
免疫原性組成物およびその使用
本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物を提供する。荷電基が樹状細胞リガンドに共有結合しており、樹状細胞ターゲッティング成分と静電気的に会合している。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、静電的相互作用を通して抗原が樹状細胞リガンドと会合している免疫原性組成物に関する。本発明は、対象で免疫応答を誘導するための、これらの組成物の使用にさらに関する。
【背景技術】
【0002】
特に疾患から保護するために、ヒトおよび他の動物で免疫応答を起こすために用いることができる組成物の開発に対して、益々関心が高まっている。免疫応答を起こす際には、抗原が樹状細胞に遭遇することが必要である。樹状細胞は免疫細胞であって、哺乳動物の免疫系の一部を形成する。それらの主機能は、抗原物質をプロセシングして、免疫系の他の細胞の表面にそれを提示し、それによって抗原提示細胞として機能することである。
【0003】
樹状細胞への抗原のターゲッティング、および樹状細胞上のToll様受容体が強い免疫応答を誘発することが示されている脂質を含むリポペプチドは、以前に企図されている(国際公開第2004/014956号と国際公開第2004/014957号、それらの開示は参照により本明細書に組み込まれる)。
【0004】
いかなるペプチドも、有効な抗体応答を誘導できるためには、それは、免疫系によって認識されるエピトープとして知られる特定のアミノ酸配列を含まなければならない。詳細には、エピトープは、抗体応答をもたらすのに、Bリンパ球の表面に存在する特異的免疫グロブリン(Ig)受容体によって認識される必要がある。そのエピトープに特異的な抗体を産生できる形質細胞に最終的に分化するのは、これらの細胞である。これらのB細胞エピトープに加えて、免疫原は、B細胞が抗体産生細胞に分化するのに必要とされるシグナルを提供するのに必要な細胞、ヘルパーTリンパ球上に存在する特異的受容体に対して抗原提示細胞(APC)によって提示されるエピトープも含まなければならない。
【0005】
ウイルス感染の場合および癌の多くの場合では、回復における抗体の利点は限定され、免疫系は、ウイルス感染細胞または癌細胞を死滅させることができる細胞傷害性T細胞(CTL)で応答する。ヘルパーT細胞のように、CTLは、今度はクラスII分子ではなくMHCクラスIと会合して、それらの特異的ペプチドエピトープを表面に提示しているAPCとの相互作用によって先ず活性化される。活性化されると、CTLは同じペプチド/クラスI複合体を有する標的細胞にエンゲージし、その溶解を引き起こすことができる。ヘルパーT細胞がこの過程で役割を果たすことも、次第に明らかになってきている;APCがCTLを活性化することができるには、それは、ヘルパーT細胞からシグナルを先ず受け取り必要な共刺激分子の発現を上方制御しなければばらない。
【0006】
ヘルパーT細胞エピトープは、主要組織適合性複合体(MHC)のクラスII遺伝子によってコードされる、APCの表面に存在する分子により結合される。クラスII分子およびペプチドエピトープの複合体は、次にTヘルパーリンパ球の表面の特異的T細胞受容体(TCR)によって認識される。このように、MHC分子との関連において抗原エピトープで提示されるT細胞は活性化され、Bリンパ球が分化するのに必要なシグナルを提供することができる。
【0007】
その場合一般に、免疫原は、B細胞または細胞傷害性T細胞によって認識されるエピトープに加えて、ヘルパーT細胞が認識することができるエピトープを含まなければならない。この種のエピトープは非常に異なることがあることを理解するべきである。B細胞エピトープについては、B細胞受容体は本来の免疫原に直接に結合するので、立体構造が重要である。対照的に、T細胞によって認識されるエピトープは、エピトープの立体構造完全性に依存せず、CTLについては約9アミノ酸の短い配列、およびヘルパーT細胞については、長さの制限はより少ないが、わずかにより長い配列からなる。これらのエピトープのための唯一の要件は、それらがそれぞれクラスIまたはクラスII分子の結合クレフトに収まることができ、その結果複合体がT細胞受容体にエンゲージできることである。クラスII分子の結合部位は両端がオープンであり、結合ペプチドの長さのかなりより大きな変動を可能にしているが(Brown、J. H.、T. S. Jardetzky、J. C. Gorga、L. J. Stern、R. G. Urban、J. L. StromingerおよびD. C. Wiley. 1993. Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1. Nature 364: 33頁)、8アミノ酸残基と短いエピトープが報告されている(Fahrer, A. M.、Geysen, H. M.、White, D. O.、Jackson, D. C.およびBrown, L. E. Analysis of the requirements for class II-restricted T-cell recognition of a single determinant reveals considerable diversity in the T-cell response and degeneracy of peptide binding to I-ED J. Immunol. 1995. 155: 2849〜2857頁)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公開第2004/014956号
【特許文献2】国際公開第2004/014957号
【非特許文献】
【0009】
【非特許文献1】Brown、J. H.、T. S. Jardetzky、J. C. Gorga、L. J. Stern、R. G. Urban、J. L. StromingerおよびD. C. Wiley. 1993. Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1. Nature 364: 33頁
【非特許文献2】Fahrer, A. M.、Geysen, H. M.、White, D. O.、Jackson, D. C.およびBrown, L. E. Analysis of the requirements for class II-restricted T-cell recognition of a single determinant reveals considerable diversity in the T-cell response and degeneracy of peptide binding to I-ED J. Immunol. 1995. 155: 2849〜2857頁
【非特許文献3】Metzger, J. W.ら1995. J Pept Sci 1:184頁
【非特許文献4】Sacht, G.ら1998. Eur J Immunol 28:4207頁
【非特許文献5】Muhiradt, P. F.ら1998. Infect Immun 66: 4804頁
【非特許文献6】Muhiradt, P. F.ら1997. J Exp Med 185: 1951頁
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明者は、抗原が樹状細胞ターゲッティング成分と静電気的に会合している、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物を開発した。
【課題を解決するための手段】
【0011】
第1の態様では、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は負荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している正荷電基を含み、抗原の負荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0012】
第2の態様では、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は正荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している負荷電基を含み、抗原の正荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0013】
第3態様では、本発明は、対象に本発明の第1または第2の態様の免疫原性組成物を投与する段階を含む、対象で免疫応答を起こす方法を提供する。
【0014】
以下では、OVA=オボアルブミン;HEL=ニワトリ卵白リゾチーム;R4(S2Pam2Cys)=図5の1に示す構築物;E4(S2Pam2Cys)=図5の2に示す構築物;CFA=完全フロイントアジュバント。
【図面の簡単な説明】
【0015】
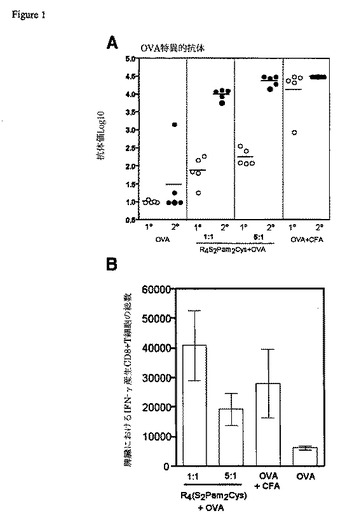
【図1】R4(S2Pam2Cys-OVA)複合体によるワクチン接種によって誘発された抗体および細胞媒介性応答を示す図である。(A)BALB/cマウスの群に、皮下に25μgのOVA単独、CFAに乳濁させたOVA、または等モルもしくは5倍モル過剰のR4(S2Pam2Cys)に混合したOVAを接種した。動物は、26日後に、抗原の第2の類似した用量が投与された。抗原の一次接種の27日後(○)、および二次接種の13日後(●)に採取された血液から血清を得た。抗体レベルをELISAで決定し、個々の抗体価を水平バーによって表される平均値と示した。(B)マウスに50日目に3回目の接種をし、脾臓を7日後に得た。SIINFEKL特異的IFN-γ産生CD8+T細胞の存在を検出するために、細胞内のサイトカイン染色を脾細胞で実施した。データは、3反復の試料の平均および標準偏差で示される。
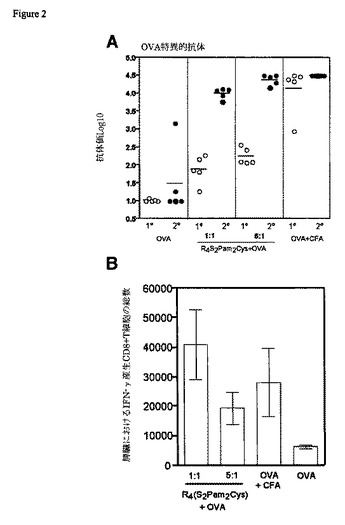
