
光学活性なアミン誘導体の製造方法
【課題】光学活性なアミン誘導体の製造方法の提供。
【解決手段】イミン誘導体に、当該化合物を立体選択的に還元しうる能力を有する酵素または該酵素を生産する微生物もしくはその処理物を作用させ、生成する光学活性なアミン誘導体を回収する、光学活性なS体のアミン誘導体の製造方法。得られる光学活性なアミン誘導体は、医薬の合成原料として有用である。たとえば下記式(IV)で示される光学活性な化合物を製造することができる。式(IV)

(式中、R基は炭素数1〜3個のアルキル基を、nは1〜4の整数を表す)
【解決手段】イミン誘導体に、当該化合物を立体選択的に還元しうる能力を有する酵素または該酵素を生産する微生物もしくはその処理物を作用させ、生成する光学活性なアミン誘導体を回収する、光学活性なS体のアミン誘導体の製造方法。得られる光学活性なアミン誘導体は、医薬の合成原料として有用である。たとえば下記式(IV)で示される光学活性な化合物を製造することができる。式(IV)
(式中、R基は炭素数1〜3個のアルキル基を、nは1〜4の整数を表す)
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、イミン誘導体を立体選択的に還元することによって各種医薬品の原料あるいは合成中間体として利用されている光学活性なアミン誘導体を製造する方法に関する。
【背景技術】
【0002】
イミン誘導体の立体選択還元による光学活性なアミン誘導体の製造方法としては、次のような報告を除いてほとんど知られていない。
−非特許文献1;Tetrahedron Asymmetry, 19, 93,96(2008)−
キャンディダ・パラプシロシスによるN−フェニル−1−フェニルエチルアミン誘導体の還元
ただし、これらの公知技術においては、N−ベンジリデンベンジルアミンの還元生成物がキラルな構造ではないため、反応の立体選択性は不明である。また、N−((E)−N−(1−フェニルエチリデン)アニリン誘導体のキャンディダ・パラプシロシスによる還元反応は極めて弱いため、工業的な利用には適さない。また、該イミン還元反応を行う酵素の実態は全く不明である。
【0003】
また、2−メチルピロリジンの製法としては、次のような方法が知られている。
−非特許文献2;Acta. Pharm. Suec., 1978, 15, 255-263−
2−メチル−1−ピロリンを水素化ホウ素ナトリウムによって還元し、ラセミ2−メチルピロリジンとした後、酒石酸を用いた光学分割により、キラル−2−メチルピロリジンを得る方法
−非特許文献3;J. Org. Chem., 1989, 54, 209-216−
L−プロリンから誘導されるL−プロリノールの水酸基を塩化チオニルでクロロ基に変換し、窒素原子をベンジルオキシカルボニル基により保護した後、水素化トリブチル錫を用いてラジカル的に還元することにより、(R)−1−ベンジルオキシカルボニル−2−メチルピロリジンを製造し、さらに脱保護する方法
【0004】
−特許文献1;WO2005/073388号公報−
光学活性1,4−ペンダンジオールをジスルホナートとした後、アミンと反応させて環化し、2−メチルピロリジンを得る方法
しかし、いずれの方法にも、次のような問題点が伴う。
−工程が長い
−危険な水素化試薬を使用する
−分割方法によって得るため収率が50%を超えない
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Tetrahedron Asymmetry, 19, 93,96(2008)
【非特許文献2】Acta. Pharm. Suec., 1978, 15, 255-263
【非特許文献3】J. Org. Chem., 1989, 54, 209-216
【特許文献】
【0006】
【特許文献1】WO2005/073388号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の課題は、イミン誘導体から光学活性なアミン誘導体を製造するための方法を提供することである。更に本発明は、イミン誘導体を基質として光学活性なアミン誘導体を生成することができる酵素、あるいは該酵素を発現する微生物の提供を課題とする。更に本発明は、目的とする性状を備えた該酵素をコードするDNAを単離してベクターに組み込み、組換え体として得ることを課題とする。加えて、組換え体による光学活性アミン誘導体の製造方法の提供をも課題とするものである。
【課題を解決するための手段】
【0008】
本発明者らは、このような高度な要求を満足する方法を鋭意検討した。そして微生物の有する立体選択的な還元能力を利用して、イミン誘導体を立体選択的に還元することにより、光学活性なアミン誘導体を生成させうることを見出した。
【0009】
既に述べたように、イミン誘導体を立体選択的に還元する生合成に関与する酵素は知られている。しかし公知の酵素はいずれも代謝に関与する酵素である。そのため医薬品の中間体などの製造に応用した場合には、工業的な生産に求められる基質特異性や活性は期待できない。特に2−メチル−1−ピロリンなどの環状アルキルイミン誘導体を還元して、光学活性環状アルキルアミン誘導体を得る方法は、まったく知られていなかった。
【0010】
したがって、本出願人が先の国際出願(PCT/JP2009/065244)において開示したように、医薬品の中間体などの製造に好適な基質特異性や触媒活性を備えた微生物が見出されたことは、非常に意外な成果であった。更に驚くべきことに、微生物が有する立体選択的なイミンの還元能力は、十分に高く、工業的な利用に際し問題の無いレベルであった。たとえば、ストレプトマイセス・エスピー GF3546(Streptomyces sp. GF3546)は前記の課題を解決するための有用な性質を備えた微生物であることが見出された。更に該国際出願(PCT/JP2009/065244)には、GF3546を大量培養し、無細胞抽出液を調製した後、次の工程を経てイミン還元酵素(SsSIR)を高度に精製することに成功したことが開示されている。
−Phenyl Sepharoseを用いた疎水クロマトグラフィー、および
−MonoQを用いたイオン交換クロマトグラフィー
また、該国際出願(PCT/JP2009/065244)においては、イミン還元酵素をコードするDNA(配列番号:6)を単離し、当該酵素を高発現する組換え菌を造成したことも開示されている。
【0011】
本発明者らは、その後さらに、Streptomyces sp. GF 3546由来イミン還元酵素(SsSIR)のアミノ酸配列(配列番号:5)を用いて同一性検索を行い、EMBLに登録されている機能不明のアミノ酸配列が、当該イミン還元酵素に57%の同一性を有していることを見出した。具体的には、ストレプトスポランジウム・ロセウム(Streptosporangium roseum)のゲノム解析の結果より得られたC4EPL6_STRRSと命名された予想タンパク質であり、機能はDDBJ(DNA Data Bank of Japan)では、C4EPL6_STRRSはbeta-hydroxyacid dehydrogenase, 3-hydroxyisobutyrate dehydrogenaseであると予測されている。本発明者らは、C4EPL6_STRRSを高発現する組換え菌を造成し、性質を調べた結果、上記DDBJの予測とまったく異なるイミン還元活性を有する事を見出し、本発明を完成した。
【0012】
すなわち、本発明は、以下の光学活性アミンの製造方法を提供する。あるいは本発明は、当該方法に有用な微生物、あるいはイミン還元酵素、当該酵素をコードするDNAを含むベクター、当該酵素を含有する立体選択的イミン還元剤に関する。
〔1〕
式(I)で示されるイミン誘導体に、下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素、当該イミン還元酵素を生産する微生物、およびその処理物からなる群から選択される、少なくとも一つの酵素活性物質を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
式(I)
【化1】
式(II)
【化2】
(式中、R1、R2基は炭素数1〜3個のアルキル基を、R3基は、炭素数1〜3個のアルキル基もしくは水素を表し、R1とR3は環を巻いていてもよい)
〔2〕
式(I)の化合物が下記式(III)で表されるイミン誘導体であり、式(II)の化合物が式(IV)で表されるアミン誘導体である〔1〕に記載の光学活性なS体のアミン誘導体の製造方法。
式(III)
【化3】
式(IV)
【化4】
(式中R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)
〔3〕
式(III)の化合物が2−メチル−1−ピロリンであり、式(IV)で表される化合物が、2−メチルピロリジンである、〔2〕に記載の光学活性なS体のアミン誘導体の製造方法。
〔4〕
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
〔5〕
前記イミン還元酵素が、配列番号:1に記載のアミノ酸配列において、1個以上かつ50個以下のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質である、〔4〕に記載のイミン還元酵素。
〔6〕
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元活性を有するタンパク質を有効成分として含む、立体選択的イミン還元剤。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
〔7〕
下記(a)から(e)のいずれかに記載のイミン還元酵素をコードするポリヌクレオチドと、NADP+を補酵素とする酸化還元反応を触媒することができる脱水素酵素をコードするポリヌクレオチドとが一緒に挿入された組換えベクター。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
〔8〕
脱水素酵素がグルコース脱水素酵素である、〔7〕に記載のベクター。
〔9〕
グルコース脱水素酵素がバシラス・サブチルス(Bacillus subtilis)に由来する、〔8〕に記載の組換えベクター。
〔10〕
〔7〕〜〔9〕のいずれかに記載のベクターを発現可能に保持した形質転換体。
〔11〕
〔10〕に記載の形質転換体を培養する工程を含む、〔1〕〜〔3〕のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
〔12〕
〔1〕記載のイミン還元酵素を生産する微生物がストレプトスポランジウム(Streptosporangium)属に属する微生物である〔1〕〜〔3〕のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
〔13〕
式(I)で示されるイミン誘導体に、ストレプトスポランジウム(Streptosporangium)属に属する微生物またはその処理物を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
【発明の効果】
【0013】
本発明により、イミン還元酵素もしくは該酵素を発現する微生物等を使用したイミンの立体選択的還元により、(S)体光学活性アミンを効率的に製造する方法が提供された。また、該反応を担うイミン還元酵素が提供された。さらに、イミン還元酵素をコードするDNAを単離し、当該酵素を高発現する組換え菌が提供された。本発明の方法は、安価に合成可能なイミン誘導体を基質とし、微生物の高い立体選択性を利用して、光学活性アミンが合成できるため、工業的に有利である。本発明は、特に、(S)−2−メチルピロリジンの製造に有用である。(S)−2−メチルピロリジンは、医薬品を合成するための中間体として有用な化合物である。本発明によって、当該化合物を微生物や酵素の作用によって簡便にかつ効率的に製造し、さらに非常に高い光学純度を達成することが可能となった。本発明は、97.7%eeという高い光学純度で化合物を生成するという、顕著な効果を奏するものである。
【図面の簡単な説明】
【0014】
【図1】C4EPL6_STRRSイミン還元酵素遺伝子が挿入されたプラスミド(pSU-SRI1)の構築の過程を示した図である。
【図2】C4EPL6_STRRSイミン還元酵素遺伝子と枯草菌由来のグルコース脱水素酵素遺伝子が挿入されたプラスミド(pSG-SRI1)の構築の過程を示した図である。
【図3】SsSIRイミン還元酵素遺伝子が挿入されたプラスミド(pSUS2MP1)の構築の過程を示した図である。
【図4】SsSIRイミン還元酵素遺伝子と枯草菌由来のグルコース脱水素酵素遺伝子が挿入されたプラスミド(pSG-S2MP)の構築の過程を示した図である。
【発明を実施するための形態】
【0015】
本発明は、前記式(I)で示されるイミン誘導体に、当該化合物を立体選択的に還元しうる能力を有する酵素タンパク質(配列番号:1のタンパク質もしくはそのバリアント)、該タンパク質を発現する微生物の培養物、菌体およびその処理物からなる群から選択される、少なくとも一つの酵素活性物質を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法に関する。
式(I)
【化5】
式(II)
【化6】
(式中、R1、R2基は炭素数1〜3個のアルキル基を、R3基は、炭素数1〜3個のアルキル基もしくは水素を表し、R1とR3は環を巻いていてもよい)。
【0016】
本発明において、環状イミン化合物は式(I)の化合物として好ましい。本発明において、環状イミン化合物は、たとえば下記式(III)で表されるイミン誘導体を含む。
式(III)
【化7】
(式中、R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)。
【0017】
上記式(III)の化合物を基質として用いた場合、光学活性アミン誘導体として式(IV)に示される化合物を得ることができる。当該化合物は、本発明によって製造することができる好ましい光学活性アミン誘導体である。
式(IV)
【化8】
(式中、R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)。
【0018】
本発明における好ましい化合物は、前記式(III)において、たとえばRがメチル基、エチル基、あるいはプロピル基であり、nが2〜3の化合物である。より具体的には、本発明における好ましいイミン誘導体として次の化合物を示すことができる。
2−メチル−1−ピロリン
2−メチル−1−ピペリデイン
2−エチル−1−ピロリン
2−エチル−1−ピペリデイン
【化9】
【0019】
これらの化合物を基質として、それぞれ以下の光学活性アミン誘導体を製造することができる。
2−メチル−1−ピロリン → (S)−2−メチルピロリジン
2−メチル−1−ピペリデイン → (S)−2−メチルピペリジン
2−エチル−1−ピロリン → (S)−2−エチルピロリジン
2−エチル−1−ピペリデイン → (S)−2−エチルピペリジン
【化10】
【0020】
本発明において、イミン誘導体を立体選択的に還元しうる能力とは、前記式(I)望ましくは式(III)で示されるイミン誘導体を基質として、S体のアミン誘導体を生成する能力を言う。本発明の微生物は、配列番号:1のイミン還元酵素もしくはそのバリアントを発現し、S体のアミン誘導体を生成しうる微生物であれば、任意の微生物を用いることができる。本発明に用いる微生物は、たとえば以下に示すような微生物について、配列番号:1のイミン還元酵素もしくはそのバリアントの有無とS体のアミン誘導体を生成する能力を調べることにより得ることができる。
【0021】
たとえば、化合物(I)望ましくは化合物(III)で示されるイミン誘導体を含む培地中で被験微生物を培養し、その培養物中に生成した光学活性アミン誘導体の光学純度を測定する。その結果、光学活性な(S)−アミン誘導体の生成が確認できれば、当該微生物を本発明に用いることができる。
【0022】
あるいは予め培地中で増殖させた被験微生物を集菌し、適当な緩衝液中に懸濁させる。更に化合物(I)望ましくは化合物(III)で示されるイミン誘導体と接触、反応させて、この緩衝液中に生成される光学活性アミン誘導体の光学純度を測定する。光学活性なアミン誘導体の生成が確認できれば、当該微生物を本発明に用いることができる。このとき、被験微生物の培養時に、培地中に化合物(I)望ましくは化合物(III)を加えておけば、この化合物を還元するための酵素の誘導が期待できる。更に反応時に還元エネルギー源を加えておけば、より効果的な生成物の蓄積が期待できる。還元エネルギー源としては、通常、グルコースなどを用いることができる。
【0023】
例えば、微生物を必要に応じて2−メチル−1−ピロリンを含む適当な培地で培養し、得られた微生物菌体を、10mM 2−メチル−1−ピロリン、25mM リン酸カリウム緩衝液(pH7.0)、及び必要に応じてグルコースを含む反応液に懸濁し、25℃で24時間振盪し、生成物をTLCもしくはHPLCにより分析することにより、微生物の2−メチル−1−ピロリン還元能を確認することができる。
TLCの条件としては、例えば、Silica gel 60 F254 プレートを用い、1−ブタノール/酢酸/水=2/1/1の展開液で展開し、ニンヒドリンにより呈色する方法が挙げられる。
【0024】
生成したアミン誘導体の光学純度は、例えば以下の方法により測定することができる。
被検サンプル50μLに以下の成分を添加して、40℃、1時間反応することにより生成したアミンをGITC化し、HPLCにより分離し定量する。
2mg/mL トリエチルアミン 50μL、および
8mg/mL 2,3,4,6−テトラ−O−アセチル−β−D−グルコピラノシルイソチオシアネート(GITC) 100μL
【0025】
HPLCの条件としては、例えば、以下のような条件を用いることができる。
−HPLCカラム:和光純薬株式会社製 Wakosil II 5C18(4.6mm x 150mm)
−溶離液:10mMリン酸バッファー(pH2.5):メタノール=55:45
−流速:1mL/min
−検出:254nmにおけるUV吸収
上記条件下で、(R)−2−メチルピロリジンは26.3分に、(S)−2−メチルピロリジン誘導体は24.9分に溶出される。
同様な方法を用いて、単離した配列番号:1のイミン還元酵素もしくはそのバリアントタンパク質が有するS体のアミン誘導体を生成する能力を調べることもできる。
【0026】
光学純度の高い生成物を得るには、より選択性の高い微生物を用いることが有利であることは言うまでも無い。具体的には、その立体選択性は、たとえば60%、通常70%以上、好ましくは80%以上、更に好ましくは90%以上の、光学活性なS体のアミン誘導体を生成しうる微生物を用いることができる。本願の実施例に開示されるように、配列番号:1の酵素を発現する微生物を利用すると、97.7%eeの光学純度(enantiomeric excess;ee)を達成することができる。
【0027】
本発明における「光学活性なアミン誘導体」とは、ある光学異性体が別の光学異性体より多く含まれるアミン誘導体を言う。本発明において、好ましい光学活性なアミン誘導体は、たとえば60%、通常70%以上、好ましくは80%以上、更に好ましくは90%以上の光学純度(enantiomeric excess;ee)を有する。光学活性なアミン誘導体の光学純度は、たとえば光学分割カラムなどを用いて確認することができる。本発明の「光学異性体」は、一般的に「光学活性体」および「鏡像異性体」(enantiomeric)と呼ばれる場合もある。
【0028】
本出願人による先の国際出願(PCT/JP2009/065244)に開示のように、たとえばストレプトマイセス(Streptomyces)属に属する微生物は、前記式(I)で示されるイミン誘導体を還元し、S体のアミン誘導体を生成する。このような微生物は、本発明のイミン還元酵素の性能を比較分析する際などに利用できる。また、このような微生物に配列番号:1の酵素をコードする遺伝子を導入することも考えられる。
さらに具体的には、ストレプトマイセス(Streptomyces)属に属する微生物として、次の微生物を示すことができる。この微生物は、高い光学純度の(S)アミン誘導体を生成する微生物である。
ストレプトマイセス・エスピーGF3546(Streptomyces sp. GF3546)
【0029】
上記の微生物は、次のとおり寄託されている(受託日;2008年6月24日)。
(1)寄託機関名:独立行政法人製品評価技術基盤機構
(2)連絡先:〒292−0818千葉県木更津市かずさ鎌足2−5−8
(3)識別のための表示および受託番号:
ストレプトマイセス・エスピー GF3546(Streptomyces sp. GF3546);受託番号 NITE P-593
【0030】
