
免疫原性マラリア原虫周辺スポロゾイトタンパク質エピトープによる組換えアデノウイルスの修飾
本開示は、組換えアデノウイルスの導入遺伝子に対する免疫応答を増強するため、かつ既存する抗アデノウイルス免疫を避けるためのアデノウイルスタンパク質の修飾に関する。一部の実施形態は、組換えアデノウイルスプラスミドベクターに由来する組換えアデノウイルスに関し、該組換えアデノウイルプラスミドベクターは、異種プロモーターに作用可能に連結したマラリア原虫スポロゾイト周囲タンパク質またはその抗原部分と、修飾型カプシドタンパク質またはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトーブがカプシドタンパク質またはコアタンパク質の少なくとも一部に挿入されるか、または一部を置換する。別の実施形態は、上述の実施形態による組換えアデノウイルスを含む医薬組成物またはマラリアワクチン組成物に関する。さらなる実施形態は、上述の実施形態による医薬組成物またはマラリアワクチン組成物の治療量を投与することを含む、マラリアの治療法、予防法、または診断法を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、医学および生物工学の分野に関する。より詳しくは、本発明はマラリア原虫(Plasmodium)スポロゾイト周囲タンパク質などのマラリア寄生虫抗原に対して強力な免疫応答を誘発するためのカプシド修飾型アデノウイルスベクターの使用に関する。該ベクターはマラリアに対するワクチンに適している。
優先権主張
本出願は、2009年8月18日に提出された国際出願PCT/US09/054212に対する優先権を主張し、かつ一部継続出願である。該国際出願の全体を完全に記述するように参照によって本明細書に組み入れる。
米国連邦政府による委託研究に関する記載
本発明は、米国国立衛生研究所の一部である国立アレルギー・感染症研究所(NIAID)によって与えられた助成金番号1R01A1081510−01A1の下で政府支援によってなされた。米国政府は、本発明において特定の権利を有する。
【背景技術】
【0002】
マラリアは、世界中の熱帯で最も蔓延している感染症に入る重篤な疾患である。毎年、およそ3億〜5億の人々が感染し、罹患率と死亡率が比較的高い。重度の罹患率と死亡率は、特に幼児と、これまでにマラリアに曝露されたことがないマラリア流行地に移住した成人に起こっている。世界保健機構(WHO)では、アフリカだけでマラリアによる小児の死亡数が毎年200万〜300万人と推定している。多くの国では、薬剤耐性寄生虫(熱帯熱マラリア原虫(Plasmodium falciparum)、最近では三日熱マラリア原虫(Plasmodium vivax)も)および殺虫剤耐性ベクター(ハマダラカ属(Anopheles)蚊)によってもたらされるマラリアの広範な発生および発生率の増加のため、この疾患を制御する新規の方法を開発する必要性が強調されているる(NussenzweigおよびLong 1994)。
【0003】
マラリア寄生虫は、前赤内期、赤血球期および生殖期の寄生形態からなる複雑なライフサイクルを有し、これらの形態はマラリアワクチンの開発のための潜在的標的を表す。前赤内期、赤血球期の形態は宿主内で見られるが、生殖形態はベクター内で起こる。弱毒化照射したスポロゾイト(IrSp)の生ワクチンによる免疫化は、マウスにおいて(Nussenzweigら 1967)、非ヒト霊長類(Gwadzら 1979)およびヒト((Clydeら 1973、Edelmanら 1993)において無菌の防御作用(すなわち、寄生体の攻撃に対する完全な耐性)を誘発することが示された。IrSpによって与えられる防御作用は、体液性(B細胞)および細胞性(T細胞)の両免疫応答によるスポロゾイト中和によって媒介される(Tsujiら 2001)。IrSpワクチン接種は魅力ある解決策ではあるが、スポロゾイトを採取する唯一の方法は蚊唾液腺を詳細に分析することにより、かつ大量のスポロゾイトをin vitroで増殖させる技術で現在知られているものはない。したがって、マラリアに対して同様に強い防御免疫を誘発することができる代替ワクチンのベクターが必要とされる。
【0004】
かかるワクチンベクターの有望な標的の1つは、スポロゾイト周囲(CS)タンパクであり、該タンパク質はスポロゾイトの表面に発現される。有効な中和抗体は、スポロゾイト周囲(CS)タンパク質の免疫優性で、種特異的な反復ドメインに向けられる。熱帯熱マラリア原虫(ヒトマラリア寄生虫)において、反復(NANP)nは、世界のすべての領域に由来する分離株の間で保存されている。この中心反復はB細胞エピトープの複数の反復を含み、したがってCSタンパク質はB細胞を誘発することによって強い体液性免疫反応を誘発することができる(Tsujiら 2001)。CSタンパク質のC末端領域では、いくつかのT細胞エピトープが存在し、該エピトープが細胞性免疫応答を著しく誘発することができる(Tsujiら 2001)。体液性(抗体)応答は、肝細胞に入る前にスポロゾイト(細胞外寄生体)の感染力と相互作用して、中和することによって寄生虫を除去することができるが、一方細胞性(T細胞)応答は、インターフェロンγを分泌することによってEEF(細胞内寄生体)を攻撃することができる。これらの免疫応答は、EEFが成熟し、急速に分裂して、血液に再度浸入して赤血球に感染する何千ものメロゾイトを形成して、我々がマラリアと認識する疾患を引き起こすことを防ぐ。
【0005】
現在ヒト臨床試験を受けているCSをベースにしたマラリアワクチンの1種は、GlaxoSmithKline社のRTS、S、B型肝炎表面抗原の融合タンパク質、およびウイルス様の粒子の形態での熱帯熱マラリア原虫スポロゾイト周囲タンパク質(PfCSP)の一部(1992年11月11日に、SmithKline Beecham Biologicals S.A.に対して出願された国際特許出願第PCT/EP1992/002591号)であり、臨床試験においてマラリア感染症を減少させることが示されている(Alonsoら 2004, Alonsoら 2005, Bejonら2008)。RTS、Sは、抗PfCSP体液性免疫応答を誘発するが、PfCSP特異的細胞(CD8+)応答は比較的弱い(Kesteら 2008)。これは、RTS、Sによる防御作用が比較的弱いことが理由でありうる。対照的に、アデノウイルスベースのマラリアワクチンは、防御細胞性免疫応答を誘発することができる(2003年12月16日に出願された国際特許出願第PCT/EP2003/051019号、Rodriguesら 1997)。しかし、現在のところマラリアワクチンとしてアデノウイルスベースのプラットフォームの利用を限定する障害が2つある:(1)導入遺伝子産物に対する強力な体液性免疫応答を誘発する能力の欠如と、(2)アデノウイルス、特に、アデノウイルスベースのワクチンの免疫原性を妨害するアデノウイルス血清型5に対する既存免疫との2つである。
【0006】
アデノウイルスによって誘発される体液性免疫応答を増強する試みの中で最近なされている1つのアプローチは、B細胞抗原エピトープ(たとえば、細菌エピトープまたはウイルスエピトープ)をHexon、Fiber、PentonおよびpIXなどのアデノウイルスカプシドタンパク質に挿入することである(Worgallら 2005, McConnellら 2006、Krauseら2006、Worgallら 2007)。
【0007】
加えて、アデノウイルス血清型5(Ad5)に対する既存免疫を避けるために、アデノウイルス血清型11、35、26、48、49および50などの低い血清有病率を有する他のアデノウイルス血清型が、ワクチンプラットフォームとして評価され、抗Ad5免疫が存在するにもかかわらず導入遺伝子に対する免疫応答を誘発することが示された(2005年10月12日にCrucell Holland B.V.に対して出願された国際特許出願第PCT/EP2005/055183号)。既存の抗Ad5免疫を回避するために、中和抗体の標的カプシドタンパク質であるAd5 Hexonと他の血清型のそれとの置換が構築された(Wuら 2002、Robertsら 2006)。
【0008】
しかし、アデノウイルスベクターを上述のマラリアワクチンに適用する際に同時に前述の2つの障害を克服したと報告される改良型アデノウイルスベクターはない。マラリア流行地においてAd5に対する血清有病率が高い(Ophorstら 2006)ことを考慮すると、アデノウイルスに対する既存免疫の存在下で、防御体液性免疫応答および細胞性免疫応答の双方を誘発するアデノウイルスベースのマラリアワクチンが求められている。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本開示は、組換えアデノウイルスワクチンの導入遺伝子に対する免疫応答を増強し、かつ既存の抗アデノウイルス免疫を回避するための種々のアデノウイルスタンパク質修飾に関する。
【0010】
より具体的には、一実施形態は、組換えアデノウイルスプラスミドベクターに由来する組換えアデノウイルスに関し、該組換えアデノウイルスプラスミドベクターは、(i)異種プロモーターに作用可能に連結した、マラリア原虫スポロゾイト周囲タンパク質、またはその抗原部分と、(ii)修飾型カプシドタンパク質もしくはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープはカプシドタンパク質もしくはコアタンパク質に挿入されているまたはその少なくとも一部を置換する。
【0011】
一部の実施形態では、マラリア原虫スポロゾイト周囲タンパク質は、熱帯熱マラリア原虫またはネズミマラリア原虫(Plasmodium yoelii)スポロゾイト周囲タンパク質をさらに含む。該スポロゾイト周囲タンパク質は、コドン最適化した熱帯熱マラリア原虫またはネズミマラリア原虫のスポロゾイト周囲タンパク質をさらに含むこともあり、一部の態様では、配列番号2または配列番号1それぞれののヌクレオチド配列によってコードされうる。
【0012】
別の実施形態では、該免疫原性エピトープは、マラリア原虫スポロゾイト周囲タンパク質のB細胞エピトープをさら含む。該B細胞エピトープは、修飾型カプシドタンパク質に取り込まれることもあり、かつ一部の態様では、カプシドタンパク質はHexon高頻度可変領域(HVR)を含みうる。該HVRは、HVR1またはHVR5の一部がB細胞エピトープに置換されているHVR1またはHVR5をさらに含みうる。別の態様では、カプシドタンパク質は、B細胞エピトープがFiberタンパク質に挿入されているカプシドFiberタンパク質をさらに含みうる。一部の態様では、B細胞エピトープは、熱帯熱マラリア原虫スポロゾイト周囲タンパク質B細胞エピトープであり、該B細胞エピトープが、たとえば(NANP)n(配列番号60)の反復配列であり、該反復配列が(NANP)4、(NANP)6、(NANP)8、(NANP)10、(NANP)12、(NANP)14、(NANP)16、(NANP)18、(NANP)20、(NANP)22、または(NANP)28でありうる。別の態様では、B細胞エピトープはネズミマラリア原虫スポロゾイト周囲タンパク質B細胞エピトープであり、B細胞エピトープが、たとえば(QGPGAP)n(配列番号59)の反復配列であり、該反復配列が(QGPGAP)3、(QGPGAP)4(QGPGAP)5、(QGPGAP)6、(QGPGAP)7、(QGPGAP)8、(QGPGAP)9、(QGPGAP)11、または(QGPGAP)12でありうる。
【0013】
さらに別の実施形態では、免疫原性エピトープは、マラリア原虫スポロゾイト周囲タンパク質のCD4+またはCD8+のT細胞エピトープをさらに含む。該CD4+ T細胞エピトープまたはCD8+ T細胞エピトープは、修飾型カプシドタンパク質またはコアタンパク質に取り込まれることもある。一部の態様では、該カプシドタンパク質は、Hexon高頻度可変領域(HVR)を含みうる。該HVRは、HVR1の一部がCD4+またはCD8+のT細胞エピトープに置換されているHVR1をさらに含みうる。別の態様では、該コアタンパク質は、pVIIタンパク質をさらに含み、かつCD4+ T細胞エピトープが該pVIIタンパク質に挿入されている。一部の態様では、該CD4+ T細胞エピトープは、熱帯熱マラリア原虫スポロゾイト周囲CD4+ T細胞エピトープであり、該CD4+ T細胞エピトープがEYLNKIQNSLSTEWSPCSVT(配列番号62)である。別の態様では、該CD4+ T細胞エピトープは、ネズミマラリア原虫スポロゾイト周囲CD4+ T細胞エピトープであり、該CD4+ T細胞エピトープがYNRNIVNRLLGDALNGKPEEK(配列番号61)である。
【0014】
別の実施形態は、上記の実施形態による組換えアデノウイルスを含む医薬組成物またはマラリアワクチン組成物に関する。さらなる実施形態は、上記の実施形態による該医薬組成物またはマラリアワクチン組成物の治療量を投与することを含むマラリアを治療する、予防する、または診断する方法を含む。
【0015】
別の実施形態では、治療法は、初回刺激−追加免疫のワクチン接種の投与を含み、ここで被験者は、所定の時間間隔で用量を増やしてまたは同じ用量で一連の投与を与えられる。所定の時間間隔は、体液性および/または細胞性の免疫応答を生成するのに十分な任意の長さであってよい。たとえば、後述するように、該時間間隔は、3週間ごとであってもよいが、これに限定されるものではない。
【課題を解決するための手段】
【0016】
本発明者らは、組換えアデノウイルスプラスミドベクターに由来する新しい、カプシド修飾型構造を有する新規の組換えアデノウイルスを見出した。新規の組換えアデノウイルスは哺乳類細胞を感染させ、哺乳類細胞にマラリア原虫スポロゾイト周囲タンパク質を発現させることができる。該組換えアデノウイルスは、所望の免疫原性の抗原、たとえばマラリア原虫スポロゾイト周囲タンパク質のB細胞エピトープ、T細胞エピトープを有することによって修飾された1つまたは複数のカプシドタンパク質も有する。該組換えアデノウイルスは、直線化組換えアデノウイルスプラスミドベクターで細胞をトランスフェクトする方法によって得られる。得られた組換えアデノウイルスを用いて、本発明者らは、有効成分としてマラリア感染予防効果および治療効果がある組換えアデノウイルスを含有する医薬品について広範囲の研究を行った。その結果、発明者らは、得られた組換えアデノウイルスが所望の薬学的効果を有することを見出した。
【図面の簡単な説明】
【0017】
【図1】本開示の実施形態によるカプシド修飾型組換えアデノウイルスの模式図である。
【図2】HVR1修飾型組換えアデノウイルスプラスミドベクターのHVR1修飾型アデノウイルスDNAの構築を示す模式図である。
【図3】HVR5修飾型組換えアデノウイルスプラスミドベクターのHVR5修飾型アデノウイルスDNAの構築を示す模式図である。
【図4】Fiber修飾型組換えアデノウイルスプラスミドベクターのFiber修飾型アデノウイルスDNAの構築を示す模式図である。
【図5】HVR1とFiberとの修飾型組換えアデノウイルスプラスミドベクターの、HVR1とFiberとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図6】FiberとpVIIとの修飾型組換えアデノウイルスプラスミドベクターの、FiberとpVIIとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図7】HVR1とpVIIとの修飾型組換えアデノウイルスプラスミドベクターの、HVR1とpVIIとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図8】HVR1とFiberとpVIIとの修飾型組換えアデノウイルスプラスミドベクターの、HVR1とFiberとpVIIとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図9】コドン最適化ネズミマラリア原虫スポロゾイト周囲タンパク質の核酸配列(PyCS、配列番号1)および対応するアミノ酸配列(配列番号30)を示す。
【図10】コドン最適化熱帯熱マラリア原虫スポロゾイト周囲タンパク質の核酸配列(PfCSP、配列番号2)および対応するアミノ酸配列(配列番号43)を示す。
【図11−1】HVR1(配列番号3、核酸;配列番号31、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図11−2】HVR1(配列番号3、核酸;配列番号31、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図11−3】HVR1(配列番号3、核酸;配列番号31、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図12−1】HVR1(配列番号4、核酸;配列番号32、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの4の反復(配列番号59;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)4の配列は、下線で示す。
【図12−2】HVR1(配列番号4、核酸;配列番号32、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの4の反復(配列番号59;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)4の配列は、下線で示す。
【図12−3】HVR1(配列番号4、核酸;配列番号32、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの4の反復(配列番号59;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)4の配列は、下線で示す。
【図13−1】HVR1(配列番号5、核酸;配列番号33、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの5の反復(配列番号59;n=5)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)5の配列は、下線で示す。
【図13−2】HVR1(配列番号5、核酸;配列番号33、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの5の反復(配列番号59;n=5)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)5の配列は、下線で示す。
【図13−3】HVR1(配列番号5、核酸;配列番号33、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの5の反復(配列番号59;n=5)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)5の配列は、下線で示す。
【図14−1】HVR1(配列番号6、核酸;配列番号34、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの6の反復(配列番号59;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)6の配列は、下線で示す。
【図14−2】HVR1(配列番号6、核酸;配列番号34、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの6の反復(配列番号59;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)6の配列は、下線で示す。
【図14−3】HVR1(配列番号6、核酸;配列番号34、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの6の反復(配列番号59;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)6の配列は、下線で示す。
【図15−1】HVR1(配列番号7、核酸;配列番号35、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの7の反復(配列番号59;n=7)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)7の配列は、下線で示す。
【図15−2】HVR1(配列番号7、核酸;配列番号35、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの7の反復(配列番号59;n=7)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)7の配列は、下線で示す。
【図15−3】HVR1(配列番号7、核酸;配列番号35、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの7の反復(配列番号59;n=7)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)7の配列は、下線で示す。
【図16−1】HVR1(配列番号8、核酸;配列番号36、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの8の反復(配列番号59;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)8の配列は、下線で示す。
【図16−2】HVR1(配列番号8、核酸;配列番号36、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの8の反復(配列番号59;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)8の配列は、下線で示す。
【図16−3】HVR1(配列番号8、核酸;配列番号36、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの8の反復(配列番号59;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)8の配列は、下線で示す。
【図17−1】HVR1(配列番号9、核酸;配列番号37、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの9の反復(配列番号59;n=9)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)9の配列は、下線で示す。
【図17−2】HVR1(配列番号9、核酸;配列番号37、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの9の反復(配列番号59;n=9)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)9の配列は、下線で示す。
【図17−3】HVR1(配列番号9、核酸;配列番号37、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの9の反復(配列番号59;n=9)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)9の配列は、下線で示す。
【図18−1】HVR1(配列番号10、核酸;配列番号38、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの11の反復(配列番号59;n=11)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)11の配列は、下線で示す。
【図18−2】HVR1(配列番号10、核酸;配列番号38、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの11の反復(配列番号59;n=11)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)11の配列は、下線で示す。
【図18−3】HVR1(配列番号10、核酸;配列番号38、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの11の反復(配列番号59;n=11)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)11の配列は、下線で示す。
【図19−1】HVR1(配列番号11、核酸;配列番号39、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの12の反復(配列番号59;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)12の配列は、下線で示す。
【図19−2】HVR1(配列番号11、核酸;配列番号39、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの12の反復(配列番号59;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)12の配列は、下線で示す。
【図19−3】HVR1(配列番号11、核酸;配列番号39、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの12の反復(配列番号59;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)12の配列は、下線で示す。
【図20−1】HVR1(配列番号12、核酸;配列番号44、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)4の配列は、下線で示す。
【図20−2】HVR1(配列番号12、核酸;配列番号44、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)4の配列は、下線で示す。
【図20−3】HVR1(配列番号12、核酸;配列番号44、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)4の配列は、下線で示す。
【図21−1】HVR1(配列番号13、核酸;配列番号45、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの6の反復(配列番号60;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)6の配列は、下線で示す。
【図21−2】HVR1(配列番号13、核酸;配列番号45、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの6の反復(配列番号60;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)6の配列は、下線で示す。
【図21−3】HVR1(配列番号13、核酸;配列番号45、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの6の反復(配列番号60;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)6の配列は、下線で示す。
【図22−1】HVR1(配列番号14、核酸;配列番号46、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの8の反復(配列番号60;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)8の配列は、下線で示す。
【図22−2】HVR1(配列番号14、核酸;配列番号46、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの8の反復(配列番号60;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)8の配列は、下線で示す。
【図22−3】HVR1(配列番号14、核酸;配列番号46、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの8の反復(配列番号60;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)8の配列は、下線で示す。
【図23−1】HVR1(配列番号15、核酸;配列番号47、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの10の反復(配列番号60;n=10)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)10の配列は、下線で示す。
【図23−2】HVR1(配列番号15、核酸;配列番号47、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの10の反復(配列番号60;n=10)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)10の配列は、下線で示す。
【図23−3】HVR1(配列番号15、核酸;配列番号47、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの10の反復(配列番号60;n=10)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)10の配列は、下線で示す。
【図24−1】HVR1(配列番号16、核酸;配列番号48、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの12の反復(配列番号60;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)12の配列は、下線で示す。
【図24−2】HVR1(配列番号16、核酸;配列番号48、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの12の反復(配列番号60;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)12の配列は、下線で示す。
【図24−3】HVR1(配列番号16、核酸;配列番号48、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの12の反復(配列番号60;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)12の配列は、下線で示す。
【図25−1】HVR1(配列番号17、核酸;配列番号49、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの14の反復(配列番号60;n=14)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)14の配列は、下線で示す。
【図25−2】HVR1(配列番号17、核酸;配列番号49、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの14の反復(配列番号60;n=14)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)14の配列は、下線で示す。
【図25−3】HVR1(配列番号17、核酸;配列番号49、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの14の反復(配列番号60;n=14)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)14の配列は、下線で示す。
【図26−1】HVR1(配列番号18、核酸;配列番号50、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの16の反復(配列番号60;n=16)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)16の配列は、下線で示す。
【図26−2】HVR1(配列番号18、核酸;配列番号50、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの16の反復(配列番号60;n=16)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)16の配列は、下線で示す。
【図26−3】HVR1(配列番号18、核酸;配列番号50、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの16の反復(配列番号60;n=16)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)16の配列は、下線で示す。
【図27−1】HVR1(配列番号19、核酸;配列番号51、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの18の反復(配列番号60;n=18)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)18の配列は、下線で示す。
【図27−2】HVR1(配列番号19、核酸;配列番号51、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの18の反復(配列番号60;n=18)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)18の配列は、下線で示す。
【図27−3】HVR1(配列番号19、核酸;配列番号51、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの18の反復(配列番号60;n=18)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)18の配列は、下線で示す。
【図28−1】HVR1(配列番号20、核酸;配列番号52、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの20の反復(配列番号60;n=20)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)20の配列は、下線で示す。
【図28−2】HVR1(配列番号20、核酸;配列番号52、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの20の反復(配列番号60;n=20)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)20の配列は、下線で示す。
【図28−3】HVR1(配列番号20、核酸;配列番号52、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの20の反復(配列番号60;n=20)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)20の配列は、下線で示す。
【図29−1】HVR1(配列番号21、核酸;配列番号53、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの22の反復(配列番号60;n=22)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)22の配列は、下線で示す。
【図29−2】HVR1(配列番号21、核酸;配列番号53、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの22の反復(配列番号60;n=22)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)22の配列は、下線で示す。
【図29−3】HVR1(配列番号21、核酸;配列番号53、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの22の反復(配列番号60;n=22)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)22の配列は、下線で示す。
【図30−1】HVR1(配列番号22、核酸;配列番号54、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの28の反復(配列番号60;n=28)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)28の配列は、下線で示す。
【図30−2】HVR1(配列番号22、核酸;配列番号54、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの28の反復(配列番号60;n=28)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)28の配列は、下線で示す。
【図30−3】HVR1(配列番号22、核酸;配列番号54、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの28の反復(配列番号60;n=28)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)28の配列は、下線で示す。
【図31−1】HVR5(配列番号23、核酸;配列番号40、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図31−2】HVR5(配列番号23、核酸;配列番号40、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図31−3】HVR5(配列番号23、核酸;配列番号40、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図32−1】Fiber(配列番号24、核酸;配列番号41、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図32−2】Fiber(配列番号24、核酸;配列番号41、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図33−1】Fiber(配列番号25、核酸;配列番号55、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(NANP)3の配列は、下線で示す。
【図33−2】Fiber(配列番号25、核酸;配列番号55、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(NANP)3の配列は、下線で示す。
【図34】N末端pVII(配列番号26、核酸;配列番号42、アミノ酸)においてPyCS CD4+エピトープ配列YNRNIVNRLLGDALNGKPEEK(配列番号61)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたYNRNIVNRLLGDALNGKPEEKの配列は、下線で示す。
【図35】C末端pVII(pVII−1;配列番号27、核酸;配列番号56、アミノ酸)においてPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたEYLNKIQNSLSTEWSPCSVT配列は、下線で示す。
【図36】pVII(pVII−2;配列番号28、核酸;配列番号57、アミノ酸)の第1の核移行シグナル(NLS)の前にPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたEYLNKIQNSLSTEWSPCSVT配列は、下線で示す。
【図37】pVII(pVII−3;配列番号29、核酸;配列番号58、アミノ酸)の2つNLSの間にPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたEYLNKIQNSLSTEWSPCSVT配列は、下線で示す。
【図38】PyCS−GFP/pShuttle−CMVによる一過性トランスフェクションの後、AD293細胞におけるPyCSタンパク質発現を示す。PyCSタンパク質は、マウスモノクローナル抗PyCS抗体(9D3)を用いるウエスタンブロット法によって検出された。
【図39】(QGPGAP)3エピトープ(配列番号59;n=3)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPyCS−GFPアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。ELISAアッセイでは、ELISAプレートは精製アデノウイルスで直接コーティングされ、次いでアデノウイルス粒子に挿入されたエピトープは抗PyCS抗体で検出された。
【図40】(QGPGAP)nエピトープ(配列番号59)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPyCSアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。ELISAアッセイでは、ELISAプレートは精製アデノウイルスで直接コーティングされ、次いでアデノウイルス粒子に挿入されたエピトープは抗PyCS抗体で検出された。
【図41】(QGPGAP)n反復(配列番号59、n=3、4、5、6、9、12)を有するカプシド修飾型PyCSアデノウイルスによる単回免疫レジメン(A)と、免疫化から2週間後のPyCS特異的CD8+応答(B)とを示す。
【図42】(QGPGAP)n反復(配列番号59、n=3)を有するカプシド修飾型PyCSアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、10週目のPyCS特異的体液性免疫応答(B)と、スポロゾイト抗原投与から42時間後の肝臓でのマラリア寄生虫の負荷(C)とを示す。
【図43】図42でのレジメンを与えられたマウスから調製され、プールした血清試料についての間接的免疫蛍光アッセイ(IFA)によって測定された抗スポロゾイト抗体価(A)と、in vitroスポロゾイト中和活性(B)とを示す。
【図44】HVR1において(QGPGAP)n反復(配列番号59、n=4、6)を有するカプシド修飾型PyCSアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPyCS特異的体液性免疫応答(B)と、スポロゾイト抗原投与から42時間後の肝臓でのマラリア寄生虫負荷(C)と、プールした血清「試料のin vitroスポロゾイト中和活性(D)とを示す。マウスにアジュバントと共に、またはアジュバントを含まずに免疫した。
【図45】HVR1において(QGPGAP)n反復(配列番号59、n=6、9、12)を有するカプシド修飾型PyCSアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPyCS特異的体液性免疫応答(B)と、9週目のPyCS特異的CD8+ T細胞応答(C)と、スポロゾイト抗原投与から42時間後の肝臓でのマラリア寄生虫負荷(D)とを示す。マウスにアジュバントと共に、またはアジュバントを含まずに免疫した。
【図46】PfCSP/pShuttle−CMVによる一過性トランスフェクションの後のAD293細胞におけるPfCSPタンパク質の発現を示す。PfCSPは、マウスモノクローナル抗NANP抗体(2A10)を用いるウエスタンブロット法によって検出された。
【図47】(NANP)4エピトープ(配列番号60、n=4)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPfCSPアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。挿入された(NANP)4のエピトープ(配列番号60、n=4)は、マウスモノクローナル抗NANP抗体(2A10)によって検出された。ELISAアッセイでは、ELISAプレートは、精製アデノウイルスで直接コーティングされた。
【図48】(NANP)nエピトープ(配列番号60、n=4、6、8、10、12、14、16、18、20、22)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPfCSPアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。ELISAアッセイでは、ELISAプレートは、精製アデノウイルスで直接コーティングされ、次いでアデノウイルス粒子に挿入されたエピトープは抗PfCSP抗体(2A10)で検出された。
【図49】(NANP)4(配列番号60、n=4)を有するカプシド修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPfCSP特異的体液性免疫応答(B)とを示す。
【図50】HVR1において(NANP)n(配列番号60、n=4、6、8、10)を有するカプシド修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPfCSP特異的体液性免疫応答(B)とを示す。
【図51】HVR1において(NANP)n(配列番号60、n=10、16、22)を有するカプシド修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPfCSP特異的体液性免疫応答(B)とを示す。マウスにアジュバントと共に、またはアジュバントを含まずに免疫した。
【図52】精製した(QGPGAP)修飾型FiberおよびpVII−1((QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFP)のアデノウイルスの銀染色分析(A)の結果と、図49に記述したように(QGPGAP)3−Fib/PyCS−GFPまたは(QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFPで免疫したマウスにおいて10週目の抗QGPGAP抗体価(B)とを示す。2つの独立した実験の結果を、B−Fib/PyCS−GFPで免疫した群において抗体価の中央値で規準化した後、図にプロットした。
【図53】pVII(PfCD4−pVII−2;配列番号28、核酸;配列番号57、アミノ酸)の第1の核移行シグナル(NLS)の前に、およびpVII(PfCD4−pVII−3;配列番号29、核酸;配列番号58、アミノ酸)の2つのNLSの間に挿入されたPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)(A)と、pVIIへのエピトープの挿入を確認するための銀染色の結果(B)とによるアデノウイルスpVIIタンパク質の構造の模式図を示す。
【図54】HVR1およびpVII修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、6週目のPfCSP特異的体液性免疫応答(B)と、9週目のPfCSP特異的CD4+(EYLNKIQNSLSTEWSPCSVT;配列番号62)の応答(C)とを示す。
【図55】ヒト血清試料による組換えアデノウイルスのin vitro組換え中和を示す。希釈ヒト血清の存在下でAD293細胞を組換えアデノウイルスに終夜感染させ、次いでGFP発現を感染のマーカーとして測定した。
【図56】抗アデノウイルス免疫の、カプシド修飾型PyCS−GFPアデノウイルスin vivoによるPyCS特異的T細胞応答の誘発への影響を示す。(A)は、研究デザインの簡単な説明である。(B)は、野生型(wt)/空のアデノウイルスで2回免疫して、続いてカプシド修飾型PyCS−GFPアデノウイルスで初回刺激したマウスにおけるPyCS特異的CD8+ T細胞応答を示す。
【図57】抗アデノウイルス免疫の、カプシド修飾型PyCS−GFPアデノウイルスin vivoによるPyCS特異的体液性免疫応答の誘発への影響を示す。(A)は、研究デザインの簡単な説明である。(B)は、野生型(wt)/空のアデノウイルスで2回、続いて2用量のカプシド修飾型PyCS−GFPアデノウイルスで免疫したマウスにおけるPyCS特異的体液性免疫応答を示す。
【発明を実施するための形態】
【0018】
以下の記述は、本開示の記述、実施形態を十分に理解するため、および可能にするための具体的な詳細を提供する。しかし、当業者は、該開示がこれらの詳細なしでも実行されうることを理解するであろう。別の例では、周知の構造および機能は、本開示の実施形態の説明を不必要に不明瞭にしないように詳細に示されていない、または記述されていない。
【0019】
本開示においてアミノ酸、ペプチド、塩基配列、および核酸のために使用された略語は、IUPAC−IUB Communication on Biochemical Nomenclature, Eur. J. Biochem., 138:9(1984), ”Guideline for Preparing Specifications Including Base Sequences and Amino Acid Sequences”(米国特許商標庁)に特定された略語に基づいており、かつ本技術分野で一般に使用されているものである。
【0020】
本開示によって意図される「ヌクレオチド配列」、「ポリヌクレオチド」または「DNA分子」は、二本鎖DNAまたは一本鎖DNA(すなわち、二本鎖DNAを構成するセンス鎖およびアンチセンス鎖)を含みうるが、その完全長に限定されるものではない。たとえば本明細書で以下に開示する免疫原性外来遺伝子をコードするヌクレオチド配列は、特に明記しない限り、ゲノムDNAを含む二本鎖DNA;cDNAを含む一本鎖DNA(センス鎖);センス鎖、合成DNA、およびその断片に相補的な配列を有する一本鎖DNA(アンチセンス鎖)を包含する。
【0021】
本明細書で用いる場合、ヌクレオチド配列、ポリヌクレオチドまたはDNA分子は、機能的な領域に限定されるものではなく、かつ発現抑制領域、コード領域、リーダー配列、エキソンおよびイントロンのうちの少なくとも1つを含みうる。さらに、ヌクレオチド配列またはポリヌクレオチドの例としては、RNAまたはDNAを含みうる。特異的アミノ酸配列を含むポリペプチドおよび特異的DNA配列を含むポリヌクレオチドは、該ポリヌクレオチドの断片、相同体、誘導体および変異体を含みうる。ヌクレオチド配列またはポリヌクレオチドの変異体の例(たとえば変異体DNA)としては、天然に存在する対立遺伝子変異体;人工変異体;および欠失、置換、追加、および/または挿入を有する変異体が挙げられる。かかる変異体は、元の非変異型ポリペプチドによってコードされるポリペプチドと同じ機能を実質的に有するポリペプチドをコードすることを理解すべきである。
【0022】
本開示は、マラリア原虫の抗原決定基を発現させることができ、および1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質を含む組換えアデノウイルスに関する。組換えアデノウイルスは、組換えアデノウイルスプラスミドベクターに由来し、この産生は以下の実施例で記述される。ベクターとしてのアデノウイルスの使用を、さらに以下で考察する。本明細書に記述する組換えアデノウイルスプラスミドベクターは、マラリア原虫に対する体液性および細胞性の両免疫応答が誘発されるマラリアワクチンまたは医薬組成物として用いられてもよい。
【0023】
マラリア原虫は、既知のマラリア原虫(P.)種、たとえば、熱帯熱マラリア原虫、四日熱マラリア原虫(P.malariae)、卵形マラリア原虫(P.ovale)、三日熱マラリア原虫、二日熱マラリア原虫(P.knowlesi)、マウスマラリア原虫(P.berghei)、ネズミマラリア原虫(P.chabaudi)およびネズミマラリア原虫(P.yoelii)のいずれかから選択されうる。一部の実施形態では、抗原決定基は齧歯動物特異的ネズミマラリア原虫またはヒト特異的熱帯熱マラリア原虫に由来する。
【0024】
一実施形態では、組換えアデノウイルスカプシド修飾型プラスミドベクター(本明細書では組換えアデノウイルスプラスミドベクターとも記述される)は、カプシドをコードして、生成するプラスミドであり、および/または1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質を含む構造を有するコア修飾型組換えアデノウイルスである。本開示の実施形態によれば、カプシドタンパク質および/またはコアタンパク質の修飾は、マラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープを挿入することによって達成されうる。あるいは、カプシドタンパク質および/またはコアタンパク質の少なくとも一部は、除去されてマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープと置換されうる。一部の実施形態では、該免疫原性エピトープは、マラリア原虫スポロゾイト周囲タンパク質のB細胞および/またはT細胞のエピトープである。B細胞またはT細胞のピトープを追加することで、CSタンパク質に対する液体性免疫応答を確立する、または増強することによってマラリアワクチンとして用いられるアデノウイルスベクターの有効性を増強するのに役立ちうる。修飾型カプシドタンパク質と、コアタンパク質と、本明細書に記述する組換えアデノウイルスでのこれらのタンパク質の使用に関するこれらの有意性とを以下でさらに考察する。
【0025】
1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質は、修飾型Hexonタンパク質、修飾型Fiberタンパク質、修飾型pVIIタンパク質またはそれらの組み合わせであってもよい。一実施形態では、Hexon高頻度可変領域(HVR)の一部および/またはFiberタンパク質の一部は、マラリア原虫スポロゾイト周囲タンパク質の少なくとも1つのB細胞および/またはT細胞のエピトープと置換される。あるいは、マラリア原虫スポロゾイト周囲タンパク質の1つもしくは複数のB細胞および/またはT細胞のエピトープは、Fiberタンパク質またはHexonのHVRに挿入されうる。一部の態様では、修飾型HVRは、HVR1、HVR2、HVR3、HVR4、HVR5、HVR6またはHVR7でありうる。別の態様では、修飾型HVRは、HVR1またはHVR5でありうる。一部の実施形態では、HVR修飾型Hexonは、以下の配列番号の核酸配列を有すこともありうる:配列番号3(図11−1〜図11−3)、配列番号4(図12−1〜図12−3)、配列番号5(図13−1〜図13−3)、配列番号6(図14−1〜図14−3)、配列番号7(図15−1〜図15−3)、配列番号8(図16−1〜図16−3)、配列番号9(図17−1〜図17−3)、配列番号10(図18−1〜図18−3)、配列番号11(図19−1〜図19−3)、配列番号12(図20−1〜図20−3)、配列番号13(図21−1〜図21−3)、配列番号14(図22−1〜図22−3)、配列番号15(図23−1〜図23−3)、配列番号16(図24−1〜図24−3)、配列番号17(図25−1〜図25−3)、配列番号18(図26−1〜図26−3)、配列番号19(図27−1〜図27−3)、配列番号20(図28−1〜図28−3)、配列番号21(図29−1〜図29−3)、配列番号22(図30−1〜図30−3)、または配列番号23(図31−1〜図31−3)。別の実施形態では、修飾型Fiberタンパク質は、配列番号24(図32−1〜図32−2)または配列番号25(図33−1〜図33−2)の核酸配列を有すこともありうる。
【0026】
別の実施形態では、マラリア原虫スポロゾイト周囲タンパク質のT細胞エピトープは、以下のいずれかの部位で、アデノウイルスコアpVIIタンパク質に挿入されうる:C末端、第1の核移行シグナル(NLS)の前、または2つのNLSの間。あるいは、マラリア原虫スポロゾイト周囲タンパク質のT細胞エピトープは、pVIIタンパク質の一部を置換しうる。一部の実施形態では、修飾型pVIIタンパク質は、配列番号26(図34)、配列番号27(図35)、配列番号28(図36)、または配列番号29(図37)の核酸配列を有することもある。
【0027】
組換えアデノウイルスでは、トランスジェニックタンパク質または組換えトランスジェニックタンパク質が発現しうる。一部の実施形態では、トランスジェニックタンパク質または組換えトランスジェニックタンパク質は、本明細書に記述する組換えアデノウイルスプラスミドベクターによってコードされるマラリア原虫スポロゾイト周囲タンパク質または抗原決定基であり、かつ1つまたは複数の宿主細胞の感染の後、前述の組換えアデノウイルスプラスミドベクターによって生成される組換えアデノウイルスによって発現される。
【0028】
したがって、一部の実施形態では、組換えアデノウイルスプラスミドベクターは、組換えトランスジェニックタンパク質をコードするヌクレオチド配列を含む。一実施形態では、組換えトランスジェニックタンパク質は、齧歯類特異的寄生虫であるネズミマラリア原虫の抗原決定基を含むこともあり、該抗原決定基が、ネズミマラリア原虫スポロゾイト周囲(CS)タンパク質遺伝子またはその抗原部分を含む。別の実施例形態では、組換えトランスジェニックタンパク質は、ヒト特異的寄生虫である熱帯熱マラリア原虫の抗原決定基を含むこともあり、該抗原決定基が熱帯熱マラリア原虫スポロゾイト周囲遺伝子(CS)タンパク質またはその抗原部分を含む。蚊媒介感染症に対するヒトにおける能動免疫の基礎として用いられるとき、熱帯熱マラリア原虫CSタンパク質は、マラリアの予防を示した。抗原決定基は、B細胞エピトープおよび/またはT細胞エピトープなどの免疫原性エピトープをさらに含みうる。
【0029】
一部の実施形態では、CSタンパク質は、被験者において発現の増強のためにコドン最適化される。コドン最適化は、必要なアミノ酸含有量、関心の対象において一般的な最適コドンの使用、ならびに適切な発現を確実にするために回避されるべきいずれの態様に基づく。かかる態様は、スプライス供与部位または受容部位、終止コドン、ポリアデニル化(pA)シグナル、GCリッチ配列とATリッチ配列、内部TATAボックス、または当技術分野で既知の他のいかなる態様でありうる。一部の実施形態では、コドン最適化CS導入遺伝子のDNA配列は、図9(配列番号1、ネズミマラリア原虫)および図10(配列番号2、熱帯熱マラリア原虫)に示される。
【0030】
一部の実施形態では、組換えアデノウイルスプラスミドベクターは、以下の修飾型熱帯熱マラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つでありうる:(NANP)n(配列番号60)のB細胞エピトープコード配列を用いて、図2に示すように構築されたHVR1修飾型アデノウイルスベクター(NANP−HVR1/PfCSP);(NANP)n(配列番号60)のB細胞エピトープコード配列を用いて、図4に示すように構築されたFiber修飾型アデノウイルスベクター(NANP−Fib/PfCSP);(NANP)n(配列番号60)のB細胞エピトープコード配列を用いて、図5に示すように構築されたHVR1とFiberとの修飾型アデノウイルスベクター(NANP−HVR1/B−Fib/PfCSP);(NANP)n(配列番号60)のB細胞エピトープコード配列およびEYLNKIQNSLSTEWSPCSVT(配列番号62)のCD4エピトープコード配列を用いて、図7に示すように構築されたHVR1とpVIIとの修飾型アデノウイルスベクター(NANP−HVR1/CD4−pVII/PfCSP);(NANP)n(配列番号60)のB細胞エピトープおよびEYLNKIQNSLSTEWSPCSVT(配列番号62)のCD4エピトープコード配列を用いて、図6に示すように構築されたFiberとpVIIとの修飾型アデノウイルスベクター(NANP−Fib/CD4−pVII/PfCSP);ならびに(NANP)n(配列番号60)のB細胞エピトープおよびEYLNKIQNSLSTEWSPCSVT(配列番号62)のCD4エピトープコード配列を用いて、図8に示すように構築されたHVR1とFiberとpVIIとの修飾型アデノウイルスベクター(NANP−HVR1/Fib/CD4−pVII/PfCSP)。