【図2】陽イオンおよび陰イオンのリポペプチド-タンパク質複合体によるワクチン接種によって誘発されたOVAおよびHEL-特異的抗体応答を示す図である。BALB/cマウスに、皮下に25μgのHEL(A)またはOVA(B)を単独で、CFAで乳濁させて、または抗原と混合したR4(S2Pam2Cys)もしくはE4(S2Pam2Cys)の同等量と一緒に接種した。マウスは一次接種の28日後(○)に採血し、32日目に追加免疫し、46日目に再度採血した(●)。その後抗体レベルをELISAで決定した。個々の動物の力価を、水平バーによって表される平均値と示す。
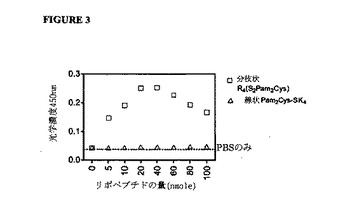
【図3】オボアルブミン-リポペプチド複合体の沈殿を示す図である。分枝状R4(S2Pam2Cys)または線状Pam2Cys-SK4リポペプチドの漸増量を、平底96ウェルプレート中の100μl PBSの総容量中の1nmoleのオボアルブミン(OVA)と混合した。450nmで溶液の光学濃度を決定することによって、次に溶液の混濁度を測定した。
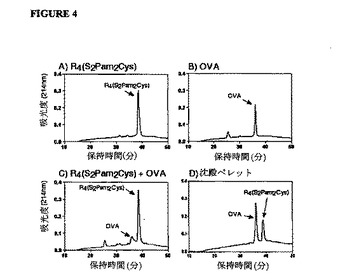
【図4】オボアルブミン- R4(S2Pam2Cys)リポペプチド溶液のHPLC分析を示す図である。遠心分離(1.2×105G)の後、100μl PBSの総容量に(A)100nmoleの分枝状R4(S2Pam2Cys)リポペプチド、(B)1nmoleのオボアルブミン(OVA)または(C)R4(S2Pam2Cys)リポペプチドおよびオボアルブミンの混合物のいずれかを含む溶液の上清で、HPLC分析を実施した。(D)リポペプチドおよびOVAの両方を含む混合物からの沈殿物を50%アセトニトリル水溶液に溶解し、その後HPLCによって分析した。R4(S2Pam2Cys)を含むすべての試料で、リポペプチドに対応するピークのアイデンティティーを質量分析によって検証した。
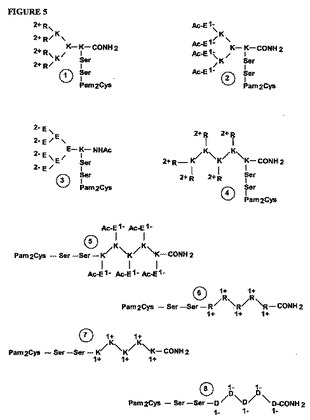
【図5】それらのそれぞれの静電的電荷が環境に提示されるように、末端位置に正荷電(アルギニン、R;リジン、K)または負荷電(アスパラギン酸、D;グルタミン酸、E)アミノ酸を含む分枝状(構造1〜5)および線状(構造6〜8)免疫原性組成物の一部の例の模式図である。各免疫原性組成物は、Toll様受容体2のリガンドであるジパルミトイル-S-グリセリルシステイン(Pam2Cys)も含む。2つのセリン残基(Ser)も組み込まれる。構築物2の場合、ペプチド構造は方向N→Cで組み立てられ、図示されるすべての他の構造はC→Nで組み立てられた。正負の静電的電荷は、電荷のサイズによって2-、2+、1-、1+などで示される。Ac=N末端にあるグルタミン酸の場合にアルファアミノ基の正電荷を抑制するために用いたアセチル基。
【発明を実施するための形態】
【0016】
本発明者は、樹状細胞ターゲッティング基に共有結合している荷電部分が静電気的に抗原と会合して、免疫応答を起こすために用いることができる免疫原性複合体を形成することを見出した。
【0017】
したがって、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は負荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している正荷電基を含み、抗原の負荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0018】
本発明のこの態様では、荷電したターゲッティング成分と静電気的に相互作用することができる負荷電領域または負荷電ドメインを抗原が含むことが重要である。しかし、好ましいことではあるが、抗原が総負電荷を有することは必須でない。負荷電基を加えることによって抗原の負電荷を増加させることも可能である。例えば、ポリペプチド抗原で、アスパラギン酸またはグルタミン酸残基の鎖をポリペプチドに加えることができよう。
【0019】
正荷電基は、少なくとも1つの正荷電アミノ酸を含むことが好ましい。正荷電基が、分枝状または線状のペプチド、好ましくは分枝状のペプチドであることも好ましい。様々な実施形態では、ペプチドは、少なくとも1つのアルギニン、ヒスチジン、オルニチンまたはリジン残基、またはそれらの組合せを含む。ペプチドが、少なくとも4つのアルギニン残基および/または少なくとも4つのリジン残基を含むことが好ましい。正荷電基が、少なくとも4つのアルギニン残基を含む分枝状のペプチドを含むことが特に好ましい。
【0020】
第2の態様では、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は正荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している負荷電基を含み、抗原の正荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0021】
本発明のこの態様では、荷電したターゲッティング成分と静電気的に相互作用することができる正荷電領域または正荷電ドメインを抗原が含むことが重要である。しかし、好ましいことではあるが、抗原が総正電荷を有することは必須でない。正荷電基を加えることによって、抗原の正電荷を増加させることも可能である。例えば、ポリペプチド抗原で、リジン、アルギニンまたはヒスチジン残基の鎖をポリペプチドに加えることができよう。
【0022】
負荷電基は、少なくとも1つの負荷電アミノ酸を含むことが好ましい。負荷電基が、分枝状または線状のペプチド、好ましくは分枝状のペプチドであることも好ましい。様々な実施形態では、ペプチドは、少なくとも1つのアスパラギン酸またはグルタミン酸残基、またはそれらの組合せを含む。ペプチドが、少なくとも4つのアスパラギン酸残基および/または少なくとも4つのグルタミン酸残基を含むことが好ましい。正荷電基が、少なくとも4つのグルタミン酸残基を含む分枝状のペプチドを含むことが特に好ましい。
【0023】
本発明の好ましい実施形態では、抗原は核酸でない。静電的相互作用だけによって抗原が樹状細胞ターゲッティング成分と結合していることも好ましい。
【0024】
本発明で用いることができる様々な樹状細胞リガンドを、表1に提示する。しかし、樹状細胞リガンドはTLRリガンドであることが好ましい。TLRリガンドは、脂質またはペプチドグリカンまたはリポタンパク質またはリポ多糖を含むことができる。詳細には、TLRリガンドは、パルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイルまたはデカノイルを含むことができる。好ましくは、TLRリガンドは、Pam2Cys、Pam3Cys、Ste2Cys、Lau2CysおよびOct2Cysからなる群から選択される。
【0025】
特定の実施形態では、TLRリガンドは、TLR-1またはTLR-6と会合していてもよいTLR-2に結合する。
【0026】
第3態様では、本発明は、対象に本発明の第1または第2の態様による免疫原性組成物を投与する段階を含む、対象で免疫応答を起こす方法を提供する。
【0027】
【表1】

【0028】
本発明の例示的な樹状細胞ターゲッティング化合物は、リポペプチド「Pam2Cys」である。当業者は、用語「リポペプチド」が、コンジュゲートされている1つまたは複数の脂質部分および1つまたは複数のアミノ酸配列を含む任意の組成物を意味することを理解しよう。「Pam2Cys」(ジパルミトイル-S-グリセリル-システインまたはS-[2,3ビス(パルミトイルオキシ)プロピル]システインとしても知られる)が合成されており(Metzger, J. W.ら1995. J Pept Sci 1:184頁)、マイコプラズマ・ファーメンタンス(Mycoplasma fermentans)から単離されたマクロファージ活性化リポペプチドであるMALP-2の脂質部分に対応する(Sacht, G.ら1998. Eur J Immunol 28:4207頁;Muhiradt, P. F.ら1998. Infect Immun 66: 4804頁;Muhiradt, P. F.ら1997. J Exp Med 185: 1951頁)。Pam2Cysは、TLR-2のリガンドであることが知られている。
【0029】
Pam2Cysは、下記式(I)の構造を有する:
【0030】
【化1】
【0031】
細胞表面TLRを標的にするために用いることができる他の脂質部分には、パルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイルまたはデカノイルが含まれる。好ましい基には、Pam2Cys、Pam3Cys、Ste2Cys、Lau2CysおよびOct2Cysが含まれる。
【0032】
本明細書で用いるように、「免疫応答」への言及は、侵入病原体、病原体に感染した細胞もしくは組織、癌細胞、または自己免疫もしくは病理学的炎症の場合には正常なヒト細胞もしくは組織に対する選択的な損傷、破壊、またはヒト体内からの除去をもたらす、リンパ球、抗原提示細胞、食細胞、顆粒球、および上記の細胞または肝臓によって産生される溶解性高分子(抗体、サイトカインおよび補体を含む)の協調作用への言及を意味する。
【0033】
本明細書全体を通して、単語「含む(comprise)」、または「含む(comprises)」もしくは「含んでいる(comprising)」などの変形形態は、明示された段階もしくは要素もしくは整数、または段階もしくは要素もしくは整数の群が含まれることを意味するが、いかなる他の段階もしくは要素もしくは整数または要素もしくは整数の群の除外を意味するものではないと理解される。
【0034】
本出願で引用されるすべての参考文献は、参照により本明細書に具体的に組み込まれている。
【0035】
本明細書での任意の先行技術への言及は、この先行技術がオーストラリアで一般常識の一部を形成することの肯定またはいかなる形の示唆ではなく、かつそのように解釈されるべきでない。
【0036】
当業者が、所望の用途に適するように、正荷電基およびTLRリガンドの任意の組合せを導入することができることが理解されよう。
【0037】
本発明は、次に、以下の非限定的実施例によってさらに記載される。
【0038】
材料および方法
リポペプチドの合成
