
免疫抑制作用を有するヒト化抗CD4抗体
本発明は、マウスのモノクローナル抗CD4抗体B−F5由来のヒト化抗体に関する。該抗体は、CD25陽性CD4陽性制御性T細胞を活性化させることができ、免疫抑制組成物の製造に有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒト化抗CD4抗体、および免疫調整のためのヒト化抗CD4抗体の使用に関する。
自己免疫疾患や移植片拒絶反応は、組織抗原、すなわち自己免疫疾患の場合は自己抗原、移植片拒絶反応の場合は同種移植片抗原に対する不適切な免疫応答から生じる。
自己免疫疾患には、例えば、関節リウマチ、タイプI型糖尿病、多発性硬化症、クローン氏病、潰瘍性大腸炎、アトピー性皮膚炎などが含まれる。
これらの免疫疾患に対する従来の治療法には免疫抑制薬が含まれる。しかしながら、これらの薬剤は、全身的な免疫抑制を引き起こし、免疫系の有害な機能だけでなく有益な機能をも阻害することになる。その結果として、そうした薬剤は、日和見感染のような副作用を引き起こす。
【背景技術】
【0002】
代替アプローチとして、特定のリンパ球サブセット(T細胞を減少させるタイプの抗体)を除去するために、あるいは抗体(T細胞を減少させないタイプの抗体)を有する細胞を殺さずに、標的表面分子の機能を阻害するために、細胞表面分子に対する免疫抑制性のモノクローナル抗体(mAbs)を使用することが提案されている。
CD4陽性T細胞が、自己免疫を開始し、維持することに主要な役割を果たしていることは一般に認められている。したがって、CD4陽性T細胞表面分子に対するモノクローナル抗体、特に抗CD4陽性モノクローナル抗体を免疫抑制剤として使用することが提案されてきている。多数の臨床研究が、この種のアプローチの可能性に関心を示しているが、これらの研究はまた、抗CD4モノクローナル抗体を定常的な臨床実務で使用するのにより適したものとするために取り組むべき幾つかの問題点を提起したのである。
【0003】
具体的には、B−F5抗体(ネズミIgG1抗ヒトCD4)が、種々の自己免疫疾患で試験された:
− 関節リウマチ患者において、いくつかの公開試験で、B−F5の少なくとも20mgの一日服用量が臨床的に有効であることが示唆された(Racadotら、Clin Exp Rheumatol;10(4)、365−74、1992;Wendlingら、Clin Rheumatol;11(4)、542−7、1992)。しかしながら、偽薬管理下の臨床試験で観察された結果では、20mg/日の服用では、有意な改善を示さなかった(Wendlingら、J Rheumatol;25(8)、1457−61、1998)。
【0004】
− 乾癬では、0.2〜0.8 mg/kg/日の投与量で7日間または8日間治療することにより乾癬傷害の改善が観察された(Morelら、J Autoimmun;5巻、4号、465−77頁、1992年)。
【0005】
− 多発性硬化症(MS)患者では、再発−寛解の病態の患者を10日間、治療した結果では若干有効であり、患者の幾人かは治療後6ケ月の時点で再発しなかった(Racadotら、J Autoimmun;6巻、6号、771−86頁、1993年)。同様の効果が、Rumbachらによって観察された(Mult Scler;1巻、4号、207−12頁、1996年)。
【0006】
− 重度のクローン氏病においては、B−F5の0.5mg/日/kgを7日間、連続投与、あるいは最初の日(ゼロ日)に0.5mg/日/kgを、そして2〜7日目は、1mg/日/kgを投与した患者では、有意ある改善は観察されなかった(Canva−Delcambreら、Pharmacol Ther;10巻、5号、721−7頁、1996年)。
【0007】
− 同種移植片拒絶反応の予防において、生物学的パラメーターの変更、すなわち30mg/日の服用量によるインビボにおけるB−F5の作用について報告された。しかしながら、B−F5の生物学的利用能が充分でなく、同種移植片拒絶反応の予防に使用することができないと報告された(Dantalら、Transplantation;27;62(10):1502−6頁、1996年)。
【0008】
上に述べた事項から、第一に解決しなければならない問題は、臨床的改善を達成するためには高用量のモノクローナル抗体を使用する必要があるという点であるように思われる。これは、標的組織におけるリンパ球のモノクローナル抗体の入手が困難であることに起因する。より高用量の使用は、血液リンパ球に対する過剰反応を引き起こし、望ましくない副作用をもたらすかもしれない。
【0009】
ヒトに対してモノクローナル抗体を使用する治療法の別の欠点としては、これらの抗体が一般にマウス細胞から得られるので、それを投与されたヒト患者は抗マウス応答が刺激されることである。これは治療効果の減少とさらには将来のいかなるマウスモノクローナル抗体による治療効果の減少のみならず、アナフィラキシーの危険性をも増大させるものである。
【0010】
このような欠点は、原則としてヒト化抗体を使用することにより回避できる。このヒト化抗体は、抗原結合特異性を決定するものであり、マウスモノクローナル抗体の相補性決定領域(CDR)を、ヒト免疫グロブリン分子のフレームワーク領域(FR)に移植することにより得られる。ヒト化の目的は、マウスモノクローナル抗体に由来する相補性決定領域と同じ抗原結合性を有し、ヒトに対してより少ない免疫原性を有する組み換え抗体を得ることにある。
【0011】
ある場合には、マウス抗体からのCDRをヒトのフレームワーク中のヒトCDRに置き換えることにより、抗原結合性の性質(抗原に対する特異性のみならず親和性)を改変させるのに充分である。しかしながら、多くの抗体では、いくつかのFR残基が抗原との結合にとって重要である。その理由は、それらのFR残基が抗原抗体複合体における抗原と直接接触する、あるいはそれらがCDRの立体構造に影響を及ぼし、そのために抗原との結合能に影響を及ぼすからである。
【0012】
したがって、ほとんどの場合、マウス抗体の1個あるいは数個のフレームワーク残基を、ヒトの対応するFR残基と置き換えることもまた、必要である。抗マウス反応を防ぐためには、置換された残基の数はできるだけ小さくなければならないので、問題はどのアミノ酸残基が抗原結合性を保持するのに必須であるかを決定することである。より適切な置換部位を予測するために、種々の方法が提案されてきている。これらの方法は、ヒト化の最初の工程で少し役に立つかも知れない一般的な原理を提供しているけれども、最終的な結果は個々の抗体によって変わり得る。したがって、与えられた抗体について、どのような置換が望ましい結果をもたらすかを予測するのは非常に困難である。
【発明の開示】
【発明が解決しようとする課題】
【0013】
しかしながら、本発明者らは、マウスB−F5のヒト化を試みて、親マウスB−F5と
