
全細胞の生体触媒
本発明は、ニトリラーゼによって触媒された反応の産物を生産する方法に関し、その方法は、(i)その表面に位置された前記ニトリラーゼを含む微生物および/または、前記微生物の膜標本を用意し、(ii)ニトリラーゼ活性と適合性をもつ条件の下、微生物および/またはそれの膜標本を1つ以上のニトリラーゼの基質に接触させる、のステップを含む。本発明は、さらに、ラセミのマンデロニトリルの転換のため、ニトリラーゼを表示する全細胞生体触媒あるいはそれの膜標本を使用する、鏡像異性的に純粋な(R)-マンデル酸を生産する方法に関する。

【発明の詳細な説明】
【説明】
【0001】
本発明は、ニトリラーゼ(nitrilase)によって触媒作用を及ぼされた反応の産物を生産する方法に関し、その方法は、(i)その表面に置かれた前記ニトリラーゼを含む微生物、および/または、前記微生物の膜標本(membrane preparation)を用意し、そして(ii)ニトリラーゼ活性と適合性をもつ条件の下で、微生物および/またはそれの膜標本を1つ以上のニトリラーゼ基質と接触させる、ステップを含む。特に、本発明は、(i)その表面上のニトリラーゼを含む微生物および/またはそれの膜標本を用意し、(ii)ニトリラーゼがニトリルのカルボン酸への転換に触媒するように、微生物および/またはそれの膜標本をニトリルと接触させる、のステップを含むカルボン酸を生産する方法に関する。本発明は、さらに、ラセミ(racemic)のマンデロニトリルの処理に使用する、ニトリラーゼを提示する全細胞生体触媒あるいはそれの膜標本を使用して、純粋な(R)-マンデル酸を鏡像異性的に生産する方法に関する。
ニトリルは、様々な産物(例えばカルボン酸、アミド(Banerjee et al.、2002年))を生産するための重要な前駆体分子である。ニトリルの高い安定性により、しかしながら、ニトリルの化学的転換のために高温でそれらを強い塩基性条件あるいは酸性条件にさらすことが必要である(Banerjee et al.、2002年、Nagasawa et al、1990年)。さらに、それぞれの反応の混合物からのこれらの産物を精製することは非常に長時間かかりうんざりするものである。
【0002】
ニトリラーゼ (例えばEC 3.5.5.1)は、ニトリルを一つのステップ(ThimannおよびMahadevan、1964年)で対応するカルボン酸およびアンモニアに変換することができる酵素である。一方では、カルボキシリック(carboxylic)な産物が種々様々の化学の生産工程の中で中間体としてしばしば使用され、他方では、ニトリラーゼが、廃液(waste)の解毒に使用されてもよい(Banerjee et al.、2002年; Thuku et al.、2009年)ので、ニトリラーゼ酵素の使用は本質的に産業的な興味を引きつけた。更に、従来の化学(Banerjee et al.、2002年)において必要な、集中的な(intensive)ブロッキングおよび脱ブロッキングステップおよび触媒を必要としないで、穏やかな条件の下でニトリラーゼは高度に選択的で、鏡像異性的に純粋なカルボン酸を産出する。前記選択性の高さは、従来の方法を使用して達成しなかった、或いは困難とともに達成された産物合成を可能にする。
【0003】
量に関するニトリラーゼの産業適用で最も重要なものは、ラセミのマンデロニトリルを鏡像異性的に純粋な(R)-マンデル酸に転換することである(Rey et al.、2004年)。(R)-マンデル酸は、ジアステレオマー塩を使用して、ラセミ化合物中の鏡像異性体を分離するために使用された。更に、それは、新規の活動的な薬剤の化学合成中での重要なキラル(chiral)中間生成物である。
【0004】
A.faecalisからのニトリラーゼがマンデロニトリルを高い鏡像異性選択性を備えたマンデル酸に変換することができることは文献から有名である。マンデル酸のようなキラルのカルボン酸は、それらが多くの薬学的薬剤か病虫害防除薬剤のための基礎製品として役立つので、有機化学の中で望まれた化合物を表す。したがって、例えば(R)−あるいは(S)−マンデル酸、ラセミのアミンのラセミ溶液のために使用されます。さらに(R)−マンデル酸は中間体として合成に使用されます。
