許出願は、全ての目的でその全体が本明細書に参照により組み込まれる。
【技術分野】
【0001】
本発明は、非相同性シグナル配列を有する代替軽鎖配列を含む代替軽鎖(SURROBODY)(商標)構造体に関する。
【背景技術】
【0002】
Bリンパ球により産生される抗体(Ig)分子は、重(H)鎖および軽(L)鎖で構築されている。HおよびL鎖のアミノ末端ドメインのアミノ酸配列は、特に抗原結合部位を形成する3つの高度可変領域(CDR1、CDR2、CDR3)に可変性(VHおよびVL)がある。HおよびL鎖のアッセンブリは、L鎖の定常領域(CL)と、重鎖の第一の定常領域(CH1)の間のジスルフィド結合により、そしてVHドメインとVLドメインの間の非共有結合的相互作用により安定化されている。
【0003】
ヒトおよび多くの動物、例えばマウスにおいて、抗体のHおよびL鎖をコードする遺伝子は、V領域の遺伝子フラグメントコード化部分の段階的な体細胞再配列により組立てられる。Bリンパ球発達の様々な段階が、Ig遺伝子lociの再配列状態により特徴づけられる(例えば、Melchers,F.&Rolink,A.,B−Lymphocyte Development and Biology,Paul,W.E.ed.,1999,Lippincott,Philadephia参照)。
【0004】
B細胞の前駆体(プレB細胞)は、完全に発達した軽鎖の代わりにVpreB(1〜3)およびλ5と呼ばれる遺伝子組が生成されること、ならびにμ重鎖が同時発現されることにより、骨髄において同定された。
【0005】
ヒトVpreB1(CAG30495)の主なアイソフォームは、145aa長ポリペプチド(配列番号1)である。それは、IgVドメイン様構造を有するが、典型的なVドメインの最後のβ鎖(β7)が欠如しており、任意の別のタンパク質に対して配列相同性を示さないカルボキシル末端を有する。VpreB2は、142アミノ酸のマウスVpreB2ポリペプチド(P13373;配列番号2)およびマウスVpreB2配列の171アミノ酸長スプライスバリアント(CAA019641;配列番号3)をはじめとする複数のアイソフォームを有する。VpreB1およびVpreB2配列は、EP0269127および米国特許第5,182,205号;Collins et al.,Genome Biol.5(10):R84(2004);およびHollins et al.,Proc.Natl.Acad.Sci.USA 86(14):5552−5556(1989)に開示されている。ヒトVpreB3(配列番号4)の主なアイソフォームは、Collins et al.,Genome Biol.5(10):R84(2004)に開示された123aa長タンパク質(CAG30496)である。
【0006】
VpreB(1〜3)は、もう一方タンパク質λ5とは非共有結合的に会合されている。ヒトλ5は、209アミノ酸ポリペプチド(CAA01962:配列番号5)であり、抗体軽鎖と強力な相同性を有するIgCドメイン様構造と、そのアミノ酸末端に向かって、2つの機能的に異なる領域とを含み、その領域の一方は、Vλドメインのβ7鎖に対して強力な相同性を示す。ヒトλ5様タンパク質は、213アミノ酸(NP_064455;配列番号6)を有し、抗体λ軽鎖定常領域に対して約84%の配列同一性を示す。
【0007】
更なる詳細については、以下のレビュー雑誌を参照されたい:Karasuyama et al.,Adv.Immunol.63:1−41(1996);Melchers et al.,Immunology Today 14:60−68(1993);およびMelchers,Proc.Natl.Acad.Sci.USA 96:2571−2573(1999)。
【0008】
VpreBとλ5のポリペプチドは、一緒になって、代替軽鎖または偽軽鎖と呼ばれる非共有結合的に会合されたIg軽鎖様構造を形成している。初期のプレB細胞の表面では、代替軽鎖が、シグナルトランスデューサーCD79a/CD79bヘテロ二量体と会合された膜結合Igμ重鎖にジスルフィド結合して、B細胞受容体様構造、いわゆるプレB細胞受容体(プレBCR)を形成している。
【0009】
Surrobodyは、抗体レパートリーの正常な発達の間に生成されるプレB細胞受容体(プレBCR)に基づいている。抗体とは異なり、プレBCRは、2つの代替軽鎖成分、VpreBおよびλ5と対になった抗体重鎖で構成された三量体である。VpreBおよびλ5は両者とも、遺伝子再配列を受けず、V(D)J組換えの開始前に初期プレB細胞内で発現される遺伝子によりコードされている。プレBCRは、重鎖と、2つの非共有結合的に会合されたタンパク質:VpreBおよびλ5とで構成されている成熟免疫グロブリンと構造的に異なっており、即ちそれらは、抗体内の2つとは対照的に3つの成分を有する。更に、VpreBは、Vλ Igドメインと相同性があり、λ5は、抗体のCλドメインと相同性があるが、それぞれは、非標準のペプチド伸長を有しており、つまりVpreB1は、C末端に追加の21残基を有し、λ5は、N末端に50アミノ酸の伸長を有する。
【0010】
κ様代替軽鎖(κ様SLC)を用いて、κ様β細胞受容体(κ様BCR)が同定された(Frances et al.,EMBO J 13:5937−43(1994);Thompson et al.,Immunogenetics 48:305−11(1998);Rangel et al.,J Biol Chem 280:17807−14(2005))。
【0011】
Rangel et al.,J Biol Chem 280(18):17807−17814(2005)には、Thompson et al.,Immunogenetics 48:308−311(1998)により過去に報告されたcDNA配列と同一であることが判明された、再配列されていないVκ遺伝子の生成物であるVκ様タンパク質の同定または分子特徴づけが報告された。ところが、Fnances et al.,EMBO J 13:5937−43(1994)には、B細胞前駆体の表面でμ重鎖と会合し、それによりB細胞発達のためのλ5経路の代替法を提供する能力を有する、再配列された生殖細胞系JCkの同定および特徴づけが報告されている。
【0012】
κ様およびλ様のプレBCRが協調的に作用して軽鎖再配列を促進し、B細胞始原を確実に成熟させることが提案された。レビューについては、McKeller and Martinez−Valdez Seminars in Immunology 18:4043(2006)を参照されたい。
【0013】
Surrobodyの設計および生成の更なる詳細は、全体の開示が本明細書に参照により明確に組み込まれる、Xu et al.,Proc.Natl.Acad.Sci.USA 2008,105(31):10756−61、2008年10月2日公開のPCT公開第WO2008/118970号、2008年7月11日出願の米国特許仮出願第61/134,929号、およびXu et al.,J.Mol.Biol.2010,397,352−360に提供されている。
【0014】
代替軽鎖は、プレB細胞上でのタンパク質産生および細胞外提示を可能にするリーダー配列を有する。しかし、典型的には、設計された代替軽鎖構造体の組換え発現は、同一の重鎖を用いた抗体よりも低いことが見出されている。それ故、代替軽鎖構造体の組換え発現の効率を改善することが求められている。
【発明の概要】
【発明が解決しようとする課題】
【0015】
本発明は、少なくとも一部は、非相同性リーダー配列を用いることにより、代替軽鎖構造体の組換え発現の効率を顕著に改善しうるという実験的発見に基づく。
【課題を解決するための手段】
【0016】
一態様において、本発明は、代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、SLCポリペプチドまたはSLCポリペプチドを含有するSLC構造体をコードする単離された核酸分子を提供する。一実施形態において、SLCポリペプチドとしては、VpreBポリペプチド、λ5ポリペプチド、またはそれらのフラグメントもしくはバリアントが挙げられる。別の実施形態において、VpreBポリペプチドは、天然型VpreB1配列、天然型VpreB2配列、天然型VpreB3配列、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。幾つかの実施形態において、天然型VpreB配列は、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3、配列番号5のヒトVpreB様ポリペプチド、配列番号6のヒトVpreB dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の一実施形態において、λ5ポリペプチドは、配列番号7のヒトλ5様、配列番号8のヒトλ5ポリペプチド、配列番号9のヒトλ5 dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の実施形態において、SLCポリペプチドとしては、Vκ様ポリペプチド、JCκポリペプチド、またはそれらのフラグメントもしくはバリアントが挙げられる。別の一実施形態において、Vκ様ポリペプチド配列は、配列番号12〜24、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。幾つかの実施形態において、JCκポリペプチド配列は、配列番号26〜39、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。
【0017】
別の態様において、本発明は、代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、SLCポリペプチドをコードした単離された核酸分子を提供し、SLCポリペプチドとしては、SLCポリペプチド融合体、またはそれらのフラグメントもしくはバリアントが挙げられる。一実施形態において、SLC融合体としては、VpreB−λ5ポリペプチド融合体、またはそれらのフラグメントもしくはバリアントが挙げられる。別の実施形態において、VpreBポリペプチド配列およびλ5ポリペプチド配列の融合は、それぞれVpreB配列およびλ5配列のCDR3類似領域で、またはその周辺で起こる。別の一実施形態において、VpreBポリペプチド配列は、カルボキシ末端で、λ5ポリペプチド配列のアミノ末端に連結されている。一実施形態において、SLC融合体としては、Vκ様−JCκポリペプチド融合体、またはそれらのフラグメントもしくはバリアントが挙げられる。別の実施形態において、Vκ様ポリペプチド配列およびJCκポリペプチド配列の融合は、それぞれVκ様配列およびJCκ配列のCDR3類似領域で、またはその周辺で起こる。別の一実施形態において、Vκ様ポリペプチド配列は、カルボキシ末端で、JCκポリペプチド配列のアミノ末端に融合されている。
【0018】
別の一態様において、本発明は、非SLC分子を含有するSLC融合体を提供する。一実施形態において、SLC融合体は、非SLC分子と、VpreB、λ5、Vκ様、およびJCκ配列の少なくとも1つとを含有する。別の実施形態において、非SLC分子は、非SLCポリペプチドであってもよい。一実施形態において、融合体は、非SLCポリペプチドに融合されたλ5配列またはVpreB配列を含む。別の一実施形態において、融合は、VpreB配列またはλ5配列のCDR3類似領域で、またはその周辺で起こる。幾つかの実施形態において、λ5配列のN末端が、非SLCポリペプチドのC末端に融合されているか、またはVpreB配列のV末端が、非SLCポリペプチドのN末端に融合されている。別の一実施形態において、融合体は、非SLCポリペプチドに融合されたVκ様またはJCκ配列を含む。別の一実施形態において、融合は、Vκ様配列またはJCκ配列のCDR3類似領域で、またはその周辺で起こる。幾つかの実施形態において、JCκ配列のN末端が、非SLCポリペプチドのC末端に融合されているか、またはVκ様配列のC末端が、非SLCポリペプチドのN末端に融合されている。一実施形態において、本発明は、SLCポリペプチドが非SLC分子を含有するSLC融合ポリペプチドを含む、SLCポリペプチドをコードする単離された核酸分子を提供する。
【0019】
全ての実施形態において、非相同性分泌リーダー配列は、抗体、サイトカイン、リンフォカイン、モノカイン、ケモカイン、ポリペプチドホルモン、消化酵素、および細胞外マトリックスの成分からなる群より選択される分泌されたポリペプチドのリーダー配列であってもよい。一実施形態において、サイトカインは、成長ホルモン、例えば、ヒト成長ホルモン、N−メチオニルヒト成長ホルモン、およびウシ成長ホルモン;副甲状腺ホルモン;チロキシン;インスリン;プロインスリン;リラキシン;プロリラキシン;糖タンパク質ホルモン、例えば、卵胞刺激ホルモン(FSH)、甲状腺刺激ホルモン(TSH)、および黄体形成ホルモン(LH);肝細胞増殖因子;線維芽細胞増殖因子;プロラクチン;胎盤性ラクトゲン;腫瘍壊死因子−αおよび−β(TNF−αおよび−β);ミュラー管抑制物質;マウスゴナドトロピン関連ペプチド;インヒビン;アクチビン;血管内皮増殖因子;インテグリン;トロンボポエチン(TPO);神経成長因子、例えば、NGF−β;血小板成長因子;トランスフォーミング増殖因子(TGF)、例えば、TGF−αおよびTGF−β;インスリン様成長因子−Iおよび−II;エリスロポエチン(EPO);骨誘導因子;インターフェロン、例えば、インターフェロン−α、−βおよび−γ;コロニー刺激因子(CSF)、例えば、マクロファージ−CSF(M−CSF)、顆粒球−マクロファージ−CSF(GM−CSF)、および顆粒球−CSF(G−CSF);インターロイキン(IL)、例えば、IL−1、IL−1a、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12;腫瘍壊死因子、例えば、TNF−αまたはTNF−β;MIP−1α;MIP−1β;ならびにLIFおよびkitリガンド(KL)をはじめとする他のポリペプチド因子からなる群より選択される分泌された他のポリペプチドからなる群より選択されてもよい。
【0020】
全ての実施形態において、分泌リーダー配列は、ヒトおよび非ヒトホ乳類アルブミン、トランスフェリン、CD36、成長ホルモン、組織プラスミノーゲン活性化因子(t−PA)、エリスロポエチン(EPO)、ならびにニューブラスチンのリーダー配列からなる群より選択されてもよい。
【0021】
全ての実施形態において、分泌リーダー配列は、合成配列であってもよい。
【0022】
全ての実施形態において、分泌リーダー配列は、天然型分泌リーダー配列のコンセンサス配列であってもよい。
【0023】
全ての実施形態において、非相同性シグナル配列は、配列番号36(METDTLLLWVLLLWVPGSTG)であってもよい。
【0024】
全ての実施形態において、本発明は、代替軽鎖(SLC)構造体をコードする単離された核酸分子を提供する。
【0025】
一態様において、本発明は、ベクターおよび組換え宿主細胞を提供する。全ての実施形態において、ベクターは、本明細書に記載された核酸分子を含有していてもよい。全ての実施形態において、組換え宿主細胞は、本明細書に記載された核酸で形質転換されていてもよい。
【0026】
別の態様において、本発明は、組換え宿主細胞内での代替軽鎖(SLC)ポリペプチドまたはSLC構造体の発現方法を提供する。一実施形態において、その方法は、SLCポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、組換え宿主細胞を、SLCポリペプチドまたはSLC構造体をコードする核酸分子で形質転換するステップを含む。別の一実施形態において、組換え宿主細胞は、真核細胞である。別の一実施形態において、組換え宿主細胞は、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞である。幾つかの実施形態において、SLCポリペプチドまたはSLC構造体は、VpreBポリペプチド、λ5ポリペプチド、VpreB−λ5ポリペプチド融合体、Vκ様ポリペプチド、JCκポリペプチド、およびVκ様−JCκポリペプチド融合体の1種以上を含むSLCポリペプチドからなる群より選択される。
【図面の簡単な説明】
【0027】
【図1】天然型リーダー配列を有する配列番号1のヒトVpreB1アミノ酸配列;配列番号2および3のマウスVpreB2配列;配列番号4のヒトVpreB3様配列;図11で「VpreB dTail」(配列番号5)と称された「三量体」内のトランケート型VpreB1の配列;ならびにマウスIgκリーダー配列を有する配列番号6のヒトVpreB1アミノ酸配列を示す。下線は、VpreBアミノ酸配列内のリーダー配列を示す。
【図2】配列番号7のヒトλ5様配列;配列番号8のヒトλ5配列;図11で「λ5 dTail」(配列番号9)と称された「三量体」内のトランケート型λ5配列;ならびにマウスIgκリーダー配列を有する配列番号10のヒトλ5 dTail配列を示す。下線は、λ5アミノ酸配列内のリーダー配列を示す。
