
免疫モジュレーション用のヘルペスウイルス
【課題】樹状細胞に効率よく感染する能力がある弱毒化単純ヘルペスウイルスの免疫応答を刺激するための薬剤を提供する。
【解決手段】機能性vhs遺伝子またはその機能性等価体を欠失している弱毒化ヘルペスウイルスの、樹状細胞に該ウイルスを感染させることを含む免疫療法またはワクチン接種法において使用する医薬の製造における使用。
【解決手段】機能性vhs遺伝子またはその機能性等価体を欠失している弱毒化ヘルペスウイルスの、樹状細胞に該ウイルスを感染させることを含む免疫療法またはワクチン接種法において使用する医薬の製造における使用。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、樹状細胞に効率よく感染する能力がある弱毒化単純ヘルペスウイルスに関する。本発明はまた、疾患の処置に対する免疫療法アプローチにおける前記ウイルスの使用に関する。
【背景技術】
【0002】
樹状細胞(dendritic cell: DC)は最も効力のある抗原提示細胞であって、免疫系が寛容になっている抗原に対してさえも応答を効果的に引き出すことができる。かくして、腫瘍に対する免疫応答を引き出すための腫瘍免疫療法の場合、DCの使用は、それらが腫瘍特異的抗原を提示するように作られたならば、理想的であると考えられる。また、細菌、ウイルス、寄生虫などの病原体に由来する抗原を提示させるためにDCを使用することができ、そのような疾患に対する防御用または治療用ワクチンが提供される可能性がある。しかしながら、これらの標的のいずれについても、抗原をDCに効率よく移入することが、このアプローチに関しての最大の問題点であることが判っている。
【0003】
腫瘍抗原または他の疾患関連抗原に対する治療免疫応答を生じさせる実際の機会を与えるためには、いくつかの条件が満たされねばならない。まず第一に、分子の発現が腫瘍または疾患に特異的(少なくとも選択的)であり、それゆえに、免疫応答の標的となりうる、そのような分子を同定することが必要である。この作業は、大多数の一般的な腫瘍にとっては非常に難しいとされているが、例えば子宮頸癌の場合には、ほとんどがウイルスの癌遺伝子E6およびE7の存在により解決され、他の腫瘍の場合にも、良好な候補抗原が同定されつつある。例えば、MUC-1遺伝子産物は、90%の卵巣癌を含めて、いくつかの腫瘍で過剰発現されている。種々の他の腫瘍関連抗原も同定されており、そのうちのどれかを癌の免疫療法処置に使用できる可能性がある。今後もおそらく、追加の腫瘍関連抗原が発見され続けるであろう。第二に、1つ以上の抗原が同定された後には、免疫原性がある形態の抗原を免疫系に送達することが必要となる。腫瘍拒絶にとって最も重要な細胞性免疫応答を生起させるために、これは、タンパク質を宿主細胞の細胞質の中に送達させるか(高分子量のタンパク質抗原の場合には難しい)、あるいは、遺伝子送達またはDNA免疫化の後で宿主細胞自体にタンパク質を合成させるか、のいずれかが必要であることを意味する。この目的のために開発されたウイルスベクターとして、ワクシニア、アデノウイルス、またはレトロウイルスが挙げられる。
【0004】
現在、最適な免疫刺激を提供すると広く認められている細胞型は樹状細胞である(DC; 例えば、GirolomoniおよびRicciardi-Castagnoli, 1997を参照のこと)。事実、DCはin vivoで一次免疫応答を刺激することができる唯一の細胞型であると考えられており、さらに、ある状況下では確立された免疫寛容を破壊する能力があることさえも示されている。いくつかのグループは、DCが治療効果を示すという希望を抱いて、腫瘍に対する免疫応答を刺激するための自己養子免疫療法プロトコルでのDCの使用を検討している。そのようなプロトコルには、末梢血由来のDCの培養および/または富化、DCへの抗原のin vitroローディングおよび患者への該DCの再導入、またはDCへの抗原の直接in vivoローディングが必要である。しかし、このアプローチは、樹状細胞に抗原をローディングするための効率のよい手段が存在しないことで、これまで阻まれてきた。ところが、最近の研究により、ペプチドパルス化DCによる抗原の提示は、in vivoにおいて抗腫瘍応答を生み出すことが示された(Celluzziら, 1996; Zitvogelら, 1996)。ウイルスベクターに関しては、レトロウイルスは樹状細胞への効率のよい遺伝子送達をもたらさず(Reevesら, 1996; Aicherら, 1997)、我々の研究では、他のグループ(Arthurら, 1997)が報告した研究と相違して、アデノウイルスのみが低効率の遺伝子送達を可能にするにすぎない。
【0005】
我々は、以前に、単純ヘルペスウイルス(HSV)が樹状細胞に効率よく感染して遺伝子を送達することができることを試験し、報告した(Coffinら, 1998; WO 00/08191)。HSVは、他のベクター系と比べてこの目的に対していくつかの利点を有する。HSVは、様々な細胞型(他のベクター系では感染させるのが非常に難しいものを含み、例えば、Dillooら, 1997; Coffinら, 1998)に効率よく感染する能力があり、操作するのが簡単であり、しかも、大きなDNA挿入物を受け入れて複数の遺伝子の発現を可能にする(CoffinおよびLatchman 1996に概説)という利点である。樹状細胞への複数の抗原のex vivo送達とその後の体内への再導入、または樹状細胞への抗原の直接in vivo投与は、いくつかの癌および感染症の治療に対する特に有望なアプローチであると考えられる。
【0006】
国際公開WO 00/08191には、野生型の単純ヘルペスウイルスが、感染した樹状細胞内で抗原プロセッシングが起こるのを妨げること、および、機能性UL43およびvhs両遺伝子を欠失しているヘルペスウイルス、または前初期遺伝子の発現を最小限に抑える突然変異を含むヘルペスウイルスは、感染細胞内で抗原プロセッシングが起こるのを妨げることなく、樹状細胞に効率よく感染する能力がある、と教示されている。
【発明の概要】
【0007】
本発明者らは、このたび、驚いたことに、HSVベクター内のビリオン宿主シャットオフ(virion host shut-off: vhs)タンパク質をコードする遺伝子を破壊すると、HSV感染細胞において効率的な樹状細胞活性化を起こさせることができることを見出した。UL43遺伝子の破壊は必要でない。以前には、HSVに感染した樹状細胞は通常、感染それ自体によっても、他の刺激によっても活性化されることはない、ということが明らかにされている(Salioら, 1999; Kruseら, 2000)。
【0008】
本発明者らは、これまで知られていない、樹状細胞活性化を妨げるというvhsタンパク質の機能を確認した。樹状細胞活性化は、非活性化状態と比べたときの、特定の細胞表面マーカーのアップレギュレーションとして定義される。これらのマーカーにはCD83およびCD86が含まれる。樹状細胞活性化はリポ多糖(LPS)による処理で刺激することができる。HSVに感染した樹状細胞のLPS処理は、CD83またはCD86のアップレギュレーションをもたらさない。本発明者らは、変異型HSV(vhsが不活性化されているが、機能性UL43遺伝子をもつ)に感染した樹状細胞をLPSで処理すると、CD83とCD86の双方がアップレギュレートされることを見出した。機能性vhs遺伝子をもつウイルスに感染した樹状細胞をLPSで処理してもCD83とCD86のアップレギュレーションは認められない。したがって、これらの結果は、ヘルペスウイルス感染後に形質導入樹状細胞が免疫応答を最大限刺激するには、vhsをコードする遺伝子を破壊する必要があるが、UL43をコードする遺伝子はその必要がない、ことを示している。
【0009】
本発明者らの結果は、さらに、野生型の単純ヘルペスウイルスの病因におけるvhsの役割を実証するものである。HSVは高効率で樹状細胞に感染するが、HSVがその天然の生活環の一部としてそれを行うように進化した理由は、細胞性免疫応答(潜伏HSV感染が効率よく確立されるのを妨げるか、または潜伏と再活性化の反復サイクル中にそのウイルスのクリアランスをもたらす可能性がある)を最小限にしうるようにするためであろう。樹状細胞活性化は効果的な細胞性免疫応答の刺激において重要である。vhsはビリオンタンパク質であり、そうしてHSV遺伝子は一般に樹状細胞内で高レベルには発現されないが、vhsタンパク質は侵入してくるウイルスと共に樹状細胞に送達されるであろう。したがって、HSVに感染した樹状細胞の活性化を妨げるというvhsの新規な機能は、ヒトに感染した後のHSV生活環において重要なvhsの機能であるらしい。
【0010】
かくして、本発明は、下記ウイルスを樹状細胞に感染させることを含む免疫応答の刺激方法において使用するための医薬の製造における、
(i) 機能性vhs遺伝子またはその機能性等価体を欠失しており、
(ii) 機能性UL43遺伝子またはその機能性等価体を含んでいる、
弱毒化ヘルペスウイルスの使用を提供する。好ましくは、前記ウイルスはヒトの単純ヘルペスウイルスである。より好ましくは、前記ウイルスはHSV1またはHSV2である。樹状細胞にはin vitroで感染させてもin vivoで感染させてもよい。
【0011】
前記ウイルスは1つ以上の追加の突然変異を含むことができる。追加の突然変異はウイルスの毒性を最小限に抑えるものが好ましい。典型的には、そのような突然変異は前初期(immediate early: IE)遺伝子の発現を低下させるか、最小限にするものである。IE遺伝子の発現を阻止または低下させると、ウイルスの複製が妨げられるか、または減少する。そのような突然変異として、例えば、ICP4、ICP27、ICP0および/またはICP22(好ましくは、ICP27および/またはICP4)をコードする遺伝子を不活性化する突然変異が挙げられる。さらに、vmw65をコードする遺伝子の、トランス活性化機能を排除する不活性化突然変異も含まれる(例えば、Aceら, 1989またはSmileyら, 1997に記載されるようなvmw65突然変異)。追加の突然変異はまた、そのウイルスの免疫応答阻害活性を最小限に抑えることが好ましい。そのような突然変異には、ICP47をコードする遺伝子の不活性化が含まれる。
【図面の簡単な説明】
【0012】
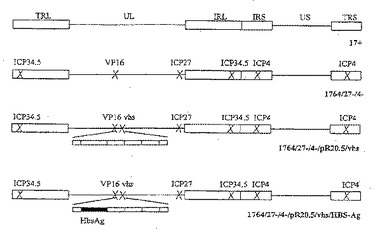
【図1】ウイルス株 1764/27-/4-、1764/27-/4-/pR20.5/vhs、1764/27-/4-/pR20.5/vhsHBS-Ag、および野生型HSV 17+株を示した図である。
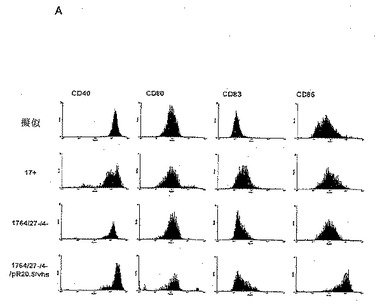
【図2A】刺激してない、擬似感染細胞ならびに17+、1764/27-/4-および1764/27-/4-/pR20.5/vhsウイルス株を感染させた細胞上のCD40、CD80、CD83およびCD86の細胞表面発現レベルを測定するFACS分析の結果を示した図である。
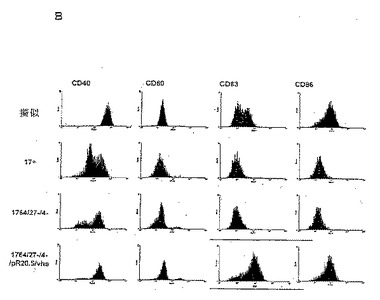
【図2B】LPSで刺激した、擬似感染細胞ならびに17+、1764/27-/4-および1764/27-/4-/pR20.5/vhsウイルス株を感染させた細胞上のCD40、CD80、CD83およびCD86の細胞表面発現レベルを測定するFACS分析の結果を示した図である。
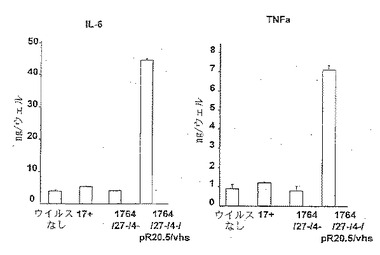
【図3】未感染の樹状細胞ならびに17+、1764/27-/4-および1764/27-/4-/pR20.5/vhsウイルス株を感染させた樹状細胞からのIL-6およびTNFα分泌に及ぼすウイルス感染の影響を調べるためのELISA分析の結果を示した図である。
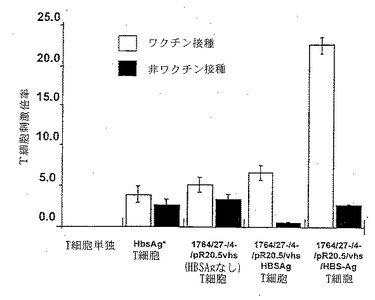
【図4】HBS-Agに応答した、B型肝炎ワクチン接種個体および非接種個体から調製したT細胞の増殖応答を示した図である。各個体から採取した樹状細胞は、処理しなかったか、組換えHBS-Agタンパク質(HBSAg*)と混合したか、対照ベクター(1764/27-/4-/pR20.5/vhs HBSAg)を感染させたか、あるいは、HBS-Agを発現するベクター(1764/27-/4-/pR20.5/vhs/HBS-Ag)を感染させたか、のいずれかであり、その後T細胞と混合した。
【発明を実施するための形態】
【0013】
A.ウイルス
本発明のウイルスは、感染した樹状細胞が活性化されるのを妨げることなく、樹状細胞に感染する能力がある。好ましくは、本発明のウイルスを感染多重度(MOI)1で感染させた樹状細胞は、LPSまたは他の活性化刺激で処理することにより活性化させることが可能である。
【0014】
本発明のウイルスは樹状細胞の活性化を妨げることがない。ウイルスがいつ樹状細胞の活性化を起こさせるかを調べるには、樹状細胞にウイルスをMOI≧1で感染させ、感染した樹状細胞をLPSで処理する。樹状細胞の活性化時にアップレギュレートされるCD83および/またはCD86などの細胞表面マーカーのレベルをモニターして、例えばFACS分析により、樹状細胞の活性化を判定する。細胞表面上のマーカーのレベルは、細胞に感染したウイルスが樹状細胞の活性化を起こさせるならば、LPS処理を行わなかった細胞よりもLPS処理を行った樹状細胞において顕著に高くなるだろう。また、樹状細胞の活性化を起こさせるウイルスを感染させた樹状細胞では、未感染の樹状細胞と比較して、これらのマーカーの一部または全部がより高くなるだろう。vhsに不活性化突然変異を含まない単純ヘルペスウイルスを樹状細胞に感染させる場合は、これらのマーカーのアップレギュレーションの著しい低下が観察される。
【0015】
感染樹状細胞の活性化を起こさせるために、本発明のウイルスは、vhs(HSVに含まれる)をコードする機能性遺伝子、または他のウイルス種に含まれるその相同性もしくは機能性等価体を欠失している。その上、本発明のウイルスは機能性UL43遺伝子をもっている。さらなる突然変異を行わせて、例えばICP47をコードする遺伝子に突然変異を導入して、ウイルスの免疫応答阻害作用をさらに低減させることも可能である。
【0016】
好ましくは、本発明の弱毒化ウイルスは、毒性が最小限となるように樹状細胞に感染させることができる。感染後の細胞生存率は、好ましくは、感染後1日目で少なくとも50%、より好ましくは、感染後1日目で少なくとも60、70、80または90%である。毒性低下を達成するために、ウイルス複製を減少させる突然変異を本発明のウイルスに導入することができる。例えば、VMW65をコードする遺伝子の突然変異(この突然変異はそのタンパク質のトランス活性化作用を最小にする)および/またはICP4、ICP27、ICP0およびICP22などの1つ以上の調節性前初期遺伝子の突然変異を含んでいてもよい。こうして、本発明のウイルスは典型的には、機能性vhs、ICP27、ICP4遺伝子を欠失しており、かつ転写活性化作用を示さないタンパク質をコードするVMW65遺伝子を含んでいてもよく、あるいは、典型的には、機能性vhs、ICP47およびICP4遺伝子を欠失していてもよい。
【0017】
in vivoで直接使用する場合には、ある程度の複製能力は一般に、誘導された免疫応答をブースト(増強)するのに有益でありうる。したがって、こうした状況下では、本発明のウイルスは、機能性vhs遺伝子を欠き、さらに、ウイルスの十分な病原性には必要であるがウイルスの複製には必要でない1つ以上の機能性遺伝子を欠くことが好ましい。そのような遺伝子として、例えば、ICP34.5、ICP6、チミジンキナーゼ、gHなどの糖タンパク質をコードするものが含まれる。しかし、チミジンキナーゼをコードする遺伝子は、この遺伝子の突然変異がアシクロビルのような抗ウイルス薬に対してウイルスを不感受性にするので、機能性であることが好ましい。
【0018】
