
免疫系刺激のための材料および方法
本発明は、新規多糖に関する。RR1は、(1→4)結合主鎖および(1→6)結合分枝から構成されるα-D-グルカン多糖であり、薬草のティノスポラ・コルディフォリア(Tinospora cordifolia)から単離されている。RR1は、固有の免疫刺激特性を示し、正常リンパ球および腫瘍細胞株に対して無毒であり、かつ非増殖性である。本発明はまた、RR1化合物を含有する組成物、およびRR1化合物を用いた被験体の免疫応答を調節するための方法にも関する。本発明はまた、免疫応答を刺激する抗原と共にRR1化合物を使用するための方法を提供し、このRR1化合物は、抗原に対するTh1型免疫応答の生成においてアジュバント様の活性を提供する。
【発明の詳細な説明】
【背景技術】
【0001】
関連出願の相互参照
本出願は、いかなる図、表、核酸配列、アミノ酸配列、および図面も含み、全体として参照により本願明細書に組み入れられる、2004年7月9日出願の米国仮特許出願第60/586,548号の恩典を主張する。
【0002】
発明の背景
バイオテロ、重症急性呼吸器症候群(SARS)および鳥インフルエンザウイルスの大発生、HIV/AIDSの持続的拡大、ならびに現行治療薬剤に耐性である病原株の出現により、これら脅威に対するバイオ防御戦略を含めた新たな防衛的手段の探索が研究者に切実に求められている。免疫活性化は、新興感染症に対するバイオ防御戦略のための、有効的かつ予防的取り組みである(Hackett, C.J. J. Allergy and Clin. Immunol., 2003, 112:686-694)。ヒト免疫系は幅広い微生物検出および宿主防御メカニズムを有しているが、免疫監視および破壊システムの病原体回避が一般的である。この状況は、ほとんどの腫瘍細胞が検出されないまま免疫監視を免れている癌の場合と異なりはしない(Wajchman, H.J. et al. Cancer Res., 2004, 64:1171-1180)。
【0003】
特異的受容体の助けを借りて、病原体または非自己侵入物を検出すること、かつ免疫コンピテント細胞の活性化、サイトカインおよびケモカインの合成ならびに侵入物を排除または封じ込めるために炎症性メディエイターの放出により直ちに応答することが、自然免疫系の構成要素である。自然免疫の活性化はまた、抗原特異的TおよびBリンパ球による適応的免疫応答を誘発し、道を開く。ナチュラルキラー(NK)細胞は、異常な細胞を認識して破壊するその能力を活かして、適応免疫が未発生の感染したウイルスおよび悪性細胞に対する自然免疫防御に決定的な役割を果たしている(Smith, H.R. et al. Proc. Natl. Acad. Sci. USA, 2002, 99:8826-8831; Moser, J.M. et al. Curr. Opin. Immunol., 2002, 14:509-516)。
【0004】
複数の化合物が免疫系を活性化し、微生物のリポ多糖類(LPS)、非メチル化CpGモチーフを含有している2本鎖RNAおよびDNAオリゴヌクレオチドなどの化合物が早期に報告されている(Hackett, C.J. J. Allergy and Clin. Immunol., 2003, 112:686-694; Kandimalla, E.R. et al. Proc. Natl. Acad. Sci. USA, 2003, 100:14303-14308)。複数の受容体刺激が、サイトカイン産生に相乗的な効果を示すことが報告されている(Gao, J.J. et al. J. Immunol., 2001, 166:6855-6860)。補体活性カスケードは、自然免疫システムのもう一つの重要な部分であり、そこでは細胞内細菌などの細胞性病原体が補体化合物によって覆われて(オプソニン化)食作用を受けやすくなる。したがって、補体活性化は、微生物の殺滅に重要な役割を果たし、かつ免疫複合体の移送およびクリアランスにとって必須である。
【0005】
感染またその他抗原性の攻撃に対し生ずるタイプの適応免疫応答(特異的または獲得免疫応答としても知られている)は、応答に関与するTヘルパー(Th)細胞のサブセットによって一般的に区別できる。Th1応答(細胞性または細胞媒介応答)は、遅延型過敏症および細胞傷害性Tリンパ球(CTL)の活性化などの古典的な細胞媒介機能を担っているのに対し、Th2応答(体液性免疫)は、B細胞活性化のためのヘルパーとしてより効果的に機能する。抗原に対する免疫応答のタイプは、一般的には、細胞が抗原に応答して産生するサイトカインによって決定される。Th1およびTh2細胞が分泌するサイトカインの違いは、これら2つサブセットの生物学的機能の差に反映されていると考えられている。
【0006】
サイトカインが、免疫応答の様々な局面の制御において重要な役割を果すことが明らかになってきた。サイトカインの中でもインターロイキン(IL)-12は、自然免疫と細胞媒介適応免疫との協調について中心的な役割を果たしている(Watford, W.T. et al. Cytokine Growth Factor Rev., 2003, 14:361-368)。免疫刺激による予防的防御および曝露後防御が証明されている(Walker, P.S. et al. Proc. Natl. Acad. Sci. USA, 1999, 96:6970-6975; Juffermans, N.P. et al. Infect. Immunol, 2002, 70:147-152)。通常、これら防御的手段は、適応免疫系に関係するT細胞分化のTh1経路のサイトカインである、IL-12とインターフェロン(IFN)-γとの合成と相関している(Gramzinski, A.M. et al. Infect. Immunol., 2001, 69:1643-1649)。
【0007】
Th1サブセットは、IL-2およびINF-γを分泌し、これがCTLを活性化することから、特にウイルス感染および細胞内病原体に対する応答に適しているであろう。Th2サブセットは、IL-4およびIL-5がそれぞれIgE産生および好酸球の活性化を誘発することが知られていることから、自由生活細菌および寄生虫に対する応答により好適であり、アレルギー応答を仲介することもある。一般的に、Th1およびTh2細胞は、明らかに異なるパターンのサイトカインを分泌しており、それゆえに一方のタイプの応答が他方のタイプの応答の活性を弱めることがある。Th1/Th2のバランスのシフトは、例えばアレルギー応答、またはCTL応答の増大をもたらし得る。
【0008】
特定抗原に対する宿主動物の免疫化は、伝統的には免疫原性の形をした抗原を宿主に、繰り返しワクチン接種することによって成し遂げられている。最近のワクチンは、効果的な体液性(抗体または「Th2型」)応答を引き出すが、それらは細胞性応答(特に主要組織適合性複合体(MHC)クラスI拘束CTLまたは「Th1型」応答)は引き出すことができず、応答は一般的には存在しないかまたは弱い。Th2型応答は、結核およびマラリアなどの多くの感染症について、感染に対する防御的な価値をほとんど有していない。さらには、抗体応答は、特定の適応症では不適当であり、特にアレルギーで顕著であり、この場合抗体応答はアナフィラキシーショックを起こすことがある。標的抗原に由来する小量ペプチドを利用する提案されているワクチン、および感染の可能性があるインタクトなウイルス粒子の使用を回避するその他現在用いられている抗原性作用物質は、治療効果を達成するのに必要な免疫応答を必ずしも引き出すわけではない。治療に有効なヒト免疫不全症ウイルス(HIV)ワクチンが存在しないことは、この失敗の不幸な例である。
【0009】
マクロファージ、好中球およびナチュラルキラー(NK)細胞が媒介する自然免疫は、微生物侵入に対する宿主防御機構の第一線である。自然免疫系は、構造的に保存された病原体関連分子パターン(PAMP)を、パターン認識受容体(PRR)と呼ばれる特異的な生殖系列がコードする受容体を介して標的としている(Aderem, A. and Ulevitch, R.J. Nature, 2000, 406:782-787)。自然および合成免疫促進剤による免疫系の強化は、特に免疫系が弱体化し、抗生物質耐性である場合には、従来の治療法に比べはるかに優れている。
【0010】
いくつかの前臨床および臨床研究は、生物学的応答調整物質(BRM)の一分類であるβ-グルカンの、創傷の治癒の加速、ならびに全身性炎症応答症候群および敗血症ショックの編成にとっての有用性を示している(Ross, G.D. et al. Immunopharmacology, 1999, 42:61-74; Williams, D.L. Mediators Inflamm., 1997, 6:247-250; Hetland, G. Curr. Med. Chem., 2003, 2:135-146)。β-グルカンは、無脊椎動物の自然免疫系の強力な刺激因子であるが、哺乳動物では、それらは補体系の強力な活性化因子である。これらのポリマーは、抗腫瘍活性および抗感染活性、ならびに真菌、細菌、ウイルスおよび原虫感染に対する防御を含めた免疫系に対するそれらの効果から、治療可能性を有している。可溶性および粒状のβ-グルカンは、マクロファージにある起源を同じくする受容体と相互作用し、サイトカイン、ケモカインおよび反応性酸素中間体の合成を刺激する(Ganter, B.N. et al. J. Exp. Med., 2003, 197:1107-1117)。マクロファージ上でのβ-グルカン認識/結合について報告されている主要受容体は、補体受容体3(CD11b/CD18またはCR3)、デクチン-1、ならびにトール様受容体(TLR)2および6である。β-グルカン認識においては、ラクトシルセラミドおよびスカベンジャー受容体も確認されているが、それらの機能はよく分かっていない(Willment, J.A. et al. J. Biol. Chem., 2001, 276:43813-43823)。
【0011】
トール様受容体(TLR)は、細胞内シグナル伝達機能に関する細胞質モチーフを持つトール-インターロイキン(IL)-1受容体(TIR)の大きなスーパーファミリーの一部である。これらの分子は、第一線の宿主防御を提供し、ハエから哺乳動物に至る様々な生物において、感染および自己免疫疾患に関係している。現在TLRは、それにより哺乳動物が感染を感知する、第一のシグナル伝達分子であることが受け入れられている(Beutler, B. et al. J Leukocyte Biol., 2003, 74:479-485)。哺乳動物では、12種類のTLRが同定されており、それぞれが異なるPAMPを認識している(Akira, S. and Sato, S. Scand. J. Infect. Dis., 2003, 35:555-562)。TLR3を除く全てのTLR、IL-1受容体およびその他TIRドメイン含有受容体は、アダプター骨髄分化因子88(MyD88)に依存した、共通のシグナル伝達経路を共有している(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117; Mukhopadhyay, S. et al. Immunol., 2004, 112:521-530)。MyD88以外にも、いくつかのアダプター分子が最近報告されており、これらアダプター分子の差別的な利用が、TLRシグナル伝達のために特異性を提供しているかもしれない(Akira, S. J. Biol. Chem., 2003, 278:38105-38108)。TLR間、ならびにTLRとその他表面受容体との間の物理的、および/または機能的相互作用の証拠が得られている。Gantnerらは、β-グルカン刺激によるデクチン-1とTLRの共同誘導、ならびにこれら2つの受容体間のNFκB活性化に関する相乗的相互作用について報告している(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117)。
【0012】
TLR媒介サイトカイン産生は、JNKおよび核因子(NF)-κBを活性化するIL-1R関連キナーゼ(IRAK)-4およびTNF受容体関連因子-6(TRAF-6)といった、その下流のメディエーターに依存している(Akira, S. J. Biol. Chem., 2003, 278:38105-38108)。NF-κBは、多くの免疫エフェクター細胞でのサイトカイン遺伝子発現を制御する普遍的な転写因子である。多くの細胞において、NF-κBは、通常、細胞質内に潜伏的、不活性な状態で、阻害タンパク質κB(I-κB)と結合して存在している(Ghosh, S and Karin, M., Cell, 2002, 109:S81-96)。それは、炎症性サイトカイン、ウイルス産物、リポ多糖類、タキソールなどの植物由来化合物、ならびに病原菌および非病原菌由来のβ-グルカンのような様々な刺激物によって活性化される(Akira, S. and Sato, S. Scand. J. Infect. Dis., 2003, 35:555-562; Young, S.H. et al. J. Biol Chem., 2001, 276:20781-20787; Lebron, F. et al. J Biol Chem., 2003, 278:25001-25008)。刺激を受けるとI-κBαは、直ちにリン酸化されて、プロテアソーム機構により急速に分解され、次に活動性NF-κBを放出して核に移行し、DNAに結合してサイトカイン/ケモカイン遺伝子の転写を開始させる(Ghosh, S, and Karin, M., Cell, 2002, 109:S81-96; Auphan, N. et al. Science, 1995, 270:232-233)。
【0013】
グルカンの免疫刺激特性は、β-グリコシド結合、枝分れ度、および溶液のコンホメーションに拠るとされている(Muller, A et al. Glycobiology, 2000, 10:339-346)。本発明は、薬用植物のティノスポラ・コルディフォリア(Tinospora cordifolia)から得ることができる新規の多糖、(1,4)-α-D-グルカン、RR1の免疫刺激特性の特徴を明らかにし報告している(Nair, P.K. et al. Int. Immunopharmacol., 2004, 4:1645-1659)。本新規α-グルカンは水溶性であり、かつ6〜7グルコース単位の間隔で(1,6)-α-D-グリコシド結合側鎖を持つ主鎖の中に(1,4)-α-D-グリコシド結合を有している。それは正常細胞ならびに腫瘍細胞株(CEM、CEM/VLB)に対しては、1000μg/mlまで非細胞傷害性であり、ヒトリンパ球サブセットを様々なレベルで活性化する。自然免疫の主要細胞の一つであるNK細胞の活性化は、機能的アッセイにおけるRR1処理リンパ球による標的細胞の殺滅レベルの増大によって証明された。RR1刺激を受けたサイトカインプロフィールは、Th1応答の過剰産生の自己制御機構と思われる、高レベルの調節サイトカイン誘導を伴った、Tヘルパー細胞分化のより望ましいTh1経路を証明している。その水溶性、非細胞傷害性、および草本起源であることは、RR1の免疫刺激に関する臨床上の可能性を示している。
【発明の開示】
【0014】
発明の概要
本発明は、新規の免疫系刺激多糖に関する。本明細書に例示する、一つのこの様な化合物は、薬用植物のティノスポラ・コルディフォリアから単離され、特徴付けされている。一つの局面では、本発明は、図3に示す構造を有する化合物(本明細書ではRR1とも呼ぶ)を提供し、この化合物は、(1→4)結合主鎖および(1→6)結合分枝から構成され、550kDaより大きな分子量を持ち、固有の免疫刺激特性を示すα-D-グルカン多糖である。
【0015】
RR1の免疫刺激剤特性は、RR1誘発のヒトリンパ球活性化、NK細胞機能活性、食作用活性、補体活性化、サイトカインおよびケモカイン合成、誘導性酸化窒素合成酵素(iNOS)の誘発、酸化窒素産生、ならびに酸化ストレス測定の分析によって確認された。結果は、自然免疫系コンポーネントの強化を明瞭に証明し、一方サイトカインプロフィールは、抗原特異的細胞媒介(適応)免疫でのTヘルパー細胞分化のTh1経路を証明した。RR1による刺激は、自然免疫系の主要エフェクター細胞であるナチュラルキラー(NK)細胞の活性化、ならびに自己増幅しかつ抗体非存在時の病原体認識および排除において重要である、第2経路での補体活性化を数倍高めた。RR1は、マクロファージによるザイモサン粒子の食作用も高める。本明細書中でさらに詳しく記載するように、RR1はまた、Th1サイトカイン、IL-12、IL-18、IL-1βおよびIFN-γの合成をアップレギュレートした。RR1はまた、抗腫瘍サイトカインのTNF-αおよびMCP-1の産生を誘発したが、iNOSの有意な誘発は起こさなかった。このサイトカイン合成のアップレギュレーションは、iκBタンパク質のリン酸化を通じたNF-κB転写因子の活性化によって促進される。RR1免疫刺激のプロセスは、トール様受容体(TLR)6およびデクチン-1などの受容体が関係する。
【0016】
RR1の投与によって強化された自然免疫系は、病原体の侵入から被験体を守ることができ、一方ブーストされたTh1細胞は、細胞内病原体、ウイルスおよび悪性細胞を見つけ出して破壊する。それゆえにRR1は、様々なヒトおよび動物の病気の免疫賦活性剤、化学予防剤および治療薬剤として用いることができる。RR1はまた、癌患者の化学療法薬剤治療のための抗癌剤(例えばモノクローナル抗体)など、その他の薬物と組み合わせることもできる。
【0017】
したがって、本発明の別の局面は、被験体の免疫応答を調節するための方法を提供する。一つの態様では、RR1化合物(RR1または薬学的に許容されるその塩または類似体)は、被験体に対し、RR1化合物のアジュバント様効果を通して、免疫応答の全般的刺激を提供するのに有効である量が投与される。本発明はまた、本発明のRR1化合物を抗原と共に使用し、免疫応答を刺激するための方法も提供する。好ましくは、このような方法で使用する場合、RR1化合物は、抗原に対しTh1型の免疫応答を作り出すアジュバント様活性を提供する。
【0018】
別の局面では、本発明は、少なくとも一つのRR1化合物および薬学的に許容される担体を含有する、免疫調節組成物を提供する。
【0019】
本発明のさらなる局面は、RR1をティノスポラ・コルディフォリア植物材料から得るためのプロセスを提供する。
【0020】
発明の詳細な説明
本発明は、固有の免疫ブースト特性を示す新規多糖である単離化合物(RR1)を提供する。RR1は、図3に示す構造を有し、(1→4)結合主鎖および(1→6)結合分枝から構成され、550kDaより大きな分子量を持つα-D-グルカン多糖である。RR1は、薬草のティノスポラ・コルディフォリア(ツツラフジ(Minispermaceae)科)から単離されており、その薬効特性が研究の主題となってきた(Singh, S.S. et al. Ind. J. Pharm., 2003, 35:83-91; Chintalwar, G. et al. Phytochemistry, 1999, 52:1089-1094; Manjerakar, P.N. et al. Fitotherapia, 2000, 71:254-257; Desai, V.R. et al. Proc. Indian Acad. Sci, 2002, 114:713-719; Subramanyan, M. et al., Redox. Rep., 2002, 7:137-143)。
【0021】
RR1は、正常リンパ球ならびに1000μg/mlの腫瘍細胞株に対してでさえ非細胞傷害性および非増殖性である。RR1は、100μg/mlの濃度で、NK細胞(331%)、T細胞(102%)およびB細胞(39%)などの様々なリンパ球のサブセットを活性化した。NK細胞の有意な活性化は、機能的アッセイにおいて、活性化された正常リンパ球による用量依存的な腫瘍細胞の殺滅を導く。正常リンパ球でのRR1による免疫活性化は、100μg/ml濃度でIL-1β(1080pg/ml)、IL-6(21833pg/ml)、IL-12 p70(50.19pg/ml)、IL-12 p40(918.23pg/ml)、IL-18(27.47pg/ml)、IFN-γ(90.16pg/ml)、TNF-α(2225pg/ml)およびMCP-1(2307pg/ml)の合成を引き出したが、IL-2、IL-4、IL-10、IFN-αおよびTNF-βの産生は誘発しなかった。このサイトカインプロフィールは、Tヘルパー細胞分化のTh1経路が、その過剰産生を制御する自己調節機構と共に、細胞媒介免疫にとって必須であることを証明している。
【0022】
RR1はまた、C3a des Arg成分が段階的に増加することで証明されたように第2経路の補体成分も活性化した。偶然に、RR1刺激はリンパ球にいかなる酸化的ストレスも起こさず、iNOSの誘発が有意でないことから証明されるように、酸化窒素産生増大を極わずかに引き起こしたに過ぎない。RR1は水溶性であり、高分子量を有し、リンパ球を活性化(特にNK細胞)し、補体を活性化し、Th1経路に関係するサイトカインプロフィールを誘発し、低レベルの酸化窒素合成を伴い、および酸化的ストレスがないという事実は、本新規α-D-グルカンに可能な生体防御特性を付与している。
【0023】
RR1のシグナル伝達機構がマクロファージで研究され、その免疫刺激特性の理解はさらに進んだ。RAW264.7マクロファージをRR1と4℃でインキュベートすると、RR1は、ザイモサンA生体粒子の非オプソニン結合および食作用を用量依存的な方法で阻害した。しかしながら、それはザイモサンA生体粒子のオプソニン結合および内在化に対してはほとんど影響しなかった。マクロファージを抗CD11b mAbとインキュベーションしてからRR1とインキュベーションすると、RR1誘発のTNF-α合成に対する任意の阻害作用は見られず、このことからCR3が、ザイモサンとは異なり、マクロファージでのRR1のオプソニン結合および内在化に関係していないことが確認された。抗CD11b mAbは、ザイモサンAが誘発するTNF-α合成に対し有意な阻害作用を有している。RR1は、マクロファージでのTNF-α合成を用量依存的な方法で誘発したが、これはNF-κBインヒビターであるカフェ酸フェネチルエステル(CAPE)によって完全に阻害され得る。RR1はNF-κBを、時間および用量依存的な方法で活性化し、この核NF-κB活性の調節に伴ってI-κBαが分解し、これがNF-κBの核内への移行を促進する。RR-1が誘発するNF-κB活性は、RR1刺激8時間目にピークに達するが、一方I-κB-αの分解は刺激1時間以内に起こった。RR1が誘発するNF-κBの活性化は、TLR6シグナル伝達を介して起こった。なぜならば、RR1は、TLR6をトランスフェクトしたHEK293細胞でIL-8合成を誘発したが、他のトランスフェクタントでは誘発しなかった。これらの結果は、RR1が、TLR6シグナル伝達、NF-κBの移行、および免疫タンパク質の産生を通して起こるマクロファージの活性化を通じて免疫系を活性化することを示している。単球/マクロファージにおけるRR1による免疫刺激のシグナル伝達経路の概略図を図20に示す。
【0024】
本発明のRR1化合物によって刺激された免疫応答は、Th1型の表現系に向かって偏り、Th2型の表現系とは疎遠であることが好ましい。本発明との関係では、Th1型免疫応答への刺激は、RR1化合物を用いて処理した細胞のサイトカイン産生を、RR1化合物なしで処理した細胞の産生と比較して測定することによって、インビトロまたエクスビボで決定できる。細胞のサイトカイン産生を決定する方法としては、本明細書に記載の方法ならびに当技術分野で公知のその他の方法を挙げることができるが、これらに限定されない。RR1処理に応答して産生されるサイトカインは、細胞によるTh1型またはTh2型に偏った免疫応答を示している。
【0025】
本明細書で使用する、用語「Th1型に偏った」サイトカインの産生とは、刺激物存在下におけるTh1型応答関連サイトカイン産生と、刺激物非存在下におけるTh1型応答関連サイトカイン産生とを比べ測定可能に増加した産生を指す。このようなTh1型に偏ったサイトカインの例としては、IL-2、IL-12およびIFN-γが挙げられるが、これらに限定されない。これに対し「Th2型に偏ったサイトカイン」とは、Th2型免疫応答に関係するサイトカインを指し、IL-4、IL-5、IL-10およびIL-13を挙げることができるが、これらに限定されない。RR1活性の決定に有用な細胞としては、免疫系細胞、被験体より単離された一次細胞、および/または細胞株、好ましくはAPCおよびリンパ球、よりさらに好ましくはマクロファージならびにT細胞を挙げることができる。
【0026】
Th1型免疫応答への刺激は、RR1化合物で処理された被験体で測定することができ、下記の方法を含む任意の公知の方法によって決定することができるが、これらに限定されない:(1)抗原に曝露された前後で測定されたIL-4レベルの低下;あるいは、RR1処理被験体と、抗原により刺激され、または刺激および曝露されたRR1なしで処理された対照とを比較した時の、IL-4のより低いレベル(もしくは非存在)を検出する方法;(2)抗原に曝露された前後でのIL-12、IL-18および/またはIFN(α、β、もしくはγ)レベルの増大;あるいは、RR1処理被験体と、抗原により刺激され、または刺激および曝露されたRR1なしで処理された対照とを比較した時の、より高いレベルのIL-12、IL-18および/またはIFN(α、β、もしくはγ)を検出する方法;(3)RR1処理被験体と、RR1なしで処理された対照とを比較した時の、IgG2a抗体産生による方法;かつ/または(4)抗原に曝露された前後で測定された抗原特異的IgEレベルの低下;あるいは、RR1処理被験体と、抗原により刺激され、または刺激および曝露されたRR1なしで処理された対照とを比較した時の、抗原特異的IgEのより低いレベル(もしくは非存在)を検出する方法。これら各種決定は、APCおよび/またはリンパ球、好ましくはマクロファージおよび/またはT細胞が作るサイトカインを、本明細書に記載されている方法、または当技術分野公知のいずれかの方法を用いてインビトロまたはエクスビボで測定することによって、実施できる。
【0027】
RR1投与の結果として起こるTh1に偏ったサイトカインの誘発は、NK細胞、細胞傷害性キラー細胞、Th1ヘルパーおよびメモリー細胞が実行するような強化された細胞性免疫応答を生ずる。これらの応答は、ウイルス、真菌、寄生原虫、細菌、アレルギー性疾患および喘息、ならびに腫瘍に対する予防または治療ワクチン接種での使用のために特に有益である。
【0028】
本発明のRR1化合物は、免疫抑制を伴う状態などの、免疫刺激を必要とする、ヒトまたはヒト以外の動物の病理的状態を治療するために有用である。免疫刺激が望まれる状態の例としては、骨髄炎、慢性気管支炎、結核、下気道感染症、扁桃炎、中耳炎、肝炎、AIDS、糖尿病、糖尿病性潰瘍、および小児疾患が挙げられるが、これらに限定されない。本発明のRR1化合物は、他の推定免疫賦活剤の活性を評価するための標準物質としても有用である。
【0029】
RR1の免疫賦活性特性のおかげで、本発明の治療法、化合物および組成物を用いて、急性リンパ性白血病、急性非リンパ性白血病、慢性リンパ性白血病、慢性骨髄性白血病、ホジキン病、非ホジキン性リンパ腫、および多発性骨髄腫などの白血病およびリンパ腫、脳腫瘍、神経芽腫、網膜芽腫、ウィルムス腫瘍、骨腫瘍、および軟組織肉腫などの小児固形腫瘍、肺癌、結腸および直腸癌、乳癌、前立腺癌、泌尿器癌、子宮癌、膀胱癌、口腔癌、膵臓癌、メラノーマおよびその他皮膚癌、胃癌、卵巣癌、脳腫瘍、肝臓癌、咽頭癌、甲状腺癌、食道癌、および精巣癌などの成人の一般的固形癌を含むが、これらに限定されない癌などの、様々な細胞増殖性の障害を治療できる。本発明の方法は、インビボまたはインビトロで実施して、ヒトまたはヒト以外の哺乳動物における癌性細胞の増殖を阻止できる。
【0030】
β-グルカンに起因する幾つかの特異的作用はまた、RR-1にも関係していることがある。これには、リポ多糖類(LPS)またはその他エンドトキシン/エキソトキシンによって誘発されるNF-κBの活性化の阻害が包含される。NF-κBの活性化およびその他経路は、敗血症および敗血症ショックの発生に重要な役割を果たしている。(1→3)-β-D-グルカンは、NF-κBの活性化を減少させ、敗血症のマウスの長期生存期間を延長することが証明されている(Williams, D.L. et al. Shock, 2000, 13:446-452; Williams, D.L. et al. Surgery, 1999, 126:54-65)。したがって、本発明は、少なくとも一つのRR1化合物を投与することによって、ヒトまたはヒト以外の哺乳動物の敗血症または敗血症ショックを治療する方法を含む。別の局面では、本発明は、少なくとも一つの本発明のRR1化合物を投与することによって、LPSまたはその他エンドトキシンおよび/またはエキソトキシンによってヒトまたはヒト以外の哺乳動物に誘発されたNF-κBの活性化を低下させるための方法を含む。
【0031】
生物学的応答調節因子としてのその作用のおかげで、本発明のRR1化合物はまた急性および慢性の肺疾患等のその他の障害、または自然もしくは適応免疫系の異常に関係し得るその他の障害の治療を目的として、被験体に投与してもよい。本発明は、このような障害を治療するための方法を含む。抗腫瘍効果以外にも、植物多糖類が癌の化学的予防に関して役割を果たすことを裏付ける証拠がある(Kim, H.S. et al. Carcinogenesis, 1999, 20:1637-1640)。本発明は、癌の化学的予防のための方法を含むが、ここで、少なくとも一つの、本発明のRR1化合物が被験体に投与される。
【0032】
本発明の治療法は、細胞増殖性障害(例えば癌)、または免疫賦活もしくは免疫抑制によって治療もしくは管理され得るその他の病理的状態(例えば疾患またはその他の障害)の治療または管理について、当業者に公知である少なくとも一つの追加的治療法または療法と、都合良く組み合わせることができる。このような補助療法の例としては、化学療法、放射線療法、および抗癌剤もしくはその他抗癌性作用物質の投与が挙げられるが、これらに限定されない。補助療法は、鍼、マッサージ療法、エネルギー療法のように、逆症療法でも非逆症療法でもよい。病理的状態は、急性でも慢性でも、疾患でも障害でもよい。
【0033】
RR1およびRR1類似体は、単独化合物として投与することができるが、これら化合物は薬学的組成物として投与することが好ましい。それゆえに本発明は、RR1、またはその類似体、または生理学的に許容されるその塩を、活性成分として、少なくとも一つの薬学的に許容される担体と共に含む薬学的組成物も提供する。薬学的組成物は、経腸的、非経口的、静脈内、筋肉内、局所、皮下等、各種投与経路に合わせることができる。投与は、当業者によって決定できる場合は、連続的または特定の間隔でよい。
【0034】
本発明のRR1化合物は、医薬として有用な組成物の調製について公知である方法に従って製剤化できる。製剤は、当業者に周知であり、かつ容易に入手できる様々な情報源に記載されている。例えば、Remington's Pharmaceutical Science (Martin EW, Easton Pennsylvania, Mack Publishing Company, 19th ed., 1995)は、本発明と関連して用いることができる製剤を記載している。投与に適した製剤としては、例えば、酸化防止剤、バッファー、抗菌剤、および製剤を目的のレシピエントの血液と等張にする溶液を含むことがある、水性滅菌注射液;ならびに懸濁化剤および増粘剤を含む水性および非水性無菌懸濁液が挙げられる。製剤は、単位投与量または複数回投与量容器、例えば密封されたアンプルおよびバイアルの中に存在してよく、使用前に無菌溶液担体、例えば注射用の水の状態のみを必要とするフリーズドライ(凍結乾燥)状態で保存することができる。準備を必要としない注射液および懸濁液を、無菌粉末、顆粒、錠剤から調製してもよい。本発明の組成物は、上記具体的に記載した成分に加えて、問題の製剤のタイプを考慮し、当技術分野で通常であるその他作用物質を含むことができる。
【0035】
本発明のRR1化合物は、当業者が調製できるRR1のあらゆる水和物および塩を含む。本発明の化合物が十分塩基性または酸性であり、無毒の酸または塩基性塩を形成できる条件では、塩としての化合物の投与が適切であろう。薬学的に許容される塩の例は、生理学的に許容されるアニオン、例えばトシラート、メタンスルホナート、アセタート、シトラート、マロナート、タルトラート、スクシナート、ベンゾアート、アスコルベート、α-ケトグルタラートおよびα-グリセロホスフェートを形成する、酸と一緒に形成した有機酸付加塩である。塩酸塩、硫酸塩、硝酸塩、重炭酸塩、および炭酸塩を含む、好適な無機塩も形成できる。
【0036】
薬学的に許容できるRR1の塩は、当技術分野で周知である標準的な方法を用いて、例えばアミンなどの十分に塩基性である化合物を好適な酸と反応させて、生理学的に許容できるアニオンを供給することによって、得ることができる。カルボン酸のアルカリ金属(例えばナトリウム、カリウムもしくはリチウム)またはアルカリ土金属(例えばカルシウム)塩も作ることができる。
【0037】
本明細書で使用する「類似体」という用語は、実質的にRR1(図3に示すような)と同一であるが、親化合物のαコンホメーションおよび(1→4)および(1→6)グルコピラノシル結合が未変性のままで、かつ親化合物の免疫賦活性特性の一つまたは複数が実質的に保持されている限りにおいて、例えば側基の付加、親構造の酸化もしくは還元により修飾されてもよい化合物を指す。例示の化合物の類似体は、公知の標準的な反応を用いて容易に調製できる。これら標準の反応には、水素化、アルキル化、アセチル化、および酸性化反応が挙げられるが、これらに限定されない。
【0038】
それらを含有するRR1化合物および組成物の治療応用は、現在または将来、当業者にとって公知となる任意の好適な治療方法および技術によって達成できる。さらに、本発明のRR1化合物は、他の有用な化合物および組成物の調製のための出発材料または中間体として有用である。
【0039】
本発明のRR1化合物およびこのような化合物を含有する免疫調節組成物は、不活性希釈液または同化性食用担体などの薬学的に許容される担体と一緒に全身投与(例えば経口または静脈内)できる。RR1化合物は、硬質または軟質シェルゼラチンカプセルに封入してもよく、錠剤に圧縮してもよく、または患者食の食材に直接組み込んでもよい。経口治療投与の場合には、RR1化合物は、一つまたは複数の賦形剤と組み合わせ、摂取可能な錠剤、バッカル剤、トローチ剤、カプセル剤、エリキシル剤、懸濁剤、シロップ剤、ウェーハ等の形で用いることができる。
【0040】
錠剤、トローチ剤、丸剤、カプセル剤等は、下記のものを含有してもよい:トラガントゴム、アカシア、コーンスターチ、またはゼラチンなどの結合剤;リン酸二カルシウムなどの賦形剤;コーンスターチ、ポテトスターチ、アルギン酸等などの崩壊剤;ステアリン酸マグネシウムなどの潤滑剤;およびショ糖、フルクトース、ラクトース、もしくはアスパルテームのような甘味料、またはペパーミント、ウインターグリーン油、もしくはチェリーフレーバーのような香味剤を加えてもよい。単位投与形態がカプセル剤の場合、上記物質以外に、植物油またはポリエチレングリコールなどの液体担体を含有してもよい。様々なその他材料が、コーティング剤として存在してもよく、そうでなければ固形の単位投与形態の物理形態に変更してもよい。例えば、錠剤、丸剤、またはカプセル剤は、ゼラチン、ワックス、セラック、または糖等でコーティングしてもよい。シロップまたはエリキシルは、活性化合物、甘味料としてのショ糖またはフルクトース、保存剤としてのメチルおよびプロピルパラベン、着色料、ならびにチェリーまたはオレンジフレーバーのような香味料を含有してよい。もちろん、いずれの単位投与形態の調製に用いられるいずれの材料も、使用する量において薬学的に許容されるものであり、実質的に無毒でなければならない。これに加えて、本発明のRR1化合物は、徐放性調製物および装置に組み入れてもよい。
【0041】
本発明のRR1化合物(即ち、RR1または、RR1の薬学的に許容される塩もしくは類似体)である活性作用物質はまた、輸液または注射によって静脈内または腹腔内投与してもよい。活性作用物質またはその塩の溶液は、水を用いて、任意で無毒の界面活性剤と混合して調製できる。分散液は、グリセロール、液体ポリエチレングリコール、トリアセチン、およびそれらの混合物、ならびに油において調製できる。保存および使用の通常の条件下では、これら調製物は、保存剤を含有し、微生物の増殖を阻止する。
【0042】
注射または輸液に好適な薬学的投与形態としては、無菌の注射可能もしくは輸液可能な溶液もしくは分散液の即時調製物に適応し、リポソームに封入されていてもよい活性成分を含む、無菌の水溶液もしくは分散液または無菌の粉末が挙げられる。いずれの場合も、最終的な投与形態は、無菌の流体であり、かつ製造および保管状態下で安定でなければならない。液体担体もしくは媒質は、例えば水、エタノール、ポリオール(例えばグリセロール、プロピレングリコール、液体ポリエチレングリコール等)、植物油、無毒のグリセリルエステルおよびそれらの好適混合物を含む、溶媒または液体分散媒体であり得る。適当な流動性は、例えば、リポソームの形成により、分散体の場合には必要な粒子サイズを維持することにより、または界面活性剤の使用により維持できる。微生物の活動は、様々な抗菌剤および抗真菌剤、例えばパラベン、クロロブタノール、フェノール、ソルビン酸、チメロサール等によって阻止できる。多くの例では、等張剤、例えば糖、緩衝剤または塩化ナトリウムを含めることが好ましいだろう。注射可能な組成物の長期吸収は、吸収遅延剤、例えばモノステアリン酸アルミニウムおよびゼラチンを組成物に用いることで実現できる。
【0043】
無菌注射液は、適切な溶媒に、必要に応じて、上に列挙した各種のその他成分と一緒にRR1化合物(即ちRR1、またはRR1の薬学的に許容される塩もしくは類似体)を組み入れ、続いて濾過滅菌して調製される。無菌注射液調製用の無菌粉末の場合は、好ましい調製方法は、真空乾燥およびフリーズドライ技術であり、これら技術により、事前に無菌濾過した溶液中に存在することが望ましい任意の追加成分を加えた活性成分の粉末が生じる。
【0044】
局所投与では、RR1化合物は純粋な形、即ち液体の場合投与できる。しかしながら、それらは、固体または液体でもよい、皮膚科学的に許容される担体と組み合わせた組成物として、皮膚に投与することが一般的には望ましいであろう。
【0045】
