
体細胞から多能性細胞を生成する方法
本明細書において、発達的再プログラミングの際にマウスおよびヒト多能性幹細胞の生成に関して選択するための方法が開示される。本明細書において記述される方法は、誘導多能性幹細胞、すなわち遺伝子選択を必要とすることなく、分化した細胞から生成または誘導された多能性幹細胞の選択に関する。本明細書において1)コロニーの形態、または2)雌性細胞におけるX染色体の再活性化に基づいて、再プログラムされた細胞を選択するための特定の態様が記述される。

【発明の詳細な説明】
【技術分野】
【0001】
本出願は、35 U.S.C. §119(e)の下で、その全内容物が参照により本明細書に組み入れられる2007年5月30日に提出された米国特許仮出願第60/932,267号に対する優先権を主張する。
【背景技術】
【0002】
発明の背景
核の移入(Wakayama, T., Perry, A. C., Zuccotti, M., Johnson, K. R., and Yanagimachi, R. (1998) Nature 394, 369-374(非特許文献1);Wilmut, I., Schnieke, A. E., McWhir, J., Kind, A. J., and Campbell, K. H. (1997) Nature 385, 810-813(非特許文献2))および細胞融合(Cowan, C. A., Atienza, J., Melton, D. A., and Eggan, K. (2005) Science 309, 1369-1373(非特許文献3);Tada, M., Takahama, Y., Abe, K., Nakatsuji, N., and Tada, T. (2001) Curr Biol 11, 1553-1558(非特許文献4))による細胞の再プログラミングによって、体細胞核において多能性状態は再度確立される(Hochedlinger, K., and Jaenisch, R. (2006) Nature 441, 1061-1067(非特許文献5))。核再プログラミングの分子メカニズムは完全に解明されていないが、細胞融合実験から、再プログラミング因子をES細胞において同定することができ、そしてこれを用いて体細胞において再プログラミングを直接誘導できることが暗示されている。実際に、合理的アプローチによって最近、その発現が成体線維芽細胞における多能性状態の誘導を可能にする4つの転写因子が同定された(Takahashi, K., and Yamanaka, S. (2006) Cell 126, 663-676(非特許文献6))。Yamanakaと共同研究者らは、転写因子Oct4、Sox2、c-Myc、およびKlf4のレトロウイルス発現をFbx15発現に関する遺伝子選択と組み合わせると、線維芽細胞培養物からiPS細胞を直接生じることを証明した。Fbx15選択iPS細胞は、妊娠中期胚における多様な組織に関与したが、これらの胚は妊娠中期で死亡し、ES細胞と比較してiPS細胞の発達能が限定的であることを指し示している。この所見と一致して、iPS細胞において発現されたのはES細胞のトランスクリプトームのごく一部に過ぎず、Oct4およびNanogプロモーターの染色質状態のメチル化分析から、線維芽細胞とES細胞の中間のエピジェネティックパターンが証明された。
【0003】
これらの所見から、直接再プログラムされた細胞の分子的および機能的性質に関して3つの根本的な疑問が持ち上がった:(i)ES細胞状態にとって必須である遺伝子に関して選択することによって、既に記述されたFbx15選択iPS細胞よりES細胞に類似する多能性細胞を生成することができるか;(ii)iPS細胞の多能性状態は、外因性の因子の持続的発現に依存するか;および(iii)転写因子誘導再プログラミングは、線維芽細胞ゲノムの後成的風景を多能性細胞の後成的風景へとリセットするか。
【0004】
核移入または細胞融合による体細胞の再プログラミングが成功するためには、DNAのメチル化、ヒストン修飾、および雌性細胞における沈黙X染色体の再活性化などの後成的修飾の忠実なリモデリングが必要であると考えられている(Rideout, W. M., 3rd, Eggan, K., and Jaenisch, R. (2001) Science 293, 1093-1098(非特許文献7))。異常な後成的再プログラミングは、核移入によってクローン化された動物において認められる発達不全および異常の主要な理由であると仮定されている。このように、後成的異常によって癌などの病理的状態が起こりうることから、後成的再プログラミングに関する疑問は、iPS細胞に関する将来的な治療応用から見て特に意味がある(Gaudet, F., Hodgson, J. G., Eden, A., Jackson-Grusby, L., Dausman, J., Gray, J. W., Leonhardt, H., and Jaenisch, R. (2003) Science 300, 489-492(非特許文献8))。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Wakayama, T., Perry, A. C., Zuccotti, M., Johnson, K. R., and Yanagimachi, R. (1998) Nature 394, 369-374
【非特許文献2】Wilmut, I., Schnieke, A. E., McWhir, J., Kind, A. J., and Campbell, K. H. (1997) Nature 385, 810-813
【非特許文献3】Cowan, C. A., Atienza, J., Melton, D. A., and Eggan, K. (2005) Science 309, 1369-1373
【非特許文献4】Tada, M., Takahama, Y., Abe, K., Nakatsuji, N., and Tada, T. (2001) Curr Biol 11, 1553-1558
【非特許文献5】Hochedlinger, K., and Jaenisch, R. (2006) Nature 441, 1061-1067
【非特許文献6】Takahashi, K., and Yamanaka, S. (2006) Cell 126, 663-676
【非特許文献7】Rideout, W. M., 3rd, Eggan, K., and Jaenisch, R. (2001) Science 293, 1093-1098
【非特許文献8】Gaudet, F., Hodgson, J. G., Eden, A., Jackson-Grusby, L., Dausman, J., Gray, J. W., Leonhardt, H., and Jaenisch, R. (2003) Science 300, 489-492
【発明の概要】
【0006】
本明細書において記述される方法は、誘導多能性幹細胞、すなわち、たとえば成体線維芽細胞が含まれる分化した細胞から生成または誘導された多能性幹細胞の選択に関する。限られた数の転写因子の発現を誘導することによる多能性の誘導は、当技術分野において証明されており、これを任意の哺乳動物細胞、非ヒト哺乳動物細胞、またはヒト細胞に応用することができる。
【0007】
本明細書において記述される方法によって、発達的再プログラミングの際に哺乳動物(たとえば、マウスおよびヒト)多能性細胞を生成するように選択することができる。定義された組の転写因子の過剰発現により、成体の体細胞を胚幹(ES)細胞様の細胞に変換することができるが、このプロセスは一般的に、ES細胞特異的遺伝子を再活性化するための遺伝子選択を必要とし;選択を行わなければES様細胞に加えて多くの非ES様細胞が生成される。そのような遺伝子選択技術は、一般的にヒト細胞において実現可能ではなく、一般的に、細胞をヒト患者に導入するために望ましくない。この問題に取り組むために、本明細書において、1)コロニーの形態のみ、および2)雌性細胞における染色体の再活性化、に基づいて再プログラムされた細胞に関して選択することができる新規選択戦略が記述される。それは、遺伝子選択、化学的選択、またはそのいずれも行わないことである。
【0008】
形態に基づく選択は、既存の選択アプローチと比較して再プログラミングのためにより長い期間、再プログラミング因子を加えた後におよそ1〜2ヶ月を必要とする。この期間のあと、ES様コロニーを採取して拡大させることができる。採取の時点では多くの非ES様細胞が残っているが、たとえばクローン密度で細胞を継代すると、ES様コロニーを容易に回収することができ、細胞株を生成することができる。
【0009】
X染色体の再活性化に基づく選択は、Hprt座における変異に関してヘテロ接合である雌性細胞株を利用する。X染色体の再活性化は、定義された因子による再プログラミングの際に起こり、この事象は再プログラミングプロセスの後期に起こることが本明細書において示される(およそ3〜4週間)。雌性の体細胞では、1つのみのX染色体が活性であり、他は沈黙である。1つの局面において、Hprtヘテロ接合細胞において、活性なX染色体上に変異体Hprt遺伝子を潜在的に有する細胞は6-チオグアニンに対して耐性であろう。再プログラミングおよびX染色体再活性化の際に、これらの細胞は、正常なHprt遺伝子を発現して、HAT培地に対する耐性を獲得するが、6-チオグアニンに対する耐性を失う。
【0010】
本明細書において記述される方法の1つの局面によって、以下の段階を含む、誘導多能性幹細胞の選択を行うことができる:a)RT-PCRによって測定した場合にNanog mRNAを発現していない分化した初代培養細胞を、多能性表現型へと再プログラムする段階;b)再プログラミング後、段階(a)において再プログラムされた細胞を選択物質の非存在下で培養する段階;c)段階(b)の培養物を顕微鏡下で観察して、培養物においてなめらかで丸い外観となった細胞のクローンを単離する段階;およびd)幹細胞マーカーの発現が検出されれば、細胞が誘導多能性幹細胞であることが指し示される、幹細胞マーカーの発現に関してクローンの細胞を試験する段階。
【0011】
この局面および本明細書において記述される他の全ての局面の1つの態様において、再プログラミング段階は、以下の1つを含む:因子を発現させるための調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-Myc、およびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;ならびに細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【0012】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、幹細胞マーカーを発現するクローンの細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階をさらに含み、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0013】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、培養する段階は、細胞を継代する段階をさらに含む。
【0014】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、分化した体細胞は、ES細胞とは明確に異なる形態を有する。
【0015】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、分化した初代培養細胞は線維芽細胞であり、線維芽細胞は再プログラミングの前は平坦で不規則な形状を有する。
【0016】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される。
【0017】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、分化した初代培養細胞が雌性の個体に由来する場合、不活性なX染色体の再活性化に関してクローンの細胞を試験する段階をさらに含む。
【0018】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、核酸配列はウイルスベクターまたはプラスミドに含まれる。
【0019】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、ウイルスベクターは、レトロウイルスベクター、レンチウイルスベクター、またはアデノウイルスベクターである。
【0020】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、外因性のOct4、Sox2、c-Myc、および/またはKlf4の発現に関してクローンの細胞を試験する段階をさらに含む。
【0021】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、初代培養細胞はヒト細胞を含む。
【0022】
本明細書において記述されるもう1つの局面は、以下の段階を含む、誘導多能性幹細胞を選択する方法である:a)選択可能マーカーが活性なX染色体上で変異体であり、不活性なX染色体上では野生型である、およびRT-PCRによって測定した場合に細胞がNanog mRNAを発現しない、X染色体上の選択可能マーカーに関してヘテロ接合である雌性細胞を提供する段階;b)多能性表現型となるように細胞を再プログラムする段階;ならびにc)不活性なX染色体の再活性化によって、野生型選択可能マーカーを発現させることができ、選択物質の存在下で細胞を生存させることができ、それによって生存する細胞が誘導多能性幹細胞である、細胞を選択物質と共に培養する段階。
【0023】
この局面の1つの態様において、方法は、選択物質の存在下で生存する細胞を幹細胞マーカーの発現に関して試験する段階をさらに含む。
【0024】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される。
【0025】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、再プログラムする段階は、因子を発現させるための調節エレメントに機能的に連結している、転写因子Oct4、Sox2、c-Myc、およびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;ならびに細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階、の1つを含む。
【0026】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、選択物質の存在下で生存する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階を含み、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0027】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は細胞株の細胞である。
【0028】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は、X染色体上の変異体Hprt遺伝子に関してヘテロ接合である。
【0029】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は、核酸の導入前に不活性である野生型Hprt遺伝子をX染色体上に持ち、再プログラミング前に活性である変異体の非機能的Hprt遺伝子をX染色体上に持つ。
【0030】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は、再プログラミング前に6-チオグアニンに対して耐性である。
【0031】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、選択物質はHAT培地を含む。
【0032】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞はヒト細胞を含む。
【0033】
本明細書において記述されるもう1つの局面は、以下の段階を含む、誘導多能性幹細胞を選択する方法である:a)RT-PCRによって測定した場合にNanog mRNAを発現しない、X不活化による沈黙化に供されるX染色体連鎖レポーター遺伝子を持つ雌性細胞を提供する段階;b)細胞を多能性表現型へと再プログラムする段階;c)再プログラミング後に細胞を培養する段階;およびd)レポーターが発現すれば、クローンが誘導多能性幹細胞を含むことが指し示される、X染色体連鎖レポーターを発現する細胞のクローンを培養物から単離する段階。
【0034】
この局面および本明細書において記述される他の全ての局面の1つの態様において、方法は、クローンの細胞を幹細胞マーカーの発現に関して試験する段階をさらに含む。
【0035】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される。
【0036】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、レポーターを発現する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に対して組織学的検査を行う段階をさらに含む。
【0037】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞はヒト細胞を含む。
【0038】
定義
本明細書において用いられる「多能性」という用語は、異なる条件下で1つより多い分化した細胞タイプに分化する、好ましくは3つ全ての胚細胞層に特徴的な細胞タイプに分化する能力を有する細胞を指す。多能性細胞は、たとえばヌードマウス奇形腫形成アッセイ(本明細書において記述される実施例を参照されたい)を用いて、1つより多い細胞タイプに分化する能力、好ましくは、3つ全ての胚細胞層に分化する能力によって主に特徴付けされる。多能性はまた、胚幹(ES)細胞マーカーの発現も証拠となるが、多能性に関する好ましい試験は、3つの胚細胞層のそれぞれの細胞への分化能を証明することである。
【0039】
本明細書において用いられるように「再プログラミング」という用語は、最終分化した体細胞の分化状態を多能性表現型へと変更するプロセスを指す。
【0040】
「分化した初代培養細胞」とは、その本来の型において、本明細書において定義される多能性ではない任意の初代培養細胞を意味する。多くの初代培養細胞を培養すると、完全に分化した特徴のいくつかが失われうることに注意すべきである。しかし、そのような細胞を単純に培養しても、ひとりでにそれらが多能性となるわけではない。多能性への移行は、培養における分化特徴の部分的喪失に至る刺激を超える再プログラミング刺激を必要とする。再プログラムされた多能性細胞はまた、一般的に培養においてごく限られた回数の分裂能を有する初代培養細胞の親と比較して、生育能を失うことなく、長期の継代能の特徴を有する。
【0041】
「ベクター」という用語は、それが複製される宿主細胞に導入するために、その中にDNA配列を挿入することができる低分子の担体DNA分子を指す。「発現ベクター」は、宿主細胞における発現にとって必要とされる必要な調節領域を有する遺伝子を含有する特殊なベクターである。「機能的に連結した」という用語は、コード配列の発現にとって必要な調節配列が、コード配列の発現を行うために、コード配列に対して適当な位置でDNA分子内に置かれていることを意味する。この同じ定義は時に、発現ベクターにおけるコード配列および転写制御エレメント(たとえば、プロモーター、エンハンサー、および終止エレメント)の整列にも当てはまる。この定義はまた、ハイブリッド核酸分子が生成される、第一および第二の核酸分子の核酸配列の整列にも当てはまる。
【図面の簡単な説明】
【0042】
【図1】Nanog-選択iPS細胞のES細胞様特性(A)Nanog-GFP(NGiP)ES細胞、ならびに持続的ピューロマイシン選択によっておよびピューロマイシン選択を行わずに生育させた2つのiPS細胞株と共に、追加の基準点としての野生型ES細胞(V6.5)およびMEFsにおけるES細胞マーカー遺伝子発現のRT-PCR分析。Oct4およびSox2のプライマーは、それぞれの内因性の座からの転写物に対して特異的である。Nat1はローディング対照として用いた。(B)iPS細胞株、MEF、およびNGiP-ES細胞におけるNanog、Oct4、Sox2、c-myc、およびKlf4の発現に関するウェスタンブロット分析。抗チューブリンおよび抗アクチン抗体をローディングの対照とするために用いた。(C)それぞれのpMXウイルスを感染させた、1)野生型MEF、2)野生型ES細胞、3)ソーティングおよびサブクローニングの前の不均一なiPS株1A2からの細胞、4)1D4 iPS、5)2D4 iPS、および6)MEF、におけるpMXレトロウイルス転写の定量的PCR分析。転写物レベルをβ-アクチンに対して標準化した。2D4 iPS株におけるレトロウイルスは、完全に沈黙しているように思われるが、不均一な1A2株はなおも外因性の因子の豊富な発現を示すことに注意すべきである。
【図2】iPS細胞と体細胞との融合(A)2D4 iPS細胞と、Oct4Neo選択可能な対立遺伝子を持つハイグロマイシン耐性MEFとの細胞融合の略図。(B)ピューロマイシン/ハイグロマイシン選択またはピューロマイシン/G418選択のいずれかの下で維持された2D4 iPS細胞、MEF、および2D4/MEF細胞ハイブリッドのDNA含有量分析。
【図3】iPS細胞を維持するための外因性のOct4に関する必要条件。(A)Oct4-誘導可能な線維芽細胞を用いたiPS細胞生成の略図。(B)ドキシサイクリン誘導可能なOct4発現の非存在下または存在下でSox2、c-MYC、およびKlf4に感染したMEF。アルカリホスファターゼ染色したプレートを示す。(C)Oct4-誘導可能なiPS細胞におけるOct4レベルの定量的PCR分析。内因性および誘導可能な対立遺伝子からの転写物のレベルを、非分化iPS細胞(+LIF、-dox)、分化したiPS細胞(-LIF、-dox)、およびLIF除去後5日目に再誘導した分化iPS細胞(-LIF、+dox)において測定した。転写物のレベルをβ-アクチンに対して標準化した。誘導可能なOct4対立遺伝子を持つES細胞;および野生型MEFを対照とした。
【図4】iPS細胞における遺伝子特異的および全体的なDNAメチル化状態(A)ES細胞、2D4 iPS細胞、およびMEFにおけるOct4およびNanogプロモーター領域のバイサルファイトシークエンシング。異なるようにメチル化したCpGを含有するプロモーター領域を転写開始部位(矢印)に関して示す。白丸は、非メチル化CpGを表す;黒丸はメチル化CpGを表示する。(B)iPS 2D4細胞とMEFとの融合を通して生成された細胞ハイブリッドにおけるNanogプロモーターのバイサルファイトシークエンシング。図2Cと同様にピューロマイシン/ハイグロマイシン耐性ハイブリッドに関するデータを示す。(C)サテライトリピートプローブを用いた全体的なDNAメチル化のサザンブロット分析。MEF、雄性Nanog-GFP ES細胞、雌性ES細胞、iPS 2D4親細胞および3つのサブクローンからのゲノムDNAを、メチル化感受性制限酵素HpaIIによって消化して、より小さいサテライトリピートプローブにハイブリダイズさせた。非メチル化感受性イソシゾマーMspIによって消化した雄性ES細胞DNAを対照とした。より低い分子量のバンドは、低メチル化を指し示している。
【図5】iPS細胞におけるX染色体の動力学(A)iPS細胞株2D4、NGiP MEF、および雄性対照ES細胞におけるXite遺伝子間転写物のRT-PCR分析。Xite座に沿って異なる位置での転写物を検出した(領域5〜7)。陽性対照Rrm2は、ハウスキーピング遺伝子である。雌性ES細胞と同様に、雄性ES細胞はXite転写物を発現する。(B)分化しつつある2D4 iPSにおけるXi上でのEzh2およびH3me3K27の濃縮。グラフは、2D4 iPS細胞のレチノイン酸誘導分化の際の異なる時点で、Xi上でEzh2またはH3me3K27との同時局在を示すXist RNAコーティングを有する細胞の百分率を示す(各時点に関してn>100)。
【図6】分化しつつあるTTF由来iPS細胞における無作為なX-不活化(A)XGFPX TTFからiPS細胞を得るためのおよびX-不活化をその後分析するためのフロースキーム。Oct4-Neo対立遺伝子を持つXGFPX TTFを、2回連続して通過させることによってソーティングして、GFP陰性集団(XiGFPXa;<0.05%緑色細胞)を得た。再プログラムされた細胞を、ES細胞の形態およびGFP再活性化に基づいて選択した。G418による薬物選択を、iPS細胞の再プログラム状態をレトロスペクティブに確かめるために使用したが、iPS細胞の確立に関して選択するためには使用しなかった。iPS細胞をサブクローニングして、分化させ、FACSおよびXistFISHによって分析した。FACSによって決定されたGFP+またはGFP-細胞の数をオレンジ色で与え、青色で与えた数値はそれぞれ、GFP+およびGFP-分化iPS細胞内のXiのXist RNAコーティングを有する細胞の百分率を指し示している。
【図7】iPS細胞におけるH3K4およびH3K27トリメチル化の全体的な分析(A)全ての細胞タイプのあいだでのK4およびK27トリメチル化データの全体的な相関。表は、細胞タイプの起こりうる全ての対のあいだおよびアレイにおける全ての遺伝子(〜16500個)に関してそれぞれ、K4およびK27トリメチル化の2成分の全体的相関を示す。(B)全ての細胞タイプのあいだでのEクラス遺伝子内でのK4およびK27トリメチル化の相関。細胞タイプのそれぞれの2つの対に関するK4またはK27メチル化に関する相関値を、500 bpずつ増加させた転写開始部位からの距離の関数としてプロットした。
【図8】Nanog-選択可能なiPS細胞のインビボ発達能。(A)無作為に組み入れられたGFPトランスジーンを持つiPS株2D4からの細胞を胚盤胞に注入した。代理母は、GFP-陽性の仔を産んだ。GFPを発現しない非キメラの仔を示す。(B)iPS細胞由来新生仔キメラマウスの脾臓および胸腺から単離された造血細胞のフローサイトメトリー分析。ヒストグラムは、系列特異的マーカーに関してゲートを設定した集団におけるGFP-陽性細胞の百分率を表示する。(C)野生型の同腹子の横に示される、胚盤胞注入2D4 iPS細胞に由来する10日齢のキメラマウス(iPS由来細胞は、アグーチ外皮色の原因である)。
【図9】iPS細胞株1A2および2D4におけるレトロウイルス組み込みDNA刷り込み状態の分析。(A)iPS細胞におけるレトロウイルス組み込み部位の分析。レトロウイルス組み込みをサザンブロット分析によって決定した。DNAをBamHI(Oct4およびKlf4に関して)、HindIII(Sox2に関して)、またはBglII(c-MYCに関して)によって消化して、それぞれのcDNAプローブとハイブリダイズさせた。組み込みをV6.5 ES細胞(wt)および2つのIPS株1A2および2D4に関して示す。(B)組み込み部位分析のために用いられる内部制限部位が含まれる個々のウイルス構築物の概略図。cDNAプローブは、ウイルスが組み入れられるゲノム領域における1つのpMX内部切断と1つの外部切断とによって生成された制限断片を検出することに注意すべきである。(C)Igf2rの異なるようにメチル化された領域でのメチル化状態。異なる細胞タイプからのDNAをPvuIIおよびMIUI制限酵素を用いて消化して、サザンブロッティングによって分析した。メチル化(M)および非メチル化(U)対立遺伝子を指し示す。Dnmt1を欠損するES細胞、またはE 12.5胚に由来する胚生殖(EG)細胞では、非メチル化対立遺伝子のみが検出された。刷り込みがiPS細胞において維持されるという事実は、iPS細胞が、線維芽細胞培養物に混入する可能性があるまれな生殖細胞に由来するのではないことを示唆する。
