
免疫治療組成物調製用のレンチウイルスベクター
【課題】免疫治療組成物調製用のレンチウイルスベクターを提供する。
【解決手段】シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を備えたレトロウイルス又はレトロウイルス様の組換えベクターからなる。該ベクターは、所定のヌクレオチド配列(目的の導入遺伝子又は配列)及びレトロウイルス又はレトロウイルス様の逆転写、発現及びウイルス粒子形成の調節遺伝子を含み、細胞性応答、例えば前記ベクターに存在する導入遺伝子によってコードされた1又は数個のエピトープに対するCTL(細胞障害性リンパ球)応答又はCD4応答を誘導又は促進することが可能である。
【解決手段】シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を備えたレトロウイルス又はレトロウイルス様の組換えベクターからなる。該ベクターは、所定のヌクレオチド配列(目的の導入遺伝子又は配列)及びレトロウイルス又はレトロウイルス様の逆転写、発現及びウイルス粒子形成の調節遺伝子を含み、細胞性応答、例えば前記ベクターに存在する導入遺伝子によってコードされた1又は数個のエピトープに対するCTL(細胞障害性リンパ球)応答又はCD4応答を誘導又は促進することが可能である。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
本発明は、インビトロで、及び好ましい実施態様ではインビボで、ベクター中に存在するヌクレオチド配列によってコードされたエピトープに対する免疫反応の発生若しくは増強を誘導し、又はこれらに寄与することが可能な組成物を調製するためのレトロウイルスベクター、特にレンチウイルスベクターの使用に関する。
【0002】
本発明者等は、本発明に従って調製したベクターによって、細胞性免疫応答、特にエピトープに対する細胞障害性Tリンパ球(CTL)反応を得ることが可能であることを示した。
【0003】
さらに、この細胞性免疫応答がベクターに含まれるヌクレオチド配列によってコードされた1又は数個のエピトープに対して得られた特異的応答であり得ることを示すデータが得られた。
【0004】
従って、本発明は腫瘍及び癌に対する治療プロトコールに使用でき、特に腫瘍に対する免疫療法又は予防接種療法のプロトコールに使用できる手段を提供する。
【0005】
本発明は、感染症、特にウイルス感染、例えばレトロウイルス感染に関連した疾患の治療また予防に使用できる手段も開示する。
【0006】
本発明者らは、さらに、本発明の組成物による治療に関連した細胞性免疫応答、特にCTL応答は腫瘍若しくはウイルスの抗原、又はウイルス感染細胞の抗原に特異的であることが可能で、MHC(主要組織適合複合体)の特定の分子に限定され得ることを示す結果を得た。
【0007】
特に本発明は、MHC複合体のクラスI分子、例えばHLA−A2又はB7分子に限定した細胞性免疫応答を得るために、免疫原組成物に前記ベクターを使用することに関する。
【0008】
従って、本発明はシス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を包含するポリヌクレオチドを含む組換えベクターを含む免疫原組成物に関する。cPPT及びCTS配列は、逆転写中に3本鎖DNA構造(本明細書ではDNA3重鎖と称する)の形成を誘導する。このDNA3重鎖は、ベクターDNAゲノムの核移入を刺激し、これらの領域の起源はレトロウイルス又はレトロウイルス様で、前記ベクターは所定のヌクレオチド配列(目的の導入遺伝子又は配列)に加えて、レトロウイルス又はレトロウイルス様起源の逆転写(retrotranscription)、発現、及びウイルス粒子形成の調節シグナルを含み、この組成物はベクター内に存在する導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)又はCD4の応答を誘導又は刺激することが可能である。
【0009】
本発明の好ましい実施態様では、細胞性免疫応答、特に1又は数個のエピトープに対するCTL応答又はCD4応答は記憶CTL又はCD4応答である。
【0010】
ベクター内にcPPT及びCTS領域が存在すると、3重鎖DNA構造が形成されることが可能で、それによって前記ベクターで組換えられた細胞でのベクターゲノムの核移入が影響を受け、特に増強され得ることが強調される。
【0011】
本発明による免疫原組成物は記憶CTL応答の発生を誘導、改良し、又は一般にその発生に関与することができるので、(前記免疫原組成物の投与後、前記免疫応答が長期間にわたって誘導されなければならない場合、又は少なくとも応答が必要とされる期間に(前記応答の長期誘導であってもよい)誘導可能でなければならない場合を含む)抗腫瘍治療又は抗ウイルス治療又は抗病原性治療のプロトコールにおいて前記免疫原組成物の使用を企図することが可能である。言い換えると、前記免疫原組成物は、細胞性免疫応答、特にCTL応答又は記憶応答の発生を誘導又は刺激し、又はそれに寄与することによって、腫瘍疾患又は感染性疾患(例えば細菌又はウイルスによる)を治療するための治療用組成物の調製に使用することができる。
【0012】
本発明の免疫原組成物は、ベクターの配列中に3重鎖構造が存在する結果、ベクター及びベクター粒子内にcPPT及びCTS領域が存在するため、標的細胞においてベクターゲノムの核移入を刺激させ得ることが注目される。ベクター内で誘導されるエピトープは細胞自身のものであってもよいし、細胞以外のものであってもよい。
【0013】
本発明は、標的細胞又は受容細胞の核への核酸配列又はペプチド配列の移入を増加させるために、レトロウイルス由来又は合成によるcPPT及びCTS配列を含むヌクレオチド配列の使用も含む。
【0014】
例えば、前記3重鎖配列には受容細胞の外来配列又は自己配列が含まれることが理解されよう。
【0015】
従って、本発明は、腫瘍治療、特に抗癌治療又は抗感染性疾患治療のための予防接種プロトコールと類似した治療プロトコールにおいて使用可能な組成物を開示する。
【0016】
本発明では、目的の該導入遺伝子又は配列が、1又は数個の腫瘍細胞の1又は数個のエピトープ、例えば腫瘍に対する細胞性免疫を誘導することが可能な標的抗原に同定されたエピトープをコードする配列である得ることに注目すると興味深い。
【0017】
ポリペプチドを形成する数個のエピトープは、本発明の導入遺伝子によってコードすることができる。特定の実施態様では、それらは腫瘍で確認された標的抗原から得ることができ、コーディング配列が一緒になって導入遺伝子を形成するとき、細胞性免疫応答がエピトープ全て又はエピトープのほとんどに対して得られるような方法で選択され得る。細胞性免疫応答はインビトロ、又は好ましい実施態様ではインビボでアッセイすることができる。このようなアッセイの実施を可能にするプロトコールは実施例で説明する。
【0018】
標的抗原はいくつかの種類の腫瘍、特にメラノーマ又は腎癌、膀胱癌、大腸癌、肺癌、乳癌、白血病及びリンパ腫などを含む癌腫で同定されている。
【0019】
本発明の他の態様では、前記免疫原組成物はウイルス関連感染症、又はミコバクテリア、例えばM tuberculosisを含む任意の種類の病原体に関連する感染症を含めた感染性疾患において、細胞性免疫応答を得るために使用することができる。
【0020】
この場合、細胞性免疫応答を引き起こすことが可能な特定の抗原を同定して、そのコーディング配列をベクターに挿入して前記免疫原組成物で使用することができる。例えば、M tuberculosisのdes遺伝子が使用できる。
【0021】
本発明では、前記免疫原組成物で使用されるベクターは、エピトープを発現するか、又は腫瘍細胞若しくはウイルス感染細胞上の標的抗原として同定された(糖タンパク質又はその他のタンパク質由来化合物を含む)タンパク質上に提示することが可能であることも付け加えなければならない。
【0022】
さらに、エピトープ、エピトープを提供するために使用されたポリペプチド又はタンパク質は、例えば変異、欠失又は挿入によって変更することが可能で、例えば安定性を高めるために変更することが可能であることにも留意すべきである。
【0023】
本発明は、
1)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成の調節シグナルの制御下に置かれた所定のヌクレオチド配列(導入遺伝子)を含有する組換えヌクレオチド配列と、
2)シス作用性中心開始領域(CTS)及びシス作用性終止領域(CTS)を包含するポリヌクレオチドであって、これらの領域の起源がレトロウイルス又はレトロウイルス様であるか、又はトランスポゾン由来であり、レトロウイルス又はレトロウイルス様起源の逆転写調節シグナル又はトランスポゾン調節シグナルとともに、機能的な方向及び位置に挿入されたポリヌクレオチドと
を備えた組換えレトロウイルス粒子を含む免疫原組成物であって、
ベクターに存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は刺激することができる免疫原組成物に関する。
【0024】
cPPT及びCTSシス作用性配列を包含するDNA配列は、逆転写後に3本鎖DNA構造「DNA3重鎖」を採り、ベクターDNAの核移入を刺激することができる。
【0025】
特定の実施態様では、ベクターに存在する導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は刺激することが可能な前記免疫原組成物は、
a)レンチウイルスの核タンパク質又は機能的に得られたポリペプチドに対応するgagポリペプチド(GAGポリペプチド)と、
b)レンチウイルスのRT、PRO、INタンパク質又は機能的に得られたポリペプチドによって構成されたpolポリペプチド(POLポリペプチド)と、
c)エンベロープポリペプチド又は機能的に得られたポリペプチド(ENVポリペプチド)と、
d)転写及び発現の調節シグナルの制御下に置かれ、1又は数個のエピトープをコードする所定のヌクレオチド配列(目的の導入遺伝子又は配列)と、レトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の調節シグナルを含有する配列と、並びにシス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含有するポリヌクレオチドであって、これらの領域がレトロウイルス又はレトロウイルス様の起源であり、上記レトロウイルス又はレトロウイルス様起源の調節シグナルとともに機能的な方向に挿入されているポリヌクレオチドとを備えた組換えヌクレオチド配列と
を具備する組換えレトロウイルスベクター粒子を含む。
【0026】
他の実施態様では、ベクターに存在する導入遺伝子によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は促進することが可能な免疫原組成物は、
a)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域がレトロトランスポゾン由来で、レトロトランスポゾン調節シグナルと共に機能的な方向挿入されたポリヌクレオチドと、
b)レトロトランスポゾンの核タンパク質又は又は機能的に得られたポリペプチドに対応するポリペプチド(GAGポリヌクレオチド)と、
c)レトロトランスポゾンのRT、PRO、INタンパク質に対応するpolポリペプチド又は機能的に得られたポリペプチド(POLポリペプチド)と、
d)ウイルスエンベロープポリペプチドと、
e)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成のレトロトランスポゾン調節シグナルの制御下に置かれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)を備えた組換えヌクレオチド配列を具備する組換えレトロウイルス様粒子と
を含む。
【0027】
好ましい実施態様では、前記の定義の何れか1つに応じた免疫原組成物に存在する組換えレトロウイルスベクター粒子は、記憶細胞性免疫応答、特に記憶CTL応答の発生を誘導、増強するか、又は関連することが可能である。
【0028】
前記で開示した定義に従って、ベクター又はベクター粒子を含む免疫組成物は、いくつか可能な実施態様によって調製することができる。
【0029】
本発明の好ましい実施態様では、レトロウイルス起源の配列をレンチウイルスゲノムから得る方法で免疫原組成物は調製される。
【0030】
他の実施態様では、これらの配列の起源はレトロウイルス様であり、レトロトランスポゾンからも得られる。
【0031】
本発明の他の実施態様では、又は前記で定義した特徴に加えて、組換えベクターに含有される目的の導入遺伝子又は配列は、転写及び発現の調節シグナルを含有する発現カセットに含まれる。
【0032】
あるいは、ベクターの逆転写、発現及びウイルス粒子形成の調節シグナルはレンチウイルス起源であり、cPPT及びCTS領域を備えたポリヌクレオチドもレンチウイルス起源である。
【0033】
他の実施態様では、ベクター内の逆転写、発現、及びウイルス粒子形成の調節シグナル並びにcPPT及びCTS領域を含むポリヌクレオチドは、HIV型、特にHIV−2又はHIV−2のレトロウイルスから得られる。
【0034】
他のウイルス、特にレンチウイルスは、逆転写発現及びウイルス粒子形成の調節シグナルを設計するために、さらにはcPPT及びCTS領域を含むポリヌクレオチドを得るために使用することができる。従って、特にレンチウイルスCAEV、EIAV、VISNA、HIV、SIV又はFIVを使用することができる。
【0035】
本発明の免疫原組成物の組換えレトロウイルス粒子を獲得するため、ベクターのトランス相補性に必要なポリペプチド又はタンパク質をコードする配列は、例えばレンチウイルス、特にHIV−1及びHIV−2レトロウイルスを含むHIV由来のGAG、POL及びENVタンパク質である。
【0036】
あるいは、GAG及びPOL配列は、ENV配列とは異なるウイルスに由来してもよい。例えば、GAG及びPOL配列はHIVレトロウイルス由来であり、ENV配列は他のウイルス又はレトロウイルス由来し、且つアンフォトロピック又はエコトロピックENV配列の何れであってもよい。
【0037】
他の実施態様では、ENV配列は水疱性口炎ウイルス(VSV)から得られる。
【0038】
本発明の特定の実施態様では、ベクター中に存在する導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は促進することが可能な免疫原組成物は、
a)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域がレトロトランスポゾンから得られ、レトロトランスポゾン調節シグナルと共に機能的な方向に挿入されたポリヌクレオチドと、
b)レトロトランスポゾンの核タンパク質又は機能的に得られたポリペプチドに対応するポリペプチド(GAGポリペプチド)と、
c)レトロトランスポゾンのRT、PRO、INタンパク質に対応するpolポリペプチド又は機能的に得られたポリペプチド(POLポリペプチド)と、
d)ウイルスエンベロープポリペプチドと、
e)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成のレトロトランスポゾン調節シグナルの制御下に置かれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)を含む組換えヌクレオチド配列と
を備えた組換えレトロウイルス様粒子を含む。
【0039】
好ましい実施態様では、組換えレトロウイルス様粒子を含む本発明の免疫原組成物は、前記で開示した特徴によって、記憶細胞性応答、特に記憶CTL応答を引き起こすことが可能である。
【0040】
本発明は、1999年10月11日にCNCM(Collection Nationale de Culture de Microorganismes at Institut Pasteur in Paris、France)に寄託されたベクター構築物にも関する。
【0041】