同様のCD4結合能を有するヒト化B−F5(以下、hB−F5と称する)の産生に成功した。
さらに、本発明者らは驚くべきことに、hB−F5が、親B−F5で使用された用量よりも、あるいは他の抗CD4モノクローナル抗体で現在使用されている用量よりも遥かに低用量で最適のインビボ免疫抑制効果を示すことを見出した。
【課題を解決するための手段】
【0014】
実際、本発明者らは、hB−F5を1mg/日の低用量で10日間、好ましくは5mgの用量で2日おきに10日間、使用して治療すると、関節リウマチ患者で臨床的に効果が認められているように、効果的な免疫抑制作用を示すことを観察した。
【0015】
本発明では、マウスB−F5モノクローナル抗体由来の、ヒト化抗体(hB−F5)が提供され、この抗体は次に示すポリペプチド配列で示される可変領域を有する:
− 重鎖可変領域:EEQLVESGGGLVKPGGSLRLSCAASGFSFSDCRMYWLRQAPGKGLEWIGVISVKSENYGANYAESVRGRFTISRDDSKNTVYLQMNSLKTEDTAVYYCSASYYRYDVGAWFAYWGQGTLVTVSS(配列番号:1)
− 軽鎖可変領域:DIVMTQSPDSLAVSLGERATINCRASKSVSTSGYSYIYWYQQKPGQPPKLLIYLASILESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQHSRELPWTFGQGTKVEIK(配列番号:2)
【0016】
一般に、本発明のhB−F5抗体は、さらにヒト定常領域(Fc)を含むものである。この定常領域は、定常ドメインの中で、IgM、 IgG、 IgD、 IgAおよびIgEなどのいずれのクラスの免疫グロブリンからも、そしてIgG1、IgG2、IgG3およびIgG4などのいずれのアイソタイプからも選ぶことができる。好ましい定常領域は、IgG、特にIgG1の定常ドメインから選択される。
【0017】
本発明はまた、可変領域を含むhB−F5抗体の如何なる断片をも包含する。このような断片としては、特に、Fab、Fab’、F(ab)’2、FvおよびscFv断片を挙げることができる。
【0018】
本発明はまた、配列番号:1のポリペプチドをコードするポリヌクレオチド、および配列番号2のポリペプチドをコードするポリヌクレオチドから選択されるポリヌクレオチドを包含する。好ましくは、配列番号:3のポリヌクレオチド、および配列番号:4のポリヌクレオチドから選ばれるポリヌクレオチドである。
本発明のポリヌクレオチドは、組み換えDNA技術および/または化学的DNA合成の公知の方法により容易に得ることができる。
【0019】
hB−F5抗体の重鎖あるいは軽鎖の可変領域をコードするポリヌクレオチドは、このようにして得られる完全な重鎖あるいは軽鎖を発現させるために、ヒト重鎖あるいは軽鎖の定常領域をコードするポリヌクレオチドと融合させてもよい;すなわち、タンパクの分泌ができるようにするシグナルペプチドをコードする配列も附加することができる。 こうした組み換えポリヌクレオチド類もまた本発明の一部を構成する。
【0020】
本発明はまた、本発明のポリヌクレオチドが適当な制御配列と結合してその転写および翻訳を選択された宿主中で可能にする発現カセット、および本発明のポリヌクレオチドあるいは発現カセットを含む組み換えベクターを提供する。
これらの組み換えDNA構築物は、DNAの組み換えおよび遺伝子工学の公知の組み換え手法により得ることができ、宿主細胞に導入することができる。
【0021】
本発明はまた、本発明のポリヌクレオチドで形質転換された宿主細胞を包含する。
本発明の構成に包含される有用な宿主細胞としては、原核細胞あるいは真核細胞である。適当な真核細胞としては、具体的には植物細胞、サッカロマイセスなどの酵母細胞、ショウジョウバエ(Drosophila)あるいはヨトウ(Spodoptera)などの昆虫細胞、およびHeLa、CHO、3T3、C127、BHK、COSなどの動物細胞が挙げられる。
【0022】
本発明の発現ベクターの構築および宿主細胞の形質転換は分子生物学の標準的な手法により達成することができる。
本発明のhB−F5抗体は、当該抗体をコードする核酸配列を含有する発現ベクターを含む宿主細胞を、その発現に適した条件下で培養し、宿主細胞の培養液から回収して得ることができる。
本発明はまた、本発明のhB−F5抗体あるいは上記したそれらの断片からなる治療用組成物をも包含する。
【0023】
好ましくは、そのような組成物は、非経口投与用組成物であって、0.1〜10mg、好都合には1〜5mgのhB−F5の用量で投与することができるように製剤化される。
さらに詳しく述べると、本発明は、免疫抑制組成物の製造のための本発明のhB−F5抗体あるいはその断片の使用を包含するものである。そのような免疫抑制組成物は、同種移植片拒絶反応、移植片対宿主病、宿主対移植片拒絶反応、あるいは例えば、心筋炎、糖尿病、乾癬、紅斑性狼瘡、クローン氏病、多発性硬化症、関節リウマチなどの自己免疫疾患等の治療あるいは予防に特に有用である。
【0024】
さらに、本発明者らは、hB−F5は、CD4陽性T細胞の特定サブセット、すなわちCD4陽性CD25陽性細胞を活性化することができることを見出した。
CD25陽性CD4陽性制御性T細胞(Treg細胞)は、末梢性のCD4陽性T細胞5〜10%を構成成分とする。これらのTreg細胞は、1995年、Sakaguchiらにより、マウスにおける制御細胞として最初に報告された(J.Immunol.,155:1151−1164)。これらのTreg細胞は、活性化されると、CD4陽性およびCD8陽性のT細胞の活性化および増殖を共に抑制することができる。その後、CD25陽性CD4陽性サプレッサーT細胞は、ヒトでも発見された(Jonuleitら,J.Exp.Med.193,1285−1294,2001;Levingsら,J.Exp.Med.193,1295−1302,2001;Dieckmannら,J.Exp.Med.193,1303−1310,2001)。数多くの論文が公開され、これらの細胞が免疫抑制の役割を果たしていることを、種々の自己免疫疾患のモデルやインビトロの系で報告している(総説:例えばShevachら,J.Exp.Med.193,11,41−46,2001)。Ex vivoで活性化されたCD4陽性CD25陽性制御性T細胞(Treg細胞)は、移植片対宿主病の予防に効果的であることが証明された(Taylorら,Blood,99,3493−3499、2002; Cohenら,J.Exp.Med.196,401−406,2002; Hoffmannら,J.Exp.Med.196,389−399,2002)。したがって、CD4陽性CD25陽性制御性T細胞(Treg細胞)を活性化する手段を提供することは、非常に興味あることである。
【0025】
本発明はまた、インビトロでCD25陽性CD4陽性制御性T細胞を活性化するための本発明のhB−F5抗体の使用、またはその親抗体B−F5の使用に関する。
