
低免疫原性のタンパク質製剤のための組成物および方法
低免疫原性のタンパク質、ポリペプチドまたはペプチド製剤にかかる方法および組成物に関する。前記方法は、タンパク質、ポリペプチドまたはペプチドと、セリン含有化合物または他のホスファチジル化合物もしくはリン脂質でありうる結合剤との複合体の形成を含む。形成された複合体は、単純複合体、ミセル、渦巻形構造、リポソーム、非二重層、および新規な脂質構造を有する。タンパク質、ポリペプチドまたはペプチドの抗原性および免疫原性はこのような複合体の形成によって減弱される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、低い免疫原性を有するタンパク質複合体およびこれを作るための方法に関する。
【背景技術】
【0002】
血友病は、ファクターVIII(抗血友病ファクター,AHFあるいはFVIII)の欠損によってもたらされる出血性疾患である。FVIIIは6つのドメインA1,A2,A3,B,C1およびC2からなるマルチドメインタンパク質であり、トロンビンによるこのタンパク質の活性化は、重鎖(A1およびA2)と軽鎖(A3,C1およびC2)内に生じる[1,2]。血液濃縮物、組換えファクターVIIIおよびファクターVIIIの変異体を用いた補充療法が、血友病のための一次治療である。しかしながら、15〜35%の患者では中和抗体が発現し、 このような免疫応答は血友病の治療を損なう。中和抗体の発現を克服するための最新の治療計画が存在するが、コスト効率が良くない。免疫原性のより少ないファクターVIII製剤の開発は、代替的なクリニカル・アプローチを提供しうる。
【0003】
一般に、治療用タンパク質への免疫応答は、以下の理由が原因とされている;(i)投与経路、(ii)凝集物の存在、(iii)頻回投与、および(iv)特定のエピトープ領域[3]。最近、FVIIIが凝集物を形成する傾向をもつことが明らかになったが[4]、免疫応答の進行におけるこれらの凝集物の役割はよく理解されていない。さらに、FVIII分子上には主に2つのエピトープ領域、C2およびA2ドメインが存在する。スカンデラら[5]は、抗体価が軽鎖のC2ドメインに対して最も高いことを明らかにした。C2ドメインは膜結合ドメインでもあり、それはその凝固カスケードの一環として血小板膜上のホスファチジルセリン(PS)に結合する[6,7]。C2ドメインに対する抗体の抗凝固作用は、リン脂質へのファクターVIIIの結合阻害に起因する。C2ドメインに対するモノクローナル抗体が、リン脂質含有膜へのファクターVIIIの結合を妨げることが明らかにされ、そしてこれらの観察に基づいて、エピトープと膜結合領域が重複することが結論づけられた[10,11]。
【0004】
FVIIIのリポソーム封入に関する従来の研究は、従来のリポソーム内へのFVIII(VIIIファクター)の封入効率を増加することを目的とし、さらに生体内安定性および経口バイオアベイラビリティの増加を目的とする[12,13]。その分子構造に起因して、脂質分子は、リポソーム、ミセル、非二重層構造および渦巻形構造などの複数の分子アセンブリを形成する傾向がある。これらの分子アセンブリを、タンパク質およびペプチドを含むいくつかの薬物の治療特性を改良するための薬物送達媒体として使用するため、多数の研究が行われてきた。これらの治療の特徴は、循環時間の延長、毒性の低減、免疫応答の亢進および生体内分解の減少である[14-25]。したがって、リポソームは免疫応答を増加させるためのアジュバント(adjuvants)として使用されてきた[26-31]。
【0005】
【非特許文献1】Fay, P.J. Factor VIII structure and function. Thromb Haemost 70, 63-7 (1993).
【非特許文献2】Foster, P.A. & Zimmerman, T.S. Factor VIII structure and function. Blood Rev 3, 180-91 (1989).
【非特許文献3】Braun, A., Kwee, L., Labow, M.A. & Alsenz, J. Protein aggregates seem to play a key role among the parameters influencing the antigenicity of interferon alpha (IFN-alpha) in normal and transgenic mice. Pharmaceutical Research 14, 1472-8 (1997).
【非特許文献4】Grillo, A.O. et al. Conformational origin of the aggregation of recombinant human factor VIII. Biochemistry 40, 586-95. (2001).
【非特許文献5】Scandella, D.H. et al. In hemophilia a and autoantibody inhibitor patients: the factor viii a2 domain and light chain are most immunogenic. Thromb Res 101, 377-85. (2001).
【非特許文献6】Lenting, P.J. et al. The light chain of factor VIII comprises a binding site for low density lipoprotein receptor-related protein. Journal of Biological Chemistry. 274, 23734-9 (1999).
【非特許文献7】Saenko, E.L., Yakhyaev, A.V., Mikhailenko, I., Strickland, D.K. & Sarafanov, A.G. Role of the low density lipoprotein-related protein receptor in mediation of factor VIII catabolism. Journal of Biological Chemistry. 274, 37685-92 (1999).
【非特許文献8】Gilbert, G.E., Furie, B.C. & Furie, B. Binding of human factor VIII to phospholipid vesicles. J Biol Chem 265, 815-22 (1990).
【非特許文献9】Stoilova-McPhie, S., Villoutreix, B.O., Mertens, K., Kemball-Cook, G. & Holzenburg, A. 3-Dimensional structure of membrane-bound coagulation factor VIII: modeling of the factor VIII heterodimer within a 3-dimensional density map derived by electron crystallography. Blood 99, 1215-23. (2002).
【非特許文献10】Scandella, D. et al. Some factor VIII inhibitor antibodies recognize a common epitope corresponding to C2 domain amino acids 2248 through 2312, which overlap a phospholipid-binding site. Blood 86, 1811-9. (1995).
【非特許文献11】Barrow, R.T., Healey, J.F., Jacquemin, M.G., Saint-Remy, J.M. & Lollar, P. Antigenicity of putative phospholipid membrane-binding residues in factor VIII. Blood 97, 169-74. (2001).
【非特許文献12】Kirby, C.J. & Gregoriadis, G. Preparation of liposomes containing factor VIII for oral treatment of haemophilia. J Microencapsul 1, 33-45. (1984).
【非特許文献13】Hemker, H.C., Hermens, W.T., Muller, A.D. & Zwaal, R.F. Oral treatment of haemophilia A by gastrointestinal absorption of factor VIII entrapped in liposomes. Lancet 1, 70-1. (1980).
【非特許文献14】Martin, F.J., Hubbell, W.L. & Papahadjopoulos, D. Immunospecific targeting of liposomes to cells: a novel and efficient method for covalent attachment of Fab' fragments via disulfide bonds. Biochemistry 20, 4229-38. (1981).
【非特許文献15】Matthay, K.K., Heath, T.D., Badger, C.C., Bernstein, I.D. & Papahadjopoulos, D. Antibody-directed liposomes: comparison of various ligands for association, endocytosis, and drug delivery. Cancer Res 46, 4904-10. (1986).
【非特許文献16】Olson, F., Hunt, C.A., Szoka, F.C., Vail, W.J. & Papahadjopoulos, D. Preparation of liposomes of defined size distribution by extrusion through polycarbonate membranes. Biochim Biophys Acta 557, 9-23. (1979).
【非特許文献17】Papahadjopoulos, D. et al. Sterically stabilized liposomes: improvements in pharmacokinetics and antitumor therapeutic efficacy. Proc Natl Acad Sci USA 88, 11460-4 (1991).
【非特許文献18】Phillips, N.C. & Emili, A. Enhanced antibody response to liposome-associated protein antigens: preferential stimulation of IgG2a/b production. Vaccine 10, 151-8. (1992).
【非特許文献19】Storm, G., Wilms, H.P. & Crommelin, D.J. Liposomes and biotherapeutics. Biotherapy 3, 25-42 (1991).
【非特許文献20】Szoka, F., Jr. & Papahadjopoulos, D. Procedure for preparation of liposomes with large internal aqueous space and high capture by reverse-phase evaporation. Proc Natl Acad Sci USA 75, 4194-8. (1978).
【非特許文献21】Szoka, F. et al. Preparation of unilamellar liposomes of intermediate size (0.1-0.2 mumol) by a combination of reverse phase evaporation and extrusion through polycarbonate membranes. Biochim Biophys Acta 601, 559-71. (1980).
【非特許文献22】Van Slooten, M.L. et al. Liposomes as sustained release system for human interferon-gamma: biopharmaceutical aspects. Biochim Biophys Acta 1530, 134-45. (2001).
【非特許文献23】Papahadjopoulos, D., Vail, W.J., Jacobson, K. & Poste, G. Cochleate lipid cylinders: formation by fusion of unilamellar lipid vesicles. Biochim Biophys Acta 394, 483-91. (1975).
【非特許文献24】Coorssen, J.R. & Rand, R.P. Structural effects of neutral lipids on divalent cation-induced interactions of phosphatidylserine-containing bilayers. Biophys J 68, 1009-18. (1995).
【非特許文献25】Duzgunes, N. et al. Calcium- and magnesium-induced fusion of mixed phosphatidylserine/phosphatidylcholine vesicles: effect of ion binding. J Membr Biol 59, 115-25. (1981).
【非特許文献26】Duzgunes, N., Wilschut, J., Fraley, R. & Papahadjopoulos, D. Studies on the mechanism of membrane fusion. Role of head-group composition in calcium- and magnesium-induced fusion of mixed phospholipid vesicles. Biochim Biophys Acta 642, 182-95. (1981).
【非特許文献27】Alving, C.R., Liposomes as carriers of antigens and adjuvants. J. Immunol. Methods, 140:1-13, (1991).
【非特許文献28】Van Rooijen, N., Liposomes as carrier and immunoadjuvant of vaccine antigens. Adv. Biotechnolo Processes, 13: 255-279 (1990).
【非特許文献29】Huong, T.M., Ishida, T., Harashima, H. & Kiwada, H. The complement system enhances the clearance of phosphatidylserine (PS)-liposomes in rat and guinea pig. Int J Pharm 215, 197-205. (2001).
【非特許文献30】Dal Monte, P. & Szoka, F.C., Jr. Effect of liposome encapsulation on antigen presentation in vitro. Comparison of presentation by peritoneal macrophages and B cell tumors. J Immunol 142, 1437-43. (1989).
【非特許文献31】Heath, T.D., Edwards, D.C. & Ryman, B.E. The adjuvant properties of liposomes. Biochemical Society Transactions 4, 129-33 (1976).
【非特許文献32】Rosenblum et al., Modified Proteins, designer toxins, and methods of making thereof., U.S. patent application publication 20030176331, 2003.
