
健康的な老化および長寿を予測および促進するFOXO3Aの多型およびハプロタイプを使用する方法
本発明は、個々の寿命の可能性、物理的および認知機能を加齢中の保持を分析し増加させ、冠状動脈(心臓)疾患、脳卒中、癌、慢性肺疾患、糖尿病、パーキンソン病および認知症を含有する心血管,代謝,年齢関連性疾患を進行する個々の可能性を分析し減少させるために使用することができるFOXO3A遺伝子からの、遺伝子情報の同定および使用に関する方法および組成物を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、診断において予測し、または、治療計画および関与において健康的な老化および長寿を促進するために、FOXO3Aの多型およびハロタイプを使用する方法に関する。
【0002】
関連出願の説明
本出願は、本願明細書に引用した2008年8月10日に出願した米国特許出願公開第61/087,722号明細書に、部分的に基づき、その利益を要求する。
【0003】
連邦支援の研究または開発に関する記載
本発明を、アメリカ国立老化研究所からの1 R01 AG027060−01 (Defining the Healthy Aging Phenotype)の研究費で、政府支援のもと行った。更なる資金提供は、国立心臓肺臓血液学会からの契約書N01−HC−05102、国立老化研究所からの契約書N01−AG4−2149および研究費5 U01 AG019349−05およびK08 AG22788−02に基づいて、米国政府の支援によって提供された。米国政府は、本発明における特定の権利を有する。更なる支援は、ハワイコミュニティー財団からの研究費2004−0463に基づいて行われた。
【背景技術】
【0004】
FOXO3A遺伝子は、明瞭なフォークヘッドドメインによって特徴づけられるフォークヘッド型転写因子に属する。この遺伝子は、細胞死に必要な遺伝子の発現によって、アポトーシスの誘因として機能するらしい。この遺伝子とMLL遺伝子の転座は、二次急性白血病と関連する。同じタンパク質をコードする選択的なスプライスされた転写変異体を観測した。
【0005】
FOXO3A遺伝子は、DAF−16のヒト相同体の1つであり、モデル生物であるシノラブディスエレガンスで寿命を延ばすと記載されていた遺伝子である(Murphy CT (2006)The search for DAF−16/FOXO transcriptional targets: approaches and discoveries. Exp Gerontol 41 :910−921) and D. melanogaster. (Giannakou ME et al. (2007) Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging Cell 6:429−438)。
【0006】
FOXO3A遺伝子は、ヒト染色体6q21の位置108,987,719から109112664(NCBI ver.36)までに配置され、選択的に発現できる4つのエクソンから構成され、同じタンパク質を生じる(異型#1は、ファイルNM 001455.3に記載され;異型#2は、ファイルNM 201559.2に記載される)。FOXO3Aタンパク質は、アミノ酸673から構成され、71,277Daサイズである。FOXO3Aのアミノ酸配列は、NCBIでファイル「NP 963853」によって規定され、SEQ ID NO.1として同定し、以下の通りである:
【0007】
【0008】
FOXO3Aは、YWHAB/14−3−3−betaおよびYWHAZ/14−3−3−zeta、UniProt:the Universal Protein Resource (www.uniprot.org)と相互作用し、細胞質の分泌で必要である。酸化性ストレスに応じて、STKと相互作用し、YWHAB/14−3−3βとの相互作用を妨害して、核移動に導く。FOXO3Aの細胞内局在は、細胞質およびサイトソルである。それは、酸化ストレスに応じておよび生存因子の欠損で、核に移行する。生存因子、IGF−1が存在する場合において、FOXO3Aは、Thr−32およびSer−253でAKT1/PKBによってリン酸化される。それからこのリン酸化された形態は、14−3−3タンパク質と相互作用し、細胞質にて保持される。生存因子の脱離は脱リン酸化を誘導し、脱リン酸化されたタンパク質が、標的遺伝子の転写を誘導する場合には核への移行を促進し、アポトーシスを誘発する。AKT1/PKBは、直接Ser−315をリン酸化するようではないが、それはこの残基でリン酸化を誘発する他のキナーゼを活性化することができる。FOXO3Aは、STK4によってSer−209に酸化性ストレスに応じてリン酸化され、YWHAB/14−3−3−βおよび核移行からの脱離を導く。
【0009】
ヒトの寿命は、多くの決定因子を有する複雑な表現型である。食事、物理的活動、健康習慣および社会心理的因子を含む非遺伝的な因子が重要である一方、ヒト寿命において最高50%の変異が遺伝子の相違点によって説明される可能性がある1−5。いくつかの研究は、平均的なヒトの寿命において、約25%の変異を示唆することをできるが、例外的な生存者の多くの人において、寿命に対する遺伝的貢献は非常に高い可能性がある。例えば、90歳代および100歳代の家系調査では、兄弟間の相対的危険度は複雑な表現型への潜在的遺伝子の寄与を評価する一般的な方法6で、特に高く、家系発端者の年齢の増加と共に増大することを示した7〜10。しかしながら、ヒトにおける「長寿に関連する」遺伝子候補の研究は、以下「長寿遺伝子」といい、一般的に期待はずれである。ほとんどの複製は、ApoE遺伝子を除いて、集団全体に観測されなかった。
【0010】
対照的に、老化のモデル生物において、いくつかの強力な遺伝子の発見があった11〜13。例えば、単一の遺伝子の変異体は、モデル生物における寿命で、特にインスリン/IGF−1(IIS)シグナル経路の一部と考えられる遺伝子14〜18で、実質的な相違を生じることができる。
【0011】
SIR2活性を向上させるか、またはインスリン/IGF−1シグナルを減少させる突然変異は、DAF−16/FOXOタンパク質を活性することによって、シノラブディスエレガンスの寿命を共に上昇させる19、20。哺乳類の細胞において、Sir2相同体物である「SIRT1」は、ストレスへの細胞反応を含む、寿命に影響するいくつかの下流転写現象に影響を及ぼしている。SIRT1は、これを、FOXO(フォークヘッドボックス転写)因子、IIS経路でセンサーとして機能し、いくつかの哺乳類において寿命の調節因子ともなるタンパク質のファミリーを調整することによって達成する17。
【0012】
哺乳類(および他の種)における、遺伝子ノックアウトモデルは、同様にIIS仮説を支持する。例えば、脂肪細胞に特異的なインスリン受容体ノックアウトのマウス(FIRKO)は、脂肪の質量、加齢に関連した肥満に対する予防を減少し、寿命を延長する。IIS経路における多くの他の突然変異体は、マウスにおいて長寿に影響を与えると考えられる。これらは、IGF−1受容体22、IRS−122、IRS−223、PAPP−A24の突然変異体およびエイムス矮小マウス突然変異を含む。
【0013】
インスリンシグナルの基本的な分子経路は、酵母、ハエ、虫、齧歯動物およびヒトに示される証拠があるように、進化にわたって保存されている。虫における前記経路の重要な調節因子は、転写制御因子DAF−16(異常なDAuer形成16)であり、シノラブディスエレガンスにもたらされる広範な寿命延長に、インスリン/IGF−1シグナルを阻害することに必要である。多くの因子は、daf−16に依存した方法、例えばAMPキナーゼ26、14−3−3のタンパク質27、lin−4マイクロRNA28および熱ショック因子において、シノラブディスエレガンスの寿命を延ばすと考えられる。いくつかの種のDAF−16の相同体は、老化表現型および長寿に関連していた30。例えば、ストレス応答Junアミノ末端キナーゼ(JNK)経路は、FOXOが必要であると考え、ショウジョウバエの寿命を延長し31、ハエがdFOXO、DAF−16オーソログを過剰に発現しているとき、それは寿命を顕著に上昇することができる32。DAF−16/FOXOでの前記多様な配列シグナルの著しい収束は、このタンパク質が、老化および長寿に影響を与えるシグナルネットワークにおいて、重要で、進化的に保存されている「ノード」であることを示唆する。
【0014】
DAF−16のヒトの相同体は、4つのFOXO:FOXO1、FOXO3A、FOXO4およびFOXO6を含有する。従って、FOXOの単一の塩基多型(SNP)形態における、一般的な、野生の変異体および関連した遺伝子は、ヒトの長寿に影響する可能性があることを仮定することを試みる。「FOXO3」は、FOXO3Bが染色体17上の偽遺伝子であるため、「F0X03A」と同義である。
【0015】
これは興味深い仮説である。インスリン、FOXO、酸化性ストレスとヒトの寿命との間の関係は、酸化ストレスが、好ましい一般的に考えられる加齢のメカニズムであるため、特に関連性がある。1956年以来、加齢の遊離基説は、加齢がDNA、反応性酸素分子の累積的な照射によって細胞および組織に損傷を部分的に生じることを仮定し、普遍的にまだ受け入れられないにも関わらず、有効な証拠が長年にわたって蓄積した34,35。従って、FOXOは、潜在的な分岐点または橋かけを、インスリンシグナル、遊離基とヒトの老化/寿命との間で提供することができる。
【0016】
Suh et alによる関連性のある最近の報告38を含むヒトの寿命で、IIS経路における、いくつかの従来の研究と関連のある遺伝子があり36、37、機能的に重要なIGF−I受容体の突然変異体と特別な寿命とを関連するが、FOXO遺伝子およびヒト寿命の間の関連性の発表報告は全く見当たらなかった。従来の研究は、FOXO遺伝子と、4年の生存率、脳卒中の危険性39および早発閉経40を含む他の老化表現型との関連が明らかになった。
【0017】
しかしながら、ヒトの寿命は、疾患特定の危険性ならびに個々の加齢の割合を包含する複雑な表現型である。その遺伝子の先例の研究は、困難である。寿命の研究は、小さい遺伝子影響によるサイズ、人為的な層化産物、人口異質、長寿研究の参加者の充分な人数の不足および他の課題3、4,41によって影響される。従って、ヒトの寿命への遺伝子寄与の可能性を、IISシグナルに関連する遺伝子から評価するために、加齢表現型としてよく特徴付けられる人の多く、均質な長寿の集団を選択し、IIS経路に関連のある5つの候補の寿命遺伝子のコホート内症例対照研究(nested case-control study)を実行した。前記遺伝子は、加齢表現型の従来の関連物に基づいて、主に加齢の遺伝子ノックアウト、トランスジェニック、突然変異および他のモデル生物から選択された3、4、14〜17、36,42。優先順位は、インスリン感受性およびグルコース(エネルギー)ホメオスタシスに関連する遺伝子で行った。
【0018】
集団の急激な老化は、慢性疾患の罹患率および障害を増大することにより、先例のない難題を社会にもたらすであろう。ヒトが齢をとる方法に関して、広く影響することができる生物学的経路を包含する、加齢のメカニズムの良好な理解は、加齢に関連した疾患および障害への低下で重要な意味を有する。ヒトの寿命で多くの生物学的に信頼できる遺伝子候補があるが、1つの発見のみが、ApoE遺伝子のものより、今まで多くの集団において広く繰り返されてきた。前記遺伝子は、加齢表現型、特に心血管疾患および認知症、例えば長く健康な寿命を達成する能力に影響するようなものに広く達成した。
【発明の概要】
【課題を解決するための手段】
【0019】
ヒトの加齢表現型および寿命への広く達成した遺伝子を発見する際の課題は、ヒトにおける研究を実施する前、モデル生物を使用して先見的で潜在的な候補を同定することに役立つことができることを示唆する。従って、ヒトのインスリン/IGF−1シグナル経路および/または配列に基づく酸化ストレス反応システムおよび/または加齢のモデル生物または従来のヒト研究で機能的相同性の中で、いくつかの候補遺伝子の研究するために選択する。前記シグナル経路からヒトの候補遺伝子のリストを構成し、日本人の集団の約10%以上の頻度にて発生する前記候補遺伝子において変異体を評価した。限られた資源のため、3つのSNPのみを、分析でそれぞれの遺伝子から選択した。SNPを、可能であれば、それぞれの遺伝子の最大範囲を提供するために、連鎖不平衡(LD)で領域から選択した。
【0020】
一般的に、本発明は、FOXO3A「GCC」ハプロタイプ(例えば、FOXO3Aハロタイプは増加した寿命と関連があり、本明細書において更に15年以上生存するヒトの対象の可能性として規定する)を検出する組成物および方法を提供する。好ましい実施形態において、検出されたFOXO3Aハプロタイプは、長く生きる増加した可能性または減少した可能性のいずれかと関連するが、本発明は、長い寿命の増加も減少の可能性と関連がなくおよび/または加齢に関連した疾患(例えば、「正常」または「wt」の遺伝子型)の危険性を最少にするFOXO3Aハプロタイプを検出する物質および方法を必然的に包含する。加齢に関連した疾患とは、疾患冠動脈疾患(CHD)であり、冠状動脈疾患、脳卒中、癌、慢性閉塞性肺疾患(COPD)、他の慢性的な肺疾患、パーキンソン病、糖尿病、肥満、認知症(一般的な認識機能)、虚弱(歩行能力)もしくは他の加齢と関連性の疾患、または身体および/または認識障害と規定する。またヒトの肥満との関連性が存在する。
【0021】
「GCC」ハプロタイプは、何十キロベースのDNAを包含する。この領域における他のSNPは、本願明細書で記載される3つのSNPを有する連鎖不平衡を示す。更なるSNPが、同様に寿命および健康的な老化に関連のあるこのGCCハロタイプで同定され、予測される加齢と関連する疾患に実用的であることができると予期される。「GCCハロタイプ」は、特別な長寿および/または健康的な加齢の予測へと導く“機能的な変異体”であることを最終的に見出すDNA変性の他の種の代用物として、FOXO3A遺伝子の範囲または隣接したいずれかでもたらすことができる。前記他の変性は、転移、複製、欠失の形態であることができ、例えば“LOC100130966”といった依然知られていない他の遺伝子または転写物を含有することができる。LOC100130966は、mif2のSMT3制御因子3ホモログ2と同類であり、FOXO3A遺伝子のエクソン2の範囲内であることが同定され、“GCC”ハロタイプ内である。LOC100130966に関するDNA配列は、GenBank取得のID#「XM 00 1725519」として記載され、LOC100130966の予想アミノ酸配列は、GenBankファイル「XP 00 1725571」として記載されている。
【0022】
ハプロタイプ分析を使用して、患者が積極的な健康または疾患の予防/治療介入によって利益があるものを潜在的に予測することができる。ハプロタイプ分析は、キットの様式で提供することができる。危険性の測定は、このような情報を、疾患、障害もしくは死亡の可能性の評価、またはヒトがどれほど生存または無病生存であるかを確定する目的に使用することができる。前記情報は患者、健康保険会社、長期介護保険会社および医師または他の医療供給者への、多少の指導を患者の長期ニーズとして提供するために重要である。医薬を、加齢に関連した健康または疾患に影響を与えるために、FOXO3A遺伝子の作用の改善、FOXO3Aタンパク質および/または他のタンパク質との相互作用の細胞局在の改善、遺伝子によって産出されるタンパク質の量または種類の改善するものを開発することができる。
【0023】
マウスにおいて相同配列は、早期卵巣機能不全と関連することができる。Castrillon DH, Miao L, Kollipara R, Horner JW, DePinho RA. Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science. 2003 Jul 11;301(5630):215−8。その結果、同類のハプロタイプ分析は、獣医学的応用に実用的であることができる。
【0024】
更なる本発明の特徴は、以下の記載から、例えば添付した図面および表を参照して、明らかになるだろう。
【図面の簡単な説明】
【0025】
【図1】表10に記載のプライマーおよび条件を使用してFOXO3A G/T変異体を検出するARMS−PCR定量法の結果を示す図である。
【図2】表10に記載のプライマーを使用してFOXO3AのG/T変異体を検出するARMS−PCR分析の模式的に概略を示す図である。
【0026】
以下の表は、本明細書の一部である:
表1:HHP/HAAS Cohortの1991−93における基本特性(n=3,741)
表2:ケースコントロール状態による基本特性
表3:ケースおよびコントロールにおけるヒトの寿命およびMAFの候補遺伝子
表4:ケースコントロール状況によるFOXO3A3遺伝子型
表5:基準での遺伝子型集団間の健康状況の相違点
表6:FOX03A遺伝子型によるインスリン感受性表現型
表7:FOXO3A3遺伝子型に関する加齢に関連した表現型の優位性
表8:最高到達年齢による遺伝子型分布
表9:rs2802292 G−T多型の同定のためのプライマー
表10:rs2802292 G−T多型の同定のためのPCR条件
【発明を実施するための形態】
【0027】
A.ハワイでの寿命の研究
調査集団
コホート内症例対照研究は、ハワイ寿命調査の一部で、ホノルル心臓計画(HHP)およびホノルルアジア加齢研究(HAAS)の当初の集団から集められた健康的な加齢の配属されたコホート研究として実施された。HHPは、1965年に開始した日系米国人8,006人の中での、心血管疾患の集団に基づく潜在的な調査である。HHP参加者は、正式な連絡先を持ち、1900年から1919年の間に生まれ、オアフ島に1965年に居住していた9,877人の男性から募った。
【0028】
試験参加者は、日本、通常日本の西部および南部からの両親を有し(中央領域またはさらに西部および南部から94%);全体の49%は、親の出身が広島および山口の隣接した県からである61、62。大半の参加者はハワイ生まれ(88%)であるが、対立遺伝子頻度のケースコントロール条件の混絡の理論的な可能性が、地理的起点による。従って、特定の分析、ケースおよびコントロールに関して、親の出身の県によって、条件付きロジスティック回帰モデルを使用して、階層化した。分析は、データセットにおける、集団階層化で全く証拠を示さなかった(資料未記載)。
【0029】
HHPコホートの補充、設計および方法を、詳細に他で概説する62。簡潔に、調査登録時(1965〜1968)、参加者は、45〜68までの歳であった(平均年齢54歳)。HHPの開始から、冠動脈疾患および脳卒中の発生の進行、ならびに全ての原因による死亡率に関する情報が、地方新聞(英語および日本語)の死亡記事の観測および退院記録の査察から得られた61。1991〜1993年検査における追跡調査は、5人のみが死亡情報をたどることができなかったことが分かった63。
【0030】
現在の調査に関する全ての参加者は、2007年8月までに更新された調査参加者の記録から導いた。HHP(1991−1993)の試験4から達成された表現型データおよび血液サンプルは、ホノルルアジア加齢研究(HAAS)の開始と一致し、コホート内症例対照研究のための基礎調査として使用された。HAASは、高齢者における神経変性疾患、認識機能および他の加齢表現型の調査のためのHHPの展開として開始された。64参加者は、試験4での71〜93歳の3,741人(平均年齢77.9±4.7歳)、最初のHHPの約半数を含有する64。
【0031】
現在のコホート内症例対照研究に関して、「ケース」(寿命表現型)を、1910年アメリカ出生コホートの特異的生存率の少なくとも上位1%(最小95歳)を募集時期から生存した全てのHHP参加者として規定した。2007年8月時点にて、少なくとも95歳まで生存した合計213人の個人を調査した。176人のこれら個人が死亡し(平均年齢97.5歳、SD2.1;範囲95〜106歳)、37人が依然生存している(平均年齢98.7歳、SD2.1;範囲97〜106歳)。
【0032】
コントロールは、中年男性で、1910年のアメリカ出生コホートの特異的生存に関する平均死亡年齢の間近に死亡したHHP/HAASコホートからの402人からなる。症例を達成するために:約1:2のコントロール比率において、81歳までに死亡したコントロールに関するHHP/HAAS調査集団をサンプルとして採取した。コントロール集団における死亡平均年齢は、78.5歳であった(SD1.8、範囲73〜81歳)。これはアメリカの男性集団より若干高いが、ハワイにおける日系米国人の男性の高い平均寿命に合致し、最後の報告で白人男性より3.5年長かった67。全てのケースおよびコントロールは、家系が主に日本の中央西部を出身とする日本人人種である61、62。
【0033】
方法は、組織のガイドラインに従って実行され、Institutional Review Board of Kuakini Medical Centerによって承認された。書面に基づくインフォームドコンセントを、参加者が同意を提供することができない場合、全ての試験参加者または家族代表者から得た。
【0034】
遺伝子型
3つのSNPを5つの候補遺伝子の各々から選択する。モデル生物において、加齢経路で影響を十分に記載された遺伝子を選択する。全ての遺伝子を、図1を参照するように、IIS経路に仮定的な関連、ならびにエネルギー恒常性、グルコースおよび/または脂質代謝に潜在的な関連に基づいて選択した。SNPを、HapMapまたはJSNPデータベースに報告された、それらの少数の対立遺伝子頻度に基づいて選択した(snp.ims.u−tokyo.ac.jp)。
【0035】
総細胞DNAを、PureGeneシステム(Gentra Systems, Inc.)を使用して分離し、PicoGreen染色(Molecular Probes, Eugene, OR)を使用して定量化し、候補遺伝子からSNPは、対立遺伝子識別分析を使用して遺伝子型を求めた。TaqMan(登録商標)(Applied Biosystems, Inc.)試薬をABIから購入し、SNPを頻度≧〜0.1で日本人集団において選択した(http://www.ncbi.nlm.nih.gov/projects/SNP/)。PCRは、Taq Gold(Perkin −Elmer, Corp)およびTaq Man(登録商標)分析でPCR産物の検出を使用し、1つの対立遺伝子および他の対立遺伝子に対してVICラベルされたプローブで6−FAMラベルされたFRETプローブを使用し、分析の検出を向上させるために副溝結合(MGB)クエンチャーを使用して、標準の条件下にて増幅した。PCR産物を、ABI Prism 7000 Sequence Detection Systemで測定した。
【0036】
遺伝子型データを、統合されたデータベースシステムによって管理した(MS Excel, Microsoft, Inc)。それぞれの遺伝子型プレート上の全ての正のコントロールを、同様に一貫して評価した。陽性マーカーをハーディワインベルグ平衡からの偏差で検出した。細胞頻度は98%を超えた。
【0037】
統計学的分析
SNPを、ハーディワインベルグ平衡からの偏差で評価した。ピアソンカイ2乗検定を使用して、ソフトウエアプログラムStatXact68を用いて同様の遺伝子型頻度に対するケースおよびコントロールを比較した。関連の強度評価するために、確率比率をSAS69のロジスティック回帰モデルを使用して評価した。一般化線形モデル(GLM)および共分散分析(ANCOVA)を更に使用して、健康な試験参加者の比率をFOXO3Aの遺伝子型によって比較した。ケースおよびコントロールにおける加齢表現型の分析のためには、連続変数の分布を比較するスチューデントt検定、および比例変数でのカイ二乗である。
【0038】
結果
1991〜1993の調査でHHP/HAAS調査集団の基本的特徴を表1に示す。平均年齢は、77.9歳であり、集団の100%は男性および日本人人種であった。生物学的特徴、公衆衛生状態、疾病の罹患および基本的な状態を示す。
【0039】
【表1】
【0040】
前記1991〜93年の基本的集団から、「長寿」のケースとして2007年までに95歳以上生存した全ての参加者を選択した(n=213)。その後「平均的な」生存率のコントロールとして81歳以前に死亡した全ての参加者を選択した(n=402)。ケースおよびコントロールの基本特徴を表2に示す。生物学的特徴において、高齢者のケースは、比較的老年で、脂肪が少なく(低い腰周り:尻周りの比率)、低いトリグリセリド(ボーダーライン)、低いグルコース、低いインスリンレベルおよび高いFOXO3A3対立遺伝子の傾向を、基礎調査で有する。ケースはまた、自己評価による健康、ならびに心血管疾患(CHDおよび脳卒中)および癌の低い傾向を改善した。機能的に、彼等は十分に歩行できるようであるが、低い握力を有した。認識スコアに関する有意差は存在しなかった。
【0041】
【表2−1】
【表2−2】
【0042】
5つの遺伝子を調査した(ADIPOQ、FOXOlA、FOXO3A、SIRTl、およびCOQ7)。ケースおよびコントロールに関して、少数の対立遺伝子頻度および他の関連する遺伝子情報を表3に示す。しかしながら、FOXO3A遺伝子型のみが、p<0.05の初期カットオフ値を使用して寿命と関連した。
【0043】
【表3】
【0044】
変異体“rs2764264”を、以下に“rs 12524491”として規定する。SEQ ID NO.2として同定されたSNP rs2764264(“F0X03A1”)のDNA配列は、以下である:
【0045】
【0046】
SEQ ID NO.3として同定されたSNP rsl3217795(“FOXO3A2”)のDNA配列は、以下である:
【0047】
【0048】
SEQ ID NO.4として同定されたSNP rs2802292(“FOXO3A3”)のDNA配列は、以下である:
【0049】
【0050】
「GCCハプロタイプ」は、SNP rs2764264、rs13217795およびrs2802292を使用して記載することができ、遺伝子型の以下の組み合わせを含む対立遺伝子である:
【0051】
【表4】
【0052】
これらの変異体を、染色体の最上部(遺伝子地図において低いヌクレオチド位置)から最下部(遺伝子地図において高いヌクレオチド位置)まで観測するとき、NCBI命名法によると「GCCハプロタイプ」を、SNPrs2802292、rs2764264およびrsl3217795を使用して記載することができ、以下の遺伝子型の組み合わせを含有する対立遺伝子である:
【0053】
【表5】
【0054】
上記のデータは、Database of Single Nucleotide Polymorphisms (dbSNP). Bethesda (MD): National Center for Biotechnology Information, National Library of Medicine. (dbSNP Build ID: 129, NCBI genome build 36.3)からである。http://www.ncbi.nlm.nih.gov/SNP/70から入手可能である。