【図3】配列番号35としてのヒトVpreB1−λ5キメラアミノ酸配列を示す(マウスIgκリーダー配列に下線)。
【図4A】および
【図4B】(A)配列番号11のヒトVκ様ヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(AJ004956;配列番号12)(天然型リーダー配列に下線)、ならびに(B)それぞれがAJ004956Vκ様プロトタイプ配列(配列番号12)に直線状に並んだ、異なる長さの伸長(配列番号13〜24)を含む、Vκファミリー全てからの可能なVκ様タンパク質の予測される成熟アミノ酸配列を示す。
【図5A】(A)配列番号25のヒトJCκヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(配列番号26)(予測される成熟JCkタンパク質と比較したユニーク配列には2本の下線を、潜在的リーダー開裂配列には1本の下線を付す)、(B)残りのκJ定常領域再配列(J1〜J5Cκ)からの予測されるJCκ様アミノ酸配列(配列番号27〜31)、ならびに(C)追加されたマウスIgκリーダー配列に下線を付したJCκ(配列番号32)、追加されたマウスIgκリーダー配列のみに下線を付した組換えJCκ(配列番号33)、および追加されたマウスIgκリーダー配列に下線を付した予測されるプロセッシングJCκ(配列番号34)、を含むJCkを設計された分泌最適化バリアント示す。
【図5B】(A)配列番号25のヒトJCκヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(配列番号26)(予測される成熟JCkタンパク質と比較したユニーク配列には2本の下線を、潜在的リーダー開裂配列には1本の下線を付す)、(B)残りのκJ定常領域再配列(J1〜J5Cκ)からの予測されるJCκ様アミノ酸配列(配列番号27〜31)、ならびに(C)追加されたマウスIgκリーダー配列に下線を付したJCκ(配列番号32)、追加されたマウスIgκリーダー配列のみに下線を付した組換えJCκ(配列番号33)、および追加されたマウスIgκリーダー配列に下線を付した予測されるプロセッシングJCκ(配列番号34)、を含むJCkを設計された分泌最適化バリアント示す。
【図5C】(A)配列番号25のヒトJCκヌクレオチド配列およびそのコード化タンパク質のアミノ酸配列(配列番号26)(予測される成熟JCkタンパク質と比較したユニーク配列には2本の下線を、潜在的リーダー開裂配列には1本の下線を付す)、(B)残りのκJ定常領域再配列(J1〜J5Cκ)からの予測されるJCκ様アミノ酸配列(配列番号27〜31)、ならびに(C)追加されたマウスIgκリーダー配列に下線を付したJCκ(配列番号32)、追加されたマウスIgκリーダー配列のみに下線を付した組換えJCκ(配列番号33)、および追加されたマウスIgκリーダー配列に下線を付した予測されるプロセッシングJCκ(配列番号34)、を含むJCkを設計された分泌最適化バリアント示す。
【図6】VpreBおよびλ5配列により形成された代替軽鎖の略図、代替軽鎖配列を含む例示的な融合ポリペプチド、ならびにV−J結合から得られた抗体軽鎖構造である。
【図7】様々な代替軽鎖欠失および一本鎖構造体の略図である。
【図8】代替軽鎖構造体への組み合わせ的機能多様性(combinatorial functional diversity)の組込みを概略的に図示している。
【図9】様々な例示的代替軽鎖構造体の遺伝子およびタンパク質構造を示す。
【図10】代替軽鎖(SLC)成分に機能性を加える様々な代表的方法を示す。
【図11】様々な三量体および二量体代替軽鎖(SLC)構造体を示す。
【図12】様々なヘテロ二量体代替κ軽鎖欠失バリアントの略図である。「完全長」構造体において、Vκ様およびJCκ配列は両者とも、それぞれC−末端およびN−末端の伸長(テール)を保持している。dJバリアントでは、JCκのN−末端伸長が欠失されている。dVκテールバリアントでは、Vκ様配列のC−末端伸長が、除去されているが、JCκのN−末端伸長は保持されている。「短κ」バリアントでは、Vκ様配列のC末端テールおよびJCκ配列のN−末端伸長は両者とも、保持されている。
【図13】個別に、または別のタンパク質、例えば、抗体重鎖またはそのフラグメントと一緒に用いることができる、κ様軽鎖欠失および一本鎖構造体。
【図14】κ様代替軽鎖構造体への組み合わせ的機能多様性の組込み。赤線は、追加された多様性、例えば、ペプチドライブラリーを示す。
【図15】軽鎖は、遺伝子再配列およびRNAプロセッシングの生成物である。
【図16A】Vκ様タンパク質が再配列されていないVκIV−遺伝子転写および翻訳から得られることを示す。VκIVは、71のVL生殖細胞系遺伝子のうちの1つである。Vκ様タンパク質を作製することが可能なVL生殖細胞系遺伝子は更に70存在するため、κV遺伝子は更に39、そしてλV遺伝子は更に31存在する。
【図16B】JCκが、再配列されていないJおよびC生殖細胞系からのプロセッシングRNAの生成物であることを示す。JCκは、45のJC生殖細胞系の組み合わせのうちの1つである。JCκ様タンパク質を作製することが可能なVL生殖細胞系遺伝子が更に44存在し、つまりCκを組み合わせるJκ遺伝子が更に4、そして10のCλ遺伝子と組み合わせるJλ遺伝子が更に4存在する(合計40)。
【図17】κ様代替軽鎖成分に機能性を加えた略図を示す。二機能性および三機能性構造を示す。A:scFvを制約された融合体;B:Vκ様−scFv融合体;C:JCκ−scFv融合体;SLC二重融合。
【図18】代替軽鎖の機能的テール伸長のタイプを示す。
【図19】κ様およびλ様代替軽鎖機能的キメラを示す。
【図20】(A)Surrobody構成、(B)二機能性および二重特異性Surrobody構成、ならびに(C)(A)および(B)に表わされた分子のクローニング方策を示す。
【発明を実施するための形態】
【0028】
発明の詳細な説明
A.定義
他に断りがなければ、本明細書で用いられる技術的および科学的用語は、当業者により一般に理解されるものと同じ意味を有する。Singleton et al.,Dictionary of Microbiology and Molecular Biology 2nd ed.,J.Wiley&Sons(New York,NY 1994)は、本願で用いられる多くの用語の概説を当業者に提供する。
【0029】
当業者には、本発明の実践で用いられうる、本明細書に記載されたものと類似または同一の多くの方法および材料が認識されよう。事実、本発明は、記載された方法および材料に限定されない。本発明の目的では、次の用語を以下の通り定義する。
【0030】
本願全体を通して、単数形の使用は、他に明記されない限り、複数形を包含する。
【0031】
本願において、「または」の使用は、他に明記されない限り、「および/または」を包含する。
【0032】
更に用語「包含する」、「はじめとする」、および「包含された」は、限定されていない。
【0033】
本発明の文脈において、用語「抗体(Ab)」は、V(D)J遺伝子組換えから得られた伝統的組換え重鎖およびVJ遺伝子組換えから得られた伝統的組換え軽鎖、またはそれらのフラグメントからの天然型抗体を指すために用いられる。
【0034】
「天然型抗体」は、2つの同一軽(L)鎖および2つの同一重(H)鎖で構成された約150,000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は、ジスルフィド共有結合により重鎖に連結されているが、ジスルフィド結合の数は、異なる免疫グロブリンアイソタイプの重鎖間で変動する。各重鎖および軽鎖は、規則的な間隔の鎖内ジスルフィド架橋も有する。各重鎖は、一方の末端に、可変ドメイン(VH)と、それに続く多数の定常ドメインを有する。各軽鎖は、一方の末端の可変ドメイン(VL)および他方の末端の定常ドメインを有し、軽鎖の定常ドメインは、重鎖の第一の定常ドメインと直線状に並べられ、軽鎖の可変ドメインは、重鎖の可変ドメインと直線状に並べられている。特定のアミノ酸残基が、軽鎖可変ドメインと重鎖可変ドメインの間に境界を形成すると考えられている(Chothia et al.,J.Mol.Biol.186:651(1985);Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0035】
抗体鎖に関連する用語「可変」は、抗体の間で配列が広範囲にわたって異なり、特定抗原に対する各特定抗体の結合および特異性に関与する、抗体鎖の部分を指すために用いられる。そのような可変性は、軽鎖可変ドメインと重鎖可変ドメインの両方の高度可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存された部分は、フレームワーク領域(FR)と呼ばれる。天然型重鎖および軽鎖の可変領域はそれぞれ、4つのFR(それぞれFR1、FR2、FR3およびFR4)を含み、3つの高度可変領域により接続されたβシート配置を大きく採用し、それがループ接続を形成し、幾つかの例ではβシート構造の一部を形成している。各鎖の高度可変領域は、FRによって接近してまとめられていて、別の鎖の高度可変領域と共に抗体の抗原結合部位の形成に寄与する(Kabat et al.,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.(1991),p647−669参照)。定常ドメインは、抗原への抗体の結合に直接関与せず、様々なエフェクター機能、例えば、抗体依存性細胞毒性における抗体の関与などを示す。
【0036】
本明細書で用いられる用語「高度可変領域」は、抗原結合を担う抗体のアミノ酸残基を指す。高度可変領域は、「相補性決定領域」または「CDR」からのアミノ酸残基(即ち、軽鎖可変ドメイン内の30〜36(L1)、46〜55(L2)および86〜96(L3)の残基、ならびに重鎖可変ドメイン内の30〜35(HI)、47〜58(H2)、および93〜101(H3)の残基;MacCallum et al.,J Mol Biol.262(5):732−45(1996))を含む。
【0037】
用語「フレームワーク領域」は、より異なるCDR領域の間に存在する、当該技術分野で認識された抗体可変領域の部分を指す。そのようなフレームワーク領域は、典型的には、フレームワーク1〜4(FR1、FR2、FR3、およびFR4)と呼ばれ、重鎖または軽鎖抗体可変領域内に見出される3つのCDRを、三次元空間に保持するための足場を提供することで、CDRが抗原結合表面を形成することができる。
【0038】
それらの重鎖の定常ドメインのアミノ酸配列に応じて、抗体を異なる分類に割り付けることができる。抗体には5つの主要な分類、IgA、IgD、IgE、IgG、およびIgMが存在し、これらの複数は、亜分類(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgAおよびIgA2に更に分別される場合がある。好ましい実施形態において、本発明のイムノアドヘシンの構築に用いられる免疫グロブリン配列は、IgG免疫グロブリン重鎖ドメインのものである。ヒトイムノアドヘシンでは、ヒトIgG1およびIgG3免疫グロブリン配列の使用が好ましい。IgG1を用いることの主な利点は、IgG1イムノアドヘシンが固定化プロテインA上で効率的に精製されうることである。しかし、特定のイムノアドヘシン構築用にIg融合パートナーを選択する場合には、別の構造的および機能的性質を考慮すべきである。例えば、IgG3ヒンジは、より長く、より可撓性があるため、より大きな「アドヘシン」ドメインを収容することができ、IgG1に融合されると適切にフォールディングまたは機能しない可能性がある。別の考慮すべき事柄は、結合価かもしれず、IgGイムノアドヘシンは、二価ホモ二量体であるが、IgAおよびIgMのようなIgサブタイプは、基本的なIgホモ二量体単位の、それぞれ二量体または五量体構造をもたらす可能性がある。インビボ適用のために設計されたVEGF受容体Ig様ドメイン/免疫グロブリンキメラでは、Fc領域に特有の薬物動態性およびエフェクター機能も重要となる。IgG1、IgG2およびIgG4は全て、インビボ半減期21日を有するが、補体系を活性化させる際のそれらの相対的能力は異なる。その上、様々な免疫グロブリンが、様々な数のアロタイピックアイソタイプ(allotypic isotypes)を有する。
【0039】
異なる分類の免疫グロブリンに対応する重鎖定常ドメインは、それぞれα、δ、ε、γ、およびμと呼ばれる。
【0040】
任意の脊椎動物種からの抗体の「軽鎖」は、定常ドメインのアミノ酸配列に基づいて、カッパ(κ)およびラムダ(λ)と呼ばれる2種の明確に異なる型の一方に割り付けることができる。本明細書における抗体軽鎖の任意の参照は、κおよびλの両方の軽鎖を包含する。
【0041】
「抗体フラグメント」は、一般に抗原が結合した、完全長抗体の一部、またはその可変ドメインを含む。抗体フラグメントの例としては、非限定的に、Fab、Fab’、F(ab’)2、scFv、および(scFv)2フラグメントが挙げられる。
【0042】
本明細書で用いられる用語「抗体結合領域」は、抗原に結合しうる免疫グロブリンまたは抗体可変領域の1部分以上を指す。典型的には、抗体結合領域は、例えば、抗体軽鎖(VL)(またはその可変領域)、抗体重鎖(VH)(またはその可変領域)、重鎖Fd領域、抗体軽鎖と重鎖の混合体(またはその可変領域)、例えば、Fab、F(ab’)2、単一ドメイン、もしくは一本鎖抗体(scFv)、または完全長抗体、例えば、IgG(例えば、IgG1、IgG2、IgG3、またはIgG4サブタイプ)、IgA1、IgA2、IgD、IgE、またはIgM抗体である。
【0043】
本明細書で用いられる用語「エピトープ」は、アミノ酸が少なくとも約3〜5、好ましくは少なくとも約5〜10、または少なくとも約5〜15、典型的には約500以下または約1000以下の配列を指し、単独で、またはより大きな配列の一部として、そのような配列に応答して生成された抗体に結合している配列を定義する。エピトープは、由来する親タンパク質の部分と同一の配列を有するポリペプチドに限定されない。事実、ウイルスゲノムは、不断に変化する状態にあり、単離物の間に比較的高度の変動性が示される。したがって用語「エピトープ」は、天然型配列と同一の配列に加え、天然型配列への修飾、例えば欠失、置換および/または挿入を包含する。一般にそのような修飾は、本質的に保存的であるが、非保存的修飾も企図される。その用語は、具体的には「ミモトープ」、即ち、連続した直線状天然型配列を画定しない配列、または天然型タンパク質中には必ずしも生じないが、天然型タンパク質上のエピトープを機能的に模倣している配列を包含する。用語「エピトープ」は、具体的には線形および立体配座エピトープを包含する。
【0044】
用語「代替軽鎖ポリペプチド」または「SLCポリペプチド」は、本明細書では、VpreBポリペプチド、λ5ポリペプチド、Vκ様ポリペプチド、JCκポリペプチド、またはそれらのバリアントを指すために用いられる。
【0045】
用語「非代替軽鎖分子」または「非SLC分子」は、本明細書では、SLCポリペプチドでない分子を指すために用いられる。非SLC分子は、ポリペプチド、例えば、サイトカインまたは抗体フラグメントであってもよい。
【0046】
用語「VpreB」は、本明細書では最も広い意味で用いられ、詳細には、非限定的に、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3様配列、配列番号5のヒトVpreB dTをはじめとする任意の天然型配列またはバリアントVpreBポリペプチド、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、そのような天然型配列ポリペプチドのバリアントをはじめとするアイソフォームを指す。
【0047】
用語「λ5」は、本明細書では最も広い意味で用いられ、詳細には、非限定的に、配列番号6のヒトλ5、配列番号7のヒトλ5様タンパク質、配列番号9として示されるヒトλ5 dT、配列番号10のヒトVpreB1アミノ酸配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、そのような天然型配列λ5ポリペプチドのバリアントをはじめとするアイソフォームを指す。
【0048】