分枝状および線状リポペプチドの合成を、PEG-S RAM樹脂(Rapp Polymere、Tubingen、Germany;置換因子0.27mmol/g)上で行った。分枝状リポペプチドR4(S2Pam2Cys)(構築物1)およびE4(S2Pam2Cys)(構築物2;模式図を参照されたい)の合成には、最初にFmoc-リジン(Mtt)-OH(Novabiochem、Laufelfingen、Switzerland)を4倍過剰で、等モルのO-ベンゾトリアゾール-N,N,N,N’,N’-テトラメチル(tetamethyl)-ウロニウム-ヘキサフルオロリン酸(HBTU; Novabiochem、Darmstadt、Germany)、1-ヒドロキシベンゾトリアゾール(hydoxybenzotriazole)(HOBt)および1.5倍モル過剰のジイソプロピルエチルアミン(DIPEA; Sigma、Castle Hill、Australia)を用いて、樹脂にカップリングさせた。アシル化は、40分間行った。α-アミノ上のFmoc保護基を次に除去し、これらのFmoc基の除去の後に、2つの一次アミノ基が分岐点の役目をするために曝露されるように、Fmoc-リジン-(Fmoc)-OHをカップリングさせた。それに続く別の1ラウンドの8倍過剰Fmoc-リジン(Fmoc)-OHのカップリングは、4つの分岐点を与え、それらに、4つのアルギニン(R)またはグルタミン酸(E)残基が16倍過剰でカップリングされた。
【0039】
分枝状構築物の脂質化のために、N末端アルギニン酸残基のアミノ基を、DIPEA存在下で10倍モル過剰のジ-tert-ブチルジカーボネート(Fluka Chemika、Switzerland)を用いて保護した。脂質化および切断に成功の後、Boc保護基を除去した。a-アミノ基をブロックして、最終生成物のa-アミノ基の正電荷も抑制するために、グルタミン酸残基を無水酢酸でアセチル化する。C末端リジンのεアミノ基に存在するMtt保護基を次に除去し、2つのセリンをカップリングさせた。次にPam2Cys脂質部分をZengら(Zeng 2003)に従ってカップリングさせ、分枝状R4(S2Pam2Cys)または分枝状E4(S2Pam2Cys)を生成した。
【0040】
線状リポペプチドPam2CyS-SK4の場合、Fmoc-リジン(Boc)-OHおよびFmoc-セリン(tBu)-OHを用いて線状のペプチドを合成し、続いてPam2Cys脂質部分をカップリングさせた。
【0041】
組立ての後に、88%TFA、5%フェノール、2%TIPS、5%水により室温で3時間、リポペプチドを固相支持体から切断し、Waters HPLC系に設置されたVydac C4カラム(4.6×300mm)を用いる逆相高圧液体クロマトグラフィー(RP-HPLC)によって分析した。制限溶媒としてH2O中の0.1%TFAおよびアセトニトリル中の0.1%TFAを用いて、クロマトグラムを1ml/分の流速で展開した。生成物を必要に応じて精製し、分析的RP-HPLC上の単一の主ピークとして提示し、Agilentシリーズ1100イオントラップ質量分析計を用いて分析すると、予想質量を有した。
【0042】
免疫化プロトコル
5匹の8〜12週齢BALB/c雌マウスの群に、特に明記しない限り0日目および28日目に再び、尾の基部の皮下に25μgのオボアルブミン(OVA; Sigma Aldrich、USA)またはニワトリ卵白リゾチーム(HEL; Sigma Aldrich、USA)のいずれかを生理食塩水で、またはCFAに乳濁させて、または生理食塩水中の異なる量のR4(S2Pam2Cys)もしくはE4(S2Pam2Cys)と混合して接種した。血清は、特に明記しない限り一次接種の約4週後および二次接種の2週後に採取された血液から調製された。
【0043】
酵素結合免疫吸着検定法(ELISA)
平底ウェルポリビニルプレート(Thermo、USA)を、加湿雰囲気下の室温で18〜20時間、0.1w/v%アジ化ナトリウムを含むPBS中のOVAまたはHEL(5μg/ml)(Chem Supply、Australia)のいずれかでコーティングした。抗原を除去し、10mg/mlウシ血清アルブミンを含有するPBS(BSA10PBS)を1時間加え、その後0.05v/v%Tween-20を含有するPBS(Sigma Aldrich、Milwaukee、USA)(PBST)で洗浄した。免疫化したマウスから得られた血清の連続希釈をウェルに加え、室温で一晩インキュベートした。PBSTによる洗浄の後、西洋ワサビペルオキシダーゼコンジュゲートウサギ抗マウスIgG抗体(Dako、Glostrup、Denmark)を、酵素基質(0.004%過酸化水素[Ajax Chemicals、Australia]を含む50mMクエン酸[M&B、England]中の0.2mM 2,2’-アジノ-ビス3-エチルベンズチアゾリン-スルホン酸[Sigma Aldrich、Milwaukee、USA])と一緒に用いて結合抗体を検出した。酵素基質反応によって誘導される発色を15分間進行させ、50mMフッ化ナトリウム(BDH Chemicals、Australia)の添加によって停止した。Labsystems Multiscan Multisoftマイクロプレートリーダー(Pathtech Diagnostics、Australia)を用いて、405nm(450nmで波長補正)における吸光度読取値を判定した。抗体力価は、0.2の光学濃度を達成するのに必要とされた血清の最も高い希釈の逆数で表す。
【0044】
細胞内サイトカイン染色によるIFN-γ産生の検出
特異的CD8+T細胞媒介サイトカイン産生の検出のために、マウスに第3の用量のOVA(25μg)を生理食塩水で、またはCFAに乳濁させて、または異なる量のR4(S2Pam2Cys)と混合して投与した。7日後、脾臓を接種マウスから得、圧力で金属篩を通して単一細胞懸濁液を得た。次に、組換えIL-2(10U/ml; Roche、Mannheim、Germany)の存在下で、OVA258〜265ペプチドSIINFEKL(2μg/ml)の有り無しで、同系照射(2200ラド、60Co源)を受けたナイーブな脾細胞(5×105)と一緒に脾細胞(1×106)を培養した。Cytofix/Cytoperm Plus Kit(Becton Dickinson、USA)からのBD GolgiPlugの形のブレフェルジンA(1μg/ml)も、この培養に含まれた。6時間後に、リンパ球をFAC洗浄液で洗浄し、4℃で30分間、PerCPコンジュゲートラット抗マウスCD8抗体(クローン53-6.7; Becton Dickinson、USA)で染色した。次に、製造業者の指示によりCytofix/Cytoperm溶液(Cytofix/Cytoperm Plus Kit、Becton Dickinson、USA)を用いて4℃で20分間、固定および透過化処理を実施した。キットによって供給されたPerm/Wash溶液で細胞を一度洗浄し、4℃で30分間、FITCコンジュゲートラット抗マウスIFN-γ抗体(クローンXMG1.2; Becton Dickinson、USA)で染色し、その後フローサイトメトリーによって分析した。FlowJoソフトウェア(Treestar Inc、USA)を用いてデータ分析を実施した。生きている生存細胞をそれらの前方および側方拡散特性に基づいてゲーティングし、合計1×105 CD8+細胞を計数した。
【0045】
OVA-リポペプチド複合体の沈澱
OVA(1nmole)を、平底96ウェルプレート中の100μl PBSの総容量で、分枝状R4(S2Pam2Cys)または線状Pam2Cys-SK4リポペプチドの漸増量と混合した。次に各ウェル中の溶液の混濁度を、Labsystems Multiscan Multisoftマイクロプレートリーダーで450nmでのその光学濃度を測定することによって決定した。
【0046】
100nmoleのR4(S2Pam2Cys)、1nmoleのOVA、またはR4(S2Pam2Cys)およびOVAの混合物のいずれかを含む溶液も遠心分離(1.2×105G)し、上清に対してHPLC分析を実施した。リポペプチドおよびOVAの両方を含む混合物からの沈殿物も50%アセトニトリル水溶液に溶解し、その後HPLCによって分析した。すべての試料で、リポペプチドに対応するピークのアイデンティティーを質量分析で検証し、OVAのそれは、OVA単独を含有する溶液でのその保持時間に基づいた。
【実施例1】
【0047】
序論
タンパク質抗原、特に組換えタンパク質はしばしば免疫原性でないことがあり、それらの免疫原性を増強するために、アジュバントを用いた製剤化が必要であるが、それらがヒトでの使用のために認可される前にアジュバント毒性に対する懸念およびそれらの作用機構が解決される必要がある。したがって、樹状細胞などの抗原提示細胞を直接標的にし、同時に活性化することによってタンパク質抗原の送達を促進することができる、新規系の開発が有利であることが判明し得る。
【0048】
この実施例では、タンパク質抗原に静電気的に会合することおよび樹状細胞にそれらを送達することができる、荷電した分枝状リポペプチド構造の使用を記載する。リポペプチドの電荷は、送達モジュールにそれぞれ総正電荷または総負電荷を付与する、4つのN末端アルギニン(R4)またはグルタミン酸残基(E4)の存在によって媒介される。それゆえに、R4は負に荷電したタンパク質に結合するために、E4は正に荷電したもののために用いることができる。分枝状リポペプチドの陽イオン性(R4(S2Pam2Cys))または陰イオン性バージョン(E4(S2Pam2Cys))へのTLR-2ターゲッティング脂質部分Pam2Cysの組込みは、樹状細胞にターゲッティングされた結合タンパク質の送達を保証する。
【0049】
結果
リポペプチドR4(S2Pam2Cys)は、4つのN末端アルギニン残基(各アルギニンは+2電荷を有する)の存在のために、+8の総電荷を有する。R4(S2Pam2Cys)が静電的相互作用を通してタンパク質免疫原性を増強することができるかを判断するために、11の総負電荷を有するオボアルブミン(OVA)をモデルタンパク質抗原として用いた。
【0050】
