
免疫疾患治療用ラパマイシン及びIL−10
本発明は、免疫抑制及び抗原特異免疫耐性を誘発することができるIL−10及びラパマイシンの組み合わせ及びT細胞によって媒介される、亢進、機能不全又は管理されていない免疫反応を含む疾患の治療におけるその用途を開示する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、それらを必要としている患者における免疫抑制及び/又は抗原−特異免疫耐性を誘発する方法及び組成物に関する。より詳細には、本発明は、T細胞によって媒介される、亢進、機能不全又は制御されていない自己又は非自己免疫応答を含む疾患の治療に用いるためのラパマイシン及びIL−10の組み合わせ製剤を提供する。また、本発明は、IL−10及びラパマイシンを含有する医薬組成物及び免疫応答のモデュレータとしてのそれらの使用に指向する。
【背景技術】
【0002】
移植及び免疫抑制薬
移植は、末期の腎不全、ハース(hearth)又は肝疾患、自己免疫タイプI糖尿病のほとんどの患者に対する代案治療であり、それは、患者に小腸及び肺機能における欠乏性疾患を発現させる可能性がある。移植片生着は、多くの因子に依存するが、これらの最も顕著なものは強力な免疫抑制剤の投与である。遺伝的に共通点のない個体間の移植は、もし制御できないままであれば、移植された臓器の完全な破壊を招き得る、急速な及び場合によっては破壊的な同種異系反応性免疫応答を誘発する。免疫抑制剤の投与はこの応答を和らげ、よって、急性組織不適合を防止する。しかし、免疫抑制剤を中止すると、拒否反応の再活性化を招き、急速な移植破壊を導くため、移植片生着を継続するためには終身の免疫抑制に依存する。
【0003】
最近、免疫抑制剤のなかで、シクロスポリンA(CsA)、FK506及びラパマイシンを含む選択的T細胞阻害剤が開発されている。CsA及びFK506の双方は、カルシニュリン機能をブロックすることによってT細胞活性化を阻害し、よって、活性化T細胞の核因子(NFAT)能の発現を防止する。この工程は、IL−2を含むいくつかのサイトカインのmRNAを調節(増加)するために不可欠である。CsA及びFK506の主な制約は、それらの種々の毒性である。さらに、CsA及びFK506の双方は、T細胞アポチーシスを防止する(Yuら、2001年、参照)。
【0004】
これに反して、ラパマイシンは、細胞質蛋白(FKBP−12)に結合し、かつIL−2信号経路をブロックすることによって、T細胞の増殖を阻害する免疫抑制能がある(Sehgal、1998)。その複合体はラパマイシンの哺乳類標的(mTOR)に結合し、かつブロックし、サイトカイン誘発T細胞増殖の阻害をもたらす。重要なことは、CsA及びFK506に反して、ラパマイシンは、TCR−媒介T細胞活性(Blahaら、2003)及び活性誘導細胞死(AICD)に対するIL−2細胞プライミングをブロックしないことである。後者は、末梢移植耐性の誘発の役割を果たすと思われるT細胞アポトーシスの一形態である(Wellsら、1999)。樹状細胞(DC)に作用しないCsAとは異なり、ラパマイシンは、DCの表現型及び機能に大いに影響を及ぼす(Hacksteinら、2002)。それは、それらの抗原取り込み能を著しく減少させ、よって、免疫寛容原性表現型を有するDCの分化に有利である。この作用は、低濃度で現われ、ラパマイシンが生理学的に適当な濃度(1ng/ml)ではDC成熟とは無関係であり、インビトロ及びインビボの双方で証明されている(Hacksteinら、2002)。
【0005】
最近の入手可能な免疫抑制剤は、非常に短期間では非常に有効であるが、実質的な問題は、移植の拒絶を防止する代替の又は高性能な方法を開発することの差し迫った必要を示している。主な障害物は、感染性の病原体に対する有効な免疫応答と、移植に対する破壊的免疫応答との間の区別ができないことである。よって、免疫抑制の治療は、日和見感染のリスクの増大を招くかも知れない。いくつかの研究は、非特異的免疫応答は、移植患者におけるガンの発生率を増加することを導くであろうことを示す(Hojoら、1999)。したがって、非特異免疫抑制の代替が見出された場合にのみ、移植の最大の可能性が履行されるであろう。移植免疫の主なねらいは、移植に対する免疫応答を防止するが、残りの免疫システムを完全に損なわれないまま残すプロトコルを開発することである。これを達成することにより、移植耐性がもたらされるであろう。
【0006】
自己免疫
自己免疫疾患において、自己抗原に対する望まない免疫応答は末梢組織の破壊を招く。自己免疫疾患の治療は、現在、炎症の抑制的調節及び非抗原(Ag)特異免疫抑制に基づいている。同種異系移植片拒絶の防止のために、この戦略は、一旦薬物の投与を中止すると再発の高い危険及び感染及び腫瘍を含む過度の免疫抑制の危険を伴うため、長期においてはしばしば有効ではない。代替のアプローチは、一時的な免疫抑制及び/又は特異免疫耐性の誘導に基づいており、宿主の防衛メカニズムを完全に保全しながら、自己Agに対する「沈黙」の病原性応答を目的としている。
【0007】
免疫システムは、自己又は有害でない抗原に対する耐性を誘発する2つの異なるメカニズムを引き出している。これらは、中枢及び末梢T細胞耐性と称される。中枢耐性は、胎児の発育中及び非常に初期の出生時期に実現され、胸腺の発現中に自己反応性T細胞の特定クローン欠損によって媒介される。末梢メカニズムは、成熟T細胞における耐性を誘発し、一生の間末梢で起こる。これらのメカニズムは、抗原特異性リンパ球の機能的不活性化(アネルギーと呼ばれる)及び抑制及び調節能を有するT細胞のサブセットの活性化(T regulatory cells、Battagliaらにおいて概説されたT調節細胞、2002参照)を含む。
【0008】
耐性及びT調節細胞
近年、移植特異耐性を達成するための戦略として、T調節(Tr)細胞の誘導における興味が増加しつつある。CD8+、D8+CD28- 及びTCR+CD4-CD8-のような他のT細胞サブセットが調節能を有する細胞に含まれることが示されているが、今日までに確認されたTr細胞のほとんどは、CD4+集団内に見出されている。CD4+集団では、抑制的な特性を有する種々の分画が確認されている。我々のグループは、タイプ1調節T細胞(Tr1)として定義されるTr細胞のサブセットを特徴づけた。それは、Th1及びTh2細胞のそれと区別できるサイトカイン産生プロファイルを有する。ヒト及びマウスのTr1細胞は、高レベルのIL−10、かなりの量のIL−5、TGF−β及びIFN−γを産生するが、IL−2は低レベルであり、IL−4は全く産生しない(Grouxら、1997)。IL−10は、Tr1細胞の分化及びエフェクター機能にとって重要なサイトカインである。抗原及びIL−10の存在下でCD4+T細胞を培養すると、インビトロでは、抗原特異T細胞応答を抑制することができるTr1細胞の発現を導き、インビボでは、自己免疫大腸炎の発現を導く(Grouxら、1997)。また、Tr1細胞は、インビボで発現することができる。Tr1細胞は、実際、SCID−再構成患者(高レベルのIL−10が成功した同種異系幹細胞移植を伴う)の末梢血から単離される(Bacchettaら、1994)。
【0009】
耐性及びIL−10
IL−10は、免疫調節の解決の鍵となる役割を果たす(Mooreらにおいて概説、2001)。それは、Tリンパ球の増殖及びIL−2産生を阻害する。IL−10は、活性化された単球/マクロファージ、好中球、好酸球及び肥満細胞によるTNF−α、IL−1、IL−6等の前炎症性サイトカイン及びIL−8、MIPlα及びMIPlβ等のケモカインの産生を阻害することによって、強力な抗炎症作用を有する。さらに、IL−10は、MHCII及び補刺激性分子を抑制することによる単球/マクロファージ/DC等の抗原提示細胞の抗原提示能を抑制する。T細胞媒介および抗炎症免疫応答の誘発及びエフェクター機能を抑制するIL−10の能力は、骨髄及び臓器移植におけるIL−10形質発現、機能及び潜在的有用性の多くの研究を導いた。マウスにおける血管新生化心臓同種異形移植の研究では、移植の前の被移植動物のIL−10処置が移植片生着を高め、一方、移植時またはその後にIL−10を与えても、ほとんど有用な効果はないか、拒絶を高めるのみであった(Liら、1999)。BMT前のIL−10産生レベルの上昇を示した患者は、GVHDの発生率が低く、生存を向上させた(Bakerら、1999)。これに対して、ポスト−BMT GNHD患者における高IL−10レベルは、生存に対する予後ははかばかしくなかった(Hempel、1997)。しかし、Blazar及びその同僚は、少量のIL−10(死亡率を増加させる10-3、10-4の量)でのマウスの処置は、GVHD関連致死率を防ぐことを示した(Blazarら、1998)。
【0010】
免疫抑制剤とIL−10との組み合わせ
現在の臨床上の使用における免疫抑制剤の大部分は、T細胞の活性化を抑制することによって作用し、これにより、移植拒絶を防止する。しかし、これは、適切な状況下としては逆効果かもしれず、T細胞の活性化は、移植特異耐性の発現を促進するプロセスの誘発を招くかもしれない。したがって、その目的が耐性誘発である場合には、免疫抑制剤の使用は、最適でないかもしれない。この現象の明確な証明は、いかなる免疫抑制治療も行うことなしに、同種造血幹細胞移植術後に耐性を獲得したSCID患者によってもたらされる(Bacchettaら、1994)。これらの患者において、ホストアロ抗原に対するTr1細胞特異性を誘導したドナーの存在は、安定な混合キメラ現象、インビボにおける高レベルのIL−10産生及びいかなる免疫抑制治療も受けない正常免疫機能と相関関係がある。これに対して、急性GVHDを管理するために免疫抑制の養生法を受けたBMT患者では、ホストアロ抗原に対するT細胞特異性が誘導されたドナーが検出できたとして、Tr1細胞は末梢血から単離されないであろう(Bacchettaら、1995)。
【0011】
ラパマイシンは、興味深い免疫調節特性を有する新規な化合物を意味する。この理由のため、我々は、同種移植片拒絶を防止し又はタイプ1糖尿病を和らげ、ならびにインビボにおいてTr細胞を発現させるために、ラパマイシンとIL−10とのインビボにおける投与を組み合わせた。
【0012】
従来技術
米国特許第6,277, 635号は、移植による拒絶反応を抑制するためのIL−10の使用に関連する。この特許は、組織の拒絶反応を治療し、阻害する方法を教示し、GVHD及び抗原特異応答を抑制する。さらに、特定の抗原に対するアネルギーを示すT細胞について述べている。
米国特許第6,428, 985号は、ヒトを含む哺乳動物の、IL−10ポリペプチドと少なくとも1つの野生型配列における突然変異体(MutIL−10)とを含む免疫抑制組成物であって、その単独又は他の剤との組み合わせのいずれかで、インビトロ及びインビボで、そのような組成物及びそれらの組み合わせを用いる種々の方法について述べている。用途は、免疫抑制剤及び多くの疾患のための組み合わせ治療、ならびに炎症、移植、線維症、傷に関連する障害、及び腫瘍治療を含む。MutIL−10の作用は、動物実験において示されているが、ヒトの臨床上の設定においては示されていない。
【0013】
米国特許第5,624, 823号は、豚IL−10をコードするDNA、及び異質遺伝子型移植を受ける被移植動物(例えば、霊長類)における耐性の誘導方法を示している。ラパマイシン、サイクロスポリン及びFK506は、「剤を減少させる助け」、例えば、サイトカイン放出を減少する剤として記載されている。豚のIL−10は、胸腺移植のみの文脈で用いられている。
米国特許第6,022, 536号は、自己免疫疾患及びGVHDの治療のための免疫抑制治療として、IL−10及びサイクロスポリンを組み合わせた使用が示されている。低容量のIL−10及びサイクロスポリンならびに医薬担体の相乗効果のある組み合わせが提案されている。
【0014】
米国特許第6,403, 562号は、自己免疫疾患に罹患した又は感受性のある人に対して、IL−10とともにTGF−βを一緒に投与することによって、多発性硬化症のような自己免疫関連疾患の治療方法が示されている。これらのサイトカインは、相乗効果のある方法において、自己免疫疾患に含まれる自己反応性T細胞の活性化を阻害するための抑制因子として作用する。
【発明の開示】
【0015】
発明の要旨
本発明は、T細胞媒介免疫応答の調節における使用のための、特に、それが必要な患者における免疫抑制及び抗原−特異免疫耐性を誘発するためのIL−10及びラパマイシンを含有する医薬製剤を提供する。Tr1及びCD4+CD25+Tr細胞−媒介抗原−特異免疫耐性の誘発は、亢進、機能不全、規制されていない又は管理されていない自己−又は非自己−T細胞−媒介免疫応答を含む病理学的症状の治療に有用である。
【0016】
本発明の好ましい実施形態では、IL−10及びラパマイシンは、同種異系の臓器拒絶;タイプ1糖尿病;乾癬、多発硬化症、炎症性腸疾患、クローン病、慢性関節リウマチを含む自己免疫及び慢性炎症疾患;又はGVHD、喘息、アトピー性皮膚炎のような他のT−細胞媒介疾患;慢性閉塞性肺疾患;ならびに遺伝子治療誘発生成物に対する免疫反応の予防又は治療上の処置における同時の、別々の又は連続的な使用のための組み合わせ製剤の形態である。さらに、肝臓及び肺繊維症を含む繊維増多疾患の治療が認識される。
好ましくは、その組み合わせ製剤は、堅固な同種異系臓器拒絶、特に、同種移植片β−島拒絶及び自己免疫疾患、特にタイプ1糖尿病の予防及び治療上の処置に使用される。
【0017】
その組み合わせ製剤は、ヒト又はウィルス性のIL−10、それらのアナログ、誘導体又ポリエチレングリコール(PEG)抱合IL−10のような、天然分子の生物学的利用率又は生物学的有効性を改善する、それらの抱合体を含んでもよい。IL−10機能的アナログは、IL−10作用を模倣する小分子及びIL−10レセプター又はIL−10融合蛋白に対するモノクローナル抗体(mAbs)を含み、それらはIL−10情報伝達系を誘引する。
【0018】