第1のベクターは、1999年10月11日にl−2326号として寄託されたpTRIP.TEL/AML−IRES−GFPであり、第2のベクターはpTRIP−ILKE−IRES−GFPと称し、1999年10月11日にl−2327号として寄託された。
【0042】
第3のベクター、pTRIP.DES−IRES−GFPは、1999年10月11日にl−2331号としてCNCMに寄託された。
【0043】
前記構築物に存在する抗原をコードする配列は、前記で引用したM tuberculosisの完全なDES遺伝子を含む、他の任意の目的の抗原又はエピトープによって置き換えることができる。
【0044】
本発明の他の態様では、ベクター、ベクター粒子及び同物を含む免疫原組成物は、cPPT及びCTS領域がベクター配列の中心に位置するように設計される。
【0045】
「中心に位置する」とは、cPPT及びCTS領域がベクター配列の中心、又はこの配列のほぼ中心にあることを意味する。特に、cPPT及びCTS領域は逆転写された直鎖状ベクターDNAの中央の3分の1以内にあることが可能である。
【0046】
cPPT及びCTS配列が存在する結果としてウイルス逆転写中に形成された3重鎖配列が中央に位置することによって、そのベクター又はベクター粒子と接触する細胞の形質導入量を増大させることが可能である。
【0047】
あるいは、前記ベクターの変種によっては、導入遺伝子を含むベクターの転写単位はLTR領域のU3領域内に挿入することができる。従って、逆転写の後、導入遺伝子は複製され、従って3重鎖配列のそれぞれの鎖に現れ、従って導入遺伝子の大きさにかかわらず、3重鎖配列がベクターの中心部分に位置することが可能となる。
【0048】
本発明は、本発明による免疫原組成物と接触するように置かれた細胞に関し、特に免疫原組成物のベクター又はベクター粒子によって形質導入された組換え細胞にも関する。
【0049】
これらの細胞は、有利には、抗原提示細胞である。例えば、これらの細胞は、肺細胞、脳細胞、上皮細胞、星状細胞、マイクログリア、乏突起神経膠細胞、ニューロン、筋細胞、肝細胞、樹状細胞、神経細胞、骨髄の細胞株、マクロファージ、繊維芽細胞、造血細胞から選択することができる。
【0050】
従って、本発明の免疫原組成物は、一次細胞性免疫応答、特に、有利には記憶CTL応答であるCTL応答を発生させ、腫瘍、特に癌の治療方法又は治療処置のための治療組成物の調製に使用することができる。あるいは、その他公知の抗癌治療の補助処置として使用することができる。
【0051】
例えば、本発明の免疫原組成物は、化学療法又は免疫化学療法又は抗癌治療のその他のアプローチと関連させて使用することができる。
【0052】
「抗癌治療」という表現は、本発明では、腫瘍の増殖又は腫瘍が増殖する可能性を阻害すること、若しくは転移巣形成の制御を含めた悪性細胞の拡散を阻害することを意味するか、又はその両者を意味するものである。
【0053】
従って、本発明では、「抗癌治療」という表現は、特にこの免疫原組成物が記憶細胞性応答を誘導又は増強し、一般的には関与することが可能であるという事実を考慮すると、疾患の治癒を期待して、悪性細胞の増殖及び腫瘍の拡散を制御するために使用する方法に関している。
【0054】
本発明の組成物で治療し得る腫瘍は、例えば、メラノーマ又は(肺、膀胱、腎臓、大腸)含めた癌腫及びリンパ滲出である。
【0055】
治療可能な腫瘍も、変異した自己タンパク質及び/又は過剰発現した自己タンパク質を含む腫瘍特異的抗原を発現している全ての腫瘍である。
【0056】
エキソビボ段階を含む投与方法、例えば、標的細胞にエキソビボ導入した後、標的細胞を治療する患者に投与することを含めて、受容可能な本発明の免疫原組成物の投与方法はいずれも興味深い。
【0057】
あるいは、本発明の免疫原組成物は、全身(IV)、局所、又は皮膚、皮下、例えば筋肉内投与経路を含めた通常の投与経路によって患者に直接投与し得る。
【0058】
特定の実施態様では、本発明の免疫原組成物は、細胞性免疫応答、特にCTL性免疫応答の発生をインビボで誘導、増強、又は関与する方法で患者に直接投与し得る。
【0059】
他の実施態様では、前記免疫原組成物は、長期間記憶細胞性応答を発生させるために使用される。
【0060】
本発明の前記免疫原組成物は、ベクター又はベクター粒子に存在するcPPT及びCTS配列の特性によって、標的細胞内でベクターゲノムの核移入を誘導又は促進するので特に興味深いことが強調される。
【0061】
本発明の免疫原組成物が特に優位なのは、これらがベクター又はベクター粒子に存在する関心のあるヌクレオチド配列又は導入遺伝子によってコードされた複数のエピトープに対する細胞性免疫応答を惹起又は促進するために使用できること、及び完全な遺伝子配列、例えば病原性物質の遺伝子の産物又は少なくとも8から15アミノ酸、好ましくは9から12アミノ酸をコードし得る前記遺伝子の断片の産物に対する細胞性免疫応答を惹起又は促進するためにも使用できることである。本発明はまた、細胞性応答を誘導する前記アミノ酸配列の複数反復(少なくとも2回の同一配列)及び/又は異なる病原性抗原又は腫瘍抗原の2個のエピトープに対応する少なくとも2種の異なる配列を含むアミノ酸配列をコードするヌクレオチド配列を含むヌクレオチド配列を含む。
【0062】
本発明のその他の特性又は利点を、以下の実施例及び図面で開示する。
【0063】
実施例
レンチウイルスベクターは、非分裂細胞を含む細胞を形質導入する能力を有しており、遺伝子治療にとってますます重要となっている。最近、ポリプリン広域シス作用性配列(中心DNA3重鎖)を含むレンチウイルスベクターが中心DNA3重鎖を欠失したものよりもヒト及びマウスをより効果的に形質導入することが示された(Charneau P.et.al.J.Mol.Biol.1994、241、651〜662)。中心DNA3重鎖を含むか、又は含まず、同様のHLA−A2.1限定メラノーマCTLポリエピトープをコードするレンチウイルスベクターのTL応答を誘導する能力を本明細書で試験した。HHD(HLA−A2.1純系)マウスにおいて、中心DNAを含むレンチウイルスベクターを直接インビボ投与すると、ポリエピトープ配列によってコードされたエピトープペプチド全てに対する強いCTL応答が誘導された。レンチウイルスベクターはDNA3重鎖を含むと明らかに有利であることが示された。さらに、DNA3重鎖を含むレンチウイルスが3重鎖を含まないベクターよりもヒト樹状細胞を7倍も効果的に形質導入することが示された。これらのエキソビボ導入した樹状細胞は、メラノーマエピトープペプチドのほとんどに対して効果的かつ特異的な1次CTL応答を引き起こす。変更したレンチウイルスの安全性の問題がほぼ解決されたので、このベクターをインビボにおけるヒトの遺伝子治療だけでなく、癌患者の免疫療法にも使用することを提案する。
【0064】
緒言
レトロウイルスの亜種レンチウイルスは、非分裂細胞を含めたほとんどの細胞種に感染することができる。この特性があるため、レンチウイルスは遺伝子治療にとって魅力的な存在である。いくつかの複製欠失組換えレンチウイルスベクターは、既に様々なグループによって構築されている(Naldini PNAS 93、11382〜8、Science、1996)。再操作し、無毒化したこれらのレンチウイルスベクターは最も効果的かつ安全な遺伝子治療用ベクターとして提案されている(Zufferey R、& Kim V.N.J Virol、72、9873〜80、1998)。本発明者等は、水疱性口炎ウイルスGタンパク質(VSVG)偽型レンチウイルス(HIV)をベースとしたベクターを開発した(Burns J.C.、1993 PNAS 90、8033〜7)。このベクターは非必須なウイルスタンパク質をほとんど欠失しているが、ポリプリン領域シス作用性配列(cPPT、中心DNA3重鎖)を含む。この中心DNA3重鎖は、逆転写されたHIV−DNA分子の核移入をかなり促進する。さらに、中心DNA3重鎖を含むレンチウイルスは、この3重鎖を含まないベクターよりも効果的にマウス及びヒト細胞を形質導入させることが認められた。
【0065】
HHD(HLA−A2.1純系)トランスジェニックマウス(Pascolo et al.、J.Exp.Med.、1997、185:2043〜2051)を用いると、実験的に制御したエピトープペプチドの免疫原性の評価及び様々な免疫戦略の評価が可能である。これらのHHDマウスを使用して、異なる組換えベクターによってコードされたメラノーマポリペプチドが単一の動物個体内で同時にCTL応答を誘導する能力が報告された。中心DNA3重鎖を含むか、又は含まず、同様のメラノーマポリエピトープをコードするレンチウイルスベクターが、HHDトランスジェニックマウスにおいてインビボCTLを誘導する能力について最初に調べた。さらに同様の組換えレンチウイルスによって形質導入されたヒト樹状細胞(hDC)がエキソビボでメラノーマポリエピトープモチーフに対して1次CTL応答を誘導するかどうかについても調べた。本発明の結果によって、組換えベクターを直接投与するか、又はエキソビボで形質導入した樹状細胞を使用すると、DNA3重鎖によってレンチウイルスベクターがインビボで特異的なCTL応答を誘導する能力が著しく増強されることが示された。
【0066】
結果
実施例1:メラノーマポリエピトープの免疫
メラノーマから得られたポリエピトープモチーフをコードする組換えDNAをHHDマウスに注射することによって、いくつかのメラノーマエピトープに対して同時にCTL応答を惹起することができることを証明した後、同様のメラノーマポリペプチドモチーフ(図1及び表1)をコードするレンチウイルスの免疫原能を試験した。TRIP−mel−IRES−GFPベクター(CNCM 1−2185として1999年4月20日に寄託)をマウス当たり1.25μg/p24で静脈内、腹腔内、又は皮下内に投与した。1群当たり少なくとも3匹のマウスを使用した。
【0067】
10個のメラノーマエピトープのほとんどに対して特異的な複数のCTL応答が同時に誘導された。投与経路に関わらず、負荷したペプチド(表2)及びTRIP−mel−IRES−GFPで形質導入したHHDトランスフェクトHeLa細胞(データは示していない)の両者に対して同様のCTL応答が認められた。しかし、腹腔内注射によって、わずかに良好なCTL応答が誘導された。強い応答がNA17−A.nt38及びgp100.154エピトープペプチドに対して引き起こされた。gp100.457、MART.1.27、Mage−3、及びチロシナーゼ368−Dに対してまた、著しい応答が認められた。gp.100.209、gp.100.280、MART−1.32、及びチロシナーゼ1に対するCTL応答は弱かった。TRIP−mel−IRES−GFPベクターで免疫した後に弱いCTL応答が引き起こされたエピトープは全て、その他のレンチウイルスベクター以外を投与したときのCTL非誘導群に分類された。
【0068】
インビボで検出可能なCTLを誘導するために必要なレンチウイルスベクターの最小用量
著しいCTL応答を引き起こすレンチウイルスベクターの最小用量は、6種の異なる用量のTRIP−mel−IRES−GFPベクターを腹腔内投与して(1用量あたり4匹のHHDマウス)決定した。51Cr試験の直前に2匹のマウスのエフェクター細胞を混合して同一のE/T比を得る実験を2回実施した。ほとんどの実験において、メラノーマエピトープペプチド全てに対するCTL応答を試験した。結果は非常に類似しており、極めて同質であったので、「線量−効果」関係を明確かつ簡潔に比較するためにNA17/Aエピトープペプチドに対するCTL応答のみを考慮した。マウス当たり500ngと2500ng/p24との間の用量を使用して、最良のCTL応答が得られた。いくつかのメラノーマエピトープに対して検出可能なCTL応答が得られたが、レンチウイルスの用量が高くても用量の低い場合よりも良好なCTL応答が誘導されるわけではなかった。いくつかのメラノーマペプチドに対するいくつかの特異的なCTL応答が実証されたとしても、マウス当たり500ng/p24未満の用量では、効果的なCTL応答を誘導するには不十分であった。マウス当たり1250ng/p24の用量で、何匹かのマウスにおいてポリエピトープモチーフに含まれる10個のエピトープ全てに対してCTL応答が生じたことは注目に値する(図3)。
【0069】
長期の記憶CTL誘導
マウス8匹にTRIP−mel−IRES−GFPベクターを注射した。免疫後、このマウスを12日目又は5ヶ月目に殺処分した。5日間はメラノーマエピトープペプチドで、さらに2日間は10%TCGFでインビトロ誘発した後、マウス4匹のエフェクター細胞を混合してペプチドを負荷したHHDトランスフェクトRMAS細胞に対して試験した。12日前に免疫したマウスでは、gp100.29及びMart1.32以外のメラノーマエピトープペプチド全てについてCTL応答が証明された。TRIP−mel−IRES−GFPを注射して5ヶ月経つと、1次CTL誘導エピトープ全てがまだ強いCTL応答を誘導した(図2)。マウス免疫後、12日目又は5ヶ月目のCTL応答のレベルは、驚くほど同等であった。レンチウイルスベクターによってインビボ導入された細胞は、免疫系によって破壊されずにコードされたメラノーマポリエピトープを産生し続けることが示唆された。
【0070】
インビボ免疫における中心DNA3重鎖の役割
HHDマウスそれぞれに、同時にTRIP−mel−IRES−GFP及びHR−mel−IRES−GFPベクターをマウス当たり800ng、200ng、50ng、12ng、及び3ng/p24の用量で腹腔内投与して免疫した。別個の2回の実験において、各用量について少なくとも4匹のマウスを試験した。合成ペプチドでインビトロ誘発した後、ペプチドを加えたRMAS−HHD標的細胞を使用して脾細胞の細胞溶解能を試験した。結果は極めて似通っていたので、明確かつ簡潔にするためにNA17/A、Mart−1.27、gp100.154、及びチロシナーゼ.368−Dエピトープペプチドに対するCTL応答のみを試験した。
【0071】
結果を表3に示す。一般に、マウス当たり800ng、200ng、50ng/p24の用量のTRIP−LVベクターで免疫したマウスはHR−LVベクターで免疫したマウスよりも強いCTL応答を引き起こした。使用したベクターにかかわらず、マウス当たりの12ng/p24未満の用量では検出可能なCTL応答は認められなかった(データは示さず)。これによって、中心DNA複合体を含むレンチウイルスベクターの形質導入能はこの複合体を欠失したものよりも増強されていることが確認された。
【0072】
レンチウイルスベクターによる樹状細胞の形質導入
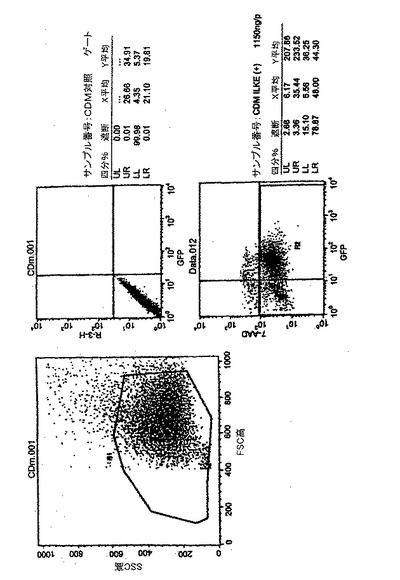
MT4及びHeLa細胞などの腫瘍細胞が中心DNA3重鎖を含むレンチウイルスによって中心DNA3重鎖を持たないベクターよりも30倍まで効果的に形質導入されることが最初に観察された。次に、様々な濃度でこれら2種のレンチウイルスベクターの形質導入能を健常提供者又はHHDマウスのDCについて試験した。GFPを発現するDCのパーセント及び平均蛍光強度をFACSによって測定し、2種のレンチウイルスベクターによるDCの形質導入濃度と見なした。
【0073】
TRIP−GFPベクターは、どのベクター濃度でもHR−GFPよりも効果的に細胞を形質導入した。TRIP−GFPベクターによって形質導入されたマウス及びヒトDCのGFP発現は、それぞれHR−GFPベクターによって形質導入されたGFPの発現の3倍及び7倍に達した(図4)。
【0074】
TRIP mel IRES−GFPベクターによって形質導入されたヒト樹状細胞を使用した1次CTL誘発
健常なHLA−A2.1提供者から得られた単核細胞(MNC)を、同提供者から得られたTRIP mel−IRES−GFPによって形質導入したDCを使用して週に1回インビトロで誘発した。樹状細胞におけるGFP発現の存在をFACSによって分析し、形質導入の効果を確かめた。3週間後、MNC細胞障害性は、FCSを含まない培養条件で標的としてペプチドを付加したT2細胞を使用した51Crアッセイによって試験した。
【0075】
TRIP mel−IRES−GFPベクターで形質導入したhDCは、Mart1.31以外のメラノーマエピトープペプチド全てに対して著しいCTL応答を誘発したが、形質導入しなかったhDCでは通常のバックグランドの応答しか誘発しなかった(図5)。
【0076】