【0055】
FOXO3 A3の遺伝子頻度をケースおよびコントロールの間で比較する更なる調査は、ピアソンのカイ二乗統計の置換分布を用いて、正確なp値0.00009で、非常に重要な相違点を示した。これらの結果を表4に示す。この研究において、それぞれの対立遺伝子内に5箇所の遺伝子座と3つのSNPが存在するため(表3)、多重比較に関するボンフェローニ補正は、15×0.00009=0.00135の補正されたp値を生じた。FOXO3Aの3SNPの間で高い関連性のLDのため、FOXO3A3のSNPのみに関して、更に調査した(rs2802292)。ケースおよびコントロールの間でFOXO3A3に関する対立遺伝子の同型劣性(homozygous minor)対同型優性(homozygous major)の確立比率(OR)は、2.75(95%CI:1.51−5.02、p=0.0007)であり、ケースおよびコントロールの間で異型対同系の優性対立遺伝子は、1.91(95%CI:1.34−2.72、p=0.0003)であった。これらの結果は、寿命に関して相加効果を示唆する。
【0056】
【表6】
【0057】
若い年齢で長寿表現型についてさらに理解するために、Willcox et al.(2006)の健康な生存者の定義を用いて3つのFOXO3A遺伝子型群のそれぞれに関する基礎調査(1991)にて、健康である人の比率を比較した。相違点は、非常に重要であった(表5)。1つ以上のG対立遺伝子を有する人々は、優性(TT)対立遺伝子に関して同型である人々より、正常にて健康であるようである。劣性対立遺伝子に関して同型な人の約75%は、優性対立遺伝子に関して同型な人々の約57%のみに対して、基礎調査にて健康的である。ケースコントロール状態を調整した後も、相違点は依然わずかながら重要であった。これは、長期生存者(ケース)およびコントロールの部類の範囲内で、対立遺伝子と健康状態の関連性が存続することを示唆する。
【0058】
【表7】
【0059】
インスリン感受性、寿命の潜在的な中間の表現型、および遺伝子型の間の関係があるかどうかを評価するため、空腹時インスリン、グルコース、HOMAおよび遺伝子型の間の関係を試験した(表6)。非正規分布変数のため、対数変換を正規分布に使用した。優位な関係がインスリン、インスリンの対数、HOMAおよび遺伝子型に存在する。同型G対立遺伝子は、コントロールのみにおいてであるが、インスリン、インスリンの対数およびHOMAスコアの顕著な低下と関連している。
【0060】
【表8】
【0061】
同様に、いくつかの慢性疾患およびFOXO3A遺伝子型の生涯有病率の間の関係に関して試験参加者にて調査した(表7)。
【0062】
【表9】
【0063】
優位な保護関係が、CHDの有病率に関する同型G対立遺伝子、並びに癌および認識機能に関する境界関係で同型接合性が明らかになった。最後に、統合した全ての参加者(ケースおよびコントロール)において最高到達年齢によって、FOXO3A3劣性対立遺伝子頻度(MAF)分布を評価した。MAFは、以前のケースコントロール分析によって予測したように、年齢と共に著しく上昇した(表8)。
【0064】
【表10】
【0065】
5つの候補遺伝子の分析は、1つの遺伝子が、潜在的なヒト長寿遺伝子であるFOXO3Aに関して、他の遺伝子より明確に卓越していることを示した。前記遺伝子が、ヒトの寿命に重要でありえることが、いくつかの証拠によって支持されている。第一に、コホート内症例対照分析において、前記遺伝子内における変異体は、寿命と強く関連している。ケースおよびコントロールの間のFOXO3A3の対立遺伝子の同型劣性(homozygous minor)対立遺伝子の同型優性(homozygous major)であるオッズ比(OR)は、2.75(95%CI:1.51−5.02、p=0.0007)であり、ケースおよびコントロールの間の異型対同系の優性でのORは、1.91(95%CI:1.34−2.72、p=0.0003)であった。これらの結果は、寿命におけるFOXO3A3 G対立遺伝子の相加効果を示唆する(即ち、G対立遺伝子の2つのコピーは、約2倍の保護作用を与えた)。一貫して、劣性対立遺伝子頻度は、70歳代から100歳代までの年代の試験参加者の年齢と共に、著しく上昇した(表8)。
【0066】
第二に、FOXO3A遺伝子において評価された全3つのSNPは、密接した連鎖不均衡(LD)であり、長寿表現型と強い相関関係があった。これは、調査結果が可能性によるものでないことを示す。第三に、1つ以上の劣勢(G)対立遺伝子を有する人は、同型優性(TT)対立遺伝子での人と比較して、約15年前の基礎調査にてより健康である可能性があった。劣性対立遺伝子に関して同型な人の約75%は、優性(TT)対立遺伝子に関して同型な人々約57%のみに対して、基礎調査にて健康的である(表5)。
【0067】
実際に、基礎調査は、ケースが、平均して11歳老年であるにも関わらず、ケースがコントロールより顕著に健康的であることを示唆した。ケースは、CHD、脳卒中および癌の低下した有病率を含有する著しく少ない加齢関連疾患を有する。同様に、彼等は優れた自己評価で健康的であり、低い歩行障害を含む、高い身体機能を一般的に有した。興味深いことに、コントロールより10歳を超える年にも関わらず、長寿のケースは同様のレベルの認識機能を有した。これは、個々が主な臨床疾患および障害を人生の末期まで何とか遅らせるか、回避する場合に、「健康的な加齢」の表現型の存在を支持する。ケースで観測された健康的な加齢表現型は、彼等の年齢に適合する出生コホート46〜48および100歳以上の子孫と比較した若い年代で、100歳以上の人々において報告された健康的な加齢表現型に類似する49。同様に長命のケースは、若い年代で比較的高いインスリン感受性と、低い腰周りと尻周りの比率、低いグルコースレベル、低いインスリンレベルおよび低いHOMA値である代謝プロフィールを有する。いくつかの表現型が、FOXO3A遺伝子型において変異体と関連した。
【0068】
意外にも、ケースおよびコントロールの間で糖尿病の有病率で優位な違いは全く存在しなかった。しかしながら、ケースはコントロールより10歳を超えて、糖尿病は年齢と共に顕著に増加するため、それは糖尿病の有病率が非常に独特でないことに注目すべきである。実際に、ケースおよびコントロールの両方は、高い糖尿病の有病率(約60%)を、比較的低いBMIに関わらず有した。2型糖尿病が、日本人において比較的低いBMIで高い有病率の傾向がある理由は、完全には解明されていない50。しかしながら、白人および黒人より低いBMIでアジア人において、高い内臓脂肪を有する日本人(多少の他のアジア人)で、代謝の相違点が存在する可能性がある51、52。実際には、日本国ガイドラインが、このような集団の違いを反映し、日本人の肥満をBMI25とみなす。HHP/HAASコホートにおける糖尿病の高い有病率への他の原因要因は、全ての参加者を、いくつかの異なった臨床試験、および更に可能性がある検出するいくつかの事前検査で糖尿病を試験する事実を包含する。
【0069】
当然、FOXO3A遺伝子型は、血漿インスリンレベル並びにCHD、癌および2型糖尿病の有病率に著しい関連があった。これは、細胞増殖、アポトーシスおよび代謝を包含する多様な生理的機能で、インスリンおよびインスリン様成長因子の効果のメディエーターとしてのFOXOでの知られている役割と一致する。シノラブディスエレガンスおよびショウジョウバエにおける遺伝学の研究は、FOXOタンパク質が、代謝および寿命を調節するインスリン様シグナルの従来の標的であることを示した。哺乳類細胞における更なる研究は、FOXOタンパク質が、タンパク質キナーゼの標的であり、細胞周期の進行に影響し、生体外において酸化ストレスに対する抵抗を制御することを示した54。生体内研究は、FOXOが、インスリンに反応して肝臓グルコース産物を転換し、他の代謝活性を媒介することを示した。これは、FOXOタンパク質が、インスリン効果を代謝で媒介して、ヒトの寿命に影響することができるという根拠を支援する。
【0070】
概して、根拠の全体は、FOXO3Aの潜在的役割を、ヒトの健康、加齢および寿命で支持する。インスリン感受性、CHD、癌、2型糖尿病および寿命を含有する多様な長寿表現型と、FOXOとの関連は、「ゲートキーパー」の役割を、IIS経路において示唆する。FOXO3Aがヒトの老化に影響する重要な下流のメカニズムは、酸化ストレスの修正による−かねてのヒトの老化に関する仮説であるが、これに対する直接的な証拠は現在研究中で全くない。しかしながら、FOXO遺伝子が、シノラブディスエレガンスDAF−16の、最も密接なヒトの相同物であり、細胞を酸化ストレスから保護するため、これはヒトの加齢の変化に対して優れた作用のメカニズムである。シノラブディスエレガンスにおいて、DAF−16は、マンガンスーパーオキシドジスムターゼ(SOD2)の発現を増加させ、これは他の「加齢防止」効果の中で、過酸化物を比較的低い損傷で過酸化物に転換し、遊離基に対して有力な内因性保護作用がある。生体内研究は、DNA、タンパク質および他の組織における酸化障害が、年齢と共に蓄積し、カロリー的に制限された食事を、齧歯動物56およびヒト57に供給することが、この障害を暖和することを示した。
【0071】
FOXOが明らかに寿命と関連している一方、インスリン感受性で遺伝子型の強い効果は、ケースには観測されず、コントロールであった。しかしながら、GG遺伝子型は、同様に低血漿インスリンレベルをケースおよびコントロールの両方において示し、遺伝子型の調節作用とインスリンレベルで両方の群において一致する。ケースが比較的強いインスリン感受性を示すため、それらの遺伝子型に関わらず、それらはインスリン感受性をFOXO以外に維持する複数のメカニズムを有することを推測する傾向がある。これは、大半の長寿遺伝子が適度または小さい効果サイズを有する仮説と一致した。小さいサンプル数が、ケースにおける相違点を検出する能力を制限することがまた可能である。一方では、突然変異をIRS−158またはIRS−223のいずれでもたらす長命のマウスは、実際インスリン抵抗性であるため、インスリン感受性は比較的長い寿命を与えることができるIIS経路における突然変異で、必要条件でない。
【0072】
しかしながら、シノラブディスエレガンスにおいて、単独で寿命に小さい影響を有することができるいくつかの遺伝子が、転写調節“マスター遺伝子”DAF−1659によって影響することに留意する関連性がある。さもなくば検出が困難な可能性があるFOXO3Aにおけるわずかな差異が、論理的にDNA結合、タンパク質間相互作用、細胞周期進行、アポトーシスおよび代謝に関するいくつかの下流遺伝子を修飾される。このように、FOXO3Aによる小さい改良する効果が、加齢表現型および寿命に、大きく付加的下流効果を潜在的に有する。
【0073】
裏づけになる証拠は、ヒトの加齢および寿命において、インスリンシグナルの役割が集積し始めたが、これらの効果を媒介する可能性がある遺伝子は知られていない。従来の研究は、長命のイタリア36、日本37,42、ドイツ60およびアシュケナージユダヤ人38の民族において、いくつかの加齢表現型と結びついて、インスリン−IGF−1シグナル経路から一塩基多型(SNPs)の過多または過少の発現を見出した。これらの発見のいくつかが、小さい効果サイズおよび重要でない統計的優位性によって制限される一方、Suh et al.38の研究は、同様にIGF−1受容体において機能的に重要な突然変異が、多少の長命のヒト、例えば100歳以上のヒトに存在することを示した。
【0074】
今日までに、ヒトにおけるFOXO遺伝子および加齢の表現型に関しては、ほとんど研究がされていない。二つの最近の研究は、FOXO遺伝子が更なる詳細な調査に値することを示唆する。第一に、年配のオランダ人の男性および女性の長期に渡る研究は、FOXO1Aハプロタイプが4年の生存率を予測し、FOXO3Aハプロタイプが、脳卒中の危険性を予測したことを明らかにした39。第二に、フラミンガム研究は、ゲノムの広範囲にわたる分析において、FOXO3A SNPが、女性において正常な閉経期で、年齢と強い関連性があることを見出した(p=0.00003)。しかしながら、オランダの調査結果は、多重比較を相当して、両方の研究が複製を必要とするとき、統計学的に有意でなかった。本研究は、FOXO3Aの関連性をヒトの寿命およびインスリン感受性に関して支持し、拡張する。
【0075】
現在の研究の主な利点の1つは、それがコホート内症例対照設計を使用することである。この調査設計は、ケースおよびコントロールを、進行中のコホート研究から長期に渡って収集したデータで選択する。従って、関連のあるいくつかの表現型(例えば、疾病の罹患、健康状態、機能)を、参加者がリコールバイアスし難いデータを作成する比較的若いときに、直接的な臨床検診によって得た。リコールバイアスは、調査結果が過去の事象を覚えることが困難なため正確でなく、高齢者にとっては大きな問題となる可能性がある。
【0076】
実際、比較的高齢の加齢を示唆する表現型の証拠を見出した特別な生存者の調査、例えば100歳以上の人は、重大なリコールバイアスを潜在的に被るおそれがある。即ち、高齢の参加者は、彼らの過去の病歴および彼らの過去の機能状態を、正確に思い出すことができない。しかしながら、現在の調査において、主な疾患は死亡率および罹病率の受託者に解決され、身体および認識機能の能力に基づく測定を自己評価の補充に使用され、証拠はこのような健康な加齢の表現型にて観測された。これは従来の遡及研究に潜在的な支持を与える。
【0077】
いくつかの他の長所が、この調査に存在する。第一に、分析で選択された候補遺伝子は、仮説に基づく基準に基づいて選択された。即ち、様々な方法を試用する加齢のモデル生物、特にノックアウトの研究は、IIS経路が加齢および寿命に重要であることを示した。多くの機能が、進化的に保存されていると考えられる。第二に、調査結果は強力で、著しく重要であり、いくつかの隣接したSNAPをFOXO3A遺伝子内に含有する。第三に、調査結果は生物学的に妥当で、従来の調査結果を老化動物モデルにて支持し、また制限された従来のヒトの研究をも支持する。第四に、寿命とケースコントロールの関係性は、コホート内症例対照分析を高度な事象の頻度(死亡率)で、検証の長期間を使用して検出した。第五に、HHPコホートは、高度に同種のコホートであり、研究の参加者において人口層化を全く検出されなかった。
【0078】
可能性のある欠点は、ケースおよびコントロールが、11歳の平均年齢の違いがあるため、誕生コホートを交絡因子として除くことができないことである。しかし、これは、参加者の間に最高19歳の違いが誕生年にあったため、疑わしい。同様に、部分分析は、ケースおよびコントロールにおいて、教育および職業(資料未記載)の違いが全く存在しないことを明らかにした。さらに、それは、95歳以上生存して、従って長寿の表現型を得る可能性のある基準にて年上である参加者であった。大抵のコホートの効果は、健康な利点を比較的若いコホートで示す。他の可能性のある欠点は、本研究を1つの集団のみで実施し、従って他の集団において、その総括を評価するために繰り返すべきである。
【0079】
要約すると、FOXO3A遺伝子内の一般的で、正常な遺伝子変異体は、ヒトの寿命と密接して関連したことを見出した。保護対立遺伝子の有病率が、年齢と共に著しく上昇した。長命のケースは、同様に癌および心血管疾患の低下した有病率、優れた自己報告の健康、高い機能状態を含有する健康的な加齢と関連のあるいくつかの付加的な表現型をおそらく保持し、それらが高いインスリン感受性のいくつかの生物学的マーカーの示唆を基礎調査にて示した。最後に、FOXO3A遺伝子内の特定の変異体は、同様にインスリン感受性、寿命の推定の中間表現型を含有するこれら加齢表現型のいくつかと関連した。
【0080】
B.対立遺伝子の患者(ヒトおよびヒト以外)の検出
多くの方法が、特定の対立遺伝子を、多型遺伝子座で検出するために利用できる。特異的な多型対立遺伝子の検出のための好適な方法は、多型の分子性質に部分的に依存するだろう。例えば、多型遺伝子座の様々な対立遺伝子形状は、DNAの一塩基対と異なる可能性がある。このような一塩基多型(またはSNP)は、全ての知られている多型の80%程を含有する遺伝子変異体の主な要因であり、ヒトゲノムにおけるそれらの濃度は平均1/1,000塩基対と推定する。SNPは最も頻繁な2対立遺伝子である―2つの形態のみで生じる(しかしDNAにて生じる4つの異なるヌクレオチド塩基に対応するSNPの最大4つの異なった形状が、理論的に可能である)。それにもかかわらず、SNPは他の多形体より突然変異的に安定していて、それらがマーカーおよび未知の変異体の間の連鎖不均衡を使用して、病原性突然変異体をマップする関連性の研究に適合できる。さらにSNPは二つの対立遺伝子のみを通常有するため、それらは長時間の測定よりはむしろ単純なプラス/マイナス分析によって遺伝子型を同定することができ、それらが自動的に従わせる。
【0081】
種々の方法が、特定の一塩基多型の対立遺伝子の存在を個々に検出するために利用可能である。本分野の促進は、正確、簡単および安価で大規模なSNP遺伝子型を同定で提供される。つい最近、例えば、いくつかの新技術は、ダイナミックアレルスペシフィックハイブリダイゼーション法(DASH)、ミクロプレートアレイ対角ゲル電気泳動(MADGE)、パイロシークエンス法、オリゴヌクレオチド特異的ライゲーション、TaqManシステム並びに様々なDNA「チップ」技術、例えばAffymetrix SNPチップを含有するものが記載された。これらの方法は、標的遺伝子領域の増幅を、通常PCRによって必要とする。さらに他の新しく開発された方法は、小さいシグナル分子の生成に基づく侵入切断(invasive cleavage)の後に、質量分析または固定されたパッドロックプローブおよびローリングサークル増幅によって小さいシグナル分子の生成に基づき、最終的にPCRの必要性を排除することがある。特異的な一塩基多型を検出する当業者に知られたいくつかの方法を以下に要約する。本発明の方法は、全ての利用可能な方法を包含することを理解される。
【0082】
いくつかの方法は、一塩基多型の分析法を容易にするため、開発された。一の実施形態において、一塩基多型を、例えばMundy,C.R.(米国特許第4,656,127号明細書)に開示されたような、特異的なエキソヌクレアーゼ抵抗性ヌクレオチドを使用して検出することができる。この方法によると、多型サイトの3’周辺の対立遺伝子に相補的なプライマーは、特定の動物またはヒトから得られた標的分子にハイブリッド形成することを許容する。標的分子での多型部位が、存在する特定のエキソヌクレアーゼ抵抗性ヌクレオチド誘導体に相補的であるヌクレオチドを含む場合、その誘導体をハイブリッド形成したプライマーの末端に組む込まれる。前記組み込みが、エキソヌクレアーゼに対して抵抗性のプライマーを与え、それによってそれの検出を可能にする。サンプルのエキソヌクレアーゼ抵抗性誘導体の同定が知られているため、プライマーはヌクレオチドが標的分子の多型部位に存在するエキソヌクレアーゼの曝露に抵抗性になった結果は、反応において使用されるヌクレオチド誘導体のものに相補的であった。この方法は、それが大量の無関係の配列データの測定を必要としない利点がある。
【0083】
本発明の他の実施形態において、溶液に基づく方法を、多型サイトのヌクレオチドの同一性の同定を決定するために使用する。Cohen, D. et al.(仏国特許発明第2,650,840号明細書;国際公開第91/02087号)。米国特許第4,656,127号明細書のMundyの方法のように、プライマーは多型サイトの3’周辺の対立遺伝子の配列に相補的であるものを用いた。この方法は、そのサイトのヌクレオチドの同一性を、ラベルされたジデオキシヌクレオチド誘導体を使用して決定し、多型のヌクレオチドに相補的な場合は、プライマーの末端に組み込まれる。
【0084】
遺伝子ビット分析(Genetic Bit Analysis)またはGBATMとして知られている代わりの方法が、Goelet,P et al.に記載されている(国際公開第92/15712号)。Goelet,P. et al.の方法は、ラベルを付けられたターミネータと、多型サイトの3’配列に相補的なおよびプライマーとの混合物を使用する。従って組み込まれてラベルを付けられたターミネータは、評価される標的分子の多型サイトに存在するヌクレオチドおよびその相補物によって決定される。Cohn et al.(仏国特許発明第2,650,840号明細書;国際公開第91/02087号)の方法とは対照的に、Goelet,P. et al.の方法は、好ましくは異質の相分析であり、プライマーまたは標的分子を固体相に固定する。
【0085】
近年では、DNAにおいて多型サイトの分析のためのいくつかのプライマーによって誘導されたヌクレオチドの組込方法を記載した(Komher, J. S. et al., Nucl. Acids. Res.17:7779−7784(1989); Sokolov, B. P., Nucl. Acids Res.18:3671(1990); Syvanen, A.−C, et al., Genomics8:684−692(1990); Kuppuswamy, M. N. et al., Proc. Natl. Acad. Sci. (U.S.A.)88:1143−1147(1991); Prezant, T. R. et al., Hum. Mutat. 1:159−164(1992); Ugozzoli, L. et al., GATA 9:107−112(1992); Nyren, P. et al., Anal. Biochem. 208:171−175(1993)。これらの方法は、これら全てがラベルされたデオキシヌクレオチドの組み合わせに依存して、多型サイトで塩基間を区別する点でGBATMと異なる。このようなフォーマットにおいて、シグナルは組み込まれたデオキシヌクレオチドの数に比例するため、同じヌクレオチドの流れを生じる多型は、流れの長さと比例するシグナルを生じることができる(Syvanen, A. −C, et al., Amer. J. Hum. Genet.52:46−59(1993))。
【0086】
タンパク質翻訳の早い終結を引き起こす突然変異体に関して、プロテイントランケーションテスト(PTT)は、効率的な診断方法を提供する(Roest, et. al., (1993) Hum. MoI Genet. 2:1719−21; van der Luijt, et. al., (1994) Genomics 20:1 −4)。PTTに関して、RNAは利用可能な組織から最初に分離、逆転写され、関連する断片をPCRによって増幅された。その後、逆転写PCRの生成物を、ネステッドPCR増幅により、RNAポリメラーゼプロモーターを含有するプライマーおよび真核生物の翻訳を開始する配列と共に使用された。関連領域の増幅後、プライマー内に組み込まれた固有のモチーフは、一連の生体外におけるPCR産物の転写および翻訳を可能にする。翻訳の生産物のドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動で、短縮型ポリペプチドの出現は、翻訳の早い終結の原因となる突然変異の存在を示す。この技術の多様化において、DNA(RNAに対して)を、関連のある標的領域が単一のエクソンに由来するとき、PCRの鋳型として使用する。
【0087】
あらゆる細胞種または組織を利用して、本明細書に記載の診断における使用で核酸サンプルを得ることができる。好ましい実施形態において、DNAサンプルを、既知の技術(例えば、静脈穿刺)によって得られる体液、例えば血液または唾液から得る。選択的に、核酸試験を乾式サンプルで実施することができる(例えば、髪の毛または皮膚)。RNAまたはタンパク質を使用するとき、用いることができる細胞または組織は、FOXO3A遺伝子を発現する必要がある。
【0088】
診断法をまた、生検または切除から得られる患者の組織の(固定および/または凍結されている)組織断片で、核酸精製が全く必要とすることないようにその場にて直接実施することができる。核酸試薬を、プローブおよび/またはプライマーとして、前記その場の方法で使用することができる(例えば、Nuovo, G. J., 1992, PCR in situ hybridization: protocols and applications, Raven Press, N.Y.参照)。
【0089】
一つの核酸配列の検出で主に焦点を合わせる方法に加えて、プロフィールをまたこのような検出方式において評価することができる。指紋プロフィールを、例えばディファレンシャルディスプレイ法、ノーザン分析および/またはRT−PCRを利用することによって生成することができる。
【0090】
好適な検出方法は、FOXO3Aハプロタイプの少なくとも一つの対立遺伝子の重なり合った領域のプローブを使用し、約5、10、20、25または30ヌクレオチドを突然変異または多型の領域付近にて有する対立遺伝子特異的なハイブリダイゼーション形成である。本発明の好ましい実施形態において、他の対立遺伝子変異体に特異的にハイブリッド形成することができるいくつかのプローブが、固相担体、例えば「チップ」(最高約250,000オリゴヌクレオチドを適用することができる)に付着される。オリゴヌクレオチドは、リソグラフィーを含む様々な工程によって固体支持体に結合することができる。オリゴヌクレオチドを含有するこれらのチップを用いる突然変異検出分析は、また「DNAプローブアレイ」といい、例えばin Cronin et al.(1996) Human Mutation7:244に記載される。一実施形態において、チップは遺伝子の多型領域の少なくとも1つの、全ての対立遺伝子変異体を含有する。その後固相担体を、テスト核酸と接触させ、特定のプローブに対するハイブリッド形成を検出する。従って、一つ以上の遺伝子の多くの対立遺伝子の変異体の同定は、単純なハイブリッド実験で同定することができる。
【0091】
これらの技術は、また核酸の増幅する工程を分析前に含有することができる。増幅技術は、当業者に知られ、クローニング、ポリメラーゼ連鎖反応(PCR)、対立遺伝子特異的増幅法(ASA)、リガーゼ連鎖反応(LCR)、自己持続型の核反応(Guatelli, J. C. et al., 1990, Proc. Natl. Acad. Sci. USA 87:1874−1878)、転写増幅システム(Kwoh, D. Y. et al., 1989, Proc. Natl. Acad. Sci. USA 86:1173−1177)、およびQβレプリカーゼを含有するが、これらに制限されない。
【0092】
増幅産物は様々な方法によって検出することができ、これはサイズ分析、サイズ分析後の制限分解作用による切断、反応生成物の特異的タグを付けられたオリゴヌクレオチドプライマーの検出、対立遺伝子の特異的オリゴヌクレオチド(ASO)ハイブリッド形成、対立遺伝子の特異的5’エキソヌクレアーゼ検出、塩基配列決定、ハイブリッド形成等を含有する。
