
低免疫原性蛋白質−脂質複合体に関する組成物および方法
本発明により、第VIII因子等の治療用蛋白質の免疫原性を低減させるための、および、前記蛋白質の循環半減期を増加させるための組成物および方法が提供される。前記組成物は、ネガティブに荷電した脂質およびポリエチレングリコール誘導化ホスファチジルエタノールアミンを含むリポソーム、ミセルおよびコクリエート等の脂質構造を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本願は、2006年6月29日に出願された米国仮出願第60/695,080号に対する優先権を主張し、その内容を出典明示により本明細書中に組込む。
本発明は、ザ・ナショナル・インスティテュート・オブ・ヘルスからの交付番号第R01HL-70227下に政府基金により支持されたものである。当該政府が本発明に関する所定の権利を有する。
本発明は概して、治療に関して免疫原性を低減するための手段に関するものであり、より詳細には、第VIII因子の免疫原性を低減するための組成物および方法を提供するものである。
【背景技術】
【0002】
血友病Aは、第VIII因子(FVIII)の欠損または機能障害により特徴づけられる遺伝性の出血性疾患である。FVIIIは、凝固カスケードの本質的経路における決定的な補因子として作用する。組換えヒトFVIII(rFVIII)または血漿由来のFVIIIでの補充療法は、出血発現を制御するのに用いられる最も一般的な治療である。しかし、投与される前記蛋白質に対する中和抗体の導入は、患者の約15〜30%において、治療における主要問題である[1-3]。中和抗体は、インビボでリン脂質への結合にも関与するC2ドメインをしばしば標的する。
【0003】
FVIIIは、ドメインA1、A2、B、A3、C1およびC2から成る巨大なマルチドメイン糖蛋白質である[4、5]。系統的なエピトープマッピング研究により、抗-FVIII抗体が、FVIIIのA2(重鎖)、A3およびC2ドメイン(軽鎖)における規定領域を主に標的することが明らかになった[6、7]。A2ドメイン内のエピトープ決定因子は、残基Arg484-Ile-508にマップされている[8、9]。この領域を標的する抗体は、A2ドメインの第IXa因子(FIXa)との相互作用を遮断することによりFVIIIの活性化形態(FVIIIa)を阻害することが示されている[10]。A3ドメイン内の主要なエピトープ決定因子は、残基1811〜1818から成り、この領域に対する抗体も、FVIIIのFIXaとの相互作用を防いで補因子の活性を減損させる[11]。C2ドメイン内のエピトープ決定因子は残基2181〜2312にマップされており[12、13]、これは免疫優性、ユニバーサルCD4+エピトープ、2191〜2210、2241〜2290、2291〜2330を含む[14、15]。C2ドメインに対する抗体は、凝固カスケードの増幅に必須の、ホスファチジルセリン(PS)に富む血小板膜表面へのFVIIIの結合を妨害する。
【0004】
投与される第VIII因子に対して生じる免疫反応のために、第VIII因子の免疫原性を、好ましくは循環半減期に悪影響を及ぼすことなく、低減させる製剤の同定が必要とされている。
【発明の開示】
【発明が解決しようとする課題】
【0005】
発明者等は、ネガティブに荷電した脂質(ホスファチジルセリンを含むリン脂質等)およびPEG誘導化リン脂質を含むリポソームおよび他の脂質構造が、第VIII因子等の蛋白質の免疫原性を高め得るか否かを研究した。
一例では、PSおよびPEG誘導化PEを含むリポソームに結合したおよび/または当該リポソームに組込まれたrFVIIIの免疫原性を、血友病Aのネズミモデルで評価した。かかる組成物で処置された動物は、rFVIII単独で処置された動物と比較して、全-および阻害抗-rFVIII抗体の両方に関して低いタイターを有した。本発明の組成物を受容した動物から単離した脾臓細胞の平均刺激指数は、rFVIIIのみを受容した動物よりも低かった。サイトカイン分析により、これらリポソーム組成物の存在下で投与されるrFVIIIの免疫原性の低減には、部分的に、低減したIL-10の生成が介在する可能性があることが示唆された。静脈(i.v.)投与後の薬物動態研究は、これらの組成物を用いてrFVIIIの循環半減期が増加することを示した。
【0006】
従って、蛋白質の免疫原性が、循環半減期を有意に損なうことなく低減された組成物が本願において提供される。この組成物は、ネガティブに荷電した脂質、PEGで誘導化された両親媒性脂質、およびFVIII等の蛋白質を含むリポソームおよび/または他の脂質構造を含む。本明細書中に記載されるPEGを含むリポソームおよび他の脂質構造は、「PEG化」されていると本願において記載される。前記組成物の調製および使用のための方法も提供される。
【0007】
本明細書中に用いる省略は次のとおりである。:APTT、活性化部分トロンボプラスチン時間;ACD、クエン酸デキストロース;BPS、脳ホスファチジルセリン;BSA、牛血清アルブミン;DMPC、ジミリストイルホスファチジルコリン;DMPE-PEG2000、1,2-ジミリストイル-sn-グリセロ-3-ホスホエタノールアミン-N-[メトキシ(ポリエチレングリコール)-2000];ELISA、酵素結合免疫吸着アッセイ;FVIIIa、活性化されたFVIII;FIXa、第IXa因子;Ig、免疫グロブリン;KO、ノックアウト;PB、ホスフェートバッファー;PBA、アルブミンを含むホスフェートバッファー;PBT、トゥイーンを含むホスフェートバッファー;PA、ホスファチジン酸;PC、ホスファチジルコリン;PS、ホスファチジルセリン;rFVIIa、組換え第VIIIa因子;rFVIII、組換えヒト第VIII因子;RES、細網内皮系;TB、トリスバッファー。
【課題を解決するための手段】
【0008】
本発明により、rFVIII製剤が提供される。この製剤は、PSまたはPA等のネガティブに荷電した脂質を含むリポソームおよび/または他の脂質構造(ミセルまたはコクリエート等)を含む。リポソームは、PEGで誘導化された第一の両親媒性脂質(PE等)と、PC、PE(PEGで誘導化されていないもの)またはPG等の第二の両親媒性脂質とを含む。ネガティブに荷電した脂質に加えて、ミセルはPCおよび/または非-PEG誘導化PEを含んでよい。ネガティブに荷電した脂質に加えて、コクリエートはPCを含んでもよい。
【0009】
本発明の組成物は、第VIII因子が低免疫原性であり、遊離状態の第VIII因子よりも長い循環半減期を有することを特徴とするものである。特に、本発明により、免疫優性エピトープが遮蔽された脂質-rFVIII調製物が提供される。低い免疫原性および長い循環半減期のために、前記蛋白質の投与の頻度を減らすことができる。
【0010】
本発明の組成物は、ネガティブに荷電した脂質、PEG誘導化両親媒性脂質を含む脂質構造を含む。第VIII因子または他の蛋白質またはポリペプチドはかかる構造と結合でき(即ち、表面吸着され得)、またはかかる構造に導入され得る。前記蛋白質は、PSまたはPA等のネガティブに荷電した脂質と結合すると考えられる。
【0011】
両親媒性脂質の例は、PC、PEおよびPGである。ネガティブに荷電した脂質の例は、PSおよびPAである。PEGで誘導化され得る脂質の例はPEである。PEは単独で、および./またはPEGで誘導化されるように、脂質構造に用いられ得ることに注意すべきである。
一態様では、前記蛋白質はFVIIIである。インビボデータを、血友病Aのネズミモデルにおいて示す。データは、PSを含むPEG化リポソームrFVIIIの投与が当該蛋白質の免疫原性を低減し、rFVIIIのt1/2の増加を生じることを示す。
【0012】
本発明のリポソームに関し、ネガティブに荷電した脂質の量は30〜50モル%の範囲である。両親媒性脂質の量は50〜70モル%の範囲である。PEGで誘導化されたPEは1〜15モル%の範囲である。任意に、前記リポソームはコレステロールを0〜30モル%の範囲で含んでいてもよい。一態様では、PCのPSに対する比は、50:50〜90:10の範囲内である。一態様では、前記比は70:30である。20%までのPSまたはPCを、非-PEG誘導化PEにより置き換えることができる。
【0013】
本発明のリン脂質は2つのアシル鎖を有する。グリセロール骨格に結合する前記アシル鎖の長さは12〜22個の炭素原子長で変化する。前記グリセロール骨格に結合する2つのアシル基は同じであっても異なっていてもよい。前記アシル鎖は飽和したものであっても不飽和のものであってもよい。12〜22個の炭素原子の飽和および不飽和アシル鎖のいくつかの非限定的な例を表1Aと1Bに示す。
【0014】
【表1】

【0015】
【表1B】
【0016】
短鎖(6〜12個の炭素原子)のホスファチジルセリンは、臨界ミセル濃度以上の濃度でミセルとして存在し得るユニークな水溶性脂質である。短鎖ホスファチジルセリンはrFVIIIと相互作用し、rFVIIIの安定性、免疫原性および薬物動態パラメータに影響する。PEG誘導化PEはミセルに用いることもできる。
【0017】
追加的に、ネガティブに荷電した脂質およびPEG誘導化PEを含むコクリエート構造またはシリンダーを調製することもできる。これらの茎は薬物送達システムとして有用である。コクリエートの調製のためには、長鎖(12〜22個の炭素原子)のリン脂質を用いる。
ミセルは100モル%のPSおよび1〜15モル%のPEG誘導化PEを含んでよい。任意に50%までのPSをPCで置き換えてもよく、および/または5%までのPSをPE(PEGで誘導化されていないもの)で置き換えてもよい。ミセルに関し、50%までのPSをPCで置き換えてよく、および/または5%までPEで置き換えてよい。
コクリエートは100モル%のPSを含んでもよい。30モル%までのPSをPCで置き換えてもよい。
【0018】
本発明の組成物はいくつかの方法により調製できる。例えば、一態様では、前記方法は、PS、PCおよび/またはPEを含むリポソームを調製し、このリポソームにFVIIIを結合および/または組込み、次いでPEG誘導化PEを、FVIII結合/組込みリポソームに添加することを含む。PEG誘導化PEのリポソームへの組込みのためには、好ましくはミセルが形成されないように、PEG誘導化PEをCMCより低い濃度で用いることが好ましい。概して、ミセルの形成はPEG誘導化PEのリポソームへの組込みのプロセスを遅延させる。
【0019】
他の態様では、PC(および任意にPE)、PSおよびPEG誘導化PEを、調製されたリポソームに対して使用し、次いでFVIIIを、リポソームと結合するようにおよび/またはリポソームに組込まれるように、添加する。
他の態様では、多様な量のPEGの組込みを、多様な量の、活性化されたPE(アミノ基、カルボキシル基またはチオール基により活性化されたもの)を含めることにより達成し得、FVIIIの組込み後、活性化されたPEを、当該活性化されたPEGに共有結合することができる。PEの存在は、FVIIIのPSとの結合特性を改善することが示されている。本態様の変法では、スペーサーをPEとPEGの間に用いることができる。適当なスペーサーは6〜12個の炭素原子を有する。6〜12個の炭素原子と同じ長さを有する他のスペーサーを用いることができる。
【0020】
さらなる態様では、脂質構造のリポソームはコレステロールを含んでもよい。この態様では、コレステロールを、リポソームまたは他の脂質構造を作成するステップにおいて添加する。
本発明のリポソームは80〜500nmである。一態様では、前記リポソームは100〜200nmの直径である。蛋白質の脂質に対するモル比は1:1000〜1:20,000である。コレステロールは、概して、高粘度のバッファー中で形成され、150〜300nmの平均範囲を有する。ミセルは70〜90nmの範囲内である。
【0021】
本発明に有用なポリエチレングリコールは、700〜30,000の分子量を有し得る。PEGに関して有用な分子量の例は、750、1000、2000、3000、5000、20000、30000Daである。PEGを脂質へ誘導化するために、種々の方法が公知である。例えば、前記誘導化は、塩化シアヌル基により、またはカルボニルジイミダゾールカップリング試薬を用いることにより、為すことができる。より詳細は、米国特許第5,013,556号に見出すことができる。種々のPEG誘導化PE脂質が市場入手可能である。例には、制限するものではないが、DMPE-PEG、DPPE-PEGおよびDSPE-PEGが含まれる。PEGでのPEの誘導化は共有結合による。
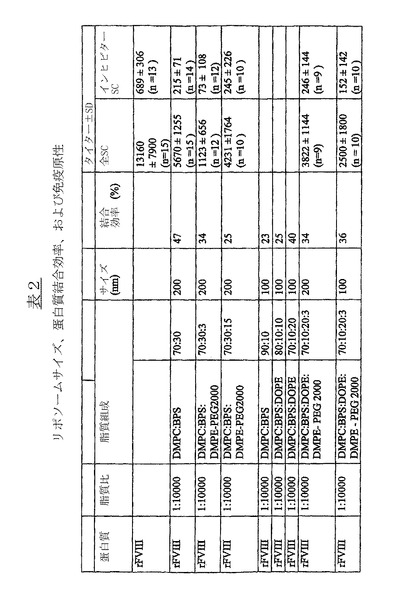
有用なリポソーム組成物の例およびその特性を表2(図7)に示す。
【実施例】
【0022】
以下の実施例を、本発明を説明するために示す。これは、限定を意図するものではない。
[実施例1]
本実施例は、PCを含むリポソームの調製を記載するものである。本実施例では、蛋白質をまず、PS含有リポソームと結合させ、次いでPEGをそれに添加した。
材料