考察
複製欠失レンチウイルス由来のベクターが様々な非分裂細胞を形質導入できることは、近年報告で示されている(Maldini、1996、Kafri、1997)。レンチウイルスは非分裂細胞の完全な核膜を通過して移入することができる(Case、1999)。本明細書では、この方法と中心DNA3重鎖と称したポリプリン領域シス作用性配列を組み合わせると、大いに可能性が広がることを実証した。この配列をレンチウイルス由来ベクターに導入すると、ニューロン、肝細胞、及び造血幹細胞/始原細胞を含めた数種の細胞の安定したインビボ及びエキソビボ形質導入を30倍高めることが可能であった。
【0077】
最近、レンチウイルス由来のベクターの安全性及び遺伝子治療での使用に関する取り組みで、進歩が認められた(Narry Kim V et al 1988、Zufferey、1999)。しかし、古典的なマウスのレトロウイルスベクター及びレトロウイルスではないベクターの使用が成功した免疫治療で(Condon、1996、Song、1997、Specht、1997)、レンチウイルスベクターを適用することについては研究されなかった。有望なワクチン戦略を開発するために、中心DNA3重鎖を含み、メラノーマポリエピトープをコードするレンチウイルスベクターの免疫原能を試験した。最初に、このベクターを直接注射して、メラノーマエピトープに対してインビボで特異的なCTL応答が引き起こされ得るかどうかについて調べた。今までに、HHD HLA−A2.1(純系)トランスジェニックマウスにおいていくつかの免疫戦略の比較が行われた。組換えpCMV−B10(HBs)DNA又は同様のメラノーマポリエピトープをコードする組換えワクチンを使用すると、1個体マウスにおいて、メラノーマポリエピトープに含まれる4から6個の異なるペプチドに対するCTL応答が同時に誘導されることが可能であった。同様のメラノーマポリエピトープをコードするTRIP mel IRES−GFPベクターでは、特異的細胞溶解についてだけでなく、CTL応答を誘発するポリペプチドの数についても、同様のポリエピトープモチーフをコードするその他のベクターよりも顕著に優れた結果が得られた。特に、ベクターを腹腔内及び皮下注射すると、TRIP mel IRES−GFPベクターによってコードされたエピトープ全てに対してCTL応答が誘導されるマウスがいた。
【0078】
形質導入又はペプチド付加された樹状細胞は、様々なガン細胞に対して有望な免疫方法として使用できることがいくつかのグループによって報告された(Mayardomo、JI、1995、Song W、1997、Specht、JM、1997)。従って、本発明の目的のために、マウス及びヒトの樹状細胞が効果的に形質導入され、インビトロで1次CTL応答を誘導するかどうかを調べた。本発明の結果によって、これらの細胞が容易に形質導入され得ることが示唆された。さらに、これらの形質導入細胞は組換えレンチウイルスによってコードされたエピトープ全てを提示した。興味深いことに、ヒト樹状細胞は最も容易に形質導入された。細胞をインビトロ形質導入することによって、レンチウイルスベクターが様々な宿主細胞にゲノムを有害な方法で組み込むことを回避できる。この理由によって、インビトロ形質導入された細胞は、最初の臨床応用で適切かつ安全な送達方法となるだろう。
【0079】
Warner等は、マウスのレトロウイルスベクターによって形質導入したHIV−タンパク質を発現する繊維芽細胞を使用して、レトロウイルスベクターがいくつかの動物モデルだけでなく、HIV患者のCTL応答を誘導する能力を初めて示した(1998年にWarnerによって概説されている)。マウスに直接注射すると、プロウイルスのDNAが効果的に抗原を提示する樹状細胞で検出された(Song、1997)。しかし、レンチウイルスとは対照的に、マウスのレトロウイルスベクターはDCなどの非分裂細胞を形質導入することができず、コードされた遺伝子のインビボ発現は「遮断される」ことが多い。結果的に、これらのレトロウイルスベクターはヒトの免疫治療にとって有力な候補とはならない(Kafri)。
【0080】
我々の結果によって、中心DNA3重鎖を含むレンチウイルスベクターは、HHDマウスにおいて中心DNA3重鎖を含まないベクターよりも強くインビボでCTL応答を誘導することが示唆される。さらに、中心DNA3重鎖を含むレンチウイルスベクターはインビトロで容易にhDCを形質導入することができ、ひいては臨床的免疫治療に使用することができよう。
【0081】
材料及び方法
レンチウイルスベクター構造
ベクタープラスミド:HR′CMVLacZ由来のpTRIP−EGFP(Naldini et al、PNAS 93、11382〜8、1996)。LacZリポーター遺伝子は、EGFP遺伝子で置き換えた(Clontech)。TRIP−EGFPでは、EGFP遺伝子をcPPT及びCTS配列を含むHIV−1LAIの中心断片のClal部位に挿入した。
【0082】
EGFP遺伝子は、pEGFP−N1プラスミドからPfuポリメラーゼ(Stratagene)を使用してPCRによって増幅し、5′及び3′それぞれにBamHI及びXhoI制限部位を付加した。PCRプライマは以下の通りであった。
【0083】
Bam GFP 5′CC GGATCC CCA CCG GTC GCC ACC 3′
Xho GFP 5′CC CTCGAG CTA GAG TCG CGG CCG 3′
HR GFPベクターは、このPCR断片をpHR′CMVLacZのBamHI及びXhoI部位にクローニングし直して、LacZ ORFをEGFPに置き換えることによって構築した。
【0084】
cPPT及びCTSを含むpLAI3の178bp断片(4793から4971)をPCRによって増幅した。この断片をHR GFPの特有のClaI部位に挿入するために、NarI制限部位をプライマの5′に付加した。
【0085】
Nar TRIP+: 5′ GTC GTC GGCGCC GAATTC ACA AAT GGC AGT ATT CAT CC 3′
Nar TRIP−: 5′GTC GTC GGCGCC CCA AAG TGG ATC TCT GCT GTC C 3′
正方向でこの3重鎖配列を挿入すると、TRIP GFPプラスミドベクターが生じ、逆向きだとTRIPinv GFPが生じた。あるいは、同様の3重鎖断片をpcPPT−AG、pcPPT−D、pcPPT−225及びpCTSプラスミドから増幅して、対応するウイルスのようにcPPT又はCTSに同様の変異を含むベクターを作製した。
【0086】
まず、TRIP GFPベクター又はTRIP−EGFP(CNCMに1998年4月15日I−2005号として寄託)に存在するEcoRI部位を充填して、TRIP GFPΔEベクターを作製した。次に、TRIP GFP ΔEベクターの0.7kb BamHI/XhoI EGFPの代わりに、ピコナウイルスの内部リボゾーム結合部位(IRES)を含む1.2kb BamHI/XhoI断片をEGFPの上流にクローニングして、TRIP ΔE IRES GFPベクターを作製した。CMVプロモータとIRES GFP配列の間に存在する部位は、BamHI−BstXI−SnaBI及びEcoRIである。CTLポリエピトープメラノーマ(mel)断片をPCRによってpBS mel poly上に作製し、プライマ5BglMlu Mel:
5′CCAGATCTACGCGTGCCACCATGGCTGCTGGT 3′
3RIMel: 5′CGGAATTCGACCTAAACGCAACGGATG3′
の内部にkozacコンセンサス配列を挿入した。
【0087】
mel PCR断片をBglII/EcoRIによって消化し、TRIP ΔE IRES GFPのBamHI/EcoRI部位に挿入して、TRIP mel IRES GFPとも称するTRIP ΔE mel IRES GFPを作製した。
【0088】
HR mel IRES GFPは、メラノーマポリエピトープ及びTRIP mel IRES GFPのIRES GFPを含むNdeI/XhoIフラグメントをHR GFPのものと交換して作製した。NdeI部位はCMVプロモータの末端に位置している。
【0089】
ウイルスベクターの産生
レンチウイルスベクターは、既に記載されたように(Naldini、I.M.PNAS 1996 及びscience 1996)、リン酸−カルシウム法を用いて、293T細胞を3種のプラスミドによって一過性にトランスフェクトすることによって作製した。簡単に説明すると、293T細胞にVSVエンベローププラスミド(pMDG)20μg及び様々なパッケージング(8.2又は8.91)プラスミド及びレンチウイルスベクタープラスミド40μgをトランスフェクトした。トランスフェクトして60h後及び84h後に、調整培地を収集した。次いで、ウイルスを濃縮して、dNTPを既に記載されたように処理した(Naldini science 1996)。HeLa P4.2細胞及びMT3細胞に対するウイルス力価は、系列希釈及びp24 ELISAアッセイによって測定した(Naldini Science 1996)。
【0090】
DEAE−デキストラン20μM存在下で、96ウェルプレートに接種したP4細胞に等量の粒子(ウェル当たりp24ウイルス抗原1ng)を感染させることによって、ウイルスの1サイクル力価測定を実施した。実験全体を通して、プロテアーゼ阻害剤サキナビル(Roche)を1μMで添加して、感染1回に限定した分析を行った。細胞の有糸分裂は、感染前日にアフィコリン(aphicolin)(8μM)処理して阻害した。β−ガラクトシダーゼ活性は、化学ルミネセンスβ−Galリポーター遺伝子アッセイ(Boehringer)を使用して、感染48時間後に測定した。
【0091】
HeLa細胞を等量のベクター粒子(ウェル当たりp24 5ng)で3回感染させた。形質導入後48時間して、培地をTNB(Tris 50mM、pH7.5、NaCl 50mM)200μlに置換して、生細胞の蛍光をマイクロプレート蛍光計(Victor2、Wallac)及びEGFP対応フィルター(励起:485nm、発光:520nm)を使用して定量した。
【0092】
マウス
HHDマウスについては既に説明されている(Pascolo、1997)。このマウスは、ヒトb2mのC末端がペプチドのアーム(GGGGS)x3によってキメラ重鎖(HLA−A2.1、a1−a2、H−2Db a3−膜間ドメイン、及び細胞質内ドメイン)のN末端に共有結合したトランスジェニック1本鎖組織適合クラスI分子を発現する。これらのマウスのH−2Db及びマウスb2m遺伝子はさらに相同的組換えによって分断され、マウス組織適合クラス1分子の細胞表面での発現が完全に欠如しており、血清学的に検出されなくなっている。
【0093】
hDC及び1次CTL誘導の発生
ヒト樹状細胞は、HLA−A2.1ハプロタイプの健常提供者のサイタフェレシス生成物(IDM、パリ、フランス)から得られた。CD3、CD14、CD80、CD83、HLA−ABC、及びHLA−DRに対するmAbを使用したこれらのDCのFACS分析によって、未熟DC表現型が示された。レンチウイルスベクターを1.106細胞当たり600ng、300ng、150ng、及び150ng/p24の濃度で含むAMV−5培地中で、このhDCを10日間形質導入した。2種のレンチウイルスベクターで形質導入したhDCにおけるGFP発現のパーセント及び平均蛍光強度をFACSによって測定した(Becton Dickinson、BD、USA)。
【0094】
同一提供者の単核細胞(MNC)をhDC又は形質導入したhDCで1hDCに対して4MNCの比でインビトロ誘発した。このMNCを同一の低温保存した形質導入hDCを使用して2回再誘発して、次いで関連ペプチド又は陰性対照(lnf.m.58)ペプチドを付加したT2細胞を標的として使用した4h51Cr−放出アッセイで細胞溶解活性を試験した(10μg/ml、5.106細胞/ml、FCSを含まないRPMI培地、室温で2時間)。
【0095】
マウス樹状細胞の生成
骨髄由来樹状細胞を既に記載されたように生成した[43、51]。骨髄の単核細胞を10%FCS、Lグルタミン 2mM、ペニシリン 50U/ml、ストレプトマイシン 50μg/ml、メルカプトエタノール 5.105Mを補足したRPMI(完全RPMI培地)に、さらに組換えマウスGM−CSF 20ng/ml及び組換えマウスIL4 100ng/ml(いずれもGENZYME(ケンブリッジ、MA)製)を補足して培養した。2日目及び6日目に、付着しなかった細胞を注意深く除去して、マウスGN−CSF 10ng/ml及びマウスIL4 50ngを補足した新鮮な完全RPMI培地を添加した。7日目に、培養培地を1.106細胞当たり600ng、300ng、150ng、及び150ng/p24の濃度のレンチウイルスベクターを補足したRPMI完全培地1mlで置換した。9日目に集めた樹状細胞を適切なmAbで評価したところ、純度は95%を上回っていた(IAb+、HHD+、CD−3、33D1+、NDL145+、及びCD11c+)。次いで、マウスDCにおけるGFP発現のパーセント及び平均蛍光強度をFACSで9日目及び12日目に測定した。
【0096】
ベクターによる免疫及びインビトロ再誘導及び細胞溶解アッセイ
HHDマウスにレンチウイルスベクターを12日間腹腔内、静脈内又は皮下内注射した。次いで感作されたマウスの脾細胞を各エピトープペプチドで独立してRPMI完全培地中で7日間再誘導した。最後の2日間に、培養細胞を10%TCGFによって再誘導した。7日目に、培養した細胞の細胞溶解活性は、既に記載されたように(Pascolo、1997)、関連ペプチド又は陰性対照ペプチド(lnf.m.58)を付加したHHDトランスフェクトTAP−RMA−S細胞を標的として使用して、4h 51Cr−放出アッセイで試験した(FCSを含まないRPMI培地中10μg/ml、5.106細胞/ml、室温で2時間)。TRIP−mel−ITES−GFPで形質導入した、又は形質導入していないHHDトランスフェクトHeLa細胞を並行して標的細胞として使用した。特異的溶解のパーセントを以下の通りに算出した:(実験上の放出−自発放出)(総放出−自発放出)x100。
【0097】
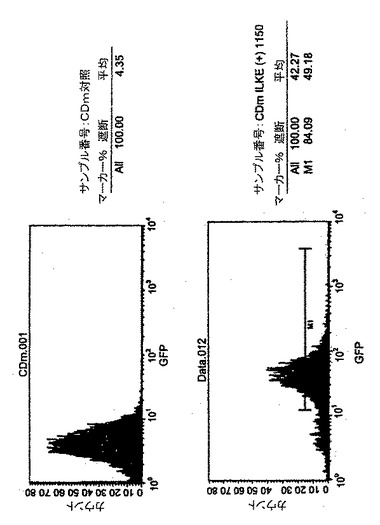
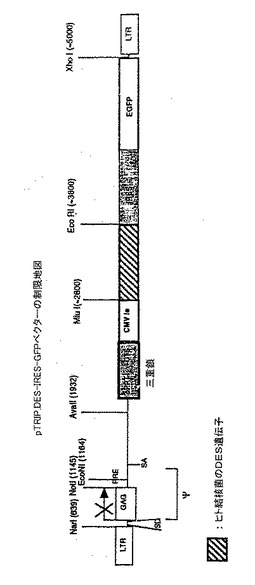
実施例II:Mycobacterium tuberculosisのDES遺伝子をコードするTRIP−des−IRES−GFPベクターで免疫したHHDマウスのCTL応答の評価
DES遺伝子についてはWO98/04711で開示している。
【0098】
実験方法
第1段階では、TRIP−des−IRES−GFPベクターを使用してHeLa−HDD細胞を形質導入した。これらの細胞を形質導入して、限界希釈によってクローニングした。GFPを高濃度で発現しているクローンを選択して、古典的な51CrCTL試験の標的細胞として使用した。
【0099】
第2段階では、HDDマウスにTRIP−des−IRES−GFPベクター粒子をマウス当たり1.2micg/p24で腹腔内注射した。注射後12日目に、これらのマウスの脾細胞を、ベクター粒子0.2micg、又は1micg/p24/ml(ml当たり2106細胞)、あるいはTRIP−des−IRES−GFPで形質導入し、LPSで誘導した同系繊維芽細胞1micg/p24/ml/2106細胞/mlで誘発した。インビトロ誘発後6日目に、細胞の細胞溶解活性をdes形質導入HeLa−HDD標的細胞を使用した51Cr試験で試験した。対照の標的細胞はメラノーマポリエピトープによって形質導入した(TRIP−mel−IRES−GFP)HeLa−HDD細胞とした。
【0100】
結果
これらの実験は、全遺伝子をコードするHIV由来の3重鎖陽性ベクターが細胞を形質導入したり、同遺伝子を発現する標的細胞に対して特異的なCTL応答を誘導したりできるかどうかを調べるために実施した。図9に例示したように、特異的な細胞溶解が全ての条件で認められる。形質導入したLPS−繊維芽細胞では弱いCTL応答が誘導された。ベクター粒子1micg/p24/mlを使用すると最も良い結果が得られた。これらの結果によって、完全な遺伝子が宿主のゲノムに導入され、その産物が処理されて、提示されて著しいCTL応答が誘導され得ることが示される。これらの結果によってまた、des遺伝子は少なくとも1個又は複数のHLA−A2.1に限定されたCTLエピトープペプチドを含んでいることが示唆される。
