
光学活性1−置換テトラヒドロイソキノリン誘導体の製造方法
【課題】酵素反応による光学活性1−置換テトラヒドロイソキノリン誘導体の効率的な製造方法を提供する。
【解決手段】アルキル側鎖の長い芳香族アルデヒドやアルキルアルデヒドと2−(3−ヒドロキシフェニル)エチルアミン誘導体に酵素を作用させることで、立体選択的な縮合反応による光学活性1−置換テトラヒドロイソキノリン誘導体の効率的な製造方法。
【解決手段】アルキル側鎖の長い芳香族アルデヒドやアルキルアルデヒドと2−(3−ヒドロキシフェニル)エチルアミン誘導体に酵素を作用させることで、立体選択的な縮合反応による光学活性1−置換テトラヒドロイソキノリン誘導体の効率的な製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は光学活性1−置換テトラヒドロイソキノリン誘導体の製造方法に関する。
【背景技術】
【0002】
光学活性1−置換テトラヒドロイソキノリン誘導体には、(S)-ラウダノシン、(S)-シナクチン、(S)-カルネギン、(S)-カリコトミン等、生理活性を示す化合物が多く、医薬品や農薬の合成中間体として有用な化合物群である(非特許文献1)。
【0003】
1−置換テトラヒドロイソキノリン誘導体の製造方法としては、2−(3−ヒドロキシフェニル)エチルアミン誘導体と、ケトンやアルデヒド化合物を、リン酸やカルシウム存在下で縮合する方法(Pictet−Spengler反応)が知られている。しかし、この方法では、ラセミ体の1−置換テトラヒドロイソキノリン誘導体しか得られない(非特許文献2、非特許文献3)。
【0004】
一方、光学活性な1−置換テトラヒドロイソキノリン誘導体の製造方法としては、プロキラルな1−置換3,4−ジヒドロイソキノリンの不斉水素化反応や、3,4−ジヒドロイソキノリン類の不斉アルキル化、3−(2−フェニルエチルアミン)−2−プロペン−1−オール誘導体の不斉分子内アリル位アミノ化反応による方法が知られている。しかし、これらの方法では、高価な希少金属錯体の触媒や多段階の反応が必要であり、効率的、経済的な製造方法とは言えない(非特許文献4、非特許文献5、特許文献1)。
【0005】
また、1−置換テトラヒドロイソキノリン誘導体の生合成メカニズムが植物において明らかになっており、ノルコクラウリンシンターゼ(以下、NCS)が初発反応を触媒することが知られている。しかし、NCSの作用する基質は、ドーパミンと4−ヒドロキシフェニルアセトアルデヒド、及び一部の限られた代替基質のみであり(非特許文献6、非特許文献7、非特許文献8)、基質特異性が非常に厳密で、非天然の基質に対する寛容性は低いと考えられていた(非特許文献2)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2005−41814
【非特許文献】
【0007】
【非特許文献1】Nat.Prod.Rep.(1989),6,405−432
【非特許文献2】Chem.Commun. (2011), 47, 3242
【非特許文献3】J.Org.Chem.(2010),75,8542
【非特許文献4】J.Am.Chem.Soc.(1986),108,7117
【非特許文献5】Chem.Lett.(1997),59
【非特許文献6】J.Biol.Chem.(2002),277,33878
【非特許文献7】Biochemistry(2007),46,10153
【非特許文献8】J.Biol.Chem.(2007),282,6274
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の課題は、酵素反応により光学活性1−置換テトラヒドロイソキノリン誘導体の効率的な製造方法を提供することである。
【課題を解決するための手段】
【0009】
本発明者らは、NCSの基質としては困難と考えられていた、アルキル側鎖の長い芳香族アルデヒドやアルキルアルデヒドと、2−(3−ヒドロキシフェニル)エチルアミン誘導体にNCSを作用させることで、立体選択的に光学活性1−置換テトラヒドロイソキノリン誘導体を製造できることを見出し、本発明を完成するに至った。
【0010】
すなわち、本発明は
一般式(1):
【0011】
【化1】

【0012】
(式中、R1〜R5は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、ヒドロキシル基、アミノ基、またはニトロ基である。)で表される2−(3−ヒドロキシフェニル)エチルアミン誘導体と一般式(2):
【0013】
【化2】
【0014】
(式中、R6〜R8は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、アミノ基、またはニトロ基であるか、もしくは、R6〜R10が互いに結合し環を形成しても良い。ただし、R6〜R8のいずれか2つが水素原子であり、1つが置換されていても良いフェニル基である場合は除く)で表されるアルデヒド化合物に酵素を作用させることを特徴とする、一般式(3):
【0015】
【化3】
【0016】
(式中R1〜R8は前記と同じ、*は不斉炭素を表す)で表される光学活性1−置換テトラヒドロイソキノリン誘導体の製造方法である。
【発明の効果】
【0017】
本発明により、光学活性1−置換テトラヒドロイソイソキノリン誘導体の効率的、な製造方法を提供することができる。
【図面の簡単な説明】
【0018】
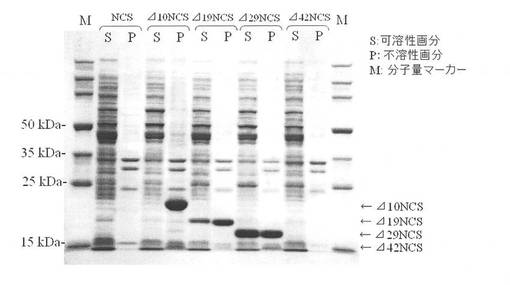
【図1】本発明の実施例6に係る遺伝子組換え生産したN末端削除NCSの電気泳動写真を示す図である。
【発明を実施するための形態】
【0019】
以下、本発明について詳述する。
【0020】
本発明の酵素反応の基質となる2−(3−ヒドロキシフェニル)エチルアミン誘導体は一般式(1):
【0021】
【化4】
【0022】
(式中、R1〜R5は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、ヒドロキシル基、アミノ基、またはニトロ基である。)である。好ましくは前記R1〜R3及びR5が水素であり、R4が水素原子、メチル基、水酸基、メトキシ基のいずれかである。最も好ましくはR1〜R3及びR5が水素であり、R4が水酸基である。
【0023】
また、本発明の酵素反応のもう1つの基質であるアルデヒド化合物は、
一般式(2):
【0024】
【化5】
【0025】
(式中、R6〜R8は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、アミノ基、またはニトロ基であるか、もしくは、R6〜R10が互いに結合し環を形成しても良い。ただし、R6〜R8のいずれか2つが水素原子であり、1つがフェニル基である場合は除く)である。好ましくは前記R6が水素原子、R7が水素原子または直鎖状または分岐鎖状の炭素数1〜10のアルキル基、R8が直鎖状または分岐鎖状の炭素数2〜10のアルキル基、置換されていても良い炭素数7〜17のアラルキル基、置換されていても良い炭素数1〜10のアルコキシ基である。さらに好ましくは下記(1)〜(10)のいずれかで表される化合物である。
(1)前記R6及びR7が水素原子、R8がエチル基;
(2)前記R6が水素原子、R7がメチル基、R8がメチル基;
(3)前記R6及びR7が水素原子、R8がn−プロピル基;
(4)前記R6及びR7が水素原子、R8がn−ブチル基;
(5)前記R6及びR7が水素原子、R8がn−ペンチル基;
(6)前記R6及びR7が水素原子、R8がn−プロピルアルデヒド基;
(7)前記R6が水素原子、R7がメチル基、R8がフェニル基;
(8)前記R6及びR7が水素原子、R8がベンジル基;
(9)前記R6及びR7が水素原子、R8が4−(トリフルオロメチル)ベンジル基;
本発明では酵素を触媒とした不斉Pictet−Spengler反応により、前記一般式(1):
【0026】
【化6】
【0027】
で表される2−(3−ヒドロキシフェニル)エチルアミン誘導体と前記一般式(2):
【0028】
【化7】
【0029】
で表されるアルデヒド化合物を縮合し、一般式(3):
【0030】
【化8】
【0031】
で表される光学活性1−置換テトラヒドロイソイソキノリン誘導体を合成する。
【0032】
Pictet−Spengler反応は、β−アリールエチルアミンとアルデヒド化合物を縮合閉環させる反応である。有機合成反応では補助剤やキラルな酸触媒を利用した不斉Pictet−Spengler反応が知られている(J.Org.Lett.(2000), 2,1955、J.Am.Chem.Soc.(2006)128,1086)。
【0033】
なお、ここでいう「光学活性」とは、考えられる各種光学異性体のうち、ひとつの光学異性体の光学純度が高いことをいう。
【0034】
光学純度は高速液体クロマトグラフィーやガスクロマトグラフィーによって光学異性体を分析し、そのピーク面積から下記式より求めることができる。
【0035】
一般式(3)で示される光学異性体の(S)体が主生成物である場合:
((S)体の光学純度(%e.e.)={((S)体のピーク面積)−((R)体ピーク面積)}÷{(S)体のピーク面積)+((R)体ピーク面積)}×100
本発明では前記一般式(3)で表される光学活性1−置換テトラヒドロイソイソキノリン誘導体においては、光学純度は50%e.e.以上が好ましく、より好ましくは75%e.e.以上、さらに好ましくは90%e.e.以上である。
【0036】
なお、J.Biol.Chem.(2009)284,897に示される触媒機構から、NCSによって生成される光学活性1−置換テトラヒドロイソキノリン誘導体の絶対立体配置はS体と予想される。
【0037】
前記酵素としては、前記反応を触媒する酵素であればいかなる酵素を用いても良いが、好ましくは国際生化学・分子生物学連合の酵素分類法によりEC4.2.1.78に分類されるノルコクラウリンシンターゼ(NCS)、もしくはPathogenesis−Related 10/Bet V1 Protein Familyに属する酵素(以下、PR10タンパク質)である。PR10タンパク質は生体防御タンパク質ファミリーの1つであり、アミノ酸配列の相同性や血清学的・免疫学的な相関性、酵素活性の特徴に基づいて17種類に分類されたうちの1つである(Plant Mol.Biol.Report.(1994),12,245、Physiol.Mol. Plant Pathol. (1999),55,85、Curr.Opin.Immunol.(1998) ,10,16)。PR10タンパク質にはアレルギー抗原であるBet V1やリボヌクレアーゼ様タンパク質の構造類似タンパク質が含まれる。
【0038】
NCS及びPR10タンパク質は植物中から単離することが可能である。NCS及びPR10タンパク質の由来となる植物は特に限定されないが、アルカロイドを体内に蓄積するアマリリダセエ(Amaryllidaceae)科、アリストロチセエ(Aristolochiaceae)科、ベルベリダセエ(Berberidaceae)科、ユーホルビアセエ(Euphorbiaceae)科、ファバセエ(Fabaceae)科、リリアセエ(Liliaceae)科、パパベラセエ(Papaveraceae)科、ラヌンチュラセエ(Ranunculaceae)科、ソラナセエ(Solanaceae)科の植物が好ましい。特に好ましくはラヌンチュラセエ(Ranunculaceae)科のコプティス・ジャポニカ(Coptis japonica)、タリクトラム・フラバム(Thalictrum flavum)及びパパベラセエ(Papaveraceae)科のパパヴェラ・ソムニフェラム(Papaver somniferum)パパヴェラ・ブラクテアタム(Papaver bracteatum)である。また、酵素を単離せずに、植物体もしくは植物培養細胞をそのまま利用することも可能である。具体的にはJ.Biol.Chem.(2007),282,6274に記載の方法で植物培養細胞を得ることができる。
【0039】
ここで、NCS及びPR10タンパク質の酵素活性は以下のようにして検出することが可能である。例えば、ドーパミン塩酸塩10mM、ジメチルホルムアミドに溶解したヒドロキシシンナムアルデヒド10mMを添加した100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に、NCSもしくはPR10が含まれる溶液(例えば、該酵素を生産した組換え生物や植物の精製物など)を加え、30℃で15分間反応を行う。反応後、下記溶離液を添加して反応を停止し、遠心後、上清を下記分析条件で分析して1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの生成量を定量する。
【0040】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 555ml、蒸留水 945ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 11.5分
【0041】
NCS及びPR10タンパク質のアミノ酸配列は、その上記の方法で検出している酵素活性を有している限り特に限定されないが、好ましくは配列表の配列番号1、2、3、4、5のいずれかで表されるポリペプチドと好ましくは60%以上、さらに好ましくは80%以上、90%以上、95%以上、97%以上、98%以上、最も好ましくは99%以上の配列同一性を示すタンパク質である。
【0042】
配列表の配列番号1、2、3、4、5のアミノ酸配列をコードするDNA配列を利用して、前記の植物から配列相同性の高いDNAを取得し、その生産物を本発明に利用することもできる。具体的にはJ.Biol.Chem.(2007),282,6274に記載の方法で取得することができる。また、ゲノム情報が解析され、塩基配列が明らかになっている場合は、DNAを人工合成することも可能である。
【0043】
