
光合成組織における遺伝子発現制御に関わるDNA
【課題】成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAを提供する。
【解決手段】以下の(a)〜(d)のDNAを含む、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA:(a)特定の塩基配列からなるDNA;(b)特定の塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;(c)特定の塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;(d)特定の塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【解決手段】以下の(a)〜(d)のDNAを含む、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA:(a)特定の塩基配列からなるDNA;(b)特定の塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;(c)特定の塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;(d)特定の塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、光合成組織、特に成葉、に特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA及びその利用に関する。
【背景技術】
【0002】
生物の遺伝子情報は、機能性遺伝子のDNAの情報がmRNAに転写され、mRNAの情報を翻訳して機能性タンパク質を合成する、セントラルドグマと呼ばれる一連のプロセスを経て情報伝達がなされ、生物機能を発現する。植物体においても、一般的に両親の半数体の会合によって形成された遺伝子情報の集合体であるゲノムDNAが、それを含む細胞及び当該細胞に由来する植物体の運命を規定している。
【0003】
その中で、細胞が有するゲノムDNAの機能性遺伝子情報が正確に情報伝達され生物機能を発現するためには、適当な遺伝子が適当な時期や場所に適当な強度で発現されなければならず、特定の遺伝子の発現が厳密に制御されていることが必要とされる。例えば、植物の葉に存在する気孔形成に関与する遺伝子は、気孔が形成されるべき組織、時期にて適切に発現されることが必要とされる。
【0004】
機能性遺伝子の発現は、当該遺伝子の5’上流域に存在する遺伝子発現制御DNAによりその発現時期、部位および強度が制御されている。
【0005】
近年、シロイヌナズナやイネなどを用いたゲノムDNAの解読により、これら植物における各種機能性遺伝子の遺伝子発現制御DNAが容易に取得可能となった(非特許文献1,2)。
【0006】
一方、実用作物であるサトウキビについては、ゲノムDNA解読が実施されていないことから、各種機能性遺伝子の遺伝子発現制御DNAを取得することは容易ではない。そのため従来的に、サトウキビに遺伝子導入する際には、その導入遺伝子の発現制御を、他の植物に由来する遺伝子発現制御DNAを用いて行なっていた(非特許文献3,4)。
【0007】
しかしながら、サトウキビにおいて他の植物に由来する遺伝子発現制御DNAを用いて機能性遺伝子を発現させた場合、その発現時期、部位および強度などが厳密に制御されない場合が観察され、当該分野においてはサトウキビに由来する遺伝子発現制御DNA、特に組織特異的な遺伝子発現制御DNAの取得が切望されていた。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Nature 2000 408:769−815
【非特許文献2】Science 2002 296:92−100
【非特許文献3】Plant Mol Biol.1992 Feb;18(4):675−89
【非特許文献4】Planta 1998 206:20−27
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、本発明の目的は、光合成組織特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAを提供することである。
【課題を解決するための手段】
【0010】
本発明者は上記課題を解決すべく鋭意研究を行なっていたところ、サトウキビの成葉において強度に発現している遺伝子の5’上流領域より、成葉に特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAを見出し、本発明を完成させるに至った。
すなわち、本発明は以下[1]〜[4]の特徴を包含する。
[1] 以下の(a)〜(d)のDNAを含む、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA:
(a)配列番号1に表される塩基配列からなるDNA;
(b)配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(c)配列番号1に表わされる塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(d)配列番号1に表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【0011】
[2] [1]の遺伝子発現制御DNAを含む、組換えベクター。
[3] [2]の組換えベクターで形質転換された形質転換植物体。
[4] [2]の組換えベクターで植物細胞を形質転換し、形質転換された植物細胞から得られた形質転換植物体。
【発明の効果】
【0012】
本発明によれば、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA、当該DNAを含有する、機能性遺伝子を成葉で特異的に発現させることを可能にする組換えベクター、当該組換えベクターを導入した形質転換植物体を提供することができる。
【図面の簡単な説明】
【0013】
【図1】図1は、Saccharum officinarumに由来するecc0002 ESTの塩基配列を示す(DFCI Sugarcane Gene Indexで検索)。
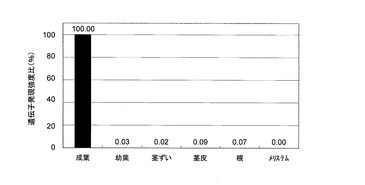
【図2】図2は、Saccharum spp. cv. NiF8の各組織におけるecc0002 ESTの発現量の解析結果を示す。最も発現の強い成葉の発現量を100%として示す。
【図3】図3は、5’末端にHindIII制限酵素認識配列および3’末端にBlnI制限酵素認識配列を導入したecc0002遺伝子の遺伝子発現制御領域の塩基配列を示す。
【図4−1】図4−1は、プロモーター解析ツールを用いて同定されたecc0002遺伝子の遺伝子発現制御領域における推定プロモーター領域を示す。
【図4−2】(図4−1の続き)
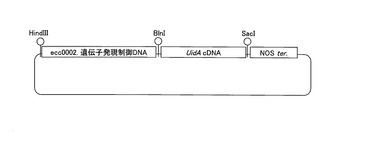
【図5】図5は、ecc0002遺伝子の遺伝子発現制御領域とβ−グルクロニダーゼ遺伝子を連結して含む遺伝子発現ベクターの模式図を示す。
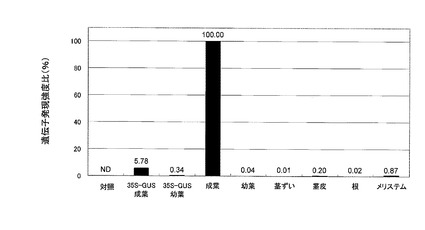
【図6】図6は、β−グルクロニダーゼ遺伝子の発現をecc0002遺伝子の発現制御DNAによって制御する遺伝子組換えサトウキビの各組織におけるGUS遺伝子の発現量の解析結果を示す。最も強い発現量が観察された当該遺伝子組換えサトウキビの成葉の発現量を100%として示す。
【発明を実施するための形態】
【0014】
以下、本発明を詳細に説明する。
先ず、本発明に係る成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAについて説明する。本発明に係る遺伝子発現制御DNAは、以下の(a)〜(d)のいずれかのDNAを含む。
(a)配列番号1に表される塩基配列からなるDNA
(b)配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA
(c)配列番号1に表わされる塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA
(d)配列番号1に表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【0015】
本発明に係る遺伝子発現制御DNAは、サトウキビの各組織(例えば、茎、成葉、幼葉)に由来する総RNA又はそれに由来するcDNAを利用した遺伝子発現解析により、成葉特異的に発現している候補遺伝子を取得し、当該候補遺伝子について発現特性を評価し、これにより成葉特異的に発現していると評価された遺伝子を特定し、特定した遺伝子のcDNA又はゲノムDNAをもとに当該候補遺伝子の5’上流領域の塩基配列を同定することによって取得することができる。ここで遺伝子発現解析は、DNAチップ、ディファレンシャルディスプレイ法など、当業者に公知である遺伝子発現の網羅的な解析手法を利用することができる。
【0016】
具体的に、配列番号1の塩基配列は、サトウキビの成葉において、特異的に発現している遺伝子(以下、「ecc0002」と記載する)の5’上流領域に存在する。本明細書において「サトウキビ」には、サトウキビ属に属する植物、例えば、Saccharum officinarum、Saccharum sinense、Saccharum barberi、Saccharum robustum、Saccharum spontaneum、Saccharum edule、Saccharum spp. hybrids cv. NiF8など(特にこれらに限定されない)、およびその近縁属種、例えば、ソルガム等が含まれる。好ましくは、Saccharum spp. hybrids cv. NiF8である。