【参照文献】
【0101】

【図面の簡単な説明】
【0102】
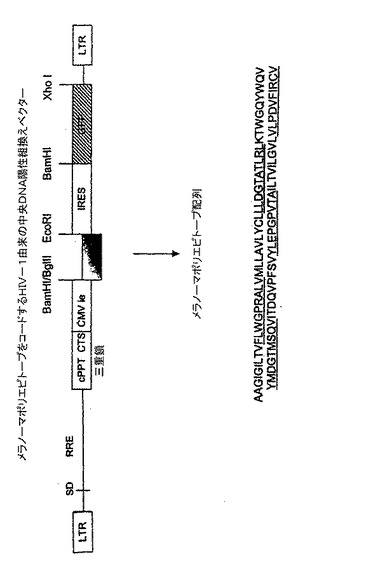
【図1】メラノーマポリペプチドをコードするHIV−1由来の3重鎖DNA陽性の組換えベクター。
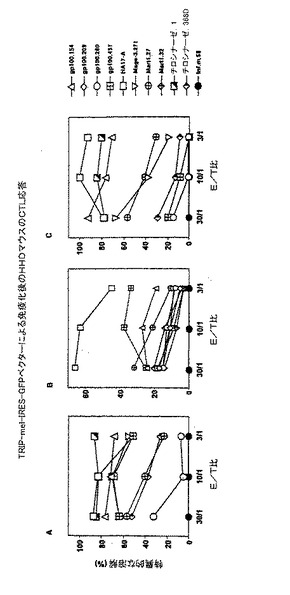
【図2】TRIP−mel−IRES−GFPベクターで免疫した後のHHDマウスのCTL応答。
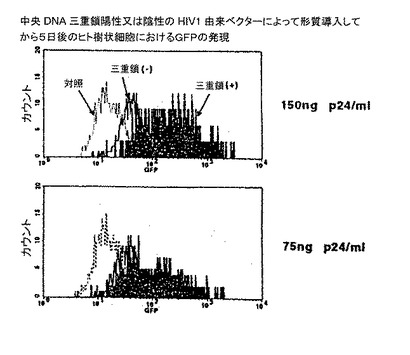
【図3】中心DNA3重鎖陽性又は陰性HIV由来ベクターによる形質導入後5日目のヒト細胞でのGFP発現。
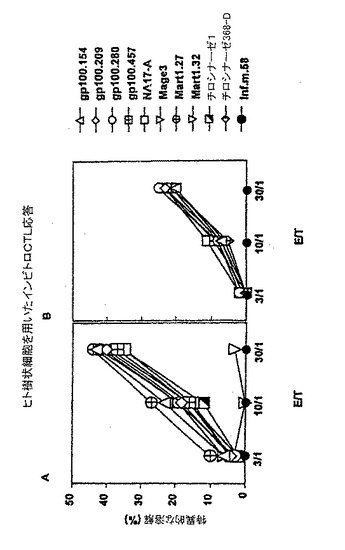
【図4】ヒト樹状細胞を使用したインビトロCTL応答。
【図5】pHR.MEL−IRES−GFPベクターの制限カード。メラノーマ特異的CTLモノ又はポリエピトープ配列HR.MEL−IRES−GFPの構築 HR.MEL−IRES−GFPプラスミドを構築するために、1999年4月20日に1−2185号としてCNCMに寄託されたCMVプロモータの一部、ポリペプチドMEL及びIRES−GFPを含むTRIP.MEL−IRES−GFPのNdeI/XhoIフラグメントをCMVプロモータの一部及びGFPを含むHR GFPのNdeI/XhoIフラグメントと置き換えた。
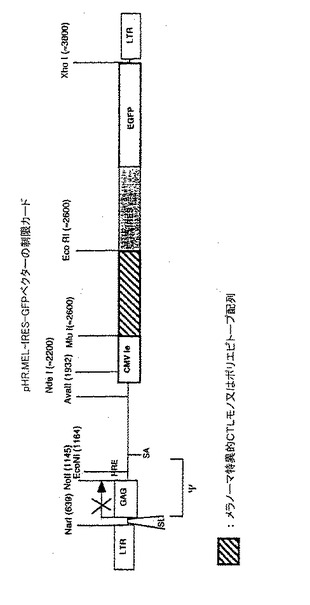
【図6】pTRIP.ILKE−IRES−GFPベクターの制限カードHIV特異的CTLモノ又はポリエピトープ配列、そのエピトープはI9V.(ILKE);(RT476−484)TRIP.ILKE−IRES−GFP(1999年10月6日、_号としてCNCM(フランス、パリ)に寄託)の構築 TRIP.ILKE−IRES−GFPは、TRIPΔE IRES−GFPのCMVプロモータとIRESの間にILKEのPCR産物を挿入することによって構築した。ILKEによって始まるCTLエピトープの周りの領域は、プライマ5 ILKE:5′TCAGATCTGCCACCATGGCACTAACAGAAGTAATACCAC 3′ 3 RIILKE:5′CGGAATTCTTATTGGCCTTGCCCCTGCTTC 3′を用いてマトリックスpLAI上にPCRによって増幅した。 Kozak配列は、上流プライマに挿入し、終止コドンは下流プライマに挿入した。 BglII/EcoRIで消化した後、PCR産物をTRIPΔE IRES−GFPのBamHI/EcoRI部位に挿入した。 このベクターは、GFP及びエピトープのクラスタに対応し、特にHLA.A2.1に限定されたI9Vエピトープ(RT476〜484)を含むVIHのRT遺伝子の領域をコードするバイシストロン性情報を発現する(Walker B.D.、1989 PNAS 86 p.9514〜9518)。
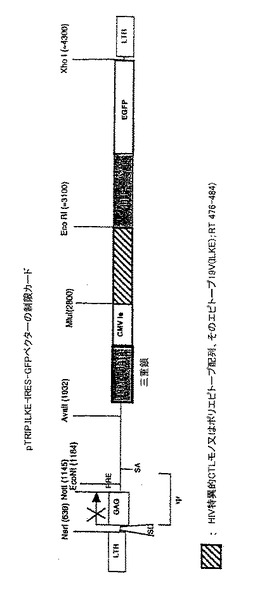
【図7】pTRIP.TEL/AML−IRES−GFPベクターの制限地図TEL/AML配列の転座 TRIP.TEL/AML−IRES−GFPは、TEL/AMLのPCR産物をTRIPΔE IRES−GFPのCMVプロモータとIRESの間に挿入することによって構築した。TELとAMLとの間の転座周囲の領域は、プライマ5 BgI TA: 5′GAAGATCTGCCACCATGAAGCCCATCAACCTCTCTCAT 3′ 3 RITA: 5′CGGAATTCTTACCCAGCGCAACGCCTC 3′ を用いてPCRによって増幅した。 Kozak配列は、上流プライマに挿入し、終止コドンは下流プライマに挿入した。 BglII/EcoRIで消化した後、PCR産物をTRIPΔE IRES−GFPのBamHI/EcoRI部位に挿入した。
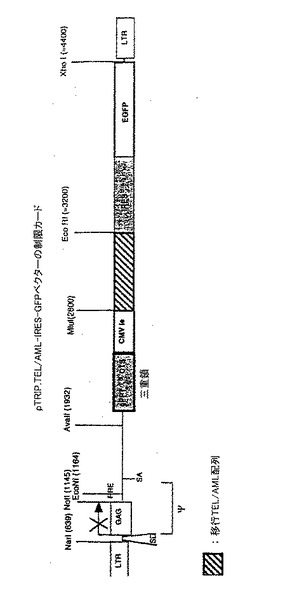
【図8A】(HIV1polから得られた)I9Vエピトープペプチド及びHHDトランスジェニックマウスの骨髄細胞を使用して産生したマウス樹状細胞のGFPをコードするHIV由来DNA3重鎖陽性ベクターの形質導入能。 細胞内GFP発現のFACS分析によって実証されたように、マウス樹状細胞の約80%がHIV由来3重鎖陽性組換えベクターによって形質導入された。
【図8B】(HIV1polから得られた)I9Vエピトープペプチド及びHHDトランスジェニックマウスの骨髄細胞を使用して産生したマウス樹状細胞のGFPをコードするHIV由来DNA3重鎖陽性ベクターの形質導入能。 細胞内GFP発現のFACS分析によって実証されたように、マウス樹状細胞の約80%がHIV由来3重鎖陽性組換えベクターによって形質導入された。
【図9】Mycobacterium tuberculosisのDES遺伝子をコードするTRIP−des−IRES−GFPベクターで免疫したHHDマウスのCTL応答の評価。
【図10】1999年10月11日にCNCMに寄託されたpTRIP.DES−IRES−GFPの制限地図。
【発明の詳細な説明】
【0001】
本発明は、インビトロで、及び好ましい実施態様ではインビボで、ベクター中に存在するヌクレオチド配列によってコードされたエピトープに対する免疫反応の発生若しくは増強を誘導し、又はこれらに寄与することが可能な組成物を調製するためのレトロウイルスベクター、特にレンチウイルスベクターの使用に関する。
【0002】
本発明者等は、本発明に従って調製したベクターによって、細胞性免疫応答、特にエピトープに対する細胞障害性Tリンパ球(CTL)反応を得ることが可能であることを示した。
【0003】
さらに、この細胞性免疫応答がベクターに含まれるヌクレオチド配列によってコードされた1又は数個のエピトープに対して得られた特異的応答であり得ることを示すデータが得られた。
【0004】
従って、本発明は腫瘍及び癌に対する治療プロトコールに使用でき、特に腫瘍に対する免疫療法又は予防接種療法のプロトコールに使用できる手段を提供する。
【0005】
本発明は、感染症、特にウイルス感染、例えばレトロウイルス感染に関連した疾患の治療また予防に使用できる手段も開示する。
【0006】
本発明者らは、さらに、本発明の組成物による治療に関連した細胞性免疫応答、特にCTL応答は腫瘍若しくはウイルスの抗原、又はウイルス感染細胞の抗原に特異的であることが可能で、MHC(主要組織適合複合体)の特定の分子に限定され得ることを示す結果を得た。
【0007】
特に本発明は、MHC複合体のクラスI分子、例えばHLA−A2又はB7分子に限定した細胞性免疫応答を得るために、免疫原組成物に前記ベクターを使用することに関する。
【0008】
従って、本発明はシス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を包含するポリヌクレオチドを含む組換えベクターを含む免疫原組成物に関する。cPPT及びCTS配列は、逆転写中に3本鎖DNA構造(本明細書ではDNA3重鎖と称する)の形成を誘導する。このDNA3重鎖は、ベクターDNAゲノムの核移入を刺激し、これらの領域の起源はレトロウイルス又はレトロウイルス様で、前記ベクターは所定のヌクレオチド配列(目的の導入遺伝子又は配列)に加えて、レトロウイルス又はレトロウイルス様起源の逆転写(retrotranscription)、発現、及びウイルス粒子形成の調節シグナルを含み、この組成物はベクター内に存在する導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)又はCD4の応答を誘導又は刺激することが可能である。
【0009】
本発明の好ましい実施態様では、細胞性免疫応答、特に1又は数個のエピトープに対するCTL応答又はCD4応答は記憶CTL又はCD4応答である。
【0010】
ベクター内にcPPT及びCTS領域が存在すると、3重鎖DNA構造が形成されることが可能で、それによって前記ベクターで組換えられた細胞でのベクターゲノムの核移入が影響を受け、特に増強され得ることが強調される。
【0011】
本発明による免疫原組成物は記憶CTL応答の発生を誘導、改良し、又は一般にその発生に関与することができるので、(前記免疫原組成物の投与後、前記免疫応答が長期間にわたって誘導されなければならない場合、又は少なくとも応答が必要とされる期間に(前記応答の長期誘導であってもよい)誘導可能でなければならない場合を含む)抗腫瘍治療又は抗ウイルス治療又は抗病原性治療のプロトコールにおいて前記免疫原組成物の使用を企図することが可能である。言い換えると、前記免疫原組成物は、細胞性免疫応答、特にCTL応答又は記憶応答の発生を誘導又は刺激し、又はそれに寄与することによって、腫瘍疾患又は感染性疾患(例えば細菌又はウイルスによる)を治療するための治療用組成物の調製に使用することができる。
【0012】
本発明の免疫原組成物は、ベクターの配列中に3重鎖構造が存在する結果、ベクター及びベクター粒子内にcPPT及びCTS領域が存在するため、標的細胞においてベクターゲノムの核移入を刺激させ得ることが注目される。ベクター内で誘導されるエピトープは細胞自身のものであってもよいし、細胞以外のものであってもよい。
【0013】
本発明は、標的細胞又は受容細胞の核への核酸配列又はペプチド配列の移入を増加させるために、レトロウイルス由来又は合成によるcPPT及びCTS配列を含むヌクレオチド配列の使用も含む。
【0014】
例えば、前記3重鎖配列には受容細胞の外来配列又は自己配列が含まれることが理解されよう。
【0015】
従って、本発明は、腫瘍治療、特に抗癌治療又は抗感染性疾患治療のための予防接種プロトコールと類似した治療プロトコールにおいて使用可能な組成物を開示する。
【0016】
本発明では、目的の該導入遺伝子又は配列が、1又は数個の腫瘍細胞の1又は数個のエピトープ、例えば腫瘍に対する細胞性免疫を誘導することが可能な標的抗原に同定されたエピトープをコードする配列である得ることに注目すると興味深い。
【0017】
ポリペプチドを形成する数個のエピトープは、本発明の導入遺伝子によってコードすることができる。特定の実施態様では、それらは腫瘍で確認された標的抗原から得ることができ、コーディング配列が一緒になって導入遺伝子を形成するとき、細胞性免疫応答がエピトープ全て又はエピトープのほとんどに対して得られるような方法で選択され得る。細胞性免疫応答はインビトロ、又は好ましい実施態様ではインビボでアッセイすることができる。このようなアッセイの実施を可能にするプロトコールは実施例で説明する。
【0018】
標的抗原はいくつかの種類の腫瘍、特にメラノーマ又は腎癌、膀胱癌、大腸癌、肺癌、乳癌、白血病及びリンパ腫などを含む癌腫で同定されている。
【0019】
本発明の他の態様では、前記免疫原組成物はウイルス関連感染症、又はミコバクテリア、例えばM tuberculosisを含む任意の種類の病原体に関連する感染症を含めた感染性疾患において、細胞性免疫応答を得るために使用することができる。
【0020】
この場合、細胞性免疫応答を引き起こすことが可能な特定の抗原を同定して、そのコーディング配列をベクターに挿入して前記免疫原組成物で使用することができる。例えば、M tuberculosisのdes遺伝子が使用できる。
【0021】
本発明では、前記免疫原組成物で使用されるベクターは、エピトープを発現するか、又は腫瘍細胞若しくはウイルス感染細胞上の標的抗原として同定された(糖タンパク質又はその他のタンパク質由来化合物を含む)タンパク質上に提示することが可能であることも付け加えなければならない。
【0022】
さらに、エピトープ、エピトープを提供するために使用されたポリペプチド又はタンパク質は、例えば変異、欠失又は挿入によって変更することが可能で、例えば安定性を高めるために変更することが可能であることにも留意すべきである。
【0023】
本発明は、
1)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成の調節シグナルの制御下に置かれた所定のヌクレオチド配列(導入遺伝子)を含有する組換えヌクレオチド配列と、
2)シス作用性中心開始領域(CTS)及びシス作用性終止領域(CTS)を包含するポリヌクレオチドであって、これらの領域の起源がレトロウイルス又はレトロウイルス様であるか、又はトランスポゾン由来であり、レトロウイルス又はレトロウイルス様起源の逆転写調節シグナル又はトランスポゾン調節シグナルとともに、機能的な方向及び位置に挿入されたポリヌクレオチドと
を備えた組換えレトロウイルス粒子を含む免疫原組成物であって、
ベクターに存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は刺激することができる免疫原組成物に関する。
【0024】
cPPT及びCTSシス作用性配列を包含するDNA配列は、逆転写後に3本鎖DNA構造「DNA3重鎖」を採り、ベクターDNAの核移入を刺激することができる。
【0025】
特定の実施態様では、ベクターに存在する導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は刺激することが可能な前記免疫原組成物は、
a)レンチウイルスの核タンパク質又は機能的に得られたポリペプチドに対応するgagポリペプチド(GAGポリペプチド)と、
b)レンチウイルスのRT、PRO、INタンパク質又は機能的に得られたポリペプチドによって構成されたpolポリペプチド(POLポリペプチド)と、
c)エンベロープポリペプチド又は機能的に得られたポリペプチド(ENVポリペプチド)と、
d)転写及び発現の調節シグナルの制御下に置かれ、1又は数個のエピトープをコードする所定のヌクレオチド配列(目的の導入遺伝子又は配列)と、レトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の調節シグナルを含有する配列と、並びにシス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含有するポリヌクレオチドであって、これらの領域がレトロウイルス又はレトロウイルス様の起源であり、上記レトロウイルス又はレトロウイルス様起源の調節シグナルとともに機能的な方向に挿入されているポリヌクレオチドとを備えた組換えヌクレオチド配列と
を具備する組換えレトロウイルスベクター粒子を含む。
【0026】
他の実施態様では、ベクターに存在する導入遺伝子によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は促進することが可能な免疫原組成物は、
a)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域がレトロトランスポゾン由来で、レトロトランスポゾン調節シグナルと共に機能的な方向挿入されたポリヌクレオチドと、
b)レトロトランスポゾンの核タンパク質又は又は機能的に得られたポリペプチドに対応するポリペプチド(GAGポリヌクレオチド)と、
c)レトロトランスポゾンのRT、PRO、INタンパク質に対応するpolポリペプチド又は機能的に得られたポリペプチド(POLポリペプチド)と、
d)ウイルスエンベロープポリペプチドと、
e)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成のレトロトランスポゾン調節シグナルの制御下に置かれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)を備えた組換えヌクレオチド配列を具備する組換えレトロウイルス様粒子と
を含む。
【0027】
好ましい実施態様では、前記の定義の何れか1つに応じた免疫原組成物に存在する組換えレトロウイルスベクター粒子は、記憶細胞性免疫応答、特に記憶CTL応答の発生を誘導、増強するか、又は関連することが可能である。
【0028】
前記で開示した定義に従って、ベクター又はベクター粒子を含む免疫組成物は、いくつか可能な実施態様によって調製することができる。
【0029】
本発明の好ましい実施態様では、レトロウイルス起源の配列をレンチウイルスゲノムから得る方法で免疫原組成物は調製される。