有用な固体担体としては、タルク、クレー、微結晶セルロース、シリカ、アルミナ等の微細分割固体が挙げられる。有用な液体担体としては、水、アルコールもしくはグリコール、または水-アルコール/グリコール混合物が挙げられ、これらの中にRR1化合物は、任意で無毒の界面活性剤の助けを借りて、有効レベルで溶解もしくは分散することができる。芳香剤のような補助剤および追加の抗菌剤を加えて、所与の使用に合わせて特性を最適化できる。得られた液体組成物は、吸収性パッドから作用させること、絆創膏および包帯剤の含浸に用いること、またはポンプタイプもしくはエアロゾルスプレーを用いて患部に噴霧することができる。
【0046】
合成ポリマー、脂肪酸、脂肪酸塩およびエステル、脂肪アルコール、変性セルロースもしくは変性ミネラル物質などの増粘剤も、液体担体と一緒に用いることができ、使用者の皮膚に直接適用するための、塗布可能なペースト、ゲル、軟膏、石けん等を形成できる。皮膚にRR1化合物を送達するのに用いることができる有用な皮膚科学的組成物の例は、Jacquetら(米国特許第4,608,392号)、Geria(米国特許第4,992,478号)、Smithら(米国特許第4,559,157号)およびWoltzman(米国特許第4,820,508号)に開示されている。
【0047】
本発明の組成物の有用投与量は、それらのインビトロ活性と、動物モデルでのインビボ活性とを比較することで決定できる。マウスおよびその他動物での有効投与量をヒトに外挿する方法は、当技術分野で公知である:例えば、米国特許第4,938,949号を参照されたい。
【0048】
したがって、本発明は、RR1化合物(即ち、RR1、またはその薬学的に許容されるその塩もしくは類似体)を薬学的に許容される担体と組み合わせて含む、免疫調節剤である薬学的組成物を含む。経口、局所、または非経口投与に適した、ある量のRR1化合物を含む免疫調節組成物は、本発明の好ましい態様を構成する。患者、特にヒトに、本発明に即して投与される量は、合理的な時間枠の中で患者に治療応答を起こさせるのに十分なものでなければならない。投与量が被験体の状態(健康)、被験体の体重、該当する場合には併用する治療の種類、治療頻度、治療比、ならびに病理的状態の重篤度およびステージを含む様々な要因に依存するだろうということを、当業者は認識するであろう。
【0049】
癌との関係では、好適投与量は、所望の応答を達成することが知られている活性剤(本発明のRR1化合物)の濃度を腫瘍組織内に生ずる投与量である。好ましい投与量は、管理不能な副作用なしに、癌細胞増殖の最大阻止を生じる量である。RR1化合物、およびこのような化合物を含有する組成物の投与は、当業者が決めることができる場合には、連続的でも一定間隔でもよい。
【0050】
所望する治療処置のためのそのような投与量を提供するために、本発明の薬学的組成物は、担体または希釈液を含めた全組成物重量を基にして、一つまたは複数の新規化合物の総量を約0.1重量%〜45重量%、特には1〜15重量%を有利に含むであろう。例示的には、投与される活性成分の投与量レベルは、下記である:静脈投与の場合、0.01〜約20mg/kg;腹腔内投与の場合、0.01〜約100mg/kg;皮下投与の場合、0.01〜約100mg/kg;筋肉内投与の場合、0.01〜約100mg/kg;経口投与の場合、0.01〜約200mg/kg、および好ましくは約1〜100mg/kg;点鼻注入の場合、0.01〜約20mg/kg;ならびにエアロゾルの場合、0.01〜約20mg/kg動物(体)重。
【0051】
開示の方法、化合物および組成物から便益を受ける哺乳動物種としては、無尾猿、チンパンジー、オラウータン、ヒト、有尾猿などの霊長類;イヌ、ネコ、モルモット、ハムスター、ベトナム産ポットベリーブタ(Vietnamese pot-bellied pigs)、ウサギおよびフェレットなどの家庭用動物(例えばペット);ウシ、バッファロー、バイソン、ロバ、ブタ、ヒツジ、およびヤギなどの家畜動物;クマ、ライオン、トラ、パンサー、ゾウ、カバ、サイ、キリン、レイヨウ、ナマケモノ、ガゼル、シマウマ、ワイルドビースト、プレーリードッグ、コアラ、カンガルー、オポッサム、アライグマ、パンダ、ハイエナ、アザラシ、アシカ、ゾウアザラシ、オットセイ、ネズミイルカ、イルカ、およびクジラなどの、典型的には動物園で見られる珍しい動物種が挙げられるが、これらに限定されない。開示の方法から便益を受けることができる他の種としては、魚類、両生類、鳥類、および爬虫類が挙げられる。本明細書で使用する「患者」および「被験体」という用語は、互換的に用いられ、このようなヒトおよびヒト以外の種を含むことを意図している。同様に、本発明のインビトロの方法は、このような種の細胞に実施できる。
【0052】
本発明の方法を用いた処置を必要とする患者は、医学専門家に公知である標準的な技術を用いて特定できる。
【0053】
本明細書で使用する「癌」および「癌性」という用語は、未制御の細胞増殖を典型的な特徴とする、哺乳動物の病理的状態を指すか、または表す。癌の例としては、癌腫、リンパ腫、芽腫、肉腫、および白血病が挙げられるが、これらに限定されない。このような癌のより具体的な例としては、乳癌、前立腺癌、結腸癌、扁平上皮癌、小細胞肺癌、非小細胞肺癌、消化管癌、膵臓癌、子宮頚癌、卵巣癌、肝臓癌、例えば肝癌、膀胱癌、結腸直腸癌、子宮内膜癌、腎臓癌、および甲状腺癌が挙げられる。
【0054】
癌のその他の非限定例は、基底細胞癌、胆道癌;骨癌;脳およびCNS癌;絨毛上皮腫;結合組織癌;食道癌;眼癌;頭部および頚部の癌;胃癌;上皮内癌;咽頭癌;ホジキンおよび非ホジキンリンパ腫を含むリンパ腫;メラノーマ;骨髄腫;神経芽腫;口腔癌(例えば唇、舌、口および咽頭);膵臓癌;網膜芽腫;黄紋筋肉腫;直腸癌;呼吸器癌;肉腫;皮膚癌;胃癌;精巣癌;子宮癌;泌尿器系の癌、ならびにその他の癌腫および肉腫である。
【0055】
本発明の方法によれば、RR1またはその薬学的に許容される塩もしくは類似体は、それ自体を患者に投与でき、または他の作用物質と共投与できる。共投与は、同時(同一または異なる調合物として)または連続的に行うことができる。さらには、本発明の方法によれば、RR1、またはその薬学的に許容される塩もしくは類似体は、補助治療として患者に投与できる。例えば、RR1、またはその薬学的に許容される塩もしくは類似体は、化学療法と共に、患者に投与できる。
【0056】
したがって、本発明のRR1化合物(RR1、またはその薬学的に許容される塩もしくは類似体)は、別々に投与するか、あるいは薬学的組成物として投与するかを問わず、添加物として各種のその他の成分を含むことができる。関連する状況において用いることができる許容される成分または補助剤としては、酸化防止剤、フリーラジカルスカベンジャー剤、ペプチド、成長因子、抗生物質、静菌剤、免疫抑制剤、他の免疫賦活性剤、抗凝固剤、緩衝化剤、抗炎症剤、抗発熱剤、時間放出結合剤、麻酔剤、ステロイド、およびコルチコステロイドが挙げられる。このような成分は、追加の治療便益を提供でき、RR1化合物の治療作用に作用して影響を及ぼし、またはRR1化合物投与の結果として生ずるかもしれない任意の潜在的な副作用を阻止するように働くことができる。本発明のRR1化合物は、同様に、治療薬剤とコンジュゲートさせることもできる。
【0057】
同一の調合物または別々の調合物として、連続的または同時に、患者に共投与できるさらなる作用物質としては、免疫調節剤のような、所与の生物学的応答を調整する作用物質が挙げられる。例えば、腫瘍壊死因子(TNF)、インターフェロン(α-インターフェロンおよびβ-インターフェロンなど)、神経成長因子(NGF)、血小板由来成長因子(PDGF)、および組織プラスミノーゲン活性化因子などのタンパク質が投与できる。リンフォカイン、インターロイキン(インターロイキン-1(IL-1)、インターロイキン-2(IL-2)、およびインターロイキン-6(IL-6)など)、顆粒球マクロファージコロニー刺激因子(GM-CSF)、顆粒球コロニー刺激因子(G-CSF)、またはその他の成長因子などの生物学的応答調整物質を投与できる。抗癌剤、細胞傷害性薬剤、および/または化学療法薬剤は、本発明のRR1化合物と一緒に、連続的または同時に、同一調合物中でまたは別々の調合物として患者に共投与できる。一つの態様では、本発明のRR1化合物は、抗原と共投与される。好ましくは、抗原はワクチンの活性成分である。
【0058】
一つの態様では、本発明は、唯一の免疫学的活性物質としてRR1化合物を含む組成物を提供する。投与されると、直ちにRR1化合物は免疫系刺激を誘発する。
【0059】
別の態様では、上記のように、RR1化合物は、抗原(タンパク質、糖タンパク質、多糖および脂質を含むが、これらに限定されない)、および/または共刺激分子(サイトカイン、ケモカイン、標的化タンパク質リガンド、トランス活性化因子、ペプチド、および修飾アミノ酸を含むペプチドが挙げられるが、これらに限定されない)などの免疫調節刺激剤、ならびにアジュバント(ミョウバン、脂質乳剤およびポリラクチド/ポリグリコリド微小粒子を含むが、これらに限定されない)などのその他の免疫調節分子の一つ、または複数と共に投与することができる。本明細書で使用する「免疫調節」という用語は、免疫賦活性作用だけでなく免疫抑制作用も含む。免疫賦活性作用は、細胞性または液性免疫応答を直接または間接的に高める作用を含むが、これらに限定されない。免疫賦活性作用の例としては、抗原特異的抗体産生の増進;NK細胞、CD4+Tリンパ球、CD8+Tリンパ球、マクロファージ等のリンパ球集団の活性化または増殖;IL-1、IL-2、IL-4、IL-5、IL-6、IL-12、IFN-γ、TNF-αを含むが、これらに限定されない免疫賦活性サイトカインの合成増加等が挙げられるが、これらに限定されない。免疫抑制作用としては、細胞性または液性免疫応答を直接または間接的に減少させる作用を含む。免疫抑制作用の例としては、IgE産生低下などの抗原特異的抗体産生の低下;免疫寛容性を生ずるような免疫抑制活性を有するリンパ球またはその他細胞集団の活性化;ならびに特定の細胞機能に対し抑制作用を有するサイトカインの合成増加が挙げられるが、これらに限定されない。この一例は、IFN-γであり、これはIL-4が誘導するIgEおよびIgG1へのクラス切り替えを阻止し、それによってこれら抗体サブクラスのレベルを下げると考えられている。
【0060】
RR1化合物ならびに抗原および/または免疫調節促進剤は、コンジュゲートの形で一緒に投与でき、または免疫応答を調節できるように、十分近接した時間で、混合物として共投与することができる。好ましくは、RR1化合物および免疫調節分子は、同時に投与される。本明細書で用いられる「共投与」という用語は、この意味においては、少なくとも2つの物質を、免疫応答を調節するのに十分近い時間的関係で投与することを指す。好ましくは、共投与は、少なくとも2つの物質の同時投与を指す。
【0061】
本明細書で使用する「コンジュゲート」という用語は、その中でRR1化合物および免疫調節分子が結合している複合体を指す。このようなコンジュゲートの結合には、共有結合および/または非共有結合が含まれる。
【0062】
本明細書で使用する「抗原」という用語は、抗体またはT細胞抗原受容体が特異的に認識および結合する物質を意味する。抗原には、ペプチド、タンパク質、糖タンパク質、多糖、ガングリオシドおよび脂質;それらの一部およびそれらの組み合わせを含めることができる。抗原は、自然界に見いだされるものでも、または合成のものであり得る。ハプテンは、「抗原」の範囲内に含まれる。ハプテンは、それ自体は免疫原性でないが、免疫原性決定因子含有免疫原性分子とコンジュゲートすると免疫原性が付与される低分子化合物である。
【0063】
本明細書で使用する「アジュバント」という用語は、免疫原性作用物質に加えると、混合物に曝露したレシピエント宿主における、その作用物質に対する免疫応答を非特異的に高めるか、または増強する物質を指す。
【0064】
別の態様では、本発明は、RR1化合物および抗原を含む組成物を提供する。RR1化合物と一緒の投与に好適な抗原は、B細胞またはT細胞抗原特異的応答を引き出すことができる任意の分子を含む。好ましくは、抗原は、抗原に特異的な抗体応答を引き出す。広範な分子が抗原である。これら抗原としては、糖、脂質およびポリペプチド、ならびに複合炭化水素のような高分子、ならびにリン脂質が挙げられるが、これらに限定されない。小分子は、抗原性を付与するためにハプテン化が必要な場合がある。好ましくは、本発明の抗原としては、ペプチド、脂質(例えばステロール、脂肪酸、およびリン脂質)、ヘモフィラス・インフルエンザワクチンで用いられるような多糖、ガングリオシドおよび糖タンパク質が挙げられる。
【0065】
本明細書で使用する「ペプチド」という用語は、ペプチドがハプテンであるかないかに関係なく、生物学的応答、例えば抗体産生もしくはサイトカイン活性を達成する十分な長さおよび組成を有するペプチドおよびタンパク質を含む。典型的には、ペプチドは、少なくとも6アミノ酸残基長である。用語「ペプチド」はさらに、修飾アミノ酸を包含し、その場合の修飾としては、リン酸化、グリコシル化、ペジル化、脂質化およびメチル化が挙げられるが、これらに限定されない。
【0066】
一つの態様では、本発明は、RR1化合物および抗原性ペプチドを含む組成物を提供する。抗原性ペプチドとしては、精製された未変性ペプチド、合成ペプチド、組換えタンパク質、粗タンパク質抽出物、弱毒化もしくは不活性化ウイルス、細胞、微生物またはそのようなペプチドの断片を包含できる。
【0067】
多くの抗原性ペプチドおよびタンパク質が公知であり、当技術分野で利用可能である;その他のものは、通常技術を用いて特定できる。免疫賦活性促進剤としての要求を満たすことができるタンパク質抗原としては、以下の例が挙げられるが、これらに限定されない。単離された未変性または組換え抗原は、植物の花粉(例えば、Rafnar et al. (1991) J. Biol. Chem. 266:1229-1236; Breiteneder et al. (1989) EMBO J. 8:1935-1938; Elsayed et al. (1991) Scand. J. Clin. Lab. Invest. Suppl. 204:17-31; およびMalley (1989) J. Reprod. Immunol. 16:173-186を参照されたい)、イエダニ(例えば、Chua et al. (1988) J. Exp. Med. 167:175-182; Chua et al. (1990) Int. Arch. Allergy Appl. Immunol. 91:124-129;およびJoost van Neerven et al. (1993) J. Immunol. 151:2326-2335)、動物のフケ(例えば、Rogers et al. (1993) Mol. Immunol. 30:559-568を参照されたい)、動物の唾液、ミツバチの毒、ならびにカビの胞子から得ることができる。HIV-1、HIV-2、単純ヘルペスウイルス、A型肝炎ウイルス(Bradley et al. (1984) J. Med. Virol. 14:373-386)、ロタウイルス、ポリオウイルス(Jiang et al. (1986) J. Biol. Stand. 14:103-109)、B型肝炎ウイルス、麻疹ウイルス(James et al. (1995) N. Engl. J. Med. 332:1262-1266)、ヒトおよびウシパピローマウイルス、およびスローブレイン(slow brain)ウイルスなどの、生きている弱毒化および不活性化された微生物は、ペプチド抗原を提供する。腫瘍形成に対する免疫化については、免疫賦活性ペプチドとして、腫瘍細胞(生きているか、または放射線処理したもの)、腫瘍細胞抽出物または腫瘍抗原のタンパク質サブユニットを挙げることができる。免疫に基づく避妊向けワクチンは、RR1化合物と一緒に投与する精子タンパク質を含ませることによって形成できる(Lea et al. (1996) Biochim. Biophys. Acta 1307:263)。
【0068】
本発明の組成物および方法で使用する抗原は、決定的ではない。各種抗原の混合物を、本発明に従って用いることができる。好ましくは、ウイルスまたは細菌病原体、あるいは真菌または寄生虫に由来するタンパク質またはペプチドを、このような抗原として使用する(誘導体化抗原またはグリコシル化もしくは脂質化抗原もしくは多糖もしくは脂質を含む)。別の好ましい抗原供給源は、腫瘍抗原である。好ましい病原体は、ヒト免疫不全症ウイルス(HIV)、A型およびB型肝炎ウイルス、C型肝炎ウイルス(HCV)、ラウス肉腫ウイルス(RSV)、エプスタインバーウイルス(EBV)、インフルエンザウイルス、ロタウイルス、黄色ブドウ球菌、クラミジア肺炎、クラミジア・トラコマチス、ヒト型結核菌、肺炎連鎖球菌、炭疽菌、ビブリオ・コレラ、プラスモディウム属種(熱帯熱マラリア原虫、三日熱マラリア原虫、その他)、アスペルギルス属種またはカンジダ・アルビカンスから選択される。抗原はまた、癌細胞が発現する分子でもよい(例えば腫瘍抗原)。誘導体化のプロセスは、病原体/癌細胞からの特異的タンパク質の精製、病原体の不活性化、ならびにそのようなタンパク質のタンパク質分解もしくは化学的誘導体化もしくは安定化を含んでよい。同様にして、腫瘍抗原(癌ワクチン)または自己免疫抗原も本発明の組成物および方法に用いることができる。このようにして、腫瘍ワクチン接種または自己免疫疾患の治療を行うことができる。
【0069】
RR1化合物および抗原は、RR1-抗原コンジュゲートとして投与でき、かつ/またはそれらは乳剤などの、混合物の形をした複合体として共投与できる。RR1-抗原コンジュゲート内でのRR1化合物と抗原分子との会合は、高親和性および/または低親和性相互作用を含む、共有結合形の互作用、および/または非共有結合形の相互作用を通して可能となる。RR1-抗原コンジュゲート内でRR1化合物と抗原とをカップリングできる非共有結合形の相互作用の例としては、イオン結合、疎水性相互作用、水素結合、およびファン・デール・ワールス力が挙げられるが、これらに限定されない。
【0070】
別の態様では、RR1化合物は、一つまたは複数の免疫調節促進剤と共に投与できる。したがって本発明は、RR1化合物および免疫調節促進剤を含む組成物を提供する。本明細書で使用する「免疫調節促進剤」という用語は、RR1化合物の免疫調節活性を支持および/または強化する分子を指す。免疫調節促進剤の例としては、サイトカイン、および/またはアジュバントのような共刺激分子を挙げることができる。RR1化合物および促進剤は、RR1-促進剤コンジュゲートとして投与でき、かつ/またはそれらは乳剤などの、混合物の形をした複合体として共投与できる。RR1-促進コンジュゲート内のRR1化合物と促進剤分子とは、高親和性および/または低親和性相互作用を含む、共有結合形の相互作用、および/または非共有結合形の相互作用を通じて結びつくことができる。RR1-促進コンジュゲート内でRR1化合物と促進剤とをカップリングできる非共有結合形の相互作用の例としては、イオン結合、疎水性相互作用、水素結合、およびファン・デール・ワールス引力が挙げられるが、これらに限定されない。
【0071】
免疫調節促進剤としては、共刺激分子(サイトカイン、ケモカイン、標的化タンパク質リガンド、トランス活性化因子、ペプチド、および修飾アミノ酸を含むペプチドなど)ならびにアジュバント(ミョウバン、脂質乳剤、およびポリラクチド/ポリグリコリド微小粒子など)が挙げられるが、これらに限定されない。
【0072】
なかでも、RR1化合物と共に投与するのに好適な免疫調節サイトカインペプチドは、インターロイキン(例えばIL-1、IL-2、IL-3等)、インターフェロン(例えばIFN-α、INF-β、IFN-γ)、エリスロポイエチン、コロニー刺激因子(例えばG-CSF、M-CSF、GM-CSF)およびTNF-αである。好ましくは、RR1化合物と共に使用するための免疫賦活性ペプチドは、IL-12(Bliss et al. (1996) J. Immunol. 156:887-894)、IL-18、TNF-α、β、およびγ、ならびに/または形質転換成長因子(TGF)-αなどTh1型免疫応答を刺激するペプチドである。
【0073】
RR1化合物と共に投与されるペプチドは、特異的受容体とのタンパク質との結合を媒介するか、または特異タイプの細胞もしくは組織への標的化を媒介するアミノ酸配列を含むことができる。例としては、抗体もしくは抗体断片、ヒト成長ホルモンなどのペプチドホルモン、および酵素が挙げられるが、これらに限定されない。免疫調節ペプチドもまた、ペプチドホルモン、ペプチド神経伝達物質およびペプチド成長因子を含む。B7(CD80)などの共刺激分子、転写因子などのトランス活性化タンパク質、マクロファージ走化性タンパク質(MCP)および他の化学誘引物質または走化性ペプチドなどのケモカインもまた、本発明のRR1化合物との共投与に有用なペプチドである。
【0074】
RR1化合物は、脂質、多糖、ガングリオシドなどの他の抗原と、ペプチドなどの連結基を介してコンジュゲートさせることもできる。
【0075】
本発明はまた、アジュバントと併せてRR1化合物の投与も提供する。抗原を、RR1化合物およびアジュバントと一緒に投与すると、抗原に対する免疫応答は強化され、それによってRR1化合物および抗原を単独で含む組成物で得られる応答よりも高い免疫応答を起こすことができる。したがって、別の態様では、本発明は、RR1化合物、抗原およびアジュバントを含む組成物を提供し、これによってRR1化合物/抗原/アジュバントを共投与できる。好ましくは、免疫原性組成物は、免疫原に対する免疫応答を強化するのに十分な量のアジュバントを含有する。好ましくは、アジュバントとしては、水中油型乳剤、油中水乳剤、ミョウバン(アルミニウム塩)、リポソーム、ならびにポリスチレン、澱粉、ポリホスファゼンおよびポリラクチド/ポリグリコシドを含むが、これらに限定されない微小粒子を挙げることができるが、これらに限定されない。より好ましくは、RR1化合物および抗原は、ミョウバンと一緒に共投与される。より好ましくは、RR1化合物および抗原は、リポソームと一緒に共投与される。よりさらに好ましくは、RR1化合物および抗原は、水中油型乳剤と共に共投与される。
【0076】
好適なアジュバントとしては、スクアレン混合物(SAF-1)、ムラミルペプチド、サポニン誘導体、マイコバクテリウム細胞壁調製物、モノホスホリル脂質A、ミコール酸誘導体、非イオン系コポリマー界面活性剤、Quil A、コレラトキシンBサブユニット、ポリホスファゼンおよび誘導体、ならびにTakahashiら(1990) Nature 344:873-875が記載したような免疫刺激複合体(ISCOM)ならびに脂質をベースとしたアジュバントおよび本明細書に記載されている他のアジュバントを挙げることができるが、これらに限定されない。獣医分野での使用、および動物での抗体産生での使用では、フロイントのアジュバント(完全および不完全の両方)の分裂誘発成分を用いることができる。
【0077】
全ての免疫原性組成物と同様に、成分の免疫学的有効量は、経験的に決めることができる。考慮すべき要素としては、RR1化合物および/または抗原が、免疫調節促進剤、アジュバントもしくは担体タンパク質もしくは他の担体と複合体化しているか、または共有結合しているか否かの抗原性、投与経路かつ投与する免疫投与の回数が挙げられる。このような要素は、ワクチン分野では公知であり、過度の実験なしにこのような決定を行うことは、免疫学者の通常の技量の範囲内である。
【0078】
本発明はさらに、RR1化合物および免疫調節分子を混合物として投与した場合に比べ、発生する免疫応答を高めるのに有効である距離まで、RR1化合物および免疫調節分子が近接して結びついている組成物を提供する。したがって、本発明は、複合体が標的に作用するまで、RR1化合物と免疫調節分子の近接した結びつきを維持できる、封入化剤を含む組成物およびその使用方法を提供する。好ましくは、RR1成分、免疫調節分子、および封入化剤を含む組成物は、アジュバント水中油型乳剤、微小粒子および/またはリポソームの形態である。より好ましくは、RR1-免疫調節分子を封入しているアジュバント水中油型乳剤、微小粒子および/またはリポソームは、約0.04μm〜約100μmの大きさ、より好ましくは約0.1μm〜約20μm、よりさらに好ましくは約0.15μm〜約10μmの粒子の形態である。
【0079】
マイクロスフェア、ビーズ、高分子複合体、ナノカプセルのようなコロイド分散系、ならびに水中油型乳剤、ミセル、混合ミセル、およびリポソームなどの脂質をベースとした系は、RR1化合物含有組成物の有効な封入を提供する。
【0080】
本発明のさらなる局面は、ティノスポラ・コルディフォリア植物材料からRR1を得るためのプロセスを提供する。好ましい態様によれば、RR1は植物材料の空中部分(茎および葉)から単離される。乾燥し、粉末化した植物材料500gを、ソックスレー(sox let)抽出器の中で、純粋メタノールを用いて72時間抽出する。残留物を熱した蒸留水に取り、遠心分離し、上清から集める。再度遠心分離を行い、80%アルコールを用いて上清から粗多糖を沈殿させる。沈殿物を濾過し、アルコールで洗浄し、蒸留水に溶解し、透明な溶液を15%トリクロロ酢酸で処理する。溶液を遠心分離にかけ、いかなる残留物も取り除き、アセトンを用いて透明な上清を再度沈殿させる。沈殿物を濾過して、アセトンで数回洗浄し、蒸留水に再溶解する。透明な溶液を流水中で72時間透析し(メンブレン、MWCO 60000Da)、チューブに入れてスピードバック(speedvac)で凍結乾燥する。本プロセスの例示的態様は、図1のフローチャートに示されている。天然産物精製分野の科学者は、記載の方法を、本発明の好ましい態様に記載されているものに容易に適合させ、各種溶媒および固定相を交換できる。ティノスポラ・コルディフォリア抽出物の水性画分は、水切り、濾過、デカンテーションによって、または溶液もしくは混合物の水性部分を分離に関する分野において公知である他の任意の方法によって分離できる。クロマトグラフィーに加えて、所望の化合物は、結晶化および分配などの方法を用いても精製できる。RR1化合物の単離には、様々な代替試薬が利用できる。例えば、エタノールに代わって一価アルコールを用いてもよい。アセトンは、単離手順では、エタノールに替えることができる。例えば、全体として参照により本明細書に組み入れられる、Brown G.G., Unit Operations, John Wiley & Sons, 1956;McCabe, W.L. and J.C. Smith, Unit Operations of Chemical Engineering, McGraw-hill, 1956;およびPerry, R.H., and D. Green, Perry's Chemical Engineers' Handbook, 7th Edition, McGraw-Hill, 1997を参照されたい。
【0081】
任意で、ひとたび得られた単離RR1化合物を出発材料として用い、本発明のRR1類似体を合成することができる。任意で、プロセスは、単離RR1化合物または類似体を、本明細書に記載のようなインビトロまたはインビボアッセイを用いて、免疫調節活性について評価する段階をさらに含むことができる。
【0082】
植物からRR1を抽出するための方法に関しては、特に制限はない。例えば、各種溶媒を用いた抽出または超臨界液体抽出が利用できる。植物からのRR1抽出に用いる溶媒に関して、特に制限はない。好適な溶媒の例としては、水、無機塩水溶液、および緩衝液などの水性媒体、ならびにアルコール、ヘキサン、トルエン、石油、エーテル、ベンゼン、酢酸エチル、クロロホルム、ジクロロメタン、1,1,2-トリクロロエタン、ジ-メチルスルホキシド、およびアセトンなどの有機溶媒が挙げられ、とりわけアルコールが好ましい。水は未蒸留水、蒸留水、脱イオン水、または純水でよい。使用できる緩衝液の例は、リン酸緩衝液およびクエン酸緩衝液が挙げられる。
【0083】
植物材料からRR1を抽出する場合、水、水含有エタノールまたは無水エタノールなどの治療薬剤に適した溶媒を使用することが適正である。使用してもよいアルコールの例としては、メタノール、エタノール、プロパノールおよびブタノールなどの一価アルコール、ならびにプロピレングリコールおよびグリセロールなどの多価アルコールが挙げられ、なかでも一価アルコールが好ましく、特にエタノールが好ましい。これらの溶媒は、単独または混合物として用いてもよい。混合溶媒として用いる場合、水含有アルコールが好ましい。水含有一価アルコールがより好ましく、水含有エタノールが特に好ましい。
【0084】
抽出に関しては、溶媒を、例えば植物1重量部を基にして、0.1〜10,000重量部、好ましくは1〜100重量部の量を用いることができる。抽出温度に関して特に制限はないが、抽出は0〜100℃、より好ましくは20〜90℃で実施するのが好ましい。抽出時間に関しては特に制限はないが、例えば1分間〜1週間、より好ましくは30分間〜1日の間に行うことが好ましいであろう。
【0085】
抽出に用いる装置に関しては特に制限はないが、効率的な抽出用に設計された容器、撹拌機、還流冷却器、ソックスレー抽出器、ホモジェナイザー、シェーカー、超音波発生装置等を用いてもよい。液体抽出物は、沈殿物、脱水濾過、透明濾過、遠心分離濾過、遠心分離沈殿、圧縮分離または圧搾濾過などの、各種固体-液体分離の手段によって処理できる。
【0086】
植物粉末を調製する場合は、葉、枝、茎、樹皮、根、種、培養細胞または器官、あるいはカルスなどの、RR1を含む植物の全てまたは一部を用いてよい。空中部分(茎および葉)が好ましい。植物材料は、そのまま用いても、または物理学的もしくは化学的もしくは生物学的に処理した後に用いてもよい。物理学的もしくは化学的処理の方法の例は、乾燥、フリーズドライ、破壊、および抽出である。物理学的もしくは化学的に処理された物体としては、乾燥物、フリーズドライ物、破壊物および抽出物が挙げられる。抽出物は、抽出後に得た植物残留物を含む。
【0087】
植物粉末を調製するために、好ましく乾燥したティノスポラ・コルディフォリア植物材料は、下記を用いて破砕できる:ジョークラッシャ、ジャイレートリークラッシャ、もしくはコーンクラッシャなどの圧縮破砕機;カッターミルもしくはシュレッダーなどの剪断機;ハンマークラッシャなどのインパクトクラッシャ;ロールクラッシャなどのロールミル;ディスインテグレータ、もしくはケージミルなどのロータリーミル;コーヒーミルなどのスクリューミル;エッジランナーなどのローリングミル;スタンプミルなどのハンマリングミル;遠心ローラーミル、ボールベアリングミル、ボールミル、もしくはゼゴミルなどのローラーミル;スウイングハンマーミル、ピンミル、ケージミル、ターボ型ミル、もしくは遠心ミルなどの高速ロータリーミル;ローリングボールミル、振動ボールミル、遊星ボールミル、もしくはCFミルなどの容器振動ミル;フローパイプ型ミル、撹拌タンクミル、管型ミル、空気吸込み型ミル、インパクトプレートミラー、もしくは流動床ミルなどのジェットミル;超音波シュレッダーなどの破砕機;ストーンモーターもしくはモーター。上記の方法で得た生成物を、さらに物理的または化学的に処理し、植物粉末を得ることができる。
【0088】
植物粉末は荒くとも細かくともよい。植物粉末の平均粒子サイズは、乾燥状態において、好ましくは0.1μm〜1mm、より好ましくは1〜100μm、特には2〜50μmである。乾燥状態の植物粉末の平均粒子サイズは、例えば、レーザー分散粒子分布アナライザーによって決定できる。または、植物粉末をグリセロールと水の1:1の混合液で膨潤させた場合、粉末の平均粒子サイズは、好ましくは1μm〜10mm、より好ましくは10μm〜1mm、特には20〜500μmである。膨潤状態の植物粉末の平均粒子サイズは、例えば顕微鏡を用いた観察によって決定できる。
【0089】
乾燥方法は、当技術分野で公知である。例えば、乾燥方法としては、加熱および減圧下での乾燥、加熱および大気圧下での乾燥、またはスプレー乾燥器もしくはドラム乾燥器を用いた乾燥、またはフリーズドライが挙げられるが、とりわけ加熱および減圧下での乾燥またはフリーズドライが好ましい。
【0090】
本明細書で使用する「腫瘍」という用語は、悪性であるか良性であるかにかかわらず、全ての新生物細胞の生育および増殖を指し、全ての前癌性および癌性細胞および組織を指す。例えば、特定の癌は、固体の塊状の腫瘍を特徴とする。固体腫瘍の塊は、存在する場合、原発腫瘍塊のことがある。原発腫瘍塊は、その組織の正常細胞が形質転換した結果生じた癌細胞が組織内で生育したものを指す。ほとんどの例では、原発腫瘍塊は、視覚法または触診法によって見つけることができる嚢胞の存在によって、あるいは組織の形状、質感または重量の異常によって同定する。しかしながら、いくつかの原発腫瘍は、触診できず、X線などの医学的画像技術(例えばマンモグラフィー)を用いるか、または針吸引によってのみ検出できる。これら後者の技術の使用は、早期発見においてより一般的である。本明細書で使用する「腫瘍」という用語は、白血病のような、非固形性の新生物も含む。組織内の癌細胞の分子および表現形の分析は通常、癌が組織に対し内因性であるか、または疾患が別の場所からの転移によるものか確認するであろう。
【0091】
本明細書で使用する「治療する」または「治療」という用語は、治療的処置および予防もしくは防御的処置の両方を指し、この場合の目的は、癌などの細胞増殖障害、または他の生理学的状態(例えば疾患もしくは他の障害)の発生もしくは拡大といった、望ましくない生理学的変化もしくは障害を予防するか、遅らせる(軽減する)ことである。好ましくは、病理学的状態は、免疫刺激または免疫抑制のいずれかによる治療または管理ができるものである。本発明の目的に関して、有利または望ましい臨床結果とは、検出可能であるか否かにかかわらず、症状の軽減、疾患の広がりの縮小、疾患の状態の安定化(即ち悪化しない)、疾患の進行の遅延または減速、疾患状態の改善もしくは緩和および寛解(部分的または完全を問わず)を含むが、これらに限定されない。「治療」はまた、治療を受けていない場合に予想される生存期間に比べ、生存期間が延長することも意味する。治療が必要な対象としては、既に状態または障害を持つ対象だけでなく、状態または障害を持つ傾向がある対象、もしくは防止しなければならない状態および疾患を持つ対象が挙げられる。
【0092】
本明細書で使用する「(治療)有効量」という用語は、哺乳動物における、細胞増殖性障害などの病理的状態(例えば疾患または他の障害)を治療するのに有効な、本発明のRR1化合物または他の作用物質などの、作用物質の量を指す。癌の場合、作用物質の治療有効量は、癌細胞の数を減らし;腫瘍の大きさを縮小し;癌細胞の周辺臓器への浸潤を阻止し(即ち、ある程度遅延させる、および好ましくは停止させる);腫瘍の転移を阻止し(即ち、ある程度まで遅延させる、および好ましくは停止させる);腫瘍の増殖をある程度まで阻止し;ならびに/または、癌に関係する一つまたは複数の症状を、ある程度まで軽減する。作用物質が増殖を阻止できるおよび/または既存の癌細胞を殺すことができる程度まで、それは細胞増殖抑制性および/または細胞傷害性でよい。癌治療に関しては、効力は、例えば、疾患が進行するまでの時間(TTP)を評価すること、および/または応答率(RR)を決定することによって測定できる。
【0093】
本明細書で使用する「抗癌剤」という用語は、癌細胞の機能を阻止し、それらの形成を阻止し、および/またはそれらのインビトロまたはインビボでの破壊を引き起こす物質または治療を指す。例としては、細胞傷害性剤(例えば5-フルオロウラシル、タキソール)および抗シグナル伝達剤(例えばPI3KインヒビターLY)が挙げられるが、これらに限定されない。
【0094】
本発明細書で使用する「細胞傷害性剤」という用語は、細胞の機能を阻止または妨害、および/または細胞のインビトロまたはインビボでの破壊を引き起こす物質を指す。用語は、放射性同位元素(例えばAt211、I131、I125、Y90、Re186、Re188、Sm153、Bi212、P32、およびLuの放射性同位元素)、化学療法薬剤、毒素、例えば細菌、真菌、植物もしくは動物起源の低分子毒素もしくは酵素活性毒素、それらの断片および/もしくは変種を含む抗体を含むことを意図する。
【0095】
本明細書で使用する「化学療法薬剤」という用語は、例えば、パクリタキセル(TAXOL、BRISTOL-MYERS SQUIBB Oncology, Princeton, N.J.)およびドキセタキセル(TAXOTERE、Rhone-Poulenc Rorer, Antony, France)などのタキセン、クロラムブシル、ビンクリスチン、ビンブラスチン、例えばタモキシフェン、ラロキシフェン、アロマターゼ阻害4(5)-イミダゾール、4-ヒドロキシタモキシフェン、トリオキシフェン、ケオキシフェン、LY117018、オナプリストン、およびトレミフェン(Fareston)などの抗エストロゲン剤、ならびにフルタミド、ニルタミド、ビカルタミド、ロイプロリド、およびガセレリン等などの抗アンドロゲン剤の、癌治療に有用な化合物である。
【0096】
本明細書で使用する「RR1化合物」という用語は、図3に示すようなRR1、またはRR1の薬学的に許容される塩または類似体を指す。本明細書で使用する、RR1またはRR1化合物に関連する「単離された」という用語は、自然に生ずる、例えばティノスポラ・コルディフォリア植物材料もしくは植物抽出物から生ずる、媒体を実質的に含まない化合物を指す。しかしながら、単離RR1化合物は、当業者に公知である適切な化学合成反応によっても得られる(Greene, T.W. and Wuts, P.G.M.「Protective Groups in Organic Synthesis」John Wiley & Sons, Inc. New York. 3rd Ed. pg. 819, 1999; Honda, T. et al. Bioorg. Med. Chem. Lett., 1997, 7:1623-1628; Honda, T. et al. Bioorg. Med. Chem. Lett, 1998, 8:2711-2714; Konoike, T. et al. J. Org. Chem., 1997, 62:960-966; Honda, T. et al. J. Med. Chem., 2000, 43:4233-4246)。
【0097】
「含む」、「からなる」および「本質的にからなる」という用語は、その標準的な意味に従って定義される。これら用語は、本明細書を通じて、各用語に関連する具体的な意味を与えるために相互に代用できる。
【0098】
本明細書で使用する場合、単数形の「一つの(a)」、「一つの(an)」、および「その(the)」は、文脈が特に明瞭に指図しない限り複数形も含む。したがって、例えば、「一つのRR1化合物」という言及は、一つより多いこのようなRR1化合物を含む。「一つの免疫調節分子」という言及は、一つより多いこのような免疫調節分子、または一つより多い免疫調節分子のタイプを含む。「一つの抗原」という言及は、一つより多い抗原、または一つより多い抗原のタイプを指すのに用いられるといった具合である。
【0099】