【0017】
5’上流領域のDNAを単離する方法は特に限定されず、当業者に公知の一般的な手法を用いて行うことができる。例えば、上記ecc0002遺伝子の塩基配列(配列番号2)に基づいて、未知の領域(この場合、5’上流領域)をクローニングする公知方法を用いて単離することができる。例えば、その一つとして、ecc0002遺伝子の5’上流領域を含むゲノムDNAを制限酵素処理をして所定の塩基配列からなるアダプターを結合させ、ecc0002遺伝子の塩基配列とアダプターにプライマーを設定してPCRを行う。これにより、ecc0002遺伝子の塩基配列に5’上流で隣接する未知の塩基配列を増幅することができる。そして増幅した塩基配列の配列決定を行った後、決定した塩基配列に基づいて、さらに一対のプライマーを設計し、決定した塩基配列に隣接するさらなる未知の塩基配列を同様にして増幅することができる。当該方法においては、市販のクローニングキット、例えば、RightWalk(登録商標)キット(ベックス)を用いて行うことができる。また、上記以外の方法としては、インバースPCRによる方法が挙げられる。この場合、ecc0002遺伝子の塩基配列情報に基づいて一対のプライマーを設計し、これら一対のプライマーと、所定の制限酵素で処理した後にセルフライゲーションさせたゲノムDNA断片とを用いてPCRを行うことによって、ecc0002遺伝子の上流領域を増幅することができる。さらに別の方法として、ecc0002遺伝子の上流領域をゲノムDNAライブラリーから単離する方法が挙げられる。この場合には、ecc0002遺伝子を含むcDNAをプローブとして、定法に従って調製したゲノムDNAライブラリーからecc0002遺伝子を含むゲノムDNAをスクリーニングする。その後、スクリーニングしたゲノムDNAの塩基配列を決定することによってecc0002遺伝子の上流領域に存在する5’上流領域を特定することができ、さらに、5’上流領域のみをPCR等によって増幅することができる。
【0018】
このように、ecc0002遺伝子の上流に位置する未知の塩基配列を順次増幅またはスクリーニングし、定法に従って塩基配列を決定することによって、配列番号1で表される塩基配列を得ることができる。一旦、配列番号1で表される塩基配列が決定されると、その後は、配列番号1で表される塩基配列に基づいて設計したプライマーを用い、サトウキビから抽出したゲノムDNAを鋳型としたPCRによって、配列番号1に表される塩基配列を得ることができる。
【0019】
配列番号1に表される塩基配列は、成葉において特異的に遺伝子発現を誘導する遺伝子発現制御領域として機能する。遺伝子発現制御領域には、プロモーター領域、エンハンサー領域、TATAボックスおよび/またはCATボックスなど(特にこれらに限定されない)、遺伝子の転写調節に関与する塩基配列が含まれる。
【0020】
本明細書において「特異的」とは、植物体を構成する各種組織のなかで成葉においてのみ遺伝子発現誘導機能を有することに加えて、成葉における遺伝子発現誘導機能が成葉以外の組織(例えば、幼葉、茎ずい、茎皮、根、メリステムなど)における遺伝子発現誘導機能と比較して顕著に高いこと、または統計的に有意に高いこと(例えば、およそ2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、又はそれ以上)を意味する。
【0021】
本明細書において「成葉」とは、光合成を行うために葉緑体を細胞内に蓄積し、緑色を帯びた葉を意味し、光合成のための葉緑体を有さない葉である幼葉以外の葉組織を意味する。
【0022】
遺伝子発現誘導機能は、当業者に公知であるレポーターアッセイ等によって確認することができる。レポーターアッセイは、種々のレポーター遺伝子(例えば、β−グルクロニダーゼ遺伝子(GUS)、ルシフェラーゼ遺伝子(LUC)、グリーンフルオレッセントプロテイン遺伝子(GFP)等)を遺伝子発現誘導機能を検討する塩基配列の制御下(下流域)に連結したベクターを作製し、当該ベクターを用いて宿主のゲノムに導入又は一過的に導入した後、当該レポーター遺伝子の発現レベルを測定することにより確認することができる。当該レポーター遺伝子としては、その発現が検出可能なものであれば特に制限されず、例えば、当業者において一般的に使用されるCAT遺伝子、lacZ遺伝子、ルシフェラーゼ遺伝子(以下、LUC)、β−グルクロニダーゼ(以下、GUS)遺伝子、及びグリーンフルオレッセントプロテイン(以下、GFP)遺伝子等を挙げることができる。
【0023】
レポーター遺伝子の発現レベルは、当該レポーター遺伝子の種類に応じて、当業者に公知の方法により測定することができる。例えば、レポーター遺伝子がCAT遺伝子である場合には、当該遺伝子産物によるクロラムフェニコールのアセチル化を検出することによって、レポーター遺伝子の発現レベルを測定することができる。各レポーター遺伝子の発現レベルは以下の手法で測定することができる。レポーター遺伝子がlacZ遺伝子である場合には、当該遺伝子発現産物の触媒作用による色素化合物の発色を検出する。LUC遺伝子である場合には、当該遺伝子発現産物の触媒作用による蛍光化合物の蛍光を検出する。GFP遺伝子である場合には、GFPタンパク質による蛍光を検出する。例えば、レポーター遺伝子がGUSの場合には、宿主細胞内でのプロモーター活性は、(i)ヒストケミカルなGUS染色による方法(EMBO J. 6, 3901−3907 (1987))により、及び/又は(ii)蛍光基質を用いるCastle&Morrisの方法(Plant Molecular Biology Manual, B5, 1−16 (1994); S.B.Gelvin & R.A.Schilperoort, Kluwer Academic Publishers)に従ってGUS活性を測定し、さらにBradfordの方法(Anal. Biochem. 72, 248−254(1976))に従ってタンパク質量を測定して、GUS活性をタンパク量当たりに換算する(nmole 4−MU/min/mg proteinとして算出する)ことにより、それぞれ確認することができる。
【0024】
また、上記以外の遺伝子をレポーターとして使用する場合には、当該遺伝子の転写レベルをノーザンハイブリダイゼーション法、RT−PCR法、DNAアレイ技術などを用いて測定することによって、又は当該遺伝子にコードされるタンパク質の発現量をSDS−PAGE等の電気泳動法、ウェスタンブロッティング法などを用いて測定することによって行うことができる。
【0025】
本発明に係る遺伝子発現制御DNAは、配列番号1に表される塩基配列からなるものに限定されず、上記(b)に記載したように、成葉に特異的に遺伝子発現を促進する活性を有する限り、配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列であってもよい。
【0026】
例えば、配列番号1に表される塩基配列において、1〜100個の塩基、好ましくは1〜50個の塩基、より好ましくは1〜10個の塩基が欠失、置換、付加もしくは挿入された塩基配列であっても、成葉に特異的に遺伝子発現を促進する活性を示す限り本発明に係る遺伝子発現制御DNAに含まれる。
【0027】
また、本発明に係る遺伝子発現制御DNAは、配列番号1に表される塩基配列からなるものに限定されず、上記(c)に記載したように、成葉に特異的に遺伝子発現を促進する活性を示す限り、配列番号1に表わされる塩基配列と80%以上、より好ましくは90%以上、さらに好ましくは95%以上、最も好ましくは99%以上の配列同一性を有する塩基配列であってもよい。塩基配列の比較は公知の手法によって行うことができ、例えば、BLAST(Basic Local Alignment Search Tool at the National Center for Biological Information(米国国立生物学情報センターの基本ローカルアラインメント検索ツール))等を例えば、デフォルトの設定で用いて実施できる。
【0028】
さらに、本発明に係る遺伝子発現制御DNAは、配列番号1に表される塩基配列からなるものに限定されず、上記(d)に記載したように、成葉に特異的に遺伝子発現を促進する活性を示す限り、配列番号1で表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズする塩基配列であってもよい。
【0029】
本発明において「ストリンジェントな条件」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいい、例えば、2〜6×SSC(1×SSCの組成:0.15M NaCl, 0.015M クエン酸ナトリウム, pH 7.0)及び0.1〜0.5%SDSを含有する溶液中42〜55℃にてハイブリダイズを行い、0.1〜0.2×SSC及び0.1〜0.5%SDSを含有する溶液中55〜65℃にて洗浄を行う条件をいう。
【0030】