しかしながら、ニトリラーゼは悪評高いほどに不安定な酵素(Buchholz、KascheおよびBornscheuer、2005年)であると見なされており、従って、産業の中で精製された酵素としてめったに使用されないか、全体細胞の標本(preparation)として使用されます。ニトリラーゼの活性中心は、自動酸化(Kobayashi et al.、1992年)に弱く、チオール含んでいる試薬のような多くの化学薬品の方へ反応的である、保存された活性部位のシステイン残基(Kobayashi et al.、1992年)を含む3つのアミノ酸残基(触媒現象のトリオ)の触媒配列を含んでいる。
【0005】
細胞の内部に存在する間、ニトリラーゼはバクテリアの細胞質ゾルの酸化還元反応の環境によって保護され、それは減少され(ChoiおよびLee、2004年)、そして、チオール基(thiol groups)の酸化を防ぐ。しかしながら、ニトリラーゼは酸素へ接触すると活動を失うことが示され、また、この所見は、活性中心(Mateo et al.、2006年)中の重要な酸化還元反応感受性のトリオ・システインの感受性に起因した。
【0006】
いくつかの戦略は不十分な活性および/または安定性の点からニトリラーゼの欠点を克服するために使用された。
【0007】
1つのアプローチでは、精製した酵素製剤ではなくむしろニトリラーゼを発現する全細胞が使用され、それは系統特異的か或いは組み換えの形式で(Kaul et al.、2007年)ニトリラーゼを細胞内で提供される。例えば、研究者は、細胞の内部で置いたニトリラーゼを含む全細胞触媒として、ニトリラーゼを発現するAlcaligenes faecalisの細胞を使用した(Kaul, A、BanerjeeおよU. C. Banerjee、2006年)。97%の鏡像異性体過剰率(enantiomeric)が達成された。このアプローチの基礎となる論理的根拠は、酵素がそれらの自然環境、すなわちバクテリアの細胞質ゾルに残るということであり、そこでは内発的な折り畳み装置類および連続的な転覆(turnover)および蛋白質生合成が、安定で活性のあるタンパク質を一定のレベルの維持する。しかしながら、時間を消費し労力の大きいの手順を使用して、全細胞触媒は調製されるに違いないし、適切なマトリックス系(アルギン酸塩)に付けられるに違いない。さらに、ひっくり返される基質は、選択的な細菌細胞の細胞膜を横切るとともに、選択的な活性を有するとともに基質競合的な酵素の射程範囲内である細胞質ゾル内に一度入るように対向する必要がある。最後に、全細胞触媒は、望ましい産物の調製を汚染するので、したがって、敏感な適用、例えば飲料および食料と同様に医学、化粧品および衛生的製品の調製、のためのそのような製品への使用を除外される。
【0008】
工業的用途用の機能的なニトリラーゼの高いレベルの生産への別のアプローチは、シャペロン(chaperones)の使用に関係する。GroELファミリーのシャペロンは細胞質ゾルにある不可欠なタンパク質で、それは、ATP依存性の結合の多数の輪(rounds)、カプセル化および基質タンパク質の放出を経由した、正確に折り重ねられたタンパク質の生産に必要である(Hartl, F.U.およびHayer-Hartl, M、2002年)。GroELの削除は、結局、機能的な基質タンパク質の低いレベルだけでなく、大量の折り畳まれないで蓄積されたタンパク質に起因する細菌細胞の死に導かれる。シャペロン、より明確に、GroEL系の構成要素は、大腸菌の細胞質ゾル中で組み換えのニトリラーゼとともに共発現(co-expressed)された(米国特許5,629,190番および図9)。このアプローチは、内発的なレベルのシャペロンだけをもつ細胞に比して、より高いレベルの可溶のタンパク質に至った。しかしながら、そのような折り畳み装置が設けられた全細胞の使用、あるいは単独に精製されたシャペロンおよびそれらの共同因子(cofactors)およびニトリラーゼ調製のために追加的な基質の付加は、そのようなアプローチの必要条件である。
米国特許5,629,190番は、ニトリラーゼを組換え的(recombinantly)に発現する全(whole)大腸菌細胞の使用を教示する。しかしながら、たとえGroELのようなタンパク質薬剤、微生物が合成するポリペプチドを折り畳むことを支援するための、例えば、組み換え的に発現されたニトリラーゼ、を組み換え的に発現する場合であっても、細胞の特定の細胞内酵素設備のため、生じる反応は複雑で、それらの結果は予測不能である。米国特許5,629,190番に示されるように、ニトリラーゼが触媒作用するニトリルのカルボン酸への転換は、産物の減少および除去するのが難しい望まれない副産物に結びつく、ニトリルヒドラターゼ(hydratase)が触媒作用する反応(酸アミド形成に帰着する)と競合する。細胞の中にある他の酵素はニトリル基あるいは他の半分(moiety)でもニトリルを修飾するかもしれない。
