
免疫応答に関与する新規ポリペプチド
【課題】T細胞の活性化に関与する受容体−リガンド対を含む新規ポリペプチド、ポリペプチドをコードする核酸、ならびにこれらを発現するベクターおよび宿主細胞を提供する。本ポリペプチド、またはそれらのアゴニストおよびアンタゴニストはT細胞媒介疾患の治療に使用される。
【解決手段】CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアンタゴニスト。自己免疫疾患(リウマチ様関節炎、乾癬、多発性硬化病、糖尿病、および全身性エリテマトーデスなど)、トキシックショック症候群、骨髄または臓器移植、炎症性腸疾患、輸血による異種感作、および移植片対宿主疾患の治療を含む多くの徴候の免疫抑制剤として使用することができる。
【解決手段】CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアンタゴニスト。自己免疫疾患(リウマチ様関節炎、乾癬、多発性硬化病、糖尿病、および全身性エリテマトーデスなど)、トキシックショック症候群、骨髄または臓器移植、炎症性腸疾患、輸血による異種感作、および移植片対宿主疾患の治療を含む多くの徴候の免疫抑制剤として使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、Tリンパ球の活性化に関与するポリペプチドに関する。特に、本発明は、Tリンパ球共刺激ポリペプチド、該ポリペプチドをコードする核酸、該ポリペプチドを産生するための発現ベクターと宿主細胞、ならびに免疫抑制および免疫活性化に関連する疾患の治療用組成物および方法に関する。
【背景技術】
【0002】
発明の背景
適切なTリンパ球(T細胞)の免疫応答を得るためには、抗原提示細胞(APC)によって2種類のシグナルがT細胞に送られる必要がある。第1に、特異性を決定する場合に、抗原は主要組織適合抗原複合体(MHC)によってT細胞受容体(TCR)に提示される必要がある。第2に、抗原依存性の共刺激シグナルは、APC上のB7ファミリーに属する要素がT細胞上のCD28タンパク質と連結することによって送り出される必要がある。増殖性の免疫応答は、増殖、分化、クローン性増殖、および作用または機能につながる。第2の共刺激シグナルがない場合は、T細胞はアネルギーと称される持続性抗原特異的不応答性の状態になる。
【0003】
T細胞は、免疫応答を開始し、抗原特異的エフェクター機能を媒介し、サイトカインの分泌によって他の白血球の活性を制限する。T細胞受容体(TCR)は、T細胞を他のリンパ球と区別するものであり、APCのMHCによって抗原が提示された場合にのみその抗原と結合可能である。特定のT細胞の機能は、CD4やCD8などの膜抗原の発現と関連づけることができる。例えば、一般にCD4+T細胞はヘルパーT細胞(TH)として機能しMHCクラスIIに限定され、一般にCD8+細胞は細胞障害性T細胞(TC)として機能しMHCクラスIに限定される。
【0004】
従来同定された有効なT細胞共刺激ポリペプチドとしては、B7.1と称されるポリペプチド(フリーマン(Freeman)らJ.Immunology 143,2714−2722(1989)、フリーマン(Freeman)らJour.Expt.Med.174,625−31(1991))、およびB7.2と称される(フリーマン(Freeman)らScience 262,909−911(1993)、およびフリーマン(Freeman)らJour.Expt.Med.178,2185−2192(1993))(あるいはそれぞれCD80およびCD86と称される)が挙げられる。これらのポリペプチドは、種々のAPC上において誘発的または恒常的のいずれかで発現され、それぞれT細胞上のCD28およびCTLA−4に対する膜結合リガンドである。CD28(アルフォ(Aruffo)およびシード(Seed)Proc.Natl.Acad.Sci.84,8573−8577(1987)、およびグロス(Gross)らJ.Immun.144,3201−3210(1990))は、休止T細胞上で発現され、陽性共刺激シグナルを媒介する。CTLA−4(ブルネット(Brunet)らNature 328,267−270(1987)、およびダリアバク(Dariavach)らEur.Jour.Immun.18,1901−1905(1988))の発現は、T細胞の活性化によって誘発され、B7.1およびB7.2に対する結合親和性がより高いために、CD28シグナルを負の方向に制御する。CTLA−4遺伝子をもたないマウスはCTLA−4がない状態では増殖シグナルのスイッチオフ機構が損なわれているため、T細胞量が非常に多く見られる。この表現型は、T細胞の増殖に対してCTLA−4共刺激タンパク質が有する主要な阻害作用を明確に示している。CD28またはB7.1またはB7.2の欠如したマウスでは、これほど過度な表現型は見られないが、これはT細胞共刺激に関する別の経路が存在しうることを示している。
【0005】
T細胞の活性化および増殖を調節する手段としてCD28/CTLA−4経路について大きな関心が示されてきている。ヒトFcと融合したCTLA−4の細胞外部分を含有するキメラタンパク質は強力な免疫抑制効果を有し、種々の臨床状況において研究が行われてきた。B7.1タンパク質とB7.2タンパク質に対する抗体についても、免疫抑制の分野で同様の徴候について評価されている。抗CTLA−4抗体は、T細胞活性化の促進に有用であることが示されている。さらに、B7.1およびB7.2の遺伝子治療が、癌免疫療法の分野で非常に有用であることが示されている。
【0006】
今までのところ、CD28、CTLA−4、B7.1、およびB7.2は、1つのT細胞共刺激経路と関連している。T細胞共刺激を調節することによって免疫応答を変化できるのであれば、宿主T細胞の機能および免疫応答の調節に関して好都合な性質を有する可能性のある同一または別のT細胞共刺激経路の他の要素を同定することが望ましい。
【0007】
したがって本発明の目的の1つは、T細胞活性および/または増殖を刺激する新規ポリペプチドを提供することである。本発明のさらなる目的は、T細胞媒介免疫疾患の予防および治療のために該新規ポリペプチドを使用することである。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】フリーマン(Freeman)らJ.Immunology 143,2714−2722(1989)
【非特許文献2】フリーマン(Freeman)らJour.Expt.Med.174,625−31(1991)
【非特許文献3】フリーマン(Freeman)らScience 262,909−911(1993)
【非特許文献4】フリーマン(Freeman)らJour.Expt.Med.178,2185−2192(1993)
【非特許文献5】アルフォ(Aruffo)およびシード(Seed)Proc.Natl.Acad.Sci.84,8573−8577(1987)
【非特許文献6】グロス(Gross)らJ.Immun.144,3201−3210(1990)
【非特許文献7】ブルネット(Brunet)らNature 328,267−270(1987)
【非特許文献8】ダリアバク(Dariavach)らEur.Jour.Immun.18,1901−1905(1988)
【発明の概要】
【0009】
発明の要約
驚くべきことに、T細胞共刺激経路の2種類の新規ポリペプチドがこれまでに同定されている。これらのポリペプチドは、前述のタンパク質CD28、CTLA−4、B7.1、およびB7.2からなる経路とは異なるものと思われるただ1つの共刺激経路におけるリガンド−受容体対を表している。これらのポリペプチドは、CD28関連タンパク質−1またはCRP1、およびB7関連タンパク質−1またはB7RP1と称される。
【0010】
本発明は、CRP1およびB7RP1ポリペプチド、ならびに関連ポリペプチドをコードする核酸分子を提供する。本発明の単離核酸分子は、
a)図1A(配列番号1)に記載のヌクレオチド配列;
b)図1A(配列番号1)に記載の残基1〜200または残基21〜200のポリペプチドをコードするヌクレオチド配列;
c)図1A(配列番号1)に記載のポリペプチドと少なくとも約70%が同一であるポリペプチドをコードするヌクレオチド配列;
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体;
e)(a)、(b)、または(c)のいずれかと相補的なヌクレオチド配列;
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリペプチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列;
g)少なくとも約10、15、20、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列;および
h)ストリンジェンシー条件下で(a)〜(g)のいずれかとハイブリッド形成するヌクレオチド配列;
からなる群より選択されるヌクレオチド配列を含む。
【0011】
さらに、本発明は、
a)図2A(配列番号6)または図3A(配列番号11)または図12A(配列番号16)に記載されるヌクレオチド配列;
b)図2A(配列番号6)に記載の残基1〜322または残基47〜322のポリペプチド、または図3A(配列番号11)に記載の残基1〜288、あるいは残基19〜288、20〜288、21〜288、22〜288、24〜288、または28〜288のポリペプチド、または図12Aに記載の残基1〜302、あるいは残基19〜302、20〜302、21〜302、22〜302、24〜302、または28〜302のポリペプチド、をコードするヌクレオチド配列;
c)図2A(配列番号6)または図3A(配列番号11)または図12A(配列番号16)に記載のポリペプチドと少なくとも約70%が同一であるポリペプチドをコードするヌクレオチド配列;
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体;
e)(a)、(b)、または(c)のいずれかと相補的なヌクレオチド配列;
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリペプチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列;
g)少なくとも約10個、15個、20個、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列;および
h)ストリンジェンシー条件下で(a)〜(g)のいずれかとハイブリット形成するヌクレオチド配列;
からなる群より選択されるヌクレオチド配列を含む単離核酸分子を提供する。
【0012】
さらに本発明の主題は、CRP1ポリペプチドおよびB7RP1ポリペプチド、ならびに関連ポリペプチドにも関する。本発明は、
a)図1A(配列番号2)に記載のアミノ酸配列;
b)残基21に成熟アミノ末端を含む図1A(配列番号2)に記載の成熟アミノ酸配列;
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む図1A(配列番号2)に記載のアミノ酸配列断片;
d)(a)、(b)、または(c)のオルソログ;および
e)(a)、(b)、または(d)の対立遺伝子変異体または選択的スプライス変異体;
からなる群より選択されるアミノ酸配列を含む単離ポリペプチドを提供する。
【0013】
さらに、本発明は、
a)図2A(配列番号7)または図3A(配列番号12)または図12A(配列番号17)に記載のアミノ酸配列;
b)残基47に成熟アミノ末端を含む図2A(配列番号7)に記載の成熟アミノ酸配列、あるいは残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図3A(配列番号12)に記載の成熟アミノ酸配列、あるいは残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図12A(配列番号17)に記載の成熟アミノ酸配列;
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む図2A(配列番号7)または図3A(配列番号12)または図12A(配列番号17)に記載のアミノ酸配列断片;
d)(a)、(b)、または(c)のオルソログ;および
e)(a)、(b)、(c)、または(d)の対立遺伝子変異体または選択的スプライス変異体;
からなる群より選択されるアミノ酸配列を含む単離ポリペプチドを提供する。
【0014】
また、本発明のポリペプチドを産生する発現ベクターおよび宿主細胞、CRP1ポリペプチドおよびB7RP1ポリペプチドならびに関連ポリペプチドと結合する抗体、ならびにB7RP1ポリペプチドおよびB7RP1関連ポリペプチドと、CRP1ポリペプチドおよびCRP1関連ポリペプチドとの結合を検出するアッセイも本発明に包含される。CRP1ポリペプチドまたはCRP1関連ポリペプチドと、B7RP1ポリペプチドまたはB7RP1関連ポリペプチドとを含む医薬組成物も本発明に包含される。CRP1またはB7RP1と相互作用する化合物の同定方法も、これらの化合物がCRP1とB7RP1の活性に対するアゴニストとアンタゴニストのいずれとなるかを決定するためのアッセイとして提供される。
【0015】
CRP1ポリペプチドとB7RP1ポリペプチドは、T細胞の共刺激および増殖に関与する。CRP1ポリペプチドとB7RP1ポリペプチド、それらの選択的結合剤、ならびにそれらのアゴニストおよびアンタゴニストは、T細胞応答の調節に関連する疾患の診断、予防、および治療に有用となりうる。
【0016】
CRP1ポリペプチドとB7RP1ポリペプチド、それらの選択的結合剤、ならびにそれらのアゴニストおよびアンタゴニストは、不充分な免疫応答の刺激、あるいは異常に大きいまたは不適当な免疫応答の軽減または抑制のいずれかによる免疫疾患の診断、予防、および治療に有用となりうる。免疫疾患は、T細胞が直接的または間接的に媒介しうる。
【0017】
本発明は、動物にCRP1ポリペプチドまたはB7RP1ポリペプチドを投与することを含むT細胞媒介疾患の治療、予防、または回復のために提供される。本発明は、CRP1ポリペプチドまたはB7RP1ポリペプチドの存在または発現量を決定することを含む動物のT細胞媒介疾患またはT細胞媒介疾患に対する感受性の診断方法;ならびに該ポリペプチドの存在または発現量に基づくT細胞媒介疾患またはT細胞媒介疾患に対する感受性の診断方法も提供する。通常、T細胞媒介疾患は、T細胞が直接的または間接的に媒介しうる免疫疾患である。動物は哺乳動物が好ましく、ヒトがより好ましい。本発明は、CRP1ポリペプチドまたはB7RP1ポリペプチドと結合する試験分子の同定方法であって、該ポリペプチドを試験化合物と接触させ、ポリペプチドと試験化合物の結合程度を求めることを含む同定方法も提供する。この方法は、CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアゴニストおよびアンタゴニストの同定に使用可能である。
【0018】
CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアンタゴニストは、自己免疫疾患(リウマチ様関節炎、乾癬、多発性硬化病、糖尿病、および全身性エリテマトーデスなど)、トキシックショック症候群、骨髄または臓器移植、炎症性腸疾患、輸血による異種感作、および移植片対宿主疾患の治療を含む多くの徴候の免疫抑制剤として使用することができる。さらに、アンタゴニストは、喘息およびアレルギーなどのT細胞依存性B細胞の媒介する徴候、および抗体媒介自己免疫疾患の抑制剤として使用することができる。CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアゴニストは、腫瘍のサーベイランスおよび除去のためのT細胞活性化に有用となりうるが、これに限定されるものではない。
【0019】
本発明のアンタゴニストは、B7RP1またはB7RP1の細胞外ドメインと反応性であるかそれらと結合する抗体またはその断片を含み、この抗体はB7RP1とCRP1の結合を減少させるか結合しなくする。実施態様の1つでは、この抗体は、ヒトB7RP1またはその細胞外ドメインと選択的に結合する。アンタゴニストであるこの抗体またはその断片は、B7RP1の免疫共刺激活性を部分的または完全に阻害する。好ましい実施態様では、この抗体はモノクローナル抗体であり、マウス抗体、ヒト抗体、キメラ抗体、またはヒト化抗体であってよい。
【0020】
本発明は、CRP1の選択的結合剤またはB7RP1の選択的結合剤またはその療法を動物に投与することを含む、B7RP1とCRP1の相互作用を調節する方法もさらに提供する。実施態様の1つでは、この選択的結合剤は、B7RP1と結合して、B7RP1のCRP1に対する結合を減少させたりなくしたりする抗体である。本発明は、B7RP1の選択的結合剤を動物に投与することを含む、B7RP1が媒介する免疫共刺激を調節する方法も提供する。この選択的結合剤は、B7RP1と結合して、B7RP1が媒介する免疫共刺激を部分的または完全に抑制する抗体であることが好ましい。
【0021】
本発明は、CRP1ポリペプチドまたはB7RP1ポリペプチドをコードする核酸分子を動物に投与することを含む、動物のT細胞の活性化または増殖を調節する方法も提供する。例えば、B7RP1ポリペプチドをコードする核酸分子を、種々の腫瘍に対するT細胞の活性化を促進する遺伝子治療に使用することができる。
【0022】
また、CRP1ポリペプチドまたはB7RP1ポリペプチドを含む非ヒトのトランスジェニック哺乳動物も本発明に包含される。CRP1ポリペプチドまたはB7RP1ポリペプチドを発現してその循環量を増大させる方法で、CRP1またはB7RP1核酸が哺乳動物に導入される。非ヒトのトランスジェニック哺乳動物は、囓歯類が好ましく、マウスまたはラットがより好ましい。
【図面の簡単な説明】
【0023】
【図1A】マウスCRP1(mCRP1)のDNA配列およびアミノ酸配列。CRP1の予測シグナル配列にはアミノ末端で下線が引かれており、実験的に求めたプロペプチド開裂部位はアスタリスクで示され、予測膜貫通配列はカルボキシ末端に向かって下線が引かれている。
【図1B】マウスCRP1タンパク質配列(mCRP1)とマウスCD28(mCD28)のアミノ酸配列。
【図2A−1】マウスB7RP1(mB7RP1)のDNA配列およびアミノ酸配列。B7RP1の予測シグナル配列にはアミノ末端で下線が引かれており、実験的に求めたプロペプチド開裂部位はアスタリスクで示される。予測膜貫通配列はカルボキシ末端に向かって下線が引かれている。
【図2A−2】マウスB7RP1(mB7RP1)のDNA配列およびアミノ酸配列。B7RP1の予測シグナル配列にはアミノ末端で下線が引かれており、実験的に求めたプロペプチド開裂部位はアスタリスクで示される。予測膜貫通配列はカルボキシ末端に向かって下線が引かれている。
【図2B】B7RP1タンパク質配列(mB7RP1)とマウスCD80(mCD80)のアミノ酸配列。
【図3A−1】推定ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図3A−2】推定ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図3B】推定成熟hB7RP1タンパク質と成熟マウスB7RP1(mB7Rp1)タンパク質のアミノ酸配列。
【図4A】pcDNA3/CRP1−Fcをトランスフェクションした293T細胞の可溶性CRP1−Fc融合タンパク質の発現。図に示すように規格化体積の細胞溶解物または馴化培地を10%PAGEゲルに加えて分離させる。細胞関連(細胞溶解物)および分泌された(培地)のFc融合タンパク質の発現についての、細胞溶解物および細胞培地上清のウエスタン(Western)分析。1次抗体はヤギ抗ヒトFc抗体(ピアス・ケミカルカンパニー(Pierce Chemical Company)、ロックフォード(Rockford),IL)であった。
【図4B】pcDNA3/B7RP1−Fcをトランスフェクションした293T細胞の可溶性B7RP1−Fc融合タンパク質の発現。10%PAGEゲルに20μlの規格化細胞溶解物または培地上清を加えて分離させた。(a)と同様にウエスタン分析を行った。
【図5】CRP1−Fc融合タンパク質およびB7RP1−Fc融合タンパク質と、COS−7細胞の発現した膜結合タンパク質の相互作用。COS−7細胞には一時的にpcDNA3/CRP1、pcDNA3/B7RP1、またはpcDNA3ベクター単独をトランスフェクションした。CHOのD細胞にpsDRα/hCD28をトランスフェクションすると、ヒトCD28(hCD28)を安定に発現した。細胞の発現する膜結合CRP1、B7RP1、またはhCD28は、パネルの左側の列に示される。Fc融合タンパク質は、パネル上部の列に示されるプレート結合細胞とともにインキュベートした。インキュベート後、細胞を洗い流し、結合したFc融合タンパク質を、抗ヒトFc抗体とACAS(付着細胞分析および選別(Adherent Cell Analysis and Sorting);ACASアルティマ(ACAS Ultima),メリディアン・インストルメンツ(Meridian Instruments,Inc.),オケモス(Okemos),MI)分析を使用して検出した。
【図6】活性化CD4+T細胞およびCD8+T細胞上のB7RP1受容体(CRP1と推定される)の発現のFACS(蛍光活性化細胞選別器(Fluorescence−Activated Cell Sorter))分析。マウス脾細胞はPMAとイオノマイシンで12時間活性化させた。各パネル底部に示されるように、B7RP1−Fc融合タンパク質、対照Fcタンパク質(Mock−Fc)、またはPBS(染色なし)に細胞を加えてインキュベートし、洗浄し、次にヤギ抗ヒトFc−FITC複合抗体(GaHuFc−FITC)を加えてインキュベートした。各パネルの左側に示されるように、細胞マーカー抗体(T細胞マーカーCD4およびCD8の)PE複合体、またはアイソタイプ対照抗体(ラットアイソタイプ)PE複合体、またはPBS(染色なし)を加えた。
【図7】B細胞のB7RP1発現のFACS(蛍光活性化細胞選別器)分析。マウス脾細胞のCRP1に対するリガンド(B7RP1と推定される)の発現の蛍光細胞数分析。各パネル底部に示されるように、CRP1−Fc融合タンパク質、対照Fcタンパク質(Mock−Fc)、またはPBS(染色なし)を、細胞を加えてインキュベートし、洗浄し、次にヤギ抗ヒトFc−FITC複合抗体(GaHuFc−FITC)を加えてインキュベートした。各パネルの左側に示されるように、PEを複合化したCD45Rに対する細胞マーカー抗体(CD45RはB細胞マーカーである)またはアイソタイプ対照抗体(ラットアイソタイプ)、またはPBS(染色なし)を加えた。
【図8】腹腔マクロファージのmCRP1リガンドの発現のFACS分析。最初に腹腔細胞は、光散乱特性によってサブセットに分類した(パネルA)。前方(FSC)および側方(SSC)に強く光を散乱することができ、マクロファージのマーカーであるF4/80抗原に対して陽性染色されるため、領域5(R5)にマクロファージを識別した(パネルB)。領域6(R6)のマクロファージは、F4/80抗原による染色強度が低いことに基づいて選択したものであり、CRP1−Fc融合タンパク質によって染色されることが分かった(B7RP1を発現するためと推定される)。
【図9】B7RP1−FC融合タンパク質を使用したT細胞増殖の抑制。マウス脾細胞からのT細胞を、グラフ底部に示されるようにコンカナバリン(Concanavalin)A(Con A)濃度を増加させることによって活性化させた。ConAの非存在下(添加せず)または存在下で脾細胞のT細胞を増加させるために、mCRP1−Fc、mB7RP1、およびmB7.2−Fc融合タンパク質を加えた。96ウェルプレートでのT細胞増殖アッセイに200,000細胞を使用した。グラフ凡例に示されるように、細胞を培地(添加物なし)またはFc融合タンパク質でインキュベートした。42時間後、細胞にH−チミジンを6時間加え、続いて回収し、放射活性を測定した。三重反復試料の平均CPMおよび標準偏差を示している。
【図10】A)リンパ節皮質、傍皮質、および髄質を示している対照マウス#10の正常腸間膜リンパ節。ヘマトキシリン−エオジン(H&E)染色、40倍。B)顕著な濾胞過形成(FH)、傍皮質の膨張、および髄索過形成(MH)を示すWX11マウス#40の顕著に肥大した腸間膜リンパ節。H&E、40倍。C)対照マウス#10の腸間膜リンパ節の髄索(MC)および髄洞(MS)の拡大図。新しいマクロファージを有する髄洞の近傍に大部分が小リンパ球で構成される小さな軸索が注目される。H&E、400倍。D)WX11マウス#40の腸間膜リンパ節の髄索(MC)および髄洞(MS)の拡大図。点在するラッセル小体細胞(矢印)を有する多数の形質細胞で構成される顕著に太い髄索が注目される。H&E、400倍。E)動脈周囲リンパ鞘(PALS)を有する赤脾髄領域および白脾髄領域を示している対照マウス#10の正常脾臓、100倍。挿入図:小リンパ球、マクロファージ、および時々の形質細胞を有する白脾髄の周縁領域の拡大図、400倍。F)PALSおよび濾胞(矢印)を含む肥大した白脾髄領域を有するMX11マウス#6の脾臓、100倍。挿入図:多数の形質細胞および時々のラッセル小体を有する周縁領域の拡大図、400倍。G)2つの2次濾胞が隣接する濾胞間領域(矢印)を有する対照マウス#25のパイエル板を有する回腸、40倍。H)顕著な胚中心と濾胞間組織(矢印)を有する顕著に肥大した濾胞を有するWX11マウス#32のパイエル板を有する回腸、40倍。
【図11】A)リンパ節皮質、傍皮質、および髄質を示している対照マウス#5の正常腸間膜リンパ節。ヘマトキシリン−エオジン(H&E)染色、40倍。B)顕著な濾胞過形成(上部:外部皮質の第2濾胞の列)、膨張した傍皮質(中央)および髄索過形成(下部)が見られるWX11マウス#33の顕著に肥大した腸間膜リンパ節。H&E、40倍。C)抗B220抗体(B細胞マーカー)による対照マウス#10の腸間膜リンパ節の免疫組織化学的染色。強く(褐色)染色された軸索領域と薄い髄索に注目されたい。アビジン−ビオチン複合体(ABC)免疫ペルオキシダーゼ法(DAB色原体、ヘマトキシリン中心染色)を使用して免疫染色を行った。40倍。D)抗B220抗体によるWX11マウス#33の腸間膜リンパ節の免疫組織化学的染色。強く染色された皮質濾胞および髄索(索の成熟形質細胞はB220に対して陰性である)に注目されたい。40倍。E)抗CD3抗体(T細胞マーカー)による対照マウス#10のリンパ節の免疫組織化学的染色。リンパ節の傍皮質領域の免疫染色に注目されたい。40倍。F)抗CD3抗体によるWX11マウス#33のリンパ節の免疫組織化学的染色。リンパ節の肥大し強く染色された傍皮質領域に注目されたい。40倍。
【図12A−1】ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図12A−2】ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図12B】推定成hB7RP1タンパク質と成熟マウスB7RP1(mB7RP1)タンパク質のアミノ酸配列。
【図13A−1】ヒトCRP1(hCRP1)のタンパク質コード領域の構造および配列。hCRP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位はアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図13A−2】ヒトCRP1(hCRP1)のタンパク質コード領域の構造および配列。hCRP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位はアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図13B】hCRP1タンパク質とマウスCRP1(mB7RP1)タンパク質のアミノ酸配列。
【図14】CRP−1は休止記憶T細胞上にある。6〜7月齢のマウスの休止脾細胞は、FITC複合化抗ヒトFc抗体と、CD44(図14A)、CD45RB(図14B)、またはCD69(図14C)のいずれかに対する抗体とPEの複合体によるB7RP−1−Fc標識を使用して二重染色した。
【図15】B7RP−1−Fc融合タンパク質によるT細胞の共刺激。A)異なる量のB7RP−1−Fc(黒四角)、B7.2−Fc(黒丸)、またはOPG−Fc融合タンパク質対照試料(白四角)を抗CD3抗体と併用してT細胞の増殖を誘発させた。種々の濃度の融合タンパク質を使用して、抗ヒトFcFAb2(12.5μg/ml)および抗CD3抗体(0.9μg/ml)で予備コーティングした96ウェルプレートにコーティングした。B7RP−1−FcおよびB7.2−Fcは、最大効果がえられる0.3μg/mlを最大量として用量依存的にT細胞を共刺激する。B)B7RP−1−Fc(黒四角)、B7.2−Fc(黒丸)、非融合Fc(白四角)、またはFcなし(白丸)を、抗CD3抗体(0.85μg/ml)および種々の濃度のウサギ抗B7RP−1−Fcポリクローナル抗体と併用してT細胞の増殖を誘発させた。Fc融合タンパク質は濃度0.3μg/mlで使用し、上記プレートに結合させた。アジュバントに乳化させた抗原の精製B7RP−1−Fcを皮下注射して抗B7RP−1−Fc抗体を産生させ、次にアフィニティ精製した。この抗体にFc融合タンパク質を加えて30分間インキュベートした後に細胞に加えた。抗B7RP−1−Fc抗体は用量依存的に、B7RP−1−Fcにより誘発されるT細胞増殖を特異的に阻害する。
【図16】マウスのコラーゲン誘発関節炎の(A)発生率および(B)重度に対するCRP−1−Fcタンパク質およびB7RP−1−Fcタンパク質の効果。コラーゲン誘発関節炎感受性のB10.RIIIマウスの尾の基部にCFA中の10μgのブタコラーゲンII型を免疫した。100μgの融合タンパク質をマウスに1週間当り2回与えた。Fc融合タンパク質および対照PBSによる処理は図の凡例に示している。
【図17】B7RP−1−Fcトランスジェニックマウスの近位結腸。(A)粘膜、粘膜下組織、筋層、および漿膜を有する腸壁を示す対照マウス#53F(メス)の正常近位結腸。ヘマトキシリン−エオジン(H&E)染色、倍率40倍。(b)顕著な腺肥大、裂傷潰瘍形成、および経壁炎症を有するB7RP−1−Fcトランスジェニックマウス#111Fのびまん性肥厚化近位結腸。H&E、40倍。(C)多病巣性裂傷潰瘍形成および経壁炎症を有するB7RP−1−Fcトランスジェニックマウス#111Fの近位結腸(パネルBと同じ)のより低倍率での図。H&E、20倍。(D)B7RP−1−Fcトランスジェニックマウス#111F裂傷潰瘍および肥大性直腸腺(上述のパネルBおよびCに示される)の拡大図。粘液化膿性滲出物を有する内腔に注意されたい。H&E、100倍。(E)マクロファージ、リンパ球、およびより少数の好中球に囲まれる多核巨細胞が見られるB7RP−1−Fcトランスジェニックマウス#112Fの粘膜下組織の肉芽腫性炎の拡大図。H&E、400倍。(F)粘膜腺の直下のリンパ球、形質細胞、およびより少数の好中球と混合された類上皮マクロファージが見られるB7RP−1−Fcトランスジェニックマウス#112Fの粘膜の肉芽腫性炎の拡大図。H&E、400倍。
【図18】B7RP−1−Fcトランスジェニックマウスの遠位結腸。(A)粘膜、粘膜下組織、筋層、および漿膜を有する腸壁の層が示される対照マウス#53F(メス)の正常遠位結腸。ヘマトキシリン−エオジン(H&E)染色、倍率40倍。(b)顕著な腺肥大および過形成、ならびに散在する腺窩膿瘍が見られるB7RP−1−Fcトランスジェニックマウス#111F(メス)のびまん性肥厚化遠位結腸。H&E、40倍。(C)顕著な腺肥大および過形成の見られるB7RP−1−Fcトランスジェニックマウス#55M(オス)のびまん性肥厚化遠位結腸。H&E、40倍。(D)肥大性結腸腺、限局性リンパ様凝集体、および多くの腺窩膿瘍が見られるB7RP−1−Fcトランスジェニックマウス#112F(メス)のびまん性肥厚化遠位結腸。H&E、40倍。(E)抗CD3抗体(T細胞マーカー)によるB7RP−1−Fcトランスジェニックマウス#112Fの遠位結腸の免疫組織化学的染色。表在粘膜および結腸リンパ様斑の免疫染色に注目されたい。H&E、40倍。(F)腺窩膿瘍(矢印)とB220+B細胞で構成されるリンパ様凝集体(差し込み)とが見られるB7RP−1−Fcトランスジェニックマウス#112Fの結腸粘膜の拡大図。H&E、100倍。
【図19】B7RP−1−Fcトランスジェニックマウスの小腸。(a)粘膜の内腔、絨毛、および腺窩が見られ、粘膜下組織、筋層、および漿膜がその下にある対照マウス#53F(メス)の正常十二指腸。ヘマトキシリン−エオジン(H&E)染色、倍率40倍。(b)顕著な腺窩肥大および過形成、ならびに粘膜固有層の軽度のリンパ形質細胞性浸潤が見られるB7RP−1−Fcトランスジェニックマウス#51F(メス)のびまん性肥厚化十二指腸。H&E、40倍。(c)空腸粘膜に正常な長さの絨毛および腺窩が見られる対照マウス#53F(メス)の正常空腸。H&E、倍率40倍。(d)局所的に高度の腺窩肥大および過形成が見られるB7RP−1−Fcトランスジェニックマウス#51F(メス)の顕著に 肥厚化した空腸粘膜。H&E、40倍。(E)回腸粘膜で正常な長さの絨毛および腺窩が見られる対照マウス#53F(メス)の正常回腸。H&E、倍率40倍。(F)絨毛が局所的に失われ平滑化したB7RP−1−Fcトランスジェニックマウス#231M(オス)の軽度に萎縮した回腸粘膜。H&E、40倍。
【図20】B7RP−1−Fc融合タンパク質はマウスの腫瘍増殖を抑制する。Meth A肉腫細胞をBalb/Cマウスの腹部の皮内に移植した。移植の7日後、10日後、14日後、および17日後に、マウスを媒体(黒ひし形)またはマウスB7RP−1−Fc(灰色三角、実施例7)で処理した。移植後の指定の日に、実施例20に記載のように腫瘍体積を測定した。腫瘍増殖を最長28日まで観察した。各群は8匹のマウスで構成された。
【図21A】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。A)異なる濃度の抗CD3の1次刺激における、抗CD3のみ(黒丸)、0.5μg/mlのB7RP−1 Fc(黒三角)、0.5μg/mlのOPG−Fc(白丸)、および5μg/mlの抗CD28(白三角)による共刺激。データは、抗CD3で1次刺激したT細胞のB7RP−1−Fcによる共刺激が、抗CD28抗体を使用した共刺激と同程度であることを示している。示されるデータは、3つの正常ドナーから単離したT細胞で行った数回の実験の代表的な1つについての三重反復ウェルの平均[3H]TdR±SDが含まれている。
【図21B】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。B)CRP−1−FcによるB7RP−1−FC共刺激の用量依存性阻害。0.3μg/mlおよび0.5μg/mlのB7RP−1−Fcの両方で抗CD3をコーティングしたウェルでT細胞を培養した。連続的に希釈した濃度のCRP−1−Fc(黒丸)またはOPG−Fc(白丸)に、B7RP−1−Fcを加えて30分間予備インキュベートした後に、T細胞を加えた。データは、CRP−1−FcがB7RP−1誘発共刺激を用量依存的に抑制することを示している。%阻害をCRP−1−FcまたはOPG−Fcタンパク質濃度に対してプロットする。示されるデータは、三重反復ウェルで行った3回の実験の平均[3H]TdR±SDであり、2つの正常ドナーで行った実験を代表している。
【図21C】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。C)CHOヒトB7RP−1細胞による共刺激。実施例22に記載されるように、T細胞を末梢血から精製し、抗CD3単独(黒丸)、1×104CHOベクター対照細胞(白丸)、または1×104CHO B7RP−1細胞(黒三角)の存在下で種々の濃度の抗CD3を加えて培養した。データは、膜結合B7RP−1によるT細胞増殖の共刺激が、B7RP−1−Fc融合タンパク質を使用した場合に観察される共刺激都道程度であることを示している。示されるデータは三重反復培養の平均±SDであり、2つの正常ドナーで得られる結果を代表している。
【図21D】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。D)サイトカイン産生。(図21A)に記載のようにT細胞を培養し、48時間(黒棒)および72時間(灰色棒)で上清を回収した。データは、B7RP−1−Fc共刺激細胞によって産生されるIL−2量(一番上のグラフ)は、抗CD3および対照Fcによって刺激された細胞の産生する量と同様であるが、抗CD28共刺激細胞の産生する量よりは有意に少ないことを示している。またデータは、B7RP−1−Fc共刺激が、IL−10(中央のグラフ)およびIFN−γ(一番下のグラフ)産生を向上させることも示している。
【0024】
発明の詳細な説明
本発明は、本明細書においてCRP1およびB7RP1と称される新規ポリペプチドを提供し、これらはT細胞活性化に関与する受容体−リガンド対を構成している。本発明のポリペプチドをコードするcDNAは、マウス腸上皮内細胞から作製したライブラリーから同定され、CD28ポリペプチドおよびCTLA−4ポリペプチド(CRP1の場合)、またはB7.1ポリペプチドおよびB7.2ポリペプチド(B7RP1の場合)との相同性を基準にしてスクリーニングを行った。
【0025】
CD28関連タンパク質−1、すなわちCRP1は、アミノ末端のシグナル配列と細胞外ドメインと、膜貫通ドメインと、カルボキシ末端細胞内ドメインを有するI型膜貫通タンパク質であると予測される(図1)。CRP1タンパク質の全長は、成熟形態で180アミノ酸である。予測リーダー配列はアミノ酸残基で1〜20付近(開始メチオニンに対して)に及び、成熟タンパク質の細胞外ドメインは残基21〜145付近を含む(実施例1)。予測膜貫通ドメインは残基146〜163付近にわたり、細胞内ドメインは残基164〜200付近を含む。アミノ末端細胞外ドメインはIgループと同様に、推定細胞内および細胞間結合システインが保存される。さらに、B7.1およびB7.2がCD28およびCTLA−4と結合するために重要であると従来知られていた「MYPPPY」モチーフも部分的に保存される。
【0026】
マウスCD28とCTLA−4の間でアミノ酸同一性が26%であると例示されるように、CD28とCTLA−4は相同性が弱い。CRP1とマウスCD28ではアミノ酸同一性が19%であり、CRP1とマウスCTLA−4では同一性が14%である。しかし、残基42、63、83、109、および137(CRP1タンパク質の開始メチオニンが基準。図1A参照)においてマウスCD28、CTLA−4、およびCRP1の間で重要なシステイン残基が保存されている。おおよその成熟タンパク質長さ、およびカルボキシル末端に対する膜貫通領域の位置も、CRP1、CD28、およびCTLA−4で同様である。
【0027】
ヒトCRP1は、図13Aに示されるヌクレオチド配列およびアミノ酸配列を有する膜貫通タンパク質である。予測リーダー配列は残基1〜19付近、または残基1〜20付近にわたる。予測成熟アミノ末端は残基20または21である。成熟アミノ末端は位置21が好ましい。細胞外ドメインは任意の予測成熟アミノ末端からアミノ酸残基140付近に広がり、膜貫通ドメインは残基141〜161付近に及び、細胞内ドメインは残基162〜199付近にわたる。ヒトCRP1タンパク質はマウスタンパク質と69%同一であり、対応するヌクレオチド配列は77%が同一である。ヒトCRP1の配列は、ハトロフ(Hutloff)らNature 397,263−266(1999)に報告されている。
【0028】
B7関連タンパク質−1、すなわちB7RP1は、アミノ末端のシグナル配列および細胞外ドメインと、膜貫通ドメインと、カルボキシ末端細胞内ドメインとを有するI型膜貫通タンパク質であると予測される(図2A)。B7RP1タンパク質の全長は、成熟形態で276アミノ酸である。予測リーダー配列はアミノ酸残基1〜46付近(開始メチオニンに対して)であり、成熟タンパク質の細胞外ドメインは残基47〜279を含む(実施例3)。予測膜貫通ドメインは残基280〜298にひろがり、細胞内ドメインは残基299〜322を含む。B7.1およびB7.2と同様に、B7RP1の細胞外ドメインは2つのIgループを含む。
【0029】
マウスB7.1とB7.2の間のアミノ酸同一性が24%であることで示されるように、B7.1とB7.2は相同性が弱い。B7RP1とマウスB7.1の間のアミノ酸同一性は20%であり、B7RP1とマウスB7.2の間の同一性は19%である。しかし、マウスB7.1、B7.2、およびB7RP1の間で重要なシステイン残基は、残基62、138、185、および242(B7RP1タンパク質の開始メチオニンが基準、図2A)において保存されている。おおよその成熟タンパク質長、およびカルボキシ末端に他意汁膜貫通領域の位置もmB7RP1、B7.1、およびB7.2で同様である。
【0030】
ヒトB7RP1も、Igループ構造を必要とする細胞外ドメインにシステイン残基が保存される膜貫通タンパク質である。図3Aに示されるように予測リーダー配列は、残基1〜18、1〜19、1〜20、1〜21、1〜23、または1〜27の付近を含む。予測成熟アミノ末端は、残基19、20、21、22、24、または28のいずれであってもよい。好ましくはアミノ末端は位置19である。細胞外ドメインは任意の成熟アミノ末端からアミノ酸残基259付近に広がる。予測膜貫通ドメインは残基259〜274に及ぶ。細胞内ドメインは残基275〜302を含む。ヒトB7RP1の全長のヌクレオチド配列およびアミノ酸配列は図12Aに示される。ヒトB7RP1はマウスタンパク質と約43%が同一である。
【0031】
CRP1とB7RP1は互いに結合するが、CRP1はB7RP1関連タンパク質B7.2とは検出可能な程度では結合せず;B7RP1はCRP1関連CD28またはCTLA−4との検出可能な結合を示さない(実施例8)。B7RP1はT細胞増殖を調節することが示されたが、これはB7RP1とCRP1受容体の相互作用のためと推測されている(実施例11)。従ってCRP1とB7RP1は、T細胞の増殖と活性化を調節する新規な経路を表している。
【0032】
B7RP1とCRP1の相互作用は、免疫共刺激、ならびにT細胞の増殖および活性化を増加または減少させることができるような方法で調節することができる。例として、マウスB7RP1に対して産生される抗B7RP1モノクローナル抗体およびポリクローナル抗体は、B7RP1/CRP1相互作用を遮断し、B7RP1−Fc融合タンパク質に誘発されるT細胞増殖も遮断した(実施例17参照)。ヒトCRP−1−Fc融合タンパク質は、ヒトB7RP−1−Fcによって誘発されるヒトT細胞の増殖を遮断した(実施例21)。さらに、CRP1−Fc融合タンパク質を加えることによって、リウマチ様関節炎のマウスモデルの関節炎症状の発生が遅れた(実施例18参照)。B7RP−1/CRP−1共刺激は、B7RP−1−Fc融合タンパク質またはこの経路の他の活性化物質を加えることによって増大させることができる(実施例20)。
【0033】
核酸分子
用語「単離核酸分子」は、必然的に関連する少なくとも1種類の汚染核酸分子がない核酸分子を意味し、好ましくは他のあらゆる汚染哺乳鵜動物核酸分子が実質的にない核酸分子を意味する。
【0034】
用語「対立遺伝子変異体」は、生物の染色体の所与の位置を占める遺伝子の数種類の可能な自然発生的別形態のうちの1つを意味する。
【0035】
用語「スプライス変異体」は、RNA転写において別のイントロン配列のプロセシングによって生成する核酸分子(通常はRNA)を意味する。
【0036】
用語「高ストリンジェンシー条件」は、(1)洗浄に低イオン強度および高温を使用、例えば50℃で0.1×SSC(0.015MのNaCl/0.0015Mのクエン酸ナトリウム)0.1%のNaDodSO4(SDS)、または(2)ハイブリダイゼーション中にホルムアミド、例えば50%(v/v)ホルムアミドなどの変性剤と0.1%ウシ血清アルブミン/0.1%を使用、などの条件を意味する。フィコール(Ficoll)/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム緩衝液(pH6.5)と5×SSC(750mMのNaCl、75mMのクエン酸ナトリウム)を42℃で使用。高ストリンジェンシー条件の別の例は、50%ホルムアミド、5×SSC、50mMのリン酸ナトリウム(pH6.8)、0.1%のピロリン酸ナトリウム、5×デンハート(Denhardt)液、超音波処理サケ精子DNA(50μg/ml)、0.1%のSDS、および10%の硫酸デキストランを42℃で使用し、42℃において0.2×SSCおよび0.1%のSDSで洗浄する。
【0037】
用語「中ストリンジェンシー条件」は、上記よりもストリンジェンシー性の低い洗浄溶液およびハイブリダイゼーション条件(例えば、温度やイオン強度)の使用を含む条件を意味する。中ストリンジェンシー条件の一例は、20%のホルムアミド、5×SSC、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%の硫酸デキストラン、および20μl/mlの変性剪断サケ精子DNAを含む溶液中37℃で終夜インキュベートした後、約37〜50℃で1×SSCを使用してフィルターをろ過する、などの条件である。当業者であれば、プローブ長などの因子の適合に必要な温度、イオン強度、および他の要因の調節方法が分かるであろう。
【0038】
組換えDNA技術は、サムブルック(Sambrook)ら(分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual),コールド・スプリング・パーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press),コールド・スプリング・パーバー(Cold Spring Harbor),NY(1989))および/またはオースベル(Ausubel)ら編著(分子生物学の最新プロトコル(Current Protocols in Molecular Biology),グリーン・パブリッシャー(Green Publishers Inc.)およびワイリー・アンド・サンズ(Wiley and Sons),NY(1994))に記載されており、これらの記載内容全体を本明細書に引用する。
【0039】
本発明は、CRP1ポリペプチドおよびB7RP1ポリペプチドをコードする単離核酸分子を提供する。また本発明は、CRP1ポリペプチドおよびB7RP1ポリペプチドをコードする分子の配列の断片、対立遺伝子変異体、スプライス変異体、または相補的なものである核酸分子も提供する。CRP1またはB7RP1をコードする分子と少なくとも約70%が同一である核酸分子、または中または高ストリンジェンシー条件下でCRP1またはB7RP1をコードする分子とハイブリッド形成する核酸分子も本発明に含まれる。これらの核酸分子は、cDNA、ゲノムDNA、RNA、あるいは部分的または完全に合成した核酸分子であってもよい。好ましい実施態様では、本発明の核酸分子は、CRP1またはB7RP1をコードする核酸分子と少なくとも約75%、80%、85%、90%、または95%が同一である。
【0040】
CRP1またはB7RP1のポリペプチドまたはその断片をコードする遺伝子またはcDNAは、ゲノムライブラリーまたはcDNAライブラリーのハイブリッド形成スクリーングまたはPCR増幅などによって得ることができる。ライブラリーのスクリーニングに有用なプローブまたはプライマーは、同一ファミリーまたは関連ファミリーの遺伝子の他の公知の遺伝子または遺伝子断片の配列情報、例えばシステイン残基の保存配列などのCRP1またはB7RP1関連ポリペプチドに見られる保存モチーフの配列情報を基準にして生成することができる。さらに、CRP1ポリペプチドまたはB7RP1ポリペプチドをコードする遺伝子がある種について同定された場合は、その遺伝子の全体または一部を他の種の相同性遺伝子の同定のプローブとして使用することができる。プローブまたはプライマーは、CRP1遺伝子またはB7RP1遺伝子を発現すると考えられている種々の組織源からcDNAライブラリーをスクリーニングするために使用することができる。
【0041】
オリゴヌクレオチドプローブがcDNAライブラリーまたはゲノムライブラリーのスクリーニングに使用される場合は、以下の2種類の高ストリンジェンシー溶液のうちの1つを使用することができる。これらの溶液の第1は、6×SSCと0.05%のピロリン酸ナトリウムを35℃〜62℃で使用し、この温度はオリゴヌクレオチドプローブの長さに依存する。例えば、14塩基対プローブは35〜40℃で洗浄され、17塩基対プローブは45〜50℃で洗浄され、20塩基対プローブは52〜57℃で洗浄され、23塩基対プローブは57〜63℃で洗浄される。バックグラウンドの非特異的結合が大きい場合にはこの温度を2〜3℃上昇させることができる。第2の高ストリンジェンシー溶液では、オリゴヌクレオチドプローブの洗浄に塩化テトラメチルアンモニウム(TMAC)を使用する。ストリンジェンシー洗浄溶液の1つは、3MのTMAC、50mMのTris−HCl(pH8.0)、および0.2%のSDSである。この溶液が使用される洗浄温度は、プローブの長さの関数である。例えば、17塩基対プローブは約45〜50℃で洗浄される。CRP1またはB7RP1のポリペプチドまたはそれらの断片をコードする遺伝子を作製する別の手段は、エンジェル(Engels)ら(Agnew.Chem.Intl.Ed.,28:716−734(1989))に記載されるものなどの当業者には公知の方法を用いる化学合成を使用することである。これらの方法としては、特に、リン酸トリエステル法、ホスホロアミダイト法、およびH−ホスホネート法による核酸合成が挙げられる。このような化学合成の好ましい方法は、標準的ホスホロアミダイト化合物を使用するポリマー支持合成である。通常、CRP1ポリペプチドまたはB7RP1ポリペプチドをコードするDNAは長さが数百ヌクレオチドとなる。約100ヌクレオチドを超える核酸は、これらの方法を使用して数個の断片として合成することができる。次にこれらの断片を結合させて、CRP1ポリペプチドまたはB7RP1ポリペプチドの全長を形成することができる。通常、ポリペプチドのアミノ末端をコードするDNA断片はメチオニン残基をコードするATGを有する。このメチオニンは、成熟形態のCRP1ポリペプチドまたはB7RP1ポリペプチドに存在してもしなくてもよく、これは宿主細胞で産生されるこのポリペプチドが細胞から分泌されるように計画されるかどうかに依存する。