【発明の開示】
【発明が解決しようとする課題】
【0006】
しかしながら、免疫応答および抗原性を減じるための脂質複合体の開発は研究されておらず、それゆえ、治療用タンパク質の免疫原性を減少させるためのアプローチを開発する必要性が引き続き存在している。免疫毒性を減少させるために特定のエピトープの配列を変更する別のアプローチがある[32]。しかしながら、そのようなアミノ酸置換は生物活性の損失をもたらす可能性がある。
【課題を解決するための手段】
【0007】
本発明は、低い抗原性と免疫原性を有する組成物およびその製造方法を開示する。したがって、タンパク質、ポリペプチドあるいはペプチドのような治療薬、およびその免疫原性と抗原性を減じるように前記タンパク質に結合できる一以上の分子(本明細書中で結合剤と呼ぶ)を含む組成物が開示される。このような結合剤には、ホスホセリン、ホスファチジルセリンもしくはホスファチジルセリン(PS)を含むリン脂質等のセリン化合物;ホスファチジルコリン(PC),ホスファチジン酸(PA),もしくはホスホエタノールアミン(PE);またはPA,PCもしくはPEを含むリン脂質が含まれる。
【0008】
前記のタンパク質-結合剤複合体は、その免疫応答を減じる(1)タンパク質-結合剤複合体を含む液体またはこの液体の凍結乾燥剤、(2)新規な非リポソーム構造、(3)リポソーム、(4)ミセル、(5)渦巻形、(6)非二重層構造の形で存在しうる。
【0009】
本発明はまた、結合剤(セリン含有化合物など)と複合体を形成することによってタンパク質の免疫原性および/または抗原性を減じる方法を開示する。前記タンパク質-結合剤複合体は、適切な緩衝剤を用いて安定化しうる。
【0010】
ある実施形態では、ジミリスチルホスファチジルコリン(DMPC)および脳ホスファチジルセリン(bPS)を含有する乾燥脂質膜は、各種の緩衝系中でタンパク質(FVIIIなど)を用いて水和させられる。新規の、非リポソーム構造物は、300mMのNaClと5mMのCaCl2中のDMPC、bPSを用いて形成される。従来のリポソームは、水に変化する緩衝系あるいはリン酸緩衝生理食塩水として形成される。これはカルシウムあるいはPS濃度を減じることによって達成することもできる。DMPCの除去および100%PSと超音波処理もしくは押出ステップの使用は渦巻形構造を導き、中間アシル鎖長のPSの使用はミセル構造をもたらす。より低濃度でのより短いアシル鎖長の使用は、溶液中にタンパク質-脂質複合体を産生する。
【発明を実施するための最良の形態】
【0011】
AHF、ファクターVIII およびFVIIIという語は、同じ分子を意味するために互換的に使用される。
本発明は、タンパク質の抗原性および免疫原性を低減するための方法を提供する。本出願を通して「タンパク質」という語が使用されるが、これはポリペプチド(一般的にはアミノ酸が50より多いものを指す)のみならずペプチド(一般的にはアミノ酸50以下を指す)を含む意味で用いられる。
本発明の方法は、リン脂質(好ましくはセリンを含むリン脂質)と一以上のタンパク質、ポリペプチドまたはペプチドの複合体を形成するステップを含む。様々な種類のタンパク質-脂質構造が、特定のリン脂質、濃度、リン脂質の組み合わせによって形成されうる。
【0012】
一般に、「リポソーム」という語は、広く球形のもしくは球状の脂質化合物のクラスターまたは凝集物であって、典型的には一以上の同心円性の層、例えば単層、二重層、多層の形態をとるものを意味する。それらは脂質小胞(lipid vesicles)とも呼ばれる。リポソームは、例えばイオン性脂質および/または非イオン性脂質から形成することができる。
「渦巻形」または「渦巻形構造」という語は通常、一般的にスパイラルまたは細管の形で存在する多重膜の脂質小胞を意味する。
「ミセル」という語は、脂質から形成されるコロイド状の存在を意味する。ミセルは単層、二重層、または六方相構造を構成しうる。
【0013】
本発明では、結合剤に応じて、タンパク質/ポリペプチド複合体により形成される構造が異なることが観察された。一般的に、セリン含有剤を使用した場合、単純複合体、ミセル、リポソーム、渦巻形構造および新規な凝縮物相が観察された。他のホスファチジル化合物またはリン脂質を使用した場合(例えばPC、PG、PAもしくはPEまたはPC、PGもしくはPE含有リン脂質)、ミセルまたはリポソームのみが観察された。さらに、PE含有リン脂質だけが、二重層および非二重層(六角形)構造を形成することが観察された。
【0014】
ホスホセリン、ホスファチジルセリン、もしくは短アシル鎖長(すなわち、4以下のアシル鎖炭素原子を有する)のPS含有リン脂質の使用は、脂質分子アセンブリの形成をもたらさなかった。形成された構造は単純複合体である。これらの単純複合体は、主にイオン性結合によって特徴付けられる。中間アシル鎖長(すなわち、5〜12アシル鎖炭素原子)のセリン含有リン脂質の使用は、その臨界ミセル濃度以上ではミセルを形成し、臨界ミセル濃度未満では単なる複合体(単純複合体)を形成する。長アシル鎖長(12〜18炭素原子)のリン脂質は、その分子構造に起因して、リポソーム、非二重層構造および渦巻形構造などの複数の分子アセンブリを形成する傾向がある。
【0015】
PC、PG、PAおよびPEなどの他のホスファチジル化合物およびPC、PG、PAおよびPE含有リン脂質の使用は、中間長のアシル鎖炭素原子ではミセルの形成をもたらし、より長いアシル鎖炭素原子ではリポソームの形成をもたらす。
【0016】
全てのリン脂質において、アシル鎖は、ジアシル鎖または単アシル鎖であってもよい。ホスファチジルセリン、ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルグリセロールという語は、化合物中にアシル鎖を持たないものを意味する。
【0017】
脂質組成物は、リポソーム、非二重層構造および渦巻型相を調製するために変更することができる。高PC含有量を有する、ホスファチジルコリン(PC):ホスファチジルセリン(PS)もしくはホスホセリンの脂質組成物は、Ca2+とNa+を含む緩衝液を用いた水和によりリポソームを形成するだろう。ホスファチジルエタノールアミンPEおよびPSの存在は、非二重層構造の形成を促進する。Na+を含まないか低濃度(100mM)で含むCa2+の存在下では、PS(90mol%以上)を伴う処方は、渦巻形シリンダーの形成を促進する。温度、Ca2+/Na+などの条件は、タンパク質-脂質複合体を含むナノ粒子における渦巻形シリンダーのサイズを減じるために変更できる。
【0018】
本発明にとって有用なリン脂質には、セリン含有化合物を含むリン脂質が含まれる。
このようなセリン含有リン脂質の例として、O-ホスホ-L-セリン(OPLS)、ジカプロイルホスファチジルセリン、ジオクタノイルホスファチジルセリン、ジミリストイル、ジパルミトイル、ジオレオイル-,ジステロイル-ホスファチジルセリンが挙げられる。セリン含有リン脂質は、他の分子を含む他のリン脂質と併用してもよい。例えば、ホスファチジルセリンを、ホスファチジルコリンまたはホスファチジルエタノールアミンと併用してもよい。ある実施形態では、PS、PC、PAおよびPEはアシル化またはジアシル化される。ホスファチジルセリンは、天然の(脳)あるいは合成起源などの何らかの原料から得ることができる。ホスファチジルセリンは、ホスファチジルコリン(PC)と併用されてもよい。ホスファチジルコリンはジミリストイルホスファチジルコリンであってもよい。PSとPCの比は、1:9〜9:1で変化しうる。一実施形態では、その比は3:7である。
【0019】
本発明のタンパク質-脂質組成物は、好ましくは適切な緩衝系中で安定化され保管される。このような緩衝剤には、TRISバッファー並びにHEPESバッファー及びナトリウム並びにカルシウム塩が含まれる。任意にアルコール(例えば10%エタノール)を添加してもよい。
【0020】
本発明のタンパク質-脂質複合体は、標準的な方法によって特徴づけられることができる。例えば、励起および発光スリットとして4nmを使用してSLM AMINCO 8000シリーズ装置またはPTI 380装置で蛍光研究を行うことができる。サンプルは280nmで発光させることができ、その発光スペクトルは300〜400nmの範囲でスキャンされる。遊離ファクターVIIIの発光スペクトルは、335nm付近で観察され、OPLSの添加は蛍光発光の強度を減じ、タンパク質の三次構造がわずかに変化したことを示した。
【0021】
さらに、前記の脂質結合タンパク質の粒子サイズは、標準的な粒子サイズ測定器(例えばNICOMP 315モデル)を使用して測定することができる。粒度分布は、単峰性および二峰性分布のためのガウス分布とNICOMP分析の両方を使用して分析することができる。ラテックスビーズのサイズは、各測定でスタンダード・コントロールとして使用できる。
【0022】
脂質の構造は、ネガティブ染色電子顕微鏡法によって分析することも可能である。このような方法は当該技術において一般的であり、凝集物がないことを確認するため、およびその構造をリポソーム、非二重層あるいは渦巻形に分類するために使用することができる。非二重層および渦巻形構造の形成は、Laurdan蛍光を使用して研究することもできる。前記脂質構造は、脂質含有溶液とプローブの水溶液(0.01%エタノール含有)を混合することによって、プローブで標識することもできる。そのサンプルは340nmで励起することができ、その発光スペクトルは440nmでモニターされる。励起スペクトルは440nmの発光モノクロメータを用いて、320および420nmの範囲で得ることができる。
【0023】
本発明のタンパク質-脂質複合体は、筋肉内、鼻腔内、腹腔内、静脈内、経口、直腸、皮下、局所投与などの標準的な投与手段によって、個体(例えばヒトを含む動物)に送達することができる。前記複合体は、標的部位に直接もしくはその付近に送達されてもよく、または循環中に直接もしくは間接的に送達されてもよい。前記複合体は、当該技術分野で周知の薬学的に許容されるキャリアーを用いて送達されてもよい。
【0024】
本発明のタンパク質-脂質複合体は、減弱された抗原性のみならず減弱された免疫原性をも示す。したがって、このような組成物は、個体において治療薬に対する免疫応答を減じるために使用することができる。本発明の組成物は、そのタンパク質への免疫反応がすでに発生した個体に治療薬を送達するために使用することもできる。したがって、これらの組成物は免疫反応の発生前あるいは後に使用することができる。
【0025】
ある実施形態では、本発明は特異的FVIII-脂質複合体を提供する。前記タンパク質-脂質複合体は、リポソーム、渦巻形、ミセルおよび非二重層構造などの構造だけでなく新規な脂質構造を形成し、免疫応答および抗原性を減弱しうる。その方法は、バッファーコンディションによって適切に安定化された特異的FVIII-脂質複合体を構築することを含む。特定の理論に制約されることを意図しないが、この抗原性および免疫原性の低下は、タンパク質-脂質複合体に起因し、リポソームなどの分子アセンブリにおいて、疎水性シールドおよび粒状物質の細胞(抗原提示細胞 APCs)内取り込みなどの担体特性を含むと信じられる。これらの複合体は、抗体の存在下で凝血活性を有し、ELISAアッセイにおいてモノクローナル抗体への結合能力を測定した際に減弱した抗原性を示した。前記複合体はまた動物モデルにおいて減弱した免疫応答を示した。本発明は、いまだFVIIIで治療されていない患者における免疫応答の発達を減じるだけでなく、すでに治療され抗体が構築された患者におけるFVIII凝血活性を保持するのにも役立つ。
【0026】
したがって、一実施形態では、本発明はFVIIIに対する免疫応答を減じる方法を提供する。ここに開示した組成物の使用によって、免疫原性は低減されるが、凝血活性は維持される。免疫原性の低下はリン脂質とC2およびA2ドメインの複合体形成によって達成されると考えられる。これは、(1)凝集物形成を低下させる、(2)投与回数を減少させる(複合体がクリアランス機構を変え、これにより循環時間がより長くなる)および/または (3)エピトープ領域の立体構造を遮蔽および変えるおよび/または(4)優先的な細胞取り込みを含む担体特性、によって免疫原性を低下させると考えられる。安定性、循環時間を増加させるクリアランス機構の変化、および抗原性並びに免疫原性の低下などの、複合体の改善された薬学的性質は、予想外の観察結果である。
【0027】
以下の実施例は本発明をより詳細に説明するために存在し、なんら制限を意図しない。
【実施例1】
【0028】
本実施例は、本発明にかかるタンパク質-脂質複合体の調製を説明する。この実施形態を説明するため、FVIIIが使用された。25mMのTRIS、300mMのNaClおよび5mMのCaCl2中で20ugの前記タンパク質と5および20mMのOPLSを混合することによって、FVIII-O-ホスホ-L-セリン(OPLS)複合体を形成した。
【実施例2】
【0029】
本実施例は、本発明のタンパク質-脂質複合体(実施例1)の安定性について説明するものである。遊離FVIIIにとって前記タンパク質のアンフォールディングはタンパク質の凝集をもたらし、これは次にアンフォールディングの不可逆性を引き起こす。凝集はC2ドメイン中のわずかな立体構造変化によって開始される。タンパク質、およびFVIIIのC2ドメインに結合すると信じられるO-ホスホ-L-セリンを含有する製剤の安定性を測定するために、アンフォールディング/リフォールディングの調査を、遊離FVIIIおよび実施例1に記載したPSと複合体を形成したFVIIIを用いて行った。
【0030】
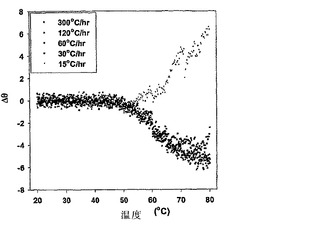
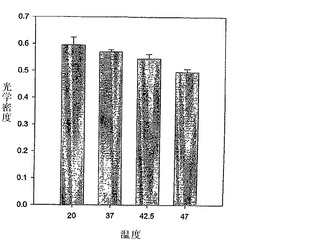
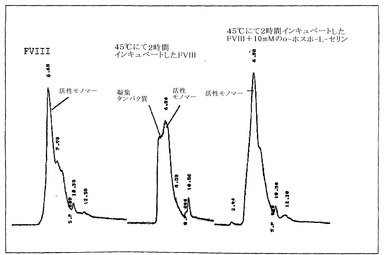
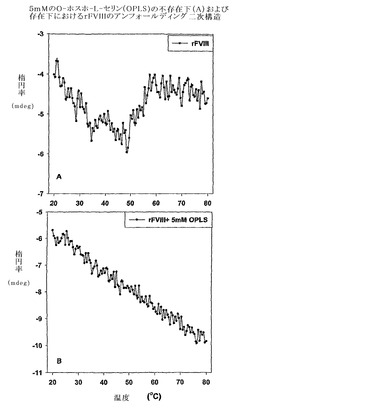