本発明のhB−F5抗体は、1〜10μg/mlの濃度でCD25陽性CD4陽性制御性T細胞に添加される。
【発明を実施するための最良の形態】
【0026】
本発明は、以下の記載によってさらに詳細に説明される。これらは、本発明のhB−F5抗体の性質を説明する実施例である。しかしながら、これらの実施例は、本発明の説明のためにだけされるものであり、本発明の限定を決して構成するものではないことを理解すべきである。
実施例1 ヒト化B−F5の構築
【0027】
ヒト化B−F5のVHおよびVκ領域の設計
マウスB−F5のVHおよびVκ領域をコードするDNA配列は、配列番号:5および配列番号:6としてそれぞれ図1および図2に示されている。マウスCDRが移植されるヒトVHおよびVκは、元のマウスB−F5のVHおよびVκに近似のヒトVHに対するデータベースを探索して選択された。ヒト抗体(M26;受付番号A36006)のVH領域は、B−F5のVHと最も高い相同性を有していた。別のヒト抗体[FK−001;NAKATANIら,Biotechnology,7(1989),805−810)]のVκ領域は、B−F5のVκと最も高い相同性を示した。
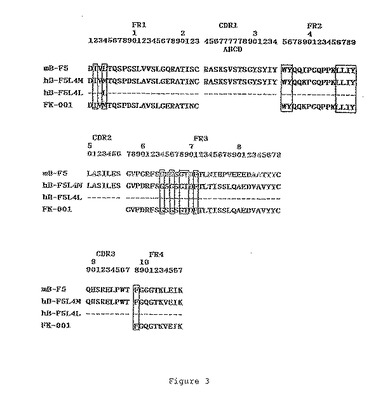
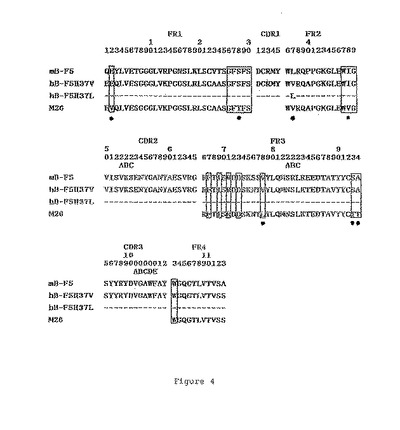
4番目の残基がロイシンあるいはメチオニンであることに違いを有するVκの二つのタイプが構築され、それぞれL4LおよびL4Mと命名された。37番目の残基がロイシンあるいはバリンであることに違いを有するVHの二つのタイプが構築され、それぞれH37LおよびH37Vと命名された。B−F5、FK−001、L4LおよびL4Mのポリペプチド配列のアライメントが、図3に示される。B−F5、M26、H37LおよびH37Vのポリペプチド配列のアライメントが図4に示される。CDRを包み込むために重要であると先に報告(Chothiaら,Nature,342,(1989),877;Footeら,J.Mol.Biol.,224(1992),487)されたフレームワークの残基は、囲んで示されている。
これらのVHおよびVκを組み合わせることにより、4種類のV領域が設計された。
【0028】
ヒト化B−F5の発現
ヒト化B−F5の製造のための次の工程は、米国特許5,886、152号でヒト化B−B10に対して記載されたものと同様であった。
要約すると、ヒト化B−F5の重鎖[ヒトγ−1鎖の定常領域(TAKAHASHIら,Cell,29(1982),671−679)と融合したヒト化VH領域]、および軽鎖[FK−0001のκ鎖の定常領域と融合したヒト化Vκ領域]に対する発現プラスミドが別々に構築された。これらのプラスミドにおいて、ヒト化B−F5の発現は、ヒトモノクローナル抗体IgMの遺伝子、すなわちFK−0001のプロモーター/エンハンサーによって推進される。図5および図6は、ヒト化BF−5のVH領域およびVκ領域をコードするプラスミド断片を、それぞれ示している。可変領域をコードする配列には、下線を付してあり、相当するポリペプチド配列は、ヌクレオチド配列の下に示されている。プラスミドおよびpSV2neoの両方を、Lipofectin(登録商標)を使用して同時にマウス骨髄細胞Sp2/0(ATCC CRL−1581)に導入した。ヒトIgGを産生する形質転換体は、抗ヒトIgG(γ鎖)抗体および抗ヒトIgGのκ鎖抗体を用いたELISA法によって選択された。
実施例2 ヒト化B−F5の種々の変異体の特性化
【0029】
CD4結合活性の評価
hB−F5の4つの変異体を産生する形質転換体の培養上澄み液を採取し、濃縮した。この種々の抗体は、プロテインAセファロースの親和性クロマトグラフィーで培養上澄み液から精製し、競合的ELISA法によりビオチン化mB−F5のマイクロタイタープレート上に被覆された可溶性CD4への結合に対する阻害活性を測定して評価した。インキュベーションは、37℃で2時間、4℃で一晩、実施した。
hB−F5(mB−F5の結合活性を100%に設定)の相対的な結合活性を、以下の表Iに示す。
【0030】
【表1】

【0031】
表Iに示された結果から、VHの37番目の残基であるロイシンは、hB−F5のCD4結合活性を維持するのに重要であるように思われる。それというのも、CD4の結合活性は、37番目のロイシンをバリンに変換することにより、数倍減少しているからである。これに対して、Vκの4番目の残基は、CD4結合活性にとってそれほど重要でないことが理解される。VHの37番目のロイシンとバリンの間の構造的な差異は、分子モデルによっては明確には証明されないので、CD4結合活性におけるH37LのH37Vに対する優位性は、予想外であった。
H37L/L4LおよびH37L/L4Mが、インビトロでの生物学的活性評価のために選択された。
インビトロでのヒト化B−F5の生物学的活性の検討
【0032】
インビトロでのマウスB−F5およびヒト化B−F5(H37L/L4M IgG1およびH37L/L4L IgG1)の生物学的活性を評価した。IgG2タイプのヒト化B−F5(H37L/L4M IgG2およびH37L/L4L IgG2)も試験された。
mB−F5および4種のhB−F5のインビトロでの生物学的活性が、健康なドナーからの末梢血単核球(PBMC)を用いて評価された。PBMCは、マウスB−F5またはhB−F5の存在下、ConA(2.5μg/ml、3日間)あるいはPPD(10μg/ml、4日間)により活性化され、[3H]−チミジンの取り込みによる増殖反応が追跡された。
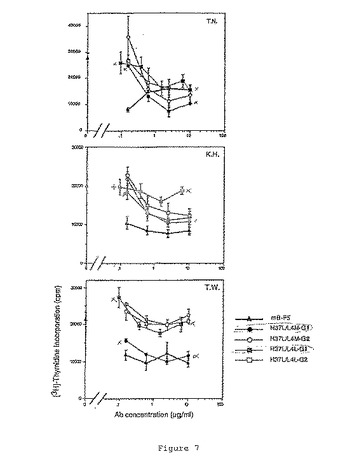
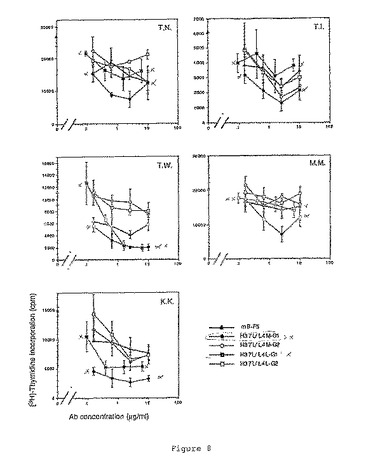
その結果を図7および図8に示す。マウスB−F5およびhB−F5は、ConA誘導性増殖を穏やかに阻害したが、その活性は抗体および/またはドナーにより変化した(図7参照)。また、マウスB−F5およびhB−F5は、PPDで誘導された抗原特異的PBMC増殖を阻害することができた(図8参照)。
hB−F5のIgG1タイプは、mB−F5よりも、PPD誘導性増殖をより効果的に阻害した(70%阻害、図7および図8参照)。IgG1タイプは、阻害活性が殆どmB−F5と同じであるIgG2タイプよりもより有効であるように思われる。IgG1タイプでは、H37L/L4MがH37L/L4Lより有効であった。H37L/L4MおよびH37L/L4LのIgG2タイプでは、殆ど同じ阻害活性を示した。要約すると、PPD誘導性PBMC増殖に対するB−F5の阻害活性は以下のようであった:H37L/L4M IgG1>H37L/L4L IgG1>H37L/L4M IgG2=H37L/L4L IgG2=mB−F5。
インビトロでの生物学的活性の効果およびマウスのアミノ酸数がより少ないことを考慮して、H37L/L4M IgG1がさらに評価するために選択された。