さらに、本発明に係る成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAは、成葉に特異的に遺伝子発現を促進する活性を示す限り、配列番号1に表わされる塩基配列において5’末端及び/又は3’末端より、連続する100個、200個、300個、400個、500個、600個、700個、800個、900個、1000個、1100個、1200個、1300個、1400個、1500個、又はそれ以上の塩基が欠失されたDNA断片であってもよい。塩基の欠失は当業者に公知の一般的な手法(例えば、PCR法、制限酵素処理など)によって行うことができる。当該DNA断片は、本発明に係る遺伝子発現制御DNAにおけるプロモーター領域であっても良い。所定の遺伝子発現制御DNAにおけるプロモーター領域は当業者に公知のプロモーター解析ツール(例えば、BioInformatics and Molecular Analysis Section(http://www−bimas.cit.nih.gov/molbio/proscan/);Prestridge, D.S. (1995). Predicting Pol II Promoter Sequences Using Transcription Factor Binding Sites. J. Mol. Biol. 249: 923−32)を使用して検索することができる。このような配列番号1に表わされる塩基配列の断片として、配列番号1の868〜1118番目の塩基からなるDNAが挙げられる。得られた当該断片は、上記レポーターアッセイ等によって、その遺伝子発現誘導機能について確認することができる。
【0031】
一旦、本発明に係る遺伝子発現制御DNAの塩基配列が確定されると、その後は化学合成によって、又はゲノムDNAを鋳型としたPCRによって、あるいは当該塩基配列を有するDNA断片をプローブとしてハイブリダイズさせることによって、本発明に係る遺伝子発現制御DNAを得ることができる。さらに、部位特定変異誘発等によって、変異を有する配列番号1に表される塩基配列を合成することもできる。なお、配列番号1に表される塩基配列に変異を導入するには、Kunkel法、Gapped duplex法等の公知の手法又はこれに準ずる方法を採用することができる。例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant−K(TAKARA社製)やMutant−G(TAKARA社製))などを用いて、あるいは、TAKARA社のLA PCR in vitro Mutagenesis シリーズキットを用いて変異の導入が行われる。
【0032】
次に、上述した成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAを用いた組換えベクターについて説明する。
【0033】
本発明の組換えベクターは、上述した遺伝子発現制御DNAに所望の機能性遺伝子を機能しうるかたちで連結したDNAを適当なベクターに導入することにより構築することができる。本明細書において、「機能しうるかたちで連結した」、「機能しうるかたちで連結された」とは、当該ベクターが導入された宿主細胞内において、当該遺伝子発現制御DNAの制御の下、機能性遺伝子が正確に発現されるように、遺伝子発現制御DNAと機能性遺伝子とを連結して含むことを意味する。ここで「連結」は直接連結されても良いし、適当な長さ及び配列のスペーサーを介して間接的に連結されても良い。本発明に用いるベクターとしては、アグロバクテリウムを介して植物に機能性遺伝子を導入することができる、pBI系、pBII系、pPZP系(Hajdukiewicz P, Svab Z, Maliga P.: The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation., Plant Mol Biol., 25: 989−94, 1994)、pCAMBIA系(http://www.cambia.org/main/r_et_camvec.htm)、pSMA系のベクターなどが好適に用いられる。特にpBII系およびpBI系のバイナリーベクター又は中間ベクター系が好適に用いられ、例えば、pBII221、pBI121、pBI101、pBI101.2、pBI101.3、pIG121などが挙げられる。バイナリーベクターとは大腸菌(Escherichia coli)及びアグロバクテリウムにおいて複製可能なシャトルベクターで、バイナリーベクターを保持するアグロバクテリムを植物に感染させると、ベクター上にあるLB配列とRB配列より成るボーダー配列で囲まれた部分のDNAを植物核DNAに組み込むことが可能である(EMBO Journal, 10(3), 697−704 (1991))。一方、pUC系のベクターは、植物に遺伝子を直接導入することができ、例えば、pUC18、pUC19、pUC9などが挙げられる。また、カリフラワーモザイクウイルス(CaMV)、インゲンマメモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)などの植物ウイルスベクターも用いることができる。
【0034】
連結及び/又はベクターへの挿入を容易にするべく、上述した遺伝子発現制御DNA及び/又は機能性遺伝子をコードするDNAには制限酵素認識配列を適宜、置換、挿入または付加することができる。ベクターへの挿入に際しては、まず、上述した遺伝子発現制御DNAおよび機能性遺伝子をコードする精製されたDNAを適当な制限酵素で切断し、適当なベクターDNAの制限酵素認識部位又はマルチクローニングサイトに挿入してベクターに連結する方法などを用いることができる。
【0035】
機能性遺伝子としては、対象となる植物における内因性遺伝子、又は外来遺伝子であって、その遺伝子産物の発現が成葉において所望される任意の遺伝子をいう。かかる遺伝子としては、光合成関連遺伝子、転流関連遺伝子、有用物質(医薬、色素、芳香成分など)生産遺伝子、糖代謝関連遺伝子、耐病虫害性〔昆虫食害抵抗性、カビ(菌類)及び細菌病抵抗性、ウイルス(病)抵抗性など〕遺伝子、環境ストレス(低温、高温、乾燥、光障害、紫外線)耐性関連遺伝子、植物生長制御(促進/抑制)遺伝子などが挙げられるが、これらに限定はされない。
【0036】
ベクターにはさらに、必要に応じて、遺伝子発現制御DNA及び/又は機能性遺伝子の上流、内部、あるいは下流に、エンハンサー、イントロン、ポリA付加シグナル、5’−UTR配列、選択マーカー遺伝子などを連結することができる。
【0037】
エンハンサーとしては、例えば、機能性遺伝子の発現効率を高めるために用いられ、例えば、CaMV35Sプロモーター内の上流側の配列を含むエンハンサー領域などが挙げられる。
【0038】
ターミネーターとしては、前記プロモーターにより転写された遺伝子の転写を終結できる配列であればよく、例えば、ノパリン合成酵素遺伝子のターミネーター、オクトピン合成酵素遺伝子のターミネーター、CaMV 35S RNA遺伝子のターミネーターなどが挙げられる。
【0039】
選択マーカー遺伝子としては、例えば、ハイグロマイシン耐性遺伝子、カナマイシン耐性遺伝子、ビアラホス耐性遺伝子、ブラストサイジンS耐性遺伝子、アセト乳酸合成酵素(Acetolactate synthase)遺伝子などが挙げられる。選択マーカー遺伝子は、上述のように機能性遺伝子とともに同一のプラスミドに連結させて組換えベクターを調製してもよいが、あるいは、選択マーカー遺伝子をプラスミドに連結して得られる組換えベクターと、機能性遺伝子をプラスミドに連結して得られる組換えベクターとを別々に調製してもよい。別々に調製した場合は、各ベクターを宿主にコトランスフェクト(共導入)する。
【0040】
このように作製された組換えベクターを用いて形質転換体を作製することができる。
形質転換植物体を調製する際には、既に報告され、確立されている種々の方法を適宜利用することができ、その好ましい例として、アグロバクテリウム法、PEG−リン酸カルシウム法、エレクトロポレーション法、リポソーム法、パーティクルガン法、マイクロインジェクション法等が挙げられる。アグロバクテリウム法を用いる場合は、プロトプラストを用いる場合、組織片を用いる場合、及び植物体そのものを用いる場合(in planta法)がある。プロトプラストを用いる場合は、Tiプラスミドをもつアグロバクテリウムと共存培養する方法、スフェロプラスト化したアグロバクテリウムと融合する方法(スフェロプラスト法)、組織片を用いる場合は、対象植物の無菌培養葉片(リーフディスク)に感染させる方法やカルスに感染させる等により行うことができる。また種子あるいは植物体を用いるin planta法を適用する場合、すなわち植物ホルモン添加の組織培養を介さない系では、吸水種子、幼植物(幼苗)、鉢植え植物などへのアグロバクテリウムの直接処理等にて実施可能である。
【0041】
遺伝子が植物体に組み込まれたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法等により行うことができる。例えば、形質転換植物体からDNAを調製し、DNA特異的プライマーを設計してPCRを行う。PCRを行った後は、増幅産物についてアガロースゲル電気泳動、ポリアクリルアミドゲル電気泳動又はキャピラリー電気泳動等を行い、臭化エチジウム、SYBR Green液等により染色し、そして増幅産物を1本のバンドとして検出することにより、形質転換されたことを確認することができる。また、予め蛍光色素等により標識したプライマーを用いてPCRを行い、増幅産物を検出することもできる。さらに、マイクロプレート等の固相に増幅産物を結合させ、蛍光又は酵素反応等により増幅産物を確認する方法でもよい。
【0042】
本発明において形質転換に用いられる植物としては、例えば、イネ科、ナス科、アブラナ科、マメ科、バラ科、キク科、ユリ科、セリ科、ナデシコ科、ウリ科、ヒルガオ科、アカザ科などに属する植物が挙げられるが、特にこれらの植物に限定されない。好ましくは、上述の遺伝子発現制御DNAが単離されたイネ科の植物、例えば、サトウキビ、イネ、オオムギ、コムギ、トウモロコシ、シバ、ソルガム、アワ、ヒエ、ネピアグラス及びスイッチグラスなどの植物が挙げられる。
【0043】