本明細書におけるポリペプチドやポリヌクレオチドの「配列同一性」とは、対比される2つのポリペプチドまたはポリヌクレオチドを最適に整列させ、核酸塩基(例えば、A、T、C、G、U、またはI)が両方の配列で一致した位置の数を比較塩基総数で除し、そして、この結果に100を乗じた数値で表される。
【0044】
配列同一性は、例えば、以下の配列分析用ツールを用いて算出し得る;GCG Wisconsin Package(ウィスコンシン大学)、the ExPASy World Wide Web分子生物学用サーバー(スイスバイオインフォマティックス研究所)、BLAST(米国生物工学情報センター)、GENETYX(ゼネティックス社)。
【0045】
また、本発明に利用するNCS及びPR10タンパク質は、目的のNCS活性を有している限りにおいては、そのアミノ酸配列(例えば、配列表の配列番号1〜5)の1若しくは複数個(例えば、40個、好ましくは20個、より好ましくは15個、さらに好ましくは10個、さらに好ましくは5個、4個、3個、または2個以下)のアミノ酸が置換、挿入、欠失及び/または付加されたポリペプチドであってもよい。また、目的のNCS活性を有する限り、本発明に利用するNCS及びPR10のアミノ酸配列に、付加的なアミノ酸配列を結合してもよい。たとえば、ヒスチジンタグやHAタグのような、タグ配列を付加することができる。あるいは、他のタンパク質との融合タンパク質とすることもできる。また、目的のNCS活性を有する限り、ペプチド断片であってもよい。
【0046】
また、本発明に利用するNCS及び/またはPR10タンパク質は下記に示す方法で、適当な宿主生物内で本発明に利用するNCS及び/またはPR10タンパク質をコードする遺伝子を発現し、遺伝子組換え生産して利用することもできる。
【0047】
NCS及び/またはPR10タンパク質のN末端部分にはシグナル配列が存在する場合があるため、宿主生物内で遺伝子組換え生産する場合はこのシグナル配列を一部もしくは全て削除することが好ましい。
【0048】
NCS及び/またはPR10タンパク質を遺伝子組換え生産する場合、宿主生物中で不溶化し、酵素活性が低下することがある。本発明では配列表の配列番号1に示すアミノ酸配列のうちN末端のアミノ酸を10個削除し、メチオニンを付加した場合(実施例4のE. coli HB101(pNΔ10NCS)が生産するΔ10NCS)はNCSの大部分が不溶化するが、N末端をさらに削除し、メチオニンを付加したポリペプチドを遺伝子組換え生産することで、不溶化が低減され、酵素活性が向上することを見出した(例えば実施例4のE. coli HB101(pNΔ19NCS)が生産するΔ19NCS)。削除するアミノ酸の数としては、好ましくは11〜41個、さらに好ましくは19〜29個である。最も好ましくは19個もしくは29個である。
【0049】
本発明においては、NCS及びPR10タンパク質をコードするDNAを含む形質転換体を使用すると、より効率的に光学活性1−置換テトラヒドロイソキノリンを製造することができる。なお、本明細書において記述されている、DNAの単離、ベクターの調製、形質転換等の遺伝子操作は、特に明記しない限り、Molecular Cloning 2nd Edition(Cold Spring Harbor Laboratory Press,1989)、Current Protocols in Molecular Biology(Greene Publishing Associates and Wiley−Interscience)等の成書に記載されている方法により実施できる。
【0050】
上記の形質転換体に用いるベクターとしては、適当な宿主生物内で本発明に利用するNCS及びPR10タンパク質をコードする遺伝子を発現できるものであれば、特に限定されない。このようなベクターとしては、例えば、プラスミドベクター、ファージベクター、コスミドベクターなどが挙げられ、さらに、他の宿主株との間での遺伝子交換が可能なシャトルベクターも使用できる。
【0051】
このようなベクターは、例えば大腸菌の場合では、通常、lacUV5プロモーター、trpプロモーター、trcプロモーター、tacプロモーター、lppプロモーター、tufBプロモーター、recAプロモーター、pLプロモーター等の制御因子を含み、本発明のDNAと作動可能に連結された発現単位を含む発現ベクターとして好適に使用できる。例えば、pSTV28(タカラバイオ社製)、pUCNT(国際特許公報WO94/03613)などが挙げられる。
【0052】
なお、「制御因子」とは、機能的プロモーター、及び、任意の関連する転写要素(例えばエンハンサー、CCAATボックス、TATAボックス、SPI部位など)を有する塩基配列をいう。
【0053】
また、「作動可能に連結」とは、遺伝子の発現を調節するプロモーター、エンハンサー等の種々の調節エレメントと遺伝子が、宿主細胞中で作動し得る状態で連結されることをいう。なお、制御因子のタイプ及び種類が宿主に応じて変わり得ることは、当業者に周知の事項である。
【0054】
各種生物において利用可能なベクター、プロモーターなどについては、「微生物学基礎講座8遺伝子工学」(共立出版、1987)などに詳細に記述されている。
【0055】
各酵素を発現させるために用いる宿主生物は、各酵素をコードするDNAを含む酵素発現ベクターにより形質転換され、DNAを導入した酵素を発現することができる生物であれば、特に制限するものではない。利用可能な微生物としては、例えば、エシェリヒア(Escherichia)属、バシラス(Bacillus)属、シュードモナス(Pseudomonas)属、セラチア(Serratia)属、ブレビバクテリウム(Brevibacterium)属、コリネバクテリイウム(Corynebacterium)属、ストレプトコッカス(Streptococcus)属、及びラクトバシラス(Lactobacillus)属など宿主ベクター系の開発されている細菌、ロドコッカス(Rhodococcus)属及びストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌、サッカロマイセス(Saccharomyces)属、クライベロマイセス(Kluyveromyces)属、シゾサッカロマイセス(Schizosaccharomyces)属、チゴサッカロマイセス(Zygosaccharomyces)属、ヤロウイア(Yarrowia)属、トリコスポロン(Trichosporon)属、ロドスポリジウム(Rhodosporidium)属、ピキア(Pichia)属、及びキャンディダ(Candida)属などの宿主ベクター系の開発されている酵母、ノイロスポラ(Neurospora)属、アスペルギルス(Aspergillus)属、セファロスポリウム(Cephalosporium)属、及びトリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ、などが挙げられる。また、微生物以外でも、植物、動物において様々な宿主・ベクター系が開発されており、特に蚕を用いた昆虫(Nature,315,592−594(1985))や菜種、トウモロコシ、ジャガイモなどの植物中に大量に異種タンパク質を発現させる系が開発されており、好適に利用できる。これらのうち、導入及び発現効率から細菌が好ましく、大腸菌(エシェリヒア・コリ(Escherichia coli)が特に好ましい。
【0056】
本発明に利用するNCS及び/またはPR10をコードするDNAを含むベクターは、公知の方法により宿主微生物に導入することができる。例えば、宿主微生物として大腸菌を用いる場合は、市販のエシェリヒア・コリ(Escherichia coli HB101(以下、E. coli HB101)コンピテントセル(タカラバイオ社製)を用いることにより、当該ベクターを宿主細胞に導入することができる。
【0057】
NCS及び/またはPR10をコードするDNAを含むベクターとしては、実施例3に記載のpNNCSが挙げられる。また、NCS及び/またはPR10をコードするDNAを含む形質転換体の例としては、ベクターpNNCSでE.coli HB101を形質転換して得られる、実施例4に記載のE.coli HB101(pNNCS)が挙げられる。
【0058】
形質転換体の培養には、通常、これらの宿主微生物が資化しうる栄養源を含む培地であればどのようなものでも使用しうる。例えば、グルコース、シュークロース、マルトース等の糖類、乳酸、酢酸、クエン酸、プロピオン酸等の有機酸類、エタノール、グリセリン等のアルコール類、パラフィン等の炭化水素類、大豆油、菜種油等の油脂類、またはこれらの混合物等の炭素源;硫酸アンモニウム、リン酸アンモニウム、尿素、酵母エキス、肉エキス、ペプトン、コーンスチープリカー等の窒素源;更に、その他の無機塩、ビタミン類等の栄養源;を適宜混合・配合した通常の培地を用いることが出来る。これら培地は用いる微生物の種類によって適宜選択すればよい。
【0059】
微生物の培養は、一般的な条件により行なうことができる。例えば、pH4.0〜9.5、温度範囲20℃〜45℃の範囲で、好気的に10〜96時間培養するのが好ましい。
【0060】
次に、酵素を用いた2−(3−ヒドロキシフェニル)エチルアミン誘導体とアルデヒド化合物の縮合反応について説明する。
【0061】
酵素を用いた縮合反応の際には、適当な溶媒と基質の2−(3−ヒドロキシフェニル)エチルアミン誘導体とアルデヒド化合物、前記酵素、前記形質転換体、その培養物、またはその処理物等を混合し、pH調整下に攪拌する。
【0062】
反応溶媒としては、通常、水や緩衝液等の水性媒体を用いる。緩衝液としては、リン酸カリウム緩衝液やトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液が挙げられる。本反応においては、非酵素的な縮合反応によりラセミ体の1−置換テトラヒドロイソキノリンが生成するため、酵素反応によって得られる光学活性1−置換テトラヒドロイソキノリンの光学純度が低下する。Chem.Commun. (2011), 47, 3242に記載されているようにリン酸緩衝液中ではこの非酵素的な縮合反応が促進されるため、不適である。一方、トリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液中では非酵素的な縮合反応の影響が少ないため、好適である。
【0063】
酵素は、通常、上記微生物の菌体を含んだ培養液をそのまま反応に使用する。培養液を濃縮して用いてもよい。また、培養液中の成分が反応に悪影響を与える場合には、培養液を遠心分離等により処理して得られる菌体または菌体処理物を使用するとよい。
【0064】
基質である2−(3−ヒドロキシフェニル)エチルアミン誘導体とアルデヒド化合物は、反応の初期に一括添加してもよく、反応の進行にあわせて逐次分割して添加してもよい。高濃度の基質存在下では非酵素的な縮合反応が促進されるため、基質の逐次分割添加が特に好適である。
【0065】
反応時の温度は、通常、10〜60℃、好ましくは20〜40℃とする。
【0066】
反応時のpHは2.5〜9、好ましくはpH5〜9の範囲である。
【0067】
反応液中の微生物の量は、これらの基質を縮合する能力に応じ適宜決定すればよく、菌体湿重量として、0.01〜50%(W/V)が好ましく、より好ましくは0.1〜10%(W/V)である。
【0068】
反応液中の基質濃度は0.01〜50%(W/V)が好ましく、より好ましくは、0.1〜30%(W/V)である。
【0069】
反応は、通常、振とうまたは通気攪拌しながら行なう。
【0070】
反応時間は、基質濃度、微生物の量及びその他の反応条件により適宜決定される。通常、2〜168時間で反応が終了するように各条件を設定することが好ましい。
【0071】
更に、トリトン(ナカライテスク株式会社製)、スパン(関東化学株式会社製)、ツイーン(ナカライテスク株式会社製)などの界面活性剤を反応液に添加することも効果的である。
【0072】
更に、基質及び/または縮合反応の生成物による反応の阻害を回避する目的で、酢酸エチル、酢酸ブチル、イソプロピルエーテル、トルエン、ヘキサンなどの水に不溶な有機溶媒を反応液に添加してもよい。
【0073】
更に、基質の溶解度を高める目的で、メタノール、エタノール、アセトン、テトラヒドロフラン、ジメチルスルホキシドなどの水に可溶な有機溶媒を添加することもできる。
【0074】
縮合反応により生成した光学活性1−置換テトラヒドロイソキノリンを取り出す方法は特に限定されないが、反応液から直接、あるいは菌体等を分離後に、酢酸エチル、トルエン、t−ブチルメチルエーテル、ヘキサン、n−ブタノール、ジクロロメタン等の溶剤で抽出し、脱水後、蒸留や結晶化操作、シリカゲルカラムクロマトグラフィー等により精製することにより、高純度の光学活性1−置換テトラヒドロイソキノリンを容易に得ることができる。
【実施例】
【0075】
以下、実施例で本発明を詳細に説明するが、本発明はこれらにより限定されるものではない。なお、以下の記載において、「%」は特に断らない限り「重量%」を意味する。また、以下の実施例において用いた組み換えDNA技術に関する詳細な操作方法などは、次の成書に記載されている:
Molecular Cloning 2nd Edition(Joseph Sambrook,Cold Spring Harbor Laboratory Press(1989))、Current Protocols in Molecular Biology(Frederick M.Ausubel,Greene Publishing Associates and Wiley−Interscience(1989))。
【0076】
(参考例1)1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの合成