【図10】シグナチャー遺伝子分析の統計学的有意性そのメチル化パターンに基づいて2D4 iPS細胞におけるほとんどのシグナチャー遺伝子をES-様(Eクラス)として分類することは、非常に有意である。上のパネルは、2D4座のE、NおよびMクラスへの観察された分布を示す(図7Aにおいて提示されたデータから)。2D4座のE、M、およびN遺伝子への分類を確証するために、2D4メチル化データを100回入れ替えて、ES-MEF対に無作為に割付して、シグナチャー遺伝子を異なるストリンジェンシー(p=0.01、p=0.05、p=0.1)で再分類した(下のパネル)。
【図11】iPS細胞におけるシグナチャー遺伝子の発現(A)ピアソン相関によって決定した、V6.5 ES細胞(ES)、ピューロマイシン選択Nanog GFP ires Puro ES細胞(ESpuro)、Nanog GFP ires Puro MEF(MEF)、および2D4 iPS細胞(iPS)のあいだの全発現データセット(Agilentマイクロアレイから)の全体的な相関。(B)ES細胞に対して2倍より大きい発現の変化を示した、(A)において記述されたESpuro、MEF、およびiPSの完全な発現データセットにおける遺伝子の数。(C)2D4 iPS細胞、雌性MEF、およびV6.5 ES細胞における選択されたシグナチャー遺伝子13個の転写物レベルのリアルタイムPCR分析。相対的発現レベルを決定するために、Qiagen RNA easyキットを用いてRNAを調製して、1μgをOmniscript RTキット(Qiagen)およびランダムプライマーを用いて逆転写した。転写物レベルをリアルタイムPCRによって定量して、△△Ct法を用いてGapdh対照に対して標準化した。ES細胞における発現を任意で1に設定して、誤差のバーは、1試料あたり3個の反応の標準偏差を表す。プライマー配列を表3に与える。Y-軸の異なる尺度に注意されたい。本発明者らのゲノム全体の発現データと一致して、Mクラスに属する試験した遺伝子3個中2個(Vgl14、HoxD10)は、そのヒストン修飾パターンに基づいてそれらがMEF様遺伝子であると分類されたにもかかわらず、2D4 iPS細胞においてES様発現パターン(MEFよりESおよびiPS細胞においてより低い発現)を証明した。このため、これらの座でのヒストンメチル化をより厳密に手動で検分することにより、MEFと比較して2D4 iPS細胞において認められた抑制が2D4 iPS細胞におけるこれらのプロモーターでのK4メチル化の低減と相関することが判明した。他の遺伝子は全て、K4メチル化のみと相対的により高い発現、K27メチル化のみと相対的により低い発現、ならびにヒストンH3 K4およびK27メチル化の二価性とより低い発現とのあいだに良好な相関を示した。
【図12】iPS細胞の造血系列へのインビトロ分化(A、B)iPS細胞株2D4および野生型V6.5 ES細胞に由来する7日胚様体を、造血マーカーCD41およびc-kitマーク未成熟造血細胞(A)と共にCD45およびc-kitマーク成熟造血細胞(B)に関してフローサイトメトリーによって分析した。二重陽性細胞の百分率を与える。EBを生成する場合、iPS株ではV6.5 ES細胞株より多数の投入細胞を用いたことに注意されたく、このことは分化細胞の百分率における定量的差を説明する可能性がある。(C)iPS細胞から作出された解離された7日EBのメチルセルロース培養物から得られた成熟造血細胞。骨髄芽球(i)、マクロファージ(ii)、肥満細胞(iii、iv)、および初期赤血球細胞(v、vi)が含まれる多数のタイプの造血細胞が存在した。血球の生成に関して、予め播種することによってフィーダー細胞を消失させた後にハンギングドロップ法を用いてEBを生成した(Geijsen, N., Horoschak, M., Kim, K., Gribnau, J., Eggan, K., and Daley, G. Q. (2004) Nature 427, 148-154)。3日後、EBを播種して、7日目に造血マーカーのFACS分析のために(別紙の表3において記述される抗体によって)、またはインビトロでさらに分化させるために、コラゲナーゼIVによってEBを単細胞浮遊液に解離した。メチルセルロース培養物に関して、7日EBの単細胞浮遊液を、造血増殖因子(M3434、Stem Cell Technologies)を補足したメチルセルロースと混合して、培養物あたり細胞1×105個で接種した。培養10日後、代表的な造血コロニーを採取して、サイトスピンを調製し、これをメイ-グリュンヴァルド-ギムザによって対比染色した。
【発明を実施するための形態】
【0043】
詳細な説明
選択物質の非存在下での誘導多能性幹細胞の単離
1つの局面において、本明細書において記述される方法は、多能性となった細胞を同定または濃縮するために選択物質を用いることに頼らず、その代わりに細胞があまり分化していないES様多能性表現型を獲得する場合に起こる当初の細胞の形態の変化を頼りとする、誘導多能性幹細胞の選択に関する。
【0044】
この局面において、本発明は、以下のような段階を有する、誘導多能性幹細胞を選択する方法に関する。第一の段階は、分化した初代培養細胞をより分化していないまたは多能性状態に再プログラミングする段階を伴う。再プログラミングは、たとえば細胞の核を卵母細胞に移入することによって(たとえば、Wilmut et al., 1997, Nature 385: 810-813を参照されたい)、または既存の胚幹細胞との融合によって(たとえば、Cowan et al., 2005, Science 309: 1369-1373、およびTada et al, 2001, Curr. Biol. 11: 1553-1558)達成されうる。そのような再プログラミングはまた、たとえば因子を発現させるための調節エレメントに機能的に連結された、転写因子Oct4、Sox2、c-Myc、およびKlf4をコードする核酸配列を、たとえば線維芽細胞に導入することによって行われうる。これらの因子が好ましいが、他の転写因子またはこれらの因子のサブセットも同様に使用することができる(たとえば、参照により本明細書に組み入れられる、Takahashi & Yamanaka, 2006, Cell 126: 663-676を参照されたい)。
【0045】
1つの態様において、転写因子はウイルスベクターまたはプラスミドによってコードされる。ウイルスベクターは、たとえばレトロウイルスベクター、レンチウイルスベクター、またはアデノウイルスベクターでありうる。当業者に公知である核酸の導入に対する非ウイルスアプローチも同様に、本明細書において記述される方法と共に用いることができる。
【0046】
または、そのような転写因子をコードする内因性の遺伝子の活性化を用いることができる。
【0047】
もう1つの代用において、細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を細胞に導入することができる。たとえば、HIV-TAT融合体を用いることを通して、タンパク質因子(たとえば、中でもc-Myc、Oct4、Sox2および/またはKlf4)を細胞に導入することができる。TATポリペプチドは、それを細胞に浸透させる特徴を有し、外因性の因子を細胞に導入するために用いられている(たとえば、Peitz et al., 2002, Proc. Natl. Acad. Sci. U S A. 99:4489-94を参照されたい)。このアプローチは、細胞の分化状態を再プログラミングするための因子を導入するために使用することができる。最後に、再プログラミングは、細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させることによって成就されうる(たとえば、Sato et al., 2004, Nature Med. 10:55-63を参照されたい)。
【0048】
線維芽細胞が好ましいが、他の初代培養細胞タイプも同様に用いることができる。親細胞は、形態的変化に基づく変化を容易にするために、ES細胞とは明確に異なる形態を有することが好ましい。「明確に異なる」とは、少なくとも、接着細胞の場合、親細胞の形状が、培養において生育した場合に丸いよりむしろ不規則であることを意味する。非接着性の初代培養細胞の場合、接着性に関して最初に選択して、次に、丸いESの形態に関して選択することができる。当業者は、位相差顕微鏡下で見た場合に、平坦であるよりむしろ丸くなる傾向があり、粗面であるよりむしろなめらかな傾向があるES細胞の形態的特徴を承知している。
【0049】
さらに、親細胞は、任意の哺乳動物種に由来しえて、非制限的な例には、ネズミ、ウシ、サル、ブタ、ウマ、ヒツジ、またはヒト細胞が含まれる。親細胞はES細胞マーカー、たとえばNanog mRNAまたは他のESマーカーを発現しない。明快かつ単純にするために、本明細書における方法の記述は、親細胞として線維芽細胞を指すが、本明細書において記述される方法は全て、他の初代培養親細胞タイプに容易に応用可能であることを理解すべきである。
【0050】
線維芽細胞を用いる場合、線維芽細胞は、再プログラミングの前は平坦で不規則な形状を有し、Nanog mRNAを発現しない。開始線維芽細胞は好ましくは他の胚幹細胞マーカーを発現しないであろう。ES細胞マーカーの発現は、たとえばRT-PCRによって測定することができる。または、測定はたとえば、ES表現型の特徴であるポリペプチドの存在を検出する免疫蛍光または他の免疫学的検出アプローチによって行うことができる。
【0051】
次の段階において、核酸配列の導入後に、線維芽細胞を選択物質の非存在下で培養する。「選択物質の非存在下」という用語は、誘導多能性幹細胞表現型に関して選択する選択物質の非存在、たとえば1つまたは複数のES細胞マーカーを発現するように脱分化した細胞に関して選択する選択物質の非存在を指す。いかなる種類の選択物質も存在しないことが好ましいが、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸の存在に関する選択物質は存在しうるものの、これらの因子の持続的な発現は、多能性表現型の維持にとって絶対的に必要ではない(以下を参照されたい)。方法には、単離されたクローンにおける導入された転写因子の存在または発現に関して試験する段階が含まれうる。
【0052】
次の段階において、選択物質の非存在下で培養される細胞を顕微鏡下で観察して(たとえば、通常の位相差光学顕微鏡または他の適当な光学手段の下で)、親細胞の不規則な形態的特徴、たとえば線維芽細胞の平坦で不規則な形態を失って、外観がなめらかで丸くなった細胞を培養において同定する。細胞は、多能性への移行を経験すると丸くなるがなおも生存している。形態による選択を容易にするために細胞を継代することができる。丸い形態を示す生存細胞のクローンを、たとえばマルチウェルプレートでの限界希釈および培養によって、または当業者に公知の他のアプローチによって単離する。
【0053】
さらなる段階において、単離されたクローンを幹細胞マーカーの発現に関して試験する。そのような発現により、細胞は誘導多能性幹細胞であると同定される。幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびNat1が含まれる非制限的な群から選択されうる。そのようなマーカーの発現を検出するための方法には、たとえばRT-PCRおよびコードされるポリペプチドの存在を検出する免疫学的方法が含まれうる。
【0054】
単離細胞の多能性幹細胞特徴を、ESマーカーの発現、および3つの胚細胞層のそれぞれの細胞への分化能を評価する多数の任意の試験によって確認することができる。1つの例として、ヌードマウスにおける奇形腫形成を用いて、単離されたクローンの多能性特徴を評価することができる。細胞をヌードマウスに導入して、細胞から発生した腫瘍に関する組織学的検査を行う。3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0055】
もう1つの態様において、細胞が雌性である場合、脱分化および多能性の測定として不活性なX染色体の再活性化を評価することができる。
【0056】
X-再活性化をモニターすることによる選択
雌性におけるX染色体の1つの不活化は、多能性から分化したことの特徴である。細胞を、たとえばOct4、Sox2、c-MycおよびKlf4の発現によって多能性状態に誘導する場合、不活性なX染色体は再活性化される。
【0057】
本明細書において記述される方法のもう1つの局面は、誘導多能性幹細胞に関して選択するために、分化した雌性細胞の不活性なX染色体の再活性化を用いる。
【0058】
この局面において、以下の段階を含む、誘導多能性幹細胞を選択するための方法が提供される。第一に、選択可能マーカーが、活性なX染色体上で変異体であり、不活性なX染色体上では野生型である、X染色体上で選択可能マーカーに関してヘテロ接合である雌性細胞を提供する。雌性細胞は、Nanog mRNAを発現せず、好ましくは他のES細胞マーカーを発現しない。または、Xが再活性化された場合に限ってマーカーの発現が観察されるように、選択可能マーカーは、たとえばトランスジェニック動物または細胞の不活性なX染色体に組み入れられるマーカーでありうる。そのようなマーカーには、たとえば任意の陽性選択可能マーカーが含まれうる。この代用物の好ましい態様はGFPを用いる(本明細書における以下の実施例を参照されたい)。
【0059】
他の好ましい態様において、選択可能マーカーは、たとえばヒポキサンチンホスホリボシルトランスフェラーゼ(Hprt)である。Hprtに関してヘテロ接合である雌性細胞株には、たとえばDR4マウス細胞(ATCC SCRC-1045を参照されたい)、ヒトTK6リンパ芽球細胞株(ECACC 87020507)、そのそれぞれが参照により本明細書に組み入れられる、Rinat et al, 2006, Mol. Genet. Metab. 87: 249-252によって記述される線維芽細胞、ならびにRivero et al., 2001, Am. J. Med. Genet. 103: 48-55およびHakoda et al., 1995, Hum. Genet. 96: 674-680によって記述されるリンパ球が含まれる。
【0060】
次の段階において、雌性細胞は、本発明の他の局面に関して本明細書において記述されるように多能性表現型となるように再プログラムされる。
【0061】
再プログラムされた細胞を選択物質と共に培養して、不活性なX染色体が再活性化されると、野生型選択可能マーカーを発現させることができ、選択物質の存在下で細胞が生存することができる。生存する細胞は誘導多能性幹細胞である。
【0062】
この局面の1つの態様において、方法はさらに、選択物質の存在下で生存する細胞を幹細胞マーカーの発現に関して試験する段階を含む。幹細胞マーカーは、たとえばSSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択されうる。
【0063】
もう1つの態様において、再プログラミングは、以下の1つを含む:因子を発現させるために調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸配列を細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;および細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【0064】
もう1つの態様において、方法は、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、選択物質の存在下で生存する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階をさらに含む。
【0065】
もう1つの態様において、細胞は細胞株に由来する。
【0066】
もう1つの態様において、細胞は、X染色体上の変異体Hprt遺伝子に関してヘテロ接合である。
【0067】
もう1つの態様において、細胞は、再プログラミングの前に不活性である野生型Hprt遺伝子をX染色体上で持ち、および再プログラミング前に活性である変異体の非機能的Hprt遺伝子をX染色体上で持つ。
【0068】
もう1つの態様において、細胞は、再プログラミングの前に6-チオグアニンに対して耐性である。
【0069】
もう1つの態様において、選択物質は、HAT培地を含む。
【0070】
もう1つの局面において、誘導多能性幹細胞を選択する方法が提供される。方法は、以下の段階を含む:(a)RT-PCRによって測定した場合にNanog mRNAを発現しない、X不活化による沈黙化に供されるX染色体連鎖レポーター遺伝子を持つ雌性細胞を提供する段階;(b)細胞が多能性表現型に再プログラムされる段階;(c)再プログラミング段階後に細胞を培養する段階;および(d)細胞のクローンがX染色体連鎖レポーターを発現する培養物から単離される段階。レポーターが発現されれば、クローンが誘導多能性幹細胞を含むことが指し示される。
【0071】
1つの態様において、方法はさらに、幹細胞マーカーの発現に関してクローンの細胞を試験する段階を含む。幹細胞マーカーは、たとえばSSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択されうる。
【0072】
もう1つの態様において、方法はさらに、レポーターを発現する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階を含む。3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0073】
X再活性化を受けた細胞に関して選択することによる多能性幹細胞の選択は、たとえば再プログラム段階の低分子調節物質、たとえば再プログラミングを容易にする低分子に関してスクリーニングするためのシステムを提供することができる。または、このように誘導化された多能性幹細胞は、幹細胞を所望の表現型に再分化させる低分子のまたは他の調節物質に関するスクリーニングアッセイを提供する。
【0074】
本発明を、制限的に解釈されてはならない以下の実施例によってさらに説明する。
【0075】
実施例
転写因子Oct4、Sox2、c-MycおよびKlf4の異所発現は、線維芽細胞ゲノムに多能性状態を付与するために十分であり、誘導多能性幹(iPS)細胞を生成する。これらの4つの因子によって誘導される核の再プログラミングが、分化した細胞と多能性細胞とのあいだの後成的な差を全体的にリセットすることができるか否かはなおも不明である。本明細書において、新規選択アプローチを用いて、iPS細胞を線維芽細胞から生成してその後成的状態を特徴付けする。雌性iPS細胞は、体細胞沈黙化X染色体の再活性化を示し、分化の際に無作為なX不活化を経験した。2つの重要なヒストン修飾に関してゲノム全体の分析を行ったところ、iPS細胞がES細胞に非常に類似していることが指し示された。これらの所見と一貫して、iPS細胞は、生殖系列に関与する生存する高度のキメラを生じた。これらのデータは、転写因子誘導再プログラミングにより、ES様状態への体細胞エピゲノムの全体的な復帰が起こることを示している。これらの結果は、核の再プログラミングを伴う後成的修飾を研究するためのパラダイムを提供し、かつ、正常でない後成的再プログラミングであっても、iPS細胞の治療応用に関する問題をもたらさないことを示唆している。これらのデータは、その全内容物が参照により本明細書に組み入れられる、Maherali, N., et al (2007) Cell-Stem Cell 1:55-70によって現在公表されている。
【0076】
実験技法
線維芽細胞の誘導化
Nanog-GFP-iresPuro構築物(Hatano et al., 2005)を雄性V6.5 ES細胞に標的化して、正確に標的化されたクローンを、標準的なサザンブロット分析によって確認し、マウスを作成した。Oct4-ネオマイシン/ハイグロマイシン選択可能なMEFをOct4-ネオマイシンマウスとpgk-ハイグロマイシンマウスとの異種交配から得た。XGFPとOct4-neo対立遺伝子とを持つTTFを、Oct4-NeoとX-連鎖GFPマウスとの異種交配から得た(Hadjantonakis et al., 1998)。誘導可能なOct4マウスは既に記述されている(Hochedlinger et al., 2005)。MEFは14.5日胚の胚に由来し、TTFは1週齢までのマウスに由来した。
【0077】
MEFのレトロウイルス産生および感染
Oct4、Sox2、c-MYC(T58A変異体)、およびKlf4に関するcDNAをレトロウイルスpMXベクターにクローニングして、Fugene(Roche)を用いてPlatEパッケージング細胞株にトランスフェクトした(Morita, S., Kojima, T., and Kitamura, T. (2000) Gene Ther 7, 1063-1066)。トランスフェクション後48時間で、ウイルス上清を用いてES培地において培養した標的MEFに感染させた。終夜感染の2〜3ラウンドを行って、細胞を放射線照射したフィーダー層において7日後に分割して、指し示された時間に1μg/mLピューロマイシン(Sigma)または300μg/mL G418(Roche)によって選択した。
【0078】
細胞培養およびインビトロ分化
iPS細胞およびES細胞を、放射線照射ネズミ胚線維芽細胞(フィーダー)および標準的なES培地(15%FBS、非必須アミノ酸、L-グルタミン、ペニシリン-ストレプトマイシン、β-メルカプトエタノール、および1000 U/mL LIFを補足したDMEM)において生育させた。胚盤胞注入のための2D4 iPS細胞を標識するために、細胞をRosa-GFP-Neoターゲティングベクターによって電気穿孔して、サザンブロット分析によって確かめた。XGFP/X Oct4-Neo iPS細胞からのサブクローンを生成するために、細胞を直線状pgk-Hygroプラスミドによって電気穿孔した。選択をそれぞれ、G418(300μg/mL)またはハイグロマイシン(140μg/mL)によってパルス後24時間目に開始した。X染色体の状態を試験するために、iPS細胞をゼラチンコーティング皿のES培地において1回継代して、フィーダー細胞の数を低減させて、LIFを欠損するES培地において40 ng/mLオールトランスレチノイン酸によって分化を誘導した。X不活化の無作為性を分析するために、EB形成時に分化を誘導した。
【0079】
卵母細胞を単離するために、雌性のキメラをPMSおよびhCGによって過剰排卵させて、卵母細胞をhCG注射後13時間で単離した。単為生殖活性化を誘導するために、卵母細胞を、10 mM塩化ストロンチウムおよび5μgml-1サイトカラシンBを補足した無カルシウムCZB培地において5時間インキュベートした後、KSOM培地において37℃で5%CO2で培養した。
【0080】
全体的なDNAメチル化に関するサザンブロット分析
ゲノムDNA 10μgをHpaIIまたはMspIによって消化して、断片を0.8%アガロースゲルにおいて分離した。DNAをHybondXLメンブレン(Amersham Biosciences)にブロットして、既に記述されているようにpMR150プローブとハイブリダイズさせた(Meissner, A., Gnirke, A., Bell, G. W., Ramsahoye, B., Lander, E. S., and Jaenisch, R. (2005). Nucleic Acids Res 33, 5868-5877)。
【0081】
バイサルファイトシークエンシング
EpiTect Bisulfiteキット(Qiagen)を用いて、製造共の説明書に従ってDNAの亜硫酸水素処置を行った。プライマー配列は既に記述されたとおりであった;Oct4(Blelloch, R., Wang, Z., Meissner, A., Pollard, S., Smith, A., and Jaenisch, R, (2006). Stem Cells 24(9):2007-13)およびNanog(Takahashi and Yamanaka, 2006)。増幅された産物をゲル濾過カラムを用いて精製してpCR2.1-TOPOベクター(Invitrogen)にクローニングして、M13フォワードおよびリバースプライマーによってシークエンシングした。
【0082】
RT-PCR分析
内因性の座からの多能性遺伝子の発現を試験するために、総RNAをDNA-freeキット(Ambion, Austin, TX)によって処置して、SuperScript First-Strand Synthesisシステム(Invitrogen)によってオリゴdTプライマーを用いて製造元の説明書に従って逆転写した。全てのプライマー配列を表3に示す。
【0083】
ウェスタン分析、免疫染色およびAP染色
本明細書において記述される方法において用いられる抗体を表3に記載する。Vector Red基質キット(Vector Labs)を用いてアルカリホスファターゼ染色を行った。免疫染色はPlath et al(2003)に従って行った。
【0084】
FISH分析
FISHは既に記述されたように行った(Panning, B., Dausman, J., and Jaenisch, R. (1997) Cell 90, 907-916)。Xist、Tsix、およびPgkI二本鎖DNAプローブを、Xist cDNA鋳型およびPgkI配列の17 kbを含有するゲノムクローンからそれぞれ、Cy3-dUTP(Perkin Elmer)、またはFITC-dUTP(Amersham)およびBioprimeキット試薬(Invitrogen)を用いてランダムプライミングによって生成した。TsixおよびXistのいずれかを特異的に検出するための鎖特異的RNAプローブを、Xistエキソン1およびエキソン6鋳型からFITC UTPの存在下でインビトロ転写によって生成した。FISHによって免疫蛍光を追跡する場合、細胞を4%PFAによって固定した後、FISH技法を開始して、ブロッキング緩衝液は1 mg/mL tRNAおよびRNアーゼ阻害剤を含有した。
【0085】
細胞融合
iPS細胞400万個をMEF 400万個と合わせて、PEG-1500(Roche)によって製造元の指示に従って融合した。融合後24時間目にピューロマイシン(1μg/mL)およびハイグロマイシン(140μg/mL)を用いて選択を開始した。Neo選択を伴う実験の場合、G418を300μg/mLで用いた。細胞周期分析を、ヨウ化プロピジウムを用いてFACS Calibur(BD)において行った;シグナル領域をDNA含有量の測定として用いた。
【0086】
染色質の免疫沈降(ChIP)およびマイクロアレイハイブリダイゼーション
ゲノム全体の染色質分析ChIPをwww.upstate.com.でのプロトコールに従って細胞約100万個について行った。各免疫沈降試料10 ngおよび対応する投入量をWhole Genome Amplificationキット(Sigma)を用いて増幅して、増幅材料2μgをBioprimeキット(Invitrogen)を用いてCy3またはCy5(Perkin Elmer)によって標識した。マウスプロモーターアレイ(Agilent -G4490)へのハイブリダイゼーション、洗浄、および走査を、製造元の説明書に従って行った。プローブシグナル(対数比)をフィーチャー抽出ソフトウェアを用いて抽出して、Chip AnalyticsソフトウェアのLowess標準化を用いて標準化して、本明細書において記述されるように統計学的に分析した。
【0087】
全ゲノム発現分析
V6.5 ES細胞、雌性NGiP MEF、ピューロマイシン選択2D4 iPS細胞、およびピューロマイシン選択対照NGiP ES細胞からのRNA 500 ngの1試料あたり2個ずつの試料を増幅して、Agilent low RNA増幅および1色標識キットを用いて製造元の説明書に従ってCy3によって標識した。標識RNAをAgilent Mouse全ゲノムアレイ(G4122F)にハイブリダイズさせて、分析した。
【0088】
フローサイトメトリー