等モルまたは5倍モル過剰のR4(S2Pam2Cys)とプレインキュベートしたOVAによるワクチン接種は、一次および二次応答で、OVA単独のワクチン接種より有意に高い抗OVA抗体の力価を誘発したが(図1A)、これは、陽イオン性リポペプチドとタンパク質との結合がその免疫原性を増強することができることを示している。等量のR4(S2Pam2Cys)が用いられたときに達成されたものと比較して、5倍過剰のR4(S2Pam2Cys)がより高い抗体レベルをもたらしたので、誘発された抗体レベルは用いられたR4(S2Pam2Cys)の量に比例するようである。これらの群のマウスの一次応答での抗体レベルは、完全フロイントアジュバント(CFA)に乳濁されたOVAをマウスが接種されたときに得られたものよりも低かったが、R4(S2Pam2Cys)中の抗原の第2の用量の後に得られた抗体レベルは、FCAによって誘導されたものとほとんど同じ高さであった。
【0051】
R4(S2Pam2Cys)+OVAによるワクチン接種の後に、細胞媒介応答も分析した。CD8+T細胞による炎症誘発性サイトカインインターフェロンγ(IFN-γ)の分泌は、T細胞活性化の指標として測定した。R4(S2Pam2Cys)+OVAによるワクチン接種は、接種されたマウスの脾臓で、OVA単独による接種よりも著しく多くの特異的IFN-γ産生CD8+T細胞を誘導し、類似したレベルのIFN-γ産生CD8+T細胞がR4(S2Pam2Cys)およびCFAで誘発された(図1B)。用いたR4(S2Pam2Cys)の量の増加がより高い抗体応答に関連していた抗体の結果と対照的に、細胞媒介応答の場合にはその反対のようである;等モル量のR4(S2Pam2Cys)が用いられた場合、5倍過剰が用いられた場合に比較してより高いサイトカイン分泌性T細胞が検出された。タンパク質抗原に対するR4(S2Pam2Cys)の比を変化させることによって、特定の病原体のクリアランスのために必要とされる免疫応答の適当な型(細胞性対体液性)を選択することが可能であり得ることを、この観察は示唆する。
【0052】
負荷電リポペプチドと正荷電抗原間の静電的相互作用も同じ免疫原性増強を提供することができるかどうかを判定するために、リポペプチドE4(S2Pam2Cys)(それは、4つのN末端グルタミン酸残基の存在のために、+4の総電荷を有する)を、+8の総電荷を有するニワトリ卵白リゾチーム(HEL)と一緒にインキュベートした。これらの複合体によるマウスの接種は、HEL単独で達成されたものよりも高い抗体価をもたらしたが、これは、それが一緒に投与される抗原の反対の電荷を組み込まれたPam2Cysを用いる戦略を、反対の電荷のタンパク質抗原を収容するために応用することもできることを示している(図2A)。
【0053】
その両方は正電荷を有するR4(S2Pam2Cys)およびHELによるマウスの接種は、抗体産生をもたらさなかったことも見出されたが、これは、増強が電荷特異的であることを示している。しかし、E4(S2Pam2Cys)および負荷電OVAで見られる予想外の抗体の結果が、OVAの上に存在する正荷電パッチへのE4(S2Pam2Cys)の結合によるものかどうかは、まだ分かっていない。
【0054】
考察
これらの知見は、正または負に荷電した分岐状リポペプチドを、逆荷電のタンパク質の免疫原性を増強するために用いることができることを示す。これは、OVA特異的応答を誘導することができるが、HEL特異的応答を増大することができないR4(S2Pam2Cys)を用いることにより、特に鮮明にされる。このリポペプチドの正電荷および調べたタンパク質のそれらを考慮すると、観察される効果は、おそらくPam2Cys部分と抗原との間の静電的相互作用、またはそこでの欠落に起因する。この仮説を確認するさらなる研究は、同じかまたは反対に荷電したタンパク質抗原、ならびに中性電荷を有する分枝状リポペプチドの組入れを用いるクロマトグラフィーの分析法およびさらなるインビボ試験を通して達成することができる。
【0055】
これらの実験結果も、誘導することができる免疫応答の型、すなわち体液性または細胞性が、用いられる荷電リポペプチドおよびタンパク質の比に依存する可能性を示唆する。特定の病原体の成功裡のクリアランスは、誘発される免疫応答の型に依存する可能性があるので、これは特に重要な知見であり得る。したがって、本発明者らがここに記載するリポペプチドは、疾患によって特定の抗原に対して所望の免疫応答を調整する手段を提供することができる。
【実施例2】
【0056】
モデル抗原オボアルブミン(OVA)と複合体を形成する分枝状R4(S2Pam2Cys)および線状Pam2Cys-SK4リポペプチドの能力を測定するために、各リポペプチドおよびOVAの漸増量を含む溶液の吸光度を、450nmでの光学濃度を測定することによって決定した(図3)。したがって、OVAとの複合体を形成させる各リポペプチドの能力は、2つの化合物間の結合を示すことになる。
【0057】
1nmoleのOVAを含有する溶液への5nmoles分枝状R4(S2Pam2Cys)の添加が、乳濁または乳白色の溶液として現れる複合体の形成をもたらすことが判明した。分枝状リポペプチドの量を増加させることは、光学濃度の読取値の上昇をもたらし、20〜40nmoleのリポペプチドを用いたときに最大値が達成された。対照的に、調査したすべての濃度で線状Pam2Cys-SK4リポペプチドを加えたときには、光学濃度のごくわずかな増加しか観察されなかったが、これは、抗原への結合において分枝状R4(S2Pam2Cys)が線状リポペプチドよりも優れていることを示している。
【0058】
OVAへのR4(S2Pam2Cys)の結合を確認するために、これらの化合物のいずれかを個々に、または両方の混合物を含む溶液を遠心分離して、あらゆる複合体を沈殿させた。上清および沈殿物に対して次にHPLC分析を実施した。
【0059】
OVAまたはR4(S2Pam2Cys)を含む溶液の上清では(図4A)、生じたクロマトグラムは、溶液中の各個々の化合物に対応する主ピークを明らかにした。両化合物を混合した後、溶液中のOVAに対応するピークのサイズの劇的な低下が明らかであったが、これは、R4(S2Pam2Cys)リポペプチドとの結合の結果、タンパク質が溶液から沈殿分離したことを示唆している。これは再構築物質のHPLC分析(図4B)で確認され、このHPLC分析はOVAおよびリポペプチドの両方に対応する2つの主ピークの存在を明らかにしたが、これは沈殿物がリポペプチドおよびタンパク質の複合体であることを確認している。
(参考文献)
【技術分野】
【0001】
本発明は、静電的相互作用を通して抗原が樹状細胞リガンドと会合している免疫原性組成物に関する。本発明は、対象で免疫応答を誘導するための、これらの組成物の使用にさらに関する。
【背景技術】
【0002】
特に疾患から保護するために、ヒトおよび他の動物で免疫応答を起こすために用いることができる組成物の開発に対して、益々関心が高まっている。免疫応答を起こす際には、抗原が樹状細胞に遭遇することが必要である。樹状細胞は免疫細胞であって、哺乳動物の免疫系の一部を形成する。それらの主機能は、抗原物質をプロセシングして、免疫系の他の細胞の表面にそれを提示し、それによって抗原提示細胞として機能することである。
【0003】
樹状細胞への抗原のターゲッティング、および樹状細胞上のToll様受容体が強い免疫応答を誘発することが示されている脂質を含むリポペプチドは、以前に企図されている(国際公開第2004/014956号と国際公開第2004/014957号、それらの開示は参照により本明細書に組み込まれる)。
【0004】
いかなるペプチドも、有効な抗体応答を誘導できるためには、それは、免疫系によって認識されるエピトープとして知られる特定のアミノ酸配列を含まなければならない。詳細には、エピトープは、抗体応答をもたらすのに、Bリンパ球の表面に存在する特異的免疫グロブリン(Ig)受容体によって認識される必要がある。そのエピトープに特異的な抗体を産生できる形質細胞に最終的に分化するのは、これらの細胞である。これらのB細胞エピトープに加えて、免疫原は、B細胞が抗体産生細胞に分化するのに必要とされるシグナルを提供するのに必要な細胞、ヘルパーTリンパ球上に存在する特異的受容体に対して抗原提示細胞(APC)によって提示されるエピトープも含まなければならない。
【0005】
ウイルス感染の場合および癌の多くの場合では、回復における抗体の利点は限定され、免疫系は、ウイルス感染細胞または癌細胞を死滅させることができる細胞傷害性T細胞(CTL)で応答する。ヘルパーT細胞のように、CTLは、今度はクラスII分子ではなくMHCクラスIと会合して、それらの特異的ペプチドエピトープを表面に提示しているAPCとの相互作用によって先ず活性化される。活性化されると、CTLは同じペプチド/クラスI複合体を有する標的細胞にエンゲージし、その溶解を引き起こすことができる。ヘルパーT細胞がこの過程で役割を果たすことも、次第に明らかになってきている;APCがCTLを活性化することができるには、それは、ヘルパーT細胞からシグナルを先ず受け取り必要な共刺激分子の発現を上方制御しなければばらない。
【0006】
ヘルパーT細胞エピトープは、主要組織適合性複合体(MHC)のクラスII遺伝子によってコードされる、APCの表面に存在する分子により結合される。クラスII分子およびペプチドエピトープの複合体は、次にTヘルパーリンパ球の表面の特異的T細胞受容体(TCR)によって認識される。このように、MHC分子との関連において抗原エピトープで提示されるT細胞は活性化され、Bリンパ球が分化するのに必要なシグナルを提供することができる。
【0007】
