
代謝リズムおよび概日リズムを調整する方法
概日(arcadian)リズムにおけるAMPKの役割およびこのようなリズムを調整する作用物質をスクリーニングする方法が開示される。このようなリズムを調整するために有用な組成物およびその使用も開示される。本開示は、代謝リズムまたは概日リズムの疾患または障害を決定する方法であって、CRY1またはCRY2の安定性を、組織において24時間周期の間に測定するステップを含み、正常または過剰ATP濃度の存在下でのCRY1またはCRY2の長期間安定性の周期が、代謝リズムまたは概日リズムの疾患または障害を示す方法も提供する。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願への相互参照)
本願は、2009年3月20日に出願された米国仮特許出願第61/162,219号の利益を主張し、本明細書において参考として援用される。
【0002】
(政府支援の認可)
本研究は、国立衛生研究所助成金番号DK057978、DK062434、CA104838、DK080425、およびEY016807により支援された。合衆国政府は、本発明において一定の権利を有する。
【0003】
本開示は、概日リズムを調整するためのAMP活性化プロテインキナーゼ(AMPK)のアゴニストおよびアンタゴニストの使用に関する。より具体的には、本開示は、睡眠行動をスクリーニングし、調整するための組成物および方法を提供する。
【背景技術】
【0004】
概日時計は、哺乳動物の組織における何千もの遺伝子のリズム性の転写を駆動することにより、行動プロセスおよび生理的プロセスを毎日の明暗サイクルに調和させる。
【発明の概要】
【課題を解決するための手段】
【0005】
本開示は、AMPKが、転写リプレッサーCRY1およびCRY2をリン酸化し、それらのプロテアソーム分解を刺激することを証明する。さらに、本開示は、クリプトクロムが、哺乳動物概日時計におけるそれらの確立された機能に加えていくつかの核ホルモン受容体と結合し、その転写活性を調節することを証明する。本開示は、クリプトクロムタンパク質が、AMPK活性化薬物を用いる治療に対する転写応答の部分集合にとって必要であることも証明する。よって、クリプトクロムの薬理学的調整は、代謝障害の治療において有用である。
【0006】
クリプトクロム転写同時制御因子機能を調整する小分子薬物の使用は、それらに限定されないが、ペルオキシソーム増殖因子活性化受容体PPARアルファ、ベータ、デルタおよびガンマを含む確立された代謝的に重要な転写因子の転写活性をクリプトクロムが調節することが証明されているので、代謝障害の治療において有用である。クリプトクロムは、天然の小分子補助因子(触媒性補助因子であるフラビンアデニンジヌクレオチドまたはFAD、および集光性補助因子である5,10−メテニルテトラヒドロフォリルポリグルタメートまたはMTHF)と結合してそれにより調節されるので、クリプトクロムは、合成小分子による調節のための良好な標的である。
【0007】
本開示は、エネルギーセンサであるAMPKが、CRY1(そのリン酸化は、CRY1−FBXL3相互作用およびCRY1のプロテアソーム分解を媒介する)中の2つのセリンを修飾することを証明する。つまり、CRY1は、元来、光受容体として進化したが、翻訳後修飾により、これは重要なシグナル伝達メディエーターとなり得る。in vivoでのAMPKの遺伝子操作または薬理学的操作は、クリプトクロム安定性と概日リズムとを共に変更し、このことは、それにより栄養調節シグナルが哺乳動物末梢器官における概日時計をリセットできる新規な同調化機構を示唆する。
【0008】
本開示は、ヒトのような哺乳動物被験体において概日リズムを改変するための方法および組成物を提供する。本開示は、AMPKが、哺乳動物被験体の概日サイクルの間に、脳および体内の他の組織において修飾されることを証明する。1つの実施形態において、本開示は、被験体における概日リズムを調整するための医薬品の製造のための、AMPキナーゼアゴニストまたはアンタゴニストの使用を提供する。1つの実施形態において、AMPKアゴニストは、AICARである。別の実施形態において、AMPKアンタゴニストは、抗体もしくはコンパウンドCまたはそれらの類似体もしくは誘導体である。さらに別の実施形態において、AMPKアゴニストは、血液脳関門を通過することができる製剤または誘導化を含む。まださらなる実施形態において、AMPKアゴニストは、経口投与、静脈内注射、筋肉内注射、硬膜外送達、頭蓋内または皮下注射用に製剤化される。
【0009】
本開示は、概日リズムを改変する第2活性成分と組み合わせて製剤化されたAMPKアゴニストを含む組成物も提供する。1つの実施形態において、第2活性成分は、睡眠補助剤である。さらなる実施形態において、組成物は、経口投与、静脈内注射、筋肉内注射、硬膜外送達、頭蓋内送達または皮下注射用に製剤化される。
【0010】
本開示は、哺乳類における睡眠を調整する方法であって、哺乳類における概日リズムを調整するのに有効な量のAMPKアゴニストまたはアンタゴニストを哺乳類に投与するステップを含む方法を提供する。
【0011】
本開示は、被験体における概日リズムまたは睡眠を調整する作用物質を同定する方法であって、(a)AMPK経路を備える試料を少なくとも1つの被検作用物質と接触させるステップと、(b)AMPKまたはAMPK経路の活性を、被検作用物質の存在下および非存在下で比較するステップであって、前記活性を変化させる被検作用物質が、概日リズム調整活性を有する作用物質を示すステップとを含む方法も提供する。
【0012】
本開示は、代謝リズムまたは概日リズムの調整において用いるための作用物質を同定する方法であって、前記作用物質をCry1またはCry2タンパク質と接触させるステップと、Cry1もしくはCry2をリン酸化もしくは脱リン酸化するか、またはCry1もしくはCry2の安定性もしくは発現を改変する前記作用物質の能力を測定するステップとを含み、Cry1またはCry2を改変する作用物質が、代謝リズムまたは概日リズムを調整するために有用な作用物質である、方法も提供する。1つの実施形態において、作用物質は、Cry1またはCry2の安定性を低下させる。
【0013】
本開示は、上記の方法により同定される、Cry1またはCry2の安定性を低下させる作用物質を含む組成物も提供する。
【0014】
本開示は、代謝性または概日性の疾患または障害を治療する方法であって、被験体を、Cry1および/またはCry2のリン酸化または脱リン酸化を促進する本開示の作用物質または組成物と接触させるステップを含む方法も提供する。1つの実施形態において、作用物質または組成物は、クリプトクロム転写同時制御因子機能を調整する。別の実施形態において、作用物質または組成物は、ペルオキシソーム増殖因子活性化受容体(PPAR)アルファ、ベータ(デルタ)およびガンマを調整する。さらに別の実施形態において、作用物質は、ビグアナイド誘導体、AICAR、メトホルミンまたはその誘導体、フェンホルミンまたはその誘導体、レプチン、アディポネクチン、AICAR(5−アミノイミダゾール−4−カルボキサミド)、ZMP、DRL−16536、BG800化合物(Betagenon)およびフラン−2−カルボン酸誘導体からなる群から選択されるAMPKアゴニストである。
【0015】
本開示は、代謝リズムまたは概日リズムの疾患または障害を決定する方法であって、CRY1またはCRY2の安定性を、組織において24時間周期の間に測定するステップを含み、正常または過剰ATP濃度の存在下でのCRY1またはCRY2の長期間安定性の周期が、代謝リズムまたは概日リズムの疾患または障害を示す方法も提供する。1つの実施形態において、方法は、mCRY1のS71またはS280を含むエピトープと特異的に結合する抗体を用いる。
【0016】
本開示は、安静および脂質異化反応を促進する方法であって、概日サイクルの夜間期の間にAMPKアゴニストを投与するステップを含み、前記AMPKアゴニストが、CRY1またはCRY2の安定性を低下させる方法も提供する。
【0017】
本開示は、代謝リズムまたは概日リズムの障害を治療する方法であって、概日サイクルの安静周期の間にAMPKアゴニストを投与するステップを含む方法も提供する。
【0018】
上記のおよびその他の特徴は、添付の図面を参照して進む以下のいくつかの実施形態の詳細な記載からより明確になる。
【図面の簡単な説明】
【0019】
【図1】図1A〜Eは、S71またはS280のリン酸化が、FBXL3およびPER2との相互作用を変更することによりmCRY1を不安定化することを示す。(A)記載される変異を有するFlagタグ付加mCRY1を発現するAD293細胞を、100μg/mlのシクロヘキシミド(CHX)で、記載される時間処理した。Flag−mCRY1は、ウェスタンブロッティングにより検出した。β−アクチンについてのイムノブロットを、負荷対照(loading control)として用いた。(B)CLOCK、BMAL1、Per1−ルシフェラーゼならびにmCRY1の記載される量および対立遺伝子を発現するAD293細胞を、トランスフェクションの48時間後にルシフェラーゼ活性について調べた。(C)Flag−mCRY1を、記載されるプラスミドを一過的に発現するAD293細胞から免疫沈降させた。CRY1と結合したFBXL3は、v5エピトープタグについてのイムノブロッティングにより検出した。(D)記載されるようにFBXL3を同時発現するかまたはしない、CLOCK、BMAL1、Per1−ルシフェラーゼおよびmCRY1の記載される対立遺伝子を一過的に発現するAD293細胞を、トランスフェクションの48時間後にルシフェラーゼ活性について調べた。**AAに対してp<0.01;## FBXL3を発現しない等価な試料に対してp<0.01。(E)Flag−mCRY1を、PER2を同時発現するかまたはしない、CRY1の記載される対立遺伝子を一過的に発現するAD293細胞から免疫沈降させた。CRY1と結合したPER2は、イムノブロッティングにより検出した。
【図2】図2A〜Gは、AMPKが、Ser71、Ser280リン酸化によりmCRY1を不安定化することを示す。(A)クリプトクロム概日転写リプレッサー(赤色のフォントでの種の名称)および青色光光受容体(青色のフォントでの種の名称)中のmCRY1のS71の周囲の領域の進化的保存を示す配列アラインメント。配列の上の強調した数字は、AMPキナーゼによるリン酸化のための標的セリンに対する、これらの位置での好まれるアミノ酸を示す。赤色は、酸性残基(K/R)への好みを示し、緑色は疎水性残基(L/I/V/F)への好みを示す。(B)上:ホスホペプチドの配列アラインメントであって、これらに対して、mCRY1−pS71およびmACC1−pS79に対する抗体を産生させた。下:抗mCRY1−pS71抗体および抗ACC1−pS79抗体は共にWTを認識するが、AD293細胞から免疫沈降させたS71A Flag−mCRY1は認識しない。(C)抗mCRY1−pS71を用いて、AMPKα1(CAα1)またはAMPKα2(CAα2)の活性化対立遺伝子の非存在下または存在下で野生型CRY1(WT)またはCRY1S71A(S71A)を一過的に発現するAD293細胞から免疫沈降させたFlag−CRY1中のSer71のリン酸化を検出した。一過的に発現されたmyc−CAα1およびmyc−CAα2は、mycエピトープタグに対して産生されたポリクローナルウサギ抗体と抗ウサギAF488(緑色)とを用いて免疫染色した。核は、DAPI(青色)を用いて対比染色した。(D)野生型(WT)またはキナーゼ非機能性(kinase dead)(KD)LKB1と共にFlag−CRY1を一過的に発現するHeLa細胞を、ビヒクル(−)または2mMのAICAR(+)で2時間処理した。(E)Flag−mCRY1の記載される対立遺伝子を一過的に発現するAD293細胞を、25mMまたは0.5mMのグルコースを含有する培地で処理した。(F)Flagタグ付加野生型CRY1(WT)またはCRY1S71A/S280A(AA)を安定的に発現する、対での野生型(AMPK+/+)およびampkα1−/−;ampkα2−/−(AMPK−/−)マウス胚線維芽細胞を、ビヒクル(−)または2mMのAICAR(+)で2時間処理した。(G)(F)に記載されるMEFを、100μg/mlのシクロヘキシミド(CHX)で、記載される時間処理した。CRY1は、(D〜G)におけるFlagエピトープについての免疫沈降およびイムノブロッティングにより検出した。
【図3】図3A〜Dは、AMPKシグナル伝達の破壊が、MEFにおける概日リズムを変更することを示す。(A)同期化していない対での野生型(AMPK+/+)またはampkα1−/−;ampkα2−/−(AMPK−/−)マウス胚線維芽細胞を、50%ウマ血清に2時間曝露することにより刺激し、その後、25mMグルコース、0.5mMグルコースまたは1mMのAICARを補った25mMグルコースを含有する培地に移した。定量的PCR分析を、刺激後の記載される時間に回収したcDNA試料を用いて行った。データは、それぞれ3重で分析した2つの独立する実験の平均値を示す。(B)Bmal1−ルシフェラーゼを安定的に発現する線維芽細胞を、記載される量のグルコースを2mMのAICARと共にまたはなしで含有する培地中で培養した。ルシフェラーゼ活性の連続モニタリングの代表的な結果を示す。(CおよびD)(B)に記載するようにして行った実験からのBmal1駆動型ルシフェラーゼ活性の概日周期(C)および振幅(D)の定量。(C)および(D)におけるデータは、条件あたり4つの試料についての平均値±標準偏差を表す。ANOVA分析は、カテゴリー間での有意差を示した。**シェッフェの事後分析において25mMのグルコースで培養した試料に対してP<0.01。
【図4】図4A〜Cは、AMPK活性および核局在化が概日調節を受けることを示す。(A)ホスホ−Raptor−S792(pRaptor)、Raptor、ホスホ−ACC1−S79(pACC1)およびACC1についてのイムノブロッティングを、記載される概日時間に回収したマウス肝臓から調製した全細胞溶解産物で行った。ブロットは、3回の独立した実験の代表である。(B)記載される概日時間に回収したマウス肝臓から調製したcDNAの定量的PCR分析。各データ点は、それぞれ独特の動物から採取され、4重で分析した3つの試料の平均値±標準偏差を表す。(C)核抽出物を、記載される概日時間のそれぞれで2匹のマウスの肝臓から調製した。AMPKα1、AMPKα2、PER2、CRY1およびREVERBαのタンパク質レベルを、イムノブロッティングにより分析した。記載される概日時間に回収した対での野生型(α1+/+)およびampkα1−/−(α1−/−)または野生型(α2+/+)およびampkα2−/−(α2−/−)マウスからの核抽出物を、抗体特異性についての対照として用いた。
【図5】図5A〜Cは、AMPK活性化が、マウス肝臓におけるCRY安定性および概日リズムを変更することを示す。(A)マウスに、生理食塩水または体重1kgあたり500mgのAICARを注射し、肝臓試料を1時間後のツァイトゲーバー時間(ZT、光点灯後の時間)6またはZT18に回収した。内因性CRY1を、肝臓核抽出物におけるイムノブロッティングにより検出した。n.s.は、試料負荷を評価するための非特異的バンドを示す。野生型(CRY+/+)およびcry1−/−;cry2−/−(CRY−/−)マウスから回収した試料を、抗体特異性についての対照として用いた。データは、2つの独立した実験からの代表的な結果を表す。(B)LKB1+/+およびLKB1fl/flマウスに、Creリコンビナーゼを発現するアデノウイルス(Ad−Cre)を尾静脈から注射した。Ad−Cre注射の1〜2週間後に、マウスを定常的な暗所に移し、記載される概日時間に肝臓を回収した。CRY1、PER2およびREVERBαをイムノブロッティングにより検出した。(C)(B)に記載される肝臓から調製したcDNA試料を、dbp、reverbα、cry1およびper2発現の定量的PCR分析により分析した。全ての転写産物を、内部対照としてのu36b4に対して正規化した。各データ点は、4重で分析した3つの試料の平均値±標準偏差を表す。
【図6】図6は、AMPKが、末梢時計の代謝同調化に寄与することを示す。マウスにおける末梢概日時計の代謝同調化におけるAMPKの役割を示すモデル。昼間、AMPKの核局在化は、食物および循環グルコースの低減によるその可能性のある活性化と連係して増加する。活性核AMPKは、クリプトクロムをリン酸化して、FBXL3とのその相互作用を増加し、プロテアソーム分解を導き、時計により制御される遺伝子(ccg)の活性化をもたらす。夜間、核AMPK活性が低下することにより、クリプトクロムが核に蓄積し、ccgが抑制される。
【図7】図7A〜Dは、mCRY1リン酸化部位の同定を示す。(A)一過性にトランスフェクトしたAD293細胞から精製されたFlag−mCRY1を、LC−MS/MSにより、リン酸化セリン、スレオニンおよびチロシン残基の存在について分析した。観察されたリン酸化を触媒すると予測されるキナーゼを、文献検索およびScansiteプログラム(http:(//)scansite.mit.edu)との組合せを用いて予測した。配列保存を、MegAlignを用いて決定した。(B)CRY1タンパク質の模式図の下のオレンジ色のバーの厚さは、タンパク質配列の各領域についてLCMS/MSにより観察されたペプチドの相対数を示す。(C)Bに示すペプチドのカバー範囲に基づく本発明者らのLC−MS/MS分析では観察可能でない場合があるScansiteにより予測されるリン酸化部位。(D)記載される変異を有するFlag−mCRY1を、100μg/mlのシクロヘキシミドで記載される時間処理したAD293細胞において発現させた。CRY1タンパク質は、Flagタグについてのイムノブロットにより検出された。
【図8】図8は、mCRY1 S280配列保存を示す。mCRY1 S280は、保存されたAMPK基質モチーフにより囲まれる。クリプトクロム概日転写リプレッサー(赤色フォントでの種の名称)および青色光光受容体(青色フォントでの種の名称)におけるmCRY1のS280の周囲の領域の進化的保存を示す配列アラインメント。配列の上の強調した数字は、AMPキナーゼによるリン酸化についての標的セリンに対するこれらの位置での好まれるアミノ酸を示す。赤色は、示される位置での酸性残基(K/R)への好みを示し、緑色は疎水性残基(L/I/V/F)への好みを示す。
【図9】図9は、精製AMPKが、mCRY1をin vitroでリン酸化することを示す。AD293細胞から精製したFlagタグ付加mCRY1を、30分間、#32P−ATPと、記載されるようにAMPキナーゼおよび300μMのAMPの非存在下または存在下でインキュベートした。mCRY1のリン酸化は、オートラジオグラフィにより検出した。合計mCRY1レベルを、Flagエピトープについてのイムノブロットにより決定した。精製AMPKは、精製mCRY1を効率的にリン酸化し、このリン酸化は、AMPの存在により強く活性化され、このことにより、精製混合物中の関連するキナーゼが、他の関連キナーゼではなくAMPKであることが確認された。
【図10】図10A〜Dは、AMPKの破壊が、MEFにおける概日リズムを変更することを示す。3T3不死化マウス胚線維芽細胞(A)または対での野生型(AMPK+/+)もしくはampkα1−/−;ampkα2−/−(AMPK−/−)線維芽細胞(B)を、50%ウマ血清に2時間曝露することにより刺激し、その後、25mMグルコース(黒色の記号)、0.5mMグルコース(灰色の記号)または1mMのAICARを補った25mMグルコース(赤色の記号)を含有する培地に移した。定量的PCR分析を、刺激後の記載される時間に回収した溶解産物から調製したcDNA試料を用いて行った。データは、それぞれ3重で分析した2または3つの独立する実験の平均値±標準偏差を表す。
【図11】図11は、mCRY1が核ホルモン受容体と相互作用することを示す。Flagタグ付加mCRY1を種々のv5タグ付加核ホルモン受容体と同時発現するAD293細胞を溶解し、mCRY1を含有するタンパク質複合体を、Flagタグの免疫沈降により単離した。mCRY1含有タンパク質複合体中の個別の核ホルモン受容体の存在を、v5タグについてのイムノブロットにより検出した(上)。免疫沈降複合体中のmCRY1の量を、Flagタグについてのイムノブロットにより示す(中)。溶解産物中に存在するそれぞれの核ホルモン受容体の量を、免疫沈降前の溶解産物から採取した試料におけるv5タグのイムノブロットにより示す(下)。RORa、b、g(レチノイン酸受容体関連オーファン受容体a、b、g)、RXRa、b(レチノイドX受容体a、b)、PPARd、g(ペルオキシソーム増殖因子活性化受容体d、g)、VDR(ビタミンD受容体)、PXR(プレグナンX受容体)、CAR(構成的アンドロスタン受容体)、ERb(エストロゲン受容体b)、ERRa、b、g(エストロゲン関連受容体a、b、g)、GR(グルココルチコイド受容体)、MR(ミネラロコルチコイド受容体)、PR(プロゲステロン受容体)、AR(アンドロゲン受容体)。データは、2または3つの独立した実験の代表的な結果を表す。
【図12】図12は、クリプトクロムが、AMPK活性化に対するいくつかの転写応答にとって必要であることを示す。野生型(WT)またはCry1−/−;Cry2−/−(CRY−/−)マウスに、生理食塩水(黒色のバー)または体重1kgあたり500mgのAICAR(赤色のバー)を、午後6時に注射した。cDNAを、4時間後の午後10時に回収した肝臓から調製し、遺伝子発現を、Sybr GreenER化学を用いる定量的PCRにより分析した。Fas(脂肪酸合成)を、Cry1およびCry2遺伝子型に関係なくAICARにより活性化される遺伝子の例として示す。Por(p450酸化還元酵素)を、そのAICARにより誘導される活性化がクリプトクロムを必要とする遺伝子の例として示す。データは、条件あたり3〜5匹のマウスについての平均値±s.e.m.を表す。
【図13】図13は、クリプトクロムの喪失が、マウスにおける代謝機能を変更することを示す。10週齢の雄の野生型(WT、黒色のバー)およびCry1−/−;Cry2−/−(CRY−/−、灰色のバー)マウスの体重測定を行い、それらの安静時血中グルコースを、尾静脈の切れ目(nick)により午後1時に測定した。データは、遺伝子型あたり10匹の動物についての平均値±s.e.m.を表す。
【発明を実施するための形態】
【0020】
本明細書においてそうでないと特に記載しない限り、用いられる用語の定義は、薬学技術において用いられる標準的な定義である。明細書および添付の特許請求の範囲において用いる場合、単数形「a」、「an」および「the」は、文脈がそうでないと明確に示さない限り、複数の引用物も含む。よって、例えば、「薬学的キャリア」への言及は、そのようなキャリアの2以上の混合物などを含む。
【0021】
また、「または」の使用は、そうでないと記載しない限り「および/または」を意味する。同様に、「含む(comprise)」、「含む(comprises)」、「含み(comprising)」、「含む(include)」、「含む(includes)」および「含み(including)」は、交換可能であり、限定することを意図しない。
【0022】
種々の実施形態の記載が「含み」の用語を用いる場合、いくつかの特定の場合において、実施形態を、「から本質的になる」または「からなる」の言葉を用いて代わりに記載できることを当業者が理解するとさらに理解される。
【0023】
そうでないと明示しない限り、本明細書において用いられる全ての技術的および科学的用語は、本開示が属する当業者に通常理解されるのと同じ意味を有する。本明細書に記載されるのと同様または等価ないずれの方法および試薬も、開示される方法および組成物の実施において用いることができるが、例示的な方法および材料をここで記載する。
【0024】
本明細書において言及する全ての出版物は、本明細書の記載と関連して用いられる可能性があるその出版物に記載される方法を記載して開示する目的のために、全体が参照により本明細書に組み込まれる。上で論じた本文全体での出版物は、本開示の出願日に先んじるそれらの開示のためにのみ提供される。本明細書中のいずれも、本発明者らが、先の開示により、そのような開示に先行する権利が与えられないことを認めると解釈されない。
【0025】
概日リズムは、限定されないが、睡眠サイクルの調整、エクササイズおよびカロリー低減に関連するエネルギー調整、ならびに摂食/栄養補給(nourishment)行動のような生理的プロセス、内分泌プロセスおよび行動プロセスの適切なタイミングを調和させることにより生物学的効率を最適化する。概日リズムは、少なくとも3つの要素を含有すると考えられる:(a)環境情報を概日ペースメーカー(時計)に中継する入力経路(複数可);(b)変動(oscillation)を生み出す概日ペースメーカー;および(c)それを介してペースメーカーが種々の出力リズムを調節する出力経路(複数可)。
【0026】
哺乳動物の視床下部視交叉上核(SCN)は、行動リズムおよび生理的リズムを明暗サイクルに整合させる主要ペースメーカーとして作用する。当初、SCNは、哺乳類における自立性分子ペースメーカーの唯一の部位であると考えられていたが、その後、複数の論文が、このような分子時計がほぼ遍在的であることを示した。SCN時計とは異なって、光非感受性末梢器官における概日時計は、摂食の毎日のリズムにより同調され、末梢組織が、毎日の食物消費を予期し、代謝プロセスのタイミングを最適化することを理論的に可能にする。いくつかの論文が、重要な代謝酵素の転写の調節および代謝生理における哺乳動物概日時計についての役割を支持している。
【0027】
本明細書で用いる場合、「概日リズム」との用語は、約24時間の過程にわたって生じる生理的および行動的パラメータにおける規則的な変動を意味することを意図する。このような活動は、睡眠サイクルおよび栄養補給サイクルなどを含む。
【0028】
本明細書で用いる場合、概日リズムに関して用いる場合の「調整する」との用語は、動物の概日タイミングシステムにより調節される生理的機能、内分泌機能もしくは行動を変更すること、または概日リズム性を示す細胞機能を変更することを意味することを意図する。動物の概日タイミングシステムにより調節される例示的な生理機能は、体温、自律神経調節、代謝および睡眠覚醒サイクルを含む。例示的な代謝機能は、体重の増加または減少および体脂肪率の増加または減少を含む増量ならびに減量の制御、持久行動の改変、減量などを含む。動物の概日タイミングシステムにより調節される例示的な内分泌機能は、松果体のメラトニン分泌、ACTHコルチゾール分泌、甲状腺刺激ホルモン分泌、成長ホルモン分泌、ニューロペプチドY分泌、セロトニン分泌、インスリン様成長因子I型分泌、副腎皮質刺激ホルモン分泌、プロラクチン分泌、ガンマ−アミノ酪酸分泌およびカテコールアミン分泌を含む。動物の概日タイミングシステムにより調節される例示的な行動は、運動(運動リズム)、精神的俊敏さ、記憶、感覚運動統合、摂食、REM睡眠、NREM睡眠および情動を含む。
【0029】
AMP活性化プロテインキナーゼ(AMPK)は、系統発生全体でよく保存されている代謝シグナルの中心的なメディエーターとして認識されている。AMPKは、触媒(α)サブユニットと2つの調節性(β、γ)サブユニットとを含むヘテロ3量体プロテインキナーゼである。これは、高いAMP/ATP比率の存在下でLKB1により、または上昇した細胞内カルシウムの存在下でCAMKKβによりリン酸化されると活性化される。生化学的研究および生物情報学的研究により、AMPKによるリン酸化が起こりそうな最適アミノ酸配列の関係が確立されている。