さらにこれらの微生物は、ブダペスト条約に基づく国際寄託に、次のとおり移管されている。
(1)国際寄託当局:独立行政法人製品評価技術基盤機構特許微生物寄託センター
(2)住所:〒292−0818 千葉県木更津市かずさ鎌足2−5−8
(3)識別の表示および受託番号
ストレプトマイセス・エスピー GF3546(Streptomyces sp. GF3546)
;受託番号 NITE BP-593 (移管日2009年8月4日)
【0031】
前記式(I)あるいは(III)で示されるイミン誘導体を立体選択的に還元する能力を有する微生物は、これらの寄託菌株のみならず、種々の微生物資源(microbiological resource)からも見出すことができる。微生物資源には、自然環境から単離される微生物群、微生物寄託機関に保存された微生物菌株などが含まれる。中でも、ストレプトマイセス属に属する微生物は、前記式(I)あるいは(III)で示されるイミン誘導体を立体選択的に還元する能力を有する微生物として好ましい。したがって、たとえば、微生物寄託機関に保存されたストレプトマイセス属微生物を対象として、先に述べたような選択方法によって、目的の作用を有する微生物を選択することができる。寄託された微生物は、たとえば次のような寄託機関から分譲を受けることができる。
生物遺伝資源センター (NBRC)
理化学研究所 (JCM)
東京農業大学菌株保存室 (IAM)
American Type Culture Collection (ATCC)
Deutsche Sammlung von Mikroorganismen (DSM)
【0032】
また、有用な微生物をリボソーマルDNA(rDNA)の塩基配列情報に基づいて、選択することもできる。一般にrDNAの塩基配列情報は、生物間の遺伝学的な近さを表す指標として有用であることが広く知られている。特に16S rRNAをコードするDNA(以下、「16S rDNA」と記載する)は、微生物の遺伝的な同一性あるいは類似性を決定するためのツールとして活用されている。
16S rRNAは、全生物に存在しており、生物間で構造の違いは見られるが、アライメント可能な程度に配列が保存されている。16S rRNAの生物間水平伝播の頻度は低いので、分類に使用する方法が汎用されている。
具体的には、小サブユニットリボソームRNA(SSU rRNA)は、タンパク質合成の場であるリボソームを構成する核酸分子の1つである。その塩基配列の長さは、多少の違いはあるものの、細菌からヒトまで起源は同じものと見なされている。したがって、rRNAの塩基配列は進化的保存性が高く、生物の系統解析において最も頻繁に利用されている(Microbiol. Rev., 58, 1-9 (1994))。原核生物におけるSSU rRNA は約1500塩基の16S rRNAである。原核生物の分類と同定に16S rRNAの塩基配列を利用する系統分類が一般に用いられている(ASM News, 65, 752-757 (1999)、生物工学実験書p113日本生物工学会編、培風館)。
現在の放線菌の分類体系は、16S rDNA塩基配列の類似性に基づいて構成されている(Stackebrandt, et al. (1997) Int. J. Syst. Bacteriol., 47, 479-491)。菌株の同定においても、属レベルだけではなく種レベルにおいても16S rDNA塩基配列の相同性に基づく同定が、細菌系統分類学の主流となっている(「放線菌の分類と同定」(日本放線菌学会)p19)。
【0033】
塩基配列情報データベースを対象として、これらの塩基配列情報を検索すれば、既に16S rDNAが決定された微生物の中から、16S rDNA中に同一性の高い塩基配列を含む菌株を見つけ出すことができる。たとえばDNA Data Bank of Japan(DDBJ)などの公共データベースサービス機関では、16S rRNA(Prokaryotes)の塩基配列情報を対象とする検索サービスが提供されている。こうして見出された微生物のうち、特にストレプトマイセス属に属する微生物を選択することによって、本発明のイミン還元酵素との比較分析や形質転換等に利用しうる微生物とすることができる。必要に応じ、こうして選択された微生物のイミン誘導体に対する作用を予め確認することもできる。
【0034】
使用する微生物は、醗酵学の分野で公知の方法に従って培養することができる。培地としては炭素源、窒素源、無機物およびその他の栄養素を適量含有する培地ならば、合成培地または天然培地のいずれでも使用することができる。培地は、液体培地または固体培地を使用することができる。
具体的には、炭素源として、次に示すような一般的な炭素源より、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
糖類: 天然炭水化物:
グルコース、 澱粉、
フルクトース、 澱粉加水分解物、
マルトース、 糖蜜、
ガラクトース、 廃糖蜜、
麦、
とうもろこしなど
アルコール類: 有機酸類:
グリセロール、 酢酸、
メタノール、 グルコン酸、
エタノールなど ピルビン酸、
ケトグルタル酸、
クエン酸など
炭化水素類:
ノルマルパラフィンなど
脂肪類:
パーム核油、
大豆油、
オリーブ油など
【0035】
窒素源としては、次に示すような一般的な窒素源の中から、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
有機窒素化合物:
肉エキス、 ペプトン、 酵母エキス、
大豆加水分解物、 ミルクカゼイン、 カザミノ酸、
各種アミノ酸、 コーンスティープリカー、
その他の動物、植物、微生物の加水分解物など
無機窒素化合物:
アンモニア、 硝酸アンモニウム、 硫酸アンモニウム、
塩化ナトリウムなどのアンモニウム塩、
硝酸ナトリウムなどの硝酸塩、尿素など
【0036】
また、微生物のイミン誘導体の立体選択的還元能力を高めるために、誘導物質を用いることができる。誘導物質としては、イミン誘導体を、使用する微生物に応じて使用することができる。
さらに、無機塩として微量のマグネシウム、マンガン、カリウム、カルシウム、ナトリウム、銅、亜鉛などのリン酸塩、塩酸塩、硝酸塩、酢酸塩等より適宜一種または二種以上を選択して使用することができる。また、必要に応じて植物油、界面活性剤、シリコンなどの消泡剤を培養液中に添加してもよい。
【0037】
培養は前記培地成分を含有する液体培地中で通常の培養方法を用いて行うことができる。たとえば次のような培養方法を利用することができる。
振とう培養 (shaking culture)
通気攪拌培養 (aeration-agitation culture)
連続培養 (continuous culture)
流加培養(feeding culture, fed batch culture)
【0038】
培養条件は、微生物の種類、培養の種類、培養方法により適宜選択することができる。利用する菌株が増殖し、イミン誘導体の立体選択的還元能力を有しうる条件であれば特に制限はない。一般的な培養条件として次のような条件を示すことができる。
培養開始時のpHを4から10、好ましくは6から8に調節
15から70℃、好ましくは20から40℃の温度条件下で培養
培養時間はイミン誘導体の還元能力を有する菌体が得ることができれば特に制限されない。通常は1日から14日、好ましくは1日から7日培養する。
【0039】
本発明において、微生物菌体の処理物とは、菌体を目的とする酵素活性が維持される条件で処理したものを意味する。酵素活性の維持とは、生菌体で得られる酵素活性の一般的には20%以上、通常30%以上、好ましくは50%以上、より好ましくは70%以上の活性を維持することを言う。生菌体とその処理物の酵素活性は、生菌体の酵素活性と、同量の生菌体から得られた処理物の酵素活性を比較することによって定量的に比較することができる。本発明における処理物は、生菌体よりも高い酵素活性を有する場合もある。
【0040】
このような処理物を得る方法としては、凍結乾燥、有機溶媒による脱水乾燥、菌体の自己消化、酵素活性画分の抽出、超音波処理などを利用することができる。このような処理方法によって、例えば、凍結乾燥菌体、アセトン乾燥菌体、菌体自己消化物、菌体抽出物、菌体磨砕物、菌体の超音波処理物等を得ることができる。
【0041】
これらの処理は、単独で、あるいは複数の処理を重複して適用することもできる。たとえば適当な培養液で微生物を培養後、菌体を破砕して、遠心分離等により無細胞抽出液を調製することができる。菌体は、機械的な破砕、超音波、高圧処理、酵素消化、自己消化などにより破砕することができる。無細胞抽出液は本発明における菌体の処理物に含まれる。
【0042】
本発明においては、配列番号:1のイミン還元酵素もしくはそのバリアントを発現する微生物から当該酵素を部分精製したものや完全に精製したものなどを用いることができる。国際出願(PCT/JP2009/065244)に開示のイミン還元酵素と同様に、本発明のイミン還元酵素も、次の(1)〜(2)に示す理化学的性状を有する。
(1)作用:NADPH依存的に、2−メチル−1−ピロリンを還元し、(S)−2−メチルピロリジンを生成する。NADP+依存的に、(S)−2−メチルピロリジンを酸化し、2−メチル−1−ピロリンを生成する;、および
(2)補酵素依存性:還元反応の補酵素としてNADPHを利用する。酸化反応の補酵素としてNADP+を利用する。
【0043】
本発明のイミン還元酵素は、配列番号:1のイミン還元酵素もしくはそのバリアントを発現する微生物、あるいはその無細胞抽出液などから精製することができる。具体的には、次のような精製工程を適宜組み合わせることによって、無細胞抽出液から実質的に純粋なイミン還元酵素を得ることができる。
硫安、アセトンなどを用いた溶解度による分割、
イオン交換クロマトグラフィー、
疎水クロマトグラフィー、
2’,5’−ADPセファロースやブルーもしくはレッドセファロースなどを用いたアフィニティクロマトグラフィー、
ゲル濾過
【0044】
例えば、微生物を集菌後、次のような精製工程を経て、本発明のイミン還元酵素を実質的に純粋な酵素として単離することができる。
超音波あるいはガラスビーズによる菌体の破砕と無細胞抽出液の調製、
Phenyl Sepharoseを用いた疎水クロマトグラフィー、および
MonoQを用いたイオン交換クロマトクロマトグラフィーによるイミン還元酵素活性分画の単離
【0045】
本発明において、「実質的に純粋」とは、当該酵素が、当該酵素以外の生物学的な高分子化合物や化学物質を実質的に含まないことを言う。当該酵素以外の生物学的な高分子化合物とは、たとえば菌体や培養液に由来するタンパク、糖類、脂質、核酸類が含まれる。本発明における実質的に純粋な酵素は、少なくとも75%以上、通常80%以上、好ましくは85%以上、より好ましくは90%以上、更に好ましくは95%以上、あるいは99%以上の純度を有する。酵素の純度を決定する方法は公知である。たとえば、各種のクロマトグラフィーや、ポリアクリルアミドゲル電気泳動などによって蛋白質の純度を決定する方法が周知である。
【0046】
あるいは本発明によって、上記理化学的性状(1)(2)を有する単離されたイミン還元酵素が提供される。本発明において、単離されたイミン還元酵素とは、それが存在する天然の環境から分離されて存在していることを意味する。したがって、菌体から分離されたイミン還元酵素は、本発明における単離されたイミン還元酵素に含まれる。
【0047】
本発明における実質的に純粋なイミン還元酵素、あるいは単離されたイミン還元酵素は、精製、あるいは単離された後に、酵素組成物とすることもできる。たとえば、精製、あるいは単離されたイミン還元酵素を適当な担体と配合することによって、本発明のイミン還元酵素を含む酵素組成物を調製することができる。担体としては、アルブミンなどの不活性蛋白質、ショ糖などの糖類あるいは糖アルコールなどを利用することができる。酵素組成物は、液状であることもできるし、乾燥状態であることもできる。酵素と担体を含む水溶液を凍結乾燥することによって得られる酵素組成物は、本発明における好ましい組成物の一つである。このようなイミン還元活性を有するタンパク質を含む組成物は、立体選択的なイミン還元剤として用いることができる。すなわち、本発明の立体選択的イミン還元剤は、有効成分として、配列番号:1に記載のアミノ酸配列からなるタンパク質および/もしくはそのバリアントを含有する。
【0048】
本発明のイミン還元酵素は、以下の方法によりその活性を定量することができる。すなわち、次の組成の反応液中(1mL)で、30℃で反応を行い、NADPHの減少に由来する340nmの吸光度の減少を測定する。
10mM 2−メチル−1−ピロリン、
100mM リン酸カリウム緩衝液(pH7.5)、
0.1mM NADPH及び
酵素
1Uは、上記条件下、1分間に1μmolのNADPHの減少を触媒する酵素量とした。
【0049】
本発明は、配列番号:1に記載のアミノ酸配列からなるタンパク質に関する。本発明はまた、配列番号:1に記載のアミノ酸配列からなるタンパク質のホモログ(本願ではバリアントとも言う)を含む。本発明のイミン還元酵素のホモログ(バリアント)とは、配列番号:1に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、配列番号:1に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質を意味する。配列番号:1に記載のアミノ酸配列において、たとえば100以下、通常50以下、好ましくは30以下、より好ましくは15以下、更に好ましくは10以下、あるいは5以下のアミノ酸残基の変異は許容される。すなわち、本発明のイミン還元酵素は、配列番号:1に記載のアミノ酸配列に対して、たとえば1以上、5以上、10以上、30以上、50以上、あるいは100以上のアミノ酸残基の変異を含みうる。
【0050】
本発明は、イミン還元酵素をコードするポリヌクレオチドおよびそのホモログ(バリアント)に関する。本発明において、ポリヌクレオチドは、DNAやRNA等の天然のポリヌクレオチドに加え、人工的なヌクレオチド誘導体を含む人工的な分子であることもできる。また本発明のポリヌクレオチドは、DNA−RNAのキメラ分子であることもできる。本発明のイミン還元酵素をコードするポリヌクレオチドは、たとえば配列番号:2に示す塩基配列を含む。配列番号:2に示す塩基配列は、配列番号:1に示すアミノ酸配列を含むタンパク質をコードしており、このアミノ酸配列を含むタンパク質は、本発明によるイミン還元酵素の好ましい態様を構成する。
【0051】
本発明のイミン還元酵素をコードするポリヌクレオチドのホモログ(バリアント)とは、配列番号:1に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列を含み、かつ、前記理化学的性質(1)〜(2)を有するタンパク質をコードするポリヌクレオチドを含む。当業者であれば、配列番号:2に記載のポリヌクレオチドに部位特異的変異導入法(Nucleic Acid Res. 10,pp.6487 (1982) , Methods in Enzymol.100,pp.448 (1983), Molecular Cloning 2ndEdt., Cold Spring Harbor Laboratory Press (1989) , PCR A Practical Approach IRL Press pp.200 (1991))などを用いて、適宜置換、欠失、挿入、および/または付加変異を導入することによりポリヌクレオチドのホモログを得ることが可能である。
【0052】
また、本発明におけるポリヌクレオチドのホモログ(バリアント)は、配列番号:2に示される塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズできるポリヌクレオチドであって、かつ、前記理化学的性質(1)〜(2)を有するタンパク質をコードするポリヌクレオチドも含む。ストリンジェントな条件でハイブリダイズできるポリヌクレオチドとは、配列番号:2に記載中の任意の少なくとも20個、好ましくは少なくとも30個、たとえば40、60または100個の連続した配列を一つまたは複数選択したDNAをプローブDNAとし、たとえばECL direct nucleic acid labeling and detection system (GEヘルスケア社製)を用いて、マニュアルに記載の条件(wash:42℃、0.5 x SSCを含むprimary wash buffer)においてハイブリダイズするポリヌクレオチドを指す。
【0053】
さらに、本発明におけるポリヌクレオチドのホモログ(バリアント)は、配列番号:1に示されるアミノ酸配列と少なくとも70%、あるいは少なくとも75%、好ましくは少なくとも80%、あるいは少なくとも85%、より好ましくは90%以上、あるいは95%以上、の同一性を有するタンパク質をコードするポリヌクレオチドを含む。タンパク質の同一性検索は、たとえばSWISS−PROT、PIR、DADなどのタンパク質のアミノ酸配列に関するデータベースやDDBJ、EMBL、あるいはGene−BankなどのDNA配列に関するデータベース、DNA配列を基にした予想アミノ酸配列に関するデータベースなどを対象に、BLAST、FASTAなどのプログラムを利用して、例えば、インターネットを通じて行うことができる。
【0054】
本発明は、配列番号:1に記載のアミノ酸配列からなるタンパク質に関する。本発明はまた、配列番号:1に記載のアミノ酸配列からなるタンパク質のホモログ(バリアント)を含む。本発明のイミン還元酵素のホモログとは、配列番号:1に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、配列番号:1に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質を意味する。本発明において、配列番号:1に記載のアミノ酸配列からなるタンパク質と機能的に同等とは、当該タンパク質が前記(1)〜(2)に示した物理化学的性状を有することを意味する。当業者であれば、配列番号:2記載のDNAに部位特異的変異導入法(Nucleic Acid Res. 10,pp.6487 (1982) , Methods in Enzymol.100,pp.448 (1983), Molecular Cloning 2ndEdt., Cold Spring Harbor Laboratory Press (1989) , PCR A Practical Approach IRL Press pp.200 (1991))などを用いて、適宜置換、欠失、挿入、および/または付加変異を導入することによりイミン還元酵素のホモログをコードするポリヌクレオチドを得ることができる。そのイミン還元酵素のホモログをコードするポリヌクレオチドを宿主に導入して発現させることにより、配列番号:1に記載のイミン還元酵素のホモログを得ることが可能である。
【0055】
さらに、本発明のイミン還元酵素のホモログ(バリアント)とは、配列番号:1に示されるアミノ酸配列と少なくとも70%、あるいは少なくとも75%、好ましくは少なくとも80%、あるいは少なくとも85%、より好ましくは90%以上、あるいは95%以上の同一性を有するタンパク質をいう。タンパク質の同一性検索は、たとえばSWISS−PROT、PIR、DADなどのタンパク質のアミノ酸配列に関するデータベースやDDBJ、EMBL、あるいはGene−BankなどのDNA配列に関するデータベース、DNA配列を基にした予想アミノ酸配列に関するデータベースなどを対象に、BLAST、FASTAなどのプログラムを利用して、例えば、インターネットを通じて行うことができる。
【0056】
本発明のイミン還元酵素をコードするポリヌクレオチドは、たとえば、以下のような方法によって単離することができる。
【0057】
配列番号:2に記載の塩基配列を基にPCR用のプライマーを設計し、酵素生産株(ストレプトスポランジウム・ロセウムなどのストレプトスポランジウム属の菌株。たとえば、ストレプトスポランジウム・ロセウムDSM 43021)の染色体DNAもしくは、cDNAライブラリーを鋳型としてPCRを行うことにより本発明のDNAを得ることができる。
【0058】
さらに、得られたDNA断片をプローブとして、酵素生産株の染色体DNAの制限酵素消化物をファージ、プラスミドなどに導入し、大腸菌を形質転換して得られたライブラリーやcDNAライブラリーを利用して、コロニーハイブリダイゼーション、プラークハイブリダイゼーションなどにより、本発明のポリヌクレオチドを得ることができる。
【0059】