実施例3 関節リウマチ患者(RA)に対するhB−F5の有効性の予備的評価
【0033】
hB−F5(H37L/L4M IgG1)の効果について、RA患者で試験した。
アッセイ条件は、以下の通りである:各患者は、hB−F5の5mgを5回注射、10日間に渡る治療を受けた(2日おきに注射)。
3人の異なる患者の結果を下記の表II〜IVに示す。
患者1(表II)
診断:関節リウマチ、活動度2
リウマチ因子: 2; 重症度: 2
性別: 女性; 年齢: 65歳; 発病: 1965年
付加的治療: ジクロフェナック(150mg/日)
【0034】
【表2】
【0035】
患者2(表III)
診断:関節リウマチ、活動度3
リウマチ因子: 2; 重症度: 2
性別: 女性; 年齢: 48歳; 発病: 2000年
付加的治療: ジクロフェナック(150mg/日)
【0036】
【表3】
【0037】
患者3(表IV)
診断:関節リウマチ、活動度3
リウマチ因子: 3; 重症度: 2
性別: 女性; 年齢: 49歳; 発病: 1989年
付加的治療: ジクロフェナック(150mg/日)
【0038】
【表4】
実施例4 hB−F5によるCD4陽性CD25陽性制御性T細胞の活性化
【0039】
T細胞の単離
1)制御性T細胞(Ttreg):
− CD25陽性細胞は、CD25マイクロビーズを用いて単離される;
− 夾雑物のCD14、CD8、CD19陽性細胞の除去はCD14/CD8/CD19 DYNALビーズを用いて実施される;
− CD45RA陽性細胞の除去は、CD45RA mAb + 抗マウスDYNALビーズを用いて実施される;CD4陽性CD25陽性Tregの純度:>95%
2)エフェクター細胞
− CD4陽性T細胞は、CD4マイクロビーズを用いて単離される;
− CD45RO陽性細胞の除去は、CD45RO陽性mAb + 抗マウスDYNALビーズを用いて実施される;CD25エフェクターT細胞であるCD4/CD45RA陽性細胞の純度:>98%
3)試験系
ドナーAからのCD25陽性Tregを、0.5μg/mlの抗CD3(OKT−3=陽性コントロール=Tregの全活性化)を加えることなく(陰性コントロール=非活性化=非抑制活性)、またはその存在下に、あるいは5μg/mlまたは30μg/mlのhB−F5の存在下に、同質遺伝子型のCD2消失PBMC中で2日間、共培養する。
前培養した細胞をよく洗って、CD25陽性Treg細胞を単離し、γ−線照射処理(3000ラド)する。
4)抑制活性試験
前培養CD25陽性Treg細胞と、ドナーB[エフェクターT細胞に対して異質遺伝子型]からの新鮮な単離CD4陽性エフェクターT細胞とを1:1の割合で、ドナーA[前培養T細胞(さらに活性化されていない)に対して同質遺伝子型]からのAPC(抗原提示細胞)(CD2消失PBMC)の存在下、4日間、共培養する(=異質遺伝子型の混合リンパ球反応)。次いで、細胞を16時間、[3H]−チミジンとインキュベーションして、エフェクターT細胞の増殖を検出する。
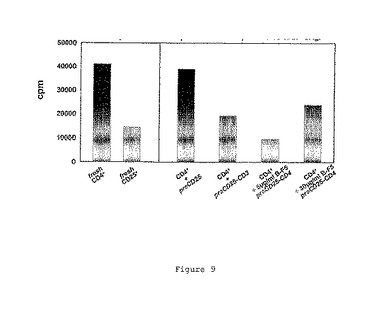
結果を図9に示す。
【0040】
図9の説明:
− 陰性コントロール(非活性化)= 前培養CD25細胞;
− 0.5μg/ml OKT−3(陽性コントロール、全活性化)=
前培養CD25−CD3細胞;
− 5μg/ml hB−F5(試験−1)=前培養CD25−CD4細胞:
− 30μg/ml hB−F5(試験−2)=前培養CD25−CD4細胞。
【図面の簡単な説明】
【0041】
【図1】は、マウスB−F5(mB−F5)のVH領域をコードするDNA配列を示す。
【図2】は、マウスB−F5(mB−F5)のVκ領域をコードするDNA配列を示す。
【図3】は、B−F5、FK−001、L4LおよびL4Mのポリペプチド配列のアライメントを示す。
【図4】は、B−F5、M26、H37LおよびH37Vのポリペプチド配列のアライメントを示す。
【図5】は、ヒト化BF−5のVH領域をコードするプラスミド断片のDNA配列およびポリペプチド配列を示す。可変領域をコードするDNA配列には、下線が付してあり、相当するポリペプチド配列は、当該DNA配列の下に示されている。
【図6】ヒト化BF−5のVκ領域をコードするプラスミド断片のDNA配列およびポリペプチド配列を示す。可変領域をコードするDNA配列には、下線が付してあり、相当するポリペプチド配列は、当該DNA配列の下に示されている。
【図7】は、ConA誘導性のPBMC増殖に対するマウスB−F5およびhB−F5の[3H]−チミジン取り込み阻害を示す。
【図8】は、PPD誘導性のPBMC増殖に対するマウスB−F5およびhB−F5の[3H]−チミジン取り込み阻害を示す。
【図9】は、各種B−F5のエフェクターT細胞増殖に対する抑制作用を示す。
【技術分野】
【0001】
本発明は、ヒト化抗CD4抗体、および免疫調整のためのヒト化抗CD4抗体の使用に関する。
自己免疫疾患や移植片拒絶反応は、組織抗原、すなわち自己免疫疾患の場合は自己抗原、移植片拒絶反応の場合は同種移植片抗原に対する不適切な免疫応答から生じる。
自己免疫疾患には、例えば、関節リウマチ、タイプI型糖尿病、多発性硬化症、クローン氏病、潰瘍性大腸炎、アトピー性皮膚炎などが含まれる。
これらの免疫疾患に対する従来の治療法には免疫抑制薬が含まれる。しかしながら、これらの薬剤は、全身的な免疫抑制を引き起こし、免疫系の有害な機能だけでなく有益な機能をも阻害することになる。その結果として、そうした薬剤は、日和見感染のような副作用を引き起こす。
【背景技術】
【0002】
代替アプローチとして、特定のリンパ球サブセット(T細胞を減少させるタイプの抗体)を除去するために、あるいは抗体(T細胞を減少させないタイプの抗体)を有する細胞を殺さずに、標的表面分子の機能を阻害するために、細胞表面分子に対する免疫抑制性のモノクローナル抗体(mAbs)を使用することが提案されている。
CD4陽性T細胞が、自己免疫を開始し、維持することに主要な役割を果たしていることは一般に認められている。したがって、CD4陽性T細胞表面分子に対するモノクローナル抗体、特に抗CD4陽性モノクローナル抗体を免疫抑制剤として使用することが提案されてきている。多数の臨床研究が、この種のアプローチの可能性に関心を示しているが、これらの研究はまた、抗CD4モノクローナル抗体を定常的な臨床実務で使用するのにより適したものとするために取り組むべき幾つかの問題点を提起したのである。
【0003】
具体的には、B−F5抗体(ネズミIgG1抗ヒトCD4)が、種々の自己免疫疾患で試験された:
− 関節リウマチ患者において、いくつかの公開試験で、B−F5の少なくとも20mgの一日服用量が臨床的に有効であることが示唆された(Racadotら、Clin Exp Rheumatol;10(4)、365−74、1992;Wendlingら、Clin Rheumatol;11(4)、542−7、1992)。しかしながら、偽薬管理下の臨床試験で観察された結果では、20mg/日の服用では、有意な改善を示さなかった(Wendlingら、J Rheumatol;25(8)、1457−61、1998)。
【0004】
− 乾癬では、0.2〜0.8 mg/kg/日の投与量で7日間または8日間治療することにより乾癬傷害の改善が観察された(Morelら、J Autoimmun;5巻、4号、465−77頁、1992年)。
【0005】
− 多発性硬化症(MS)患者では、再発−寛解の病態の患者を10日間、治療した結果では若干有効であり、患者の幾人かは治療後6ケ月の時点で再発しなかった(Racadotら、J Autoimmun;6巻、6号、771−86頁、1993年)。同様の効果が、Rumbachらによって観察された(Mult Scler;1巻、4号、207−12頁、1996年)。
【0006】
− 重度のクローン氏病においては、B−F5の0.5mg/日/kgを7日間、連続投与、あるいは最初の日(ゼロ日)に0.5mg/日/kgを、そして2〜7日目は、1mg/日/kgを投与した患者では、有意ある改善は観察されなかった(Canva−Delcambreら、Pharmacol Ther;10巻、5号、721−7頁、1996年)。
【0007】
− 同種移植片拒絶反応の予防において、生物学的パラメーターの変更、すなわち30mg/日の服用量によるインビボにおけるB−F5の作用について報告された。しかしながら、B−F5の生物学的利用能が充分でなく、同種移植片拒絶反応の予防に使用することができないと報告された(Dantalら、Transplantation;27;62(10):1502−6頁、1996年)。
【0008】
上に述べた事項から、第一に解決しなければならない問題は、臨床的改善を達成するためには高用量のモノクローナル抗体を使用する必要があるという点であるように思われる。これは、標的組織におけるリンパ球のモノクローナル抗体の入手が困難であることに起因する。より高用量の使用は、血液リンパ球に対する過剰反応を引き起こし、望ましくない副作用をもたらすかもしれない。
【0009】
ヒトに対してモノクローナル抗体を使用する治療法の別の欠点としては、これらの抗体が一般にマウス細胞から得られるので、それを投与されたヒト患者は抗マウス応答が刺激されることである。これは治療効果の減少とさらには将来のいかなるマウスモノクローナル抗体による治療効果の減少のみならず、アナフィラキシーの危険性をも増大させるものである。
【0010】
このような欠点は、原則としてヒト化抗体を使用することにより回避できる。このヒト化抗体は、抗原結合特異性を決定するものであり、マウスモノクローナル抗体の相補性決定領域(CDR)を、ヒト免疫グロブリン分子のフレームワーク領域(FR)に移植することにより得られる。ヒト化の目的は、マウスモノクローナル抗体に由来する相補性決定領域と同じ抗原結合性を有し、ヒトに対してより少ない免疫原性を有する組み換え抗体を得ることにある。
【0011】
ある場合には、マウス抗体からのCDRをヒトのフレームワーク中のヒトCDRに置き換えることにより、抗原結合性の性質(抗原に対する特異性のみならず親和性)を改変させるのに充分である。しかしながら、多くの抗体では、いくつかのFR残基が抗原との結合にとって重要である。その理由は、それらのFR残基が抗原抗体複合体における抗原と直接接触する、あるいはそれらがCDRの立体構造に影響を及ぼし、そのために抗原との結合能に影響を及ぼすからである。
【0012】
したがって、ほとんどの場合、マウス抗体の1個あるいは数個のフレームワーク残基を、ヒトの対応するFR残基と置き換えることもまた、必要である。抗マウス反応を防ぐためには、置換された残基の数はできるだけ小さくなければならないので、問題はどのアミノ酸残基が抗原結合性を保持するのに必須であるかを決定することである。より適切な置換部位を予測するために、種々の方法が提案されてきている。これらの方法は、ヒト化の最初の工程で少し役に立つかも知れない一般的な原理を提供しているけれども、最終的な結果は個々の抗体によって変わり得る。したがって、与えられた抗体について、どのような置換が望ましい結果をもたらすかを予測するのは非常に困難である。
【発明の開示】
【発明が解決しようとする課題】
【0013】
しかしながら、本発明者らは、マウスB−F5のヒト化を試みて、親マウスB−F5と
同様のCD4結合能を有するヒト化B−F5(以下、hB−F5と称する)の産生に成功した。
さらに、本発明者らは驚くべきことに、hB−F5が、親B−F5で使用された用量よりも、あるいは他の抗CD4モノクローナル抗体で現在使用されている用量よりも遥かに低用量で最適のインビボ免疫抑制効果を示すことを見出した。
【課題を解決するための手段】
【0014】
実際、本発明者らは、hB−F5を1mg/日の低用量で10日間、好ましくは5mgの用量で2日おきに10日間、使用して治療すると、関節リウマチ患者で臨床的に効果が認められているように、効果的な免疫抑制作用を示すことを観察した。
【0015】
本発明では、マウスB−F5モノクローナル抗体由来の、ヒト化抗体(hB−F5)が提供され、この抗体は次に示すポリペプチド配列で示される可変領域を有する:
− 重鎖可変領域:EEQLVESGGGLVKPGGSLRLSCAASGFSFSDCRMYWLRQAPGKGLEWIGVISVKSENYGANYAESVRGRFTISRDDSKNTVYLQMNSLKTEDTAVYYCSASYYRYDVGAWFAYWGQGTLVTVSS(配列番号:1)
− 軽鎖可変領域:DIVMTQSPDSLAVSLGERATINCRASKSVSTSGYSYIYWYQQKPGQPPKLLIYLASILESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQHSRELPWTFGQGTKVEIK(配列番号:2)
【0016】
一般に、本発明のhB−F5抗体は、さらにヒト定常領域(Fc)を含むものである。この定常領域は、定常ドメインの中で、IgM、 IgG、 IgD、 IgAおよびIgEなどのいずれのクラスの免疫グロブリンからも、そしてIgG1、IgG2、IgG3およびIgG4などのいずれのアイソタイプからも選ぶことができる。好ましい定常領域は、IgG、特にIgG1の定常ドメインから選択される。
【0017】
本発明はまた、可変領域を含むhB−F5抗体の如何なる断片をも包含する。このような断片としては、特に、Fab、Fab’、F(ab)’2、FvおよびscFv断片を挙げることができる。
【0018】
本発明はまた、配列番号:1のポリペプチドをコードするポリヌクレオチド、および配列番号2のポリペプチドをコードするポリヌクレオチドから選択されるポリヌクレオチドを包含する。好ましくは、配列番号:3のポリヌクレオチド、および配列番号:4のポリヌクレオチドから選ばれるポリヌクレオチドである。