rFVIII(Baxter Biosciences、Carlsband、CA)を抗原として用いた。正常な凝固コントロール血漿および活性アッセイのためのFVIII欠損血漿はTrinity Biotech(Co Wicklow、Ireland)から購入した。脳ホスファチジルセリン(BPS)、ジミリストイルホスファチジルコリン(DMPC)およびクロロホルムに溶解した1,2-ジミリストイル-sn-グリセロ-3-ホスホエタノールアミン-N-[メトキシ(ポリエチレングリコール)-2000](DMPE-PEG2000)をAvanti Polar Lipids(Alabaster、AL)から入手し、-70℃で保存し、さらに精製することなく用いた。滅菌したパイロジェンフリーの水はHenry Schein Inc.(Melville、NY)から購入した。アルカリホスファターゼに結合させたヤギ抗-マウス免疫グロブリン(Ig、IgM+IgG+IgA、H+L)は Southern Biotechnology Associates, Inc. (Birmingham、Alabama)から入手した。モノクローナル抗体ESH8はAmerican Diagnostica Inc, (Greenwich、CT)から入手した。IgG不含牛血清アルブミン(BSA)、ジエタノールアミンおよびアセトンはSigma(Saint Louis、MO)から入手した。p-ニトロフェニルホスフェート二ナトリウム塩はPierce(Rockford、IL)から入手した。1,6-ジフェニル-1,3,5-ヘキサトリエン(DPH)、RPMI-1640培養培地、ペニシリン、ストレプトマイシン、L-グルタミン、2-メルカプトエタノールおよびポリミキシンBはすべてInvitrogen Corp.(Carlsband、CA)から入手した。3H-チミジンはParkin Elmer Inc.(Boston、MA)から入手した。本試験で使用した全ての他のバッファー塩はFisher Scientific(Fairlawn、NJ)から入手し、精製なしで使用した。
【0023】
DMPE-PEG2000のミセル形成臨界濃度(CMC)の決定
DMPE-PEG2000のCMCを、既に記載されているように[16]、蛍光プローブDPHを用いて決定した。簡単には、DPH溶液(2μl、[DPH]=アセトン中30μM)を、種々の濃度の1mlのDMPE-PEG2000に添加後、37℃で2時間インキュべートした。ディスパーションの蛍光強度を、キセノン・アーク・ランプを備えたPTIフルオロメーター(Photon Technology International、Lawrenceville、NJ)を用いて測定した。プローブの励起波長(Ex)を360nmに設定し、発光(Em)を430nmで測定した。蛍光強度が突然増加したディスパーションの濃度をCMCと定義し、これは〜100μMであることが認められた。
【0024】
ポリエチレングリコール-(PEG化)PSリポソームの調製
PEGトランスファー法
所望量のDMPC(Tc〜23℃)およびBPS(Tc〜6-8℃)をクロロホルムに溶解し、溶媒をロータエバポレータ(Buchi-R200、Fisher Scientific)を用いて蒸発させて、丸底フラスコの壁面に薄膜を形成した。あらゆる残存溶媒を乾燥窒素流下にサンプルから除去した。トリスバッファー(TB、300mM NaCl、25mMトリス、5mM CaCl2.2H2O、pH=7.0、滅菌されたパイロジェンフリーの水中で調製されたもの)中37℃で、当該薄脂質膜を補水することによりリポソームを形成した。本試験で用いた脂質のモル比はDMPC:BPS(70:30モル%)であった。リポソームを高圧押出機(Mico、Inc. Middleton、WI)を〜200-250psiの圧力で用いて、3枚の重なった200nmのポリカーボネート膜を通して数回押し出した。リポソームを0.22μmのmillex(登録商標)-GPフィルターユニット(Millipore Corporation、Bedford、MA)を通して滅菌濾過した。脂質の回収を、Bartlettの方法[17]によるリン含量の決定により算定した。リポソームのサイズ分布を、Nicomp Model CW380粒子サイズアナライザー(Particle Sizing Systems、Santa Barbara、CA)を用いて既に記載されているように[18]決定した。サイズを揃えたリポソームを適量のrFVIIIと、37℃で穏やかに攪拌しながら〜30分間インキュべートすることにより結合させた。蛋白質-リポソーム混合物のPEG化は、蛋白質-リポソーム混合物をDMPE-PEG2000の乾燥膜に添加することにより達成した。乾燥PEG膜に添加された蛋白質-リポソーム混合物の当該体積がPEGミセルの形成を生じないことを保証した。PEGの導入をMALDI-TOFにより確認した(データは示さず)。調製物中のPEGの最終モル%は全脂質の4モル%であった。蛋白質と脂質のモル比は、全実験に関して1:10,000に維持した。PEG化されたリポソームに結合した蛋白質の量を算出するために、遊離蛋白質を、PEG化されたリポソーム結合蛋白質から、不連続デキストラン密度勾配遠心分離法[19]を用いて分離した。結合している活性蛋白質の割合を、1段階(one-stage)APTTアッセイ[20]により決定した。結合の割合は、〜27.6±9.6%(±S.E.M、n=5)であると算出された。調製物は、調製後すぐに使用した。
【0025】
リポソームにおけるポリエチレングリコール(PEG)の導入に関する理論的考察
本実施例では、脂質膜の調製中にPEGを導入するのではなく、リポソームの表面に蛋白質を結合後、PEGをリポソームに組込んだ。PEG導入のこの方法は、構造上の妨害の結果として、蛋白質がリポソームと結合する能力がPEGにより妨害される可能性を小さくすると考えられる。発明者等は、以下の理論的考察が、リポソームの表面における蛋白質の存在がPEGの挿入の能率を損ねるという可能性に反対の議論をなすものと信ずる。
【0026】
用いたリポソームの平均直径は200nmであった。二重層の厚さが40Åで、各リン脂質分子が占める面積が70Å2であるという仮定の下、小胞数/リン脂質のμモルは〜1.8×1012小胞であると算定された。免疫研究のために、動物毎に2μgの蛋白質を投与し、使用した蛋白質対脂質のモル過剰(1:10,000)に基づき、各動物は〜71.4nmolの脂質を受容した。電子クリスタログラフィーにより誘導された膜結合FVIIIの三次元構造により、FVIIIドメインがコンパクトな配置を有し、その中で当該蛋白質のC2ドメインがリン脂質と相互作用することが明らかにされた[21]。200nmの平均直径のリポソームの二次元マップのためのユニットセルディメンジョン(the unit cell dimension)および全表面面積に基づき、発明者等は、リポソームの表面に充填され得るFVIII分子の最大数は〜2400であると算定した。しかし、蛋白質分子/小胞の最大数が、発明者等の研究で用いられた蛋白質の脂質に対する比に基づき〜33となるように計画されたとすれば、前記理論的評価により、リポソームの表面の大部分が未だ非占有状態にあり、PEGによるコーティングに利用できることが明らかである。
【0027】
[実施例2]
本実施例は、実施例1で調製したリポソームの特性を記載するものである。
蛍光スペクトロスコピー
rFVIII、およびPEG化リポソームと結合したrFVIIIの発光スペクトルを、PTIフルオロメーター(Quanta Master、Photon Technology International、Lawrenceville、NJ)を用いて得た。サンプルを280nmで励起し、発光スペクトルを300〜400nmから得た。4nmのスリット幅を、励起および発光光路の両方に関して用いた。蛋白質濃度は〜4μg/mlであり、内部フィルター効果を抑えるために、可変光路長キュベットを用いた。
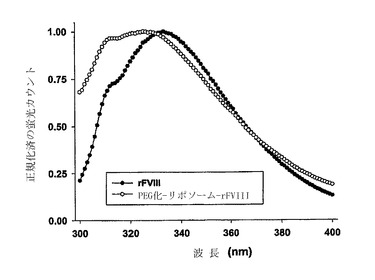
蛋白質における三次元構造の変化を蛍光スペクトロスコピーにより調べた(図1)。遊離状態のFVIIIの発光スペクトルは、333nmの最大発光を示した。PEG化されたリポソームと結合した蛋白質は、最大発光に関して325nmへの有意なブルーシフトを示し、強度の大幅な増加を伴った(データは示さず)。最大発光の明らかなブルーシフトは、rFVIIIの疎水性ドメインのリポソーム二重膜への実質的な層間挿入またはカプセル化の可能性を示唆する。これは、PEGを欠くリポソームへの蛋白質の結合後に観察された発光スペクトルにおける僅かな変化と対照的であった。PEGの不在下では、データは、最小限度の構造変化しか示さず、膜結合形態の蛋白質の大部分はリポソームの表面に存在した。発明者等は、波長シフトが、膜結合に関与するトリプトファン(Trp)残基の、リポソームの表面におけるPEGの存在によるミクロ環境における誘電率の変化、または、PEGの構造的効果による、Trp残基に対する溶媒分子の接近能力の低下の結果であると信じる。周囲の溶媒の誘電率の変化は、励起状態のフルオロフォア付近の溶媒分子の組成を変化させることにより、フルオロフォアのストロークシフトに影響を及ぼすことが示されている[25]。
【0028】
[実施例3]
本実施例は、PS含有ミセルの調製を記載するものである。ジカプロイルホスファチジルセリン(DCPS)およびジカプロイルホスファチジルチオエタノール(DCPSE)(97:3モル比、トータル脂質5μモル)から成る脂質膜を、クロロホルムストック溶液から、溶媒をロータ・エバポレータ中で蒸発させることにより調製した。膜を、1mLのトリスバッファー(5mM CaCl2、25mMトリスおよび300mM NaCl、pH=7)を用いて攪拌することにより再構築し、5mMの脂質溶液を得た。濃縮したrFVIIIストックを5mMの脂質溶液で希釈し、37℃にて30分間インキュべートした。PEG化法は、予め形成されたリポソームの、活性化されたPEG分子を用いるPEG化と同様である。PEG化は、活性化されたPEG分子(鎖状または分枝状mPEGマレイミド)を、リン脂質頭基上に存在する遊離チオール基(DCPSE)に結合することにより達成した。
【0029】
[実施例4]
本実施例は、PS含有コクリエート構造またはシリンダーの調製を記載するものである。純粋な脳ホスファチジルセリン(BPS)およびジオレオイルホスファチジルチオエタノールアミン(DOPSE)(モル比99:1)を含む100nm以下のサイズを揃えたリポソームを、Ca2+-不含トリスバッファー中で調製した。rFVIII-リポソーム複合体を、濃縮されたrFVIII溶液を、サイズを揃えたリポソームの存在下で30分間37℃でインキュべートすることにより作成した。rFVIIIリポソーム複合体の粘度を、デキストラン溶液(20%w/v)を、5または10%w/vの最終デキストラン濃度を達成するまで添加することにより増した。コクリエートシリンダーの制御成長を、前記溶液中にCa2+イオン(5mMの終濃度)を添加し、次いで、混合物をより低い温度で30分間インキュべートすることにより開始する。PEG化を、活性化されたPEG分子(鎖状または分枝状のmPEGマレイミド)をリン脂質頭基上に存在する遊離チオール基(DOPSE)に結合させることにより達成した。加えて、ナノコクリエートシリンダーのPEG化は、活性化されたPEG分子(PEGカルボン酸のN-ヒドロキシスクシンイミドエステル(PEG-NHS))とBPS頭基に存在する遊離アミノ基の間に共有結合を設計することにより行われ得る。PEG-NHS試薬によるrFVIIIの直接PEG化は、PS頭基に存在するアミノ基のかなりの過剰に基づき、殆ど起こりそうにない。
【0030】
[実施例5]
本実施例は、活性化PEG技術による、PEGとのrFVIIIリポソーム複合体の複合化を記載するものである。DMPC:BPS:ジオレオイルホスファチジルチオエタノール(DOPSE)リポソーム(モル比70:25:5)を以下に記載のように調製した。必要量のDMPC、BPSおよびDOPSEをクロロホルムに溶解した。薄い脂質膜を、Buchi-R200ロトエバポレータ(Fisher Scientific)中で溶媒を除去することにより、ガラスチューブの壁面上に形成した。リポソームを、脂質膜をトリスバッファー(TB25mm トリス、300mM NaCl、5mM CaCl2、pH=7.4)で37℃で補水することにより調製した。リポソームを、高圧押出機(Lipex Biomembranes、Inc.)を〜200psiの圧力で用いて二枚の重なった100nmのポリカーボネート膜を通して8回押出した。粒子のサイズ分布をNicompモデルCW380サイズアナライザー(Particle Sizing System)を用いて測定した。
【0031】
リポソーム蛋白質の調製
予め形成したリポソームとの蛋白質の結合を、蛋白質をリポソームの存在下で37℃で30分間、時々穏やかに攪拌しながらインキュべートすることにより達成した。蛋白質の脂質に対するモル比は、全調製物に関して同様に維持した(1:10,000)。
PEG化は、DOPSE脂質の頭基に存在する遊離チオール基と活性化PEG誘導体との間に共有結合を設計することにより達成した。そのような誘導体は、mPEG-マレイミドまたは分枝PEGマレイミドにより表記できる。遊離チオール基を標的する他の活性化PEG誘導体も、リポソームとPEG部の間の共有結合を形成するのに同様に適している。
本方法の有利な点は、チオール基が蛋白質分子の表面あまり存在しないことである。つまり、脂質の大過剰(蛋白質:脂質比は1:10000、脂質の5%はDOPSEである)が、活性化されたPEGのrFVIIIへの結合を減らし、その活性を減らすと予想される。