本発明は単純ヘルペスウイルスを用いて説明されるが、感染した樹状細胞の活性化の阻止を低下させるようにヘルペスウイルス科の他のウイルスを改変しうることが理解されよう。特に、そのようなウイルスとして、水痘-帯状疱疹ウイルス、仮性狂犬病ウイルス、またはウシヘルペスウイルスが挙げられる。
【0019】
本発明のウイルスが単純ヘルペスウイルスである場合、このウイルスは例えばHSV1株もしくはHSV2株またはその誘導体に由来するものであってよく、好ましくはHSV1である。誘導体には、HSV1株およびHSV2株由来のDNAを含有するタイプ間組換え体が含まれる。そのようなタイプ間組換え体は当技術分野で公知であり、例えば、Thompsonら(1988)およびMeignierら(1988)に記載されている。誘導体は、典型的には本明細書に記載の方法で測定したとき、好ましくはHSV1またはHSV2のいずれかのゲノムに対して、少なくとも70%の配列相同性、より好ましくは少なくとも80%、より一層好ましくは少なくとも90%または95%の相同性を有する。さらに好ましくは、誘導体は、HSV1またはHSV2のいずれかのゲノムに対して、少なくとも70%の配列同一性、より好ましくは少なくとも80%の同一性、より一層好ましくは少なくとも90%、95%または98%の同一性を有する。
【0020】
誘導体は、ヌクレオチド置換(例えば、1、2または3個から10、25、50または100個までの置換)により改変されたHSV1またはHSV2ゲノムの配列をもつことができる。これとは別に、あるいは追加的に、HSV1またはHSV2ゲノムが1以上の挿入および/または欠失および/またはいずれか一方の末端または両方の末端での伸長により改変されていてもよい。
【0021】
本発明のウイルスを取得するために使用できる誘導体には、本発明のウイルスにおいて機能的に不活性化することが望ましい遺伝子中にすでに突然変異が存在するウイルス株が含まれ、例えば、vhs不活性化株(Jonesら, 1995に記載)、ICP47不活性化株(Goldsmithら, 1998に記載)、ICP4中に欠失があるd120株(DeLucaら, 1985)、ICP27中に欠失があるd27-1株(RiceおよびKnipe, 1990)、またはICP27とICP4の両方に欠失があるd92株(Samaniegoら, 1995)などである。こうした株を使用すると、本発明の変異型HSV株を作出するのに必要なステップ数を減らすことができる。
【0022】
各種HSV遺伝子を説明する際に用いる学術用語は、CoffinおよびLatchman, 1996に見出せるとおりである。
【0023】
上記HSV遺伝子の遺伝子相同体が他のヘルペスウイルス種に存在する場合は、これらの相同体が改変されるだろう。「相同体」とは、HSV遺伝子と機能的に等しい遺伝子を意味し、典型的には、相同体は、対応するHSV遺伝子に対してアミノ酸配列相同性または核酸配列相同性のいずれかを示す。典型的には、HSV遺伝子の相同体は、対応するHSV遺伝子に対して、アミノ酸レベルで、少なくとも15%、好ましくは少なくとも20%、より好ましくは少なくとも30%、40%または50%の同一性を示すであろう。
【0024】
vhsをコードする遺伝子は、HSV1およびHSV2中のUL41遺伝子である。HSV1の17+株(EMBL登録番号HE1CG)では、UL41遺伝子はヌクレオチド91,170からヌクレオチド92,637までである。HSV2のHG52株(EMBL登録番号z86099)では、UL41遺伝子はヌクレオチド91,800からヌクレオチド93,275までである。
【0025】
核酸およびタンパク質の相同性を決定するための方法は当技術分野で周知である。例えば、UWGCGパッケージは、相同性を算出するために使用できるBESTFITプログラムを提供する(例えば、そのデフォルトセッティングで使用)(Devereuxら(1984) Nucleic Acids Research 12, p387-395)。PILEUPおよびBLASTアルゴリズムを使用すると、相同性を算出したり、配列を整列させたりすることができ(典型的には、そのデフォルトセッティングで使用)、これらは例えば、Altschul (1993) J. Mol. Evol. 36:290-300; Altschulら(1990) J. Mol. Biol. 215:403-10に記載されている。
【0026】
BLAST解析を行うためのソフトウェアは、国立バイオテクノロジー情報センター(http://www.ncbi.nlm.nih.gov/)から利用可能である。このアルゴリズムでは、最初に、データベース配列中の同じ長さのワードとアライメントするとき、ある正値の閾値スコアTに一致するかまたはそれを満たすクエリー配列中の長さWの短いワードを同定することによって、高スコアを与える配列対(high scoring sequence pair: HSP)を同定することが必要である。Tは近隣ワードスコア閾値と呼ばれている(Altschulら, 1990)。これらの初期近隣ワードヒットは、それらを含むHSPを見つける検索を開始するためのシードとして働く。累積アライメントスコアが増加するかぎり、ワードヒットを、各配列に沿って両方向に延長させる。各方向でのワードヒットを中止するのは、累積アライメントスコアがその最大達成値から量Xだけ低下するとき;累積スコアが1以上の負スコアを与える残基アライメントの蓄積のためゼロまたはそれ以下になるとき;または、いずれかの配列の末端に達したときである。BLASTアルゴリズムパラメーターW、TおよびXはアライメントの感度と速度を決定する。BLASTプログラムは、デフォルトとして、ワード長(W) 11、BLOSUM62スコアリングマトリックス(HenikoffおよびHenikoff (1992) Proc. Natl. Acad. Sci. USA 89: 10915-10919を参照)アライメント(B) 50、期待値 10、M=5、N=4、および両鎖の比較を使用する。
【0027】
BLASTアルゴリズムは2つの配列間の類似性の統計的分析を実行する。例えば、KarlinおよびAltschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-5787を参照されたい。BLASTアルゴリズムにより与えられた類似性の程度は、最小の合計確率(P(N))であり、これは2つのヌクレオチドまたはアミノ酸配列間のマッチが偶然に出現する確率を示すものである。例えば、第1配列と第2配列との比較において最小合計確率が約1より小、好ましくは約0.1より小、より好ましくは約0.01より小、最も好ましくは約0.001より小である場合、一方の配列は他方の配列と類似していると見なされる。
【0028】
HSVゲノムの相同体はいくつかの方法で同定することができ、例えば、他のウイルスから作製したゲノムまたはcDNAライブラリーを、HSV遺伝子の全部または一部を含むプローブを用いて、中程度から高度のストリンジェンシー条件(例えば、0.03M 塩化ナトリウム、0.03M クエン酸ナトリウム、約50〜60℃)下で検索することにより行なう。これとは別に、縮重PCR(degenerate PCR)を使って、種相同体を取得することもでき、縮重PCRでは、保存されたアミノ酸配列をコードする変異体または相同体内の配列を標的とするように設計されたプライマーを使用する。これらのプライマーは、1以上の縮重位置を含んでおり、既知配列に対して単一配列プライマーを用いて配列をクローニングするのに用いた条件よりも低いストリンジェンシー条件(例えば、0.03M 塩化ナトリウム、0.03M クエン酸ナトリウム、約40℃)で使用する。
【0029】
ヘルペスウイルス中の相同体は、それがHSVタンパク質と1以上の機能的特徴を共有するならば、HSVタンパク質の機能性等価体となる。例えば、vhsタンパク質は、mRNAの安定性を弱めることによって感染細胞のタンパク質発現レベルを下げるという役割を果たしている。したがって、vhsタンパク質の機能性等価体は、mRNAの安定性を弱めることで宿主細胞の遺伝子発現を抑える役割を果たすことが好ましい。さらに、vhsの機能性等価体は、未感染樹状細胞を活性化する刺激に応答して樹状細胞の活性化を妨げることが好ましい。
【0030】
安全性の理由から、本発明のウイルスは、典型的には病気を引き起こす能力がなくなるように、弱毒化される。弱毒化のために改変されるウイルス領域は、完全にまたは部分的に除去されても、非機能的にされても、あるいは、他の配列(特に、異種遺伝子配列)で置換されてもよい。弱毒化をもたらす突然変異は、ウイルスベクターとして用いられる全てのウイルスグループについて、すでに記載されている。例えば、HSVはICP34.5および/または必須の遺伝子(例えば、ICP4、ICP27および/またはvhs遺伝子それ自体)の突然変異により無毒性にすることができる。
【0031】
特に好ましい弱毒化ウイルスとしては、vhsをコードする機能性遺伝子を欠失しており、かつ場合により機能性ICP47遺伝子を欠失していることに加えて、機能性ICP34.5と機能性ICP27遺伝子を欠失しており、また、機能性ICP4遺伝子および/またはVMW65遺伝子(転写活性化作用をもつタンパク質をコードする)を欠失していてもよいウイルス、および、機能性ICP27遺伝子をもつが機能性ICP4遺伝子と機能性ICP34.5遺伝子を欠失しており、また、VMW65遺伝子(転写活性化作用をもつタンパク質をコードする)を欠失していてもよいウイルスが挙げられる。そのようなウイルスは国際公開WO 98/04726およびWO 99/60145に記載されており、それらの開示内容を参照により本明細書に含めるものとする。
【0032】
本発明の単純ヘルペスウイルスが、特定の機能性必須遺伝子、例えば、ICP4またはICP27をコードする遺伝子を欠失している場合は、そのウイルスを、必須遺伝子を発現している細胞系を使って増殖させる。例えば、ウイルスが機能性ICP27遺伝子を欠く場合は、V27細胞(RiceおよびKnipe, 1990)、2-2細胞(Smithら, 1992)、またはB130/2細胞(WO 98/30707)を使って増殖させることができ、好ましくはB130/2細胞を用いる。ウイルスが機能性ICP4遺伝子を欠く場合は、ICP4を発現している細胞系、例えば、E5細胞(DeLucaら, 1985)を使ってウイルスを増殖させることができる。ウイルスが機能性ICP4遺伝子と機能性ICP27遺伝子を欠く場合は、そのウイルスを、ICP4とICP27の両方を発現している細胞系(例えば、E26細胞;Samaniegoら, 1995)を使って増殖させ、このウイルスがさらに機能性vmw65遺伝子を欠く場合には、vmw65の非HSV相同体(例えば、ウマヘルペスウイルス遺伝子12またはウシヘルペスウイルス由来のBTIF)をも含む細胞系を使って増殖させる。
【0033】
B.突然変異の方法
言及した各種ウイルス遺伝子は、当技術分野でよく知られているいくつかの技法により機能的に不活性化することができる。例えば、それらは欠失、置換または挿入により機能的に不活性化することができ、欠失による不活性化が好ましい。欠失は遺伝子の一部分を除去しても、遺伝子全体を除去してもよい。例えば、1個のヌクレオチドだけを欠失させて、フレームシフトを起こさせることができる。しかし、好ましくは、より大きな欠失を行ない、例えば、2、3または5個から10、20、30、50、100または200個のヌクレオチドの置換である。コードおよび非コード配列全体の好ましくは25%以上、より好ましくは50%以上(あるいはまた、絶対的な言い方では、10ヌクレオチド以上、より好ましくは100ヌクレオチド以上、最も好ましくは1000ヌクレオチド以上)を欠失または置換する。特に、遺伝子全体とそのフランキング配列の一部を除去することが好適である。挿入配列には、以下で述べる異種遺伝子が含まれる。突然変異は欠失と挿入の両方を含んでいてもよい。例えば、挿入を欠失部位に行なうことができる。こうして、ウイルス遺伝子への異種遺伝子の挿入はそのウイルス遺伝子の一部または全部に取って代わることができる。特に、異種遺伝子をvhs、ICP47、ICP27またはICP4に挿入することが好適である。VMW65遺伝子の場合には、それが必須の構造タンパク質をコードしているので、その遺伝子全体を欠失させることはできないが、一般的には、IE遺伝子を転写的に活性化するVMW65の能力を消失させる(例えば、Aceら, 1989またはSmileyら, 1997に記載されるような)不活性化突然変異が可能である。
【0034】
当業者には周知の相同組換え法によりヘルペスウイルスに突然変異を導入することができる。例えば、HSVゲノムDNAを、相同HSV配列に挟まれた突然変異配列を含むベクター(好ましくは、プラスミドベクター)と共にトランスフェクトする。この突然変異配列は欠失、挿入または置換を含んでいてよく、これらはいずれもルーチンな技法で構築することができる。組換えウイルスを、例えばβ-ガラクトシダーゼ活性または蛍光により、スクリーニングするために、挿入物に選択マーカー遺伝子、例えばlacZまたはGFP、を含めてもよい。
【0035】
C.異種遺伝子およびプロモーター
本発明のウイルスは1つ以上の異種遺伝子をもつように改変することができる。「異種遺伝子」という用語はあらゆる遺伝子を包含する。異種遺伝子は一般的にはヘルペスウイルスのゲノム中に存在しない遺伝子であるが、ヘルペス遺伝子を用いてもよい。ただし、そのコード配列が、もともとそれと関連しているウイルスの制御配列に機能的に連結されていないことを条件とする。異種遺伝子は野生型遺伝子のどのような対立遺伝子変異体であってもよいし、突然変異遺伝子であってもよい。「遺伝子」という用語は、RNA分子を産生するように少なくとも転写され得る核酸配列を包含し、そのRNA分子は翻訳可能であってポリペプチドを産生するか、またはアンチセンス効果により遺伝子発現レベルをダウンレギュレートすることが好ましい。本発明のウイルスは、天然の、あるいは異種遺伝子の翻訳コード配列に結合された、5'および/または3'転写されるが翻訳されないフランキング配列の一部または全部を含んでいてもよい。さらに、本発明のウイルスは、転写される配列に通常結合された転写制御配列、例えば、転写終止シグナル、ポリアデニル化部位および下流エンハンサーエレメントを含んでいてもよい。
【0036】
異種遺伝子は、HSV株と、例えばHSV配列により挟まれた異種遺伝子を含むプラスミドベクターと、の相同組換えによってウイルスのゲノムに挿入することができる。当技術分野で周知のクローニング技法を用いて、ヘルペスウイルス配列を含む適切なプラスミドベクターに1つ以上の異種遺伝子を導入することが可能である。ウイルスが依然として増殖可能であるかぎり、ウイルスゲノムのどの位置に異種遺伝子を挿入してもよい。好ましくは、ウイルスの弱毒化をもたらす遺伝子に異種遺伝子を挿入する。複数の異種遺伝子をウイルスゲノム内の複数の部位に挿入することも可能である。
【0037】
異種遺伝子の転写配列は、樹状細胞、好ましくは哺乳動物の樹状細胞、より好ましくはヒトの樹状細胞内での異種遺伝子の発現を可能にする制御配列と機能的に連結されていることが好ましい。「機能的に連結される」という用語は、記載した成分同士がそれらの意図された様式で機能することを可能にする関係にある並置をさす。コード配列と「機能的に連結される」制御配列は、コード配列の発現が制御配列と適合しうる条件下で達成されるような方法で連結される。
【0038】
制御配列は、異種遺伝子の発現を可能にするプロモーターおよび転写を終結させるためのシグナルを含む。プロモーターは哺乳動物、好ましくはヒトの樹状細胞において機能するプロモーターから選択される。プロモーターは真核生物遺伝子のプロモーター配列に由来するものでもよい。例えば、プロモーターは、異種遺伝子の発現を起こさせようとする細胞、好ましくは哺乳動物の樹状細胞、より好ましくはヒトの樹状細胞から誘導することができる。真核細胞のプロモーターに関しては、それらは普遍的なやり方で(β-アクチン、チューブリンのプロモーターなど)あるいは樹状細胞に特異的なやり方で機能するプロモーターでありうる。ウイルスプロモーターを使用してもよく、例えば、モロニーマウス白血病ウイルスの長い末端反復配列(MMLV LTR)プロモーターまたは他のレトロウイルスプロモーター、ヒトまたはマウスサイトメガロウイルス(CMV)IEプロモーターがある。