【0042】
それにも拘らず、生物活性であるポリペプチドをそれ自身コードしない、CRP1またはB7RP1核酸分子、断片、及び/または誘導体は、哺乳類の組織または体液試料中のCRP1あるいはB7RP1のDNAあるいは対応するRNAの存在に対して、定性的あるいは定量的に試験する診断アッセイにおける交雑プローブとして有用である。
【0043】
ある場合には、天然のCRP1あるいはB7RP1ポリペプチドの核酸及び/またはアミノ酸変異体を製造することが望ましい。核酸変異体は、プライマーが所望の点突然変異を持つ、部位特異的突然変異誘発、PCR増幅、または他の適切な方法を用いて産生されてよい(Sambrookら,上記,及び突然変異誘発の手法の記述については、Ausubelら,上記を参照)。上記のEngelsらにより述べられた方法を用いての化学合成を使用して、このような変異体を製造してもよい。当業者に知られている他の方法も使用してもよい。
好ましい核酸変異体は、一定の宿主細胞におけるCRP1とB7RP1ポリペプチドの最適発現に対して変更されたコドンを含むものを含んでいる。特定のコドン変更は、タンパク質と宿主細胞の選択に依存する。一つの場合には、一定の宿主細胞の高度に発現された遺伝子において優先的に使用されるコドンを選択することにより、このような「コドン最適化」を行うことができる。高度に発現された細菌の遺伝子のコドン選択に対する「Ecohigh.Cod」等のコドン頻度表を組込むコンピューターアルゴリズムが使用されてよく、これはUniversity of Wisconsin Packageバージョン9.0、Genetics Computer Group、Madison、WIにより提供される。他の有用なコドン頻度表は、「Celegans_high.cod」、「Celegans_lowcod」、「Drosophila_high.cod」、「Human_high.cod」、「Maize_high.cod」、及び「酵母_high.cod」を含む。他の好ましい変異体は、野生のタイプに比べて上記に述べたような保存アミノ酸変化(例えば、天然のアミノ酸の側鎖の荷電または極性は異なるアミノ酸による置換により実質的に変えられない)をコードするもの、新規なグリコシル化及び/またはリン酸化部位を生成するように設計されたもの、または現存のグリコシル化及び/またはリン酸化部位を削除するように設計されたものである。
【0044】
標準連結法を用いて、CRP1またはB7RP1ポリペプチドをコードする遺伝子、cDNA、またはこれらの断片を適切な発現または増幅ベクター中に挿入することができる。このベクターは、通常、使用される特別な宿主細胞において機能的であるように選択される(すなわち、このベクターは、この遺伝子の増幅及び/またはこの遺伝子の発現が起こることができるように、宿主細胞仕組みと適合している)。CRP1またはB7RP1ポリペプチドをコードする遺伝子、cDNA、またはこれらの断片は、原核、酵母、昆虫(バキュロウイルス系)及び/または真核の宿主細胞において増幅/発現されてよい。この宿主細胞の選択は、CRP1またはB7RP1ポリペプチドまたはこれらの断片がグリコシル化及び/またはリン酸化されるかどうかに一部依存する。そうであるとすれば、酵母、昆虫、または哺乳類の宿主細胞が好ましい。
【0045】
通常、宿主細胞のいずれかで使用される発現ベクターは、プラスミド維持及び挿入ヌクレオチド配列のクローニングと発現のための配列を含む。「隣接配列」とまとめて呼ばれるこのような配列は、プロモーター及びエンハンサー、複製エレメントの起源、転写停止エレメント、供与体と受容体またはスプライス部位を含む完全なイントロン配列、シグナルペプチド配列、リボソーム結合部位エレメント、ポリアデニル化配列、発現されるポリペプチドをコードする核酸を挿入するためのポリリンカー領域及び選択マーカーエレメント等の他の調節エレメントを含む。これらのエレメントの各々は下記に述べられる。場合によっては、このベクターは、「タグ」配列、CRP1またはB7RP1ポリペプチドコーディング配列の5’または3’末端に位置するすなわちオリゴヌクレオチド分子を含んでよい。このオリゴヌクレオチド分子は、ポリヒス(ヘキサヒス等の)、または市販の抗体が存在する、FLAG、HA(血球凝集素インフルエンザウイルス)またはmyc等の他の「タグ」をコードする。このタグは、通常、このポリペプチドの発現時にこのポリペプチドに融合され、宿主細胞からのCRP1またはB7RP1ポリペプチドをアフィニティ精製する手段として機能することができる。例えば、このタグに対する抗体を親和マトリックスとして用いるカラムクロマトグラフィにより、アフィニティ精製を行うことができる。場合によっては、引き続いて、このタグは、しかるべきペプチダーゼ用いる等の種々の手段により精製されたCRP1またはB7RP1ポリペプチドから除去され得る。
【0046】
当業者によれば、ヒト免疫グロブリンヒンジとFc領域は、CRP1またはB7RP1ポリペプチドのN−末端またはC−末端のいずれかに融合されてよい。引き続くFc融合タンパク質は、タンパク質A親和カラムを使用することにより精製され得る。免疫グロブリンFc領域は、インビトロで長い薬物動力学的半減期を示すことが知られていて、Fc領域に融合されたタンパク質は、非融合品と比較して、インビトロで実質的に大きい半減期を示すことが見出されている。また、このFc領域への融合により、一部の分子の生物活性に有用であるこの分子の二量化及び/または多量化が可能になる。
【0047】
この隣接配列は、同種(すなわち、宿主細胞と同じ種及び/または菌株からの)、異種(すなわち、宿主細胞種または菌株と別の種からの)、雑種(すなわち、一つの以上のソースからの隣接配列の組み合わせ)、合成によるものであってよく、またはこれは、未変性のCRP1またはB7RP1核酸隣接配列であってよい。従って、この隣接配列が宿主細胞仕組みで機能的であり、活性化され得るという前提であれば、隣接配列のソースは、いかなる単一細胞の原核または真核生物体、いかなる脊椎あるいは無脊椎生物体、またはいかなる植物であってよい。
【0048】
本発明のベクターにおいて有用である隣接配列は、当分野においてよく知られている方法のいずれによって得られてもよい。通常、CRP1またはB7RP1核酸隣接配列以外の、ここにおいて有用な隣接配列は、マッピングにより及び/または制限エンドヌクレアーゼ消化により以前に同定されていて、このようにして、適切な制限エンドヌクレアーゼを用いて適当な組織ソースから単離され得る。いくつかの場合には、この隣接配列の完全なヌクレオチド配列は知られているかもしれない。ここで、この隣接配列は、核酸合成またはクローニングについて上述した方法を用いて合成されてよい。
【0049】
この隣接配列のすべてまたは一部のみが知られている場合には、PCRを用いて及び/または同じあるいはもう一つの種からの好適なオリゴヌクレオチド及び/または隣接配列断片によりゲノムライブラリーをスクリーニングすることによりこれを得てよい。
【0050】
この隣接配列が知られていない場合には、隣接配列を含むDNAの断片は、例えば、コーディング配列または更にもう一つの遺伝子を含む更に大きな片のDNAから単離されてよい。単離は、一つあるいはそれ以上の慎重に選ばれた酵素を用いて、適当なDNA断片を単離する、制限エンドヌクレアーゼ消化により行われてよい。消化の後、所望の断片は、アガロースゲル精製、Qiagen(登録商標)カラムまたは当業者に知られた他の方法により単離されてよい。この目的を果たすための好適な酵素の選択は、当分野分野の通常の技術を持つ者には容易に明白になるであろう。
【0051】
複製エレメントの起源は、通常、購入される原核の発現ベクターの一部であり、宿主細胞においてこのベクターの増幅を助ける。しかるべきコピー数迄のこのベクターの増幅は、ある場合には、CRP1あるいはB7RP1ポリペプチドの最適な発現に重要であり得る。選択したベクターが複製部位の起源を含まないならば、これは既知の配列に基づいて化学的に合成され、このベクター中に連結されてよい。
【0052】
この転写停止エレメントは、通常、CRP1あるいはB7RP1ポリペプチドのコーディング配列の末端の3’に位置し、CRP1またはB7RP1ポリペプチドの転写を停止させる機能をする。通常、原核細胞中の転写停止エレメントは、G−Cに富む断片とそれに続くポリT配列である。このエレメントはライブラリーから容易にクローンされるか、あるいは更にベクターの一部として購入される一方、また、上述の方法等の核酸合成の方法を用いて、容易に合成され得る。
【0053】
選択マーカー遺伝子エレメントは、選択的培養基中で育成された宿主細胞の生き残りと成長に必要なタンパク質をコードする。典型的な選択マーカー遺伝子は、(a)原核の宿主細胞に対する抗生物質または他の毒素、例えば、アンピシリン、テトラサイクリン、またはカナマイシンに耐性を賦与し、(b)細胞の栄養要求性欠乏症を補い、または(c)複雑な培地から得られない重要な栄養物を供給するタンパク質をコードする。好ましい選択マーカーは、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、及びテトラサイクリン耐性遺伝子である。
【0054】
普通、シャイン−ダルガルノ配列(原核細胞)またはKozak配列(真核細胞)と呼ばれるリボソーム結合エレメントは、mRNAの翻訳開始に通常必要である。このエレメントは、このプロモーターに3’に、また合成されるべきCRP1またはB7RP1ポリペプチドのコーディング配列に5’に通常位置する。シャイン−ダルガルノ配列は変るが、通常、ポリプリン(すなわち、高A−G含量を有する)である。多数のシャイン−ダルガルノ配列が同定されており、その各々は、上記に示し、そして原核のベクターに使用される方法を用いて、容易に合成され得る。
【0055】
CRP1またはB7RP1ポリペプチドが宿主細胞から分泌されるのが望ましい場合には、宿主細胞からポリペプチドの移行を方向付けするために、シグナル配列が使用されてよい。CRP1またはB7RP1膜貫通ドメインは、また、宿主膜への結合を防ぐために、突然変異または欠失により不活性化される。通常、このシグナル配列は、CRP1またはB7RP1遺伝子またはcDNAのコーディング領域に、あるいはCRP1またはB7RP1遺伝子コーディング領域の5’末端に直接に位置される。多数のシグナル配列が同定されており、選ばれた宿主細胞中で機能性であるシグナル配列のいずれもCRP1あるいはB7RP1遺伝子またはcDNAと一緒に使用されてよい。それゆえ、シグナル配列は、CRP1あるいはB7RP1遺伝子またはcDNAに同種あるいは異種であってよく、CRP1あるいはB7RP1ポリペプチド遺伝子またはcDNAに同種あるいは異種であってよい。加えて、このシグナル配列は、上記に示した方法を用いて化学的に合成されてよい。
【0056】
大多数の場合において、シグナルペプチドの存在によるこの宿主細胞からのポリペプチドの分泌は、結果としてこのポリペプチドからアミノ末端メチオニンを生成する。
【0057】
多くの場合、CRP1あるいはB7RP1遺伝子またはcDNAの転写は、このベクター中の一つあるいはそれ以上のイントロンの存在により増大する。これは、CRP1あるいはB7RP1ポリペプチドが真核の宿主細胞、特に哺乳類の宿主細胞中で産生する場合に特に当てはまる。使用されるイントロンは、使用される遺伝子が完全な長さのゲノム配列またはこれの断片である、CRP1あるいはB7RP1遺伝子内で天然に生じるものであってよい。このイントロンは、この遺伝子(または大多数のcDNAに関して)内で天然に生じるものでない場合には、このイントロンは他の源から得られてよい。このイントロンは転写されて有効になるので、5’隣接配列とCRP1あるいはB7RP1遺伝子に対するこのイントロンの位置は、一般に重要である。従って、発現ベクター中に挿入されるCRP1あるいはB7RP1遺伝子がcDNA分子である場合には、このイントロンに好ましい位置は、転写スタート部位の3’及びポリA転写停止配列の5’である。好ましくは、このイントロンは、このコーディング配列を妨害しないように、cDNAの一方または他方(すなわち、5’または3’)に位置する。本発明を実施するするのに、挿入される宿主細胞と適合するという前提ならば、いかなるウイルス、原核及び真核の(植物または動物)生物体をも含む、いかなる源からのいかなるイントロンも使用しうる。また、ここには合成イントロンも含まれる。場合によっては、一つ以上のイントロンがベクターに使用しうる。
【0058】
上記に示した一つあるいはそれ以上のエレメントが使用されるべきベクター中に既に存在しない場合には、これらを個別に得て、このベクターに連結してよい。このエレメントの各々を得るために使用される方法は、当業者によく知られている。
【0059】
本発明を実施するための好ましいベクターは、細菌、昆虫、及び哺乳類の宿主細胞と適合するものである。このようなベクターは、とりわけ、pCRII、pCR3、及びpcDNA3.1(Invitrogen Company,San Diego,CA)、pBSII(Stratagene Company,LaJalla,CA)、pETl5b(Novagen,Madison、WI)、pGEX(Pharmacia Biotech,Piscataway,NJ)、pEGFP−N2(Clontech,Palo Alto,CA)、pETL(BlueBacII Invitrogen)、及びpFastBacDual(Gibco/BRL,Grand Island,NY)を含む。
【0060】
このベクターが構築され、完全な長さをコードする核酸分子または切断されたCRP1あるいはB7RP1ポリペプチドがこのベクターの適当な部位に挿入された後、増幅及び/またはポリペプチド発現に好適な宿主細胞にできあがったベクターを挿入してよい。
【0061】
宿主細胞は、原核宿主細胞(大腸菌等の)または真核宿主細胞(酵母細胞、昆虫細胞、または脊椎動物の細胞等の)であってよい。この宿主細胞は、適切な条件下で培養した場合、CRP1あるいはB7RP1ポリペプチドを合成することができ、引き続いて、培養基(宿主細胞が培地中にそれを分泌するならば)から、あるいはそれを産生する宿主細胞からそれを直接に収集(それが分泌されないならば)することができる。収集後、CRP1あるいはB7RP1ポリペプチドは、モレキュラーシーブクロマトグラフィー、親和クロマトグラフィー等の方法を用いて、精製され得る。
【0062】
CRP1あるいはB7RP1ポリペプチドの産生に適切な宿主細胞の選択は、所望の発現レベル、グリコシル化またはリン酸化等の活性に必要とされるポリペプチド変性、または生物活性分子への折り畳みの容易さ等の種々の因子に依存する。
【0063】
好適な細胞または細胞株は、チャイニーズハムスター卵巣細胞(CR0)、ヒト胎児腎臓(HEK)293または293T細胞、または3T3細胞等の哺乳類の細胞であってよい。好適な哺乳類の宿主細胞の選択及び転換、培養、増幅、スクリーニング及び産物生産及び精製のための方法は、当分野で知られている。他の好適な哺乳類の細胞株は、サルCOS−1及びCOS−7細胞株とCV−1細胞株である。更なる例示の哺乳類の宿主細胞は、転換細胞株を含む、霊長類細胞株及び齧歯類細胞株を含む。正常な2倍体細胞、1次組織、並びに1次外植体のインビトロ培養に由来する細胞菌株もまた好適である。候補細胞は、遺伝子型として選択遺伝子欠乏であっても、あるいは主に作用する選択遺伝子を含んでよい。他の好適な哺乳類の細胞株は、限定ではないがマウス神経芽細胞腫N2A細胞、HeLa、マウスL−929細胞、Swiss、Balb−c由来の3T3株、またはNIHマウス、BHKまたはHaKハムスター細胞株を含む。
【0064】
宿主細胞として同様に有用なのは、細菌細胞である。例えば、大腸菌の種々の菌株(例えば、HB101、DH5α、DH10、及びMC1061)がバイオテクノロジーの分野で宿主細胞としてよく知られている。B.subtilis、Pseudomonas spp.、他のBacillus spp.、Streptomyces spp.などの種々の菌株もこの方法で使用されてよい。
【0065】
当業者に知られている酵母細胞の多数の菌株もまた本発明のポリペプチドを発現する宿主細胞として入手できる。
【0066】
加えて、所望の場合には、昆虫細胞系が本発明の方法で利用されてよい。このような系は、例えばKittsら(Biotechniques、14:810−817(1993))、Lucklow(Curr.Opin.Biotechnol.,4:564−572(1993))及びLucklowら(J.Virol.,12:4566−4579(1993))に記述されている。好ましい昆虫細胞は、Sf−9とHi5(Invitrogen,Carlsbad,CA)である。
【0067】
塩化カルシウム、電気穿孔法、マイクロインジェクション、リポフェクションまたはDEAE−デキストラン法等の方法を用いて、発現ベクターの選ばれた宿主細胞中への「転換」または「トランスフェクション」を行ってよい。選ばれた方法は、一部、使用されるべき宿主細胞のタイプの機能である。これらの方法及び他の好適な方法は、当業者によく知られていて、例えば、上記のSambrookらに示されている。
【0068】
発現ベクターにより転換あるいはトランスフェクションされる宿主細胞は、当業者によく知られた標準培地を用いて培養されてよい。この培地は、普通、この細胞の成長と生き残りに必要なすべての栄養素を含有する。大腸菌細胞を培養するための好適な培地は、例えば、Luria Broth(LB)及び/またはTerrific Broth(TB)である。真核細胞を培養するのに好適な培地は、RPMI 1640、MEM、DMEMであり、これらのすべては、培養される特別な細胞株にように必要とされるのに従って、血清及び/または成長因子により補われる。昆虫培養に好適な培地は、必要に従って、酵母オレエート、ラクトアルブミン加水分解物、及び/またはウシ胎仔血清により補われるグレース(Grace)の培地である。
【0069】
通常、転換された細胞の選択成長に有用な抗生物質または他の化合物のみがサプルメントとして培地に添加される。使用される化合物は、宿主細胞を転換したプラスミド上に存在する選択マーカーエレメントにより決定付けられる。例えば、この選択マーカーエレメントがカナマイシン耐性である場合には、培養基に添加される化合物はカナマイシンである。
【0070】
この宿主細胞中に産生されるCRP1あるいはB7RP1ポリペプチドの量は、当分野において知られている標準的な方法を用いて評価され得る。このような方法は、限定ではないが、ウエスタンブロット分析、SDS−ポリアクリルアミドゲル電気泳動、非変性ゲル電気泳動、HPLC分離、免疫沈澱、及び/またはDNA結合ゲルシフトアッセイ等の活性アッセイを含む。
【0071】
宿主細胞から分泌されるようにCRP1またはB7RP1ポリペプチドを設計した場合には、大部分のポリペプチドが細胞培養基中に見出されてよい。このように製造されたポリペプチドは、この細胞からの分泌時に除去されるので、通常、アミノ末端メチオニンを保持しない。しかしながら、CRP1またはB7RP1ポリペプチドが宿主細胞から分泌されない場合には、それは、細胞質及び/または核(真核宿主細胞に対して)中に、またはサイトソル(グラム陰性バクテリア宿主細胞に対して)中に存在し、アミノ末端メチオニンを有してよい。
【0072】
種々の手法を用いて溶液からのCRP1またはB7RP1ポリペプチドの精製を行うことができる。ヘキサヒスチジン(CRP1またはB7RP1/ヘキサヒス)等のタグ、またはFLAG(Eastman Kodak Co.,NewHaven,CT)またはmyc(Invitrogen,Carlsbad,CA)等の他の小ペプチドをそのカルボキシルまたはアミノ末端で含むように、このポリペプチドを合成した場合、このカラムマトリックスがタグに対して、あるいは直接的にポリペプチドに対して高親和性を持つならば(すなわち、CRP1またはB7RP1ポリペプチドを特異的に認識するモノクローナル抗体)、タグを付けたポリペプチドを親和カラムに通すことにより、1段階プロセスで本質的に精製してよい。例えば、ポリヒスチジンは、大きな親和性と特異性でニッケルに結合し、このようにしてニッケルの親和カラム(Qiagen(登録商標)ニッケルカラム等の)をCRP1またはB7RP1/ポリヒスの精製に使用することができる(例えば、Ausubelら編、Current Protocols in Molecular Biology,Section10.11.8,John Wiley & Sons,New York(1993)を参照)。
【0073】
タグを付けずにCRP1またはB7RP1ポリペプチドを製造し、抗体が入手できない場合には、精製によく知られた方法を使用することができる。このような方法は、限定ではないが、イオン交換クロマトグラフィー、モレキュラーシーブクロマトグラフィー、HPLC、ゲル溶離と組み合わせた未変性ゲル電気泳動、及び合成的等電点電気泳動(「Isoprime」マシン/手法、Hoefer Scientific)を含む。ある場合には、2つあるいはそれ以上のこれらの方法を組み合わせて、純度の増大を達成しうる。
【0074】
CRP1あるいはB7RP1ポリペプチドが主として細胞内に見出されることが期待される場合には、当業者に知られた任意の標準的な手法を用いて、この細胞内材料(グラム陰性バクテリア用の封入体を含む)を宿主細胞から抽出することができる。例えば、フレンチプレス、均質化、及び/または高周波による分解とそれに続く遠心分離により、この宿主細胞を溶解して、ペリプラズム/細胞質の内容物を放出させることができる。
【0075】
CRP1またはB7RP1ポリペプチドがサイトソル中に封入体を形成した場合には、この封入体は、時には内及び/または外の細胞膜に結合することができ、このようにして遠心分離後主としてペレット材料中に見出される。次に、このペレット材料は、pHの両極端で、あるいは洗浄剤、グアニジン、グアニジン誘導体、尿素、または尿素誘導体等のカオトロピック剤により、アルカリ性pHでジチオトレイトールまたは酸性pHでトリスカルボキシエチルホスフィン等の還元剤の存在下で処理され、この封入体を放出、分割、及び可溶化することができる。次に、ゲル電気泳動、免疫沈澱などを用いて、溶解した形のこのポリペプチドを分析することができる。CRP1またはB7RP1ポリペプチドを単離することを所望する場合には、下記及びMarstonら、(Meth.Enz.,182:264−275(1990))に示すような標準方法を用いて、単離を行ってよい。ある場合には、CRP1あるいはB7RP1ポリペプチドは単離時に生物活性でなくてよい。「再折りたたみ」あるいはこのポリペプチドを3級構造に変換し、そしてジスルフィド結合を生成する種々の方法を使用して、生物活性を取り戻すことができる。このような方法は、可溶化ポリペプチドを通常7以上のpHを持つ溶液と特別な濃度の適切なカオトロープの存在下で接触することを含む。また、大多数の場合、再折りたたみ/酸化溶液は、還元剤またはタンパク質のシステイン架橋の形成においてジスルフィド混合を起こさせる特別なレドックス電位を生じる還元剤と対応する酸化形を特定の比で含む。普通に使用されるレドックス対の一部は、システイン/シスタミン、グルタチオン(GSH)/ジチオビスGSH、塩化第2銅、ジチオトレイトール(DTT)/ジチアンDTT、2−メルカプトエタノール(bME)/ジチオ−b(ME)を含む。多数の例において、再折りたたみの効率を増大するために、共溶媒が必要であり、この目的に使用される更に普通の試剤は、グリセロール、種々の分子量のポリエチレングリコール、及びアルギニンを含む。
【0076】
また、化学合成方法(固相ペプチド合成等の)により、Merrifieldらにより示されたもの(J.Am.Chem.Soc.,85:2149(1963))、Houghtenら(Proc Natl Acad.Sci.USA,82:5132(1985))、及びStewart及びYoung(Solid Phase Peptide Synthesis,Pierce Chemical Co.,Rockford,IL(1984))等の当分野において知られた手法を用いて、CRP1またはB7RP1ポリペプチド、断片、及び/またはこれらの誘導体を製造してもよい。このようなポリペプチドは、アミノ末端上にメチオニンの有無のいずれで合成されてよい。これらの文献に示される方法を用いて、化学合成されたCRP1あるいはB7RP1ポリペプチドまたは断片を酸化して、ダイサルファイド架橋を形成してよい。CRP1あるいはB7RP1ポリペプチドまたは断片は、組み換えで製造された、あるいは天然源から精製されたCRP1あるいはB7RP1ポリペプチドに匹敵する生物活性を有すると予期され、このように、組み換えあるいは天然のCRP1あるいはB7RP1ポリペプチドと入れ替えて使用してもよい。
【0077】
ポリペプチド
「CRP1あるいはB7RP1ポリペプチド」という用語は、図1A(配列番号2)、図2A(配列番号7)または図3A(配列番号12)及びここで説明されたすべての関連ポリペプチドのアミノ酸配列を持つポリペプチドを指す。関連ポリペプチドは、対立変異体、スプライス変異体、断片、誘導体、置換、欠失、及び挿入変異体、融合ポリペプチド、及びオルトログを含む。このような関連ポリペプチドは、成熟ポリペプチド、すなわち、シグナルペプチドを欠いたポリペプチドであってよい。CRP1あるいはB7RP1ポリペプチドは、これらを製造する方法に依って、アミノ末端メチオニンであってよく、あるいはそうでなくてもよい。
【0078】
「CRP1あるいはB7RP1ポリペプチド断片」という用語は、図1A(配列番号2)、図2A(配列番号7)または図3A(配列番号12)に示すような、CRP1あるいはB7RP1ポリペプチドの完全長さ未満であるアミノ酸配列のペプチドまたはポリペプチドを指す。このような断片は、アミノ末端での切断、カルボキシ末端での切断、及び/またはポリペプチド配列への内部の欠失の結果から生じうる。このようなCRP1あるいはB7RP1ポリペプチド断片は、アミノ末端メチオニンの有無のいずれで製造されてよい。加えて、CRP1あるいはB7RP1ポリペプチド断片は、天然由来のスプライス変異体、他のスプライス変異体、及び天然に生じるインビボ活性の結果生じる断片であってよい。好ましいCRP1あるいはB7RP1ポリペプチド断片は、機能性膜貫通ドメインを欠き、CRP1あるいはB7RP1のいずれかの細胞外ドメインの一部または全部を構成するCRP1あるいはB7RP1の可溶性の形態を含む。
【0079】
「CRP1あるいはB7RP1ポリペプチド変異体」という用語は、図1A(配列番号2)、図2A(配列番号7)または図3A(配列番号12)に示すCRP1またはB7RP1ポリペプチドアミノ酸配列に比較して、アミノ酸配列が一つあるいはそれ以上のアミノ酸配列置換、欠失、及び/または付加を含むCRP1あるいはB7RP1ポリペプチドを指す。従って、CRP1あるいはB7RP1ポリペプチドに対するDNA配列から変るDNA配列を持つ対応するCRP1及びB7RP1ポリペプチド核酸分子変異体から、このようなCRP1あるいはB7RP1ポリペプチド変異体を製造することができる
ここで使用される時、「CRP1あるいはB7RP1ポリペプチド誘導体」という用語は、例えば、一つあるいはそれ以上の水溶性ポリマー、N−結合あるいはO−結合炭水化物、糖、リン酸塩、及び/または分子が野生型のCRP1あるいはB7RP1ポリペプチドに天然に結合していない、他のこのような分子の添加により化学的に変性したCRP1あるいはB7RPlポリペプチド、変異体、またはこれらの断片を指す。誘導体は、CRP1あるいはB7RP1ポリペプチドに天然に結合した一つあるいはそれ以上の化学基の欠失を更に含む。
【0080】
ここで使用される時、「生物活性のCRP1あるいはB7RP1ポリペプチド」、「生物活性のCRP1あるいはB7RP1ポリペプチド断片」、「生物活性のCRP1あるいはB7RP1ポリペプチド変異体」、及び「生物活性のCRP1あるいはB7RP1ポリペプチド誘導体」という用語は、CRP1あるいはB7RP1特有の少なくとも一つの活性を持つCRP1あるいはB7RP1ポリペプチドを指す。一つの活性はCRP1へのB7RP1の結合である。他の活性は、CRP1あるいはB7RP1がT−細胞増殖及び/または活性化を刺激する能力である。
【0081】
「オルトログ」という用語は、種から同定されるポリペプチドに対応するポリペプチドを指す。例えば、マウス及びヒトB7RP1ポリペプチドはオルトログと考えられる。
「成熟したアミノ酸配列」という用語は、リーダー配列を欠くポリペプチドを指す。
【0082】
「単離されたポリペプチド」という用語は、その自然な環境中で見出される少なくとも一つの汚染性のポリペプチドを含まない、そして好ましくはいかなる他の汚染性の哺乳類のポリペプチドも実質的に含まないポリペプチを指す。
【0083】
「同一性」という用語は、当分野において知られているように、配列を比較することにより決定される2つあるいはそれ以上の核酸分子または2つあるいはそれ以上のポリペプチドの配列間の関係である。当分野においては、「同一性」は、また、場合に応じて、ヌクレオチドあるいはアミノ酸配列の列間の一致により決定されるような、ポリペプチドまたは核酸分子配列間の配列関連性の程度を意味する。「同一性」は、特別なコンピュータープログラム(すなわち、「アルゴリズム」)により扱われるギャップ配列により2つあるいはそれ以上の配列の間の同一性一致のパーセントを測定する。
【0084】
「類似性」という用語は、関連する概念を指すが、対照的に、「同一性」とは、同一性一致と保存的置換一致の双方を含む。保存的置換はポリペプチドに当てはまり、核酸分子には当てはまらないので、類似性は、ポリペプチド配列の比較を扱うのみである。2つのポリペプチド配列が例えば、10/20同一のアミノ酸を持ち、残りがすべて非保存的置換である場合には、同一性と類似性のパーセントは双方とも50%である。同じ試料において、保存的置換がある、更に5つの位置がある場合には、パーセント同一性は50%のままであるが、パーセント類似性は75%である(15/20)。それゆえ、保存的置換がある場合には、2つのポリペプチド配列の間の類似性の程度は、これらの2つの配列の間のパーセント同一性よりも高い。「保存的」アミノ酸置換はここでは下記に表Iを参照して記述されている。表Iに基づくと、保存アミノ酸置換は、同じグループ、例えば、塩基性、酸性、非帯電極性、及び非極性から選ばれる交互のアミノ酸である。例えば、アルギニンに対する保存アミノ酸置換は、リジン及びヒスチジンである。
【0085】
同一性と類似性は、限定ではないが、Computational Molecular Biology,Lesk,A.M編,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.編,Academic Press,New York,1993;Computer Analysis of Sequence Data,Part1,Griffin,A.M.,及びGriffin,H.G.編,Humana Press,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,Academic Press,1987;及びSequence Analysis Primer,Gribskov,M.及び Devereux,J.編,M.Stockton Press,New York,1991;及びCarillo,H.,及びLipman,D.,SIAM J.Applied Math.,48:l073(1988)に記述されているものを含む、既知の方法により容易に計算できる。
【0086】
同一性及び/または類似性を求める好ましい方法は、試験する配列の間の最大一致を与えるように設計される。同一性と類似性を求める方法は、公に入手できるコンピュータープログラムにコード化されている。2つの配列間の同一性と類似性を求める好ましいコンピュータープログラムの方法は、限定ではないが、GAP(Devereux,J.ら,Nucleic Acids, Research 12(1):387(1984);Genetics Computer Group,University of Wisconsin,Madison,WI)、BLASTP,BLASTN,及びFASTA(Atschul,S.F.ら,J.Molec.Biol.215:403−410(1990)を含む、GCGプログラムパッケージを含む。BLAST Xプログラムは、National Center for Biotechnology Information(NCBI)及び他の源(BLAST Manual,Altschul,S.ら、NCB NLM NIH Bethesda,MD 20894;Altschul,S.ら,J.Mol.Biol.215:403−410(1990))から公に入手できる。また、同一性を求めるのに、よく知られたSmith Watermanアルゴリズムを使用してもよい。
【0087】
例として、コンピューターアルゴリズムギャップ(Genetics Computer Group,University of Wisconsin,Madison,WI)を用いて、パーセント配列同一性を求めるべき2つのポリペプチドは、各アミノ酸(このアルゴリズムにより求められるような「一致したスパン」)の最適マッチングに対して並べられる。ギャップオープニングペナルティ(3X平均対角線として計算される;「平均対角線」は、使用されている比較マトリックスの対角線の平均である。「対角線」は、特別な比較マトリックスにより各々の完全なアミノ酸一致に割り当てられた得点または数であり、そして、ギャップエクステンションペナルティ(通常、ギャップオープニングペナルティの1/10倍である)、並びにPAM 250またはBLOSUM 62等の比較マトリックスがアルゴリズムと一緒に使用される。標準比較マトリックス(PAM 250比較マトリックスに対しては、Dayhoffら,Atlas of Protein Sequence and Structure,5巻,補遺3(1978)を参照;BLOSUM 62比較マトリックスに対しては、Henikoffら,Proc.Nati.Acad.Sci USA,89:10915−10919(1992)を参照)もまたアルゴリズムにより使用される。
【0088】
ポリペプチド配列比較に対する好ましいパラメーターは、次を含む。
【0089】
アルゴリズム:Needleman及びWunsch、J.Mol.Biol.48:443453(1970)
比較マトリックス:HenikoffからのBLOSUM 62及びHenikoff,Proc.Natl.Acad.Sci.USA 89:10915−10919(1992)
ギャップペナルティ:12
ギャップ長さペナルティ:4
類似性のしきい値:0
GAPプログラムは上記パラメーターにより有用である。この上述のパラメーターは、ギャップアルゴリズムを用いたポリペプチド比較(末端ギャップに対する無ペナルティと一緒に)のデフォルトパラメーターである。
【0090】
核酸分子配列比較の好ましいパラメーターは次を含む。
【0091】
アルゴリズム:Needleman及びWunsch,J.Mol Biol.48:443−453(1970)
比較マトリックス:一致=+10、ミスマッチ=0
ギャップペナルティ:50
ギャップ長さペナルティ:3
また、GAPプログラムは上記パラメーターによっても有用である。この上述のパラメーターは、核酸分子比較のデフォルトパラメーターである。
【0092】
他の例示のアルゴリズム、ギャップオープニングペナルティ、ギャップエクステンションペナルティ、比較マトリックス、類似性のしきい値などを当業者によりProgram Manual,Wisconsin Package,バージョン9,1997年9月に示したものを含めて使用してよい。行われるべき特別な選択は、DNA対DNA、タンパク質対タンパク質、タンパク質対DNA等の行われるべき特定の比較、及び加えて、この比較が一対の配列(GAPまたはBestFitが一般的に好ましい)の間であるか、あるいはまたは一つの配列と配列の大きなデータベース(FASTAまたはBLASTAが好ましい)の間であるかどうかにに依存する。
【0093】
少なくとも約70パーセント同一であるポリペプチドは、野生型のCRP1あるいはB7RP1ポリペプチドと比較して、一つあるいはそれ以上のアミノ酸置換、欠失、及び/または付加を通常有する。好ましい実施形態においては、ポリペプチドは、CRP1あるいはB7RP1ポリペプチドに対して約75%、80%、85%、90%または95%の同一性を有する。通常、未変性の残渣の置換は、ポリペプチドの正味の全帯電、極性、または疎水性に殆ど、あるいは全く影響を及ぼさないように、アラニン、または保存アミノ酸のいずれかである。保存的置換は下記の表Iに示される。
【0094】
【表1】

【0095】
CRP1あるいはB7RP1ポリペプチド誘導体が本発明により提供される。一つの実施形態においては、CRP1あるいはB7RP1ポリペプチドがポリマーに結合している化学変性されたCRP1あるいはB7RP1ポリペプチド組成物は、本発明の範囲内に含まれている。選ばれたポリマーは、通常水溶性であり、そこで、ポリマーが結合しているタンパク質は生理的環境等の水性環境において沈殿しない。選ばれたポリマーは、通常、アシル化のための活性エステルまたはアルキル化のためのアルデヒド等の単一の反応性基を持つように変性されて、重合度は本発明の方法に提供されるように制御されてよい。このポリマーは、いかなる分子量であってよく、分岐あるいは非分岐であってよい。本発明の範囲内に入れられているのは、ポリマーの混合物である。好ましくは、最終製品製造の治療への使用のためには、このポリマーは医薬として許容されるものである。
【0096】
水溶性ポリマーまたはこれらの混合物は、例えば、ポリエチレングリコール(PEG)、モノメトキシ−ポリエチレングリコール、デキストラン、セルロース、または他の炭水化物ベースのポリマー、ポリ−(N−ビニルピロリドン)ポリエチレングリコール、プロピレングリコールホモポリマー、ポリプロピレンオキサイド/エチレンオキサイドコポリマー、ポリオキシエチル化ポリオール(例えば、グリセロール)及びポリビニルアルコールからなる群から選ばれてよい。
【0097】
このアシル化反応のためには、選ばれたポリマーは単一の反応性エステル基を有しなければならない。還元アルキル化のためには、選ばれたポリマーは単一の反応性アルデヒド基を有しなければならない。好ましい反応性アルデヒドは、水安定であるポリエチレングリコールプロピオンアルデヒド、またはモノC1−C10アルコキシまたはこれらのアリールオキシ誘導体(米国特許第5,252,714号を参照)である。
【0098】
CRP1あるいはB7RP1ポリペプチドのペギル化(Pegylation)は、例えば次の文献に記述されているような、当分野において知られている任意のペギル化反応により行われてよい。Focuson Growth Factors 3:4−10(1992);EP 0 154 316;及びEP 0 401 384。好ましくは、このペギル化は、アシル化反応またはアルキル化反応により反応性ポリエチレングリコール分子(または同族の反応性水溶性ポリマー)により下記に述べるように行われる。
【0099】
ここでの使用に特に好ましい水溶性ポリマーは、PEGと略記されるポリエチレングリコールである。ここで使用される時、ポリエチレングリコールは、モノ(C1−C10)アルコキシ−またはアリールオキシ−ポリエチレングリコール等の他のタンパク質を誘導体化するのに使用されたいかなる形のPEGを包含するとの意味である。
【0100】
一般に、生物活性物質と活性化されたポリマー分子とを反応するのに使用されるいかなる好適な条件下でも、化学的誘導体化を行ってよい。ペギル化されたCRP1及びB7RP1ポリペプチドを製造する方法は、一般に(a)ポリペプチドとポリエチレングリコール(PEGの反応性エステルあるいはアルデヒド誘導体)とを、CRP1あるいはB7RP1ポリペプチドが一つあるいはそれ以上のPEG基に結合される条件下で反応し、そして(b)反応生成物を得るステップを含んでなる。一般に、アシル化反応の最適な反応条件は、既知のパラメーターと所望の結果をベースにして決められる。例えば、PEG:タンパク質の比が大きい程、ポリペギル化された製品のパーセンテージは大きい。
【0101】
一般に、CRP1あるいはB7RP1ポリマーコンジュゲートの投与により軽減あるいは調節される状態は、非共役のCRP1あるいはB7RP1ポリペプチドについてここで述べたものを含む。しかしながら、ここに開示されているコンジュゲートは、追加の活性、増進あるいは低下した生物活性、または非誘導体化された分子に比較して、増加あるいは減少した半減期等の他の特性を有してよい。
【0102】
CRP1あるいはB7RP1ポリペプチド、断片変異体、及び誘導体は、単独で、一緒に、あるいは他の医薬組成物と組み合わせて使用されてよい。CRP1あるいはB7RP1ポリペプチド、断片、変異体、及び誘導体は、治療される適応に適切であるように、サイトカイン、成長因子、抗生物質、抗炎症剤、及び/または化学治療剤と組み合わせて使用されてよい。
【0103】
本発明は、CRP1あるいはB7RP1の選択的結合剤を提供する。選択的結合剤は、CRP1あるいはB7RP1に特異性を持つ分子を指し、タンパク質、ペプチド、核酸、炭水化物、リピッドまたは小分子量化合物を含んでよい。選択的結合剤は、CRP1またはB7RP1のいずれかと相互作用し、次には、CRP1のB7RP1への結合を調節する。一つの実施形態においては、選択的結合剤は、CRP1のB7RP1への結合を部分的あるいは完全に阻止し、免疫同時刺激活性等のCRP1またはB7RP1の少なくとも一つの生物活性を部分的あるいは完全に抑制する。もう一つの実施形態においては、選択的結合剤は抗体である。この抗体は、CRP1またはB7RP1のいずれかと免疫反応性であってよく、好ましくはB7RP1と免疫反応性である。本発明の更にもう一つの実施形態においては、B7RP1と反応性である抗体は、B7RP1上のエプチオープ(eptiope)に結合して、CRP1への結合が部分的あるいは完全に阻止され、免疫同時刺激活性等のB7RP1の少なくとも一つの生物活性が部分的あるいは完全に抑制される。部分的に抑制されるという用語は、少なくとも検出し得るレベルの抑制が起こったことを意味する。完全に抑制されるという用語は、抑制の更なる増加が起こらなかったことを意味する。
【0104】
当分野で知られている方法を用いて抗体を製造するのに、CRP1あるいはB7RP1ポリペプチド、断片、変異体、及び/または誘導体を使用してよい。このように、CRP1あるいはB7RP1ポリペプチド、並びにこのような抗体の反応性断片と反応する抗体もまた本発明の範囲内として考慮される。この抗体は、ポリクローナル、モノクローナル、組み換え、キメラ、単一鎖及び/または両特異的であってよい。通常、この抗体またはこの断片は、ヒト起源であるかあるいは、「ヒト化されている」、すなわち、または患者に投与した場合に、この抗体への免疫反応を防止あるいは最少化するように製造される。この抗体断片は、Fab、Fab’等の本発明のCRP1及びB7RP1ポリペプチドと反応性であるいかなる断片であってよい。また、本発明により提供されるのは、任意のCRP1あるいはB7RP1ポリペプチドまたはこれらの断片を抗原として選ばれた哺乳類に与え、続いて哺乳類の細胞(例えば、脾臓細胞)をしかるべきガン細胞と融合して、不死化された細胞株を知られた手法により作り出すことにより生じるヒドリドマス(hydridomas)である。本発明のヒトCRP1あるいはB7RP1ポリペプチドの全部または一部に特異的なこのような細胞株及び抗体を生じるのに使用される方法も本発明により包含される。
【0105】
本発明のモノクローナル抗体は、所望の生物活性を示す限り(米国特許第4,816,567;Morrisonら,Proc.Natl.Acad.Sci.81,6851−6855(1985))、重及び/または軽鎖の一部が特別な種に由来する、あるいは特別な抗体クラスまたはサブクラスに属する抗体における対応する配列と同一あるいは相同であり、一方、鎖の残りがもう一つの種に由来する、あるいはもう一つの抗体クラスまたはサブクラス、並びにこのような抗体の断片に属する抗体における対応する配列と同一あるいは相同である、「キメラ」抗体を含む。
【0106】
好ましい実施形態においては、キメラ抗−CRP1あるいはB7RP1抗体は「ヒト化された」抗体である。非ヒト抗体をヒト化する方法は当分野においてよく知られている。一般に、ヒト化された抗体は、非ヒトである源からその中に導入される、一つあるいはそれ以上のアミノ酸残渣を有する。当分野において知られている方法(Jonesら,Nature 321,522−525(1986);Riechmannら、Nature,332,323−327(1988);Verhoeyenら、Science 2391534−1536(1988))に従って、齧歯類の相補性決定領域(CDR)にヒト抗体の対応する領域を置き換えることにより、ヒト化を行うことができる。
【0107】
また、本発明により包含されているのは、完全にヒトの抗−CRP1または抗−B7RP1抗体である。内生免疫グロブリン産生の不存在下でヒト抗体のレパートリーを産生する能力のあるトランスジェニック動物(例えば、マウス)のCRP1あるいはB7RP1抗原による免疫化により、このような抗体を製造してよい。例えば、Jakobovitsら,Proc.Natl.Acad.Sci.90、2551−2555(1993);Jakobovitsら,Nature 362,255−258(1993)を参照のこと。また、ファージディスプレイライブラリ(phage−display libraries)においてヒト抗体を製造することもできる(Hoogenboomら,J.Mol.Biol.227,381(1991);Marksら,J.Mol.Biol.222.,581(1991))。
【0108】
本発明の選択的結合剤を使用して、CRP1のB7RP1への結合を調節し、CRP1及びB7RP1により媒介される、免疫同時刺激等の少なくとも一つの生物活性を調節してよい。このような選択的結合剤の例は、CRP1またはB7RP1との抗体免疫反応性である。CRP1とB7RP1ポリペプチドのその結合パートナーへの結合を抑制する等、この抗体を治療的に使用してよい。インビトロ及びインビトロ診断の目的でラベルされた形などでこの抗体を更に使用して、体液または細胞試料中のCRP1及びB7RP1ポリペプチドの存在を検出してよい。
【0109】
医薬組成物と投与
CRP1あるいはB7RP1ポリペプチドの医薬組成物は、本発明の範囲内である。このような組成物は、医薬として許容し得るキャリアとの混合物での治療として有効な量のポリペプチドまたは断片、変異体、または誘導体を含んでなってよい。好ましい実施形態においては、医薬組成物は、CRP1あるいはB7RP1ポリペプチドを、CRP1あるいはB7RP1細胞外ドメインの一部または全部を含んでなる、可溶性の形態として含んでなる。通常、CRP1及びB7RP1ポリペプチド治療用化合物は、一つあるいはそれ以上の生理学的に許容し得るキャリア、賦形剤、または希釈剤と一緒に、精製されたポリペプチド、断片、変異体、または誘導体を含んでなる、組成物の形で投与される。このキャリア材料は、好ましくは哺乳類への投与用に溶液中の普通の他の材料と共に補われる、注射用の水であってよい。中性の緩衝された生理食塩水または血清アルブミンと混合した生理食塩水が例示の適切なキャリアである。好ましくは、この製品は、適切な賦形剤(例えば、スクロース)を用いて凍結乾燥品として配合される。他の標準キャリア、希釈剤、及び賦形剤は、所望に応じて入れられてよい。他の例示的組成物は、ソルビトールまたはこれの好適な代替品を更に含む、約pH7.0−8.5のトリスバッファー、または約pH4.0−5.5の酢酸塩バッファーを含んでなる。
【0110】
CRP1あるいはB7RP1医薬組成物は、非経口で投与され得る。あるいは、この組成物は、静脈注射であるいは皮下注射で投与されてよい。全身的に投与する場合には、本発明の使用での治療用組成物は、発熱物質を含まない、非経口で許容し得る水溶液の形であってよい。pH、アイソトニック性、安定性などに関するこのような医薬として許容し得るタンパク質溶液の製造は、当分野の技術内である。
【0111】
所望の程度の純度を持つ選ばれた組成物を、オプションの生理的に許容し得るキャリア、賦形剤、または安定剤(Remington’s Pharmaceutical Sciences、18版Edition,A.R.Gennaro編,Mack Publishing Company(1990))と凍結乾燥されたケーキまたは水溶液の形で混合することにより、本発明を実施するのに有用なCRP1あるいはB7RP1ポリペプチド組成物の治療用配合物を貯蔵用に製造してよい。許容し得るキャリア、賦形剤または安定剤は、服用者に非毒性であり、好ましくは使用される投与量と濃度で不活性であり、そしてリン酸塩、クエン酸塩、または他の有機酸等のバッファー;アスコルビン酸等の酸化防止剤;低分子量ポリペプチド;、血清アルブミン、ゼラチン、または免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニンまたはリジン等のアミノ酸;モノサッカライド、ジサッカライド、及びグルコース、マンノース、またはデキストリンを含む他の炭水化物;EDTA等のキレート化剤;マニトールまたはソルビトール等のシュガーアルコール;ナトリウム等の対イオンを形成する塩;及び/またはトウイーン、プルロニックスまたはポリエチレングリコール(PEG)等の非イオン性界面活性剤を含む。