本発明において、形質転換の対象とする植物材料としては、例えば、根、茎、葉、種子、胚、胚珠、子房、茎頂(植物の芽の先端の生長点)、葯、花粉等の植物組織やその切片、未分化のカルス、それを酵素処置して細胞壁を除いたプロプラスト等の植物培養細胞が挙げられる。またin planta法適用の場合、吸水種子や植物体全体を利用し得る。
【0044】
また、本発明において形質転換植物体とは、植物体全体、植物器官(例えば根、茎、葉、花弁、種子、種子、実等)植物組織(例えば表皮、師部、柔組織、木部、維管束等)、植物培養細胞のいずれをも意味するものである。
【0045】
植物培養細胞を対象とする場合において、得られた形質転換細胞から形質転換植物体を再生させるためには既知の組織培養法により器官又は個体を再生させればよい。このような操作は、植物細胞から植物体への再生方法として一般的に知られている方法により、当業者であれば容易に行うことができる。植物細胞から植物体への再生については、例えば、以下のように行うことができる。
【0046】
まず、形質転換の対象とする植物材料して植物組織又はプロトプラストを用いた場合、これらを無機要素、ビタミン、炭素源、エネルギー源としての糖類、植物生長調節物質(オーキシン、サイトカイニン等の植物ホルモン)等を加えて滅菌したカルス形成用培地中で培養し、不定形に増殖する脱分化したカルスを形成させる(以下「カルス誘導」という)。このように形成されたカルスをオーキシン等の植物生長調節物質を含む新しい培地に移しかえて更に増殖(継代培養)させる。
【0047】
カルス誘導は寒天等の固型培地で行い、継代培養は例えば液体培養で行うと、それぞれの培養を効率良くかつ大量に行うことができる。次に、上記の継代培養により増殖したカルスを適当な条件下で培養することにより器官の再分化を誘導し(以下、「再分化誘導」という)、最終的に完全な植物体を再生させる。再分化誘導は、培地におけるオーキシンやサイトカイニン等の植物生長調節物質、炭素源等の各種成分の種類や量、光、温度等を適切に設定することにより行うことができる。かかる再分化誘導により、不定胚、不定根、不定芽、不定茎葉等が形成され、更に完全な植物体へと育成させる。あるいは、完全な植物体になる前の状態(例えばカプセル化された人工種子、乾燥胚、凍結乾燥細胞及び組織等)で貯蔵等を行ってもよい。
【0048】
本発明において「形質転換植物体」には、形質転換を施した再分化当代である「T1世代」のほか、その植物の種子から得られた後代である「T2世代」、薬剤選抜あるいはサザン法等による解析によりトランスジェニックであることが判明した「T2世代」植物の花を自家受粉して得られる次世代(T3世代)などの後代植物をも含む。
【0049】
このようにして作出された形質転換植物体は、導入された機能性遺伝子を成葉特異的に発現する。
【0050】
本発明の遺伝子発現制御DNAは、植物、特にサトウキビ、の成葉特異的に遺伝子発現を促進する活性を有する。本発明の遺伝子発現制御DNAに所望される機能性遺伝子を連結して、植物に導入することによって、当該植物の成葉特異的に機能性遺伝子を発現させ、当該植物に所望の特性を付与することができる。本発明の遺伝子発現制御DNAは、従来、遺伝子発現制御系が多く開発されていないサトウキビにおける使用に適しており、バイオマス資源をはじめとする各種の利用に適した性質を有するトランスジェニックサトウキビの作出を可能とする。
【実施例】
【0051】
以下、実施例を用いて本発明をより詳細に説明する。しかしながら、本発明の技術的範囲はこれら実施例に限定されるものではない。
〔実施例1〕成葉特異的遺伝子のクローニング
サトウキビ(Saccharum spp. cv. NiF8)の成葉、幼葉及び茎の各組織からRNeasy Plant Mini Kit (QIAGEN)を用いて総RNAを抽出精製し、定法に従ってcDNAライブラリーを構築して遺伝子発現解析に用いた。遺伝子発現解析は、Sugarcane Genome Array (Affimetrix)を利用して、製造元の指示書に従って行った。
【0052】
遺伝子発現解析の結果、Saccharum spp. cv. NiF8の成葉において特に発現量の高い遺伝子を見出し、当該遺伝子を「ecc0002」とした。なお、Saccharum spp. cv. NiF8に由来するecc0002遺伝子の塩基配列は、図1に示すSaccharum officinarumに由来するecc0002 ESTの塩基配列に一致する。Saccharum spp. cv. NiF8の成葉、幼葉、茎ずい、茎皮、根およびメリステムから総RNAを抽出精製し、定法に従ってcDNAを作製し、ABI7500リアルタイムPCR装置(アプライドバイオシステムズ)を使用したSYBRGreen法によって各組織におけるecc0002遺伝子の発現量を解析した。
【0053】
結果を図2に示す(最も発現の強い成葉の発現量を100%として示す)。この結果より、Saccharum spp. cv. NiF8においてecc0002遺伝子が成葉において、強く遺伝子発現が誘導されていることが明らかとなった。
【0054】
〔実施例2〕成葉特異的プロモーターの取得
サトウキビ(Saccharum spp. cv. NiF8)の成葉組織(0.5g)から、DNeasy Plant Mini Kit (QIAGEN)を用いてゲノムDNAを約300ng抽出精製した。実施例1で得たecc0002遺伝子の塩基配列をベースに、RightWallk(登録商標)Kit(BEX)を用いて、上記ゲノムDNAよりecc0002遺伝子の5’上流に存在する遺伝子発現制御領域を得た。取得した遺伝子発現制御領域の5’末端にHindIII制限酵素認識配列(AAGCTT)を、3’末端に存在するecc0002遺伝子の翻訳開始点(ATG)の3’側にBlnI制限酵素認識配列(CCTAGG)をリンカー配列として、それぞれ導入してecc0002遺伝子の発現制御領域をコードするDNAとして調製した(図3)(配列番号3)。
【0055】
上記DNAの配列について、公知のプロモーター解析ツール(BioInformatics and Molecular Analysis Section(http://www−bimas.cit.nih.gov/molbio/proscan/)を利用して解析した結果、875〜1125番目の塩基領域にプロモーターとして機能する領域が推定された(図4−1および図4−2)。
【0056】
〔実施例3〕遺伝子発現ベクターの構築
(1)β−グルクロニダーゼ遺伝子発現ベクター
実施例2で得た遺伝子発現制御領域をコードするDNAを、β−グルクロニダーゼ(GUS)遺伝子をコードするUidAcDNAと連結して含む、遺伝子発現ベクターを構築した。当該遺伝子発現ベクターには、植物形質転換用ベクター(pBII221)を用いた。遺伝子発現制御領域をコードするDNAとUidAcDNAとの連結に際しては、UidAcDNAの5’末端側に存在するファーストメチオニンをコードするATG配列と遺伝子発現制御領域をコードするDNAの3’末端側に存在するecc0002遺伝子のファーストメチオニンをコードするATG配列を一致させて連結した(翻訳融合型)。当該遺伝子発現ベクターの模式図を図5に示す。
【0057】
〔実施例4〕遺伝子組換え植物の作出
実施例3で作製した遺伝子発現ベクターを、アグロバクテリウム法を用いて宿主植物体(Saccharum spp. cv. NiF8)に導入し、GUS遺伝子の発現をecc0002遺伝子の発現制御DNAによって制御する遺伝子組換えサトウキビを作出した。
【0058】
当該遺伝子組換えサトウキビの成葉、幼葉、茎ずい、茎皮、根およびメリステムから総RNAを抽出精製し、定法に従ってcDNAを作製し、ABI7500リアルタイムPCR装置(アプライドバイオシステムズ)を使用したSYBRGreen法によって各組織におけるGUS遺伝子の発現量を解析した。
【0059】
比較として、非遺伝子組換えサトウキビのメリステム(対照)、ならびにGUS遺伝子の発現をカリフラワーモザイクウイルス(CaMV)35S プロモーターによって制御する遺伝子組換えサトウキビに由来する成葉および幼葉における、GUS遺伝子の発現量を上記と同様に解析した。
【0060】
結果を図6に示す。各組織の発現量は、解析した組織の中で最も高い発現を示した、GUS遺伝子の発現をecc0002遺伝子の発現制御DNAによって制御する遺伝子組換えサトウキビの成葉におけるGUS遺伝子の発現量を100%とする相対値で示した。
【0061】
この結果より、ecc0002遺伝子の発現制御DNAが成葉において、CaMV35Sプロモーターによって制御するGUS遺伝子の発現量と比較して約17倍と、特異的に遺伝子発現を誘導できることが明らかとなった。
【産業上の利用可能性】
【0062】
本発明における遺伝子発現制御DNAは、成葉に特異的な遺伝子発現を促進する活性を有する。この特徴から、所望の遺伝子を成葉において特異的に発現させ、所望の性質を有する植物体の作出に利用することができる。
【技術分野】
【0001】
本発明は、光合成組織、特に成葉、に特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA及びその利用に関する。
【背景技術】
【0002】
生物の遺伝子情報は、機能性遺伝子のDNAの情報がmRNAに転写され、mRNAの情報を翻訳して機能性タンパク質を合成する、セントラルドグマと呼ばれる一連のプロセスを経て情報伝達がなされ、生物機能を発現する。植物体においても、一般的に両親の半数体の会合によって形成された遺伝子情報の集合体であるゲノムDNAが、それを含む細胞及び当該細胞に由来する植物体の運命を規定している。
【0003】