また、PCRにより得られたDNA断片の塩基配列を解析し、得られた配列から、既知のDNAの外側に伸長させるためのPCRプライマーを設計し、酵素生産株の染色体DNAを適当な制限酵素で消化後、自己環化反応によりDNAを鋳型として逆PCRを行うことにより(Genetics 120, 621-623 (1988))、また、RACE法(Rapid Amplification of cDNA End、「PCR実験マニュアル」p25−33、HBJ出版局)などにより本発明のポリヌクレオチドを得ることも可能である。
【0060】
なお本発明において利用可能なポリヌクレオチドには、以上のような方法によってクローニングされたゲノムDNA、あるいはcDNAの他、合成によって得られたDNAが含まれる。
このようにして単離された、本発明によるイミン還元酵素をコードするポリヌクレオチドを公知の発現ベクターに挿入することにより、イミン還元酵素発現ベクターが提供される。
【0061】
また、この発現ベクターで形質転換した形質転換体を培養することにより、本発明のイミン還元酵素を組換え体から得ることができる。
【0062】
本発明においてイミン還元酵素を発現させるために、形質転換の対象となる微生物は、イミン還元酵素を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターにより形質転換され、イミン還元酵素活性を発現することができる生物であれば特に制限はない。利用可能な微生物としては、たとえば以下のような微生物を示すことができる。
【0063】
エシェリヒア(Escherichia)属
バチルス(Bacillus)属
シュードモナス(Pseudomonas)属
セラチア(Serratia)属
ブレビバクテリウム(Brevibacterium)属
コリネバクテリイウム(Corynebacterium)属
ストレプトコッカス(Streptococcus)属
ラクトバチルス(Lactobacillus)属など宿主ベクター系の開発されている細菌
ロドコッカス(Rhodococcus)属
ストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌
サッカロマイセス(Saccharomyces)属
クライベロマイセス(Kluyveromyces)属
シゾサッカロマイセス(Schizosaccharomyces)属
チゴサッカロマイセス(Zygosaccharomyces)属
ヤロウイア(Yarrowia)属
トリコスポロン(Trichosporon)属
ロドスポリジウム(Rhodosporidium)属
ピキア(Pichia)属
キャンディダ(Candida)属などの宿主ベクター系の開発されている酵母
ノイロスポラ(Neurospora)属
アスペルギルス(Aspergillus)属
セファロスポリウム(Cephalosporium)属
トリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ
【0064】
形質転換体の作製のための手順および宿主に適合した組み換えベクターの構築は、分子生物学、生物工学、遺伝子工学の分野において慣用されている技術に準じて行うことができる(例えば、Sambrookら、モレキュラー・クローニング、Cold Spring Harbor Laboratories)。微生物中などにおいて、本発明のNADPHを電子供与体とするイミン還元酵素遺伝子を発現させるためには、まず微生物中において安定に存在するプラスミドベクターやファージベクター中にこのDNAを導入し、その遺伝情報を転写・翻訳させる必要がある。
【0065】
そのためには、転写・翻訳を制御するユニットにあたるプロモーターを本発明のDNA鎖の5’−側上流に、より好ましくはターミネーターを3’−側下流に、それぞれ組み込めばよい。このプロモーター、ターミネーターとしては、宿主として利用する微生物中において機能することが知られているプロモーター、ターミネーターを用いる必要がある。これら各種微生物において利用可能なベクター、プロモーター、ターミネーターなどに関して「微生物学基礎講座8遺伝子工学・共立出版」、特に酵母に関しては、Adv. Biochem. Eng. 43, 75-102 (1990)、Yeast 8, 423-488 (1992)、などに詳細に記述されている。
【0066】
例えば、エシェリヒア属、特に大腸菌エシェリヒア・コリ(Escherichia coli)においては、プラスミドベクターとして、pBR、pUC系プラスミドを利用でき、lac(β−ガラクトシダーゼ)、trp(トリプトファンオペロン)、tac、trc(lac、trpの融合)、λファージPL、PRなどに由来するプロモーターなどが利用できる。また、ターミネーターとしては、trpA由来、ファージ由来、rrnBリボソーマルRNA由来のターミネーターなどを用いることができる。これらの中で、市販のpSE420(Invitrogen製)のマルチクローニングサイトを一部改変したベクターpSE420D(特開2000−189170に記載)が好適に利用できる。
【0067】
バチルス属においては、ベクターとしてpUB110系プラスミド、pC194系プラスミドなどが利用可能であり、染色体にインテグレートすることもできる。また、プロモーター、ターミネーターとしてapr(アルカリプロテアーゼ)、npr(中性プロテアーゼ)、amy(α−アミラーゼ)などが利用できる。
【0068】
シュードモナス属においては、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・セパシア(Pseudomonas cepacia)などで宿主ベクター系が開発されている。トルエン化合物の分解に関与するプラスミドTOLプラスミドを基本にした広宿主域ベクター(RSF1010などに由来する自律的複製に必要な遺伝子を含む)pKT240などが利用可能であり、プロモーター、ターミネーターとして、リパーゼ(特開平5−284973)遺伝子などが利用できる。
【0069】
ブレビバクテリウム属特に、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)においては、pAJ43(Gene 39, 281 (1985))などのプラスミドベクターが利用可能である。プロモーター、ターミネーターとしては、大腸菌で使用されているプロモーター、ターミネーターがそのまま利用可能である。
【0070】
コリネバクテリウム属、特にコリネバクテリウム・グルタミカム(Corynebacterium glutamicum)においては、pCS11(特開昭57−183799)、pCB101(Mol. Gen. Genet. 196, 175 (1984)などのプラスミドベクターが利用可能である。
【0071】
ストレプトコッカス(Streptococcus)属においては、pHV1301(FEMS Microbiol. Lett. 26, 239 (1985)、pGK1(Appl. Environ. Microbiol. 50, 94 (1985))などがプラスミドベクターとして利用可能である。
【0072】
ラクトバチルス(Lactobacillus)属においては、ストレプトコッカス属用に開発されたpAMβ1(J. Bacteriol. 137, 614 (1979))などが利用可能であり、プロモーターとして大腸菌で利用されているものが利用可能である。
【0073】
ロドコッカス(Rhodococcus)属においては、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)から単離されたプラスミドベクターが使用可能である(J. Gen. Microbiol. 138,1003 (1992))。
【0074】
ストレプトマイセス(Streptomyces)属においては、HopwoodらのGenetic Manipulation of Streptomyces: A Laboratory Manual Cold Spring Harbor Laboratories (1985)に記載の方法に従って、プラスミドを構築することができる。特に、ストレプトマイセス・リビダンス(Streptomyces lividans)においては、pIJ486(Mol. Gen. Genet. 203, 468-478, 1986)、pKC1064(Gene 103,97-99 (1991))、pUWL−KS(Gene 165,149-150 (1995))が使用できる。また、ストレプトマイセス・バージニア(Streptomyces virginiae)においても、同様のプラスミドを使用することができる(Actinomycetol. 11, 46-53 (1997))。
【0075】
サッカロマイセス(Saccharomyces)属、特にサッカロマイセス・セレビジアエ(Saccharomyces cerevisiae)においては、YRp系、YEp系、YCp系、YIp系プラスミドが利用可能であり、染色体内に多コピー存在するリボソームDNAとの相同組み換えを利用したインテグレーションベクター(EP537456など)は、多コピーで遺伝子を導入でき、かつ安定に遺伝子を保持できるため極めて有用である。また、ADH(アルコール脱水素酵素)、GAPDH(グリセルアルデヒド−3−リン酸脱水素酵素)、PHO(酸性フォスファターゼ)、GAL(β−ガラクトシダーゼ)、PGK(ホスホグリセレートキナーゼ)、ENO(エノラーゼ)などのプロモーター、ターミネーターが利用可能である。
【0076】
クライベロマイセス属、特にクライベロマイセス・ラクティス(Kluyveromyces lactis)においては、サッカロマイセス・セレビジアエ由来2μm系プラスミド、pKD1系プラスミド(J. Bacteriol. 145, 382-390 (1981))、キラー活性に関与するpGKl1由来プラスミド、クライベロマイセス属における自律増殖遺伝子KARS系プラスミド、リボソームDNAなどとの相同組み換えにより染色体中にインテグレート可能なベクタープラスミド(EP 537456など)などが利用可能である。また、ADH、PGKなどに由来するプロモーター、ターミネーターが利用可能である。
【0077】
シゾサッカロマイセス(Schizosaccharomyces)属においては、シゾサッカロマイセス・ポンベ由来のARS(自律複製に関与する遺伝子)及びサッカロマイセス・セレビジアエ由来の栄養要求性を相補する選択マーカーを含むプラスミドベクターが利用可能である(Mol. Cell. Biol. 6, 80 (1986))。また、シゾサッカロマイセス・ポンベ由来のADHプロモーターなどが利用できる(EMBO J. 6, 729 (1987))。特に、pAUR224は、タカラバイオから市販されており容易に利用できる。
【0078】
チゴサッカロマイセス属(Zygosaccharomyces)においては、チゴサッカロマイセス・ロウキシ(Zygosaccharomyces rouxii)由来のpSB3(Nucleic Acids Res. 13, 4267 (1985))などに由来するプラスミドベクターが利用可能であり、サッカロマイセス・セレビジアエ由来PHO5プロモーターや、チゴサッカロマイセス・ロウキシ由来GAP−Zr(グリセルアルデヒド−3−リン酸脱水素酵素)のプロモーター(Agri. Biol. Chem. 54, 2521 (1990))などが利用可能である。
【0079】
ピキア(Pichia)属においては、ピキア・アンガスタ(旧名:ハンゼヌラ・ポリモルファ(Hansenula polymorpha))において宿主ベクター系が開発されている。ベクターとしては、ピキア・アンガスタ由来自律複製に関与する遺伝子(HARS1、HARS2)も利用可能であるが、比較的不安定であるため、染色体への多コピーインテグレーションが有効である(Yeast 7, 431-443 (1991))。また、メタノールなどで誘導されるAOX(アルコールオキシダーゼ)、FDH(ギ酸脱水素酵素)のプロモーターなどが利用可能である。また、ピキア・パストリス(Pichia pastoris)などにピキア由来自律複製に関与する遺伝子(PARS1、PARS2)などを利用した宿主ベクター系が開発されており(Mol. Cell. Biol. 5, 3376 (1985))、高濃度培養とメタノールで誘導可能なAOXなど強いプロモーターが利用できる(Nucleic Acids Res. 15, 3859 (1987))。
【0080】
キャンディダ(Candida)属においては、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・アルビカンス(Candida albicans)、キャンディダ・トロピカリス(Candida tropicalis)、キャンディダ・ウチルス(Candida utilis)などにおいて宿主ベクター系が開発されている。キャンディダ・マルトーサにおいてはキャンディダ・マルトーサ由来ARSがクローニングされ(Agri. Biol. Chem. 51, 51, 1587 (1987))、これを利用したベクターが開発されている。また、キャンディダ・ウチルスにおいては、染色体インテグレートタイプのベクターは強力なプロモーターが開発されている(特開平08−173170)。
【0081】
アスペルギルス(Aspergillus)属においては、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・オリジー(Aspergillus oryzae)などがカビの中で最もよく研究されており、プラスミドや染色体へのインテグレーションが利用可能であり、菌体外プロテアーゼやアミラーゼ由来のプロモーターが利用可能である(Trends in Biotechnology 7, 283-287 (1989))。
【0082】
トリコデルマ(Trichoderma)属においては、トリコデルマ・リーゼイ(Trichoderma reesei)を利用したホストベクター系が開発され、菌体外セルラーゼ遺伝子由来プロモーターなどが利用できる(Biotechnology 7, 596-603 (1989))。
【0083】
また、微生物以外でも、植物、動物において様々な宿主・ベクター系が開発されており、特に蚕を用いた昆虫(Nature 315, 592-594 (1985))や菜種、トウモロコシ、ジャガイモなどの植物中に大量に異種タンパク質を発現させる系が開発されており、好適に利用できる。
【0084】
本発明において使用するイミン還元酵素生産能を有する微生物は、イミン還元酵素生産能を有するストレプトスポランジウム属に属するすべての菌株、突然変異株、変種、遺伝子操作技術の利用により作成された本発明の酵素生産能を獲得した形質転換株を含む。イミン還元酵素生産能を有するストレプトスポランジウム属に属する菌株として例えば以下の菌株をあげることが出来る。
ストレプトスポランジウム ロセウム(Streptosporangium roseum) DSM 43021
ストレプトスポランジウム アメチストゲネス サブsp.アメチストゲネス(Streptosporangium amethystogenes subsp. amethystogenes) NBRC 13986
ストレプトスポランジウム カヌム(Streptosporangium canum) NBRC 106320
ストレプトスポランジウム ロンギスポラム(Streptosporangium longisporum) NBRC 13141
ストレプトスポランジウム ブルガレ(Streptosporangium vulgare) NBRC 13985
特に好ましくは、ストレプトスポランジウム ロセウム(Streptosporangium roseum) DSM 43021をあげることが出来る。
なお、DSM番号が記載されている菌株はDSMZ(Deutsche Sammlung von Mikroorganismen und Zellkulturen)より入手することができ、NBRC番号が記載されている菌株は独立行政法人製品評価技術基盤機構、生物遺伝資源部門より入手することが出来る。
【0085】
本発明の光学活性アミン誘導体の製造方法は、式(I)あるいは式(III)で示される基質化合物を立体選択的に還元する能力を有するイミン還元酵素活性を有する酵素活性物質を、この酵素活性物質によって当該基質化合物の還元が可能な条件下で当該基質化合物に接触させる工程を含む。本発明において、酵素活性物質(enzymatic active materials)とは、式(I)あるいは式(III)で示される基質化合物を立体選択的に還元する能力を有する酵素タンパク質(配列番号:1のタンパク質もしくはそのバリアント)、または該タンパク質を発現する微生物に由来する物質であって、その能力が維持された物質を意味する。具体的には、たとえば次のような要素は、イミン還元酵素活性を維持する限り、本発明における酵素活性物質に含まれる。
i) 式(I)あるいは式(III)で示される基質化合物を立体選択的に還元する能力を有する配列番号:1のタンパク質もしくはそのバリアント
ii) i)のタンパク質を発現する微生物
iii) ii)の微生物菌体の培養物
iv) ii)の微生物菌体の処理物
【0086】
本発明において、微生物は、培養物であることもできるし、菌体であることもできる。培養物は、微生物の生存と増殖を支持する培地とともに当該微生物が存在する状態を言う。たとえば微生物を含む液体培地は、培養物である。一方、本発明においては、微生物そのものを指す用語として「菌体」(microbial cell)を用いる。菌体は、培養物から回収された微生物細胞を指す。たとえば遠心分離によって液体培地から微生物の菌体を回収することができる。培養物から回収された菌体を洗浄することによって、菌体表面の培地成分を除くこともできる。
【0087】
本発明において酵素活性物質は、そのまま、あるいは固定化して基質化合物と接触させることができる。微生物菌体あるいはその処理物を固定化するための種々の方法が公知である。たとえば、以下のような方法が微生物やその処理物の固定化方法として公知である。
ポリアクリルアミド法
含硫多糖ゲル法(κ−カラギーナンゲル法など)
アルギン酸ゲル法
寒天ゲル法
イオン交換樹脂法
あるいは、限外濾過膜等を用いたメンブレンバイオリアクター中で、微生物、その処理物、を基質化合物と接触させて反応させることもできる。
【0088】
本発明における微生物による立体選択的なイミンの還元反応は、微生物を酵素が誘導される適切な前記培養条件において培養を行い、得られた培養液、あるいは培養液から採取した菌体や該菌体処理物に反応基質であるイミン誘導体を加えて、インキュベートすることにより行うことができる。また、前記微生物の培養開始時、もしくは、培養途中に基質となるイミン誘導体を添加して、培養しながらイミンの立体選択的還元反応を行うこともできる。
【0089】
本発明における微生物による立体選択的還元方法は、微生物を酵素が誘導される適切な前記培養条件において培養を行い、得られた培養液、あるいは培養液から採取した菌体や該菌体処理物に反応基質を加えて、立体選択的還元反応を行うことができる。また、前記培養条件と同じpH、温度範囲で、1日から7日間、培養と並行しても行うことができる。
反応条件としては、たとえば以下の条件を示すことができる。
pH4.0から9.0、好ましくはpH6.0から8.0、
温度15から50℃、好ましくは20から40℃、
反応時間は、4時間から7日間接触させる。
一般に、微生物の培養と反応とを、別に行った方が、培地中の不純物の反応系への持ち込みを最小限に抑えることができる。その結果、反応後の生成物の精製において、有利な結果を生むことが期待できる。
【0090】
また酵素的還元反応においては、還元エネルギー源として、糖類などの存在下で反応を行うことにより、さらに効率的な反応が期待できる。還元エネルギー源として添加する化合物は、反応基質であるイミン誘導体量に応じて添加することができる。より具体的には、次のような還元エネルギー源を利用することができる。
糖類:グルコース、グルコース−6−リン酸、スクロースなど
有機酸:リンゴ酸、クエン酸、ギ酸など
アミノ酸:アラニン、グルタミン酸、リジンなど
アルコール:エタノール、イソプロパノールなど
その他、亜リン酸など無機化合物を還元エネルギー源として利用できる場合もある。
【0091】
本発明における還元反応は、水素供与体の存在下で進行する。したがって、本発明における基質化合物の還元が可能な条件は、水素供与体の存在下で、基質化合物と酵素活性物質とをインキュベートすることによって提供することができる。本発明における還元反応を助けるものであれば、任意の水素供与体を利用することができる。本発明における好ましい水素供与体は、NADPHあるいはNADHである。本発明者らが単離に成功したストレプトスポランジウム・ロセウムDSM 43021に由来するイミン還元酵素、およびそのホモログ、あるいは酵素活性物質を利用する場合には、NADPHを利用するのが好ましい。
【0092】