【0112】
治療に使用されるべき有効な量のCRP1あるいはB7RP1ポリペプチド組成物は、例えば、CRP1及びB7RP1ポリペプチドが使用される適応、投与経路、及び患者の状態等の治療目標に依存する。従って、療法士が最適な治療効果を得るのに必要とされるように、投与量を求めて、投与経路を調節することが必要である。通常の1日の投与量は、上述の因子に依って、約0.1μg/kgから100mg/kgあるいはそ以上迄の範囲であってよい。通常、臨床医は、所望の効果を得る投与量に到達する迄この組成物を投与する。この組成物は、それゆえ、時間にわたって、単一投与として、あるいは2あるいはそれ以上の投与(同じ量のCRP1あるいはB7RP1ポリペプチドを含んでも、あるいは含まなくともよい)として、あるいインプランテーション器具またはカテーテルにより連続的な輸液として投与されてよい。
【0113】
更なる試験を行うのに従って、種々の患者における種々の状態を治療するための適切な投与量レベルに関する情報が出てきて、通常の熟練作業者は、治療の状況、治療を受けている疾患のタイプ、服用者の年齢と全般的な健康を考慮して、適当な投与を確定することが可能であろう。
【0114】
インビボ投与に使用されるべきCRP1あるいはB7RP1ポリペプチド組成物は無菌でなければならない。これは、無菌の濾過膜により濾過することにより容易に行われる。この組成物を凍結乾燥された場合には、これらの方法を用いる殺菌は、凍結乾燥と再構成に先立って、あるいは続いて行われてよい。非経口投与用のこの組成物は、通常、凍結乾燥された形で、あるいは溶液で貯蔵される。
【0115】
この治療用組成物は、一般に無菌アクセスポートを持つ容器、例えば、皮下注射針で突き刺すことができるストッパーを持つ静脈注射溶液バックまたはバイアル中に入れられる。
【0116】
この組成物の投与経路は、知られている方法、例えば、皮下、腹腔内、大脳内(柔細胞内)、大脳室内、筋肉内、眼球内、動脈内、あるいは病変内(intralesional)の経路による、あるいは持続放出系または場合によってはカテーテルの使用を含む埋め込み器具による経口注入または輸液と一致する。所望の場合には、この組成物は、輸液、丸薬注入により、またはインプランテーション器具により連続的に投与されてよい。
【0117】
あるいはまたはそれに加えて、この組成物は、埋め込みを経て、CRP1及びB7RP1ポリペプチドを吸収した膜、スポンジ、または他の適切な材料の患部中に局所的に投与されてよい。埋め込み器具を使用する場合には、この器具はいかなる好適な組織または器官中にも埋め込まれ、CRP1あるいはB7RP1ポリペプチドの送達は、丸薬を経た、または連続投与を経た、または連続的な輸液を用いるカテーテルを経た器具を直接的に通されてよい。CRP1あるいはB7RP1ポリペプチドは、持続放出配合物または調合物で投与されてよい。持続放出調合物の好適な例は、半透過性ポリマーマトリックスを成形された物品、例えば、フィルム、またはマイクロカプセルの形で含む。持続放出マトリックスは、ポリエステル、ハイドロゲル、ポリラクチド(U.S.3,773,919、EP58,481)、L−グルタミン酸のコポリマー及びガンマエチルL−グルタメート(Sidmanら,Biopolymers,22:547−556(1983))、ポリ(2−ヒドロキシエチル−メタアクリレート)(LangerらJ.Biomed.Mater.Res,15:167−277(1981)]及びLanger、Chem.Tech.,12:98−105(1982))、エチレンビニルアセテート(Langerら、上記)またはポリ−D(−)−3−ヒドロキシ酪酸(EP133,988)を含む。持続放出組成物は、また、当分野において知られているいくつかの方法(例えば、Eppsteinら,Proc.Nati.Acad.Sci.USA,82:3688−3692(1985);EP36,676;EP88,046;EP143,949)のいずれかにより製造することができる、リポソームも含む。
【0118】
ある場合には、CRP1あるいはB7RP1ポリペプチド組成物をエクスビボ(ex vivo)の方法で使用することが望ましいかもしれない。ここで、患者から取り出した細胞、組織、または器官がCRP1あるいはB7RP1ポリペプチド組成物を曝らし、その後、この細胞、組織、または器官を患者に埋め込みにより戻す。
【0119】
他の場合には、CRP1あるいはB7RP1ポリペプチドを、ここで述べたもの等の方法を用いて、遺伝子的に設計されたしかるべき細胞を患者の中に埋め込むことにより送達して、ポリペプチド、断片、変異体、または誘導体を発現し、分泌してよい。このような細胞は、動物あるいはヒト細胞であってよく、患者自身の組織から、あるいはまたはヒトまたは非ヒトのいずれかのもう一つの源から送達されてよい。場合によっては、この細胞は不死化されてよい。しかしながら、免疫応答の機会を減少させるために、この細胞をカプセル化して、取巻く組織の浸潤を避けることが好ましい。カプセル化材料は、タンパク質生成物を放出させるが、患者の免疫系により、あるいは取巻く組織からの他の有害因子により細胞の破壊を防止する通常生体適合性の、半透過性ポリマーの囲いまたは膜である。
【0120】
細胞の膜被包のために使用される方法は当業者に熟知されており、被包細胞の調製と患者への移植は多大の実験を必要とせずに実現できる。例えば、米国特許第4,892,538号、同第5,011,472号及び同第5,106,627号参照。生存細胞を被包するための系がPCT WO91/10425号(Aebischerら)に述べられている。リポソーム担体、生体内腐食性粒子又はビーズのような様々な他の持続性又は制御された送達方法を製剤するための手法も当業者に既知であり、例えば米国特許第5,653,975号に記述されている。被包された又は被包されていない細胞を患者の適当な体組織又は器官に移植することができる。
【0121】
上述したように、細胞製剤を1個又はそれ以上のCRP1又はB7RP1ポリペプチド、変異体、誘導体及び/又は断片で処理することが望ましいと考えられる。これは、例えば骨髄細胞のようなT細胞を含む細胞を、細胞膜透過性の形態の当該ポリペプチド、変異体、誘導体又は断片に直接接触させることによって実施できる。例えば、T細胞の機能を活性化するためにT細胞を含む細胞をB7RP1ポリペプチドに接触させることができ、そのように処理した細胞を患者に移植する。
【0122】
その代わりに、遺伝子治療を用いることもできる。遺伝子治療が適用できる1つの方法は、「遺伝子治療DNA構築物」を形成するために構成的又は誘導性プロモーターに機能的に連結されうるCRP1又はB7RP1遺伝子(CRP1又はB7RP1ポリペプチドをコードするゲノムDNA、cDNA、及び/又は合成DNA、あるいはその断片、変異体又は誘導体)を使用して、「遺伝子治療DNA構築物」を形成することである。プロモーターは、構築物を挿入する細胞又は組織型において活性であることを条件として、内因性CRP1又はB7RP1遺伝子に相同性あるいは非相同性のいずれでもよい。遺伝子治療DNA構築物の他の成分は、任意に、必要に応じて、位置特異的組込みのために設計されたDNA分子(例えば相同的組換えのために有用な内因性隣接配列)、組織特異的プロモーター、エンハンサー又はサイレンサー、親細胞に比べて選択的優越性を提供することができるDNA分子、形質転換細胞を同定するための標識として有用なDNA分子、負の選択系、細胞特異的結合物質(例えば、細胞標的に関して)、細胞特異的インターナリゼーション因子、及びベクターによる発現を高めるための転写因子ならびにベクターの製造を可能にする因子を含みうる。
【0123】
次にこの遺伝子治療DNA構築物を患者の細胞に挿入することができる(エクスビボ又はインビボで)。遺伝子治療DNA構築物を挿入するための1つの方法は、ウイルスベクターによるものである。遺伝子治療DNA構築物の送達のために遺伝子治療において典型的に使用される適当なウイルスベクターは、限定されることなく、アデノウイルス、アデノ関連ウイルス、単純ヘルペスウイルス、レンチウイルス、乳頭腫ウイルス、及びレトロウイルスベクターを含む。レトロウイルスベクターのようなこれらのベクターの一部は、遺伝子治療DNA構築物を患者の細胞の染色体DNAに送達し、遺伝子治療DNA構築物は染色体DNAに入り込むことができる;他のベクターはエピソームとして機能し、遺伝子治療DNA構築物は細胞質中にとどまる。遺伝子治療ベクターの使用は、例えば米国特許第5,672,344号、同第5,399,346号に述べられている。
【0124】
ウイルスベクターを使用せずに遺伝子治療DNA構築物を患者の細胞に送達するための代替的手段は、限定されることなく、リポソームを介した移入、裸のDNAの直接注入、レセプタを介した移入(リガンド−DNA複合体)、エレクトロポレーション、リン酸カルシウム沈殿法、及び微粒子撃ち込み(例えば「遺伝子ガン」)を含む。例えば米国特許第4,970,154号、WO96/40958号、5,679,559号、5,676,954号及び5,593,875号参照。
【0125】
遺伝子治療を通して細胞における内因性CRP1又はB7RP1ポリペプチドの発現を高めるためのもう1つの方法は、1つ又はそれ以上のエンハンサー要素をCRP1又はB7RP1ポリペプチドプロモーター内に挿入することであり、エンハンサー要素はCRP1又はB7RP1ポリペプチド遺伝子の転写活性を高めるために働くことができる。使用されるエンハンサー要素は、遺伝子を活性化することを所望する組織に基づいて選択され、その組織においてプロモーターの活性化をもたらすことが知られているエンハンサー要素が選択される。例えば、CPR1又はB7RP1ポリペプチドをT細胞において「発動(turned on)」させようとする場合は、lckプロモーターエンハンサー要素が使用できる。この場合には、標準的なクローニング手法を使用して、付加する転写要素の機能的部分を、CRP1又はB7RP1ポリペプチドプロモーター(及び任意に、ベクター、5’及び/又は3’隣接配列、等々)を含むDNAの断片に挿入することができる。その後、「相同的組換え構築物」として知られるこの構築物をエクスビボ又はインビボのいずれかで所望する細胞に挿入することができる。
【0126】
内因性プロモーターのヌクレオチド配列を改変することにより、遺伝子治療を使用してCRP1又はB7RP1ポリペプチドの発現を低下させることができる。そのような改変は、典型的には相同的組換え法を通して実施される。例えば、不活性化のために選択したCPR1又はB7RP1遺伝子のプロモーターの全部又は一部を含むDNA分子を構築して、転写を調節するプロモーターの断片を取り除く及び/又は置き換えることができる。ここでは、標準的な分子生物学手法を使用して、プロモーターのTATAボックス又は転写活性化因子の結合部位が欠失されうる。そのような欠失はプロモーター活性を阻害し、それによって対応するCRP1またはB7RP1遺伝子の転写を抑制することができる。プロモーターにおけるTATAボックス又は転写活性化因子結合部位の欠失は、TATAボックス及び/又は活性化因子結合部位の活性が低下する又は完全に不活性となるように、1個又はそれ以上のヌクレオチドの置換、欠失及び/又は挿入を通して1個又はそれ以上のTATAボックス及び/又は転写活性化因子結合部位のヌクレオチドが突然変異を起こす、CRP1又はB7RP1ポリペプチドプロモーターの全部又は該当部分(調節されるCRP1又はB7RP1遺伝子と同じ又は関連種からの)を含むDNA構築物を作製することによって実施できる。典型的には、改変されたプロモーターセグメントの天然(内因性)5’及び3’隣接領域に対応する、少なくとも約500塩基のDNAを同時に含むこの構築物は、直接又は上述したようなウイルスベクターを通して、適切な細胞内に挿入されうる(エクスビボ又はインビボのいずれかで)。典型的には、細胞のゲノムDNAへの構築物の組込みは相同的組換えを通して行われ、プロモーター構築物内の5’及び3’隣接DNA配列は、内因性染色体DNAへのハイブリダイゼーションを通して改変されたプロモーター領域の組込みを助けるように働くことができる。
【0127】
1個又はそれ以上のCRP1又はB7RP1ポリペプチドを阻害することを所望する場合、他の遺伝子治療法も使用しうる。例えば、選択したCRP1又はB7RP1ポリペプチドの少なくとも一部に相補的な配列を持つアンチセンスDNA又はRNA分子を細胞に挿入することができる。典型的には、そのような各々のアンチセンス分子は、各々の選択したCRP1又はB7RP1遺伝子の開始部位(5’末端)に相補的である。その後アンチセンス分子が対応するCRP1又はB7RP1ポリペプチドmRNAにハイブリダイズするときに、このmRNAの翻訳が妨げられる。
【0128】
その代わりに、遺伝子治療を使用して、1個又はそれ以上のCRP1又はB7RP1ポリペプチドの優性の負の阻害因子を創造することができる。この状況では、各々の選択したCRP1又はB7RP1ポリペプチドの突然変異体完全長又はトランケートポリペプチドをコードするDNAを調製し、上述したようなウイルス又は非ウイルス法を用いて患者の細胞に挿入することができる。そのような各々の突然変異体は、典型的にはその生物学的役割において内因性ポリペプチドと競合するように設計される。
【0129】
作用物質と拮抗物質
本発明はまた、CRP1とB7RP1のいずれか又は両方の分子の活性を調節する、CRP1又はB7RP1の作用物質と拮抗物質を提供する。作用物質と拮抗物質は、CRP1へのB7RP1の結合を変化させる被験分子から同定されうる。
【0130】
「被験分子」という用語は、CRP1又はB7RP1ポリペプチドに結合し、それによってCRP1へのB7RP1の結合を変化させる能力についての評価の対象である分子を指す。好ましくは、被験分子は少なくとも約106Mの親和定数で結合する。
【0131】
CRP1へのB7RP1の結合を測定するためには様々なアッセイが使用できる。これらのアッセイを使用して、CRP1へのB7RP1の結合の速度又は度合を上昇させる又は低下させる能力に関して被験分子をスクリーニングすることができる。1つの種類のアッセイでは、CRP1ポリペプチド、好ましくは細胞外ドメインのような可溶性形態のCRP1を、マイクロタイタープレートのウエルの底に付着させることによって固定する。次に放射標識B7RP1と被験分子を1つずつ(いずれかの順序で)又は同時にウエルに加えることができる。インキュベーション後、ウエルを洗浄し、シンチレーション計数器を使用してB7RP1のCRP1タンパク質への結合の度合を調べるために放射能を測定することができる。典型的には、分子を一定の濃度範囲にわたって試験し、結果の評価の正確さを確認するために試験アッセイの1つ又はそれ以上の要素を欠く一連の対照ウエルが使用できる。これに代わる方法は、タンパク質の「位置」を逆にする、すなわちB7RP1をマイクロタイタープレートのウエルに固定し、被験分子と放射標識CRP1をインキュベーションして、CRP1の結合の度合を測定することを含む(例えば、Current Protocols in Molecular Biology、Ausubelら編集、John Wiley & Sons,New York,NY[1995]の18章参照)。
【0132】
放射標識に代わる方法として、CRP1又はB7RP1をビオチンに結合し、比色定量によって又はストレプタビジンの蛍光標識によって検出することができる、ホースラディシュペルオキシダーゼ[HRP]又はアルカリホスファターゼ[AP]のような酵素に連結されたストレプタビジンを使用してビオチニル化タンパク質の存在を検出することができる。ビオチンに結合したCRP1又はB7RP1に対して特異的な抗体も使用でき、AP又はHRPに連結された酵素連結ストレプタビジンとのインキュベーション後、検出することができる。
【0133】
CRP1及びB7RP1はまた、アガロースビーズ、アクリルビーズ又は他の種類のそのような不活性物質への接着によって固定しうる。基質−タンパク質複合体を相補的タンパク質と被験化合物を含む溶液に入れる;インキュベーション後、遠心分離によってビーズを沈澱させ、上述した方法を用いてCRP1とB7RP1間の結合の量を評価することができる。その代わりに、基質−タンパク質複合体をカラムに固定し、被験分子と相補的タンパク質をカラムに通過させることができる。次に上記に述べた手法、すなわち放射標識、抗体結合、等のいずれかを用いてCRP1とB7RP1間の複合体の形成を評価することができる。
【0134】
CRP1/B7RP1複合体の形成を増加させる又は減少させる被験分子を同定するために有用なもう1つのタイプのインビトロアッセイは、Biacoreアッセイ系(Pharmacia,Piscataway,NJ)のような表面プラスモン共鳴検出器システムである。Biacoreシステムは製造者のプロトコールを使用して実施しうる。このアッセイは基本的に、検出器内に位置するデキストラン被覆センサーチップへのCRP1又はB7RP1のいずれかの共有結合を含む。次に、被験化合物と他の相補的タンパク質を同時に又は連続的にセンサーチップが入ったチェンバー内に注入し、結合する相補的タンパク質の量を、センサーチップのデキストラン被覆側と物理的に関連する分子量の変化に基づいて評価することができる;分子量の変化は検出器システムによって測定できる。
【0135】
一部の場合には、CRP1/B7RP1複合体の形成を増加させる又は減少させるのに使用するため、2個又はそれ以上の被験化合物を一緒に評価することが望ましいと考えられる。これらの場合、そのような付加被験化合物を最初の被験化合物と同時に又は最初の被験化合物に続いて加えることにより、上記に述べたアッセイを容易に修正することができる。アッセイの残りの段階は上述した通りである。
【0136】
上に述べたようなインビトロアッセイは、CRP1又はB7RP1による複合体形成への作用に関して多数の化合物を速やかにスクリーニングするために好都合に使用しうる。ファージディスプレイ、合成ペプチド及び化学合成ライブラリーにおいて生成される化合物をスクリーニングするためにアッセイを自動化することができる。
【0137】
CRP1及びB7RP1の複合体形成を上昇させる又は低下させる化合物はまた、いずれかのポリペプチドを発現する細胞及び細胞系統を使用した細胞培養においてもスクリーニングしうる。細胞と細胞系統はどのような哺乳類からも入手しうるが、好ましくはヒト又は他の霊長類、イヌ、げっ歯類ソースからのものである。表面上にCRP1を発現する細胞へのB7RP1の結合を被験化合物の存在下又は不在下で評価し、結合の度合を、例えばB7RP1に対するビオチニル化抗体を使用するフローサイトメトリーによって測定することができる。細胞培養アッセイは、上述したタンパク結合アッセイにおいて陽性と判定された化合物をさらに評価するために好都合に使用しうる。
【0138】
治療用途
本発明のポリペプチド、及びその作用物質と拮抗物質は、T細胞の機能を調節するために使用しうる。作用物質と拮抗物質は、CRP1及び/又はB7RP1活性を調節し、T細胞機能に関連する活性、例えばT細胞の活性化のような、CRP1又はB7RP1タンパク質の少なくとも1つの活性を上昇させる又は低下させる分子を含む。作用物質又は拮抗物質は、CRP1又はB7RP1のいずれかと相互作用し、それによってそれらの活性を調節する、タンパク質、ペプチド、炭水化物、脂質、又は低分子量分子のような補因子でありうる。潜在的なポリペプチド作用物質又は拮抗物質は、当該タンパク質の細胞外ドメインの一部又は全部を含む、可溶性又は膜結合形態のCRP1又はB7RP1と相互作用する抗体を含む。CRP1又はB7RP1発現を調節する分子は、典型的には発現のアンチセンス調節因子として働きうる、CRP1又はB7RP1タンパク質をコードする核酸を含む。
【0139】
CRP1又はB7RP1ポリペプチド及びその作用物質と拮抗物質は、自己免疫疾患、移植片の生存、腫瘍細胞の増殖を抑制するための免疫細胞の活性化、T細胞依存性B細胞仲介疾患、及び癌遺伝子免疫療法の治療において使用しうる。1つの実施態様では、CRP1及び/又はB7RP1機能の拮抗物質又は阻害因子は、慢性免疫細胞機能不全を有する疾患において症状を軽減するために有益であると考えられる。全身性エリテマトーデス、慢性関節リウマチ、免疫性血小板減少性紫斑病(ITP)、及び乾癬のような自己免疫疾患は、CRP−1/B7RP−1の拮抗物質又は阻害因子で治療しうる。さらに、炎症性腸疾患(クローン病及び潰瘍性大腸炎)、グレーヴズ病、橋本甲状腺炎、及び糖尿病のような慢性炎症性疾患も、CRP−1/B7RP−1に対する阻害因子で治療できる。実施例18の中で述べるように、げっ歯類の慢性関節リウマチ疾患モデルにおいて、CRP−1−Fcは疾患の発症を抑制し、B7RP−1−Fcは発症を促進する。このモデルにおけるこれら正反対の作用は、B7RP−1−Fcタンパク質に関する作用性の役割とCRP−1−Fcタンパク質についての拮抗性の役割を裏付ける。その結果はまた、この経路の操作によってT細胞応答をどのように調節することができるか、そして慢性関節リウマチの進行におけるこの経路の重要性を示している。さらに、実施例19で述べるように、インビボでのB7RP−1−Fcの発現はトランスジェニックマウスにおいて炎症性腸疾患(IBD)表現型を刺激する。この実施例は、腸の炎症発生におけるB7RP−1/CRP−1の役割を裏付ける。それ故、B7RP−1/CRP−1経路の拮抗物質はヒトIBDを治療するために使用しうる。
【0140】
CRP1又はB7RP1の拮抗物質は、骨髄及び臓器移植のための免疫抑制剤として使用でき、移植片の生存を延長させるために使用しうる。そのような拮抗物質は既存の治療に比べて重要な利益を提供しうる。骨髄及び臓器移植療法は、宿主によるT細胞仲介の異種細胞又は異種組織拒絶反応と戦わねばならない。T細胞仲介の拒絶反応抑制するための現在の治療レジメンは、シクロスポリン又はFK506薬剤による治療を含む。薬剤は有効であるが、患者は肝毒性、腎毒性及び神経毒性を含めた深刻な副作用に悩まされる。シクロスポリン/FK506クラスの治療薬の標的は、遍在発現するホスファターゼ、カルシニューリンである。CRP1の発現はT細胞に限定されるので、CRP1又はB7RP1の阻害因子は現在の免疫治療薬の使用に関して認められる重篤な副作用がないと考えられる。
【0141】
CRP1又はB7RP1の拮抗物質は、慢性関節リウマチ、乾癬、多発性硬化症、糖尿病、及び全身性エリテマトーデスのような自己免疫疾患のための免疫抑制剤として使用しうる。
【0142】
CRP1/B7RP1を介する共同刺激経路の拮抗物質はまた、トキシックショック症候群、炎症性腸疾患、輸血によるアロ感作、T細胞依存性B細胞仲介疾患を軽減するため、及び対宿主性移植片病の治療のために使用しうる。
【0143】
抗体、例えば細胞外ドメインを含む可溶性タンパク質、及びT細胞活性化の延長又は促進をもたらすCRP1又はB7RP1の他の調節因子が、腫瘍に対する免疫応答を高めるために使用できる。実施例20は、B7RP−1−Fcがマウスにおいて腫瘍細胞の増殖を阻害しうることを示している。同様に、ヒトB7RP−1−Fc又はB7RP−1/CRP−1経路の他の活性化因子は、ヒト腫瘍に対する免疫応答を増強するために使用しうる。抗腫瘍活性は一般に、強力な細胞溶解性Tリンパ球成分を持つとみなされる。実際に、B7−Fc融合タンパク質の抗腫瘍作用(Sturmhoefelら、Cancer Res.59:4964−4972,1999)は、細胞溶解性CD8+T細胞によって仲介された。CRP−1も細胞溶解性CD8+T細胞上で発現されるので(実施例9)、実施例20で明らかにされた抗腫瘍作用はCD8+細胞へのB7RP−1−Fcの作用によるものであったと考えられる。B7RP−1/CRP−1経路はまた、同種移植片移植、対宿主性移植片病、及び自己免疫疾患を含めた多くの他の臨床状況においてCTL応答を調節するために操作できる。
【0144】
本発明のB7RP1遺伝子を使用する遺伝子治療は、癌免疫療法において使用しうる。癌細胞内に挿入されたB7RP1遺伝子は、動物の体内にもどされたときに免疫系のT細胞によって認識されうる抗原呈示細胞へと癌細胞を形質転換することができる。T細胞によるトランスフェクションされた腫瘍細胞の認識は、B7RP1遺伝子を発現する又は発現しない腫瘍細胞の根絶をもたらす。この免疫療法アプローチは、様々な白血病、肉腫、黒色腫、腺癌、乳癌、前立腺腫瘍、肺癌、結腸癌及び他の腫瘍のために使用しうる。本発明は、様々な腫瘍に対する応答においてT細胞の活性化を高めるためにB7RP1遺伝子を同様に使用することを包含する。
【0145】
実施例14で述べるように、B7RP1を発現するトランスジェニックマウスの表現型は、B7RP1が抗体産生の制御において重要であることを示唆する。B7RP1タンパク質活性の作用物質及び拮抗物質は、抗体産生の抑制又は強化を必要とする治療適応症において有用であると考えられる。
【0146】
例えば、多くのワクチンは有効で特異的な抗体応答を惹起することによって作用する。一部のワクチン、特に腸内微生物(例えばA型肝炎ウイルス及びサルモネラ属(Salmonellas))に対するものは、短命な抗体応答を惹起する。ワクチンの有効性を高めるためにはこの応答を増強し、延長させることが望ましい。それ故、可溶性B7RP1又はCRP1に対する活性化抗体は、ワクチンアジュバントとして役立つと考えられる。
【0147】
抗ウイルス応答はまた、B7RP−1/CRP−1経路の活性化因子又は作用物質によっても増強されうる。実施例20のデータは、細胞性免疫がB7RP−1−Fcによって高められることを示している。B7RP−1−Fc又はB7RP−1/CPR−1経路の他の活性化因子による細胞性免疫機能の増強はまた、ウイルス感染細胞を排除する上でも有益であると考えられる。補足的に、B7RP−1−Fcは、体内から遊離ウイルスを清掃するのを助ける働きをすると考えられる、実施例13で認められたような抗体仲介応答を増強しうる対液性免疫機能に影響を及ぼす。
【0148】
逆に、抗体産生の抑制によって改善される多くの臨床条件がある。過敏症は、誇張された又は不適切な、常態では有益な免疫応答であり、炎症反応と組織損傷を導く。抗体が仲介する過敏症反応は、B7RP1活性の阻害因子による拮抗作用を特に受けやすいと考えられる。アレルギー、枯草熱、喘息、及び急性浮腫はI型過敏症反応を引き起こすが、これらの反応はB7RP1活性のタンパク質、抗体又は小分子阻害因子によって抑制されうる。 全身性エリテマトーデス、関節炎(慢性関節リウマチ、反応性関節炎、乾癬性関節炎)、ネフロパシー(膜性、脈管膜毛細管性、巣状分節性、巣状壊死性、半月性、増殖性−糸球体腎炎、尿細管症)、皮膚疾患(天疱瘡及び類天疱瘡、結節性紅斑)、内分泌障害(甲状腺炎−グレーヴズ病、橋本甲状腺炎、インスリン依存性糖尿病)、種々の肺疾患(特に外因性肺胞炎)、種々の血管症、IgAの異常産生を伴う小児脂肪便症、多くの貧血及び血小板減少症、ギヤン−バレー症候群、及び重症筋無力症を含めて、抗体仲介の過敏症反応を引き起こす疾患がB7RP1拮抗物質によって治療できる。
【0149】
さらに、多発性骨髄腫、ヴァルデンストレームマクログロブリン血症、及びクリオグロブリン血症のようなリンパ球増殖性疾患が、B7RP1のタンパク質、抗体、又は小分子拮抗物質によって抑制しうる。
【0150】
最後に、「人為的」免疫疾患である対宿主性移植片病は、B7RP1拮抗物質による抗体産生の抑制によって恩恵を受けるであろう。
【0151】
下記の実施例は本発明をさらに詳しく説明するために提示するものであるが、本発明の範囲を制限するものとみなされるべきではない。
【実施例】
【0152】
実施例1
CRP1 cDNAおよびアミノ酸配列
雌C57/ブラック6マウスを犠牲にして、小腸を切除し、パイエル板を除去した。小腸組織を切開し、洗浄して、粘膜および他の壊死組織片を除去した。1mMのジチオトレイトール(DTT)を補充したRPMI−1640中で、20分間、37℃で穏やかに撹拌することによって、腸上皮内細胞(iIEL)を含有する上皮層を解離させた。解離した細胞を100μフィルターに通して、50mLのRPMI−1640中で洗浄し、混合して細胞の凝集塊をさらに破壊し、その後、40μストレーナーを通して、単一細胞集団を得た。その後、これらの細胞を、体積50mLのRPMI−1640で再び洗浄して、残留DTTを確実に除去した。その後、その組織を撹拌し、前のように洗浄して、残留iIELを採集した。3段階のパーコール勾配を用いて、iIELを40%〜80%界面でバンディングしながら、iIELを脂肪細胞および最上皮細胞から分離した。その後、これらの細胞をRPMI−1640で2回洗浄して極微量のパーコールを除去し、CD103(インテグリンαIEL)抗体で免疫染色して、FACs Star セルソーターを用いて分離した。その後、これらの選別細胞は、Trizol(Gibco BRL,Gaithersburg,MD)を直接用いて全RNAを準備するために用いるか、あるいはγ−/δ−TCR、α−/β−TCR、またはCD3を架橋するプレート結合活性化抗体を用いて一晩活性化するかした。RNAを以上のように準備し、EST cDNAライブラリを造るためにプールした。
【0153】
smi12−00082−a1で表わされるcDNAクローンは、CD28(図1B)と相同のヌクレオチド配列を含んでいた(図1B)。配列の翻訳および公開データベース中の既知タンパク質とのその後の比較は、19%のアミノ酸がマウスCD28と同一であることを示した。マウスCD28は、マウスのCTLA−4と同一のアミノ酸を26%しか共有しないため、この低相同性は有意である。CD28/CTLA−4系統における分子内および分子間システイン結合にとって重要であると考えられる推定システインのすべてが、保存されることが分かった(アミノ酸残基83、109および137;開始メチオニンに関して)。さらに、推定オープンリーディングフレームの全長および膜内外領域の相対位置は、CD28およびCRLA−4の両方のものと類似していた。遺伝子CRP1をCD28関連プロテイン−1と名づけた。
【0154】
実施例2
ヒトCRP1 cDNAのクローニング
ヒトCRP1プロテインをコードする核酸配列は、以下の手順によって同定する。ヒトcDNAライブラリは、健康なボランティアから得たヒト抹梢血からの濃縮リンパ球から製造した。リンパ球を精製し、赤血球をリンパ球分離培地(INC Pharmaceuticals,INC.,Costa Mesa,CA)によって除去した。その後、10ng/mLのPMA、500ng/mLのイノマイシン、およびCD3に対するプレート結合活性化抗体を含有する培地中で、一晩、その細胞を活性化させた。Trizol法によって活性化された細胞(Gibco/BRL)から全RNAを準備し、ダイナルビード精製によってポリA RNAを分離した。cDNAは、分離したポリA RNAから製造し、サイズは最大cDNA断片を選択した。その後、サイズ選択cDNAは、プラスミドpSPORT(Gibco/BRL)にライゲーションさせた。ヒトCRP1タンパク質をコードするDNAは、組換えバクテリオファージプラークまたは形質転換細菌コロニーハイブリダゼーションプロトコル(Sambrookら,Supra)のいずれかにより活性化リンパ球cDNAライブラリをスクリーニングすることによって得る。ファージまたはプラスミドcDNAライブラリは、実施例1および図1に記載のマウスCRP1遺伝子クローンから誘導される放射性標識したプローブを用いてスクリーニングする。プローブを用いて、プレーティングしたライブラリから切り離したナイロンフィルタをスクリーニングする。これらのフィルタは、50%ホルムアミド、5X SSPE、2X デンハート溶液、0.5%SDSおよび100μg/mLのサケ精子DNA中、42℃で4時間予備ハイブリダイズし、その後、50%ホルムアミド、5X SSPE、2X デンハート溶液、0.5%SDS、100μg/mLのサケ精子DNAおよび5ng/mLのmB7RP1プローブ中、42℃で24時間ハイブリダイズする。ブロットは、2X SSC、0.1%SDS中、室温で10分間、1X SSC、0.1%SDS中、50℃で10分間、0.2X SSC、0.1%SDS中、50℃で10分間洗浄し、その後、0.2X SSC中、50℃で10分間、再び洗浄する。一切のヒトCRP1クローンから得られた挿入物を配列し、実施例1に記載したように分析する。
【0155】
実施例3
B7RP1 DNAおよびアミノ酸配列
smi11−00003−g5で表わされるcDNAクローンは、B7.1(CD80)およびB7.2(CD86)と相同のヌクレオチド配列を含んでいた。配列の翻訳(図2A)および公開データベース中の既知タンパク質とのその後の比較は、20%のアミノ酸がマウスB7.1と同一であることを示した(図2B)。マウスB7.1は、マウスB7.2と同一のアミノ酸を24%しか共有しないため、この低相同性は有意であった。この低相同性にもかかわらず、臨界システイン残基は、このクローンのオープンリーディングフレームとマウスB7.1およびB7.1との間、残基62、138、185および242(開始メチオニンに関して、図2B)において保存される。B7.1およびB7.1と比較すると、このクローンの推定ORFおける適切な成熟タンパク質長およびカルボキシ末端に対する膜内外領域の位置も類似している。我々は遺伝子B7RP1をB7関連タンパク質−1と名づけた。
【0156】
実施例4
ヒトB7RP1 cDNAのクローニング
マウスB7RP1配列(図2参照)を用いるGenbank芽細胞相同性調査(GCG,University of Wisconsin)は、1679bpのORFを有する4358bpの配列を含むクローン(AB014553)を検索した。PCRクローニングプライマーは、この配列にしたがって設計した。Human Lymph Node Marathon−ReadyTMcDNA(Clontech,Palo Alto,CA)を用い、製造業者の指示した手順に従って、5’および3’RACEにより1313bpのDNA断片を得た。
【0157】
ヒトB7RP1全長に用いたプライマー:
【0158】
【化1】
プライマー2083−75および2083−76は、RACEプロトコルを用いて遺伝子の5’末端を増幅させるために用いた。プライマー2083−77、2083−78、2113−29、2113−30および2113−31は、RACEプロトコルを用いて遺伝子の3’末端を増幅させるために用いた。
【0159】
得られたヌクレオチド配列は、メチオニンで開始する288のアミノ酸残基のORFを含んでいた。その後、予測成熟ヒトB7RP1アミノ酸配列を成熟マウスB7RP1アミノ酸配列と比較して(図3B)、同一のアミノ酸を48%共有することがわかった。CD80(B7.1)を用いると化学種間の相同性が低く、実際に、マウスおよびヒトのCD80は、同一のアミノ酸を41%しか共有しないため、この相同性は有意である。重要なことに、ヒトB7RP1タンパク質は、Igループ構造(成熟タンパク質に関するアミノ酸残基16、92、138、194および195、図3B)に必要な臨界システイン残基を保存する。さらに、膜内外領域の全長および位置は、ヒトB7RP1ホモログと一致する。
【0160】
実施例5
B7RP1 RNAの発現
B7RP1遺伝子に対してRNAプローブ用いるRNAインシチューハイブリダイゼーション。成体マウス組織を4%パラホルムアルデヒド中に固定し、パラフィン包理して、5μmの薄切りにした。インシチューハイブリダイゼーションの前に、組織を0.2MのHClで易透化し、続いて、プロテイナーゼKを用いて消化して、トリエタノールアミンおよび無水酢酸を用いてアセチル化した。一晩、55℃で、マウスB7RP1配列のヌクレオチド1〜96に対応する969塩基の33P標識リボプローブを用いて、切片をハイブリダイズした。RNアーゼ消化によって過剰のプローブを除去し、続いて、漸減塩濃度の緩衝液中で一連の洗浄を行い、その後、0.1X SSC中、55℃で高ストリンジェンシー洗浄を行った。スライドをKodak NTB2エマルジョンに浸漬し、4℃で2〜3週間露光して、現像し、ヘマトキシリンおよびエオシンを用いて対比染色した。切片標本を暗視野および透過光照明法を用いて試験することによって、組織形態学およびハイブリダイゼーションシグナルを同時に評価することができた。
【0161】
インシチューハイブリダイゼーションによるB7RP1 RNAの分析は、B7RP1 RNAがリンパ系成熟領域において非常に発現することを示した。B7RP1 RNAは、胸腺のリンパ系組織、腸、脾臓およびリンパ節のパイエル板において発現した。これらのリンパ系組織内の発現は、B7RP1 RNAが、一般に、B細胞領域およびその他のAPC関与領域において発現することを実証していた。これらの領域には、胸腺の髄質領域、リンパ節の1次濾胞、およびパイエル板の濾胞およびドーム領域が挙げられる。B7RP1 RNAの発現は、リンパ系組織におけるAPC関与領に極めて特異的なものである。
【0162】
一部の非リンパ系組織の分析も、APC関与領域におけるB7RP1発現を示した。肺において、B7RP1発現は、抗原処理に関する機能と一致する粘膜下領域において見られた。小腸において、B7RP1 RNAは、粘膜固有層において見られた。特に、B7RP1 RNAの発現と重複するリンパ球湿潤を示す損傷肝臓部分を発見した。B7RP1発現の組織損傷に応じたリンパ球蓄積との一致は、B7RP1がリンパ球活性化に関与することを強く示している。
【0163】
実施例6
CRP1 RNAの発現
CRP1遺伝子に対してRNAプローブを用いるRNAインシチューハイブリダイゼーション。マウスの組織は、実施例5の場合のように準備した。組織の易透化、プローブハイブリダイゼーション、スライド処理、および組織染色は、実施例5に記載したとおりであった。切片は、一晩、55℃で、マウスCRP1配列のヌクレオチド1〜603に対応する603塩基の33P標識リボプローブを用いて、ハイブリダイズした。切片標本を暗視野および透過光照明法を用いて試験することによって、組織形態学およびハイブリダイゼーションシグナルを同時に評価することができた。
【0164】
正常なマウスまたはオキサゾロンで処理したマウスからのリンパ節を薄切りにし、CRP1 RNA発現について分析した。感作マウスリンパ節は、正常なマウスのリンパ節より大きいCRP1 RNA発現を示した。CRP1の発現は、T細胞活性領域である傍皮質にあった。従って、CRP1 RNAの発現は、Tリンパ球発現のものと一致し、T細胞活性次第で向上調節される。
【0165】
実施例7
CRP1−FcおよびB7RP1−Fc融合タンパク質の発現および精製
CRP1−Fc融合タンパク質についてのDNA発現ベクターを構成するために、CRP1の第一アミノ末端147アミノ酸についてのコード配列は、ヒトFc遺伝子のカルボキシ末端235アミノ酸(アイソタイプIgG1)についてのコード配列、フレーム内に融合させ、pcDNA3のポリリンカー配列内にライゲーションした(pcDNA3/CRP1−Fc)。B7RP1−Fc融合タンパク質につてのDNA発現ベクターを構成するために、B7RP1の第一アミノ末端269アミノ酸についてのコード配列は、ヒトFc遺伝子のカルボキシ末端235アミノ酸(アイソタイプIgG1)についてのコード配列、フレーム内に融合させ、pcDNA3のポリリンカー配列内にライゲーションした(pcDNA3/B7RP1−Fc)。CRP1およびB7RP1、両方のコード配列は、各タンパク質のN−末端から、そこは含まないが、各タンパク質の推定膜内外領域までの配列を含んでいた。293T細胞には、FuGene6形質移入試薬(Roche Molecular Biochmicals,Indianapolis,IN)を用いて、pcDNA3/CRP1−FcまたはpcDNA3/B7RP1−Fcのいずれかを形質移入した。4日後、調整培地を回収し、Protein A Sepharose(Pharmacia)を用いるバッチクロマトグラフィーによって、Fc融合タンパク質を精製した。カラムに結合したFc融合タンパク質を3カラム容量のImmunopure Gentle Elution Buffer(Pierce)を用いて溶離し、その後、体積150の20mMのHEPES、100mMのNaCl(pH7.5)に透析させた。Macrosep遠心コンセントレート、30kd MWCO(Pall Filtron)を用いて、透析したタンパク質を濃縮し、各タンパク質のアミノ酸配列から誘導される吸光係数を用いてタンパク質濃度を計算した。CRP1−Fc融合タンパク質の発現を図4Aに示し、B7CRP1−Fc融合タンパク質の発現を図4Bに示す。
【0166】
実施例8
レセプター−リガンド対としてのCRP1およびB7RP1の同定
新規タンパク質が、CD28、CTLA−4、B7.1およびB7.2を含むものと同じ共同刺激性経路であるかを決定するために、細胞表面画像分析を用いた。この分析は、ACAS(Adherent Cell Analysis and Sorting/粘着細胞分析および選別)分析を用いて、細胞中で発現した膜結合タンパク質が種々のFc融合タンパク質と相互作用するかを分析するものである。膜結合タンパク質を発現する細胞(図5の左側に示す)をFc融合タンパク質(同図の上に示す)と共にインキュベートした。
【0167】
10%FBSを含有するDMEM培地中で増殖させたCos−7細胞を24ウエルのプレートに500,000細胞/ウエルでプレーティングした。FuGene6試薬(Roche Molecular Biochemicals,Indianapolis,IN)を用いて細胞に形質移入した。各形質移入について、3μLのFuGene6試薬を血清を含有しない47μLのDMEM培地に添加した。室温で10分間インキュベートした後、混合物を0.25μgのプラスミドに一滴ずつ添加し、その後、15分間インキュベートした。その後、10%FBSを含有する0.5mLのDMEMと共に上の混合物を細胞に添加した。細胞を37℃、5%CO2雰囲気のもとでインキュベートした。対照として、ヒトCD28についてのcDNAを含む発現プラスミドで安定的に形質移入したCHO D細胞も、24ウエルのプレートに500,000細胞/ウエルでプレーティングした。
【0168】
48時間後、形質移入試薬を含有する培地を除去し、細胞をRPMIと5%FBSで2回洗浄した。1mLの培地中、10〜20ngの精製Fc融合タンパク質を細胞に添加し、これを30分間氷上でインキュベートした。細胞をRPMIと5%FBSで3回洗浄し、その後、2μLのFITC結合抗ヒトFc抗体(1mg/mL)と共に、さらに30分間氷上でインキュベートした。RPMIを用いて3回連続洗浄した後、細胞は、ACAS分析用のために、フェノールレッドを含有しない250μLのRPMI培地を用いて転化させた。
【0169】
種々のFc融合タンパク質を結合した細胞のACAS分析は、B7RP1タンパク質がCRP1に結合していることを示したが、既知の共同刺激性経路におけるタンパク質、CD28またはCTLA−4は示さなかった。逆に言えば、CRP1はB7RP1と相互作用したが、既知経路における成分、B7.2とは相互作用しなかった。(図5参照)。これらの結果は、CRP1およびB7RP1が、CD28およびB7.2のアナログである新規レセプター−リガンド対を表わすことを強く示している。しかし、CRP1およびB7RP1は、B7.2、CTLA−4、またはCD28と相互作用しないため、それらは離れており、既知の共同刺激性経路から独立している。
【0170】
実施例9
B7RP1レセプターを発現する細胞の同定
B7RP1−Fc融合タンパク質を用いて、CRP1タンパク質を含有すると推定されるB7RP1に対するレセプターを発現する細胞(図6参照)をFACS分析によって検出した。雌C57/ブラック6マウスから脾臓を除去して、100μメッシュフィルターを用いて磨砕し、リンパ球を解離させて、70μフィルターを通し、その後、50mLのRPMI−1640中で洗浄した。それらを1500rpmでペレット化して、新しいRPMIに再懸濁させ、混合して凝集細胞を破壊し、40μフィルターを通した。活性化させるべきT細胞をRPMI−1640、5%FBS、1XPSG、PMA、イノマイシン中で6ウエルのプレートに播種し、37℃、5%CO2で、一晩インキュベートした。12時間後、T細胞の活性化を視覚的確認によりチェックした。
【0171】
免疫染色用の活性化脾臓細胞をPBS、0.5%BSA(Path−ocyte 4、ICN Pharmaceuticals)洗浄緩衝液中で洗浄して、再懸濁させ、その後、体積100μLで分取した。CRP1−Fc融合タンパク質またはB7RP1−Fc融合タンパク質、いずれかの15μg/mLを適切に添加し(1.5μg/サンプル)、その後、その混合物を氷上で30分間、時折混合しながらインキュベートした。細胞を5.0mLの洗浄緩衝液中で2回洗浄した。細胞染色用の100μL容積中、2μgのヤギ抗ヒト(GaHuFc−FITC)複合第2抗体を用いて融合タンパク質の結合を視覚化した。PEと複合した細胞マーカー抗体をGaHUFc−FITC、ならびに指示された抗体対照(ラットアイソタイプ)と複合した対照アイソタイプ−PEと共に添加した。そのサンプルを氷上でインキュベートし、前のごとく洗浄した。リンパ球集団にゲートを設けるFACScan分析によって、視覚化を行った。CD4+抗体およびB7RP1−Fc融合タンパク質を用いた二重染色は、細胞がCD4マーカーとB7RP1に対するレセプター、おそらくCRP1との両方を発現することを示した(図6)。同様に、CD8+抗体およびB7RP1−Fc融合タンパク質を用いた二重染色は、細胞がCD8レセプターおよびB7RP1レセプターの両方を発現することを実証した(図6)。非活性化脾細胞試料中では、こうした二重染色細胞を確実に検出することはできなかった。CD4およびCD8はTリンパ球マーカーであるので、CRP1は活性化CD4+およびCD8+T細胞上で発現されると仮定することができる。これらのデータは、正常なマウスと比較して、感作マウスからのリンパ節のT細胞領域において増加したCRP1 RNAの発現と一致する(実施例6)。
【0172】
実施例10
CRP1リガンドを発現する細胞の同定
CRP1−Fc融合タンパク質を用いて、B7RP1タンパク質を含有すると推定されるCRP1に対するリガンドを発現する細胞(実施例8参照)をFACS分析によって検出した(図7)。脾細胞は、実施例8の場合のように準備したが、12時間のT細胞活性化段階を省略し、細胞を直接分析した。脾細胞は、CD45R(B220)マーカー抗体およびCRP1−Fc融合タンパク質を用いて二重染色した。CD45R B細胞マーカーとB7RP1を含有すると推定されるCRP1に対する推定リガンド(実施例8)との両方を発現する細胞を検出した。従って、B7RP1は、抗原提示細胞タイプのB細胞上で発現されると推断する。これらのデータは、種々のリンパ系組織のB細胞領域におけるB7RP1 RNAの発現(実施例5)と一致した。
【0173】
腹腔マクロファージ上でのB7RP1の発現のFACS分析(図8)。腹腔細胞を局所洗浄によって正常なマウスから採集し、CRP1−Fc融合タンパク質または対照としてのFcタンパク質と共に、あるいはF4/80モノクローナル抗体(マクロファージに対して特異的な抗原を検出する)または無関係なアイソタイプ対応モノクロナール抗体と共にインキュベートする前に洗浄した。その後、再び細胞を洗浄し、ヤギ抗ヒトFc−FITC複合抗体と共にインキュベートした。さらなる洗浄の後、細胞は、FACS分析機で光拡散および蛍光染色性について評価した。腹腔細胞は、それらの光拡散性によってサブセット中で最初に識別された(図8A)。マクロファージは、光を前方(FSC)および横向き(SSC)に強く拡散させる能力のため、およびマクロファージに対するマーカーであるF4/80抗原に対して染色陽性であるため、領域5(R5)内で確認した(図8B)。領域6(R6)内のマクロファージは、F4/80抗原に対するさほど強くない染色性をもとに選抜され、またCRP1−Fc融合タンパク質によって染色されることが分かった(図8C)。これらのデータは、B7RP1を含有することができるCRP1に対するリガンドが、プロフェッショナル抗原提示細胞であるマクロファージ上で発現されることを示している。これは、Tリンパ球活性化におけるCRP1およびB7RP1機能と一致する。
【0174】
実施例11
Con A刺激Tリンパ球上のB7RP1−Fc融合タンパク質のインヴィトロ抑制作用
マウス脾細胞を実施例8の場合のように準備し、ネガティブ選択によってTリンパ球を濃縮した(R and D Systems,Inc.,Minneapolis,MN)。200,000の脾細胞を96ウエルの丸底プレートにおけるT細胞増殖分析に用いた。細胞は、培地(添加なし)、図9に示したCRP1−Fc、B7PR1−FcまたはB7.2−Fc融合タンパク質と共に1時間インキュベートした。培地(添加なし)、または種々の濃度でのCon Aを図9の下に示したとおり添加した。その後、細胞を37℃、5%CO2でインキュベートした。42時間後、細胞に3H−チミジンを6時間適用し、回収して、組込まれた放射活性を決定した。3つの同じサンプルからの平均CPMおよび標準偏差を図9に示す。
【0175】
Fc融合タンパク質は、それ自体では有意なT細胞刺激活性または抑制活性を示さなかったが、1μg/mLおよび3μg/mLのCon Aの存在のもとで、B7RP1−Fcおよび既知のB7.2―Fc融合タンパク質は、有意な抑制活性を示した(図9)。高濃度(10μg/mL)では、Con A刺激作用は、おそらくT細胞の過剰活性化によって細胞死をもたらす。B7RP1−FcまたはB7.2−Fc、いずれかを添加すると、細胞は高濃度のCon Aの有害作用から著しく保護された。抑制および保護機能の両方において、B7PR1−Fcタンパク質によるCon A刺激細胞への影響は、B7.2−Fcタンパク質より大きかった。これらのデータは、B7RP1タンパク質が機能して、T細胞増殖を調整することを示している。
【0176】
実施例12
トランスジェニックマウスにおけるB7RP1−Fc融合タンパク質の全身送達