その場合一般に、免疫原は、B細胞または細胞傷害性T細胞によって認識されるエピトープに加えて、ヘルパーT細胞が認識することができるエピトープを含まなければならない。この種のエピトープは非常に異なることがあることを理解するべきである。B細胞エピトープについては、B細胞受容体は本来の免疫原に直接に結合するので、立体構造が重要である。対照的に、T細胞によって認識されるエピトープは、エピトープの立体構造完全性に依存せず、CTLについては約9アミノ酸の短い配列、およびヘルパーT細胞については、長さの制限はより少ないが、わずかにより長い配列からなる。これらのエピトープのための唯一の要件は、それらがそれぞれクラスIまたはクラスII分子の結合クレフトに収まることができ、その結果複合体がT細胞受容体にエンゲージできることである。クラスII分子の結合部位は両端がオープンであり、結合ペプチドの長さのかなりより大きな変動を可能にしているが(Brown、J. H.、T. S. Jardetzky、J. C. Gorga、L. J. Stern、R. G. Urban、J. L. StromingerおよびD. C. Wiley. 1993. Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1. Nature 364: 33頁)、8アミノ酸残基と短いエピトープが報告されている(Fahrer, A. M.、Geysen, H. M.、White, D. O.、Jackson, D. C.およびBrown, L. E. Analysis of the requirements for class II-restricted T-cell recognition of a single determinant reveals considerable diversity in the T-cell response and degeneracy of peptide binding to I-ED J. Immunol. 1995. 155: 2849〜2857頁)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公開第2004/014956号
【特許文献2】国際公開第2004/014957号
【非特許文献】
【0009】
【非特許文献1】Brown、J. H.、T. S. Jardetzky、J. C. Gorga、L. J. Stern、R. G. Urban、J. L. StromingerおよびD. C. Wiley. 1993. Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1. Nature 364: 33頁
【非特許文献2】Fahrer, A. M.、Geysen, H. M.、White, D. O.、Jackson, D. C.およびBrown, L. E. Analysis of the requirements for class II-restricted T-cell recognition of a single determinant reveals considerable diversity in the T-cell response and degeneracy of peptide binding to I-ED J. Immunol. 1995. 155: 2849〜2857頁
【非特許文献3】Metzger, J. W.ら1995. J Pept Sci 1:184頁
【非特許文献4】Sacht, G.ら1998. Eur J Immunol 28:4207頁
【非特許文献5】Muhiradt, P. F.ら1998. Infect Immun 66: 4804頁
【非特許文献6】Muhiradt, P. F.ら1997. J Exp Med 185: 1951頁
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明者は、抗原が樹状細胞ターゲッティング成分と静電気的に会合している、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物を開発した。
【課題を解決するための手段】
【0011】
第1の態様では、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は負荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している正荷電基を含み、抗原の負荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0012】
第2の態様では、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は正荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している負荷電基を含み、抗原の正荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0013】
第3態様では、本発明は、対象に本発明の第1または第2の態様の免疫原性組成物を投与する段階を含む、対象で免疫応答を起こす方法を提供する。
【0014】
以下では、OVA=オボアルブミン;HEL=ニワトリ卵白リゾチーム;R4(S2Pam2Cys)=図5の1に示す構築物;E4(S2Pam2Cys)=図5の2に示す構築物;CFA=完全フロイントアジュバント。
【図面の簡単な説明】
【0015】
【図1】R4(S2Pam2Cys-OVA)複合体によるワクチン接種によって誘発された抗体および細胞媒介性応答を示す図である。(A)BALB/cマウスの群に、皮下に25μgのOVA単独、CFAに乳濁させたOVA、または等モルもしくは5倍モル過剰のR4(S2Pam2Cys)に混合したOVAを接種した。動物は、26日後に、抗原の第2の類似した用量が投与された。抗原の一次接種の27日後(○)、および二次接種の13日後(●)に採取された血液から血清を得た。抗体レベルをELISAで決定し、個々の抗体価を水平バーによって表される平均値と示した。(B)マウスに50日目に3回目の接種をし、脾臓を7日後に得た。SIINFEKL特異的IFN-γ産生CD8+T細胞の存在を検出するために、細胞内のサイトカイン染色を脾細胞で実施した。データは、3反復の試料の平均および標準偏差で示される。
【図2】陽イオンおよび陰イオンのリポペプチド-タンパク質複合体によるワクチン接種によって誘発されたOVAおよびHEL-特異的抗体応答を示す図である。BALB/cマウスに、皮下に25μgのHEL(A)またはOVA(B)を単独で、CFAで乳濁させて、または抗原と混合したR4(S2Pam2Cys)もしくはE4(S2Pam2Cys)の同等量と一緒に接種した。マウスは一次接種の28日後(○)に採血し、32日目に追加免疫し、46日目に再度採血した(●)。その後抗体レベルをELISAで決定した。個々の動物の力価を、水平バーによって表される平均値と示す。
【図3】オボアルブミン-リポペプチド複合体の沈殿を示す図である。分枝状R4(S2Pam2Cys)または線状Pam2Cys-SK4リポペプチドの漸増量を、平底96ウェルプレート中の100μl PBSの総容量中の1nmoleのオボアルブミン(OVA)と混合した。450nmで溶液の光学濃度を決定することによって、次に溶液の混濁度を測定した。
【図4】オボアルブミン- R4(S2Pam2Cys)リポペプチド溶液のHPLC分析を示す図である。遠心分離(1.2×105G)の後、100μl PBSの総容量に(A)100nmoleの分枝状R4(S2Pam2Cys)リポペプチド、(B)1nmoleのオボアルブミン(OVA)または(C)R4(S2Pam2Cys)リポペプチドおよびオボアルブミンの混合物のいずれかを含む溶液の上清で、HPLC分析を実施した。(D)リポペプチドおよびOVAの両方を含む混合物からの沈殿物を50%アセトニトリル水溶液に溶解し、その後HPLCによって分析した。R4(S2Pam2Cys)を含むすべての試料で、リポペプチドに対応するピークのアイデンティティーを質量分析によって検証した。
【図5】それらのそれぞれの静電的電荷が環境に提示されるように、末端位置に正荷電(アルギニン、R;リジン、K)または負荷電(アスパラギン酸、D;グルタミン酸、E)アミノ酸を含む分枝状(構造1〜5)および線状(構造6〜8)免疫原性組成物の一部の例の模式図である。各免疫原性組成物は、Toll様受容体2のリガンドであるジパルミトイル-S-グリセリルシステイン(Pam2Cys)も含む。2つのセリン残基(Ser)も組み込まれる。構築物2の場合、ペプチド構造は方向N→Cで組み立てられ、図示されるすべての他の構造はC→Nで組み立てられた。正負の静電的電荷は、電荷のサイズによって2-、2+、1-、1+などで示される。Ac=N末端にあるグルタミン酸の場合にアルファアミノ基の正電荷を抑制するために用いたアセチル基。
【発明を実施するための形態】
【0016】
本発明者は、樹状細胞ターゲッティング基に共有結合している荷電部分が静電気的に抗原と会合して、免疫応答を起こすために用いることができる免疫原性複合体を形成することを見出した。
【0017】
したがって、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は負荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している正荷電基を含み、抗原の負荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0018】
本発明のこの態様では、荷電したターゲッティング成分と静電気的に相互作用することができる負荷電領域または負荷電ドメインを抗原が含むことが重要である。しかし、好ましいことではあるが、抗原が総負電荷を有することは必須でない。負荷電基を加えることによって抗原の負電荷を増加させることも可能である。例えば、ポリペプチド抗原で、アスパラギン酸またはグルタミン酸残基の鎖をポリペプチドに加えることができよう。
【0019】
正荷電基は、少なくとも1つの正荷電アミノ酸を含むことが好ましい。正荷電基が、分枝状または線状のペプチド、好ましくは分枝状のペプチドであることも好ましい。様々な実施形態では、ペプチドは、少なくとも1つのアルギニン、ヒスチジン、オルニチンまたはリジン残基、またはそれらの組合せを含む。ペプチドが、少なくとも4つのアルギニン残基および/または少なくとも4つのリジン残基を含むことが好ましい。正荷電基が、少なくとも4つのアルギニン残基を含む分枝状のペプチドを含むことが特に好ましい。
【0020】
