
免疫性を有するペプチド(TheireliaorientalisToMRP抗原)
【課題】
T.orientalis抗原に応答してIFN-γ及びIL-10を産生する各T細胞数の増減を評価する、タイレリア感染症の診断システムの利便性をより向上させるために、新たなタイレリア抗原を提供する。
【解決手段】
T.orientalisのToMRP抗原の部分ペプチドを合成し、該ペプチドを上記診断システムにおけるT細胞免疫応答の刺激剤として使用する。該ペプチドは、T.orientalis感染ウシにおいてToMRP抗原特異的に産生するIFN-γ分泌細胞及びIL-10を産生する分泌細胞を刺激し、IFN-γ分泌及びIL-10を分泌させる。これらの量を測定することにより、T.orientalis感染症の診断を有利に行う。
T.orientalis抗原に応答してIFN-γ及びIL-10を産生する各T細胞数の増減を評価する、タイレリア感染症の診断システムの利便性をより向上させるために、新たなタイレリア抗原を提供する。
【解決手段】
T.orientalisのToMRP抗原の部分ペプチドを合成し、該ペプチドを上記診断システムにおけるT細胞免疫応答の刺激剤として使用する。該ペプチドは、T.orientalis感染ウシにおいてToMRP抗原特異的に産生するIFN-γ分泌細胞及びIL-10を産生する分泌細胞を刺激し、IFN-γ分泌及びIL-10を分泌させる。これらの量を測定することにより、T.orientalis感染症の診断を有利に行う。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、タイレリア・オリエンターリス(Theirelia orientalis以下T. orientalis)抗原ToMRPに由来するペプチド、及び該抗原ペプチドを用いたタイレリア感染症の診断手段に関する。
【背景技術】
【0002】
タイレリア症はダニ媒介性の寄生虫感染症で、肥育効率を損ない畜産業における生産性が損なわれる原因となる。タイレリア感染症は、その病態病理像の違いからlymphoproliferative groupとnonlymphoproliferative groupに分類される。
Lymphoproliferatative group に含まれるTheileria parva(以下T.parva)やTheileria annulata(以下T.annulata)は、アフリカをはじめとする高温地域を中心に感染が確認されている。
一方、nonlymphoproliferative groupに分類されるT.orientalis (=T.selgenti)は、我が国を始め、東アジアからヨーロッパまで広範囲にその感染が確認されている。T.orientalis感染は、ダニによる吸血によってウシ体内へ送り込まれたスポロゾイトが、細網内皮系細胞に侵入してシゾントとなって増殖し、その後、血中へ出たメロゾイトが赤血球へ寄生することによって成立すると考えられている。
Lymphproliferative groupのT.parvaやT.annulataでは、寄生するシゾントがJNK1/2の恒常的活性化をもたらしてTやBリンパ球をtransformするため、リンパ系細胞の異常増殖とそれに関連すると考えられる症状lymphoproliferative disease(リンパ球増殖症)が出現する。一方、nonlymphoproliferative groupのT.orientalisには、リンパ球に対するtransform活性は確認されていない。一方で免疫系細胞から生産される活性酸素による赤血球の膜変性が生じ、脾臓の網内皮系による変性赤血球除去が亢進することで生じると考えられる貧血と、それに関連した慢性消耗疾患(Chronic Wasting Disease)が出現する事が知られている。但し、T.orientalisの感染動向を調査した結果、抗ダニ剤が普及した現在では、原虫が感染した赤血球が末梢血中に観察されるものの、貧血等の明らかな臨床所見を呈する事は稀で、大部分は不顕性感染となる。(以上、非特許文献1〜5参照)
【0003】
感染原因に対する免疫応答メカニズムを解明する事は、それぞれに特有の病態を克服する手掛かりとなる。Lymphproliferative groupのタイレリア感染について、T.parvaやT.annulataでは、その感染によってCD4+T細胞とCD8+T細胞による免疫応答が惹起され、同CD8+T細胞=細胞障害性T細胞(cytotoxic T lymphocyte=CTL)が主たる排除エフェクターとして作用する事、そしてその誘導維持にはCD4+(ヘルパーT細胞=helper T cell(Th))が必要であることが明らかにされている。またγδT cellは、MHC非拘束性に感染細胞によって活性化される事、そしてそれによって感染細胞を障害する事が知られている。
したがって、lymphoproliferative groupに属するTheileria感染症においては、原虫抗原特異的なCD8陽性T細胞の増加を検出する事は、感染状態である事を意味する。(以上、非特許文献6〜9参照)
【0004】
一方、nonlymphoproliferative groupのT.orientalis感染において、本発明以前に明らかとなっている事象としては、次のようなものが挙げられる。1)T.orientalis感染では末梢血中のリンパ球やマクロファージなどの単核球細胞が活性化するがその特異性は明らかでない。2)マラリア症、バベシア症やアナプラズマ症と言った赤血球寄生性原虫と同様、感染・変性赤血球は脾臓で除去される。3)血中のIFN-γ値が高い程、全身症状は穏やかである。4)T.orientalis感染末梢血をPHAで刺激すると、IFN-γ産生が検出される。
しかしながら、これらの事象のいずれにおいても、タイレリア抗原特異的にT細胞が存在して、免疫システムを活性化しているのかどうか、もしくは、どのような免疫応答をもたらしているのかは十分に明らかにされていない。(以上、特許文献10〜14参照)
【0005】
抗原特異的なT細胞を検出する方法が幾つか開発されている。なかでもELISpot(Enzyme-linked immunospot)technologyは、刺激よって産生される特定のサイトカインを分泌する細胞の数を評価する方法として広く活用されている。ELISpotアッセイは最初、抗体を産生する細胞を検出する方法として報告されており、IFN-γの検出に関しては、同じグループから1988年の「Reverse ELISPOT assay for clonal analysis of cytokine production I.Enumeration of gamma-interferon-secreting cells」に報告されている(非特許文献14参照)。
【0006】
一方、本発明者は、T.orientalis感染ウシにおいて免疫応答の進展(サイトカイン産生変化)を詳細に解析したところ、感染後、速やかにMPSP(major piroplasm surface protein)ペプチド特異的にIFN-γを産生する細胞が増加し(Th1免疫応答)、時間経過と伴に減少して消失したが、このIFN-γ産生する細胞の減少に従い、MPSPペプチド特異的に反応してIL-10を産生する細胞が増加する(免疫抑制性T細胞免疫応答)という知見を得たうえで、同ペプチドと一連の解析手法について特許出願している(特許文献1)。さらには、MPSPに加えて、特異的T細胞検出ペプチドの多様化を図るべく、同じくT.orientalisに由来する抗原p23においても、鋭意研究の結果、T.orientalisに感染したウシ体内において、p23に由来するペプチドに特異的に応答するT細胞が発生することを証明し、同ペプチドと一連の解析手法についても特許出願している。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2010−75121号公報