【0031】
別の実施形態では、組換えアデノウイルスプラスミドベクターは、以下の修飾型ネズミマラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つでありうる:(QGPGAP)n(配列番号59)のB細胞エピトープコード配列を用いて、図2に示すように構築されたHVR1修飾型アデノウイルスベクター(QGPGAP−HVR1/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープコード配列を用いて、図4に示すように構築されたFiber修飾型アデノウイルスベクター(QGPGAP−Fib/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープコード配列を用いて、図5に示すように構築されたHVR1とFiberとの修飾型アデノウイルスベクター(QGPGAP−HVR1/B−Fib/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープおよびYNRNIVNRLLGDALNGKPEEK(配列番号61)のCD4エピトープコード配列を用いて、図7に示すように構築されたHVR1とpVIIとの修飾型アデノウイルスベクター(QGPGAP−HVR1/CD4−pVII/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープおよびYNRNIVNRLLGDALNGKPEEK(配列番号61)のCD4エピトープコード配列を用いて、図6に示すように構築されたFiber1とpVIIとの修飾型アデノウイルスベクター(QGPGAP−Fib/CD4−pVII/PyCS);ならびに(QGPGAP)n(配列番号59)のB細胞エピトープおよびYNRNIVNRLLGDALNGKPEEK(配列番号61)のCD4エピトープコード配列を用いて、図8に示すように構築されたHVR1とFiberとpVIIとの修飾型アデノウイルスベクター(QGPGAP−HVR1/Fib/CD4−pVII/PfCSP)。
【0032】
別の実施形態では、組換えアデノウイルスは、以下の修飾型熱帯熱マラリア原虫またはネズミマラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つによって生成されうる:NANP−HVR1/PfCSPまたはQGPGAP−HVR1/PyCS(図2)、NANP−Fib/PfCSPまたはQGPGAP−Fib/PyCS(図4)、NANP−HVR1/B−Fib/PfCSPまたはQGPGAP−HVR1/B−Fib/PyCS(図5)、NANP−HVR1/CD4−pVII/PfCSPまたはQGPGAP−HVR1/CD4−pVII/PyCS(図7)、NANP−Fib/CD4−pVII/PfCSPまたはQGPGAP−Fib/CD4−pVII/PyCS(図6)、(NANP−HVR1/Fib/CD4−pVII/PfCSPまたはQGPGAP−HVR1/Fib/CD4−pVII/PyCS(図8))。組換えアデノウイルスは、哺乳類宿主細胞で組換えトランスジェニックタンパク質(たとえばマラリア原虫CSタンパク質)を発現させる能力を有する組換えアデノウイルスプラスミドベクターを生成するための本明細書に記述する方法によって生成されうる。
【0033】
組換えアデノウイルスの精製は、既知のウイルス精製方法を用いて行われてもよい。たとえば、精製する0.5〜1.0mLのストックウイルスは、AD293細胞などの昆虫細胞を播種する(1×107細胞/10cmシャーレ)ことによる組換えアデノウイスタンパク質を生成する方法によって得た。次いで、感染から数日後に培養上清を収集し、次いで遠心によって得たウイルスペレットをPBS(リン酸緩衝食塩水)で験濁する。得られた懸濁液を10〜60%のショ糖密度勾配にかけ、次いで遠心させて(4℃で、25,000rpmで60分間)、ウイルスバンドを収集する。収集したウイルスをPBSでさらに験濁させ、続いて上述と同じ条件下で遠心する。得られた精製組換えウイルスペレットをPBSなどの緩衝液中、4℃で貯蔵する。
【0034】
別の実施形態は、基本的に少なくとも1つの有効成分を含む医薬組成物に関する。一実施形態では、該医薬組成物の有効成分は、本明細書に記述する遺伝子工学技術によって得られうる組換えアデノウイルスを含みうる。より具体的には、該有効成分は、Hexon高頻度可変領域(HVR)の一部、Fiberタンパク質の一部、pVIIタンパク質の一部、またはそれらの組み合わせがマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープと置換されている、修飾型カプシドタンパク質および/またはコアタンパク質を含む組換えアデノウイルでありうる。あるいは、マラリア原虫スポロゾイト周囲タンパク質の1つもしくは複数のB細胞エピトープおよび/またはT細胞エピトープが、Fiberタンパク質、Hexon HVRまたはpVIIタンパク質に挿入されうる。組換えアデノウイルスプラスミドベクターは、1つもしくは複数の宿主細胞に感染させた後に組換えアデノウイルスによって発現されるトランスジェニックタンパク質または組換えトランスジェニックタンパク質をさらに含む。トランスジェニックタンパク質または組換えトランスジェニックタンパク質は、マラリア原虫スポロゾイト周囲タンパク質またはマラリア原虫スポロゾイト周囲タンパク質のマラリア抗原でありえ、ここで該マラリア抗原はマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープ(たとえばB細胞エピトープまたはT細胞エピトープ)を含む。
【0035】
一部の実施形態では、医薬組成物の有効成分は、組換えアデノウイルスプラスミドベクターに由来する組換えアデノウイルスプラスミドベクターであり、該組換えアデノウイルスプラスミドベクターは、以下の修飾型熱帯熱マラリア原虫またはネズミマラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つである:NANP−HVFM/PfCSPまたはQGPGAP−HVFM/PyCS(図2)、NANP−Fib/PfCSPまたはQGPGAP−Fib/PyCS(図4)、NANP−HVR1/B−Fib/PfCSPまたはQGPGAP−HVR1/B−Fib/PyCS(図5)、NANP−HVR1/CD4−pVII/PfCSPまたはQGPGAP−HVR1/CD4−pVII/PyCS(図7)、NANP−Fib/CD4−pVII/PfCSPまたはQGPGAP−Fib/CD4−pVII/PyCS(図6)、(NANP−HVR1/Fib/CD4−pVII/PfCSPまたはQGPGAP−HVR1/Fib/CD4−pVII/PyCS(図8)。これらの組換えアデノウイルスプラスミドベクターは、細胞(たとえばAD293細胞)にトランスフェクトされると、組換えアデノウイルスを生成することができ、かつ該組換えトランスジェニックタンパク質がヒト細胞を含む哺乳類細胞で発現されうる。
【0036】
下記の実施例でさらに説明するように、有効成分を含む医薬組成物は、本明細書に記述する組換えアデノウイルスであり、被験者に投与されると、マラリア感染性抗原に対するマラリア感染症予防効果を増強し、感染価を低下させる。したがって、組換えアデノウイルスは、標的細胞および組織の感染に付随したマラリア感染症の治療で用いられうる。かかるマラリア感染症に影響を受ける標的細胞の例としては、血球、肝細胞、腎細胞、脳細胞、肺細胞、上皮細胞、および筋細胞が挙げられる。かかる細胞を含む組織の例としては、肺、肝臓、腎臓、脳、動脈と静脈、胃、腸、尿道、皮膚、および筋肉が挙げられる。
【0037】
一部の態様では、医薬組成物は、感染抗原、たとえば、マラリア寄生虫のスポロゾイト表面抗原(スポロゾイト周囲タンパク質(CSP)およびトロンボスポンジン関連接着タンパク質(TRAP))などのマラリア抗原、メロゾイト表面膜タンパク質(MSPI)、マラリアに感染した赤血球から分泌されるマラリアスポロゾイト抗原、およびマラリアに感染した赤血球のノブ内に存在する熱帯熱マラリア原虫赤血球膜タンパク質−1(PfMP1)に対するマラリア感染症予防効果を増強しうる。医薬組成物は、いずれの既知のマラリア原虫種、たとえば、熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫、二日熱マラリア原虫、マウスマラリア原虫、ネズミマラリア原虫(P.chabaudi)およびネズミマラリア原虫(P.yoelii)から選択されるマラリア原虫に対して、感染価を低下させることによってマラリア感染症予防効果を増強しうる。被験者に投与されるとき、医薬組成物による感染価の低下は、該医薬組成物を投与されなかった被験者と比較すると、生存、無病生存、または無感染生存期間と生存、無病生存、または無感染生存率の増加をもたらしうる。したがって、一部の態様では、医薬組成物は、マラリア原虫などの病原体によってもたらされるマラリア感染症のための予防薬または治療薬として有用である。さらなる態様では、医薬組成物は、マラリア原虫などの病原体によってもたらされるマラリア感染症に起因する合併症のための予防薬または治療薬として有用である。
【0038】
被験者における本発明の組換えアデノウイルスの感染症予防効果は、たとえば、本発明のカプシド修飾型組換えアデノウイルスと、医薬投与用の添加剤類とを含有する医薬組成物を、脊椎動物、特にヒトを含む哺乳類に、筋肉内(i.m.)、皮下(s.c.)、皮内(i.c.)、皮内(i.d.)、腹腔内(i.p.)、経鼻、または呼吸経路によって投与し、次いで有効成分として本明細書に記述する組換えアデノウイルスを含有する医薬組成物で脊椎動物に数回免疫することによって、もたらされうる。感染症予防効果を評価するために、該医薬組成物で数回免疫され、続いて標的病原体(たとえば選択されたマラリア原虫種)により感染した被験者の生存率、無病生存、または無感染生存を、該医薬組成物を投与されなかって被験者の生存率、無病生存、または無感染生存と比較してもよい。
【0039】
一部の実施形態では、医薬組成物は、本明細書に記述する薬学的に有効量のカプシドまたはコア修飾型組換えアデノウイルスと、以下にさらに記述する薬学的に許容される担体とをさらに含みうる。
【0040】
別の実施形態は、少なくとも1つの有効成分を基本的に含むワクチン組成物に関する。一実施形態では、該ワクチン組成物の有効成分は、本明細書に記述する組換えアデノウイルスプラスミドベクターに由来する組換えアデノウイルスを含みうる。より具体的には、該有効成分は、Hexon高頻度可変領域(HVR)の一部、Fiberタンパク質の一部、pVIIタンパク質の一部、またはそれらの組み合わせがマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープと置換されている、修飾型カプシドタンパク質および/またはコアタンパク質を含む組換えアデノウイルでありうる。あるいは、マラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープが、pV11タンパク質、Fiberタンパク質もしくはHexon HVR、またはそれらの組み合わせに挿入されうる。一部の実施形態では、ワクチン組成物の有効成分は、図2〜8に示した組換えアデノウイルスプラスミドベクター、たとえば、NANP−HVR1/PfCSPまたはQGPGAP−HVR1/PyCS(図2)、NANP−Fib/PfCSPまたはQGPGAP−Fib/PyCS(図4)、NANP−HVR1/B−Fib/PfCSPまたはQGPGAP−HVR1/B−Fib/PyCS(図5)、NANP−HVR1/CD4−pVII/PfCSPまたはQGPGAP−HVR1/CD4−pVII/PyCS(図7)、NANP−Fib/CD4−pVII/PfCSPまたはQGPGAP−Fib/CD4−pVII/PyCS(図6)、NANP−HVR1/Fib/CD4−pVII/PfCSPまたはQGPGAP−HVR1/Fib/CD4−pVII/PyCS(図8)に由来してもよい。
【0041】
一部の態様では、被験者に投与される場合、ワクチン組成物は、カプシドタンパク質またはコアタンパク質の少なくとも一部に挿入されたまたは該一部を置換する、マラリア原虫CSタンパク質(すなわち、B細胞エピトープ、T細胞エピトープまたは両方)の1つまたは複数の抗原部分を有する組換えアデノウイルスをまず含んでいる。次いで、該ワクチン組成物は、組換えトランスジェニックタンパク質がB細胞エピトープ、T細胞エピトープまたは両方を含むマラリア原虫CSタンパク質である組換えトランスジェニックタンパク質を発現しうる。マラリア原虫CSタンパク質の抗原部分は、組換えトランスジェニックタンパク質に見出され、および下記の実施例に記述するように、修飾型カプシドタンパク質またはコアタンパク質は後天性の体液性免疫、細胞性免疫または両方を促進する、または増強する。このように、一部の態様では、本明細書に記述する組換えアデノウイルスは、体液性免疫、細胞性免疫または両方を促進する、または増強するワクチンとして有用である。
【0042】
一部の態様では、ワクチン組成物は、感染抗原、たとえば、マラリア寄生虫のスポロゾイト表面抗原(CSPおよびTRAP)などのマラリア抗原、メロゾイト表面膜タンパク質(MSPI)、マラリアに感染した赤血球から分泌されるマラリアスポロゾイト抗原、マラリアに感染した赤血球のノブ内に存在するPfEMP1、セリンリッチ抗原(SERA)タンパク質、チロシンリッチ酸性マトリクスタンパク質(TRAMP)、および頂端膜抗原−1(AMA1)に対するマラリア感染症予防効果を増強しうる。さらに、本明細書に記述するワクチン組成物の投与によって生ずる感染価(たとえばウイルス感染価)の低下は、該ワクチン組成物を投与されなかった被験者と比較すると、生存増加、無病生存期間もしくは無感染生存期間と生存、無病生存率もしくは無感染生存率をもたらしうる。したがって、一部の態様では、該ワクチン組成物は、マラリア原虫などの病原体によってもたらされるマラリア感染症のための予防薬または治療薬としても有用である。さらなる態様では、該ワクチン組成物は、マラリア原虫などの病原体によるマラリア感染症に起因する合併症のための予防薬または治療薬としても有用もある。
【0043】
本明細書に記述するワクチン組成物は、本明細書に記述する治療有効量の組換えアデノウイルスを含むこともあり、かつ標準方法によって薬学的に許容される担体をさらに含む。許容される担体の例としては、生理学的に許容される溶液、たとえば無菌生理食塩水および無菌緩衝生理食塩水が含まれる。
【0044】
一部の実施形態では、ワクチン組成物または医薬組成物を薬学的に有効量のアジュバントと組み合わせて用いて、抗マラリア効果を増強しうる。免疫系を刺激して、ワクチンに対する応答を増加させうるが、それ自体は何ら特定の抗原効果がない、任意の免疫アジュバントはアジュバントとして用いられうる。多くの免疫アジュバントは、病原体関連分子パターン(PAMP)として知られている保存分子を進化的に模倣して、トール様受容体(TLR)として知られている一連の免疫受容体によって認識される。本明細書に記述する実施形態によって用いられるうるアジュバントの例としては、フロイント完全アジュバント;フロイント不完全アジュバント;二本鎖RNA(TLR3リガンド);LPS;モノホスホリルリピドA(MPL)(TLR4リガンド)などのLPS類似体;フラゲリン(TLR5リガンド);リポプロテイン;リポペプチド;一本鎖RNA;一本鎖DNA;イミダゾキノリン類似体(TLR7およびTLRのリガンド);CpG DNA(TLR9リガンド);Ribiアジュバント(モノホスホリルリピドA/トレハロースジコリノミコラート);糖脂質(α−GalCer類似体);非メチル化CpGアイランド;油乳剤;リポソーム;ビロゾーム;サポニン類(QS21などのサポニンの活性画分);ムラミールジペプチド;ミョウバン;水酸化アルミニウム;スクアレン;BCG;GM−CSFおよびIL−12などのサイトカイン;MIP 1−αおよびRANTESなどのケモカイン;N−アセチルムラミル−L−アラニル−D−イソグルタミン(MDP);チモシンα1、およびMF59が挙げられる。用いられるアジュバントの量は、皮膚の軟化、疼痛、紅斑、発熱、頭痛、および筋肉痛などの症状の程度に応じて適宜選択されることができる。これらの症状は、ヒトまたは動物においてこの種のワクチン投与後の免疫応答の一部として発現されることもある。
【0045】
一部の実施形態では、本明細書に記述するワクチン組成物または医薬組成物は、他の既知の医薬品、たとえば免疫応答促進ペプチドおよび抗菌剤(合成抗菌剤)と組み合わせて用いられうる。該ワクチン組成物または医薬組成物は、他の薬物および添加剤をさらに含みうる。本明細書に記述するワクチン組成物または医薬組成物と組み合わせて用いられうる薬物ま添加剤の例としては、本発明の組換えアデノウイルスまたは組換えトランスジェニックタンパク質の細胞内取り込みを補助する薬物、トランスフェクションを促進するリポソームおよび他の薬物ならびに/または添加剤(たとえば、フルオロカーボン乳化剤、コキレート、チューブル、金粒子、生分解性マイクロスフェア、およびカチオン性ポリマー)が挙げられる。
【0046】
一部の実施形態では、本明細書に記述するワクチン組成物または医薬組成物に含有される有効成分の量は、それが治療的にもしくは薬学的に有効な量である限り、広範囲の濃度、ウイルス粒子単位(VPU)、プラーク形成単位(PFU)、質量対容量百分率(w/v%)または有効成分量の他の定量的測度から選択されうる。ワクチン組成物または医薬組成物の投与量は、所望の治療効果、投与方法(投与経路)、治療期間、患者の年齢、性別、および他の条件などによって広範囲から適切に選択されうる。
【0047】
一部の態様では、組換えアデノウイルスがワクチン組成物または医薬組成物の有効成分としてヒト被験者に投与されるとき、組換えアデノウイルスの投与量は、組換えウイルスのPFUとして算出され、患者一人あたりおよそ102〜1014PFU、好ましくは105〜1012PFU、より好ましくは106〜1010PFUに対応する量で投与されうる。
【0048】
さらなる態様では、組換えアデノウイルスがワクチン組成物または医薬組成物の有効成分として被験者に投与されるとき、投与量は、ワクチンの宿主に導入される発現可能なDNAの量または転写RNAの量に関して広範囲から選択されうる。該投与量は、使用されるいずれのトランスファーベクターで用いられる転写プロモーターおよび翻訳プロモーターの強度にも依存する。
【0049】
一部の実施形態では、本明細書に記述するワクチン組成物または医薬組成物は、組換えアデノウイルスをPBS(リン酸緩衝生理食塩水)または生理食塩水中で験濁させることで調製される組換えアデノウイルス験濁剤を直接局所部位に(たとえば肺組織、肝臓、筋肉もしくは脳内に)注射することによって、鼻腔急吸入もしくは呼吸吸入によって、または血管内(i.v.)(たとえば動脈内、静脈内、および門脈内)、皮下(s.c.)、皮内(i.e.)、皮内(i.d.)、もしくは腹腔内(i.p.)への投与によって投与されうる。本発明のワクチン組成物または医薬組成物は、複数回投与されうる。より具体的には、最初の投与の後、1回または複数回の追加接種が、追加免疫として投与されうる。1回または複数回の追加免疫投与によって、所望の効果を増強することができる。ワクチン組成物または医薬組成物の投与の後、本明細書に記述する組換えアデノウイルスを含有する医薬組成物による追加免疫が実行されうる。
【0050】
さらなる実施形態では、上述のように、本発明のワクチンと共に種々の他のアジュバント、薬物もしくは添加剤を使用することで、該ワクチン組成物または医薬組成物の投与によって達成される治療効果を増強しうる。薬学的に許容される担体は、等張性および化学安定性を増強する物質などの微量の添加剤を含んでもよい。かかる添加剤は、使用される投与量および濃度においてヒトまたは他の哺乳類の被験者に対して非毒性でなければならない。その例としてはリン酸、クエン酸、コハク酸、酢酸、および他の有機酸、ならびにその塩類;アスコルビン酸などの抗酸化剤;低分子量(たとえば約10未満の残基)ポリペプチド(たとえポリアルギニンおよびトリペプチド)タンパク質(たとえば血清アルブミン、ゼラチン、および免疫グロブリン);アミノ酸(たとえばグリシン、グルタミン酸、アスパラギン酸、およびアルギニン);単糖、二糖、および他の糖質(たとえばセルロースおよびその誘導体、グルコース、マンノース、およびデキストリン)、キレート化剤(たとえばEDTA);糖アルコール(たとえばマンニトールおよびソルビトール);対イオン(たとえばナトリウム);非イオン界面活性剤(たとえばポリソルベートおよびポロキサマー);ならびにPEGなどの緩衝剤が挙げられる。
【0051】
本明細書に記述する組換えアデノウイルスを含有するワクチン組成物または医薬組成物は、密封アンプルまたはバイアルなどの単位用量または複数用量の容器に水溶液剤または凍結乾燥製剤として保存されうる。
【0052】
さらなる別の実施形態は、有効量の組換えアデノウイルスワクチン、製剤または医薬組成物を投与することを含むマラリア感染症を予防する方法、またはマラリアを治療する方法を提供する。本発明は、有効量の組換えアデノウイルスワクチンの組成物、製剤、医薬組成物またはその組み合わせを被験者に投与することを含む免疫活性化の方法をさらに提供する。被験者は、ヒト、動物(たとえば哺乳類、鳥類、爬虫類、魚類および両生類)、またはマラリア寄生虫に感染しうる他のいずれの被験者も含みうる。マラリア寄生虫は、既知のマラリア原虫種、たとえば、熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫、二日熱マラリア原虫、マウスマラリア原虫、ネズミマラリア原虫(P.chabaudi)およびネズミマラリア原虫(P.yoelii)のいずれかから選択されマラリア原虫を含みうる。
【0053】
一部の実施形態では、本明細書に記述する組換えアデノウイルスは、単独で形成されることもあり、または薬学的に許容される担体と共にワクチン組成物、製剤、または医薬組成物に形成されることもあり、かつ前述の被験者に投与されうる。投与経路は、たとえば、上記のいずれの投与経路であってもよい。本発明で使用する薬学的に許容される担体は、生成されるべき医薬組成物の形状によって、本技術分野で一般的に用いられる担体から適宜選択されることができる。たとえば、薬理組成物が水溶液に形成されるとき、純水(滅菌水)または生理的緩衝液が担体として使用されることができる。医薬組成物が他の適切な溶液に形成されるとき、注射されることができる有機エステル、たとえばグリコール、グリセロール、およびオリーブ油が担体として用いられうる。該組成物は、安定剤、賦形剤、および本技術分野で、特にワクチン製剤分野で一般的に使用される他の物質を含みうる。
【0054】
さらなる実施形態では、ワクチン組成物、製剤、または医薬組成物で用いられる組換えアデノウイルスの量は、広範囲の濃度、VPU、PFU、質量対容量百分率(w/v%)または有効成分量の他の定量的測度から選択されうる。一部の態様では、組成物の組換えアデノウイルスの適切な範囲は、好ましくは約0.0002〜約0.2(w/v %)、より好ましくは0.001〜0.1(w/v %)ある。一部の実施形態による組換えアデノウイルスワクチン組成物、製剤または医薬組成物の投与方法は、剤形、患者の年齢、性別、および疾患の重症度など他の条件によって適宜選択されうる。適切な剤形は、注射剤、滴下剤、点鼻薬および吸入剤などの非経口的投与のための形状である。該組成物が注射剤または滴下剤に形成される場合、該注射剤は、必要に応じてグルコース溶液またはアミノ酸溶液などの代替液と混合されて静脈内に投与されることができ、または筋肉内(i.m.)、皮内(i.e.)、皮下(s.c.)、皮内(i.d.)、もしくは腹腔内(i.p.)に投与されることができる。
【0055】
別の実施態様では、組換えアデノウイルスワクチン組成物、製剤、または医薬組成物の1日投与量は、被験者の状態、体重、年齢、性別などに応じて異なりうる。一部の態様では、組換えアデノウイルスの投与量は、1日あたり約0.001〜100mg/kg体重の量で投与される。本発明のワクチン、製剤、または組成物は、1日あたり1回または複数回の投与で投与されうる。
【0056】
さらなる実施形態では、組換えアデノウイルスがワクチン組成物、製剤または医薬組成物の有効成分としてヒト被験者に投与されるとき、組換えアデノウイルスの投与量は、組換えウイルス粒子のPFUとして算出され、患者一人あたりおよそ102〜1014PFU、好ましくは105〜1012PFU、より好ましくは106〜1010PFUに対応する量で投与される。本発明のワクチンの組成物は、各患者の臨床症状(たとえば、予防または治療されるべき症状)、組換えアデノウイルスを含有するワクチン組成物の送達部位、標的組織、投与方法、投与量レジメン、および当業者にとって既知の他の因子を考慮して、良質の医療のための原則に従って投与されるべきである。したがって、本明細書のワクチン組成物の適当な投与量は、上記を考慮して決定される。
【0057】
本開示のさらに別の実施形態は、被験者においてマラリア感染症を治療する、または予防する方法に関し、該方法は、組換えアデノウイルスを含むマラリアワクチン組成物の免疫量または治療量を投与することを含む。マラリアワクチンの組換えアデノウイルスは、マラリア原虫の抗原決定基を含んでもよく、かつ1つもしくは複数の修飾型カプシドタンパク質またはコアタンパク質をさらに含んでもよい。免疫量、薬理量、または治療量は、マラリア感染症が予防されるか、または重症度が低減されるように、強力な免疫応答が(CS)タンパク質(すなわち導入遺伝子、B細胞エピトープ、またはCD4+ T細胞エピトープ)の1つまたは複数の抗原部分に対して生成される、任意の適切な量であってよい。
【0058】
被験者がアデノウイルスベクターに最初に曝露される、または「初回刺激される」とき、免疫系は、その特定のベクターに対して中和抗体を産生する。アデノウイルスに対する免疫応答は、通常カプシドタンパク質対して作られる。したがって、同じアデノウイルスベクターに対する次の曝露、すなわち「追加免疫」は、導入遺伝子発現の効力を低下させることができる。したがって、一部の実施形態では、上記のマラリア感染症を治療する、または予防する方法は、第1の組換えアデノウイルスベクターを用いる初回刺激ステップと、続いて1種または複数種の異なる組換えアデノウイルスベクターを用いる1回もしくは複数回の追加免疫のステップとを含みうる。野生型アデノウイルスにまだ曝露されたことのない被験者において、またはこれまでに野生型アデノウイルスベクターに曝露されたことのある被験者においてこの方法は、用いられてもよい。後述の被験者の場合、初回刺激ステップの組換えアデノウイルスベクターを用いて既存のアデノウイルス免疫を回避する。さらなる実施形態および実施例を後述する。
【0059】
<ベクターとしてのアデノウイルス>
アデノウイルスは、一連のウイルスカプシドタンパク質(以下に記述する)と、1つのウイルスゲノムを含む非エンベロープ被覆型DNAウイルスであり、1種もしくは複数種の治療用導入遺伝子または抗原導入遺伝子をin vitroおよびin vivoで種々の細胞に送達するために広範に用いられてきた。アデノウイルス血清型は数多く存在する。既知のアデノウイルス血清型のうち、血清型5(Ad5)は、in vivoでその高い感染性のために、外来遺伝子の導入のためのベクターとして好ましくは用いられている(Abbinkら 2007)。抗原導入遺伝子の発現は、当技術分野で既知のいずれのプロモーターまたはエンハンサーのエレメントによって制御されうる。遺伝子発現を制御するのに用いられてもよいプロモーターはとしては、サイトメガロウイルス前初期プロモーター(CMV)、シミアンウイルス40(SV40)初期プロモーター、細胞ポリペプチド鎖伸長因子1α(EF1)プロモーター、Rous肉腫ウイルス(RSV)プロモーター、およびテトラサイクリン調節(TR)プロモーターが挙げられるがこれらに限定されない。コード配列の後のポリアデニル化(pA)シグナルは、効果的な転写および翻訳のためにも用いられうる。本明細書に記述する組換えアデノウイルスベクターは、アデノウイルゲノムの少なくともE1領域において欠失を有する複製欠損でありうる。これはE1領域が複製、転写、翻訳およびパッケージングのプロセスに必要とされるからである。一部の態様では、E2、E3および/またはE4の領域も欠失しうる。さらなる態様において、Kozak共通配列は、より効果的な翻訳のために用いられうる(Kozak 1987)。
【0060】
アデノウイルス(Ad)系は、いくつかの理由で組換えワクチンの開発のための魅力あるベクターである。1つの理由は、組換えアデノウイルスベクターは、これらに限定されないがマウスおよびヒトの細胞型を含む、大部分の哺乳類細胞型(複製型および非複製型の双方)を感染させるからである。したがって、同じベクターを、マウスモデルおよびヒト臨床試験でうまく用いることが可能である。別の理由は、転移されたいずれの遺伝情報がエピ染色体にとどまり、細胞遺伝子型の挿入変異および変化を回避するからである(Crystal 1995)。さらに別の理由は、ウイルス複製のラウンドを連続した後でも、導入遺伝子が不変のままであるからである。アデノウイルスを用いる他の利点は、以下が挙げられる:組換えアデノウイルスは1)高いビリオン安定性を有する、2)耐容性がよい、3)高力価で増殖しうる、4)大きな導入遺伝子を収容することができる,5)長年にわたり広範囲に研究されたゲノムを有しており、その結果いくつかの血清型の全DNA配列が分かり、組換えDNA技術によるAdゲノムの操作が容易になっている(GrahamとPrevec 1992)。
【0061】
一実施形態では、前赤内期のマラリア寄生虫を標的にするワクチンの開発のためのウイルスベクターとしてアデノウイルスワクチンプラットフォームを用いて、マラリア感染からの防御を提供する。既知の組換えウイルスベクター(Rodriguesら 1997、Bruna−Romeroら 2001、Andersonら 2004、Taoら 2005)のなかで、アデノウイルスは、前赤内期のマラリア寄生虫に対して強い防御の細胞性免疫応答を誘発することができる(Rodriguesら 1997)。マラリア寄生虫は、マラリア原虫ファミリーのうちのいずれか1つで有りうる。一部の実施形態では、標的とされる該寄生虫は、ネズミマラリア原虫(P.yoelii)または熱帯熱マラリア原虫でありうる。
【0062】
<導入遺伝子としてPyCSを発現するアデノウイルスベクターは、マラリア特異的CD8+ T細胞応答を誘発する>
アデノウイルスは、マラリアに対して重要なCD8+ T細胞介在防御免疫を誘発するための魅力あるベクターである(Rodriguesら 1997、Rodriguesら 1998)。ネズミマラリア原虫(齧歯目のマラリア寄生虫)CSタンパク質(AdPyCS)を発現する組換えアデノウイルスの免疫原性は、齧歯目のマラリアモデルを使用して決定された。AdPyCSをマウスに接種すると、かなりの割合のマウスにおいて、完全免疫が誘発され、寄生虫血症の発生を予防する(Rodriguesら 1997)。T細胞集団を枯渇させることによって明らかになり、かつAdPyCSがマラリア寄生虫に対して高力価の抗体応答を誘発することができなかった事実によって実証されているように、この防御効果は、主にCD8+ T細胞によって介在される。
【0063】
カプシド修飾型アデノウイルスの感染性を定量的に測定するために、シャトルベクターにGFP発現カセットと導入遺伝子のクローニング部位とを含ませてもよい。結果として生じるシャトルベクター(GFP/pShuttle−CMV)は、デュアルpCMVプロモーターと、導入遺伝子のためのSV40pAと、pmaxGFP(Amaxa,Germany)からのGFPとを有する。最適化PyCS断片をGFP/pShuttle−CMVのKpnIとHindIIIの部位に挿入した。
【0064】
Ad(PyCS+GFP)の免疫原性は、CS特異的CD8+ T細胞応答の大きさと、マラリア原虫肝臓内期に対する防御免疫のレベルとを測定することによって決定された。異なる経路を介して、最適用量でAd(PyCS+GFP)を投与することで、1010ウイルス粒子(v.p.)は、AdPyCSが誘発することを示した同じパターンの抗マラリア防御応答を誘発し、s.c.およびi.m.経路で、最も強い応答を誘発し、生ネズミマラリア原虫スポロゾイトを投与されたマウスにおいて肝臓内期での最も高い程度の抑制をもたらした。これは、ワクチンとしてAd(PyCS+GFP)がAdPyCSと同等に作用し(Rodriguesら 1997)、かつaAdPyCSのin vivo向性を決定する潜在的に有用なツールであることを示している。
【0065】
<アデノウイルスのカプシドタンパク質およびコアタンパク質>
CSタンパク質を発現する組換えアデノウイルスベクターがCD8+ T細胞によって強力な細胞性免疫応答を誘発するが、はっきりと認識できる体液性応答は誘発されないことを上述の研究で確認された。したがって、野生型アデノウイルスに対する体液性応答がカプシドタンパク質に起因しうることが多いので、修飾型カプシドタンパク質およびコアタンパク質で組換えアデノウイルスベクターを構築することで、1)B細胞活性を介する体液性免疫を増強し、2)Tヘルパー細胞活性を介する体液性免疫を増強し、および3)既存するアデノウイルス免疫を回避する。
【0066】
アデノウイルスは、正三角形の20面を有する正二十面体の形状の非エンベロープ被覆型ネイキッド二重鎖DNAウイルスである。アデノウイルスカプシドは、252カプソメアからなり、そのうち240がHexon三量体であり、12がPenton五量体である。各Penton塩基から突き出ているFiberタンパク質は、細胞受容体との相互作用によって宿主細胞への接着を媒介する。二次的相互作用は、Penton塩基内のRGD(Asp−Arg−Gly)モチーフ間でαvβ3、αvβ5および同様のインテグリンによって生じ、その後の細胞へのアデノウイルスの内部移行を容易にする(Mathiasら 1994、Wickhamら 1993)。ほとんどのアデノウイルスは、細胞受容体として、コクサッキー−アデノウイルス受容体(CAR)を使用する(Bergelsonら 1997)。加えて、MHCクラス1分子、VCAM、およびヘパラン硫酸は、Ad5の接着と侵入を媒介することが示されている(Chuら 2001、Hongら 1997)。エンドサイトーシスを介する侵入に続いて、Ad5はエンドサイトーシス区画からサイトゾルに迅速に逃避する(MeierとGreber 2003、LeopoldとCrystal 2007)。次いで、ビリオンは、微小管を用いて核に移動する。Fiberタンパク質は、プロセスにおいて最も初期のカプシドタンパク質として脱落される(Nakanoら 2000、Hongら 2003)。異なる血清型のアデノウイルスは、異なる輸送パターンを示す(Miyazawaら 1999、Miyazawaら 2001)。Fiberタンパク質を変えるまたは修飾することで、輸送に影響を与えることができる。これは、抗原提示細胞(APC)の感染に続いて、抗原プロセシングおよび提示に関して特に重要になりうる。
【0067】
アデノウイルスFiberは、Fiber尾部、シャフト、およびノブのドメインに分けられる三量体である(Henryら 1994、RuxとBurnett 2004、Chroboczekら 1995)。ノブドメインの三次元構造は知られおり、変異原性研究と共に、これらの研究は、CAR相互作用と三量体形成に関与する領域を視覚化させることを可能にする(Kirbgyら 1999、Xiaら 1995)。Fiberシャフトは、ビリオンから突き出ており、Fiberノブはコサッキーおよびアデノウイルス受容体(CAR)相互作用ドメインを含む(Roelvinkら 1999、Bewleyら 1999)。FiberノブのCAR結合部位は、一次的にABループとCDループからの残基からなり、二次的にFGとHIループ、次いでB、EおよびFのβシートに伸長する(Roelvinkら 1999、Bewleyら 1999)。HIループは、最も研究されたFiberノブ上の挿入部位(Worgallら 2004、MizuguchiとHayakawa 2004、Koizumiら 2003、Belousovaら 2002、NoureddiniとCuriel 2005、Nicklinら 2001)であり、HIループ(残基543と544)へのエピトープの組み込みは、強力な抗エピトープ免疫をもたらす(Krauseら 2006)。したがって、免疫優性CS由来のB細胞エピトープは、Fiberタンパク質のHIループに最初に挿入された。
【0068】
Hexonは、1ビリオンあたり720コピーを有する、アデノウイルスカプシドで最も豊富なタンパク質である。成熟したウイルスでは、Hexonは、正二十面体ビリオンの小面から構成されるホモ三量体のカプソメアとして存在する(RuxとBurnett 2004)。アデノウイルス血清型2および5(Ad2およびAd5)のHexonのの結晶構造は解析されており、複合分子構造が明らかになった(Athapillyら 1994、Robertsら 1986、RuxとBurnett 2000)。各単量体サブユニットのベースは、多くの正二十面体のウイルスのカプシドタンパク質に存在する2つのβ−バレルモチーフからなる。3つの長いループ(DE1、FG1、およびFG2)は、ベース構造から伸びて、各分子のタワー領域を形成する(RuxとBurnett 2004)。これらのループドメイン内の配列は、カプシドの表面から突出して、ビリオンの外側を形成する。異なるアデノウイルス血清型からの配列比較は、カプシド外部上に位置する配列が長さとアミノ酸配列との両方で不完全に保存されていることを示している(Crawford−MikszaとSchnurr 1996)。さらに、これらの不完全に保存されたドメイン(高頻度可変領域(HVR)と名付けられる)に位置する配列が決定基を含み、それらの決定基に対して血清型特有の抗体が産生されることが示された(Top 1975、RuxとBurnett 2000、Topら 1971)。
【0069】
初期の配列比較に基づいて、Hexon分子全体を通して7つのHVRが同定された(Crawford−MikszaとSchnurr 1996、Robertsら 2006)。HVRは血清型間で不完全に保存されており、かつHexonの構造的完全性を維持することに関与していないと思われるので、ウイルスの生存能に影響を及ぼすことなく少量の変化がこれらのドメインになされることができた(RuxとBurnett 2000)。ととえば、ヘキサヒスチジンタグは、ウイルス生存能を損なうことなく、HVR2、HVR3、HVR5、HVR6、およびHVR7に挿入されることができる(Wuら 2005)。このようにして、Hexon HVRを標的として用いて、Hexon HVR内に位置するペプチドに対する抗体応答を効率的に誘発させることが多い(Worgallら 2005、Cromptonら 1994)。血清型間でその長さが完全に保存されないこと、およびアデノウイルスカプシドの最外側表面上のその位置のために(RuxとBurnett 2000、Crawford−MikszaとSchnurr 1996)、Hexon HVR5は、最初はエピトープ挿入のための部位として選択された。さらに、HVR5がカプシド表面上の柔軟なループであることをHexonの結晶構造が示しており、HVR5がカプシドの構造的完全性を損なうことなく比較的大きなペプチドを収容することができることを示唆している(Robertsら 1986)。Hexon特異的CD4+エピトープおよびCD6+エピトープが最近同定されて(Leenら 2008)、アデノウイルスへのCD4+ T細胞応答は、ヒトにおいてHexonタンパク質内に保存された残基に対して集中する(Onionら 2007、Heemskertら 2006)ことが判明した。
【0070】
アデノウイルスのコアは、ウイルスゲノムと4種類のコアタンパク質で構成されている。末端タンパク質(TP)は、1ビリオンあたり2つのコピーで各直鎖状ウイルスDNA鎖の5’末端に共有結合される。他の3種類のコアタンパク質、mu(μ)、V(pV)およびVII(pVII)は、アルギニンリッチタンパク質を介して非共有結合的かつ非特異的にウイルスDNAに結合している。pVIIは、1ビリオンあたりおおよそ700〜800コピーを提供する主要なコアタンパク質であり、かつその周りをウイルスDNAで包まれてヌクレオソーム構造を形成している、ヒストン様中枢として役立っている。
【0071】
<体液性免疫を増強するためのアデノウイルスカプシドタンパク質の修飾>
一部の実施形態では、アデノウイルスカプシドタンパク質(HexonまたはFiberに挿入された)内に免疫優性CSタンパク質Bエピトープを有する、スポロゾイト周囲(CS)アデノウイルスベクターが記述されている。導入遺伝子は、CSタンパク質に対する細胞介在免疫応答および体液性免疫応答を増強するために、CMVなどのプロモーター下に存在しうる。
【0072】
中心反復領域は、マラリア原虫種間でCSタンパク質の保存された構造であり、かつこの反復配列に対する抗体は、スポロゾイト中和活性を有することが示されている。マラリア原虫CSタンパク質内の反復配列の例として、(NANP)n反復(熱帯熱マラリア原虫;配列番号60)、ANGAGNQPG反復(三日熱マラリア原虫;配列番号63)、およびNAAG反復(四日熱マラリア原虫;配列番号64)があり、これらはアデノウイルスカプシドタンパク質に挿入されることができる。一部の実施形態では、PfCSPの4以上の(NANP)n反復(配列番号60)は、アデノウイルス血清型5HexonのHVR1に挿入されうる。一部の実施形態では、PfCSPの2、4、6、8、10、14、16、18、20、22、24、26、または28の(NANP)n反復(配列番号60;n=2、4、6、8、10、12、14、16、18、20、22、24、26、28)は、アデノウイルス血清型5HexonのHVR1に挿入される。(NANP)n反復配列は、FiberのHIループにさらに挿入されうる。
【0073】
一部の実施形態では、改良型CSタンパク質アデノウイルスワクチンを開発するために、CSに対する免疫優性中和B細胞エピトープがマッピングされた。組換えネズミマラリア原虫CSタンパク質(PyCS)で免疫されたマウスは、2つの主要免疫優性Bエピトープ、すなわちQGPGAP(配列番号59)とQQPP(配列番号65)に対して高力価を生成したが、in vitro中和アッセイでは、(QGPGAP)3のペプチド(配列番号59;n=3)を培地に加えることによって中和が逆転されうるために、QGPGAPエピトープ(配列番号59)それ自体に対する体液性免疫応答が中和活性の主な原因となりうることを示した。一部の実施形態では、PyCSの3以上の(QGPGAP)n反復(配列番号59)は、アデノウイルス血清型5HexonのHVR1に挿入される。一部の実施形態では、PyCSの3、4、5、6、7、8、9、10、11、または12の(QGPGAP)n反復(配列番号59;n=3、4、5、6、7、8、9、10、11、12)は、アデノウイルス血清型5HexonのHVR1に挿入されうる。(QGPGAP)n反復配列は、FiberのHIループにさらに挿入されうる。
【0074】
B細胞エピトープペプチドは、免疫系がエピトープを効率的に認識することができるように、アデノウイルスビリオンの表面に提示されなければならない。かかる挿入部位は、HexonのHVRおよびFiber内のループ構造でありえ、かつ異なる挿入部位が組み合わせられてもよい。
【0075】
<Tヘルパー細胞活性を増強するためのアデノウイルスカプシドタンパク質とコアタンパク質の修飾>
別の実施例では、アデノウイルスベクターで用いられる導入遺伝子に特異的なCD4+エピトープを、pVII、pVおよびHexonなどのアデノウイルスタンパク質に取り込んでアデノウイルスベースワクチンの免疫原性を増強させうる。樹状細胞(DC)およびB細胞などの専門の抗原提示細胞(APC)は、エンドサイトーシスを介して微粒子状の病原体様ウイルス粒子を取り込むことができ、次いで該病原体中のCD4+エピトープをCD4+ T細胞に提示し、それによって体液性免疫応答および/または細胞性免疫応答のヘルパー細胞として作用する。1ビリオンあたりのコピー数がpVII(700〜800コピー)およびHexon(720コピー)と高いため、pVIIおよびHexonをアデノウイルス標的タンパク質として用いて、抗原CD4+ペプチドに容易に挿入することができる。
【0076】
<既存のアデノウイルス免疫を増強するためのアデノウイルスカプシドタンパク質の修飾>
一部の実施形態では、アデノウイルスに対する既存の免疫を克服するため、および/またはアデノウイルスワクチンに対する体液性応答を増強するために、アデノウイルスFiberおよびHexonのカプシドタンパク質は、B細胞エピトープまたはTヘルパー細胞エピトープを挿入するよう修飾されうる。ヒト集団において若年成人の推定80%がアデノウイルスに対する循環中和抗体(Douglas 2007)、特に血清型5(Ad5)を有する。遺伝子治療ベクターとしてアデノウイルスを利用する研究では、動物内の中和抗体の存在が、アデノウイルスによって送達される導入遺伝子の発現を限定していることが判明した。中和抗体に加えて、CD8+ T細胞応答も、組換え遺伝子発現を限定する一因となっている(Yangら 1995、Yangら 1996)。アデノウイルスに対するかかる既存の免疫は、組換えアデノウイルスワクチンの有効性を阻害することがいままでに報告されており(Pappら 1999)、また臨床試験において、アデノウイルスベースワクチンの免疫原性を低下させている(Priddyら 2008)。
【0077】
Hexonは抗Adカプシド免疫応答の主要な標的であり(Royら 2005、Wohlfart 1988)、CD4+とCD8+のT細胞応答の誘発を含むアデノウイルスの強力なアジュバント効果の原因であると思われる。したがって、既存の抗アデノウイルス免疫を回避するために使用されてきた1つの戦略は、Hexon全体または一部を異なるタンパク質、たとえばアデノウイルス11、24、26および35などのまれな血清型と置換することである。Hexonは抗アデノウイルス中和抗体の主要な標的である(Youilら2002、Sumidaら2005)ために、全HexonまたはHexonのHVRはまれな血清型と交換されうる(Wuら 2002、Robertsら 2006)。
【0078】
本明細書の一実施形態で記述した別の戦略では、既存の抗アデノウウイルス免疫またはアデノウウイルスベクターによる以前のワクチン接種によって誘発された抗アデノウウイルス中和抗体を増強するために、アデノウイルスHexonは、HVR1またはHVR5と抗原ペプチドとの置換によって修飾されうる。一部の実施形態では、抗原ペプチドはマラリア原虫CSタンパク質の免疫原性エピトープであってもよく、特定の態様では、該エピトープは中心反復配列、CD4+エピトープ配列またはCD8+エピトープ配列を含みうる。
【0079】
同じ血清型のAdベクターによる反復投与は、免疫化後の坑Ad免疫のために阻止される。したがって、多くのAdワクチンは、導入遺伝子によってコードされた抗原の発現と提示を防ぐことでワクチンの追加免疫を妨げる(Yangら 1995、Hackettら 2000、Harveyら 1999、Mastrangeliら 1996)。後述の実施例に記述するように特異的エピトープをAdカプシドに追加することで、一部の実施形態によるこの妨害を低下しうる、または排除しうる。
【0080】
実施形態をよりよく説明するため、およびクレームするいずれの実施形態の範囲を限定すると解釈されないようにするために以下の実施例を提供する。特定の材料が記載される範囲は、説明の目的だけであり、本発明を限定することを意図するものではない。当業者は、発明能力を行使することなく、および本発明の範囲から逸脱することなく、同等の方法または反応物を開発しうる。本発明の範囲内に依然として留まりつつ、多くのバリエーションが本明細書に記述する方法でなされうることを理解されるであろう。かかるバリエーションが本発明の範囲内に含まれることは、本発明者らの意思である。
【実施例】
【0081】
実施例1:カプシド修飾型マラリア原虫スポロゾイト周囲タンパク質アデノウイルスプラスミドベクターおよび組換えアデノウイルス粒子の構築
【0082】
エピトープマッピング
まず、PyCSの免疫優性中和B細胞エピトープを選択した。ナイーブBALB/cマウスにフロイント不完全アジュバントと共に組換えPyCSタンパク質を3回免疫し、プールした血清を用いて、中和抗体の重要なエピトープを決定した。
【0083】
簡単に言うと、中和アッセイでは、ヒトCD81発現HepG2細胞を標的細胞として用いた。CD81は、マラリア寄生虫が増殖して、シゾントになる肝細胞においてパラシトホーラスバキュオールを形成するために必要な分子であり(Silvieら 2006)、したがってスポロゾイトのin vitro感染力が極めて高い。
【0084】
本アッセイでは、PyCS合成ペプチドをウェルに加えて、ペプチド特異的抗体を遮断した。エピトープマッピングの結果は、PyCS中心反復配列(QGPGAP;配列番号59)がPyCSにおいてQQPP(配列番号65)よりもより強力な中和エピトープであることを示した。したがって、(QGPGAP)nエピトープ(配列番号59)の挿入を用いて、アデノウイルスカプシドタンパク質、Hexonおよび/またはFiberを修飾した。
【0085】
カプシド修飾型プラスミドベクターの構築
クローニング部位下にGFP発現カセットを挿入することでアデノウイルスシャトルベクターpShuttle−CMV(STRATAGENE)を修飾した。まず、pmaxGFP(Lonza,Cologne,Germany)のBsmBI−SacI断片(pCMV+GFP)とSacI−BsmBI断片(SV40ポリAshigunaru)を平滑末端化して、pUC19の平滑末端化Sall部位とKpnI部位にそれぞれ挿入した。もたらされたpCMV−GFP/pUC19のBamHI−EcoRI断片をSV40pA/pUC19の同じ部位に挿入して、SV40pA−pCMV−GFP断片を作製した。該断片を平滑末端化して、pShuttle−CMVのEcoRV部位に挿入した。結果として生じるシャトルベクター(GFP/pShuttle−CMV)は、導入遺伝子とGFPのためのデュアルpCMVプロモーターとSV40pAを有する。
【0086】
アデノウイルスシャトルベクターpShuttle−CMVの別の修飾を行って、CMVプロモーター領域をpQBI−AdCMV5(QBIOgene)からのCMV5プロモーターと置換した。pShuttle−CMV内でpShuttle−CMVのSgrAI−KpnI断片をCMV5プロモーター配列と、CMVプロモーターからの上流配列とを含む断片と置換して、pShuttle−CMV5ベクターを構築した。
【0087】
ネズミマラリア原虫CS(PyCS)遺伝子は、JCatコドン最適化アルゴリズム(http://www jcat de/)に基づいてPCR反応を重複させることによって、(QGPGAP)n反復(配列番号59)を除いて、コドン最適化した。
【0088】
熱帯熱マラリア原虫3D7株のPfCSPアミノ酸配列をコドン最適化のための鋳型配列として用いた。ヒトにおいてタンパク質発現のためのコドン最適化は、Integrated DNA Technologies(Coralville,IA,USA)の最適化ソフトウェアによって行った。C末端でGPIアンカードモチーフを除く全PfCSPをコードするDNA断片(図10;配列番号2)をIntegrated DNA Technologiesで合成した。
【0089】
コドン最適化PyCS遺伝子(図9;配列番号1)またはPfCSP遺伝子(図10;配列番号2)を、pShuttle−CMV、pShuttle−CMV5、またはGFP/pShuttle−CMVのKpnIおよびHindIIIに挿入した。AdEasy−1による相同組換えのために、もたらされたマラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターを用いて、マラリア原虫スポロゾイト周囲抗原遺伝子と、インタクトなアデノウイルスタンパク質コード配列とを有するアデノウイルスゲノムを構築した。簡単に言うと、マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターをPmeI消化によって直線化し、次いで相同組換えのために大腸菌(E.coli)BJ5183細胞を直線化シャトルベクターおよびpAdEasy−1ベクター(Bruna−Romeroら 2003)と同時形質転換させた。
【0090】
アデノウイルスカプシドタンパク質の修飾を要約して、図1に示す。アデノウイルスゲノムDNAのHVR1配列の修飾を図2に示す。簡単に言うと、AdEasy−1をSfiIで消化させて、6.4kbp断片を、EcoRI−SfiIおよびPstI−SfiIリンカーオリゴマーを用いて、pUC19のEcoRI部位とPstI部位にサブクローニングさせた。HVR1をマラリア原虫スポロゾイト周囲タンパク質B細胞エピトープと置換するために、AgeI部位とNdeI部位を含む領域を、HVR1配列の代わりにエピトープ配列を有するプライマーを用いる2ステップPCRによって増幅させた。PCR産物をAgeIとNdeIで消化させ、次いでこれを用いてSfiI/pUC19ベクター中のSfiI断片のナイーブAgeI−NdeI領域を置換した。配列を確認した後に、アデノウイルスゲノムDNAのSfiI断片を、スポロゾイト周囲エピトープ配列を含むSfiI断片と置換して、HVR1修飾型Hexonを生成した。一部の実施形態では、HVR修飾型Hexonは、配列番号3(図11−1〜図11−3)、配列番号4(図12−1〜図12−3)、配列番号5(図13−1〜図13−3)、配列番号6(図14−1〜図14−3)、配列番号7(図15−1〜図15−3)、配列番号8(図16−1〜図16−3)、配列番号9(図17−1〜図17−3)、配列番号10(図18−1〜図18−3)、配列番号11(図19−1〜図19−3)、配列番号12(図20−1〜図20−3)、配列番号13(図21−1〜図21−3)、配列番号14(図22−1〜図22−3)、配列番号15(図23−1〜図23−3)、配列番号16(図24−1〜図24−3)、配列番号17(図25−1〜図25−3)、配列番号18(図26−1〜図26−3)、配列番号19(図27−1〜図27−3)、配列番号20(図28−1〜図28−3)、配列番号21(図29−1〜図29−3)、配列番号22(図30−1〜図30−3)、または配列番号23(図31−1〜図31−3)の核酸配列を有することもある。
【0091】
HVR1に(NANP)28(配列番号60;n=28)を挿入するために、コドン最適化PfCSPの中心反復領域の一部を、Hexon特異的配列を5’に、NANP特異的配列を3’に有するプライマーを用いてPCRによって増幅させた。もたらされたDNA断片を第2のPCRによってAgeI−NdeI領域に挿入した。
【0092】
HVR5修飾の場合、図3に示すように、XbaI部位を、AdEasy−1内のHexonのL1ループ中のHVR5に導入し、次いで合成し、マラリア原虫スポロゾイト周囲タンパク質エピトープをコードするリン酸化二本鎖オリゴマーをXbaI部位に挿入した。配列決定(図31;配列番号23)によってこの挿入を確認した。
【0093】
Fiber修飾の場合、図4に示すように、AdEasy−1のSpeI−PacI断片を、EcoRI−PacIおよびPstI−SpeIのリンカーオリゴマーを用いてpUC19のEcoRI部位とPstI部位にサブクローニングした。マラリア原虫スポロゾイト周囲タンパク質B細胞エピトープ配列をFiberノブのHIループに挿入するために、EcoNI(またはNhel)部位とMfel部位を含む領域を、エピトープ配列を有するプライマーを用いる2ステップPCRによって増幅させた。PCR産物をEcoNI(またはNheI)とMfeIで消化させ、次いでこれを用いてSpeI−PacI/pUC19ベクター内のFiberのナイーブEcoNI(またはNheI)−MfeI領域を置換した。配列(図32−1〜図32−2、配列番号24;図33−1〜図32−2、配列番号25)を確認した後、AdEasy−1のSpeI−PacI断片を、エピトープ配列を含むSpeI−PacI断片と置換した。マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターによる相同的組換えのために、もたらされたFiber修飾型アデノウイルスDNAを用いて、Fiber修飾型マラリア原虫スポロゾイト周囲タンパク質アデノウイルスDNAを生成した。
【0094】
2つのエピトープ挿入を有するHVR1およびFiber修飾型アデノウイルスDNAを構築するために、Fiber修飾型アデノウイルスDNAのSfiI−SfiI断片を、図5に示すようにHVR1内にスポロゾイト周囲タンパク質エピトープを有するSfiI−SfiI断片と置換した。
【0095】
pVIIのC末端を修飾するために、SfiI部位とSalI部位を含む領域を、スポロゾイト周囲タンパク質エピトープ配列を有するプライマーを用いて2ステップPCRによって増幅させた。PCR産物をSfiIとSaiIで消化させ、次いでこれを用いて、SfiI/pUC19ベクターのナイーブSfiI−SalI領域を置換した。配列(図34、配列番号26;図35、配列番号27)を確認した後、HVR1のSfiI−SfiI断片および/またはFiber修飾型スポロゾイト周囲タンパク質アデノウイルスDNAを、pVII内にスポロゾイト周囲タンパク質エピトープを有するSfiI−SfiI断片と置換した。
【0096】
スポロゾイト周囲タンパク質CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)をpVIIの中央に挿入するために、pAdEasy−1の約7.7kb断片をRsrII消化によって調製して、RsrIIリンカー(RsrII/pUC19)を用いて、pUC19プラスミドのEcoRI部位とHindiII部位との間にクローニングした。RsrII/pUC19内にAscI部位とBgIII部位を含む領域を、エピトープ配列を有するプライマーを用いて2ステップPCRによって増幅させた。PCR産物をAscIとBgIIIで消化させ、次いでこれを用いて、RsrII/pUC19プラスミド内のナイーブAscIとBgIII領域を置換した。置換した領域(図36、配列番号28;図37、配列番号29)の配列を確認した後、HVR1修飾型アデノウイルスDNAのRsrII断片を、エピトープ配列を含むRsrII断片と置換した。
【0097】
以下の表1(ネズミマラリア原虫)および表2(熱帯熱マラリア原虫)に記載する組換えアデノウイルスを作製して、エピトープ挿入の感染性、免疫原性、および既存の抗アデノウウイルス免疫に対する感受性への効果を評価した。使用した組換えアデノウイルスベクターは、複製欠損、E1およびE3欠失アデノウイルス血清型5(STRATAGENE)であった。図1は、カプシド修飾型マラリア原虫スポロゾイト周囲タンパク質組換えアデノウイルスの構造の模式図を示す。
【0098】
【表1】