その組み合わせ製剤は、ラパマイシンアナログ又は誘導体を含んでいてもよい。ラパマイシン及びIL−10、それらの誘導体又はアナログに加えて、その組み合わせ製剤は、さらに免疫抑制剤又は免疫調整剤、モノクローナル抗体又はサイトカインを含んでいてもよい。IL−10及びラパマイシンの組み合わせにおいて使用することができる好ましい生物学的に活性な物質は、a)サイクロスポリン、FK506(タクロリムス)、ピメクロリムスのようなカルシニュリン阻害剤、b)ミコフェノラートのような他の免疫抑制剤、c)CD45の種々のイソ型に対する抗体又はLFA−1のような接着分子、及びd)IL−2受容体α、β及びγ鎖に対する抗体を含む。適当な免疫抑制剤は、IL−2情報伝達系によって作用するそれらを含む(例えば、JAK1及びJAK3ならびにSTAT5阻害剤)。ラパマイシン+抗Tac(IL−2受容体α鎖に対するヒト化抗体)+IL−10の組み合わせは、従来の治療プロトコルによって発生した持続性免疫抑制の代わりに耐性の状態を含むことによって、異質遺伝子型拒絶、特に、同種移植片β−島拒絶のマウスモデルの予防において特に有効であることが示された。さらに、ラパマイシン+IL−10の組み合わせは、自己免疫糖尿病の治療及びNODマウスにおける長期にわたる免疫調節の誘発に有効となることが示された。耐性は、ラパマイシン+IL−10誘発拡大ならびにタイプ1T調節(Tr1)及びCD4+CD25+Tr細胞の区別化の結果として獲得され、それらは、抑制サイトカイン(IL−10及びTGF−β)の生成を含む種々のメカニズム及びT細胞活性化の阻害によって、抗原−特異耐性を媒介する。
【0019】
本発明のラパマイシン+IL−10組み合わせ製剤は、T細胞免疫能力の回復にかかわらず、長期間の予防に能力を発揮し、それは、薬物の使用中止後に維持することができる。
本発明のさらなる実施形態は、IL−10及びラパマイシン、ならびに任意に、免疫抑制剤又は免疫調節剤、モノクローナル抗体及びサイトカインから選択されたさらなる活性成分を、医薬的に許容される賦形剤とともに含む医薬組成物を提供する。適当な医薬組成物は、経口、静脈内、非経口又は皮下経路によって投与され、好ましくは、溶液、懸濁液、注射物質、錠剤又はカプセルの形態である。ラパマイシンの有効量は、0.001mg/kgから100mg/kgの範囲とすることができ、IL−10の有効量は、0.001μg/kgから1000μg/kgの範囲とすることができる。
本発明を、以下の実施例及び添付の図面によってさらに詳細に説明する。
【0020】
図面の説明
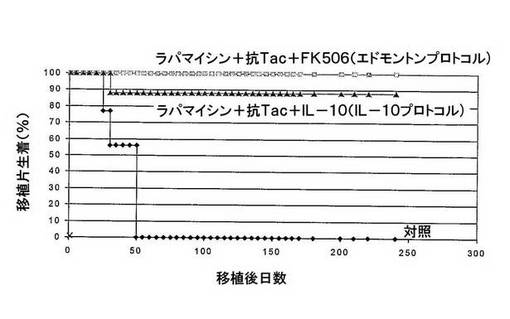
図1 IL−10プロトコル及びエドモントンプロトコルで処置したマウスは、同程度の移植片生着を示す。
ストレプトゾトシン注入によって糖尿病にされたBalb/cマウスに、腎臓被膜の下、純粋な異質遺伝子型C57BL/6β−島を移植した。マウスは、処置なし(対照、n=13マウス)か、30日間、ラパマイシン+抗Tac+IL−10で処置された(IL−10プロトコル、n=16マウス)か、ラパマイシン+抗Tac+FK506で処置(エドモントンプロトコル、n=4マウス)された。移植片生着を、血糖レベルによってモニターした。移植片は、血糖が250mg/dlより高いときに拒絶されているとみなした。
FK506(エドモントンプロトコル)をIL−10(IL−10プロトコル)に置き換えることにより、同等の移植片生着を示した。IL−10プロトコルで処置したマウスにおいて、移植片生着は89%であり、一方、100%の生存がエドモントンプロトコルで処置したマウスで観察された。
【0021】
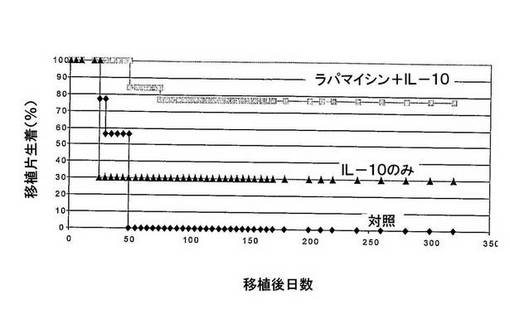
図2 IL−10プロトコルで抗Tacなしとした場合、わずかに異種遺伝子型β−島拒絶が増加した。
ストレプトゾトシン注入によって糖尿病にされたBalb/cマウスに、腎臓被膜下、純粋な異質遺伝子型C57BL/6β−島を移植した。マウスは、処置なし(対照、n=8マウス)か、ラパマイシンとIL−10との組み合わせ(ラパ+IL−10、n=8マウス)又はIL−10のみ(n=4マウス)で処置された。移植片生着を、血糖レベルによってモニターした。
IL−10プロトコルで抗Tacなしとした場合(図1参照)、わずかに移植片生着に影響を及ぼした。ラパマイシンをIL−10と組み合わせた場合、移植片生着は動物の78%で認められた。IL−10単独での処置は、移植片拒絶の予防に有効でなかった。
これらのデータは、抗Tacが同種異系移植拒絶を予防するのに必要でなかったことを示唆する。
【0022】
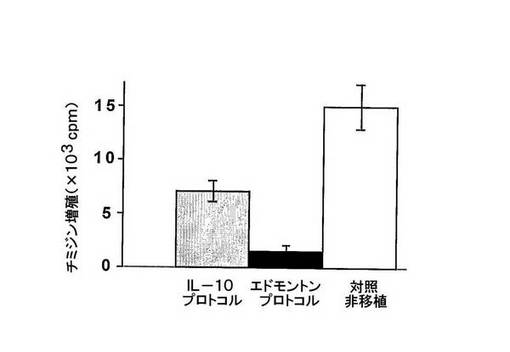
図3 IL−10プロトコルで処置したマウスからのT細胞はインビトロで増殖能を維持する。
対照の非移植マウス(白棒)及びIL−10プロトコル(グレー棒)又はエドモントンプロトコル(黒棒)で処置したマウスからのT細胞を脾臓から単離し、インビトロで、抗CD3及び抗CD28mAbsで多クローン的に刺激した。エドモントンプロトコルで処置したマウスからの細胞は、インビトロでの増殖能においてそれらを強く減少させ、一方、増殖におけるマイルドな減少のみが、IL−10プロトコルで処置したマウスから単離したT細胞において観察された。
これらのデータは、IL−10プロトコルで処置したマウスからのものはそうでないが、エドモントンプロトコルで処置したマウスから単離したT細胞において、免疫抑制が意味深い状態であることを示唆する。
【0023】
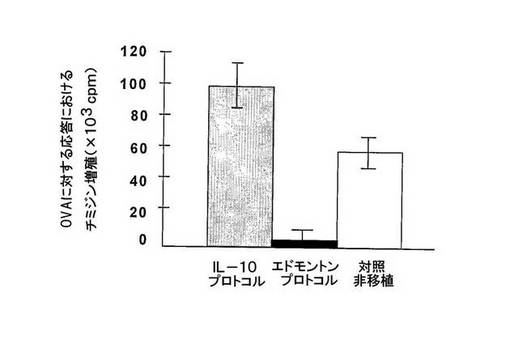
図4 IL−10プロトコルで処置したマウスからのT細胞は、抗原−特異増殖能を防止する。
移植280日前およびIL−10プロトコル(グレー棒)又はエドモントンプロトコル(黒棒)で30日間のみ処置したマウスに対して、CFA+OVAで、後足裏のやわらかい部分にインビボにおいて免疫化した。リンパ節流を採取し、OVA及び自己APCによりインビトロで再刺激した。
OVA−特異T細胞増殖は、エドモントンプロトコルで処置したマウスにおいて非常に減少し、一方、IL−10プロトコルで処置したマウスにおけるOVA−応答は、非移植免疫化マウスで観察されたのと同等であった。
これらのデータは、さらに、IL−10プロトコルではそうではないが、エドモントンプロトコルで処置したマウスにおいて、免疫抑制が一般状態であることを証明する。
【0024】
図5 IL−10プロトコルで処置したマウスから単離したT細胞はIL−10を産生する。
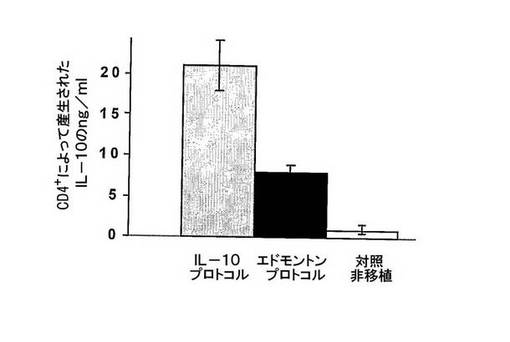
対照の非移植マウス又はIL−10プロトコル(グレー棒)及びエドモントンプロトコル(黒棒)で処置したマウスの腎臓から単離したCD4+T細胞を、抗CD3及び抗CD28mAbsで、インビトロで刺激した。刺激後96時間に上清を採取し、IL−10産生をELISAで評価した。
IL−10プロトコルで処置したマウスからのT細胞は、エドモントンプロトコルで処置したマウス及び対照の非移植マウスと比較して、高いレベルのIL−10を産生した。
【0025】
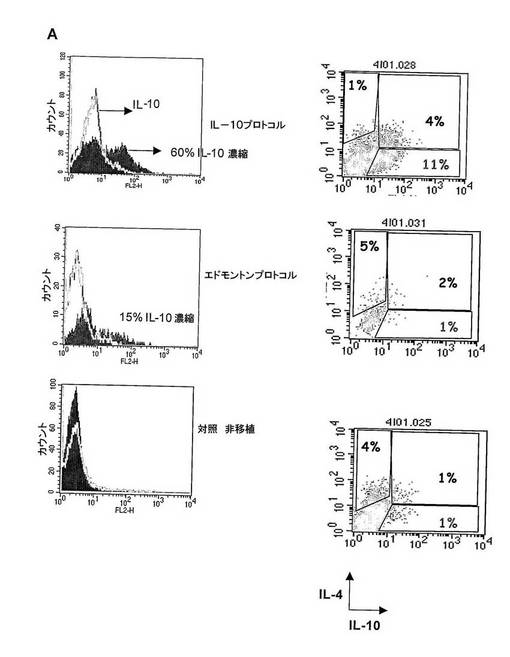
図6 IL−10産生T細胞の特異集団は、IL−10プロトコルで処置したマウスから単離することができる。
(A)IL−10プロトコル、エドモントンプロトコルで処置したマウス及び対照の非移植マウスからのT細胞を単離し、インビトロで多クローン的に刺激し、サイトカイン産生を誘導した。3時間後、細胞を、偏在細胞表面マーカーに結合する一つのmAb及びIL−10を捕捉できる他のmAbとからなるダイアボディで標識した。次いで、多クローン刺激の間に蓄積されたサイトカインを放出させるために、標識細胞を37℃でさらに1時間インキュベートした。標識細胞によって産生されたIL−10をダイアボディによって捕獲した。さらに、細胞を、PEで標識された抗IL−10mAbで標識した。IL−10+濃縮(密ヒストグラム)及びIL10-(空ヒストグラム)細胞を磁気で分離するために、抗−PEマイクロビーズを用いた。IL−10+濃縮T細胞の特異集団を、IL−10プロトコルで処置した移植マウスからのみ単離した。
【0026】
(B) 細胞質内の染色を、この特異IL−10+濃縮T細胞に行い、Tr1サイトカインプロファイル(つまり、IL−10+、IL−4-)を有する細胞の有意な集団を、エドモントンプロトコルで処置したマウス又は対照の非移植マウスではなく、IL−10プロトコルで処置したマウスにおいて確認した。
【0027】
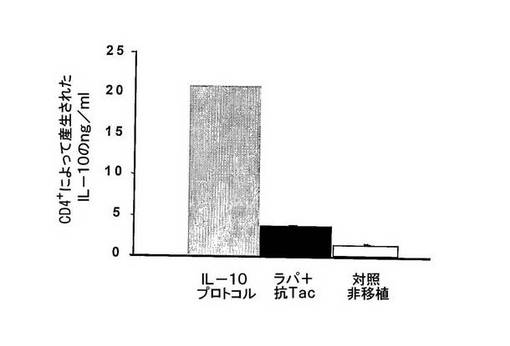
図7 IL−10は、インビボでのIL−10産生T細胞を誘発するために必要である。
インビボでIL−10+Trl細胞を誘発するためにIL−10投与が必要であることを理解するために、移植マウスを、IL−10プロトコル(ラパマイシン+抗Tac+IL−10)又はラパマイシン+抗Tacのみで処置した。次いで、マウスの2つの群におけるCD4+脾臓T細胞によるIL−10の産生を評価した。
IL−10プロトコル(グレー棒)又はラパマイシン+抗Tac(黒棒)で処置したマウス又は対照の非移植(白棒)のマウスからのT細胞を単離し、インビトロで、多クローン的に、抗CD3及び抗CD28mAbsで刺激した。
IL−10のかなりのレベルが、IL−10プロトコルで処置されたマウスから単離された細胞によってのみ産生した。
これらのデータは、IL−10が、インビボでIL−10産生細胞を誘発するために必要であることを示唆する。
【0028】
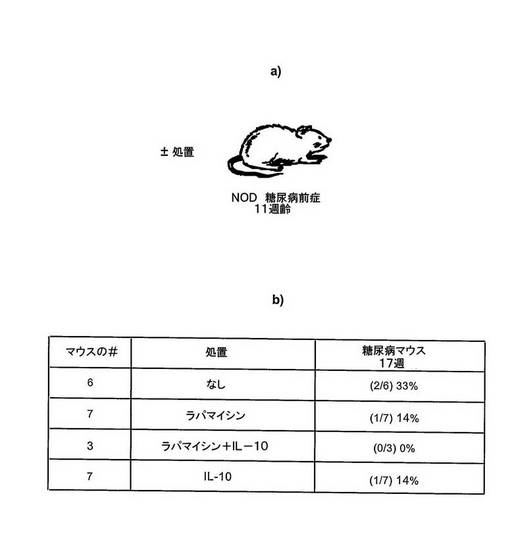
図8 NODマウスにおけるタイプI糖尿病の治療のためのラパマイシン+IL−10での予備的結果
11週齢のNODマウスは、糖尿病前症の段階である。これらのマウスはインスリン炎及びすい臓における浸潤性自己免疫T細胞を有しているが、それらは、いまだ血糖正常とする十分なインスリンを産生することができるの十分に正常なβ−島を有している。
糖尿病前症マウスを、11週齢からはじめて毎日、ラパマイシン、ラパマイシン+IL−10又はIL−10単独のいずれかで処置した。処置後6週間で、対照マウスの33%が糖尿病を発症したが、ラパマイシン及びIL−10で処置したマウスは、全て血糖正常のままであった。
これらの予備的結果は、ラパマイシン+IL−10は、その初期段階において糖尿病をブロックするために使用することができ、さらに進行した自己免疫糖尿病の自発性の発症を予防することを示唆する。
【0029】
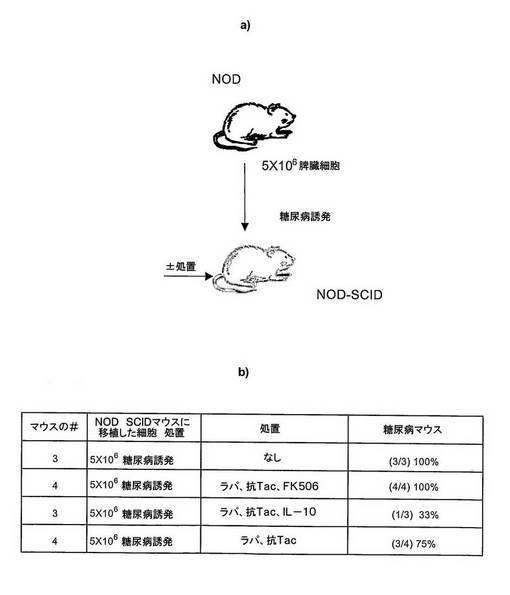
図9 IL−10プロトコルは、糖尿病誘発性T細胞の移植後のNOD.SCIDマウスにおいて誘発した糖尿病を阻害する。
NOD糖尿病マウスからの脾細胞5×106をNOD.SCIDマウスに静脈内移植した。被移植マウスを、移植後40日間、処置しないか、エドモントンプロトコル(ラパマイシン+抗Tac+FK506)、IL−10プロトコル(ラパマイシン+抗Tac+IL−10)又はラパマイシン+抗Tacのいずれかで処置した。
移植後50日で、全ての対照の被処置マウスは糖尿病になった。
エドモントンプロトコルで処置した全てのマウス及びラパマイシン+抗Tacで処置した75%のマウスは、糖尿病になった。興味深いことに、IL−10プロトコルで処置した33%のみのマウスが糖尿病になった。
これらの予備的データは、IL−10プロトコルは、自己免疫糖尿病誘発性NODT細胞の移植によってNOD.SCIDマウスにおいて誘発されたタイプI糖尿病を阻害する。
【0030】
図10 タイプI糖尿病の治療のためのラパマイシン+IL−10の使用
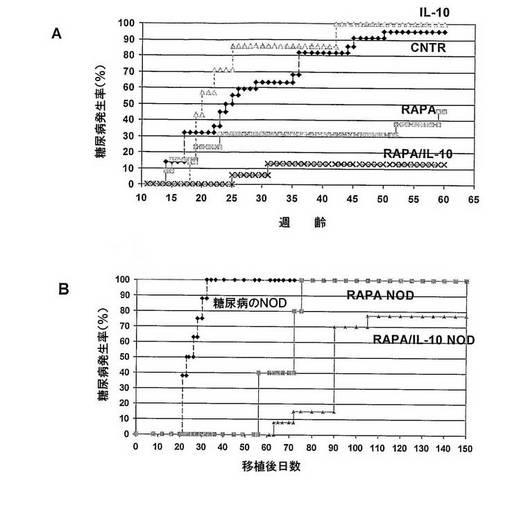
A)NODマウスにおける糖尿病の治療
NODマウスを11週齢から31週齢まで、IL−10(IL−10、n=7マウス)又はラパマイシン(RAPA、n=13マウス)又はラパマイシン+IL−10(RAPA/IL−10、n=16マウス)又は賦形剤(CNTR、n=22マウス)で処置した。血糖レベルによってモニターした糖尿病発生率は、少なくとも60週齢まで安定した。ラパマイシン単独投与では、糖尿病発生率が95%から46%に減少した。IL−10投与では、糖尿病の発症に顕著な効果はなかった。ラパマイシンの予防効果は、IL−10を処置に加えた場合に、顕著に改善され、糖尿病発生率を13%に減少した。
【0031】
B)NOD.SCIDマウスにおける糖尿病の移植に対する処置NODマウスからの脾細胞の能力
非処置糖尿病NODマウス(DIABETIC NOD、n=8)又はラパマイシンで処理したマウス(RAPA NOD、n=5)又はラパマイシン+IL−10(RAPA/IL−10 NOD、n=13)からの総脾細胞5×106を、NOD.SCIDマウスに移植し、糖尿病発生率を血糖レベルによってモニターした。
ラパマイシン処置マウスからの脾細胞の移植は、糖尿病NODマウスからの脾細胞を注入されたマウスに比べて、病気の発症を顕著に遅延させた。重要なことに、ラパマイシン+IL−10で処置されたマウスからの脾細胞も同様に、さらに糖尿病伝達を遅らせた。
【0032】
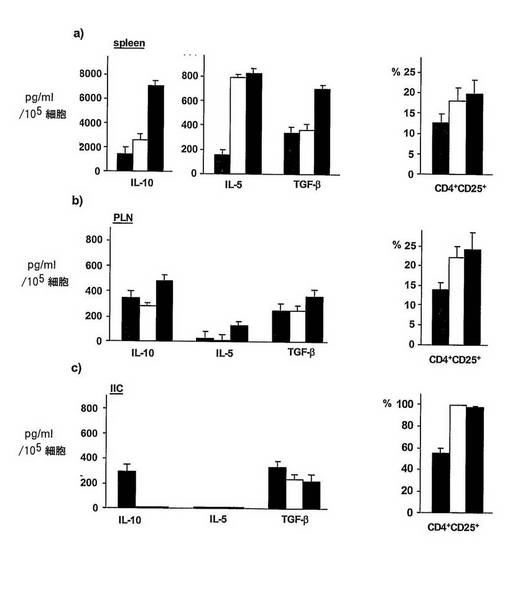
図11 ラパマイシン±IL−10で処置したマウスにおけるTr細胞の濃度
CD4+T細胞によって産生されたサイトカイン(左パネル)及びCD4+CD25+T細胞のパーセント(右パネル)を、非処置糖尿病NODマウス(グレー線)、ラパマイシン処置マウス(白棒)又はラパマイシン+IL−10処置マウス(黒棒)の脾臓(上部パネル)、膵臓のリンパ節(PLN)(中段パネル)及び島浸潤性細胞(IIC)(下部パネル)において、それぞれCBA及びFACによって評価した。それらのサイトカイン産生プロファイル(つまり、IL−10++IL−15+TGF−β+)によって測定したように、CD4+Tr1細胞の高い割合が、ラパマイシン+IL−10(黒棒)で処置した耐性マウスの脾臓にのみ存在した。CD4+CD25+T細胞のパーセントは、ラパマイシン単独又はラパマイシン+IL−10(黒棒)で処置したマウスの双方の脾臓、PLM及びIICにおいて高かった。したがって、ラパマイシン+IL−10処置マウスにおいて、Tr1細胞は脾臓において存在し、CD4+CD25+Tr細胞は、脾臓、リンパ節及び膵臓において存在した。
【0033】
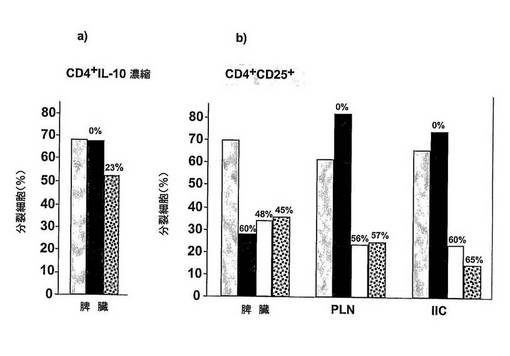
図12 インビトロにおける免疫応答を抑制するTr細胞の能力