【0093】
PCRに基づく検出手段は、同時に複数の標識の多重増幅を含有することができる。例えば、サイズにおいて重なり合わず、同時に分析することができるPCR産物を生成するPCRプライマーを選択することは、よく知られた技術である。選択的に、特異的にラベルして、従ってそれぞれ特異的に検出することができるプライマーと、異なった標識を増幅することが可能である。当然、ハイブリッド形成に基づく検出方法は、複数のPCR産物の異なった検出をサンプルにおいて可能とする。他の技術では、複数の標識の多数の分析を可能にする従来技術で知られている。
【0094】
単に例証となる実施形態において、前記方法は、(i)細胞サンプルを患者(唾液、頬の内側の粘膜、血液、その他の体液または成分)から収集する工程と、(ii)核酸(ゲノム、mRNAまたはその両方)をサンプルの細胞から分離する工程と、(iii)核酸サンプルと、FOXO3Aハプロタイプの少なくとも1つの対立遺伝子の5’および3’に、対立遺伝子のハイブリッド形成および増幅を生じるような条件下において、特異的にハイブリッド形成する一つ以上のプライマーを接触させる工程と、(iv)増幅産物を検出する工程とを含有する。これらの検出方法は、このような分子が比較的低い数で存在する場合、核酸分子の検出に特に実用的である。
【0095】
対象となる分析の好ましい実施形態において、FOXO3Aハプロタイプの対立遺伝子を、制限酵素切断パターンにおける変化によって同定する。例えば、サンプルおよびコントロールのDNAを分離し、(任意に)増幅し、1つ以上の制限エンドヌクレアーゼで切断し、断片の長さのサイズを、ゲル電気泳動によって決定する。
【0096】
さらにもう一つの実施形態において、従来技術で知られる様々な塩基配列決定反応のいずれかを、対立遺伝子配列に直接使用することができる。例示的な塩基配列決定反応は、Maxim and Gilbert ((1977) Proc. Natl Acad Sci USA74:560) or Sanger (Sanger et al (1977) Proc. Nat. Acad. Sci USA 74:5463)によって開発された技術に基づくものを含有する。自動塩基配列決定の様々な方法のいずれかは、質量分析による塩基配列決定を含有する(例えば、国際公開第94/16101号参考;Cohen et al.(1996)Adv Chromatogr36:127−162;およびGriffm et al.(1993)Appl Biochem Biotechnol38:147−159)対象分析(例えば、Biotechniques(1995)19:448参考)を実行するときに、使用することができることを、同様に予測される。特定の実施形態で、1、2または3の核酸塩基のみの発生を、配列反応において決定擦る必要があることが、当業者に明らかであるだろう。例えば、Aトラック等を、1つの核酸のみを検出する場合に、実施することができる。
【0097】
更なる実施形態において、切断剤からの保護を使用して、RNA/RNA、RNA/DNAまたはDNA/DNAのヘテロ2本鎖に基づくミスマッチ塩基を検出することができる(Myers, et al.(1985)Science230:1242)。一般的に、従来技術の「ミスマッチ切断」は、野生型対立遺伝子をサンプルに含む(ラベルされた)RNAまたはDNAにハイブリッド形成することによって、ヘテロ二本鎖の形成を提供することによって開始する。二本鎖デュプレックスを、例えばコントロールおよびサンプル鎖の間の塩基対ミスマッチのために存在するデュプレックスの一本鎖領域を切断する薬剤で処理する。例えば、RNA/DNAデュプレックスを、SIヌクレアーゼで処理してミスマッチ領域を酵素で切断するRNaseおよびDNA/DNAハイブリッドで処理することができる。他の実施形態において、DNA/DNAまたはRNA/DNAデュプレックスのいずれかを、ヒドロキシルアミンまたは四酸化オスミウムおよびピペリジンで処理して、ミスマッチ領域を切断することができる。その後ミスマッチ領域の切断後、結果として生じる物質を、サイズによって変性ポリアクリルアミドゲルにて分離し、突然変異サイトを断定する。例えば、Cotton et al(1988)Proc. Natl Acad Sci USA85:4397;およびSaleeba et al (1992) Methods Enzymol.217:286−295参考。好ましい実施形態において、コントロールDNAまたはRNAを、検出のためにラベルすることができる。
【0098】
さらに別の実施形態において、ミスマッチ切断反応は、ミスマッチ塩基対を二重鎖DNAにて認識する1つ以上のタンパク質(「DNAミスマッチ修復」酵素と称す)を使用する。例えば、大腸菌のmutY酵素は、AをG/Aミスマッチで切断し、HeLa細胞からチミジンDNAグリコシラーゼはTをG/Tミスマッチで切断する(Hsu et al.(1994)Carcinogenesis 15:1657−1662)。例示的実施形態の記載のように、FOXO3A遺伝子座ハプロタイプの対立遺伝子に基づくプローブは、テスト細胞からcDNAまたは他のDNA産物にハイブリッドを形成する。デュプレックスを、DNAミスマッチ修復酵素で処理し、切断生成物は、必要である場合、それを電気泳動プロトコール等から検出することができる。例えば、米国特許第5,459,039号明細書を参照とし、本明細書において、完全に引用したものとする。
【0099】
他の実施形態において、電気泳動移動度の違いを使用して、FOXO3A遺伝子座の対立遺伝子を同定するだろう。例えば、一本鎖高次構造多型(SSCP)を使用して、電気泳動移動度における、突然変異体および野生型核酸の間の相違を検出することができる(Orita et al.(1989)Proc Natl. Acad. Sci USA86:2766、またCotton(1993)Mutat Res285:125−144参考; and Hayashi(1992)Genet Anal Tech Appl9:73−79)。サンプルおよびコントロールFOXO3A遺伝子座の対立遺伝子の一本鎖DNA断片を、変性し、復元を可能にした。一本鎖核酸の2次構造は、配列により異なり、電気泳動移動度における結果として生じる変性は、一塩基の変化さえ検出することが可能である。DNA断片を、ラベルされたプローブで、ラベルするか検出することができる。分析の感度はRNAを使用することによって向上することができ、2次構造が、配列の変異に比較的感度がある。好ましい実施形態において、対象となる方法は、ヘテロ二本鎖分析を利用して、電気泳動移動度の変化に基づいて、二重鎖へテロ二本鎖分子を分離する(Keen et al.(1991)Trends Genet7:5)。
【0100】
さらにもう一つの実施形態において、変性剤の濃度勾配を含むポリアクリルアミドゲルにおける対立遺伝子の変動を、変性剤濃度勾配ゲル電気泳動法(DGGE)を使用して分析した(Myers et al.(1985)Nature313:495)。DGGEを分析方法として使用するとき、DNAはそれが例えばPCRで高い融解のGCに富んだDNAの約40bpのGCクランプを添加することによって完全に変性しないことを確実にするために修飾するだろう。更なる実施形態において、温度勾配を、変性剤の勾配の代わりに使用して、対象およびサンプルのDNAの移動度における相違を同定することができる(Rosenbaum and Reissner(1987)Biophys Chem265:12753)。
【0101】
対立遺伝子の検出する他の技術の例は、選択的なオリゴヌクレオチドハイブリッド形成、選択的な増幅または選択的なプライマー伸長を含有するが、これらに制限されない。例えば、オリゴヌクレオチドプライマーを、既知の突然変異またはヌクレオチド(例えば、対立遺伝子変異体)の相違を中央に配置し、その後、完全に一致を発見するときのみにハイブリッド形成を可能にする状態で標的DNAにハイブリッド形成することで調整することができる(Saiki et al. (1986) Nature 324:163); Saiki et al (1989) Proc. Natl Acad. Sci USA 86:6230)。このような対立遺伝子に特異的なオリゴヌクレオチドのハイブリッド形成技術を使用して、オリゴヌクレオチドが、PCR増幅された標的DNAにハイブリッド形成したとき1つの突然変異または多型態領域について1つの反応を、またはオリゴヌクレオチドが、ハイブリッド膜に付着し、ラベルされた標的DNAにハイブリッド形成したとき数々の異なった突然変異または多型体の領域を試験する。
【0102】
選択的に、対立遺伝子特異的な増幅技術は、選択的なPCR増幅に依存し、本発明と共に使用することができる。特異的な増幅のために使用されるオリゴヌクオチドは、(増幅が異なったハイブリッド形成に依存するために)分子の中心における関連のある突然変異または多型領域(Gibbs et al (1989) Nucleic Acids Res.17:2437−2448)、または適切な条件に基づいて、ミスマッチがポリメラーゼの伸長を防止または減少することができる場合に1つのプライマーの3’の最終末端で担持することができる(Prossner(1993)Tibtech11:238)。さらに、新規な制限酵素サイトを突然変異の領域に導入して切断に基づく検出を生成することが望ましい(Gasparini et al(1992)Mol. Cell Probes6:1)。特定の実施形態において、増幅が同様に増幅でTaqリガーゼを使用して実施することができることが予想される(Barany(1991)Proc. Natl. Acad. Sci USA88:189)。そのような場合、ライゲーションは、完全な一致が、5’配列の3’末端に存在し、増幅の有無を調べることによって、特定のサイトにて既知の突然変異の存在を検出することが可能となる場合のみに生じる。
【0103】
他の実施形態において、対立遺伝子変異体の同定を、例えば米国特許第4,998,617号明細書およびLandegren, U. et al.((1988) Science 241 :1077−1080)に記載されたように、オリゴヌクレオチドのライゲーション分析(OLA)を使用して実施する。OLAプロトコールは、標的の一本鎖の接触配列にハイブリッド形成することができるように設計される2つのオリゴヌクレオチドを使用する。オリゴヌクレオチドの1つは、分離標識、例えばビオチン化で結合し、他は検出が可能なようにラベルされる。正確な相補的配列が、標的分子に見出す場合、オリゴヌクレオチドが、それらの末端が隣接するようにハイブリッドを形成し、ライゲーション基質を生成する。その後ライゲーションは、ラベルされたオリゴヌクレオチドがアビジンまたは他のビオチンリガンドを使用して回復されることを可能にする。Nickerson, D. A. et al.は、PCRおよびOLAの性質を組み合わせる核酸検出分析を記載する(Nickerson, D. A. et al.(1990)Proc. Natl. Acad. Sci. USA87:8923−27)。この方法において、PCRを使用して、標的DNAの指数関数的増幅を達成し、その後OLAを用いて検出される。
【0104】
このOLA方法に基づくいくつかの技術が開発され、FOXO3a遺伝子座ハプロタイプの対立遺伝子を検出するために使用することができる。例えば、米国特許第5,593,826号明細書は、3’アミノ基を有するオリゴヌクレオチドおよび5’リン酸化オリゴヌクレオチドを使用して、アミド亜リン酸エステル結合を有する結合体を形成するOLAを開示する。Tobe et al.((1996)Nucleic Acids Res24:3728)に記載のOLAの他の変異体において、PCRと組み合わせたOLAは、2つの対立遺伝子の分類化を、一つのマイクロタイターウェルにて可能にする。それぞれの対立遺伝子を固有のハプテン、即ちジゴキシゲニンおよびフルオレセインでマークすることによって、それぞれのOLA反応を、異なった酵素レポーター、アルカリホスファターゼまたはセイヨウワサビペルオキシダーゼでラベルされたハプテン特異的な抗体を使用することによって検出することができる。このシステムは、2つの異なる色の製造を導くハイスループットフォーマットを使用する2つの対立遺伝子の検出を可能にする。
【0105】
本発明の他の実施形態は、近い将来における長命の可能性、または健康または診断の介入の必要性を検出するためのキットを対象とする。このキットは、FOXO3A遺伝子座のハプロタイプの少なくとも1つの対立遺伝子に5’および3’をハイブリッド形成する5’および3’オリゴヌクレオチドを含む、一つ以上のオリゴヌクレオチドを含有することができる。PCR増幅オリゴヌクレオチドは、その後の分析で適切なサイズのPCR産物を生成するために、25から2500の独自の塩基対の間、好ましくは約100から500の独自の塩基とハイブリッドを形成する必要がある。
【0106】
特に好ましいプライマーは、SEQ ID NO.2〜9に記載のヌクレオチド配列を含有する。これら遺伝子においてヒトの多型の検出のために適切なプライマーを、従来技術において既知であるこの配列情報および通常の技術を使用して、プライマー配列の設計および最適化のために容易に設計することができる。上記プライマー配列の最適な設計を、例えば、市販のプライマー選択プログラム、例えばPrimer2.1、Primer3またはGeneFisherの使用によって達成することができる。
【0107】
「GCCハプロタイプ」の検出のための簡単な方法の例は、関連のある特異的なヌクレオチドを増幅し、段落[0081]に記載のものと同様に対立遺伝子特異的なプライマーの使用を含む。この方法は、オリゴヌクレオチドプライマーが、それらの3’末端にて完全にアニールして、DNAポリメラーゼがこれらプライマーをPCR時に伸長する必要がある事実を利用する。特異的なDNAの点相違のみと一致するオリゴヌクレオチドプライマーを設計することによって、rs2802292多型にて見られるようなもの−T種対立遺伝子に結合しないプライマー−このようなプライマーは多型体対立遺伝子と区別することができる。コントロール反応を(ARMS:amplification refractory mutation system)のような同じチューブ内で設定し、所定のサンプルからの生成物の生成の欠如が、分析を確かめる「G」変異体の欠損よりむしろ、PCR反応の失敗によるように単純でないことを確実にすることが必要である。
【0108】
このために使用されるオリゴヌクレオチドは、フォワードアウター(「rs2802292_FO」)で、SEQ ID NO.5として同定される5’―GAAACTGAGGCTAACAGCTGGGTCTGGCCC―3’、リバースアウター(「rs2802292_RO」)で、SEQ ID NO.6として同定される5’―AGCTGATGCTCCTCAACGAAACCACCTTAC―3’、リバースG特異的(「rs2802292_RG」)で、SEQ ID NO.7として同定される5’―GGACCCCTTCATCTGTCACACAGAGGCTCC―3’、フォワード特異的(「rs2802292_FT」)で、SEQ ID NO.8として同定される5’―CTGTTGCTCACAAGAGCTCAGGGCTGGGcT―3’を含み、最後の2つのプライマーにおいて、下線の引かれた末端塩基は、G−Tの相違のサイトでアニールし、3’末端から2番目のbp(小文字)は、対立遺伝子の特異性を最大限にするために意図的にミスマッチされる。この例証における4つのプライマーを、表No.9に記載する。
【0109】
【表11】
表9は、出現の順において、それぞれSEQ ID NOS5〜8を開示する。
【0110】
プライマーおよびG/T変異体を示すPCR産物のDNA配列(ソースGenbank AL391646.12)は、以下の通りである:
【0111】
【0112】
このように発生した増幅物をアガロースゲルで分離したとき、Gタイププライマーは186bp生成物を発生することを示すことができ、Tタイププライマーは132bp生成物を生じる。アウトサイドプライマーは、反応が正確に進んだことを保証するために、全ての反応で存在すべき288bp生成物を生成する。
【0113】
例示的な、増幅の試薬および条件を、表10に示す。
【0114】
【表12】
【0115】
増幅された断片は、図1に示すような3%アガロースゲルに分離することができる。図1はARMS−PCR定量分析の結果を示し、上記のプライマーおよび条件を使用してFOXO3A G/T変異体を検出する。レーン1は「T」対立遺伝子(132bp)の対象となる同型を示す;レーン2および3は、「G」対立遺伝子(186bp)に同型である対象を示す;レーン4および5は、「T」および「G」の対立遺伝子(132+186bp)に異型である対象を示し、Mは100bpDNAラダー(Invitrogen, Paisley, United Kingdom)である。
【0116】
要約すると、図2は分析の模式的な概略を示す。プライマー「rs2802292_FO」および「rs2802292_RO」は、多型体遺伝子座rs2802292の隣接し、コントロール288bpバンドを全ての場合にて発生する。プライマー「rs2802292_OF」および「rs2802292_RG」は、186bpのG特異的生成物を発生し、プライマー「rs2802292_FT」および「rs2802292_OR」は、132bpのT特異的生成物を発生する。
【0117】
キットの使用のために、オリゴヌクレオチドは様々な天然および/または合成の組成物のいずれか、例えば合成オリゴヌクレオチド、制限断片、cDNA、合成ペプチド核酸(PNA)等であることができる。分析キットおよび方法は、同様にラベルされたオリゴヌクレオチドを使用して、分析において同定を容易にすることを可能にすることができる。用いることができるラベルの例は、放射性同位元素識別、酵素、蛍光化合物、ストレプトアビジン、アビジン、ビオチン、磁性部位、金属結合部位、抗原または抗体の部位等を含有する。
【0118】
キットは、任意に、またDNAサンプリング方法を含有することができる。DNAサンプリング方法は当業者によく知られる一つであり、基質、例えば濾紙、AmpliCardTM(University of Sheffield, Sheffield, England SlO 2 J F; Tarlow, J W, et al, J of Invest. Dermatol.103:387−389(1994))等;DNA精製試薬、例えばNucleonTMキット、リシスバッファー、タンパク質分解酵素溶液等;PCR試薬、例えば反応バッファー、熱安定性ポリメラーゼ、dNTP等;および対立遺伝子検出方法、例えばHinfI制限酵素、対立遺伝子特異的オリゴヌクレオチド、乾燥血痕からのネステッドPCRのための縮重オリゴヌクレオチドプライマーを含有するが、これらに限定されない。
【0119】
C.薬理ゲノミクス
特定の対立遺伝子の知識は、特定の疾患または状態の進行する感受性と関連し、特定の疾患または状態に関与する他の遺伝子異常に関する情報と単独または組み合わせては、「薬理ゲノミクス」の目標である個々の遺伝子プロフィールに従う予防または治療のカスタム化を可能とする。従って、健康的な加齢で個々のFOXO3Aプロフィールと集団的プロフィールの比較は、特定の患者または患者集団で安全且つ効果的であると期待する薬物もしくは他の治療療法の選択または設計を可能とする(即ち、同じ遺伝子の変性を有する患者の一群)。
【0120】
本発明に記載の特定の対立遺伝子の知識を使用して、細胞の性質の違いを細胞培養および組織システムにて観測し、細胞または組織培養システムに添加された化学的または生物学的薬剤への細胞の反応を測定することができる。細胞性質および反応の相違点を遺伝子型間で比較して、健康を改善または寿命を延ばす要求のため実施することができる薬物または他の薬理的物質を同定するか、または遺伝子もしくは遺伝子発現で毒性または潜在的な効果で新規化合物を試験することができる。
【0121】
さらに標的集団に対する可能性は、最も高い臨床利点を示すことを期待して、遺伝子プロフィールに基づいて:1)既にマークされた薬物の再配置;2)臨床開発が安全性または効率性の制限の結果として中断され、患者のサブグループに特有である候補薬の救済;および3)候補治療および比較的最適な薬物のラベリングで、急速な、比較的費用が高くない開発を可能とすることができる(すなわち、薬剤の様々な服用量の効果を、原因となる突然変異で測定することは、有効量の最適化に実用的である)。
【0122】
特定の治療薬で個々の治療は、長寿と関連のある遺伝子の発現レベルを測定することによって観測することができる。発現レベルを、タンパク質(例えば、FOXO3A)、mRNAおよび/または転写レベルを同定することによって、測定することができる。その後検出されるレベルに依存して、治療療法を、維持または調整することができる(増幅量を増加または減少)。好ましい実施形態において、対象を薬剤で治療する効果は以下の工程を含有する:(i)薬剤を投与する前に前投与サンプルを対象から得る;(ii)タンパク質、mRNAまたはゲノムDNAのレベルまたは量を前投与サンプルにおいて検出する;(iii)治療薬の投与後、1つ以上の投与後のサンプルを対象から得る;(iv)タンパク質、mRNAまたはゲノムDNAの発現または活性のレベルを、投与後のサンプルにおいて検出する;(v)前投与サンプルにおけるタンパク質、mRNAまたはゲノムDNAの発現または活性のレベルと、投与後のサンプルにおける対応するタンパク質、mRNAまたはゲノムDNAを、それぞれ比較する;(iv)薬剤の投与を対象に応じて変更する。
【0123】
対象の細胞をまた、FOXO3A遺伝子以外の遺伝子発現のレベルを検出するため治療的の投与前後で得て、治療が有害である可能性がある遺伝子の発現を増加または減少しないことを確認することができる。これは、例えば転写プロファイルの方法を使用して実施することができる。従って、生体内において治療に露出された細胞からのmRNA、治療に露出されていない同じ種類の細胞のmRNAは、逆転写され、多くの遺伝子からのDNAを含むチップにハイブリッド形成することができ、この結果、治療で処理または未処理の細胞の遺伝子の発現を比較する。
【0124】
「GCC」ハプロタイプを、危険率の算出において、死亡および年齢に関連する疾患(心臓病、脳卒中、癌、COPDまたはその他慢性の肺疾患、パーキンソン病、糖尿病および認知症)および将来的な身体機能(歩行能力、認識機能)の予測の補助に使用することができる。この情報は、市民、医師、健康管理会社および保険会社にとって関心があるものである。既知の危険性算出の例は、Perls、米国特許出願公開第2007/0118398A1号明細書、2007年5月24日に公開されたシステムおよび方法を含有し、本明細書において完全に引用する。危険率の算出を、例えば医師局にて、携帯端末またはオンラインにて提供することができる。人がどれくらい生存するかを予想するのに興味がある個人、健康介護専門家、保険会社、健康管理組織は、コンピューターに男/女の遺伝子型を入力して、老化関連の疾患の危険性のスコア、健康的な残余寿命の年数および残余寿命の合計年数を得ることができる。
【0125】
特定のスコアに基づいて、医師または保険専門家は、上記疾患および死亡、特にFOXO3A遺伝子の比較的保護的でない変異を有するヒトで、健康的な生活または危険性の減少に関して、患者に知らせることができる。いくつかの例示的な選択肢は、食物の選択(例えば、赤ワイン、大豆製品およびFOXO3A遺伝子の活性に影響を及ぼすことができる化合物を含む他の食品)、または強い危険性要因の改善、例えば体重減少または増加の身体活性に関するアドバイスを含む。
【0126】
FOXO3Aおよび特にGCCハプロタイプの健康な老化および長寿の予測手段の同定は、実用的な生物学および標的の有望な情報源を、製薬のスクリーンおよび試験に提供する。例えば、一つは、FOXO3A遺伝子によって年齢関連疾患を減少で予期される健康的利益のために産出される遺伝子産物またはタンパク質もしくは他の活性化合物を利用することができる。遺伝子産物を利用する方法は、製薬技術において知られている摂取、注射、経皮的投与および他の方法を含有する。化合物をスクリーンして、FOXO3A、特にGCCハプロタイプによって産出される遺伝子産物の種類、活性または量に影響するものを見つけることができる。
【0127】
本発明は、年齢関連性疾患の予防または治療するFOXO3Aの調製方法を含有する。本発明はまた、FOXO3Aが例えば年齢に関連のある疾患または対象において寿命の向上と関連する疾患または状態を治療または予防する方法を含有する。本明細書において使用される「対象」とは、ヒトまたはヒトでない動物とする。「ヒトでない動物」という用語は、哺乳類、人間以外の霊長類(特に比較的高度な霊長類)のような全ての脊髄動物、ウマ、ウシ、バイソン、バッファロ、ヤギ、ブタ、ヒツジ、ニワトリ、カモおよびガチョウのような飼育哺乳類、イヌ、ネコ、ウサギ、モルモットのようなペット、齧歯動物、爬虫類および実験動物を含有する。好ましい実施形態において、対象はヒトである。他の実施形態において、対象は、疾患モデルとして適切な実験動物または遺伝形質転換動物である。調整および治療の方法は、Geesaman et al.の2007年5月10日に公開された米国特許出願公開第2007/0105109Al号明細書において当業者に知られ、完全に本明細書に引用する。
【0128】
本発明によって教示した対立遺伝子の変異体の配列または遺伝子産物の、多くの他の診断および治療への使用は、当業者に明らかであるだろう。小分子スクリーン、アンチセンスオリゴヌクレオチド、低分子二本鎖RNA(siRNA)の使用を含有するいくつかの例が、当業者に明らかである。FOXO活性に関する診断および治療の使用を開発するいくつかの方法は、Goldberg et al.の2006年3月30日に公開された米国特許出願公開第2006/0069049 A1号明細書に記載され、関連する経路にはTissenbaum et al.の2006年11月30日に公開された米国特許出願公開第2006/0272039号明細書に関係があり、共に完全に本明細書に引用する。
【0129】
前述の明細書を、当業者が本発明を実施することを可能にするため、充分であると考慮する。当業者は、単に通常の方法を用いて、多くの本発明の特定の実施形態と均等物を理解するか、または同定することができる。これら均等物は、添付の特許請求項に含有することを意図する。実際に、本明細書に示しまたは記載されるものに加えて本発明の様々な修正は、前述の記載から当業者に明らかになり、添付の特許請求の範囲に含まれる。
【0130】
本文に記載されていない参考文献
1. Herskind AM et al. (1996). The heritability of human longevity: a population−based study of 2872 Danish twin pairs born 1870−1900. Hum Genet 97:319−323.