本発明の実施は、特に指示がない限り、分子生物学(組換え技術を含む)、微生物学、細胞生物学、生化学および免疫学の通常の技術を使用するが、それらは当技術分野の範囲内である。このような技術は、「Molecular Cloning: A Laboratory Manual」、第2版のような文献に十分に説明されている(Sambrook et al., 1989);「Oligonucleotide Synthesis」(M. J. Gait, ed., 1984);「Animal Cell Culture」(R. I. Freshney, ed., 1987);「Methods in Enzymology」(Academic Press, Inc.);「Handbook of Experimental Immunology」(D. M. Weir & C. C. Blackwell, eds.);「Gene Transfer Vectors for Mammalian Cells」(J. M. Miller & M. P. Calos, eds., 1987);「Current Protocols in Molecular Biology」(F. M. Ausubel et al., eds., 1987);「PCR: The Polymerase Chain Reaction」(Mullis et al., eds., 1994);および「Current Protocols in Immunology」(J. E. Coligan et al., eds., 1991)。
【0100】
以下は、本発明を実施するための材料、方法、および手順を例示する実施例である。実施例は例示であり、限定的に解釈してはならない。
【0101】
材料および方法
RR1の単離および特徴付け
ティノスポラ・コルディフォリア粉末(GARRY and SUN, Inc., Reno, Nevadaから市販されている)の抽出物の予備的研究により、免疫刺激の原則は、水性画分中に限定されることが明らかとなった。図1に、単離に適用された手順をフローチャートにて示す。図3に示す最終的なRR1化合物は、抽出に使用した全乾燥材料の約0.1%収率で、水に溶解した膨れた固体として単離された。最終生成物の最初の分析、および13C NMRスペクトルは、それが多糖であることを明らかにした。したがって、Complex Carbohydrate Research Center, University of Georgia, Athensにおいて、グリコシル成分、結合、分子量、およびアノマー中心におけるグルコース単位のコンホメーションなどの、多糖の詳細な分析を行った。グリコシル成分の分析は、Yorkら(York, W.S. et al. Methods Enzymol., 1985, 118:3-40)の方法に従った酸性メタノリシスによりRR1から得られた単糖メチルグリコシドのパー-O-トリメチルシリル誘導体のガスクロマトグラフィー/質量分析の組み合わせ(GC/MS)により行った。この分析では、イノシトールを内部基準として使用した。単糖誘導体は、それらの特徴的な保持時間により同定され、さらにそれらの質量スペクトルにより確認された。グリコシル結合分析を行うには、試料をCikanuおよびKereck(Ciucanu, I. and F. Kereck Carbohydr. Res., 1984, 131:209-217)の方法により3回パーメチル化し、2Mトリフルオロ酢酸で加水分解し、NaBD4で還元し、無水酢酸/ピリジンでアセチル化して、得られた一部メチル化アルジトールアセテート(PMAA)をGC/MSにより分析した。糖残基は、それらの特徴的な保持時間と、質量スペクトルデータにより同定した。グリコピラノシル単位のアノマー中心におけるコンホメーションは、D2O中で記録した500MHzプロトンNMRスペクトルに基づき、かつ基準試料のアノマープロトンシグナルとの比較により得た。分子量は、50mMギ酸アンモニウム緩衝液により速度0.5ml/分で溶出するSUPELCOシリカカラム(1.0×30cm)を使用したサイズ排除クロマトグラフィーから得て、屈折率により検出した。デキストラン試料を基準として使用して、溶出したピークの保持時間と基準との比較により分子量を得た。
【0102】
免疫刺激特性
RR1の免疫刺激特性を測定するために、リンパ球の異なるサブセットの活性化、例えばインターロイキン(IL)-1β、IL-2、IL-4、IL-6、IL-10、IL-12、p70およびp40、IL-18、インターフェロン(IFN)-αおよびγ、腫瘍壊死因子(TNF)-αおよびβ、単球遊走因子(MCP)-1などのサイトカインの合成、一酸化窒素(NO)の合成、ならびにヒトリンパ球内で誘発される酸化ストレスの程度を分析した。健康な志願者から採取した新鮮血から、histopaque密度勾配法により正常なリンパ球を単離して、様々なアッセイに使用した。ヒト白血病(CEM)および多剤耐性(CEM/VLB)細胞株を、10%ウシ胎児血清および抗生物質を補充したRPMI培地中、5%CO2インキュベーター内にて37℃で増殖させた。
【0103】
リンパ球の活性化
正常なリンパ球(106/ml)を、RR1 0〜100μgで、37℃に保持したCO2インキュベーター内にて、RPMI培地中で24時間処理した。次いで、細胞を特定の蛍光色素結合モノクローナル抗体で30分間、室温にて染色し、Coulter Elite Flow Cytometer内で、AlamoおよびMelnick(Alamo, A.L. and S.J. Melnick Cytometry, 2000, 42:363-370)の4色または5色イムノタイピング(immunotyping)アッセイプロトコルにより分析し、例えばNK、TおよびB細胞等のリンパ球のサブセットの活性化は百分率を計算した。
【0104】
RR1処理リンパ球の細胞傷害性
活性化したリンパ球の高められた細胞傷害性を評価するために、RR1活性化リンパ球をエフェクター細胞とし、ヒト白血病細胞(CEM)を標的として使用する機能的アッセイを採用した(Liu, L. et al. Nat. Med., 2002, 8:185-189; Jerome, K.R. et al. Nat. Med., 2003, 9:4-5)。簡単に言えば、正常なリンパ球(106/ml)を、RPMI培地中の異なる濃度のRR1で、CO2インキュベーター内にて37℃で24時間処理した。翌日、標的細胞(CEM 1×106/ml)を、1ml PBS中の4.6μM膜標識色素PKH26(SIGMA)により室温で3分間標識した。等容積のウシ胎児血清(GIBCO, Life Sciences, MD)を1分間加えて、標識を停止した。次いで、標識した腫瘍細胞を、エフェクターと標的との比を1:1としてRR1処理リンパ球と共に4時間インキュベートし、未処理リンパ球を対照として使用した。活性化NK細胞により殺傷された細胞の百分率を、氷上にて2%パラホルムアルデヒド溶液1mlで混合物を30分間固定し、続いてPBS中0.5%ツイーン20中に懸濁させることにより測定した。細胞混合物を抗活性カスパーゼ3-FITC抗体(BD Biosciences, CA)7.5μlで、室温にて30分間染色し、PBSで洗浄し、Coulter Elite Flow Cytometer内で分析した。
【0105】
サイトカインおよびケモカイン合成の定量
IL-1β、IL-2、IL-4、IL-6、IL-10、IL-12p40、IL-12p70、IL-18、IFN-γおよびTNF-α、βなどのサイトカイン、ならびにケモカイン、MCP-1を、ELISA法により、BD Biosciencesのキットを使用して、製造業者のプロトコルにしたがって定量した。IFN-αは、RESEARCH DIAGNOSTICS Inc.(New Jersey)のELISAキットを使用してアッセイした。簡単に言えば、ELISA希釈液50μlをピペットで、96ウェルプレートの抗体コートウェル内に入れ、続いて各標準試料および試験試料100μlを入れて、5秒間振とうしてウェル内の内容物を混合し、プレートシーラーで覆い、室温で2時間インキュベートした。インキュベーション後、ウェルの内容物を吸引し、洗浄溶液で5回洗浄した。最終的な洗浄の洗浄溶液を完全に除去した後、検出溶液100μlを加え、プレートシーラーで覆い、1時間インキュベートした。ウェルを洗浄溶液で7回洗浄し、一段階基質試薬100μlを加えて、暗闇で30分間インキュベートした。停止溶液50μlを加えて着色を停止し、BIORAD Benchmarkプレートリーダー内で、参照波長570nmと共に450nmにて吸収を記録した。
【0106】
補体活性化経路
ヒト補体C3a des ArgおよびC4a des Arg関連のEIAキット(ASSAY DESIGN Inc., Ann Arbor, MI)を使用して、製造業者のプロトコルにしたがって、切断された補体成分(C3a des ArgおよびC4a des Arg)を測定した。簡単に言えば、健康な志願者から採取した正常血液1mlを、CO2インキュベーター内で37℃にて24時間、RR1 0〜100μg/mlと共にインキュベートした。処理した血液試料を4℃にて2000×gで遠心分離にかけ、補体試薬「A」225μlを等容積の試料上澄み液に加え、徹底的にボルテックスで撹拌した。この混合物に10N HCl 50μlを加えて、再度ボルテックスで撹拌し、室温で1時間インキュベートした。試料を室温で5分間、ミクロ遠心分離機内で10,000rpmにて遠心分離し、上澄み液180μlを15ml管に移し、9N NaOH 20μlを加えて、徹底的にボルテックスで撹拌した。この混合物に、補体試薬「B」600μlを加えた後、アッセイ緩衝液10.7mlを加え、ボルテックスで撹拌して、分析に使用した。アッセイ試料(100μl)を96ウェルのマイクロプレート内のウェル内にピペットで入れ、次いで青色コンジュゲート50μlおよび黄色抗体50μlを入れた。プレートをプラットホームシェーカー上で、500rpmにて2時間振とうした。ウェルを吸引して非結合材料を除去し、洗浄溶液200μlで3回洗浄し、p-Npp基質溶液200μlを加えた。プレートを37℃で1時間、振とうせずにインキュベートし、停止溶液50μlを加え、BIO-RAD Bench topプレートリーダー内で、参照波長570nmと共に405nmの吸収を測定した。
【0107】
誘発可能な一酸化窒素シンターゼ(iNOS)誘導、および一酸化窒素(NO)合成
NOの増大した、および持続的な放出は、アルギニン刺激による、酵素iNOSの刺激を原因とする。RR1刺激によるiNOS誘導を、サンドイッチ酵素イムノアッセイ法を用いるQUANTIKINE iNOSイムノアッセイキット(R&D systems, Minneapolis, MN)によりアッセイした。異なる濃度のRR1で処理した細胞の細胞質抽出物から、iNOSをアッセイした。試料および標準を96ウェルプレートのウェル内にピペットで入れ、存在する任意のiNOSは固定化した抗体に結合した。未結合物質を洗浄除去した後、iNOSに特異的な酵素結合モノクローナル抗体を加えた。未結合抗体酵素試薬を洗浄除去した後、着色物質溶液を加え、停止溶液で着色を停止して、Bio-Radプレートリーダー内で、参照波長570nmと共に450nmの吸収を読み取った。
【0108】
NOの量を、一酸化窒素定量キット(ACTIVE MOTIF, Carlsbad, CA)により、製造業者のプロトコルにしたがってアッセイした。簡単に言えば、リンパ球(106/ml)を様々な濃度のRR1(0〜100μg/ml)と共に、硝酸を含まない培地(DULBECO Minimum Essential Medium)中にて37℃で24時間インキュベートした。細胞培養物の上澄み液(70μl)を、再構成した補因子20μl、および硝酸還元酵素溶液10μlと共に96ウェルプレート内にピペットで入れた。プレートをプレートシェーカー上で、室温にて30分間、150rpmにて振とうした。Griess試薬AおよびB(各50μl)を各ウェルに加え、20分間着色させて、BIO-RADプレートリーダー内で参照波長620nmと共に540nmの吸収を測定した。
【0109】
酸化ストレス(GSH/GSSGレベル)
グルタチオンの還元(GSH)およびグルタチオンの酸化(GSSG)レベルの測定値、ならびにそれらの比は、酸化ストレスの有用な指標である。Biotech GSH/GSSG-412キット(Oxis Research, Portland, OR)を使用して、製造業者のプロトコルにしたがって、比色法によりGSHおよびGSSGのレベルをアッセイした。簡単に言えば、健康なドナーからの正常血液試料を、異なる濃度のRR1と共に、5% CO2インキュベーター内で37℃にて24時間インキュベートした。GSSGに関しては、各処理試料100μlを-70℃で4時間冷凍し、解凍して、5%メタリン酸(MPA)290μlを加えた。試料を15〜20秒間ボルテックスで撹拌し、1000×gで10分間遠心分離した。MPA抽出物(50μl)を比色アッセイで使用したGSSG緩衝液700μlに加えた。GSH測定に関しては、全血50μlを-70℃で冷凍し、解凍して、5% MPA 350μlを加え、15〜20秒間ボルテックスで撹拌し、10,000×gで10分間遠心分離した。MPA抽出物(50μl)をアッセイ緩衝液3mlに加え、更なる分析にて使用した。各標準および試料のMPA緩衝液混合物(200μl)を分光光度計のキュベットに移して、色素原200μlを加えた後、酵素200μlを順に加え、室温で5分間インキュベートした。その後、キュベット内にNADPH 200μlを加え、BECKMAN分光光度計内で412nmの吸収における変化を記録した。吸収値を使用して反応速度をプロットし、GSHおよびGSSGレベルを決定した。
【0110】
統計的分析
データ分析に一要因分散分析(ANOVA)を使用した。
【0111】
細胞株
マウス単球(RAW 264.7)、およびヒト胎児腎臓(HEK293)細胞株を、研究のためにAmerican Type Culture collection, Manassas, VAから得た。TLR2、TLR2/6およびTLR4/CD14/MD2遺伝子をトランスフェクトしたHEK293細胞を、INVIVOGEN Corporation, San Diego, CAより購入した。細胞は、5%CO2インキュベーター内の組織培養フラスコ内で、10%ウシ胎児血清、L-グルタミン(2mM)、および抗生物質(100単位/mlペニシリンおよび100μg/mlストレプトマイシン)を補充したDMEM培地中で増殖させた。Nair, P.K.ら(Nair, P.K. et al. Int. Immunopharmacol., 2004, 4:1645-1659)に記載されている抽出手順にしたがって、GARRY and SUNS(Reno, Nevada, USA)から購入したティノスポラ・コルディフォリア粉末からRR1を単離した。試料の内毒素含有量をLymulus Amoebocyte Lysate Assay(Cambrex, MA)により試験し、有意なレベル(0.0008ng/ml)が示された。研究全体を通して、内毒素を含まない緩衝液、試薬、および無菌水を使用することにより、必要な予防策をとって内毒素汚染を回避した。
【0112】
試薬
ザイモサンA、およびFITC標識ザイモサンA生体粒子を、Molecular Probes, Inc., Eugene, ORより購入した。ラミナリンおよびカフェ酸フェネチルエステル(CAPE)を、SIGMA Chemical Co., St. Louis, MOより購入した。抗マウスCD11bモノクローナル抗体(mAb)を、BD BIOSCIENCES, San Jose, CAより購入した。FITC標識TLR2およびPE標識TLR4抗体を、eBiosciences, San Diego, CAより購入した。
【0113】
RR1の非オプソニン化結合
ザイモサンA生体粒子の非オプソニン化結合に対するRR1の阻害効果を測定するために、マウス単球/マクロファージ細胞(0.5×106)を、0、100、500および1000μg/mlのRR1と共に4℃でインキュベートして、補体を含むオプソニンの局所放出を防止した(Ezekowitz, R. A. et al. J. Clin. Invest., 1985, 76:2368-2376; Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)。氷上で1時間インキュベーション後、細胞を予備冷却した培養液で3回洗浄し、FITCコンジュゲートザイモサンA粒子を25粒子/細胞の比で加えて、氷上でさらに1時間保持した。細胞をPBSで2回洗浄し、CO2インキュベーター内にてDMEM中で37℃にて30分間インキュベートした。未結合ザイモサン粒子を大量の培地で洗浄して除去し、ザイモサン-FITCの細胞内蛍光をCoulter Eliteフローサイトメーターで分析した。
【0114】
オプソニン化結合および食作用
RR1およびラミナリンの、ザイモサンA粒子のオプソニン化結合および食作用に対する効果を測定するために、RAW264.7細胞を、DMEM中で37℃にて1時間、RR1またはラミナリン(0および500μg/ml)で処理した。このことは、オプソニンの放出とオプソニン結合の促進とを確実にするであろう。細胞をPBSで2回洗浄し、25粒子/細胞の比でザイモサンA-FITC生体粒子と共に、37℃で1時間インキュベートした。大量の培地での洗浄により未結合ザイモサン粒子を除去した後、細胞をPBS中に懸濁させ、細胞内蛍光をCoulter Eliteフローサイトメーターで分析した。蛍光強度を対照の百分率で表した。
【0115】
RR1誘導によるTNF-α合成
0.5% FBS(飢餓)を含有するDMEM培地を有する6ウェルプレート上に、RAW264.7細胞(0.5×106/ml)をプレーティングした。ひとたび細胞が付着したら、培地を交換し、0、100、500および1000μg/ml用量のRR1を加え、37℃で24時間インキュベートした。ELISAキット(BD BIOSCIENCES kit, San Jose, CA)を使用して、製造業者のプロトコルにしたがって、分泌されたTNF-αに関して培地を分析した。
【0116】
TNF-α合成に対するNF-κB阻害剤CAPEの効果を測定するために、インキュベーター内で、RAW264.7細胞を10μg/mlのCAPEで37℃にて1時間処理した後、RR1(0〜1000μg/ml)で一夜処理した。培地中の分泌されたTNF-αを、ELIZA(BD BIOSCIENCES kit, San Jose, CA)を使用して分析した。
【0117】
NF-κB活性
RAW264.7細胞を、0、100、500および1000μg/mlのRR1と共に8時間インキュベートし、Active Motif, CAのNF-κB活性キットを使用して、製造業者のELISA説明書にしたがって、核タンパク質を抽出した。NF-κB活性化をさらに視覚化するために、核タンパク質を用いて電気泳動移動度シフトアッセイ(EMSA)を行った(Ghosh, S, and Karin, M., Cell, 2002, 109:S81-96, Young, S.H. et al. J. Biol Chem., 2001, 276:20781-20787; Lebron, F. et al. J Biol Chem., 2003, 278:25001-25008)。NF-κBに結合する二本鎖プローブ

を32P-ATPで放射標識し、核抽出物5μgを用いてEMSAを実施した。EMSA反応混合物を室温で30分間インキュベートし、DNA-タンパク質複合体を6%ポリアクリルアミドゲル上で分離して、オートラジオグラフィーで視覚化した。NF-κBの時間依存的活性化を測定するために、異なる期間にて(刺激0、4、8、14、20時間後)RR1(100μg/ml)処理細胞から核抽出物を調製した。ACTIVE MOTIF, Carlsbad, CAのELISAキットを使用して、核抽出物のタンパク質含有量と、NF-κB活性を分析した。
【0118】
I-κBαの分解
IκB-αの分解は、NF-κB活性化の必須条件である。細胞のI-κBαレベルおよびその分解に対するRR1の効果を測定するために、異なる期間にて(0、15分、30分、1時間)、RR1処理RAW264.7マクロファージ由来の細胞質ゾル溶解物を用いてイムノブロット分析を実施した。溶解物を12%ゲル上でSDS-PAGEにより分離して、TransBlot装置(BIO-RAD, Hercules, CA)を使用してニトロセルロース膜に転写した。ウエスタンブロットハイブリダイゼーションプロトコル(Lebron, F.et al. J Biol Chem., 2003, 278:25001-25008)により、抗マウスIκB-α抗体(CELL SIGNALING TECHNOLOGY, Inc, Beverly, MA)を用いて、色検出システム(BIO-RAD, Hercules, CA)を使用して、膜をIκB-α特異的モノクローナル抗体(1:1000)でハイブリダイズした。
【0119】
オプソニン化結合に関するCR3のCD11b mAbによる遮断効果
RR1のマクロファージに対するオプソニン化認識/結合にグルカン特異的受容体CR3が関与しているか否かを試験するために、CR3に特異的なモノクローナル抗体(mAbs)(BD BIOSCIENCES, San Jose, CAのラット抗マウスCD11b)を使用して、CR3を遮断した。RR1(0〜100μg/ml)と共に3時間インキュベートするのに先だって、RAW264.7細胞(0.5×106)をCO2インキュベーター内で5μg/mlのモノクローナル抗体により37℃で1時間処理した。培地中にマクロファージにより放出されたTNF-αを、ELISAキット(BD Biosciences, San Jose, CA)を用いて分析した(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Brown, G.D. et al. J. Exp. Med., 2003, 197:1119-1124)。
【0120】
TLRシグナル伝達
TLRシグナル伝達の関与を測定するために、親のHEK293、ならびにヒトTLR2、TLR6、TLR2/6またはTLR4/CD14/MD2遺伝子をトランスフェクトした(pDUOプラスミド内にクローン化した)HEK293細胞を使用した。これらの細胞(0.5×106/ml)を、RR1 0〜100μg/mlと共に24時間インキュベートした。これらの細胞は、活性化により非常に低いレベルのTNF-αおよび検出レベルのIL-8を産生するため、培地中に分泌されたIL-8はELISA法により分析した。偶然にも、IL-8合成も転写因子NF-κBの制御下にある。
【0121】
TLR mRNAおよびタンパク質レベル
RR1処理細胞から抽出した全RNAを使用して、RT-PCRによりTLR2、4、および6 mRNAレベルを分析した。全RNAをRR1処理および未処理RAW264.7細胞から抽出し、0.5μg/ml RNAを逆転写して、マウスTLR2、4、および6に特異的なPCRプライマー(R&D SYSTEMS, Minneapolis, MN)を使用して増幅した。対照としてマウスGAPDH遺伝子も増幅した。PCR生成物をアガロースゲル上で分離した。
【0122】
実施例1 RR1の特徴付け
グリコシル組成および結合分析の結果は、最初の発見、およびRR1の炭水化物組成に関する13C NMRスペクトルデータを裏付けるものであった。グリコシル組成の分析は、グルコースがRR1の唯一の成分であることを示したが、一方、結合分析は、結合の3つのタイプに対応するグルコピラノシル残基の3つのタイプを明らかにした:4-結合グルコピラノシル残基(80%)、4,6-結合グルコピラノシル残基(12%)および末端グルコピラノシル残基(8%)。13C NMRスペクトル(図2A)は、グルコピラノシル部分内の炭素原子の良好に分離したシグナルを示した-C1(δ99.97ppm)、C2(δ73.68ppm)、C3(δ71.89ppm)、C4(δ77.09ppm)、C5(δ69.67ppm)、およびC6(δ60.83ppm)。C1およびC4シグナルの低磁場シフトは(1→4)結合を裏付けるが、一方、δ71.53ppmのシグナルは、(1→6)結合のC6に起因し得る。500MHzプロトンNMRスペクトル(図2B)におけるδ5.44ppmのシグナル(良好に分離せず)およびδ5.00ppmのシグナルは、グリコピラノシル単位のアノマー炭素に関連したαプロトンに起因する。他方、δ4.66ppmの非常に弱いシグナルは、βアノマーに起因する可能性がある。しかしながら、α-D-グルコースとβ-D-グルコースのシグナルの比は約99.9:1であり、それ故殆ど全てのグルコース単位がα立体配置を有すると想定される。したがって、RR1は、主鎖内の(1→4)結合したグルコピラノシル単位、(1→6)結合したグルコピラノシル単位の分岐、および0.15の分岐度を有するα-D-グルカンである(図3)。サイズ排除クロマトグラフィーでは、RR1は12.32分において単一ピークとして溶出し、これは511kDaデキストラン試料(保持時間:12.72分)のピークに非常に近かった。したがって、RR1は>550kDaの分子量が割り当てられた。
【0123】
実施例2 非細胞傷害性/細胞増殖効果
図4Aおよび図4Bに示すMTTアッセイの結果は、RR1が1000μg/mlの高濃度においても、正常リンパ球または腫瘍細胞株(CEMおよびCEM/VLB)のいずれに対しても、直接的な細胞傷害性または細胞増殖効果を有さないことを示す。
【0124】
実施例3 リンパ球活性化
リンパ球は、哺乳動物免疫系の重要なエフェクター細胞であり、これらの試験は、異なるリンパ球亜集団が、RR1により多様なレベルで活性化されることを示す。RR1 100μg/mlにより、B細胞は39%、T細胞は102%およびNK細胞は331%活性化される(図5)。NK細胞は、適応免疫系による抗体産生および認識前に、抗原/マイトジェンと接触する自然免疫系の主なエフェクターであるため、NK細胞の>3倍の活性化は非常に意義がある。RR1によるNK細胞の増大された活性化は、図6に示す機能的細胞傷害性アッセイの結果から全く明らかである。RR1処理された正常リンパ球は、未処理細胞と比較して高い割合の腫瘍細胞を殺すことができ、かつ活性化リンパ球の細胞傷害性の用量依存的増強が明らかであった。
【0125】
実施例4 補体活性化経路
図7に示すように、RR1濃度の上昇に伴う、代替的経路のC3a des Argレベルの段階的増大が注目された。この観察結果は、他の多糖による補体活性化に関するいくつかの報告と類似する。C3aおよびC4aは、代替的および古典的経路における補体活性化カスケード中に、血漿成分C3およびC4から放出される生理活性切断生成物であり(Ember, J.A. et al. The Human complement system in Health and Disease, Ed. Volnakis, J.E. and M.M. Frank, Marcel Dekker Inc., New York, 1998, pp. 241-248)、これらはより活性が低いC3a-des ArgおよびC4a-des Arg形態に急速に転換されて、細胞免疫応答の仲介に関与する。代替的経路は自己増幅性であり、抗体不在下での病原の排除および認識に重要である(Stahl, G.M. et al. Am. J. Pathol., 2003, 162:449-455)。β-グルカンは代替的経路を活性化することが報告されており、これらのポリマーが示す宿主仲介性抗腫瘍活性は、補体系の活性化と相関していた(Hamuro, J. et al. Am. J. Pathol., 1978, 93:526-617)。リポ多糖は、代替的および古典的経路を経由して補体系を活性化し;即ち、脂質部分が古典的経路を活性化し、一方、多糖部分が代替的経路を活性化する(Morrison, D. C. and R. J. Ulevitch Am. J. Pathol., 1978, 93:526-617)。
【0126】
実施例5 サイトカインの合成
図8A〜図8Hに示すように、RR1は、IL-1β(1080pg)、IL-6(21833pg)、IL-12p40(918.23pg)、IL-12p70(50.19pg)、IL-18(27.47pg)、IFN-γ(90.16pg)、MCP-1(2307pg)およびTNF-α(2225pg)の合成を誘導したが、IL-2、IL-4、IL-10、TNF-βおよびIFN-αの産生を誘導しなかった。一般に、図8Cに示すように、最大値が10μg/mlにて記録されかつRR1の増加により低下傾向が示されたIL-12(p40)を除いて、RR1によりサイトカイン産生の用量依存的増大が観察された。図8Hに示すように、RR1 10μg/mlまでのMCP-1産生は非常に有意であり、より高いRR1濃度では増大は僅かであった。一般に、プロ炎症性サイトカインIL-1β、IL-6、およびTNF-α、ならびに調節サイトカインIL-12p(40)は、他のサイトカインと比較してより高いレベルで産生された。
【0127】
哺乳動物の免疫系は、抗原、病原および非自己分子を認識して、免疫コンピテント細胞の活性化、化学的メッセンジャー(サイトカインおよびケモカイン)の産生、補体カスケード経路の活性化および一酸化窒素(NO)の合成を介した防御機構を引き起こす。サイトカインは、刺激に応答して免疫成分細胞により分泌される、低分子量の調節性非抗体タンパク質の群である。これらは、標的細胞の特異的受容体に結合し、最終的に標的細胞内の遺伝子発現に繋がるシグナル伝達経路を引き起こす。サイトカインは、様々な細胞の活性化、増殖、および/または分化を刺激または抑制することによって、かつ抗体または他のサイトカインの分泌を調節することによって、免疫応答の強度および持続時間を調節する。サイトカインが示す多面発現性、重複性、相乗作用および拮抗作用により、サイトカインは細胞活性を調節および制御することができる。炎症応答は、病原体の制御および排除、ならびに組織の完全性を回復する創傷治癒の促進に重要である。
【0128】
IL-1βおよび/またはTNF-αは、互いの産生を誘導すると共にIL-6の産生を誘導し、相乗的に作用して数個の生物学的作用を調節し、その上IL-1は自己誘導性である(Horai, R. et al. J. Exp. Med., 2000, 191:313-320)。RR1処理リンパ球内でのIL-1βおよびTNF-αの産生は、LPS刺激ヒト単球細胞株(THP-1)内でのそれと一致する(Baqui, A.A. et al. Clin. Diagn. Lab. Immunol., 1998, 5:341-347)。IL-12(p70)は、IL-12の生理活性を有するイソ型であり、ナイーブT細胞が、IFN-γを分泌するエフェクターTヘルパータイプ1(Th1)CD4+リンパ球に分化する際の重要な因子である(Trinchieri, G. et al. Prog. Growth Factor Res., 1992, 4:355-368)。加えて、NK細胞上に刺激効果を有することも報告されている(Wajchman, H.J. et al. Cancer Res., 2004, 64:1171-1180)。最近、IL-12(p70)は、そのTh1応答を引き出す能力により、有効で毒性の少ない抗腫瘍サイトカインとして浮上している(Hiscox, S. and Jiang, W.G. In Vivo, 1997, 11:125-132)。免疫系を越えて、このサイトカインは、UV誘導アポトーシスを抑制する能力、および究極的には細胞を悪性腫瘍から保護するUV損傷ケラチノサイト内のDNA修復を開始させる能力を有することも報告されている(Schwarz, A. et al. Nat. Cell Biol., 2002, 4:26-31)。ホモ二量体イソ型であるIL-12(p40)は、マウスにおいて生理活性ヘテロ二量体イソ型p70の受容体拮抗剤である(Gillessen, S. et al. Eur. J. Immunol., 1995, 25:200-206)。RR1処理細胞内では、調節サイトカインであるIL-12(p40)は、生理活性型の何倍かの過剰量で産生され、これは生理活性型p70の過剰産生を制御するための天然の機構であろう。IL-18は、IFN-γ、および明らかにNK細胞のもう一つの強力な誘導物質である(Micallef, M.J. et al. Eur. J. Immunol., 1996, 26:1647-1651)。IL-18産生のRR1誘導は、IL-12およびIFN-γと調和して作用する、Th1応答の発達における早期の応答であろう。
【0129】
MCP-1は、単球、および活性化CD4およびCD8 T細胞の強力な化学誘引物質であり、NKおよびCD8+細胞からの顆粒放出を誘導し、CD56+細胞内のNK機能を活性化し、かつ好塩基球からのヒスタミンの強力な放出因子として作用することが報告されている(Rollins, B.J. Blood, 1997, 90:909-928)。さらに、MCP-1は、おそらくT細胞依存的に、腫瘍特異的免疫を高めることにより、抗腫瘍効果を示すことが報告されている(Laning, J. et al., J. Immunol, 1994, 153:4625-4635)。本発明者らの観察によれば、RR1は、10μg/mlにおいて、このサイトカインの産生を有意に誘導する。RR1刺激によるTNF-αの有意な合成は、感染および悪性腫瘍の増殖に対する宿主抵抗力に重要な役割を果たしているだろう。TNF-αおよびその受容体は、結核に対する保護、および感染の早期におけるマクロファージ内でのNO合成に不可欠である(Lipton, S.A. Neurochem. Int., 1996, 29:111-114)。RR1誘導によるIFN-γ産生は、一部には、例えば細胞傷害性Tリンパ球(CTL)、マクロファージおよびNK細胞のような細胞仲介免疫システムの構成要素の活性を促進するよう機能し、またTh2応答において抑制的役割を果たすだろう。これは食細胞の殺菌活性を刺激し、したがって自然免疫応答を向上させる(Watford, W.T. et al. Cytokine Growth Factor Rev., 2003, 14:361-368)。さらに、これはLPS内と同様にマクロファージ内でMCP-1合成を調節し得る(Munder, M. et al. J. Exp. Med, 1998, 187:2103-2108)。
【0130】
CD4+T細胞は、抗原の認識と、その結果としてのサイトカイン産生を介した抗原特異的(適応)免疫系の調節に寄与する。CD4+細胞によるサイトカイン産生の異なるパターンは、IFN-γ産生により特徴付けられかつ細胞内病原の排除を促進するタイプ1(Th1)と、IL-4産生により特徴付けられかつ細胞外病原の排除に適切なIgEおよび好酸球に関与するタイプ2(Th2)とに対生を形成する。サイトカインは、最初の活性中にT細胞上に直接作用し、最終分化に影響する因子の中で最も直接的なメディエーターであると思われる。IL-12の存在はTh1応答を導き、一方、IL-4はTh2発達を導き、これら二つの経路は互いに抑制する作用も発現する。IL-1はIL-12の誘導物質として同定されており(Rollins, B.J. Blood, 1997, 90:909-928)、一方、IL-18は、IFN-γ誘導によるTh1細胞の発達における早期の応答である。RR1刺激によるサイトカインプロファイル、IL-12、IL-18、IFN-γおよびIL-1、ならびにこれらサイトカインの用量依存的合成は、細胞免疫ならびに細胞内病原および悪性腫瘍細胞の殺傷に不可欠なTh1経路を明白に示している。この観察は、IL-12誘導による、IFN-γ依存的なT細胞のTh1およびCD8+細胞傷害性エフェクター細胞への発達と一致する(Russo, D.M. et al. Expt. Parasitol., 1999, 93:161-170; Yoshimoto, T. et al. J. Immunol., 1998, 161:3400-3407; Russo, D.M. et al. Infect. Immun., 2000, 68:6826-6832)。IL-12およびIL-18の同時発生的なシグナル伝達ならびに相乗作用は、T細胞内で、延長したIFN-γ産生と、連続するIL-18R mRNAの強力な発現とを誘導する(Yoshimoto, T. et al. J. Immunol., 1998, 161:3400-3407)。IL-12およびIL-18に関しては、T細胞の炎症部位に対する接着など、早期の炎症性事象も報告された(Ariel, A. et al. J. Leuk. Biol., 2002, 72:192-198)。調節性イソ型IL-12(p40)の高いレベルの合成は、Th1応答の過剰な生成を含有する天然の機構であろう。
【0131】
NOレベルもまた、Th細胞分化および極性化の調節に重要な役割を果たすことが報告されている。Yamasakiら(Yamasaki, A. et al. Yonago Acta Medica, 1998, 44:7-15)およびTaylor-Robinsonら(Taylor-Robinson, A.W. et al. Eur. J. Immunol., 1994, 24:980-984)は、Th1サイトカインに対するNOの抑制的役割を報告したが、一方、Bauerら(Bauer, H. et al., Immunol., 1997, 90:205-211)は、活性化T細胞からのNOによるTh1およびTh2サイトカインに対する同一の役割を報告した。本研究において、RR1処理は、NOの増加を<21%だけ誘導した(図9)。酵素iNOSは、刺激によるマクロファージからのNOの高産生量を生じた。本研究は、RR1がiNOSの誘導を有意に刺激しないことを明らかにした。RR1によるiNOS誘導の不在は、NO産生の有意な増加が存在しないという結論を支持している。Monocyte IsolationキットII(Miltenyi Biotec, Germany)による末梢血単核細胞から単離された単球を用いた、またはヒト単球細胞株THP-1(ATCC)を用いた本研究でも、RR1処理によってより高いレベルのNOは全く誘導されなかった。RR1による低いレベルのNO合成は、Th1経路を全く阻害しないと思われる。IFN-α、IFN-γ、およびTNF-αは、NO合成の公知の上方調節因子である(Shin, J.Y. et al. Immunopharmcol. Immunotoxicol., 2002, 24:469-482; Hirazumi, A. and Furusawa, E. Phytother Res., 1999, 13:380-387)。RR1刺激により、IFN-γおよびTNF-α、ならびに非常に少量のIFN-αが合成されたために、多糖によるより高いレベルのNO産生を引き起こすためには、IFN-αプライミングが必要ステップであろうと想定することが正当である(Ring, A. et al. J. Infect. Dis., 2002, 186:1518-1521)。