上記還元反応に付随してNADPHから生成するNADP+のNADPHへの再生は、微生物の持つNADP+還元能(解糖系、メチロトローフのC1化合物資化経路など)を用いて行うことができる。これらNADP+還元能は、反応系にグルコースやエタノール、などを添加することにより増強することが可能である。また、NADP+からNADPHを生成する能力を有する微生物やその処理物、酵素を反応系に添加することによっても行うことができる。たとえば、グルコース脱水素酵素、アルコール脱水素酵素、アミノ酸脱水素酵素、有機酸脱水素酵素(リンゴ酸脱水素酵素など)などを含む微生物、その処理物、ならびに部分精製もしくは精製酵素を用いてNADPHの再生を行うことができる。これらのNADPH再生に必要な反応を構成する成分は、本発明による光学活性アミンの製造のための反応系に添加する、固定化したものを添加する、あるいはNADPHの交換が可能な膜を介して接触させることができる。
【0093】
また、本発明のDNAを含む組換えベクターで形質転換した微生物の生菌体を前記光学活性アミンの製造方法に利用する場合には、NADPH再生のための付加的な反応系を不要とできる場合がある。すなわち、NADPH再生活性の高い微生物を用いることにより、形質転換体を用いた還元反応において、NADPH再生用の酵素を添加することなく効率的な反応が行える。さらに、NADPH再生に利用可能なグルコース脱水素酵素、アルコール脱水素酵素、アミノ酸脱水素酵素、有機酸脱水素酵素(リンゴ酸脱水素酵素など)などの遺伝子を、本発明のイミン還元酵素をコードするDNAと同時に宿主に導入することによって、より効率的なNADPH再生酵素とイミン還元酵素の同時発現、イミン還元反応を行うことも可能である。これらの2つもしくはそれ以上の遺伝子の宿主への導入には、不和合性を避けるために複製起源のことなる複数のベクターに別々に遺伝子を導入した組換えベクターにより宿主を形質転換する方法や、単一のベクターに両遺伝子を導入する方法、両方、もしくは、片方の遺伝子を染色体中に導入する方法などを利用することができる。
【0094】
単一のベクター中に複数の遺伝子を導入する場合には、プロモーター、ターミネーターなど発現制御に関わる領域をそれぞれの遺伝子に連結する方法やラクトースオペロンのような複数のシストロンを含むオペロンとして発現させることも可能である。
例えば、NADPH再生用酵素として、バシラス・サブチルス(Bacillus subtilis)やサーモプラズマ・アシドフィラム(Thermoplasma acidophilum)に由来するグルコース脱水素酵素が利用可能であり、具体的には、イミン還元酵素とバシラス・サブチリス由来のグルコース脱水素酵素の遺伝子を導入した組換えベクターであるpSG-SRI1などが好適に利用される。
【0095】
アミン誘導体の分解に伴って、反応液のpHに変化が見られる場合には、適当な酸、アルカリを用いてpHを一定範囲に調節することで、さらに良好な反応性を維持して、反応を継続することもできる。
【0096】
本発明において生菌体を使用した場合、反応液中に界面活性剤を添加しておけば反応時間の短縮を期待できる場合がある。この目的に用いられる界面活性剤としては、生菌体の細胞壁の透過性をあげるものであれば特に制限はなく、臭化セチルピリミジウム、臭化セチルトリメチルアンモニウム、トリトンX−100、パライソオクチルフェニルエーテル、トゥイーン80、スパン60等があげられ、反応液に対して、0.0001〜1%程度使用することが好ましい。
また、反応液中に有機溶媒を添加することによっても、同様の効果を期待できる場合がある。この目的に用いられる界面活性剤としては、生菌体の細胞壁の透過性をあげるものであれば特に制限はなく、トルエン、キシレンなどの有機溶媒があげられ、反応液に対して、0.0001〜1%程度使用することが好ましい。
また、反応液に添加せずとも、菌体を集菌後、界面活性剤および有機溶媒を含む水もしくはバッファーで前処理することにより、細胞壁の透過性を上げた菌体を用いてもよい。
【0097】
本発明における反応基質であるイミン誘導体は、目的とする生成物を効率的に生成できるように、適切な濃度で用いることができる。イミン誘導体は、菌体および当該反応を触媒する酵素に対して毒性を有することがあり、必ずしも高濃度反応が効率的な生産に結びつかないことがあり、注意を要する。イミン誘導体の反応液中における濃度として、たとえば0.05から50%w/v、好ましくは0.1から10%w/vを示すことができる。イミン誘導体は、一括(バッチ法)、分割添加(フェドバッチ法)あるいは連続添加(フィード法)等の任意の方法で添加することができる。更に、基質となるイミンを添加せず、イミンの原料となるケトンとアミンを反応液に添加し、反応液中でイミンを生成させ、基質として供給することもできる。
【0098】
添加されるイミン誘導体は、不活性ガス雰囲気下で扱うことによって、着色が抑制され、製品品質の向上が期待できる。このような効果を期待して、反応液を不活性ガス雰囲気下で原料を反応液に添加することもできる。更に酵素反応をも不活性化ガス雰囲気下で行うことによって、着色を防ぐことができる。ここで使用する不活性ガスとしては、たとえば窒素、アルゴン、ヘリウムなどが使用できる。特に窒素は、容易に入手することができる不活性ガスである。あるいは、アルゴンは空気よりも比重が大きいため、容易に拡散しない。したがって、開放作業においても、不活性化ガス雰囲気下環境を維持できることから、操作上有利である。
【0099】
本発明は、基質化合物と酵素活性物質との接触によって得られた光学活性アミン誘導体を回収する工程を含む。具体的には、イミン誘導体の立体選択的還元により、光学活性なアミン誘導体は、各種クロマトグラフィーや結晶化などを組み合わせることにより、反応液から単離することができる。
反応液中に蓄積された光学活性アミン誘導体は、たとえば次のような溶媒で抽出することができる。
酢酸エチル、
酢酸ブチル、
トルエン、
キシレン、
ヘキサン、
ヘプタン、
メチルイソブチルケトン、
メチル−t−ブチルエーテル、
ハロゲン系などの有機溶媒
【0100】
そして抽出されたアミン誘導体は、以下のようなクロマトグラフィーによって更に精製することができる。精製に先立って、必要に応じて、酸やアルカリによる洗浄、脱水剤による脱水などの処理を組み合わせることもできる。
イオン交換クロマトグラフィー、
吸着クロマトグラフィー
【0101】
たとえば、反応液から遠心分離によって、菌体などの不溶性物質を除去した後、アルカリ条件下で溶媒抽出後、減圧濃縮することにより、光学活性アミン誘導体を採取することができる。
菌体などの不溶性物質を除去した液を、アミン誘導体を吸着できるイオン交換樹脂などに吸着させ、アンモニア水や水酸化ナトリウム水溶液、水酸化ナトリウムメタノール溶液、水酸化ナトリウムエタノール溶液などのアルカリで溶出させれば、簡便に高濃度溶液を得ることができる。
また、菌体などの不溶性物質を除去した液を、アミン誘導体を吸着できるイオン交換樹脂などに吸着させ、アンモニア水や水酸化ナトリウム水溶液、水酸化ナトリウムメタノール溶液、水酸化ナトリウムエタノール溶液などのアルカリで溶出させれば、簡便に高濃度溶液を得ることができる。
また、溶媒抽出後、塩酸で塩酸塩化することにより、光学活性アミン誘導体を結晶として採取することができる。回収率を高めるために、溶媒を減圧濃縮しておいてもよく、あるいは溶媒を結晶性のよい溶媒に置換しておくこともできる。
また、回収率を高めるために、溶媒を減圧濃縮することもできるあるいは溶媒を結晶性のよい溶媒に置換しておくこともできる。
【0102】
あるいは、光学活性アミン誘導体を、酸との塩の結晶として採取することにより、その光学純度を高めることができる場合がある。溶媒抽出後、塩酸で塩酸塩化することにより、光学活性アミン誘導体を結晶として採取することができる。塩を精製する酸としては、塩酸、硫酸などの鉱酸、あるいはマンデル酸、クエン酸、酒石酸などの有機酸を利用することができる。こうして得られたアミン誘導体の塩を、強酸あるいは強アルカリに溶解した後、適当なアルカリもしくは酸で、pHを中性付近とし、中和晶析することで、容易に他の不純物と分離することができる。アミン誘導体の塩を溶解する強酸性水溶液には、たとえば塩酸、硫酸などの鉱酸を利用することができる。また、強アルカリ性水溶液には、NaOH水溶液やアンモニア水を利用することができる。
【0103】
また、加熱下水溶液とし、冷却し、再結晶することで容易に他の不純物と分離することができる。
更に得られた結晶を少量の水に溶解し、適当な展開相で溶出するシリカゲルカラムクロマトグラフィーを行うことで更に高度に精製することができる。シリカゲルゲルクロマトグラフィーの展開相には、たとえばブタノール、酢酸、あるいはそれらと水の混合溶剤などを利用することができる。
なお本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0104】
以下、実施例により本発明を詳細に説明するが、本発明はこれに限定されない。
[実施例1] イミン還元酵素のホモログC4EPL6_STRRSのクローニング
EMBLに登録されている予想タンパク質C4EPL6_STRRS(配列番号:1)に対応するDNA配列 (配列番号:2)を基にしてPCR用プライマー(配列番号:3および配列番号:4)を合成した。プライマーを各50 pmol、dNTP 10 nmol、Streptosporangium roseum DSM 43021由来染色体DNA (DSMZより購入) 50 ng、Prime STAR DNA polymerase用緩衝液 (TAKARA製)、Prime STAR DNA polymerase 2.5 U (TAKARA製)を含む50 μLの反応液を用い、変性(98℃、10秒) 、アニール(58℃、5秒)、伸長(72℃、1分)を30サイクル、GeneAmp PCR System 9700 (アプライド・バイオシステムズ製)を用いて行った。得られたDNA断片をGFX PCR DNA and Gel Band Purification Kit(GEヘルスケア製)により精製して回収した。
回収したDNA断片をBspH I、Xba Iで2重消化し、Nco I、Xba Iで2重消化したpSE420D(Invitrogen製のプラスミドベクターpSE420のマルチクローニングサイトを改変したプラスミド、特開2000-189170)とTakara Ligation Kitを用いて、ライゲーションし、大腸菌JM109株を形質転換した。
形質転換株をアンピシリンを含むLB培地で生育させ、生育した形質転換体からPlasmidを抽出し、イミン還元酵素遺伝子が挿入されている事が確認できたプラスミドをpSU-SRI1とした。プラスミド構築の過程を図1に示した。
【0105】
[実施例2] C4EPL6_STRRSの活性評価
C4EPL6_STRRSを発現するプラスミドpSU-SRI1で形質転換された大腸菌JM109株をアンピシリンを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに4時間培養を行った。
菌体を遠心分離により集菌した後、1 mM DTTを含む100 mM リン酸カリウム緩衝液(pH7.5)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収した。10 mM 2-メチル-1-ピロリンと、10 mM NADPHを含む 100 mM リン酸バッファー(pH 7.5)に菌体抽出液 100μLを加え、30℃で保持しながら340 nmの吸光度を測定してNADPHの減少を追跡することで、酵素活性を測定した。1Uは、上記反応条件において1分間に1μmolのNADPH消費を触媒する酵素量とした。結果、イミン還元活性は11.4mU/mgだった。
【0106】
[実施例3] C4EPL6_STRRSと枯草菌由来のグルコース脱水素酵素を共発現するプラスミドpSG-SRI1の構築
枯草菌由来のグルコース脱水素酵素遺伝子を発現するプラスミドpSE-BSG1(特開2000-189170)をEcoR I、HindIIIの2つの制限酵素で二重消化し、切出した枯草菌由来のグルコース脱水素酵素遺伝子を、同酵素で二重消化したpSU-SRI1とTakara Ligation Kitを用いてライゲーションし、C4EPL6_STRRSとグルコース脱水素酵素を同時に発現可能なプラスミドであるpSG-SRI1を得た。プラスミド構築の過程を図2に示した。
【0107】
[実施例4] C4EPL6_STRRSとグルコース脱水素酵素の大腸菌による同時発現
pSG-SRI1で形質転換された大腸菌JM109株をアンピシリンを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに4時間培養を行った。
菌体を遠心分離により集菌した後、1 mM DTTを含む100 mM リン酸カリウム緩衝液(pH 7.0)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収し、2-メチル-1-ピロリンに対するイミン還元活性とグルコース脱水素活性を測定した。なお、グルコース脱水素活性の測定は、100mMリン酸カリウム緩衝液(pH6.5)、2.5mM NADP+、100mMグルコースおよび菌体抽出液を含む反応液中で、30℃で行った。1Uは、上記反応条件において1分間に1μmolのNADPH生成を触媒する酵素量とした。結果、イミン還元活性5.13mU/mg、グルコース脱水素活性9.36mU/mgだった。
【0108】
[実施例5] 組換え大腸菌による(S)-2-メチルピロリジンの製造
pSG-SRI1で形質転換された大腸菌JM109株をアンピシリンを含む10 mLの液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに4時間培養を行った。
菌体を遠心分離により集菌した後、42.0 mM 2-メチル-1-ピロリン、84.0 mM グルコースを含む100 mMリン酸カリウム緩衝液(pH 7.5)10 mLに懸濁し、終夜25℃で振とう反応を行った。反応後、生成した2-メチルピロリジンの光学純度、及び、反応液中における2-メチル-1-ピロリンと2-メチルピロリジンの濃度を分析した。その結果、生成した(S)-2-メチルピロリジンの光学純度は97.7%ee、反応収率は99.4%であった。
光学純度の測定方法は以下のとおりである。
反応液50μLに以下の成分を添加して、40℃, 1 時間反応することにより生成したアミンをGITC化し、HPLCにより分離し定量した。
2 mg/mL トリエチルアミン 50μL および
8 mg/mL 2,3,4,6-テトラ-O-アセチル-β-D-グルコピラノシル イソチオシアネート (GITC) 100μL
HPLCの条件は、以下のとおりである。
− HPLCカラム: 和光純薬株式会社製 Wakosil II 5C18 (4.6mm x 150 mm)
− 溶離液: 10 mM リン酸バッファー(pH 2.5):メタノール=55:45
− 流速: 1 mL/min
− 検出: 254 nmにおけるUV吸収
上記条件下で、(R)-2-メチルピロリジンは 26.3 分に、(S)-2-メチルピロリジンは 24.9 分に溶出された。各溶出フラクションの254 nmにおけるUV吸収に基づいて光学純度を決定した。
【0109】
[実施例6]
SsSIRとC4EPL6_STRRSの酵素活性および反応結果を比較した表を以下に示す。
[表1]
ここで、pSUS2MP1およびpSG-S2MPはそれぞれ、イミン還元酵素SsSIRを発現するプラスミド(図3)、イミン還元酵素SsSIRと枯草菌由来のグルコース脱水素酵素を共発現するプラスミド(図4)である。
なお、SsSIRの酵素活性、光学純度の測定は以下のようにして行った。
pSG-S2MPで形質転換された大腸菌JM109株をアンピシリンを含む5mLの液体LB培地で終夜30℃培養し、0.1mM IPTGを加え、さらに4時間培養を行った。菌体を遠心分離により集菌した後、20mM 2−メチル−1−ピロリン、40mM グルコースを含む100mMリン酸カリウム緩衝液(pH7.0)1mLに懸濁し、終夜25℃で振とう反応を行った。反応後、生成した2−メチルピロリジンの光学純度、及び、反応液中における2−メチル−1−ピロリンと2−メチルピロリジンの濃度を実施例5と同様にして分析した。その結果、生成した(S)−2−メチルピロリジンの光学純度は92%eeであった。
上記の表から明らかなように、C4EPL6_STRRSの使用により、SsSIRを上回る光学純度を達成することができた。
【産業上の利用可能性】
【0110】
本発明は、微生物由来のイミン還元酵素を使用した立体選択的還元により、光学活性なアミン誘導体を効率的に製造する方法を提供する。本発明の方法は、高い立体選択性と光学純度(97.7%ee)を期待できるので、工業的にも有利である。本発明によって製造される光学活性なアミン誘導体は、光学活性な医薬の合成原料として有用である。たとえば、(S)−2−メチルピロリジンは、医薬品原料として有用な化合物である。
【技術分野】
【0001】
本発明は、イミン誘導体を立体選択的に還元することによって各種医薬品の原料あるいは合成中間体として利用されている光学活性なアミン誘導体を製造する方法に関する。
【背景技術】
【0002】
イミン誘導体の立体選択還元による光学活性なアミン誘導体の製造方法としては、次のような報告を除いてほとんど知られていない。
−非特許文献1;Tetrahedron Asymmetry, 19, 93,96(2008)−
キャンディダ・パラプシロシスによるN−フェニル−1−フェニルエチルアミン誘導体の還元
ただし、これらの公知技術においては、N−ベンジリデンベンジルアミンの還元生成物がキラルな構造ではないため、反応の立体選択性は不明である。また、N−((E)−N−(1−フェニルエチリデン)アニリン誘導体のキャンディダ・パラプシロシスによる還元反応は極めて弱いため、工業的な利用には適さない。また、該イミン還元反応を行う酵素の実態は全く不明である。
【0003】
また、2−メチルピロリジンの製法としては、次のような方法が知られている。
−非特許文献2;Acta. Pharm. Suec., 1978, 15, 255-263−
2−メチル−1−ピロリンを水素化ホウ素ナトリウムによって還元し、ラセミ2−メチルピロリジンとした後、酒石酸を用いた光学分割により、キラル−2−メチルピロリジンを得る方法
−非特許文献3;J. Org. Chem., 1989, 54, 209-216−
L−プロリンから誘導されるL−プロリノールの水酸基を塩化チオニルでクロロ基に変換し、窒素原子をベンジルオキシカルボニル基により保護した後、水素化トリブチル錫を用いてラジカル的に還元することにより、(R)−1−ベンジルオキシカルボニル−2−メチルピロリジンを製造し、さらに脱保護する方法
【0004】
−特許文献1;WO2005/073388号公報−
光学活性1,4−ペンダンジオールをジスルホナートとした後、アミンと反応させて環化し、2−メチルピロリジンを得る方法
しかし、いずれの方法にも、次のような問題点が伴う。
−工程が長い
−危険な水素化試薬を使用する
−分割方法によって得るため収率が50%を超えない
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Tetrahedron Asymmetry, 19, 93,96(2008)
【非特許文献2】Acta. Pharm. Suec., 1978, 15, 255-263
【非特許文献3】J. Org. Chem., 1989, 54, 209-216
【特許文献】
【0006】
【特許文献1】WO2005/073388号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の課題は、イミン誘導体から光学活性なアミン誘導体を製造するための方法を提供することである。更に本発明は、イミン誘導体を基質として光学活性なアミン誘導体を生成することができる酵素、あるいは該酵素を発現する微生物の提供を課題とする。更に本発明は、目的とする性状を備えた該酵素をコードするDNAを単離してベクターに組み込み、組換え体として得ることを課題とする。加えて、組換え体による光学活性アミン誘導体の製造方法の提供をも課題とするものである。
【課題を解決するための手段】
【0008】
本発明者らは、このような高度な要求を満足する方法を鋭意検討した。そして微生物の有する立体選択的な還元能力を利用して、イミン誘導体を立体選択的に還元することにより、光学活性なアミン誘導体を生成させうることを見出した。
【0009】
既に述べたように、イミン誘導体を立体選択的に還元する生合成に関与する酵素は知られている。しかし公知の酵素はいずれも代謝に関与する酵素である。そのため医薬品の中間体などの製造に応用した場合には、工業的な生産に求められる基質特異性や活性は期待できない。特に2−メチル−1−ピロリンなどの環状アルキルイミン誘導体を還元して、光学活性環状アルキルアミン誘導体を得る方法は、まったく知られていなかった。