【0030】
他の実施態様では、これらの配列の起源はレトロウイルス様であり、レトロトランスポゾンからも得られる。
【0031】
本発明の他の実施態様では、又は前記で定義した特徴に加えて、組換えベクターに含有される目的の導入遺伝子又は配列は、転写及び発現の調節シグナルを含有する発現カセットに含まれる。
【0032】
あるいは、ベクターの逆転写、発現及びウイルス粒子形成の調節シグナルはレンチウイルス起源であり、cPPT及びCTS領域を備えたポリヌクレオチドもレンチウイルス起源である。
【0033】
他の実施態様では、ベクター内の逆転写、発現、及びウイルス粒子形成の調節シグナル並びにcPPT及びCTS領域を含むポリヌクレオチドは、HIV型、特にHIV−2又はHIV−2のレトロウイルスから得られる。
【0034】
他のウイルス、特にレンチウイルスは、逆転写発現及びウイルス粒子形成の調節シグナルを設計するために、さらにはcPPT及びCTS領域を含むポリヌクレオチドを得るために使用することができる。従って、特にレンチウイルスCAEV、EIAV、VISNA、HIV、SIV又はFIVを使用することができる。
【0035】
本発明の免疫原組成物の組換えレトロウイルス粒子を獲得するため、ベクターのトランス相補性に必要なポリペプチド又はタンパク質をコードする配列は、例えばレンチウイルス、特にHIV−1及びHIV−2レトロウイルスを含むHIV由来のGAG、POL及びENVタンパク質である。
【0036】
あるいは、GAG及びPOL配列は、ENV配列とは異なるウイルスに由来してもよい。例えば、GAG及びPOL配列はHIVレトロウイルス由来であり、ENV配列は他のウイルス又はレトロウイルス由来し、且つアンフォトロピック又はエコトロピックENV配列の何れであってもよい。
【0037】
他の実施態様では、ENV配列は水疱性口炎ウイルス(VSV)から得られる。
【0038】
本発明の特定の実施態様では、ベクター中に存在する導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は促進することが可能な免疫原組成物は、
a)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域がレトロトランスポゾンから得られ、レトロトランスポゾン調節シグナルと共に機能的な方向に挿入されたポリヌクレオチドと、
b)レトロトランスポゾンの核タンパク質又は機能的に得られたポリペプチドに対応するポリペプチド(GAGポリペプチド)と、
c)レトロトランスポゾンのRT、PRO、INタンパク質に対応するpolポリペプチド又は機能的に得られたポリペプチド(POLポリペプチド)と、
d)ウイルスエンベロープポリペプチドと、
e)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成のレトロトランスポゾン調節シグナルの制御下に置かれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)を含む組換えヌクレオチド配列と
を備えた組換えレトロウイルス様粒子を含む。
【0039】
好ましい実施態様では、組換えレトロウイルス様粒子を含む本発明の免疫原組成物は、前記で開示した特徴によって、記憶細胞性応答、特に記憶CTL応答を引き起こすことが可能である。
【0040】
本発明は、1999年10月11日にCNCM(Collection Nationale de Culture de Microorganismes at Institut Pasteur in Paris、France)に寄託されたベクター構築物にも関する。
【0041】
第1のベクターは、1999年10月11日にl−2326号として寄託されたpTRIP.TEL/AML−IRES−GFPであり、第2のベクターはpTRIP−ILKE−IRES−GFPと称し、1999年10月11日にl−2327号として寄託された。
【0042】
第3のベクター、pTRIP.DES−IRES−GFPは、1999年10月11日にl−2331号としてCNCMに寄託された。
【0043】
前記構築物に存在する抗原をコードする配列は、前記で引用したM tuberculosisの完全なDES遺伝子を含む、他の任意の目的の抗原又はエピトープによって置き換えることができる。
【0044】
本発明の他の態様では、ベクター、ベクター粒子及び同物を含む免疫原組成物は、cPPT及びCTS領域がベクター配列の中心に位置するように設計される。
【0045】
「中心に位置する」とは、cPPT及びCTS領域がベクター配列の中心、又はこの配列のほぼ中心にあることを意味する。特に、cPPT及びCTS領域は逆転写された直鎖状ベクターDNAの中央の3分の1以内にあることが可能である。
【0046】
cPPT及びCTS配列が存在する結果としてウイルス逆転写中に形成された3重鎖配列が中央に位置することによって、そのベクター又はベクター粒子と接触する細胞の形質導入量を増大させることが可能である。
【0047】
あるいは、前記ベクターの変種によっては、導入遺伝子を含むベクターの転写単位はLTR領域のU3領域内に挿入することができる。従って、逆転写の後、導入遺伝子は複製され、従って3重鎖配列のそれぞれの鎖に現れ、従って導入遺伝子の大きさにかかわらず、3重鎖配列がベクターの中心部分に位置することが可能となる。
【0048】
本発明は、本発明による免疫原組成物と接触するように置かれた細胞に関し、特に免疫原組成物のベクター又はベクター粒子によって形質導入された組換え細胞にも関する。
【0049】
これらの細胞は、有利には、抗原提示細胞である。例えば、これらの細胞は、肺細胞、脳細胞、上皮細胞、星状細胞、マイクログリア、乏突起神経膠細胞、ニューロン、筋細胞、肝細胞、樹状細胞、神経細胞、骨髄の細胞株、マクロファージ、繊維芽細胞、造血細胞から選択することができる。
【0050】
従って、本発明の免疫原組成物は、一次細胞性免疫応答、特に、有利には記憶CTL応答であるCTL応答を発生させ、腫瘍、特に癌の治療方法又は治療処置のための治療組成物の調製に使用することができる。あるいは、その他公知の抗癌治療の補助処置として使用することができる。
【0051】
例えば、本発明の免疫原組成物は、化学療法又は免疫化学療法又は抗癌治療のその他のアプローチと関連させて使用することができる。
【0052】
「抗癌治療」という表現は、本発明では、腫瘍の増殖又は腫瘍が増殖する可能性を阻害すること、若しくは転移巣形成の制御を含めた悪性細胞の拡散を阻害することを意味するか、又はその両者を意味するものである。
【0053】
従って、本発明では、「抗癌治療」という表現は、特にこの免疫原組成物が記憶細胞性応答を誘導又は増強し、一般的には関与することが可能であるという事実を考慮すると、疾患の治癒を期待して、悪性細胞の増殖及び腫瘍の拡散を制御するために使用する方法に関している。
【0054】
本発明の組成物で治療し得る腫瘍は、例えば、メラノーマ又は(肺、膀胱、腎臓、大腸)含めた癌腫及びリンパ滲出である。
【0055】
治療可能な腫瘍も、変異した自己タンパク質及び/又は過剰発現した自己タンパク質を含む腫瘍特異的抗原を発現している全ての腫瘍である。
【0056】
エキソビボ段階を含む投与方法、例えば、標的細胞にエキソビボ導入した後、標的細胞を治療する患者に投与することを含めて、受容可能な本発明の免疫原組成物の投与方法はいずれも興味深い。
【0057】
あるいは、本発明の免疫原組成物は、全身(IV)、局所、又は皮膚、皮下、例えば筋肉内投与経路を含めた通常の投与経路によって患者に直接投与し得る。
【0058】
特定の実施態様では、本発明の免疫原組成物は、細胞性免疫応答、特にCTL性免疫応答の発生をインビボで誘導、増強、又は関与する方法で患者に直接投与し得る。
【0059】
他の実施態様では、前記免疫原組成物は、長期間記憶細胞性応答を発生させるために使用される。
【0060】
本発明の前記免疫原組成物は、ベクター又はベクター粒子に存在するcPPT及びCTS配列の特性によって、標的細胞内でベクターゲノムの核移入を誘導又は促進するので特に興味深いことが強調される。
【0061】
本発明の免疫原組成物が特に優位なのは、これらがベクター又はベクター粒子に存在する関心のあるヌクレオチド配列又は導入遺伝子によってコードされた複数のエピトープに対する細胞性免疫応答を惹起又は促進するために使用できること、及び完全な遺伝子配列、例えば病原性物質の遺伝子の産物又は少なくとも8から15アミノ酸、好ましくは9から12アミノ酸をコードし得る前記遺伝子の断片の産物に対する細胞性免疫応答を惹起又は促進するためにも使用できることである。本発明はまた、細胞性応答を誘導する前記アミノ酸配列の複数反復(少なくとも2回の同一配列)及び/又は異なる病原性抗原又は腫瘍抗原の2個のエピトープに対応する少なくとも2種の異なる配列を含むアミノ酸配列をコードするヌクレオチド配列を含むヌクレオチド配列を含む。
【0062】
本発明のその他の特性又は利点を、以下の実施例及び図面で開示する。
【0063】
実施例
レンチウイルスベクターは、非分裂細胞を含む細胞を形質導入する能力を有しており、遺伝子治療にとってますます重要となっている。最近、ポリプリン広域シス作用性配列(中心DNA3重鎖)を含むレンチウイルスベクターが中心DNA3重鎖を欠失したものよりもヒト及びマウスをより効果的に形質導入することが示された(Charneau P.et.al.J.Mol.Biol.1994、241、651〜662)。中心DNA3重鎖を含むか、又は含まず、同様のHLA−A2.1限定メラノーマCTLポリエピトープをコードするレンチウイルスベクターのTL応答を誘導する能力を本明細書で試験した。HHD(HLA−A2.1純系)マウスにおいて、中心DNAを含むレンチウイルスベクターを直接インビボ投与すると、ポリエピトープ配列によってコードされたエピトープペプチド全てに対する強いCTL応答が誘導された。レンチウイルスベクターはDNA3重鎖を含むと明らかに有利であることが示された。さらに、DNA3重鎖を含むレンチウイルスが3重鎖を含まないベクターよりもヒト樹状細胞を7倍も効果的に形質導入することが示された。これらのエキソビボ導入した樹状細胞は、メラノーマエピトープペプチドのほとんどに対して効果的かつ特異的な1次CTL応答を引き起こす。変更したレンチウイルスの安全性の問題がほぼ解決されたので、このベクターをインビボにおけるヒトの遺伝子治療だけでなく、癌患者の免疫療法にも使用することを提案する。
【0064】
緒言
レトロウイルスの亜種レンチウイルスは、非分裂細胞を含めたほとんどの細胞種に感染することができる。この特性があるため、レンチウイルスは遺伝子治療にとって魅力的な存在である。いくつかの複製欠失組換えレンチウイルスベクターは、既に様々なグループによって構築されている(Naldini PNAS 93、11382〜8、Science、1996)。再操作し、無毒化したこれらのレンチウイルスベクターは最も効果的かつ安全な遺伝子治療用ベクターとして提案されている(Zufferey R、& Kim V.N.J Virol、72、9873〜80、1998)。本発明者等は、水疱性口炎ウイルスGタンパク質(VSVG)偽型レンチウイルス(HIV)をベースとしたベクターを開発した(Burns J.C.、1993 PNAS 90、8033〜7)。このベクターは非必須なウイルスタンパク質をほとんど欠失しているが、ポリプリン領域シス作用性配列(cPPT、中心DNA3重鎖)を含む。この中心DNA3重鎖は、逆転写されたHIV−DNA分子の核移入をかなり促進する。さらに、中心DNA3重鎖を含むレンチウイルスは、この3重鎖を含まないベクターよりも効果的にマウス及びヒト細胞を形質導入させることが認められた。
【0065】
HHD(HLA−A2.1純系)トランスジェニックマウス(Pascolo et al.、J.Exp.Med.、1997、185:2043〜2051)を用いると、実験的に制御したエピトープペプチドの免疫原性の評価及び様々な免疫戦略の評価が可能である。これらのHHDマウスを使用して、異なる組換えベクターによってコードされたメラノーマポリペプチドが単一の動物個体内で同時にCTL応答を誘導する能力が報告された。中心DNA3重鎖を含むか、又は含まず、同様のメラノーマポリエピトープをコードするレンチウイルスベクターが、HHDトランスジェニックマウスにおいてインビボCTLを誘導する能力について最初に調べた。さらに同様の組換えレンチウイルスによって形質導入されたヒト樹状細胞(hDC)がエキソビボでメラノーマポリエピトープモチーフに対して1次CTL応答を誘導するかどうかについても調べた。本発明の結果によって、組換えベクターを直接投与するか、又はエキソビボで形質導入した樹状細胞を使用すると、DNA3重鎖によってレンチウイルスベクターがインビボで特異的なCTL応答を誘導する能力が著しく増強されることが示された。
【0066】
結果
実施例1:メラノーマポリエピトープの免疫
メラノーマから得られたポリエピトープモチーフをコードする組換えDNAをHHDマウスに注射することによって、いくつかのメラノーマエピトープに対して同時にCTL応答を惹起することができることを証明した後、同様のメラノーマポリペプチドモチーフ(図1及び表1)をコードするレンチウイルスの免疫原能を試験した。TRIP−mel−IRES−GFPベクター(CNCM 1−2185として1999年4月20日に寄託)をマウス当たり1.25μg/p24で静脈内、腹腔内、又は皮下内に投与した。1群当たり少なくとも3匹のマウスを使用した。
【0067】
10個のメラノーマエピトープのほとんどに対して特異的な複数のCTL応答が同時に誘導された。投与経路に関わらず、負荷したペプチド(表2)及びTRIP−mel−IRES−GFPで形質導入したHHDトランスフェクトHeLa細胞(データは示していない)の両者に対して同様のCTL応答が認められた。しかし、腹腔内注射によって、わずかに良好なCTL応答が誘導された。強い応答がNA17−A.nt38及びgp100.154エピトープペプチドに対して引き起こされた。gp100.457、MART.1.27、Mage−3、及びチロシナーゼ368−Dに対してまた、著しい応答が認められた。gp.100.209、gp.100.280、MART−1.32、及びチロシナーゼ1に対するCTL応答は弱かった。TRIP−mel−IRES−GFPベクターで免疫した後に弱いCTL応答が引き起こされたエピトープは全て、その他のレンチウイルスベクター以外を投与したときのCTL非誘導群に分類された。
【0068】
インビボで検出可能なCTLを誘導するために必要なレンチウイルスベクターの最小用量
著しいCTL応答を引き起こすレンチウイルスベクターの最小用量は、6種の異なる用量のTRIP−mel−IRES−GFPベクターを腹腔内投与して(1用量あたり4匹のHHDマウス)決定した。51Cr試験の直前に2匹のマウスのエフェクター細胞を混合して同一のE/T比を得る実験を2回実施した。ほとんどの実験において、メラノーマエピトープペプチド全てに対するCTL応答を試験した。結果は非常に類似しており、極めて同質であったので、「線量−効果」関係を明確かつ簡潔に比較するためにNA17/Aエピトープペプチドに対するCTL応答のみを考慮した。マウス当たり500ngと2500ng/p24との間の用量を使用して、最良のCTL応答が得られた。いくつかのメラノーマエピトープに対して検出可能なCTL応答が得られたが、レンチウイルスの用量が高くても用量の低い場合よりも良好なCTL応答が誘導されるわけではなかった。いくつかのメラノーマペプチドに対するいくつかの特異的なCTL応答が実証されたとしても、マウス当たり500ng/p24未満の用量では、効果的なCTL応答を誘導するには不十分であった。マウス当たり1250ng/p24の用量で、何匹かのマウスにおいてポリエピトープモチーフに含まれる10個のエピトープ全てに対してCTL応答が生じたことは注目に値する(図3)。
【0069】
長期の記憶CTL誘導