【0009】
別のアプローチは、および架橋された酵素集合体(つまり可溶の組み換えの酵素、およびグルタルアルデヒドまたはポリ(エチレンイミン)のような反応性の二価性の化学薬品を使用する二次的架橋)の調製および使用に関係する。しかしながらKaul et al. (2007)は、そのような固定が反応速度の著しい減少および一定の特異性の減少に帰着したことを実証した。さらに、化学的架橋は、変更された基質特異性(特定の形態に「固定された」タンパク質分子を容易に産出した架橋プロセスに関連された観察)に結びついた。さらに、それらの分解された産物および残骸と同様に動けなくされた酵素の一群は、望ましい反応生成物から分離するには簡単ではないかもしれない。
【0010】
Yamamoto et al.(1991)は、マンデロニトリルを(R)-(-)-マンデル酸に変換するそれらの能力に関して、様々な分離物(野生種(wildtype strains))をテストした。A.faecalis ATCC 8750は、ラセミのマンデロニトリルからの(R)-(-)-マンデル酸を生産するために最高レベルの活性と鏡像異性選択性とを持つことが発見された(Yamamoto et al.、1991年)。特に、休眠細胞(resting cells)によってマンデロニトリルから作られた(R)-(-)-マンデル酸は、100%の鏡像体過剰率にあった。A.faecalis ATCC 8750は、マンデロニトリル(マンデロニトリル)およびマンデルアミド(mandelamide)、をそれぞれ処理するために使用される1つの(R)−鏡像異性選択的なニトリラーゼおよびアミダーゼを持っている。(R)-(-)-マンデル酸は、91%の収量でのラセミのマンデロニトリルから生まれるが、しかしながら、反応で残る(S)-マンデロニトリルは、一方では(R)―および(S)―異性体のマンデロニトリルの化学平衡、および他方ではベンズアルデヒド/HCNの化学平衡のために、(S)-マンデロニトリルは残されず自発的にラセミ化される。従って、マンデロニトリルはほとんどすべて消費され、(R)-(-)-マンデル酸に転換される。Yamamoto et al. (1991)によれば、(R)―マンデル酸の生産のために使用されたA.faecalis細胞は、特に最適化された培養条件を要することが注目される。
【0011】
バクテリアの起源の異なるニトリラーゼは、クローン化され、大腸菌の中で組み換え的に発現された(Kiziak et al.、2005年; Luo et al.、2008年; Rustler et al.、2008年; 米国特許6,180,359番; Ress-Loschke et al.、1998年)。ニトリラーゼは生化学的な特徴が述べられた(Kiziak et al.、2005年)。さらに、大腸菌のA.faecalis ATCC8750のニトリラーゼ遺伝子を組み換え的に発現させることが可能で、それは、精製後に「架橋した酵素集合体」として、動けなくされた実験の中でその後使用された(リイ、2004年ら)。この方法では、宿主微生物、つまり大腸菌、からニトリラーゼを分離し、浄化しなければならない。このアプローチは全く長時間かかりうんざりするものである。さらに、ニトリラーゼは手順(procedure)の間に非活性化されるかもしれない。
【0012】
Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750からのニトリラーゼは、1つのサブユニット当たりの分子量32 kDを備えたおよそ14のサブユニットから構成されるホモオリゴマー(homooligomeric)の酵素で、よって、天然タンパク質の合計の結合した分子量はおよそ460 kDであるとして、記述された(Yamamoto et al.、1992年)。一般に、ニトリラーゼは不活発な二量体として存在し、自己会合を通じて活発なオリゴマーを形成する。しかしながら、活動に必要とされたサブユニットの数はニトリラーゼごとに異なる(Thuku et al.、2009年)。
要約すると、ニトリラーゼには有機化合物の生産でのニトリラーゼの使用を妨げるいくつかの欠点がある:
ニトリラーゼは、酸化に対して敏感であり、時間とともにニトリラーゼ活動の縮小に帰着する。
