
光駆動カルシウムイオン制御タンパク質

【課題】本発明の目的は、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中において、光刺激によりカルシウムイオン濃度を制御することができる化合物及び方法を提供することである。
【解決手段】前記課題は、(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;を含む、光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体により解決することができる。
【解決手段】前記課題は、(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;を含む、光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体により解決することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、光駆動カルシウムイオン制御タンパク及びカルシウムイオン濃度の制御方法に関する。本発明によれば、カルシウムイオン濃度を制御することができ、例えば、細胞内におけるカルシウムイオンの濃度を上昇させること、又は低下させることができる。
【背景技術】
【0002】
カルシウムイオンは、通常、細胞内液にはほとんど存在せず、細胞外からのカルシウムイオンの流入、又は細胞内の小胞体等に蓄えられたカルシウムイオンの放出などによって、細胞内の濃度が上昇し、細胞内の様々な生理活性のシグナルとしての役割をはたしている。この細胞内のカルシウムイオン濃度の変化によって、筋肉収縮、神経の情報伝達、短期記憶、長期記憶、神経の成長、免疫系細胞の活性化、胃酸・胃液・腸粘液・汗等の分泌、酵素の活性化、アポトーシス、及び細胞分裂など、生体の多くの生理現象がコントロールされている。
【0003】
従って、細胞内で起きるカルシウムイオンの濃度変化と、生体の生理現象との関係は、正常な生体の機能ばかりではなく、多くの疾患との関連性も指摘されている。そのため、細胞内で起きるカルシウムイオンの濃度変化を解析することは、非常に重要である。特に、カルシウムイオン濃度の変化は、細胞内の特定の領域又は細胞内小器官付近で起きることもあり、細胞内の任意の位置でカルシウムイオン濃度を制御する実験方法の開発が期待されている。
【0004】
従来、カルシウムイオンによる細胞又は生体への影響を調べる方法として、光照射によりカルシウムイオンを放出するカルシウムイオンキレート剤である光駆動ケージドカルシウムイオン化合物(例えば、nitrophenyl−EGTA:以下、NP−EGTAと称する)を用いる方法が知られている(非特許文献1)。NP−EGTAは、マイクロインジェクションにより、細胞内に注入し、光照射によりカルシウムイオンを放出させ、細胞内のカルシウムイオン濃度を上昇させることができる。しかしながら、マイクロインジェクションによる細胞への導入は煩雑な手技を要し、また導入したNP−EGTAを細胞内の任意の位置にとどめることは困難であり、細胞内の特定の領域のカルシウムイオン濃度を制御することは不可能であった。更に、励起時に紫外光を用いなければならず、細胞に対して害を与える可能性があった。
一方、光照射によって細胞外からカルシウムイオンを流入させる光駆動カルシウムイオントランスポーターを用いる方法が開発された(非特許文献2)。しかしながら、この方法も、カルシウムイオンを細胞外から流入させるため、カルシウムイオン濃度の変化が、細胞全体で起こるものであった。従って、細胞内の任意の領域又は位置でのカルシウムイオン濃度の変化に対する応答を調べることは、原理上不可能であった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】「プロシーディング・オブ・ザ・ナショナル・アカデミー・オブ・サイエンス・ユーエスエー(Proceedings of the National Academy of Sciences)」(米国)1994年、第91巻、p.187−91
【非特許文献2】「ネイチャー(Nature)」(英国)2002年、第28巻、p.398−401
【発明の概要】
【発明が解決しようとする課題】
【0006】
前記の光駆動ケージドカルシウムイオン化合物(例えば、NP−EGTA)には、マイクロインジェクションが不要な膜透過性のものも開発されているが、膜透過性の化合物でも、細胞内の局所におけるカルシウムイオンの濃度の制御は困難である。また、前記のように、光駆動カルシウムイオントランスポーターは、細胞外からのカルシウムイオンの流入を行うものであり、やはり細胞内におけるカルシウムイオンの濃度の制御は不可能である。
従って、本発明の目的は、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中において、光刺激によりカルシウムイオン濃度を制御することができる化合物及び方法を提供することである。具体的には、カルシウムイオン濃度を上昇又は低下させることが可能な化合物及び方法を提供することである。
【課題を解決するための手段】
【0007】
本発明者らは、カルシウムイオン濃度を制御することができる実験方法について、鋭意研究した結果、驚くべきことに、カルシウムイオン結合タンパク質、光励起によって立体構造が変化する光受容体ペプチド、カルシウムイオン結合タンパク質に結合するカルシウムイオン結合タンパク質結合ペプチドを含む融合タンパク質を用いることにより、カルシウムイオン濃度を低下させること、又は上昇させることが可能であることを見出した。すなわち、光刺激により光受容体ペプチドの構造が変化することを利用して、カルシウムイオン結合タンパク質及びカルシウムイオン結合タンパク質結合ペプチドの構造を調整し、融合タンパク質にカルシウムイオンを結合させること、又は融合タンパク質からカルシウムイオンを放出させることが可能であることを見出した。従って、これらの融合タンパク質を用いることによって、細胞内などのカルシウムイオン濃度を制御することが可能である。
前記カルシウムイオン結合タンパク質として、カルモジュリンが最もよく研究されているが、従来、カルモジュリンを用いた融合タンパク質としては、細胞内のカルシウムイオン濃度を測定することのできる融合タンパク質が、本発明者らによって報告されている(Nature Methods, vol.7, No.9, p.729-732)。この融合タンパク質は、カルモジュリン、カルモジュリンと結合するM13ペプチド、及び蛍光タンパク質からなる融合タンパク質であり、カルモジュリンへのカルシウムイオンの結合により、蛍光タンパク質の構造が変化し、カルシウムイオンの濃度を測定することができるプローブ(融合タンパク質)である。しかしながら、このカルシウムイオン濃度測定のプローブ(融合タンパク質)は、カルモジュリン及びM13の複合体にカルシウムイオンが結合することにより、蛍光タンパク質の構造を変化させ、それによって蛍光タンパク質の蛍光強度が変化することを利用するものである。一方、本発明の融合タンパク質は、光刺激により光受容体ペプチドの構造が変化することを利用して、カルモジュリン及びM13の複合体と、カルシウムイオンとの結合を制御するものであり、カルモジュリン、M13ペプチド及び蛍光タンパク質からなる融合タンパク質とは、全く異なるものである。更に、本発明者らは、カルシウムイオンと結合する融合タンパク質と、カルシウムイオンを放出するタンパク質とを調製することができたが、この2種類の融合タンパク質は、カルモジュリン、M13ペプチド、及び光受容体ペプチドの3つのタンパク質の配置を変更することのみによって、得られたものであり、驚くべきことである。
本発明は、こうした知見に基づくものである。
【0008】
すなわち、本発明は、
[1](a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;を含む、光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[2](a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムと結合する、[1]に記載の光駆動カルシウムイオン制御タンパク質(以下、カルシウムイオン吸収融合タンパク質と称することがある)、又はその機能的等価改変体(以下、まとめてカルシウムイオン吸収融合タンパク質等と称することがある)、
[3](a1)カルシウムイオン結合タンパク質N末側ペプチド、又はその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;(a2)カルシウムイオン結合タンパク質C末側ペプチド、又はその改変体;及び(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムを放出する、[1]に記載の光駆動カルシウムイオン制御タンパク質(以下、カルシウムイオン吸収融合タンパク質と称することがある)、又はその機能的等価改変体(以下、まとめてカルシウムイオン吸収融合タンパク質等と称することがある)、
[4]前記光受容体ペプチドが、フォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、及びFKF−1のLOVドメインからなる群から選択される光受容体ペプチドであり;
前記カルシウムイオン結合タンパク質及びカルシウムイオン結合タンパク質結合ペプチドの組み合わせが、カルモジュリン及びカルモジュリン結合ペプチド、トロポニンC及びトロポニンI、ミオシン軽鎖及びミオシンII重鎖、並びにレコベリン及びロドプシンキナーゼからなる群から選択される、[1]〜[3]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[5](a)カルモジュリン、又はカルシウムイオン結合能を有するカルモジュリン改変体;(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;をN末端から順に含み、光刺激によりカルシウムと結合する、[2]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[6]配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、又は配列番号7で表されるアミノ酸配列からなる、[5]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[7](a1)カルモジュリンN末側ペプチド、又はその改変体;(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;(a2)カルモジュリンC末側ペプチド、又はその改変体;及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;をN末端から順に含み、光刺激によりカルシウムを放出する、[3]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[8]配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、又は配列番号15で表されるアミノ酸配列からなる、[7]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[9]シグナルペプチドを含む、[1]〜[8]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[10][1]〜[9]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体をコードするDNA、
[11]配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、又は配列番号30で表される塩基配列からなる、[10]に記載のDNA、
[12][10]又は[11]に記載のDNAを含むプラスミド、
[13][1]〜[9]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、若しくはその機能的等価改変体、又は[10]若しくは[11]に記載のDNAを用いて発現するタンパク質に、励起光を照射することを特徴とする、カルシウムイオン濃度の制御方法、
[14]前記励起光の励起波長が、300〜500nmである[13]に記載のカルシウムイオン濃度の制御方法、
に関する。
【発明の効果】
【0009】
本発明の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体によれば、カルシウムイオン濃度を制御すること、すなわちカルシウムイオン濃度を上昇、又は低下させることができる。
また、本発明のDNA又はプラスミドを用いることにより、細胞内において、容易に本発明の光駆動カルシウムイオン制御タンパク質を発現させることが可能である。従って、従来の光駆動ケージドカルシウムイオン化合物又は光駆動カルシウムイオントランスポーターと比較して、本発明の光駆動カルシウムイオン制御タンパク質は、細胞内での取り扱いが容易である。
更に、本発明の細胞内小器官ターゲット配列を有する光駆動カルシウムイオン制御タンパク質は、細胞内の特定の領域に移行(存在)させることが可能であり、細胞内の任意の位置でカルシウムイオン濃度を制御することができる。すなわち、本発明のカルシウムイオン制御タンパク質、又はその機能的等価改変体は、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中のいずれにおいても、使用することが可能であり、生体内等のカルシウムイオン濃度を容易に制御することができる。
【図面の簡単な説明】
【0010】
【図1】本発明のPacman(カルシウムイオン吸収融合タンパク質)及びCARBY(カルシウムイオン放出融合タンパク質)の吸収スペクトルを、LOV2ドメインの吸収スペクトルと比較したグラフである。
【図2】本発明のPacman(カルシウムイオン吸収融合タンパク質)又はCARBY(カルシウムイオン放出融合タンパク質)を用いて、光励起によりカルシウムイオン濃度が変化すること示したグラフである。
【図3】本発明のPacman(カルシウムイオン吸収融合タンパク質)の構造の模式図である。
【図4】本発明のCARBY(カルシウムイオン放出融合タンパク質)の構造の模式図である。
【発明を実施するための形態】
【0011】
[1]光駆動カルシウムイオン制御タンパク質
(光駆動カルシウムイオン制御タンパク質)
本発明の光駆動カルシウムイオン制御タンパク質は、(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体(以下、カルシウムイオン結合タンパク質等(a)と称することがある);(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体(以下、光受容体ペプチド等(b)と称することがある);及び(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質結合能を有するその改変体(以下、カルシウムイオン結合タンパク質結合ペプチド等(c)と称することがある);を含む。また、光駆動カルシウムイオン制御タンパク質は、カルシウムイオン結合タンパク質等(a)、光受容体ペプチド等(b)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)からなってもよく、更に後述のペプチドリンカー等、及び/又はシグナルペプチドを含んでもよい。。
【0012】
(カルシウムイオン結合タンパク質)
本発明のカルシウムイオン制御タンパク質に用いるカルシウムイオン結合タンパク質は、カルシウムイオン結合能を有する限り、特に限定されるものはないが、例えばカルモジュリン、トロポニンC、ミオシン軽鎖、又はレコベリンを挙げることができる。これらのカルシウムイオン結合タンパク質は、後述のカルシウムイオン結合タンパク質結合ペプチドとの複合体を形成することにより、カルシウムイオンとの親和性が上昇するタンパク質であり、本発明のカルシウムイオン制御タンパク質に好ましく用いることができる。また、カルシウムイオン結合タンパク質の由来も特に限定されるものではなく、すべての生物由来のカルシウムイオン結合タンパク質を使用することが可能である。
【0013】
前記カルシウムイオン結合タンパク質のうち、カルモジュリンは酸性のタンパク質で、約148残基のアミノ酸からなる。4つのEFハンドモチーフを有しており、それぞれにカルシウムイオンが結合する。カルモジュリンは、様々な細胞で発現し、細胞における存在部位も、細胞内小器官内及び膜上など様々であり、カルシウムイオンによるシグナル伝達に重要な役割を果たしているタンパク質である。
本発明のカルシウムイオン制御タンパク質に用いるカルモジュリンは、カルシウムイオン結合能を有する限り、特に限定されるものはなく、すべての生物由来のカルモジュリンを使用することが可能であり、例えば配列番号31で表されたアミノ酸配列からなるタンパク質であるカルモジュリンを用いることができる。
【0014】
(カルシウムイオン結合タンパク質改変体)
カルシウムイオン結合タンパク質の改変体とは、カルシウムイオン結合タンパク質の本来有しているカルシウムイオン結合能を有している改変体を意味する。具体的には、そのアミノ酸配列が、元となるカルシウムイオン結合タンパク質のアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるカルシウムイオン結合タンパク質と実質的に同じ活性(カルシウムイオン結合能)を有しているタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。例えば、N末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体、C末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体を挙げることができる。
【0015】
カルシウムイオン結合タンパク質の改変体のうち、カルモジュリンの改変体とは、カルモジュリンの本来有しているカルシウムイオン結合能を有している改変体を意味する。例えば、そのアミノ酸配列が、配列番号31で表されるアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるカルモジュリンと実質的に同じ活性(カルシウムイオン結合能)を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。例えば、N末側の1〜30個のアミノ酸の欠失したカルモジュリン改変体、より好ましくは1〜20個のアミノ酸の欠失したカルモジュリン改変体、更に好ましくは1〜10個のアミノ酸欠失したカルモジュリン改変体、C末側の1〜30個のアミノ酸の欠失したカルモジュリン改変体、より好ましくは1〜20個のアミノ酸の欠失したカルモジュリン改変体、更に好ましくは1〜10個のアミノ酸欠失したカルモジュリン改変体を挙げることができる。
【0016】
(光受容体ペプチド)
本発明の光駆動生カルシウムイオン制御タンパク質に用いる光受容体ペプチドは、光刺激により構造の変化するペプチドであり、例えばフォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、FKF−1のLOVドメイン、AppAのBLUFドメイン、及びTePixD−I66CのBLUFドメインを挙げることができ、好ましくはフォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、又はFKF−1のLOVドメインであり、より好ましくはフォトトロピン1のLOV2ドメイン、又はフォトトロピン2のLOV2ドメインである。
【0017】
前記光受容体ペプチドの由来も、特に限定されるものではなく、すべての生物由来(例えば、動物由来、又は植物由来)の光受容体ペプチドを用いることができる。すなわち、多くの生物が、光受容体ペプチドを含むタンパク質であるフォトトロピン1、フォトトロピン2、オーレオクロム1、オーレオクロム2、FKF−1、AppA、及びTePixD−I66Cを有しており、それぞれのタンパク質に含まれる光受容体ペプチドを用いることができる。
【0018】
なお、フォトトロピンは、植物特有の青色光受容体であり、光屈性、葉緑体の光定位運動、気孔の開口、葉の平坦化、葉の光定位運動などを誘導する。フォトトロピンには、前記のようにフォトトロピン1及びフォトトロピン2があり、フォトトロピン1は弱光から中高強光域の光センサーとして、フォトトロピン2は強光域の光センサーとして機能する。フォトトロピン1及びフォトトロピン2は、ともにLOV1ドメイン及びLOV2ドメインを有しており、LOV1ドメインは光感受性制御に関連した機能を有しており、LOV2ドメインがキナーゼ活性の光制御分子スイッチとして機能すると考えられている。また、オーレオクロムは、黄色植物(ヒバマタ、コンブ、及びケイ藻など)が有する青色光受容体であり、LOVドメインを有しており、青色光を吸収して構造が変化することによって、光センサーとして機能する。
【0019】
例えば、前記フォトトロピン1は、燕麦、シロイヌナズナ、タバコ、トウモロコシ、イネ又はシダ類などが有しており、これらの植物のフォトトロピン1のLOV2ドメインを本発明の光駆動生カルシウムイオン制御タンパク質に用いることができるが、特には配列番号32で表されるアミノ酸配列からなるペプチドである燕麦由来のLOV2ドメインが好ましい。
【0020】
光受容体ペプチドは、光刺激によりその立体構造が変化するが、立体構造が変化する励起波長は、光受容体ペプチドの種類により、適宜選択することができる。例えば、フォトトロピン1、フォトトロピン2、オーレオクロム1、及びオーレオクロム2は青色光受容体であり、青色周辺の励起波長を用いることが好ましい。従って、フォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、及びフォトトロピン2のLOV2ドメイン、及びオーレオクロム1のLOVドメイン、オーレオクロム2LOVドメインの励起波長は、300〜500nmを用いることが可能であり、350〜490nmが好ましく、430〜470nmがより好ましい。
【0021】
(光受容体ペプチド改変体)
前記光受容体ペプチドの改変体とは、その光受容体ペプチドの本来有している性質、すなわち光刺激により構造の変化することのできる改変体を意味する。具体的には、そのアミノ酸配列が、元となる光受容体ペプチドのアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となる光受容体ペプチドと実質的に同じ活性(光刺激による構造変化)を示すペプチドを意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。例えば、N末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体、C末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体を挙げることができる。