用語「バリアントVpreBポリペプチド」および「VpreBポリペプチドのバリアント」は、互換的に用いられ、本明細書ではアミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列VpreBポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントVpreBポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントVpreBポリペプチド」は、好ましくは天然型配列VpreBポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントVpreBポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントVpreBポリペプチドとしては、非限定的に、VpreB配列のC末端の非Ig様ユニークテールが部分的または完全に除去されているVpreBポリペプチドが挙げられる。
【0049】
用語「バリアントλ5ポリペプチド」および「λ5ポリペプチドのバリアント」は、互換的に用いられ、本明細書では、アミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列λ5ポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントλ5ポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントλ5ポリペプチド」は、好ましくは天然型配列λ5ポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントλ5ポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントλ5ポリペプチドとしては、非限定的に、λ5配列のN末端のユニークテールが部分的または完全に除去されているλ5ポリペプチドが挙げられる。
【0050】
用語「バリアントVκ様ポリペプチド」および「Vκ様ポリペプチドのバリアント」は、互換的に用いられ、本明細書では、アミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列Vκ様ポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントVκ様ポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントVκ様ポリペプチド」は、好ましくは天然型配列Vκ様ポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントVκ様ポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントVκ様ポリペプチドとしては、非限定的に、Vκ様配列のC末端の非Ig様ユニークテールが部分的または完全に除去されているVκ様ポリペプチドが挙げられる。
【0051】
用語「バリアントJCκポリペプチド」および「JCκポリペプチドのバリアント」は、互換的に用いられ、本明細書では、アミノ酸修飾の結果、1ヶ所以上のアミノ酸位置が天然型配列JCκポリペプチドと異なっているポリペプチドと定義される。本明細書で定義される「バリアントJCκポリペプチド」は、天然型抗体λもしくはκ軽鎖配列、またはそれらのフラグメントとは異なる。「バリアントJCκポリペプチド」は、好ましくは天然型配列JCκポリペプチドと、少なくとも約65%、または少なくとも約70%、または少なくとも約75%、または少なくとも約80%、または少なくとも約85%、または少なくとも約90%、または少なくとも約95%、または少なくとも約98%の配列同一性を保持する。別の好ましい実施形態において、「バリアントJCκポリペプチド」は、天然型抗体λまたはκ軽鎖配列と95%未満、または90%未満、または85%未満、または80%未満、または75%未満、または70%未満、または65%未満、または60%未満のアミノ酸配列が同一である。具体的には、バリアントJCκポリペプチドとしては、非限定的に、JCκ配列のN末端のユニークテールが部分的または完全に除去されているJCκポリペプチドが挙げられる。
【0052】
アミノ酸配列同一性の百分率は、配列比較プログラムNCBI−BLAST2を用いて決定してもよい(Altschul et al.,Nucleic Acids Res.25:3389−3402(1997))。NCBI−BLAST2配列比較プログラムは、からダウンロードしてもよく、さもなければメリーランド州ベセスダのアメリカ国立衛生研究所から得てもよい。NCBI−BLAST2は、複数の研究パラメータを利用しており、それらの研究パラメータの全てが、例えば、unmask=yes、strand=all、expected occurrences=10、minimum low complexity length=15/5、multi−pass e−value=0.01、constant for multi−pass=25、dropoff for final gapped alignment=25、scoring matrix=BLOSUM62、をはじめとするデフォルト値に設定される。
【0053】
用語「VpreB配列」は、本明細書では、本明細書の先に定義された「VpreB」またはそのフラグメントの配列を指すために用いられる。
【0054】
用語「λ5配列」は、本明細書では、本明細書の先に定義された「λ5」またはそのフラグメントの配列を指すために用いられる。
【0055】
用語「Vκ様配列」は、本明細書では、本明細書の先に定義された「Vκ様」またはそのフラグメントの配列を指すために用いられる。
【0056】
用語「JCκ配列」は、本明細書では、本明細書の先に定義された「JCκ」またはそのフラグメントの配列を指すために用いられる。
【0057】
本明細書で用いられる用語「λ様代替軽鎖」は、VpreBとλ5タンパク質との非共有結合的会合により形成される二量体を指す。
【0058】
本明細書で用いられる用語「κ様代替軽鎖」は、Vκ様とJCκタンパク質との非共有結合的会合により形成される二量体を指す。
【0059】
本明細書に定義される用語「λ様代替軽鎖」は、本明細書の先に定義された「VpreB配列」および/または「λ5配列」を含む任意のポリペプチド配列を意味する。具体的には、本明細書に定義される「λ様代替軽鎖配列」としては、非限定的に、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3配列、配列番号5のヒトVpreB dT;および配列番号6のヒトVpreB1アミノ酸配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、それらのフラグメントおよびバリアントをはじめとする様々なアイソフォームが挙げられる。加えて用語「λ様代替軽鎖配列」としては、非限定的に、配列番号7のヒトλ5配列、配列番号8のヒトλ5様配列、配列番号9として示されるヒトλ5 dTail、配列番号10のヒトλ5 dTail、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、それらのフラグメントおよびバリアントをはじめとするアイソフォームが挙げられる。加えて用語「λ様代替軽鎖配列」としては、本明細書の先に定義されたVpreBおよびλ5の両方の配列を含む配列が挙げられる。
【0060】
本明細書に定義される用語「κ様代替軽鎖配列」は、本明細書の先に定義された「Vκ様配列」および/または「JCκ」を含む任意のポリペプチド配列を意味する。具体的には、本明細書に定義される「κ様代替軽鎖配列」としては、非限定的に、配列番号12〜24のいずれかのヒトVκ様配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、それらのフラグメントおよびバリアントをはじめとする様々なアイソフォームが挙げられる。加えて「κ様代替軽鎖配列」としては、非限定的に、配列番号12〜24のいずれかのヒトVκ様配列、配列番号25〜35のいずれかのヒトJCκ配列、ならびにスプライスバリアントおよび翻訳後修飾により形成されたバリアント、別のホ乳類のその相同物に加えて、そのフラグメントおよびバリアントをはじめとするアイソフォームが挙げられる。加えて用語「κ様代替軽鎖配列」としては、本明細書の先に定義されたVκ様およびJCκの両方の配列を含む配列が挙げられる。
【0061】
用語「代替軽鎖構造体」は、最も広い意味で用いられ、異種アミノ酸配列、核酸、および代替軽鎖配列にコンジュゲートされた別の分子をはじめとする任意の、そして全ての追加の異種成分を包含し、「コンジュゲーション」は、以下に定義する。
【0062】
「代替軽鎖構造体」は、本明細書では、「Surrobody(商標)」または「Surrobody」とも呼ばれ、その2つの用語は、互換的に用いられる。特定のSurrobody(商標)λ様代替軽鎖構造体は、Xu et al.,Proc.Natl.Acad.Sci.USA 2008,105(31):10756−61および2008年10月2日公開のPCT公開第WO2008/118970号に開示されている。同じく企図されるのは、米国特許公開第2010−0062950号、およびXu et al.,J.Mol.Biol.2010,397,352−360に記載されたκ様代替軽鎖構造体であり、それらの開示の全体は、本明細書に参照により明確に組み込まれる。
【0063】
本発明のポリペプチドの文脈において、第一のアミノ酸配列に関する用語「異種アミノ酸配列」は、第一のアミノ酸配列に自然に会合されておらず、少なくとも第一のアミノ酸配列が本明細書の代替軽鎖構造体内に存在する形態ではないアミノ酸配列を指すために用いられる。したがって、VpreB、λ5、Vκ様、またはJCκに関する「異種アミノ酸配列」は、その天然の環境において、天然型VpreB、λ5、Vκ様、またはJCκと会合されていない任意のアミノ酸配列である。これらには、非限定的に、i)VpreBと共に発達中のB細胞上に代替軽鎖を形成するそれらのλ5配列とは異なるλ5配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたλ5配列;ii)λ5と共に発達中のB細胞上に代替軽鎖を形成するそれらのVpreB配列とは異なるVpreB配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたVpreB配列;iii)JCκ様と共に発達中のB細胞上にκ様代替軽鎖を形成するそれらのVκ様配列とは異なるVκ様配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたVκ様配列;ならびにiv)Vκ様と共に発達中のB細胞上にκ様代替軽鎖を形成するそれらのJCκ配列とは異なるJCκ配列、例えば、アミノ酸配列バリアント、例えば、トランケートおよび/または誘導体化されたJCκ配列が挙げられる。
【0064】
VpreBまたはλ5に関する「異種アミノ酸配列」は、天然の環境では、VpreBまたはλ5配列は互いに共有結合的に会合、例えば融合されていないため、天然型配列VpreBまたはλ5をはじめとする対応するVpreBまたはλ5と共有結合的に会合、例えば融合されている、VpreBまたはλ5配列も包含する。同様に、Vκ様またはJCκに関する「異種アミノ酸配列」は、天然の環境では、Vκ様またはJCκ配列は互いに共有的に会合、例えば融合されていないため、天然型配列Vκ様またはJCκをはじめとする対応するVκ様またはJCκと共有結合的に会合、例えば融合されている、Vκ様またはJCκ配列も包含する。異種アミノ酸配列としては、非限定的に、抗体および重鎖配列をはじめとする抗体配列ならびにそれらのフラグメント、例えば、抗体軽鎖および重鎖可変領域配列、ならびに抗体軽鎖および重鎖定常領域配列も挙げられる。
【0065】
用語「コンジュゲート」、「コンジュゲートされた」および「コンジュゲーション」は、共有結合または非共有結合の任意の、そして全ての形態を指し、非限定的に、直接遺伝子融合または化学的融合、リンカーまたは架橋剤を介したカップリング、そして例えば、ファンデルワールス力を介した、またはロイシンジッパーを用いた非共有結合的会合が挙げられる。
【0066】
用語「可撓性リンカー」は、本明細書では、化学構造に基づけば、意図された文脈および環境において三次元空間に固定されていると予測されない任意のリンカーを指すために用いられる。
【0067】
用語「融合」は、本明細書では、1本のポリペプチド鎖内の異なる起点のアミノ酸配列をコードするヌクレオチド配列をインフレームで組み合わせることによって、それらのアミノ酸配列を組み合わせることを指すのに用いられる。用語「融合」は、その末端の1つと融合することに加えて、内部融合、即ちポリペプチド鎖内の異なる起点の配列を挿入することを明示的に包含する。
【0068】
本明細書で用いられる用語「ペプチド」、「ポリペプチド」および「タンパク質」は全て、共有結合的「ペプチド結合」によって連結されたアミノ酸の一次配列を指す。一般に、ペプチドは、少数のアミノ酸、典型的には約2〜約50のアミノ酸からなり、タンパク質よりも短い。本明細書で定義される用語「ポリペプチド」は、ペプチドおよびタンパク質を包含する。
【0069】
用語「アミノ酸」または「アミノ酸残基」は、典型的には、当該技術分野で認識された定義を有するアミノ酸、例えば、アラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Gln)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ile)、ロイシン(Leu)、リシン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、トレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)およびバリン(Val)からなる群より選択されるアミノ酸を指すが、修飾アミノ酸、合成アミノ酸または希アミノ酸を所望どおり使用してもよい。したがって、具体的には37 CFR 1.822(b)(4)に列挙された修飾および異常アミノ酸は、この定義に含まれ、本明細書に参照により明確に組み込まれる。アミノ酸は、様々な亜群に細分することができる。したがってアミノ酸は、非極性側鎖(例えば、Ala、Cys、Ile、Leu、Met、Phe、Pro、Val)、負電荷の側鎖(例えば、Asp、Glu)、正電荷の側鎖(例えば、Arg、His、Lys)、または非荷電の極性側鎖(例えば、Asn、Cys、Gln、Gly、His、Met、Phe、Ser、Thr、TrpおよびTyr)を有するとして分類することができる。アミノ酸は、小アミノ酸(Gly、Ala)、求核性アミノ酸(Ser、His、Thr、Cys)、疎水性アミノ酸(Val、Leu、Ile、Met、Pro)、芳香族アミノ酸(Phe、Tyr、Trp、Asp、Glu)、アミド(Asp、Glu)および塩基性アミノ酸(Lys、Arg)として分類することもできる。
【0070】
用語「ポリヌクレオチド」は、核酸、例えば、DNA分子およびRNA分子、ならびにそれらの類似体(例えば、ヌクレオチド類似体または核酸化学を利用して生成されるDNAまたはRNA)を指す。ポリヌクレオチドは、所望通り、例えば当該技術分野で認識された核酸化学を利用して、合成的に生成し、または例えばポリメラーゼを用いて、酵素的に生成し、そして所望なら修飾してもよい。典型的な修飾としては、メチル化、ビオチン化、および当該技術分野で公知の別の修飾が挙げられる。加えて核酸分子は、一本鎖または二本鎖であってもよく、所望ならば、検出可能な部分と連結させることができる。
【0071】
参照ポリペプチドに関する用語「バリアント」は、天然型ポリペプチドと比較して、少なくとも1つのアミノ酸変異または修飾(すなわち、改変)を有するポリペプチドを指す。「アミノ酸修飾」により生成されたバリアントは、例えば、天然型アミノ酸配列中の少なくとも1つのアミノ酸を置換、欠失、挿入および/または化学修飾することによって生成することができる。
【0072】
「アミノ酸修飾」は、所定のアミノ酸配列のアミノ酸配列における変化を指す。例示的な修飾としては、アミノ酸置換、挿入および/または欠失が挙げられる
【0073】
特定の位置「でのアミノ酸修飾」は、特定の残基の置換もしくは欠失、または特定の残基に隣接する少なくとも1つのアミノ酸残基の挿入を指す。特定の残基に「隣接する」挿入は、その1つまたは2つの残基の内部の挿入を意味する。挿入は、N末端またはC末端から特定の残基までに存在してもよい。
【0074】
「アミノ酸置換」は、所定のアミノ酸配列内の少なくとも1つの既存のアミノ酸残基を、別の異なる「交換」アミノ酸残基で交換することを指す。交換残基は、「天然アミノ酸残基」(即ち、遺伝子コードによりコードされている)であってもよく、アラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リシン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);トレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)からなる群より選択されてもよい。1つ以上の非天然アミノ酸残基での置換も、本明細書でのアミノ酸置換の定義に包含される。