2. Yashin AI, Iachine IA, Harris JR (1999) Half of variation in susceptibility to mortality is genetic: findings from Swedish twin survival data. Behav Genet 29:11−19.
3. Christensen K, Johnson TE, Vaupel JW (2006) The quest for genetic determinants of human longevity: challenges and insights. Nat Rev Genet 7:436−448.
4. Willcox DC, Willcox BJ, Hsueh WC, Suzuki M (2006) Genetic determinants of exceptional human longevity: insights from the Okinawa Centenarian Study. AGE 28:313−332.
5. Bishop NA, Guarente L (2007) Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nat Rev Genet 8:835−844.
6. Risch N, Zhang H (1995) Extreme discordant sib pairs for mapping quantitative trait loci in humans. Science 268: 1584−1589.
7. Gundmundsson H et al. (2000) Inheritance of human longevity in Iceland. Eur J Hum Genet 8:743−749.
8. Kerber RA, O´Brien E, Smith KR, Cawthon RM (2001) Familial excess longevity in Utah genealogies. J Gerontol A Biol Sci MedSci 56:B130−B139.
9. Perls TT et al. (2002) Life− long sustained mortality advantage of siblings of centenarians. Proc Natl Acad Sci USA 99:8442−8447.
10. Willcox BJ, Willcox DC, He Q, Curb JD, Suzuki M (2006) Siblings of Okinawan centenarians exhibit lifelong mortality advantages. J Gerontol A Biol Med Sci 61 :345− 354.
11. Partridge L (2007) Some highlights of research on aging with invertebrates, 2006−2007. Aging Cell 6:595−598.
12. Ghazi A, Henis−Korenblit S, Kenyon C (2007) Regulation of Caenorhabditis elegans lifespan by a proteasomal E3 ligase complex. Proc Natl Acad Sci USA 104:5947−5952.
13. Bartke A (2008) New findings in gene knockout, mutant and transgenic mice. Exp Gerontol 43:11 −14.
14. Lin K, Dorman JB, Rodan A, Kenyon C (1997) daf−16: An HNF−3/forkhead family member that can function to double the life−span of Caenorhabditis elegans. Science
278:1319−1322.
15. Brunet A et al. (2004) Stress−dependent regulation of FOXO transcription factors by the SIRTl deacetylase. Science 303:2011−2015.
16. Kenyon C, Murphy CT (2006) Enrichment of regulatory motifs upstream of predicted DAF−16 targets. Nat Genet 38:397−398.
17. van der Horst A, Burgering BM (2007) Stressing the role of FOXO proteins in lifespan and disease. Nat Rev MoI Cell Biol 8:440−450.
18. Russell SJ, Kahn CR (2007) Endocrine regulation of ageing. Nat Rev MoI Cell Biol 8:681−691.
19. Sauve AA, Wolberger C, Schramm VL, Boeke JD (2006) The biochemistry of Sirtuins. Ann Rev Biochem 75:435−465.
20. Tissenbaum HA, Guarente L (2001) Increased dosage of a sir −2 gene extends lifespan in Caenorhabditis elegans. Nature 410:227−230.
21. Blvher M, Kahn BB, Kahn CR (2003) Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 299:572−574.
22. Bartke A (2008) Impact of reduced insulin−like growth factor− 1 /insulin signaling on aging in mammals: novel findings. Aging Cell 7:285−290.
23. White MF (2003) Insulin signaling in health and disease. Science 302:1710−1711.24. Conover CA, Bale LK (2007) Loss of pregnancy −associated plasma protein A extends lifespan in mice. Aging Cell 6:727−729.
25. Curran SP, Ruvkun G (2007) Lifespan regulation by evolutionarily conserved genes essential for viability. PLoS Genet 3:e56.
26. Greer EL et al. (2007) An AMPK−FOXO pathway mediates the extension of lifespan induced by a novel method of dietary restriction in C. elegans. Curr Biol 17:1646− 1656.
27. Berdichevsky A, Viswanathan M, Horvitz HR, Guarente L (2006) C. elegans SIR −2.1 interacts with 14−3−3 proteins to activate DAF−16 and extend life span. Cell 125:1165− 1177.
28. Boehm M, Slack F (2005) A developmental timing MicroRNA and its target regulate life span in C. elegans. Science 310:1954−1957.
29. Hsu AL, Murphy CT, Kenyon C (2003) Regulation of aging and age−related disease by DAF−16 and heat−shock factor. Science 300:1142−1145.
30. Murphy CT (2006) The search for DAF−16/FOXO transcriptional targets: approaches and discoveries. Exp Gerontol 41 :910−921.
31. Wang MC, Bohmann D, Jasper H (2005) JNK extends lifespan and limits growth by
antagonizing cellular and organism −wide responses to insulin signaling. Cell 121 :115− 125.
32. Giannakou ME et al. (2007) Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging Cell 6:429438.
33. Harman D (1956) Aging: a theory based on free radical and radiation chemistry. J Gerontol 11 :298−300.
34. Beckman KB, Ames BN (1998) The free radical theory of aging matures. Physiol Rev 78:547−581.
35. Fusco D, Colloca G, Lo Monaco MR, Cesari M (2007) Effects of antioxidant supplementation on the aging process. Clin Interv Aging 2:377−387.
36. Bonafe M et al. (2003) Polymorphic variants of IGF−I receptor and phosphoinositide 3− kinase genes affect IGF−I plasma levels and human longevity: cues for an evolutionarily conserved mechanism of life span control. J Clin Endocrinol Metab 88:3299−3304.
37. Kojima et al. (2004) Association analysis between longevity in the Japanese population and polymorphic variants of genes involved in insulin and insulin−like growth factor 1 signaling pathways. Exp Gerontol 39:1595−1598.
38. Suh Y et al. (2008) Functionally significant insulin−like growth factor I receptor mutations in centenarians. Proc Natl Acad Sci USA 105:3438−3439.
39. Kuningas M et al. (2007) Haplotypes in the human FOXOIa and FOXO3a genes: impact on disease and mortality at old age. Eur J Hum Genet 15:294−301.
40. Lunetta KL et al. (2007) Genetic correlates of longevity and selected age−related phenotypes: a genome −wide association study in the Framingham Study. BMC Med Genet 8 Supple 1 : S13.
41. Nebel A et al. (2005) No association between microsomal triglyceride transfer protein (MTP) haplotype and longevity in humans. Proc Natl Acad Sci USA 102:7906−7909.
42. Arai Y et al. (2001) Serum insulin like growth factor 1 (IGF−I) in centenarians: implication of IGF− 1 as a turnover protein. J Gerontol A Biol Sci Med Sci 56A:M79−M82.
43. Teng EL et al. (1994) The Cognitive Abilities Screening Instrument (CASI): a practical test for cross−cultural epidemiological studies of dementia. Int Psychogeriatr 6:45−58.
44. Willcox BJ et al. (2006) Midlife risk factors and survival in men. JAMA 296:2343−2350.
45. Kramarow E, Lubitz J, Lentzner H, Gorina Y (2007) Trends in the health of older Americans, 1970−2005. Health AfJ (Millwood) 26:1417−1425.
46. Evert J, Lawler E, Bogan H, Perls T (2003) Morbidity profiles of centenarians: survivors, delayers, and escapers. J Gerontol A Biol Sci Med Sci 58:232−237.
47. Bernstein AM et al. (2004) First autopsy study of an Okinawan centenarian: absence of many age−related diseases. J Gerontol A Biol Sci Med Sci 59:1195− 1199.
48. Willcox DC et al. (2007) Aging gracefully: a retrospective analysis of functional status in Okinawan centenarians. Am J Geriatr Psychiatry 15:252−256.
49. Terry DF, Wilcox MA, McCormick MA, Perls TT (2004) Cardiovascular disease delay in centenarian offspring. J Gerontol A Biol Sci Med Sci 59:385−389.
50. Rodriguez BL et al. (2002) The A.D.A and W.H.O. classifications for diabetes: their impact on diabetes prevalence and total and cardiovascular disease mortality in elderly Japanese −American men. Diabetes Care 25:951−955.
51. McNeely MJ, Boyko EJ (2004) Type 2 diabetes prevalence in Asian Americans: results of a national health survey. Diabetes Care 27:66−69.
52. Fujimoto WY et al. (1995) Susceptibility to development of central adiposity among populations. Obes Res 3Suppl2:179S−186S.
53. Examination Committee of Criteria for Obesity Disease´ in Japan, et al. (2002). New criteria for Obesity disease´ in Japan. Circ J 66:987−992.
54. Barthel A, Schmoll D, Unterman TG (2005). FoxO proteins in insulin action and metabolism. Trends Endocrinol Metab 16:183−189.
55. Kops JG et al. (2002) Forkhead transcription factor FOXO3a protects quiescent cells from oxidative stress. Nature 419:316−321.
56. Youngman LD, Park JY, Ames BN (1992) Protein oxidation associated with aging is reduced by dietary restriction of protein or calories. Proc Natl Acad Sci USA 89:9112− 9116.
57. Heilbronn LK et al. (2006) Effect of 6−month calorie restriction on biomarkers of longevity, metabolic adaptation, and oxidative stress in overweight individuals: a randomized controlled trial. JAMA 295:1539−1548.
58. Selman C et al. (2008) Evidence for lifespan extension and delayed age−related biomarkers in insulin receptor substrate 1 null mice. FJffi´´5 J22:807−818.
59. Murphy CT et al. (2003) Genes that act downstream of DAF−16 to influence the lifespan of Caenorhabditis elegans. Nature 424:277−283.
60. Kuningas M et al. (2007) SIRTl gene, age−related diseases, and mortality: the Leiden 85− plus study. J Gerontol A Biol Sci Med Sci 62:960−965.
61. Kagan A. Ed (1996) The Honolulu Heart Program: An Epidemiological Study of Coronary Heart Disease and Stroke (Harwood Academic Publishers, the Netherlands).
62. Worth RM, Kagan A (1970) Ascertainment of men of Japanese ancestry in Hawaii
through World War II Selective Service registration. J Chronic Dis 23:389−397.
63. Rodriguez BL, Curb JD (1998) Cardiovascular risk factors in the elderly: the Honolulu Heart Program. Cardiovasc Risk Factors 8:99− 103.
64. White L, et al. (1996) Prevalence of dementia in Japanese− American men living in Hawaii: the Honolulu− Asia Aging Study. JAMA 276:955−960.
65. National Institutes of Health (NIH): U.S. Department of Health and Human Services, NIH, National Institute on Aging (2001) Report of the National Institute on Aging Advisory Panel on Exceptional Longevity (APEL). NIH Pub 01−4951 (NIH, Washington, DC).
66. Arias E (2006) United States life tables, 2003 (National Center for Health Statistics, Hyattsville, MD), vol. 54, no. 14.
67. Nordyke EC, Lee R, Gardner RW (1984) A profile of Hawaii´s elderly population. Papers East W1 est Popul Inst 91 :13−14.
68. Mehta C, Patel N (2001) StatXact 5 (CYTEL Software Corporation, Cambridge, MA).
69. SAS Institute, Inc. (1990) SAS/STAT user´s guide, version 6 (SAS Institute, Cary, NC).
70. Sherry ST, Ward MH, Kholodov M, Baker J, Phan L, Smigielski EM, Sirotkin K. dbSNP: the NCBI database of genetic variation. Nucleic Acids Res. 2001 Jan l;29(l):308−l l.)