【0099】
【表2】
【0100】
カプシド修飾型アデノウイルスゲノムDNAプラスミドを精製し、Pad消化によって直線化して、Ad293細胞のトランスフェクションのために用いた。
【0101】
凍結/解凍を4回繰り返すことでトランスフェクトされたAD293細胞からアデノウイルス粒子を調製して、さらなるウイルス増幅のために用いた。最後の増幅を終えてから、アデノウイルス粒子をCsCl勾配遠心分離によって精製した。次いで、バンドを収集して、透析緩衝液に対して透析して、CsClを除去した。ウイルス粒子(v.p.)は、O.D.260(1 O.D.260=1.25×1012v.p./mL)(Bruna−Romeroら 2003)に基づいて算出した。
【0102】
アデノウイルス増幅処置の間、アデノウイルス増殖でのカプシド修飾型アデノウイルス間で観察された差異は小さく、アデノウイルスの感染力および産生力が、修飾によって悪影響を受けていなかったことを示していた。
【0103】
実施例2:ネズミマラリア原虫スポロゾイト周囲タンパク質特的免疫応答
【0104】
ネズミマラリア原虫組換えアデノウイルスの検証
マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターを一過性トランスフェクションのために用いて、AD293細胞を用いてマラリア原虫スポロゾイト周囲タンパク質発現を確認した(図38)。トランスフェクションから24時間後に、細胞をSDS試料緩衝液中で溶解し、続いてSDS PAGE電気泳動および抗PyCSモノクローナル抗体(9D3)によるウエスタンブロットを行った。
【0105】
アデノウイルスカプシドタンパク質へのエピトープ挿入を確認するために、精製した組換えアデノウイルスをSDS−PAGE(2×109v.p./レーン)によって分析し、図39Aおよび40Aに示すように(QGPGAP)n(配列番号59)反復を認識する、抗スポロゾイト抗体によるウエスタンブロット(1×109v.p./レーン)を行った。図39Aでのバンドの強度は、アデノウイルスビリオン中のカプシドタンパク質のコピー数と相関した:Fiberのコピー数(1ビリオンあたり36コピー)は、Hexonのそれ(1ビリオンあたり720コピー)の20分の1に満たない。図39Aのレーン4の下位のバンドは、分解したHexonと思われる。図40Aのバンドの強度は、HVR1に挿入された(QGPGAP)n(配列番号59)の反復数と相関した。
【0106】
PyCS−Bエピトープがアデノウイルスビリオンの外側に曝露されたかどうかを評価するために、連続希釈、精製した組換えアデノウイルス粒子を、酵素結合免疫吸着測定(ELISA)プレート上にコーティングして、(QGPGAP)n(配列番号59)反復を認識する抗PyCS抗体で検出した。該抗体は、すべてのカプシド修飾型アデノウイルスを認識した(図39Bおよび40B)。ELISA分析の結果は、カプシドタンパク質に取り込まれたPyCS−Bエピトープがアデノウイルスビリオンの外側に十分に曝露されたことを示唆する。
【0107】
カプシド修飾型PyCSアデノウイルスによる免疫化後のネズミマラリア原虫スポロゾイト周囲タンパク質特異的免疫応答
6〜8週齢の雌BALB/cマウスをTaconic(Hudson,NY, USA)から購入して,Rockefeller UniversityのLaboratory Animal Research Centerにおいて標準条件下で維持した。免疫化のために、アデノウイルスをPBSで希釈して、表示された用量で筋肉内に注射した。
【0108】
単回免疫後に組換えアデノウイルスの免疫原性を評価するために、ナイーブBALB/cマウスの群(1群あたり5匹)に、種々の組換えPyCSアデノウイルスを1×109v.p.で筋肉内に免疫し、次いで免疫化から2週間後にPyCS特異的細胞媒介免疫応答(CMI)をELISPOTで測定した(図41A).
【0109】
免疫したマウスの脾臓のPyCS特異的、予防接種IFN−γ分泌CD8+ T細胞の数を、PyCSタンパク質内のCD8+ T細胞エピトープ(SYVPSAEQI;配列番号66)に対応する合成ペプチドを用いてELISPOTによって決定した。簡単に言うと、96ウェルニトロセルロースプレート(Milititer HA,Millipore)を抗マウスインターフェロンγモノクローナル抗体、R4で終夜コーティングした。室温での終夜インキュベーションの後、ウェルを培地で繰り返し洗浄し、次いで4時間培地で遮断した。免疫したマウスからの5×105脾細胞を、10μg/mLのCD8+ T細胞エピトープペプチドの存在下または非存在下でELISPOTに加えて、37℃、5%CO2で24時間インキュベートした。0.05%Tween20(PBST)を含有するPBSで、該プレートを十分に洗浄した後、PBST中のビオチン化抗マウスインターフェロンγモノクーナル抗体、XMG1.2を加えて、40℃で終夜インキュベートした。PBSTで洗浄した後に、該プレートをペルオキシダーゼ標識アビジン(eBiosciences)と共にインキュベートすることになる。AEC基質(BD Biosciences)を加えることによって複数のスポットが発現した。
【0110】
カプシド修飾型アデノウイルスのすべては、この用量で、インタクトなカプシドタンパク質を有するアデノウイルスに対して同程度のCMIを誘発した(図41B)。
【0111】
次に、図42Aに示すように、ナイーブBALB/cマウスに増加した用量(すなわち、1×108、1×109、および1×1010v.p.)の組換えアデノウイルスを3週間間隔で複数回投与した。PyCS特異的体液性応答をELISAで決定した。免疫したマウスの尾静脈から5マイクロリットルの血液を採取して、495μLのPBSで希釈し、次いでこれらの試料を5,000rpmで5分間遠心して、希釈血漿試料(×100)を調製した。Maxisorp ELISAプレートを、0.1Mの炭酸ナトリウム緩衝液中の5μg/mLのCS特異的ペプチド((QGPGAP)3;配列番号59、n=3)で、40℃で終夜コーティングした。プレートを洗浄して、室温で、1×希釈液で2時間遮断した。プレートを再度洗浄して、1×希釈液中の100μLの連続2倍希釈した血漿または血清をプレートに加え、次いで該プレートを室温で1時間インキュベートした。プレートを洗浄して、100μLのHRP標識ヤギ抗マウスIgG抗体と共にインキュベートした。ペプチドのすべては、Biosyntheis(Lewisville,TX,USA)で合成されたものであった。
【0112】
この免疫レジメンを使用して、10週目にすべてのカプシド修飾型アデノウイルスは、野生型PyCS−GFPよりも高いレベルの抗−(QGPGAP)3抗体応答を有意に誘発した(図42B)。
【0113】
カプシド修飾型アデノウイルスのワクチン有効性を決定するために、10週目に、免疫したマウスに2×104感染性ネズミマラリア原虫スポロゾイトを尾静脈注で投与した。スポロゾイト投与から42時間後、マウスの肝臓内の寄生虫特異的リボソームRNAの量を定量化することによって寄生虫負荷を決定し、次いで寄生虫リボソームRNAの絶対コピー数対マウスGAPDH mRNAの絶対コピー数の比として表した。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて差異を決定した。
【0114】
(QGPGAP)3−HVR1/PyCS−GFP、(QGPGAP)3−Fib/PyCS−GFPまたは(QGPGAP)3−HVR1/Fib/PyCS−GFPによるワクチン接種は、野生型PyCS−GFPよりも高いレベルの予防を誘発し、マラリアを投与されたマウスにおいて有意に低い寄生虫負荷をもたらした(図42C)。
【0115】
次に、カプシド修飾型アデノウイルスによって誘発されたPyCS特異的抗体の機能性を評価した。第1に、免疫されたマウスのアデノウイルスの血清が10週目に、インタクトなスポロゾイトを認識することができるかどうかを調べるために、間接的免疫蛍光アッセイ(IFA)を行った。IFAでは、マルチスポットガラススライド上の風乾したスポロゾイトを、PBS中3%のウシ血清アルブミン(BSA)と共に1時間インキュベートし、次いで希釈血清と共に1時間インキュベートした。洗浄後、該スライドを蛍光標識二次抗体と共に1時間インキュベートした。該スライドを洗浄して、蛍光顕微鏡下でIFA力価を、蛍光を発する最高希釈として決定した。(QGPGAP)3−HVR1/PyCS−GFPと(QGPGAP)3−HVFM/Fib/PyCS−GFPは共に、スポロゾイトに対して最高のIFA力価を誘発し(図43A)、アデノウイルスHexonのHVR1への(QGPGAP)3のエピトープの挿入によってPyCSアデノウイルスが合成ペプチドだけでなく、マラリア寄生虫に存在するナイーブエピトープに対しても強い抗体応答を誘発することを可能にすることを示した。
【0116】
第2に、カプシド修飾型アデノウイルスで免疫したマウス(図42A)が、スポロゾイトの感染力の効力を中和することができうる「機能的」抗体を発現したかどうか決定するために、in vitroスポロゾイト中和アッセイを行った。
【0117】
in vitro中和アッセイでは、アデノウイルス免疫マウスに由来するプールした血清を30倍に希釈して添加した96ウェルプレート中のCD81/He−pG2にネズミマラリア原虫スポロゾイトを加えた。2時間のインキュベーションの後、非感染スポロゾイトを培地で洗い落とし、次いで該細胞を42時間培養した。ヒトGAPDH mRNAに対する寄生虫リボソームRNAの相対量を、リアルタイムPCR(Ophorstら 2006)で測定した。
【0118】
カプシド修飾型アデノウイルス、特に(QGPGAP)3−HVR1/PyCS−GFPおよび(QGPGAP)3−HVR1/Fib/PyCS−GFP)で免疫したマウスに由来するプールした血清は、in vitroでスポロゾイト感染力をほぼ完全に(99%)阻害した(図43B)。このアッセイでの阻害の程度は、図43Aに示すIFA力価と逆相関したことが注目される。
【0119】
赤血球期マラリア感染からの防御。
次に、スポロゾイト投与の後、カプシド修飾型rAdによる免疫化が赤血球期マラリア感染症の発現からマウスを防御するかどうかを決定した。実験を2回行った。各実験では、各群で20匹のBALB/cマウスに、図42Aに示すように、野生型PyCS−GFPまたは(QGPGAP)3−HVR1/PyCS−GFPで3回免疫した。最後の免疫化から4週目に、マウスに50ネズミマラリア原虫スポロゾイトを静脈内投与した。抗原投与から3〜12日後に、Giemsa染色した血液塗沫標本を分析して、赤血球期マラリア寄生虫感染を検出した。野生型CS−GFP免疫群では、40匹中30匹(75%)のマウスが感染し、一方ナイーブ群では40匹中35匹(87.5%)が感染した(以下の表3)。(QGPGAP)3−HVRI/CS−GFP予免疫マウスは、野生型CS−GFPよりもより防御された;40匹中わずかに15匹(37.5%)が感染した。これは肝臓での寄生虫負荷によって測定された防御実験(図42C)の結果と一致する。
【0120】
【表3】
【0121】
初回刺激−追加免疫による免疫化1
次に、図44Aに示すように、ナイーブBALB/cマウスに、(QGPGAP)nの4または6の反復(配列番号59;n=4、6)を有するHVR1−修飾型PyCSアデノウイルスの「追加免疫」をアジュバントと共にまたはアジュバントを含まずに、用量を増加して(すなわち、1×108、1×109、および1×1010v.p.)3週間間隔で投与した。この実験で使用したアジュバントは、200μg/mLのSaponin(Sigma−Aldrich)を含有するSigma Adjuvant System(Sigma−Aldrich)である。Sigma Adjuvant System(1mL)のバイアルは、Salmonellaミネソタ由来のモノホスホリルリピドA(解毒化内毒素)を0.5mgおよび水中2%オイル(スクアレン)−Tween80中の合成トレハロースジコリノミコラートを0.5mg含有する。免疫の前に、アデノウイルス溶液を同量のアジュバントと混合した。100マイクロリットルのアデノウイルス−アジュバント混合液を筋肉内に注射した。PyCS特異的体液性免疫応答および細胞介在性免疫応答を上述のように測定した。6反復を有するHVR1修飾型アデノウイルスは、4反復を有する該修飾型アデノウイルスよりも高い抗体価を誘発し、かつアジュバントの使用が抗体価を増強する傾向が観察された(図44B)。対照的に、CMIへのアジュバントの効果はみられなかった(データ図示せず)。
【0122】
HVR1修飾型PyCSアデノウイルスのワクチン有効性を決定するために、9週目に各群の5匹のマウスに2×104の感染性ネズミマラリア原虫スポロゾイトを尾静脈から投与した。スポロゾイト投与から42時間後の寄生虫負荷を上述のように決定した。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて差異を決定した。6反復を有するHVR1修飾型アデノウイルスは、4反復を有する該修飾型アデノウイルスよりも寄生虫負荷を減少させ、かつアジュバントの使用が防御を増強する傾向がみられた(図44C)。
【0123】
HVR1修飾型PyCSアデノウイルスによって誘発される抗体の機能性を評価するために、我々は上述のようにin vitroスポロゾイト中和分析を行った。HVR1修飾型PyCSアデノウイルスで免疫したマウス由来のプールした血清試料を9週目に、50倍希釈でスポロゾイト浸潤を中和させた(図44D)。
【0124】
初回刺激−追加免疫による免疫化2
図45Aに示すように、ナイーブBALB/cマウスに、(QGPGAP)nの反復を6、9、または12有する(配列番号59;n=6、9、12)HVR1修飾型PyCSアデノウイルスの「追加免疫」を、アジュバント共に、またはアジュバントを含まずに、1×1010v.p.の用量で、3週間間隔で3回、投与した。この実験で使用したアジュバントは、200μg/mLのSaponin(Sigma−Aldrich)を含有するSigma Adjuvant System(Sigma−Aldrich)である。免疫の前に、アデノウイルス溶液を同量のアジュバントと混合した。PyCS特異的体液性免疫応答および細胞介在性免疫応答を上述のように測定した。(QGPGAP)nの反復を12有する(配列番号59;n=12)HVR1修飾型PyCSアデノウイルスとアジュバントは、9週目に群の間で最も高い抗体価を誘発した(図45B)。PyCS特異的CMIに関して、群の間で差はみられず、最高12の長いエピトープ挿入がin vivoでアデノウイルス感染力を損なわないことを示した(図45C)。さらに、アジュバンントはCMIを誘発するるアデノウイルスの能力に影響を及ぼさなかった(図45C)。
【0125】
HVR1修飾型PyCSアデノウイルスのワクチン有効性を決定するために、9週目に各群の5匹のマウスに2×104の感染性ネズミマラリア原虫スポロゾイトを尾静脈から投与した。スポロゾイト投与から42時間後の寄生虫負荷を上述のように決定した。(QGPGAP)n反復(配列番号59;n=6、9、12)を有するすべてのHVR−1修飾型PyCSアデノウイルスは、アジュバントの有無にかかわらず防御の増加を示した。しかし、(QGPGAP)nの反復を12有する(配列番号59;n=12)HVR1修飾型PyCSアデノウイルスとアジュバントは、最良の防御を示した(図45D)。これは他のいずれの処置よりも有意により防御的であった。
【0126】
実施例3:熱帯熱マラリア原虫スポロゾイト周囲タンパク質特異的免疫応答
【0127】
熱帯熱マラリア原虫組換えアデノウイルスの検証
一過性トランスフェクションのために、マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターを用いて、AD293細胞を用いるマラリア原虫スポロゾイト周囲タンパク質発現を確認した(図46A)。
トランスフェクションから24時間後に、細胞をSDS試料緩衝液中で溶解し、続いてSDS PAGE電気泳動および抗NANPモノクローナル抗体(2A10)によるウエスタンブロットを行った。
【0128】
アデノウイルスカプシドタンパク質へのエピトープ挿入を確認するために、精製した組換えアデノウイルスをSDS−PAGE(2×109v.p./レーン)によって分析し、図47Aおよび48Aに示すように(NANP)n(配列番号60)反復を認識する、抗スポロゾイト抗体によるウエスタンブロット(1×109v.p./レーン)を行った。図47Aのバンドの強度は、アデノウイルスビリオン中のカプシドタンパク質のコピー数と相関した;Fiberのコピー数(1ビリオンあたり36コピー)は、Hexonのそれ(1ビリオンあたり720コピー)の20分の1に満たない。図48Aのバンドの強度は、NANP反復HVR1の数と相関した。
【0129】
PfCSP−Bエピトープがアデノウイルスビリオンの外側に曝露されたかどうか評価するために、連続希釈し、精製した組換えアデノウイルス粒子を、酵素結合免疫吸着測定(ELISA)プレート上にコーティングし、次いで(NANP)n(配列番号60)反復を認識する抗PyCS抗体で検出した。該抗体は、すべてのカプシド修飾型アデノウイルスを認識した(図47Bおよび48B)。ELISA分析の結果は、カプシドタンパク質に取り込まれたPfSP−Bエピトープがアデノウイルスビリオンの外側に十分に曝露されたことを示唆する。
【0130】
初回刺激−追加免疫による免疫化3
図49Aに示すように、ナイーブBALB/cマウスに 、組換えPfCSPアデノウイルスを複数回、用量を増加して(すなわち、1×108、1×109、および1×1010v.p.)、3週間間隔で投与した。上述のように、PfCSP特異的体液性応答を、(NANP)n反復配列(配列番号60)を含有する1μg/mL(T1B)4、CS反復ペプチドでコーティングしたELISAプレートを用いるELISAによって決定した(Calvo−Calleら 2006)。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて、野生型PfCSPとカプシド修飾型アデノウイルスとの差異を決定した。すべてのカプシド修飾型アデノウイルスは、野生型PfCSPよりも統計学的に高い抗NANP抗体価を誘発した。
【0131】
初回刺激−追加免疫による免疫化4
次に、図50Aに示すように、ナイーブBALB/cマウスに、(NANP)nの反復を4、6、8、または10有する(配列番号60;n=4、6、8、10)HVR1修飾型PfCSPアデノウイルスの「追加免疫」を、用量を増加して複数回(すなわち、1×108、1×109、および1×1010v.p.)、3週間間隔で投与した。PfCSP特異的体液体性免疫応答を上述のように測定した。9週目に、すべてのHVR1修飾型アデノウイルスは、野生型PfCSPよりも有意に高い抗NANP抗体価を誘発した(図50B)。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて、差異を決定した。
【0132】
初回刺激-追加免疫による免疫化5
次に、図51Aに示すように、ナイーブBALB/cマウスに、(NANP)nの反復を10、16、または22有する(配列番号60;n=10、16、22)HVR1修飾型アデノウイルスの「追加免疫」を、アジュバントと共に、またはアジュバントを含まずに、1×1010v.p.の用量で、3週間間隔で3回投与した。この実験で使用したアジュバントは、200μg/mLのSaponin(Sigma−Aldrich)を含有するSigma Adjuvant System(Sigma−Aldrich)である。免疫の前に、アデノウイルス溶液を同量のアジュバントと混合した。PfCSP特異的液体性免疫応答を上述のように測定し、次いで長いBエピトープを有するHVR1修飾型アデノウイルスがより高い抗体価を誘発したことを決定した(図51B)。
【0133】
実施例4:アデノウイルスコアタンパク質pVIIへのPyCS CD4エピトープの挿入
抗原特異的CD4 T細胞は、抗原特異的B細胞の発現と増殖のために必要とされる。したがって、PyCS CD4エピトープのアデノウイルスタンパク質への挿入によるカプシド修飾型アデノウイルスによってPyCS特異的体液性免疫応答を増強することが可能であるかどうかを決定するために、pVII内にPyCS CD4エピトープを有する(QGPGAP)3−Fib/PyCS−GFP、((QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFP)を構築した。pVIIは、アデノウイルスコアタンパク質のうちの1つであり、1ビリオンあたりのコピー数は700〜800である。これは、MHCクラスII分子上への効率的なCD4エピトープ提示に理想的である。図52Aに示すように、PyCS CD4エピトープをSDS−PAGEゲル上でpVIIに挿入することによってpVIIバンドを移動させる。
【0134】
PyCS CD4エピトープのpVIIへの挿入の効果を試験するために、図42Aに示すように、ナイーブBALB/cマウスを(QGPGAP)3−Fib/PyCS−GFPまたは(QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFPで免疫して、10週目に抗QGPGAP抗体価をELISAで決定した。(QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFPは、(QGPGAP)3−Fib/PyCS−GFPよりも有意に高い抗QGPGAP抗体価を誘発し(図52B)、pVIIへのPyCS CD4エピトープの挿入が、カプシド修飾型アデノウイルスによって誘発された体液性免疫応答を増強したことを示した。
【0135】
アデノウイルスコアタンパク質pVIIへのPfCSP CD4エピトープ挿入
アデノウイルスコアタンパク質内の異なる位置へのPfCSP CD4+エピトープ挿入の、アデノウイルス誘発免疫応答への効果を評価するために、第1の核移行シグナル(NLS)の前、または2つのNLSの間にPfCSP CD4+エピトープを有するHVR1修飾型PfCSPアデノウイルスを構築した(図53A)。
【0136】
pVIIへのエピトープ挿入を確認するために、上述のように精製組換えアデノウイルスをSDS−PAGEで分析した。図53Bに示すように、(NANP)4−HVR1/CD4−pVII−2/PfCSPおよび(NANP)4−HVR1/CD4−pVII−3/PfCSPのpVIIバンドを、エピトープを挿入することによって上方に移動させた。
【0137】
HVR1およびpVIIの修飾型PfCSPアデノウイルスによって誘発されたPfCSP特異的免疫応答
次に、図54Aに示すように、ナイーブBALB/cマウスに、HVR1にNANPの4反復と、pVIIにPfCD4+エピトープとを有するHVR1およびpVIIの修飾型PfCSPアデノウイルスの「追加免疫」を、用量を増加して複数回(すなわち、1×108、1×109、および1×1010v.p.)、3週間間隔で投与した。PfCSP特異的体液性免疫応答を上述のように測定した。6週目に、(NANP)4−HVR1/CD4−pVII−2/PfCSPおよび(NANP)4−HVRI/CD4−pVII−3/PfCSPは、(NANP)4−HVR1/PfCSPより有意に高い抗NANP抗体価を誘発した。CMIに関して、(NANP)4−HVR1/CD4−pVII−3/PfCSPは、(NANP)4−HVR1/PfCSPより有意に高いIFNγおよびIL−4分泌PfCSP特異的CD4+ T細胞を誘発した(図54C)。
【0138】
実施例5:カプシド修飾の抗アデノウイルス免疫への効果
in vitroアデノウイルス中和実験のために、アデノウイルス感染の前に表示した希釈の血清をAD293細胞に加えた。Caucasian血清試料は、Innovative Research(Novi,MI,USA)から入手した。すべてのフローサイトメトリーデータは、FlowJo v8.8ソフトウェア(Tree Star,Inc.Ashland,OR,USA)で分析した。表示した希釈のヒトアデノウイルス中和血清試料の存在下で、AD293細胞に各カプシド修飾型アデノウイルスを感染させ、続いてフローサイトメトリーによってGFP発現を測定した。HVR1のPyCS−Bエピトープとの置換によって、アデノウイルスは抗アデノウイルス血清型5血清に対して明らかに回復力があるようになり、一方HVR5またはFiberの修飾は効果がなかった(図55)。
【0139】
次に、HVR1がin vivo中和にとって重要な分子であるかどうかを決定した。この目的のために、既存の抗アデノウイルス免疫を十分に開始させるために、マウスに1×1010v.p.野生型空のアデノウイルスを2回感染させて、(図56A)、次いでELISAで決定したそれらの抗アデノウイルス抗体価に基づいてランダム化した。次いで、マウスにカプシド修飾型アデノウイルスまたは非修飾型アデノウイルスを単回免疫量で投与して、上述のようにPyCS−特異的CD8+ T細胞応答のレベルを測定した。他のカプシド修飾型または非修飾型アデノウイルスによって誘発されたワクチン接種と比較すると、(QGPGAP)3−HVR1/PyCS−GFPまたは(QGPGAP)3−HVR1/Fib/PyCS−GFPによるワクチン接種だけが有意により強力なCS特異的CD8+ T細胞応答を誘発することができた(図56B)。
【0140】
(QGPGAP)3エピトープに対する抗体応答のレベルも、測定した。抗体応答は、野生型空のアデノウイルスに感染させ、続いてカプシド修飾型rAdでワクチン接種したマウスにおいて、rAdのカプシドタンパク質上で発現される(図57A)。(QGPGAP)3−HVR1/PyCS−GFPおよび(QGPGAP)3−HVR1/Fib/PyCS−GFPをワクチン接種したマウスだけが、野生型PyCS−GFPをワクチン接種したマウスよりも有意に高い抗QGPGAP抗体価を開始することができた。
【0141】
上述の実施例は、実施形態をより十分に説明するように意図され、クレームされるいずれの実施形態の範囲を限定すると解釈されるものではない。加えて、本開示内に引用した参照文献、および後述するすべての参照文献は、それら全体を参照することによって本明細書に十分に記載されているかのように本明細書によって組み込まれる。
【0142】
参考文献
Abbink,P., Lemckert,A.A., Ewald,B.A., Lynch,D.M., Denholtz,M.,ら 2007. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J Virol. 81:4654−4663.
Alonso,P.L., Sacarlal,J., Aponte,J.J., Leach,A., Macete,E.,ら 2004. Efficacy of the RTS,S/AS02A vaccine against Plasmodium falciparum infection and disease in young African children: randomised controlled trial. Lancet. 364:1411−1420.
Alonso,P.L., Sacarlal,J., Aponte,J.J., Leach,A., Macete,E.,ら 2005. Duration of protection with RTS,S/AS02A malaria vaccine in prevention of Plasmodium falciparum disease in Mozambican children:single−blind extended follow−up of a randomised controlled trial. Lancet. 366:2012−2018.
Anderson,R.J., Hannan,CM., Gilbert,S.C., Laidlaw,S.M., Sheu,E.G.,ら 2004. Enhanced CD8+ T cell immune responses and protection elicited against Plasmodium berghei malaria by prime boost immunization regimens using a novel attenuated fowlpox virus. J Immunol. 172:3094−3100.
Athappilly,F.K., Murali,R., Rux,J.J., Cai, Z. & Burnett,R.M. The refined crystal structure of Hexon, the major coat protein of adenovirus type 2, at 2.9 A resolution. Journal of molecular biology 242,430−455(1994).
Barrat,F.J., Meeker,T,, Gregorio,J,, Chan, J.H., Uematsu,S.,ら. Nucleic acids of mammalian origin can act as endogenous ligands for Toll−like receptors and may promote systemic lupus erythematosus.J Exp Med. 202, 1131−1139(2005).
Bejon,P., Lusingu,J., Olotu,A., Leach,A., Lievens,M.,ら 2008. Efficacy of RTS,S/AS01 E vaccine against malaria in children 5 to 17 months of age. N Engl J Med. 359:2521−2532.
Belousova,N., Krendelchtchikova,V., Curiel,DT. & Krasnykh,V. Modulation of adenovirus vector tropism via incorporation of polypeptide ligands into the Fiber protein. Journal of virology 76, 8621−8631(2002).
Bergelson,J.M.ら. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science (New York, N.Y275, 1320−1323(1997).
Bewley,M.C, Springer,K., Zhang,Y.B., Freimuth,P. & Flanagan,J.M. Structural analysis of the mechanism of adenovirus binding to its human cellular receptor, CAR. Science (New York, N.Y286, 1579−1583(1999).
Bruna−Romero,O., Schmieg,J., Del Val,M., Buschle,M. & Tsuji,M. The dendritic cell−specific chemokine, dendritic cell−derived CC chemokine 1, enhances protective cell−mediated immunity to murine malaria. J Immunol 170, 3195−3203(2003).Hong,S.S., Karayan,L, Tournier,J., Curiel,DT. & Boulanger,P.A. Adenovirus type 5 Fiber knob binds to MHC class I alpha2 domain at the surface of human epithelial and B lymphoblastoid cells. The EMBO journal 16, 2294−2306(1997).
Bruna−Romero,O., Gonzalez−Aseguinolaza,G., Hafalla,J.C, Tsuji,M.,およびNussenzweig,R.S. 2001. Complete, long−lasting protection against malaria of mice primed and boosted with two distinct viral vectors expressing the same plasmodial antigen.Proc Natl Acad Sci U S A. 98:11491−11496.
Bruna−Romero,O., Rocha,CD., Tsuji,M. & Gazzinelli,RT. Enhanced protective immunity against malaria by vaccination with a recombinant adenovirus encoding the circumsporozoite protein of Plasmodium lacking the GPI−anchoring motif. Vaccine 22, 3575−3584(2004).
Calvo−Calle,J.M., Oliveira,G.A., Watta,CO., Soverow,J., Parra−Lopez,C,ら 2006. A linear peptide containing minimal T− and B−cell epitopes of Plasmodium falciparum circumsporozoite protein elicits protection against transgenic sporozoite challenge.Infect Immun. 74:6929−6939.
Clyde,D.F., Most,H., McCarthy,V.C,およびVanderberg,J.P. 1973. Immunization of man against sporozite−induced falciparum malaria. Am J Med Sci. 266:169−177.
Chroboczek,J., Ruigrok,R.W.& Cusack,S. Adenovirus Fiber.Current topics in microbiology and immunology 199(Pt1), 163−200(1995).
Chu,Y., Heistad,D., Cybulsky,M.I.& Davidson,B.L. Vascular cell adhesion molecule−1 augments adenovirus−mediated gene transfer.Arterioscler Thromb Vasc Biol 21, 238−242(2001).
Crawford−Miksza,L. & Schnurr,D.P. Analysis of 15 adenovirus Hexon proteins reveals the location and structure of seven hypervariable regions containing serotype−specific residues.Journal of virology 70, 1836−1844(1996).
Crompton,J., Toogood,Cl., Wallis,N. & Hay, RT. Expression of a foreign epitope on the surface of the adenovirus Hexon.The Journal of general virology75(Pt1), 133−139(1994).
Crystal,R.G. Transfer of genes to humans:early lessons and obstacles to success.
Science (New York, N.Y270, 404−410(1995).
Douglas,JT. Adenoviral vectors for gene therapy.Molecular biotechnology 36, 71−80(2007).
Edelman,R., Hoffman,S.L., Davis,J.R., Beier,M., Sztein,M.B.,ら 1993. Long−term persistence of sterile immunity in a volunteer immunized with X−irradiated Plasmodium falciparum sporozoites.J Infect Dis. 168: 1066−1070.
Grillot,D., Valmori,D., Lambert,P.H., Corradin,G.,およびDel Giudice,G. 1993. Presentation of T cell epitopes assembled as multiple−antigen peptides to murine and human T lymphocytes.Infect Immun. 61:3064−3067.
Graham,F.L.& Prevec,L. Adenovirus−based expression vectors and recombinant vaccines.Biotechnology (Reading, Mass 20, 363−390(1992).
Gwadz,R.W., Cochrane,A.H., Nussenzweig,V.,およびNussenzweig,R.S. 1979. Preliminary studies on vaccination of rhesus monkeys with irradiated sporozoites of Plasmodium knowlesi and characterization of surface antigens of these parasites.Bull World Health Organ.57 Suppl 1:165−173.
Hackett,N.R.ら. Use of quantitative TaqMan real−time PCR to track the time−dependent distribution of gene transfer vectors in vivo. Mol Ther2, 649−656(2000).
Harvey,B.G.ら. Airway epithelial CFTR mRNA expression in cystic fibrosis patients after repetitive administration of a recombinant adenovirus.The Journal of clinical investigation 104, 1245−1255(1999).
Heemskerk,B.ら. Adenovirus−specific CD4+ T cell clones recognizing endogenous antigen inhibit viral replication in vitro through cognate interaction. J Immunol 177, 8851−8859(2006).
Henry,L.J., Xia,D., Wilke,M.E., Deisenhofer,J. & Gerard,R.D. Characterization of the knob domain of the adenovirus type 5 Fiber protein expressed in Escherichia coli.Journal of virology 68, 5239−5246(1994).
Hong,S.S., Habib,N.A., Franqueville,L, Jensen,S. & Boulanger,P.A. Identification of adenovirus (ad) penton base neutralizing epitopes by use of sera from patients who had received conditionally replicative ad (addl1520) for treatment of liver tumors.Journal of virology 77, 10366−10375(2003).
Kester,K.E., Cummings,J.F., Ockenhouse,C.F., Nielsen,R., Hall,B.T.,ら 2008. Phase 2a trial of 0, 1, and 3 month and 0, 7, and 28 day immunization schedules of malaria vaccine RTS,S/AS02 in malaria−naive adults at the Walter Reed Army Institute of Research.Vaccine. 26:2191−2202.
Kirby,I.ら. Mutations in the DG loop of adenovirus type 5 Fiber knob protein abolish highaffinity binding to its cellular receptor CAR.Journal of virology 73, 9508−9514(1999).
Koizumi,N., Mizuguchi,H., Utoguchi,., Watanabe,Y. & Hayakawa,T. Generation of Fiber−modified adenovirus vectors containing heterologous peptides in both the HI loop and C terminus of the Fiber knob.The journal of gene medicine 5, 267−276(2003).
Kozak M. 1987.An analysis of 5’−noncoding sequences from 699 vertebrate messenger RNAs.Nucleic Acids Res. 15:8125−148.
Krause,A., Joh,J.H., Hackett,N.R., Roelvink,P.W., Bruder,J.T.,ら 2006. Epitopes expressed in different adenovirus capsid proteins induce different levels of epitope−specific immunity.J Virol. 80:5523−5530.
Labow,D., Lee,S., Ginsberg,R.J., Crystal,R.G. & Korst, R.J. Adenovirus vector−mediated gene transfer to regional lymph nodes.Human gene therapy11,759−769(2000).
Leen,A.M.ら. Identification of Hexon−specific CD4 and CD8 T cell epitopes for vaccine and immunotherapy.Journal of virology 82, 546−554(2008).
Leopold,P.L. & Crystal,R.G. Intracellular trafficking of adenovirus:many means to many ends.Advanced drug delivery reviews 59, 810−821(2007).
Mastrangeli,A.ら. ”Sero−switch” adenovirus−mediated in v/Vo gene transfer:circumvention of anti−adenovirus humoral immune defenses against repeat adenovirus vector administration by changing the adenovirus serotype.Human gene therapy 7, 79−87(1996).
Mathias,P., Wickham,T., Moore,M. & Nemerow,G. Multiple adenovirus serotypes use alpha v integrins for infection.Journal of virology 68, 6811−6814(1994).
McConnell,M.J., Danthinne,X.,およびImperiale,M.J. 2006.Characterization of a permissive epitope insertion site in adenovirus Hexon.J Virol. 80:5361−5370.
Meier,O. & Greber,U.F. Adenovirus endocytosis.The journal of gene medicine 5, 451−462(2003).
Miyazawa,N.ら. Fiber swap between adenovirus subgroups B and C alters intracellular trafficking of adenovirus gene transfer vectors.Journal of virology 73, 6056−6065(1999).
Miyazawa,N., Crystal,R.G. & Leopold,P.L. Adenovirus serotype 7 retention in a late endosomal compartment prior to cytosol escape is modulated by Fiber protein.Journal of virology75, 1387−1400(2001).
Mizuguchi,H. & Hayakawa,T. Targeted adenovirus vectors.Human gene therapy 15, 1034−1044(2004).
Nakano,M.Y., Boucke,K., Suomalainen,M., Stidwill,R.P. & Greber,U.F. The first step of adenovirus type 2 disassembly occurs at the cell surface, independently of endocytosis and escape to the cytosol.Journal of virology 74, 7085−7095(2000).
Nicklin,S.A.ら. Ablating adenovirus type 5 Fiber−CAR binding and HI loop insertion of the SIGYPLP peptide generate an endothelial cell−selective adenovirus.Mol Ther4, 534−542(2001).
Noureddini,S.C. & Curiel,DT. Genetic targeting strategies for adenovirus.Molecular pharmaceutics 2, 341−347(2005).
Nussenzweig,R.S., Vanderberg,J., Most,H.,およびOrton,C. 1967. Protective immunity produced by the injection of x−irradiated sporozoites of Plasmodium berghei.Nature. 216:160−162.
Nussenzweig,R.S. & Long, CA.Malaria vaccines: multiple targets.Science (New York, N.Y 265, 1381−1383(1994).
Onion,D.ら. The CD4+ T cell response to adenovirus is focused against conserved residues within the Hexon protein.The Journal of general virology 88, 2417−2425(2007).
Ophorst,O.J., Radosevic,K., Havenga,M.J., Pau,M.G., Holterman,L.,ら 2006. lmmunogenicity and protection of a recombinant human adenovirus serotype 35−based malaria vaccine against Plasmodium yoelii in mice.Infect Immun. 74:313−320.
Oualikene,W., Gonin,P. & Eloit,M. Short and long term dissemination of deletion mutants of adenovirus in permissive (cotton rat) and non−permissive (mouse) species.The Journal of general virology 75(Pt10), 2765−2768(1994).
Priddy,F.H., Brown,D., Kublin,J., Monahan,K., Wright,D.P.,ら 2008. Safety and immunogenicity of a replication−incompetent adenovirus type 5 HIV−1 clade B gag/pol/nef vaccine in healthy adults.Clin Infect Dis. 46: 1769−1781.
Roberts,M.M., White,J.L., Grutter,M.G. & Burnett,R.M. Three−dimensional structure of the adenovirus major coat protein Hexon.Science (New York, N.Y 232, 1148−1151(1986).
Roberts,D.M., Nanda,A., Havenga,M.J., Abbink,P., Lynch,D.M.,ら 2006. Hexon−chimaeric adenovirus serotype 5 vectors circumvent pre−existing anti−vector immunity.Nature. 441 : 239−243.
Rodrigues,E.G., Zavala,F., Eichinger,D., Wilson,J.M.,およびTsuji,M. 1997. Single immunizing dose of recombinant adenovirus efficiently induces CD8+ T cell−mediated protective immunity against malaria.J.Immunol. 158:1268−1274.
Rodrigues,E.G., Zavala,F., Nussenzweig,R.S., Wilson,J.M. & Tsuji,M. Efficient induction of protective anti−malaria immunity by recombinant adenovirus.Vaccine 16, 1812−1817(1998).
Roelvink,P.W., Mi Lee,G., Einfeld,D.A., Kovesdi,I. & Wickham, T.J. Identification of a conserved receptor−binding site on the Fiber proteins of CAR−recognizing adenoviridae.Science (New York, N.Y 286, 1568−1571(1999).
Rux,J.J. & Burnett,R.M. Type−specific epitope locations revealed by X−ray crystallographic study of adenovirus type 5 Hexon.Mol Ther 1,18−30(2000).
Rux,J.J.& Burnett,R.M. Adenovirus structure.Human gene therapy f(C)5, 1167−1176(2004).
Roy,S.ら. Use of chimeric adenoviral vectors to assess capsid neutralization determinants.Virology 333, 207−214(2005).
Silvie,O., Greco,C, Franetich,J.F., Dubart−Kupperschmitt,A., Hannoun,L,ら 2006. Expression of human CD81 differently affects host cell susceptibility to malaria sporozoites depending on the Plasmodium species.Cell Microbiol. 8:1134−1146.
Sumida,S.M., Truitt,D.M., Lemckert,A.A., Vogels,R., Custers,J.H.,ら 2005. Neutralizing antibodies to adenovirus serotype 5 vaccine vectors are directed primarily against the adenovirus Hexon protein.J Immunol. 174:7179−7185.
Sun,P., Schwenk,R., White,K., Stoute,J.A., Cohen,J.,ら 2003. Protective immunity induced with malaria vaccine, RTS, S, is linked to Plasmodium falciparum circumsporozoite protein−specific CD4+ and CD8+ T cells producing IFN−gamma.J Immunol. 171:6961−6967.
Tao,D., Barba−Spaeth,G., Rai,U., Nussenzweig,V., Rice,CM.,およびNussenzweig,R.S. 2005. Yellow fever 17D as a vaccine vector for microbial CTL epitopes:protection in a rodent malaria model.J Exp Med. 201:201−209.
Teramoto,S.ら. Investigation of effects of anesthesia and age on aspiration in mice through LacZ gene transfer by recombinant E1 −deleted adenovirus vectors.American journal of respiratory and critical care medicine 158, 1914−1919(1998).
Top,F.H.,Jr., Dudding,B.A., Russell,P.K. & Buescher,E.L. Control of respiratory disease in recruits with types 4 and 7 adenovirus vaccines.American journal of epidemiology 94, 142−146(1971).
Top,F.H.,Jr. Control of adenovirus acute respiratory disease in U.S. Army trainees.Yale J Biol Med 48, 185−195(1975).
Tsuji,M., Romero,P., Nussenzweig,R.S.,およびZavala,F. 1990. CD4+ cytolytic T cell clone confers protection against murine malaria.J Exp Med. 172:1353−1357.
Tsuji,M., Rodrigues,E.G.& Nussenzweig,S. Progress toward a malaria vaccine:efficient induction of protective anti−malaria immunity.Biol Chem 382, 553−570(2001).
Wickham,T.J., Mathias,P., Cheresh,D.A. & Nemerow,G.R. lntegrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment.Cell 73, 309−319(1993).
Wohlfart,C. Neutralization of adenoviruses:kinetics, stoichiometry, and mechanisms.Journal of virology 62, 2321−2328(1988).
Worgall,S.ら. Modification to the capsid of the adenovirus vector that enhances dendritic cell infection and transgene−specific cellular immune responses.Journal of virology 78, 2572−2580(2004).
Worgall,S., Krause,A., Rivara,M., Hee,K.K., Vintayen,E.V.,ら 2005. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid.J Clin Invest.115:1281−1289.
Worgall,S., Krause,A., Qiu,J., Joh,J., Hackett,N.R.,およびCrystal,R.G. 2007. Protective immunity to pseudomonas aeruginosa induced with a capsid−modified adenovirus expressing P. aeruginosa OprF.J Virol. 81:13801−13808.
Wu,H., Dmitriev,I., Kashentseva,E., Seki,T., Wang,M.,ら 2002. Construction and characterization of adenovirus serotype 5 packaged by serotype 3 Hexon.J Virol. 76:12775−12782.
Wu,H.ら. Identification of sites in adenovirus Hexon for foreign peptide incorporation.Journal of virol, 78:3382−3390(2005).
Xia,D., Henry,L., Gerard,R.D. & Deisenhofer,J. Structure of the receptor binding domain of adenovirus type 5 Fiber protein.Current topics in microbiology and immunology 199(Pt1), 39−46(1995).
Yang,Y., Li,Q., Ertl,H.C. & Wilson,J.M. Cellular and humoral immune responses to viral antigens create barriers to lung−directed gene therapy with recombinant adenoviruses.Journal of virology 69, 2004−2015(1995).
Youil,R., Toner,T.J., Su,Q., Chen,M., Tang,A.,ら 2002. Hexon gene switch strategy for the generation of chimeric recombinant adenovirus.Hum Gene Ther. 13:311−320.
【配列表フリーテキスト】
【0143】
配列番号1:コドン最適化ネズミマラリア原虫スポロゾイト周囲タンパク質
配列番号2:コドン最適化熱帯熱マラリア原虫スポロゾイト周囲タンパク質
配列番号3:(QGPGAP)3−HVR1修飾型Hexonタンパク質
配列番号4:(QGPGAP)4−HVR1修飾型Hexonタンパク質
配列番号5:(QGPGAP)5−HVR1修飾型Hexonタンパク質
配列番号6:(QGPGAP)6−HVR1修飾型Hexonタンパク質
配列番号7:(QGPGAP)7−HVR1修飾型Hexonタンパク質
配列番号8:(QGPGAP)8−HVR1修飾型Hexon配列
配列番号9:(QGPGAP)9−HVR1修飾型Hexonタンパク質
配列番号10:(QGPGAP)11−HVR1修飾型Hexonタンパク質
配列番号11:(QGPGAP)12−HVR1修飾型Hexonタンパク質
配列番号12:(NANP)4−HVR1修飾型Hexonタンパク質
配列番号13:(NANP)6−HVR1修飾型Hexonタンパク質
配列番号14:(NANP)8−HVR1修飾型Hexonタンパク質
配列番号15:(NANP)10−HVR1修飾型Hexonタンパク質
配列番号16:(NANP)12−HVR1修飾型Hexonタンパク質
配列番号17:(NANP)14−HVR1修飾型Hexonタンパク質
配列番号18:(NANP)16−HVR1修飾型Hexonタンパク質
配列番号19:(NANP)18−HVR1修飾型Hexonタンパク質
配列番号20:(NANP)20−HVR1修飾型Hexonタンパク質
配列番号21:(NANP)22−HVR1修飾型Hexonタンパク質
配列番号22:(NANP)28−HVR1修飾型Hexonタンパク質
配列番号23:(QGPGAP)3−HVR5修飾型Hexonタンパク質
配列番号24:(QGPGAP)3修飾型Fiberタンパク質
配列番号25:(NANP)4修飾型Fiberタンパク質
配列番号26:ネズミマラリア原虫CD4+エピトープ修飾型pVII配列
配列番号27:熱帯熱マラリア原虫CD4+エピトープ修飾型pVII−1配列
配列番号28:熱帯熱マラリア原虫CD4+エピトープ修飾型pVII−2配列
配列番号29:熱帯熱マラリア原虫CD4+エピトープ修飾型pVII−3配列
配列番号30:合成コンストラクト
配列番号31:合成コンストラクト
配列番号32:合成コンストラクト
配列番号33:合成コンストラクト
配列番号34:合成コンストラクト
配列番号35:合成コンストラクト
配列番号36:合成コンストラクト
配列番号37:合成コンストラクト
配列番号38:合成コンストラクト
配列番号39:合成コンストラクト
配列番号40:合成コンストラクト
配列番号41:合成コンストラクト
配列番号42:合成コンストラクト
配列番号43:合成コンストラクト
配列番号44:合成コンストラクト
配列番号45:合成コンストラクト
配列番号46:合成コンストラクト
配列番号47:合成コンストラクト
配列番号48:合成コンストラクト
配列番号49:合成コンストラクト
配列番号50:合成コンストラクト
配列番号51:合成コンストラクト
配列番号52:合成コンストラクト
配列番号53:合成コンストラクト
配列番号54:合成コンストラクト
配列番号55:合成コンストラクト
配列番号56:合成コンストラクト
配列番号57:合成コンストラクト
配列番号58:合成コンストラクト
配列番号59:ネズミマラリア原虫
配列番号60:熱帯熱マラリア原虫
配列番号61:ネズミマラリア原虫
配列番号62:熱帯熱マラリア原虫
配列番号63:三日熱マラリア原虫
配列番号64:四日熱マラリア原虫
配列番号65:ネズミマラリア原虫
配列番号66:ネズミマラリア原虫
【技術分野】
【0001】
本発明は、医学および生物工学の分野に関する。より詳しくは、本発明はマラリア原虫(Plasmodium)スポロゾイト周囲タンパク質などのマラリア寄生虫抗原に対して強力な免疫応答を誘発するためのカプシド修飾型アデノウイルスベクターの使用に関する。該ベクターはマラリアに対するワクチンに適している。
優先権主張
本出願は、2009年8月18日に提出された国際出願PCT/US09/054212に対する優先権を主張し、かつ一部継続出願である。該国際出願の全体を完全に記述するように参照によって本明細書に組み入れる。
米国連邦政府による委託研究に関する記載
本発明は、米国国立衛生研究所の一部である国立アレルギー・感染症研究所(NIAID)によって与えられた助成金番号1R01A1081510−01A1の下で政府支援によってなされた。米国政府は、本発明において特定の権利を有する。
【背景技術】
【0002】
マラリアは、世界中の熱帯で最も蔓延している感染症に入る重篤な疾患である。毎年、およそ3億〜5億の人々が感染し、罹患率と死亡率が比較的高い。重度の罹患率と死亡率は、特に幼児と、これまでにマラリアに曝露されたことがないマラリア流行地に移住した成人に起こっている。世界保健機構(WHO)では、アフリカだけでマラリアによる小児の死亡数が毎年200万〜300万人と推定している。多くの国では、薬剤耐性寄生虫(熱帯熱マラリア原虫(Plasmodium falciparum)、最近では三日熱マラリア原虫(Plasmodium vivax)も)および殺虫剤耐性ベクター(ハマダラカ属(Anopheles)蚊)によってもたらされるマラリアの広範な発生および発生率の増加のため、この疾患を制御する新規の方法を開発する必要性が強調されているる(NussenzweigおよびLong 1994)。