組換えヒトFVIII(rFVIII)の温度依存性の物理的および機能的変化を研究するために、円偏光二色性(CD)、蛍光偏光測定法、サイズ排除クロマトグラフィー(SEC)、ドメイン特異的抗体結合、および凝血活性の調査を行った。フォールディングおよびアンフォールディングを測定するために、CDスペクトルの調査を行った。CDスペクトルはd10ショウノウスルホン酸で較正したJASCO-715分光偏光計を用いて得た。サンプルは二次構造分析のために255〜208nmの範囲で調査され、そして典型的には、使用したタンパク質濃度は20〜22μg/mlであった。タンパク質のCDスペクトルは、バッファー・ベースラインのスペクトルを差し引くことによって補正した。シグナルのクオリティを高めるために複数の調査を行い、平均化した。タンパク質の融解は、2.5℃ごとに2分の保持時間で20〜80℃の温度範囲にわたって行った。温度調査は、Peltier 300RTSユニットを用いて行い、その分析結果は、製造業者によって提供されるソフトウェアを用いて作成した。アンフォールディング特性の加熱速度依存は、キネティック・コントロール下でタンパク質の熱変性が少なくともある程度存在することを示した(図1)。フォールディングモデルは、ファクターVIIIの凝集キネティクスを説明するために提唱された。このモデルに基づき、ファクターVIIIのアンフォールディングは、単なる二状態のキネティックモデル[凝集性(A)→未変性(N)、ここでkはアレニウス式によって与えられるように、温度に伴って変化する一次速度定数である]において解釈される。上記遷移に関する活性化エネルギーは、〜127.98 Kcal/Mole(〜534.97 KJ/Mole)と算出された。抗体結合の研究は、C2ドメインの脂質-結合領域(2303-2332)中の立体構造変化が、少なくともある程度、凝集の開始に関与しうることを示した(図2)。OPLSの存在下および不存在下におけるFVIIIのSEC特性の分析は、単量体の集団がPSの存在下の凝集性タンパク質のものより有意に高いことを明確に示し、これはファクターVIIIの凝集キネティクスにおけるOPLSの干渉が原因かもしれない(図3)。前記データは、複合体がFVIIIの安定性を改善し、凝集物を減らすことによって免疫原性を減じるのに役立ちうることを示す。
【0031】
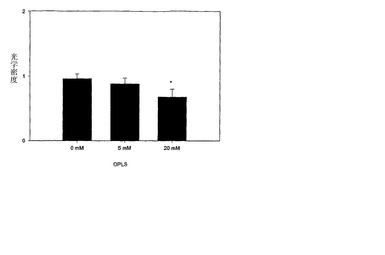
本実施例のさらなる実例では、O-ホスホ-L-セリンは前記タンパク質と複合体を形成する(C2ドメインを遮蔽すると思われる)。20ug/mlのタンパク質と5および20mMのホスホ-L-セリンとを、25mMのTRIS、300mMのNaClおよび5mMのCaCl2を含む別のバッファー中で混合した。モノクローナル抗体への結合は、サンドイッチELISAによって調査した。図4に示すように、添加無しのタンパク質では凝集が原因で未変性様構造の回復は不可能であり、一方、O-ホスホ-L-セリンの存在は未変性構造の実質的な回復をもたらした。
【実施例3】
【0032】
本実施例は、本発明のタンパク質-脂質組成物(実施例1)が、Sprague-Dawleyラットにおいて、前記タンパク質に対し免疫原性を減弱することを実証する。この実施形態を説明するために、OPLS-ファクターVIII複合体を、Sprague-Dawleyラットに投与した。このラットモデルは、FVIIIへの抗体開発を研究するのに好適であることが実証されている。ELISAによって、遊離FVIIIおよびFVIII-PS複合体に対する抗体価を測定した。投与から2週間、ファクターVIII-OPLS複合体に対する抗体価の分析が非免疫毒性であることが見い出された。遊離FVIIIに対する抗体価は563.72±916.15であり、FVIII-OPLS複合体に対する検出可能な抗体価は観察されなかった。
【実施例4】
【0033】
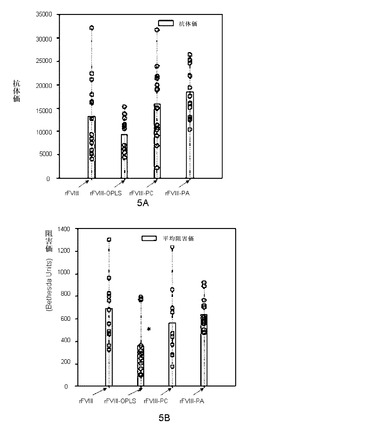
本実施例は、ホスホセリンが、免疫原性を減少させる単純複合体において必須であることを実証する。本実施例を説明するために、ホスホセリン、ホスホコリンまたはホスファチジン酸を用いて単純複合体を調製した。免疫原性は、血友病Aマウスにおいて複合体の皮下投与によってテストした。総抗体価(ELISAによって測定)または特異的阻害力価(Bethesda Units)を図5Aおよび5Bに示す。効果はホスホセリンでは統計的に有意であったが、ホスホコリンまたはホスファチジン酸では統計的に有意ではなかった。
【実施例5】
【0034】
本実施例は、本発明の組成物を小さな単層ベシクル(vesicles)として作ることができることを実証する。0.3mg/mlのDMPCおよび0.15mg/mlのbPSを丸底フラスコ中で溶解し、溶媒を蒸発させて薄膜を形成した。その後この薄膜を水和してMLV’sを形成し、そしてこのMLVsを200nmのポリカーボネートフィルターを通して押し出し、160nmのサイズ範囲内のSUV’sを形成した。この製剤(ファクターVIIIを含む)の免疫応答を実施例5で説明する。
【0035】
全ての上述した研究において、PS無しの状態を使用して行った実験、および脂質と結び付いたタンパク質の濃度並びに脂質構造は、これら全ての脂質製剤において特異的な脂質-FVIII複合体の存在を示唆している。さらに、C2ドメインに特異的なモノクローナル抗体が、セリンの存在下でFVIIIに結合しなかったことは、調査した脂質構造における複合体の存在を明確に示唆する。
【実施例6】
【0036】
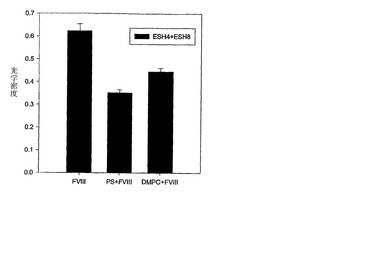
本実施例は、抗原性および、複合体の形成におけるタンパク質の特定のエピトープ領域の関与を研究するためのELISA分析の一般的な方法論を実証する。タンパク質-脂質複合体の抗体結合は、抗体捕獲ELISAとサンドイッチ ELISAを用いて調べた。サンドイッチELISAでは、炭酸緩衝液(0.2M、pH9.4)中5μg/mlの濃度にて50μl/ウェルの抗体溶液を4℃で一晩インキュベートすることによって、96ウェルプレート(Nunc-Maxisorb)を抗-C2ドメイン抗体(ESH4)でコートした。前記プレートをその後、0.05%Tween20(10mM Na2HPO4,1.8mM KH2PO4,0.14mM NaCl,2.7mM KCl,および0.02% NaN3からなるPBT)を含む100μlのリン酸緩衝液で10回洗浄した。プラスチックの吸着性表面上の残りの非特異的タンパク質結合サイトを、リン酸緩衝液(10mM Na2HPO4,1.8mM KH2PO4,0.14mM NaCl,および2.7mM KClからなるPB)中の1%ウシ血清アルブミンからなるブロッキングバッファー200μlを室温で2時間インキュベートすることによってブロックした。プレートをPBTで10回洗浄し、ブロッキングバッファー中の100ng/mlのrFVIIIもしくはrFVIII/OPLS(実施例1および3)またはリポソーム付随rFVIII(実施例5)50μlを添加して37℃で1時間インキュベートした。プレートをPBTで10回洗浄し、そして、50μlのビオチン化ESH8−別の抗-C2抗体を用いて、1μg/ml濃度および50μlの1:1000希釈のアビジン-アルカリホスファターゼ複合物にて、どちらもブロッキングバッファー中、室温にて1時間インキュベートした。プレートをPBTおよびジエタノールアミン緩衝液(1Mジエタノールアミン,0.5mM MgCl2および0.02% NaN3からなる)中の1mg/ml p-ニトロフェニルリン酸塩溶液100μlで10回洗浄した。プレートを室温で30分間インキュベートし、3NのNaOHを100μl添加することによって反応をクエンチした。吸光度を405nmでプレートリーダーを用いて読み取った。このELISA研究は、より少ないC2ドメイン特異的抗体、ESH8またはESH4がPS(OPLSまたはリポソーム 図6)の存在下でタンパク質に結合することを示した。その結果は、これらのモノクローナル抗体の結合が脂質によって阻害されることを示し、エピトープ領域がタンパク質-脂質複合体中に遮蔽されることを示唆する。さらに、前記特異的複合体FVIII-PSが、前記エピトープ遮蔽の原因であることが、さらに対照実験によって確認された。この際、抗原性はDMPC/PS混合物を用いたときのみ観察され、DMPC単独では観察されなかった(図7)。
【実施例7】
【0037】
本実施例は、本発明のタンパク質-脂質組成物(実施例5)が、動物モデル、Sprague-DawleyラットおよびファクターVIIIノックアウトマウス,血友病Aモデルにおいて、前記タンパク質に対し免疫原性を減じることを実証する。
【0038】
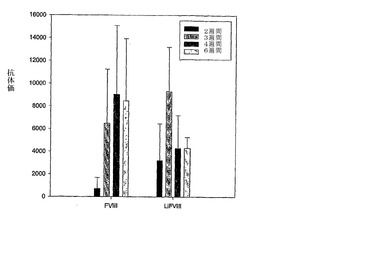
タンパク質-リポソーム複合体の投与後4および6週間目にSprague-Dawleyラットで評価された抗体価は、遊離FVIIIに比べてFVIII-PS複合体でより低いことが見いだされた(図8A)。
【0039】
本実施形態の別の実例では、免疫原性に対する本発明のタンパク質-脂質複合体の効果を、ファクターVIIIノックアウトマウスモデルでテストした。この表現型のマウスは重度の血友病である(ネオ・カセットを用いる標的破壊によるエクソン16ノックアウト)。前記タンパク質-リポソーム複合体(DMPC+bPS)は皮下に投与し、その抗体価はELISA分析を用いて測定した。図8Bから明らかなように、抗体の濃度は投与後6週間、リポソーム付随タンパク質でより低かった。このデータは明確に、リポソーム結合ファクターVIIIが遊離ファクターVIIIに比べてより少ない抗体を誘発することを実証する。特定の理論に拘束されることを意図しないが、この観察結果は、一部、変化した(1)遊離FVIIIの立体構造および凝集キネティクス、(2)粒状物質のデコイ効果および免疫系によるプロセシングおよび (3)クリアランス機構、に起因するかもしれない。
【実施例8】
【0040】
以下の実施例は、タンパク質-脂質複合体のミセル、非二重層構造、渦巻形構造、および新規な非リポソーム脂質粒子の形成を実証する。本実施例は、ここに記載された組成物によるミセルの形成を説明する。タンパク質溶液を、低濃度および高濃度(臨界ミセル濃度の上下)の短鎖アシル鎖脂質(ジヘキサノイル・ホスファチジルセリン[0.3mM前後])と混合し、その結果生じたミセル粒子を特性化した。ミセルの構造は、光散乱、円二色性および蛍光実験によって特性化した。活性や抗体結合などの機能分析を行った。
【実施例9】
【0041】
本実施例は、ここに記載された組成物による渦巻形構造の形成について記述する。0.15mg/mlのbPSをクロロホルム中に溶解し、溶媒を蒸発させて薄い脂質膜を形成した。それからこのフィルムをpH7.0の複数の緩衝系中で水和させ、前記MLVsを押し出すか超音波処理するかのいずれかによりSUVsを形成した。この結果生じたSUVsを、5mMのCaCl2を含有する緩衝系中でタンパク質と混合し、渦巻形構造を形成した。その脂質構造を、光散乱、微分干渉顕微鏡法、ネガティブ染色電子顕微鏡法によって及び蛍光研究によって分析した。これらの研究により、この手順によって形成された脂質構造は事実上渦巻形であることが分かった。
【実施例10】
【0042】
本実施例は、本発明のタンパク質-脂質複合体調製の別の実例である。0.15mg/mlのbPSおよび0.3mg/mlのジオレオイル・ホスファチジルエタノールアミン(DOPE)をクロロホルム中に溶解し、溶媒を蒸発して薄い脂質膜を形成した。結果物である脂質膜をリン酸緩衝生理食塩水を用いて水和し、六方相を形成した。前記非二重層構造は蛍光研究によって特性化した。
【実施例11】
【0043】
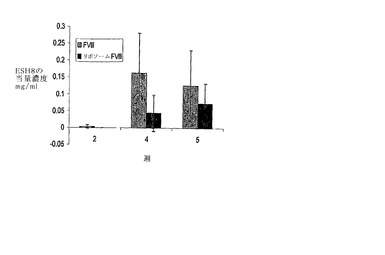
本実施例は、新規な非リポソーム構造をもつ、本発明のタンパク質-脂質複合体の調製の別の実例である。0.3mg/mlのDMPC、0.15mg/mlのbPSをクロロホルム中に溶解し、溶媒を蒸発させ、丸底フラスコ中で薄い脂質膜を形成した。この脂質を、FVIII、25mM TRIS、300mM NaClおよび5mM CaCl2を含有する緩衝系を用いて水和させ、この溶液を室温あるいは37℃のいずれかにて穏やかに回転した。その後前記フィルムを、穏やかに回旋させながら適切な緩衝液(25mM TRIS、300mM NaClおよび5mM CaCl2)中で水和させた。このようにして形成したMLV’sを、MLV’sと結びついたタンパク質から遊離タンパク質を分離するために、デキストラン遠心分離勾配にかけた。0.5mlの脂質付随タンパク質を1mlの20%w/vデキストランと混合し、そして3mlの10%w/vデキストランを上記溶液上に層状に重ねた。その後0.5mlの緩衝液をトップに層状に重ねた。前記勾配を、ベックマン社のSW50.1ローターを用いて35分間、45K PRMで遠心分離した。遠心分離研究の結果を図9に示す。図から明らかなように、浮かなかった脂質分画がある(図中では分画3として表示する)。この分画は、14%と10%のデキストランの界面で観察された。この分画が浮かなかったという事実は、この脂質分画が十分な浮力あるいは被包された水を持たないことを示す。従来のリポソームは一般的に封入された水を有するため勾配の最高部に浮く。それゆえ、浮かない分画は、脂質粒子を含有する非リポソームタンパク質の可能性がある。分画を採取して、質量分析で脂質含量を、活量でタンパク質含量をテストした。質量分析研究により、この分画が、単なるPS-Ca+複合体ではないことを示唆する高PC含量の脂質を含むことが明らかになった。活量分析は、この分画内に最初のタンパク質が約40%封入されていることを示した。
【0044】