【0030】
AMP活性化プロテインキナーゼ(AMPK)およびAMPKキナーゼ(AMPKK)は、プロテインキナーゼカスケードと関連する。AMPKカスケードは、燃料の生成および利用を細胞内で調節する。例えば、細胞での燃料が低い(例えばAMP濃度の増加)と、AMPK活性が増加する。一旦活性化されると、AMPKは、ATPを保存するか、またはATP生成の代替法を促進するように機能する。よって、その活性を調整することにより、エネルギー貯蔵の異化反応を増加でき、脂質含量を低減させてATPを増加させるか、または体を安静状態にして、ATP使用を保存する。
【0031】
AMPKは、肝臓、脳および骨格筋を含むいくつかの組織において発現される。AMPKの活性化は、肝臓の脂肪酸酸化およびケトン体生成を活性化し、コレステロール合成、脂肪生成およびトリグリセリド合成を阻害し、脂肪細胞の脂肪分解および脂肪生成を阻害し、骨格筋の脂肪酸酸化および筋肉グルコース摂取を刺激し、膵臓ベータ細胞によるインスリン分泌を調整することが示されている。
【0032】
AMPKの活性化は、AMPの濃度の増加により誘引できる。AMPKのγサブユニットは、αサブユニット上に活性部位(Thr−172)を露出するように立体構造変化を受ける。AMPKのγサブユニットの立体構造変化は、AMPの濃度が増加している場合に達成され得る。AMPの濃度の増加により、AMPKのγサブユニットの立体構造変化が、このサブユニットにある2つのBatemanドメインを2つのAMPが結合させることにより生じる。AMPのこの役割は、5−アミノ−4−イミダゾールカルボキサミドリボシド(AICAR)に由来するAMP類似体である5−アミノ−4−イミダゾールカルボキサミドリボチド(ZMP)によるAMPK活性化を示す実験において証明される。同様に、AMPのアンタゴニストは、AMPKによる下流のキナーゼの活性化を阻害する阻害抗体の使用を含む。
【0033】
断眠(SD)は、神経活性を増加させる。持続性の神経活性は、細胞エネルギー充足率を減少させる(AMPレベルが増加し、ATPが減少する)。これは、次に、細胞エネルギーセンサAMPKにおける変化を引き起こす。上で論じたように、AMPKは、ATPの保存を導くカスケードを含む種々のキナーゼカスケードを調整する。
【0034】
CLOCKおよびBMAL1は、ヘテロ2量体の形成により、概日リズムと関連する遺伝子の転写を誘導するポリペプチドである。典型的な概日サイクルの間に、分子機構は、2つの相互に連結された転写/翻訳フィードバックループを有する内部時計を形成する2つのサイクルの間で変動する。フィードバックループのポジティブアームは、塩基性ヘリックス−ループ−ヘリックス−PAS(Per−Arnt−Sim)ドメイン含有転写因子であるCLOCKおよびBMAL1により駆動される。CLOCK/BMAL1ヘテロ2量体は、時計遺伝子であるクリプトクロム(Cry1およびCry2)、period(Per1およびPer2)、およびRev−Erbαの転写を活性化する。PERおよびCRYタンパク質は、核に移動し、そこでこれらはCLOCK/BMAL1と相互作用して転写を下方調節し、主要フィードバックループのネガティブアームを生成する。
【0035】
上記の概日転写プログラムの堅固な変動は、コア時計タンパク質の翻訳後修飾を必要とする。3つの研究により、最近、Fボックスタンパク質FBXL3が、クリプトクロムのユビキチン化および分解のメディエーターとして同定された。Fボックスタンパク質がそれらの同族基質に結合することは、基質タンパク質中の1つ以上のアミノ酸のリン酸化により頻繁に調節されるが、このような調節の改変は、CRY:FBXL3相互作用について記載されなかった。
【0036】
時計タンパク質の翻訳後修飾(例えばリン酸化および脱リン酸化)は、タンパク質の局在化、分子間相互作用および安定性を決定し、よって、概日時計の周期を調節する。本開示は、この翻訳後調節が、AMPK活性により調整され得、よって、AMPKアゴニストおよびアンタゴニストが、概日時計の調節において役割を有し得ることを証明する。
【0037】
クリプトクロム(Cry1およびCry2)は、ほとんどの植物において概日光受容体として機能する。クリプトクロムは、全ての組織において発現することが見出されている。しかし、発現は、網膜においてより高く、マウスおよびヒトの両方において網膜内層に限定される。脳において、Cry1はSCNにおいて発現し、発現は、毎日の変動を示し、午後2時付近にピークになり、午前2時付近にその最低に達する。
【0038】
ヒトのクリプトクロムは共に、Cry遺伝子を異所的に発現するHeLa細胞から、および組換えタンパク質としてE.coliから精製されている。両方の供給源から単離されたタンパク質は、FADおよびプテリンを含有する。
【0039】
クリプトクロムは、光センサとして進化したが、これは、光非感受性組織においてさえ、コアとなる概日時計の重要な成分として保持されている。本開示は、AMPKが、栄養シグナルを時計に変換するように、クリプトクロムに再度目的を持たせることを証明する。概日システムと代謝システムとの間の逆調節についての証拠は、過去10年間で増加しており、出現している理論は、概日時計が、代謝プロセスの一時的な分離を可能にすることを示唆している。代謝シグナルは、哺乳動物の末梢器官において概日時計のタイミングを設定することが示されているが、このようなシグナルを伝達する分子機構は、まだ明らかになっていない。
【0040】
本開示は、AMPKによるクリプトクロムのリン酸化が、FBXL3との結合により分解を促進し、CLOCK:BMAL1抑制を軽減することを証明する。このプロセスは、過剰グルコースにより抑制され、AICARのようなAMPK活性化因子、およびampkβ2調節性サブユニットの核移動により増強される。よって、本開示は、細胞内生物エネルギーの状態が、末梢組織における概日時計に直接影響を与えることができる新規な生化学的経路を提供する。
【0041】
AMPKの概日性活性化は、CRY1のリン酸化を駆動し、そのFBXL3媒介分解を刺激することにより、リズムの維持に寄与する。AMPKは、CRY1を2つのセリン残基(マウスCRY1においてS71およびS280)上でリン酸化する。セリン71および周囲の配列は、全ての光非依存性クリプトクロム転写リプレッサーに存在し、このことは、この経路が、光に曝露されない概日時計の代謝の同調化を可能にするように進化したことを示唆する。
【0042】
AMPK活性は、グルコース利用力によりLKB1依存的な様式で調節されることができ、栄養利用力を変化させるか、またはAMPK活性は、培養線維芽細胞における時計の振幅および周期を変更する。in vivoでは、AMPK基質であるACC1およびRaptorは、リン酸化の概日性の変化を示し、このことは、AMPKの下流の細胞質経路および核での経路がリズム性に調節されることを示唆する。AMPKが代謝プロセスの中心的な制御因子であることに鑑みて、これは、代謝の概日性の調節に重要な関わりを有する。遍在的または組織特異的な様式のいずれかでの概日時計機能の遺伝子的な変更は、摂食行動、体重、ランニング持久力およびグルコース恒常性を変化させ、これらはそれぞれ、AMPKの操作によっても変更される。一括して、これらのデータは、AMPKが、細胞レベルおよび生物全体レベルの両方での概日生理的調節の重要なメディエーターであり得るという考えを支持する。
【0043】
興味深いことに、それぞれのAMPKサブユニットの転写、核局在化および活性化は、マウス肝細胞において概日リズムを示し、クリプトクロムタンパク質の量が最小の時間にピークになる。ampkβ2転写は、堅固に概日性であり、真昼には夜より8倍高い。AMPKβ2は、AMPKの核局在化と、それに対応してAMPKα1のリズム性の核蓄積を駆動する。よって、AMPKサブユニットは、概日時計の調節に寄与するだけでなく、それら自体も概日性の様式で転写調節される。
【0044】
栄養状態と時計との連絡は複雑であり、付加的な経路がそれらの同調化にin vivoで寄与する。2つの最近の研究は、SIRT1が、肝細胞においてリズム的に発現され、線維芽細胞における概日リズム性に寄与することを証明した。SIRT1は、NAD+/NADH比率によるその脱アセチル化酵素活性の調節により概日時計の代謝同調化において役割を果たすと見られる。複数の論文が、種々の時計成分の調節におけるヘムの役割を示唆し、3価鉄および2価鉄ヘムによる異なる調節が、細胞酸化還元状態についての情報を概日時計に伝達することを示唆している。これらの機構の1もしくは複数、および/またはSCNから発せられる昼間液性シグナルもしくはニューロンシグナルは、おそらく、LKB1L/Lマウスの肝臓において観察された残存概日リズムに寄与する。
【0045】
Fbxl3またはLkb1における変異は、ヒト腫瘍において頻繁に行われる。クリプトクロムのLKB1およびAMPKにより媒介されるリン酸化が、それらのFBXL3により媒介される分解を刺激することの証明は、2つの腫瘍抑制因子がクリプトクロムの不安定化において協力することを示し、このことは、クリプトクロムの異常に高いレベルが、細胞周期調節解除(deregulation)または腫瘍形成に寄与し得ることを示唆する。肝臓再生の概日性調節について記載した論文において、Matsuoおよび共同研究者らは、cry1−/−;cry2−/−マウスの肝臓が、野生型同腹子のものよりも遅く再生されたことを示し、このことは、CRYタンパク質が、細胞成長または増殖において刺激性の役割を果たすとの考えを支持する。CRY:FBXL3相互作用を媒介するLKB1およびAMPKに依存するリン酸化部位の本明細書における同定は、これらの疑問を明確にする。
【0046】
哺乳動物クリプトクロムにおける他のリン酸化部位は、さらなる入力シグナルを概日時計へ媒介し得るが、本開示は、セリン71および280のAMPKにより媒介されるリン酸化が、FBXL3とのその相互作用を増加することによりCRY1プロテアソーム分解を刺激することを証明する。さらに、グルコース欠損は、クリプトクロム安定性を低下させ、概日転写産物を変更し、培養細胞における概日周期の長さを増加させ、これらの効果はAMPKにより媒介される。さらに、マウスにおけるAMPKの遺伝子破壊は、クリプトクロム安定性および概日リズムを乱す。これらのデータはまとめて、AMPKによるクリプトクロムリン酸化が、代謝シグナルによる末梢器官時計の同調化を可能にするように進化したことを証明する。
【0047】
本開示は、睡眠または他の概日プロセスに影響するための、AMP活性化プロテインキナーゼ(AMPK)と結合するか、またはそうでなければそれを活性化もしくは不活性化する化合物(これらのいくつかは、糖尿病の治療のために現在用いられている)の使用を提供する。本開示は、AMP活性化プロテインキナーゼ活性の遺伝子操作または薬理学的操作が、培養細胞およびインタクトな動物の肝臓における概日リズムを変更することを証明する。本開示は、AMPキナーゼが、睡眠覚醒サイクルおよび他の生理的リズムのタイミングを支配するいわゆる「主要ペースメーカー」の場所である視交叉上核(SCN)で発現されることも証明する。現在利用可能な療法は、血液脳関門を通過せず、よって、睡眠障害の調整のために有用でない。
【0048】
AMPKによる概日リズムの調節は、血液脳関門を通過するAMPK調節因子が、概日リズムと関連する下流のキナーゼ活性を調節することにより、それに限定されないが不眠症を含む睡眠障害の治療に有用であることを示唆する。さらに、それらに限定されないが、CLOCK、BMAL1、PERならびにCRY−1および−2を含むある特定の概日ポリペプチドは、リン酸化および脱リン酸化により調節され、脳の外側の組織に存在する。よって、非神経組織においてAMPK活性を調整することは、キナーゼカスケードにより概日リズムを設定し、最終的に下流のポリペプチドのリン酸化および脱リン酸化を調節するためにも重要であり得る。
【0049】
さらに、本開示は、Cry1およびCry2のリン酸化および脱リン酸化が、概日性の作用を有し、よって、睡眠状態およびエネルギー代謝の調整のための有用な標的であることを証明する。例えば、CRY1のセリン71および280のリン酸化または脱リン酸化を特異的に調整することにより、FBXL3とのその相互作用を増加させることによるプロテアソーム分解を促進できる。
【0050】
AMPKを活性化するいくつかの薬理学的作用物質は、糖尿病の治療のために現在臨床使用されており、いくつかの型の癌について臨床試験されている。
【0051】
AICARのようなAMPキナーゼアゴニストは、インスリン調節、糖尿病および肥満について研究されている。しかし、AMPキナーゼは、概日リズムや睡眠行動を調整すると以前に証明されていなかった。本開示は、AMPK活性を調整することが、概日リズムに関連するタンパク質の翻訳後修飾を含む下流のプロセスに対する作用を有し得ることを証明する。1つの実施形態において、本開示は、AMPKアゴニストおよびアンタゴニストを用いて、被験体における概日リズムを調整できることを示す。例えば、AMPKは、本開示により、ヘテロ2量体CLOCK/BMAL1を活性化する転写の調整において役割を果たすことが証明される。
【0052】
種々のAMPKアゴニストが、当該技術において公知である。このようなAMPKアゴニストを含む方法および組成物が、本明細書において提供される。このようなAMPKアゴニストの使用は、概日リズムを調整する方法をもたらすことができる。種々のAMPKアゴニストが本明細書に記載され、当該技術において公知である。1つの実施形態において、AMPKアゴニストは、AICAR化合物を含む。本開示の方法において有用な他の化合物は、ビグアナイド誘導体、AICARの類似体(例えば米国特許第5,777,100号(本明細書に参照により組み込まれる)に開示されるもの)およびAICARのバイオアベイラビリティーを増加させるAICARのプロドラッグまたは前駆体(例えば米国特許第5,082,829号(本明細書に参照により組み込まれる)に開示されるもの)を含み、これらは全て当業者に周知である。AMPKの他の活性化因子は、Iyengarらへの米国特許出願公開第20060287356号(その開示は、参照により本明細書に組み込まれる)に記載されるものを含む。従来公知のAMPK活性化化合物は、上記のレプチン、アディポネクチンおよびメトホルミンに加えて、AICAR(5−アミノイミダゾール−4−カルボキサミド)を含む。他のAMPKアゴニストは、それらに限定されないが、DRL−16536(Dr.Reddy’s/Perlecan Pharma)、BG800化合物(Betagenon)、フラン−2−カルボン酸誘導体(Hanall、KR;国際出願公開第WO/2008/016278号(参照により本明細書に組み込まれる)も参照されたい)、A−769662(Abbott)(構造I;Coolら、Cell Metabol.3巻:403〜416頁、2006年も参照されたい);国際公開第WO/2006/033709号に記載されるようなMetabasisにより開発中のAMPKアゴニスト;MT−39シリーズの化合物(Mercury Therapeutics);およびTransTech Pharmaにより開発中のAMPKアゴニストを含む。
【0053】
【化1】
例えば、AICARは、細胞に取り込まれ、AMPKを活性化することが示されているAMP類似体であるZMPに変換される。ZMPは、細胞内AMP模倣物として作用し、十分に高いレベルまで蓄積されると、AMPK活性を刺激できる(Corton, J. M.ら、Eur. J. Biochem.、229巻:558頁(1995年))。しかし、ZMPは、他の酵素の調節においてもAMP模倣物として作用し、よって、特異的AMPK活性化因子ではない(Musi, N.およびGoodyear, L. J. Current Drug Targets−−Immune, Endocrine and Metabolic Disorders 2巻:119頁(2002年))。
【0054】
本開示は、AMPKアゴニストまたはAMPKアンタゴニストのいずれかを用いることにより被験体における概日時計の特定のサイクルを刺激する方法を提供する。1つの実施形態において、AMPKアゴニストを用いて、CLOCK/BMAL1転写活性の増加と関連する概日サイクルを促進する。1つの実施形態において、AMPKアゴニストは、エネルギー保存のシグナル伝達による睡眠効果を、対応するキナーゼカスケードを通して促進する。この方法は、被験体におけるエネルギー欠乏状態を刺激するのに十分な量のAMPKアゴニストを被験体に投与するステップを含む。「エネルギー欠乏状態」により、AMPKのγサブユニットが立体構造変化を受ける状態のことをいう。睡眠効果を促進するとは、そのような効果が、被験体において、AMPKアゴニストの非存在下で生じるよりも多く改善されることを意味する。
【0055】
以下により詳細に記載するように、AMPKアゴニストは、経口で、非経口で、筋肉内に、脈管内に、または任意の適切な経路により投与してよい。1つの実施形態において、AMPKアゴニストは、硬膜外投与される。1つの実施形態において、AMPKアゴニストは、血液脳関門の通過が促進されるように製剤化される。
【0056】
本開示は、AMPK活性と拮抗することにより代謝および活性を「覚醒」または「活動」サイクルに設定する作用物質を投与するステップを含む、活動状態を促進する方法も提供する。1つの実施形態において、AMPKアンタゴニストは、阻害抗体である。1つの実施形態において、AMPKアンタゴニストは、コンパウンドC(ドルソモルフィン、6−[4−(2−ピペリジン−1−イル−エトキシ)−フェニル)]−3−ピリジン−4−イル−ピラゾロ[1,5−a]−ピリミジン)、それらの類似体、誘導体または塩のような小分子阻害物質である。
【0057】
開示される方法は、限定されないが、薬学技術の当業者に周知の投与方法、投与量および製剤を含む、AMPKアゴニスト単独、またはその製剤を受容する被験体における所望の概日サイクルの状態を誘導する所望の結果を有する他の概日調節剤もしくは睡眠補助剤との組合せでの任意の投与方法、投与量および/または製剤の使用を想定する。
【0058】
本開示のAMPKアゴニストは、薬物の形態でヒトまたは動物に投与してよい。代わりに、AMPKアゴニストは、ヒトまたは動物により消費されるように、多様な食品および飲料またはペットフードに組み込んでよい。AMPKアゴニストは、一般的な食品もしくは飲料に用いてよいか、あるいは機能性食品もしくは飲料、疾患に罹患している被験体のための食品、または特定の健康用途のための食品(この食品(または飲料)は、それが生理機能を有することを記載したラベルを有する);例えば睡眠補助剤に用いてよい。
【0059】
AMPKアゴニスト単独、または他の睡眠補助剤もしくは活性成分との組合せは、薬品、例えば、錠剤もしくは顆粒剤のような経口固体製品、または液剤もしくはシロップ剤のような経口液体製品に製剤化してよい。
【0060】
開示される方法におけるAMPKアゴニストまたは製剤の投与の方式は、それらに限定されないが、くも膜下内(intrathecal)、皮内、筋肉内、腹腔内(ip)、静脈内(iv)、皮下、鼻内、硬膜外、硬膜内、頭蓋内、脳室内および経口経路を含む。具体的な例において、AMPKアゴニストは、経口投与される。AMPKアゴニストの投与のための他の簡便な経路は、例えば、注入もしくはボーラス投与、局所、上皮もしくは皮膚粘膜層を通しての吸収(例えば口腔粘膜、直腸および腸粘膜など)、眼、鼻および経皮を含む。投与は、全身性または局所的であり得る。肺投与も(例えば吸入器またはネブライザにより)、例えばエアロゾル剤を含有する製剤を用いることにより、用いることができる。
【0061】
いくつかの実施形態において、AMPKアゴニストを投与またはAMPKアゴニストを局所的に投与することが望ましい場合がある。このことは、例えば、局所的もしくは領域的な注入または灌流、局所塗布(例えば創傷包帯材)、注射、カテーテル、坐剤またはインプラント(例えば、シアラスティック(sialastic)膜または繊維のような、膜を含む多孔質、非多孔質またはゼラチン質の材料から形成されるインプラント)などにより達成してよい。
【0062】
他の実施形態において、ポンプ(例えば移植されたミニポンプ)を用いてAMPKアゴニストまたは製剤を送達してよい(例えば、Langer Science 249巻、1527頁、1990年;Sefton Crit. Rev. Biomed. Eng. 14巻、201頁、1987年;Buchwaldら、Surgery 88巻、507頁、1980年;Saudekら、N. Engl. J. Med. 321巻、574頁、1989年を参照されたい)。別の実施形態において、AMPKアゴニストまたは製剤は、小胞、特にリポソームで送達される(例えば、Langer、Science 249巻、1527頁、1990年;Liposomes in the Therapy of Infectious Disease and Cancer中のTreatら、Lopez−BeresteinおよびFidler(編)、Liss, N. Y.、353〜365頁、1989年を参照されたい)。
【0063】
さらに別の方法実施形態において、AMPKアゴニストは、制御放出製剤で送達できる。Langer(Science 249巻、1527頁、1990年)による総説で論じられているもののような制御放出系が公知である。同様に、制御放出製剤において有用なポリマー材料が公知である(例えば、Rangerら、Macromol. ScL Rev. Macromol. Chem. 23巻、61頁、1983年;Levyら、Science 228巻、190頁、1985年;Duringら、Ann. Neurol. 25巻、351頁、1989年;Howardら、J. Neurosurg. 71巻、105頁、1989年を参照されたい)。例えば、アゴニストは、ポリ乳酸、ポリグリコール酸、ポリ乳酸とポリグリコール酸とのコポリマー、ポリイプシロンカプロラクトン、ポリヒドロキシ酪酸、ポリオルトエステル、ポリアセタール、ポリジヒドロピラン、ポリシアノアクリレート、およびヒドロゲルの架橋または両親媒性ブロックコポリマーを含む、化合物の制御放出を達成するのに有用なクラスの生分解性ポリマーと結合させてよい。
【0064】
開示される方法は、アゴニスト(複数可)を送達して所望の結果を達成するAMPKアゴニストの任意の剤形またはその製剤の使用を意図する。剤形は、一般的に公知であり、例えばAllenら、Ansel’s Pharmaceutical Dosage Forms and Drug Delivery Systems、第8版、Philadelphia, PA: Lippincott Williams & Wilkins、2005年、全738頁を含む多様な参考書で教示されている。開示される方法で用いるための剤形は、それらに限定されないが、固体剤形および固体改変放出薬物送達系(例えば散剤および顆粒剤、カプセル剤ならびに/または錠剤);半固体剤形および経皮系(例えば軟膏剤、クリームおよび/またはゲル);経皮薬物送達系;薬学的挿入物(例えば坐剤および/または挿入物(insert));液体剤形(例えば液剤および分散系);ならびに/または滅菌剤形および送達系(例えば非経口製剤(parenterals)および/または生物製剤(biologies))を含む。具体的な例示的剤形は、エアロゾル(定量噴霧(metered dose)、粉末、溶液および/または噴霧剤なしを含む);ビーズ;カプセル(従来の、制御送達、制御放出、腸溶被覆および/または持続放出を含む);キャプレット;濃縮物;クリーム;結晶;ディスク(持続放出を含む);点滴剤;エリキシル;乳剤;フォーム;ゲル(ゼリーおよび/または制御放出を含む);小球;顆粒剤;ガム;インプラント;吸入薬;注射剤;挿入物(徐放(extended release)を含む);リポソーム;液体(制御放出を含む);ローション;ロゼンジ;定量噴霧(例えばポンプ);ミスト;うがい薬;噴霧化用溶液;眼用の系;油;軟膏;膣坐剤(ovule);粉末(パケット、発泡剤(effervescent)、懸濁用粉末、懸濁持続放出用粉末および/または溶液用粉末);ペレット;ペースト;液剤(長時間作用性および/または再構成されたものを含む);細片;坐剤(持続放出を含む);懸濁剤(レンテ、ウルトラレンテ(ultre lente)、再構成されたものを含む);シロップ(持続放出を含む);錠剤(咀嚼、舌下、持続放出、制御放出、遅延作用、遅延放出、腸溶被覆、発泡剤、フィルム被覆、即時溶解、緩慢放出を含む);経皮系;チンキ;および/またはカシェ剤を含む。典型的には、剤形は、AMPKアゴニストを含む少なくとも1つの薬学的活性成分の有効量(例えば治療有効量)と、薬学的に許容される賦形剤および/または他の成分(例えば1つ以上の他の活性成分)との製剤である。薬物製剤化の目的は、被験体に活性成分(例えばAMPKアゴニストまたはAMPKアンタゴニスト)を適切に投与することである。製剤は、投与の方式に適するものである。「薬学的に許容される」との用語は、連邦政府もしくは州政府の規制機関により承認されたか、または米国薬局方もしくは他の一般的に認識されている動物およびより具体的にはヒトにおける使用についての薬局方に列挙されていることを意味する。例示的な製剤において用いるための賦形剤は、例えば、1つ以上の以下のものを含む:結合剤、増量剤(filler)、崩壊剤、滑沢剤、コーティング、甘味料、香料、着色料、防腐剤、希釈剤、アジュバントおよび/またはビヒクル。賦形剤が、一括して、具体的な剤形の全重量(および/または容量)の約5%〜95%を構成するいくつかの場合がある。
【0065】
薬学的賦形剤は、例えば、水および/またはピーナツ油、大豆油、鉱油、ごま油のような石油、動物油、植物油または合成起源のものなどを含む油のような滅菌液体であり得る。水は、製剤を静脈内投与する場合の例示的なキャリアである。生理食塩水、血漿媒体、デキストロース水溶液およびグリセロール溶液を、特に注射用溶液用の液体キャリアとして用いることもできる。経口製剤は、それらに限定されないが、薬学的グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどを含み得る。非経口の薬学的賦形剤のより完全な説明は、Remington、The Science and Practice of Pharmacy、第19版、Philadelphia, PA: Lippincott Williams & Wilkins、1995年、第95章に見出すことができる。賦形剤は、例えば、浸透圧を調整するための薬学的に許容される塩、シクロデキストリンのような脂質キャリア、血清アルブミンのようなタンパク質、メチルセルロースのような親水性作用物質、界面活性剤、緩衝剤、防腐剤なども含んでよい。薬学的賦形剤の他の例は、デンプン、グルコース、ラクトース、スクロース、ゼラチン、麦芽、コメ、小麦粉、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリン酸グリセロール、タルク、塩化ナトリウム、脱脂粉乳、グリセロール、プロピレン、グリコール(propylene, glycol)、水、エタノールなどを含む。所望により、製剤は、少量の湿潤剤もしくは乳化剤またはpH緩衝剤も含有できる。