【0013】
全細胞生体触媒の形で使用された時、細胞の中にある他の酵素(ニトリルヒドラターゼのような)によって触媒作用を及ぼされた望まれない副反応は、ニトリラーゼによって触媒作用を及ぼされた反応を妨害するかもしれない。
ニトリラーゼは反応混合物から回復(recovered)することができない。
自動表示(Autodisplay)は、組換え型タンパク質のバクテリアの表面の表示用の簡潔なツールを表わす。この表現系は、タイプV分泌系に属するタンパク質の自動トランスポーター・ファミリー(autotransporter family)の分泌メカニズムに基づく。
グラム陰性菌では、自動トランスポーター経路は、細胞表面へのタンパク質の輸送、および細胞外の環境の中へのタンパク質の分泌の両方のために発展した(Jose and Meyer、2007年)。自動トランスポータータンパク質は、細胞表面への輸送のための構造の必要条件をすべて含んでいる前駆体タンパク質として合成される(Jose、2006年)。それらは、内部の薄膜の交差を可能にするSec 経路には典型的なN-末端のシグナル・ペプチドで合成される。一旦、周辺質(periplasm)では、シグナル・ペプチドの切断の後、前駆体のC-終末部がポーリン(porine)状の構造、いわゆるβ-バレルとしての外膜内へ折り畳まれる。この孔によって、N-末端に付属した乗客ドメインは、表面へ移動させられる(Jose、2002年)。そこで、それは切断(自己タンパク分解的に(autoproteolytically)あるいは追加のプロテアーゼによって)されるかもしれないし、あるいはトランスポータードメインによって細胞外被にアンカーされ続けるかもしれない。元の乗客を組換え型タンパク質に取り替えることはその適切な表面の転置(translocation)に帰着する。この目的のために、シグナル・ペプチド、組み換えの乗客、β−バレルおよびリンク地域、それらの中間にある連結位置(linking region)(それは全表面へのアクセスを達成するために必要です)、をからなる、人工の前駆体が遺伝子工学によって構築されなければならない。AIDA-I自動トランスポーターは、様々な乗客ドメインの効率的な表面への表示するため、このように使用されて成功した(Henderson et al.、2004年)。
【0014】
自己ディスプレイ(autodisplay)系では、活性のある酵素へのサブユニットの自己会合は、例えば二量体酵素ソルビトール・デヒドロゲナーゼの中で観察された(Jose (2002年); Jose and von Schwichow、2004年)。
特に、自動表示技術は、大腸菌および他のグラム陰性菌の外膜の表面への前もって定義したタンパク質のための発現方法である。そこでは自動ディスプレイ系は、自動トランスポータータンパク質の自然分泌メカニズムに基づく(Jose and Meyer、2007年)。このプロセスでは、組み換えの乗客タンパク質は、遺伝子工学の標準的方法を使用して、単にシグナル・ペプチドと自動表示ベクターの移動(translocating)ドメインの間で翻訳領域内(in-frame)にそのコード配列を導入することにより輸送されるかもしれない。 シグナル・ペプチドはコレラ毒素サブユニット(CTB)から得られるかもしれないし、人工プロモータと結合されるかもしれない。したがって、自動トランスポータータンパクと呼ばれる別のタンパク質を備えた組み換え融合タンパク質として、外膜を横切る移動が意図された乗客タンパク質が大腸菌(AIDA-I)の外膜で発現される(Jose、2006年)。自動トランスポータータンパク質のC-末端部は大腸菌の外膜内にポーリン(porine)状の構造(β-バレル)を形成する。このポーリン状の構造は、大腸菌の外膜の表面への組み換え乗客タンパク質の移動を促進する(Jose、1995年、2006年、2007年)。
【0015】
本発明の基礎となる問題は、基質分子に容易にアクセス可能な活性状態のニトリラーゼの生産および調製を可能にする方法を提供することである。更に、他の酵素の存在は望まれない副反応を防ぐために回避されるべきである。本発明のさらなる基礎となる問題は、組み換えのニトリラーゼを使用した、ニトリラーゼによって触媒作用を及ぼされた反応の産物(例えばカルボン酸)を生産する方法を提供することであり、そこでは時間とともに活性が低下する問題は少なくとも部分的に解決される。本発明の基礎となる別の問題は方法を提供することであり、そこではニトリラーゼによって触媒作用を及ぼされた反応の産物(例えばカルボン酸)は、容易に取り出されるかもしれない。