1−(2−フェニルエチル)−6,7−ジメトキシ−3,4−ジヒドロイソキノリン1.48g(5mmol)のメタノール溶液10mlを5℃に冷却した。ここに、水素化ホウ素ナトリウム189mg(5mmol)を添加して1時間撹拌し、更に25℃まで昇温して2時間撹拌した。別の容器に濃塩酸2.61g(25mmol)と水10mlを加えて5℃に冷却し、ここに反応液をゆっくりと加えた。30wt%水酸化ナトリウム水溶液でpH=12に調整後、減圧下にメタノールを留去した。残渣に酢酸エチル20mlを加えて抽出し、有機層を更に水10mlで洗浄した。減圧濃縮することにより、1−(2−フェニルエチル)−6,7−ジメトキシ−1,2,3,4−テトラヒドロイソキノリンを淡黄色油状物として得た(1.58g、含量:94.2%、収率は定量的)。
【0077】
続いて、前記化合物1.06g(3.37mmol)に48wt%臭化水素酸10g(17.6当量)を加えて、90℃、16時間撹拌した。室温まで冷却後、30wt%水酸化ナトリウム水溶液でpH=7に調整し、水20mLと酢酸エチル10mlを加えた。5℃まで冷却して30分撹拌後、析出している結晶を減圧濾別した。結晶を水10gに懸濁して25℃、30分撹拌後、減圧濾別し、水10gで洗浄、50℃で真空乾燥することにより、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン520.6mgを得た(淡褐色結晶、含量:92.1%)下記分析条件で分析し、ラセミ体のそれぞれのピークを確認した。
1H NMR(CDCl3):δ1.86(2H,m)、2.1−2.7(4H,m)、3.02(1H,m)、3.27(1H,m)、3.5−4.4(2H,brs)、3.91(1H,m)、6.57(1H,s)、6.60(1H,s)、7.1−7.3(5H,m)
【0078】
<高速液体クロマトグラフィー分析条件> 光学純度分析
カラム:ダイセル化学工業株式会社製CHIRALPAK AD−H(250mm×4.6mm)
溶離液:n−ヘキサン/エタノール/ジエチルアミン=80/20/0.1
流速:1ml/min
検出:230nm
カラム温度:30℃
検出時間:1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 11.5分、14.4分
【0079】
(実施例1)NCSをコードするDNAの調製
配列表の配列番号1のアミノ酸配列で示されるコプティス・ジャポニカ(Coptis japonica)由来のNCS(CjPR10A)をコードする配列番号6のDNAを人工合成した。この合成DNAを鋳型としてプライマー1:5’-GGAGTCCATATGCGTATGGAAGTGGTTCT-3’(配列表の配列番号7)、プライマー2:5’-TATATACTGCAGTTATTCGGAAGATTTGTG -3’(配列表の配列番号8)を用いてPCRを行った。その結果、配列表の配列番号6に示す塩基配列からなる遺伝子の開始コドン部分にNdeI認識部位が付加され、かつ終始コドンの直後にPstI認識部位が付加された二本鎖DNA(NCS遺伝子)が得られた。PCRは、DNAポリメラ−ゼとして、PrimeSTAR HS DNA polymerase(タカラバイオ社製)を用いて行い、反応条件はその取り扱い説明書に従った。
【0080】
(実施例2)N末端配列を削除したNCSをコードするDNAの調製
NCSのN末端アミノ酸配列を10、19、29、42個削除し、N末端にメチオニンを付加したポリペプチドをコードするDNAを調製した(以下、それぞれΔ10NCS、Δ19NCS、Δ29NCS、Δ42NCS遺伝子と呼ぶ)。実施例1で増幅したDNA(NCS遺伝子)を鋳型としてプライマー3:5’-GGAGTCCATATGCTGATGTTCATTGGCACC-3’(配列表の配列番号9、Δ10NCS増幅用)、プライマー4:5’-GGAGTCCATATGGAACGCCTGATTTTTAA-3’(配列表の配列番号10、Δ19NCS増幅用)、プライマー5:5’-GGAGTCCATATGCTGCATCGCGTTACGAAA-3’(配列表の配列番号11、Δ29NCS増幅用)、プライマー6:5’- GGAGTCCATATGCACGAACTGGAAGTGGCG -3’(配列表の配列番号12、Δ43NCS増幅用)のいずれか1つとプライマー2:5’- TATATACTGCAGTTATTCGGAAGATTTGTG -3’(配列表の配列番号8)を用いてPCRを行った。その結果、配列表の配列番号1に示すアミノ酸配列からなるポリペプチドのN末端部分のアミノ酸から10、19、29、42残基削除されて、かつN末端にメチオニンが付加されたポリペプチドをコードする二本鎖DNA(配列開始コドン部分にNdeI認識部位が付加され、かつ終始コドンの直後にPstI認識部位が付加された)が4種類(Δ10NCS、Δ19NCS、Δ29NCS、Δ43NCS遺伝子)得られた。
【0081】
(実施例3)組換えベクターの調製
実施例1及び2で得られた5種類のNCS遺伝子をNdeI及びPstIで消化し、プラスミドpUCN18(PCR法によりpUC18(タカラバイオ社製)の185番目のTをAに改変してNdeIサイトを破壊し、更に471−472番目のGCをTGに改変することにより新たにNdeIサイトを導入したプラスミド)のlacプロモーターの下流のNdeI認識部位とPstI認識部位の間に挿入し、組換えベクターpNNCS、pNΔ10NCS、pNΔ19NCS、pNΔ29NCS、pNΔ42NCSを構築した。
【0082】
(実施例4)NCSを生産する組換え大腸菌の育種
実施例3で構築した5種類の組換えベクターを用いて、E. coli HB101コンピテントセル(タカラバイオ社製)を形質転換し、5種類の組換え生物E. coli HB101(pNNCS)、E. coli HB101(pNΔ10NCS)、E. coli HB101(pNΔ19NCS)、E. coli HB101(pNΔ29NCS)、E. coli HB101(pNΔ42NCS)を得た。また、pUCN18を用いてE. coli HB101コンピテントセル(タカラバイオ社製)を形質転換し、組換え生物E. coli HB101(pUCN18)を得た。
【0083】
(実施例5) 組換え生物におけるDNAの発現
実施例4で得た6種類の組換え生物(E. coli HB101(pUCN18)、E. coli HB101(pNNCS)、E. coli HB101(pNΔ10NCS)、E. coli HB101(pNΔ19NCS)、E. coli HB101(pNΔ29NCS)、E. coli HB101(pNΔ42NCS)を、200μg/mlのアンピシリンを含む2×YT培地(トリプトン1.6%、イーストエキス1.0%、塩化ナトリウム 0.5%、pH7.0)5mlに接種し、30℃で24時間振盪培養した。上記の培養で得られたそれぞれ培養液1mlについて、遠心分離により菌体を集め、1mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁した。これを、UH−50型超音波ホモゲナイザー(SMT社製)を用いて破砕した後、遠心分離により菌体残渣を除去し、無細胞抽出液を得た。これらの無細胞抽出液のNCS活性を測定した。
【0084】
NCS活性は、100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に、ドーパミン塩酸塩10mM、ジメチルホルムアミドに溶解したヒドロキシシンナムアルデヒド10mM、および無細胞抽出液を添加して30℃で15分間反応を行い、下記溶離液を添加して反応を停止し、遠心後、上清を下記分析条件で分析し、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの生成量を定量した。この反応条件において、1分間に1μmolの1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを生成する酵素活性(NCS活性)を1Uと定義した。
【0085】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 555ml、蒸留水 945ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 11.5分
それぞれの組換え生物のNCS活性を以下に示す。 E. coli HB101(pUCN18)については、NCS活性は0.01U/mg以下であった。一方、NCSを発現させた組換え生物のNCS活性はそれぞれ、E. coli HB101(pNNCS):0.03U/mg、E. coli HB101(pNΔ10NCS):0.1U/mg、E. coli HB101(pNΔ19NCS):0.3U/mg、E. coli HB101(pNΔ29NCS):1.2U/mg、E. coli HB101(pNΔ42NCS):0.1U/mgであった。N末端アミノ酸を19または29残基削除したNCSでは高いNCS活性を示した。
【0086】
(実施例6) N末端アミノ酸を削除したNCSの可溶化の確認
実施例4で作製したNCSを生産する組換え大腸菌5種類について、実施例5と同様の方法で無細胞抽出液を調製した。これを可溶性画分とした。また、遠心分離後の菌体残渣を1mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁し、これを不溶性画分とした。各組換え生物の可溶性、不溶性画分についてドデシル硫酸ナトリウム(SDS)ポリアクリルアミドゲル電気泳動を行った。泳動後、クマシーブリリアントブルー染色によって可溶性画分のNCSを確認した。結果を図1に示す。
【0087】
シグナル配列を含むNCS全長を生産するE. coli HB101(pNNCS)では高生産されているタンパク質が確認できなかった。一方、E. coli HB101(pNΔ10NCS)ではΔ10NCSが高生産されいていることが確認できた。しかし、大部分のΔ10NCSは不溶性画分に確認された。E. coli HB101(pNΔ19NCS)及びE. coli HB101(pNΔ29NCS)ではE. coli HB101(pNΔ10NCS)に比べ可溶性画分のN末端削除NCSが増加していた。E. coli HB101(pNΔ42NCS)では高生産されているタンパク質が確認できなかった。
【0088】
N末端アミノ酸を19もしくは29個削除したNCSでは遺伝子組換え生産時の不溶化が低減できた。
【0089】
(実施例7)光学活性1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ10NCS)について実施例5と同様の方法で無細胞抽出液1mlを調製した。この無細胞抽出液にドーパミン塩酸塩10mM、ヒドロシンナムアルデヒド10mMを添加し、30℃で30分反応した。また、対照実験として、無細胞抽出液の代わりに100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)でも同様に反応を実施した。反応液から酢酸エチルにて生成した1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを抽出し、参考例1及び実施例5に示した分析条件で分析した。その結果、1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンへの変換率は11%、であり、参考例1の分析条件において検出時間11.5分の光学活性体が生成しており、その光学純度は96.1%e.e.であった。一方、対照実験では1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンへの変換率は1%であり、ラセミ体が生成していた。
【0090】
E. coli HB101(pNΔ10NCS)が高い立体選択性で光学活性1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを生成することが分かった。
【0091】
(実施例8)光学活性−1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ10NCS)について実施例5と同様の方法で無細胞抽出液20mlを調製した。この無細胞抽出液にドーパミン塩酸塩0.25gを添加し、攪拌しながら、ヒドロシンナムアルデヒド0.21gを逐次添加した。16時間後の反応液を実施例5及び実施例7記載の分析条件で分析したところ、変換率は58%、光学純度は76.3%e.e.(検出時間11.5分の光学活性体)であった。
【0092】
(実施例9)光学活性−1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ29NCS)について実施例5と同様の方法で無細胞抽出液20mlを調製した。この無細胞抽出液にドーパミン塩酸塩0.25gを添加し、攪拌しながら、ヒドロシンナムアルデヒド0.21gを逐次添加した。16時間後の反応液を実施例5及び実施例7記載の分析条件で分析したところ、変換率は86%、(S)−1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの光学純度は95.3%e.e.(検出時間11.5分の光学活性体)であった。
【0093】
(実施例10)3−(4−トリフルオロメチルフェニル)プロピルアルデヒドの製造
3−(4−トリフルオロメチルフェニル)プロピオン酸8.33g(37.4mmol)にTHF 40mlを加え、−10℃に冷却し、1.03M BH3−THF 42.3ml(43.6mmol)を添加した(内温推移−7〜0℃)。室温まで昇温し、13時間撹拌した。反応液に水40mlと飽和炭酸水素ナトリウム水溶液20ml、酢酸エチル40ml、飽和食塩水20mlを添加し、分液した。有機層に水40mlと飽和食塩水20mlを加え、撹拌後、分液した。有機層を減圧濃縮することで3−(4−トリフルオロメチルフェニル)プロパノール8.27gを得た。ここで得た3−(4−トリフルオロメチルフェニル)プロパノール、炭酸水素ナトリウム 9.38g(0.112mol)、NaBr 3.83g(37.2mmol)、水80ml、酢酸エチル80ml、Tempo 0.291g(1.86mmol)を氷冷し、13.02%次亜塩素酸ナトリウム 21.28g(37.2mmol)を35分間で添加した。添加終了後、1.5時間撹拌し、水120mlを添加し、分液した。有機層を水40mlで2回洗浄し、減圧濃縮することで3−(4−トリフルオロメチルフェニル)プロピルアルデヒド8.08gを得た。ガスクロマトグラフィーで分析したところ純度89.7wt%、収率95.8mol%であった。