【0010】
したがって、本出願人が先の国際出願(PCT/JP2009/065244)において開示したように、医薬品の中間体などの製造に好適な基質特異性や触媒活性を備えた微生物が見出されたことは、非常に意外な成果であった。更に驚くべきことに、微生物が有する立体選択的なイミンの還元能力は、十分に高く、工業的な利用に際し問題の無いレベルであった。たとえば、ストレプトマイセス・エスピー GF3546(Streptomyces sp. GF3546)は前記の課題を解決するための有用な性質を備えた微生物であることが見出された。更に該国際出願(PCT/JP2009/065244)には、GF3546を大量培養し、無細胞抽出液を調製した後、次の工程を経てイミン還元酵素(SsSIR)を高度に精製することに成功したことが開示されている。
−Phenyl Sepharoseを用いた疎水クロマトグラフィー、および
−MonoQを用いたイオン交換クロマトグラフィー
また、該国際出願(PCT/JP2009/065244)においては、イミン還元酵素をコードするDNA(配列番号:6)を単離し、当該酵素を高発現する組換え菌を造成したことも開示されている。
【0011】
本発明者らは、その後さらに、Streptomyces sp. GF 3546由来イミン還元酵素(SsSIR)のアミノ酸配列(配列番号:5)を用いて同一性検索を行い、EMBLに登録されている機能不明のアミノ酸配列が、当該イミン還元酵素に57%の同一性を有していることを見出した。具体的には、ストレプトスポランジウム・ロセウム(Streptosporangium roseum)のゲノム解析の結果より得られたC4EPL6_STRRSと命名された予想タンパク質であり、機能はDDBJ(DNA Data Bank of Japan)では、C4EPL6_STRRSはbeta-hydroxyacid dehydrogenase, 3-hydroxyisobutyrate dehydrogenaseであると予測されている。本発明者らは、C4EPL6_STRRSを高発現する組換え菌を造成し、性質を調べた結果、上記DDBJの予測とまったく異なるイミン還元活性を有する事を見出し、本発明を完成した。
【0012】
すなわち、本発明は、以下の光学活性アミンの製造方法を提供する。あるいは本発明は、当該方法に有用な微生物、あるいはイミン還元酵素、当該酵素をコードするDNAを含むベクター、当該酵素を含有する立体選択的イミン還元剤に関する。
〔1〕
式(I)で示されるイミン誘導体に、下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素、当該イミン還元酵素を生産する微生物、およびその処理物からなる群から選択される、少なくとも一つの酵素活性物質を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
式(I)
【化1】
式(II)
【化2】
(式中、R1、R2基は炭素数1〜3個のアルキル基を、R3基は、炭素数1〜3個のアルキル基もしくは水素を表し、R1とR3は環を巻いていてもよい)
〔2〕
式(I)の化合物が下記式(III)で表されるイミン誘導体であり、式(II)の化合物が式(IV)で表されるアミン誘導体である〔1〕に記載の光学活性なS体のアミン誘導体の製造方法。
式(III)
【化3】
式(IV)
【化4】
(式中R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)
〔3〕
式(III)の化合物が2−メチル−1−ピロリンであり、式(IV)で表される化合物が、2−メチルピロリジンである、〔2〕に記載の光学活性なS体のアミン誘導体の製造方法。
〔4〕
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
〔5〕
前記イミン還元酵素が、配列番号:1に記載のアミノ酸配列において、1個以上かつ50個以下のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質である、〔4〕に記載のイミン還元酵素。
〔6〕
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元活性を有するタンパク質を有効成分として含む、立体選択的イミン還元剤。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
〔7〕
下記(a)から(e)のいずれかに記載のイミン還元酵素をコードするポリヌクレオチドと、NADP+を補酵素とする酸化還元反応を触媒することができる脱水素酵素をコードするポリヌクレオチドとが一緒に挿入された組換えベクター。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
〔8〕
脱水素酵素がグルコース脱水素酵素である、〔7〕に記載のベクター。
〔9〕
グルコース脱水素酵素がバシラス・サブチルス(Bacillus subtilis)に由来する、〔8〕に記載の組換えベクター。
〔10〕
〔7〕〜〔9〕のいずれかに記載のベクターを発現可能に保持した形質転換体。
〔11〕
〔10〕に記載の形質転換体を培養する工程を含む、〔1〕〜〔3〕のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
〔12〕
〔1〕記載のイミン還元酵素を生産する微生物がストレプトスポランジウム(Streptosporangium)属に属する微生物である〔1〕〜〔3〕のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
〔13〕
式(I)で示されるイミン誘導体に、ストレプトスポランジウム(Streptosporangium)属に属する微生物またはその処理物を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
【発明の効果】
【0013】
本発明により、イミン還元酵素もしくは該酵素を発現する微生物等を使用したイミンの立体選択的還元により、(S)体光学活性アミンを効率的に製造する方法が提供された。また、該反応を担うイミン還元酵素が提供された。さらに、イミン還元酵素をコードするDNAを単離し、当該酵素を高発現する組換え菌が提供された。本発明の方法は、安価に合成可能なイミン誘導体を基質とし、微生物の高い立体選択性を利用して、光学活性アミンが合成できるため、工業的に有利である。本発明は、特に、(S)−2−メチルピロリジンの製造に有用である。(S)−2−メチルピロリジンは、医薬品を合成するための中間体として有用な化合物である。本発明によって、当該化合物を微生物や酵素の作用によって簡便にかつ効率的に製造し、さらに非常に高い光学純度を達成することが可能となった。本発明は、97.7%eeという高い光学純度で化合物を生成するという、顕著な効果を奏するものである。
【図面の簡単な説明】
【0014】
【図1】C4EPL6_STRRSイミン還元酵素遺伝子が挿入されたプラスミド(pSU-SRI1)の構築の過程を示した図である。
【図2】C4EPL6_STRRSイミン還元酵素遺伝子と枯草菌由来のグルコース脱水素酵素遺伝子が挿入されたプラスミド(pSG-SRI1)の構築の過程を示した図である。
【図3】SsSIRイミン還元酵素遺伝子が挿入されたプラスミド(pSUS2MP1)の構築の過程を示した図である。
【図4】SsSIRイミン還元酵素遺伝子と枯草菌由来のグルコース脱水素酵素遺伝子が挿入されたプラスミド(pSG-S2MP)の構築の過程を示した図である。
【発明を実施するための形態】
【0015】
本発明は、前記式(I)で示されるイミン誘導体に、当該化合物を立体選択的に還元しうる能力を有する酵素タンパク質(配列番号:1のタンパク質もしくはそのバリアント)、該タンパク質を発現する微生物の培養物、菌体およびその処理物からなる群から選択される、少なくとも一つの酵素活性物質を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法に関する。
式(I)
【化5】
式(II)
【化6】
(式中、R1、R2基は炭素数1〜3個のアルキル基を、R3基は、炭素数1〜3個のアルキル基もしくは水素を表し、R1とR3は環を巻いていてもよい)。
【0016】
本発明において、環状イミン化合物は式(I)の化合物として好ましい。本発明において、環状イミン化合物は、たとえば下記式(III)で表されるイミン誘導体を含む。
式(III)
【化7】
(式中、R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)。
【0017】
上記式(III)の化合物を基質として用いた場合、光学活性アミン誘導体として式(IV)に示される化合物を得ることができる。当該化合物は、本発明によって製造することができる好ましい光学活性アミン誘導体である。
式(IV)
【化8】
(式中、R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)。
【0018】
本発明における好ましい化合物は、前記式(III)において、たとえばRがメチル基、エチル基、あるいはプロピル基であり、nが2〜3の化合物である。より具体的には、本発明における好ましいイミン誘導体として次の化合物を示すことができる。
2−メチル−1−ピロリン
2−メチル−1−ピペリデイン
2−エチル−1−ピロリン
2−エチル−1−ピペリデイン
【化9】
【0019】
これらの化合物を基質として、それぞれ以下の光学活性アミン誘導体を製造することができる。
2−メチル−1−ピロリン → (S)−2−メチルピロリジン
2−メチル−1−ピペリデイン → (S)−2−メチルピペリジン
2−エチル−1−ピロリン → (S)−2−エチルピロリジン
2−エチル−1−ピペリデイン → (S)−2−エチルピペリジン
【化10】
【0020】
本発明において、イミン誘導体を立体選択的に還元しうる能力とは、前記式(I)望ましくは式(III)で示されるイミン誘導体を基質として、S体のアミン誘導体を生成する能力を言う。本発明の微生物は、配列番号:1のイミン還元酵素もしくはそのバリアントを発現し、S体のアミン誘導体を生成しうる微生物であれば、任意の微生物を用いることができる。本発明に用いる微生物は、たとえば以下に示すような微生物について、配列番号:1のイミン還元酵素もしくはそのバリアントの有無とS体のアミン誘導体を生成する能力を調べることにより得ることができる。
【0021】
たとえば、化合物(I)望ましくは化合物(III)で示されるイミン誘導体を含む培地中で被験微生物を培養し、その培養物中に生成した光学活性アミン誘導体の光学純度を測定する。その結果、光学活性な(S)−アミン誘導体の生成が確認できれば、当該微生物を本発明に用いることができる。
【0022】
あるいは予め培地中で増殖させた被験微生物を集菌し、適当な緩衝液中に懸濁させる。更に化合物(I)望ましくは化合物(III)で示されるイミン誘導体と接触、反応させて、この緩衝液中に生成される光学活性アミン誘導体の光学純度を測定する。光学活性なアミン誘導体の生成が確認できれば、当該微生物を本発明に用いることができる。このとき、被験微生物の培養時に、培地中に化合物(I)望ましくは化合物(III)を加えておけば、この化合物を還元するための酵素の誘導が期待できる。更に反応時に還元エネルギー源を加えておけば、より効果的な生成物の蓄積が期待できる。還元エネルギー源としては、通常、グルコースなどを用いることができる。
【0023】
例えば、微生物を必要に応じて2−メチル−1−ピロリンを含む適当な培地で培養し、得られた微生物菌体を、10mM 2−メチル−1−ピロリン、25mM リン酸カリウム緩衝液(pH7.0)、及び必要に応じてグルコースを含む反応液に懸濁し、25℃で24時間振盪し、生成物をTLCもしくはHPLCにより分析することにより、微生物の2−メチル−1−ピロリン還元能を確認することができる。
TLCの条件としては、例えば、Silica gel 60 F254 プレートを用い、1−ブタノール/酢酸/水=2/1/1の展開液で展開し、ニンヒドリンにより呈色する方法が挙げられる。
【0024】
生成したアミン誘導体の光学純度は、例えば以下の方法により測定することができる。
被検サンプル50μLに以下の成分を添加して、40℃、1時間反応することにより生成したアミンをGITC化し、HPLCにより分離し定量する。
2mg/mL トリエチルアミン 50μL、および
8mg/mL 2,3,4,6−テトラ−O−アセチル−β−D−グルコピラノシルイソチオシアネート(GITC) 100μL
【0025】
HPLCの条件としては、例えば、以下のような条件を用いることができる。
−HPLCカラム:和光純薬株式会社製 Wakosil II 5C18(4.6mm x 150mm)
−溶離液:10mMリン酸バッファー(pH2.5):メタノール=55:45
−流速:1mL/min
−検出:254nmにおけるUV吸収
上記条件下で、(R)−2−メチルピロリジンは26.3分に、(S)−2−メチルピロリジン誘導体は24.9分に溶出される。
同様な方法を用いて、単離した配列番号:1のイミン還元酵素もしくはそのバリアントタンパク質が有するS体のアミン誘導体を生成する能力を調べることもできる。
【0026】
光学純度の高い生成物を得るには、より選択性の高い微生物を用いることが有利であることは言うまでも無い。具体的には、その立体選択性は、たとえば60%、通常70%以上、好ましくは80%以上、更に好ましくは90%以上の、光学活性なS体のアミン誘導体を生成しうる微生物を用いることができる。本願の実施例に開示されるように、配列番号:1の酵素を発現する微生物を利用すると、97.7%eeの光学純度(enantiomeric excess;ee)を達成することができる。
【0027】
本発明における「光学活性なアミン誘導体」とは、ある光学異性体が別の光学異性体より多く含まれるアミン誘導体を言う。本発明において、好ましい光学活性なアミン誘導体は、たとえば60%、通常70%以上、好ましくは80%以上、更に好ましくは90%以上の光学純度(enantiomeric excess;ee)を有する。光学活性なアミン誘導体の光学純度は、たとえば光学分割カラムなどを用いて確認することができる。本発明の「光学異性体」は、一般的に「光学活性体」および「鏡像異性体」(enantiomeric)と呼ばれる場合もある。
【0028】
本出願人による先の国際出願(PCT/JP2009/065244)に開示のように、たとえばストレプトマイセス(Streptomyces)属に属する微生物は、前記式(I)で示されるイミン誘導体を還元し、S体のアミン誘導体を生成する。このような微生物は、本発明のイミン還元酵素の性能を比較分析する際などに利用できる。また、このような微生物に配列番号:1の酵素をコードする遺伝子を導入することも考えられる。
さらに具体的には、ストレプトマイセス(Streptomyces)属に属する微生物として、次の微生物を示すことができる。この微生物は、高い光学純度の(S)アミン誘導体を生成する微生物である。
ストレプトマイセス・エスピーGF3546(Streptomyces sp. GF3546)
【0029】
上記の微生物は、次のとおり寄託されている(受託日;2008年6月24日)。
(1)寄託機関名:独立行政法人製品評価技術基盤機構
(2)連絡先:〒292−0818千葉県木更津市かずさ鎌足2−5−8
(3)識別のための表示および受託番号:
ストレプトマイセス・エスピー GF3546(Streptomyces sp. GF3546);受託番号 NITE P-593
【0030】
さらにこれらの微生物は、ブダペスト条約に基づく国際寄託に、次のとおり移管されている。
(1)国際寄託当局:独立行政法人製品評価技術基盤機構特許微生物寄託センター
(2)住所:〒292−0818 千葉県木更津市かずさ鎌足2−5−8
(3)識別の表示および受託番号
ストレプトマイセス・エスピー GF3546(Streptomyces sp. GF3546)
;受託番号 NITE BP-593 (移管日2009年8月4日)
【0031】
前記式(I)あるいは(III)で示されるイミン誘導体を立体選択的に還元する能力を有する微生物は、これらの寄託菌株のみならず、種々の微生物資源(microbiological resource)からも見出すことができる。微生物資源には、自然環境から単離される微生物群、微生物寄託機関に保存された微生物菌株などが含まれる。中でも、ストレプトマイセス属に属する微生物は、前記式(I)あるいは(III)で示されるイミン誘導体を立体選択的に還元する能力を有する微生物として好ましい。したがって、たとえば、微生物寄託機関に保存されたストレプトマイセス属微生物を対象として、先に述べたような選択方法によって、目的の作用を有する微生物を選択することができる。寄託された微生物は、たとえば次のような寄託機関から分譲を受けることができる。
生物遺伝資源センター (NBRC)
理化学研究所 (JCM)
東京農業大学菌株保存室 (IAM)
American Type Culture Collection (ATCC)
Deutsche Sammlung von Mikroorganismen (DSM)
【0032】
また、有用な微生物をリボソーマルDNA(rDNA)の塩基配列情報に基づいて、選択することもできる。一般にrDNAの塩基配列情報は、生物間の遺伝学的な近さを表す指標として有用であることが広く知られている。特に16S rRNAをコードするDNA(以下、「16S rDNA」と記載する)は、微生物の遺伝的な同一性あるいは類似性を決定するためのツールとして活用されている。
16S rRNAは、全生物に存在しており、生物間で構造の違いは見られるが、アライメント可能な程度に配列が保存されている。16S rRNAの生物間水平伝播の頻度は低いので、分類に使用する方法が汎用されている。
具体的には、小サブユニットリボソームRNA(SSU rRNA)は、タンパク質合成の場であるリボソームを構成する核酸分子の1つである。その塩基配列の長さは、多少の違いはあるものの、細菌からヒトまで起源は同じものと見なされている。したがって、rRNAの塩基配列は進化的保存性が高く、生物の系統解析において最も頻繁に利用されている(Microbiol. Rev., 58, 1-9 (1994))。原核生物におけるSSU rRNA は約1500塩基の16S rRNAである。原核生物の分類と同定に16S rRNAの塩基配列を利用する系統分類が一般に用いられている(ASM News, 65, 752-757 (1999)、生物工学実験書p113日本生物工学会編、培風館)。
現在の放線菌の分類体系は、16S rDNA塩基配列の類似性に基づいて構成されている(Stackebrandt, et al. (1997) Int. J. Syst. Bacteriol., 47, 479-491)。菌株の同定においても、属レベルだけではなく種レベルにおいても16S rDNA塩基配列の相同性に基づく同定が、細菌系統分類学の主流となっている(「放線菌の分類と同定」(日本放線菌学会)p19)。
【0033】
塩基配列情報データベースを対象として、これらの塩基配列情報を検索すれば、既に16S rDNAが決定された微生物の中から、16S rDNA中に同一性の高い塩基配列を含む菌株を見つけ出すことができる。たとえばDNA Data Bank of Japan(DDBJ)などの公共データベースサービス機関では、16S rRNA(Prokaryotes)の塩基配列情報を対象とする検索サービスが提供されている。こうして見出された微生物のうち、特にストレプトマイセス属に属する微生物を選択することによって、本発明のイミン還元酵素との比較分析や形質転換等に利用しうる微生物とすることができる。必要に応じ、こうして選択された微生物のイミン誘導体に対する作用を予め確認することもできる。
【0034】
使用する微生物は、醗酵学の分野で公知の方法に従って培養することができる。培地としては炭素源、窒素源、無機物およびその他の栄養素を適量含有する培地ならば、合成培地または天然培地のいずれでも使用することができる。培地は、液体培地または固体培地を使用することができる。
具体的には、炭素源として、次に示すような一般的な炭素源より、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
糖類: 天然炭水化物:
グルコース、 澱粉、
フルクトース、 澱粉加水分解物、
マルトース、 糖蜜、
ガラクトース、 廃糖蜜、
麦、
とうもろこしなど
アルコール類: 有機酸類:
グリセロール、 酢酸、
メタノール、 グルコン酸、