インビトロにおいて、CFSEで標識し、抗CD3mAbの存在中で培養したCD4+の実験に未使用のNODT細胞の増殖におけるTr細胞の抑制活性を試験した。CD4+IL−10濃縮脾臓T細胞(純度40%以下)(左パネル)又はCD4+CD25+T細胞(MAC精製、純度75%以上)(右パネル)のいずれかを、実験に未使用のT細胞と同じ数で加えた抑制細胞として用いた。何も加えない細胞(グレー棒)に区分した実験に未使用のT細胞を対照として用いた。非処置糖尿病NODマウス(黒棒)又はラパマイシン処置マウス(白棒)又はラパマイシン+IL−10処置マウス(点線)から単離されたCD4+IL−10濃縮Tr1細胞又はCD4+CD25+Tr細胞の存在における細胞区分を評価し、対照に対する抑制パーセントを測定した(各ヒストグラムの上部における数)。ラパマイシン+IL−10処置マウスの脾臓からのTr1細胞は、NODマウスから得られたCD4+T細胞の増殖応答を緩やかに抑制する。強力な抑制が、ラパマイシン及びラパマイシン+IL−10処置マウスの双方のPLN及びIICから単離されたCD4+CD25+T細胞で観察された。
【実施例】
【0034】
実施例1
1.同種異系β−島移植
完全不一致のマウス島同種異系移植のモデル(Balb/CへのC57BL/6)を用いた。同種異系すい臓β−島移植は、インスリン置換療法又はタイプ1糖尿病の治療のためのすい臓移植への有効な代替となっている。過去数年のうちに、ヒトβ細胞の単離及び保存のための改善された方法及び新たな免疫抑制剤の開発が、これらの移植の臨床成果を顕著に改善している。特に、ラパマイシン+抗Tac+FK506(エドモントンプロトコル)に基づく新たなステロイドフリーの免疫抑制処方が、近年、移植後1年で、患者の80%において、インスリン非依存をもたらすことが示されている(Shapiroら、2000)。これらの結果は、全ての先の免疫抑制剤の組み合わせ治療で得られたものをはるかにしのいでいる。しかし、この処方が耐性を誘発し得るという証明は行われていない。
【0035】
重要なことは、アポトーシスの防止及びT細胞発現の阻害のために、FK506の作用メカニズムが耐性誘発の状態を防止するかもしれないことである。
寛容原性プロトコルの開発の努力において、我々は、エドモントンプロトコルにおけるFK506をIL−10によって置換した(つまり、IL−10プロトコル、ラパマイシン+抗Tac+IL−10)処方を設計した。
ストレプトゾトシン注入によって糖尿病にされたBalb/cマウスに、純粋な異質遺伝子型C57BL/6β−島を、腎臓被膜下、移植した。細胞片生着は、IL−10プロトコル及びエドモントンプロトコルで処置したマウスにおいて、それぞれ同様であった(移植後240日で89%及び100%)(図1)。
【0036】
抗Tacのインビボにおける使用は、高いIL2Rα鎖発現を伴う活性T細胞をブロックするための必要性によって、過去数年、強く支持されている。
しかし、Tr細胞のサブセットは、IL2Rα鎖(つまり、CD4+CD25+Tr細胞)を構成的に発現させ、このT細胞集団が同種異系移植拒絶を抑制し得ることは広く証明されている(Taylorら、2002)。
【0037】
したがって、CD25+T細胞集団をブロックするmAbの使用は、インビボにおいて、Tr細胞によって媒介される耐性誘導が求められる場合には、逆指示(counterindicated)されるかもしれない。この理由に対して、抗Tacを治療プロトコルからの除外することを評価した。我々は、島拒絶を十分に防止し得るか否かを測定するために、ラパマイシン+IL−10又はIL−10単独で30日間、移植マウスを処置した(図2)。長期間の移植片生着が、IL−10単独で処置したマウスの30%で観察され、ラパマイシン+IL−10で処置したマウスにおいては78%に増加した。この移植片生着のレベルは、さらに抗Tacで処置したマウスのそれよりもわずかに低いのみであった(図1に示す)。これらのデータは、IL−10プロトコルから抗Tac mAbを不在とすることにより、同種異系移植片生着を可能にし、一方、CD4+CD25+Tr細胞は影響を受けず、インビボにおいてこれら細胞による耐性誘発を保つことを示す。
【0038】
所望の成果の一つが免疫抑制よりもむしろ耐性であるため、我々は、IL−10及びエドモントンプロトコルで処置したマウスからのT細胞が多クローナル及び抗原特異刺激に対して応答したかどうかを試験した。まず、T細胞を移植後240日(治療の中断後210日)でマウス脾臓から単離し、抗CD3及び抗CD28mAbで刺激した(図3)。エドモントンプロトコルで処置したマウスからのT細胞の増殖は、対照の非移植マウスからのT細胞のそれと比較した場合、強く抑制された。T細胞増殖の抑制は、IL−10プロトコルで処置したマウスでは、それほど強くなかった(図3)。次いで、移植後280日(治療の中断後250日)のマウスを、後足裏の柔らかいところに、CFA+OVAで免疫化し、リンパ節流から単離されたT細胞の増殖応答を測定した(図4)。エドモントンプロトコルで処置したマウスからのT細胞は、OVAに対する応答において増殖しなかったが、一方、IL−10プロトコルで処置したマウスからのT細胞は、非移植免疫化対照マウスと同様の応答であった(図4)。
【0039】
FK506のIL−10による置換が潜在的にTr1細胞拡大を促進するかどうかを測定するために、島移植の部位に浸潤するCD4+T細胞を、移植後200日のマウスから単離し、それらのサイトカイン産生を調べた。IL−10プロトコルで処置したマウスから単離されたCD4+T細胞は、エドモントンプロトコルで処置されたマウスよりも、抗CD3及び抗CD28mAbで刺激した後、IL−10のかなり高い量を産生した(図5)。次いで、精製脾臓T細胞を、抗CD3及び抗CD28mAbで刺激し、IL−10分泌細胞を、IL−10捕捉ビーズを用いて濃縮した(図6A)。IL−10/IL−4サイトカインプロファイルを、細胞質内染色によって試験した(図6B)。興味深いことに、IL−10+IL−4-細胞の特異集団(つまり、Tr1細胞のサイトカインプロファイルを反映する)が、IL−10プロトコルで処置したマウスにおいてのみ確認された。
【0040】
IL−10を含まない、ラパマイシン+抗Tacで処置されたマウスからの浸潤細胞は、IL−10を産生しなかった。これは、インビトロにおいて増加したIL−10産生は、インビボにおけるIL−10の投与のためであったことを示す(図7)。
【0041】
まとめると、これらのデータは、
1.ラパマイシン+抗Tac+FK506の組み合わせ(エドモントンプロトコル)は、同種異系拒絶からマウスを保護するが、長期の慢性免疫抑制の状態を誘発する、
2.ラパマイシン+抗Tac+IL−10の組み合わせ(IL−10プロトコル)は、同種異系拒絶に対する長期保護を与える。この処置は、Tr1細胞と一致するサイトカインプロファイルを伴うT細胞の特異集団の拡大をもたらし、保護は、T細胞免疫能力の回復にもかかわらず、薬物の中断後まで維持する。
【0042】
実施例2
1.タイプI糖尿病における予備的な結果
ラパマイシン+IL−10の寛容原性作用も、タイプI糖尿病の設定において評価した。
我々は、β細胞破壊の防止(それは、タイプI糖尿病への進行を伴い、疾患の発症において見られる)が、
1.すい臓内での一般的な「バイスタンダー」炎症の抑制、
2.島特異Tエフェクター細胞拡大のブロック、
3.抗原特異Tr細胞の誘発及び拡大
によって防止することができると考えている。
【0043】
我々の提案するプロトコルでは、炎症の抑制は、IL−10によって達成すべきであり、Tエフェクター細胞拡大のブロックは、ラパマイシンによって達成すべきである。IL−10及びラパマイシンも、T細胞プライミングを防止せず、したがってそれらは抗原特異T調節細胞の誘導を可能にするはずであり、以下に説明するように、IL−10はTr1細胞の誘導及び拡大を促進するはずである。我々は、タイプI糖尿病のNODマウスのモデルにおける自己免疫を処置することにおいて、ラパマイシン単独又はIL−10との組み合わせの効果を調査した。NODマウスは、島β−細胞の破壊及び血中グルコースの上昇を伴って、15から30週齢で明白な疾患を発現し、多くの主要な特性をヒトの疾患と共有している(Tischら、1996、Delovitchら、1997)。タイプI糖尿病の阻害を、ラパマイシン、ラパマイシン+IL−10又はIL−10単独で、11週齢(つまり、ペリ膵島炎(periinsultis)を伴う糖尿病前症のマウス)から毎日、NODマウスを処置することによって評価した。処置後6週間で、非処置対照マウスの33%が糖尿病を発症しはじめ、一方、ラパマイシン+IL−10で処置したマウスはなおも全て正常血糖であった(図8)。
【0044】
また、我々のプロトコルの有効性を、養子免疫細胞移入NOD.SCIDマウスのモデルでも試験し、それは、内因性のT及びB細胞を欠いており、したがって、自発性の糖尿病を発現せず、糖尿病のNODマウスからの5×106脾細胞の移入後15から20日で糖尿病を発現する。NOD.SCID被移植マウスを、処置しないか、あるいはラパマイシン+抗Tac+FK506(エドモントンプロトコル)、ラパマイシン+抗Tac+IL−10(IL−10プロトコル)及びラパマイシン+抗Tacで糖尿病の移入後40日間処置した(図9)。対照マウスは、移入後15日で糖尿病を発症し始めた。エドモントンプロトコルで処置した全てのマウス及びラパマイシン+抗Tacで処置した75%のマウスは、移入後33日以内に糖尿病になった。興味深いことに、IL−10プロトコルで処置したうちの1匹のマウスのみ(33%)が、移入後35日で糖尿病になった。
これらの予備的データの全体から、ラパマイシン+IL−10の使用によって、タイプI糖尿病の発現を完全に阻害することは当然のことである。
【0045】
2.ラパマイシン+IL−10の処置は、自己免疫糖尿病を阻害し、長期間の耐性を誘発する。NODマウスモデルにおいて得られた有望な予備的結果に基づいて(図8)、我々は、ラパマイシン±IL−10で、11週齢(膵臓細胞自己免疫が、膵島炎及び自己インスリン抗体によって判断されるように明らかに確立された時点)から、20週間NODマウスを処置した。ラパマイシン単独の投与は、糖尿病の発生率を95%から46%に減少させた(図10A)。先の観察は、NODマウスにおけるIL−10処置の効果は、経路、用量及び投与のタイミングに依存して変化することを示す(Roncaroloら、2003)。しかし、ここで、我々は、同じ期間にわたるIL−10単独の投与は、糖尿病の発現において顕著に作用しなかったことを示す。ラパマイシンの防御効果は、IL−10が治療に加えられた際、顕著に改善され、さらに、糖尿病の発生率を13%に減少させた(図10A)。興味深いことに、防御は、治療を中止した後30週間さらに維持され、長期免疫調節が確立されたことを示す。
【0046】
さらに、ラパマイシン又はラパマイシン+IL−10が自己免疫糖尿病の発現を防止することによるメカニズムを、耐性マウスからの細胞の移植実験で調査した。免疫不全NOD.SCIDマウスにおける非処置糖尿病NODマウスからの脾細胞を移植することによって、糖尿病が迅速に誘発され、一方、ラパマイシン処置マウスからの脾細胞を移植することによって、疾患の発症は顕著に遅延した。興味深いことに、ラパマイシン+IL−10の組み合わせで処置したマウスからの脾細胞を移植することでも、さらに、糖尿病移植を遅延した(図10B)。これらのデータは、ラパマイシンで処置することは、脾臓の自己反応性T細胞における糖尿病を伝達する能力を抑制し、この作用は、IL−10が処置に加えられた際に強力に高められることを示す。
【0047】
長期耐性に横たわるメカニズムを、50週齢以上の耐性マウスで分析した。非処置糖尿病NODマウス又はラパマイシン+IL−10処置マウスからの脾臓細胞は、同程度の細胞数を含み、CD4+及びCD8+T細胞と同じ割合であるが、それらのサイトカイン産生プロファイルは制限される。CD4+Tr1細胞の高い割合は、それらのサイトカイン産生プロファイル(つまり、IL−10++IL−5+TGF−β+)によって示されるように、ラパマイシン+IL−10で処置された耐性マウスの脾臓において存在するが、ラパマイシン単独で処置されたマウス又は非処置糖尿病NODマウスの脾臓において存在しない(図11)。しかし、IL−4を産生する脾臓のCD4+T細胞の割合は、非処置及び処置マウスの双方において同じである。さらに、脾臓CD4+CD25+T細胞のパーセントは、非処置糖尿病NODマウスに比較して、ラパマイシン単独又はラパマイシン+IL−10で処置したマウスの双方において高い(図11)。反対に、Tr1細胞は、膵臓リンパ節(PLN)及び島浸潤性細胞(IIC)において検出することができず(図11)、Tr1細胞は自己免疫部位に存在しないことを示す。一方、CD4+CD25+T細胞は、PLNにおいて多数観察され、ラパマイシン単独又はラパマイシン+IL−10のいずれかで処置されたマウスのIICから単離されたCD4+T細胞のほぼ100%を示すが、非処置糖尿病マウスでは示さない(図11)。IICからのこれらCD4+CD25+T細胞は、アレルギー性であり、TGF−βは別として、サイトカインを有意なレベルで産生しない。
【0048】
次いで、我々は、ラパマイシン+IL−10処置マウスにおいて存在するTr1細胞及びラパマイシン及びラパマイシン+IL−10処置マウスの脾臓、PLN及びIICからのCD4+CD25+T細胞が、インビトロにおいて抑制活性を有するか否かを測定した。ラパマイシン+IL−10処置マウスの脾臓からのTr1細胞は、NODマウスから得られたCD4+T細胞の増殖応答を緩やかに抑制する(図12)。また、抑制は、処置及び非処置糖尿病NODマウスの双方の脾臓から精製されたCD4+CD25+T細胞で観察され(図12)、それは、CD4+CD25+Tr細胞は、また糖尿病NODマウスの脾臓において存在するが、非常に低い頻度であることを示す(図11に示す)。興味深いことに、強い抑制は、ラパマイシン又はラパマイシン+IL−10処置マウスの双方のPLN及びIICから単離されたCD4+CD25+T細胞で観察された。反対に、非処置糖尿病NODマウスから単離されたCD4+CD25+T細胞は、測定可能ないかなる抑制活性を有さなかった(図12)。これらのデータは、糖尿病NODの膵臓組織は、Tr細胞よりもむしろ、活性化Tエフェクター細胞を主に含み、一方、処置マウスのPLN及びIICは、主にCD4+CD25+サブセットのうちTr細胞を含むことを示す。
【0049】
概して、これらのデータは、ラパマイシン+IL−10処置の後に観察される定常状態の耐性は、脾臓におけるTr1細胞ならびにリンパ節及び膵臓におけるCD4+CD25+Tr細胞の蓄積に関連することを示す。