【非特許文献】
【0008】
【非特許文献1】A.D.Irvin.Parasitology Today Vol.1 No.5 p124-128,1985
【非特許文献2】Yves Galley et al,Proc.Natl,Acad,Sci.USA Vol 94,p5119-5124,1997
【非特許文献3】Regina Lizundia et al,Cancer Res:66(12)p6105-6110,2006
【非特許文献4】Wendy C.Brown Veterinary Parasitology,1001 p233-248,2001
【非特許文献5】Misao Onuma,et al Comparative Immunology,Micrology & Infectious Disease 21,p165-177,1998)
【非特許文献6】Declan McKeever,Proc.Natl,Acad,Sci.USA Vol91,p1959-1963,1994
【非特許文献7】Evans Taracha,Journal of Immunology,Vol 159,p4539-4545
【非特許文献8】Simon Graham,Proc.Natl,Acad,Sci.USA,Vol.103,p3286-3291,2006
【非特許文献9】Claudia Daubenberger et al Infection and Immunity,p2241-2249,1999
【非特許文献10】Yasutomi,Y.et al,Vet Parasitology,Vol39, p33-41,1991
【非特許文献11】Asaoka H.,et al,Res vet Sci,Vol50,p23-28,1991
【非特許文献12】Hagiwara K.et al,Vet Parasitology,Vol127,p105-110,2005
【非特許文献13】Kakuda T.,et al.,J.Vet.Med.Sci.,vol63,p895-901,2001
【非特許文献14】Czerkinsky CC et.al.,J Immunol Methods.1983 Dec 16;65(1-2):109-21
【発明の概要】
【発明が解決しようとする課題】
【0009】
上記のとおり、本発明者は、牛タイレリア感染症の診断手段として、T.orientalis抗原MPSPおよびp23に応答してIFN-γ及びIL-10を産生する各T細胞数の増減を評価する新たな診断システムの構築しており、本願発明の課題は、このような診断システム使用の利便性をより向上させるために、使用しうるタイレリア抗原の多様化を図るとともに、T細胞免疫応答を誘導する能力が高く、より使い勝手のよいタイレリア抗原を提供する点にある。
【0010】
本発明者は、上記課題を解決するため鋭意研究の結果、タイレリア抗原の一種であるToMRP抗原にT細胞免疫応答能があるとの知見を得て、これをさらに一歩進め、ToMRP抗原の各種部分ペプチドを合成し、T細胞免疫応答能を調べた結果、ToMRP抗原の部分ペプチドにおいてT細胞免疫応答を誘導する能力を有するものがあることを見いだし、本発明を完成させるに至った。
【0011】
すなわち、本発明は以下のとおりである。
(1)配列番号3〜5、10及び12のいずれかに示されるアミノ酸配列を有するペプチド。
(2)上記(1)に記載のペプチドからなる、ウシのT.orientalis感染症診断剤。
(3)上記(1)に記載のペプチド及びIFN-γに対する抗体及び/又はIL-10に対する抗体を少なくとも含むことを特徴とする、ウシのT.orientalis感染症診断キット。
【発明の効果】
【0012】
Theileriosisの原因の一つで、lymphoproliferative groupに属するT.parvaについては、すでに、T細胞免疫応の全容がほぼ解明されている。これに対してnonlymphoproliferative groupに属するT.orientalis感染症に関しては、免疫システムで極めて重要な役割を果たす病原体由来抗原特異的T細胞がジェネレーションされるかどうか詳しくはわかっていなかった。
【0013】
このような状況下、本発明は、T.orientalis感染ウシ個体中に同病原体に由来するMPSPやp23とは異なる抗原、すなわち、新たに、T.orientalis由来ToMRP抗原に特異的に応答してIFN-γ及びIL-10を産生するT細胞が存在し、これを検出することに成功したことに基づく。このことは、先のMPSP(特許文献1)およびp23に由来する抗原ペプチドを用いた解析法に加えて、別の抗原であるToMRPによる解析法を可能にしたことを意味し、より多角的なT.orientalis感染ウシに対する病態診断手法をもたらすものである。
【0014】
本発明のペプチドは、上記ToMRP抗原特異的なT細胞免疫応答の誘導能が高く、T.orientalisに感染したウシが産生するToMRP抗原特異的なIFN-γ分泌T細胞及びIL-10分泌T細胞を刺激し、IFN-γ及びIL-10を分泌させ、このIFN-γ及びIL-10を感度よく検出、測定することが可能となる。このIFN-γ及びIL-10はT.orientalis感染に基づくToMRP抗原特異的に産生されたT細胞の量を反映するから、これにより、T.orientalisの感染の有無、該感染に基づく病態の程度を診断することができる。
以上のとおり、本発明のペプチド抗原はT.orientalis感染症の診断剤として、極めて有用である。
【図面の簡単な説明】
【0015】
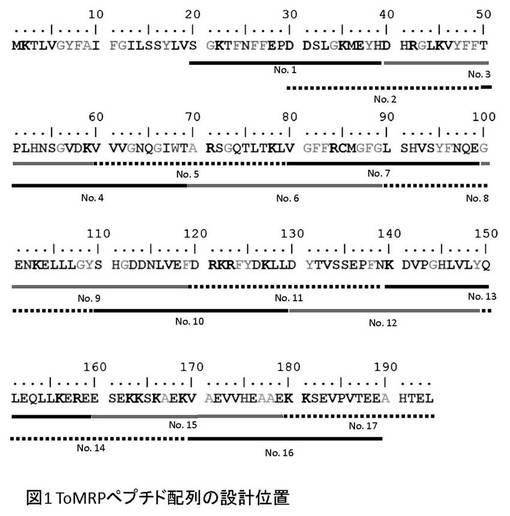
【図1】T.orientalis 由来抗原ToMRPに特異的なT細胞を検出すために合成した各ペプチドのToMRP上の位置を示す図。
【図2】T.orientalis自然感染ウシのPBMCsに対し、本発明の抗原ペプチド(全17種類)を用いて刺激し、IFN-γ ELISpot解析を行った結果を示す図。
【図3】T.orientalis人工感染ウシのPBMCsに対し、本発明の抗原ペプチド(全17種類)を用いて刺激し、IFN-γ ELISpot解析を行った結果を示す図。
【発明を実施するための最良の形態】
【0016】
本発明の抗原ペプチドは、T.orientalis原虫のToMRP(T.orientalis microneme-rhoptry protein)の部分ペプチドであって、配列番号3〜5、10、12に示される。
ToMRPは、T.orientalisのpiroplasmステージのcDNAライブラリーからクローニングされ、T.parvaのMRP(=p104)と相同性が高いことから、その名がつけられたものである。ToMRPの機能の多くは不明であるが、T.orientalisと同様に赤血球に寄生することが知られるPlasmodium(=マラリア)のmicroneme-rhoptry細胞内小器官に関連する遺伝子群は赤血球に侵入する際に重要な役割を果たすことが知られている。実際、ToMRPは赤血球膜を構成するBand3と結合することも知られているため、赤血球への侵入(もしくは脱出)に利用される可能性が考られる。