【技術分野】
【0001】
本発明は、診断において予測し、または、治療計画および関与において健康的な老化および長寿を促進するために、FOXO3Aの多型およびハロタイプを使用する方法に関する。
【0002】
関連出願の説明
本出願は、本願明細書に引用した2008年8月10日に出願した米国特許出願公開第61/087,722号明細書に、部分的に基づき、その利益を要求する。
【0003】
連邦支援の研究または開発に関する記載
本発明を、アメリカ国立老化研究所からの1 R01 AG027060−01 (Defining the Healthy Aging Phenotype)の研究費で、政府支援のもと行った。更なる資金提供は、国立心臓肺臓血液学会からの契約書N01−HC−05102、国立老化研究所からの契約書N01−AG4−2149および研究費5 U01 AG019349−05およびK08 AG22788−02に基づいて、米国政府の支援によって提供された。米国政府は、本発明における特定の権利を有する。更なる支援は、ハワイコミュニティー財団からの研究費2004−0463に基づいて行われた。
【背景技術】
【0004】
FOXO3A遺伝子は、明瞭なフォークヘッドドメインによって特徴づけられるフォークヘッド型転写因子に属する。この遺伝子は、細胞死に必要な遺伝子の発現によって、アポトーシスの誘因として機能するらしい。この遺伝子とMLL遺伝子の転座は、二次急性白血病と関連する。同じタンパク質をコードする選択的なスプライスされた転写変異体を観測した。
【0005】
FOXO3A遺伝子は、DAF−16のヒト相同体の1つであり、モデル生物であるシノラブディスエレガンスで寿命を延ばすと記載されていた遺伝子である(Murphy CT (2006)The search for DAF−16/FOXO transcriptional targets: approaches and discoveries. Exp Gerontol 41 :910−921) and D. melanogaster. (Giannakou ME et al. (2007) Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging Cell 6:429−438)。
【0006】
FOXO3A遺伝子は、ヒト染色体6q21の位置108,987,719から109112664(NCBI ver.36)までに配置され、選択的に発現できる4つのエクソンから構成され、同じタンパク質を生じる(異型#1は、ファイルNM 001455.3に記載され;異型#2は、ファイルNM 201559.2に記載される)。FOXO3Aタンパク質は、アミノ酸673から構成され、71,277Daサイズである。FOXO3Aのアミノ酸配列は、NCBIでファイル「NP 963853」によって規定され、SEQ ID NO.1として同定し、以下の通りである:
【0007】
【0008】
FOXO3Aは、YWHAB/14−3−3−betaおよびYWHAZ/14−3−3−zeta、UniProt:the Universal Protein Resource (www.uniprot.org)と相互作用し、細胞質の分泌で必要である。酸化性ストレスに応じて、STKと相互作用し、YWHAB/14−3−3βとの相互作用を妨害して、核移動に導く。FOXO3Aの細胞内局在は、細胞質およびサイトソルである。それは、酸化ストレスに応じておよび生存因子の欠損で、核に移行する。生存因子、IGF−1が存在する場合において、FOXO3Aは、Thr−32およびSer−253でAKT1/PKBによってリン酸化される。それからこのリン酸化された形態は、14−3−3タンパク質と相互作用し、細胞質にて保持される。生存因子の脱離は脱リン酸化を誘導し、脱リン酸化されたタンパク質が、標的遺伝子の転写を誘導する場合には核への移行を促進し、アポトーシスを誘発する。AKT1/PKBは、直接Ser−315をリン酸化するようではないが、それはこの残基でリン酸化を誘発する他のキナーゼを活性化することができる。FOXO3Aは、STK4によってSer−209に酸化性ストレスに応じてリン酸化され、YWHAB/14−3−3−βおよび核移行からの脱離を導く。
【0009】
ヒトの寿命は、多くの決定因子を有する複雑な表現型である。食事、物理的活動、健康習慣および社会心理的因子を含む非遺伝的な因子が重要である一方、ヒト寿命において最高50%の変異が遺伝子の相違点によって説明される可能性がある1−5。いくつかの研究は、平均的なヒトの寿命において、約25%の変異を示唆することをできるが、例外的な生存者の多くの人において、寿命に対する遺伝的貢献は非常に高い可能性がある。例えば、90歳代および100歳代の家系調査では、兄弟間の相対的危険度は複雑な表現型への潜在的遺伝子の寄与を評価する一般的な方法6で、特に高く、家系発端者の年齢の増加と共に増大することを示した7〜10。しかしながら、ヒトにおける「長寿に関連する」遺伝子候補の研究は、以下「長寿遺伝子」といい、一般的に期待はずれである。ほとんどの複製は、ApoE遺伝子を除いて、集団全体に観測されなかった。
【0010】
対照的に、老化のモデル生物において、いくつかの強力な遺伝子の発見があった11〜13。例えば、単一の遺伝子の変異体は、モデル生物における寿命で、特にインスリン/IGF−1(IIS)シグナル経路の一部と考えられる遺伝子14〜18で、実質的な相違を生じることができる。
【0011】
SIR2活性を向上させるか、またはインスリン/IGF−1シグナルを減少させる突然変異は、DAF−16/FOXOタンパク質を活性することによって、シノラブディスエレガンスの寿命を共に上昇させる19、20。哺乳類の細胞において、Sir2相同体物である「SIRT1」は、ストレスへの細胞反応を含む、寿命に影響するいくつかの下流転写現象に影響を及ぼしている。SIRT1は、これを、FOXO(フォークヘッドボックス転写)因子、IIS経路でセンサーとして機能し、いくつかの哺乳類において寿命の調節因子ともなるタンパク質のファミリーを調整することによって達成する17。
【0012】
哺乳類(および他の種)における、遺伝子ノックアウトモデルは、同様にIIS仮説を支持する。例えば、脂肪細胞に特異的なインスリン受容体ノックアウトのマウス(FIRKO)は、脂肪の質量、加齢に関連した肥満に対する予防を減少し、寿命を延長する。IIS経路における多くの他の突然変異体は、マウスにおいて長寿に影響を与えると考えられる。これらは、IGF−1受容体22、IRS−122、IRS−223、PAPP−A24の突然変異体およびエイムス矮小マウス突然変異を含む。
【0013】
インスリンシグナルの基本的な分子経路は、酵母、ハエ、虫、齧歯動物およびヒトに示される証拠があるように、進化にわたって保存されている。虫における前記経路の重要な調節因子は、転写制御因子DAF−16(異常なDAuer形成16)であり、シノラブディスエレガンスにもたらされる広範な寿命延長に、インスリン/IGF−1シグナルを阻害することに必要である。多くの因子は、daf−16に依存した方法、例えばAMPキナーゼ26、14−3−3のタンパク質27、lin−4マイクロRNA28および熱ショック因子において、シノラブディスエレガンスの寿命を延ばすと考えられる。いくつかの種のDAF−16の相同体は、老化表現型および長寿に関連していた30。例えば、ストレス応答Junアミノ末端キナーゼ(JNK)経路は、FOXOが必要であると考え、ショウジョウバエの寿命を延長し31、ハエがdFOXO、DAF−16オーソログを過剰に発現しているとき、それは寿命を顕著に上昇することができる32。DAF−16/FOXOでの前記多様な配列シグナルの著しい収束は、このタンパク質が、老化および長寿に影響を与えるシグナルネットワークにおいて、重要で、進化的に保存されている「ノード」であることを示唆する。
【0014】
DAF−16のヒトの相同体は、4つのFOXO:FOXO1、FOXO3A、FOXO4およびFOXO6を含有する。従って、FOXOの単一の塩基多型(SNP)形態における、一般的な、野生の変異体および関連した遺伝子は、ヒトの長寿に影響する可能性があることを仮定することを試みる。「FOXO3」は、FOXO3Bが染色体17上の偽遺伝子であるため、「F0X03A」と同義である。
【0015】
これは興味深い仮説である。インスリン、FOXO、酸化性ストレスとヒトの寿命との間の関係は、酸化ストレスが、好ましい一般的に考えられる加齢のメカニズムであるため、特に関連性がある。1956年以来、加齢の遊離基説は、加齢がDNA、反応性酸素分子の累積的な照射によって細胞および組織に損傷を部分的に生じることを仮定し、普遍的にまだ受け入れられないにも関わらず、有効な証拠が長年にわたって蓄積した34,35。従って、FOXOは、潜在的な分岐点または橋かけを、インスリンシグナル、遊離基とヒトの老化/寿命との間で提供することができる。
【0016】
Suh et alによる関連性のある最近の報告38を含むヒトの寿命で、IIS経路における、いくつかの従来の研究と関連のある遺伝子があり36、37、機能的に重要なIGF−I受容体の突然変異体と特別な寿命とを関連するが、FOXO遺伝子およびヒト寿命の間の関連性の発表報告は全く見当たらなかった。従来の研究は、FOXO遺伝子と、4年の生存率、脳卒中の危険性39および早発閉経40を含む他の老化表現型との関連が明らかになった。
【0017】
しかしながら、ヒトの寿命は、疾患特定の危険性ならびに個々の加齢の割合を包含する複雑な表現型である。その遺伝子の先例の研究は、困難である。寿命の研究は、小さい遺伝子影響によるサイズ、人為的な層化産物、人口異質、長寿研究の参加者の充分な人数の不足および他の課題3、4,41によって影響される。従って、ヒトの寿命への遺伝子寄与の可能性を、IISシグナルに関連する遺伝子から評価するために、加齢表現型としてよく特徴付けられる人の多く、均質な長寿の集団を選択し、IIS経路に関連のある5つの候補の寿命遺伝子のコホート内症例対照研究(nested case-control study)を実行した。前記遺伝子は、加齢表現型の従来の関連物に基づいて、主に加齢の遺伝子ノックアウト、トランスジェニック、突然変異および他のモデル生物から選択された3、4、14〜17、36,42。優先順位は、インスリン感受性およびグルコース(エネルギー)ホメオスタシスに関連する遺伝子で行った。
【0018】
集団の急激な老化は、慢性疾患の罹患率および障害を増大することにより、先例のない難題を社会にもたらすであろう。ヒトが齢をとる方法に関して、広く影響することができる生物学的経路を包含する、加齢のメカニズムの良好な理解は、加齢に関連した疾患および障害への低下で重要な意味を有する。ヒトの寿命で多くの生物学的に信頼できる遺伝子候補があるが、1つの発見のみが、ApoE遺伝子のものより、今まで多くの集団において広く繰り返されてきた。前記遺伝子は、加齢表現型、特に心血管疾患および認知症、例えば長く健康な寿命を達成する能力に影響するようなものに広く達成した。
【発明の概要】
【課題を解決するための手段】
【0019】
ヒトの加齢表現型および寿命への広く達成した遺伝子を発見する際の課題は、ヒトにおける研究を実施する前、モデル生物を使用して先見的で潜在的な候補を同定することに役立つことができることを示唆する。従って、ヒトのインスリン/IGF−1シグナル経路および/または配列に基づく酸化ストレス反応システムおよび/または加齢のモデル生物または従来のヒト研究で機能的相同性の中で、いくつかの候補遺伝子の研究するために選択する。前記シグナル経路からヒトの候補遺伝子のリストを構成し、日本人の集団の約10%以上の頻度にて発生する前記候補遺伝子において変異体を評価した。限られた資源のため、3つのSNPのみを、分析でそれぞれの遺伝子から選択した。SNPを、可能であれば、それぞれの遺伝子の最大範囲を提供するために、連鎖不平衡(LD)で領域から選択した。
【0020】
一般的に、本発明は、FOXO3A「GCC」ハプロタイプ(例えば、FOXO3Aハロタイプは増加した寿命と関連があり、本明細書において更に15年以上生存するヒトの対象の可能性として規定する)を検出する組成物および方法を提供する。好ましい実施形態において、検出されたFOXO3Aハプロタイプは、長く生きる増加した可能性または減少した可能性のいずれかと関連するが、本発明は、長い寿命の増加も減少の可能性と関連がなくおよび/または加齢に関連した疾患(例えば、「正常」または「wt」の遺伝子型)の危険性を最少にするFOXO3Aハプロタイプを検出する物質および方法を必然的に包含する。加齢に関連した疾患とは、疾患冠動脈疾患(CHD)であり、冠状動脈疾患、脳卒中、癌、慢性閉塞性肺疾患(COPD)、他の慢性的な肺疾患、パーキンソン病、糖尿病、肥満、認知症(一般的な認識機能)、虚弱(歩行能力)もしくは他の加齢と関連性の疾患、または身体および/または認識障害と規定する。またヒトの肥満との関連性が存在する。
【0021】
「GCC」ハプロタイプは、何十キロベースのDNAを包含する。この領域における他のSNPは、本願明細書で記載される3つのSNPを有する連鎖不平衡を示す。更なるSNPが、同様に寿命および健康的な老化に関連のあるこのGCCハロタイプで同定され、予測される加齢と関連する疾患に実用的であることができると予期される。「GCCハロタイプ」は、特別な長寿および/または健康的な加齢の予測へと導く“機能的な変異体”であることを最終的に見出すDNA変性の他の種の代用物として、FOXO3A遺伝子の範囲または隣接したいずれかでもたらすことができる。前記他の変性は、転移、複製、欠失の形態であることができ、例えば“LOC100130966”といった依然知られていない他の遺伝子または転写物を含有することができる。LOC100130966は、mif2のSMT3制御因子3ホモログ2と同類であり、FOXO3A遺伝子のエクソン2の範囲内であることが同定され、“GCC”ハロタイプ内である。LOC100130966に関するDNA配列は、GenBank取得のID#「XM 00 1725519」として記載され、LOC100130966の予想アミノ酸配列は、GenBankファイル「XP 00 1725571」として記載されている。
【0022】
ハプロタイプ分析を使用して、患者が積極的な健康または疾患の予防/治療介入によって利益があるものを潜在的に予測することができる。ハプロタイプ分析は、キットの様式で提供することができる。危険性の測定は、このような情報を、疾患、障害もしくは死亡の可能性の評価、またはヒトがどれほど生存または無病生存であるかを確定する目的に使用することができる。前記情報は患者、健康保険会社、長期介護保険会社および医師または他の医療供給者への、多少の指導を患者の長期ニーズとして提供するために重要である。医薬を、加齢に関連した健康または疾患に影響を与えるために、FOXO3A遺伝子の作用の改善、FOXO3Aタンパク質および/または他のタンパク質との相互作用の細胞局在の改善、遺伝子によって産出されるタンパク質の量または種類の改善するものを開発することができる。
【0023】
マウスにおいて相同配列は、早期卵巣機能不全と関連することができる。Castrillon DH, Miao L, Kollipara R, Horner JW, DePinho RA. Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science. 2003 Jul 11;301(5630):215−8。その結果、同類のハプロタイプ分析は、獣医学的応用に実用的であることができる。
【0024】
更なる本発明の特徴は、以下の記載から、例えば添付した図面および表を参照して、明らかになるだろう。
【図面の簡単な説明】
【0025】
【図1】表10に記載のプライマーおよび条件を使用してFOXO3A G/T変異体を検出するARMS−PCR定量法の結果を示す図である。
【図2】表10に記載のプライマーを使用してFOXO3AのG/T変異体を検出するARMS−PCR分析の模式的に概略を示す図である。
【0026】
以下の表は、本明細書の一部である:
表1:HHP/HAAS Cohortの1991−93における基本特性(n=3,741)
表2:ケースコントロール状態による基本特性
表3:ケースおよびコントロールにおけるヒトの寿命およびMAFの候補遺伝子
表4:ケースコントロール状況によるFOXO3A3遺伝子型
表5:基準での遺伝子型集団間の健康状況の相違点
表6:FOX03A遺伝子型によるインスリン感受性表現型
表7:FOXO3A3遺伝子型に関する加齢に関連した表現型の優位性
表8:最高到達年齢による遺伝子型分布
表9:rs2802292 G−T多型の同定のためのプライマー
表10:rs2802292 G−T多型の同定のためのPCR条件
【発明を実施するための形態】
【0027】
A.ハワイでの寿命の研究
調査集団
コホート内症例対照研究は、ハワイ寿命調査の一部で、ホノルル心臓計画(HHP)およびホノルルアジア加齢研究(HAAS)の当初の集団から集められた健康的な加齢の配属されたコホート研究として実施された。HHPは、1965年に開始した日系米国人8,006人の中での、心血管疾患の集団に基づく潜在的な調査である。HHP参加者は、正式な連絡先を持ち、1900年から1919年の間に生まれ、オアフ島に1965年に居住していた9,877人の男性から募った。
【0028】
試験参加者は、日本、通常日本の西部および南部からの両親を有し(中央領域またはさらに西部および南部から94%);全体の49%は、親の出身が広島および山口の隣接した県からである61、62。大半の参加者はハワイ生まれ(88%)であるが、対立遺伝子頻度のケースコントロール条件の混絡の理論的な可能性が、地理的起点による。従って、特定の分析、ケースおよびコントロールに関して、親の出身の県によって、条件付きロジスティック回帰モデルを使用して、階層化した。分析は、データセットにおける、集団階層化で全く証拠を示さなかった(資料未記載)。
【0029】
HHPコホートの補充、設計および方法を、詳細に他で概説する62。簡潔に、調査登録時(1965〜1968)、参加者は、45〜68までの歳であった(平均年齢54歳)。HHPの開始から、冠動脈疾患および脳卒中の発生の進行、ならびに全ての原因による死亡率に関する情報が、地方新聞(英語および日本語)の死亡記事の観測および退院記録の査察から得られた61。1991〜1993年検査における追跡調査は、5人のみが死亡情報をたどることができなかったことが分かった63。
【0030】
現在の調査に関する全ての参加者は、2007年8月までに更新された調査参加者の記録から導いた。HHP(1991−1993)の試験4から達成された表現型データおよび血液サンプルは、ホノルルアジア加齢研究(HAAS)の開始と一致し、コホート内症例対照研究のための基礎調査として使用された。HAASは、高齢者における神経変性疾患、認識機能および他の加齢表現型の調査のためのHHPの展開として開始された。64参加者は、試験4での71〜93歳の3,741人(平均年齢77.9±4.7歳)、最初のHHPの約半数を含有する64。
【0031】
現在のコホート内症例対照研究に関して、「ケース」(寿命表現型)を、1910年アメリカ出生コホートの特異的生存率の少なくとも上位1%(最小95歳)を募集時期から生存した全てのHHP参加者として規定した。2007年8月時点にて、少なくとも95歳まで生存した合計213人の個人を調査した。176人のこれら個人が死亡し(平均年齢97.5歳、SD2.1;範囲95〜106歳)、37人が依然生存している(平均年齢98.7歳、SD2.1;範囲97〜106歳)。
【0032】
コントロールは、中年男性で、1910年のアメリカ出生コホートの特異的生存に関する平均死亡年齢の間近に死亡したHHP/HAASコホートからの402人からなる。症例を達成するために:約1:2のコントロール比率において、81歳までに死亡したコントロールに関するHHP/HAAS調査集団をサンプルとして採取した。コントロール集団における死亡平均年齢は、78.5歳であった(SD1.8、範囲73〜81歳)。これはアメリカの男性集団より若干高いが、ハワイにおける日系米国人の男性の高い平均寿命に合致し、最後の報告で白人男性より3.5年長かった67。全てのケースおよびコントロールは、家系が主に日本の中央西部を出身とする日本人人種である61、62。
【0033】
方法は、組織のガイドラインに従って実行され、Institutional Review Board of Kuakini Medical Centerによって承認された。書面に基づくインフォームドコンセントを、参加者が同意を提供することができない場合、全ての試験参加者または家族代表者から得た。
【0034】
遺伝子型
3つのSNPを5つの候補遺伝子の各々から選択する。モデル生物において、加齢経路で影響を十分に記載された遺伝子を選択する。全ての遺伝子を、図1を参照するように、IIS経路に仮定的な関連、ならびにエネルギー恒常性、グルコースおよび/または脂質代謝に潜在的な関連に基づいて選択した。SNPを、HapMapまたはJSNPデータベースに報告された、それらの少数の対立遺伝子頻度に基づいて選択した(snp.ims.u−tokyo.ac.jp)。
【0035】
総細胞DNAを、PureGeneシステム(Gentra Systems, Inc.)を使用して分離し、PicoGreen染色(Molecular Probes, Eugene, OR)を使用して定量化し、候補遺伝子からSNPは、対立遺伝子識別分析を使用して遺伝子型を求めた。TaqMan(登録商標)(Applied Biosystems, Inc.)試薬をABIから購入し、SNPを頻度≧〜0.1で日本人集団において選択した(http://www.ncbi.nlm.nih.gov/projects/SNP/)。PCRは、Taq Gold(Perkin −Elmer, Corp)およびTaq Man(登録商標)分析でPCR産物の検出を使用し、1つの対立遺伝子および他の対立遺伝子に対してVICラベルされたプローブで6−FAMラベルされたFRETプローブを使用し、分析の検出を向上させるために副溝結合(MGB)クエンチャーを使用して、標準の条件下にて増幅した。PCR産物を、ABI Prism 7000 Sequence Detection Systemで測定した。
【0036】
遺伝子型データを、統合されたデータベースシステムによって管理した(MS Excel, Microsoft, Inc)。それぞれの遺伝子型プレート上の全ての正のコントロールを、同様に一貫して評価した。陽性マーカーをハーディワインベルグ平衡からの偏差で検出した。細胞頻度は98%を超えた。
【0037】
統計学的分析
SNPを、ハーディワインベルグ平衡からの偏差で評価した。ピアソンカイ2乗検定を使用して、ソフトウエアプログラムStatXact68を用いて同様の遺伝子型頻度に対するケースおよびコントロールを比較した。関連の強度評価するために、確率比率をSAS69のロジスティック回帰モデルを使用して評価した。一般化線形モデル(GLM)および共分散分析(ANCOVA)を更に使用して、健康な試験参加者の比率をFOXO3Aの遺伝子型によって比較した。ケースおよびコントロールにおける加齢表現型の分析のためには、連続変数の分布を比較するスチューデントt検定、および比例変数でのカイ二乗である。
【0038】
結果
1991〜1993の調査でHHP/HAAS調査集団の基本的特徴を表1に示す。平均年齢は、77.9歳であり、集団の100%は男性および日本人人種であった。生物学的特徴、公衆衛生状態、疾病の罹患および基本的な状態を示す。
【0039】
【表1】
【0040】
前記1991〜93年の基本的集団から、「長寿」のケースとして2007年までに95歳以上生存した全ての参加者を選択した(n=213)。その後「平均的な」生存率のコントロールとして81歳以前に死亡した全ての参加者を選択した(n=402)。ケースおよびコントロールの基本特徴を表2に示す。生物学的特徴において、高齢者のケースは、比較的老年で、脂肪が少なく(低い腰周り:尻周りの比率)、低いトリグリセリド(ボーダーライン)、低いグルコース、低いインスリンレベルおよび高いFOXO3A3対立遺伝子の傾向を、基礎調査で有する。ケースはまた、自己評価による健康、ならびに心血管疾患(CHDおよび脳卒中)および癌の低い傾向を改善した。機能的に、彼等は十分に歩行できるようであるが、低い握力を有した。認識スコアに関する有意差は存在しなかった。
【0041】
【表2−1】
【表2−2】
【0042】
5つの遺伝子を調査した(ADIPOQ、FOXOlA、FOXO3A、SIRTl、およびCOQ7)。ケースおよびコントロールに関して、少数の対立遺伝子頻度および他の関連する遺伝子情報を表3に示す。しかしながら、FOXO3A遺伝子型のみが、p<0.05の初期カットオフ値を使用して寿命と関連した。
【0043】
【表3】
【0044】
変異体“rs2764264”を、以下に“rs 12524491”として規定する。SEQ ID NO.2として同定されたSNP rs2764264(“F0X03A1”)のDNA配列は、以下である:
【0045】
【0046】
SEQ ID NO.3として同定されたSNP rsl3217795(“FOXO3A2”)のDNA配列は、以下である:
【0047】
【0048】
SEQ ID NO.4として同定されたSNP rs2802292(“FOXO3A3”)のDNA配列は、以下である:
【0049】
【0050】
「GCCハプロタイプ」は、SNP rs2764264、rs13217795およびrs2802292を使用して記載することができ、遺伝子型の以下の組み合わせを含む対立遺伝子である:
【0051】
【表4】
【0052】
これらの変異体を、染色体の最上部(遺伝子地図において低いヌクレオチド位置)から最下部(遺伝子地図において高いヌクレオチド位置)まで観測するとき、NCBI命名法によると「GCCハプロタイプ」を、SNPrs2802292、rs2764264およびrsl3217795を使用して記載することができ、以下の遺伝子型の組み合わせを含有する対立遺伝子である:
【0053】
【表5】
【0054】
上記のデータは、Database of Single Nucleotide Polymorphisms (dbSNP). Bethesda (MD): National Center for Biotechnology Information, National Library of Medicine. (dbSNP Build ID: 129, NCBI genome build 36.3)からである。http://www.ncbi.nlm.nih.gov/SNP/70から入手可能である。
【0055】