【0003】
マラリア寄生虫は、前赤内期、赤血球期および生殖期の寄生形態からなる複雑なライフサイクルを有し、これらの形態はマラリアワクチンの開発のための潜在的標的を表す。前赤内期、赤血球期の形態は宿主内で見られるが、生殖形態はベクター内で起こる。弱毒化照射したスポロゾイト(IrSp)の生ワクチンによる免疫化は、マウスにおいて(Nussenzweigら 1967)、非ヒト霊長類(Gwadzら 1979)およびヒト((Clydeら 1973、Edelmanら 1993)において無菌の防御作用(すなわち、寄生体の攻撃に対する完全な耐性)を誘発することが示された。IrSpによって与えられる防御作用は、体液性(B細胞)および細胞性(T細胞)の両免疫応答によるスポロゾイト中和によって媒介される(Tsujiら 2001)。IrSpワクチン接種は魅力ある解決策ではあるが、スポロゾイトを採取する唯一の方法は蚊唾液腺を詳細に分析することにより、かつ大量のスポロゾイトをin vitroで増殖させる技術で現在知られているものはない。したがって、マラリアに対して同様に強い防御免疫を誘発することができる代替ワクチンのベクターが必要とされる。
【0004】
かかるワクチンベクターの有望な標的の1つは、スポロゾイト周囲(CS)タンパクであり、該タンパク質はスポロゾイトの表面に発現される。有効な中和抗体は、スポロゾイト周囲(CS)タンパク質の免疫優性で、種特異的な反復ドメインに向けられる。熱帯熱マラリア原虫(ヒトマラリア寄生虫)において、反復(NANP)nは、世界のすべての領域に由来する分離株の間で保存されている。この中心反復はB細胞エピトープの複数の反復を含み、したがってCSタンパク質はB細胞を誘発することによって強い体液性免疫反応を誘発することができる(Tsujiら 2001)。CSタンパク質のC末端領域では、いくつかのT細胞エピトープが存在し、該エピトープが細胞性免疫応答を著しく誘発することができる(Tsujiら 2001)。体液性(抗体)応答は、肝細胞に入る前にスポロゾイト(細胞外寄生体)の感染力と相互作用して、中和することによって寄生虫を除去することができるが、一方細胞性(T細胞)応答は、インターフェロンγを分泌することによってEEF(細胞内寄生体)を攻撃することができる。これらの免疫応答は、EEFが成熟し、急速に分裂して、血液に再度浸入して赤血球に感染する何千ものメロゾイトを形成して、我々がマラリアと認識する疾患を引き起こすことを防ぐ。
【0005】
現在ヒト臨床試験を受けているCSをベースにしたマラリアワクチンの1種は、GlaxoSmithKline社のRTS、S、B型肝炎表面抗原の融合タンパク質、およびウイルス様の粒子の形態での熱帯熱マラリア原虫スポロゾイト周囲タンパク質(PfCSP)の一部(1992年11月11日に、SmithKline Beecham Biologicals S.A.に対して出願された国際特許出願第PCT/EP1992/002591号)であり、臨床試験においてマラリア感染症を減少させることが示されている(Alonsoら 2004, Alonsoら 2005, Bejonら2008)。RTS、Sは、抗PfCSP体液性免疫応答を誘発するが、PfCSP特異的細胞(CD8+)応答は比較的弱い(Kesteら 2008)。これは、RTS、Sによる防御作用が比較的弱いことが理由でありうる。対照的に、アデノウイルスベースのマラリアワクチンは、防御細胞性免疫応答を誘発することができる(2003年12月16日に出願された国際特許出願第PCT/EP2003/051019号、Rodriguesら 1997)。しかし、現在のところマラリアワクチンとしてアデノウイルスベースのプラットフォームの利用を限定する障害が2つある:(1)導入遺伝子産物に対する強力な体液性免疫応答を誘発する能力の欠如と、(2)アデノウイルス、特に、アデノウイルスベースのワクチンの免疫原性を妨害するアデノウイルス血清型5に対する既存免疫との2つである。
【0006】
アデノウイルスによって誘発される体液性免疫応答を増強する試みの中で最近なされている1つのアプローチは、B細胞抗原エピトープ(たとえば、細菌エピトープまたはウイルスエピトープ)をHexon、Fiber、PentonおよびpIXなどのアデノウイルスカプシドタンパク質に挿入することである(Worgallら 2005, McConnellら 2006、Krauseら2006、Worgallら 2007)。
【0007】
加えて、アデノウイルス血清型5(Ad5)に対する既存免疫を避けるために、アデノウイルス血清型11、35、26、48、49および50などの低い血清有病率を有する他のアデノウイルス血清型が、ワクチンプラットフォームとして評価され、抗Ad5免疫が存在するにもかかわらず導入遺伝子に対する免疫応答を誘発することが示された(2005年10月12日にCrucell Holland B.V.に対して出願された国際特許出願第PCT/EP2005/055183号)。既存の抗Ad5免疫を回避するために、中和抗体の標的カプシドタンパク質であるAd5 Hexonと他の血清型のそれとの置換が構築された(Wuら 2002、Robertsら 2006)。
【0008】
しかし、アデノウイルスベクターを上述のマラリアワクチンに適用する際に同時に前述の2つの障害を克服したと報告される改良型アデノウイルスベクターはない。マラリア流行地においてAd5に対する血清有病率が高い(Ophorstら 2006)ことを考慮すると、アデノウイルスに対する既存免疫の存在下で、防御体液性免疫応答および細胞性免疫応答の双方を誘発するアデノウイルスベースのマラリアワクチンが求められている。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本開示は、組換えアデノウイルスワクチンの導入遺伝子に対する免疫応答を増強し、かつ既存の抗アデノウイルス免疫を回避するための種々のアデノウイルスタンパク質修飾に関する。
【0010】
より具体的には、一実施形態は、組換えアデノウイルスプラスミドベクターに由来する組換えアデノウイルスに関し、該組換えアデノウイルスプラスミドベクターは、(i)異種プロモーターに作用可能に連結した、マラリア原虫スポロゾイト周囲タンパク質、またはその抗原部分と、(ii)修飾型カプシドタンパク質もしくはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープはカプシドタンパク質もしくはコアタンパク質に挿入されているまたはその少なくとも一部を置換する。
【0011】
一部の実施形態では、マラリア原虫スポロゾイト周囲タンパク質は、熱帯熱マラリア原虫またはネズミマラリア原虫(Plasmodium yoelii)スポロゾイト周囲タンパク質をさらに含む。該スポロゾイト周囲タンパク質は、コドン最適化した熱帯熱マラリア原虫またはネズミマラリア原虫のスポロゾイト周囲タンパク質をさらに含むこともあり、一部の態様では、配列番号2または配列番号1それぞれののヌクレオチド配列によってコードされうる。
【0012】
別の実施形態では、該免疫原性エピトープは、マラリア原虫スポロゾイト周囲タンパク質のB細胞エピトープをさら含む。該B細胞エピトープは、修飾型カプシドタンパク質に取り込まれることもあり、かつ一部の態様では、カプシドタンパク質はHexon高頻度可変領域(HVR)を含みうる。該HVRは、HVR1またはHVR5の一部がB細胞エピトープに置換されているHVR1またはHVR5をさらに含みうる。別の態様では、カプシドタンパク質は、B細胞エピトープがFiberタンパク質に挿入されているカプシドFiberタンパク質をさらに含みうる。一部の態様では、B細胞エピトープは、熱帯熱マラリア原虫スポロゾイト周囲タンパク質B細胞エピトープであり、該B細胞エピトープが、たとえば(NANP)n(配列番号60)の反復配列であり、該反復配列が(NANP)4、(NANP)6、(NANP)8、(NANP)10、(NANP)12、(NANP)14、(NANP)16、(NANP)18、(NANP)20、(NANP)22、または(NANP)28でありうる。別の態様では、B細胞エピトープはネズミマラリア原虫スポロゾイト周囲タンパク質B細胞エピトープであり、B細胞エピトープが、たとえば(QGPGAP)n(配列番号59)の反復配列であり、該反復配列が(QGPGAP)3、(QGPGAP)4(QGPGAP)5、(QGPGAP)6、(QGPGAP)7、(QGPGAP)8、(QGPGAP)9、(QGPGAP)11、または(QGPGAP)12でありうる。
【0013】
さらに別の実施形態では、免疫原性エピトープは、マラリア原虫スポロゾイト周囲タンパク質のCD4+またはCD8+のT細胞エピトープをさらに含む。該CD4+ T細胞エピトープまたはCD8+ T細胞エピトープは、修飾型カプシドタンパク質またはコアタンパク質に取り込まれることもある。一部の態様では、該カプシドタンパク質は、Hexon高頻度可変領域(HVR)を含みうる。該HVRは、HVR1の一部がCD4+またはCD8+のT細胞エピトープに置換されているHVR1をさらに含みうる。別の態様では、該コアタンパク質は、pVIIタンパク質をさらに含み、かつCD4+ T細胞エピトープが該pVIIタンパク質に挿入されている。一部の態様では、該CD4+ T細胞エピトープは、熱帯熱マラリア原虫スポロゾイト周囲CD4+ T細胞エピトープであり、該CD4+ T細胞エピトープがEYLNKIQNSLSTEWSPCSVT(配列番号62)である。別の態様では、該CD4+ T細胞エピトープは、ネズミマラリア原虫スポロゾイト周囲CD4+ T細胞エピトープであり、該CD4+ T細胞エピトープがYNRNIVNRLLGDALNGKPEEK(配列番号61)である。
【0014】
別の実施形態は、上記の実施形態による組換えアデノウイルスを含む医薬組成物またはマラリアワクチン組成物に関する。さらなる実施形態は、上記の実施形態による該医薬組成物またはマラリアワクチン組成物の治療量を投与することを含むマラリアを治療する、予防する、または診断する方法を含む。
【0015】
別の実施形態では、治療法は、初回刺激−追加免疫のワクチン接種の投与を含み、ここで被験者は、所定の時間間隔で用量を増やしてまたは同じ用量で一連の投与を与えられる。所定の時間間隔は、体液性および/または細胞性の免疫応答を生成するのに十分な任意の長さであってよい。たとえば、後述するように、該時間間隔は、3週間ごとであってもよいが、これに限定されるものではない。
【課題を解決するための手段】
【0016】
本発明者らは、組換えアデノウイルスプラスミドベクターに由来する新しい、カプシド修飾型構造を有する新規の組換えアデノウイルスを見出した。新規の組換えアデノウイルスは哺乳類細胞を感染させ、哺乳類細胞にマラリア原虫スポロゾイト周囲タンパク質を発現させることができる。該組換えアデノウイルスは、所望の免疫原性の抗原、たとえばマラリア原虫スポロゾイト周囲タンパク質のB細胞エピトープ、T細胞エピトープを有することによって修飾された1つまたは複数のカプシドタンパク質も有する。該組換えアデノウイルスは、直線化組換えアデノウイルスプラスミドベクターで細胞をトランスフェクトする方法によって得られる。得られた組換えアデノウイルスを用いて、本発明者らは、有効成分としてマラリア感染予防効果および治療効果がある組換えアデノウイルスを含有する医薬品について広範囲の研究を行った。その結果、発明者らは、得られた組換えアデノウイルスが所望の薬学的効果を有することを見出した。
【図面の簡単な説明】
【0017】
【図1】本開示の実施形態によるカプシド修飾型組換えアデノウイルスの模式図である。
【図2】HVR1修飾型組換えアデノウイルスプラスミドベクターのHVR1修飾型アデノウイルスDNAの構築を示す模式図である。
【図3】HVR5修飾型組換えアデノウイルスプラスミドベクターのHVR5修飾型アデノウイルスDNAの構築を示す模式図である。
【図4】Fiber修飾型組換えアデノウイルスプラスミドベクターのFiber修飾型アデノウイルスDNAの構築を示す模式図である。
【図5】HVR1とFiberとの修飾型組換えアデノウイルスプラスミドベクターの、HVR1とFiberとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図6】FiberとpVIIとの修飾型組換えアデノウイルスプラスミドベクターの、FiberとpVIIとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図7】HVR1とpVIIとの修飾型組換えアデノウイルスプラスミドベクターの、HVR1とpVIIとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図8】HVR1とFiberとpVIIとの修飾型組換えアデノウイルスプラスミドベクターの、HVR1とFiberとpVIIとの修飾型アデノウイルスDNAの構築を示す模式図である。
【図9】コドン最適化ネズミマラリア原虫スポロゾイト周囲タンパク質の核酸配列(PyCS、配列番号1)および対応するアミノ酸配列(配列番号30)を示す。
【図10】コドン最適化熱帯熱マラリア原虫スポロゾイト周囲タンパク質の核酸配列(PfCSP、配列番号2)および対応するアミノ酸配列(配列番号43)を示す。
【図11−1】HVR1(配列番号3、核酸;配列番号31、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図11−2】HVR1(配列番号3、核酸;配列番号31、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図11−3】HVR1(配列番号3、核酸;配列番号31、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図12−1】HVR1(配列番号4、核酸;配列番号32、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの4の反復(配列番号59;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)4の配列は、下線で示す。
【図12−2】HVR1(配列番号4、核酸;配列番号32、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの4の反復(配列番号59;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)4の配列は、下線で示す。
【図12−3】HVR1(配列番号4、核酸;配列番号32、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの4の反復(配列番号59;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)4の配列は、下線で示す。
【図13−1】HVR1(配列番号5、核酸;配列番号33、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの5の反復(配列番号59;n=5)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)5の配列は、下線で示す。
【図13−2】HVR1(配列番号5、核酸;配列番号33、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの5の反復(配列番号59;n=5)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)5の配列は、下線で示す。
【図13−3】HVR1(配列番号5、核酸;配列番号33、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの5の反復(配列番号59;n=5)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)5の配列は、下線で示す。
【図14−1】HVR1(配列番号6、核酸;配列番号34、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの6の反復(配列番号59;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)6の配列は、下線で示す。
【図14−2】HVR1(配列番号6、核酸;配列番号34、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの6の反復(配列番号59;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)6の配列は、下線で示す。
【図14−3】HVR1(配列番号6、核酸;配列番号34、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの6の反復(配列番号59;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)6の配列は、下線で示す。
【図15−1】HVR1(配列番号7、核酸;配列番号35、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの7の反復(配列番号59;n=7)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)7の配列は、下線で示す。
【図15−2】HVR1(配列番号7、核酸;配列番号35、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの7の反復(配列番号59;n=7)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)7の配列は、下線で示す。
【図15−3】HVR1(配列番号7、核酸;配列番号35、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの7の反復(配列番号59;n=7)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)7の配列は、下線で示す。
【図16−1】HVR1(配列番号8、核酸;配列番号36、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの8の反復(配列番号59;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)8の配列は、下線で示す。
【図16−2】HVR1(配列番号8、核酸;配列番号36、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの8の反復(配列番号59;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)8の配列は、下線で示す。
【図16−3】HVR1(配列番号8、核酸;配列番号36、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの8の反復(配列番号59;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)8の配列は、下線で示す。
【図17−1】HVR1(配列番号9、核酸;配列番号37、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの9の反復(配列番号59;n=9)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)9の配列は、下線で示す。
【図17−2】HVR1(配列番号9、核酸;配列番号37、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの9の反復(配列番号59;n=9)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)9の配列は、下線で示す。
【図17−3】HVR1(配列番号9、核酸;配列番号37、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの9の反復(配列番号59;n=9)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)9の配列は、下線で示す。
【図18−1】HVR1(配列番号10、核酸;配列番号38、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの11の反復(配列番号59;n=11)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)11の配列は、下線で示す。
【図18−2】HVR1(配列番号10、核酸;配列番号38、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの11の反復(配列番号59;n=11)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)11の配列は、下線で示す。
【図18−3】HVR1(配列番号10、核酸;配列番号38、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの11の反復(配列番号59;n=11)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)11の配列は、下線で示す。
【図19−1】HVR1(配列番号11、核酸;配列番号39、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの12の反復(配列番号59;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)12の配列は、下線で示す。
【図19−2】HVR1(配列番号11、核酸;配列番号39、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの12の反復(配列番号59;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)12の配列は、下線で示す。
【図19−3】HVR1(配列番号11、核酸;配列番号39、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの12の反復(配列番号59;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)12の配列は、下線で示す。
【図20−1】HVR1(配列番号12、核酸;配列番号44、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)4の配列は、下線で示す。
【図20−2】HVR1(配列番号12、核酸;配列番号44、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)4の配列は、下線で示す。
【図20−3】HVR1(配列番号12、核酸;配列番号44、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)4の配列は、下線で示す。
【図21−1】HVR1(配列番号13、核酸;配列番号45、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの6の反復(配列番号60;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)6の配列は、下線で示す。
【図21−2】HVR1(配列番号13、核酸;配列番号45、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの6の反復(配列番号60;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)6の配列は、下線で示す。
【図21−3】HVR1(配列番号13、核酸;配列番号45、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの6の反復(配列番号60;n=6)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)6の配列は、下線で示す。
【図22−1】HVR1(配列番号14、核酸;配列番号46、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの8の反復(配列番号60;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)8の配列は、下線で示す。
【図22−2】HVR1(配列番号14、核酸;配列番号46、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの8の反復(配列番号60;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)8の配列は、下線で示す。
【図22−3】HVR1(配列番号14、核酸;配列番号46、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの8の反復(配列番号60;n=8)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)8の配列は、下線で示す。
【図23−1】HVR1(配列番号15、核酸;配列番号47、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの10の反復(配列番号60;n=10)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)10の配列は、下線で示す。
【図23−2】HVR1(配列番号15、核酸;配列番号47、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの10の反復(配列番号60;n=10)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)10の配列は、下線で示す。
【図23−3】HVR1(配列番号15、核酸;配列番号47、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの10の反復(配列番号60;n=10)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)10の配列は、下線で示す。
【図24−1】HVR1(配列番号16、核酸;配列番号48、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの12の反復(配列番号60;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)12の配列は、下線で示す。
【図24−2】HVR1(配列番号16、核酸;配列番号48、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの12の反復(配列番号60;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)12の配列は、下線で示す。
【図24−3】HVR1(配列番号16、核酸;配列番号48、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの12の反復(配列番号60;n=12)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)12の配列は、下線で示す。
【図25−1】HVR1(配列番号17、核酸;配列番号49、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの14の反復(配列番号60;n=14)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)14の配列は、下線で示す。
【図25−2】HVR1(配列番号17、核酸;配列番号49、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの14の反復(配列番号60;n=14)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)14の配列は、下線で示す。
【図25−3】HVR1(配列番号17、核酸;配列番号49、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの14の反復(配列番号60;n=14)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)14の配列は、下線で示す。
【図26−1】HVR1(配列番号18、核酸;配列番号50、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの16の反復(配列番号60;n=16)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)16の配列は、下線で示す。
【図26−2】HVR1(配列番号18、核酸;配列番号50、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの16の反復(配列番号60;n=16)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)16の配列は、下線で示す。
【図26−3】HVR1(配列番号18、核酸;配列番号50、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの16の反復(配列番号60;n=16)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)16の配列は、下線で示す。
【図27−1】HVR1(配列番号19、核酸;配列番号51、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの18の反復(配列番号60;n=18)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)18の配列は、下線で示す。
【図27−2】HVR1(配列番号19、核酸;配列番号51、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの18の反復(配列番号60;n=18)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)18の配列は、下線で示す。
【図27−3】HVR1(配列番号19、核酸;配列番号51、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの18の反復(配列番号60;n=18)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)18の配列は、下線で示す。
【図28−1】HVR1(配列番号20、核酸;配列番号52、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの20の反復(配列番号60;n=20)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)20の配列は、下線で示す。
【図28−2】HVR1(配列番号20、核酸;配列番号52、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの20の反復(配列番号60;n=20)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)20の配列は、下線で示す。
【図28−3】HVR1(配列番号20、核酸;配列番号52、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの20の反復(配列番号60;n=20)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)20の配列は、下線で示す。
【図29−1】HVR1(配列番号21、核酸;配列番号53、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの22の反復(配列番号60;n=22)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)22の配列は、下線で示す。
【図29−2】HVR1(配列番号21、核酸;配列番号53、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの22の反復(配列番号60;n=22)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)22の配列は、下線で示す。
【図29−3】HVR1(配列番号21、核酸;配列番号53、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの22の反復(配列番号60;n=22)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)22の配列は、下線で示す。
【図30−1】HVR1(配列番号22、核酸;配列番号54、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの28の反復(配列番号60;n=28)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)28の配列は、下線で示す。
【図30−2】HVR1(配列番号22、核酸;配列番号54、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの28の反復(配列番号60;n=28)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)28の配列は、下線で示す。
【図30−3】HVR1(配列番号22、核酸;配列番号54、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの28の反復(配列番号60;n=28)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(NANP)28の配列は、下線で示す。
【図31−1】HVR5(配列番号23、核酸;配列番号40、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図31−2】HVR5(配列番号23、核酸;配列番号40、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図31−3】HVR5(配列番号23、核酸;配列番号40、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Hexonの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図32−1】Fiber(配列番号24、核酸;配列番号41、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図32−2】Fiber(配列番号24、核酸;配列番号41、アミノ酸)においてPyCS B細胞エピトープ配列(QGPGAP)nの3の反復(配列番号59;n=3)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(QGPGAP)3の配列は、下線で示す。
【図33−1】Fiber(配列番号25、核酸;配列番号55、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(NANP)3の配列は、下線で示す。
【図33−2】Fiber(配列番号25、核酸;配列番号55、アミノ酸)においてPfCSP B細胞エピトープ配列(NANP)nの4の反復(配列番号60;n=4)を有する修飾型Fiberの核酸配列およびアミノ酸配列を示す。挿入された(NANP)3の配列は、下線で示す。
【図34】N末端pVII(配列番号26、核酸;配列番号42、アミノ酸)においてPyCS CD4+エピトープ配列YNRNIVNRLLGDALNGKPEEK(配列番号61)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたYNRNIVNRLLGDALNGKPEEKの配列は、下線で示す。
【図35】C末端pVII(pVII−1;配列番号27、核酸;配列番号56、アミノ酸)においてPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたEYLNKIQNSLSTEWSPCSVT配列は、下線で示す。
【図36】pVII(pVII−2;配列番号28、核酸;配列番号57、アミノ酸)の第1の核移行シグナル(NLS)の前にPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたEYLNKIQNSLSTEWSPCSVT配列は、下線で示す。
【図37】pVII(pVII−3;配列番号29、核酸;配列番号58、アミノ酸)の2つNLSの間にPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)を有する修飾型pVIIの核酸配列およびアミノ酸配列を示す。挿入されたEYLNKIQNSLSTEWSPCSVT配列は、下線で示す。
【図38】PyCS−GFP/pShuttle−CMVによる一過性トランスフェクションの後、AD293細胞におけるPyCSタンパク質発現を示す。PyCSタンパク質は、マウスモノクローナル抗PyCS抗体(9D3)を用いるウエスタンブロット法によって検出された。
【図39】(QGPGAP)3エピトープ(配列番号59;n=3)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPyCS−GFPアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。ELISAアッセイでは、ELISAプレートは精製アデノウイルスで直接コーティングされ、次いでアデノウイルス粒子に挿入されたエピトープは抗PyCS抗体で検出された。
【図40】(QGPGAP)nエピトープ(配列番号59)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPyCSアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。ELISAアッセイでは、ELISAプレートは精製アデノウイルスで直接コーティングされ、次いでアデノウイルス粒子に挿入されたエピトープは抗PyCS抗体で検出された。
【図41】(QGPGAP)n反復(配列番号59、n=3、4、5、6、9、12)を有するカプシド修飾型PyCSアデノウイルスによる単回免疫レジメン(A)と、免疫化から2週間後のPyCS特異的CD8+応答(B)とを示す。
【図42】(QGPGAP)n反復(配列番号59、n=3)を有するカプシド修飾型PyCSアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、10週目のPyCS特異的体液性免疫応答(B)と、スポロゾイト抗原投与から42時間後の肝臓でのマラリア寄生虫の負荷(C)とを示す。
【図43】図42でのレジメンを与えられたマウスから調製され、プールした血清試料についての間接的免疫蛍光アッセイ(IFA)によって測定された抗スポロゾイト抗体価(A)と、in vitroスポロゾイト中和活性(B)とを示す。
【図44】HVR1において(QGPGAP)n反復(配列番号59、n=4、6)を有するカプシド修飾型PyCSアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPyCS特異的体液性免疫応答(B)と、スポロゾイト抗原投与から42時間後の肝臓でのマラリア寄生虫負荷(C)と、プールした血清「試料のin vitroスポロゾイト中和活性(D)とを示す。マウスにアジュバントと共に、またはアジュバントを含まずに免疫した。
【図45】HVR1において(QGPGAP)n反復(配列番号59、n=6、9、12)を有するカプシド修飾型PyCSアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPyCS特異的体液性免疫応答(B)と、9週目のPyCS特異的CD8+ T細胞応答(C)と、スポロゾイト抗原投与から42時間後の肝臓でのマラリア寄生虫負荷(D)とを示す。マウスにアジュバントと共に、またはアジュバントを含まずに免疫した。
【図46】PfCSP/pShuttle−CMVによる一過性トランスフェクションの後のAD293細胞におけるPfCSPタンパク質の発現を示す。PfCSPは、マウスモノクローナル抗NANP抗体(2A10)を用いるウエスタンブロット法によって検出された。
【図47】(NANP)4エピトープ(配列番号60、n=4)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPfCSPアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。挿入された(NANP)4のエピトープ(配列番号60、n=4)は、マウスモノクローナル抗NANP抗体(2A10)によって検出された。ELISAアッセイでは、ELISAプレートは、精製アデノウイルスで直接コーティングされた。
【図48】(NANP)nエピトープ(配列番号60、n=4、6、8、10、12、14、16、18、20、22)のアデノウイルウスカプシドタンパク質への挿入を確認するために、精製したカプシド修飾型組換えPfCSPアデノウイルスの銀染色とウエスタンブロット法(A)およびELISAアッセイ(B)による結果を示す。ELISAアッセイでは、ELISAプレートは、精製アデノウイルスで直接コーティングされ、次いでアデノウイルス粒子に挿入されたエピトープは抗PfCSP抗体(2A10)で検出された。
【図49】(NANP)4(配列番号60、n=4)を有するカプシド修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPfCSP特異的体液性免疫応答(B)とを示す。
【図50】HVR1において(NANP)n(配列番号60、n=4、6、8、10)を有するカプシド修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPfCSP特異的体液性免疫応答(B)とを示す。
【図51】HVR1において(NANP)n(配列番号60、n=10、16、22)を有するカプシド修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、9週目のPfCSP特異的体液性免疫応答(B)とを示す。マウスにアジュバントと共に、またはアジュバントを含まずに免疫した。
【図52】精製した(QGPGAP)修飾型FiberおよびpVII−1((QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFP)のアデノウイルスの銀染色分析(A)の結果と、図49に記述したように(QGPGAP)3−Fib/PyCS−GFPまたは(QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFPで免疫したマウスにおいて10週目の抗QGPGAP抗体価(B)とを示す。2つの独立した実験の結果を、B−Fib/PyCS−GFPで免疫した群において抗体価の中央値で規準化した後、図にプロットした。
【図53】pVII(PfCD4−pVII−2;配列番号28、核酸;配列番号57、アミノ酸)の第1の核移行シグナル(NLS)の前に、およびpVII(PfCD4−pVII−3;配列番号29、核酸;配列番号58、アミノ酸)の2つのNLSの間に挿入されたPfCSP CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)(A)と、pVIIへのエピトープの挿入を確認するための銀染色の結果(B)とによるアデノウイルスpVIIタンパク質の構造の模式図を示す。
【図54】HVR1およびpVII修飾型組換えPfCSPアデノウイルスによる初回刺激および追加免疫の免疫レジメン(A)と、6週目のPfCSP特異的体液性免疫応答(B)と、9週目のPfCSP特異的CD4+(EYLNKIQNSLSTEWSPCSVT;配列番号62)の応答(C)とを示す。
【図55】ヒト血清試料による組換えアデノウイルスのin vitro組換え中和を示す。希釈ヒト血清の存在下でAD293細胞を組換えアデノウイルスに終夜感染させ、次いでGFP発現を感染のマーカーとして測定した。
【図56】抗アデノウイルス免疫の、カプシド修飾型PyCS−GFPアデノウイルスin vivoによるPyCS特異的T細胞応答の誘発への影響を示す。(A)は、研究デザインの簡単な説明である。(B)は、野生型(wt)/空のアデノウイルスで2回免疫して、続いてカプシド修飾型PyCS−GFPアデノウイルスで初回刺激したマウスにおけるPyCS特異的CD8+ T細胞応答を示す。
【図57】抗アデノウイルス免疫の、カプシド修飾型PyCS−GFPアデノウイルスin vivoによるPyCS特異的体液性免疫応答の誘発への影響を示す。(A)は、研究デザインの簡単な説明である。(B)は、野生型(wt)/空のアデノウイルスで2回、続いて2用量のカプシド修飾型PyCS−GFPアデノウイルスで免疫したマウスにおけるPyCS特異的体液性免疫応答を示す。
【発明を実施するための形態】
【0018】
以下の記述は、本開示の記述、実施形態を十分に理解するため、および可能にするための具体的な詳細を提供する。しかし、当業者は、該開示がこれらの詳細なしでも実行されうることを理解するであろう。別の例では、周知の構造および機能は、本開示の実施形態の説明を不必要に不明瞭にしないように詳細に示されていない、または記述されていない。
【0019】
本開示においてアミノ酸、ペプチド、塩基配列、および核酸のために使用された略語は、IUPAC−IUB Communication on Biochemical Nomenclature, Eur. J. Biochem., 138:9(1984), ”Guideline for Preparing Specifications Including Base Sequences and Amino Acid Sequences”(米国特許商標庁)に特定された略語に基づいており、かつ本技術分野で一般に使用されているものである。
【0020】
本開示によって意図される「ヌクレオチド配列」、「ポリヌクレオチド」または「DNA分子」は、二本鎖DNAまたは一本鎖DNA(すなわち、二本鎖DNAを構成するセンス鎖およびアンチセンス鎖)を含みうるが、その完全長に限定されるものではない。たとえば本明細書で以下に開示する免疫原性外来遺伝子をコードするヌクレオチド配列は、特に明記しない限り、ゲノムDNAを含む二本鎖DNA;cDNAを含む一本鎖DNA(センス鎖);センス鎖、合成DNA、およびその断片に相補的な配列を有する一本鎖DNA(アンチセンス鎖)を包含する。
【0021】
本明細書で用いる場合、ヌクレオチド配列、ポリヌクレオチドまたはDNA分子は、機能的な領域に限定されるものではなく、かつ発現抑制領域、コード領域、リーダー配列、エキソンおよびイントロンのうちの少なくとも1つを含みうる。さらに、ヌクレオチド配列またはポリヌクレオチドの例としては、RNAまたはDNAを含みうる。特異的アミノ酸配列を含むポリペプチドおよび特異的DNA配列を含むポリヌクレオチドは、該ポリヌクレオチドの断片、相同体、誘導体および変異体を含みうる。ヌクレオチド配列またはポリヌクレオチドの変異体の例(たとえば変異体DNA)としては、天然に存在する対立遺伝子変異体;人工変異体;および欠失、置換、追加、および/または挿入を有する変異体が挙げられる。かかる変異体は、元の非変異型ポリペプチドによってコードされるポリペプチドと同じ機能を実質的に有するポリペプチドをコードすることを理解すべきである。
【0022】
本開示は、マラリア原虫の抗原決定基を発現させることができ、および1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質を含む組換えアデノウイルスに関する。組換えアデノウイルスは、組換えアデノウイルスプラスミドベクターに由来し、この産生は以下の実施例で記述される。ベクターとしてのアデノウイルスの使用を、さらに以下で考察する。本明細書に記述する組換えアデノウイルスプラスミドベクターは、マラリア原虫に対する体液性および細胞性の両免疫応答が誘発されるマラリアワクチンまたは医薬組成物として用いられてもよい。
【0023】
マラリア原虫は、既知のマラリア原虫(P.)種、たとえば、熱帯熱マラリア原虫、四日熱マラリア原虫(P.malariae)、卵形マラリア原虫(P.ovale)、三日熱マラリア原虫、二日熱マラリア原虫(P.knowlesi)、マウスマラリア原虫(P.berghei)、ネズミマラリア原虫(P.chabaudi)およびネズミマラリア原虫(P.yoelii)のいずれかから選択されうる。一部の実施形態では、抗原決定基は齧歯動物特異的ネズミマラリア原虫またはヒト特異的熱帯熱マラリア原虫に由来する。
【0024】
一実施形態では、組換えアデノウイルスカプシド修飾型プラスミドベクター(本明細書では組換えアデノウイルスプラスミドベクターとも記述される)は、カプシドをコードして、生成するプラスミドであり、および/または1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質を含む構造を有するコア修飾型組換えアデノウイルスである。本開示の実施形態によれば、カプシドタンパク質および/またはコアタンパク質の修飾は、マラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープを挿入することによって達成されうる。あるいは、カプシドタンパク質および/またはコアタンパク質の少なくとも一部は、除去されてマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープと置換されうる。一部の実施形態では、該免疫原性エピトープは、マラリア原虫スポロゾイト周囲タンパク質のB細胞および/またはT細胞のエピトープである。B細胞またはT細胞のピトープを追加することで、CSタンパク質に対する液体性免疫応答を確立する、または増強することによってマラリアワクチンとして用いられるアデノウイルスベクターの有効性を増強するのに役立ちうる。修飾型カプシドタンパク質と、コアタンパク質と、本明細書に記述する組換えアデノウイルスでのこれらのタンパク質の使用に関するこれらの有意性とを以下でさらに考察する。
【0025】
1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質は、修飾型Hexonタンパク質、修飾型Fiberタンパク質、修飾型pVIIタンパク質またはそれらの組み合わせであってもよい。一実施形態では、Hexon高頻度可変領域(HVR)の一部および/またはFiberタンパク質の一部は、マラリア原虫スポロゾイト周囲タンパク質の少なくとも1つのB細胞および/またはT細胞のエピトープと置換される。あるいは、マラリア原虫スポロゾイト周囲タンパク質の1つもしくは複数のB細胞および/またはT細胞のエピトープは、Fiberタンパク質またはHexonのHVRに挿入されうる。一部の態様では、修飾型HVRは、HVR1、HVR2、HVR3、HVR4、HVR5、HVR6またはHVR7でありうる。別の態様では、修飾型HVRは、HVR1またはHVR5でありうる。一部の実施形態では、HVR修飾型Hexonは、以下の配列番号の核酸配列を有すこともありうる:配列番号3(図11−1〜図11−3)、配列番号4(図12−1〜図12−3)、配列番号5(図13−1〜図13−3)、配列番号6(図14−1〜図14−3)、配列番号7(図15−1〜図15−3)、配列番号8(図16−1〜図16−3)、配列番号9(図17−1〜図17−3)、配列番号10(図18−1〜図18−3)、配列番号11(図19−1〜図19−3)、配列番号12(図20−1〜図20−3)、配列番号13(図21−1〜図21−3)、配列番号14(図22−1〜図22−3)、配列番号15(図23−1〜図23−3)、配列番号16(図24−1〜図24−3)、配列番号17(図25−1〜図25−3)、配列番号18(図26−1〜図26−3)、配列番号19(図27−1〜図27−3)、配列番号20(図28−1〜図28−3)、配列番号21(図29−1〜図29−3)、配列番号22(図30−1〜図30−3)、または配列番号23(図31−1〜図31−3)。別の実施形態では、修飾型Fiberタンパク質は、配列番号24(図32−1〜図32−2)または配列番号25(図33−1〜図33−2)の核酸配列を有すこともありうる。
【0026】
別の実施形態では、マラリア原虫スポロゾイト周囲タンパク質のT細胞エピトープは、以下のいずれかの部位で、アデノウイルスコアpVIIタンパク質に挿入されうる:C末端、第1の核移行シグナル(NLS)の前、または2つのNLSの間。あるいは、マラリア原虫スポロゾイト周囲タンパク質のT細胞エピトープは、pVIIタンパク質の一部を置換しうる。一部の実施形態では、修飾型pVIIタンパク質は、配列番号26(図34)、配列番号27(図35)、配列番号28(図36)、または配列番号29(図37)の核酸配列を有することもある。
【0027】
組換えアデノウイルスでは、トランスジェニックタンパク質または組換えトランスジェニックタンパク質が発現しうる。一部の実施形態では、トランスジェニックタンパク質または組換えトランスジェニックタンパク質は、本明細書に記述する組換えアデノウイルスプラスミドベクターによってコードされるマラリア原虫スポロゾイト周囲タンパク質または抗原決定基であり、かつ1つまたは複数の宿主細胞の感染の後、前述の組換えアデノウイルスプラスミドベクターによって生成される組換えアデノウイルスによって発現される。
【0028】
したがって、一部の実施形態では、組換えアデノウイルスプラスミドベクターは、組換えトランスジェニックタンパク質をコードするヌクレオチド配列を含む。一実施形態では、組換えトランスジェニックタンパク質は、齧歯類特異的寄生虫であるネズミマラリア原虫の抗原決定基を含むこともあり、該抗原決定基が、ネズミマラリア原虫スポロゾイト周囲(CS)タンパク質遺伝子またはその抗原部分を含む。別の実施例形態では、組換えトランスジェニックタンパク質は、ヒト特異的寄生虫である熱帯熱マラリア原虫の抗原決定基を含むこともあり、該抗原決定基が熱帯熱マラリア原虫スポロゾイト周囲遺伝子(CS)タンパク質またはその抗原部分を含む。蚊媒介感染症に対するヒトにおける能動免疫の基礎として用いられるとき、熱帯熱マラリア原虫CSタンパク質は、マラリアの予防を示した。抗原決定基は、B細胞エピトープおよび/またはT細胞エピトープなどの免疫原性エピトープをさらに含みうる。
【0029】
一部の実施形態では、CSタンパク質は、被験者において発現の増強のためにコドン最適化される。コドン最適化は、必要なアミノ酸含有量、関心の対象において一般的な最適コドンの使用、ならびに適切な発現を確実にするために回避されるべきいずれの態様に基づく。かかる態様は、スプライス供与部位または受容部位、終止コドン、ポリアデニル化(pA)シグナル、GCリッチ配列とATリッチ配列、内部TATAボックス、または当技術分野で既知の他のいかなる態様でありうる。一部の実施形態では、コドン最適化CS導入遺伝子のDNA配列は、図9(配列番号1、ネズミマラリア原虫)および図10(配列番号2、熱帯熱マラリア原虫)に示される。
【0030】
一部の実施形態では、組換えアデノウイルスプラスミドベクターは、以下の修飾型熱帯熱マラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つでありうる:(NANP)n(配列番号60)のB細胞エピトープコード配列を用いて、図2に示すように構築されたHVR1修飾型アデノウイルスベクター(NANP−HVR1/PfCSP);(NANP)n(配列番号60)のB細胞エピトープコード配列を用いて、図4に示すように構築されたFiber修飾型アデノウイルスベクター(NANP−Fib/PfCSP);(NANP)n(配列番号60)のB細胞エピトープコード配列を用いて、図5に示すように構築されたHVR1とFiberとの修飾型アデノウイルスベクター(NANP−HVR1/B−Fib/PfCSP);(NANP)n(配列番号60)のB細胞エピトープコード配列およびEYLNKIQNSLSTEWSPCSVT(配列番号62)のCD4エピトープコード配列を用いて、図7に示すように構築されたHVR1とpVIIとの修飾型アデノウイルスベクター(NANP−HVR1/CD4−pVII/PfCSP);(NANP)n(配列番号60)のB細胞エピトープおよびEYLNKIQNSLSTEWSPCSVT(配列番号62)のCD4エピトープコード配列を用いて、図6に示すように構築されたFiberとpVIIとの修飾型アデノウイルスベクター(NANP−Fib/CD4−pVII/PfCSP);ならびに(NANP)n(配列番号60)のB細胞エピトープおよびEYLNKIQNSLSTEWSPCSVT(配列番号62)のCD4エピトープコード配列を用いて、図8に示すように構築されたHVR1とFiberとpVIIとの修飾型アデノウイルスベクター(NANP−HVR1/Fib/CD4−pVII/PfCSP)。
【0031】
別の実施形態では、組換えアデノウイルスプラスミドベクターは、以下の修飾型ネズミマラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つでありうる:(QGPGAP)n(配列番号59)のB細胞エピトープコード配列を用いて、図2に示すように構築されたHVR1修飾型アデノウイルスベクター(QGPGAP−HVR1/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープコード配列を用いて、図4に示すように構築されたFiber修飾型アデノウイルスベクター(QGPGAP−Fib/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープコード配列を用いて、図5に示すように構築されたHVR1とFiberとの修飾型アデノウイルスベクター(QGPGAP−HVR1/B−Fib/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープおよびYNRNIVNRLLGDALNGKPEEK(配列番号61)のCD4エピトープコード配列を用いて、図7に示すように構築されたHVR1とpVIIとの修飾型アデノウイルスベクター(QGPGAP−HVR1/CD4−pVII/PyCS);(QGPGAP)n(配列番号59)のB細胞エピトープおよびYNRNIVNRLLGDALNGKPEEK(配列番号61)のCD4エピトープコード配列を用いて、図6に示すように構築されたFiber1とpVIIとの修飾型アデノウイルスベクター(QGPGAP−Fib/CD4−pVII/PyCS);ならびに(QGPGAP)n(配列番号59)のB細胞エピトープおよびYNRNIVNRLLGDALNGKPEEK(配列番号61)のCD4エピトープコード配列を用いて、図8に示すように構築されたHVR1とFiberとpVIIとの修飾型アデノウイルスベクター(QGPGAP−HVR1/Fib/CD4−pVII/PfCSP)。
【0032】
別の実施形態では、組換えアデノウイルスは、以下の修飾型熱帯熱マラリア原虫またはネズミマラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つによって生成されうる:NANP−HVR1/PfCSPまたはQGPGAP−HVR1/PyCS(図2)、NANP−Fib/PfCSPまたはQGPGAP−Fib/PyCS(図4)、NANP−HVR1/B−Fib/PfCSPまたはQGPGAP−HVR1/B−Fib/PyCS(図5)、NANP−HVR1/CD4−pVII/PfCSPまたはQGPGAP−HVR1/CD4−pVII/PyCS(図7)、NANP−Fib/CD4−pVII/PfCSPまたはQGPGAP−Fib/CD4−pVII/PyCS(図6)、(NANP−HVR1/Fib/CD4−pVII/PfCSPまたはQGPGAP−HVR1/Fib/CD4−pVII/PyCS(図8))。組換えアデノウイルスは、哺乳類宿主細胞で組換えトランスジェニックタンパク質(たとえばマラリア原虫CSタンパク質)を発現させる能力を有する組換えアデノウイルスプラスミドベクターを生成するための本明細書に記述する方法によって生成されうる。
【0033】
組換えアデノウイルスの精製は、既知のウイルス精製方法を用いて行われてもよい。たとえば、精製する0.5〜1.0mLのストックウイルスは、AD293細胞などの昆虫細胞を播種する(1×107細胞/10cmシャーレ)ことによる組換えアデノウイスタンパク質を生成する方法によって得た。次いで、感染から数日後に培養上清を収集し、次いで遠心によって得たウイルスペレットをPBS(リン酸緩衝食塩水)で験濁する。得られた懸濁液を10〜60%のショ糖密度勾配にかけ、次いで遠心させて(4℃で、25,000rpmで60分間)、ウイルスバンドを収集する。収集したウイルスをPBSでさらに験濁させ、続いて上述と同じ条件下で遠心する。得られた精製組換えウイルスペレットをPBSなどの緩衝液中、4℃で貯蔵する。
【0034】
別の実施形態は、基本的に少なくとも1つの有効成分を含む医薬組成物に関する。一実施形態では、該医薬組成物の有効成分は、本明細書に記述する遺伝子工学技術によって得られうる組換えアデノウイルスを含みうる。より具体的には、該有効成分は、Hexon高頻度可変領域(HVR)の一部、Fiberタンパク質の一部、pVIIタンパク質の一部、またはそれらの組み合わせがマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープと置換されている、修飾型カプシドタンパク質および/またはコアタンパク質を含む組換えアデノウイルでありうる。あるいは、マラリア原虫スポロゾイト周囲タンパク質の1つもしくは複数のB細胞エピトープおよび/またはT細胞エピトープが、Fiberタンパク質、Hexon HVRまたはpVIIタンパク質に挿入されうる。組換えアデノウイルスプラスミドベクターは、1つもしくは複数の宿主細胞に感染させた後に組換えアデノウイルスによって発現されるトランスジェニックタンパク質または組換えトランスジェニックタンパク質をさらに含む。トランスジェニックタンパク質または組換えトランスジェニックタンパク質は、マラリア原虫スポロゾイト周囲タンパク質またはマラリア原虫スポロゾイト周囲タンパク質のマラリア抗原でありえ、ここで該マラリア抗原はマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープ(たとえばB細胞エピトープまたはT細胞エピトープ)を含む。
【0035】
一部の実施形態では、医薬組成物の有効成分は、組換えアデノウイルスプラスミドベクターに由来する組換えアデノウイルスプラスミドベクターであり、該組換えアデノウイルスプラスミドベクターは、以下の修飾型熱帯熱マラリア原虫またはネズミマラリア原虫組換えアデノウイルスプラスミドベクターのうちの1つである:NANP−HVFM/PfCSPまたはQGPGAP−HVFM/PyCS(図2)、NANP−Fib/PfCSPまたはQGPGAP−Fib/PyCS(図4)、NANP−HVR1/B−Fib/PfCSPまたはQGPGAP−HVR1/B−Fib/PyCS(図5)、NANP−HVR1/CD4−pVII/PfCSPまたはQGPGAP−HVR1/CD4−pVII/PyCS(図7)、NANP−Fib/CD4−pVII/PfCSPまたはQGPGAP−Fib/CD4−pVII/PyCS(図6)、(NANP−HVR1/Fib/CD4−pVII/PfCSPまたはQGPGAP−HVR1/Fib/CD4−pVII/PyCS(図8)。これらの組換えアデノウイルスプラスミドベクターは、細胞(たとえばAD293細胞)にトランスフェクトされると、組換えアデノウイルスを生成することができ、かつ該組換えトランスジェニックタンパク質がヒト細胞を含む哺乳類細胞で発現されうる。
【0036】
下記の実施例でさらに説明するように、有効成分を含む医薬組成物は、本明細書に記述する組換えアデノウイルスであり、被験者に投与されると、マラリア感染性抗原に対するマラリア感染症予防効果を増強し、感染価を低下させる。したがって、組換えアデノウイルスは、標的細胞および組織の感染に付随したマラリア感染症の治療で用いられうる。かかるマラリア感染症に影響を受ける標的細胞の例としては、血球、肝細胞、腎細胞、脳細胞、肺細胞、上皮細胞、および筋細胞が挙げられる。かかる細胞を含む組織の例としては、肺、肝臓、腎臓、脳、動脈と静脈、胃、腸、尿道、皮膚、および筋肉が挙げられる。
【0037】
一部の態様では、医薬組成物は、感染抗原、たとえば、マラリア寄生虫のスポロゾイト表面抗原(スポロゾイト周囲タンパク質(CSP)およびトロンボスポンジン関連接着タンパク質(TRAP))などのマラリア抗原、メロゾイト表面膜タンパク質(MSPI)、マラリアに感染した赤血球から分泌されるマラリアスポロゾイト抗原、およびマラリアに感染した赤血球のノブ内に存在する熱帯熱マラリア原虫赤血球膜タンパク質−1(PfMP1)に対するマラリア感染症予防効果を増強しうる。医薬組成物は、いずれの既知のマラリア原虫種、たとえば、熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫、二日熱マラリア原虫、マウスマラリア原虫、ネズミマラリア原虫(P.chabaudi)およびネズミマラリア原虫(P.yoelii)から選択されるマラリア原虫に対して、感染価を低下させることによってマラリア感染症予防効果を増強しうる。被験者に投与されるとき、医薬組成物による感染価の低下は、該医薬組成物を投与されなかった被験者と比較すると、生存、無病生存、または無感染生存期間と生存、無病生存、または無感染生存率の増加をもたらしうる。したがって、一部の態様では、医薬組成物は、マラリア原虫などの病原体によってもたらされるマラリア感染症のための予防薬または治療薬として有用である。さらなる態様では、医薬組成物は、マラリア原虫などの病原体によってもたらされるマラリア感染症に起因する合併症のための予防薬または治療薬として有用である。