マウス8匹にTRIP−mel−IRES−GFPベクターを注射した。免疫後、このマウスを12日目又は5ヶ月目に殺処分した。5日間はメラノーマエピトープペプチドで、さらに2日間は10%TCGFでインビトロ誘発した後、マウス4匹のエフェクター細胞を混合してペプチドを負荷したHHDトランスフェクトRMAS細胞に対して試験した。12日前に免疫したマウスでは、gp100.29及びMart1.32以外のメラノーマエピトープペプチド全てについてCTL応答が証明された。TRIP−mel−IRES−GFPを注射して5ヶ月経つと、1次CTL誘導エピトープ全てがまだ強いCTL応答を誘導した(図2)。マウス免疫後、12日目又は5ヶ月目のCTL応答のレベルは、驚くほど同等であった。レンチウイルスベクターによってインビボ導入された細胞は、免疫系によって破壊されずにコードされたメラノーマポリエピトープを産生し続けることが示唆された。
【0070】
インビボ免疫における中心DNA3重鎖の役割
HHDマウスそれぞれに、同時にTRIP−mel−IRES−GFP及びHR−mel−IRES−GFPベクターをマウス当たり800ng、200ng、50ng、12ng、及び3ng/p24の用量で腹腔内投与して免疫した。別個の2回の実験において、各用量について少なくとも4匹のマウスを試験した。合成ペプチドでインビトロ誘発した後、ペプチドを加えたRMAS−HHD標的細胞を使用して脾細胞の細胞溶解能を試験した。結果は極めて似通っていたので、明確かつ簡潔にするためにNA17/A、Mart−1.27、gp100.154、及びチロシナーゼ.368−Dエピトープペプチドに対するCTL応答のみを試験した。
【0071】
結果を表3に示す。一般に、マウス当たり800ng、200ng、50ng/p24の用量のTRIP−LVベクターで免疫したマウスはHR−LVベクターで免疫したマウスよりも強いCTL応答を引き起こした。使用したベクターにかかわらず、マウス当たりの12ng/p24未満の用量では検出可能なCTL応答は認められなかった(データは示さず)。これによって、中心DNA複合体を含むレンチウイルスベクターの形質導入能はこの複合体を欠失したものよりも増強されていることが確認された。
【0072】
レンチウイルスベクターによる樹状細胞の形質導入
MT4及びHeLa細胞などの腫瘍細胞が中心DNA3重鎖を含むレンチウイルスによって中心DNA3重鎖を持たないベクターよりも30倍まで効果的に形質導入されることが最初に観察された。次に、様々な濃度でこれら2種のレンチウイルスベクターの形質導入能を健常提供者又はHHDマウスのDCについて試験した。GFPを発現するDCのパーセント及び平均蛍光強度をFACSによって測定し、2種のレンチウイルスベクターによるDCの形質導入濃度と見なした。
【0073】
TRIP−GFPベクターは、どのベクター濃度でもHR−GFPよりも効果的に細胞を形質導入した。TRIP−GFPベクターによって形質導入されたマウス及びヒトDCのGFP発現は、それぞれHR−GFPベクターによって形質導入されたGFPの発現の3倍及び7倍に達した(図4)。
【0074】
TRIP mel IRES−GFPベクターによって形質導入されたヒト樹状細胞を使用した1次CTL誘発
健常なHLA−A2.1提供者から得られた単核細胞(MNC)を、同提供者から得られたTRIP mel−IRES−GFPによって形質導入したDCを使用して週に1回インビトロで誘発した。樹状細胞におけるGFP発現の存在をFACSによって分析し、形質導入の効果を確かめた。3週間後、MNC細胞障害性は、FCSを含まない培養条件で標的としてペプチドを付加したT2細胞を使用した51Crアッセイによって試験した。
【0075】
TRIP mel−IRES−GFPベクターで形質導入したhDCは、Mart1.31以外のメラノーマエピトープペプチド全てに対して著しいCTL応答を誘発したが、形質導入しなかったhDCでは通常のバックグランドの応答しか誘発しなかった(図5)。
【0076】
考察
複製欠失レンチウイルス由来のベクターが様々な非分裂細胞を形質導入できることは、近年報告で示されている(Maldini、1996、Kafri、1997)。レンチウイルスは非分裂細胞の完全な核膜を通過して移入することができる(Case、1999)。本明細書では、この方法と中心DNA3重鎖と称したポリプリン領域シス作用性配列を組み合わせると、大いに可能性が広がることを実証した。この配列をレンチウイルス由来ベクターに導入すると、ニューロン、肝細胞、及び造血幹細胞/始原細胞を含めた数種の細胞の安定したインビボ及びエキソビボ形質導入を30倍高めることが可能であった。
【0077】
最近、レンチウイルス由来のベクターの安全性及び遺伝子治療での使用に関する取り組みで、進歩が認められた(Narry Kim V et al 1988、Zufferey、1999)。しかし、古典的なマウスのレトロウイルスベクター及びレトロウイルスではないベクターの使用が成功した免疫治療で(Condon、1996、Song、1997、Specht、1997)、レンチウイルスベクターを適用することについては研究されなかった。有望なワクチン戦略を開発するために、中心DNA3重鎖を含み、メラノーマポリエピトープをコードするレンチウイルスベクターの免疫原能を試験した。最初に、このベクターを直接注射して、メラノーマエピトープに対してインビボで特異的なCTL応答が引き起こされ得るかどうかについて調べた。今までに、HHD HLA−A2.1(純系)トランスジェニックマウスにおいていくつかの免疫戦略の比較が行われた。組換えpCMV−B10(HBs)DNA又は同様のメラノーマポリエピトープをコードする組換えワクチンを使用すると、1個体マウスにおいて、メラノーマポリエピトープに含まれる4から6個の異なるペプチドに対するCTL応答が同時に誘導されることが可能であった。同様のメラノーマポリエピトープをコードするTRIP mel IRES−GFPベクターでは、特異的細胞溶解についてだけでなく、CTL応答を誘発するポリペプチドの数についても、同様のポリエピトープモチーフをコードするその他のベクターよりも顕著に優れた結果が得られた。特に、ベクターを腹腔内及び皮下注射すると、TRIP mel IRES−GFPベクターによってコードされたエピトープ全てに対してCTL応答が誘導されるマウスがいた。
【0078】
形質導入又はペプチド付加された樹状細胞は、様々なガン細胞に対して有望な免疫方法として使用できることがいくつかのグループによって報告された(Mayardomo、JI、1995、Song W、1997、Specht、JM、1997)。従って、本発明の目的のために、マウス及びヒトの樹状細胞が効果的に形質導入され、インビトロで1次CTL応答を誘導するかどうかを調べた。本発明の結果によって、これらの細胞が容易に形質導入され得ることが示唆された。さらに、これらの形質導入細胞は組換えレンチウイルスによってコードされたエピトープ全てを提示した。興味深いことに、ヒト樹状細胞は最も容易に形質導入された。細胞をインビトロ形質導入することによって、レンチウイルスベクターが様々な宿主細胞にゲノムを有害な方法で組み込むことを回避できる。この理由によって、インビトロ形質導入された細胞は、最初の臨床応用で適切かつ安全な送達方法となるだろう。
【0079】
Warner等は、マウスのレトロウイルスベクターによって形質導入したHIV−タンパク質を発現する繊維芽細胞を使用して、レトロウイルスベクターがいくつかの動物モデルだけでなく、HIV患者のCTL応答を誘導する能力を初めて示した(1998年にWarnerによって概説されている)。マウスに直接注射すると、プロウイルスのDNAが効果的に抗原を提示する樹状細胞で検出された(Song、1997)。しかし、レンチウイルスとは対照的に、マウスのレトロウイルスベクターはDCなどの非分裂細胞を形質導入することができず、コードされた遺伝子のインビボ発現は「遮断される」ことが多い。結果的に、これらのレトロウイルスベクターはヒトの免疫治療にとって有力な候補とはならない(Kafri)。
【0080】
我々の結果によって、中心DNA3重鎖を含むレンチウイルスベクターは、HHDマウスにおいて中心DNA3重鎖を含まないベクターよりも強くインビボでCTL応答を誘導することが示唆される。さらに、中心DNA3重鎖を含むレンチウイルスベクターはインビトロで容易にhDCを形質導入することができ、ひいては臨床的免疫治療に使用することができよう。
【0081】
材料及び方法
レンチウイルスベクター構造
ベクタープラスミド:HR′CMVLacZ由来のpTRIP−EGFP(Naldini et al、PNAS 93、11382〜8、1996)。LacZリポーター遺伝子は、EGFP遺伝子で置き換えた(Clontech)。TRIP−EGFPでは、EGFP遺伝子をcPPT及びCTS配列を含むHIV−1LAIの中心断片のClal部位に挿入した。
【0082】
EGFP遺伝子は、pEGFP−N1プラスミドからPfuポリメラーゼ(Stratagene)を使用してPCRによって増幅し、5′及び3′それぞれにBamHI及びXhoI制限部位を付加した。PCRプライマは以下の通りであった。
【0083】
Bam GFP 5′CC GGATCC CCA CCG GTC GCC ACC 3′
Xho GFP 5′CC CTCGAG CTA GAG TCG CGG CCG 3′
HR GFPベクターは、このPCR断片をpHR′CMVLacZのBamHI及びXhoI部位にクローニングし直して、LacZ ORFをEGFPに置き換えることによって構築した。
【0084】
cPPT及びCTSを含むpLAI3の178bp断片(4793から4971)をPCRによって増幅した。この断片をHR GFPの特有のClaI部位に挿入するために、NarI制限部位をプライマの5′に付加した。
【0085】
Nar TRIP+: 5′ GTC GTC GGCGCC GAATTC ACA AAT GGC AGT ATT CAT CC 3′
Nar TRIP−: 5′GTC GTC GGCGCC CCA AAG TGG ATC TCT GCT GTC C 3′
正方向でこの3重鎖配列を挿入すると、TRIP GFPプラスミドベクターが生じ、逆向きだとTRIPinv GFPが生じた。あるいは、同様の3重鎖断片をpcPPT−AG、pcPPT−D、pcPPT−225及びpCTSプラスミドから増幅して、対応するウイルスのようにcPPT又はCTSに同様の変異を含むベクターを作製した。
【0086】
まず、TRIP GFPベクター又はTRIP−EGFP(CNCMに1998年4月15日I−2005号として寄託)に存在するEcoRI部位を充填して、TRIP GFPΔEベクターを作製した。次に、TRIP GFP ΔEベクターの0.7kb BamHI/XhoI EGFPの代わりに、ピコナウイルスの内部リボゾーム結合部位(IRES)を含む1.2kb BamHI/XhoI断片をEGFPの上流にクローニングして、TRIP ΔE IRES GFPベクターを作製した。CMVプロモータとIRES GFP配列の間に存在する部位は、BamHI−BstXI−SnaBI及びEcoRIである。CTLポリエピトープメラノーマ(mel)断片をPCRによってpBS mel poly上に作製し、プライマ5BglMlu Mel:
5′CCAGATCTACGCGTGCCACCATGGCTGCTGGT 3′
3RIMel: 5′CGGAATTCGACCTAAACGCAACGGATG3′
の内部にkozacコンセンサス配列を挿入した。
【0087】
mel PCR断片をBglII/EcoRIによって消化し、TRIP ΔE IRES GFPのBamHI/EcoRI部位に挿入して、TRIP mel IRES GFPとも称するTRIP ΔE mel IRES GFPを作製した。
【0088】
HR mel IRES GFPは、メラノーマポリエピトープ及びTRIP mel IRES GFPのIRES GFPを含むNdeI/XhoIフラグメントをHR GFPのものと交換して作製した。NdeI部位はCMVプロモータの末端に位置している。
【0089】
ウイルスベクターの産生
レンチウイルスベクターは、既に記載されたように(Naldini、I.M.PNAS 1996 及びscience 1996)、リン酸−カルシウム法を用いて、293T細胞を3種のプラスミドによって一過性にトランスフェクトすることによって作製した。簡単に説明すると、293T細胞にVSVエンベローププラスミド(pMDG)20μg及び様々なパッケージング(8.2又は8.91)プラスミド及びレンチウイルスベクタープラスミド40μgをトランスフェクトした。トランスフェクトして60h後及び84h後に、調整培地を収集した。次いで、ウイルスを濃縮して、dNTPを既に記載されたように処理した(Naldini science 1996)。HeLa P4.2細胞及びMT3細胞に対するウイルス力価は、系列希釈及びp24 ELISAアッセイによって測定した(Naldini Science 1996)。
【0090】
DEAE−デキストラン20μM存在下で、96ウェルプレートに接種したP4細胞に等量の粒子(ウェル当たりp24ウイルス抗原1ng)を感染させることによって、ウイルスの1サイクル力価測定を実施した。実験全体を通して、プロテアーゼ阻害剤サキナビル(Roche)を1μMで添加して、感染1回に限定した分析を行った。細胞の有糸分裂は、感染前日にアフィコリン(aphicolin)(8μM)処理して阻害した。β−ガラクトシダーゼ活性は、化学ルミネセンスβ−Galリポーター遺伝子アッセイ(Boehringer)を使用して、感染48時間後に測定した。
【0091】
HeLa細胞を等量のベクター粒子(ウェル当たりp24 5ng)で3回感染させた。形質導入後48時間して、培地をTNB(Tris 50mM、pH7.5、NaCl 50mM)200μlに置換して、生細胞の蛍光をマイクロプレート蛍光計(Victor2、Wallac)及びEGFP対応フィルター(励起:485nm、発光:520nm)を使用して定量した。
【0092】
マウス
HHDマウスについては既に説明されている(Pascolo、1997)。このマウスは、ヒトb2mのC末端がペプチドのアーム(GGGGS)x3によってキメラ重鎖(HLA−A2.1、a1−a2、H−2Db a3−膜間ドメイン、及び細胞質内ドメイン)のN末端に共有結合したトランスジェニック1本鎖組織適合クラスI分子を発現する。これらのマウスのH−2Db及びマウスb2m遺伝子はさらに相同的組換えによって分断され、マウス組織適合クラス1分子の細胞表面での発現が完全に欠如しており、血清学的に検出されなくなっている。
【0093】
hDC及び1次CTL誘導の発生
ヒト樹状細胞は、HLA−A2.1ハプロタイプの健常提供者のサイタフェレシス生成物(IDM、パリ、フランス)から得られた。CD3、CD14、CD80、CD83、HLA−ABC、及びHLA−DRに対するmAbを使用したこれらのDCのFACS分析によって、未熟DC表現型が示された。レンチウイルスベクターを1.106細胞当たり600ng、300ng、150ng、及び150ng/p24の濃度で含むAMV−5培地中で、このhDCを10日間形質導入した。2種のレンチウイルスベクターで形質導入したhDCにおけるGFP発現のパーセント及び平均蛍光強度をFACSによって測定した(Becton Dickinson、BD、USA)。
【0094】
同一提供者の単核細胞(MNC)をhDC又は形質導入したhDCで1hDCに対して4MNCの比でインビトロ誘発した。このMNCを同一の低温保存した形質導入hDCを使用して2回再誘発して、次いで関連ペプチド又は陰性対照(lnf.m.58)ペプチドを付加したT2細胞を標的として使用した4h51Cr−放出アッセイで細胞溶解活性を試験した(10μg/ml、5.106細胞/ml、FCSを含まないRPMI培地、室温で2時間)。
【0095】
マウス樹状細胞の生成
骨髄由来樹状細胞を既に記載されたように生成した[43、51]。骨髄の単核細胞を10%FCS、Lグルタミン 2mM、ペニシリン 50U/ml、ストレプトマイシン 50μg/ml、メルカプトエタノール 5.105Mを補足したRPMI(完全RPMI培地)に、さらに組換えマウスGM−CSF 20ng/ml及び組換えマウスIL4 100ng/ml(いずれもGENZYME(ケンブリッジ、MA)製)を補足して培養した。2日目及び6日目に、付着しなかった細胞を注意深く除去して、マウスGN−CSF 10ng/ml及びマウスIL4 50ngを補足した新鮮な完全RPMI培地を添加した。7日目に、培養培地を1.106細胞当たり600ng、300ng、150ng、及び150ng/p24の濃度のレンチウイルスベクターを補足したRPMI完全培地1mlで置換した。9日目に集めた樹状細胞を適切なmAbで評価したところ、純度は95%を上回っていた(IAb+、HHD+、CD−3、33D1+、NDL145+、及びCD11c+)。次いで、マウスDCにおけるGFP発現のパーセント及び平均蛍光強度をFACSで9日目及び12日目に測定した。