驚いたことに、本発明者らは、微生物のような宿主細胞の表面上でニトリラーゼが発現されるかもしれないことを発見し、そこではニトリラーゼは完全に機能的(つまり、カルボン酸の形成に帰着するニトリルの転換に触媒作用を及ぼすことができる。)である。
【0016】
本発明の最初の側面は、ニトリラーゼによって触媒作用を及ぼされた反応の産物を生産する方法で、次のステップを含む、
(i)その表面に置かれた前記ニトリラーゼを含む微生物、および/または前記微生物の膜標本を用意し、
(ii)ニトリラーゼ活性と適合性をもつ条件の下で、前記微生物および/またはそれの膜標本と1つ以上のニトリラーゼ基質とを接触させる。
【0017】
本発明の方法では、ニトリラーゼのどんな既知の基質が使用されてもよい。産物は、ニトリラーゼによって触媒作用を及ぼされた反応の産物であるあらゆる化合物であってもよい。ここに使用されるような「ニトリラーゼ活性と適合性をもつ条件」は、ニトリラーゼが活発な条件を含んでおり、つまり、そこではニトリラーゼは1つ以上の基質を産物に変換することができる。ニトリラーゼが活発になるあらゆる既知の条件が使用されてもよい。
【0018】
本発明の方法の中で使用された1あるいは少なくとも1個のニトリラーゼ基質は、ニトリルであることが好ましい。本発明の方法によって得られた産物がカルボン酸であることはさらに好まれます。「ニトリラーゼ活性と適合性をもつ条件」は、ニトリラーゼがカルボン酸の形成に帰着するニトリルの転換に触媒作用を及ぼすような条件であってよい。本発明の好ましい実施形態はカルボン酸を生産する方法であり、その方法は以下のステップを含む、
(i)その表面に置かれたニトリラーゼを含む微生物および/またはそれの膜標本を用意し、
(ii)カルボン酸の形成に帰着するニトリルの転換に前記ニトリラーゼが触媒作用を及ぼすように、前記微生物および/またはそれの前記膜標本をニトリルに接触させる。
【0019】
任意に、本発明の方法は、ステップ(ii)で使用された微生物を取り出すステップ(iii)をさらに含む。
【0020】
ここに使用されるように、「ステップ」(i)、「ステップ」(ii)あるいは「ステップ」(iii)への言及は、ここに記述されるように、ニトリラーゼによって触媒作用を及ぼされた反応の産物を生産する方法、およびカルボン酸を生産する好ましい方法のそれぞれのステップへの言及である。
【0021】
本発明は、組み換え法によって適切な自動トランスポーター系に融合された時、細菌のニトリラーゼが、それらが数日に対する触媒能力を保持したままで、産業上適切な反応に触媒作用を及ぼすために使用されるように、細菌細胞の表面へ移動させられ、細菌の細胞膜中への活性のある折りたたみ形態に統合されるという驚くべき発見に基づく。
【0022】
細菌細胞の外膜は、細胞質ゾル中のタンパク質に適切なタンパク質折り畳み装置類を欠いているので、これは特に驚くべきことである。更に、本発明者らは、もっとも驚いたことに、還元性のあるバクテリアの細胞質ゾルから、酸素が高いレベルで存在する(非常に酸化しやすい環境を形成する)外膜まで、酸化還元反応に敏感な酵素を転送した後或いは転送中に、酵素が失活しないことを見つけ出した。
【0023】
任意の理論に結合される意図なしで、本発明者らは、ニトリラーゼは、自動ディスプレイ経由で細菌細胞の表面上で動けなくされた時、大量の細菌細胞を育てるために使用されるルーチン方法の一部としての、高い酸素レベルおよび活発な揺れに関連した強い機械的な力にさらされた時さえ、反応性のある触媒アミノ酸残基を非活性化から保護する集合体あるいは3次元構造を形成すると仮定する。
【0024】
ここに記述された全細胞生体触媒は、大腸菌のようなグラム陰性細胞の表面上に活性のあるニトリラーゼを配置することを初めて可能にし、その細胞は、ニトリル転換および対応するカルボン酸の生産に使用されてもよい。特に、ここに記述されたニトリラーゼを表示している全細胞生体触媒は、ニトリルの生物工学的な転換に使用されてもよい。
【0025】
重要なことは、前記反応は、穏やかな条件の下で、よい生産速度で、および有毒な副産物の生成なしで遂行されることである。特に、微生物の表面上で組み換え的に発現されたニトリラーゼは、pH7.0前後のように穏やかな条件の下でニトリルの転換に触媒作用を及ぼすことができる。
【0026】