エタノールなど ピルビン酸、
ケトグルタル酸、
クエン酸など
炭化水素類:
ノルマルパラフィンなど
脂肪類:
パーム核油、
大豆油、
オリーブ油など
【0035】
窒素源としては、次に示すような一般的な窒素源の中から、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
有機窒素化合物:
肉エキス、 ペプトン、 酵母エキス、
大豆加水分解物、 ミルクカゼイン、 カザミノ酸、
各種アミノ酸、 コーンスティープリカー、
その他の動物、植物、微生物の加水分解物など
無機窒素化合物:
アンモニア、 硝酸アンモニウム、 硫酸アンモニウム、
塩化ナトリウムなどのアンモニウム塩、
硝酸ナトリウムなどの硝酸塩、尿素など
【0036】
また、微生物のイミン誘導体の立体選択的還元能力を高めるために、誘導物質を用いることができる。誘導物質としては、イミン誘導体を、使用する微生物に応じて使用することができる。
さらに、無機塩として微量のマグネシウム、マンガン、カリウム、カルシウム、ナトリウム、銅、亜鉛などのリン酸塩、塩酸塩、硝酸塩、酢酸塩等より適宜一種または二種以上を選択して使用することができる。また、必要に応じて植物油、界面活性剤、シリコンなどの消泡剤を培養液中に添加してもよい。
【0037】
培養は前記培地成分を含有する液体培地中で通常の培養方法を用いて行うことができる。たとえば次のような培養方法を利用することができる。
振とう培養 (shaking culture)
通気攪拌培養 (aeration-agitation culture)
連続培養 (continuous culture)
流加培養(feeding culture, fed batch culture)
【0038】
培養条件は、微生物の種類、培養の種類、培養方法により適宜選択することができる。利用する菌株が増殖し、イミン誘導体の立体選択的還元能力を有しうる条件であれば特に制限はない。一般的な培養条件として次のような条件を示すことができる。
培養開始時のpHを4から10、好ましくは6から8に調節
15から70℃、好ましくは20から40℃の温度条件下で培養
培養時間はイミン誘導体の還元能力を有する菌体が得ることができれば特に制限されない。通常は1日から14日、好ましくは1日から7日培養する。
【0039】
本発明において、微生物菌体の処理物とは、菌体を目的とする酵素活性が維持される条件で処理したものを意味する。酵素活性の維持とは、生菌体で得られる酵素活性の一般的には20%以上、通常30%以上、好ましくは50%以上、より好ましくは70%以上の活性を維持することを言う。生菌体とその処理物の酵素活性は、生菌体の酵素活性と、同量の生菌体から得られた処理物の酵素活性を比較することによって定量的に比較することができる。本発明における処理物は、生菌体よりも高い酵素活性を有する場合もある。
【0040】
このような処理物を得る方法としては、凍結乾燥、有機溶媒による脱水乾燥、菌体の自己消化、酵素活性画分の抽出、超音波処理などを利用することができる。このような処理方法によって、例えば、凍結乾燥菌体、アセトン乾燥菌体、菌体自己消化物、菌体抽出物、菌体磨砕物、菌体の超音波処理物等を得ることができる。
【0041】
これらの処理は、単独で、あるいは複数の処理を重複して適用することもできる。たとえば適当な培養液で微生物を培養後、菌体を破砕して、遠心分離等により無細胞抽出液を調製することができる。菌体は、機械的な破砕、超音波、高圧処理、酵素消化、自己消化などにより破砕することができる。無細胞抽出液は本発明における菌体の処理物に含まれる。
【0042】
本発明においては、配列番号:1のイミン還元酵素もしくはそのバリアントを発現する微生物から当該酵素を部分精製したものや完全に精製したものなどを用いることができる。国際出願(PCT/JP2009/065244)に開示のイミン還元酵素と同様に、本発明のイミン還元酵素も、次の(1)〜(2)に示す理化学的性状を有する。
(1)作用:NADPH依存的に、2−メチル−1−ピロリンを還元し、(S)−2−メチルピロリジンを生成する。NADP+依存的に、(S)−2−メチルピロリジンを酸化し、2−メチル−1−ピロリンを生成する;、および
(2)補酵素依存性:還元反応の補酵素としてNADPHを利用する。酸化反応の補酵素としてNADP+を利用する。
【0043】
本発明のイミン還元酵素は、配列番号:1のイミン還元酵素もしくはそのバリアントを発現する微生物、あるいはその無細胞抽出液などから精製することができる。具体的には、次のような精製工程を適宜組み合わせることによって、無細胞抽出液から実質的に純粋なイミン還元酵素を得ることができる。
硫安、アセトンなどを用いた溶解度による分割、
イオン交換クロマトグラフィー、
疎水クロマトグラフィー、
2’,5’−ADPセファロースやブルーもしくはレッドセファロースなどを用いたアフィニティクロマトグラフィー、
ゲル濾過
【0044】
例えば、微生物を集菌後、次のような精製工程を経て、本発明のイミン還元酵素を実質的に純粋な酵素として単離することができる。
超音波あるいはガラスビーズによる菌体の破砕と無細胞抽出液の調製、
Phenyl Sepharoseを用いた疎水クロマトグラフィー、および
MonoQを用いたイオン交換クロマトクロマトグラフィーによるイミン還元酵素活性分画の単離
【0045】
本発明において、「実質的に純粋」とは、当該酵素が、当該酵素以外の生物学的な高分子化合物や化学物質を実質的に含まないことを言う。当該酵素以外の生物学的な高分子化合物とは、たとえば菌体や培養液に由来するタンパク、糖類、脂質、核酸類が含まれる。本発明における実質的に純粋な酵素は、少なくとも75%以上、通常80%以上、好ましくは85%以上、より好ましくは90%以上、更に好ましくは95%以上、あるいは99%以上の純度を有する。酵素の純度を決定する方法は公知である。たとえば、各種のクロマトグラフィーや、ポリアクリルアミドゲル電気泳動などによって蛋白質の純度を決定する方法が周知である。
【0046】
あるいは本発明によって、上記理化学的性状(1)(2)を有する単離されたイミン還元酵素が提供される。本発明において、単離されたイミン還元酵素とは、それが存在する天然の環境から分離されて存在していることを意味する。したがって、菌体から分離されたイミン還元酵素は、本発明における単離されたイミン還元酵素に含まれる。
【0047】
本発明における実質的に純粋なイミン還元酵素、あるいは単離されたイミン還元酵素は、精製、あるいは単離された後に、酵素組成物とすることもできる。たとえば、精製、あるいは単離されたイミン還元酵素を適当な担体と配合することによって、本発明のイミン還元酵素を含む酵素組成物を調製することができる。担体としては、アルブミンなどの不活性蛋白質、ショ糖などの糖類あるいは糖アルコールなどを利用することができる。酵素組成物は、液状であることもできるし、乾燥状態であることもできる。酵素と担体を含む水溶液を凍結乾燥することによって得られる酵素組成物は、本発明における好ましい組成物の一つである。このようなイミン還元活性を有するタンパク質を含む組成物は、立体選択的なイミン還元剤として用いることができる。すなわち、本発明の立体選択的イミン還元剤は、有効成分として、配列番号:1に記載のアミノ酸配列からなるタンパク質および/もしくはそのバリアントを含有する。
【0048】
本発明のイミン還元酵素は、以下の方法によりその活性を定量することができる。すなわち、次の組成の反応液中(1mL)で、30℃で反応を行い、NADPHの減少に由来する340nmの吸光度の減少を測定する。
10mM 2−メチル−1−ピロリン、
100mM リン酸カリウム緩衝液(pH7.5)、
0.1mM NADPH及び
酵素
1Uは、上記条件下、1分間に1μmolのNADPHの減少を触媒する酵素量とした。
【0049】
本発明は、配列番号:1に記載のアミノ酸配列からなるタンパク質に関する。本発明はまた、配列番号:1に記載のアミノ酸配列からなるタンパク質のホモログ(本願ではバリアントとも言う)を含む。本発明のイミン還元酵素のホモログ(バリアント)とは、配列番号:1に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、配列番号:1に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質を意味する。配列番号:1に記載のアミノ酸配列において、たとえば100以下、通常50以下、好ましくは30以下、より好ましくは15以下、更に好ましくは10以下、あるいは5以下のアミノ酸残基の変異は許容される。すなわち、本発明のイミン還元酵素は、配列番号:1に記載のアミノ酸配列に対して、たとえば1以上、5以上、10以上、30以上、50以上、あるいは100以上のアミノ酸残基の変異を含みうる。
【0050】
本発明は、イミン還元酵素をコードするポリヌクレオチドおよびそのホモログ(バリアント)に関する。本発明において、ポリヌクレオチドは、DNAやRNA等の天然のポリヌクレオチドに加え、人工的なヌクレオチド誘導体を含む人工的な分子であることもできる。また本発明のポリヌクレオチドは、DNA−RNAのキメラ分子であることもできる。本発明のイミン還元酵素をコードするポリヌクレオチドは、たとえば配列番号:2に示す塩基配列を含む。配列番号:2に示す塩基配列は、配列番号:1に示すアミノ酸配列を含むタンパク質をコードしており、このアミノ酸配列を含むタンパク質は、本発明によるイミン還元酵素の好ましい態様を構成する。
【0051】
本発明のイミン還元酵素をコードするポリヌクレオチドのホモログ(バリアント)とは、配列番号:1に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列を含み、かつ、前記理化学的性質(1)〜(2)を有するタンパク質をコードするポリヌクレオチドを含む。当業者であれば、配列番号:2に記載のポリヌクレオチドに部位特異的変異導入法(Nucleic Acid Res. 10,pp.6487 (1982) , Methods in Enzymol.100,pp.448 (1983), Molecular Cloning 2ndEdt., Cold Spring Harbor Laboratory Press (1989) , PCR A Practical Approach IRL Press pp.200 (1991))などを用いて、適宜置換、欠失、挿入、および/または付加変異を導入することによりポリヌクレオチドのホモログを得ることが可能である。
【0052】
また、本発明におけるポリヌクレオチドのホモログ(バリアント)は、配列番号:2に示される塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズできるポリヌクレオチドであって、かつ、前記理化学的性質(1)〜(2)を有するタンパク質をコードするポリヌクレオチドも含む。ストリンジェントな条件でハイブリダイズできるポリヌクレオチドとは、配列番号:2に記載中の任意の少なくとも20個、好ましくは少なくとも30個、たとえば40、60または100個の連続した配列を一つまたは複数選択したDNAをプローブDNAとし、たとえばECL direct nucleic acid labeling and detection system (GEヘルスケア社製)を用いて、マニュアルに記載の条件(wash:42℃、0.5 x SSCを含むprimary wash buffer)においてハイブリダイズするポリヌクレオチドを指す。
【0053】
さらに、本発明におけるポリヌクレオチドのホモログ(バリアント)は、配列番号:1に示されるアミノ酸配列と少なくとも70%、あるいは少なくとも75%、好ましくは少なくとも80%、あるいは少なくとも85%、より好ましくは90%以上、あるいは95%以上、の同一性を有するタンパク質をコードするポリヌクレオチドを含む。タンパク質の同一性検索は、たとえばSWISS−PROT、PIR、DADなどのタンパク質のアミノ酸配列に関するデータベースやDDBJ、EMBL、あるいはGene−BankなどのDNA配列に関するデータベース、DNA配列を基にした予想アミノ酸配列に関するデータベースなどを対象に、BLAST、FASTAなどのプログラムを利用して、例えば、インターネットを通じて行うことができる。
【0054】
本発明は、配列番号:1に記載のアミノ酸配列からなるタンパク質に関する。本発明はまた、配列番号:1に記載のアミノ酸配列からなるタンパク質のホモログ(バリアント)を含む。本発明のイミン還元酵素のホモログとは、配列番号:1に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、配列番号:1に記載のアミノ酸配列からなるタンパク質と機能的に同等なタンパク質を意味する。本発明において、配列番号:1に記載のアミノ酸配列からなるタンパク質と機能的に同等とは、当該タンパク質が前記(1)〜(2)に示した物理化学的性状を有することを意味する。当業者であれば、配列番号:2記載のDNAに部位特異的変異導入法(Nucleic Acid Res. 10,pp.6487 (1982) , Methods in Enzymol.100,pp.448 (1983), Molecular Cloning 2ndEdt., Cold Spring Harbor Laboratory Press (1989) , PCR A Practical Approach IRL Press pp.200 (1991))などを用いて、適宜置換、欠失、挿入、および/または付加変異を導入することによりイミン還元酵素のホモログをコードするポリヌクレオチドを得ることができる。そのイミン還元酵素のホモログをコードするポリヌクレオチドを宿主に導入して発現させることにより、配列番号:1に記載のイミン還元酵素のホモログを得ることが可能である。
【0055】
さらに、本発明のイミン還元酵素のホモログ(バリアント)とは、配列番号:1に示されるアミノ酸配列と少なくとも70%、あるいは少なくとも75%、好ましくは少なくとも80%、あるいは少なくとも85%、より好ましくは90%以上、あるいは95%以上の同一性を有するタンパク質をいう。タンパク質の同一性検索は、たとえばSWISS−PROT、PIR、DADなどのタンパク質のアミノ酸配列に関するデータベースやDDBJ、EMBL、あるいはGene−BankなどのDNA配列に関するデータベース、DNA配列を基にした予想アミノ酸配列に関するデータベースなどを対象に、BLAST、FASTAなどのプログラムを利用して、例えば、インターネットを通じて行うことができる。
【0056】
本発明のイミン還元酵素をコードするポリヌクレオチドは、たとえば、以下のような方法によって単離することができる。
【0057】
配列番号:2に記載の塩基配列を基にPCR用のプライマーを設計し、酵素生産株(ストレプトスポランジウム・ロセウムなどのストレプトスポランジウム属の菌株。たとえば、ストレプトスポランジウム・ロセウムDSM 43021)の染色体DNAもしくは、cDNAライブラリーを鋳型としてPCRを行うことにより本発明のDNAを得ることができる。
【0058】
さらに、得られたDNA断片をプローブとして、酵素生産株の染色体DNAの制限酵素消化物をファージ、プラスミドなどに導入し、大腸菌を形質転換して得られたライブラリーやcDNAライブラリーを利用して、コロニーハイブリダイゼーション、プラークハイブリダイゼーションなどにより、本発明のポリヌクレオチドを得ることができる。
【0059】
また、PCRにより得られたDNA断片の塩基配列を解析し、得られた配列から、既知のDNAの外側に伸長させるためのPCRプライマーを設計し、酵素生産株の染色体DNAを適当な制限酵素で消化後、自己環化反応によりDNAを鋳型として逆PCRを行うことにより(Genetics 120, 621-623 (1988))、また、RACE法(Rapid Amplification of cDNA End、「PCR実験マニュアル」p25−33、HBJ出版局)などにより本発明のポリヌクレオチドを得ることも可能である。
【0060】
なお本発明において利用可能なポリヌクレオチドには、以上のような方法によってクローニングされたゲノムDNA、あるいはcDNAの他、合成によって得られたDNAが含まれる。
このようにして単離された、本発明によるイミン還元酵素をコードするポリヌクレオチドを公知の発現ベクターに挿入することにより、イミン還元酵素発現ベクターが提供される。
【0061】
また、この発現ベクターで形質転換した形質転換体を培養することにより、本発明のイミン還元酵素を組換え体から得ることができる。
【0062】
本発明においてイミン還元酵素を発現させるために、形質転換の対象となる微生物は、イミン還元酵素を有するポリペプチドをコードするポリヌクレオチドを含む組換えベクターにより形質転換され、イミン還元酵素活性を発現することができる生物であれば特に制限はない。利用可能な微生物としては、たとえば以下のような微生物を示すことができる。
【0063】
エシェリヒア(Escherichia)属
バチルス(Bacillus)属
シュードモナス(Pseudomonas)属
セラチア(Serratia)属
ブレビバクテリウム(Brevibacterium)属
コリネバクテリイウム(Corynebacterium)属
ストレプトコッカス(Streptococcus)属
ラクトバチルス(Lactobacillus)属など宿主ベクター系の開発されている細菌
ロドコッカス(Rhodococcus)属
ストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌
サッカロマイセス(Saccharomyces)属
クライベロマイセス(Kluyveromyces)属
シゾサッカロマイセス(Schizosaccharomyces)属
チゴサッカロマイセス(Zygosaccharomyces)属
ヤロウイア(Yarrowia)属
トリコスポロン(Trichosporon)属
ロドスポリジウム(Rhodosporidium)属
ピキア(Pichia)属
キャンディダ(Candida)属などの宿主ベクター系の開発されている酵母
ノイロスポラ(Neurospora)属
アスペルギルス(Aspergillus)属
セファロスポリウム(Cephalosporium)属
トリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ
【0064】
形質転換体の作製のための手順および宿主に適合した組み換えベクターの構築は、分子生物学、生物工学、遺伝子工学の分野において慣用されている技術に準じて行うことができる(例えば、Sambrookら、モレキュラー・クローニング、Cold Spring Harbor Laboratories)。微生物中などにおいて、本発明のNADPHを電子供与体とするイミン還元酵素遺伝子を発現させるためには、まず微生物中において安定に存在するプラスミドベクターやファージベクター中にこのDNAを導入し、その遺伝情報を転写・翻訳させる必要がある。
【0065】
そのためには、転写・翻訳を制御するユニットにあたるプロモーターを本発明のDNA鎖の5’−側上流に、より好ましくはターミネーターを3’−側下流に、それぞれ組み込めばよい。このプロモーター、ターミネーターとしては、宿主として利用する微生物中において機能することが知られているプロモーター、ターミネーターを用いる必要がある。これら各種微生物において利用可能なベクター、プロモーター、ターミネーターなどに関して「微生物学基礎講座8遺伝子工学・共立出版」、特に酵母に関しては、Adv. Biochem. Eng. 43, 75-102 (1990)、Yeast 8, 423-488 (1992)、などに詳細に記述されている。
【0066】
例えば、エシェリヒア属、特に大腸菌エシェリヒア・コリ(Escherichia coli)においては、プラスミドベクターとして、pBR、pUC系プラスミドを利用でき、lac(β−ガラクトシダーゼ)、trp(トリプトファンオペロン)、tac、trc(lac、trpの融合)、λファージPL、PRなどに由来するプロモーターなどが利用できる。また、ターミネーターとしては、trpA由来、ファージ由来、rrnBリボソーマルRNA由来のターミネーターなどを用いることができる。これらの中で、市販のpSE420(Invitrogen製)のマルチクローニングサイトを一部改変したベクターpSE420D(特開2000−189170に記載)が好適に利用できる。
【0067】
バチルス属においては、ベクターとしてpUB110系プラスミド、pC194系プラスミドなどが利用可能であり、染色体にインテグレートすることもできる。また、プロモーター、ターミネーターとしてapr(アルカリプロテアーゼ)、npr(中性プロテアーゼ)、amy(α−アミラーゼ)などが利用できる。