【0096】
ベクターによる免疫及びインビトロ再誘導及び細胞溶解アッセイ
HHDマウスにレンチウイルスベクターを12日間腹腔内、静脈内又は皮下内注射した。次いで感作されたマウスの脾細胞を各エピトープペプチドで独立してRPMI完全培地中で7日間再誘導した。最後の2日間に、培養細胞を10%TCGFによって再誘導した。7日目に、培養した細胞の細胞溶解活性は、既に記載されたように(Pascolo、1997)、関連ペプチド又は陰性対照ペプチド(lnf.m.58)を付加したHHDトランスフェクトTAP−RMA−S細胞を標的として使用して、4h 51Cr−放出アッセイで試験した(FCSを含まないRPMI培地中10μg/ml、5.106細胞/ml、室温で2時間)。TRIP−mel−ITES−GFPで形質導入した、又は形質導入していないHHDトランスフェクトHeLa細胞を並行して標的細胞として使用した。特異的溶解のパーセントを以下の通りに算出した:(実験上の放出−自発放出)(総放出−自発放出)x100。
【0097】
実施例II:Mycobacterium tuberculosisのDES遺伝子をコードするTRIP−des−IRES−GFPベクターで免疫したHHDマウスのCTL応答の評価
DES遺伝子についてはWO98/04711で開示している。
【0098】
実験方法
第1段階では、TRIP−des−IRES−GFPベクターを使用してHeLa−HDD細胞を形質導入した。これらの細胞を形質導入して、限界希釈によってクローニングした。GFPを高濃度で発現しているクローンを選択して、古典的な51CrCTL試験の標的細胞として使用した。
【0099】
第2段階では、HDDマウスにTRIP−des−IRES−GFPベクター粒子をマウス当たり1.2micg/p24で腹腔内注射した。注射後12日目に、これらのマウスの脾細胞を、ベクター粒子0.2micg、又は1micg/p24/ml(ml当たり2106細胞)、あるいはTRIP−des−IRES−GFPで形質導入し、LPSで誘導した同系繊維芽細胞1micg/p24/ml/2106細胞/mlで誘発した。インビトロ誘発後6日目に、細胞の細胞溶解活性をdes形質導入HeLa−HDD標的細胞を使用した51Cr試験で試験した。対照の標的細胞はメラノーマポリエピトープによって形質導入した(TRIP−mel−IRES−GFP)HeLa−HDD細胞とした。
【0100】
結果
これらの実験は、全遺伝子をコードするHIV由来の3重鎖陽性ベクターが細胞を形質導入したり、同遺伝子を発現する標的細胞に対して特異的なCTL応答を誘導したりできるかどうかを調べるために実施した。図9に例示したように、特異的な細胞溶解が全ての条件で認められる。形質導入したLPS−繊維芽細胞では弱いCTL応答が誘導された。ベクター粒子1micg/p24/mlを使用すると最も良い結果が得られた。これらの結果によって、完全な遺伝子が宿主のゲノムに導入され、その産物が処理されて、提示されて著しいCTL応答が誘導され得ることが示される。これらの結果によってまた、des遺伝子は少なくとも1個又は複数のHLA−A2.1に限定されたCTLエピトープペプチドを含んでいることが示唆される。
【参照文献】
【0101】
【図面の簡単な説明】
【0102】
【図1】メラノーマポリペプチドをコードするHIV−1由来の3重鎖DNA陽性の組換えベクター。
【図2】TRIP−mel−IRES−GFPベクターで免疫した後のHHDマウスのCTL応答。
【図3】中心DNA3重鎖陽性又は陰性HIV由来ベクターによる形質導入後5日目のヒト細胞でのGFP発現。
【図4】ヒト樹状細胞を使用したインビトロCTL応答。
【図5】pHR.MEL−IRES−GFPベクターの制限カード。メラノーマ特異的CTLモノ又はポリエピトープ配列HR.MEL−IRES−GFPの構築 HR.MEL−IRES−GFPプラスミドを構築するために、1999年4月20日に1−2185号としてCNCMに寄託されたCMVプロモータの一部、ポリペプチドMEL及びIRES−GFPを含むTRIP.MEL−IRES−GFPのNdeI/XhoIフラグメントをCMVプロモータの一部及びGFPを含むHR GFPのNdeI/XhoIフラグメントと置き換えた。
【図6】pTRIP.ILKE−IRES−GFPベクターの制限カードHIV特異的CTLモノ又はポリエピトープ配列、そのエピトープはI9V.(ILKE);(RT476−484)TRIP.ILKE−IRES−GFP(1999年10月6日、_号としてCNCM(フランス、パリ)に寄託)の構築 TRIP.ILKE−IRES−GFPは、TRIPΔE IRES−GFPのCMVプロモータとIRESの間にILKEのPCR産物を挿入することによって構築した。ILKEによって始まるCTLエピトープの周りの領域は、プライマ5 ILKE:5′TCAGATCTGCCACCATGGCACTAACAGAAGTAATACCAC 3′ 3 RIILKE:5′CGGAATTCTTATTGGCCTTGCCCCTGCTTC 3′を用いてマトリックスpLAI上にPCRによって増幅した。 Kozak配列は、上流プライマに挿入し、終止コドンは下流プライマに挿入した。 BglII/EcoRIで消化した後、PCR産物をTRIPΔE IRES−GFPのBamHI/EcoRI部位に挿入した。 このベクターは、GFP及びエピトープのクラスタに対応し、特にHLA.A2.1に限定されたI9Vエピトープ(RT476〜484)を含むVIHのRT遺伝子の領域をコードするバイシストロン性情報を発現する(Walker B.D.、1989 PNAS 86 p.9514〜9518)。
【図7】pTRIP.TEL/AML−IRES−GFPベクターの制限地図TEL/AML配列の転座 TRIP.TEL/AML−IRES−GFPは、TEL/AMLのPCR産物をTRIPΔE IRES−GFPのCMVプロモータとIRESの間に挿入することによって構築した。TELとAMLとの間の転座周囲の領域は、プライマ5 BgI TA: 5′GAAGATCTGCCACCATGAAGCCCATCAACCTCTCTCAT 3′ 3 RITA: 5′CGGAATTCTTACCCAGCGCAACGCCTC 3′ を用いてPCRによって増幅した。 Kozak配列は、上流プライマに挿入し、終止コドンは下流プライマに挿入した。 BglII/EcoRIで消化した後、PCR産物をTRIPΔE IRES−GFPのBamHI/EcoRI部位に挿入した。
【図8A】(HIV1polから得られた)I9Vエピトープペプチド及びHHDトランスジェニックマウスの骨髄細胞を使用して産生したマウス樹状細胞のGFPをコードするHIV由来DNA3重鎖陽性ベクターの形質導入能。 細胞内GFP発現のFACS分析によって実証されたように、マウス樹状細胞の約80%がHIV由来3重鎖陽性組換えベクターによって形質導入された。
【図8B】(HIV1polから得られた)I9Vエピトープペプチド及びHHDトランスジェニックマウスの骨髄細胞を使用して産生したマウス樹状細胞のGFPをコードするHIV由来DNA3重鎖陽性ベクターの形質導入能。 細胞内GFP発現のFACS分析によって実証されたように、マウス樹状細胞の約80%がHIV由来3重鎖陽性組換えベクターによって形質導入された。
【図9】Mycobacterium tuberculosisのDES遺伝子をコードするTRIP−des−IRES−GFPベクターで免疫したHHDマウスのCTL応答の評価。
【図10】1999年10月11日にCNCMに寄託されたpTRIP.DES−IRES−GFPの制限地図。
【特許請求の範囲】
【請求項1】
組換えベクターを含む免疫原組成物であって、
前記ベクターがシス作用性中心開始領域(cPPT)とシス作用性終止領域(CTS)とを備えたポリヌクレオチドを含み、該領域はレトロウイルス又はレトロウイルス様の起源であり、前記ベクターはさらに所定のヌクレオチド配列(目的の導入遺伝子又は配列)とレトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の制御シグナルとを含み、前記組成物は細胞性応答、例えば前記ベクター中に存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は刺激することができることを特徴とする免疫原組成物。
【請求項2】
発生したCTL応答が記憶CTL応答である請求項1に記載の免疫原組成物。
【請求項3】
レトロウイルス起源の前記配列がレンチウイルスのゲノムから得られたことを特徴とする請求項1に記載の免疫原組成物。
【請求項4】
前記目的の導入遺伝子又は配列が転写及び発現の調節シグナルを含む発現カセットに含有されることを特徴とする請求項1から3の何れか1項に記載の免疫原組成物。
【請求項5】
1)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成の調節シグナルの制御下に置かれた所定のヌクレオチド配列(導入遺伝子)を含有する組換えヌクレオチド配列と、2)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域の起源がレトロウイルス又はレトロウイルス様であるか、トランスポゾン由来であり、レトロウイルス又はレトロウイルス様起源の逆転写調節シグナル又はトランスポゾン調節シグナルとともに、機能的な方向及び位置に挿入されているポリヌクレオチドとを備えた組換えレトロウイルス粒子を含む免疫原組成物であって、
前記免疫原組成物が細胞性応答、例えば前記ベクター中に存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は刺激することができる免疫組成物。
【請求項6】
a)レンチウイルスの核タンパク質又は機能的に得られたポリペプチドに対応するgagポリペプチド(GAGポリペプチド)と、
b)レンチウイルスのRT、PRO、INタンパク質又は機能的に得られたポリペプチドによって構成されたpolポリペプチド(POLポリペプチド)と、
c)エンベロープポリペプチド又は機能的に得られたポリペプチド(ENVポリペプチド)と、
d)1又は数個のエピトープをコードし、転写及び発現の調節シグナルの制御下におかれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)と、レトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の調節シグナルを含有する配列と、シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)とを備えたポリヌクレオチドを含む組換えヌクレオチド配列であって、これらの領域の起源がレトロウイルス又はレトロウイルス様であり、レトロウイルス又はレトロウイルス様起源の上記調節シグナルとともに、機能的な方向及び位置に挿入された組換えヌクレオチド配列と
を備えた組換えレトロウイルスベクター粒子を含む免疫原組成物であって、
前記組成物が、前記ベクターに存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は促進することができる免疫原組成物。
【請求項7】
前記組換えレトロウイルス様粒子が、
a)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域がレトロトランスポゾン由来であり、レトロトランスポゾン調節シグナルとともに機能的な方向に挿入されているポリヌクレオチドと、
b)レトロトランスポゾンの核タンパク質又は機能的に得られたポリペプチドに対応するポリペプチド(GAGポリヌクレオチド)と、
c)レトロトランスポゾンのRT、PRO、INタンパク質に対応するpolポリペプチド又は機能的に得られたポリペプチド(POLポリペプチド)と、
d)ウイルスエンベロープポリペプチドと、
e)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成のレトロトランスポゾン調節シグナルの制御下に置かれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)を備えた組換えヌクレオチド配列と
を含む免疫原組成物であって、
前記組成物は細胞性応答、例えば前記ベクターに存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は促進することができ、前記組成物は前記導入遺伝子配列によってコードされた1又は数個のエピトープに対して細胞性応答CTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は促進することができる免疫原組成物。
【請求項8】
発生したCTL応答が記憶CTL応答又はCD4応答である請求項5〜7の何れか1項に記載の免疫原組成物。
【請求項9】
前記逆転写、発現及びウイルス粒子形成の調節シグナルの起源がレンチウイルスであり、cPPT及びCTSを含む前記ポリヌクレオチドの起源がレンチウイルスであることを特徴とする請求項1〜8の何れか1項に記載の免疫原組成物。
【請求項10】
前記逆転写、発現及びウイルス粒子形成の調節シグナル及び前記ベクター内のcPPT及びCTSを含む前記ポリヌクレオチドがHIV型レトロウイルス、特にHIV−1又はHIV−2由来であることを特徴とする請求項1〜9の何れか1項に記載の免疫原組成物。
【請求項11】
前記逆転写、発現及びウイルス粒子形成の調節シグナル及び前記ベクター内のcPPT及びCTSを含む前記ポリヌクレオチドが、レンチウイルスCAEV、EIAV、VISNA、HIV、SIV又はFIVから選択されたウイルス由来であることを特徴とする請求項10に記載の免疫原組成物。
【請求項12】
前記ベクターの前記ポリヌクレオチドがHIV−1レトロウイルスゲノムのシス作用性中心開始領域(cPPT)及び終止領域(CTS)を含むDNA配列であることを特徴とする請求項1〜11の何れか1項に記載の免疫原組成物。
【請求項13】
前記ベクター粒子のgag、pol及びenv配列がHIVレトロウイルス、特にHIV−1又はHIV−2の配列由来であることを特徴とする請求項5〜12の何れか1項に記載の免疫原組成物。
【請求項14】
gag及びpol配列がHIVレトロウイルスの配列由来であり、env配列が異なるHIVレトロウイルス又はウイルス由来であることを特徴とする請求項5〜12の何れか1項に記載の免疫原組成物。
【請求項15】
前記env配列がアンフォトロピックENVポリペプチドをコードすることを特徴とする請求項14に記載の免疫原組成物。
【請求項16】
前記env配列がエコトロピックENVポリペプチドをコードすることを特徴とする請求項14に記載の免疫原組成物。
【請求項17】
前記env配列が水疱性口内炎ウイルス(VSV)から得られたことを特徴とする請求項14に記載の免疫原組成物。
【請求項18】
CNCMに1999年10月11日、I−2326号として寄託されたプラスミドpTRIP.TEL/AML−IRES−EGFP、又はCNCMに1999年10月11日、I−2331号として寄託されたPtrip.DES−IRES−GFPであることを特徴とするベクター。
【請求項19】
CNCMに1999年10月11日、I−2337号として寄託されたプラスミドpTRIP.ILKE−IRES−GFPであることを特徴とするベクター。
【請求項20】
請求項1〜17の何れか1項に記載の免疫原組成物と接触した組換え細胞。
【請求項21】
抗原提示細胞又は肺細胞、脳細胞、上皮細胞、星状細胞、マイクログリア、乏突起神経膠細胞、ニューロン、筋細胞、肝細胞、樹状細胞、神経細胞、骨髄の細胞株、マクロファージ、繊維芽細胞、造血細胞から選択された細胞である請求項20に記載の組換え細胞。
【請求項22】