【0066】
経口投与を含むいくつかの実施形態において、AMPKアゴニストの経口投与量は、通常、1日あたり体重1kgあたり約0.001mg(mg/kg/日)〜約100mg/kg/日、例えば約0.01〜10mg/kg/日の範囲である(そうでないと記載しない限り、活性成分の量は、遊離の酸または遊離の塩基であり得る中性分子に基づく)。例えば、80kgの被験体は、約0.08mg/日〜8g/日、例えば約0.8mg/日〜800mg/日を受容する。1日1回の投与のための適切に調製された医薬品は、よって、0.08mg〜8g、例えば0.8mg〜800mgを含有する。いくつかの場合において、AMPKアゴニストまたはアンタゴニストを含む製剤は、1日2回、3回または4回の分割用量で投与してよい。1日2回の投与のために、上記のような適切に調製された医薬品は、0.04mg〜4g、例えば0.4mg〜400mgを含有する。上記の範囲外の投与量が必要となる場合があり得る。1日あたり0.08mg〜8gの範囲で与えられ得る1日投与量の例は、0.1mg、0.5mg、1mg、2.5mg、5mg、10mg、25mg、50mg、100mg、200mg、300mg、400mg、500mg、600mg、800mg、1g、2g、4gおよび8gを含む。これらの量は、1日に1回より多く投与する場合は、より少ない用量に分けることができる(例えば、薬物を1日2回摂取する場合は、各投与において半分の量)。
【0067】
注射による投与(例えば静脈内または皮下注射)を含むいくつかの方法実施形態において、被験体は、ほぼ上記の量で活性成分を送達する注射量を受容する。量は、消化系を回避する注射薬物形態に起因する送達効率の差を考慮して調整してよい。このような量は、いくつかの適切な方法、例えば1回の長期間または1日に数回の間での低濃度で高容量の活性成分、短期間、例えば1日1回の間での高濃度で低容量の活性成分で投与してよい。典型的には、約0.01〜1.0mg/mlの間の、例えば0.1mg/ml、0.3mg/mlまたは0.6mg/mlなどの活性成分の濃度を含有する従来の静脈内製剤を調製し、上記の1日あたりの量に等しい1日あたりの量で投与してよい。例えば、0.5mg/mlの活性成分の濃度を有する静脈内製剤を1日2回8ml受容する80kgの被験体は、1日あたり8mgの活性成分を受容する。
【0068】
他の方法実施形態において、AMPKアゴニストまたはアンタゴニスト(またはその製剤)は、治療期間を通してほぼ同じ用量、漸増用量計画または負荷用量計画(例えば負荷用量が維持用量の約2〜5倍である)で投与できる。いくつかの実施形態において、用量は、使用期間の間に、組成物を受容する被験体の状態、組成物に対する見かけの応答および/または当業者により判断される他の因子に基づいて変動する。いくつかの実施形態において、例えば慢性不眠症または睡眠覚醒サイクル障害を管理するために、AMPKアゴニストまたはアンタゴニストの長期間の投与が意図される。
【0069】
本開示は、AMPK活性化または阻害を測定することによる、概日リズムを調整する作用物質のスクリーニング方法も提供する。概日リズムを調整する化合物をスクリーニングするための本開示の方法は、AMPK経路を備える細胞、組織または被験体(例えば動物)を提供するステップと、前記被験体を、概日リズム調整活性を有すると推測される作用物質と接触させるステップと、AMPK活性に対する作用を、直接、または下流のキナーゼ活性により測定するステップとを含む。被検作用物質は、概日リズム機能の少なくとも1つの観察可能な指標を有し、かつAMPKを発現する細胞調製物、組織、器官、生物または動物に対して提供することができる。概日リズムを調整する作用物質の能力は、概日リズム機能の徴候を示す多様な動物種、ならびにそのような動物から得られる器官、組織および細胞、およびそれらに由来する細胞調製物で試験できる。AMPK活性を調整する作用物質は、次いで、推定概日リズム調整活性を有する作用物質として同定できる。
【0070】
多様なin vitroスクリーニング方法が、概日リズムを調整する化合物を同定するための本開示の方法において提供されるアンタゴニストまたはアゴニストを同定するために有用である。AMPKを調整する化合物の能力は、例えば、AMPKと結合してAMPKを活性化もしくは不活性化するか、下流のキナーゼ活性を遮断するか、リン酸化および脱リン酸化を調整するか(例えばCry1またはCry2のリン酸化、脱リン酸化)、またはAMPKにより生成される予め決定されたシグナルを調整する化合物の能力により示され得る。よって、シグナル伝達および結合アッセイを用いて、概日リズムを調整する化合物を同定するための本開示の方法において提供されるAMPKのアンタゴニストまたはアゴニストを同定できる。
【0071】
「作用物質」は、目的または結果を達成するために有用である、任意の物質または物質の任意の組合せ、例えば、AMPK活性化カスケードと関連するタンパク質活性を調整するために有用であり(例えばAMPK依存性リン酸化事象)、またはタンパク質−タンパク質相互作用もしくはATP代謝を改変するかまたはそれに影響するために有用である物質あるいは物質の組合せである。
【0072】
例示的な作用物質は、それらに限定されないが、それらに限定されないがランダムペプチドライブラリーのメンバー(例えば、Lamら、Nature、354巻:82〜84頁、1991年;Houghtenら、Nature、354巻:84〜86頁、1991年を参照されたい)を含む例えば可溶性ペプチドのようなペプチド、ならびにDおよび/またはL立体配置のアミノ酸で作製されたコンビナトリアル化学由来分子ライブラリー、ホスホペプチド(それらに限定されないが、ランダムまたは部分的に縮重された、定向ホスホペプチドライブラリーのメンバーを含む;例えば、Songyangら、Cell、72巻:767〜778頁、1993年を参照されたい)、抗体(それらに限定されないが、ポリクローナル、モノクローナル、ヒト化、抗イディオタイプ、キメラまたは単鎖抗体、ならびにFab、F(ab’)2およびFab発現ライブラリーフラグメント、およびそれらのエピトープ結合フラグメントを含む)、有機または無機の小分子(例えば、いわゆる天然生成物または化学コンビナトリアルライブラリーのメンバー)、分子複合体(例えばタンパク質複合体)、あるいは核酸を含む。
【0073】
本開示の方法において有用なライブラリー(例えばコンビナトリアル化学ライブラリー)は、それらに限定されないが、ペプチドライブラリー(例えば米国特許第5,010,175号;Furka、Int. J. Pept. Prot. Res.、37巻:487〜493頁、1991年;Houghtonら、Nature、354巻:84〜88頁、1991年;PCT公報第WO91/19735号を参照されたい)、コードされたペプチド(例えばPCT公報第WO93/20242号)、ランダムバイオオリゴマー(例えばPCT公報第WO92/00091号)、ベンゾジアゼピン(例えば米国特許第5,288,514号)、ヒダントイン、ベンゾジアゼピンおよびジペプチドのようなダイバーソマー(diversomer)(Hobbsら、Proc. Natl. Acad. Sci. USA、90巻:6909〜6913頁、1993年)、ビニローグ(vinylogous)ポリペプチド(Hagiharaら、J. Am. Chem. Soc、114巻:6568頁、1992年)、グルコース足場を有する非ペプチド性ペプチド模倣物(Hirschmannら、J. Am. Chem. Soc、114巻:9217〜9218頁、1992年)、小化合物ライブラリーの類似した有機合成(Chenら、J. Am. Chem. Soc、116巻:2661頁、1994年)、オリゴカルバメート(Choら、Science、261巻:1303頁、1003年)および/またはペプチジルホスホネート(Campbellら、J. Org. Chem.、59巻:658頁、1994年)、核酸ライブラリー(Sambrookら、 Molecular Cloning, A Laboratory Manual、Cold Springs Harbor Press, N. Y.、1989年;Ausubelら、Current Protocols in Molecular Biology、Green Publishing Associates and Wiley Interscience, N. Y.、1989年を参照されたい)、ペプチド核酸ライブラリー(例えば米国特許第5,539,083号を参照されたい)、抗体ライブラリー(例えばVaughnら、Nat. Biotechnol、14巻:309〜314頁、1996年;PCT出願第PCT/US96/10287号を参照されたい)、炭水化物ライブラリー(例えばLiangら、Science、274巻:1520〜1522頁、1996年;米国特許第5,593,853号を参照されたい)、有機小分子ライブラリー(例えばベンゾジアゼピン、Baum、C&EN、1月18日、33頁、1993年;イソプレノイド、米国特許第5,569,588号;チアゾリジオノンおよびメタチアゾン、米国特許第5,549,974号;ピロリジン、米国特許第5,525,735号および第5,519,134号;モルホリノ化合物、米国特許第5,506,337号;ベンゾジアゼピン、第5,288,514号を参照されたい)などを含む。
【0074】
開示されるスクリーニング方法のために有用なライブラリーは、それらに限定されないが、空間的に整列させたマルチピンペプチド合成(Geysenら、Proc Natl. Acad. Sci.、81巻(13号):3998〜4002頁、1984年)、「ティーバッグ」ペプチド合成(Houghten、Proc Natl. Acad. Sci.、82巻(15号):5131〜5135頁、1985年)、ファージディスプレイ(ScottおよびSmith、Science、249巻:386〜390頁、1990年)、スポットまたはディスク合成(Dittrichら、Bioorg. Med. Chem. Lett.、8巻(17号):2351〜2356頁、1998年)、あるいはビーズ上での分割および混合固相合成(Furkaら、Int. J. Pept. Protein Res.、37巻(6号):487〜493頁、1991年;Lamら、Chem. Rev.、97巻(2号):411〜448頁、1997年)を含む多様な方法で生成できる。ライブラリーは、種々の数の構成成分(composition)(メンバー)、例えば約100メンバーまで、例えば約1000メンバーまで、例えば約5000メンバーまで、例えば約10,000メンバーまで、例えば約100,000メンバーまで、例えば約500,000メンバーまでまたは500,000メンバーより多くさえ含んでよい。
【0075】
1つの実施形態において、ハイスループットスクリーニング法は、多数の可能性のある治療化合物(例えば、AMPKタンパク質−タンパク質相互作用の影響因子)を含有するコンビナトリアル化学またはペプチドライブラリーを提供するステップを含む。このようなコンビナトリアルライブラリーを、次いで、本明細書に記載されるような1つ以上のアッセイでスクリーニングして、所望の特徴的な活性を示す(例えばAMPKタンパク質−タンパク質相互作用を増加または減少させる)ライブラリーメンバー(特に化学種またはサブクラス)を同定する。このようにして同定された化合物は、通常の「リード化合物」として用い得るか、またはそれら自体を可能性のあるもしくは実際の治療剤として用いることができる。いくつかの場合において、候補作用物質のプールを同定し、さらにスクリーニングして、どの個別の作用物質または集合的な作用物質のより小さいプールが所望の活性を有するか決定してよい。AMPK相互作用またはプロセスのAMP依存性リン酸化に影響する(例えば増加または減少させる)作用物質は、被験体において概日リズム(例えば睡眠行動)を調整する効果を有する場合があり、よって、同定することが望まれる。
【0076】
本明細書に記載されるスクリーニング法において、組織試料、単離細胞、単離ポリペプチドおよび/または被検作用物質は、ハイスループットスクリーニングに適する様式で提示できる。例えば、1または複数の単離組織試料、単離細胞または単離ポリペプチドを、マイクロタイタープレートのウェルに入れ、1または複数の被検作用物質をマイクロタイタープレートのウェルに加えることができる。代わりに、1または複数の被検作用物質を、マイクロタイタープレートのウェル(溶液でまたはプレート表面に接着した)のようなハイスループットフォーマットで提示でき、1または複数の単離組織試料、単離細胞および/または単離ポリペプチドと、組織試料もしくは単離細胞もしくは所望のポリペプチド機能および/または構造物を少なくとも維持する条件下で接触させる。被検作用物質は、組織試料、単離細胞または単離ポリペプチドに、組織もしくは細胞に致死的でなく、ポリペプチド構造物および/または機能に有害な影響を与えない任意の濃度で加えることができる。異なる被検作用物質は、異なる有効濃度を有することが予想される。よって、いくつかの方法において、被検作用物質濃度の範囲を試験することが有利である。
【0077】
タンパク質リン酸化を検出するための方法は、従来からあり(例えばGloffke、The Scientist、16巻(19号):52頁、2002年;Screatonら、Cell、119巻:61〜74頁、2004年を参照されたい)、検出キットは、多様な商業的供給源から入手可能である(例えばUpstate(Charlottesville、VA、USA)、Bio−Rad(Hercules、CA、USA)、Marligen Biosciences,Inc.(Ijamsville、MD、USA)、Calbiochem(San Diego、CA、USA)を参照されたい)。簡単に述べると、リン酸化タンパク質は、ゲル中のリン酸化タンパク質に特異的な染色を用いて検出できる。代わりに、リン酸化タンパク質に特異的な抗体を作製できるか、または商業的に入手できる。リン酸化タンパク質に特異的な抗体は、なかでも、ビーズに繋留されているか(特定の色のサインを有するビーズを含む)、またはELISAもしくはウェスタンブロットアッセイにおいて用いることができる。
【0078】
特定の方法において、ポリペプチドのリン酸化は、このような翻訳後修飾が検出可能に測定される場合、またはこのような翻訳後修飾が対照測定(例えば被検作用物質を加える前の同じ試験系、または被検作用物質の非存在下での同様の試験系、またはAMPKの非存在下での同様の試験系)よりも少なくとも20%、少なくとも30%、少なくとも50%、少なくとも100%または少なくとも250%高い場合に、増加している。
【0079】
AMPKサブユニット(例えばAMPKα1および/またはAMPKα2)の基本型のアミノ酸配列(および基本型のAMPKサブユニット(例えばAMPKα1および/またはAMPKα2)をコードする核酸配列)は、周知である。例示的なAMPKα1アミノ酸配列および対応する核酸配列は、例えば、GenBank受託番号NM_206907.3(GI:94557298)(アミノ酸配列および核酸配列を含むHomo sapiens転写産物バリアント2 REFSEQ);NM_006251.5(GI:94557300)(アミノ酸および核酸配列を含むHomo sapiens転写産物バリアント1 REFSEQ);NM_001013367.3(GI:94681060)(アミノ酸および核酸配列を含むMus musculus REFSEQ);NMJ)01039603.1(GI:88853844)(アミノ酸および核酸配列を含むGallus gallus REFSEQ);およびNM_019142.1(GI:11862979XRaJfWSアミノ酸配列および核酸配列を含むnorvegicus REFSEQ)に記載される。例示的なAMPKα2アミノ酸配列および対応する核酸配列は、例えば、GenBank受託番号NM_006252.2(GI:46877067)(アミノ酸配列および核酸配列を含むHomo sapiens REFSEQ);NM_178143.1(GI:54792085)(アミノ酸配列および核酸配列を含むMus musculus REFSEQ);NM_001039605.1(GI:88853850)(アミノ酸配列および核酸配列を含むGallus gallus REFSEQ);およびNM_214266.1(GI:47523597)(アミノ酸配列および核酸配列を含むMus musculus REFSEQ)に記載される。
【0080】
いくつかの方法実施形態において、AMPKサブユニットのホモログまたは機能的バリアントは、基本型のAMPKα1および/またはAMPKα2ポリペプチドと少なくとも60%のアミノ酸配列同一性、例えばGenBank受託番号NM_206907.3;NM_006251.5;NMJ)01013367.3;NM_001039603.1;NM_019142.1;NM_006252.2;NM_178143.1;NM_001039605.1;またはNM_214266.1に記載されるアミノ酸配列と、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%または少なくとも98%のアミノ酸配列同一性を共有する。他の方法実施形態において、AMPKサブユニットのホモログまたは機能的バリアントは、基本型のAMPKα1および/またはAMPKα2ポリペプチドと比較して1つ以上の保存的アミノ酸置換、例えばGenBank受託番号NM_206907.3;NM_006251.5;NM_001013367.3;NM_001039603.1;NM_019142.1;NM_006252.2;NM_178143.1;NM_001039605.1;またはNM_214266.1に記載されるアミノ酸配列と比較して、3以下、5以下、10以下、15以下、20以下、25以下、30以下、40以下または50以下の保存的アミノ酸の変化を有する。例示的な保存的アミノ酸置換は、本明細書において先に記載した。
【0081】
いくつかの方法実施形態は、AMPKまたはそのサブユニット(例えばAMPKα1および/またはAMPKα2)の機能的フラグメントを含む。AMPKまたはそのサブユニット(例えばAMPKα1および/またはAMPKα2)の機能的フラグメントは、例えばその約20、約30、約40、約50、約75、約100、約150または約200連続アミノ酸残基を含む、全長もしくはインタクトなAMPKポリペプチド複合体またはそのサブユニット(例えばAMPKα1および/またはAMPKα2)の任意の部分であり得るが、ただし、フラグメントが、対象とする少なくとも1つのAMPK(またはAMPKα1および/もしくはAMPKα2)機能を保持することを条件とする。AMPK経路におけるポリペプチド間のタンパク質−タンパク質相互作用は、少なくともAMPKαサブユニット(例えばAMPKα1および/またはAMPKα2)を含むと考えられる。
【0082】
「単離」生体成分(例えばポリヌクレオチド、ポリペプチドまたは細胞)は、混合試料(例えば細胞または組織の抽出物)中の他の生体成分から分けて精製されている。例えば、「単離」ポリペプチドまたはポリヌクレオチドは、ポリペプチドまたはポリヌクレオチドが存在していた細胞(例えば組換えポリペプチドまたはポリヌクレオチドについての発現宿主細胞)の他の成分から分けられたポリペプチドまたはポリヌクレオチドである。
【0083】
「精製された」との用語は、試料から1つ以上の無関係の成分を除去することをいう。例えば、組換えポリペプチドを宿主細胞において発現させる場合、ポリペプチドは、例えば、宿主細胞タンパク質を除去し、それにより試料中の組換えポリペプチドの割合が増加することにより精製される。同様に、組換えポリヌクレオチドが宿主細胞中に存在する場合、ポリヌクレオチドは、例えば、宿主細胞ポリヌクレオチドを除去し、それにより試料中の組換えポリヌクレオチドの割合が増加することにより精製される。
【0084】
単離ポリペプチドまたは核酸分子は、典型的には、試料の少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%または99%(w/wまたはw/v)超をさらに含む。
【0085】
ポリペプチドおよび核酸分子は、当該技術において一般的に公知であり本明細書に記載される方法により単離される。ポリペプチドまたは核酸分子の純度は、ポリペプチドについてポリアクリルアミドゲル電気泳動、または核酸分子についてアガロースゲル電気泳動のようないくつかの周知の方法により決定してよい。
【0086】
2つの核酸配列間または2つのアミノ酸配列間の類似性は、これらの配列間で共有される配列同一性のレベルの点で表される。配列同一性は、典型的には、パーセント同一性に関して表される。パーセンテージが高いほど、2つの配列はより類似する。
【0087】
比較のために配列を整列させる方法は、当該技術において周知である。種々のプログラムおよびアラインメントアルゴリズムが、SmithおよびWaterman、Adv. Appl. Math.2巻:482頁、1981年;NeedlemanおよびWunsch、J. Mol. Biol.48巻:443頁、1970年;PearsonおよびLipman、Proc. Natl. Acad. ScL USA85巻:2444頁、1988年;HigginsおよびSharp、Gene73巻:237〜244頁、1988年;HigginsおよびSharp、CABIOS5巻:151〜153頁、1989年;Corpetら、Nucleic Acids Research16巻:10881〜10890頁、1988年;Huangら、Computer Applications in the Biosciences8巻:155〜165頁、1992年;Pearsonら、Methods in Molecular Biology24巻:307〜331頁、1994年;Tatianaら(1999年)、FEMS Microbiol. Lett.、174巻:247〜250頁、1999年に記載されている。Altschulらは、配列アラインメント法および相同性算出について詳細な考察を示している(J. Mol. Biol.215巻:403〜410頁、1990年)。国立バイオテクノロジー情報センター(NCBI)のBasic Local Alignment Search Tool(BLAST(商標)、Altschulら、J. Mol. Biol.215巻:403〜410頁、1990年)は、blastp、blastn、blastx、tblastnおよびtblastxの配列分析プログラムと関連して用いるために、国立バイオテクノロジー情報センター(NCBI、Bethesda、MD)およびインターネット上を含む種々の供給源から利用可能である。このプログラムを用いてどのようにして配列同一性を決定するかについての記載は、インターネット上で、BLAST(商標)についてのヘルプ部分から利用可能である。
【0088】
約30アミノ酸より大きいアミノ酸配列の比較のために、BLAST(商標)(Blastp)プログラムの「Blast2配列」機能を、デフォルトパラメータ(ギャップ開始コスト[デフォルト=5];ギャップ伸長コスト[デフォルト=2];ミスマッチペナルティ[デフォルト=−3];マッチ報酬[デフォルト=1];期待値(E)[デフォルト=10.0];ワードサイズ[デフォルト=3];1行の記載の数(V)[デフォルト=100];表示するアラインメントの数(B)[デフォルト=100])に設定されたデフォルトBLOSUM62行列を用いて行う。短いペプチド(30アミノ酸くらいより少ない)を整列させる場合、アラインメントは、デフォルトパラメータ(開始ギャップ9、伸長ギャップ1ペナルティ)に設定されたPAM30行列を用いるBlast2配列機能を用いて行う。参照配列に対してさらにより大きい類似性を有するタンパク質は、この方法により評価する場合に、パーセント同一性の増加を示す。
【0089】
核酸配列の比較のために、BLAST(商標)(Blastn)プログラムの「Blast2配列」機能を、デフォルトパラメータ(ギャップ開始コスト[デフォルト=11];ギャップ伸長コスト[デフォルト=1];期待値(E)[デフォルト=10.0];ワードサイズ[デフォルト=11];1行の記載の数(V)[デフォルト=100];表示するアラインメントの数(B)[デフォルト=100])に設定されたデフォルトBLOSUM62行列を用いて行う。参照配列に対してさらにより大きい類似性を有する核酸配列は、この方法により評価する場合に、パーセント同一性の増加を示す。
【0090】
特異的結合とは、一方の結合パートナー(例えば結合剤)と他方の結合パートナー(例えば標的)との間の特定の相互作用のことをいう。このような相互作用は、結合パートナー間(またはより頻繁には、各結合パートナーの特定の領域または部分間)の1または典型的にはそれより多い非共有結合により媒介される。非特異的結合部位とは対照的に、特異的結合部位は、飽和可能である。よって、特異的結合を特徴づける1つの例示的な方法は、特異的結合曲線による。特異的結合曲線は、例えば、一定量の一方の結合パートナーと結合した他方の結合パートナー(第1結合パートナー)の量を、第1結合パートナーの濃度の関数として示す。第1結合パートナーの濃度がこれらの条件下で増加すると、結合した第1結合パートナーの量が飽和する。非特異的結合部位とはまた別に対照的に、互いの直接の会合(例えばタンパク質−タンパク質相互作用)に関与する特異的結合パートナーは、このような会合(例えばタンパク質複合体)から、過剰量のいずれかの特異的結合パートナーにより競合的に除去(または置換)され得る。このような競合アッセイ(または置換アッセイ)は、当該技術において周知である。
【0091】
本開示は、概日リズムおよび睡眠行動をもたらすために有用な作用物質を同定する方法および作用物質も提供する。
【実施例】
【0092】
以下の実施例は、ある特定の特徴および/または実施形態を例示するために提供される。これらの実施例は、記載される特定の特徴または実施形態に本発明を限定すると解釈されない。
【0093】
(実施例1)
CRY1−S71またはCRY1−S280のリン酸化は、CRY1:FBXL3相互作用を増加させる。末梢時計のリセットにおける機構としての翻訳後修飾の役割を調べるために、質量分析および生物情報学分析の組合せを用いて、調節されたリン酸化の部位であると予測されるmCRY1およびmCRY2中の8つのセリンまたはスレオニン残基を同定した。リン酸化し得ない変異体(non−phosphorylatable mutant)をそれぞれについて作製し、セリン71のアラニンへの変異がmCRY1を安定化するが、残りの変異は安定性に対してほとんどまたは全く効果を有さないことが分かった(図7)。
【0094】