【0022】
例えば、LOV2ドメイン改変体とは、LOV2ドメインの本来有している性質を有する改変体、すなわち光刺激により構造の変化することのできる改変体を意味する。例えば、そのアミノ酸配列が、配列番号32で表されるアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるLOV2ドメインと実質的に同じ活性(光刺激による構造変化)を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。特に、例えば、N末側の1〜30個のアミノ酸の欠失したLOV2ドメイン改変体、より好ましくは1〜20個のアミノ酸の欠失したLOV2ドメイン改変体、更に好ましくは1〜10個のアミノ酸欠失したLOV2ドメイン改変体、C末側の1〜30個のアミノ酸の欠失したLOV2ドメイン改変体、より好ましくは1〜20個のアミノ酸の欠失したLOV2ドメイン改変体、更に好ましくは1〜10個のアミノ酸欠失したLOV2ドメイン改変体を挙げることができる。具体的には、後述の実施例に示すように、前記フォトトロピン1のLOV2ドメインの改変体としては、C末側アミノ酸が17個、16個、15個、14個、12個、9個、又は3個欠失した改変体を挙げることができ、これらの改変体はLOV2ドメインと同じように、光刺激により構造が変化する。
【0023】
(カルシウムイオン結合タンパク質結合ペプチド)
本発明のカルシウムイオン制御タンパク質は、更にカルシウムイオン結合タンパク質結合ペプチドを含む。このペプチドは、カルシウムイオン結合タンパク質への結合能を有する限り、特に限定されるものはないが、例えばカルモジュリン結合ペプチド、トロポニンI、ミオシンII重鎖、又はロドプシンキナーゼを挙げることができる。これらのカルシウムイオン結合タンパク質結合ペプチドは、前記カルシウムイオン結合タンパク質との複合体を形成することにより、カルシウムイオン結合タンパク質のカルシウムイオンとの親和性を上昇させることができる。具体的には、カルモジュリン結合ペプチドはカルモジュリンと、トロポニンIはトロポニンCと、ミオシンII重鎖はミオシン軽鎖と、及びロドプシンキナーゼはレコベリンとそれぞれ複合体を形成する。
【0024】
前記カルモジュリン結合ペプチドは、いくつかのキナーゼなどに存在しており、例えば骨格筋ミオシン軽鎖キナーゼ、平滑筋ミオシン軽鎖キナーゼ、カルモジュリン依存キナーゼII、カルモジュリン依存キナーゼキナーゼ等のカルモジュリン結合領域のペプチドを挙げることができる。
【0025】
本発明のカルシウムイオン制御タンパク質に用いるカルモジュリン結合ペプチドの生物種の由来も、カルモジュリンに結合するものである限り、特に限定されるものではなく、すべての生物由来のカルモジュリン結合ペプチドを用いることが可能である。すなわち、すべての生物由来に骨格筋ミオシン軽鎖キナーゼのカルモジュリン結合領域のペプチド、平滑筋ミオシン軽鎖キナーゼのカルモジュリン結合領域のペプチド、カルモジュリン依存キナーゼIIのカルモジュリン結合領域のペプチド、カルモジュリン依存キナーゼキナーゼのカルモジュリン結合領域のペプチド等を用いることができる。例えば、配列番号33で表されるアミノ酸配列からなる、アフリカツメガエルの骨格筋ミオシン軽鎖キナーゼのカルモジュリン結合領域であるM13ペプチドを用いることが可能である。
前記M13ペプチドは、骨格筋ミオシン軽鎖キナーゼのカルモジュリン結合領域に由来する26残基ペプチド(KRRWKKNFIAVSAANRFKKISSSGAL)(Blumenthal, D. K., & Krebs, E. G. (1987) Methods Enzymol. 139,115-126)であり、カルシウムイオンが結合したカルモジュリン(カルシウムイオン−カルモジュリン)に結合し、安定した複合体を形成する。従って、本発明のカルシウムイオン制御タンパク質はM13ペプチドを含むことにより、カルモジュリンがカルシウムイオンと結合した場合、強固な結合を形成することが可能である。
【0026】
更に、M13ペプチド以外の具体的なカルモジュリン結合ペプチドを以下の表1に挙げる。
【0027】
【表1】

【0028】
本発明のカルシウムイオン制御タンパク質において、前記表1に記載の23種類のカルモジェリン結合ペプチドを、M13ペプチドに代えて用いることが可能である。それぞれのカルモジェリン結合ペプチドは、20アミノ酸からなるペプチド又は16アミノ酸からなるペプチドのみでもカルモジュリンと結合可能であり、そのまま本発明のカルシウムイオン制御タンパク質に用いることが可能である。しかしながら、M13ペプチドと同じように、それぞれのペプチドのC末側に続く1〜10個のアミノ酸(好ましくは4〜8アミノ酸、より好ましくは6アミノ酸)を含んだペプチドを用いることもできる。更に、それらのペプチドの改変体を用いることも可能である。
【0029】
(カルシウムイオン結合タンパク質結合ペプチド改変体)
カルシウムイオン結合タンパク質結合ペプチド改変体とは、カルシウムイオン結合タンパク質結合ペプチド改変体の本来有しているカルシウムイオン結合タンパク質への結合能を有している改変体を意味する。具体的には、そのアミノ酸配列が、元となるペプチドのアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるペプチドと実質的に同じ活性(カルシウムイオン結合タンパク質結合能)を有しているペプチドを意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜3個である。例えば、N末側の1〜10個のアミノ酸の欠失したペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したペプチド改変体、C末側の1〜10個のアミノ酸の欠失したペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したペプチド改変体を挙げることができる。
【0030】
カルシウムイオン結合タンパク質結合ペプチド改変体のうち、M13ペプチド改変体とは、M13ペプチドの本来有している性質を有する改変体、すなわちカルモジュリンへの結合能を有する改変体を意味する。例えば、そのアミノ酸配列が、配列番号33で表されるアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるM13ペプチドと実質的に同じ活性(カルモジュリンへの結合能)を示すペプチドを意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜3個である。特に、例えば、N末側の1〜10個のアミノ酸の欠失したM13ペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したM13ペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したM13ペプチド改変体、C末側の1〜10個のアミノ酸の欠失したM13ペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したM13ペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したM13ペプチド改変体を挙げることができる。例えば、後述の実施例に示すように、前記M13ペプチド改変体としては、N末側アミノ酸が1個又は2個欠失した改変体を挙げることができ、この改変体はM13ペプチドと同じように、カルモジュリンへの結合能を示す。
【0031】
(シグナルペプチド)
また、本発明の光駆動カルシウムイオン制御タンパク質を、細胞内の任意の位置で発現させるため、光駆動カルシウムイオン制御タンパク質は、シグナルペプチドを含んでもよい。シグナルペプチドを含むことによって、細胞内の特定の領域又は細胞内小器官内若しくは細胞内小器官の膜上で光駆動カルシウムイオン制御タンパク質を発現させることができる。
シグナルペプチドは、約3〜60アミノ酸からなるペプチドであり、局在シグナル、輸送シグナル、又は移行シグナルと呼ばれることもある。シグナルペプチドは、その種類に応じて、N末端、C末端、又はタンパク質内部のアミノ酸配列に存在し、細胞質内で生合成されたタンパク質の細胞内の局所への輸送および局在化を誘導する。具体的には、シグナルペプチドとして、ミトコンドリア膜間スペース局在化シグナル、ゴルジ体局在化シグナル、小胞体局在化シグナル、ペルオキシソーム局在化シグナル、核局在シグナル、ミトコンドリアのマトリックス局在シグナル、細胞膜局在シグナル及び葉緑体局在シグナルなどを挙げることができる。
例えば、N末端に存在するシグナルペプチドとしては、小胞体移行シグナル(H2N-Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-Gly-Ile-Leu-Phe-Trp-Ala-Thr-Glu-Ala-Glu-Gln-Leu-Thr-Lys-Cys-Glu-Val-Phe-Gln-)、ミトコンドリアマトリックスシグナル(H2N-Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-Phe-Lys-Pro-Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-Leu-Leu-)、及びペルオキシソームシグナル(PTS2)(H2N-----Arg-Leu-X5-His-Leu)を挙げることができ、C末端に存在するシグナルペプチドとしては、小胞体保留シグナル(-Lys-Asp-Glu-Leu-COOH)、及びペルオキシソームシグナル(PTS1)(-Ser-Lys-Leu-COOH)を挙げることができ、タンパク質内部に存在するシグナルペプチドとしては、核移行シグナル(NLS)(-Pro-Pro-Lys-Lys-Lys-Arg-Lys-Val)を挙げることができる。
これらのシグナルペプチドを、本発明の光駆動カルシウムイオン制御タンパク質のN末端、C末端、又はタンパク質内部に含ませることにより、光駆動カルシウムイオン制御タンパク質を細胞内の特定の領域に局在させ、カルシウムイオン濃度の変化による細胞の動態を解析することが可能である。
【0032】
(機能的等価改変体)
本発明の機能的等価改変体は、前記光駆動カルシウムイオン制御タンパク質の機能的等価改変体である。本明細書において「機能的等価改変体」とは、元となるタンパク質のアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列からなるタンパク質であって、しかも、元となるタンパク質と実質的に同じ活性を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜2個である。例えば、配列番号1〜15で表されたアミノ酸配列のそれぞれにおいて、1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列からなるタンパク質であって、しかも、光駆動カルシウムイオン制御タンパク質と実質的に同じ活性を有するタンパク質、すなわち、光照射によりカルシウムイオンの濃度を制御可能であり、そしてその光照射の励起波長が実質的に同じであるタンパク質を意味する。
【0033】
(リンカー及び制限酵素サイト)
本発明のカルシウムイオン制御タンパク質における、カルシウムイオン結合タンパク質等(a)と光受容体ペプチド等(b)との間、及び光受容体ペプチド等(b)とカルシウムイオン結合タンパク質結合ペプチド等(c)との間には、カルシウムイオン制御タンパク質の機能を阻害しない限りにおいて、適当な長さのペプチドリンカー又は制限酵素サイトのヌクレオチド配列から翻訳される2〜数アミノ酸の配列を含むことができる。
【0034】
本発明のカルシウムイオン制御タンパク質は、カルシウムイオン吸収融合タンパク質及びカルシウムイオン放出融合タンパク質を含む。カルシウムイオン吸収融合タンパク質は、光刺激により周囲のカルシウムを吸収(結合)し、周囲の環境のカルシウム濃度を低下させることができる。カルシウムイオン放出融合タンパク質は、保持しているカルシウムイオンを光刺激により放出し、周囲の環境のカルシウムイオン濃度を上昇させることができる。
【0035】
(カルシウムイオン吸収融合タンパク質)
本発明のカルシウムイオン吸収融合タンパク質は、(a)カルシウム結合タンパク質、又はカルシウム結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び(c)カルシウム結合タンパク質結合ペプチド、又はカルシウム結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムと結合するものである。また、光駆動カルシウムイオン吸収融合タンパク質は、カルシウムイオン結合タンパク質等(a)、光受容体ペプチド等(b)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)からなってもよく、更に前記ペプチドリンカー等、及び/又はシグナルペプチドを含んでもよい。
【0036】
カルシウムイオン吸収融合タンパク質の1つの態様として、(a)カルモジュリン、又はカルシウムイオン結合能を有するカルモジュリン改変体(以下、カルモジュリン等(a)と称する);(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体(以下、LOV2ドメイン等(b)と称する);及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体(以下、M13ペプチド等(c)と称する);をN末端から順に含み、光刺激によりカルシウムイオンと結合する融合タンパク質を挙げることができる。具体的には、カルシウムイオン吸収融合タンパク質として、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、又は配列番号7で表されるアミノ酸配列からなる融合タンパク質を挙げることができる。
【0037】
本発明のカルシウムイオン吸収融合タンパク質の機能的等価改変体は、実質的にカルシウムイオン吸収融合タンパク質と同じ活性を有するものである。「カルシウムイオン吸収融合タンパク質と同じ活性」とは、光照射によりカルシウムイオンと結合し、そしてその光照射の励起波長が実質的に同じであることを意味する。例えば、カルモジュリン又はカルモジュリン改変体のカルシウムイオンと結合する機能が維持されており、そしてM13ペプチド又はM13ペプチド改変体のカルモジュリンに結合する機能が維持されていることを意味する。そして、励起波長として300〜500nmを用いることによって、機能的等価改変体がカルシウムイオンと結合できることを意味する。
【0038】
本発明のカルシウムイオン吸収融合タンパク質は、例えば、カルモジュリン又はカルモジュリン改変体(a)、LOV2ドメイン又はLOV2ドメイン改変体(b)、及びM13ペプチド又はM13ペプチド改変体(c)が、N末端から順に連結された構造をとっている。カルモジュリンは、単体ではカルシウムイオンとの結合はそれほど強くないが、M13ペプチドと結合することにより、解離定数が小さくなり、カルモジュリン及びM13の複合体は、カルシウムイオンを強固に結合させることが可能になる。本発明のカルシウムイオン吸収融合タンパク質は、光照射されない状態では、LOV2ドメイン等(b)が、カルモジュリン等(a)と、M13ペプチド等(c)との間に組み込まれていることにより、カルモジュリン等(a)とM13ペプチド等(c)とが結合できない状態であり、カルモジュリンの解離定数は大きいと推定される。一方、光照射された場合、LOV2ドメイン等(b)の構造が変化し、カルモジュリン等(a)と、M13ペプチド等(c)とが結合可能な状態になり、カルモジュリン等(a)とM13ペプチド等(c)との複合体は、カルシウムイオン吸収融合タンパク質の周囲のカルシウムイオン濃度より小さな解離定数を有する状態になる。そのため、本発明のカルシウムイオン吸収融合タンパク質周囲のカルシウムイオンを吸収することができると推定される。また、カルシウムイオン結合タンパク質等(a)、光受容体ペプチド等(b)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)との関係も、それぞれカルモジュリン等(a)、LOV2ドメイン等(b)、及びM13ペプチド等(c)との関係と同じであると推定される。しかしながら、本発明は前記の推定により限定されるものではない。
【0039】
(リンカー及び制限酵素サイト)
本発明のカルシウムイオン吸収融合タンパク質において、カルシウムイオン結合タンパク質等(a)(例えば、カルモジュリン等(a))と光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))との間、及び光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))とカルシウムイオン結合タンパク質結合ペプチド等(c)(例えば、M13ペプチド等(c))との間には、カルシウムイオン吸収融合タンパク質の機能を阻害しない限りにおいて、適当な長さのペプチドリンカー又は制限酵素サイトのヌクレオチド配列から翻訳される2〜数アミノ酸の配列を含むことができる。
ペプチドリンカーしては、例えば「G」、「GG」、「GGS」、「GGGS」、「GGGGS」、「GGGSK」、「GGSGG」、又は「GGSGGSGGS」を挙げることができる。また、制限酵素サイトのヌクレオチド配列から翻訳されるアミノ酸配列としては、LE及びGTなどを含むことができる。
後述の実施例に示すように、カルモジュリン等(a)とLOV2ドメイン等(b)との間に制限酵素サイトXhoIから翻訳されるLE、及びペプチドリンカー「G」、「GGS」、「GGGGS」を含んでも、本発明のカルシウムイオン吸収融合タンパク質を発揮することができる。
【0040】
(カルシウムイオン放出融合タンパク質)
本発明のカルシウムイオン放出融合タンパク質は、(a1)カルシウム結合タンパク質N末側ペプチド、又はその改変体(以下、カルシウム結合タンパク質N末側ペプチド等(a1)と称することがある);(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;(a2)カルシウム結合タンパク質C末側ペプチド、又はその改変体(以下、カルシウム結合タンパク質C末側ペプチド等(a2)と称することがある);及び(c)カルシウム結合タンパク質結合ペプチド、又はカルシウム結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムを放出するものである。また、光駆動カルシウムイオン放出収融合タンパク質は、カルシウム結合タンパク質N末側ペプチド等(a1)、光受容体ペプチド等(b)、カルシウム結合タンパク質C末側ペプチド等(a2)、及びカルシウム結合タンパク質結合ペプチド(c)からなってもよく、更に前記ペプチドリンカー等、及び/又はシグナルペプチドを含んでもよい。
【0041】
カルシウムイオン吸収融合タンパク質の1つの態様として、(a1)カルモジュリンN末側ペプチド、又はその改変体;(b)(以下、カルモジュリンN末側ペプチド等(a1)と称することがある);LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体(以下、LOV2ドメイン等(b)と称する);(a2)カルモジュリンC末側ペプチド、又はその改変体(以下、カルモジュリンC末側ペプチド等(a2)と称することがある);及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体(以下、M13ペプチド等(c)と称する)、をN末端から順に含み、光刺激によりカルシウムを放出する融合タンパク質を挙げることができる。具体的には、カルシウムイオン放出融合タンパク質として、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、又は配列番号15で表されるアミノ酸配列からなる融合タンパク質を挙げることができる。
【0042】
LOV2ドメインが挿入される、カルモジュリンN末側ペプチド等(a1)と、カルモジュリンC末側ペプチド等(a2)との切断位置は、カルモジュリンN末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体のカルシウムイオン結合能が阻害されない限り、限定されるものではなく、例えばカルモジュリンの20〜130番に存在する連続する2アミノ酸の間であればよい。例えば、25〜55番に存在する任意の2アミノ酸の間で切断してよく、30〜50番に存在する任意の2アミノ酸の間で切断してよく、35〜55番に存在する任意の2アミノ酸の間で切断してもよい。
【0043】
従って、カルモジュリンN末側ペプチド等(a1)と、カルモジュリンC末側ペプチド等(a2)のアミノ酸の鎖長も限定されるものはないが、例えばカルモジュリンN末側ペプチド等(a1)は、1〜20番のアミノ酸からなるペプチドから1〜130番のアミノ酸からなるペプチドであってよく、1〜20番のアミノ酸からなるペプチドから1〜60番のアミノ酸からなるペプチドであってよく、1〜30番のアミノ酸からなるペプチドから1〜50番のアミノ酸からなるペプチドであってよく、1〜40番のアミノ酸からなるペプチドであってよい。また、カルモジュリンC末側ペプチド等(a2)は、21〜150番のアミノ酸からなるペプチドから131〜150番のアミノ酸からなるペプチドであってよく、21〜150番のアミノ酸からなるペプチドから61〜150番のアミノ酸からなるペプチドであってよく、31〜150番のアミノ酸からなるペプチドから51〜150番のアミノ酸からなるペプチドであってよく、41〜150番のアミノ酸からなるペプチドであってよい。但し、カルモジュリンN末側ペプチド等(a1)のC末端と、カルモジュリンC末側ペプチド等(a2)のN末端は、実質的に元のカルモジュリンのアミノ酸配列におけるアミノ酸配列の重複及び欠失を含まないものする。ここで、実質的にアミノ酸配列の重複及び欠失を含まないとは、カルモジュリンN末側ペプチド等(a1)、カルシウムイオン吸収融合タンパク質におけるカルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体の形成に影響を与えないものであればよいが、具体的には、10アミノ酸以下であり、好ましくは5アミノ酸以下である。