【0094】
3−(4−トリフルオロメチルフェニル)プロパノール
1H NMR(CDCl3):δ=1.87−1.94(m、2H)、2.78(t、2H、J=7.8Hz)、3.67−3.77(m、2H)、7.31(d、2H、J=8.0Hz)、7.53(d、2H、J=8.1Hz)
3−(4−トリフルオロメチルフェニル)プロピルアルデヒド
1H NMR(CDCl3):δ=2.81(d、2H、J=7.3Hz)、3.01(t、2H、J=7.6Hz)、7.30(d、2H、J=8.0Hz)、7.53(d、2H、J=8.1Hz)、9.81(s、1H)
<ガスクロマトグラフィー分析系>
カラム:Restek Corporation製Rtx−35A(30mx0.25mm)
検出器:FID
カラム温度:50℃→(17分)→220℃→(13分)→220℃
キャリアー:He 150Kpa
【0095】
(参考例2)1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
1.94mlの100mMリン酸緩衝液(pH6.5)にドーパミン塩酸塩194mg(1.0mmol)、メタノール0.5mlを添加し40℃で攪拌した。3−(4−トリフルオロメチルフェニル)プロピルアルデヒド207mg(1.0mmol)を分割添加し、4時間反応した。反応液にNaCl添加後、生成した1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンをn−ブタノールで抽出した。有機層に硫酸ナトリウムを添加後、ろ過により除去した。ろ液から溶媒を留去した後、濃縮物をトルエンで洗浄した後、有機層を除去した。これを真空乾燥し、51mgの1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン(淡黄色の固体)を得た。さらに結晶化により精製した後、下記分析条件で分析し、ラセミ体のそれぞれのピークを確認した。
1H NMR(CD3OD):δ2.07−2.19(m、2H)、2.74−2.84(m、4H)、3.09−3.12(m、1H)、3.32−3.38(m、1H)、4.12−4.13(m、1H)、6.51(s、1H)、6.57(s,1H)、7.40−7.41(d,2H)、7.54−7.59(d、2H)
【0096】
<高速液体クロマトグラフィー分析条件> 光学純度分析
カラム:ダイセル化学工業株式会社製CHIRALPAK AD−H(250mm×4.6mm)
溶離液:n−ヘキサン/エタノール/ジエチルアミン=80/20/0.1
流速:1ml/min
検出:230nm
カラム温度:30℃
検出時間:1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 9.2分、11.5分
【0097】
(実施例11)光学活性1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ10NCS)について実施例5と同様の方法で無細胞抽出液120mlを調製した。この無細胞抽出液にドーパミン塩酸塩1.51gを添加し、攪拌しながら、4−トリフルオロメチルヒドロシンナムアルデヒド2gを逐次添加した。28時間後の反応液を下記分析条件で分析したところ、変換率は45%であった。また、参考例2に記載の分析条件で光学純度を求めたところ、検出時間9.2分の光学活性1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの光学純度が82.0%e.e.であった。
【0098】
<高速液体クロマトグラフィー分析条件> 変換率分析
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 600ml、蒸留水 900ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 28.1分
【0099】
(実施例12)NCSの基質特異性の評価
組換え生物E. coli HB101(pNΔ29NCS)及びE. coliHB101(pUCN18)を実施例5と同様の方法で培養した。各培養液から遠心分離により菌体を集め、5mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁した。これを、UH−50型超音波ホモゲナイザー(SMT社製)を用いて破砕した後、遠心分離により菌体残渣を除去し、無細胞抽出液を得た。これらの無細胞抽出液1mlにドーパミン塩酸塩10mM、表1に示すアルデヒド化合物10mM(ジメチルホルムアミドに溶解して添加)を添加し、30℃で15分反応した(対照実験としてE. coli HB101(pUCN18)の無細胞抽出液についても同様に反応した)。反応終了後、下記溶離液を添加して反応を終了し、遠心後、上清を下記分析条件で分析し、ドーパミンの減少量を定量した。
【0100】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 600ml、蒸留水 900ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分
E. coli HB101(pNΔ29NCS)を用いた反応でのドーパミン減少量からE. coli HB101(pUCN18)の反応液でのドーパミン減少量を差し引いた値をNCSの触媒作用によって減少したドーパミン量と見なした。本条件において、1分間に1μmolのドーパミンが消費される酵素活性を1Uと定義した。また、フェニルアセトアルデヒドをアルデヒド化合物の標準基質とし、フェニルアセトアルデヒドに対する酵素活性を100%としたときの相対活性で各アルデヒド化合物に対する酵素活性を示した。結果を表1に示す。
NCSはフェニルアセトアルデヒドや、ヒドロシンナムアルデヒド以外にもアルキルアルデヒドに対して活性を示した。
【0101】
【表1】
【0102】
(参考例3)1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
1.94mlの100mMリン酸緩衝液(pH6.5)にドーパミン塩酸塩194mg(1.0mmol)、n−ブチルアルデヒド111mg(1.5mmol)を添加し、40℃で2時間攪拌した。薄層クロマトグラフィーによりドーパミンの消失を確認した後、生成した1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを4mlのn−ブタノールで2回抽出した。有機層に硫酸ナトリウムを添加後、ろ過により除去した。ろ液から溶媒を留去し、241mgの1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを得た(黄色の固体)。下記分析条件で分析し、ラセミ体であることを確認した。
1H NMR(CD3OD):δ1.02(t、J=7.45Hz、3H)、1.42−1.65(m、2H)、1.76−2.09(m、2H)、2.79−3.04(m、2H)、3.21(m、1H)、3.48(dt,J=12.46,6.09Hz、1H)、4.25−4.42(m,1H)、6.59(s,1H)、6.71(s、1H)
【0103】
<高速液体クロマトグラフィー分析条件>
ダイセル化学工業株式会社製CHIRALPAK AD−H(250mm×4.6mm)
溶離液:n−ヘキサン/エタノール/ジエチルアミン=80/20/0.1
流速:0.5ml/min
検出:230nm
カラム温度:30℃
検出時間:1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 33.9分、39.1分
【0104】
(実施例13)光学活性1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
組換え生物E. coli HB101(pNΔ29NCS)を実施例5と同様の方法で培養した。培養液から遠心分離により菌体を集め、1mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁した。これを、UH−50型超音波ホモゲナイザー(SMT社製)を用いて破砕した後、遠心分離により菌体残渣を除去し、無細胞抽出液を得た。この無細胞抽出液1mlに19.4mgのドーパミン塩酸塩を加え、攪拌しながらn−ブチルアルデヒド21mgを分割添加した(反応時間1.5時間)。反応液を下記分析条件で分析したところ、変換率は99.6%であった。
【0105】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 600ml、蒸留水 900ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 7.1分
反応終了後、n−ブタノールで生成物を抽出し、溶媒を留去して20mgの1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを得た。参考例3に示した分析方法で分析したところ、検出時間33.9分の光学活性体の光学純度が98.0%e.e.であった。
【0106】
NCSを用いて光学活性1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを合成できた。
【技術分野】
【0001】
本発明は光学活性1−置換テトラヒドロイソキノリン誘導体の製造方法に関する。
【背景技術】
【0002】
光学活性1−置換テトラヒドロイソキノリン誘導体には、(S)-ラウダノシン、(S)-シナクチン、(S)-カルネギン、(S)-カリコトミン等、生理活性を示す化合物が多く、医薬品や農薬の合成中間体として有用な化合物群である(非特許文献1)。
【0003】
1−置換テトラヒドロイソキノリン誘導体の製造方法としては、2−(3−ヒドロキシフェニル)エチルアミン誘導体と、ケトンやアルデヒド化合物を、リン酸やカルシウム存在下で縮合する方法(Pictet−Spengler反応)が知られている。しかし、この方法では、ラセミ体の1−置換テトラヒドロイソキノリン誘導体しか得られない(非特許文献2、非特許文献3)。
【0004】
一方、光学活性な1−置換テトラヒドロイソキノリン誘導体の製造方法としては、プロキラルな1−置換3,4−ジヒドロイソキノリンの不斉水素化反応や、3,4−ジヒドロイソキノリン類の不斉アルキル化、3−(2−フェニルエチルアミン)−2−プロペン−1−オール誘導体の不斉分子内アリル位アミノ化反応による方法が知られている。しかし、これらの方法では、高価な希少金属錯体の触媒や多段階の反応が必要であり、効率的、経済的な製造方法とは言えない(非特許文献4、非特許文献5、特許文献1)。
【0005】
また、1−置換テトラヒドロイソキノリン誘導体の生合成メカニズムが植物において明らかになっており、ノルコクラウリンシンターゼ(以下、NCS)が初発反応を触媒することが知られている。しかし、NCSの作用する基質は、ドーパミンと4−ヒドロキシフェニルアセトアルデヒド、及び一部の限られた代替基質のみであり(非特許文献6、非特許文献7、非特許文献8)、基質特異性が非常に厳密で、非天然の基質に対する寛容性は低いと考えられていた(非特許文献2)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2005−41814
【非特許文献】
【0007】
【非特許文献1】Nat.Prod.Rep.(1989),6,405−432
【非特許文献2】Chem.Commun. (2011), 47, 3242
【非特許文献3】J.Org.Chem.(2010),75,8542
【非特許文献4】J.Am.Chem.Soc.(1986),108,7117
【非特許文献5】Chem.Lett.(1997),59
【非特許文献6】J.Biol.Chem.(2002),277,33878
【非特許文献7】Biochemistry(2007),46,10153
【非特許文献8】J.Biol.Chem.(2007),282,6274
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の課題は、酵素反応により光学活性1−置換テトラヒドロイソキノリン誘導体の効率的な製造方法を提供することである。
【課題を解決するための手段】
【0009】
本発明者らは、NCSの基質としては困難と考えられていた、アルキル側鎖の長い芳香族アルデヒドやアルキルアルデヒドと、2−(3−ヒドロキシフェニル)エチルアミン誘導体にNCSを作用させることで、立体選択的に光学活性1−置換テトラヒドロイソキノリン誘導体を製造できることを見出し、本発明を完成するに至った。
【0010】
すなわち、本発明は
一般式(1):
【0011】
【化1】
【0012】
(式中、R1〜R5は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、ヒドロキシル基、アミノ基、またはニトロ基である。)で表される2−(3−ヒドロキシフェニル)エチルアミン誘導体と一般式(2):
【0013】
【化2】
【0014】
(式中、R6〜R8は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、アミノ基、またはニトロ基であるか、もしくは、R6〜R10が互いに結合し環を形成しても良い。ただし、R6〜R8のいずれか2つが水素原子であり、1つが置換されていても良いフェニル基である場合は除く)で表されるアルデヒド化合物に酵素を作用させることを特徴とする、一般式(3):