【0038】
被験者における本発明の組換えアデノウイルスの感染症予防効果は、たとえば、本発明のカプシド修飾型組換えアデノウイルスと、医薬投与用の添加剤類とを含有する医薬組成物を、脊椎動物、特にヒトを含む哺乳類に、筋肉内(i.m.)、皮下(s.c.)、皮内(i.c.)、皮内(i.d.)、腹腔内(i.p.)、経鼻、または呼吸経路によって投与し、次いで有効成分として本明細書に記述する組換えアデノウイルスを含有する医薬組成物で脊椎動物に数回免疫することによって、もたらされうる。感染症予防効果を評価するために、該医薬組成物で数回免疫され、続いて標的病原体(たとえば選択されたマラリア原虫種)により感染した被験者の生存率、無病生存、または無感染生存を、該医薬組成物を投与されなかって被験者の生存率、無病生存、または無感染生存と比較してもよい。
【0039】
一部の実施形態では、医薬組成物は、本明細書に記述する薬学的に有効量のカプシドまたはコア修飾型組換えアデノウイルスと、以下にさらに記述する薬学的に許容される担体とをさらに含みうる。
【0040】
別の実施形態は、少なくとも1つの有効成分を基本的に含むワクチン組成物に関する。一実施形態では、該ワクチン組成物の有効成分は、本明細書に記述する組換えアデノウイルスプラスミドベクターに由来する組換えアデノウイルスを含みうる。より具体的には、該有効成分は、Hexon高頻度可変領域(HVR)の一部、Fiberタンパク質の一部、pVIIタンパク質の一部、またはそれらの組み合わせがマラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープと置換されている、修飾型カプシドタンパク質および/またはコアタンパク質を含む組換えアデノウイルでありうる。あるいは、マラリア原虫スポロゾイト周囲タンパク質の少なくとも1つの免疫原性エピトープが、pV11タンパク質、Fiberタンパク質もしくはHexon HVR、またはそれらの組み合わせに挿入されうる。一部の実施形態では、ワクチン組成物の有効成分は、図2〜8に示した組換えアデノウイルスプラスミドベクター、たとえば、NANP−HVR1/PfCSPまたはQGPGAP−HVR1/PyCS(図2)、NANP−Fib/PfCSPまたはQGPGAP−Fib/PyCS(図4)、NANP−HVR1/B−Fib/PfCSPまたはQGPGAP−HVR1/B−Fib/PyCS(図5)、NANP−HVR1/CD4−pVII/PfCSPまたはQGPGAP−HVR1/CD4−pVII/PyCS(図7)、NANP−Fib/CD4−pVII/PfCSPまたはQGPGAP−Fib/CD4−pVII/PyCS(図6)、NANP−HVR1/Fib/CD4−pVII/PfCSPまたはQGPGAP−HVR1/Fib/CD4−pVII/PyCS(図8)に由来してもよい。
【0041】
一部の態様では、被験者に投与される場合、ワクチン組成物は、カプシドタンパク質またはコアタンパク質の少なくとも一部に挿入されたまたは該一部を置換する、マラリア原虫CSタンパク質(すなわち、B細胞エピトープ、T細胞エピトープまたは両方)の1つまたは複数の抗原部分を有する組換えアデノウイルスをまず含んでいる。次いで、該ワクチン組成物は、組換えトランスジェニックタンパク質がB細胞エピトープ、T細胞エピトープまたは両方を含むマラリア原虫CSタンパク質である組換えトランスジェニックタンパク質を発現しうる。マラリア原虫CSタンパク質の抗原部分は、組換えトランスジェニックタンパク質に見出され、および下記の実施例に記述するように、修飾型カプシドタンパク質またはコアタンパク質は後天性の体液性免疫、細胞性免疫または両方を促進する、または増強する。このように、一部の態様では、本明細書に記述する組換えアデノウイルスは、体液性免疫、細胞性免疫または両方を促進する、または増強するワクチンとして有用である。
【0042】
一部の態様では、ワクチン組成物は、感染抗原、たとえば、マラリア寄生虫のスポロゾイト表面抗原(CSPおよびTRAP)などのマラリア抗原、メロゾイト表面膜タンパク質(MSPI)、マラリアに感染した赤血球から分泌されるマラリアスポロゾイト抗原、マラリアに感染した赤血球のノブ内に存在するPfEMP1、セリンリッチ抗原(SERA)タンパク質、チロシンリッチ酸性マトリクスタンパク質(TRAMP)、および頂端膜抗原−1(AMA1)に対するマラリア感染症予防効果を増強しうる。さらに、本明細書に記述するワクチン組成物の投与によって生ずる感染価(たとえばウイルス感染価)の低下は、該ワクチン組成物を投与されなかった被験者と比較すると、生存増加、無病生存期間もしくは無感染生存期間と生存、無病生存率もしくは無感染生存率をもたらしうる。したがって、一部の態様では、該ワクチン組成物は、マラリア原虫などの病原体によってもたらされるマラリア感染症のための予防薬または治療薬としても有用である。さらなる態様では、該ワクチン組成物は、マラリア原虫などの病原体によるマラリア感染症に起因する合併症のための予防薬または治療薬としても有用もある。
【0043】
本明細書に記述するワクチン組成物は、本明細書に記述する治療有効量の組換えアデノウイルスを含むこともあり、かつ標準方法によって薬学的に許容される担体をさらに含む。許容される担体の例としては、生理学的に許容される溶液、たとえば無菌生理食塩水および無菌緩衝生理食塩水が含まれる。
【0044】
一部の実施形態では、ワクチン組成物または医薬組成物を薬学的に有効量のアジュバントと組み合わせて用いて、抗マラリア効果を増強しうる。免疫系を刺激して、ワクチンに対する応答を増加させうるが、それ自体は何ら特定の抗原効果がない、任意の免疫アジュバントはアジュバントとして用いられうる。多くの免疫アジュバントは、病原体関連分子パターン(PAMP)として知られている保存分子を進化的に模倣して、トール様受容体(TLR)として知られている一連の免疫受容体によって認識される。本明細書に記述する実施形態によって用いられるうるアジュバントの例としては、フロイント完全アジュバント;フロイント不完全アジュバント;二本鎖RNA(TLR3リガンド);LPS;モノホスホリルリピドA(MPL)(TLR4リガンド)などのLPS類似体;フラゲリン(TLR5リガンド);リポプロテイン;リポペプチド;一本鎖RNA;一本鎖DNA;イミダゾキノリン類似体(TLR7およびTLRのリガンド);CpG DNA(TLR9リガンド);Ribiアジュバント(モノホスホリルリピドA/トレハロースジコリノミコラート);糖脂質(α−GalCer類似体);非メチル化CpGアイランド;油乳剤;リポソーム;ビロゾーム;サポニン類(QS21などのサポニンの活性画分);ムラミールジペプチド;ミョウバン;水酸化アルミニウム;スクアレン;BCG;GM−CSFおよびIL−12などのサイトカイン;MIP 1−αおよびRANTESなどのケモカイン;N−アセチルムラミル−L−アラニル−D−イソグルタミン(MDP);チモシンα1、およびMF59が挙げられる。用いられるアジュバントの量は、皮膚の軟化、疼痛、紅斑、発熱、頭痛、および筋肉痛などの症状の程度に応じて適宜選択されることができる。これらの症状は、ヒトまたは動物においてこの種のワクチン投与後の免疫応答の一部として発現されることもある。
【0045】
一部の実施形態では、本明細書に記述するワクチン組成物または医薬組成物は、他の既知の医薬品、たとえば免疫応答促進ペプチドおよび抗菌剤(合成抗菌剤)と組み合わせて用いられうる。該ワクチン組成物または医薬組成物は、他の薬物および添加剤をさらに含みうる。本明細書に記述するワクチン組成物または医薬組成物と組み合わせて用いられうる薬物ま添加剤の例としては、本発明の組換えアデノウイルスまたは組換えトランスジェニックタンパク質の細胞内取り込みを補助する薬物、トランスフェクションを促進するリポソームおよび他の薬物ならびに/または添加剤(たとえば、フルオロカーボン乳化剤、コキレート、チューブル、金粒子、生分解性マイクロスフェア、およびカチオン性ポリマー)が挙げられる。
【0046】
一部の実施形態では、本明細書に記述するワクチン組成物または医薬組成物に含有される有効成分の量は、それが治療的にもしくは薬学的に有効な量である限り、広範囲の濃度、ウイルス粒子単位(VPU)、プラーク形成単位(PFU)、質量対容量百分率(w/v%)または有効成分量の他の定量的測度から選択されうる。ワクチン組成物または医薬組成物の投与量は、所望の治療効果、投与方法(投与経路)、治療期間、患者の年齢、性別、および他の条件などによって広範囲から適切に選択されうる。
【0047】
一部の態様では、組換えアデノウイルスがワクチン組成物または医薬組成物の有効成分としてヒト被験者に投与されるとき、組換えアデノウイルスの投与量は、組換えウイルスのPFUとして算出され、患者一人あたりおよそ102〜1014PFU、好ましくは105〜1012PFU、より好ましくは106〜1010PFUに対応する量で投与されうる。
【0048】
さらなる態様では、組換えアデノウイルスがワクチン組成物または医薬組成物の有効成分として被験者に投与されるとき、投与量は、ワクチンの宿主に導入される発現可能なDNAの量または転写RNAの量に関して広範囲から選択されうる。該投与量は、使用されるいずれのトランスファーベクターで用いられる転写プロモーターおよび翻訳プロモーターの強度にも依存する。
【0049】
一部の実施形態では、本明細書に記述するワクチン組成物または医薬組成物は、組換えアデノウイルスをPBS(リン酸緩衝生理食塩水)または生理食塩水中で験濁させることで調製される組換えアデノウイルス験濁剤を直接局所部位に(たとえば肺組織、肝臓、筋肉もしくは脳内に)注射することによって、鼻腔急吸入もしくは呼吸吸入によって、または血管内(i.v.)(たとえば動脈内、静脈内、および門脈内)、皮下(s.c.)、皮内(i.e.)、皮内(i.d.)、もしくは腹腔内(i.p.)への投与によって投与されうる。本発明のワクチン組成物または医薬組成物は、複数回投与されうる。より具体的には、最初の投与の後、1回または複数回の追加接種が、追加免疫として投与されうる。1回または複数回の追加免疫投与によって、所望の効果を増強することができる。ワクチン組成物または医薬組成物の投与の後、本明細書に記述する組換えアデノウイルスを含有する医薬組成物による追加免疫が実行されうる。
【0050】
さらなる実施形態では、上述のように、本発明のワクチンと共に種々の他のアジュバント、薬物もしくは添加剤を使用することで、該ワクチン組成物または医薬組成物の投与によって達成される治療効果を増強しうる。薬学的に許容される担体は、等張性および化学安定性を増強する物質などの微量の添加剤を含んでもよい。かかる添加剤は、使用される投与量および濃度においてヒトまたは他の哺乳類の被験者に対して非毒性でなければならない。その例としてはリン酸、クエン酸、コハク酸、酢酸、および他の有機酸、ならびにその塩類;アスコルビン酸などの抗酸化剤;低分子量(たとえば約10未満の残基)ポリペプチド(たとえポリアルギニンおよびトリペプチド)タンパク質(たとえば血清アルブミン、ゼラチン、および免疫グロブリン);アミノ酸(たとえばグリシン、グルタミン酸、アスパラギン酸、およびアルギニン);単糖、二糖、および他の糖質(たとえばセルロースおよびその誘導体、グルコース、マンノース、およびデキストリン)、キレート化剤(たとえばEDTA);糖アルコール(たとえばマンニトールおよびソルビトール);対イオン(たとえばナトリウム);非イオン界面活性剤(たとえばポリソルベートおよびポロキサマー);ならびにPEGなどの緩衝剤が挙げられる。
【0051】
本明細書に記述する組換えアデノウイルスを含有するワクチン組成物または医薬組成物は、密封アンプルまたはバイアルなどの単位用量または複数用量の容器に水溶液剤または凍結乾燥製剤として保存されうる。
【0052】
さらなる別の実施形態は、有効量の組換えアデノウイルスワクチン、製剤または医薬組成物を投与することを含むマラリア感染症を予防する方法、またはマラリアを治療する方法を提供する。本発明は、有効量の組換えアデノウイルスワクチンの組成物、製剤、医薬組成物またはその組み合わせを被験者に投与することを含む免疫活性化の方法をさらに提供する。被験者は、ヒト、動物(たとえば哺乳類、鳥類、爬虫類、魚類および両生類)、またはマラリア寄生虫に感染しうる他のいずれの被験者も含みうる。マラリア寄生虫は、既知のマラリア原虫種、たとえば、熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫、二日熱マラリア原虫、マウスマラリア原虫、ネズミマラリア原虫(P.chabaudi)およびネズミマラリア原虫(P.yoelii)のいずれかから選択されマラリア原虫を含みうる。
【0053】
一部の実施形態では、本明細書に記述する組換えアデノウイルスは、単独で形成されることもあり、または薬学的に許容される担体と共にワクチン組成物、製剤、または医薬組成物に形成されることもあり、かつ前述の被験者に投与されうる。投与経路は、たとえば、上記のいずれの投与経路であってもよい。本発明で使用する薬学的に許容される担体は、生成されるべき医薬組成物の形状によって、本技術分野で一般的に用いられる担体から適宜選択されることができる。たとえば、薬理組成物が水溶液に形成されるとき、純水(滅菌水)または生理的緩衝液が担体として使用されることができる。医薬組成物が他の適切な溶液に形成されるとき、注射されることができる有機エステル、たとえばグリコール、グリセロール、およびオリーブ油が担体として用いられうる。該組成物は、安定剤、賦形剤、および本技術分野で、特にワクチン製剤分野で一般的に使用される他の物質を含みうる。
【0054】
さらなる実施形態では、ワクチン組成物、製剤、または医薬組成物で用いられる組換えアデノウイルスの量は、広範囲の濃度、VPU、PFU、質量対容量百分率(w/v%)または有効成分量の他の定量的測度から選択されうる。一部の態様では、組成物の組換えアデノウイルスの適切な範囲は、好ましくは約0.0002〜約0.2(w/v %)、より好ましくは0.001〜0.1(w/v %)ある。一部の実施形態による組換えアデノウイルスワクチン組成物、製剤または医薬組成物の投与方法は、剤形、患者の年齢、性別、および疾患の重症度など他の条件によって適宜選択されうる。適切な剤形は、注射剤、滴下剤、点鼻薬および吸入剤などの非経口的投与のための形状である。該組成物が注射剤または滴下剤に形成される場合、該注射剤は、必要に応じてグルコース溶液またはアミノ酸溶液などの代替液と混合されて静脈内に投与されることができ、または筋肉内(i.m.)、皮内(i.e.)、皮下(s.c.)、皮内(i.d.)、もしくは腹腔内(i.p.)に投与されることができる。
【0055】
別の実施態様では、組換えアデノウイルスワクチン組成物、製剤、または医薬組成物の1日投与量は、被験者の状態、体重、年齢、性別などに応じて異なりうる。一部の態様では、組換えアデノウイルスの投与量は、1日あたり約0.001〜100mg/kg体重の量で投与される。本発明のワクチン、製剤、または組成物は、1日あたり1回または複数回の投与で投与されうる。
【0056】
さらなる実施形態では、組換えアデノウイルスがワクチン組成物、製剤または医薬組成物の有効成分としてヒト被験者に投与されるとき、組換えアデノウイルスの投与量は、組換えウイルス粒子のPFUとして算出され、患者一人あたりおよそ102〜1014PFU、好ましくは105〜1012PFU、より好ましくは106〜1010PFUに対応する量で投与される。本発明のワクチンの組成物は、各患者の臨床症状(たとえば、予防または治療されるべき症状)、組換えアデノウイルスを含有するワクチン組成物の送達部位、標的組織、投与方法、投与量レジメン、および当業者にとって既知の他の因子を考慮して、良質の医療のための原則に従って投与されるべきである。したがって、本明細書のワクチン組成物の適当な投与量は、上記を考慮して決定される。
【0057】
本開示のさらに別の実施形態は、被験者においてマラリア感染症を治療する、または予防する方法に関し、該方法は、組換えアデノウイルスを含むマラリアワクチン組成物の免疫量または治療量を投与することを含む。マラリアワクチンの組換えアデノウイルスは、マラリア原虫の抗原決定基を含んでもよく、かつ1つもしくは複数の修飾型カプシドタンパク質またはコアタンパク質をさらに含んでもよい。免疫量、薬理量、または治療量は、マラリア感染症が予防されるか、または重症度が低減されるように、強力な免疫応答が(CS)タンパク質(すなわち導入遺伝子、B細胞エピトープ、またはCD4+ T細胞エピトープ)の1つまたは複数の抗原部分に対して生成される、任意の適切な量であってよい。
【0058】
被験者がアデノウイルスベクターに最初に曝露される、または「初回刺激される」とき、免疫系は、その特定のベクターに対して中和抗体を産生する。アデノウイルスに対する免疫応答は、通常カプシドタンパク質対して作られる。したがって、同じアデノウイルスベクターに対する次の曝露、すなわち「追加免疫」は、導入遺伝子発現の効力を低下させることができる。したがって、一部の実施形態では、上記のマラリア感染症を治療する、または予防する方法は、第1の組換えアデノウイルスベクターを用いる初回刺激ステップと、続いて1種または複数種の異なる組換えアデノウイルスベクターを用いる1回もしくは複数回の追加免疫のステップとを含みうる。野生型アデノウイルスにまだ曝露されたことのない被験者において、またはこれまでに野生型アデノウイルスベクターに曝露されたことのある被験者においてこの方法は、用いられてもよい。後述の被験者の場合、初回刺激ステップの組換えアデノウイルスベクターを用いて既存のアデノウイルス免疫を回避する。さらなる実施形態および実施例を後述する。
【0059】
<ベクターとしてのアデノウイルス>
アデノウイルスは、一連のウイルスカプシドタンパク質(以下に記述する)と、1つのウイルスゲノムを含む非エンベロープ被覆型DNAウイルスであり、1種もしくは複数種の治療用導入遺伝子または抗原導入遺伝子をin vitroおよびin vivoで種々の細胞に送達するために広範に用いられてきた。アデノウイルス血清型は数多く存在する。既知のアデノウイルス血清型のうち、血清型5(Ad5)は、in vivoでその高い感染性のために、外来遺伝子の導入のためのベクターとして好ましくは用いられている(Abbinkら 2007)。抗原導入遺伝子の発現は、当技術分野で既知のいずれのプロモーターまたはエンハンサーのエレメントによって制御されうる。遺伝子発現を制御するのに用いられてもよいプロモーターはとしては、サイトメガロウイルス前初期プロモーター(CMV)、シミアンウイルス40(SV40)初期プロモーター、細胞ポリペプチド鎖伸長因子1α(EF1)プロモーター、Rous肉腫ウイルス(RSV)プロモーター、およびテトラサイクリン調節(TR)プロモーターが挙げられるがこれらに限定されない。コード配列の後のポリアデニル化(pA)シグナルは、効果的な転写および翻訳のためにも用いられうる。本明細書に記述する組換えアデノウイルスベクターは、アデノウイルゲノムの少なくともE1領域において欠失を有する複製欠損でありうる。これはE1領域が複製、転写、翻訳およびパッケージングのプロセスに必要とされるからである。一部の態様では、E2、E3および/またはE4の領域も欠失しうる。さらなる態様において、Kozak共通配列は、より効果的な翻訳のために用いられうる(Kozak 1987)。
【0060】
アデノウイルス(Ad)系は、いくつかの理由で組換えワクチンの開発のための魅力あるベクターである。1つの理由は、組換えアデノウイルスベクターは、これらに限定されないがマウスおよびヒトの細胞型を含む、大部分の哺乳類細胞型(複製型および非複製型の双方)を感染させるからである。したがって、同じベクターを、マウスモデルおよびヒト臨床試験でうまく用いることが可能である。別の理由は、転移されたいずれの遺伝情報がエピ染色体にとどまり、細胞遺伝子型の挿入変異および変化を回避するからである(Crystal 1995)。さらに別の理由は、ウイルス複製のラウンドを連続した後でも、導入遺伝子が不変のままであるからである。アデノウイルスを用いる他の利点は、以下が挙げられる:組換えアデノウイルスは1)高いビリオン安定性を有する、2)耐容性がよい、3)高力価で増殖しうる、4)大きな導入遺伝子を収容することができる,5)長年にわたり広範囲に研究されたゲノムを有しており、その結果いくつかの血清型の全DNA配列が分かり、組換えDNA技術によるAdゲノムの操作が容易になっている(GrahamとPrevec 1992)。
【0061】
一実施形態では、前赤内期のマラリア寄生虫を標的にするワクチンの開発のためのウイルスベクターとしてアデノウイルスワクチンプラットフォームを用いて、マラリア感染からの防御を提供する。既知の組換えウイルスベクター(Rodriguesら 1997、Bruna−Romeroら 2001、Andersonら 2004、Taoら 2005)のなかで、アデノウイルスは、前赤内期のマラリア寄生虫に対して強い防御の細胞性免疫応答を誘発することができる(Rodriguesら 1997)。マラリア寄生虫は、マラリア原虫ファミリーのうちのいずれか1つで有りうる。一部の実施形態では、標的とされる該寄生虫は、ネズミマラリア原虫(P.yoelii)または熱帯熱マラリア原虫でありうる。
【0062】
<導入遺伝子としてPyCSを発現するアデノウイルスベクターは、マラリア特異的CD8+ T細胞応答を誘発する>
アデノウイルスは、マラリアに対して重要なCD8+ T細胞介在防御免疫を誘発するための魅力あるベクターである(Rodriguesら 1997、Rodriguesら 1998)。ネズミマラリア原虫(齧歯目のマラリア寄生虫)CSタンパク質(AdPyCS)を発現する組換えアデノウイルスの免疫原性は、齧歯目のマラリアモデルを使用して決定された。AdPyCSをマウスに接種すると、かなりの割合のマウスにおいて、完全免疫が誘発され、寄生虫血症の発生を予防する(Rodriguesら 1997)。T細胞集団を枯渇させることによって明らかになり、かつAdPyCSがマラリア寄生虫に対して高力価の抗体応答を誘発することができなかった事実によって実証されているように、この防御効果は、主にCD8+ T細胞によって介在される。
【0063】
カプシド修飾型アデノウイルスの感染性を定量的に測定するために、シャトルベクターにGFP発現カセットと導入遺伝子のクローニング部位とを含ませてもよい。結果として生じるシャトルベクター(GFP/pShuttle−CMV)は、デュアルpCMVプロモーターと、導入遺伝子のためのSV40pAと、pmaxGFP(Amaxa,Germany)からのGFPとを有する。最適化PyCS断片をGFP/pShuttle−CMVのKpnIとHindIIIの部位に挿入した。
【0064】
Ad(PyCS+GFP)の免疫原性は、CS特異的CD8+ T細胞応答の大きさと、マラリア原虫肝臓内期に対する防御免疫のレベルとを測定することによって決定された。異なる経路を介して、最適用量でAd(PyCS+GFP)を投与することで、1010ウイルス粒子(v.p.)は、AdPyCSが誘発することを示した同じパターンの抗マラリア防御応答を誘発し、s.c.およびi.m.経路で、最も強い応答を誘発し、生ネズミマラリア原虫スポロゾイトを投与されたマウスにおいて肝臓内期での最も高い程度の抑制をもたらした。これは、ワクチンとしてAd(PyCS+GFP)がAdPyCSと同等に作用し(Rodriguesら 1997)、かつaAdPyCSのin vivo向性を決定する潜在的に有用なツールであることを示している。
【0065】
<アデノウイルスのカプシドタンパク質およびコアタンパク質>
CSタンパク質を発現する組換えアデノウイルスベクターがCD8+ T細胞によって強力な細胞性免疫応答を誘発するが、はっきりと認識できる体液性応答は誘発されないことを上述の研究で確認された。したがって、野生型アデノウイルスに対する体液性応答がカプシドタンパク質に起因しうることが多いので、修飾型カプシドタンパク質およびコアタンパク質で組換えアデノウイルスベクターを構築することで、1)B細胞活性を介する体液性免疫を増強し、2)Tヘルパー細胞活性を介する体液性免疫を増強し、および3)既存するアデノウイルス免疫を回避する。
【0066】
アデノウイルスは、正三角形の20面を有する正二十面体の形状の非エンベロープ被覆型ネイキッド二重鎖DNAウイルスである。アデノウイルスカプシドは、252カプソメアからなり、そのうち240がHexon三量体であり、12がPenton五量体である。各Penton塩基から突き出ているFiberタンパク質は、細胞受容体との相互作用によって宿主細胞への接着を媒介する。二次的相互作用は、Penton塩基内のRGD(Asp−Arg−Gly)モチーフ間でαvβ3、αvβ5および同様のインテグリンによって生じ、その後の細胞へのアデノウイルスの内部移行を容易にする(Mathiasら 1994、Wickhamら 1993)。ほとんどのアデノウイルスは、細胞受容体として、コクサッキー−アデノウイルス受容体(CAR)を使用する(Bergelsonら 1997)。加えて、MHCクラス1分子、VCAM、およびヘパラン硫酸は、Ad5の接着と侵入を媒介することが示されている(Chuら 2001、Hongら 1997)。エンドサイトーシスを介する侵入に続いて、Ad5はエンドサイトーシス区画からサイトゾルに迅速に逃避する(MeierとGreber 2003、LeopoldとCrystal 2007)。次いで、ビリオンは、微小管を用いて核に移動する。Fiberタンパク質は、プロセスにおいて最も初期のカプシドタンパク質として脱落される(Nakanoら 2000、Hongら 2003)。異なる血清型のアデノウイルスは、異なる輸送パターンを示す(Miyazawaら 1999、Miyazawaら 2001)。Fiberタンパク質を変えるまたは修飾することで、輸送に影響を与えることができる。これは、抗原提示細胞(APC)の感染に続いて、抗原プロセシングおよび提示に関して特に重要になりうる。
【0067】
アデノウイルスFiberは、Fiber尾部、シャフト、およびノブのドメインに分けられる三量体である(Henryら 1994、RuxとBurnett 2004、Chroboczekら 1995)。ノブドメインの三次元構造は知られおり、変異原性研究と共に、これらの研究は、CAR相互作用と三量体形成に関与する領域を視覚化させることを可能にする(Kirbgyら 1999、Xiaら 1995)。Fiberシャフトは、ビリオンから突き出ており、Fiberノブはコサッキーおよびアデノウイルス受容体(CAR)相互作用ドメインを含む(Roelvinkら 1999、Bewleyら 1999)。FiberノブのCAR結合部位は、一次的にABループとCDループからの残基からなり、二次的にFGとHIループ、次いでB、EおよびFのβシートに伸長する(Roelvinkら 1999、Bewleyら 1999)。HIループは、最も研究されたFiberノブ上の挿入部位(Worgallら 2004、MizuguchiとHayakawa 2004、Koizumiら 2003、Belousovaら 2002、NoureddiniとCuriel 2005、Nicklinら 2001)であり、HIループ(残基543と544)へのエピトープの組み込みは、強力な抗エピトープ免疫をもたらす(Krauseら 2006)。したがって、免疫優性CS由来のB細胞エピトープは、Fiberタンパク質のHIループに最初に挿入された。
【0068】
Hexonは、1ビリオンあたり720コピーを有する、アデノウイルスカプシドで最も豊富なタンパク質である。成熟したウイルスでは、Hexonは、正二十面体ビリオンの小面から構成されるホモ三量体のカプソメアとして存在する(RuxとBurnett 2004)。アデノウイルス血清型2および5(Ad2およびAd5)のHexonのの結晶構造は解析されており、複合分子構造が明らかになった(Athapillyら 1994、Robertsら 1986、RuxとBurnett 2000)。各単量体サブユニットのベースは、多くの正二十面体のウイルスのカプシドタンパク質に存在する2つのβ−バレルモチーフからなる。3つの長いループ(DE1、FG1、およびFG2)は、ベース構造から伸びて、各分子のタワー領域を形成する(RuxとBurnett 2004)。これらのループドメイン内の配列は、カプシドの表面から突出して、ビリオンの外側を形成する。異なるアデノウイルス血清型からの配列比較は、カプシド外部上に位置する配列が長さとアミノ酸配列との両方で不完全に保存されていることを示している(Crawford−MikszaとSchnurr 1996)。さらに、これらの不完全に保存されたドメイン(高頻度可変領域(HVR)と名付けられる)に位置する配列が決定基を含み、それらの決定基に対して血清型特有の抗体が産生されることが示された(Top 1975、RuxとBurnett 2000、Topら 1971)。
【0069】
初期の配列比較に基づいて、Hexon分子全体を通して7つのHVRが同定された(Crawford−MikszaとSchnurr 1996、Robertsら 2006)。HVRは血清型間で不完全に保存されており、かつHexonの構造的完全性を維持することに関与していないと思われるので、ウイルスの生存能に影響を及ぼすことなく少量の変化がこれらのドメインになされることができた(RuxとBurnett 2000)。ととえば、ヘキサヒスチジンタグは、ウイルス生存能を損なうことなく、HVR2、HVR3、HVR5、HVR6、およびHVR7に挿入されることができる(Wuら 2005)。このようにして、Hexon HVRを標的として用いて、Hexon HVR内に位置するペプチドに対する抗体応答を効率的に誘発させることが多い(Worgallら 2005、Cromptonら 1994)。血清型間でその長さが完全に保存されないこと、およびアデノウイルスカプシドの最外側表面上のその位置のために(RuxとBurnett 2000、Crawford−MikszaとSchnurr 1996)、Hexon HVR5は、最初はエピトープ挿入のための部位として選択された。さらに、HVR5がカプシド表面上の柔軟なループであることをHexonの結晶構造が示しており、HVR5がカプシドの構造的完全性を損なうことなく比較的大きなペプチドを収容することができることを示唆している(Robertsら 1986)。Hexon特異的CD4+エピトープおよびCD6+エピトープが最近同定されて(Leenら 2008)、アデノウイルスへのCD4+ T細胞応答は、ヒトにおいてHexonタンパク質内に保存された残基に対して集中する(Onionら 2007、Heemskertら 2006)ことが判明した。
【0070】
アデノウイルスのコアは、ウイルスゲノムと4種類のコアタンパク質で構成されている。末端タンパク質(TP)は、1ビリオンあたり2つのコピーで各直鎖状ウイルスDNA鎖の5’末端に共有結合される。他の3種類のコアタンパク質、mu(μ)、V(pV)およびVII(pVII)は、アルギニンリッチタンパク質を介して非共有結合的かつ非特異的にウイルスDNAに結合している。pVIIは、1ビリオンあたりおおよそ700〜800コピーを提供する主要なコアタンパク質であり、かつその周りをウイルスDNAで包まれてヌクレオソーム構造を形成している、ヒストン様中枢として役立っている。
【0071】
<体液性免疫を増強するためのアデノウイルスカプシドタンパク質の修飾>
一部の実施形態では、アデノウイルスカプシドタンパク質(HexonまたはFiberに挿入された)内に免疫優性CSタンパク質Bエピトープを有する、スポロゾイト周囲(CS)アデノウイルスベクターが記述されている。導入遺伝子は、CSタンパク質に対する細胞介在免疫応答および体液性免疫応答を増強するために、CMVなどのプロモーター下に存在しうる。
【0072】
中心反復領域は、マラリア原虫種間でCSタンパク質の保存された構造であり、かつこの反復配列に対する抗体は、スポロゾイト中和活性を有することが示されている。マラリア原虫CSタンパク質内の反復配列の例として、(NANP)n反復(熱帯熱マラリア原虫;配列番号60)、ANGAGNQPG反復(三日熱マラリア原虫;配列番号63)、およびNAAG反復(四日熱マラリア原虫;配列番号64)があり、これらはアデノウイルスカプシドタンパク質に挿入されることができる。一部の実施形態では、PfCSPの4以上の(NANP)n反復(配列番号60)は、アデノウイルス血清型5HexonのHVR1に挿入されうる。一部の実施形態では、PfCSPの2、4、6、8、10、14、16、18、20、22、24、26、または28の(NANP)n反復(配列番号60;n=2、4、6、8、10、12、14、16、18、20、22、24、26、28)は、アデノウイルス血清型5HexonのHVR1に挿入される。(NANP)n反復配列は、FiberのHIループにさらに挿入されうる。
【0073】
一部の実施形態では、改良型CSタンパク質アデノウイルスワクチンを開発するために、CSに対する免疫優性中和B細胞エピトープがマッピングされた。組換えネズミマラリア原虫CSタンパク質(PyCS)で免疫されたマウスは、2つの主要免疫優性Bエピトープ、すなわちQGPGAP(配列番号59)とQQPP(配列番号65)に対して高力価を生成したが、in vitro中和アッセイでは、(QGPGAP)3のペプチド(配列番号59;n=3)を培地に加えることによって中和が逆転されうるために、QGPGAPエピトープ(配列番号59)それ自体に対する体液性免疫応答が中和活性の主な原因となりうることを示した。一部の実施形態では、PyCSの3以上の(QGPGAP)n反復(配列番号59)は、アデノウイルス血清型5HexonのHVR1に挿入される。一部の実施形態では、PyCSの3、4、5、6、7、8、9、10、11、または12の(QGPGAP)n反復(配列番号59;n=3、4、5、6、7、8、9、10、11、12)は、アデノウイルス血清型5HexonのHVR1に挿入されうる。(QGPGAP)n反復配列は、FiberのHIループにさらに挿入されうる。
【0074】
B細胞エピトープペプチドは、免疫系がエピトープを効率的に認識することができるように、アデノウイルスビリオンの表面に提示されなければならない。かかる挿入部位は、HexonのHVRおよびFiber内のループ構造でありえ、かつ異なる挿入部位が組み合わせられてもよい。
【0075】
<Tヘルパー細胞活性を増強するためのアデノウイルスカプシドタンパク質とコアタンパク質の修飾>
別の実施例では、アデノウイルスベクターで用いられる導入遺伝子に特異的なCD4+エピトープを、pVII、pVおよびHexonなどのアデノウイルスタンパク質に取り込んでアデノウイルスベースワクチンの免疫原性を増強させうる。樹状細胞(DC)およびB細胞などの専門の抗原提示細胞(APC)は、エンドサイトーシスを介して微粒子状の病原体様ウイルス粒子を取り込むことができ、次いで該病原体中のCD4+エピトープをCD4+ T細胞に提示し、それによって体液性免疫応答および/または細胞性免疫応答のヘルパー細胞として作用する。1ビリオンあたりのコピー数がpVII(700〜800コピー)およびHexon(720コピー)と高いため、pVIIおよびHexonをアデノウイルス標的タンパク質として用いて、抗原CD4+ペプチドに容易に挿入することができる。
【0076】
<既存のアデノウイルス免疫を増強するためのアデノウイルスカプシドタンパク質の修飾>
一部の実施形態では、アデノウイルスに対する既存の免疫を克服するため、および/またはアデノウイルスワクチンに対する体液性応答を増強するために、アデノウイルスFiberおよびHexonのカプシドタンパク質は、B細胞エピトープまたはTヘルパー細胞エピトープを挿入するよう修飾されうる。ヒト集団において若年成人の推定80%がアデノウイルスに対する循環中和抗体(Douglas 2007)、特に血清型5(Ad5)を有する。遺伝子治療ベクターとしてアデノウイルスを利用する研究では、動物内の中和抗体の存在が、アデノウイルスによって送達される導入遺伝子の発現を限定していることが判明した。中和抗体に加えて、CD8+ T細胞応答も、組換え遺伝子発現を限定する一因となっている(Yangら 1995、Yangら 1996)。アデノウイルスに対するかかる既存の免疫は、組換えアデノウイルスワクチンの有効性を阻害することがいままでに報告されており(Pappら 1999)、また臨床試験において、アデノウイルスベースワクチンの免疫原性を低下させている(Priddyら 2008)。
【0077】
Hexonは抗Adカプシド免疫応答の主要な標的であり(Royら 2005、Wohlfart 1988)、CD4+とCD8+のT細胞応答の誘発を含むアデノウイルスの強力なアジュバント効果の原因であると思われる。したがって、既存の抗アデノウイルス免疫を回避するために使用されてきた1つの戦略は、Hexon全体または一部を異なるタンパク質、たとえばアデノウイルス11、24、26および35などのまれな血清型と置換することである。Hexonは抗アデノウイルス中和抗体の主要な標的である(Youilら2002、Sumidaら2005)ために、全HexonまたはHexonのHVRはまれな血清型と交換されうる(Wuら 2002、Robertsら 2006)。
【0078】
本明細書の一実施形態で記述した別の戦略では、既存の抗アデノウウイルス免疫またはアデノウウイルスベクターによる以前のワクチン接種によって誘発された抗アデノウウイルス中和抗体を増強するために、アデノウイルスHexonは、HVR1またはHVR5と抗原ペプチドとの置換によって修飾されうる。一部の実施形態では、抗原ペプチドはマラリア原虫CSタンパク質の免疫原性エピトープであってもよく、特定の態様では、該エピトープは中心反復配列、CD4+エピトープ配列またはCD8+エピトープ配列を含みうる。
【0079】
同じ血清型のAdベクターによる反復投与は、免疫化後の坑Ad免疫のために阻止される。したがって、多くのAdワクチンは、導入遺伝子によってコードされた抗原の発現と提示を防ぐことでワクチンの追加免疫を妨げる(Yangら 1995、Hackettら 2000、Harveyら 1999、Mastrangeliら 1996)。後述の実施例に記述するように特異的エピトープをAdカプシドに追加することで、一部の実施形態によるこの妨害を低下しうる、または排除しうる。
【0080】
実施形態をよりよく説明するため、およびクレームするいずれの実施形態の範囲を限定すると解釈されないようにするために以下の実施例を提供する。特定の材料が記載される範囲は、説明の目的だけであり、本発明を限定することを意図するものではない。当業者は、発明能力を行使することなく、および本発明の範囲から逸脱することなく、同等の方法または反応物を開発しうる。本発明の範囲内に依然として留まりつつ、多くのバリエーションが本明細書に記述する方法でなされうることを理解されるであろう。かかるバリエーションが本発明の範囲内に含まれることは、本発明者らの意思である。
【実施例】
【0081】
実施例1:カプシド修飾型マラリア原虫スポロゾイト周囲タンパク質アデノウイルスプラスミドベクターおよび組換えアデノウイルス粒子の構築
【0082】
エピトープマッピング
まず、PyCSの免疫優性中和B細胞エピトープを選択した。ナイーブBALB/cマウスにフロイント不完全アジュバントと共に組換えPyCSタンパク質を3回免疫し、プールした血清を用いて、中和抗体の重要なエピトープを決定した。
【0083】
簡単に言うと、中和アッセイでは、ヒトCD81発現HepG2細胞を標的細胞として用いた。CD81は、マラリア寄生虫が増殖して、シゾントになる肝細胞においてパラシトホーラスバキュオールを形成するために必要な分子であり(Silvieら 2006)、したがってスポロゾイトのin vitro感染力が極めて高い。
【0084】
本アッセイでは、PyCS合成ペプチドをウェルに加えて、ペプチド特異的抗体を遮断した。エピトープマッピングの結果は、PyCS中心反復配列(QGPGAP;配列番号59)がPyCSにおいてQQPP(配列番号65)よりもより強力な中和エピトープであることを示した。したがって、(QGPGAP)nエピトープ(配列番号59)の挿入を用いて、アデノウイルスカプシドタンパク質、Hexonおよび/またはFiberを修飾した。
【0085】
カプシド修飾型プラスミドベクターの構築
クローニング部位下にGFP発現カセットを挿入することでアデノウイルスシャトルベクターpShuttle−CMV(STRATAGENE)を修飾した。まず、pmaxGFP(Lonza,Cologne,Germany)のBsmBI−SacI断片(pCMV+GFP)とSacI−BsmBI断片(SV40ポリAshigunaru)を平滑末端化して、pUC19の平滑末端化Sall部位とKpnI部位にそれぞれ挿入した。もたらされたpCMV−GFP/pUC19のBamHI−EcoRI断片をSV40pA/pUC19の同じ部位に挿入して、SV40pA−pCMV−GFP断片を作製した。該断片を平滑末端化して、pShuttle−CMVのEcoRV部位に挿入した。結果として生じるシャトルベクター(GFP/pShuttle−CMV)は、導入遺伝子とGFPのためのデュアルpCMVプロモーターとSV40pAを有する。
【0086】
アデノウイルスシャトルベクターpShuttle−CMVの別の修飾を行って、CMVプロモーター領域をpQBI−AdCMV5(QBIOgene)からのCMV5プロモーターと置換した。pShuttle−CMV内でpShuttle−CMVのSgrAI−KpnI断片をCMV5プロモーター配列と、CMVプロモーターからの上流配列とを含む断片と置換して、pShuttle−CMV5ベクターを構築した。
【0087】
ネズミマラリア原虫CS(PyCS)遺伝子は、JCatコドン最適化アルゴリズム(http://www jcat de/)に基づいてPCR反応を重複させることによって、(QGPGAP)n反復(配列番号59)を除いて、コドン最適化した。
【0088】
熱帯熱マラリア原虫3D7株のPfCSPアミノ酸配列をコドン最適化のための鋳型配列として用いた。ヒトにおいてタンパク質発現のためのコドン最適化は、Integrated DNA Technologies(Coralville,IA,USA)の最適化ソフトウェアによって行った。C末端でGPIアンカードモチーフを除く全PfCSPをコードするDNA断片(図10;配列番号2)をIntegrated DNA Technologiesで合成した。
【0089】
コドン最適化PyCS遺伝子(図9;配列番号1)またはPfCSP遺伝子(図10;配列番号2)を、pShuttle−CMV、pShuttle−CMV5、またはGFP/pShuttle−CMVのKpnIおよびHindIIIに挿入した。AdEasy−1による相同組換えのために、もたらされたマラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターを用いて、マラリア原虫スポロゾイト周囲抗原遺伝子と、インタクトなアデノウイルスタンパク質コード配列とを有するアデノウイルスゲノムを構築した。簡単に言うと、マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターをPmeI消化によって直線化し、次いで相同組換えのために大腸菌(E.coli)BJ5183細胞を直線化シャトルベクターおよびpAdEasy−1ベクター(Bruna−Romeroら 2003)と同時形質転換させた。
【0090】
アデノウイルスカプシドタンパク質の修飾を要約して、図1に示す。アデノウイルスゲノムDNAのHVR1配列の修飾を図2に示す。簡単に言うと、AdEasy−1をSfiIで消化させて、6.4kbp断片を、EcoRI−SfiIおよびPstI−SfiIリンカーオリゴマーを用いて、pUC19のEcoRI部位とPstI部位にサブクローニングさせた。HVR1をマラリア原虫スポロゾイト周囲タンパク質B細胞エピトープと置換するために、AgeI部位とNdeI部位を含む領域を、HVR1配列の代わりにエピトープ配列を有するプライマーを用いる2ステップPCRによって増幅させた。PCR産物をAgeIとNdeIで消化させ、次いでこれを用いてSfiI/pUC19ベクター中のSfiI断片のナイーブAgeI−NdeI領域を置換した。配列を確認した後に、アデノウイルスゲノムDNAのSfiI断片を、スポロゾイト周囲エピトープ配列を含むSfiI断片と置換して、HVR1修飾型Hexonを生成した。一部の実施形態では、HVR修飾型Hexonは、配列番号3(図11−1〜図11−3)、配列番号4(図12−1〜図12−3)、配列番号5(図13−1〜図13−3)、配列番号6(図14−1〜図14−3)、配列番号7(図15−1〜図15−3)、配列番号8(図16−1〜図16−3)、配列番号9(図17−1〜図17−3)、配列番号10(図18−1〜図18−3)、配列番号11(図19−1〜図19−3)、配列番号12(図20−1〜図20−3)、配列番号13(図21−1〜図21−3)、配列番号14(図22−1〜図22−3)、配列番号15(図23−1〜図23−3)、配列番号16(図24−1〜図24−3)、配列番号17(図25−1〜図25−3)、配列番号18(図26−1〜図26−3)、配列番号19(図27−1〜図27−3)、配列番号20(図28−1〜図28−3)、配列番号21(図29−1〜図29−3)、配列番号22(図30−1〜図30−3)、または配列番号23(図31−1〜図31−3)の核酸配列を有することもある。
【0091】
HVR1に(NANP)28(配列番号60;n=28)を挿入するために、コドン最適化PfCSPの中心反復領域の一部を、Hexon特異的配列を5’に、NANP特異的配列を3’に有するプライマーを用いてPCRによって増幅させた。もたらされたDNA断片を第2のPCRによってAgeI−NdeI領域に挿入した。
【0092】
HVR5修飾の場合、図3に示すように、XbaI部位を、AdEasy−1内のHexonのL1ループ中のHVR5に導入し、次いで合成し、マラリア原虫スポロゾイト周囲タンパク質エピトープをコードするリン酸化二本鎖オリゴマーをXbaI部位に挿入した。配列決定(図31;配列番号23)によってこの挿入を確認した。
【0093】
Fiber修飾の場合、図4に示すように、AdEasy−1のSpeI−PacI断片を、EcoRI−PacIおよびPstI−SpeIのリンカーオリゴマーを用いてpUC19のEcoRI部位とPstI部位にサブクローニングした。マラリア原虫スポロゾイト周囲タンパク質B細胞エピトープ配列をFiberノブのHIループに挿入するために、EcoNI(またはNhel)部位とMfel部位を含む領域を、エピトープ配列を有するプライマーを用いる2ステップPCRによって増幅させた。PCR産物をEcoNI(またはNheI)とMfeIで消化させ、次いでこれを用いてSpeI−PacI/pUC19ベクター内のFiberのナイーブEcoNI(またはNheI)−MfeI領域を置換した。配列(図32−1〜図32−2、配列番号24;図33−1〜図32−2、配列番号25)を確認した後、AdEasy−1のSpeI−PacI断片を、エピトープ配列を含むSpeI−PacI断片と置換した。マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターによる相同的組換えのために、もたらされたFiber修飾型アデノウイルスDNAを用いて、Fiber修飾型マラリア原虫スポロゾイト周囲タンパク質アデノウイルスDNAを生成した。
【0094】
2つのエピトープ挿入を有するHVR1およびFiber修飾型アデノウイルスDNAを構築するために、Fiber修飾型アデノウイルスDNAのSfiI−SfiI断片を、図5に示すようにHVR1内にスポロゾイト周囲タンパク質エピトープを有するSfiI−SfiI断片と置換した。
【0095】
pVIIのC末端を修飾するために、SfiI部位とSalI部位を含む領域を、スポロゾイト周囲タンパク質エピトープ配列を有するプライマーを用いて2ステップPCRによって増幅させた。PCR産物をSfiIとSaiIで消化させ、次いでこれを用いて、SfiI/pUC19ベクターのナイーブSfiI−SalI領域を置換した。配列(図34、配列番号26;図35、配列番号27)を確認した後、HVR1のSfiI−SfiI断片および/またはFiber修飾型スポロゾイト周囲タンパク質アデノウイルスDNAを、pVII内にスポロゾイト周囲タンパク質エピトープを有するSfiI−SfiI断片と置換した。
【0096】
スポロゾイト周囲タンパク質CD4+エピトープ配列EYLNKIQNSLSTEWSPCSVT(配列番号62)をpVIIの中央に挿入するために、pAdEasy−1の約7.7kb断片をRsrII消化によって調製して、RsrIIリンカー(RsrII/pUC19)を用いて、pUC19プラスミドのEcoRI部位とHindiII部位との間にクローニングした。RsrII/pUC19内にAscI部位とBgIII部位を含む領域を、エピトープ配列を有するプライマーを用いて2ステップPCRによって増幅させた。PCR産物をAscIとBgIIIで消化させ、次いでこれを用いて、RsrII/pUC19プラスミド内のナイーブAscIとBgIII領域を置換した。置換した領域(図36、配列番号28;図37、配列番号29)の配列を確認した後、HVR1修飾型アデノウイルスDNAのRsrII断片を、エピトープ配列を含むRsrII断片と置換した。
【0097】
以下の表1(ネズミマラリア原虫)および表2(熱帯熱マラリア原虫)に記載する組換えアデノウイルスを作製して、エピトープ挿入の感染性、免疫原性、および既存の抗アデノウウイルス免疫に対する感受性への効果を評価した。使用した組換えアデノウイルスベクターは、複製欠損、E1およびE3欠失アデノウイルス血清型5(STRATAGENE)であった。図1は、カプシド修飾型マラリア原虫スポロゾイト周囲タンパク質組換えアデノウイルスの構造の模式図を示す。
【0098】
【表1】
【0099】
【表2】
【0100】
カプシド修飾型アデノウイルスゲノムDNAプラスミドを精製し、Pad消化によって直線化して、Ad293細胞のトランスフェクションのために用いた。
【0101】
凍結/解凍を4回繰り返すことでトランスフェクトされたAD293細胞からアデノウイルス粒子を調製して、さらなるウイルス増幅のために用いた。最後の増幅を終えてから、アデノウイルス粒子をCsCl勾配遠心分離によって精製した。次いで、バンドを収集して、透析緩衝液に対して透析して、CsClを除去した。ウイルス粒子(v.p.)は、O.D.260(1 O.D.260=1.25×1012v.p./mL)(Bruna−Romeroら 2003)に基づいて算出した。
【0102】
アデノウイルス増幅処置の間、アデノウイルス増殖でのカプシド修飾型アデノウイルス間で観察された差異は小さく、アデノウイルスの感染力および産生力が、修飾によって悪影響を受けていなかったことを示していた。
【0103】
実施例2:ネズミマラリア原虫スポロゾイト周囲タンパク質特的免疫応答
【0104】
ネズミマラリア原虫組換えアデノウイルスの検証
マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターを一過性トランスフェクションのために用いて、AD293細胞を用いてマラリア原虫スポロゾイト周囲タンパク質発現を確認した(図38)。トランスフェクションから24時間後に、細胞をSDS試料緩衝液中で溶解し、続いてSDS PAGE電気泳動および抗PyCSモノクローナル抗体(9D3)によるウエスタンブロットを行った。
【0105】
アデノウイルスカプシドタンパク質へのエピトープ挿入を確認するために、精製した組換えアデノウイルスをSDS−PAGE(2×109v.p./レーン)によって分析し、図39Aおよび40Aに示すように(QGPGAP)n(配列番号59)反復を認識する、抗スポロゾイト抗体によるウエスタンブロット(1×109v.p./レーン)を行った。図39Aでのバンドの強度は、アデノウイルスビリオン中のカプシドタンパク質のコピー数と相関した:Fiberのコピー数(1ビリオンあたり36コピー)は、Hexonのそれ(1ビリオンあたり720コピー)の20分の1に満たない。図39Aのレーン4の下位のバンドは、分解したHexonと思われる。図40Aのバンドの強度は、HVR1に挿入された(QGPGAP)n(配列番号59)の反復数と相関した。
【0106】
PyCS−Bエピトープがアデノウイルスビリオンの外側に曝露されたかどうかを評価するために、連続希釈、精製した組換えアデノウイルス粒子を、酵素結合免疫吸着測定(ELISA)プレート上にコーティングして、(QGPGAP)n(配列番号59)反復を認識する抗PyCS抗体で検出した。該抗体は、すべてのカプシド修飾型アデノウイルスを認識した(図39Bおよび40B)。ELISA分析の結果は、カプシドタンパク質に取り込まれたPyCS−Bエピトープがアデノウイルスビリオンの外側に十分に曝露されたことを示唆する。
【0107】
カプシド修飾型PyCSアデノウイルスによる免疫化後のネズミマラリア原虫スポロゾイト周囲タンパク質特異的免疫応答
6〜8週齢の雌BALB/cマウスをTaconic(Hudson,NY, USA)から購入して,Rockefeller UniversityのLaboratory Animal Research Centerにおいて標準条件下で維持した。免疫化のために、アデノウイルスをPBSで希釈して、表示された用量で筋肉内に注射した。
【0108】
単回免疫後に組換えアデノウイルスの免疫原性を評価するために、ナイーブBALB/cマウスの群(1群あたり5匹)に、種々の組換えPyCSアデノウイルスを1×109v.p.で筋肉内に免疫し、次いで免疫化から2週間後にPyCS特異的細胞媒介免疫応答(CMI)をELISPOTで測定した(図41A).
【0109】
免疫したマウスの脾臓のPyCS特異的、予防接種IFN−γ分泌CD8+ T細胞の数を、PyCSタンパク質内のCD8+ T細胞エピトープ(SYVPSAEQI;配列番号66)に対応する合成ペプチドを用いてELISPOTによって決定した。簡単に言うと、96ウェルニトロセルロースプレート(Milititer HA,Millipore)を抗マウスインターフェロンγモノクローナル抗体、R4で終夜コーティングした。室温での終夜インキュベーションの後、ウェルを培地で繰り返し洗浄し、次いで4時間培地で遮断した。免疫したマウスからの5×105脾細胞を、10μg/mLのCD8+ T細胞エピトープペプチドの存在下または非存在下でELISPOTに加えて、37℃、5%CO2で24時間インキュベートした。0.05%Tween20(PBST)を含有するPBSで、該プレートを十分に洗浄した後、PBST中のビオチン化抗マウスインターフェロンγモノクーナル抗体、XMG1.2を加えて、40℃で終夜インキュベートした。PBSTで洗浄した後に、該プレートをペルオキシダーゼ標識アビジン(eBiosciences)と共にインキュベートすることになる。AEC基質(BD Biosciences)を加えることによって複数のスポットが発現した。
【0110】
カプシド修飾型アデノウイルスのすべては、この用量で、インタクトなカプシドタンパク質を有するアデノウイルスに対して同程度のCMIを誘発した(図41B)。
【0111】
次に、図42Aに示すように、ナイーブBALB/cマウスに増加した用量(すなわち、1×108、1×109、および1×1010v.p.)の組換えアデノウイルスを3週間間隔で複数回投与した。PyCS特異的体液性応答をELISAで決定した。免疫したマウスの尾静脈から5マイクロリットルの血液を採取して、495μLのPBSで希釈し、次いでこれらの試料を5,000rpmで5分間遠心して、希釈血漿試料(×100)を調製した。Maxisorp ELISAプレートを、0.1Mの炭酸ナトリウム緩衝液中の5μg/mLのCS特異的ペプチド((QGPGAP)3;配列番号59、n=3)で、40℃で終夜コーティングした。プレートを洗浄して、室温で、1×希釈液で2時間遮断した。プレートを再度洗浄して、1×希釈液中の100μLの連続2倍希釈した血漿または血清をプレートに加え、次いで該プレートを室温で1時間インキュベートした。プレートを洗浄して、100μLのHRP標識ヤギ抗マウスIgG抗体と共にインキュベートした。ペプチドのすべては、Biosyntheis(Lewisville,TX,USA)で合成されたものであった。
【0112】
この免疫レジメンを使用して、10週目にすべてのカプシド修飾型アデノウイルスは、野生型PyCS−GFPよりも高いレベルの抗−(QGPGAP)3抗体応答を有意に誘発した(図42B)。
【0113】
カプシド修飾型アデノウイルスのワクチン有効性を決定するために、10週目に、免疫したマウスに2×104感染性ネズミマラリア原虫スポロゾイトを尾静脈注で投与した。スポロゾイト投与から42時間後、マウスの肝臓内の寄生虫特異的リボソームRNAの量を定量化することによって寄生虫負荷を決定し、次いで寄生虫リボソームRNAの絶対コピー数対マウスGAPDH mRNAの絶対コピー数の比として表した。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて差異を決定した。
【0114】
(QGPGAP)3−HVR1/PyCS−GFP、(QGPGAP)3−Fib/PyCS−GFPまたは(QGPGAP)3−HVR1/Fib/PyCS−GFPによるワクチン接種は、野生型PyCS−GFPよりも高いレベルの予防を誘発し、マラリアを投与されたマウスにおいて有意に低い寄生虫負荷をもたらした(図42C)。
【0115】
次に、カプシド修飾型アデノウイルスによって誘発されたPyCS特異的抗体の機能性を評価した。第1に、免疫されたマウスのアデノウイルスの血清が10週目に、インタクトなスポロゾイトを認識することができるかどうかを調べるために、間接的免疫蛍光アッセイ(IFA)を行った。IFAでは、マルチスポットガラススライド上の風乾したスポロゾイトを、PBS中3%のウシ血清アルブミン(BSA)と共に1時間インキュベートし、次いで希釈血清と共に1時間インキュベートした。洗浄後、該スライドを蛍光標識二次抗体と共に1時間インキュベートした。該スライドを洗浄して、蛍光顕微鏡下でIFA力価を、蛍光を発する最高希釈として決定した。(QGPGAP)3−HVR1/PyCS−GFPと(QGPGAP)3−HVFM/Fib/PyCS−GFPは共に、スポロゾイトに対して最高のIFA力価を誘発し(図43A)、アデノウイルスHexonのHVR1への(QGPGAP)3のエピトープの挿入によってPyCSアデノウイルスが合成ペプチドだけでなく、マラリア寄生虫に存在するナイーブエピトープに対しても強い抗体応答を誘発することを可能にすることを示した。
【0116】
第2に、カプシド修飾型アデノウイルスで免疫したマウス(図42A)が、スポロゾイトの感染力の効力を中和することができうる「機能的」抗体を発現したかどうか決定するために、in vitroスポロゾイト中和アッセイを行った。
【0117】
in vitro中和アッセイでは、アデノウイルス免疫マウスに由来するプールした血清を30倍に希釈して添加した96ウェルプレート中のCD81/He−pG2にネズミマラリア原虫スポロゾイトを加えた。2時間のインキュベーションの後、非感染スポロゾイトを培地で洗い落とし、次いで該細胞を42時間培養した。ヒトGAPDH mRNAに対する寄生虫リボソームRNAの相対量を、リアルタイムPCR(Ophorstら 2006)で測定した。
【0118】
カプシド修飾型アデノウイルス、特に(QGPGAP)3−HVR1/PyCS−GFPおよび(QGPGAP)3−HVR1/Fib/PyCS−GFP)で免疫したマウスに由来するプールした血清は、in vitroでスポロゾイト感染力をほぼ完全に(99%)阻害した(図43B)。このアッセイでの阻害の程度は、図43Aに示すIFA力価と逆相関したことが注目される。
【0119】
赤血球期マラリア感染からの防御。
次に、スポロゾイト投与の後、カプシド修飾型rAdによる免疫化が赤血球期マラリア感染症の発現からマウスを防御するかどうかを決定した。実験を2回行った。各実験では、各群で20匹のBALB/cマウスに、図42Aに示すように、野生型PyCS−GFPまたは(QGPGAP)3−HVR1/PyCS−GFPで3回免疫した。最後の免疫化から4週目に、マウスに50ネズミマラリア原虫スポロゾイトを静脈内投与した。抗原投与から3〜12日後に、Giemsa染色した血液塗沫標本を分析して、赤血球期マラリア寄生虫感染を検出した。野生型CS−GFP免疫群では、40匹中30匹(75%)のマウスが感染し、一方ナイーブ群では40匹中35匹(87.5%)が感染した(以下の表3)。(QGPGAP)3−HVRI/CS−GFP予免疫マウスは、野生型CS−GFPよりもより防御された;40匹中わずかに15匹(37.5%)が感染した。これは肝臓での寄生虫負荷によって測定された防御実験(図42C)の結果と一致する。
【0120】
【表3】
【0121】
初回刺激−追加免疫による免疫化1
次に、図44Aに示すように、ナイーブBALB/cマウスに、(QGPGAP)nの4または6の反復(配列番号59;n=4、6)を有するHVR1−修飾型PyCSアデノウイルスの「追加免疫」をアジュバントと共にまたはアジュバントを含まずに、用量を増加して(すなわち、1×108、1×109、および1×1010v.p.)3週間間隔で投与した。この実験で使用したアジュバントは、200μg/mLのSaponin(Sigma−Aldrich)を含有するSigma Adjuvant System(Sigma−Aldrich)である。Sigma Adjuvant System(1mL)のバイアルは、Salmonellaミネソタ由来のモノホスホリルリピドA(解毒化内毒素)を0.5mgおよび水中2%オイル(スクアレン)−Tween80中の合成トレハロースジコリノミコラートを0.5mg含有する。免疫の前に、アデノウイルス溶液を同量のアジュバントと混合した。100マイクロリットルのアデノウイルス−アジュバント混合液を筋肉内に注射した。PyCS特異的体液性免疫応答および細胞介在性免疫応答を上述のように測定した。6反復を有するHVR1修飾型アデノウイルスは、4反復を有する該修飾型アデノウイルスよりも高い抗体価を誘発し、かつアジュバントの使用が抗体価を増強する傾向が観察された(図44B)。対照的に、CMIへのアジュバントの効果はみられなかった(データ図示せず)。
【0122】
HVR1修飾型PyCSアデノウイルスのワクチン有効性を決定するために、9週目に各群の5匹のマウスに2×104の感染性ネズミマラリア原虫スポロゾイトを尾静脈から投与した。スポロゾイト投与から42時間後の寄生虫負荷を上述のように決定した。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて差異を決定した。6反復を有するHVR1修飾型アデノウイルスは、4反復を有する該修飾型アデノウイルスよりも寄生虫負荷を減少させ、かつアジュバントの使用が防御を増強する傾向がみられた(図44C)。
【0123】
HVR1修飾型PyCSアデノウイルスによって誘発される抗体の機能性を評価するために、我々は上述のようにin vitroスポロゾイト中和分析を行った。HVR1修飾型PyCSアデノウイルスで免疫したマウス由来のプールした血清試料を9週目に、50倍希釈でスポロゾイト浸潤を中和させた(図44D)。
【0124】
初回刺激−追加免疫による免疫化2
図45Aに示すように、ナイーブBALB/cマウスに、(QGPGAP)nの反復を6、9、または12有する(配列番号59;n=6、9、12)HVR1修飾型PyCSアデノウイルスの「追加免疫」を、アジュバント共に、またはアジュバントを含まずに、1×1010v.p.の用量で、3週間間隔で3回、投与した。この実験で使用したアジュバントは、200μg/mLのSaponin(Sigma−Aldrich)を含有するSigma Adjuvant System(Sigma−Aldrich)である。免疫の前に、アデノウイルス溶液を同量のアジュバントと混合した。PyCS特異的体液性免疫応答および細胞介在性免疫応答を上述のように測定した。(QGPGAP)nの反復を12有する(配列番号59;n=12)HVR1修飾型PyCSアデノウイルスとアジュバントは、9週目に群の間で最も高い抗体価を誘発した(図45B)。PyCS特異的CMIに関して、群の間で差はみられず、最高12の長いエピトープ挿入がin vivoでアデノウイルス感染力を損なわないことを示した(図45C)。さらに、アジュバンントはCMIを誘発するるアデノウイルスの能力に影響を及ぼさなかった(図45C)。
【0125】
HVR1修飾型PyCSアデノウイルスのワクチン有効性を決定するために、9週目に各群の5匹のマウスに2×104の感染性ネズミマラリア原虫スポロゾイトを尾静脈から投与した。スポロゾイト投与から42時間後の寄生虫負荷を上述のように決定した。(QGPGAP)n反復(配列番号59;n=6、9、12)を有するすべてのHVR−1修飾型PyCSアデノウイルスは、アジュバントの有無にかかわらず防御の増加を示した。しかし、(QGPGAP)nの反復を12有する(配列番号59;n=12)HVR1修飾型PyCSアデノウイルスとアジュバントは、最良の防御を示した(図45D)。これは他のいずれの処置よりも有意により防御的であった。
【0126】
実施例3:熱帯熱マラリア原虫スポロゾイト周囲タンパク質特異的免疫応答
【0127】
熱帯熱マラリア原虫組換えアデノウイルスの検証
一過性トランスフェクションのために、マラリア原虫スポロゾイト周囲タンパク質をコードするアデノウイルスシャトルベクターを用いて、AD293細胞を用いるマラリア原虫スポロゾイト周囲タンパク質発現を確認した(図46A)。