【0032】
[実施例6]
本実施例は、実施例1に記載される組成物を用いるインビボ研究を記載するものである。血友病のマウス(FVIII遺伝子のエキソン16に標的欠損を有するもの)のコロニー[22]。18〜12週齢の大人の雄および雌のマウスの同数を、rFVIIIに対するその免疫反応の特性が同等であることが示されている[23]ので、本試験のために使用した。
血液サンプルを心臓穿刺により得、10:1(v/v)比でクエン酸デキストロース(ACD、85mMのクエン酸ナトリウム、110mMのD-グルコースおよび71mMのクエン酸を含むもの)に添加した。血漿を遠心分離により分離し、サンプルを-80℃で分析まで保存した。全試験は、バッファローの大学で、動物実験委員会(Institutional Animal Care and Use Committee)(IACUC)のガイドラインに従って行った。
FVIIIノックアウトマウス(n=12)の免疫化は、rFVIIIまたはrFVIII-PEG化リポソーム(2μg)の隔週毎4回の皮下(s.c.)注射から成るものであった。血液サンプルを、6週の終時に得た。
【0033】
抗体測定
全抗-rFVIII抗体の検出
全抗-rFVIII抗体のタイターをELISAにより決定した。簡単には、Nunc-Maxisorbの96ウェルプレートをカーボネートバッファー(0.2M、pH=9.4)中2.5μg/mlのrFVIIIの50μlで覆い、4℃で一晩インキュべートした。プレートを次いで6回、0.05%のトゥイーン20を含むホスフェートバッファー(PB;10mM Na2HPO4、1.8mM KH2PO4、14mM NaCl、2.7mM KCl)(PBT)の100μlで洗浄した。当該プラスチックの吸着表面上の非特異的蛋白質結合部位を、1%のウシ血清アルブミンを含むPBバッファー(PBA)の200μlを2時間室温でインキュべートすることによりブロックした。プレートをPBTで6回洗浄し、次いで50μlの、マウス血漿サンプルのPBA中での種々の希釈物を添加し、37℃で1時間インキュベートした。プレートをPBTで6回洗浄し、アルカリホスファターゼ結合ヤギ抗-マウスIgのPBA中での1:1000希釈の50μlと共に室温で1時間インキュベートした。プレートをPBTと、ジエタノールアミンバッファー(1Mジエタノールアミン、0.5mM MgCl2から成る)中1mg/mlのリン酸p-ニトロフェニルホスフェート溶液の100μlで6回洗浄した。プレートを室温で30分間インキュベートし、反応を、3NのNaOHの100μlを添加することにより停止した。アルカリホスファターゼ反応生成物を、スペクトラマックスプレートリーダー(Molecular Devices Corporation、Sunnyvale、CA)を用いて405nmでの吸光度(absorbance)により決定した。免疫原性の結果を、以下のように表現した。:モノクローナルネズミIgG抗-ヒトFVIII抗体、C2ドメインに結合するESH8を用いて得られた吸光値に関して、線形回帰を行った。最大および最小の予測された吸光度の差の半分を、プレート特異的ファクター(PSF)として算出した。種々の希釈(1:100〜1:40,000)の吸光度の値の、希釈のlogに対するプロットの線形回帰を、PSFに等しい光学密度を与える希釈を算出するのに用いた。このように得られた希釈をサンプルの抗体タイターと見なした。
【0034】
阻害抗-rFVIII抗体の検出
阻害(中和)抗-rFVIII抗体を、ベセスダアッセイ(Bethesda assay)のニジメゲン(Nijmegen)変更[24]を用いて検出した。残るrFVIII活性を、1段階APTTアッセイ[20]を用いて測定した。各希釈を二対で試験した。1ベセスダユニット(BU)は、rFVIII活性の50%阻害を生ずる阻害活性である。50%阻害のポイントを、少なくとも20〜80%阻害の範囲内に収まるデータポイントの線形回帰により決定した。
【0035】
T-細胞増殖試験
8〜12週齢の雌の血友病マウスを、rFVIIIまたはPEG化リポソーム-rFVIIIの、隔週毎2回の皮下(s.c)注射(注射毎2μg蛋白質)を用いて免疫化した。コントロールマウスはrFVIIIを受容しなかった。2回目の注射の3日後に動物を屠殺し、脾臓をT-細胞の源として回収した。脾臓細胞から、CD8細胞上に発現されたLyt2膜抗原(Dynal Biotech、Oslo、Norway)に関するラット抗-マウスモノクローナル抗体でコートした磁気ビーズを用いて、製造業者のプロトコルを用いて、CD8+を除去した。残る細胞(2×105細胞/200μl)を96ウェルの平底プレートにてrFVIII(100ng/ウェルまたは1000ng/ウェル)と共に、10,000U/mlのペニシリン、10mg/mlのストレプトマイシン、2.5mMのピルビン酸ナトリウム、4mMのL-グルタミン、0.05mMの2-メルカプトエタノール、2mg/mlのポリミキシンBおよび0.5%の熱不活化血友病マウス血清を含む完全RPMI-1640培養培地中で培養した。1μCi/ウェルの3H-チミジン(6.7Ci/mmol)を培養の72時間後37℃で添加した。16時間の終時に、細胞をミクロメート・ハーベスター(Packard、Meriden、CT)を用いて回収し、次いで、3H-チミジンの導入をTopCount(登録商標)ミクロプレートシンチレーションおよびルミネセンスカウンター(Packard Instrument Company、Meriden、CT)を用いて測定した。処置群は3匹の同型の動物から成り、各々のマウスからの細胞を、抗原依存性の増幅に関して4ツ組で試験した。データを、抗原の存在下での平均3H-チミジン導入の、抗原の不在下での平均導入に対する比である刺激指標(SI)として示す。この方法により、各試験のデータが正規化され、異なる時間に行われた実験の比較が可能となる。
【0036】
サイトカイン分析
インキュベーションの72時間後、抗原-刺激性T細胞の上清を回収し、さらなる分析のために-70℃で保存した。上清を抗体捕捉ELISA(R&Dシステム、Minneapolis、MN)により分析した。IFN-γを典型的なTh1サイトカインとして測定し、IL-10を典型的なTh2サイトカインとして測定した。
【0037】
薬物動態試験
27匹の雄の血友病マウス(20〜26g、8〜12週齢)に、400IU/kgのrFVIIIまたはPEG化リポソーム-rFVIIIを単一のi.v.ボーラス注射として陰茎静脈経由で受容させた。血液サンプル(〜600μl)を、心臓穿刺により投与後.08、0.5、1、2、4、8、16、24、36および48時間で回収し(n=2〜3匹のマウス/時間点)、ACDに添加した。血漿を分離し、-70℃で分析まで保存した。血漿サンプルを、色原性アッセイ(chromogenic assay)(Coamatic FVIII、DaiPharma Group、West Chaster、OH)により蛋白質の活性に関して分析した。各時点で算出された活性を次いで用いて、Win Nonlin(Pharsight Corporation、Mountainview、CA)を用いて非区画分析(non-compartmental analysis)により基本的な薬物動態パラメータを算出した。
統計学的分析
データをSAS(SAS Institute Inc.、Cary、NC)またはMinitab(Minitab Inc.、State College、PA)のアナリスト・アプリケーション(Analyst Application)を用いて分散分析(ANOVA)により分析した。Dunnetteのpost-hocマルチプル比較試験を用いて有意な差(p<0.05)を検出した。
【0038】
結果
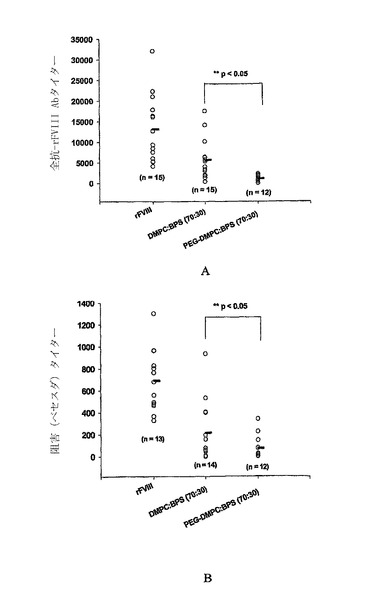
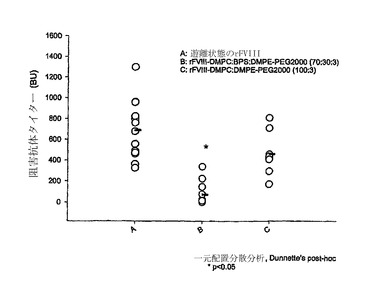
図2のAに、DMPCおよびBPSから成るPEG化リポソームの不在下および存在下での全抗-rFVIII抗体タイターを示す。PEG化リポソーム-rFVIIIで処置した動物は、rFVIIIで処置した動物(13,166.7±2042.2、±S.E.M、n=15)と比較して、有意に低い抗体タイター(1123.1±189.5、±S.E.M、n=12、p-値<0.05)を示した。これらの結果は、抗体形成がPEG化リポソームの存在下で低減されることを示す。
【0039】
蛋白質の活性を妨害する中和抗体(即ち、第VIII因子に対して特異的な抗体)をベセスダアッセイを用いて検出した。図2のBは、rFVIIIおよびPEG化リポソーム-rFVIII処置後6週間の終時にベセスダユニット(BU)にて表現される阻害抗体タイターを示す。データは、中和抗体が、rFVIII単独(689.7±78.1BU/ml、±S.E.M、n=13)と比較して、PEG化リポソームの存在下で有意に低いこと(73.65±31.25BU/ml、±S.E.M.、n=12、p-値<0.05)を示した。かかる結果は、PEG化されたリポソームが全抗-FVIII抗体タイターを低減させるだけでなく、蛋白質を不活性化する抗体のタイターも低下させることを示す。比較目的のために、非-PEG化PC/PSリポソームの投与後の全抗体および阻害タイターも示す。データは、差は統計学的に差はないが(p>0.05)、PEG化リポソームの存在下での平均全抗体および阻害タイターが、非-PEG化リポソームのものよりも低いことを示した。
【0040】
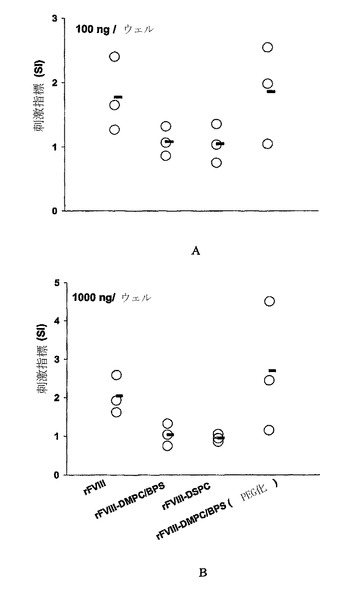
FVIII特異的T細胞が、インビボで、PEG化リポソーム-rFVIIIでの免疫化後に刺激されるかどうかを決定するために、rFVIIIチャレンジに対するT-細胞増殖反応をインビボで評価した。PEG化リポソーム-rFVIII処置を受容した動物から単離された脾臓細胞の平均刺激指標は、rFVIII処置のみを受容した動物と比較して低かった(図3)。データは、動物がPEG化PS含有リポソームの存在下および不在下でrFVIIIに曝されたかどうかにより、クローナル細胞増殖(clonal expansion)のために活性化されたT-細胞クローンにおいて生じ得る差を示唆する。
【0041】
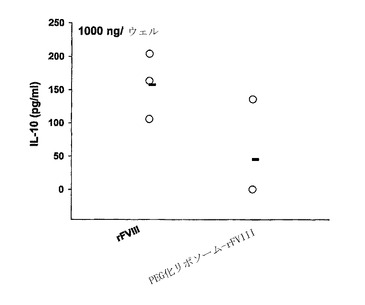
PEG化されたPS含有リポソームの存在下でのrFVIIIの免疫原性の低下が、低下したIL-10の分泌の結果かどうかを決定するために、抗原-刺激性T-細胞のサイトカイン分析を、遊離状態の-またはリポソーム-rFVIIIでの動物の免疫化後に行った。図4に示すように、PEG化リポソームと結合したrFVIIIを与えられた動物のT-細胞により分泌された平均IL-10レベルは、rFVIIIのみ与えられた動物よりも低かった。微量レベルのIFN-γが、全処置群に関して培養培地中で検出された(データは示さず)。総じて、データは、PEG化されたPS-含有リポソームの存在下で投与されたrFVIIIの免疫原性の低下には、部分的には、低下したIL-10生成が介在する可能性があることを示唆する。さらに、データは、免疫原性の低下がTh1/Th2反応の偏光の結果ではないことを示唆する。
【0042】
いかなる特定の理論に拘束されることを意図するものではないが、リポソームにおけるPSの含有が、免疫調節に寄与すると考えられる。rFVIIIに対する抗体反応はT-細胞依存性のプロセスであることを考慮すると、PEG化されたPS含有リポソームの存在下でのrFVIIIの免疫原性の低下は、インビボでのrFVIII特異的T-細胞クローンの抑制の結果である可能性がある。
rFVIIIの免疫原性の低下に加えて、rFVIIIのPA含有リポソームとの結合は、また、インビボでのrFVIIIの循環時間を延長し得、こうして、コントロールの血友病Aに対し、必要とされる蛋白質の投与頻度を減らし得る。薬物動態(PK)研究は、PEG化リポソーム-rFVIIIの循環半減期(t1/2)が、rFVIII単独に対して〜35%まで増加することを示唆した(表1)。全身暴露は、処置間で同様であった(図5および表2)。
【0043】
【表2】
【0044】
[実施例6]
本実施例は、ネガティブに荷電したリン脂質を用いてまたは用いずに調製したPEG結合リポソームの比較分析を提供するものである。阻害タイターを、遊離状態のrFVIII;PASを用いて調製されたリポソームと結合したないしこれに組込まれたFVIII;および、PSを用いずに調製したリポソームと結合したないしこれに組込まれたrFVIIIに関して、実施例5に記載したように決定した。図6に示すように、PSを用いて調製されたリポソームと結合したないしこれに組込まれたrFVIIIに関する阻害抗体タイターは、遊離状態のrFVIIIおよびPSを用いずに調製されたリポソームと結合したないしこれに組込まれたrFVIIIに関するタイターよりも有意に低かった。
当業者は個々の調製を最適化することができる。加えて、PS含有PEG化リポソーム-rFVIIIの免疫原性がrFVIII単独よりもかなり低いという所見は、低免疫原性製剤の開発への有意な進展を示す。
【0045】
引用文献:
[1] J.Klinge、N.M.Ananyeva、C.A. Hauser、E.L. Saenko、血友病A--基礎科学から臨床実施まで、Semin Thromb Hemost 28 (2002) 309-322.