【0015】
【化3】
【0016】
(式中R1〜R8は前記と同じ、*は不斉炭素を表す)で表される光学活性1−置換テトラヒドロイソキノリン誘導体の製造方法である。
【発明の効果】
【0017】
本発明により、光学活性1−置換テトラヒドロイソイソキノリン誘導体の効率的、な製造方法を提供することができる。
【図面の簡単な説明】
【0018】
【図1】本発明の実施例6に係る遺伝子組換え生産したN末端削除NCSの電気泳動写真を示す図である。
【発明を実施するための形態】
【0019】
以下、本発明について詳述する。
【0020】
本発明の酵素反応の基質となる2−(3−ヒドロキシフェニル)エチルアミン誘導体は一般式(1):
【0021】
【化4】
【0022】
(式中、R1〜R5は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、ヒドロキシル基、アミノ基、またはニトロ基である。)である。好ましくは前記R1〜R3及びR5が水素であり、R4が水素原子、メチル基、水酸基、メトキシ基のいずれかである。最も好ましくはR1〜R3及びR5が水素であり、R4が水酸基である。
【0023】
また、本発明の酵素反応のもう1つの基質であるアルデヒド化合物は、
一般式(2):
【0024】
【化5】
【0025】
(式中、R6〜R8は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、アミノ基、またはニトロ基であるか、もしくは、R6〜R10が互いに結合し環を形成しても良い。ただし、R6〜R8のいずれか2つが水素原子であり、1つがフェニル基である場合は除く)である。好ましくは前記R6が水素原子、R7が水素原子または直鎖状または分岐鎖状の炭素数1〜10のアルキル基、R8が直鎖状または分岐鎖状の炭素数2〜10のアルキル基、置換されていても良い炭素数7〜17のアラルキル基、置換されていても良い炭素数1〜10のアルコキシ基である。さらに好ましくは下記(1)〜(10)のいずれかで表される化合物である。
(1)前記R6及びR7が水素原子、R8がエチル基;
(2)前記R6が水素原子、R7がメチル基、R8がメチル基;
(3)前記R6及びR7が水素原子、R8がn−プロピル基;
(4)前記R6及びR7が水素原子、R8がn−ブチル基;
(5)前記R6及びR7が水素原子、R8がn−ペンチル基;
(6)前記R6及びR7が水素原子、R8がn−プロピルアルデヒド基;
(7)前記R6が水素原子、R7がメチル基、R8がフェニル基;
(8)前記R6及びR7が水素原子、R8がベンジル基;
(9)前記R6及びR7が水素原子、R8が4−(トリフルオロメチル)ベンジル基;
本発明では酵素を触媒とした不斉Pictet−Spengler反応により、前記一般式(1):
【0026】
【化6】
【0027】
で表される2−(3−ヒドロキシフェニル)エチルアミン誘導体と前記一般式(2):
【0028】
【化7】
【0029】
で表されるアルデヒド化合物を縮合し、一般式(3):
【0030】
【化8】
【0031】
で表される光学活性1−置換テトラヒドロイソイソキノリン誘導体を合成する。
【0032】
Pictet−Spengler反応は、β−アリールエチルアミンとアルデヒド化合物を縮合閉環させる反応である。有機合成反応では補助剤やキラルな酸触媒を利用した不斉Pictet−Spengler反応が知られている(J.Org.Lett.(2000), 2,1955、J.Am.Chem.Soc.(2006)128,1086)。
【0033】
なお、ここでいう「光学活性」とは、考えられる各種光学異性体のうち、ひとつの光学異性体の光学純度が高いことをいう。
【0034】
光学純度は高速液体クロマトグラフィーやガスクロマトグラフィーによって光学異性体を分析し、そのピーク面積から下記式より求めることができる。
【0035】
一般式(3)で示される光学異性体の(S)体が主生成物である場合:
((S)体の光学純度(%e.e.)={((S)体のピーク面積)−((R)体ピーク面積)}÷{(S)体のピーク面積)+((R)体ピーク面積)}×100
本発明では前記一般式(3)で表される光学活性1−置換テトラヒドロイソイソキノリン誘導体においては、光学純度は50%e.e.以上が好ましく、より好ましくは75%e.e.以上、さらに好ましくは90%e.e.以上である。
【0036】
なお、J.Biol.Chem.(2009)284,897に示される触媒機構から、NCSによって生成される光学活性1−置換テトラヒドロイソキノリン誘導体の絶対立体配置はS体と予想される。
【0037】
前記酵素としては、前記反応を触媒する酵素であればいかなる酵素を用いても良いが、好ましくは国際生化学・分子生物学連合の酵素分類法によりEC4.2.1.78に分類されるノルコクラウリンシンターゼ(NCS)、もしくはPathogenesis−Related 10/Bet V1 Protein Familyに属する酵素(以下、PR10タンパク質)である。PR10タンパク質は生体防御タンパク質ファミリーの1つであり、アミノ酸配列の相同性や血清学的・免疫学的な相関性、酵素活性の特徴に基づいて17種類に分類されたうちの1つである(Plant Mol.Biol.Report.(1994),12,245、Physiol.Mol. Plant Pathol. (1999),55,85、Curr.Opin.Immunol.(1998) ,10,16)。PR10タンパク質にはアレルギー抗原であるBet V1やリボヌクレアーゼ様タンパク質の構造類似タンパク質が含まれる。
【0038】
NCS及びPR10タンパク質は植物中から単離することが可能である。NCS及びPR10タンパク質の由来となる植物は特に限定されないが、アルカロイドを体内に蓄積するアマリリダセエ(Amaryllidaceae)科、アリストロチセエ(Aristolochiaceae)科、ベルベリダセエ(Berberidaceae)科、ユーホルビアセエ(Euphorbiaceae)科、ファバセエ(Fabaceae)科、リリアセエ(Liliaceae)科、パパベラセエ(Papaveraceae)科、ラヌンチュラセエ(Ranunculaceae)科、ソラナセエ(Solanaceae)科の植物が好ましい。特に好ましくはラヌンチュラセエ(Ranunculaceae)科のコプティス・ジャポニカ(Coptis japonica)、タリクトラム・フラバム(Thalictrum flavum)及びパパベラセエ(Papaveraceae)科のパパヴェラ・ソムニフェラム(Papaver somniferum)パパヴェラ・ブラクテアタム(Papaver bracteatum)である。また、酵素を単離せずに、植物体もしくは植物培養細胞をそのまま利用することも可能である。具体的にはJ.Biol.Chem.(2007),282,6274に記載の方法で植物培養細胞を得ることができる。
【0039】
ここで、NCS及びPR10タンパク質の酵素活性は以下のようにして検出することが可能である。例えば、ドーパミン塩酸塩10mM、ジメチルホルムアミドに溶解したヒドロキシシンナムアルデヒド10mMを添加した100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に、NCSもしくはPR10が含まれる溶液(例えば、該酵素を生産した組換え生物や植物の精製物など)を加え、30℃で15分間反応を行う。反応後、下記溶離液を添加して反応を停止し、遠心後、上清を下記分析条件で分析して1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの生成量を定量する。
【0040】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 555ml、蒸留水 945ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 11.5分
【0041】
NCS及びPR10タンパク質のアミノ酸配列は、その上記の方法で検出している酵素活性を有している限り特に限定されないが、好ましくは配列表の配列番号1、2、3、4、5のいずれかで表されるポリペプチドと好ましくは60%以上、さらに好ましくは80%以上、90%以上、95%以上、97%以上、98%以上、最も好ましくは99%以上の配列同一性を示すタンパク質である。
【0042】
配列表の配列番号1、2、3、4、5のアミノ酸配列をコードするDNA配列を利用して、前記の植物から配列相同性の高いDNAを取得し、その生産物を本発明に利用することもできる。具体的にはJ.Biol.Chem.(2007),282,6274に記載の方法で取得することができる。また、ゲノム情報が解析され、塩基配列が明らかになっている場合は、DNAを人工合成することも可能である。
【0043】
本明細書におけるポリペプチドやポリヌクレオチドの「配列同一性」とは、対比される2つのポリペプチドまたはポリヌクレオチドを最適に整列させ、核酸塩基(例えば、A、T、C、G、U、またはI)が両方の配列で一致した位置の数を比較塩基総数で除し、そして、この結果に100を乗じた数値で表される。
【0044】
配列同一性は、例えば、以下の配列分析用ツールを用いて算出し得る;GCG Wisconsin Package(ウィスコンシン大学)、the ExPASy World Wide Web分子生物学用サーバー(スイスバイオインフォマティックス研究所)、BLAST(米国生物工学情報センター)、GENETYX(ゼネティックス社)。
【0045】
また、本発明に利用するNCS及びPR10タンパク質は、目的のNCS活性を有している限りにおいては、そのアミノ酸配列(例えば、配列表の配列番号1〜5)の1若しくは複数個(例えば、40個、好ましくは20個、より好ましくは15個、さらに好ましくは10個、さらに好ましくは5個、4個、3個、または2個以下)のアミノ酸が置換、挿入、欠失及び/または付加されたポリペプチドであってもよい。また、目的のNCS活性を有する限り、本発明に利用するNCS及びPR10のアミノ酸配列に、付加的なアミノ酸配列を結合してもよい。たとえば、ヒスチジンタグやHAタグのような、タグ配列を付加することができる。あるいは、他のタンパク質との融合タンパク質とすることもできる。また、目的のNCS活性を有する限り、ペプチド断片であってもよい。
【0046】
また、本発明に利用するNCS及び/またはPR10タンパク質は下記に示す方法で、適当な宿主生物内で本発明に利用するNCS及び/またはPR10タンパク質をコードする遺伝子を発現し、遺伝子組換え生産して利用することもできる。
【0047】
NCS及び/またはPR10タンパク質のN末端部分にはシグナル配列が存在する場合があるため、宿主生物内で遺伝子組換え生産する場合はこのシグナル配列を一部もしくは全て削除することが好ましい。
【0048】
NCS及び/またはPR10タンパク質を遺伝子組換え生産する場合、宿主生物中で不溶化し、酵素活性が低下することがある。本発明では配列表の配列番号1に示すアミノ酸配列のうちN末端のアミノ酸を10個削除し、メチオニンを付加した場合(実施例4のE. coli HB101(pNΔ10NCS)が生産するΔ10NCS)はNCSの大部分が不溶化するが、N末端をさらに削除し、メチオニンを付加したポリペプチドを遺伝子組換え生産することで、不溶化が低減され、酵素活性が向上することを見出した(例えば実施例4のE. coli HB101(pNΔ19NCS)が生産するΔ19NCS)。削除するアミノ酸の数としては、好ましくは11〜41個、さらに好ましくは19〜29個である。最も好ましくは19個もしくは29個である。
【0049】
本発明においては、NCS及びPR10タンパク質をコードするDNAを含む形質転換体を使用すると、より効率的に光学活性1−置換テトラヒドロイソキノリンを製造することができる。なお、本明細書において記述されている、DNAの単離、ベクターの調製、形質転換等の遺伝子操作は、特に明記しない限り、Molecular Cloning 2nd Edition(Cold Spring Harbor Laboratory Press,1989)、Current Protocols in Molecular Biology(Greene Publishing Associates and Wiley−Interscience)等の成書に記載されている方法により実施できる。
【0050】
上記の形質転換体に用いるベクターとしては、適当な宿主生物内で本発明に利用するNCS及びPR10タンパク質をコードする遺伝子を発現できるものであれば、特に限定されない。このようなベクターとしては、例えば、プラスミドベクター、ファージベクター、コスミドベクターなどが挙げられ、さらに、他の宿主株との間での遺伝子交換が可能なシャトルベクターも使用できる。
【0051】
このようなベクターは、例えば大腸菌の場合では、通常、lacUV5プロモーター、trpプロモーター、trcプロモーター、tacプロモーター、lppプロモーター、tufBプロモーター、recAプロモーター、pLプロモーター等の制御因子を含み、本発明のDNAと作動可能に連結された発現単位を含む発現ベクターとして好適に使用できる。例えば、pSTV28(タカラバイオ社製)、pUCNT(国際特許公報WO94/03613)などが挙げられる。
【0052】
なお、「制御因子」とは、機能的プロモーター、及び、任意の関連する転写要素(例えばエンハンサー、CCAATボックス、TATAボックス、SPI部位など)を有する塩基配列をいう。
【0053】
また、「作動可能に連結」とは、遺伝子の発現を調節するプロモーター、エンハンサー等の種々の調節エレメントと遺伝子が、宿主細胞中で作動し得る状態で連結されることをいう。なお、制御因子のタイプ及び種類が宿主に応じて変わり得ることは、当業者に周知の事項である。
【0054】
各種生物において利用可能なベクター、プロモーターなどについては、「微生物学基礎講座8遺伝子工学」(共立出版、1987)などに詳細に記述されている。