また、カルモジュリンN末側ペプチド等(a1)及びカルモジュリンC末側ペプチド等(a2)の関係は、カルシウム結合タンパク質N末側ペプチド等(a1)及びカルシウム結合タンパク質C末側ペプチド等(a2)の関係にも当てはまるものである。
【0044】
本発明のカルシウムイオン放出融合タンパク質の機能的等価改変体は、実質的にカルシウムイオン放出融合タンパク質と同じ活性を有するものである。「カルシウムイオン放出融合タンパク質と同じ活性」とは、光照射によりカルシウムイオンを放出し、そしてその光照射の励起波長が実質的に同じであることを意味する。例えば、カルモジュリンC末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体のカルシウムイオン結合能が維持されており、そして光刺激により複合体のカルシウムイオンとの結合能が消失することを意味する。そして、励起波長として300〜500nmを用いることによって、機能的等価改変体がカルシウムイオン放出できることを意味する。
【0045】
本発明のカルシウムイオン放出融合タンパク質は、前記のように(a1)カルモジュリンN末側ペプチド、又はその改変体、(b)LOV2ドメイン、又はLOV2ドメイン改変体、(a2)カルモジュリンC末側ペプチド、又はその改変体、及び(c)M13ペプチド、又はM13ペプチド改変体が、N末端から順に連結された構造をとっている。このカルシウムイオン放出融合タンパク質は、光照射されない状態では、カルモジュリンN末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)及びM13ペプチド等(c)が複合体を形成し、解離定数が小さくなりカルシウムイオンと強固に結合できる構造となっていると推定される。一方、光照射された場合、カルモジュリンN末側ペプチド等(a1)及びカルモジュリンC末側ペプチド等(a2)の間に組み込まれたLOV2ドメイン等(b)の構造が変化し、カルモジュリンN末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体の結合が維持できなくなり、解離定数が大きくなることによって、カルシウムイオンを放出するものと推定される。また、カルシウム結合タンパク質N末側ペプチド等(a1)、光受容体ペプチド等(b)、カルシウム結合タンパク質C末側ペプチド等(a2)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)との関係も、それぞれカルモジュリンN末側ペプチド等(a1)、LOV2ドメイン等(b)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)との関係と同じであると推定される。しかしながら、本発明は前記の推定により限定されるものではない。
【0046】
(リンカー及び制限酵素サイト)
本発明のカルシウムイオン吸収融合タンパク質において、カルシウム結合タンパク質N末側ペプチド等(a1)(例えば、カルモジュリンN末側ペプチド等(a1))と光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))との間、光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))とカルシウム結合タンパク質C末側ペプチド等(a2)(例えば、カルモジュリンC末側ペプチド等(a2))との間、及びカルシウム結合タンパク質C末側ペプチド等(a2)(例えば、カルモジュリンC末側ペプチド等(a2))とカルシウムイオン結合タンパク質結合ペプチド等(c)(例えば、M13ペプチド等(c))との間には、カルシウムイオン吸収融合タンパク質の機能を阻害しない限りにおいて、適当な長さのペプチドリンカー又は制限酵素サイトのヌクレオチド配列から翻訳される2〜数アミノ酸の配列を含むことができる。
ペプチドリンカーしては、例えば「G」、「GG」、「GGS」、「GGGS」、「GGGGS」、「GGGSK」、「GGSGG」、「GGGGSK」、又は「GGSGGSGGS」を挙げることができる。また、制限酵素サイトのヌクレオチド配列から翻訳されるアミノ酸配列としては、LE及びGTなどを含むことができる。
例えば、後述の実施例に示すように、カルモジュリンN末側ペプチド等とLOV2ドメイン等(b)との間に、「G」、「GG」、「GGS」、又は「GGGS」のペプチドリンカーを含んでもよい。また、カルモジュリンC末側ペプチド等(a2)とM13ペプチド等(c)との間に、「GGGGS」のペプチドリンカーを含んでもよい。
【0047】
本発明によるDNAは、本発明の光駆動カルシウムイオン制御タンパク質又はその機能的等価改変体をコードする限り、特に限定されるものではなく、例えば、配列表の配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、又は配列番号30で表される塩基配列からなるDNAを挙げることができる。
配列表の配列番号16で表される塩基配列からなる前記DNAは、配列表の配列番号1で表されるアミノ酸配列からなるPacman(カルシウムイオン吸収融合タンパク質)をコードする。また、配列表の配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、及び配列番号22で表される塩基配列からなる前記DNAは、それぞれ、配列表の配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、及び配列番号7で表されるアミノ酸配列からなるカルシウムイオン吸収融合タンパク質をコードする。
配列表の配列番号23で表される塩基配列からなる前記DNAは、配列表の配列番号8で表されるアミノ酸配列からなるCARBY(カルシウムイオン放出融合タンパク質)をコードする。また、配列表の配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、及び配列番号30で表される塩基配列からなる前記DNAは、それぞれ、配列表の配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、及び配列番号15で表されるアミノ酸配列からなるカルシウムイオン放出融合タンパク質をコードする。
【0048】
本発明によるプラスミドは、本発明による前記DNAを含む限り、特に限定されるものではなく、例えば、用いる宿主細胞に応じて適宜選択した公知の発現ベクターに、本発明による前記DNAを挿入することにより得られるプラスミドを挙げることができる。
得られたプラスミドを所望の宿主細胞に形質転換することにより、形質転換体を得ることができる。前記形質転換体も、本発明による前記プラスミドを含む限り、特に限定されるものではない。
【0049】
前記宿主細胞としては、例えば、通常使用される公知の微生物、例えば、大腸菌又は酵母(Saccharomyces cerevisiae)、あるいは、公知の培養細胞、例えば、動物細胞(例えば、CHO細胞、HEK−293細胞、又はCOS細胞)又は昆虫細胞(例えば、BmN4細胞)を挙げることができる。
【0050】
また、公知の前記発現ベクターとしては、例えば、大腸菌に対しては、pUC、pTV、pGEX、pKK、又はpTrcHisを;酵母に対しては、pEMBLY又はpYES2を;CHO細胞に対してはpcDNA3又はpMAMneoを;HEK−293細胞に対してはpcDNA3を;COS細胞に対してはpcDNA3を;BmN4細胞に対しては、カイコ核多角体ウイルス(BmNPV)のポリヘドリンプロモーターを有するベクター(例えば、pBK283)を挙げることができる。
【0051】
[2]カルシウムイオン濃度の制御方法
本発明のカルシウムイオン濃度の制御方法は、前記光駆動カルシウムイオン制御タンパク質若しくはその機能的等価改変体、又は前記DNAを用いる。そして光駆動カルシウムイオン制御タンパク質又はその機能的等価改変体に、励起光を照射することにより、カルシウムイオンの濃度を制御する方法である。
本発明のカルシウムイオンの濃度制御方法において、用いる励起波長は光駆動カルシウムイオン制御タンパク質に使用されている光受容体ペプチドの種類により、適宜選択することができる。すなわち、光刺激により立体構造が変化する励起波長を用いることができる。例えば、青色光受容体であるフォトトロピン1、又はフォトトロピン2のLOV1ドメイン又はLOV2ドメイン、又はオーレオクロム1、又はオーレオクロム2のLOVドメインを光受容体ペプチドとして用いた場合は、300〜500nmの励起波長を用いることが可能であり、350〜490nmが好ましく、430〜470nmがより好ましい。
前記カルシウムイオン吸収融合タンパク質を用いることにより、周囲のカルシウムイオン濃度を低下させることができる。また、カルシウムイオン放出融合タンパク質を用いることにより、周囲のカルシウムイオン濃度を上昇させることができる。
更に、前記シグナルペプチドを含む光駆動カルシウムイオン制御タンパク質を用いることによって、細胞内の特定の領域又は細胞内小器官内若しくは細胞内小器官の膜上でカルシウムイオン濃度を制御することが可能である。具体的には、ミトコンドリア膜間スペース、ゴルジ体小胞体、ペルオキシソーム、核、ミトコンドリアのマトリックス、細胞膜、及び葉緑体などにおいて、カルシウムイオン濃度を制御することができる。
【0052】
[3]光駆動カルシウムイオン制御タンパク質等の利用
本発明の光駆動カルシウムイオン制御タンパク質若しくはその機能的等価改変体、又は前記DNAは、カルシウムイオン代謝異常を伴う疾患の治療又は予防用医薬組成物として用いることができる。カルシウムイオン代謝異常を伴う疾患としては、中枢神経系疾患(例えば、脳卒中、頭部外傷、脊髄損傷、外傷性ショック、アルツハイマー病、パーキンソン病、ハンチントン病、筋萎縮側索硬化症、てんかん、発作、痙攣性疾患、低酸素誘因神経細胞損傷、疼痛、うつ病、不安神経症、パニック障害、脅迫神経症、外傷後ストレス障害、麻酔後認識衰退、オピオイド耐性、薬物乱用、アルコール依存症、統合失調症)、又は循環器系疾患(例えば、高血圧、狭心症、不整脈)を挙げることができる。
【実施例】
【0053】
以下、実施例によって本発明を具体的に説明するが、これらは本発明の範囲を限定するものではない。
【0054】
《実施例1》
本実施例1では、カルシウムイオン吸収融合タンパク質であるPacmanを製造した。
(1)LOV2遺伝子の調製
融合タンパク質の構築に用いるLOV2ドメインをコードするDNAを調製した。Leeらの報告(Lee J. et al. Surface sites for engineering allosteric control in proteins.Science.17. 438-42 2008)を参考にして、以下のLOV2の骨格オリゴヌクレオチド、アダプターオリゴヌクレオチド、及びPCRに用いるプライマーを合成した。
【0055】
(骨格オリゴヌクレオチド)
LOV2-skel1 5’-TCGAGCTGGCCACCACTCTAGAGCGCATCGAGAAGAACTTCGTGATCACC-3’(配列番号37)
LOV2-skel2 5’-GACCCCCGCCTGCCCGACAACCCCATCATCTTCGCCTCCGACTCC-3’(配列番号38)
LOV2-skel3 5’-TTCCTGCAGCTGACCGAGTACTCCCGCGAGGAGATCCTGGGCCGCAAC-3’(配列番号39)
LOV2-skel4 5’-TGCCGCTTCCTGCAGGGCCCCGAGACCGACCGCGCCACCGTGCGC-3’(配列番号40)
LOV2-skel5 5’-AAGATCCGCGACGCCATCGACAACCAGACCGAGGTGACCGTG-3’(配列番号41)
LOV2-skel6 5’-CAGCTGATCAACTACACCAAGTCCGGCAAGAAGTTCTGGAACCTGTTC-3’(配列番号42)
LOV2-skel7 5’-CACCTGCAGCCCATGCGCGACCAGAAGGGCGACGTGCAGTACTTCATC-3’(配列番号43)
LOV2-skel8 5’-GGCGTGCAGCTGGACGGCACCGAGCACGTGCGCGACGCCGCC-3’(配列番号44)
LOV2-skel9 5’-GAGCGCGAGGGCGTGATGCTGATCAAGAAGACCGCCGAGAAC-3’(配列番号45)
LOV2-skel10 5’-ATCGACGAGGCCGCCAAGGAGCTGCCCGACGCCAACCTGCGC-3’(配列番号46)
LOV2-skel11 5’-CCCGAAGATCTGTGGGCCAACCACGGTAC-3’(配列番号47)
【0056】
(アダプターオリゴヌクレオチド)
LOV2-adop1 5’-GGGGTCGGTGAT-3’(配列番号48)
LOV2-adop2 5’-CAGGAAGGAGTC-3’(配列番号49)
LOV2-adop3 5’-GCGGCAGTTGCG-3’(配列番号50)
LOV2-adop4 5’-GATCTTGCGCAC-3’(配列番号51)
LOV2-adop5 5’-CAGCTGCACGGT-3’(配列番号52)
LOV2-adop6 5’-CAGGTGGAACAG-3’(配列番号53)
LOV2-adop7 5’-CACGCCGATGAA-3’(配列番号54)
LOV2-adop8 5’-GCGCTCGGCGGC-3’(配列番号55)
LOV2-adop9 5’-GTCGATGTTCTC-3’(配列番号56)
LOV2-adop10 5’-TTCGGGGCGCAG-3’(配列番号57)
【0057】
(プライマー)
LOV2-fw 5’-TTT CTCGAG CTG GCC ACC ACT CTA-3’(配列番号58)
LOV2-rv 5’-TT GGTACC GTG GTT GGC CCA CA-3’(配列番号59)
【0058】
前記骨格オリゴヌクレオチドLOV2−skel1〜11を、1μMモル濃度に調整し、5’末端をT4ポリヌクレオチドキナーゼによってリン酸化した。その後、T4ポリヌクレオチドキナーゼを70℃/10minにて失活した。得られたオリゴヌクレオチドのリン酸化処理産物に、アダプターオリゴヌクレオチドLOV2−adop1〜10とT4DNALigaseを加え、16℃でO/Nのライゲーションを行った。すなわち、LOV2−skel1〜11は、それぞれ終濃度0.5μMになるように調製し、LOV2−adop1〜10は、それぞれ終濃度5μMになるように調整し、混合した。この混合物10.5μLに2XRapid Ligation Buffer(NEB)を12μL及びT4 DNAライゲースを1.5μLとを加え、16℃でO/Nのライゲーションを行った。得られたライゲーション産物を鋳型とし、フォワードプライマーLOV2−fwと、リバースプライマーLOV2−rvを用いてPCRを行った。PCRサイクルは94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間、保持した。ポリメラーゼは、KOD−Plus(TOYOBO)を用いた。PCR産物をフェノール/クロロホルム抽出にて精製し、XhoI/KpnIで制限酵素処理を行った。制限酵素処理産物を電気泳動した後、目的のバンドを切り出し、QIAEXIIGel extract kit(QIAGEN)を用いて精製を行った。得られたフラグメントを、試料1とする。プラスミドベクターpRSETB(インビトロジェン社製(Invitrogen))を、同様にXhoI/KpnIで制限酵素処理を行い、電気泳動により精製した。得られたXhoI/KpnI切断ベクターを、試料2とする。試料1と試料2を混和し、T4DNAライゲースでライゲーションし、LOV2遺伝子を導入したpRSETBを得た。このプラスミドベクターを、「pRSETB/LOV2」と称する。
【0059】
(2)pRSETB/CaM-LOV2の構築
CaM及びLOV2を含むベクターpRSETB/CaM-LOV2の構築を行った。
用いたプライマーを以下に示す。
BHI-CaM-fw 5’-CG GGATCCC CGC ATG CAT GAC CAA CTG-3’(配列番号60)
CaM-ERI-rv 5’-G GAATTC TTA CTT TGC TGT CAT CAT TTG-3’(配列番号61)
CaM及びM13ペプチドを含むベクターpRSETB/YC3.6(Nagai T. et al Expanded dynamic range of fluorescent indicators for Ca(2+) by circularly permuted yellow fluorescent proteins. Proc Natl Acad Sci U S A. 20. 10554-9 2004)を鋳型として、フォワードプライマーBHI−CaM−fw及びリバースプライマーCaM−XhoI−rvを用いてPCRを行った。PCRサイクルは94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間である。PCR産物を、フェノール/クロロホルム抽出し、BamHI/XhoIで制限酵素処理し、電気泳動によって、ゲル精製を行った。得られたCaMのフラグメントを試料3とする。前記(1)の工程で得られたpRSETB/LOV2を、BamHI/XhoIで制限酵素処理し、電気泳動によりゲル精製を行った。得られたLOV2を含むベクターを試料4とする。試料3と試料4を混和し、T4DNAライゲースで、ライゲーションしてカルモジュリンをLOV2の5’側に持つ融合遺伝子を得た、このプラスミドベクターを、「pRSETB/CaM−LOV2」と称する。
【0060】
(3)Pacman遺伝子の構築
図3の模式図に示したPacman遺伝子を構築するために、LOV2ドメインを含む前記(1)の工程で得られたベクターpRSETB/LOV2、及びCaM及びM13ペプチドを含むベクターpRSETB/YC3.6(Nagai T. et al Expanded dynamic range of fluorescent indicators for Ca(2+) by circularly permuted yellow fluorescent proteins. Proc Natl Acad Sci U S A. 20. 10554-9 2004)を鋳型として、PCRを行い、得られたフラグメントをpRSETB/CaM−LOV2に組み込んだ。使用したプライマーを以下に示す。
LOV2-fw-GGGGS-XhoI 5’-TTT CTCGAG GGAGGTGGAGGTAGT CTG GCC ACC ACT CT-3’(配列番号62)
LOV2-Cd17-M13 5’-GAC GAG GCC GCC AGG CGC TGG AAG -3’(配列番号63)
M13-ERI-rv 5’-G GAATTC TTA GAG CTC CAG TGC CCC GGA-3’(配列番号64)
【0061】
PCRの鋳型として、pRSETB/LOV2及びpRSETB/YC3.6を用いた。プライマーとして、LOV2−fw−GGGGS−XhoI及びM13−ERI−rvを終濃度0.3μMになるように添加し、LOV2−Cd17−M13を終濃度0.03μMとなるように添加した。PCRのサイクルは、94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間、保持した。
なお、本PCRでは、初期段階において、主にpRSETB/YC3.6を鋳型にして、フォワードプライマーLOV2−Cd17−M13及びリバースプライマーM13−ERI−rvによって、一段階目のM13ペプチドを含む断片の増幅が起こる。この断片の5’端側にはLOV2−CD17のC末端側の配列がコードされている。この断片が蓄積すると、pRSETB/LOV2を鋳型にして、フォワードプライマーLOV2−fw−GGGGS−XhoI3、及びリバースプライマーとして一段階目のM13ペプチドを含む断片の相補鎖を用いた二段階目の増幅が起き、LOV2ドメイン及びM13ペプチドを含む全長の配列が得られる。さらに、この全長の配列を鋳型にしてフォワードプライマーLOV2−fw−GGGGS−XhoI3及びリバースプライマーM13−ER−rvを用いた増幅により、目的の増幅断片が得られる。
得られたPCR産物をフェノール/クロロホルム抽出し、XhoI/EcoRIで制限酵素処理し、電気泳動により、ゲル精製を行った。得られたフラグメントを、試料5とした。次に、pRSETB/CaM−LOV2を、XhoI/EcoRIで制限酵素処理し、電気泳動により、ゲル精製を行った。得られた切断ベクターを試料6とする。試料5と試料6を混和し、T4DNAライゲースで、ライゲーションしてカルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子を得た。得られたベクターを「pRSETB/Pacman」と称する。
なお、この融合遺伝子から得られる融合タンパク質は、カルモジュリンとLOV2△C17との間にXhoIサイトから翻訳される2個のアミノ酸LEを含む。
【0062】
《実施例2》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を14個欠失したLOV2(LOV2△C14)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd14-M13を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd14-M13 5’-GAG GCC GCC AAG GAG CTG AGG CGC TGG AGG-3’(配列番号65)
【0063】
《実施例3》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を15個欠失したLOV2(LOV2△C15)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd15-M13を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd15-M13 5’-GAG GCC GCC AAG GAG AGG CGC TGG AAG-3’(配列番号66)
【0064】
《実施例4》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を16個欠失したLOV2(LOV2△C16)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd16-M13を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd16-M13 5’-GAG GCC GCC AAG AGG CGC TGG AAG-3’(配列番号67)
【0065】