セリン71に影響するCRY1安定化変異は、AMPKによりリン酸化された最適配列によく順応するので、特に興味が持たれる。哺乳動物クリプトクロムは、mCRY1の280位に別のセリンを含有し、これも、AMPK基質モチーフによく順応する(図8)。セリン71またはセリン280のいずれかをリン酸化し得ないアミノ酸(アラニン)に変異させることは、mCRY1を安定化するために十分であったが、セリン71またはセリン280のいずれかをホスホ模倣アミノ酸(アスパルテート)に変異することは、mCRY1を不安定化するのに十分であり、これらの両方の残基を一緒に変異させると、安定性に対する影響が増加した(図1)。両方の場合において、全ての光非感受性昆虫クリプトクロムおよび高等生物において進化的に保存されているセリン71(図2)の変異は、S280の変異よりも強い効果を有した(図1A)。S71およびS280の両方のアスパラギン酸へのホスホ模倣変異を有するmCRY1は、イムノブロットにより検出できなかった。セリン71および/またはセリン280でリン酸化されているmCRY1の安定性の低下と一致して、これらの部位のホスホ模倣変異体は、CLOCK:BMAL1転写活性のより効果が低いリプレッサーでもあった(図1B)。クリプトクロムは、元来、植物において青色光光受容体として同定され、後に、動物の概日時計の成分として認識された。多くの昆虫が、クリプトクロムのそれぞれの型のうちの1つを発現する:露光により分解される青色光光受容体(「1型」)および概日転写調節に関与するが、光により誘導される不安定化に対して感受性でない転写リプレッサー(「2型」)。昆虫「2型」クリプトクロムタンパク質は、それらの哺乳動物での対応物と同様に、一昼夜の経過のうちで変動し、このことは、それらの安定化が、非光(non−light)シグナルにより、FBXL3(そのオルソログは昆虫に存在する(GenBank参照番号XM_001120533.1))が関与する保存された機構をおそらく用いて調節されるはずであることを示す。
【0095】
セリン71および280のリン酸化を模倣する変異を有するmCRY1変異体の不安定性が、FBXL3との相互作用の増加を反映するかを決定するために、v5タグ付加FBXL3を有するFlagタグ付加野生型または変異体mCRY1を発現させ、それらの結合親和性を、Flagタグ付加mCRY1の免疫沈降と、その後のv5タグ付加FBXL3のイムノブロットにより分析した。S71またはS280のいずれかのアスパラギン酸への変異は、FBXL3へのmCRY1の結合親和性を増加させ(図1C)、このことは、これらの部位のリン酸化が、mCRY1とFBXL3との間の相互作用の増加を媒介することを示唆した。2重変異体は不安定すぎて、FBXL3とのその相互作用を生化学的に決定できなかった。
【0096】
これらの変異体を、同時トランスフェクトしたCLOCKおよびBMAL1の転写活性を抑制する能力について試験した場合、CRY1のリン酸化し得ないAA変異体は、効果的なリプレッサーであることが示され、この抑制は、FBXL3の同時トランスフェクションによって変更されなかった。予測されたように、2重ホスホ模倣物CRY1 DDは、CLOCK:BMAL1活性のより効果が低い阻害物質であり、これは、この変異体のより低い安定性を反映し得る。CRY1 AA抑制に対するFBXL3媒介の作用の欠如とは対照的に、CLOCK:BMAL1駆動型転写を抑制するCRY1 DDの弱い能力は、FBXL3の同時発現により喪失した(図1D)。
【0097】
S71、S280およびS281変異の影響を、mCRY1とその公知の結合パートナーであるPER2との相互作用に対して調べた。セリン71のホスホ模倣変異(S71D)は、CRY1とPER2との間の相互作用を遮断したが、S280D変異体はPER2結合を保持し、調べた他の変異体も同様であった(図1E)。この差は、CRY1 S280Dを上回るCRY1 S71Dの分解の増進が一因であり得る。よって、S71D変異体は、PER2との結合の減少と、FBXL3との結合の増加を示し、これらはそれぞれCRY1を不安定化すると予想され、これらは一緒にCRY1 S71Dの観察された不安定性の原因であると見られる。
【0098】
AMPKは、リン酸化依存性クリプトクロム分解を媒介する。mCRY1のセリン71の周囲の配列の関係は、これが、近くの好ましい配列特異性(標的セリンに対して−4位および−3位での正に荷電された残基、ならびに−5位および+4位での疎水性残基)だけでなく、標的セリンに対して−16位および−9位での好ましい遠位ロイシン残基さえ含み、AMPKによるリン酸化の優れた候補であることを示唆する(図2A)。S280の周囲のアミノ酸配列の関係も、近傍の好ましい配列特異性に従ってAMPKリン酸化を示唆する(図8)。
【0099】
ホスホ特異抗体を、mCRY1 S71の配列の関係を周囲に有するホスホセリンを含有するペプチド抗原に対して作製し、この抗体を用いて、外因的に発現させた野生型mCRY1のリン酸化を観察したが、リン酸化し得ないS71A変異体では観察されなかった(図2B)。mCRY1のセリン71の周囲の配列は、アセチル補酵素Aカルボキシラーゼ1(ACC1)のセリン79の周囲のものと類似し、これは、AMPKの最もよく研究された基質の1つである(図2B)。実際に、セリン79でリン酸化されたACC1の残基73〜85に相当するペプチドに対して産生された抗体は、野生型mCRY1を検出できたが、セリン71をアラニンに置き換える変異を有するmCRY1は検出できず(図2B)、このことは、mCRY1のセリン71が、in vivoでリン酸化されることができることのさらなる証拠を提供し、このリン酸化事象がAMPKにより媒介され得ることをさらに示唆する。AMPKa2触媒ドメインの構成的に活性な変異体(CAa2)をmCRY1と発現させた場合、セリン71のリン酸化の増加(図2C)が観察され、このことにより、AMPKがCRY1をin vivoにてセリン71上でリン酸化できることが確認された。AMPKa1の構成的に活性な変異体(CAa1)は、核から排除されたが、セリン71のリン酸化を認識可能には増加しなかった(図2C)。AMPKは、精製された成分を用いるin vitroキナーゼアッセイにおいて、mCRY1を直接リン酸化することもできた(図9)。
【0100】
内因性AMPKの活性化は、クリプトクロムを不安定化する。いくつかの相補的ストラテジーを用いて、S71およびS280のリン酸化ならびにmCRY1の不安定化に対する内因性AMPKの寄与を分析した。HeLa細胞は、AMPK活性化キナーゼLKB1のプロモーターのメチル化によるエネルギーストレスに応答して、内因性AMPKの活性化を低減する。野生型(WT)LKB1の導入は、外因的に発現されるmCRY1のレベルを低減させ、不活性(KD)LKB1の導入では低減させなかった。この低減は、AMPK活性化AMP模倣AICARを、WT LKB1の存在下に加えることにより増強し、KD変異体の存在下では増強しなかった(図2D)。同様に、グルコース欠損によるAD293細胞におけるAMPKの活性化は、トランスフェクトした野生型mCRY1(WT)発現を低減させ、予測されるAMPKリン酸化部位を欠く変異体mCRY1(AA)ではそうではなかった(図2E)。
【0101】
クリプトクロム安定性の調節におけるAMPKの役割をさらに調べるために、遺伝子的に野生型(WT)またはAMPKの触媒サブユニットについてヌル(ampka1−/−;ampka2−/−)(AMPK−/−)であるマウス胚線維芽細胞(MEF)を用いた。flagタグ付加野生型(WT)または2重リン酸化し得ない(AA)のmCRY1をこれらの細胞において安定的に発現するレトロウイルスを用いて、AA CRY1ではなく野生型が、野生型細胞においてのみAMPKアゴニストAICARを用いる処理により急速に分解されることを示した。機能的AMPKの非存在下では、AICARは、WTまたはAA CRY1のいずれに対しても作用を有さなかった(図2F)。S71およびS280のAMPKリン酸化によるCRY1安定性の調節は、これらの細胞を4時間の過程のAMPK活性化AICARの存在下でのシクロヘキシミド処理に供することによりさらに確認された(図2G)。AICAR処理は、野生型の安定性の低減をもたらしたが、CRY1のリン酸化し得ない変異体の安定性の低減はもたらさなかった。
【0102】
AMPKは、線維芽細胞における概日リズムの代謝的変更に寄与する。概日時計リセットのための摂食由来シグナルの重要性、グルコース利用力によるAMPKの調節、およびクリプトクロム不安定化におけるAMPKの役割についての蓄積している証拠に鑑みて、AMPK発現およびグルコース利用力の影響を、線維芽細胞における概日リズム性に対して調べた。野生型線維芽細胞を、制限されたグルコースを含有する培地で培養した場合、概日性reverbαおよびdbp発現の振幅は、著しく増進され(図3Aおよび図10)、これは、グルコース欠損がAMPKを活性化し、CRY安定性を低減させるモデル(これは、CLOCK:BMAL1標的であるreverbαおよびdbpの脱抑制を導く)と一致した。予想されたように、培養培地にAICARを加えることは、グルコース欠損の効果を模倣した。目ざましいことに、グルコース欠損もAICAR処理も、AMPKを欠くMEF(ampkα1−/−;ampkα2−/−、「AMPK−/−」)におけるreverbαおよびdbpの発現に影響せず(図3Aおよび図10)、このことは、線維芽細胞概日リズムに対するグルコース制限の影響が、AMPKにより媒介されることを示した。
【0103】
Bmal1プロモーターは、REVERBαにより抑制される。よって、グルコース利用力の低減が概日リズムに与える影響を、Bmal1プロモーターの制御下でルシフェラーゼを安定的に発現する線維芽細胞を用いて調べた。標準的(高グルコース)培養条件下では、Bmal1−ルシフェラーゼの発現の高振幅概日リズムが、25.3時間の周期で観察された(図3B、C)。培地中のグルコース量の低下は、概日周期を30.7時間まで増加させた。Bmal1−ルシフェラーゼ発現細胞を、AICARを補った高グルコース培地で培養した場合、概日周期は、低グルコースで観察されたものと同様であり、グルコース欠損の概日性の影響が、AMPKにより媒介されるという考えを補強した。制限されたグルコース条件のもとで観察されたREVERBαの発現の増加は、Bmal1を含むREVERBαにより抑制される遺伝子の発現の低下をもたらすと予想される。実際に、グルコース濃度の低下またはAICAR処理のいずれかによるAMPKの活性化は、Bmal1−ルシフェラーゼ発現の振幅を低下させた(図3D)。これらの結果はまとめて、培養線維芽細胞の概日リズムが、グルコース利用力の変更を担い、これらの効果が、AMPK誘導リン酸化により媒介されることを示す。
【0104】
in vivoでのAMPKの概日性の調節。AMPKの日周期性調節を調べるために、AMPK転写、局在化および基質リン酸化を、インタクトな動物の末梢器官において調べた。全ての実験は、標準的な明暗サイクルへの同調化の後に定常的な暗所に維持した動物を用いて行って、観察された効果が、外部の環境における変化に対する日周期性の応答よりもむしろ概日性であることを確実にした。
【0105】
調べた両方のAMPK基質、ACC1−Ser79およびRaptor−Ser792のリン酸化は、主体的な昼において夜よりも再現可能により高く(図4A)、ネガティブフィードバックタンパク質が不安定な昼の時間にほぼ対応し、これは、リズム性のAMPK活性化が分解に寄与するモデルと一致した。マウス肝臓におけるAMPKの概日性の調節を探索する一方、最小限の核クリプトクロムタンパク質の時間と一致するピーク発現(図4C)を有する調節性ampkβ2サブユニットの堅固な概日性の発現(図4B)。AMPKβ2は、AMPK複合体の核局在化を駆動すると報告されているが、AMPKβ1含有複合体は、形質膜を標的にする。よって、ampkβ2の概日転写は、変動するAMPKβ2が、AMPKα1およびAMPKα2の核局在化を日周期性に調節することを示唆する。この仮説を試験するために、概日サイクル全体にわたって回収した肝臓核(liver nucleus)におけるAMPKα1およびAMPKα2のタンパク質レベルを測定し(図4C)、ampkβ2発現と同期してピークになる核AMPKα1のリズム性を観察した。AMPKα2は、核局在化シグナルを含有し、核に常に存在した。AMPKα1核局在化がピークになる時間は、肝臓核における最小CRY1タンパク質の時間でもあり、このことは、AMPKの核へのリズム性の移入が、AMPK媒介リン酸化およびクリプトクロムの分解に寄与し得ることを示唆する。
【0106】
AMPKは、概日時計をin vivoにて変更する。マウスにおいてAMPKα1およびAMPKα2の両方を遺伝子欠失させることにより、早期の胚致死性が導かれる。よって、肝臓概日時計におけるAMPKの役割をさらに調べるために、概日タンパク質および転写産物を、明暗サイクルへの同調化の後に定常的な暗所で飼育した対照マウス(LKB1+/+)または肝細胞におけるlkb1が喪失した同腹子(LKB1L/L)の肝臓において24時間にわたって調べた。lkb1の肝臓特異的欠失は、この器官におけるAMPK活性化を廃止し、概日サイクル全体で、特に、変更されていないマウスにおいてAMPKが最も活性であることが見出された昼間の時間中に、肝臓核に存在するCRY1およびCRY2タンパク質の量を著しく増加させた(図5B)。この増加は、昼光に対応する期間におけるREVERBα発現の減少(図5B)および概日サイクルを通しての概日転写産物の振幅の減少(図5C)と関連した。よって、AMPKシグナル伝達のin vivoにおける喪失は、クリプトクロムを安定化し、概日リズムを混乱させ、光非依存性末梢概日時計についての同期化の機構を確立する。
【0107】
材料および方法
細胞および細胞培養物 − AMPK+/+およびAMPK−/−マウス胚線維芽細胞は、Benoit Viollet博士からの贈物であった。HeLa細胞およびAD293細胞は、American Type Culture Collection(ATCC)から購入した。3T3不死化MEFは、以前に記載された。そうでないと記載しない限り、細胞は、10%ウシ胎仔血清、ペニシリンおよびストレプトマイシンを補った完全ダルベッコ改変イーグル培地(DMEM)(Invitrogen cat#11995またはcat#11965)で、5%CO2に維持した37℃インキュベータ中で成長させた。グルコース濃度を操作した実験において、細胞は、グルタミン、非必須アミノ酸、ペニシリン、ストレプトマイシンおよび記載される量のD−グルコースを補った最小DMEM(Sigma cat#D5030)、またはペニシリン、ストレプトマイシン、L−グルタミンおよび記載される量のD−グルコースを補ったグルコース非含有DMEM(Invitrogen cat#11966)で成長させた。0.5mMグルコースを用いる実験は、浸透圧の影響を制御するためにD−マンニトールを補った。細胞刺激は、50%ウマ血清(Invitrogen cat#26050)を含む完全DMEMを用いて行い、以前に記載されたようにして行った。
【0108】
プラスミドおよびトランスフェクション − pDONR221およびpcDNA3.1/v5−His−TOPOは、Invitrogenから購入した。pcDNA3−2xFlag−mCRY1(WT)およびpcDNA3−PER2は、Charles Weitz博士からの贈物であった。pCMV−SPORT6−Fbxl3は、Open Biosystemsから購入し、FBXL3は、pcDNA3.1/v5−His−TOPO中に、標準的なプロトコルによりクローニングした。flag−LKB1、myc−AMPKa1およびmyc−AMPKa2構築物は、以前に記載され、構成的に活性な対立遺伝子(CAa1およびCAa2)は、残基T312の後に停止コドンを挿入することにより作製した。全ての変異は、Stratagene部位特異的変異誘発プロトコルを用いて作製した。トランスフェクションは、FuGene HD(Roche)を用いて行った。
【0109】
ウイルスおよび安定細胞株の作製 − AD293細胞にpLXSP3puro発現クローンをpCL−Amphoと共にトランスフェクトしてウイルスを生成した。トランスフェクションの48時間後にウイルス上清を回収し、0.45μmフィルターでろ過し、6μg/mlポリブレンを添加し、親細胞株に加えた。4時間後に、さらなる培地を加えてポリブレンを3μg/ml未満に希釈した。ウイルス形質導入の48時間後に、感染細胞を、1〜5μg/mlピューロマイシンを含有する選択培地中に分配した。選択培地は、選択が完了するまで2〜3日毎に置き換えた。
【0110】
質量分析 − Flag−mCRY1をトランスフェクトしたAD293細胞を、10μMのMG132で6時間処理し、1%のTx−100を含有する緩衝液中で溶解させた。Flag−mCRY1を、M2−アガロース(Sigma)上で精製し、SDS−PAGEにより夾雑物から分けた。クーマシー染色されたバンドを切り出し、HPLCグレードの50%アセトニトリル中で2回すすぎ、Beth Israel Deaconess Medical Center質量分析施設に送付した。
【0111】
タンパク質抽出物の調製、免疫沈降およびイムノブロッティング − 全細胞抽出物は、以前に記載されたように1%のTritonX−100を含有する溶解緩衝液中で調製し、肝臓核抽出物を、NUN手順により調製した。用いた抗体は、Sigmaからの抗Flag M2アガロース、抗v5アガロース、抗Flagポリクローナル、抗v5ポリクローナルおよび抗βアクチン;Alpha Diagnostics InternationalからのCRY11A、CRY21AおよびPER21A;Cell Signaling Technologiesからの抗ホスホACC1(S79)、抗ACC1、抗ホスホ−AMPKa、抗ホスホ−Raptor、抗Raptorおよび抗REVERBa;Upstate Biotechologyからの抗AMPKa1および抗AMPKa2;ならびにMilliporeと共同で作製したホスホ−CRY1(S71)および周囲の残基を含有するホスホペプチドに対して産生されたポリクローナル抗血清であった。
【0112】
in vitroリン酸化アッセイ − Flag−mCRY1を、トランスフェクトしたAD293細胞から精製し、32P−ATPおよび精製AMPK(Upstate Biotechnologyから)と、300μMのAMPの存在下または非存在下で30分間室温にて組み合わせた。反応混合物をSDS−PAGEにより分離し、ニトロセルロースに移した。ホスホイメージャーによる放射活性の視覚化の後に、ニトロセルロースをFlagタグについてイムノブロットした。
【0113】
リアルタイム生物発光モニタリング − Bmal1プロモーター駆動型ルシフェラーゼを安定的に発現するヒト骨肉腫U2OSレポーター細胞株は、記載されている。2×104細胞を35mmディッシュに播種し、10%血清を補ったDMEM中で集密まで3日間にわたって成長させた。集密細胞を、50%ウマ血清で2時間刺激し、次いで、0.1%透析血清および上記のように種々の量のグルコースを含有する培地に移した。生物発光を、Actimetrics,Inc.からのLumiCycle装置により連続的に記録した。
【0114】
遺伝子発現 − RNAを、肝臓または培養線維芽細胞から、TrizolでまたはQiagen RNeasy精製システムを用いて抽出した。cDNAは、SuperscriptII逆転写酵素(Invitrogen)を用いて調製し、遺伝子発現について、SYBRグリーン(Invitrogen)またはTaqMan(Applied Biosystems)化学のいずれかを用いる定量的リアルタイムPCRを用いて分析した。プライマー配列は、要請に応じて入手可能である。
【0115】
マウス − LKB1fl/flマウスは、Ronald De Pinho博士からの贈物であり、Cry1−/−;Cry2−/−マウスは、Aziz Sancar博士からの贈物であった。Creリコンビナーゼを発現するアデノウイルスは、University of Iowa Transgenic Core施設からであった。全ての動物の世話および処置は、動物の世話および使用についてのSalk Instituteガイドラインに準拠した。
【0116】
本開示は、mCRY1が、実際に、今まで調べた47の核ホルモン受容体のうち20と相互作用し、PPARdと特に良好に相互作用することを証明する(図11)。さらに、生理食塩水またはAMPK活性化薬物であるAICARを注射した野生型およびクリプトクロム欠損マウスの肝臓における遺伝子発現は、クリプトクロムが、遺伝子のサブセットのAICARにより誘導される活性化に必要であることを証明する(図12)。さらに、マウスにおけるCry1およびCry2の両方の遺伝子破壊の代謝生理に対する影響を調べる。データは、Cry1−/−;Cry2−/−マウスが、野生型対照よりも著しく低い体重と、著しく低減された安静時血中グルコースとを有することを示す(図13)。一括して、これらのデータは、哺乳動物のクリプトクロムが、細胞のエネルギー状態の以前には認識されていなかったセンサとして機能すること、これらが生物のエネルギー恒常性において役割を果たすこと、およびクリプトクロムの薬理学的調整が、代謝障害の治療に有用であり得ることを示唆する。
【0117】
本開示は、具体的な実施形態に重点をおいて記載しているが、具体的な実施形態の変形を用いることができることが当業者に明らかであり、本開示を、本明細書に具体的に記載する以外の別な方法で行うことができることを意図する。よって、本開示は、以下の特許請求の範囲により定義される本開示の精神および範囲内に包含される全ての改変を含む。
【技術分野】
【0001】
(関連出願への相互参照)
本願は、2009年3月20日に出願された米国仮特許出願第61/162,219号の利益を主張し、本明細書において参考として援用される。
【0002】
(政府支援の認可)
本研究は、国立衛生研究所助成金番号DK057978、DK062434、CA104838、DK080425、およびEY016807により支援された。合衆国政府は、本発明において一定の権利を有する。
【0003】
本開示は、概日リズムを調整するためのAMP活性化プロテインキナーゼ(AMPK)のアゴニストおよびアンタゴニストの使用に関する。より具体的には、本開示は、睡眠行動をスクリーニングし、調整するための組成物および方法を提供する。
【背景技術】
【0004】
概日時計は、哺乳動物の組織における何千もの遺伝子のリズム性の転写を駆動することにより、行動プロセスおよび生理的プロセスを毎日の明暗サイクルに調和させる。
【発明の概要】
【課題を解決するための手段】
【0005】
本開示は、AMPKが、転写リプレッサーCRY1およびCRY2をリン酸化し、それらのプロテアソーム分解を刺激することを証明する。さらに、本開示は、クリプトクロムが、哺乳動物概日時計におけるそれらの確立された機能に加えていくつかの核ホルモン受容体と結合し、その転写活性を調節することを証明する。本開示は、クリプトクロムタンパク質が、AMPK活性化薬物を用いる治療に対する転写応答の部分集合にとって必要であることも証明する。よって、クリプトクロムの薬理学的調整は、代謝障害の治療において有用である。
【0006】
クリプトクロム転写同時制御因子機能を調整する小分子薬物の使用は、それらに限定されないが、ペルオキシソーム増殖因子活性化受容体PPARアルファ、ベータ、デルタおよびガンマを含む確立された代謝的に重要な転写因子の転写活性をクリプトクロムが調節することが証明されているので、代謝障害の治療において有用である。クリプトクロムは、天然の小分子補助因子(触媒性補助因子であるフラビンアデニンジヌクレオチドまたはFAD、および集光性補助因子である5,10−メテニルテトラヒドロフォリルポリグルタメートまたはMTHF)と結合してそれにより調節されるので、クリプトクロムは、合成小分子による調節のための良好な標的である。
【0007】
本開示は、エネルギーセンサであるAMPKが、CRY1(そのリン酸化は、CRY1−FBXL3相互作用およびCRY1のプロテアソーム分解を媒介する)中の2つのセリンを修飾することを証明する。つまり、CRY1は、元来、光受容体として進化したが、翻訳後修飾により、これは重要なシグナル伝達メディエーターとなり得る。in vivoでのAMPKの遺伝子操作または薬理学的操作は、クリプトクロム安定性と概日リズムとを共に変更し、このことは、それにより栄養調節シグナルが哺乳動物末梢器官における概日時計をリセットできる新規な同調化機構を示唆する。
【0008】
本開示は、ヒトのような哺乳動物被験体において概日リズムを改変するための方法および組成物を提供する。本開示は、AMPKが、哺乳動物被験体の概日サイクルの間に、脳および体内の他の組織において修飾されることを証明する。1つの実施形態において、本開示は、被験体における概日リズムを調整するための医薬品の製造のための、AMPキナーゼアゴニストまたはアンタゴニストの使用を提供する。1つの実施形態において、AMPKアゴニストは、AICARである。別の実施形態において、AMPKアンタゴニストは、抗体もしくはコンパウンドCまたはそれらの類似体もしくは誘導体である。さらに別の実施形態において、AMPKアゴニストは、血液脳関門を通過することができる製剤または誘導化を含む。まださらなる実施形態において、AMPKアゴニストは、経口投与、静脈内注射、筋肉内注射、硬膜外送達、頭蓋内または皮下注射用に製剤化される。
【0009】
本開示は、概日リズムを改変する第2活性成分と組み合わせて製剤化されたAMPKアゴニストを含む組成物も提供する。1つの実施形態において、第2活性成分は、睡眠補助剤である。さらなる実施形態において、組成物は、経口投与、静脈内注射、筋肉内注射、硬膜外送達、頭蓋内送達または皮下注射用に製剤化される。
【0010】
本開示は、哺乳類における睡眠を調整する方法であって、哺乳類における概日リズムを調整するのに有効な量のAMPKアゴニストまたはアンタゴニストを哺乳類に投与するステップを含む方法を提供する。
【0011】
本開示は、被験体における概日リズムまたは睡眠を調整する作用物質を同定する方法であって、(a)AMPK経路を備える試料を少なくとも1つの被検作用物質と接触させるステップと、(b)AMPKまたはAMPK経路の活性を、被検作用物質の存在下および非存在下で比較するステップであって、前記活性を変化させる被検作用物質が、概日リズム調整活性を有する作用物質を示すステップとを含む方法も提供する。
【0012】
本開示は、代謝リズムまたは概日リズムの調整において用いるための作用物質を同定する方法であって、前記作用物質をCry1またはCry2タンパク質と接触させるステップと、Cry1もしくはCry2をリン酸化もしくは脱リン酸化するか、またはCry1もしくはCry2の安定性もしくは発現を改変する前記作用物質の能力を測定するステップとを含み、Cry1またはCry2を改変する作用物質が、代謝リズムまたは概日リズムを調整するために有用な作用物質である、方法も提供する。1つの実施形態において、作用物質は、Cry1またはCry2の安定性を低下させる。
【0013】
本開示は、上記の方法により同定される、Cry1またはCry2の安定性を低下させる作用物質を含む組成物も提供する。
【0014】
本開示は、代謝性または概日性の疾患または障害を治療する方法であって、被験体を、Cry1および/またはCry2のリン酸化または脱リン酸化を促進する本開示の作用物質または組成物と接触させるステップを含む方法も提供する。1つの実施形態において、作用物質または組成物は、クリプトクロム転写同時制御因子機能を調整する。別の実施形態において、作用物質または組成物は、ペルオキシソーム増殖因子活性化受容体(PPAR)アルファ、ベータ(デルタ)およびガンマを調整する。さらに別の実施形態において、作用物質は、ビグアナイド誘導体、AICAR、メトホルミンまたはその誘導体、フェンホルミンまたはその誘導体、レプチン、アディポネクチン、AICAR(5−アミノイミダゾール−4−カルボキサミド)、ZMP、DRL−16536、BG800化合物(Betagenon)およびフラン−2−カルボン酸誘導体からなる群から選択されるAMPKアゴニストである。