【0068】
シュードモナス属においては、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・セパシア(Pseudomonas cepacia)などで宿主ベクター系が開発されている。トルエン化合物の分解に関与するプラスミドTOLプラスミドを基本にした広宿主域ベクター(RSF1010などに由来する自律的複製に必要な遺伝子を含む)pKT240などが利用可能であり、プロモーター、ターミネーターとして、リパーゼ(特開平5−284973)遺伝子などが利用できる。
【0069】
ブレビバクテリウム属特に、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)においては、pAJ43(Gene 39, 281 (1985))などのプラスミドベクターが利用可能である。プロモーター、ターミネーターとしては、大腸菌で使用されているプロモーター、ターミネーターがそのまま利用可能である。
【0070】
コリネバクテリウム属、特にコリネバクテリウム・グルタミカム(Corynebacterium glutamicum)においては、pCS11(特開昭57−183799)、pCB101(Mol. Gen. Genet. 196, 175 (1984)などのプラスミドベクターが利用可能である。
【0071】
ストレプトコッカス(Streptococcus)属においては、pHV1301(FEMS Microbiol. Lett. 26, 239 (1985)、pGK1(Appl. Environ. Microbiol. 50, 94 (1985))などがプラスミドベクターとして利用可能である。
【0072】
ラクトバチルス(Lactobacillus)属においては、ストレプトコッカス属用に開発されたpAMβ1(J. Bacteriol. 137, 614 (1979))などが利用可能であり、プロモーターとして大腸菌で利用されているものが利用可能である。
【0073】
ロドコッカス(Rhodococcus)属においては、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)から単離されたプラスミドベクターが使用可能である(J. Gen. Microbiol. 138,1003 (1992))。
【0074】
ストレプトマイセス(Streptomyces)属においては、HopwoodらのGenetic Manipulation of Streptomyces: A Laboratory Manual Cold Spring Harbor Laboratories (1985)に記載の方法に従って、プラスミドを構築することができる。特に、ストレプトマイセス・リビダンス(Streptomyces lividans)においては、pIJ486(Mol. Gen. Genet. 203, 468-478, 1986)、pKC1064(Gene 103,97-99 (1991))、pUWL−KS(Gene 165,149-150 (1995))が使用できる。また、ストレプトマイセス・バージニア(Streptomyces virginiae)においても、同様のプラスミドを使用することができる(Actinomycetol. 11, 46-53 (1997))。
【0075】
サッカロマイセス(Saccharomyces)属、特にサッカロマイセス・セレビジアエ(Saccharomyces cerevisiae)においては、YRp系、YEp系、YCp系、YIp系プラスミドが利用可能であり、染色体内に多コピー存在するリボソームDNAとの相同組み換えを利用したインテグレーションベクター(EP537456など)は、多コピーで遺伝子を導入でき、かつ安定に遺伝子を保持できるため極めて有用である。また、ADH(アルコール脱水素酵素)、GAPDH(グリセルアルデヒド−3−リン酸脱水素酵素)、PHO(酸性フォスファターゼ)、GAL(β−ガラクトシダーゼ)、PGK(ホスホグリセレートキナーゼ)、ENO(エノラーゼ)などのプロモーター、ターミネーターが利用可能である。
【0076】
クライベロマイセス属、特にクライベロマイセス・ラクティス(Kluyveromyces lactis)においては、サッカロマイセス・セレビジアエ由来2μm系プラスミド、pKD1系プラスミド(J. Bacteriol. 145, 382-390 (1981))、キラー活性に関与するpGKl1由来プラスミド、クライベロマイセス属における自律増殖遺伝子KARS系プラスミド、リボソームDNAなどとの相同組み換えにより染色体中にインテグレート可能なベクタープラスミド(EP 537456など)などが利用可能である。また、ADH、PGKなどに由来するプロモーター、ターミネーターが利用可能である。
【0077】
シゾサッカロマイセス(Schizosaccharomyces)属においては、シゾサッカロマイセス・ポンベ由来のARS(自律複製に関与する遺伝子)及びサッカロマイセス・セレビジアエ由来の栄養要求性を相補する選択マーカーを含むプラスミドベクターが利用可能である(Mol. Cell. Biol. 6, 80 (1986))。また、シゾサッカロマイセス・ポンベ由来のADHプロモーターなどが利用できる(EMBO J. 6, 729 (1987))。特に、pAUR224は、タカラバイオから市販されており容易に利用できる。
【0078】
チゴサッカロマイセス属(Zygosaccharomyces)においては、チゴサッカロマイセス・ロウキシ(Zygosaccharomyces rouxii)由来のpSB3(Nucleic Acids Res. 13, 4267 (1985))などに由来するプラスミドベクターが利用可能であり、サッカロマイセス・セレビジアエ由来PHO5プロモーターや、チゴサッカロマイセス・ロウキシ由来GAP−Zr(グリセルアルデヒド−3−リン酸脱水素酵素)のプロモーター(Agri. Biol. Chem. 54, 2521 (1990))などが利用可能である。
【0079】
ピキア(Pichia)属においては、ピキア・アンガスタ(旧名:ハンゼヌラ・ポリモルファ(Hansenula polymorpha))において宿主ベクター系が開発されている。ベクターとしては、ピキア・アンガスタ由来自律複製に関与する遺伝子(HARS1、HARS2)も利用可能であるが、比較的不安定であるため、染色体への多コピーインテグレーションが有効である(Yeast 7, 431-443 (1991))。また、メタノールなどで誘導されるAOX(アルコールオキシダーゼ)、FDH(ギ酸脱水素酵素)のプロモーターなどが利用可能である。また、ピキア・パストリス(Pichia pastoris)などにピキア由来自律複製に関与する遺伝子(PARS1、PARS2)などを利用した宿主ベクター系が開発されており(Mol. Cell. Biol. 5, 3376 (1985))、高濃度培養とメタノールで誘導可能なAOXなど強いプロモーターが利用できる(Nucleic Acids Res. 15, 3859 (1987))。
【0080】
キャンディダ(Candida)属においては、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・アルビカンス(Candida albicans)、キャンディダ・トロピカリス(Candida tropicalis)、キャンディダ・ウチルス(Candida utilis)などにおいて宿主ベクター系が開発されている。キャンディダ・マルトーサにおいてはキャンディダ・マルトーサ由来ARSがクローニングされ(Agri. Biol. Chem. 51, 51, 1587 (1987))、これを利用したベクターが開発されている。また、キャンディダ・ウチルスにおいては、染色体インテグレートタイプのベクターは強力なプロモーターが開発されている(特開平08−173170)。
【0081】
アスペルギルス(Aspergillus)属においては、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・オリジー(Aspergillus oryzae)などがカビの中で最もよく研究されており、プラスミドや染色体へのインテグレーションが利用可能であり、菌体外プロテアーゼやアミラーゼ由来のプロモーターが利用可能である(Trends in Biotechnology 7, 283-287 (1989))。
【0082】
トリコデルマ(Trichoderma)属においては、トリコデルマ・リーゼイ(Trichoderma reesei)を利用したホストベクター系が開発され、菌体外セルラーゼ遺伝子由来プロモーターなどが利用できる(Biotechnology 7, 596-603 (1989))。
【0083】
また、微生物以外でも、植物、動物において様々な宿主・ベクター系が開発されており、特に蚕を用いた昆虫(Nature 315, 592-594 (1985))や菜種、トウモロコシ、ジャガイモなどの植物中に大量に異種タンパク質を発現させる系が開発されており、好適に利用できる。
【0084】
本発明において使用するイミン還元酵素生産能を有する微生物は、イミン還元酵素生産能を有するストレプトスポランジウム属に属するすべての菌株、突然変異株、変種、遺伝子操作技術の利用により作成された本発明の酵素生産能を獲得した形質転換株を含む。イミン還元酵素生産能を有するストレプトスポランジウム属に属する菌株として例えば以下の菌株をあげることが出来る。
ストレプトスポランジウム ロセウム(Streptosporangium roseum) DSM 43021
ストレプトスポランジウム アメチストゲネス サブsp.アメチストゲネス(Streptosporangium amethystogenes subsp. amethystogenes) NBRC 13986
ストレプトスポランジウム カヌム(Streptosporangium canum) NBRC 106320
ストレプトスポランジウム ロンギスポラム(Streptosporangium longisporum) NBRC 13141
ストレプトスポランジウム ブルガレ(Streptosporangium vulgare) NBRC 13985
特に好ましくは、ストレプトスポランジウム ロセウム(Streptosporangium roseum) DSM 43021をあげることが出来る。
なお、DSM番号が記載されている菌株はDSMZ(Deutsche Sammlung von Mikroorganismen und Zellkulturen)より入手することができ、NBRC番号が記載されている菌株は独立行政法人製品評価技術基盤機構、生物遺伝資源部門より入手することが出来る。
【0085】
本発明の光学活性アミン誘導体の製造方法は、式(I)あるいは式(III)で示される基質化合物を立体選択的に還元する能力を有するイミン還元酵素活性を有する酵素活性物質を、この酵素活性物質によって当該基質化合物の還元が可能な条件下で当該基質化合物に接触させる工程を含む。本発明において、酵素活性物質(enzymatic active materials)とは、式(I)あるいは式(III)で示される基質化合物を立体選択的に還元する能力を有する酵素タンパク質(配列番号:1のタンパク質もしくはそのバリアント)、または該タンパク質を発現する微生物に由来する物質であって、その能力が維持された物質を意味する。具体的には、たとえば次のような要素は、イミン還元酵素活性を維持する限り、本発明における酵素活性物質に含まれる。
i) 式(I)あるいは式(III)で示される基質化合物を立体選択的に還元する能力を有する配列番号:1のタンパク質もしくはそのバリアント
ii) i)のタンパク質を発現する微生物
iii) ii)の微生物菌体の培養物
iv) ii)の微生物菌体の処理物
【0086】
本発明において、微生物は、培養物であることもできるし、菌体であることもできる。培養物は、微生物の生存と増殖を支持する培地とともに当該微生物が存在する状態を言う。たとえば微生物を含む液体培地は、培養物である。一方、本発明においては、微生物そのものを指す用語として「菌体」(microbial cell)を用いる。菌体は、培養物から回収された微生物細胞を指す。たとえば遠心分離によって液体培地から微生物の菌体を回収することができる。培養物から回収された菌体を洗浄することによって、菌体表面の培地成分を除くこともできる。
【0087】
本発明において酵素活性物質は、そのまま、あるいは固定化して基質化合物と接触させることができる。微生物菌体あるいはその処理物を固定化するための種々の方法が公知である。たとえば、以下のような方法が微生物やその処理物の固定化方法として公知である。
ポリアクリルアミド法
含硫多糖ゲル法(κ−カラギーナンゲル法など)
アルギン酸ゲル法
寒天ゲル法
イオン交換樹脂法
あるいは、限外濾過膜等を用いたメンブレンバイオリアクター中で、微生物、その処理物、を基質化合物と接触させて反応させることもできる。
【0088】
本発明における微生物による立体選択的なイミンの還元反応は、微生物を酵素が誘導される適切な前記培養条件において培養を行い、得られた培養液、あるいは培養液から採取した菌体や該菌体処理物に反応基質であるイミン誘導体を加えて、インキュベートすることにより行うことができる。また、前記微生物の培養開始時、もしくは、培養途中に基質となるイミン誘導体を添加して、培養しながらイミンの立体選択的還元反応を行うこともできる。
【0089】
本発明における微生物による立体選択的還元方法は、微生物を酵素が誘導される適切な前記培養条件において培養を行い、得られた培養液、あるいは培養液から採取した菌体や該菌体処理物に反応基質を加えて、立体選択的還元反応を行うことができる。また、前記培養条件と同じpH、温度範囲で、1日から7日間、培養と並行しても行うことができる。
反応条件としては、たとえば以下の条件を示すことができる。
pH4.0から9.0、好ましくはpH6.0から8.0、
温度15から50℃、好ましくは20から40℃、
反応時間は、4時間から7日間接触させる。
一般に、微生物の培養と反応とを、別に行った方が、培地中の不純物の反応系への持ち込みを最小限に抑えることができる。その結果、反応後の生成物の精製において、有利な結果を生むことが期待できる。
【0090】
また酵素的還元反応においては、還元エネルギー源として、糖類などの存在下で反応を行うことにより、さらに効率的な反応が期待できる。還元エネルギー源として添加する化合物は、反応基質であるイミン誘導体量に応じて添加することができる。より具体的には、次のような還元エネルギー源を利用することができる。
糖類:グルコース、グルコース−6−リン酸、スクロースなど
有機酸:リンゴ酸、クエン酸、ギ酸など
アミノ酸:アラニン、グルタミン酸、リジンなど
アルコール:エタノール、イソプロパノールなど
その他、亜リン酸など無機化合物を還元エネルギー源として利用できる場合もある。
【0091】
本発明における還元反応は、水素供与体の存在下で進行する。したがって、本発明における基質化合物の還元が可能な条件は、水素供与体の存在下で、基質化合物と酵素活性物質とをインキュベートすることによって提供することができる。本発明における還元反応を助けるものであれば、任意の水素供与体を利用することができる。本発明における好ましい水素供与体は、NADPHあるいはNADHである。本発明者らが単離に成功したストレプトスポランジウム・ロセウムDSM 43021に由来するイミン還元酵素、およびそのホモログ、あるいは酵素活性物質を利用する場合には、NADPHを利用するのが好ましい。
【0092】
上記還元反応に付随してNADPHから生成するNADP+のNADPHへの再生は、微生物の持つNADP+還元能(解糖系、メチロトローフのC1化合物資化経路など)を用いて行うことができる。これらNADP+還元能は、反応系にグルコースやエタノール、などを添加することにより増強することが可能である。また、NADP+からNADPHを生成する能力を有する微生物やその処理物、酵素を反応系に添加することによっても行うことができる。たとえば、グルコース脱水素酵素、アルコール脱水素酵素、アミノ酸脱水素酵素、有機酸脱水素酵素(リンゴ酸脱水素酵素など)などを含む微生物、その処理物、ならびに部分精製もしくは精製酵素を用いてNADPHの再生を行うことができる。これらのNADPH再生に必要な反応を構成する成分は、本発明による光学活性アミンの製造のための反応系に添加する、固定化したものを添加する、あるいはNADPHの交換が可能な膜を介して接触させることができる。
【0093】
また、本発明のDNAを含む組換えベクターで形質転換した微生物の生菌体を前記光学活性アミンの製造方法に利用する場合には、NADPH再生のための付加的な反応系を不要とできる場合がある。すなわち、NADPH再生活性の高い微生物を用いることにより、形質転換体を用いた還元反応において、NADPH再生用の酵素を添加することなく効率的な反応が行える。さらに、NADPH再生に利用可能なグルコース脱水素酵素、アルコール脱水素酵素、アミノ酸脱水素酵素、有機酸脱水素酵素(リンゴ酸脱水素酵素など)などの遺伝子を、本発明のイミン還元酵素をコードするDNAと同時に宿主に導入することによって、より効率的なNADPH再生酵素とイミン還元酵素の同時発現、イミン還元反応を行うことも可能である。これらの2つもしくはそれ以上の遺伝子の宿主への導入には、不和合性を避けるために複製起源のことなる複数のベクターに別々に遺伝子を導入した組換えベクターにより宿主を形質転換する方法や、単一のベクターに両遺伝子を導入する方法、両方、もしくは、片方の遺伝子を染色体中に導入する方法などを利用することができる。
【0094】
単一のベクター中に複数の遺伝子を導入する場合には、プロモーター、ターミネーターなど発現制御に関わる領域をそれぞれの遺伝子に連結する方法やラクトースオペロンのような複数のシストロンを含むオペロンとして発現させることも可能である。
例えば、NADPH再生用酵素として、バシラス・サブチルス(Bacillus subtilis)やサーモプラズマ・アシドフィラム(Thermoplasma acidophilum)に由来するグルコース脱水素酵素が利用可能であり、具体的には、イミン還元酵素とバシラス・サブチリス由来のグルコース脱水素酵素の遺伝子を導入した組換えベクターであるpSG-SRI1などが好適に利用される。
【0095】
アミン誘導体の分解に伴って、反応液のpHに変化が見られる場合には、適当な酸、アルカリを用いてpHを一定範囲に調節することで、さらに良好な反応性を維持して、反応を継続することもできる。
【0096】
本発明において生菌体を使用した場合、反応液中に界面活性剤を添加しておけば反応時間の短縮を期待できる場合がある。この目的に用いられる界面活性剤としては、生菌体の細胞壁の透過性をあげるものであれば特に制限はなく、臭化セチルピリミジウム、臭化セチルトリメチルアンモニウム、トリトンX−100、パライソオクチルフェニルエーテル、トゥイーン80、スパン60等があげられ、反応液に対して、0.0001〜1%程度使用することが好ましい。
また、反応液中に有機溶媒を添加することによっても、同様の効果を期待できる場合がある。この目的に用いられる界面活性剤としては、生菌体の細胞壁の透過性をあげるものであれば特に制限はなく、トルエン、キシレンなどの有機溶媒があげられ、反応液に対して、0.0001〜1%程度使用することが好ましい。
また、反応液に添加せずとも、菌体を集菌後、界面活性剤および有機溶媒を含む水もしくはバッファーで前処理することにより、細胞壁の透過性を上げた菌体を用いてもよい。
【0097】
本発明における反応基質であるイミン誘導体は、目的とする生成物を効率的に生成できるように、適切な濃度で用いることができる。イミン誘導体は、菌体および当該反応を触媒する酵素に対して毒性を有することがあり、必ずしも高濃度反応が効率的な生産に結びつかないことがあり、注意を要する。イミン誘導体の反応液中における濃度として、たとえば0.05から50%w/v、好ましくは0.1から10%w/vを示すことができる。イミン誘導体は、一括(バッチ法)、分割添加(フェドバッチ法)あるいは連続添加(フィード法)等の任意の方法で添加することができる。更に、基質となるイミンを添加せず、イミンの原料となるケトンとアミンを反応液に添加し、反応液中でイミンを生成させ、基質として供給することもできる。
【0098】
添加されるイミン誘導体は、不活性ガス雰囲気下で扱うことによって、着色が抑制され、製品品質の向上が期待できる。このような効果を期待して、反応液を不活性ガス雰囲気下で原料を反応液に添加することもできる。更に酵素反応をも不活性化ガス雰囲気下で行うことによって、着色を防ぐことができる。ここで使用する不活性ガスとしては、たとえば窒素、アルゴン、ヘリウムなどが使用できる。特に窒素は、容易に入手することができる不活性ガスである。あるいは、アルゴンは空気よりも比重が大きいため、容易に拡散しない。したがって、開放作業においても、不活性化ガス雰囲気下環境を維持できることから、操作上有利である。