【0132】
RR1刺激は、GSH、GSSGのレベル、およびそれらの比に示されるように(表1)、リンパ球内で酸化ストレスを全く引き起こさず、これはNO合成データと一致する。過酸化水素量の有意な増加も全く観察されなかった(データは示さず)。NO合成の低いレベルは、NOに対する免疫調節的役割を示唆する。RR1の非細胞傷害性の性質は、かなり高濃度(1000μg/ml)においても、この多糖が細胞内で酸化ストレスを誘導しない能力とともに低いレベルのNO産生に寄与する可能性がある。
【0133】
(表1)RR1で処理した正常リンパ球のGSH、GSSG、およびGSH/GSSG比
【0134】
サイトカインの誘導およびNOの合成による免疫刺激、マクロファージの活性化、食作用、細胞傷害性および抗腫瘍活性の誘導が、最近、オタネニンジン(Phanax ginseng)、ヤエヤマアオキ(Morinda citrifolia)およびエキナセア(Echinacea)の多糖または多糖含有画分内に報告されている(Shin, J.Y. et al. Immunopharmcol. Immunotoxicol., 2002, 24:469-482; Hirazumi, A. and Furusawa, E. Phytother Res., 1999, 13:380-387; Goel, V. et al. J. Nutri. Biochem, 2002, 13:487-492)。RR1は、アミロペクチンが免疫刺激を全く誘導できないことから、アミロペクチンとは構造的に区別されるα-D-グルカンである。RR1構造の、真菌β-グルカンの細胞壁成分の「保存された分子形態」との類似性が、免疫系の活性化の理由であり、一方、相違(即ち、αコンホメーション、および(1→4)結合)が、低いレベルのNO産生、およびその結果としての非細胞傷害性の原因である可能性がある。この水溶性を有する中性α-グルカンは、有意な免疫刺激性を示す多糖の範囲内に十分含まれる分子量および分岐の連続を有している(Bohn, J.A. and N. BeMiller Carbohydr. Polymers, 1995, 28:3-14; Brown, G.D. and S. Gordon Immunity, 2003:19:311-315; Kulicke, W.M. et al. Carbohydr. Res., 1997, 295:135-143; Bao, X. et al Carbohyd. Res., 2001, 336:127-140)。水溶性は粒子β-グルカンが示す肉芽腫形成を克服でき、一方、高い分子量は、最も強力な範囲内にあるので分解することなく宿主の器官により長期間維持されることを可能にする(Williams, D.L. et al. Clin. Immunotherapy, 1996, 5:392-396; Nono, I. et al. Pharmacobiodyn., 1991, 14:9-19; Suda, M. et al. FEMS Immunol. Med. Microbiol, 1996, 15:93-100)。
【0135】
非細胞傷害性の性質、リンパ球、特にNK細胞の有意な活性化および補体活性化の代替的経路は、自然免疫系の刺激を明らかに示し、一方、活性化の結果としてのサイトカインプロファイルは、その過剰産生に対する自己調節機構と共に抗原特異的な細胞仲介(適応)免疫性(Th1経路)の刺激を明白に示す。RR1刺激によるIL-4合成(Th1抑制因子サイトカイン)およびIL-10(IL-12の阻害剤)の不在は、T細胞分化のTh1経路と十分一致している。IL-12およびIL-18の同時発生シグナル伝達によるIFN-γ産生は、強力な抗炭疽菌剤として働き、外因性のIFN処置の副作用も、もし存在したとしても、回避するであろう。刺激を受けた自然免疫系は、宿主内への病原のいかなる侵入とも戦うことができ、一次感染が実際の疾病を引き起こすことを防止する能力を有する(Parham, P. Nature, 2003, 423:20)。ここまで説明したように、RR1の独特な免疫刺激特性は、酸化ストレスおよび直接的な細胞傷害性効果を全く与えることなく、RR1を多数の病原およびヒト悪性腫瘍に対する強力な生体防御剤とし得る。
【0136】
実施例6 ザイモサンA-FITC生体粒子の非オプソニン化結合に対するRR1の効果
図10に、非オプソニン化認識およびFITC標識ザイモサンA生体粒子のマクロファージへの結合に対する、RR1の阻害効果を示す。RR1の濃度の増加に伴い蛍光強度の用量依存的阻害が観察され、1000μg/mlのRR1濃度において約65%の阻害を示す。実験中、細胞を4℃でインキュベートするとオプソニンの放出が防止され、それにより非オプソニン化結合が促進されるであろう。
【0137】
実施例7 オプソニン化結合および食作用に対するRR1の効果
ザイモサンA生体粒子の食作用に対するRR1およびラミナリンの37℃での阻害効果を、図11に示す。構造的に定義されたβ-グルカンであるラミナリンは、食作用に対して完全な阻害作用を有するが、RR1は有意な効果を全く示さなかった。食作用は、マクロファージによる粒子の結合および内在化を含む。
【0138】
実施例8 RR1効果に対するCR3の関与
CR3は、マクロファージ活性化および炎症応答に繋がる多糖の単球に対するオプソニン化結合に関与する、β-グルカン特異的受容体である。単球を5μg/mlのCR3(CD11b)特異的モノクローナル抗体と共にインキュベートした後、RR1でインキュベートした結果、RAW264.7細胞のTNF-α合成は阻害または低下されなかった。しかしながら、RAW264.7マクロファージをCD11b抗体で処理した後、可溶性ザイモサンで処理した場合、TNF-α合成の有意な低下(41.31%)が観察された(表2)。
【0139】
(表2)グルカンのオプソニン化結合の抗CD11b(CR3特異的)阻害後の、RAW264.7マクロファージ内のTNF-α合成
細胞をCR3(5μg/ml)と共に2時間インキュベートした後、37℃で1時間グルカン処理した。
**p<0.05
【0140】
実施例9 TNF-αの合成
図12に、NF-κB阻害剤、カフェ酸フェネチルエステル(CAPE)を伴うおよび伴わないRR1誘導によるTNF-α合成を示す。RR1用量の漸増に伴うTNF-α合成の用量依存的増大が明らかであった。単球のRR1処理に先立って、CAPEを10μg/mlで1時間インキュベートした場合、RR1の全濃度にてTNF-α合成は完全に阻害された。
【0141】
NF-κB活性化の動態を測定するために、ELISAプロトコルを使用して、RR1処理マクロファージ内のNF-κB活性を様々な時間間隔において分析した(図13)。刺激時間に伴なってNF-κB活性が増大し、8時間でピークとなり、その後活性は減少した。これらの結果に基づいて、8時間の刺激による更なるNF-κB活性化試験の全てを行い、RR1濃度の増加に伴うNF-κB活性の用量依存的増大を観察し、RR1用量500〜1000μg/mlの間でプラトーに達した(図14)。EMSAによるNF-κBのヘテロ二量体バンドにおけるシフトの視覚化は、上方のバンド(p65ヘテロ二量体バンド)の強度の濃度依存的な増大から明らかであるように、ELISA実験による定量を実証する(図15)。
【0142】
実施例10 IκB-αの分解
IκB-αのリン酸化および分解は、NF-κB活性化の必須条件であり、これがNF-κBの細胞質から核への移行を促進する。IκB-αウエスタンブロットは、RR1処理後の刺激の様々な間隔におけるIκB-αレベルを示す。IκB-αの着実な減少がRR1インキュベーションの30分にて明らかであり、その発現は100μg/mlのRR1処理の60分後に完全に除去された(図16)。
【0143】
実施例11 TLR mRNAおよびタンパク質
TLR2、4、および6(図17)遺伝子のmRNAレベルは、RR1濃度0〜1000μg/mlにおいて同様のままであった。したがって、TLRの転写は、RR1処理により変化しないものと思われた。
【0144】
RR1シグナル伝達におけるTLRの役割
RR1シグナル伝達に対する異なるTLRがなす役割を試験するために、親HEK293細胞を、受容体トランスフェクタント(TLR2、TLR4/CD14/MD2、TLR6/またはTLR2/TLR6)およびRR1と共にインキュベートし、24時間後にIL-8が産生した。RR1は、親HEK、およびTLR2またはTLR4/CD14/MD2遺伝子をトランスフェクトしたHEK細胞内でIL-8産生を誘導しなかった。しかしながら、TLR6およびTLR2/6遺伝子をトランスフェクトしたHEK細胞は、大量のIL-8を誘導し、その合成において用量依存的誘導が明らかであった(図18および図19)。TLR2をトランスフェクトした細胞は、RR1処理(0〜100μM)によりIL-8を全く生産しなかったことも興味深い。
【0145】
ティノスポラ・コルディフォリアは、アーユルヴェーダ(Ayurvedic)(インド)系医学にて広く使用されている免疫刺激性の草本である(Chopra, R.N., I.C. Chopra, K.D. Handa, L.D Kanpur (Editors), 1982, Glossary of Indian Medicinal Plants; Council of Scientific an Industrial Research, New Delhi, Dhar VN & Sons, Kolkata, India)。本発明者らは、免疫刺激特性を有することが見出されたこの薬草から、新規な1,4-α-D-グルカン(RR1)を単離および特徴付けしたことを説明した。この水溶性の多糖は、主鎖に(1,4)-α-D-グリコシド結合を有し、(1,6)-α-D-グリコシド結合した側鎖を6、7グルコース単位の区間に有する。免疫系刺激の更なる機序を理解するために、本研究を行った。本明細書に記載する結果は、RR1が、オプソニン化ザイモサンA生体粒子の結合および内在化(食作用)に対して阻害効果を殆ど有さないことを示す。これは、ザイモサン生体粒子の内在化を約90%阻害した、水溶性の、真菌由来のβ-グルカンであるラミナリンの効果と対照的であった(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Czop, J.K. and Austen, K.F. J. Immunol., 1985, 134:2588-2593; Goldman, R. Exp. Cell Res., 1988, 174:481-490)。
【0146】
補体受容体3(CR3)は、β2-インテグリンファミリーに属するヘテロ二量体膜貫通糖タンパク質であり、CD18に非共有結合的に結合したCD11bからなる。CR3のCD11bサブユニットの細胞外部分に、明確な機能的ドメインが確認されている(Diamond, M.S. et al. J. Cell Biol., 1993, 120:1031-1043; Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246; Lu, C. et al. J. Biol. Chem., 1998, 273:15138-15147; Dana, N. et al. J. Immunol., 1986, 137:3259-3263)。β-グルカンシグナル伝達において、CR3は、粒子(Ross, G.D. et al. Complement, 1987, 4:61-74)および可溶性グルカン(Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246)のための白血球受容体として働く。この受容体は、受容体をプライミングするよう作用するCOOH末端レクチン部位を介して、グルカン受容体として機能する。C3bi被覆粒子の結合および食作用に、CR3のIまたはAドメインが必須であり(Diamond, M.S. et al. J. Cell Biol., 1993, 120:1031-1043; Dana, N. et al. J. Immunol., 1986, 137:3259-3263)、IドメインのC末端に位置するレクチンドメイン(Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246)は、CR3の非オプソニン化結合特性に関与することが報告されている(Balsam, L.B. et al. J. Immunol., 1998, 160:5058-5065; Ross, G.D. et al. J. Immunol., 1985, 134:3307-3315)。多糖のCR3をプライミングする能力は、レクチン部位の近傍に結合する抗体により模倣され得る(Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246)。Cabecら(Cabec, V.L. et al. Infect. Immunity, 2000, 68:4736-4745)は、非オプソニン条件下で、ザイモサンの食作用が、受容体の区別された分子決定因子に関連すること、かつ非オプソニン化食作用はC末端レクチン部位と無関係に起きることを示した。Brownら(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)は、CR3特異的mAb 5C6を用いた、オプソニン化ザイモサンのマクロファージに対する結合の阻害を示し、阻害のレベルは、以前の報告と同様であった(Xia, Y. et al. J. Immunol., 1999, 162:2281-2290)。しかしながら、5C6 mAbによるCR3の阻害は、ザイモサン粒子の非オプソニン化結合を阻害しなかった。抗CD11bモノクローナル抗体を使用したRAW264.7細胞によるCR3阻害実験では、オプソニン化条件下(37℃)で、ザイモサン誘導TNF-α合成の41%の低下が示された。しかしながら、抗体のインキュベーションは、オプソニンおよび非オプソニン条件下で、RAW264.7細胞のRR1誘導TNF-α合成を阻害しなかった。したがって、マクロファージ上のCR3受容体は、サイトカイン合成に繋がるRR1の認識およびプライミングに関与しないであろう。
【0147】
ザイモサンは酵母由来の粒子であり、主としてβ-グルカンおよびマンナンが主成分の多糖からなる。ザイモサンは、受容体に特異的な選択的遮断剤を利用した、マクロファージ上の受容体結合研究のモデルとして使用されている。マウスマクロファージに対するザイモサンの非オプソニン化結合にβ-グルカン受容体、CR3およびマンノース受容体が関与しないことは、各々、ジメチルグリコシドおよびマンナンに対する阻害効果が全く欠乏していたことから既に証明された(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)。一方、構造的に定義された、水溶性のβ-グルカンであるラミナリンは、マクロファージに対する非オプソニン化結合を非常に強力に阻害し、β-グルカン受容体の関与を証明した(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Ross, G.D. et al. Complement, 1987, 4:61-74)。RR1の、非オプソニン化ザイモサン-FITC粒子の蛍光に対する強力かつ濃度依存的な阻害効果は、TNF-α産生と矛盾しない非オプソニン化認識および結合におけるザイモサンおよびRR1内の同一タイプの受容体の関与を示し、そのリン酸化グルカンによる遮断は、マクロファージ活性化を阻害することができる(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117)。オプソニン化結合およびその結果のTNF-α産生に対するラミナリンの強力な阻害効果は、ザイモサン粒子およびラミナリンの双方が同様の分子形態およびコンホメーション(β-グリコシド結合による直鎖状グルコースポリマー)を有するために、これらの結合および内在化に関して同一タイプの受容体が関与することを原因とし得る。また一方、RR1は、異なる分子コンホメーションを有する(α-グリコシド結合による螺旋状)α-グルカンであり、したがって結合/内在化に関しては異なるタイプの受容体が関与してもよい。
【0148】
Brownら(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Brown, G.D. et al. J. Exp. Med., 2003, 197:1119-1124; Brown, G.D. and Gordon, S. Nature, 2001, 413:36-37)は、第一マクロファージによるザイモサンの非オプソニン化認識を独占的に担うデクチン-1受容体を記載した。この著者らは、デクチン-1受容体に特異的なデクチン-1 mAb 2A11が、外因性β-グルカン、リン酸化グルカンまたはラミナリンにより得られる阻害に匹敵するレベルで、非オプソニン化ザイモサンの結合を阻害することができ、mAbはβ-グルカン結合部位にまたはその近傍に結合することを示した。また、Adachiら(Adachi, Y. et al. Infect Immun., 2004, 72:4159-71)は、最近、マウスデクチン-1 cDNAをトランスフェクトしたHEK293細胞が、ゲル化β-グルカン、シゾフィランに結合することができ、この結合は、他のβ-グルカンによる前処理により阻害し得るが、α-グルカンでは阻害されないことを示し、デクチン-1のβ-グルカンに対する特異性を示した。
【0149】
RR1のプライミングを担うさらなる受容体を同定するために、様々なTLRをトランスフェクトしたHEK293細胞内でのRR1誘導サイトカイン合成を研究した。HEK293細胞は、多糖刺激下でTNF-αを産生せず;したがって他のサイトカインを分析した。これらの細胞は、刺激によりIL-8を産生し、またIL-8産生はNF-κB活性化の制御下にあるという事実に基づき、HEK293トランスフェクタント内のIL-8を分析した(Wang, Q. et al. Infect Immun., 2001, 69:2270-2276; Torok, A.M. et al. Infect Immun., 2005, 73:1523-1531)。TLRは、タイプIの膜貫通タンパク質であり、今日までヒトにおけるTLRの12メンバーが記述されている。HEK293細胞は、これら受容体を欠いており、TLR1、TLR2、およびTLR6は全て、ファゴソーム含有ザイモサンA粒子に補充されることが報告されている(Underhill, D.M. et al. Nature, 1999, 401:811-815; Ozinsky, A. et al. Proc. Natl. Acad. Sci. USA., 2000, 97:13766-13771)。TLR2およびTLR6の双方が、ザイモサンA粒子によるNF-κBの活性化、およびTNF-αなどの炎症性サイトカインの産生に必要である。本研究では、RR1は、TLR6およびTLR2/6トランスフェクトHEK293細胞内でIL-8産生を誘導したが、他のトランスフェクタント内では誘導しなかった。IL-8合成の用量依存的増加も観察された。RR1処理は、TLR2またはTLR4遺伝子トランスフェクトHEK293細胞内でIL-8産生を全く誘導しなかった。
【0150】
自然免疫系は、TLRおよび他の受容体を介して多様な微生物生成物および非自己分子を認識しかつ応答し、高度に特異的な応答を引き出す(Netea, M.G. et al. Trends Microbiol., 2004, 12:484-488)。少数の受容体のリガンド特異性は、ホモ/ヘテロ二量体化などの受容体間の協力、ならびにCD14、MD2およびデクチン-1などの非TLRとの共同による攻撃の特異性の達成を提示する(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117; Wright, S.D. et al. Science, 1990, 252:1321-1322; Medzhitov, R. Nature Reviews Immunol., 2001, 1:135-145; Brown, G.D. and Gordon, S. Cell. Microbiol., 2005, 7:471-479; Shimazu, R. et al. J Exp Med., 1999, 189:1777-1782)。Underhillら(Underhill, D.M. et al. Nature, 1999, 401:811-815)は、粒子の内在化は、応答がTLRにより仲介されるため、ザイモサン誘導マクロファージ活性化における炎症応答には必要ないことを証明した。
【0151】
β-グルカンの研究は、免疫刺激に繋がるサイトカインおよびケモカイン合成に、マクロファージ内のNF-κB活性化が重要なステップの一つであることを示している(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Brown, G.D. et al. J. Exp. Med., 2003, 197:1119-1124)。非活性化NF-κBは、細胞質内に位置し、阻害性タンパク質IκB-αに結合している。IκB-αは、炎症性刺激に応答してリン酸化および分解して、NF-κBの活性化を誘導する。活性化NKκBは細胞質から核へ移行して、そこで標的遺伝子のプロモーター領域に結合し、それらの転写を調節する。標的遺伝子がNF-κBにより作動されると、mRNA合成が起こり、次いでタンパク質が発現される。本研究にて実施したNF-κB活性化の試験は、NF-κB活性化が、RR1の免疫系刺激特性の根底にある機構であることを確認した。
【0152】
RelおよびNFκBタンパク質は、癌、関節炎、慢性炎症、喘息、神経変成および心疾患などの多数の疾病状態の持続的活性化に加えて、免疫および炎症応答、ならびにアポトーシスを含む大多数の細胞および組織プロセスの制御に関与する、構造的に関連する真核生物転写因子のファミリーを含む(Beyaert, R. (ed.), 2004,「Nuclear Factor-κB: Regulation and Role in Disease」Kluwer Academy Publishers Dordrecht, The Netherlands; Ghosh, S. et al. Annu Rev Immunol, 1998, 16:225-260)。これらタンパク質は、Relホモロジーと称される、高度に保存されたDNA結合/二量体化ドメインにより関連付けられる。NF-κBは、p50-RelA(p65)ヘテロ二量体を指し、ほとんどの細胞内で主要なRel/NF-κB複合体である。Rel-NF-κB転写因子は、二量体の状態で、κBと称される9〜10塩基対のDNA部位に結合する。個々の二量体は、関連するκB部位の収集のために、明確なDNA結合特異性を有する。マクロファージのCAPEとの、および続くRR1とのプレインキュベーションによるTNF-α合成の完全な阻害は、RR1の細胞シグナル伝達経路にNF-κBが関与していることを明白に示している。CAPEは、p65サブユニットの核への移行を防止することによってNF-κBを阻害し、該阻害はp65ヘテロ二量体移行に特異的であることを示している(Natarajan, K. et al. Proc. Natl. Acad. Sci. USA, 1999, 199693:9090-9095)。ELISA測定における濃度依存的なNF-κBの発現およびEMSAゲル中のNF-κBヘテロ二量体バンドは、サイトカイン産生の転写調節にNF-κBが積極関与していることをさらに確認する。NF-κB発現の発現前のIκB-αの分解は、シグナル伝達経路においてNF-κB活性に不可欠な条件であり、RR1刺激の最初の60分以内の細胞質ゾルIκB-αの分解は、NF-κB経路を正当化する。時間経過実験は、IκB-α分解後にNF-κBの活性化が開始し、刺激の8時間目にピークに到達した後、減少することを示した。NF-κB活性化の一過性の性質は、次にNF-κBの連続的かつ持続的な活性化、およびその結果の炎症性サイトカインの過剰産出を防止する時間経過実験から明らかである。
【0153】
NF-κBにより活性化される標的遺伝子の一つは、IκB-αをコードする遺伝子であり得る。新たに合成されたIκB-αは核に入り、DNAからNF-κBを除去して、複合体を細胞質に輸送し戻し、元の潜在的状態を回復し、それによって連続的な活性化を防止する。8時間後の核抽出物中のNFκB活性化の減少は、この逆輸送に起因する可能性があり、したがって、NF-κB活性化は、数時間のみ継続する一過性プロセスとして維持される。
【0154】
ニューモシスチス・カリニ(Pneumocystis carinii)由来の細胞壁β-グルカンは、p65 NF-κBヘテロ二量体の核内への移行の誘導を介してマウスのマクロファージを活性化することが報告されている。このプロセスは時間依存的に起こり、早くも刺激の1時間目から検出可能であり、2〜4時間目にピークに到達し、連続的刺激の6時間目以降に減少を開始した(Lebron, F. et al. J Biol Chem., 2003, 278:25001-25008)。LPSを用いた同様の試験では、p65 NF-κBは10〜30分間で急速に移行し、先の報告と一致した(O'Connell, M.A. et al. J. Biol. Chem., 1998, 273:30410-30414)。しかしながら、IκB-αの分解は、4時間で完了した刺激のうちの2時間後に初めて認められた。ザイモサン刺激マクロファージ内では、Youngらが、刺激の2時間目のNF-κB活性化を報告し、該活性化は8時間目にピークに到達した後、減少したことを報告した(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)。本発明者らは、RR1刺激についてNF-κB移行の動態にほとんど同様の傾向を観察し、これは4時間目に検出され、8時間目にピークに到達し、より長時間の刺激により減少した。細胞質ゾル抽出物中のIκB-α分解は、早くも刺激の30分目に観察され、1時間後に完了した。LPSまたはニューモシスチス・カリニのグルカンと比較した、NF-κBの核内への移行の程度および時間の差異は、RR1およびβ-グルカンの防御特性に寄与する要素の一つであろう。LPSと比較した、IκB-α分解のより遅い動態およびより長い過渡期を伴うNF-κBのさらに遅い核移行は、RR1およびβ-グルカンの宿主防御特性に寄与するだろう。さらに、ヒト白血球内のRR1刺激は、高いレベルの調節サイトカイン、IL-12p(40)、ならびに抗炎症性サイトカインIL-10およびMCP-1も産出する(Nair, P.K. et al. Int. Immunopharmacol., 2004, 4:1645-1659)。すなわち、これらの結果は、RR1が、TLR6シグナル伝達およびNF-κB活性化機構を介してマクロファージを活性化することにより免疫システムを刺激して、免疫タンパク質の産生を誘導することを示した。突然のIκB-α分解を伴うNF-κBの潜在的なおよび延長した活性化は、LPSおよび真菌細胞壁グルカンなどの他の活性化分子から、その活性化を区別できるであろう。
【0155】
本明細書に言及または引用した全特許、特許出願、仮出願、および刊行物は、それらが本明細書の明白な開示と矛盾しない限り、その図面および表を含む全体が参照により本明細書に組み入れられる。
【0156】
本明細書に記載した実施例および態様は、例示のみを目的とし、当業者にはそれらに照らした様々な修正または変更が示唆されるであろう。それらは、本明細書の趣旨および範囲内に含まれるものとする。
【図面の簡単な説明】
【0157】
【図1】RR1の単離を表すフローチャートを示す。
【図2】図2A〜2Bは、(a)(1-4)および(1-6)グリコシド結合を示すD2Oにおける13CNMRスペクトル、(b)α-コンホメーションを示すD2Oにおける500 MHzプロトンNMRスペクトルを示す。
【図3】RR1((1-4)-α-D-グルカン)の化学構造を示す。
【図4】図4A〜4Bは、正常リンパ球(a)および腫瘍細胞株(b)でのRR1の細胞傷害性分析を示す。細胞(106)をRR1で24時間処理してから、MTT細胞増殖キット(Roche Biochemicals)を用いて細胞傷害性アッセイを行った。対照に対する生存細胞のパーセンテージを、RR1濃度に対しプロットした。
【図5】RR1によるNK、T、およびB細胞の活性化のフローサイトメトリー分析を示す。正常リンパ球を、CO2インキュベーター中、37℃において、RR1(0〜100μg/ml)で24時間処理した。細胞を、各種蛍光色素とコンジュゲートした細胞特異的抗体のパネルで染色した:CD3-FITC、CD16/56-PE、CD19-ECD、CD69-PC5またはCD8-FITC、CD4-PE、CD3-ECD、CD69-ECD。染色した細胞は、Coulter Eliteフローサイトメトリーで、4色アッセイで分析した。***p<0.001。
【図6】RR1活性化ヒトリンパ球の細胞傷害性についてのフローサイトメトリーアッセイを示す。正常リンパ球をRR1(0〜100μg/ml)で24時間、RPMI培地中で処理した。細胞を洗浄し、PKH-26標識ヒト白血病CEM細胞とともに12時間、共インキュベートし、NK細胞を活性化させて、腫瘍細胞を溶解した。細胞混合液を、FITC標識し活性化したカスパーゼ3モノクローナル抗体で染色した。PKH26+およびカスパーゼ3+細胞のパーセンテージを、Beckman-Coulter Eliteフローサイトメーターで分析し、定量化した(**p=0.01)。
【図7】RR1が誘発した補体経路活性化の分析を示す。健康な志願者から得た正常血液試料を、RR1(0〜100μg)で24時間インキュベートした。ヒト補体C3a des ArgおよびC4a des Arg相関EIAキット(Assay Design Inc., Ann Arbor, MI)を用いて、ELISA法を用いた補体活性化の古典的経路および第2経路の定量化を行った。***p<0.001。
【図8】図8A〜8Hは、RR1誘発正常リンパ球におけるサイトカインおよびケモカインの合成を示す。健康な志願者から得た正常血液試料を、CO2インキュベーター内、37℃にて、RR1(0〜100μg)でRPMI培地中24時間処理した。上清培地を、試薬キット(BD Biosciences)を利用して、ELISA法でサイトカインおよびケモカイン産生について分析した。RR1は、IL-1β、IL-6、IL12p40、IL-12p70、およびIL-18、TNF-α、IFN-γ、およびMCP-1の合成を有意に誘発した(***p<0.001)。
【図9】正常リンパ球でのRR1誘発NO合成を示す。健康な志願者から得た正常リンパ球を、CO2インキュベーター内、37℃にて、RR1(0〜100μg/ml)でMEM培地中24時間処理した。上清培地を、Active Motif, Carlsbad, CA社製のキットを用いてNO産生について分析し、データはANOVAを用いて分析した(p=0.109)。
【図10】RAW264.7マクロファージに対するFITC標識ザイモサンA生体粒子の非オプソニン認識/結合へのRR1の阻害作用を示す。RR1は、用量依存的にザイモサンA生体粒子の非オプソニン結合の結合および内在化を阻害する。細胞を0.5%FBSを含有するDMEM中、1時間、氷上(4℃)でRR1とインキュベートした後、ザイモサンA生体粒子と氷上で1時間インキュベートした。細胞を37℃で7分間温め、ザイモサンA生体粒子を内在化させ、Coulter Eliteフローサイトメーターで細胞内FITC蛍光を分析した。
【図11】RAW264.7マクロファージ内への、FITC標識ザイモサンA生体粒子のオプソニン化結合および内在化(食作用)に対するRR1およびラミナリンの阻害作用を示す。単球をRR1で37℃にて1時間インキュベートした後、FITC標識ザイモサンA生体粒子とさらに1時間、37℃でインキュベートした。細胞を洗浄し、Coulter Eliteフローサイトメーターで細胞内の蛍光を測定した。
【図12】NF-κBインヒビターのカフェ酸フェネチルエステル(CAPE)存在下および非存在下での、RR1が誘発したRAW264.7細胞のTNF-α合成を示す。細胞を、0.5%のFBSを含有するEMEM培地中、10μg/mlのCAPEと1時間予備インキュベートしてから、培地を様々な濃度のRR1を含有する培地に交換し37℃で4時間インキュべートした。培地を、ELISAプロトコルを用いて、分泌されたTNF-αについて分析した。
【図13】RR1処理したRAW264.7マクロファージにおける転写因子NF-κBのDNA結合活性の経時変化を示す。マクロファージを100μg/mlのRR1で処理し、核抽出物のNF-κBの活性を、ELISAプロトコルを用いて、インキュベーション後2、4、8、14、および20時間目について分析した。
【図14】RR1処理したRAW264.7マクロファージにおける転写因子NF-κBの用量依存的活性化を示す。NF-κB活性は、RR1用量の漸増に伴って増大し、500μg/mlでプラトーに達した。
【図15】EMSAゲルシフトアッセイにより決定した、RAW264.7細胞の核抽出物における転写因子NF-κBのDNA結合活性を示す。非特異的バンドの強度を基に、NF-κB特異的バンドの用量依存的な強度増加が見られる。
【図16】RR1によるRAW264.7マクロファージ刺激が、細胞質ゾルのI-κBα分解を誘発することを示す。細胞を15、30、および60分の間隔でRR1処理した。細胞質の溶解物中のI-κBα分解レベルは、抗マウスI-κBαmAbを用いたイムノブロット分析で決定した。
【図17】RR1で刺激したRAW264.7細胞におけるTLR2、TLR4、およびTLR6 mRNAのRT-PCRを示す。細胞(1×106)を、CO2インキュベーター内、37℃で、RR1(0、10、100、500、および1000μg/ml)で24時間処理し、マウスTLR特異的プライマーを用いてmRNAを増幅した。
【図18】TLR2およびTLR6遺伝子を安定にトランスフェクトしたHEK293細胞における、RR1誘発IL-8合成の分析を示す。HEK293/TLR2/TLR6細胞(1×106)を、37℃で24時間、RR1(0、10、50、および100μg/ml)で処理し、培地中に分泌されたIL-8をELISAプロトコルで分析した。
【図19】TLR6遺伝子を安定にトランスフェクトしたHEK293細胞における、RR1誘発IL-8合成の分析を示す。HEK293/TLR6細胞(1×106)を、37℃で24時間、RR1(0、10、50、および100μg/ml)で処理し、培地中に分泌されたIL-8をELISAプロトコルで分析した。
【図20】マクロファージにおけるRR1の免疫系刺激経路を示す。
【背景技術】
【0001】
関連出願の相互参照
本出願は、いかなる図、表、核酸配列、アミノ酸配列、および図面も含み、全体として参照により本願明細書に組み入れられる、2004年7月9日出願の米国仮特許出願第60/586,548号の恩典を主張する。
【0002】
発明の背景
バイオテロ、重症急性呼吸器症候群(SARS)および鳥インフルエンザウイルスの大発生、HIV/AIDSの持続的拡大、ならびに現行治療薬剤に耐性である病原株の出現により、これら脅威に対するバイオ防御戦略を含めた新たな防衛的手段の探索が研究者に切実に求められている。免疫活性化は、新興感染症に対するバイオ防御戦略のための、有効的かつ予防的取り組みである(Hackett, C.J. J. Allergy and Clin. Immunol., 2003, 112:686-694)。ヒト免疫系は幅広い微生物検出および宿主防御メカニズムを有しているが、免疫監視および破壊システムの病原体回避が一般的である。この状況は、ほとんどの腫瘍細胞が検出されないまま免疫監視を免れている癌の場合と異なりはしない(Wajchman, H.J. et al. Cancer Res., 2004, 64:1171-1180)。
【0003】
特異的受容体の助けを借りて、病原体または非自己侵入物を検出すること、かつ免疫コンピテント細胞の活性化、サイトカインおよびケモカインの合成ならびに侵入物を排除または封じ込めるために炎症性メディエイターの放出により直ちに応答することが、自然免疫系の構成要素である。自然免疫の活性化はまた、抗原特異的TおよびBリンパ球による適応的免疫応答を誘発し、道を開く。ナチュラルキラー(NK)細胞は、異常な細胞を認識して破壊するその能力を活かして、適応免疫が未発生の感染したウイルスおよび悪性細胞に対する自然免疫防御に決定的な役割を果たしている(Smith, H.R. et al. Proc. Natl. Acad. Sci. USA, 2002, 99:8826-8831; Moser, J.M. et al. Curr. Opin. Immunol., 2002, 14:509-516)。
【0004】
複数の化合物が免疫系を活性化し、微生物のリポ多糖類(LPS)、非メチル化CpGモチーフを含有している2本鎖RNAおよびDNAオリゴヌクレオチドなどの化合物が早期に報告されている(Hackett, C.J. J. Allergy and Clin. Immunol., 2003, 112:686-694; Kandimalla, E.R. et al. Proc. Natl. Acad. Sci. USA, 2003, 100:14303-14308)。複数の受容体刺激が、サイトカイン産生に相乗的な効果を示すことが報告されている(Gao, J.J. et al. J. Immunol., 2001, 166:6855-6860)。補体活性カスケードは、自然免疫システムのもう一つの重要な部分であり、そこでは細胞内細菌などの細胞性病原体が補体化合物によって覆われて(オプソニン化)食作用を受けやすくなる。したがって、補体活性化は、微生物の殺滅に重要な役割を果たし、かつ免疫複合体の移送およびクリアランスにとって必須である。
【0005】
感染またその他抗原性の攻撃に対し生ずるタイプの適応免疫応答(特異的または獲得免疫応答としても知られている)は、応答に関与するTヘルパー(Th)細胞のサブセットによって一般的に区別できる。Th1応答(細胞性または細胞媒介応答)は、遅延型過敏症および細胞傷害性Tリンパ球(CTL)の活性化などの古典的な細胞媒介機能を担っているのに対し、Th2応答(体液性免疫)は、B細胞活性化のためのヘルパーとしてより効果的に機能する。抗原に対する免疫応答のタイプは、一般的には、細胞が抗原に応答して産生するサイトカインによって決定される。Th1およびTh2細胞が分泌するサイトカインの違いは、これら2つサブセットの生物学的機能の差に反映されていると考えられている。
【0006】