本発明のポリヌクレオチドは、組み換えDNA技術および/または化学的DNA合成の公知の方法により容易に得ることができる。
【0019】
hB−F5抗体の重鎖あるいは軽鎖の可変領域をコードするポリヌクレオチドは、このようにして得られる完全な重鎖あるいは軽鎖を発現させるために、ヒト重鎖あるいは軽鎖の定常領域をコードするポリヌクレオチドと融合させてもよい;すなわち、タンパクの分泌ができるようにするシグナルペプチドをコードする配列も附加することができる。 こうした組み換えポリヌクレオチド類もまた本発明の一部を構成する。
【0020】
本発明はまた、本発明のポリヌクレオチドが適当な制御配列と結合してその転写および翻訳を選択された宿主中で可能にする発現カセット、および本発明のポリヌクレオチドあるいは発現カセットを含む組み換えベクターを提供する。
これらの組み換えDNA構築物は、DNAの組み換えおよび遺伝子工学の公知の組み換え手法により得ることができ、宿主細胞に導入することができる。
【0021】
本発明はまた、本発明のポリヌクレオチドで形質転換された宿主細胞を包含する。
本発明の構成に包含される有用な宿主細胞としては、原核細胞あるいは真核細胞である。適当な真核細胞としては、具体的には植物細胞、サッカロマイセスなどの酵母細胞、ショウジョウバエ(Drosophila)あるいはヨトウ(Spodoptera)などの昆虫細胞、およびHeLa、CHO、3T3、C127、BHK、COSなどの動物細胞が挙げられる。
【0022】
本発明の発現ベクターの構築および宿主細胞の形質転換は分子生物学の標準的な手法により達成することができる。
本発明のhB−F5抗体は、当該抗体をコードする核酸配列を含有する発現ベクターを含む宿主細胞を、その発現に適した条件下で培養し、宿主細胞の培養液から回収して得ることができる。
本発明はまた、本発明のhB−F5抗体あるいは上記したそれらの断片からなる治療用組成物をも包含する。
【0023】
好ましくは、そのような組成物は、非経口投与用組成物であって、0.1〜10mg、好都合には1〜5mgのhB−F5の用量で投与することができるように製剤化される。
さらに詳しく述べると、本発明は、免疫抑制組成物の製造のための本発明のhB−F5抗体あるいはその断片の使用を包含するものである。そのような免疫抑制組成物は、同種移植片拒絶反応、移植片対宿主病、宿主対移植片拒絶反応、あるいは例えば、心筋炎、糖尿病、乾癬、紅斑性狼瘡、クローン氏病、多発性硬化症、関節リウマチなどの自己免疫疾患等の治療あるいは予防に特に有用である。
【0024】
さらに、本発明者らは、hB−F5は、CD4陽性T細胞の特定サブセット、すなわちCD4陽性CD25陽性細胞を活性化することができることを見出した。
CD25陽性CD4陽性制御性T細胞(Treg細胞)は、末梢性のCD4陽性T細胞5〜10%を構成成分とする。これらのTreg細胞は、1995年、Sakaguchiらにより、マウスにおける制御細胞として最初に報告された(J.Immunol.,155:1151−1164)。これらのTreg細胞は、活性化されると、CD4陽性およびCD8陽性のT細胞の活性化および増殖を共に抑制することができる。その後、CD25陽性CD4陽性サプレッサーT細胞は、ヒトでも発見された(Jonuleitら,J.Exp.Med.193,1285−1294,2001;Levingsら,J.Exp.Med.193,1295−1302,2001;Dieckmannら,J.Exp.Med.193,1303−1310,2001)。数多くの論文が公開され、これらの細胞が免疫抑制の役割を果たしていることを、種々の自己免疫疾患のモデルやインビトロの系で報告している(総説:例えばShevachら,J.Exp.Med.193,11,41−46,2001)。Ex vivoで活性化されたCD4陽性CD25陽性制御性T細胞(Treg細胞)は、移植片対宿主病の予防に効果的であることが証明された(Taylorら,Blood,99,3493−3499、2002; Cohenら,J.Exp.Med.196,401−406,2002; Hoffmannら,J.Exp.Med.196,389−399,2002)。したがって、CD4陽性CD25陽性制御性T細胞(Treg細胞)を活性化する手段を提供することは、非常に興味あることである。
【0025】
本発明はまた、インビトロでCD25陽性CD4陽性制御性T細胞を活性化するための本発明のhB−F5抗体の使用、またはその親抗体B−F5の使用に関する。
本発明のhB−F5抗体は、1〜10μg/mlの濃度でCD25陽性CD4陽性制御性T細胞に添加される。
【発明を実施するための最良の形態】
【0026】
本発明は、以下の記載によってさらに詳細に説明される。これらは、本発明のhB−F5抗体の性質を説明する実施例である。しかしながら、これらの実施例は、本発明の説明のためにだけされるものであり、本発明の限定を決して構成するものではないことを理解すべきである。
実施例1 ヒト化B−F5の構築
【0027】
ヒト化B−F5のVHおよびVκ領域の設計
マウスB−F5のVHおよびVκ領域をコードするDNA配列は、配列番号:5および配列番号:6としてそれぞれ図1および図2に示されている。マウスCDRが移植されるヒトVHおよびVκは、元のマウスB−F5のVHおよびVκに近似のヒトVHに対するデータベースを探索して選択された。ヒト抗体(M26;受付番号A36006)のVH領域は、B−F5のVHと最も高い相同性を有していた。別のヒト抗体[FK−001;NAKATANIら,Biotechnology,7(1989),805−810)]のVκ領域は、B−F5のVκと最も高い相同性を示した。
4番目の残基がロイシンあるいはメチオニンであることに違いを有するVκの二つのタイプが構築され、それぞれL4LおよびL4Mと命名された。37番目の残基がロイシンあるいはバリンであることに違いを有するVHの二つのタイプが構築され、それぞれH37LおよびH37Vと命名された。B−F5、FK−001、L4LおよびL4Mのポリペプチド配列のアライメントが、図3に示される。B−F5、M26、H37LおよびH37Vのポリペプチド配列のアライメントが図4に示される。CDRを包み込むために重要であると先に報告(Chothiaら,Nature,342,(1989),877;Footeら,J.Mol.Biol.,224(1992),487)されたフレームワークの残基は、囲んで示されている。
これらのVHおよびVκを組み合わせることにより、4種類のV領域が設計された。
【0028】
ヒト化B−F5の発現
ヒト化B−F5の製造のための次の工程は、米国特許5,886、152号でヒト化B−B10に対して記載されたものと同様であった。