【0075】
「非天然アミノ酸残基」は、先に列挙された天然アミノ酸残基以外の、ポリペプチド鎖内の隣接アミノ酸残基と共有結合できる残基を指す。非天然アミノ酸残基の例としては、ノルロイシン、オルニチン、ノルバリン、ホモセリンおよびEllman et al.,Meth.Enzym.202:301 336(1991)に記載されるような別のアミノ酸残基類似体が挙げられる。そのような非天然アミノ酸残基を生成するために、Noren et al.Science 224:182(1989)およびEllman et al.の上記文献の手順を用いることができる。簡潔に述べると、これらの手順は、非天然アミノ酸残基によりサプレッサーtRNAを化学的に活性化させて、RNAのインビトロ転写および翻訳を行うことを含む。
【0076】
「アミノ酸挿入」は、少なくとも1つのアミノ酸を所定のアミノ酸配列に組込むことを指す。挿入は通常、1つまたは2つのアミノ酸残基の挿入からなるが、本願は、より大きな「ペプチド挿入」、例えば、約3〜約5または最大約10のアミノ酸残基の挿入を企図する。挿入された残基は、先に開示された通り天然または非天然のものであってもよい。
【0077】
「アミノ酸欠失」は、所定のアミノ酸配列からの少なくとも1つのアミノ酸残基の除去を指す。
【0078】
用語「突然変異誘発」は、他に明記されていなければ、ポリヌクレオチドまたはポリペプチド配列を改変するための、任意の当該技術分野で認識された技術を指す。突然変異誘発の好ましいタイプとしては、エラープローンPCR突然変異誘発、飽和突然変異誘発、または別の部位特異的突然変異誘発が挙げられる。
【0079】
「部位特異的突然変異誘発」は、当該技術分野における技術標準であり、一本鎖ファージDNAに相補的な合成オリゴヌクレオチドプライマーを用いて実施して、限定されたミスマッチ以外の変異を誘発させると、所望の突然変異が現れる。簡潔に述べると、合成オリゴヌクレオチドを、一本鎖ファージDNAに相補的な鎖の合成を導くプライマーとして用い、得られた二本鎖DNAをファージ支持宿主細菌(phage−supporting host bacterium)内に形質変換させる。形質転換された細菌の培養物を上層寒天に塗布し、ファージを含む単細胞からのプラーク形成を可能にする。理論的には、新しいプラークの50%が、突然変異形態を一本鎖として有するファージを含有し、50%が本来の配列を有する。完全一致のハイブリダイゼーションが可能であるが、本来の鎖とのミスマッチがハイブリダイゼーションを防止するのに十分となる温度で、該当するプラークを、キナーゼ処理した合成プライマーとハイブリダイズすることによって選択する。その後、プローブとハイブリダイズするプラークが選択され、配列決定および培養されて、DNAが回収される。
【0080】
用語「ベクター」は、細胞内での自己複製が可能で、DNAセグメント、例えば遺伝子またはポリヌクレオチドが作動可能に連結されることで、結合したセグメントの複製をもたらすことができるrDNA分子を指すために用いられる。1種以上のポリペプチドをコードする遺伝子の発現を導くことが可能なベクターを、本明細書では「発現ベクター」と呼ぶ。用語「制御配列(control sequences)」は、特定の宿主生物中の作動可能に連結されたコード配列の発現に必要なDNA配列を指す。例えば、原核生物に適した制御配列としては、プロモータ、場合によりオペレータ配列、およびリボソーム結合部位が挙げられる。真核細胞は、プロモータ、ポリアデニル化シグナル、およびエンハンサーを用いることが公知である。ベクターは、追加のDNAセグメントにライゲートされている場合がある環状二本鎖DNAループを指す「プラスミド」であってもよい。ベクターは、追加のDNAセグメントがウイルスゲノムにライゲートされている場合があるファージベクターまたはウイルスベクターであってもよい。適切なベクター、例えば、細菌起源を有する細菌ベクターまたは複製およびエピソームホ乳類ベクターは、導入される宿主細胞内で自己複製が可能である。ベクターは、宿主細胞に導入する際、宿主細胞ゲノム、例えば、非エピソームホ乳類ベクターに合体させて、宿主ゲノムと一緒に複製してもよい。
【0081】
核酸は、別の核酸配列と機能的関係にある場合に、「作動可能に連結」されている。例えば、プレ配列または分泌リーダーのDNAは、ポリペプチドの分泌に関与するプレタンパク質として発現される場合、ポリペプチドのDNAに作動可能に連結されており、プロモータもしくはエンハンサーは、コード配列の転写に影響を及ぼす場合、コード配列に作動可能に連結されており、またはリボソーム結合部位は、翻訳を促進するように位置する場合、コード配列に作動可能に連結されている。一般に、「作動可能に連結された」とは、連結されているDNA配列が隣接すること、そして分泌リーダーの場合、隣接し、かつ読み取り期(reading phase)にあることを意味する。しかし、エンハンサーは、隣接している必要はない。連結は、簡便な制限部位でのライゲーションによって果される。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーを従来の実践法に従って使用する。
【0082】
「ファージディスプレイライブラリー」は、クローン化されたタンパク質配列の回収物をファージコートタンパク質との融合体として発現するタンパク質発現ライブラリーである。したがって、語句「ファージディスプレイライブラリー」は、本明細書では、外来(一般には非相同性)タンパク質を発現するファージ(例えば、繊維状ファージ)の回収物を指す。外来タンパク質は、ファージと接触している別の部分と自由に相互作用(結合)することができる。外来タンパク質を提示する各ファージは、ファージディスプレイライブラリーの「メンバー」である。
【0083】
用語「繊維状ファージ」は、異種ポリペプチドを表面に提示することができるウイルス粒子を指し、非限定的に、fl、fd、Pf1、およびM13が挙げられる。繊維状ファージは、選択マーカー、例えば、テトラサイクリン(例えば、「fd−tet」)を含有していてもよい。様々な繊維状ファージディスプレイシステムが、当業者に周知である(例えば、Zacher et al.,Gene 9:127−140(1980),Smith et al.,Science 228:1315−1317(1985);およびParmley and Smith Gene 73:305−318(1988)参照)。
【0084】
用語「パンニング」は、標的に対する親和力および特異性が高い、抗体などの化合物を運搬する化合物を同定および単離する際の複数回のスクリーニング工程を指すために用いられる。
【0085】
「リーダー配列」、「シグナルペプチド」、または「分泌リーダー」は、互換的に用いられ、一部であるポリペプチドの細胞内輸送を導くアミノ酸残基を含む配列を含有する。ポリペプチドは、典型的にはN末端に、分泌リーダー、シグナルペプチド、またはリーダー配列を含有する。これらのポリペプチドは、リーダー配列がシグナルエンドペプチターゼによりポリペプチドの残り部分から開裂されうる開裂部位を含んでいてもよい。そのような開裂により、成熟ポリペプチドが生成する。開裂は、典型的には、分泌の間に、またはインタクトポリペプチドが適切な細胞内コンパートメントに導かれた後に起こる。
【0086】
「宿主細胞」は、本明細書に記載された分子をコードする核酸および/または核酸含有ベクターの形質転換のレシピエントとなりうる、またはレシピエントになった個々の細胞または細胞培養物を包含する。本発明の方法において、宿主細胞は、真核細胞、例えば、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞であってもよい。別の適切な宿主細胞は、当業者に公知である。
【0087】
B.詳細な説明
本発明の方法を実施する技術は、当該技術分野で周知であり、例えば、Ausubel et al.,Current Protocols of Molecular Biology,John Wiley and Sons(1997);Molecular Cloning:A Laboratory Manual,Third Edition,J.Sambrook and D.W.Russell,eds,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;O’Brian et al.,Analytical Chemistry of Bacillus Thuringiensis,Hickle and Fitch,eds,Am.Chem.Soc.,1990;Bacillus thuringiensis:biology,ecology and safety,T.R.Glare and M.O’Callaghan,eds,John Wiley,2000;Antibody Phage Display,Methods and Protocols,Humana Press,2001;およびAntibodies,G.Subramanian,ed,Kluwer Academic,2004をはじめとする標準的な実験教書に記載されている。突然変異誘発は、例えば、部位特異的突然変異誘発(Kunkel et al.,Proc.Natl.Acad.Sci.USA 82:488−492(1985))を利用して実施することができる。PCR増幅法は、米国特許第4,683,192号、同第4,683,202号、同第4,800,159号、および同第4,965,188号に、ならびに「PCR Technology:Principles and Applications for DNA Amplification」,H.Erlich,ed,Stockton Press,New York(1989);およびPCR Protocols:A Guide to Methods and Applications,Innis et al.,eds,Academic Press,San Diego,Calif.(1990)をはじめとする複数の教書に記載されている。
【0088】
非相同性リーダー配列
ヒトVpreB1(CAG30495)の主なアイソフォームは、19アミノ酸のリーダー配列を含む145アミノ酸長のポリペプチド(図1の配列番号1)である。類似のリーダー配列が、別のVpreBポリペプチド内に存在する。ヒトトランケート型VpreB1配列(天然型VpreB1のC末端にある特徴的「テール」が欠如)は、「VpreB1 dTail配列」とも呼ばれ、配列番号5として示される。
【0089】
ヒトλ5(CAA10962)の主なアイソフォームは、30アミノ酸のリーダー配列を含む209アミノ酸長のポリペプチド(配列番号7)である。類似のリーダー配列が、別のλ5ポリペプチド内に存在する。ヒトトランケート型λ5配列(天然型λ5のN末端にある特徴的「テール」が欠如)は、「λ5 dTail配列」とも呼ばれ、配列番号9として示される。
【0090】
具体的には、天然型ヒトVκ様ポリペプチド配列としては、非限定的に、20アミノ酸のリーダー配列を含む配列番号11として示されるAJ004956のポリヌクレオチドによりコードされた、ヒトκ様ポリペプチド(配列番号12)が挙げられる。類似のリーダー配列が、別のVκ様ポリペプチド内に存在する。
【0091】
天然型配列JCκ様ポリペプチドとしては、非限定的に、潜在的な22アミノ酸のリーダー配列を含み、そのうちの15のアミノ酸が伝統的に組換えられたJCκ配列に特有に付加されている、プロトタイプリーダー配列(配列番号26)が欠如した、図5Aに示されたAAB32987 ヒトJCκポリペプチドが挙げられる。類似の組換えリーダー配列が、別のJCκポリペプチド内に存在する。
【0092】
本発明は、代替軽鎖構造体が、内在性リーダーVpreBリーダー配列および/もしくはλ5リーダー配列、または内在性Vκ様リーダー配列および/もしくはJCκリーダー配列を含む配列から生成された場合よりも高い収率で、代替軽鎖構造体を生成するための核酸およびポリペプチド構造体を提供する。本発明は、代替軽鎖構造体が、VpreB配列および/もしくはλ5の内在性リーダー、またはVκ様および/もしくはJCκの内在性リーダーのコード配列を含むか、あるいは内在性リーダー配列を含まずにDNA配列から生成された場合よりも高い収率で、代替軽鎖構造体を生成するためのベクター、宿主細胞および方法も提供する。より高い収率は、少なくとも1種の内在性分泌リーダー配列を、本発明の非相同性リーダー配列と交換することにより実現される。したがって本発明は、非相同性リーダー配列を含む、代替軽鎖および代替軽鎖構造体を提供する。
【0093】
好ましくは、非相同性リーダーペプチドにより実現される発現レベルは、発現が本質的に同じ条件下で実施される場合に、相同性リーダー配列を用いて実現される発現レベルよりも、少なくとも約5%高い、少なくとも約10%高い、少なくとも約20%高い、少なくとも約30%高い、少なくとも約40%高い、または少なくとも約50%高い。
【0094】
本発明において、非相同性リーダー配列は、天然型VpreBリーダー配列および/もしくは天然型λ5リーダー配列の代わりに代替軽鎖ポリペプチドのアミノ末端に融合されるか、または天然型Vκ様リーダー配列および/もしくは天然型JCκリーダー配列の代わりにκ様代替軽鎖ポリペプチドのアミノ末端に融合される。本発明人は、代替軽鎖配列(一緒に融合されるか、または非共有結合的に会合されている、VpreB/λ5配列またはVκ様/JCκ配列)および抗体重鎖配列を含む代替軽鎖配列構造体の生成時に、代替軽鎖の天然型リーダー配列とは対照的に、特定の非相同性リーダー配列が意外にも良好に機能することを発見した。
【0095】
本発明によれば、非相同性リーダー配列は、抗体軽鎖ならびにヒトおよび非ヒトホ乳類分泌タンパク質のリーダー配列をはじめとする、高度翻訳タンパク質からの任意のリーダー配列であってもよい。分泌タンパク質が含まれており、それらの配列は、一般のデータベース、例えばSwiss−Prot、UniProt、TrEMBL、RefSeq、EnsemblおよびCBI−Geneから入手できる。加えて、SPD、つまりウェブに基づく分泌タンパク質データベース、http://spd.cbi.pku.edu.cn.で入手できる、そのような配列の供給源である(Chen et al.,Nucleic Acids Res.,2005,33:D169−D173)。そのような分泌タンパク質としては、非限定的に、抗体、サイトカイン、リンフォカイン、モノカイン、ケモカイン、ポリペプチドホルモン、消化酵素、および細胞外マトリックスの成分が挙げられる。
【0096】
サイトカインのうちで含まれるのは、成長ホルモン、例えば、ヒト成長ホルモン、N−メチオニルヒト成長ホルモン、およびウシ成長ホルモン;副甲状腺ホルモン;チロキシン;インスリン;プロインスリン;リラキシン;プロリラキシン;糖タンパク質ホルモン、例えば、卵胞刺激ホルモン(FSH)、甲状腺刺激ホルモン(TSH)、および黄体形成ホルモン(LH);肝細胞増殖因子;線維芽細胞増殖因子;プロラクチン;胎盤性ラクトゲン;腫瘍壊死因子−αおよび−β(TNF−αおよび−β);ミュラー管抑制物質;マウスゴナドトロピン関連ペプチド;インヒビン;アクチビン;血管内皮増殖因子;インテグリン;トロンボポエチン(TPO);神経成長因子、例えば、NGF−β;血小板成長因子;トランスフォーミング増殖因子(TGF)、例えば、TGF−αおよびTGF−β;インスリン様成長因子−Iおよび−II;エリスロポエチン(EPO);骨誘導因子;インターフェロン、例えば、インターフェロン−α、−βおよび−γ;コロニー刺激因子(CSF)、例えば、マクロファージ−CSF(M−CSF);顆粒球−マクロファージ−CSF(GM−CSF);ならびに顆粒球−CSF(G−CSF);インターロイキン(IL)、例えば、IL−1、IL−1a、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12;腫瘍壊死因子、例えば、TNF−αまたはTNF−β;MIP−1α;MIP−1β;ならびにLIFおよびkitリガンド(KL)をはじめとする他のポリペプチド因子である。
【0097】
本発明の構造体での使用に適した追加のリーダー配列は、一般に入手可能なシグナルペプチドデータベース、例えば、http://proline.bis.nus.edu.sq/spdbでアクセス可能なSPdbシグナルペプチドデータベースに含まれる(Choo et al.,BMC Bioinformatics 2005,6:249参照)。
【0098】
適切な非相同性リーダー配列の具体的な例としては、非限定的に、ヒトおよび非ヒトホ乳類アルブミン、トランスフェリン、CD36、成長ホルモン、組織プラスミノーゲン活性化因子(t−PA)、エリスロポエチン(EPO)、ニューブラスチンのリーダー配列、ならびに別の分泌ヒトおよび非ヒトタンパク質からのリーダーペプチドが挙げられる。
【0099】