実施例7に記載したB7RP1−Fc融合タンパク質は、ApoE肝臓特異的発現ベクターにさらにクローニングした(Simonetら,J.Clin.Invest.94,1310−1319(1994)およびPCT出願番号US94/11675)。制限酵素、Spe IおよびNot Iを用いてコード領域をpCEP4/B7RP1−Fcから切除して、破片を前述のApoE肝臓特異的発現ベクター中の同じ部位にさらにクローニングした。得られたプラスミド、HE−B7RP1−Fcをそのタンパク質コード領域およびコード領域の側面に位置する配列を通して配列し、突然変異体が確実に含まれないようにした。
【0177】
プラスミドを増幅させ、CsC1密度勾配遠心法を2回行って精製した。精製したプラスミドDNAを制限酵素、Cla IおよびAse Iを用いて消化し、1.5kbのトランスジーン挿入物をアガロースゲル電気泳動法によって精製した。精製した破片を希釈して、5mMのTris(pH7.4)および0.2mMのEDTA中1μg/mLの保存注射溶液にした。使用前に注射針を斜めに切り、シリコーン処理したことを除いて、本質的には(Brinster et al.,Proc.Natl.Acad.Sci.USA 82,4338(1985))に記載されているとおりにBDF1 X BDF1−交配マウスからの単個細胞胚を注入した。胚をCO2インキュベータ内で一晩培養し、15〜20の2−細胞胚を偽妊娠CD1雌マウスの卵管に移入した。
【0178】
満期妊娠の後、微量注入した胚に基づく体内移植から56の子孫を得た。ゲノムDNAサンプルに組込まれたトランスジーンのPCR増幅によって、その子孫をスクリーニングした。増幅の標的領域は、発現ベクター中に含まれるヒトApo Eイントロンの369bp領域であった。PCR増幅に用いたオリゴは:
【0179】
【化2】
であった。
【0180】
PCRについての条件は、94℃で2分間を1サイクル、94℃で1分間、63℃で20秒間、そして72℃で30秒間を30サイクルであった。56の原子孫のうち7匹をPCR陽性トランスジェニックファウンダーマウスとして確認した。
【0181】
12週齢において、9匹のトランスジェニックファウンダー(マウス#1、2、4、6、8、30、32、33、40)および5匹の対照(マウス#5、9、10、25、28)を屍検および病理学分析のために犠牲にした。全細胞性RNAをファウンダー動物および陰性対照同腹子の肝臓から、(McDonald et al.,Meth.Enzymol.152,219(1987))に記載されているように分離した。これらのサンプルを用いてノーザン法分析を行い、トランスジーン発現のレベルを評価した。アガロース電気泳動変性ゲル(Ogden et al.,Meth.Enzymol.152,61(1987))によって、各動物から約10μgの全RNAを分離し、その後、HYBOND−Nナイロン膜(Amersham)に移送して、32P dCTP標識mB7RP1−Fc挿入DNAを用いてプローブした。ExpressHyb溶液(Clonetech)およびハイブリダイゼーション緩衝液1mLあたり2〜4 X 106CPMの標識プローブ中、63℃で1時間ハイブリダイゼーションを行った。ハイブリダイゼーションに続いて、ブロットを2X SSC、0.1%SDS中、室温で2回、各5分間洗浄し、その後、0.1X SSC、0.1%SDS中、55℃で2回、各15〜20分間洗浄した。ファウンダーおよび対照同腹子におけるトランスジーンの発現は、オートラジオグラフ法に従って決定した。
【0182】
ノーザン法分析は、7匹のトランスジェニックファウンダーが検出可能レベルのトランスジーンRNAを発現することを示した(マウス#1、2、6、8、32、33および40)。陰性対照マウスおよび3匹のファウンダー(#4、30および31)は、検出可能レベルのRNAを発現しなかった。B7RP1−Fc融合タンパク質は、培養状態の哺乳動物細胞から分泌されるものと決めたので(図4Bおよび実施例7)、トランスジーンmRNAの発現は、全身送達遺伝子産物レベルを示すはずである。
【0183】
実施例13
B7RP1−Fc融合タンパク質の生物活性
7匹のトランスジェニックマウス(マウス#1、2、6、8、32、33および40)および5匹の対照同腹子(#5、9、10、25および28)を以下の手順を用いる屍検および病理学分析のために犠牲にした。安楽死させる前に、すべての動物のID番号を確認し、その後、体重を計り、麻酔をかけて、血液を取った。血液は、全血清化学および血液学パネルのために、血清および全血の両方として保管した。致死性CO2吸入による終末麻酔の直後、グロス解剖の前にX線撮影を行った。その後、組織試験のために組織を除去し、10%Zn−ホルマリン緩衝液中に固定した。回収した組織には、肝臓、脾臓、膵臓、胃、十二指腸、回腸、パイエル板、結腸、腎臓、生殖器官、皮膚、乳腺、骨、脳、心臓、肺、胸腺、気管、食道、甲状腺/副甲状腺、空腸、盲腸、直腸、副臓、白色および褐色脂肪、坐骨神経、骨髄、膀胱、および骨格筋が挙げられる。固定前に、肝臓、心臓、胃、腎臓、副腎、脾臓、および胸腺について全器官重量を測定した。固定後、組織をパラフィンブロックに加工して、3μmの切片を得た。
【0184】
Bリンパ球マーカー、B220およびTリンパ球マーカー、CD3についての免疫組織化学法を実施した。B220またはCD3発現を検出するために、ホルマリン固定し、パラフィン包理した4μm切片を脱パラフィン化し、脱イオン水に水和した。切片は、3%過酸化水素を用いて反応停止させ、Protein Block(Lipshaw,Pittsburgh,PA)を用いてブロックして、B220に対するラットモノクローナル抗体(Pharmingen,San Diego,CA)またはCD3に対するウサギポリクローナル抗体(Dako,Carpinteria,CA)中でインキュベートした。クロマーゲンとしてDAB(Biotek,Santa Barbara,CA)を用いて、ビオチニル化ウサギ抗ラットまたはヤギ抗ウサギ免疫グロブリン、ペルオキシダーゼ複合ストレプタビジン、(BioGenex,San Ramon,CA)により、抗体を検出した。切片をヘマトキシリンで対比染色した。
【0185】
この研究では、寿命がある段階での研究中に、正常な臨床症状が報告された。トランスジェニックマウスの全身放射線写真を対照マウスのものと比較した。トランスジェニックマウスの全血液学的パラメータを陰性対照群ののものと比較したが、個々のマウスに孤立の変化が存在した。トランスジーン系#8および#40は、増大した血清グロブリンレベル(グロブレン過剰血)を有し、そして#32および#33は、免疫系の慢性抗原刺激により一般に見られるパターンである低い正常範囲のアルブミンレベルを随伴する高い正常範囲のグロブリンレベルを有した。その他のトランスジェニックマウスの器官重量は、対照群のものとさほど差がなかった。
【0186】
トランスジェニックマウスには以下の病理組織学的変化が存在した。トランスジェニックB7RP1−Fcマウスの腸間膜リンパ節は、対照マウスと比較して中等度〜著しく肥大した(図10A〜10D;図11A〜11E)。皮質は、主としてB220+ B細胞(図11D)、そして少数の散在CD3+ T細胞(図11F)を含む大きな胚中心を有する肥大した第二濾胞(図10B〜11B)のように見える隆起濾胞過形成を有した。傍皮質(CD3+ T細胞)領域も中等度に肥大化し(図11B〜11F)、髄洞は、わずかに増加した数の肉マクロファージ(洞組織球増殖)を有した。節における最も顕著な変化は、B7RP1−Fcトランスジェニックマウスにおける多数の分化型プラズマ細胞によって穏やか〜著しく膨張した髄索に存在した(図10D)。トランスジェニックマウス#40において、少数の散在ラッセル体(すなわち、免疫グロブリンを含有する隆起した大きく丸い細胞質内小疱)も髄索内に見られた(図10D)。興味深いことに、その他の内部および抹消リンパ節(例えば、頚部、鼠径部)は、全身応答を思わせる類似の形態学的特徴の活性リンパ細網細胞増生を有した。これらの発見は、B細胞増殖およびプラズマ細胞への終末分化を導く体液性免疫反応を強化させる慢性進行性免疫刺激と一致する。
【0187】
B7RP1−Fcトランスジェニックマウスの脾臓は、対照マウスと比較して、特に、胚中心隆起を伴うB細胞第二濾胞および細動脈周囲T細胞鞘が関係する中程度活性リンパ組織過形成により可変的に肥大した白色脾髄領域を有した(図10E〜10F)。B7RP1−Fcトランスジェニックマウスにおけるもう一つの顕著な発見は、白色脾髄領域周囲の帯域および隣接赤色脾髄における最小限〜軽度のプラズマ細胞増加であった。トランスジェニックマウス#6は、少数の散在ラッセル体を有した(図10F、挿入)。赤色脾髄は、対照マウスに見られるものと比較して、軽度〜中程度の骨髄外造血を有した(図10E)。
【0188】
小腸パイエル板は、B7RP1−Fcトランスジェニックマウスにおいて、対照マウスのものに比べ軽度〜著しく肥大し(図10G)、また、特にトランスジェニックマウス#40および#32において、胚中心が突起した非常に大きな濾胞を有した(図10H)。さらに、小腸内に存在する粘膜であるが、トランスジェニックマウスの結腸内ではより大きく突起した粘膜の濃化粘膜固有層中のリンパ球およびプラズマ細胞の数(マウス#32の回腸における軽度好酸性浸潤と混合)について、最小限(#32において)〜軽度(#8および#33)の増加があった。大腸リンパ球系凝集体(GALT)も、対照群中に比べ、一部のB7RP1−Fcマウス(特に、マウス#8および#2)中でわずかに大きく突起していた。
【0189】
胸腺、骨髄、肝臓、肺、心臓、膵臓、腎臓、副臓、甲状腺、副甲状腺、気管、生殖器官、膀胱、乳腺、皮膚、骨格筋、抹消神経、脳、食道、胃、小および大腸、骨(大腿骨/頚骨)、後膝関節、正常に見え、対照マウスにおいて検出された背景変化と比較できる白色および褐色脂肪を含むその他の組織をあまねく試験した。
【0190】
この研究からのデータは、トランスジェニックマウスにおけるB7関連タンパク質Fcキメラ(B7RP1−Fc)の過剰発現が、脾臓、抹消および内部リンパ節、および消化管関連リンパ系組織において検出される突起した反応性リンパ組織過形成によって特徴づけられる、濾胞過形成、T細胞領域の膨張、および一部の動物においてグロブリン過剰血を随伴する顕著なプラズマ細胞増加などの表現型を誘導することを実証している。プラズマ細胞増加は、より高レベルの循環IgG(±SD = トランスジェニックマウスにおける597±298mg/mL 対 対照同腹子における209±80mg/mL、n=7、P<0.05、t testを意味する)、特に、IgG2a(217±100mg/mL 対 75±29mg/mL、n=7、P<0.01、t test)を随伴する。IgG2aの誘導は、通常、IFN−gなどのTh1サイトカインに関連する。従って、B7RP−1は、BおよびT細胞増殖を誘導し、B細胞を刺激してプラズマ細胞に分化させ、免疫グロブリンを生成する。
【0191】
これらの変化は、試験したリンパ器官全体にわたって、抗体生産プラズマ細胞にB細胞刺激、増殖および分化をもたらす体液性上肢免疫系の過剰刺激に対する持続性全身免疫応答と一致する。
【0192】
B7RP1−Fcトランスジェニックマウスにおいて実証される著しいリンパ組織過形成から、我々は、B7RP1タンパク質が免疫系刺激に関連する有意な生体内生物活性を有するものと推断する。
【0193】
実施例14
ヒトB7RP1のクローニング
正常なヒト循環抹消リンパ球をリンパ球分離培地(ICN Pharmaceuticals)を用いて赤血球から分離した。その後、10μg/mLのプレート結合抗CD3抗体(Immunotech,Westbrook,ME)、10ng/mLのPMA、および500ng/mLのイノマイシンを用いて、一晩(16時間)、37℃、5%CO2で、T細胞を活性化した。その後、TRIzol試薬(Gibco BRL)を用いて全RNAを細胞から準備した。遠心分離によって細胞をペレット化し、その細胞ペレットを各々5 x 106の細胞に対して1mLのTRIzol試薬に再懸濁させて、室温で5分間インキュベートした。その後、原TRIzol試薬1mLあたり0.2mLのクロロホルムを添加した。試験管を手で15秒間激しく振り、3分間室温でインキュベートして、13,000rpmで15分間、4℃で遠心分離した。遠心分離後、RNAを含有する水性上澄み相を回収し、イソプロピルアルコールの添加によってサンプルRNAを沈殿させた。その後、溶液を室温で10分間インキュベートし、RNAをペレット化して、75%のエタノールで洗浄し、その後、15,000rpmで5分間、4℃で遠心分離した。ペレットを空気乾燥して、RNアーゼを含有しない水に再懸濁させ、その後、分取して、後で用いるまで−80℃で保管した。
【0194】
cDNA合成用のSuperScript Plasmid SystemおよびPlasmid Cloning(Gibco BRL)を用いてライブラリを構築した。簡単に言えば、平均サイズ2kbのcDNA挿入物をSal1/Not1クローニング部位においてpSportベクターにライゲーションした。ライゲーションしたプラスミドは、Electromax 形質転換担当E.コリ(Gibco BRL)へのエレクトロポレーションを施し、滴定して、LBプレートあたり15,000コロニー(アンピシリン100μg/mL)でプレーティングした。300,000のコロニーを切離してコロニー/プラークスクリーンハイブリダイゼーション伝達膜(NEN Life Scinece)に送り、0.5NのNaOH、1.5MのNaCl中で5分間変性させ、その後、次の緩衝液、1MのTris HCl(pH8.0)、0.5MのTris HCl(pH8.0)および1.5MのNaClならびに2X SSC中で各5分間、連続して中和した。その後、フィルターを紫外線照射によって架橋し、真空オーブン内で30分間80℃で焼いた。フィルターを2X SSC中、42℃で大いに予備洗浄して壊死組織片を除去し、その後、42℃で、50%ホルムアミド、5X SSPE、5X デンハート溶液、0.5%SDS、100μg/mLのサケ精液DNA中で、2時間予備ハイブリダイズした。
【0195】
図3Aに示すヌクレオチド1〜711を有する895bpのDNA断片、167bpsの5’直から図3中〜開始メチオニンコドン、および17bpsの3’直〜図3A中の位置711を用いてヒトリンパ球cDNAライブラリをスクリーニングした。167塩基対のこの上流5’配列は、HuB7RP1 cDNAの5’RACEによって得られ(実施例4)、Eco RI 制限酵素切断部位においてTOPO TAベクター(Invitrogen,Carlsbad,CA)から解離させた。この挿入物を0.8%アガロースTAEゲルを用いて2回精製した。DNAゲル精製キット(Qiagen)を用いて、アガロースからDNA挿入物を分離した。
【0196】
Redi−Prime2(Amersham)ランダム第一標識システムプロトコルに従って、125ngのDNA断片を32P dCTP(Amersham)を用いて標識した。その後、コロニーリフトフィルターは、プローブを用いて42℃でハイブリダイズさせた。次の緩衝液、50%ホルムアミド、5X SSPE、2X デンハート溶液、0.5%SDS、100mg/mLのssDNA中で、一晩、42℃。プローブの比活性は、ハイブリダイゼーション緩衝液1mLにつき約2ngの標識プローブ中、2.38x109cpm/μgのDNAであった。プローブを除去し、次回のスクリーニングのために保管した。その後、フィルターを2X SSC、0.1%SDS RT中で15分間、続いて1X SSC、0.1%SDS中、55℃で15分間、そして1X SSC、0.1%SDS中、60℃で10分間洗浄した。フィルターをプラスチック中にラップし、一晩、−80℃で、2つの増強スクリーンを用いてオートラジオグラフィーフィルムに曝した。3つの独立した陽性クローンを同定した。暴露したものを細菌プレートに配列して、陽性クローンをスクラップし、アンピシリン100μg/mLを用いて希釈してLBプレート上に再びプレーティングし、前のように一晩増殖させて、コロニーを切り離し、準備して、前に記載したようにプローブした。3つの独立したクローンコロニーを分離し、DNAを分離して、同じ3つのクローンそれぞれについてDNAを配列した。
【0197】
ヒトB7RP1タンパク質の全長は、302アミノ酸である。ポリペプチド長および膜内外領域の関係位置は、B7系列の他の構成員と一致した。ヒトB7RP1遺伝子は、マウスクローンと一致するアミノ酸を43%有する。マウスおよびヒトCD80タンパク質は41%しか一致しないので、この相同性度は有意である。マウスとヒトの間で特に保存される遺伝子は、アミノ酸位置37、113、158、215および216におけるシステイン残基である。
【0198】
実施例15
ヒトCRP1のクローニング
マウスB7RP1配列(図2参照)を使用したGenbank芽細胞相同性検索(GCG,University of Wisconsin)により、マウスCRP1遺伝子と高い相同性を示す104bp配列を含むゲノムクローン(Gen Bankアクセス番号AQ022676)を取り出した。この配列とオーバーラップするPCRクローニングプライマーを設計した。
【0199】
【化3】
【0200】
上記のプライマーを使用し、図1と実施例1に示したマウスCRP1プラスミドを鋳型としてマウスCRP1の151bp DNA断片をPCR増幅した。かかるDNA 125ngを、Redi−Prime 2(Amersham)ランダムプライム標識システムプロトコールに従って32P dCTP(Amersham)で標識した。実施例15で述べるヒト末梢血ライブラリーからのコロニーリフトフィルターを下記のハイブリダイゼーション緩衝液中41℃でひと晩(15時間)、プローブとハイブリダイズさせた;50%ホルムアミド、5×SSPE、2×デンハルト溶液、0.5%SDS、100μg/ml ssDNA。プローブの比放射能は3.52×109cpm/μg DNA、1.5ng標識プローブ/mlハイブリダイゼーション緩衝液であった。プローブを取り出し、次回のスクリーニングのために保存した。次にフィルターを2×SSC、0.1%SDSにおいて室温で10分間洗浄し、その後1×SSC、0.1%SDSにより37℃で7分間、40℃で7分間、44℃で7分間、さらに50℃で7分間、フィルターが標識プローブを放出する速度を絶えずモニターしながら洗浄した。フィルターをプラスチックに包み、2枚の増感紙を用いて−80℃でひと晩フィルムに暴露した。この方法は、可能性のある9個の独立した陽性クローンを明らかにした。暴露したものを細菌プレートに整列し、陽性クローンを削りとり、200μl SOCに入れて、1:10の2回の連続希釈を行い、2回目の希釈液から70μlを100μg/mlのアンピシリンを含むLBプレートに接種して上記のようにひと晩増殖させた。コロニーを採取し、調製して、上記のようにプローブした。8個の独立クローンを分離し、Qiagenミニプレップ法によってDNAを調製した。
【0201】
199個のアミノ酸のオープンリーディングフレームを含むcDNAクローンを得た(図13A)。このcDNAクローンは、実施例1と図1に示したマウスCRP1クローンとヌクレオチド及びアミノ酸の相同性を有していた。このヒトクローンのオープンリーディングフレームに対応するヌクレオチドは、マウスCRP1遺伝子と77%同一であった。ヒト配列の翻訳及びそれに続くマウスCRP1タンパク質との比較は、マウスタンパク質と69%のアミノ酸同一性を明らかにした(図13B)。さらに、アミノ酸114から119の間のモチーフ、「FDPPPF」は、マウスとヒトCRP1遺伝子間で保存されていた。このモチーフは、マウスにおける「MYPPPY」モチーフ及びB7タンパク質の相互作用にとって必須のヒトCD28に対応する。さらに、アミノ酸42、109及び141位のシステインも保存されていた。これらのシステインはCD28及びCTLA−4におけるシステインに対応し、Igループの形成と分子間ジスルフィド二量体化に関与する。マウスCRP1との緊密な類似性及びCD28ホモロジーファミリーとの構造的類似性は、これがヒトCRP1の同族体であることを示唆している。
【0202】
実施例16
CRP−1は休止記憶Tリンパ球上に発現される
記憶T細胞上のCRP−1発現を調べるため、6−7ヵ月齢のマウスから脾T細胞を採集した。これらの細胞を、FITC複合抗ヒトFc抗体及びCD44、CD45RB、又はCD69のいずれかに対するPE複合抗体によって標識したB7RP−1−Fcを使用して二重染色した。B7RP−1−Fc融合タンパク質による染色は、これらのT細胞でのCRP−1タンパク質の発現を検出する。より高齢のマウスは若齢マウスよりも多くのCRP−1+脾T細胞を示す。興味深いことに、顕著な数のこれらの細胞が、記憶T細胞に典型的なプロフィールである、CD44が高く(図14a)、CD45RBが低い(図14b)。これらのCRP−1+記憶T細胞は、活性化マーカーであるCD69を発現しないため(図14c)、休止状態にある。記憶T細胞上のCRP−1発現は、CRP−1が記憶T細胞への共同刺激機能を持つことを示唆する。
【0203】
実施例17
B7RP−1に対する抗体によって阻害される、インビトロでのT細胞の共同刺激
B7RP−1タンパク質がT細胞と機能的関連性を持つかどうかを調べるため、インビトロ増殖アッセイにおいてCD3+T細胞をB7RP−1−Fc融合タンパク質及び抗CD3抗体と共にインキュベーションした。次にウサギ抗マウスB7RP1ポリクローナル抗体又はラット抗マウスB7RP1モノクローナル抗体を使用して、B7RP1−Fcが共同刺激するインビトロでの増殖を特異的に阻害した。
【0204】
B7RP−1ウサギポリクローナル抗血清の調製
3匹のニュージーランド白色ウサギ(初期重量5−8ポンド)にマウスB7RP1タンパク質を筋肉内注射した。各々のウサギを1日目に、等量のHunters Titer Max完全アジュバントに乳化したマウスB7RP1タンパク質150μgで免疫した。さらに同じ手順で追加免疫(14日目と28日目)を実施した。EIAによって抗体価をモニターした。2回目の追加免疫後、抗血清は中等度の抗体価を示した。各々の動物から30mlの採血を行った。これを週に1回6週間にわたって繰り返した。その後ポリクローナル抗体をプロテインAアガロースクロマトグラフィーによって精製し、次いでFcタンパクアフィニティークロマトグラフィーによる陰性選択及びB7RP−1−Fcアフィニティークロマトグラフィーによる陽性選択を実施した。
【0205】
ラット抗マウスB7RP1モノクローナル抗体の調製
ラット抗マウスB7RP1モノクローナル抗体を、Practical Immunology,第2版(1980;L.Hudson and F.C.Hay;Blackwell Scientific Publications;St.Louis,MO)に述べられているように作製した。簡単に述べると、Louラット(Harlan;Indianapolis,IN)に、フロイントアジュバントに乳化したmuB7RP1−Fc融合タンパクを4週間隔で腹腔内注入した。融合の3日前に、ラットを可溶性muB7RP1により静脈内経路で追加免疫した。融合の当日、動物を二酸化炭素下で犠牲屠殺し、無菌的に脾を切除した。組織ストマカーを用いて単細胞懸濁液を調製した。脾細胞とY3−Ag1.2.3.骨髄腫細胞(American Type Culture Collection;Rockville,MD)の両方を無血清培地中で洗浄し、ポリエチレングリコール(PEG 1500;Boehringer Mannheim Biochemicals;Indianapolis,IN)を加えて融合した。細胞を1回すすぎ、血清含有培地に再懸濁して、96穴の組織培養プレートに接種した。10−12日後、各々の穴からの培地を、直接酵素免疫測定法(EIA)によりB7RP1に対する特異性抗体に関して試験した。潜在的な結合を示す穴からの細胞を10ml培養物に増殖させ、液体窒素中で冷凍した。各培養からの培地をフローサイトメトリー及び機能的T細胞増殖アッセイにおいてさらに試験した。これらの方法によって興味深いと判定されたものを単細胞コロニーに接種し、再びEIAによって選択して、最終的な細胞系統を抗体作製のために保持した。プロテインAアガロースクロマトグラフィーによって細胞培地から抗体を精製した。
【0206】
T細胞の調製及びT細胞増殖アッセイ
C57B1/6マウス(8−12週齢の雌性、Charles River Laboratories)の脾からのT細胞を、マウスT細胞強化カラム(R&D Systems)を通して陰性選択によって精製した。その後T細胞を直接使用するか、若しくは次のような抗体と補体の溶解によってさらに精製した。細胞を、マウスCD11b(クローンM1/70)、NK−1.1(クローンPK136)、CD8a(クローン53−6.7)、I−Ab(クローンM5/114.15.2)、CD11c(クローンHL3)、及びB220抗原(クローンRA3−6B2)に対する抗体(すべて10μg/ml、Pharmingenより)を含むRPMI培地に懸濁した(2.5×106細胞/ml)。次に細胞を氷上で30分間インキュベーションし、1200rpmでペレット化して、4:1v/vのRPMI:ウサギ補体(Sigma,No.S−7764)中に懸濁し、37℃でさらに30分間インキュベーションした。再び細胞をペレット化し、補体処理を繰り返した。平板培養の前に、10%FCSを含むRPMIで細胞を洗浄した。U底96穴プレートを、0から1.2μg/mlの濃度範囲の抗CD3抗体(クローン145−2C11,Pharmingen)及び抗ヒトIgG Fab2(Sigma,12.5μg/ml)により4℃でひと晩被覆し、次いで37℃で6−9時間インキュベーションした。T細胞(1×105/穴)を種々のFc融合タンパクの不在下又は存在下で48時間培養し、最後の18時間は1μCiの3H−チミジンでパルスした。対照Fcタンパクは、OPGとFcの融合タンパク及び非融合Fcタンパク断片を含んだ。その後細胞を採集し、取り込まれた放射能を測定した。B7RP−1−FcはT細胞を用量依存的に共同刺激して増殖させ(図15a)、抗B7RP−1−Fc抗体はこの共同刺激を用量依存的に阻害する(図15b)。
【0207】
実施例18
CRP−1/B7RP−1経路の阻害因子は、コラーゲンによって誘発される慢性関節リウマチの発症を低下させる
コラーゲン誘発の関節炎(CIA)は、ヒトにおける慢性関節リウマチと多くの類似性を持つ、げっ歯類及び霊長類での自己免疫性多発性関節炎の動物モデルである。異種のII型コラーゲン(CII)による免疫はCIIに対する自己免疫応答を誘発し、感受性のあるマウス系統においてCIAの発症を導く。H−2r及びH−2qを有する類似遺伝子型系統のマウスはCIAに対して極めて感受性が高い。CIAは、CII反応性T細胞と抗体の共力作用によって仲介される。ブタCII(Naboznyら、Autoimmunity 20,51−58(1995))を2mg/mlの濃度で0.01N酢酸に溶解し、次に1:1の比率でCFA(Difco)により乳化した。関節炎感受性B10.RIII(H−2r)マウス(Jackson Laboratories,Bar Harbor,ME)を、尾の基部において乳剤100μlにより皮内経路で免疫した。マウスを関節炎の発症に関して週に2−3回モニターした。関節炎の重症度は、各々の足について次のような採点システムを用いて判定した:0:関節炎なし;1:1−3本の足指に発赤又は腫脹;足の重篤な腫脹;3:関節強直。各々の肢の評点を合計し、各動物について0から12の重症度範囲を決定した。
【0208】
マウスにタンパク質100μg(200μL中)を週に2回、腹腔内経路で注入した。処置は、ブタCIIによる免疫の1日後に開始し、免疫後52日目に中止した。10匹のマウスから成る処置群において実験を行い、1又はそれ以上の評点を有する動物を陽性と判定した。結果を図16及び表1に示す。
【0209】
【表2】
【0210】
CRP1−Fc融合タンパクで処置したマウスでは、PBS処置マウスと比較して関節炎症状の発現が約10日間遅延した。これは、CRP−1/B7RP−1経路の阻害がこの慢性関節リウマチのマウスモデルにおいて疾患の症状を軽減しうることを示している。
【0211】
B7RP−1−Fc又はB7.2−Fcで処置したマウスは、PBS処置した対照と比較すると、関節炎の重症度の上昇(図16b)を伴って疾患の早期発症(表1及び図16a)を示した。これは、B7RP−1−Fc融合タンパクがT細胞免疫応答を増強することを示唆している。そのような活性は、インビボで抗腫瘍免疫を生じさせる上で有用と考えられる。
【0212】
慢性関節リウマチのこのマウスモデルにおけるCRP−1−Fc及びB7RP−1−Fcの反対の作用は、疾患の進行を促進する又は阻害するように経路を操作できることを示唆する。可溶性B7RP−1−FcによるCRP−1タンパクのターゲティングは疾患を促進するが、B7RP−1と可溶性CRP−1−Fcの相互作用は疾患の症状を抑制する。
【0213】
実施例19
B7RP−1−Fcはトランスジェニックマウスにおいて炎症性腸疾患表現型を誘導する。
【0214】
22−25週齢のトランスジェニックマウスにおけるB7関連タンパク(B7RP1−Fc)の持続的な過剰発現は、一部の動物において小腸及び大腸の著しい肥厚と慢性炎症(腸炎)及び体重減少を伴った炎症性腸疾患(IBD)の著明な表現型を誘導する。組織学的には、近位及び遠位結腸において最も重症の炎症性変化が認められ、小腸ではより軽度の変化が認められた。近位結腸は、裂溝状潰瘍、経壁炎症、及び結腸粘膜の肥厚を伴って著しく肥厚していたが、遠位結腸は、潰瘍形成を伴わない散在性粘膜肥厚(又は限局性びらん及び腺萎縮)を有していた。近位小腸は、より軽度の炎症性変化を伴って軽度から著明な粘膜肥厚を示し、一方遠位小腸(回腸)は、一部の動物では軽度の粘膜肥厚、また別の動物では萎縮を示した。腸の変化は、雌性B7RP−1トランスジェニックマウスにおいて最も重症であり、一貫して認められたが、本試験では雄性トランスジェニックマウスの一部においても認められた。
【0215】
裂溝状潰瘍及び多核巨細胞を伴った経壁性慢性肉芽腫性炎症を含めて、近位結腸で認められた組織学的特徴が、ヒトにおける潰瘍性結腸炎よりもクローン病で認められるものにより密接に類似していることは興味深い。形態学的には、この結腸炎はまた、インターロイキン−10欠損のマウスにおいて記述されているIBDに類似しており、この疾患は消耗、貧血、及び腸管全体を冒す腸炎を発現する(Kuhnら、1993;Sartor,1995;Leachら、1999)。IL−10ノックアウトマウスにおけるように、B7RP−1−Fcトランスジェニックマウスでの初期変化は、結腸上皮肥厚を伴わない粘膜固有層における炎症細胞の軽度で限局性の浸潤から成る(実施例13)。より高齢のマウスでは、罹患した結腸セグメントは腺肥大/過形成と慢性炎症のため肥厚する。B7RP1マウスの近位及び遠位結腸は、炎症性腸疾患(IBD)の組織学的特徴を持つ中等度から重度の結腸炎を有していた。近位結腸の罹患セグメント(図17B−17D)は、粘液腺の伸長と拡大を伴った顕著な腺上皮肥厚と過形成のため、広汎に肥厚しており(図17B)、粘液腺は有糸分裂像の数の増大とまれな陰窩膿瘍を有していたが、ムチンを含む杯細胞は保持していた(図17D)。粘膜は、固有層に広汎な慢性炎症があり、一部の動物では膜を越えて拡大し、粘膜下組織、筋層、漿液、及び隣接する腸間膜脂肪組織を含めて腸管壁の下層にまで及んでいた(図17B−17C)。炎症性浸潤は、慢性肉芽腫性炎症の特徴である、いくつかの好中球及び時として多核巨細胞が混じった(図17E)、リンパ球(主としてCD3+、CD44+T細胞)、プラスマ細胞、及び類上皮マクロファージ(図17F)から成った。リンパ球様凝集物(少数のCD3+細胞が混じった、主にB220+細胞)も、粘膜中及び腸間膜脂肪を含めた粘膜下組織と深層のより微小な血管の周囲に存在した(図17C)。管腔は粘液膿性又は粘液性の滲出物を含んでいた(図17D)。これらのB7RP−1−Fcトランスジェニックマウスにおいて、粘膜の多病巣性裂溝状潰瘍と経壁性炎症(図17B−17C)を伴う結腸炎の深刻な証拠が認められた。
【0216】
B7RP−1−Fcトランスジェニックマウスの遠位結腸も、結腸腺の伸長、好塩基球増加及び拡大を伴って広汎に肥厚し、増殖しており(図18B−18G)、その内の一部は陰窩膿瘍(図18D及び18F)及び粘液を含んでいた。粘膜固有層は、リンパ球(主としてCD3+、CD44+細胞、特に浅粘膜において;図18E)の軽度の散在性炎症性浸潤、ならびにプラスマ細胞と一部好中球が混じった類上皮マクロファージの巣状凝集物を有していた。リンパ球様凝集物(主としてB220+細胞;図18D及び18F)も粘膜全体に散らばっていた。B7RP−1−Fcトランスジェニックマウスの小腸は、粘膜固有層の主としてリンパプラスマ細胞性浸潤を伴った、軽度から局所的に著明な粘膜及び陰窩の肥厚と過形成(図19B及び19D、陰窩/絨毛比は、対照マウスでの1:10と比較して、1:4から1.5:1)を含めて、より多様な変化を有していた。粘膜過形成は、十二指腸(図19B)、そして特に空腸(図19D)を含めた近位小腸において最も顕著であった。最も重症のマウスでは、陰窩構造が局所的に混乱し、形成異常であった(図19D)。これに対し、一部のマウスの遠位小腸(回腸)は、絨毛の鈍化、肥厚又は局所的喪失(陰窩:絨毛比は、正常比1:2に対して1:1又はそれ以下)を伴った回腸粘膜の軽度で斑状の絨毛萎縮を有しており、また他の一部のマウスは軽度の回腸粘膜肥厚を有していた。
【0217】
B7RP−1−Fc融合タンパクは、ヒトクローン病のものに非常に類似した表現型を誘発する役割を担う細胞を活性化するように働く。これは、クローン病における炎症の原因と考えられる細胞がB7RP−1−Fc融合タンパクによって活性化されることを示唆する。B7RP−1の可溶性タンパク、抗体又は小分子阻害因子は、それ故、IBDを抑制する上で有用と考えられる。
【0218】
実施例20
B7RP−1−Fc融合タンパクはマウスにおける腫瘍増殖を抑制する
免疫原性マウスMeth A肉腫の増殖へのB7RP−1及びCRP−1の作用を調べるため、可溶性B7RP−1−FcがBalb/cマウスの確認されたMeth A肉腫の増殖に影響を及ぼすかどうかを検討した。
【0219】
対数増殖するMeth A肉腫細胞を、0日目にBalb/cマウスの腹部に50万個の細胞を皮内注入して移植した。7日目に腫瘍が〜100mm3に達し、7、10、14及び17日目にマウスを賦形剤(PBS)又はB7RP−1−Fc(8mg/kg)のいずれかにより皮下経路で処置した。腫瘍の二次元直径をカリパスで測定し、次の式を用いて腫瘍容積(mm3)を算定した:腫瘍容積=[{(幅)2×長さ}/2]。28日目まで腫瘍増殖をモニターした。各々の群は8匹のマウスで構成された。
【0220】
対照腫瘍のMeth A肉腫増殖パターンは二相性であった:緩慢な初期相の後に比較的迅速な対数期が続いた。B7RP−1−Fc処置マウスでは、腫瘍の増殖は迅速な対数期において有意に緩慢であった。28日目に、対照とB7RP−1−Fc処置マウスの平均容積はそれぞれ1410mm3と580mm3であった(図20)。それ故、B7RP−1−Fc処置はこのモデルにおいて腫瘍の増殖を有意に抑制した。データは、免疫原性腫瘍の治療における可溶性B7RP−1−Fcタンパク及びB7RP−1/CRP−1経路の他の活性化因子の有益な治療上の有用性を強く示唆している。
【0221】
免疫学的抗腫瘍活性は、細胞溶解性Tリンパ球(CTL)の機能と緊密に結びついている。一貫して、B7RP−1−Fcタンパクは細胞溶解性CD8+T細胞上で発現される(実施例9、図6)。これらのデータは、細胞溶解性CD8+T細胞へのB7RP−1の機能を強く裏付けるものである。B7RP−1−Fc又はB7RP−1/CRP−1経路の他の刺激因子は、それ故、多くの非癌関連適応症に関して細胞溶解性T細胞及び細胞性免疫機能を増強するために使用しうるであろう。
【0222】
実施例21
in vitroでのヒトB7RP−1活性の阻害
ヒトB7RP−1が正の同時刺激性を有するかどうかを同定するために、我々は、T細胞増殖アッセイにおけるヒトB7RP−1およびヒトB7RP−1−Fc融合タンパク質を発現する細胞を試験した。ヒトB7RP−1−Fc融合タンパク質を、部分的ヒトIgG1遺伝子配列へのアミノ酸1〜247に対応する遺伝子配列の融合によって構築した(実施例14)。ヒトCRP−1−Fc融合タンパク質を、部分的ヒトIgG1遺伝子配列へのアミノ酸1〜146に対応する遺伝子配列の融合によって構築した(実施例14)。両融合タンパク質の構築法、発現法、および精製法を、実施例7に記載のように行った。B7RP−1−Fcは、抗CD3刺激に依存する同時刺激活性を示した(図21a)。さらに、この活性を、溶解性CRP−1−Fcタンパク質で特異的に阻害することができる(図21b)。全コード配列を含む、膜結合ヒトB7RP−1を発現するCHO細胞を用いて、類似の同時刺激効果が得られた(図21c)。
【0223】
上記のin vitro増殖条件下でのヒトT細胞によるサイトカインの産生を同定した。48時間および72時間刺激したT細胞培養物由来の上清を、製造者の仕様書(BioSource International)に従ったELISAによってIL−2、IL−10、およびIFN−γについて分析した。IFN−γおよびIL−10レベルは有意に増加したが、CD28同時刺激の場合と異なり、IL−2は特に誘導されなかった(図21d)。実施例13に記載のように、IFN−γ(Th1サイトカイン)レベルの増加は、IgG2aを増加させるB7RP−1の機能と相関する。
【0224】
in vitroT細胞同時刺激アッセイを、以下のように行った。高度に精製したヒトT細胞(>98% CD3+)を、mAb標識磁性ビーズ(Miltenyi Biotec)を用いた新鮮または解凍した接着減少PBMCのネガティブ選択によって単離した。T細胞(1×105細胞/ウェル)を、96ウェルの3連のウェルにおいて200μl/ウェル RPMI+10% FCS中で培養した。B7RP−1−Fc同時刺激を評価するために、種々の濃度の抗CD−3(Pharmingen)および10μg/ml抗ヒトIgG Fc(Sigma)の100μl 1×PBS溶液を4℃で一晩のインキュベーションによってU底プレート上にプレコートした。非結合抗CD3および抗ヒトIgG Fcを取り除き、細胞を種々の濃度のB7RP−1−Fc、OPG−Fcコントロール、または抗CD28(Pharmingen)の存在下または非存在下で培養した。B7RP−1−Fc同時刺激のCRP−1−Fc阻害用に、連続希釈したCRP−1−FcまたはOPG−Fcの存在下で、0.5μg/mlB7RP−1−Fcを含む0.33μg/ml抗CD−3および10μg/ml抗ヒトIgG Fcプレコートウェル中で、10μg/mlから開始してT細胞を培養した。B7RP−1を発現するCHO細胞による同時刺激を評価するために、種々の量のマイトマイシン−C処理CHO B7RP−1細胞またはCHOベクター細胞の存在下または非存在下で、種々の濃度の溶解性抗CD3を含む平底プレート中でT細胞を培養した。T細胞増殖について試験するために、72時間の培養の最後の18時間に1 uCi/ウェル [3H]TdRで培養物をパルスした。T細胞増殖を、[3H]TdR組み込みによって同定した。3つの無作為なドナー由来の1つの代表的な実験結果を、平均CPM組み込み+/−SDで示す。サイトカイン産生分析用に、細胞を48時間および72時間培養し、ELISA用に上清を回収した。
【0225】
3つの実験は、実施例14に記載するように、ヒトFc断片に融合した場合、ヒトB7RP−1の細胞外部分は、in vitroでT細胞を同時刺激することができることを示す。この同時刺激は、CRP−1−Fcで阻害されるので、これは、ヒトB7RP−1の溶解性インヒビターがどのように機能し得るのかを示す。in vitroアッセイ(本明細書中に記載のヒトB7RP−1およびCRP−1を用いたアッセイ)を使用して、B7RP−1/CRP−1活性の抗体、溶解性タンパク質、ペプチ体、または小分子インヒビターをスクリーニングすることができる。
【0226】
本発明は好ましい実施形態に関して記載しているが、当業者により変形形態および修正形態が実施されることが理解される。したがって、添付の特許請求の範囲は、特許請求の範囲に記載の本発明の範囲内で実施されるこのような全ての等価な変形形態を対象とすることが意図される。
【技術分野】
【0001】
発明の分野
本発明は、Tリンパ球の活性化に関与するポリペプチドに関する。特に、本発明は、Tリンパ球共刺激ポリペプチド、該ポリペプチドをコードする核酸、該ポリペプチドを産生するための発現ベクターと宿主細胞、ならびに免疫抑制および免疫活性化に関連する疾患の治療用組成物および方法に関する。
【背景技術】
【0002】
発明の背景
適切なTリンパ球(T細胞)の免疫応答を得るためには、抗原提示細胞(APC)によって2種類のシグナルがT細胞に送られる必要がある。第1に、特異性を決定する場合に、抗原は主要組織適合抗原複合体(MHC)によってT細胞受容体(TCR)に提示される必要がある。第2に、抗原依存性の共刺激シグナルは、APC上のB7ファミリーに属する要素がT細胞上のCD28タンパク質と連結することによって送り出される必要がある。増殖性の免疫応答は、増殖、分化、クローン性増殖、および作用または機能につながる。第2の共刺激シグナルがない場合は、T細胞はアネルギーと称される持続性抗原特異的不応答性の状態になる。
【0003】
T細胞は、免疫応答を開始し、抗原特異的エフェクター機能を媒介し、サイトカインの分泌によって他の白血球の活性を制限する。T細胞受容体(TCR)は、T細胞を他のリンパ球と区別するものであり、APCのMHCによって抗原が提示された場合にのみその抗原と結合可能である。特定のT細胞の機能は、CD4やCD8などの膜抗原の発現と関連づけることができる。例えば、一般にCD4+T細胞はヘルパーT細胞(TH)として機能しMHCクラスIIに限定され、一般にCD8+細胞は細胞障害性T細胞(TC)として機能しMHCクラスIに限定される。
【0004】
従来同定された有効なT細胞共刺激ポリペプチドとしては、B7.1と称されるポリペプチド(フリーマン(Freeman)らJ.Immunology 143,2714−2722(1989)、フリーマン(Freeman)らJour.Expt.Med.174,625−31(1991))、およびB7.2と称される(フリーマン(Freeman)らScience 262,909−911(1993)、およびフリーマン(Freeman)らJour.Expt.Med.178,2185−2192(1993))(あるいはそれぞれCD80およびCD86と称される)が挙げられる。これらのポリペプチドは、種々のAPC上において誘発的または恒常的のいずれかで発現され、それぞれT細胞上のCD28およびCTLA−4に対する膜結合リガンドである。CD28(アルフォ(Aruffo)およびシード(Seed)Proc.Natl.Acad.Sci.84,8573−8577(1987)、およびグロス(Gross)らJ.Immun.144,3201−3210(1990))は、休止T細胞上で発現され、陽性共刺激シグナルを媒介する。CTLA−4(ブルネット(Brunet)らNature 328,267−270(1987)、およびダリアバク(Dariavach)らEur.Jour.Immun.18,1901−1905(1988))の発現は、T細胞の活性化によって誘発され、B7.1およびB7.2に対する結合親和性がより高いために、CD28シグナルを負の方向に制御する。CTLA−4遺伝子をもたないマウスはCTLA−4がない状態では増殖シグナルのスイッチオフ機構が損なわれているため、T細胞量が非常に多く見られる。この表現型は、T細胞の増殖に対してCTLA−4共刺激タンパク質が有する主要な阻害作用を明確に示している。CD28またはB7.1またはB7.2の欠如したマウスでは、これほど過度な表現型は見られないが、これはT細胞共刺激に関する別の経路が存在しうることを示している。
【0005】
T細胞の活性化および増殖を調節する手段としてCD28/CTLA−4経路について大きな関心が示されてきている。ヒトFcと融合したCTLA−4の細胞外部分を含有するキメラタンパク質は強力な免疫抑制効果を有し、種々の臨床状況において研究が行われてきた。B7.1タンパク質とB7.2タンパク質に対する抗体についても、免疫抑制の分野で同様の徴候について評価されている。抗CTLA−4抗体は、T細胞活性化の促進に有用であることが示されている。さらに、B7.1およびB7.2の遺伝子治療が、癌免疫療法の分野で非常に有用であることが示されている。
【0006】
今までのところ、CD28、CTLA−4、B7.1、およびB7.2は、1つのT細胞共刺激経路と関連している。T細胞共刺激を調節することによって免疫応答を変化できるのであれば、宿主T細胞の機能および免疫応答の調節に関して好都合な性質を有する可能性のある同一または別のT細胞共刺激経路の他の要素を同定することが望ましい。
【0007】
したがって本発明の目的の1つは、T細胞活性および/または増殖を刺激する新規ポリペプチドを提供することである。本発明のさらなる目的は、T細胞媒介免疫疾患の予防および治療のために該新規ポリペプチドを使用することである。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】フリーマン(Freeman)らJ.Immunology 143,2714−2722(1989)
【非特許文献2】フリーマン(Freeman)らJour.Expt.Med.174,625−31(1991)
【非特許文献3】フリーマン(Freeman)らScience 262,909−911(1993)
【非特許文献4】フリーマン(Freeman)らJour.Expt.Med.178,2185−2192(1993)
【非特許文献5】アルフォ(Aruffo)およびシード(Seed)Proc.Natl.Acad.Sci.84,8573−8577(1987)
【非特許文献6】グロス(Gross)らJ.Immun.144,3201−3210(1990)
【非特許文献7】ブルネット(Brunet)らNature 328,267−270(1987)
【非特許文献8】ダリアバク(Dariavach)らEur.Jour.Immun.18,1901−1905(1988)
【発明の概要】
【0009】
発明の要約
驚くべきことに、T細胞共刺激経路の2種類の新規ポリペプチドがこれまでに同定されている。これらのポリペプチドは、前述のタンパク質CD28、CTLA−4、B7.1、およびB7.2からなる経路とは異なるものと思われるただ1つの共刺激経路におけるリガンド−受容体対を表している。これらのポリペプチドは、CD28関連タンパク質−1またはCRP1、およびB7関連タンパク質−1またはB7RP1と称される。
【0010】
本発明は、CRP1およびB7RP1ポリペプチド、ならびに関連ポリペプチドをコードする核酸分子を提供する。本発明の単離核酸分子は、
a)図1A(配列番号1)に記載のヌクレオチド配列;
b)図1A(配列番号1)に記載の残基1〜200または残基21〜200のポリペプチドをコードするヌクレオチド配列;
c)図1A(配列番号1)に記載のポリペプチドと少なくとも約70%が同一であるポリペプチドをコードするヌクレオチド配列;
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体;
e)(a)、(b)、または(c)のいずれかと相補的なヌクレオチド配列;
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリペプチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列;
g)少なくとも約10、15、20、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列;および
h)ストリンジェンシー条件下で(a)〜(g)のいずれかとハイブリッド形成するヌクレオチド配列;
からなる群より選択されるヌクレオチド配列を含む。
【0011】
さらに、本発明は、
a)図2A(配列番号6)または図3A(配列番号11)または図12A(配列番号16)に記載されるヌクレオチド配列;
b)図2A(配列番号6)に記載の残基1〜322または残基47〜322のポリペプチド、または図3A(配列番号11)に記載の残基1〜288、あるいは残基19〜288、20〜288、21〜288、22〜288、24〜288、または28〜288のポリペプチド、または図12Aに記載の残基1〜302、あるいは残基19〜302、20〜302、21〜302、22〜302、24〜302、または28〜302のポリペプチド、をコードするヌクレオチド配列;
c)図2A(配列番号6)または図3A(配列番号11)または図12A(配列番号16)に記載のポリペプチドと少なくとも約70%が同一であるポリペプチドをコードするヌクレオチド配列;
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体;
e)(a)、(b)、または(c)のいずれかと相補的なヌクレオチド配列;
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリペプチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列;
g)少なくとも約10個、15個、20個、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列;および
h)ストリンジェンシー条件下で(a)〜(g)のいずれかとハイブリット形成するヌクレオチド配列;
からなる群より選択されるヌクレオチド配列を含む単離核酸分子を提供する。
【0012】
さらに本発明の主題は、CRP1ポリペプチドおよびB7RP1ポリペプチド、ならびに関連ポリペプチドにも関する。本発明は、
a)図1A(配列番号2)に記載のアミノ酸配列;
b)残基21に成熟アミノ末端を含む図1A(配列番号2)に記載の成熟アミノ酸配列;