【0039】
異種遺伝子と制御配列を含む発現カセットおよび他の適当な構築物は、当業者には公知である通常のクローニング法を用いて作製することができる(例えば、Sambrookら, 1989, Molecular Cloning - a laboratory manual; Cold Spring Harbor Press を参照されたい)。
【0040】
さらに、これらのプロモーターはどれも、さらなる調節配列、例えばエンハンサー配列(HSV LAT領域を含む)を付加することによって改変することができる。上記の2種以上のプロモーター由来の配列エレメントを含むキメラプロモーター、例えば、MMLV LTR/LAT融合プロモーター(Lokensgardら, 1994)またはLAT領域のエレメントを含むプロモーター(WO98/30707)を用いてもよい。
【0041】
異種遺伝子は典型的には治療用のポリペプチドをコードする。例えば、特定の腫瘍に対する免疫応答を特異的に促進させるには、1以上の腫瘍抗原の発現を指令する本発明のウイルスで樹状細胞をトランスフェクトすることが望ましい。腫瘍抗原は腫瘍細胞に特異的なもの(すなわち、腫瘍細胞に存在するが、非腫瘍細胞には存在しないもの)であっても、同じタイプの非腫瘍細胞よりも(例えば、抗原発現のアップレギュレーションのため)腫瘍細胞に高レベルで存在するものであってもよい。これは癌の治療に有効であろう。なんとなれば、本発明の感染樹状細胞を用いることにより、宿主の免疫系が刺激されて腫瘍特異的抗原または腫瘍普遍的抗原に反応し、腫瘍の縮小/後退をもたらすからである。特に、腫瘍抗原は腫瘍細胞の表面に発現されることが好ましく、例えば、細胞表面受容体または細胞接着タンパク質でありうる。腫瘍抗原の例として、卵巣癌をはじめとする幾つかの腫瘍において過剰発現されるMUC-1遺伝子産物(Gendlerら, 1990)、子宮頸癌と関連があるヒト乳頭腫ウイルスタンパク質E6およびE7、メラノーマにおけるMART-I、MAGE-I、gp100およびチロシナーゼ、前立腺癌におけるPSA、様々なタイプの幾つかの腫瘍におけるCEA、および乳癌を含めた各種の癌におけるHer2neuが挙げられる。
【0042】
また、異種遺伝子は免疫応答を改変することができるポリペプチドをコードしてもよく、かかるポリペプチドには、例えば、サイトカイン(α-、β-またはγ-インターフェロン、IL-1、IL-2などのインターロイキン、腫瘍壊死因子、またはインスリン様増殖因子IもしくはIIなど)または他の免疫調節タンパク質(RANTESなどのケモカイン、およびCD80、CD86、CD40およびCD40リガンドのような共刺激分子など)が含まれる。
【0043】
異種遺伝子はまた、病原体由来のポリペプチドをコードしてもよく、かくして、例えば、本発明のウイルスを感染させた樹状細胞を用いることにより、宿主の免疫系を刺激して病原体に対する免疫応答を、該病原体への宿主の感染前または感染後に、生じさせることができる。ワクチンに用いるウイルスは、典型的には抗原性ポリペプチドをコードする異種遺伝子を含みうる。好ましくは、そのような病原体由来のポリペプチドは、寄生虫、細菌またはウイルスなどの病原性の生物に由来するものである。かかる抗原性ポリペプチドの例として、C型肝炎ウイルス抗原、B型肝炎表面またはコア抗原、乳頭腫ウイルス抗原、HIV抗原およびマラリア抗原が挙げられる。病原性生物由来の異種遺伝子を含むウイルスは、治療処置または予防処置のいずれかまたは両方に使用することができる。
【0044】
治療用途には、おそらく複数の遺伝子の投与が必要である。複数の遺伝子の発現はさまざまな症状を治療するのに有利であろう。ヘルペスウイルスは、他のウイルスベクター系のようにパッケージング能力が限られていないので、唯一適切なものである。したがって、ヘルペスウイルスのゲノム内には複数の異種遺伝子を収容することができる。例えば、2〜6個の遺伝子をそのゲノムに挿入することができる。
【0045】
これを達成するための方法は、少なくとも2つある。例えば、2個以上の異種遺伝子と関連制御配列を、特定のHSV株のウイルスゲノムの単一部位または複数の部位に導入することである。また、互いに離れて反対の方向に向いているプロモーター(同一のまたは異なるプロモーター)の対を使用することも可能であり、これらのプロモーターはそれぞれが上記のような異種遺伝子(同一のまたは異なる異種遺伝子)の発現を駆動する。
【0046】
D.樹状細胞
樹状細胞はいくつかの手段により単離/調製することができ、例えば、それらは末梢血から直接精製されるか、例えばG-CSF処理により末梢血に移動した後の、または直接骨髄からの、CD34+前駆細胞から生成される。末梢血からの付着前駆細胞はGM-CSF/IL-4混合物で処理し(Inabaら, 1992)、骨髄からの非付着CD34+細胞はGM-CSFとTNF-αで処理することができる(Cauxら, 1992)。樹状細胞をヒトボランティアの末梢血からルーチンに調製するには、SallustoおよびLanzavecchia, 1994に記載の方法と同様にして、精製した末梢血単核細胞(PBMC)を使用し、そして付着細胞をGM-CSFとIL-4で2時間処理する。次に、磁性ビーズを使ってこれらからCD19+B細胞とCD3+, CD2+T細胞を枯渇する(Coffinら, 1998)。樹状細胞の調製には他の方法も使用することができる。
【0047】
E.治療用途
本発明のウイルスおよび本発明のウイルスを感染させた樹状細胞は、治療方法において使用することができる。特に、腫瘍抗原を発現する本発明のウイルス、および腫瘍抗原を発現する本発明のウイルスを感染させた樹状細胞は、癌の治療方法に使用される。特定すると、本発明のウイルスおよび本発明のウイルスを感染させた樹状細胞は、ヒトを含む哺乳動物の各種腫瘍の増殖を抑制するために使用することができ、例えば、卵巣癌、子宮頸癌および子宮内膜腫瘍、ならびに、乳癌、肺癌、膀胱癌、大腸癌などの癌の抑制である。増殖抑制が可能なその他の新生物としては、肉腫(例えば、軟組織および骨の肉腫)、血液の悪性腫瘍(例えば、白血病)などがある。腫瘍抗原を発現する本発明のウイルスおよび/または該ウイルスを感染させた樹状細胞を用いて治療しうる癌の例として、特に、メラノーマ、白血病、子宮頸癌および卵巣癌が挙げられる。癌治療用のウイルスは典型的には腫瘍抗原をコードする異種遺伝子を含む。かかるウイルスまたはウイルスを感染させた樹状細胞の投与により、一般的には、腫瘍抗原に対する免疫応答が引き出されるであろう。
【0048】
本発明のウイルスおよび本発明のウイルスを感染させた樹状細胞は、病原体による感染症(例えば、寄生虫、細菌またはウイルスによる感染症)の治療または予防方法において使用することができる。病原体による感染症の治療に用いるウイルスは、典型的には、病原性生物に由来する抗原をコードする異種遺伝子を含む。そのようなウイルスまたはウイルスを感染させた樹状細胞を投与することにより、一般的には、病原性生物由来の抗原に対する免疫応答が引き出されるであろう。かかるウイルス感染症として、ヘルペスウイルス感染症が含まれる。こうして、本発明のウイルスを使って、例えば、HSV1またはHSV2感染症の治療またはワクチン接種において、該ウイルス自体に対する免疫応答を誘発させることが可能である。ウイルスがHSV1またはHSV2感染の治療に用いられるものである場合、そのウイルスはHSV抗原(その天然のプロモーターの制御下にないもの)または免疫調節分子をコードする異種遺伝子を含んでいてもよい。ウイルス/樹状細胞は、宿主の防御免疫応答を刺激するために感染前に、あるいは宿主の免疫系を刺激して感染症を撲滅するために感染後に、投与することができる。
【0049】
F.投与
こうして、本発明のヘルペスウイルスを用いて、処置を必要とするヒトまたは動物に治療用遺伝子を送達することができる。本発明のヘルペスウイルスを用いた治療用遺伝子の送達は、例えば、悪性疾患および/または病原体による感染症の治療に使用することができる。
【0050】
本発明のウイルスは、治療を必要とする患者(好ましくは、ヒト患者)に使用される。治療を必要とする患者は、癌を患う個体または感染症のある患者である。治療処置の目的は患者の状態を改善することにある。典型的には、本発明のウイルスを用いた治療処置は癌の症状を軽減する。本発明に従う癌の治療方法は、機能性UL43遺伝子をもつが機能性vhs遺伝子を欠失しているウイルスを、治療に有効な量で癌患者に投与して、該ウイルスが該患者の樹状細胞中に存在するようにすることを含んでなる。腫瘍を患う個体に本発明のウイルスを投与すると、典型的には、腫瘍細胞が殺傷される結果、腫瘍の大きさが小さくなったり、その腫瘍由来の悪性細胞の広がりがくい止められたりするだろう。
【0051】
一般的に、本発明のウイルスを用いた感染症の治療処置は、感染症の症状を軽減し、さらには病原性生物を死滅させることが好ましい。本発明に従う感染症の治療方法は、機能性vhs遺伝子を欠失しているウイルスを、治療に有効な量で感染症の患者に投与することを含んでなる。好ましくは、該ウイルスを患者の樹状細胞に侵入させるか、あるいはex vivoで該ウイルスを感染させた樹状細胞を患者に投与する。本発明のウイルスを用いた予防処置は、典型的には、癌や感染症のリスクがある患者において、腫瘍抗原に対する抗体または病原性生物由来の抗原に対する抗体の産生をうながす。一般的に、癌のリスクがある患者とは、遺伝的にその傾向があるか、癌原物質に暴露されてしまったか、癌原物質に暴露される危険がある者でありうる。一般的に、感染症のリスクがある患者とは、病原性生物にさらされる可能性がある者でありうる。
【0052】
治療を実施するための一つの方法は、1以上の治療用遺伝子を、上記のような本発明のヘルペスウイルスのゲノムに挿入し、次に、得られた組換えウイルスを製薬上許容される担体または希釈剤と組み合わせて医薬組成物を調製することを含む。適当な担体および希釈剤として、等張食塩水、例えばリン酸緩衝食塩水が挙げられる。医薬組成物は非経口、筋肉内、静脈内、腹腔内、皮下または経皮投与用に製剤化することができる。皮下または腹腔内投与が好適である。経皮または皮内投与が特に好適でありうる。
【0053】
本発明のウイルスを樹状細胞に感染させるには、ウイルスを含む組成物を患者に投与することによってin vivoで実施する。治療用遺伝子を含むウイルスが樹状細胞に感染できるようなやり方で医薬組成物を投与する。投与されるウイルスの量は、104〜1010 pfu、好ましくは105〜108または105〜109 pfu、より好ましくは約106〜108 pfuの範囲とする。皮内に注射する場合、あるいは、例えば針のない器具を使って、経皮投与する場合、製薬上許容される適当な担体もしくは希釈剤中または特定の組成物中のウイルスを、一般的には10μl〜1ml、好ましくは100μl〜1mlの量で投与する。
【0054】
別の方法では、樹状細胞を末梢血または骨髄から単離/調製し、その細胞にin vitroで本発明のウイルスを感染させる。次に、形質導入された樹状細胞を、一般的には、筋肉内、腹腔内、皮下もしくは静脈内注射によって、あるいは、患者のリンパ節への直接注入によって、好ましくは皮下、腹腔内注射またはリンパ節への直接注入により、患者に投与する。形質導入された樹状細胞は、一般的には105〜109個、好ましくは106〜108個、より好ましくは約107個を患者に投与する。
【0055】
記載した投与経路および投与量は単なる指針であって、当業者であれば、個々の患者に最適な投与経路および投与量を容易に決定することができよう。投与量はさまざまなパラメーターに従って、特に患者の年齢、体重および症状に応じて決定される。
以下の実施例は本発明を例示するためのものである。
【0056】
実施例
材料および方法
ウイルス株の構築および増殖
全てのウイルス株はHSV1 17+株に由来し、そのヌクレオチド配列はGenBankに登録されている(登録番号 HE1CG)。ウイルス株は、BHK C-21細胞(ECACC番号 8501143)またはHSV1 ICP27、ICP4およびウマヘルペスウイルス遺伝子12をコードする遺伝子で安定にトランスフェクトされたBHK細胞(Thomasら, 1999)を使って産生し、増殖させた。
【0057】
VMW65に突然変異があるウイルスの場合は、ウイルス増殖用の培地に3mMのヘキサメチレン-ビスアセトアミド(HMBA)を加えた(McFarlaneら, 1992)。以下のウイルス株を使用した。
【0058】
(i) 17+ (野生型HSV1)
(ii) 17+/pR20.5/UL43
HSV LAT領域配列により隔離されたRSV/lacZ/pA配列とCMV/GFP/pA配列(相反する背中合わせの方向にある)からなるプラスミドpR20.5(Thomasら, 1999b)由来のカセット(ヌクレオチド118,866-120,219)を、標準的な方法で、精製したHSV1 17+株のゲノムDNAとの相同組換えにより、UL43遺伝子座に挿入した。最初に、pR20.5カセットを、UL43フランキング領域を含むプラスミド(Coffinら, 1996)にユニークなNsiI部位で挿入して、プラスミドpR20.5/43を得た。20.5カセットは、SrfIをコードするオリゴヌクレオチドがそのカセットの両側に挿入されたので、そのpGEM5(Promega)プラスミド骨格からSrfIで切り出すことができる。プラスミドpR20.5の構築のために、RSVプロモーターをpRc/RSV(Invitrogen)から、lacZ/pAをpCHI10(Pharmacia)から、CMV/pAをpcDNA3(Invitrogen)から、そしてGFPをpEGFP-N1(Clontech)から切り出した。
【0059】
(iii) 1764/27-/4-
1764/27-/4-ウイルス株は、1764/27-/4-/pR20.5ウイルス株DNAの空のICP4フランキング領域による組換え、およびGFPまたはlacZを発現しないウイルスプラークの選択により構築した。1764/27-/4-/pR20.5ウイルス株は、Thomasら, 1999bに記載されており、ウイルス(ICP27およびICP34.5も欠失されていて、VMW65をコードする遺伝子に不活性化突然変異がある)のICP4をコードする遺伝子を置換するように、ICP4遺伝子にpR20.5カセットが挿入されている。
【0060】
(iv) 1764/27-/4-/pR20.5/vhs
1764/27-/4-/pR20.5/vhsウイルス株を構築するために、pR20.5カセットを、HSV1 17+株のvhsコード遺伝子中のユニークなNruI部位でvhsフランキング領域に挿入し、得られたプラスミド(pR20.5/vhs)をHSV 1764/27-/4-株のDNAに組換えにより挿入した。したがって、1764/27-/4-/pR20.5/vhsウイルス株は、ICP4、ICP27およびICP34.5をコードする遺伝子が欠失しており、vmw65およびvhsをコードする遺伝子には不活性化突然変異が存在する。
【0061】
(v) 1764/27-/4-/pR19lacZ
1764/27-/4-/pR19lacZウイルス株は上記ウイルス(iv)と同様に構築したが、pR19lacZカセット(Wagstaffら, 1998)を、vhsにpR20.5カセットを挿入するのではなく、1764/27-/4-ウイルス株の潜伏関連転写物(latency associated transcript: LAT)領域に挿入した。
【0062】
(vi) 1764/27-/4-/pR20.5/vhs/HBS-Ag