含水量が無いかわずかである高密度な分画3に関し、いくつかの考えられる解釈が存在する。この分画は以下のことを意味するかもしれない:(1)わずかな封入水量を有する非常に小さな単層ベシクル ごく小さなベシクル(200nm未満)が同一のコンディション下で浮くことができるかどうかを確認するために、事前にサイズを揃えたリポソームを、ポリカーボネートフィルターを通して押し出すことによって調製した。前記押し出しを3回繰り返し、粒子のサイズを約160nmとした。結果物であるSUVsをFVIIIと混合し、デキストラン遠心分離勾配を適用した。この対照研究は、これらのSUV’sが分画3のバンドを示さないことを明らかにし、これにより、このようなバンドの観察が小さなリポソームの形成に起因しないことが示された。さらにこの実験を、デキストラン勾配における人為的な影響を排除するために、同一の緩衝液および実験条件の下で行った。(2)第二の可能性は、分画3が、含水量がわずかな、渦巻形構造の形成を意味するかもしれないということである[26]。しかしながら、渦巻形構造の形成は、非常に高いPS含量(>50%)を必要とする。分画3バンドの形成において、使われるPS含量は約30%であり、これらの条件の下で渦巻形構造の形成は見られなかった。(3)脱水構造[27]を有する崩壊したCa(PS)2複合体の形成は、浮かない可能性があり、高密度なバンドを形成しうる。このような崩壊した構造の形成には、非常に高いPS含量(>50%)と、Na+が存在しないことが必要である。しかしながら、本実施例では、組成物は低いPS含量と、非常に高い濃度のNa+を有する、言い換えると、30%のPSおよび300mMのNaClが使用されており、それゆえ、Ca(PS)2 形成の可能性は除外される。(4)PSおよびカルシウム系は、ベシクル融合を促進することが示されている。しかしながら、Ca2+などの二価カチオンによるベシクルの融合は、脂質結合サイトでカルシウムと競合するNa+の存在によって阻害される。このPC/PS比7:3における、および300mMのNaClと5mMのCaCl2の存在下におけるPSあたりのカルシウム結合の推定量は、0.22(500mM NaCl)〜0.35(100mM NaCl)の間である[28, 29]。PSあたりの推定結合カルシウムは、小さな単層PS/PCベシクル系において融合に必要とされる臨界比0.35〜0.39より小さい。したがって、ベシクルの融合が、高密度なバンドを示すとは考えられない。これは、より大きなPC分画(>50%)は結果的により少ないPS-二価カチオン複合体をもたらすかもしれないため、および、大きなドメインへクラスター形成して融合を誘導するその能力のためである[28,29]。これらの理由に基き、高密度なバンドは、新規な非リポソーム脂質粒子の形成に起因する可能性が高いと思われる。このバンドは、封入水量がわずかであるため、非リポソームである。この新規な脂質構造の構造を理解するため、ネガティブ染色電子顕微鏡法、微分干渉コントラスト光学顕微鏡法、光散乱、円二色性および蛍光測定(データは示さず)を行った。
【0045】
複数のコンディション下で行われた遠心分離の研究は、緩衝剤条件、バルクタンパク質濃度、アルコールの使用(例えば10%エタノール)およびタンパク質-脂質比を、所望の収量の複合体を得るために変化させることができることを示す。これらのパラメーターにおけるこのようなバリエーションは、当業者の範囲内であると見なされる。視覚的な検査で確認できるように、一般に、より高いタンパク質濃度はより高密度の分画をもたらすことが確認された。
【実施例12】
【0046】
本実施例は、本発明のタンパク質-脂質組成物が生物活性を保持することを実証する。この実施形態を実証するために、ファクターVIIIを含むタンパク質-脂質複合体(OLPS,DCPS,およびDCPC)の凝固に対する効果をテストした。rFVIII凝血活性は、活性化剤として微粉化シリカおよび基質としてFVIII欠損プラズマを用いた一段階活性化部分トロンボプラスチン時間(APTT)分析によって測定した。APTT分析は、COAG-A-MATEモデルの凝固分析器(Organon Teknika社、ダラム,NC)を用いて行った。手短に言えば、rFVIIIをFVIII欠損プラズマに添加し、凝固時間をモニターした。その後、既知の活性を持つ凍結乾燥参考濃縮物の様々な希釈物から測定された凝固時間を利用して作成した較正曲線から、rFVIIIの活性を求めた。タンパク質の濃度は、ビシンコニン酸(BCA)分析を用いて独立的に測定して、活性と比較した。例えば、タンパク質の20〜22μg/mlは全て87〜95.6IUの比活性に対応する。サンプルを調製するために使用された原液は、2174IU/0.5mg/mlの比活性を有していた。
【実施例13】
【0047】
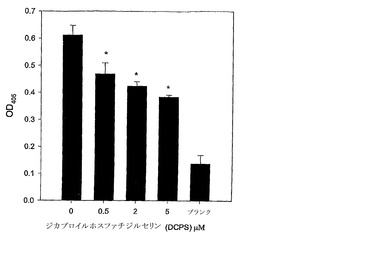
本実施例は、PS含有リン脂質の存在下におけるタンパク質の減弱した抗原性を実証する。本実施形態を説明するために、ESH4抗体へのrFVIIIの結合に対するジカプロイルホスファチジルセリン(DCPS)の効果をテストした。2ugのタンパク質と0.5,2および5mMのDCPSを、25mM TRIS、300mM NaClおよび5mM CaCl2中で混合することによって、FVIII-(DCPS)複合体を形成した。この複合体の抗原性をELISAを用いて研究した。図10に示すように、試験した濃度(すなわち、0.5,2および5.0uM)にて、DCPSはESH4抗体へのrFVIIIの結合を阻害した。これらの濃度はこのリン脂質のCMCの上および下である。
【実施例14】
【0048】
本実施例は、タンパク質の免疫原性に対するPS含有リン脂質の効果を説明する。2ugのタンパク質と5mMのDCPSを、25mM TRIS、300mM NaClおよび1mM CaCl2中で混合することによって、FVIII-(DCPS)複合体を形成した。この組成物はミセル構造を形成し、複合体の免疫原性は血友病Aマウスにおいて研究した。総抗体価は遊離FVIIIでは13,167(SD=7909,n=16)およびFVIII-DCPS複合体では3506(SD=1150.7,n=12)である。
【実施例15】
【0049】
2ugのタンパク質と20mMのDCPCを、25mM TRIS、300mM NaClおよび1mM CaCl2中で混合することによって、FVIII-ジカプロイル・ホスファチジルコリン(DCPC)複合体を形成した。この組成物はミセル構造を形成し、複合体の免疫原性は血友病Aマウスにおいて研究した。総抗体価は遊離FVIIIでは13,167(SD=7909,n=16)およびFVIII-DCPC複合体では1293(SD=946.57,n=12)である。
【実施例16】
【0050】
本実施例は、本発明の組成物をリポソーム小胞として作ることができることを実証する。クロロホルム中に溶解した11.25μmolのDMPCと4.83μmolのBPSを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発しフラスコ壁上に薄膜を形成した。その後この薄膜を、3mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)で水和させ、MLV’sを形成し、このMLVsを80nmもしくは200nmもしくは400nmのいずれかのポリカーボネートフィルターを通して押し出し、80nm〜400nmのサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。その後このリポソームを以下のように免疫化で使用した。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。
【0051】
別のサンプルでは、クロロホルム中に溶解した11.25μmolのDMPCを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発し、フラスコ壁上に薄膜を形成した。その後この薄膜を、3mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)で水和させ、MLV’sを形成し、このMLVsを80nmもしくは200nmもしくは400nmのいずれかのポリカーボネートフィルターを通して押し出し、80nm〜400nmのサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。高い相転移温度を持つDSPC(固体状態)はネガティブ・コントロールに使用された。
【0052】
別のサンプルでは、クロロホルム中に溶解した7.5μmolのDMPCと3.22μmolのDOPAを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発し、フラスコ壁上に薄膜を形成した。その後この薄膜を、2mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)で水和させ、MLV’sを形成し、このMLVsを200nmのポリカーボネートフィルターを通して押し出し、200nm以下のサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。
【0053】
別のサンプルでは、クロロホルム中に溶解した7.5μmolのDMPCと3.22μmolのDOPGを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発し、フラスコ壁上に薄膜を形成した。その後この薄膜を、2mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)中で水和させ、MLV’sを形成し、このMLVsを200nmのポリカーボネートフィルターを通して押し出し、200nm以下のサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。この製剤の免疫応答を表1に示す。
【0054】
【表1】

【0055】
平均総抗-rFVIII抗体価および阻害価は、リポソーム不存在下および存在下でのrFVIIIを用いたFVIII-KOマウスの免疫化に続く6週間後の値である。*はリポソームを押し出すのに使用したポリカーボネート膜の孔サイズを示し、**P<0.05,統計分析は、実験手順において記載したようにして行った。
【0056】
本発明をここに記載した実施例を用いて説明したが、本発明の精神を逸脱することなく本発明について一般的な修正を行うことができる。このような修正は本願の特許請求の範囲内に含まれる。
【図面の簡単な説明】
【0057】
【図1】図1は、様々な加熱速度でのFVIIIの融解特性を示す。
【図2】図2は、C2ドメインにおける立体構造変化を示す抗体結合アッセイを示す。モノクローナル抗体ESH4およびESH8のrFVIIIへの結合は、60℃および15℃/hrでrFVIIIを表示の温度まで加熱した後サンドイッチERISAによって測定した。
【図3】図3は、O-ホスホ-L-セリンの存在下または不存在下でのファクターVIIIのサイズ排除クロマトグラフィー(SEC)特性を示す。
【図4】図4は、サンドイッチELISAによって研究されたFVIII-O-ホスホ-L-セリンの抗原性を示す。
【図5】図5Aおよび5Bは、rFVIIIの免疫原性に対するOPLS、ホスホコリンおよびホスファチジン酸の影響を示す。平均総抗体価(5A)および阻害価(5B)は、rFVIIIと比較した表記の結合剤-rFVIII複合体を示す。
【図6】図6は、O-ホスホ-L-セリン中のFVIIIのフォールディング研究を示す。
【図7】図7は、サンドイッチELISAによって研究されたリポソーム中のFVIII-PS複合体の抗原性を示す。FVIII:遊離FVIII、本発明のFVIII:本発明で使用される組成物、およびDMPC+FVIII:DMPCリポソームおよびFVIII欠損特異的タンパク質(FVIII)脂質(PS)複合体の物理的混合物。
【図8A】図8Aは、遊離FVIIIおよびFVIII-PS複合体に対する動物モデルにおける免疫応答を示す。FVIII:遊離FVIII、本発明のFVIII:本発明で使用される組成物
【図8B】図8Bは、ファクターVIII欠損血友病AマウスモデルにおけるFVIIIとFVIII-PS複合体リポソームを示す。
【図9】図9は、FVIII-PS複合体を含む、非リポソームで低水分量フラクションを示すデキストラン密度勾配の写真である。
【図10】図10は、rFVIIIの抗原性に対するDCPSの影響を示す。
【技術分野】
【0001】
本発明は、低い免疫原性を有するタンパク質複合体およびこれを作るための方法に関する。
【背景技術】
【0002】
血友病は、ファクターVIII(抗血友病ファクター,AHFあるいはFVIII)の欠損によってもたらされる出血性疾患である。FVIIIは6つのドメインA1,A2,A3,B,C1およびC2からなるマルチドメインタンパク質であり、トロンビンによるこのタンパク質の活性化は、重鎖(A1およびA2)と軽鎖(A3,C1およびC2)内に生じる[1,2]。血液濃縮物、組換えファクターVIIIおよびファクターVIIIの変異体を用いた補充療法が、血友病のための一次治療である。しかしながら、15〜35%の患者では中和抗体が発現し、 このような免疫応答は血友病の治療を損なう。中和抗体の発現を克服するための最新の治療計画が存在するが、コスト効率が良くない。免疫原性のより少ないファクターVIII製剤の開発は、代替的なクリニカル・アプローチを提供しうる。
【0003】
一般に、治療用タンパク質への免疫応答は、以下の理由が原因とされている;(i)投与経路、(ii)凝集物の存在、(iii)頻回投与、および(iv)特定のエピトープ領域[3]。最近、FVIIIが凝集物を形成する傾向をもつことが明らかになったが[4]、免疫応答の進行におけるこれらの凝集物の役割はよく理解されていない。さらに、FVIII分子上には主に2つのエピトープ領域、C2およびA2ドメインが存在する。スカンデラら[5]は、抗体価が軽鎖のC2ドメインに対して最も高いことを明らかにした。C2ドメインは膜結合ドメインでもあり、それはその凝固カスケードの一環として血小板膜上のホスファチジルセリン(PS)に結合する[6,7]。C2ドメインに対する抗体の抗凝固作用は、リン脂質へのファクターVIIIの結合阻害に起因する。C2ドメインに対するモノクローナル抗体が、リン脂質含有膜へのファクターVIIIの結合を妨げることが明らかにされ、そしてこれらの観察に基づいて、エピトープと膜結合領域が重複することが結論づけられた[10,11]。
【0004】
FVIIIのリポソーム封入に関する従来の研究は、従来のリポソーム内へのFVIII(VIIIファクター)の封入効率を増加することを目的とし、さらに生体内安定性および経口バイオアベイラビリティの増加を目的とする[12,13]。その分子構造に起因して、脂質分子は、リポソーム、ミセル、非二重層構造および渦巻形構造などの複数の分子アセンブリを形成する傾向がある。これらの分子アセンブリを、タンパク質およびペプチドを含むいくつかの薬物の治療特性を改良するための薬物送達媒体として使用するため、多数の研究が行われてきた。これらの治療の特徴は、循環時間の延長、毒性の低減、免疫応答の亢進および生体内分解の減少である[14-25]。したがって、リポソームは免疫応答を増加させるためのアジュバント(adjuvants)として使用されてきた[26-31]。
【0005】
【非特許文献1】Fay, P.J. Factor VIII structure and function. Thromb Haemost 70, 63-7 (1993).