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む図1A(配列番号2)に記載のアミノ酸配列断片;
d)(a)、(b)、または(c)のオルソログ;および
e)(a)、(b)、または(d)の対立遺伝子変異体または選択的スプライス変異体;
からなる群より選択されるアミノ酸配列を含む単離ポリペプチドを提供する。
【0013】
さらに、本発明は、
a)図2A(配列番号7)または図3A(配列番号12)または図12A(配列番号17)に記載のアミノ酸配列;
b)残基47に成熟アミノ末端を含む図2A(配列番号7)に記載の成熟アミノ酸配列、あるいは残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図3A(配列番号12)に記載の成熟アミノ酸配列、あるいは残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図12A(配列番号17)に記載の成熟アミノ酸配列;
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む図2A(配列番号7)または図3A(配列番号12)または図12A(配列番号17)に記載のアミノ酸配列断片;
d)(a)、(b)、または(c)のオルソログ;および
e)(a)、(b)、(c)、または(d)の対立遺伝子変異体または選択的スプライス変異体;
からなる群より選択されるアミノ酸配列を含む単離ポリペプチドを提供する。
【0014】
また、本発明のポリペプチドを産生する発現ベクターおよび宿主細胞、CRP1ポリペプチドおよびB7RP1ポリペプチドならびに関連ポリペプチドと結合する抗体、ならびにB7RP1ポリペプチドおよびB7RP1関連ポリペプチドと、CRP1ポリペプチドおよびCRP1関連ポリペプチドとの結合を検出するアッセイも本発明に包含される。CRP1ポリペプチドまたはCRP1関連ポリペプチドと、B7RP1ポリペプチドまたはB7RP1関連ポリペプチドとを含む医薬組成物も本発明に包含される。CRP1またはB7RP1と相互作用する化合物の同定方法も、これらの化合物がCRP1とB7RP1の活性に対するアゴニストとアンタゴニストのいずれとなるかを決定するためのアッセイとして提供される。
【0015】
CRP1ポリペプチドとB7RP1ポリペプチドは、T細胞の共刺激および増殖に関与する。CRP1ポリペプチドとB7RP1ポリペプチド、それらの選択的結合剤、ならびにそれらのアゴニストおよびアンタゴニストは、T細胞応答の調節に関連する疾患の診断、予防、および治療に有用となりうる。
【0016】
CRP1ポリペプチドとB7RP1ポリペプチド、それらの選択的結合剤、ならびにそれらのアゴニストおよびアンタゴニストは、不充分な免疫応答の刺激、あるいは異常に大きいまたは不適当な免疫応答の軽減または抑制のいずれかによる免疫疾患の診断、予防、および治療に有用となりうる。免疫疾患は、T細胞が直接的または間接的に媒介しうる。
【0017】
本発明は、動物にCRP1ポリペプチドまたはB7RP1ポリペプチドを投与することを含むT細胞媒介疾患の治療、予防、または回復のために提供される。本発明は、CRP1ポリペプチドまたはB7RP1ポリペプチドの存在または発現量を決定することを含む動物のT細胞媒介疾患またはT細胞媒介疾患に対する感受性の診断方法;ならびに該ポリペプチドの存在または発現量に基づくT細胞媒介疾患またはT細胞媒介疾患に対する感受性の診断方法も提供する。通常、T細胞媒介疾患は、T細胞が直接的または間接的に媒介しうる免疫疾患である。動物は哺乳動物が好ましく、ヒトがより好ましい。本発明は、CRP1ポリペプチドまたはB7RP1ポリペプチドと結合する試験分子の同定方法であって、該ポリペプチドを試験化合物と接触させ、ポリペプチドと試験化合物の結合程度を求めることを含む同定方法も提供する。この方法は、CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアゴニストおよびアンタゴニストの同定に使用可能である。
【0018】
CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアンタゴニストは、自己免疫疾患(リウマチ様関節炎、乾癬、多発性硬化病、糖尿病、および全身性エリテマトーデスなど)、トキシックショック症候群、骨髄または臓器移植、炎症性腸疾患、輸血による異種感作、および移植片対宿主疾患の治療を含む多くの徴候の免疫抑制剤として使用することができる。さらに、アンタゴニストは、喘息およびアレルギーなどのT細胞依存性B細胞の媒介する徴候、および抗体媒介自己免疫疾患の抑制剤として使用することができる。CRP1ポリペプチドおよび/またはB7RP1ポリペプチドのアゴニストは、腫瘍のサーベイランスおよび除去のためのT細胞活性化に有用となりうるが、これに限定されるものではない。
【0019】
本発明のアンタゴニストは、B7RP1またはB7RP1の細胞外ドメインと反応性であるかそれらと結合する抗体またはその断片を含み、この抗体はB7RP1とCRP1の結合を減少させるか結合しなくする。実施態様の1つでは、この抗体は、ヒトB7RP1またはその細胞外ドメインと選択的に結合する。アンタゴニストであるこの抗体またはその断片は、B7RP1の免疫共刺激活性を部分的または完全に阻害する。好ましい実施態様では、この抗体はモノクローナル抗体であり、マウス抗体、ヒト抗体、キメラ抗体、またはヒト化抗体であってよい。
【0020】
本発明は、CRP1の選択的結合剤またはB7RP1の選択的結合剤またはその療法を動物に投与することを含む、B7RP1とCRP1の相互作用を調節する方法もさらに提供する。実施態様の1つでは、この選択的結合剤は、B7RP1と結合して、B7RP1のCRP1に対する結合を減少させたりなくしたりする抗体である。本発明は、B7RP1の選択的結合剤を動物に投与することを含む、B7RP1が媒介する免疫共刺激を調節する方法も提供する。この選択的結合剤は、B7RP1と結合して、B7RP1が媒介する免疫共刺激を部分的または完全に抑制する抗体であることが好ましい。
【0021】
本発明は、CRP1ポリペプチドまたはB7RP1ポリペプチドをコードする核酸分子を動物に投与することを含む、動物のT細胞の活性化または増殖を調節する方法も提供する。例えば、B7RP1ポリペプチドをコードする核酸分子を、種々の腫瘍に対するT細胞の活性化を促進する遺伝子治療に使用することができる。
【0022】
また、CRP1ポリペプチドまたはB7RP1ポリペプチドを含む非ヒトのトランスジェニック哺乳動物も本発明に包含される。CRP1ポリペプチドまたはB7RP1ポリペプチドを発現してその循環量を増大させる方法で、CRP1またはB7RP1核酸が哺乳動物に導入される。非ヒトのトランスジェニック哺乳動物は、囓歯類が好ましく、マウスまたはラットがより好ましい。
【図面の簡単な説明】
【0023】
【図1A】マウスCRP1(mCRP1)のDNA配列およびアミノ酸配列。CRP1の予測シグナル配列にはアミノ末端で下線が引かれており、実験的に求めたプロペプチド開裂部位はアスタリスクで示され、予測膜貫通配列はカルボキシ末端に向かって下線が引かれている。
【図1B】マウスCRP1タンパク質配列(mCRP1)とマウスCD28(mCD28)のアミノ酸配列。
【図2A−1】マウスB7RP1(mB7RP1)のDNA配列およびアミノ酸配列。B7RP1の予測シグナル配列にはアミノ末端で下線が引かれており、実験的に求めたプロペプチド開裂部位はアスタリスクで示される。予測膜貫通配列はカルボキシ末端に向かって下線が引かれている。
【図2A−2】マウスB7RP1(mB7RP1)のDNA配列およびアミノ酸配列。B7RP1の予測シグナル配列にはアミノ末端で下線が引かれており、実験的に求めたプロペプチド開裂部位はアスタリスクで示される。予測膜貫通配列はカルボキシ末端に向かって下線が引かれている。
【図2B】B7RP1タンパク質配列(mB7RP1)とマウスCD80(mCD80)のアミノ酸配列。
【図3A−1】推定ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図3A−2】推定ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図3B】推定成熟hB7RP1タンパク質と成熟マウスB7RP1(mB7Rp1)タンパク質のアミノ酸配列。
【図4A】pcDNA3/CRP1−Fcをトランスフェクションした293T細胞の可溶性CRP1−Fc融合タンパク質の発現。図に示すように規格化体積の細胞溶解物または馴化培地を10%PAGEゲルに加えて分離させる。細胞関連(細胞溶解物)および分泌された(培地)のFc融合タンパク質の発現についての、細胞溶解物および細胞培地上清のウエスタン(Western)分析。1次抗体はヤギ抗ヒトFc抗体(ピアス・ケミカルカンパニー(Pierce Chemical Company)、ロックフォード(Rockford),IL)であった。
【図4B】pcDNA3/B7RP1−Fcをトランスフェクションした293T細胞の可溶性B7RP1−Fc融合タンパク質の発現。10%PAGEゲルに20μlの規格化細胞溶解物または培地上清を加えて分離させた。(a)と同様にウエスタン分析を行った。
【図5】CRP1−Fc融合タンパク質およびB7RP1−Fc融合タンパク質と、COS−7細胞の発現した膜結合タンパク質の相互作用。COS−7細胞には一時的にpcDNA3/CRP1、pcDNA3/B7RP1、またはpcDNA3ベクター単独をトランスフェクションした。CHOのD細胞にpsDRα/hCD28をトランスフェクションすると、ヒトCD28(hCD28)を安定に発現した。細胞の発現する膜結合CRP1、B7RP1、またはhCD28は、パネルの左側の列に示される。Fc融合タンパク質は、パネル上部の列に示されるプレート結合細胞とともにインキュベートした。インキュベート後、細胞を洗い流し、結合したFc融合タンパク質を、抗ヒトFc抗体とACAS(付着細胞分析および選別(Adherent Cell Analysis and Sorting);ACASアルティマ(ACAS Ultima),メリディアン・インストルメンツ(Meridian Instruments,Inc.),オケモス(Okemos),MI)分析を使用して検出した。
【図6】活性化CD4+T細胞およびCD8+T細胞上のB7RP1受容体(CRP1と推定される)の発現のFACS(蛍光活性化細胞選別器(Fluorescence−Activated Cell Sorter))分析。マウス脾細胞はPMAとイオノマイシンで12時間活性化させた。各パネル底部に示されるように、B7RP1−Fc融合タンパク質、対照Fcタンパク質(Mock−Fc)、またはPBS(染色なし)に細胞を加えてインキュベートし、洗浄し、次にヤギ抗ヒトFc−FITC複合抗体(GaHuFc−FITC)を加えてインキュベートした。各パネルの左側に示されるように、細胞マーカー抗体(T細胞マーカーCD4およびCD8の)PE複合体、またはアイソタイプ対照抗体(ラットアイソタイプ)PE複合体、またはPBS(染色なし)を加えた。
【図7】B細胞のB7RP1発現のFACS(蛍光活性化細胞選別器)分析。マウス脾細胞のCRP1に対するリガンド(B7RP1と推定される)の発現の蛍光細胞数分析。各パネル底部に示されるように、CRP1−Fc融合タンパク質、対照Fcタンパク質(Mock−Fc)、またはPBS(染色なし)を、細胞を加えてインキュベートし、洗浄し、次にヤギ抗ヒトFc−FITC複合抗体(GaHuFc−FITC)を加えてインキュベートした。各パネルの左側に示されるように、PEを複合化したCD45Rに対する細胞マーカー抗体(CD45RはB細胞マーカーである)またはアイソタイプ対照抗体(ラットアイソタイプ)、またはPBS(染色なし)を加えた。
【図8】腹腔マクロファージのmCRP1リガンドの発現のFACS分析。最初に腹腔細胞は、光散乱特性によってサブセットに分類した(パネルA)。前方(FSC)および側方(SSC)に強く光を散乱することができ、マクロファージのマーカーであるF4/80抗原に対して陽性染色されるため、領域5(R5)にマクロファージを識別した(パネルB)。領域6(R6)のマクロファージは、F4/80抗原による染色強度が低いことに基づいて選択したものであり、CRP1−Fc融合タンパク質によって染色されることが分かった(B7RP1を発現するためと推定される)。
【図9】B7RP1−FC融合タンパク質を使用したT細胞増殖の抑制。マウス脾細胞からのT細胞を、グラフ底部に示されるようにコンカナバリン(Concanavalin)A(Con A)濃度を増加させることによって活性化させた。ConAの非存在下(添加せず)または存在下で脾細胞のT細胞を増加させるために、mCRP1−Fc、mB7RP1、およびmB7.2−Fc融合タンパク質を加えた。96ウェルプレートでのT細胞増殖アッセイに200,000細胞を使用した。グラフ凡例に示されるように、細胞を培地(添加物なし)またはFc融合タンパク質でインキュベートした。42時間後、細胞にH−チミジンを6時間加え、続いて回収し、放射活性を測定した。三重反復試料の平均CPMおよび標準偏差を示している。
【図10】A)リンパ節皮質、傍皮質、および髄質を示している対照マウス#10の正常腸間膜リンパ節。ヘマトキシリン−エオジン(H&E)染色、40倍。B)顕著な濾胞過形成(FH)、傍皮質の膨張、および髄索過形成(MH)を示すWX11マウス#40の顕著に肥大した腸間膜リンパ節。H&E、40倍。C)対照マウス#10の腸間膜リンパ節の髄索(MC)および髄洞(MS)の拡大図。新しいマクロファージを有する髄洞の近傍に大部分が小リンパ球で構成される小さな軸索が注目される。H&E、400倍。D)WX11マウス#40の腸間膜リンパ節の髄索(MC)および髄洞(MS)の拡大図。点在するラッセル小体細胞(矢印)を有する多数の形質細胞で構成される顕著に太い髄索が注目される。H&E、400倍。E)動脈周囲リンパ鞘(PALS)を有する赤脾髄領域および白脾髄領域を示している対照マウス#10の正常脾臓、100倍。挿入図:小リンパ球、マクロファージ、および時々の形質細胞を有する白脾髄の周縁領域の拡大図、400倍。F)PALSおよび濾胞(矢印)を含む肥大した白脾髄領域を有するMX11マウス#6の脾臓、100倍。挿入図:多数の形質細胞および時々のラッセル小体を有する周縁領域の拡大図、400倍。G)2つの2次濾胞が隣接する濾胞間領域(矢印)を有する対照マウス#25のパイエル板を有する回腸、40倍。H)顕著な胚中心と濾胞間組織(矢印)を有する顕著に肥大した濾胞を有するWX11マウス#32のパイエル板を有する回腸、40倍。
【図11】A)リンパ節皮質、傍皮質、および髄質を示している対照マウス#5の正常腸間膜リンパ節。ヘマトキシリン−エオジン(H&E)染色、40倍。B)顕著な濾胞過形成(上部:外部皮質の第2濾胞の列)、膨張した傍皮質(中央)および髄索過形成(下部)が見られるWX11マウス#33の顕著に肥大した腸間膜リンパ節。H&E、40倍。C)抗B220抗体(B細胞マーカー)による対照マウス#10の腸間膜リンパ節の免疫組織化学的染色。強く(褐色)染色された軸索領域と薄い髄索に注目されたい。アビジン−ビオチン複合体(ABC)免疫ペルオキシダーゼ法(DAB色原体、ヘマトキシリン中心染色)を使用して免疫染色を行った。40倍。D)抗B220抗体によるWX11マウス#33の腸間膜リンパ節の免疫組織化学的染色。強く染色された皮質濾胞および髄索(索の成熟形質細胞はB220に対して陰性である)に注目されたい。40倍。E)抗CD3抗体(T細胞マーカー)による対照マウス#10のリンパ節の免疫組織化学的染色。リンパ節の傍皮質領域の免疫染色に注目されたい。40倍。F)抗CD3抗体によるWX11マウス#33のリンパ節の免疫組織化学的染色。リンパ節の肥大し強く染色された傍皮質領域に注目されたい。40倍。
【図12A−1】ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図12A−2】ヒトB7RP1(hB7RP1)のタンパク質コード領域の構造および配列。hB7RP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位にはアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図12B】推定成hB7RP1タンパク質と成熟マウスB7RP1(mB7RP1)タンパク質のアミノ酸配列。
【図13A−1】ヒトCRP1(hCRP1)のタンパク質コード領域の構造および配列。hCRP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位はアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図13A−2】ヒトCRP1(hCRP1)のタンパク質コード領域の構造および配列。hCRP1の予測シグナル配列にはアミノ末端で下線が引かれている。予測シグナルペプチド開裂部位はアスタリスクが付けられている。予測膜貫通配列にはカルボキシ末端に向かって下線が引かれている。
【図13B】hCRP1タンパク質とマウスCRP1(mB7RP1)タンパク質のアミノ酸配列。
【図14】CRP−1は休止記憶T細胞上にある。6〜7月齢のマウスの休止脾細胞は、FITC複合化抗ヒトFc抗体と、CD44(図14A)、CD45RB(図14B)、またはCD69(図14C)のいずれかに対する抗体とPEの複合体によるB7RP−1−Fc標識を使用して二重染色した。
【図15】B7RP−1−Fc融合タンパク質によるT細胞の共刺激。A)異なる量のB7RP−1−Fc(黒四角)、B7.2−Fc(黒丸)、またはOPG−Fc融合タンパク質対照試料(白四角)を抗CD3抗体と併用してT細胞の増殖を誘発させた。種々の濃度の融合タンパク質を使用して、抗ヒトFcFAb2(12.5μg/ml)および抗CD3抗体(0.9μg/ml)で予備コーティングした96ウェルプレートにコーティングした。B7RP−1−FcおよびB7.2−Fcは、最大効果がえられる0.3μg/mlを最大量として用量依存的にT細胞を共刺激する。B)B7RP−1−Fc(黒四角)、B7.2−Fc(黒丸)、非融合Fc(白四角)、またはFcなし(白丸)を、抗CD3抗体(0.85μg/ml)および種々の濃度のウサギ抗B7RP−1−Fcポリクローナル抗体と併用してT細胞の増殖を誘発させた。Fc融合タンパク質は濃度0.3μg/mlで使用し、上記プレートに結合させた。アジュバントに乳化させた抗原の精製B7RP−1−Fcを皮下注射して抗B7RP−1−Fc抗体を産生させ、次にアフィニティ精製した。この抗体にFc融合タンパク質を加えて30分間インキュベートした後に細胞に加えた。抗B7RP−1−Fc抗体は用量依存的に、B7RP−1−Fcにより誘発されるT細胞増殖を特異的に阻害する。
【図16】マウスのコラーゲン誘発関節炎の(A)発生率および(B)重度に対するCRP−1−Fcタンパク質およびB7RP−1−Fcタンパク質の効果。コラーゲン誘発関節炎感受性のB10.RIIIマウスの尾の基部にCFA中の10μgのブタコラーゲンII型を免疫した。100μgの融合タンパク質をマウスに1週間当り2回与えた。Fc融合タンパク質および対照PBSによる処理は図の凡例に示している。
【図17】B7RP−1−Fcトランスジェニックマウスの近位結腸。(A)粘膜、粘膜下組織、筋層、および漿膜を有する腸壁を示す対照マウス#53F(メス)の正常近位結腸。ヘマトキシリン−エオジン(H&E)染色、倍率40倍。(b)顕著な腺肥大、裂傷潰瘍形成、および経壁炎症を有するB7RP−1−Fcトランスジェニックマウス#111Fのびまん性肥厚化近位結腸。H&E、40倍。(C)多病巣性裂傷潰瘍形成および経壁炎症を有するB7RP−1−Fcトランスジェニックマウス#111Fの近位結腸(パネルBと同じ)のより低倍率での図。H&E、20倍。(D)B7RP−1−Fcトランスジェニックマウス#111F裂傷潰瘍および肥大性直腸腺(上述のパネルBおよびCに示される)の拡大図。粘液化膿性滲出物を有する内腔に注意されたい。H&E、100倍。(E)マクロファージ、リンパ球、およびより少数の好中球に囲まれる多核巨細胞が見られるB7RP−1−Fcトランスジェニックマウス#112Fの粘膜下組織の肉芽腫性炎の拡大図。H&E、400倍。(F)粘膜腺の直下のリンパ球、形質細胞、およびより少数の好中球と混合された類上皮マクロファージが見られるB7RP−1−Fcトランスジェニックマウス#112Fの粘膜の肉芽腫性炎の拡大図。H&E、400倍。
【図18】B7RP−1−Fcトランスジェニックマウスの遠位結腸。(A)粘膜、粘膜下組織、筋層、および漿膜を有する腸壁の層が示される対照マウス#53F(メス)の正常遠位結腸。ヘマトキシリン−エオジン(H&E)染色、倍率40倍。(b)顕著な腺肥大および過形成、ならびに散在する腺窩膿瘍が見られるB7RP−1−Fcトランスジェニックマウス#111F(メス)のびまん性肥厚化遠位結腸。H&E、40倍。(C)顕著な腺肥大および過形成の見られるB7RP−1−Fcトランスジェニックマウス#55M(オス)のびまん性肥厚化遠位結腸。H&E、40倍。(D)肥大性結腸腺、限局性リンパ様凝集体、および多くの腺窩膿瘍が見られるB7RP−1−Fcトランスジェニックマウス#112F(メス)のびまん性肥厚化遠位結腸。H&E、40倍。(E)抗CD3抗体(T細胞マーカー)によるB7RP−1−Fcトランスジェニックマウス#112Fの遠位結腸の免疫組織化学的染色。表在粘膜および結腸リンパ様斑の免疫染色に注目されたい。H&E、40倍。(F)腺窩膿瘍(矢印)とB220+B細胞で構成されるリンパ様凝集体(差し込み)とが見られるB7RP−1−Fcトランスジェニックマウス#112Fの結腸粘膜の拡大図。H&E、100倍。
【図19】B7RP−1−Fcトランスジェニックマウスの小腸。(a)粘膜の内腔、絨毛、および腺窩が見られ、粘膜下組織、筋層、および漿膜がその下にある対照マウス#53F(メス)の正常十二指腸。ヘマトキシリン−エオジン(H&E)染色、倍率40倍。(b)顕著な腺窩肥大および過形成、ならびに粘膜固有層の軽度のリンパ形質細胞性浸潤が見られるB7RP−1−Fcトランスジェニックマウス#51F(メス)のびまん性肥厚化十二指腸。H&E、40倍。(c)空腸粘膜に正常な長さの絨毛および腺窩が見られる対照マウス#53F(メス)の正常空腸。H&E、倍率40倍。(d)局所的に高度の腺窩肥大および過形成が見られるB7RP−1−Fcトランスジェニックマウス#51F(メス)の顕著に 肥厚化した空腸粘膜。H&E、40倍。(E)回腸粘膜で正常な長さの絨毛および腺窩が見られる対照マウス#53F(メス)の正常回腸。H&E、倍率40倍。(F)絨毛が局所的に失われ平滑化したB7RP−1−Fcトランスジェニックマウス#231M(オス)の軽度に萎縮した回腸粘膜。H&E、40倍。
【図20】B7RP−1−Fc融合タンパク質はマウスの腫瘍増殖を抑制する。Meth A肉腫細胞をBalb/Cマウスの腹部の皮内に移植した。移植の7日後、10日後、14日後、および17日後に、マウスを媒体(黒ひし形)またはマウスB7RP−1−Fc(灰色三角、実施例7)で処理した。移植後の指定の日に、実施例20に記載のように腫瘍体積を測定した。腫瘍増殖を最長28日まで観察した。各群は8匹のマウスで構成された。
【図21A】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。A)異なる濃度の抗CD3の1次刺激における、抗CD3のみ(黒丸)、0.5μg/mlのB7RP−1 Fc(黒三角)、0.5μg/mlのOPG−Fc(白丸)、および5μg/mlの抗CD28(白三角)による共刺激。データは、抗CD3で1次刺激したT細胞のB7RP−1−Fcによる共刺激が、抗CD28抗体を使用した共刺激と同程度であることを示している。示されるデータは、3つの正常ドナーから単離したT細胞で行った数回の実験の代表的な1つについての三重反復ウェルの平均[3H]TdR±SDが含まれている。
【図21B】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。B)CRP−1−FcによるB7RP−1−FC共刺激の用量依存性阻害。0.3μg/mlおよび0.5μg/mlのB7RP−1−Fcの両方で抗CD3をコーティングしたウェルでT細胞を培養した。連続的に希釈した濃度のCRP−1−Fc(黒丸)またはOPG−Fc(白丸)に、B7RP−1−Fcを加えて30分間予備インキュベートした後に、T細胞を加えた。データは、CRP−1−FcがB7RP−1誘発共刺激を用量依存的に抑制することを示している。%阻害をCRP−1−FcまたはOPG−Fcタンパク質濃度に対してプロットする。示されるデータは、三重反復ウェルで行った3回の実験の平均[3H]TdR±SDであり、2つの正常ドナーで行った実験を代表している。
【図21C】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。C)CHOヒトB7RP−1細胞による共刺激。実施例22に記載されるように、T細胞を末梢血から精製し、抗CD3単独(黒丸)、1×104CHOベクター対照細胞(白丸)、または1×104CHO B7RP−1細胞(黒三角)の存在下で種々の濃度の抗CD3を加えて培養した。データは、膜結合B7RP−1によるT細胞増殖の共刺激が、B7RP−1−Fc融合タンパク質を使用した場合に観察される共刺激都道程度であることを示している。示されるデータは三重反復培養の平均±SDであり、2つの正常ドナーで得られる結果を代表している。
【図21D】ヒトB7RP−1−FcによるT細胞共刺激。実施例21に記載のように、抗CD3およびヒトB7RP−1 Fcを使用して96ウェルプレートをコーティングし、1×105T細胞/ウェル(>98%CD3+)を培養して回収した。D)サイトカイン産生。(図21A)に記載のようにT細胞を培養し、48時間(黒棒)および72時間(灰色棒)で上清を回収した。データは、B7RP−1−Fc共刺激細胞によって産生されるIL−2量(一番上のグラフ)は、抗CD3および対照Fcによって刺激された細胞の産生する量と同様であるが、抗CD28共刺激細胞の産生する量よりは有意に少ないことを示している。またデータは、B7RP−1−Fc共刺激が、IL−10(中央のグラフ)およびIFN−γ(一番下のグラフ)産生を向上させることも示している。
【0024】
発明の詳細な説明
本発明は、本明細書においてCRP1およびB7RP1と称される新規ポリペプチドを提供し、これらはT細胞活性化に関与する受容体−リガンド対を構成している。本発明のポリペプチドをコードするcDNAは、マウス腸上皮内細胞から作製したライブラリーから同定され、CD28ポリペプチドおよびCTLA−4ポリペプチド(CRP1の場合)、またはB7.1ポリペプチドおよびB7.2ポリペプチド(B7RP1の場合)との相同性を基準にしてスクリーニングを行った。
【0025】
CD28関連タンパク質−1、すなわちCRP1は、アミノ末端のシグナル配列と細胞外ドメインと、膜貫通ドメインと、カルボキシ末端細胞内ドメインを有するI型膜貫通タンパク質であると予測される(図1)。CRP1タンパク質の全長は、成熟形態で180アミノ酸である。予測リーダー配列はアミノ酸残基で1〜20付近(開始メチオニンに対して)に及び、成熟タンパク質の細胞外ドメインは残基21〜145付近を含む(実施例1)。予測膜貫通ドメインは残基146〜163付近にわたり、細胞内ドメインは残基164〜200付近を含む。アミノ末端細胞外ドメインはIgループと同様に、推定細胞内および細胞間結合システインが保存される。さらに、B7.1およびB7.2がCD28およびCTLA−4と結合するために重要であると従来知られていた「MYPPPY」モチーフも部分的に保存される。
【0026】
マウスCD28とCTLA−4の間でアミノ酸同一性が26%であると例示されるように、CD28とCTLA−4は相同性が弱い。CRP1とマウスCD28ではアミノ酸同一性が19%であり、CRP1とマウスCTLA−4では同一性が14%である。しかし、残基42、63、83、109、および137(CRP1タンパク質の開始メチオニンが基準。図1A参照)においてマウスCD28、CTLA−4、およびCRP1の間で重要なシステイン残基が保存されている。おおよその成熟タンパク質長さ、およびカルボキシル末端に対する膜貫通領域の位置も、CRP1、CD28、およびCTLA−4で同様である。
【0027】
ヒトCRP1は、図13Aに示されるヌクレオチド配列およびアミノ酸配列を有する膜貫通タンパク質である。予測リーダー配列は残基1〜19付近、または残基1〜20付近にわたる。予測成熟アミノ末端は残基20または21である。成熟アミノ末端は位置21が好ましい。細胞外ドメインは任意の予測成熟アミノ末端からアミノ酸残基140付近に広がり、膜貫通ドメインは残基141〜161付近に及び、細胞内ドメインは残基162〜199付近にわたる。ヒトCRP1タンパク質はマウスタンパク質と69%同一であり、対応するヌクレオチド配列は77%が同一である。ヒトCRP1の配列は、ハトロフ(Hutloff)らNature 397,263−266(1999)に報告されている。
【0028】
B7関連タンパク質−1、すなわちB7RP1は、アミノ末端のシグナル配列および細胞外ドメインと、膜貫通ドメインと、カルボキシ末端細胞内ドメインとを有するI型膜貫通タンパク質であると予測される(図2A)。B7RP1タンパク質の全長は、成熟形態で276アミノ酸である。予測リーダー配列はアミノ酸残基1〜46付近(開始メチオニンに対して)であり、成熟タンパク質の細胞外ドメインは残基47〜279を含む(実施例3)。予測膜貫通ドメインは残基280〜298にひろがり、細胞内ドメインは残基299〜322を含む。B7.1およびB7.2と同様に、B7RP1の細胞外ドメインは2つのIgループを含む。
【0029】
マウスB7.1とB7.2の間のアミノ酸同一性が24%であることで示されるように、B7.1とB7.2は相同性が弱い。B7RP1とマウスB7.1の間のアミノ酸同一性は20%であり、B7RP1とマウスB7.2の間の同一性は19%である。しかし、マウスB7.1、B7.2、およびB7RP1の間で重要なシステイン残基は、残基62、138、185、および242(B7RP1タンパク質の開始メチオニンが基準、図2A)において保存されている。おおよその成熟タンパク質長、およびカルボキシ末端に他意汁膜貫通領域の位置もmB7RP1、B7.1、およびB7.2で同様である。
【0030】
ヒトB7RP1も、Igループ構造を必要とする細胞外ドメインにシステイン残基が保存される膜貫通タンパク質である。図3Aに示されるように予測リーダー配列は、残基1〜18、1〜19、1〜20、1〜21、1〜23、または1〜27の付近を含む。予測成熟アミノ末端は、残基19、20、21、22、24、または28のいずれであってもよい。好ましくはアミノ末端は位置19である。細胞外ドメインは任意の成熟アミノ末端からアミノ酸残基259付近に広がる。予測膜貫通ドメインは残基259〜274に及ぶ。細胞内ドメインは残基275〜302を含む。ヒトB7RP1の全長のヌクレオチド配列およびアミノ酸配列は図12Aに示される。ヒトB7RP1はマウスタンパク質と約43%が同一である。
【0031】
CRP1とB7RP1は互いに結合するが、CRP1はB7RP1関連タンパク質B7.2とは検出可能な程度では結合せず;B7RP1はCRP1関連CD28またはCTLA−4との検出可能な結合を示さない(実施例8)。B7RP1はT細胞増殖を調節することが示されたが、これはB7RP1とCRP1受容体の相互作用のためと推測されている(実施例11)。従ってCRP1とB7RP1は、T細胞の増殖と活性化を調節する新規な経路を表している。
【0032】
B7RP1とCRP1の相互作用は、免疫共刺激、ならびにT細胞の増殖および活性化を増加または減少させることができるような方法で調節することができる。例として、マウスB7RP1に対して産生される抗B7RP1モノクローナル抗体およびポリクローナル抗体は、B7RP1/CRP1相互作用を遮断し、B7RP1−Fc融合タンパク質に誘発されるT細胞増殖も遮断した(実施例17参照)。ヒトCRP−1−Fc融合タンパク質は、ヒトB7RP−1−Fcによって誘発されるヒトT細胞の増殖を遮断した(実施例21)。さらに、CRP1−Fc融合タンパク質を加えることによって、リウマチ様関節炎のマウスモデルの関節炎症状の発生が遅れた(実施例18参照)。B7RP−1/CRP−1共刺激は、B7RP−1−Fc融合タンパク質またはこの経路の他の活性化物質を加えることによって増大させることができる(実施例20)。
【0033】
核酸分子
用語「単離核酸分子」は、必然的に関連する少なくとも1種類の汚染核酸分子がない核酸分子を意味し、好ましくは他のあらゆる汚染哺乳鵜動物核酸分子が実質的にない核酸分子を意味する。
【0034】
用語「対立遺伝子変異体」は、生物の染色体の所与の位置を占める遺伝子の数種類の可能な自然発生的別形態のうちの1つを意味する。
【0035】
用語「スプライス変異体」は、RNA転写において別のイントロン配列のプロセシングによって生成する核酸分子(通常はRNA)を意味する。
【0036】
用語「高ストリンジェンシー条件」は、(1)洗浄に低イオン強度および高温を使用、例えば50℃で0.1×SSC(0.015MのNaCl/0.0015Mのクエン酸ナトリウム)0.1%のNaDodSO4(SDS)、または(2)ハイブリダイゼーション中にホルムアミド、例えば50%(v/v)ホルムアミドなどの変性剤と0.1%ウシ血清アルブミン/0.1%を使用、などの条件を意味する。フィコール(Ficoll)/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム緩衝液(pH6.5)と5×SSC(750mMのNaCl、75mMのクエン酸ナトリウム)を42℃で使用。高ストリンジェンシー条件の別の例は、50%ホルムアミド、5×SSC、50mMのリン酸ナトリウム(pH6.8)、0.1%のピロリン酸ナトリウム、5×デンハート(Denhardt)液、超音波処理サケ精子DNA(50μg/ml)、0.1%のSDS、および10%の硫酸デキストランを42℃で使用し、42℃において0.2×SSCおよび0.1%のSDSで洗浄する。
【0037】
用語「中ストリンジェンシー条件」は、上記よりもストリンジェンシー性の低い洗浄溶液およびハイブリダイゼーション条件(例えば、温度やイオン強度)の使用を含む条件を意味する。中ストリンジェンシー条件の一例は、20%のホルムアミド、5×SSC、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%の硫酸デキストラン、および20μl/mlの変性剪断サケ精子DNAを含む溶液中37℃で終夜インキュベートした後、約37〜50℃で1×SSCを使用してフィルターをろ過する、などの条件である。当業者であれば、プローブ長などの因子の適合に必要な温度、イオン強度、および他の要因の調節方法が分かるであろう。
【0038】
組換えDNA技術は、サムブルック(Sambrook)ら(分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual),コールド・スプリング・パーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press),コールド・スプリング・パーバー(Cold Spring Harbor),NY(1989))および/またはオースベル(Ausubel)ら編著(分子生物学の最新プロトコル(Current Protocols in Molecular Biology),グリーン・パブリッシャー(Green Publishers Inc.)およびワイリー・アンド・サンズ(Wiley and Sons),NY(1994))に記載されており、これらの記載内容全体を本明細書に引用する。
【0039】
本発明は、CRP1ポリペプチドおよびB7RP1ポリペプチドをコードする単離核酸分子を提供する。また本発明は、CRP1ポリペプチドおよびB7RP1ポリペプチドをコードする分子の配列の断片、対立遺伝子変異体、スプライス変異体、または相補的なものである核酸分子も提供する。CRP1またはB7RP1をコードする分子と少なくとも約70%が同一である核酸分子、または中または高ストリンジェンシー条件下でCRP1またはB7RP1をコードする分子とハイブリッド形成する核酸分子も本発明に含まれる。これらの核酸分子は、cDNA、ゲノムDNA、RNA、あるいは部分的または完全に合成した核酸分子であってもよい。好ましい実施態様では、本発明の核酸分子は、CRP1またはB7RP1をコードする核酸分子と少なくとも約75%、80%、85%、90%、または95%が同一である。
【0040】
CRP1またはB7RP1のポリペプチドまたはその断片をコードする遺伝子またはcDNAは、ゲノムライブラリーまたはcDNAライブラリーのハイブリッド形成スクリーングまたはPCR増幅などによって得ることができる。ライブラリーのスクリーニングに有用なプローブまたはプライマーは、同一ファミリーまたは関連ファミリーの遺伝子の他の公知の遺伝子または遺伝子断片の配列情報、例えばシステイン残基の保存配列などのCRP1またはB7RP1関連ポリペプチドに見られる保存モチーフの配列情報を基準にして生成することができる。さらに、CRP1ポリペプチドまたはB7RP1ポリペプチドをコードする遺伝子がある種について同定された場合は、その遺伝子の全体または一部を他の種の相同性遺伝子の同定のプローブとして使用することができる。プローブまたはプライマーは、CRP1遺伝子またはB7RP1遺伝子を発現すると考えられている種々の組織源からcDNAライブラリーをスクリーニングするために使用することができる。
【0041】
オリゴヌクレオチドプローブがcDNAライブラリーまたはゲノムライブラリーのスクリーニングに使用される場合は、以下の2種類の高ストリンジェンシー溶液のうちの1つを使用することができる。これらの溶液の第1は、6×SSCと0.05%のピロリン酸ナトリウムを35℃〜62℃で使用し、この温度はオリゴヌクレオチドプローブの長さに依存する。例えば、14塩基対プローブは35〜40℃で洗浄され、17塩基対プローブは45〜50℃で洗浄され、20塩基対プローブは52〜57℃で洗浄され、23塩基対プローブは57〜63℃で洗浄される。バックグラウンドの非特異的結合が大きい場合にはこの温度を2〜3℃上昇させることができる。第2の高ストリンジェンシー溶液では、オリゴヌクレオチドプローブの洗浄に塩化テトラメチルアンモニウム(TMAC)を使用する。ストリンジェンシー洗浄溶液の1つは、3MのTMAC、50mMのTris−HCl(pH8.0)、および0.2%のSDSである。この溶液が使用される洗浄温度は、プローブの長さの関数である。例えば、17塩基対プローブは約45〜50℃で洗浄される。CRP1またはB7RP1のポリペプチドまたはそれらの断片をコードする遺伝子を作製する別の手段は、エンジェル(Engels)ら(Agnew.Chem.Intl.Ed.,28:716−734(1989))に記載されるものなどの当業者には公知の方法を用いる化学合成を使用することである。これらの方法としては、特に、リン酸トリエステル法、ホスホロアミダイト法、およびH−ホスホネート法による核酸合成が挙げられる。このような化学合成の好ましい方法は、標準的ホスホロアミダイト化合物を使用するポリマー支持合成である。通常、CRP1ポリペプチドまたはB7RP1ポリペプチドをコードするDNAは長さが数百ヌクレオチドとなる。約100ヌクレオチドを超える核酸は、これらの方法を使用して数個の断片として合成することができる。次にこれらの断片を結合させて、CRP1ポリペプチドまたはB7RP1ポリペプチドの全長を形成することができる。通常、ポリペプチドのアミノ末端をコードするDNA断片はメチオニン残基をコードするATGを有する。このメチオニンは、成熟形態のCRP1ポリペプチドまたはB7RP1ポリペプチドに存在してもしなくてもよく、これは宿主細胞で産生されるこのポリペプチドが細胞から分泌されるように計画されるかどうかに依存する。
【0042】
それにも拘らず、生物活性であるポリペプチドをそれ自身コードしない、CRP1またはB7RP1核酸分子、断片、及び/または誘導体は、哺乳類の組織または体液試料中のCRP1あるいはB7RP1のDNAあるいは対応するRNAの存在に対して、定性的あるいは定量的に試験する診断アッセイにおける交雑プローブとして有用である。
【0043】
ある場合には、天然のCRP1あるいはB7RP1ポリペプチドの核酸及び/またはアミノ酸変異体を製造することが望ましい。核酸変異体は、プライマーが所望の点突然変異を持つ、部位特異的突然変異誘発、PCR増幅、または他の適切な方法を用いて産生されてよい(Sambrookら,上記,及び突然変異誘発の手法の記述については、Ausubelら,上記を参照)。上記のEngelsらにより述べられた方法を用いての化学合成を使用して、このような変異体を製造してもよい。当業者に知られている他の方法も使用してもよい。
好ましい核酸変異体は、一定の宿主細胞におけるCRP1とB7RP1ポリペプチドの最適発現に対して変更されたコドンを含むものを含んでいる。特定のコドン変更は、タンパク質と宿主細胞の選択に依存する。一つの場合には、一定の宿主細胞の高度に発現された遺伝子において優先的に使用されるコドンを選択することにより、このような「コドン最適化」を行うことができる。高度に発現された細菌の遺伝子のコドン選択に対する「Ecohigh.Cod」等のコドン頻度表を組込むコンピューターアルゴリズムが使用されてよく、これはUniversity of Wisconsin Packageバージョン9.0、Genetics Computer Group、Madison、WIにより提供される。他の有用なコドン頻度表は、「Celegans_high.cod」、「Celegans_lowcod」、「Drosophila_high.cod」、「Human_high.cod」、「Maize_high.cod」、及び「酵母_high.cod」を含む。他の好ましい変異体は、野生のタイプに比べて上記に述べたような保存アミノ酸変化(例えば、天然のアミノ酸の側鎖の荷電または極性は異なるアミノ酸による置換により実質的に変えられない)をコードするもの、新規なグリコシル化及び/またはリン酸化部位を生成するように設計されたもの、または現存のグリコシル化及び/またはリン酸化部位を削除するように設計されたものである。
【0044】
標準連結法を用いて、CRP1またはB7RP1ポリペプチドをコードする遺伝子、cDNA、またはこれらの断片を適切な発現または増幅ベクター中に挿入することができる。このベクターは、通常、使用される特別な宿主細胞において機能的であるように選択される(すなわち、このベクターは、この遺伝子の増幅及び/またはこの遺伝子の発現が起こることができるように、宿主細胞仕組みと適合している)。CRP1またはB7RP1ポリペプチドをコードする遺伝子、cDNA、またはこれらの断片は、原核、酵母、昆虫(バキュロウイルス系)及び/または真核の宿主細胞において増幅/発現されてよい。この宿主細胞の選択は、CRP1またはB7RP1ポリペプチドまたはこれらの断片がグリコシル化及び/またはリン酸化されるかどうかに一部依存する。そうであるとすれば、酵母、昆虫、または哺乳類の宿主細胞が好ましい。
【0045】
通常、宿主細胞のいずれかで使用される発現ベクターは、プラスミド維持及び挿入ヌクレオチド配列のクローニングと発現のための配列を含む。「隣接配列」とまとめて呼ばれるこのような配列は、プロモーター及びエンハンサー、複製エレメントの起源、転写停止エレメント、供与体と受容体またはスプライス部位を含む完全なイントロン配列、シグナルペプチド配列、リボソーム結合部位エレメント、ポリアデニル化配列、発現されるポリペプチドをコードする核酸を挿入するためのポリリンカー領域及び選択マーカーエレメント等の他の調節エレメントを含む。これらのエレメントの各々は下記に述べられる。場合によっては、このベクターは、「タグ」配列、CRP1またはB7RP1ポリペプチドコーディング配列の5’または3’末端に位置するすなわちオリゴヌクレオチド分子を含んでよい。このオリゴヌクレオチド分子は、ポリヒス(ヘキサヒス等の)、または市販の抗体が存在する、FLAG、HA(血球凝集素インフルエンザウイルス)またはmyc等の他の「タグ」をコードする。このタグは、通常、このポリペプチドの発現時にこのポリペプチドに融合され、宿主細胞からのCRP1またはB7RP1ポリペプチドをアフィニティ精製する手段として機能することができる。例えば、このタグに対する抗体を親和マトリックスとして用いるカラムクロマトグラフィにより、アフィニティ精製を行うことができる。場合によっては、引き続いて、このタグは、しかるべきペプチダーゼ用いる等の種々の手段により精製されたCRP1またはB7RP1ポリペプチドから除去され得る。
【0046】
当業者によれば、ヒト免疫グロブリンヒンジとFc領域は、CRP1またはB7RP1ポリペプチドのN−末端またはC−末端のいずれかに融合されてよい。引き続くFc融合タンパク質は、タンパク質A親和カラムを使用することにより精製され得る。免疫グロブリンFc領域は、インビトロで長い薬物動力学的半減期を示すことが知られていて、Fc領域に融合されたタンパク質は、非融合品と比較して、インビトロで実質的に大きい半減期を示すことが見出されている。また、このFc領域への融合により、一部の分子の生物活性に有用であるこの分子の二量化及び/または多量化が可能になる。
【0047】
この隣接配列は、同種(すなわち、宿主細胞と同じ種及び/または菌株からの)、異種(すなわち、宿主細胞種または菌株と別の種からの)、雑種(すなわち、一つの以上のソースからの隣接配列の組み合わせ)、合成によるものであってよく、またはこれは、未変性のCRP1またはB7RP1核酸隣接配列であってよい。従って、この隣接配列が宿主細胞仕組みで機能的であり、活性化され得るという前提であれば、隣接配列のソースは、いかなる単一細胞の原核または真核生物体、いかなる脊椎あるいは無脊椎生物体、またはいかなる植物であってよい。
【0048】
本発明のベクターにおいて有用である隣接配列は、当分野においてよく知られている方法のいずれによって得られてもよい。通常、CRP1またはB7RP1核酸隣接配列以外の、ここにおいて有用な隣接配列は、マッピングにより及び/または制限エンドヌクレアーゼ消化により以前に同定されていて、このようにして、適切な制限エンドヌクレアーゼを用いて適当な組織ソースから単離され得る。いくつかの場合には、この隣接配列の完全なヌクレオチド配列は知られているかもしれない。ここで、この隣接配列は、核酸合成またはクローニングについて上述した方法を用いて合成されてよい。
【0049】
この隣接配列のすべてまたは一部のみが知られている場合には、PCRを用いて及び/または同じあるいはもう一つの種からの好適なオリゴヌクレオチド及び/または隣接配列断片によりゲノムライブラリーをスクリーニングすることによりこれを得てよい。
【0050】
この隣接配列が知られていない場合には、隣接配列を含むDNAの断片は、例えば、コーディング配列または更にもう一つの遺伝子を含む更に大きな片のDNAから単離されてよい。単離は、一つあるいはそれ以上の慎重に選ばれた酵素を用いて、適当なDNA断片を単離する、制限エンドヌクレアーゼ消化により行われてよい。消化の後、所望の断片は、アガロースゲル精製、Qiagen(登録商標)カラムまたは当業者に知られた他の方法により単離されてよい。この目的を果たすための好適な酵素の選択は、当分野分野の通常の技術を持つ者には容易に明白になるであろう。
【0051】
複製エレメントの起源は、通常、購入される原核の発現ベクターの一部であり、宿主細胞においてこのベクターの増幅を助ける。しかるべきコピー数迄のこのベクターの増幅は、ある場合には、CRP1あるいはB7RP1ポリペプチドの最適な発現に重要であり得る。選択したベクターが複製部位の起源を含まないならば、これは既知の配列に基づいて化学的に合成され、このベクター中に連結されてよい。
【0052】
この転写停止エレメントは、通常、CRP1あるいはB7RP1ポリペプチドのコーディング配列の末端の3’に位置し、CRP1またはB7RP1ポリペプチドの転写を停止させる機能をする。通常、原核細胞中の転写停止エレメントは、G−Cに富む断片とそれに続くポリT配列である。このエレメントはライブラリーから容易にクローンされるか、あるいは更にベクターの一部として購入される一方、また、上述の方法等の核酸合成の方法を用いて、容易に合成され得る。
【0053】
選択マーカー遺伝子エレメントは、選択的培養基中で育成された宿主細胞の生き残りと成長に必要なタンパク質をコードする。典型的な選択マーカー遺伝子は、(a)原核の宿主細胞に対する抗生物質または他の毒素、例えば、アンピシリン、テトラサイクリン、またはカナマイシンに耐性を賦与し、(b)細胞の栄養要求性欠乏症を補い、または(c)複雑な培地から得られない重要な栄養物を供給するタンパク質をコードする。好ましい選択マーカーは、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、及びテトラサイクリン耐性遺伝子である。
【0054】
普通、シャイン−ダルガルノ配列(原核細胞)またはKozak配列(真核細胞)と呼ばれるリボソーム結合エレメントは、mRNAの翻訳開始に通常必要である。このエレメントは、このプロモーターに3’に、また合成されるべきCRP1またはB7RP1ポリペプチドのコーディング配列に5’に通常位置する。シャイン−ダルガルノ配列は変るが、通常、ポリプリン(すなわち、高A−G含量を有する)である。多数のシャイン−ダルガルノ配列が同定されており、その各々は、上記に示し、そして原核のベクターに使用される方法を用いて、容易に合成され得る。
【0055】