【0015】
本開示は、代謝リズムまたは概日リズムの疾患または障害を決定する方法であって、CRY1またはCRY2の安定性を、組織において24時間周期の間に測定するステップを含み、正常または過剰ATP濃度の存在下でのCRY1またはCRY2の長期間安定性の周期が、代謝リズムまたは概日リズムの疾患または障害を示す方法も提供する。1つの実施形態において、方法は、mCRY1のS71またはS280を含むエピトープと特異的に結合する抗体を用いる。
【0016】
本開示は、安静および脂質異化反応を促進する方法であって、概日サイクルの夜間期の間にAMPKアゴニストを投与するステップを含み、前記AMPKアゴニストが、CRY1またはCRY2の安定性を低下させる方法も提供する。
【0017】
本開示は、代謝リズムまたは概日リズムの障害を治療する方法であって、概日サイクルの安静周期の間にAMPKアゴニストを投与するステップを含む方法も提供する。
【0018】
上記のおよびその他の特徴は、添付の図面を参照して進む以下のいくつかの実施形態の詳細な記載からより明確になる。
【図面の簡単な説明】
【0019】
【図1】図1A〜Eは、S71またはS280のリン酸化が、FBXL3およびPER2との相互作用を変更することによりmCRY1を不安定化することを示す。(A)記載される変異を有するFlagタグ付加mCRY1を発現するAD293細胞を、100μg/mlのシクロヘキシミド(CHX)で、記載される時間処理した。Flag−mCRY1は、ウェスタンブロッティングにより検出した。β−アクチンについてのイムノブロットを、負荷対照(loading control)として用いた。(B)CLOCK、BMAL1、Per1−ルシフェラーゼならびにmCRY1の記載される量および対立遺伝子を発現するAD293細胞を、トランスフェクションの48時間後にルシフェラーゼ活性について調べた。(C)Flag−mCRY1を、記載されるプラスミドを一過的に発現するAD293細胞から免疫沈降させた。CRY1と結合したFBXL3は、v5エピトープタグについてのイムノブロッティングにより検出した。(D)記載されるようにFBXL3を同時発現するかまたはしない、CLOCK、BMAL1、Per1−ルシフェラーゼおよびmCRY1の記載される対立遺伝子を一過的に発現するAD293細胞を、トランスフェクションの48時間後にルシフェラーゼ活性について調べた。**AAに対してp<0.01;## FBXL3を発現しない等価な試料に対してp<0.01。(E)Flag−mCRY1を、PER2を同時発現するかまたはしない、CRY1の記載される対立遺伝子を一過的に発現するAD293細胞から免疫沈降させた。CRY1と結合したPER2は、イムノブロッティングにより検出した。
【図2】図2A〜Gは、AMPKが、Ser71、Ser280リン酸化によりmCRY1を不安定化することを示す。(A)クリプトクロム概日転写リプレッサー(赤色のフォントでの種の名称)および青色光光受容体(青色のフォントでの種の名称)中のmCRY1のS71の周囲の領域の進化的保存を示す配列アラインメント。配列の上の強調した数字は、AMPキナーゼによるリン酸化のための標的セリンに対する、これらの位置での好まれるアミノ酸を示す。赤色は、酸性残基(K/R)への好みを示し、緑色は疎水性残基(L/I/V/F)への好みを示す。(B)上:ホスホペプチドの配列アラインメントであって、これらに対して、mCRY1−pS71およびmACC1−pS79に対する抗体を産生させた。下:抗mCRY1−pS71抗体および抗ACC1−pS79抗体は共にWTを認識するが、AD293細胞から免疫沈降させたS71A Flag−mCRY1は認識しない。(C)抗mCRY1−pS71を用いて、AMPKα1(CAα1)またはAMPKα2(CAα2)の活性化対立遺伝子の非存在下または存在下で野生型CRY1(WT)またはCRY1S71A(S71A)を一過的に発現するAD293細胞から免疫沈降させたFlag−CRY1中のSer71のリン酸化を検出した。一過的に発現されたmyc−CAα1およびmyc−CAα2は、mycエピトープタグに対して産生されたポリクローナルウサギ抗体と抗ウサギAF488(緑色)とを用いて免疫染色した。核は、DAPI(青色)を用いて対比染色した。(D)野生型(WT)またはキナーゼ非機能性(kinase dead)(KD)LKB1と共にFlag−CRY1を一過的に発現するHeLa細胞を、ビヒクル(−)または2mMのAICAR(+)で2時間処理した。(E)Flag−mCRY1の記載される対立遺伝子を一過的に発現するAD293細胞を、25mMまたは0.5mMのグルコースを含有する培地で処理した。(F)Flagタグ付加野生型CRY1(WT)またはCRY1S71A/S280A(AA)を安定的に発現する、対での野生型(AMPK+/+)およびampkα1−/−;ampkα2−/−(AMPK−/−)マウス胚線維芽細胞を、ビヒクル(−)または2mMのAICAR(+)で2時間処理した。(G)(F)に記載されるMEFを、100μg/mlのシクロヘキシミド(CHX)で、記載される時間処理した。CRY1は、(D〜G)におけるFlagエピトープについての免疫沈降およびイムノブロッティングにより検出した。
【図3】図3A〜Dは、AMPKシグナル伝達の破壊が、MEFにおける概日リズムを変更することを示す。(A)同期化していない対での野生型(AMPK+/+)またはampkα1−/−;ampkα2−/−(AMPK−/−)マウス胚線維芽細胞を、50%ウマ血清に2時間曝露することにより刺激し、その後、25mMグルコース、0.5mMグルコースまたは1mMのAICARを補った25mMグルコースを含有する培地に移した。定量的PCR分析を、刺激後の記載される時間に回収したcDNA試料を用いて行った。データは、それぞれ3重で分析した2つの独立する実験の平均値を示す。(B)Bmal1−ルシフェラーゼを安定的に発現する線維芽細胞を、記載される量のグルコースを2mMのAICARと共にまたはなしで含有する培地中で培養した。ルシフェラーゼ活性の連続モニタリングの代表的な結果を示す。(CおよびD)(B)に記載するようにして行った実験からのBmal1駆動型ルシフェラーゼ活性の概日周期(C)および振幅(D)の定量。(C)および(D)におけるデータは、条件あたり4つの試料についての平均値±標準偏差を表す。ANOVA分析は、カテゴリー間での有意差を示した。**シェッフェの事後分析において25mMのグルコースで培養した試料に対してP<0.01。
【図4】図4A〜Cは、AMPK活性および核局在化が概日調節を受けることを示す。(A)ホスホ−Raptor−S792(pRaptor)、Raptor、ホスホ−ACC1−S79(pACC1)およびACC1についてのイムノブロッティングを、記載される概日時間に回収したマウス肝臓から調製した全細胞溶解産物で行った。ブロットは、3回の独立した実験の代表である。(B)記載される概日時間に回収したマウス肝臓から調製したcDNAの定量的PCR分析。各データ点は、それぞれ独特の動物から採取され、4重で分析した3つの試料の平均値±標準偏差を表す。(C)核抽出物を、記載される概日時間のそれぞれで2匹のマウスの肝臓から調製した。AMPKα1、AMPKα2、PER2、CRY1およびREVERBαのタンパク質レベルを、イムノブロッティングにより分析した。記載される概日時間に回収した対での野生型(α1+/+)およびampkα1−/−(α1−/−)または野生型(α2+/+)およびampkα2−/−(α2−/−)マウスからの核抽出物を、抗体特異性についての対照として用いた。
【図5】図5A〜Cは、AMPK活性化が、マウス肝臓におけるCRY安定性および概日リズムを変更することを示す。(A)マウスに、生理食塩水または体重1kgあたり500mgのAICARを注射し、肝臓試料を1時間後のツァイトゲーバー時間(ZT、光点灯後の時間)6またはZT18に回収した。内因性CRY1を、肝臓核抽出物におけるイムノブロッティングにより検出した。n.s.は、試料負荷を評価するための非特異的バンドを示す。野生型(CRY+/+)およびcry1−/−;cry2−/−(CRY−/−)マウスから回収した試料を、抗体特異性についての対照として用いた。データは、2つの独立した実験からの代表的な結果を表す。(B)LKB1+/+およびLKB1fl/flマウスに、Creリコンビナーゼを発現するアデノウイルス(Ad−Cre)を尾静脈から注射した。Ad−Cre注射の1〜2週間後に、マウスを定常的な暗所に移し、記載される概日時間に肝臓を回収した。CRY1、PER2およびREVERBαをイムノブロッティングにより検出した。(C)(B)に記載される肝臓から調製したcDNA試料を、dbp、reverbα、cry1およびper2発現の定量的PCR分析により分析した。全ての転写産物を、内部対照としてのu36b4に対して正規化した。各データ点は、4重で分析した3つの試料の平均値±標準偏差を表す。
【図6】図6は、AMPKが、末梢時計の代謝同調化に寄与することを示す。マウスにおける末梢概日時計の代謝同調化におけるAMPKの役割を示すモデル。昼間、AMPKの核局在化は、食物および循環グルコースの低減によるその可能性のある活性化と連係して増加する。活性核AMPKは、クリプトクロムをリン酸化して、FBXL3とのその相互作用を増加し、プロテアソーム分解を導き、時計により制御される遺伝子(ccg)の活性化をもたらす。夜間、核AMPK活性が低下することにより、クリプトクロムが核に蓄積し、ccgが抑制される。
【図7】図7A〜Dは、mCRY1リン酸化部位の同定を示す。(A)一過性にトランスフェクトしたAD293細胞から精製されたFlag−mCRY1を、LC−MS/MSにより、リン酸化セリン、スレオニンおよびチロシン残基の存在について分析した。観察されたリン酸化を触媒すると予測されるキナーゼを、文献検索およびScansiteプログラム(http:(//)scansite.mit.edu)との組合せを用いて予測した。配列保存を、MegAlignを用いて決定した。(B)CRY1タンパク質の模式図の下のオレンジ色のバーの厚さは、タンパク質配列の各領域についてLCMS/MSにより観察されたペプチドの相対数を示す。(C)Bに示すペプチドのカバー範囲に基づく本発明者らのLC−MS/MS分析では観察可能でない場合があるScansiteにより予測されるリン酸化部位。(D)記載される変異を有するFlag−mCRY1を、100μg/mlのシクロヘキシミドで記載される時間処理したAD293細胞において発現させた。CRY1タンパク質は、Flagタグについてのイムノブロットにより検出された。
【図8】図8は、mCRY1 S280配列保存を示す。mCRY1 S280は、保存されたAMPK基質モチーフにより囲まれる。クリプトクロム概日転写リプレッサー(赤色フォントでの種の名称)および青色光光受容体(青色フォントでの種の名称)におけるmCRY1のS280の周囲の領域の進化的保存を示す配列アラインメント。配列の上の強調した数字は、AMPキナーゼによるリン酸化についての標的セリンに対するこれらの位置での好まれるアミノ酸を示す。赤色は、示される位置での酸性残基(K/R)への好みを示し、緑色は疎水性残基(L/I/V/F)への好みを示す。
【図9】図9は、精製AMPKが、mCRY1をin vitroでリン酸化することを示す。AD293細胞から精製したFlagタグ付加mCRY1を、30分間、#32P−ATPと、記載されるようにAMPキナーゼおよび300μMのAMPの非存在下または存在下でインキュベートした。mCRY1のリン酸化は、オートラジオグラフィにより検出した。合計mCRY1レベルを、Flagエピトープについてのイムノブロットにより決定した。精製AMPKは、精製mCRY1を効率的にリン酸化し、このリン酸化は、AMPの存在により強く活性化され、このことにより、精製混合物中の関連するキナーゼが、他の関連キナーゼではなくAMPKであることが確認された。
【図10】図10A〜Dは、AMPKの破壊が、MEFにおける概日リズムを変更することを示す。3T3不死化マウス胚線維芽細胞(A)または対での野生型(AMPK+/+)もしくはampkα1−/−;ampkα2−/−(AMPK−/−)線維芽細胞(B)を、50%ウマ血清に2時間曝露することにより刺激し、その後、25mMグルコース(黒色の記号)、0.5mMグルコース(灰色の記号)または1mMのAICARを補った25mMグルコース(赤色の記号)を含有する培地に移した。定量的PCR分析を、刺激後の記載される時間に回収した溶解産物から調製したcDNA試料を用いて行った。データは、それぞれ3重で分析した2または3つの独立する実験の平均値±標準偏差を表す。
【図11】図11は、mCRY1が核ホルモン受容体と相互作用することを示す。Flagタグ付加mCRY1を種々のv5タグ付加核ホルモン受容体と同時発現するAD293細胞を溶解し、mCRY1を含有するタンパク質複合体を、Flagタグの免疫沈降により単離した。mCRY1含有タンパク質複合体中の個別の核ホルモン受容体の存在を、v5タグについてのイムノブロットにより検出した(上)。免疫沈降複合体中のmCRY1の量を、Flagタグについてのイムノブロットにより示す(中)。溶解産物中に存在するそれぞれの核ホルモン受容体の量を、免疫沈降前の溶解産物から採取した試料におけるv5タグのイムノブロットにより示す(下)。RORa、b、g(レチノイン酸受容体関連オーファン受容体a、b、g)、RXRa、b(レチノイドX受容体a、b)、PPARd、g(ペルオキシソーム増殖因子活性化受容体d、g)、VDR(ビタミンD受容体)、PXR(プレグナンX受容体)、CAR(構成的アンドロスタン受容体)、ERb(エストロゲン受容体b)、ERRa、b、g(エストロゲン関連受容体a、b、g)、GR(グルココルチコイド受容体)、MR(ミネラロコルチコイド受容体)、PR(プロゲステロン受容体)、AR(アンドロゲン受容体)。データは、2または3つの独立した実験の代表的な結果を表す。
【図12】図12は、クリプトクロムが、AMPK活性化に対するいくつかの転写応答にとって必要であることを示す。野生型(WT)またはCry1−/−;Cry2−/−(CRY−/−)マウスに、生理食塩水(黒色のバー)または体重1kgあたり500mgのAICAR(赤色のバー)を、午後6時に注射した。cDNAを、4時間後の午後10時に回収した肝臓から調製し、遺伝子発現を、Sybr GreenER化学を用いる定量的PCRにより分析した。Fas(脂肪酸合成)を、Cry1およびCry2遺伝子型に関係なくAICARにより活性化される遺伝子の例として示す。Por(p450酸化還元酵素)を、そのAICARにより誘導される活性化がクリプトクロムを必要とする遺伝子の例として示す。データは、条件あたり3〜5匹のマウスについての平均値±s.e.m.を表す。
【図13】図13は、クリプトクロムの喪失が、マウスにおける代謝機能を変更することを示す。10週齢の雄の野生型(WT、黒色のバー)およびCry1−/−;Cry2−/−(CRY−/−、灰色のバー)マウスの体重測定を行い、それらの安静時血中グルコースを、尾静脈の切れ目(nick)により午後1時に測定した。データは、遺伝子型あたり10匹の動物についての平均値±s.e.m.を表す。
【発明を実施するための形態】
【0020】
本明細書においてそうでないと特に記載しない限り、用いられる用語の定義は、薬学技術において用いられる標準的な定義である。明細書および添付の特許請求の範囲において用いる場合、単数形「a」、「an」および「the」は、文脈がそうでないと明確に示さない限り、複数の引用物も含む。よって、例えば、「薬学的キャリア」への言及は、そのようなキャリアの2以上の混合物などを含む。
【0021】
また、「または」の使用は、そうでないと記載しない限り「および/または」を意味する。同様に、「含む(comprise)」、「含む(comprises)」、「含み(comprising)」、「含む(include)」、「含む(includes)」および「含み(including)」は、交換可能であり、限定することを意図しない。
【0022】
種々の実施形態の記載が「含み」の用語を用いる場合、いくつかの特定の場合において、実施形態を、「から本質的になる」または「からなる」の言葉を用いて代わりに記載できることを当業者が理解するとさらに理解される。
【0023】
そうでないと明示しない限り、本明細書において用いられる全ての技術的および科学的用語は、本開示が属する当業者に通常理解されるのと同じ意味を有する。本明細書に記載されるのと同様または等価ないずれの方法および試薬も、開示される方法および組成物の実施において用いることができるが、例示的な方法および材料をここで記載する。
【0024】
本明細書において言及する全ての出版物は、本明細書の記載と関連して用いられる可能性があるその出版物に記載される方法を記載して開示する目的のために、全体が参照により本明細書に組み込まれる。上で論じた本文全体での出版物は、本開示の出願日に先んじるそれらの開示のためにのみ提供される。本明細書中のいずれも、本発明者らが、先の開示により、そのような開示に先行する権利が与えられないことを認めると解釈されない。
【0025】
概日リズムは、限定されないが、睡眠サイクルの調整、エクササイズおよびカロリー低減に関連するエネルギー調整、ならびに摂食/栄養補給(nourishment)行動のような生理的プロセス、内分泌プロセスおよび行動プロセスの適切なタイミングを調和させることにより生物学的効率を最適化する。概日リズムは、少なくとも3つの要素を含有すると考えられる:(a)環境情報を概日ペースメーカー(時計)に中継する入力経路(複数可);(b)変動(oscillation)を生み出す概日ペースメーカー;および(c)それを介してペースメーカーが種々の出力リズムを調節する出力経路(複数可)。
【0026】
哺乳動物の視床下部視交叉上核(SCN)は、行動リズムおよび生理的リズムを明暗サイクルに整合させる主要ペースメーカーとして作用する。当初、SCNは、哺乳類における自立性分子ペースメーカーの唯一の部位であると考えられていたが、その後、複数の論文が、このような分子時計がほぼ遍在的であることを示した。SCN時計とは異なって、光非感受性末梢器官における概日時計は、摂食の毎日のリズムにより同調され、末梢組織が、毎日の食物消費を予期し、代謝プロセスのタイミングを最適化することを理論的に可能にする。いくつかの論文が、重要な代謝酵素の転写の調節および代謝生理における哺乳動物概日時計についての役割を支持している。
【0027】
本明細書で用いる場合、「概日リズム」との用語は、約24時間の過程にわたって生じる生理的および行動的パラメータにおける規則的な変動を意味することを意図する。このような活動は、睡眠サイクルおよび栄養補給サイクルなどを含む。
【0028】
本明細書で用いる場合、概日リズムに関して用いる場合の「調整する」との用語は、動物の概日タイミングシステムにより調節される生理的機能、内分泌機能もしくは行動を変更すること、または概日リズム性を示す細胞機能を変更することを意味することを意図する。動物の概日タイミングシステムにより調節される例示的な生理機能は、体温、自律神経調節、代謝および睡眠覚醒サイクルを含む。例示的な代謝機能は、体重の増加または減少および体脂肪率の増加または減少を含む増量ならびに減量の制御、持久行動の改変、減量などを含む。動物の概日タイミングシステムにより調節される例示的な内分泌機能は、松果体のメラトニン分泌、ACTHコルチゾール分泌、甲状腺刺激ホルモン分泌、成長ホルモン分泌、ニューロペプチドY分泌、セロトニン分泌、インスリン様成長因子I型分泌、副腎皮質刺激ホルモン分泌、プロラクチン分泌、ガンマ−アミノ酪酸分泌およびカテコールアミン分泌を含む。動物の概日タイミングシステムにより調節される例示的な行動は、運動(運動リズム)、精神的俊敏さ、記憶、感覚運動統合、摂食、REM睡眠、NREM睡眠および情動を含む。
【0029】
AMP活性化プロテインキナーゼ(AMPK)は、系統発生全体でよく保存されている代謝シグナルの中心的なメディエーターとして認識されている。AMPKは、触媒(α)サブユニットと2つの調節性(β、γ)サブユニットとを含むヘテロ3量体プロテインキナーゼである。これは、高いAMP/ATP比率の存在下でLKB1により、または上昇した細胞内カルシウムの存在下でCAMKKβによりリン酸化されると活性化される。生化学的研究および生物情報学的研究により、AMPKによるリン酸化が起こりそうな最適アミノ酸配列の関係が確立されている。
【0030】
AMP活性化プロテインキナーゼ(AMPK)およびAMPKキナーゼ(AMPKK)は、プロテインキナーゼカスケードと関連する。AMPKカスケードは、燃料の生成および利用を細胞内で調節する。例えば、細胞での燃料が低い(例えばAMP濃度の増加)と、AMPK活性が増加する。一旦活性化されると、AMPKは、ATPを保存するか、またはATP生成の代替法を促進するように機能する。よって、その活性を調整することにより、エネルギー貯蔵の異化反応を増加でき、脂質含量を低減させてATPを増加させるか、または体を安静状態にして、ATP使用を保存する。
【0031】
AMPKは、肝臓、脳および骨格筋を含むいくつかの組織において発現される。AMPKの活性化は、肝臓の脂肪酸酸化およびケトン体生成を活性化し、コレステロール合成、脂肪生成およびトリグリセリド合成を阻害し、脂肪細胞の脂肪分解および脂肪生成を阻害し、骨格筋の脂肪酸酸化および筋肉グルコース摂取を刺激し、膵臓ベータ細胞によるインスリン分泌を調整することが示されている。
【0032】
AMPKの活性化は、AMPの濃度の増加により誘引できる。AMPKのγサブユニットは、αサブユニット上に活性部位(Thr−172)を露出するように立体構造変化を受ける。AMPKのγサブユニットの立体構造変化は、AMPの濃度が増加している場合に達成され得る。AMPの濃度の増加により、AMPKのγサブユニットの立体構造変化が、このサブユニットにある2つのBatemanドメインを2つのAMPが結合させることにより生じる。AMPのこの役割は、5−アミノ−4−イミダゾールカルボキサミドリボシド(AICAR)に由来するAMP類似体である5−アミノ−4−イミダゾールカルボキサミドリボチド(ZMP)によるAMPK活性化を示す実験において証明される。同様に、AMPのアンタゴニストは、AMPKによる下流のキナーゼの活性化を阻害する阻害抗体の使用を含む。
【0033】
断眠(SD)は、神経活性を増加させる。持続性の神経活性は、細胞エネルギー充足率を減少させる(AMPレベルが増加し、ATPが減少する)。これは、次に、細胞エネルギーセンサAMPKにおける変化を引き起こす。上で論じたように、AMPKは、ATPの保存を導くカスケードを含む種々のキナーゼカスケードを調整する。
【0034】
CLOCKおよびBMAL1は、ヘテロ2量体の形成により、概日リズムと関連する遺伝子の転写を誘導するポリペプチドである。典型的な概日サイクルの間に、分子機構は、2つの相互に連結された転写/翻訳フィードバックループを有する内部時計を形成する2つのサイクルの間で変動する。フィードバックループのポジティブアームは、塩基性ヘリックス−ループ−ヘリックス−PAS(Per−Arnt−Sim)ドメイン含有転写因子であるCLOCKおよびBMAL1により駆動される。CLOCK/BMAL1ヘテロ2量体は、時計遺伝子であるクリプトクロム(Cry1およびCry2)、period(Per1およびPer2)、およびRev−Erbαの転写を活性化する。PERおよびCRYタンパク質は、核に移動し、そこでこれらはCLOCK/BMAL1と相互作用して転写を下方調節し、主要フィードバックループのネガティブアームを生成する。
【0035】
上記の概日転写プログラムの堅固な変動は、コア時計タンパク質の翻訳後修飾を必要とする。3つの研究により、最近、Fボックスタンパク質FBXL3が、クリプトクロムのユビキチン化および分解のメディエーターとして同定された。Fボックスタンパク質がそれらの同族基質に結合することは、基質タンパク質中の1つ以上のアミノ酸のリン酸化により頻繁に調節されるが、このような調節の改変は、CRY:FBXL3相互作用について記載されなかった。
【0036】
時計タンパク質の翻訳後修飾(例えばリン酸化および脱リン酸化)は、タンパク質の局在化、分子間相互作用および安定性を決定し、よって、概日時計の周期を調節する。本開示は、この翻訳後調節が、AMPK活性により調整され得、よって、AMPKアゴニストおよびアンタゴニストが、概日時計の調節において役割を有し得ることを証明する。
【0037】
クリプトクロム(Cry1およびCry2)は、ほとんどの植物において概日光受容体として機能する。クリプトクロムは、全ての組織において発現することが見出されている。しかし、発現は、網膜においてより高く、マウスおよびヒトの両方において網膜内層に限定される。脳において、Cry1はSCNにおいて発現し、発現は、毎日の変動を示し、午後2時付近にピークになり、午前2時付近にその最低に達する。
【0038】
ヒトのクリプトクロムは共に、Cry遺伝子を異所的に発現するHeLa細胞から、および組換えタンパク質としてE.coliから精製されている。両方の供給源から単離されたタンパク質は、FADおよびプテリンを含有する。
【0039】
クリプトクロムは、光センサとして進化したが、これは、光非感受性組織においてさえ、コアとなる概日時計の重要な成分として保持されている。本開示は、AMPKが、栄養シグナルを時計に変換するように、クリプトクロムに再度目的を持たせることを証明する。概日システムと代謝システムとの間の逆調節についての証拠は、過去10年間で増加しており、出現している理論は、概日時計が、代謝プロセスの一時的な分離を可能にすることを示唆している。代謝シグナルは、哺乳動物の末梢器官において概日時計のタイミングを設定することが示されているが、このようなシグナルを伝達する分子機構は、まだ明らかになっていない。
【0040】
本開示は、AMPKによるクリプトクロムのリン酸化が、FBXL3との結合により分解を促進し、CLOCK:BMAL1抑制を軽減することを証明する。このプロセスは、過剰グルコースにより抑制され、AICARのようなAMPK活性化因子、およびampkβ2調節性サブユニットの核移動により増強される。よって、本開示は、細胞内生物エネルギーの状態が、末梢組織における概日時計に直接影響を与えることができる新規な生化学的経路を提供する。
【0041】
AMPKの概日性活性化は、CRY1のリン酸化を駆動し、そのFBXL3媒介分解を刺激することにより、リズムの維持に寄与する。AMPKは、CRY1を2つのセリン残基(マウスCRY1においてS71およびS280)上でリン酸化する。セリン71および周囲の配列は、全ての光非依存性クリプトクロム転写リプレッサーに存在し、このことは、この経路が、光に曝露されない概日時計の代謝の同調化を可能にするように進化したことを示唆する。
【0042】
AMPK活性は、グルコース利用力によりLKB1依存的な様式で調節されることができ、栄養利用力を変化させるか、またはAMPK活性は、培養線維芽細胞における時計の振幅および周期を変更する。in vivoでは、AMPK基質であるACC1およびRaptorは、リン酸化の概日性の変化を示し、このことは、AMPKの下流の細胞質経路および核での経路がリズム性に調節されることを示唆する。AMPKが代謝プロセスの中心的な制御因子であることに鑑みて、これは、代謝の概日性の調節に重要な関わりを有する。遍在的または組織特異的な様式のいずれかでの概日時計機能の遺伝子的な変更は、摂食行動、体重、ランニング持久力およびグルコース恒常性を変化させ、これらはそれぞれ、AMPKの操作によっても変更される。一括して、これらのデータは、AMPKが、細胞レベルおよび生物全体レベルの両方での概日生理的調節の重要なメディエーターであり得るという考えを支持する。