トランスフェクションから24時間後に、細胞をSDS試料緩衝液中で溶解し、続いてSDS PAGE電気泳動および抗NANPモノクローナル抗体(2A10)によるウエスタンブロットを行った。
【0128】
アデノウイルスカプシドタンパク質へのエピトープ挿入を確認するために、精製した組換えアデノウイルスをSDS−PAGE(2×109v.p./レーン)によって分析し、図47Aおよび48Aに示すように(NANP)n(配列番号60)反復を認識する、抗スポロゾイト抗体によるウエスタンブロット(1×109v.p./レーン)を行った。図47Aのバンドの強度は、アデノウイルスビリオン中のカプシドタンパク質のコピー数と相関した;Fiberのコピー数(1ビリオンあたり36コピー)は、Hexonのそれ(1ビリオンあたり720コピー)の20分の1に満たない。図48Aのバンドの強度は、NANP反復HVR1の数と相関した。
【0129】
PfCSP−Bエピトープがアデノウイルスビリオンの外側に曝露されたかどうか評価するために、連続希釈し、精製した組換えアデノウイルス粒子を、酵素結合免疫吸着測定(ELISA)プレート上にコーティングし、次いで(NANP)n(配列番号60)反復を認識する抗PyCS抗体で検出した。該抗体は、すべてのカプシド修飾型アデノウイルスを認識した(図47Bおよび48B)。ELISA分析の結果は、カプシドタンパク質に取り込まれたPfSP−Bエピトープがアデノウイルスビリオンの外側に十分に曝露されたことを示唆する。
【0130】
初回刺激−追加免疫による免疫化3
図49Aに示すように、ナイーブBALB/cマウスに 、組換えPfCSPアデノウイルスを複数回、用量を増加して(すなわち、1×108、1×109、および1×1010v.p.)、3週間間隔で投与した。上述のように、PfCSP特異的体液性応答を、(NANP)n反復配列(配列番号60)を含有する1μg/mL(T1B)4、CS反復ペプチドでコーティングしたELISAプレートを用いるELISAによって決定した(Calvo−Calleら 2006)。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて、野生型PfCSPとカプシド修飾型アデノウイルスとの差異を決定した。すべてのカプシド修飾型アデノウイルスは、野生型PfCSPよりも統計学的に高い抗NANP抗体価を誘発した。
【0131】
初回刺激−追加免疫による免疫化4
次に、図50Aに示すように、ナイーブBALB/cマウスに、(NANP)nの反復を4、6、8、または10有する(配列番号60;n=4、6、8、10)HVR1修飾型PfCSPアデノウイルスの「追加免疫」を、用量を増加して複数回(すなわち、1×108、1×109、および1×1010v.p.)、3週間間隔で投与した。PfCSP特異的体液体性免疫応答を上述のように測定した。9週目に、すべてのHVR1修飾型アデノウイルスは、野生型PfCSPよりも有意に高い抗NANP抗体価を誘発した(図50B)。統計分析のために、値を対数変換し、次いで一元配置分散分析の後にDunnett検定を用いて、差異を決定した。
【0132】
初回刺激-追加免疫による免疫化5
次に、図51Aに示すように、ナイーブBALB/cマウスに、(NANP)nの反復を10、16、または22有する(配列番号60;n=10、16、22)HVR1修飾型アデノウイルスの「追加免疫」を、アジュバントと共に、またはアジュバントを含まずに、1×1010v.p.の用量で、3週間間隔で3回投与した。この実験で使用したアジュバントは、200μg/mLのSaponin(Sigma−Aldrich)を含有するSigma Adjuvant System(Sigma−Aldrich)である。免疫の前に、アデノウイルス溶液を同量のアジュバントと混合した。PfCSP特異的液体性免疫応答を上述のように測定し、次いで長いBエピトープを有するHVR1修飾型アデノウイルスがより高い抗体価を誘発したことを決定した(図51B)。
【0133】
実施例4:アデノウイルスコアタンパク質pVIIへのPyCS CD4エピトープの挿入
抗原特異的CD4 T細胞は、抗原特異的B細胞の発現と増殖のために必要とされる。したがって、PyCS CD4エピトープのアデノウイルスタンパク質への挿入によるカプシド修飾型アデノウイルスによってPyCS特異的体液性免疫応答を増強することが可能であるかどうかを決定するために、pVII内にPyCS CD4エピトープを有する(QGPGAP)3−Fib/PyCS−GFP、((QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFP)を構築した。pVIIは、アデノウイルスコアタンパク質のうちの1つであり、1ビリオンあたりのコピー数は700〜800である。これは、MHCクラスII分子上への効率的なCD4エピトープ提示に理想的である。図52Aに示すように、PyCS CD4エピトープをSDS−PAGEゲル上でpVIIに挿入することによってpVIIバンドを移動させる。
【0134】
PyCS CD4エピトープのpVIIへの挿入の効果を試験するために、図42Aに示すように、ナイーブBALB/cマウスを(QGPGAP)3−Fib/PyCS−GFPまたは(QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFPで免疫して、10週目に抗QGPGAP抗体価をELISAで決定した。(QGPGAP)3−Fib/CD4−pVII−1/PyCS−GFPは、(QGPGAP)3−Fib/PyCS−GFPよりも有意に高い抗QGPGAP抗体価を誘発し(図52B)、pVIIへのPyCS CD4エピトープの挿入が、カプシド修飾型アデノウイルスによって誘発された体液性免疫応答を増強したことを示した。
【0135】
アデノウイルスコアタンパク質pVIIへのPfCSP CD4エピトープ挿入
アデノウイルスコアタンパク質内の異なる位置へのPfCSP CD4+エピトープ挿入の、アデノウイルス誘発免疫応答への効果を評価するために、第1の核移行シグナル(NLS)の前、または2つのNLSの間にPfCSP CD4+エピトープを有するHVR1修飾型PfCSPアデノウイルスを構築した(図53A)。
【0136】
pVIIへのエピトープ挿入を確認するために、上述のように精製組換えアデノウイルスをSDS−PAGEで分析した。図53Bに示すように、(NANP)4−HVR1/CD4−pVII−2/PfCSPおよび(NANP)4−HVR1/CD4−pVII−3/PfCSPのpVIIバンドを、エピトープを挿入することによって上方に移動させた。
【0137】
HVR1およびpVIIの修飾型PfCSPアデノウイルスによって誘発されたPfCSP特異的免疫応答
次に、図54Aに示すように、ナイーブBALB/cマウスに、HVR1にNANPの4反復と、pVIIにPfCD4+エピトープとを有するHVR1およびpVIIの修飾型PfCSPアデノウイルスの「追加免疫」を、用量を増加して複数回(すなわち、1×108、1×109、および1×1010v.p.)、3週間間隔で投与した。PfCSP特異的体液性免疫応答を上述のように測定した。6週目に、(NANP)4−HVR1/CD4−pVII−2/PfCSPおよび(NANP)4−HVRI/CD4−pVII−3/PfCSPは、(NANP)4−HVR1/PfCSPより有意に高い抗NANP抗体価を誘発した。CMIに関して、(NANP)4−HVR1/CD4−pVII−3/PfCSPは、(NANP)4−HVR1/PfCSPより有意に高いIFNγおよびIL−4分泌PfCSP特異的CD4+ T細胞を誘発した(図54C)。
【0138】
実施例5:カプシド修飾の抗アデノウイルス免疫への効果
in vitroアデノウイルス中和実験のために、アデノウイルス感染の前に表示した希釈の血清をAD293細胞に加えた。Caucasian血清試料は、Innovative Research(Novi,MI,USA)から入手した。すべてのフローサイトメトリーデータは、FlowJo v8.8ソフトウェア(Tree Star,Inc.Ashland,OR,USA)で分析した。表示した希釈のヒトアデノウイルス中和血清試料の存在下で、AD293細胞に各カプシド修飾型アデノウイルスを感染させ、続いてフローサイトメトリーによってGFP発現を測定した。HVR1のPyCS−Bエピトープとの置換によって、アデノウイルスは抗アデノウイルス血清型5血清に対して明らかに回復力があるようになり、一方HVR5またはFiberの修飾は効果がなかった(図55)。
【0139】
次に、HVR1がin vivo中和にとって重要な分子であるかどうかを決定した。この目的のために、既存の抗アデノウイルス免疫を十分に開始させるために、マウスに1×1010v.p.野生型空のアデノウイルスを2回感染させて、(図56A)、次いでELISAで決定したそれらの抗アデノウイルス抗体価に基づいてランダム化した。次いで、マウスにカプシド修飾型アデノウイルスまたは非修飾型アデノウイルスを単回免疫量で投与して、上述のようにPyCS−特異的CD8+ T細胞応答のレベルを測定した。他のカプシド修飾型または非修飾型アデノウイルスによって誘発されたワクチン接種と比較すると、(QGPGAP)3−HVR1/PyCS−GFPまたは(QGPGAP)3−HVR1/Fib/PyCS−GFPによるワクチン接種だけが有意により強力なCS特異的CD8+ T細胞応答を誘発することができた(図56B)。
【0140】
(QGPGAP)3エピトープに対する抗体応答のレベルも、測定した。抗体応答は、野生型空のアデノウイルスに感染させ、続いてカプシド修飾型rAdでワクチン接種したマウスにおいて、rAdのカプシドタンパク質上で発現される(図57A)。(QGPGAP)3−HVR1/PyCS−GFPおよび(QGPGAP)3−HVR1/Fib/PyCS−GFPをワクチン接種したマウスだけが、野生型PyCS−GFPをワクチン接種したマウスよりも有意に高い抗QGPGAP抗体価を開始することができた。
【0141】
上述の実施例は、実施形態をより十分に説明するように意図され、クレームされるいずれの実施形態の範囲を限定すると解釈されるものではない。加えて、本開示内に引用した参照文献、および後述するすべての参照文献は、それら全体を参照することによって本明細書に十分に記載されているかのように本明細書によって組み込まれる。
【0142】
参考文献
Abbink,P., Lemckert,A.A., Ewald,B.A., Lynch,D.M., Denholtz,M.,ら 2007. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J Virol. 81:4654−4663.
Alonso,P.L., Sacarlal,J., Aponte,J.J., Leach,A., Macete,E.,ら 2004. Efficacy of the RTS,S/AS02A vaccine against Plasmodium falciparum infection and disease in young African children: randomised controlled trial. Lancet. 364:1411−1420.
Alonso,P.L., Sacarlal,J., Aponte,J.J., Leach,A., Macete,E.,ら 2005. Duration of protection with RTS,S/AS02A malaria vaccine in prevention of Plasmodium falciparum disease in Mozambican children:single−blind extended follow−up of a randomised controlled trial. Lancet. 366:2012−2018.
Anderson,R.J., Hannan,CM., Gilbert,S.C., Laidlaw,S.M., Sheu,E.G.,ら 2004. Enhanced CD8+ T cell immune responses and protection elicited against Plasmodium berghei malaria by prime boost immunization regimens using a novel attenuated fowlpox virus. J Immunol. 172:3094−3100.
Athappilly,F.K., Murali,R., Rux,J.J., Cai, Z. & Burnett,R.M. The refined crystal structure of Hexon, the major coat protein of adenovirus type 2, at 2.9 A resolution. Journal of molecular biology 242,430−455(1994).
Barrat,F.J., Meeker,T,, Gregorio,J,, Chan, J.H., Uematsu,S.,ら. Nucleic acids of mammalian origin can act as endogenous ligands for Toll−like receptors and may promote systemic lupus erythematosus.J Exp Med. 202, 1131−1139(2005).
Bejon,P., Lusingu,J., Olotu,A., Leach,A., Lievens,M.,ら 2008. Efficacy of RTS,S/AS01 E vaccine against malaria in children 5 to 17 months of age. N Engl J Med. 359:2521−2532.
Belousova,N., Krendelchtchikova,V., Curiel,DT. & Krasnykh,V. Modulation of adenovirus vector tropism via incorporation of polypeptide ligands into the Fiber protein. Journal of virology 76, 8621−8631(2002).
Bergelson,J.M.ら. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science (New York, N.Y275, 1320−1323(1997).
Bewley,M.C, Springer,K., Zhang,Y.B., Freimuth,P. & Flanagan,J.M. Structural analysis of the mechanism of adenovirus binding to its human cellular receptor, CAR. Science (New York, N.Y286, 1579−1583(1999).
Bruna−Romero,O., Schmieg,J., Del Val,M., Buschle,M. & Tsuji,M. The dendritic cell−specific chemokine, dendritic cell−derived CC chemokine 1, enhances protective cell−mediated immunity to murine malaria. J Immunol 170, 3195−3203(2003).Hong,S.S., Karayan,L, Tournier,J., Curiel,DT. & Boulanger,P.A. Adenovirus type 5 Fiber knob binds to MHC class I alpha2 domain at the surface of human epithelial and B lymphoblastoid cells. The EMBO journal 16, 2294−2306(1997).
Bruna−Romero,O., Gonzalez−Aseguinolaza,G., Hafalla,J.C, Tsuji,M.,およびNussenzweig,R.S. 2001. Complete, long−lasting protection against malaria of mice primed and boosted with two distinct viral vectors expressing the same plasmodial antigen.Proc Natl Acad Sci U S A. 98:11491−11496.
Bruna−Romero,O., Rocha,CD., Tsuji,M. & Gazzinelli,RT. Enhanced protective immunity against malaria by vaccination with a recombinant adenovirus encoding the circumsporozoite protein of Plasmodium lacking the GPI−anchoring motif. Vaccine 22, 3575−3584(2004).
Calvo−Calle,J.M., Oliveira,G.A., Watta,CO., Soverow,J., Parra−Lopez,C,ら 2006. A linear peptide containing minimal T− and B−cell epitopes of Plasmodium falciparum circumsporozoite protein elicits protection against transgenic sporozoite challenge.Infect Immun. 74:6929−6939.
Clyde,D.F., Most,H., McCarthy,V.C,およびVanderberg,J.P. 1973. Immunization of man against sporozite−induced falciparum malaria. Am J Med Sci. 266:169−177.
Chroboczek,J., Ruigrok,R.W.& Cusack,S. Adenovirus Fiber.Current topics in microbiology and immunology 199(Pt1), 163−200(1995).
Chu,Y., Heistad,D., Cybulsky,M.I.& Davidson,B.L. Vascular cell adhesion molecule−1 augments adenovirus−mediated gene transfer.Arterioscler Thromb Vasc Biol 21, 238−242(2001).
Crawford−Miksza,L. & Schnurr,D.P. Analysis of 15 adenovirus Hexon proteins reveals the location and structure of seven hypervariable regions containing serotype−specific residues.Journal of virology 70, 1836−1844(1996).
Crompton,J., Toogood,Cl., Wallis,N. & Hay, RT. Expression of a foreign epitope on the surface of the adenovirus Hexon.The Journal of general virology75(Pt1), 133−139(1994).
Crystal,R.G. Transfer of genes to humans:early lessons and obstacles to success.
Science (New York, N.Y270, 404−410(1995).
Douglas,JT. Adenoviral vectors for gene therapy.Molecular biotechnology 36, 71−80(2007).
Edelman,R., Hoffman,S.L., Davis,J.R., Beier,M., Sztein,M.B.,ら 1993. Long−term persistence of sterile immunity in a volunteer immunized with X−irradiated Plasmodium falciparum sporozoites.J Infect Dis. 168: 1066−1070.
Grillot,D., Valmori,D., Lambert,P.H., Corradin,G.,およびDel Giudice,G. 1993. Presentation of T cell epitopes assembled as multiple−antigen peptides to murine and human T lymphocytes.Infect Immun. 61:3064−3067.
Graham,F.L.& Prevec,L. Adenovirus−based expression vectors and recombinant vaccines.Biotechnology (Reading, Mass 20, 363−390(1992).
Gwadz,R.W., Cochrane,A.H., Nussenzweig,V.,およびNussenzweig,R.S. 1979. Preliminary studies on vaccination of rhesus monkeys with irradiated sporozoites of Plasmodium knowlesi and characterization of surface antigens of these parasites.Bull World Health Organ.57 Suppl 1:165−173.
Hackett,N.R.ら. Use of quantitative TaqMan real−time PCR to track the time−dependent distribution of gene transfer vectors in vivo. Mol Ther2, 649−656(2000).
Harvey,B.G.ら. Airway epithelial CFTR mRNA expression in cystic fibrosis patients after repetitive administration of a recombinant adenovirus.The Journal of clinical investigation 104, 1245−1255(1999).
Heemskerk,B.ら. Adenovirus−specific CD4+ T cell clones recognizing endogenous antigen inhibit viral replication in vitro through cognate interaction. J Immunol 177, 8851−8859(2006).
Henry,L.J., Xia,D., Wilke,M.E., Deisenhofer,J. & Gerard,R.D. Characterization of the knob domain of the adenovirus type 5 Fiber protein expressed in Escherichia coli.Journal of virology 68, 5239−5246(1994).
Hong,S.S., Habib,N.A., Franqueville,L, Jensen,S. & Boulanger,P.A. Identification of adenovirus (ad) penton base neutralizing epitopes by use of sera from patients who had received conditionally replicative ad (addl1520) for treatment of liver tumors.Journal of virology 77, 10366−10375(2003).
Kester,K.E., Cummings,J.F., Ockenhouse,C.F., Nielsen,R., Hall,B.T.,ら 2008. Phase 2a trial of 0, 1, and 3 month and 0, 7, and 28 day immunization schedules of malaria vaccine RTS,S/AS02 in malaria−naive adults at the Walter Reed Army Institute of Research.Vaccine. 26:2191−2202.
Kirby,I.ら. Mutations in the DG loop of adenovirus type 5 Fiber knob protein abolish highaffinity binding to its cellular receptor CAR.Journal of virology 73, 9508−9514(1999).
Koizumi,N., Mizuguchi,H., Utoguchi,., Watanabe,Y. & Hayakawa,T. Generation of Fiber−modified adenovirus vectors containing heterologous peptides in both the HI loop and C terminus of the Fiber knob.The journal of gene medicine 5, 267−276(2003).
Kozak M. 1987.An analysis of 5’−noncoding sequences from 699 vertebrate messenger RNAs.Nucleic Acids Res. 15:8125−148.
Krause,A., Joh,J.H., Hackett,N.R., Roelvink,P.W., Bruder,J.T.,ら 2006. Epitopes expressed in different adenovirus capsid proteins induce different levels of epitope−specific immunity.J Virol. 80:5523−5530.
Labow,D., Lee,S., Ginsberg,R.J., Crystal,R.G. & Korst, R.J. Adenovirus vector−mediated gene transfer to regional lymph nodes.Human gene therapy11,759−769(2000).
Leen,A.M.ら. Identification of Hexon−specific CD4 and CD8 T cell epitopes for vaccine and immunotherapy.Journal of virology 82, 546−554(2008).
Leopold,P.L. & Crystal,R.G. Intracellular trafficking of adenovirus:many means to many ends.Advanced drug delivery reviews 59, 810−821(2007).
Mastrangeli,A.ら. ”Sero−switch” adenovirus−mediated in v/Vo gene transfer:circumvention of anti−adenovirus humoral immune defenses against repeat adenovirus vector administration by changing the adenovirus serotype.Human gene therapy 7, 79−87(1996).
Mathias,P., Wickham,T., Moore,M. & Nemerow,G. Multiple adenovirus serotypes use alpha v integrins for infection.Journal of virology 68, 6811−6814(1994).
McConnell,M.J., Danthinne,X.,およびImperiale,M.J. 2006.Characterization of a permissive epitope insertion site in adenovirus Hexon.J Virol. 80:5361−5370.
Meier,O. & Greber,U.F. Adenovirus endocytosis.The journal of gene medicine 5, 451−462(2003).
Miyazawa,N.ら. Fiber swap between adenovirus subgroups B and C alters intracellular trafficking of adenovirus gene transfer vectors.Journal of virology 73, 6056−6065(1999).
Miyazawa,N., Crystal,R.G. & Leopold,P.L. Adenovirus serotype 7 retention in a late endosomal compartment prior to cytosol escape is modulated by Fiber protein.Journal of virology75, 1387−1400(2001).
Mizuguchi,H. & Hayakawa,T. Targeted adenovirus vectors.Human gene therapy 15, 1034−1044(2004).
Nakano,M.Y., Boucke,K., Suomalainen,M., Stidwill,R.P. & Greber,U.F. The first step of adenovirus type 2 disassembly occurs at the cell surface, independently of endocytosis and escape to the cytosol.Journal of virology 74, 7085−7095(2000).
Nicklin,S.A.ら. Ablating adenovirus type 5 Fiber−CAR binding and HI loop insertion of the SIGYPLP peptide generate an endothelial cell−selective adenovirus.Mol Ther4, 534−542(2001).
Noureddini,S.C. & Curiel,DT. Genetic targeting strategies for adenovirus.Molecular pharmaceutics 2, 341−347(2005).
Nussenzweig,R.S., Vanderberg,J., Most,H.,およびOrton,C. 1967. Protective immunity produced by the injection of x−irradiated sporozoites of Plasmodium berghei.Nature. 216:160−162.
Nussenzweig,R.S. & Long, CA.Malaria vaccines: multiple targets.Science (New York, N.Y 265, 1381−1383(1994).
Onion,D.ら. The CD4+ T cell response to adenovirus is focused against conserved residues within the Hexon protein.The Journal of general virology 88, 2417−2425(2007).
Ophorst,O.J., Radosevic,K., Havenga,M.J., Pau,M.G., Holterman,L.,ら 2006. lmmunogenicity and protection of a recombinant human adenovirus serotype 35−based malaria vaccine against Plasmodium yoelii in mice.Infect Immun. 74:313−320.
Oualikene,W., Gonin,P. & Eloit,M. Short and long term dissemination of deletion mutants of adenovirus in permissive (cotton rat) and non−permissive (mouse) species.The Journal of general virology 75(Pt10), 2765−2768(1994).
Priddy,F.H., Brown,D., Kublin,J., Monahan,K., Wright,D.P.,ら 2008. Safety and immunogenicity of a replication−incompetent adenovirus type 5 HIV−1 clade B gag/pol/nef vaccine in healthy adults.Clin Infect Dis. 46: 1769−1781.
Roberts,M.M., White,J.L., Grutter,M.G. & Burnett,R.M. Three−dimensional structure of the adenovirus major coat protein Hexon.Science (New York, N.Y 232, 1148−1151(1986).
Roberts,D.M., Nanda,A., Havenga,M.J., Abbink,P., Lynch,D.M.,ら 2006. Hexon−chimaeric adenovirus serotype 5 vectors circumvent pre−existing anti−vector immunity.Nature. 441 : 239−243.
Rodrigues,E.G., Zavala,F., Eichinger,D., Wilson,J.M.,およびTsuji,M. 1997. Single immunizing dose of recombinant adenovirus efficiently induces CD8+ T cell−mediated protective immunity against malaria.J.Immunol. 158:1268−1274.
Rodrigues,E.G., Zavala,F., Nussenzweig,R.S., Wilson,J.M. & Tsuji,M. Efficient induction of protective anti−malaria immunity by recombinant adenovirus.Vaccine 16, 1812−1817(1998).
Roelvink,P.W., Mi Lee,G., Einfeld,D.A., Kovesdi,I. & Wickham, T.J. Identification of a conserved receptor−binding site on the Fiber proteins of CAR−recognizing adenoviridae.Science (New York, N.Y 286, 1568−1571(1999).
Rux,J.J. & Burnett,R.M. Type−specific epitope locations revealed by X−ray crystallographic study of adenovirus type 5 Hexon.Mol Ther 1,18−30(2000).
Rux,J.J.& Burnett,R.M. Adenovirus structure.Human gene therapy f(C)5, 1167−1176(2004).
Roy,S.ら. Use of chimeric adenoviral vectors to assess capsid neutralization determinants.Virology 333, 207−214(2005).
Silvie,O., Greco,C, Franetich,J.F., Dubart−Kupperschmitt,A., Hannoun,L,ら 2006. Expression of human CD81 differently affects host cell susceptibility to malaria sporozoites depending on the Plasmodium species.Cell Microbiol. 8:1134−1146.
Sumida,S.M., Truitt,D.M., Lemckert,A.A., Vogels,R., Custers,J.H.,ら 2005. Neutralizing antibodies to adenovirus serotype 5 vaccine vectors are directed primarily against the adenovirus Hexon protein.J Immunol. 174:7179−7185.
Sun,P., Schwenk,R., White,K., Stoute,J.A., Cohen,J.,ら 2003. Protective immunity induced with malaria vaccine, RTS, S, is linked to Plasmodium falciparum circumsporozoite protein−specific CD4+ and CD8+ T cells producing IFN−gamma.J Immunol. 171:6961−6967.
Tao,D., Barba−Spaeth,G., Rai,U., Nussenzweig,V., Rice,CM.,およびNussenzweig,R.S. 2005. Yellow fever 17D as a vaccine vector for microbial CTL epitopes:protection in a rodent malaria model.J Exp Med. 201:201−209.
Teramoto,S.ら. Investigation of effects of anesthesia and age on aspiration in mice through LacZ gene transfer by recombinant E1 −deleted adenovirus vectors.American journal of respiratory and critical care medicine 158, 1914−1919(1998).
Top,F.H.,Jr., Dudding,B.A., Russell,P.K. & Buescher,E.L. Control of respiratory disease in recruits with types 4 and 7 adenovirus vaccines.American journal of epidemiology 94, 142−146(1971).
Top,F.H.,Jr. Control of adenovirus acute respiratory disease in U.S. Army trainees.Yale J Biol Med 48, 185−195(1975).
Tsuji,M., Romero,P., Nussenzweig,R.S.,およびZavala,F. 1990. CD4+ cytolytic T cell clone confers protection against murine malaria.J Exp Med. 172:1353−1357.
Tsuji,M., Rodrigues,E.G.& Nussenzweig,S. Progress toward a malaria vaccine:efficient induction of protective anti−malaria immunity.Biol Chem 382, 553−570(2001).
Wickham,T.J., Mathias,P., Cheresh,D.A. & Nemerow,G.R. lntegrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment.Cell 73, 309−319(1993).
Wohlfart,C. Neutralization of adenoviruses:kinetics, stoichiometry, and mechanisms.Journal of virology 62, 2321−2328(1988).
Worgall,S.ら. Modification to the capsid of the adenovirus vector that enhances dendritic cell infection and transgene−specific cellular immune responses.Journal of virology 78, 2572−2580(2004).
Worgall,S., Krause,A., Rivara,M., Hee,K.K., Vintayen,E.V.,ら 2005. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid.J Clin Invest.115:1281−1289.
Worgall,S., Krause,A., Qiu,J., Joh,J., Hackett,N.R.,およびCrystal,R.G. 2007. Protective immunity to pseudomonas aeruginosa induced with a capsid−modified adenovirus expressing P. aeruginosa OprF.J Virol. 81:13801−13808.
Wu,H., Dmitriev,I., Kashentseva,E., Seki,T., Wang,M.,ら 2002. Construction and characterization of adenovirus serotype 5 packaged by serotype 3 Hexon.J Virol. 76:12775−12782.
Wu,H.ら. Identification of sites in adenovirus Hexon for foreign peptide incorporation.Journal of virol, 78:3382−3390(2005).
Xia,D., Henry,L., Gerard,R.D. & Deisenhofer,J. Structure of the receptor binding domain of adenovirus type 5 Fiber protein.Current topics in microbiology and immunology 199(Pt1), 39−46(1995).
Yang,Y., Li,Q., Ertl,H.C. & Wilson,J.M. Cellular and humoral immune responses to viral antigens create barriers to lung−directed gene therapy with recombinant adenoviruses.Journal of virology 69, 2004−2015(1995).
Youil,R., Toner,T.J., Su,Q., Chen,M., Tang,A.,ら 2002. Hexon gene switch strategy for the generation of chimeric recombinant adenovirus.Hum Gene Ther. 13:311−320.
【配列表フリーテキスト】
【0143】
配列番号1:コドン最適化ネズミマラリア原虫スポロゾイト周囲タンパク質
配列番号2:コドン最適化熱帯熱マラリア原虫スポロゾイト周囲タンパク質
配列番号3:(QGPGAP)3−HVR1修飾型Hexonタンパク質
配列番号4:(QGPGAP)4−HVR1修飾型Hexonタンパク質
配列番号5:(QGPGAP)5−HVR1修飾型Hexonタンパク質
配列番号6:(QGPGAP)6−HVR1修飾型Hexonタンパク質
配列番号7:(QGPGAP)7−HVR1修飾型Hexonタンパク質
配列番号8:(QGPGAP)8−HVR1修飾型Hexon配列
配列番号9:(QGPGAP)9−HVR1修飾型Hexonタンパク質
配列番号10:(QGPGAP)11−HVR1修飾型Hexonタンパク質
配列番号11:(QGPGAP)12−HVR1修飾型Hexonタンパク質
配列番号12:(NANP)4−HVR1修飾型Hexonタンパク質
配列番号13:(NANP)6−HVR1修飾型Hexonタンパク質
配列番号14:(NANP)8−HVR1修飾型Hexonタンパク質
配列番号15:(NANP)10−HVR1修飾型Hexonタンパク質
配列番号16:(NANP)12−HVR1修飾型Hexonタンパク質
配列番号17:(NANP)14−HVR1修飾型Hexonタンパク質
配列番号18:(NANP)16−HVR1修飾型Hexonタンパク質
配列番号19:(NANP)18−HVR1修飾型Hexonタンパク質
配列番号20:(NANP)20−HVR1修飾型Hexonタンパク質
配列番号21:(NANP)22−HVR1修飾型Hexonタンパク質
配列番号22:(NANP)28−HVR1修飾型Hexonタンパク質
配列番号23:(QGPGAP)3−HVR5修飾型Hexonタンパク質
配列番号24:(QGPGAP)3修飾型Fiberタンパク質
配列番号25:(NANP)4修飾型Fiberタンパク質
配列番号26:ネズミマラリア原虫CD4+エピトープ修飾型pVII配列
配列番号27:熱帯熱マラリア原虫CD4+エピトープ修飾型pVII−1配列
配列番号28:熱帯熱マラリア原虫CD4+エピトープ修飾型pVII−2配列
配列番号29:熱帯熱マラリア原虫CD4+エピトープ修飾型pVII−3配列
配列番号30:合成コンストラクト
配列番号31:合成コンストラクト
配列番号32:合成コンストラクト
配列番号33:合成コンストラクト
配列番号34:合成コンストラクト
配列番号35:合成コンストラクト
配列番号36:合成コンストラクト
配列番号37:合成コンストラクト
配列番号38:合成コンストラクト
配列番号39:合成コンストラクト
配列番号40:合成コンストラクト
配列番号41:合成コンストラクト
配列番号42:合成コンストラクト
配列番号43:合成コンストラクト
配列番号44:合成コンストラクト
配列番号45:合成コンストラクト
配列番号46:合成コンストラクト
配列番号47:合成コンストラクト
配列番号48:合成コンストラクト
配列番号49:合成コンストラクト
配列番号50:合成コンストラクト
配列番号51:合成コンストラクト
配列番号52:合成コンストラクト
配列番号53:合成コンストラクト
配列番号54:合成コンストラクト
配列番号55:合成コンストラクト
配列番号56:合成コンストラクト
配列番号57:合成コンストラクト
配列番号58:合成コンストラクト
配列番号59:ネズミマラリア原虫
配列番号60:熱帯熱マラリア原虫
配列番号61:ネズミマラリア原虫
配列番号62:熱帯熱マラリア原虫
配列番号63:三日熱マラリア原虫
配列番号64:四日熱マラリア原虫
配列番号65:ネズミマラリア原虫
配列番号66:ネズミマラリア原虫
【特許請求の範囲】
【請求項1】
組換えアデノウイルスプラスミドベクター由来の組換えアデノウイルスであって、前記組換えアデノウイルスプラスミドベクターが、
異種プロモーター配列に作用可能に連結されたマラリア原虫(Plasmodium)スポロゾイト周囲タンパク質遺伝子またはその抗原部分をコードするヌクレオチド配列と、
1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープ配列が前記1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質の少なくとも一部に挿入されている、または一部を置換する組換えアデノウイルス。
【請求項2】
前記マラリア原虫スポロゾイト周囲タンパク質遺伝子が、熱帯熱マラリア原虫(Plasmodium falciparum)スポロゾイト周囲タンパク質遺伝子またはネズミマラリア原虫(Plasmodium yoelii)スポロゾイト周囲タンパク質遺伝子をさらに含む請求項1に記載のアデノウイルス。
【請求項3】
前記マラリア原虫スポロゾイト周囲タンパク質遺伝子が、コドン最適化熱帯熱マラリア原虫スポロゾイト周囲タンパク質遺伝子またはコドン最適化ネズミマラリア原虫スポロゾイト周囲タンパク質遺伝子をさらに含む、請求項1に記載のアデノウイルス。
【請求項4】
前記コドン最適化タンパク質遺伝子が、配列番号1または配列番号2によってコードされる請求項3に記載のアデノウイルス。
【請求項5】
前記免疫原性エピトープ配列が、マラリア原虫スポロゾイト周囲タンパク質遺伝子のB細胞エピトープ配列および/またはT細胞エピトープ配列をさらに含む請求項1に記載のアデノウイルス。
【請求項6】
前記カプシドタンパク質遺伝子がHexon高頻度可変領域(HVR)配列をさらに含む請求項5に記載のアデノウイルス。
【請求項7】
前記HVR配列がHVR1またはHVR5の配列をさらに含み、および前記B細胞エピトープが、
a)HVR1配列またはHVR5配列に挿入される;または
b)HVR1配列またはHVR5配列の一部を置換する
請求項6に記載のアデノウイルス。
【請求項8】
前記修飾型カプシドタンパク質遺伝子が、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、または配列番号23からなる群から選択される核酸配列によってコードされる請求項7に記載のアデノウイルス。
【請求項9】
前記HVR配列がHVR1配列もしくはHVR5配列を含み、CD4+またはCD8+のT細胞エピトープが、
a)HVR1配列またはHVR5配列に挿入される;または
b)HVR1配列またはHVR5配列の一部を置換する
請求項6に記載のアデノウイルス。
【請求項10】
前記カプシドタンパク質遺伝子がカプシドFiberタンパク質遺伝子をさらに含み、前記B細胞エピトープ配列が該Fiberタンパク質遺伝子に挿入される請求項5に記載のアデノウイルス。
【請求項11】
前記修飾型カプシドタンパク質遺伝子が配列番号24または配列番号25によってコードされる請求項10に記載のアデノウイルス。
【請求項12】
前記B細胞エピトープ配列が熱帯熱マラリア原虫スポロゾイト周囲タンパク質遺伝子B細胞エピトープ配列である請求項5から11のいずれかに記載のアデノウイルス。
【請求項13】
前記B細胞エピトープ配列が(NANP)n(配列番号60)であり、nが4以上である請求項12に記載のアデノウイルス。
【請求項14】
前記B細胞エピトープ配列が(NANP)4、(NANP)6、(NANP)8、(NANP)10、(NANP)12、(NANP)14、(NANP)16、(NANP)18、(NANP)20、(NANP)22または(NANP)28である請求項12に記載のアデノウイルス。
【請求項15】
前記免疫原性エピトープ配列がマラリア原虫スポロゾイト周囲タンパク質遺伝子のCD4+ T細胞エピトープ配列をさらに含む請求項1に記載のアデノウイルス。
【請求項16】
前記コアタンパク質遺伝子がpVIIタンパク質遺伝子をさらに含み、および前記CD4+ T細胞エピトープ配列が前記pVIIタンパク質遺伝子に挿入される請求項15に記載のアデノウイルス。
【請求項17】
前記修飾型コアタンパク質遺伝子が配列番号26、配列番号27、配列番号28または配列番号29によってコードされる請求項16に記載のアデノウイルス。
【請求項18】
前記CD4+ T細胞エピトープ配列が熱帯熱マラリア原虫スポロゾイト周囲タンパク質遺伝子CD4+ T細胞エピトープ配列である請求項15から17のいずれかに記載のアデノウイルス。
【請求項19】
前記CD4+ T細胞エピトープ配列がEYLNKIQNSLSTEWSPCSVT(配列番号62)である請求項16に記載のアデノウイルス。
【請求項20】
前記アデノウイルスが、HVR1−修飾型アデノウイルスベクター、Fiber−修飾型アデノウイルスベクター、HVR1とFiberとの修飾型アデノウイルスベクター、FiberとpVIIとの修飾型アデノウイルスベクター、HVR1とpVIIとの修飾型アデノウイルスベクター、およびHVR1とFiberとpVIIとの修飾型アデノウイルスベクターからなる群から選択されるいずれか1つの組換えアデノウイルスプラスミドベクターから生成される請求項1に記載のアデノウイルス。
【請求項21】
被験者においてマラリア原虫スポロゾイト周囲タンパク質に対する細胞性および体液性の免疫応答を誘発する方法であって、前記被験者に組換えアデノウイルスの少なくとも1用量を投与することを含み、前記組換えアデノウイルスが組換えアデノウイルスプラスミドベクターに由来し、前記組換えアデノウイルスプラスミドベクターが、
異種プロモーターに作用可能に連結されたマラリア原虫スポロゾイト周囲タンパク質またはその抗原部分と、
修飾型カプシドタンパク質またはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープがカプシドタンパク質もしくはコアタンパク質に挿入れているか、または一部を置換する方法。
【請求項22】
前記組換えアデノウイルスと共にアジュバントを投与することをさらに含む請求項21に記載の方法。
【請求項23】
アデノウイルス血清型に対する既存の中和抗体が欠乏している被験者においてマラリア原虫スポロゾイト周囲タンパク質に対する細胞性および体液性の免疫応答を誘発する方法であって、前記被験者に
第1の組換えアデノウイルスの第1の初回刺激用量と、
続いて第2の組換えアデノウイルスの追加免疫の用量とを投与することを含み、
前記第1の組換えアデノウイルスが組換えアデノウイルスプラスミドベクターに由来し、および前記組換えアデノウイルスプラスミドベクターが、異種プロモーターに作用可能に連結したマラリア原虫スポロゾイト周囲タンパク質またはその抗原部分をコードするヌクレオチド配列を含み、および
前記第2の組換えアデノウイルスが組換えアデノウイルスプラスミドベクターに由来し、および前記組換えアデノウイルスプラスミドベクターが、異種プロモーターに作用可能に連結したマラリア原虫スポロゾイト周囲タンパク質またはその抗原部分と、修飾型カプシドタンパク質またはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープがカプシドタンパク質もしくはコアタンパク質の少なくとも一部に挿入されているか、または一部を置換する方法。
【請求項24】
前記組換えアデノウイルスと共にアジュバントを投与することをさらに含む請求項23に記載の方法。
【請求項25】
組換えアデノウイルスを含む医療組成物であって、前記組換えアデノウイルスがHVR1−修飾型アデノウイルスベクター、Fiber−修飾型アデノウイルスベクター、HVR1とFiberとの修飾型アデノウイルスベクター、FiberとpVIIとの修飾型アデノウイルスベクター、HVR1とpVIIとの修飾型アデノウイルスベクター、およびHVR1とFiberとpVIIとの修飾型アデノウイルスベクターからなる群から選択されるいずれか1つの組換えアデノウイルスプラスミドベクターから生成される医療組成物。
【請求項26】
組換えアデノウイルスを含むマラリア感染症用のワクチンであって、前記組換えアデノウイルスがHVR1−修飾型アデノウイルスベクター、Fiber−修飾型アデノウイルスベクター、HVR1とFiberとの修飾型アデノウイルスベクター、FiberとpVIIとの修飾型アデノウイルスベクター、HVR1とpVIIとの修飾型アデノウイルスベクター、およびHVR1とFiberとpVIIとの修飾型アデノウイルスベクターからなる群から選択されるいずれか1つの組換えアデノウイルスプラスミドベクターから生成されるマラリア感染症用のワクチン。
【請求項27】
前記ワクチンが前記被験者の筋肉内に、皮内に、または皮下に投与される請求項26に記載のマラリア感染症用のワクチン。
【請求項28】
前記ワクチンが前記被験者にアジュバントと共に投与される請求項26に記載のマラリア感染症用のワクチン。
【請求項1】
組換えアデノウイルスプラスミドベクター由来の組換えアデノウイルスであって、前記組換えアデノウイルスプラスミドベクターが、
異種プロモーター配列に作用可能に連結されたマラリア原虫(Plasmodium)スポロゾイト周囲タンパク質遺伝子またはその抗原部分をコードするヌクレオチド配列と、
1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープ配列が前記1つもしくは複数の修飾型カプシドタンパク質および/またはコアタンパク質の少なくとも一部に挿入されている、または一部を置換する組換えアデノウイルス。
【請求項2】
前記マラリア原虫スポロゾイト周囲タンパク質遺伝子が、熱帯熱マラリア原虫(Plasmodium falciparum)スポロゾイト周囲タンパク質遺伝子またはネズミマラリア原虫(Plasmodium yoelii)スポロゾイト周囲タンパク質遺伝子をさらに含む請求項1に記載のアデノウイルス。
【請求項3】
前記マラリア原虫スポロゾイト周囲タンパク質遺伝子が、コドン最適化熱帯熱マラリア原虫スポロゾイト周囲タンパク質遺伝子またはコドン最適化ネズミマラリア原虫スポロゾイト周囲タンパク質遺伝子をさらに含む、請求項1に記載のアデノウイルス。
【請求項4】
前記コドン最適化タンパク質遺伝子が、配列番号1または配列番号2によってコードされる請求項3に記載のアデノウイルス。
【請求項5】
前記免疫原性エピトープ配列が、マラリア原虫スポロゾイト周囲タンパク質遺伝子のB細胞エピトープ配列および/またはT細胞エピトープ配列をさらに含む請求項1に記載のアデノウイルス。
【請求項6】
前記カプシドタンパク質遺伝子がHexon高頻度可変領域(HVR)配列をさらに含む請求項5に記載のアデノウイルス。
【請求項7】
前記HVR配列がHVR1またはHVR5の配列をさらに含み、および前記B細胞エピトープが、
a)HVR1配列またはHVR5配列に挿入される;または
b)HVR1配列またはHVR5配列の一部を置換する
請求項6に記載のアデノウイルス。
【請求項8】
前記修飾型カプシドタンパク質遺伝子が、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、または配列番号23からなる群から選択される核酸配列によってコードされる請求項7に記載のアデノウイルス。
【請求項9】
前記HVR配列がHVR1配列もしくはHVR5配列を含み、CD4+またはCD8+のT細胞エピトープが、
a)HVR1配列またはHVR5配列に挿入される;または
b)HVR1配列またはHVR5配列の一部を置換する
請求項6に記載のアデノウイルス。
【請求項10】
前記カプシドタンパク質遺伝子がカプシドFiberタンパク質遺伝子をさらに含み、前記B細胞エピトープ配列が該Fiberタンパク質遺伝子に挿入される請求項5に記載のアデノウイルス。
【請求項11】
前記修飾型カプシドタンパク質遺伝子が配列番号24または配列番号25によってコードされる請求項10に記載のアデノウイルス。
【請求項12】
前記B細胞エピトープ配列が熱帯熱マラリア原虫スポロゾイト周囲タンパク質遺伝子B細胞エピトープ配列である請求項5から11のいずれかに記載のアデノウイルス。
【請求項13】
前記B細胞エピトープ配列が(NANP)n(配列番号60)であり、nが4以上である請求項12に記載のアデノウイルス。
【請求項14】
前記B細胞エピトープ配列が(NANP)4、(NANP)6、(NANP)8、(NANP)10、(NANP)12、(NANP)14、(NANP)16、(NANP)18、(NANP)20、(NANP)22または(NANP)28である請求項12に記載のアデノウイルス。
【請求項15】
前記免疫原性エピトープ配列がマラリア原虫スポロゾイト周囲タンパク質遺伝子のCD4+ T細胞エピトープ配列をさらに含む請求項1に記載のアデノウイルス。
【請求項16】
前記コアタンパク質遺伝子がpVIIタンパク質遺伝子をさらに含み、および前記CD4+ T細胞エピトープ配列が前記pVIIタンパク質遺伝子に挿入される請求項15に記載のアデノウイルス。
【請求項17】
前記修飾型コアタンパク質遺伝子が配列番号26、配列番号27、配列番号28または配列番号29によってコードされる請求項16に記載のアデノウイルス。
【請求項18】
前記CD4+ T細胞エピトープ配列が熱帯熱マラリア原虫スポロゾイト周囲タンパク質遺伝子CD4+ T細胞エピトープ配列である請求項15から17のいずれかに記載のアデノウイルス。
【請求項19】
前記CD4+ T細胞エピトープ配列がEYLNKIQNSLSTEWSPCSVT(配列番号62)である請求項16に記載のアデノウイルス。
【請求項20】
前記アデノウイルスが、HVR1−修飾型アデノウイルスベクター、Fiber−修飾型アデノウイルスベクター、HVR1とFiberとの修飾型アデノウイルスベクター、FiberとpVIIとの修飾型アデノウイルスベクター、HVR1とpVIIとの修飾型アデノウイルスベクター、およびHVR1とFiberとpVIIとの修飾型アデノウイルスベクターからなる群から選択されるいずれか1つの組換えアデノウイルスプラスミドベクターから生成される請求項1に記載のアデノウイルス。
【請求項21】
被験者においてマラリア原虫スポロゾイト周囲タンパク質に対する細胞性および体液性の免疫応答を誘発する方法であって、前記被験者に組換えアデノウイルスの少なくとも1用量を投与することを含み、前記組換えアデノウイルスが組換えアデノウイルスプラスミドベクターに由来し、前記組換えアデノウイルスプラスミドベクターが、
異種プロモーターに作用可能に連結されたマラリア原虫スポロゾイト周囲タンパク質またはその抗原部分と、
修飾型カプシドタンパク質またはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープがカプシドタンパク質もしくはコアタンパク質に挿入れているか、または一部を置換する方法。
【請求項22】
前記組換えアデノウイルスと共にアジュバントを投与することをさらに含む請求項21に記載の方法。
【請求項23】
アデノウイルス血清型に対する既存の中和抗体が欠乏している被験者においてマラリア原虫スポロゾイト周囲タンパク質に対する細胞性および体液性の免疫応答を誘発する方法であって、前記被験者に
第1の組換えアデノウイルスの第1の初回刺激用量と、
続いて第2の組換えアデノウイルスの追加免疫の用量とを投与することを含み、
前記第1の組換えアデノウイルスが組換えアデノウイルスプラスミドベクターに由来し、および前記組換えアデノウイルスプラスミドベクターが、異種プロモーターに作用可能に連結したマラリア原虫スポロゾイト周囲タンパク質またはその抗原部分をコードするヌクレオチド配列を含み、および
前記第2の組換えアデノウイルスが組換えアデノウイルスプラスミドベクターに由来し、および前記組換えアデノウイルスプラスミドベクターが、異種プロモーターに作用可能に連結したマラリア原虫スポロゾイト周囲タンパク質またはその抗原部分と、修飾型カプシドタンパク質またはコアタンパク質とをコードするヌクレオチド配列を含み、マラリア原虫スポロゾイト周囲の免疫原性エピトープがカプシドタンパク質もしくはコアタンパク質の少なくとも一部に挿入されているか、または一部を置換する方法。
【請求項24】
前記組換えアデノウイルスと共にアジュバントを投与することをさらに含む請求項23に記載の方法。
【請求項25】
組換えアデノウイルスを含む医療組成物であって、前記組換えアデノウイルスがHVR1−修飾型アデノウイルスベクター、Fiber−修飾型アデノウイルスベクター、HVR1とFiberとの修飾型アデノウイルスベクター、FiberとpVIIとの修飾型アデノウイルスベクター、HVR1とpVIIとの修飾型アデノウイルスベクター、およびHVR1とFiberとpVIIとの修飾型アデノウイルスベクターからなる群から選択されるいずれか1つの組換えアデノウイルスプラスミドベクターから生成される医療組成物。
【請求項26】
組換えアデノウイルスを含むマラリア感染症用のワクチンであって、前記組換えアデノウイルスがHVR1−修飾型アデノウイルスベクター、Fiber−修飾型アデノウイルスベクター、HVR1とFiberとの修飾型アデノウイルスベクター、FiberとpVIIとの修飾型アデノウイルスベクター、HVR1とpVIIとの修飾型アデノウイルスベクター、およびHVR1とFiberとpVIIとの修飾型アデノウイルスベクターからなる群から選択されるいずれか1つの組換えアデノウイルスプラスミドベクターから生成されるマラリア感染症用のワクチン。
【請求項27】
前記ワクチンが前記被験者の筋肉内に、皮内に、または皮下に投与される請求項26に記載のマラリア感染症用のワクチン。
【請求項28】
前記ワクチンが前記被験者にアジュバントと共に投与される請求項26に記載のマラリア感染症用のワクチン。
【図1】