CRP1またはB7RP1ポリペプチドが宿主細胞から分泌されるのが望ましい場合には、宿主細胞からポリペプチドの移行を方向付けするために、シグナル配列が使用されてよい。CRP1またはB7RP1膜貫通ドメインは、また、宿主膜への結合を防ぐために、突然変異または欠失により不活性化される。通常、このシグナル配列は、CRP1またはB7RP1遺伝子またはcDNAのコーディング領域に、あるいはCRP1またはB7RP1遺伝子コーディング領域の5’末端に直接に位置される。多数のシグナル配列が同定されており、選ばれた宿主細胞中で機能性であるシグナル配列のいずれもCRP1あるいはB7RP1遺伝子またはcDNAと一緒に使用されてよい。それゆえ、シグナル配列は、CRP1あるいはB7RP1遺伝子またはcDNAに同種あるいは異種であってよく、CRP1あるいはB7RP1ポリペプチド遺伝子またはcDNAに同種あるいは異種であってよい。加えて、このシグナル配列は、上記に示した方法を用いて化学的に合成されてよい。
【0056】
大多数の場合において、シグナルペプチドの存在によるこの宿主細胞からのポリペプチドの分泌は、結果としてこのポリペプチドからアミノ末端メチオニンを生成する。
【0057】
多くの場合、CRP1あるいはB7RP1遺伝子またはcDNAの転写は、このベクター中の一つあるいはそれ以上のイントロンの存在により増大する。これは、CRP1あるいはB7RP1ポリペプチドが真核の宿主細胞、特に哺乳類の宿主細胞中で産生する場合に特に当てはまる。使用されるイントロンは、使用される遺伝子が完全な長さのゲノム配列またはこれの断片である、CRP1あるいはB7RP1遺伝子内で天然に生じるものであってよい。このイントロンは、この遺伝子(または大多数のcDNAに関して)内で天然に生じるものでない場合には、このイントロンは他の源から得られてよい。このイントロンは転写されて有効になるので、5’隣接配列とCRP1あるいはB7RP1遺伝子に対するこのイントロンの位置は、一般に重要である。従って、発現ベクター中に挿入されるCRP1あるいはB7RP1遺伝子がcDNA分子である場合には、このイントロンに好ましい位置は、転写スタート部位の3’及びポリA転写停止配列の5’である。好ましくは、このイントロンは、このコーディング配列を妨害しないように、cDNAの一方または他方(すなわち、5’または3’)に位置する。本発明を実施するするのに、挿入される宿主細胞と適合するという前提ならば、いかなるウイルス、原核及び真核の(植物または動物)生物体をも含む、いかなる源からのいかなるイントロンも使用しうる。また、ここには合成イントロンも含まれる。場合によっては、一つ以上のイントロンがベクターに使用しうる。
【0058】
上記に示した一つあるいはそれ以上のエレメントが使用されるべきベクター中に既に存在しない場合には、これらを個別に得て、このベクターに連結してよい。このエレメントの各々を得るために使用される方法は、当業者によく知られている。
【0059】
本発明を実施するための好ましいベクターは、細菌、昆虫、及び哺乳類の宿主細胞と適合するものである。このようなベクターは、とりわけ、pCRII、pCR3、及びpcDNA3.1(Invitrogen Company,San Diego,CA)、pBSII(Stratagene Company,LaJalla,CA)、pETl5b(Novagen,Madison、WI)、pGEX(Pharmacia Biotech,Piscataway,NJ)、pEGFP−N2(Clontech,Palo Alto,CA)、pETL(BlueBacII Invitrogen)、及びpFastBacDual(Gibco/BRL,Grand Island,NY)を含む。
【0060】
このベクターが構築され、完全な長さをコードする核酸分子または切断されたCRP1あるいはB7RP1ポリペプチドがこのベクターの適当な部位に挿入された後、増幅及び/またはポリペプチド発現に好適な宿主細胞にできあがったベクターを挿入してよい。
【0061】
宿主細胞は、原核宿主細胞(大腸菌等の)または真核宿主細胞(酵母細胞、昆虫細胞、または脊椎動物の細胞等の)であってよい。この宿主細胞は、適切な条件下で培養した場合、CRP1あるいはB7RP1ポリペプチドを合成することができ、引き続いて、培養基(宿主細胞が培地中にそれを分泌するならば)から、あるいはそれを産生する宿主細胞からそれを直接に収集(それが分泌されないならば)することができる。収集後、CRP1あるいはB7RP1ポリペプチドは、モレキュラーシーブクロマトグラフィー、親和クロマトグラフィー等の方法を用いて、精製され得る。
【0062】
CRP1あるいはB7RP1ポリペプチドの産生に適切な宿主細胞の選択は、所望の発現レベル、グリコシル化またはリン酸化等の活性に必要とされるポリペプチド変性、または生物活性分子への折り畳みの容易さ等の種々の因子に依存する。
【0063】
好適な細胞または細胞株は、チャイニーズハムスター卵巣細胞(CR0)、ヒト胎児腎臓(HEK)293または293T細胞、または3T3細胞等の哺乳類の細胞であってよい。好適な哺乳類の宿主細胞の選択及び転換、培養、増幅、スクリーニング及び産物生産及び精製のための方法は、当分野で知られている。他の好適な哺乳類の細胞株は、サルCOS−1及びCOS−7細胞株とCV−1細胞株である。更なる例示の哺乳類の宿主細胞は、転換細胞株を含む、霊長類細胞株及び齧歯類細胞株を含む。正常な2倍体細胞、1次組織、並びに1次外植体のインビトロ培養に由来する細胞菌株もまた好適である。候補細胞は、遺伝子型として選択遺伝子欠乏であっても、あるいは主に作用する選択遺伝子を含んでよい。他の好適な哺乳類の細胞株は、限定ではないがマウス神経芽細胞腫N2A細胞、HeLa、マウスL−929細胞、Swiss、Balb−c由来の3T3株、またはNIHマウス、BHKまたはHaKハムスター細胞株を含む。
【0064】
宿主細胞として同様に有用なのは、細菌細胞である。例えば、大腸菌の種々の菌株(例えば、HB101、DH5α、DH10、及びMC1061)がバイオテクノロジーの分野で宿主細胞としてよく知られている。B.subtilis、Pseudomonas spp.、他のBacillus spp.、Streptomyces spp.などの種々の菌株もこの方法で使用されてよい。
【0065】
当業者に知られている酵母細胞の多数の菌株もまた本発明のポリペプチドを発現する宿主細胞として入手できる。
【0066】
加えて、所望の場合には、昆虫細胞系が本発明の方法で利用されてよい。このような系は、例えばKittsら(Biotechniques、14:810−817(1993))、Lucklow(Curr.Opin.Biotechnol.,4:564−572(1993))及びLucklowら(J.Virol.,12:4566−4579(1993))に記述されている。好ましい昆虫細胞は、Sf−9とHi5(Invitrogen,Carlsbad,CA)である。
【0067】
塩化カルシウム、電気穿孔法、マイクロインジェクション、リポフェクションまたはDEAE−デキストラン法等の方法を用いて、発現ベクターの選ばれた宿主細胞中への「転換」または「トランスフェクション」を行ってよい。選ばれた方法は、一部、使用されるべき宿主細胞のタイプの機能である。これらの方法及び他の好適な方法は、当業者によく知られていて、例えば、上記のSambrookらに示されている。
【0068】
発現ベクターにより転換あるいはトランスフェクションされる宿主細胞は、当業者によく知られた標準培地を用いて培養されてよい。この培地は、普通、この細胞の成長と生き残りに必要なすべての栄養素を含有する。大腸菌細胞を培養するための好適な培地は、例えば、Luria Broth(LB)及び/またはTerrific Broth(TB)である。真核細胞を培養するのに好適な培地は、RPMI 1640、MEM、DMEMであり、これらのすべては、培養される特別な細胞株にように必要とされるのに従って、血清及び/または成長因子により補われる。昆虫培養に好適な培地は、必要に従って、酵母オレエート、ラクトアルブミン加水分解物、及び/またはウシ胎仔血清により補われるグレース(Grace)の培地である。
【0069】
通常、転換された細胞の選択成長に有用な抗生物質または他の化合物のみがサプルメントとして培地に添加される。使用される化合物は、宿主細胞を転換したプラスミド上に存在する選択マーカーエレメントにより決定付けられる。例えば、この選択マーカーエレメントがカナマイシン耐性である場合には、培養基に添加される化合物はカナマイシンである。
【0070】
この宿主細胞中に産生されるCRP1あるいはB7RP1ポリペプチドの量は、当分野において知られている標準的な方法を用いて評価され得る。このような方法は、限定ではないが、ウエスタンブロット分析、SDS−ポリアクリルアミドゲル電気泳動、非変性ゲル電気泳動、HPLC分離、免疫沈澱、及び/またはDNA結合ゲルシフトアッセイ等の活性アッセイを含む。
【0071】
宿主細胞から分泌されるようにCRP1またはB7RP1ポリペプチドを設計した場合には、大部分のポリペプチドが細胞培養基中に見出されてよい。このように製造されたポリペプチドは、この細胞からの分泌時に除去されるので、通常、アミノ末端メチオニンを保持しない。しかしながら、CRP1またはB7RP1ポリペプチドが宿主細胞から分泌されない場合には、それは、細胞質及び/または核(真核宿主細胞に対して)中に、またはサイトソル(グラム陰性バクテリア宿主細胞に対して)中に存在し、アミノ末端メチオニンを有してよい。
【0072】
種々の手法を用いて溶液からのCRP1またはB7RP1ポリペプチドの精製を行うことができる。ヘキサヒスチジン(CRP1またはB7RP1/ヘキサヒス)等のタグ、またはFLAG(Eastman Kodak Co.,NewHaven,CT)またはmyc(Invitrogen,Carlsbad,CA)等の他の小ペプチドをそのカルボキシルまたはアミノ末端で含むように、このポリペプチドを合成した場合、このカラムマトリックスがタグに対して、あるいは直接的にポリペプチドに対して高親和性を持つならば(すなわち、CRP1またはB7RP1ポリペプチドを特異的に認識するモノクローナル抗体)、タグを付けたポリペプチドを親和カラムに通すことにより、1段階プロセスで本質的に精製してよい。例えば、ポリヒスチジンは、大きな親和性と特異性でニッケルに結合し、このようにしてニッケルの親和カラム(Qiagen(登録商標)ニッケルカラム等の)をCRP1またはB7RP1/ポリヒスの精製に使用することができる(例えば、Ausubelら編、Current Protocols in Molecular Biology,Section10.11.8,John Wiley & Sons,New York(1993)を参照)。
【0073】
タグを付けずにCRP1またはB7RP1ポリペプチドを製造し、抗体が入手できない場合には、精製によく知られた方法を使用することができる。このような方法は、限定ではないが、イオン交換クロマトグラフィー、モレキュラーシーブクロマトグラフィー、HPLC、ゲル溶離と組み合わせた未変性ゲル電気泳動、及び合成的等電点電気泳動(「Isoprime」マシン/手法、Hoefer Scientific)を含む。ある場合には、2つあるいはそれ以上のこれらの方法を組み合わせて、純度の増大を達成しうる。
【0074】
CRP1あるいはB7RP1ポリペプチドが主として細胞内に見出されることが期待される場合には、当業者に知られた任意の標準的な手法を用いて、この細胞内材料(グラム陰性バクテリア用の封入体を含む)を宿主細胞から抽出することができる。例えば、フレンチプレス、均質化、及び/または高周波による分解とそれに続く遠心分離により、この宿主細胞を溶解して、ペリプラズム/細胞質の内容物を放出させることができる。
【0075】
CRP1またはB7RP1ポリペプチドがサイトソル中に封入体を形成した場合には、この封入体は、時には内及び/または外の細胞膜に結合することができ、このようにして遠心分離後主としてペレット材料中に見出される。次に、このペレット材料は、pHの両極端で、あるいは洗浄剤、グアニジン、グアニジン誘導体、尿素、または尿素誘導体等のカオトロピック剤により、アルカリ性pHでジチオトレイトールまたは酸性pHでトリスカルボキシエチルホスフィン等の還元剤の存在下で処理され、この封入体を放出、分割、及び可溶化することができる。次に、ゲル電気泳動、免疫沈澱などを用いて、溶解した形のこのポリペプチドを分析することができる。CRP1またはB7RP1ポリペプチドを単離することを所望する場合には、下記及びMarstonら、(Meth.Enz.,182:264−275(1990))に示すような標準方法を用いて、単離を行ってよい。ある場合には、CRP1あるいはB7RP1ポリペプチドは単離時に生物活性でなくてよい。「再折りたたみ」あるいはこのポリペプチドを3級構造に変換し、そしてジスルフィド結合を生成する種々の方法を使用して、生物活性を取り戻すことができる。このような方法は、可溶化ポリペプチドを通常7以上のpHを持つ溶液と特別な濃度の適切なカオトロープの存在下で接触することを含む。また、大多数の場合、再折りたたみ/酸化溶液は、還元剤またはタンパク質のシステイン架橋の形成においてジスルフィド混合を起こさせる特別なレドックス電位を生じる還元剤と対応する酸化形を特定の比で含む。普通に使用されるレドックス対の一部は、システイン/シスタミン、グルタチオン(GSH)/ジチオビスGSH、塩化第2銅、ジチオトレイトール(DTT)/ジチアンDTT、2−メルカプトエタノール(bME)/ジチオ−b(ME)を含む。多数の例において、再折りたたみの効率を増大するために、共溶媒が必要であり、この目的に使用される更に普通の試剤は、グリセロール、種々の分子量のポリエチレングリコール、及びアルギニンを含む。
【0076】
また、化学合成方法(固相ペプチド合成等の)により、Merrifieldらにより示されたもの(J.Am.Chem.Soc.,85:2149(1963))、Houghtenら(Proc Natl Acad.Sci.USA,82:5132(1985))、及びStewart及びYoung(Solid Phase Peptide Synthesis,Pierce Chemical Co.,Rockford,IL(1984))等の当分野において知られた手法を用いて、CRP1またはB7RP1ポリペプチド、断片、及び/またはこれらの誘導体を製造してもよい。このようなポリペプチドは、アミノ末端上にメチオニンの有無のいずれで合成されてよい。これらの文献に示される方法を用いて、化学合成されたCRP1あるいはB7RP1ポリペプチドまたは断片を酸化して、ダイサルファイド架橋を形成してよい。CRP1あるいはB7RP1ポリペプチドまたは断片は、組み換えで製造された、あるいは天然源から精製されたCRP1あるいはB7RP1ポリペプチドに匹敵する生物活性を有すると予期され、このように、組み換えあるいは天然のCRP1あるいはB7RP1ポリペプチドと入れ替えて使用してもよい。
【0077】
ポリペプチド
「CRP1あるいはB7RP1ポリペプチド」という用語は、図1A(配列番号2)、図2A(配列番号7)または図3A(配列番号12)及びここで説明されたすべての関連ポリペプチドのアミノ酸配列を持つポリペプチドを指す。関連ポリペプチドは、対立変異体、スプライス変異体、断片、誘導体、置換、欠失、及び挿入変異体、融合ポリペプチド、及びオルトログを含む。このような関連ポリペプチドは、成熟ポリペプチド、すなわち、シグナルペプチドを欠いたポリペプチドであってよい。CRP1あるいはB7RP1ポリペプチドは、これらを製造する方法に依って、アミノ末端メチオニンであってよく、あるいはそうでなくてもよい。
【0078】
「CRP1あるいはB7RP1ポリペプチド断片」という用語は、図1A(配列番号2)、図2A(配列番号7)または図3A(配列番号12)に示すような、CRP1あるいはB7RP1ポリペプチドの完全長さ未満であるアミノ酸配列のペプチドまたはポリペプチドを指す。このような断片は、アミノ末端での切断、カルボキシ末端での切断、及び/またはポリペプチド配列への内部の欠失の結果から生じうる。このようなCRP1あるいはB7RP1ポリペプチド断片は、アミノ末端メチオニンの有無のいずれで製造されてよい。加えて、CRP1あるいはB7RP1ポリペプチド断片は、天然由来のスプライス変異体、他のスプライス変異体、及び天然に生じるインビボ活性の結果生じる断片であってよい。好ましいCRP1あるいはB7RP1ポリペプチド断片は、機能性膜貫通ドメインを欠き、CRP1あるいはB7RP1のいずれかの細胞外ドメインの一部または全部を構成するCRP1あるいはB7RP1の可溶性の形態を含む。
【0079】
「CRP1あるいはB7RP1ポリペプチド変異体」という用語は、図1A(配列番号2)、図2A(配列番号7)または図3A(配列番号12)に示すCRP1またはB7RP1ポリペプチドアミノ酸配列に比較して、アミノ酸配列が一つあるいはそれ以上のアミノ酸配列置換、欠失、及び/または付加を含むCRP1あるいはB7RP1ポリペプチドを指す。従って、CRP1あるいはB7RP1ポリペプチドに対するDNA配列から変るDNA配列を持つ対応するCRP1及びB7RP1ポリペプチド核酸分子変異体から、このようなCRP1あるいはB7RP1ポリペプチド変異体を製造することができる
ここで使用される時、「CRP1あるいはB7RP1ポリペプチド誘導体」という用語は、例えば、一つあるいはそれ以上の水溶性ポリマー、N−結合あるいはO−結合炭水化物、糖、リン酸塩、及び/または分子が野生型のCRP1あるいはB7RP1ポリペプチドに天然に結合していない、他のこのような分子の添加により化学的に変性したCRP1あるいはB7RPlポリペプチド、変異体、またはこれらの断片を指す。誘導体は、CRP1あるいはB7RP1ポリペプチドに天然に結合した一つあるいはそれ以上の化学基の欠失を更に含む。
【0080】
ここで使用される時、「生物活性のCRP1あるいはB7RP1ポリペプチド」、「生物活性のCRP1あるいはB7RP1ポリペプチド断片」、「生物活性のCRP1あるいはB7RP1ポリペプチド変異体」、及び「生物活性のCRP1あるいはB7RP1ポリペプチド誘導体」という用語は、CRP1あるいはB7RP1特有の少なくとも一つの活性を持つCRP1あるいはB7RP1ポリペプチドを指す。一つの活性はCRP1へのB7RP1の結合である。他の活性は、CRP1あるいはB7RP1がT−細胞増殖及び/または活性化を刺激する能力である。
【0081】
「オルトログ」という用語は、種から同定されるポリペプチドに対応するポリペプチドを指す。例えば、マウス及びヒトB7RP1ポリペプチドはオルトログと考えられる。
「成熟したアミノ酸配列」という用語は、リーダー配列を欠くポリペプチドを指す。
【0082】
「単離されたポリペプチド」という用語は、その自然な環境中で見出される少なくとも一つの汚染性のポリペプチドを含まない、そして好ましくはいかなる他の汚染性の哺乳類のポリペプチドも実質的に含まないポリペプチを指す。
【0083】
「同一性」という用語は、当分野において知られているように、配列を比較することにより決定される2つあるいはそれ以上の核酸分子または2つあるいはそれ以上のポリペプチドの配列間の関係である。当分野においては、「同一性」は、また、場合に応じて、ヌクレオチドあるいはアミノ酸配列の列間の一致により決定されるような、ポリペプチドまたは核酸分子配列間の配列関連性の程度を意味する。「同一性」は、特別なコンピュータープログラム(すなわち、「アルゴリズム」)により扱われるギャップ配列により2つあるいはそれ以上の配列の間の同一性一致のパーセントを測定する。
【0084】
「類似性」という用語は、関連する概念を指すが、対照的に、「同一性」とは、同一性一致と保存的置換一致の双方を含む。保存的置換はポリペプチドに当てはまり、核酸分子には当てはまらないので、類似性は、ポリペプチド配列の比較を扱うのみである。2つのポリペプチド配列が例えば、10/20同一のアミノ酸を持ち、残りがすべて非保存的置換である場合には、同一性と類似性のパーセントは双方とも50%である。同じ試料において、保存的置換がある、更に5つの位置がある場合には、パーセント同一性は50%のままであるが、パーセント類似性は75%である(15/20)。それゆえ、保存的置換がある場合には、2つのポリペプチド配列の間の類似性の程度は、これらの2つの配列の間のパーセント同一性よりも高い。「保存的」アミノ酸置換はここでは下記に表Iを参照して記述されている。表Iに基づくと、保存アミノ酸置換は、同じグループ、例えば、塩基性、酸性、非帯電極性、及び非極性から選ばれる交互のアミノ酸である。例えば、アルギニンに対する保存アミノ酸置換は、リジン及びヒスチジンである。
【0085】
同一性と類似性は、限定ではないが、Computational Molecular Biology,Lesk,A.M編,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.編,Academic Press,New York,1993;Computer Analysis of Sequence Data,Part1,Griffin,A.M.,及びGriffin,H.G.編,Humana Press,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,Academic Press,1987;及びSequence Analysis Primer,Gribskov,M.及び Devereux,J.編,M.Stockton Press,New York,1991;及びCarillo,H.,及びLipman,D.,SIAM J.Applied Math.,48:l073(1988)に記述されているものを含む、既知の方法により容易に計算できる。
【0086】
同一性及び/または類似性を求める好ましい方法は、試験する配列の間の最大一致を与えるように設計される。同一性と類似性を求める方法は、公に入手できるコンピュータープログラムにコード化されている。2つの配列間の同一性と類似性を求める好ましいコンピュータープログラムの方法は、限定ではないが、GAP(Devereux,J.ら,Nucleic Acids, Research 12(1):387(1984);Genetics Computer Group,University of Wisconsin,Madison,WI)、BLASTP,BLASTN,及びFASTA(Atschul,S.F.ら,J.Molec.Biol.215:403−410(1990)を含む、GCGプログラムパッケージを含む。BLAST Xプログラムは、National Center for Biotechnology Information(NCBI)及び他の源(BLAST Manual,Altschul,S.ら、NCB NLM NIH Bethesda,MD 20894;Altschul,S.ら,J.Mol.Biol.215:403−410(1990))から公に入手できる。また、同一性を求めるのに、よく知られたSmith Watermanアルゴリズムを使用してもよい。
【0087】
例として、コンピューターアルゴリズムギャップ(Genetics Computer Group,University of Wisconsin,Madison,WI)を用いて、パーセント配列同一性を求めるべき2つのポリペプチドは、各アミノ酸(このアルゴリズムにより求められるような「一致したスパン」)の最適マッチングに対して並べられる。ギャップオープニングペナルティ(3X平均対角線として計算される;「平均対角線」は、使用されている比較マトリックスの対角線の平均である。「対角線」は、特別な比較マトリックスにより各々の完全なアミノ酸一致に割り当てられた得点または数であり、そして、ギャップエクステンションペナルティ(通常、ギャップオープニングペナルティの1/10倍である)、並びにPAM 250またはBLOSUM 62等の比較マトリックスがアルゴリズムと一緒に使用される。標準比較マトリックス(PAM 250比較マトリックスに対しては、Dayhoffら,Atlas of Protein Sequence and Structure,5巻,補遺3(1978)を参照;BLOSUM 62比較マトリックスに対しては、Henikoffら,Proc.Nati.Acad.Sci USA,89:10915−10919(1992)を参照)もまたアルゴリズムにより使用される。
【0088】
ポリペプチド配列比較に対する好ましいパラメーターは、次を含む。
【0089】
アルゴリズム:Needleman及びWunsch、J.Mol.Biol.48:443453(1970)
比較マトリックス:HenikoffからのBLOSUM 62及びHenikoff,Proc.Natl.Acad.Sci.USA 89:10915−10919(1992)
ギャップペナルティ:12
ギャップ長さペナルティ:4
類似性のしきい値:0
GAPプログラムは上記パラメーターにより有用である。この上述のパラメーターは、ギャップアルゴリズムを用いたポリペプチド比較(末端ギャップに対する無ペナルティと一緒に)のデフォルトパラメーターである。
【0090】
核酸分子配列比較の好ましいパラメーターは次を含む。
【0091】
アルゴリズム:Needleman及びWunsch,J.Mol Biol.48:443−453(1970)
比較マトリックス:一致=+10、ミスマッチ=0
ギャップペナルティ:50
ギャップ長さペナルティ:3
また、GAPプログラムは上記パラメーターによっても有用である。この上述のパラメーターは、核酸分子比較のデフォルトパラメーターである。
【0092】
他の例示のアルゴリズム、ギャップオープニングペナルティ、ギャップエクステンションペナルティ、比較マトリックス、類似性のしきい値などを当業者によりProgram Manual,Wisconsin Package,バージョン9,1997年9月に示したものを含めて使用してよい。行われるべき特別な選択は、DNA対DNA、タンパク質対タンパク質、タンパク質対DNA等の行われるべき特定の比較、及び加えて、この比較が一対の配列(GAPまたはBestFitが一般的に好ましい)の間であるか、あるいはまたは一つの配列と配列の大きなデータベース(FASTAまたはBLASTAが好ましい)の間であるかどうかにに依存する。
【0093】
少なくとも約70パーセント同一であるポリペプチドは、野生型のCRP1あるいはB7RP1ポリペプチドと比較して、一つあるいはそれ以上のアミノ酸置換、欠失、及び/または付加を通常有する。好ましい実施形態においては、ポリペプチドは、CRP1あるいはB7RP1ポリペプチドに対して約75%、80%、85%、90%または95%の同一性を有する。通常、未変性の残渣の置換は、ポリペプチドの正味の全帯電、極性、または疎水性に殆ど、あるいは全く影響を及ぼさないように、アラニン、または保存アミノ酸のいずれかである。保存的置換は下記の表Iに示される。
【0094】
【表1】
【0095】
CRP1あるいはB7RP1ポリペプチド誘導体が本発明により提供される。一つの実施形態においては、CRP1あるいはB7RP1ポリペプチドがポリマーに結合している化学変性されたCRP1あるいはB7RP1ポリペプチド組成物は、本発明の範囲内に含まれている。選ばれたポリマーは、通常水溶性であり、そこで、ポリマーが結合しているタンパク質は生理的環境等の水性環境において沈殿しない。選ばれたポリマーは、通常、アシル化のための活性エステルまたはアルキル化のためのアルデヒド等の単一の反応性基を持つように変性されて、重合度は本発明の方法に提供されるように制御されてよい。このポリマーは、いかなる分子量であってよく、分岐あるいは非分岐であってよい。本発明の範囲内に入れられているのは、ポリマーの混合物である。好ましくは、最終製品製造の治療への使用のためには、このポリマーは医薬として許容されるものである。
【0096】
水溶性ポリマーまたはこれらの混合物は、例えば、ポリエチレングリコール(PEG)、モノメトキシ−ポリエチレングリコール、デキストラン、セルロース、または他の炭水化物ベースのポリマー、ポリ−(N−ビニルピロリドン)ポリエチレングリコール、プロピレングリコールホモポリマー、ポリプロピレンオキサイド/エチレンオキサイドコポリマー、ポリオキシエチル化ポリオール(例えば、グリセロール)及びポリビニルアルコールからなる群から選ばれてよい。
【0097】
このアシル化反応のためには、選ばれたポリマーは単一の反応性エステル基を有しなければならない。還元アルキル化のためには、選ばれたポリマーは単一の反応性アルデヒド基を有しなければならない。好ましい反応性アルデヒドは、水安定であるポリエチレングリコールプロピオンアルデヒド、またはモノC1−C10アルコキシまたはこれらのアリールオキシ誘導体(米国特許第5,252,714号を参照)である。
【0098】
CRP1あるいはB7RP1ポリペプチドのペギル化(Pegylation)は、例えば次の文献に記述されているような、当分野において知られている任意のペギル化反応により行われてよい。Focuson Growth Factors 3:4−10(1992);EP 0 154 316;及びEP 0 401 384。好ましくは、このペギル化は、アシル化反応またはアルキル化反応により反応性ポリエチレングリコール分子(または同族の反応性水溶性ポリマー)により下記に述べるように行われる。
【0099】
ここでの使用に特に好ましい水溶性ポリマーは、PEGと略記されるポリエチレングリコールである。ここで使用される時、ポリエチレングリコールは、モノ(C1−C10)アルコキシ−またはアリールオキシ−ポリエチレングリコール等の他のタンパク質を誘導体化するのに使用されたいかなる形のPEGを包含するとの意味である。
【0100】
一般に、生物活性物質と活性化されたポリマー分子とを反応するのに使用されるいかなる好適な条件下でも、化学的誘導体化を行ってよい。ペギル化されたCRP1及びB7RP1ポリペプチドを製造する方法は、一般に(a)ポリペプチドとポリエチレングリコール(PEGの反応性エステルあるいはアルデヒド誘導体)とを、CRP1あるいはB7RP1ポリペプチドが一つあるいはそれ以上のPEG基に結合される条件下で反応し、そして(b)反応生成物を得るステップを含んでなる。一般に、アシル化反応の最適な反応条件は、既知のパラメーターと所望の結果をベースにして決められる。例えば、PEG:タンパク質の比が大きい程、ポリペギル化された製品のパーセンテージは大きい。
【0101】
一般に、CRP1あるいはB7RP1ポリマーコンジュゲートの投与により軽減あるいは調節される状態は、非共役のCRP1あるいはB7RP1ポリペプチドについてここで述べたものを含む。しかしながら、ここに開示されているコンジュゲートは、追加の活性、増進あるいは低下した生物活性、または非誘導体化された分子に比較して、増加あるいは減少した半減期等の他の特性を有してよい。
【0102】
CRP1あるいはB7RP1ポリペプチド、断片変異体、及び誘導体は、単独で、一緒に、あるいは他の医薬組成物と組み合わせて使用されてよい。CRP1あるいはB7RP1ポリペプチド、断片、変異体、及び誘導体は、治療される適応に適切であるように、サイトカイン、成長因子、抗生物質、抗炎症剤、及び/または化学治療剤と組み合わせて使用されてよい。
【0103】
本発明は、CRP1あるいはB7RP1の選択的結合剤を提供する。選択的結合剤は、CRP1あるいはB7RP1に特異性を持つ分子を指し、タンパク質、ペプチド、核酸、炭水化物、リピッドまたは小分子量化合物を含んでよい。選択的結合剤は、CRP1またはB7RP1のいずれかと相互作用し、次には、CRP1のB7RP1への結合を調節する。一つの実施形態においては、選択的結合剤は、CRP1のB7RP1への結合を部分的あるいは完全に阻止し、免疫同時刺激活性等のCRP1またはB7RP1の少なくとも一つの生物活性を部分的あるいは完全に抑制する。もう一つの実施形態においては、選択的結合剤は抗体である。この抗体は、CRP1またはB7RP1のいずれかと免疫反応性であってよく、好ましくはB7RP1と免疫反応性である。本発明の更にもう一つの実施形態においては、B7RP1と反応性である抗体は、B7RP1上のエプチオープ(eptiope)に結合して、CRP1への結合が部分的あるいは完全に阻止され、免疫同時刺激活性等のB7RP1の少なくとも一つの生物活性が部分的あるいは完全に抑制される。部分的に抑制されるという用語は、少なくとも検出し得るレベルの抑制が起こったことを意味する。完全に抑制されるという用語は、抑制の更なる増加が起こらなかったことを意味する。
【0104】
当分野で知られている方法を用いて抗体を製造するのに、CRP1あるいはB7RP1ポリペプチド、断片、変異体、及び/または誘導体を使用してよい。このように、CRP1あるいはB7RP1ポリペプチド、並びにこのような抗体の反応性断片と反応する抗体もまた本発明の範囲内として考慮される。この抗体は、ポリクローナル、モノクローナル、組み換え、キメラ、単一鎖及び/または両特異的であってよい。通常、この抗体またはこの断片は、ヒト起源であるかあるいは、「ヒト化されている」、すなわち、または患者に投与した場合に、この抗体への免疫反応を防止あるいは最少化するように製造される。この抗体断片は、Fab、Fab’等の本発明のCRP1及びB7RP1ポリペプチドと反応性であるいかなる断片であってよい。また、本発明により提供されるのは、任意のCRP1あるいはB7RP1ポリペプチドまたはこれらの断片を抗原として選ばれた哺乳類に与え、続いて哺乳類の細胞(例えば、脾臓細胞)をしかるべきガン細胞と融合して、不死化された細胞株を知られた手法により作り出すことにより生じるヒドリドマス(hydridomas)である。本発明のヒトCRP1あるいはB7RP1ポリペプチドの全部または一部に特異的なこのような細胞株及び抗体を生じるのに使用される方法も本発明により包含される。
【0105】
本発明のモノクローナル抗体は、所望の生物活性を示す限り(米国特許第4,816,567;Morrisonら,Proc.Natl.Acad.Sci.81,6851−6855(1985))、重及び/または軽鎖の一部が特別な種に由来する、あるいは特別な抗体クラスまたはサブクラスに属する抗体における対応する配列と同一あるいは相同であり、一方、鎖の残りがもう一つの種に由来する、あるいはもう一つの抗体クラスまたはサブクラス、並びにこのような抗体の断片に属する抗体における対応する配列と同一あるいは相同である、「キメラ」抗体を含む。
【0106】
好ましい実施形態においては、キメラ抗−CRP1あるいはB7RP1抗体は「ヒト化された」抗体である。非ヒト抗体をヒト化する方法は当分野においてよく知られている。一般に、ヒト化された抗体は、非ヒトである源からその中に導入される、一つあるいはそれ以上のアミノ酸残渣を有する。当分野において知られている方法(Jonesら,Nature 321,522−525(1986);Riechmannら、Nature,332,323−327(1988);Verhoeyenら、Science 2391534−1536(1988))に従って、齧歯類の相補性決定領域(CDR)にヒト抗体の対応する領域を置き換えることにより、ヒト化を行うことができる。
【0107】
また、本発明により包含されているのは、完全にヒトの抗−CRP1または抗−B7RP1抗体である。内生免疫グロブリン産生の不存在下でヒト抗体のレパートリーを産生する能力のあるトランスジェニック動物(例えば、マウス)のCRP1あるいはB7RP1抗原による免疫化により、このような抗体を製造してよい。例えば、Jakobovitsら,Proc.Natl.Acad.Sci.90、2551−2555(1993);Jakobovitsら,Nature 362,255−258(1993)を参照のこと。また、ファージディスプレイライブラリ(phage−display libraries)においてヒト抗体を製造することもできる(Hoogenboomら,J.Mol.Biol.227,381(1991);Marksら,J.Mol.Biol.222.,581(1991))。
【0108】
本発明の選択的結合剤を使用して、CRP1のB7RP1への結合を調節し、CRP1及びB7RP1により媒介される、免疫同時刺激等の少なくとも一つの生物活性を調節してよい。このような選択的結合剤の例は、CRP1またはB7RP1との抗体免疫反応性である。CRP1とB7RP1ポリペプチドのその結合パートナーへの結合を抑制する等、この抗体を治療的に使用してよい。インビトロ及びインビトロ診断の目的でラベルされた形などでこの抗体を更に使用して、体液または細胞試料中のCRP1及びB7RP1ポリペプチドの存在を検出してよい。
【0109】
医薬組成物と投与
CRP1あるいはB7RP1ポリペプチドの医薬組成物は、本発明の範囲内である。このような組成物は、医薬として許容し得るキャリアとの混合物での治療として有効な量のポリペプチドまたは断片、変異体、または誘導体を含んでなってよい。好ましい実施形態においては、医薬組成物は、CRP1あるいはB7RP1ポリペプチドを、CRP1あるいはB7RP1細胞外ドメインの一部または全部を含んでなる、可溶性の形態として含んでなる。通常、CRP1及びB7RP1ポリペプチド治療用化合物は、一つあるいはそれ以上の生理学的に許容し得るキャリア、賦形剤、または希釈剤と一緒に、精製されたポリペプチド、断片、変異体、または誘導体を含んでなる、組成物の形で投与される。このキャリア材料は、好ましくは哺乳類への投与用に溶液中の普通の他の材料と共に補われる、注射用の水であってよい。中性の緩衝された生理食塩水または血清アルブミンと混合した生理食塩水が例示の適切なキャリアである。好ましくは、この製品は、適切な賦形剤(例えば、スクロース)を用いて凍結乾燥品として配合される。他の標準キャリア、希釈剤、及び賦形剤は、所望に応じて入れられてよい。他の例示的組成物は、ソルビトールまたはこれの好適な代替品を更に含む、約pH7.0−8.5のトリスバッファー、または約pH4.0−5.5の酢酸塩バッファーを含んでなる。
【0110】
CRP1あるいはB7RP1医薬組成物は、非経口で投与され得る。あるいは、この組成物は、静脈注射であるいは皮下注射で投与されてよい。全身的に投与する場合には、本発明の使用での治療用組成物は、発熱物質を含まない、非経口で許容し得る水溶液の形であってよい。pH、アイソトニック性、安定性などに関するこのような医薬として許容し得るタンパク質溶液の製造は、当分野の技術内である。
【0111】
所望の程度の純度を持つ選ばれた組成物を、オプションの生理的に許容し得るキャリア、賦形剤、または安定剤(Remington’s Pharmaceutical Sciences、18版Edition,A.R.Gennaro編,Mack Publishing Company(1990))と凍結乾燥されたケーキまたは水溶液の形で混合することにより、本発明を実施するのに有用なCRP1あるいはB7RP1ポリペプチド組成物の治療用配合物を貯蔵用に製造してよい。許容し得るキャリア、賦形剤または安定剤は、服用者に非毒性であり、好ましくは使用される投与量と濃度で不活性であり、そしてリン酸塩、クエン酸塩、または他の有機酸等のバッファー;アスコルビン酸等の酸化防止剤;低分子量ポリペプチド;、血清アルブミン、ゼラチン、または免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニンまたはリジン等のアミノ酸;モノサッカライド、ジサッカライド、及びグルコース、マンノース、またはデキストリンを含む他の炭水化物;EDTA等のキレート化剤;マニトールまたはソルビトール等のシュガーアルコール;ナトリウム等の対イオンを形成する塩;及び/またはトウイーン、プルロニックスまたはポリエチレングリコール(PEG)等の非イオン性界面活性剤を含む。
【0112】
治療に使用されるべき有効な量のCRP1あるいはB7RP1ポリペプチド組成物は、例えば、CRP1及びB7RP1ポリペプチドが使用される適応、投与経路、及び患者の状態等の治療目標に依存する。従って、療法士が最適な治療効果を得るのに必要とされるように、投与量を求めて、投与経路を調節することが必要である。通常の1日の投与量は、上述の因子に依って、約0.1μg/kgから100mg/kgあるいはそ以上迄の範囲であってよい。通常、臨床医は、所望の効果を得る投与量に到達する迄この組成物を投与する。この組成物は、それゆえ、時間にわたって、単一投与として、あるいは2あるいはそれ以上の投与(同じ量のCRP1あるいはB7RP1ポリペプチドを含んでも、あるいは含まなくともよい)として、あるいインプランテーション器具またはカテーテルにより連続的な輸液として投与されてよい。
【0113】
更なる試験を行うのに従って、種々の患者における種々の状態を治療するための適切な投与量レベルに関する情報が出てきて、通常の熟練作業者は、治療の状況、治療を受けている疾患のタイプ、服用者の年齢と全般的な健康を考慮して、適当な投与を確定することが可能であろう。
【0114】
インビボ投与に使用されるべきCRP1あるいはB7RP1ポリペプチド組成物は無菌でなければならない。これは、無菌の濾過膜により濾過することにより容易に行われる。この組成物を凍結乾燥された場合には、これらの方法を用いる殺菌は、凍結乾燥と再構成に先立って、あるいは続いて行われてよい。非経口投与用のこの組成物は、通常、凍結乾燥された形で、あるいは溶液で貯蔵される。
【0115】
この治療用組成物は、一般に無菌アクセスポートを持つ容器、例えば、皮下注射針で突き刺すことができるストッパーを持つ静脈注射溶液バックまたはバイアル中に入れられる。
【0116】
この組成物の投与経路は、知られている方法、例えば、皮下、腹腔内、大脳内(柔細胞内)、大脳室内、筋肉内、眼球内、動脈内、あるいは病変内(intralesional)の経路による、あるいは持続放出系または場合によってはカテーテルの使用を含む埋め込み器具による経口注入または輸液と一致する。所望の場合には、この組成物は、輸液、丸薬注入により、またはインプランテーション器具により連続的に投与されてよい。
【0117】
あるいはまたはそれに加えて、この組成物は、埋め込みを経て、CRP1及びB7RP1ポリペプチドを吸収した膜、スポンジ、または他の適切な材料の患部中に局所的に投与されてよい。埋め込み器具を使用する場合には、この器具はいかなる好適な組織または器官中にも埋め込まれ、CRP1あるいはB7RP1ポリペプチドの送達は、丸薬を経た、または連続投与を経た、または連続的な輸液を用いるカテーテルを経た器具を直接的に通されてよい。CRP1あるいはB7RP1ポリペプチドは、持続放出配合物または調合物で投与されてよい。持続放出調合物の好適な例は、半透過性ポリマーマトリックスを成形された物品、例えば、フィルム、またはマイクロカプセルの形で含む。持続放出マトリックスは、ポリエステル、ハイドロゲル、ポリラクチド(U.S.3,773,919、EP58,481)、L−グルタミン酸のコポリマー及びガンマエチルL−グルタメート(Sidmanら,Biopolymers,22:547−556(1983))、ポリ(2−ヒドロキシエチル−メタアクリレート)(LangerらJ.Biomed.Mater.Res,15:167−277(1981)]及びLanger、Chem.Tech.,12:98−105(1982))、エチレンビニルアセテート(Langerら、上記)またはポリ−D(−)−3−ヒドロキシ酪酸(EP133,988)を含む。持続放出組成物は、また、当分野において知られているいくつかの方法(例えば、Eppsteinら,Proc.Nati.Acad.Sci.USA,82:3688−3692(1985);EP36,676;EP88,046;EP143,949)のいずれかにより製造することができる、リポソームも含む。
【0118】
ある場合には、CRP1あるいはB7RP1ポリペプチド組成物をエクスビボ(ex vivo)の方法で使用することが望ましいかもしれない。ここで、患者から取り出した細胞、組織、または器官がCRP1あるいはB7RP1ポリペプチド組成物を曝らし、その後、この細胞、組織、または器官を患者に埋め込みにより戻す。
【0119】
他の場合には、CRP1あるいはB7RP1ポリペプチドを、ここで述べたもの等の方法を用いて、遺伝子的に設計されたしかるべき細胞を患者の中に埋め込むことにより送達して、ポリペプチド、断片、変異体、または誘導体を発現し、分泌してよい。このような細胞は、動物あるいはヒト細胞であってよく、患者自身の組織から、あるいはまたはヒトまたは非ヒトのいずれかのもう一つの源から送達されてよい。場合によっては、この細胞は不死化されてよい。しかしながら、免疫応答の機会を減少させるために、この細胞をカプセル化して、取巻く組織の浸潤を避けることが好ましい。カプセル化材料は、タンパク質生成物を放出させるが、患者の免疫系により、あるいは取巻く組織からの他の有害因子により細胞の破壊を防止する通常生体適合性の、半透過性ポリマーの囲いまたは膜である。
【0120】
細胞の膜被包のために使用される方法は当業者に熟知されており、被包細胞の調製と患者への移植は多大の実験を必要とせずに実現できる。例えば、米国特許第4,892,538号、同第5,011,472号及び同第5,106,627号参照。生存細胞を被包するための系がPCT WO91/10425号(Aebischerら)に述べられている。リポソーム担体、生体内腐食性粒子又はビーズのような様々な他の持続性又は制御された送達方法を製剤するための手法も当業者に既知であり、例えば米国特許第5,653,975号に記述されている。被包された又は被包されていない細胞を患者の適当な体組織又は器官に移植することができる。
【0121】
上述したように、細胞製剤を1個又はそれ以上のCRP1又はB7RP1ポリペプチド、変異体、誘導体及び/又は断片で処理することが望ましいと考えられる。これは、例えば骨髄細胞のようなT細胞を含む細胞を、細胞膜透過性の形態の当該ポリペプチド、変異体、誘導体又は断片に直接接触させることによって実施できる。例えば、T細胞の機能を活性化するためにT細胞を含む細胞をB7RP1ポリペプチドに接触させることができ、そのように処理した細胞を患者に移植する。
【0122】
その代わりに、遺伝子治療を用いることもできる。遺伝子治療が適用できる1つの方法は、「遺伝子治療DNA構築物」を形成するために構成的又は誘導性プロモーターに機能的に連結されうるCRP1又はB7RP1遺伝子(CRP1又はB7RP1ポリペプチドをコードするゲノムDNA、cDNA、及び/又は合成DNA、あるいはその断片、変異体又は誘導体)を使用して、「遺伝子治療DNA構築物」を形成することである。プロモーターは、構築物を挿入する細胞又は組織型において活性であることを条件として、内因性CRP1又はB7RP1遺伝子に相同性あるいは非相同性のいずれでもよい。遺伝子治療DNA構築物の他の成分は、任意に、必要に応じて、位置特異的組込みのために設計されたDNA分子(例えば相同的組換えのために有用な内因性隣接配列)、組織特異的プロモーター、エンハンサー又はサイレンサー、親細胞に比べて選択的優越性を提供することができるDNA分子、形質転換細胞を同定するための標識として有用なDNA分子、負の選択系、細胞特異的結合物質(例えば、細胞標的に関して)、細胞特異的インターナリゼーション因子、及びベクターによる発現を高めるための転写因子ならびにベクターの製造を可能にする因子を含みうる。
【0123】
次にこの遺伝子治療DNA構築物を患者の細胞に挿入することができる(エクスビボ又はインビボで)。遺伝子治療DNA構築物を挿入するための1つの方法は、ウイルスベクターによるものである。遺伝子治療DNA構築物の送達のために遺伝子治療において典型的に使用される適当なウイルスベクターは、限定されることなく、アデノウイルス、アデノ関連ウイルス、単純ヘルペスウイルス、レンチウイルス、乳頭腫ウイルス、及びレトロウイルスベクターを含む。レトロウイルスベクターのようなこれらのベクターの一部は、遺伝子治療DNA構築物を患者の細胞の染色体DNAに送達し、遺伝子治療DNA構築物は染色体DNAに入り込むことができる;他のベクターはエピソームとして機能し、遺伝子治療DNA構築物は細胞質中にとどまる。遺伝子治療ベクターの使用は、例えば米国特許第5,672,344号、同第5,399,346号に述べられている。
【0124】
ウイルスベクターを使用せずに遺伝子治療DNA構築物を患者の細胞に送達するための代替的手段は、限定されることなく、リポソームを介した移入、裸のDNAの直接注入、レセプタを介した移入(リガンド−DNA複合体)、エレクトロポレーション、リン酸カルシウム沈殿法、及び微粒子撃ち込み(例えば「遺伝子ガン」)を含む。例えば米国特許第4,970,154号、WO96/40958号、5,679,559号、5,676,954号及び5,593,875号参照。
【0125】
遺伝子治療を通して細胞における内因性CRP1又はB7RP1ポリペプチドの発現を高めるためのもう1つの方法は、1つ又はそれ以上のエンハンサー要素をCRP1又はB7RP1ポリペプチドプロモーター内に挿入することであり、エンハンサー要素はCRP1又はB7RP1ポリペプチド遺伝子の転写活性を高めるために働くことができる。使用されるエンハンサー要素は、遺伝子を活性化することを所望する組織に基づいて選択され、その組織においてプロモーターの活性化をもたらすことが知られているエンハンサー要素が選択される。例えば、CPR1又はB7RP1ポリペプチドをT細胞において「発動(turned on)」させようとする場合は、lckプロモーターエンハンサー要素が使用できる。この場合には、標準的なクローニング手法を使用して、付加する転写要素の機能的部分を、CRP1又はB7RP1ポリペプチドプロモーター(及び任意に、ベクター、5’及び/又は3’隣接配列、等々)を含むDNAの断片に挿入することができる。その後、「相同的組換え構築物」として知られるこの構築物をエクスビボ又はインビボのいずれかで所望する細胞に挿入することができる。
【0126】