【0050】
材料及び方法
マウス Balb/c、C57BL/6、NOD/Lt及びNOD.SCIDの雌性マウスをCharles River Laboratories (Calco, Italy)から購入した。全てのマウスは、特定の病原体フリーの状態で保持した。尾の静脈血におけるグルコースレベルをGlucometer Elite system (Bayer, Wuppertal, Germany)を用いて定量した。糖尿病を、ストレプトゾトシン(シグマ、セントルイス、MO)170mg/kgの静脈内注入によって、Balb/cマウスで誘発した。糖尿病の診断を、250mg/dlより高い2回の連続的なグルコース測定の後に行った。
【0051】
島移植 厳選されたC57BL/6の膵臓島を、37℃にて一晩培養した後、先に述べたように、Balb/c糖尿病マウスの腎臓膜の下に移植した(300島/マウス)(Davalliら、1996)。
【0052】
移植マウスの処置 移植されたBalb/cマウスの処置を、移植して30日後の日に始めた。ラパマイシン(Rapamune, Wyeth-Ayerst Research, Pearl River, NY)をピーナツ油(シグマ)で希釈し、胃管栄養法によって1mg/kgの用量で一日1回投与した。ヒトIL−10(BD Biosciences, Mountain View, CA)をPBSで希釈し、0.05mg/kgの用量で、腹腔内に1日2回投与した。FK506(Prograf, Fujisawa, Milano)を生理食塩水で希釈し、0.3mg/kgの用量で、腹腔内に1日1回投与した。抗IL−2Rα鎖mAb(抗Tac)(クローン7D4、BD)を生理食塩水で希釈し、1mg/マウスの最終用量になるように、移植後0及び4日目に、腹腔内に投与した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0053】
糖尿病阻害研究 雌性NODマウスを、11週齢から31週齢まで、移植マウスで用いたのと同じ用量で用いて、ラパマイシン、ラパマイシン+IL−10又はIL−10単独で処置した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0054】
糖尿病移植研究 糖尿病NOD雌性マウスから脾細胞を採取し、NOD.SCIDにおいて5×106/マウスの用量で、静脈内に注入した。移植マウスにおいて使用されたのと同じ用量で、被移植マウスを、処置しないか、移植後40日間、ラパマイシン+抗Tac+FK506又はラパマイシン+抗Tac+IL−10、又はラパマイシン+抗Tacで処置した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0055】
NOD.SCIDマウスにおける養子免疫細胞移入 対照又は処置NODマウスからの脾臓を、治療の中止後採取した。500万の総脾細胞を、NOD.SCIDマウスに静脈内注入によって養子免疫細胞移入した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0056】
インビボにおける免疫化 CFA(Difco, Detroit, MI)に乳化したオバアルブミン(OVA)ペプチド323−339(Primm, Milano, Italy)を移植Balb/cマウスの後足裏の柔らかい部位の皮下に、100μg/マウスの用量で、一回注入した。リンパ節流を採取し、インビトロアッセイに用いた。
【0057】
細胞選別 浸潤性膵臓の細胞を、説明されたように単離した(Gregoriら、2003)。得られた細胞集団を、抗CD90mAB−被覆マイクロビーズとともにインキュベートし、精製T細胞を得るためにMiniMacs カラム(Miltenyi Biotec, Bergisch Gladbach, Germany)に付した。CD4+CD25+T細胞を、Multisortキット(Miltenyi)で選別した(平均精製度≧75%)。いくつかの実験において、CD4+CD25+T細胞を、無菌でFACStar細胞ソーター(BD)で選別した(平均精製度99%)。IL−10産生細胞を、ネズミIL−10分泌アッセイ濃縮及び検出キット(Miltenyi)で選別した(平均精製度≧40%)。
【0058】
IL−10陽性細胞の濃縮 IL−10産生細胞を、市販のキット(Miltenyi)によって濃縮した。精製T細胞を、不動状態の抗CD3及び溶解抗CD28の存在下、106/mlの濃度で培養した。培養の3時間後、細胞を回収し、CD45及び他のmAb捕獲ネズミIL−10に対して指向するmAbからなるダイアボディで、4℃にて10分間標識した。次いで、細胞を105/mlの最終濃度で希釈し、37℃で45分間サイトカインを分泌させた。サイトカイン捕捉期間の後、細胞を回収し、0.5%のBSA及び5nMのEDTA(バッファ)を含有するPBSに108/モル再懸濁し、PE抱合αIL−10mAb(BD)で4℃にて10分間染色した。細胞を1回バッファで洗浄し、108/モル再懸濁し、抗−PEマイクロビーズで4℃にて10分間染色した。IL−10濃縮細胞集団を磁気カラムで単離した。細胞サンプルをFACSカリバー・フローサイトメトリー(BD)で分析した。
【0059】
細胞培養 抑制実験のために、実験に使用していないCD4+NODのT細胞を、他で述べたように(Lyonsら、1994)、CFSE(Molecular Probes、Eugene、OR)で染色し、96ウェルプレート(1×105ウェル)中にて、抗CD3mAb(BD)10μg/mlの存在下で培養した。ラパマイシン又はラパマイシン+IL−10で20週間処置したNODマウスから得られたCD4+T細胞を、1:1の割合で培養液に加え、分裂した実験に使用していない細胞のパーセントを評価し、なんら加えられていない細胞における分裂細胞のパーセントと比較した。分裂細胞を、総事象CFSE+によって増殖集団に含有された分裂事象によって評価した。媒体中に放出されたサイトカインの測定のために、精製T細胞(1×105/ウェル)を、96ウェルプレートで培養し、10μg/mlの免疫化抗CD3(BD)及び1μg/mlの溶解抗CD28(BD)で刺激した。培養の48時間(IL−5検出のために)及び96時間(IL−10及びTGF−β検出のために)後に上清を採取した。
【0060】
フローサイトメトリー 説明したAbs(全てBDから)で細胞を染色し、CellQuest ソフトウェア (BD)を装備したFACスキャンフローサイトメータで分析した。
【0061】
サイトカイン測定 採取された上清中に存在するサイトカインを、標準的な市販のキット(BD)を用いて、サンドイッチERISA又はフローサイトメトリー原理のアッセイ(CBA)によって定量した。特定のサイトカインを産生する細胞のパーセントを細胞内染色によって測定した。精製T細胞を6時間、10μg/mlの免疫化抗CD3及び1μg/mlの溶解抗CD28(BD)で、1×106/mlの濃度で、染色した。ブレフェドリンAを培養の最後の3時間添加した。細胞内染色を、先に述べたように(Trembleauら、2000)行った。
【図面の簡単な説明】
【0062】
【図1】IL−10プロトコル及びエドモントンプロトコルで処置したマウスの移植片生着を示す。
【図2】IL−10プロトコルで抗Tacなしとした場合の異種遺伝子型β−島拒絶の増加を示す。
【図3】IL−10プロトコルで処置したマウスからのT細胞について、インビトロでの増殖能を示す。
【図4】IL−10プロトコルで処置したマウスからのT細胞についての抗原−特異増殖能を示す。
【図5】IL−10プロトコルで処置したマウスから単離したT細胞について、IL−10の産生を示す。
【図6】IL−10産生T細胞の特異集団について、IL−10プロトコルで処置したマウスからの単離を示す。
【図7】IL−10によるインビボでのIL−10産生T細胞の誘発を示す。
【図8】NODマウスにおけるタイプI糖尿病の治療のためのラパマイシン+IL−10での予備的結果を示す。
【図9】IL−10プロトコルが、糖尿病誘発性T細胞の移植後のNOD.SCIDマウスにおいて誘発した糖尿病を阻害した実験結果を示す。
【図10】タイプI糖尿病の治療のためのラパマイシン+IL−10の使用効果を示す。
【図11】ラパマイシン±IL−10で処置したマウスにおけるTr細胞の濃度を示す。
【図12】インビトロにおける免疫応答を抑制するTr細胞の能力を示す。
【0063】
参考文献
Bacchetta, R. , M. Biglerら (1994)."High levels of interleukin 10 production in vivo are associated with tolerance in SCID patients transplanted with HLA mismatched hematopoietic stem cells. Growth and expansion of human T regulatory type 1 cells are independent from TCR activation but require exogenous cytokines. "Journal of Experimental Medicine 179 (2): 493- 502.
Bacchetta R. , Parkman R. ら(1995) "Dysfunctional cytokine production by host-reactive T-cell clones isolated from a chimeric severe combined immunodeficiency patient transplanted with haploidentical bone marrow." Blood 85: 1944-1953.
Baker, K. S. , M. G. Roncaroloら(1999). "High spontaneous IL-10 production in unrelated bone marrow transplant recipients is associated with fewer transplant-related complications and early deaths. "Bone Marrow Transplant 23 (11): 1123-9.
Battaglia, M. , B. R. Blazarら(2002). "The puzzling world of murine T regulatory cells. "Microbes Infect 4 (5): 559-66.
Blaha, P. Bigenzahn, S. ら(2003). "The influence of immunosuppressive drugs on tolerance induction through bone marrow transplantation with costimulation blockade. "Blood 101: 2886-2893.
Blazar, B. R., P. A. Taylorら(1998). "Interleukin-10 dose-dependent regulation of CD4+and CD8+T cell-mediated graft-versus-host disease." Transplantation 66 (9): 1220-9.
Davalli, A. M. , L. Scagliaら(1996). "Vulnerability of islets in the immediate posttransplantation period. Dynamic changes in structure and function. "Diabetes 45 (9): 1161-7.
Delovitch, T. L. , B. Singh. (1997) "The nonobese diabetic mouse as a model of autoimmune diabetes: immune dysregulation gets the NOD"Immunity 6 (7): 727-738.
Gregori S. , Giarratana N. ら(2003) "Dynamics of pathogenic and suppressor T cells in development of autoimmune diabetes"J. Immunol 171: 4040-4047.
Groux, H. , A. O'Garraら(1997). "A CD4+T-cell subset inhibits antigenspecific T-cell responses and prevents colitis. "Nature 389 (6652): 737-42.
Hackstein, H. , T. Tanerら(2002). "Rapamycin inhibits macropinocytosis and mannose receptor-mediated endocytosis by bone marrow-derived dendritic cells. "Blood 100 (3): 1084-7.
Hempel, L. , D. Korholzら(1997). "High interleukin-10 serum levels are associated with fatal outcome in patients after bone marrow transplantation." Bone Marrow Transplant 20 (5): 365-8.
Hojo, M. , T. Morimotoら(1999). "Cyclosporine induces cancer progression by a cell-autonomous mechanism. "Nature 397 (6719): 530-4.
Li, W. , F. Fuら(1999). "Recipient pretreatment with mammalian IL-10 prolongs mouse cardiac allograft survival by inhibition of anti-donor T cell responses. "Transplant Proc 31 (1-2): 115.