サイトカインが、免疫応答の様々な局面の制御において重要な役割を果すことが明らかになってきた。サイトカインの中でもインターロイキン(IL)-12は、自然免疫と細胞媒介適応免疫との協調について中心的な役割を果たしている(Watford, W.T. et al. Cytokine Growth Factor Rev., 2003, 14:361-368)。免疫刺激による予防的防御および曝露後防御が証明されている(Walker, P.S. et al. Proc. Natl. Acad. Sci. USA, 1999, 96:6970-6975; Juffermans, N.P. et al. Infect. Immunol, 2002, 70:147-152)。通常、これら防御的手段は、適応免疫系に関係するT細胞分化のTh1経路のサイトカインである、IL-12とインターフェロン(IFN)-γとの合成と相関している(Gramzinski, A.M. et al. Infect. Immunol., 2001, 69:1643-1649)。
【0007】
Th1サブセットは、IL-2およびINF-γを分泌し、これがCTLを活性化することから、特にウイルス感染および細胞内病原体に対する応答に適しているであろう。Th2サブセットは、IL-4およびIL-5がそれぞれIgE産生および好酸球の活性化を誘発することが知られていることから、自由生活細菌および寄生虫に対する応答により好適であり、アレルギー応答を仲介することもある。一般的に、Th1およびTh2細胞は、明らかに異なるパターンのサイトカインを分泌しており、それゆえに一方のタイプの応答が他方のタイプの応答の活性を弱めることがある。Th1/Th2のバランスのシフトは、例えばアレルギー応答、またはCTL応答の増大をもたらし得る。
【0008】
特定抗原に対する宿主動物の免疫化は、伝統的には免疫原性の形をした抗原を宿主に、繰り返しワクチン接種することによって成し遂げられている。最近のワクチンは、効果的な体液性(抗体または「Th2型」)応答を引き出すが、それらは細胞性応答(特に主要組織適合性複合体(MHC)クラスI拘束CTLまたは「Th1型」応答)は引き出すことができず、応答は一般的には存在しないかまたは弱い。Th2型応答は、結核およびマラリアなどの多くの感染症について、感染に対する防御的な価値をほとんど有していない。さらには、抗体応答は、特定の適応症では不適当であり、特にアレルギーで顕著であり、この場合抗体応答はアナフィラキシーショックを起こすことがある。標的抗原に由来する小量ペプチドを利用する提案されているワクチン、および感染の可能性があるインタクトなウイルス粒子の使用を回避するその他現在用いられている抗原性作用物質は、治療効果を達成するのに必要な免疫応答を必ずしも引き出すわけではない。治療に有効なヒト免疫不全症ウイルス(HIV)ワクチンが存在しないことは、この失敗の不幸な例である。
【0009】
マクロファージ、好中球およびナチュラルキラー(NK)細胞が媒介する自然免疫は、微生物侵入に対する宿主防御機構の第一線である。自然免疫系は、構造的に保存された病原体関連分子パターン(PAMP)を、パターン認識受容体(PRR)と呼ばれる特異的な生殖系列がコードする受容体を介して標的としている(Aderem, A. and Ulevitch, R.J. Nature, 2000, 406:782-787)。自然および合成免疫促進剤による免疫系の強化は、特に免疫系が弱体化し、抗生物質耐性である場合には、従来の治療法に比べはるかに優れている。
【0010】
いくつかの前臨床および臨床研究は、生物学的応答調整物質(BRM)の一分類であるβ-グルカンの、創傷の治癒の加速、ならびに全身性炎症応答症候群および敗血症ショックの編成にとっての有用性を示している(Ross, G.D. et al. Immunopharmacology, 1999, 42:61-74; Williams, D.L. Mediators Inflamm., 1997, 6:247-250; Hetland, G. Curr. Med. Chem., 2003, 2:135-146)。β-グルカンは、無脊椎動物の自然免疫系の強力な刺激因子であるが、哺乳動物では、それらは補体系の強力な活性化因子である。これらのポリマーは、抗腫瘍活性および抗感染活性、ならびに真菌、細菌、ウイルスおよび原虫感染に対する防御を含めた免疫系に対するそれらの効果から、治療可能性を有している。可溶性および粒状のβ-グルカンは、マクロファージにある起源を同じくする受容体と相互作用し、サイトカイン、ケモカインおよび反応性酸素中間体の合成を刺激する(Ganter, B.N. et al. J. Exp. Med., 2003, 197:1107-1117)。マクロファージ上でのβ-グルカン認識/結合について報告されている主要受容体は、補体受容体3(CD11b/CD18またはCR3)、デクチン-1、ならびにトール様受容体(TLR)2および6である。β-グルカン認識においては、ラクトシルセラミドおよびスカベンジャー受容体も確認されているが、それらの機能はよく分かっていない(Willment, J.A. et al. J. Biol. Chem., 2001, 276:43813-43823)。
【0011】
トール様受容体(TLR)は、細胞内シグナル伝達機能に関する細胞質モチーフを持つトール-インターロイキン(IL)-1受容体(TIR)の大きなスーパーファミリーの一部である。これらの分子は、第一線の宿主防御を提供し、ハエから哺乳動物に至る様々な生物において、感染および自己免疫疾患に関係している。現在TLRは、それにより哺乳動物が感染を感知する、第一のシグナル伝達分子であることが受け入れられている(Beutler, B. et al. J Leukocyte Biol., 2003, 74:479-485)。哺乳動物では、12種類のTLRが同定されており、それぞれが異なるPAMPを認識している(Akira, S. and Sato, S. Scand. J. Infect. Dis., 2003, 35:555-562)。TLR3を除く全てのTLR、IL-1受容体およびその他TIRドメイン含有受容体は、アダプター骨髄分化因子88(MyD88)に依存した、共通のシグナル伝達経路を共有している(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117; Mukhopadhyay, S. et al. Immunol., 2004, 112:521-530)。MyD88以外にも、いくつかのアダプター分子が最近報告されており、これらアダプター分子の差別的な利用が、TLRシグナル伝達のために特異性を提供しているかもしれない(Akira, S. J. Biol. Chem., 2003, 278:38105-38108)。TLR間、ならびにTLRとその他表面受容体との間の物理的、および/または機能的相互作用の証拠が得られている。Gantnerらは、β-グルカン刺激によるデクチン-1とTLRの共同誘導、ならびにこれら2つの受容体間のNFκB活性化に関する相乗的相互作用について報告している(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117)。
【0012】
TLR媒介サイトカイン産生は、JNKおよび核因子(NF)-κBを活性化するIL-1R関連キナーゼ(IRAK)-4およびTNF受容体関連因子-6(TRAF-6)といった、その下流のメディエーターに依存している(Akira, S. J. Biol. Chem., 2003, 278:38105-38108)。NF-κBは、多くの免疫エフェクター細胞でのサイトカイン遺伝子発現を制御する普遍的な転写因子である。多くの細胞において、NF-κBは、通常、細胞質内に潜伏的、不活性な状態で、阻害タンパク質κB(I-κB)と結合して存在している(Ghosh, S and Karin, M., Cell, 2002, 109:S81-96)。それは、炎症性サイトカイン、ウイルス産物、リポ多糖類、タキソールなどの植物由来化合物、ならびに病原菌および非病原菌由来のβ-グルカンのような様々な刺激物によって活性化される(Akira, S. and Sato, S. Scand. J. Infect. Dis., 2003, 35:555-562; Young, S.H. et al. J. Biol Chem., 2001, 276:20781-20787; Lebron, F. et al. J Biol Chem., 2003, 278:25001-25008)。刺激を受けるとI-κBαは、直ちにリン酸化されて、プロテアソーム機構により急速に分解され、次に活動性NF-κBを放出して核に移行し、DNAに結合してサイトカイン/ケモカイン遺伝子の転写を開始させる(Ghosh, S, and Karin, M., Cell, 2002, 109:S81-96; Auphan, N. et al. Science, 1995, 270:232-233)。
【0013】
グルカンの免疫刺激特性は、β-グリコシド結合、枝分れ度、および溶液のコンホメーションに拠るとされている(Muller, A et al. Glycobiology, 2000, 10:339-346)。本発明は、薬用植物のティノスポラ・コルディフォリア(Tinospora cordifolia)から得ることができる新規の多糖、(1,4)-α-D-グルカン、RR1の免疫刺激特性の特徴を明らかにし報告している(Nair, P.K. et al. Int. Immunopharmacol., 2004, 4:1645-1659)。本新規α-グルカンは水溶性であり、かつ6〜7グルコース単位の間隔で(1,6)-α-D-グリコシド結合側鎖を持つ主鎖の中に(1,4)-α-D-グリコシド結合を有している。それは正常細胞ならびに腫瘍細胞株(CEM、CEM/VLB)に対しては、1000μg/mlまで非細胞傷害性であり、ヒトリンパ球サブセットを様々なレベルで活性化する。自然免疫の主要細胞の一つであるNK細胞の活性化は、機能的アッセイにおけるRR1処理リンパ球による標的細胞の殺滅レベルの増大によって証明された。RR1刺激を受けたサイトカインプロフィールは、Th1応答の過剰産生の自己制御機構と思われる、高レベルの調節サイトカイン誘導を伴った、Tヘルパー細胞分化のより望ましいTh1経路を証明している。その水溶性、非細胞傷害性、および草本起源であることは、RR1の免疫刺激に関する臨床上の可能性を示している。
【発明の開示】
【0014】
発明の概要
本発明は、新規の免疫系刺激多糖に関する。本明細書に例示する、一つのこの様な化合物は、薬用植物のティノスポラ・コルディフォリアから単離され、特徴付けされている。一つの局面では、本発明は、図3に示す構造を有する化合物(本明細書ではRR1とも呼ぶ)を提供し、この化合物は、(1→4)結合主鎖および(1→6)結合分枝から構成され、550kDaより大きな分子量を持ち、固有の免疫刺激特性を示すα-D-グルカン多糖である。
【0015】
RR1の免疫刺激剤特性は、RR1誘発のヒトリンパ球活性化、NK細胞機能活性、食作用活性、補体活性化、サイトカインおよびケモカイン合成、誘導性酸化窒素合成酵素(iNOS)の誘発、酸化窒素産生、ならびに酸化ストレス測定の分析によって確認された。結果は、自然免疫系コンポーネントの強化を明瞭に証明し、一方サイトカインプロフィールは、抗原特異的細胞媒介(適応)免疫でのTヘルパー細胞分化のTh1経路を証明した。RR1による刺激は、自然免疫系の主要エフェクター細胞であるナチュラルキラー(NK)細胞の活性化、ならびに自己増幅しかつ抗体非存在時の病原体認識および排除において重要である、第2経路での補体活性化を数倍高めた。RR1は、マクロファージによるザイモサン粒子の食作用も高める。本明細書中でさらに詳しく記載するように、RR1はまた、Th1サイトカイン、IL-12、IL-18、IL-1βおよびIFN-γの合成をアップレギュレートした。RR1はまた、抗腫瘍サイトカインのTNF-αおよびMCP-1の産生を誘発したが、iNOSの有意な誘発は起こさなかった。このサイトカイン合成のアップレギュレーションは、iκBタンパク質のリン酸化を通じたNF-κB転写因子の活性化によって促進される。RR1免疫刺激のプロセスは、トール様受容体(TLR)6およびデクチン-1などの受容体が関係する。
【0016】
RR1の投与によって強化された自然免疫系は、病原体の侵入から被験体を守ることができ、一方ブーストされたTh1細胞は、細胞内病原体、ウイルスおよび悪性細胞を見つけ出して破壊する。それゆえにRR1は、様々なヒトおよび動物の病気の免疫賦活性剤、化学予防剤および治療薬剤として用いることができる。RR1はまた、癌患者の化学療法薬剤治療のための抗癌剤(例えばモノクローナル抗体)など、その他の薬物と組み合わせることもできる。
【0017】
したがって、本発明の別の局面は、被験体の免疫応答を調節するための方法を提供する。一つの態様では、RR1化合物(RR1または薬学的に許容されるその塩または類似体)は、被験体に対し、RR1化合物のアジュバント様効果を通して、免疫応答の全般的刺激を提供するのに有効である量が投与される。本発明はまた、本発明のRR1化合物を抗原と共に使用し、免疫応答を刺激するための方法も提供する。好ましくは、このような方法で使用する場合、RR1化合物は、抗原に対しTh1型の免疫応答を作り出すアジュバント様活性を提供する。
【0018】
別の局面では、本発明は、少なくとも一つのRR1化合物および薬学的に許容される担体を含有する、免疫調節組成物を提供する。
【0019】
本発明のさらなる局面は、RR1をティノスポラ・コルディフォリア植物材料から得るためのプロセスを提供する。
【0020】
発明の詳細な説明
本発明は、固有の免疫ブースト特性を示す新規多糖である単離化合物(RR1)を提供する。RR1は、図3に示す構造を有し、(1→4)結合主鎖および(1→6)結合分枝から構成され、550kDaより大きな分子量を持つα-D-グルカン多糖である。RR1は、薬草のティノスポラ・コルディフォリア(ツツラフジ(Minispermaceae)科)から単離されており、その薬効特性が研究の主題となってきた(Singh, S.S. et al. Ind. J. Pharm., 2003, 35:83-91; Chintalwar, G. et al. Phytochemistry, 1999, 52:1089-1094; Manjerakar, P.N. et al. Fitotherapia, 2000, 71:254-257; Desai, V.R. et al. Proc. Indian Acad. Sci, 2002, 114:713-719; Subramanyan, M. et al., Redox. Rep., 2002, 7:137-143)。
【0021】
RR1は、正常リンパ球ならびに1000μg/mlの腫瘍細胞株に対してでさえ非細胞傷害性および非増殖性である。RR1は、100μg/mlの濃度で、NK細胞(331%)、T細胞(102%)およびB細胞(39%)などの様々なリンパ球のサブセットを活性化した。NK細胞の有意な活性化は、機能的アッセイにおいて、活性化された正常リンパ球による用量依存的な腫瘍細胞の殺滅を導く。正常リンパ球でのRR1による免疫活性化は、100μg/ml濃度でIL-1β(1080pg/ml)、IL-6(21833pg/ml)、IL-12 p70(50.19pg/ml)、IL-12 p40(918.23pg/ml)、IL-18(27.47pg/ml)、IFN-γ(90.16pg/ml)、TNF-α(2225pg/ml)およびMCP-1(2307pg/ml)の合成を引き出したが、IL-2、IL-4、IL-10、IFN-αおよびTNF-βの産生は誘発しなかった。このサイトカインプロフィールは、Tヘルパー細胞分化のTh1経路が、その過剰産生を制御する自己調節機構と共に、細胞媒介免疫にとって必須であることを証明している。
【0022】
RR1はまた、C3a des Arg成分が段階的に増加することで証明されたように第2経路の補体成分も活性化した。偶然に、RR1刺激はリンパ球にいかなる酸化的ストレスも起こさず、iNOSの誘発が有意でないことから証明されるように、酸化窒素産生増大を極わずかに引き起こしたに過ぎない。RR1は水溶性であり、高分子量を有し、リンパ球を活性化(特にNK細胞)し、補体を活性化し、Th1経路に関係するサイトカインプロフィールを誘発し、低レベルの酸化窒素合成を伴い、および酸化的ストレスがないという事実は、本新規α-D-グルカンに可能な生体防御特性を付与している。
【0023】
RR1のシグナル伝達機構がマクロファージで研究され、その免疫刺激特性の理解はさらに進んだ。RAW264.7マクロファージをRR1と4℃でインキュベートすると、RR1は、ザイモサンA生体粒子の非オプソニン結合および食作用を用量依存的な方法で阻害した。しかしながら、それはザイモサンA生体粒子のオプソニン結合および内在化に対してはほとんど影響しなかった。マクロファージを抗CD11b mAbとインキュベーションしてからRR1とインキュベーションすると、RR1誘発のTNF-α合成に対する任意の阻害作用は見られず、このことからCR3が、ザイモサンとは異なり、マクロファージでのRR1のオプソニン結合および内在化に関係していないことが確認された。抗CD11b mAbは、ザイモサンAが誘発するTNF-α合成に対し有意な阻害作用を有している。RR1は、マクロファージでのTNF-α合成を用量依存的な方法で誘発したが、これはNF-κBインヒビターであるカフェ酸フェネチルエステル(CAPE)によって完全に阻害され得る。RR1はNF-κBを、時間および用量依存的な方法で活性化し、この核NF-κB活性の調節に伴ってI-κBαが分解し、これがNF-κBの核内への移行を促進する。RR-1が誘発するNF-κB活性は、RR1刺激8時間目にピークに達するが、一方I-κB-αの分解は刺激1時間以内に起こった。RR1が誘発するNF-κBの活性化は、TLR6シグナル伝達を介して起こった。なぜならば、RR1は、TLR6をトランスフェクトしたHEK293細胞でIL-8合成を誘発したが、他のトランスフェクタントでは誘発しなかった。これらの結果は、RR1が、TLR6シグナル伝達、NF-κBの移行、および免疫タンパク質の産生を通して起こるマクロファージの活性化を通じて免疫系を活性化することを示している。単球/マクロファージにおけるRR1による免疫刺激のシグナル伝達経路の概略図を図20に示す。
【0024】
本発明のRR1化合物によって刺激された免疫応答は、Th1型の表現系に向かって偏り、Th2型の表現系とは疎遠であることが好ましい。本発明との関係では、Th1型免疫応答への刺激は、RR1化合物を用いて処理した細胞のサイトカイン産生を、RR1化合物なしで処理した細胞の産生と比較して測定することによって、インビトロまたエクスビボで決定できる。細胞のサイトカイン産生を決定する方法としては、本明細書に記載の方法ならびに当技術分野で公知のその他の方法を挙げることができるが、これらに限定されない。RR1処理に応答して産生されるサイトカインは、細胞によるTh1型またはTh2型に偏った免疫応答を示している。
【0025】
本明細書で使用する、用語「Th1型に偏った」サイトカインの産生とは、刺激物存在下におけるTh1型応答関連サイトカイン産生と、刺激物非存在下におけるTh1型応答関連サイトカイン産生とを比べ測定可能に増加した産生を指す。このようなTh1型に偏ったサイトカインの例としては、IL-2、IL-12およびIFN-γが挙げられるが、これらに限定されない。これに対し「Th2型に偏ったサイトカイン」とは、Th2型免疫応答に関係するサイトカインを指し、IL-4、IL-5、IL-10およびIL-13を挙げることができるが、これらに限定されない。RR1活性の決定に有用な細胞としては、免疫系細胞、被験体より単離された一次細胞、および/または細胞株、好ましくはAPCおよびリンパ球、よりさらに好ましくはマクロファージならびにT細胞を挙げることができる。
【0026】
Th1型免疫応答への刺激は、RR1化合物で処理された被験体で測定することができ、下記の方法を含む任意の公知の方法によって決定することができるが、これらに限定されない:(1)抗原に曝露された前後で測定されたIL-4レベルの低下;あるいは、RR1処理被験体と、抗原により刺激され、または刺激および曝露されたRR1なしで処理された対照とを比較した時の、IL-4のより低いレベル(もしくは非存在)を検出する方法;(2)抗原に曝露された前後でのIL-12、IL-18および/またはIFN(α、β、もしくはγ)レベルの増大;あるいは、RR1処理被験体と、抗原により刺激され、または刺激および曝露されたRR1なしで処理された対照とを比較した時の、より高いレベルのIL-12、IL-18および/またはIFN(α、β、もしくはγ)を検出する方法;(3)RR1処理被験体と、RR1なしで処理された対照とを比較した時の、IgG2a抗体産生による方法;かつ/または(4)抗原に曝露された前後で測定された抗原特異的IgEレベルの低下;あるいは、RR1処理被験体と、抗原により刺激され、または刺激および曝露されたRR1なしで処理された対照とを比較した時の、抗原特異的IgEのより低いレベル(もしくは非存在)を検出する方法。これら各種決定は、APCおよび/またはリンパ球、好ましくはマクロファージおよび/またはT細胞が作るサイトカインを、本明細書に記載されている方法、または当技術分野公知のいずれかの方法を用いてインビトロまたはエクスビボで測定することによって、実施できる。
【0027】
RR1投与の結果として起こるTh1に偏ったサイトカインの誘発は、NK細胞、細胞傷害性キラー細胞、Th1ヘルパーおよびメモリー細胞が実行するような強化された細胞性免疫応答を生ずる。これらの応答は、ウイルス、真菌、寄生原虫、細菌、アレルギー性疾患および喘息、ならびに腫瘍に対する予防または治療ワクチン接種での使用のために特に有益である。
【0028】
本発明のRR1化合物は、免疫抑制を伴う状態などの、免疫刺激を必要とする、ヒトまたはヒト以外の動物の病理的状態を治療するために有用である。免疫刺激が望まれる状態の例としては、骨髄炎、慢性気管支炎、結核、下気道感染症、扁桃炎、中耳炎、肝炎、AIDS、糖尿病、糖尿病性潰瘍、および小児疾患が挙げられるが、これらに限定されない。本発明のRR1化合物は、他の推定免疫賦活剤の活性を評価するための標準物質としても有用である。
【0029】
RR1の免疫賦活性特性のおかげで、本発明の治療法、化合物および組成物を用いて、急性リンパ性白血病、急性非リンパ性白血病、慢性リンパ性白血病、慢性骨髄性白血病、ホジキン病、非ホジキン性リンパ腫、および多発性骨髄腫などの白血病およびリンパ腫、脳腫瘍、神経芽腫、網膜芽腫、ウィルムス腫瘍、骨腫瘍、および軟組織肉腫などの小児固形腫瘍、肺癌、結腸および直腸癌、乳癌、前立腺癌、泌尿器癌、子宮癌、膀胱癌、口腔癌、膵臓癌、メラノーマおよびその他皮膚癌、胃癌、卵巣癌、脳腫瘍、肝臓癌、咽頭癌、甲状腺癌、食道癌、および精巣癌などの成人の一般的固形癌を含むが、これらに限定されない癌などの、様々な細胞増殖性の障害を治療できる。本発明の方法は、インビボまたはインビトロで実施して、ヒトまたはヒト以外の哺乳動物における癌性細胞の増殖を阻止できる。
【0030】
β-グルカンに起因する幾つかの特異的作用はまた、RR-1にも関係していることがある。これには、リポ多糖類(LPS)またはその他エンドトキシン/エキソトキシンによって誘発されるNF-κBの活性化の阻害が包含される。NF-κBの活性化およびその他経路は、敗血症および敗血症ショックの発生に重要な役割を果たしている。(1→3)-β-D-グルカンは、NF-κBの活性化を減少させ、敗血症のマウスの長期生存期間を延長することが証明されている(Williams, D.L. et al. Shock, 2000, 13:446-452; Williams, D.L. et al. Surgery, 1999, 126:54-65)。したがって、本発明は、少なくとも一つのRR1化合物を投与することによって、ヒトまたはヒト以外の哺乳動物の敗血症または敗血症ショックを治療する方法を含む。別の局面では、本発明は、少なくとも一つの本発明のRR1化合物を投与することによって、LPSまたはその他エンドトキシンおよび/またはエキソトキシンによってヒトまたはヒト以外の哺乳動物に誘発されたNF-κBの活性化を低下させるための方法を含む。
【0031】
生物学的応答調節因子としてのその作用のおかげで、本発明のRR1化合物はまた急性および慢性の肺疾患等のその他の障害、または自然もしくは適応免疫系の異常に関係し得るその他の障害の治療を目的として、被験体に投与してもよい。本発明は、このような障害を治療するための方法を含む。抗腫瘍効果以外にも、植物多糖類が癌の化学的予防に関して役割を果たすことを裏付ける証拠がある(Kim, H.S. et al. Carcinogenesis, 1999, 20:1637-1640)。本発明は、癌の化学的予防のための方法を含むが、ここで、少なくとも一つの、本発明のRR1化合物が被験体に投与される。
【0032】
本発明の治療法は、細胞増殖性障害(例えば癌)、または免疫賦活もしくは免疫抑制によって治療もしくは管理され得るその他の病理的状態(例えば疾患またはその他の障害)の治療または管理について、当業者に公知である少なくとも一つの追加的治療法または療法と、都合良く組み合わせることができる。このような補助療法の例としては、化学療法、放射線療法、および抗癌剤もしくはその他抗癌性作用物質の投与が挙げられるが、これらに限定されない。補助療法は、鍼、マッサージ療法、エネルギー療法のように、逆症療法でも非逆症療法でもよい。病理的状態は、急性でも慢性でも、疾患でも障害でもよい。
【0033】
RR1およびRR1類似体は、単独化合物として投与することができるが、これら化合物は薬学的組成物として投与することが好ましい。それゆえに本発明は、RR1、またはその類似体、または生理学的に許容されるその塩を、活性成分として、少なくとも一つの薬学的に許容される担体と共に含む薬学的組成物も提供する。薬学的組成物は、経腸的、非経口的、静脈内、筋肉内、局所、皮下等、各種投与経路に合わせることができる。投与は、当業者によって決定できる場合は、連続的または特定の間隔でよい。
【0034】
本発明のRR1化合物は、医薬として有用な組成物の調製について公知である方法に従って製剤化できる。製剤は、当業者に周知であり、かつ容易に入手できる様々な情報源に記載されている。例えば、Remington's Pharmaceutical Science (Martin EW, Easton Pennsylvania, Mack Publishing Company, 19th ed., 1995)は、本発明と関連して用いることができる製剤を記載している。投与に適した製剤としては、例えば、酸化防止剤、バッファー、抗菌剤、および製剤を目的のレシピエントの血液と等張にする溶液を含むことがある、水性滅菌注射液;ならびに懸濁化剤および増粘剤を含む水性および非水性無菌懸濁液が挙げられる。製剤は、単位投与量または複数回投与量容器、例えば密封されたアンプルおよびバイアルの中に存在してよく、使用前に無菌溶液担体、例えば注射用の水の状態のみを必要とするフリーズドライ(凍結乾燥)状態で保存することができる。準備を必要としない注射液および懸濁液を、無菌粉末、顆粒、錠剤から調製してもよい。本発明の組成物は、上記具体的に記載した成分に加えて、問題の製剤のタイプを考慮し、当技術分野で通常であるその他作用物質を含むことができる。
【0035】
本発明のRR1化合物は、当業者が調製できるRR1のあらゆる水和物および塩を含む。本発明の化合物が十分塩基性または酸性であり、無毒の酸または塩基性塩を形成できる条件では、塩としての化合物の投与が適切であろう。薬学的に許容される塩の例は、生理学的に許容されるアニオン、例えばトシラート、メタンスルホナート、アセタート、シトラート、マロナート、タルトラート、スクシナート、ベンゾアート、アスコルベート、α-ケトグルタラートおよびα-グリセロホスフェートを形成する、酸と一緒に形成した有機酸付加塩である。塩酸塩、硫酸塩、硝酸塩、重炭酸塩、および炭酸塩を含む、好適な無機塩も形成できる。
【0036】
薬学的に許容できるRR1の塩は、当技術分野で周知である標準的な方法を用いて、例えばアミンなどの十分に塩基性である化合物を好適な酸と反応させて、生理学的に許容できるアニオンを供給することによって、得ることができる。カルボン酸のアルカリ金属(例えばナトリウム、カリウムもしくはリチウム)またはアルカリ土金属(例えばカルシウム)塩も作ることができる。
【0037】
本明細書で使用する「類似体」という用語は、実質的にRR1(図3に示すような)と同一であるが、親化合物のαコンホメーションおよび(1→4)および(1→6)グルコピラノシル結合が未変性のままで、かつ親化合物の免疫賦活性特性の一つまたは複数が実質的に保持されている限りにおいて、例えば側基の付加、親構造の酸化もしくは還元により修飾されてもよい化合物を指す。例示の化合物の類似体は、公知の標準的な反応を用いて容易に調製できる。これら標準の反応には、水素化、アルキル化、アセチル化、および酸性化反応が挙げられるが、これらに限定されない。
【0038】
それらを含有するRR1化合物および組成物の治療応用は、現在または将来、当業者にとって公知となる任意の好適な治療方法および技術によって達成できる。さらに、本発明のRR1化合物は、他の有用な化合物および組成物の調製のための出発材料または中間体として有用である。
【0039】
本発明のRR1化合物およびこのような化合物を含有する免疫調節組成物は、不活性希釈液または同化性食用担体などの薬学的に許容される担体と一緒に全身投与(例えば経口または静脈内)できる。RR1化合物は、硬質または軟質シェルゼラチンカプセルに封入してもよく、錠剤に圧縮してもよく、または患者食の食材に直接組み込んでもよい。経口治療投与の場合には、RR1化合物は、一つまたは複数の賦形剤と組み合わせ、摂取可能な錠剤、バッカル剤、トローチ剤、カプセル剤、エリキシル剤、懸濁剤、シロップ剤、ウェーハ等の形で用いることができる。
【0040】
錠剤、トローチ剤、丸剤、カプセル剤等は、下記のものを含有してもよい:トラガントゴム、アカシア、コーンスターチ、またはゼラチンなどの結合剤;リン酸二カルシウムなどの賦形剤;コーンスターチ、ポテトスターチ、アルギン酸等などの崩壊剤;ステアリン酸マグネシウムなどの潤滑剤;およびショ糖、フルクトース、ラクトース、もしくはアスパルテームのような甘味料、またはペパーミント、ウインターグリーン油、もしくはチェリーフレーバーのような香味剤を加えてもよい。単位投与形態がカプセル剤の場合、上記物質以外に、植物油またはポリエチレングリコールなどの液体担体を含有してもよい。様々なその他材料が、コーティング剤として存在してもよく、そうでなければ固形の単位投与形態の物理形態に変更してもよい。例えば、錠剤、丸剤、またはカプセル剤は、ゼラチン、ワックス、セラック、または糖等でコーティングしてもよい。シロップまたはエリキシルは、活性化合物、甘味料としてのショ糖またはフルクトース、保存剤としてのメチルおよびプロピルパラベン、着色料、ならびにチェリーまたはオレンジフレーバーのような香味料を含有してよい。もちろん、いずれの単位投与形態の調製に用いられるいずれの材料も、使用する量において薬学的に許容されるものであり、実質的に無毒でなければならない。これに加えて、本発明のRR1化合物は、徐放性調製物および装置に組み入れてもよい。
【0041】
本発明のRR1化合物(即ち、RR1または、RR1の薬学的に許容される塩もしくは類似体)である活性作用物質はまた、輸液または注射によって静脈内または腹腔内投与してもよい。活性作用物質またはその塩の溶液は、水を用いて、任意で無毒の界面活性剤と混合して調製できる。分散液は、グリセロール、液体ポリエチレングリコール、トリアセチン、およびそれらの混合物、ならびに油において調製できる。保存および使用の通常の条件下では、これら調製物は、保存剤を含有し、微生物の増殖を阻止する。
【0042】
注射または輸液に好適な薬学的投与形態としては、無菌の注射可能もしくは輸液可能な溶液もしくは分散液の即時調製物に適応し、リポソームに封入されていてもよい活性成分を含む、無菌の水溶液もしくは分散液または無菌の粉末が挙げられる。いずれの場合も、最終的な投与形態は、無菌の流体であり、かつ製造および保管状態下で安定でなければならない。液体担体もしくは媒質は、例えば水、エタノール、ポリオール(例えばグリセロール、プロピレングリコール、液体ポリエチレングリコール等)、植物油、無毒のグリセリルエステルおよびそれらの好適混合物を含む、溶媒または液体分散媒体であり得る。適当な流動性は、例えば、リポソームの形成により、分散体の場合には必要な粒子サイズを維持することにより、または界面活性剤の使用により維持できる。微生物の活動は、様々な抗菌剤および抗真菌剤、例えばパラベン、クロロブタノール、フェノール、ソルビン酸、チメロサール等によって阻止できる。多くの例では、等張剤、例えば糖、緩衝剤または塩化ナトリウムを含めることが好ましいだろう。注射可能な組成物の長期吸収は、吸収遅延剤、例えばモノステアリン酸アルミニウムおよびゼラチンを組成物に用いることで実現できる。
【0043】
無菌注射液は、適切な溶媒に、必要に応じて、上に列挙した各種のその他成分と一緒にRR1化合物(即ちRR1、またはRR1の薬学的に許容される塩もしくは類似体)を組み入れ、続いて濾過滅菌して調製される。無菌注射液調製用の無菌粉末の場合は、好ましい調製方法は、真空乾燥およびフリーズドライ技術であり、これら技術により、事前に無菌濾過した溶液中に存在することが望ましい任意の追加成分を加えた活性成分の粉末が生じる。
【0044】
局所投与では、RR1化合物は純粋な形、即ち液体の場合投与できる。しかしながら、それらは、固体または液体でもよい、皮膚科学的に許容される担体と組み合わせた組成物として、皮膚に投与することが一般的には望ましいであろう。
【0045】
有用な固体担体としては、タルク、クレー、微結晶セルロース、シリカ、アルミナ等の微細分割固体が挙げられる。有用な液体担体としては、水、アルコールもしくはグリコール、または水-アルコール/グリコール混合物が挙げられ、これらの中にRR1化合物は、任意で無毒の界面活性剤の助けを借りて、有効レベルで溶解もしくは分散することができる。芳香剤のような補助剤および追加の抗菌剤を加えて、所与の使用に合わせて特性を最適化できる。得られた液体組成物は、吸収性パッドから作用させること、絆創膏および包帯剤の含浸に用いること、またはポンプタイプもしくはエアロゾルスプレーを用いて患部に噴霧することができる。
【0046】
合成ポリマー、脂肪酸、脂肪酸塩およびエステル、脂肪アルコール、変性セルロースもしくは変性ミネラル物質などの増粘剤も、液体担体と一緒に用いることができ、使用者の皮膚に直接適用するための、塗布可能なペースト、ゲル、軟膏、石けん等を形成できる。皮膚にRR1化合物を送達するのに用いることができる有用な皮膚科学的組成物の例は、Jacquetら(米国特許第4,608,392号)、Geria(米国特許第4,992,478号)、Smithら(米国特許第4,559,157号)およびWoltzman(米国特許第4,820,508号)に開示されている。
【0047】
本発明の組成物の有用投与量は、それらのインビトロ活性と、動物モデルでのインビボ活性とを比較することで決定できる。マウスおよびその他動物での有効投与量をヒトに外挿する方法は、当技術分野で公知である:例えば、米国特許第4,938,949号を参照されたい。
【0048】
したがって、本発明は、RR1化合物(即ち、RR1、またはその薬学的に許容されるその塩もしくは類似体)を薬学的に許容される担体と組み合わせて含む、免疫調節剤である薬学的組成物を含む。経口、局所、または非経口投与に適した、ある量のRR1化合物を含む免疫調節組成物は、本発明の好ましい態様を構成する。患者、特にヒトに、本発明に即して投与される量は、合理的な時間枠の中で患者に治療応答を起こさせるのに十分なものでなければならない。投与量が被験体の状態(健康)、被験体の体重、該当する場合には併用する治療の種類、治療頻度、治療比、ならびに病理的状態の重篤度およびステージを含む様々な要因に依存するだろうということを、当業者は認識するであろう。
【0049】
癌との関係では、好適投与量は、所望の応答を達成することが知られている活性剤(本発明のRR1化合物)の濃度を腫瘍組織内に生ずる投与量である。好ましい投与量は、管理不能な副作用なしに、癌細胞増殖の最大阻止を生じる量である。RR1化合物、およびこのような化合物を含有する組成物の投与は、当業者が決めることができる場合には、連続的でも一定間隔でもよい。
【0050】
所望する治療処置のためのそのような投与量を提供するために、本発明の薬学的組成物は、担体または希釈液を含めた全組成物重量を基にして、一つまたは複数の新規化合物の総量を約0.1重量%〜45重量%、特には1〜15重量%を有利に含むであろう。例示的には、投与される活性成分の投与量レベルは、下記である:静脈投与の場合、0.01〜約20mg/kg;腹腔内投与の場合、0.01〜約100mg/kg;皮下投与の場合、0.01〜約100mg/kg;筋肉内投与の場合、0.01〜約100mg/kg;経口投与の場合、0.01〜約200mg/kg、および好ましくは約1〜100mg/kg;点鼻注入の場合、0.01〜約20mg/kg;ならびにエアロゾルの場合、0.01〜約20mg/kg動物(体)重。
【0051】
開示の方法、化合物および組成物から便益を受ける哺乳動物種としては、無尾猿、チンパンジー、オラウータン、ヒト、有尾猿などの霊長類;イヌ、ネコ、モルモット、ハムスター、ベトナム産ポットベリーブタ(Vietnamese pot-bellied pigs)、ウサギおよびフェレットなどの家庭用動物(例えばペット);ウシ、バッファロー、バイソン、ロバ、ブタ、ヒツジ、およびヤギなどの家畜動物;クマ、ライオン、トラ、パンサー、ゾウ、カバ、サイ、キリン、レイヨウ、ナマケモノ、ガゼル、シマウマ、ワイルドビースト、プレーリードッグ、コアラ、カンガルー、オポッサム、アライグマ、パンダ、ハイエナ、アザラシ、アシカ、ゾウアザラシ、オットセイ、ネズミイルカ、イルカ、およびクジラなどの、典型的には動物園で見られる珍しい動物種が挙げられるが、これらに限定されない。開示の方法から便益を受けることができる他の種としては、魚類、両生類、鳥類、および爬虫類が挙げられる。本明細書で使用する「患者」および「被験体」という用語は、互換的に用いられ、このようなヒトおよびヒト以外の種を含むことを意図している。同様に、本発明のインビトロの方法は、このような種の細胞に実施できる。
【0052】
本発明の方法を用いた処置を必要とする患者は、医学専門家に公知である標準的な技術を用いて特定できる。
【0053】
本明細書で使用する「癌」および「癌性」という用語は、未制御の細胞増殖を典型的な特徴とする、哺乳動物の病理的状態を指すか、または表す。癌の例としては、癌腫、リンパ腫、芽腫、肉腫、および白血病が挙げられるが、これらに限定されない。このような癌のより具体的な例としては、乳癌、前立腺癌、結腸癌、扁平上皮癌、小細胞肺癌、非小細胞肺癌、消化管癌、膵臓癌、子宮頚癌、卵巣癌、肝臓癌、例えば肝癌、膀胱癌、結腸直腸癌、子宮内膜癌、腎臓癌、および甲状腺癌が挙げられる。