その中で、細胞が有するゲノムDNAの機能性遺伝子情報が正確に情報伝達され生物機能を発現するためには、適当な遺伝子が適当な時期や場所に適当な強度で発現されなければならず、特定の遺伝子の発現が厳密に制御されていることが必要とされる。例えば、植物の葉に存在する気孔形成に関与する遺伝子は、気孔が形成されるべき組織、時期にて適切に発現されることが必要とされる。
【0004】
機能性遺伝子の発現は、当該遺伝子の5’上流域に存在する遺伝子発現制御DNAによりその発現時期、部位および強度が制御されている。
【0005】
近年、シロイヌナズナやイネなどを用いたゲノムDNAの解読により、これら植物における各種機能性遺伝子の遺伝子発現制御DNAが容易に取得可能となった(非特許文献1,2)。
【0006】
一方、実用作物であるサトウキビについては、ゲノムDNA解読が実施されていないことから、各種機能性遺伝子の遺伝子発現制御DNAを取得することは容易ではない。そのため従来的に、サトウキビに遺伝子導入する際には、その導入遺伝子の発現制御を、他の植物に由来する遺伝子発現制御DNAを用いて行なっていた(非特許文献3,4)。
【0007】
しかしながら、サトウキビにおいて他の植物に由来する遺伝子発現制御DNAを用いて機能性遺伝子を発現させた場合、その発現時期、部位および強度などが厳密に制御されない場合が観察され、当該分野においてはサトウキビに由来する遺伝子発現制御DNA、特に組織特異的な遺伝子発現制御DNAの取得が切望されていた。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Nature 2000 408:769−815
【非特許文献2】Science 2002 296:92−100
【非特許文献3】Plant Mol Biol.1992 Feb;18(4):675−89
【非特許文献4】Planta 1998 206:20−27
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、本発明の目的は、光合成組織特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAを提供することである。
【課題を解決するための手段】
【0010】
本発明者は上記課題を解決すべく鋭意研究を行なっていたところ、サトウキビの成葉において強度に発現している遺伝子の5’上流領域より、成葉に特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAを見出し、本発明を完成させるに至った。
すなわち、本発明は以下[1]〜[4]の特徴を包含する。
[1] 以下の(a)〜(d)のDNAを含む、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA:
(a)配列番号1に表される塩基配列からなるDNA;
(b)配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(c)配列番号1に表わされる塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(d)配列番号1に表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【0011】
[2] [1]の遺伝子発現制御DNAを含む、組換えベクター。
[3] [2]の組換えベクターで形質転換された形質転換植物体。
[4] [2]の組換えベクターで植物細胞を形質転換し、形質転換された植物細胞から得られた形質転換植物体。
【発明の効果】
【0012】
本発明によれば、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA、当該DNAを含有する、機能性遺伝子を成葉で特異的に発現させることを可能にする組換えベクター、当該組換えベクターを導入した形質転換植物体を提供することができる。
【図面の簡単な説明】
【0013】
【図1】図1は、Saccharum officinarumに由来するecc0002 ESTの塩基配列を示す(DFCI Sugarcane Gene Indexで検索)。
【図2】図2は、Saccharum spp. cv. NiF8の各組織におけるecc0002 ESTの発現量の解析結果を示す。最も発現の強い成葉の発現量を100%として示す。
【図3】図3は、5’末端にHindIII制限酵素認識配列および3’末端にBlnI制限酵素認識配列を導入したecc0002遺伝子の遺伝子発現制御領域の塩基配列を示す。
【図4−1】図4−1は、プロモーター解析ツールを用いて同定されたecc0002遺伝子の遺伝子発現制御領域における推定プロモーター領域を示す。
【図4−2】(図4−1の続き)
【図5】図5は、ecc0002遺伝子の遺伝子発現制御領域とβ−グルクロニダーゼ遺伝子を連結して含む遺伝子発現ベクターの模式図を示す。
【図6】図6は、β−グルクロニダーゼ遺伝子の発現をecc0002遺伝子の発現制御DNAによって制御する遺伝子組換えサトウキビの各組織におけるGUS遺伝子の発現量の解析結果を示す。最も強い発現量が観察された当該遺伝子組換えサトウキビの成葉の発現量を100%として示す。
【発明を実施するための形態】
【0014】
以下、本発明を詳細に説明する。
先ず、本発明に係る成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAについて説明する。本発明に係る遺伝子発現制御DNAは、以下の(a)〜(d)のいずれかのDNAを含む。
(a)配列番号1に表される塩基配列からなるDNA
(b)配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA
(c)配列番号1に表わされる塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA
(d)配列番号1に表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【0015】
本発明に係る遺伝子発現制御DNAは、サトウキビの各組織(例えば、茎、成葉、幼葉)に由来する総RNA又はそれに由来するcDNAを利用した遺伝子発現解析により、成葉特異的に発現している候補遺伝子を取得し、当該候補遺伝子について発現特性を評価し、これにより成葉特異的に発現していると評価された遺伝子を特定し、特定した遺伝子のcDNA又はゲノムDNAをもとに当該候補遺伝子の5’上流領域の塩基配列を同定することによって取得することができる。ここで遺伝子発現解析は、DNAチップ、ディファレンシャルディスプレイ法など、当業者に公知である遺伝子発現の網羅的な解析手法を利用することができる。
【0016】
具体的に、配列番号1の塩基配列は、サトウキビの成葉において、特異的に発現している遺伝子(以下、「ecc0002」と記載する)の5’上流領域に存在する。本明細書において「サトウキビ」には、サトウキビ属に属する植物、例えば、Saccharum officinarum、Saccharum sinense、Saccharum barberi、Saccharum robustum、Saccharum spontaneum、Saccharum edule、Saccharum spp. hybrids cv. NiF8など(特にこれらに限定されない)、およびその近縁属種、例えば、ソルガム等が含まれる。好ましくは、Saccharum spp. hybrids cv. NiF8である。
【0017】
5’上流領域のDNAを単離する方法は特に限定されず、当業者に公知の一般的な手法を用いて行うことができる。例えば、上記ecc0002遺伝子の塩基配列(配列番号2)に基づいて、未知の領域(この場合、5’上流領域)をクローニングする公知方法を用いて単離することができる。例えば、その一つとして、ecc0002遺伝子の5’上流領域を含むゲノムDNAを制限酵素処理をして所定の塩基配列からなるアダプターを結合させ、ecc0002遺伝子の塩基配列とアダプターにプライマーを設定してPCRを行う。これにより、ecc0002遺伝子の塩基配列に5’上流で隣接する未知の塩基配列を増幅することができる。そして増幅した塩基配列の配列決定を行った後、決定した塩基配列に基づいて、さらに一対のプライマーを設計し、決定した塩基配列に隣接するさらなる未知の塩基配列を同様にして増幅することができる。当該方法においては、市販のクローニングキット、例えば、RightWalk(登録商標)キット(ベックス)を用いて行うことができる。また、上記以外の方法としては、インバースPCRによる方法が挙げられる。この場合、ecc0002遺伝子の塩基配列情報に基づいて一対のプライマーを設計し、これら一対のプライマーと、所定の制限酵素で処理した後にセルフライゲーションさせたゲノムDNA断片とを用いてPCRを行うことによって、ecc0002遺伝子の上流領域を増幅することができる。さらに別の方法として、ecc0002遺伝子の上流領域をゲノムDNAライブラリーから単離する方法が挙げられる。この場合には、ecc0002遺伝子を含むcDNAをプローブとして、定法に従って調製したゲノムDNAライブラリーからecc0002遺伝子を含むゲノムDNAをスクリーニングする。その後、スクリーニングしたゲノムDNAの塩基配列を決定することによってecc0002遺伝子の上流領域に存在する5’上流領域を特定することができ、さらに、5’上流領域のみをPCR等によって増幅することができる。
【0018】