キメラ分析に関して、脾臓、胸腺、および骨髄を、既に記述されているように単離した(Ye, M., Iwasaki, H., Laiosa, C. V., Stadtfeld, M., Xie, H., Heck, S., Clausen, B., Akashi, K., and Graf, T. (2003) Immunity 19, 689-699);細胞を抗体によって染色して、FACSによって分析した。Oct4-Neo XGFP/X尾部線維芽細胞を2回連続して通過させることによってソーティングして、純粋なGFP陰性集団を確かめるために再分析した。EB分化の際に、細胞をGFP+/GFP-集団にソーティングして、FISH分析のために用いた。細胞をBD FACS ARIA(BD Pharmingen)において獲得してFlow Joソフトウェア(Tree Star, Inc.)を用いてデータを分析した。
【0089】
奇形腫形成
各株の細胞200万個を、イソフルラン麻酔SCIDマウスの背側脇腹に皮下注射した。注射の3〜4週間後に奇形腫を回収して、10%ホルマリンにおいて終夜固定して、パラフィン抱埋して、ヘマトキシリン・エオジンによってまたは特異的抗体によって処理した。
【0090】
キメラマウスにおける組織学的検査およびGFP発現の免疫組織学的分析
4%PFAおよび20%蔗糖において組織を続いてインキュベートした後にOCT化合物に抱埋して低温槽において切片(厚さ10μm)を作製する段階によって凍結切片を生成した。切片をVectashield封入培地およびDAPIと共にカバーガラスで覆い、GFPシグナルに関して直接可視化した。
【0091】
iPS細胞の生成効率
ウイルスパッケージングPlatE細胞に、4つの因子12μg(各因子3μg)、またはGFPベクター:空のベクターの1:3混合物の全12μgのいずれかをトランスフェクトした。Nanog-GFP MEFを50%コンフルエンスで接種して、パッケージング細胞からの上清を感染させた。感染後7日目に、4つの因子を感染させた細胞を、放射線照射フィーダーにおいて1:2に分割して、選択(1μg/mLピューロマイシン)または非選択条件のいずれかに置いた。GFP感染細胞を計数して(5.3×106個)、FACSによって分析した。GFP+細胞の百分率(15%)を1つの因子による感染頻度であるとすると、このように4つ全ての因子の頻度は0.15-4であり、理論的にはコロニー〜2700個をもたらした。選択条件下で4週間後、AP陽性ピューロマイシン耐性コロニー20個が出現して、効率は〜0.74%であった。非選択条件ではコロニー〜240個が出現して、効率は〜9%であった。
【0092】
染色質の免疫沈降
フィーダー依存的雄性ES細胞株V6.5(129/B16)、フィーダー非依存的雄性ES細胞株E14(129/ola)、ならびに129/B16マウスに由来する初代培養雄性および雌性MEFを、ピューロマイシンの存在下で生育させた2D4 iPS株と共に用いた。V6.5および2D4細胞の場合、線維芽細胞の混入を低減させるために、細胞の最後の継代は、追加のフィーダー細胞を加えずに行った。細胞は、これらの条件下でその非分化状態を維持した(図1およびデータは示していない)。細胞をホルムアルデヒドによって室温で10分間クロスリンクさせた後、1%SDSを有する10 mM Tris-EDTA、pH 8.0において溶解して、氷中で45秒間の休止をはさんで15秒間のパルス6回の超音波処理を行った。浄化された剪断染色質をH3me3K4(Abcam 8580)またはH3me3K27(Upstate 07-449)に対する抗体によって4℃で終夜免疫沈降させて、プロテインAビーズによって2時間収集して、5分間の洗浄を2回行い、緩衝液(Upstateウェブサイト上でのレシピ)によって溶出した。溶出液を逆クロスリンクして、RNアーゼおよびプロテナーゼK処置を行い、Qiagen PCR精製キットを用いてDNAを精製した。ウサギIgG抗体によるChIPは、いかなる濃縮も見いださなかった(データは示していない)。
【0093】
ヒストンメチル化データと共にゲノムの分析のための統計学的方法
平均プローブシグナルを500 bpのウィンドウで段階的に抽出した。遺伝子16339個を、領域の少なくとも50%が500 bpのウィンドウでプローブによって覆われるという基準に基づいて選択した。ES細胞とMEF細胞のあいだでH3me3K4およびH3me3K27パターンの有意な差を有する遺伝子をシグナチャー遺伝子としてフィルタリングした。各遺伝子に関して、2つの細胞タイプのあいだのヒストン修飾パターンの差を16ウィンドウのシグナルベクトルのユークリッド距離によって定義した。2つのES細胞株(E14対V6.5の距離)および2つの初代培養MEF細胞株(雄性対雌性の距離)の自己距離をプールして、2つのES細胞株または2つのMEF細胞株のあいだの差が小さいと仮定して無効分布を生成した。XおよびY染色体上でコードされる遺伝子を分析から除外した。全てのシグナチャー遺伝子に関して、任意のES-MEF対の距離(E14対Mの距離;E14対Fの距離、V6.5対Mの距離;V6.5対Fの距離)は、ヌル分布の99%分位点である(p値0.01に対応する)既定のシグナチャー-遺伝子閾値(SigT)より大きくなければならない。
【0094】
2D4株におけるシグナチャー遺伝子のEs様遺伝子(Eクラス)、MEF様遺伝子(M-クラス)およびNeutral遺伝子(Nクラス;ES細胞またはMEFのいずれに対しても有意により強い優先性を示さない遺伝子)へのメチル化パターンを明快にするために、2D4とES細胞との平均距離(2D4対ESの距離)および2D4とMEFとの平均距離(2D4対MEFの距離)をコンピューターによって計算した。優先性スコアPS_2D4=(2D4対ESの距離−2D4対MEFの距離)を、2D4細胞における特定の遺伝子のヒストンメチル化パターンが、ES細胞のパターンをどれほど強く「優先して」おそらく模倣するかに関する指標として用いた。この場合も、PSのヌル分布を以下のようにして生成した。各ES細胞株のデータセットを他のES株およびMEFのデータセットと比較した。全ての遺伝子16339個からのPS_ES(E14対V6.5の距離−E 14対MEFの距離)および(V6.5対E14の距離−V6.5対MEFの距離)をコンピューターによって計算してプールした。95%分位点をEクラス閾値(ET)として用いた。ETより大きいPS_2D4を有する任意のシグナチャー遺伝子を「Mクラス」と呼んだ。M閾値(MT)およびEクラスを類似のように定義した。PS-2D4がMTとETの間に入る遺伝子を「Nクラス」と呼んだ。異なる細胞タイプのあいだの8 kb領域内での各500 bpウィンドウに関するメチル化データのピアソン相関係数を、MS Excelにおいて相関関数を用いて計算した。
【0095】
遺伝子発現分析
発現データをFeature Extractionソフトウェア(Agilent)を用いて抽出した。未加工データをlog2変換して、同じ遺伝子に関する多数のプローブからのシグナルを平均した。平均値が0であり、標準偏差が1であるようにそれぞれのアレイを標準化した。再現実験からのデータを平均した。MEFとES細胞のあいだで発現が2倍変化した遺伝子を選択して、それによってこれらの2つの細胞タイプのあいだで最も異なるように発現される遺伝子2473個が同定された(全遺伝子33376個中)。偏りのない階層的クラスタリングを使用して、これらの遺伝子2473個の発現パターンをES細胞、MEF、ピューロマイシン選択NGiP ES細胞およびiPS細胞に対して群分けした。加えて、シグナチャー遺伝子に関する発現パターンをESとMEFの比、またはiPSとMEFの比としてコンピューターにより計算して、メチル化データと共にプロットした。
【0096】
実施例1:Nanog選択可能な線維芽細胞を用いるiPS細胞の生成
Nanog-GFP-puro(Hatano, S. Y., Tada, M., Kimura, H., Yamaguchi, S., Kono, T., Nakano, T., Suemori, H., Nakatsuji, N., and Tada, T. (2005) Mech Dev 122, 67-79)と呼ばれる内因性のNanog座においてGFP-IRES-Puroカセットを持つ雌性マウス胚線維芽細胞(MEF)に、Oct4、Sox2、タンパク質を安定化させる(Sears, R., Nuckolls, F., Haura, E., Taya, Y., Tamai, K., and Nevins, J. R. (2000) Genes Dev 14, 2501-2514)c-MYC-T58A変異体、およびKlf4をコードするcDNAをレトロウイルスによって感染させた。先に報告された、感染後3日目に適用されたFbx15選択(Takahashi and Yamanaka, 2006)とは対照的に、感染後3日目でのNanog発現に関する選択によってコロニーは得られず、Fbx15およびNanog遺伝子の再活性化速度論が異なることを示唆している。感染後7日またはそれより後の日に選択を適用した場合、耐性コロニーが再現性よく出現した。拡大した株5株のうち(表1を参照されたい)、2つの株は、ES細胞と同一であるように思われ、ES細胞表面マーカーSSEA1およびCD9を発現する均一な培養を維持した(データは示していない)。対照的に、他の3つのクローンは、多数回の継代後不均一な培養物を生じ、これは、ES様集団と小さく丸い急速に分裂する細胞の個別の集団の双方を含有した。Nanog-GFP、SSEA-1、およびCD9に関するFACSソーティングの後にサブクローニングを行うと、これらの丸い細胞を消失させるために十分であり、この集団が、ES様細胞とは別個であることを示唆した。興味深いことに、2つの均一な細胞株に関する選択の開始は感染後3週間で起こったが、不均一な株は、感染後1週間で選択を受け、選択の遅れがiPS細胞のより純粋な集団を得るために都合がよい可能性があることを示唆している。
【0097】
その後の研究は、均一なES様細胞株2D4、および本明細書においてiPS細胞と呼ばれる、再ソーティングされて再クローニングされた株1A2に重点を置いた。レトロウイルス組み込み部位のサザンブロット分析により、双方のiPS株に4つ全てのレトロウイルスコード遺伝子が存在することが判明し、ゲノム刷り込みに関する試験により、iPS細胞が、線維芽細胞培養物に存在する可能性があるまれな始原生殖細胞に由来するのではないことを確認した(図9)。Fbx15選択iPS細胞(Takahashi and Yamanaka, 2006)とは対照的に、Nanog選択可能なiPS細胞は、フィーダーの非存在下およびピューロマイシン選択下でES様形態、Nanog発現、およびアルカリホスファターゼ活性(AP)を維持したことから(データは示していない)、フィーダー非依存的生育を示した。LIFを除去すると、始原内胚葉に類似するGATA-4発現細胞への予想される分化が起こり(データは示していない)、分化はNanog発現の喪失を伴った(データは示していない)。RT-PCR分析は、他のES細胞マーカー、Nanog、ERas、およびCriptoと共に内因性の座からのOct4およびSox2の発現を指し示した(図1A)。4つのレトロウイルス発現遺伝子に関する定量的PCR分析は、個々のレトロウイルスに感染した線維芽細胞における強い発現を示したが、均一なiPS細胞では効率よい沈黙化を示した(図1C)。Oct4、Sox2、c-MycおよびKlf4のタンパク質レベルは、iPS細胞と対照ES細胞のあいだで類似であり(図1B)、免疫蛍光は、Oct4およびSox2がレチノイン酸誘導分化の際に効率よくダウンレギュレートされることを示し、ウイルスコード転写因子遺伝子が分化した細胞において有効に沈黙化されたままであることを証明した(データは示していない)。2D4 iPS細胞をSCIDマウスに注射すると、3つの胚細胞層を代表する細胞タイプを含有する奇形腫を生じ、その多能性を確認した(データは示していない)。これらのデータは、レトロウイルス発現Oct4、Sox-2、c-MYCおよびKlf4が、Nanog再活性化に関する選択と組み合わせて、ES細胞と多くの特性を共有するiPS細胞をもたらしうることを指し示している。
【0098】
実施例2:Nanog-選択可能なiPS細胞は、ES細胞様表現型を体細胞に付与する
Nanog-選択可能なiPS細胞がES細胞と類似の機能的属性を保有するか否かを決定するために、細胞融合のコンテクストにおいて体細胞にES-様表現型を押しつける能力を試験した。ピューロマイシン耐性2D4 iPS細胞株からの細胞と共にハイグロマイシン耐性MEF(図2A)。融合後2週間目、ES細胞様形態を有してNanog-GFPを発現し続ける二重耐性4倍体ハイブリッドクローン7個(図2Bおよびデータは示していない)を回収した。対照Nanog-GFP-puro ES細胞をハイグロマイシン耐性MEFと融合させると、1つのハイブリッドコロニーが回収された。多能性を試験するために、ハイブリッド細胞を免疫無防備マウスに注入したところ、4週間後に、3つ全ての胚細胞層を代表する細胞タイプを含有する奇形腫が単離された(データは示していない)。
【0099】
体細胞ゲノムの再プログラミングに関する試験として、内因性のOct4座の制御下で構成的ハイグロマイシン耐性遺伝子とネオマイシン選択可能マーカーの双方を含有するMEF(Oct4-Neo対立遺伝子と呼ばれる)について融合実験を繰り返した。G418を初回選択プロセスにおいて用いた場合にはクローンは得られず、体細胞Oct4座の再プログラミングは、内因性のNanog座の再プログラミングと同様に、逐次的プロセスであることを示唆した。ゆえに、ピューロマイシン/ハイグロマイシン耐性ハイブリッドを拡大した後、それらをピューロマイシン/G418選択に供して、体細胞Oct4遺伝子の再活性化に関して試験した。ピューロマイシン/ハイグロマイシン耐性コロニーは全てピューロマイシン/G418選択下で生存しており、体細胞ゲノムが内因性Oct4座で再プログラムされていることを指し示している(データは示していない)。これらの結果は、Nanog-選択細胞がES細胞と同様に、再プログラミング活性を持ち、体細胞ゲノムに対してES様状態を付与できることを示している。
【0100】
実施例3:異所Oct4発現はiPS細胞の維持にとって不可欠である
Fbx15-選択2D4 iPS細胞は、Oct4およびSox2の永続的なレトロウイルス発現を示し、それぞれの内因性の座からの発現は無視できる程度であり、iPS細胞の自己再生および多能性を維持するために外から提供された因子が持続的に必要であることを示唆した(Takahashi and Yamanaka, 2006)。iPS細胞における効率的なレトロウイルス遺伝子沈黙化を示唆する遺伝子発現データを補強するために、そのゲノムにおいてドキシサイクリン誘導可能なOct4トランスジーンを持つ線維芽細胞を用いることによって、持続的なOct4発現がiPS細胞の維持にとって必要であるか否かを遺伝的に試験することに決定した(Hochedlinger, K., Yamada, Y., Beard, C, and Jaenisch, R. (2005) Cell 121, 465-477)(図3A)。
【0101】
コロニーがOct4誘導可能なシステムを用いて得られうるか否かを最初に決定するために、Oct4誘導可能なMEFに、いかなる選択も行わずにSox2、c-MYCおよびKlf4レトロウイルスを感染させた。ドキシサイクリンの非存在下では、AP陽性コロニーは回収されなかったが、ドキシサイクリンの存在下では、数百個のAP陽性コロニーが出現し、AP陽性コロニーを確立するためにトランスジェニックOct4発現に厳密に依存することを指し示している(図3B)。その後、得られた細胞の再プログラム状態を確かめるために、iPS細胞を、Oct4誘導可能な対立遺伝子とOct4-Neo対立遺伝子の双方を持つ尾部線維芽細胞(TTF)から生成した(図3A)。ドキシサイクリンの存在下で、標的細胞にSox2、c-MYCおよびKlf4を感染させた。薬物選択の開始が遅いことは有利であるというこれまでの所見に基づいて、初回選択を行わずに、ES細胞様形態のみに基づいてiPSコロニーを確立しようと試みた。個々のES様コロニー48個を感染後3週目に採取して、うち2つは、ドキシサイクリンの持続的存在下で安定なES細胞様株に生育した。G418培地に再度播種した後、いずれの細胞株も生存し、内因性のOct4遺伝子が再活性化されて、iPS細胞が生成されていることを指し示した。重要なことに、ドキシサイクリンを培地から除去すると、その生育挙動または形態が変化することなく(データは示していない)、これらの細胞をG418の存在下で何回も継代することが可能であった。ドキシサイクリンの非存在下でのウイルス挿入および異常なOct4トランスジーン活性化の可能性を除外するために、内因性および誘導Oct4発現の定量的PCR分析を行って、分化および誘導の際の発現レベルを分析した(図3C)。非分化iPS細胞は、高レベルの内因性のOct4発現およびトランスジーン発現の完全な非存在を示した。Oct4レベルは、LIFの非存在下で下落して、ドキシサイクリンを投与すると再度出現し、内因性のOct4発現の分化依存的ダウンレギュレーションおよびドキシサイクリンに対する細胞の継続的応答性をそれぞれ示している(図3C)。十分に分化した奇形腫の形成能は、これらの細胞の多能性を証明した(データは示していない)。このように、内因性のOct4座は、外因性のOct4発現の非存在下で多能性状態でiPS細胞を維持するために4つの転写因子によって十分に再プログラムされた。
【0102】
実施例4:遺伝子特異的および全体的DNAメチル化はiPS細胞とES細胞のあいだで類似である
再プログラムされた線維芽細胞のES細胞様特性に基づいて、iPS細胞が、ES細胞と類似の後成的状態を獲得したか否かを調べた。核移入または細胞融合による体細胞ゲノムの再プログラミングは、そのプロモーター領域での多能性遺伝子のDNA脱メチル化などの後成的変化を伴う(Cowan et al., 2005;Tada et al., 2001)。バイサルファイトシークエンシングを用いて、Fbx15-選択iPS細胞において不完全に脱メチル化されていることが既に示されている(Takahashi and Yamanaka, 2006)Oct4およびNanogプロモーターのメチル化状態を査定した。MEFにおいてメチル化されているプロモーターエレメントはいずれも、Nanog選択iPS細胞およびES細胞において脱メチル化を示し、これらの2つの多能性遺伝子が適切に後成的に再プログラミングされたことを示唆している(図4A)。さらに、Nanogプロモーターの脱メチル化は、iPS細胞とMEFの融合を通して生成された細胞ハイブリッドにおいて起こり(図4B、図2を参照されたい)、iPS細胞が、再プログラミング活性を潜在的に持ち、分化した細胞において後成的変化を誘導できることを確認した。
【0103】
雌性ES細胞は、雄性ES細胞および分化した細胞とは対照的に、ゲノムの全体的なDNA低メチル化を示し、これは2つの活性なX染色体(Xa)の存在に帰因しうる(Zvetkova, I., Apedaile, A., Ramsahoye, B., Mermoud, J. E., Crompton, L. A., John, R., Feil, R., and Brockdorff, N. (2005) Nat Genet 37, 1274-1279)。メチル化感受性制限酵素アッセイを用いて、雌性対照ES細胞と同様に、小さいサテライトリピートの全体的な低メチル化が、2D4 iPS細胞株において検出された(図4C)。これらの結果は、iPS細胞が、雌性ES細胞と類似の後成的状態を得たことを示唆している。
【0104】
実施例5:雌性Nanog-選択可能なiPS細胞におけるX-不活化
iPS細胞における全体的なDNA低メチル化は、不活性なX染色体(Xi)が雌性iPS細胞において再活性化されることを指し示している。X-不活化は、哺乳動物細胞における異質染色質形成の最も劇的な例の1つであり、相反して発現される2つの非コードRNA、すなわちXistとそのアンチセンス転写物Tsixによって調節される(Thorvaldsen, J. L., Verona, R. I., and Bartolomei, M. S. (2006) Dev Biol 298, 344-353)。非分化雌性ES細胞は、2つのXaを持ち、両方のX染色体からのTsixを発現して、Xist発現を抑制する。分化すると、Xistは将来のXiにおいて強くアップレギュレートされるようになり、沈黙化を誘導するが、Tsixは消失して体細胞には存在しない。Tsixの下流に位置するX-不活化にとって重要な第3の座であるXite座は、Tsix-様パターンで発現される(Ogawa, Y., and Lee, J. T. (2003) Mol Cell 11, 731-743)。
【0105】
雌性Nanog-GFP-puro MEFにおけるX不活化状態を、蛍光インサイチューハイブリダイゼーション(FISH)を用いて最初に査定して、Xist RNA局在およびX-連鎖遺伝子発現を分析した。Xiの存在と一致して、線維芽細胞の96%がXist RNAコーティングX染色体を持ち、他のX染色体からのPgk1遺伝子の発現を示した(データは示していない)。2D4 iPS細胞株は、非分化ES細胞を強く暗示するXist、Tsix、およびPgk1発現パターンを示した(データは示していない)。すなわち、TsixおよびPgk1は、高レベルで二対立遺伝子で発現されたが、Xist RNAを検出することができず、2つのXaが存在することを証明した。加えて、RT-PCR分析は、ES細胞および2D4 iPS細胞の双方においてXite座からの転写物を検出したが、親線維芽細胞集団では検出しなかった(図5A)。
【0106】
X不活化が開始されると、特徴的な染色質の修飾が将来のXiに押しつけられて、これは染色体の安定な沈黙化を確実にする(Heard, E. (2005) Curr Opin Genet Dev 15, 482-489;Ng, K., Pullirsch, D., Leeb, M., and Wutz, A. (2007) EMBO Rep 8, 34-39)。免疫蛍光を用いて、iPS細胞におけるXi連鎖染色質修飾の存在を分析した。雌性Nanog-GFP-puro MEFは、リジン27位でトリメチル化されたヒストンH3、ヒストンH4リジン20モノメチル化に関する、およびH3K27トリメチル化の媒介の原因であるPolycomb群(PcG)タンパク質Ezh2に関するXi様濃縮の予想された頻度を示した。対照的に、ES細胞と同様にiPS細胞は、これらの染色質のマークに関して豊富で均一な核染色を示し、Xi様濃縮を示さなかった(データは示していない)。併せて、これらのデータは、Xiの転写再活性化を誘導するために、X不活化の調節にとって必須である3つの非コード転写物の発現パターンをリセットするために、およびXiに対して特異的である染色質修飾を消すために、Nanog選択と組み合わせて4つの転写因子が十分であることを指し示している。
【0107】
次に、2D4細胞が分化の際にX不活化を受けるか否かを試験した。iPS細胞がそのX染色体の1つを沈黙化させる能力と一貫して、レチノイン酸誘導分化を受けつつある2D4 iPS細胞においてXist RNAコーティング染色体が検出された(データは示していない)。Xistコーティング染色体は、そのXの沈黙状態と一致してRNAポリメラーゼIIとのオーバーラップを示さなかった(データは示していない)。さらに、分化しつつある雌性ES細胞と同様に、iPS細胞におけるXist RNAコーティングX染色体は、X-不活化の開始時にほとんど常にH3me3K27およびそのメチルトランスフェラーゼEzh2の濃縮領域と一致した(図5B)。XiにおけるEzh2蓄積およびH3me3K27濃縮の一致は、X不活化の初期相のみの特徴である(Plath, K., Fang, J., Mlynarczyk-Evans, S. K., Cao, R., Worringer, K. A., Wang, H., de la Cruz, C. C, Otte, A. P., Panning, B., and Zhang, Y. (2003) Science 300, 131-135;Silva, J., Mak, W., Zvetkova, L, Appanah, R., Nesterova, T. B., Webster, Z., Peters, A. H., Jenuwein, T., Otte, A. P., and Brockdorff, N. (2003) Dev Cell 4, 481-495)。このように、雌性iPS細胞におけるX染色体の不活化は、雌性ES細胞と同じ動力学を表す。
【0108】
実施例6:分化しつつあるiPS細胞における無作為なX不活化
X染色体不活化は、胚外系列および初期着床前胚において非無作為に起こるが、胚盤葉上層および分化しつつあるES細胞においては無作為である。クローニングしたマウス胚におけるX不活化の分析から、体細胞Xiが胚細胞における無作為なX不活化を可能にするために核移入の際に再プログラムされるが、Xiの記憶は、それが配偶子刷り込みを交換する胚外組織において維持されることが示されている(Eggan et al., 2000)。ゆえに、転写因子誘導再プログラミングが体細胞不活化Xiの記憶を消して、このように、分化しつつあるiPS細胞において無作為なX不活化を可能にするか否かを試験した。Nanog-選択可能な2D4 iPS細胞において2つのX染色体を区別することは不可能であったことから、iPS細胞を、緑色蛍光タンパク質(GFP)の発現を駆動するサイトメガロウイルスプロモーターと共にX-連鎖レポータートランスジーン(XGFP)を持つ雌性線維芽細胞から生成した(Hadjantonakis, A. K., Gertsenstein, M., Ikawa, M., Okabe, M., and Nagy, A. (1998) Nat Genet 19, 220-222)(図6A)。このレポーターを、X不活化による沈黙化に供すると、分化しつつあるiPS細胞において沈黙化したX染色体を決定することができる。TTFを、GFPトランスジーンに関してヘテロ接合であり、Oct4-Neo対立遺伝子を持つ雌性マウスから単離した。線維芽細胞集団における無作為なX-不活化と一貫して、TTF細胞の34%がGFP陽性であり(XaGFP/Xi)、細胞の66%がGFP陰性であった(XiGFP/Xa)(図6A、データは示していない)。X不活化の何らかの非対称性が予想され、おそらく、2つのX染色体の遺伝的バックグラウンドの差を反映した。2ラウンドのFACSソーティングによって単離されたGFP陰性細胞に4つの転写因子をコードするレトロウイルスを感染させて、得られたES様コロニーをGFP再発現に基づいてXiGFPの再活性化に関してスクリーニングした。完全に緑色の4つのコロニーが単離されたが、これは再播種した場合に、G418に対して耐性であることが同様に見いだされ、このように沈黙X染色体の再活性化に加えてOct4座が活性化されたことを指し示している。XistおよびTsix発現のES細胞様パターンは、X再プログラミングを確認した(データは示していない)。
【0109】
これらの雌性iPS細胞が、ES細胞と同様に、培養において持続的に維持した場合にXを失う傾向を有したと仮定して、iPS細胞の純粋なクローン集団がX不活化の無作為性に関して確実に分析されるように、XaGFPXa iPS細胞をサブクローニングした。サブクローンの分化を胎芽様体の形成によって誘導して、分化した細胞をFACSによって、GFP陽性およびGFP陰性集団にソーティングして、FISHによって分析した(図6A)。X不活化の無作為なパターンと一貫して、平均で細胞の38%がGFP陽性であり、細胞の62%がGFP陰性であり、双方の集団の大多数が、XiのXist RNAコーティングと一貫するXistシグナルを有した(データは示していない)。無作為なX-不活化により、体細胞においてXaとXiとを区別する後成的なマークがインビトロ再プログラミングの際に除去されうること、およびその後のインビトロ分化の際にいずれかのXにおいて再確立されることが確認される。
【0110】
実施例7:iPS細胞におけるヒストンメチル化パターンの全体的な再プログラミング