《実施例5》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。また、カルモジュリンとLOV2△C17との間のリンカーを、GGGGSからGGSに変更した。
プライマーLOV2-fw-GGGGS-XhoIに代えて、以下のLOV2-fw-GGS-XhoIを用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-fw-GGS-XhoI 5’-TTT CTCGAG GGAGGTAGT CTG GCC ACC ACT CT-3’(配列番号68)
【0066】
《実施例6》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。また、カルモジュリンとLOV2△C17との間のリンカーを、GGGGSからGに変更した。
プライマーLOV2-fw-GGGGS-XhoIに代えて、以下のLOV2-fw-G-XhoIを用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-fw-G-XhoI 5’-TTT CTCGAG GGT CTG GCC ACC ACT CT-3’(配列番号69)
【0067】
《実施例7》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を2個欠失したM13ペプチド(M13△N2)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd17-M13△N1を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd17-M13△N1 5’-GAC GAG GCC GCC CGC TGG AAG AAA-3’(配列番号70)
【0068】
《実施例8》
本実施例8では、カルシウムイオン放出融合タンパク質であるCARBYを製造した。
(1)pRSETB/CaM40−XhoI−KpnI−41CaM−M13の作製
CaM及びM13ペプチドが、GGGGSのリンカーで結合されているベクターpRSETB/YC−nano15(Horikawa K. et al. Spontaneous network activity visualized by ultrasensitive Ca(2+) indicators, yellow Cameleon-Nano.. Nat Methods.7.729-32. 2010)を用いて、カルモジュリンの第40番のアミノ酸と41番のアミノ酸との間に、XhoI及びKpnIの制限酵素サイトを導入したベクターpRSETB/CaM40−XhoI−KpnI−41CaM−M13を作製した。
【0069】
変異を導入するために以下のプライマーを合成した。
QC-CaM41-XhoI-TAA-KpnI 5’-G TCG CTT GGA CTCGAG TAA GGTACC CAA CCA ACG-3(配列番号71)
プライマーQC-CaM41-XhoI-TAA-KpnIの5’端を、T4ポリヌクレオチドキナーゼによってリン酸化した。得られたプライマーを用いて、PCR法により変異を含むPCR産物を調製した。pRSETB/YC−nano15を50ng、QC−CaM41−XhoI−TAA−KpnIを10pmol、dNTP3.75nmol、Pfu DNAポリメラーゼ(STRATAGENE社製)1.25U、Pfu DNA リガーゼ(STRATAGENE社製)を20Uを含む反応溶液40μLを準備した。まず、65℃、5分のプレインキュベーションを行ってPfu DNA リガーゼにより、鋳型DNAのニックを修復し、その後94℃、1分の最初の変性を行った。PCRのサイクルは、94℃、10秒のDNA変性、55℃、30秒のアニーリング反応及び65℃、10分の伸長・連結反応を1サイクルとして35サイクル行った。最後に75℃、10分のポストインキュベーションを行った。
【0070】
得られたPCR産物のうち、変異が含まれていない断片を、DpnIを用いて切断した。すなわち、前記PCR後の反応溶液40μLに1μL(20U)のDpnI(New England BioLab社製)を加え、37℃で2時間インキュベートすることによって、メチル化またはヘミメチル化した鋳型プラスミドDNAを選択的に消化した。DpnIは[5’−Gm6ATC−3’]を認識して2本鎖DNAを切断するエンドヌクレアーゼである。
【0071】
次に、得られた反応液を用いて、メガプライマーを用いた突然変異が導入されたDNAの相補鎖の合成を行った。
前記反応溶液41μLを94℃、30秒で変性させ、次いで95℃で30秒、55℃で1分、65℃で10分の3サイクルのPCRを行った。このとき反応溶液に含まれるDNA断片のうちのいくつかが、メガプライマーとして作用する。この2本鎖突然変異導入環状DNA鎖反応溶液41μLを、エタノール沈殿し、沈殿物を乾燥させた。
【0072】
前記2本鎖突然変異導入環状DNA鎖を鋳型とし、フォワードプライマーBHI−CaM−fw及びリバースプライマーM13−ERI−rvを用いてPCRを行った。PCR産物を、フェノール/クロロホルム抽出し、BamHI/EcoRIで制限酵素処理し、電気泳動によりゲル精製を行った。得られたフラグメントを、試料7とする。ベクターpRSETBを、BamHI/EcoRIで制限酵素処理し、電気泳動によりゲル精製を行った。得られた切断ベクターを、試料8とする。試料7と試料8を混和し、T4 DNAライゲースで、ライゲーションしてカルモジュリンの40番目のアミノ酸と41番目のアミノ酸の間にXhoI制限酵素サイト、終始コドン、KpnI制限酵素サイトを持ち、カルモジュリンのC末端側にリンカーとしてGGGGSを持ち、更にそのC末端側にM13を持つ融合遺伝子を得た、このプラスミドベクターを「pRSETB/CaM40−XhoI−KpnI−41CaM−M13」と称する。
【0073】
(2)CARBYの作製
ベクターpRSETB/CaM40−XhoI−KpnI−41CaM−M13を、XhoI/KpnIで制限酵素処理し、電気泳動によってゲル精製を行った。得られた切断ベクターを試料9とする。試料9と前記実施例1の工程(1)で得られた試料1を混和し、T4 DNAライゲースで、ライゲーションしてpRSETB/CaM40−XhoI−KpnI−41CaM−M13のカルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入された融合遺伝子を得た。すなわち、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを持つ、融合遺伝子を得た。得られたベクターを「pRSETB/CARBY」と称する。
なお、この融合遺伝子から得られる融合タンパク質は、カルモジュリン(1〜40番目のアミノ酸)とLOV2との間のXhoI制限酵素サイトから翻訳される2個のアミノ酸LE、及びLOV2とカルモジュリン(41〜150番目のアミノ酸)との間のKpnI制限酵素サイトから翻訳される2個のアミノ酸GTとを含む。
【0074】
《実施例9》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、C末端アミノ酸を3個欠失したLOV2(LOV2△C3)、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。
前記pRSETB/LOV2を鋳型として、以下のプライマーによってPCRを行った。
LOV2−fw
3'LOV2-Nd3-KpnI 5’-GG GGTACC CCA CAG ATC TTC GGG GCG-3’(配列番号72)
pRSETB/LOV2を鋳型として、フォワードプライマーLOV2−fw及びリバースプライマー3'LOV2-Nd3-KpnIを用いてPCRを行った。PCRサイクルは94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間である。PCR産物を、フェノール/クロロホルム抽出し、XhoI/KpnIで制限酵素処理し、電気泳動によって、ゲル精製を行った。得られたLOV2△3のフラグメントを試料10とする。
前記試料9と試料10を混和し、T4 DNAライゲースで、ライゲーションしてpRSETB/CaM40−XhoI−KpnI−41CaM−M13のカルモジュリンの40番目と41番目のアミノ酸の間にLOV2△3が挿入された融合遺伝子を得た。
【0075】
《実施例10》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、C末端アミノ酸を9個欠失したLOV2(LOV2△C9)、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。
リバースプライマー3'LOV2-Nd3-KpnIに代えて、3'LOV2-Nd9-KpnI 5’-GG GGTACC CAG GTT GGC GTC GGG CAG-3’(配列番号73)を用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2△9が挿入された融合遺伝子を得た。
【0076】
《実施例11》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、C末端アミノ酸を12個欠失したLOV2(LOV2△C12)、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。
リバースプライマー3'LOV2-Nd3-KpnIに代えて、3'LOV2-Nd12-KpnI 5’-GG GGTACC GTC GGG CAG CTC CTT GGC-3’(配列番号74)を用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2△12が挿入された融合遺伝子を得た。
【0077】
《実施例12》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-G-XhoIを用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGを有する融合遺伝子を得た。
【0078】
《実施例13》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-GG-XhoI 5’-TTT CTCGAG GGTAGT CTG GCC ACC ACT CT-3’(配列番号75)を用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGを有する融合遺伝子を得た。
【0079】
《実施例14》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGSを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-GGS-XhoIを用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGSを有する融合遺伝子を得た。
【0080】
《実施例15》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGGSを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-GGGS-XhoIを用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGGSを有する融合遺伝子を得た。
【0081】
《融合タンパク質の発現及び精製》
実施例1〜15において得られたベクターを用いて、ヒートショック法により大腸菌コンピテントセル(JM109(DE3))を形質転換した。形質転換した大腸菌細胞を、100μg/mLのアンピシリンを含むLB固体培地で、37℃にて培養し、単一コロニーをピックアップして100μg/mLのアンピシリンを含むLB培養液2mL中へ移し、37℃で一晩培養して増殖させた。融合タンパク質は、T7発現系(pRSETB/JM109(DE3))を用いて発現させた。すなわち、培養液の100倍希釈物を作製し、更に23℃で3日間培養した。得られた細胞をフレンチプレスにより破砕し、得られた上清液から、ニッケルキレートカラム(QIAGEN社製)を用いてポリヒスチジン標識化タンパク質を精製した。さらに、この溶出液(100mMイミダゾール、50mMTris−Cl pH7.4、300mMNaCl)中のタンパク質サンプルをPD10脱塩・バッファー交換カラム(GE healthcare Bio−Sciences社製)により、HEPES10mM(pH7.5)を用いて精製を行った。
【0082】
《融合タンパク質の吸収スペクトルの測定》
得られた融合タンパク質に含まれているLOV2が、LOV2本来の構造を維持していることを、吸収スペクトルから確認した。LOV2は、447nm、473nm、及び370nmに吸収極大が存在する。
実施例1〜15において得られたベクターを発現させて得られた融合タンパク質の吸収スペクトルのうち、実施例1のPacman及び実施例8のCARBYの吸収スペクトルを規格化したものを図1に示す。また、陽性コントロールとして、LOV2の吸収スペクトルも記載した(Imaizumi T. et al FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis. Nature 20. 302-6 2003 Nov 20;426(6964):302-6.)。図1中、(A)がPacman、(B)がCARBY、(C)がLOV2の規格化吸収スペクトルである。
本発明の光駆動カルシウムイオン制御タンパク質は、LOV2と同じ吸収スペクトルを示しており、融合したLOV2がその機能を維持していることを示している。
【0083】
更に、LOV2の吸収極大ε447=13,800 M−1cm−1(Strickland D. et al Light-activated DNA binding in a designed allosteric protein. Proc Natl Acad Sci U S A. 105. 10709-14 2008)を用いてタンパク質のモル濃度を決定した、その後タンパク質のモル濃度をPacmanは100μMに合わせ、CARBYは200μMに合わせてサンプル溶液とした。
【0084】
《光照射によるカルシウムイオン濃度変化の測定》
本発明の光駆動カルシウムイオン制御タンパク質が、光照射により周囲のカルシウムイオン濃度を制御可能であることを確認した。
カルシウムイオン濃度の測定は、測定用試薬としてカルシウムイオン指示薬を用いて行った。カルシウムイオン指示薬は、Rhod−2−AM(同仁堂)を、エステラーゼ・ブタ肝臓由来(Sigma)で処理した化合物であるRhod−2を用いた。終濃度100μMになるようにRhod−2−AMを10mMのHEPES(pH8.0)に溶解し、エステラーゼを10U加え、室温でO/N放置した後、10mMのHEPES(pH7.5)で10倍希釈し、10μMのRhod−2を得た。
【0085】
前記精製した融合タンパク質を含むサンプル溶液を、以下の測定条件に従って、測定した。10mMのHEPES(pH7.5)を80μL、サンプル溶液を10μL、10μMのRhod−2を10μLを石英セルに滴下し、分光蛍光光度計であるF2500(HITACHI)を用いて測定を行った。測定条件は蛍光スペクトルモード、励起波長は550nm、スキャンスピードは1500nm/min、初期待ち時間は0秒、励起側/蛍光側スリットは5nm/5nm、ホトマル電圧は700V、レスポンスは0.04秒である。最初に、前記の条件で測定したのち、励起波長を450nmに変更し10秒光刺激した後、励起波長を550nmに戻して、再度蛍光スペクトルを測定した、その後シャッターを閉め、1分間待ち、シャッターをあけて、再度蛍光スペクトルを測定した。
Pacmanでは570nmにおける蛍光強度が40%程度減少した後にその蛍光強度が回復することが観測され(図2A)、CARBYでは570nmにおける蛍光強度が15%程度増加した後にその蛍光強度が回復することが観測された(図2B)。これはPacmanでは溶液中のカルシウムイオン濃度が光励起により低下し、時間経過と共に回復したことを示しており、CARBYでは溶液中のカルシウムイオン濃度が光励起により上昇し、時間経過と共に低下したことを示している。Rhod−2の570nmにおける蛍光強度から計算したカルシウムイオン濃度は、Pacmanの場合は約400nmが約200nmに、CARBYの場合は約1.3μMが約2μMに変化していた。また、実施例2〜7、及び実施例9〜15の融合タンパク質のカルシウムの吸収(結合)又はカルシウムの放出については、表2及び表3に示す。
【0086】
カルモジュリン、M13ペプチド、及びLOV2ドメインを、適切な順番で結合させることにより、カルシウムイオン指示薬であるRhod−2の励起光/蛍光を、550nm/570nmで測定した時の蛍光強度が、450nmの光刺激により蛍光強度が低下すること、又は蛍光強度が増加することが確認できた。すなわち、本発明の光駆動カルシウムイオン制御タンパク質は光刺激によってカルシウムイオン濃度を低下、又は上昇させることができる。
【0087】
【表2】
【0088】
【表3】
【産業上の利用可能性】
【0089】
本発明の光駆動カルシウムイオン制御タンパク質は、カルシウムイオンの生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中における濃度を制御することが可能であり、カルシウムイオンの細胞等における機能の解析に用いることができる。また、カルシウムイオンの濃度を制御することにより、それらの濃度異常を原因とする疾患の治療、又は予防に用いることも可能である。
【技術分野】
【0001】
本発明は、光駆動カルシウムイオン制御タンパク及びカルシウムイオン濃度の制御方法に関する。本発明によれば、カルシウムイオン濃度を制御することができ、例えば、細胞内におけるカルシウムイオンの濃度を上昇させること、又は低下させることができる。
【背景技術】
【0002】
カルシウムイオンは、通常、細胞内液にはほとんど存在せず、細胞外からのカルシウムイオンの流入、又は細胞内の小胞体等に蓄えられたカルシウムイオンの放出などによって、細胞内の濃度が上昇し、細胞内の様々な生理活性のシグナルとしての役割をはたしている。この細胞内のカルシウムイオン濃度の変化によって、筋肉収縮、神経の情報伝達、短期記憶、長期記憶、神経の成長、免疫系細胞の活性化、胃酸・胃液・腸粘液・汗等の分泌、酵素の活性化、アポトーシス、及び細胞分裂など、生体の多くの生理現象がコントロールされている。
【0003】
従って、細胞内で起きるカルシウムイオンの濃度変化と、生体の生理現象との関係は、正常な生体の機能ばかりではなく、多くの疾患との関連性も指摘されている。そのため、細胞内で起きるカルシウムイオンの濃度変化を解析することは、非常に重要である。特に、カルシウムイオン濃度の変化は、細胞内の特定の領域又は細胞内小器官付近で起きることもあり、細胞内の任意の位置でカルシウムイオン濃度を制御する実験方法の開発が期待されている。
【0004】
従来、カルシウムイオンによる細胞又は生体への影響を調べる方法として、光照射によりカルシウムイオンを放出するカルシウムイオンキレート剤である光駆動ケージドカルシウムイオン化合物(例えば、nitrophenyl−EGTA:以下、NP−EGTAと称する)を用いる方法が知られている(非特許文献1)。NP−EGTAは、マイクロインジェクションにより、細胞内に注入し、光照射によりカルシウムイオンを放出させ、細胞内のカルシウムイオン濃度を上昇させることができる。しかしながら、マイクロインジェクションによる細胞への導入は煩雑な手技を要し、また導入したNP−EGTAを細胞内の任意の位置にとどめることは困難であり、細胞内の特定の領域のカルシウムイオン濃度を制御することは不可能であった。更に、励起時に紫外光を用いなければならず、細胞に対して害を与える可能性があった。
一方、光照射によって細胞外からカルシウムイオンを流入させる光駆動カルシウムイオントランスポーターを用いる方法が開発された(非特許文献2)。しかしながら、この方法も、カルシウムイオンを細胞外から流入させるため、カルシウムイオン濃度の変化が、細胞全体で起こるものであった。従って、細胞内の任意の領域又は位置でのカルシウムイオン濃度の変化に対する応答を調べることは、原理上不可能であった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】「プロシーディング・オブ・ザ・ナショナル・アカデミー・オブ・サイエンス・ユーエスエー(Proceedings of the National Academy of Sciences)」(米国)1994年、第91巻、p.187−91
【非特許文献2】「ネイチャー(Nature)」(英国)2002年、第28巻、p.398−401
【発明の概要】
【発明が解決しようとする課題】
【0006】
前記の光駆動ケージドカルシウムイオン化合物(例えば、NP−EGTA)には、マイクロインジェクションが不要な膜透過性のものも開発されているが、膜透過性の化合物でも、細胞内の局所におけるカルシウムイオンの濃度の制御は困難である。また、前記のように、光駆動カルシウムイオントランスポーターは、細胞外からのカルシウムイオンの流入を行うものであり、やはり細胞内におけるカルシウムイオンの濃度の制御は不可能である。
従って、本発明の目的は、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中において、光刺激によりカルシウムイオン濃度を制御することができる化合物及び方法を提供することである。具体的には、カルシウムイオン濃度を上昇又は低下させることが可能な化合物及び方法を提供することである。
【課題を解決するための手段】
【0007】
本発明者らは、カルシウムイオン濃度を制御することができる実験方法について、鋭意研究した結果、驚くべきことに、カルシウムイオン結合タンパク質、光励起によって立体構造が変化する光受容体ペプチド、カルシウムイオン結合タンパク質に結合するカルシウムイオン結合タンパク質結合ペプチドを含む融合タンパク質を用いることにより、カルシウムイオン濃度を低下させること、又は上昇させることが可能であることを見出した。すなわち、光刺激により光受容体ペプチドの構造が変化することを利用して、カルシウムイオン結合タンパク質及びカルシウムイオン結合タンパク質結合ペプチドの構造を調整し、融合タンパク質にカルシウムイオンを結合させること、又は融合タンパク質からカルシウムイオンを放出させることが可能であることを見出した。従って、これらの融合タンパク質を用いることによって、細胞内などのカルシウムイオン濃度を制御することが可能である。