【非特許文献2】Foster, P.A. & Zimmerman, T.S. Factor VIII structure and function. Blood Rev 3, 180-91 (1989).
【非特許文献3】Braun, A., Kwee, L., Labow, M.A. & Alsenz, J. Protein aggregates seem to play a key role among the parameters influencing the antigenicity of interferon alpha (IFN-alpha) in normal and transgenic mice. Pharmaceutical Research 14, 1472-8 (1997).
【非特許文献4】Grillo, A.O. et al. Conformational origin of the aggregation of recombinant human factor VIII. Biochemistry 40, 586-95. (2001).
【非特許文献5】Scandella, D.H. et al. In hemophilia a and autoantibody inhibitor patients: the factor viii a2 domain and light chain are most immunogenic. Thromb Res 101, 377-85. (2001).
【非特許文献6】Lenting, P.J. et al. The light chain of factor VIII comprises a binding site for low density lipoprotein receptor-related protein. Journal of Biological Chemistry. 274, 23734-9 (1999).
【非特許文献7】Saenko, E.L., Yakhyaev, A.V., Mikhailenko, I., Strickland, D.K. & Sarafanov, A.G. Role of the low density lipoprotein-related protein receptor in mediation of factor VIII catabolism. Journal of Biological Chemistry. 274, 37685-92 (1999).
【非特許文献8】Gilbert, G.E., Furie, B.C. & Furie, B. Binding of human factor VIII to phospholipid vesicles. J Biol Chem 265, 815-22 (1990).
【非特許文献9】Stoilova-McPhie, S., Villoutreix, B.O., Mertens, K., Kemball-Cook, G. & Holzenburg, A. 3-Dimensional structure of membrane-bound coagulation factor VIII: modeling of the factor VIII heterodimer within a 3-dimensional density map derived by electron crystallography. Blood 99, 1215-23. (2002).
【非特許文献10】Scandella, D. et al. Some factor VIII inhibitor antibodies recognize a common epitope corresponding to C2 domain amino acids 2248 through 2312, which overlap a phospholipid-binding site. Blood 86, 1811-9. (1995).
【非特許文献11】Barrow, R.T., Healey, J.F., Jacquemin, M.G., Saint-Remy, J.M. & Lollar, P. Antigenicity of putative phospholipid membrane-binding residues in factor VIII. Blood 97, 169-74. (2001).
【非特許文献12】Kirby, C.J. & Gregoriadis, G. Preparation of liposomes containing factor VIII for oral treatment of haemophilia. J Microencapsul 1, 33-45. (1984).
【非特許文献13】Hemker, H.C., Hermens, W.T., Muller, A.D. & Zwaal, R.F. Oral treatment of haemophilia A by gastrointestinal absorption of factor VIII entrapped in liposomes. Lancet 1, 70-1. (1980).
【非特許文献14】Martin, F.J., Hubbell, W.L. & Papahadjopoulos, D. Immunospecific targeting of liposomes to cells: a novel and efficient method for covalent attachment of Fab' fragments via disulfide bonds. Biochemistry 20, 4229-38. (1981).
【非特許文献15】Matthay, K.K., Heath, T.D., Badger, C.C., Bernstein, I.D. & Papahadjopoulos, D. Antibody-directed liposomes: comparison of various ligands for association, endocytosis, and drug delivery. Cancer Res 46, 4904-10. (1986).
【非特許文献16】Olson, F., Hunt, C.A., Szoka, F.C., Vail, W.J. & Papahadjopoulos, D. Preparation of liposomes of defined size distribution by extrusion through polycarbonate membranes. Biochim Biophys Acta 557, 9-23. (1979).
【非特許文献17】Papahadjopoulos, D. et al. Sterically stabilized liposomes: improvements in pharmacokinetics and antitumor therapeutic efficacy. Proc Natl Acad Sci USA 88, 11460-4 (1991).
【非特許文献18】Phillips, N.C. & Emili, A. Enhanced antibody response to liposome-associated protein antigens: preferential stimulation of IgG2a/b production. Vaccine 10, 151-8. (1992).
【非特許文献19】Storm, G., Wilms, H.P. & Crommelin, D.J. Liposomes and biotherapeutics. Biotherapy 3, 25-42 (1991).
【非特許文献20】Szoka, F., Jr. & Papahadjopoulos, D. Procedure for preparation of liposomes with large internal aqueous space and high capture by reverse-phase evaporation. Proc Natl Acad Sci USA 75, 4194-8. (1978).
【非特許文献21】Szoka, F. et al. Preparation of unilamellar liposomes of intermediate size (0.1-0.2 mumol) by a combination of reverse phase evaporation and extrusion through polycarbonate membranes. Biochim Biophys Acta 601, 559-71. (1980).
【非特許文献22】Van Slooten, M.L. et al. Liposomes as sustained release system for human interferon-gamma: biopharmaceutical aspects. Biochim Biophys Acta 1530, 134-45. (2001).
【非特許文献23】Papahadjopoulos, D., Vail, W.J., Jacobson, K. & Poste, G. Cochleate lipid cylinders: formation by fusion of unilamellar lipid vesicles. Biochim Biophys Acta 394, 483-91. (1975).
【非特許文献24】Coorssen, J.R. & Rand, R.P. Structural effects of neutral lipids on divalent cation-induced interactions of phosphatidylserine-containing bilayers. Biophys J 68, 1009-18. (1995).
【非特許文献25】Duzgunes, N. et al. Calcium- and magnesium-induced fusion of mixed phosphatidylserine/phosphatidylcholine vesicles: effect of ion binding. J Membr Biol 59, 115-25. (1981).
【非特許文献26】Duzgunes, N., Wilschut, J., Fraley, R. & Papahadjopoulos, D. Studies on the mechanism of membrane fusion. Role of head-group composition in calcium- and magnesium-induced fusion of mixed phospholipid vesicles. Biochim Biophys Acta 642, 182-95. (1981).
【非特許文献27】Alving, C.R., Liposomes as carriers of antigens and adjuvants. J. Immunol. Methods, 140:1-13, (1991).
【非特許文献28】Van Rooijen, N., Liposomes as carrier and immunoadjuvant of vaccine antigens. Adv. Biotechnolo Processes, 13: 255-279 (1990).
【非特許文献29】Huong, T.M., Ishida, T., Harashima, H. & Kiwada, H. The complement system enhances the clearance of phosphatidylserine (PS)-liposomes in rat and guinea pig. Int J Pharm 215, 197-205. (2001).
【非特許文献30】Dal Monte, P. & Szoka, F.C., Jr. Effect of liposome encapsulation on antigen presentation in vitro. Comparison of presentation by peritoneal macrophages and B cell tumors. J Immunol 142, 1437-43. (1989).
【非特許文献31】Heath, T.D., Edwards, D.C. & Ryman, B.E. The adjuvant properties of liposomes. Biochemical Society Transactions 4, 129-33 (1976).
【非特許文献32】Rosenblum et al., Modified Proteins, designer toxins, and methods of making thereof., U.S. patent application publication 20030176331, 2003.
【発明の開示】
【発明が解決しようとする課題】
【0006】
しかしながら、免疫応答および抗原性を減じるための脂質複合体の開発は研究されておらず、それゆえ、治療用タンパク質の免疫原性を減少させるためのアプローチを開発する必要性が引き続き存在している。免疫毒性を減少させるために特定のエピトープの配列を変更する別のアプローチがある[32]。しかしながら、そのようなアミノ酸置換は生物活性の損失をもたらす可能性がある。
【課題を解決するための手段】
【0007】
本発明は、低い抗原性と免疫原性を有する組成物およびその製造方法を開示する。したがって、タンパク質、ポリペプチドあるいはペプチドのような治療薬、およびその免疫原性と抗原性を減じるように前記タンパク質に結合できる一以上の分子(本明細書中で結合剤と呼ぶ)を含む組成物が開示される。このような結合剤には、ホスホセリン、ホスファチジルセリンもしくはホスファチジルセリン(PS)を含むリン脂質等のセリン化合物;ホスファチジルコリン(PC),ホスファチジン酸(PA),もしくはホスホエタノールアミン(PE);またはPA,PCもしくはPEを含むリン脂質が含まれる。
【0008】
前記のタンパク質-結合剤複合体は、その免疫応答を減じる(1)タンパク質-結合剤複合体を含む液体またはこの液体の凍結乾燥剤、(2)新規な非リポソーム構造、(3)リポソーム、(4)ミセル、(5)渦巻形、(6)非二重層構造の形で存在しうる。
【0009】
本発明はまた、結合剤(セリン含有化合物など)と複合体を形成することによってタンパク質の免疫原性および/または抗原性を減じる方法を開示する。前記タンパク質-結合剤複合体は、適切な緩衝剤を用いて安定化しうる。
【0010】