次に、Oct4およびNanogプロモーターのDNA脱メチル化ならびにXiの再活性化に加えて、完全な線維芽細胞ゲノムがiPS細胞誘導化の際にES様状態に後成的に再プログラムされていたか否かを調べた。ヒストンメチル化は、哺乳動物の発達および細胞分化の際の遺伝子発現の後成的調節において重要な役割を果たす。一般的に、転写された遺伝子は、H3K4トリメチル化に関連しているが(Bernstein, B. E., Kamal, M., Lindblad-Toh, K., Bekiranov, S., Bailey, D. K., Huebert, D. J., McMahon, S., Karlsson, E. K., Kulbokas, E. J., 3rd, Gingeras, T. R., et al. (2005) Cell 120, 169-181;Kim, T. H., Barrera, L. O., Zheng, M., Qu, C, Singer, M. A., Richmond, T. A., Wu, Y., Green, R. D., and Ren, B. (2005) Nature 436, 876-880)、沈黙化された多くの遺伝子はH3K27トリメチル化に関連している(Boyer, L. A., Plath, K., Zeitlinger, J., Brambrink, T., Medeiros, L. A., Lee, T. I., Levine, S. S., Wernig, M., Tajonar, A., Ray, M. K., et al. (2006). Nature 441(7091):349-53;Lee, T. I, Jenner, R. G., Boyer, L. A., Guenther, M. G., Levine, S. S., Kumar, R. M., Chevalier, B., Johnstone, S. E., Cole, M. F., Isono, K., et al. (2006) Cell 125(2):301-13)。Nanog-選択2D4 iPS株、雄性および雌性MEF、ならびに2つの雄性ES細胞株におけるK4およびK27トリメチル化に関するゲノム全体の位置分析を、染色質の免疫沈降の後に、マウスプロモーターアレイとのハイブリダイゼーションを用いて行った。このアレイ上のプロモーターは、遺伝子約16,500個に関して転写開始部位の-5.5 kb上流から+2.5 kb下流までの領域を範囲に持つ。2D4 iPS株がES細胞またはMEFにより類似しているか否かを決定するために、ES細胞とMEFのあいだでヒストンメチル化パターンが有意に異なる一組の遺伝子を定義した。高ストリンジェンシー(p=0.01)では、ES細胞とMEFのあいだで遺伝子957個が異なると同定され、「シグナチャー」遺伝子として分類された(実験技法を参照されたい)。特に、2D4 iPS細胞では、シグナチャー遺伝子の94.4%がES細胞と実質的に同一であるメチル化パターン(Eクラス遺伝子)を持ったが、よりMEF様パターン(Mクラス遺伝子)では遺伝子の0.7%がメチル化されたに過ぎなかった。座の残りの4.9%は、差が小さすぎて有意ではなかったために(データは示していない)Nクラス遺伝子(中性)であると分類された。iPS座の大部分(91%)は、より大きい組のシグナチャー遺伝子を含めるためにたとえストリンジェンシーをp=0.05まで低下させてもEクラスに留まった(データは示していない)。E、M、およびN遺伝子への分布は、無作為入れ替え試験によって確認したところ、非常に有意である(図10)。非シグナチャークラスに属する遺伝子は、MEF、ES細胞、およびiPS細胞のあいだでメチル化パターンにほとんどまたは全く差を示さず(データは示していない)、iPS株がES細胞またはMEFのいずれにおいても見いだされない完全に新規の後成的同一性を獲得したことを指し示している。集合的に、これらの結果は、インビトロ再プログラミングが、線維芽細胞ゲノムの後成的記憶をES細胞と非常に類似の記憶に逆転させることができることを指し示している。
【0111】
K4およびK27メチル化パターンが再プログラミングの際に異なる程度にリセットされたか否かを決定する試みにおいて、アレイ上の遺伝子16,500個全てに関してそれぞれのメチル化のマークに関するピアソン相関を個々に計算した(図7A)。この分析から、iPS細胞およびES細胞が、そのK27メチル化パターンにおいて2つのES株が互いに対して類似であるのと同程度に類似であるが、MEFはiPSおよびES細胞の双方から同程度に明らかに異なることが判明した。興味深いことに、K4メチル化は、全ての細胞タイプのあいだでより類似であり、再プログラミングが、K4トリメチル化よりむしろK27の変化に主に関連することを示唆している。この全体的分析からの1つの予測は、K27メチル化の変化が、シグナチャー遺伝子のEクラスにおいて顕著であるはずであるという点である。これを試験するために、8 kbのプロモーター領域にわたって500 bpの間隔で起こりうる全ての細胞タイプのあいだで対応のある相関分析を行い、各比較に関して相関値16個を得た(図7B)。E遺伝子として分類された遺伝子は実際に、全分析領域にわたってES細胞と2D4 iPS細胞のあいだでそのK4およびK27メチル化パターンが非常に類似であったが、MEFは全体を通していずれの細胞タイプとも劇的に異なった。全体的相関とさらに一致して、K27メチル化は、MEFとES/iPS細胞のあいだでK4メチル化より劇的に異なった。発達遺伝子は、ネズミES細胞におけるPcG媒介K27メチル化の最も重要な標的群であるというこれまでの所見(Boyer et al., 2006)に基づいて、これらの座がシグナチャー遺伝子内で濃縮されるか否かを試験することに決定した。実際に、遺伝子オントロジー分析により、発達遺伝子はシグナチャー遺伝子のEクラスにおける最も有意に濃縮された遺伝子群であることが判明した(p=8xe-10)。これらの知見は、MEFからiPS細胞への再プログラミングにとって、K27メチル化の変化がK4メチル化の変化より重要であることを示唆しており、再プログラミングにおけるPcGタンパク質の重要な役割を示唆している。
【0112】
iPSとES細胞のヒストンメチル化パターンの相関が、iPS細胞の転写状態の変化を忠実に捕らえているか否かを試験するために、全ゲノムレベルでAgilentマイクロアレイを用いて、ES細胞、2D4 iPS細胞、およびMEF細胞について発現分析を行った。ESおよびiPS細胞は、ピアソン相関によって決定した場合に、全体的レベルで発現パターンに非常に高い相関を示した(図11Aおよび11B)。ES細胞とMEFのあいだで発現が2倍より大きい差を有する遺伝子は、ESとiPS細胞のあいだでほとんど同一に発現された(データは示していない)。ゆえに、これらのデータは、後成的データから予想されるように、iPS細胞が、転写的にES細胞と非常に同等であることを指し示している。シグナチャー遺伝子13個の無作為に選ばれたサブセットのレベルをリアルタイムRT-PCRによって確認した(図11C)。試験した遺伝子は全て、iPS細胞とES細胞において類似のレベルで発現された。ES、iPS細胞、およびMEFのあいだのシグナチャー遺伝子の発現の差は、ヒストンメチル化パターンにおいて観察された差と良好に相関し(データは示していない)、K4およびK27メチル化がそれらの遺伝子の発現状態の重要な決定因子であることを示唆している。併せて、これらのデータは、4つの転写因子による核の再プログラミングが、線維芽細胞ゲノムの全体的な転写および後成的リセットを誘導できることを証明している。
【0113】
実施例8:MEFおよびTTF由来iPS細胞は胚細胞が含まれる多数の細胞タイプに分化する
iPS細胞の忠実な後成的再プログラミングによって、ES細胞と同等の発達能が得られるであろうと推論された。GFPマークMEF由来2D4 iPS細胞を2倍体胚盤胞に注入すると、明白なGFP蛍光を有する新生仔キメラ3例を生じた(図8A、表2)。新生仔からの組織切片は、軟骨、腺構造、肝臓、心臓、および肺に対するiPS細胞の広いクローン性の関与を示した(データは示していない)。新生仔に由来する造血細胞のFACS分析により、脾臓B細胞およびマクロファージと共に胸腺CD4+およびCD8+ T細胞の18〜28%がiPS細胞に由来することが判明した(図8B)。その上、iPS細胞由来の尾部線維芽細胞およびニューロスフェア培養物をこのキメラ仔から単離することが可能であり、これらは宿主由来の線維芽細胞およびニューロスフェアと比較して類似の生育速度およびサイトカイン依存性を示した(データは示していない)。成体まで発達したキメラ1例は、外皮色キメラ現象を示し、iPS細胞が機能的メラノサイトへと分化したことを指し示した(図8C)。
【0114】
次に、MEF由来iPS細胞に加えて、雌性TTF由来iPS細胞が同様に発達を支持しうるか否かを調べた。XiGFPトランスジーンの再発現に基づいて選択されている異なる2つのiPSクローンの胚盤胞注射によって、株あたり1例の生後動物(postnatal animal)を生じた(表2を参照されたい)。キメラ動物は、健康であるように見え、成体マウスまで正常に生育した。これらの結果は、TTFに由来するiPS細胞が、胎児線維芽細胞に由来するiPS細胞と同様に、正常な外観の生後キメラを生じたことを示している。
【0115】
生殖系列伝搬は、細胞の多能性に関する最もストリンジェントな試験の1つであると見なされている。XiGFP/X TTF由来iPS細胞が、生殖系列に寄与しうるか否かを査定するために、過剰排卵させたiPSキメラ1例から卵母細胞16個を単離したところ、その4個が明らかにGFP陽性であり、iPS細胞が雌性生殖系列に関与することを支持している(データは示していない)。これらの卵母細胞を塩化ストロンチウムおよびサイトカラシンBによって処置すると、単為生殖活性化に成功して、その後胚盤胞段階へと卵割して、卵母細胞の機能性を証明した(データは示していない)。
【0116】
ES細胞の成熟細胞タイプへの方向性の分化は、明らかに治療的将来性を有する。iPS細胞がインビトロで成熟細胞を生じるか否かを決定するために、EBを生成して、造血細胞の運命を誘導するために培養において拡大させた。実際に、未成熟および成熟血球のマーカーを発現する細胞タイプが検出され、このように再生医学におけるiPS細胞の将来的な使用を強調する(図12)。
【0117】
線維芽細胞培養物から直接多能性細胞を生成することは、核の再プログラミングを支配するメカニズムの理解に向けて主要な進歩を表した(Takahashi and Yamanaka, 2006)。ここに、ゲノムの忠実な後成的リセットは転写因子誘導再プログラミングを伴う、という第一の証拠を提供する。そのエピゲノムにおいてES細胞と顕著に類似であるiPS細胞を回収した。たとえば、雌性iPS細胞は、重要な多能性遺伝子のプロモーターで適切な脱メチル化を示し、それらは分化の際に無作為なX不活化を受ける体細胞沈黙化X染色体を再活性化して、ES細胞とほとんど同一である全体的なヒストンメチル化パターンを有した。iPS細胞はまた、増殖因子応答性、細胞融合における再プログラミングドナーとしての作用能と共に、インビトロおよびインビボの双方でES様分化を受ける能力が含まれる他のES様性質も示し、1つの生殖系列キメラが含まれる高グレード生後キメラに関与した。
【0118】
トランスジェニックOct4発現がiPS細胞の維持にとって必要ではないという知見は、多能性を確実に維持するために、内因性の遺伝子発現プログラムが十分に再活性化されていることを指し示している。このことは、Oct4の外因性の発現、ならびにおそらくSox2、c-MycおよびKlf4の外因性の発現が、多能性に至る転写の変化および後成的変化を誘発するために再プログラミングの最初の段階の際に限って必要である可能性があることを指し示している。この考え方を支持して、4つの因子のレトロウイルス発現は、感染したドナー線維芽細胞において高く、iPS細胞において沈黙化した。このように、体細胞に4つの因子を一過性に供給することが実現可能であり、それによって、それぞれ挿入突然変異または遺伝子発現のアーチファクトを生じる可能性のあるレトロウイルスまたはトランスジェニックエレメントを含有しない、安定に再プログラムされた細胞が生成される。
【0119】
意外にも、Nanog選択iPS細胞は、既に報告されているFbx15選択iPS細胞とは、表現型的および分子的に異なっていた。Nanogは、胚の発達にとって必須であり、始原内胚葉への分化を抑制することによって多能性を維持するために必要である(Chambers, I., Colby, D., Robertson, M., Nichols, J., Lee, S., Tweedie, S., and Smith, A. (2003) Cell 113, 643-655.;Mitsui, K., Tokuzawa, Y., Itoh, H., Segawa, K., Murakami, M., Takahashi, K., Maruyama, M., Maeda, M., and Yamanaka, S. (2003) Cell 113, 631-642)。対照的に、Fbx15は、ES細胞におけるその独占的発現にもかかわらず、多能性または発達にとって必須ではない(Tokuzawa, Y., Kaiho, E., Maruyama, M., Takahashi, K., Mitsui, K., Maeda, M., Niwa, H., and Yamanaka, S. (2003) Mol Cell Biol 23, 2699-2708)。理論に拘束されることを望むものではないが、Fbx15選択iPS細胞と本明細書において記述されるiPS細胞のあいだの性質の差に関して、いくつかの可能性のある説明がある。1つの可能性は、Nanog選択が、Fbx15選択と比較してより大きい発達能を有する異なる多能性細胞タイプを生じることである。これと一致して、ほとんどのFbx15選択iPS細胞は、Nanogを発現しなかったが(Takahashi and Yamanaka, 2006)、このことは、MEFの非存在下でそれらが不適当に分化し、満期産のキメラを生じなかった理由を説明する可能性がある。この考え方をさらに支持するのは、正常なES細胞培養において必ずしも全てのOct4発現細胞がNanog陽性ではないという所見であり、このことは、ES細胞集団内の不均一性を示唆している(Hatano et al., 2005)。興味深いことに、ES細胞が由来する胚盤胞の内細胞塊は、Oct4およびNanogに関して類似の不均一な発現パターンを示す(Chazaud, C, Yamanaka, Y., Pawson, T., and Rossant, J. (2006) Dev Cell 10, 615-624.)。
【0120】
再び、理論に拘束されることを望むものではないが、得られたiPSの品質に関するNanog選択の効果に関する別の説明は、Nanogタンパク質それ自身が忠実な後成的再プログラミングにおいて重大な役割を果たしていることでありうるあろう。この考え方と一致して、ES細胞と体細胞との細胞融合実験により、NanogがES細胞において過剰発現されている場合に、200倍多くのコロニーが得られることが示されている(Silva, J., Chambers, I, Pollard, S., and Smith, A. (2006) Nature 441, 997-1001)。Nanogは体細胞において多能性を誘導する場合には必要ないが、再プログラミングプロセスの際のその過剰発現が、iPS細胞を得る効率を増強するか否か、およびiPS細胞の発達効能に影響を及ぼすか否かを査定することは有益である。
【0121】
再び、理論に拘束されることを望むものではないが、既に報告されたiPS細胞と本明細書において記述されたiPS細胞のあいだに観察された差のもう1つの可能性は、選択の時期である可能性がある。感染後3日目に選択を適用した場合には、Nanog-GFP-puro MEFからiPS細胞を誘導化することは不可能であったが、これはYamanakaとその共同研究者による知見とは対照的であり、彼らはこの時期にFbx15発現に関して選択することが可能であった。よって、選択を感染後1週目に開始するか、またはES細胞の形態もしくは沈黙化したX-連鎖GFPトランスジーンの再活性化のみに基づいてiPS細胞を単離した後、Oct4-Neo対立遺伝子を用いて多能性をレトロスペクティブに確認した。初回薬物選択を行わずに誘導化されたiPS細胞は全て、キメラの関与およびES細胞様後成的特色に関して既に報告されたFbx15-選択iPS細胞より良好であるように思われた。再プログラミングは、数日または数週間を要する逐次的プロセスであり、再活性化される必要がある遺伝子のカスケードに依存するという仮説が立てられる。このシナリオにおいて、Nanog再活性化は、Fbx15再活性化より核の再プログラミングの際の後期に起こるであろう。このように、Fbx15の初期選択は、核の再プログラミングを完了していない細胞集団を拡大して、その結果、再プログラミングプロセスの後期に現れるであろう潜在的によりよく再プログラムされる細胞を消失させる可能性がある;Nanogに関して後期に選択することは、再プログラミングがより完了している段階を捕える可能性がある。この仮説を探る1つの方法は、Fbx15発現に関して後期に選択した場合に、ES細胞により類似するiPS細胞を生成するか否かを試験することであろう。薬物選択の代わりにES様コロニーの形態的選択がiPS細胞を得るために十分でありうるという所見は、ヒト細胞にレポータートランスジーンを導入する段階が技術的に難題であり、挿入突然変異を引き起こす可能性があることから、ヒトにおける直接再プログラミングに関して重要な意味を有する。
【0122】
多能性への細胞の直接再プログラミングは、明らかな治療的意味を有し、ゆえにiPS細胞がES細胞と同じ後成的状態で存在するか否かを確かめることは非常に重要であった。これらのデータは、異常な後成的再プログラミングが直接再プログラムされた細胞の治療的有用性を損なわないことを指し示している。
【0123】
実施例9:ヒトiPS細胞は選択の非存在下で生成されうる
患者特異的線維芽細胞またはケラチノサイト細胞に、テトラサイクリン誘導可能なレンチウイルスシステムによって発現された、4つ(OCT4、SOX2、CMYC、KLF4)または5つ(4 + NANOG)の再プログラミング因子を感染させた。ウイルスを、リバーステトラサイクリントランスアクチベータ(rtTA)を発現するレンチウイルスと共に同時感染させた。細胞をフィーダー細胞上で継代して、ドキシサイクリンによって誘導した;細胞を、最初の3日間線維芽細胞培地において維持した後、ヒトES細胞条件に切り換えた。小さいコロニー様構造が4日以内に目に見えるようになり、30日までに、別個のhES様の丸い石のような外観を有するヒトES細胞形態を有するコロニーが存在した(データは示していない)。非hES細胞形態を有するコロニーもまた存在したが、hES様コロニーの生成を妨害しなかった。
【0124】
hES様コロニーを採取して、拡大させ、特徴付けを行った。ヒトES細胞と同様に、それらは多能性であり(奇形腫を生成した)、重要な多能性遺伝子を発現し、後成的修飾の適切なリセットを示した。加えて、それらは同様にレンチウイルストランスジーンを沈黙化した。
【0125】
本明細書において引用された任意の特許、または特許出願が含まれる全ての参考文献は、図面および表と共に参照により本明細書に組み入れられる。いかなる参考文献も先行技術を構成すると認めたわけではない。参考文献の考察は、その著者が主張する内容を述べているのであり、本出願者らは、引用された文書の精度および適当性に異議を申し立てる権利を保有する。多数の先行技術の出版物が本明細書において引用されるが、このように引用することが、これらの文書のいかなるものも、アメリカ、または任意の他の国における当技術分野において共通の一般的知識の一部を形成すると認めているわけではないと明らかに理解されるであろう。
【0126】
(表1)得られたiPS細胞株の要約
* 細胞をGFP、SSEA-1およびCD9に関して三重にソーティングした後、サブクローニングして均一で安定なES様集団を得た;
** 三重ソーティング後に行った分析;ND=決定していない
【0127】
(表2)iPS細胞由来マウスにおける満期発達効率および推定されるキメラ現象の程度
【0128】
(表3)RT-PCR分析のためのプライマー配列と抗体
【図1A】
【図1B】
【図1C】
【図2A】
【図2B】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【技術分野】
【0001】
本出願は、35 U.S.C. §119(e)の下で、その全内容物が参照により本明細書に組み入れられる2007年5月30日に提出された米国特許仮出願第60/932,267号に対する優先権を主張する。
【背景技術】
【0002】
発明の背景
核の移入(Wakayama, T., Perry, A. C., Zuccotti, M., Johnson, K. R., and Yanagimachi, R. (1998) Nature 394, 369-374(非特許文献1);Wilmut, I., Schnieke, A. E., McWhir, J., Kind, A. J., and Campbell, K. H. (1997) Nature 385, 810-813(非特許文献2))および細胞融合(Cowan, C. A., Atienza, J., Melton, D. A., and Eggan, K. (2005) Science 309, 1369-1373(非特許文献3);Tada, M., Takahama, Y., Abe, K., Nakatsuji, N., and Tada, T. (2001) Curr Biol 11, 1553-1558(非特許文献4))による細胞の再プログラミングによって、体細胞核において多能性状態は再度確立される(Hochedlinger, K., and Jaenisch, R. (2006) Nature 441, 1061-1067(非特許文献5))。核再プログラミングの分子メカニズムは完全に解明されていないが、細胞融合実験から、再プログラミング因子をES細胞において同定することができ、そしてこれを用いて体細胞において再プログラミングを直接誘導できることが暗示されている。実際に、合理的アプローチによって最近、その発現が成体線維芽細胞における多能性状態の誘導を可能にする4つの転写因子が同定された(Takahashi, K., and Yamanaka, S. (2006) Cell 126, 663-676(非特許文献6))。Yamanakaと共同研究者らは、転写因子Oct4、Sox2、c-Myc、およびKlf4のレトロウイルス発現をFbx15発現に関する遺伝子選択と組み合わせると、線維芽細胞培養物からiPS細胞を直接生じることを証明した。Fbx15選択iPS細胞は、妊娠中期胚における多様な組織に関与したが、これらの胚は妊娠中期で死亡し、ES細胞と比較してiPS細胞の発達能が限定的であることを指し示している。この所見と一致して、iPS細胞において発現されたのはES細胞のトランスクリプトームのごく一部に過ぎず、Oct4およびNanogプロモーターの染色質状態のメチル化分析から、線維芽細胞とES細胞の中間のエピジェネティックパターンが証明された。
【0003】
これらの所見から、直接再プログラムされた細胞の分子的および機能的性質に関して3つの根本的な疑問が持ち上がった:(i)ES細胞状態にとって必須である遺伝子に関して選択することによって、既に記述されたFbx15選択iPS細胞よりES細胞に類似する多能性細胞を生成することができるか;(ii)iPS細胞の多能性状態は、外因性の因子の持続的発現に依存するか;および(iii)転写因子誘導再プログラミングは、線維芽細胞ゲノムの後成的風景を多能性細胞の後成的風景へとリセットするか。
【0004】
核移入または細胞融合による体細胞の再プログラミングが成功するためには、DNAのメチル化、ヒストン修飾、および雌性細胞における沈黙X染色体の再活性化などの後成的修飾の忠実なリモデリングが必要であると考えられている(Rideout, W. M., 3rd, Eggan, K., and Jaenisch, R. (2001) Science 293, 1093-1098(非特許文献7))。異常な後成的再プログラミングは、核移入によってクローン化された動物において認められる発達不全および異常の主要な理由であると仮定されている。このように、後成的異常によって癌などの病理的状態が起こりうることから、後成的再プログラミングに関する疑問は、iPS細胞に関する将来的な治療応用から見て特に意味がある(Gaudet, F., Hodgson, J. G., Eden, A., Jackson-Grusby, L., Dausman, J., Gray, J. W., Leonhardt, H., and Jaenisch, R. (2003) Science 300, 489-492(非特許文献8))。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Wakayama, T., Perry, A. C., Zuccotti, M., Johnson, K. R., and Yanagimachi, R. (1998) Nature 394, 369-374
【非特許文献2】Wilmut, I., Schnieke, A. E., McWhir, J., Kind, A. J., and Campbell, K. H. (1997) Nature 385, 810-813
【非特許文献3】Cowan, C. A., Atienza, J., Melton, D. A., and Eggan, K. (2005) Science 309, 1369-1373
【非特許文献4】Tada, M., Takahama, Y., Abe, K., Nakatsuji, N., and Tada, T. (2001) Curr Biol 11, 1553-1558
【非特許文献5】Hochedlinger, K., and Jaenisch, R. (2006) Nature 441, 1061-1067
【非特許文献6】Takahashi, K., and Yamanaka, S. (2006) Cell 126, 663-676
【非特許文献7】Rideout, W. M., 3rd, Eggan, K., and Jaenisch, R. (2001) Science 293, 1093-1098
【非特許文献8】Gaudet, F., Hodgson, J. G., Eden, A., Jackson-Grusby, L., Dausman, J., Gray, J. W., Leonhardt, H., and Jaenisch, R. (2003) Science 300, 489-492
【発明の概要】
【0006】
本明細書において記述される方法は、誘導多能性幹細胞、すなわち、たとえば成体線維芽細胞が含まれる分化した細胞から生成または誘導された多能性幹細胞の選択に関する。限られた数の転写因子の発現を誘導することによる多能性の誘導は、当技術分野において証明されており、これを任意の哺乳動物細胞、非ヒト哺乳動物細胞、またはヒト細胞に応用することができる。
【0007】
本明細書において記述される方法によって、発達的再プログラミングの際に哺乳動物(たとえば、マウスおよびヒト)多能性細胞を生成するように選択することができる。定義された組の転写因子の過剰発現により、成体の体細胞を胚幹(ES)細胞様の細胞に変換することができるが、このプロセスは一般的に、ES細胞特異的遺伝子を再活性化するための遺伝子選択を必要とし;選択を行わなければES様細胞に加えて多くの非ES様細胞が生成される。そのような遺伝子選択技術は、一般的にヒト細胞において実現可能ではなく、一般的に、細胞をヒト患者に導入するために望ましくない。この問題に取り組むために、本明細書において、1)コロニーの形態のみ、および2)雌性細胞における染色体の再活性化、に基づいて再プログラムされた細胞に関して選択することができる新規選択戦略が記述される。それは、遺伝子選択、化学的選択、またはそのいずれも行わないことである。
【0008】