【0055】
各酵素を発現させるために用いる宿主生物は、各酵素をコードするDNAを含む酵素発現ベクターにより形質転換され、DNAを導入した酵素を発現することができる生物であれば、特に制限するものではない。利用可能な微生物としては、例えば、エシェリヒア(Escherichia)属、バシラス(Bacillus)属、シュードモナス(Pseudomonas)属、セラチア(Serratia)属、ブレビバクテリウム(Brevibacterium)属、コリネバクテリイウム(Corynebacterium)属、ストレプトコッカス(Streptococcus)属、及びラクトバシラス(Lactobacillus)属など宿主ベクター系の開発されている細菌、ロドコッカス(Rhodococcus)属及びストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌、サッカロマイセス(Saccharomyces)属、クライベロマイセス(Kluyveromyces)属、シゾサッカロマイセス(Schizosaccharomyces)属、チゴサッカロマイセス(Zygosaccharomyces)属、ヤロウイア(Yarrowia)属、トリコスポロン(Trichosporon)属、ロドスポリジウム(Rhodosporidium)属、ピキア(Pichia)属、及びキャンディダ(Candida)属などの宿主ベクター系の開発されている酵母、ノイロスポラ(Neurospora)属、アスペルギルス(Aspergillus)属、セファロスポリウム(Cephalosporium)属、及びトリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ、などが挙げられる。また、微生物以外でも、植物、動物において様々な宿主・ベクター系が開発されており、特に蚕を用いた昆虫(Nature,315,592−594(1985))や菜種、トウモロコシ、ジャガイモなどの植物中に大量に異種タンパク質を発現させる系が開発されており、好適に利用できる。これらのうち、導入及び発現効率から細菌が好ましく、大腸菌(エシェリヒア・コリ(Escherichia coli)が特に好ましい。
【0056】
本発明に利用するNCS及び/またはPR10をコードするDNAを含むベクターは、公知の方法により宿主微生物に導入することができる。例えば、宿主微生物として大腸菌を用いる場合は、市販のエシェリヒア・コリ(Escherichia coli HB101(以下、E. coli HB101)コンピテントセル(タカラバイオ社製)を用いることにより、当該ベクターを宿主細胞に導入することができる。
【0057】
NCS及び/またはPR10をコードするDNAを含むベクターとしては、実施例3に記載のpNNCSが挙げられる。また、NCS及び/またはPR10をコードするDNAを含む形質転換体の例としては、ベクターpNNCSでE.coli HB101を形質転換して得られる、実施例4に記載のE.coli HB101(pNNCS)が挙げられる。
【0058】
形質転換体の培養には、通常、これらの宿主微生物が資化しうる栄養源を含む培地であればどのようなものでも使用しうる。例えば、グルコース、シュークロース、マルトース等の糖類、乳酸、酢酸、クエン酸、プロピオン酸等の有機酸類、エタノール、グリセリン等のアルコール類、パラフィン等の炭化水素類、大豆油、菜種油等の油脂類、またはこれらの混合物等の炭素源;硫酸アンモニウム、リン酸アンモニウム、尿素、酵母エキス、肉エキス、ペプトン、コーンスチープリカー等の窒素源;更に、その他の無機塩、ビタミン類等の栄養源;を適宜混合・配合した通常の培地を用いることが出来る。これら培地は用いる微生物の種類によって適宜選択すればよい。
【0059】
微生物の培養は、一般的な条件により行なうことができる。例えば、pH4.0〜9.5、温度範囲20℃〜45℃の範囲で、好気的に10〜96時間培養するのが好ましい。
【0060】
次に、酵素を用いた2−(3−ヒドロキシフェニル)エチルアミン誘導体とアルデヒド化合物の縮合反応について説明する。
【0061】
酵素を用いた縮合反応の際には、適当な溶媒と基質の2−(3−ヒドロキシフェニル)エチルアミン誘導体とアルデヒド化合物、前記酵素、前記形質転換体、その培養物、またはその処理物等を混合し、pH調整下に攪拌する。
【0062】
反応溶媒としては、通常、水や緩衝液等の水性媒体を用いる。緩衝液としては、リン酸カリウム緩衝液やトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液が挙げられる。本反応においては、非酵素的な縮合反応によりラセミ体の1−置換テトラヒドロイソキノリンが生成するため、酵素反応によって得られる光学活性1−置換テトラヒドロイソキノリンの光学純度が低下する。Chem.Commun. (2011), 47, 3242に記載されているようにリン酸緩衝液中ではこの非酵素的な縮合反応が促進されるため、不適である。一方、トリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液中では非酵素的な縮合反応の影響が少ないため、好適である。
【0063】
酵素は、通常、上記微生物の菌体を含んだ培養液をそのまま反応に使用する。培養液を濃縮して用いてもよい。また、培養液中の成分が反応に悪影響を与える場合には、培養液を遠心分離等により処理して得られる菌体または菌体処理物を使用するとよい。
【0064】
基質である2−(3−ヒドロキシフェニル)エチルアミン誘導体とアルデヒド化合物は、反応の初期に一括添加してもよく、反応の進行にあわせて逐次分割して添加してもよい。高濃度の基質存在下では非酵素的な縮合反応が促進されるため、基質の逐次分割添加が特に好適である。
【0065】
反応時の温度は、通常、10〜60℃、好ましくは20〜40℃とする。
【0066】
反応時のpHは2.5〜9、好ましくはpH5〜9の範囲である。
【0067】
反応液中の微生物の量は、これらの基質を縮合する能力に応じ適宜決定すればよく、菌体湿重量として、0.01〜50%(W/V)が好ましく、より好ましくは0.1〜10%(W/V)である。
【0068】
反応液中の基質濃度は0.01〜50%(W/V)が好ましく、より好ましくは、0.1〜30%(W/V)である。
【0069】
反応は、通常、振とうまたは通気攪拌しながら行なう。
【0070】
反応時間は、基質濃度、微生物の量及びその他の反応条件により適宜決定される。通常、2〜168時間で反応が終了するように各条件を設定することが好ましい。
【0071】
更に、トリトン(ナカライテスク株式会社製)、スパン(関東化学株式会社製)、ツイーン(ナカライテスク株式会社製)などの界面活性剤を反応液に添加することも効果的である。
【0072】
更に、基質及び/または縮合反応の生成物による反応の阻害を回避する目的で、酢酸エチル、酢酸ブチル、イソプロピルエーテル、トルエン、ヘキサンなどの水に不溶な有機溶媒を反応液に添加してもよい。
【0073】
更に、基質の溶解度を高める目的で、メタノール、エタノール、アセトン、テトラヒドロフラン、ジメチルスルホキシドなどの水に可溶な有機溶媒を添加することもできる。
【0074】
縮合反応により生成した光学活性1−置換テトラヒドロイソキノリンを取り出す方法は特に限定されないが、反応液から直接、あるいは菌体等を分離後に、酢酸エチル、トルエン、t−ブチルメチルエーテル、ヘキサン、n−ブタノール、ジクロロメタン等の溶剤で抽出し、脱水後、蒸留や結晶化操作、シリカゲルカラムクロマトグラフィー等により精製することにより、高純度の光学活性1−置換テトラヒドロイソキノリンを容易に得ることができる。
【実施例】
【0075】
以下、実施例で本発明を詳細に説明するが、本発明はこれらにより限定されるものではない。なお、以下の記載において、「%」は特に断らない限り「重量%」を意味する。また、以下の実施例において用いた組み換えDNA技術に関する詳細な操作方法などは、次の成書に記載されている:
Molecular Cloning 2nd Edition(Joseph Sambrook,Cold Spring Harbor Laboratory Press(1989))、Current Protocols in Molecular Biology(Frederick M.Ausubel,Greene Publishing Associates and Wiley−Interscience(1989))。
【0076】
(参考例1)1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの合成
1−(2−フェニルエチル)−6,7−ジメトキシ−3,4−ジヒドロイソキノリン1.48g(5mmol)のメタノール溶液10mlを5℃に冷却した。ここに、水素化ホウ素ナトリウム189mg(5mmol)を添加して1時間撹拌し、更に25℃まで昇温して2時間撹拌した。別の容器に濃塩酸2.61g(25mmol)と水10mlを加えて5℃に冷却し、ここに反応液をゆっくりと加えた。30wt%水酸化ナトリウム水溶液でpH=12に調整後、減圧下にメタノールを留去した。残渣に酢酸エチル20mlを加えて抽出し、有機層を更に水10mlで洗浄した。減圧濃縮することにより、1−(2−フェニルエチル)−6,7−ジメトキシ−1,2,3,4−テトラヒドロイソキノリンを淡黄色油状物として得た(1.58g、含量:94.2%、収率は定量的)。
【0077】
続いて、前記化合物1.06g(3.37mmol)に48wt%臭化水素酸10g(17.6当量)を加えて、90℃、16時間撹拌した。室温まで冷却後、30wt%水酸化ナトリウム水溶液でpH=7に調整し、水20mLと酢酸エチル10mlを加えた。5℃まで冷却して30分撹拌後、析出している結晶を減圧濾別した。結晶を水10gに懸濁して25℃、30分撹拌後、減圧濾別し、水10gで洗浄、50℃で真空乾燥することにより、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン520.6mgを得た(淡褐色結晶、含量:92.1%)下記分析条件で分析し、ラセミ体のそれぞれのピークを確認した。
1H NMR(CDCl3):δ1.86(2H,m)、2.1−2.7(4H,m)、3.02(1H,m)、3.27(1H,m)、3.5−4.4(2H,brs)、3.91(1H,m)、6.57(1H,s)、6.60(1H,s)、7.1−7.3(5H,m)
【0078】
<高速液体クロマトグラフィー分析条件> 光学純度分析
カラム:ダイセル化学工業株式会社製CHIRALPAK AD−H(250mm×4.6mm)
溶離液:n−ヘキサン/エタノール/ジエチルアミン=80/20/0.1
流速:1ml/min
検出:230nm
カラム温度:30℃
検出時間:1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 11.5分、14.4分
【0079】
(実施例1)NCSをコードするDNAの調製
配列表の配列番号1のアミノ酸配列で示されるコプティス・ジャポニカ(Coptis japonica)由来のNCS(CjPR10A)をコードする配列番号6のDNAを人工合成した。この合成DNAを鋳型としてプライマー1:5’-GGAGTCCATATGCGTATGGAAGTGGTTCT-3’(配列表の配列番号7)、プライマー2:5’-TATATACTGCAGTTATTCGGAAGATTTGTG -3’(配列表の配列番号8)を用いてPCRを行った。その結果、配列表の配列番号6に示す塩基配列からなる遺伝子の開始コドン部分にNdeI認識部位が付加され、かつ終始コドンの直後にPstI認識部位が付加された二本鎖DNA(NCS遺伝子)が得られた。PCRは、DNAポリメラ−ゼとして、PrimeSTAR HS DNA polymerase(タカラバイオ社製)を用いて行い、反応条件はその取り扱い説明書に従った。
【0080】
(実施例2)N末端配列を削除したNCSをコードするDNAの調製
NCSのN末端アミノ酸配列を10、19、29、42個削除し、N末端にメチオニンを付加したポリペプチドをコードするDNAを調製した(以下、それぞれΔ10NCS、Δ19NCS、Δ29NCS、Δ42NCS遺伝子と呼ぶ)。実施例1で増幅したDNA(NCS遺伝子)を鋳型としてプライマー3:5’-GGAGTCCATATGCTGATGTTCATTGGCACC-3’(配列表の配列番号9、Δ10NCS増幅用)、プライマー4:5’-GGAGTCCATATGGAACGCCTGATTTTTAA-3’(配列表の配列番号10、Δ19NCS増幅用)、プライマー5:5’-GGAGTCCATATGCTGCATCGCGTTACGAAA-3’(配列表の配列番号11、Δ29NCS増幅用)、プライマー6:5’- GGAGTCCATATGCACGAACTGGAAGTGGCG -3’(配列表の配列番号12、Δ43NCS増幅用)のいずれか1つとプライマー2:5’- TATATACTGCAGTTATTCGGAAGATTTGTG -3’(配列表の配列番号8)を用いてPCRを行った。その結果、配列表の配列番号1に示すアミノ酸配列からなるポリペプチドのN末端部分のアミノ酸から10、19、29、42残基削除されて、かつN末端にメチオニンが付加されたポリペプチドをコードする二本鎖DNA(配列開始コドン部分にNdeI認識部位が付加され、かつ終始コドンの直後にPstI認識部位が付加された)が4種類(Δ10NCS、Δ19NCS、Δ29NCS、Δ43NCS遺伝子)得られた。
【0081】
(実施例3)組換えベクターの調製
実施例1及び2で得られた5種類のNCS遺伝子をNdeI及びPstIで消化し、プラスミドpUCN18(PCR法によりpUC18(タカラバイオ社製)の185番目のTをAに改変してNdeIサイトを破壊し、更に471−472番目のGCをTGに改変することにより新たにNdeIサイトを導入したプラスミド)のlacプロモーターの下流のNdeI認識部位とPstI認識部位の間に挿入し、組換えベクターpNNCS、pNΔ10NCS、pNΔ19NCS、pNΔ29NCS、pNΔ42NCSを構築した。