このように、ecc0002遺伝子の上流に位置する未知の塩基配列を順次増幅またはスクリーニングし、定法に従って塩基配列を決定することによって、配列番号1で表される塩基配列を得ることができる。一旦、配列番号1で表される塩基配列が決定されると、その後は、配列番号1で表される塩基配列に基づいて設計したプライマーを用い、サトウキビから抽出したゲノムDNAを鋳型としたPCRによって、配列番号1に表される塩基配列を得ることができる。
【0019】
配列番号1に表される塩基配列は、成葉において特異的に遺伝子発現を誘導する遺伝子発現制御領域として機能する。遺伝子発現制御領域には、プロモーター領域、エンハンサー領域、TATAボックスおよび/またはCATボックスなど(特にこれらに限定されない)、遺伝子の転写調節に関与する塩基配列が含まれる。
【0020】
本明細書において「特異的」とは、植物体を構成する各種組織のなかで成葉においてのみ遺伝子発現誘導機能を有することに加えて、成葉における遺伝子発現誘導機能が成葉以外の組織(例えば、幼葉、茎ずい、茎皮、根、メリステムなど)における遺伝子発現誘導機能と比較して顕著に高いこと、または統計的に有意に高いこと(例えば、およそ2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、又はそれ以上)を意味する。
【0021】
本明細書において「成葉」とは、光合成を行うために葉緑体を細胞内に蓄積し、緑色を帯びた葉を意味し、光合成のための葉緑体を有さない葉である幼葉以外の葉組織を意味する。
【0022】
遺伝子発現誘導機能は、当業者に公知であるレポーターアッセイ等によって確認することができる。レポーターアッセイは、種々のレポーター遺伝子(例えば、β−グルクロニダーゼ遺伝子(GUS)、ルシフェラーゼ遺伝子(LUC)、グリーンフルオレッセントプロテイン遺伝子(GFP)等)を遺伝子発現誘導機能を検討する塩基配列の制御下(下流域)に連結したベクターを作製し、当該ベクターを用いて宿主のゲノムに導入又は一過的に導入した後、当該レポーター遺伝子の発現レベルを測定することにより確認することができる。当該レポーター遺伝子としては、その発現が検出可能なものであれば特に制限されず、例えば、当業者において一般的に使用されるCAT遺伝子、lacZ遺伝子、ルシフェラーゼ遺伝子(以下、LUC)、β−グルクロニダーゼ(以下、GUS)遺伝子、及びグリーンフルオレッセントプロテイン(以下、GFP)遺伝子等を挙げることができる。
【0023】
レポーター遺伝子の発現レベルは、当該レポーター遺伝子の種類に応じて、当業者に公知の方法により測定することができる。例えば、レポーター遺伝子がCAT遺伝子である場合には、当該遺伝子産物によるクロラムフェニコールのアセチル化を検出することによって、レポーター遺伝子の発現レベルを測定することができる。各レポーター遺伝子の発現レベルは以下の手法で測定することができる。レポーター遺伝子がlacZ遺伝子である場合には、当該遺伝子発現産物の触媒作用による色素化合物の発色を検出する。LUC遺伝子である場合には、当該遺伝子発現産物の触媒作用による蛍光化合物の蛍光を検出する。GFP遺伝子である場合には、GFPタンパク質による蛍光を検出する。例えば、レポーター遺伝子がGUSの場合には、宿主細胞内でのプロモーター活性は、(i)ヒストケミカルなGUS染色による方法(EMBO J. 6, 3901−3907 (1987))により、及び/又は(ii)蛍光基質を用いるCastle&Morrisの方法(Plant Molecular Biology Manual, B5, 1−16 (1994); S.B.Gelvin & R.A.Schilperoort, Kluwer Academic Publishers)に従ってGUS活性を測定し、さらにBradfordの方法(Anal. Biochem. 72, 248−254(1976))に従ってタンパク質量を測定して、GUS活性をタンパク量当たりに換算する(nmole 4−MU/min/mg proteinとして算出する)ことにより、それぞれ確認することができる。
【0024】
また、上記以外の遺伝子をレポーターとして使用する場合には、当該遺伝子の転写レベルをノーザンハイブリダイゼーション法、RT−PCR法、DNAアレイ技術などを用いて測定することによって、又は当該遺伝子にコードされるタンパク質の発現量をSDS−PAGE等の電気泳動法、ウェスタンブロッティング法などを用いて測定することによって行うことができる。
【0025】
本発明に係る遺伝子発現制御DNAは、配列番号1に表される塩基配列からなるものに限定されず、上記(b)に記載したように、成葉に特異的に遺伝子発現を促進する活性を有する限り、配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列であってもよい。
【0026】
例えば、配列番号1に表される塩基配列において、1〜100個の塩基、好ましくは1〜50個の塩基、より好ましくは1〜10個の塩基が欠失、置換、付加もしくは挿入された塩基配列であっても、成葉に特異的に遺伝子発現を促進する活性を示す限り本発明に係る遺伝子発現制御DNAに含まれる。
【0027】
また、本発明に係る遺伝子発現制御DNAは、配列番号1に表される塩基配列からなるものに限定されず、上記(c)に記載したように、成葉に特異的に遺伝子発現を促進する活性を示す限り、配列番号1に表わされる塩基配列と80%以上、より好ましくは90%以上、さらに好ましくは95%以上、最も好ましくは99%以上の配列同一性を有する塩基配列であってもよい。塩基配列の比較は公知の手法によって行うことができ、例えば、BLAST(Basic Local Alignment Search Tool at the National Center for Biological Information(米国国立生物学情報センターの基本ローカルアラインメント検索ツール))等を例えば、デフォルトの設定で用いて実施できる。
【0028】
さらに、本発明に係る遺伝子発現制御DNAは、配列番号1に表される塩基配列からなるものに限定されず、上記(d)に記載したように、成葉に特異的に遺伝子発現を促進する活性を示す限り、配列番号1で表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズする塩基配列であってもよい。
【0029】
本発明において「ストリンジェントな条件」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいい、例えば、2〜6×SSC(1×SSCの組成:0.15M NaCl, 0.015M クエン酸ナトリウム, pH 7.0)及び0.1〜0.5%SDSを含有する溶液中42〜55℃にてハイブリダイズを行い、0.1〜0.2×SSC及び0.1〜0.5%SDSを含有する溶液中55〜65℃にて洗浄を行う条件をいう。
【0030】
さらに、本発明に係る成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAは、成葉に特異的に遺伝子発現を促進する活性を示す限り、配列番号1に表わされる塩基配列において5’末端及び/又は3’末端より、連続する100個、200個、300個、400個、500個、600個、700個、800個、900個、1000個、1100個、1200個、1300個、1400個、1500個、又はそれ以上の塩基が欠失されたDNA断片であってもよい。塩基の欠失は当業者に公知の一般的な手法(例えば、PCR法、制限酵素処理など)によって行うことができる。当該DNA断片は、本発明に係る遺伝子発現制御DNAにおけるプロモーター領域であっても良い。所定の遺伝子発現制御DNAにおけるプロモーター領域は当業者に公知のプロモーター解析ツール(例えば、BioInformatics and Molecular Analysis Section(http://www−bimas.cit.nih.gov/molbio/proscan/);Prestridge, D.S. (1995). Predicting Pol II Promoter Sequences Using Transcription Factor Binding Sites. J. Mol. Biol. 249: 923−32)を使用して検索することができる。このような配列番号1に表わされる塩基配列の断片として、配列番号1の868〜1118番目の塩基からなるDNAが挙げられる。得られた当該断片は、上記レポーターアッセイ等によって、その遺伝子発現誘導機能について確認することができる。
【0031】
一旦、本発明に係る遺伝子発現制御DNAの塩基配列が確定されると、その後は化学合成によって、又はゲノムDNAを鋳型としたPCRによって、あるいは当該塩基配列を有するDNA断片をプローブとしてハイブリダイズさせることによって、本発明に係る遺伝子発現制御DNAを得ることができる。さらに、部位特定変異誘発等によって、変異を有する配列番号1に表される塩基配列を合成することもできる。なお、配列番号1に表される塩基配列に変異を導入するには、Kunkel法、Gapped duplex法等の公知の手法又はこれに準ずる方法を採用することができる。例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant−K(TAKARA社製)やMutant−G(TAKARA社製))などを用いて、あるいは、TAKARA社のLA PCR in vitro Mutagenesis シリーズキットを用いて変異の導入が行われる。
【0032】
次に、上述した成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNAを用いた組換えベクターについて説明する。
【0033】