形態に基づく選択は、既存の選択アプローチと比較して再プログラミングのためにより長い期間、再プログラミング因子を加えた後におよそ1〜2ヶ月を必要とする。この期間のあと、ES様コロニーを採取して拡大させることができる。採取の時点では多くの非ES様細胞が残っているが、たとえばクローン密度で細胞を継代すると、ES様コロニーを容易に回収することができ、細胞株を生成することができる。
【0009】
X染色体の再活性化に基づく選択は、Hprt座における変異に関してヘテロ接合である雌性細胞株を利用する。X染色体の再活性化は、定義された因子による再プログラミングの際に起こり、この事象は再プログラミングプロセスの後期に起こることが本明細書において示される(およそ3〜4週間)。雌性の体細胞では、1つのみのX染色体が活性であり、他は沈黙である。1つの局面において、Hprtヘテロ接合細胞において、活性なX染色体上に変異体Hprt遺伝子を潜在的に有する細胞は6-チオグアニンに対して耐性であろう。再プログラミングおよびX染色体再活性化の際に、これらの細胞は、正常なHprt遺伝子を発現して、HAT培地に対する耐性を獲得するが、6-チオグアニンに対する耐性を失う。
【0010】
本明細書において記述される方法の1つの局面によって、以下の段階を含む、誘導多能性幹細胞の選択を行うことができる:a)RT-PCRによって測定した場合にNanog mRNAを発現していない分化した初代培養細胞を、多能性表現型へと再プログラムする段階;b)再プログラミング後、段階(a)において再プログラムされた細胞を選択物質の非存在下で培養する段階;c)段階(b)の培養物を顕微鏡下で観察して、培養物においてなめらかで丸い外観となった細胞のクローンを単離する段階;およびd)幹細胞マーカーの発現が検出されれば、細胞が誘導多能性幹細胞であることが指し示される、幹細胞マーカーの発現に関してクローンの細胞を試験する段階。
【0011】
この局面および本明細書において記述される他の全ての局面の1つの態様において、再プログラミング段階は、以下の1つを含む:因子を発現させるための調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-Myc、およびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;ならびに細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【0012】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、幹細胞マーカーを発現するクローンの細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階をさらに含み、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0013】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、培養する段階は、細胞を継代する段階をさらに含む。
【0014】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、分化した体細胞は、ES細胞とは明確に異なる形態を有する。
【0015】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、分化した初代培養細胞は線維芽細胞であり、線維芽細胞は再プログラミングの前は平坦で不規則な形状を有する。
【0016】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される。
【0017】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、分化した初代培養細胞が雌性の個体に由来する場合、不活性なX染色体の再活性化に関してクローンの細胞を試験する段階をさらに含む。
【0018】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、核酸配列はウイルスベクターまたはプラスミドに含まれる。
【0019】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、ウイルスベクターは、レトロウイルスベクター、レンチウイルスベクター、またはアデノウイルスベクターである。
【0020】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、外因性のOct4、Sox2、c-Myc、および/またはKlf4の発現に関してクローンの細胞を試験する段階をさらに含む。
【0021】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、初代培養細胞はヒト細胞を含む。
【0022】
本明細書において記述されるもう1つの局面は、以下の段階を含む、誘導多能性幹細胞を選択する方法である:a)選択可能マーカーが活性なX染色体上で変異体であり、不活性なX染色体上では野生型である、およびRT-PCRによって測定した場合に細胞がNanog mRNAを発現しない、X染色体上の選択可能マーカーに関してヘテロ接合である雌性細胞を提供する段階;b)多能性表現型となるように細胞を再プログラムする段階;ならびにc)不活性なX染色体の再活性化によって、野生型選択可能マーカーを発現させることができ、選択物質の存在下で細胞を生存させることができ、それによって生存する細胞が誘導多能性幹細胞である、細胞を選択物質と共に培養する段階。
【0023】
この局面の1つの態様において、方法は、選択物質の存在下で生存する細胞を幹細胞マーカーの発現に関して試験する段階をさらに含む。
【0024】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される。
【0025】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、再プログラムする段階は、因子を発現させるための調節エレメントに機能的に連結している、転写因子Oct4、Sox2、c-Myc、およびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;ならびに細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階、の1つを含む。
【0026】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、選択物質の存在下で生存する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階を含み、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0027】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は細胞株の細胞である。
【0028】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は、X染色体上の変異体Hprt遺伝子に関してヘテロ接合である。
【0029】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は、核酸の導入前に不活性である野生型Hprt遺伝子をX染色体上に持ち、再プログラミング前に活性である変異体の非機能的Hprt遺伝子をX染色体上に持つ。
【0030】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞は、再プログラミング前に6-チオグアニンに対して耐性である。
【0031】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、選択物質はHAT培地を含む。
【0032】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞はヒト細胞を含む。
【0033】
本明細書において記述されるもう1つの局面は、以下の段階を含む、誘導多能性幹細胞を選択する方法である:a)RT-PCRによって測定した場合にNanog mRNAを発現しない、X不活化による沈黙化に供されるX染色体連鎖レポーター遺伝子を持つ雌性細胞を提供する段階;b)細胞を多能性表現型へと再プログラムする段階;c)再プログラミング後に細胞を培養する段階;およびd)レポーターが発現すれば、クローンが誘導多能性幹細胞を含むことが指し示される、X染色体連鎖レポーターを発現する細胞のクローンを培養物から単離する段階。
【0034】
この局面および本明細書において記述される他の全ての局面の1つの態様において、方法は、クローンの細胞を幹細胞マーカーの発現に関して試験する段階をさらに含む。
【0035】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される。
【0036】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、方法は、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、レポーターを発現する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に対して組織学的検査を行う段階をさらに含む。
【0037】
この局面および本明細書において記述される他の全ての局面のもう1つの態様において、細胞はヒト細胞を含む。
【0038】
定義
本明細書において用いられる「多能性」という用語は、異なる条件下で1つより多い分化した細胞タイプに分化する、好ましくは3つ全ての胚細胞層に特徴的な細胞タイプに分化する能力を有する細胞を指す。多能性細胞は、たとえばヌードマウス奇形腫形成アッセイ(本明細書において記述される実施例を参照されたい)を用いて、1つより多い細胞タイプに分化する能力、好ましくは、3つ全ての胚細胞層に分化する能力によって主に特徴付けされる。多能性はまた、胚幹(ES)細胞マーカーの発現も証拠となるが、多能性に関する好ましい試験は、3つの胚細胞層のそれぞれの細胞への分化能を証明することである。
【0039】
本明細書において用いられるように「再プログラミング」という用語は、最終分化した体細胞の分化状態を多能性表現型へと変更するプロセスを指す。
【0040】
「分化した初代培養細胞」とは、その本来の型において、本明細書において定義される多能性ではない任意の初代培養細胞を意味する。多くの初代培養細胞を培養すると、完全に分化した特徴のいくつかが失われうることに注意すべきである。しかし、そのような細胞を単純に培養しても、ひとりでにそれらが多能性となるわけではない。多能性への移行は、培養における分化特徴の部分的喪失に至る刺激を超える再プログラミング刺激を必要とする。再プログラムされた多能性細胞はまた、一般的に培養においてごく限られた回数の分裂能を有する初代培養細胞の親と比較して、生育能を失うことなく、長期の継代能の特徴を有する。
【0041】
「ベクター」という用語は、それが複製される宿主細胞に導入するために、その中にDNA配列を挿入することができる低分子の担体DNA分子を指す。「発現ベクター」は、宿主細胞における発現にとって必要とされる必要な調節領域を有する遺伝子を含有する特殊なベクターである。「機能的に連結した」という用語は、コード配列の発現にとって必要な調節配列が、コード配列の発現を行うために、コード配列に対して適当な位置でDNA分子内に置かれていることを意味する。この同じ定義は時に、発現ベクターにおけるコード配列および転写制御エレメント(たとえば、プロモーター、エンハンサー、および終止エレメント)の整列にも当てはまる。この定義はまた、ハイブリッド核酸分子が生成される、第一および第二の核酸分子の核酸配列の整列にも当てはまる。
【図面の簡単な説明】
【0042】
【図1】Nanog-選択iPS細胞のES細胞様特性(A)Nanog-GFP(NGiP)ES細胞、ならびに持続的ピューロマイシン選択によっておよびピューロマイシン選択を行わずに生育させた2つのiPS細胞株と共に、追加の基準点としての野生型ES細胞(V6.5)およびMEFsにおけるES細胞マーカー遺伝子発現のRT-PCR分析。Oct4およびSox2のプライマーは、それぞれの内因性の座からの転写物に対して特異的である。Nat1はローディング対照として用いた。(B)iPS細胞株、MEF、およびNGiP-ES細胞におけるNanog、Oct4、Sox2、c-myc、およびKlf4の発現に関するウェスタンブロット分析。抗チューブリンおよび抗アクチン抗体をローディングの対照とするために用いた。(C)それぞれのpMXウイルスを感染させた、1)野生型MEF、2)野生型ES細胞、3)ソーティングおよびサブクローニングの前の不均一なiPS株1A2からの細胞、4)1D4 iPS、5)2D4 iPS、および6)MEF、におけるpMXレトロウイルス転写の定量的PCR分析。転写物レベルをβ-アクチンに対して標準化した。2D4 iPS株におけるレトロウイルスは、完全に沈黙しているように思われるが、不均一な1A2株はなおも外因性の因子の豊富な発現を示すことに注意すべきである。
【図2】iPS細胞と体細胞との融合(A)2D4 iPS細胞と、Oct4Neo選択可能な対立遺伝子を持つハイグロマイシン耐性MEFとの細胞融合の略図。(B)ピューロマイシン/ハイグロマイシン選択またはピューロマイシン/G418選択のいずれかの下で維持された2D4 iPS細胞、MEF、および2D4/MEF細胞ハイブリッドのDNA含有量分析。
【図3】iPS細胞を維持するための外因性のOct4に関する必要条件。(A)Oct4-誘導可能な線維芽細胞を用いたiPS細胞生成の略図。(B)ドキシサイクリン誘導可能なOct4発現の非存在下または存在下でSox2、c-MYC、およびKlf4に感染したMEF。アルカリホスファターゼ染色したプレートを示す。(C)Oct4-誘導可能なiPS細胞におけるOct4レベルの定量的PCR分析。内因性および誘導可能な対立遺伝子からの転写物のレベルを、非分化iPS細胞(+LIF、-dox)、分化したiPS細胞(-LIF、-dox)、およびLIF除去後5日目に再誘導した分化iPS細胞(-LIF、+dox)において測定した。転写物のレベルをβ-アクチンに対して標準化した。誘導可能なOct4対立遺伝子を持つES細胞;および野生型MEFを対照とした。
【図4】iPS細胞における遺伝子特異的および全体的なDNAメチル化状態(A)ES細胞、2D4 iPS細胞、およびMEFにおけるOct4およびNanogプロモーター領域のバイサルファイトシークエンシング。異なるようにメチル化したCpGを含有するプロモーター領域を転写開始部位(矢印)に関して示す。白丸は、非メチル化CpGを表す;黒丸はメチル化CpGを表示する。(B)iPS 2D4細胞とMEFとの融合を通して生成された細胞ハイブリッドにおけるNanogプロモーターのバイサルファイトシークエンシング。図2Cと同様にピューロマイシン/ハイグロマイシン耐性ハイブリッドに関するデータを示す。(C)サテライトリピートプローブを用いた全体的なDNAメチル化のサザンブロット分析。MEF、雄性Nanog-GFP ES細胞、雌性ES細胞、iPS 2D4親細胞および3つのサブクローンからのゲノムDNAを、メチル化感受性制限酵素HpaIIによって消化して、より小さいサテライトリピートプローブにハイブリダイズさせた。非メチル化感受性イソシゾマーMspIによって消化した雄性ES細胞DNAを対照とした。より低い分子量のバンドは、低メチル化を指し示している。
【図5】iPS細胞におけるX染色体の動力学(A)iPS細胞株2D4、NGiP MEF、および雄性対照ES細胞におけるXite遺伝子間転写物のRT-PCR分析。Xite座に沿って異なる位置での転写物を検出した(領域5〜7)。陽性対照Rrm2は、ハウスキーピング遺伝子である。雌性ES細胞と同様に、雄性ES細胞はXite転写物を発現する。(B)分化しつつある2D4 iPSにおけるXi上でのEzh2およびH3me3K27の濃縮。グラフは、2D4 iPS細胞のレチノイン酸誘導分化の際の異なる時点で、Xi上でEzh2またはH3me3K27との同時局在を示すXist RNAコーティングを有する細胞の百分率を示す(各時点に関してn>100)。
【図6】分化しつつあるTTF由来iPS細胞における無作為なX-不活化(A)XGFPX TTFからiPS細胞を得るためのおよびX-不活化をその後分析するためのフロースキーム。Oct4-Neo対立遺伝子を持つXGFPX TTFを、2回連続して通過させることによってソーティングして、GFP陰性集団(XiGFPXa;<0.05%緑色細胞)を得た。再プログラムされた細胞を、ES細胞の形態およびGFP再活性化に基づいて選択した。G418による薬物選択を、iPS細胞の再プログラム状態をレトロスペクティブに確かめるために使用したが、iPS細胞の確立に関して選択するためには使用しなかった。iPS細胞をサブクローニングして、分化させ、FACSおよびXistFISHによって分析した。FACSによって決定されたGFP+またはGFP-細胞の数をオレンジ色で与え、青色で与えた数値はそれぞれ、GFP+およびGFP-分化iPS細胞内のXiのXist RNAコーティングを有する細胞の百分率を指し示している。
【図7】iPS細胞におけるH3K4およびH3K27トリメチル化の全体的な分析(A)全ての細胞タイプのあいだでのK4およびK27トリメチル化データの全体的な相関。表は、細胞タイプの起こりうる全ての対のあいだおよびアレイにおける全ての遺伝子(〜16500個)に関してそれぞれ、K4およびK27トリメチル化の2成分の全体的相関を示す。(B)全ての細胞タイプのあいだでのEクラス遺伝子内でのK4およびK27トリメチル化の相関。細胞タイプのそれぞれの2つの対に関するK4またはK27メチル化に関する相関値を、500 bpずつ増加させた転写開始部位からの距離の関数としてプロットした。
【図8】Nanog-選択可能なiPS細胞のインビボ発達能。(A)無作為に組み入れられたGFPトランスジーンを持つiPS株2D4からの細胞を胚盤胞に注入した。代理母は、GFP-陽性の仔を産んだ。GFPを発現しない非キメラの仔を示す。(B)iPS細胞由来新生仔キメラマウスの脾臓および胸腺から単離された造血細胞のフローサイトメトリー分析。ヒストグラムは、系列特異的マーカーに関してゲートを設定した集団におけるGFP-陽性細胞の百分率を表示する。(C)野生型の同腹子の横に示される、胚盤胞注入2D4 iPS細胞に由来する10日齢のキメラマウス(iPS由来細胞は、アグーチ外皮色の原因である)。
【図9】iPS細胞株1A2および2D4におけるレトロウイルス組み込みDNA刷り込み状態の分析。(A)iPS細胞におけるレトロウイルス組み込み部位の分析。レトロウイルス組み込みをサザンブロット分析によって決定した。DNAをBamHI(Oct4およびKlf4に関して)、HindIII(Sox2に関して)、またはBglII(c-MYCに関して)によって消化して、それぞれのcDNAプローブとハイブリダイズさせた。組み込みをV6.5 ES細胞(wt)および2つのIPS株1A2および2D4に関して示す。(B)組み込み部位分析のために用いられる内部制限部位が含まれる個々のウイルス構築物の概略図。cDNAプローブは、ウイルスが組み入れられるゲノム領域における1つのpMX内部切断と1つの外部切断とによって生成された制限断片を検出することに注意すべきである。(C)Igf2rの異なるようにメチル化された領域でのメチル化状態。異なる細胞タイプからのDNAをPvuIIおよびMIUI制限酵素を用いて消化して、サザンブロッティングによって分析した。メチル化(M)および非メチル化(U)対立遺伝子を指し示す。Dnmt1を欠損するES細胞、またはE 12.5胚に由来する胚生殖(EG)細胞では、非メチル化対立遺伝子のみが検出された。刷り込みがiPS細胞において維持されるという事実は、iPS細胞が、線維芽細胞培養物に混入する可能性があるまれな生殖細胞に由来するのではないことを示唆する。
【図10】シグナチャー遺伝子分析の統計学的有意性そのメチル化パターンに基づいて2D4 iPS細胞におけるほとんどのシグナチャー遺伝子をES-様(Eクラス)として分類することは、非常に有意である。上のパネルは、2D4座のE、NおよびMクラスへの観察された分布を示す(図7Aにおいて提示されたデータから)。2D4座のE、M、およびN遺伝子への分類を確証するために、2D4メチル化データを100回入れ替えて、ES-MEF対に無作為に割付して、シグナチャー遺伝子を異なるストリンジェンシー(p=0.01、p=0.05、p=0.1)で再分類した(下のパネル)。
【図11】iPS細胞におけるシグナチャー遺伝子の発現(A)ピアソン相関によって決定した、V6.5 ES細胞(ES)、ピューロマイシン選択Nanog GFP ires Puro ES細胞(ESpuro)、Nanog GFP ires Puro MEF(MEF)、および2D4 iPS細胞(iPS)のあいだの全発現データセット(Agilentマイクロアレイから)の全体的な相関。(B)ES細胞に対して2倍より大きい発現の変化を示した、(A)において記述されたESpuro、MEF、およびiPSの完全な発現データセットにおける遺伝子の数。(C)2D4 iPS細胞、雌性MEF、およびV6.5 ES細胞における選択されたシグナチャー遺伝子13個の転写物レベルのリアルタイムPCR分析。相対的発現レベルを決定するために、Qiagen RNA easyキットを用いてRNAを調製して、1μgをOmniscript RTキット(Qiagen)およびランダムプライマーを用いて逆転写した。転写物レベルをリアルタイムPCRによって定量して、△△Ct法を用いてGapdh対照に対して標準化した。ES細胞における発現を任意で1に設定して、誤差のバーは、1試料あたり3個の反応の標準偏差を表す。プライマー配列を表3に与える。Y-軸の異なる尺度に注意されたい。本発明者らのゲノム全体の発現データと一致して、Mクラスに属する試験した遺伝子3個中2個(Vgl14、HoxD10)は、そのヒストン修飾パターンに基づいてそれらがMEF様遺伝子であると分類されたにもかかわらず、2D4 iPS細胞においてES様発現パターン(MEFよりESおよびiPS細胞においてより低い発現)を証明した。このため、これらの座でのヒストンメチル化をより厳密に手動で検分することにより、MEFと比較して2D4 iPS細胞において認められた抑制が2D4 iPS細胞におけるこれらのプロモーターでのK4メチル化の低減と相関することが判明した。他の遺伝子は全て、K4メチル化のみと相対的により高い発現、K27メチル化のみと相対的により低い発現、ならびにヒストンH3 K4およびK27メチル化の二価性とより低い発現とのあいだに良好な相関を示した。
【図12】iPS細胞の造血系列へのインビトロ分化(A、B)iPS細胞株2D4および野生型V6.5 ES細胞に由来する7日胚様体を、造血マーカーCD41およびc-kitマーク未成熟造血細胞(A)と共にCD45およびc-kitマーク成熟造血細胞(B)に関してフローサイトメトリーによって分析した。二重陽性細胞の百分率を与える。EBを生成する場合、iPS株ではV6.5 ES細胞株より多数の投入細胞を用いたことに注意されたく、このことは分化細胞の百分率における定量的差を説明する可能性がある。(C)iPS細胞から作出された解離された7日EBのメチルセルロース培養物から得られた成熟造血細胞。骨髄芽球(i)、マクロファージ(ii)、肥満細胞(iii、iv)、および初期赤血球細胞(v、vi)が含まれる多数のタイプの造血細胞が存在した。血球の生成に関して、予め播種することによってフィーダー細胞を消失させた後にハンギングドロップ法を用いてEBを生成した(Geijsen, N., Horoschak, M., Kim, K., Gribnau, J., Eggan, K., and Daley, G. Q. (2004) Nature 427, 148-154)。3日後、EBを播種して、7日目に造血マーカーのFACS分析のために(別紙の表3において記述される抗体によって)、またはインビトロでさらに分化させるために、コラゲナーゼIVによってEBを単細胞浮遊液に解離した。メチルセルロース培養物に関して、7日EBの単細胞浮遊液を、造血増殖因子(M3434、Stem Cell Technologies)を補足したメチルセルロースと混合して、培養物あたり細胞1×105個で接種した。培養10日後、代表的な造血コロニーを採取して、サイトスピンを調製し、これをメイ-グリュンヴァルド-ギムザによって対比染色した。
【発明を実施するための形態】
【0043】
詳細な説明
選択物質の非存在下での誘導多能性幹細胞の単離
1つの局面において、本明細書において記述される方法は、多能性となった細胞を同定または濃縮するために選択物質を用いることに頼らず、その代わりに細胞があまり分化していないES様多能性表現型を獲得する場合に起こる当初の細胞の形態の変化を頼りとする、誘導多能性幹細胞の選択に関する。
【0044】
この局面において、本発明は、以下のような段階を有する、誘導多能性幹細胞を選択する方法に関する。第一の段階は、分化した初代培養細胞をより分化していないまたは多能性状態に再プログラミングする段階を伴う。再プログラミングは、たとえば細胞の核を卵母細胞に移入することによって(たとえば、Wilmut et al., 1997, Nature 385: 810-813を参照されたい)、または既存の胚幹細胞との融合によって(たとえば、Cowan et al., 2005, Science 309: 1369-1373、およびTada et al, 2001, Curr. Biol. 11: 1553-1558)達成されうる。そのような再プログラミングはまた、たとえば因子を発現させるための調節エレメントに機能的に連結された、転写因子Oct4、Sox2、c-Myc、およびKlf4をコードする核酸配列を、たとえば線維芽細胞に導入することによって行われうる。これらの因子が好ましいが、他の転写因子またはこれらの因子のサブセットも同様に使用することができる(たとえば、参照により本明細書に組み入れられる、Takahashi & Yamanaka, 2006, Cell 126: 663-676を参照されたい)。
【0045】
1つの態様において、転写因子はウイルスベクターまたはプラスミドによってコードされる。ウイルスベクターは、たとえばレトロウイルスベクター、レンチウイルスベクター、またはアデノウイルスベクターでありうる。当業者に公知である核酸の導入に対する非ウイルスアプローチも同様に、本明細書において記述される方法と共に用いることができる。
【0046】
または、そのような転写因子をコードする内因性の遺伝子の活性化を用いることができる。
【0047】
もう1つの代用において、細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を細胞に導入することができる。たとえば、HIV-TAT融合体を用いることを通して、タンパク質因子(たとえば、中でもc-Myc、Oct4、Sox2および/またはKlf4)を細胞に導入することができる。TATポリペプチドは、それを細胞に浸透させる特徴を有し、外因性の因子を細胞に導入するために用いられている(たとえば、Peitz et al., 2002, Proc. Natl. Acad. Sci. U S A. 99:4489-94を参照されたい)。このアプローチは、細胞の分化状態を再プログラミングするための因子を導入するために使用することができる。最後に、再プログラミングは、細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させることによって成就されうる(たとえば、Sato et al., 2004, Nature Med. 10:55-63を参照されたい)。
【0048】