【0082】
(実施例4)NCSを生産する組換え大腸菌の育種
実施例3で構築した5種類の組換えベクターを用いて、E. coli HB101コンピテントセル(タカラバイオ社製)を形質転換し、5種類の組換え生物E. coli HB101(pNNCS)、E. coli HB101(pNΔ10NCS)、E. coli HB101(pNΔ19NCS)、E. coli HB101(pNΔ29NCS)、E. coli HB101(pNΔ42NCS)を得た。また、pUCN18を用いてE. coli HB101コンピテントセル(タカラバイオ社製)を形質転換し、組換え生物E. coli HB101(pUCN18)を得た。
【0083】
(実施例5) 組換え生物におけるDNAの発現
実施例4で得た6種類の組換え生物(E. coli HB101(pUCN18)、E. coli HB101(pNNCS)、E. coli HB101(pNΔ10NCS)、E. coli HB101(pNΔ19NCS)、E. coli HB101(pNΔ29NCS)、E. coli HB101(pNΔ42NCS)を、200μg/mlのアンピシリンを含む2×YT培地(トリプトン1.6%、イーストエキス1.0%、塩化ナトリウム 0.5%、pH7.0)5mlに接種し、30℃で24時間振盪培養した。上記の培養で得られたそれぞれ培養液1mlについて、遠心分離により菌体を集め、1mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁した。これを、UH−50型超音波ホモゲナイザー(SMT社製)を用いて破砕した後、遠心分離により菌体残渣を除去し、無細胞抽出液を得た。これらの無細胞抽出液のNCS活性を測定した。
【0084】
NCS活性は、100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に、ドーパミン塩酸塩10mM、ジメチルホルムアミドに溶解したヒドロキシシンナムアルデヒド10mM、および無細胞抽出液を添加して30℃で15分間反応を行い、下記溶離液を添加して反応を停止し、遠心後、上清を下記分析条件で分析し、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの生成量を定量した。この反応条件において、1分間に1μmolの1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを生成する酵素活性(NCS活性)を1Uと定義した。
【0085】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 555ml、蒸留水 945ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 11.5分
それぞれの組換え生物のNCS活性を以下に示す。 E. coli HB101(pUCN18)については、NCS活性は0.01U/mg以下であった。一方、NCSを発現させた組換え生物のNCS活性はそれぞれ、E. coli HB101(pNNCS):0.03U/mg、E. coli HB101(pNΔ10NCS):0.1U/mg、E. coli HB101(pNΔ19NCS):0.3U/mg、E. coli HB101(pNΔ29NCS):1.2U/mg、E. coli HB101(pNΔ42NCS):0.1U/mgであった。N末端アミノ酸を19または29残基削除したNCSでは高いNCS活性を示した。
【0086】
(実施例6) N末端アミノ酸を削除したNCSの可溶化の確認
実施例4で作製したNCSを生産する組換え大腸菌5種類について、実施例5と同様の方法で無細胞抽出液を調製した。これを可溶性画分とした。また、遠心分離後の菌体残渣を1mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁し、これを不溶性画分とした。各組換え生物の可溶性、不溶性画分についてドデシル硫酸ナトリウム(SDS)ポリアクリルアミドゲル電気泳動を行った。泳動後、クマシーブリリアントブルー染色によって可溶性画分のNCSを確認した。結果を図1に示す。
【0087】
シグナル配列を含むNCS全長を生産するE. coli HB101(pNNCS)では高生産されているタンパク質が確認できなかった。一方、E. coli HB101(pNΔ10NCS)ではΔ10NCSが高生産されいていることが確認できた。しかし、大部分のΔ10NCSは不溶性画分に確認された。E. coli HB101(pNΔ19NCS)及びE. coli HB101(pNΔ29NCS)ではE. coli HB101(pNΔ10NCS)に比べ可溶性画分のN末端削除NCSが増加していた。E. coli HB101(pNΔ42NCS)では高生産されているタンパク質が確認できなかった。
【0088】
N末端アミノ酸を19もしくは29個削除したNCSでは遺伝子組換え生産時の不溶化が低減できた。
【0089】
(実施例7)光学活性1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ10NCS)について実施例5と同様の方法で無細胞抽出液1mlを調製した。この無細胞抽出液にドーパミン塩酸塩10mM、ヒドロシンナムアルデヒド10mMを添加し、30℃で30分反応した。また、対照実験として、無細胞抽出液の代わりに100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)でも同様に反応を実施した。反応液から酢酸エチルにて生成した1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを抽出し、参考例1及び実施例5に示した分析条件で分析した。その結果、1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンへの変換率は11%、であり、参考例1の分析条件において検出時間11.5分の光学活性体が生成しており、その光学純度は96.1%e.e.であった。一方、対照実験では1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンへの変換率は1%であり、ラセミ体が生成していた。
【0090】
E. coli HB101(pNΔ10NCS)が高い立体選択性で光学活性1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを生成することが分かった。
【0091】
(実施例8)光学活性−1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ10NCS)について実施例5と同様の方法で無細胞抽出液20mlを調製した。この無細胞抽出液にドーパミン塩酸塩0.25gを添加し、攪拌しながら、ヒドロシンナムアルデヒド0.21gを逐次添加した。16時間後の反応液を実施例5及び実施例7記載の分析条件で分析したところ、変換率は58%、光学純度は76.3%e.e.(検出時間11.5分の光学活性体)であった。
【0092】
(実施例9)光学活性−1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ29NCS)について実施例5と同様の方法で無細胞抽出液20mlを調製した。この無細胞抽出液にドーパミン塩酸塩0.25gを添加し、攪拌しながら、ヒドロシンナムアルデヒド0.21gを逐次添加した。16時間後の反応液を実施例5及び実施例7記載の分析条件で分析したところ、変換率は86%、(S)−1−(2−フェニルエチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの光学純度は95.3%e.e.(検出時間11.5分の光学活性体)であった。
【0093】
(実施例10)3−(4−トリフルオロメチルフェニル)プロピルアルデヒドの製造
3−(4−トリフルオロメチルフェニル)プロピオン酸8.33g(37.4mmol)にTHF 40mlを加え、−10℃に冷却し、1.03M BH3−THF 42.3ml(43.6mmol)を添加した(内温推移−7〜0℃)。室温まで昇温し、13時間撹拌した。反応液に水40mlと飽和炭酸水素ナトリウム水溶液20ml、酢酸エチル40ml、飽和食塩水20mlを添加し、分液した。有機層に水40mlと飽和食塩水20mlを加え、撹拌後、分液した。有機層を減圧濃縮することで3−(4−トリフルオロメチルフェニル)プロパノール8.27gを得た。ここで得た3−(4−トリフルオロメチルフェニル)プロパノール、炭酸水素ナトリウム 9.38g(0.112mol)、NaBr 3.83g(37.2mmol)、水80ml、酢酸エチル80ml、Tempo 0.291g(1.86mmol)を氷冷し、13.02%次亜塩素酸ナトリウム 21.28g(37.2mmol)を35分間で添加した。添加終了後、1.5時間撹拌し、水120mlを添加し、分液した。有機層を水40mlで2回洗浄し、減圧濃縮することで3−(4−トリフルオロメチルフェニル)プロピルアルデヒド8.08gを得た。ガスクロマトグラフィーで分析したところ純度89.7wt%、収率95.8mol%であった。
【0094】
3−(4−トリフルオロメチルフェニル)プロパノール
1H NMR(CDCl3):δ=1.87−1.94(m、2H)、2.78(t、2H、J=7.8Hz)、3.67−3.77(m、2H)、7.31(d、2H、J=8.0Hz)、7.53(d、2H、J=8.1Hz)
3−(4−トリフルオロメチルフェニル)プロピルアルデヒド
1H NMR(CDCl3):δ=2.81(d、2H、J=7.3Hz)、3.01(t、2H、J=7.6Hz)、7.30(d、2H、J=8.0Hz)、7.53(d、2H、J=8.1Hz)、9.81(s、1H)
<ガスクロマトグラフィー分析系>
カラム:Restek Corporation製Rtx−35A(30mx0.25mm)
検出器:FID
カラム温度:50℃→(17分)→220℃→(13分)→220℃
キャリアー:He 150Kpa
【0095】
(参考例2)1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
1.94mlの100mMリン酸緩衝液(pH6.5)にドーパミン塩酸塩194mg(1.0mmol)、メタノール0.5mlを添加し40℃で攪拌した。3−(4−トリフルオロメチルフェニル)プロピルアルデヒド207mg(1.0mmol)を分割添加し、4時間反応した。反応液にNaCl添加後、生成した1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンをn−ブタノールで抽出した。有機層に硫酸ナトリウムを添加後、ろ過により除去した。ろ液から溶媒を留去した後、濃縮物をトルエンで洗浄した後、有機層を除去した。これを真空乾燥し、51mgの1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン(淡黄色の固体)を得た。さらに結晶化により精製した後、下記分析条件で分析し、ラセミ体のそれぞれのピークを確認した。
1H NMR(CD3OD):δ2.07−2.19(m、2H)、2.74−2.84(m、4H)、3.09−3.12(m、1H)、3.32−3.38(m、1H)、4.12−4.13(m、1H)、6.51(s、1H)、6.57(s,1H)、7.40−7.41(d,2H)、7.54−7.59(d、2H)
【0096】
<高速液体クロマトグラフィー分析条件> 光学純度分析
カラム:ダイセル化学工業株式会社製CHIRALPAK AD−H(250mm×4.6mm)
溶離液:n−ヘキサン/エタノール/ジエチルアミン=80/20/0.1
流速:1ml/min
検出:230nm
カラム温度:30℃
検出時間:1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 9.2分、11.5分
【0097】
(実施例11)光学活性1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
E. coli HB101(pNΔ10NCS)について実施例5と同様の方法で無細胞抽出液120mlを調製した。この無細胞抽出液にドーパミン塩酸塩1.51gを添加し、攪拌しながら、4−トリフルオロメチルヒドロシンナムアルデヒド2gを逐次添加した。28時間後の反応液を下記分析条件で分析したところ、変換率は45%であった。また、参考例2に記載の分析条件で光学純度を求めたところ、検出時間9.2分の光学活性1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの光学純度が82.0%e.e.であった。
【0098】
<高速液体クロマトグラフィー分析条件> 変換率分析
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 600ml、蒸留水 900ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−(2−(4−トリフルオロメチルフェニル)エチル)−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 28.1分
【0099】
(実施例12)NCSの基質特異性の評価