【0099】
本発明は、基質化合物と酵素活性物質との接触によって得られた光学活性アミン誘導体を回収する工程を含む。具体的には、イミン誘導体の立体選択的還元により、光学活性なアミン誘導体は、各種クロマトグラフィーや結晶化などを組み合わせることにより、反応液から単離することができる。
反応液中に蓄積された光学活性アミン誘導体は、たとえば次のような溶媒で抽出することができる。
酢酸エチル、
酢酸ブチル、
トルエン、
キシレン、
ヘキサン、
ヘプタン、
メチルイソブチルケトン、
メチル−t−ブチルエーテル、
ハロゲン系などの有機溶媒
【0100】
そして抽出されたアミン誘導体は、以下のようなクロマトグラフィーによって更に精製することができる。精製に先立って、必要に応じて、酸やアルカリによる洗浄、脱水剤による脱水などの処理を組み合わせることもできる。
イオン交換クロマトグラフィー、
吸着クロマトグラフィー
【0101】
たとえば、反応液から遠心分離によって、菌体などの不溶性物質を除去した後、アルカリ条件下で溶媒抽出後、減圧濃縮することにより、光学活性アミン誘導体を採取することができる。
菌体などの不溶性物質を除去した液を、アミン誘導体を吸着できるイオン交換樹脂などに吸着させ、アンモニア水や水酸化ナトリウム水溶液、水酸化ナトリウムメタノール溶液、水酸化ナトリウムエタノール溶液などのアルカリで溶出させれば、簡便に高濃度溶液を得ることができる。
また、菌体などの不溶性物質を除去した液を、アミン誘導体を吸着できるイオン交換樹脂などに吸着させ、アンモニア水や水酸化ナトリウム水溶液、水酸化ナトリウムメタノール溶液、水酸化ナトリウムエタノール溶液などのアルカリで溶出させれば、簡便に高濃度溶液を得ることができる。
また、溶媒抽出後、塩酸で塩酸塩化することにより、光学活性アミン誘導体を結晶として採取することができる。回収率を高めるために、溶媒を減圧濃縮しておいてもよく、あるいは溶媒を結晶性のよい溶媒に置換しておくこともできる。
また、回収率を高めるために、溶媒を減圧濃縮することもできるあるいは溶媒を結晶性のよい溶媒に置換しておくこともできる。
【0102】
あるいは、光学活性アミン誘導体を、酸との塩の結晶として採取することにより、その光学純度を高めることができる場合がある。溶媒抽出後、塩酸で塩酸塩化することにより、光学活性アミン誘導体を結晶として採取することができる。塩を精製する酸としては、塩酸、硫酸などの鉱酸、あるいはマンデル酸、クエン酸、酒石酸などの有機酸を利用することができる。こうして得られたアミン誘導体の塩を、強酸あるいは強アルカリに溶解した後、適当なアルカリもしくは酸で、pHを中性付近とし、中和晶析することで、容易に他の不純物と分離することができる。アミン誘導体の塩を溶解する強酸性水溶液には、たとえば塩酸、硫酸などの鉱酸を利用することができる。また、強アルカリ性水溶液には、NaOH水溶液やアンモニア水を利用することができる。
【0103】
また、加熱下水溶液とし、冷却し、再結晶することで容易に他の不純物と分離することができる。
更に得られた結晶を少量の水に溶解し、適当な展開相で溶出するシリカゲルカラムクロマトグラフィーを行うことで更に高度に精製することができる。シリカゲルゲルクロマトグラフィーの展開相には、たとえばブタノール、酢酸、あるいはそれらと水の混合溶剤などを利用することができる。
なお本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0104】
以下、実施例により本発明を詳細に説明するが、本発明はこれに限定されない。
[実施例1] イミン還元酵素のホモログC4EPL6_STRRSのクローニング
EMBLに登録されている予想タンパク質C4EPL6_STRRS(配列番号:1)に対応するDNA配列 (配列番号:2)を基にしてPCR用プライマー(配列番号:3および配列番号:4)を合成した。プライマーを各50 pmol、dNTP 10 nmol、Streptosporangium roseum DSM 43021由来染色体DNA (DSMZより購入) 50 ng、Prime STAR DNA polymerase用緩衝液 (TAKARA製)、Prime STAR DNA polymerase 2.5 U (TAKARA製)を含む50 μLの反応液を用い、変性(98℃、10秒) 、アニール(58℃、5秒)、伸長(72℃、1分)を30サイクル、GeneAmp PCR System 9700 (アプライド・バイオシステムズ製)を用いて行った。得られたDNA断片をGFX PCR DNA and Gel Band Purification Kit(GEヘルスケア製)により精製して回収した。
回収したDNA断片をBspH I、Xba Iで2重消化し、Nco I、Xba Iで2重消化したpSE420D(Invitrogen製のプラスミドベクターpSE420のマルチクローニングサイトを改変したプラスミド、特開2000-189170)とTakara Ligation Kitを用いて、ライゲーションし、大腸菌JM109株を形質転換した。
形質転換株をアンピシリンを含むLB培地で生育させ、生育した形質転換体からPlasmidを抽出し、イミン還元酵素遺伝子が挿入されている事が確認できたプラスミドをpSU-SRI1とした。プラスミド構築の過程を図1に示した。
【0105】
[実施例2] C4EPL6_STRRSの活性評価
C4EPL6_STRRSを発現するプラスミドpSU-SRI1で形質転換された大腸菌JM109株をアンピシリンを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに4時間培養を行った。
菌体を遠心分離により集菌した後、1 mM DTTを含む100 mM リン酸カリウム緩衝液(pH7.5)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収した。10 mM 2-メチル-1-ピロリンと、10 mM NADPHを含む 100 mM リン酸バッファー(pH 7.5)に菌体抽出液 100μLを加え、30℃で保持しながら340 nmの吸光度を測定してNADPHの減少を追跡することで、酵素活性を測定した。1Uは、上記反応条件において1分間に1μmolのNADPH消費を触媒する酵素量とした。結果、イミン還元活性は11.4mU/mgだった。
【0106】
[実施例3] C4EPL6_STRRSと枯草菌由来のグルコース脱水素酵素を共発現するプラスミドpSG-SRI1の構築
枯草菌由来のグルコース脱水素酵素遺伝子を発現するプラスミドpSE-BSG1(特開2000-189170)をEcoR I、HindIIIの2つの制限酵素で二重消化し、切出した枯草菌由来のグルコース脱水素酵素遺伝子を、同酵素で二重消化したpSU-SRI1とTakara Ligation Kitを用いてライゲーションし、C4EPL6_STRRSとグルコース脱水素酵素を同時に発現可能なプラスミドであるpSG-SRI1を得た。プラスミド構築の過程を図2に示した。
【0107】
[実施例4] C4EPL6_STRRSとグルコース脱水素酵素の大腸菌による同時発現
pSG-SRI1で形質転換された大腸菌JM109株をアンピシリンを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに4時間培養を行った。
菌体を遠心分離により集菌した後、1 mM DTTを含む100 mM リン酸カリウム緩衝液(pH 7.0)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収し、2-メチル-1-ピロリンに対するイミン還元活性とグルコース脱水素活性を測定した。なお、グルコース脱水素活性の測定は、100mMリン酸カリウム緩衝液(pH6.5)、2.5mM NADP+、100mMグルコースおよび菌体抽出液を含む反応液中で、30℃で行った。1Uは、上記反応条件において1分間に1μmolのNADPH生成を触媒する酵素量とした。結果、イミン還元活性5.13mU/mg、グルコース脱水素活性9.36mU/mgだった。
【0108】
[実施例5] 組換え大腸菌による(S)-2-メチルピロリジンの製造
pSG-SRI1で形質転換された大腸菌JM109株をアンピシリンを含む10 mLの液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに4時間培養を行った。
菌体を遠心分離により集菌した後、42.0 mM 2-メチル-1-ピロリン、84.0 mM グルコースを含む100 mMリン酸カリウム緩衝液(pH 7.5)10 mLに懸濁し、終夜25℃で振とう反応を行った。反応後、生成した2-メチルピロリジンの光学純度、及び、反応液中における2-メチル-1-ピロリンと2-メチルピロリジンの濃度を分析した。その結果、生成した(S)-2-メチルピロリジンの光学純度は97.7%ee、反応収率は99.4%であった。
光学純度の測定方法は以下のとおりである。
反応液50μLに以下の成分を添加して、40℃, 1 時間反応することにより生成したアミンをGITC化し、HPLCにより分離し定量した。
2 mg/mL トリエチルアミン 50μL および
8 mg/mL 2,3,4,6-テトラ-O-アセチル-β-D-グルコピラノシル イソチオシアネート (GITC) 100μL
HPLCの条件は、以下のとおりである。
− HPLCカラム: 和光純薬株式会社製 Wakosil II 5C18 (4.6mm x 150 mm)
− 溶離液: 10 mM リン酸バッファー(pH 2.5):メタノール=55:45
− 流速: 1 mL/min
− 検出: 254 nmにおけるUV吸収
上記条件下で、(R)-2-メチルピロリジンは 26.3 分に、(S)-2-メチルピロリジンは 24.9 分に溶出された。各溶出フラクションの254 nmにおけるUV吸収に基づいて光学純度を決定した。
【0109】
[実施例6]
SsSIRとC4EPL6_STRRSの酵素活性および反応結果を比較した表を以下に示す。
[表1]
ここで、pSUS2MP1およびpSG-S2MPはそれぞれ、イミン還元酵素SsSIRを発現するプラスミド(図3)、イミン還元酵素SsSIRと枯草菌由来のグルコース脱水素酵素を共発現するプラスミド(図4)である。
なお、SsSIRの酵素活性、光学純度の測定は以下のようにして行った。
pSG-S2MPで形質転換された大腸菌JM109株をアンピシリンを含む5mLの液体LB培地で終夜30℃培養し、0.1mM IPTGを加え、さらに4時間培養を行った。菌体を遠心分離により集菌した後、20mM 2−メチル−1−ピロリン、40mM グルコースを含む100mMリン酸カリウム緩衝液(pH7.0)1mLに懸濁し、終夜25℃で振とう反応を行った。反応後、生成した2−メチルピロリジンの光学純度、及び、反応液中における2−メチル−1−ピロリンと2−メチルピロリジンの濃度を実施例5と同様にして分析した。その結果、生成した(S)−2−メチルピロリジンの光学純度は92%eeであった。
上記の表から明らかなように、C4EPL6_STRRSの使用により、SsSIRを上回る光学純度を達成することができた。
【産業上の利用可能性】
【0110】
本発明は、微生物由来のイミン還元酵素を使用した立体選択的還元により、光学活性なアミン誘導体を効率的に製造する方法を提供する。本発明の方法は、高い立体選択性と光学純度(97.7%ee)を期待できるので、工業的にも有利である。本発明によって製造される光学活性なアミン誘導体は、光学活性な医薬の合成原料として有用である。たとえば、(S)−2−メチルピロリジンは、医薬品原料として有用な化合物である。
【特許請求の範囲】
【請求項1】
式(I)で示されるイミン誘導体に、下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素、当該イミン還元酵素を生産する微生物、およびその処理物からなる群から選択される、少なくとも一つの酵素活性物質を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
式(I)
【化1】
式(II)
【化2】
(式中、R1、R2基は炭素数1〜3個のアルキル基を、R3基は、炭素数1〜3個のアルキル基もしくは水素を表し、R1とR3は環を巻いていてもよい)
【請求項2】
式(I)の化合物が下記式(III)で表されるイミン誘導体であり、式(II)の化合物が式(IV)で表されるアミン誘導体である、請求項1に記載の光学活性なS体のアミン誘導体の製造方法。
式(III)
【化3】
式(IV)
【化4】
(式中R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)
【請求項3】
式(III)の化合物が2−メチル−1−ピロリンであり、式(IV)で表される化合物が、2−メチルピロリジンである、請求項2に記載の光学活性なS体のアミン誘導体の製造方法。
【請求項4】
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
【請求項5】
前記イミン還元酵素が、配列番号:1に記載のアミノ酸配列において、1個以上かつ50個以下のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質である、請求項4に記載のイミン還元酵素。
【請求項6】
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元活性を有するタンパク質を有効成分として含む、立体選択的イミン還元剤。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
【請求項7】
下記(a)から(e)のいずれかに記載のイミン還元酵素をコードするポリヌクレオチドと、NADP+を補酵素とする酸化還元反応を触媒することができる脱水素酵素をコードするポリヌクレオチドとが一緒に挿入された組換えベクター。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
【請求項8】
脱水素酵素がグルコース脱水素酵素である、請求項7に記載のベクター。
【請求項9】
グルコース脱水素酵素がバシラス・サブチルス(Bacillus subtilis)に由来する、請求項8に記載の組換えベクター。
【請求項10】
請求項7〜9のいずれかに記載のベクターを発現可能に保持した形質転換体。
【請求項11】
請求項10に記載の形質転換体を培養する工程を含む、請求項1〜3のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
【請求項12】
請求項1記載のイミン還元酵素を生産する微生物がストレプトスポランジウム(Streptosporangium)属に属する微生物である請求項1〜3のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
【請求項13】
式(I)で示されるイミン誘導体に、ストレプトスポランジウム(Streptosporangium)属に属する微生物またはその処理物を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
【請求項1】
式(I)で示されるイミン誘導体に、下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素、当該イミン還元酵素を生産する微生物、およびその処理物からなる群から選択される、少なくとも一つの酵素活性物質を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
式(I)
【化1】
式(II)
【化2】
(式中、R1、R2基は炭素数1〜3個のアルキル基を、R3基は、炭素数1〜3個のアルキル基もしくは水素を表し、R1とR3は環を巻いていてもよい)
【請求項2】
式(I)の化合物が下記式(III)で表されるイミン誘導体であり、式(II)の化合物が式(IV)で表されるアミン誘導体である、請求項1に記載の光学活性なS体のアミン誘導体の製造方法。
式(III)
【化3】
式(IV)
【化4】
(式中R基は炭素数1〜3個のアルキル基を、nは、1〜4の整数を表す)
【請求項3】
式(III)の化合物が2−メチル−1−ピロリンであり、式(IV)で表される化合物が、2−メチルピロリジンである、請求項2に記載の光学活性なS体のアミン誘導体の製造方法。
【請求項4】
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元酵素。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
【請求項5】
前記イミン還元酵素が、配列番号:1に記載のアミノ酸配列において、1個以上かつ50個以下のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質である、請求項4に記載のイミン還元酵素。
【請求項6】
下記(a)から(e)のいずれかに記載のポリヌクレオチドによりコードされるイミン還元活性を有するタンパク質を有効成分として含む、立体選択的イミン還元剤。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
【請求項7】
下記(a)から(e)のいずれかに記載のイミン還元酵素をコードするポリヌクレオチドと、NADP+を補酵素とする酸化還元反応を触媒することができる脱水素酵素をコードするポリヌクレオチドとが一緒に挿入された組換えベクター。
(a)配列番号:2に記載された塩基配列を含むポリヌクレオチド、
(b)配列番号:1に記載のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(c)配列番号:1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(d)配列番号:2に記載された塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするポリヌクレオチド、
(e)配列番号:1に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列をコードするポリヌクレオチド。
【請求項8】
脱水素酵素がグルコース脱水素酵素である、請求項7に記載のベクター。
【請求項9】
グルコース脱水素酵素がバシラス・サブチルス(Bacillus subtilis)に由来する、請求項8に記載の組換えベクター。
【請求項10】
請求項7〜9のいずれかに記載のベクターを発現可能に保持した形質転換体。
【請求項11】
請求項10に記載の形質転換体を培養する工程を含む、請求項1〜3のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
【請求項12】
請求項1記載のイミン還元酵素を生産する微生物がストレプトスポランジウム(Streptosporangium)属に属する微生物である請求項1〜3のいずれかに記載の光学活性なS体のアミン誘導体の製造方法。
【請求項13】
式(I)で示されるイミン誘導体に、ストレプトスポランジウム(Streptosporangium)属に属する微生物またはその処理物を接触させる工程と、式(II)で示される光学活性なS体のアミン誘導体を回収する工程を含む、光学活性なS体のアミン誘導体の製造方法。
【図1】

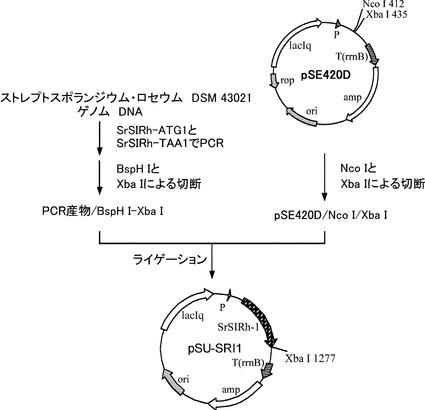
【図2】
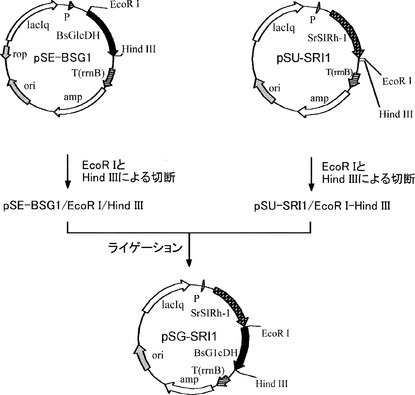
【図3】
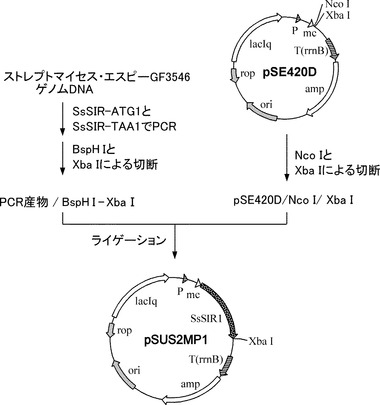
【図4】
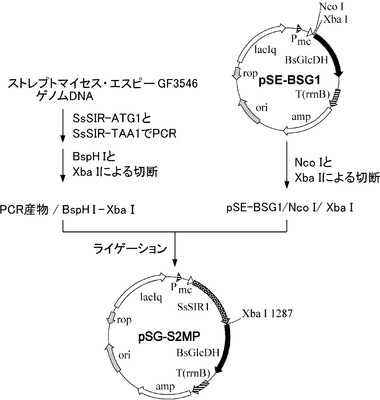
【図2】
【図3】
【図4】
【公開番号】特開2011−177029(P2011−177029A)
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【出願番号】特願2010−41378(P2010−41378)
【出願日】平成22年2月26日(2010.2.26)
【出願人】(000002901)ダイセル化学工業株式会社 (1,236)
【Fターム(参考)】
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【出願日】平成22年2月26日(2010.2.26)
【出願人】(000002901)ダイセル化学工業株式会社 (1,236)
【Fターム(参考)】
[ Back to top ]