pR20.5/vhsプラスミド中のlacZ遺伝子は、pHBV130(GoughおよびMurray, 1982)をXhoIとNsiIで消化し、放出された断片をpSP72(Promega)のSalI部位とSmaI部位の間に挿入することにより、肝炎表面抗原(HBS-Ag)をコードする遺伝子で置換した。pR20.5/vhsをXbaIとEcoRIで消化してlacZ遺伝子を放出させ、HindIIIとEcoRIを用いてpSP72から切り出したHBS-Ag遺伝子と置き換えた。得られたプラスミドを1764/27-/4-/pR20.5/vhsウイルスDNAに挿入し、lacZを発現しないプラークを選択し、精製した。その後ゲノムの構造をサザンブロットで確認した。
【0063】
樹状細胞の調製
樹状細胞は以前に記載されたとおりに末梢血から調製した(Coffinら, 1998)。簡単に説明すると、末梢血単核細胞(PBMC)を、健康でB型肝炎ワクチンを接種したドナー血液60mlからリンフォプレップ(lymphoprep; Nycomed)を使って調製した。赤血球の除去後、非付着細胞(主にT細胞とB細胞)を分離し、HBSSで洗浄し、室温、1400rpmで5分間遠心した。細胞ペレットを2mlの90% FCS:10% ジメチルスルホキシド(DMSO)混合物中に再懸濁し、アリコートに分けて、その後のT細胞分離のために-80℃で保存した。付着細胞は、GM-CSF(0.1μg/ml)とIL-4(0.05μg/ml)を補充したRPMI培地で培養し、37℃、5% CO2で7日間インキュベートした。さらなるリンフォプレップ精製の後、抗CD19抗体、抗CD2抗体(Harlan)、および抗CD3抗体(Harlan)を用いて磁気的に細胞を枯渇させ、即使用のために樹状細胞を完全RPMI培地に再懸濁した。
【0064】
CD4+T細胞の分離
上記のように凍結させたT細胞とB細胞をすばやく解凍し、HBSS中で洗浄し、1400rpmで5分間遠心した。細胞を2mlの完全RPMI培地に再懸濁し、カウントし、抗CD19(BU12 - 200μl正味、Immunology dept, UCL)、抗CD14(HB246 - 200μl正味、Immunology dept, UCL)、および抗HLA-DR(L243 - 100μl正味、Immunology dept, UCL)モノクローナル抗体と共にインキュベートし、氷上に30分間放置した。細胞をHBSS中で洗浄し、2mlの完全RPMI培地に再懸濁し、磁性ビーズに結合させたヒツジ抗マウス抗体(Dynabeads, Dynal)と、汚染細胞106個あたり10μlのビーズの割合で混合して、ローターミキサー上で4℃、45分間インキュベートした。その後、細胞懸濁液/磁性ビーズの混合物を磁石と接触させた状態で氷上に10分間置いた後上清を分離することにより、CD4+T細胞を枯渇させた。CD4+T細胞をカウントし、完全RPMI培地に適切な濃度で再懸濁し、氷上に配置するか、またはその後の使用のために37℃、5% CO2で培養した。
【0065】
樹状細胞の感染
樹状細胞は1400rpm、室温で5分間ペレット化した。次に、ウイルスを含むRPMI培地中に37℃、5% CO2で1時間再懸濁することにより、感染多重度(MOI)1で樹状細胞にウイルスを感染させた。GM-CSF(0.1μg/ml)とIL-4(0.05μg/ml)を補充したRPMI培地 1mlを加え、樹状細胞を37℃、5% CO2でインキュベートした。LPS刺激のためには、100ng/mlのLPSを追加的に含むRPMI培地を使用した。
【0066】
サイトカイン分析
市販のELISAキット(R&D Systems)を使って樹状細胞の培養上清中に含まれるIL-6およびTNF-αを測定した。ELISAに先立って、先に示したウイルスを樹状細胞に感染させてから42時間後に上清を集めて、使用前に-20℃で保存した。
【0067】
T細胞増殖アッセイ
B型肝炎ワクチンを接種したヒト個体と接種してないヒト個体から樹状細胞とCD4+T細胞を分離し、上記のように処理した。樹状細胞は1×105個/ml〜1×104個/mlの希釈物として使用し、CD4+T細胞は1×106個/mlで使用した。それぞれの樹状細胞(DC)濃度での実験を3回反復して行なった。各アッセイウェルに100μlの樹状細胞と100μlのCD4+T細胞を加えた。表示した場合は、組換えB型肝炎表面抗原(Austral)を1μg/ウェルの最終濃度でウェルに添加した。HSV-1を感染させた樹状細胞と未感染の樹状細胞をCD4+T細胞と共に37℃、5% CO2で6日間培養した。その後、1μCu/ウェルの[3H]チミジン(Amersham)を添加し、18時間後細胞を回収して、[3H]チミジンの取込みをカウントした。
【0068】
実施例1
予備的データは、機能性vhs遺伝子を含まないHSV株がウイルス感染後の樹状細胞の活性化を増大させる、ことを示す
ここでは、それぞれの場合に、1×105個の樹状細胞に各ウイルスを感染させるために、樹状細胞を穏やかにペレット化し、DMEM中の約100μlのウイルス懸濁液に再懸濁させ、37℃で1時間インキュベートし、そして2mlのRPMI/10% FCS+100ng/mlのGM-CSF、50ng/mlのIL-4を含む24ウェルプレートに移した。次いで、これらのプレートを37℃、5% CO2で一夜インキュベートした。また、公知の樹状細胞活性化剤であるリポ多糖(LPS)で樹状細胞を処理し、未処理の樹状細胞を対照として用いた。
【0069】
その後、これらの感染物および対照物から得られた上清をELISA試験で使用して、分泌されたサイトカインのレベルを検出した。また、感染させた樹状細胞および対照の樹状細胞の表面上に発現されたCD86のレベルを検出するために、蛍光活性化セルソーティング(FACS)を使用した。樹状細胞の培養物には、CD86発現のレベルに関して2つの細胞集団が存在していた。これらはFACS分析で2つのピークとして観察され、第1ピークはCD86発現のレベルが比較的低い細胞を示し、第2ピークはCD86発現のレベルが比較的高い細胞を示す。例えばLPSで活性化すると、より多くの細胞がより高レベルのCD86を発現し、こうして、第2ピーク中に見出せる細胞がより多くなる。
【0070】
【表1】

表1: 表示したウイルスを感染多重度1で感染させてから24時間後の培養上清中または対照の上清中に含まれるサイトカイン濃度。ELISAで測定。
【0071】
【表2】
表2: 対照の細胞上および表示したウイルスを感染させた細胞上のCD86の発現。
【0072】
結論
上記の結果から、未処理の樹状細胞は試験したサイトカインを最低レベルで分泌しており、しかもその表面上に予想された「休止」レベルのCD86を有する、ことがわかる。LPS処理後に、サイトカインレベルは著しく刺激され、CD86の表面発現のレベルが大いに増加する。上記の実験では、LPS処理細胞の平均蛍光強度は、抗CD86抗体を用いたFACS分析により約1×103である。これらの結果は、樹状細胞が活性化された状態にあることを示している。
【0073】
表示したウイルスを樹状細胞に感染させた後で、これらのアッセイにより樹状細胞の活性化が生じるためには、ウイルスがvhsをコードする遺伝子に不活性化突然変異を含まねばならない、ことが明らかに見てとれる。さまざまな無能化レベルのウイルスを使用しているが、vhs突然変異を含むウイルスのみがこれらのアッセイで樹状細胞の顕著な活性化を与える。また、vhs突然変異を含まないならば、表示のウイルスを細胞に感染させて、さらにLPSで処理しても、LPS単独で処理した場合ほど多くの細胞においてCD86レベルが増加しない、ことが見てとれる。さらに、vhs突然変異を含まないならば、ウイルス感染後にFACSで測定したCD86発現細胞の平均蛍光強度は、細胞をLPSで処理した場合に見られる強度よりも低下している。したがって、樹状細胞による最大免疫刺激を得るためには、vhsをコードする遺伝子に不活性化突然変異を含めるべきである、と結論付けることができる。
【0074】
実施例2
機能性vhsタンパク質を含まないHSV株は、樹状細胞の活性化を阻止しない
蛍光活性化セルソーティング(FACS)を用いて、感染樹状細胞と対照樹状細胞の表面上のCD86、CD80、CD83およびCD40の発現レベルを検出した。感染細胞からの上清を用いてELISAによりサイトカインのレベルを評価した。
【0075】
結果
ELISAの結果(図3)から、樹状細胞にHSVを高頻度で感染させることができるが、樹状細胞活性化の指標となるサイトカインは野生型(17+株)ウイルスまたは無能化(1764/27-/4-株)ウイルスのいずれの場合にも産生されない、ことがわかる。しかしながら、vhsを1764/27-/4-株から不活性化させると、得られる1764/27-/4-/pR20.5/vhs株では、樹状細胞の活性化を示すサイトカインが産生される。非LPS刺激樹状細胞に対するFACS分析(図2)からは、本質的に野生型HSV(17+株)または複製能のないHSVベクター(1764/27-/4-株)による感染は、CD86発現の増加を妨げることがわかる。上述したように、樹状細胞が感染プロセスによって活性化されると、CD86発現の増加が期待される。また、CD40レベルはHSV感染細胞において変化/低下する。しかし、vhsが不活性化されると(1764/27-/4-/pR20.5/vhs株)、CD86レベルは活性化を示して増加し、CD40レベルは変化しない。CD80とCD83は非刺激のHSV感染樹状細胞では大きく変化しない。CD83(B7.1)およびCD86(B7.2)はともに重要なT細胞共刺激分子であり、CD40は重要なT細胞活性化剤であり、そしてCD83は樹状細胞の成熟化および活性化の間にアップレギュレートされる樹状細胞のマーカーである。
【0076】
感染時に樹状細胞をLPSで刺激する場合には、野生型(17+株)ウイルスまたは無能化(1764/27-/4-株)ウイルスのいずれにおいてもCD40レベルに対する影響がより顕著である。しかし、vhsが不活性化されると、CD40に対するこれらの影響は妨げられる。LPSで刺激した樹状細胞は通常、CD83およびCD86発現を著しくアップレギュレートするが、これは、vhsが不活性化されない限り、HSV(17+株および1764/27-/4-株)によって阻止される。vhsが不活性化されると、CD83およびCD86レベルはどちらも、LPSで刺激した未感染細胞と同程度またはそれ以上に増加する。
【0077】
結論
予備実験と同様に、樹状細胞がHSV感染またはHSV感染とLPS刺激に応答して活性化される(表面マーカーの発現レベルにより測定)ためには、vhsをコードする遺伝子が不活性化されねばならないことが明らかである、と明確に見てとれる。機能性vhsをコードするウイルスは、試験した表面マーカーの発現レベルにより測定したとき、樹状細胞を顕著に活性化させることができない。
【0078】
実施例3
vhs不活性化HSVで形質導入された樹状細胞は、in vitroで抗原特異的T細胞応答を引き出す
上記の結果から、vhsが不活性化されているHSVベクターは、樹状細胞におけるHSVの不活性化作用が妨げられているので、樹状細胞用の効果的なベクターとして使用できることが示唆された。実際、そのようなHSV変異体を感染させた樹状細胞は、CD86のアップレギュレーションおよび特定のサイトカインの分泌により測定したとき、感染に応答して特異的に活性化されるようである。vhs不活性化HSV変異体が、樹状細胞への抗原コード化遺伝子の送達に続いて、抗原特異的免疫応答を引き出すことができるか否かを試験するために、B型肝炎ワクチンを接種した個体と接種しなかった個体から調製した樹状細胞およびT細胞を用いて実験を行なった。ここでは、最初に、B型肝炎表面抗原(HBS-Ag)発現カセットがIE遺伝子欠損ウイルスのvhsコード化遺伝子に挿入されているウイルスを構築した。次にT細胞増殖アッセイを実施したが、ここにおいて、ワクチン接種個体または非接種個体からの樹状細胞は、処理しなかったか、組換えHBS-Agタンパク質と混合して抗原「負荷」を行なったか、対照のマーカー遺伝子含有ベクター(1764/27-/4-/pR20.5/vhs、MOI=1)を感染させたか、対照のベクター(1764/27-/4-/pR20.5/vhs、MOI=1)を感染させかつ組換えHBS-Agタンパク質とも混合したか、あるいは、HBS-Agを発現するベクター(1764/27-/4-/pR20.5/vhs/HBS-Ag、MOI=1)を感染させたか、のいずれかである。その後、樹状細胞を、それぞれ同一のワクチン接種個体または非接種個体に由来するT細胞と混合し、T細胞増殖に及ぼす効果を標準的なT細胞増殖アッセイで観察した。
【0079】
これらの実験から明らかなように(図4)、HBS-Ag組換えタンパク質および対照のHSVベクターはワクチン接種個体において小さなT細胞増殖応答を引き出すことができ(HSV応答はおそらくHSV構造タンパク質に特異的なT細胞の増殖を示す)、また、組換えHBS-Agと混合された対照のベクターはやや大きな応答を引き出すことができたが、HBS-Agを発現するベクターはこれらのいずれよりも格別に大きな応答を与えた。こうして、HSVベクターを用いて樹状細胞にHBS-Agを直接送達した後に、顕著な特異的T細胞増殖応答が誘導されたが、かかる応答は組換え抗原単独と混合した後では生じなかった。したがって、vhsが不活性化されているHSVベクターは、抗原特異的T細胞増殖応答を刺激する樹状細胞の能力を保持したままで、抗原コード化遺伝子を樹状細胞に送達することを可能にする。
【0080】
参考文献
【技術分野】
【0001】
本発明は、樹状細胞に効率よく感染する能力がある弱毒化単純ヘルペスウイルスに関する。本発明はまた、疾患の処置に対する免疫療法アプローチにおける前記ウイルスの使用に関する。
【背景技術】
【0002】
樹状細胞(dendritic cell: DC)は最も効力のある抗原提示細胞であって、免疫系が寛容になっている抗原に対してさえも応答を効果的に引き出すことができる。かくして、腫瘍に対する免疫応答を引き出すための腫瘍免疫療法の場合、DCの使用は、それらが腫瘍特異的抗原を提示するように作られたならば、理想的であると考えられる。また、細菌、ウイルス、寄生虫などの病原体に由来する抗原を提示させるためにDCを使用することができ、そのような疾患に対する防御用または治療用ワクチンが提供される可能性がある。しかしながら、これらの標的のいずれについても、抗原をDCに効率よく移入することが、このアプローチに関しての最大の問題点であることが判っている。
【0003】
腫瘍抗原または他の疾患関連抗原に対する治療免疫応答を生じさせる実際の機会を与えるためには、いくつかの条件が満たされねばならない。まず第一に、分子の発現が腫瘍または疾患に特異的(少なくとも選択的)であり、それゆえに、免疫応答の標的となりうる、そのような分子を同定することが必要である。この作業は、大多数の一般的な腫瘍にとっては非常に難しいとされているが、例えば子宮頸癌の場合には、ほとんどがウイルスの癌遺伝子E6およびE7の存在により解決され、他の腫瘍の場合にも、良好な候補抗原が同定されつつある。例えば、MUC-1遺伝子産物は、90%の卵巣癌を含めて、いくつかの腫瘍で過剰発現されている。種々の他の腫瘍関連抗原も同定されており、そのうちのどれかを癌の免疫療法処置に使用できる可能性がある。今後もおそらく、追加の腫瘍関連抗原が発見され続けるであろう。第二に、1つ以上の抗原が同定された後には、免疫原性がある形態の抗原を免疫系に送達することが必要となる。腫瘍拒絶にとって最も重要な細胞性免疫応答を生起させるために、これは、タンパク質を宿主細胞の細胞質の中に送達させるか(高分子量のタンパク質抗原の場合には難しい)、あるいは、遺伝子送達またはDNA免疫化の後で宿主細胞自体にタンパク質を合成させるか、のいずれかが必要であることを意味する。この目的のために開発されたウイルスベクターとして、ワクシニア、アデノウイルス、またはレトロウイルスが挙げられる。
【0004】