線維芽細胞が好ましいが、他の初代培養細胞タイプも同様に用いることができる。親細胞は、形態的変化に基づく変化を容易にするために、ES細胞とは明確に異なる形態を有することが好ましい。「明確に異なる」とは、少なくとも、接着細胞の場合、親細胞の形状が、培養において生育した場合に丸いよりむしろ不規則であることを意味する。非接着性の初代培養細胞の場合、接着性に関して最初に選択して、次に、丸いESの形態に関して選択することができる。当業者は、位相差顕微鏡下で見た場合に、平坦であるよりむしろ丸くなる傾向があり、粗面であるよりむしろなめらかな傾向があるES細胞の形態的特徴を承知している。
【0049】
さらに、親細胞は、任意の哺乳動物種に由来しえて、非制限的な例には、ネズミ、ウシ、サル、ブタ、ウマ、ヒツジ、またはヒト細胞が含まれる。親細胞はES細胞マーカー、たとえばNanog mRNAまたは他のESマーカーを発現しない。明快かつ単純にするために、本明細書における方法の記述は、親細胞として線維芽細胞を指すが、本明細書において記述される方法は全て、他の初代培養親細胞タイプに容易に応用可能であることを理解すべきである。
【0050】
線維芽細胞を用いる場合、線維芽細胞は、再プログラミングの前は平坦で不規則な形状を有し、Nanog mRNAを発現しない。開始線維芽細胞は好ましくは他の胚幹細胞マーカーを発現しないであろう。ES細胞マーカーの発現は、たとえばRT-PCRによって測定することができる。または、測定はたとえば、ES表現型の特徴であるポリペプチドの存在を検出する免疫蛍光または他の免疫学的検出アプローチによって行うことができる。
【0051】
次の段階において、核酸配列の導入後に、線維芽細胞を選択物質の非存在下で培養する。「選択物質の非存在下」という用語は、誘導多能性幹細胞表現型に関して選択する選択物質の非存在、たとえば1つまたは複数のES細胞マーカーを発現するように脱分化した細胞に関して選択する選択物質の非存在を指す。いかなる種類の選択物質も存在しないことが好ましいが、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸の存在に関する選択物質は存在しうるものの、これらの因子の持続的な発現は、多能性表現型の維持にとって絶対的に必要ではない(以下を参照されたい)。方法には、単離されたクローンにおける導入された転写因子の存在または発現に関して試験する段階が含まれうる。
【0052】
次の段階において、選択物質の非存在下で培養される細胞を顕微鏡下で観察して(たとえば、通常の位相差光学顕微鏡または他の適当な光学手段の下で)、親細胞の不規則な形態的特徴、たとえば線維芽細胞の平坦で不規則な形態を失って、外観がなめらかで丸くなった細胞を培養において同定する。細胞は、多能性への移行を経験すると丸くなるがなおも生存している。形態による選択を容易にするために細胞を継代することができる。丸い形態を示す生存細胞のクローンを、たとえばマルチウェルプレートでの限界希釈および培養によって、または当業者に公知の他のアプローチによって単離する。
【0053】
さらなる段階において、単離されたクローンを幹細胞マーカーの発現に関して試験する。そのような発現により、細胞は誘導多能性幹細胞であると同定される。幹細胞マーカーは、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびNat1が含まれる非制限的な群から選択されうる。そのようなマーカーの発現を検出するための方法には、たとえばRT-PCRおよびコードされるポリペプチドの存在を検出する免疫学的方法が含まれうる。
【0054】
単離細胞の多能性幹細胞特徴を、ESマーカーの発現、および3つの胚細胞層のそれぞれの細胞への分化能を評価する多数の任意の試験によって確認することができる。1つの例として、ヌードマウスにおける奇形腫形成を用いて、単離されたクローンの多能性特徴を評価することができる。細胞をヌードマウスに導入して、細胞から発生した腫瘍に関する組織学的検査を行う。3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0055】
もう1つの態様において、細胞が雌性である場合、脱分化および多能性の測定として不活性なX染色体の再活性化を評価することができる。
【0056】
X-再活性化をモニターすることによる選択
雌性におけるX染色体の1つの不活化は、多能性から分化したことの特徴である。細胞を、たとえばOct4、Sox2、c-MycおよびKlf4の発現によって多能性状態に誘導する場合、不活性なX染色体は再活性化される。
【0057】
本明細書において記述される方法のもう1つの局面は、誘導多能性幹細胞に関して選択するために、分化した雌性細胞の不活性なX染色体の再活性化を用いる。
【0058】
この局面において、以下の段階を含む、誘導多能性幹細胞を選択するための方法が提供される。第一に、選択可能マーカーが、活性なX染色体上で変異体であり、不活性なX染色体上では野生型である、X染色体上で選択可能マーカーに関してヘテロ接合である雌性細胞を提供する。雌性細胞は、Nanog mRNAを発現せず、好ましくは他のES細胞マーカーを発現しない。または、Xが再活性化された場合に限ってマーカーの発現が観察されるように、選択可能マーカーは、たとえばトランスジェニック動物または細胞の不活性なX染色体に組み入れられるマーカーでありうる。そのようなマーカーには、たとえば任意の陽性選択可能マーカーが含まれうる。この代用物の好ましい態様はGFPを用いる(本明細書における以下の実施例を参照されたい)。
【0059】
他の好ましい態様において、選択可能マーカーは、たとえばヒポキサンチンホスホリボシルトランスフェラーゼ(Hprt)である。Hprtに関してヘテロ接合である雌性細胞株には、たとえばDR4マウス細胞(ATCC SCRC-1045を参照されたい)、ヒトTK6リンパ芽球細胞株(ECACC 87020507)、そのそれぞれが参照により本明細書に組み入れられる、Rinat et al, 2006, Mol. Genet. Metab. 87: 249-252によって記述される線維芽細胞、ならびにRivero et al., 2001, Am. J. Med. Genet. 103: 48-55およびHakoda et al., 1995, Hum. Genet. 96: 674-680によって記述されるリンパ球が含まれる。
【0060】
次の段階において、雌性細胞は、本発明の他の局面に関して本明細書において記述されるように多能性表現型となるように再プログラムされる。
【0061】
再プログラムされた細胞を選択物質と共に培養して、不活性なX染色体が再活性化されると、野生型選択可能マーカーを発現させることができ、選択物質の存在下で細胞が生存することができる。生存する細胞は誘導多能性幹細胞である。
【0062】
この局面の1つの態様において、方法はさらに、選択物質の存在下で生存する細胞を幹細胞マーカーの発現に関して試験する段階を含む。幹細胞マーカーは、たとえばSSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択されうる。
【0063】
もう1つの態様において、再プログラミングは、以下の1つを含む:因子を発現させるために調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸配列を細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;および細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【0064】
もう1つの態様において、方法は、3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、選択物質の存在下で生存する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階をさらに含む。
【0065】
もう1つの態様において、細胞は細胞株に由来する。
【0066】
もう1つの態様において、細胞は、X染色体上の変異体Hprt遺伝子に関してヘテロ接合である。
【0067】
もう1つの態様において、細胞は、再プログラミングの前に不活性である野生型Hprt遺伝子をX染色体上で持ち、および再プログラミング前に活性である変異体の非機能的Hprt遺伝子をX染色体上で持つ。
【0068】
もう1つの態様において、細胞は、再プログラミングの前に6-チオグアニンに対して耐性である。
【0069】
もう1つの態様において、選択物質は、HAT培地を含む。
【0070】
もう1つの局面において、誘導多能性幹細胞を選択する方法が提供される。方法は、以下の段階を含む:(a)RT-PCRによって測定した場合にNanog mRNAを発現しない、X不活化による沈黙化に供されるX染色体連鎖レポーター遺伝子を持つ雌性細胞を提供する段階;(b)細胞が多能性表現型に再プログラムされる段階;(c)再プログラミング段階後に細胞を培養する段階;および(d)細胞のクローンがX染色体連鎖レポーターを発現する培養物から単離される段階。レポーターが発現されれば、クローンが誘導多能性幹細胞を含むことが指し示される。
【0071】
1つの態様において、方法はさらに、幹細胞マーカーの発現に関してクローンの細胞を試験する段階を含む。幹細胞マーカーは、たとえばSSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択されうる。
【0072】
もう1つの態様において、方法はさらに、レポーターを発現する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階を含む。3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される。
【0073】
X再活性化を受けた細胞に関して選択することによる多能性幹細胞の選択は、たとえば再プログラム段階の低分子調節物質、たとえば再プログラミングを容易にする低分子に関してスクリーニングするためのシステムを提供することができる。または、このように誘導化された多能性幹細胞は、幹細胞を所望の表現型に再分化させる低分子のまたは他の調節物質に関するスクリーニングアッセイを提供する。
【0074】
本発明を、制限的に解釈されてはならない以下の実施例によってさらに説明する。
【0075】
実施例
転写因子Oct4、Sox2、c-MycおよびKlf4の異所発現は、線維芽細胞ゲノムに多能性状態を付与するために十分であり、誘導多能性幹(iPS)細胞を生成する。これらの4つの因子によって誘導される核の再プログラミングが、分化した細胞と多能性細胞とのあいだの後成的な差を全体的にリセットすることができるか否かはなおも不明である。本明細書において、新規選択アプローチを用いて、iPS細胞を線維芽細胞から生成してその後成的状態を特徴付けする。雌性iPS細胞は、体細胞沈黙化X染色体の再活性化を示し、分化の際に無作為なX不活化を経験した。2つの重要なヒストン修飾に関してゲノム全体の分析を行ったところ、iPS細胞がES細胞に非常に類似していることが指し示された。これらの所見と一貫して、iPS細胞は、生殖系列に関与する生存する高度のキメラを生じた。これらのデータは、転写因子誘導再プログラミングにより、ES様状態への体細胞エピゲノムの全体的な復帰が起こることを示している。これらの結果は、核の再プログラミングを伴う後成的修飾を研究するためのパラダイムを提供し、かつ、正常でない後成的再プログラミングであっても、iPS細胞の治療応用に関する問題をもたらさないことを示唆している。これらのデータは、その全内容物が参照により本明細書に組み入れられる、Maherali, N., et al (2007) Cell-Stem Cell 1:55-70によって現在公表されている。
【0076】
実験技法
線維芽細胞の誘導化
Nanog-GFP-iresPuro構築物(Hatano et al., 2005)を雄性V6.5 ES細胞に標的化して、正確に標的化されたクローンを、標準的なサザンブロット分析によって確認し、マウスを作成した。Oct4-ネオマイシン/ハイグロマイシン選択可能なMEFをOct4-ネオマイシンマウスとpgk-ハイグロマイシンマウスとの異種交配から得た。XGFPとOct4-neo対立遺伝子とを持つTTFを、Oct4-NeoとX-連鎖GFPマウスとの異種交配から得た(Hadjantonakis et al., 1998)。誘導可能なOct4マウスは既に記述されている(Hochedlinger et al., 2005)。MEFは14.5日胚の胚に由来し、TTFは1週齢までのマウスに由来した。
【0077】
MEFのレトロウイルス産生および感染
Oct4、Sox2、c-MYC(T58A変異体)、およびKlf4に関するcDNAをレトロウイルスpMXベクターにクローニングして、Fugene(Roche)を用いてPlatEパッケージング細胞株にトランスフェクトした(Morita, S., Kojima, T., and Kitamura, T. (2000) Gene Ther 7, 1063-1066)。トランスフェクション後48時間で、ウイルス上清を用いてES培地において培養した標的MEFに感染させた。終夜感染の2〜3ラウンドを行って、細胞を放射線照射したフィーダー層において7日後に分割して、指し示された時間に1μg/mLピューロマイシン(Sigma)または300μg/mL G418(Roche)によって選択した。
【0078】
細胞培養およびインビトロ分化
iPS細胞およびES細胞を、放射線照射ネズミ胚線維芽細胞(フィーダー)および標準的なES培地(15%FBS、非必須アミノ酸、L-グルタミン、ペニシリン-ストレプトマイシン、β-メルカプトエタノール、および1000 U/mL LIFを補足したDMEM)において生育させた。胚盤胞注入のための2D4 iPS細胞を標識するために、細胞をRosa-GFP-Neoターゲティングベクターによって電気穿孔して、サザンブロット分析によって確かめた。XGFP/X Oct4-Neo iPS細胞からのサブクローンを生成するために、細胞を直線状pgk-Hygroプラスミドによって電気穿孔した。選択をそれぞれ、G418(300μg/mL)またはハイグロマイシン(140μg/mL)によってパルス後24時間目に開始した。X染色体の状態を試験するために、iPS細胞をゼラチンコーティング皿のES培地において1回継代して、フィーダー細胞の数を低減させて、LIFを欠損するES培地において40 ng/mLオールトランスレチノイン酸によって分化を誘導した。X不活化の無作為性を分析するために、EB形成時に分化を誘導した。
【0079】
卵母細胞を単離するために、雌性のキメラをPMSおよびhCGによって過剰排卵させて、卵母細胞をhCG注射後13時間で単離した。単為生殖活性化を誘導するために、卵母細胞を、10 mM塩化ストロンチウムおよび5μgml-1サイトカラシンBを補足した無カルシウムCZB培地において5時間インキュベートした後、KSOM培地において37℃で5%CO2で培養した。
【0080】
全体的なDNAメチル化に関するサザンブロット分析
ゲノムDNA 10μgをHpaIIまたはMspIによって消化して、断片を0.8%アガロースゲルにおいて分離した。DNAをHybondXLメンブレン(Amersham Biosciences)にブロットして、既に記述されているようにpMR150プローブとハイブリダイズさせた(Meissner, A., Gnirke, A., Bell, G. W., Ramsahoye, B., Lander, E. S., and Jaenisch, R. (2005). Nucleic Acids Res 33, 5868-5877)。
【0081】
バイサルファイトシークエンシング
EpiTect Bisulfiteキット(Qiagen)を用いて、製造共の説明書に従ってDNAの亜硫酸水素処置を行った。プライマー配列は既に記述されたとおりであった;Oct4(Blelloch, R., Wang, Z., Meissner, A., Pollard, S., Smith, A., and Jaenisch, R, (2006). Stem Cells 24(9):2007-13)およびNanog(Takahashi and Yamanaka, 2006)。増幅された産物をゲル濾過カラムを用いて精製してpCR2.1-TOPOベクター(Invitrogen)にクローニングして、M13フォワードおよびリバースプライマーによってシークエンシングした。
【0082】
RT-PCR分析
内因性の座からの多能性遺伝子の発現を試験するために、総RNAをDNA-freeキット(Ambion, Austin, TX)によって処置して、SuperScript First-Strand Synthesisシステム(Invitrogen)によってオリゴdTプライマーを用いて製造元の説明書に従って逆転写した。全てのプライマー配列を表3に示す。
【0083】
ウェスタン分析、免疫染色およびAP染色
本明細書において記述される方法において用いられる抗体を表3に記載する。Vector Red基質キット(Vector Labs)を用いてアルカリホスファターゼ染色を行った。免疫染色はPlath et al(2003)に従って行った。
【0084】
FISH分析
FISHは既に記述されたように行った(Panning, B., Dausman, J., and Jaenisch, R. (1997) Cell 90, 907-916)。Xist、Tsix、およびPgkI二本鎖DNAプローブを、Xist cDNA鋳型およびPgkI配列の17 kbを含有するゲノムクローンからそれぞれ、Cy3-dUTP(Perkin Elmer)、またはFITC-dUTP(Amersham)およびBioprimeキット試薬(Invitrogen)を用いてランダムプライミングによって生成した。TsixおよびXistのいずれかを特異的に検出するための鎖特異的RNAプローブを、Xistエキソン1およびエキソン6鋳型からFITC UTPの存在下でインビトロ転写によって生成した。FISHによって免疫蛍光を追跡する場合、細胞を4%PFAによって固定した後、FISH技法を開始して、ブロッキング緩衝液は1 mg/mL tRNAおよびRNアーゼ阻害剤を含有した。
【0085】
細胞融合
iPS細胞400万個をMEF 400万個と合わせて、PEG-1500(Roche)によって製造元の指示に従って融合した。融合後24時間目にピューロマイシン(1μg/mL)およびハイグロマイシン(140μg/mL)を用いて選択を開始した。Neo選択を伴う実験の場合、G418を300μg/mLで用いた。細胞周期分析を、ヨウ化プロピジウムを用いてFACS Calibur(BD)において行った;シグナル領域をDNA含有量の測定として用いた。
【0086】
染色質の免疫沈降(ChIP)およびマイクロアレイハイブリダイゼーション
ゲノム全体の染色質分析ChIPをwww.upstate.com.でのプロトコールに従って細胞約100万個について行った。各免疫沈降試料10 ngおよび対応する投入量をWhole Genome Amplificationキット(Sigma)を用いて増幅して、増幅材料2μgをBioprimeキット(Invitrogen)を用いてCy3またはCy5(Perkin Elmer)によって標識した。マウスプロモーターアレイ(Agilent -G4490)へのハイブリダイゼーション、洗浄、および走査を、製造元の説明書に従って行った。プローブシグナル(対数比)をフィーチャー抽出ソフトウェアを用いて抽出して、Chip AnalyticsソフトウェアのLowess標準化を用いて標準化して、本明細書において記述されるように統計学的に分析した。
【0087】
全ゲノム発現分析
V6.5 ES細胞、雌性NGiP MEF、ピューロマイシン選択2D4 iPS細胞、およびピューロマイシン選択対照NGiP ES細胞からのRNA 500 ngの1試料あたり2個ずつの試料を増幅して、Agilent low RNA増幅および1色標識キットを用いて製造元の説明書に従ってCy3によって標識した。標識RNAをAgilent Mouse全ゲノムアレイ(G4122F)にハイブリダイズさせて、分析した。
【0088】
フローサイトメトリー
キメラ分析に関して、脾臓、胸腺、および骨髄を、既に記述されているように単離した(Ye, M., Iwasaki, H., Laiosa, C. V., Stadtfeld, M., Xie, H., Heck, S., Clausen, B., Akashi, K., and Graf, T. (2003) Immunity 19, 689-699);細胞を抗体によって染色して、FACSによって分析した。Oct4-Neo XGFP/X尾部線維芽細胞を2回連続して通過させることによってソーティングして、純粋なGFP陰性集団を確かめるために再分析した。EB分化の際に、細胞をGFP+/GFP-集団にソーティングして、FISH分析のために用いた。細胞をBD FACS ARIA(BD Pharmingen)において獲得してFlow Joソフトウェア(Tree Star, Inc.)を用いてデータを分析した。
【0089】
奇形腫形成
各株の細胞200万個を、イソフルラン麻酔SCIDマウスの背側脇腹に皮下注射した。注射の3〜4週間後に奇形腫を回収して、10%ホルマリンにおいて終夜固定して、パラフィン抱埋して、ヘマトキシリン・エオジンによってまたは特異的抗体によって処理した。
【0090】
キメラマウスにおける組織学的検査およびGFP発現の免疫組織学的分析
4%PFAおよび20%蔗糖において組織を続いてインキュベートした後にOCT化合物に抱埋して低温槽において切片(厚さ10μm)を作製する段階によって凍結切片を生成した。切片をVectashield封入培地およびDAPIと共にカバーガラスで覆い、GFPシグナルに関して直接可視化した。
【0091】
iPS細胞の生成効率
ウイルスパッケージングPlatE細胞に、4つの因子12μg(各因子3μg)、またはGFPベクター:空のベクターの1:3混合物の全12μgのいずれかをトランスフェクトした。Nanog-GFP MEFを50%コンフルエンスで接種して、パッケージング細胞からの上清を感染させた。感染後7日目に、4つの因子を感染させた細胞を、放射線照射フィーダーにおいて1:2に分割して、選択(1μg/mLピューロマイシン)または非選択条件のいずれかに置いた。GFP感染細胞を計数して(5.3×106個)、FACSによって分析した。GFP+細胞の百分率(15%)を1つの因子による感染頻度であるとすると、このように4つ全ての因子の頻度は0.15-4であり、理論的にはコロニー〜2700個をもたらした。選択条件下で4週間後、AP陽性ピューロマイシン耐性コロニー20個が出現して、効率は〜0.74%であった。非選択条件ではコロニー〜240個が出現して、効率は〜9%であった。
【0092】
染色質の免疫沈降
フィーダー依存的雄性ES細胞株V6.5(129/B16)、フィーダー非依存的雄性ES細胞株E14(129/ola)、ならびに129/B16マウスに由来する初代培養雄性および雌性MEFを、ピューロマイシンの存在下で生育させた2D4 iPS株と共に用いた。V6.5および2D4細胞の場合、線維芽細胞の混入を低減させるために、細胞の最後の継代は、追加のフィーダー細胞を加えずに行った。細胞は、これらの条件下でその非分化状態を維持した(図1およびデータは示していない)。細胞をホルムアルデヒドによって室温で10分間クロスリンクさせた後、1%SDSを有する10 mM Tris-EDTA、pH 8.0において溶解して、氷中で45秒間の休止をはさんで15秒間のパルス6回の超音波処理を行った。浄化された剪断染色質をH3me3K4(Abcam 8580)またはH3me3K27(Upstate 07-449)に対する抗体によって4℃で終夜免疫沈降させて、プロテインAビーズによって2時間収集して、5分間の洗浄を2回行い、緩衝液(Upstateウェブサイト上でのレシピ)によって溶出した。溶出液を逆クロスリンクして、RNアーゼおよびプロテナーゼK処置を行い、Qiagen PCR精製キットを用いてDNAを精製した。ウサギIgG抗体によるChIPは、いかなる濃縮も見いださなかった(データは示していない)。
【0093】
ヒストンメチル化データと共にゲノムの分析のための統計学的方法
平均プローブシグナルを500 bpのウィンドウで段階的に抽出した。遺伝子16339個を、領域の少なくとも50%が500 bpのウィンドウでプローブによって覆われるという基準に基づいて選択した。ES細胞とMEF細胞のあいだでH3me3K4およびH3me3K27パターンの有意な差を有する遺伝子をシグナチャー遺伝子としてフィルタリングした。各遺伝子に関して、2つの細胞タイプのあいだのヒストン修飾パターンの差を16ウィンドウのシグナルベクトルのユークリッド距離によって定義した。2つのES細胞株(E14対V6.5の距離)および2つの初代培養MEF細胞株(雄性対雌性の距離)の自己距離をプールして、2つのES細胞株または2つのMEF細胞株のあいだの差が小さいと仮定して無効分布を生成した。XおよびY染色体上でコードされる遺伝子を分析から除外した。全てのシグナチャー遺伝子に関して、任意のES-MEF対の距離(E14対Mの距離;E14対Fの距離、V6.5対Mの距離;V6.5対Fの距離)は、ヌル分布の99%分位点である(p値0.01に対応する)既定のシグナチャー-遺伝子閾値(SigT)より大きくなければならない。
【0094】
2D4株におけるシグナチャー遺伝子のEs様遺伝子(Eクラス)、MEF様遺伝子(M-クラス)およびNeutral遺伝子(Nクラス;ES細胞またはMEFのいずれに対しても有意により強い優先性を示さない遺伝子)へのメチル化パターンを明快にするために、2D4とES細胞との平均距離(2D4対ESの距離)および2D4とMEFとの平均距離(2D4対MEFの距離)をコンピューターによって計算した。優先性スコアPS_2D4=(2D4対ESの距離−2D4対MEFの距離)を、2D4細胞における特定の遺伝子のヒストンメチル化パターンが、ES細胞のパターンをどれほど強く「優先して」おそらく模倣するかに関する指標として用いた。この場合も、PSのヌル分布を以下のようにして生成した。各ES細胞株のデータセットを他のES株およびMEFのデータセットと比較した。全ての遺伝子16339個からのPS_ES(E14対V6.5の距離−E 14対MEFの距離)および(V6.5対E14の距離−V6.5対MEFの距離)をコンピューターによって計算してプールした。95%分位点をEクラス閾値(ET)として用いた。ETより大きいPS_2D4を有する任意のシグナチャー遺伝子を「Mクラス」と呼んだ。M閾値(MT)およびEクラスを類似のように定義した。PS-2D4がMTとETの間に入る遺伝子を「Nクラス」と呼んだ。異なる細胞タイプのあいだの8 kb領域内での各500 bpウィンドウに関するメチル化データのピアソン相関係数を、MS Excelにおいて相関関数を用いて計算した。
【0095】
遺伝子発現分析
発現データをFeature Extractionソフトウェア(Agilent)を用いて抽出した。未加工データをlog2変換して、同じ遺伝子に関する多数のプローブからのシグナルを平均した。平均値が0であり、標準偏差が1であるようにそれぞれのアレイを標準化した。再現実験からのデータを平均した。MEFとES細胞のあいだで発現が2倍変化した遺伝子を選択して、それによってこれらの2つの細胞タイプのあいだで最も異なるように発現される遺伝子2473個が同定された(全遺伝子33376個中)。偏りのない階層的クラスタリングを使用して、これらの遺伝子2473個の発現パターンをES細胞、MEF、ピューロマイシン選択NGiP ES細胞およびiPS細胞に対して群分けした。加えて、シグナチャー遺伝子に関する発現パターンをESとMEFの比、またはiPSとMEFの比としてコンピューターにより計算して、メチル化データと共にプロットした。
【0096】
実施例1:Nanog選択可能な線維芽細胞を用いるiPS細胞の生成