[2] K.Fijnvandraat、W.S. Bril、J.Voorberg、血友病Aにおけるインヒビター開発に関する免疫生物学、Semin Thromb Hemost 29 (2003) 61-68.
[3] P. Lollar、第VIII因子に対する免疫反応の分子特定、Vox Sang 83 Suppl 1 (2002) 403-408.
[4] P. A. Foster、T.S. Zimmerman、第VIII因子の構造および機能、Blood Rev 3 (1989) 180-191.
[5] P.J.Fay、第VIII因子の構造および機能、Thromb Haemost 70 (1993) 63-67.
[6] D. Scandella、M. Mattingly、S. de Graaf、C.A. Fulcher、ヒト第VIII因子インヒビター抗体に関するエピトープの、イムノブロッティングおよび抗体中和による同定、
Blood 74 (1989) 1618-1626.
[7] P. Lollar、ハイブリッドヒト/ブタ第VIII因子を用いる第VIII因子インヒビターの分析、Thromb Haemost 78 (1997) 647-651.
[8] J.F. Healey、I.M. Lubin、H. Nakai、E.L. Saenko、L.W. Hoyer、D. Scandella、P. Lollar、残基484-508は、ヒト第VIII因子のA2ドメインにおける阻害エピトープの主要決定基を含む、J Biol Chem 270 (1995) 14505-14509.
[9] I.M. Lubin、J.F. Healey、R.T. Barrow、D. Scandella、P. Lollar、第VIII因子A2インヒビターエピトープの、アラニンスキャニング変異形成による分析、J Biol Chem 272 (1997) 30191-30195.
[10] P.J.Fay、D. Scandella、第VIII因子A2ドメインに特異的なヒトインヒビター抗体は、サブユニットと第IXa因子の間の相互作用を破壊する、J Biol Chem 274 (1999) 29826-29830.
[11] D. Zhong、E.L. Saenko、M. Shima、M. Felch、D.scandella、いくつかのヒトインヒビター抗体は、第IX因子への第VIII因子の結合を妨害する、Blood 92 (1998) 136-142.
[12] J.F. Healey、R.T. Barrow、H.M. Tamim、I.M.Lubin、M. Shima、D. Scandella、P. Lollar、残基Glu2181-Val2243は、ヒト第VIII因子のC2ドメインに阻害エピトープの主要決定基を含む、Blood 92(1998) 3701-3709.
[13] D. Scandella、G.E. Gilbert、M. Shima、H. Nakai、C. Eagleson、M. Felch、R.Prescott、K.J. Rajalakshmi、L.W. Hoyer、E. Saenko、いくつかの第VIII因子インヒビター抗体は、リン脂質結合部位に重なるC2ドメインアミノ酸2248〜2312に対応する共通のエピトープを認識する、Blood 86 (1995) 1811-1819.
[14] M.T. Reding、D.K. Okita、B.M. Diethelm-Okita、T.A. Anderson、B.M. Conti-Fine、凝固第VIII因子のC2ドメインにおけるヒトCE4+T-細胞エピトープレパートリー、J Thromb Haemost 1(2003) 1777-1784.
[15] K.P. Pratt、J. Qian、E. Ellaban、D.K. Okita、B.M.Diethelm-Okita、B. Conti-Fine、D.W. Scott、第VIII因子C2ドメインにおける免疫優性T-細胞エピトープは、阻害抗体結合部位に位置する、Thromb Haemost 92 (2004) 522-528.
[16] K. Sou、T. Endo、S. Takeoka、E. Tsuchida、ポリ(エチレングリコール)-脂質の、小胞への自発的組込みを用いることによる、リン脂質小胞のポリ(エチレングリコール)-修飾、Bioconjug Chem 11(2000) 372-379.
[17] G.R.Bartlett、カラムクロマトグラフィーにおけるリンアッセイ、J Biol Chem 234(1959) 466-468.
[18] V.S. Purohit、K. Ramani、R.S. Kashi、M.J. Durrani、T.J. Kreiger、S.V. Balasubramanian、ホスファチジルセリン-含有モデル膜に結合した第VIII因子のトポロジー、Biochim Biophys Acta 1617 (2003) 31-38.
[19] T.D. Heath、B.A. Macher、D. Papahadjopoulos、グリコスフィンド脂質によるリポソームへのイムノグロブリンの共有結合、Biochim Biophys Acta 640 (1981) 66-81.
[20] J. Over、第VIII因子(VIII:C)の1段階アッセイの方法、Scand J Haematol Suppl 41 (1984) 13-24.
[21] S. Stoilova-Mcphie、B.O. Villoutreix、K. Mertens、G. Kemball-Cook、A. Holzenburg、膜結合凝固第VIII因子の三次元構造:電子クリスタログラフィーにより誘導された三次元密度マップ内の第VIII因子ヘテロダイマーのモデリング、Blood 99(2002) 1215-1223.
[22] L. Bi、A.M. Lawler、S.E. Antonarakis、K.A. High、J.D. Gearhart、H.H. Kazazian, Jr.、マウス第VIII因子遺伝子の標的破壊は、血友病Aのモデルを生じる、Nat Genet 10 (1995) 119-121.
[23] J. Qian、M. Borovok、L. Bi、H.H. Kazazian, Jr.、L.W, Hoyer、ネズミ血友病Aにおけるヒト第VIII因子に対するインヒビター抗体発生およびT細胞反応、Thromb Haemost 81 (1999) 240-244.
[24] B. Verbruggen、I. Novakova、H. Wessels、J. Boezeman、M. van den Berg、E. Mauser-Bunschoten、第VIII因子に関するベセスダアッセイのニジメゲン変更:Cインヒビター:改良された特異性および信頼性、Thromb Haemost 73 (1995) 247-251.
[25] J.R. Lakowicz、蛍光スペクトロスコピーの原理、Kluwer Academic/Plenum.
【図面の簡単な説明】
【0046】
【図1】PEG化リポソームの存在下および不在下でのrFVIIIの三次元構造。蛍光発光スペクトルを300〜400nmの範囲で取得した。励起モノクロメータは280nmで設定した。用いた蛋白質濃度は〜4μg/mlであった。
【図2】DMPC:BPS(70:30)から成るPEG化リポソームの存在下および不在下でのrFVIIIの投与後6週の終時での、血友病マウスにおける全抗-FVIII抗体タイター(A)および阻害抗-rFVIII抗体(B)。各ポイントは、処置を受けた個々のマウスからの値を示し、水平バーは、全抗体または阻害タイターの平均を示す。比較目的のために、非-PEG化DMPC:BPSリポソームの存在下でrFVIIIの投与後に得られたデータも示す。血液サンプルを、4回目の注射の2週間後に得た。全抗FVIII抗体タイターはELISAにより決定し、阻害タイターはベセスダアッセイにより決定した。統計学的分析は、実施例に記載するように行った。
【図3】2μgのrFVIII、非-PEG化リポソーム-rFVIII、PEG化リポソーム-rFVIIIまたはPS不含リポソーム-rFVIIIの2回の皮下投与後の、多様な免疫優性エピトープを運搬している完全なrFVIII(100ng/ウェル(A)または1000ng/ウェル(B))に対する血友病マウスのCD4+T細胞増殖反応を刺激指標として表記したもの。刺激指標の算出および統計学的分析は実施例に記載されている。各ポイントは個々の動物からの値を示し、水平バーは刺激指標の平均を示す。
【図4】2μgの遊離状態のrFVIIIまたはPEG化リポソーム-rFVIIIの2回の皮下投与製剤の投与を受けた動物からの、抗原でチャレンジされたCD4+T細胞によるIL-10の分泌。CD4+に富むT細胞をrFVIII(1000ng/ウェル)でチャレンジした。各ポイントは個々の動物からの値を示し、水平バーは培養培地中に分泌された平均IL-10レベルを示す。統計学的分析は、実施例に記載されているように行った。
【図5】rFVIII、PEG化または非-PEG化リポソーム-rFVIII投与後の血友病マウスにおける、血漿rFIII活性の時間に対する特性
【図6】種々の脂質組成のPEG化-リポソームの不在および存在下でのrFVIIIの投与後6週の終時での、血友病マウスにおける阻害抗-rFVIII抗体。各ポイントは処置を受けた個々のマウスからの値を示し、水平バーは、全抗体または阻害抗体タイターの平均を示す。血液サンプルを4回目の注射の2週間後に得た。阻害タイターはベセスダアッセイにより決定した。統計学的分析は実施例に記載されているように行った。
【図7】本発明のいくつかのリポソーム組成物の例およびそれらのリポソームサイズ、蛋白質結合効率および免疫原性。
【技術分野】
【0001】
本願は、2006年6月29日に出願された米国仮出願第60/695,080号に対する優先権を主張し、その内容を出典明示により本明細書中に組込む。
本発明は、ザ・ナショナル・インスティテュート・オブ・ヘルスからの交付番号第R01HL-70227下に政府基金により支持されたものである。当該政府が本発明に関する所定の権利を有する。
本発明は概して、治療に関して免疫原性を低減するための手段に関するものであり、より詳細には、第VIII因子の免疫原性を低減するための組成物および方法を提供するものである。
【背景技術】
【0002】
血友病Aは、第VIII因子(FVIII)の欠損または機能障害により特徴づけられる遺伝性の出血性疾患である。FVIIIは、凝固カスケードの本質的経路における決定的な補因子として作用する。組換えヒトFVIII(rFVIII)または血漿由来のFVIIIでの補充療法は、出血発現を制御するのに用いられる最も一般的な治療である。しかし、投与される前記蛋白質に対する中和抗体の導入は、患者の約15〜30%において、治療における主要問題である[1-3]。中和抗体は、インビボでリン脂質への結合にも関与するC2ドメインをしばしば標的する。
【0003】
FVIIIは、ドメインA1、A2、B、A3、C1およびC2から成る巨大なマルチドメイン糖蛋白質である[4、5]。系統的なエピトープマッピング研究により、抗-FVIII抗体が、FVIIIのA2(重鎖)、A3およびC2ドメイン(軽鎖)における規定領域を主に標的することが明らかになった[6、7]。A2ドメイン内のエピトープ決定因子は、残基Arg484-Ile-508にマップされている[8、9]。この領域を標的する抗体は、A2ドメインの第IXa因子(FIXa)との相互作用を遮断することによりFVIIIの活性化形態(FVIIIa)を阻害することが示されている[10]。A3ドメイン内の主要なエピトープ決定因子は、残基1811〜1818から成り、この領域に対する抗体も、FVIIIのFIXaとの相互作用を防いで補因子の活性を減損させる[11]。C2ドメイン内のエピトープ決定因子は残基2181〜2312にマップされており[12、13]、これは免疫優性、ユニバーサルCD4+エピトープ、2191〜2210、2241〜2290、2291〜2330を含む[14、15]。C2ドメインに対する抗体は、凝固カスケードの増幅に必須の、ホスファチジルセリン(PS)に富む血小板膜表面へのFVIIIの結合を妨害する。
【0004】
投与される第VIII因子に対して生じる免疫反応のために、第VIII因子の免疫原性を、好ましくは循環半減期に悪影響を及ぼすことなく、低減させる製剤の同定が必要とされている。
【発明の開示】
【発明が解決しようとする課題】
【0005】
発明者等は、ネガティブに荷電した脂質(ホスファチジルセリンを含むリン脂質等)およびPEG誘導化リン脂質を含むリポソームおよび他の脂質構造が、第VIII因子等の蛋白質の免疫原性を高め得るか否かを研究した。
一例では、PSおよびPEG誘導化PEを含むリポソームに結合したおよび/または当該リポソームに組込まれたrFVIIIの免疫原性を、血友病Aのネズミモデルで評価した。かかる組成物で処置された動物は、rFVIII単独で処置された動物と比較して、全-および阻害抗-rFVIII抗体の両方に関して低いタイターを有した。本発明の組成物を受容した動物から単離した脾臓細胞の平均刺激指数は、rFVIIIのみを受容した動物よりも低かった。サイトカイン分析により、これらリポソーム組成物の存在下で投与されるrFVIIIの免疫原性の低減には、部分的に、低減したIL-10の生成が介在する可能性があることが示唆された。静脈(i.v.)投与後の薬物動態研究は、これらの組成物を用いてrFVIIIの循環半減期が増加することを示した。
【0006】
従って、蛋白質の免疫原性が、循環半減期を有意に損なうことなく低減された組成物が本願において提供される。この組成物は、ネガティブに荷電した脂質、PEGで誘導化された両親媒性脂質、およびFVIII等の蛋白質を含むリポソームおよび/または他の脂質構造を含む。本明細書中に記載されるPEGを含むリポソームおよび他の脂質構造は、「PEG化」されていると本願において記載される。前記組成物の調製および使用のための方法も提供される。