マウスIgκリーダー配列は、非相同性リーダー配列として用いられてもよい(METDTLLLWVLLLWVPGSTG−配列番号36)。
【0100】
非相同性リーダー配列が、i)VpreBおよびλ5の両方の代替軽鎖構造体、またはii)Vκ様およびJCκの両方の代替軽鎖構造体、に存在する場合、i)またはii)内の各非相同性リーダー配列は、他方と同一であってもよく、または他方と異なっていてもよい。
【0101】
天然型タンパク質からのシグナルペプチドに加えて、本発明の非相同性リーダー配列としては、消化されて、本質的に生じるリーダー配列の性能を更に改善することができ、本発明の代替軽鎖構造体の発現で用いられる宿主生物中での最良の性能のために特別に適合させることができる、合成およびコンセンサスリーダー配列が挙げられる。
【0102】
代替軽鎖構造体
本明細書の代替軽鎖(SLC)構造体は、抗体レパートリーの通常の発達時に生成されるプレB細胞受容体(プレBCR)に基づいている。抗体とは異なり、プレBCRは、2つの代替軽鎖成分、VpreBおよびλ5と対になった抗体重鎖で構成された三量体である。VpreBおよびλ5は両者とも、遺伝子再配列を受けない遺伝子によりコードされ、V(D)J組換えの開始前に初期のプロB細胞内で発現される。プレBCRは、1つの重鎖と、2つの非共有結合的に会合されたタンパク質:VpreBおよびλ5とで構成された成熟免疫グロブリンとは、構造的に異なっており、即ちそれらは、抗体内の2つの成分とは対照的に3つの成分を有する。更に、VpreBはVλIgドメインと相同性があり、λ5は抗体のCλドメインと相同性があるが、それぞれは、非標準のペプチド伸長を有し、VpreB1は、C末端に追加の21の残基を有し;λ5は、N末端に50アミノ酸の伸長を有する。Surrobodyの設計および生成の更なる詳細については、Xu et al.,Proc.Natl.Acad.Sci.USA 2008,105(31):10756−61、2008年10月2日公開のPCT公開第WO2008/118970号に提供されている。
【0103】
同様に、本明細書に記載されたκ様代替軽鎖構造体は、プレB細胞受容体(プレBCR)に基づいている。κ様軽鎖は、JCκ融合遺伝子と組になった生殖細胞系VκIV遺伝子である。これらの遺伝子のそれぞれにおいて、ペプチド伸長は、CDR3と類似した部位の付近に存在する。これらの2種のタンパク質は、ゲノムレベルで組換えられていないようであるため、重鎖への会合が相互に排他的で、λ様代替軽鎖に記載された会合と類似していると思われる。κ様代替軽鎖構造体の設計および生成の更なる詳細については、全体の開示が本明細書に参照により明確に組み込まれる、米国特許公開第2010−0062950号、およびXu et al.,J.Mol.Biol.2010,397,352−360に見出すことができる。
【0104】
本発明は、代替軽鎖(SLC)ポリペプチドと、非相同性シグナル配列を含む代替軽鎖配列を有するSLCポリペプチドを含有するSLC構造体とを企図する。一実施形態において、SLC構造体は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、λ5配列にコンジュゲートされたVpreB配列を含んでいてもよい。別の実施形態において、VpreB配列は、天然型VpreB1配列、天然型VpreB2配列、天然型VpreB3配列ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の一実施形態において、天然型VpreB配列は、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3、配列番号5のヒトVpreB様ポリペプチド、配列番号6のヒトVpreB dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。別の一実施形態において、λ5配列は、配列番号7のヒトλ5様、配列番号8のヒトλ5ポリペプチド、または配列番号9のヒトλ5 dTailポリペプチドの全てまたは一部を含む。
【0105】
本発明は、λ5配列およびVpreB配列が共有結合リンカーにより結合されている、SLC構造体も企図する。一実施形態において、本発明は、λ5配列がVpreB配列と非共有結合的に会合されている、SLC構造体を提供する。別の一実施形態において、本発明は、前記VpreB配列とλ5配列とのコンジュゲートが抗体重鎖配列と非共有結合的に会合されている、SLC構造体を企図する。
【0106】
本明細書に記載された通り、本発明は、SLCポリペプチドをコードする単離された核酸分子、およびSLCポリペプチドを含むSLC構造体に関する。一実施形態において、本発明は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、λ5配列に融合されたVpreB配列を含む代替軽鎖をコードする核酸を提供する。別の実施形態において、本発明は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、ペプチドまたはペプチドリンカーによりλ5配列に結合されたVpreB配列を含む代替軽鎖をコードする核酸を提供する。別の一実施形態において、本発明は、核酸を含むベクターを提供する。別の実施形態において、本発明は、核酸で形質転換された組換え宿主細胞を提供する。
【0107】
別の態様において、本発明は、代替軽鎖構造体のライブラリーを提供する。別の実施形態において、ライブラリーは、SLCをコードする核酸を含む。別の一実施形態において、ライブラリーは、ディスプレイの形態であってもよい。
【0108】
別の一態様において、本発明は、κ様代替軽鎖ポリペプチドと、κ様SLCポリペプチドを含むSLC構造体とを企図する。一実施形態において、本発明は、前記Vκ配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列にコンジュゲートされたVκ様配列を含むκ様SLC構造体に関する。別の実施形態において、Vκ様配列は、配列番号12〜24、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。他の一実施形態において、JCκ配列は、配列番号26〜39、ならびにそれらのフラグメントおよびバリアントからなる群より選択される。
【0109】
一実施形態において、本発明は、Vκ様配列が前記JCκ配列に融合されている、κ様SLC構造体を企図する。別の実施形態において、融合は、それぞれ前記Vκ様配列および前記JCκ配列のCDR3類似領域で、またはその周辺で起こる。一実施形態において、本発明は、前記Vκ様配列および前記JCκ配列が共有結合リンカーにより結合されている、κ様SLC構造体を企図する。
【0110】
一実施形態において、本発明は、前記Vκ様配列が前記JCκと非共有結合的に会合されている、κ様SLC構造体を提供する。一実施形態において、本発明は、前記Vκ様配列と前記JCκ配列とのコンジュゲートが抗体重鎖配列と非共有結合的に会合されている、κ様SLC構造体を提供する。
【0111】
一実施形態において、本発明は、前記分泌リーダー配列が合成配列であってもよい、κ様SLC構造体を提供する。一実施形態において、本発明は、前記分泌リーダー配列が天然型分泌リーダー配列のコンセンサス配列であってもよい、κ様SLC構造体を提供する。
【0112】
別の態様において、本発明は、κ様SLC構造体をコードする、単離された核酸を提供する。一実施形態において、本発明は、前記Vκ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列に融合されたVκ様配列を含むκ様代替軽鎖をコードする核酸を提供する。別の実施形態において、本発明は、前記Vκ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、ペプチドまたはペプチドリンカーによりJCκ配列に結合されたVκ様配列を含むκ様代替軽鎖をコードする核酸を提供する。別の一実施形態において、本発明は、核酸を含むベクターを提供する。別の実施形態において、本発明は、核酸で形質転換された組換え宿主細胞を提供する。
【0113】
一実施形態において、本発明は、κ様代替軽鎖構造体のライブラリーを提供する。別の実施形態において、ライブラリーは、κ様SLCをコードする核酸を含む。別の一実施形態において、ライブラリーは、ディスプレイの形態であってもよい。
【0114】
別の一態様において、本発明は、κ様SLCの発現方法を提供する。一実施形態において、本発明は、前記Vκ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列に共有結合されたVκ様配列を含むキメラ分子をコードする核酸で前記組換え宿主細胞を形質転換することを含む、組換え宿主細胞におけるκ様代替軽鎖の発現方法を提供する。一実施形態において、Vκ様配列は、JCκ配列に融合されている。別の実施形態において、Vκ様配列は、ペプチドまたはペプチドリンカーを介してJCκ配列に結合されている。別の実施形態において、組換え宿主細胞は、真核細胞である。一実施形態において、組換え宿主細胞は、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞である。
【0115】
別の一実施形態において、本発明は、配列番号6として示されたVpreB配列を含むSLC構造体を提供する。別の実施形態において、本発明は、配列番号10として示されたλ5配列を含むSLC構造体を提供する。一実施形態において、本発明は、配列番号35として示されたポリペプチドを含むSLC構造体を提供する。
【0116】
λ様Surrobodyの具体的な例としては、天然型配列のフラグメントおよびバリアントを含むVpreB配列、例えば、VpreB1、VpreB2、またはVpreB3配列が、天然型配列のフラグメントおよびバリアントを含むλ5配列にコンジュゲートされている、ポリペプチドが挙げられる。このタイプの代表的融合は、全体の開示が本明細書に参照により明確に組み込まれる、2008年10月2日公開のPCT公開第WO2008/118970号に提供されている。非相同性リーダー配列での融合の例は、図3(配列番号35)に示されている。
【0117】
直接の融合において、典型的にはVpreB配列のC末端(例えば、VpreB1、VpreB2、またはVpreB3配列)は、λ5配列のN末端に融合されている。完全長の天然型VpreB配列を完全長λ5配列に融合することは可能であるが(例えば、図7の最初の図解を参照)、典型的には融合は、2つのポリペプチドそれぞれのCDR3類似部位で、またはその周辺で起こる。VpreB1およびλ5の類似CDR3部位に基づく代表的な融合構造体を、図6に示す。この実施形態において、融合は、CDR3類似領域の片側の約10のアミノ酸残基の内部で、またはその内部のある位置で起こってもよい。好ましい実施形態において、融合は、天然型ヒトVpreB1配列(配列番号1)の約116アミノ酸残基から126アミノ酸残基の間、および天然型ヒトλ5配列(配列番号7)の約82アミノ酸残基から93アミノ酸残基の間で起こる。
【0118】
VpreB配列またはλ5配列を、それぞれ抗体λ軽鎖のCDR3領域または抗体軽鎖の可変領域に融合することもできる。VpreBおよびλ5の一方だけがトランケートされている追加の構造体も、図7に示す。類似の構造体を、抗体κ軽鎖配列を用いて調製することができる。κ様代替軽鎖構造体の例示を、図12〜19に見出すことができる。
【0119】
更なる直接融合構造を、図11の右側に示す。「SLC融合1」と称される構造は、トランケート型VpreB1配列(天然型VpreB1配列のC末端にある特徴的「テール」が欠如)の、同様にトランケートされたλ5配列への融合体が、抗体重鎖と非共有結合的に会合されている、2つの二量体で構成された四量体である。「SLC融合2」と称される構造は、トランケート型VpreB1配列(天然型VpreB1配列のC末端にある特徴的「テール」が欠如)の、抗体軽鎖構造体領域への融合体が、抗体重鎖と非共有結合的に会合されている、2つの二量体で構成された四量体である。「SLC融合3」と称される構造は、抗体軽鎖可変領域のトランケート型λ5配列(天然型λ5のN末端にある特徴的「テール」が欠如)への融合体が、抗体重鎖と非共有結合的に会合されている、2つの二量体で構成された四量体である。
【0120】
先に注記された通り、本発明のポリペプチド構造体は、直接融合に加えて、VpreB配列(天然型配列のフラグメントまたはバリアントを含む)と、異種配列、例えばλ5配列(天然型配列のフラグメントまたはバリアントを含む)および/または抗体配列との非共有結合的会合を包含する。したがって、例えば完全長VpreB配列が、トランケート型λ5配列と非共有結合的に会合されていてもよい。あるいはトランケート型VpreB配列が、完全長λ5配列と非共有結合的に会合されていてもよい。
【0121】
抗体重鎖との非共有結合的会合において、非共有結合的に会合されたVpreB1およびλ5配列を含む代替軽鎖構造体を、図11の左側に示す。様々な例示を示したが、構造が、例えば完全長VpreB1およびλ5配列、トランケート型λ5配列と会合された完全長VpreB1配列(「λ5 dT」)、完全長λ5配列と会合されたトランケート型VpreB1配列(「VpreB dT」)およびトランケート型λ5配列と会合されたトランケート型VpreB1配列(「短」)を包含してもよい。
【0122】
図11は、特定の具体的構造体を示しているが、様々な他の構造体を類似の様式で製造および使用しうることは、当業者には理解されよう。例えば構造は、図11に示された構造とは対照的に、非対称で、各アームに異なる代替軽鎖配列を含んでいても、そして/または三量体もしくは五量体構造を有していてもよい。
【0123】
本明細書の代替軽鎖構造体(Surrobody)の全てが、抗体配列と会合されていてもよい。例えば図9に示す通り、VpreB−λ5融合体は、ペプチドリンカーにより、抗体重鎖の可変領域配列に連結することができる。別の実施形態において、VpreB−λ5融合体は、抗体重鎖、または可変領域配列を含むそのフラグメントと非共有結合的に会合されて、二量体複合体を形成している。更に別の実施形態において、VpreBおよびλ5配列は、互いに、そして抗体重鎖または可変領域配列を含むそのフラグメントに、非共有結合的に会合されており、それにより三量体複合体を形成している。抗体重鎖を含む実例の構造体を、図11に示す。
【0124】
κ様Surrobodyの具体的例としては、天然型配列のフラグメントまたはバリアントを含むVκ様配列が、天然型配列のフラグメントまたはバリアントを含むJCκ配列にコンジュゲートされているポリペプチドが挙げられる。この型の代表的な融合体は、全体の開示が本明細書に参照により明確に組み込まれる、米国特許公開第2001−0062950号、およびXu et al.,J.Mol.Biol.2010,397,352−360に示されている。
【0125】
本明細書のポリペプチド構造体の具体的例としては、Vκ様および/またはJCκ配列が抗体重鎖、またはそのフラグメントに会合されているポリペプチドが挙げられる。Vκ様配列およびJCκ配列の両方を含む具体的なヘテロ二量体構造体を、図12に示す。図12に示す通り、本発明のκ様代替軽鎖構造体では、Vκ様ポリペプチドおよび/またはJCκポリペプチドは、類似の抗体配列内に存在しない、それぞれC−およびN−末端伸長を含有していてもよい。あるいは伸長の一部または全体が、本明細書のκ様代替軽鎖構造体から除去されていてもよい。
【0126】
個別に用いられうる、または追加の異種配列、例えば抗体重鎖配列、例えば、完全長抗体重鎖またはそのフラグメントにより、更に誘導体化および/または会合されうる他のκ様代替軽鎖構造体。
【0127】
Vκ様ポリペプチドおよび/またはJCκポリペプチドのC−およびN−末端伸長は、本発明の構造体に存在する必要はないが、図14に示す通り、ループライブラリーをスクリーニングした結果、それらは直線伸長、または例えば多様性に制約のある形態のいずれかにより、組み合わせ的機能多様性を創り出すための特有の好機を提供するため、少なくとも1種のそのような付加物の少なくとも一部を保持することが有益となる。加えて、Vκ様ポリペプチドおよび/またはJCκポリペプチドの「テール」部分が、他のペプチドおよび/またはポリペプチドに融合されて、様々な所望の特性、例えば、結合の強化、追加の結合特異性、pKの上昇、半減期の改善、半減期の短縮、細胞表面への固定、細胞内トランスロケーションの増加、ドミナントネガティブ活性などを提供することができる。具体的な機能的テール伸長を、図18に列挙する。
【0128】
所望なら、本発明の構造体を、例えば、公知の治療抗体をはじめとする抗体のCDR1、CDR2および/またはCDR3領域の公知配列または配列モチーフを、κ様代替軽鎖配列のCDR1、CDR2および/またはCDR3類似領域に組込む、または付加することにより、設計することができる。これにより、抗体ではないが、公知の治療抗体と同様の、またはそれを超えた結合特異性または親和力を示す分子を作製することができる。