前記カルシウムイオン結合タンパク質として、カルモジュリンが最もよく研究されているが、従来、カルモジュリンを用いた融合タンパク質としては、細胞内のカルシウムイオン濃度を測定することのできる融合タンパク質が、本発明者らによって報告されている(Nature Methods, vol.7, No.9, p.729-732)。この融合タンパク質は、カルモジュリン、カルモジュリンと結合するM13ペプチド、及び蛍光タンパク質からなる融合タンパク質であり、カルモジュリンへのカルシウムイオンの結合により、蛍光タンパク質の構造が変化し、カルシウムイオンの濃度を測定することができるプローブ(融合タンパク質)である。しかしながら、このカルシウムイオン濃度測定のプローブ(融合タンパク質)は、カルモジュリン及びM13の複合体にカルシウムイオンが結合することにより、蛍光タンパク質の構造を変化させ、それによって蛍光タンパク質の蛍光強度が変化することを利用するものである。一方、本発明の融合タンパク質は、光刺激により光受容体ペプチドの構造が変化することを利用して、カルモジュリン及びM13の複合体と、カルシウムイオンとの結合を制御するものであり、カルモジュリン、M13ペプチド及び蛍光タンパク質からなる融合タンパク質とは、全く異なるものである。更に、本発明者らは、カルシウムイオンと結合する融合タンパク質と、カルシウムイオンを放出するタンパク質とを調製することができたが、この2種類の融合タンパク質は、カルモジュリン、M13ペプチド、及び光受容体ペプチドの3つのタンパク質の配置を変更することのみによって、得られたものであり、驚くべきことである。
本発明は、こうした知見に基づくものである。
【0008】
すなわち、本発明は、
[1](a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;を含む、光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[2](a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムと結合する、[1]に記載の光駆動カルシウムイオン制御タンパク質(以下、カルシウムイオン吸収融合タンパク質と称することがある)、又はその機能的等価改変体(以下、まとめてカルシウムイオン吸収融合タンパク質等と称することがある)、
[3](a1)カルシウムイオン結合タンパク質N末側ペプチド、又はその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;(a2)カルシウムイオン結合タンパク質C末側ペプチド、又はその改変体;及び(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムを放出する、[1]に記載の光駆動カルシウムイオン制御タンパク質(以下、カルシウムイオン吸収融合タンパク質と称することがある)、又はその機能的等価改変体(以下、まとめてカルシウムイオン吸収融合タンパク質等と称することがある)、
[4]前記光受容体ペプチドが、フォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、及びFKF−1のLOVドメインからなる群から選択される光受容体ペプチドであり;
前記カルシウムイオン結合タンパク質及びカルシウムイオン結合タンパク質結合ペプチドの組み合わせが、カルモジュリン及びカルモジュリン結合ペプチド、トロポニンC及びトロポニンI、ミオシン軽鎖及びミオシンII重鎖、並びにレコベリン及びロドプシンキナーゼからなる群から選択される、[1]〜[3]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[5](a)カルモジュリン、又はカルシウムイオン結合能を有するカルモジュリン改変体;(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;をN末端から順に含み、光刺激によりカルシウムと結合する、[2]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[6]配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、又は配列番号7で表されるアミノ酸配列からなる、[5]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[7](a1)カルモジュリンN末側ペプチド、又はその改変体;(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;(a2)カルモジュリンC末側ペプチド、又はその改変体;及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;をN末端から順に含み、光刺激によりカルシウムを放出する、[3]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[8]配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、又は配列番号15で表されるアミノ酸配列からなる、[7]に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[9]シグナルペプチドを含む、[1]〜[8]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体、
[10][1]〜[9]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体をコードするDNA、
[11]配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、又は配列番号30で表される塩基配列からなる、[10]に記載のDNA、
[12][10]又は[11]に記載のDNAを含むプラスミド、
[13][1]〜[9]のいずれかに記載の光駆動カルシウムイオン制御タンパク質、若しくはその機能的等価改変体、又は[10]若しくは[11]に記載のDNAを用いて発現するタンパク質に、励起光を照射することを特徴とする、カルシウムイオン濃度の制御方法、
[14]前記励起光の励起波長が、300〜500nmである[13]に記載のカルシウムイオン濃度の制御方法、
に関する。
【発明の効果】
【0009】
本発明の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体によれば、カルシウムイオン濃度を制御すること、すなわちカルシウムイオン濃度を上昇、又は低下させることができる。
また、本発明のDNA又はプラスミドを用いることにより、細胞内において、容易に本発明の光駆動カルシウムイオン制御タンパク質を発現させることが可能である。従って、従来の光駆動ケージドカルシウムイオン化合物又は光駆動カルシウムイオントランスポーターと比較して、本発明の光駆動カルシウムイオン制御タンパク質は、細胞内での取り扱いが容易である。
更に、本発明の細胞内小器官ターゲット配列を有する光駆動カルシウムイオン制御タンパク質は、細胞内の特定の領域に移行(存在)させることが可能であり、細胞内の任意の位置でカルシウムイオン濃度を制御することができる。すなわち、本発明のカルシウムイオン制御タンパク質、又はその機能的等価改変体は、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中のいずれにおいても、使用することが可能であり、生体内等のカルシウムイオン濃度を容易に制御することができる。
【図面の簡単な説明】
【0010】
【図1】本発明のPacman(カルシウムイオン吸収融合タンパク質)及びCARBY(カルシウムイオン放出融合タンパク質)の吸収スペクトルを、LOV2ドメインの吸収スペクトルと比較したグラフである。
【図2】本発明のPacman(カルシウムイオン吸収融合タンパク質)又はCARBY(カルシウムイオン放出融合タンパク質)を用いて、光励起によりカルシウムイオン濃度が変化すること示したグラフである。
【図3】本発明のPacman(カルシウムイオン吸収融合タンパク質)の構造の模式図である。
【図4】本発明のCARBY(カルシウムイオン放出融合タンパク質)の構造の模式図である。
【発明を実施するための形態】
【0011】
[1]光駆動カルシウムイオン制御タンパク質
(光駆動カルシウムイオン制御タンパク質)
本発明の光駆動カルシウムイオン制御タンパク質は、(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体(以下、カルシウムイオン結合タンパク質等(a)と称することがある);(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体(以下、光受容体ペプチド等(b)と称することがある);及び(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質結合能を有するその改変体(以下、カルシウムイオン結合タンパク質結合ペプチド等(c)と称することがある);を含む。また、光駆動カルシウムイオン制御タンパク質は、カルシウムイオン結合タンパク質等(a)、光受容体ペプチド等(b)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)からなってもよく、更に後述のペプチドリンカー等、及び/又はシグナルペプチドを含んでもよい。。
【0012】
(カルシウムイオン結合タンパク質)
本発明のカルシウムイオン制御タンパク質に用いるカルシウムイオン結合タンパク質は、カルシウムイオン結合能を有する限り、特に限定されるものはないが、例えばカルモジュリン、トロポニンC、ミオシン軽鎖、又はレコベリンを挙げることができる。これらのカルシウムイオン結合タンパク質は、後述のカルシウムイオン結合タンパク質結合ペプチドとの複合体を形成することにより、カルシウムイオンとの親和性が上昇するタンパク質であり、本発明のカルシウムイオン制御タンパク質に好ましく用いることができる。また、カルシウムイオン結合タンパク質の由来も特に限定されるものではなく、すべての生物由来のカルシウムイオン結合タンパク質を使用することが可能である。
【0013】
前記カルシウムイオン結合タンパク質のうち、カルモジュリンは酸性のタンパク質で、約148残基のアミノ酸からなる。4つのEFハンドモチーフを有しており、それぞれにカルシウムイオンが結合する。カルモジュリンは、様々な細胞で発現し、細胞における存在部位も、細胞内小器官内及び膜上など様々であり、カルシウムイオンによるシグナル伝達に重要な役割を果たしているタンパク質である。
本発明のカルシウムイオン制御タンパク質に用いるカルモジュリンは、カルシウムイオン結合能を有する限り、特に限定されるものはなく、すべての生物由来のカルモジュリンを使用することが可能であり、例えば配列番号31で表されたアミノ酸配列からなるタンパク質であるカルモジュリンを用いることができる。
【0014】
(カルシウムイオン結合タンパク質改変体)
カルシウムイオン結合タンパク質の改変体とは、カルシウムイオン結合タンパク質の本来有しているカルシウムイオン結合能を有している改変体を意味する。具体的には、そのアミノ酸配列が、元となるカルシウムイオン結合タンパク質のアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるカルシウムイオン結合タンパク質と実質的に同じ活性(カルシウムイオン結合能)を有しているタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。例えば、N末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体、C末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体を挙げることができる。
【0015】
カルシウムイオン結合タンパク質の改変体のうち、カルモジュリンの改変体とは、カルモジュリンの本来有しているカルシウムイオン結合能を有している改変体を意味する。例えば、そのアミノ酸配列が、配列番号31で表されるアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるカルモジュリンと実質的に同じ活性(カルシウムイオン結合能)を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。例えば、N末側の1〜30個のアミノ酸の欠失したカルモジュリン改変体、より好ましくは1〜20個のアミノ酸の欠失したカルモジュリン改変体、更に好ましくは1〜10個のアミノ酸欠失したカルモジュリン改変体、C末側の1〜30個のアミノ酸の欠失したカルモジュリン改変体、より好ましくは1〜20個のアミノ酸の欠失したカルモジュリン改変体、更に好ましくは1〜10個のアミノ酸欠失したカルモジュリン改変体を挙げることができる。
【0016】
(光受容体ペプチド)
本発明の光駆動生カルシウムイオン制御タンパク質に用いる光受容体ペプチドは、光刺激により構造の変化するペプチドであり、例えばフォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、FKF−1のLOVドメイン、AppAのBLUFドメイン、及びTePixD−I66CのBLUFドメインを挙げることができ、好ましくはフォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、又はFKF−1のLOVドメインであり、より好ましくはフォトトロピン1のLOV2ドメイン、又はフォトトロピン2のLOV2ドメインである。
【0017】
前記光受容体ペプチドの由来も、特に限定されるものではなく、すべての生物由来(例えば、動物由来、又は植物由来)の光受容体ペプチドを用いることができる。すなわち、多くの生物が、光受容体ペプチドを含むタンパク質であるフォトトロピン1、フォトトロピン2、オーレオクロム1、オーレオクロム2、FKF−1、AppA、及びTePixD−I66Cを有しており、それぞれのタンパク質に含まれる光受容体ペプチドを用いることができる。
【0018】
なお、フォトトロピンは、植物特有の青色光受容体であり、光屈性、葉緑体の光定位運動、気孔の開口、葉の平坦化、葉の光定位運動などを誘導する。フォトトロピンには、前記のようにフォトトロピン1及びフォトトロピン2があり、フォトトロピン1は弱光から中高強光域の光センサーとして、フォトトロピン2は強光域の光センサーとして機能する。フォトトロピン1及びフォトトロピン2は、ともにLOV1ドメイン及びLOV2ドメインを有しており、LOV1ドメインは光感受性制御に関連した機能を有しており、LOV2ドメインがキナーゼ活性の光制御分子スイッチとして機能すると考えられている。また、オーレオクロムは、黄色植物(ヒバマタ、コンブ、及びケイ藻など)が有する青色光受容体であり、LOVドメインを有しており、青色光を吸収して構造が変化することによって、光センサーとして機能する。
【0019】
例えば、前記フォトトロピン1は、燕麦、シロイヌナズナ、タバコ、トウモロコシ、イネ又はシダ類などが有しており、これらの植物のフォトトロピン1のLOV2ドメインを本発明の光駆動生カルシウムイオン制御タンパク質に用いることができるが、特には配列番号32で表されるアミノ酸配列からなるペプチドである燕麦由来のLOV2ドメインが好ましい。
【0020】
光受容体ペプチドは、光刺激によりその立体構造が変化するが、立体構造が変化する励起波長は、光受容体ペプチドの種類により、適宜選択することができる。例えば、フォトトロピン1、フォトトロピン2、オーレオクロム1、及びオーレオクロム2は青色光受容体であり、青色周辺の励起波長を用いることが好ましい。従って、フォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、及びフォトトロピン2のLOV2ドメイン、及びオーレオクロム1のLOVドメイン、オーレオクロム2LOVドメインの励起波長は、300〜500nmを用いることが可能であり、350〜490nmが好ましく、430〜470nmがより好ましい。
【0021】
(光受容体ペプチド改変体)
前記光受容体ペプチドの改変体とは、その光受容体ペプチドの本来有している性質、すなわち光刺激により構造の変化することのできる改変体を意味する。具体的には、そのアミノ酸配列が、元となる光受容体ペプチドのアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となる光受容体ペプチドと実質的に同じ活性(光刺激による構造変化)を示すペプチドを意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。例えば、N末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体、C末側の1〜30個のアミノ酸の欠失した改変体、より好ましくは1〜20個のアミノ酸の欠失した改変体、更に好ましくは1〜10個のアミノ酸欠失した改変体を挙げることができる。
【0022】
例えば、LOV2ドメイン改変体とは、LOV2ドメインの本来有している性質を有する改変体、すなわち光刺激により構造の変化することのできる改変体を意味する。例えば、そのアミノ酸配列が、配列番号32で表されるアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるLOV2ドメインと実質的に同じ活性(光刺激による構造変化)を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば30個であり、好ましくは1〜30個、より好ましくは1〜20個、更に好ましくは1〜10個である。特に、例えば、N末側の1〜30個のアミノ酸の欠失したLOV2ドメイン改変体、より好ましくは1〜20個のアミノ酸の欠失したLOV2ドメイン改変体、更に好ましくは1〜10個のアミノ酸欠失したLOV2ドメイン改変体、C末側の1〜30個のアミノ酸の欠失したLOV2ドメイン改変体、より好ましくは1〜20個のアミノ酸の欠失したLOV2ドメイン改変体、更に好ましくは1〜10個のアミノ酸欠失したLOV2ドメイン改変体を挙げることができる。具体的には、後述の実施例に示すように、前記フォトトロピン1のLOV2ドメインの改変体としては、C末側アミノ酸が17個、16個、15個、14個、12個、9個、又は3個欠失した改変体を挙げることができ、これらの改変体はLOV2ドメインと同じように、光刺激により構造が変化する。
【0023】
(カルシウムイオン結合タンパク質結合ペプチド)
本発明のカルシウムイオン制御タンパク質は、更にカルシウムイオン結合タンパク質結合ペプチドを含む。このペプチドは、カルシウムイオン結合タンパク質への結合能を有する限り、特に限定されるものはないが、例えばカルモジュリン結合ペプチド、トロポニンI、ミオシンII重鎖、又はロドプシンキナーゼを挙げることができる。これらのカルシウムイオン結合タンパク質結合ペプチドは、前記カルシウムイオン結合タンパク質との複合体を形成することにより、カルシウムイオン結合タンパク質のカルシウムイオンとの親和性を上昇させることができる。具体的には、カルモジュリン結合ペプチドはカルモジュリンと、トロポニンIはトロポニンCと、ミオシンII重鎖はミオシン軽鎖と、及びロドプシンキナーゼはレコベリンとそれぞれ複合体を形成する。
【0024】
前記カルモジュリン結合ペプチドは、いくつかのキナーゼなどに存在しており、例えば骨格筋ミオシン軽鎖キナーゼ、平滑筋ミオシン軽鎖キナーゼ、カルモジュリン依存キナーゼII、カルモジュリン依存キナーゼキナーゼ等のカルモジュリン結合領域のペプチドを挙げることができる。
【0025】
本発明のカルシウムイオン制御タンパク質に用いるカルモジュリン結合ペプチドの生物種の由来も、カルモジュリンに結合するものである限り、特に限定されるものではなく、すべての生物由来のカルモジュリン結合ペプチドを用いることが可能である。すなわち、すべての生物由来に骨格筋ミオシン軽鎖キナーゼのカルモジュリン結合領域のペプチド、平滑筋ミオシン軽鎖キナーゼのカルモジュリン結合領域のペプチド、カルモジュリン依存キナーゼIIのカルモジュリン結合領域のペプチド、カルモジュリン依存キナーゼキナーゼのカルモジュリン結合領域のペプチド等を用いることができる。例えば、配列番号33で表されるアミノ酸配列からなる、アフリカツメガエルの骨格筋ミオシン軽鎖キナーゼのカルモジュリン結合領域であるM13ペプチドを用いることが可能である。
前記M13ペプチドは、骨格筋ミオシン軽鎖キナーゼのカルモジュリン結合領域に由来する26残基ペプチド(KRRWKKNFIAVSAANRFKKISSSGAL)(Blumenthal, D. K., & Krebs, E. G. (1987) Methods Enzymol. 139,115-126)であり、カルシウムイオンが結合したカルモジュリン(カルシウムイオン−カルモジュリン)に結合し、安定した複合体を形成する。従って、本発明のカルシウムイオン制御タンパク質はM13ペプチドを含むことにより、カルモジュリンがカルシウムイオンと結合した場合、強固な結合を形成することが可能である。
【0026】
更に、M13ペプチド以外の具体的なカルモジュリン結合ペプチドを以下の表1に挙げる。
【0027】
【表1】
【0028】
本発明のカルシウムイオン制御タンパク質において、前記表1に記載の23種類のカルモジェリン結合ペプチドを、M13ペプチドに代えて用いることが可能である。それぞれのカルモジェリン結合ペプチドは、20アミノ酸からなるペプチド又は16アミノ酸からなるペプチドのみでもカルモジュリンと結合可能であり、そのまま本発明のカルシウムイオン制御タンパク質に用いることが可能である。しかしながら、M13ペプチドと同じように、それぞれのペプチドのC末側に続く1〜10個のアミノ酸(好ましくは4〜8アミノ酸、より好ましくは6アミノ酸)を含んだペプチドを用いることもできる。更に、それらのペプチドの改変体を用いることも可能である。
【0029】
(カルシウムイオン結合タンパク質結合ペプチド改変体)
カルシウムイオン結合タンパク質結合ペプチド改変体とは、カルシウムイオン結合タンパク質結合ペプチド改変体の本来有しているカルシウムイオン結合タンパク質への結合能を有している改変体を意味する。具体的には、そのアミノ酸配列が、元となるペプチドのアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるペプチドと実質的に同じ活性(カルシウムイオン結合タンパク質結合能)を有しているペプチドを意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜3個である。例えば、N末側の1〜10個のアミノ酸の欠失したペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したペプチド改変体、C末側の1〜10個のアミノ酸の欠失したペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したペプチド改変体を挙げることができる。