【0054】
癌のその他の非限定例は、基底細胞癌、胆道癌;骨癌;脳およびCNS癌;絨毛上皮腫;結合組織癌;食道癌;眼癌;頭部および頚部の癌;胃癌;上皮内癌;咽頭癌;ホジキンおよび非ホジキンリンパ腫を含むリンパ腫;メラノーマ;骨髄腫;神経芽腫;口腔癌(例えば唇、舌、口および咽頭);膵臓癌;網膜芽腫;黄紋筋肉腫;直腸癌;呼吸器癌;肉腫;皮膚癌;胃癌;精巣癌;子宮癌;泌尿器系の癌、ならびにその他の癌腫および肉腫である。
【0055】
本発明の方法によれば、RR1またはその薬学的に許容される塩もしくは類似体は、それ自体を患者に投与でき、または他の作用物質と共投与できる。共投与は、同時(同一または異なる調合物として)または連続的に行うことができる。さらには、本発明の方法によれば、RR1、またはその薬学的に許容される塩もしくは類似体は、補助治療として患者に投与できる。例えば、RR1、またはその薬学的に許容される塩もしくは類似体は、化学療法と共に、患者に投与できる。
【0056】
したがって、本発明のRR1化合物(RR1、またはその薬学的に許容される塩もしくは類似体)は、別々に投与するか、あるいは薬学的組成物として投与するかを問わず、添加物として各種のその他の成分を含むことができる。関連する状況において用いることができる許容される成分または補助剤としては、酸化防止剤、フリーラジカルスカベンジャー剤、ペプチド、成長因子、抗生物質、静菌剤、免疫抑制剤、他の免疫賦活性剤、抗凝固剤、緩衝化剤、抗炎症剤、抗発熱剤、時間放出結合剤、麻酔剤、ステロイド、およびコルチコステロイドが挙げられる。このような成分は、追加の治療便益を提供でき、RR1化合物の治療作用に作用して影響を及ぼし、またはRR1化合物投与の結果として生ずるかもしれない任意の潜在的な副作用を阻止するように働くことができる。本発明のRR1化合物は、同様に、治療薬剤とコンジュゲートさせることもできる。
【0057】
同一の調合物または別々の調合物として、連続的または同時に、患者に共投与できるさらなる作用物質としては、免疫調節剤のような、所与の生物学的応答を調整する作用物質が挙げられる。例えば、腫瘍壊死因子(TNF)、インターフェロン(α-インターフェロンおよびβ-インターフェロンなど)、神経成長因子(NGF)、血小板由来成長因子(PDGF)、および組織プラスミノーゲン活性化因子などのタンパク質が投与できる。リンフォカイン、インターロイキン(インターロイキン-1(IL-1)、インターロイキン-2(IL-2)、およびインターロイキン-6(IL-6)など)、顆粒球マクロファージコロニー刺激因子(GM-CSF)、顆粒球コロニー刺激因子(G-CSF)、またはその他の成長因子などの生物学的応答調整物質を投与できる。抗癌剤、細胞傷害性薬剤、および/または化学療法薬剤は、本発明のRR1化合物と一緒に、連続的または同時に、同一調合物中でまたは別々の調合物として患者に共投与できる。一つの態様では、本発明のRR1化合物は、抗原と共投与される。好ましくは、抗原はワクチンの活性成分である。
【0058】
一つの態様では、本発明は、唯一の免疫学的活性物質としてRR1化合物を含む組成物を提供する。投与されると、直ちにRR1化合物は免疫系刺激を誘発する。
【0059】
別の態様では、上記のように、RR1化合物は、抗原(タンパク質、糖タンパク質、多糖および脂質を含むが、これらに限定されない)、および/または共刺激分子(サイトカイン、ケモカイン、標的化タンパク質リガンド、トランス活性化因子、ペプチド、および修飾アミノ酸を含むペプチドが挙げられるが、これらに限定されない)などの免疫調節刺激剤、ならびにアジュバント(ミョウバン、脂質乳剤およびポリラクチド/ポリグリコリド微小粒子を含むが、これらに限定されない)などのその他の免疫調節分子の一つ、または複数と共に投与することができる。本明細書で使用する「免疫調節」という用語は、免疫賦活性作用だけでなく免疫抑制作用も含む。免疫賦活性作用は、細胞性または液性免疫応答を直接または間接的に高める作用を含むが、これらに限定されない。免疫賦活性作用の例としては、抗原特異的抗体産生の増進;NK細胞、CD4+Tリンパ球、CD8+Tリンパ球、マクロファージ等のリンパ球集団の活性化または増殖;IL-1、IL-2、IL-4、IL-5、IL-6、IL-12、IFN-γ、TNF-αを含むが、これらに限定されない免疫賦活性サイトカインの合成増加等が挙げられるが、これらに限定されない。免疫抑制作用としては、細胞性または液性免疫応答を直接または間接的に減少させる作用を含む。免疫抑制作用の例としては、IgE産生低下などの抗原特異的抗体産生の低下;免疫寛容性を生ずるような免疫抑制活性を有するリンパ球またはその他細胞集団の活性化;ならびに特定の細胞機能に対し抑制作用を有するサイトカインの合成増加が挙げられるが、これらに限定されない。この一例は、IFN-γであり、これはIL-4が誘導するIgEおよびIgG1へのクラス切り替えを阻止し、それによってこれら抗体サブクラスのレベルを下げると考えられている。
【0060】
RR1化合物ならびに抗原および/または免疫調節促進剤は、コンジュゲートの形で一緒に投与でき、または免疫応答を調節できるように、十分近接した時間で、混合物として共投与することができる。好ましくは、RR1化合物および免疫調節分子は、同時に投与される。本明細書で用いられる「共投与」という用語は、この意味においては、少なくとも2つの物質を、免疫応答を調節するのに十分近い時間的関係で投与することを指す。好ましくは、共投与は、少なくとも2つの物質の同時投与を指す。
【0061】
本明細書で使用する「コンジュゲート」という用語は、その中でRR1化合物および免疫調節分子が結合している複合体を指す。このようなコンジュゲートの結合には、共有結合および/または非共有結合が含まれる。
【0062】
本明細書で使用する「抗原」という用語は、抗体またはT細胞抗原受容体が特異的に認識および結合する物質を意味する。抗原には、ペプチド、タンパク質、糖タンパク質、多糖、ガングリオシドおよび脂質;それらの一部およびそれらの組み合わせを含めることができる。抗原は、自然界に見いだされるものでも、または合成のものであり得る。ハプテンは、「抗原」の範囲内に含まれる。ハプテンは、それ自体は免疫原性でないが、免疫原性決定因子含有免疫原性分子とコンジュゲートすると免疫原性が付与される低分子化合物である。
【0063】
本明細書で使用する「アジュバント」という用語は、免疫原性作用物質に加えると、混合物に曝露したレシピエント宿主における、その作用物質に対する免疫応答を非特異的に高めるか、または増強する物質を指す。
【0064】
別の態様では、本発明は、RR1化合物および抗原を含む組成物を提供する。RR1化合物と一緒の投与に好適な抗原は、B細胞またはT細胞抗原特異的応答を引き出すことができる任意の分子を含む。好ましくは、抗原は、抗原に特異的な抗体応答を引き出す。広範な分子が抗原である。これら抗原としては、糖、脂質およびポリペプチド、ならびに複合炭化水素のような高分子、ならびにリン脂質が挙げられるが、これらに限定されない。小分子は、抗原性を付与するためにハプテン化が必要な場合がある。好ましくは、本発明の抗原としては、ペプチド、脂質(例えばステロール、脂肪酸、およびリン脂質)、ヘモフィラス・インフルエンザワクチンで用いられるような多糖、ガングリオシドおよび糖タンパク質が挙げられる。
【0065】
本明細書で使用する「ペプチド」という用語は、ペプチドがハプテンであるかないかに関係なく、生物学的応答、例えば抗体産生もしくはサイトカイン活性を達成する十分な長さおよび組成を有するペプチドおよびタンパク質を含む。典型的には、ペプチドは、少なくとも6アミノ酸残基長である。用語「ペプチド」はさらに、修飾アミノ酸を包含し、その場合の修飾としては、リン酸化、グリコシル化、ペジル化、脂質化およびメチル化が挙げられるが、これらに限定されない。
【0066】
一つの態様では、本発明は、RR1化合物および抗原性ペプチドを含む組成物を提供する。抗原性ペプチドとしては、精製された未変性ペプチド、合成ペプチド、組換えタンパク質、粗タンパク質抽出物、弱毒化もしくは不活性化ウイルス、細胞、微生物またはそのようなペプチドの断片を包含できる。
【0067】
多くの抗原性ペプチドおよびタンパク質が公知であり、当技術分野で利用可能である;その他のものは、通常技術を用いて特定できる。免疫賦活性促進剤としての要求を満たすことができるタンパク質抗原としては、以下の例が挙げられるが、これらに限定されない。単離された未変性または組換え抗原は、植物の花粉(例えば、Rafnar et al. (1991) J. Biol. Chem. 266:1229-1236; Breiteneder et al. (1989) EMBO J. 8:1935-1938; Elsayed et al. (1991) Scand. J. Clin. Lab. Invest. Suppl. 204:17-31; およびMalley (1989) J. Reprod. Immunol. 16:173-186を参照されたい)、イエダニ(例えば、Chua et al. (1988) J. Exp. Med. 167:175-182; Chua et al. (1990) Int. Arch. Allergy Appl. Immunol. 91:124-129;およびJoost van Neerven et al. (1993) J. Immunol. 151:2326-2335)、動物のフケ(例えば、Rogers et al. (1993) Mol. Immunol. 30:559-568を参照されたい)、動物の唾液、ミツバチの毒、ならびにカビの胞子から得ることができる。HIV-1、HIV-2、単純ヘルペスウイルス、A型肝炎ウイルス(Bradley et al. (1984) J. Med. Virol. 14:373-386)、ロタウイルス、ポリオウイルス(Jiang et al. (1986) J. Biol. Stand. 14:103-109)、B型肝炎ウイルス、麻疹ウイルス(James et al. (1995) N. Engl. J. Med. 332:1262-1266)、ヒトおよびウシパピローマウイルス、およびスローブレイン(slow brain)ウイルスなどの、生きている弱毒化および不活性化された微生物は、ペプチド抗原を提供する。腫瘍形成に対する免疫化については、免疫賦活性ペプチドとして、腫瘍細胞(生きているか、または放射線処理したもの)、腫瘍細胞抽出物または腫瘍抗原のタンパク質サブユニットを挙げることができる。免疫に基づく避妊向けワクチンは、RR1化合物と一緒に投与する精子タンパク質を含ませることによって形成できる(Lea et al. (1996) Biochim. Biophys. Acta 1307:263)。
【0068】
本発明の組成物および方法で使用する抗原は、決定的ではない。各種抗原の混合物を、本発明に従って用いることができる。好ましくは、ウイルスまたは細菌病原体、あるいは真菌または寄生虫に由来するタンパク質またはペプチドを、このような抗原として使用する(誘導体化抗原またはグリコシル化もしくは脂質化抗原もしくは多糖もしくは脂質を含む)。別の好ましい抗原供給源は、腫瘍抗原である。好ましい病原体は、ヒト免疫不全症ウイルス(HIV)、A型およびB型肝炎ウイルス、C型肝炎ウイルス(HCV)、ラウス肉腫ウイルス(RSV)、エプスタインバーウイルス(EBV)、インフルエンザウイルス、ロタウイルス、黄色ブドウ球菌、クラミジア肺炎、クラミジア・トラコマチス、ヒト型結核菌、肺炎連鎖球菌、炭疽菌、ビブリオ・コレラ、プラスモディウム属種(熱帯熱マラリア原虫、三日熱マラリア原虫、その他)、アスペルギルス属種またはカンジダ・アルビカンスから選択される。抗原はまた、癌細胞が発現する分子でもよい(例えば腫瘍抗原)。誘導体化のプロセスは、病原体/癌細胞からの特異的タンパク質の精製、病原体の不活性化、ならびにそのようなタンパク質のタンパク質分解もしくは化学的誘導体化もしくは安定化を含んでよい。同様にして、腫瘍抗原(癌ワクチン)または自己免疫抗原も本発明の組成物および方法に用いることができる。このようにして、腫瘍ワクチン接種または自己免疫疾患の治療を行うことができる。
【0069】
RR1化合物および抗原は、RR1-抗原コンジュゲートとして投与でき、かつ/またはそれらは乳剤などの、混合物の形をした複合体として共投与できる。RR1-抗原コンジュゲート内でのRR1化合物と抗原分子との会合は、高親和性および/または低親和性相互作用を含む、共有結合形の互作用、および/または非共有結合形の相互作用を通して可能となる。RR1-抗原コンジュゲート内でRR1化合物と抗原とをカップリングできる非共有結合形の相互作用の例としては、イオン結合、疎水性相互作用、水素結合、およびファン・デール・ワールス力が挙げられるが、これらに限定されない。
【0070】
別の態様では、RR1化合物は、一つまたは複数の免疫調節促進剤と共に投与できる。したがって本発明は、RR1化合物および免疫調節促進剤を含む組成物を提供する。本明細書で使用する「免疫調節促進剤」という用語は、RR1化合物の免疫調節活性を支持および/または強化する分子を指す。免疫調節促進剤の例としては、サイトカイン、および/またはアジュバントのような共刺激分子を挙げることができる。RR1化合物および促進剤は、RR1-促進剤コンジュゲートとして投与でき、かつ/またはそれらは乳剤などの、混合物の形をした複合体として共投与できる。RR1-促進コンジュゲート内のRR1化合物と促進剤分子とは、高親和性および/または低親和性相互作用を含む、共有結合形の相互作用、および/または非共有結合形の相互作用を通じて結びつくことができる。RR1-促進コンジュゲート内でRR1化合物と促進剤とをカップリングできる非共有結合形の相互作用の例としては、イオン結合、疎水性相互作用、水素結合、およびファン・デール・ワールス引力が挙げられるが、これらに限定されない。
【0071】
免疫調節促進剤としては、共刺激分子(サイトカイン、ケモカイン、標的化タンパク質リガンド、トランス活性化因子、ペプチド、および修飾アミノ酸を含むペプチドなど)ならびにアジュバント(ミョウバン、脂質乳剤、およびポリラクチド/ポリグリコリド微小粒子など)が挙げられるが、これらに限定されない。
【0072】
なかでも、RR1化合物と共に投与するのに好適な免疫調節サイトカインペプチドは、インターロイキン(例えばIL-1、IL-2、IL-3等)、インターフェロン(例えばIFN-α、INF-β、IFN-γ)、エリスロポイエチン、コロニー刺激因子(例えばG-CSF、M-CSF、GM-CSF)およびTNF-αである。好ましくは、RR1化合物と共に使用するための免疫賦活性ペプチドは、IL-12(Bliss et al. (1996) J. Immunol. 156:887-894)、IL-18、TNF-α、β、およびγ、ならびに/または形質転換成長因子(TGF)-αなどTh1型免疫応答を刺激するペプチドである。
【0073】
RR1化合物と共に投与されるペプチドは、特異的受容体とのタンパク質との結合を媒介するか、または特異タイプの細胞もしくは組織への標的化を媒介するアミノ酸配列を含むことができる。例としては、抗体もしくは抗体断片、ヒト成長ホルモンなどのペプチドホルモン、および酵素が挙げられるが、これらに限定されない。免疫調節ペプチドもまた、ペプチドホルモン、ペプチド神経伝達物質およびペプチド成長因子を含む。B7(CD80)などの共刺激分子、転写因子などのトランス活性化タンパク質、マクロファージ走化性タンパク質(MCP)および他の化学誘引物質または走化性ペプチドなどのケモカインもまた、本発明のRR1化合物との共投与に有用なペプチドである。
【0074】
RR1化合物は、脂質、多糖、ガングリオシドなどの他の抗原と、ペプチドなどの連結基を介してコンジュゲートさせることもできる。
【0075】
本発明はまた、アジュバントと併せてRR1化合物の投与も提供する。抗原を、RR1化合物およびアジュバントと一緒に投与すると、抗原に対する免疫応答は強化され、それによってRR1化合物および抗原を単独で含む組成物で得られる応答よりも高い免疫応答を起こすことができる。したがって、別の態様では、本発明は、RR1化合物、抗原およびアジュバントを含む組成物を提供し、これによってRR1化合物/抗原/アジュバントを共投与できる。好ましくは、免疫原性組成物は、免疫原に対する免疫応答を強化するのに十分な量のアジュバントを含有する。好ましくは、アジュバントとしては、水中油型乳剤、油中水乳剤、ミョウバン(アルミニウム塩)、リポソーム、ならびにポリスチレン、澱粉、ポリホスファゼンおよびポリラクチド/ポリグリコシドを含むが、これらに限定されない微小粒子を挙げることができるが、これらに限定されない。より好ましくは、RR1化合物および抗原は、ミョウバンと一緒に共投与される。より好ましくは、RR1化合物および抗原は、リポソームと一緒に共投与される。よりさらに好ましくは、RR1化合物および抗原は、水中油型乳剤と共に共投与される。
【0076】
好適なアジュバントとしては、スクアレン混合物(SAF-1)、ムラミルペプチド、サポニン誘導体、マイコバクテリウム細胞壁調製物、モノホスホリル脂質A、ミコール酸誘導体、非イオン系コポリマー界面活性剤、Quil A、コレラトキシンBサブユニット、ポリホスファゼンおよび誘導体、ならびにTakahashiら(1990) Nature 344:873-875が記載したような免疫刺激複合体(ISCOM)ならびに脂質をベースとしたアジュバントおよび本明細書に記載されている他のアジュバントを挙げることができるが、これらに限定されない。獣医分野での使用、および動物での抗体産生での使用では、フロイントのアジュバント(完全および不完全の両方)の分裂誘発成分を用いることができる。
【0077】
全ての免疫原性組成物と同様に、成分の免疫学的有効量は、経験的に決めることができる。考慮すべき要素としては、RR1化合物および/または抗原が、免疫調節促進剤、アジュバントもしくは担体タンパク質もしくは他の担体と複合体化しているか、または共有結合しているか否かの抗原性、投与経路かつ投与する免疫投与の回数が挙げられる。このような要素は、ワクチン分野では公知であり、過度の実験なしにこのような決定を行うことは、免疫学者の通常の技量の範囲内である。
【0078】
本発明はさらに、RR1化合物および免疫調節分子を混合物として投与した場合に比べ、発生する免疫応答を高めるのに有効である距離まで、RR1化合物および免疫調節分子が近接して結びついている組成物を提供する。したがって、本発明は、複合体が標的に作用するまで、RR1化合物と免疫調節分子の近接した結びつきを維持できる、封入化剤を含む組成物およびその使用方法を提供する。好ましくは、RR1成分、免疫調節分子、および封入化剤を含む組成物は、アジュバント水中油型乳剤、微小粒子および/またはリポソームの形態である。より好ましくは、RR1-免疫調節分子を封入しているアジュバント水中油型乳剤、微小粒子および/またはリポソームは、約0.04μm〜約100μmの大きさ、より好ましくは約0.1μm〜約20μm、よりさらに好ましくは約0.15μm〜約10μmの粒子の形態である。
【0079】
マイクロスフェア、ビーズ、高分子複合体、ナノカプセルのようなコロイド分散系、ならびに水中油型乳剤、ミセル、混合ミセル、およびリポソームなどの脂質をベースとした系は、RR1化合物含有組成物の有効な封入を提供する。
【0080】
本発明のさらなる局面は、ティノスポラ・コルディフォリア植物材料からRR1を得るためのプロセスを提供する。好ましい態様によれば、RR1は植物材料の空中部分(茎および葉)から単離される。乾燥し、粉末化した植物材料500gを、ソックスレー(sox let)抽出器の中で、純粋メタノールを用いて72時間抽出する。残留物を熱した蒸留水に取り、遠心分離し、上清から集める。再度遠心分離を行い、80%アルコールを用いて上清から粗多糖を沈殿させる。沈殿物を濾過し、アルコールで洗浄し、蒸留水に溶解し、透明な溶液を15%トリクロロ酢酸で処理する。溶液を遠心分離にかけ、いかなる残留物も取り除き、アセトンを用いて透明な上清を再度沈殿させる。沈殿物を濾過して、アセトンで数回洗浄し、蒸留水に再溶解する。透明な溶液を流水中で72時間透析し(メンブレン、MWCO 60000Da)、チューブに入れてスピードバック(speedvac)で凍結乾燥する。本プロセスの例示的態様は、図1のフローチャートに示されている。天然産物精製分野の科学者は、記載の方法を、本発明の好ましい態様に記載されているものに容易に適合させ、各種溶媒および固定相を交換できる。ティノスポラ・コルディフォリア抽出物の水性画分は、水切り、濾過、デカンテーションによって、または溶液もしくは混合物の水性部分を分離に関する分野において公知である他の任意の方法によって分離できる。クロマトグラフィーに加えて、所望の化合物は、結晶化および分配などの方法を用いても精製できる。RR1化合物の単離には、様々な代替試薬が利用できる。例えば、エタノールに代わって一価アルコールを用いてもよい。アセトンは、単離手順では、エタノールに替えることができる。例えば、全体として参照により本明細書に組み入れられる、Brown G.G., Unit Operations, John Wiley & Sons, 1956;McCabe, W.L. and J.C. Smith, Unit Operations of Chemical Engineering, McGraw-hill, 1956;およびPerry, R.H., and D. Green, Perry's Chemical Engineers' Handbook, 7th Edition, McGraw-Hill, 1997を参照されたい。
【0081】
任意で、ひとたび得られた単離RR1化合物を出発材料として用い、本発明のRR1類似体を合成することができる。任意で、プロセスは、単離RR1化合物または類似体を、本明細書に記載のようなインビトロまたはインビボアッセイを用いて、免疫調節活性について評価する段階をさらに含むことができる。
【0082】
植物からRR1を抽出するための方法に関しては、特に制限はない。例えば、各種溶媒を用いた抽出または超臨界液体抽出が利用できる。植物からのRR1抽出に用いる溶媒に関して、特に制限はない。好適な溶媒の例としては、水、無機塩水溶液、および緩衝液などの水性媒体、ならびにアルコール、ヘキサン、トルエン、石油、エーテル、ベンゼン、酢酸エチル、クロロホルム、ジクロロメタン、1,1,2-トリクロロエタン、ジ-メチルスルホキシド、およびアセトンなどの有機溶媒が挙げられ、とりわけアルコールが好ましい。水は未蒸留水、蒸留水、脱イオン水、または純水でよい。使用できる緩衝液の例は、リン酸緩衝液およびクエン酸緩衝液が挙げられる。
【0083】
植物材料からRR1を抽出する場合、水、水含有エタノールまたは無水エタノールなどの治療薬剤に適した溶媒を使用することが適正である。使用してもよいアルコールの例としては、メタノール、エタノール、プロパノールおよびブタノールなどの一価アルコール、ならびにプロピレングリコールおよびグリセロールなどの多価アルコールが挙げられ、なかでも一価アルコールが好ましく、特にエタノールが好ましい。これらの溶媒は、単独または混合物として用いてもよい。混合溶媒として用いる場合、水含有アルコールが好ましい。水含有一価アルコールがより好ましく、水含有エタノールが特に好ましい。
【0084】
抽出に関しては、溶媒を、例えば植物1重量部を基にして、0.1〜10,000重量部、好ましくは1〜100重量部の量を用いることができる。抽出温度に関して特に制限はないが、抽出は0〜100℃、より好ましくは20〜90℃で実施するのが好ましい。抽出時間に関しては特に制限はないが、例えば1分間〜1週間、より好ましくは30分間〜1日の間に行うことが好ましいであろう。
【0085】
抽出に用いる装置に関しては特に制限はないが、効率的な抽出用に設計された容器、撹拌機、還流冷却器、ソックスレー抽出器、ホモジェナイザー、シェーカー、超音波発生装置等を用いてもよい。液体抽出物は、沈殿物、脱水濾過、透明濾過、遠心分離濾過、遠心分離沈殿、圧縮分離または圧搾濾過などの、各種固体-液体分離の手段によって処理できる。
【0086】
植物粉末を調製する場合は、葉、枝、茎、樹皮、根、種、培養細胞または器官、あるいはカルスなどの、RR1を含む植物の全てまたは一部を用いてよい。空中部分(茎および葉)が好ましい。植物材料は、そのまま用いても、または物理学的もしくは化学的もしくは生物学的に処理した後に用いてもよい。物理学的もしくは化学的処理の方法の例は、乾燥、フリーズドライ、破壊、および抽出である。物理学的もしくは化学的に処理された物体としては、乾燥物、フリーズドライ物、破壊物および抽出物が挙げられる。抽出物は、抽出後に得た植物残留物を含む。
【0087】
植物粉末を調製するために、好ましく乾燥したティノスポラ・コルディフォリア植物材料は、下記を用いて破砕できる:ジョークラッシャ、ジャイレートリークラッシャ、もしくはコーンクラッシャなどの圧縮破砕機;カッターミルもしくはシュレッダーなどの剪断機;ハンマークラッシャなどのインパクトクラッシャ;ロールクラッシャなどのロールミル;ディスインテグレータ、もしくはケージミルなどのロータリーミル;コーヒーミルなどのスクリューミル;エッジランナーなどのローリングミル;スタンプミルなどのハンマリングミル;遠心ローラーミル、ボールベアリングミル、ボールミル、もしくはゼゴミルなどのローラーミル;スウイングハンマーミル、ピンミル、ケージミル、ターボ型ミル、もしくは遠心ミルなどの高速ロータリーミル;ローリングボールミル、振動ボールミル、遊星ボールミル、もしくはCFミルなどの容器振動ミル;フローパイプ型ミル、撹拌タンクミル、管型ミル、空気吸込み型ミル、インパクトプレートミラー、もしくは流動床ミルなどのジェットミル;超音波シュレッダーなどの破砕機;ストーンモーターもしくはモーター。上記の方法で得た生成物を、さらに物理的または化学的に処理し、植物粉末を得ることができる。
【0088】
植物粉末は荒くとも細かくともよい。植物粉末の平均粒子サイズは、乾燥状態において、好ましくは0.1μm〜1mm、より好ましくは1〜100μm、特には2〜50μmである。乾燥状態の植物粉末の平均粒子サイズは、例えば、レーザー分散粒子分布アナライザーによって決定できる。または、植物粉末をグリセロールと水の1:1の混合液で膨潤させた場合、粉末の平均粒子サイズは、好ましくは1μm〜10mm、より好ましくは10μm〜1mm、特には20〜500μmである。膨潤状態の植物粉末の平均粒子サイズは、例えば顕微鏡を用いた観察によって決定できる。
【0089】
乾燥方法は、当技術分野で公知である。例えば、乾燥方法としては、加熱および減圧下での乾燥、加熱および大気圧下での乾燥、またはスプレー乾燥器もしくはドラム乾燥器を用いた乾燥、またはフリーズドライが挙げられるが、とりわけ加熱および減圧下での乾燥またはフリーズドライが好ましい。
【0090】
本明細書で使用する「腫瘍」という用語は、悪性であるか良性であるかにかかわらず、全ての新生物細胞の生育および増殖を指し、全ての前癌性および癌性細胞および組織を指す。例えば、特定の癌は、固体の塊状の腫瘍を特徴とする。固体腫瘍の塊は、存在する場合、原発腫瘍塊のことがある。原発腫瘍塊は、その組織の正常細胞が形質転換した結果生じた癌細胞が組織内で生育したものを指す。ほとんどの例では、原発腫瘍塊は、視覚法または触診法によって見つけることができる嚢胞の存在によって、あるいは組織の形状、質感または重量の異常によって同定する。しかしながら、いくつかの原発腫瘍は、触診できず、X線などの医学的画像技術(例えばマンモグラフィー)を用いるか、または針吸引によってのみ検出できる。これら後者の技術の使用は、早期発見においてより一般的である。本明細書で使用する「腫瘍」という用語は、白血病のような、非固形性の新生物も含む。組織内の癌細胞の分子および表現形の分析は通常、癌が組織に対し内因性であるか、または疾患が別の場所からの転移によるものか確認するであろう。
【0091】
本明細書で使用する「治療する」または「治療」という用語は、治療的処置および予防もしくは防御的処置の両方を指し、この場合の目的は、癌などの細胞増殖障害、または他の生理学的状態(例えば疾患もしくは他の障害)の発生もしくは拡大といった、望ましくない生理学的変化もしくは障害を予防するか、遅らせる(軽減する)ことである。好ましくは、病理学的状態は、免疫刺激または免疫抑制のいずれかによる治療または管理ができるものである。本発明の目的に関して、有利または望ましい臨床結果とは、検出可能であるか否かにかかわらず、症状の軽減、疾患の広がりの縮小、疾患の状態の安定化(即ち悪化しない)、疾患の進行の遅延または減速、疾患状態の改善もしくは緩和および寛解(部分的または完全を問わず)を含むが、これらに限定されない。「治療」はまた、治療を受けていない場合に予想される生存期間に比べ、生存期間が延長することも意味する。治療が必要な対象としては、既に状態または障害を持つ対象だけでなく、状態または障害を持つ傾向がある対象、もしくは防止しなければならない状態および疾患を持つ対象が挙げられる。
【0092】
本明細書で使用する「(治療)有効量」という用語は、哺乳動物における、細胞増殖性障害などの病理的状態(例えば疾患または他の障害)を治療するのに有効な、本発明のRR1化合物または他の作用物質などの、作用物質の量を指す。癌の場合、作用物質の治療有効量は、癌細胞の数を減らし;腫瘍の大きさを縮小し;癌細胞の周辺臓器への浸潤を阻止し(即ち、ある程度遅延させる、および好ましくは停止させる);腫瘍の転移を阻止し(即ち、ある程度まで遅延させる、および好ましくは停止させる);腫瘍の増殖をある程度まで阻止し;ならびに/または、癌に関係する一つまたは複数の症状を、ある程度まで軽減する。作用物質が増殖を阻止できるおよび/または既存の癌細胞を殺すことができる程度まで、それは細胞増殖抑制性および/または細胞傷害性でよい。癌治療に関しては、効力は、例えば、疾患が進行するまでの時間(TTP)を評価すること、および/または応答率(RR)を決定することによって測定できる。
【0093】
本明細書で使用する「抗癌剤」という用語は、癌細胞の機能を阻止し、それらの形成を阻止し、および/またはそれらのインビトロまたはインビボでの破壊を引き起こす物質または治療を指す。例としては、細胞傷害性剤(例えば5-フルオロウラシル、タキソール)および抗シグナル伝達剤(例えばPI3KインヒビターLY)が挙げられるが、これらに限定されない。
【0094】
本発明細書で使用する「細胞傷害性剤」という用語は、細胞の機能を阻止または妨害、および/または細胞のインビトロまたはインビボでの破壊を引き起こす物質を指す。用語は、放射性同位元素(例えばAt211、I131、I125、Y90、Re186、Re188、Sm153、Bi212、P32、およびLuの放射性同位元素)、化学療法薬剤、毒素、例えば細菌、真菌、植物もしくは動物起源の低分子毒素もしくは酵素活性毒素、それらの断片および/もしくは変種を含む抗体を含むことを意図する。
【0095】
本明細書で使用する「化学療法薬剤」という用語は、例えば、パクリタキセル(TAXOL、BRISTOL-MYERS SQUIBB Oncology, Princeton, N.J.)およびドキセタキセル(TAXOTERE、Rhone-Poulenc Rorer, Antony, France)などのタキセン、クロラムブシル、ビンクリスチン、ビンブラスチン、例えばタモキシフェン、ラロキシフェン、アロマターゼ阻害4(5)-イミダゾール、4-ヒドロキシタモキシフェン、トリオキシフェン、ケオキシフェン、LY117018、オナプリストン、およびトレミフェン(Fareston)などの抗エストロゲン剤、ならびにフルタミド、ニルタミド、ビカルタミド、ロイプロリド、およびガセレリン等などの抗アンドロゲン剤の、癌治療に有用な化合物である。
【0096】
本明細書で使用する「RR1化合物」という用語は、図3に示すようなRR1、またはRR1の薬学的に許容される塩または類似体を指す。本明細書で使用する、RR1またはRR1化合物に関連する「単離された」という用語は、自然に生ずる、例えばティノスポラ・コルディフォリア植物材料もしくは植物抽出物から生ずる、媒体を実質的に含まない化合物を指す。しかしながら、単離RR1化合物は、当業者に公知である適切な化学合成反応によっても得られる(Greene, T.W. and Wuts, P.G.M.「Protective Groups in Organic Synthesis」John Wiley & Sons, Inc. New York. 3rd Ed. pg. 819, 1999; Honda, T. et al. Bioorg. Med. Chem. Lett., 1997, 7:1623-1628; Honda, T. et al. Bioorg. Med. Chem. Lett, 1998, 8:2711-2714; Konoike, T. et al. J. Org. Chem., 1997, 62:960-966; Honda, T. et al. J. Med. Chem., 2000, 43:4233-4246)。
【0097】
「含む」、「からなる」および「本質的にからなる」という用語は、その標準的な意味に従って定義される。これら用語は、本明細書を通じて、各用語に関連する具体的な意味を与えるために相互に代用できる。
【0098】
本明細書で使用する場合、単数形の「一つの(a)」、「一つの(an)」、および「その(the)」は、文脈が特に明瞭に指図しない限り複数形も含む。したがって、例えば、「一つのRR1化合物」という言及は、一つより多いこのようなRR1化合物を含む。「一つの免疫調節分子」という言及は、一つより多いこのような免疫調節分子、または一つより多い免疫調節分子のタイプを含む。「一つの抗原」という言及は、一つより多い抗原、または一つより多い抗原のタイプを指すのに用いられるといった具合である。
【0099】
本発明の実施は、特に指示がない限り、分子生物学(組換え技術を含む)、微生物学、細胞生物学、生化学および免疫学の通常の技術を使用するが、それらは当技術分野の範囲内である。このような技術は、「Molecular Cloning: A Laboratory Manual」、第2版のような文献に十分に説明されている(Sambrook et al., 1989);「Oligonucleotide Synthesis」(M. J. Gait, ed., 1984);「Animal Cell Culture」(R. I. Freshney, ed., 1987);「Methods in Enzymology」(Academic Press, Inc.);「Handbook of Experimental Immunology」(D. M. Weir & C. C. Blackwell, eds.);「Gene Transfer Vectors for Mammalian Cells」(J. M. Miller & M. P. Calos, eds., 1987);「Current Protocols in Molecular Biology」(F. M. Ausubel et al., eds., 1987);「PCR: The Polymerase Chain Reaction」(Mullis et al., eds., 1994);および「Current Protocols in Immunology」(J. E. Coligan et al., eds., 1991)。
【0100】
以下は、本発明を実施するための材料、方法、および手順を例示する実施例である。実施例は例示であり、限定的に解釈してはならない。
【0101】
材料および方法
RR1の単離および特徴付け
ティノスポラ・コルディフォリア粉末(GARRY and SUN, Inc., Reno, Nevadaから市販されている)の抽出物の予備的研究により、免疫刺激の原則は、水性画分中に限定されることが明らかとなった。図1に、単離に適用された手順をフローチャートにて示す。図3に示す最終的なRR1化合物は、抽出に使用した全乾燥材料の約0.1%収率で、水に溶解した膨れた固体として単離された。最終生成物の最初の分析、および13C NMRスペクトルは、それが多糖であることを明らかにした。したがって、Complex Carbohydrate Research Center, University of Georgia, Athensにおいて、グリコシル成分、結合、分子量、およびアノマー中心におけるグルコース単位のコンホメーションなどの、多糖の詳細な分析を行った。グリコシル成分の分析は、Yorkら(York, W.S. et al. Methods Enzymol., 1985, 118:3-40)の方法に従った酸性メタノリシスによりRR1から得られた単糖メチルグリコシドのパー-O-トリメチルシリル誘導体のガスクロマトグラフィー/質量分析の組み合わせ(GC/MS)により行った。この分析では、イノシトールを内部基準として使用した。単糖誘導体は、それらの特徴的な保持時間により同定され、さらにそれらの質量スペクトルにより確認された。グリコシル結合分析を行うには、試料をCikanuおよびKereck(Ciucanu, I. and F. Kereck Carbohydr. Res., 1984, 131:209-217)の方法により3回パーメチル化し、2Mトリフルオロ酢酸で加水分解し、NaBD4で還元し、無水酢酸/ピリジンでアセチル化して、得られた一部メチル化アルジトールアセテート(PMAA)をGC/MSにより分析した。糖残基は、それらの特徴的な保持時間と、質量スペクトルデータにより同定した。グリコピラノシル単位のアノマー中心におけるコンホメーションは、D2O中で記録した500MHzプロトンNMRスペクトルに基づき、かつ基準試料のアノマープロトンシグナルとの比較により得た。分子量は、50mMギ酸アンモニウム緩衝液により速度0.5ml/分で溶出するSUPELCOシリカカラム(1.0×30cm)を使用したサイズ排除クロマトグラフィーから得て、屈折率により検出した。デキストラン試料を基準として使用して、溶出したピークの保持時間と基準との比較により分子量を得た。
【0102】
免疫刺激特性
RR1の免疫刺激特性を測定するために、リンパ球の異なるサブセットの活性化、例えばインターロイキン(IL)-1β、IL-2、IL-4、IL-6、IL-10、IL-12、p70およびp40、IL-18、インターフェロン(IFN)-αおよびγ、腫瘍壊死因子(TNF)-αおよびβ、単球遊走因子(MCP)-1などのサイトカインの合成、一酸化窒素(NO)の合成、ならびにヒトリンパ球内で誘発される酸化ストレスの程度を分析した。健康な志願者から採取した新鮮血から、histopaque密度勾配法により正常なリンパ球を単離して、様々なアッセイに使用した。ヒト白血病(CEM)および多剤耐性(CEM/VLB)細胞株を、10%ウシ胎児血清および抗生物質を補充したRPMI培地中、5%CO2インキュベーター内にて37℃で増殖させた。