【0129】
Vκ様およびJCκ遺伝子は、独立したタンパク質として、そして代替軽鎖として機能することができるポリペプチドをコードしているため、代替様の軽鎖を、真の軽鎖から設計することができ、そして設計された真の代替軽鎖のために提案されたこれまでの適用例に用いることができる。これは、VpreBまたはVκ様遺伝子のいずれかに類似したペプチド伸長を含む可変軽領域を発現することにより、果たすことができる。同様に、定常領域を、λ5またはJCκ遺伝子のいずれかおよびそれらのペプチド伸長と似せるように設計することができる。更に任意のキメラまたはヘテロ二量体の組み合わせは、本明細書の範囲内である。
【0130】
幾つかの実施形態において、SLC構造体は、異種アミノ酸配列または非SLCポリペプチドを含む。特定の実施形態において、異種アミノ酸配列は、1種以上の追加の機能性を、本発明の構造体に付加する。SLC構造体は、非SLCポリペプチドを含むように設計されてもよい。一実施形態において、非SLCポリペプチドは、第一のSLC成分および/または第二のSLC成分に融合されている。非SLCポリペプチドのアミノ末端が、第一のSLC成分のカルボキシ末端に融合されていてもよく、そして/または非SLCポリペプチドのカルボキシ末端が、第二のSLC成分のアミノ末端に融合されていてもよい。別の実施形態において、第一のSLC成分は、VpreBポリペプチドまたはVκ様ポリペプチドである。別の一実施形態において、第二のSLC成分は、λ5ポリペプチドまたはJCκポリペプチドである。
【0131】
所望の結合特異性を有する抗体可変領域配列を含む、追加の機能性を有するλ様SLC構造体を、図10に示す。詳細には図10は、VpreBおよびλ5に連結する融合タンパク質を作製するための抗VEGF一本鎖Fv(scFv)の挿入を示している(図10A)。こうして得られた、設計されたSLC制約scFvは、抗TNF−α抗体の重鎖と対をなしている。図10Bは、VpreBのC末端への抗VEGF scFvの融合を表わしている。図10Cは、λ5のアミノ末端への抗オボアルブミンscFvの融合を表わしている。TNF−αおよびオボアルブミンの両方に結合する能力を有する3部のタンパク質複合体を形成することができる。図10Dは、三特異性分子を作製するための、2つの融合構造体(VpreB−抗VEGF scFvおよびλ5−抗オボアルブミン)と抗TNF−α抗体の重鎖との組み合わせを表わしている。VpreBおよびλ5ポリペプチド配列を含む様々な二機能性および三機能性構造体を、そのような方策を用いて構築してもよい。加えて図8に表わされる通り、組み合わせ的機能多様性を、λ様SLC構造体に組込んでもよい。
【0132】
所望の結合特異性を有する抗体可変領域配列を含む追加の機能性を有するκ様SLC構造体を、図17に示す。詳細には図17は、本明細書の先に記載された、Vκ様およびJCκポリペプチド配列を含む様々な二機能性および三機能性構造体を示している。
【0133】
本発明の代替軽鎖(SLC)構造体は、二量体または2部構成で提供されてもよい。この構成の例を図9に提供し、それは、2部構成に対応するVpreB−λ5融合体および抗体重鎖(右側、下から二番目の図)のタンパク質構造を示している。SLC構造体は、三量体または3部構成で提供されてもよい。図9は、3部構成に対応するVpreB、λ5、および抗体重鎖(右側、一番下の図)のタンパク質構造を示している。
【0134】
本発明の代替軽鎖(SLC)構造体は、二機能または二特異性形態で提供されてもよい。図20は、この例を示し、(A)は、Surrobody構成を表しており、(B)は、二機能性および二特異性Surrobody構成を表わしている。図20(B)に示される通り、SLC構造体は、SLCポリペプチド成分(例えば、VpreB、λ5、Vκ様、JCκポリペプチド、またはそれらのフラグメントもしくはバリアント)、および非SLC分子を有するSLC融合ポリペプチドを含んでいてもよい。一実施形態において、非SLC分子は、特定の機能を有する任意のポリペプチドであってもよい。別の実施形態において、ポリペプチドは、追加の機能性を提供しうるサイトカインであってもよい。別の実施形態において、非SLCポリペプチドは、追加の特異性を提供しうる抗体フラグメントであってもよい。図20(C)は、実例のSLC融合クローニング方策およびその各アミノ酸を表わしている。斜線のエリアは、VpreB(アミノ酸120〜145)およびλ5(アミノ酸38〜92)の非免疫グロブリンテール領域を表す。「L」は、内在性リーダー配列を示し、「mL」は、合成Igκリーダー配列を示す。「融合」は、非SLC分子など、該当する遺伝子の融合部位を示す。
【0135】
代替軽鎖構造体の調製
核酸コード代替軽鎖、例えば、VpreBおよびλ5ポリペプチド、またはVκ様もしくはJCκポリペプチドを、天然供給源、例えば、発達中のB細胞から単離することができ、そして/または合成もしくは半合成法により得ることができる。このDNAが同定され単離されるか、さもなければ生成されれば、更なるクローニングまたは発現のために複製可能なベクターにライゲートすることができる。
【0136】
本明細書のポリペプチドのコード配列を発現するために用いうるクローニングおよび発現ベクターは、当該技術分野で周知であり、市販されている。一般にベクター成分は、非限定的に、以下のものの1つ以上が挙げられる:シグナル配列、複製起点、1つ以上のマーカー遺伝子、エンハンサー要素、プロモータ、および転写終結配列。本明細書のベクター内の代替軽鎖構造体をコードするDNAをクローニングまたは発現するのに適した宿主細胞は、原核生物、酵母、またはより高等な真核生物(ホ乳類)細胞であり、ホ乳類細胞が好ましい。
【0137】
適切なホ乳類宿主細胞系の例としては、非限定的に、SV40により形質転換されたサル腎臓CV1系(COS−7、ATCC CRL 1651);懸濁培養での増殖用にサブクローニングされたヒト胚性腎(HEK)細胞系293(HEK293細胞)、(Graham et al.J.Gen Virol.36:59(1977));ベビーハムスター腎臓細胞(BHK、ATCC CCL 10);チャイニーズハムスター卵巣細胞/−DHFR(CHO、Urlaub et al.,Proc.Natl.Acad.Sci.USA 77:4216(1980));マウスセルトリ細胞(TM4、Mather,Biol.Reprod.23:243−251(1980));サル腎臓細胞(CV1 ATCC CCL 70);アフリカミドリザル腎臓細胞(VERO−76、ATCC CRL−1587);ヒト子宮頸癌細胞(HELA、ATCC CCL 2);イヌ腎臓細胞(MDCK、ATCC CCL 34);バッファロー系ラット肝臓細胞(BRL 3A、ATCC CRL 1442);ヒト肺細胞(W138、ATCC CCL 75);ヒト肝臓細胞(Hep G2、HB 8065);マウス乳腺腫(MMT 060562、ATCC CCL 51);TRI細胞(Mather et al.,Annals N.Y.Acad.Sci.383:44−68(1982));MRC5細胞;FS4細胞;およびヒトヘパトーマ系(Hep G2)が挙げられる。
【0138】
ホ乳類細胞内での使用に関しては、発現ベクターでの制御機能が、多くの場合、ウイルス材料により提供される。したがって、一般に用いられるプロモータは、ポリオーマ、アデノウイルス2、レトロウイルス、サイトメガロウイルス、およびシミアンウイルス40(SV40)のゲノムから得ることができる。他のプロモータ、例えば、β−アクチンプロモータは、非相同性供給源から生成させる。適切なプロモータの例としては、非限定的に、SV40ウイルスの初期または後期プロモータ(Fiers et al.,Nature,273:113(1978))、ヒトサイトメガロウイルスの最初期プロモータ(Greenaway et al.,Gene,18;355−360(1982))、ならびに通常は所望の遺伝子配列に関連するプロモータおよび/または制御配列が挙げられるが、但し、そのような制御配列が、宿主細胞系と適合することが前提である。
【0139】
高等な真核生物による、所望の非相同性ポリペプチドをコードするDNAの転写は、エンハンサー配列をベクターに挿入することにより増加する。エンハンサーは、プロモータ上で作用してその転写開始活性を高める、通常は10〜300bpのDNAのシス作用性要素である。エンハンサーは、相対的に配列および位置非依存性であるが、好ましくは発現ベクター内に存在するプロモータ配列の上流に位置する。エンハンサーは、プロモータと同じ供給源、例えば真核細胞ウイルスから、例えば、複製起点の後期側のSV40エンハンサー(bp100〜270)、サイトメガロウイルス初期プロモータエンハンサー、複製起点の後期側のポリオーマエンハンサー、およびアデノウイルスエンハンサーから生成させてもよい。
【0140】
ホ乳類宿主細胞内で用いられる発現ベクターは、ポリアデニル化部位、例えば、SV40(初期および後期)またはHBVなどのウイルス由来のものも含んでいる。
【0141】
複製起点は、外来起源、例えばSV40または他のウイルス(例えば、ポリオーマ、アデノ、VSV、BPV)供給源から得ることができる、または宿主細胞により提供することができる、ベクターの構築により提供されてよい。
【0142】
発現ベクターは、通常、ベクターで形質転換された宿主細胞の生存または発育に必要なタンパク質をコードする選択マーカーを含む。ホ乳類細胞に適した選択マーカーの例としては、ジヒドロ葉酸還元酵素(DHFR)、チミジンキナーゼ(TK)、およびネオマイシンが挙げられる。
【0143】
適切なホ乳類発現ベクターは、当該技術分野で周知であり、市販されている。したがって例えば、本発明の代替軽鎖構造体は、ヒトサイトメガロウイルス(CMV)最初期エンハンサー/プロモータ領域を含むpCI発現ベクター(プロメガ)を用いてホ乳類宿主細胞内で産生させて、DNA挿入部の構成的発現を促進することができる。ベクターは、pTT5発現ベクター(カナダ国立研究機構)であってもよい。ベクターは、ネオマイシンホスホトランスフェラーゼ遺伝子を選択マーカーとして含むことができる。
【0144】
本発明の代替軽鎖構造体は、細菌宿主細胞内で産生することもできる。細菌系内で使用される制御要素としては、場合によりオペレータ−配列を含むプロモータ、およびリボソーム結合部位が挙げられる。適切なプロモータとしては、非限定的に、ガラクトース(gal)、ラクトース(lac)、マルトース、トリプトファン(trp)、β−ラクタマーゼプロモータ、バクテリオファージλ、およびT7プロモータが挙げられる。加えて合成プロモータ、例えばtacプロモータを用いることができる。細菌系内で使用されるプロモータは、一般に、Fab分子をコードするDNAに作動可能に連結されたシャイン・ダルガルノ(SD)配列も含有する。プラスミドpBR322からの複製起点は、ほとんどのグラム陰性菌に適している。
【0145】
抗体代替軽鎖配列を含む多重鎖構造体内の各鎖のコード配列は、真核生物および原核生物の宿主をはじめとする所望の宿主細胞をコトランスフェクトするのに用いられる、別個の調節配列の制御下では、または別個の発現ベクター内では、同じ発現ベクター内に存在することができる。したがって、ノバゲン(Novagen)から市販されるDuet(商標)ベクターを用いて、同義遺伝子を共発現させることができる。
【0146】
形質転換された宿主細胞は、様々な培地で培養させてもよい。ホ乳類宿主細胞を培養するための市販の培地としては、Ham’s F10(シグマ)、最小必須培地(MEM)(シグマ)、RPMI−1640(シグマ)、およびダルベッコ改変イーグル培地((DMEM),(シグマ)が挙げられる。加えてHam et al.,Meth.Enz.58:44(1979)およびBarnes et al.,Anal.Biochem.102:255(1980)に記載された培地のいずれも、宿主細胞用の培地として用いることができる。培養条件、例えば温度、pHなどは、発現用に選択される宿主細胞によって過去に利用されたものであり、製造業者の説明書に含まれるか、さもなければ当業者には自明であろう。
【0147】
ホ乳類、細菌(例えば大腸菌)、または他の宿主細胞を培養するのに更に適した培地は、標準の教書、例えば、Sambrook et al.の上記文献またはAusubel et al.の上記文献にも記載されている。
【0148】
一態様において、本発明は、組換え宿主細胞内での代替軽鎖の発現方法を提供する。一実施形態において、その方法は、SLCポリペプチドまたはSLC融合ポリペプチドをコードする核酸を提供するステップを含む。別の実施形態において、その方法は、組換え宿主細胞を、SLCポリペプチドまたはSLC融合ポリペプチドをコードする核酸で形質転換またはトランスフェクトするステップを含む。一実施形態において、SLC融合ポリペプチドをコードする核酸は、第一のSLC配列および/または第二のSLC配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、第二のSLC配列に共有結合された第一のSLC配列を含むキメラ分子である。第一のSLC配列は、VpreB配列、Vκ様配列、またはそれらの融合ポリペプチドであってもよい。第二のSLC配列は、λ5配列、JCκ配列、またはそれらの融合ポリペプチドであってもよい。
【0149】
一実施形態において、VpreB配列は、前記VpreB配列および/または前記λ5配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、λ5配列に共有結合されている。別の実施形態において、VpreB配列は、λ5配列に融合されている。別の一実施形態において、VpreB配列は、ペプチドまたはペプチドリンカーを介して、λ5配列に結合されている。別の一実施形態において、Vκ様配列は、前記κ様配列および/または前記JCκ配列の天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、JCκ配列に共有結合されている。別の医師実施形態において、Vκ様配列は、JCκ配列に融合されている。別の実施形態において、Vκ様配列は、ペプチドまたはペプチドリンカーを介して、JCκ配列に結合されている。
【0150】
全ての実施形態において、発現方法は、代替軽鎖ポリペプチドおよび/または代替軽鎖融合ポリペプチドをはじめとする、代替軽鎖ポリペプチドをコードする1つ以上の核酸で宿主細胞を形質転換またはトランスフェクトするステップを含んでいてもよい。
【0151】
全ての実施形態において、その方法は、抗体重鎖をコードする核酸で宿主細胞を形質転換またはトランスフェクトするステップを更に含んでいてもよい。
【0152】
一態様において、本発明は、改善された収率を有する代替軽鎖ポリペプチドおよび/または代替軽鎖融合ポリペプチドの発現方法を提供する。一実施形態において、天然型リーダー配列の代わりに非相同性リーダー配列を用いる本発明の方法は、天然型リーダー配列を非相同性リーダー配列で交換されていない方法よりも、大きなポリペプチド発現および収率を特徴とする。
【0153】
一実施形態において、組換え宿主細胞は、細菌細胞である。別の実施形態において、宿主細胞は、真核細胞である。一実施形態において、組換え宿主細胞は、チャイニーズハムスター卵巣(CHO)細胞、またはヒト胚性腎(HEK)293細胞である。
【0154】
一態様において、本発明は、本明細書に記載された核酸を含有する宿主細胞を提供する。一実施形態において、本発明は、本明細書に記載された少なくとも1種の核酸で形質転換された組換え宿主細胞を提供する。別の一実施形態において、宿主細胞は、非SLC分子を含んでいても、または含んでいなくてもよい、SLC融合をコードする核酸で形質転換されている。
【0155】
全ての実施形態において、宿主細胞は、抗体重鎖をコードする核酸で更に形質転換されている。
【0156】
全ての実施形態において、本発明は、本明細書に記載された核酸を含有するベクターを提供する。全ての実施形態において、宿主細胞は、本明細書に記載された核酸を含有する少なくとも1種のベクターで形質転換されている。
【0157】
精製は、当該技術分野で公知の方法により実施することができる。好ましい実施形態において、代替軽鎖構造体は、Ni−NTA精製システム(インビトロジェン)を用いて6×Hisタグ形態で精製される。
【0158】
κ様SLC分子は、既存の軽鎖V遺伝子および軽鎖定常遺伝子から設計することができる。図15に示す通り、軽鎖は、遺伝子再配列およびRNAプロセッシングの生成物である。κ様SLC分子の成分は、再配列されていない軽鎖V遺伝子および再配列された軽鎖JC遺伝子からの別の機能を提供するため、残りのκおよびλ軽鎖V遺伝子全てから類似の翻訳タンパク質を設計して、Vκ様分子(図16A)、ならびに残りのκJC再配列(4JCκ様)(図16B)と、λJC再配列(4“J”×10“定数”=40JCλ様)(図16B)との全ての組み合わせを作製することが実行可能である。これらの設計分子のそれぞれ1つは、VpreBおよびλ5、ならびにそれらの組み合わせおよびキメラを用いて、Vκ様およびJCκを用いたもの、および2008年10月2日公開のPCT公開WO2008/118970に含まれるものと類似の目的を果たすことができる。