【0030】
カルシウムイオン結合タンパク質結合ペプチド改変体のうち、M13ペプチド改変体とは、M13ペプチドの本来有している性質を有する改変体、すなわちカルモジュリンへの結合能を有する改変体を意味する。例えば、そのアミノ酸配列が、配列番号33で表されるアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるM13ペプチドと実質的に同じ活性(カルモジュリンへの結合能)を示すペプチドを意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜3個である。特に、例えば、N末側の1〜10個のアミノ酸の欠失したM13ペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したM13ペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したM13ペプチド改変体、C末側の1〜10個のアミノ酸の欠失したM13ペプチド改変体、より好ましくは1〜5個のアミノ酸の欠失したM13ペプチド改変体、更に好ましくは1〜3個のアミノ酸欠失したM13ペプチド改変体を挙げることができる。例えば、後述の実施例に示すように、前記M13ペプチド改変体としては、N末側アミノ酸が1個又は2個欠失した改変体を挙げることができ、この改変体はM13ペプチドと同じように、カルモジュリンへの結合能を示す。
【0031】
(シグナルペプチド)
また、本発明の光駆動カルシウムイオン制御タンパク質を、細胞内の任意の位置で発現させるため、光駆動カルシウムイオン制御タンパク質は、シグナルペプチドを含んでもよい。シグナルペプチドを含むことによって、細胞内の特定の領域又は細胞内小器官内若しくは細胞内小器官の膜上で光駆動カルシウムイオン制御タンパク質を発現させることができる。
シグナルペプチドは、約3〜60アミノ酸からなるペプチドであり、局在シグナル、輸送シグナル、又は移行シグナルと呼ばれることもある。シグナルペプチドは、その種類に応じて、N末端、C末端、又はタンパク質内部のアミノ酸配列に存在し、細胞質内で生合成されたタンパク質の細胞内の局所への輸送および局在化を誘導する。具体的には、シグナルペプチドとして、ミトコンドリア膜間スペース局在化シグナル、ゴルジ体局在化シグナル、小胞体局在化シグナル、ペルオキシソーム局在化シグナル、核局在シグナル、ミトコンドリアのマトリックス局在シグナル、細胞膜局在シグナル及び葉緑体局在シグナルなどを挙げることができる。
例えば、N末端に存在するシグナルペプチドとしては、小胞体移行シグナル(H2N-Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-Gly-Ile-Leu-Phe-Trp-Ala-Thr-Glu-Ala-Glu-Gln-Leu-Thr-Lys-Cys-Glu-Val-Phe-Gln-)、ミトコンドリアマトリックスシグナル(H2N-Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-Phe-Lys-Pro-Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-Leu-Leu-)、及びペルオキシソームシグナル(PTS2)(H2N-----Arg-Leu-X5-His-Leu)を挙げることができ、C末端に存在するシグナルペプチドとしては、小胞体保留シグナル(-Lys-Asp-Glu-Leu-COOH)、及びペルオキシソームシグナル(PTS1)(-Ser-Lys-Leu-COOH)を挙げることができ、タンパク質内部に存在するシグナルペプチドとしては、核移行シグナル(NLS)(-Pro-Pro-Lys-Lys-Lys-Arg-Lys-Val)を挙げることができる。
これらのシグナルペプチドを、本発明の光駆動カルシウムイオン制御タンパク質のN末端、C末端、又はタンパク質内部に含ませることにより、光駆動カルシウムイオン制御タンパク質を細胞内の特定の領域に局在させ、カルシウムイオン濃度の変化による細胞の動態を解析することが可能である。
【0032】
(機能的等価改変体)
本発明の機能的等価改変体は、前記光駆動カルシウムイオン制御タンパク質の機能的等価改変体である。本明細書において「機能的等価改変体」とは、元となるタンパク質のアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列からなるタンパク質であって、しかも、元となるタンパク質と実質的に同じ活性を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜2個である。例えば、配列番号1〜15で表されたアミノ酸配列のそれぞれにおいて、1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列からなるタンパク質であって、しかも、光駆動カルシウムイオン制御タンパク質と実質的に同じ活性を有するタンパク質、すなわち、光照射によりカルシウムイオンの濃度を制御可能であり、そしてその光照射の励起波長が実質的に同じであるタンパク質を意味する。
【0033】
(リンカー及び制限酵素サイト)
本発明のカルシウムイオン制御タンパク質における、カルシウムイオン結合タンパク質等(a)と光受容体ペプチド等(b)との間、及び光受容体ペプチド等(b)とカルシウムイオン結合タンパク質結合ペプチド等(c)との間には、カルシウムイオン制御タンパク質の機能を阻害しない限りにおいて、適当な長さのペプチドリンカー又は制限酵素サイトのヌクレオチド配列から翻訳される2〜数アミノ酸の配列を含むことができる。
【0034】
本発明のカルシウムイオン制御タンパク質は、カルシウムイオン吸収融合タンパク質及びカルシウムイオン放出融合タンパク質を含む。カルシウムイオン吸収融合タンパク質は、光刺激により周囲のカルシウムを吸収(結合)し、周囲の環境のカルシウム濃度を低下させることができる。カルシウムイオン放出融合タンパク質は、保持しているカルシウムイオンを光刺激により放出し、周囲の環境のカルシウムイオン濃度を上昇させることができる。
【0035】
(カルシウムイオン吸収融合タンパク質)
本発明のカルシウムイオン吸収融合タンパク質は、(a)カルシウム結合タンパク質、又はカルシウム結合能を有するその改変体;(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び(c)カルシウム結合タンパク質結合ペプチド、又はカルシウム結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムと結合するものである。また、光駆動カルシウムイオン吸収融合タンパク質は、カルシウムイオン結合タンパク質等(a)、光受容体ペプチド等(b)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)からなってもよく、更に前記ペプチドリンカー等、及び/又はシグナルペプチドを含んでもよい。
【0036】
カルシウムイオン吸収融合タンパク質の1つの態様として、(a)カルモジュリン、又はカルシウムイオン結合能を有するカルモジュリン改変体(以下、カルモジュリン等(a)と称する);(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体(以下、LOV2ドメイン等(b)と称する);及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体(以下、M13ペプチド等(c)と称する);をN末端から順に含み、光刺激によりカルシウムイオンと結合する融合タンパク質を挙げることができる。具体的には、カルシウムイオン吸収融合タンパク質として、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、又は配列番号7で表されるアミノ酸配列からなる融合タンパク質を挙げることができる。
【0037】
本発明のカルシウムイオン吸収融合タンパク質の機能的等価改変体は、実質的にカルシウムイオン吸収融合タンパク質と同じ活性を有するものである。「カルシウムイオン吸収融合タンパク質と同じ活性」とは、光照射によりカルシウムイオンと結合し、そしてその光照射の励起波長が実質的に同じであることを意味する。例えば、カルモジュリン又はカルモジュリン改変体のカルシウムイオンと結合する機能が維持されており、そしてM13ペプチド又はM13ペプチド改変体のカルモジュリンに結合する機能が維持されていることを意味する。そして、励起波長として300〜500nmを用いることによって、機能的等価改変体がカルシウムイオンと結合できることを意味する。
【0038】
本発明のカルシウムイオン吸収融合タンパク質は、例えば、カルモジュリン又はカルモジュリン改変体(a)、LOV2ドメイン又はLOV2ドメイン改変体(b)、及びM13ペプチド又はM13ペプチド改変体(c)が、N末端から順に連結された構造をとっている。カルモジュリンは、単体ではカルシウムイオンとの結合はそれほど強くないが、M13ペプチドと結合することにより、解離定数が小さくなり、カルモジュリン及びM13の複合体は、カルシウムイオンを強固に結合させることが可能になる。本発明のカルシウムイオン吸収融合タンパク質は、光照射されない状態では、LOV2ドメイン等(b)が、カルモジュリン等(a)と、M13ペプチド等(c)との間に組み込まれていることにより、カルモジュリン等(a)とM13ペプチド等(c)とが結合できない状態であり、カルモジュリンの解離定数は大きいと推定される。一方、光照射された場合、LOV2ドメイン等(b)の構造が変化し、カルモジュリン等(a)と、M13ペプチド等(c)とが結合可能な状態になり、カルモジュリン等(a)とM13ペプチド等(c)との複合体は、カルシウムイオン吸収融合タンパク質の周囲のカルシウムイオン濃度より小さな解離定数を有する状態になる。そのため、本発明のカルシウムイオン吸収融合タンパク質周囲のカルシウムイオンを吸収することができると推定される。また、カルシウムイオン結合タンパク質等(a)、光受容体ペプチド等(b)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)との関係も、それぞれカルモジュリン等(a)、LOV2ドメイン等(b)、及びM13ペプチド等(c)との関係と同じであると推定される。しかしながら、本発明は前記の推定により限定されるものではない。
【0039】
(リンカー及び制限酵素サイト)
本発明のカルシウムイオン吸収融合タンパク質において、カルシウムイオン結合タンパク質等(a)(例えば、カルモジュリン等(a))と光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))との間、及び光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))とカルシウムイオン結合タンパク質結合ペプチド等(c)(例えば、M13ペプチド等(c))との間には、カルシウムイオン吸収融合タンパク質の機能を阻害しない限りにおいて、適当な長さのペプチドリンカー又は制限酵素サイトのヌクレオチド配列から翻訳される2〜数アミノ酸の配列を含むことができる。
ペプチドリンカーしては、例えば「G」、「GG」、「GGS」、「GGGS」、「GGGGS」、「GGGSK」、「GGSGG」、又は「GGSGGSGGS」を挙げることができる。また、制限酵素サイトのヌクレオチド配列から翻訳されるアミノ酸配列としては、LE及びGTなどを含むことができる。
後述の実施例に示すように、カルモジュリン等(a)とLOV2ドメイン等(b)との間に制限酵素サイトXhoIから翻訳されるLE、及びペプチドリンカー「G」、「GGS」、「GGGGS」を含んでも、本発明のカルシウムイオン吸収融合タンパク質を発揮することができる。
【0040】
(カルシウムイオン放出融合タンパク質)
本発明のカルシウムイオン放出融合タンパク質は、(a1)カルシウム結合タンパク質N末側ペプチド、又はその改変体(以下、カルシウム結合タンパク質N末側ペプチド等(a1)と称することがある);(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;(a2)カルシウム結合タンパク質C末側ペプチド、又はその改変体(以下、カルシウム結合タンパク質C末側ペプチド等(a2)と称することがある);及び(c)カルシウム結合タンパク質結合ペプチド、又はカルシウム結合タンパク質への結合能を有するその改変体;をN末端から順に含み、光刺激によりカルシウムを放出するものである。また、光駆動カルシウムイオン放出収融合タンパク質は、カルシウム結合タンパク質N末側ペプチド等(a1)、光受容体ペプチド等(b)、カルシウム結合タンパク質C末側ペプチド等(a2)、及びカルシウム結合タンパク質結合ペプチド(c)からなってもよく、更に前記ペプチドリンカー等、及び/又はシグナルペプチドを含んでもよい。
【0041】
カルシウムイオン吸収融合タンパク質の1つの態様として、(a1)カルモジュリンN末側ペプチド、又はその改変体;(b)(以下、カルモジュリンN末側ペプチド等(a1)と称することがある);LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体(以下、LOV2ドメイン等(b)と称する);(a2)カルモジュリンC末側ペプチド、又はその改変体(以下、カルモジュリンC末側ペプチド等(a2)と称することがある);及び(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体(以下、M13ペプチド等(c)と称する)、をN末端から順に含み、光刺激によりカルシウムを放出する融合タンパク質を挙げることができる。具体的には、カルシウムイオン放出融合タンパク質として、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、又は配列番号15で表されるアミノ酸配列からなる融合タンパク質を挙げることができる。
【0042】
LOV2ドメインが挿入される、カルモジュリンN末側ペプチド等(a1)と、カルモジュリンC末側ペプチド等(a2)との切断位置は、カルモジュリンN末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体のカルシウムイオン結合能が阻害されない限り、限定されるものではなく、例えばカルモジュリンの20〜130番に存在する連続する2アミノ酸の間であればよい。例えば、25〜55番に存在する任意の2アミノ酸の間で切断してよく、30〜50番に存在する任意の2アミノ酸の間で切断してよく、35〜55番に存在する任意の2アミノ酸の間で切断してもよい。
【0043】
従って、カルモジュリンN末側ペプチド等(a1)と、カルモジュリンC末側ペプチド等(a2)のアミノ酸の鎖長も限定されるものはないが、例えばカルモジュリンN末側ペプチド等(a1)は、1〜20番のアミノ酸からなるペプチドから1〜130番のアミノ酸からなるペプチドであってよく、1〜20番のアミノ酸からなるペプチドから1〜60番のアミノ酸からなるペプチドであってよく、1〜30番のアミノ酸からなるペプチドから1〜50番のアミノ酸からなるペプチドであってよく、1〜40番のアミノ酸からなるペプチドであってよい。また、カルモジュリンC末側ペプチド等(a2)は、21〜150番のアミノ酸からなるペプチドから131〜150番のアミノ酸からなるペプチドであってよく、21〜150番のアミノ酸からなるペプチドから61〜150番のアミノ酸からなるペプチドであってよく、31〜150番のアミノ酸からなるペプチドから51〜150番のアミノ酸からなるペプチドであってよく、41〜150番のアミノ酸からなるペプチドであってよい。但し、カルモジュリンN末側ペプチド等(a1)のC末端と、カルモジュリンC末側ペプチド等(a2)のN末端は、実質的に元のカルモジュリンのアミノ酸配列におけるアミノ酸配列の重複及び欠失を含まないものする。ここで、実質的にアミノ酸配列の重複及び欠失を含まないとは、カルモジュリンN末側ペプチド等(a1)、カルシウムイオン吸収融合タンパク質におけるカルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体の形成に影響を与えないものであればよいが、具体的には、10アミノ酸以下であり、好ましくは5アミノ酸以下である。
また、カルモジュリンN末側ペプチド等(a1)及びカルモジュリンC末側ペプチド等(a2)の関係は、カルシウム結合タンパク質N末側ペプチド等(a1)及びカルシウム結合タンパク質C末側ペプチド等(a2)の関係にも当てはまるものである。
【0044】
本発明のカルシウムイオン放出融合タンパク質の機能的等価改変体は、実質的にカルシウムイオン放出融合タンパク質と同じ活性を有するものである。「カルシウムイオン放出融合タンパク質と同じ活性」とは、光照射によりカルシウムイオンを放出し、そしてその光照射の励起波長が実質的に同じであることを意味する。例えば、カルモジュリンC末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体のカルシウムイオン結合能が維持されており、そして光刺激により複合体のカルシウムイオンとの結合能が消失することを意味する。そして、励起波長として300〜500nmを用いることによって、機能的等価改変体がカルシウムイオン放出できることを意味する。
【0045】
本発明のカルシウムイオン放出融合タンパク質は、前記のように(a1)カルモジュリンN末側ペプチド、又はその改変体、(b)LOV2ドメイン、又はLOV2ドメイン改変体、(a2)カルモジュリンC末側ペプチド、又はその改変体、及び(c)M13ペプチド、又はM13ペプチド改変体が、N末端から順に連結された構造をとっている。このカルシウムイオン放出融合タンパク質は、光照射されない状態では、カルモジュリンN末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)及びM13ペプチド等(c)が複合体を形成し、解離定数が小さくなりカルシウムイオンと強固に結合できる構造となっていると推定される。一方、光照射された場合、カルモジュリンN末側ペプチド等(a1)及びカルモジュリンC末側ペプチド等(a2)の間に組み込まれたLOV2ドメイン等(b)の構造が変化し、カルモジュリンN末側ペプチド等(a1)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)の複合体の結合が維持できなくなり、解離定数が大きくなることによって、カルシウムイオンを放出するものと推定される。また、カルシウム結合タンパク質N末側ペプチド等(a1)、光受容体ペプチド等(b)、カルシウム結合タンパク質C末側ペプチド等(a2)、及びカルシウムイオン結合タンパク質結合ペプチド等(c)との関係も、それぞれカルモジュリンN末側ペプチド等(a1)、LOV2ドメイン等(b)、カルモジュリンC末側ペプチド等(a2)、及びM13ペプチド等(c)との関係と同じであると推定される。しかしながら、本発明は前記の推定により限定されるものではない。
【0046】
(リンカー及び制限酵素サイト)
本発明のカルシウムイオン吸収融合タンパク質において、カルシウム結合タンパク質N末側ペプチド等(a1)(例えば、カルモジュリンN末側ペプチド等(a1))と光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))との間、光受容体ペプチド等(b)(例えば、LOV2ドメイン等(b))とカルシウム結合タンパク質C末側ペプチド等(a2)(例えば、カルモジュリンC末側ペプチド等(a2))との間、及びカルシウム結合タンパク質C末側ペプチド等(a2)(例えば、カルモジュリンC末側ペプチド等(a2))とカルシウムイオン結合タンパク質結合ペプチド等(c)(例えば、M13ペプチド等(c))との間には、カルシウムイオン吸収融合タンパク質の機能を阻害しない限りにおいて、適当な長さのペプチドリンカー又は制限酵素サイトのヌクレオチド配列から翻訳される2〜数アミノ酸の配列を含むことができる。
ペプチドリンカーしては、例えば「G」、「GG」、「GGS」、「GGGS」、「GGGGS」、「GGGSK」、「GGSGG」、「GGGGSK」、又は「GGSGGSGGS」を挙げることができる。また、制限酵素サイトのヌクレオチド配列から翻訳されるアミノ酸配列としては、LE及びGTなどを含むことができる。
例えば、後述の実施例に示すように、カルモジュリンN末側ペプチド等とLOV2ドメイン等(b)との間に、「G」、「GG」、「GGS」、又は「GGGS」のペプチドリンカーを含んでもよい。また、カルモジュリンC末側ペプチド等(a2)とM13ペプチド等(c)との間に、「GGGGS」のペプチドリンカーを含んでもよい。
【0047】
本発明によるDNAは、本発明の光駆動カルシウムイオン制御タンパク質又はその機能的等価改変体をコードする限り、特に限定されるものではなく、例えば、配列表の配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、又は配列番号30で表される塩基配列からなるDNAを挙げることができる。
配列表の配列番号16で表される塩基配列からなる前記DNAは、配列表の配列番号1で表されるアミノ酸配列からなるPacman(カルシウムイオン吸収融合タンパク質)をコードする。また、配列表の配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、及び配列番号22で表される塩基配列からなる前記DNAは、それぞれ、配列表の配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、及び配列番号7で表されるアミノ酸配列からなるカルシウムイオン吸収融合タンパク質をコードする。
配列表の配列番号23で表される塩基配列からなる前記DNAは、配列表の配列番号8で表されるアミノ酸配列からなるCARBY(カルシウムイオン放出融合タンパク質)をコードする。また、配列表の配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、及び配列番号30で表される塩基配列からなる前記DNAは、それぞれ、配列表の配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、及び配列番号15で表されるアミノ酸配列からなるカルシウムイオン放出融合タンパク質をコードする。