第2の態様では、本発明は、抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は正荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している負荷電基を含み、抗原の正荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物を提供する。
【0021】
本発明のこの態様では、荷電したターゲッティング成分と静電気的に相互作用することができる正荷電領域または正荷電ドメインを抗原が含むことが重要である。しかし、好ましいことではあるが、抗原が総正電荷を有することは必須でない。正荷電基を加えることによって、抗原の正電荷を増加させることも可能である。例えば、ポリペプチド抗原で、リジン、アルギニンまたはヒスチジン残基の鎖をポリペプチドに加えることができよう。
【0022】
負荷電基は、少なくとも1つの負荷電アミノ酸を含むことが好ましい。負荷電基が、分枝状または線状のペプチド、好ましくは分枝状のペプチドであることも好ましい。様々な実施形態では、ペプチドは、少なくとも1つのアスパラギン酸またはグルタミン酸残基、またはそれらの組合せを含む。ペプチドが、少なくとも4つのアスパラギン酸残基および/または少なくとも4つのグルタミン酸残基を含むことが好ましい。正荷電基が、少なくとも4つのグルタミン酸残基を含む分枝状のペプチドを含むことが特に好ましい。
【0023】
本発明の好ましい実施形態では、抗原は核酸でない。静電的相互作用だけによって抗原が樹状細胞ターゲッティング成分と結合していることも好ましい。
【0024】
本発明で用いることができる様々な樹状細胞リガンドを、表1に提示する。しかし、樹状細胞リガンドはTLRリガンドであることが好ましい。TLRリガンドは、脂質またはペプチドグリカンまたはリポタンパク質またはリポ多糖を含むことができる。詳細には、TLRリガンドは、パルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイルまたはデカノイルを含むことができる。好ましくは、TLRリガンドは、Pam2Cys、Pam3Cys、Ste2Cys、Lau2CysおよびOct2Cysからなる群から選択される。
【0025】
特定の実施形態では、TLRリガンドは、TLR-1またはTLR-6と会合していてもよいTLR-2に結合する。
【0026】
第3態様では、本発明は、対象に本発明の第1または第2の態様による免疫原性組成物を投与する段階を含む、対象で免疫応答を起こす方法を提供する。
【0027】
【表1】
【0028】
本発明の例示的な樹状細胞ターゲッティング化合物は、リポペプチド「Pam2Cys」である。当業者は、用語「リポペプチド」が、コンジュゲートされている1つまたは複数の脂質部分および1つまたは複数のアミノ酸配列を含む任意の組成物を意味することを理解しよう。「Pam2Cys」(ジパルミトイル-S-グリセリル-システインまたはS-[2,3ビス(パルミトイルオキシ)プロピル]システインとしても知られる)が合成されており(Metzger, J. W.ら1995. J Pept Sci 1:184頁)、マイコプラズマ・ファーメンタンス(Mycoplasma fermentans)から単離されたマクロファージ活性化リポペプチドであるMALP-2の脂質部分に対応する(Sacht, G.ら1998. Eur J Immunol 28:4207頁;Muhiradt, P. F.ら1998. Infect Immun 66: 4804頁;Muhiradt, P. F.ら1997. J Exp Med 185: 1951頁)。Pam2Cysは、TLR-2のリガンドであることが知られている。
【0029】
Pam2Cysは、下記式(I)の構造を有する:
【0030】
【化1】
【0031】
細胞表面TLRを標的にするために用いることができる他の脂質部分には、パルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイルまたはデカノイルが含まれる。好ましい基には、Pam2Cys、Pam3Cys、Ste2Cys、Lau2CysおよびOct2Cysが含まれる。
【0032】
本明細書で用いるように、「免疫応答」への言及は、侵入病原体、病原体に感染した細胞もしくは組織、癌細胞、または自己免疫もしくは病理学的炎症の場合には正常なヒト細胞もしくは組織に対する選択的な損傷、破壊、またはヒト体内からの除去をもたらす、リンパ球、抗原提示細胞、食細胞、顆粒球、および上記の細胞または肝臓によって産生される溶解性高分子(抗体、サイトカインおよび補体を含む)の協調作用への言及を意味する。
【0033】
本明細書全体を通して、単語「含む(comprise)」、または「含む(comprises)」もしくは「含んでいる(comprising)」などの変形形態は、明示された段階もしくは要素もしくは整数、または段階もしくは要素もしくは整数の群が含まれることを意味するが、いかなる他の段階もしくは要素もしくは整数または要素もしくは整数の群の除外を意味するものではないと理解される。
【0034】
本出願で引用されるすべての参考文献は、参照により本明細書に具体的に組み込まれている。
【0035】
本明細書での任意の先行技術への言及は、この先行技術がオーストラリアで一般常識の一部を形成することの肯定またはいかなる形の示唆ではなく、かつそのように解釈されるべきでない。
【0036】
当業者が、所望の用途に適するように、正荷電基およびTLRリガンドの任意の組合せを導入することができることが理解されよう。
【0037】
本発明は、次に、以下の非限定的実施例によってさらに記載される。
【0038】
材料および方法
リポペプチドの合成
分枝状および線状リポペプチドの合成を、PEG-S RAM樹脂(Rapp Polymere、Tubingen、Germany;置換因子0.27mmol/g)上で行った。分枝状リポペプチドR4(S2Pam2Cys)(構築物1)およびE4(S2Pam2Cys)(構築物2;模式図を参照されたい)の合成には、最初にFmoc-リジン(Mtt)-OH(Novabiochem、Laufelfingen、Switzerland)を4倍過剰で、等モルのO-ベンゾトリアゾール-N,N,N,N’,N’-テトラメチル(tetamethyl)-ウロニウム-ヘキサフルオロリン酸(HBTU; Novabiochem、Darmstadt、Germany)、1-ヒドロキシベンゾトリアゾール(hydoxybenzotriazole)(HOBt)および1.5倍モル過剰のジイソプロピルエチルアミン(DIPEA; Sigma、Castle Hill、Australia)を用いて、樹脂にカップリングさせた。アシル化は、40分間行った。α-アミノ上のFmoc保護基を次に除去し、これらのFmoc基の除去の後に、2つの一次アミノ基が分岐点の役目をするために曝露されるように、Fmoc-リジン-(Fmoc)-OHをカップリングさせた。それに続く別の1ラウンドの8倍過剰Fmoc-リジン(Fmoc)-OHのカップリングは、4つの分岐点を与え、それらに、4つのアルギニン(R)またはグルタミン酸(E)残基が16倍過剰でカップリングされた。
【0039】
分枝状構築物の脂質化のために、N末端アルギニン酸残基のアミノ基を、DIPEA存在下で10倍モル過剰のジ-tert-ブチルジカーボネート(Fluka Chemika、Switzerland)を用いて保護した。脂質化および切断に成功の後、Boc保護基を除去した。a-アミノ基をブロックして、最終生成物のa-アミノ基の正電荷も抑制するために、グルタミン酸残基を無水酢酸でアセチル化する。C末端リジンのεアミノ基に存在するMtt保護基を次に除去し、2つのセリンをカップリングさせた。次にPam2Cys脂質部分をZengら(Zeng 2003)に従ってカップリングさせ、分枝状R4(S2Pam2Cys)または分枝状E4(S2Pam2Cys)を生成した。
【0040】
線状リポペプチドPam2CyS-SK4の場合、Fmoc-リジン(Boc)-OHおよびFmoc-セリン(tBu)-OHを用いて線状のペプチドを合成し、続いてPam2Cys脂質部分をカップリングさせた。
【0041】
組立ての後に、88%TFA、5%フェノール、2%TIPS、5%水により室温で3時間、リポペプチドを固相支持体から切断し、Waters HPLC系に設置されたVydac C4カラム(4.6×300mm)を用いる逆相高圧液体クロマトグラフィー(RP-HPLC)によって分析した。制限溶媒としてH2O中の0.1%TFAおよびアセトニトリル中の0.1%TFAを用いて、クロマトグラムを1ml/分の流速で展開した。生成物を必要に応じて精製し、分析的RP-HPLC上の単一の主ピークとして提示し、Agilentシリーズ1100イオントラップ質量分析計を用いて分析すると、予想質量を有した。
【0042】
免疫化プロトコル
5匹の8〜12週齢BALB/c雌マウスの群に、特に明記しない限り0日目および28日目に再び、尾の基部の皮下に25μgのオボアルブミン(OVA; Sigma Aldrich、USA)またはニワトリ卵白リゾチーム(HEL; Sigma Aldrich、USA)のいずれかを生理食塩水で、またはCFAに乳濁させて、または生理食塩水中の異なる量のR4(S2Pam2Cys)もしくはE4(S2Pam2Cys)と混合して接種した。血清は、特に明記しない限り一次接種の約4週後および二次接種の2週後に採取された血液から調製された。
【0043】
酵素結合免疫吸着検定法(ELISA)