Lyons, A. B. Parish, C. R. (1994)"Determination of lymphocyte division by flow cytometry"J Immunol Methods 171: 131-137.
Moore, K. W. , R. de Waal Malefytら(2001). "Interleukin-10 and the interleukin-10 receptor. "Annu Rev Immunol 19: 683-765.
Roncarolo M. G. , Battaglia M. ら(2003) "The role of interleukin 10 in the control of autoimmunity"J. Autoimm 4: 269-272 Sehgal, S. N. (1998). "Rapamune (RAPA, Rapamycin, sirolimus): mechanism of action immunosuppressive effect results from blockade of signal transduction and inhibition of cell cycle progression. "Clin Biochem 31 (5): 335-40.
Shapiro, A. M. , J. R. Lakeyら(2000). "Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen."N Engl J Med 343 (4): 230-8.
Taylor, P. A. , C. J. Leesら(2002). "The infusion of ex vivo activated and expanded CD4 (+) CD25 (+) immune regulatory cells inhibits graft-versus-host disease lethality. "Blood 99 (10): 3493-9.
Tisch, R. and H. McDevitt (1996). "Insulin-dependent diabetes mellitus."Cell 85 (3) : 291-7.
Trembleau, S. , G. Pennaら(2000)."Early Thl response in unprimed nonobese diabetic mice to the tyrosine phosphatase-like insulinoma-associated protein 2, an autoantigen in type 1 diabetes."3 Immunol 165 (12): 6748-55.
Wells, A. D. , X. C. Liら(1999). "Requirement for T-cell apoptosis in the induction of peripheral transplantation tolerance. " Nat Med 5 (11): 1303-7.
Yu, X. , P. Carpenterら(2001). "Advances in transplantation tolerance." Lancet 357 (9272): 1959-63.
【技術分野】
【0001】
本発明は、それらを必要としている患者における免疫抑制及び/又は抗原−特異免疫耐性を誘発する方法及び組成物に関する。より詳細には、本発明は、T細胞によって媒介される、亢進、機能不全又は制御されていない自己又は非自己免疫応答を含む疾患の治療に用いるためのラパマイシン及びIL−10の組み合わせ製剤を提供する。また、本発明は、IL−10及びラパマイシンを含有する医薬組成物及び免疫応答のモデュレータとしてのそれらの使用に指向する。
【背景技術】
【0002】
移植及び免疫抑制薬
移植は、末期の腎不全、ハース(hearth)又は肝疾患、自己免疫タイプI糖尿病のほとんどの患者に対する代案治療であり、それは、患者に小腸及び肺機能における欠乏性疾患を発現させる可能性がある。移植片生着は、多くの因子に依存するが、これらの最も顕著なものは強力な免疫抑制剤の投与である。遺伝的に共通点のない個体間の移植は、もし制御できないままであれば、移植された臓器の完全な破壊を招き得る、急速な及び場合によっては破壊的な同種異系反応性免疫応答を誘発する。免疫抑制剤の投与はこの応答を和らげ、よって、急性組織不適合を防止する。しかし、免疫抑制剤を中止すると、拒否反応の再活性化を招き、急速な移植破壊を導くため、移植片生着を継続するためには終身の免疫抑制に依存する。
【0003】
最近、免疫抑制剤のなかで、シクロスポリンA(CsA)、FK506及びラパマイシンを含む選択的T細胞阻害剤が開発されている。CsA及びFK506の双方は、カルシニュリン機能をブロックすることによってT細胞活性化を阻害し、よって、活性化T細胞の核因子(NFAT)能の発現を防止する。この工程は、IL−2を含むいくつかのサイトカインのmRNAを調節(増加)するために不可欠である。CsA及びFK506の主な制約は、それらの種々の毒性である。さらに、CsA及びFK506の双方は、T細胞アポチーシスを防止する(Yuら、2001年、参照)。
【0004】
これに反して、ラパマイシンは、細胞質蛋白(FKBP−12)に結合し、かつIL−2信号経路をブロックすることによって、T細胞の増殖を阻害する免疫抑制能がある(Sehgal、1998)。その複合体はラパマイシンの哺乳類標的(mTOR)に結合し、かつブロックし、サイトカイン誘発T細胞増殖の阻害をもたらす。重要なことは、CsA及びFK506に反して、ラパマイシンは、TCR−媒介T細胞活性(Blahaら、2003)及び活性誘導細胞死(AICD)に対するIL−2細胞プライミングをブロックしないことである。後者は、末梢移植耐性の誘発の役割を果たすと思われるT細胞アポトーシスの一形態である(Wellsら、1999)。樹状細胞(DC)に作用しないCsAとは異なり、ラパマイシンは、DCの表現型及び機能に大いに影響を及ぼす(Hacksteinら、2002)。それは、それらの抗原取り込み能を著しく減少させ、よって、免疫寛容原性表現型を有するDCの分化に有利である。この作用は、低濃度で現われ、ラパマイシンが生理学的に適当な濃度(1ng/ml)ではDC成熟とは無関係であり、インビトロ及びインビボの双方で証明されている(Hacksteinら、2002)。
【0005】
最近の入手可能な免疫抑制剤は、非常に短期間では非常に有効であるが、実質的な問題は、移植の拒絶を防止する代替の又は高性能な方法を開発することの差し迫った必要を示している。主な障害物は、感染性の病原体に対する有効な免疫応答と、移植に対する破壊的免疫応答との間の区別ができないことである。よって、免疫抑制の治療は、日和見感染のリスクの増大を招くかも知れない。いくつかの研究は、非特異的免疫応答は、移植患者におけるガンの発生率を増加することを導くであろうことを示す(Hojoら、1999)。したがって、非特異免疫抑制の代替が見出された場合にのみ、移植の最大の可能性が履行されるであろう。移植免疫の主なねらいは、移植に対する免疫応答を防止するが、残りの免疫システムを完全に損なわれないまま残すプロトコルを開発することである。これを達成することにより、移植耐性がもたらされるであろう。
【0006】
自己免疫
自己免疫疾患において、自己抗原に対する望まない免疫応答は末梢組織の破壊を招く。自己免疫疾患の治療は、現在、炎症の抑制的調節及び非抗原(Ag)特異免疫抑制に基づいている。同種異系移植片拒絶の防止のために、この戦略は、一旦薬物の投与を中止すると再発の高い危険及び感染及び腫瘍を含む過度の免疫抑制の危険を伴うため、長期においてはしばしば有効ではない。代替のアプローチは、一時的な免疫抑制及び/又は特異免疫耐性の誘導に基づいており、宿主の防衛メカニズムを完全に保全しながら、自己Agに対する「沈黙」の病原性応答を目的としている。
【0007】
免疫システムは、自己又は有害でない抗原に対する耐性を誘発する2つの異なるメカニズムを引き出している。これらは、中枢及び末梢T細胞耐性と称される。中枢耐性は、胎児の発育中及び非常に初期の出生時期に実現され、胸腺の発現中に自己反応性T細胞の特定クローン欠損によって媒介される。末梢メカニズムは、成熟T細胞における耐性を誘発し、一生の間末梢で起こる。これらのメカニズムは、抗原特異性リンパ球の機能的不活性化(アネルギーと呼ばれる)及び抑制及び調節能を有するT細胞のサブセットの活性化(T regulatory cells、Battagliaらにおいて概説されたT調節細胞、2002参照)を含む。
【0008】
耐性及びT調節細胞
近年、移植特異耐性を達成するための戦略として、T調節(Tr)細胞の誘導における興味が増加しつつある。CD8+、D8+CD28- 及びTCR+CD4-CD8-のような他のT細胞サブセットが調節能を有する細胞に含まれることが示されているが、今日までに確認されたTr細胞のほとんどは、CD4+集団内に見出されている。CD4+集団では、抑制的な特性を有する種々の分画が確認されている。我々のグループは、タイプ1調節T細胞(Tr1)として定義されるTr細胞のサブセットを特徴づけた。それは、Th1及びTh2細胞のそれと区別できるサイトカイン産生プロファイルを有する。ヒト及びマウスのTr1細胞は、高レベルのIL−10、かなりの量のIL−5、TGF−β及びIFN−γを産生するが、IL−2は低レベルであり、IL−4は全く産生しない(Grouxら、1997)。IL−10は、Tr1細胞の分化及びエフェクター機能にとって重要なサイトカインである。抗原及びIL−10の存在下でCD4+T細胞を培養すると、インビトロでは、抗原特異T細胞応答を抑制することができるTr1細胞の発現を導き、インビボでは、自己免疫大腸炎の発現を導く(Grouxら、1997)。また、Tr1細胞は、インビボで発現することができる。Tr1細胞は、実際、SCID−再構成患者(高レベルのIL−10が成功した同種異系幹細胞移植を伴う)の末梢血から単離される(Bacchettaら、1994)。
【0009】
耐性及びIL−10
IL−10は、免疫調節の解決の鍵となる役割を果たす(Mooreらにおいて概説、2001)。それは、Tリンパ球の増殖及びIL−2産生を阻害する。IL−10は、活性化された単球/マクロファージ、好中球、好酸球及び肥満細胞によるTNF−α、IL−1、IL−6等の前炎症性サイトカイン及びIL−8、MIPlα及びMIPlβ等のケモカインの産生を阻害することによって、強力な抗炎症作用を有する。さらに、IL−10は、MHCII及び補刺激性分子を抑制することによる単球/マクロファージ/DC等の抗原提示細胞の抗原提示能を抑制する。T細胞媒介および抗炎症免疫応答の誘発及びエフェクター機能を抑制するIL−10の能力は、骨髄及び臓器移植におけるIL−10形質発現、機能及び潜在的有用性の多くの研究を導いた。マウスにおける血管新生化心臓同種異形移植の研究では、移植の前の被移植動物のIL−10処置が移植片生着を高め、一方、移植時またはその後にIL−10を与えても、ほとんど有用な効果はないか、拒絶を高めるのみであった(Liら、1999)。BMT前のIL−10産生レベルの上昇を示した患者は、GVHDの発生率が低く、生存を向上させた(Bakerら、1999)。これに対して、ポスト−BMT GNHD患者における高IL−10レベルは、生存に対する予後ははかばかしくなかった(Hempel、1997)。しかし、Blazar及びその同僚は、少量のIL−10(死亡率を増加させる10-3、10-4の量)でのマウスの処置は、GVHD関連致死率を防ぐことを示した(Blazarら、1998)。
【0010】
免疫抑制剤とIL−10との組み合わせ
現在の臨床上の使用における免疫抑制剤の大部分は、T細胞の活性化を抑制することによって作用し、これにより、移植拒絶を防止する。しかし、これは、適切な状況下としては逆効果かもしれず、T細胞の活性化は、移植特異耐性の発現を促進するプロセスの誘発を招くかもしれない。したがって、その目的が耐性誘発である場合には、免疫抑制剤の使用は、最適でないかもしれない。この現象の明確な証明は、いかなる免疫抑制治療も行うことなしに、同種造血幹細胞移植術後に耐性を獲得したSCID患者によってもたらされる(Bacchettaら、1994)。これらの患者において、ホストアロ抗原に対するTr1細胞特異性を誘導したドナーの存在は、安定な混合キメラ現象、インビボにおける高レベルのIL−10産生及びいかなる免疫抑制治療も受けない正常免疫機能と相関関係がある。これに対して、急性GVHDを管理するために免疫抑制の養生法を受けたBMT患者では、ホストアロ抗原に対するT細胞特異性が誘導されたドナーが検出できたとして、Tr1細胞は末梢血から単離されないであろう(Bacchettaら、1995)。
【0011】
ラパマイシンは、興味深い免疫調節特性を有する新規な化合物を意味する。この理由のため、我々は、同種移植片拒絶を防止し又はタイプ1糖尿病を和らげ、ならびにインビボにおいてTr細胞を発現させるために、ラパマイシンとIL−10とのインビボにおける投与を組み合わせた。
【0012】
従来技術
米国特許第6,277, 635号は、移植による拒絶反応を抑制するためのIL−10の使用に関連する。この特許は、組織の拒絶反応を治療し、阻害する方法を教示し、GVHD及び抗原特異応答を抑制する。さらに、特定の抗原に対するアネルギーを示すT細胞について述べている。
米国特許第6,428, 985号は、ヒトを含む哺乳動物の、IL−10ポリペプチドと少なくとも1つの野生型配列における突然変異体(MutIL−10)とを含む免疫抑制組成物であって、その単独又は他の剤との組み合わせのいずれかで、インビトロ及びインビボで、そのような組成物及びそれらの組み合わせを用いる種々の方法について述べている。用途は、免疫抑制剤及び多くの疾患のための組み合わせ治療、ならびに炎症、移植、線維症、傷に関連する障害、及び腫瘍治療を含む。MutIL−10の作用は、動物実験において示されているが、ヒトの臨床上の設定においては示されていない。
【0013】
米国特許第5,624, 823号は、豚IL−10をコードするDNA、及び異質遺伝子型移植を受ける被移植動物(例えば、霊長類)における耐性の誘導方法を示している。ラパマイシン、サイクロスポリン及びFK506は、「剤を減少させる助け」、例えば、サイトカイン放出を減少する剤として記載されている。豚のIL−10は、胸腺移植のみの文脈で用いられている。
米国特許第6,022, 536号は、自己免疫疾患及びGVHDの治療のための免疫抑制治療として、IL−10及びサイクロスポリンを組み合わせた使用が示されている。低容量のIL−10及びサイクロスポリンならびに医薬担体の相乗効果のある組み合わせが提案されている。
【0014】
米国特許第6,403, 562号は、自己免疫疾患に罹患した又は感受性のある人に対して、IL−10とともにTGF−βを一緒に投与することによって、多発性硬化症のような自己免疫関連疾患の治療方法が示されている。これらのサイトカインは、相乗効果のある方法において、自己免疫疾患に含まれる自己反応性T細胞の活性化を阻害するための抑制因子として作用する。
【発明の開示】
【0015】
発明の要旨
本発明は、T細胞媒介免疫応答の調節における使用のための、特に、それが必要な患者における免疫抑制及び抗原−特異免疫耐性を誘発するためのIL−10及びラパマイシンを含有する医薬製剤を提供する。Tr1及びCD4+CD25+Tr細胞−媒介抗原−特異免疫耐性の誘発は、亢進、機能不全、規制されていない又は管理されていない自己−又は非自己−T細胞−媒介免疫応答を含む病理学的症状の治療に有用である。
【0016】
本発明の好ましい実施形態では、IL−10及びラパマイシンは、同種異系の臓器拒絶;タイプ1糖尿病;乾癬、多発硬化症、炎症性腸疾患、クローン病、慢性関節リウマチを含む自己免疫及び慢性炎症疾患;又はGVHD、喘息、アトピー性皮膚炎のような他のT−細胞媒介疾患;慢性閉塞性肺疾患;ならびに遺伝子治療誘発生成物に対する免疫反応の予防又は治療上の処置における同時の、別々の又は連続的な使用のための組み合わせ製剤の形態である。さらに、肝臓及び肺繊維症を含む繊維増多疾患の治療が認識される。
好ましくは、その組み合わせ製剤は、堅固な同種異系臓器拒絶、特に、同種移植片β−島拒絶及び自己免疫疾患、特にタイプ1糖尿病の予防及び治療上の処置に使用される。
【0017】
その組み合わせ製剤は、ヒト又はウィルス性のIL−10、それらのアナログ、誘導体又ポリエチレングリコール(PEG)抱合IL−10のような、天然分子の生物学的利用率又は生物学的有効性を改善する、それらの抱合体を含んでもよい。IL−10機能的アナログは、IL−10作用を模倣する小分子及びIL−10レセプター又はIL−10融合蛋白に対するモノクローナル抗体(mAbs)を含み、それらはIL−10情報伝達系を誘引する。
【0018】