現在、最適な免疫刺激を提供すると広く認められている細胞型は樹状細胞である(DC; 例えば、GirolomoniおよびRicciardi-Castagnoli, 1997を参照のこと)。事実、DCはin vivoで一次免疫応答を刺激することができる唯一の細胞型であると考えられており、さらに、ある状況下では確立された免疫寛容を破壊する能力があることさえも示されている。いくつかのグループは、DCが治療効果を示すという希望を抱いて、腫瘍に対する免疫応答を刺激するための自己養子免疫療法プロトコルでのDCの使用を検討している。そのようなプロトコルには、末梢血由来のDCの培養および/または富化、DCへの抗原のin vitroローディングおよび患者への該DCの再導入、またはDCへの抗原の直接in vivoローディングが必要である。しかし、このアプローチは、樹状細胞に抗原をローディングするための効率のよい手段が存在しないことで、これまで阻まれてきた。ところが、最近の研究により、ペプチドパルス化DCによる抗原の提示は、in vivoにおいて抗腫瘍応答を生み出すことが示された(Celluzziら, 1996; Zitvogelら, 1996)。ウイルスベクターに関しては、レトロウイルスは樹状細胞への効率のよい遺伝子送達をもたらさず(Reevesら, 1996; Aicherら, 1997)、我々の研究では、他のグループ(Arthurら, 1997)が報告した研究と相違して、アデノウイルスのみが低効率の遺伝子送達を可能にするにすぎない。
【0005】
我々は、以前に、単純ヘルペスウイルス(HSV)が樹状細胞に効率よく感染して遺伝子を送達することができることを試験し、報告した(Coffinら, 1998; WO 00/08191)。HSVは、他のベクター系と比べてこの目的に対していくつかの利点を有する。HSVは、様々な細胞型(他のベクター系では感染させるのが非常に難しいものを含み、例えば、Dillooら, 1997; Coffinら, 1998)に効率よく感染する能力があり、操作するのが簡単であり、しかも、大きなDNA挿入物を受け入れて複数の遺伝子の発現を可能にする(CoffinおよびLatchman 1996に概説)という利点である。樹状細胞への複数の抗原のex vivo送達とその後の体内への再導入、または樹状細胞への抗原の直接in vivo投与は、いくつかの癌および感染症の治療に対する特に有望なアプローチであると考えられる。
【0006】
国際公開WO 00/08191には、野生型の単純ヘルペスウイルスが、感染した樹状細胞内で抗原プロセッシングが起こるのを妨げること、および、機能性UL43およびvhs両遺伝子を欠失しているヘルペスウイルス、または前初期遺伝子の発現を最小限に抑える突然変異を含むヘルペスウイルスは、感染細胞内で抗原プロセッシングが起こるのを妨げることなく、樹状細胞に効率よく感染する能力がある、と教示されている。
【発明の概要】
【0007】
本発明者らは、このたび、驚いたことに、HSVベクター内のビリオン宿主シャットオフ(virion host shut-off: vhs)タンパク質をコードする遺伝子を破壊すると、HSV感染細胞において効率的な樹状細胞活性化を起こさせることができることを見出した。UL43遺伝子の破壊は必要でない。以前には、HSVに感染した樹状細胞は通常、感染それ自体によっても、他の刺激によっても活性化されることはない、ということが明らかにされている(Salioら, 1999; Kruseら, 2000)。
【0008】
本発明者らは、これまで知られていない、樹状細胞活性化を妨げるというvhsタンパク質の機能を確認した。樹状細胞活性化は、非活性化状態と比べたときの、特定の細胞表面マーカーのアップレギュレーションとして定義される。これらのマーカーにはCD83およびCD86が含まれる。樹状細胞活性化はリポ多糖(LPS)による処理で刺激することができる。HSVに感染した樹状細胞のLPS処理は、CD83またはCD86のアップレギュレーションをもたらさない。本発明者らは、変異型HSV(vhsが不活性化されているが、機能性UL43遺伝子をもつ)に感染した樹状細胞をLPSで処理すると、CD83とCD86の双方がアップレギュレートされることを見出した。機能性vhs遺伝子をもつウイルスに感染した樹状細胞をLPSで処理してもCD83とCD86のアップレギュレーションは認められない。したがって、これらの結果は、ヘルペスウイルス感染後に形質導入樹状細胞が免疫応答を最大限刺激するには、vhsをコードする遺伝子を破壊する必要があるが、UL43をコードする遺伝子はその必要がない、ことを示している。
【0009】
本発明者らの結果は、さらに、野生型の単純ヘルペスウイルスの病因におけるvhsの役割を実証するものである。HSVは高効率で樹状細胞に感染するが、HSVがその天然の生活環の一部としてそれを行うように進化した理由は、細胞性免疫応答(潜伏HSV感染が効率よく確立されるのを妨げるか、または潜伏と再活性化の反復サイクル中にそのウイルスのクリアランスをもたらす可能性がある)を最小限にしうるようにするためであろう。樹状細胞活性化は効果的な細胞性免疫応答の刺激において重要である。vhsはビリオンタンパク質であり、そうしてHSV遺伝子は一般に樹状細胞内で高レベルには発現されないが、vhsタンパク質は侵入してくるウイルスと共に樹状細胞に送達されるであろう。したがって、HSVに感染した樹状細胞の活性化を妨げるというvhsの新規な機能は、ヒトに感染した後のHSV生活環において重要なvhsの機能であるらしい。
【0010】
かくして、本発明は、下記ウイルスを樹状細胞に感染させることを含む免疫応答の刺激方法において使用するための医薬の製造における、
(i) 機能性vhs遺伝子またはその機能性等価体を欠失しており、
(ii) 機能性UL43遺伝子またはその機能性等価体を含んでいる、
弱毒化ヘルペスウイルスの使用を提供する。好ましくは、前記ウイルスはヒトの単純ヘルペスウイルスである。より好ましくは、前記ウイルスはHSV1またはHSV2である。樹状細胞にはin vitroで感染させてもin vivoで感染させてもよい。
【0011】
前記ウイルスは1つ以上の追加の突然変異を含むことができる。追加の突然変異はウイルスの毒性を最小限に抑えるものが好ましい。典型的には、そのような突然変異は前初期(immediate early: IE)遺伝子の発現を低下させるか、最小限にするものである。IE遺伝子の発現を阻止または低下させると、ウイルスの複製が妨げられるか、または減少する。そのような突然変異として、例えば、ICP4、ICP27、ICP0および/またはICP22(好ましくは、ICP27および/またはICP4)をコードする遺伝子を不活性化する突然変異が挙げられる。さらに、vmw65をコードする遺伝子の、トランス活性化機能を排除する不活性化突然変異も含まれる(例えば、Aceら, 1989またはSmileyら, 1997に記載されるようなvmw65突然変異)。追加の突然変異はまた、そのウイルスの免疫応答阻害活性を最小限に抑えることが好ましい。そのような突然変異には、ICP47をコードする遺伝子の不活性化が含まれる。
【図面の簡単な説明】
【0012】
【図1】ウイルス株 1764/27-/4-、1764/27-/4-/pR20.5/vhs、1764/27-/4-/pR20.5/vhsHBS-Ag、および野生型HSV 17+株を示した図である。
【図2A】刺激してない、擬似感染細胞ならびに17+、1764/27-/4-および1764/27-/4-/pR20.5/vhsウイルス株を感染させた細胞上のCD40、CD80、CD83およびCD86の細胞表面発現レベルを測定するFACS分析の結果を示した図である。
【図2B】LPSで刺激した、擬似感染細胞ならびに17+、1764/27-/4-および1764/27-/4-/pR20.5/vhsウイルス株を感染させた細胞上のCD40、CD80、CD83およびCD86の細胞表面発現レベルを測定するFACS分析の結果を示した図である。
【図3】未感染の樹状細胞ならびに17+、1764/27-/4-および1764/27-/4-/pR20.5/vhsウイルス株を感染させた樹状細胞からのIL-6およびTNFα分泌に及ぼすウイルス感染の影響を調べるためのELISA分析の結果を示した図である。
【図4】HBS-Agに応答した、B型肝炎ワクチン接種個体および非接種個体から調製したT細胞の増殖応答を示した図である。各個体から採取した樹状細胞は、処理しなかったか、組換えHBS-Agタンパク質(HBSAg*)と混合したか、対照ベクター(1764/27-/4-/pR20.5/vhs HBSAg)を感染させたか、あるいは、HBS-Agを発現するベクター(1764/27-/4-/pR20.5/vhs/HBS-Ag)を感染させたか、のいずれかであり、その後T細胞と混合した。
【発明を実施するための形態】
【0013】
A.ウイルス
本発明のウイルスは、感染した樹状細胞が活性化されるのを妨げることなく、樹状細胞に感染する能力がある。好ましくは、本発明のウイルスを感染多重度(MOI)1で感染させた樹状細胞は、LPSまたは他の活性化刺激で処理することにより活性化させることが可能である。
【0014】
本発明のウイルスは樹状細胞の活性化を妨げることがない。ウイルスがいつ樹状細胞の活性化を起こさせるかを調べるには、樹状細胞にウイルスをMOI≧1で感染させ、感染した樹状細胞をLPSで処理する。樹状細胞の活性化時にアップレギュレートされるCD83および/またはCD86などの細胞表面マーカーのレベルをモニターして、例えばFACS分析により、樹状細胞の活性化を判定する。細胞表面上のマーカーのレベルは、細胞に感染したウイルスが樹状細胞の活性化を起こさせるならば、LPS処理を行わなかった細胞よりもLPS処理を行った樹状細胞において顕著に高くなるだろう。また、樹状細胞の活性化を起こさせるウイルスを感染させた樹状細胞では、未感染の樹状細胞と比較して、これらのマーカーの一部または全部がより高くなるだろう。vhsに不活性化突然変異を含まない単純ヘルペスウイルスを樹状細胞に感染させる場合は、これらのマーカーのアップレギュレーションの著しい低下が観察される。
【0015】
感染樹状細胞の活性化を起こさせるために、本発明のウイルスは、vhs(HSVに含まれる)をコードする機能性遺伝子、または他のウイルス種に含まれるその相同性もしくは機能性等価体を欠失している。その上、本発明のウイルスは機能性UL43遺伝子をもっている。さらなる突然変異を行わせて、例えばICP47をコードする遺伝子に突然変異を導入して、ウイルスの免疫応答阻害作用をさらに低減させることも可能である。
【0016】
好ましくは、本発明の弱毒化ウイルスは、毒性が最小限となるように樹状細胞に感染させることができる。感染後の細胞生存率は、好ましくは、感染後1日目で少なくとも50%、より好ましくは、感染後1日目で少なくとも60、70、80または90%である。毒性低下を達成するために、ウイルス複製を減少させる突然変異を本発明のウイルスに導入することができる。例えば、VMW65をコードする遺伝子の突然変異(この突然変異はそのタンパク質のトランス活性化作用を最小にする)および/またはICP4、ICP27、ICP0およびICP22などの1つ以上の調節性前初期遺伝子の突然変異を含んでいてもよい。こうして、本発明のウイルスは典型的には、機能性vhs、ICP27、ICP4遺伝子を欠失しており、かつ転写活性化作用を示さないタンパク質をコードするVMW65遺伝子を含んでいてもよく、あるいは、典型的には、機能性vhs、ICP47およびICP4遺伝子を欠失していてもよい。
【0017】
in vivoで直接使用する場合には、ある程度の複製能力は一般に、誘導された免疫応答をブースト(増強)するのに有益でありうる。したがって、こうした状況下では、本発明のウイルスは、機能性vhs遺伝子を欠き、さらに、ウイルスの十分な病原性には必要であるがウイルスの複製には必要でない1つ以上の機能性遺伝子を欠くことが好ましい。そのような遺伝子として、例えば、ICP34.5、ICP6、チミジンキナーゼ、gHなどの糖タンパク質をコードするものが含まれる。しかし、チミジンキナーゼをコードする遺伝子は、この遺伝子の突然変異がアシクロビルのような抗ウイルス薬に対してウイルスを不感受性にするので、機能性であることが好ましい。
【0018】
本発明は単純ヘルペスウイルスを用いて説明されるが、感染した樹状細胞の活性化の阻止を低下させるようにヘルペスウイルス科の他のウイルスを改変しうることが理解されよう。特に、そのようなウイルスとして、水痘-帯状疱疹ウイルス、仮性狂犬病ウイルス、またはウシヘルペスウイルスが挙げられる。
【0019】
本発明のウイルスが単純ヘルペスウイルスである場合、このウイルスは例えばHSV1株もしくはHSV2株またはその誘導体に由来するものであってよく、好ましくはHSV1である。誘導体には、HSV1株およびHSV2株由来のDNAを含有するタイプ間組換え体が含まれる。そのようなタイプ間組換え体は当技術分野で公知であり、例えば、Thompsonら(1988)およびMeignierら(1988)に記載されている。誘導体は、典型的には本明細書に記載の方法で測定したとき、好ましくはHSV1またはHSV2のいずれかのゲノムに対して、少なくとも70%の配列相同性、より好ましくは少なくとも80%、より一層好ましくは少なくとも90%または95%の相同性を有する。さらに好ましくは、誘導体は、HSV1またはHSV2のいずれかのゲノムに対して、少なくとも70%の配列同一性、より好ましくは少なくとも80%の同一性、より一層好ましくは少なくとも90%、95%または98%の同一性を有する。
【0020】
誘導体は、ヌクレオチド置換(例えば、1、2または3個から10、25、50または100個までの置換)により改変されたHSV1またはHSV2ゲノムの配列をもつことができる。これとは別に、あるいは追加的に、HSV1またはHSV2ゲノムが1以上の挿入および/または欠失および/またはいずれか一方の末端または両方の末端での伸長により改変されていてもよい。
【0021】
本発明のウイルスを取得するために使用できる誘導体には、本発明のウイルスにおいて機能的に不活性化することが望ましい遺伝子中にすでに突然変異が存在するウイルス株が含まれ、例えば、vhs不活性化株(Jonesら, 1995に記載)、ICP47不活性化株(Goldsmithら, 1998に記載)、ICP4中に欠失があるd120株(DeLucaら, 1985)、ICP27中に欠失があるd27-1株(RiceおよびKnipe, 1990)、またはICP27とICP4の両方に欠失があるd92株(Samaniegoら, 1995)などである。こうした株を使用すると、本発明の変異型HSV株を作出するのに必要なステップ数を減らすことができる。
【0022】
各種HSV遺伝子を説明する際に用いる学術用語は、CoffinおよびLatchman, 1996に見出せるとおりである。
【0023】
上記HSV遺伝子の遺伝子相同体が他のヘルペスウイルス種に存在する場合は、これらの相同体が改変されるだろう。「相同体」とは、HSV遺伝子と機能的に等しい遺伝子を意味し、典型的には、相同体は、対応するHSV遺伝子に対してアミノ酸配列相同性または核酸配列相同性のいずれかを示す。典型的には、HSV遺伝子の相同体は、対応するHSV遺伝子に対して、アミノ酸レベルで、少なくとも15%、好ましくは少なくとも20%、より好ましくは少なくとも30%、40%または50%の同一性を示すであろう。
【0024】
vhsをコードする遺伝子は、HSV1およびHSV2中のUL41遺伝子である。HSV1の17+株(EMBL登録番号HE1CG)では、UL41遺伝子はヌクレオチド91,170からヌクレオチド92,637までである。HSV2のHG52株(EMBL登録番号z86099)では、UL41遺伝子はヌクレオチド91,800からヌクレオチド93,275までである。
【0025】