要約すると、ヒト化B−F5の重鎖[ヒトγ−1鎖の定常領域(TAKAHASHIら,Cell,29(1982),671−679)と融合したヒト化VH領域]、および軽鎖[FK−0001のκ鎖の定常領域と融合したヒト化Vκ領域]に対する発現プラスミドが別々に構築された。これらのプラスミドにおいて、ヒト化B−F5の発現は、ヒトモノクローナル抗体IgMの遺伝子、すなわちFK−0001のプロモーター/エンハンサーによって推進される。図5および図6は、ヒト化BF−5のVH領域およびVκ領域をコードするプラスミド断片を、それぞれ示している。可変領域をコードする配列には、下線を付してあり、相当するポリペプチド配列は、ヌクレオチド配列の下に示されている。プラスミドおよびpSV2neoの両方を、Lipofectin(登録商標)を使用して同時にマウス骨髄細胞Sp2/0(ATCC CRL−1581)に導入した。ヒトIgGを産生する形質転換体は、抗ヒトIgG(γ鎖)抗体および抗ヒトIgGのκ鎖抗体を用いたELISA法によって選択された。
実施例2 ヒト化B−F5の種々の変異体の特性化
【0029】
CD4結合活性の評価
hB−F5の4つの変異体を産生する形質転換体の培養上澄み液を採取し、濃縮した。この種々の抗体は、プロテインAセファロースの親和性クロマトグラフィーで培養上澄み液から精製し、競合的ELISA法によりビオチン化mB−F5のマイクロタイタープレート上に被覆された可溶性CD4への結合に対する阻害活性を測定して評価した。インキュベーションは、37℃で2時間、4℃で一晩、実施した。
hB−F5(mB−F5の結合活性を100%に設定)の相対的な結合活性を、以下の表Iに示す。
【0030】
【表1】
【0031】
表Iに示された結果から、VHの37番目の残基であるロイシンは、hB−F5のCD4結合活性を維持するのに重要であるように思われる。それというのも、CD4の結合活性は、37番目のロイシンをバリンに変換することにより、数倍減少しているからである。これに対して、Vκの4番目の残基は、CD4結合活性にとってそれほど重要でないことが理解される。VHの37番目のロイシンとバリンの間の構造的な差異は、分子モデルによっては明確には証明されないので、CD4結合活性におけるH37LのH37Vに対する優位性は、予想外であった。
H37L/L4LおよびH37L/L4Mが、インビトロでの生物学的活性評価のために選択された。
インビトロでのヒト化B−F5の生物学的活性の検討
【0032】
インビトロでのマウスB−F5およびヒト化B−F5(H37L/L4M IgG1およびH37L/L4L IgG1)の生物学的活性を評価した。IgG2タイプのヒト化B−F5(H37L/L4M IgG2およびH37L/L4L IgG2)も試験された。
mB−F5および4種のhB−F5のインビトロでの生物学的活性が、健康なドナーからの末梢血単核球(PBMC)を用いて評価された。PBMCは、マウスB−F5またはhB−F5の存在下、ConA(2.5μg/ml、3日間)あるいはPPD(10μg/ml、4日間)により活性化され、[3H]−チミジンの取り込みによる増殖反応が追跡された。
その結果を図7および図8に示す。マウスB−F5およびhB−F5は、ConA誘導性増殖を穏やかに阻害したが、その活性は抗体および/またはドナーにより変化した(図7参照)。また、マウスB−F5およびhB−F5は、PPDで誘導された抗原特異的PBMC増殖を阻害することができた(図8参照)。
hB−F5のIgG1タイプは、mB−F5よりも、PPD誘導性増殖をより効果的に阻害した(70%阻害、図7および図8参照)。IgG1タイプは、阻害活性が殆どmB−F5と同じであるIgG2タイプよりもより有効であるように思われる。IgG1タイプでは、H37L/L4MがH37L/L4Lより有効であった。H37L/L4MおよびH37L/L4LのIgG2タイプでは、殆ど同じ阻害活性を示した。要約すると、PPD誘導性PBMC増殖に対するB−F5の阻害活性は以下のようであった:H37L/L4M IgG1>H37L/L4L IgG1>H37L/L4M IgG2=H37L/L4L IgG2=mB−F5。
インビトロでの生物学的活性の効果およびマウスのアミノ酸数がより少ないことを考慮して、H37L/L4M IgG1がさらに評価するために選択された。
実施例3 関節リウマチ患者(RA)に対するhB−F5の有効性の予備的評価
【0033】
hB−F5(H37L/L4M IgG1)の効果について、RA患者で試験した。
アッセイ条件は、以下の通りである:各患者は、hB−F5の5mgを5回注射、10日間に渡る治療を受けた(2日おきに注射)。
3人の異なる患者の結果を下記の表II〜IVに示す。
患者1(表II)
診断:関節リウマチ、活動度2
リウマチ因子: 2; 重症度: 2
性別: 女性; 年齢: 65歳; 発病: 1965年
付加的治療: ジクロフェナック(150mg/日)
【0034】
【表2】
【0035】
患者2(表III)
診断:関節リウマチ、活動度3
リウマチ因子: 2; 重症度: 2
性別: 女性; 年齢: 48歳; 発病: 2000年
付加的治療: ジクロフェナック(150mg/日)
【0036】
【表3】
【0037】
患者3(表IV)
診断:関節リウマチ、活動度3
リウマチ因子: 3; 重症度: 2
性別: 女性; 年齢: 49歳; 発病: 1989年
付加的治療: ジクロフェナック(150mg/日)
【0038】
【表4】
実施例4 hB−F5によるCD4陽性CD25陽性制御性T細胞の活性化
【0039】
T細胞の単離
1)制御性T細胞(Ttreg):
− CD25陽性細胞は、CD25マイクロビーズを用いて単離される;
− 夾雑物のCD14、CD8、CD19陽性細胞の除去はCD14/CD8/CD19 DYNALビーズを用いて実施される;
− CD45RA陽性細胞の除去は、CD45RA mAb + 抗マウスDYNALビーズを用いて実施される;CD4陽性CD25陽性Tregの純度:>95%
2)エフェクター細胞
− CD4陽性T細胞は、CD4マイクロビーズを用いて単離される;
− CD45RO陽性細胞の除去は、CD45RO陽性mAb + 抗マウスDYNALビーズを用いて実施される;CD25エフェクターT細胞であるCD4/CD45RA陽性細胞の純度:>98%
3)試験系
ドナーAからのCD25陽性Tregを、0.5μg/mlの抗CD3(OKT−3=陽性コントロール=Tregの全活性化)を加えることなく(陰性コントロール=非活性化=非抑制活性)、またはその存在下に、あるいは5μg/mlまたは30μg/mlのhB−F5の存在下に、同質遺伝子型のCD2消失PBMC中で2日間、共培養する。
前培養した細胞をよく洗って、CD25陽性Treg細胞を単離し、γ−線照射処理(3000ラド)する。
4)抑制活性試験