本発明のペプチドは、ウシのT.orientalis原虫感染の有無あるいは、その病態を診断するための診断剤として用いられる。以下に、本発明のペプチドを用いたT.orientalisの感染症の診断手法について説明する。
ウシがT.orientalis原虫に感染した場合のT細胞免疫応答は、IFN-γの分泌により特徴づけられる細胞性免疫促進型のT細胞免疫応答と、IL-10の分泌により特徴づけられる免疫抑制型のT細胞免疫応答で構成される。これらT.orientalis原虫抗原特異的なIFN-γ分泌T細胞とIL-10分泌T細胞の数は、T.orientalis感染の進行を反映し、IFN-γ分泌T細胞とIL-10分泌細胞の数がともに一定値を超えるウシは、T.orientalis感染による、貧血等の特徴的な症状が顕在化することが、MPSPペプチドを用いた解析によって明らかになっている(特許文献1)。
【0017】
本発明を構成するT.orientalis抗原ToMRP由来ペプチドは、ウシ末梢血に含まれる精製されたPBMCs懸濁液中に含まれるT細胞のうち、T.orientalis抗原ToMRPに特異的に応答するIFN-γ分泌T細胞及びIL-10分泌T細胞を刺激し、活性化する。
T.orientalis抗原ToMRP特異的に応答してIFN-γを分泌するT細胞もしくは、IL-10を分泌するT細胞は、それぞれIFN-γ ELISpot法およびIL-10 ELISpot法によって検出される(最終的にT.orientalis抗原ToMRP特異的に応答するIFN-γ分泌T細胞及びIL-10分泌T細胞数がスポット数として計測される)(特許文献1)。
【0018】
このToMRPペプチドによる刺激は、複数のToMRPペプチド抗原が混和されたものでも遂行可能である。
本発明のペプチドを使用するウシT.orientalis感染症の診断に使用される被験試料のPBMCs懸濁液は、ウシから採取された末梢血から精製されたものをそのまま用いてもよいし、また、凍結保存したものを用いてもよい。
【0019】
本発明における細胞の具体的な調製法は次のとおりである。ウシより採取された抗凝固処理した末梢血液に対して、HISTOPAQE-1077(登録商標SIGMA-ALDRICH社)などを用いた密度遠心分離作業を施すことによって単離・精製されたPBMCsをペプチド刺激対象細胞として使用する。このように精製されたPBMCを、精製直後に解析に使用しない場合は、細胞凍結保護液CP-1(極東製薬工業株式会社)などを用いて凍結保存してもよい。凍結保存されたPBMCsは解凍することでペプチドに応答可能な細胞群として取り扱うことができる。また、ELISpot解析時に使用される細胞懸濁液の調製に使用される培地は、特に限定されるものではなく、通常の生命科学の細胞培養実験に使用される培地等が使用されるが、可能な限りRPMI1640培地へ最終濃度10%非働済ウシ胎仔血清(FCS)、50μM2-メルカプトエタノール、100units/ml penicillin、100μg/ml streptomycinを添加したRPMIコンプリートメディウムが望ましい。また、使用するPBMCsはRPMIコンプリートメディウムへ懸濁した際に、市販の細胞数検出機(例:chemometec社製NucleoCassette)もしくは通常の細胞計算に使われる血球計算盤を用いて細胞数を算出した後に測定に用いる。
【0020】
同ELISpot解析に使用する抗IFN-γ抗体および抗体IL-10抗体は、ウシ由来のIFN-γ及びIL-10を認識・結合することできるものであればよい。又、抗体の種類は、モノクローナル抗体およびポリクローナル抗体のどちらであってもよい。
サイトカイン分泌T細胞の数を測定する方法は、抗体による細胞内染色を行ないフローサイトメーターで検出する等いくつか知られているが、本願発明においては、感度の高さおよび抗原特異的T細胞を検出する目的からELISpot(Enzyme-Linked ImmunoSpot)法が好ましく用いられる。
【0021】
このように求められたT.orientalis由来抗原ToMRPペプチドに特異的に応答するIFN-γ分泌T細胞及びIL-10分泌T細胞数は、T.orientalis感染の状態を反映し、診断対象のウシにおいてIFN-γ分泌T細胞及びIL-10分泌T細胞数がともに検出されない場合には、診断対象のウシはT.orientalisに感染しておらず、また、IFN-γ分泌T細胞及びIL-10分泌T細胞数が単独もしくは両方ともに一定基準値を超える場合には、T.orientalisに感染した上で、同感染に特徴的な病態を発症しているか、あるいは発症する可能性がある状態であるものと判定することができる。この基準値は、IFN-γ分泌T細胞及びIL-10分泌T細胞数とT.orientalis感染症の発症との関係を照らし合わせることにより、求めることが可能である。
【0022】
例えば、IFN-γ検出ウエルにおいて1×10E6cells、IL-10検出ウエルにおいて1×10E5cellsのPBMCを播種して測定する場合、それぞれの検出されたサイトカインスポット数が「10」とすると、PBMCs1×10E6個あたりIFN-γ分泌T細胞が10個、IL-10分泌T細胞が100個含まれていることを意味する。これらの1ウエル当たりのスポット数についての有効値は、同システムの利用者が用いる実験条件(例えば、1ウェル当たりのPBMCsの数や培養時間)や、研究者の求める病態・精度によって最適化される可能性がある。
【0023】
上記ELISpot法に対して、既存のPCR法、血液スメア解析、既存の一般血液解析を組み合わせる事で、免疫応答のバランスと病態の診断をより確実に行うことも可能である。すなわち、PCR法および血液スメア解析によって、血液内に存在するT.orientalisをELISpot結果に遅れて検出することでELISpot結果を補強することが可能であるし、一般血液解析によって貧血等の病態の発症の有無を直接示すことでELISpot結果との相関性を示すことが可能である。
【実施例】
【0024】
以下に、本発明を実施例として説明するが、本発明は該実施例に何ら限定・制約されるものではない。
【0025】
実施例1
本実施例において用いた手法及び抗原ペプチドは以下に示される。
1)ELISpotアッセイの準備
下記の免疫活性化反応の培養が終了するまでの操作は安全キャビネットもしくはクリーンベンチ内で無菌的に行った。
底面がPolyVinylidine DiFluoride(PVDF)メンブランで構成されている96ウエルプレート(Millipore社)を準備し、各ウエルへ70%EtOHを50μlずつ加え30秒−2分間処理することにより疎水性PVDFメンブランの親水処理を行った。ウエル内のEtOHをアスピレーターにより吸引除去後、Phosphate Buffered Saline(Ca2+ Mg2+ 無添加)(PBS)を各ウエルあたり150μl加えた後にアスピレーターで取り除く洗浄作業を5回繰り返す。洗浄操作が終了した後、抗bovine IFN-γキャプチャー抗体(MABTECH code 3115-3)または、抗IL-10キャプチャー抗体(AbD serotec,code MCA2110,clone CC318)をPBSにより7.5μg/mlに希釈し、80μlずつ加え4℃、一晩処理することにより底面のPVDF メンブランへキャプチャー抗体をコーティングさせた。翌日に余剰なキャプチャー抗体はアスピレーターにより吸引除去後、各ウエルを150μlのPBSで洗浄作業を行った。洗浄終了後のウエルにはRPMIコンプリートメディウム(RPMIメディウムへ最終濃度10%FCS(本実験系では非働化処理したFCSを使用)、50μM2-メルカプトエタノール、100units/ml penicillin、100μg/ml streptomycinになるように添加したもの)を100μlずつ加えて室温30分間もしくは4℃、1時間以上のブロッキング処理を行った。また、処理温度が4℃でのブロッキング処理を行った場合はELISpot assayを行う前にプレートを室温へ戻した。