内因性プロモーターのヌクレオチド配列を改変することにより、遺伝子治療を使用してCRP1又はB7RP1ポリペプチドの発現を低下させることができる。そのような改変は、典型的には相同的組換え法を通して実施される。例えば、不活性化のために選択したCPR1又はB7RP1遺伝子のプロモーターの全部又は一部を含むDNA分子を構築して、転写を調節するプロモーターの断片を取り除く及び/又は置き換えることができる。ここでは、標準的な分子生物学手法を使用して、プロモーターのTATAボックス又は転写活性化因子の結合部位が欠失されうる。そのような欠失はプロモーター活性を阻害し、それによって対応するCRP1またはB7RP1遺伝子の転写を抑制することができる。プロモーターにおけるTATAボックス又は転写活性化因子結合部位の欠失は、TATAボックス及び/又は活性化因子結合部位の活性が低下する又は完全に不活性となるように、1個又はそれ以上のヌクレオチドの置換、欠失及び/又は挿入を通して1個又はそれ以上のTATAボックス及び/又は転写活性化因子結合部位のヌクレオチドが突然変異を起こす、CRP1又はB7RP1ポリペプチドプロモーターの全部又は該当部分(調節されるCRP1又はB7RP1遺伝子と同じ又は関連種からの)を含むDNA構築物を作製することによって実施できる。典型的には、改変されたプロモーターセグメントの天然(内因性)5’及び3’隣接領域に対応する、少なくとも約500塩基のDNAを同時に含むこの構築物は、直接又は上述したようなウイルスベクターを通して、適切な細胞内に挿入されうる(エクスビボ又はインビボのいずれかで)。典型的には、細胞のゲノムDNAへの構築物の組込みは相同的組換えを通して行われ、プロモーター構築物内の5’及び3’隣接DNA配列は、内因性染色体DNAへのハイブリダイゼーションを通して改変されたプロモーター領域の組込みを助けるように働くことができる。
【0127】
1個又はそれ以上のCRP1又はB7RP1ポリペプチドを阻害することを所望する場合、他の遺伝子治療法も使用しうる。例えば、選択したCRP1又はB7RP1ポリペプチドの少なくとも一部に相補的な配列を持つアンチセンスDNA又はRNA分子を細胞に挿入することができる。典型的には、そのような各々のアンチセンス分子は、各々の選択したCRP1又はB7RP1遺伝子の開始部位(5’末端)に相補的である。その後アンチセンス分子が対応するCRP1又はB7RP1ポリペプチドmRNAにハイブリダイズするときに、このmRNAの翻訳が妨げられる。
【0128】
その代わりに、遺伝子治療を使用して、1個又はそれ以上のCRP1又はB7RP1ポリペプチドの優性の負の阻害因子を創造することができる。この状況では、各々の選択したCRP1又はB7RP1ポリペプチドの突然変異体完全長又はトランケートポリペプチドをコードするDNAを調製し、上述したようなウイルス又は非ウイルス法を用いて患者の細胞に挿入することができる。そのような各々の突然変異体は、典型的にはその生物学的役割において内因性ポリペプチドと競合するように設計される。
【0129】
作用物質と拮抗物質
本発明はまた、CRP1とB7RP1のいずれか又は両方の分子の活性を調節する、CRP1又はB7RP1の作用物質と拮抗物質を提供する。作用物質と拮抗物質は、CRP1へのB7RP1の結合を変化させる被験分子から同定されうる。
【0130】
「被験分子」という用語は、CRP1又はB7RP1ポリペプチドに結合し、それによってCRP1へのB7RP1の結合を変化させる能力についての評価の対象である分子を指す。好ましくは、被験分子は少なくとも約106Mの親和定数で結合する。
【0131】
CRP1へのB7RP1の結合を測定するためには様々なアッセイが使用できる。これらのアッセイを使用して、CRP1へのB7RP1の結合の速度又は度合を上昇させる又は低下させる能力に関して被験分子をスクリーニングすることができる。1つの種類のアッセイでは、CRP1ポリペプチド、好ましくは細胞外ドメインのような可溶性形態のCRP1を、マイクロタイタープレートのウエルの底に付着させることによって固定する。次に放射標識B7RP1と被験分子を1つずつ(いずれかの順序で)又は同時にウエルに加えることができる。インキュベーション後、ウエルを洗浄し、シンチレーション計数器を使用してB7RP1のCRP1タンパク質への結合の度合を調べるために放射能を測定することができる。典型的には、分子を一定の濃度範囲にわたって試験し、結果の評価の正確さを確認するために試験アッセイの1つ又はそれ以上の要素を欠く一連の対照ウエルが使用できる。これに代わる方法は、タンパク質の「位置」を逆にする、すなわちB7RP1をマイクロタイタープレートのウエルに固定し、被験分子と放射標識CRP1をインキュベーションして、CRP1の結合の度合を測定することを含む(例えば、Current Protocols in Molecular Biology、Ausubelら編集、John Wiley & Sons,New York,NY[1995]の18章参照)。
【0132】
放射標識に代わる方法として、CRP1又はB7RP1をビオチンに結合し、比色定量によって又はストレプタビジンの蛍光標識によって検出することができる、ホースラディシュペルオキシダーゼ[HRP]又はアルカリホスファターゼ[AP]のような酵素に連結されたストレプタビジンを使用してビオチニル化タンパク質の存在を検出することができる。ビオチンに結合したCRP1又はB7RP1に対して特異的な抗体も使用でき、AP又はHRPに連結された酵素連結ストレプタビジンとのインキュベーション後、検出することができる。
【0133】
CRP1及びB7RP1はまた、アガロースビーズ、アクリルビーズ又は他の種類のそのような不活性物質への接着によって固定しうる。基質−タンパク質複合体を相補的タンパク質と被験化合物を含む溶液に入れる;インキュベーション後、遠心分離によってビーズを沈澱させ、上述した方法を用いてCRP1とB7RP1間の結合の量を評価することができる。その代わりに、基質−タンパク質複合体をカラムに固定し、被験分子と相補的タンパク質をカラムに通過させることができる。次に上記に述べた手法、すなわち放射標識、抗体結合、等のいずれかを用いてCRP1とB7RP1間の複合体の形成を評価することができる。
【0134】
CRP1/B7RP1複合体の形成を増加させる又は減少させる被験分子を同定するために有用なもう1つのタイプのインビトロアッセイは、Biacoreアッセイ系(Pharmacia,Piscataway,NJ)のような表面プラスモン共鳴検出器システムである。Biacoreシステムは製造者のプロトコールを使用して実施しうる。このアッセイは基本的に、検出器内に位置するデキストラン被覆センサーチップへのCRP1又はB7RP1のいずれかの共有結合を含む。次に、被験化合物と他の相補的タンパク質を同時に又は連続的にセンサーチップが入ったチェンバー内に注入し、結合する相補的タンパク質の量を、センサーチップのデキストラン被覆側と物理的に関連する分子量の変化に基づいて評価することができる;分子量の変化は検出器システムによって測定できる。
【0135】
一部の場合には、CRP1/B7RP1複合体の形成を増加させる又は減少させるのに使用するため、2個又はそれ以上の被験化合物を一緒に評価することが望ましいと考えられる。これらの場合、そのような付加被験化合物を最初の被験化合物と同時に又は最初の被験化合物に続いて加えることにより、上記に述べたアッセイを容易に修正することができる。アッセイの残りの段階は上述した通りである。
【0136】
上に述べたようなインビトロアッセイは、CRP1又はB7RP1による複合体形成への作用に関して多数の化合物を速やかにスクリーニングするために好都合に使用しうる。ファージディスプレイ、合成ペプチド及び化学合成ライブラリーにおいて生成される化合物をスクリーニングするためにアッセイを自動化することができる。
【0137】
CRP1及びB7RP1の複合体形成を上昇させる又は低下させる化合物はまた、いずれかのポリペプチドを発現する細胞及び細胞系統を使用した細胞培養においてもスクリーニングしうる。細胞と細胞系統はどのような哺乳類からも入手しうるが、好ましくはヒト又は他の霊長類、イヌ、げっ歯類ソースからのものである。表面上にCRP1を発現する細胞へのB7RP1の結合を被験化合物の存在下又は不在下で評価し、結合の度合を、例えばB7RP1に対するビオチニル化抗体を使用するフローサイトメトリーによって測定することができる。細胞培養アッセイは、上述したタンパク結合アッセイにおいて陽性と判定された化合物をさらに評価するために好都合に使用しうる。
【0138】
治療用途
本発明のポリペプチド、及びその作用物質と拮抗物質は、T細胞の機能を調節するために使用しうる。作用物質と拮抗物質は、CRP1及び/又はB7RP1活性を調節し、T細胞機能に関連する活性、例えばT細胞の活性化のような、CRP1又はB7RP1タンパク質の少なくとも1つの活性を上昇させる又は低下させる分子を含む。作用物質又は拮抗物質は、CRP1又はB7RP1のいずれかと相互作用し、それによってそれらの活性を調節する、タンパク質、ペプチド、炭水化物、脂質、又は低分子量分子のような補因子でありうる。潜在的なポリペプチド作用物質又は拮抗物質は、当該タンパク質の細胞外ドメインの一部又は全部を含む、可溶性又は膜結合形態のCRP1又はB7RP1と相互作用する抗体を含む。CRP1又はB7RP1発現を調節する分子は、典型的には発現のアンチセンス調節因子として働きうる、CRP1又はB7RP1タンパク質をコードする核酸を含む。
【0139】
CRP1又はB7RP1ポリペプチド及びその作用物質と拮抗物質は、自己免疫疾患、移植片の生存、腫瘍細胞の増殖を抑制するための免疫細胞の活性化、T細胞依存性B細胞仲介疾患、及び癌遺伝子免疫療法の治療において使用しうる。1つの実施態様では、CRP1及び/又はB7RP1機能の拮抗物質又は阻害因子は、慢性免疫細胞機能不全を有する疾患において症状を軽減するために有益であると考えられる。全身性エリテマトーデス、慢性関節リウマチ、免疫性血小板減少性紫斑病(ITP)、及び乾癬のような自己免疫疾患は、CRP−1/B7RP−1の拮抗物質又は阻害因子で治療しうる。さらに、炎症性腸疾患(クローン病及び潰瘍性大腸炎)、グレーヴズ病、橋本甲状腺炎、及び糖尿病のような慢性炎症性疾患も、CRP−1/B7RP−1に対する阻害因子で治療できる。実施例18の中で述べるように、げっ歯類の慢性関節リウマチ疾患モデルにおいて、CRP−1−Fcは疾患の発症を抑制し、B7RP−1−Fcは発症を促進する。このモデルにおけるこれら正反対の作用は、B7RP−1−Fcタンパク質に関する作用性の役割とCRP−1−Fcタンパク質についての拮抗性の役割を裏付ける。その結果はまた、この経路の操作によってT細胞応答をどのように調節することができるか、そして慢性関節リウマチの進行におけるこの経路の重要性を示している。さらに、実施例19で述べるように、インビボでのB7RP−1−Fcの発現はトランスジェニックマウスにおいて炎症性腸疾患(IBD)表現型を刺激する。この実施例は、腸の炎症発生におけるB7RP−1/CRP−1の役割を裏付ける。それ故、B7RP−1/CRP−1経路の拮抗物質はヒトIBDを治療するために使用しうる。
【0140】
CRP1又はB7RP1の拮抗物質は、骨髄及び臓器移植のための免疫抑制剤として使用でき、移植片の生存を延長させるために使用しうる。そのような拮抗物質は既存の治療に比べて重要な利益を提供しうる。骨髄及び臓器移植療法は、宿主によるT細胞仲介の異種細胞又は異種組織拒絶反応と戦わねばならない。T細胞仲介の拒絶反応抑制するための現在の治療レジメンは、シクロスポリン又はFK506薬剤による治療を含む。薬剤は有効であるが、患者は肝毒性、腎毒性及び神経毒性を含めた深刻な副作用に悩まされる。シクロスポリン/FK506クラスの治療薬の標的は、遍在発現するホスファターゼ、カルシニューリンである。CRP1の発現はT細胞に限定されるので、CRP1又はB7RP1の阻害因子は現在の免疫治療薬の使用に関して認められる重篤な副作用がないと考えられる。
【0141】
CRP1又はB7RP1の拮抗物質は、慢性関節リウマチ、乾癬、多発性硬化症、糖尿病、及び全身性エリテマトーデスのような自己免疫疾患のための免疫抑制剤として使用しうる。
【0142】
CRP1/B7RP1を介する共同刺激経路の拮抗物質はまた、トキシックショック症候群、炎症性腸疾患、輸血によるアロ感作、T細胞依存性B細胞仲介疾患を軽減するため、及び対宿主性移植片病の治療のために使用しうる。
【0143】
抗体、例えば細胞外ドメインを含む可溶性タンパク質、及びT細胞活性化の延長又は促進をもたらすCRP1又はB7RP1の他の調節因子が、腫瘍に対する免疫応答を高めるために使用できる。実施例20は、B7RP−1−Fcがマウスにおいて腫瘍細胞の増殖を阻害しうることを示している。同様に、ヒトB7RP−1−Fc又はB7RP−1/CRP−1経路の他の活性化因子は、ヒト腫瘍に対する免疫応答を増強するために使用しうる。抗腫瘍活性は一般に、強力な細胞溶解性Tリンパ球成分を持つとみなされる。実際に、B7−Fc融合タンパク質の抗腫瘍作用(Sturmhoefelら、Cancer Res.59:4964−4972,1999)は、細胞溶解性CD8+T細胞によって仲介された。CRP−1も細胞溶解性CD8+T細胞上で発現されるので(実施例9)、実施例20で明らかにされた抗腫瘍作用はCD8+細胞へのB7RP−1−Fcの作用によるものであったと考えられる。B7RP−1/CRP−1経路はまた、同種移植片移植、対宿主性移植片病、及び自己免疫疾患を含めた多くの他の臨床状況においてCTL応答を調節するために操作できる。
【0144】
本発明のB7RP1遺伝子を使用する遺伝子治療は、癌免疫療法において使用しうる。癌細胞内に挿入されたB7RP1遺伝子は、動物の体内にもどされたときに免疫系のT細胞によって認識されうる抗原呈示細胞へと癌細胞を形質転換することができる。T細胞によるトランスフェクションされた腫瘍細胞の認識は、B7RP1遺伝子を発現する又は発現しない腫瘍細胞の根絶をもたらす。この免疫療法アプローチは、様々な白血病、肉腫、黒色腫、腺癌、乳癌、前立腺腫瘍、肺癌、結腸癌及び他の腫瘍のために使用しうる。本発明は、様々な腫瘍に対する応答においてT細胞の活性化を高めるためにB7RP1遺伝子を同様に使用することを包含する。
【0145】
実施例14で述べるように、B7RP1を発現するトランスジェニックマウスの表現型は、B7RP1が抗体産生の制御において重要であることを示唆する。B7RP1タンパク質活性の作用物質及び拮抗物質は、抗体産生の抑制又は強化を必要とする治療適応症において有用であると考えられる。
【0146】
例えば、多くのワクチンは有効で特異的な抗体応答を惹起することによって作用する。一部のワクチン、特に腸内微生物(例えばA型肝炎ウイルス及びサルモネラ属(Salmonellas))に対するものは、短命な抗体応答を惹起する。ワクチンの有効性を高めるためにはこの応答を増強し、延長させることが望ましい。それ故、可溶性B7RP1又はCRP1に対する活性化抗体は、ワクチンアジュバントとして役立つと考えられる。
【0147】
抗ウイルス応答はまた、B7RP−1/CRP−1経路の活性化因子又は作用物質によっても増強されうる。実施例20のデータは、細胞性免疫がB7RP−1−Fcによって高められることを示している。B7RP−1−Fc又はB7RP−1/CPR−1経路の他の活性化因子による細胞性免疫機能の増強はまた、ウイルス感染細胞を排除する上でも有益であると考えられる。補足的に、B7RP−1−Fcは、体内から遊離ウイルスを清掃するのを助ける働きをすると考えられる、実施例13で認められたような抗体仲介応答を増強しうる対液性免疫機能に影響を及ぼす。
【0148】
逆に、抗体産生の抑制によって改善される多くの臨床条件がある。過敏症は、誇張された又は不適切な、常態では有益な免疫応答であり、炎症反応と組織損傷を導く。抗体が仲介する過敏症反応は、B7RP1活性の阻害因子による拮抗作用を特に受けやすいと考えられる。アレルギー、枯草熱、喘息、及び急性浮腫はI型過敏症反応を引き起こすが、これらの反応はB7RP1活性のタンパク質、抗体又は小分子阻害因子によって抑制されうる。 全身性エリテマトーデス、関節炎(慢性関節リウマチ、反応性関節炎、乾癬性関節炎)、ネフロパシー(膜性、脈管膜毛細管性、巣状分節性、巣状壊死性、半月性、増殖性−糸球体腎炎、尿細管症)、皮膚疾患(天疱瘡及び類天疱瘡、結節性紅斑)、内分泌障害(甲状腺炎−グレーヴズ病、橋本甲状腺炎、インスリン依存性糖尿病)、種々の肺疾患(特に外因性肺胞炎)、種々の血管症、IgAの異常産生を伴う小児脂肪便症、多くの貧血及び血小板減少症、ギヤン−バレー症候群、及び重症筋無力症を含めて、抗体仲介の過敏症反応を引き起こす疾患がB7RP1拮抗物質によって治療できる。
【0149】
さらに、多発性骨髄腫、ヴァルデンストレームマクログロブリン血症、及びクリオグロブリン血症のようなリンパ球増殖性疾患が、B7RP1のタンパク質、抗体、又は小分子拮抗物質によって抑制しうる。
【0150】
最後に、「人為的」免疫疾患である対宿主性移植片病は、B7RP1拮抗物質による抗体産生の抑制によって恩恵を受けるであろう。
【0151】
下記の実施例は本発明をさらに詳しく説明するために提示するものであるが、本発明の範囲を制限するものとみなされるべきではない。
【実施例】
【0152】
実施例1
CRP1 cDNAおよびアミノ酸配列
雌C57/ブラック6マウスを犠牲にして、小腸を切除し、パイエル板を除去した。小腸組織を切開し、洗浄して、粘膜および他の壊死組織片を除去した。1mMのジチオトレイトール(DTT)を補充したRPMI−1640中で、20分間、37℃で穏やかに撹拌することによって、腸上皮内細胞(iIEL)を含有する上皮層を解離させた。解離した細胞を100μフィルターに通して、50mLのRPMI−1640中で洗浄し、混合して細胞の凝集塊をさらに破壊し、その後、40μストレーナーを通して、単一細胞集団を得た。その後、これらの細胞を、体積50mLのRPMI−1640で再び洗浄して、残留DTTを確実に除去した。その後、その組織を撹拌し、前のように洗浄して、残留iIELを採集した。3段階のパーコール勾配を用いて、iIELを40%〜80%界面でバンディングしながら、iIELを脂肪細胞および最上皮細胞から分離した。その後、これらの細胞をRPMI−1640で2回洗浄して極微量のパーコールを除去し、CD103(インテグリンαIEL)抗体で免疫染色して、FACs Star セルソーターを用いて分離した。その後、これらの選別細胞は、Trizol(Gibco BRL,Gaithersburg,MD)を直接用いて全RNAを準備するために用いるか、あるいはγ−/δ−TCR、α−/β−TCR、またはCD3を架橋するプレート結合活性化抗体を用いて一晩活性化するかした。RNAを以上のように準備し、EST cDNAライブラリを造るためにプールした。
【0153】
smi12−00082−a1で表わされるcDNAクローンは、CD28(図1B)と相同のヌクレオチド配列を含んでいた(図1B)。配列の翻訳および公開データベース中の既知タンパク質とのその後の比較は、19%のアミノ酸がマウスCD28と同一であることを示した。マウスCD28は、マウスのCTLA−4と同一のアミノ酸を26%しか共有しないため、この低相同性は有意である。CD28/CTLA−4系統における分子内および分子間システイン結合にとって重要であると考えられる推定システインのすべてが、保存されることが分かった(アミノ酸残基83、109および137;開始メチオニンに関して)。さらに、推定オープンリーディングフレームの全長および膜内外領域の相対位置は、CD28およびCRLA−4の両方のものと類似していた。遺伝子CRP1をCD28関連プロテイン−1と名づけた。
【0154】
実施例2
ヒトCRP1 cDNAのクローニング
ヒトCRP1プロテインをコードする核酸配列は、以下の手順によって同定する。ヒトcDNAライブラリは、健康なボランティアから得たヒト抹梢血からの濃縮リンパ球から製造した。リンパ球を精製し、赤血球をリンパ球分離培地(INC Pharmaceuticals,INC.,Costa Mesa,CA)によって除去した。その後、10ng/mLのPMA、500ng/mLのイノマイシン、およびCD3に対するプレート結合活性化抗体を含有する培地中で、一晩、その細胞を活性化させた。Trizol法によって活性化された細胞(Gibco/BRL)から全RNAを準備し、ダイナルビード精製によってポリA RNAを分離した。cDNAは、分離したポリA RNAから製造し、サイズは最大cDNA断片を選択した。その後、サイズ選択cDNAは、プラスミドpSPORT(Gibco/BRL)にライゲーションさせた。ヒトCRP1タンパク質をコードするDNAは、組換えバクテリオファージプラークまたは形質転換細菌コロニーハイブリダゼーションプロトコル(Sambrookら,Supra)のいずれかにより活性化リンパ球cDNAライブラリをスクリーニングすることによって得る。ファージまたはプラスミドcDNAライブラリは、実施例1および図1に記載のマウスCRP1遺伝子クローンから誘導される放射性標識したプローブを用いてスクリーニングする。プローブを用いて、プレーティングしたライブラリから切り離したナイロンフィルタをスクリーニングする。これらのフィルタは、50%ホルムアミド、5X SSPE、2X デンハート溶液、0.5%SDSおよび100μg/mLのサケ精子DNA中、42℃で4時間予備ハイブリダイズし、その後、50%ホルムアミド、5X SSPE、2X デンハート溶液、0.5%SDS、100μg/mLのサケ精子DNAおよび5ng/mLのmB7RP1プローブ中、42℃で24時間ハイブリダイズする。ブロットは、2X SSC、0.1%SDS中、室温で10分間、1X SSC、0.1%SDS中、50℃で10分間、0.2X SSC、0.1%SDS中、50℃で10分間洗浄し、その後、0.2X SSC中、50℃で10分間、再び洗浄する。一切のヒトCRP1クローンから得られた挿入物を配列し、実施例1に記載したように分析する。
【0155】
実施例3
B7RP1 DNAおよびアミノ酸配列
smi11−00003−g5で表わされるcDNAクローンは、B7.1(CD80)およびB7.2(CD86)と相同のヌクレオチド配列を含んでいた。配列の翻訳(図2A)および公開データベース中の既知タンパク質とのその後の比較は、20%のアミノ酸がマウスB7.1と同一であることを示した(図2B)。マウスB7.1は、マウスB7.2と同一のアミノ酸を24%しか共有しないため、この低相同性は有意であった。この低相同性にもかかわらず、臨界システイン残基は、このクローンのオープンリーディングフレームとマウスB7.1およびB7.1との間、残基62、138、185および242(開始メチオニンに関して、図2B)において保存される。B7.1およびB7.1と比較すると、このクローンの推定ORFおける適切な成熟タンパク質長およびカルボキシ末端に対する膜内外領域の位置も類似している。我々は遺伝子B7RP1をB7関連タンパク質−1と名づけた。
【0156】
実施例4
ヒトB7RP1 cDNAのクローニング
マウスB7RP1配列(図2参照)を用いるGenbank芽細胞相同性調査(GCG,University of Wisconsin)は、1679bpのORFを有する4358bpの配列を含むクローン(AB014553)を検索した。PCRクローニングプライマーは、この配列にしたがって設計した。Human Lymph Node Marathon−ReadyTMcDNA(Clontech,Palo Alto,CA)を用い、製造業者の指示した手順に従って、5’および3’RACEにより1313bpのDNA断片を得た。
【0157】
ヒトB7RP1全長に用いたプライマー:
【0158】
【化1】
プライマー2083−75および2083−76は、RACEプロトコルを用いて遺伝子の5’末端を増幅させるために用いた。プライマー2083−77、2083−78、2113−29、2113−30および2113−31は、RACEプロトコルを用いて遺伝子の3’末端を増幅させるために用いた。
【0159】
得られたヌクレオチド配列は、メチオニンで開始する288のアミノ酸残基のORFを含んでいた。その後、予測成熟ヒトB7RP1アミノ酸配列を成熟マウスB7RP1アミノ酸配列と比較して(図3B)、同一のアミノ酸を48%共有することがわかった。CD80(B7.1)を用いると化学種間の相同性が低く、実際に、マウスおよびヒトのCD80は、同一のアミノ酸を41%しか共有しないため、この相同性は有意である。重要なことに、ヒトB7RP1タンパク質は、Igループ構造(成熟タンパク質に関するアミノ酸残基16、92、138、194および195、図3B)に必要な臨界システイン残基を保存する。さらに、膜内外領域の全長および位置は、ヒトB7RP1ホモログと一致する。
【0160】
実施例5
B7RP1 RNAの発現
B7RP1遺伝子に対してRNAプローブ用いるRNAインシチューハイブリダイゼーション。成体マウス組織を4%パラホルムアルデヒド中に固定し、パラフィン包理して、5μmの薄切りにした。インシチューハイブリダイゼーションの前に、組織を0.2MのHClで易透化し、続いて、プロテイナーゼKを用いて消化して、トリエタノールアミンおよび無水酢酸を用いてアセチル化した。一晩、55℃で、マウスB7RP1配列のヌクレオチド1〜96に対応する969塩基の33P標識リボプローブを用いて、切片をハイブリダイズした。RNアーゼ消化によって過剰のプローブを除去し、続いて、漸減塩濃度の緩衝液中で一連の洗浄を行い、その後、0.1X SSC中、55℃で高ストリンジェンシー洗浄を行った。スライドをKodak NTB2エマルジョンに浸漬し、4℃で2〜3週間露光して、現像し、ヘマトキシリンおよびエオシンを用いて対比染色した。切片標本を暗視野および透過光照明法を用いて試験することによって、組織形態学およびハイブリダイゼーションシグナルを同時に評価することができた。
【0161】
インシチューハイブリダイゼーションによるB7RP1 RNAの分析は、B7RP1 RNAがリンパ系成熟領域において非常に発現することを示した。B7RP1 RNAは、胸腺のリンパ系組織、腸、脾臓およびリンパ節のパイエル板において発現した。これらのリンパ系組織内の発現は、B7RP1 RNAが、一般に、B細胞領域およびその他のAPC関与領域において発現することを実証していた。これらの領域には、胸腺の髄質領域、リンパ節の1次濾胞、およびパイエル板の濾胞およびドーム領域が挙げられる。B7RP1 RNAの発現は、リンパ系組織におけるAPC関与領に極めて特異的なものである。
【0162】
一部の非リンパ系組織の分析も、APC関与領域におけるB7RP1発現を示した。肺において、B7RP1発現は、抗原処理に関する機能と一致する粘膜下領域において見られた。小腸において、B7RP1 RNAは、粘膜固有層において見られた。特に、B7RP1 RNAの発現と重複するリンパ球湿潤を示す損傷肝臓部分を発見した。B7RP1発現の組織損傷に応じたリンパ球蓄積との一致は、B7RP1がリンパ球活性化に関与することを強く示している。
【0163】
実施例6
CRP1 RNAの発現
CRP1遺伝子に対してRNAプローブを用いるRNAインシチューハイブリダイゼーション。マウスの組織は、実施例5の場合のように準備した。組織の易透化、プローブハイブリダイゼーション、スライド処理、および組織染色は、実施例5に記載したとおりであった。切片は、一晩、55℃で、マウスCRP1配列のヌクレオチド1〜603に対応する603塩基の33P標識リボプローブを用いて、ハイブリダイズした。切片標本を暗視野および透過光照明法を用いて試験することによって、組織形態学およびハイブリダイゼーションシグナルを同時に評価することができた。
【0164】
正常なマウスまたはオキサゾロンで処理したマウスからのリンパ節を薄切りにし、CRP1 RNA発現について分析した。感作マウスリンパ節は、正常なマウスのリンパ節より大きいCRP1 RNA発現を示した。CRP1の発現は、T細胞活性領域である傍皮質にあった。従って、CRP1 RNAの発現は、Tリンパ球発現のものと一致し、T細胞活性次第で向上調節される。
【0165】
実施例7
CRP1−FcおよびB7RP1−Fc融合タンパク質の発現および精製
CRP1−Fc融合タンパク質についてのDNA発現ベクターを構成するために、CRP1の第一アミノ末端147アミノ酸についてのコード配列は、ヒトFc遺伝子のカルボキシ末端235アミノ酸(アイソタイプIgG1)についてのコード配列、フレーム内に融合させ、pcDNA3のポリリンカー配列内にライゲーションした(pcDNA3/CRP1−Fc)。B7RP1−Fc融合タンパク質につてのDNA発現ベクターを構成するために、B7RP1の第一アミノ末端269アミノ酸についてのコード配列は、ヒトFc遺伝子のカルボキシ末端235アミノ酸(アイソタイプIgG1)についてのコード配列、フレーム内に融合させ、pcDNA3のポリリンカー配列内にライゲーションした(pcDNA3/B7RP1−Fc)。CRP1およびB7RP1、両方のコード配列は、各タンパク質のN−末端から、そこは含まないが、各タンパク質の推定膜内外領域までの配列を含んでいた。293T細胞には、FuGene6形質移入試薬(Roche Molecular Biochmicals,Indianapolis,IN)を用いて、pcDNA3/CRP1−FcまたはpcDNA3/B7RP1−Fcのいずれかを形質移入した。4日後、調整培地を回収し、Protein A Sepharose(Pharmacia)を用いるバッチクロマトグラフィーによって、Fc融合タンパク質を精製した。カラムに結合したFc融合タンパク質を3カラム容量のImmunopure Gentle Elution Buffer(Pierce)を用いて溶離し、その後、体積150の20mMのHEPES、100mMのNaCl(pH7.5)に透析させた。Macrosep遠心コンセントレート、30kd MWCO(Pall Filtron)を用いて、透析したタンパク質を濃縮し、各タンパク質のアミノ酸配列から誘導される吸光係数を用いてタンパク質濃度を計算した。CRP1−Fc融合タンパク質の発現を図4Aに示し、B7CRP1−Fc融合タンパク質の発現を図4Bに示す。
【0166】
実施例8
レセプター−リガンド対としてのCRP1およびB7RP1の同定
新規タンパク質が、CD28、CTLA−4、B7.1およびB7.2を含むものと同じ共同刺激性経路であるかを決定するために、細胞表面画像分析を用いた。この分析は、ACAS(Adherent Cell Analysis and Sorting/粘着細胞分析および選別)分析を用いて、細胞中で発現した膜結合タンパク質が種々のFc融合タンパク質と相互作用するかを分析するものである。膜結合タンパク質を発現する細胞(図5の左側に示す)をFc融合タンパク質(同図の上に示す)と共にインキュベートした。
【0167】
10%FBSを含有するDMEM培地中で増殖させたCos−7細胞を24ウエルのプレートに500,000細胞/ウエルでプレーティングした。FuGene6試薬(Roche Molecular Biochemicals,Indianapolis,IN)を用いて細胞に形質移入した。各形質移入について、3μLのFuGene6試薬を血清を含有しない47μLのDMEM培地に添加した。室温で10分間インキュベートした後、混合物を0.25μgのプラスミドに一滴ずつ添加し、その後、15分間インキュベートした。その後、10%FBSを含有する0.5mLのDMEMと共に上の混合物を細胞に添加した。細胞を37℃、5%CO2雰囲気のもとでインキュベートした。対照として、ヒトCD28についてのcDNAを含む発現プラスミドで安定的に形質移入したCHO D細胞も、24ウエルのプレートに500,000細胞/ウエルでプレーティングした。
【0168】
48時間後、形質移入試薬を含有する培地を除去し、細胞をRPMIと5%FBSで2回洗浄した。1mLの培地中、10〜20ngの精製Fc融合タンパク質を細胞に添加し、これを30分間氷上でインキュベートした。細胞をRPMIと5%FBSで3回洗浄し、その後、2μLのFITC結合抗ヒトFc抗体(1mg/mL)と共に、さらに30分間氷上でインキュベートした。RPMIを用いて3回連続洗浄した後、細胞は、ACAS分析用のために、フェノールレッドを含有しない250μLのRPMI培地を用いて転化させた。
【0169】
種々のFc融合タンパク質を結合した細胞のACAS分析は、B7RP1タンパク質がCRP1に結合していることを示したが、既知の共同刺激性経路におけるタンパク質、CD28またはCTLA−4は示さなかった。逆に言えば、CRP1はB7RP1と相互作用したが、既知経路における成分、B7.2とは相互作用しなかった。(図5参照)。これらの結果は、CRP1およびB7RP1が、CD28およびB7.2のアナログである新規レセプター−リガンド対を表わすことを強く示している。しかし、CRP1およびB7RP1は、B7.2、CTLA−4、またはCD28と相互作用しないため、それらは離れており、既知の共同刺激性経路から独立している。
【0170】
実施例9
B7RP1レセプターを発現する細胞の同定
B7RP1−Fc融合タンパク質を用いて、CRP1タンパク質を含有すると推定されるB7RP1に対するレセプターを発現する細胞(図6参照)をFACS分析によって検出した。雌C57/ブラック6マウスから脾臓を除去して、100μメッシュフィルターを用いて磨砕し、リンパ球を解離させて、70μフィルターを通し、その後、50mLのRPMI−1640中で洗浄した。それらを1500rpmでペレット化して、新しいRPMIに再懸濁させ、混合して凝集細胞を破壊し、40μフィルターを通した。活性化させるべきT細胞をRPMI−1640、5%FBS、1XPSG、PMA、イノマイシン中で6ウエルのプレートに播種し、37℃、5%CO2で、一晩インキュベートした。12時間後、T細胞の活性化を視覚的確認によりチェックした。
【0171】
免疫染色用の活性化脾臓細胞をPBS、0.5%BSA(Path−ocyte 4、ICN Pharmaceuticals)洗浄緩衝液中で洗浄して、再懸濁させ、その後、体積100μLで分取した。CRP1−Fc融合タンパク質またはB7RP1−Fc融合タンパク質、いずれかの15μg/mLを適切に添加し(1.5μg/サンプル)、その後、その混合物を氷上で30分間、時折混合しながらインキュベートした。細胞を5.0mLの洗浄緩衝液中で2回洗浄した。細胞染色用の100μL容積中、2μgのヤギ抗ヒト(GaHuFc−FITC)複合第2抗体を用いて融合タンパク質の結合を視覚化した。PEと複合した細胞マーカー抗体をGaHUFc−FITC、ならびに指示された抗体対照(ラットアイソタイプ)と複合した対照アイソタイプ−PEと共に添加した。そのサンプルを氷上でインキュベートし、前のごとく洗浄した。リンパ球集団にゲートを設けるFACScan分析によって、視覚化を行った。CD4+抗体およびB7RP1−Fc融合タンパク質を用いた二重染色は、細胞がCD4マーカーとB7RP1に対するレセプター、おそらくCRP1との両方を発現することを示した(図6)。同様に、CD8+抗体およびB7RP1−Fc融合タンパク質を用いた二重染色は、細胞がCD8レセプターおよびB7RP1レセプターの両方を発現することを実証した(図6)。非活性化脾細胞試料中では、こうした二重染色細胞を確実に検出することはできなかった。CD4およびCD8はTリンパ球マーカーであるので、CRP1は活性化CD4+およびCD8+T細胞上で発現されると仮定することができる。これらのデータは、正常なマウスと比較して、感作マウスからのリンパ節のT細胞領域において増加したCRP1 RNAの発現と一致する(実施例6)。
【0172】
実施例10
CRP1リガンドを発現する細胞の同定
CRP1−Fc融合タンパク質を用いて、B7RP1タンパク質を含有すると推定されるCRP1に対するリガンドを発現する細胞(実施例8参照)をFACS分析によって検出した(図7)。脾細胞は、実施例8の場合のように準備したが、12時間のT細胞活性化段階を省略し、細胞を直接分析した。脾細胞は、CD45R(B220)マーカー抗体およびCRP1−Fc融合タンパク質を用いて二重染色した。CD45R B細胞マーカーとB7RP1を含有すると推定されるCRP1に対する推定リガンド(実施例8)との両方を発現する細胞を検出した。従って、B7RP1は、抗原提示細胞タイプのB細胞上で発現されると推断する。これらのデータは、種々のリンパ系組織のB細胞領域におけるB7RP1 RNAの発現(実施例5)と一致した。
【0173】
腹腔マクロファージ上でのB7RP1の発現のFACS分析(図8)。腹腔細胞を局所洗浄によって正常なマウスから採集し、CRP1−Fc融合タンパク質または対照としてのFcタンパク質と共に、あるいはF4/80モノクローナル抗体(マクロファージに対して特異的な抗原を検出する)または無関係なアイソタイプ対応モノクロナール抗体と共にインキュベートする前に洗浄した。その後、再び細胞を洗浄し、ヤギ抗ヒトFc−FITC複合抗体と共にインキュベートした。さらなる洗浄の後、細胞は、FACS分析機で光拡散および蛍光染色性について評価した。腹腔細胞は、それらの光拡散性によってサブセット中で最初に識別された(図8A)。マクロファージは、光を前方(FSC)および横向き(SSC)に強く拡散させる能力のため、およびマクロファージに対するマーカーであるF4/80抗原に対して染色陽性であるため、領域5(R5)内で確認した(図8B)。領域6(R6)内のマクロファージは、F4/80抗原に対するさほど強くない染色性をもとに選抜され、またCRP1−Fc融合タンパク質によって染色されることが分かった(図8C)。これらのデータは、B7RP1を含有することができるCRP1に対するリガンドが、プロフェッショナル抗原提示細胞であるマクロファージ上で発現されることを示している。これは、Tリンパ球活性化におけるCRP1およびB7RP1機能と一致する。
【0174】
実施例11
Con A刺激Tリンパ球上のB7RP1−Fc融合タンパク質のインヴィトロ抑制作用
マウス脾細胞を実施例8の場合のように準備し、ネガティブ選択によってTリンパ球を濃縮した(R and D Systems,Inc.,Minneapolis,MN)。200,000の脾細胞を96ウエルの丸底プレートにおけるT細胞増殖分析に用いた。細胞は、培地(添加なし)、図9に示したCRP1−Fc、B7PR1−FcまたはB7.2−Fc融合タンパク質と共に1時間インキュベートした。培地(添加なし)、または種々の濃度でのCon Aを図9の下に示したとおり添加した。その後、細胞を37℃、5%CO2でインキュベートした。42時間後、細胞に3H−チミジンを6時間適用し、回収して、組込まれた放射活性を決定した。3つの同じサンプルからの平均CPMおよび標準偏差を図9に示す。
【0175】
Fc融合タンパク質は、それ自体では有意なT細胞刺激活性または抑制活性を示さなかったが、1μg/mLおよび3μg/mLのCon Aの存在のもとで、B7RP1−Fcおよび既知のB7.2―Fc融合タンパク質は、有意な抑制活性を示した(図9)。高濃度(10μg/mL)では、Con A刺激作用は、おそらくT細胞の過剰活性化によって細胞死をもたらす。B7RP1−FcまたはB7.2−Fc、いずれかを添加すると、細胞は高濃度のCon Aの有害作用から著しく保護された。抑制および保護機能の両方において、B7PR1−Fcタンパク質によるCon A刺激細胞への影響は、B7.2−Fcタンパク質より大きかった。これらのデータは、B7RP1タンパク質が機能して、T細胞増殖を調整することを示している。
【0176】
実施例12
トランスジェニックマウスにおけるB7RP1−Fc融合タンパク質の全身送達
実施例7に記載したB7RP1−Fc融合タンパク質は、ApoE肝臓特異的発現ベクターにさらにクローニングした(Simonetら,J.Clin.Invest.94,1310−1319(1994)およびPCT出願番号US94/11675)。制限酵素、Spe IおよびNot Iを用いてコード領域をpCEP4/B7RP1−Fcから切除して、破片を前述のApoE肝臓特異的発現ベクター中の同じ部位にさらにクローニングした。得られたプラスミド、HE−B7RP1−Fcをそのタンパク質コード領域およびコード領域の側面に位置する配列を通して配列し、突然変異体が確実に含まれないようにした。
【0177】
プラスミドを増幅させ、CsC1密度勾配遠心法を2回行って精製した。精製したプラスミドDNAを制限酵素、Cla IおよびAse Iを用いて消化し、1.5kbのトランスジーン挿入物をアガロースゲル電気泳動法によって精製した。精製した破片を希釈して、5mMのTris(pH7.4)および0.2mMのEDTA中1μg/mLの保存注射溶液にした。使用前に注射針を斜めに切り、シリコーン処理したことを除いて、本質的には(Brinster et al.,Proc.Natl.Acad.Sci.USA 82,4338(1985))に記載されているとおりにBDF1 X BDF1−交配マウスからの単個細胞胚を注入した。胚をCO2インキュベータ内で一晩培養し、15〜20の2−細胞胚を偽妊娠CD1雌マウスの卵管に移入した。
【0178】
満期妊娠の後、微量注入した胚に基づく体内移植から56の子孫を得た。ゲノムDNAサンプルに組込まれたトランスジーンのPCR増幅によって、その子孫をスクリーニングした。増幅の標的領域は、発現ベクター中に含まれるヒトApo Eイントロンの369bp領域であった。PCR増幅に用いたオリゴは:
【0179】
【化2】
であった。
【0180】
PCRについての条件は、94℃で2分間を1サイクル、94℃で1分間、63℃で20秒間、そして72℃で30秒間を30サイクルであった。56の原子孫のうち7匹をPCR陽性トランスジェニックファウンダーマウスとして確認した。
【0181】
12週齢において、9匹のトランスジェニックファウンダー(マウス#1、2、4、6、8、30、32、33、40)および5匹の対照(マウス#5、9、10、25、28)を屍検および病理学分析のために犠牲にした。全細胞性RNAをファウンダー動物および陰性対照同腹子の肝臓から、(McDonald et al.,Meth.Enzymol.152,219(1987))に記載されているように分離した。これらのサンプルを用いてノーザン法分析を行い、トランスジーン発現のレベルを評価した。アガロース電気泳動変性ゲル(Ogden et al.,Meth.Enzymol.152,61(1987))によって、各動物から約10μgの全RNAを分離し、その後、HYBOND−Nナイロン膜(Amersham)に移送して、32P dCTP標識mB7RP1−Fc挿入DNAを用いてプローブした。ExpressHyb溶液(Clonetech)およびハイブリダイゼーション緩衝液1mLあたり2〜4 X 106CPMの標識プローブ中、63℃で1時間ハイブリダイゼーションを行った。ハイブリダイゼーションに続いて、ブロットを2X SSC、0.1%SDS中、室温で2回、各5分間洗浄し、その後、0.1X SSC、0.1%SDS中、55℃で2回、各15〜20分間洗浄した。ファウンダーおよび対照同腹子におけるトランスジーンの発現は、オートラジオグラフ法に従って決定した。
【0182】
ノーザン法分析は、7匹のトランスジェニックファウンダーが検出可能レベルのトランスジーンRNAを発現することを示した(マウス#1、2、6、8、32、33および40)。陰性対照マウスおよび3匹のファウンダー(#4、30および31)は、検出可能レベルのRNAを発現しなかった。B7RP1−Fc融合タンパク質は、培養状態の哺乳動物細胞から分泌されるものと決めたので(図4Bおよび実施例7)、トランスジーンmRNAの発現は、全身送達遺伝子産物レベルを示すはずである。
【0183】
実施例13
B7RP1−Fc融合タンパク質の生物活性
7匹のトランスジェニックマウス(マウス#1、2、6、8、32、33および40)および5匹の対照同腹子(#5、9、10、25および28)を以下の手順を用いる屍検および病理学分析のために犠牲にした。安楽死させる前に、すべての動物のID番号を確認し、その後、体重を計り、麻酔をかけて、血液を取った。血液は、全血清化学および血液学パネルのために、血清および全血の両方として保管した。致死性CO2吸入による終末麻酔の直後、グロス解剖の前にX線撮影を行った。その後、組織試験のために組織を除去し、10%Zn−ホルマリン緩衝液中に固定した。回収した組織には、肝臓、脾臓、膵臓、胃、十二指腸、回腸、パイエル板、結腸、腎臓、生殖器官、皮膚、乳腺、骨、脳、心臓、肺、胸腺、気管、食道、甲状腺/副甲状腺、空腸、盲腸、直腸、副臓、白色および褐色脂肪、坐骨神経、骨髄、膀胱、および骨格筋が挙げられる。固定前に、肝臓、心臓、胃、腎臓、副腎、脾臓、および胸腺について全器官重量を測定した。固定後、組織をパラフィンブロックに加工して、3μmの切片を得た。
【0184】
Bリンパ球マーカー、B220およびTリンパ球マーカー、CD3についての免疫組織化学法を実施した。B220またはCD3発現を検出するために、ホルマリン固定し、パラフィン包理した4μm切片を脱パラフィン化し、脱イオン水に水和した。切片は、3%過酸化水素を用いて反応停止させ、Protein Block(Lipshaw,Pittsburgh,PA)を用いてブロックして、B220に対するラットモノクローナル抗体(Pharmingen,San Diego,CA)またはCD3に対するウサギポリクローナル抗体(Dako,Carpinteria,CA)中でインキュベートした。クロマーゲンとしてDAB(Biotek,Santa Barbara,CA)を用いて、ビオチニル化ウサギ抗ラットまたはヤギ抗ウサギ免疫グロブリン、ペルオキシダーゼ複合ストレプタビジン、(BioGenex,San Ramon,CA)により、抗体を検出した。切片をヘマトキシリンで対比染色した。
【0185】
この研究では、寿命がある段階での研究中に、正常な臨床症状が報告された。トランスジェニックマウスの全身放射線写真を対照マウスのものと比較した。トランスジェニックマウスの全血液学的パラメータを陰性対照群ののものと比較したが、個々のマウスに孤立の変化が存在した。トランスジーン系#8および#40は、増大した血清グロブリンレベル(グロブレン過剰血)を有し、そして#32および#33は、免疫系の慢性抗原刺激により一般に見られるパターンである低い正常範囲のアルブミンレベルを随伴する高い正常範囲のグロブリンレベルを有した。その他のトランスジェニックマウスの器官重量は、対照群のものとさほど差がなかった。
【0186】
トランスジェニックマウスには以下の病理組織学的変化が存在した。トランスジェニックB7RP1−Fcマウスの腸間膜リンパ節は、対照マウスと比較して中等度〜著しく肥大した(図10A〜10D;図11A〜11E)。皮質は、主としてB220+ B細胞(図11D)、そして少数の散在CD3+ T細胞(図11F)を含む大きな胚中心を有する肥大した第二濾胞(図10B〜11B)のように見える隆起濾胞過形成を有した。傍皮質(CD3+ T細胞)領域も中等度に肥大化し(図11B〜11F)、髄洞は、わずかに増加した数の肉マクロファージ(洞組織球増殖)を有した。節における最も顕著な変化は、B7RP1−Fcトランスジェニックマウスにおける多数の分化型プラズマ細胞によって穏やか〜著しく膨張した髄索に存在した(図10D)。トランスジェニックマウス#40において、少数の散在ラッセル体(すなわち、免疫グロブリンを含有する隆起した大きく丸い細胞質内小疱)も髄索内に見られた(図10D)。興味深いことに、その他の内部および抹消リンパ節(例えば、頚部、鼠径部)は、全身応答を思わせる類似の形態学的特徴の活性リンパ細網細胞増生を有した。これらの発見は、B細胞増殖およびプラズマ細胞への終末分化を導く体液性免疫反応を強化させる慢性進行性免疫刺激と一致する。
【0187】
B7RP1−Fcトランスジェニックマウスの脾臓は、対照マウスと比較して、特に、胚中心隆起を伴うB細胞第二濾胞および細動脈周囲T細胞鞘が関係する中程度活性リンパ組織過形成により可変的に肥大した白色脾髄領域を有した(図10E〜10F)。B7RP1−Fcトランスジェニックマウスにおけるもう一つの顕著な発見は、白色脾髄領域周囲の帯域および隣接赤色脾髄における最小限〜軽度のプラズマ細胞増加であった。トランスジェニックマウス#6は、少数の散在ラッセル体を有した(図10F、挿入)。赤色脾髄は、対照マウスに見られるものと比較して、軽度〜中程度の骨髄外造血を有した(図10E)。
【0188】
小腸パイエル板は、B7RP1−Fcトランスジェニックマウスにおいて、対照マウスのものに比べ軽度〜著しく肥大し(図10G)、また、特にトランスジェニックマウス#40および#32において、胚中心が突起した非常に大きな濾胞を有した(図10H)。さらに、小腸内に存在する粘膜であるが、トランスジェニックマウスの結腸内ではより大きく突起した粘膜の濃化粘膜固有層中のリンパ球およびプラズマ細胞の数(マウス#32の回腸における軽度好酸性浸潤と混合)について、最小限(#32において)〜軽度(#8および#33)の増加があった。大腸リンパ球系凝集体(GALT)も、対照群中に比べ、一部のB7RP1−Fcマウス(特に、マウス#8および#2)中でわずかに大きく突起していた。
【0189】
胸腺、骨髄、肝臓、肺、心臓、膵臓、腎臓、副臓、甲状腺、副甲状腺、気管、生殖器官、膀胱、乳腺、皮膚、骨格筋、抹消神経、脳、食道、胃、小および大腸、骨(大腿骨/頚骨)、後膝関節、正常に見え、対照マウスにおいて検出された背景変化と比較できる白色および褐色脂肪を含むその他の組織をあまねく試験した。
【0190】
この研究からのデータは、トランスジェニックマウスにおけるB7関連タンパク質Fcキメラ(B7RP1−Fc)の過剰発現が、脾臓、抹消および内部リンパ節、および消化管関連リンパ系組織において検出される突起した反応性リンパ組織過形成によって特徴づけられる、濾胞過形成、T細胞領域の膨張、および一部の動物においてグロブリン過剰血を随伴する顕著なプラズマ細胞増加などの表現型を誘導することを実証している。プラズマ細胞増加は、より高レベルの循環IgG(±SD = トランスジェニックマウスにおける597±298mg/mL 対 対照同腹子における209±80mg/mL、n=7、P<0.05、t testを意味する)、特に、IgG2a(217±100mg/mL 対 75±29mg/mL、n=7、P<0.01、t test)を随伴する。IgG2aの誘導は、通常、IFN−gなどのTh1サイトカインに関連する。従って、B7RP−1は、BおよびT細胞増殖を誘導し、B細胞を刺激してプラズマ細胞に分化させ、免疫グロブリンを生成する。
【0191】