ある実施形態では、ジミリスチルホスファチジルコリン(DMPC)および脳ホスファチジルセリン(bPS)を含有する乾燥脂質膜は、各種の緩衝系中でタンパク質(FVIIIなど)を用いて水和させられる。新規の、非リポソーム構造物は、300mMのNaClと5mMのCaCl2中のDMPC、bPSを用いて形成される。従来のリポソームは、水に変化する緩衝系あるいはリン酸緩衝生理食塩水として形成される。これはカルシウムあるいはPS濃度を減じることによって達成することもできる。DMPCの除去および100%PSと超音波処理もしくは押出ステップの使用は渦巻形構造を導き、中間アシル鎖長のPSの使用はミセル構造をもたらす。より低濃度でのより短いアシル鎖長の使用は、溶液中にタンパク質-脂質複合体を産生する。
【発明を実施するための最良の形態】
【0011】
AHF、ファクターVIII およびFVIIIという語は、同じ分子を意味するために互換的に使用される。
本発明は、タンパク質の抗原性および免疫原性を低減するための方法を提供する。本出願を通して「タンパク質」という語が使用されるが、これはポリペプチド(一般的にはアミノ酸が50より多いものを指す)のみならずペプチド(一般的にはアミノ酸50以下を指す)を含む意味で用いられる。
本発明の方法は、リン脂質(好ましくはセリンを含むリン脂質)と一以上のタンパク質、ポリペプチドまたはペプチドの複合体を形成するステップを含む。様々な種類のタンパク質-脂質構造が、特定のリン脂質、濃度、リン脂質の組み合わせによって形成されうる。
【0012】
一般に、「リポソーム」という語は、広く球形のもしくは球状の脂質化合物のクラスターまたは凝集物であって、典型的には一以上の同心円性の層、例えば単層、二重層、多層の形態をとるものを意味する。それらは脂質小胞(lipid vesicles)とも呼ばれる。リポソームは、例えばイオン性脂質および/または非イオン性脂質から形成することができる。
「渦巻形」または「渦巻形構造」という語は通常、一般的にスパイラルまたは細管の形で存在する多重膜の脂質小胞を意味する。
「ミセル」という語は、脂質から形成されるコロイド状の存在を意味する。ミセルは単層、二重層、または六方相構造を構成しうる。
【0013】
本発明では、結合剤に応じて、タンパク質/ポリペプチド複合体により形成される構造が異なることが観察された。一般的に、セリン含有剤を使用した場合、単純複合体、ミセル、リポソーム、渦巻形構造および新規な凝縮物相が観察された。他のホスファチジル化合物またはリン脂質を使用した場合(例えばPC、PG、PAもしくはPEまたはPC、PGもしくはPE含有リン脂質)、ミセルまたはリポソームのみが観察された。さらに、PE含有リン脂質だけが、二重層および非二重層(六角形)構造を形成することが観察された。
【0014】
ホスホセリン、ホスファチジルセリン、もしくは短アシル鎖長(すなわち、4以下のアシル鎖炭素原子を有する)のPS含有リン脂質の使用は、脂質分子アセンブリの形成をもたらさなかった。形成された構造は単純複合体である。これらの単純複合体は、主にイオン性結合によって特徴付けられる。中間アシル鎖長(すなわち、5〜12アシル鎖炭素原子)のセリン含有リン脂質の使用は、その臨界ミセル濃度以上ではミセルを形成し、臨界ミセル濃度未満では単なる複合体(単純複合体)を形成する。長アシル鎖長(12〜18炭素原子)のリン脂質は、その分子構造に起因して、リポソーム、非二重層構造および渦巻形構造などの複数の分子アセンブリを形成する傾向がある。
【0015】
PC、PG、PAおよびPEなどの他のホスファチジル化合物およびPC、PG、PAおよびPE含有リン脂質の使用は、中間長のアシル鎖炭素原子ではミセルの形成をもたらし、より長いアシル鎖炭素原子ではリポソームの形成をもたらす。
【0016】
全てのリン脂質において、アシル鎖は、ジアシル鎖または単アシル鎖であってもよい。ホスファチジルセリン、ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルグリセロールという語は、化合物中にアシル鎖を持たないものを意味する。
【0017】
脂質組成物は、リポソーム、非二重層構造および渦巻型相を調製するために変更することができる。高PC含有量を有する、ホスファチジルコリン(PC):ホスファチジルセリン(PS)もしくはホスホセリンの脂質組成物は、Ca2+とNa+を含む緩衝液を用いた水和によりリポソームを形成するだろう。ホスファチジルエタノールアミンPEおよびPSの存在は、非二重層構造の形成を促進する。Na+を含まないか低濃度(100mM)で含むCa2+の存在下では、PS(90mol%以上)を伴う処方は、渦巻形シリンダーの形成を促進する。温度、Ca2+/Na+などの条件は、タンパク質-脂質複合体を含むナノ粒子における渦巻形シリンダーのサイズを減じるために変更できる。
【0018】
本発明にとって有用なリン脂質には、セリン含有化合物を含むリン脂質が含まれる。
このようなセリン含有リン脂質の例として、O-ホスホ-L-セリン(OPLS)、ジカプロイルホスファチジルセリン、ジオクタノイルホスファチジルセリン、ジミリストイル、ジパルミトイル、ジオレオイル-,ジステロイル-ホスファチジルセリンが挙げられる。セリン含有リン脂質は、他の分子を含む他のリン脂質と併用してもよい。例えば、ホスファチジルセリンを、ホスファチジルコリンまたはホスファチジルエタノールアミンと併用してもよい。ある実施形態では、PS、PC、PAおよびPEはアシル化またはジアシル化される。ホスファチジルセリンは、天然の(脳)あるいは合成起源などの何らかの原料から得ることができる。ホスファチジルセリンは、ホスファチジルコリン(PC)と併用されてもよい。ホスファチジルコリンはジミリストイルホスファチジルコリンであってもよい。PSとPCの比は、1:9〜9:1で変化しうる。一実施形態では、その比は3:7である。
【0019】
本発明のタンパク質-脂質組成物は、好ましくは適切な緩衝系中で安定化され保管される。このような緩衝剤には、TRISバッファー並びにHEPESバッファー及びナトリウム並びにカルシウム塩が含まれる。任意にアルコール(例えば10%エタノール)を添加してもよい。
【0020】
本発明のタンパク質-脂質複合体は、標準的な方法によって特徴づけられることができる。例えば、励起および発光スリットとして4nmを使用してSLM AMINCO 8000シリーズ装置またはPTI 380装置で蛍光研究を行うことができる。サンプルは280nmで発光させることができ、その発光スペクトルは300〜400nmの範囲でスキャンされる。遊離ファクターVIIIの発光スペクトルは、335nm付近で観察され、OPLSの添加は蛍光発光の強度を減じ、タンパク質の三次構造がわずかに変化したことを示した。
【0021】
さらに、前記の脂質結合タンパク質の粒子サイズは、標準的な粒子サイズ測定器(例えばNICOMP 315モデル)を使用して測定することができる。粒度分布は、単峰性および二峰性分布のためのガウス分布とNICOMP分析の両方を使用して分析することができる。ラテックスビーズのサイズは、各測定でスタンダード・コントロールとして使用できる。
【0022】
脂質の構造は、ネガティブ染色電子顕微鏡法によって分析することも可能である。このような方法は当該技術において一般的であり、凝集物がないことを確認するため、およびその構造をリポソーム、非二重層あるいは渦巻形に分類するために使用することができる。非二重層および渦巻形構造の形成は、Laurdan蛍光を使用して研究することもできる。前記脂質構造は、脂質含有溶液とプローブの水溶液(0.01%エタノール含有)を混合することによって、プローブで標識することもできる。そのサンプルは340nmで励起することができ、その発光スペクトルは440nmでモニターされる。励起スペクトルは440nmの発光モノクロメータを用いて、320および420nmの範囲で得ることができる。
【0023】
本発明のタンパク質-脂質複合体は、筋肉内、鼻腔内、腹腔内、静脈内、経口、直腸、皮下、局所投与などの標準的な投与手段によって、個体(例えばヒトを含む動物)に送達することができる。前記複合体は、標的部位に直接もしくはその付近に送達されてもよく、または循環中に直接もしくは間接的に送達されてもよい。前記複合体は、当該技術分野で周知の薬学的に許容されるキャリアーを用いて送達されてもよい。
【0024】
本発明のタンパク質-脂質複合体は、減弱された抗原性のみならず減弱された免疫原性をも示す。したがって、このような組成物は、個体において治療薬に対する免疫応答を減じるために使用することができる。本発明の組成物は、そのタンパク質への免疫反応がすでに発生した個体に治療薬を送達するために使用することもできる。したがって、これらの組成物は免疫反応の発生前あるいは後に使用することができる。
【0025】
ある実施形態では、本発明は特異的FVIII-脂質複合体を提供する。前記タンパク質-脂質複合体は、リポソーム、渦巻形、ミセルおよび非二重層構造などの構造だけでなく新規な脂質構造を形成し、免疫応答および抗原性を減弱しうる。その方法は、バッファーコンディションによって適切に安定化された特異的FVIII-脂質複合体を構築することを含む。特定の理論に制約されることを意図しないが、この抗原性および免疫原性の低下は、タンパク質-脂質複合体に起因し、リポソームなどの分子アセンブリにおいて、疎水性シールドおよび粒状物質の細胞(抗原提示細胞 APCs)内取り込みなどの担体特性を含むと信じられる。これらの複合体は、抗体の存在下で凝血活性を有し、ELISAアッセイにおいてモノクローナル抗体への結合能力を測定した際に減弱した抗原性を示した。前記複合体はまた動物モデルにおいて減弱した免疫応答を示した。本発明は、いまだFVIIIで治療されていない患者における免疫応答の発達を減じるだけでなく、すでに治療され抗体が構築された患者におけるFVIII凝血活性を保持するのにも役立つ。
【0026】
したがって、一実施形態では、本発明はFVIIIに対する免疫応答を減じる方法を提供する。ここに開示した組成物の使用によって、免疫原性は低減されるが、凝血活性は維持される。免疫原性の低下はリン脂質とC2およびA2ドメインの複合体形成によって達成されると考えられる。これは、(1)凝集物形成を低下させる、(2)投与回数を減少させる(複合体がクリアランス機構を変え、これにより循環時間がより長くなる)および/または (3)エピトープ領域の立体構造を遮蔽および変えるおよび/または(4)優先的な細胞取り込みを含む担体特性、によって免疫原性を低下させると考えられる。安定性、循環時間を増加させるクリアランス機構の変化、および抗原性並びに免疫原性の低下などの、複合体の改善された薬学的性質は、予想外の観察結果である。
【0027】
以下の実施例は本発明をより詳細に説明するために存在し、なんら制限を意図しない。
【実施例1】
【0028】
本実施例は、本発明にかかるタンパク質-脂質複合体の調製を説明する。この実施形態を説明するため、FVIIIが使用された。25mMのTRIS、300mMのNaClおよび5mMのCaCl2中で20ugの前記タンパク質と5および20mMのOPLSを混合することによって、FVIII-O-ホスホ-L-セリン(OPLS)複合体を形成した。
【実施例2】
【0029】
本実施例は、本発明のタンパク質-脂質複合体(実施例1)の安定性について説明するものである。遊離FVIIIにとって前記タンパク質のアンフォールディングはタンパク質の凝集をもたらし、これは次にアンフォールディングの不可逆性を引き起こす。凝集はC2ドメイン中のわずかな立体構造変化によって開始される。タンパク質、およびFVIIIのC2ドメインに結合すると信じられるO-ホスホ-L-セリンを含有する製剤の安定性を測定するために、アンフォールディング/リフォールディングの調査を、遊離FVIIIおよび実施例1に記載したPSと複合体を形成したFVIIIを用いて行った。
【0030】
組換えヒトFVIII(rFVIII)の温度依存性の物理的および機能的変化を研究するために、円偏光二色性(CD)、蛍光偏光測定法、サイズ排除クロマトグラフィー(SEC)、ドメイン特異的抗体結合、および凝血活性の調査を行った。フォールディングおよびアンフォールディングを測定するために、CDスペクトルの調査を行った。CDスペクトルはd10ショウノウスルホン酸で較正したJASCO-715分光偏光計を用いて得た。サンプルは二次構造分析のために255〜208nmの範囲で調査され、そして典型的には、使用したタンパク質濃度は20〜22μg/mlであった。タンパク質のCDスペクトルは、バッファー・ベースラインのスペクトルを差し引くことによって補正した。シグナルのクオリティを高めるために複数の調査を行い、平均化した。タンパク質の融解は、2.5℃ごとに2分の保持時間で20〜80℃の温度範囲にわたって行った。温度調査は、Peltier 300RTSユニットを用いて行い、その分析結果は、製造業者によって提供されるソフトウェアを用いて作成した。アンフォールディング特性の加熱速度依存は、キネティック・コントロール下でタンパク質の熱変性が少なくともある程度存在することを示した(図1)。フォールディングモデルは、ファクターVIIIの凝集キネティクスを説明するために提唱された。このモデルに基づき、ファクターVIIIのアンフォールディングは、単なる二状態のキネティックモデル[凝集性(A)→未変性(N)、ここでkはアレニウス式によって与えられるように、温度に伴って変化する一次速度定数である]において解釈される。上記遷移に関する活性化エネルギーは、〜127.98 Kcal/Mole(〜534.97 KJ/Mole)と算出された。抗体結合の研究は、C2ドメインの脂質-結合領域(2303-2332)中の立体構造変化が、少なくともある程度、凝集の開始に関与しうることを示した(図2)。OPLSの存在下および不存在下におけるFVIIIのSEC特性の分析は、単量体の集団がPSの存在下の凝集性タンパク質のものより有意に高いことを明確に示し、これはファクターVIIIの凝集キネティクスにおけるOPLSの干渉が原因かもしれない(図3)。前記データは、複合体がFVIIIの安定性を改善し、凝集物を減らすことによって免疫原性を減じるのに役立ちうることを示す。
【0031】
本実施例のさらなる実例では、O-ホスホ-L-セリンは前記タンパク質と複合体を形成する(C2ドメインを遮蔽すると思われる)。20ug/mlのタンパク質と5および20mMのホスホ-L-セリンとを、25mMのTRIS、300mMのNaClおよび5mMのCaCl2を含む別のバッファー中で混合した。モノクローナル抗体への結合は、サンドイッチELISAによって調査した。図4に示すように、添加無しのタンパク質では凝集が原因で未変性様構造の回復は不可能であり、一方、O-ホスホ-L-セリンの存在は未変性構造の実質的な回復をもたらした。
【実施例3】
【0032】
本実施例は、本発明のタンパク質-脂質組成物(実施例1)が、Sprague-Dawleyラットにおいて、前記タンパク質に対し免疫原性を減弱することを実証する。この実施形態を説明するために、OPLS-ファクターVIII複合体を、Sprague-Dawleyラットに投与した。このラットモデルは、FVIIIへの抗体開発を研究するのに好適であることが実証されている。ELISAによって、遊離FVIIIおよびFVIII-PS複合体に対する抗体価を測定した。投与から2週間、ファクターVIII-OPLS複合体に対する抗体価の分析が非免疫毒性であることが見い出された。遊離FVIIIに対する抗体価は563.72±916.15であり、FVIII-OPLS複合体に対する検出可能な抗体価は観察されなかった。