核酸およびタンパク質の相同性を決定するための方法は当技術分野で周知である。例えば、UWGCGパッケージは、相同性を算出するために使用できるBESTFITプログラムを提供する(例えば、そのデフォルトセッティングで使用)(Devereuxら(1984) Nucleic Acids Research 12, p387-395)。PILEUPおよびBLASTアルゴリズムを使用すると、相同性を算出したり、配列を整列させたりすることができ(典型的には、そのデフォルトセッティングで使用)、これらは例えば、Altschul (1993) J. Mol. Evol. 36:290-300; Altschulら(1990) J. Mol. Biol. 215:403-10に記載されている。
【0026】
BLAST解析を行うためのソフトウェアは、国立バイオテクノロジー情報センター(http://www.ncbi.nlm.nih.gov/)から利用可能である。このアルゴリズムでは、最初に、データベース配列中の同じ長さのワードとアライメントするとき、ある正値の閾値スコアTに一致するかまたはそれを満たすクエリー配列中の長さWの短いワードを同定することによって、高スコアを与える配列対(high scoring sequence pair: HSP)を同定することが必要である。Tは近隣ワードスコア閾値と呼ばれている(Altschulら, 1990)。これらの初期近隣ワードヒットは、それらを含むHSPを見つける検索を開始するためのシードとして働く。累積アライメントスコアが増加するかぎり、ワードヒットを、各配列に沿って両方向に延長させる。各方向でのワードヒットを中止するのは、累積アライメントスコアがその最大達成値から量Xだけ低下するとき;累積スコアが1以上の負スコアを与える残基アライメントの蓄積のためゼロまたはそれ以下になるとき;または、いずれかの配列の末端に達したときである。BLASTアルゴリズムパラメーターW、TおよびXはアライメントの感度と速度を決定する。BLASTプログラムは、デフォルトとして、ワード長(W) 11、BLOSUM62スコアリングマトリックス(HenikoffおよびHenikoff (1992) Proc. Natl. Acad. Sci. USA 89: 10915-10919を参照)アライメント(B) 50、期待値 10、M=5、N=4、および両鎖の比較を使用する。
【0027】
BLASTアルゴリズムは2つの配列間の類似性の統計的分析を実行する。例えば、KarlinおよびAltschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-5787を参照されたい。BLASTアルゴリズムにより与えられた類似性の程度は、最小の合計確率(P(N))であり、これは2つのヌクレオチドまたはアミノ酸配列間のマッチが偶然に出現する確率を示すものである。例えば、第1配列と第2配列との比較において最小合計確率が約1より小、好ましくは約0.1より小、より好ましくは約0.01より小、最も好ましくは約0.001より小である場合、一方の配列は他方の配列と類似していると見なされる。
【0028】
HSVゲノムの相同体はいくつかの方法で同定することができ、例えば、他のウイルスから作製したゲノムまたはcDNAライブラリーを、HSV遺伝子の全部または一部を含むプローブを用いて、中程度から高度のストリンジェンシー条件(例えば、0.03M 塩化ナトリウム、0.03M クエン酸ナトリウム、約50〜60℃)下で検索することにより行なう。これとは別に、縮重PCR(degenerate PCR)を使って、種相同体を取得することもでき、縮重PCRでは、保存されたアミノ酸配列をコードする変異体または相同体内の配列を標的とするように設計されたプライマーを使用する。これらのプライマーは、1以上の縮重位置を含んでおり、既知配列に対して単一配列プライマーを用いて配列をクローニングするのに用いた条件よりも低いストリンジェンシー条件(例えば、0.03M 塩化ナトリウム、0.03M クエン酸ナトリウム、約40℃)で使用する。
【0029】
ヘルペスウイルス中の相同体は、それがHSVタンパク質と1以上の機能的特徴を共有するならば、HSVタンパク質の機能性等価体となる。例えば、vhsタンパク質は、mRNAの安定性を弱めることによって感染細胞のタンパク質発現レベルを下げるという役割を果たしている。したがって、vhsタンパク質の機能性等価体は、mRNAの安定性を弱めることで宿主細胞の遺伝子発現を抑える役割を果たすことが好ましい。さらに、vhsの機能性等価体は、未感染樹状細胞を活性化する刺激に応答して樹状細胞の活性化を妨げることが好ましい。
【0030】
安全性の理由から、本発明のウイルスは、典型的には病気を引き起こす能力がなくなるように、弱毒化される。弱毒化のために改変されるウイルス領域は、完全にまたは部分的に除去されても、非機能的にされても、あるいは、他の配列(特に、異種遺伝子配列)で置換されてもよい。弱毒化をもたらす突然変異は、ウイルスベクターとして用いられる全てのウイルスグループについて、すでに記載されている。例えば、HSVはICP34.5および/または必須の遺伝子(例えば、ICP4、ICP27および/またはvhs遺伝子それ自体)の突然変異により無毒性にすることができる。
【0031】
特に好ましい弱毒化ウイルスとしては、vhsをコードする機能性遺伝子を欠失しており、かつ場合により機能性ICP47遺伝子を欠失していることに加えて、機能性ICP34.5と機能性ICP27遺伝子を欠失しており、また、機能性ICP4遺伝子および/またはVMW65遺伝子(転写活性化作用をもつタンパク質をコードする)を欠失していてもよいウイルス、および、機能性ICP27遺伝子をもつが機能性ICP4遺伝子と機能性ICP34.5遺伝子を欠失しており、また、VMW65遺伝子(転写活性化作用をもつタンパク質をコードする)を欠失していてもよいウイルスが挙げられる。そのようなウイルスは国際公開WO 98/04726およびWO 99/60145に記載されており、それらの開示内容を参照により本明細書に含めるものとする。
【0032】
本発明の単純ヘルペスウイルスが、特定の機能性必須遺伝子、例えば、ICP4またはICP27をコードする遺伝子を欠失している場合は、そのウイルスを、必須遺伝子を発現している細胞系を使って増殖させる。例えば、ウイルスが機能性ICP27遺伝子を欠く場合は、V27細胞(RiceおよびKnipe, 1990)、2-2細胞(Smithら, 1992)、またはB130/2細胞(WO 98/30707)を使って増殖させることができ、好ましくはB130/2細胞を用いる。ウイルスが機能性ICP4遺伝子を欠く場合は、ICP4を発現している細胞系、例えば、E5細胞(DeLucaら, 1985)を使ってウイルスを増殖させることができる。ウイルスが機能性ICP4遺伝子と機能性ICP27遺伝子を欠く場合は、そのウイルスを、ICP4とICP27の両方を発現している細胞系(例えば、E26細胞;Samaniegoら, 1995)を使って増殖させ、このウイルスがさらに機能性vmw65遺伝子を欠く場合には、vmw65の非HSV相同体(例えば、ウマヘルペスウイルス遺伝子12またはウシヘルペスウイルス由来のBTIF)をも含む細胞系を使って増殖させる。
【0033】
B.突然変異の方法
言及した各種ウイルス遺伝子は、当技術分野でよく知られているいくつかの技法により機能的に不活性化することができる。例えば、それらは欠失、置換または挿入により機能的に不活性化することができ、欠失による不活性化が好ましい。欠失は遺伝子の一部分を除去しても、遺伝子全体を除去してもよい。例えば、1個のヌクレオチドだけを欠失させて、フレームシフトを起こさせることができる。しかし、好ましくは、より大きな欠失を行ない、例えば、2、3または5個から10、20、30、50、100または200個のヌクレオチドの置換である。コードおよび非コード配列全体の好ましくは25%以上、より好ましくは50%以上(あるいはまた、絶対的な言い方では、10ヌクレオチド以上、より好ましくは100ヌクレオチド以上、最も好ましくは1000ヌクレオチド以上)を欠失または置換する。特に、遺伝子全体とそのフランキング配列の一部を除去することが好適である。挿入配列には、以下で述べる異種遺伝子が含まれる。突然変異は欠失と挿入の両方を含んでいてもよい。例えば、挿入を欠失部位に行なうことができる。こうして、ウイルス遺伝子への異種遺伝子の挿入はそのウイルス遺伝子の一部または全部に取って代わることができる。特に、異種遺伝子をvhs、ICP47、ICP27またはICP4に挿入することが好適である。VMW65遺伝子の場合には、それが必須の構造タンパク質をコードしているので、その遺伝子全体を欠失させることはできないが、一般的には、IE遺伝子を転写的に活性化するVMW65の能力を消失させる(例えば、Aceら, 1989またはSmileyら, 1997に記載されるような)不活性化突然変異が可能である。
【0034】
当業者には周知の相同組換え法によりヘルペスウイルスに突然変異を導入することができる。例えば、HSVゲノムDNAを、相同HSV配列に挟まれた突然変異配列を含むベクター(好ましくは、プラスミドベクター)と共にトランスフェクトする。この突然変異配列は欠失、挿入または置換を含んでいてよく、これらはいずれもルーチンな技法で構築することができる。組換えウイルスを、例えばβ-ガラクトシダーゼ活性または蛍光により、スクリーニングするために、挿入物に選択マーカー遺伝子、例えばlacZまたはGFP、を含めてもよい。
【0035】
C.異種遺伝子およびプロモーター
本発明のウイルスは1つ以上の異種遺伝子をもつように改変することができる。「異種遺伝子」という用語はあらゆる遺伝子を包含する。異種遺伝子は一般的にはヘルペスウイルスのゲノム中に存在しない遺伝子であるが、ヘルペス遺伝子を用いてもよい。ただし、そのコード配列が、もともとそれと関連しているウイルスの制御配列に機能的に連結されていないことを条件とする。異種遺伝子は野生型遺伝子のどのような対立遺伝子変異体であってもよいし、突然変異遺伝子であってもよい。「遺伝子」という用語は、RNA分子を産生するように少なくとも転写され得る核酸配列を包含し、そのRNA分子は翻訳可能であってポリペプチドを産生するか、またはアンチセンス効果により遺伝子発現レベルをダウンレギュレートすることが好ましい。本発明のウイルスは、天然の、あるいは異種遺伝子の翻訳コード配列に結合された、5'および/または3'転写されるが翻訳されないフランキング配列の一部または全部を含んでいてもよい。さらに、本発明のウイルスは、転写される配列に通常結合された転写制御配列、例えば、転写終止シグナル、ポリアデニル化部位および下流エンハンサーエレメントを含んでいてもよい。
【0036】
異種遺伝子は、HSV株と、例えばHSV配列により挟まれた異種遺伝子を含むプラスミドベクターと、の相同組換えによってウイルスのゲノムに挿入することができる。当技術分野で周知のクローニング技法を用いて、ヘルペスウイルス配列を含む適切なプラスミドベクターに1つ以上の異種遺伝子を導入することが可能である。ウイルスが依然として増殖可能であるかぎり、ウイルスゲノムのどの位置に異種遺伝子を挿入してもよい。好ましくは、ウイルスの弱毒化をもたらす遺伝子に異種遺伝子を挿入する。複数の異種遺伝子をウイルスゲノム内の複数の部位に挿入することも可能である。
【0037】
異種遺伝子の転写配列は、樹状細胞、好ましくは哺乳動物の樹状細胞、より好ましくはヒトの樹状細胞内での異種遺伝子の発現を可能にする制御配列と機能的に連結されていることが好ましい。「機能的に連結される」という用語は、記載した成分同士がそれらの意図された様式で機能することを可能にする関係にある並置をさす。コード配列と「機能的に連結される」制御配列は、コード配列の発現が制御配列と適合しうる条件下で達成されるような方法で連結される。
【0038】
制御配列は、異種遺伝子の発現を可能にするプロモーターおよび転写を終結させるためのシグナルを含む。プロモーターは哺乳動物、好ましくはヒトの樹状細胞において機能するプロモーターから選択される。プロモーターは真核生物遺伝子のプロモーター配列に由来するものでもよい。例えば、プロモーターは、異種遺伝子の発現を起こさせようとする細胞、好ましくは哺乳動物の樹状細胞、より好ましくはヒトの樹状細胞から誘導することができる。真核細胞のプロモーターに関しては、それらは普遍的なやり方で(β-アクチン、チューブリンのプロモーターなど)あるいは樹状細胞に特異的なやり方で機能するプロモーターでありうる。ウイルスプロモーターを使用してもよく、例えば、モロニーマウス白血病ウイルスの長い末端反復配列(MMLV LTR)プロモーターまたは他のレトロウイルスプロモーター、ヒトまたはマウスサイトメガロウイルス(CMV)IEプロモーターがある。
【0039】
異種遺伝子と制御配列を含む発現カセットおよび他の適当な構築物は、当業者には公知である通常のクローニング法を用いて作製することができる(例えば、Sambrookら, 1989, Molecular Cloning - a laboratory manual; Cold Spring Harbor Press を参照されたい)。
【0040】
さらに、これらのプロモーターはどれも、さらなる調節配列、例えばエンハンサー配列(HSV LAT領域を含む)を付加することによって改変することができる。上記の2種以上のプロモーター由来の配列エレメントを含むキメラプロモーター、例えば、MMLV LTR/LAT融合プロモーター(Lokensgardら, 1994)またはLAT領域のエレメントを含むプロモーター(WO98/30707)を用いてもよい。
【0041】
異種遺伝子は典型的には治療用のポリペプチドをコードする。例えば、特定の腫瘍に対する免疫応答を特異的に促進させるには、1以上の腫瘍抗原の発現を指令する本発明のウイルスで樹状細胞をトランスフェクトすることが望ましい。腫瘍抗原は腫瘍細胞に特異的なもの(すなわち、腫瘍細胞に存在するが、非腫瘍細胞には存在しないもの)であっても、同じタイプの非腫瘍細胞よりも(例えば、抗原発現のアップレギュレーションのため)腫瘍細胞に高レベルで存在するものであってもよい。これは癌の治療に有効であろう。なんとなれば、本発明の感染樹状細胞を用いることにより、宿主の免疫系が刺激されて腫瘍特異的抗原または腫瘍普遍的抗原に反応し、腫瘍の縮小/後退をもたらすからである。特に、腫瘍抗原は腫瘍細胞の表面に発現されることが好ましく、例えば、細胞表面受容体または細胞接着タンパク質でありうる。腫瘍抗原の例として、卵巣癌をはじめとする幾つかの腫瘍において過剰発現されるMUC-1遺伝子産物(Gendlerら, 1990)、子宮頸癌と関連があるヒト乳頭腫ウイルスタンパク質E6およびE7、メラノーマにおけるMART-I、MAGE-I、gp100およびチロシナーゼ、前立腺癌におけるPSA、様々なタイプの幾つかの腫瘍におけるCEA、および乳癌を含めた各種の癌におけるHer2neuが挙げられる。
【0042】
また、異種遺伝子は免疫応答を改変することができるポリペプチドをコードしてもよく、かかるポリペプチドには、例えば、サイトカイン(α-、β-またはγ-インターフェロン、IL-1、IL-2などのインターロイキン、腫瘍壊死因子、またはインスリン様増殖因子IもしくはIIなど)または他の免疫調節タンパク質(RANTESなどのケモカイン、およびCD80、CD86、CD40およびCD40リガンドのような共刺激分子など)が含まれる。
【0043】
異種遺伝子はまた、病原体由来のポリペプチドをコードしてもよく、かくして、例えば、本発明のウイルスを感染させた樹状細胞を用いることにより、宿主の免疫系を刺激して病原体に対する免疫応答を、該病原体への宿主の感染前または感染後に、生じさせることができる。ワクチンに用いるウイルスは、典型的には抗原性ポリペプチドをコードする異種遺伝子を含みうる。好ましくは、そのような病原体由来のポリペプチドは、寄生虫、細菌またはウイルスなどの病原性の生物に由来するものである。かかる抗原性ポリペプチドの例として、C型肝炎ウイルス抗原、B型肝炎表面またはコア抗原、乳頭腫ウイルス抗原、HIV抗原およびマラリア抗原が挙げられる。病原性生物由来の異種遺伝子を含むウイルスは、治療処置または予防処置のいずれかまたは両方に使用することができる。