【0103】
リンパ球の活性化
正常なリンパ球(106/ml)を、RR1 0〜100μgで、37℃に保持したCO2インキュベーター内にて、RPMI培地中で24時間処理した。次いで、細胞を特定の蛍光色素結合モノクローナル抗体で30分間、室温にて染色し、Coulter Elite Flow Cytometer内で、AlamoおよびMelnick(Alamo, A.L. and S.J. Melnick Cytometry, 2000, 42:363-370)の4色または5色イムノタイピング(immunotyping)アッセイプロトコルにより分析し、例えばNK、TおよびB細胞等のリンパ球のサブセットの活性化は百分率を計算した。
【0104】
RR1処理リンパ球の細胞傷害性
活性化したリンパ球の高められた細胞傷害性を評価するために、RR1活性化リンパ球をエフェクター細胞とし、ヒト白血病細胞(CEM)を標的として使用する機能的アッセイを採用した(Liu, L. et al. Nat. Med., 2002, 8:185-189; Jerome, K.R. et al. Nat. Med., 2003, 9:4-5)。簡単に言えば、正常なリンパ球(106/ml)を、RPMI培地中の異なる濃度のRR1で、CO2インキュベーター内にて37℃で24時間処理した。翌日、標的細胞(CEM 1×106/ml)を、1ml PBS中の4.6μM膜標識色素PKH26(SIGMA)により室温で3分間標識した。等容積のウシ胎児血清(GIBCO, Life Sciences, MD)を1分間加えて、標識を停止した。次いで、標識した腫瘍細胞を、エフェクターと標的との比を1:1としてRR1処理リンパ球と共に4時間インキュベートし、未処理リンパ球を対照として使用した。活性化NK細胞により殺傷された細胞の百分率を、氷上にて2%パラホルムアルデヒド溶液1mlで混合物を30分間固定し、続いてPBS中0.5%ツイーン20中に懸濁させることにより測定した。細胞混合物を抗活性カスパーゼ3-FITC抗体(BD Biosciences, CA)7.5μlで、室温にて30分間染色し、PBSで洗浄し、Coulter Elite Flow Cytometer内で分析した。
【0105】
サイトカインおよびケモカイン合成の定量
IL-1β、IL-2、IL-4、IL-6、IL-10、IL-12p40、IL-12p70、IL-18、IFN-γおよびTNF-α、βなどのサイトカイン、ならびにケモカイン、MCP-1を、ELISA法により、BD Biosciencesのキットを使用して、製造業者のプロトコルにしたがって定量した。IFN-αは、RESEARCH DIAGNOSTICS Inc.(New Jersey)のELISAキットを使用してアッセイした。簡単に言えば、ELISA希釈液50μlをピペットで、96ウェルプレートの抗体コートウェル内に入れ、続いて各標準試料および試験試料100μlを入れて、5秒間振とうしてウェル内の内容物を混合し、プレートシーラーで覆い、室温で2時間インキュベートした。インキュベーション後、ウェルの内容物を吸引し、洗浄溶液で5回洗浄した。最終的な洗浄の洗浄溶液を完全に除去した後、検出溶液100μlを加え、プレートシーラーで覆い、1時間インキュベートした。ウェルを洗浄溶液で7回洗浄し、一段階基質試薬100μlを加えて、暗闇で30分間インキュベートした。停止溶液50μlを加えて着色を停止し、BIORAD Benchmarkプレートリーダー内で、参照波長570nmと共に450nmにて吸収を記録した。
【0106】
補体活性化経路
ヒト補体C3a des ArgおよびC4a des Arg関連のEIAキット(ASSAY DESIGN Inc., Ann Arbor, MI)を使用して、製造業者のプロトコルにしたがって、切断された補体成分(C3a des ArgおよびC4a des Arg)を測定した。簡単に言えば、健康な志願者から採取した正常血液1mlを、CO2インキュベーター内で37℃にて24時間、RR1 0〜100μg/mlと共にインキュベートした。処理した血液試料を4℃にて2000×gで遠心分離にかけ、補体試薬「A」225μlを等容積の試料上澄み液に加え、徹底的にボルテックスで撹拌した。この混合物に10N HCl 50μlを加えて、再度ボルテックスで撹拌し、室温で1時間インキュベートした。試料を室温で5分間、ミクロ遠心分離機内で10,000rpmにて遠心分離し、上澄み液180μlを15ml管に移し、9N NaOH 20μlを加えて、徹底的にボルテックスで撹拌した。この混合物に、補体試薬「B」600μlを加えた後、アッセイ緩衝液10.7mlを加え、ボルテックスで撹拌して、分析に使用した。アッセイ試料(100μl)を96ウェルのマイクロプレート内のウェル内にピペットで入れ、次いで青色コンジュゲート50μlおよび黄色抗体50μlを入れた。プレートをプラットホームシェーカー上で、500rpmにて2時間振とうした。ウェルを吸引して非結合材料を除去し、洗浄溶液200μlで3回洗浄し、p-Npp基質溶液200μlを加えた。プレートを37℃で1時間、振とうせずにインキュベートし、停止溶液50μlを加え、BIO-RAD Bench topプレートリーダー内で、参照波長570nmと共に405nmの吸収を測定した。
【0107】
誘発可能な一酸化窒素シンターゼ(iNOS)誘導、および一酸化窒素(NO)合成
NOの増大した、および持続的な放出は、アルギニン刺激による、酵素iNOSの刺激を原因とする。RR1刺激によるiNOS誘導を、サンドイッチ酵素イムノアッセイ法を用いるQUANTIKINE iNOSイムノアッセイキット(R&D systems, Minneapolis, MN)によりアッセイした。異なる濃度のRR1で処理した細胞の細胞質抽出物から、iNOSをアッセイした。試料および標準を96ウェルプレートのウェル内にピペットで入れ、存在する任意のiNOSは固定化した抗体に結合した。未結合物質を洗浄除去した後、iNOSに特異的な酵素結合モノクローナル抗体を加えた。未結合抗体酵素試薬を洗浄除去した後、着色物質溶液を加え、停止溶液で着色を停止して、Bio-Radプレートリーダー内で、参照波長570nmと共に450nmの吸収を読み取った。
【0108】
NOの量を、一酸化窒素定量キット(ACTIVE MOTIF, Carlsbad, CA)により、製造業者のプロトコルにしたがってアッセイした。簡単に言えば、リンパ球(106/ml)を様々な濃度のRR1(0〜100μg/ml)と共に、硝酸を含まない培地(DULBECO Minimum Essential Medium)中にて37℃で24時間インキュベートした。細胞培養物の上澄み液(70μl)を、再構成した補因子20μl、および硝酸還元酵素溶液10μlと共に96ウェルプレート内にピペットで入れた。プレートをプレートシェーカー上で、室温にて30分間、150rpmにて振とうした。Griess試薬AおよびB(各50μl)を各ウェルに加え、20分間着色させて、BIO-RADプレートリーダー内で参照波長620nmと共に540nmの吸収を測定した。
【0109】
酸化ストレス(GSH/GSSGレベル)
グルタチオンの還元(GSH)およびグルタチオンの酸化(GSSG)レベルの測定値、ならびにそれらの比は、酸化ストレスの有用な指標である。Biotech GSH/GSSG-412キット(Oxis Research, Portland, OR)を使用して、製造業者のプロトコルにしたがって、比色法によりGSHおよびGSSGのレベルをアッセイした。簡単に言えば、健康なドナーからの正常血液試料を、異なる濃度のRR1と共に、5% CO2インキュベーター内で37℃にて24時間インキュベートした。GSSGに関しては、各処理試料100μlを-70℃で4時間冷凍し、解凍して、5%メタリン酸(MPA)290μlを加えた。試料を15〜20秒間ボルテックスで撹拌し、1000×gで10分間遠心分離した。MPA抽出物(50μl)を比色アッセイで使用したGSSG緩衝液700μlに加えた。GSH測定に関しては、全血50μlを-70℃で冷凍し、解凍して、5% MPA 350μlを加え、15〜20秒間ボルテックスで撹拌し、10,000×gで10分間遠心分離した。MPA抽出物(50μl)をアッセイ緩衝液3mlに加え、更なる分析にて使用した。各標準および試料のMPA緩衝液混合物(200μl)を分光光度計のキュベットに移して、色素原200μlを加えた後、酵素200μlを順に加え、室温で5分間インキュベートした。その後、キュベット内にNADPH 200μlを加え、BECKMAN分光光度計内で412nmの吸収における変化を記録した。吸収値を使用して反応速度をプロットし、GSHおよびGSSGレベルを決定した。
【0110】
統計的分析
データ分析に一要因分散分析(ANOVA)を使用した。
【0111】
細胞株
マウス単球(RAW 264.7)、およびヒト胎児腎臓(HEK293)細胞株を、研究のためにAmerican Type Culture collection, Manassas, VAから得た。TLR2、TLR2/6およびTLR4/CD14/MD2遺伝子をトランスフェクトしたHEK293細胞を、INVIVOGEN Corporation, San Diego, CAより購入した。細胞は、5%CO2インキュベーター内の組織培養フラスコ内で、10%ウシ胎児血清、L-グルタミン(2mM)、および抗生物質(100単位/mlペニシリンおよび100μg/mlストレプトマイシン)を補充したDMEM培地中で増殖させた。Nair, P.K.ら(Nair, P.K. et al. Int. Immunopharmacol., 2004, 4:1645-1659)に記載されている抽出手順にしたがって、GARRY and SUNS(Reno, Nevada, USA)から購入したティノスポラ・コルディフォリア粉末からRR1を単離した。試料の内毒素含有量をLymulus Amoebocyte Lysate Assay(Cambrex, MA)により試験し、有意なレベル(0.0008ng/ml)が示された。研究全体を通して、内毒素を含まない緩衝液、試薬、および無菌水を使用することにより、必要な予防策をとって内毒素汚染を回避した。
【0112】
試薬
ザイモサンA、およびFITC標識ザイモサンA生体粒子を、Molecular Probes, Inc., Eugene, ORより購入した。ラミナリンおよびカフェ酸フェネチルエステル(CAPE)を、SIGMA Chemical Co., St. Louis, MOより購入した。抗マウスCD11bモノクローナル抗体(mAb)を、BD BIOSCIENCES, San Jose, CAより購入した。FITC標識TLR2およびPE標識TLR4抗体を、eBiosciences, San Diego, CAより購入した。
【0113】
RR1の非オプソニン化結合
ザイモサンA生体粒子の非オプソニン化結合に対するRR1の阻害効果を測定するために、マウス単球/マクロファージ細胞(0.5×106)を、0、100、500および1000μg/mlのRR1と共に4℃でインキュベートして、補体を含むオプソニンの局所放出を防止した(Ezekowitz, R. A. et al. J. Clin. Invest., 1985, 76:2368-2376; Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)。氷上で1時間インキュベーション後、細胞を予備冷却した培養液で3回洗浄し、FITCコンジュゲートザイモサンA粒子を25粒子/細胞の比で加えて、氷上でさらに1時間保持した。細胞をPBSで2回洗浄し、CO2インキュベーター内にてDMEM中で37℃にて30分間インキュベートした。未結合ザイモサン粒子を大量の培地で洗浄して除去し、ザイモサン-FITCの細胞内蛍光をCoulter Eliteフローサイトメーターで分析した。
【0114】
オプソニン化結合および食作用
RR1およびラミナリンの、ザイモサンA粒子のオプソニン化結合および食作用に対する効果を測定するために、RAW264.7細胞を、DMEM中で37℃にて1時間、RR1またはラミナリン(0および500μg/ml)で処理した。このことは、オプソニンの放出とオプソニン結合の促進とを確実にするであろう。細胞をPBSで2回洗浄し、25粒子/細胞の比でザイモサンA-FITC生体粒子と共に、37℃で1時間インキュベートした。大量の培地での洗浄により未結合ザイモサン粒子を除去した後、細胞をPBS中に懸濁させ、細胞内蛍光をCoulter Eliteフローサイトメーターで分析した。蛍光強度を対照の百分率で表した。
【0115】
RR1誘導によるTNF-α合成
0.5% FBS(飢餓)を含有するDMEM培地を有する6ウェルプレート上に、RAW264.7細胞(0.5×106/ml)をプレーティングした。ひとたび細胞が付着したら、培地を交換し、0、100、500および1000μg/ml用量のRR1を加え、37℃で24時間インキュベートした。ELISAキット(BD BIOSCIENCES kit, San Jose, CA)を使用して、製造業者のプロトコルにしたがって、分泌されたTNF-αに関して培地を分析した。
【0116】
TNF-α合成に対するNF-κB阻害剤CAPEの効果を測定するために、インキュベーター内で、RAW264.7細胞を10μg/mlのCAPEで37℃にて1時間処理した後、RR1(0〜1000μg/ml)で一夜処理した。培地中の分泌されたTNF-αを、ELIZA(BD BIOSCIENCES kit, San Jose, CA)を使用して分析した。
【0117】
NF-κB活性
RAW264.7細胞を、0、100、500および1000μg/mlのRR1と共に8時間インキュベートし、Active Motif, CAのNF-κB活性キットを使用して、製造業者のELISA説明書にしたがって、核タンパク質を抽出した。NF-κB活性化をさらに視覚化するために、核タンパク質を用いて電気泳動移動度シフトアッセイ(EMSA)を行った(Ghosh, S, and Karin, M., Cell, 2002, 109:S81-96, Young, S.H. et al. J. Biol Chem., 2001, 276:20781-20787; Lebron, F. et al. J Biol Chem., 2003, 278:25001-25008)。NF-κBに結合する二本鎖プローブ
を32P-ATPで放射標識し、核抽出物5μgを用いてEMSAを実施した。EMSA反応混合物を室温で30分間インキュベートし、DNA-タンパク質複合体を6%ポリアクリルアミドゲル上で分離して、オートラジオグラフィーで視覚化した。NF-κBの時間依存的活性化を測定するために、異なる期間にて(刺激0、4、8、14、20時間後)RR1(100μg/ml)処理細胞から核抽出物を調製した。ACTIVE MOTIF, Carlsbad, CAのELISAキットを使用して、核抽出物のタンパク質含有量と、NF-κB活性を分析した。
【0118】
I-κBαの分解
IκB-αの分解は、NF-κB活性化の必須条件である。細胞のI-κBαレベルおよびその分解に対するRR1の効果を測定するために、異なる期間にて(0、15分、30分、1時間)、RR1処理RAW264.7マクロファージ由来の細胞質ゾル溶解物を用いてイムノブロット分析を実施した。溶解物を12%ゲル上でSDS-PAGEにより分離して、TransBlot装置(BIO-RAD, Hercules, CA)を使用してニトロセルロース膜に転写した。ウエスタンブロットハイブリダイゼーションプロトコル(Lebron, F.et al. J Biol Chem., 2003, 278:25001-25008)により、抗マウスIκB-α抗体(CELL SIGNALING TECHNOLOGY, Inc, Beverly, MA)を用いて、色検出システム(BIO-RAD, Hercules, CA)を使用して、膜をIκB-α特異的モノクローナル抗体(1:1000)でハイブリダイズした。
【0119】
オプソニン化結合に関するCR3のCD11b mAbによる遮断効果
RR1のマクロファージに対するオプソニン化認識/結合にグルカン特異的受容体CR3が関与しているか否かを試験するために、CR3に特異的なモノクローナル抗体(mAbs)(BD BIOSCIENCES, San Jose, CAのラット抗マウスCD11b)を使用して、CR3を遮断した。RR1(0〜100μg/ml)と共に3時間インキュベートするのに先だって、RAW264.7細胞(0.5×106)をCO2インキュベーター内で5μg/mlのモノクローナル抗体により37℃で1時間処理した。培地中にマクロファージにより放出されたTNF-αを、ELISAキット(BD Biosciences, San Jose, CA)を用いて分析した(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Brown, G.D. et al. J. Exp. Med., 2003, 197:1119-1124)。
【0120】
TLRシグナル伝達
TLRシグナル伝達の関与を測定するために、親のHEK293、ならびにヒトTLR2、TLR6、TLR2/6またはTLR4/CD14/MD2遺伝子をトランスフェクトした(pDUOプラスミド内にクローン化した)HEK293細胞を使用した。これらの細胞(0.5×106/ml)を、RR1 0〜100μg/mlと共に24時間インキュベートした。これらの細胞は、活性化により非常に低いレベルのTNF-αおよび検出レベルのIL-8を産生するため、培地中に分泌されたIL-8はELISA法により分析した。偶然にも、IL-8合成も転写因子NF-κBの制御下にある。
【0121】
TLR mRNAおよびタンパク質レベル
RR1処理細胞から抽出した全RNAを使用して、RT-PCRによりTLR2、4、および6 mRNAレベルを分析した。全RNAをRR1処理および未処理RAW264.7細胞から抽出し、0.5μg/ml RNAを逆転写して、マウスTLR2、4、および6に特異的なPCRプライマー(R&D SYSTEMS, Minneapolis, MN)を使用して増幅した。対照としてマウスGAPDH遺伝子も増幅した。PCR生成物をアガロースゲル上で分離した。
【0122】
実施例1 RR1の特徴付け
グリコシル組成および結合分析の結果は、最初の発見、およびRR1の炭水化物組成に関する13C NMRスペクトルデータを裏付けるものであった。グリコシル組成の分析は、グルコースがRR1の唯一の成分であることを示したが、一方、結合分析は、結合の3つのタイプに対応するグルコピラノシル残基の3つのタイプを明らかにした:4-結合グルコピラノシル残基(80%)、4,6-結合グルコピラノシル残基(12%)および末端グルコピラノシル残基(8%)。13C NMRスペクトル(図2A)は、グルコピラノシル部分内の炭素原子の良好に分離したシグナルを示した-C1(δ99.97ppm)、C2(δ73.68ppm)、C3(δ71.89ppm)、C4(δ77.09ppm)、C5(δ69.67ppm)、およびC6(δ60.83ppm)。C1およびC4シグナルの低磁場シフトは(1→4)結合を裏付けるが、一方、δ71.53ppmのシグナルは、(1→6)結合のC6に起因し得る。500MHzプロトンNMRスペクトル(図2B)におけるδ5.44ppmのシグナル(良好に分離せず)およびδ5.00ppmのシグナルは、グリコピラノシル単位のアノマー炭素に関連したαプロトンに起因する。他方、δ4.66ppmの非常に弱いシグナルは、βアノマーに起因する可能性がある。しかしながら、α-D-グルコースとβ-D-グルコースのシグナルの比は約99.9:1であり、それ故殆ど全てのグルコース単位がα立体配置を有すると想定される。したがって、RR1は、主鎖内の(1→4)結合したグルコピラノシル単位、(1→6)結合したグルコピラノシル単位の分岐、および0.15の分岐度を有するα-D-グルカンである(図3)。サイズ排除クロマトグラフィーでは、RR1は12.32分において単一ピークとして溶出し、これは511kDaデキストラン試料(保持時間:12.72分)のピークに非常に近かった。したがって、RR1は>550kDaの分子量が割り当てられた。
【0123】
実施例2 非細胞傷害性/細胞増殖効果
図4Aおよび図4Bに示すMTTアッセイの結果は、RR1が1000μg/mlの高濃度においても、正常リンパ球または腫瘍細胞株(CEMおよびCEM/VLB)のいずれに対しても、直接的な細胞傷害性または細胞増殖効果を有さないことを示す。
【0124】
実施例3 リンパ球活性化
リンパ球は、哺乳動物免疫系の重要なエフェクター細胞であり、これらの試験は、異なるリンパ球亜集団が、RR1により多様なレベルで活性化されることを示す。RR1 100μg/mlにより、B細胞は39%、T細胞は102%およびNK細胞は331%活性化される(図5)。NK細胞は、適応免疫系による抗体産生および認識前に、抗原/マイトジェンと接触する自然免疫系の主なエフェクターであるため、NK細胞の>3倍の活性化は非常に意義がある。RR1によるNK細胞の増大された活性化は、図6に示す機能的細胞傷害性アッセイの結果から全く明らかである。RR1処理された正常リンパ球は、未処理細胞と比較して高い割合の腫瘍細胞を殺すことができ、かつ活性化リンパ球の細胞傷害性の用量依存的増強が明らかであった。
【0125】
実施例4 補体活性化経路
図7に示すように、RR1濃度の上昇に伴う、代替的経路のC3a des Argレベルの段階的増大が注目された。この観察結果は、他の多糖による補体活性化に関するいくつかの報告と類似する。C3aおよびC4aは、代替的および古典的経路における補体活性化カスケード中に、血漿成分C3およびC4から放出される生理活性切断生成物であり(Ember, J.A. et al. The Human complement system in Health and Disease, Ed. Volnakis, J.E. and M.M. Frank, Marcel Dekker Inc., New York, 1998, pp. 241-248)、これらはより活性が低いC3a-des ArgおよびC4a-des Arg形態に急速に転換されて、細胞免疫応答の仲介に関与する。代替的経路は自己増幅性であり、抗体不在下での病原の排除および認識に重要である(Stahl, G.M. et al. Am. J. Pathol., 2003, 162:449-455)。β-グルカンは代替的経路を活性化することが報告されており、これらのポリマーが示す宿主仲介性抗腫瘍活性は、補体系の活性化と相関していた(Hamuro, J. et al. Am. J. Pathol., 1978, 93:526-617)。リポ多糖は、代替的および古典的経路を経由して補体系を活性化し;即ち、脂質部分が古典的経路を活性化し、一方、多糖部分が代替的経路を活性化する(Morrison, D. C. and R. J. Ulevitch Am. J. Pathol., 1978, 93:526-617)。
【0126】
実施例5 サイトカインの合成
図8A〜図8Hに示すように、RR1は、IL-1β(1080pg)、IL-6(21833pg)、IL-12p40(918.23pg)、IL-12p70(50.19pg)、IL-18(27.47pg)、IFN-γ(90.16pg)、MCP-1(2307pg)およびTNF-α(2225pg)の合成を誘導したが、IL-2、IL-4、IL-10、TNF-βおよびIFN-αの産生を誘導しなかった。一般に、図8Cに示すように、最大値が10μg/mlにて記録されかつRR1の増加により低下傾向が示されたIL-12(p40)を除いて、RR1によりサイトカイン産生の用量依存的増大が観察された。図8Hに示すように、RR1 10μg/mlまでのMCP-1産生は非常に有意であり、より高いRR1濃度では増大は僅かであった。一般に、プロ炎症性サイトカインIL-1β、IL-6、およびTNF-α、ならびに調節サイトカインIL-12p(40)は、他のサイトカインと比較してより高いレベルで産生された。
【0127】
哺乳動物の免疫系は、抗原、病原および非自己分子を認識して、免疫コンピテント細胞の活性化、化学的メッセンジャー(サイトカインおよびケモカイン)の産生、補体カスケード経路の活性化および一酸化窒素(NO)の合成を介した防御機構を引き起こす。サイトカインは、刺激に応答して免疫成分細胞により分泌される、低分子量の調節性非抗体タンパク質の群である。これらは、標的細胞の特異的受容体に結合し、最終的に標的細胞内の遺伝子発現に繋がるシグナル伝達経路を引き起こす。サイトカインは、様々な細胞の活性化、増殖、および/または分化を刺激または抑制することによって、かつ抗体または他のサイトカインの分泌を調節することによって、免疫応答の強度および持続時間を調節する。サイトカインが示す多面発現性、重複性、相乗作用および拮抗作用により、サイトカインは細胞活性を調節および制御することができる。炎症応答は、病原体の制御および排除、ならびに組織の完全性を回復する創傷治癒の促進に重要である。
【0128】
IL-1βおよび/またはTNF-αは、互いの産生を誘導すると共にIL-6の産生を誘導し、相乗的に作用して数個の生物学的作用を調節し、その上IL-1は自己誘導性である(Horai, R. et al. J. Exp. Med., 2000, 191:313-320)。RR1処理リンパ球内でのIL-1βおよびTNF-αの産生は、LPS刺激ヒト単球細胞株(THP-1)内でのそれと一致する(Baqui, A.A. et al. Clin. Diagn. Lab. Immunol., 1998, 5:341-347)。IL-12(p70)は、IL-12の生理活性を有するイソ型であり、ナイーブT細胞が、IFN-γを分泌するエフェクターTヘルパータイプ1(Th1)CD4+リンパ球に分化する際の重要な因子である(Trinchieri, G. et al. Prog. Growth Factor Res., 1992, 4:355-368)。加えて、NK細胞上に刺激効果を有することも報告されている(Wajchman, H.J. et al. Cancer Res., 2004, 64:1171-1180)。最近、IL-12(p70)は、そのTh1応答を引き出す能力により、有効で毒性の少ない抗腫瘍サイトカインとして浮上している(Hiscox, S. and Jiang, W.G. In Vivo, 1997, 11:125-132)。免疫系を越えて、このサイトカインは、UV誘導アポトーシスを抑制する能力、および究極的には細胞を悪性腫瘍から保護するUV損傷ケラチノサイト内のDNA修復を開始させる能力を有することも報告されている(Schwarz, A. et al. Nat. Cell Biol., 2002, 4:26-31)。ホモ二量体イソ型であるIL-12(p40)は、マウスにおいて生理活性ヘテロ二量体イソ型p70の受容体拮抗剤である(Gillessen, S. et al. Eur. J. Immunol., 1995, 25:200-206)。RR1処理細胞内では、調節サイトカインであるIL-12(p40)は、生理活性型の何倍かの過剰量で産生され、これは生理活性型p70の過剰産生を制御するための天然の機構であろう。IL-18は、IFN-γ、および明らかにNK細胞のもう一つの強力な誘導物質である(Micallef, M.J. et al. Eur. J. Immunol., 1996, 26:1647-1651)。IL-18産生のRR1誘導は、IL-12およびIFN-γと調和して作用する、Th1応答の発達における早期の応答であろう。
【0129】
MCP-1は、単球、および活性化CD4およびCD8 T細胞の強力な化学誘引物質であり、NKおよびCD8+細胞からの顆粒放出を誘導し、CD56+細胞内のNK機能を活性化し、かつ好塩基球からのヒスタミンの強力な放出因子として作用することが報告されている(Rollins, B.J. Blood, 1997, 90:909-928)。さらに、MCP-1は、おそらくT細胞依存的に、腫瘍特異的免疫を高めることにより、抗腫瘍効果を示すことが報告されている(Laning, J. et al., J. Immunol, 1994, 153:4625-4635)。本発明者らの観察によれば、RR1は、10μg/mlにおいて、このサイトカインの産生を有意に誘導する。RR1刺激によるTNF-αの有意な合成は、感染および悪性腫瘍の増殖に対する宿主抵抗力に重要な役割を果たしているだろう。TNF-αおよびその受容体は、結核に対する保護、および感染の早期におけるマクロファージ内でのNO合成に不可欠である(Lipton, S.A. Neurochem. Int., 1996, 29:111-114)。RR1誘導によるIFN-γ産生は、一部には、例えば細胞傷害性Tリンパ球(CTL)、マクロファージおよびNK細胞のような細胞仲介免疫システムの構成要素の活性を促進するよう機能し、またTh2応答において抑制的役割を果たすだろう。これは食細胞の殺菌活性を刺激し、したがって自然免疫応答を向上させる(Watford, W.T. et al. Cytokine Growth Factor Rev., 2003, 14:361-368)。さらに、これはLPS内と同様にマクロファージ内でMCP-1合成を調節し得る(Munder, M. et al. J. Exp. Med, 1998, 187:2103-2108)。
【0130】
CD4+T細胞は、抗原の認識と、その結果としてのサイトカイン産生を介した抗原特異的(適応)免疫系の調節に寄与する。CD4+細胞によるサイトカイン産生の異なるパターンは、IFN-γ産生により特徴付けられかつ細胞内病原の排除を促進するタイプ1(Th1)と、IL-4産生により特徴付けられかつ細胞外病原の排除に適切なIgEおよび好酸球に関与するタイプ2(Th2)とに対生を形成する。サイトカインは、最初の活性中にT細胞上に直接作用し、最終分化に影響する因子の中で最も直接的なメディエーターであると思われる。IL-12の存在はTh1応答を導き、一方、IL-4はTh2発達を導き、これら二つの経路は互いに抑制する作用も発現する。IL-1はIL-12の誘導物質として同定されており(Rollins, B.J. Blood, 1997, 90:909-928)、一方、IL-18は、IFN-γ誘導によるTh1細胞の発達における早期の応答である。RR1刺激によるサイトカインプロファイル、IL-12、IL-18、IFN-γおよびIL-1、ならびにこれらサイトカインの用量依存的合成は、細胞免疫ならびに細胞内病原および悪性腫瘍細胞の殺傷に不可欠なTh1経路を明白に示している。この観察は、IL-12誘導による、IFN-γ依存的なT細胞のTh1およびCD8+細胞傷害性エフェクター細胞への発達と一致する(Russo, D.M. et al. Expt. Parasitol., 1999, 93:161-170; Yoshimoto, T. et al. J. Immunol., 1998, 161:3400-3407; Russo, D.M. et al. Infect. Immun., 2000, 68:6826-6832)。IL-12およびIL-18の同時発生的なシグナル伝達ならびに相乗作用は、T細胞内で、延長したIFN-γ産生と、連続するIL-18R mRNAの強力な発現とを誘導する(Yoshimoto, T. et al. J. Immunol., 1998, 161:3400-3407)。IL-12およびIL-18に関しては、T細胞の炎症部位に対する接着など、早期の炎症性事象も報告された(Ariel, A. et al. J. Leuk. Biol., 2002, 72:192-198)。調節性イソ型IL-12(p40)の高いレベルの合成は、Th1応答の過剰な生成を含有する天然の機構であろう。
【0131】
NOレベルもまた、Th細胞分化および極性化の調節に重要な役割を果たすことが報告されている。Yamasakiら(Yamasaki, A. et al. Yonago Acta Medica, 1998, 44:7-15)およびTaylor-Robinsonら(Taylor-Robinson, A.W. et al. Eur. J. Immunol., 1994, 24:980-984)は、Th1サイトカインに対するNOの抑制的役割を報告したが、一方、Bauerら(Bauer, H. et al., Immunol., 1997, 90:205-211)は、活性化T細胞からのNOによるTh1およびTh2サイトカインに対する同一の役割を報告した。本研究において、RR1処理は、NOの増加を<21%だけ誘導した(図9)。酵素iNOSは、刺激によるマクロファージからのNOの高産生量を生じた。本研究は、RR1がiNOSの誘導を有意に刺激しないことを明らかにした。RR1によるiNOS誘導の不在は、NO産生の有意な増加が存在しないという結論を支持している。Monocyte IsolationキットII(Miltenyi Biotec, Germany)による末梢血単核細胞から単離された単球を用いた、またはヒト単球細胞株THP-1(ATCC)を用いた本研究でも、RR1処理によってより高いレベルのNOは全く誘導されなかった。RR1による低いレベルのNO合成は、Th1経路を全く阻害しないと思われる。IFN-α、IFN-γ、およびTNF-αは、NO合成の公知の上方調節因子である(Shin, J.Y. et al. Immunopharmcol. Immunotoxicol., 2002, 24:469-482; Hirazumi, A. and Furusawa, E. Phytother Res., 1999, 13:380-387)。RR1刺激により、IFN-γおよびTNF-α、ならびに非常に少量のIFN-αが合成されたために、多糖によるより高いレベルのNO産生を引き起こすためには、IFN-αプライミングが必要ステップであろうと想定することが正当である(Ring, A. et al. J. Infect. Dis., 2002, 186:1518-1521)。
【0132】
RR1刺激は、GSH、GSSGのレベル、およびそれらの比に示されるように(表1)、リンパ球内で酸化ストレスを全く引き起こさず、これはNO合成データと一致する。過酸化水素量の有意な増加も全く観察されなかった(データは示さず)。NO合成の低いレベルは、NOに対する免疫調節的役割を示唆する。RR1の非細胞傷害性の性質は、かなり高濃度(1000μg/ml)においても、この多糖が細胞内で酸化ストレスを誘導しない能力とともに低いレベルのNO産生に寄与する可能性がある。
【0133】
(表1)RR1で処理した正常リンパ球のGSH、GSSG、およびGSH/GSSG比
【0134】
サイトカインの誘導およびNOの合成による免疫刺激、マクロファージの活性化、食作用、細胞傷害性および抗腫瘍活性の誘導が、最近、オタネニンジン(Phanax ginseng)、ヤエヤマアオキ(Morinda citrifolia)およびエキナセア(Echinacea)の多糖または多糖含有画分内に報告されている(Shin, J.Y. et al. Immunopharmcol. Immunotoxicol., 2002, 24:469-482; Hirazumi, A. and Furusawa, E. Phytother Res., 1999, 13:380-387; Goel, V. et al. J. Nutri. Biochem, 2002, 13:487-492)。RR1は、アミロペクチンが免疫刺激を全く誘導できないことから、アミロペクチンとは構造的に区別されるα-D-グルカンである。RR1構造の、真菌β-グルカンの細胞壁成分の「保存された分子形態」との類似性が、免疫系の活性化の理由であり、一方、相違(即ち、αコンホメーション、および(1→4)結合)が、低いレベルのNO産生、およびその結果としての非細胞傷害性の原因である可能性がある。この水溶性を有する中性α-グルカンは、有意な免疫刺激性を示す多糖の範囲内に十分含まれる分子量および分岐の連続を有している(Bohn, J.A. and N. BeMiller Carbohydr. Polymers, 1995, 28:3-14; Brown, G.D. and S. Gordon Immunity, 2003:19:311-315; Kulicke, W.M. et al. Carbohydr. Res., 1997, 295:135-143; Bao, X. et al Carbohyd. Res., 2001, 336:127-140)。水溶性は粒子β-グルカンが示す肉芽腫形成を克服でき、一方、高い分子量は、最も強力な範囲内にあるので分解することなく宿主の器官により長期間維持されることを可能にする(Williams, D.L. et al. Clin. Immunotherapy, 1996, 5:392-396; Nono, I. et al. Pharmacobiodyn., 1991, 14:9-19; Suda, M. et al. FEMS Immunol. Med. Microbiol, 1996, 15:93-100)。
【0135】
非細胞傷害性の性質、リンパ球、特にNK細胞の有意な活性化および補体活性化の代替的経路は、自然免疫系の刺激を明らかに示し、一方、活性化の結果としてのサイトカインプロファイルは、その過剰産生に対する自己調節機構と共に抗原特異的な細胞仲介(適応)免疫性(Th1経路)の刺激を明白に示す。RR1刺激によるIL-4合成(Th1抑制因子サイトカイン)およびIL-10(IL-12の阻害剤)の不在は、T細胞分化のTh1経路と十分一致している。IL-12およびIL-18の同時発生シグナル伝達によるIFN-γ産生は、強力な抗炭疽菌剤として働き、外因性のIFN処置の副作用も、もし存在したとしても、回避するであろう。刺激を受けた自然免疫系は、宿主内への病原のいかなる侵入とも戦うことができ、一次感染が実際の疾病を引き起こすことを防止する能力を有する(Parham, P. Nature, 2003, 423:20)。ここまで説明したように、RR1の独特な免疫刺激特性は、酸化ストレスおよび直接的な細胞傷害性効果を全く与えることなく、RR1を多数の病原およびヒト悪性腫瘍に対する強力な生体防御剤とし得る。
【0136】
実施例6 ザイモサンA-FITC生体粒子の非オプソニン化結合に対するRR1の効果
図10に、非オプソニン化認識およびFITC標識ザイモサンA生体粒子のマクロファージへの結合に対する、RR1の阻害効果を示す。RR1の濃度の増加に伴い蛍光強度の用量依存的阻害が観察され、1000μg/mlのRR1濃度において約65%の阻害を示す。実験中、細胞を4℃でインキュベートするとオプソニンの放出が防止され、それにより非オプソニン化結合が促進されるであろう。
【0137】
実施例7 オプソニン化結合および食作用に対するRR1の効果