【0007】
本明細書中に用いる省略は次のとおりである。:APTT、活性化部分トロンボプラスチン時間;ACD、クエン酸デキストロース;BPS、脳ホスファチジルセリン;BSA、牛血清アルブミン;DMPC、ジミリストイルホスファチジルコリン;DMPE-PEG2000、1,2-ジミリストイル-sn-グリセロ-3-ホスホエタノールアミン-N-[メトキシ(ポリエチレングリコール)-2000];ELISA、酵素結合免疫吸着アッセイ;FVIIIa、活性化されたFVIII;FIXa、第IXa因子;Ig、免疫グロブリン;KO、ノックアウト;PB、ホスフェートバッファー;PBA、アルブミンを含むホスフェートバッファー;PBT、トゥイーンを含むホスフェートバッファー;PA、ホスファチジン酸;PC、ホスファチジルコリン;PS、ホスファチジルセリン;rFVIIa、組換え第VIIIa因子;rFVIII、組換えヒト第VIII因子;RES、細網内皮系;TB、トリスバッファー。
【課題を解決するための手段】
【0008】
本発明により、rFVIII製剤が提供される。この製剤は、PSまたはPA等のネガティブに荷電した脂質を含むリポソームおよび/または他の脂質構造(ミセルまたはコクリエート等)を含む。リポソームは、PEGで誘導化された第一の両親媒性脂質(PE等)と、PC、PE(PEGで誘導化されていないもの)またはPG等の第二の両親媒性脂質とを含む。ネガティブに荷電した脂質に加えて、ミセルはPCおよび/または非-PEG誘導化PEを含んでよい。ネガティブに荷電した脂質に加えて、コクリエートはPCを含んでもよい。
【0009】
本発明の組成物は、第VIII因子が低免疫原性であり、遊離状態の第VIII因子よりも長い循環半減期を有することを特徴とするものである。特に、本発明により、免疫優性エピトープが遮蔽された脂質-rFVIII調製物が提供される。低い免疫原性および長い循環半減期のために、前記蛋白質の投与の頻度を減らすことができる。
【0010】
本発明の組成物は、ネガティブに荷電した脂質、PEG誘導化両親媒性脂質を含む脂質構造を含む。第VIII因子または他の蛋白質またはポリペプチドはかかる構造と結合でき(即ち、表面吸着され得)、またはかかる構造に導入され得る。前記蛋白質は、PSまたはPA等のネガティブに荷電した脂質と結合すると考えられる。
【0011】
両親媒性脂質の例は、PC、PEおよびPGである。ネガティブに荷電した脂質の例は、PSおよびPAである。PEGで誘導化され得る脂質の例はPEである。PEは単独で、および./またはPEGで誘導化されるように、脂質構造に用いられ得ることに注意すべきである。
一態様では、前記蛋白質はFVIIIである。インビボデータを、血友病Aのネズミモデルにおいて示す。データは、PSを含むPEG化リポソームrFVIIIの投与が当該蛋白質の免疫原性を低減し、rFVIIIのt1/2の増加を生じることを示す。
【0012】
本発明のリポソームに関し、ネガティブに荷電した脂質の量は30〜50モル%の範囲である。両親媒性脂質の量は50〜70モル%の範囲である。PEGで誘導化されたPEは1〜15モル%の範囲である。任意に、前記リポソームはコレステロールを0〜30モル%の範囲で含んでいてもよい。一態様では、PCのPSに対する比は、50:50〜90:10の範囲内である。一態様では、前記比は70:30である。20%までのPSまたはPCを、非-PEG誘導化PEにより置き換えることができる。
【0013】
本発明のリン脂質は2つのアシル鎖を有する。グリセロール骨格に結合する前記アシル鎖の長さは12〜22個の炭素原子長で変化する。前記グリセロール骨格に結合する2つのアシル基は同じであっても異なっていてもよい。前記アシル鎖は飽和したものであっても不飽和のものであってもよい。12〜22個の炭素原子の飽和および不飽和アシル鎖のいくつかの非限定的な例を表1Aと1Bに示す。
【0014】
【表1】
【0015】
【表1B】
【0016】
短鎖(6〜12個の炭素原子)のホスファチジルセリンは、臨界ミセル濃度以上の濃度でミセルとして存在し得るユニークな水溶性脂質である。短鎖ホスファチジルセリンはrFVIIIと相互作用し、rFVIIIの安定性、免疫原性および薬物動態パラメータに影響する。PEG誘導化PEはミセルに用いることもできる。
【0017】
追加的に、ネガティブに荷電した脂質およびPEG誘導化PEを含むコクリエート構造またはシリンダーを調製することもできる。これらの茎は薬物送達システムとして有用である。コクリエートの調製のためには、長鎖(12〜22個の炭素原子)のリン脂質を用いる。
ミセルは100モル%のPSおよび1〜15モル%のPEG誘導化PEを含んでよい。任意に50%までのPSをPCで置き換えてもよく、および/または5%までのPSをPE(PEGで誘導化されていないもの)で置き換えてもよい。ミセルに関し、50%までのPSをPCで置き換えてよく、および/または5%までPEで置き換えてよい。
コクリエートは100モル%のPSを含んでもよい。30モル%までのPSをPCで置き換えてもよい。
【0018】
本発明の組成物はいくつかの方法により調製できる。例えば、一態様では、前記方法は、PS、PCおよび/またはPEを含むリポソームを調製し、このリポソームにFVIIIを結合および/または組込み、次いでPEG誘導化PEを、FVIII結合/組込みリポソームに添加することを含む。PEG誘導化PEのリポソームへの組込みのためには、好ましくはミセルが形成されないように、PEG誘導化PEをCMCより低い濃度で用いることが好ましい。概して、ミセルの形成はPEG誘導化PEのリポソームへの組込みのプロセスを遅延させる。
【0019】
他の態様では、PC(および任意にPE)、PSおよびPEG誘導化PEを、調製されたリポソームに対して使用し、次いでFVIIIを、リポソームと結合するようにおよび/またはリポソームに組込まれるように、添加する。
他の態様では、多様な量のPEGの組込みを、多様な量の、活性化されたPE(アミノ基、カルボキシル基またはチオール基により活性化されたもの)を含めることにより達成し得、FVIIIの組込み後、活性化されたPEを、当該活性化されたPEGに共有結合することができる。PEの存在は、FVIIIのPSとの結合特性を改善することが示されている。本態様の変法では、スペーサーをPEとPEGの間に用いることができる。適当なスペーサーは6〜12個の炭素原子を有する。6〜12個の炭素原子と同じ長さを有する他のスペーサーを用いることができる。
【0020】
さらなる態様では、脂質構造のリポソームはコレステロールを含んでもよい。この態様では、コレステロールを、リポソームまたは他の脂質構造を作成するステップにおいて添加する。
本発明のリポソームは80〜500nmである。一態様では、前記リポソームは100〜200nmの直径である。蛋白質の脂質に対するモル比は1:1000〜1:20,000である。コレステロールは、概して、高粘度のバッファー中で形成され、150〜300nmの平均範囲を有する。ミセルは70〜90nmの範囲内である。
【0021】
本発明に有用なポリエチレングリコールは、700〜30,000の分子量を有し得る。PEGに関して有用な分子量の例は、750、1000、2000、3000、5000、20000、30000Daである。PEGを脂質へ誘導化するために、種々の方法が公知である。例えば、前記誘導化は、塩化シアヌル基により、またはカルボニルジイミダゾールカップリング試薬を用いることにより、為すことができる。より詳細は、米国特許第5,013,556号に見出すことができる。種々のPEG誘導化PE脂質が市場入手可能である。例には、制限するものではないが、DMPE-PEG、DPPE-PEGおよびDSPE-PEGが含まれる。PEGでのPEの誘導化は共有結合による。
有用なリポソーム組成物の例およびその特性を表2(図7)に示す。
【実施例】
【0022】
以下の実施例を、本発明を説明するために示す。これは、限定を意図するものではない。
[実施例1]
本実施例は、PCを含むリポソームの調製を記載するものである。本実施例では、蛋白質をまず、PS含有リポソームと結合させ、次いでPEGをそれに添加した。
材料
rFVIII(Baxter Biosciences、Carlsband、CA)を抗原として用いた。正常な凝固コントロール血漿および活性アッセイのためのFVIII欠損血漿はTrinity Biotech(Co Wicklow、Ireland)から購入した。脳ホスファチジルセリン(BPS)、ジミリストイルホスファチジルコリン(DMPC)およびクロロホルムに溶解した1,2-ジミリストイル-sn-グリセロ-3-ホスホエタノールアミン-N-[メトキシ(ポリエチレングリコール)-2000](DMPE-PEG2000)をAvanti Polar Lipids(Alabaster、AL)から入手し、-70℃で保存し、さらに精製することなく用いた。滅菌したパイロジェンフリーの水はHenry Schein Inc.(Melville、NY)から購入した。アルカリホスファターゼに結合させたヤギ抗-マウス免疫グロブリン(Ig、IgM+IgG+IgA、H+L)は Southern Biotechnology Associates, Inc. (Birmingham、Alabama)から入手した。モノクローナル抗体ESH8はAmerican Diagnostica Inc, (Greenwich、CT)から入手した。IgG不含牛血清アルブミン(BSA)、ジエタノールアミンおよびアセトンはSigma(Saint Louis、MO)から入手した。p-ニトロフェニルホスフェート二ナトリウム塩はPierce(Rockford、IL)から入手した。1,6-ジフェニル-1,3,5-ヘキサトリエン(DPH)、RPMI-1640培養培地、ペニシリン、ストレプトマイシン、L-グルタミン、2-メルカプトエタノールおよびポリミキシンBはすべてInvitrogen Corp.(Carlsband、CA)から入手した。3H-チミジンはParkin Elmer Inc.(Boston、MA)から入手した。本試験で使用した全ての他のバッファー塩はFisher Scientific(Fairlawn、NJ)から入手し、精製なしで使用した。
【0023】
DMPE-PEG2000のミセル形成臨界濃度(CMC)の決定
DMPE-PEG2000のCMCを、既に記載されているように[16]、蛍光プローブDPHを用いて決定した。簡単には、DPH溶液(2μl、[DPH]=アセトン中30μM)を、種々の濃度の1mlのDMPE-PEG2000に添加後、37℃で2時間インキュべートした。ディスパーションの蛍光強度を、キセノン・アーク・ランプを備えたPTIフルオロメーター(Photon Technology International、Lawrenceville、NJ)を用いて測定した。プローブの励起波長(Ex)を360nmに設定し、発光(Em)を430nmで測定した。蛍光強度が突然増加したディスパーションの濃度をCMCと定義し、これは〜100μMであることが認められた。
【0024】
ポリエチレングリコール-(PEG化)PSリポソームの調製
PEGトランスファー法
所望量のDMPC(Tc〜23℃)およびBPS(Tc〜6-8℃)をクロロホルムに溶解し、溶媒をロータエバポレータ(Buchi-R200、Fisher Scientific)を用いて蒸発させて、丸底フラスコの壁面に薄膜を形成した。あらゆる残存溶媒を乾燥窒素流下にサンプルから除去した。トリスバッファー(TB、300mM NaCl、25mMトリス、5mM CaCl2.2H2O、pH=7.0、滅菌されたパイロジェンフリーの水中で調製されたもの)中37℃で、当該薄脂質膜を補水することによりリポソームを形成した。本試験で用いた脂質のモル比はDMPC:BPS(70:30モル%)であった。リポソームを高圧押出機(Mico、Inc. Middleton、WI)を〜200-250psiの圧力で用いて、3枚の重なった200nmのポリカーボネート膜を通して数回押し出した。リポソームを0.22μmのmillex(登録商標)-GPフィルターユニット(Millipore Corporation、Bedford、MA)を通して滅菌濾過した。脂質の回収を、Bartlettの方法[17]によるリン含量の決定により算定した。リポソームのサイズ分布を、Nicomp Model CW380粒子サイズアナライザー(Particle Sizing Systems、Santa Barbara、CA)を用いて既に記載されているように[18]決定した。サイズを揃えたリポソームを適量のrFVIIIと、37℃で穏やかに攪拌しながら〜30分間インキュべートすることにより結合させた。蛋白質-リポソーム混合物のPEG化は、蛋白質-リポソーム混合物をDMPE-PEG2000の乾燥膜に添加することにより達成した。乾燥PEG膜に添加された蛋白質-リポソーム混合物の当該体積がPEGミセルの形成を生じないことを保証した。PEGの導入をMALDI-TOFにより確認した(データは示さず)。調製物中のPEGの最終モル%は全脂質の4モル%であった。蛋白質と脂質のモル比は、全実験に関して1:10,000に維持した。PEG化されたリポソームに結合した蛋白質の量を算出するために、遊離蛋白質を、PEG化されたリポソーム結合蛋白質から、不連続デキストラン密度勾配遠心分離法[19]を用いて分離した。結合している活性蛋白質の割合を、1段階(one-stage)APTTアッセイ[20]により決定した。結合の割合は、〜27.6±9.6%(±S.E.M、n=5)であると算出された。調製物は、調製後すぐに使用した。
【0025】
リポソームにおけるポリエチレングリコール(PEG)の導入に関する理論的考察