その組み合わせ製剤は、ラパマイシンアナログ又は誘導体を含んでいてもよい。ラパマイシン及びIL−10、それらの誘導体又はアナログに加えて、その組み合わせ製剤は、さらに免疫抑制剤又は免疫調整剤、モノクローナル抗体又はサイトカインを含んでいてもよい。IL−10及びラパマイシンの組み合わせにおいて使用することができる好ましい生物学的に活性な物質は、a)サイクロスポリン、FK506(タクロリムス)、ピメクロリムスのようなカルシニュリン阻害剤、b)ミコフェノラートのような他の免疫抑制剤、c)CD45の種々のイソ型に対する抗体又はLFA−1のような接着分子、及びd)IL−2受容体α、β及びγ鎖に対する抗体を含む。適当な免疫抑制剤は、IL−2情報伝達系によって作用するそれらを含む(例えば、JAK1及びJAK3ならびにSTAT5阻害剤)。ラパマイシン+抗Tac(IL−2受容体α鎖に対するヒト化抗体)+IL−10の組み合わせは、従来の治療プロトコルによって発生した持続性免疫抑制の代わりに耐性の状態を含むことによって、異質遺伝子型拒絶、特に、同種移植片β−島拒絶のマウスモデルの予防において特に有効であることが示された。さらに、ラパマイシン+IL−10の組み合わせは、自己免疫糖尿病の治療及びNODマウスにおける長期にわたる免疫調節の誘発に有効となることが示された。耐性は、ラパマイシン+IL−10誘発拡大ならびにタイプ1T調節(Tr1)及びCD4+CD25+Tr細胞の区別化の結果として獲得され、それらは、抑制サイトカイン(IL−10及びTGF−β)の生成を含む種々のメカニズム及びT細胞活性化の阻害によって、抗原−特異耐性を媒介する。
【0019】
本発明のラパマイシン+IL−10組み合わせ製剤は、T細胞免疫能力の回復にかかわらず、長期間の予防に能力を発揮し、それは、薬物の使用中止後に維持することができる。
本発明のさらなる実施形態は、IL−10及びラパマイシン、ならびに任意に、免疫抑制剤又は免疫調節剤、モノクローナル抗体及びサイトカインから選択されたさらなる活性成分を、医薬的に許容される賦形剤とともに含む医薬組成物を提供する。適当な医薬組成物は、経口、静脈内、非経口又は皮下経路によって投与され、好ましくは、溶液、懸濁液、注射物質、錠剤又はカプセルの形態である。ラパマイシンの有効量は、0.001mg/kgから100mg/kgの範囲とすることができ、IL−10の有効量は、0.001μg/kgから1000μg/kgの範囲とすることができる。
本発明を、以下の実施例及び添付の図面によってさらに詳細に説明する。
【0020】
図面の説明
図1 IL−10プロトコル及びエドモントンプロトコルで処置したマウスは、同程度の移植片生着を示す。
ストレプトゾトシン注入によって糖尿病にされたBalb/cマウスに、腎臓被膜の下、純粋な異質遺伝子型C57BL/6β−島を移植した。マウスは、処置なし(対照、n=13マウス)か、30日間、ラパマイシン+抗Tac+IL−10で処置された(IL−10プロトコル、n=16マウス)か、ラパマイシン+抗Tac+FK506で処置(エドモントンプロトコル、n=4マウス)された。移植片生着を、血糖レベルによってモニターした。移植片は、血糖が250mg/dlより高いときに拒絶されているとみなした。
FK506(エドモントンプロトコル)をIL−10(IL−10プロトコル)に置き換えることにより、同等の移植片生着を示した。IL−10プロトコルで処置したマウスにおいて、移植片生着は89%であり、一方、100%の生存がエドモントンプロトコルで処置したマウスで観察された。
【0021】
図2 IL−10プロトコルで抗Tacなしとした場合、わずかに異種遺伝子型β−島拒絶が増加した。
ストレプトゾトシン注入によって糖尿病にされたBalb/cマウスに、腎臓被膜下、純粋な異質遺伝子型C57BL/6β−島を移植した。マウスは、処置なし(対照、n=8マウス)か、ラパマイシンとIL−10との組み合わせ(ラパ+IL−10、n=8マウス)又はIL−10のみ(n=4マウス)で処置された。移植片生着を、血糖レベルによってモニターした。
IL−10プロトコルで抗Tacなしとした場合(図1参照)、わずかに移植片生着に影響を及ぼした。ラパマイシンをIL−10と組み合わせた場合、移植片生着は動物の78%で認められた。IL−10単独での処置は、移植片拒絶の予防に有効でなかった。
これらのデータは、抗Tacが同種異系移植拒絶を予防するのに必要でなかったことを示唆する。
【0022】
図3 IL−10プロトコルで処置したマウスからのT細胞はインビトロで増殖能を維持する。
対照の非移植マウス(白棒)及びIL−10プロトコル(グレー棒)又はエドモントンプロトコル(黒棒)で処置したマウスからのT細胞を脾臓から単離し、インビトロで、抗CD3及び抗CD28mAbsで多クローン的に刺激した。エドモントンプロトコルで処置したマウスからの細胞は、インビトロでの増殖能においてそれらを強く減少させ、一方、増殖におけるマイルドな減少のみが、IL−10プロトコルで処置したマウスから単離したT細胞において観察された。
これらのデータは、IL−10プロトコルで処置したマウスからのものはそうでないが、エドモントンプロトコルで処置したマウスから単離したT細胞において、免疫抑制が意味深い状態であることを示唆する。
【0023】
図4 IL−10プロトコルで処置したマウスからのT細胞は、抗原−特異増殖能を防止する。
移植280日前およびIL−10プロトコル(グレー棒)又はエドモントンプロトコル(黒棒)で30日間のみ処置したマウスに対して、CFA+OVAで、後足裏のやわらかい部分にインビボにおいて免疫化した。リンパ節流を採取し、OVA及び自己APCによりインビトロで再刺激した。
OVA−特異T細胞増殖は、エドモントンプロトコルで処置したマウスにおいて非常に減少し、一方、IL−10プロトコルで処置したマウスにおけるOVA−応答は、非移植免疫化マウスで観察されたのと同等であった。
これらのデータは、さらに、IL−10プロトコルではそうではないが、エドモントンプロトコルで処置したマウスにおいて、免疫抑制が一般状態であることを証明する。
【0024】
図5 IL−10プロトコルで処置したマウスから単離したT細胞はIL−10を産生する。
対照の非移植マウス又はIL−10プロトコル(グレー棒)及びエドモントンプロトコル(黒棒)で処置したマウスの腎臓から単離したCD4+T細胞を、抗CD3及び抗CD28mAbsで、インビトロで刺激した。刺激後96時間に上清を採取し、IL−10産生をELISAで評価した。
IL−10プロトコルで処置したマウスからのT細胞は、エドモントンプロトコルで処置したマウス及び対照の非移植マウスと比較して、高いレベルのIL−10を産生した。
【0025】
図6 IL−10産生T細胞の特異集団は、IL−10プロトコルで処置したマウスから単離することができる。
(A)IL−10プロトコル、エドモントンプロトコルで処置したマウス及び対照の非移植マウスからのT細胞を単離し、インビトロで多クローン的に刺激し、サイトカイン産生を誘導した。3時間後、細胞を、偏在細胞表面マーカーに結合する一つのmAb及びIL−10を捕捉できる他のmAbとからなるダイアボディで標識した。次いで、多クローン刺激の間に蓄積されたサイトカインを放出させるために、標識細胞を37℃でさらに1時間インキュベートした。標識細胞によって産生されたIL−10をダイアボディによって捕獲した。さらに、細胞を、PEで標識された抗IL−10mAbで標識した。IL−10+濃縮(密ヒストグラム)及びIL10-(空ヒストグラム)細胞を磁気で分離するために、抗−PEマイクロビーズを用いた。IL−10+濃縮T細胞の特異集団を、IL−10プロトコルで処置した移植マウスからのみ単離した。
【0026】
(B) 細胞質内の染色を、この特異IL−10+濃縮T細胞に行い、Tr1サイトカインプロファイル(つまり、IL−10+、IL−4-)を有する細胞の有意な集団を、エドモントンプロトコルで処置したマウス又は対照の非移植マウスではなく、IL−10プロトコルで処置したマウスにおいて確認した。
【0027】
図7 IL−10は、インビボでのIL−10産生T細胞を誘発するために必要である。
インビボでIL−10+Trl細胞を誘発するためにIL−10投与が必要であることを理解するために、移植マウスを、IL−10プロトコル(ラパマイシン+抗Tac+IL−10)又はラパマイシン+抗Tacのみで処置した。次いで、マウスの2つの群におけるCD4+脾臓T細胞によるIL−10の産生を評価した。
IL−10プロトコル(グレー棒)又はラパマイシン+抗Tac(黒棒)で処置したマウス又は対照の非移植(白棒)のマウスからのT細胞を単離し、インビトロで、多クローン的に、抗CD3及び抗CD28mAbsで刺激した。
IL−10のかなりのレベルが、IL−10プロトコルで処置されたマウスから単離された細胞によってのみ産生した。
これらのデータは、IL−10が、インビボでIL−10産生細胞を誘発するために必要であることを示唆する。
【0028】
図8 NODマウスにおけるタイプI糖尿病の治療のためのラパマイシン+IL−10での予備的結果
11週齢のNODマウスは、糖尿病前症の段階である。これらのマウスはインスリン炎及びすい臓における浸潤性自己免疫T細胞を有しているが、それらは、いまだ血糖正常とする十分なインスリンを産生することができるの十分に正常なβ−島を有している。
糖尿病前症マウスを、11週齢からはじめて毎日、ラパマイシン、ラパマイシン+IL−10又はIL−10単独のいずれかで処置した。処置後6週間で、対照マウスの33%が糖尿病を発症したが、ラパマイシン及びIL−10で処置したマウスは、全て血糖正常のままであった。
これらの予備的結果は、ラパマイシン+IL−10は、その初期段階において糖尿病をブロックするために使用することができ、さらに進行した自己免疫糖尿病の自発性の発症を予防することを示唆する。
【0029】
図9 IL−10プロトコルは、糖尿病誘発性T細胞の移植後のNOD.SCIDマウスにおいて誘発した糖尿病を阻害する。
NOD糖尿病マウスからの脾細胞5×106をNOD.SCIDマウスに静脈内移植した。被移植マウスを、移植後40日間、処置しないか、エドモントンプロトコル(ラパマイシン+抗Tac+FK506)、IL−10プロトコル(ラパマイシン+抗Tac+IL−10)又はラパマイシン+抗Tacのいずれかで処置した。
移植後50日で、全ての対照の被処置マウスは糖尿病になった。
エドモントンプロトコルで処置した全てのマウス及びラパマイシン+抗Tacで処置した75%のマウスは、糖尿病になった。興味深いことに、IL−10プロトコルで処置した33%のみのマウスが糖尿病になった。
これらの予備的データは、IL−10プロトコルは、自己免疫糖尿病誘発性NODT細胞の移植によってNOD.SCIDマウスにおいて誘発されたタイプI糖尿病を阻害する。
【0030】
図10 タイプI糖尿病の治療のためのラパマイシン+IL−10の使用
A)NODマウスにおける糖尿病の治療
NODマウスを11週齢から31週齢まで、IL−10(IL−10、n=7マウス)又はラパマイシン(RAPA、n=13マウス)又はラパマイシン+IL−10(RAPA/IL−10、n=16マウス)又は賦形剤(CNTR、n=22マウス)で処置した。血糖レベルによってモニターした糖尿病発生率は、少なくとも60週齢まで安定した。ラパマイシン単独投与では、糖尿病発生率が95%から46%に減少した。IL−10投与では、糖尿病の発症に顕著な効果はなかった。ラパマイシンの予防効果は、IL−10を処置に加えた場合に、顕著に改善され、糖尿病発生率を13%に減少した。
【0031】
B)NOD.SCIDマウスにおける糖尿病の移植に対する処置NODマウスからの脾細胞の能力
非処置糖尿病NODマウス(DIABETIC NOD、n=8)又はラパマイシンで処理したマウス(RAPA NOD、n=5)又はラパマイシン+IL−10(RAPA/IL−10 NOD、n=13)からの総脾細胞5×106を、NOD.SCIDマウスに移植し、糖尿病発生率を血糖レベルによってモニターした。
ラパマイシン処置マウスからの脾細胞の移植は、糖尿病NODマウスからの脾細胞を注入されたマウスに比べて、病気の発症を顕著に遅延させた。重要なことに、ラパマイシン+IL−10で処置されたマウスからの脾細胞も同様に、さらに糖尿病伝達を遅らせた。
【0032】
図11 ラパマイシン±IL−10で処置したマウスにおけるTr細胞の濃度
CD4+T細胞によって産生されたサイトカイン(左パネル)及びCD4+CD25+T細胞のパーセント(右パネル)を、非処置糖尿病NODマウス(グレー線)、ラパマイシン処置マウス(白棒)又はラパマイシン+IL−10処置マウス(黒棒)の脾臓(上部パネル)、膵臓のリンパ節(PLN)(中段パネル)及び島浸潤性細胞(IIC)(下部パネル)において、それぞれCBA及びFACによって評価した。それらのサイトカイン産生プロファイル(つまり、IL−10++IL−15+TGF−β+)によって測定したように、CD4+Tr1細胞の高い割合が、ラパマイシン+IL−10(黒棒)で処置した耐性マウスの脾臓にのみ存在した。CD4+CD25+T細胞のパーセントは、ラパマイシン単独又はラパマイシン+IL−10(黒棒)で処置したマウスの双方の脾臓、PLM及びIICにおいて高かった。したがって、ラパマイシン+IL−10処置マウスにおいて、Tr1細胞は脾臓において存在し、CD4+CD25+Tr細胞は、脾臓、リンパ節及び膵臓において存在した。
【0033】
図12 インビトロにおける免疫応答を抑制するTr細胞の能力
インビトロにおいて、CFSEで標識し、抗CD3mAbの存在中で培養したCD4+の実験に未使用のNODT細胞の増殖におけるTr細胞の抑制活性を試験した。CD4+IL−10濃縮脾臓T細胞(純度40%以下)(左パネル)又はCD4+CD25+T細胞(MAC精製、純度75%以上)(右パネル)のいずれかを、実験に未使用のT細胞と同じ数で加えた抑制細胞として用いた。何も加えない細胞(グレー棒)に区分した実験に未使用のT細胞を対照として用いた。非処置糖尿病NODマウス(黒棒)又はラパマイシン処置マウス(白棒)又はラパマイシン+IL−10処置マウス(点線)から単離されたCD4+IL−10濃縮Tr1細胞又はCD4+CD25+Tr細胞の存在における細胞区分を評価し、対照に対する抑制パーセントを測定した(各ヒストグラムの上部における数)。ラパマイシン+IL−10処置マウスの脾臓からのTr1細胞は、NODマウスから得られたCD4+T細胞の増殖応答を緩やかに抑制する。強力な抑制が、ラパマイシン及びラパマイシン+IL−10処置マウスの双方のPLN及びIICから単離されたCD4+CD25+T細胞で観察された。
【実施例】
【0034】
実施例1
1.同種異系β−島移植
完全不一致のマウス島同種異系移植のモデル(Balb/CへのC57BL/6)を用いた。同種異系すい臓β−島移植は、インスリン置換療法又はタイプ1糖尿病の治療のためのすい臓移植への有効な代替となっている。過去数年のうちに、ヒトβ細胞の単離及び保存のための改善された方法及び新たな免疫抑制剤の開発が、これらの移植の臨床成果を顕著に改善している。特に、ラパマイシン+抗Tac+FK506(エドモントンプロトコル)に基づく新たなステロイドフリーの免疫抑制処方が、近年、移植後1年で、患者の80%において、インスリン非依存をもたらすことが示されている(Shapiroら、2000)。これらの結果は、全ての先の免疫抑制剤の組み合わせ治療で得られたものをはるかにしのいでいる。しかし、この処方が耐性を誘発し得るという証明は行われていない。
【0035】
重要なことは、アポトーシスの防止及びT細胞発現の阻害のために、FK506の作用メカニズムが耐性誘発の状態を防止するかもしれないことである。
寛容原性プロトコルの開発の努力において、我々は、エドモントンプロトコルにおけるFK506をIL−10によって置換した(つまり、IL−10プロトコル、ラパマイシン+抗Tac+IL−10)処方を設計した。
ストレプトゾトシン注入によって糖尿病にされたBalb/cマウスに、純粋な異質遺伝子型C57BL/6β−島を、腎臓被膜下、移植した。細胞片生着は、IL−10プロトコル及びエドモントンプロトコルで処置したマウスにおいて、それぞれ同様であった(移植後240日で89%及び100%)(図1)。
【0036】
抗Tacのインビボにおける使用は、高いIL2Rα鎖発現を伴う活性T細胞をブロックするための必要性によって、過去数年、強く支持されている。
しかし、Tr細胞のサブセットは、IL2Rα鎖(つまり、CD4+CD25+Tr細胞)を構成的に発現させ、このT細胞集団が同種異系移植拒絶を抑制し得ることは広く証明されている(Taylorら、2002)。
【0037】
したがって、CD25+T細胞集団をブロックするmAbの使用は、インビボにおいて、Tr細胞によって媒介される耐性誘導が求められる場合には、逆指示(counterindicated)されるかもしれない。この理由に対して、抗Tacを治療プロトコルからの除外することを評価した。我々は、島拒絶を十分に防止し得るか否かを測定するために、ラパマイシン+IL−10又はIL−10単独で30日間、移植マウスを処置した(図2)。長期間の移植片生着が、IL−10単独で処置したマウスの30%で観察され、ラパマイシン+IL−10で処置したマウスにおいては78%に増加した。この移植片生着のレベルは、さらに抗Tacで処置したマウスのそれよりもわずかに低いのみであった(図1に示す)。これらのデータは、IL−10プロトコルから抗Tac mAbを不在とすることにより、同種異系移植片生着を可能にし、一方、CD4+CD25+Tr細胞は影響を受けず、インビボにおいてこれら細胞による耐性誘発を保つことを示す。