FOXO3 A3の遺伝子頻度をケースおよびコントロールの間で比較する更なる調査は、ピアソンのカイ二乗統計の置換分布を用いて、正確なp値0.00009で、非常に重要な相違点を示した。これらの結果を表4に示す。この研究において、それぞれの対立遺伝子内に5箇所の遺伝子座と3つのSNPが存在するため(表3)、多重比較に関するボンフェローニ補正は、15×0.00009=0.00135の補正されたp値を生じた。FOXO3Aの3SNPの間で高い関連性のLDのため、FOXO3A3のSNPのみに関して、更に調査した(rs2802292)。ケースおよびコントロールの間でFOXO3A3に関する対立遺伝子の同型劣性(homozygous minor)対同型優性(homozygous major)の確立比率(OR)は、2.75(95%CI:1.51−5.02、p=0.0007)であり、ケースおよびコントロールの間で異型対同系の優性対立遺伝子は、1.91(95%CI:1.34−2.72、p=0.0003)であった。これらの結果は、寿命に関して相加効果を示唆する。
【0056】
【表6】
【0057】
若い年齢で長寿表現型についてさらに理解するために、Willcox et al.(2006)の健康な生存者の定義を用いて3つのFOXO3A遺伝子型群のそれぞれに関する基礎調査(1991)にて、健康である人の比率を比較した。相違点は、非常に重要であった(表5)。1つ以上のG対立遺伝子を有する人々は、優性(TT)対立遺伝子に関して同型である人々より、正常にて健康であるようである。劣性対立遺伝子に関して同型な人の約75%は、優性対立遺伝子に関して同型な人々の約57%のみに対して、基礎調査にて健康的である。ケースコントロール状態を調整した後も、相違点は依然わずかながら重要であった。これは、長期生存者(ケース)およびコントロールの部類の範囲内で、対立遺伝子と健康状態の関連性が存続することを示唆する。
【0058】
【表7】
【0059】
インスリン感受性、寿命の潜在的な中間の表現型、および遺伝子型の間の関係があるかどうかを評価するため、空腹時インスリン、グルコース、HOMAおよび遺伝子型の間の関係を試験した(表6)。非正規分布変数のため、対数変換を正規分布に使用した。優位な関係がインスリン、インスリンの対数、HOMAおよび遺伝子型に存在する。同型G対立遺伝子は、コントロールのみにおいてであるが、インスリン、インスリンの対数およびHOMAスコアの顕著な低下と関連している。
【0060】
【表8】
【0061】
同様に、いくつかの慢性疾患およびFOXO3A遺伝子型の生涯有病率の間の関係に関して試験参加者にて調査した(表7)。
【0062】
【表9】
【0063】
優位な保護関係が、CHDの有病率に関する同型G対立遺伝子、並びに癌および認識機能に関する境界関係で同型接合性が明らかになった。最後に、統合した全ての参加者(ケースおよびコントロール)において最高到達年齢によって、FOXO3A3劣性対立遺伝子頻度(MAF)分布を評価した。MAFは、以前のケースコントロール分析によって予測したように、年齢と共に著しく上昇した(表8)。
【0064】
【表10】
【0065】
5つの候補遺伝子の分析は、1つの遺伝子が、潜在的なヒト長寿遺伝子であるFOXO3Aに関して、他の遺伝子より明確に卓越していることを示した。前記遺伝子が、ヒトの寿命に重要でありえることが、いくつかの証拠によって支持されている。第一に、コホート内症例対照分析において、前記遺伝子内における変異体は、寿命と強く関連している。ケースおよびコントロールの間のFOXO3A3の対立遺伝子の同型劣性(homozygous minor)対立遺伝子の同型優性(homozygous major)であるオッズ比(OR)は、2.75(95%CI:1.51−5.02、p=0.0007)であり、ケースおよびコントロールの間の異型対同系の優性でのORは、1.91(95%CI:1.34−2.72、p=0.0003)であった。これらの結果は、寿命におけるFOXO3A3 G対立遺伝子の相加効果を示唆する(即ち、G対立遺伝子の2つのコピーは、約2倍の保護作用を与えた)。一貫して、劣性対立遺伝子頻度は、70歳代から100歳代までの年代の試験参加者の年齢と共に、著しく上昇した(表8)。
【0066】
第二に、FOXO3A遺伝子において評価された全3つのSNPは、密接した連鎖不均衡(LD)であり、長寿表現型と強い相関関係があった。これは、調査結果が可能性によるものでないことを示す。第三に、1つ以上の劣勢(G)対立遺伝子を有する人は、同型優性(TT)対立遺伝子での人と比較して、約15年前の基礎調査にてより健康である可能性があった。劣性対立遺伝子に関して同型な人の約75%は、優性(TT)対立遺伝子に関して同型な人々約57%のみに対して、基礎調査にて健康的である(表5)。
【0067】
実際に、基礎調査は、ケースが、平均して11歳老年であるにも関わらず、ケースがコントロールより顕著に健康的であることを示唆した。ケースは、CHD、脳卒中および癌の低下した有病率を含有する著しく少ない加齢関連疾患を有する。同様に、彼等は優れた自己評価で健康的であり、低い歩行障害を含む、高い身体機能を一般的に有した。興味深いことに、コントロールより10歳を超える年にも関わらず、長寿のケースは同様のレベルの認識機能を有した。これは、個々が主な臨床疾患および障害を人生の末期まで何とか遅らせるか、回避する場合に、「健康的な加齢」の表現型の存在を支持する。ケースで観測された健康的な加齢表現型は、彼等の年齢に適合する出生コホート46〜48および100歳以上の子孫と比較した若い年代で、100歳以上の人々において報告された健康的な加齢表現型に類似する49。同様に長命のケースは、若い年代で比較的高いインスリン感受性と、低い腰周りと尻周りの比率、低いグルコースレベル、低いインスリンレベルおよび低いHOMA値である代謝プロフィールを有する。いくつかの表現型が、FOXO3A遺伝子型において変異体と関連した。
【0068】
意外にも、ケースおよびコントロールの間で糖尿病の有病率で優位な違いは全く存在しなかった。しかしながら、ケースはコントロールより10歳を超えて、糖尿病は年齢と共に顕著に増加するため、それは糖尿病の有病率が非常に独特でないことに注目すべきである。実際に、ケースおよびコントロールの両方は、高い糖尿病の有病率(約60%)を、比較的低いBMIに関わらず有した。2型糖尿病が、日本人において比較的低いBMIで高い有病率の傾向がある理由は、完全には解明されていない50。しかしながら、白人および黒人より低いBMIでアジア人において、高い内臓脂肪を有する日本人(多少の他のアジア人)で、代謝の相違点が存在する可能性がある51、52。実際には、日本国ガイドラインが、このような集団の違いを反映し、日本人の肥満をBMI25とみなす。HHP/HAASコホートにおける糖尿病の高い有病率への他の原因要因は、全ての参加者を、いくつかの異なった臨床試験、および更に可能性がある検出するいくつかの事前検査で糖尿病を試験する事実を包含する。
【0069】
当然、FOXO3A遺伝子型は、血漿インスリンレベル並びにCHD、癌および2型糖尿病の有病率に著しい関連があった。これは、細胞増殖、アポトーシスおよび代謝を包含する多様な生理的機能で、インスリンおよびインスリン様成長因子の効果のメディエーターとしてのFOXOでの知られている役割と一致する。シノラブディスエレガンスおよびショウジョウバエにおける遺伝学の研究は、FOXOタンパク質が、代謝および寿命を調節するインスリン様シグナルの従来の標的であることを示した。哺乳類細胞における更なる研究は、FOXOタンパク質が、タンパク質キナーゼの標的であり、細胞周期の進行に影響し、生体外において酸化ストレスに対する抵抗を制御することを示した54。生体内研究は、FOXOが、インスリンに反応して肝臓グルコース産物を転換し、他の代謝活性を媒介することを示した。これは、FOXOタンパク質が、インスリン効果を代謝で媒介して、ヒトの寿命に影響することができるという根拠を支援する。
【0070】
概して、根拠の全体は、FOXO3Aの潜在的役割を、ヒトの健康、加齢および寿命で支持する。インスリン感受性、CHD、癌、2型糖尿病および寿命を含有する多様な長寿表現型と、FOXOとの関連は、「ゲートキーパー」の役割を、IIS経路において示唆する。FOXO3Aがヒトの老化に影響する重要な下流のメカニズムは、酸化ストレスの修正による−かねてのヒトの老化に関する仮説であるが、これに対する直接的な証拠は現在研究中で全くない。しかしながら、FOXO遺伝子が、シノラブディスエレガンスDAF−16の、最も密接なヒトの相同物であり、細胞を酸化ストレスから保護するため、これはヒトの加齢の変化に対して優れた作用のメカニズムである。シノラブディスエレガンスにおいて、DAF−16は、マンガンスーパーオキシドジスムターゼ(SOD2)の発現を増加させ、これは他の「加齢防止」効果の中で、過酸化物を比較的低い損傷で過酸化物に転換し、遊離基に対して有力な内因性保護作用がある。生体内研究は、DNA、タンパク質および他の組織における酸化障害が、年齢と共に蓄積し、カロリー的に制限された食事を、齧歯動物56およびヒト57に供給することが、この障害を暖和することを示した。
【0071】
FOXOが明らかに寿命と関連している一方、インスリン感受性で遺伝子型の強い効果は、ケースには観測されず、コントロールであった。しかしながら、GG遺伝子型は、同様に低血漿インスリンレベルをケースおよびコントロールの両方において示し、遺伝子型の調節作用とインスリンレベルで両方の群において一致する。ケースが比較的強いインスリン感受性を示すため、それらの遺伝子型に関わらず、それらはインスリン感受性をFOXO以外に維持する複数のメカニズムを有することを推測する傾向がある。これは、大半の長寿遺伝子が適度または小さい効果サイズを有する仮説と一致した。小さいサンプル数が、ケースにおける相違点を検出する能力を制限することがまた可能である。一方では、突然変異をIRS−158またはIRS−223のいずれでもたらす長命のマウスは、実際インスリン抵抗性であるため、インスリン感受性は比較的長い寿命を与えることができるIIS経路における突然変異で、必要条件でない。
【0072】
しかしながら、シノラブディスエレガンスにおいて、単独で寿命に小さい影響を有することができるいくつかの遺伝子が、転写調節“マスター遺伝子”DAF−1659によって影響することに留意する関連性がある。さもなくば検出が困難な可能性があるFOXO3Aにおけるわずかな差異が、論理的にDNA結合、タンパク質間相互作用、細胞周期進行、アポトーシスおよび代謝に関するいくつかの下流遺伝子を修飾される。このように、FOXO3Aによる小さい改良する効果が、加齢表現型および寿命に、大きく付加的下流効果を潜在的に有する。
【0073】
裏づけになる証拠は、ヒトの加齢および寿命において、インスリンシグナルの役割が集積し始めたが、これらの効果を媒介する可能性がある遺伝子は知られていない。従来の研究は、長命のイタリア36、日本37,42、ドイツ60およびアシュケナージユダヤ人38の民族において、いくつかの加齢表現型と結びついて、インスリン−IGF−1シグナル経路から一塩基多型(SNPs)の過多または過少の発現を見出した。これらの発見のいくつかが、小さい効果サイズおよび重要でない統計的優位性によって制限される一方、Suh et al.38の研究は、同様にIGF−1受容体において機能的に重要な突然変異が、多少の長命のヒト、例えば100歳以上のヒトに存在することを示した。
【0074】
今日までに、ヒトにおけるFOXO遺伝子および加齢の表現型に関しては、ほとんど研究がされていない。二つの最近の研究は、FOXO遺伝子が更なる詳細な調査に値することを示唆する。第一に、年配のオランダ人の男性および女性の長期に渡る研究は、FOXO1Aハプロタイプが4年の生存率を予測し、FOXO3Aハプロタイプが、脳卒中の危険性を予測したことを明らかにした39。第二に、フラミンガム研究は、ゲノムの広範囲にわたる分析において、FOXO3A SNPが、女性において正常な閉経期で、年齢と強い関連性があることを見出した(p=0.00003)。しかしながら、オランダの調査結果は、多重比較を相当して、両方の研究が複製を必要とするとき、統計学的に有意でなかった。本研究は、FOXO3Aの関連性をヒトの寿命およびインスリン感受性に関して支持し、拡張する。
【0075】
現在の研究の主な利点の1つは、それがコホート内症例対照設計を使用することである。この調査設計は、ケースおよびコントロールを、進行中のコホート研究から長期に渡って収集したデータで選択する。従って、関連のあるいくつかの表現型(例えば、疾病の罹患、健康状態、機能)を、参加者がリコールバイアスし難いデータを作成する比較的若いときに、直接的な臨床検診によって得た。リコールバイアスは、調査結果が過去の事象を覚えることが困難なため正確でなく、高齢者にとっては大きな問題となる可能性がある。
【0076】
実際、比較的高齢の加齢を示唆する表現型の証拠を見出した特別な生存者の調査、例えば100歳以上の人は、重大なリコールバイアスを潜在的に被るおそれがある。即ち、高齢の参加者は、彼らの過去の病歴および彼らの過去の機能状態を、正確に思い出すことができない。しかしながら、現在の調査において、主な疾患は死亡率および罹病率の受託者に解決され、身体および認識機能の能力に基づく測定を自己評価の補充に使用され、証拠はこのような健康な加齢の表現型にて観測された。これは従来の遡及研究に潜在的な支持を与える。
【0077】
いくつかの他の長所が、この調査に存在する。第一に、分析で選択された候補遺伝子は、仮説に基づく基準に基づいて選択された。即ち、様々な方法を試用する加齢のモデル生物、特にノックアウトの研究は、IIS経路が加齢および寿命に重要であることを示した。多くの機能が、進化的に保存されていると考えられる。第二に、調査結果は強力で、著しく重要であり、いくつかの隣接したSNAPをFOXO3A遺伝子内に含有する。第三に、調査結果は生物学的に妥当で、従来の調査結果を老化動物モデルにて支持し、また制限された従来のヒトの研究をも支持する。第四に、寿命とケースコントロールの関係性は、コホート内症例対照分析を高度な事象の頻度(死亡率)で、検証の長期間を使用して検出した。第五に、HHPコホートは、高度に同種のコホートであり、研究の参加者において人口層化を全く検出されなかった。
【0078】
可能性のある欠点は、ケースおよびコントロールが、11歳の平均年齢の違いがあるため、誕生コホートを交絡因子として除くことができないことである。しかし、これは、参加者の間に最高19歳の違いが誕生年にあったため、疑わしい。同様に、部分分析は、ケースおよびコントロールにおいて、教育および職業(資料未記載)の違いが全く存在しないことを明らかにした。さらに、それは、95歳以上生存して、従って長寿の表現型を得る可能性のある基準にて年上である参加者であった。大抵のコホートの効果は、健康な利点を比較的若いコホートで示す。他の可能性のある欠点は、本研究を1つの集団のみで実施し、従って他の集団において、その総括を評価するために繰り返すべきである。
【0079】
要約すると、FOXO3A遺伝子内の一般的で、正常な遺伝子変異体は、ヒトの寿命と密接して関連したことを見出した。保護対立遺伝子の有病率が、年齢と共に著しく上昇した。長命のケースは、同様に癌および心血管疾患の低下した有病率、優れた自己報告の健康、高い機能状態を含有する健康的な加齢と関連のあるいくつかの付加的な表現型をおそらく保持し、それらが高いインスリン感受性のいくつかの生物学的マーカーの示唆を基礎調査にて示した。最後に、FOXO3A遺伝子内の特定の変異体は、同様にインスリン感受性、寿命の推定の中間表現型を含有するこれら加齢表現型のいくつかと関連した。
【0080】
B.対立遺伝子の患者(ヒトおよびヒト以外)の検出
多くの方法が、特定の対立遺伝子を、多型遺伝子座で検出するために利用できる。特異的な多型対立遺伝子の検出のための好適な方法は、多型の分子性質に部分的に依存するだろう。例えば、多型遺伝子座の様々な対立遺伝子形状は、DNAの一塩基対と異なる可能性がある。このような一塩基多型(またはSNP)は、全ての知られている多型の80%程を含有する遺伝子変異体の主な要因であり、ヒトゲノムにおけるそれらの濃度は平均1/1,000塩基対と推定する。SNPは最も頻繁な2対立遺伝子である―2つの形態のみで生じる(しかしDNAにて生じる4つの異なるヌクレオチド塩基に対応するSNPの最大4つの異なった形状が、理論的に可能である)。それにもかかわらず、SNPは他の多形体より突然変異的に安定していて、それらがマーカーおよび未知の変異体の間の連鎖不均衡を使用して、病原性突然変異体をマップする関連性の研究に適合できる。さらにSNPは二つの対立遺伝子のみを通常有するため、それらは長時間の測定よりはむしろ単純なプラス/マイナス分析によって遺伝子型を同定することができ、それらが自動的に従わせる。
【0081】
種々の方法が、特定の一塩基多型の対立遺伝子の存在を個々に検出するために利用可能である。本分野の促進は、正確、簡単および安価で大規模なSNP遺伝子型を同定で提供される。つい最近、例えば、いくつかの新技術は、ダイナミックアレルスペシフィックハイブリダイゼーション法(DASH)、ミクロプレートアレイ対角ゲル電気泳動(MADGE)、パイロシークエンス法、オリゴヌクレオチド特異的ライゲーション、TaqManシステム並びに様々なDNA「チップ」技術、例えばAffymetrix SNPチップを含有するものが記載された。これらの方法は、標的遺伝子領域の増幅を、通常PCRによって必要とする。さらに他の新しく開発された方法は、小さいシグナル分子の生成に基づく侵入切断(invasive cleavage)の後に、質量分析または固定されたパッドロックプローブおよびローリングサークル増幅によって小さいシグナル分子の生成に基づき、最終的にPCRの必要性を排除することがある。特異的な一塩基多型を検出する当業者に知られたいくつかの方法を以下に要約する。本発明の方法は、全ての利用可能な方法を包含することを理解される。
【0082】
いくつかの方法は、一塩基多型の分析法を容易にするため、開発された。一の実施形態において、一塩基多型を、例えばMundy,C.R.(米国特許第4,656,127号明細書)に開示されたような、特異的なエキソヌクレアーゼ抵抗性ヌクレオチドを使用して検出することができる。この方法によると、多型サイトの3’周辺の対立遺伝子に相補的なプライマーは、特定の動物またはヒトから得られた標的分子にハイブリッド形成することを許容する。標的分子での多型部位が、存在する特定のエキソヌクレアーゼ抵抗性ヌクレオチド誘導体に相補的であるヌクレオチドを含む場合、その誘導体をハイブリッド形成したプライマーの末端に組む込まれる。前記組み込みが、エキソヌクレアーゼに対して抵抗性のプライマーを与え、それによってそれの検出を可能にする。サンプルのエキソヌクレアーゼ抵抗性誘導体の同定が知られているため、プライマーはヌクレオチドが標的分子の多型部位に存在するエキソヌクレアーゼの曝露に抵抗性になった結果は、反応において使用されるヌクレオチド誘導体のものに相補的であった。この方法は、それが大量の無関係の配列データの測定を必要としない利点がある。
【0083】
本発明の他の実施形態において、溶液に基づく方法を、多型サイトのヌクレオチドの同一性の同定を決定するために使用する。Cohen, D. et al.(仏国特許発明第2,650,840号明細書;国際公開第91/02087号)。米国特許第4,656,127号明細書のMundyの方法のように、プライマーは多型サイトの3’周辺の対立遺伝子の配列に相補的であるものを用いた。この方法は、そのサイトのヌクレオチドの同一性を、ラベルされたジデオキシヌクレオチド誘導体を使用して決定し、多型のヌクレオチドに相補的な場合は、プライマーの末端に組み込まれる。
【0084】
遺伝子ビット分析(Genetic Bit Analysis)またはGBATMとして知られている代わりの方法が、Goelet,P et al.に記載されている(国際公開第92/15712号)。Goelet,P. et al.の方法は、ラベルを付けられたターミネータと、多型サイトの3’配列に相補的なおよびプライマーとの混合物を使用する。従って組み込まれてラベルを付けられたターミネータは、評価される標的分子の多型サイトに存在するヌクレオチドおよびその相補物によって決定される。Cohn et al.(仏国特許発明第2,650,840号明細書;国際公開第91/02087号)の方法とは対照的に、Goelet,P. et al.の方法は、好ましくは異質の相分析であり、プライマーまたは標的分子を固体相に固定する。
【0085】
近年では、DNAにおいて多型サイトの分析のためのいくつかのプライマーによって誘導されたヌクレオチドの組込方法を記載した(Komher, J. S. et al., Nucl. Acids. Res.17:7779−7784(1989); Sokolov, B. P., Nucl. Acids Res.18:3671(1990); Syvanen, A.−C, et al., Genomics8:684−692(1990); Kuppuswamy, M. N. et al., Proc. Natl. Acad. Sci. (U.S.A.)88:1143−1147(1991); Prezant, T. R. et al., Hum. Mutat. 1:159−164(1992); Ugozzoli, L. et al., GATA 9:107−112(1992); Nyren, P. et al., Anal. Biochem. 208:171−175(1993)。これらの方法は、これら全てがラベルされたデオキシヌクレオチドの組み合わせに依存して、多型サイトで塩基間を区別する点でGBATMと異なる。このようなフォーマットにおいて、シグナルは組み込まれたデオキシヌクレオチドの数に比例するため、同じヌクレオチドの流れを生じる多型は、流れの長さと比例するシグナルを生じることができる(Syvanen, A. −C, et al., Amer. J. Hum. Genet.52:46−59(1993))。
【0086】
タンパク質翻訳の早い終結を引き起こす突然変異体に関して、プロテイントランケーションテスト(PTT)は、効率的な診断方法を提供する(Roest, et. al., (1993) Hum. MoI Genet. 2:1719−21; van der Luijt, et. al., (1994) Genomics 20:1 −4)。PTTに関して、RNAは利用可能な組織から最初に分離、逆転写され、関連する断片をPCRによって増幅された。その後、逆転写PCRの生成物を、ネステッドPCR増幅により、RNAポリメラーゼプロモーターを含有するプライマーおよび真核生物の翻訳を開始する配列と共に使用された。関連領域の増幅後、プライマー内に組み込まれた固有のモチーフは、一連の生体外におけるPCR産物の転写および翻訳を可能にする。翻訳の生産物のドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動で、短縮型ポリペプチドの出現は、翻訳の早い終結の原因となる突然変異の存在を示す。この技術の多様化において、DNA(RNAに対して)を、関連のある標的領域が単一のエクソンに由来するとき、PCRの鋳型として使用する。
【0087】
あらゆる細胞種または組織を利用して、本明細書に記載の診断における使用で核酸サンプルを得ることができる。好ましい実施形態において、DNAサンプルを、既知の技術(例えば、静脈穿刺)によって得られる体液、例えば血液または唾液から得る。選択的に、核酸試験を乾式サンプルで実施することができる(例えば、髪の毛または皮膚)。RNAまたはタンパク質を使用するとき、用いることができる細胞または組織は、FOXO3A遺伝子を発現する必要がある。
【0088】
診断法をまた、生検または切除から得られる患者の組織の(固定および/または凍結されている)組織断片で、核酸精製が全く必要とすることないようにその場にて直接実施することができる。核酸試薬を、プローブおよび/またはプライマーとして、前記その場の方法で使用することができる(例えば、Nuovo, G. J., 1992, PCR in situ hybridization: protocols and applications, Raven Press, N.Y.参照)。
【0089】
一つの核酸配列の検出で主に焦点を合わせる方法に加えて、プロフィールをまたこのような検出方式において評価することができる。指紋プロフィールを、例えばディファレンシャルディスプレイ法、ノーザン分析および/またはRT−PCRを利用することによって生成することができる。
【0090】
好適な検出方法は、FOXO3Aハプロタイプの少なくとも一つの対立遺伝子の重なり合った領域のプローブを使用し、約5、10、20、25または30ヌクレオチドを突然変異または多型の領域付近にて有する対立遺伝子特異的なハイブリダイゼーション形成である。本発明の好ましい実施形態において、他の対立遺伝子変異体に特異的にハイブリッド形成することができるいくつかのプローブが、固相担体、例えば「チップ」(最高約250,000オリゴヌクレオチドを適用することができる)に付着される。オリゴヌクレオチドは、リソグラフィーを含む様々な工程によって固体支持体に結合することができる。オリゴヌクレオチドを含有するこれらのチップを用いる突然変異検出分析は、また「DNAプローブアレイ」といい、例えばin Cronin et al.(1996) Human Mutation7:244に記載される。一実施形態において、チップは遺伝子の多型領域の少なくとも1つの、全ての対立遺伝子変異体を含有する。その後固相担体を、テスト核酸と接触させ、特定のプローブに対するハイブリッド形成を検出する。従って、一つ以上の遺伝子の多くの対立遺伝子の変異体の同定は、単純なハイブリッド実験で同定することができる。
【0091】
これらの技術は、また核酸の増幅する工程を分析前に含有することができる。増幅技術は、当業者に知られ、クローニング、ポリメラーゼ連鎖反応(PCR)、対立遺伝子特異的増幅法(ASA)、リガーゼ連鎖反応(LCR)、自己持続型の核反応(Guatelli, J. C. et al., 1990, Proc. Natl. Acad. Sci. USA 87:1874−1878)、転写増幅システム(Kwoh, D. Y. et al., 1989, Proc. Natl. Acad. Sci. USA 86:1173−1177)、およびQβレプリカーゼを含有するが、これらに制限されない。
【0092】
増幅産物は様々な方法によって検出することができ、これはサイズ分析、サイズ分析後の制限分解作用による切断、反応生成物の特異的タグを付けられたオリゴヌクレオチドプライマーの検出、対立遺伝子の特異的オリゴヌクレオチド(ASO)ハイブリッド形成、対立遺伝子の特異的5’エキソヌクレアーゼ検出、塩基配列決定、ハイブリッド形成等を含有する。
【0093】