腫瘍又は感染性疾患を治療的に処置するための請求項1〜17の何れか1項に記載の免疫原組成物又は組換え細胞。
【請求項23】
前記ベクターに含まれた導入遺伝子が腫瘍細胞又は感染性病原体から得られたエピトープ、ポリエピトープを含むポリペプチド、タンパク質である請求項1から17のいずれかに記載の免疫原組成物。
【請求項24】
前記エピトープ、ポリエピトープを含むポリペプチド又は前記タンパク質がメラノーマ細胞由来である請求項23に記載の免疫原組成物。
【請求項25】
前記cPPT及びCTS領域が前記ベクターの前記配列内の中心に位置する請求項1〜17の組成物、又は請求項1〜17のいずれかに定義されたベクター又はベクター粒子。
【請求項26】
前記導入遺伝子が逆転写の調節シグナルのU3領域に挿入された請求項1〜17の何れか1項に記載された組成物又は請求項1〜17の何れか1項に記載されたベクター又はベクター粒子。
【請求項27】
シス作用性中心開始領域(cPPT)及びシス作用性末端領域(CTS)を含むポリヌクレオチドを含む組換えベクターであって、これらの領域の起源がレトロウイルス又はレトロウイルス様であり、前記ベクターがさらに定義されたヌクレオチド配列(目的の導入遺伝子又は配列)を含み、前記ヌクレオチド配列が病原体の遺伝子配列、及びレトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の調節シグナルであり、前記組成物が細胞性応答、例えば前記ベクターに存在する前記導入遺伝子配列によってコードされた1個又はいくつかのエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は促進することが可能であることを特徴とする組換えベクター。
【請求項28】
前記目的のヌクレオチド配列又は導入遺伝子が細菌、例えばミコバクテリア、特にMycobacterium tuberculosisの抗原をコードする遺伝子である請求項27に記載の組換えベクター。
【請求項29】
前記目的のヌクレオチド配列又は導入遺伝子がウイルス又はレトロウイルスの抗原をコードする遺伝子である請求項27に記載の組換えベクター。
【請求項30】
前記TEL/AML又は前記ILKEエピトープが病原体の遺伝子、例えばM tuberculosisのDES遺伝子の一部と置換された請求項18又は19に記載のベクター。
【請求項31】
前記ベクターのゲノムの標的細胞核への移入を誘導又は促進することが可能な請求項1〜17の何れか1項に記載の免疫原組成物。
【請求項32】
請求項1〜17の何れか1項に記載の免疫原組成物における請求項18又は19又は27又は31の何れかに記載のベクターの使用。
【請求項33】
レンチウイルス又はレトロトランスポゾンベクター中で逆転写後に3本鎖DNA構造(DNA3重鎖)をとるcPPT及びCTS領域を含み、前記ベクターDNAの形質導入細胞核への侵入速度を促進するヌクレオチド配列の使用。
【請求項34】
細胞又は患者の細胞性応答を誘導するための請求項33に記載のヌクレオチド配列の使用。
【請求項35】
前記ベクターDNAの速い侵入速度を誘導するか、又は前記ベクターDNAの核移入速度を増加させることが可能なcPPT及びCTS領域によって誘導された3本鎖DNA配列。
【請求項1】
組換えベクターを含む免疫原組成物であって、
前記ベクターがシス作用性中心開始領域(cPPT)とシス作用性終止領域(CTS)とを備えたポリヌクレオチドを含み、該領域はレトロウイルス又はレトロウイルス様の起源であり、前記ベクターはさらに所定のヌクレオチド配列(目的の導入遺伝子又は配列)とレトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の制御シグナルとを含み、前記組成物は細胞性応答、例えば前記ベクター中に存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は刺激することができることを特徴とする免疫原組成物。
【請求項2】
発生したCTL応答が記憶CTL応答である請求項1に記載の免疫原組成物。
【請求項3】
レトロウイルス起源の前記配列がレンチウイルスのゲノムから得られたことを特徴とする請求項1に記載の免疫原組成物。
【請求項4】
前記目的の導入遺伝子又は配列が転写及び発現の調節シグナルを含む発現カセットに含有されることを特徴とする請求項1から3の何れか1項に記載の免疫原組成物。
【請求項5】
1)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成の調節シグナルの制御下に置かれた所定のヌクレオチド配列(導入遺伝子)を含有する組換えヌクレオチド配列と、2)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域の起源がレトロウイルス又はレトロウイルス様であるか、トランスポゾン由来であり、レトロウイルス又はレトロウイルス様起源の逆転写調節シグナル又はトランスポゾン調節シグナルとともに、機能的な方向及び位置に挿入されているポリヌクレオチドとを備えた組換えレトロウイルス粒子を含む免疫原組成物であって、
前記免疫原組成物が細胞性応答、例えば前記ベクター中に存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は刺激することができる免疫組成物。
【請求項6】
a)レンチウイルスの核タンパク質又は機能的に得られたポリペプチドに対応するgagポリペプチド(GAGポリペプチド)と、
b)レンチウイルスのRT、PRO、INタンパク質又は機能的に得られたポリペプチドによって構成されたpolポリペプチド(POLポリペプチド)と、
c)エンベロープポリペプチド又は機能的に得られたポリペプチド(ENVポリペプチド)と、
d)1又は数個のエピトープをコードし、転写及び発現の調節シグナルの制御下におかれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)と、レトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の調節シグナルを含有する配列と、シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)とを備えたポリヌクレオチドを含む組換えヌクレオチド配列であって、これらの領域の起源がレトロウイルス又はレトロウイルス様であり、レトロウイルス又はレトロウイルス様起源の上記調節シグナルとともに、機能的な方向及び位置に挿入された組換えヌクレオチド配列と
を備えた組換えレトロウイルスベクター粒子を含む免疫原組成物であって、
前記組成物が、前記ベクターに存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答を誘導又は促進することができる免疫原組成物。
【請求項7】
前記組換えレトロウイルス様粒子が、
a)シス作用性中心開始領域(cPPT)及びシス作用性終止領域(CTS)を含むポリヌクレオチドであって、これらの領域がレトロトランスポゾン由来であり、レトロトランスポゾン調節シグナルとともに機能的な方向に挿入されているポリヌクレオチドと、
b)レトロトランスポゾンの核タンパク質又は機能的に得られたポリペプチドに対応するポリペプチド(GAGポリヌクレオチド)と、
c)レトロトランスポゾンのRT、PRO、INタンパク質に対応するpolポリペプチド又は機能的に得られたポリペプチド(POLポリペプチド)と、
d)ウイルスエンベロープポリペプチドと、
e)転写及び発現の調節シグナル、逆転写、発現及びウイルス粒子形成のレトロトランスポゾン調節シグナルの制御下に置かれた所定のヌクレオチド配列(目的の導入遺伝子又は配列)を備えた組換えヌクレオチド配列と
を含む免疫原組成物であって、
前記組成物は細胞性応答、例えば前記ベクターに存在する前記導入遺伝子配列によってコードされた1又は数個のエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は促進することができ、前記組成物は前記導入遺伝子配列によってコードされた1又は数個のエピトープに対して細胞性応答CTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は促進することができる免疫原組成物。
【請求項8】
発生したCTL応答が記憶CTL応答又はCD4応答である請求項5〜7の何れか1項に記載の免疫原組成物。
【請求項9】
前記逆転写、発現及びウイルス粒子形成の調節シグナルの起源がレンチウイルスであり、cPPT及びCTSを含む前記ポリヌクレオチドの起源がレンチウイルスであることを特徴とする請求項1〜8の何れか1項に記載の免疫原組成物。
【請求項10】
前記逆転写、発現及びウイルス粒子形成の調節シグナル及び前記ベクター内のcPPT及びCTSを含む前記ポリヌクレオチドがHIV型レトロウイルス、特にHIV−1又はHIV−2由来であることを特徴とする請求項1〜9の何れか1項に記載の免疫原組成物。
【請求項11】
前記逆転写、発現及びウイルス粒子形成の調節シグナル及び前記ベクター内のcPPT及びCTSを含む前記ポリヌクレオチドが、レンチウイルスCAEV、EIAV、VISNA、HIV、SIV又はFIVから選択されたウイルス由来であることを特徴とする請求項10に記載の免疫原組成物。
【請求項12】
前記ベクターの前記ポリヌクレオチドがHIV−1レトロウイルスゲノムのシス作用性中心開始領域(cPPT)及び終止領域(CTS)を含むDNA配列であることを特徴とする請求項1〜11の何れか1項に記載の免疫原組成物。
【請求項13】
前記ベクター粒子のgag、pol及びenv配列がHIVレトロウイルス、特にHIV−1又はHIV−2の配列由来であることを特徴とする請求項5〜12の何れか1項に記載の免疫原組成物。
【請求項14】
gag及びpol配列がHIVレトロウイルスの配列由来であり、env配列が異なるHIVレトロウイルス又はウイルス由来であることを特徴とする請求項5〜12の何れか1項に記載の免疫原組成物。
【請求項15】
前記env配列がアンフォトロピックENVポリペプチドをコードすることを特徴とする請求項14に記載の免疫原組成物。
【請求項16】
前記env配列がエコトロピックENVポリペプチドをコードすることを特徴とする請求項14に記載の免疫原組成物。
【請求項17】
前記env配列が水疱性口内炎ウイルス(VSV)から得られたことを特徴とする請求項14に記載の免疫原組成物。
【請求項18】
CNCMに1999年10月11日、I−2326号として寄託されたプラスミドpTRIP.TEL/AML−IRES−EGFP、又はCNCMに1999年10月11日、I−2331号として寄託されたPtrip.DES−IRES−GFPであることを特徴とするベクター。
【請求項19】
CNCMに1999年10月11日、I−2337号として寄託されたプラスミドpTRIP.ILKE−IRES−GFPであることを特徴とするベクター。
【請求項20】
請求項1〜17の何れか1項に記載の免疫原組成物と接触した組換え細胞。
【請求項21】
抗原提示細胞又は肺細胞、脳細胞、上皮細胞、星状細胞、マイクログリア、乏突起神経膠細胞、ニューロン、筋細胞、肝細胞、樹状細胞、神経細胞、骨髄の細胞株、マクロファージ、繊維芽細胞、造血細胞から選択された細胞である請求項20に記載の組換え細胞。
【請求項22】
腫瘍又は感染性疾患を治療的に処置するための請求項1〜17の何れか1項に記載の免疫原組成物又は組換え細胞。
【請求項23】
前記ベクターに含まれた導入遺伝子が腫瘍細胞又は感染性病原体から得られたエピトープ、ポリエピトープを含むポリペプチド、タンパク質である請求項1から17のいずれかに記載の免疫原組成物。
【請求項24】
前記エピトープ、ポリエピトープを含むポリペプチド又は前記タンパク質がメラノーマ細胞由来である請求項23に記載の免疫原組成物。
【請求項25】
前記cPPT及びCTS領域が前記ベクターの前記配列内の中心に位置する請求項1〜17の組成物、又は請求項1〜17のいずれかに定義されたベクター又はベクター粒子。
【請求項26】
前記導入遺伝子が逆転写の調節シグナルのU3領域に挿入された請求項1〜17の何れか1項に記載された組成物又は請求項1〜17の何れか1項に記載されたベクター又はベクター粒子。
【請求項27】
シス作用性中心開始領域(cPPT)及びシス作用性末端領域(CTS)を含むポリヌクレオチドを含む組換えベクターであって、これらの領域の起源がレトロウイルス又はレトロウイルス様であり、前記ベクターがさらに定義されたヌクレオチド配列(目的の導入遺伝子又は配列)を含み、前記ヌクレオチド配列が病原体の遺伝子配列、及びレトロウイルス又はレトロウイルス様起源の逆転写、発現及びウイルス粒子形成の調節シグナルであり、前記組成物が細胞性応答、例えば前記ベクターに存在する前記導入遺伝子配列によってコードされた1個又はいくつかのエピトープに対するCTL(細胞障害性Tリンパ球)応答又はCD4応答を誘導又は促進することが可能であることを特徴とする組換えベクター。
【請求項28】
前記目的のヌクレオチド配列又は導入遺伝子が細菌、例えばミコバクテリア、特にMycobacterium tuberculosisの抗原をコードする遺伝子である請求項27に記載の組換えベクター。
【請求項29】
前記目的のヌクレオチド配列又は導入遺伝子がウイルス又はレトロウイルスの抗原をコードする遺伝子である請求項27に記載の組換えベクター。
【請求項30】
前記TEL/AML又は前記ILKEエピトープが病原体の遺伝子、例えばM tuberculosisのDES遺伝子の一部と置換された請求項18又は19に記載のベクター。
【請求項31】
前記ベクターのゲノムの標的細胞核への移入を誘導又は促進することが可能な請求項1〜17の何れか1項に記載の免疫原組成物。
【請求項32】
請求項1〜17の何れか1項に記載の免疫原組成物における請求項18又は19又は27又は31の何れかに記載のベクターの使用。
【請求項33】
レンチウイルス又はレトロトランスポゾンベクター中で逆転写後に3本鎖DNA構造(DNA3重鎖)をとるcPPT及びCTS領域を含み、前記ベクターDNAの形質導入細胞核への侵入速度を促進するヌクレオチド配列の使用。
【請求項34】
細胞又は患者の細胞性応答を誘導するための請求項33に記載のヌクレオチド配列の使用。
【請求項35】
前記ベクターDNAの速い侵入速度を誘導するか、又は前記ベクターDNAの核移入速度を増加させることが可能なcPPT及びCTS領域によって誘導された3本鎖DNA配列。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8B】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8B】
【図9】
【図10】
【公開番号】特開2011−78420(P2011−78420A)
【公開日】平成23年4月21日(2011.4.21)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−251771(P2010−251771)
【出願日】平成22年11月10日(2010.11.10)
【分割の表示】特願2001−530503(P2001−530503)の分割
【原出願日】平成12年10月10日(2000.10.10)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(598125981)
【氏名又は名称原語表記】Centre National de la Recherche Scientifique
【Fターム(参考)】
【公開日】平成23年4月21日(2011.4.21)
【国際特許分類】
【出願番号】特願2010−251771(P2010−251771)
【出願日】平成22年11月10日(2010.11.10)
【分割の表示】特願2001−530503(P2001−530503)の分割
【原出願日】平成12年10月10日(2000.10.10)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(598125981)
【氏名又は名称原語表記】Centre National de la Recherche Scientifique
【Fターム(参考)】
[ Back to top ]