本実施例では、脂質膜の調製中にPEGを導入するのではなく、リポソームの表面に蛋白質を結合後、PEGをリポソームに組込んだ。PEG導入のこの方法は、構造上の妨害の結果として、蛋白質がリポソームと結合する能力がPEGにより妨害される可能性を小さくすると考えられる。発明者等は、以下の理論的考察が、リポソームの表面における蛋白質の存在がPEGの挿入の能率を損ねるという可能性に反対の議論をなすものと信ずる。
【0026】
用いたリポソームの平均直径は200nmであった。二重層の厚さが40Åで、各リン脂質分子が占める面積が70Å2であるという仮定の下、小胞数/リン脂質のμモルは〜1.8×1012小胞であると算定された。免疫研究のために、動物毎に2μgの蛋白質を投与し、使用した蛋白質対脂質のモル過剰(1:10,000)に基づき、各動物は〜71.4nmolの脂質を受容した。電子クリスタログラフィーにより誘導された膜結合FVIIIの三次元構造により、FVIIIドメインがコンパクトな配置を有し、その中で当該蛋白質のC2ドメインがリン脂質と相互作用することが明らかにされた[21]。200nmの平均直径のリポソームの二次元マップのためのユニットセルディメンジョン(the unit cell dimension)および全表面面積に基づき、発明者等は、リポソームの表面に充填され得るFVIII分子の最大数は〜2400であると算定した。しかし、蛋白質分子/小胞の最大数が、発明者等の研究で用いられた蛋白質の脂質に対する比に基づき〜33となるように計画されたとすれば、前記理論的評価により、リポソームの表面の大部分が未だ非占有状態にあり、PEGによるコーティングに利用できることが明らかである。
【0027】
[実施例2]
本実施例は、実施例1で調製したリポソームの特性を記載するものである。
蛍光スペクトロスコピー
rFVIII、およびPEG化リポソームと結合したrFVIIIの発光スペクトルを、PTIフルオロメーター(Quanta Master、Photon Technology International、Lawrenceville、NJ)を用いて得た。サンプルを280nmで励起し、発光スペクトルを300〜400nmから得た。4nmのスリット幅を、励起および発光光路の両方に関して用いた。蛋白質濃度は〜4μg/mlであり、内部フィルター効果を抑えるために、可変光路長キュベットを用いた。
蛋白質における三次元構造の変化を蛍光スペクトロスコピーにより調べた(図1)。遊離状態のFVIIIの発光スペクトルは、333nmの最大発光を示した。PEG化されたリポソームと結合した蛋白質は、最大発光に関して325nmへの有意なブルーシフトを示し、強度の大幅な増加を伴った(データは示さず)。最大発光の明らかなブルーシフトは、rFVIIIの疎水性ドメインのリポソーム二重膜への実質的な層間挿入またはカプセル化の可能性を示唆する。これは、PEGを欠くリポソームへの蛋白質の結合後に観察された発光スペクトルにおける僅かな変化と対照的であった。PEGの不在下では、データは、最小限度の構造変化しか示さず、膜結合形態の蛋白質の大部分はリポソームの表面に存在した。発明者等は、波長シフトが、膜結合に関与するトリプトファン(Trp)残基の、リポソームの表面におけるPEGの存在によるミクロ環境における誘電率の変化、または、PEGの構造的効果による、Trp残基に対する溶媒分子の接近能力の低下の結果であると信じる。周囲の溶媒の誘電率の変化は、励起状態のフルオロフォア付近の溶媒分子の組成を変化させることにより、フルオロフォアのストロークシフトに影響を及ぼすことが示されている[25]。
【0028】
[実施例3]
本実施例は、PS含有ミセルの調製を記載するものである。ジカプロイルホスファチジルセリン(DCPS)およびジカプロイルホスファチジルチオエタノール(DCPSE)(97:3モル比、トータル脂質5μモル)から成る脂質膜を、クロロホルムストック溶液から、溶媒をロータ・エバポレータ中で蒸発させることにより調製した。膜を、1mLのトリスバッファー(5mM CaCl2、25mMトリスおよび300mM NaCl、pH=7)を用いて攪拌することにより再構築し、5mMの脂質溶液を得た。濃縮したrFVIIIストックを5mMの脂質溶液で希釈し、37℃にて30分間インキュべートした。PEG化法は、予め形成されたリポソームの、活性化されたPEG分子を用いるPEG化と同様である。PEG化は、活性化されたPEG分子(鎖状または分枝状mPEGマレイミド)を、リン脂質頭基上に存在する遊離チオール基(DCPSE)に結合することにより達成した。
【0029】
[実施例4]
本実施例は、PS含有コクリエート構造またはシリンダーの調製を記載するものである。純粋な脳ホスファチジルセリン(BPS)およびジオレオイルホスファチジルチオエタノールアミン(DOPSE)(モル比99:1)を含む100nm以下のサイズを揃えたリポソームを、Ca2+-不含トリスバッファー中で調製した。rFVIII-リポソーム複合体を、濃縮されたrFVIII溶液を、サイズを揃えたリポソームの存在下で30分間37℃でインキュべートすることにより作成した。rFVIIIリポソーム複合体の粘度を、デキストラン溶液(20%w/v)を、5または10%w/vの最終デキストラン濃度を達成するまで添加することにより増した。コクリエートシリンダーの制御成長を、前記溶液中にCa2+イオン(5mMの終濃度)を添加し、次いで、混合物をより低い温度で30分間インキュべートすることにより開始する。PEG化を、活性化されたPEG分子(鎖状または分枝状のmPEGマレイミド)をリン脂質頭基上に存在する遊離チオール基(DOPSE)に結合させることにより達成した。加えて、ナノコクリエートシリンダーのPEG化は、活性化されたPEG分子(PEGカルボン酸のN-ヒドロキシスクシンイミドエステル(PEG-NHS))とBPS頭基に存在する遊離アミノ基の間に共有結合を設計することにより行われ得る。PEG-NHS試薬によるrFVIIIの直接PEG化は、PS頭基に存在するアミノ基のかなりの過剰に基づき、殆ど起こりそうにない。
【0030】
[実施例5]
本実施例は、活性化PEG技術による、PEGとのrFVIIIリポソーム複合体の複合化を記載するものである。DMPC:BPS:ジオレオイルホスファチジルチオエタノール(DOPSE)リポソーム(モル比70:25:5)を以下に記載のように調製した。必要量のDMPC、BPSおよびDOPSEをクロロホルムに溶解した。薄い脂質膜を、Buchi-R200ロトエバポレータ(Fisher Scientific)中で溶媒を除去することにより、ガラスチューブの壁面上に形成した。リポソームを、脂質膜をトリスバッファー(TB25mm トリス、300mM NaCl、5mM CaCl2、pH=7.4)で37℃で補水することにより調製した。リポソームを、高圧押出機(Lipex Biomembranes、Inc.)を〜200psiの圧力で用いて二枚の重なった100nmのポリカーボネート膜を通して8回押出した。粒子のサイズ分布をNicompモデルCW380サイズアナライザー(Particle Sizing System)を用いて測定した。
【0031】
リポソーム蛋白質の調製
予め形成したリポソームとの蛋白質の結合を、蛋白質をリポソームの存在下で37℃で30分間、時々穏やかに攪拌しながらインキュべートすることにより達成した。蛋白質の脂質に対するモル比は、全調製物に関して同様に維持した(1:10,000)。
PEG化は、DOPSE脂質の頭基に存在する遊離チオール基と活性化PEG誘導体との間に共有結合を設計することにより達成した。そのような誘導体は、mPEG-マレイミドまたは分枝PEGマレイミドにより表記できる。遊離チオール基を標的する他の活性化PEG誘導体も、リポソームとPEG部の間の共有結合を形成するのに同様に適している。
本方法の有利な点は、チオール基が蛋白質分子の表面あまり存在しないことである。つまり、脂質の大過剰(蛋白質:脂質比は1:10000、脂質の5%はDOPSEである)が、活性化されたPEGのrFVIIIへの結合を減らし、その活性を減らすと予想される。
【0032】
[実施例6]
本実施例は、実施例1に記載される組成物を用いるインビボ研究を記載するものである。血友病のマウス(FVIII遺伝子のエキソン16に標的欠損を有するもの)のコロニー[22]。18〜12週齢の大人の雄および雌のマウスの同数を、rFVIIIに対するその免疫反応の特性が同等であることが示されている[23]ので、本試験のために使用した。
血液サンプルを心臓穿刺により得、10:1(v/v)比でクエン酸デキストロース(ACD、85mMのクエン酸ナトリウム、110mMのD-グルコースおよび71mMのクエン酸を含むもの)に添加した。血漿を遠心分離により分離し、サンプルを-80℃で分析まで保存した。全試験は、バッファローの大学で、動物実験委員会(Institutional Animal Care and Use Committee)(IACUC)のガイドラインに従って行った。
FVIIIノックアウトマウス(n=12)の免疫化は、rFVIIIまたはrFVIII-PEG化リポソーム(2μg)の隔週毎4回の皮下(s.c.)注射から成るものであった。血液サンプルを、6週の終時に得た。
【0033】
抗体測定
全抗-rFVIII抗体の検出
全抗-rFVIII抗体のタイターをELISAにより決定した。簡単には、Nunc-Maxisorbの96ウェルプレートをカーボネートバッファー(0.2M、pH=9.4)中2.5μg/mlのrFVIIIの50μlで覆い、4℃で一晩インキュべートした。プレートを次いで6回、0.05%のトゥイーン20を含むホスフェートバッファー(PB;10mM Na2HPO4、1.8mM KH2PO4、14mM NaCl、2.7mM KCl)(PBT)の100μlで洗浄した。当該プラスチックの吸着表面上の非特異的蛋白質結合部位を、1%のウシ血清アルブミンを含むPBバッファー(PBA)の200μlを2時間室温でインキュべートすることによりブロックした。プレートをPBTで6回洗浄し、次いで50μlの、マウス血漿サンプルのPBA中での種々の希釈物を添加し、37℃で1時間インキュベートした。プレートをPBTで6回洗浄し、アルカリホスファターゼ結合ヤギ抗-マウスIgのPBA中での1:1000希釈の50μlと共に室温で1時間インキュベートした。プレートをPBTと、ジエタノールアミンバッファー(1Mジエタノールアミン、0.5mM MgCl2から成る)中1mg/mlのリン酸p-ニトロフェニルホスフェート溶液の100μlで6回洗浄した。プレートを室温で30分間インキュベートし、反応を、3NのNaOHの100μlを添加することにより停止した。アルカリホスファターゼ反応生成物を、スペクトラマックスプレートリーダー(Molecular Devices Corporation、Sunnyvale、CA)を用いて405nmでの吸光度(absorbance)により決定した。免疫原性の結果を、以下のように表現した。:モノクローナルネズミIgG抗-ヒトFVIII抗体、C2ドメインに結合するESH8を用いて得られた吸光値に関して、線形回帰を行った。最大および最小の予測された吸光度の差の半分を、プレート特異的ファクター(PSF)として算出した。種々の希釈(1:100〜1:40,000)の吸光度の値の、希釈のlogに対するプロットの線形回帰を、PSFに等しい光学密度を与える希釈を算出するのに用いた。このように得られた希釈をサンプルの抗体タイターと見なした。
【0034】
阻害抗-rFVIII抗体の検出
阻害(中和)抗-rFVIII抗体を、ベセスダアッセイ(Bethesda assay)のニジメゲン(Nijmegen)変更[24]を用いて検出した。残るrFVIII活性を、1段階APTTアッセイ[20]を用いて測定した。各希釈を二対で試験した。1ベセスダユニット(BU)は、rFVIII活性の50%阻害を生ずる阻害活性である。50%阻害のポイントを、少なくとも20〜80%阻害の範囲内に収まるデータポイントの線形回帰により決定した。
【0035】
T-細胞増殖試験
8〜12週齢の雌の血友病マウスを、rFVIIIまたはPEG化リポソーム-rFVIIIの、隔週毎2回の皮下(s.c)注射(注射毎2μg蛋白質)を用いて免疫化した。コントロールマウスはrFVIIIを受容しなかった。2回目の注射の3日後に動物を屠殺し、脾臓をT-細胞の源として回収した。脾臓細胞から、CD8細胞上に発現されたLyt2膜抗原(Dynal Biotech、Oslo、Norway)に関するラット抗-マウスモノクローナル抗体でコートした磁気ビーズを用いて、製造業者のプロトコルを用いて、CD8+を除去した。残る細胞(2×105細胞/200μl)を96ウェルの平底プレートにてrFVIII(100ng/ウェルまたは1000ng/ウェル)と共に、10,000U/mlのペニシリン、10mg/mlのストレプトマイシン、2.5mMのピルビン酸ナトリウム、4mMのL-グルタミン、0.05mMの2-メルカプトエタノール、2mg/mlのポリミキシンBおよび0.5%の熱不活化血友病マウス血清を含む完全RPMI-1640培養培地中で培養した。1μCi/ウェルの3H-チミジン(6.7Ci/mmol)を培養の72時間後37℃で添加した。16時間の終時に、細胞をミクロメート・ハーベスター(Packard、Meriden、CT)を用いて回収し、次いで、3H-チミジンの導入をTopCount(登録商標)ミクロプレートシンチレーションおよびルミネセンスカウンター(Packard Instrument Company、Meriden、CT)を用いて測定した。処置群は3匹の同型の動物から成り、各々のマウスからの細胞を、抗原依存性の増幅に関して4ツ組で試験した。データを、抗原の存在下での平均3H-チミジン導入の、抗原の不在下での平均導入に対する比である刺激指標(SI)として示す。この方法により、各試験のデータが正規化され、異なる時間に行われた実験の比較が可能となる。
【0036】
サイトカイン分析
インキュベーションの72時間後、抗原-刺激性T細胞の上清を回収し、さらなる分析のために-70℃で保存した。上清を抗体捕捉ELISA(R&Dシステム、Minneapolis、MN)により分析した。IFN-γを典型的なTh1サイトカインとして測定し、IL-10を典型的なTh2サイトカインとして測定した。