【0048】
本発明によるプラスミドは、本発明による前記DNAを含む限り、特に限定されるものではなく、例えば、用いる宿主細胞に応じて適宜選択した公知の発現ベクターに、本発明による前記DNAを挿入することにより得られるプラスミドを挙げることができる。
得られたプラスミドを所望の宿主細胞に形質転換することにより、形質転換体を得ることができる。前記形質転換体も、本発明による前記プラスミドを含む限り、特に限定されるものではない。
【0049】
前記宿主細胞としては、例えば、通常使用される公知の微生物、例えば、大腸菌又は酵母(Saccharomyces cerevisiae)、あるいは、公知の培養細胞、例えば、動物細胞(例えば、CHO細胞、HEK−293細胞、又はCOS細胞)又は昆虫細胞(例えば、BmN4細胞)を挙げることができる。
【0050】
また、公知の前記発現ベクターとしては、例えば、大腸菌に対しては、pUC、pTV、pGEX、pKK、又はpTrcHisを;酵母に対しては、pEMBLY又はpYES2を;CHO細胞に対してはpcDNA3又はpMAMneoを;HEK−293細胞に対してはpcDNA3を;COS細胞に対してはpcDNA3を;BmN4細胞に対しては、カイコ核多角体ウイルス(BmNPV)のポリヘドリンプロモーターを有するベクター(例えば、pBK283)を挙げることができる。
【0051】
[2]カルシウムイオン濃度の制御方法
本発明のカルシウムイオン濃度の制御方法は、前記光駆動カルシウムイオン制御タンパク質若しくはその機能的等価改変体、又は前記DNAを用いる。そして光駆動カルシウムイオン制御タンパク質又はその機能的等価改変体に、励起光を照射することにより、カルシウムイオンの濃度を制御する方法である。
本発明のカルシウムイオンの濃度制御方法において、用いる励起波長は光駆動カルシウムイオン制御タンパク質に使用されている光受容体ペプチドの種類により、適宜選択することができる。すなわち、光刺激により立体構造が変化する励起波長を用いることができる。例えば、青色光受容体であるフォトトロピン1、又はフォトトロピン2のLOV1ドメイン又はLOV2ドメイン、又はオーレオクロム1、又はオーレオクロム2のLOVドメインを光受容体ペプチドとして用いた場合は、300〜500nmの励起波長を用いることが可能であり、350〜490nmが好ましく、430〜470nmがより好ましい。
前記カルシウムイオン吸収融合タンパク質を用いることにより、周囲のカルシウムイオン濃度を低下させることができる。また、カルシウムイオン放出融合タンパク質を用いることにより、周囲のカルシウムイオン濃度を上昇させることができる。
更に、前記シグナルペプチドを含む光駆動カルシウムイオン制御タンパク質を用いることによって、細胞内の特定の領域又は細胞内小器官内若しくは細胞内小器官の膜上でカルシウムイオン濃度を制御することが可能である。具体的には、ミトコンドリア膜間スペース、ゴルジ体小胞体、ペルオキシソーム、核、ミトコンドリアのマトリックス、細胞膜、及び葉緑体などにおいて、カルシウムイオン濃度を制御することができる。
【0052】
[3]光駆動カルシウムイオン制御タンパク質等の利用
本発明の光駆動カルシウムイオン制御タンパク質若しくはその機能的等価改変体、又は前記DNAは、カルシウムイオン代謝異常を伴う疾患の治療又は予防用医薬組成物として用いることができる。カルシウムイオン代謝異常を伴う疾患としては、中枢神経系疾患(例えば、脳卒中、頭部外傷、脊髄損傷、外傷性ショック、アルツハイマー病、パーキンソン病、ハンチントン病、筋萎縮側索硬化症、てんかん、発作、痙攣性疾患、低酸素誘因神経細胞損傷、疼痛、うつ病、不安神経症、パニック障害、脅迫神経症、外傷後ストレス障害、麻酔後認識衰退、オピオイド耐性、薬物乱用、アルコール依存症、統合失調症)、又は循環器系疾患(例えば、高血圧、狭心症、不整脈)を挙げることができる。
【実施例】
【0053】
以下、実施例によって本発明を具体的に説明するが、これらは本発明の範囲を限定するものではない。
【0054】
《実施例1》
本実施例1では、カルシウムイオン吸収融合タンパク質であるPacmanを製造した。
(1)LOV2遺伝子の調製
融合タンパク質の構築に用いるLOV2ドメインをコードするDNAを調製した。Leeらの報告(Lee J. et al. Surface sites for engineering allosteric control in proteins.Science.17. 438-42 2008)を参考にして、以下のLOV2の骨格オリゴヌクレオチド、アダプターオリゴヌクレオチド、及びPCRに用いるプライマーを合成した。
【0055】
(骨格オリゴヌクレオチド)
LOV2-skel1 5’-TCGAGCTGGCCACCACTCTAGAGCGCATCGAGAAGAACTTCGTGATCACC-3’(配列番号37)
LOV2-skel2 5’-GACCCCCGCCTGCCCGACAACCCCATCATCTTCGCCTCCGACTCC-3’(配列番号38)
LOV2-skel3 5’-TTCCTGCAGCTGACCGAGTACTCCCGCGAGGAGATCCTGGGCCGCAAC-3’(配列番号39)
LOV2-skel4 5’-TGCCGCTTCCTGCAGGGCCCCGAGACCGACCGCGCCACCGTGCGC-3’(配列番号40)
LOV2-skel5 5’-AAGATCCGCGACGCCATCGACAACCAGACCGAGGTGACCGTG-3’(配列番号41)
LOV2-skel6 5’-CAGCTGATCAACTACACCAAGTCCGGCAAGAAGTTCTGGAACCTGTTC-3’(配列番号42)
LOV2-skel7 5’-CACCTGCAGCCCATGCGCGACCAGAAGGGCGACGTGCAGTACTTCATC-3’(配列番号43)
LOV2-skel8 5’-GGCGTGCAGCTGGACGGCACCGAGCACGTGCGCGACGCCGCC-3’(配列番号44)
LOV2-skel9 5’-GAGCGCGAGGGCGTGATGCTGATCAAGAAGACCGCCGAGAAC-3’(配列番号45)
LOV2-skel10 5’-ATCGACGAGGCCGCCAAGGAGCTGCCCGACGCCAACCTGCGC-3’(配列番号46)
LOV2-skel11 5’-CCCGAAGATCTGTGGGCCAACCACGGTAC-3’(配列番号47)
【0056】
(アダプターオリゴヌクレオチド)
LOV2-adop1 5’-GGGGTCGGTGAT-3’(配列番号48)
LOV2-adop2 5’-CAGGAAGGAGTC-3’(配列番号49)
LOV2-adop3 5’-GCGGCAGTTGCG-3’(配列番号50)
LOV2-adop4 5’-GATCTTGCGCAC-3’(配列番号51)
LOV2-adop5 5’-CAGCTGCACGGT-3’(配列番号52)
LOV2-adop6 5’-CAGGTGGAACAG-3’(配列番号53)
LOV2-adop7 5’-CACGCCGATGAA-3’(配列番号54)
LOV2-adop8 5’-GCGCTCGGCGGC-3’(配列番号55)
LOV2-adop9 5’-GTCGATGTTCTC-3’(配列番号56)
LOV2-adop10 5’-TTCGGGGCGCAG-3’(配列番号57)
【0057】
(プライマー)
LOV2-fw 5’-TTT CTCGAG CTG GCC ACC ACT CTA-3’(配列番号58)
LOV2-rv 5’-TT GGTACC GTG GTT GGC CCA CA-3’(配列番号59)
【0058】
前記骨格オリゴヌクレオチドLOV2−skel1〜11を、1μMモル濃度に調整し、5’末端をT4ポリヌクレオチドキナーゼによってリン酸化した。その後、T4ポリヌクレオチドキナーゼを70℃/10minにて失活した。得られたオリゴヌクレオチドのリン酸化処理産物に、アダプターオリゴヌクレオチドLOV2−adop1〜10とT4DNALigaseを加え、16℃でO/Nのライゲーションを行った。すなわち、LOV2−skel1〜11は、それぞれ終濃度0.5μMになるように調製し、LOV2−adop1〜10は、それぞれ終濃度5μMになるように調整し、混合した。この混合物10.5μLに2XRapid Ligation Buffer(NEB)を12μL及びT4 DNAライゲースを1.5μLとを加え、16℃でO/Nのライゲーションを行った。得られたライゲーション産物を鋳型とし、フォワードプライマーLOV2−fwと、リバースプライマーLOV2−rvを用いてPCRを行った。PCRサイクルは94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間、保持した。ポリメラーゼは、KOD−Plus(TOYOBO)を用いた。PCR産物をフェノール/クロロホルム抽出にて精製し、XhoI/KpnIで制限酵素処理を行った。制限酵素処理産物を電気泳動した後、目的のバンドを切り出し、QIAEXIIGel extract kit(QIAGEN)を用いて精製を行った。得られたフラグメントを、試料1とする。プラスミドベクターpRSETB(インビトロジェン社製(Invitrogen))を、同様にXhoI/KpnIで制限酵素処理を行い、電気泳動により精製した。得られたXhoI/KpnI切断ベクターを、試料2とする。試料1と試料2を混和し、T4DNAライゲースでライゲーションし、LOV2遺伝子を導入したpRSETBを得た。このプラスミドベクターを、「pRSETB/LOV2」と称する。
【0059】
(2)pRSETB/CaM-LOV2の構築
CaM及びLOV2を含むベクターpRSETB/CaM-LOV2の構築を行った。
用いたプライマーを以下に示す。
BHI-CaM-fw 5’-CG GGATCCC CGC ATG CAT GAC CAA CTG-3’(配列番号60)
CaM-ERI-rv 5’-G GAATTC TTA CTT TGC TGT CAT CAT TTG-3’(配列番号61)
CaM及びM13ペプチドを含むベクターpRSETB/YC3.6(Nagai T. et al Expanded dynamic range of fluorescent indicators for Ca(2+) by circularly permuted yellow fluorescent proteins. Proc Natl Acad Sci U S A. 20. 10554-9 2004)を鋳型として、フォワードプライマーBHI−CaM−fw及びリバースプライマーCaM−XhoI−rvを用いてPCRを行った。PCRサイクルは94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間である。PCR産物を、フェノール/クロロホルム抽出し、BamHI/XhoIで制限酵素処理し、電気泳動によって、ゲル精製を行った。得られたCaMのフラグメントを試料3とする。前記(1)の工程で得られたpRSETB/LOV2を、BamHI/XhoIで制限酵素処理し、電気泳動によりゲル精製を行った。得られたLOV2を含むベクターを試料4とする。試料3と試料4を混和し、T4DNAライゲースで、ライゲーションしてカルモジュリンをLOV2の5’側に持つ融合遺伝子を得た、このプラスミドベクターを、「pRSETB/CaM−LOV2」と称する。
【0060】
(3)Pacman遺伝子の構築
図3の模式図に示したPacman遺伝子を構築するために、LOV2ドメインを含む前記(1)の工程で得られたベクターpRSETB/LOV2、及びCaM及びM13ペプチドを含むベクターpRSETB/YC3.6(Nagai T. et al Expanded dynamic range of fluorescent indicators for Ca(2+) by circularly permuted yellow fluorescent proteins. Proc Natl Acad Sci U S A. 20. 10554-9 2004)を鋳型として、PCRを行い、得られたフラグメントをpRSETB/CaM−LOV2に組み込んだ。使用したプライマーを以下に示す。
LOV2-fw-GGGGS-XhoI 5’-TTT CTCGAG GGAGGTGGAGGTAGT CTG GCC ACC ACT CT-3’(配列番号62)
LOV2-Cd17-M13 5’-GAC GAG GCC GCC AGG CGC TGG AAG -3’(配列番号63)
M13-ERI-rv 5’-G GAATTC TTA GAG CTC CAG TGC CCC GGA-3’(配列番号64)
【0061】
PCRの鋳型として、pRSETB/LOV2及びpRSETB/YC3.6を用いた。プライマーとして、LOV2−fw−GGGGS−XhoI及びM13−ERI−rvを終濃度0.3μMになるように添加し、LOV2−Cd17−M13を終濃度0.03μMとなるように添加した。PCRのサイクルは、94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間、保持した。
なお、本PCRでは、初期段階において、主にpRSETB/YC3.6を鋳型にして、フォワードプライマーLOV2−Cd17−M13及びリバースプライマーM13−ERI−rvによって、一段階目のM13ペプチドを含む断片の増幅が起こる。この断片の5’端側にはLOV2−CD17のC末端側の配列がコードされている。この断片が蓄積すると、pRSETB/LOV2を鋳型にして、フォワードプライマーLOV2−fw−GGGGS−XhoI3、及びリバースプライマーとして一段階目のM13ペプチドを含む断片の相補鎖を用いた二段階目の増幅が起き、LOV2ドメイン及びM13ペプチドを含む全長の配列が得られる。さらに、この全長の配列を鋳型にしてフォワードプライマーLOV2−fw−GGGGS−XhoI3及びリバースプライマーM13−ER−rvを用いた増幅により、目的の増幅断片が得られる。
得られたPCR産物をフェノール/クロロホルム抽出し、XhoI/EcoRIで制限酵素処理し、電気泳動により、ゲル精製を行った。得られたフラグメントを、試料5とした。次に、pRSETB/CaM−LOV2を、XhoI/EcoRIで制限酵素処理し、電気泳動により、ゲル精製を行った。得られた切断ベクターを試料6とする。試料5と試料6を混和し、T4DNAライゲースで、ライゲーションしてカルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子を得た。得られたベクターを「pRSETB/Pacman」と称する。
なお、この融合遺伝子から得られる融合タンパク質は、カルモジュリンとLOV2△C17との間にXhoIサイトから翻訳される2個のアミノ酸LEを含む。
【0062】
《実施例2》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を14個欠失したLOV2(LOV2△C14)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd14-M13を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd14-M13 5’-GAG GCC GCC AAG GAG CTG AGG CGC TGG AGG-3’(配列番号65)
【0063】
《実施例3》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を15個欠失したLOV2(LOV2△C15)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd15-M13を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd15-M13 5’-GAG GCC GCC AAG GAG AGG CGC TGG AAG-3’(配列番号66)
【0064】
《実施例4》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を16個欠失したLOV2(LOV2△C16)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd16-M13を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd16-M13 5’-GAG GCC GCC AAG AGG CGC TGG AAG-3’(配列番号67)
【0065】
《実施例5》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。また、カルモジュリンとLOV2△C17との間のリンカーを、GGGGSからGGSに変更した。
プライマーLOV2-fw-GGGGS-XhoIに代えて、以下のLOV2-fw-GGS-XhoIを用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-fw-GGS-XhoI 5’-TTT CTCGAG GGAGGTAGT CTG GCC ACC ACT CT-3’(配列番号68)
【0066】
《実施例6》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を1個欠失したM13ペプチド(M13△N1)を持つ、融合遺伝子の調製を行った。また、カルモジュリンとLOV2△C17との間のリンカーを、GGGGSからGに変更した。
プライマーLOV2-fw-GGGGS-XhoIに代えて、以下のLOV2-fw-G-XhoIを用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-fw-G-XhoI 5’-TTT CTCGAG GGT CTG GCC ACC ACT CT-3’(配列番号69)
【0067】
《実施例7》
本実施例では、カルモジュリンをLOV2の3’側に持ち、C末端アミノ酸を17個欠失したLOV2(LOV2△C17)、及びN末端アミノ酸を2個欠失したM13ペプチド(M13△N2)を持つ、融合遺伝子の調製を行った。
プライマーLOV2-Cd17-M13に代えて、以下のLOV2-Cd17-M13△N1を用いたことを除いては、実施例1の(3)の操作を繰り返し、融合遺伝子を得た。
LOV2-Cd17-M13△N1 5’-GAC GAG GCC GCC CGC TGG AAG AAA-3’(配列番号70)
【0068】
《実施例8》
本実施例8では、カルシウムイオン放出融合タンパク質であるCARBYを製造した。
(1)pRSETB/CaM40−XhoI−KpnI−41CaM−M13の作製
CaM及びM13ペプチドが、GGGGSのリンカーで結合されているベクターpRSETB/YC−nano15(Horikawa K. et al. Spontaneous network activity visualized by ultrasensitive Ca(2+) indicators, yellow Cameleon-Nano.. Nat Methods.7.729-32. 2010)を用いて、カルモジュリンの第40番のアミノ酸と41番のアミノ酸との間に、XhoI及びKpnIの制限酵素サイトを導入したベクターpRSETB/CaM40−XhoI−KpnI−41CaM−M13を作製した。
【0069】
変異を導入するために以下のプライマーを合成した。
QC-CaM41-XhoI-TAA-KpnI 5’-G TCG CTT GGA CTCGAG TAA GGTACC CAA CCA ACG-3(配列番号71)
プライマーQC-CaM41-XhoI-TAA-KpnIの5’端を、T4ポリヌクレオチドキナーゼによってリン酸化した。得られたプライマーを用いて、PCR法により変異を含むPCR産物を調製した。pRSETB/YC−nano15を50ng、QC−CaM41−XhoI−TAA−KpnIを10pmol、dNTP3.75nmol、Pfu DNAポリメラーゼ(STRATAGENE社製)1.25U、Pfu DNA リガーゼ(STRATAGENE社製)を20Uを含む反応溶液40μLを準備した。まず、65℃、5分のプレインキュベーションを行ってPfu DNA リガーゼにより、鋳型DNAのニックを修復し、その後94℃、1分の最初の変性を行った。PCRのサイクルは、94℃、10秒のDNA変性、55℃、30秒のアニーリング反応及び65℃、10分の伸長・連結反応を1サイクルとして35サイクル行った。最後に75℃、10分のポストインキュベーションを行った。
【0070】
得られたPCR産物のうち、変異が含まれていない断片を、DpnIを用いて切断した。すなわち、前記PCR後の反応溶液40μLに1μL(20U)のDpnI(New England BioLab社製)を加え、37℃で2時間インキュベートすることによって、メチル化またはヘミメチル化した鋳型プラスミドDNAを選択的に消化した。DpnIは[5’−Gm6ATC−3’]を認識して2本鎖DNAを切断するエンドヌクレアーゼである。
【0071】
次に、得られた反応液を用いて、メガプライマーを用いた突然変異が導入されたDNAの相補鎖の合成を行った。
前記反応溶液41μLを94℃、30秒で変性させ、次いで95℃で30秒、55℃で1分、65℃で10分の3サイクルのPCRを行った。このとき反応溶液に含まれるDNA断片のうちのいくつかが、メガプライマーとして作用する。この2本鎖突然変異導入環状DNA鎖反応溶液41μLを、エタノール沈殿し、沈殿物を乾燥させた。
【0072】
前記2本鎖突然変異導入環状DNA鎖を鋳型とし、フォワードプライマーBHI−CaM−fw及びリバースプライマーM13−ERI−rvを用いてPCRを行った。PCR産物を、フェノール/クロロホルム抽出し、BamHI/EcoRIで制限酵素処理し、電気泳動によりゲル精製を行った。得られたフラグメントを、試料7とする。ベクターpRSETBを、BamHI/EcoRIで制限酵素処理し、電気泳動によりゲル精製を行った。得られた切断ベクターを、試料8とする。試料7と試料8を混和し、T4 DNAライゲースで、ライゲーションしてカルモジュリンの40番目のアミノ酸と41番目のアミノ酸の間にXhoI制限酵素サイト、終始コドン、KpnI制限酵素サイトを持ち、カルモジュリンのC末端側にリンカーとしてGGGGSを持ち、更にそのC末端側にM13を持つ融合遺伝子を得た、このプラスミドベクターを「pRSETB/CaM40−XhoI−KpnI−41CaM−M13」と称する。
【0073】
(2)CARBYの作製