【図2】
【図3】
【図4】
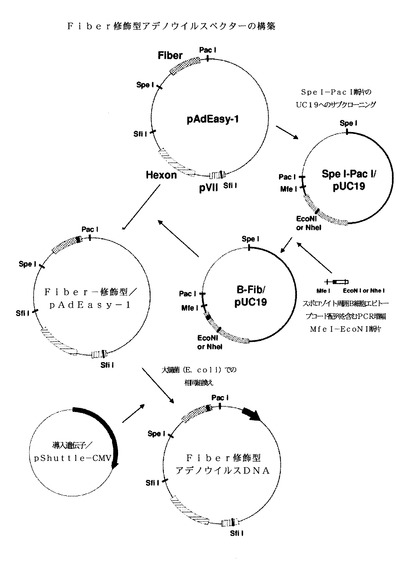
【図5】
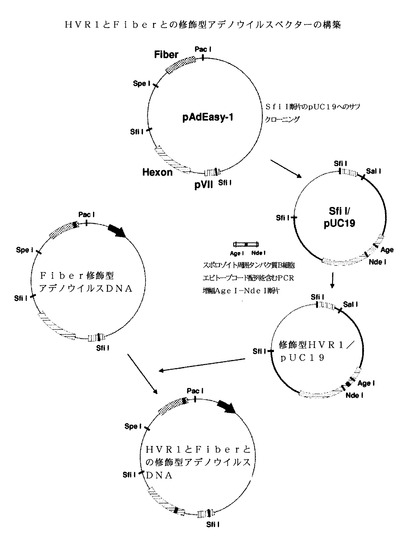
【図6】
【図7】
【図8】
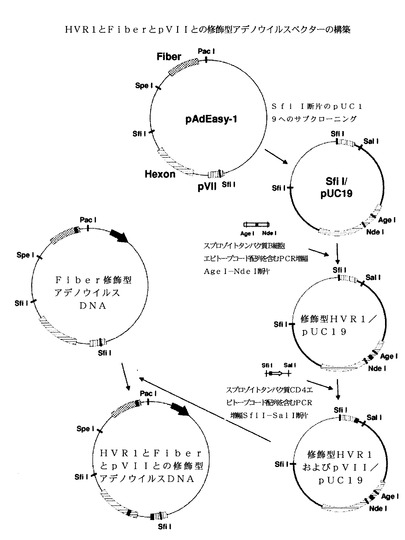
【図9】