【0026】
2)ELISpotアッセイに使用する細胞の準備
目的のウシ個体から抗凝固処理した末梢血液を遠心することでPBMCsを含むバフィーコートを形成させた。同バフィーコートを慎重に回収後、混入してきた赤血球をHISTOPAQE(登録商標)-1077(SIGMA-ALDRICH社)を使用した密度遠心分離法によって取り除くことで、精製PBMCsを得た。また、単離後すぐに使用しない場合は細胞凍結保護液CP-1(極東製薬工業株式会社)などを用いて必要になるまで保存した。精製・回収されたPBMCsはRPMIコンプリートメディウムで洗浄後、同メディウムへ再懸濁させ細胞数検出キットNucleoCassetteTM(chemometec)もしくは血球計算盤を用いて細胞数を算出しておいた。
【0027】
3)ELISpotアッセイにおける免疫活性化反応
上記1)のキャプチャー抗体がコーティングされ、加えてブロッキング処理が施された96ウエルフィルタープレートにおいて、ブロッキング処理に用いたRPMIコンプリートメディウムを除去した後に、抗bovine IFN-γキャプチャー抗体をコーティングしたウエルには3×10E5cellsの細胞を含むRPMIコンプリートメディウムを100μlずつウエルへ播種した。続けて、ToMRPに由来するT細胞刺激用抗原ペプチドをRPMIコンプリートメディウムで最終使用濃度の2倍濃度に調製し、同刺激用抗原ペプチド入りコンプリートメディウムを100μlずつPBMCs入りのウエルに加えた(この段階で、目的の最終濃度のT細胞刺激用抗原ペプチドを含む細胞懸濁液が1ウエル当たり200μl含まれることになる)。ELISPOTアッセイのコントロールとして、最終濃度1μg/ml phytohemagglutin(PHA)とPBMCsを含むウエルを用意した。最後に、ウエル内のPBMCsと刺激物質の混合液を数回ピペティングし、37℃のCO2インキュベーター内で18−42時間、振動を与えないように培養した。
【0028】
4)ELISpotアッセイにおけるスポットの検出
細胞懸濁液を除去した後に200μlPBSで5回洗浄作業をし、0.5%FCSを含むPBS(0.5%FCS/PBS)で希釈した抗bovine IFN-γビオチン化抗体(最終濃度0.25μg/ml MABTECH code 3115-6)をウエルへ80μlずつ加えて室温2時間反応させた。ビオチン化抗体溶液を除去し、200μl PBSで5回洗浄作業をした後に0.5%FCS/PBS で500〜1000倍希釈したHorseradish Peroxidase(HRP)をコンジュゲートしたstreptavidin(MABTECH code 3310-9)を室温1時間反応させた。その後、HRP 溶液を除去し、200μlPBSで5回洗浄作業をした後に3-Amino-9-ethylcarbazole(AEC)substrate kit(BD Biosciences code 551951)を用いて室温30分間、発色させた。流水により反応液を洗い流し発色反応を停止させた。プレートは風乾によって十分に乾燥させた。ウエル底面に現れたスポットは顕微鏡によってカウントした。
【0029】
5)抗原ペプチド
図1に示すように、T.orientalis由来抗原ToMRPタンパク質のアミノ酸配列(配列番号18)において、シグナルペプタイド部位以降のアミノ酸配列に基づき、C末端方向に10アミノ酸をオーバーラップさせながら20アミノ酸残基を有するペプチドを順次合成し、17種のペプチドを得た(Int.J.Parasitol.,29:593-599,1999 Sako Y et al)。なお、最後のNo.17のみ15アミノ酸で構成した。これら17種のペプチドのアミノ酸配列(配列番号1〜17)を以下に示す。
【0030】
・ToMRP No.1
SGKTFNFFEPDDSLGKMEYH(配列番号1)
・ToMRP No.2
DDSLGKMEYHDHRGLKVYFF(配列番号2)
・ToMRP No.3
DHRGLKVYFFTPLHNSGVDK(配列番号3)
・ToMRP No.4
TPLHNSGVDKVVVGNQGIWT(配列番号4)
・ToMRP No.5
VVVGNQGIWTARSGQTLTKL(配列番号5)
・ToMRP No.6
ARSGQTLTKLVGFFRCMGFG(配列番号6)
・ToMRP No.7
VGFFRCMGFGLSHVSYFNQE(配列番号7)
・ToMRP No.8
LSHVSYFNQEGENKELLLGY(配列番号8)
・ToMRP No.9
GENKELLLGYSHGDDNLVEF(配列番号9)
・ToMRP No.10
SHGDDNLVEFDRKRFYDKLL(配列番号10)
・ToMRP No.11
DRKRFYDKLLDYTVSSEPFN(配列番号11)
・ToMRP No.12
DYTVSSEPFNKDVPGHLVLY(配列番号12)
・ToMRP No.13
KDVPGHLVLYQLEQLLKERE(配列番号13)
・ToMRP No.14
QLEQLLKEREESEKKSKAEK(配列番号14)
・ToMRP No.15
ESEKKSKAEKVAEVVHEAAE(配列番号15)
・ToMRP No.16
VAEVVHEAAEKKSEVPVTEE(配列番号16)
・ToMRP No.17
KKSEVPVTEEAHTEL(配列番号17)
【0031】
6)上記5)の方法により合成したに示した17種のペプチドを用いた刺激により、IFN-γELISpotアッセイを行った。
放牧によりT.orientalis原虫に自然に感染したウシ(自然感染ウシ)2頭(#1と#2)と、タイレリア原虫を含む血液を接種したウシ(人工感染ウシ)2頭(#3と#4)から、上記2)の方法により回収したPBMCを用いた。上記3)に記載の手法(免疫活性化反応)に従い、上記1)に記載の抗bovine IFN-γ キャプチャー抗体をコーティングしたプレート(IFN-γ検出プレート)に、上記2) の方法に従い調整した標的細胞と、上記5)の方法により合成した17種の抗原ペプチド(最終濃度:1μM) を混合し42時間インキュベーションした。その後、上記4)のELISpotアッセイの操作を行ってスポットを検出した。結果を図2、3に示す。
感染が成立したウシを解析した結果であるウシ#2、#3、#4において、共通してToMRP No.3〜5、10、12のペプチド(ウシ#1ではNo.3、5)に強い反応が認められ、加えて、ウシ#1において ToMRP No.1が、ウシ#4においてToMRP No.16が独立して高い反応を示す領域であることが認められた。
【技術分野】
【0001】
本発明は、タイレリア・オリエンターリス(Theirelia orientalis以下T. orientalis)抗原ToMRPに由来するペプチド、及び該抗原ペプチドを用いたタイレリア感染症の診断手段に関する。