組換え生物E. coli HB101(pNΔ29NCS)及びE. coliHB101(pUCN18)を実施例5と同様の方法で培養した。各培養液から遠心分離により菌体を集め、5mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁した。これを、UH−50型超音波ホモゲナイザー(SMT社製)を用いて破砕した後、遠心分離により菌体残渣を除去し、無細胞抽出液を得た。これらの無細胞抽出液1mlにドーパミン塩酸塩10mM、表1に示すアルデヒド化合物10mM(ジメチルホルムアミドに溶解して添加)を添加し、30℃で15分反応した(対照実験としてE. coli HB101(pUCN18)の無細胞抽出液についても同様に反応した)。反応終了後、下記溶離液を添加して反応を終了し、遠心後、上清を下記分析条件で分析し、ドーパミンの減少量を定量した。
【0100】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 600ml、蒸留水 900ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分
E. coli HB101(pNΔ29NCS)を用いた反応でのドーパミン減少量からE. coli HB101(pUCN18)の反応液でのドーパミン減少量を差し引いた値をNCSの触媒作用によって減少したドーパミン量と見なした。本条件において、1分間に1μmolのドーパミンが消費される酵素活性を1Uと定義した。また、フェニルアセトアルデヒドをアルデヒド化合物の標準基質とし、フェニルアセトアルデヒドに対する酵素活性を100%としたときの相対活性で各アルデヒド化合物に対する酵素活性を示した。結果を表1に示す。
NCSはフェニルアセトアルデヒドや、ヒドロシンナムアルデヒド以外にもアルキルアルデヒドに対して活性を示した。
【0101】
【表1】
【0102】
(参考例3)1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
1.94mlの100mMリン酸緩衝液(pH6.5)にドーパミン塩酸塩194mg(1.0mmol)、n−ブチルアルデヒド111mg(1.5mmol)を添加し、40℃で2時間攪拌した。薄層クロマトグラフィーによりドーパミンの消失を確認した後、生成した1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを4mlのn−ブタノールで2回抽出した。有機層に硫酸ナトリウムを添加後、ろ過により除去した。ろ液から溶媒を留去し、241mgの1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを得た(黄色の固体)。下記分析条件で分析し、ラセミ体であることを確認した。
1H NMR(CD3OD):δ1.02(t、J=7.45Hz、3H)、1.42−1.65(m、2H)、1.76−2.09(m、2H)、2.79−3.04(m、2H)、3.21(m、1H)、3.48(dt,J=12.46,6.09Hz、1H)、4.25−4.42(m,1H)、6.59(s,1H)、6.71(s、1H)
【0103】
<高速液体クロマトグラフィー分析条件>
ダイセル化学工業株式会社製CHIRALPAK AD−H(250mm×4.6mm)
溶離液:n−ヘキサン/エタノール/ジエチルアミン=80/20/0.1
流速:0.5ml/min
検出:230nm
カラム温度:30℃
検出時間:1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 33.9分、39.1分
【0104】
(実施例13)光学活性1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンの製造
組換え生物E. coli HB101(pNΔ29NCS)を実施例5と同様の方法で培養した。培養液から遠心分離により菌体を集め、1mlの100mMトリス(ヒドロキシメチル)アミノメタン−塩酸緩衝液(pH7)に懸濁した。これを、UH−50型超音波ホモゲナイザー(SMT社製)を用いて破砕した後、遠心分離により菌体残渣を除去し、無細胞抽出液を得た。この無細胞抽出液1mlに19.4mgのドーパミン塩酸塩を加え、攪拌しながらn−ブチルアルデヒド21mgを分割添加した(反応時間1.5時間)。反応液を下記分析条件で分析したところ、変換率は99.6%であった。
【0105】
<高速液体クロマトグラフィー分析条件>
カラム:日本分光工業株式会社製Finepak SIL C18T−5(250mm×4.6mm)
溶離液:アセトニトリル 600ml、蒸留水 900ml、リン酸二水素カリウム 4.75g、ドデシル硫酸ナトリウム 2.16g(リン酸にてpH3.5に調整)
流速:1ml/min
検出:230nm
カラム温度:40℃
検出時間:ドーパミン4.8分、1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリン 7.1分
反応終了後、n−ブタノールで生成物を抽出し、溶媒を留去して20mgの1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを得た。参考例3に示した分析方法で分析したところ、検出時間33.9分の光学活性体の光学純度が98.0%e.e.であった。
【0106】
NCSを用いて光学活性1−プロピル−6,7−ジヒドロキシ−1,2,3,4−テトラヒドロイソキノリンを合成できた。
【特許請求の範囲】
【請求項1】
一般式(1):
【化1】
(式中、R1〜R5は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、ヒドロキシル基、アミノ基、またはニトロ基である。)で表される2−(3−ヒドロキシフェニル)エチルアミン誘導体と一般式(2):
【化2】
(式中、R6〜R8は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、アミノ基、またはニトロ基であるか、もしくは、R6〜R8が互いに結合し環を形成しても良い。ただし、R6〜R8のいずれか2つが水素原子であり、1つがフェニル基である場合は除く)で表されるアルデヒド化合物に酵素を作用させることを特徴とする、一般式(3):
【化3】
(式中R1〜R8は前記と同じ、*は不斉炭素を表す)で表される光学活性1−置換テトラヒドロイソキノリン誘導体の製造方法。
【請求項2】
前記R1〜R3及びR5が水素であり、R4が水素原子、メチル基、水酸基、メトキシ基のいずれかである、請求項1記載の製造方法。
【請求項3】
前記R6が水素原子、R7が水素原子または直鎖状または分岐鎖状の炭素数1〜10のアルキル基、R8が直鎖状または分岐鎖状の炭素数2〜10のアルキル基、置換されていても良い炭素数7〜17のアラルキル基、置換されていても良い炭素数1〜10のアルコキシ基である、請求項1または2記載の製造方法。
【請求項4】
前記R6〜R8が下記(1)〜(10):
(1)前記R6及びR7が水素原子、R8がエチル基、
(2)前記R6が水素原子、R7がメチル基、R8がメチル基、
(3)前記R6及びR7が水素原子、R8がn−プロピル基、
(4)前記R6及びR7が水素原子、R8がn−ブチル基、
(5)前記R6及びR7が水素原子、R8がn−ペンチル基、
(6)前記R6及びR7が水素原子、R8がn−プロピルアルデヒド基、
(7)前記R6が水素原子、R7がメチル基、R8がフェニル基、
(8)前記R6及びR7が水素原子、R8がベンジル基、
(9)前記R6及びR7が水素原子、R8が4−(トリフルオロメチル)ベンジル基、
(10)前記R6が水素原子、R7がメチル基、R8が 4−(イソプロピル)ベンジル基、
のいずれかである、請求項3記載の製造方法。
【請求項5】
前記酵素がノルコクラウリンシンターゼもしくはPathogenesis−Related 10/Bet V1 Protein Familyに属する酵素である、請求項1〜4のいずれかに記載の製造方法。
【請求項6】
前記酵素が、以下の(a)〜(c):
(a)配列表の配列番号1、2、3、4、又は5に示すアミノ酸配列からなるポリペプチド、
(b) 配列表の配列番号1、2、3、4、又は5に示すアミノ酸配列において1若しくは複数個のアミノ酸配列が欠失、挿入、置換及び/又は付加されたアミノ酸配列からなるポリペプチド、
(c) 配列表の配列番号1、2、3、4、又は5に示すアミノ酸配列と85%以上の配列同一性を有するアミノ酸配列からなるポリペプチド、
のいずれかである、請求項5記載の製造方法。
【請求項7】
前記酵素が、配列表の配列番号1に示すアミノ酸配列から、N末端のアミノ酸を11〜41個削除し、N末端にメチオニンを付加したアミノ酸配列からなるポリペプチドである、請求項5記載の製造方法。
【請求項8】
請求項1から7のいずれかに記載の製造方法に用いるポリペプチドであって、以下の(d)〜(f):
(d)配列表の配列番号1に示すアミノ酸配列において、1個または複数個のアミノ酸が欠失、挿入、置換及び/又は付加されている、
(e)配列表の配列番号1に示すアミノ酸配列から、N末端のアミノ酸が11〜41個削除され、N末端にメチオニンが付加されている、
(f)配列表の配列番号2のポリヌクレオチドに比べて遺伝子組換え生産時の不溶化が低減されている、
を特徴とする、ポリペプチド。
【請求項9】
一般式(4);
【化4】
(式中、R11はメチル基、エチル基、n−プロピル基、n−ブチル基、エチルアルデヒド基、フェニル基、4−(トリフルオロメチル)フェニル基のいずれかである)で表される、光学活性化合物。
【請求項1】
一般式(1):
【化1】
(式中、R1〜R5は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、ヒドロキシル基、アミノ基、またはニトロ基である。)で表される2−(3−ヒドロキシフェニル)エチルアミン誘導体と一般式(2):
【化2】
(式中、R6〜R8は水素原子、ハロゲン原子、置換されていても良いアルキル基、置換されていても良いアラルキル基、置換されていてもよいアリール基、置換されていても良いアルコキシ基、アミノ基、またはニトロ基であるか、もしくは、R6〜R8が互いに結合し環を形成しても良い。ただし、R6〜R8のいずれか2つが水素原子であり、1つがフェニル基である場合は除く)で表されるアルデヒド化合物に酵素を作用させることを特徴とする、一般式(3):
【化3】
(式中R1〜R8は前記と同じ、*は不斉炭素を表す)で表される光学活性1−置換テトラヒドロイソキノリン誘導体の製造方法。
【請求項2】
前記R1〜R3及びR5が水素であり、R4が水素原子、メチル基、水酸基、メトキシ基のいずれかである、請求項1記載の製造方法。
【請求項3】
前記R6が水素原子、R7が水素原子または直鎖状または分岐鎖状の炭素数1〜10のアルキル基、R8が直鎖状または分岐鎖状の炭素数2〜10のアルキル基、置換されていても良い炭素数7〜17のアラルキル基、置換されていても良い炭素数1〜10のアルコキシ基である、請求項1または2記載の製造方法。
【請求項4】
前記R6〜R8が下記(1)〜(10):
(1)前記R6及びR7が水素原子、R8がエチル基、
(2)前記R6が水素原子、R7がメチル基、R8がメチル基、
(3)前記R6及びR7が水素原子、R8がn−プロピル基、
(4)前記R6及びR7が水素原子、R8がn−ブチル基、
(5)前記R6及びR7が水素原子、R8がn−ペンチル基、
(6)前記R6及びR7が水素原子、R8がn−プロピルアルデヒド基、
(7)前記R6が水素原子、R7がメチル基、R8がフェニル基、
(8)前記R6及びR7が水素原子、R8がベンジル基、
(9)前記R6及びR7が水素原子、R8が4−(トリフルオロメチル)ベンジル基、
(10)前記R6が水素原子、R7がメチル基、R8が 4−(イソプロピル)ベンジル基、
のいずれかである、請求項3記載の製造方法。
【請求項5】
前記酵素がノルコクラウリンシンターゼもしくはPathogenesis−Related 10/Bet V1 Protein Familyに属する酵素である、請求項1〜4のいずれかに記載の製造方法。
【請求項6】
前記酵素が、以下の(a)〜(c):
(a)配列表の配列番号1、2、3、4、又は5に示すアミノ酸配列からなるポリペプチド、
(b) 配列表の配列番号1、2、3、4、又は5に示すアミノ酸配列において1若しくは複数個のアミノ酸配列が欠失、挿入、置換及び/又は付加されたアミノ酸配列からなるポリペプチド、
(c) 配列表の配列番号1、2、3、4、又は5に示すアミノ酸配列と85%以上の配列同一性を有するアミノ酸配列からなるポリペプチド、
のいずれかである、請求項5記載の製造方法。
【請求項7】
前記酵素が、配列表の配列番号1に示すアミノ酸配列から、N末端のアミノ酸を11〜41個削除し、N末端にメチオニンを付加したアミノ酸配列からなるポリペプチドである、請求項5記載の製造方法。
【請求項8】
請求項1から7のいずれかに記載の製造方法に用いるポリペプチドであって、以下の(d)〜(f):
(d)配列表の配列番号1に示すアミノ酸配列において、1個または複数個のアミノ酸が欠失、挿入、置換及び/又は付加されている、
(e)配列表の配列番号1に示すアミノ酸配列から、N末端のアミノ酸が11〜41個削除され、N末端にメチオニンが付加されている、
(f)配列表の配列番号2のポリヌクレオチドに比べて遺伝子組換え生産時の不溶化が低減されている、
を特徴とする、ポリペプチド。
【請求項9】
一般式(4);
【化4】
(式中、R11はメチル基、エチル基、n−プロピル基、n−ブチル基、エチルアルデヒド基、フェニル基、4−(トリフルオロメチル)フェニル基のいずれかである)で表される、光学活性化合物。
【図1】

【公開番号】特開2013−74811(P2013−74811A)
【公開日】平成25年4月25日(2013.4.25)
【国際特許分類】
【出願番号】特願2011−215407(P2011−215407)
【出願日】平成23年9月29日(2011.9.29)
【出願人】(000000941)株式会社カネカ (3,932)
【Fターム(参考)】
【公開日】平成25年4月25日(2013.4.25)
【国際特許分類】
【出願日】平成23年9月29日(2011.9.29)
【出願人】(000000941)株式会社カネカ (3,932)
【Fターム(参考)】
[ Back to top ]