【0037】
薬物動態試験
27匹の雄の血友病マウス(20〜26g、8〜12週齢)に、400IU/kgのrFVIIIまたはPEG化リポソーム-rFVIIIを単一のi.v.ボーラス注射として陰茎静脈経由で受容させた。血液サンプル(〜600μl)を、心臓穿刺により投与後.08、0.5、1、2、4、8、16、24、36および48時間で回収し(n=2〜3匹のマウス/時間点)、ACDに添加した。血漿を分離し、-70℃で分析まで保存した。血漿サンプルを、色原性アッセイ(chromogenic assay)(Coamatic FVIII、DaiPharma Group、West Chaster、OH)により蛋白質の活性に関して分析した。各時点で算出された活性を次いで用いて、Win Nonlin(Pharsight Corporation、Mountainview、CA)を用いて非区画分析(non-compartmental analysis)により基本的な薬物動態パラメータを算出した。
統計学的分析
データをSAS(SAS Institute Inc.、Cary、NC)またはMinitab(Minitab Inc.、State College、PA)のアナリスト・アプリケーション(Analyst Application)を用いて分散分析(ANOVA)により分析した。Dunnetteのpost-hocマルチプル比較試験を用いて有意な差(p<0.05)を検出した。
【0038】
結果
図2のAに、DMPCおよびBPSから成るPEG化リポソームの不在下および存在下での全抗-rFVIII抗体タイターを示す。PEG化リポソーム-rFVIIIで処置した動物は、rFVIIIで処置した動物(13,166.7±2042.2、±S.E.M、n=15)と比較して、有意に低い抗体タイター(1123.1±189.5、±S.E.M、n=12、p-値<0.05)を示した。これらの結果は、抗体形成がPEG化リポソームの存在下で低減されることを示す。
【0039】
蛋白質の活性を妨害する中和抗体(即ち、第VIII因子に対して特異的な抗体)をベセスダアッセイを用いて検出した。図2のBは、rFVIIIおよびPEG化リポソーム-rFVIII処置後6週間の終時にベセスダユニット(BU)にて表現される阻害抗体タイターを示す。データは、中和抗体が、rFVIII単独(689.7±78.1BU/ml、±S.E.M、n=13)と比較して、PEG化リポソームの存在下で有意に低いこと(73.65±31.25BU/ml、±S.E.M.、n=12、p-値<0.05)を示した。かかる結果は、PEG化されたリポソームが全抗-FVIII抗体タイターを低減させるだけでなく、蛋白質を不活性化する抗体のタイターも低下させることを示す。比較目的のために、非-PEG化PC/PSリポソームの投与後の全抗体および阻害タイターも示す。データは、差は統計学的に差はないが(p>0.05)、PEG化リポソームの存在下での平均全抗体および阻害タイターが、非-PEG化リポソームのものよりも低いことを示した。
【0040】
FVIII特異的T細胞が、インビボで、PEG化リポソーム-rFVIIIでの免疫化後に刺激されるかどうかを決定するために、rFVIIIチャレンジに対するT-細胞増殖反応をインビボで評価した。PEG化リポソーム-rFVIII処置を受容した動物から単離された脾臓細胞の平均刺激指標は、rFVIII処置のみを受容した動物と比較して低かった(図3)。データは、動物がPEG化PS含有リポソームの存在下および不在下でrFVIIIに曝されたかどうかにより、クローナル細胞増殖(clonal expansion)のために活性化されたT-細胞クローンにおいて生じ得る差を示唆する。
【0041】
PEG化されたPS含有リポソームの存在下でのrFVIIIの免疫原性の低下が、低下したIL-10の分泌の結果かどうかを決定するために、抗原-刺激性T-細胞のサイトカイン分析を、遊離状態の-またはリポソーム-rFVIIIでの動物の免疫化後に行った。図4に示すように、PEG化リポソームと結合したrFVIIIを与えられた動物のT-細胞により分泌された平均IL-10レベルは、rFVIIIのみ与えられた動物よりも低かった。微量レベルのIFN-γが、全処置群に関して培養培地中で検出された(データは示さず)。総じて、データは、PEG化されたPS-含有リポソームの存在下で投与されたrFVIIIの免疫原性の低下には、部分的には、低下したIL-10生成が介在する可能性があることを示唆する。さらに、データは、免疫原性の低下がTh1/Th2反応の偏光の結果ではないことを示唆する。
【0042】
いかなる特定の理論に拘束されることを意図するものではないが、リポソームにおけるPSの含有が、免疫調節に寄与すると考えられる。rFVIIIに対する抗体反応はT-細胞依存性のプロセスであることを考慮すると、PEG化されたPS含有リポソームの存在下でのrFVIIIの免疫原性の低下は、インビボでのrFVIII特異的T-細胞クローンの抑制の結果である可能性がある。
rFVIIIの免疫原性の低下に加えて、rFVIIIのPA含有リポソームとの結合は、また、インビボでのrFVIIIの循環時間を延長し得、こうして、コントロールの血友病Aに対し、必要とされる蛋白質の投与頻度を減らし得る。薬物動態(PK)研究は、PEG化リポソーム-rFVIIIの循環半減期(t1/2)が、rFVIII単独に対して〜35%まで増加することを示唆した(表1)。全身暴露は、処置間で同様であった(図5および表2)。
【0043】
【表2】
【0044】
[実施例6]
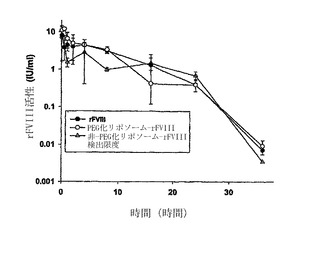
本実施例は、ネガティブに荷電したリン脂質を用いてまたは用いずに調製したPEG結合リポソームの比較分析を提供するものである。阻害タイターを、遊離状態のrFVIII;PASを用いて調製されたリポソームと結合したないしこれに組込まれたFVIII;および、PSを用いずに調製したリポソームと結合したないしこれに組込まれたrFVIIIに関して、実施例5に記載したように決定した。図6に示すように、PSを用いて調製されたリポソームと結合したないしこれに組込まれたrFVIIIに関する阻害抗体タイターは、遊離状態のrFVIIIおよびPSを用いずに調製されたリポソームと結合したないしこれに組込まれたrFVIIIに関するタイターよりも有意に低かった。
当業者は個々の調製を最適化することができる。加えて、PS含有PEG化リポソーム-rFVIIIの免疫原性がrFVIII単独よりもかなり低いという所見は、低免疫原性製剤の開発への有意な進展を示す。
【0045】
引用文献:
[1] J.Klinge、N.M.Ananyeva、C.A. Hauser、E.L. Saenko、血友病A--基礎科学から臨床実施まで、Semin Thromb Hemost 28 (2002) 309-322.
[2] K.Fijnvandraat、W.S. Bril、J.Voorberg、血友病Aにおけるインヒビター開発に関する免疫生物学、Semin Thromb Hemost 29 (2003) 61-68.
[3] P. Lollar、第VIII因子に対する免疫反応の分子特定、Vox Sang 83 Suppl 1 (2002) 403-408.
[4] P. A. Foster、T.S. Zimmerman、第VIII因子の構造および機能、Blood Rev 3 (1989) 180-191.
[5] P.J.Fay、第VIII因子の構造および機能、Thromb Haemost 70 (1993) 63-67.
[6] D. Scandella、M. Mattingly、S. de Graaf、C.A. Fulcher、ヒト第VIII因子インヒビター抗体に関するエピトープの、イムノブロッティングおよび抗体中和による同定、
Blood 74 (1989) 1618-1626.
[7] P. Lollar、ハイブリッドヒト/ブタ第VIII因子を用いる第VIII因子インヒビターの分析、Thromb Haemost 78 (1997) 647-651.
[8] J.F. Healey、I.M. Lubin、H. Nakai、E.L. Saenko、L.W. Hoyer、D. Scandella、P. Lollar、残基484-508は、ヒト第VIII因子のA2ドメインにおける阻害エピトープの主要決定基を含む、J Biol Chem 270 (1995) 14505-14509.
[9] I.M. Lubin、J.F. Healey、R.T. Barrow、D. Scandella、P. Lollar、第VIII因子A2インヒビターエピトープの、アラニンスキャニング変異形成による分析、J Biol Chem 272 (1997) 30191-30195.
[10] P.J.Fay、D. Scandella、第VIII因子A2ドメインに特異的なヒトインヒビター抗体は、サブユニットと第IXa因子の間の相互作用を破壊する、J Biol Chem 274 (1999) 29826-29830.
[11] D. Zhong、E.L. Saenko、M. Shima、M. Felch、D.scandella、いくつかのヒトインヒビター抗体は、第IX因子への第VIII因子の結合を妨害する、Blood 92 (1998) 136-142.
[12] J.F. Healey、R.T. Barrow、H.M. Tamim、I.M.Lubin、M. Shima、D. Scandella、P. Lollar、残基Glu2181-Val2243は、ヒト第VIII因子のC2ドメインに阻害エピトープの主要決定基を含む、Blood 92(1998) 3701-3709.
[13] D. Scandella、G.E. Gilbert、M. Shima、H. Nakai、C. Eagleson、M. Felch、R.Prescott、K.J. Rajalakshmi、L.W. Hoyer、E. Saenko、いくつかの第VIII因子インヒビター抗体は、リン脂質結合部位に重なるC2ドメインアミノ酸2248〜2312に対応する共通のエピトープを認識する、Blood 86 (1995) 1811-1819.
[14] M.T. Reding、D.K. Okita、B.M. Diethelm-Okita、T.A. Anderson、B.M. Conti-Fine、凝固第VIII因子のC2ドメインにおけるヒトCE4+T-細胞エピトープレパートリー、J Thromb Haemost 1(2003) 1777-1784.
[15] K.P. Pratt、J. Qian、E. Ellaban、D.K. Okita、B.M.Diethelm-Okita、B. Conti-Fine、D.W. Scott、第VIII因子C2ドメインにおける免疫優性T-細胞エピトープは、阻害抗体結合部位に位置する、Thromb Haemost 92 (2004) 522-528.
[16] K. Sou、T. Endo、S. Takeoka、E. Tsuchida、ポリ(エチレングリコール)-脂質の、小胞への自発的組込みを用いることによる、リン脂質小胞のポリ(エチレングリコール)-修飾、Bioconjug Chem 11(2000) 372-379.
[17] G.R.Bartlett、カラムクロマトグラフィーにおけるリンアッセイ、J Biol Chem 234(1959) 466-468.
[18] V.S. Purohit、K. Ramani、R.S. Kashi、M.J. Durrani、T.J. Kreiger、S.V. Balasubramanian、ホスファチジルセリン-含有モデル膜に結合した第VIII因子のトポロジー、Biochim Biophys Acta 1617 (2003) 31-38.
[19] T.D. Heath、B.A. Macher、D. Papahadjopoulos、グリコスフィンド脂質によるリポソームへのイムノグロブリンの共有結合、Biochim Biophys Acta 640 (1981) 66-81.
[20] J. Over、第VIII因子(VIII:C)の1段階アッセイの方法、Scand J Haematol Suppl 41 (1984) 13-24.
[21] S. Stoilova-Mcphie、B.O. Villoutreix、K. Mertens、G. Kemball-Cook、A. Holzenburg、膜結合凝固第VIII因子の三次元構造:電子クリスタログラフィーにより誘導された三次元密度マップ内の第VIII因子ヘテロダイマーのモデリング、Blood 99(2002) 1215-1223.
[22] L. Bi、A.M. Lawler、S.E. Antonarakis、K.A. High、J.D. Gearhart、H.H. Kazazian, Jr.、マウス第VIII因子遺伝子の標的破壊は、血友病Aのモデルを生じる、Nat Genet 10 (1995) 119-121.
[23] J. Qian、M. Borovok、L. Bi、H.H. Kazazian, Jr.、L.W, Hoyer、ネズミ血友病Aにおけるヒト第VIII因子に対するインヒビター抗体発生およびT細胞反応、Thromb Haemost 81 (1999) 240-244.
[24] B. Verbruggen、I. Novakova、H. Wessels、J. Boezeman、M. van den Berg、E. Mauser-Bunschoten、第VIII因子に関するベセスダアッセイのニジメゲン変更:Cインヒビター:改良された特異性および信頼性、Thromb Haemost 73 (1995) 247-251.
[25] J.R. Lakowicz、蛍光スペクトロスコピーの原理、Kluwer Academic/Plenum.