ここに使用されるように、「全細胞生体触媒」は、有機化合物、特にニトリルに対して化学変化を行なうための、タンパク質酵素、特にニトリラーゼのような天然の触媒を含む、全体の細胞、特に微生物を表し、そこでは天然の触媒は、好ましくは細胞の表面に置かれる。発明によれば、触媒は組み換え的に発現されてもよい。
【0027】
細菌細胞は、疎水性の細胞膜によって互いから分けられたある範囲の区画を含む。グラム陽性の細菌細胞は、細胞質ゾル(細胞の内部)を閉じ込める原形質膜を含む。 原形質膜はペプチドグリカンの層に包まれる。対照的に、グラム陰性菌には原形質膜に加えて、外膜と呼ばれた別の薄膜がある。用語「表面」は、ここに使用されるように、好ましくは環境(例えば液体培養培地)に接している微生物の層を指す。本発明の好ましい実施形態では、表面は、グラム陰性の細菌細胞の液体培養培地に接している側である。ここに使用されるように、「表面に表示され」および「表面上で発現された」の表現は、交換可能に使用され、細胞表面上にニトリラーゼを局在化させることを指す。
【0028】
ここに使用されるように、「微生物」は宿主細胞を指す。発明による宿主細胞は、1つの原核生物あるいは真核生物の微生物、好ましくは原核生物の微生物、より好ましくは細菌細胞、さらに好ましくはグラム陰性の細菌細胞であってよい。最も好ましくは、微生物は大腸菌細胞(E. coli cell)である。「微生物」、「宿主細胞」という用語はここで交換可能に使用される。
ここに使用される「ニトリラーゼ」はニトリラーゼ酵素、触媒現象的に活性のある部分、それの誘導体あるいは類似体を指し、それは、ニトリルのカルボン酸およびアンモニアへの転換に触媒作用を及ぼす。ここに使用されるように、分子の「誘導体」あるいは「類似体」は、その分子から由来された部分或いは分子の変形版(modified version)を指すことが注目されるべきである。本発明では、ニトリラーゼは、カルボン酸の形成に帰着するニトリルの転換に触媒作用を及ぼすことができるニトリラーゼのような任意のニトリラーゼでもよい。好ましいニトリラーゼはEC 3.5.5.1による酵素である(さらに「ニトリルアミノヒドラーゼ(nitrile aminohydrolase)」と呼ばれる)。
【0029】
ニトリラーゼはニトリラーゼ活性のみを表示してもよい。ニトリラーゼはさらに多機能酵素でもよく、それは、他の活性に加えて、ニトリラーゼ活性を示す。
【0030】
熟練した人は、ニトリラーゼが設けられた多くの種を知っており、その種は原核生物と真核生物を含む。例えば、本発明の中で使用されたニトリラーゼは、Bordetella, Klebsiella, Aspergillus, Alcaligenes, Saccharomyces, Burkholderia, Neurospora, Lachancea, Debaryomyces, Yarrowia, Candida, Kluyveromyces, Rhodococcus, Nocardia, and/or Rhizobium.および/またはRhizobiumから得られるものでもよい。本発明の中で使用することができるニトリラーゼを有した種の例は、Bordetella bronchiseptica, Klebsiella pneumoniae, Aspergillus niger, Alcaligenes faecalis, Saccharomyces cerevisiae, Burkholderia multivorans, Aspergillus fumigatus, Neurospora crassa, Lachancea thermotolerans, Debaryomyces hansenii, Yarrowia lipolytica, Candida glabrata, Kluyveromyces lactis, Rhodococcus rhodochrous, Nocardia sp., Rhizobium leguminosarum, および/またはNocardia farcinicaである。
【0031】
本発明のニトリラーゼは、好ましくはAlcaligenes, Klebsiella および/または Saccharomycesから得られる。より好ましいニトリラーゼは、Alcaligenes faecalis, Klebsiella pneumoniae および Saccharomyces cerevisiaeから得られたニトリラーゼから選ばれる。Alcaligenes faecalisは、Alcaligenes faecalisの亜種のfaecalisでもよい。Klebsiella pneumoniaeは、Klebsiella pneumoniaeの亜種ozaenaeでもよい。より好ましいニトリラーゼは、Alcaligenes faecalis のニトリラーゼである。