【0159】
本発明の代替軽鎖を用いて、疾患の予防および/または処置のための分子を構築することができる。そのような適用では、代替軽鎖を含有する分子は、通常、医薬組成物の形態で用いられる。技術および配合は、一般に、Remington’s Pharmaceutical Sciences,18th Edition,Mack Publishing Co.(Easton,PA.1990)に見出してもよい。Wang and Hanson「Parenteral Formulations of Proteins and Peptides:Stability and Stabilizers,」Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)も参照されたい。
【0160】
ポリペプチドを基剤とする医薬組成物は、典型的には、凍結乾燥された配合剤または水性配合剤の形態で配合される。許容されうる担体、賦形剤、または安定化剤は、用いられる投与量および濃度ではレシピエントに対して非毒性であり、緩衝剤、例えば、リン酸塩、クエン酸塩、および他の有機酸;アスコルビン酸およびメチオニンをはじめとする抗酸化剤;防腐剤(例えば、塩化オクタデシルジメチルベンジルアンモニウム;塩化ヘキサメトニウム;塩化ベンザルコニウム;塩化ベンゼトニウム;フェノール、ブチルもしくはベンジルアルコール;アルキルパラベン、例えば、メチルもしくはプロピルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;およびクレゾール);低分子量(約10残基未満)のポリペプチド;タンパク質、例えば、血清アルブミン、ゼラチン、もしくは免疫グロブリン;親水性ポリマー、例えば、ポリビニルピロリドン;アミノ酸、例えば、グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、もしくはリシン;単糖類、二糖類、およびグルコース、マンノース、もしくはデキストリンをはじめとする他の炭水化物;キレート化剤、例えば、EDTA;糖類、例えば、ショ糖、マンニトール、トレハロースもしくはソルビトール;塩形成対イオン、例えば、ナトリウム;金属錯体(例えば、Zn−タンパク質錯体);ならびに/または非イオン性界面活性剤、例えば、TWEEN(商標)、PLURONICS(商標)もしくはポリエチレングリコール(PEG)が挙げられる。
【0161】
その分子は、例えば、コロイド状薬物送達系(例えば、リポソーム、アルブミンマイクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)、またはマクロエマルジョン中で、コアセルベーション技術、または界面重合(例えば、ヒドロキシメチルセリロースまたはゼラチン−マイクロカプセルおよびポリ(メチルメタクリラート)マイクロカプセル)により、マイクロカプセルに封入してもよい。そのような技術は、上記のRemington’s Pharmaceutical Sciencesに開示されている。
【0162】
本明細書に開示された代替軽鎖を含有する分子は、免疫リポソームとして配合されてもよい。分子を含有するリポソームは、Epstein et al.,Proc.Natl.Acad.Sci.USA 82:3688;Hwang et al,Proc.Natl.Acad.Sci.USA 77:4030(1980);米国特許第4,485,045号、同第4,544,545号および1997年10月23日公開のWO97/38731に記載されたものなど、当該技術分野で公知の方法により調製される。循環時間を高めたリポソームは、米国特許第5,013,556号に開示されている。
【0163】
特に有用なリポソームは、ホスファチジルコリン、コレステロールおよびPEG誘導体化ホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を用いる逆相エバポレーション法により生成させることができる。リポソームは、画定された孔径のフィルターから押出されて、所望の口径を有するリポソームが生成される。本発明の分子のフラグメントは、ジスルフィド交換反応を介してリポソームにコンジュゲートすることができる(Martin et al.J.Biol.Chem.257:286−288(1982))。化学療法剤が、場合によりリポソーム中に含有されている。Gabizon et al.J.National Cancer Inst.81(19)1484(1989)を参照されたい。
【0164】
疾患の予防または処置では、分子の適切な投与量は、処置される感染のタイプ、疾患の重症度および経過、ならびに抗体が予防目的または治療目的で投与されるか否かに依存する。分子は、一度に、または一連の処置を通して、適切に患者に投与される。疾患のタイプおよび重症度にもよるが、例えば、1回以上の別々の投与または連続注入のいずれにしろ、抗体約1μg/kg〜約15mg/kgが、患者に投与される典型的な初期候補投与量である。
【0165】
本発明の代替軽鎖を含有する分子は、疾患の処置または予防における使用に適している。一実施形態において、本発明は、医薬として使用される、または疾患の処置に使用される代替軽鎖含有分子を提供する。別の実施形態において、本発明は、疾患を処置する医薬の製造のための、代替軽鎖含有分子の使用を提供する。その分子は、SLCペプチドまたはSLC融合体をコードする核酸であってもよい。
【0166】
一態様において、本発明は、治療上効果的な量の代替軽鎖含有分子をホ乳類に投与するステップを含む、ホ乳類における疾患を処置するのに有用な方法を提供する。治療組成物を、医師の指示通り、短期(急性)もしくは慢性で、または間欠的に投与することができる。
【0167】
本発明は、疾患の処置、予防および/または診断に有用な材料を含有する製造品のキットおよび物品も提供する。キットは、容器と、容器の表面に設置しうる、または容器と結合しうるラベルとを含んでいる。容器は、瓶、バイアル、注射器、または任意の他の適切な容器であってもよく、様々な材料、例えばガラスまたはプラスチックから形成させてもよい。容器は、本明細書に記載された代替軽鎖含有分子を有する組成物を保持しており、滅菌されたアクセスポートを有していてもよい。容器の例としては、静注液バッグ、または皮下注射針により穿孔することができるストッパー付きバイアルが挙げられる。キットは、様々な試薬、例えば希釈剤および緩衝剤を保持する追加の容器を有していてもよい。ラベルは、組成物および目的の使用のための説明書の記載を提供してもよい。分子を含有するキットは、細胞からのポリペプチドの精製または免疫沈降法のための、例えば細胞アッセイ用に、使用が見出される。例えば、タンパク質の単離および精製では、キットは、ビーズ(セファロースビーズ)に結び付いたタンパク質に結合する代替軽鎖含有分子を含有することができる。インビトロで、例えばELISAまたはウェスタンブロットにおいて、タンパク質の検出および定量用の分子を含有するキットを提供することができる。検出に有用なそのような分子は、標識、例えば蛍光または放射線標識と共に提供されてもよい。
【0168】
キットは、本明細書に記載された代替軽鎖を活性剤として含む分子を含む少なくとも1つの容器を有する。ラベルを提供して、組成物が疾患の処置に使用しうることを示してもよい。ラベルは、処置を必要とする対象へ投与するための説明書を提供してもよい。キットは、薬学的に許容しうる緩衝剤、例えば、注射用静菌水(BWFI)、リン酸緩衝生理食塩水、リンガー溶液およびデキストロース溶液を有する追加の容器を更に含んでいてもよい。最後に、キットは、別の緩衝剤、希釈剤、フィルター、針、および注射器をはじめとする任意の別の適切な材料を含有していてもよい。
【0169】
本発明の更なる詳細を、以下の非限定的実施例において提供する。
【0170】
実施例1−HEK293細胞における一過性発現
Surrobodyの上清を、ヒト胚性腎293(HEK293)細胞において一過性に生成させた。pTT5プラスミド(カナダ国立研究機構)を用いて、組換えSurrobody軽鎖構造体を提供した。非相同性リーダー配列METDTLLLWVLLLWVPGSTG(配列番号36−マウスIgκリーダー配列)での天然型リーダー配列の置換を行った、または行わなかったプラスミドの代替軽鎖核酸配列を提供した。以下の核酸配列を含有するpTT5プラスミドを用いた:(a)天然型リーダー配列(配列番号1−図1)もしくは配列番号36の非相同性マウスIgκリーダー配列を含むヒトVpreB1、(b)天然型リーダー配列(配列番号8−図2)もしくは配列番号36の非相同性マウスIgκリーダー配列を含むヒトλ5、または(c)VpreB1およびλ5と、天然型VpreB1リーダー配列もしくは配列番号36の非相同性マウスIgκリーダー配列との融合(配列番号35−図3)。(a)および(b)のプラスミドは、3部Surrobody構成に対応し、(c)のプラスミドは、3部Surrobody構成に対応する。これらのプラスミドを、抗体重鎖を含有するpTT5プラスミドでコトランスフェクトした。
【0171】
Surrobodyを、本質的には過去にXu et al.,(2008).Proc.Natl.Acad.Sci.USA,105:10756−10761;Kashyap et al.,(2008).Proc.Natl.Acad.Sci.USA,105:5986−5991に記載された通り、HEK293のFreestyleに基づく系(インビトロジェン)で一過性に生成させた。HEK293細胞を0.25〜2.0×106細胞/mlの密度で増殖培地中で増殖させ、その後、トランスフェクションの1日前に、増殖培地90mlを含む新しい振とうフラスコに0.75×106細胞/mlの密度で接種した。一晩発育させた後、細胞密度を1〜1.5×106細胞/mlであることを検証した。発現では、pTT5発現ベクターを用いた。次に、DNAトランスフェクション剤混合物を、以下の通り調製した。プラスミドDNA(pTT5−SLC分子)合計0.1mgに対応するDNA溶液を10ml遠沈管内で混合して、増殖培地で最終容量を5mLにした。2部Surrobody構成では、抗体重鎖を含有するプラスミド0.05mgを、VpreB1−λ5キメラプラスミド(Surrobody融合体)0.05mgと混合した。3部Surrobody構成では、抗体重鎖を含有するプラスミド0.033mgを、VpreB1プラスミド0.033mgおよびλ5プラスミド0.033mgと混合した。次に、増殖培地4.8mlをPEI 0.2mlと混和してプラスミドDNA溶液に添加することにより、ポリエチレンイミン(PEI)トランスフェクション溶液を調製した。混合物を力強く1〜2秒間ボルテックス処理した。室温で15分間インキュベートした後、プラスミドDNA−PEI混合物を、10mlピペットで1〜1.5×106細胞/mlの密度のHEK293細胞を含むフラスコに移した。フラスコを直ちに回転させて、振とうインキュベータに移動させた。125rpmのシェーカープラットホームの付いた、37℃、5%CO2の加湿インキュベータで、細胞を6日間発育させた。得られた培養上清におけるタンパク質生成レベルを、定量的反応速度分析(フォルテバイオのOctet:抗Fcセンサー)により決定した。下の表に示す通り、マウスIgκ軽鎖リーダー配列を置換したことで、一過性組換えSurrobody発現レベルが改善している。表1(mg/L)に示す通り、タンパク質レベルが少なくとも20倍改善された。
【0172】
表1
【0173】
3部および2部Surrobodyの複数回のトランスフェクションから得られた精製タンパク質を更に分析すれば、高レベルの収量が裏づけられる。非相同性リーダーを用いた、独立したトランスフェクションのタンパク質収量を、4ヶ月の期間にわたりモニタリングした。プロテインAまたはプロテインGのいずれかのクロマトグラフィー担体および低pHでの溶出を用いた高速タンパク質液体クロマトグラフィー(FPLC)システムで、タンパク質を精製した。表2(mg/L)に示される通り、どちらの構成においても、平均収量は、内在性代替軽鎖リーダー配列を用いて認められたものよりも実質的に高くなった。
【0174】
表2
【0175】
以下の表3は、表2で平均をとった、様々な2部および3部SLC形態で測定された各濃度を提供している。先に記載された通り、一般に2部構成は、Surrobody軽鎖融合体および1つの抗体重鎖を含み、3部構成は、2つのSurrobodyポリペプチドおよび抗体重鎖を含む。1列目の1〜47行目は、2部構成を有する47の異なる構造体に対応し、1列目の49〜63行目は、3部構成を有する15の異なる構造体に対応する。4列目は、検査されたSurrobodyの特徴の幾つかを提供している。「Surrobody」は、2つのプロペプチドおよび1つの重鎖で作製されたSurrobody構造体である。「融合体」は、2つのSLCポリペプチドおよび1つの重鎖の融合体で作製されたSurrobody構造体である。「ペプチドタグを含む融合体」は、エピトープタグが組込まれたSurrobody構造体である。「機能性ペプチド融合体」は、特定の機能を有する少なくとも1つの非SLCポリペプチドが組込まれているSurrobody構造体である。
【0176】
表3
【0177】
表3は、収率の改善が、SLCポリペプチドと、SLC融合ポリペプチドと、非SLC分子を含有するSLC融合ポリペプチドと、を含むSurrobodyをはじめとする複数のSurrobody構成で得ることができる証拠を提供している。
【0178】
Surrobody分子を、チャイニーズハムスター卵巣K1(CHO−K1)細胞中でも一過性に発現させることができる。Surrobodyの上清を、(CHO−K1)細胞中で一過性に産生してもよい。組換えSLCポリペプチドまたは非SLCポリペプチドのプラスミドを、製造業者の説明書に従って、10%ウシ胎仔血清を補充されたダルベッコ改変イーグル培地/F12培地中のLipofectamine−2000(インビトロジェン、カタログ番号11668−027)を用いて、抗体重鎖を含有するプラスミドとコトランスフェクトした。37℃、5%CO2で一晩インキュベートした後、培地をGlutamax−1(インビトロジェン、カタログ番号12362)を含む新しいOpti−MEMI血清使用量低減培地と交換した。トランスフェクトされた上清を72時間後に回収して、0.22μmフィルター装置でろ過した。
【0179】
実施例2−CHO細胞における安定発現
真核生物コドンを最適化させた可溶性分泌遺伝子(DNA2.0)として、ホ乳類で発現された、またはデノボ合成で生成された代替軽鎖構造体を、ホ乳類タンパク質発現のために、pCIプラスミド(プロメガ)にサブクローニングした。製造業者の指導書に従ってチャイニーズハムスター卵巣(CHO−K1)細胞(インビトロジェン)にトランスフェクトする前に、配列を検証した。製造業者の指導書に従って、DNAおよびLipofectamine 2000(インビトロジェン)を合計32μgとし、所望の代替軽鎖を同量用いて、T−75フラスコ中の80%コンフルエントの細胞のトランスフェクションを実施した。トランスフェクションごとにOpti−MEMI 20ml中に、細胞にタンパク質を産生させた。4日後に、ニッケルキレートクロマトグラフィー(Ni−NTAアガロース、キアゲン)を用いて、分泌されたSurrobodyを培養上清から精製した。得られた精製Surrobodyを、遠心式サイズろ過(Centricon Plus−20)を用いて滅菌PBSに緩衝液交換して、A280読み取り、SDSゲル、またはウェスタンブロット分析により、公知の標準物質と比較して、それらのタンパク質濃度を決定した。
【0180】
前述の記載では本発明は、特定の実施形態を参照して示したが、それはそのように限定されるものではない。事実、図示されたものおよび本明細書に記載されたものに加えて、本発明の様々な改変が、前述の説明から当業者に自明となり、それは添付の特許請求の範囲に含まれる。
【0181】
本明細書に引用された個々の発行物、特許および特許出願が、具体的かつ個別に参考として本明細書に組込まれていることを示されたのと同程度に、全ての発行物、特許および特許出願は、全ての目的でその全体が本明細書に参照により組み込まれる。
【特許請求の範囲】
【請求項1】
代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、SLCポリペプチドをコードする単離された核酸分子。
【請求項2】
SLCポリペプチドが、VpreBポリペプチドを含む、請求項1記載の単離された核酸分子。
【請求項3】
SLCポリペプチドが、λ5ポリペプチドである、請求項1記載の単離された核酸分子。
【請求項4】