【0044】
治療用途には、おそらく複数の遺伝子の投与が必要である。複数の遺伝子の発現はさまざまな症状を治療するのに有利であろう。ヘルペスウイルスは、他のウイルスベクター系のようにパッケージング能力が限られていないので、唯一適切なものである。したがって、ヘルペスウイルスのゲノム内には複数の異種遺伝子を収容することができる。例えば、2〜6個の遺伝子をそのゲノムに挿入することができる。
【0045】
これを達成するための方法は、少なくとも2つある。例えば、2個以上の異種遺伝子と関連制御配列を、特定のHSV株のウイルスゲノムの単一部位または複数の部位に導入することである。また、互いに離れて反対の方向に向いているプロモーター(同一のまたは異なるプロモーター)の対を使用することも可能であり、これらのプロモーターはそれぞれが上記のような異種遺伝子(同一のまたは異なる異種遺伝子)の発現を駆動する。
【0046】
D.樹状細胞
樹状細胞はいくつかの手段により単離/調製することができ、例えば、それらは末梢血から直接精製されるか、例えばG-CSF処理により末梢血に移動した後の、または直接骨髄からの、CD34+前駆細胞から生成される。末梢血からの付着前駆細胞はGM-CSF/IL-4混合物で処理し(Inabaら, 1992)、骨髄からの非付着CD34+細胞はGM-CSFとTNF-αで処理することができる(Cauxら, 1992)。樹状細胞をヒトボランティアの末梢血からルーチンに調製するには、SallustoおよびLanzavecchia, 1994に記載の方法と同様にして、精製した末梢血単核細胞(PBMC)を使用し、そして付着細胞をGM-CSFとIL-4で2時間処理する。次に、磁性ビーズを使ってこれらからCD19+B細胞とCD3+, CD2+T細胞を枯渇する(Coffinら, 1998)。樹状細胞の調製には他の方法も使用することができる。
【0047】
E.治療用途
本発明のウイルスおよび本発明のウイルスを感染させた樹状細胞は、治療方法において使用することができる。特に、腫瘍抗原を発現する本発明のウイルス、および腫瘍抗原を発現する本発明のウイルスを感染させた樹状細胞は、癌の治療方法に使用される。特定すると、本発明のウイルスおよび本発明のウイルスを感染させた樹状細胞は、ヒトを含む哺乳動物の各種腫瘍の増殖を抑制するために使用することができ、例えば、卵巣癌、子宮頸癌および子宮内膜腫瘍、ならびに、乳癌、肺癌、膀胱癌、大腸癌などの癌の抑制である。増殖抑制が可能なその他の新生物としては、肉腫(例えば、軟組織および骨の肉腫)、血液の悪性腫瘍(例えば、白血病)などがある。腫瘍抗原を発現する本発明のウイルスおよび/または該ウイルスを感染させた樹状細胞を用いて治療しうる癌の例として、特に、メラノーマ、白血病、子宮頸癌および卵巣癌が挙げられる。癌治療用のウイルスは典型的には腫瘍抗原をコードする異種遺伝子を含む。かかるウイルスまたはウイルスを感染させた樹状細胞の投与により、一般的には、腫瘍抗原に対する免疫応答が引き出されるであろう。
【0048】
本発明のウイルスおよび本発明のウイルスを感染させた樹状細胞は、病原体による感染症(例えば、寄生虫、細菌またはウイルスによる感染症)の治療または予防方法において使用することができる。病原体による感染症の治療に用いるウイルスは、典型的には、病原性生物に由来する抗原をコードする異種遺伝子を含む。そのようなウイルスまたはウイルスを感染させた樹状細胞を投与することにより、一般的には、病原性生物由来の抗原に対する免疫応答が引き出されるであろう。かかるウイルス感染症として、ヘルペスウイルス感染症が含まれる。こうして、本発明のウイルスを使って、例えば、HSV1またはHSV2感染症の治療またはワクチン接種において、該ウイルス自体に対する免疫応答を誘発させることが可能である。ウイルスがHSV1またはHSV2感染の治療に用いられるものである場合、そのウイルスはHSV抗原(その天然のプロモーターの制御下にないもの)または免疫調節分子をコードする異種遺伝子を含んでいてもよい。ウイルス/樹状細胞は、宿主の防御免疫応答を刺激するために感染前に、あるいは宿主の免疫系を刺激して感染症を撲滅するために感染後に、投与することができる。
【0049】
F.投与
こうして、本発明のヘルペスウイルスを用いて、処置を必要とするヒトまたは動物に治療用遺伝子を送達することができる。本発明のヘルペスウイルスを用いた治療用遺伝子の送達は、例えば、悪性疾患および/または病原体による感染症の治療に使用することができる。
【0050】
本発明のウイルスは、治療を必要とする患者(好ましくは、ヒト患者)に使用される。治療を必要とする患者は、癌を患う個体または感染症のある患者である。治療処置の目的は患者の状態を改善することにある。典型的には、本発明のウイルスを用いた治療処置は癌の症状を軽減する。本発明に従う癌の治療方法は、機能性UL43遺伝子をもつが機能性vhs遺伝子を欠失しているウイルスを、治療に有効な量で癌患者に投与して、該ウイルスが該患者の樹状細胞中に存在するようにすることを含んでなる。腫瘍を患う個体に本発明のウイルスを投与すると、典型的には、腫瘍細胞が殺傷される結果、腫瘍の大きさが小さくなったり、その腫瘍由来の悪性細胞の広がりがくい止められたりするだろう。
【0051】
一般的に、本発明のウイルスを用いた感染症の治療処置は、感染症の症状を軽減し、さらには病原性生物を死滅させることが好ましい。本発明に従う感染症の治療方法は、機能性vhs遺伝子を欠失しているウイルスを、治療に有効な量で感染症の患者に投与することを含んでなる。好ましくは、該ウイルスを患者の樹状細胞に侵入させるか、あるいはex vivoで該ウイルスを感染させた樹状細胞を患者に投与する。本発明のウイルスを用いた予防処置は、典型的には、癌や感染症のリスクがある患者において、腫瘍抗原に対する抗体または病原性生物由来の抗原に対する抗体の産生をうながす。一般的に、癌のリスクがある患者とは、遺伝的にその傾向があるか、癌原物質に暴露されてしまったか、癌原物質に暴露される危険がある者でありうる。一般的に、感染症のリスクがある患者とは、病原性生物にさらされる可能性がある者でありうる。
【0052】
治療を実施するための一つの方法は、1以上の治療用遺伝子を、上記のような本発明のヘルペスウイルスのゲノムに挿入し、次に、得られた組換えウイルスを製薬上許容される担体または希釈剤と組み合わせて医薬組成物を調製することを含む。適当な担体および希釈剤として、等張食塩水、例えばリン酸緩衝食塩水が挙げられる。医薬組成物は非経口、筋肉内、静脈内、腹腔内、皮下または経皮投与用に製剤化することができる。皮下または腹腔内投与が好適である。経皮または皮内投与が特に好適でありうる。
【0053】
本発明のウイルスを樹状細胞に感染させるには、ウイルスを含む組成物を患者に投与することによってin vivoで実施する。治療用遺伝子を含むウイルスが樹状細胞に感染できるようなやり方で医薬組成物を投与する。投与されるウイルスの量は、104〜1010 pfu、好ましくは105〜108または105〜109 pfu、より好ましくは約106〜108 pfuの範囲とする。皮内に注射する場合、あるいは、例えば針のない器具を使って、経皮投与する場合、製薬上許容される適当な担体もしくは希釈剤中または特定の組成物中のウイルスを、一般的には10μl〜1ml、好ましくは100μl〜1mlの量で投与する。
【0054】
別の方法では、樹状細胞を末梢血または骨髄から単離/調製し、その細胞にin vitroで本発明のウイルスを感染させる。次に、形質導入された樹状細胞を、一般的には、筋肉内、腹腔内、皮下もしくは静脈内注射によって、あるいは、患者のリンパ節への直接注入によって、好ましくは皮下、腹腔内注射またはリンパ節への直接注入により、患者に投与する。形質導入された樹状細胞は、一般的には105〜109個、好ましくは106〜108個、より好ましくは約107個を患者に投与する。
【0055】
記載した投与経路および投与量は単なる指針であって、当業者であれば、個々の患者に最適な投与経路および投与量を容易に決定することができよう。投与量はさまざまなパラメーターに従って、特に患者の年齢、体重および症状に応じて決定される。
以下の実施例は本発明を例示するためのものである。
【0056】
実施例
材料および方法
ウイルス株の構築および増殖
全てのウイルス株はHSV1 17+株に由来し、そのヌクレオチド配列はGenBankに登録されている(登録番号 HE1CG)。ウイルス株は、BHK C-21細胞(ECACC番号 8501143)またはHSV1 ICP27、ICP4およびウマヘルペスウイルス遺伝子12をコードする遺伝子で安定にトランスフェクトされたBHK細胞(Thomasら, 1999)を使って産生し、増殖させた。
【0057】
VMW65に突然変異があるウイルスの場合は、ウイルス増殖用の培地に3mMのヘキサメチレン-ビスアセトアミド(HMBA)を加えた(McFarlaneら, 1992)。以下のウイルス株を使用した。
【0058】
(i) 17+ (野生型HSV1)
(ii) 17+/pR20.5/UL43
HSV LAT領域配列により隔離されたRSV/lacZ/pA配列とCMV/GFP/pA配列(相反する背中合わせの方向にある)からなるプラスミドpR20.5(Thomasら, 1999b)由来のカセット(ヌクレオチド118,866-120,219)を、標準的な方法で、精製したHSV1 17+株のゲノムDNAとの相同組換えにより、UL43遺伝子座に挿入した。最初に、pR20.5カセットを、UL43フランキング領域を含むプラスミド(Coffinら, 1996)にユニークなNsiI部位で挿入して、プラスミドpR20.5/43を得た。20.5カセットは、SrfIをコードするオリゴヌクレオチドがそのカセットの両側に挿入されたので、そのpGEM5(Promega)プラスミド骨格からSrfIで切り出すことができる。プラスミドpR20.5の構築のために、RSVプロモーターをpRc/RSV(Invitrogen)から、lacZ/pAをpCHI10(Pharmacia)から、CMV/pAをpcDNA3(Invitrogen)から、そしてGFPをpEGFP-N1(Clontech)から切り出した。
【0059】
(iii) 1764/27-/4-
1764/27-/4-ウイルス株は、1764/27-/4-/pR20.5ウイルス株DNAの空のICP4フランキング領域による組換え、およびGFPまたはlacZを発現しないウイルスプラークの選択により構築した。1764/27-/4-/pR20.5ウイルス株は、Thomasら, 1999bに記載されており、ウイルス(ICP27およびICP34.5も欠失されていて、VMW65をコードする遺伝子に不活性化突然変異がある)のICP4をコードする遺伝子を置換するように、ICP4遺伝子にpR20.5カセットが挿入されている。
【0060】
(iv) 1764/27-/4-/pR20.5/vhs
1764/27-/4-/pR20.5/vhsウイルス株を構築するために、pR20.5カセットを、HSV1 17+株のvhsコード遺伝子中のユニークなNruI部位でvhsフランキング領域に挿入し、得られたプラスミド(pR20.5/vhs)をHSV 1764/27-/4-株のDNAに組換えにより挿入した。したがって、1764/27-/4-/pR20.5/vhsウイルス株は、ICP4、ICP27およびICP34.5をコードする遺伝子が欠失しており、vmw65およびvhsをコードする遺伝子には不活性化突然変異が存在する。
【0061】
(v) 1764/27-/4-/pR19lacZ
1764/27-/4-/pR19lacZウイルス株は上記ウイルス(iv)と同様に構築したが、pR19lacZカセット(Wagstaffら, 1998)を、vhsにpR20.5カセットを挿入するのではなく、1764/27-/4-ウイルス株の潜伏関連転写物(latency associated transcript: LAT)領域に挿入した。
【0062】
(vi) 1764/27-/4-/pR20.5/vhs/HBS-Ag
pR20.5/vhsプラスミド中のlacZ遺伝子は、pHBV130(GoughおよびMurray, 1982)をXhoIとNsiIで消化し、放出された断片をpSP72(Promega)のSalI部位とSmaI部位の間に挿入することにより、肝炎表面抗原(HBS-Ag)をコードする遺伝子で置換した。pR20.5/vhsをXbaIとEcoRIで消化してlacZ遺伝子を放出させ、HindIIIとEcoRIを用いてpSP72から切り出したHBS-Ag遺伝子と置き換えた。得られたプラスミドを1764/27-/4-/pR20.5/vhsウイルスDNAに挿入し、lacZを発現しないプラークを選択し、精製した。その後ゲノムの構造をサザンブロットで確認した。
【0063】
樹状細胞の調製
樹状細胞は以前に記載されたとおりに末梢血から調製した(Coffinら, 1998)。簡単に説明すると、末梢血単核細胞(PBMC)を、健康でB型肝炎ワクチンを接種したドナー血液60mlからリンフォプレップ(lymphoprep; Nycomed)を使って調製した。赤血球の除去後、非付着細胞(主にT細胞とB細胞)を分離し、HBSSで洗浄し、室温、1400rpmで5分間遠心した。細胞ペレットを2mlの90% FCS:10% ジメチルスルホキシド(DMSO)混合物中に再懸濁し、アリコートに分けて、その後のT細胞分離のために-80℃で保存した。付着細胞は、GM-CSF(0.1μg/ml)とIL-4(0.05μg/ml)を補充したRPMI培地で培養し、37℃、5% CO2で7日間インキュベートした。さらなるリンフォプレップ精製の後、抗CD19抗体、抗CD2抗体(Harlan)、および抗CD3抗体(Harlan)を用いて磁気的に細胞を枯渇させ、即使用のために樹状細胞を完全RPMI培地に再懸濁した。
【0064】
CD4+T細胞の分離
上記のように凍結させたT細胞とB細胞をすばやく解凍し、HBSS中で洗浄し、1400rpmで5分間遠心した。細胞を2mlの完全RPMI培地に再懸濁し、カウントし、抗CD19(BU12 - 200μl正味、Immunology dept, UCL)、抗CD14(HB246 - 200μl正味、Immunology dept, UCL)、および抗HLA-DR(L243 - 100μl正味、Immunology dept, UCL)モノクローナル抗体と共にインキュベートし、氷上に30分間放置した。細胞をHBSS中で洗浄し、2mlの完全RPMI培地に再懸濁し、磁性ビーズに結合させたヒツジ抗マウス抗体(Dynabeads, Dynal)と、汚染細胞106個あたり10μlのビーズの割合で混合して、ローターミキサー上で4℃、45分間インキュベートした。その後、細胞懸濁液/磁性ビーズの混合物を磁石と接触させた状態で氷上に10分間置いた後上清を分離することにより、CD4+T細胞を枯渇させた。CD4+T細胞をカウントし、完全RPMI培地に適切な濃度で再懸濁し、氷上に配置するか、またはその後の使用のために37℃、5% CO2で培養した。
【0065】
樹状細胞の感染
樹状細胞は1400rpm、室温で5分間ペレット化した。次に、ウイルスを含むRPMI培地中に37℃、5% CO2で1時間再懸濁することにより、感染多重度(MOI)1で樹状細胞にウイルスを感染させた。GM-CSF(0.1μg/ml)とIL-4(0.05μg/ml)を補充したRPMI培地 1mlを加え、樹状細胞を37℃、5% CO2でインキュベートした。LPS刺激のためには、100ng/mlのLPSを追加的に含むRPMI培地を使用した。
【0066】
サイトカイン分析
市販のELISAキット(R&D Systems)を使って樹状細胞の培養上清中に含まれるIL-6およびTNF-αを測定した。ELISAに先立って、先に示したウイルスを樹状細胞に感染させてから42時間後に上清を集めて、使用前に-20℃で保存した。
【0067】
T細胞増殖アッセイ