【0043】
興味深いことに、それぞれのAMPKサブユニットの転写、核局在化および活性化は、マウス肝細胞において概日リズムを示し、クリプトクロムタンパク質の量が最小の時間にピークになる。ampkβ2転写は、堅固に概日性であり、真昼には夜より8倍高い。AMPKβ2は、AMPKの核局在化と、それに対応してAMPKα1のリズム性の核蓄積を駆動する。よって、AMPKサブユニットは、概日時計の調節に寄与するだけでなく、それら自体も概日性の様式で転写調節される。
【0044】
栄養状態と時計との連絡は複雑であり、付加的な経路がそれらの同調化にin vivoで寄与する。2つの最近の研究は、SIRT1が、肝細胞においてリズム的に発現され、線維芽細胞における概日リズム性に寄与することを証明した。SIRT1は、NAD+/NADH比率によるその脱アセチル化酵素活性の調節により概日時計の代謝同調化において役割を果たすと見られる。複数の論文が、種々の時計成分の調節におけるヘムの役割を示唆し、3価鉄および2価鉄ヘムによる異なる調節が、細胞酸化還元状態についての情報を概日時計に伝達することを示唆している。これらの機構の1もしくは複数、および/またはSCNから発せられる昼間液性シグナルもしくはニューロンシグナルは、おそらく、LKB1L/Lマウスの肝臓において観察された残存概日リズムに寄与する。
【0045】
Fbxl3またはLkb1における変異は、ヒト腫瘍において頻繁に行われる。クリプトクロムのLKB1およびAMPKにより媒介されるリン酸化が、それらのFBXL3により媒介される分解を刺激することの証明は、2つの腫瘍抑制因子がクリプトクロムの不安定化において協力することを示し、このことは、クリプトクロムの異常に高いレベルが、細胞周期調節解除(deregulation)または腫瘍形成に寄与し得ることを示唆する。肝臓再生の概日性調節について記載した論文において、Matsuoおよび共同研究者らは、cry1−/−;cry2−/−マウスの肝臓が、野生型同腹子のものよりも遅く再生されたことを示し、このことは、CRYタンパク質が、細胞成長または増殖において刺激性の役割を果たすとの考えを支持する。CRY:FBXL3相互作用を媒介するLKB1およびAMPKに依存するリン酸化部位の本明細書における同定は、これらの疑問を明確にする。
【0046】
哺乳動物クリプトクロムにおける他のリン酸化部位は、さらなる入力シグナルを概日時計へ媒介し得るが、本開示は、セリン71および280のAMPKにより媒介されるリン酸化が、FBXL3とのその相互作用を増加することによりCRY1プロテアソーム分解を刺激することを証明する。さらに、グルコース欠損は、クリプトクロム安定性を低下させ、概日転写産物を変更し、培養細胞における概日周期の長さを増加させ、これらの効果はAMPKにより媒介される。さらに、マウスにおけるAMPKの遺伝子破壊は、クリプトクロム安定性および概日リズムを乱す。これらのデータはまとめて、AMPKによるクリプトクロムリン酸化が、代謝シグナルによる末梢器官時計の同調化を可能にするように進化したことを証明する。
【0047】
本開示は、睡眠または他の概日プロセスに影響するための、AMP活性化プロテインキナーゼ(AMPK)と結合するか、またはそうでなければそれを活性化もしくは不活性化する化合物(これらのいくつかは、糖尿病の治療のために現在用いられている)の使用を提供する。本開示は、AMP活性化プロテインキナーゼ活性の遺伝子操作または薬理学的操作が、培養細胞およびインタクトな動物の肝臓における概日リズムを変更することを証明する。本開示は、AMPキナーゼが、睡眠覚醒サイクルおよび他の生理的リズムのタイミングを支配するいわゆる「主要ペースメーカー」の場所である視交叉上核(SCN)で発現されることも証明する。現在利用可能な療法は、血液脳関門を通過せず、よって、睡眠障害の調整のために有用でない。
【0048】
AMPKによる概日リズムの調節は、血液脳関門を通過するAMPK調節因子が、概日リズムと関連する下流のキナーゼ活性を調節することにより、それに限定されないが不眠症を含む睡眠障害の治療に有用であることを示唆する。さらに、それらに限定されないが、CLOCK、BMAL1、PERならびにCRY−1および−2を含むある特定の概日ポリペプチドは、リン酸化および脱リン酸化により調節され、脳の外側の組織に存在する。よって、非神経組織においてAMPK活性を調整することは、キナーゼカスケードにより概日リズムを設定し、最終的に下流のポリペプチドのリン酸化および脱リン酸化を調節するためにも重要であり得る。
【0049】
さらに、本開示は、Cry1およびCry2のリン酸化および脱リン酸化が、概日性の作用を有し、よって、睡眠状態およびエネルギー代謝の調整のための有用な標的であることを証明する。例えば、CRY1のセリン71および280のリン酸化または脱リン酸化を特異的に調整することにより、FBXL3とのその相互作用を増加させることによるプロテアソーム分解を促進できる。
【0050】
AMPKを活性化するいくつかの薬理学的作用物質は、糖尿病の治療のために現在臨床使用されており、いくつかの型の癌について臨床試験されている。
【0051】
AICARのようなAMPキナーゼアゴニストは、インスリン調節、糖尿病および肥満について研究されている。しかし、AMPキナーゼは、概日リズムや睡眠行動を調整すると以前に証明されていなかった。本開示は、AMPK活性を調整することが、概日リズムに関連するタンパク質の翻訳後修飾を含む下流のプロセスに対する作用を有し得ることを証明する。1つの実施形態において、本開示は、AMPKアゴニストおよびアンタゴニストを用いて、被験体における概日リズムを調整できることを示す。例えば、AMPKは、本開示により、ヘテロ2量体CLOCK/BMAL1を活性化する転写の調整において役割を果たすことが証明される。
【0052】
種々のAMPKアゴニストが、当該技術において公知である。このようなAMPKアゴニストを含む方法および組成物が、本明細書において提供される。このようなAMPKアゴニストの使用は、概日リズムを調整する方法をもたらすことができる。種々のAMPKアゴニストが本明細書に記載され、当該技術において公知である。1つの実施形態において、AMPKアゴニストは、AICAR化合物を含む。本開示の方法において有用な他の化合物は、ビグアナイド誘導体、AICARの類似体(例えば米国特許第5,777,100号(本明細書に参照により組み込まれる)に開示されるもの)およびAICARのバイオアベイラビリティーを増加させるAICARのプロドラッグまたは前駆体(例えば米国特許第5,082,829号(本明細書に参照により組み込まれる)に開示されるもの)を含み、これらは全て当業者に周知である。AMPKの他の活性化因子は、Iyengarらへの米国特許出願公開第20060287356号(その開示は、参照により本明細書に組み込まれる)に記載されるものを含む。従来公知のAMPK活性化化合物は、上記のレプチン、アディポネクチンおよびメトホルミンに加えて、AICAR(5−アミノイミダゾール−4−カルボキサミド)を含む。他のAMPKアゴニストは、それらに限定されないが、DRL−16536(Dr.Reddy’s/Perlecan Pharma)、BG800化合物(Betagenon)、フラン−2−カルボン酸誘導体(Hanall、KR;国際出願公開第WO/2008/016278号(参照により本明細書に組み込まれる)も参照されたい)、A−769662(Abbott)(構造I;Coolら、Cell Metabol.3巻:403〜416頁、2006年も参照されたい);国際公開第WO/2006/033709号に記載されるようなMetabasisにより開発中のAMPKアゴニスト;MT−39シリーズの化合物(Mercury Therapeutics);およびTransTech Pharmaにより開発中のAMPKアゴニストを含む。
【0053】
【化1】
例えば、AICARは、細胞に取り込まれ、AMPKを活性化することが示されているAMP類似体であるZMPに変換される。ZMPは、細胞内AMP模倣物として作用し、十分に高いレベルまで蓄積されると、AMPK活性を刺激できる(Corton, J. M.ら、Eur. J. Biochem.、229巻:558頁(1995年))。しかし、ZMPは、他の酵素の調節においてもAMP模倣物として作用し、よって、特異的AMPK活性化因子ではない(Musi, N.およびGoodyear, L. J. Current Drug Targets−−Immune, Endocrine and Metabolic Disorders 2巻:119頁(2002年))。
【0054】
本開示は、AMPKアゴニストまたはAMPKアンタゴニストのいずれかを用いることにより被験体における概日時計の特定のサイクルを刺激する方法を提供する。1つの実施形態において、AMPKアゴニストを用いて、CLOCK/BMAL1転写活性の増加と関連する概日サイクルを促進する。1つの実施形態において、AMPKアゴニストは、エネルギー保存のシグナル伝達による睡眠効果を、対応するキナーゼカスケードを通して促進する。この方法は、被験体におけるエネルギー欠乏状態を刺激するのに十分な量のAMPKアゴニストを被験体に投与するステップを含む。「エネルギー欠乏状態」により、AMPKのγサブユニットが立体構造変化を受ける状態のことをいう。睡眠効果を促進するとは、そのような効果が、被験体において、AMPKアゴニストの非存在下で生じるよりも多く改善されることを意味する。
【0055】
以下により詳細に記載するように、AMPKアゴニストは、経口で、非経口で、筋肉内に、脈管内に、または任意の適切な経路により投与してよい。1つの実施形態において、AMPKアゴニストは、硬膜外投与される。1つの実施形態において、AMPKアゴニストは、血液脳関門の通過が促進されるように製剤化される。
【0056】
本開示は、AMPK活性と拮抗することにより代謝および活性を「覚醒」または「活動」サイクルに設定する作用物質を投与するステップを含む、活動状態を促進する方法も提供する。1つの実施形態において、AMPKアンタゴニストは、阻害抗体である。1つの実施形態において、AMPKアンタゴニストは、コンパウンドC(ドルソモルフィン、6−[4−(2−ピペリジン−1−イル−エトキシ)−フェニル)]−3−ピリジン−4−イル−ピラゾロ[1,5−a]−ピリミジン)、それらの類似体、誘導体または塩のような小分子阻害物質である。
【0057】
開示される方法は、限定されないが、薬学技術の当業者に周知の投与方法、投与量および製剤を含む、AMPKアゴニスト単独、またはその製剤を受容する被験体における所望の概日サイクルの状態を誘導する所望の結果を有する他の概日調節剤もしくは睡眠補助剤との組合せでの任意の投与方法、投与量および/または製剤の使用を想定する。
【0058】
本開示のAMPKアゴニストは、薬物の形態でヒトまたは動物に投与してよい。代わりに、AMPKアゴニストは、ヒトまたは動物により消費されるように、多様な食品および飲料またはペットフードに組み込んでよい。AMPKアゴニストは、一般的な食品もしくは飲料に用いてよいか、あるいは機能性食品もしくは飲料、疾患に罹患している被験体のための食品、または特定の健康用途のための食品(この食品(または飲料)は、それが生理機能を有することを記載したラベルを有する);例えば睡眠補助剤に用いてよい。
【0059】
AMPKアゴニスト単独、または他の睡眠補助剤もしくは活性成分との組合せは、薬品、例えば、錠剤もしくは顆粒剤のような経口固体製品、または液剤もしくはシロップ剤のような経口液体製品に製剤化してよい。
【0060】
開示される方法におけるAMPKアゴニストまたは製剤の投与の方式は、それらに限定されないが、くも膜下内(intrathecal)、皮内、筋肉内、腹腔内(ip)、静脈内(iv)、皮下、鼻内、硬膜外、硬膜内、頭蓋内、脳室内および経口経路を含む。具体的な例において、AMPKアゴニストは、経口投与される。AMPKアゴニストの投与のための他の簡便な経路は、例えば、注入もしくはボーラス投与、局所、上皮もしくは皮膚粘膜層を通しての吸収(例えば口腔粘膜、直腸および腸粘膜など)、眼、鼻および経皮を含む。投与は、全身性または局所的であり得る。肺投与も(例えば吸入器またはネブライザにより)、例えばエアロゾル剤を含有する製剤を用いることにより、用いることができる。
【0061】
いくつかの実施形態において、AMPKアゴニストを投与またはAMPKアゴニストを局所的に投与することが望ましい場合がある。このことは、例えば、局所的もしくは領域的な注入または灌流、局所塗布(例えば創傷包帯材)、注射、カテーテル、坐剤またはインプラント(例えば、シアラスティック(sialastic)膜または繊維のような、膜を含む多孔質、非多孔質またはゼラチン質の材料から形成されるインプラント)などにより達成してよい。
【0062】
他の実施形態において、ポンプ(例えば移植されたミニポンプ)を用いてAMPKアゴニストまたは製剤を送達してよい(例えば、Langer Science 249巻、1527頁、1990年;Sefton Crit. Rev. Biomed. Eng. 14巻、201頁、1987年;Buchwaldら、Surgery 88巻、507頁、1980年;Saudekら、N. Engl. J. Med. 321巻、574頁、1989年を参照されたい)。別の実施形態において、AMPKアゴニストまたは製剤は、小胞、特にリポソームで送達される(例えば、Langer、Science 249巻、1527頁、1990年;Liposomes in the Therapy of Infectious Disease and Cancer中のTreatら、Lopez−BeresteinおよびFidler(編)、Liss, N. Y.、353〜365頁、1989年を参照されたい)。
【0063】
さらに別の方法実施形態において、AMPKアゴニストは、制御放出製剤で送達できる。Langer(Science 249巻、1527頁、1990年)による総説で論じられているもののような制御放出系が公知である。同様に、制御放出製剤において有用なポリマー材料が公知である(例えば、Rangerら、Macromol. ScL Rev. Macromol. Chem. 23巻、61頁、1983年;Levyら、Science 228巻、190頁、1985年;Duringら、Ann. Neurol. 25巻、351頁、1989年;Howardら、J. Neurosurg. 71巻、105頁、1989年を参照されたい)。例えば、アゴニストは、ポリ乳酸、ポリグリコール酸、ポリ乳酸とポリグリコール酸とのコポリマー、ポリイプシロンカプロラクトン、ポリヒドロキシ酪酸、ポリオルトエステル、ポリアセタール、ポリジヒドロピラン、ポリシアノアクリレート、およびヒドロゲルの架橋または両親媒性ブロックコポリマーを含む、化合物の制御放出を達成するのに有用なクラスの生分解性ポリマーと結合させてよい。
【0064】
開示される方法は、アゴニスト(複数可)を送達して所望の結果を達成するAMPKアゴニストの任意の剤形またはその製剤の使用を意図する。剤形は、一般的に公知であり、例えばAllenら、Ansel’s Pharmaceutical Dosage Forms and Drug Delivery Systems、第8版、Philadelphia, PA: Lippincott Williams & Wilkins、2005年、全738頁を含む多様な参考書で教示されている。開示される方法で用いるための剤形は、それらに限定されないが、固体剤形および固体改変放出薬物送達系(例えば散剤および顆粒剤、カプセル剤ならびに/または錠剤);半固体剤形および経皮系(例えば軟膏剤、クリームおよび/またはゲル);経皮薬物送達系;薬学的挿入物(例えば坐剤および/または挿入物(insert));液体剤形(例えば液剤および分散系);ならびに/または滅菌剤形および送達系(例えば非経口製剤(parenterals)および/または生物製剤(biologies))を含む。具体的な例示的剤形は、エアロゾル(定量噴霧(metered dose)、粉末、溶液および/または噴霧剤なしを含む);ビーズ;カプセル(従来の、制御送達、制御放出、腸溶被覆および/または持続放出を含む);キャプレット;濃縮物;クリーム;結晶;ディスク(持続放出を含む);点滴剤;エリキシル;乳剤;フォーム;ゲル(ゼリーおよび/または制御放出を含む);小球;顆粒剤;ガム;インプラント;吸入薬;注射剤;挿入物(徐放(extended release)を含む);リポソーム;液体(制御放出を含む);ローション;ロゼンジ;定量噴霧(例えばポンプ);ミスト;うがい薬;噴霧化用溶液;眼用の系;油;軟膏;膣坐剤(ovule);粉末(パケット、発泡剤(effervescent)、懸濁用粉末、懸濁持続放出用粉末および/または溶液用粉末);ペレット;ペースト;液剤(長時間作用性および/または再構成されたものを含む);細片;坐剤(持続放出を含む);懸濁剤(レンテ、ウルトラレンテ(ultre lente)、再構成されたものを含む);シロップ(持続放出を含む);錠剤(咀嚼、舌下、持続放出、制御放出、遅延作用、遅延放出、腸溶被覆、発泡剤、フィルム被覆、即時溶解、緩慢放出を含む);経皮系;チンキ;および/またはカシェ剤を含む。典型的には、剤形は、AMPKアゴニストを含む少なくとも1つの薬学的活性成分の有効量(例えば治療有効量)と、薬学的に許容される賦形剤および/または他の成分(例えば1つ以上の他の活性成分)との製剤である。薬物製剤化の目的は、被験体に活性成分(例えばAMPKアゴニストまたはAMPKアンタゴニスト)を適切に投与することである。製剤は、投与の方式に適するものである。「薬学的に許容される」との用語は、連邦政府もしくは州政府の規制機関により承認されたか、または米国薬局方もしくは他の一般的に認識されている動物およびより具体的にはヒトにおける使用についての薬局方に列挙されていることを意味する。例示的な製剤において用いるための賦形剤は、例えば、1つ以上の以下のものを含む:結合剤、増量剤(filler)、崩壊剤、滑沢剤、コーティング、甘味料、香料、着色料、防腐剤、希釈剤、アジュバントおよび/またはビヒクル。賦形剤が、一括して、具体的な剤形の全重量(および/または容量)の約5%〜95%を構成するいくつかの場合がある。
【0065】
薬学的賦形剤は、例えば、水および/またはピーナツ油、大豆油、鉱油、ごま油のような石油、動物油、植物油または合成起源のものなどを含む油のような滅菌液体であり得る。水は、製剤を静脈内投与する場合の例示的なキャリアである。生理食塩水、血漿媒体、デキストロース水溶液およびグリセロール溶液を、特に注射用溶液用の液体キャリアとして用いることもできる。経口製剤は、それらに限定されないが、薬学的グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどを含み得る。非経口の薬学的賦形剤のより完全な説明は、Remington、The Science and Practice of Pharmacy、第19版、Philadelphia, PA: Lippincott Williams & Wilkins、1995年、第95章に見出すことができる。賦形剤は、例えば、浸透圧を調整するための薬学的に許容される塩、シクロデキストリンのような脂質キャリア、血清アルブミンのようなタンパク質、メチルセルロースのような親水性作用物質、界面活性剤、緩衝剤、防腐剤なども含んでよい。薬学的賦形剤の他の例は、デンプン、グルコース、ラクトース、スクロース、ゼラチン、麦芽、コメ、小麦粉、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリン酸グリセロール、タルク、塩化ナトリウム、脱脂粉乳、グリセロール、プロピレン、グリコール(propylene, glycol)、水、エタノールなどを含む。所望により、製剤は、少量の湿潤剤もしくは乳化剤またはpH緩衝剤も含有できる。
【0066】
経口投与を含むいくつかの実施形態において、AMPKアゴニストの経口投与量は、通常、1日あたり体重1kgあたり約0.001mg(mg/kg/日)〜約100mg/kg/日、例えば約0.01〜10mg/kg/日の範囲である(そうでないと記載しない限り、活性成分の量は、遊離の酸または遊離の塩基であり得る中性分子に基づく)。例えば、80kgの被験体は、約0.08mg/日〜8g/日、例えば約0.8mg/日〜800mg/日を受容する。1日1回の投与のための適切に調製された医薬品は、よって、0.08mg〜8g、例えば0.8mg〜800mgを含有する。いくつかの場合において、AMPKアゴニストまたはアンタゴニストを含む製剤は、1日2回、3回または4回の分割用量で投与してよい。1日2回の投与のために、上記のような適切に調製された医薬品は、0.04mg〜4g、例えば0.4mg〜400mgを含有する。上記の範囲外の投与量が必要となる場合があり得る。1日あたり0.08mg〜8gの範囲で与えられ得る1日投与量の例は、0.1mg、0.5mg、1mg、2.5mg、5mg、10mg、25mg、50mg、100mg、200mg、300mg、400mg、500mg、600mg、800mg、1g、2g、4gおよび8gを含む。これらの量は、1日に1回より多く投与する場合は、より少ない用量に分けることができる(例えば、薬物を1日2回摂取する場合は、各投与において半分の量)。
【0067】
注射による投与(例えば静脈内または皮下注射)を含むいくつかの方法実施形態において、被験体は、ほぼ上記の量で活性成分を送達する注射量を受容する。量は、消化系を回避する注射薬物形態に起因する送達効率の差を考慮して調整してよい。このような量は、いくつかの適切な方法、例えば1回の長期間または1日に数回の間での低濃度で高容量の活性成分、短期間、例えば1日1回の間での高濃度で低容量の活性成分で投与してよい。典型的には、約0.01〜1.0mg/mlの間の、例えば0.1mg/ml、0.3mg/mlまたは0.6mg/mlなどの活性成分の濃度を含有する従来の静脈内製剤を調製し、上記の1日あたりの量に等しい1日あたりの量で投与してよい。例えば、0.5mg/mlの活性成分の濃度を有する静脈内製剤を1日2回8ml受容する80kgの被験体は、1日あたり8mgの活性成分を受容する。
【0068】
他の方法実施形態において、AMPKアゴニストまたはアンタゴニスト(またはその製剤)は、治療期間を通してほぼ同じ用量、漸増用量計画または負荷用量計画(例えば負荷用量が維持用量の約2〜5倍である)で投与できる。いくつかの実施形態において、用量は、使用期間の間に、組成物を受容する被験体の状態、組成物に対する見かけの応答および/または当業者により判断される他の因子に基づいて変動する。いくつかの実施形態において、例えば慢性不眠症または睡眠覚醒サイクル障害を管理するために、AMPKアゴニストまたはアンタゴニストの長期間の投与が意図される。
【0069】
本開示は、AMPK活性化または阻害を測定することによる、概日リズムを調整する作用物質のスクリーニング方法も提供する。概日リズムを調整する化合物をスクリーニングするための本開示の方法は、AMPK経路を備える細胞、組織または被験体(例えば動物)を提供するステップと、前記被験体を、概日リズム調整活性を有すると推測される作用物質と接触させるステップと、AMPK活性に対する作用を、直接、または下流のキナーゼ活性により測定するステップとを含む。被検作用物質は、概日リズム機能の少なくとも1つの観察可能な指標を有し、かつAMPKを発現する細胞調製物、組織、器官、生物または動物に対して提供することができる。概日リズムを調整する作用物質の能力は、概日リズム機能の徴候を示す多様な動物種、ならびにそのような動物から得られる器官、組織および細胞、およびそれらに由来する細胞調製物で試験できる。AMPK活性を調整する作用物質は、次いで、推定概日リズム調整活性を有する作用物質として同定できる。
【0070】
多様なin vitroスクリーニング方法が、概日リズムを調整する化合物を同定するための本開示の方法において提供されるアンタゴニストまたはアゴニストを同定するために有用である。AMPKを調整する化合物の能力は、例えば、AMPKと結合してAMPKを活性化もしくは不活性化するか、下流のキナーゼ活性を遮断するか、リン酸化および脱リン酸化を調整するか(例えばCry1またはCry2のリン酸化、脱リン酸化)、またはAMPKにより生成される予め決定されたシグナルを調整する化合物の能力により示され得る。よって、シグナル伝達および結合アッセイを用いて、概日リズムを調整する化合物を同定するための本開示の方法において提供されるAMPKのアンタゴニストまたはアゴニストを同定できる。
【0071】
「作用物質」は、目的または結果を達成するために有用である、任意の物質または物質の任意の組合せ、例えば、AMPK活性化カスケードと関連するタンパク質活性を調整するために有用であり(例えばAMPK依存性リン酸化事象)、またはタンパク質−タンパク質相互作用もしくはATP代謝を改変するかまたはそれに影響するために有用である物質あるいは物質の組合せである。
【0072】
例示的な作用物質は、それらに限定されないが、それらに限定されないがランダムペプチドライブラリーのメンバー(例えば、Lamら、Nature、354巻:82〜84頁、1991年;Houghtenら、Nature、354巻:84〜86頁、1991年を参照されたい)を含む例えば可溶性ペプチドのようなペプチド、ならびにDおよび/またはL立体配置のアミノ酸で作製されたコンビナトリアル化学由来分子ライブラリー、ホスホペプチド(それらに限定されないが、ランダムまたは部分的に縮重された、定向ホスホペプチドライブラリーのメンバーを含む;例えば、Songyangら、Cell、72巻:767〜778頁、1993年を参照されたい)、抗体(それらに限定されないが、ポリクローナル、モノクローナル、ヒト化、抗イディオタイプ、キメラまたは単鎖抗体、ならびにFab、F(ab’)2およびFab発現ライブラリーフラグメント、およびそれらのエピトープ結合フラグメントを含む)、有機または無機の小分子(例えば、いわゆる天然生成物または化学コンビナトリアルライブラリーのメンバー)、分子複合体(例えばタンパク質複合体)、あるいは核酸を含む。
【0073】