本発明の組換えベクターは、上述した遺伝子発現制御DNAに所望の機能性遺伝子を機能しうるかたちで連結したDNAを適当なベクターに導入することにより構築することができる。本明細書において、「機能しうるかたちで連結した」、「機能しうるかたちで連結された」とは、当該ベクターが導入された宿主細胞内において、当該遺伝子発現制御DNAの制御の下、機能性遺伝子が正確に発現されるように、遺伝子発現制御DNAと機能性遺伝子とを連結して含むことを意味する。ここで「連結」は直接連結されても良いし、適当な長さ及び配列のスペーサーを介して間接的に連結されても良い。本発明に用いるベクターとしては、アグロバクテリウムを介して植物に機能性遺伝子を導入することができる、pBI系、pBII系、pPZP系(Hajdukiewicz P, Svab Z, Maliga P.: The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation., Plant Mol Biol., 25: 989−94, 1994)、pCAMBIA系(http://www.cambia.org/main/r_et_camvec.htm)、pSMA系のベクターなどが好適に用いられる。特にpBII系およびpBI系のバイナリーベクター又は中間ベクター系が好適に用いられ、例えば、pBII221、pBI121、pBI101、pBI101.2、pBI101.3、pIG121などが挙げられる。バイナリーベクターとは大腸菌(Escherichia coli)及びアグロバクテリウムにおいて複製可能なシャトルベクターで、バイナリーベクターを保持するアグロバクテリムを植物に感染させると、ベクター上にあるLB配列とRB配列より成るボーダー配列で囲まれた部分のDNAを植物核DNAに組み込むことが可能である(EMBO Journal, 10(3), 697−704 (1991))。一方、pUC系のベクターは、植物に遺伝子を直接導入することができ、例えば、pUC18、pUC19、pUC9などが挙げられる。また、カリフラワーモザイクウイルス(CaMV)、インゲンマメモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)などの植物ウイルスベクターも用いることができる。
【0034】
連結及び/又はベクターへの挿入を容易にするべく、上述した遺伝子発現制御DNA及び/又は機能性遺伝子をコードするDNAには制限酵素認識配列を適宜、置換、挿入または付加することができる。ベクターへの挿入に際しては、まず、上述した遺伝子発現制御DNAおよび機能性遺伝子をコードする精製されたDNAを適当な制限酵素で切断し、適当なベクターDNAの制限酵素認識部位又はマルチクローニングサイトに挿入してベクターに連結する方法などを用いることができる。
【0035】
機能性遺伝子としては、対象となる植物における内因性遺伝子、又は外来遺伝子であって、その遺伝子産物の発現が成葉において所望される任意の遺伝子をいう。かかる遺伝子としては、光合成関連遺伝子、転流関連遺伝子、有用物質(医薬、色素、芳香成分など)生産遺伝子、糖代謝関連遺伝子、耐病虫害性〔昆虫食害抵抗性、カビ(菌類)及び細菌病抵抗性、ウイルス(病)抵抗性など〕遺伝子、環境ストレス(低温、高温、乾燥、光障害、紫外線)耐性関連遺伝子、植物生長制御(促進/抑制)遺伝子などが挙げられるが、これらに限定はされない。
【0036】
ベクターにはさらに、必要に応じて、遺伝子発現制御DNA及び/又は機能性遺伝子の上流、内部、あるいは下流に、エンハンサー、イントロン、ポリA付加シグナル、5’−UTR配列、選択マーカー遺伝子などを連結することができる。
【0037】
エンハンサーとしては、例えば、機能性遺伝子の発現効率を高めるために用いられ、例えば、CaMV35Sプロモーター内の上流側の配列を含むエンハンサー領域などが挙げられる。
【0038】
ターミネーターとしては、前記プロモーターにより転写された遺伝子の転写を終結できる配列であればよく、例えば、ノパリン合成酵素遺伝子のターミネーター、オクトピン合成酵素遺伝子のターミネーター、CaMV 35S RNA遺伝子のターミネーターなどが挙げられる。
【0039】
選択マーカー遺伝子としては、例えば、ハイグロマイシン耐性遺伝子、カナマイシン耐性遺伝子、ビアラホス耐性遺伝子、ブラストサイジンS耐性遺伝子、アセト乳酸合成酵素(Acetolactate synthase)遺伝子などが挙げられる。選択マーカー遺伝子は、上述のように機能性遺伝子とともに同一のプラスミドに連結させて組換えベクターを調製してもよいが、あるいは、選択マーカー遺伝子をプラスミドに連結して得られる組換えベクターと、機能性遺伝子をプラスミドに連結して得られる組換えベクターとを別々に調製してもよい。別々に調製した場合は、各ベクターを宿主にコトランスフェクト(共導入)する。
【0040】
このように作製された組換えベクターを用いて形質転換体を作製することができる。
形質転換植物体を調製する際には、既に報告され、確立されている種々の方法を適宜利用することができ、その好ましい例として、アグロバクテリウム法、PEG−リン酸カルシウム法、エレクトロポレーション法、リポソーム法、パーティクルガン法、マイクロインジェクション法等が挙げられる。アグロバクテリウム法を用いる場合は、プロトプラストを用いる場合、組織片を用いる場合、及び植物体そのものを用いる場合(in planta法)がある。プロトプラストを用いる場合は、Tiプラスミドをもつアグロバクテリウムと共存培養する方法、スフェロプラスト化したアグロバクテリウムと融合する方法(スフェロプラスト法)、組織片を用いる場合は、対象植物の無菌培養葉片(リーフディスク)に感染させる方法やカルスに感染させる等により行うことができる。また種子あるいは植物体を用いるin planta法を適用する場合、すなわち植物ホルモン添加の組織培養を介さない系では、吸水種子、幼植物(幼苗)、鉢植え植物などへのアグロバクテリウムの直接処理等にて実施可能である。
【0041】
遺伝子が植物体に組み込まれたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法等により行うことができる。例えば、形質転換植物体からDNAを調製し、DNA特異的プライマーを設計してPCRを行う。PCRを行った後は、増幅産物についてアガロースゲル電気泳動、ポリアクリルアミドゲル電気泳動又はキャピラリー電気泳動等を行い、臭化エチジウム、SYBR Green液等により染色し、そして増幅産物を1本のバンドとして検出することにより、形質転換されたことを確認することができる。また、予め蛍光色素等により標識したプライマーを用いてPCRを行い、増幅産物を検出することもできる。さらに、マイクロプレート等の固相に増幅産物を結合させ、蛍光又は酵素反応等により増幅産物を確認する方法でもよい。
【0042】
本発明において形質転換に用いられる植物としては、例えば、イネ科、ナス科、アブラナ科、マメ科、バラ科、キク科、ユリ科、セリ科、ナデシコ科、ウリ科、ヒルガオ科、アカザ科などに属する植物が挙げられるが、特にこれらの植物に限定されない。好ましくは、上述の遺伝子発現制御DNAが単離されたイネ科の植物、例えば、サトウキビ、イネ、オオムギ、コムギ、トウモロコシ、シバ、ソルガム、アワ、ヒエ、ネピアグラス及びスイッチグラスなどの植物が挙げられる。
【0043】
本発明において、形質転換の対象とする植物材料としては、例えば、根、茎、葉、種子、胚、胚珠、子房、茎頂(植物の芽の先端の生長点)、葯、花粉等の植物組織やその切片、未分化のカルス、それを酵素処置して細胞壁を除いたプロプラスト等の植物培養細胞が挙げられる。またin planta法適用の場合、吸水種子や植物体全体を利用し得る。
【0044】
また、本発明において形質転換植物体とは、植物体全体、植物器官(例えば根、茎、葉、花弁、種子、種子、実等)植物組織(例えば表皮、師部、柔組織、木部、維管束等)、植物培養細胞のいずれをも意味するものである。
【0045】
植物培養細胞を対象とする場合において、得られた形質転換細胞から形質転換植物体を再生させるためには既知の組織培養法により器官又は個体を再生させればよい。このような操作は、植物細胞から植物体への再生方法として一般的に知られている方法により、当業者であれば容易に行うことができる。植物細胞から植物体への再生については、例えば、以下のように行うことができる。
【0046】
まず、形質転換の対象とする植物材料して植物組織又はプロトプラストを用いた場合、これらを無機要素、ビタミン、炭素源、エネルギー源としての糖類、植物生長調節物質(オーキシン、サイトカイニン等の植物ホルモン)等を加えて滅菌したカルス形成用培地中で培養し、不定形に増殖する脱分化したカルスを形成させる(以下「カルス誘導」という)。このように形成されたカルスをオーキシン等の植物生長調節物質を含む新しい培地に移しかえて更に増殖(継代培養)させる。
【0047】
カルス誘導は寒天等の固型培地で行い、継代培養は例えば液体培養で行うと、それぞれの培養を効率良くかつ大量に行うことができる。次に、上記の継代培養により増殖したカルスを適当な条件下で培養することにより器官の再分化を誘導し(以下、「再分化誘導」という)、最終的に完全な植物体を再生させる。再分化誘導は、培地におけるオーキシンやサイトカイニン等の植物生長調節物質、炭素源等の各種成分の種類や量、光、温度等を適切に設定することにより行うことができる。かかる再分化誘導により、不定胚、不定根、不定芽、不定茎葉等が形成され、更に完全な植物体へと育成させる。あるいは、完全な植物体になる前の状態(例えばカプセル化された人工種子、乾燥胚、凍結乾燥細胞及び組織等)で貯蔵等を行ってもよい。
【0048】