【背景技術】
【0002】
タイレリア症はダニ媒介性の寄生虫感染症で、肥育効率を損ない畜産業における生産性が損なわれる原因となる。タイレリア感染症は、その病態病理像の違いからlymphoproliferative groupとnonlymphoproliferative groupに分類される。
Lymphoproliferatative group に含まれるTheileria parva(以下T.parva)やTheileria annulata(以下T.annulata)は、アフリカをはじめとする高温地域を中心に感染が確認されている。
一方、nonlymphoproliferative groupに分類されるT.orientalis (=T.selgenti)は、我が国を始め、東アジアからヨーロッパまで広範囲にその感染が確認されている。T.orientalis感染は、ダニによる吸血によってウシ体内へ送り込まれたスポロゾイトが、細網内皮系細胞に侵入してシゾントとなって増殖し、その後、血中へ出たメロゾイトが赤血球へ寄生することによって成立すると考えられている。
Lymphproliferative groupのT.parvaやT.annulataでは、寄生するシゾントがJNK1/2の恒常的活性化をもたらしてTやBリンパ球をtransformするため、リンパ系細胞の異常増殖とそれに関連すると考えられる症状lymphoproliferative disease(リンパ球増殖症)が出現する。一方、nonlymphoproliferative groupのT.orientalisには、リンパ球に対するtransform活性は確認されていない。一方で免疫系細胞から生産される活性酸素による赤血球の膜変性が生じ、脾臓の網内皮系による変性赤血球除去が亢進することで生じると考えられる貧血と、それに関連した慢性消耗疾患(Chronic Wasting Disease)が出現する事が知られている。但し、T.orientalisの感染動向を調査した結果、抗ダニ剤が普及した現在では、原虫が感染した赤血球が末梢血中に観察されるものの、貧血等の明らかな臨床所見を呈する事は稀で、大部分は不顕性感染となる。(以上、非特許文献1〜5参照)
【0003】
感染原因に対する免疫応答メカニズムを解明する事は、それぞれに特有の病態を克服する手掛かりとなる。Lymphproliferative groupのタイレリア感染について、T.parvaやT.annulataでは、その感染によってCD4+T細胞とCD8+T細胞による免疫応答が惹起され、同CD8+T細胞=細胞障害性T細胞(cytotoxic T lymphocyte=CTL)が主たる排除エフェクターとして作用する事、そしてその誘導維持にはCD4+(ヘルパーT細胞=helper T cell(Th))が必要であることが明らかにされている。またγδT cellは、MHC非拘束性に感染細胞によって活性化される事、そしてそれによって感染細胞を障害する事が知られている。
したがって、lymphoproliferative groupに属するTheileria感染症においては、原虫抗原特異的なCD8陽性T細胞の増加を検出する事は、感染状態である事を意味する。(以上、非特許文献6〜9参照)
【0004】
一方、nonlymphoproliferative groupのT.orientalis感染において、本発明以前に明らかとなっている事象としては、次のようなものが挙げられる。1)T.orientalis感染では末梢血中のリンパ球やマクロファージなどの単核球細胞が活性化するがその特異性は明らかでない。2)マラリア症、バベシア症やアナプラズマ症と言った赤血球寄生性原虫と同様、感染・変性赤血球は脾臓で除去される。3)血中のIFN-γ値が高い程、全身症状は穏やかである。4)T.orientalis感染末梢血をPHAで刺激すると、IFN-γ産生が検出される。
しかしながら、これらの事象のいずれにおいても、タイレリア抗原特異的にT細胞が存在して、免疫システムを活性化しているのかどうか、もしくは、どのような免疫応答をもたらしているのかは十分に明らかにされていない。(以上、特許文献10〜14参照)
【0005】
抗原特異的なT細胞を検出する方法が幾つか開発されている。なかでもELISpot(Enzyme-linked immunospot)technologyは、刺激よって産生される特定のサイトカインを分泌する細胞の数を評価する方法として広く活用されている。ELISpotアッセイは最初、抗体を産生する細胞を検出する方法として報告されており、IFN-γの検出に関しては、同じグループから1988年の「Reverse ELISPOT assay for clonal analysis of cytokine production I.Enumeration of gamma-interferon-secreting cells」に報告されている(非特許文献14参照)。
【0006】
一方、本発明者は、T.orientalis感染ウシにおいて免疫応答の進展(サイトカイン産生変化)を詳細に解析したところ、感染後、速やかにMPSP(major piroplasm surface protein)ペプチド特異的にIFN-γを産生する細胞が増加し(Th1免疫応答)、時間経過と伴に減少して消失したが、このIFN-γ産生する細胞の減少に従い、MPSPペプチド特異的に反応してIL-10を産生する細胞が増加する(免疫抑制性T細胞免疫応答)という知見を得たうえで、同ペプチドと一連の解析手法について特許出願している(特許文献1)。さらには、MPSPに加えて、特異的T細胞検出ペプチドの多様化を図るべく、同じくT.orientalisに由来する抗原p23においても、鋭意研究の結果、T.orientalisに感染したウシ体内において、p23に由来するペプチドに特異的に応答するT細胞が発生することを証明し、同ペプチドと一連の解析手法についても特許出願している。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2010−75121号公報
【非特許文献】
【0008】
【非特許文献1】A.D.Irvin.Parasitology Today Vol.1 No.5 p124-128,1985
【非特許文献2】Yves Galley et al,Proc.Natl,Acad,Sci.USA Vol 94,p5119-5124,1997
【非特許文献3】Regina Lizundia et al,Cancer Res:66(12)p6105-6110,2006
【非特許文献4】Wendy C.Brown Veterinary Parasitology,1001 p233-248,2001
【非特許文献5】Misao Onuma,et al Comparative Immunology,Micrology & Infectious Disease 21,p165-177,1998)
【非特許文献6】Declan McKeever,Proc.Natl,Acad,Sci.USA Vol91,p1959-1963,1994
【非特許文献7】Evans Taracha,Journal of Immunology,Vol 159,p4539-4545
【非特許文献8】Simon Graham,Proc.Natl,Acad,Sci.USA,Vol.103,p3286-3291,2006
【非特許文献9】Claudia Daubenberger et al Infection and Immunity,p2241-2249,1999