【実施例4】
【0033】
本実施例は、ホスホセリンが、免疫原性を減少させる単純複合体において必須であることを実証する。本実施例を説明するために、ホスホセリン、ホスホコリンまたはホスファチジン酸を用いて単純複合体を調製した。免疫原性は、血友病Aマウスにおいて複合体の皮下投与によってテストした。総抗体価(ELISAによって測定)または特異的阻害力価(Bethesda Units)を図5Aおよび5Bに示す。効果はホスホセリンでは統計的に有意であったが、ホスホコリンまたはホスファチジン酸では統計的に有意ではなかった。
【実施例5】
【0034】
本実施例は、本発明の組成物を小さな単層ベシクル(vesicles)として作ることができることを実証する。0.3mg/mlのDMPCおよび0.15mg/mlのbPSを丸底フラスコ中で溶解し、溶媒を蒸発させて薄膜を形成した。その後この薄膜を水和してMLV’sを形成し、そしてこのMLVsを200nmのポリカーボネートフィルターを通して押し出し、160nmのサイズ範囲内のSUV’sを形成した。この製剤(ファクターVIIIを含む)の免疫応答を実施例5で説明する。
【0035】
全ての上述した研究において、PS無しの状態を使用して行った実験、および脂質と結び付いたタンパク質の濃度並びに脂質構造は、これら全ての脂質製剤において特異的な脂質-FVIII複合体の存在を示唆している。さらに、C2ドメインに特異的なモノクローナル抗体が、セリンの存在下でFVIIIに結合しなかったことは、調査した脂質構造における複合体の存在を明確に示唆する。
【実施例6】
【0036】
本実施例は、抗原性および、複合体の形成におけるタンパク質の特定のエピトープ領域の関与を研究するためのELISA分析の一般的な方法論を実証する。タンパク質-脂質複合体の抗体結合は、抗体捕獲ELISAとサンドイッチ ELISAを用いて調べた。サンドイッチELISAでは、炭酸緩衝液(0.2M、pH9.4)中5μg/mlの濃度にて50μl/ウェルの抗体溶液を4℃で一晩インキュベートすることによって、96ウェルプレート(Nunc-Maxisorb)を抗-C2ドメイン抗体(ESH4)でコートした。前記プレートをその後、0.05%Tween20(10mM Na2HPO4,1.8mM KH2PO4,0.14mM NaCl,2.7mM KCl,および0.02% NaN3からなるPBT)を含む100μlのリン酸緩衝液で10回洗浄した。プラスチックの吸着性表面上の残りの非特異的タンパク質結合サイトを、リン酸緩衝液(10mM Na2HPO4,1.8mM KH2PO4,0.14mM NaCl,および2.7mM KClからなるPB)中の1%ウシ血清アルブミンからなるブロッキングバッファー200μlを室温で2時間インキュベートすることによってブロックした。プレートをPBTで10回洗浄し、ブロッキングバッファー中の100ng/mlのrFVIIIもしくはrFVIII/OPLS(実施例1および3)またはリポソーム付随rFVIII(実施例5)50μlを添加して37℃で1時間インキュベートした。プレートをPBTで10回洗浄し、そして、50μlのビオチン化ESH8−別の抗-C2抗体を用いて、1μg/ml濃度および50μlの1:1000希釈のアビジン-アルカリホスファターゼ複合物にて、どちらもブロッキングバッファー中、室温にて1時間インキュベートした。プレートをPBTおよびジエタノールアミン緩衝液(1Mジエタノールアミン,0.5mM MgCl2および0.02% NaN3からなる)中の1mg/ml p-ニトロフェニルリン酸塩溶液100μlで10回洗浄した。プレートを室温で30分間インキュベートし、3NのNaOHを100μl添加することによって反応をクエンチした。吸光度を405nmでプレートリーダーを用いて読み取った。このELISA研究は、より少ないC2ドメイン特異的抗体、ESH8またはESH4がPS(OPLSまたはリポソーム 図6)の存在下でタンパク質に結合することを示した。その結果は、これらのモノクローナル抗体の結合が脂質によって阻害されることを示し、エピトープ領域がタンパク質-脂質複合体中に遮蔽されることを示唆する。さらに、前記特異的複合体FVIII-PSが、前記エピトープ遮蔽の原因であることが、さらに対照実験によって確認された。この際、抗原性はDMPC/PS混合物を用いたときのみ観察され、DMPC単独では観察されなかった(図7)。
【実施例7】
【0037】
本実施例は、本発明のタンパク質-脂質組成物(実施例5)が、動物モデル、Sprague-DawleyラットおよびファクターVIIIノックアウトマウス,血友病Aモデルにおいて、前記タンパク質に対し免疫原性を減じることを実証する。
【0038】
タンパク質-リポソーム複合体の投与後4および6週間目にSprague-Dawleyラットで評価された抗体価は、遊離FVIIIに比べてFVIII-PS複合体でより低いことが見いだされた(図8A)。
【0039】
本実施形態の別の実例では、免疫原性に対する本発明のタンパク質-脂質複合体の効果を、ファクターVIIIノックアウトマウスモデルでテストした。この表現型のマウスは重度の血友病である(ネオ・カセットを用いる標的破壊によるエクソン16ノックアウト)。前記タンパク質-リポソーム複合体(DMPC+bPS)は皮下に投与し、その抗体価はELISA分析を用いて測定した。図8Bから明らかなように、抗体の濃度は投与後6週間、リポソーム付随タンパク質でより低かった。このデータは明確に、リポソーム結合ファクターVIIIが遊離ファクターVIIIに比べてより少ない抗体を誘発することを実証する。特定の理論に拘束されることを意図しないが、この観察結果は、一部、変化した(1)遊離FVIIIの立体構造および凝集キネティクス、(2)粒状物質のデコイ効果および免疫系によるプロセシングおよび (3)クリアランス機構、に起因するかもしれない。
【実施例8】
【0040】
以下の実施例は、タンパク質-脂質複合体のミセル、非二重層構造、渦巻形構造、および新規な非リポソーム脂質粒子の形成を実証する。本実施例は、ここに記載された組成物によるミセルの形成を説明する。タンパク質溶液を、低濃度および高濃度(臨界ミセル濃度の上下)の短鎖アシル鎖脂質(ジヘキサノイル・ホスファチジルセリン[0.3mM前後])と混合し、その結果生じたミセル粒子を特性化した。ミセルの構造は、光散乱、円二色性および蛍光実験によって特性化した。活性や抗体結合などの機能分析を行った。
【実施例9】
【0041】
本実施例は、ここに記載された組成物による渦巻形構造の形成について記述する。0.15mg/mlのbPSをクロロホルム中に溶解し、溶媒を蒸発させて薄い脂質膜を形成した。それからこのフィルムをpH7.0の複数の緩衝系中で水和させ、前記MLVsを押し出すか超音波処理するかのいずれかによりSUVsを形成した。この結果生じたSUVsを、5mMのCaCl2を含有する緩衝系中でタンパク質と混合し、渦巻形構造を形成した。その脂質構造を、光散乱、微分干渉顕微鏡法、ネガティブ染色電子顕微鏡法によって及び蛍光研究によって分析した。これらの研究により、この手順によって形成された脂質構造は事実上渦巻形であることが分かった。
【実施例10】
【0042】
本実施例は、本発明のタンパク質-脂質複合体調製の別の実例である。0.15mg/mlのbPSおよび0.3mg/mlのジオレオイル・ホスファチジルエタノールアミン(DOPE)をクロロホルム中に溶解し、溶媒を蒸発して薄い脂質膜を形成した。結果物である脂質膜をリン酸緩衝生理食塩水を用いて水和し、六方相を形成した。前記非二重層構造は蛍光研究によって特性化した。
【実施例11】
【0043】
本実施例は、新規な非リポソーム構造をもつ、本発明のタンパク質-脂質複合体の調製の別の実例である。0.3mg/mlのDMPC、0.15mg/mlのbPSをクロロホルム中に溶解し、溶媒を蒸発させ、丸底フラスコ中で薄い脂質膜を形成した。この脂質を、FVIII、25mM TRIS、300mM NaClおよび5mM CaCl2を含有する緩衝系を用いて水和させ、この溶液を室温あるいは37℃のいずれかにて穏やかに回転した。その後前記フィルムを、穏やかに回旋させながら適切な緩衝液(25mM TRIS、300mM NaClおよび5mM CaCl2)中で水和させた。このようにして形成したMLV’sを、MLV’sと結びついたタンパク質から遊離タンパク質を分離するために、デキストラン遠心分離勾配にかけた。0.5mlの脂質付随タンパク質を1mlの20%w/vデキストランと混合し、そして3mlの10%w/vデキストランを上記溶液上に層状に重ねた。その後0.5mlの緩衝液をトップに層状に重ねた。前記勾配を、ベックマン社のSW50.1ローターを用いて35分間、45K PRMで遠心分離した。遠心分離研究の結果を図9に示す。図から明らかなように、浮かなかった脂質分画がある(図中では分画3として表示する)。この分画は、14%と10%のデキストランの界面で観察された。この分画が浮かなかったという事実は、この脂質分画が十分な浮力あるいは被包された水を持たないことを示す。従来のリポソームは一般的に封入された水を有するため勾配の最高部に浮く。それゆえ、浮かない分画は、脂質粒子を含有する非リポソームタンパク質の可能性がある。分画を採取して、質量分析で脂質含量を、活量でタンパク質含量をテストした。質量分析研究により、この分画が、単なるPS-Ca+複合体ではないことを示唆する高PC含量の脂質を含むことが明らかになった。活量分析は、この分画内に最初のタンパク質が約40%封入されていることを示した。
【0044】
含水量が無いかわずかである高密度な分画3に関し、いくつかの考えられる解釈が存在する。この分画は以下のことを意味するかもしれない:(1)わずかな封入水量を有する非常に小さな単層ベシクル ごく小さなベシクル(200nm未満)が同一のコンディション下で浮くことができるかどうかを確認するために、事前にサイズを揃えたリポソームを、ポリカーボネートフィルターを通して押し出すことによって調製した。前記押し出しを3回繰り返し、粒子のサイズを約160nmとした。結果物であるSUVsをFVIIIと混合し、デキストラン遠心分離勾配を適用した。この対照研究は、これらのSUV’sが分画3のバンドを示さないことを明らかにし、これにより、このようなバンドの観察が小さなリポソームの形成に起因しないことが示された。さらにこの実験を、デキストラン勾配における人為的な影響を排除するために、同一の緩衝液および実験条件の下で行った。(2)第二の可能性は、分画3が、含水量がわずかな、渦巻形構造の形成を意味するかもしれないということである[26]。しかしながら、渦巻形構造の形成は、非常に高いPS含量(>50%)を必要とする。分画3バンドの形成において、使われるPS含量は約30%であり、これらの条件の下で渦巻形構造の形成は見られなかった。(3)脱水構造[27]を有する崩壊したCa(PS)2複合体の形成は、浮かない可能性があり、高密度なバンドを形成しうる。このような崩壊した構造の形成には、非常に高いPS含量(>50%)と、Na+が存在しないことが必要である。しかしながら、本実施例では、組成物は低いPS含量と、非常に高い濃度のNa+を有する、言い換えると、30%のPSおよび300mMのNaClが使用されており、それゆえ、Ca(PS)2 形成の可能性は除外される。(4)PSおよびカルシウム系は、ベシクル融合を促進することが示されている。しかしながら、Ca2+などの二価カチオンによるベシクルの融合は、脂質結合サイトでカルシウムと競合するNa+の存在によって阻害される。このPC/PS比7:3における、および300mMのNaClと5mMのCaCl2の存在下におけるPSあたりのカルシウム結合の推定量は、0.22(500mM NaCl)〜0.35(100mM NaCl)の間である[28, 29]。PSあたりの推定結合カルシウムは、小さな単層PS/PCベシクル系において融合に必要とされる臨界比0.35〜0.39より小さい。したがって、ベシクルの融合が、高密度なバンドを示すとは考えられない。これは、より大きなPC分画(>50%)は結果的により少ないPS-二価カチオン複合体をもたらすかもしれないため、および、大きなドメインへクラスター形成して融合を誘導するその能力のためである[28,29]。これらの理由に基き、高密度なバンドは、新規な非リポソーム脂質粒子の形成に起因する可能性が高いと思われる。このバンドは、封入水量がわずかであるため、非リポソームである。この新規な脂質構造の構造を理解するため、ネガティブ染色電子顕微鏡法、微分干渉コントラスト光学顕微鏡法、光散乱、円二色性および蛍光測定(データは示さず)を行った。
【0045】
複数のコンディション下で行われた遠心分離の研究は、緩衝剤条件、バルクタンパク質濃度、アルコールの使用(例えば10%エタノール)およびタンパク質-脂質比を、所望の収量の複合体を得るために変化させることができることを示す。これらのパラメーターにおけるこのようなバリエーションは、当業者の範囲内であると見なされる。視覚的な検査で確認できるように、一般に、より高いタンパク質濃度はより高密度の分画をもたらすことが確認された。
【実施例12】
【0046】
本実施例は、本発明のタンパク質-脂質組成物が生物活性を保持することを実証する。この実施形態を実証するために、ファクターVIIIを含むタンパク質-脂質複合体(OLPS,DCPS,およびDCPC)の凝固に対する効果をテストした。rFVIII凝血活性は、活性化剤として微粉化シリカおよび基質としてFVIII欠損プラズマを用いた一段階活性化部分トロンボプラスチン時間(APTT)分析によって測定した。APTT分析は、COAG-A-MATEモデルの凝固分析器(Organon Teknika社、ダラム,NC)を用いて行った。手短に言えば、rFVIIIをFVIII欠損プラズマに添加し、凝固時間をモニターした。その後、既知の活性を持つ凍結乾燥参考濃縮物の様々な希釈物から測定された凝固時間を利用して作成した較正曲線から、rFVIIIの活性を求めた。タンパク質の濃度は、ビシンコニン酸(BCA)分析を用いて独立的に測定して、活性と比較した。例えば、タンパク質の20〜22μg/mlは全て87〜95.6IUの比活性に対応する。サンプルを調製するために使用された原液は、2174IU/0.5mg/mlの比活性を有していた。
【実施例13】
【0047】
本実施例は、PS含有リン脂質の存在下におけるタンパク質の減弱した抗原性を実証する。本実施形態を説明するために、ESH4抗体へのrFVIIIの結合に対するジカプロイルホスファチジルセリン(DCPS)の効果をテストした。2ugのタンパク質と0.5,2および5mMのDCPSを、25mM TRIS、300mM NaClおよび5mM CaCl2中で混合することによって、FVIII-(DCPS)複合体を形成した。この複合体の抗原性をELISAを用いて研究した。図10に示すように、試験した濃度(すなわち、0.5,2および5.0uM)にて、DCPSはESH4抗体へのrFVIIIの結合を阻害した。これらの濃度はこのリン脂質のCMCの上および下である。
【実施例14】
【0048】