B型肝炎ワクチンを接種したヒト個体と接種してないヒト個体から樹状細胞とCD4+T細胞を分離し、上記のように処理した。樹状細胞は1×105個/ml〜1×104個/mlの希釈物として使用し、CD4+T細胞は1×106個/mlで使用した。それぞれの樹状細胞(DC)濃度での実験を3回反復して行なった。各アッセイウェルに100μlの樹状細胞と100μlのCD4+T細胞を加えた。表示した場合は、組換えB型肝炎表面抗原(Austral)を1μg/ウェルの最終濃度でウェルに添加した。HSV-1を感染させた樹状細胞と未感染の樹状細胞をCD4+T細胞と共に37℃、5% CO2で6日間培養した。その後、1μCu/ウェルの[3H]チミジン(Amersham)を添加し、18時間後細胞を回収して、[3H]チミジンの取込みをカウントした。
【0068】
実施例1
予備的データは、機能性vhs遺伝子を含まないHSV株がウイルス感染後の樹状細胞の活性化を増大させる、ことを示す
ここでは、それぞれの場合に、1×105個の樹状細胞に各ウイルスを感染させるために、樹状細胞を穏やかにペレット化し、DMEM中の約100μlのウイルス懸濁液に再懸濁させ、37℃で1時間インキュベートし、そして2mlのRPMI/10% FCS+100ng/mlのGM-CSF、50ng/mlのIL-4を含む24ウェルプレートに移した。次いで、これらのプレートを37℃、5% CO2で一夜インキュベートした。また、公知の樹状細胞活性化剤であるリポ多糖(LPS)で樹状細胞を処理し、未処理の樹状細胞を対照として用いた。
【0069】
その後、これらの感染物および対照物から得られた上清をELISA試験で使用して、分泌されたサイトカインのレベルを検出した。また、感染させた樹状細胞および対照の樹状細胞の表面上に発現されたCD86のレベルを検出するために、蛍光活性化セルソーティング(FACS)を使用した。樹状細胞の培養物には、CD86発現のレベルに関して2つの細胞集団が存在していた。これらはFACS分析で2つのピークとして観察され、第1ピークはCD86発現のレベルが比較的低い細胞を示し、第2ピークはCD86発現のレベルが比較的高い細胞を示す。例えばLPSで活性化すると、より多くの細胞がより高レベルのCD86を発現し、こうして、第2ピーク中に見出せる細胞がより多くなる。
【0070】
【表1】
表1: 表示したウイルスを感染多重度1で感染させてから24時間後の培養上清中または対照の上清中に含まれるサイトカイン濃度。ELISAで測定。
【0071】
【表2】
表2: 対照の細胞上および表示したウイルスを感染させた細胞上のCD86の発現。
【0072】
結論
上記の結果から、未処理の樹状細胞は試験したサイトカインを最低レベルで分泌しており、しかもその表面上に予想された「休止」レベルのCD86を有する、ことがわかる。LPS処理後に、サイトカインレベルは著しく刺激され、CD86の表面発現のレベルが大いに増加する。上記の実験では、LPS処理細胞の平均蛍光強度は、抗CD86抗体を用いたFACS分析により約1×103である。これらの結果は、樹状細胞が活性化された状態にあることを示している。
【0073】
表示したウイルスを樹状細胞に感染させた後で、これらのアッセイにより樹状細胞の活性化が生じるためには、ウイルスがvhsをコードする遺伝子に不活性化突然変異を含まねばならない、ことが明らかに見てとれる。さまざまな無能化レベルのウイルスを使用しているが、vhs突然変異を含むウイルスのみがこれらのアッセイで樹状細胞の顕著な活性化を与える。また、vhs突然変異を含まないならば、表示のウイルスを細胞に感染させて、さらにLPSで処理しても、LPS単独で処理した場合ほど多くの細胞においてCD86レベルが増加しない、ことが見てとれる。さらに、vhs突然変異を含まないならば、ウイルス感染後にFACSで測定したCD86発現細胞の平均蛍光強度は、細胞をLPSで処理した場合に見られる強度よりも低下している。したがって、樹状細胞による最大免疫刺激を得るためには、vhsをコードする遺伝子に不活性化突然変異を含めるべきである、と結論付けることができる。
【0074】
実施例2
機能性vhsタンパク質を含まないHSV株は、樹状細胞の活性化を阻止しない
蛍光活性化セルソーティング(FACS)を用いて、感染樹状細胞と対照樹状細胞の表面上のCD86、CD80、CD83およびCD40の発現レベルを検出した。感染細胞からの上清を用いてELISAによりサイトカインのレベルを評価した。
【0075】
結果
ELISAの結果(図3)から、樹状細胞にHSVを高頻度で感染させることができるが、樹状細胞活性化の指標となるサイトカインは野生型(17+株)ウイルスまたは無能化(1764/27-/4-株)ウイルスのいずれの場合にも産生されない、ことがわかる。しかしながら、vhsを1764/27-/4-株から不活性化させると、得られる1764/27-/4-/pR20.5/vhs株では、樹状細胞の活性化を示すサイトカインが産生される。非LPS刺激樹状細胞に対するFACS分析(図2)からは、本質的に野生型HSV(17+株)または複製能のないHSVベクター(1764/27-/4-株)による感染は、CD86発現の増加を妨げることがわかる。上述したように、樹状細胞が感染プロセスによって活性化されると、CD86発現の増加が期待される。また、CD40レベルはHSV感染細胞において変化/低下する。しかし、vhsが不活性化されると(1764/27-/4-/pR20.5/vhs株)、CD86レベルは活性化を示して増加し、CD40レベルは変化しない。CD80とCD83は非刺激のHSV感染樹状細胞では大きく変化しない。CD83(B7.1)およびCD86(B7.2)はともに重要なT細胞共刺激分子であり、CD40は重要なT細胞活性化剤であり、そしてCD83は樹状細胞の成熟化および活性化の間にアップレギュレートされる樹状細胞のマーカーである。
【0076】
感染時に樹状細胞をLPSで刺激する場合には、野生型(17+株)ウイルスまたは無能化(1764/27-/4-株)ウイルスのいずれにおいてもCD40レベルに対する影響がより顕著である。しかし、vhsが不活性化されると、CD40に対するこれらの影響は妨げられる。LPSで刺激した樹状細胞は通常、CD83およびCD86発現を著しくアップレギュレートするが、これは、vhsが不活性化されない限り、HSV(17+株および1764/27-/4-株)によって阻止される。vhsが不活性化されると、CD83およびCD86レベルはどちらも、LPSで刺激した未感染細胞と同程度またはそれ以上に増加する。
【0077】
結論
予備実験と同様に、樹状細胞がHSV感染またはHSV感染とLPS刺激に応答して活性化される(表面マーカーの発現レベルにより測定)ためには、vhsをコードする遺伝子が不活性化されねばならないことが明らかである、と明確に見てとれる。機能性vhsをコードするウイルスは、試験した表面マーカーの発現レベルにより測定したとき、樹状細胞を顕著に活性化させることができない。
【0078】
実施例3
vhs不活性化HSVで形質導入された樹状細胞は、in vitroで抗原特異的T細胞応答を引き出す
上記の結果から、vhsが不活性化されているHSVベクターは、樹状細胞におけるHSVの不活性化作用が妨げられているので、樹状細胞用の効果的なベクターとして使用できることが示唆された。実際、そのようなHSV変異体を感染させた樹状細胞は、CD86のアップレギュレーションおよび特定のサイトカインの分泌により測定したとき、感染に応答して特異的に活性化されるようである。vhs不活性化HSV変異体が、樹状細胞への抗原コード化遺伝子の送達に続いて、抗原特異的免疫応答を引き出すことができるか否かを試験するために、B型肝炎ワクチンを接種した個体と接種しなかった個体から調製した樹状細胞およびT細胞を用いて実験を行なった。ここでは、最初に、B型肝炎表面抗原(HBS-Ag)発現カセットがIE遺伝子欠損ウイルスのvhsコード化遺伝子に挿入されているウイルスを構築した。次にT細胞増殖アッセイを実施したが、ここにおいて、ワクチン接種個体または非接種個体からの樹状細胞は、処理しなかったか、組換えHBS-Agタンパク質と混合して抗原「負荷」を行なったか、対照のマーカー遺伝子含有ベクター(1764/27-/4-/pR20.5/vhs、MOI=1)を感染させたか、対照のベクター(1764/27-/4-/pR20.5/vhs、MOI=1)を感染させかつ組換えHBS-Agタンパク質とも混合したか、あるいは、HBS-Agを発現するベクター(1764/27-/4-/pR20.5/vhs/HBS-Ag、MOI=1)を感染させたか、のいずれかである。その後、樹状細胞を、それぞれ同一のワクチン接種個体または非接種個体に由来するT細胞と混合し、T細胞増殖に及ぼす効果を標準的なT細胞増殖アッセイで観察した。
【0079】
これらの実験から明らかなように(図4)、HBS-Ag組換えタンパク質および対照のHSVベクターはワクチン接種個体において小さなT細胞増殖応答を引き出すことができ(HSV応答はおそらくHSV構造タンパク質に特異的なT細胞の増殖を示す)、また、組換えHBS-Agと混合された対照のベクターはやや大きな応答を引き出すことができたが、HBS-Agを発現するベクターはこれらのいずれよりも格別に大きな応答を与えた。こうして、HSVベクターを用いて樹状細胞にHBS-Agを直接送達した後に、顕著な特異的T細胞増殖応答が誘導されたが、かかる応答は組換え抗原単独と混合した後では生じなかった。したがって、vhsが不活性化されているHSVベクターは、抗原特異的T細胞増殖応答を刺激する樹状細胞の能力を保持したままで、抗原コード化遺伝子を樹状細胞に送達することを可能にする。
【0080】
参考文献
【特許請求の範囲】
【請求項1】
個体において樹状細胞にin vivoで感染して該個体の免疫応答を刺激するための薬剤であって、
(i) 機能性vhs遺伝子を欠失しており、
(ii) 機能性UL43遺伝子を含んでいる、
弱毒化単純ヘルペスウイルス(HSV)を含む、前記薬剤。
【請求項2】
前記ウイルスが単純ヘルペスウイルス1または2である、請求項1に記載の薬剤。
【請求項3】
前記ウイルスがICP47をコードする機能性遺伝子を欠失している、請求項1または2に記載の薬剤。
【請求項4】
前記ウイルスが、転写活性化活性を欠くタンパク質をコードするVMW65遺伝子を有している、請求項1〜3のいずれか1項に記載の薬剤。
【請求項5】
前記ウイルスが、ICP0、ICP4、ICP22およびICP27をコードする遺伝子から選択される少なくとも1つの機能性前初期遺伝子を欠失している、請求項1〜4のいずれか1項に記載の薬剤。
【請求項6】
前記ウイルスがICP27をコードする機能性遺伝子とICP4をコードする機能性遺伝子の両方を欠失している、請求項5に記載の薬剤。
【請求項7】
前記ウイルスがICP27をコードする機能性遺伝子とICP4をコードする機能性遺伝子の両方を欠失しており、かつ、転写活性化活性を欠くタンパク質をコードするVMW65遺伝子を有している、請求項4〜6のいずれか1項に記載の薬剤。
【請求項8】
前記ウイルスがICP0、ICP4、ICP22およびICP27をコードする機能性遺伝子を欠失している、請求項5〜7のいずれか1項に記載の薬剤。
【請求項9】
前記ウイルスが機能性ICP34.5遺伝子をさらに欠失している、請求項1〜8のいずれか1項に記載の薬剤。
【請求項10】
前記ウイルスが機能性ICP34.5遺伝子および機能性ICP47遺伝子を欠失している、請求項9に記載の薬剤。
【請求項11】
前記ウイルスが異種遺伝子を含んでいる、請求項1〜10のいずれか1項に記載の薬剤。
【請求項12】
前記異種遺伝子が治療用のポリペプチドをコードしている、請求項11に記載の薬剤。
【請求項13】
前記ウイルスが2つ以上の異種遺伝子を含んでいる、請求項11または12に記載の薬剤。
【請求項14】
前記ウイルスが、ケモカイン、サイトカインまたは共刺激分子をコードする異種遺伝子を含んでいる、請求項1〜13のいずれか1項に記載の薬剤。
【請求項15】
前記樹状細胞がヒト樹状細胞である、請求項1〜14のいずれか1項に記載の薬剤。
【請求項16】
注射、注入、皮内もしくは経皮経路、またはバイオリスティック手段により投与するためのものである、請求項1〜15のいずれか1項に記載の薬剤。
【請求項1】
個体において樹状細胞にin vivoで感染して該個体の免疫応答を刺激するための薬剤であって、
(i) 機能性vhs遺伝子を欠失しており、
(ii) 機能性UL43遺伝子を含んでいる、
弱毒化単純ヘルペスウイルス(HSV)を含む、前記薬剤。
【請求項2】
前記ウイルスが単純ヘルペスウイルス1または2である、請求項1に記載の薬剤。
【請求項3】
前記ウイルスがICP47をコードする機能性遺伝子を欠失している、請求項1または2に記載の薬剤。
【請求項4】
前記ウイルスが、転写活性化活性を欠くタンパク質をコードするVMW65遺伝子を有している、請求項1〜3のいずれか1項に記載の薬剤。
【請求項5】
前記ウイルスが、ICP0、ICP4、ICP22およびICP27をコードする遺伝子から選択される少なくとも1つの機能性前初期遺伝子を欠失している、請求項1〜4のいずれか1項に記載の薬剤。
【請求項6】
前記ウイルスがICP27をコードする機能性遺伝子とICP4をコードする機能性遺伝子の両方を欠失している、請求項5に記載の薬剤。
【請求項7】
前記ウイルスがICP27をコードする機能性遺伝子とICP4をコードする機能性遺伝子の両方を欠失しており、かつ、転写活性化活性を欠くタンパク質をコードするVMW65遺伝子を有している、請求項4〜6のいずれか1項に記載の薬剤。
【請求項8】
前記ウイルスがICP0、ICP4、ICP22およびICP27をコードする機能性遺伝子を欠失している、請求項5〜7のいずれか1項に記載の薬剤。
【請求項9】
前記ウイルスが機能性ICP34.5遺伝子をさらに欠失している、請求項1〜8のいずれか1項に記載の薬剤。
【請求項10】
前記ウイルスが機能性ICP34.5遺伝子および機能性ICP47遺伝子を欠失している、請求項9に記載の薬剤。
【請求項11】
前記ウイルスが異種遺伝子を含んでいる、請求項1〜10のいずれか1項に記載の薬剤。
【請求項12】
前記異種遺伝子が治療用のポリペプチドをコードしている、請求項11に記載の薬剤。
【請求項13】
前記ウイルスが2つ以上の異種遺伝子を含んでいる、請求項11または12に記載の薬剤。
【請求項14】
前記ウイルスが、ケモカイン、サイトカインまたは共刺激分子をコードする異種遺伝子を含んでいる、請求項1〜13のいずれか1項に記載の薬剤。
【請求項15】
前記樹状細胞がヒト樹状細胞である、請求項1〜14のいずれか1項に記載の薬剤。
【請求項16】
注射、注入、皮内もしくは経皮経路、またはバイオリスティック手段により投与するためのものである、請求項1〜15のいずれか1項に記載の薬剤。
【図1】

【図2A】
【図2B】
【図3】
【図4】
【図2A】
【図2B】
【図3】
【図4】
【公開番号】特開2012−188440(P2012−188440A)
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願番号】特願2012−125645(P2012−125645)
【出願日】平成24年6月1日(2012.6.1)
【分割の表示】特願2001−575212(P2001−575212)の分割
【原出願日】平成13年4月12日(2001.4.12)
【出願人】(500354883)バイオヴェックス リミテッド (3)
【Fターム(参考)】
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願日】平成24年6月1日(2012.6.1)
【分割の表示】特願2001−575212(P2001−575212)の分割
【原出願日】平成13年4月12日(2001.4.12)
【出願人】(500354883)バイオヴェックス リミテッド (3)
【Fターム(参考)】
[ Back to top ]