【非特許文献10】Yasutomi,Y.et al,Vet Parasitology,Vol39, p33-41,1991
【非特許文献11】Asaoka H.,et al,Res vet Sci,Vol50,p23-28,1991
【非特許文献12】Hagiwara K.et al,Vet Parasitology,Vol127,p105-110,2005
【非特許文献13】Kakuda T.,et al.,J.Vet.Med.Sci.,vol63,p895-901,2001
【非特許文献14】Czerkinsky CC et.al.,J Immunol Methods.1983 Dec 16;65(1-2):109-21
【発明の概要】
【発明が解決しようとする課題】
【0009】
上記のとおり、本発明者は、牛タイレリア感染症の診断手段として、T.orientalis抗原MPSPおよびp23に応答してIFN-γ及びIL-10を産生する各T細胞数の増減を評価する新たな診断システムの構築しており、本願発明の課題は、このような診断システム使用の利便性をより向上させるために、使用しうるタイレリア抗原の多様化を図るとともに、T細胞免疫応答を誘導する能力が高く、より使い勝手のよいタイレリア抗原を提供する点にある。
【0010】
本発明者は、上記課題を解決するため鋭意研究の結果、タイレリア抗原の一種であるToMRP抗原にT細胞免疫応答能があるとの知見を得て、これをさらに一歩進め、ToMRP抗原の各種部分ペプチドを合成し、T細胞免疫応答能を調べた結果、ToMRP抗原の部分ペプチドにおいてT細胞免疫応答を誘導する能力を有するものがあることを見いだし、本発明を完成させるに至った。
【0011】
すなわち、本発明は以下のとおりである。
(1)配列番号3〜5、10及び12のいずれかに示されるアミノ酸配列を有するペプチド。
(2)上記(1)に記載のペプチドからなる、ウシのT.orientalis感染症診断剤。
(3)上記(1)に記載のペプチド及びIFN-γに対する抗体及び/又はIL-10に対する抗体を少なくとも含むことを特徴とする、ウシのT.orientalis感染症診断キット。
【発明の効果】
【0012】
Theileriosisの原因の一つで、lymphoproliferative groupに属するT.parvaについては、すでに、T細胞免疫応の全容がほぼ解明されている。これに対してnonlymphoproliferative groupに属するT.orientalis感染症に関しては、免疫システムで極めて重要な役割を果たす病原体由来抗原特異的T細胞がジェネレーションされるかどうか詳しくはわかっていなかった。
【0013】
このような状況下、本発明は、T.orientalis感染ウシ個体中に同病原体に由来するMPSPやp23とは異なる抗原、すなわち、新たに、T.orientalis由来ToMRP抗原に特異的に応答してIFN-γ及びIL-10を産生するT細胞が存在し、これを検出することに成功したことに基づく。このことは、先のMPSP(特許文献1)およびp23に由来する抗原ペプチドを用いた解析法に加えて、別の抗原であるToMRPによる解析法を可能にしたことを意味し、より多角的なT.orientalis感染ウシに対する病態診断手法をもたらすものである。
【0014】
本発明のペプチドは、上記ToMRP抗原特異的なT細胞免疫応答の誘導能が高く、T.orientalisに感染したウシが産生するToMRP抗原特異的なIFN-γ分泌T細胞及びIL-10分泌T細胞を刺激し、IFN-γ及びIL-10を分泌させ、このIFN-γ及びIL-10を感度よく検出、測定することが可能となる。このIFN-γ及びIL-10はT.orientalis感染に基づくToMRP抗原特異的に産生されたT細胞の量を反映するから、これにより、T.orientalisの感染の有無、該感染に基づく病態の程度を診断することができる。
以上のとおり、本発明のペプチド抗原はT.orientalis感染症の診断剤として、極めて有用である。
【図面の簡単な説明】
【0015】
【図1】T.orientalis 由来抗原ToMRPに特異的なT細胞を検出すために合成した各ペプチドのToMRP上の位置を示す図。
【図2】T.orientalis自然感染ウシのPBMCsに対し、本発明の抗原ペプチド(全17種類)を用いて刺激し、IFN-γ ELISpot解析を行った結果を示す図。
【図3】T.orientalis人工感染ウシのPBMCsに対し、本発明の抗原ペプチド(全17種類)を用いて刺激し、IFN-γ ELISpot解析を行った結果を示す図。
【発明を実施するための最良の形態】
【0016】
本発明の抗原ペプチドは、T.orientalis原虫のToMRP(T.orientalis microneme-rhoptry protein)の部分ペプチドであって、配列番号3〜5、10、12に示される。
ToMRPは、T.orientalisのpiroplasmステージのcDNAライブラリーからクローニングされ、T.parvaのMRP(=p104)と相同性が高いことから、その名がつけられたものである。ToMRPの機能の多くは不明であるが、T.orientalisと同様に赤血球に寄生することが知られるPlasmodium(=マラリア)のmicroneme-rhoptry細胞内小器官に関連する遺伝子群は赤血球に侵入する際に重要な役割を果たすことが知られている。実際、ToMRPは赤血球膜を構成するBand3と結合することも知られているため、赤血球への侵入(もしくは脱出)に利用される可能性が考られる。
本発明のペプチドは、ウシのT.orientalis原虫感染の有無あるいは、その病態を診断するための診断剤として用いられる。以下に、本発明のペプチドを用いたT.orientalisの感染症の診断手法について説明する。
ウシがT.orientalis原虫に感染した場合のT細胞免疫応答は、IFN-γの分泌により特徴づけられる細胞性免疫促進型のT細胞免疫応答と、IL-10の分泌により特徴づけられる免疫抑制型のT細胞免疫応答で構成される。これらT.orientalis原虫抗原特異的なIFN-γ分泌T細胞とIL-10分泌T細胞の数は、T.orientalis感染の進行を反映し、IFN-γ分泌T細胞とIL-10分泌細胞の数がともに一定値を超えるウシは、T.orientalis感染による、貧血等の特徴的な症状が顕在化することが、MPSPペプチドを用いた解析によって明らかになっている(特許文献1)。
【0017】
本発明を構成するT.orientalis抗原ToMRP由来ペプチドは、ウシ末梢血に含まれる精製されたPBMCs懸濁液中に含まれるT細胞のうち、T.orientalis抗原ToMRPに特異的に応答するIFN-γ分泌T細胞及びIL-10分泌T細胞を刺激し、活性化する。
T.orientalis抗原ToMRP特異的に応答してIFN-γを分泌するT細胞もしくは、IL-10を分泌するT細胞は、それぞれIFN-γ ELISpot法およびIL-10 ELISpot法によって検出される(最終的にT.orientalis抗原ToMRP特異的に応答するIFN-γ分泌T細胞及びIL-10分泌T細胞数がスポット数として計測される)(特許文献1)。
【0018】
このToMRPペプチドによる刺激は、複数のToMRPペプチド抗原が混和されたものでも遂行可能である。