ザイモサンA生体粒子の食作用に対するRR1およびラミナリンの37℃での阻害効果を、図11に示す。構造的に定義されたβ-グルカンであるラミナリンは、食作用に対して完全な阻害作用を有するが、RR1は有意な効果を全く示さなかった。食作用は、マクロファージによる粒子の結合および内在化を含む。
【0138】
実施例8 RR1効果に対するCR3の関与
CR3は、マクロファージ活性化および炎症応答に繋がる多糖の単球に対するオプソニン化結合に関与する、β-グルカン特異的受容体である。単球を5μg/mlのCR3(CD11b)特異的モノクローナル抗体と共にインキュベートした後、RR1でインキュベートした結果、RAW264.7細胞のTNF-α合成は阻害または低下されなかった。しかしながら、RAW264.7マクロファージをCD11b抗体で処理した後、可溶性ザイモサンで処理した場合、TNF-α合成の有意な低下(41.31%)が観察された(表2)。
【0139】
(表2)グルカンのオプソニン化結合の抗CD11b(CR3特異的)阻害後の、RAW264.7マクロファージ内のTNF-α合成
細胞をCR3(5μg/ml)と共に2時間インキュベートした後、37℃で1時間グルカン処理した。
**p<0.05
【0140】
実施例9 TNF-αの合成
図12に、NF-κB阻害剤、カフェ酸フェネチルエステル(CAPE)を伴うおよび伴わないRR1誘導によるTNF-α合成を示す。RR1用量の漸増に伴うTNF-α合成の用量依存的増大が明らかであった。単球のRR1処理に先立って、CAPEを10μg/mlで1時間インキュベートした場合、RR1の全濃度にてTNF-α合成は完全に阻害された。
【0141】
NF-κB活性化の動態を測定するために、ELISAプロトコルを使用して、RR1処理マクロファージ内のNF-κB活性を様々な時間間隔において分析した(図13)。刺激時間に伴なってNF-κB活性が増大し、8時間でピークとなり、その後活性は減少した。これらの結果に基づいて、8時間の刺激による更なるNF-κB活性化試験の全てを行い、RR1濃度の増加に伴うNF-κB活性の用量依存的増大を観察し、RR1用量500〜1000μg/mlの間でプラトーに達した(図14)。EMSAによるNF-κBのヘテロ二量体バンドにおけるシフトの視覚化は、上方のバンド(p65ヘテロ二量体バンド)の強度の濃度依存的な増大から明らかであるように、ELISA実験による定量を実証する(図15)。
【0142】
実施例10 IκB-αの分解
IκB-αのリン酸化および分解は、NF-κB活性化の必須条件であり、これがNF-κBの細胞質から核への移行を促進する。IκB-αウエスタンブロットは、RR1処理後の刺激の様々な間隔におけるIκB-αレベルを示す。IκB-αの着実な減少がRR1インキュベーションの30分にて明らかであり、その発現は100μg/mlのRR1処理の60分後に完全に除去された(図16)。
【0143】
実施例11 TLR mRNAおよびタンパク質
TLR2、4、および6(図17)遺伝子のmRNAレベルは、RR1濃度0〜1000μg/mlにおいて同様のままであった。したがって、TLRの転写は、RR1処理により変化しないものと思われた。
【0144】
RR1シグナル伝達におけるTLRの役割
RR1シグナル伝達に対する異なるTLRがなす役割を試験するために、親HEK293細胞を、受容体トランスフェクタント(TLR2、TLR4/CD14/MD2、TLR6/またはTLR2/TLR6)およびRR1と共にインキュベートし、24時間後にIL-8が産生した。RR1は、親HEK、およびTLR2またはTLR4/CD14/MD2遺伝子をトランスフェクトしたHEK細胞内でIL-8産生を誘導しなかった。しかしながら、TLR6およびTLR2/6遺伝子をトランスフェクトしたHEK細胞は、大量のIL-8を誘導し、その合成において用量依存的誘導が明らかであった(図18および図19)。TLR2をトランスフェクトした細胞は、RR1処理(0〜100μM)によりIL-8を全く生産しなかったことも興味深い。
【0145】
ティノスポラ・コルディフォリアは、アーユルヴェーダ(Ayurvedic)(インド)系医学にて広く使用されている免疫刺激性の草本である(Chopra, R.N., I.C. Chopra, K.D. Handa, L.D Kanpur (Editors), 1982, Glossary of Indian Medicinal Plants; Council of Scientific an Industrial Research, New Delhi, Dhar VN & Sons, Kolkata, India)。本発明者らは、免疫刺激特性を有することが見出されたこの薬草から、新規な1,4-α-D-グルカン(RR1)を単離および特徴付けしたことを説明した。この水溶性の多糖は、主鎖に(1,4)-α-D-グリコシド結合を有し、(1,6)-α-D-グリコシド結合した側鎖を6、7グルコース単位の区間に有する。免疫系刺激の更なる機序を理解するために、本研究を行った。本明細書に記載する結果は、RR1が、オプソニン化ザイモサンA生体粒子の結合および内在化(食作用)に対して阻害効果を殆ど有さないことを示す。これは、ザイモサン生体粒子の内在化を約90%阻害した、水溶性の、真菌由来のβ-グルカンであるラミナリンの効果と対照的であった(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Czop, J.K. and Austen, K.F. J. Immunol., 1985, 134:2588-2593; Goldman, R. Exp. Cell Res., 1988, 174:481-490)。
【0146】
補体受容体3(CR3)は、β2-インテグリンファミリーに属するヘテロ二量体膜貫通糖タンパク質であり、CD18に非共有結合的に結合したCD11bからなる。CR3のCD11bサブユニットの細胞外部分に、明確な機能的ドメインが確認されている(Diamond, M.S. et al. J. Cell Biol., 1993, 120:1031-1043; Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246; Lu, C. et al. J. Biol. Chem., 1998, 273:15138-15147; Dana, N. et al. J. Immunol., 1986, 137:3259-3263)。β-グルカンシグナル伝達において、CR3は、粒子(Ross, G.D. et al. Complement, 1987, 4:61-74)および可溶性グルカン(Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246)のための白血球受容体として働く。この受容体は、受容体をプライミングするよう作用するCOOH末端レクチン部位を介して、グルカン受容体として機能する。C3bi被覆粒子の結合および食作用に、CR3のIまたはAドメインが必須であり(Diamond, M.S. et al. J. Cell Biol., 1993, 120:1031-1043; Dana, N. et al. J. Immunol., 1986, 137:3259-3263)、IドメインのC末端に位置するレクチンドメイン(Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246)は、CR3の非オプソニン化結合特性に関与することが報告されている(Balsam, L.B. et al. J. Immunol., 1998, 160:5058-5065; Ross, G.D. et al. J. Immunol., 1985, 134:3307-3315)。多糖のCR3をプライミングする能力は、レクチン部位の近傍に結合する抗体により模倣され得る(Thornton, B.P. et al. J. Immunol., 1996, 156:1235-1246)。Cabecら(Cabec, V.L. et al. Infect. Immunity, 2000, 68:4736-4745)は、非オプソニン条件下で、ザイモサンの食作用が、受容体の区別された分子決定因子に関連すること、かつ非オプソニン化食作用はC末端レクチン部位と無関係に起きることを示した。Brownら(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)は、CR3特異的mAb 5C6を用いた、オプソニン化ザイモサンのマクロファージに対する結合の阻害を示し、阻害のレベルは、以前の報告と同様であった(Xia, Y. et al. J. Immunol., 1999, 162:2281-2290)。しかしながら、5C6 mAbによるCR3の阻害は、ザイモサン粒子の非オプソニン化結合を阻害しなかった。抗CD11bモノクローナル抗体を使用したRAW264.7細胞によるCR3阻害実験では、オプソニン化条件下(37℃)で、ザイモサン誘導TNF-α合成の41%の低下が示された。しかしながら、抗体のインキュベーションは、オプソニンおよび非オプソニン条件下で、RAW264.7細胞のRR1誘導TNF-α合成を阻害しなかった。したがって、マクロファージ上のCR3受容体は、サイトカイン合成に繋がるRR1の認識およびプライミングに関与しないであろう。
【0147】
ザイモサンは酵母由来の粒子であり、主としてβ-グルカンおよびマンナンが主成分の多糖からなる。ザイモサンは、受容体に特異的な選択的遮断剤を利用した、マクロファージ上の受容体結合研究のモデルとして使用されている。マウスマクロファージに対するザイモサンの非オプソニン化結合にβ-グルカン受容体、CR3およびマンノース受容体が関与しないことは、各々、ジメチルグリコシドおよびマンナンに対する阻害効果が全く欠乏していたことから既に証明された(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)。一方、構造的に定義された、水溶性のβ-グルカンであるラミナリンは、マクロファージに対する非オプソニン化結合を非常に強力に阻害し、β-グルカン受容体の関与を証明した(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Ross, G.D. et al. Complement, 1987, 4:61-74)。RR1の、非オプソニン化ザイモサン-FITC粒子の蛍光に対する強力かつ濃度依存的な阻害効果は、TNF-α産生と矛盾しない非オプソニン化認識および結合におけるザイモサンおよびRR1内の同一タイプの受容体の関与を示し、そのリン酸化グルカンによる遮断は、マクロファージ活性化を阻害することができる(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117)。オプソニン化結合およびその結果のTNF-α産生に対するラミナリンの強力な阻害効果は、ザイモサン粒子およびラミナリンの双方が同様の分子形態およびコンホメーション(β-グリコシド結合による直鎖状グルコースポリマー)を有するために、これらの結合および内在化に関して同一タイプの受容体が関与することを原因とし得る。また一方、RR1は、異なる分子コンホメーションを有する(α-グリコシド結合による螺旋状)α-グルカンであり、したがって結合/内在化に関しては異なるタイプの受容体が関与してもよい。
【0148】
Brownら(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Brown, G.D. et al. J. Exp. Med., 2003, 197:1119-1124; Brown, G.D. and Gordon, S. Nature, 2001, 413:36-37)は、第一マクロファージによるザイモサンの非オプソニン化認識を独占的に担うデクチン-1受容体を記載した。この著者らは、デクチン-1受容体に特異的なデクチン-1 mAb 2A11が、外因性β-グルカン、リン酸化グルカンまたはラミナリンにより得られる阻害に匹敵するレベルで、非オプソニン化ザイモサンの結合を阻害することができ、mAbはβ-グルカン結合部位にまたはその近傍に結合することを示した。また、Adachiら(Adachi, Y. et al. Infect Immun., 2004, 72:4159-71)は、最近、マウスデクチン-1 cDNAをトランスフェクトしたHEK293細胞が、ゲル化β-グルカン、シゾフィランに結合することができ、この結合は、他のβ-グルカンによる前処理により阻害し得るが、α-グルカンでは阻害されないことを示し、デクチン-1のβ-グルカンに対する特異性を示した。
【0149】
RR1のプライミングを担うさらなる受容体を同定するために、様々なTLRをトランスフェクトしたHEK293細胞内でのRR1誘導サイトカイン合成を研究した。HEK293細胞は、多糖刺激下でTNF-αを産生せず;したがって他のサイトカインを分析した。これらの細胞は、刺激によりIL-8を産生し、またIL-8産生はNF-κB活性化の制御下にあるという事実に基づき、HEK293トランスフェクタント内のIL-8を分析した(Wang, Q. et al. Infect Immun., 2001, 69:2270-2276; Torok, A.M. et al. Infect Immun., 2005, 73:1523-1531)。TLRは、タイプIの膜貫通タンパク質であり、今日までヒトにおけるTLRの12メンバーが記述されている。HEK293細胞は、これら受容体を欠いており、TLR1、TLR2、およびTLR6は全て、ファゴソーム含有ザイモサンA粒子に補充されることが報告されている(Underhill, D.M. et al. Nature, 1999, 401:811-815; Ozinsky, A. et al. Proc. Natl. Acad. Sci. USA., 2000, 97:13766-13771)。TLR2およびTLR6の双方が、ザイモサンA粒子によるNF-κBの活性化、およびTNF-αなどの炎症性サイトカインの産生に必要である。本研究では、RR1は、TLR6およびTLR2/6トランスフェクトHEK293細胞内でIL-8産生を誘導したが、他のトランスフェクタント内では誘導しなかった。IL-8合成の用量依存的増加も観察された。RR1処理は、TLR2またはTLR4遺伝子トランスフェクトHEK293細胞内でIL-8産生を全く誘導しなかった。
【0150】
自然免疫系は、TLRおよび他の受容体を介して多様な微生物生成物および非自己分子を認識しかつ応答し、高度に特異的な応答を引き出す(Netea, M.G. et al. Trends Microbiol., 2004, 12:484-488)。少数の受容体のリガンド特異性は、ホモ/ヘテロ二量体化などの受容体間の協力、ならびにCD14、MD2およびデクチン-1などの非TLRとの共同による攻撃の特異性の達成を提示する(Ganter, B.N. et al. J. Expt. Med., 2003, 197:1107-1117; Wright, S.D. et al. Science, 1990, 252:1321-1322; Medzhitov, R. Nature Reviews Immunol., 2001, 1:135-145; Brown, G.D. and Gordon, S. Cell. Microbiol., 2005, 7:471-479; Shimazu, R. et al. J Exp Med., 1999, 189:1777-1782)。Underhillら(Underhill, D.M. et al. Nature, 1999, 401:811-815)は、粒子の内在化は、応答がTLRにより仲介されるため、ザイモサン誘導マクロファージ活性化における炎症応答には必要ないことを証明した。
【0151】
β-グルカンの研究は、免疫刺激に繋がるサイトカインおよびケモカイン合成に、マクロファージ内のNF-κB活性化が重要なステップの一つであることを示している(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412; Brown, G.D. et al. J. Exp. Med., 2003, 197:1119-1124)。非活性化NF-κBは、細胞質内に位置し、阻害性タンパク質IκB-αに結合している。IκB-αは、炎症性刺激に応答してリン酸化および分解して、NF-κBの活性化を誘導する。活性化NKκBは細胞質から核へ移行して、そこで標的遺伝子のプロモーター領域に結合し、それらの転写を調節する。標的遺伝子がNF-κBにより作動されると、mRNA合成が起こり、次いでタンパク質が発現される。本研究にて実施したNF-κB活性化の試験は、NF-κB活性化が、RR1の免疫系刺激特性の根底にある機構であることを確認した。
【0152】
RelおよびNFκBタンパク質は、癌、関節炎、慢性炎症、喘息、神経変成および心疾患などの多数の疾病状態の持続的活性化に加えて、免疫および炎症応答、ならびにアポトーシスを含む大多数の細胞および組織プロセスの制御に関与する、構造的に関連する真核生物転写因子のファミリーを含む(Beyaert, R. (ed.), 2004,「Nuclear Factor-κB: Regulation and Role in Disease」Kluwer Academy Publishers Dordrecht, The Netherlands; Ghosh, S. et al. Annu Rev Immunol, 1998, 16:225-260)。これらタンパク質は、Relホモロジーと称される、高度に保存されたDNA結合/二量体化ドメインにより関連付けられる。NF-κBは、p50-RelA(p65)ヘテロ二量体を指し、ほとんどの細胞内で主要なRel/NF-κB複合体である。Rel-NF-κB転写因子は、二量体の状態で、κBと称される9〜10塩基対のDNA部位に結合する。個々の二量体は、関連するκB部位の収集のために、明確なDNA結合特異性を有する。マクロファージのCAPEとの、および続くRR1とのプレインキュベーションによるTNF-α合成の完全な阻害は、RR1の細胞シグナル伝達経路にNF-κBが関与していることを明白に示している。CAPEは、p65サブユニットの核への移行を防止することによってNF-κBを阻害し、該阻害はp65ヘテロ二量体移行に特異的であることを示している(Natarajan, K. et al. Proc. Natl. Acad. Sci. USA, 1999, 199693:9090-9095)。ELISA測定における濃度依存的なNF-κBの発現およびEMSAゲル中のNF-κBヘテロ二量体バンドは、サイトカイン産生の転写調節にNF-κBが積極関与していることをさらに確認する。NF-κB発現の発現前のIκB-αの分解は、シグナル伝達経路においてNF-κB活性に不可欠な条件であり、RR1刺激の最初の60分以内の細胞質ゾルIκB-αの分解は、NF-κB経路を正当化する。時間経過実験は、IκB-α分解後にNF-κBの活性化が開始し、刺激の8時間目にピークに到達した後、減少することを示した。NF-κB活性化の一過性の性質は、次にNF-κBの連続的かつ持続的な活性化、およびその結果の炎症性サイトカインの過剰産出を防止する時間経過実験から明らかである。
【0153】
NF-κBにより活性化される標的遺伝子の一つは、IκB-αをコードする遺伝子であり得る。新たに合成されたIκB-αは核に入り、DNAからNF-κBを除去して、複合体を細胞質に輸送し戻し、元の潜在的状態を回復し、それによって連続的な活性化を防止する。8時間後の核抽出物中のNFκB活性化の減少は、この逆輸送に起因する可能性があり、したがって、NF-κB活性化は、数時間のみ継続する一過性プロセスとして維持される。
【0154】
ニューモシスチス・カリニ(Pneumocystis carinii)由来の細胞壁β-グルカンは、p65 NF-κBヘテロ二量体の核内への移行の誘導を介してマウスのマクロファージを活性化することが報告されている。このプロセスは時間依存的に起こり、早くも刺激の1時間目から検出可能であり、2〜4時間目にピークに到達し、連続的刺激の6時間目以降に減少を開始した(Lebron, F. et al. J Biol Chem., 2003, 278:25001-25008)。LPSを用いた同様の試験では、p65 NF-κBは10〜30分間で急速に移行し、先の報告と一致した(O'Connell, M.A. et al. J. Biol. Chem., 1998, 273:30410-30414)。しかしながら、IκB-αの分解は、4時間で完了した刺激のうちの2時間後に初めて認められた。ザイモサン刺激マクロファージ内では、Youngらが、刺激の2時間目のNF-κB活性化を報告し、該活性化は8時間目にピークに到達した後、減少したことを報告した(Brown, G.D. et al. J. Expt. Med., 2002, 196:407-412)。本発明者らは、RR1刺激についてNF-κB移行の動態にほとんど同様の傾向を観察し、これは4時間目に検出され、8時間目にピークに到達し、より長時間の刺激により減少した。細胞質ゾル抽出物中のIκB-α分解は、早くも刺激の30分目に観察され、1時間後に完了した。LPSまたはニューモシスチス・カリニのグルカンと比較した、NF-κBの核内への移行の程度および時間の差異は、RR1およびβ-グルカンの防御特性に寄与する要素の一つであろう。LPSと比較した、IκB-α分解のより遅い動態およびより長い過渡期を伴うNF-κBのさらに遅い核移行は、RR1およびβ-グルカンの宿主防御特性に寄与するだろう。さらに、ヒト白血球内のRR1刺激は、高いレベルの調節サイトカイン、IL-12p(40)、ならびに抗炎症性サイトカインIL-10およびMCP-1も産出する(Nair, P.K. et al. Int. Immunopharmacol., 2004, 4:1645-1659)。すなわち、これらの結果は、RR1が、TLR6シグナル伝達およびNF-κB活性化機構を介してマクロファージを活性化することにより免疫システムを刺激して、免疫タンパク質の産生を誘導することを示した。突然のIκB-α分解を伴うNF-κBの潜在的なおよび延長した活性化は、LPSおよび真菌細胞壁グルカンなどの他の活性化分子から、その活性化を区別できるであろう。
【0155】
本明細書に言及または引用した全特許、特許出願、仮出願、および刊行物は、それらが本明細書の明白な開示と矛盾しない限り、その図面および表を含む全体が参照により本明細書に組み入れられる。
【0156】
本明細書に記載した実施例および態様は、例示のみを目的とし、当業者にはそれらに照らした様々な修正または変更が示唆されるであろう。それらは、本明細書の趣旨および範囲内に含まれるものとする。
【図面の簡単な説明】
【0157】
【図1】RR1の単離を表すフローチャートを示す。
【図2】図2A〜2Bは、(a)(1-4)および(1-6)グリコシド結合を示すD2Oにおける13CNMRスペクトル、(b)α-コンホメーションを示すD2Oにおける500 MHzプロトンNMRスペクトルを示す。
【図3】RR1((1-4)-α-D-グルカン)の化学構造を示す。
【図4】図4A〜4Bは、正常リンパ球(a)および腫瘍細胞株(b)でのRR1の細胞傷害性分析を示す。細胞(106)をRR1で24時間処理してから、MTT細胞増殖キット(Roche Biochemicals)を用いて細胞傷害性アッセイを行った。対照に対する生存細胞のパーセンテージを、RR1濃度に対しプロットした。
【図5】RR1によるNK、T、およびB細胞の活性化のフローサイトメトリー分析を示す。正常リンパ球を、CO2インキュベーター中、37℃において、RR1(0〜100μg/ml)で24時間処理した。細胞を、各種蛍光色素とコンジュゲートした細胞特異的抗体のパネルで染色した:CD3-FITC、CD16/56-PE、CD19-ECD、CD69-PC5またはCD8-FITC、CD4-PE、CD3-ECD、CD69-ECD。染色した細胞は、Coulter Eliteフローサイトメトリーで、4色アッセイで分析した。***p<0.001。
【図6】RR1活性化ヒトリンパ球の細胞傷害性についてのフローサイトメトリーアッセイを示す。正常リンパ球をRR1(0〜100μg/ml)で24時間、RPMI培地中で処理した。細胞を洗浄し、PKH-26標識ヒト白血病CEM細胞とともに12時間、共インキュベートし、NK細胞を活性化させて、腫瘍細胞を溶解した。細胞混合液を、FITC標識し活性化したカスパーゼ3モノクローナル抗体で染色した。PKH26+およびカスパーゼ3+細胞のパーセンテージを、Beckman-Coulter Eliteフローサイトメーターで分析し、定量化した(**p=0.01)。
【図7】RR1が誘発した補体経路活性化の分析を示す。健康な志願者から得た正常血液試料を、RR1(0〜100μg)で24時間インキュベートした。ヒト補体C3a des ArgおよびC4a des Arg相関EIAキット(Assay Design Inc., Ann Arbor, MI)を用いて、ELISA法を用いた補体活性化の古典的経路および第2経路の定量化を行った。***p<0.001。
【図8】図8A〜8Hは、RR1誘発正常リンパ球におけるサイトカインおよびケモカインの合成を示す。健康な志願者から得た正常血液試料を、CO2インキュベーター内、37℃にて、RR1(0〜100μg)でRPMI培地中24時間処理した。上清培地を、試薬キット(BD Biosciences)を利用して、ELISA法でサイトカインおよびケモカイン産生について分析した。RR1は、IL-1β、IL-6、IL12p40、IL-12p70、およびIL-18、TNF-α、IFN-γ、およびMCP-1の合成を有意に誘発した(***p<0.001)。
【図9】正常リンパ球でのRR1誘発NO合成を示す。健康な志願者から得た正常リンパ球を、CO2インキュベーター内、37℃にて、RR1(0〜100μg/ml)でMEM培地中24時間処理した。上清培地を、Active Motif, Carlsbad, CA社製のキットを用いてNO産生について分析し、データはANOVAを用いて分析した(p=0.109)。
【図10】RAW264.7マクロファージに対するFITC標識ザイモサンA生体粒子の非オプソニン認識/結合へのRR1の阻害作用を示す。RR1は、用量依存的にザイモサンA生体粒子の非オプソニン結合の結合および内在化を阻害する。細胞を0.5%FBSを含有するDMEM中、1時間、氷上(4℃)でRR1とインキュベートした後、ザイモサンA生体粒子と氷上で1時間インキュベートした。細胞を37℃で7分間温め、ザイモサンA生体粒子を内在化させ、Coulter Eliteフローサイトメーターで細胞内FITC蛍光を分析した。
【図11】RAW264.7マクロファージ内への、FITC標識ザイモサンA生体粒子のオプソニン化結合および内在化(食作用)に対するRR1およびラミナリンの阻害作用を示す。単球をRR1で37℃にて1時間インキュベートした後、FITC標識ザイモサンA生体粒子とさらに1時間、37℃でインキュベートした。細胞を洗浄し、Coulter Eliteフローサイトメーターで細胞内の蛍光を測定した。
【図12】NF-κBインヒビターのカフェ酸フェネチルエステル(CAPE)存在下および非存在下での、RR1が誘発したRAW264.7細胞のTNF-α合成を示す。細胞を、0.5%のFBSを含有するEMEM培地中、10μg/mlのCAPEと1時間予備インキュベートしてから、培地を様々な濃度のRR1を含有する培地に交換し37℃で4時間インキュべートした。培地を、ELISAプロトコルを用いて、分泌されたTNF-αについて分析した。
【図13】RR1処理したRAW264.7マクロファージにおける転写因子NF-κBのDNA結合活性の経時変化を示す。マクロファージを100μg/mlのRR1で処理し、核抽出物のNF-κBの活性を、ELISAプロトコルを用いて、インキュベーション後2、4、8、14、および20時間目について分析した。
【図14】RR1処理したRAW264.7マクロファージにおける転写因子NF-κBの用量依存的活性化を示す。NF-κB活性は、RR1用量の漸増に伴って増大し、500μg/mlでプラトーに達した。
【図15】EMSAゲルシフトアッセイにより決定した、RAW264.7細胞の核抽出物における転写因子NF-κBのDNA結合活性を示す。非特異的バンドの強度を基に、NF-κB特異的バンドの用量依存的な強度増加が見られる。
【図16】RR1によるRAW264.7マクロファージ刺激が、細胞質ゾルのI-κBα分解を誘発することを示す。細胞を15、30、および60分の間隔でRR1処理した。細胞質の溶解物中のI-κBα分解レベルは、抗マウスI-κBαmAbを用いたイムノブロット分析で決定した。
【図17】RR1で刺激したRAW264.7細胞におけるTLR2、TLR4、およびTLR6 mRNAのRT-PCRを示す。細胞(1×106)を、CO2インキュベーター内、37℃で、RR1(0、10、100、500、および1000μg/ml)で24時間処理し、マウスTLR特異的プライマーを用いてmRNAを増幅した。
【図18】TLR2およびTLR6遺伝子を安定にトランスフェクトしたHEK293細胞における、RR1誘発IL-8合成の分析を示す。HEK293/TLR2/TLR6細胞(1×106)を、37℃で24時間、RR1(0、10、50、および100μg/ml)で処理し、培地中に分泌されたIL-8をELISAプロトコルで分析した。
【図19】TLR6遺伝子を安定にトランスフェクトしたHEK293細胞における、RR1誘発IL-8合成の分析を示す。HEK293/TLR6細胞(1×106)を、37℃で24時間、RR1(0、10、50、および100μg/ml)で処理し、培地中に分泌されたIL-8をELISAプロトコルで分析した。
【図20】マクロファージにおけるRR1の免疫系刺激経路を示す。
【特許請求の範囲】
【請求項1】
下記の化学構造(I)を有する単離された化合物、またはその薬学的に許容される塩もしくは類似体。
【請求項2】
下記の化学構造(I)を有する単離された化合物、またはその薬学的に許容される塩もしくは類似体;および薬学的に許容される担体を含む、薬学的組成物。
【請求項3】
免疫調節促進剤をさらに含む、請求項2記載の薬学的組成物。
【請求項4】
抗原をさらに含む、請求項2または3記載の薬学的組成物。
【請求項5】
アジュバントをさらに含む、請求項2または3記載の薬学的組成物。
【請求項6】
抗原およびアジュバントをさらに含む、請求項2または3記載の薬学的組成物。
【請求項7】
抗癌剤をさらに含む、請求項2〜6のいずれか一項記載の薬学的組成物。
【請求項8】
細胞傷害性薬剤をさらに含む、請求項2〜7のいずれか一項記載の薬学的組成物。
【請求項9】
化学療法薬剤をさらに含む、請求項2〜8のいずれか一項記載の薬学的組成物。
【請求項10】
腫瘍壊死因子(TNF)、インターフェロン、神経成長因子(NGF)、血小板由来成長因子(PDGF)、および組織プラスミノーゲン活性化因子からなる群より選択される免疫調節物をさらに含む、請求項2〜9のいずれか一項記載の薬学的組成物。
【請求項11】
リンフォカイン、インターロイキン、および成長因子からなる群より選択される生物学的応答調整物質をさらに含む、請求項2〜10のいずれか一項記載の薬学的組成物。
【請求項12】
請求項1記載の単離化合物の有効量を細胞にインビトロまたはインビボ投与する工程を含む、免疫応答を刺激するための方法。
【請求項13】
化合物が被験体に投与され、かつ被験体の免疫系が該投与によって刺激される、請求項12記載の方法。
【請求項14】
被験体の免疫系がマクロファージの活性化によって刺激される、請求項13記載の方法。
【請求項15】
被験体の免疫系が下記の様式の一つまたは複数で刺激される、請求項13または14記載の方法:
(a)ナチュラルキラー(NK)細胞の活性化の増大;
(b)マクロファージの腫瘍壊死因子(TNF)-α合成の増大;
(c)マクロファージ走化性タンパク質(MCP)-1合成の増大;
(d)NF-κBの活性化;
(e)Th1サイトカイン産生の増大;および
(f)第2補体活性化経路のC3a des Argレベルの増大。
【請求項16】
被験体が細胞増殖性障害を患っている、請求項13〜15のいずれか一項記載の方法。
【請求項17】
細胞増殖性障害が癌である、請求項16記載の方法。
【請求項18】
被験体が感染症を患っている、請求項13〜17のいずれか一項記載の方法。
【請求項19】
下記の工程を含む、ティノスポラ・コルディフォリア(Tinospora cordifolia)植物材料からRR1を得るためのプロセス:
(a)ティノスポラ・コルディフォリア植物材料を提供する工程;および
(b)請求項1記載の化合物を抽出する工程。
【請求項20】
抽出した化合物を薬学的に許容される担体と混合して薬学的組成物を形成する工程をさらに含む、請求項19記載のプロセス。
【請求項1】
下記の化学構造(I)を有する単離された化合物、またはその薬学的に許容される塩もしくは類似体。
【請求項2】
下記の化学構造(I)を有する単離された化合物、またはその薬学的に許容される塩もしくは類似体;および薬学的に許容される担体を含む、薬学的組成物。
【請求項3】
免疫調節促進剤をさらに含む、請求項2記載の薬学的組成物。
【請求項4】
抗原をさらに含む、請求項2または3記載の薬学的組成物。
【請求項5】
アジュバントをさらに含む、請求項2または3記載の薬学的組成物。
【請求項6】
抗原およびアジュバントをさらに含む、請求項2または3記載の薬学的組成物。
【請求項7】
抗癌剤をさらに含む、請求項2〜6のいずれか一項記載の薬学的組成物。
【請求項8】
細胞傷害性薬剤をさらに含む、請求項2〜7のいずれか一項記載の薬学的組成物。
【請求項9】
化学療法薬剤をさらに含む、請求項2〜8のいずれか一項記載の薬学的組成物。
【請求項10】
腫瘍壊死因子(TNF)、インターフェロン、神経成長因子(NGF)、血小板由来成長因子(PDGF)、および組織プラスミノーゲン活性化因子からなる群より選択される免疫調節物をさらに含む、請求項2〜9のいずれか一項記載の薬学的組成物。
【請求項11】
リンフォカイン、インターロイキン、および成長因子からなる群より選択される生物学的応答調整物質をさらに含む、請求項2〜10のいずれか一項記載の薬学的組成物。
【請求項12】
請求項1記載の単離化合物の有効量を細胞にインビトロまたはインビボ投与する工程を含む、免疫応答を刺激するための方法。
【請求項13】
化合物が被験体に投与され、かつ被験体の免疫系が該投与によって刺激される、請求項12記載の方法。
【請求項14】
被験体の免疫系がマクロファージの活性化によって刺激される、請求項13記載の方法。
【請求項15】
被験体の免疫系が下記の様式の一つまたは複数で刺激される、請求項13または14記載の方法:
(a)ナチュラルキラー(NK)細胞の活性化の増大;
(b)マクロファージの腫瘍壊死因子(TNF)-α合成の増大;
(c)マクロファージ走化性タンパク質(MCP)-1合成の増大;
(d)NF-κBの活性化;
(e)Th1サイトカイン産生の増大;および
(f)第2補体活性化経路のC3a des Argレベルの増大。
【請求項16】
被験体が細胞増殖性障害を患っている、請求項13〜15のいずれか一項記載の方法。
【請求項17】
細胞増殖性障害が癌である、請求項16記載の方法。
【請求項18】
被験体が感染症を患っている、請求項13〜17のいずれか一項記載の方法。
【請求項19】
下記の工程を含む、ティノスポラ・コルディフォリア(Tinospora cordifolia)植物材料からRR1を得るためのプロセス:
(a)ティノスポラ・コルディフォリア植物材料を提供する工程;および
(b)請求項1記載の化合物を抽出する工程。
【請求項20】
抽出した化合物を薬学的に許容される担体と混合して薬学的組成物を形成する工程をさらに含む、請求項19記載のプロセス。
【図1】


【図2A】

【図2B】
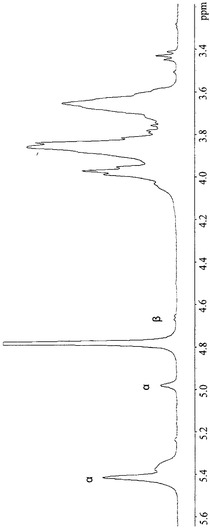
【図3】
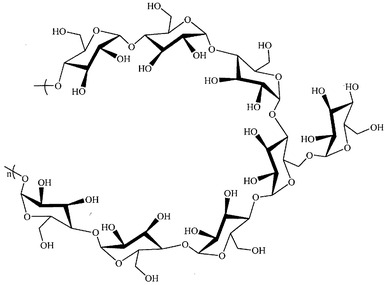
【図4】
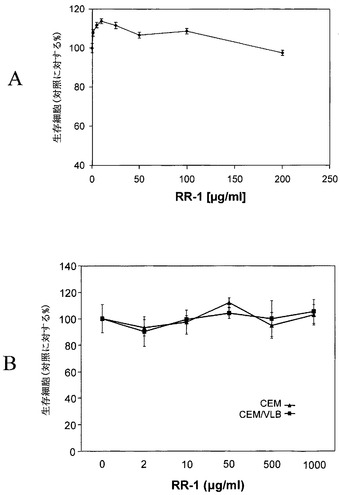
【図5】
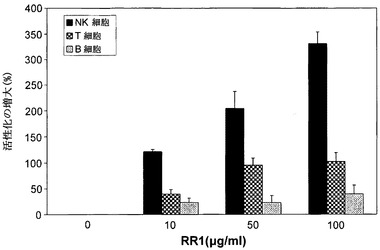
【図6】

【図7】
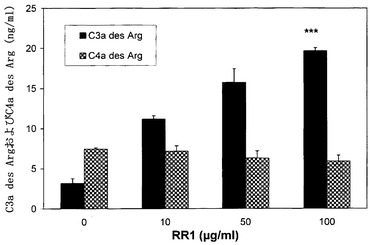
【図8A】
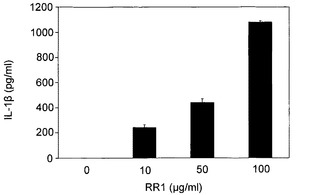
【図8B】
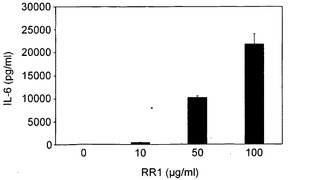
【図8C】
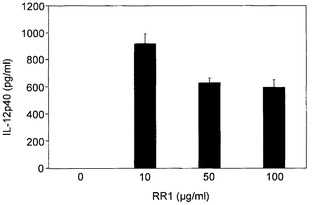
【図8D】

【図8E】
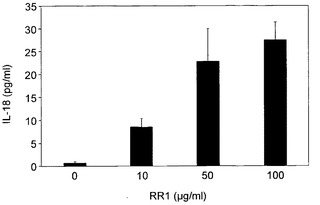
【図8F】

【図8G】
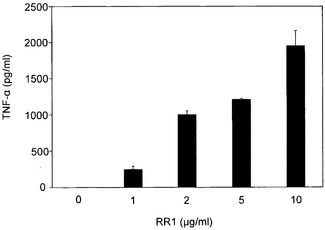
【図8H】

【図9】
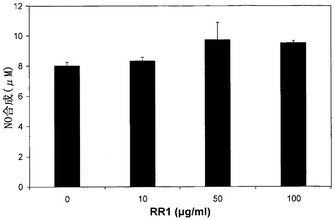
【図10】
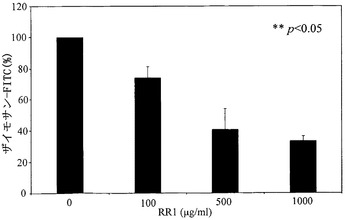
【図11】

【図12】

【図13】
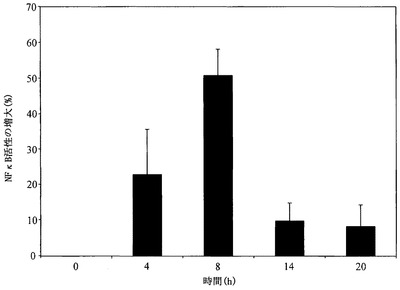
【図14】

【図15】
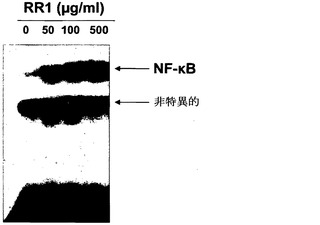
【図16】

【図17】
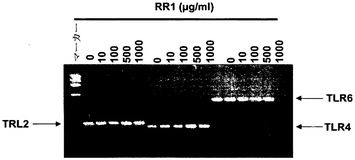
【図18】

【図19】

【図20】
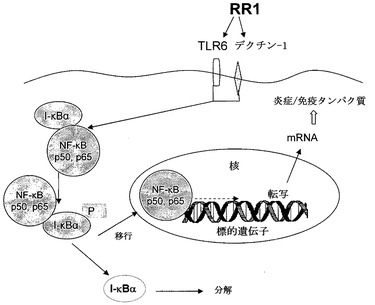
【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図8H】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【公表番号】特表2008−506024(P2008−506024A)
【公表日】平成20年2月28日(2008.2.28)
【国際特許分類】
【出願番号】特願2007−520570(P2007−520570)
【出願日】平成17年7月11日(2005.7.11)
【国際出願番号】PCT/US2005/024410
【国際公開番号】WO2006/010069
【国際公開日】平成18年1月26日(2006.1.26)
【出願人】(507007474)
【Fターム(参考)】
【公表日】平成20年2月28日(2008.2.28)
【国際特許分類】
【出願日】平成17年7月11日(2005.7.11)
【国際出願番号】PCT/US2005/024410
【国際公開番号】WO2006/010069
【国際公開日】平成18年1月26日(2006.1.26)
【出願人】(507007474)
【Fターム(参考)】
[ Back to top ]