本発明において「形質転換植物体」には、形質転換を施した再分化当代である「T1世代」のほか、その植物の種子から得られた後代である「T2世代」、薬剤選抜あるいはサザン法等による解析によりトランスジェニックであることが判明した「T2世代」植物の花を自家受粉して得られる次世代(T3世代)などの後代植物をも含む。
【0049】
このようにして作出された形質転換植物体は、導入された機能性遺伝子を成葉特異的に発現する。
【0050】
本発明の遺伝子発現制御DNAは、植物、特にサトウキビ、の成葉特異的に遺伝子発現を促進する活性を有する。本発明の遺伝子発現制御DNAに所望される機能性遺伝子を連結して、植物に導入することによって、当該植物の成葉特異的に機能性遺伝子を発現させ、当該植物に所望の特性を付与することができる。本発明の遺伝子発現制御DNAは、従来、遺伝子発現制御系が多く開発されていないサトウキビにおける使用に適しており、バイオマス資源をはじめとする各種の利用に適した性質を有するトランスジェニックサトウキビの作出を可能とする。
【実施例】
【0051】
以下、実施例を用いて本発明をより詳細に説明する。しかしながら、本発明の技術的範囲はこれら実施例に限定されるものではない。
〔実施例1〕成葉特異的遺伝子のクローニング
サトウキビ(Saccharum spp. cv. NiF8)の成葉、幼葉及び茎の各組織からRNeasy Plant Mini Kit (QIAGEN)を用いて総RNAを抽出精製し、定法に従ってcDNAライブラリーを構築して遺伝子発現解析に用いた。遺伝子発現解析は、Sugarcane Genome Array (Affimetrix)を利用して、製造元の指示書に従って行った。
【0052】
遺伝子発現解析の結果、Saccharum spp. cv. NiF8の成葉において特に発現量の高い遺伝子を見出し、当該遺伝子を「ecc0002」とした。なお、Saccharum spp. cv. NiF8に由来するecc0002遺伝子の塩基配列は、図1に示すSaccharum officinarumに由来するecc0002 ESTの塩基配列に一致する。Saccharum spp. cv. NiF8の成葉、幼葉、茎ずい、茎皮、根およびメリステムから総RNAを抽出精製し、定法に従ってcDNAを作製し、ABI7500リアルタイムPCR装置(アプライドバイオシステムズ)を使用したSYBRGreen法によって各組織におけるecc0002遺伝子の発現量を解析した。
【0053】
結果を図2に示す(最も発現の強い成葉の発現量を100%として示す)。この結果より、Saccharum spp. cv. NiF8においてecc0002遺伝子が成葉において、強く遺伝子発現が誘導されていることが明らかとなった。
【0054】
〔実施例2〕成葉特異的プロモーターの取得
サトウキビ(Saccharum spp. cv. NiF8)の成葉組織(0.5g)から、DNeasy Plant Mini Kit (QIAGEN)を用いてゲノムDNAを約300ng抽出精製した。実施例1で得たecc0002遺伝子の塩基配列をベースに、RightWallk(登録商標)Kit(BEX)を用いて、上記ゲノムDNAよりecc0002遺伝子の5’上流に存在する遺伝子発現制御領域を得た。取得した遺伝子発現制御領域の5’末端にHindIII制限酵素認識配列(AAGCTT)を、3’末端に存在するecc0002遺伝子の翻訳開始点(ATG)の3’側にBlnI制限酵素認識配列(CCTAGG)をリンカー配列として、それぞれ導入してecc0002遺伝子の発現制御領域をコードするDNAとして調製した(図3)(配列番号3)。
【0055】
上記DNAの配列について、公知のプロモーター解析ツール(BioInformatics and Molecular Analysis Section(http://www−bimas.cit.nih.gov/molbio/proscan/)を利用して解析した結果、875〜1125番目の塩基領域にプロモーターとして機能する領域が推定された(図4−1および図4−2)。
【0056】
〔実施例3〕遺伝子発現ベクターの構築
(1)β−グルクロニダーゼ遺伝子発現ベクター
実施例2で得た遺伝子発現制御領域をコードするDNAを、β−グルクロニダーゼ(GUS)遺伝子をコードするUidAcDNAと連結して含む、遺伝子発現ベクターを構築した。当該遺伝子発現ベクターには、植物形質転換用ベクター(pBII221)を用いた。遺伝子発現制御領域をコードするDNAとUidAcDNAとの連結に際しては、UidAcDNAの5’末端側に存在するファーストメチオニンをコードするATG配列と遺伝子発現制御領域をコードするDNAの3’末端側に存在するecc0002遺伝子のファーストメチオニンをコードするATG配列を一致させて連結した(翻訳融合型)。当該遺伝子発現ベクターの模式図を図5に示す。
【0057】
〔実施例4〕遺伝子組換え植物の作出
実施例3で作製した遺伝子発現ベクターを、アグロバクテリウム法を用いて宿主植物体(Saccharum spp. cv. NiF8)に導入し、GUS遺伝子の発現をecc0002遺伝子の発現制御DNAによって制御する遺伝子組換えサトウキビを作出した。
【0058】
当該遺伝子組換えサトウキビの成葉、幼葉、茎ずい、茎皮、根およびメリステムから総RNAを抽出精製し、定法に従ってcDNAを作製し、ABI7500リアルタイムPCR装置(アプライドバイオシステムズ)を使用したSYBRGreen法によって各組織におけるGUS遺伝子の発現量を解析した。
【0059】
比較として、非遺伝子組換えサトウキビのメリステム(対照)、ならびにGUS遺伝子の発現をカリフラワーモザイクウイルス(CaMV)35S プロモーターによって制御する遺伝子組換えサトウキビに由来する成葉および幼葉における、GUS遺伝子の発現量を上記と同様に解析した。
【0060】
結果を図6に示す。各組織の発現量は、解析した組織の中で最も高い発現を示した、GUS遺伝子の発現をecc0002遺伝子の発現制御DNAによって制御する遺伝子組換えサトウキビの成葉におけるGUS遺伝子の発現量を100%とする相対値で示した。
【0061】
この結果より、ecc0002遺伝子の発現制御DNAが成葉において、CaMV35Sプロモーターによって制御するGUS遺伝子の発現量と比較して約17倍と、特異的に遺伝子発現を誘導できることが明らかとなった。
【産業上の利用可能性】
【0062】
本発明における遺伝子発現制御DNAは、成葉に特異的な遺伝子発現を促進する活性を有する。この特徴から、所望の遺伝子を成葉において特異的に発現させ、所望の性質を有する植物体の作出に利用することができる。
【特許請求の範囲】
【請求項1】
以下の(a)〜(d)のDNAを含む、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA:
(a)配列番号1に表される塩基配列からなるDNA;
(b)配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(c)配列番号1に表わされる塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(d)配列番号1に表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【請求項2】
請求項1に記載の遺伝子発現制御DNAを含む、組換えベクター。
【請求項3】
請求項2に記載の組換えベクターで形質転換された形質転換植物体。
【請求項4】
請求項2に記載の組換えベクターで植物細胞を形質転換し、形質転換された植物細胞から得られた形質転換植物体。
【請求項1】
以下の(a)〜(d)のDNAを含む、成葉特異的に遺伝子発現を促進する活性を有する遺伝子発現制御DNA:
(a)配列番号1に表される塩基配列からなるDNA;
(b)配列番号1に表される塩基配列において、1〜複数個の塩基が欠失、置換、付加もしくは挿入された塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(c)配列番号1に表わされる塩基配列と90%以上の配列同一性を有する塩基配列からなり、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA;
(d)配列番号1に表される塩基配列の全部もしくは一部と相補的な配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ成葉に特異的に遺伝子発現を促進する活性を有するDNA。
【請求項2】
請求項1に記載の遺伝子発現制御DNAを含む、組換えベクター。
【請求項3】
請求項2に記載の組換えベクターで形質転換された形質転換植物体。
【請求項4】
請求項2に記載の組換えベクターで植物細胞を形質転換し、形質転換された植物細胞から得られた形質転換植物体。
【図1】



【図2】
【図3】

【図4−1】

【図4−2】

【図5】
【図6】
【図2】
【図3】
【図4−1】
【図4−2】
【図5】
【図6】
【公開番号】特開2012−139156(P2012−139156A)
【公開日】平成24年7月26日(2012.7.26)
【国際特許分類】
【出願番号】特願2010−293580(P2010−293580)
【出願日】平成22年12月28日(2010.12.28)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【Fターム(参考)】
【公開日】平成24年7月26日(2012.7.26)
【国際特許分類】
【出願日】平成22年12月28日(2010.12.28)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【Fターム(参考)】
[ Back to top ]