PCRに基づく検出手段は、同時に複数の標識の多重増幅を含有することができる。例えば、サイズにおいて重なり合わず、同時に分析することができるPCR産物を生成するPCRプライマーを選択することは、よく知られた技術である。選択的に、特異的にラベルして、従ってそれぞれ特異的に検出することができるプライマーと、異なった標識を増幅することが可能である。当然、ハイブリッド形成に基づく検出方法は、複数のPCR産物の異なった検出をサンプルにおいて可能とする。他の技術では、複数の標識の多数の分析を可能にする従来技術で知られている。
【0094】
単に例証となる実施形態において、前記方法は、(i)細胞サンプルを患者(唾液、頬の内側の粘膜、血液、その他の体液または成分)から収集する工程と、(ii)核酸(ゲノム、mRNAまたはその両方)をサンプルの細胞から分離する工程と、(iii)核酸サンプルと、FOXO3Aハプロタイプの少なくとも1つの対立遺伝子の5’および3’に、対立遺伝子のハイブリッド形成および増幅を生じるような条件下において、特異的にハイブリッド形成する一つ以上のプライマーを接触させる工程と、(iv)増幅産物を検出する工程とを含有する。これらの検出方法は、このような分子が比較的低い数で存在する場合、核酸分子の検出に特に実用的である。
【0095】
対象となる分析の好ましい実施形態において、FOXO3Aハプロタイプの対立遺伝子を、制限酵素切断パターンにおける変化によって同定する。例えば、サンプルおよびコントロールのDNAを分離し、(任意に)増幅し、1つ以上の制限エンドヌクレアーゼで切断し、断片の長さのサイズを、ゲル電気泳動によって決定する。
【0096】
さらにもう一つの実施形態において、従来技術で知られる様々な塩基配列決定反応のいずれかを、対立遺伝子配列に直接使用することができる。例示的な塩基配列決定反応は、Maxim and Gilbert ((1977) Proc. Natl Acad Sci USA74:560) or Sanger (Sanger et al (1977) Proc. Nat. Acad. Sci USA 74:5463)によって開発された技術に基づくものを含有する。自動塩基配列決定の様々な方法のいずれかは、質量分析による塩基配列決定を含有する(例えば、国際公開第94/16101号参考;Cohen et al.(1996)Adv Chromatogr36:127−162;およびGriffm et al.(1993)Appl Biochem Biotechnol38:147−159)対象分析(例えば、Biotechniques(1995)19:448参考)を実行するときに、使用することができることを、同様に予測される。特定の実施形態で、1、2または3の核酸塩基のみの発生を、配列反応において決定擦る必要があることが、当業者に明らかであるだろう。例えば、Aトラック等を、1つの核酸のみを検出する場合に、実施することができる。
【0097】
更なる実施形態において、切断剤からの保護を使用して、RNA/RNA、RNA/DNAまたはDNA/DNAのヘテロ2本鎖に基づくミスマッチ塩基を検出することができる(Myers, et al.(1985)Science230:1242)。一般的に、従来技術の「ミスマッチ切断」は、野生型対立遺伝子をサンプルに含む(ラベルされた)RNAまたはDNAにハイブリッド形成することによって、ヘテロ二本鎖の形成を提供することによって開始する。二本鎖デュプレックスを、例えばコントロールおよびサンプル鎖の間の塩基対ミスマッチのために存在するデュプレックスの一本鎖領域を切断する薬剤で処理する。例えば、RNA/DNAデュプレックスを、SIヌクレアーゼで処理してミスマッチ領域を酵素で切断するRNaseおよびDNA/DNAハイブリッドで処理することができる。他の実施形態において、DNA/DNAまたはRNA/DNAデュプレックスのいずれかを、ヒドロキシルアミンまたは四酸化オスミウムおよびピペリジンで処理して、ミスマッチ領域を切断することができる。その後ミスマッチ領域の切断後、結果として生じる物質を、サイズによって変性ポリアクリルアミドゲルにて分離し、突然変異サイトを断定する。例えば、Cotton et al(1988)Proc. Natl Acad Sci USA85:4397;およびSaleeba et al (1992) Methods Enzymol.217:286−295参考。好ましい実施形態において、コントロールDNAまたはRNAを、検出のためにラベルすることができる。
【0098】
さらに別の実施形態において、ミスマッチ切断反応は、ミスマッチ塩基対を二重鎖DNAにて認識する1つ以上のタンパク質(「DNAミスマッチ修復」酵素と称す)を使用する。例えば、大腸菌のmutY酵素は、AをG/Aミスマッチで切断し、HeLa細胞からチミジンDNAグリコシラーゼはTをG/Tミスマッチで切断する(Hsu et al.(1994)Carcinogenesis 15:1657−1662)。例示的実施形態の記載のように、FOXO3A遺伝子座ハプロタイプの対立遺伝子に基づくプローブは、テスト細胞からcDNAまたは他のDNA産物にハイブリッドを形成する。デュプレックスを、DNAミスマッチ修復酵素で処理し、切断生成物は、必要である場合、それを電気泳動プロトコール等から検出することができる。例えば、米国特許第5,459,039号明細書を参照とし、本明細書において、完全に引用したものとする。
【0099】
他の実施形態において、電気泳動移動度の違いを使用して、FOXO3A遺伝子座の対立遺伝子を同定するだろう。例えば、一本鎖高次構造多型(SSCP)を使用して、電気泳動移動度における、突然変異体および野生型核酸の間の相違を検出することができる(Orita et al.(1989)Proc Natl. Acad. Sci USA86:2766、またCotton(1993)Mutat Res285:125−144参考; and Hayashi(1992)Genet Anal Tech Appl9:73−79)。サンプルおよびコントロールFOXO3A遺伝子座の対立遺伝子の一本鎖DNA断片を、変性し、復元を可能にした。一本鎖核酸の2次構造は、配列により異なり、電気泳動移動度における結果として生じる変性は、一塩基の変化さえ検出することが可能である。DNA断片を、ラベルされたプローブで、ラベルするか検出することができる。分析の感度はRNAを使用することによって向上することができ、2次構造が、配列の変異に比較的感度がある。好ましい実施形態において、対象となる方法は、ヘテロ二本鎖分析を利用して、電気泳動移動度の変化に基づいて、二重鎖へテロ二本鎖分子を分離する(Keen et al.(1991)Trends Genet7:5)。
【0100】
さらにもう一つの実施形態において、変性剤の濃度勾配を含むポリアクリルアミドゲルにおける対立遺伝子の変動を、変性剤濃度勾配ゲル電気泳動法(DGGE)を使用して分析した(Myers et al.(1985)Nature313:495)。DGGEを分析方法として使用するとき、DNAはそれが例えばPCRで高い融解のGCに富んだDNAの約40bpのGCクランプを添加することによって完全に変性しないことを確実にするために修飾するだろう。更なる実施形態において、温度勾配を、変性剤の勾配の代わりに使用して、対象およびサンプルのDNAの移動度における相違を同定することができる(Rosenbaum and Reissner(1987)Biophys Chem265:12753)。
【0101】
対立遺伝子の検出する他の技術の例は、選択的なオリゴヌクレオチドハイブリッド形成、選択的な増幅または選択的なプライマー伸長を含有するが、これらに制限されない。例えば、オリゴヌクレオチドプライマーを、既知の突然変異またはヌクレオチド(例えば、対立遺伝子変異体)の相違を中央に配置し、その後、完全に一致を発見するときのみにハイブリッド形成を可能にする状態で標的DNAにハイブリッド形成することで調整することができる(Saiki et al. (1986) Nature 324:163); Saiki et al (1989) Proc. Natl Acad. Sci USA 86:6230)。このような対立遺伝子に特異的なオリゴヌクレオチドのハイブリッド形成技術を使用して、オリゴヌクレオチドが、PCR増幅された標的DNAにハイブリッド形成したとき1つの突然変異または多型態領域について1つの反応を、またはオリゴヌクレオチドが、ハイブリッド膜に付着し、ラベルされた標的DNAにハイブリッド形成したとき数々の異なった突然変異または多型体の領域を試験する。
【0102】
選択的に、対立遺伝子特異的な増幅技術は、選択的なPCR増幅に依存し、本発明と共に使用することができる。特異的な増幅のために使用されるオリゴヌクオチドは、(増幅が異なったハイブリッド形成に依存するために)分子の中心における関連のある突然変異または多型領域(Gibbs et al (1989) Nucleic Acids Res.17:2437−2448)、または適切な条件に基づいて、ミスマッチがポリメラーゼの伸長を防止または減少することができる場合に1つのプライマーの3’の最終末端で担持することができる(Prossner(1993)Tibtech11:238)。さらに、新規な制限酵素サイトを突然変異の領域に導入して切断に基づく検出を生成することが望ましい(Gasparini et al(1992)Mol. Cell Probes6:1)。特定の実施形態において、増幅が同様に増幅でTaqリガーゼを使用して実施することができることが予想される(Barany(1991)Proc. Natl. Acad. Sci USA88:189)。そのような場合、ライゲーションは、完全な一致が、5’配列の3’末端に存在し、増幅の有無を調べることによって、特定のサイトにて既知の突然変異の存在を検出することが可能となる場合のみに生じる。
【0103】
他の実施形態において、対立遺伝子変異体の同定を、例えば米国特許第4,998,617号明細書およびLandegren, U. et al.((1988) Science 241 :1077−1080)に記載されたように、オリゴヌクレオチドのライゲーション分析(OLA)を使用して実施する。OLAプロトコールは、標的の一本鎖の接触配列にハイブリッド形成することができるように設計される2つのオリゴヌクレオチドを使用する。オリゴヌクレオチドの1つは、分離標識、例えばビオチン化で結合し、他は検出が可能なようにラベルされる。正確な相補的配列が、標的分子に見出す場合、オリゴヌクレオチドが、それらの末端が隣接するようにハイブリッドを形成し、ライゲーション基質を生成する。その後ライゲーションは、ラベルされたオリゴヌクレオチドがアビジンまたは他のビオチンリガンドを使用して回復されることを可能にする。Nickerson, D. A. et al.は、PCRおよびOLAの性質を組み合わせる核酸検出分析を記載する(Nickerson, D. A. et al.(1990)Proc. Natl. Acad. Sci. USA87:8923−27)。この方法において、PCRを使用して、標的DNAの指数関数的増幅を達成し、その後OLAを用いて検出される。
【0104】
このOLA方法に基づくいくつかの技術が開発され、FOXO3a遺伝子座ハプロタイプの対立遺伝子を検出するために使用することができる。例えば、米国特許第5,593,826号明細書は、3’アミノ基を有するオリゴヌクレオチドおよび5’リン酸化オリゴヌクレオチドを使用して、アミド亜リン酸エステル結合を有する結合体を形成するOLAを開示する。Tobe et al.((1996)Nucleic Acids Res24:3728)に記載のOLAの他の変異体において、PCRと組み合わせたOLAは、2つの対立遺伝子の分類化を、一つのマイクロタイターウェルにて可能にする。それぞれの対立遺伝子を固有のハプテン、即ちジゴキシゲニンおよびフルオレセインでマークすることによって、それぞれのOLA反応を、異なった酵素レポーター、アルカリホスファターゼまたはセイヨウワサビペルオキシダーゼでラベルされたハプテン特異的な抗体を使用することによって検出することができる。このシステムは、2つの異なる色の製造を導くハイスループットフォーマットを使用する2つの対立遺伝子の検出を可能にする。
【0105】
本発明の他の実施形態は、近い将来における長命の可能性、または健康または診断の介入の必要性を検出するためのキットを対象とする。このキットは、FOXO3A遺伝子座のハプロタイプの少なくとも1つの対立遺伝子に5’および3’をハイブリッド形成する5’および3’オリゴヌクレオチドを含む、一つ以上のオリゴヌクレオチドを含有することができる。PCR増幅オリゴヌクレオチドは、その後の分析で適切なサイズのPCR産物を生成するために、25から2500の独自の塩基対の間、好ましくは約100から500の独自の塩基とハイブリッドを形成する必要がある。
【0106】
特に好ましいプライマーは、SEQ ID NO.2〜9に記載のヌクレオチド配列を含有する。これら遺伝子においてヒトの多型の検出のために適切なプライマーを、従来技術において既知であるこの配列情報および通常の技術を使用して、プライマー配列の設計および最適化のために容易に設計することができる。上記プライマー配列の最適な設計を、例えば、市販のプライマー選択プログラム、例えばPrimer2.1、Primer3またはGeneFisherの使用によって達成することができる。
【0107】
「GCCハプロタイプ」の検出のための簡単な方法の例は、関連のある特異的なヌクレオチドを増幅し、段落[0081]に記載のものと同様に対立遺伝子特異的なプライマーの使用を含む。この方法は、オリゴヌクレオチドプライマーが、それらの3’末端にて完全にアニールして、DNAポリメラーゼがこれらプライマーをPCR時に伸長する必要がある事実を利用する。特異的なDNAの点相違のみと一致するオリゴヌクレオチドプライマーを設計することによって、rs2802292多型にて見られるようなもの−T種対立遺伝子に結合しないプライマー−このようなプライマーは多型体対立遺伝子と区別することができる。コントロール反応を(ARMS:amplification refractory mutation system)のような同じチューブ内で設定し、所定のサンプルからの生成物の生成の欠如が、分析を確かめる「G」変異体の欠損よりむしろ、PCR反応の失敗によるように単純でないことを確実にすることが必要である。
【0108】
このために使用されるオリゴヌクレオチドは、フォワードアウター(「rs2802292_FO」)で、SEQ ID NO.5として同定される5’―GAAACTGAGGCTAACAGCTGGGTCTGGCCC―3’、リバースアウター(「rs2802292_RO」)で、SEQ ID NO.6として同定される5’―AGCTGATGCTCCTCAACGAAACCACCTTAC―3’、リバースG特異的(「rs2802292_RG」)で、SEQ ID NO.7として同定される5’―GGACCCCTTCATCTGTCACACAGAGGCTCC―3’、フォワード特異的(「rs2802292_FT」)で、SEQ ID NO.8として同定される5’―CTGTTGCTCACAAGAGCTCAGGGCTGGGcT―3’を含み、最後の2つのプライマーにおいて、下線の引かれた末端塩基は、G−Tの相違のサイトでアニールし、3’末端から2番目のbp(小文字)は、対立遺伝子の特異性を最大限にするために意図的にミスマッチされる。この例証における4つのプライマーを、表No.9に記載する。
【0109】
【表11】
表9は、出現の順において、それぞれSEQ ID NOS5〜8を開示する。
【0110】
プライマーおよびG/T変異体を示すPCR産物のDNA配列(ソースGenbank AL391646.12)は、以下の通りである:
【0111】
【0112】
このように発生した増幅物をアガロースゲルで分離したとき、Gタイププライマーは186bp生成物を発生することを示すことができ、Tタイププライマーは132bp生成物を生じる。アウトサイドプライマーは、反応が正確に進んだことを保証するために、全ての反応で存在すべき288bp生成物を生成する。
【0113】
例示的な、増幅の試薬および条件を、表10に示す。
【0114】
【表12】
【0115】
増幅された断片は、図1に示すような3%アガロースゲルに分離することができる。図1はARMS−PCR定量分析の結果を示し、上記のプライマーおよび条件を使用してFOXO3A G/T変異体を検出する。レーン1は「T」対立遺伝子(132bp)の対象となる同型を示す;レーン2および3は、「G」対立遺伝子(186bp)に同型である対象を示す;レーン4および5は、「T」および「G」の対立遺伝子(132+186bp)に異型である対象を示し、Mは100bpDNAラダー(Invitrogen, Paisley, United Kingdom)である。
【0116】
要約すると、図2は分析の模式的な概略を示す。プライマー「rs2802292_FO」および「rs2802292_RO」は、多型体遺伝子座rs2802292の隣接し、コントロール288bpバンドを全ての場合にて発生する。プライマー「rs2802292_OF」および「rs2802292_RG」は、186bpのG特異的生成物を発生し、プライマー「rs2802292_FT」および「rs2802292_OR」は、132bpのT特異的生成物を発生する。
【0117】
キットの使用のために、オリゴヌクレオチドは様々な天然および/または合成の組成物のいずれか、例えば合成オリゴヌクレオチド、制限断片、cDNA、合成ペプチド核酸(PNA)等であることができる。分析キットおよび方法は、同様にラベルされたオリゴヌクレオチドを使用して、分析において同定を容易にすることを可能にすることができる。用いることができるラベルの例は、放射性同位元素識別、酵素、蛍光化合物、ストレプトアビジン、アビジン、ビオチン、磁性部位、金属結合部位、抗原または抗体の部位等を含有する。
【0118】
キットは、任意に、またDNAサンプリング方法を含有することができる。DNAサンプリング方法は当業者によく知られる一つであり、基質、例えば濾紙、AmpliCardTM(University of Sheffield, Sheffield, England SlO 2 J F; Tarlow, J W, et al, J of Invest. Dermatol.103:387−389(1994))等;DNA精製試薬、例えばNucleonTMキット、リシスバッファー、タンパク質分解酵素溶液等;PCR試薬、例えば反応バッファー、熱安定性ポリメラーゼ、dNTP等;および対立遺伝子検出方法、例えばHinfI制限酵素、対立遺伝子特異的オリゴヌクレオチド、乾燥血痕からのネステッドPCRのための縮重オリゴヌクレオチドプライマーを含有するが、これらに限定されない。
【0119】
C.薬理ゲノミクス
特定の対立遺伝子の知識は、特定の疾患または状態の進行する感受性と関連し、特定の疾患または状態に関与する他の遺伝子異常に関する情報と単独または組み合わせては、「薬理ゲノミクス」の目標である個々の遺伝子プロフィールに従う予防または治療のカスタム化を可能とする。従って、健康的な加齢で個々のFOXO3Aプロフィールと集団的プロフィールの比較は、特定の患者または患者集団で安全且つ効果的であると期待する薬物もしくは他の治療療法の選択または設計を可能とする(即ち、同じ遺伝子の変性を有する患者の一群)。
【0120】
本発明に記載の特定の対立遺伝子の知識を使用して、細胞の性質の違いを細胞培養および組織システムにて観測し、細胞または組織培養システムに添加された化学的または生物学的薬剤への細胞の反応を測定することができる。細胞性質および反応の相違点を遺伝子型間で比較して、健康を改善または寿命を延ばす要求のため実施することができる薬物または他の薬理的物質を同定するか、または遺伝子もしくは遺伝子発現で毒性または潜在的な効果で新規化合物を試験することができる。
【0121】
さらに標的集団に対する可能性は、最も高い臨床利点を示すことを期待して、遺伝子プロフィールに基づいて:1)既にマークされた薬物の再配置;2)臨床開発が安全性または効率性の制限の結果として中断され、患者のサブグループに特有である候補薬の救済;および3)候補治療および比較的最適な薬物のラベリングで、急速な、比較的費用が高くない開発を可能とすることができる(すなわち、薬剤の様々な服用量の効果を、原因となる突然変異で測定することは、有効量の最適化に実用的である)。
【0122】
特定の治療薬で個々の治療は、長寿と関連のある遺伝子の発現レベルを測定することによって観測することができる。発現レベルを、タンパク質(例えば、FOXO3A)、mRNAおよび/または転写レベルを同定することによって、測定することができる。その後検出されるレベルに依存して、治療療法を、維持または調整することができる(増幅量を増加または減少)。好ましい実施形態において、対象を薬剤で治療する効果は以下の工程を含有する:(i)薬剤を投与する前に前投与サンプルを対象から得る;(ii)タンパク質、mRNAまたはゲノムDNAのレベルまたは量を前投与サンプルにおいて検出する;(iii)治療薬の投与後、1つ以上の投与後のサンプルを対象から得る;(iv)タンパク質、mRNAまたはゲノムDNAの発現または活性のレベルを、投与後のサンプルにおいて検出する;(v)前投与サンプルにおけるタンパク質、mRNAまたはゲノムDNAの発現または活性のレベルと、投与後のサンプルにおける対応するタンパク質、mRNAまたはゲノムDNAを、それぞれ比較する;(iv)薬剤の投与を対象に応じて変更する。
【0123】
対象の細胞をまた、FOXO3A遺伝子以外の遺伝子発現のレベルを検出するため治療的の投与前後で得て、治療が有害である可能性がある遺伝子の発現を増加または減少しないことを確認することができる。これは、例えば転写プロファイルの方法を使用して実施することができる。従って、生体内において治療に露出された細胞からのmRNA、治療に露出されていない同じ種類の細胞のmRNAは、逆転写され、多くの遺伝子からのDNAを含むチップにハイブリッド形成することができ、この結果、治療で処理または未処理の細胞の遺伝子の発現を比較する。
【0124】
「GCC」ハプロタイプを、危険率の算出において、死亡および年齢に関連する疾患(心臓病、脳卒中、癌、COPDまたはその他慢性の肺疾患、パーキンソン病、糖尿病および認知症)および将来的な身体機能(歩行能力、認識機能)の予測の補助に使用することができる。この情報は、市民、医師、健康管理会社および保険会社にとって関心があるものである。既知の危険性算出の例は、Perls、米国特許出願公開第2007/0118398A1号明細書、2007年5月24日に公開されたシステムおよび方法を含有し、本明細書において完全に引用する。危険率の算出を、例えば医師局にて、携帯端末またはオンラインにて提供することができる。人がどれくらい生存するかを予想するのに興味がある個人、健康介護専門家、保険会社、健康管理組織は、コンピューターに男/女の遺伝子型を入力して、老化関連の疾患の危険性のスコア、健康的な残余寿命の年数および残余寿命の合計年数を得ることができる。
【0125】
特定のスコアに基づいて、医師または保険専門家は、上記疾患および死亡、特にFOXO3A遺伝子の比較的保護的でない変異を有するヒトで、健康的な生活または危険性の減少に関して、患者に知らせることができる。いくつかの例示的な選択肢は、食物の選択(例えば、赤ワイン、大豆製品およびFOXO3A遺伝子の活性に影響を及ぼすことができる化合物を含む他の食品)、または強い危険性要因の改善、例えば体重減少または増加の身体活性に関するアドバイスを含む。
【0126】
FOXO3Aおよび特にGCCハプロタイプの健康な老化および長寿の予測手段の同定は、実用的な生物学および標的の有望な情報源を、製薬のスクリーンおよび試験に提供する。例えば、一つは、FOXO3A遺伝子によって年齢関連疾患を減少で予期される健康的利益のために産出される遺伝子産物またはタンパク質もしくは他の活性化合物を利用することができる。遺伝子産物を利用する方法は、製薬技術において知られている摂取、注射、経皮的投与および他の方法を含有する。化合物をスクリーンして、FOXO3A、特にGCCハプロタイプによって産出される遺伝子産物の種類、活性または量に影響するものを見つけることができる。
【0127】
本発明は、年齢関連性疾患の予防または治療するFOXO3Aの調製方法を含有する。本発明はまた、FOXO3Aが例えば年齢に関連のある疾患または対象において寿命の向上と関連する疾患または状態を治療または予防する方法を含有する。本明細書において使用される「対象」とは、ヒトまたはヒトでない動物とする。「ヒトでない動物」という用語は、哺乳類、人間以外の霊長類(特に比較的高度な霊長類)のような全ての脊髄動物、ウマ、ウシ、バイソン、バッファロ、ヤギ、ブタ、ヒツジ、ニワトリ、カモおよびガチョウのような飼育哺乳類、イヌ、ネコ、ウサギ、モルモットのようなペット、齧歯動物、爬虫類および実験動物を含有する。好ましい実施形態において、対象はヒトである。他の実施形態において、対象は、疾患モデルとして適切な実験動物または遺伝形質転換動物である。調整および治療の方法は、Geesaman et al.の2007年5月10日に公開された米国特許出願公開第2007/0105109Al号明細書において当業者に知られ、完全に本明細書に引用する。
【0128】
本発明によって教示した対立遺伝子の変異体の配列または遺伝子産物の、多くの他の診断および治療への使用は、当業者に明らかであるだろう。小分子スクリーン、アンチセンスオリゴヌクレオチド、低分子二本鎖RNA(siRNA)の使用を含有するいくつかの例が、当業者に明らかである。FOXO活性に関する診断および治療の使用を開発するいくつかの方法は、Goldberg et al.の2006年3月30日に公開された米国特許出願公開第2006/0069049 A1号明細書に記載され、関連する経路にはTissenbaum et al.の2006年11月30日に公開された米国特許出願公開第2006/0272039号明細書に関係があり、共に完全に本明細書に引用する。
【0129】
前述の明細書を、当業者が本発明を実施することを可能にするため、充分であると考慮する。当業者は、単に通常の方法を用いて、多くの本発明の特定の実施形態と均等物を理解するか、または同定することができる。これら均等物は、添付の特許請求項に含有することを意図する。実際に、本明細書に示しまたは記載されるものに加えて本発明の様々な修正は、前述の記載から当業者に明らかになり、添付の特許請求の範囲に含まれる。
【0130】
本文に記載されていない参考文献
1. Herskind AM et al. (1996). The heritability of human longevity: a population−based study of 2872 Danish twin pairs born 1870−1900. Hum Genet 97:319−323.