ベクターpRSETB/CaM40−XhoI−KpnI−41CaM−M13を、XhoI/KpnIで制限酵素処理し、電気泳動によってゲル精製を行った。得られた切断ベクターを試料9とする。試料9と前記実施例1の工程(1)で得られた試料1を混和し、T4 DNAライゲースで、ライゲーションしてpRSETB/CaM40−XhoI−KpnI−41CaM−M13のカルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入された融合遺伝子を得た。すなわち、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを持つ、融合遺伝子を得た。得られたベクターを「pRSETB/CARBY」と称する。
なお、この融合遺伝子から得られる融合タンパク質は、カルモジュリン(1〜40番目のアミノ酸)とLOV2との間のXhoI制限酵素サイトから翻訳される2個のアミノ酸LE、及びLOV2とカルモジュリン(41〜150番目のアミノ酸)との間のKpnI制限酵素サイトから翻訳される2個のアミノ酸GTとを含む。
【0074】
《実施例9》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、C末端アミノ酸を3個欠失したLOV2(LOV2△C3)、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。
前記pRSETB/LOV2を鋳型として、以下のプライマーによってPCRを行った。
LOV2−fw
3'LOV2-Nd3-KpnI 5’-GG GGTACC CCA CAG ATC TTC GGG GCG-3’(配列番号72)
pRSETB/LOV2を鋳型として、フォワードプライマーLOV2−fw及びリバースプライマー3'LOV2-Nd3-KpnIを用いてPCRを行った。PCRサイクルは94℃2分の後、94℃30秒−62℃30秒−68℃1分のサイクルを30サイクル行い、その後に68℃10分間である。PCR産物を、フェノール/クロロホルム抽出し、XhoI/KpnIで制限酵素処理し、電気泳動によって、ゲル精製を行った。得られたLOV2△3のフラグメントを試料10とする。
前記試料9と試料10を混和し、T4 DNAライゲースで、ライゲーションしてpRSETB/CaM40−XhoI−KpnI−41CaM−M13のカルモジュリンの40番目と41番目のアミノ酸の間にLOV2△3が挿入された融合遺伝子を得た。
【0075】
《実施例10》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、C末端アミノ酸を9個欠失したLOV2(LOV2△C9)、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。
リバースプライマー3'LOV2-Nd3-KpnIに代えて、3'LOV2-Nd9-KpnI 5’-GG GGTACC CAG GTT GGC GTC GGG CAG-3’(配列番号73)を用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2△9が挿入された融合遺伝子を得た。
【0076】
《実施例11》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、C末端アミノ酸を12個欠失したLOV2(LOV2△C12)、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。
リバースプライマー3'LOV2-Nd3-KpnIに代えて、3'LOV2-Nd12-KpnI 5’-GG GGTACC GTC GGG CAG CTC CTT GGC-3’(配列番号74)を用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2△12が挿入された融合遺伝子を得た。
【0077】
《実施例12》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-G-XhoIを用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGを有する融合遺伝子を得た。
【0078】
《実施例13》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-GG-XhoI 5’-TTT CTCGAG GGTAGT CTG GCC ACC ACT CT-3’(配列番号75)を用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGを有する融合遺伝子を得た。
【0079】
《実施例14》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGSを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-GGS-XhoIを用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGSを有する融合遺伝子を得た。
【0080】
《実施例15》
本実施例では、カルモジュリンの1〜40番目のアミノ酸からなるペプチド(N−CaM(1−40))、LOV2ドメイン、カルモジュリンの41〜150番目のアミノ酸からなるペプチド(C−CaM(41−150))及びM13ペプチドを、この順番に持つ融合遺伝子の調製を行った。また、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGGSを導入した。
フォワードプライマーLOV2−fwに代えて、LOV2-fw-GGGS-XhoIを用いたことを除いては実施例9の操作を繰り返して、カルモジュリンの40番目と41番目のアミノ酸の間にLOV2が挿入され、N−CaM(1−40)とLOV2ドメインとの間にリンカーGGGSを有する融合遺伝子を得た。
【0081】
《融合タンパク質の発現及び精製》
実施例1〜15において得られたベクターを用いて、ヒートショック法により大腸菌コンピテントセル(JM109(DE3))を形質転換した。形質転換した大腸菌細胞を、100μg/mLのアンピシリンを含むLB固体培地で、37℃にて培養し、単一コロニーをピックアップして100μg/mLのアンピシリンを含むLB培養液2mL中へ移し、37℃で一晩培養して増殖させた。融合タンパク質は、T7発現系(pRSETB/JM109(DE3))を用いて発現させた。すなわち、培養液の100倍希釈物を作製し、更に23℃で3日間培養した。得られた細胞をフレンチプレスにより破砕し、得られた上清液から、ニッケルキレートカラム(QIAGEN社製)を用いてポリヒスチジン標識化タンパク質を精製した。さらに、この溶出液(100mMイミダゾール、50mMTris−Cl pH7.4、300mMNaCl)中のタンパク質サンプルをPD10脱塩・バッファー交換カラム(GE healthcare Bio−Sciences社製)により、HEPES10mM(pH7.5)を用いて精製を行った。
【0082】
《融合タンパク質の吸収スペクトルの測定》
得られた融合タンパク質に含まれているLOV2が、LOV2本来の構造を維持していることを、吸収スペクトルから確認した。LOV2は、447nm、473nm、及び370nmに吸収極大が存在する。
実施例1〜15において得られたベクターを発現させて得られた融合タンパク質の吸収スペクトルのうち、実施例1のPacman及び実施例8のCARBYの吸収スペクトルを規格化したものを図1に示す。また、陽性コントロールとして、LOV2の吸収スペクトルも記載した(Imaizumi T. et al FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis. Nature 20. 302-6 2003 Nov 20;426(6964):302-6.)。図1中、(A)がPacman、(B)がCARBY、(C)がLOV2の規格化吸収スペクトルである。
本発明の光駆動カルシウムイオン制御タンパク質は、LOV2と同じ吸収スペクトルを示しており、融合したLOV2がその機能を維持していることを示している。
【0083】
更に、LOV2の吸収極大ε447=13,800 M−1cm−1(Strickland D. et al Light-activated DNA binding in a designed allosteric protein. Proc Natl Acad Sci U S A. 105. 10709-14 2008)を用いてタンパク質のモル濃度を決定した、その後タンパク質のモル濃度をPacmanは100μMに合わせ、CARBYは200μMに合わせてサンプル溶液とした。
【0084】
《光照射によるカルシウムイオン濃度変化の測定》
本発明の光駆動カルシウムイオン制御タンパク質が、光照射により周囲のカルシウムイオン濃度を制御可能であることを確認した。
カルシウムイオン濃度の測定は、測定用試薬としてカルシウムイオン指示薬を用いて行った。カルシウムイオン指示薬は、Rhod−2−AM(同仁堂)を、エステラーゼ・ブタ肝臓由来(Sigma)で処理した化合物であるRhod−2を用いた。終濃度100μMになるようにRhod−2−AMを10mMのHEPES(pH8.0)に溶解し、エステラーゼを10U加え、室温でO/N放置した後、10mMのHEPES(pH7.5)で10倍希釈し、10μMのRhod−2を得た。
【0085】
前記精製した融合タンパク質を含むサンプル溶液を、以下の測定条件に従って、測定した。10mMのHEPES(pH7.5)を80μL、サンプル溶液を10μL、10μMのRhod−2を10μLを石英セルに滴下し、分光蛍光光度計であるF2500(HITACHI)を用いて測定を行った。測定条件は蛍光スペクトルモード、励起波長は550nm、スキャンスピードは1500nm/min、初期待ち時間は0秒、励起側/蛍光側スリットは5nm/5nm、ホトマル電圧は700V、レスポンスは0.04秒である。最初に、前記の条件で測定したのち、励起波長を450nmに変更し10秒光刺激した後、励起波長を550nmに戻して、再度蛍光スペクトルを測定した、その後シャッターを閉め、1分間待ち、シャッターをあけて、再度蛍光スペクトルを測定した。
Pacmanでは570nmにおける蛍光強度が40%程度減少した後にその蛍光強度が回復することが観測され(図2A)、CARBYでは570nmにおける蛍光強度が15%程度増加した後にその蛍光強度が回復することが観測された(図2B)。これはPacmanでは溶液中のカルシウムイオン濃度が光励起により低下し、時間経過と共に回復したことを示しており、CARBYでは溶液中のカルシウムイオン濃度が光励起により上昇し、時間経過と共に低下したことを示している。Rhod−2の570nmにおける蛍光強度から計算したカルシウムイオン濃度は、Pacmanの場合は約400nmが約200nmに、CARBYの場合は約1.3μMが約2μMに変化していた。また、実施例2〜7、及び実施例9〜15の融合タンパク質のカルシウムの吸収(結合)又はカルシウムの放出については、表2及び表3に示す。
【0086】
カルモジュリン、M13ペプチド、及びLOV2ドメインを、適切な順番で結合させることにより、カルシウムイオン指示薬であるRhod−2の励起光/蛍光を、550nm/570nmで測定した時の蛍光強度が、450nmの光刺激により蛍光強度が低下すること、又は蛍光強度が増加することが確認できた。すなわち、本発明の光駆動カルシウムイオン制御タンパク質は光刺激によってカルシウムイオン濃度を低下、又は上昇させることができる。
【0087】
【表2】
【0088】
【表3】
【産業上の利用可能性】
【0089】
本発明の光駆動カルシウムイオン制御タンパク質は、カルシウムイオンの生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中における濃度を制御することが可能であり、カルシウムイオンの細胞等における機能の解析に用いることができる。また、カルシウムイオンの濃度を制御することにより、それらの濃度異常を原因とする疾患の治療、又は予防に用いることも可能である。
【特許請求の範囲】
【請求項1】
(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;
を含む、光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項2】
(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;
をN末端から順に含み、光刺激によりカルシウムと結合する、請求項1に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項3】
(a1)カルシウムイオン結合タンパク質N末側ペプチド、又はその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;
(a2)カルシウムイオン結合タンパク質C末側ペプチド、又はその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;
をN末端から順に含み、光刺激によりカルシウムを放出する、請求項1に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項4】
前記光受容体ペプチドが、フォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、及びFKF−1のLOVドメインからなる群から選択される光受容体ペプチドであり;
前記カルシウムイオン結合タンパク質及びカルシウムイオン結合タンパク質結合ペプチドの組み合わせが、カルモジュリン及びカルモジュリン結合ペプチド、トロポニンC及びトロポニンI、ミオシン軽鎖及びミオシンII重鎖、並びにレコベリン及びロドプシンキナーゼからなる群から選択される、請求項1〜3のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項5】
(a)カルモジュリン、又はカルシウムイオン結合能を有するカルモジュリン改変体;
(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;及び
(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;
をN末端から順に含み、光刺激によりカルシウムと結合する、請求項2に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項6】
配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、又は配列番号7で表されるアミノ酸配列からなる、請求項5に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項7】
(a1)カルモジュリンN末側ペプチド、又はその改変体;
(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;
(a2)カルモジュリンC末側ペプチド、又はその改変体;及び
(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;
をN末端から順に含み、光刺激によりカルシウムを放出する、請求項3に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項8】
配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、又は配列番号15で表されるアミノ酸配列からなる、請求項7に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項9】
シグナルペプチドを含む、請求項1〜8のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項10】
請求項1〜9のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体をコードするDNA。
【請求項11】
配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、又は配列番号30で表される塩基配列からなる、請求項10に記載のDNA。
【請求項12】
請求項10又は11に記載のDNAを含むプラスミド。
【請求項13】
請求項1〜9のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、若しくはその機能的等価改変体、又は請求項10若しくは11に記載のDNAを用いて発現するタンパク質に、励起光を照射することを特徴とする、カルシウムイオン濃度の制御方法。
【請求項14】
前記励起光の励起波長が、300〜500nmである請求項13に記載のカルシウムイオン濃度の制御方法。
【請求項1】
(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;
を含む、光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項2】
(a)カルシウムイオン結合タンパク質、又はカルシウムイオン結合能を有するその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;
をN末端から順に含み、光刺激によりカルシウムと結合する、請求項1に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項3】
(a1)カルシウムイオン結合タンパク質N末側ペプチド、又はその改変体;
(b)光受容体ペプチド、又は光刺激により構造の変化するその改変体;
(a2)カルシウムイオン結合タンパク質C末側ペプチド、又はその改変体;及び
(c)カルシウムイオン結合タンパク質結合ペプチド、又はカルシウムイオン結合タンパク質への結合能を有するその改変体;
をN末端から順に含み、光刺激によりカルシウムを放出する、請求項1に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項4】
前記光受容体ペプチドが、フォトトロピン1のLOV1ドメイン、フォトトロピン1のLOV2ドメイン、フォトトロピン2のLOV1ドメイン、フォトトロピン2のLOV2ドメイン、オーレオクロム1のLOVドメイン、オーレオクロム2のLOVドメイン、及びFKF−1のLOVドメインからなる群から選択される光受容体ペプチドであり;
前記カルシウムイオン結合タンパク質及びカルシウムイオン結合タンパク質結合ペプチドの組み合わせが、カルモジュリン及びカルモジュリン結合ペプチド、トロポニンC及びトロポニンI、ミオシン軽鎖及びミオシンII重鎖、並びにレコベリン及びロドプシンキナーゼからなる群から選択される、請求項1〜3のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項5】
(a)カルモジュリン、又はカルシウムイオン結合能を有するカルモジュリン改変体;
(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;及び
(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;
をN末端から順に含み、光刺激によりカルシウムと結合する、請求項2に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項6】
配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、又は配列番号7で表されるアミノ酸配列からなる、請求項5に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項7】
(a1)カルモジュリンN末側ペプチド、又はその改変体;
(b)LOV2ドメイン、又は光刺激により構造の変化するLOV2ドメイン改変体;
(a2)カルモジュリンC末側ペプチド、又はその改変体;及び
(c)M13ペプチド、又はカルモジュリン結合能を有するM13ペプチド改変体;
をN末端から順に含み、光刺激によりカルシウムを放出する、請求項3に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項8】
配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、又は配列番号15で表されるアミノ酸配列からなる、請求項7に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項9】
シグナルペプチドを含む、請求項1〜8のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体。
【請求項10】
請求項1〜9のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、又はその機能的等価改変体をコードするDNA。
【請求項11】
配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、又は配列番号30で表される塩基配列からなる、請求項10に記載のDNA。
【請求項12】
請求項10又は11に記載のDNAを含むプラスミド。
【請求項13】
請求項1〜9のいずれか一項に記載の光駆動カルシウムイオン制御タンパク質、若しくはその機能的等価改変体、又は請求項10若しくは11に記載のDNAを用いて発現するタンパク質に、励起光を照射することを特徴とする、カルシウムイオン濃度の制御方法。
【請求項14】
前記励起光の励起波長が、300〜500nmである請求項13に記載のカルシウムイオン濃度の制御方法。
【図1】

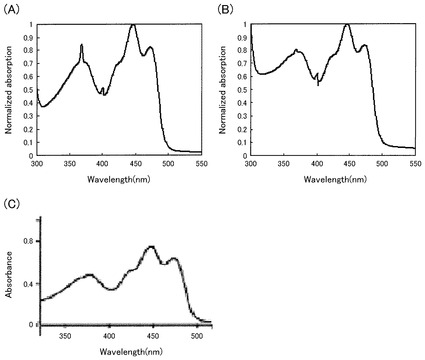
【図2】
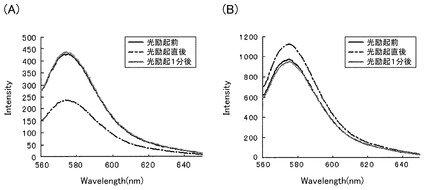
【図3】
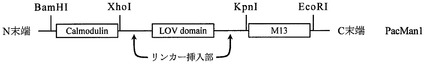
【図4】
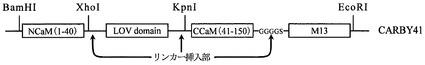
【図2】
【図3】
【図4】
【公開番号】特開2012−157306(P2012−157306A)
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2011−20234(P2011−20234)
【出願日】平成23年2月1日(2011.2.1)
【出願人】(504173471)国立大学法人北海道大学 (971)
【Fターム(参考)】
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成23年2月1日(2011.2.1)
【出願人】(504173471)国立大学法人北海道大学 (971)
【Fターム(参考)】
[ Back to top ]