Nanog-GFP-puro(Hatano, S. Y., Tada, M., Kimura, H., Yamaguchi, S., Kono, T., Nakano, T., Suemori, H., Nakatsuji, N., and Tada, T. (2005) Mech Dev 122, 67-79)と呼ばれる内因性のNanog座においてGFP-IRES-Puroカセットを持つ雌性マウス胚線維芽細胞(MEF)に、Oct4、Sox2、タンパク質を安定化させる(Sears, R., Nuckolls, F., Haura, E., Taya, Y., Tamai, K., and Nevins, J. R. (2000) Genes Dev 14, 2501-2514)c-MYC-T58A変異体、およびKlf4をコードするcDNAをレトロウイルスによって感染させた。先に報告された、感染後3日目に適用されたFbx15選択(Takahashi and Yamanaka, 2006)とは対照的に、感染後3日目でのNanog発現に関する選択によってコロニーは得られず、Fbx15およびNanog遺伝子の再活性化速度論が異なることを示唆している。感染後7日またはそれより後の日に選択を適用した場合、耐性コロニーが再現性よく出現した。拡大した株5株のうち(表1を参照されたい)、2つの株は、ES細胞と同一であるように思われ、ES細胞表面マーカーSSEA1およびCD9を発現する均一な培養を維持した(データは示していない)。対照的に、他の3つのクローンは、多数回の継代後不均一な培養物を生じ、これは、ES様集団と小さく丸い急速に分裂する細胞の個別の集団の双方を含有した。Nanog-GFP、SSEA-1、およびCD9に関するFACSソーティングの後にサブクローニングを行うと、これらの丸い細胞を消失させるために十分であり、この集団が、ES様細胞とは別個であることを示唆した。興味深いことに、2つの均一な細胞株に関する選択の開始は感染後3週間で起こったが、不均一な株は、感染後1週間で選択を受け、選択の遅れがiPS細胞のより純粋な集団を得るために都合がよい可能性があることを示唆している。
【0097】
その後の研究は、均一なES様細胞株2D4、および本明細書においてiPS細胞と呼ばれる、再ソーティングされて再クローニングされた株1A2に重点を置いた。レトロウイルス組み込み部位のサザンブロット分析により、双方のiPS株に4つ全てのレトロウイルスコード遺伝子が存在することが判明し、ゲノム刷り込みに関する試験により、iPS細胞が、線維芽細胞培養物に存在する可能性があるまれな始原生殖細胞に由来するのではないことを確認した(図9)。Fbx15選択iPS細胞(Takahashi and Yamanaka, 2006)とは対照的に、Nanog選択可能なiPS細胞は、フィーダーの非存在下およびピューロマイシン選択下でES様形態、Nanog発現、およびアルカリホスファターゼ活性(AP)を維持したことから(データは示していない)、フィーダー非依存的生育を示した。LIFを除去すると、始原内胚葉に類似するGATA-4発現細胞への予想される分化が起こり(データは示していない)、分化はNanog発現の喪失を伴った(データは示していない)。RT-PCR分析は、他のES細胞マーカー、Nanog、ERas、およびCriptoと共に内因性の座からのOct4およびSox2の発現を指し示した(図1A)。4つのレトロウイルス発現遺伝子に関する定量的PCR分析は、個々のレトロウイルスに感染した線維芽細胞における強い発現を示したが、均一なiPS細胞では効率よい沈黙化を示した(図1C)。Oct4、Sox2、c-MycおよびKlf4のタンパク質レベルは、iPS細胞と対照ES細胞のあいだで類似であり(図1B)、免疫蛍光は、Oct4およびSox2がレチノイン酸誘導分化の際に効率よくダウンレギュレートされることを示し、ウイルスコード転写因子遺伝子が分化した細胞において有効に沈黙化されたままであることを証明した(データは示していない)。2D4 iPS細胞をSCIDマウスに注射すると、3つの胚細胞層を代表する細胞タイプを含有する奇形腫を生じ、その多能性を確認した(データは示していない)。これらのデータは、レトロウイルス発現Oct4、Sox-2、c-MYCおよびKlf4が、Nanog再活性化に関する選択と組み合わせて、ES細胞と多くの特性を共有するiPS細胞をもたらしうることを指し示している。
【0098】
実施例2:Nanog-選択可能なiPS細胞は、ES細胞様表現型を体細胞に付与する
Nanog-選択可能なiPS細胞がES細胞と類似の機能的属性を保有するか否かを決定するために、細胞融合のコンテクストにおいて体細胞にES-様表現型を押しつける能力を試験した。ピューロマイシン耐性2D4 iPS細胞株からの細胞と共にハイグロマイシン耐性MEF(図2A)。融合後2週間目、ES細胞様形態を有してNanog-GFPを発現し続ける二重耐性4倍体ハイブリッドクローン7個(図2Bおよびデータは示していない)を回収した。対照Nanog-GFP-puro ES細胞をハイグロマイシン耐性MEFと融合させると、1つのハイブリッドコロニーが回収された。多能性を試験するために、ハイブリッド細胞を免疫無防備マウスに注入したところ、4週間後に、3つ全ての胚細胞層を代表する細胞タイプを含有する奇形腫が単離された(データは示していない)。
【0099】
体細胞ゲノムの再プログラミングに関する試験として、内因性のOct4座の制御下で構成的ハイグロマイシン耐性遺伝子とネオマイシン選択可能マーカーの双方を含有するMEF(Oct4-Neo対立遺伝子と呼ばれる)について融合実験を繰り返した。G418を初回選択プロセスにおいて用いた場合にはクローンは得られず、体細胞Oct4座の再プログラミングは、内因性のNanog座の再プログラミングと同様に、逐次的プロセスであることを示唆した。ゆえに、ピューロマイシン/ハイグロマイシン耐性ハイブリッドを拡大した後、それらをピューロマイシン/G418選択に供して、体細胞Oct4遺伝子の再活性化に関して試験した。ピューロマイシン/ハイグロマイシン耐性コロニーは全てピューロマイシン/G418選択下で生存しており、体細胞ゲノムが内因性Oct4座で再プログラムされていることを指し示している(データは示していない)。これらの結果は、Nanog-選択細胞がES細胞と同様に、再プログラミング活性を持ち、体細胞ゲノムに対してES様状態を付与できることを示している。
【0100】
実施例3:異所Oct4発現はiPS細胞の維持にとって不可欠である
Fbx15-選択2D4 iPS細胞は、Oct4およびSox2の永続的なレトロウイルス発現を示し、それぞれの内因性の座からの発現は無視できる程度であり、iPS細胞の自己再生および多能性を維持するために外から提供された因子が持続的に必要であることを示唆した(Takahashi and Yamanaka, 2006)。iPS細胞における効率的なレトロウイルス遺伝子沈黙化を示唆する遺伝子発現データを補強するために、そのゲノムにおいてドキシサイクリン誘導可能なOct4トランスジーンを持つ線維芽細胞を用いることによって、持続的なOct4発現がiPS細胞の維持にとって必要であるか否かを遺伝的に試験することに決定した(Hochedlinger, K., Yamada, Y., Beard, C, and Jaenisch, R. (2005) Cell 121, 465-477)(図3A)。
【0101】
コロニーがOct4誘導可能なシステムを用いて得られうるか否かを最初に決定するために、Oct4誘導可能なMEFに、いかなる選択も行わずにSox2、c-MYCおよびKlf4レトロウイルスを感染させた。ドキシサイクリンの非存在下では、AP陽性コロニーは回収されなかったが、ドキシサイクリンの存在下では、数百個のAP陽性コロニーが出現し、AP陽性コロニーを確立するためにトランスジェニックOct4発現に厳密に依存することを指し示している(図3B)。その後、得られた細胞の再プログラム状態を確かめるために、iPS細胞を、Oct4誘導可能な対立遺伝子とOct4-Neo対立遺伝子の双方を持つ尾部線維芽細胞(TTF)から生成した(図3A)。ドキシサイクリンの存在下で、標的細胞にSox2、c-MYCおよびKlf4を感染させた。薬物選択の開始が遅いことは有利であるというこれまでの所見に基づいて、初回選択を行わずに、ES細胞様形態のみに基づいてiPSコロニーを確立しようと試みた。個々のES様コロニー48個を感染後3週目に採取して、うち2つは、ドキシサイクリンの持続的存在下で安定なES細胞様株に生育した。G418培地に再度播種した後、いずれの細胞株も生存し、内因性のOct4遺伝子が再活性化されて、iPS細胞が生成されていることを指し示した。重要なことに、ドキシサイクリンを培地から除去すると、その生育挙動または形態が変化することなく(データは示していない)、これらの細胞をG418の存在下で何回も継代することが可能であった。ドキシサイクリンの非存在下でのウイルス挿入および異常なOct4トランスジーン活性化の可能性を除外するために、内因性および誘導Oct4発現の定量的PCR分析を行って、分化および誘導の際の発現レベルを分析した(図3C)。非分化iPS細胞は、高レベルの内因性のOct4発現およびトランスジーン発現の完全な非存在を示した。Oct4レベルは、LIFの非存在下で下落して、ドキシサイクリンを投与すると再度出現し、内因性のOct4発現の分化依存的ダウンレギュレーションおよびドキシサイクリンに対する細胞の継続的応答性をそれぞれ示している(図3C)。十分に分化した奇形腫の形成能は、これらの細胞の多能性を証明した(データは示していない)。このように、内因性のOct4座は、外因性のOct4発現の非存在下で多能性状態でiPS細胞を維持するために4つの転写因子によって十分に再プログラムされた。
【0102】
実施例4:遺伝子特異的および全体的DNAメチル化はiPS細胞とES細胞のあいだで類似である
再プログラムされた線維芽細胞のES細胞様特性に基づいて、iPS細胞が、ES細胞と類似の後成的状態を獲得したか否かを調べた。核移入または細胞融合による体細胞ゲノムの再プログラミングは、そのプロモーター領域での多能性遺伝子のDNA脱メチル化などの後成的変化を伴う(Cowan et al., 2005;Tada et al., 2001)。バイサルファイトシークエンシングを用いて、Fbx15-選択iPS細胞において不完全に脱メチル化されていることが既に示されている(Takahashi and Yamanaka, 2006)Oct4およびNanogプロモーターのメチル化状態を査定した。MEFにおいてメチル化されているプロモーターエレメントはいずれも、Nanog選択iPS細胞およびES細胞において脱メチル化を示し、これらの2つの多能性遺伝子が適切に後成的に再プログラミングされたことを示唆している(図4A)。さらに、Nanogプロモーターの脱メチル化は、iPS細胞とMEFの融合を通して生成された細胞ハイブリッドにおいて起こり(図4B、図2を参照されたい)、iPS細胞が、再プログラミング活性を潜在的に持ち、分化した細胞において後成的変化を誘導できることを確認した。
【0103】
雌性ES細胞は、雄性ES細胞および分化した細胞とは対照的に、ゲノムの全体的なDNA低メチル化を示し、これは2つの活性なX染色体(Xa)の存在に帰因しうる(Zvetkova, I., Apedaile, A., Ramsahoye, B., Mermoud, J. E., Crompton, L. A., John, R., Feil, R., and Brockdorff, N. (2005) Nat Genet 37, 1274-1279)。メチル化感受性制限酵素アッセイを用いて、雌性対照ES細胞と同様に、小さいサテライトリピートの全体的な低メチル化が、2D4 iPS細胞株において検出された(図4C)。これらの結果は、iPS細胞が、雌性ES細胞と類似の後成的状態を得たことを示唆している。
【0104】
実施例5:雌性Nanog-選択可能なiPS細胞におけるX-不活化
iPS細胞における全体的なDNA低メチル化は、不活性なX染色体(Xi)が雌性iPS細胞において再活性化されることを指し示している。X-不活化は、哺乳動物細胞における異質染色質形成の最も劇的な例の1つであり、相反して発現される2つの非コードRNA、すなわちXistとそのアンチセンス転写物Tsixによって調節される(Thorvaldsen, J. L., Verona, R. I., and Bartolomei, M. S. (2006) Dev Biol 298, 344-353)。非分化雌性ES細胞は、2つのXaを持ち、両方のX染色体からのTsixを発現して、Xist発現を抑制する。分化すると、Xistは将来のXiにおいて強くアップレギュレートされるようになり、沈黙化を誘導するが、Tsixは消失して体細胞には存在しない。Tsixの下流に位置するX-不活化にとって重要な第3の座であるXite座は、Tsix-様パターンで発現される(Ogawa, Y., and Lee, J. T. (2003) Mol Cell 11, 731-743)。
【0105】
雌性Nanog-GFP-puro MEFにおけるX不活化状態を、蛍光インサイチューハイブリダイゼーション(FISH)を用いて最初に査定して、Xist RNA局在およびX-連鎖遺伝子発現を分析した。Xiの存在と一致して、線維芽細胞の96%がXist RNAコーティングX染色体を持ち、他のX染色体からのPgk1遺伝子の発現を示した(データは示していない)。2D4 iPS細胞株は、非分化ES細胞を強く暗示するXist、Tsix、およびPgk1発現パターンを示した(データは示していない)。すなわち、TsixおよびPgk1は、高レベルで二対立遺伝子で発現されたが、Xist RNAを検出することができず、2つのXaが存在することを証明した。加えて、RT-PCR分析は、ES細胞および2D4 iPS細胞の双方においてXite座からの転写物を検出したが、親線維芽細胞集団では検出しなかった(図5A)。
【0106】
X不活化が開始されると、特徴的な染色質の修飾が将来のXiに押しつけられて、これは染色体の安定な沈黙化を確実にする(Heard, E. (2005) Curr Opin Genet Dev 15, 482-489;Ng, K., Pullirsch, D., Leeb, M., and Wutz, A. (2007) EMBO Rep 8, 34-39)。免疫蛍光を用いて、iPS細胞におけるXi連鎖染色質修飾の存在を分析した。雌性Nanog-GFP-puro MEFは、リジン27位でトリメチル化されたヒストンH3、ヒストンH4リジン20モノメチル化に関する、およびH3K27トリメチル化の媒介の原因であるPolycomb群(PcG)タンパク質Ezh2に関するXi様濃縮の予想された頻度を示した。対照的に、ES細胞と同様にiPS細胞は、これらの染色質のマークに関して豊富で均一な核染色を示し、Xi様濃縮を示さなかった(データは示していない)。併せて、これらのデータは、Xiの転写再活性化を誘導するために、X不活化の調節にとって必須である3つの非コード転写物の発現パターンをリセットするために、およびXiに対して特異的である染色質修飾を消すために、Nanog選択と組み合わせて4つの転写因子が十分であることを指し示している。
【0107】
次に、2D4細胞が分化の際にX不活化を受けるか否かを試験した。iPS細胞がそのX染色体の1つを沈黙化させる能力と一貫して、レチノイン酸誘導分化を受けつつある2D4 iPS細胞においてXist RNAコーティング染色体が検出された(データは示していない)。Xistコーティング染色体は、そのXの沈黙状態と一致してRNAポリメラーゼIIとのオーバーラップを示さなかった(データは示していない)。さらに、分化しつつある雌性ES細胞と同様に、iPS細胞におけるXist RNAコーティングX染色体は、X-不活化の開始時にほとんど常にH3me3K27およびそのメチルトランスフェラーゼEzh2の濃縮領域と一致した(図5B)。XiにおけるEzh2蓄積およびH3me3K27濃縮の一致は、X不活化の初期相のみの特徴である(Plath, K., Fang, J., Mlynarczyk-Evans, S. K., Cao, R., Worringer, K. A., Wang, H., de la Cruz, C. C, Otte, A. P., Panning, B., and Zhang, Y. (2003) Science 300, 131-135;Silva, J., Mak, W., Zvetkova, L, Appanah, R., Nesterova, T. B., Webster, Z., Peters, A. H., Jenuwein, T., Otte, A. P., and Brockdorff, N. (2003) Dev Cell 4, 481-495)。このように、雌性iPS細胞におけるX染色体の不活化は、雌性ES細胞と同じ動力学を表す。
【0108】
実施例6:分化しつつあるiPS細胞における無作為なX不活化
X染色体不活化は、胚外系列および初期着床前胚において非無作為に起こるが、胚盤葉上層および分化しつつあるES細胞においては無作為である。クローニングしたマウス胚におけるX不活化の分析から、体細胞Xiが胚細胞における無作為なX不活化を可能にするために核移入の際に再プログラムされるが、Xiの記憶は、それが配偶子刷り込みを交換する胚外組織において維持されることが示されている(Eggan et al., 2000)。ゆえに、転写因子誘導再プログラミングが体細胞不活化Xiの記憶を消して、このように、分化しつつあるiPS細胞において無作為なX不活化を可能にするか否かを試験した。Nanog-選択可能な2D4 iPS細胞において2つのX染色体を区別することは不可能であったことから、iPS細胞を、緑色蛍光タンパク質(GFP)の発現を駆動するサイトメガロウイルスプロモーターと共にX-連鎖レポータートランスジーン(XGFP)を持つ雌性線維芽細胞から生成した(Hadjantonakis, A. K., Gertsenstein, M., Ikawa, M., Okabe, M., and Nagy, A. (1998) Nat Genet 19, 220-222)(図6A)。このレポーターを、X不活化による沈黙化に供すると、分化しつつあるiPS細胞において沈黙化したX染色体を決定することができる。TTFを、GFPトランスジーンに関してヘテロ接合であり、Oct4-Neo対立遺伝子を持つ雌性マウスから単離した。線維芽細胞集団における無作為なX-不活化と一貫して、TTF細胞の34%がGFP陽性であり(XaGFP/Xi)、細胞の66%がGFP陰性であった(XiGFP/Xa)(図6A、データは示していない)。X不活化の何らかの非対称性が予想され、おそらく、2つのX染色体の遺伝的バックグラウンドの差を反映した。2ラウンドのFACSソーティングによって単離されたGFP陰性細胞に4つの転写因子をコードするレトロウイルスを感染させて、得られたES様コロニーをGFP再発現に基づいてXiGFPの再活性化に関してスクリーニングした。完全に緑色の4つのコロニーが単離されたが、これは再播種した場合に、G418に対して耐性であることが同様に見いだされ、このように沈黙X染色体の再活性化に加えてOct4座が活性化されたことを指し示している。XistおよびTsix発現のES細胞様パターンは、X再プログラミングを確認した(データは示していない)。
【0109】
これらの雌性iPS細胞が、ES細胞と同様に、培養において持続的に維持した場合にXを失う傾向を有したと仮定して、iPS細胞の純粋なクローン集団がX不活化の無作為性に関して確実に分析されるように、XaGFPXa iPS細胞をサブクローニングした。サブクローンの分化を胎芽様体の形成によって誘導して、分化した細胞をFACSによって、GFP陽性およびGFP陰性集団にソーティングして、FISHによって分析した(図6A)。X不活化の無作為なパターンと一貫して、平均で細胞の38%がGFP陽性であり、細胞の62%がGFP陰性であり、双方の集団の大多数が、XiのXist RNAコーティングと一貫するXistシグナルを有した(データは示していない)。無作為なX-不活化により、体細胞においてXaとXiとを区別する後成的なマークがインビトロ再プログラミングの際に除去されうること、およびその後のインビトロ分化の際にいずれかのXにおいて再確立されることが確認される。
【0110】
実施例7:iPS細胞におけるヒストンメチル化パターンの全体的な再プログラミング
次に、Oct4およびNanogプロモーターのDNA脱メチル化ならびにXiの再活性化に加えて、完全な線維芽細胞ゲノムがiPS細胞誘導化の際にES様状態に後成的に再プログラムされていたか否かを調べた。ヒストンメチル化は、哺乳動物の発達および細胞分化の際の遺伝子発現の後成的調節において重要な役割を果たす。一般的に、転写された遺伝子は、H3K4トリメチル化に関連しているが(Bernstein, B. E., Kamal, M., Lindblad-Toh, K., Bekiranov, S., Bailey, D. K., Huebert, D. J., McMahon, S., Karlsson, E. K., Kulbokas, E. J., 3rd, Gingeras, T. R., et al. (2005) Cell 120, 169-181;Kim, T. H., Barrera, L. O., Zheng, M., Qu, C, Singer, M. A., Richmond, T. A., Wu, Y., Green, R. D., and Ren, B. (2005) Nature 436, 876-880)、沈黙化された多くの遺伝子はH3K27トリメチル化に関連している(Boyer, L. A., Plath, K., Zeitlinger, J., Brambrink, T., Medeiros, L. A., Lee, T. I., Levine, S. S., Wernig, M., Tajonar, A., Ray, M. K., et al. (2006). Nature 441(7091):349-53;Lee, T. I, Jenner, R. G., Boyer, L. A., Guenther, M. G., Levine, S. S., Kumar, R. M., Chevalier, B., Johnstone, S. E., Cole, M. F., Isono, K., et al. (2006) Cell 125(2):301-13)。Nanog-選択2D4 iPS株、雄性および雌性MEF、ならびに2つの雄性ES細胞株におけるK4およびK27トリメチル化に関するゲノム全体の位置分析を、染色質の免疫沈降の後に、マウスプロモーターアレイとのハイブリダイゼーションを用いて行った。このアレイ上のプロモーターは、遺伝子約16,500個に関して転写開始部位の-5.5 kb上流から+2.5 kb下流までの領域を範囲に持つ。2D4 iPS株がES細胞またはMEFにより類似しているか否かを決定するために、ES細胞とMEFのあいだでヒストンメチル化パターンが有意に異なる一組の遺伝子を定義した。高ストリンジェンシー(p=0.01)では、ES細胞とMEFのあいだで遺伝子957個が異なると同定され、「シグナチャー」遺伝子として分類された(実験技法を参照されたい)。特に、2D4 iPS細胞では、シグナチャー遺伝子の94.4%がES細胞と実質的に同一であるメチル化パターン(Eクラス遺伝子)を持ったが、よりMEF様パターン(Mクラス遺伝子)では遺伝子の0.7%がメチル化されたに過ぎなかった。座の残りの4.9%は、差が小さすぎて有意ではなかったために(データは示していない)Nクラス遺伝子(中性)であると分類された。iPS座の大部分(91%)は、より大きい組のシグナチャー遺伝子を含めるためにたとえストリンジェンシーをp=0.05まで低下させてもEクラスに留まった(データは示していない)。E、M、およびN遺伝子への分布は、無作為入れ替え試験によって確認したところ、非常に有意である(図10)。非シグナチャークラスに属する遺伝子は、MEF、ES細胞、およびiPS細胞のあいだでメチル化パターンにほとんどまたは全く差を示さず(データは示していない)、iPS株がES細胞またはMEFのいずれにおいても見いだされない完全に新規の後成的同一性を獲得したことを指し示している。集合的に、これらの結果は、インビトロ再プログラミングが、線維芽細胞ゲノムの後成的記憶をES細胞と非常に類似の記憶に逆転させることができることを指し示している。
【0111】
K4およびK27メチル化パターンが再プログラミングの際に異なる程度にリセットされたか否かを決定する試みにおいて、アレイ上の遺伝子16,500個全てに関してそれぞれのメチル化のマークに関するピアソン相関を個々に計算した(図7A)。この分析から、iPS細胞およびES細胞が、そのK27メチル化パターンにおいて2つのES株が互いに対して類似であるのと同程度に類似であるが、MEFはiPSおよびES細胞の双方から同程度に明らかに異なることが判明した。興味深いことに、K4メチル化は、全ての細胞タイプのあいだでより類似であり、再プログラミングが、K4トリメチル化よりむしろK27の変化に主に関連することを示唆している。この全体的分析からの1つの予測は、K27メチル化の変化が、シグナチャー遺伝子のEクラスにおいて顕著であるはずであるという点である。これを試験するために、8 kbのプロモーター領域にわたって500 bpの間隔で起こりうる全ての細胞タイプのあいだで対応のある相関分析を行い、各比較に関して相関値16個を得た(図7B)。E遺伝子として分類された遺伝子は実際に、全分析領域にわたってES細胞と2D4 iPS細胞のあいだでそのK4およびK27メチル化パターンが非常に類似であったが、MEFは全体を通していずれの細胞タイプとも劇的に異なった。全体的相関とさらに一致して、K27メチル化は、MEFとES/iPS細胞のあいだでK4メチル化より劇的に異なった。発達遺伝子は、ネズミES細胞におけるPcG媒介K27メチル化の最も重要な標的群であるというこれまでの所見(Boyer et al., 2006)に基づいて、これらの座がシグナチャー遺伝子内で濃縮されるか否かを試験することに決定した。実際に、遺伝子オントロジー分析により、発達遺伝子はシグナチャー遺伝子のEクラスにおける最も有意に濃縮された遺伝子群であることが判明した(p=8xe-10)。これらの知見は、MEFからiPS細胞への再プログラミングにとって、K27メチル化の変化がK4メチル化の変化より重要であることを示唆しており、再プログラミングにおけるPcGタンパク質の重要な役割を示唆している。
【0112】
iPSとES細胞のヒストンメチル化パターンの相関が、iPS細胞の転写状態の変化を忠実に捕らえているか否かを試験するために、全ゲノムレベルでAgilentマイクロアレイを用いて、ES細胞、2D4 iPS細胞、およびMEF細胞について発現分析を行った。ESおよびiPS細胞は、ピアソン相関によって決定した場合に、全体的レベルで発現パターンに非常に高い相関を示した(図11Aおよび11B)。ES細胞とMEFのあいだで発現が2倍より大きい差を有する遺伝子は、ESとiPS細胞のあいだでほとんど同一に発現された(データは示していない)。ゆえに、これらのデータは、後成的データから予想されるように、iPS細胞が、転写的にES細胞と非常に同等であることを指し示している。シグナチャー遺伝子13個の無作為に選ばれたサブセットのレベルをリアルタイムRT-PCRによって確認した(図11C)。試験した遺伝子は全て、iPS細胞とES細胞において類似のレベルで発現された。ES、iPS細胞、およびMEFのあいだのシグナチャー遺伝子の発現の差は、ヒストンメチル化パターンにおいて観察された差と良好に相関し(データは示していない)、K4およびK27メチル化がそれらの遺伝子の発現状態の重要な決定因子であることを示唆している。併せて、これらのデータは、4つの転写因子による核の再プログラミングが、線維芽細胞ゲノムの全体的な転写および後成的リセットを誘導できることを証明している。
【0113】
実施例8:MEFおよびTTF由来iPS細胞は胚細胞が含まれる多数の細胞タイプに分化する
iPS細胞の忠実な後成的再プログラミングによって、ES細胞と同等の発達能が得られるであろうと推論された。GFPマークMEF由来2D4 iPS細胞を2倍体胚盤胞に注入すると、明白なGFP蛍光を有する新生仔キメラ3例を生じた(図8A、表2)。新生仔からの組織切片は、軟骨、腺構造、肝臓、心臓、および肺に対するiPS細胞の広いクローン性の関与を示した(データは示していない)。新生仔に由来する造血細胞のFACS分析により、脾臓B細胞およびマクロファージと共に胸腺CD4+およびCD8+ T細胞の18〜28%がiPS細胞に由来することが判明した(図8B)。その上、iPS細胞由来の尾部線維芽細胞およびニューロスフェア培養物をこのキメラ仔から単離することが可能であり、これらは宿主由来の線維芽細胞およびニューロスフェアと比較して類似の生育速度およびサイトカイン依存性を示した(データは示していない)。成体まで発達したキメラ1例は、外皮色キメラ現象を示し、iPS細胞が機能的メラノサイトへと分化したことを指し示した(図8C)。