本開示の方法において有用なライブラリー(例えばコンビナトリアル化学ライブラリー)は、それらに限定されないが、ペプチドライブラリー(例えば米国特許第5,010,175号;Furka、Int. J. Pept. Prot. Res.、37巻:487〜493頁、1991年;Houghtonら、Nature、354巻:84〜88頁、1991年;PCT公報第WO91/19735号を参照されたい)、コードされたペプチド(例えばPCT公報第WO93/20242号)、ランダムバイオオリゴマー(例えばPCT公報第WO92/00091号)、ベンゾジアゼピン(例えば米国特許第5,288,514号)、ヒダントイン、ベンゾジアゼピンおよびジペプチドのようなダイバーソマー(diversomer)(Hobbsら、Proc. Natl. Acad. Sci. USA、90巻:6909〜6913頁、1993年)、ビニローグ(vinylogous)ポリペプチド(Hagiharaら、J. Am. Chem. Soc、114巻:6568頁、1992年)、グルコース足場を有する非ペプチド性ペプチド模倣物(Hirschmannら、J. Am. Chem. Soc、114巻:9217〜9218頁、1992年)、小化合物ライブラリーの類似した有機合成(Chenら、J. Am. Chem. Soc、116巻:2661頁、1994年)、オリゴカルバメート(Choら、Science、261巻:1303頁、1003年)および/またはペプチジルホスホネート(Campbellら、J. Org. Chem.、59巻:658頁、1994年)、核酸ライブラリー(Sambrookら、 Molecular Cloning, A Laboratory Manual、Cold Springs Harbor Press, N. Y.、1989年;Ausubelら、Current Protocols in Molecular Biology、Green Publishing Associates and Wiley Interscience, N. Y.、1989年を参照されたい)、ペプチド核酸ライブラリー(例えば米国特許第5,539,083号を参照されたい)、抗体ライブラリー(例えばVaughnら、Nat. Biotechnol、14巻:309〜314頁、1996年;PCT出願第PCT/US96/10287号を参照されたい)、炭水化物ライブラリー(例えばLiangら、Science、274巻:1520〜1522頁、1996年;米国特許第5,593,853号を参照されたい)、有機小分子ライブラリー(例えばベンゾジアゼピン、Baum、C&EN、1月18日、33頁、1993年;イソプレノイド、米国特許第5,569,588号;チアゾリジオノンおよびメタチアゾン、米国特許第5,549,974号;ピロリジン、米国特許第5,525,735号および第5,519,134号;モルホリノ化合物、米国特許第5,506,337号;ベンゾジアゼピン、第5,288,514号を参照されたい)などを含む。
【0074】
開示されるスクリーニング方法のために有用なライブラリーは、それらに限定されないが、空間的に整列させたマルチピンペプチド合成(Geysenら、Proc Natl. Acad. Sci.、81巻(13号):3998〜4002頁、1984年)、「ティーバッグ」ペプチド合成(Houghten、Proc Natl. Acad. Sci.、82巻(15号):5131〜5135頁、1985年)、ファージディスプレイ(ScottおよびSmith、Science、249巻:386〜390頁、1990年)、スポットまたはディスク合成(Dittrichら、Bioorg. Med. Chem. Lett.、8巻(17号):2351〜2356頁、1998年)、あるいはビーズ上での分割および混合固相合成(Furkaら、Int. J. Pept. Protein Res.、37巻(6号):487〜493頁、1991年;Lamら、Chem. Rev.、97巻(2号):411〜448頁、1997年)を含む多様な方法で生成できる。ライブラリーは、種々の数の構成成分(composition)(メンバー)、例えば約100メンバーまで、例えば約1000メンバーまで、例えば約5000メンバーまで、例えば約10,000メンバーまで、例えば約100,000メンバーまで、例えば約500,000メンバーまでまたは500,000メンバーより多くさえ含んでよい。
【0075】
1つの実施形態において、ハイスループットスクリーニング法は、多数の可能性のある治療化合物(例えば、AMPKタンパク質−タンパク質相互作用の影響因子)を含有するコンビナトリアル化学またはペプチドライブラリーを提供するステップを含む。このようなコンビナトリアルライブラリーを、次いで、本明細書に記載されるような1つ以上のアッセイでスクリーニングして、所望の特徴的な活性を示す(例えばAMPKタンパク質−タンパク質相互作用を増加または減少させる)ライブラリーメンバー(特に化学種またはサブクラス)を同定する。このようにして同定された化合物は、通常の「リード化合物」として用い得るか、またはそれら自体を可能性のあるもしくは実際の治療剤として用いることができる。いくつかの場合において、候補作用物質のプールを同定し、さらにスクリーニングして、どの個別の作用物質または集合的な作用物質のより小さいプールが所望の活性を有するか決定してよい。AMPK相互作用またはプロセスのAMP依存性リン酸化に影響する(例えば増加または減少させる)作用物質は、被験体において概日リズム(例えば睡眠行動)を調整する効果を有する場合があり、よって、同定することが望まれる。
【0076】
本明細書に記載されるスクリーニング法において、組織試料、単離細胞、単離ポリペプチドおよび/または被検作用物質は、ハイスループットスクリーニングに適する様式で提示できる。例えば、1または複数の単離組織試料、単離細胞または単離ポリペプチドを、マイクロタイタープレートのウェルに入れ、1または複数の被検作用物質をマイクロタイタープレートのウェルに加えることができる。代わりに、1または複数の被検作用物質を、マイクロタイタープレートのウェル(溶液でまたはプレート表面に接着した)のようなハイスループットフォーマットで提示でき、1または複数の単離組織試料、単離細胞および/または単離ポリペプチドと、組織試料もしくは単離細胞もしくは所望のポリペプチド機能および/または構造物を少なくとも維持する条件下で接触させる。被検作用物質は、組織試料、単離細胞または単離ポリペプチドに、組織もしくは細胞に致死的でなく、ポリペプチド構造物および/または機能に有害な影響を与えない任意の濃度で加えることができる。異なる被検作用物質は、異なる有効濃度を有することが予想される。よって、いくつかの方法において、被検作用物質濃度の範囲を試験することが有利である。
【0077】
タンパク質リン酸化を検出するための方法は、従来からあり(例えばGloffke、The Scientist、16巻(19号):52頁、2002年;Screatonら、Cell、119巻:61〜74頁、2004年を参照されたい)、検出キットは、多様な商業的供給源から入手可能である(例えばUpstate(Charlottesville、VA、USA)、Bio−Rad(Hercules、CA、USA)、Marligen Biosciences,Inc.(Ijamsville、MD、USA)、Calbiochem(San Diego、CA、USA)を参照されたい)。簡単に述べると、リン酸化タンパク質は、ゲル中のリン酸化タンパク質に特異的な染色を用いて検出できる。代わりに、リン酸化タンパク質に特異的な抗体を作製できるか、または商業的に入手できる。リン酸化タンパク質に特異的な抗体は、なかでも、ビーズに繋留されているか(特定の色のサインを有するビーズを含む)、またはELISAもしくはウェスタンブロットアッセイにおいて用いることができる。
【0078】
特定の方法において、ポリペプチドのリン酸化は、このような翻訳後修飾が検出可能に測定される場合、またはこのような翻訳後修飾が対照測定(例えば被検作用物質を加える前の同じ試験系、または被検作用物質の非存在下での同様の試験系、またはAMPKの非存在下での同様の試験系)よりも少なくとも20%、少なくとも30%、少なくとも50%、少なくとも100%または少なくとも250%高い場合に、増加している。
【0079】
AMPKサブユニット(例えばAMPKα1および/またはAMPKα2)の基本型のアミノ酸配列(および基本型のAMPKサブユニット(例えばAMPKα1および/またはAMPKα2)をコードする核酸配列)は、周知である。例示的なAMPKα1アミノ酸配列および対応する核酸配列は、例えば、GenBank受託番号NM_206907.3(GI:94557298)(アミノ酸配列および核酸配列を含むHomo sapiens転写産物バリアント2 REFSEQ);NM_006251.5(GI:94557300)(アミノ酸および核酸配列を含むHomo sapiens転写産物バリアント1 REFSEQ);NM_001013367.3(GI:94681060)(アミノ酸および核酸配列を含むMus musculus REFSEQ);NMJ)01039603.1(GI:88853844)(アミノ酸および核酸配列を含むGallus gallus REFSEQ);およびNM_019142.1(GI:11862979XRaJfWSアミノ酸配列および核酸配列を含むnorvegicus REFSEQ)に記載される。例示的なAMPKα2アミノ酸配列および対応する核酸配列は、例えば、GenBank受託番号NM_006252.2(GI:46877067)(アミノ酸配列および核酸配列を含むHomo sapiens REFSEQ);NM_178143.1(GI:54792085)(アミノ酸配列および核酸配列を含むMus musculus REFSEQ);NM_001039605.1(GI:88853850)(アミノ酸配列および核酸配列を含むGallus gallus REFSEQ);およびNM_214266.1(GI:47523597)(アミノ酸配列および核酸配列を含むMus musculus REFSEQ)に記載される。
【0080】
いくつかの方法実施形態において、AMPKサブユニットのホモログまたは機能的バリアントは、基本型のAMPKα1および/またはAMPKα2ポリペプチドと少なくとも60%のアミノ酸配列同一性、例えばGenBank受託番号NM_206907.3;NM_006251.5;NMJ)01013367.3;NM_001039603.1;NM_019142.1;NM_006252.2;NM_178143.1;NM_001039605.1;またはNM_214266.1に記載されるアミノ酸配列と、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%または少なくとも98%のアミノ酸配列同一性を共有する。他の方法実施形態において、AMPKサブユニットのホモログまたは機能的バリアントは、基本型のAMPKα1および/またはAMPKα2ポリペプチドと比較して1つ以上の保存的アミノ酸置換、例えばGenBank受託番号NM_206907.3;NM_006251.5;NM_001013367.3;NM_001039603.1;NM_019142.1;NM_006252.2;NM_178143.1;NM_001039605.1;またはNM_214266.1に記載されるアミノ酸配列と比較して、3以下、5以下、10以下、15以下、20以下、25以下、30以下、40以下または50以下の保存的アミノ酸の変化を有する。例示的な保存的アミノ酸置換は、本明細書において先に記載した。
【0081】
いくつかの方法実施形態は、AMPKまたはそのサブユニット(例えばAMPKα1および/またはAMPKα2)の機能的フラグメントを含む。AMPKまたはそのサブユニット(例えばAMPKα1および/またはAMPKα2)の機能的フラグメントは、例えばその約20、約30、約40、約50、約75、約100、約150または約200連続アミノ酸残基を含む、全長もしくはインタクトなAMPKポリペプチド複合体またはそのサブユニット(例えばAMPKα1および/またはAMPKα2)の任意の部分であり得るが、ただし、フラグメントが、対象とする少なくとも1つのAMPK(またはAMPKα1および/もしくはAMPKα2)機能を保持することを条件とする。AMPK経路におけるポリペプチド間のタンパク質−タンパク質相互作用は、少なくともAMPKαサブユニット(例えばAMPKα1および/またはAMPKα2)を含むと考えられる。
【0082】
「単離」生体成分(例えばポリヌクレオチド、ポリペプチドまたは細胞)は、混合試料(例えば細胞または組織の抽出物)中の他の生体成分から分けて精製されている。例えば、「単離」ポリペプチドまたはポリヌクレオチドは、ポリペプチドまたはポリヌクレオチドが存在していた細胞(例えば組換えポリペプチドまたはポリヌクレオチドについての発現宿主細胞)の他の成分から分けられたポリペプチドまたはポリヌクレオチドである。
【0083】
「精製された」との用語は、試料から1つ以上の無関係の成分を除去することをいう。例えば、組換えポリペプチドを宿主細胞において発現させる場合、ポリペプチドは、例えば、宿主細胞タンパク質を除去し、それにより試料中の組換えポリペプチドの割合が増加することにより精製される。同様に、組換えポリヌクレオチドが宿主細胞中に存在する場合、ポリヌクレオチドは、例えば、宿主細胞ポリヌクレオチドを除去し、それにより試料中の組換えポリヌクレオチドの割合が増加することにより精製される。
【0084】
単離ポリペプチドまたは核酸分子は、典型的には、試料の少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%または99%(w/wまたはw/v)超をさらに含む。
【0085】
ポリペプチドおよび核酸分子は、当該技術において一般的に公知であり本明細書に記載される方法により単離される。ポリペプチドまたは核酸分子の純度は、ポリペプチドについてポリアクリルアミドゲル電気泳動、または核酸分子についてアガロースゲル電気泳動のようないくつかの周知の方法により決定してよい。
【0086】
2つの核酸配列間または2つのアミノ酸配列間の類似性は、これらの配列間で共有される配列同一性のレベルの点で表される。配列同一性は、典型的には、パーセント同一性に関して表される。パーセンテージが高いほど、2つの配列はより類似する。
【0087】
比較のために配列を整列させる方法は、当該技術において周知である。種々のプログラムおよびアラインメントアルゴリズムが、SmithおよびWaterman、Adv. Appl. Math.2巻:482頁、1981年;NeedlemanおよびWunsch、J. Mol. Biol.48巻:443頁、1970年;PearsonおよびLipman、Proc. Natl. Acad. ScL USA85巻:2444頁、1988年;HigginsおよびSharp、Gene73巻:237〜244頁、1988年;HigginsおよびSharp、CABIOS5巻:151〜153頁、1989年;Corpetら、Nucleic Acids Research16巻:10881〜10890頁、1988年;Huangら、Computer Applications in the Biosciences8巻:155〜165頁、1992年;Pearsonら、Methods in Molecular Biology24巻:307〜331頁、1994年;Tatianaら(1999年)、FEMS Microbiol. Lett.、174巻:247〜250頁、1999年に記載されている。Altschulらは、配列アラインメント法および相同性算出について詳細な考察を示している(J. Mol. Biol.215巻:403〜410頁、1990年)。国立バイオテクノロジー情報センター(NCBI)のBasic Local Alignment Search Tool(BLAST(商標)、Altschulら、J. Mol. Biol.215巻:403〜410頁、1990年)は、blastp、blastn、blastx、tblastnおよびtblastxの配列分析プログラムと関連して用いるために、国立バイオテクノロジー情報センター(NCBI、Bethesda、MD)およびインターネット上を含む種々の供給源から利用可能である。このプログラムを用いてどのようにして配列同一性を決定するかについての記載は、インターネット上で、BLAST(商標)についてのヘルプ部分から利用可能である。
【0088】
約30アミノ酸より大きいアミノ酸配列の比較のために、BLAST(商標)(Blastp)プログラムの「Blast2配列」機能を、デフォルトパラメータ(ギャップ開始コスト[デフォルト=5];ギャップ伸長コスト[デフォルト=2];ミスマッチペナルティ[デフォルト=−3];マッチ報酬[デフォルト=1];期待値(E)[デフォルト=10.0];ワードサイズ[デフォルト=3];1行の記載の数(V)[デフォルト=100];表示するアラインメントの数(B)[デフォルト=100])に設定されたデフォルトBLOSUM62行列を用いて行う。短いペプチド(30アミノ酸くらいより少ない)を整列させる場合、アラインメントは、デフォルトパラメータ(開始ギャップ9、伸長ギャップ1ペナルティ)に設定されたPAM30行列を用いるBlast2配列機能を用いて行う。参照配列に対してさらにより大きい類似性を有するタンパク質は、この方法により評価する場合に、パーセント同一性の増加を示す。
【0089】
核酸配列の比較のために、BLAST(商標)(Blastn)プログラムの「Blast2配列」機能を、デフォルトパラメータ(ギャップ開始コスト[デフォルト=11];ギャップ伸長コスト[デフォルト=1];期待値(E)[デフォルト=10.0];ワードサイズ[デフォルト=11];1行の記載の数(V)[デフォルト=100];表示するアラインメントの数(B)[デフォルト=100])に設定されたデフォルトBLOSUM62行列を用いて行う。参照配列に対してさらにより大きい類似性を有する核酸配列は、この方法により評価する場合に、パーセント同一性の増加を示す。
【0090】
特異的結合とは、一方の結合パートナー(例えば結合剤)と他方の結合パートナー(例えば標的)との間の特定の相互作用のことをいう。このような相互作用は、結合パートナー間(またはより頻繁には、各結合パートナーの特定の領域または部分間)の1または典型的にはそれより多い非共有結合により媒介される。非特異的結合部位とは対照的に、特異的結合部位は、飽和可能である。よって、特異的結合を特徴づける1つの例示的な方法は、特異的結合曲線による。特異的結合曲線は、例えば、一定量の一方の結合パートナーと結合した他方の結合パートナー(第1結合パートナー)の量を、第1結合パートナーの濃度の関数として示す。第1結合パートナーの濃度がこれらの条件下で増加すると、結合した第1結合パートナーの量が飽和する。非特異的結合部位とはまた別に対照的に、互いの直接の会合(例えばタンパク質−タンパク質相互作用)に関与する特異的結合パートナーは、このような会合(例えばタンパク質複合体)から、過剰量のいずれかの特異的結合パートナーにより競合的に除去(または置換)され得る。このような競合アッセイ(または置換アッセイ)は、当該技術において周知である。
【0091】
本開示は、概日リズムおよび睡眠行動をもたらすために有用な作用物質を同定する方法および作用物質も提供する。
【実施例】
【0092】
以下の実施例は、ある特定の特徴および/または実施形態を例示するために提供される。これらの実施例は、記載される特定の特徴または実施形態に本発明を限定すると解釈されない。
【0093】
(実施例1)
CRY1−S71またはCRY1−S280のリン酸化は、CRY1:FBXL3相互作用を増加させる。末梢時計のリセットにおける機構としての翻訳後修飾の役割を調べるために、質量分析および生物情報学分析の組合せを用いて、調節されたリン酸化の部位であると予測されるmCRY1およびmCRY2中の8つのセリンまたはスレオニン残基を同定した。リン酸化し得ない変異体(non−phosphorylatable mutant)をそれぞれについて作製し、セリン71のアラニンへの変異がmCRY1を安定化するが、残りの変異は安定性に対してほとんどまたは全く効果を有さないことが分かった(図7)。
【0094】
セリン71に影響するCRY1安定化変異は、AMPKによりリン酸化された最適配列によく順応するので、特に興味が持たれる。哺乳動物クリプトクロムは、mCRY1の280位に別のセリンを含有し、これも、AMPK基質モチーフによく順応する(図8)。セリン71またはセリン280のいずれかをリン酸化し得ないアミノ酸(アラニン)に変異させることは、mCRY1を安定化するために十分であったが、セリン71またはセリン280のいずれかをホスホ模倣アミノ酸(アスパルテート)に変異することは、mCRY1を不安定化するのに十分であり、これらの両方の残基を一緒に変異させると、安定性に対する影響が増加した(図1)。両方の場合において、全ての光非感受性昆虫クリプトクロムおよび高等生物において進化的に保存されているセリン71(図2)の変異は、S280の変異よりも強い効果を有した(図1A)。S71およびS280の両方のアスパラギン酸へのホスホ模倣変異を有するmCRY1は、イムノブロットにより検出できなかった。セリン71および/またはセリン280でリン酸化されているmCRY1の安定性の低下と一致して、これらの部位のホスホ模倣変異体は、CLOCK:BMAL1転写活性のより効果が低いリプレッサーでもあった(図1B)。クリプトクロムは、元来、植物において青色光光受容体として同定され、後に、動物の概日時計の成分として認識された。多くの昆虫が、クリプトクロムのそれぞれの型のうちの1つを発現する:露光により分解される青色光光受容体(「1型」)および概日転写調節に関与するが、光により誘導される不安定化に対して感受性でない転写リプレッサー(「2型」)。昆虫「2型」クリプトクロムタンパク質は、それらの哺乳動物での対応物と同様に、一昼夜の経過のうちで変動し、このことは、それらの安定化が、非光(non−light)シグナルにより、FBXL3(そのオルソログは昆虫に存在する(GenBank参照番号XM_001120533.1))が関与する保存された機構をおそらく用いて調節されるはずであることを示す。
【0095】
セリン71および280のリン酸化を模倣する変異を有するmCRY1変異体の不安定性が、FBXL3との相互作用の増加を反映するかを決定するために、v5タグ付加FBXL3を有するFlagタグ付加野生型または変異体mCRY1を発現させ、それらの結合親和性を、Flagタグ付加mCRY1の免疫沈降と、その後のv5タグ付加FBXL3のイムノブロットにより分析した。S71またはS280のいずれかのアスパラギン酸への変異は、FBXL3へのmCRY1の結合親和性を増加させ(図1C)、このことは、これらの部位のリン酸化が、mCRY1とFBXL3との間の相互作用の増加を媒介することを示唆した。2重変異体は不安定すぎて、FBXL3とのその相互作用を生化学的に決定できなかった。
【0096】
これらの変異体を、同時トランスフェクトしたCLOCKおよびBMAL1の転写活性を抑制する能力について試験した場合、CRY1のリン酸化し得ないAA変異体は、効果的なリプレッサーであることが示され、この抑制は、FBXL3の同時トランスフェクションによって変更されなかった。予測されたように、2重ホスホ模倣物CRY1 DDは、CLOCK:BMAL1活性のより効果が低い阻害物質であり、これは、この変異体のより低い安定性を反映し得る。CRY1 AA抑制に対するFBXL3媒介の作用の欠如とは対照的に、CLOCK:BMAL1駆動型転写を抑制するCRY1 DDの弱い能力は、FBXL3の同時発現により喪失した(図1D)。
【0097】
S71、S280およびS281変異の影響を、mCRY1とその公知の結合パートナーであるPER2との相互作用に対して調べた。セリン71のホスホ模倣変異(S71D)は、CRY1とPER2との間の相互作用を遮断したが、S280D変異体はPER2結合を保持し、調べた他の変異体も同様であった(図1E)。この差は、CRY1 S280Dを上回るCRY1 S71Dの分解の増進が一因であり得る。よって、S71D変異体は、PER2との結合の減少と、FBXL3との結合の増加を示し、これらはそれぞれCRY1を不安定化すると予想され、これらは一緒にCRY1 S71Dの観察された不安定性の原因であると見られる。
【0098】
AMPKは、リン酸化依存性クリプトクロム分解を媒介する。mCRY1のセリン71の周囲の配列の関係は、これが、近くの好ましい配列特異性(標的セリンに対して−4位および−3位での正に荷電された残基、ならびに−5位および+4位での疎水性残基)だけでなく、標的セリンに対して−16位および−9位での好ましい遠位ロイシン残基さえ含み、AMPKによるリン酸化の優れた候補であることを示唆する(図2A)。S280の周囲のアミノ酸配列の関係も、近傍の好ましい配列特異性に従ってAMPKリン酸化を示唆する(図8)。
【0099】
ホスホ特異抗体を、mCRY1 S71の配列の関係を周囲に有するホスホセリンを含有するペプチド抗原に対して作製し、この抗体を用いて、外因的に発現させた野生型mCRY1のリン酸化を観察したが、リン酸化し得ないS71A変異体では観察されなかった(図2B)。mCRY1のセリン71の周囲の配列は、アセチル補酵素Aカルボキシラーゼ1(ACC1)のセリン79の周囲のものと類似し、これは、AMPKの最もよく研究された基質の1つである(図2B)。実際に、セリン79でリン酸化されたACC1の残基73〜85に相当するペプチドに対して産生された抗体は、野生型mCRY1を検出できたが、セリン71をアラニンに置き換える変異を有するmCRY1は検出できず(図2B)、このことは、mCRY1のセリン71が、in vivoでリン酸化されることができることのさらなる証拠を提供し、このリン酸化事象がAMPKにより媒介され得ることをさらに示唆する。AMPKa2触媒ドメインの構成的に活性な変異体(CAa2)をmCRY1と発現させた場合、セリン71のリン酸化の増加(図2C)が観察され、このことにより、AMPKがCRY1をin vivoにてセリン71上でリン酸化できることが確認された。AMPKa1の構成的に活性な変異体(CAa1)は、核から排除されたが、セリン71のリン酸化を認識可能には増加しなかった(図2C)。AMPKは、精製された成分を用いるin vitroキナーゼアッセイにおいて、mCRY1を直接リン酸化することもできた(図9)。
【0100】
内因性AMPKの活性化は、クリプトクロムを不安定化する。いくつかの相補的ストラテジーを用いて、S71およびS280のリン酸化ならびにmCRY1の不安定化に対する内因性AMPKの寄与を分析した。HeLa細胞は、AMPK活性化キナーゼLKB1のプロモーターのメチル化によるエネルギーストレスに応答して、内因性AMPKの活性化を低減する。野生型(WT)LKB1の導入は、外因的に発現されるmCRY1のレベルを低減させ、不活性(KD)LKB1の導入では低減させなかった。この低減は、AMPK活性化AMP模倣AICARを、WT LKB1の存在下に加えることにより増強し、KD変異体の存在下では増強しなかった(図2D)。同様に、グルコース欠損によるAD293細胞におけるAMPKの活性化は、トランスフェクトした野生型mCRY1(WT)発現を低減させ、予測されるAMPKリン酸化部位を欠く変異体mCRY1(AA)ではそうではなかった(図2E)。
【0101】
クリプトクロム安定性の調節におけるAMPKの役割をさらに調べるために、遺伝子的に野生型(WT)またはAMPKの触媒サブユニットについてヌル(ampka1−/−;ampka2−/−)(AMPK−/−)であるマウス胚線維芽細胞(MEF)を用いた。flagタグ付加野生型(WT)または2重リン酸化し得ない(AA)のmCRY1をこれらの細胞において安定的に発現するレトロウイルスを用いて、AA CRY1ではなく野生型が、野生型細胞においてのみAMPKアゴニストAICARを用いる処理により急速に分解されることを示した。機能的AMPKの非存在下では、AICARは、WTまたはAA CRY1のいずれに対しても作用を有さなかった(図2F)。S71およびS280のAMPKリン酸化によるCRY1安定性の調節は、これらの細胞を4時間の過程のAMPK活性化AICARの存在下でのシクロヘキシミド処理に供することによりさらに確認された(図2G)。AICAR処理は、野生型の安定性の低減をもたらしたが、CRY1のリン酸化し得ない変異体の安定性の低減はもたらさなかった。