【0038】
所望の成果の一つが免疫抑制よりもむしろ耐性であるため、我々は、IL−10及びエドモントンプロトコルで処置したマウスからのT細胞が多クローナル及び抗原特異刺激に対して応答したかどうかを試験した。まず、T細胞を移植後240日(治療の中断後210日)でマウス脾臓から単離し、抗CD3及び抗CD28mAbで刺激した(図3)。エドモントンプロトコルで処置したマウスからのT細胞の増殖は、対照の非移植マウスからのT細胞のそれと比較した場合、強く抑制された。T細胞増殖の抑制は、IL−10プロトコルで処置したマウスでは、それほど強くなかった(図3)。次いで、移植後280日(治療の中断後250日)のマウスを、後足裏の柔らかいところに、CFA+OVAで免疫化し、リンパ節流から単離されたT細胞の増殖応答を測定した(図4)。エドモントンプロトコルで処置したマウスからのT細胞は、OVAに対する応答において増殖しなかったが、一方、IL−10プロトコルで処置したマウスからのT細胞は、非移植免疫化対照マウスと同様の応答であった(図4)。
【0039】
FK506のIL−10による置換が潜在的にTr1細胞拡大を促進するかどうかを測定するために、島移植の部位に浸潤するCD4+T細胞を、移植後200日のマウスから単離し、それらのサイトカイン産生を調べた。IL−10プロトコルで処置したマウスから単離されたCD4+T細胞は、エドモントンプロトコルで処置されたマウスよりも、抗CD3及び抗CD28mAbで刺激した後、IL−10のかなり高い量を産生した(図5)。次いで、精製脾臓T細胞を、抗CD3及び抗CD28mAbで刺激し、IL−10分泌細胞を、IL−10捕捉ビーズを用いて濃縮した(図6A)。IL−10/IL−4サイトカインプロファイルを、細胞質内染色によって試験した(図6B)。興味深いことに、IL−10+IL−4-細胞の特異集団(つまり、Tr1細胞のサイトカインプロファイルを反映する)が、IL−10プロトコルで処置したマウスにおいてのみ確認された。
【0040】
IL−10を含まない、ラパマイシン+抗Tacで処置されたマウスからの浸潤細胞は、IL−10を産生しなかった。これは、インビトロにおいて増加したIL−10産生は、インビボにおけるIL−10の投与のためであったことを示す(図7)。
【0041】
まとめると、これらのデータは、
1.ラパマイシン+抗Tac+FK506の組み合わせ(エドモントンプロトコル)は、同種異系拒絶からマウスを保護するが、長期の慢性免疫抑制の状態を誘発する、
2.ラパマイシン+抗Tac+IL−10の組み合わせ(IL−10プロトコル)は、同種異系拒絶に対する長期保護を与える。この処置は、Tr1細胞と一致するサイトカインプロファイルを伴うT細胞の特異集団の拡大をもたらし、保護は、T細胞免疫能力の回復にもかかわらず、薬物の中断後まで維持する。
【0042】
実施例2
1.タイプI糖尿病における予備的な結果
ラパマイシン+IL−10の寛容原性作用も、タイプI糖尿病の設定において評価した。
我々は、β細胞破壊の防止(それは、タイプI糖尿病への進行を伴い、疾患の発症において見られる)が、
1.すい臓内での一般的な「バイスタンダー」炎症の抑制、
2.島特異Tエフェクター細胞拡大のブロック、
3.抗原特異Tr細胞の誘発及び拡大
によって防止することができると考えている。
【0043】
我々の提案するプロトコルでは、炎症の抑制は、IL−10によって達成すべきであり、Tエフェクター細胞拡大のブロックは、ラパマイシンによって達成すべきである。IL−10及びラパマイシンも、T細胞プライミングを防止せず、したがってそれらは抗原特異T調節細胞の誘導を可能にするはずであり、以下に説明するように、IL−10はTr1細胞の誘導及び拡大を促進するはずである。我々は、タイプI糖尿病のNODマウスのモデルにおける自己免疫を処置することにおいて、ラパマイシン単独又はIL−10との組み合わせの効果を調査した。NODマウスは、島β−細胞の破壊及び血中グルコースの上昇を伴って、15から30週齢で明白な疾患を発現し、多くの主要な特性をヒトの疾患と共有している(Tischら、1996、Delovitchら、1997)。タイプI糖尿病の阻害を、ラパマイシン、ラパマイシン+IL−10又はIL−10単独で、11週齢(つまり、ペリ膵島炎(periinsultis)を伴う糖尿病前症のマウス)から毎日、NODマウスを処置することによって評価した。処置後6週間で、非処置対照マウスの33%が糖尿病を発症しはじめ、一方、ラパマイシン+IL−10で処置したマウスはなおも全て正常血糖であった(図8)。
【0044】
また、我々のプロトコルの有効性を、養子免疫細胞移入NOD.SCIDマウスのモデルでも試験し、それは、内因性のT及びB細胞を欠いており、したがって、自発性の糖尿病を発現せず、糖尿病のNODマウスからの5×106脾細胞の移入後15から20日で糖尿病を発現する。NOD.SCID被移植マウスを、処置しないか、あるいはラパマイシン+抗Tac+FK506(エドモントンプロトコル)、ラパマイシン+抗Tac+IL−10(IL−10プロトコル)及びラパマイシン+抗Tacで糖尿病の移入後40日間処置した(図9)。対照マウスは、移入後15日で糖尿病を発症し始めた。エドモントンプロトコルで処置した全てのマウス及びラパマイシン+抗Tacで処置した75%のマウスは、移入後33日以内に糖尿病になった。興味深いことに、IL−10プロトコルで処置したうちの1匹のマウスのみ(33%)が、移入後35日で糖尿病になった。
これらの予備的データの全体から、ラパマイシン+IL−10の使用によって、タイプI糖尿病の発現を完全に阻害することは当然のことである。
【0045】
2.ラパマイシン+IL−10の処置は、自己免疫糖尿病を阻害し、長期間の耐性を誘発する。NODマウスモデルにおいて得られた有望な予備的結果に基づいて(図8)、我々は、ラパマイシン±IL−10で、11週齢(膵臓細胞自己免疫が、膵島炎及び自己インスリン抗体によって判断されるように明らかに確立された時点)から、20週間NODマウスを処置した。ラパマイシン単独の投与は、糖尿病の発生率を95%から46%に減少させた(図10A)。先の観察は、NODマウスにおけるIL−10処置の効果は、経路、用量及び投与のタイミングに依存して変化することを示す(Roncaroloら、2003)。しかし、ここで、我々は、同じ期間にわたるIL−10単独の投与は、糖尿病の発現において顕著に作用しなかったことを示す。ラパマイシンの防御効果は、IL−10が治療に加えられた際、顕著に改善され、さらに、糖尿病の発生率を13%に減少させた(図10A)。興味深いことに、防御は、治療を中止した後30週間さらに維持され、長期免疫調節が確立されたことを示す。
【0046】
さらに、ラパマイシン又はラパマイシン+IL−10が自己免疫糖尿病の発現を防止することによるメカニズムを、耐性マウスからの細胞の移植実験で調査した。免疫不全NOD.SCIDマウスにおける非処置糖尿病NODマウスからの脾細胞を移植することによって、糖尿病が迅速に誘発され、一方、ラパマイシン処置マウスからの脾細胞を移植することによって、疾患の発症は顕著に遅延した。興味深いことに、ラパマイシン+IL−10の組み合わせで処置したマウスからの脾細胞を移植することでも、さらに、糖尿病移植を遅延した(図10B)。これらのデータは、ラパマイシンで処置することは、脾臓の自己反応性T細胞における糖尿病を伝達する能力を抑制し、この作用は、IL−10が処置に加えられた際に強力に高められることを示す。
【0047】
長期耐性に横たわるメカニズムを、50週齢以上の耐性マウスで分析した。非処置糖尿病NODマウス又はラパマイシン+IL−10処置マウスからの脾臓細胞は、同程度の細胞数を含み、CD4+及びCD8+T細胞と同じ割合であるが、それらのサイトカイン産生プロファイルは制限される。CD4+Tr1細胞の高い割合は、それらのサイトカイン産生プロファイル(つまり、IL−10++IL−5+TGF−β+)によって示されるように、ラパマイシン+IL−10で処置された耐性マウスの脾臓において存在するが、ラパマイシン単独で処置されたマウス又は非処置糖尿病NODマウスの脾臓において存在しない(図11)。しかし、IL−4を産生する脾臓のCD4+T細胞の割合は、非処置及び処置マウスの双方において同じである。さらに、脾臓CD4+CD25+T細胞のパーセントは、非処置糖尿病NODマウスに比較して、ラパマイシン単独又はラパマイシン+IL−10で処置したマウスの双方において高い(図11)。反対に、Tr1細胞は、膵臓リンパ節(PLN)及び島浸潤性細胞(IIC)において検出することができず(図11)、Tr1細胞は自己免疫部位に存在しないことを示す。一方、CD4+CD25+T細胞は、PLNにおいて多数観察され、ラパマイシン単独又はラパマイシン+IL−10のいずれかで処置されたマウスのIICから単離されたCD4+T細胞のほぼ100%を示すが、非処置糖尿病マウスでは示さない(図11)。IICからのこれらCD4+CD25+T細胞は、アレルギー性であり、TGF−βは別として、サイトカインを有意なレベルで産生しない。
【0048】
次いで、我々は、ラパマイシン+IL−10処置マウスにおいて存在するTr1細胞及びラパマイシン及びラパマイシン+IL−10処置マウスの脾臓、PLN及びIICからのCD4+CD25+T細胞が、インビトロにおいて抑制活性を有するか否かを測定した。ラパマイシン+IL−10処置マウスの脾臓からのTr1細胞は、NODマウスから得られたCD4+T細胞の増殖応答を緩やかに抑制する(図12)。また、抑制は、処置及び非処置糖尿病NODマウスの双方の脾臓から精製されたCD4+CD25+T細胞で観察され(図12)、それは、CD4+CD25+Tr細胞は、また糖尿病NODマウスの脾臓において存在するが、非常に低い頻度であることを示す(図11に示す)。興味深いことに、強い抑制は、ラパマイシン又はラパマイシン+IL−10処置マウスの双方のPLN及びIICから単離されたCD4+CD25+T細胞で観察された。反対に、非処置糖尿病NODマウスから単離されたCD4+CD25+T細胞は、測定可能ないかなる抑制活性を有さなかった(図12)。これらのデータは、糖尿病NODの膵臓組織は、Tr細胞よりもむしろ、活性化Tエフェクター細胞を主に含み、一方、処置マウスのPLN及びIICは、主にCD4+CD25+サブセットのうちTr細胞を含むことを示す。
【0049】
概して、これらのデータは、ラパマイシン+IL−10処置の後に観察される定常状態の耐性は、脾臓におけるTr1細胞ならびにリンパ節及び膵臓におけるCD4+CD25+Tr細胞の蓄積に関連することを示す。
【0050】
材料及び方法
マウス Balb/c、C57BL/6、NOD/Lt及びNOD.SCIDの雌性マウスをCharles River Laboratories (Calco, Italy)から購入した。全てのマウスは、特定の病原体フリーの状態で保持した。尾の静脈血におけるグルコースレベルをGlucometer Elite system (Bayer, Wuppertal, Germany)を用いて定量した。糖尿病を、ストレプトゾトシン(シグマ、セントルイス、MO)170mg/kgの静脈内注入によって、Balb/cマウスで誘発した。糖尿病の診断を、250mg/dlより高い2回の連続的なグルコース測定の後に行った。
【0051】
島移植 厳選されたC57BL/6の膵臓島を、37℃にて一晩培養した後、先に述べたように、Balb/c糖尿病マウスの腎臓膜の下に移植した(300島/マウス)(Davalliら、1996)。
【0052】
移植マウスの処置 移植されたBalb/cマウスの処置を、移植して30日後の日に始めた。ラパマイシン(Rapamune, Wyeth-Ayerst Research, Pearl River, NY)をピーナツ油(シグマ)で希釈し、胃管栄養法によって1mg/kgの用量で一日1回投与した。ヒトIL−10(BD Biosciences, Mountain View, CA)をPBSで希釈し、0.05mg/kgの用量で、腹腔内に1日2回投与した。FK506(Prograf, Fujisawa, Milano)を生理食塩水で希釈し、0.3mg/kgの用量で、腹腔内に1日1回投与した。抗IL−2Rα鎖mAb(抗Tac)(クローン7D4、BD)を生理食塩水で希釈し、1mg/マウスの最終用量になるように、移植後0及び4日目に、腹腔内に投与した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0053】
糖尿病阻害研究 雌性NODマウスを、11週齢から31週齢まで、移植マウスで用いたのと同じ用量で用いて、ラパマイシン、ラパマイシン+IL−10又はIL−10単独で処置した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0054】
糖尿病移植研究 糖尿病NOD雌性マウスから脾細胞を採取し、NOD.SCIDにおいて5×106/マウスの用量で、静脈内に注入した。移植マウスにおいて使用されたのと同じ用量で、被移植マウスを、処置しないか、移植後40日間、ラパマイシン+抗Tac+FK506又はラパマイシン+抗Tac+IL−10、又はラパマイシン+抗Tacで処置した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0055】
NOD.SCIDマウスにおける養子免疫細胞移入 対照又は処置NODマウスからの脾臓を、治療の中止後採取した。500万の総脾細胞を、NOD.SCIDマウスに静脈内注入によって養子免疫細胞移入した。糖尿病の発生を血中グルコースレベルによってモニターした。
【0056】
インビボにおける免疫化 CFA(Difco, Detroit, MI)に乳化したオバアルブミン(OVA)ペプチド323−339(Primm, Milano, Italy)を移植Balb/cマウスの後足裏の柔らかい部位の皮下に、100μg/マウスの用量で、一回注入した。リンパ節流を採取し、インビトロアッセイに用いた。
【0057】
細胞選別 浸潤性膵臓の細胞を、説明されたように単離した(Gregoriら、2003)。得られた細胞集団を、抗CD90mAB−被覆マイクロビーズとともにインキュベートし、精製T細胞を得るためにMiniMacs カラム(Miltenyi Biotec, Bergisch Gladbach, Germany)に付した。CD4+CD25+T細胞を、Multisortキット(Miltenyi)で選別した(平均精製度≧75%)。いくつかの実験において、CD4+CD25+T細胞を、無菌でFACStar細胞ソーター(BD)で選別した(平均精製度99%)。IL−10産生細胞を、ネズミIL−10分泌アッセイ濃縮及び検出キット(Miltenyi)で選別した(平均精製度≧40%)。
【0058】
IL−10陽性細胞の濃縮 IL−10産生細胞を、市販のキット(Miltenyi)によって濃縮した。精製T細胞を、不動状態の抗CD3及び溶解抗CD28の存在下、106/mlの濃度で培養した。培養の3時間後、細胞を回収し、CD45及び他のmAb捕獲ネズミIL−10に対して指向するmAbからなるダイアボディで、4℃にて10分間標識した。次いで、細胞を105/mlの最終濃度で希釈し、37℃で45分間サイトカインを分泌させた。サイトカイン捕捉期間の後、細胞を回収し、0.5%のBSA及び5nMのEDTA(バッファ)を含有するPBSに108/モル再懸濁し、PE抱合αIL−10mAb(BD)で4℃にて10分間染色した。細胞を1回バッファで洗浄し、108/モル再懸濁し、抗−PEマイクロビーズで4℃にて10分間染色した。IL−10濃縮細胞集団を磁気カラムで単離した。細胞サンプルをFACSカリバー・フローサイトメトリー(BD)で分析した。
【0059】
細胞培養 抑制実験のために、実験に使用していないCD4+NODのT細胞を、他で述べたように(Lyonsら、1994)、CFSE(Molecular Probes、Eugene、OR)で染色し、96ウェルプレート(1×105ウェル)中にて、抗CD3mAb(BD)10μg/mlの存在下で培養した。ラパマイシン又はラパマイシン+IL−10で20週間処置したNODマウスから得られたCD4+T細胞を、1:1の割合で培養液に加え、分裂した実験に使用していない細胞のパーセントを評価し、なんら加えられていない細胞における分裂細胞のパーセントと比較した。分裂細胞を、総事象CFSE+によって増殖集団に含有された分裂事象によって評価した。媒体中に放出されたサイトカインの測定のために、精製T細胞(1×105/ウェル)を、96ウェルプレートで培養し、10μg/mlの免疫化抗CD3(BD)及び1μg/mlの溶解抗CD28(BD)で刺激した。培養の48時間(IL−5検出のために)及び96時間(IL−10及びTGF−β検出のために)後に上清を採取した。
【0060】
フローサイトメトリー 説明したAbs(全てBDから)で細胞を染色し、CellQuest ソフトウェア (BD)を装備したFACスキャンフローサイトメータで分析した。