【0114】
次に、MEF由来iPS細胞に加えて、雌性TTF由来iPS細胞が同様に発達を支持しうるか否かを調べた。XiGFPトランスジーンの再発現に基づいて選択されている異なる2つのiPSクローンの胚盤胞注射によって、株あたり1例の生後動物(postnatal animal)を生じた(表2を参照されたい)。キメラ動物は、健康であるように見え、成体マウスまで正常に生育した。これらの結果は、TTFに由来するiPS細胞が、胎児線維芽細胞に由来するiPS細胞と同様に、正常な外観の生後キメラを生じたことを示している。
【0115】
生殖系列伝搬は、細胞の多能性に関する最もストリンジェントな試験の1つであると見なされている。XiGFP/X TTF由来iPS細胞が、生殖系列に寄与しうるか否かを査定するために、過剰排卵させたiPSキメラ1例から卵母細胞16個を単離したところ、その4個が明らかにGFP陽性であり、iPS細胞が雌性生殖系列に関与することを支持している(データは示していない)。これらの卵母細胞を塩化ストロンチウムおよびサイトカラシンBによって処置すると、単為生殖活性化に成功して、その後胚盤胞段階へと卵割して、卵母細胞の機能性を証明した(データは示していない)。
【0116】
ES細胞の成熟細胞タイプへの方向性の分化は、明らかに治療的将来性を有する。iPS細胞がインビトロで成熟細胞を生じるか否かを決定するために、EBを生成して、造血細胞の運命を誘導するために培養において拡大させた。実際に、未成熟および成熟血球のマーカーを発現する細胞タイプが検出され、このように再生医学におけるiPS細胞の将来的な使用を強調する(図12)。
【0117】
線維芽細胞培養物から直接多能性細胞を生成することは、核の再プログラミングを支配するメカニズムの理解に向けて主要な進歩を表した(Takahashi and Yamanaka, 2006)。ここに、ゲノムの忠実な後成的リセットは転写因子誘導再プログラミングを伴う、という第一の証拠を提供する。そのエピゲノムにおいてES細胞と顕著に類似であるiPS細胞を回収した。たとえば、雌性iPS細胞は、重要な多能性遺伝子のプロモーターで適切な脱メチル化を示し、それらは分化の際に無作為なX不活化を受ける体細胞沈黙化X染色体を再活性化して、ES細胞とほとんど同一である全体的なヒストンメチル化パターンを有した。iPS細胞はまた、増殖因子応答性、細胞融合における再プログラミングドナーとしての作用能と共に、インビトロおよびインビボの双方でES様分化を受ける能力が含まれる他のES様性質も示し、1つの生殖系列キメラが含まれる高グレード生後キメラに関与した。
【0118】
トランスジェニックOct4発現がiPS細胞の維持にとって必要ではないという知見は、多能性を確実に維持するために、内因性の遺伝子発現プログラムが十分に再活性化されていることを指し示している。このことは、Oct4の外因性の発現、ならびにおそらくSox2、c-MycおよびKlf4の外因性の発現が、多能性に至る転写の変化および後成的変化を誘発するために再プログラミングの最初の段階の際に限って必要である可能性があることを指し示している。この考え方を支持して、4つの因子のレトロウイルス発現は、感染したドナー線維芽細胞において高く、iPS細胞において沈黙化した。このように、体細胞に4つの因子を一過性に供給することが実現可能であり、それによって、それぞれ挿入突然変異または遺伝子発現のアーチファクトを生じる可能性のあるレトロウイルスまたはトランスジェニックエレメントを含有しない、安定に再プログラムされた細胞が生成される。
【0119】
意外にも、Nanog選択iPS細胞は、既に報告されているFbx15選択iPS細胞とは、表現型的および分子的に異なっていた。Nanogは、胚の発達にとって必須であり、始原内胚葉への分化を抑制することによって多能性を維持するために必要である(Chambers, I., Colby, D., Robertson, M., Nichols, J., Lee, S., Tweedie, S., and Smith, A. (2003) Cell 113, 643-655.;Mitsui, K., Tokuzawa, Y., Itoh, H., Segawa, K., Murakami, M., Takahashi, K., Maruyama, M., Maeda, M., and Yamanaka, S. (2003) Cell 113, 631-642)。対照的に、Fbx15は、ES細胞におけるその独占的発現にもかかわらず、多能性または発達にとって必須ではない(Tokuzawa, Y., Kaiho, E., Maruyama, M., Takahashi, K., Mitsui, K., Maeda, M., Niwa, H., and Yamanaka, S. (2003) Mol Cell Biol 23, 2699-2708)。理論に拘束されることを望むものではないが、Fbx15選択iPS細胞と本明細書において記述されるiPS細胞のあいだの性質の差に関して、いくつかの可能性のある説明がある。1つの可能性は、Nanog選択が、Fbx15選択と比較してより大きい発達能を有する異なる多能性細胞タイプを生じることである。これと一致して、ほとんどのFbx15選択iPS細胞は、Nanogを発現しなかったが(Takahashi and Yamanaka, 2006)、このことは、MEFの非存在下でそれらが不適当に分化し、満期産のキメラを生じなかった理由を説明する可能性がある。この考え方をさらに支持するのは、正常なES細胞培養において必ずしも全てのOct4発現細胞がNanog陽性ではないという所見であり、このことは、ES細胞集団内の不均一性を示唆している(Hatano et al., 2005)。興味深いことに、ES細胞が由来する胚盤胞の内細胞塊は、Oct4およびNanogに関して類似の不均一な発現パターンを示す(Chazaud, C, Yamanaka, Y., Pawson, T., and Rossant, J. (2006) Dev Cell 10, 615-624.)。
【0120】
再び、理論に拘束されることを望むものではないが、得られたiPSの品質に関するNanog選択の効果に関する別の説明は、Nanogタンパク質それ自身が忠実な後成的再プログラミングにおいて重大な役割を果たしていることでありうるあろう。この考え方と一致して、ES細胞と体細胞との細胞融合実験により、NanogがES細胞において過剰発現されている場合に、200倍多くのコロニーが得られることが示されている(Silva, J., Chambers, I, Pollard, S., and Smith, A. (2006) Nature 441, 997-1001)。Nanogは体細胞において多能性を誘導する場合には必要ないが、再プログラミングプロセスの際のその過剰発現が、iPS細胞を得る効率を増強するか否か、およびiPS細胞の発達効能に影響を及ぼすか否かを査定することは有益である。
【0121】
再び、理論に拘束されることを望むものではないが、既に報告されたiPS細胞と本明細書において記述されたiPS細胞のあいだに観察された差のもう1つの可能性は、選択の時期である可能性がある。感染後3日目に選択を適用した場合には、Nanog-GFP-puro MEFからiPS細胞を誘導化することは不可能であったが、これはYamanakaとその共同研究者による知見とは対照的であり、彼らはこの時期にFbx15発現に関して選択することが可能であった。よって、選択を感染後1週目に開始するか、またはES細胞の形態もしくは沈黙化したX-連鎖GFPトランスジーンの再活性化のみに基づいてiPS細胞を単離した後、Oct4-Neo対立遺伝子を用いて多能性をレトロスペクティブに確認した。初回薬物選択を行わずに誘導化されたiPS細胞は全て、キメラの関与およびES細胞様後成的特色に関して既に報告されたFbx15-選択iPS細胞より良好であるように思われた。再プログラミングは、数日または数週間を要する逐次的プロセスであり、再活性化される必要がある遺伝子のカスケードに依存するという仮説が立てられる。このシナリオにおいて、Nanog再活性化は、Fbx15再活性化より核の再プログラミングの際の後期に起こるであろう。このように、Fbx15の初期選択は、核の再プログラミングを完了していない細胞集団を拡大して、その結果、再プログラミングプロセスの後期に現れるであろう潜在的によりよく再プログラムされる細胞を消失させる可能性がある;Nanogに関して後期に選択することは、再プログラミングがより完了している段階を捕える可能性がある。この仮説を探る1つの方法は、Fbx15発現に関して後期に選択した場合に、ES細胞により類似するiPS細胞を生成するか否かを試験することであろう。薬物選択の代わりにES様コロニーの形態的選択がiPS細胞を得るために十分でありうるという所見は、ヒト細胞にレポータートランスジーンを導入する段階が技術的に難題であり、挿入突然変異を引き起こす可能性があることから、ヒトにおける直接再プログラミングに関して重要な意味を有する。
【0122】
多能性への細胞の直接再プログラミングは、明らかな治療的意味を有し、ゆえにiPS細胞がES細胞と同じ後成的状態で存在するか否かを確かめることは非常に重要であった。これらのデータは、異常な後成的再プログラミングが直接再プログラムされた細胞の治療的有用性を損なわないことを指し示している。
【0123】
実施例9:ヒトiPS細胞は選択の非存在下で生成されうる
患者特異的線維芽細胞またはケラチノサイト細胞に、テトラサイクリン誘導可能なレンチウイルスシステムによって発現された、4つ(OCT4、SOX2、CMYC、KLF4)または5つ(4 + NANOG)の再プログラミング因子を感染させた。ウイルスを、リバーステトラサイクリントランスアクチベータ(rtTA)を発現するレンチウイルスと共に同時感染させた。細胞をフィーダー細胞上で継代して、ドキシサイクリンによって誘導した;細胞を、最初の3日間線維芽細胞培地において維持した後、ヒトES細胞条件に切り換えた。小さいコロニー様構造が4日以内に目に見えるようになり、30日までに、別個のhES様の丸い石のような外観を有するヒトES細胞形態を有するコロニーが存在した(データは示していない)。非hES細胞形態を有するコロニーもまた存在したが、hES様コロニーの生成を妨害しなかった。
【0124】
hES様コロニーを採取して、拡大させ、特徴付けを行った。ヒトES細胞と同様に、それらは多能性であり(奇形腫を生成した)、重要な多能性遺伝子を発現し、後成的修飾の適切なリセットを示した。加えて、それらは同様にレンチウイルストランスジーンを沈黙化した。
【0125】
本明細書において引用された任意の特許、または特許出願が含まれる全ての参考文献は、図面および表と共に参照により本明細書に組み入れられる。いかなる参考文献も先行技術を構成すると認めたわけではない。参考文献の考察は、その著者が主張する内容を述べているのであり、本出願者らは、引用された文書の精度および適当性に異議を申し立てる権利を保有する。多数の先行技術の出版物が本明細書において引用されるが、このように引用することが、これらの文書のいかなるものも、アメリカ、または任意の他の国における当技術分野において共通の一般的知識の一部を形成すると認めているわけではないと明らかに理解されるであろう。
【0126】
(表1)得られたiPS細胞株の要約
* 細胞をGFP、SSEA-1およびCD9に関して三重にソーティングした後、サブクローニングして均一で安定なES様集団を得た;
** 三重ソーティング後に行った分析;ND=決定していない
【0127】
(表2)iPS細胞由来マウスにおける満期発達効率および推定されるキメラ現象の程度
【0128】
(表3)RT-PCR分析のためのプライマー配列と抗体
【図1A】
【図1B】
【図1C】
【図2A】
【図2B】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【特許請求の範囲】
【請求項1】
以下の段階を含む、誘導多能性幹細胞を選択する方法:
a)RT-PCRによって測定した場合に、Nanog mRNAを発現しない分化した初代培養細胞を、多能性表現型に再プログラムする段階;
b)再プログラミング後、選択物質の非存在下で段階(a)において再プログラムされた細胞を培養する段階;
c)段階(b)の培養物を顕微鏡下で観察して、培養物において外観がなめらかで丸くなった細胞のクローンを単離する段階;および
d)幹細胞マーカーの発現が検出されれば、細胞が誘導多能性幹細胞であることが指し示される、幹細胞マーカーの発現に関してクローンの細胞を試験する段階。
【請求項2】
再プログラミング段階が、以下の1つを含む、請求項1記載の方法:因子を発現させるための調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;および細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【請求項3】
3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、幹細胞マーカーを発現するクローンの細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に対する組織学的検査を行う段階をさらに含む、請求項1記載の方法。
【請求項4】
培養する段階が細胞を継代する段階をさらに含む、請求項1記載の方法。
【請求項5】
分化した体細胞がES細胞の形態とは明確に異なる形態を有する、請求項1記載の方法。
【請求項6】
分化した初代培養細胞が線維芽細胞であり、線維芽細胞が再プログラミング前に平坦で不規則な形状を有する、請求項1記載の方法。
【請求項7】
幹細胞マーカーが、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される、請求項1記載の方法。
【請求項8】
分化した初代培養細胞が雌性の個体に由来する場合、不活性なX染色体の再活性化に関してクローンの細胞を試験する段階をさらに含む、請求項1記載の方法。
【請求項9】
核酸配列がウイルスベクターまたはプラスミドに含まれる、請求項2記載の方法。
【請求項10】
ウイルスベクターがレトロウイルスベクター、レンチウイルスベクター、またはアデノウイルスベクターである、請求項9記載の方法。
【請求項11】
外因性のOct4、Sox2、c-Mycおよび/またはKlf4の発現に関してクローンの細胞を試験する段階をさらに含む、請求項1記載の方法。
【請求項12】
細胞がヒト細胞を含む、請求項1記載の方法。
【請求項13】
以下の段階を含む、誘導多能性幹細胞を選択する方法:
a)選択可能マーカーが活性なX染色体上で変異体であり、不活性なX染色体上で野生型であり、およびRT-PCRによって測定した場合に、細胞がNanog mRNAを発現しない、X染色体上で選択可能マーカーに関してヘテロ接合である雌性細胞を提供する段階;
b)細胞を多能性表現型に再プログラムする段階;
c)不活性なX染色体の再活性化によって、野生型選択可能マーカーを発現することができ、選択物質の存在下で細胞を生存させることができ、それによって生存細胞が誘導多能性幹細胞である、細胞を選択物質と共に培養する段階。
【請求項14】
選択物質の存在下で生存する細胞を幹細胞マーカーの発現に関して試験する段階をさらに含む、請求項13記載の方法。
【請求項15】
幹細胞マーカーが、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される、請求項14記載の方法。
【請求項16】
再プログラムする段階が、以下の1つを含む、請求項13記載の方法:因子を発現させるための調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;および細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【請求項17】
3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、選択物質の存在下で生存する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に対する組織学的検査を行う段階をさらに含む、請求項13記載の方法。
【請求項18】
細胞が細胞株の細胞である、請求項13記載の方法。
【請求項19】
細胞がX染色体上で変異体Hprt遺伝子に関してヘテロ接合である、請求項13記載の方法。
【請求項20】
細胞が、前記核酸の導入前に不活性である野生型Hprt遺伝子をX染色体上で持ち、および再プログラミング前に活性である変異体の非機能的Hprt遺伝子をX染色体上で持つ、請求項19記載の方法。
【請求項21】
細胞が再プログラミング前に6-チオグアニンに対して耐性である、請求項13記載の方法。
【請求項22】
選択物質が、HAT培地を含む、請求項13記載の方法。
【請求項23】
細胞がヒト細胞を含む、請求項13記載の方法。
【請求項24】
以下の段階を含む、誘導多能性幹細胞を選択する方法:
a)RT-PCRによって測定した場合に、Nanog mRNAを発現しない、X不活化による沈黙化に供されるX染色体連鎖レポーター遺伝子を持つ雌性細胞を提供する段階;
b)細胞を多能性表現型に再プログラムする段階;
c)再プログラミング後に細胞を培養する段階;および
d)レポーターが発現すれば、クローンが誘導多能性幹細胞を含むことが指し示される、X染色体連鎖レポーターを発現する細胞のクローンを培養物から単離する段階。
【請求項25】
幹細胞マーカーの発現に関してクローンの細胞を試験する段階をさらに含む、請求項24記載の方法。
【請求項26】
幹細胞マーカーが、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される、請求項25記載の方法。
【請求項27】
3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、レポーターを発現する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階をさらに含む、請求項24記載の方法。
【請求項28】
細胞がヒト細胞を含む、請求項24記載の方法。
【請求項1】
以下の段階を含む、誘導多能性幹細胞を選択する方法:
a)RT-PCRによって測定した場合に、Nanog mRNAを発現しない分化した初代培養細胞を、多能性表現型に再プログラムする段階;
b)再プログラミング後、選択物質の非存在下で段階(a)において再プログラムされた細胞を培養する段階;
c)段階(b)の培養物を顕微鏡下で観察して、培養物において外観がなめらかで丸くなった細胞のクローンを単離する段階;および
d)幹細胞マーカーの発現が検出されれば、細胞が誘導多能性幹細胞であることが指し示される、幹細胞マーカーの発現に関してクローンの細胞を試験する段階。
【請求項2】
再プログラミング段階が、以下の1つを含む、請求項1記載の方法:因子を発現させるための調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;および細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【請求項3】
3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、幹細胞マーカーを発現するクローンの細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に対する組織学的検査を行う段階をさらに含む、請求項1記載の方法。
【請求項4】
培養する段階が細胞を継代する段階をさらに含む、請求項1記載の方法。
【請求項5】
分化した体細胞がES細胞の形態とは明確に異なる形態を有する、請求項1記載の方法。
【請求項6】
分化した初代培養細胞が線維芽細胞であり、線維芽細胞が再プログラミング前に平坦で不規則な形状を有する、請求項1記載の方法。
【請求項7】
幹細胞マーカーが、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される、請求項1記載の方法。
【請求項8】
分化した初代培養細胞が雌性の個体に由来する場合、不活性なX染色体の再活性化に関してクローンの細胞を試験する段階をさらに含む、請求項1記載の方法。
【請求項9】
核酸配列がウイルスベクターまたはプラスミドに含まれる、請求項2記載の方法。
【請求項10】
ウイルスベクターがレトロウイルスベクター、レンチウイルスベクター、またはアデノウイルスベクターである、請求項9記載の方法。
【請求項11】
外因性のOct4、Sox2、c-Mycおよび/またはKlf4の発現に関してクローンの細胞を試験する段階をさらに含む、請求項1記載の方法。
【請求項12】
細胞がヒト細胞を含む、請求項1記載の方法。
【請求項13】
以下の段階を含む、誘導多能性幹細胞を選択する方法:
a)選択可能マーカーが活性なX染色体上で変異体であり、不活性なX染色体上で野生型であり、およびRT-PCRによって測定した場合に、細胞がNanog mRNAを発現しない、X染色体上で選択可能マーカーに関してヘテロ接合である雌性細胞を提供する段階;
b)細胞を多能性表現型に再プログラムする段階;
c)不活性なX染色体の再活性化によって、野生型選択可能マーカーを発現することができ、選択物質の存在下で細胞を生存させることができ、それによって生存細胞が誘導多能性幹細胞である、細胞を選択物質と共に培養する段階。
【請求項14】
選択物質の存在下で生存する細胞を幹細胞マーカーの発現に関して試験する段階をさらに含む、請求項13記載の方法。
【請求項15】
幹細胞マーカーが、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される、請求項14記載の方法。
【請求項16】
再プログラムする段階が、以下の1つを含む、請求項13記載の方法:因子を発現させるための調節エレメントに機能的に連結した、転写因子Oct4、Sox2、c-MycおよびKlf4をコードする核酸配列を分化した体細胞に導入する段階;細胞の分化状態を再プログラムする1つまたは複数のタンパク質因子を導入する段階;および細胞の分化状態の再プログラミングを誘導する低分子を細胞に接触させる段階。
【請求項17】
3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、選択物質の存在下で生存する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に対する組織学的検査を行う段階をさらに含む、請求項13記載の方法。
【請求項18】
細胞が細胞株の細胞である、請求項13記載の方法。
【請求項19】
細胞がX染色体上で変異体Hprt遺伝子に関してヘテロ接合である、請求項13記載の方法。
【請求項20】
細胞が、前記核酸の導入前に不活性である野生型Hprt遺伝子をX染色体上で持ち、および再プログラミング前に活性である変異体の非機能的Hprt遺伝子をX染色体上で持つ、請求項19記載の方法。
【請求項21】
細胞が再プログラミング前に6-チオグアニンに対して耐性である、請求項13記載の方法。
【請求項22】
選択物質が、HAT培地を含む、請求項13記載の方法。
【請求項23】
細胞がヒト細胞を含む、請求項13記載の方法。
【請求項24】
以下の段階を含む、誘導多能性幹細胞を選択する方法:
a)RT-PCRによって測定した場合に、Nanog mRNAを発現しない、X不活化による沈黙化に供されるX染色体連鎖レポーター遺伝子を持つ雌性細胞を提供する段階;
b)細胞を多能性表現型に再プログラムする段階;
c)再プログラミング後に細胞を培養する段階;および
d)レポーターが発現すれば、クローンが誘導多能性幹細胞を含むことが指し示される、X染色体連鎖レポーターを発現する細胞のクローンを培養物から単離する段階。
【請求項25】
幹細胞マーカーの発現に関してクローンの細胞を試験する段階をさらに含む、請求項24記載の方法。
【請求項26】
幹細胞マーカーが、SSEA1、CD9、Nanog、Fbx15、Ecat1、Esg1、Eras、Gdf3、Fgf4、Cripto、Dax1、Zpf296、Slc2a3、Rex1、Utf1、およびOct4からなる群より選択される、請求項25記載の方法。
【請求項27】
3つ全ての胚細胞層からの細胞を含む腫瘍が生育すれば、細胞が多能性幹細胞であることがさらに指し示される、レポーターを発現する細胞をヌードマウスに導入する段階、および細胞から発生した腫瘍に関する組織学的検査を行う段階をさらに含む、請求項24記載の方法。
【請求項28】
細胞がヒト細胞を含む、請求項24記載の方法。
【図5】

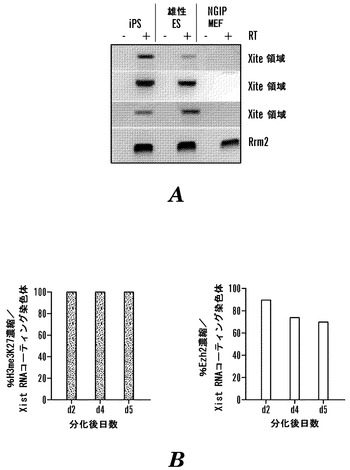
【図6】
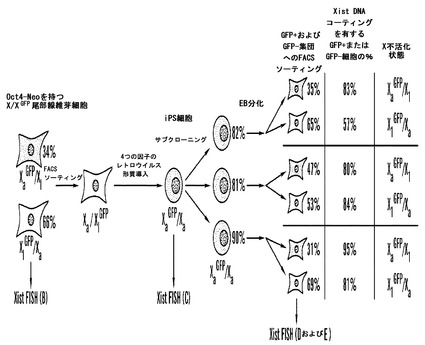
【図7A】
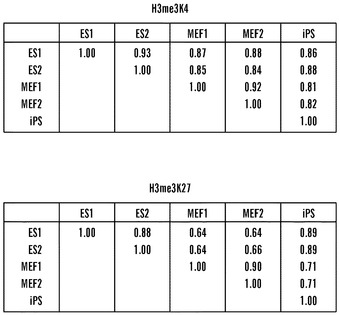
【図7B】
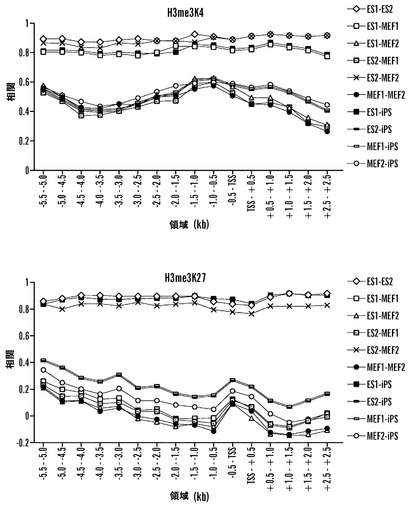
【図8A】
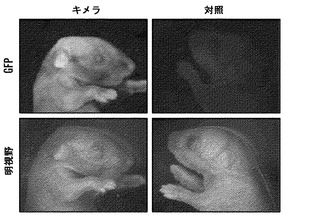
【図8B】
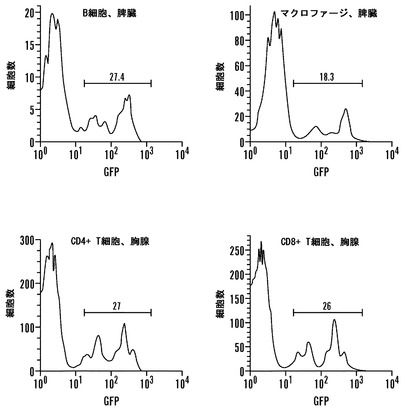
【図8C】

【図9】
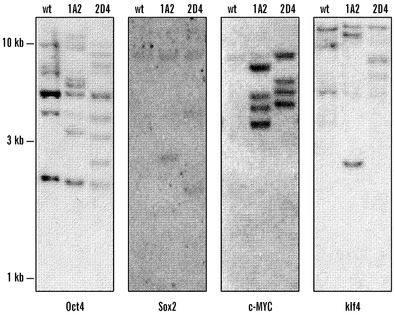
【図10】
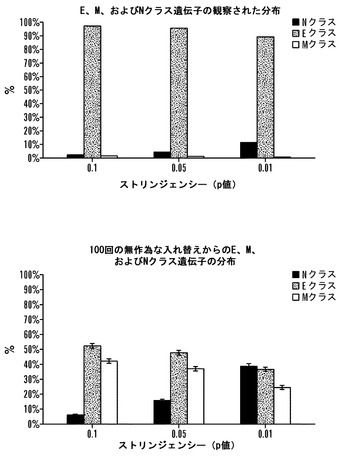
【図11A】
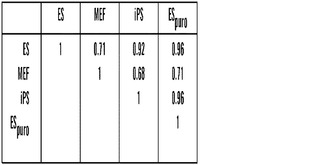
【図11B】
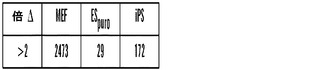
【図11C】
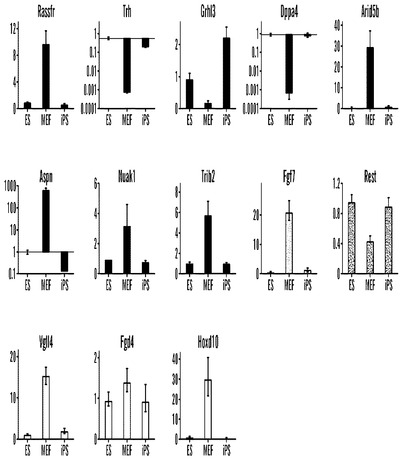
【図12A】
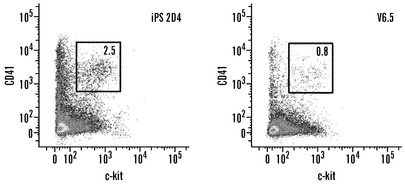
【図12B】
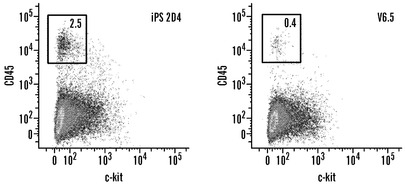
【図12C】
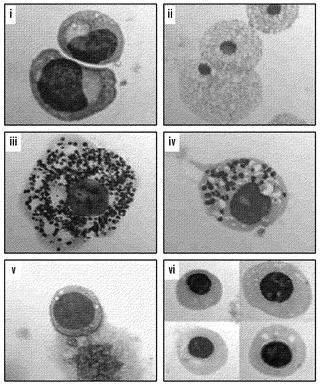
【図6】
【図7A】
【図7B】
【図8A】
【図8B】
【図8C】
【図9】
【図10】
【図11A】
【図11B】
【図11C】
【図12A】
【図12B】
【図12C】
【公表番号】特表2010−528622(P2010−528622A)
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願番号】特願2010−510528(P2010−510528)
【出願日】平成20年5月30日(2008.5.30)
【国際出願番号】PCT/US2008/065384
【国際公開番号】WO2008/151058
【国際公開日】平成20年12月11日(2008.12.11)
【出願人】(592017633)ザ ジェネラル ホスピタル コーポレイション (177)
【出願人】(507244910)プレジデント・アンド・フェロウズ・オブ・ハーバード・カレッジ (18)
【Fターム(参考)】
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願日】平成20年5月30日(2008.5.30)
【国際出願番号】PCT/US2008/065384
【国際公開番号】WO2008/151058
【国際公開日】平成20年12月11日(2008.12.11)
【出願人】(592017633)ザ ジェネラル ホスピタル コーポレイション (177)
【出願人】(507244910)プレジデント・アンド・フェロウズ・オブ・ハーバード・カレッジ (18)
【Fターム(参考)】
[ Back to top ]