平底ウェルポリビニルプレート(Thermo、USA)を、加湿雰囲気下の室温で18〜20時間、0.1w/v%アジ化ナトリウムを含むPBS中のOVAまたはHEL(5μg/ml)(Chem Supply、Australia)のいずれかでコーティングした。抗原を除去し、10mg/mlウシ血清アルブミンを含有するPBS(BSA10PBS)を1時間加え、その後0.05v/v%Tween-20を含有するPBS(Sigma Aldrich、Milwaukee、USA)(PBST)で洗浄した。免疫化したマウスから得られた血清の連続希釈をウェルに加え、室温で一晩インキュベートした。PBSTによる洗浄の後、西洋ワサビペルオキシダーゼコンジュゲートウサギ抗マウスIgG抗体(Dako、Glostrup、Denmark)を、酵素基質(0.004%過酸化水素[Ajax Chemicals、Australia]を含む50mMクエン酸[M&B、England]中の0.2mM 2,2’-アジノ-ビス3-エチルベンズチアゾリン-スルホン酸[Sigma Aldrich、Milwaukee、USA])と一緒に用いて結合抗体を検出した。酵素基質反応によって誘導される発色を15分間進行させ、50mMフッ化ナトリウム(BDH Chemicals、Australia)の添加によって停止した。Labsystems Multiscan Multisoftマイクロプレートリーダー(Pathtech Diagnostics、Australia)を用いて、405nm(450nmで波長補正)における吸光度読取値を判定した。抗体力価は、0.2の光学濃度を達成するのに必要とされた血清の最も高い希釈の逆数で表す。
【0044】
細胞内サイトカイン染色によるIFN-γ産生の検出
特異的CD8+T細胞媒介サイトカイン産生の検出のために、マウスに第3の用量のOVA(25μg)を生理食塩水で、またはCFAに乳濁させて、または異なる量のR4(S2Pam2Cys)と混合して投与した。7日後、脾臓を接種マウスから得、圧力で金属篩を通して単一細胞懸濁液を得た。次に、組換えIL-2(10U/ml; Roche、Mannheim、Germany)の存在下で、OVA258〜265ペプチドSIINFEKL(2μg/ml)の有り無しで、同系照射(2200ラド、60Co源)を受けたナイーブな脾細胞(5×105)と一緒に脾細胞(1×106)を培養した。Cytofix/Cytoperm Plus Kit(Becton Dickinson、USA)からのBD GolgiPlugの形のブレフェルジンA(1μg/ml)も、この培養に含まれた。6時間後に、リンパ球をFAC洗浄液で洗浄し、4℃で30分間、PerCPコンジュゲートラット抗マウスCD8抗体(クローン53-6.7; Becton Dickinson、USA)で染色した。次に、製造業者の指示によりCytofix/Cytoperm溶液(Cytofix/Cytoperm Plus Kit、Becton Dickinson、USA)を用いて4℃で20分間、固定および透過化処理を実施した。キットによって供給されたPerm/Wash溶液で細胞を一度洗浄し、4℃で30分間、FITCコンジュゲートラット抗マウスIFN-γ抗体(クローンXMG1.2; Becton Dickinson、USA)で染色し、その後フローサイトメトリーによって分析した。FlowJoソフトウェア(Treestar Inc、USA)を用いてデータ分析を実施した。生きている生存細胞をそれらの前方および側方拡散特性に基づいてゲーティングし、合計1×105 CD8+細胞を計数した。
【0045】
OVA-リポペプチド複合体の沈澱
OVA(1nmole)を、平底96ウェルプレート中の100μl PBSの総容量で、分枝状R4(S2Pam2Cys)または線状Pam2Cys-SK4リポペプチドの漸増量と混合した。次に各ウェル中の溶液の混濁度を、Labsystems Multiscan Multisoftマイクロプレートリーダーで450nmでのその光学濃度を測定することによって決定した。
【0046】
100nmoleのR4(S2Pam2Cys)、1nmoleのOVA、またはR4(S2Pam2Cys)およびOVAの混合物のいずれかを含む溶液も遠心分離(1.2×105G)し、上清に対してHPLC分析を実施した。リポペプチドおよびOVAの両方を含む混合物からの沈殿物も50%アセトニトリル水溶液に溶解し、その後HPLCによって分析した。すべての試料で、リポペプチドに対応するピークのアイデンティティーを質量分析で検証し、OVAのそれは、OVA単独を含有する溶液でのその保持時間に基づいた。
【実施例1】
【0047】
序論
タンパク質抗原、特に組換えタンパク質はしばしば免疫原性でないことがあり、それらの免疫原性を増強するために、アジュバントを用いた製剤化が必要であるが、それらがヒトでの使用のために認可される前にアジュバント毒性に対する懸念およびそれらの作用機構が解決される必要がある。したがって、樹状細胞などの抗原提示細胞を直接標的にし、同時に活性化することによってタンパク質抗原の送達を促進することができる、新規系の開発が有利であることが判明し得る。
【0048】
この実施例では、タンパク質抗原に静電気的に会合することおよび樹状細胞にそれらを送達することができる、荷電した分枝状リポペプチド構造の使用を記載する。リポペプチドの電荷は、送達モジュールにそれぞれ総正電荷または総負電荷を付与する、4つのN末端アルギニン(R4)またはグルタミン酸残基(E4)の存在によって媒介される。それゆえに、R4は負に荷電したタンパク質に結合するために、E4は正に荷電したもののために用いることができる。分枝状リポペプチドの陽イオン性(R4(S2Pam2Cys))または陰イオン性バージョン(E4(S2Pam2Cys))へのTLR-2ターゲッティング脂質部分Pam2Cysの組込みは、樹状細胞にターゲッティングされた結合タンパク質の送達を保証する。
【0049】
結果
リポペプチドR4(S2Pam2Cys)は、4つのN末端アルギニン残基(各アルギニンは+2電荷を有する)の存在のために、+8の総電荷を有する。R4(S2Pam2Cys)が静電的相互作用を通してタンパク質免疫原性を増強することができるかを判断するために、11の総負電荷を有するオボアルブミン(OVA)をモデルタンパク質抗原として用いた。
【0050】
等モルまたは5倍モル過剰のR4(S2Pam2Cys)とプレインキュベートしたOVAによるワクチン接種は、一次および二次応答で、OVA単独のワクチン接種より有意に高い抗OVA抗体の力価を誘発したが(図1A)、これは、陽イオン性リポペプチドとタンパク質との結合がその免疫原性を増強することができることを示している。等量のR4(S2Pam2Cys)が用いられたときに達成されたものと比較して、5倍過剰のR4(S2Pam2Cys)がより高い抗体レベルをもたらしたので、誘発された抗体レベルは用いられたR4(S2Pam2Cys)の量に比例するようである。これらの群のマウスの一次応答での抗体レベルは、完全フロイントアジュバント(CFA)に乳濁されたOVAをマウスが接種されたときに得られたものよりも低かったが、R4(S2Pam2Cys)中の抗原の第2の用量の後に得られた抗体レベルは、FCAによって誘導されたものとほとんど同じ高さであった。
【0051】
R4(S2Pam2Cys)+OVAによるワクチン接種の後に、細胞媒介応答も分析した。CD8+T細胞による炎症誘発性サイトカインインターフェロンγ(IFN-γ)の分泌は、T細胞活性化の指標として測定した。R4(S2Pam2Cys)+OVAによるワクチン接種は、接種されたマウスの脾臓で、OVA単独による接種よりも著しく多くの特異的IFN-γ産生CD8+T細胞を誘導し、類似したレベルのIFN-γ産生CD8+T細胞がR4(S2Pam2Cys)およびCFAで誘発された(図1B)。用いたR4(S2Pam2Cys)の量の増加がより高い抗体応答に関連していた抗体の結果と対照的に、細胞媒介応答の場合にはその反対のようである;等モル量のR4(S2Pam2Cys)が用いられた場合、5倍過剰が用いられた場合に比較してより高いサイトカイン分泌性T細胞が検出された。タンパク質抗原に対するR4(S2Pam2Cys)の比を変化させることによって、特定の病原体のクリアランスのために必要とされる免疫応答の適当な型(細胞性対体液性)を選択することが可能であり得ることを、この観察は示唆する。
【0052】
負荷電リポペプチドと正荷電抗原間の静電的相互作用も同じ免疫原性増強を提供することができるかどうかを判定するために、リポペプチドE4(S2Pam2Cys)(それは、4つのN末端グルタミン酸残基の存在のために、+4の総電荷を有する)を、+8の総電荷を有するニワトリ卵白リゾチーム(HEL)と一緒にインキュベートした。これらの複合体によるマウスの接種は、HEL単独で達成されたものよりも高い抗体価をもたらしたが、これは、それが一緒に投与される抗原の反対の電荷を組み込まれたPam2Cysを用いる戦略を、反対の電荷のタンパク質抗原を収容するために応用することもできることを示している(図2A)。
【0053】
その両方は正電荷を有するR4(S2Pam2Cys)およびHELによるマウスの接種は、抗体産生をもたらさなかったことも見出されたが、これは、増強が電荷特異的であることを示している。しかし、E4(S2Pam2Cys)および負荷電OVAで見られる予想外の抗体の結果が、OVAの上に存在する正荷電パッチへのE4(S2Pam2Cys)の結合によるものかどうかは、まだ分かっていない。
【0054】
考察
これらの知見は、正または負に荷電した分岐状リポペプチドを、逆荷電のタンパク質の免疫原性を増強するために用いることができることを示す。これは、OVA特異的応答を誘導することができるが、HEL特異的応答を増大することができないR4(S2Pam2Cys)を用いることにより、特に鮮明にされる。このリポペプチドの正電荷および調べたタンパク質のそれらを考慮すると、観察される効果は、おそらくPam2Cys部分と抗原との間の静電的相互作用、またはそこでの欠落に起因する。この仮説を確認するさらなる研究は、同じかまたは反対に荷電したタンパク質抗原、ならびに中性電荷を有する分枝状リポペプチドの組入れを用いるクロマトグラフィーの分析法およびさらなるインビボ試験を通して達成することができる。
【0055】
これらの実験結果も、誘導することができる免疫応答の型、すなわち体液性または細胞性が、用いられる荷電リポペプチドおよびタンパク質の比に依存する可能性を示唆する。特定の病原体の成功裡のクリアランスは、誘発される免疫応答の型に依存する可能性があるので、これは特に重要な知見であり得る。したがって、本発明者らがここに記載するリポペプチドは、疾患によって特定の抗原に対して所望の免疫応答を調整する手段を提供することができる。
【実施例2】
【0056】
モデル抗原オボアルブミン(OVA)と複合体を形成する分枝状R4(S2Pam2Cys)および線状Pam2Cys-SK4リポペプチドの能力を測定するために、各リポペプチドおよびOVAの漸増量を含む溶液の吸光度を、450nmでの光学濃度を測定することによって決定した(図3)。したがって、OVAとの複合体を形成させる各リポペプチドの能力は、2つの化合物間の結合を示すことになる。