2. Yashin AI, Iachine IA, Harris JR (1999) Half of variation in susceptibility to mortality is genetic: findings from Swedish twin survival data. Behav Genet 29:11−19.
3. Christensen K, Johnson TE, Vaupel JW (2006) The quest for genetic determinants of human longevity: challenges and insights. Nat Rev Genet 7:436−448.
4. Willcox DC, Willcox BJ, Hsueh WC, Suzuki M (2006) Genetic determinants of exceptional human longevity: insights from the Okinawa Centenarian Study. AGE 28:313−332.
5. Bishop NA, Guarente L (2007) Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nat Rev Genet 8:835−844.
6. Risch N, Zhang H (1995) Extreme discordant sib pairs for mapping quantitative trait loci in humans. Science 268: 1584−1589.
7. Gundmundsson H et al. (2000) Inheritance of human longevity in Iceland. Eur J Hum Genet 8:743−749.
8. Kerber RA, O´Brien E, Smith KR, Cawthon RM (2001) Familial excess longevity in Utah genealogies. J Gerontol A Biol Sci MedSci 56:B130−B139.
9. Perls TT et al. (2002) Life− long sustained mortality advantage of siblings of centenarians. Proc Natl Acad Sci USA 99:8442−8447.
10. Willcox BJ, Willcox DC, He Q, Curb JD, Suzuki M (2006) Siblings of Okinawan centenarians exhibit lifelong mortality advantages. J Gerontol A Biol Med Sci 61 :345− 354.
11. Partridge L (2007) Some highlights of research on aging with invertebrates, 2006−2007. Aging Cell 6:595−598.
12. Ghazi A, Henis−Korenblit S, Kenyon C (2007) Regulation of Caenorhabditis elegans lifespan by a proteasomal E3 ligase complex. Proc Natl Acad Sci USA 104:5947−5952.
13. Bartke A (2008) New findings in gene knockout, mutant and transgenic mice. Exp Gerontol 43:11 −14.
14. Lin K, Dorman JB, Rodan A, Kenyon C (1997) daf−16: An HNF−3/forkhead family member that can function to double the life−span of Caenorhabditis elegans. Science
278:1319−1322.
15. Brunet A et al. (2004) Stress−dependent regulation of FOXO transcription factors by the SIRTl deacetylase. Science 303:2011−2015.
16. Kenyon C, Murphy CT (2006) Enrichment of regulatory motifs upstream of predicted DAF−16 targets. Nat Genet 38:397−398.
17. van der Horst A, Burgering BM (2007) Stressing the role of FOXO proteins in lifespan and disease. Nat Rev MoI Cell Biol 8:440−450.
18. Russell SJ, Kahn CR (2007) Endocrine regulation of ageing. Nat Rev MoI Cell Biol 8:681−691.
19. Sauve AA, Wolberger C, Schramm VL, Boeke JD (2006) The biochemistry of Sirtuins. Ann Rev Biochem 75:435−465.
20. Tissenbaum HA, Guarente L (2001) Increased dosage of a sir −2 gene extends lifespan in Caenorhabditis elegans. Nature 410:227−230.
21. Blvher M, Kahn BB, Kahn CR (2003) Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 299:572−574.
22. Bartke A (2008) Impact of reduced insulin−like growth factor− 1 /insulin signaling on aging in mammals: novel findings. Aging Cell 7:285−290.
23. White MF (2003) Insulin signaling in health and disease. Science 302:1710−1711.24. Conover CA, Bale LK (2007) Loss of pregnancy −associated plasma protein A extends lifespan in mice. Aging Cell 6:727−729.
25. Curran SP, Ruvkun G (2007) Lifespan regulation by evolutionarily conserved genes essential for viability. PLoS Genet 3:e56.
26. Greer EL et al. (2007) An AMPK−FOXO pathway mediates the extension of lifespan induced by a novel method of dietary restriction in C. elegans. Curr Biol 17:1646− 1656.
27. Berdichevsky A, Viswanathan M, Horvitz HR, Guarente L (2006) C. elegans SIR −2.1 interacts with 14−3−3 proteins to activate DAF−16 and extend life span. Cell 125:1165− 1177.
28. Boehm M, Slack F (2005) A developmental timing MicroRNA and its target regulate life span in C. elegans. Science 310:1954−1957.
29. Hsu AL, Murphy CT, Kenyon C (2003) Regulation of aging and age−related disease by DAF−16 and heat−shock factor. Science 300:1142−1145.
30. Murphy CT (2006) The search for DAF−16/FOXO transcriptional targets: approaches and discoveries. Exp Gerontol 41 :910−921.
31. Wang MC, Bohmann D, Jasper H (2005) JNK extends lifespan and limits growth by
antagonizing cellular and organism −wide responses to insulin signaling. Cell 121 :115− 125.
32. Giannakou ME et al. (2007) Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging Cell 6:429438.
33. Harman D (1956) Aging: a theory based on free radical and radiation chemistry. J Gerontol 11 :298−300.
34. Beckman KB, Ames BN (1998) The free radical theory of aging matures. Physiol Rev 78:547−581.
35. Fusco D, Colloca G, Lo Monaco MR, Cesari M (2007) Effects of antioxidant supplementation on the aging process. Clin Interv Aging 2:377−387.
36. Bonafe M et al. (2003) Polymorphic variants of IGF−I receptor and phosphoinositide 3− kinase genes affect IGF−I plasma levels and human longevity: cues for an evolutionarily conserved mechanism of life span control. J Clin Endocrinol Metab 88:3299−3304.
37. Kojima et al. (2004) Association analysis between longevity in the Japanese population and polymorphic variants of genes involved in insulin and insulin−like growth factor 1 signaling pathways. Exp Gerontol 39:1595−1598.
38. Suh Y et al. (2008) Functionally significant insulin−like growth factor I receptor mutations in centenarians. Proc Natl Acad Sci USA 105:3438−3439.
39. Kuningas M et al. (2007) Haplotypes in the human FOXOIa and FOXO3a genes: impact on disease and mortality at old age. Eur J Hum Genet 15:294−301.
40. Lunetta KL et al. (2007) Genetic correlates of longevity and selected age−related phenotypes: a genome −wide association study in the Framingham Study. BMC Med Genet 8 Supple 1 : S13.
41. Nebel A et al. (2005) No association between microsomal triglyceride transfer protein (MTP) haplotype and longevity in humans. Proc Natl Acad Sci USA 102:7906−7909.
42. Arai Y et al. (2001) Serum insulin like growth factor 1 (IGF−I) in centenarians: implication of IGF− 1 as a turnover protein. J Gerontol A Biol Sci Med Sci 56A:M79−M82.
43. Teng EL et al. (1994) The Cognitive Abilities Screening Instrument (CASI): a practical test for cross−cultural epidemiological studies of dementia. Int Psychogeriatr 6:45−58.
44. Willcox BJ et al. (2006) Midlife risk factors and survival in men. JAMA 296:2343−2350.
45. Kramarow E, Lubitz J, Lentzner H, Gorina Y (2007) Trends in the health of older Americans, 1970−2005. Health AfJ (Millwood) 26:1417−1425.
46. Evert J, Lawler E, Bogan H, Perls T (2003) Morbidity profiles of centenarians: survivors, delayers, and escapers. J Gerontol A Biol Sci Med Sci 58:232−237.
47. Bernstein AM et al. (2004) First autopsy study of an Okinawan centenarian: absence of many age−related diseases. J Gerontol A Biol Sci Med Sci 59:1195− 1199.
48. Willcox DC et al. (2007) Aging gracefully: a retrospective analysis of functional status in Okinawan centenarians. Am J Geriatr Psychiatry 15:252−256.
49. Terry DF, Wilcox MA, McCormick MA, Perls TT (2004) Cardiovascular disease delay in centenarian offspring. J Gerontol A Biol Sci Med Sci 59:385−389.
50. Rodriguez BL et al. (2002) The A.D.A and W.H.O. classifications for diabetes: their impact on diabetes prevalence and total and cardiovascular disease mortality in elderly Japanese −American men. Diabetes Care 25:951−955.
51. McNeely MJ, Boyko EJ (2004) Type 2 diabetes prevalence in Asian Americans: results of a national health survey. Diabetes Care 27:66−69.
52. Fujimoto WY et al. (1995) Susceptibility to development of central adiposity among populations. Obes Res 3Suppl2:179S−186S.
53. Examination Committee of Criteria for Obesity Disease´ in Japan, et al. (2002). New criteria for Obesity disease´ in Japan. Circ J 66:987−992.
54. Barthel A, Schmoll D, Unterman TG (2005). FoxO proteins in insulin action and metabolism. Trends Endocrinol Metab 16:183−189.
55. Kops JG et al. (2002) Forkhead transcription factor FOXO3a protects quiescent cells from oxidative stress. Nature 419:316−321.
56. Youngman LD, Park JY, Ames BN (1992) Protein oxidation associated with aging is reduced by dietary restriction of protein or calories. Proc Natl Acad Sci USA 89:9112− 9116.
57. Heilbronn LK et al. (2006) Effect of 6−month calorie restriction on biomarkers of longevity, metabolic adaptation, and oxidative stress in overweight individuals: a randomized controlled trial. JAMA 295:1539−1548.
58. Selman C et al. (2008) Evidence for lifespan extension and delayed age−related biomarkers in insulin receptor substrate 1 null mice. FJffi´´5 J22:807−818.
59. Murphy CT et al. (2003) Genes that act downstream of DAF−16 to influence the lifespan of Caenorhabditis elegans. Nature 424:277−283.
60. Kuningas M et al. (2007) SIRTl gene, age−related diseases, and mortality: the Leiden 85− plus study. J Gerontol A Biol Sci Med Sci 62:960−965.
61. Kagan A. Ed (1996) The Honolulu Heart Program: An Epidemiological Study of Coronary Heart Disease and Stroke (Harwood Academic Publishers, the Netherlands).
62. Worth RM, Kagan A (1970) Ascertainment of men of Japanese ancestry in Hawaii
through World War II Selective Service registration. J Chronic Dis 23:389−397.
63. Rodriguez BL, Curb JD (1998) Cardiovascular risk factors in the elderly: the Honolulu Heart Program. Cardiovasc Risk Factors 8:99− 103.
64. White L, et al. (1996) Prevalence of dementia in Japanese− American men living in Hawaii: the Honolulu− Asia Aging Study. JAMA 276:955−960.
65. National Institutes of Health (NIH): U.S. Department of Health and Human Services, NIH, National Institute on Aging (2001) Report of the National Institute on Aging Advisory Panel on Exceptional Longevity (APEL). NIH Pub 01−4951 (NIH, Washington, DC).
66. Arias E (2006) United States life tables, 2003 (National Center for Health Statistics, Hyattsville, MD), vol. 54, no. 14.
67. Nordyke EC, Lee R, Gardner RW (1984) A profile of Hawaii´s elderly population. Papers East W1 est Popul Inst 91 :13−14.
68. Mehta C, Patel N (2001) StatXact 5 (CYTEL Software Corporation, Cambridge, MA).
69. SAS Institute, Inc. (1990) SAS/STAT user´s guide, version 6 (SAS Institute, Cary, NC).
70. Sherry ST, Ward MH, Kholodov M, Baker J, Phan L, Smigielski EM, Sirotkin K. dbSNP: the NCBI database of genetic variation. Nucleic Acids Res. 2001 Jan l;29(l):308−l l.)
【特許請求の範囲】
【請求項1】
対象における長寿に関する要因を診断する方法において、前記対象が遺伝子座/多型rs2802292の「G」対立遺伝子の少なくとも1つを位置109015211でFOXO3A遺伝子の染色体6に有するかどうかを、前記対象の組織サンプルで決定する工程を含む方法。
【請求項2】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項1に記載の方法。
【請求項3】
加齢と関連のある少なくとも1つの慢性疾患は、糖尿病、冠状動脈疾患および癌からなる群から選択され、請求項2に記載の方法。
【請求項4】
対象における長寿に関する要因を診断する方法において、前記対象が遺伝子座/多型の「C」対立遺伝子rs2802292の少なくとも1つを位置109015211でのFOXO3A遺伝子の染色体6に有するかどうかを、前記対象の組織サンプルで決定する工程を含む方法。
【請求項5】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項1に記載の方法。
【請求項6】
加齢と関連のある少なくとも1つの慢性疾患は、糖尿病、冠状動脈疾患および癌からなる群から選択され、請求項2に記載の方法。
【請求項7】
対象における長寿に関する要因を診断する方法において、前記対象が遺伝子座/多型rs13217795の「C」対立遺伝子の少なくとも1つを位置109080791でFOXO3A遺伝子の染色体6に有するかどうかを、前記対象の組織サンプルで決定する工程を含む方法。
【請求項8】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項7に記載の方法。
【請求項9】
加齢と関連のある少なくとも1つの慢性疾患は、糖尿病、冠状動脈疾患および癌からなる群から選択され、請求項8に記載の方法。
【請求項10】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項1に記載の方法。
【請求項11】
対象における長寿に関する要因を診断する方法において、前記対象が「GCC」ハプロタイプを有するかどうかを、前記対象の組織サンプルで決定する工程を含み、「GCC」ハプロタイプはFOXO3A遺伝子においてそれぞれ遺伝子座/多型rs2802292、rs2764264およびrs13217795である方法。
【請求項12】
ヒト以外の対象における長寿に関する要因を診断する方法において、前記対象がヒト以外の相同物「GCC」ハプロタイプを有するかどうかを、前記対象の組織サンプルで決定する工程を含み、「GCC」ハプロタイプはFOXO3A遺伝子においてそれぞれ遺伝子座/多型rs2802292、rs2764264およびrs13217795である方法。
【請求項13】
FOXO3A遺伝子の染色体6に位置109015211で遺伝子座/多型rs2802292の「G」対立遺伝子;FOXO3A遺伝子の染色体6に位置109015211で遺伝子座/多型rs2802292の「G」対立遺伝子;およびFOXO3A遺伝子の染色体6に位置109041154で遺伝子座/多型rs2764264の「C」対立遺伝子からなる群から選択される第一の標的核酸配列の位置にハイブリッドを形成する十分な長さの第一の核酸であって、前記第一の標的核酸配列の前記位置、または前記第一の標的核酸配列相補体とハイブリッドを形成する前記第一の核酸を、1つ以上の容器内に含有するものおよび使用説明書を含むキット。
【請求項14】
FOXO3Aハプロタイプの少なくとも1つの対立遺伝子の存在を試験する方法において、
a)対象から細胞サンプルを採取する工程;
b)細胞サンプルから核酸サンプルを分離する工程;
c)対立遺伝子のハイブリッド形成および増幅が生じる条件下に基づいて、FOXO3Aハプロタイプの少なくとも1つの対立遺伝子に5’および3’を特異的にハイブリッド形成する少なくとも1つのプライマーと、核酸サンプルを接触させる工程;および
d)対立遺伝子検出方法により増幅産物の検出工程
を含有する方法。
【請求項15】
少なくとも1つのプライマーは、SEQ ID NOS.5〜8からなる群から選択される、請求項14に記載の方法。
【請求項16】
寿命と関連のある遺伝子の遺伝子発現レベルを観測することによって、対象の治療を治療薬で観測する方法において、
a)治療薬の投与前に前投与サンプルを対象から得る工程;
b)前投与サンプルにおいて遺伝子発現レベルを検出する工程;
c)対象から1つ以上の投与後サンプルを得る工程;
d)投与後サンプルにおいて、遺伝子発現のレベルを検出する工程;
e)投与前後サンプルにおいて遺伝子発現のレベルを比較して、治療薬の遺伝子発現への効果を分析する工程;
f)遺伝子発現に対する治療薬の効果に基づいて対象の治療を変更する工程
を含有する方法。
【請求項17】
遺伝子発現のレベルを検出する工程は、寿命と関連のある遺伝子の、タンパク質、mRNAまたはDNA転写レベルの少なくとも1つを検出する、請求項16に記載の方法。
【請求項18】
長寿と関連のある遺伝子は、FOXO3A、LOC100130966、FOXO3Aのヒト以外の相同物およびLOC100130966のヒト以外の相同物からなる群から選択される、請求項16に記載の方法。
【請求項19】
死亡、加齢関係の疾患またはさらなる物理的機能の予想を助けるため、危険率の計算において「GCC」FOXO3Aハプロタイプの有無の使用。
【請求項1】
対象における長寿に関する要因を診断する方法において、前記対象が遺伝子座/多型rs2802292の「G」対立遺伝子の少なくとも1つを位置109015211でFOXO3A遺伝子の染色体6に有するかどうかを、前記対象の組織サンプルで決定する工程を含む方法。
【請求項2】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項1に記載の方法。
【請求項3】
加齢と関連のある少なくとも1つの慢性疾患は、糖尿病、冠状動脈疾患および癌からなる群から選択され、請求項2に記載の方法。
【請求項4】
対象における長寿に関する要因を診断する方法において、前記対象が遺伝子座/多型の「C」対立遺伝子rs2802292の少なくとも1つを位置109015211でのFOXO3A遺伝子の染色体6に有するかどうかを、前記対象の組織サンプルで決定する工程を含む方法。
【請求項5】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項1に記載の方法。
【請求項6】
加齢と関連のある少なくとも1つの慢性疾患は、糖尿病、冠状動脈疾患および癌からなる群から選択され、請求項2に記載の方法。
【請求項7】
対象における長寿に関する要因を診断する方法において、前記対象が遺伝子座/多型rs13217795の「C」対立遺伝子の少なくとも1つを位置109080791でFOXO3A遺伝子の染色体6に有するかどうかを、前記対象の組織サンプルで決定する工程を含む方法。
【請求項8】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項7に記載の方法。
【請求項9】
加齢と関連のある少なくとも1つの慢性疾患は、糖尿病、冠状動脈疾患および癌からなる群から選択され、請求項8に記載の方法。
【請求項10】
長寿とは、少なくともさらに15年間生存し、前記要因はさらに加齢と関連のある少なくとも1つの慢性疾患から独立している要因を含有する、請求項1に記載の方法。
【請求項11】
対象における長寿に関する要因を診断する方法において、前記対象が「GCC」ハプロタイプを有するかどうかを、前記対象の組織サンプルで決定する工程を含み、「GCC」ハプロタイプはFOXO3A遺伝子においてそれぞれ遺伝子座/多型rs2802292、rs2764264およびrs13217795である方法。
【請求項12】
ヒト以外の対象における長寿に関する要因を診断する方法において、前記対象がヒト以外の相同物「GCC」ハプロタイプを有するかどうかを、前記対象の組織サンプルで決定する工程を含み、「GCC」ハプロタイプはFOXO3A遺伝子においてそれぞれ遺伝子座/多型rs2802292、rs2764264およびrs13217795である方法。
【請求項13】
FOXO3A遺伝子の染色体6に位置109015211で遺伝子座/多型rs2802292の「G」対立遺伝子;FOXO3A遺伝子の染色体6に位置109015211で遺伝子座/多型rs2802292の「G」対立遺伝子;およびFOXO3A遺伝子の染色体6に位置109041154で遺伝子座/多型rs2764264の「C」対立遺伝子からなる群から選択される第一の標的核酸配列の位置にハイブリッドを形成する十分な長さの第一の核酸であって、前記第一の標的核酸配列の前記位置、または前記第一の標的核酸配列相補体とハイブリッドを形成する前記第一の核酸を、1つ以上の容器内に含有するものおよび使用説明書を含むキット。
【請求項14】
FOXO3Aハプロタイプの少なくとも1つの対立遺伝子の存在を試験する方法において、
a)対象から細胞サンプルを採取する工程;
b)細胞サンプルから核酸サンプルを分離する工程;
c)対立遺伝子のハイブリッド形成および増幅が生じる条件下に基づいて、FOXO3Aハプロタイプの少なくとも1つの対立遺伝子に5’および3’を特異的にハイブリッド形成する少なくとも1つのプライマーと、核酸サンプルを接触させる工程;および
d)対立遺伝子検出方法により増幅産物の検出工程
を含有する方法。
【請求項15】
少なくとも1つのプライマーは、SEQ ID NOS.5〜8からなる群から選択される、請求項14に記載の方法。
【請求項16】
寿命と関連のある遺伝子の遺伝子発現レベルを観測することによって、対象の治療を治療薬で観測する方法において、
a)治療薬の投与前に前投与サンプルを対象から得る工程;
b)前投与サンプルにおいて遺伝子発現レベルを検出する工程;
c)対象から1つ以上の投与後サンプルを得る工程;
d)投与後サンプルにおいて、遺伝子発現のレベルを検出する工程;
e)投与前後サンプルにおいて遺伝子発現のレベルを比較して、治療薬の遺伝子発現への効果を分析する工程;
f)遺伝子発現に対する治療薬の効果に基づいて対象の治療を変更する工程
を含有する方法。
【請求項17】
遺伝子発現のレベルを検出する工程は、寿命と関連のある遺伝子の、タンパク質、mRNAまたはDNA転写レベルの少なくとも1つを検出する、請求項16に記載の方法。
【請求項18】
長寿と関連のある遺伝子は、FOXO3A、LOC100130966、FOXO3Aのヒト以外の相同物およびLOC100130966のヒト以外の相同物からなる群から選択される、請求項16に記載の方法。
【請求項19】
死亡、加齢関係の疾患またはさらなる物理的機能の予想を助けるため、危険率の計算において「GCC」FOXO3Aハプロタイプの有無の使用。
【図1】


【図2】
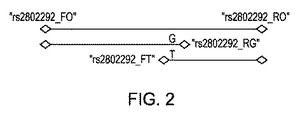
【図2】
【公表番号】特表2011−530312(P2011−530312A)
【公表日】平成23年12月22日(2011.12.22)
【国際特許分類】
【出願番号】特願2011−523071(P2011−523071)
【出願日】平成21年8月10日(2009.8.10)
【国際出願番号】PCT/US2009/053307
【国際公開番号】WO2010/019519
【国際公開日】平成22年2月18日(2010.2.18)
【出願人】(511036015)クアキニ メディカル センター (1)
【氏名又は名称原語表記】KUAKINI MEDICAL CENTER
【Fターム(参考)】
【公表日】平成23年12月22日(2011.12.22)
【国際特許分類】
【出願日】平成21年8月10日(2009.8.10)
【国際出願番号】PCT/US2009/053307
【国際公開番号】WO2010/019519
【国際公開日】平成22年2月18日(2010.2.18)
【出願人】(511036015)クアキニ メディカル センター (1)
【氏名又は名称原語表記】KUAKINI MEDICAL CENTER
【Fターム(参考)】
[ Back to top ]