これらの変化は、試験したリンパ器官全体にわたって、抗体生産プラズマ細胞にB細胞刺激、増殖および分化をもたらす体液性上肢免疫系の過剰刺激に対する持続性全身免疫応答と一致する。
【0192】
B7RP1−Fcトランスジェニックマウスにおいて実証される著しいリンパ組織過形成から、我々は、B7RP1タンパク質が免疫系刺激に関連する有意な生体内生物活性を有するものと推断する。
【0193】
実施例14
ヒトB7RP1のクローニング
正常なヒト循環抹消リンパ球をリンパ球分離培地(ICN Pharmaceuticals)を用いて赤血球から分離した。その後、10μg/mLのプレート結合抗CD3抗体(Immunotech,Westbrook,ME)、10ng/mLのPMA、および500ng/mLのイノマイシンを用いて、一晩(16時間)、37℃、5%CO2で、T細胞を活性化した。その後、TRIzol試薬(Gibco BRL)を用いて全RNAを細胞から準備した。遠心分離によって細胞をペレット化し、その細胞ペレットを各々5 x 106の細胞に対して1mLのTRIzol試薬に再懸濁させて、室温で5分間インキュベートした。その後、原TRIzol試薬1mLあたり0.2mLのクロロホルムを添加した。試験管を手で15秒間激しく振り、3分間室温でインキュベートして、13,000rpmで15分間、4℃で遠心分離した。遠心分離後、RNAを含有する水性上澄み相を回収し、イソプロピルアルコールの添加によってサンプルRNAを沈殿させた。その後、溶液を室温で10分間インキュベートし、RNAをペレット化して、75%のエタノールで洗浄し、その後、15,000rpmで5分間、4℃で遠心分離した。ペレットを空気乾燥して、RNアーゼを含有しない水に再懸濁させ、その後、分取して、後で用いるまで−80℃で保管した。
【0194】
cDNA合成用のSuperScript Plasmid SystemおよびPlasmid Cloning(Gibco BRL)を用いてライブラリを構築した。簡単に言えば、平均サイズ2kbのcDNA挿入物をSal1/Not1クローニング部位においてpSportベクターにライゲーションした。ライゲーションしたプラスミドは、Electromax 形質転換担当E.コリ(Gibco BRL)へのエレクトロポレーションを施し、滴定して、LBプレートあたり15,000コロニー(アンピシリン100μg/mL)でプレーティングした。300,000のコロニーを切離してコロニー/プラークスクリーンハイブリダイゼーション伝達膜(NEN Life Scinece)に送り、0.5NのNaOH、1.5MのNaCl中で5分間変性させ、その後、次の緩衝液、1MのTris HCl(pH8.0)、0.5MのTris HCl(pH8.0)および1.5MのNaClならびに2X SSC中で各5分間、連続して中和した。その後、フィルターを紫外線照射によって架橋し、真空オーブン内で30分間80℃で焼いた。フィルターを2X SSC中、42℃で大いに予備洗浄して壊死組織片を除去し、その後、42℃で、50%ホルムアミド、5X SSPE、5X デンハート溶液、0.5%SDS、100μg/mLのサケ精液DNA中で、2時間予備ハイブリダイズした。
【0195】
図3Aに示すヌクレオチド1〜711を有する895bpのDNA断片、167bpsの5’直から図3中〜開始メチオニンコドン、および17bpsの3’直〜図3A中の位置711を用いてヒトリンパ球cDNAライブラリをスクリーニングした。167塩基対のこの上流5’配列は、HuB7RP1 cDNAの5’RACEによって得られ(実施例4)、Eco RI 制限酵素切断部位においてTOPO TAベクター(Invitrogen,Carlsbad,CA)から解離させた。この挿入物を0.8%アガロースTAEゲルを用いて2回精製した。DNAゲル精製キット(Qiagen)を用いて、アガロースからDNA挿入物を分離した。
【0196】
Redi−Prime2(Amersham)ランダム第一標識システムプロトコルに従って、125ngのDNA断片を32P dCTP(Amersham)を用いて標識した。その後、コロニーリフトフィルターは、プローブを用いて42℃でハイブリダイズさせた。次の緩衝液、50%ホルムアミド、5X SSPE、2X デンハート溶液、0.5%SDS、100mg/mLのssDNA中で、一晩、42℃。プローブの比活性は、ハイブリダイゼーション緩衝液1mLにつき約2ngの標識プローブ中、2.38x109cpm/μgのDNAであった。プローブを除去し、次回のスクリーニングのために保管した。その後、フィルターを2X SSC、0.1%SDS RT中で15分間、続いて1X SSC、0.1%SDS中、55℃で15分間、そして1X SSC、0.1%SDS中、60℃で10分間洗浄した。フィルターをプラスチック中にラップし、一晩、−80℃で、2つの増強スクリーンを用いてオートラジオグラフィーフィルムに曝した。3つの独立した陽性クローンを同定した。暴露したものを細菌プレートに配列して、陽性クローンをスクラップし、アンピシリン100μg/mLを用いて希釈してLBプレート上に再びプレーティングし、前のように一晩増殖させて、コロニーを切り離し、準備して、前に記載したようにプローブした。3つの独立したクローンコロニーを分離し、DNAを分離して、同じ3つのクローンそれぞれについてDNAを配列した。
【0197】
ヒトB7RP1タンパク質の全長は、302アミノ酸である。ポリペプチド長および膜内外領域の関係位置は、B7系列の他の構成員と一致した。ヒトB7RP1遺伝子は、マウスクローンと一致するアミノ酸を43%有する。マウスおよびヒトCD80タンパク質は41%しか一致しないので、この相同性度は有意である。マウスとヒトの間で特に保存される遺伝子は、アミノ酸位置37、113、158、215および216におけるシステイン残基である。
【0198】
実施例15
ヒトCRP1のクローニング
マウスB7RP1配列(図2参照)を使用したGenbank芽細胞相同性検索(GCG,University of Wisconsin)により、マウスCRP1遺伝子と高い相同性を示す104bp配列を含むゲノムクローン(Gen Bankアクセス番号AQ022676)を取り出した。この配列とオーバーラップするPCRクローニングプライマーを設計した。
【0199】
【化3】
【0200】
上記のプライマーを使用し、図1と実施例1に示したマウスCRP1プラスミドを鋳型としてマウスCRP1の151bp DNA断片をPCR増幅した。かかるDNA 125ngを、Redi−Prime 2(Amersham)ランダムプライム標識システムプロトコールに従って32P dCTP(Amersham)で標識した。実施例15で述べるヒト末梢血ライブラリーからのコロニーリフトフィルターを下記のハイブリダイゼーション緩衝液中41℃でひと晩(15時間)、プローブとハイブリダイズさせた;50%ホルムアミド、5×SSPE、2×デンハルト溶液、0.5%SDS、100μg/ml ssDNA。プローブの比放射能は3.52×109cpm/μg DNA、1.5ng標識プローブ/mlハイブリダイゼーション緩衝液であった。プローブを取り出し、次回のスクリーニングのために保存した。次にフィルターを2×SSC、0.1%SDSにおいて室温で10分間洗浄し、その後1×SSC、0.1%SDSにより37℃で7分間、40℃で7分間、44℃で7分間、さらに50℃で7分間、フィルターが標識プローブを放出する速度を絶えずモニターしながら洗浄した。フィルターをプラスチックに包み、2枚の増感紙を用いて−80℃でひと晩フィルムに暴露した。この方法は、可能性のある9個の独立した陽性クローンを明らかにした。暴露したものを細菌プレートに整列し、陽性クローンを削りとり、200μl SOCに入れて、1:10の2回の連続希釈を行い、2回目の希釈液から70μlを100μg/mlのアンピシリンを含むLBプレートに接種して上記のようにひと晩増殖させた。コロニーを採取し、調製して、上記のようにプローブした。8個の独立クローンを分離し、Qiagenミニプレップ法によってDNAを調製した。
【0201】
199個のアミノ酸のオープンリーディングフレームを含むcDNAクローンを得た(図13A)。このcDNAクローンは、実施例1と図1に示したマウスCRP1クローンとヌクレオチド及びアミノ酸の相同性を有していた。このヒトクローンのオープンリーディングフレームに対応するヌクレオチドは、マウスCRP1遺伝子と77%同一であった。ヒト配列の翻訳及びそれに続くマウスCRP1タンパク質との比較は、マウスタンパク質と69%のアミノ酸同一性を明らかにした(図13B)。さらに、アミノ酸114から119の間のモチーフ、「FDPPPF」は、マウスとヒトCRP1遺伝子間で保存されていた。このモチーフは、マウスにおける「MYPPPY」モチーフ及びB7タンパク質の相互作用にとって必須のヒトCD28に対応する。さらに、アミノ酸42、109及び141位のシステインも保存されていた。これらのシステインはCD28及びCTLA−4におけるシステインに対応し、Igループの形成と分子間ジスルフィド二量体化に関与する。マウスCRP1との緊密な類似性及びCD28ホモロジーファミリーとの構造的類似性は、これがヒトCRP1の同族体であることを示唆している。
【0202】
実施例16
CRP−1は休止記憶Tリンパ球上に発現される
記憶T細胞上のCRP−1発現を調べるため、6−7ヵ月齢のマウスから脾T細胞を採集した。これらの細胞を、FITC複合抗ヒトFc抗体及びCD44、CD45RB、又はCD69のいずれかに対するPE複合抗体によって標識したB7RP−1−Fcを使用して二重染色した。B7RP−1−Fc融合タンパク質による染色は、これらのT細胞でのCRP−1タンパク質の発現を検出する。より高齢のマウスは若齢マウスよりも多くのCRP−1+脾T細胞を示す。興味深いことに、顕著な数のこれらの細胞が、記憶T細胞に典型的なプロフィールである、CD44が高く(図14a)、CD45RBが低い(図14b)。これらのCRP−1+記憶T細胞は、活性化マーカーであるCD69を発現しないため(図14c)、休止状態にある。記憶T細胞上のCRP−1発現は、CRP−1が記憶T細胞への共同刺激機能を持つことを示唆する。
【0203】
実施例17
B7RP−1に対する抗体によって阻害される、インビトロでのT細胞の共同刺激
B7RP−1タンパク質がT細胞と機能的関連性を持つかどうかを調べるため、インビトロ増殖アッセイにおいてCD3+T細胞をB7RP−1−Fc融合タンパク質及び抗CD3抗体と共にインキュベーションした。次にウサギ抗マウスB7RP1ポリクローナル抗体又はラット抗マウスB7RP1モノクローナル抗体を使用して、B7RP1−Fcが共同刺激するインビトロでの増殖を特異的に阻害した。
【0204】
B7RP−1ウサギポリクローナル抗血清の調製
3匹のニュージーランド白色ウサギ(初期重量5−8ポンド)にマウスB7RP1タンパク質を筋肉内注射した。各々のウサギを1日目に、等量のHunters Titer Max完全アジュバントに乳化したマウスB7RP1タンパク質150μgで免疫した。さらに同じ手順で追加免疫(14日目と28日目)を実施した。EIAによって抗体価をモニターした。2回目の追加免疫後、抗血清は中等度の抗体価を示した。各々の動物から30mlの採血を行った。これを週に1回6週間にわたって繰り返した。その後ポリクローナル抗体をプロテインAアガロースクロマトグラフィーによって精製し、次いでFcタンパクアフィニティークロマトグラフィーによる陰性選択及びB7RP−1−Fcアフィニティークロマトグラフィーによる陽性選択を実施した。
【0205】
ラット抗マウスB7RP1モノクローナル抗体の調製
ラット抗マウスB7RP1モノクローナル抗体を、Practical Immunology,第2版(1980;L.Hudson and F.C.Hay;Blackwell Scientific Publications;St.Louis,MO)に述べられているように作製した。簡単に述べると、Louラット(Harlan;Indianapolis,IN)に、フロイントアジュバントに乳化したmuB7RP1−Fc融合タンパクを4週間隔で腹腔内注入した。融合の3日前に、ラットを可溶性muB7RP1により静脈内経路で追加免疫した。融合の当日、動物を二酸化炭素下で犠牲屠殺し、無菌的に脾を切除した。組織ストマカーを用いて単細胞懸濁液を調製した。脾細胞とY3−Ag1.2.3.骨髄腫細胞(American Type Culture Collection;Rockville,MD)の両方を無血清培地中で洗浄し、ポリエチレングリコール(PEG 1500;Boehringer Mannheim Biochemicals;Indianapolis,IN)を加えて融合した。細胞を1回すすぎ、血清含有培地に再懸濁して、96穴の組織培養プレートに接種した。10−12日後、各々の穴からの培地を、直接酵素免疫測定法(EIA)によりB7RP1に対する特異性抗体に関して試験した。潜在的な結合を示す穴からの細胞を10ml培養物に増殖させ、液体窒素中で冷凍した。各培養からの培地をフローサイトメトリー及び機能的T細胞増殖アッセイにおいてさらに試験した。これらの方法によって興味深いと判定されたものを単細胞コロニーに接種し、再びEIAによって選択して、最終的な細胞系統を抗体作製のために保持した。プロテインAアガロースクロマトグラフィーによって細胞培地から抗体を精製した。
【0206】
T細胞の調製及びT細胞増殖アッセイ
C57B1/6マウス(8−12週齢の雌性、Charles River Laboratories)の脾からのT細胞を、マウスT細胞強化カラム(R&D Systems)を通して陰性選択によって精製した。その後T細胞を直接使用するか、若しくは次のような抗体と補体の溶解によってさらに精製した。細胞を、マウスCD11b(クローンM1/70)、NK−1.1(クローンPK136)、CD8a(クローン53−6.7)、I−Ab(クローンM5/114.15.2)、CD11c(クローンHL3)、及びB220抗原(クローンRA3−6B2)に対する抗体(すべて10μg/ml、Pharmingenより)を含むRPMI培地に懸濁した(2.5×106細胞/ml)。次に細胞を氷上で30分間インキュベーションし、1200rpmでペレット化して、4:1v/vのRPMI:ウサギ補体(Sigma,No.S−7764)中に懸濁し、37℃でさらに30分間インキュベーションした。再び細胞をペレット化し、補体処理を繰り返した。平板培養の前に、10%FCSを含むRPMIで細胞を洗浄した。U底96穴プレートを、0から1.2μg/mlの濃度範囲の抗CD3抗体(クローン145−2C11,Pharmingen)及び抗ヒトIgG Fab2(Sigma,12.5μg/ml)により4℃でひと晩被覆し、次いで37℃で6−9時間インキュベーションした。T細胞(1×105/穴)を種々のFc融合タンパクの不在下又は存在下で48時間培養し、最後の18時間は1μCiの3H−チミジンでパルスした。対照Fcタンパクは、OPGとFcの融合タンパク及び非融合Fcタンパク断片を含んだ。その後細胞を採集し、取り込まれた放射能を測定した。B7RP−1−FcはT細胞を用量依存的に共同刺激して増殖させ(図15a)、抗B7RP−1−Fc抗体はこの共同刺激を用量依存的に阻害する(図15b)。
【0207】
実施例18
CRP−1/B7RP−1経路の阻害因子は、コラーゲンによって誘発される慢性関節リウマチの発症を低下させる
コラーゲン誘発の関節炎(CIA)は、ヒトにおける慢性関節リウマチと多くの類似性を持つ、げっ歯類及び霊長類での自己免疫性多発性関節炎の動物モデルである。異種のII型コラーゲン(CII)による免疫はCIIに対する自己免疫応答を誘発し、感受性のあるマウス系統においてCIAの発症を導く。H−2r及びH−2qを有する類似遺伝子型系統のマウスはCIAに対して極めて感受性が高い。CIAは、CII反応性T細胞と抗体の共力作用によって仲介される。ブタCII(Naboznyら、Autoimmunity 20,51−58(1995))を2mg/mlの濃度で0.01N酢酸に溶解し、次に1:1の比率でCFA(Difco)により乳化した。関節炎感受性B10.RIII(H−2r)マウス(Jackson Laboratories,Bar Harbor,ME)を、尾の基部において乳剤100μlにより皮内経路で免疫した。マウスを関節炎の発症に関して週に2−3回モニターした。関節炎の重症度は、各々の足について次のような採点システムを用いて判定した:0:関節炎なし;1:1−3本の足指に発赤又は腫脹;足の重篤な腫脹;3:関節強直。各々の肢の評点を合計し、各動物について0から12の重症度範囲を決定した。
【0208】
マウスにタンパク質100μg(200μL中)を週に2回、腹腔内経路で注入した。処置は、ブタCIIによる免疫の1日後に開始し、免疫後52日目に中止した。10匹のマウスから成る処置群において実験を行い、1又はそれ以上の評点を有する動物を陽性と判定した。結果を図16及び表1に示す。
【0209】
【表2】
【0210】
CRP1−Fc融合タンパクで処置したマウスでは、PBS処置マウスと比較して関節炎症状の発現が約10日間遅延した。これは、CRP−1/B7RP−1経路の阻害がこの慢性関節リウマチのマウスモデルにおいて疾患の症状を軽減しうることを示している。
【0211】
B7RP−1−Fc又はB7.2−Fcで処置したマウスは、PBS処置した対照と比較すると、関節炎の重症度の上昇(図16b)を伴って疾患の早期発症(表1及び図16a)を示した。これは、B7RP−1−Fc融合タンパクがT細胞免疫応答を増強することを示唆している。そのような活性は、インビボで抗腫瘍免疫を生じさせる上で有用と考えられる。
【0212】
慢性関節リウマチのこのマウスモデルにおけるCRP−1−Fc及びB7RP−1−Fcの反対の作用は、疾患の進行を促進する又は阻害するように経路を操作できることを示唆する。可溶性B7RP−1−FcによるCRP−1タンパクのターゲティングは疾患を促進するが、B7RP−1と可溶性CRP−1−Fcの相互作用は疾患の症状を抑制する。
【0213】
実施例19
B7RP−1−Fcはトランスジェニックマウスにおいて炎症性腸疾患表現型を誘導する。
【0214】
22−25週齢のトランスジェニックマウスにおけるB7関連タンパク(B7RP1−Fc)の持続的な過剰発現は、一部の動物において小腸及び大腸の著しい肥厚と慢性炎症(腸炎)及び体重減少を伴った炎症性腸疾患(IBD)の著明な表現型を誘導する。組織学的には、近位及び遠位結腸において最も重症の炎症性変化が認められ、小腸ではより軽度の変化が認められた。近位結腸は、裂溝状潰瘍、経壁炎症、及び結腸粘膜の肥厚を伴って著しく肥厚していたが、遠位結腸は、潰瘍形成を伴わない散在性粘膜肥厚(又は限局性びらん及び腺萎縮)を有していた。近位小腸は、より軽度の炎症性変化を伴って軽度から著明な粘膜肥厚を示し、一方遠位小腸(回腸)は、一部の動物では軽度の粘膜肥厚、また別の動物では萎縮を示した。腸の変化は、雌性B7RP−1トランスジェニックマウスにおいて最も重症であり、一貫して認められたが、本試験では雄性トランスジェニックマウスの一部においても認められた。
【0215】
裂溝状潰瘍及び多核巨細胞を伴った経壁性慢性肉芽腫性炎症を含めて、近位結腸で認められた組織学的特徴が、ヒトにおける潰瘍性結腸炎よりもクローン病で認められるものにより密接に類似していることは興味深い。形態学的には、この結腸炎はまた、インターロイキン−10欠損のマウスにおいて記述されているIBDに類似しており、この疾患は消耗、貧血、及び腸管全体を冒す腸炎を発現する(Kuhnら、1993;Sartor,1995;Leachら、1999)。IL−10ノックアウトマウスにおけるように、B7RP−1−Fcトランスジェニックマウスでの初期変化は、結腸上皮肥厚を伴わない粘膜固有層における炎症細胞の軽度で限局性の浸潤から成る(実施例13)。より高齢のマウスでは、罹患した結腸セグメントは腺肥大/過形成と慢性炎症のため肥厚する。B7RP1マウスの近位及び遠位結腸は、炎症性腸疾患(IBD)の組織学的特徴を持つ中等度から重度の結腸炎を有していた。近位結腸の罹患セグメント(図17B−17D)は、粘液腺の伸長と拡大を伴った顕著な腺上皮肥厚と過形成のため、広汎に肥厚しており(図17B)、粘液腺は有糸分裂像の数の増大とまれな陰窩膿瘍を有していたが、ムチンを含む杯細胞は保持していた(図17D)。粘膜は、固有層に広汎な慢性炎症があり、一部の動物では膜を越えて拡大し、粘膜下組織、筋層、漿液、及び隣接する腸間膜脂肪組織を含めて腸管壁の下層にまで及んでいた(図17B−17C)。炎症性浸潤は、慢性肉芽腫性炎症の特徴である、いくつかの好中球及び時として多核巨細胞が混じった(図17E)、リンパ球(主としてCD3+、CD44+T細胞)、プラスマ細胞、及び類上皮マクロファージ(図17F)から成った。リンパ球様凝集物(少数のCD3+細胞が混じった、主にB220+細胞)も、粘膜中及び腸間膜脂肪を含めた粘膜下組織と深層のより微小な血管の周囲に存在した(図17C)。管腔は粘液膿性又は粘液性の滲出物を含んでいた(図17D)。これらのB7RP−1−Fcトランスジェニックマウスにおいて、粘膜の多病巣性裂溝状潰瘍と経壁性炎症(図17B−17C)を伴う結腸炎の深刻な証拠が認められた。
【0216】
B7RP−1−Fcトランスジェニックマウスの遠位結腸も、結腸腺の伸長、好塩基球増加及び拡大を伴って広汎に肥厚し、増殖しており(図18B−18G)、その内の一部は陰窩膿瘍(図18D及び18F)及び粘液を含んでいた。粘膜固有層は、リンパ球(主としてCD3+、CD44+細胞、特に浅粘膜において;図18E)の軽度の散在性炎症性浸潤、ならびにプラスマ細胞と一部好中球が混じった類上皮マクロファージの巣状凝集物を有していた。リンパ球様凝集物(主としてB220+細胞;図18D及び18F)も粘膜全体に散らばっていた。B7RP−1−Fcトランスジェニックマウスの小腸は、粘膜固有層の主としてリンパプラスマ細胞性浸潤を伴った、軽度から局所的に著明な粘膜及び陰窩の肥厚と過形成(図19B及び19D、陰窩/絨毛比は、対照マウスでの1:10と比較して、1:4から1.5:1)を含めて、より多様な変化を有していた。粘膜過形成は、十二指腸(図19B)、そして特に空腸(図19D)を含めた近位小腸において最も顕著であった。最も重症のマウスでは、陰窩構造が局所的に混乱し、形成異常であった(図19D)。これに対し、一部のマウスの遠位小腸(回腸)は、絨毛の鈍化、肥厚又は局所的喪失(陰窩:絨毛比は、正常比1:2に対して1:1又はそれ以下)を伴った回腸粘膜の軽度で斑状の絨毛萎縮を有しており、また他の一部のマウスは軽度の回腸粘膜肥厚を有していた。
【0217】
B7RP−1−Fc融合タンパクは、ヒトクローン病のものに非常に類似した表現型を誘発する役割を担う細胞を活性化するように働く。これは、クローン病における炎症の原因と考えられる細胞がB7RP−1−Fc融合タンパクによって活性化されることを示唆する。B7RP−1の可溶性タンパク、抗体又は小分子阻害因子は、それ故、IBDを抑制する上で有用と考えられる。
【0218】
実施例20
B7RP−1−Fc融合タンパクはマウスにおける腫瘍増殖を抑制する
免疫原性マウスMeth A肉腫の増殖へのB7RP−1及びCRP−1の作用を調べるため、可溶性B7RP−1−FcがBalb/cマウスの確認されたMeth A肉腫の増殖に影響を及ぼすかどうかを検討した。
【0219】
対数増殖するMeth A肉腫細胞を、0日目にBalb/cマウスの腹部に50万個の細胞を皮内注入して移植した。7日目に腫瘍が〜100mm3に達し、7、10、14及び17日目にマウスを賦形剤(PBS)又はB7RP−1−Fc(8mg/kg)のいずれかにより皮下経路で処置した。腫瘍の二次元直径をカリパスで測定し、次の式を用いて腫瘍容積(mm3)を算定した:腫瘍容積=[{(幅)2×長さ}/2]。28日目まで腫瘍増殖をモニターした。各々の群は8匹のマウスで構成された。
【0220】
対照腫瘍のMeth A肉腫増殖パターンは二相性であった:緩慢な初期相の後に比較的迅速な対数期が続いた。B7RP−1−Fc処置マウスでは、腫瘍の増殖は迅速な対数期において有意に緩慢であった。28日目に、対照とB7RP−1−Fc処置マウスの平均容積はそれぞれ1410mm3と580mm3であった(図20)。それ故、B7RP−1−Fc処置はこのモデルにおいて腫瘍の増殖を有意に抑制した。データは、免疫原性腫瘍の治療における可溶性B7RP−1−Fcタンパク及びB7RP−1/CRP−1経路の他の活性化因子の有益な治療上の有用性を強く示唆している。
【0221】
免疫学的抗腫瘍活性は、細胞溶解性Tリンパ球(CTL)の機能と緊密に結びついている。一貫して、B7RP−1−Fcタンパクは細胞溶解性CD8+T細胞上で発現される(実施例9、図6)。これらのデータは、細胞溶解性CD8+T細胞へのB7RP−1の機能を強く裏付けるものである。B7RP−1−Fc又はB7RP−1/CRP−1経路の他の刺激因子は、それ故、多くの非癌関連適応症に関して細胞溶解性T細胞及び細胞性免疫機能を増強するために使用しうるであろう。
【0222】
実施例21
in vitroでのヒトB7RP−1活性の阻害
ヒトB7RP−1が正の同時刺激性を有するかどうかを同定するために、我々は、T細胞増殖アッセイにおけるヒトB7RP−1およびヒトB7RP−1−Fc融合タンパク質を発現する細胞を試験した。ヒトB7RP−1−Fc融合タンパク質を、部分的ヒトIgG1遺伝子配列へのアミノ酸1〜247に対応する遺伝子配列の融合によって構築した(実施例14)。ヒトCRP−1−Fc融合タンパク質を、部分的ヒトIgG1遺伝子配列へのアミノ酸1〜146に対応する遺伝子配列の融合によって構築した(実施例14)。両融合タンパク質の構築法、発現法、および精製法を、実施例7に記載のように行った。B7RP−1−Fcは、抗CD3刺激に依存する同時刺激活性を示した(図21a)。さらに、この活性を、溶解性CRP−1−Fcタンパク質で特異的に阻害することができる(図21b)。全コード配列を含む、膜結合ヒトB7RP−1を発現するCHO細胞を用いて、類似の同時刺激効果が得られた(図21c)。
【0223】
上記のin vitro増殖条件下でのヒトT細胞によるサイトカインの産生を同定した。48時間および72時間刺激したT細胞培養物由来の上清を、製造者の仕様書(BioSource International)に従ったELISAによってIL−2、IL−10、およびIFN−γについて分析した。IFN−γおよびIL−10レベルは有意に増加したが、CD28同時刺激の場合と異なり、IL−2は特に誘導されなかった(図21d)。実施例13に記載のように、IFN−γ(Th1サイトカイン)レベルの増加は、IgG2aを増加させるB7RP−1の機能と相関する。
【0224】
in vitroT細胞同時刺激アッセイを、以下のように行った。高度に精製したヒトT細胞(>98% CD3+)を、mAb標識磁性ビーズ(Miltenyi Biotec)を用いた新鮮または解凍した接着減少PBMCのネガティブ選択によって単離した。T細胞(1×105細胞/ウェル)を、96ウェルの3連のウェルにおいて200μl/ウェル RPMI+10% FCS中で培養した。B7RP−1−Fc同時刺激を評価するために、種々の濃度の抗CD−3(Pharmingen)および10μg/ml抗ヒトIgG Fc(Sigma)の100μl 1×PBS溶液を4℃で一晩のインキュベーションによってU底プレート上にプレコートした。非結合抗CD3および抗ヒトIgG Fcを取り除き、細胞を種々の濃度のB7RP−1−Fc、OPG−Fcコントロール、または抗CD28(Pharmingen)の存在下または非存在下で培養した。B7RP−1−Fc同時刺激のCRP−1−Fc阻害用に、連続希釈したCRP−1−FcまたはOPG−Fcの存在下で、0.5μg/mlB7RP−1−Fcを含む0.33μg/ml抗CD−3および10μg/ml抗ヒトIgG Fcプレコートウェル中で、10μg/mlから開始してT細胞を培養した。B7RP−1を発現するCHO細胞による同時刺激を評価するために、種々の量のマイトマイシン−C処理CHO B7RP−1細胞またはCHOベクター細胞の存在下または非存在下で、種々の濃度の溶解性抗CD3を含む平底プレート中でT細胞を培養した。T細胞増殖について試験するために、72時間の培養の最後の18時間に1 uCi/ウェル [3H]TdRで培養物をパルスした。T細胞増殖を、[3H]TdR組み込みによって同定した。3つの無作為なドナー由来の1つの代表的な実験結果を、平均CPM組み込み+/−SDで示す。サイトカイン産生分析用に、細胞を48時間および72時間培養し、ELISA用に上清を回収した。
【0225】
3つの実験は、実施例14に記載するように、ヒトFc断片に融合した場合、ヒトB7RP−1の細胞外部分は、in vitroでT細胞を同時刺激することができることを示す。この同時刺激は、CRP−1−Fcで阻害されるので、これは、ヒトB7RP−1の溶解性インヒビターがどのように機能し得るのかを示す。in vitroアッセイ(本明細書中に記載のヒトB7RP−1およびCRP−1を用いたアッセイ)を使用して、B7RP−1/CRP−1活性の抗体、溶解性タンパク質、ペプチ体、または小分子インヒビターをスクリーニングすることができる。
【0226】
本発明は好ましい実施形態に関して記載しているが、当業者により変形形態および修正形態が実施されることが理解される。したがって、添付の特許請求の範囲は、特許請求の範囲に記載の本発明の範囲内で実施されるこのような全ての等価な変形形態を対象とすることが意図される。
【特許請求の範囲】
【請求項1】
a)図1A(配列番号1)に記載のヌクレオチド配列、
b)図1A(配列番号1)に記載の残基1〜200または残基21〜200由来のペプチドをコードするヌクレオチド配列、
c)図1A(配列番号1)に記載のポリペプチドに少なくとも約70%同一であるポリペプチドをコードするヌクレオチド配列、
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体、
e)(a)、(b)、または(c)のいずれかに相補的なヌクレオチド配列、
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリヌクレオチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列、
g)少なくとも約10個、15個、20個、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列、および
h)ストリンジェントな条件下で、(a)〜(g)いずれかとハイブリダイズするヌクレオチド配列
からなる群から選択されるヌクレオチド配列を含む、単離核酸分子。
【請求項2】
a)図2A(配列番号11)、図3A(配列番号6)、または図12A(配列番号16)に記載のヌクレオチド配列、
b)図2A(配列番号6)に記載の残基1〜322もしくは残基47〜322、図3A(配列番号11)に記載の残基1〜288、残基19〜288、20〜288、21〜288、22〜288、24〜288、もしくは28〜288、または図12Aに記載の残基1〜302、もしくは残基19〜302、20〜302、21〜302、22〜302、24〜302、もしくは28〜302由来のポリペプチドをコードするヌクレオチド配列、
c)図2A(配列番号6)、図3A(配列番号11)または図12A(配列番号6)に記載のポリペプチドに少なくとも約70%同一であるポリペプチドをコードするヌクレオチド配列、
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体、
e)(a)、(b)、または(c)のいずれかに相補的なヌクレオチド配列、
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリヌクレオチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列、
g)少なくとも約10個、15個、20個、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列、および
h)ストリンジェントな条件下で、(a)〜(g)いずれかとハイブリダイズ形成するヌクレオチド配列
からなる群から選択されるヌクレオチド配列を含む、単離核酸分子。
【請求項3】
前記ヌクレオチド配列が発現調節配列に機能的に連結された、請求項1または請求項2に記載の核酸分子。
【請求項4】
請求項2に記載の核酸配列を含む、宿主細胞。
【請求項5】
真核細胞である、請求項3に記載の宿主細胞。
【請求項6】
原核細胞である、請求項3に記載の宿主細胞。
【請求項7】
請求項3に記載の宿主細胞の培養物を適切な培養培地中で増殖させる工程と前記培養物からポリペプチドを単離する工程とを包含する、ポリペプチド産生法。
【請求項8】
請求項7に記載の方法によって産生された、ポリペプチド。
【請求項9】
請求項1に記載の核酸分子によってコードされる、ポリペプチド。
【請求項10】
請求項2に記載の核酸分子によってコードされる、ポリペプチド。
【請求項11】
a)図1A(配列番号2)に記載のアミノ酸配列、
b)残基21に成熟アミノ末端を含む図1A(配列番号2)に記載の成熟アミノ酸配列、
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む、図1A(配列番号2)に記載のアミノ酸配列の断片、
d)(a)、(b)、または(c)のオルソログ、および
e)(a)、(b)、(c)または(d)の対立遺伝子変異体または選択的スプライス変異体
からなる群から選択されるアミノ酸配列を含む、単離ポリペプチド。
【請求項12】
a)図2A(配列番号7)、図3A(配列番号12)、または図12A(配列番号17)に記載のアミノ酸配列、
b)残基47に成熟アミノ末端を含む図2A(配列番号7)に記載の成熟アミノ酸配列、残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図3A(配列番号12)に記載の成熟アミノ酸配列、または残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図12A(配列番号17)に記載の成熟アミノ酸配列、
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む、図2A(配列番号7)、図3A(配列番号12)、または図12A(配列番号17)に記載のアミノ酸配列の断片、
d)(a)、(b)、または(c)のオルソログ、および
e)(a)、(b)、(c)または(d)の対立遺伝子変異体または選択的スプライス変異体からなる群から選択されるアミノ酸配列を含む、単離ポリペプチド。
【請求項13】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドに特異的に結合する、抗体またはその断片。
【請求項14】
モノクローナル抗体である、請求項11に記載の抗体。
【請求項15】
ヒト抗体である、請求項13に記載の抗体。
【請求項16】
ヒト化またはCDR移植抗体である、請求項13に記載の抗体。
【請求項17】
B7RP1またはB7RP1細胞外ドメインに結合する、請求項13に記載の抗体または断片。
【請求項18】
B7RP1のCRP1への結合を阻害する、請求項13に記載の抗体または断片。
【請求項19】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドおよび薬学的に受容可能なキャリア、アジュバント、可溶化剤、安定剤、または抗酸化剤を含む、組成物。
【請求項20】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドの誘導体を含む、ポリペプチド。
【請求項21】
水溶性ポリマーで共有結合的に修飾された、請求項20に記載のポリペプチド。
【請求項22】
異種アミノ酸配列に融合された請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドを含む、融合ポリペプチド。
【請求項23】
前記異種アミノ酸配列がIgG定常ドメインまたはその断片である、請求項22に記載の融合ポリペプチド。
【請求項24】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドを動物に投与する工程を包含する、T細胞媒介疾患の治療、予防、または回復法。
【請求項25】
a)請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドの発現の存在または量を同定する工程と、
b)前記ポリペプチドの発現の存在または量に基づいてT細胞媒介疾患またはT細胞媒介疾患に対する感受性を診断する工程
とを包含する、動物におけるT細胞媒介疾患またはT細胞媒介疾患に対する感受性の診断法。
【請求項26】
a)請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドと試験分子とを接触させる工程と、
b)前記試験分子への前記ポリペプチドの結合範囲を同定する工程
とを包含する、ポリペプチドに結合する試験分子の同定法。
【請求項27】
前記化合物に結合する場合に、前記ポリペプチドの活性を同定する工程をさらに包含する、請求項26に記載の方法。
【請求項28】
請求項1、請求項2、または請求項3に記載の核酸分子を動物に投与する工程を包含する、動物におけるT細胞活性化または増殖の制御法。
【請求項29】
請求項3に記載の核酸分子を含む、トランスジェニック非ヒト哺乳動物。
【請求項30】
CRP−1またはB7RP−1のアンタゴニストを動物に投与する工程を包含する、動物の免疫応答の抑制法。
【請求項31】
前記アンタゴニストがB7RP−1に結合する抗体である、請求項30に記載の方法。
【請求項1】
a)図1A(配列番号1)に記載のヌクレオチド配列、
b)図1A(配列番号1)に記載の残基1〜200または残基21〜200由来のペプチドをコードするヌクレオチド配列、
c)図1A(配列番号1)に記載のポリペプチドに少なくとも約70%同一であるポリペプチドをコードするヌクレオチド配列、
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体、
e)(a)、(b)、または(c)のいずれかに相補的なヌクレオチド配列、
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリヌクレオチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列、
g)少なくとも約10個、15個、20個、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列、および
h)ストリンジェントな条件下で、(a)〜(g)いずれかとハイブリダイズするヌクレオチド配列
からなる群から選択されるヌクレオチド配列を含む、単離核酸分子。
【請求項2】
a)図2A(配列番号11)、図3A(配列番号6)、または図12A(配列番号16)に記載のヌクレオチド配列、
b)図2A(配列番号6)に記載の残基1〜322もしくは残基47〜322、図3A(配列番号11)に記載の残基1〜288、残基19〜288、20〜288、21〜288、22〜288、24〜288、もしくは28〜288、または図12Aに記載の残基1〜302、もしくは残基19〜302、20〜302、21〜302、22〜302、24〜302、もしくは28〜302由来のポリペプチドをコードするヌクレオチド配列、
c)図2A(配列番号6)、図3A(配列番号11)または図12A(配列番号6)に記載のポリペプチドに少なくとも約70%同一であるポリペプチドをコードするヌクレオチド配列、
d)(a)、(b)、または(c)のいずれかの天然の対立遺伝子変異体または選択的スプライス変異体、
e)(a)、(b)、または(c)のいずれかに相補的なヌクレオチド配列、
f)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基のポリヌクレオチド断片をコードする(b)、(c)、または(d)のヌクレオチド配列、
g)少なくとも約10個、15個、20個、25個、50個、75個、100個、または100個を超えるヌクレオチドの断片を含む(a)、(b)、または(c)のヌクレオチド配列、および
h)ストリンジェントな条件下で、(a)〜(g)いずれかとハイブリダイズ形成するヌクレオチド配列
からなる群から選択されるヌクレオチド配列を含む、単離核酸分子。
【請求項3】
前記ヌクレオチド配列が発現調節配列に機能的に連結された、請求項1または請求項2に記載の核酸分子。
【請求項4】
請求項2に記載の核酸配列を含む、宿主細胞。
【請求項5】
真核細胞である、請求項3に記載の宿主細胞。
【請求項6】
原核細胞である、請求項3に記載の宿主細胞。
【請求項7】
請求項3に記載の宿主細胞の培養物を適切な培養培地中で増殖させる工程と前記培養物からポリペプチドを単離する工程とを包含する、ポリペプチド産生法。
【請求項8】
請求項7に記載の方法によって産生された、ポリペプチド。
【請求項9】
請求項1に記載の核酸分子によってコードされる、ポリペプチド。
【請求項10】
請求項2に記載の核酸分子によってコードされる、ポリペプチド。
【請求項11】
a)図1A(配列番号2)に記載のアミノ酸配列、
b)残基21に成熟アミノ末端を含む図1A(配列番号2)に記載の成熟アミノ酸配列、
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む、図1A(配列番号2)に記載のアミノ酸配列の断片、
d)(a)、(b)、または(c)のオルソログ、および
e)(a)、(b)、(c)または(d)の対立遺伝子変異体または選択的スプライス変異体
からなる群から選択されるアミノ酸配列を含む、単離ポリペプチド。
【請求項12】
a)図2A(配列番号7)、図3A(配列番号12)、または図12A(配列番号17)に記載のアミノ酸配列、
b)残基47に成熟アミノ末端を含む図2A(配列番号7)に記載の成熟アミノ酸配列、残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図3A(配列番号12)に記載の成熟アミノ酸配列、または残基19、20、21、22、24、または28のいずれかに成熟アミノ末端を含む図12A(配列番号17)に記載の成熟アミノ酸配列、
c)少なくとも約25個、50個、75個、100個、または100個を超えるアミノ酸残基を含む、図2A(配列番号7)、図3A(配列番号12)、または図12A(配列番号17)に記載のアミノ酸配列の断片、
d)(a)、(b)、または(c)のオルソログ、および
e)(a)、(b)、(c)または(d)の対立遺伝子変異体または選択的スプライス変異体からなる群から選択されるアミノ酸配列を含む、単離ポリペプチド。
【請求項13】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドに特異的に結合する、抗体またはその断片。
【請求項14】
モノクローナル抗体である、請求項11に記載の抗体。
【請求項15】
ヒト抗体である、請求項13に記載の抗体。
【請求項16】
ヒト化またはCDR移植抗体である、請求項13に記載の抗体。
【請求項17】
B7RP1またはB7RP1細胞外ドメインに結合する、請求項13に記載の抗体または断片。
【請求項18】
B7RP1のCRP1への結合を阻害する、請求項13に記載の抗体または断片。
【請求項19】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドおよび薬学的に受容可能なキャリア、アジュバント、可溶化剤、安定剤、または抗酸化剤を含む、組成物。
【請求項20】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドの誘導体を含む、ポリペプチド。
【請求項21】
水溶性ポリマーで共有結合的に修飾された、請求項20に記載のポリペプチド。
【請求項22】
異種アミノ酸配列に融合された請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドを含む、融合ポリペプチド。
【請求項23】
前記異種アミノ酸配列がIgG定常ドメインまたはその断片である、請求項22に記載の融合ポリペプチド。
【請求項24】
請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドを動物に投与する工程を包含する、T細胞媒介疾患の治療、予防、または回復法。
【請求項25】
a)請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドの発現の存在または量を同定する工程と、
b)前記ポリペプチドの発現の存在または量に基づいてT細胞媒介疾患またはT細胞媒介疾患に対する感受性を診断する工程
とを包含する、動物におけるT細胞媒介疾患またはT細胞媒介疾患に対する感受性の診断法。
【請求項26】
a)請求項9、請求項10、請求項11、または請求項12に記載のポリペプチドと試験分子とを接触させる工程と、
b)前記試験分子への前記ポリペプチドの結合範囲を同定する工程
とを包含する、ポリペプチドに結合する試験分子の同定法。
【請求項27】
前記化合物に結合する場合に、前記ポリペプチドの活性を同定する工程をさらに包含する、請求項26に記載の方法。
【請求項28】
請求項1、請求項2、または請求項3に記載の核酸分子を動物に投与する工程を包含する、動物におけるT細胞活性化または増殖の制御法。
【請求項29】
請求項3に記載の核酸分子を含む、トランスジェニック非ヒト哺乳動物。
【請求項30】
CRP−1またはB7RP−1のアンタゴニストを動物に投与する工程を包含する、動物の免疫応答の抑制法。
【請求項31】
前記アンタゴニストがB7RP−1に結合する抗体である、請求項30に記載の方法。
【図1A】


【図1B】

【図2A−1】
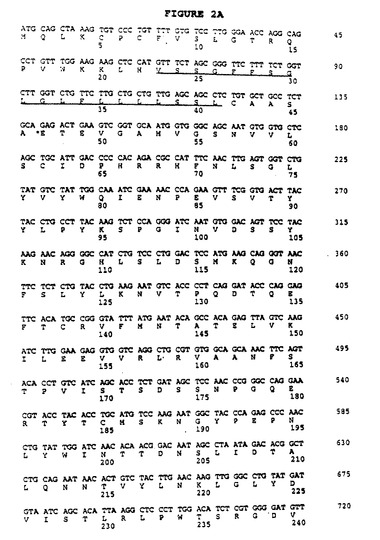
【図2A−2】

【図2B】

【図3A−1】
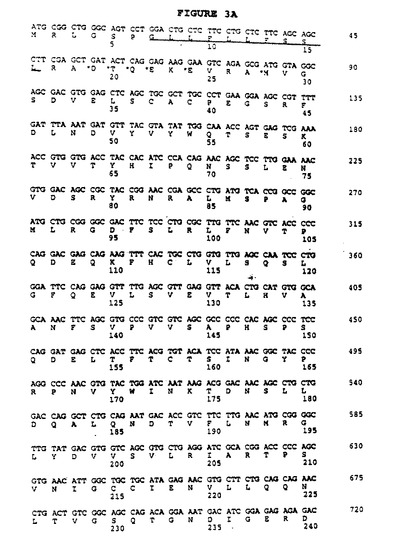
【図3A−2】

【図3B】

【図4A】
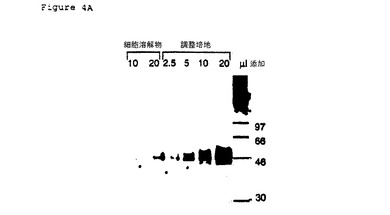
【図4B】
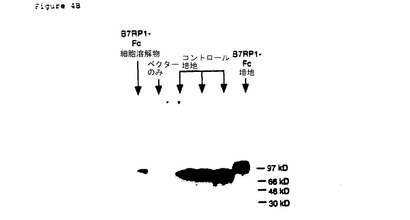
【図5】
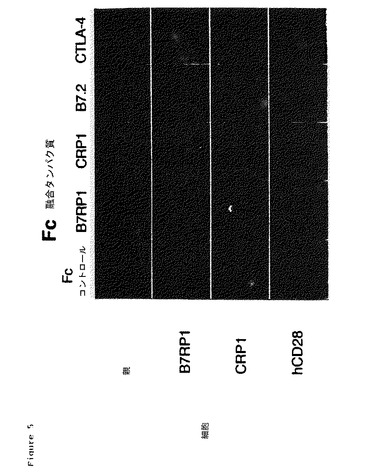
【図6】
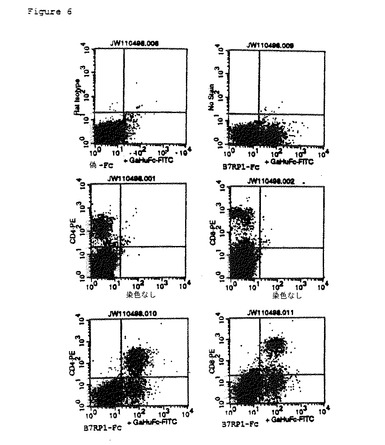
【図7】
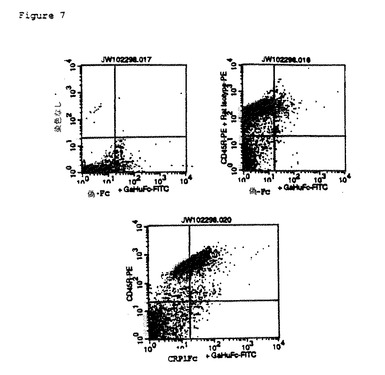
【図8】
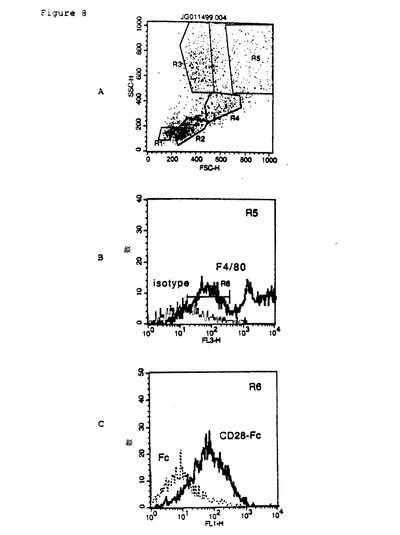
【図9】
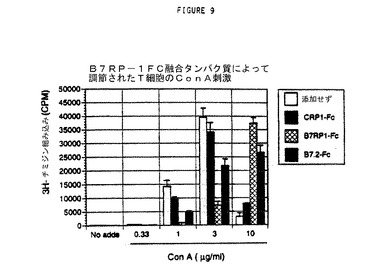
【図10】
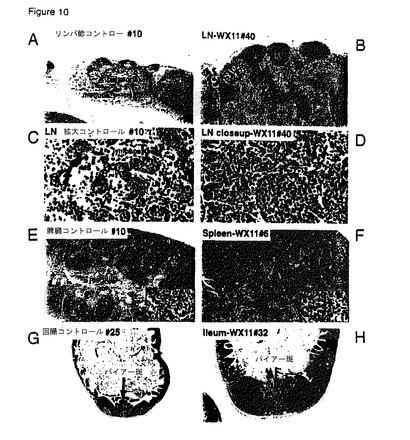
【図11】
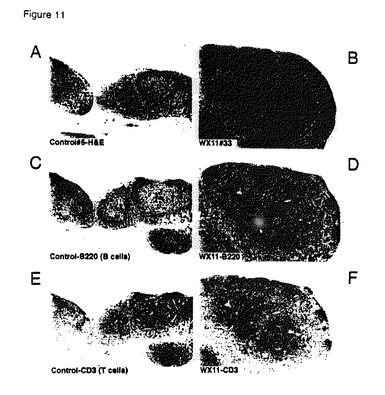
【図12A−1】

【図12A−2】

【図12B】

【図13A−1】
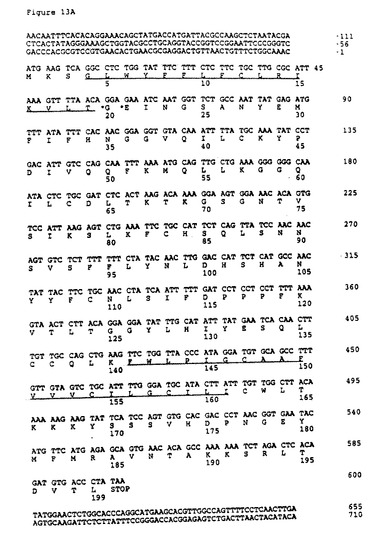
【図13A−2】

【図13B】

【図14】
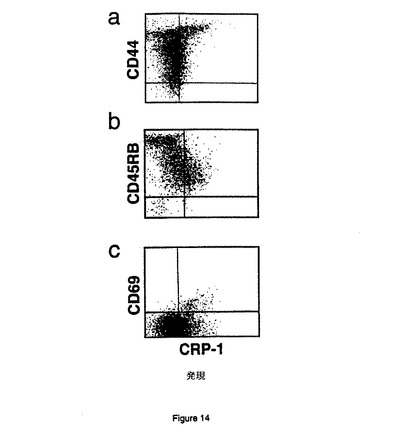
【図15】
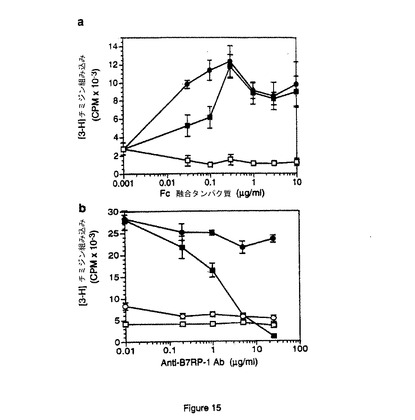
【図16】
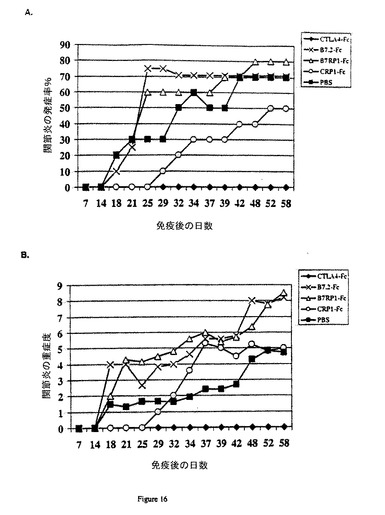
【図17】
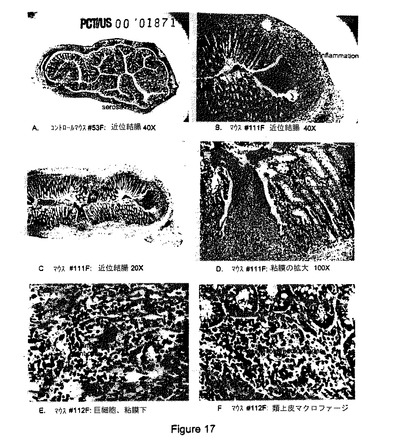
【図18】
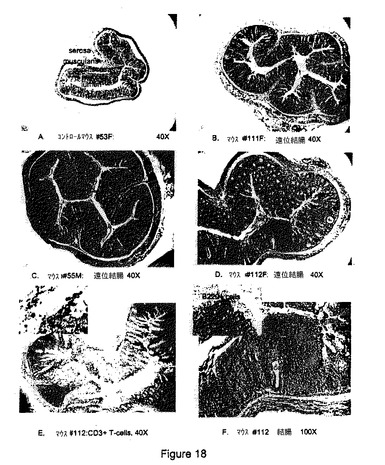
【図19】
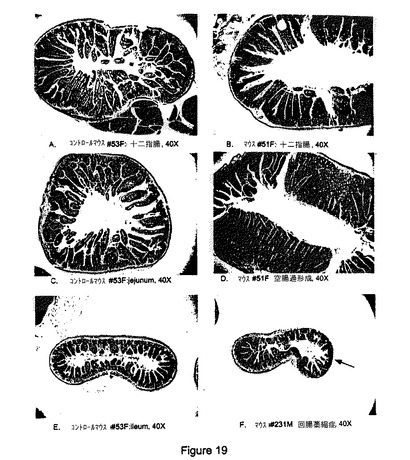
【図20】
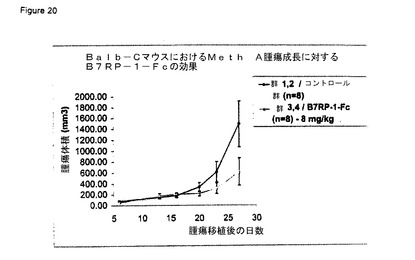
【図21A】
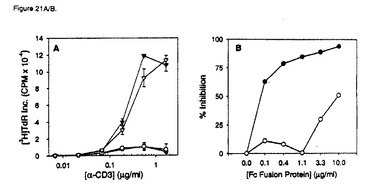
【図21B】
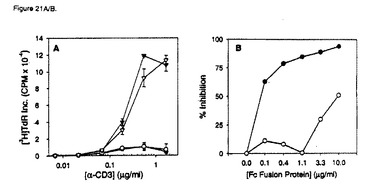
【図21C】
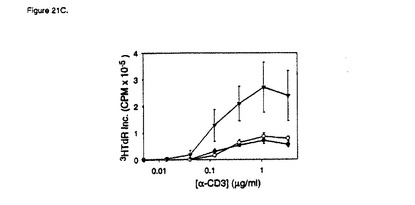
【図21D】
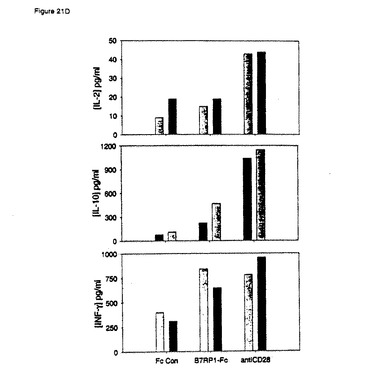
【図1B】
【図2A−1】
【図2A−2】
【図2B】
【図3A−1】
【図3A−2】
【図3B】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12A−1】
【図12A−2】
【図12B】
【図13A−1】
【図13A−2】
【図13B】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21A】
【図21B】
【図21C】
【図21D】
【公開番号】特開2012−110333(P2012−110333A)
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−749(P2012−749)
【出願日】平成24年1月5日(2012.1.5)
【分割の表示】特願2000−597310(P2000−597310)の分割
【原出願日】平成12年1月27日(2000.1.27)
【出願人】(500049716)アムジエン・インコーポレーテツド (242)
【Fターム(参考)】
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願番号】特願2012−749(P2012−749)
【出願日】平成24年1月5日(2012.1.5)
【分割の表示】特願2000−597310(P2000−597310)の分割
【原出願日】平成12年1月27日(2000.1.27)
【出願人】(500049716)アムジエン・インコーポレーテツド (242)
【Fターム(参考)】
[ Back to top ]