【図面の簡単な説明】
【0046】
【図1】PEG化リポソームの存在下および不在下でのrFVIIIの三次元構造。蛍光発光スペクトルを300〜400nmの範囲で取得した。励起モノクロメータは280nmで設定した。用いた蛋白質濃度は〜4μg/mlであった。
【図2】DMPC:BPS(70:30)から成るPEG化リポソームの存在下および不在下でのrFVIIIの投与後6週の終時での、血友病マウスにおける全抗-FVIII抗体タイター(A)および阻害抗-rFVIII抗体(B)。各ポイントは、処置を受けた個々のマウスからの値を示し、水平バーは、全抗体または阻害タイターの平均を示す。比較目的のために、非-PEG化DMPC:BPSリポソームの存在下でrFVIIIの投与後に得られたデータも示す。血液サンプルを、4回目の注射の2週間後に得た。全抗FVIII抗体タイターはELISAにより決定し、阻害タイターはベセスダアッセイにより決定した。統計学的分析は、実施例に記載するように行った。
【図3】2μgのrFVIII、非-PEG化リポソーム-rFVIII、PEG化リポソーム-rFVIIIまたはPS不含リポソーム-rFVIIIの2回の皮下投与後の、多様な免疫優性エピトープを運搬している完全なrFVIII(100ng/ウェル(A)または1000ng/ウェル(B))に対する血友病マウスのCD4+T細胞増殖反応を刺激指標として表記したもの。刺激指標の算出および統計学的分析は実施例に記載されている。各ポイントは個々の動物からの値を示し、水平バーは刺激指標の平均を示す。
【図4】2μgの遊離状態のrFVIIIまたはPEG化リポソーム-rFVIIIの2回の皮下投与製剤の投与を受けた動物からの、抗原でチャレンジされたCD4+T細胞によるIL-10の分泌。CD4+に富むT細胞をrFVIII(1000ng/ウェル)でチャレンジした。各ポイントは個々の動物からの値を示し、水平バーは培養培地中に分泌された平均IL-10レベルを示す。統計学的分析は、実施例に記載されているように行った。
【図5】rFVIII、PEG化または非-PEG化リポソーム-rFVIII投与後の血友病マウスにおける、血漿rFIII活性の時間に対する特性
【図6】種々の脂質組成のPEG化-リポソームの不在および存在下でのrFVIIIの投与後6週の終時での、血友病マウスにおける阻害抗-rFVIII抗体。各ポイントは処置を受けた個々のマウスからの値を示し、水平バーは、全抗体または阻害抗体タイターの平均を示す。血液サンプルを4回目の注射の2週間後に得た。阻害タイターはベセスダアッセイにより決定した。統計学的分析は実施例に記載されているように行った。
【図7】本発明のいくつかのリポソーム組成物の例およびそれらのリポソームサイズ、蛋白質結合効率および免疫原性。
【特許請求の範囲】
【請求項1】
a)ポリエチレングリコール(PEG)で誘導化されたホスファチジルエタノールアミン(PE)、
b)ホスファチジルコリン、ホスファチジルグリセロール、およびこれらの組合せから成る群から選択される両親媒性脂質、および任意にPE、
c)ホスファチジルセリン(PS)、ホスファチジン酸(PA)およびこれらの組合せから成る群から選択されるネガティブに荷電した脂質、および、
d)第VIII因子
を含むリポソームを含み、第VIII因子の免疫原性が、遊離状態の第VIII因子の免疫原性よりも低減されていることを特徴とする医薬組成物。
【請求項2】
前記両親媒性脂質の、前記ネガティブに荷電した脂質に対する比が50:50〜90:10であり、かつ、PEG誘導化PEの量が1〜15モル%であることを特徴とする請求項1に記載の医薬組成物。
【請求項3】
前記両親媒性脂質がPCであり、かつ、前記ネガティブに荷電した脂質がPSであることを特徴とする請求項1に記載の医薬組成物。
【請求項4】
PCのPSに対する比が70:30であることを特徴とする請求項2に記載の医薬組成物。
【請求項5】
前記両親媒性脂質がPCおよびPEであり、前記ネガティブに荷電した脂質がPSであり、PC:PS:PEの比が80:10:10または70:10:20であることを特徴とする請求項1に記載の医薬組成物。
【請求項6】
PC:PS:PEG誘導化PEの比が70:30:15であることを特徴とする請求項2に記載の医薬組成物。
【請求項7】
前記リポソームが、さらに、PEGで誘導化されていないPEを1〜10%の量で含むことを特徴とする請求項1に記載の医薬組成物。
【請求項8】
前記リン脂質の2つのアシル鎖が各鎖中に12〜22個の炭素原子を有し、前記2つのアシル鎖が同数または異数の炭素原子を有することを特徴とする請求項1に記載の医薬組成物。
【請求項9】
前記アシル鎖が、ミリスチン酸、パルミチン酸、ステアリン酸から成る群から選択されることを特徴とする請求項8に記載の医薬組成物。
【請求項10】
さらに、0.5〜30モル%のコレステロールを含むことを特徴とする請求項1に記載の医薬組成物。
【請求項11】
前記PEG誘導化PEのPE部分とPEG部分の間に6〜12個の炭素原子のスペーサーが存在することを特徴とする請求項1に記載の医薬組成物。
【請求項12】
a)両親媒性脂質およびネガティブに荷電した脂質を含むリポソームを調製するステップ、
b)第VIII因子蛋白質を前記リポソームに添加して、前記蛋白質を前記リポソームと結合させる、または前記蛋白質を前記リポソームに組込むステップ、および、
c) PEG誘導化両親媒性脂質をb)に、PEGの1〜15モル%が前記蛋白質結合/組込みリポソームと複合化するように、添加するステップ、
を含む、請求項1に記載の組成物の作成方法。
【請求項13】
前記両親媒性脂質がPCであり、前記ネガティブに荷電した脂質がPSであり、かつ、前記PEGがPEに誘導化されていることを特徴とする請求項12に記載の方法。
【請求項14】
PC:PS:PEの比が70:30:15であることを特徴とする請求項12に記載の方法。
【請求項15】
前記PEがDMPE、DPPEおよびDSPEから成る群から選択されることを特徴とする請求項12に記載の方法。
【請求項16】
ステップaで調製される前記リポソームが、両親媒性脂質、ネガティブに荷電した脂質およびコレステロールを含むことを特徴とする請求項12に記載の方法。
【請求項17】
a)ネガティブに荷電した脂質、両親媒性脂質および、PEGで誘導化されたPEを含むリポソームを調製すること、および、
b)第VIII因子を添加して、第VIII因子の免疫原性が、遊離状態の第VIII因子の免疫原性よりも低減されているリポソームを形成すること
を含む請求項1に記載の医薬組成物の作成方法。
【請求項18】
前記ネガティブに荷電した脂質がPSであり、かつ、前記両親媒性脂質がPCであることを特徴とする請求項17に記載の方法。
【請求項19】
前記PEG誘導化PEのPEが、DMPE、DPPEおよびDSPEから成る群から選択されることを特徴とする請求項17に記載の方法。
【請求項20】
ステップaにおける前記リポソームが、さらにコレステロールを含むことを特徴とする請求項17に記載の方法。
【請求項21】
a)PC、PSおよび活性化されたPEを含むリポソームを調製すること、
b)第VIII因子を添加すること、
c)活性化されたPEGを、当該活性化されたPEGが前記活性化されたPEに結合するように添加すること、
を含む、請求項1に記載の医薬組成物の作成方法。
【請求項22】
6〜12個の炭素原子を含むスペーサーアームが、PEGまたはPEに結合されることを特徴とする請求項21に記載の方法。
【請求項23】
1)PEGで誘導化されたPE、
b)PS、PAおよびこれらの組合せから成る群から選択されるネガティブに荷電した脂質、
c)任意に、PC、PE、PGおよびこれらの組合せから成る群から選択される両親媒性脂質、および、
d)第VIII因子
を含むミセルおよび/またはコクリエート構造を含み、PEG誘導化PEの量が1〜15モル%であり、第VIIIの免疫原性が遊離状態の第VIII因子の免疫原性よりも低減されていることを特徴とする医薬組成物。
【請求項24】
投与される第VIII因子蛋白質の免疫原性を低減する方法であって、
リポソーム、ミセルおよびコクリアー構造を含む脂質構造と前記蛋白質が結合されるか、または前記脂質構造に前記蛋白質が組込まれて成り、前記脂質構造がネガティブに荷電した脂質とPEG誘導化PEを含むことを特徴とする製剤を調製するステップ;
前記製剤を出血性疾患に罹患している個人に投与するステップ
を含み、前記投与される蛋白質の免疫原性が、遊離状態の第VIII因子の免疫原性よりも低く、かつ、前記投与される蛋白質の循環半減期が遊離状態の第VIII因子蛋白質の循環半減期よりも長いことを特徴とする方法。
【請求項1】
a)ポリエチレングリコール(PEG)で誘導化されたホスファチジルエタノールアミン(PE)、
b)ホスファチジルコリン、ホスファチジルグリセロール、およびこれらの組合せから成る群から選択される両親媒性脂質、および任意にPE、
c)ホスファチジルセリン(PS)、ホスファチジン酸(PA)およびこれらの組合せから成る群から選択されるネガティブに荷電した脂質、および、
d)第VIII因子
を含むリポソームを含み、第VIII因子の免疫原性が、遊離状態の第VIII因子の免疫原性よりも低減されていることを特徴とする医薬組成物。
【請求項2】
前記両親媒性脂質の、前記ネガティブに荷電した脂質に対する比が50:50〜90:10であり、かつ、PEG誘導化PEの量が1〜15モル%であることを特徴とする請求項1に記載の医薬組成物。
【請求項3】
前記両親媒性脂質がPCであり、かつ、前記ネガティブに荷電した脂質がPSであることを特徴とする請求項1に記載の医薬組成物。
【請求項4】
PCのPSに対する比が70:30であることを特徴とする請求項2に記載の医薬組成物。
【請求項5】
前記両親媒性脂質がPCおよびPEであり、前記ネガティブに荷電した脂質がPSであり、PC:PS:PEの比が80:10:10または70:10:20であることを特徴とする請求項1に記載の医薬組成物。
【請求項6】
PC:PS:PEG誘導化PEの比が70:30:15であることを特徴とする請求項2に記載の医薬組成物。
【請求項7】
前記リポソームが、さらに、PEGで誘導化されていないPEを1〜10%の量で含むことを特徴とする請求項1に記載の医薬組成物。
【請求項8】
前記リン脂質の2つのアシル鎖が各鎖中に12〜22個の炭素原子を有し、前記2つのアシル鎖が同数または異数の炭素原子を有することを特徴とする請求項1に記載の医薬組成物。
【請求項9】
前記アシル鎖が、ミリスチン酸、パルミチン酸、ステアリン酸から成る群から選択されることを特徴とする請求項8に記載の医薬組成物。
【請求項10】
さらに、0.5〜30モル%のコレステロールを含むことを特徴とする請求項1に記載の医薬組成物。
【請求項11】
前記PEG誘導化PEのPE部分とPEG部分の間に6〜12個の炭素原子のスペーサーが存在することを特徴とする請求項1に記載の医薬組成物。
【請求項12】
a)両親媒性脂質およびネガティブに荷電した脂質を含むリポソームを調製するステップ、
b)第VIII因子蛋白質を前記リポソームに添加して、前記蛋白質を前記リポソームと結合させる、または前記蛋白質を前記リポソームに組込むステップ、および、
c) PEG誘導化両親媒性脂質をb)に、PEGの1〜15モル%が前記蛋白質結合/組込みリポソームと複合化するように、添加するステップ、
を含む、請求項1に記載の組成物の作成方法。
【請求項13】
前記両親媒性脂質がPCであり、前記ネガティブに荷電した脂質がPSであり、かつ、前記PEGがPEに誘導化されていることを特徴とする請求項12に記載の方法。
【請求項14】
PC:PS:PEの比が70:30:15であることを特徴とする請求項12に記載の方法。
【請求項15】
前記PEがDMPE、DPPEおよびDSPEから成る群から選択されることを特徴とする請求項12に記載の方法。
【請求項16】
ステップaで調製される前記リポソームが、両親媒性脂質、ネガティブに荷電した脂質およびコレステロールを含むことを特徴とする請求項12に記載の方法。
【請求項17】
a)ネガティブに荷電した脂質、両親媒性脂質および、PEGで誘導化されたPEを含むリポソームを調製すること、および、
b)第VIII因子を添加して、第VIII因子の免疫原性が、遊離状態の第VIII因子の免疫原性よりも低減されているリポソームを形成すること
を含む請求項1に記載の医薬組成物の作成方法。
【請求項18】
前記ネガティブに荷電した脂質がPSであり、かつ、前記両親媒性脂質がPCであることを特徴とする請求項17に記載の方法。
【請求項19】
前記PEG誘導化PEのPEが、DMPE、DPPEおよびDSPEから成る群から選択されることを特徴とする請求項17に記載の方法。
【請求項20】
ステップaにおける前記リポソームが、さらにコレステロールを含むことを特徴とする請求項17に記載の方法。
【請求項21】
a)PC、PSおよび活性化されたPEを含むリポソームを調製すること、
b)第VIII因子を添加すること、
c)活性化されたPEGを、当該活性化されたPEGが前記活性化されたPEに結合するように添加すること、
を含む、請求項1に記載の医薬組成物の作成方法。
【請求項22】
6〜12個の炭素原子を含むスペーサーアームが、PEGまたはPEに結合されることを特徴とする請求項21に記載の方法。
【請求項23】
1)PEGで誘導化されたPE、
b)PS、PAおよびこれらの組合せから成る群から選択されるネガティブに荷電した脂質、
c)任意に、PC、PE、PGおよびこれらの組合せから成る群から選択される両親媒性脂質、および、
d)第VIII因子
を含むミセルおよび/またはコクリエート構造を含み、PEG誘導化PEの量が1〜15モル%であり、第VIIIの免疫原性が遊離状態の第VIII因子の免疫原性よりも低減されていることを特徴とする医薬組成物。
【請求項24】
投与される第VIII因子蛋白質の免疫原性を低減する方法であって、
リポソーム、ミセルおよびコクリアー構造を含む脂質構造と前記蛋白質が結合されるか、または前記脂質構造に前記蛋白質が組込まれて成り、前記脂質構造がネガティブに荷電した脂質とPEG誘導化PEを含むことを特徴とする製剤を調製するステップ;
前記製剤を出血性疾患に罹患している個人に投与するステップ
を含み、前記投与される蛋白質の免疫原性が、遊離状態の第VIII因子の免疫原性よりも低く、かつ、前記投与される蛋白質の循環半減期が遊離状態の第VIII因子蛋白質の循環半減期よりも長いことを特徴とする方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2008−544990(P2008−544990A)
【公表日】平成20年12月11日(2008.12.11)
【国際特許分類】
【出願番号】特願2008−519595(P2008−519595)
【出願日】平成18年6月29日(2006.6.29)
【国際出願番号】PCT/US2006/025519
【国際公開番号】WO2007/002886
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(506194117)ザ リサーチ ファウンデイション オブ ステイト ユニバーシティー オブ ニューヨーク (11)
【Fターム(参考)】
【公表日】平成20年12月11日(2008.12.11)
【国際特許分類】
【出願日】平成18年6月29日(2006.6.29)
【国際出願番号】PCT/US2006/025519
【国際公開番号】WO2007/002886
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(506194117)ザ リサーチ ファウンデイション オブ ステイト ユニバーシティー オブ ニューヨーク (11)
【Fターム(参考)】
[ Back to top ]