【図10】

【図11−1】

【図11−2】

【図11−3】

【図12−1】

【図12−2】

【図12−3】

【図13−1】

【図13−2】

【図13−3】

【図14−1】

【図14−2】

【図14−3】

【図15−1】

【図15−2】

【図15−3】

【図16−1】

【図16−2】

【図16−3】

【図17−1】

【図17−2】

【図17−3】

【図18−1】

【図18−2】

【図18−3】

【図19−1】

【図19−2】

【図19−3】

【図20−1】

【図20−2】

【図20−3】

【図21−1】

【図21−2】

【図21−3】

【図22−1】

【図22−2】

【図22−3】

【図23−1】

【図23−2】

【図23−3】

【図24−1】

【図24−2】

【図24−3】

【図25−1】

【図25−2】

【図25−3】

【図26−1】

【図26−2】

【図26−3】

【図27−1】

【図27−2】

【図27−3】

【図28−1】

【図28−2】

【図28−3】

【図29−1】

【図29−2】

【図29−3】

【図30−1】

【図30−2】

【図30−3】

【図31−1】

【図31−2】

【図31−3】

【図32−1】

【図32−2】

【図33−1】

【図33−2】

【図34】

【図35】

【図36】

【図37】

【図38】
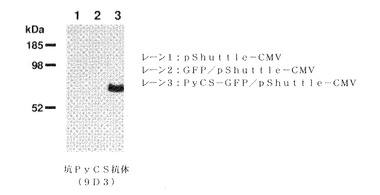
【図39】
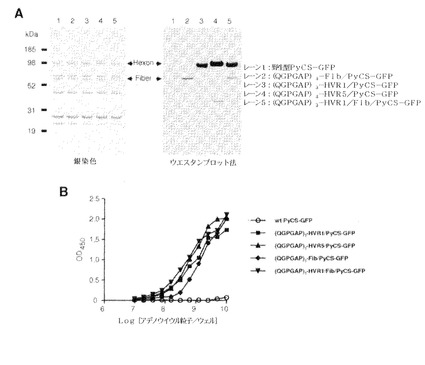
【図40】
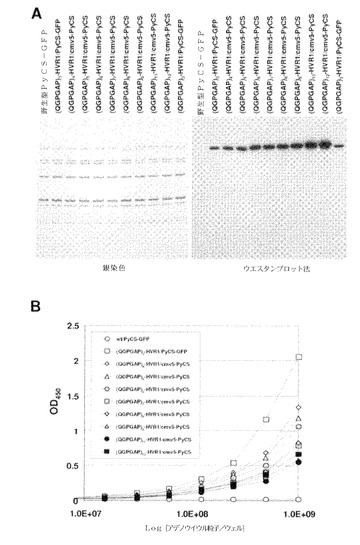
【図41】
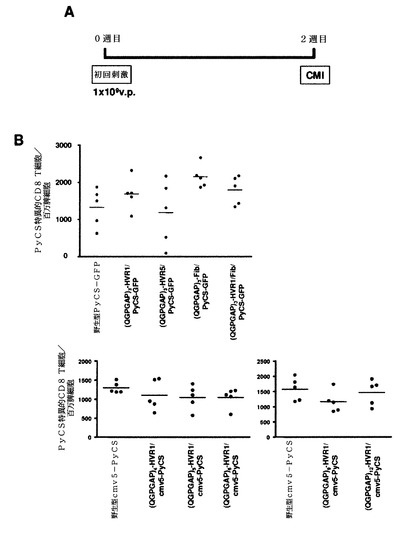
【図42】
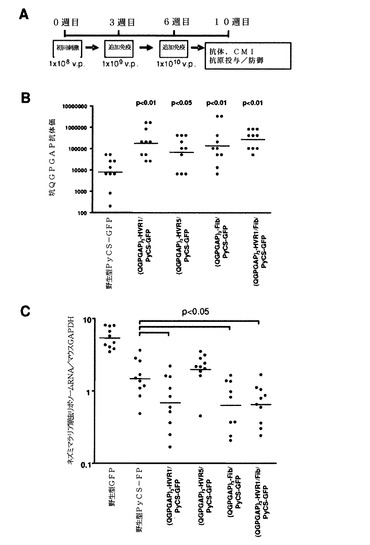
【図43】
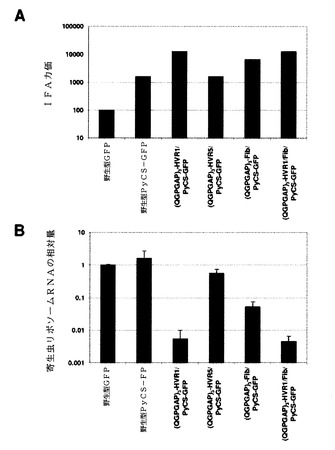
【図44】
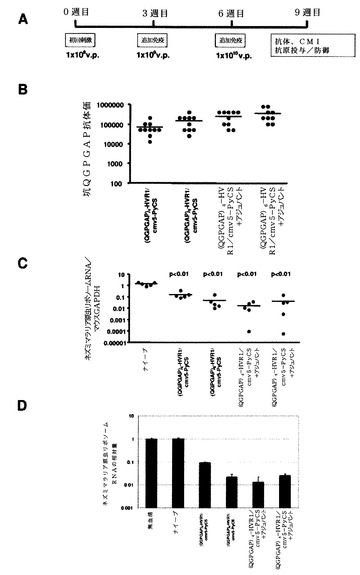
【図45】
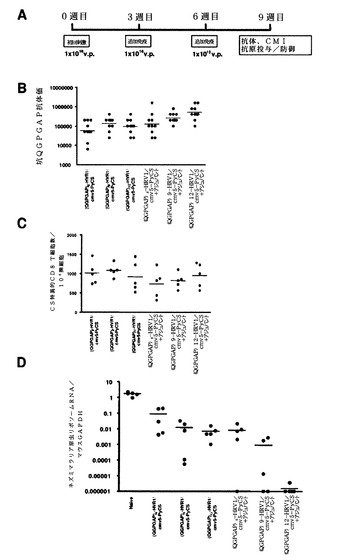
【図46】
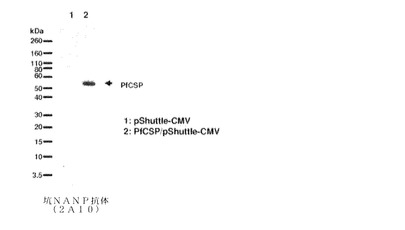
【図47】
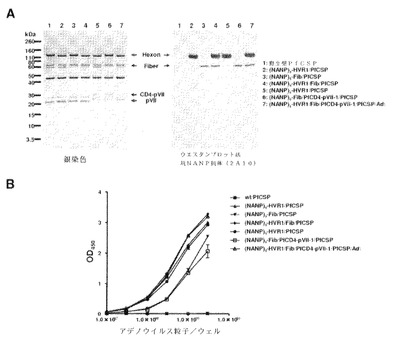
【図48】
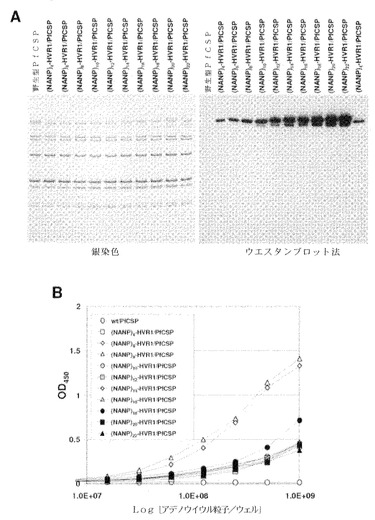
【図49】
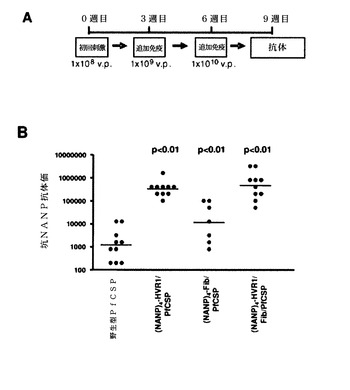
【図50】
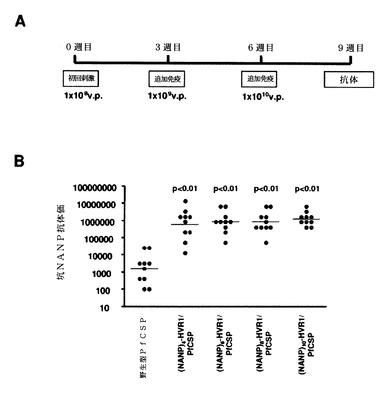
【図51】
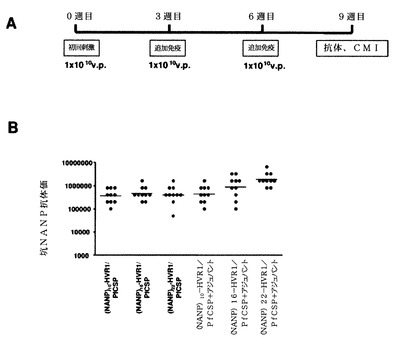
【図52】
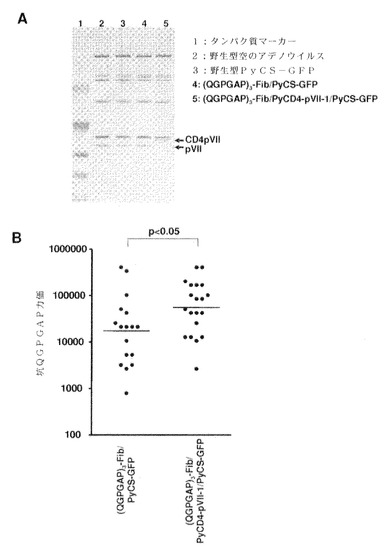
【図53】
【図54】
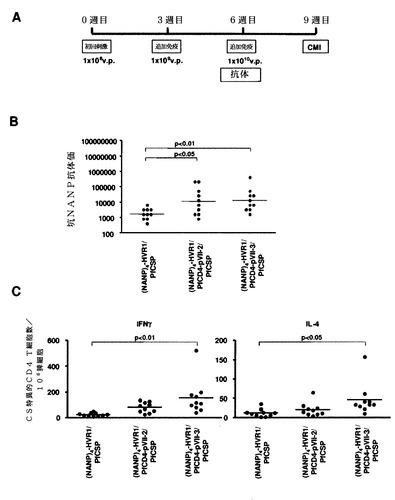
【図55】
【図56】
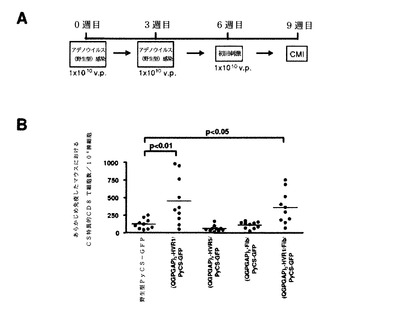
【図57】
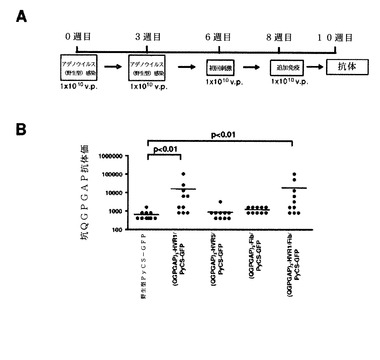
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11−1】
【図11−2】
【図11−3】
【図12−1】
【図12−2】
【図12−3】
【図13−1】
【図13−2】
【図13−3】
【図14−1】
【図14−2】
【図14−3】
【図15−1】
【図15−2】
【図15−3】
【図16−1】
【図16−2】
【図16−3】
【図17−1】
【図17−2】
【図17−3】
【図18−1】
【図18−2】
【図18−3】
【図19−1】
【図19−2】
【図19−3】
【図20−1】
【図20−2】
【図20−3】
【図21−1】
【図21−2】
【図21−3】
【図22−1】
【図22−2】
【図22−3】
【図23−1】
【図23−2】
【図23−3】
【図24−1】
【図24−2】
【図24−3】
【図25−1】
【図25−2】
【図25−3】
【図26−1】
【図26−2】
【図26−3】
【図27−1】
【図27−2】
【図27−3】
【図28−1】
【図28−2】
【図28−3】
【図29−1】
【図29−2】
【図29−3】
【図30−1】
【図30−2】
【図30−3】
【図31−1】
【図31−2】
【図31−3】
【図32−1】
【図32−2】
【図33−1】
【図33−2】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【図48】
【図49】
【図50】
【図51】
【図52】
【図53】
【図54】
【図55】
【図56】
【図57】
【公表番号】特表2013−502221(P2013−502221A)
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−525677(P2012−525677)
【出願日】平成22年8月18日(2010.8.18)
【国際出願番号】PCT/US2010/045952
【国際公開番号】WO2011/022522
【国際公開日】平成23年2月24日(2011.2.24)
【出願人】(596171834)ザ ロックフェラー ユニヴァーシティ (11)
【Fターム(参考)】
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願日】平成22年8月18日(2010.8.18)
【国際出願番号】PCT/US2010/045952
【国際公開番号】WO2011/022522
【国際公開日】平成23年2月24日(2011.2.24)
【出願人】(596171834)ザ ロックフェラー ユニヴァーシティ (11)
【Fターム(参考)】
[ Back to top ]