VpreBポリペプチドが、天然型VpreB1配列、天然型VpreB2配列、天然型VpreB3配列、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項2記載の単離された核酸分子。
【請求項5】
天然型VpreB配列が、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3、配列番号5のヒトVpreB様ポリペプチド、配列番号6のヒトVpreB dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項4記載の単離された核酸分子。
【請求項6】
λ5ポリペプチドが、配列番号7のヒトλ5様、配列番号8のヒトλ5ポリペプチド、配列番号9のヒトλ5 dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項3記載の単離された核酸分子。
【請求項7】
SLCポリペプチドが、VpreB−λ5ポリペプチド融合体を含む、請求項1記載の単離された核酸分子。
【請求項8】
VpreBポリペプチド配列およびλ5ポリペプチド配列の融合が、それぞれ前記VpreB配列および前記λ5配列のCDR3類似領域で、またはその周辺で起こる、請求項7記載の単離された核酸分子。
【請求項9】
VpreBポリペプチド配列が、カルボキシ末端で、λ5ポリペプチド配列のアミノ末端に連結されている、請求項7記載の単離された核酸分子。
【請求項10】
SLCポリペプチドが、Vκ様ポリペプチドを含む、請求項1記載の単離された核酸分子。
【請求項11】
SLCポリペプチドが、JCκポリペプチドを含む、請求項1または10記載の単離された核酸分子。
【請求項12】
Vκ様ポリペプチド配列が、配列番号12〜24、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項10記載の単離された核酸分子。
【請求項13】
JCκポリペプチド配列が、配列番号26〜39、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項11記載の単離された核酸分子。
【請求項14】
SLCポリペプチドが、Vκ様−JCκポリペプチド融合体である、請求項1記載の単離された核酸分子。
【請求項15】
Vκ様ポリペプチド配列およびJCκポリペプチド配列の融合が、それぞれ前記Vκ様配列および前記JCκ配列のCDR3類似領域で、またはその周辺で起こる、請求項14記載の単離された核酸分子。
【請求項16】
Vκ様ポリペプチド配列が、カルボキシ末端で、JCκポリペプチド配列のアミノ末端に連結されている、請求項14記載の単離された核酸分子。
【請求項17】
前記非相同性分泌リーダー配列が、抗体、サイトカイン、リンフォカイン、モノカイン、ケモカイン、ポリペプチドホルモン、消化酵素、および細胞外マトリックスの成分からなる群より選択される分泌されたポリペプチドのリーダー配列である、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項18】
前記サイトカインが、成長ホルモン、例えば、ヒト成長ホルモン、N−メチオニルヒト成長ホルモン、およびウシ成長ホルモン;副甲状腺ホルモン;チロキシン;インスリン;プロインスリン;リラキシン;プロリラキシン;糖タンパク質ホルモン、例えば、卵胞刺激ホルモン(FSH)、甲状腺刺激ホルモン(TSH)、および黄体形成ホルモン(LH);肝細胞増殖因子;線維芽細胞増殖因子;プロラクチン;胎盤性ラクトゲン;腫瘍壊死因子−αおよび−β(TNF−αおよび−β);ミュラー管抑制物質;マウスゴナドトロピン関連ペプチド;インヒビン;アクチビン;血管内皮増殖因子;インテグリン;トロンボポエチン(TPO);神経成長因子、例えば、NGF−β;血小板成長因子;トランスフォーミング増殖因子(TGF)、例えば、TGF−αおよびTGF−β;インスリン様成長因子−Iおよび−II;エリスロポエチン(EPO);骨誘導因子;インターフェロン、例えば、インターフェロン−α、−βおよび−γ;コロニー刺激因子(CSF)、例えば、マクロファージ−CSF(M−CSF)、顆粒球−マクロファージ−CSF(GM−CSF)、および顆粒球−CSF(G−CSF);インターロイキン(IL)、例えば、IL−1、IL−1a、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12;腫瘍壊死因子、例えば、TNF−αまたはTNF−β;MIP−1α;MIP−1β;ならびにLIFおよびkitリガンド(KL)をはじめとする他のポリペプチド因子からなる群より選択される、請求項17記載の単離された核酸分子。
【請求項19】
前記分泌リーダー配列が、ヒトおよび非ヒトホ乳類アルブミン、トランスフェリン、CD36、成長ホルモン、組織プラスミノーゲン活性化因子(t−PA)、エリスロポエチン(EPO)、ならびにニューブラスチンのリーダー配列からなる群より選択される、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項20】
前記分泌リーダー配列が、合成配列である、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項21】
前記分泌リーダー配列が、天然型分泌リーダー配列のコンセンサス配列である、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項22】
代替軽鎖(SLC)構造体をコードする、請求項1〜21のいずれか1項記載の単離された核酸分子。
【請求項23】
請求項1〜22のいずれか1項記載の核酸分子を含むベクター。
【請求項24】
請求項1〜22のいずれか1項記載の核酸分子または請求項23記載のベクターでトランスフェクトされた組換え宿主細胞。
【請求項25】
代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、組換え宿主細胞を、SLCポリペプチドまたはSLC構造体をコードする核酸分子で形質転換することを含む、組換え宿主細胞内でのSLCポリペプチドまたはSLC構造体の発現方法。
【請求項26】
前記組換え宿主細胞が、真核細胞である、請求項25記載の方法。
【請求項27】
前記組換え宿主細胞が、チャイニーズハムスター卵巣(CHO)細胞である、請求項25記載の方法。
【請求項28】
前記組換え宿主細胞が、ヒト胚性腎(HEK)293細胞である、請求項25記載の方法。
【請求項29】
SLCポリペプチドまたはSLC構造体が、VpreBポリペプチド、λ5ポリペプチド、VpreB−λ5ポリペプチド融合体、Vκ様ポリペプチド、JCκポリペプチド、およびVκ様−JCκポリペプチド融合体の1種以上を含むSLCポリペプチドからなる群より選択される、請求項25記載の方法。
【請求項1】
代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、SLCポリペプチドをコードする単離された核酸分子。
【請求項2】
SLCポリペプチドが、VpreBポリペプチドを含む、請求項1記載の単離された核酸分子。
【請求項3】
SLCポリペプチドが、λ5ポリペプチドである、請求項1記載の単離された核酸分子。
【請求項4】
VpreBポリペプチドが、天然型VpreB1配列、天然型VpreB2配列、天然型VpreB3配列、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項2記載の単離された核酸分子。
【請求項5】
天然型VpreB配列が、配列番号1のヒトVpreB1、配列番号2および3のマウスVpreB2、配列番号4のヒトVpreB3、配列番号5のヒトVpreB様ポリペプチド、配列番号6のヒトVpreB dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項4記載の単離された核酸分子。
【請求項6】
λ5ポリペプチドが、配列番号7のヒトλ5様、配列番号8のヒトλ5ポリペプチド、配列番号9のヒトλ5 dTailポリペプチド、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項3記載の単離された核酸分子。
【請求項7】
SLCポリペプチドが、VpreB−λ5ポリペプチド融合体を含む、請求項1記載の単離された核酸分子。
【請求項8】
VpreBポリペプチド配列およびλ5ポリペプチド配列の融合が、それぞれ前記VpreB配列および前記λ5配列のCDR3類似領域で、またはその周辺で起こる、請求項7記載の単離された核酸分子。
【請求項9】
VpreBポリペプチド配列が、カルボキシ末端で、λ5ポリペプチド配列のアミノ末端に連結されている、請求項7記載の単離された核酸分子。
【請求項10】
SLCポリペプチドが、Vκ様ポリペプチドを含む、請求項1記載の単離された核酸分子。
【請求項11】
SLCポリペプチドが、JCκポリペプチドを含む、請求項1または10記載の単離された核酸分子。
【請求項12】
Vκ様ポリペプチド配列が、配列番号12〜24、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項10記載の単離された核酸分子。
【請求項13】
JCκポリペプチド配列が、配列番号26〜39、ならびにそれらのフラグメントおよびバリアントからなる群より選択される、請求項11記載の単離された核酸分子。
【請求項14】
SLCポリペプチドが、Vκ様−JCκポリペプチド融合体である、請求項1記載の単離された核酸分子。
【請求項15】
Vκ様ポリペプチド配列およびJCκポリペプチド配列の融合が、それぞれ前記Vκ様配列および前記JCκ配列のCDR3類似領域で、またはその周辺で起こる、請求項14記載の単離された核酸分子。
【請求項16】
Vκ様ポリペプチド配列が、カルボキシ末端で、JCκポリペプチド配列のアミノ末端に連結されている、請求項14記載の単離された核酸分子。
【請求項17】
前記非相同性分泌リーダー配列が、抗体、サイトカイン、リンフォカイン、モノカイン、ケモカイン、ポリペプチドホルモン、消化酵素、および細胞外マトリックスの成分からなる群より選択される分泌されたポリペプチドのリーダー配列である、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項18】
前記サイトカインが、成長ホルモン、例えば、ヒト成長ホルモン、N−メチオニルヒト成長ホルモン、およびウシ成長ホルモン;副甲状腺ホルモン;チロキシン;インスリン;プロインスリン;リラキシン;プロリラキシン;糖タンパク質ホルモン、例えば、卵胞刺激ホルモン(FSH)、甲状腺刺激ホルモン(TSH)、および黄体形成ホルモン(LH);肝細胞増殖因子;線維芽細胞増殖因子;プロラクチン;胎盤性ラクトゲン;腫瘍壊死因子−αおよび−β(TNF−αおよび−β);ミュラー管抑制物質;マウスゴナドトロピン関連ペプチド;インヒビン;アクチビン;血管内皮増殖因子;インテグリン;トロンボポエチン(TPO);神経成長因子、例えば、NGF−β;血小板成長因子;トランスフォーミング増殖因子(TGF)、例えば、TGF−αおよびTGF−β;インスリン様成長因子−Iおよび−II;エリスロポエチン(EPO);骨誘導因子;インターフェロン、例えば、インターフェロン−α、−βおよび−γ;コロニー刺激因子(CSF)、例えば、マクロファージ−CSF(M−CSF)、顆粒球−マクロファージ−CSF(GM−CSF)、および顆粒球−CSF(G−CSF);インターロイキン(IL)、例えば、IL−1、IL−1a、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12;腫瘍壊死因子、例えば、TNF−αまたはTNF−β;MIP−1α;MIP−1β;ならびにLIFおよびkitリガンド(KL)をはじめとする他のポリペプチド因子からなる群より選択される、請求項17記載の単離された核酸分子。
【請求項19】
前記分泌リーダー配列が、ヒトおよび非ヒトホ乳類アルブミン、トランスフェリン、CD36、成長ホルモン、組織プラスミノーゲン活性化因子(t−PA)、エリスロポエチン(EPO)、ならびにニューブラスチンのリーダー配列からなる群より選択される、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項20】
前記分泌リーダー配列が、合成配列である、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項21】
前記分泌リーダー配列が、天然型分泌リーダー配列のコンセンサス配列である、請求項1〜16のいずれか1項記載の単離された核酸分子。
【請求項22】
代替軽鎖(SLC)構造体をコードする、請求項1〜21のいずれか1項記載の単離された核酸分子。
【請求項23】
請求項1〜22のいずれか1項記載の核酸分子を含むベクター。
【請求項24】
請求項1〜22のいずれか1項記載の核酸分子または請求項23記載のベクターでトランスフェクトされた組換え宿主細胞。
【請求項25】
代替軽鎖(SLC)ポリペプチドの天然型分泌リーダー配列が非相同性分泌リーダー配列により交換されている、組換え宿主細胞を、SLCポリペプチドまたはSLC構造体をコードする核酸分子で形質転換することを含む、組換え宿主細胞内でのSLCポリペプチドまたはSLC構造体の発現方法。
【請求項26】
前記組換え宿主細胞が、真核細胞である、請求項25記載の方法。
【請求項27】
前記組換え宿主細胞が、チャイニーズハムスター卵巣(CHO)細胞である、請求項25記載の方法。
【請求項28】
前記組換え宿主細胞が、ヒト胚性腎(HEK)293細胞である、請求項25記載の方法。
【請求項29】
SLCポリペプチドまたはSLC構造体が、VpreBポリペプチド、λ5ポリペプチド、VpreB−λ5ポリペプチド融合体、Vκ様ポリペプチド、JCκポリペプチド、およびVκ様−JCκポリペプチド融合体の1種以上を含むSLCポリペプチドからなる群より選択される、請求項25記載の方法。
【図1】


【図2】

【図3】

【図4A】

【図4B】

【図5A】

【図5B】

【図5C】

【図6】
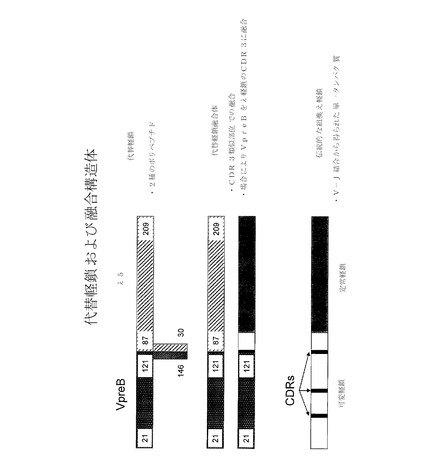
【図7】
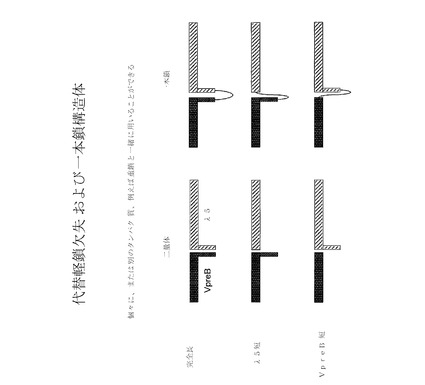
【図8】
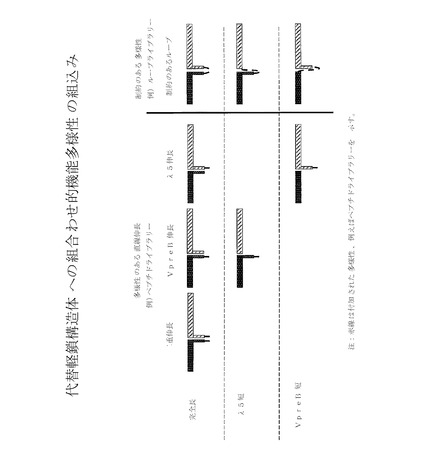
【図9】
【図10】
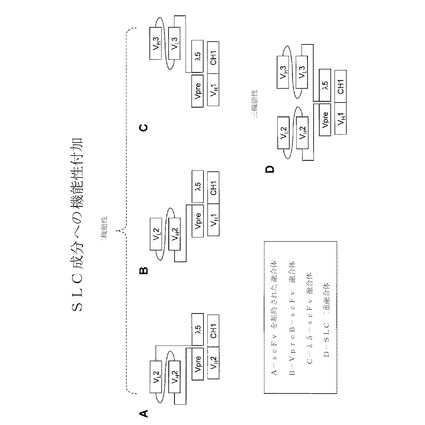
【図11】
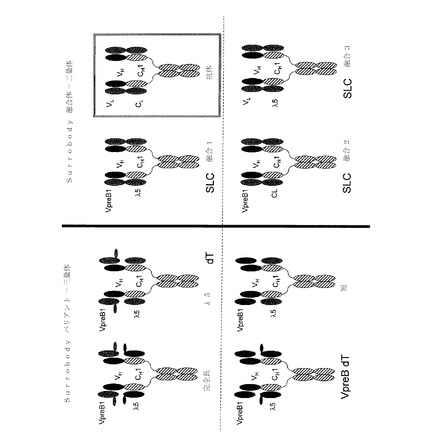
【図12】
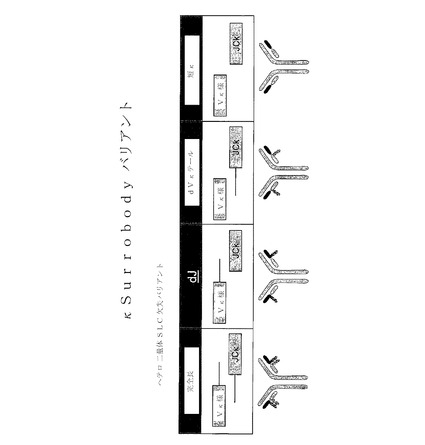
【図13】
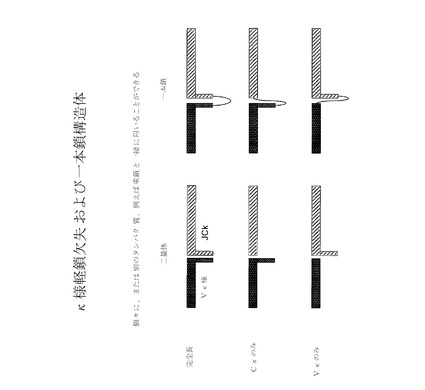
【図14】

【図15】
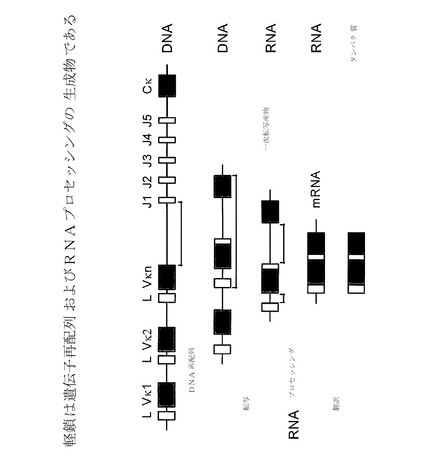
【図16A】
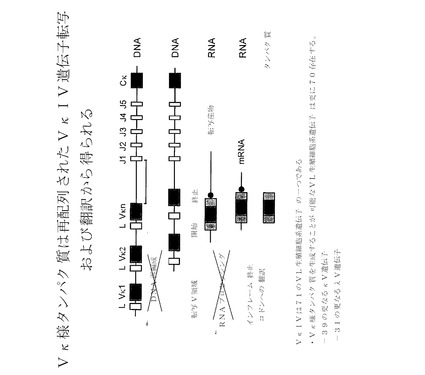
【図16B】
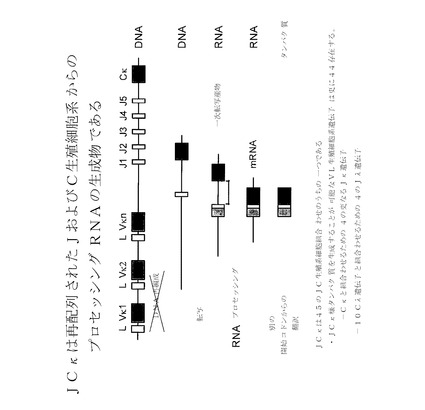
【図17】
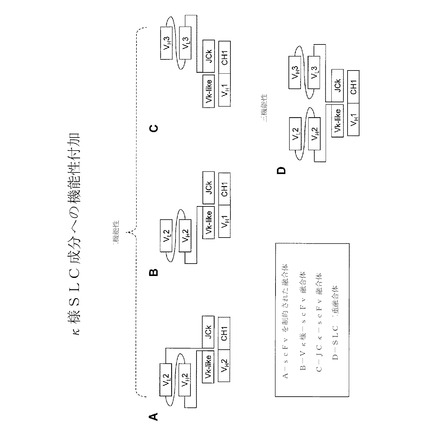
【図18】

【図19】
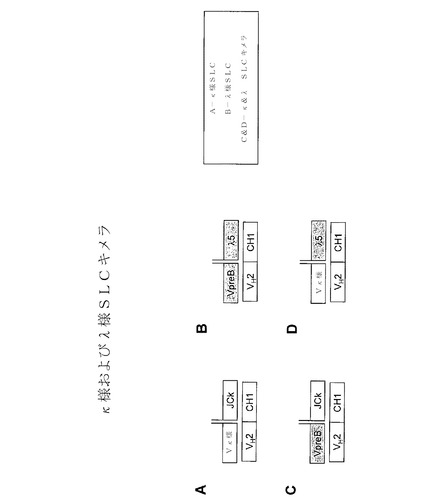
【図20】
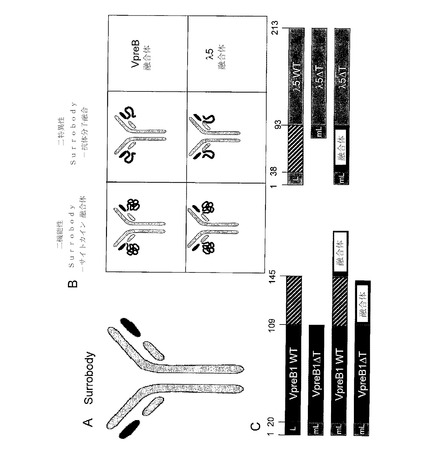
【図2】
【図3】
【図4A】
【図4B】
【図5A】
【図5B】
【図5C】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16A】
【図16B】
【図17】
【図18】
【図19】
【図20】
【公表番号】特表2012−531208(P2012−531208A)
【公表日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願番号】特願2012−517785(P2012−517785)
【出願日】平成22年6月25日(2010.6.25)
【国際出願番号】PCT/US2010/040052
【国際公開番号】WO2010/151808
【国際公開日】平成22年12月29日(2010.12.29)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
【公表日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願日】平成22年6月25日(2010.6.25)
【国際出願番号】PCT/US2010/040052
【国際公開番号】WO2010/151808
【国際公開日】平成22年12月29日(2010.12.29)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
[ Back to top ]