本実施例は、タンパク質の免疫原性に対するPS含有リン脂質の効果を説明する。2ugのタンパク質と5mMのDCPSを、25mM TRIS、300mM NaClおよび1mM CaCl2中で混合することによって、FVIII-(DCPS)複合体を形成した。この組成物はミセル構造を形成し、複合体の免疫原性は血友病Aマウスにおいて研究した。総抗体価は遊離FVIIIでは13,167(SD=7909,n=16)およびFVIII-DCPS複合体では3506(SD=1150.7,n=12)である。
【実施例15】
【0049】
2ugのタンパク質と20mMのDCPCを、25mM TRIS、300mM NaClおよび1mM CaCl2中で混合することによって、FVIII-ジカプロイル・ホスファチジルコリン(DCPC)複合体を形成した。この組成物はミセル構造を形成し、複合体の免疫原性は血友病Aマウスにおいて研究した。総抗体価は遊離FVIIIでは13,167(SD=7909,n=16)およびFVIII-DCPC複合体では1293(SD=946.57,n=12)である。
【実施例16】
【0050】
本実施例は、本発明の組成物をリポソーム小胞として作ることができることを実証する。クロロホルム中に溶解した11.25μmolのDMPCと4.83μmolのBPSを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発しフラスコ壁上に薄膜を形成した。その後この薄膜を、3mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)で水和させ、MLV’sを形成し、このMLVsを80nmもしくは200nmもしくは400nmのいずれかのポリカーボネートフィルターを通して押し出し、80nm〜400nmのサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。その後このリポソームを以下のように免疫化で使用した。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。
【0051】
別のサンプルでは、クロロホルム中に溶解した11.25μmolのDMPCを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発し、フラスコ壁上に薄膜を形成した。その後この薄膜を、3mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)で水和させ、MLV’sを形成し、このMLVsを80nmもしくは200nmもしくは400nmのいずれかのポリカーボネートフィルターを通して押し出し、80nm〜400nmのサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。高い相転移温度を持つDSPC(固体状態)はネガティブ・コントロールに使用された。
【0052】
別のサンプルでは、クロロホルム中に溶解した7.5μmolのDMPCと3.22μmolのDOPAを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発し、フラスコ壁上に薄膜を形成した。その後この薄膜を、2mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)で水和させ、MLV’sを形成し、このMLVsを200nmのポリカーボネートフィルターを通して押し出し、200nm以下のサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。
【0053】
別のサンプルでは、クロロホルム中に溶解した7.5μmolのDMPCと3.22μmolのDOPGを丸底フラスコに入れ、回転エバポレータを使用して溶媒を蒸発し、フラスコ壁上に薄膜を形成した。その後この薄膜を、2mlのトリス緩衝液(300mM NaCl,25mM Tris,5mM CaCl2含有 pH=7.0)中で水和させ、MLV’sを形成し、このMLVsを200nmのポリカーボネートフィルターを通して押し出し、200nm以下のサイズ範囲のSUV’sを形成した。脂質リカバリーはバートレット法によってリン含有量を測定することにより推定した。タンパク質を37℃で30分間インキュベートすることによってリポソームと結合させた。FVIII欠損マウスの免疫化は、一週間間隔でrFVIIIもしくはrFVIII-リポソーム(100μlのトリス緩衝液中2μg)を4回皮下(s.c.)注射して行った。タンパク質と脂質のモル比は1:10,000に維持した。この製剤の免疫応答を表1に示す。
【0054】
【表1】
【0055】
平均総抗-rFVIII抗体価および阻害価は、リポソーム不存在下および存在下でのrFVIIIを用いたFVIII-KOマウスの免疫化に続く6週間後の値である。*はリポソームを押し出すのに使用したポリカーボネート膜の孔サイズを示し、**P<0.05,統計分析は、実験手順において記載したようにして行った。
【0056】
本発明をここに記載した実施例を用いて説明したが、本発明の精神を逸脱することなく本発明について一般的な修正を行うことができる。このような修正は本願の特許請求の範囲内に含まれる。
【図面の簡単な説明】
【0057】
【図1】図1は、様々な加熱速度でのFVIIIの融解特性を示す。
【図2】図2は、C2ドメインにおける立体構造変化を示す抗体結合アッセイを示す。モノクローナル抗体ESH4およびESH8のrFVIIIへの結合は、60℃および15℃/hrでrFVIIIを表示の温度まで加熱した後サンドイッチERISAによって測定した。
【図3】図3は、O-ホスホ-L-セリンの存在下または不存在下でのファクターVIIIのサイズ排除クロマトグラフィー(SEC)特性を示す。
【図4】図4は、サンドイッチELISAによって研究されたFVIII-O-ホスホ-L-セリンの抗原性を示す。
【図5】図5Aおよび5Bは、rFVIIIの免疫原性に対するOPLS、ホスホコリンおよびホスファチジン酸の影響を示す。平均総抗体価(5A)および阻害価(5B)は、rFVIIIと比較した表記の結合剤-rFVIII複合体を示す。
【図6】図6は、O-ホスホ-L-セリン中のFVIIIのフォールディング研究を示す。
【図7】図7は、サンドイッチELISAによって研究されたリポソーム中のFVIII-PS複合体の抗原性を示す。FVIII:遊離FVIII、本発明のFVIII:本発明で使用される組成物、およびDMPC+FVIII:DMPCリポソームおよびFVIII欠損特異的タンパク質(FVIII)脂質(PS)複合体の物理的混合物。
【図8A】図8Aは、遊離FVIIIおよびFVIII-PS複合体に対する動物モデルにおける免疫応答を示す。FVIII:遊離FVIII、本発明のFVIII:本発明で使用される組成物
【図8B】図8Bは、ファクターVIII欠損血友病AマウスモデルにおけるFVIIIとFVIII-PS複合体リポソームを示す。
【図9】図9は、FVIII-PS複合体を含む、非リポソームで低水分量フラクションを示すデキストラン密度勾配の写真である。
【図10】図10は、rFVIIIの抗原性に対するDCPSの影響を示す。
【特許請求の範囲】
【請求項1】
ペプチド、ポリペプチドおよびタンパク質からなる群から選択される治療薬の免疫原性を減弱する方法であって、
前記治療薬と、セリン化合物、ホスファチジルコリン(PC)、ホスファチジルエタノールアミン(PE)、ホスファチジン酸(PA)、およびPC、PEもしくはPA含有リン脂質からなる群から選択される一以上の結合剤との複合体を形成する工程を含み、
個体に投与された前記治療薬の免疫原性が、結合剤と複合体を形成した時に減弱されることを特徴とする方法。
【請求項2】
前記セリン化合物が、ホスホセリン、ホスファチジルセリン(PS)およびPS含有リン脂質からなる群から選択される、請求項1に記載の方法。
【請求項3】
前記複合体が、単純複合体、リポソーム、ミセル、渦巻形および非リポソーム脂質構造からなる群から選択される一以上の構造を有し、前記非リポソーム脂質構造は10%と14%のデキストラン勾配の界面に存在する、請求項1に記載の方法。
【請求項4】
前記結合剤がホスホセリンであり、形成される複合体が単純複合体である、請求項3に記載の方法。
【請求項5】
前記ホスホセリンがO-ホスホ-L-セリンである、請求項4に記載の方法。
【請求項6】
前記リン脂質がアシル化またはジアシル化されている、請求項1に記載の方法。
【請求項7】
前記のアシル鎖長が炭素原子数1〜18の範囲である、請求項6に記載の方法。
【請求項8】
前記のアシル鎖長が炭素原子数1〜4の範囲である、請求項7に記載の方法。
【請求項9】
前記のアシル鎖長が炭素原子数5〜12の範囲である、請求項7に記載の方法。
【請求項10】
前記のアシル鎖長が炭素原子数13〜18の範囲である、請求項7に記載の方法。
【請求項11】
前記リン脂質がPS含有リン脂質である、請求項6に記載の方法。
【請求項12】
前記リン脂質が、さらにPC含有リン脂質および/またはPE含有リン脂質を含む、請求項11に記載の方法。
【請求項13】
PC含有リン脂質とPS含有リン脂質の比が7:3である、請求項12に記載の方法。
【請求項14】
前記アシル鎖が、カプロイル、オレオイル、オクタノイル、ミリストイル、パルミトイル、およびステロイルからなる群から選択される、請求項6に記載の方法。
【請求項15】
前記治療薬がファクターVIIIである、請求項1に記載の方法。
【請求項16】
非-脂質二重層が、前記治療薬とセリン含有リン脂質とを約100nMのナトリウム塩と約5nMのカルシウム塩の存在下でインキュベートすることによって形成される、請求項1に記載の方法。
【請求項17】
約100nMのナトリウム塩と約5nMのカルシウム塩の存在下で、タンパク質をセリン含有リン脂質とインキュベートすることによって形成された脂質-タンパク質複合体であって、前記複合体が10%と14%のデキストラン勾配の界面に移行することを特徴とする、脂質-タンパク質複合体。
【請求項18】
前記セリン含有リン脂質がホスファチジルセリンである、請求項17に記載の脂質-タンパク質複合体。
【請求項19】
ホスファチジルコリンをさらに含有し、ホスファチジルコリンとホスファチジルセリンの比が7:3である、請求項18に記載の脂質-タンパク質複合体。
【請求項20】
前記タンパク質がファクターVIIIである、請求項19に記載の脂質-タンパク質複合体。
【請求項1】
ペプチド、ポリペプチドおよびタンパク質からなる群から選択される治療薬の免疫原性を減弱する方法であって、
前記治療薬と、セリン化合物、ホスファチジルコリン(PC)、ホスファチジルエタノールアミン(PE)、ホスファチジン酸(PA)、およびPC、PEもしくはPA含有リン脂質からなる群から選択される一以上の結合剤との複合体を形成する工程を含み、
個体に投与された前記治療薬の免疫原性が、結合剤と複合体を形成した時に減弱されることを特徴とする方法。
【請求項2】
前記セリン化合物が、ホスホセリン、ホスファチジルセリン(PS)およびPS含有リン脂質からなる群から選択される、請求項1に記載の方法。
【請求項3】
前記複合体が、単純複合体、リポソーム、ミセル、渦巻形および非リポソーム脂質構造からなる群から選択される一以上の構造を有し、前記非リポソーム脂質構造は10%と14%のデキストラン勾配の界面に存在する、請求項1に記載の方法。
【請求項4】
前記結合剤がホスホセリンであり、形成される複合体が単純複合体である、請求項3に記載の方法。
【請求項5】
前記ホスホセリンがO-ホスホ-L-セリンである、請求項4に記載の方法。
【請求項6】
前記リン脂質がアシル化またはジアシル化されている、請求項1に記載の方法。
【請求項7】
前記のアシル鎖長が炭素原子数1〜18の範囲である、請求項6に記載の方法。
【請求項8】
前記のアシル鎖長が炭素原子数1〜4の範囲である、請求項7に記載の方法。
【請求項9】
前記のアシル鎖長が炭素原子数5〜12の範囲である、請求項7に記載の方法。
【請求項10】
前記のアシル鎖長が炭素原子数13〜18の範囲である、請求項7に記載の方法。
【請求項11】
前記リン脂質がPS含有リン脂質である、請求項6に記載の方法。
【請求項12】
前記リン脂質が、さらにPC含有リン脂質および/またはPE含有リン脂質を含む、請求項11に記載の方法。
【請求項13】
PC含有リン脂質とPS含有リン脂質の比が7:3である、請求項12に記載の方法。
【請求項14】
前記アシル鎖が、カプロイル、オレオイル、オクタノイル、ミリストイル、パルミトイル、およびステロイルからなる群から選択される、請求項6に記載の方法。
【請求項15】
前記治療薬がファクターVIIIである、請求項1に記載の方法。
【請求項16】
非-脂質二重層が、前記治療薬とセリン含有リン脂質とを約100nMのナトリウム塩と約5nMのカルシウム塩の存在下でインキュベートすることによって形成される、請求項1に記載の方法。
【請求項17】
約100nMのナトリウム塩と約5nMのカルシウム塩の存在下で、タンパク質をセリン含有リン脂質とインキュベートすることによって形成された脂質-タンパク質複合体であって、前記複合体が10%と14%のデキストラン勾配の界面に移行することを特徴とする、脂質-タンパク質複合体。
【請求項18】
前記セリン含有リン脂質がホスファチジルセリンである、請求項17に記載の脂質-タンパク質複合体。
【請求項19】
ホスファチジルコリンをさらに含有し、ホスファチジルコリンとホスファチジルセリンの比が7:3である、請求項18に記載の脂質-タンパク質複合体。
【請求項20】
前記タンパク質がファクターVIIIである、請求項19に記載の脂質-タンパク質複合体。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8B】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8B】
【図9】
【図10】
【公表番号】特表2008−528696(P2008−528696A)
【公表日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願番号】特願2007−554228(P2007−554228)
【出願日】平成18年2月2日(2006.2.2)
【国際出願番号】PCT/US2006/003779
【国際公開番号】WO2006/084095
【国際公開日】平成18年8月10日(2006.8.10)
【出願人】(506194117)ザ リサーチ ファウンデイション オブ ステイト ユニバーシティー オブ ニューヨーク (11)
【Fターム(参考)】
【公表日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願日】平成18年2月2日(2006.2.2)
【国際出願番号】PCT/US2006/003779
【国際公開番号】WO2006/084095
【国際公開日】平成18年8月10日(2006.8.10)
【出願人】(506194117)ザ リサーチ ファウンデイション オブ ステイト ユニバーシティー オブ ニューヨーク (11)
【Fターム(参考)】
[ Back to top ]