前培養CD25陽性Treg細胞と、ドナーB[エフェクターT細胞に対して異質遺伝子型]からの新鮮な単離CD4陽性エフェクターT細胞とを1:1の割合で、ドナーA[前培養T細胞(さらに活性化されていない)に対して同質遺伝子型]からのAPC(抗原提示細胞)(CD2消失PBMC)の存在下、4日間、共培養する(=異質遺伝子型の混合リンパ球反応)。次いで、細胞を16時間、[3H]−チミジンとインキュベーションして、エフェクターT細胞の増殖を検出する。
結果を図9に示す。
【0040】
図9の説明:
− 陰性コントロール(非活性化)= 前培養CD25細胞;
− 0.5μg/ml OKT−3(陽性コントロール、全活性化)=
前培養CD25−CD3細胞;
− 5μg/ml hB−F5(試験−1)=前培養CD25−CD4細胞:
− 30μg/ml hB−F5(試験−2)=前培養CD25−CD4細胞。
【図面の簡単な説明】
【0041】
【図1】は、マウスB−F5(mB−F5)のVH領域をコードするDNA配列を示す。
【図2】は、マウスB−F5(mB−F5)のVκ領域をコードするDNA配列を示す。
【図3】は、B−F5、FK−001、L4LおよびL4Mのポリペプチド配列のアライメントを示す。
【図4】は、B−F5、M26、H37LおよびH37Vのポリペプチド配列のアライメントを示す。
【図5】は、ヒト化BF−5のVH領域をコードするプラスミド断片のDNA配列およびポリペプチド配列を示す。可変領域をコードするDNA配列には、下線が付してあり、相当するポリペプチド配列は、当該DNA配列の下に示されている。
【図6】ヒト化BF−5のVκ領域をコードするプラスミド断片のDNA配列およびポリペプチド配列を示す。可変領域をコードするDNA配列には、下線が付してあり、相当するポリペプチド配列は、当該DNA配列の下に示されている。
【図7】は、ConA誘導性のPBMC増殖に対するマウスB−F5およびhB−F5の[3H]−チミジン取り込み阻害を示す。
【図8】は、PPD誘導性のPBMC増殖に対するマウスB−F5およびhB−F5の[3H]−チミジン取り込み阻害を示す。
【図9】は、各種B−F5のエフェクターT細胞増殖に対する抑制作用を示す。
【特許請求の範囲】
【請求項1】
マウスのマウスモノクローナル抗CD4抗体B−F5由来のヒト化抗体(hB−F5)であって、当該hB−F5抗体は以下のポリペプチド配列で示される可変領域:
− 重鎖可変領域:EEQLVESGGGLVKPGGSLRLSCAASGFSFSDCRMYWLRQAPGKGLEWIGVISVKSENYGANYAESVRGRFTISRDDSKNTVYLQMNSLKTEDTAVYYCSASYYRYDVGAWFAYWGQGTLVTVSS(配列番号:1)
− 軽鎖可変領域:DIVMTQSPDSLAVSLGERATINCRASKSVSTSGYSYIYWYQQKPGQPPKLLIYLASILESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQHSRELPWTFGQGTKVEIK(配列番号:2)
を有することを特徴とするヒト化抗体(hB−F5)。
【請求項2】
配列番号1および配列番号2の可変領域を含むことを特徴とする、請求項1に記載のhB−F5抗体の断片。
【請求項3】
配列番号1の重鎖可変領域をコードする配列を含むポリヌクレオチドおよび配列番号2の軽鎖可変領域をコードする配列を含むポリヌクレオチドから選ばれることを特徴とするポリヌクレオチド。
【請求項4】
配列番号3の配列を含むポリヌクレオチドおよび配列番号4の配列を含むポリヌクレオチドから選ばれることを特徴とする、請求項3に記載のポリヌクレオチド。
【請求項5】
請求項1に記載のヒト化抗体または請求項2に記載の断片を含むことを特徴とする、治療用組成物。
【請求項6】
免疫抑制組成物の製造のための、請求項1に記載のヒト化抗体または請求項2に記載の断片の使用。
【請求項7】
インビトロでCD25陽性CD4陽性制御性T細胞を活性化するための、請求項1に記載のヒト化抗体の使用。
【請求項1】
マウスのマウスモノクローナル抗CD4抗体B−F5由来のヒト化抗体(hB−F5)であって、当該hB−F5抗体は以下のポリペプチド配列で示される可変領域:
− 重鎖可変領域:EEQLVESGGGLVKPGGSLRLSCAASGFSFSDCRMYWLRQAPGKGLEWIGVISVKSENYGANYAESVRGRFTISRDDSKNTVYLQMNSLKTEDTAVYYCSASYYRYDVGAWFAYWGQGTLVTVSS(配列番号:1)
− 軽鎖可変領域:DIVMTQSPDSLAVSLGERATINCRASKSVSTSGYSYIYWYQQKPGQPPKLLIYLASILESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQHSRELPWTFGQGTKVEIK(配列番号:2)
を有することを特徴とするヒト化抗体(hB−F5)。
【請求項2】
配列番号1および配列番号2の可変領域を含むことを特徴とする、請求項1に記載のhB−F5抗体の断片。
【請求項3】
配列番号1の重鎖可変領域をコードする配列を含むポリヌクレオチドおよび配列番号2の軽鎖可変領域をコードする配列を含むポリヌクレオチドから選ばれることを特徴とするポリヌクレオチド。
【請求項4】
配列番号3の配列を含むポリヌクレオチドおよび配列番号4の配列を含むポリヌクレオチドから選ばれることを特徴とする、請求項3に記載のポリヌクレオチド。
【請求項5】
請求項1に記載のヒト化抗体または請求項2に記載の断片を含むことを特徴とする、治療用組成物。
【請求項6】
免疫抑制組成物の製造のための、請求項1に記載のヒト化抗体または請求項2に記載の断片の使用。
【請求項7】
インビトロでCD25陽性CD4陽性制御性T細胞を活性化するための、請求項1に記載のヒト化抗体の使用。
【図1】


【図2】

【図3】
【図4】
【図5】

【図6】

【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2007−525156(P2007−525156A)
【公表日】平成19年9月6日(2007.9.6)
【国際特許分類】
【出願番号】特願2006−504748(P2006−504748)
【出願日】平成16年3月19日(2004.3.19)
【国際出願番号】PCT/EP2004/002888
【国際公開番号】WO2004/083247
【国際公開日】平成16年9月30日(2004.9.30)
【出願人】(505348500)ビオテスト・アーゲー (1)
【Fターム(参考)】
【公表日】平成19年9月6日(2007.9.6)
【国際特許分類】
【出願日】平成16年3月19日(2004.3.19)
【国際出願番号】PCT/EP2004/002888
【国際公開番号】WO2004/083247
【国際公開日】平成16年9月30日(2004.9.30)
【出願人】(505348500)ビオテスト・アーゲー (1)
【Fターム(参考)】
[ Back to top ]