【0061】
サイトカイン測定 採取された上清中に存在するサイトカインを、標準的な市販のキット(BD)を用いて、サンドイッチERISA又はフローサイトメトリー原理のアッセイ(CBA)によって定量した。特定のサイトカインを産生する細胞のパーセントを細胞内染色によって測定した。精製T細胞を6時間、10μg/mlの免疫化抗CD3及び1μg/mlの溶解抗CD28(BD)で、1×106/mlの濃度で、染色した。ブレフェドリンAを培養の最後の3時間添加した。細胞内染色を、先に述べたように(Trembleauら、2000)行った。
【図面の簡単な説明】
【0062】
【図1】IL−10プロトコル及びエドモントンプロトコルで処置したマウスの移植片生着を示す。
【図2】IL−10プロトコルで抗Tacなしとした場合の異種遺伝子型β−島拒絶の増加を示す。
【図3】IL−10プロトコルで処置したマウスからのT細胞について、インビトロでの増殖能を示す。
【図4】IL−10プロトコルで処置したマウスからのT細胞についての抗原−特異増殖能を示す。
【図5】IL−10プロトコルで処置したマウスから単離したT細胞について、IL−10の産生を示す。
【図6】IL−10産生T細胞の特異集団について、IL−10プロトコルで処置したマウスからの単離を示す。
【図7】IL−10によるインビボでのIL−10産生T細胞の誘発を示す。
【図8】NODマウスにおけるタイプI糖尿病の治療のためのラパマイシン+IL−10での予備的結果を示す。
【図9】IL−10プロトコルが、糖尿病誘発性T細胞の移植後のNOD.SCIDマウスにおいて誘発した糖尿病を阻害した実験結果を示す。
【図10】タイプI糖尿病の治療のためのラパマイシン+IL−10の使用効果を示す。
【図11】ラパマイシン±IL−10で処置したマウスにおけるTr細胞の濃度を示す。
【図12】インビトロにおける免疫応答を抑制するTr細胞の能力を示す。
【0063】
参考文献
Bacchetta, R. , M. Biglerら (1994)."High levels of interleukin 10 production in vivo are associated with tolerance in SCID patients transplanted with HLA mismatched hematopoietic stem cells. Growth and expansion of human T regulatory type 1 cells are independent from TCR activation but require exogenous cytokines. "Journal of Experimental Medicine 179 (2): 493- 502.
Bacchetta R. , Parkman R. ら(1995) "Dysfunctional cytokine production by host-reactive T-cell clones isolated from a chimeric severe combined immunodeficiency patient transplanted with haploidentical bone marrow." Blood 85: 1944-1953.
Baker, K. S. , M. G. Roncaroloら(1999). "High spontaneous IL-10 production in unrelated bone marrow transplant recipients is associated with fewer transplant-related complications and early deaths. "Bone Marrow Transplant 23 (11): 1123-9.
Battaglia, M. , B. R. Blazarら(2002). "The puzzling world of murine T regulatory cells. "Microbes Infect 4 (5): 559-66.
Blaha, P. Bigenzahn, S. ら(2003). "The influence of immunosuppressive drugs on tolerance induction through bone marrow transplantation with costimulation blockade. "Blood 101: 2886-2893.
Blazar, B. R., P. A. Taylorら(1998). "Interleukin-10 dose-dependent regulation of CD4+and CD8+T cell-mediated graft-versus-host disease." Transplantation 66 (9): 1220-9.
Davalli, A. M. , L. Scagliaら(1996). "Vulnerability of islets in the immediate posttransplantation period. Dynamic changes in structure and function. "Diabetes 45 (9): 1161-7.
Delovitch, T. L. , B. Singh. (1997) "The nonobese diabetic mouse as a model of autoimmune diabetes: immune dysregulation gets the NOD"Immunity 6 (7): 727-738.
Gregori S. , Giarratana N. ら(2003) "Dynamics of pathogenic and suppressor T cells in development of autoimmune diabetes"J. Immunol 171: 4040-4047.
Groux, H. , A. O'Garraら(1997). "A CD4+T-cell subset inhibits antigenspecific T-cell responses and prevents colitis. "Nature 389 (6652): 737-42.
Hackstein, H. , T. Tanerら(2002). "Rapamycin inhibits macropinocytosis and mannose receptor-mediated endocytosis by bone marrow-derived dendritic cells. "Blood 100 (3): 1084-7.
Hempel, L. , D. Korholzら(1997). "High interleukin-10 serum levels are associated with fatal outcome in patients after bone marrow transplantation." Bone Marrow Transplant 20 (5): 365-8.
Hojo, M. , T. Morimotoら(1999). "Cyclosporine induces cancer progression by a cell-autonomous mechanism. "Nature 397 (6719): 530-4.
Li, W. , F. Fuら(1999). "Recipient pretreatment with mammalian IL-10 prolongs mouse cardiac allograft survival by inhibition of anti-donor T cell responses. "Transplant Proc 31 (1-2): 115.
Lyons, A. B. Parish, C. R. (1994)"Determination of lymphocyte division by flow cytometry"J Immunol Methods 171: 131-137.
Moore, K. W. , R. de Waal Malefytら(2001). "Interleukin-10 and the interleukin-10 receptor. "Annu Rev Immunol 19: 683-765.
Roncarolo M. G. , Battaglia M. ら(2003) "The role of interleukin 10 in the control of autoimmunity"J. Autoimm 4: 269-272 Sehgal, S. N. (1998). "Rapamune (RAPA, Rapamycin, sirolimus): mechanism of action immunosuppressive effect results from blockade of signal transduction and inhibition of cell cycle progression. "Clin Biochem 31 (5): 335-40.
Shapiro, A. M. , J. R. Lakeyら(2000). "Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen."N Engl J Med 343 (4): 230-8.
Taylor, P. A. , C. J. Leesら(2002). "The infusion of ex vivo activated and expanded CD4 (+) CD25 (+) immune regulatory cells inhibits graft-versus-host disease lethality. "Blood 99 (10): 3493-9.
Tisch, R. and H. McDevitt (1996). "Insulin-dependent diabetes mellitus."Cell 85 (3) : 291-7.
Trembleau, S. , G. Pennaら(2000)."Early Thl response in unprimed nonobese diabetic mice to the tyrosine phosphatase-like insulinoma-associated protein 2, an autoantigen in type 1 diabetes."3 Immunol 165 (12): 6748-55.
Wells, A. D. , X. C. Liら(1999). "Requirement for T-cell apoptosis in the induction of peripheral transplantation tolerance. " Nat Med 5 (11): 1303-7.
Yu, X. , P. Carpenterら(2001). "Advances in transplantation tolerance." Lancet 357 (9272): 1959-63.
【特許請求の範囲】
【請求項1】
T細胞媒介免疫応答の調節における使用のためのIL−10及びラパマイシン、それらのアナログ、誘導体又は抱合体を含む組み合わせ医薬製剤。
【請求項2】
亢進、機能不全又は制御されていない自己又は非自己T細胞媒介免疫応答を含む疾患の予防又は治療上の処置における同時の、別々の又は連続使用のための請求項1の医薬製剤。
【請求項3】
異質遺伝子型臓器拒絶、同種異系移植β−島拒絶、移植片対宿主拒絶反応及びタイプI糖尿病の予防又は治療上の処置における同時の、別々の又は連続使用のための請求項2の医薬製剤。
【請求項4】
関節リウマチ、多発性硬化症、全身性エリテマトーデスから選択される自己免疫疾患;慢性炎症性疾患;クローン病及び潰瘍性大腸炎から選択される炎症性腸疾患;慢性閉塞性肺疾患;喘息及びアトピー性皮膚炎から選択されるアレルギー疾患;繊維症;及び遺伝子治療誘発生成物に対する免疫反応の予防又は治療上の処置における同時の、別々の又は連続使用のための請求項2の医薬製剤。
【請求項5】
さらに免疫抑制剤又は免疫調節剤、抗体及びサイトカインから選択される活性物質を含む請求項1の医薬製剤。
【請求項6】
活性成分が、サイクロスポリン、FK506、ピメクロリムス、マイコフェノラート、CD45に対する抗体、LFA−1分子又はIL−2受容体から選択される請求項5の医薬製剤。
【請求項7】
IL−2受容体α、β及びγ鎖に対する抗体を含む請求項6の医薬製剤。
【請求項8】
医薬的に許容される賦形剤とともに、IL−10及びラパマイシンもしくはそれらのアナログ、誘導体又は抱合体を含む医薬組成物。
【請求項9】
溶液、懸濁液、錠剤又はカプセルの形態である請求項8の医薬組成物。
【請求項10】
免疫調節剤の製造のための、IL−10及びラパマイシンもしくはそれらのアナアログ、誘導体又は抱合体の組み合わせの使用。
【請求項11】
免疫抑制剤の製造のための請求項10の使用。
【請求項12】
免疫抑制剤が抗原−特異免疫耐性を誘発する請求項11の使用。
【請求項13】
免疫抑制剤、抗原−特異免疫耐性がTr1細胞及び/又はCD4+CD25+Tr細胞によって媒介される請求項11又は12の使用。
【請求項1】
T細胞媒介免疫応答の調節における使用のためのIL−10及びラパマイシン、それらのアナログ、誘導体又は抱合体を含む組み合わせ医薬製剤。
【請求項2】
亢進、機能不全又は制御されていない自己又は非自己T細胞媒介免疫応答を含む疾患の予防又は治療上の処置における同時の、別々の又は連続使用のための請求項1の医薬製剤。
【請求項3】
異質遺伝子型臓器拒絶、同種異系移植β−島拒絶、移植片対宿主拒絶反応及びタイプI糖尿病の予防又は治療上の処置における同時の、別々の又は連続使用のための請求項2の医薬製剤。
【請求項4】
関節リウマチ、多発性硬化症、全身性エリテマトーデスから選択される自己免疫疾患;慢性炎症性疾患;クローン病及び潰瘍性大腸炎から選択される炎症性腸疾患;慢性閉塞性肺疾患;喘息及びアトピー性皮膚炎から選択されるアレルギー疾患;繊維症;及び遺伝子治療誘発生成物に対する免疫反応の予防又は治療上の処置における同時の、別々の又は連続使用のための請求項2の医薬製剤。
【請求項5】
さらに免疫抑制剤又は免疫調節剤、抗体及びサイトカインから選択される活性物質を含む請求項1の医薬製剤。
【請求項6】
活性成分が、サイクロスポリン、FK506、ピメクロリムス、マイコフェノラート、CD45に対する抗体、LFA−1分子又はIL−2受容体から選択される請求項5の医薬製剤。
【請求項7】
IL−2受容体α、β及びγ鎖に対する抗体を含む請求項6の医薬製剤。
【請求項8】
医薬的に許容される賦形剤とともに、IL−10及びラパマイシンもしくはそれらのアナログ、誘導体又は抱合体を含む医薬組成物。
【請求項9】
溶液、懸濁液、錠剤又はカプセルの形態である請求項8の医薬組成物。
【請求項10】
免疫調節剤の製造のための、IL−10及びラパマイシンもしくはそれらのアナアログ、誘導体又は抱合体の組み合わせの使用。
【請求項11】
免疫抑制剤の製造のための請求項10の使用。
【請求項12】
免疫抑制剤が抗原−特異免疫耐性を誘発する請求項11の使用。
【請求項13】
免疫抑制剤、抗原−特異免疫耐性がTr1細胞及び/又はCD4+CD25+Tr細胞によって媒介される請求項11又は12の使用。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2006−511510(P2006−511510A)
【公表日】平成18年4月6日(2006.4.6)
【国際特許分類】
【出願番号】特願2004−556199(P2004−556199)
【出願日】平成15年11月27日(2003.11.27)
【国際出願番号】PCT/EP2003/013351
【国際公開番号】WO2004/050090
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(505196808)
【氏名又は名称原語表記】RONCAROLO,Maria,Grazia
【住所又は居所原語表記】Via F.IIi Cervi,Res.Andromeda,i.131,I−20090 Segrate Italy
【Fターム(参考)】
【公表日】平成18年4月6日(2006.4.6)
【国際特許分類】
【出願日】平成15年11月27日(2003.11.27)
【国際出願番号】PCT/EP2003/013351
【国際公開番号】WO2004/050090
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(505196808)
【氏名又は名称原語表記】RONCAROLO,Maria,Grazia
【住所又は居所原語表記】Via F.IIi Cervi,Res.Andromeda,i.131,I−20090 Segrate Italy
【Fターム(参考)】
[ Back to top ]