本発明のペプチドを使用するウシT.orientalis感染症の診断に使用される被験試料のPBMCs懸濁液は、ウシから採取された末梢血から精製されたものをそのまま用いてもよいし、また、凍結保存したものを用いてもよい。
【0019】
本発明における細胞の具体的な調製法は次のとおりである。ウシより採取された抗凝固処理した末梢血液に対して、HISTOPAQE-1077(登録商標SIGMA-ALDRICH社)などを用いた密度遠心分離作業を施すことによって単離・精製されたPBMCsをペプチド刺激対象細胞として使用する。このように精製されたPBMCを、精製直後に解析に使用しない場合は、細胞凍結保護液CP-1(極東製薬工業株式会社)などを用いて凍結保存してもよい。凍結保存されたPBMCsは解凍することでペプチドに応答可能な細胞群として取り扱うことができる。また、ELISpot解析時に使用される細胞懸濁液の調製に使用される培地は、特に限定されるものではなく、通常の生命科学の細胞培養実験に使用される培地等が使用されるが、可能な限りRPMI1640培地へ最終濃度10%非働済ウシ胎仔血清(FCS)、50μM2-メルカプトエタノール、100units/ml penicillin、100μg/ml streptomycinを添加したRPMIコンプリートメディウムが望ましい。また、使用するPBMCsはRPMIコンプリートメディウムへ懸濁した際に、市販の細胞数検出機(例:chemometec社製NucleoCassette)もしくは通常の細胞計算に使われる血球計算盤を用いて細胞数を算出した後に測定に用いる。
【0020】
同ELISpot解析に使用する抗IFN-γ抗体および抗体IL-10抗体は、ウシ由来のIFN-γ及びIL-10を認識・結合することできるものであればよい。又、抗体の種類は、モノクローナル抗体およびポリクローナル抗体のどちらであってもよい。
サイトカイン分泌T細胞の数を測定する方法は、抗体による細胞内染色を行ないフローサイトメーターで検出する等いくつか知られているが、本願発明においては、感度の高さおよび抗原特異的T細胞を検出する目的からELISpot(Enzyme-Linked ImmunoSpot)法が好ましく用いられる。
【0021】
このように求められたT.orientalis由来抗原ToMRPペプチドに特異的に応答するIFN-γ分泌T細胞及びIL-10分泌T細胞数は、T.orientalis感染の状態を反映し、診断対象のウシにおいてIFN-γ分泌T細胞及びIL-10分泌T細胞数がともに検出されない場合には、診断対象のウシはT.orientalisに感染しておらず、また、IFN-γ分泌T細胞及びIL-10分泌T細胞数が単独もしくは両方ともに一定基準値を超える場合には、T.orientalisに感染した上で、同感染に特徴的な病態を発症しているか、あるいは発症する可能性がある状態であるものと判定することができる。この基準値は、IFN-γ分泌T細胞及びIL-10分泌T細胞数とT.orientalis感染症の発症との関係を照らし合わせることにより、求めることが可能である。
【0022】
例えば、IFN-γ検出ウエルにおいて1×10E6cells、IL-10検出ウエルにおいて1×10E5cellsのPBMCを播種して測定する場合、それぞれの検出されたサイトカインスポット数が「10」とすると、PBMCs1×10E6個あたりIFN-γ分泌T細胞が10個、IL-10分泌T細胞が100個含まれていることを意味する。これらの1ウエル当たりのスポット数についての有効値は、同システムの利用者が用いる実験条件(例えば、1ウェル当たりのPBMCsの数や培養時間)や、研究者の求める病態・精度によって最適化される可能性がある。
【0023】
上記ELISpot法に対して、既存のPCR法、血液スメア解析、既存の一般血液解析を組み合わせる事で、免疫応答のバランスと病態の診断をより確実に行うことも可能である。すなわち、PCR法および血液スメア解析によって、血液内に存在するT.orientalisをELISpot結果に遅れて検出することでELISpot結果を補強することが可能であるし、一般血液解析によって貧血等の病態の発症の有無を直接示すことでELISpot結果との相関性を示すことが可能である。
【実施例】
【0024】
以下に、本発明を実施例として説明するが、本発明は該実施例に何ら限定・制約されるものではない。
【0025】
実施例1
本実施例において用いた手法及び抗原ペプチドは以下に示される。
1)ELISpotアッセイの準備
下記の免疫活性化反応の培養が終了するまでの操作は安全キャビネットもしくはクリーンベンチ内で無菌的に行った。
底面がPolyVinylidine DiFluoride(PVDF)メンブランで構成されている96ウエルプレート(Millipore社)を準備し、各ウエルへ70%EtOHを50μlずつ加え30秒−2分間処理することにより疎水性PVDFメンブランの親水処理を行った。ウエル内のEtOHをアスピレーターにより吸引除去後、Phosphate Buffered Saline(Ca2+ Mg2+ 無添加)(PBS)を各ウエルあたり150μl加えた後にアスピレーターで取り除く洗浄作業を5回繰り返す。洗浄操作が終了した後、抗bovine IFN-γキャプチャー抗体(MABTECH code 3115-3)または、抗IL-10キャプチャー抗体(AbD serotec,code MCA2110,clone CC318)をPBSにより7.5μg/mlに希釈し、80μlずつ加え4℃、一晩処理することにより底面のPVDF メンブランへキャプチャー抗体をコーティングさせた。翌日に余剰なキャプチャー抗体はアスピレーターにより吸引除去後、各ウエルを150μlのPBSで洗浄作業を行った。洗浄終了後のウエルにはRPMIコンプリートメディウム(RPMIメディウムへ最終濃度10%FCS(本実験系では非働化処理したFCSを使用)、50μM2-メルカプトエタノール、100units/ml penicillin、100μg/ml streptomycinになるように添加したもの)を100μlずつ加えて室温30分間もしくは4℃、1時間以上のブロッキング処理を行った。また、処理温度が4℃でのブロッキング処理を行った場合はELISpot assayを行う前にプレートを室温へ戻した。
【0026】
2)ELISpotアッセイに使用する細胞の準備
目的のウシ個体から抗凝固処理した末梢血液を遠心することでPBMCsを含むバフィーコートを形成させた。同バフィーコートを慎重に回収後、混入してきた赤血球をHISTOPAQE(登録商標)-1077(SIGMA-ALDRICH社)を使用した密度遠心分離法によって取り除くことで、精製PBMCsを得た。また、単離後すぐに使用しない場合は細胞凍結保護液CP-1(極東製薬工業株式会社)などを用いて必要になるまで保存した。精製・回収されたPBMCsはRPMIコンプリートメディウムで洗浄後、同メディウムへ再懸濁させ細胞数検出キットNucleoCassetteTM(chemometec)もしくは血球計算盤を用いて細胞数を算出しておいた。
【0027】
3)ELISpotアッセイにおける免疫活性化反応