【0057】
1nmoleのOVAを含有する溶液への5nmoles分枝状R4(S2Pam2Cys)の添加が、乳濁または乳白色の溶液として現れる複合体の形成をもたらすことが判明した。分枝状リポペプチドの量を増加させることは、光学濃度の読取値の上昇をもたらし、20〜40nmoleのリポペプチドを用いたときに最大値が達成された。対照的に、調査したすべての濃度で線状Pam2Cys-SK4リポペプチドを加えたときには、光学濃度のごくわずかな増加しか観察されなかったが、これは、抗原への結合において分枝状R4(S2Pam2Cys)が線状リポペプチドよりも優れていることを示している。
【0058】
OVAへのR4(S2Pam2Cys)の結合を確認するために、これらの化合物のいずれかを個々に、または両方の混合物を含む溶液を遠心分離して、あらゆる複合体を沈殿させた。上清および沈殿物に対して次にHPLC分析を実施した。
【0059】
OVAまたはR4(S2Pam2Cys)を含む溶液の上清では(図4A)、生じたクロマトグラムは、溶液中の各個々の化合物に対応する主ピークを明らかにした。両化合物を混合した後、溶液中のOVAに対応するピークのサイズの劇的な低下が明らかであったが、これは、R4(S2Pam2Cys)リポペプチドとの結合の結果、タンパク質が溶液から沈殿分離したことを示唆している。これは再構築物質のHPLC分析(図4B)で確認され、このHPLC分析はOVAおよびリポペプチドの両方に対応する2つの主ピークの存在を明らかにしたが、これは沈殿物がリポペプチドおよびタンパク質の複合体であることを確認している。
(参考文献)
【特許請求の範囲】
【請求項1】
抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は負荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している正荷電基を含み、抗原の負荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物。
【請求項2】
抗原が総負電荷を有する、請求項1に記載の免疫原性組成物。
【請求項3】
正荷電基が少なくとも1つの正荷電アミノ酸を含む、請求項1または請求項2に記載の免疫原性組成物。
【請求項4】
正荷電基が分枝状または線状のペプチドである、請求項1から3のいずれか一項に記載の免疫原性組成物。
【請求項5】
ペプチドが少なくとも1つのアルギニンまたはリジン残基を含む、請求項3に記載の免疫原性組成物。
【請求項6】
ペプチドが、少なくとも4つのアルギニン残基および/または少なくとも4つのリジン残基を含む、請求項3または請求項4に記載の免疫原性組成物。
【請求項7】
抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は正荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している負荷電基を含み、抗原の正荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物。
【請求項8】
抗原が総正電荷を有する、請求項7に記載の免疫原性組成物。
【請求項9】
負荷電基が少なくとも1つの負荷電アミノ酸を含む、請求項7または請求項8に記載の免疫原性組成物。
【請求項10】
負荷電基が分枝状または線状のペプチドである、請求項7から9のいずれか一項に記載の免疫原性組成物。
【請求項11】
ペプチドが少なくとも1つのアスパラギン酸またはグルタミン酸残基を含む、請求項10に記載の免疫原性組成物。
【請求項12】
ペプチドが、少なくとも4つのアスパラギン酸残基および/または少なくとも4つのグルタミン酸残基を含む、請求項10または請求項11に記載の免疫原性組成物。
【請求項13】
抗原が核酸でない、請求項1から12のいずれか一項に記載の免疫原性組成物。
【請求項14】
静電的相互作用だけによって抗原が樹状細胞ターゲッティング成分と会合している、請求項1から13のいずれか一項に記載の免疫原性組成物。
【請求項15】
樹状細胞リガンドがTLRリガンドである、請求項1から14のいずれか一項に記載の免疫原性組成物。
【請求項16】
TLRリガンドが脂質またはペプチドグリカンまたはリポタンパク質またはリポ多糖を含む、請求項15に記載の免疫原性組成物。
【請求項17】
TLRリガンドがパルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイルまたはデカノイルを含む、請求項15または請求項16に記載の免疫原性組成物。
【請求項18】
TLRリガンドがPam2Cys、Pam3Cys、Ste2Cys、Lau2CysおよびOct2Cysからなる群から選択される、請求項15から17のいずれか一項に記載の免疫原性組成物。
【請求項19】
TLRリガンドがTLR-1、TLR-2またはTLR-6のいずれかに結合する、請求項15から18のいずれか一項に記載の免疫原性組成物。
【請求項20】
TLRリガンドがTLR-2に結合する、請求項15から19のいずれか一項に記載の免疫原性組成物。
【請求項21】
対象に請求項1から20のいずれか一項に記載の免疫原性組成物を投与する段階を含む、対象で免疫応答を起こす方法。
【請求項1】
抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は負荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している正荷電基を含み、抗原の負荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物。
【請求項2】
抗原が総負電荷を有する、請求項1に記載の免疫原性組成物。
【請求項3】
正荷電基が少なくとも1つの正荷電アミノ酸を含む、請求項1または請求項2に記載の免疫原性組成物。
【請求項4】
正荷電基が分枝状または線状のペプチドである、請求項1から3のいずれか一項に記載の免疫原性組成物。
【請求項5】
ペプチドが少なくとも1つのアルギニンまたはリジン残基を含む、請求項3に記載の免疫原性組成物。
【請求項6】
ペプチドが、少なくとも4つのアルギニン残基および/または少なくとも4つのリジン残基を含む、請求項3または請求項4に記載の免疫原性組成物。
【請求項7】
抗原および樹状細胞ターゲッティング成分を含む免疫原性組成物であって、抗原は正荷電領域を含み、樹状細胞ターゲッティング成分は樹状細胞リガンドに共有結合している負荷電基を含み、抗原の正荷電領域は樹状細胞ターゲッティング成分と静電気的に会合している、免疫原性組成物。
【請求項8】
抗原が総正電荷を有する、請求項7に記載の免疫原性組成物。
【請求項9】
負荷電基が少なくとも1つの負荷電アミノ酸を含む、請求項7または請求項8に記載の免疫原性組成物。
【請求項10】
負荷電基が分枝状または線状のペプチドである、請求項7から9のいずれか一項に記載の免疫原性組成物。
【請求項11】
ペプチドが少なくとも1つのアスパラギン酸またはグルタミン酸残基を含む、請求項10に記載の免疫原性組成物。
【請求項12】
ペプチドが、少なくとも4つのアスパラギン酸残基および/または少なくとも4つのグルタミン酸残基を含む、請求項10または請求項11に記載の免疫原性組成物。
【請求項13】
抗原が核酸でない、請求項1から12のいずれか一項に記載の免疫原性組成物。
【請求項14】
静電的相互作用だけによって抗原が樹状細胞ターゲッティング成分と会合している、請求項1から13のいずれか一項に記載の免疫原性組成物。
【請求項15】
樹状細胞リガンドがTLRリガンドである、請求項1から14のいずれか一項に記載の免疫原性組成物。
【請求項16】
TLRリガンドが脂質またはペプチドグリカンまたはリポタンパク質またはリポ多糖を含む、請求項15に記載の免疫原性組成物。
【請求項17】
TLRリガンドがパルミトイル、ミリストイル、ステアロイル、ラウロイル、オクタノイルまたはデカノイルを含む、請求項15または請求項16に記載の免疫原性組成物。
【請求項18】
TLRリガンドがPam2Cys、Pam3Cys、Ste2Cys、Lau2CysおよびOct2Cysからなる群から選択される、請求項15から17のいずれか一項に記載の免疫原性組成物。
【請求項19】
TLRリガンドがTLR-1、TLR-2またはTLR-6のいずれかに結合する、請求項15から18のいずれか一項に記載の免疫原性組成物。
【請求項20】
TLRリガンドがTLR-2に結合する、請求項15から19のいずれか一項に記載の免疫原性組成物。
【請求項21】
対象に請求項1から20のいずれか一項に記載の免疫原性組成物を投与する段階を含む、対象で免疫応答を起こす方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2012−523379(P2012−523379A)
【公表日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願番号】特願2012−503830(P2012−503830)
【出願日】平成21年4月9日(2009.4.9)
【国際出願番号】PCT/AU2009/000446
【国際公開番号】WO2010/115229
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(500074981)ザ ユニバーシティー オブ メルボルン (5)
【Fターム(参考)】
【公表日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願日】平成21年4月9日(2009.4.9)
【国際出願番号】PCT/AU2009/000446
【国際公開番号】WO2010/115229
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(500074981)ザ ユニバーシティー オブ メルボルン (5)
【Fターム(参考)】
[ Back to top ]