【0102】
AMPKは、線維芽細胞における概日リズムの代謝的変更に寄与する。概日時計リセットのための摂食由来シグナルの重要性、グルコース利用力によるAMPKの調節、およびクリプトクロム不安定化におけるAMPKの役割についての蓄積している証拠に鑑みて、AMPK発現およびグルコース利用力の影響を、線維芽細胞における概日リズム性に対して調べた。野生型線維芽細胞を、制限されたグルコースを含有する培地で培養した場合、概日性reverbαおよびdbp発現の振幅は、著しく増進され(図3Aおよび図10)、これは、グルコース欠損がAMPKを活性化し、CRY安定性を低減させるモデル(これは、CLOCK:BMAL1標的であるreverbαおよびdbpの脱抑制を導く)と一致した。予想されたように、培養培地にAICARを加えることは、グルコース欠損の効果を模倣した。目ざましいことに、グルコース欠損もAICAR処理も、AMPKを欠くMEF(ampkα1−/−;ampkα2−/−、「AMPK−/−」)におけるreverbαおよびdbpの発現に影響せず(図3Aおよび図10)、このことは、線維芽細胞概日リズムに対するグルコース制限の影響が、AMPKにより媒介されることを示した。
【0103】
Bmal1プロモーターは、REVERBαにより抑制される。よって、グルコース利用力の低減が概日リズムに与える影響を、Bmal1プロモーターの制御下でルシフェラーゼを安定的に発現する線維芽細胞を用いて調べた。標準的(高グルコース)培養条件下では、Bmal1−ルシフェラーゼの発現の高振幅概日リズムが、25.3時間の周期で観察された(図3B、C)。培地中のグルコース量の低下は、概日周期を30.7時間まで増加させた。Bmal1−ルシフェラーゼ発現細胞を、AICARを補った高グルコース培地で培養した場合、概日周期は、低グルコースで観察されたものと同様であり、グルコース欠損の概日性の影響が、AMPKにより媒介されるという考えを補強した。制限されたグルコース条件のもとで観察されたREVERBαの発現の増加は、Bmal1を含むREVERBαにより抑制される遺伝子の発現の低下をもたらすと予想される。実際に、グルコース濃度の低下またはAICAR処理のいずれかによるAMPKの活性化は、Bmal1−ルシフェラーゼ発現の振幅を低下させた(図3D)。これらの結果はまとめて、培養線維芽細胞の概日リズムが、グルコース利用力の変更を担い、これらの効果が、AMPK誘導リン酸化により媒介されることを示す。
【0104】
in vivoでのAMPKの概日性の調節。AMPKの日周期性調節を調べるために、AMPK転写、局在化および基質リン酸化を、インタクトな動物の末梢器官において調べた。全ての実験は、標準的な明暗サイクルへの同調化の後に定常的な暗所に維持した動物を用いて行って、観察された効果が、外部の環境における変化に対する日周期性の応答よりもむしろ概日性であることを確実にした。
【0105】
調べた両方のAMPK基質、ACC1−Ser79およびRaptor−Ser792のリン酸化は、主体的な昼において夜よりも再現可能により高く(図4A)、ネガティブフィードバックタンパク質が不安定な昼の時間にほぼ対応し、これは、リズム性のAMPK活性化が分解に寄与するモデルと一致した。マウス肝臓におけるAMPKの概日性の調節を探索する一方、最小限の核クリプトクロムタンパク質の時間と一致するピーク発現(図4C)を有する調節性ampkβ2サブユニットの堅固な概日性の発現(図4B)。AMPKβ2は、AMPK複合体の核局在化を駆動すると報告されているが、AMPKβ1含有複合体は、形質膜を標的にする。よって、ampkβ2の概日転写は、変動するAMPKβ2が、AMPKα1およびAMPKα2の核局在化を日周期性に調節することを示唆する。この仮説を試験するために、概日サイクル全体にわたって回収した肝臓核(liver nucleus)におけるAMPKα1およびAMPKα2のタンパク質レベルを測定し(図4C)、ampkβ2発現と同期してピークになる核AMPKα1のリズム性を観察した。AMPKα2は、核局在化シグナルを含有し、核に常に存在した。AMPKα1核局在化がピークになる時間は、肝臓核における最小CRY1タンパク質の時間でもあり、このことは、AMPKの核へのリズム性の移入が、AMPK媒介リン酸化およびクリプトクロムの分解に寄与し得ることを示唆する。
【0106】
AMPKは、概日時計をin vivoにて変更する。マウスにおいてAMPKα1およびAMPKα2の両方を遺伝子欠失させることにより、早期の胚致死性が導かれる。よって、肝臓概日時計におけるAMPKの役割をさらに調べるために、概日タンパク質および転写産物を、明暗サイクルへの同調化の後に定常的な暗所で飼育した対照マウス(LKB1+/+)または肝細胞におけるlkb1が喪失した同腹子(LKB1L/L)の肝臓において24時間にわたって調べた。lkb1の肝臓特異的欠失は、この器官におけるAMPK活性化を廃止し、概日サイクル全体で、特に、変更されていないマウスにおいてAMPKが最も活性であることが見出された昼間の時間中に、肝臓核に存在するCRY1およびCRY2タンパク質の量を著しく増加させた(図5B)。この増加は、昼光に対応する期間におけるREVERBα発現の減少(図5B)および概日サイクルを通しての概日転写産物の振幅の減少(図5C)と関連した。よって、AMPKシグナル伝達のin vivoにおける喪失は、クリプトクロムを安定化し、概日リズムを混乱させ、光非依存性末梢概日時計についての同期化の機構を確立する。
【0107】
材料および方法
細胞および細胞培養物 − AMPK+/+およびAMPK−/−マウス胚線維芽細胞は、Benoit Viollet博士からの贈物であった。HeLa細胞およびAD293細胞は、American Type Culture Collection(ATCC)から購入した。3T3不死化MEFは、以前に記載された。そうでないと記載しない限り、細胞は、10%ウシ胎仔血清、ペニシリンおよびストレプトマイシンを補った完全ダルベッコ改変イーグル培地(DMEM)(Invitrogen cat#11995またはcat#11965)で、5%CO2に維持した37℃インキュベータ中で成長させた。グルコース濃度を操作した実験において、細胞は、グルタミン、非必須アミノ酸、ペニシリン、ストレプトマイシンおよび記載される量のD−グルコースを補った最小DMEM(Sigma cat#D5030)、またはペニシリン、ストレプトマイシン、L−グルタミンおよび記載される量のD−グルコースを補ったグルコース非含有DMEM(Invitrogen cat#11966)で成長させた。0.5mMグルコースを用いる実験は、浸透圧の影響を制御するためにD−マンニトールを補った。細胞刺激は、50%ウマ血清(Invitrogen cat#26050)を含む完全DMEMを用いて行い、以前に記載されたようにして行った。
【0108】
プラスミドおよびトランスフェクション − pDONR221およびpcDNA3.1/v5−His−TOPOは、Invitrogenから購入した。pcDNA3−2xFlag−mCRY1(WT)およびpcDNA3−PER2は、Charles Weitz博士からの贈物であった。pCMV−SPORT6−Fbxl3は、Open Biosystemsから購入し、FBXL3は、pcDNA3.1/v5−His−TOPO中に、標準的なプロトコルによりクローニングした。flag−LKB1、myc−AMPKa1およびmyc−AMPKa2構築物は、以前に記載され、構成的に活性な対立遺伝子(CAa1およびCAa2)は、残基T312の後に停止コドンを挿入することにより作製した。全ての変異は、Stratagene部位特異的変異誘発プロトコルを用いて作製した。トランスフェクションは、FuGene HD(Roche)を用いて行った。
【0109】
ウイルスおよび安定細胞株の作製 − AD293細胞にpLXSP3puro発現クローンをpCL−Amphoと共にトランスフェクトしてウイルスを生成した。トランスフェクションの48時間後にウイルス上清を回収し、0.45μmフィルターでろ過し、6μg/mlポリブレンを添加し、親細胞株に加えた。4時間後に、さらなる培地を加えてポリブレンを3μg/ml未満に希釈した。ウイルス形質導入の48時間後に、感染細胞を、1〜5μg/mlピューロマイシンを含有する選択培地中に分配した。選択培地は、選択が完了するまで2〜3日毎に置き換えた。
【0110】
質量分析 − Flag−mCRY1をトランスフェクトしたAD293細胞を、10μMのMG132で6時間処理し、1%のTx−100を含有する緩衝液中で溶解させた。Flag−mCRY1を、M2−アガロース(Sigma)上で精製し、SDS−PAGEにより夾雑物から分けた。クーマシー染色されたバンドを切り出し、HPLCグレードの50%アセトニトリル中で2回すすぎ、Beth Israel Deaconess Medical Center質量分析施設に送付した。
【0111】
タンパク質抽出物の調製、免疫沈降およびイムノブロッティング − 全細胞抽出物は、以前に記載されたように1%のTritonX−100を含有する溶解緩衝液中で調製し、肝臓核抽出物を、NUN手順により調製した。用いた抗体は、Sigmaからの抗Flag M2アガロース、抗v5アガロース、抗Flagポリクローナル、抗v5ポリクローナルおよび抗βアクチン;Alpha Diagnostics InternationalからのCRY11A、CRY21AおよびPER21A;Cell Signaling Technologiesからの抗ホスホACC1(S79)、抗ACC1、抗ホスホ−AMPKa、抗ホスホ−Raptor、抗Raptorおよび抗REVERBa;Upstate Biotechologyからの抗AMPKa1および抗AMPKa2;ならびにMilliporeと共同で作製したホスホ−CRY1(S71)および周囲の残基を含有するホスホペプチドに対して産生されたポリクローナル抗血清であった。
【0112】
in vitroリン酸化アッセイ − Flag−mCRY1を、トランスフェクトしたAD293細胞から精製し、32P−ATPおよび精製AMPK(Upstate Biotechnologyから)と、300μMのAMPの存在下または非存在下で30分間室温にて組み合わせた。反応混合物をSDS−PAGEにより分離し、ニトロセルロースに移した。ホスホイメージャーによる放射活性の視覚化の後に、ニトロセルロースをFlagタグについてイムノブロットした。
【0113】
リアルタイム生物発光モニタリング − Bmal1プロモーター駆動型ルシフェラーゼを安定的に発現するヒト骨肉腫U2OSレポーター細胞株は、記載されている。2×104細胞を35mmディッシュに播種し、10%血清を補ったDMEM中で集密まで3日間にわたって成長させた。集密細胞を、50%ウマ血清で2時間刺激し、次いで、0.1%透析血清および上記のように種々の量のグルコースを含有する培地に移した。生物発光を、Actimetrics,Inc.からのLumiCycle装置により連続的に記録した。
【0114】
遺伝子発現 − RNAを、肝臓または培養線維芽細胞から、TrizolでまたはQiagen RNeasy精製システムを用いて抽出した。cDNAは、SuperscriptII逆転写酵素(Invitrogen)を用いて調製し、遺伝子発現について、SYBRグリーン(Invitrogen)またはTaqMan(Applied Biosystems)化学のいずれかを用いる定量的リアルタイムPCRを用いて分析した。プライマー配列は、要請に応じて入手可能である。
【0115】
マウス − LKB1fl/flマウスは、Ronald De Pinho博士からの贈物であり、Cry1−/−;Cry2−/−マウスは、Aziz Sancar博士からの贈物であった。Creリコンビナーゼを発現するアデノウイルスは、University of Iowa Transgenic Core施設からであった。全ての動物の世話および処置は、動物の世話および使用についてのSalk Instituteガイドラインに準拠した。
【0116】
本開示は、mCRY1が、実際に、今まで調べた47の核ホルモン受容体のうち20と相互作用し、PPARdと特に良好に相互作用することを証明する(図11)。さらに、生理食塩水またはAMPK活性化薬物であるAICARを注射した野生型およびクリプトクロム欠損マウスの肝臓における遺伝子発現は、クリプトクロムが、遺伝子のサブセットのAICARにより誘導される活性化に必要であることを証明する(図12)。さらに、マウスにおけるCry1およびCry2の両方の遺伝子破壊の代謝生理に対する影響を調べる。データは、Cry1−/−;Cry2−/−マウスが、野生型対照よりも著しく低い体重と、著しく低減された安静時血中グルコースとを有することを示す(図13)。一括して、これらのデータは、哺乳動物のクリプトクロムが、細胞のエネルギー状態の以前には認識されていなかったセンサとして機能すること、これらが生物のエネルギー恒常性において役割を果たすこと、およびクリプトクロムの薬理学的調整が、代謝障害の治療に有用であり得ることを示唆する。
【0117】
本開示は、具体的な実施形態に重点をおいて記載しているが、具体的な実施形態の変形を用いることができることが当業者に明らかであり、本開示を、本明細書に具体的に記載する以外の別な方法で行うことができることを意図する。よって、本開示は、以下の特許請求の範囲により定義される本開示の精神および範囲内に包含される全ての改変を含む。
【特許請求の範囲】
【請求項1】
代謝リズムまたは概日リズムの調整において用いるための作用物質を同定する方法であって、前記作用物質をCry1またはCry2タンパク質と接触させるステップと、Cry1またはCry2をリン酸化または脱リン酸化する前記作用物質の能力を測定するステップとを含み、Cry1またはCry2を改変する作用物質が、代謝リズムまたは概日リズムを調整するために有用な小分子作用物質である方法。
【請求項2】
前記作用物質が、S71またはS280でのリン酸化に影響する、請求項1に記載の方法。
【請求項3】
AMPKの活性の変化を測定するステップをさらに含む、請求項1に記載の方法。
【請求項4】
前記作用物質が、Cry1またはCry2の安定性を低下させる、請求項1に記載の方法。
【請求項5】
前記作用物質が、安静状態を促進する、請求項4に記載の方法。
【請求項6】
前記作用物質が、ペプチド、ポリペプチド、抗体、抗体フラグメント、核酸および小分子からなる群から選択される、請求項1に記載の方法。
【請求項7】
前記作用物質が、AMPKアゴニストである、請求項1に記載の方法。
【請求項8】
請求項1に記載の方法により同定される作用物質であって、Cry1またはCry2の安定性を低下させる作用物質を含む組成物。
【請求項9】
被験体を、請求項1に記載の方法により同定される作用物質であって、Cry1および/またはCry2のリン酸化または脱リン酸化を促進する作用物質と接触させるステップを含む、代謝性または概日性の疾患または障害を治療する方法。
【請求項10】
被験体における代謝障害および概日リズムの治療用の医薬品の調製において用いるための、クリプトクロム転写同時制御因子機能を調整する、請求項1に記載の方法により同定される作用物質の使用。
【請求項11】
前記作用物質が、ペルオキシソーム増殖因子活性化受容体(PPAR)アルファ、ベータ(デルタ)およびガンマを調整する、請求項10に記載の使用。
【請求項12】
前記作用物質が、ビグアナイド誘導体、AICAR、メトホルミンまたはその誘導体、フェンホルミンまたはその誘導体、レプチン、アディポネクチン、AICAR(5−アミノイミダゾール−4−カルボキサミド)、ZMP、DRL−16536、BG800化合物(Betagenon)およびフラン−2−カルボン酸誘導体からなる群から選択されるAMPKアゴニストである、請求項10に記載の使用。
【請求項13】
前記被験体が哺乳類である、請求項10に記載の使用。
【請求項14】
有効量が、単一用量または分割用量での1日あたり約0.5mg/kg〜1日あたり約100mg/kgである、請求項10または12に記載の使用。
【請求項15】
前記作用物質が、経口投与、静脈内注射、筋肉内注射、硬膜外送達、頭蓋内、局所、眼内、坐剤または皮下注射用に製剤化される、請求項10または12に記載の使用。
【請求項16】
CRY1またはCRY2のリン酸化を調整するか、またはクリプトクロム転写同時制御因子機能を調整する作用物質と、少なくとも1つの他の概日リズム改変作用物質または代謝改変作用物質とを含む組成物。
【請求項17】
前記少なくとも1つの他の概日リズム改変作用物質が、睡眠補助剤である、請求項16に記載の組成物。
【請求項18】
ビグアナイド誘導体、AICAR、メトホルミンまたはその誘導体、フェンホルミンまたはその誘導体、レプチン、アディポネクチン、AICAR(5−アミノイミダゾール−4−カルボキサミド)、ZMP、DRL−16536、BG800化合物(Betagenon)およびフラン−2−カルボン酸誘導体からなる群から選択されるAMPKアゴニストを含む、請求項16に記載の組成物。
【請求項19】
化合物が、経口投与、静脈内注射、筋肉内注射、硬膜外送達、局所、坐剤による、眼送達、頭蓋内送達または皮下注射用に製剤化される、請求項16に記載の組成物。
【請求項20】
哺乳類における睡眠を調整する方法であって、哺乳類における概日リズムまたは代謝を調整するのに有効な量のCRY1またはCRY2を不安定化する作用物質を前記哺乳類に投与するステップを含む方法。
【請求項21】
前記哺乳類がヒトである、請求項20に記載の方法。
【請求項22】
前記概日リズムが睡眠行動である、請求項20に記載の方法。
【請求項23】
被験体における概日リズムまたは睡眠を調整する作用物質を同定する方法であって、
(a)AMPKまたはLKB1経路を備える試料を少なくとも1つの被検作用物質と接触させるステップと、
(b)CRY1またはCRY2経路の活性を、前記被検作用物質の存在下および非存在下で比較するステップであって、CRY1またはCRY2の活性または安定性を変化させる被検作用物質が、概日リズム調整活性を有する作用物質を示すステップと
を含む方法。
【請求項24】
細胞における概日サイクルまたは代謝サイクルを調整する作用物質を同定する方法であって、前記細胞を前記作用物質と接触させるステップであって、前記細胞が、Cyr1またはCry2を含むAMPK経路またはLKB1経路を備えるステップと、Cry1およびCry2活性に対する前記作用物質の影響を測定するステップであって、Cry1またはCry2の活性の変化が、概日サイクルまたは代謝サイクルを調整できる作用物質を示すステップとを含む方法。
【請求項25】
代謝リズムまたは概日リズムの疾患または障害を決定する方法であって、CRY1またはCRY2の安定性を、組織において24時間周期の間に測定するステップを含み、正常または過剰ATP濃度の存在下でのCRY1またはCRY2の長期間安定性の周期が、代謝リズムまたは概日リズムの疾患または障害を示す方法。
【請求項26】
安静および脂質異化反応を促進する方法であって、概日サイクルの夜間期の間にAMPKアゴニストを投与するステップを含み、前記AMPKアゴニストが、CRY1またはCRY2の安定性を低下させる方法。
【請求項1】
代謝リズムまたは概日リズムの調整において用いるための作用物質を同定する方法であって、前記作用物質をCry1またはCry2タンパク質と接触させるステップと、Cry1またはCry2をリン酸化または脱リン酸化する前記作用物質の能力を測定するステップとを含み、Cry1またはCry2を改変する作用物質が、代謝リズムまたは概日リズムを調整するために有用な小分子作用物質である方法。
【請求項2】
前記作用物質が、S71またはS280でのリン酸化に影響する、請求項1に記載の方法。
【請求項3】
AMPKの活性の変化を測定するステップをさらに含む、請求項1に記載の方法。
【請求項4】
前記作用物質が、Cry1またはCry2の安定性を低下させる、請求項1に記載の方法。
【請求項5】
前記作用物質が、安静状態を促進する、請求項4に記載の方法。
【請求項6】
前記作用物質が、ペプチド、ポリペプチド、抗体、抗体フラグメント、核酸および小分子からなる群から選択される、請求項1に記載の方法。
【請求項7】
前記作用物質が、AMPKアゴニストである、請求項1に記載の方法。
【請求項8】
請求項1に記載の方法により同定される作用物質であって、Cry1またはCry2の安定性を低下させる作用物質を含む組成物。
【請求項9】
被験体を、請求項1に記載の方法により同定される作用物質であって、Cry1および/またはCry2のリン酸化または脱リン酸化を促進する作用物質と接触させるステップを含む、代謝性または概日性の疾患または障害を治療する方法。
【請求項10】
被験体における代謝障害および概日リズムの治療用の医薬品の調製において用いるための、クリプトクロム転写同時制御因子機能を調整する、請求項1に記載の方法により同定される作用物質の使用。
【請求項11】
前記作用物質が、ペルオキシソーム増殖因子活性化受容体(PPAR)アルファ、ベータ(デルタ)およびガンマを調整する、請求項10に記載の使用。
【請求項12】
前記作用物質が、ビグアナイド誘導体、AICAR、メトホルミンまたはその誘導体、フェンホルミンまたはその誘導体、レプチン、アディポネクチン、AICAR(5−アミノイミダゾール−4−カルボキサミド)、ZMP、DRL−16536、BG800化合物(Betagenon)およびフラン−2−カルボン酸誘導体からなる群から選択されるAMPKアゴニストである、請求項10に記載の使用。
【請求項13】
前記被験体が哺乳類である、請求項10に記載の使用。
【請求項14】
有効量が、単一用量または分割用量での1日あたり約0.5mg/kg〜1日あたり約100mg/kgである、請求項10または12に記載の使用。
【請求項15】
前記作用物質が、経口投与、静脈内注射、筋肉内注射、硬膜外送達、頭蓋内、局所、眼内、坐剤または皮下注射用に製剤化される、請求項10または12に記載の使用。
【請求項16】
CRY1またはCRY2のリン酸化を調整するか、またはクリプトクロム転写同時制御因子機能を調整する作用物質と、少なくとも1つの他の概日リズム改変作用物質または代謝改変作用物質とを含む組成物。
【請求項17】
前記少なくとも1つの他の概日リズム改変作用物質が、睡眠補助剤である、請求項16に記載の組成物。
【請求項18】
ビグアナイド誘導体、AICAR、メトホルミンまたはその誘導体、フェンホルミンまたはその誘導体、レプチン、アディポネクチン、AICAR(5−アミノイミダゾール−4−カルボキサミド)、ZMP、DRL−16536、BG800化合物(Betagenon)およびフラン−2−カルボン酸誘導体からなる群から選択されるAMPKアゴニストを含む、請求項16に記載の組成物。
【請求項19】
化合物が、経口投与、静脈内注射、筋肉内注射、硬膜外送達、局所、坐剤による、眼送達、頭蓋内送達または皮下注射用に製剤化される、請求項16に記載の組成物。
【請求項20】
哺乳類における睡眠を調整する方法であって、哺乳類における概日リズムまたは代謝を調整するのに有効な量のCRY1またはCRY2を不安定化する作用物質を前記哺乳類に投与するステップを含む方法。
【請求項21】
前記哺乳類がヒトである、請求項20に記載の方法。
【請求項22】
前記概日リズムが睡眠行動である、請求項20に記載の方法。
【請求項23】
被験体における概日リズムまたは睡眠を調整する作用物質を同定する方法であって、
(a)AMPKまたはLKB1経路を備える試料を少なくとも1つの被検作用物質と接触させるステップと、
(b)CRY1またはCRY2経路の活性を、前記被検作用物質の存在下および非存在下で比較するステップであって、CRY1またはCRY2の活性または安定性を変化させる被検作用物質が、概日リズム調整活性を有する作用物質を示すステップと
を含む方法。
【請求項24】
細胞における概日サイクルまたは代謝サイクルを調整する作用物質を同定する方法であって、前記細胞を前記作用物質と接触させるステップであって、前記細胞が、Cyr1またはCry2を含むAMPK経路またはLKB1経路を備えるステップと、Cry1およびCry2活性に対する前記作用物質の影響を測定するステップであって、Cry1またはCry2の活性の変化が、概日サイクルまたは代謝サイクルを調整できる作用物質を示すステップとを含む方法。
【請求項25】
代謝リズムまたは概日リズムの疾患または障害を決定する方法であって、CRY1またはCRY2の安定性を、組織において24時間周期の間に測定するステップを含み、正常または過剰ATP濃度の存在下でのCRY1またはCRY2の長期間安定性の周期が、代謝リズムまたは概日リズムの疾患または障害を示す方法。
【請求項26】
安静および脂質異化反応を促進する方法であって、概日サイクルの夜間期の間にAMPKアゴニストを投与するステップを含み、前記AMPKアゴニストが、CRY1またはCRY2の安定性を低下させる方法。
【図1】

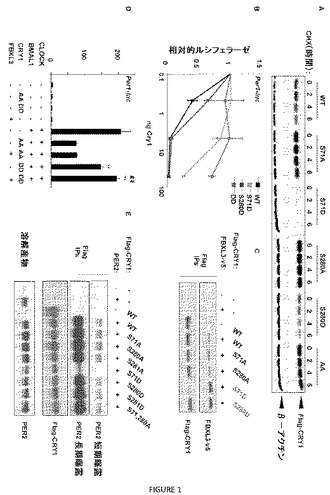
【図2】
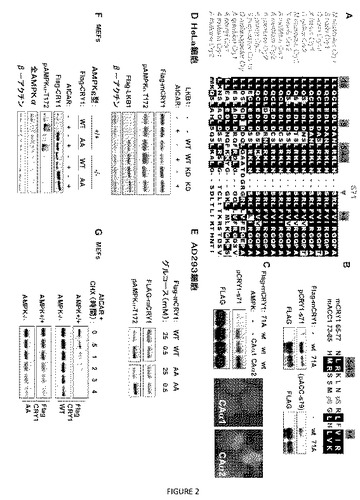
【図3】
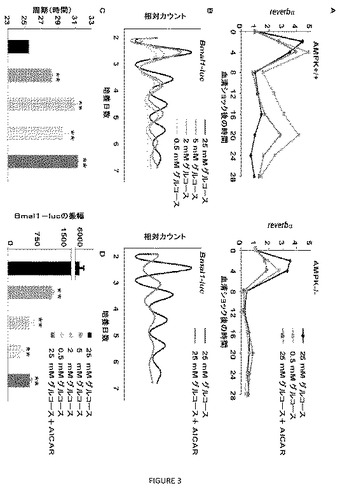
【図4】
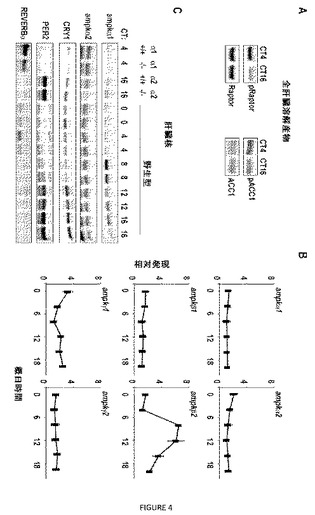
【図5】

【図6】
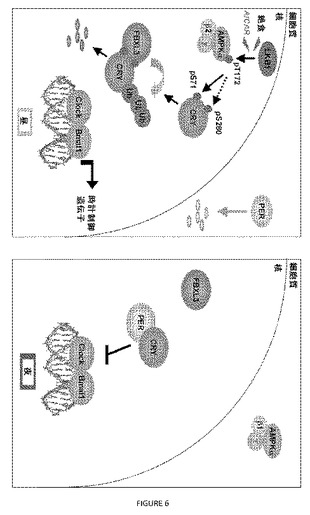
【図7】

【図8】

【図9】

【図10】
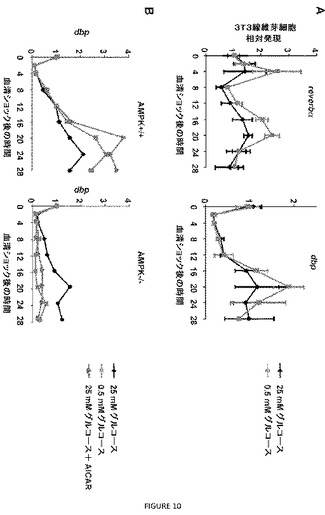
【図11】

【図12】

【図13】
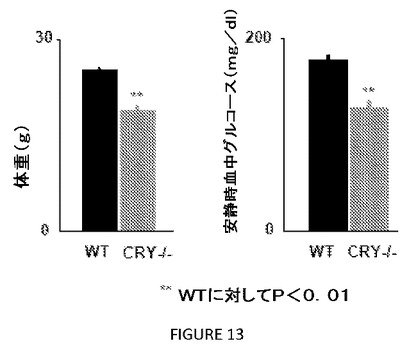
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2012−521198(P2012−521198A)
【公表日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願番号】特願2012−501034(P2012−501034)
【出願日】平成22年3月22日(2010.3.22)
【国際出願番号】PCT/US2010/028196
【国際公開番号】WO2010/108195
【国際公開日】平成22年9月23日(2010.9.23)
【出願人】(510177832)ザ ソーク インスティテュート フォー バイオロジカル スタディーズ (3)
【Fターム(参考)】
【公表日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願日】平成22年3月22日(2010.3.22)
【国際出願番号】PCT/US2010/028196
【国際公開番号】WO2010/108195
【国際公開日】平成22年9月23日(2010.9.23)
【出願人】(510177832)ザ ソーク インスティテュート フォー バイオロジカル スタディーズ (3)
【Fターム(参考)】
[ Back to top ]