上記1)のキャプチャー抗体がコーティングされ、加えてブロッキング処理が施された96ウエルフィルタープレートにおいて、ブロッキング処理に用いたRPMIコンプリートメディウムを除去した後に、抗bovine IFN-γキャプチャー抗体をコーティングしたウエルには3×10E5cellsの細胞を含むRPMIコンプリートメディウムを100μlずつウエルへ播種した。続けて、ToMRPに由来するT細胞刺激用抗原ペプチドをRPMIコンプリートメディウムで最終使用濃度の2倍濃度に調製し、同刺激用抗原ペプチド入りコンプリートメディウムを100μlずつPBMCs入りのウエルに加えた(この段階で、目的の最終濃度のT細胞刺激用抗原ペプチドを含む細胞懸濁液が1ウエル当たり200μl含まれることになる)。ELISPOTアッセイのコントロールとして、最終濃度1μg/ml phytohemagglutin(PHA)とPBMCsを含むウエルを用意した。最後に、ウエル内のPBMCsと刺激物質の混合液を数回ピペティングし、37℃のCO2インキュベーター内で18−42時間、振動を与えないように培養した。
【0028】
4)ELISpotアッセイにおけるスポットの検出
細胞懸濁液を除去した後に200μlPBSで5回洗浄作業をし、0.5%FCSを含むPBS(0.5%FCS/PBS)で希釈した抗bovine IFN-γビオチン化抗体(最終濃度0.25μg/ml MABTECH code 3115-6)をウエルへ80μlずつ加えて室温2時間反応させた。ビオチン化抗体溶液を除去し、200μl PBSで5回洗浄作業をした後に0.5%FCS/PBS で500〜1000倍希釈したHorseradish Peroxidase(HRP)をコンジュゲートしたstreptavidin(MABTECH code 3310-9)を室温1時間反応させた。その後、HRP 溶液を除去し、200μlPBSで5回洗浄作業をした後に3-Amino-9-ethylcarbazole(AEC)substrate kit(BD Biosciences code 551951)を用いて室温30分間、発色させた。流水により反応液を洗い流し発色反応を停止させた。プレートは風乾によって十分に乾燥させた。ウエル底面に現れたスポットは顕微鏡によってカウントした。
【0029】
5)抗原ペプチド
図1に示すように、T.orientalis由来抗原ToMRPタンパク質のアミノ酸配列(配列番号18)において、シグナルペプタイド部位以降のアミノ酸配列に基づき、C末端方向に10アミノ酸をオーバーラップさせながら20アミノ酸残基を有するペプチドを順次合成し、17種のペプチドを得た(Int.J.Parasitol.,29:593-599,1999 Sako Y et al)。なお、最後のNo.17のみ15アミノ酸で構成した。これら17種のペプチドのアミノ酸配列(配列番号1〜17)を以下に示す。
【0030】
・ToMRP No.1
SGKTFNFFEPDDSLGKMEYH(配列番号1)
・ToMRP No.2
DDSLGKMEYHDHRGLKVYFF(配列番号2)
・ToMRP No.3
DHRGLKVYFFTPLHNSGVDK(配列番号3)
・ToMRP No.4
TPLHNSGVDKVVVGNQGIWT(配列番号4)
・ToMRP No.5
VVVGNQGIWTARSGQTLTKL(配列番号5)
・ToMRP No.6
ARSGQTLTKLVGFFRCMGFG(配列番号6)
・ToMRP No.7
VGFFRCMGFGLSHVSYFNQE(配列番号7)
・ToMRP No.8
LSHVSYFNQEGENKELLLGY(配列番号8)
・ToMRP No.9
GENKELLLGYSHGDDNLVEF(配列番号9)
・ToMRP No.10
SHGDDNLVEFDRKRFYDKLL(配列番号10)
・ToMRP No.11
DRKRFYDKLLDYTVSSEPFN(配列番号11)
・ToMRP No.12
DYTVSSEPFNKDVPGHLVLY(配列番号12)
・ToMRP No.13
KDVPGHLVLYQLEQLLKERE(配列番号13)
・ToMRP No.14
QLEQLLKEREESEKKSKAEK(配列番号14)
・ToMRP No.15
ESEKKSKAEKVAEVVHEAAE(配列番号15)
・ToMRP No.16
VAEVVHEAAEKKSEVPVTEE(配列番号16)
・ToMRP No.17
KKSEVPVTEEAHTEL(配列番号17)
【0031】
6)上記5)の方法により合成したに示した17種のペプチドを用いた刺激により、IFN-γELISpotアッセイを行った。
放牧によりT.orientalis原虫に自然に感染したウシ(自然感染ウシ)2頭(#1と#2)と、タイレリア原虫を含む血液を接種したウシ(人工感染ウシ)2頭(#3と#4)から、上記2)の方法により回収したPBMCを用いた。上記3)に記載の手法(免疫活性化反応)に従い、上記1)に記載の抗bovine IFN-γ キャプチャー抗体をコーティングしたプレート(IFN-γ検出プレート)に、上記2) の方法に従い調整した標的細胞と、上記5)の方法により合成した17種の抗原ペプチド(最終濃度:1μM) を混合し42時間インキュベーションした。その後、上記4)のELISpotアッセイの操作を行ってスポットを検出した。結果を図2、3に示す。
感染が成立したウシを解析した結果であるウシ#2、#3、#4において、共通してToMRP No.3〜5、10、12のペプチド(ウシ#1ではNo.3、5)に強い反応が認められ、加えて、ウシ#1において ToMRP No.1が、ウシ#4においてToMRP No.16が独立して高い反応を示す領域であることが認められた。
【特許請求の範囲】
【請求項1】
配列番号3〜5、10及び12のいずれかに示されるアミノ酸配列を有するペプチド。
【請求項2】
上記(1)に記載のペプチドからなる、ウシのT.orientalis感染症診断剤。
【請求項3】
上記(1)に記載のペプチド及びIFN-γに対する抗体及び/又はIL-10に対する抗体を少なくとも含むことを特徴とする、ウシのT.orientalis感染症診断キット。
【請求項1】
配列番号3〜5、10及び12のいずれかに示されるアミノ酸配列を有するペプチド。
【請求項2】
上記(1)に記載のペプチドからなる、ウシのT.orientalis感染症診断剤。
【請求項3】
上記(1)に記載のペプチド及びIFN-γに対する抗体及び/又はIL-10に対する抗体を少なくとも含むことを特徴とする、ウシのT.orientalis感染症診断キット。
【図1】


【図2】

【図3】

【図2】
【図3】
【公開番号】特開2012−116797(P2012−116797A)
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【出願番号】特願2010−268957(P2010−268957)
【出願日】平成22年12月2日(2010.12.2)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人農業・食品産業技術総合研究機構委託研究「新技術・新分野創出のための基礎研究推進事業/病原性原虫によるTh1免疫回避機構の解明と糖鎖被覆リポソームワクチン評価技術の確立」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【出願日】平成22年12月2日(2010.12.2)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人農業・食品産業技術総合研究機構委託研究「新技術・新分野創出のための基礎研究推進事業/病原性原虫によるTh1免疫回避機構の解明と糖鎖被覆リポソームワクチン評価技術の確立」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]