【0032】
本発明のニトリラーゼは、ニトリラーゼ P_887662 [GI:33600102, 01-MAY-2009, Bordetella bronchiseptica RB50], AAA25057 [GI:149175, 26-APR-1993, Klebsiella pneumoniae], NP_943299 [GI:38639530, 30-APR-2009, Klebsiella pneumoniae], P10045 [GI:115192, 20-JAN-2009, Klebsiella pneumoniae subsp. ozaenae], XP_001389617 [GI:145230706, 28-FEB-2008, Aspergillus niger], ACS13754 [GI:239738518, 15-JUN-2009, Alcaligenes sp. ECU0401], BAA02684 [GI:216203, 16-FEB-2008, Alcaligenes faecalis], CAK46957 [GI:134083480, 24-MAR-2007, Aspergillus niger], EDV09642 [GI:190406375, 16-JUN-2008, Saccharomyces cerevisiae RM11-1a], YP_001945058 [GI:189349430, 07-MAY-2009, Burkholderia multivorans ATCC 17616], NP_012102 [GI:6322027, 16-JUN-2008, Saccharomyces cerevisiae RM11-1a], 1F89B [GI:16975400, 19-NOV-2001, Saccharomyces cerevisiae], NP_012409 [GI:6322335, 05-NOV-2009, Saccharomyces cerevisiae], EDN63257 [GI:151945002, 13-JUL-2007, Saccharomyces cerevisiae YJM789], EDN59257 [GI:151940875, 13-JUL-2007, Saccharomyces cerevisiae YJM789], CAY80342 [GI:259147089, 23-SEP-2009, Saccharomyces cerevisiae EC1118], EEU05439 [GI:256270219, 20-AUG-2009, Saccharomyces cerevisiae JAY291] P40447 [GI:731891, 03-NOV-2009, Saccharomyces cerevisiae], XP_751200 [GI:70992703, 27-FEB-2008, Aspergillus fumigatus Af293], CAD71250 [GI:28950282, 14-NOV-2006, Neurospora crassa], CAK48039 [GI:134075478, 24-MAR-2007, Aspergillus niger], CAR23067 [GI:238934886, 08-OCT-2009, Lachancea thermotolerans], CAG86637 [GI:199431326, 10-SEP-2008, Debaryomyces hansenii], XP_500602 [GI:50546150, 29-OCT-2008, Yarrowia lipolytica], CAG78819 [GI:49651877, 23-OCT-2008, Yarrowia lipolytica], CAG59341 [GI:49525722, 16-DEC-2008, Candida glabrata], XP_454637 [GI:50309261, 18-APR-2008, Kluyveromyces lactis], P20960 [GI:417386, 05-MAY-2009, Alcaligenes faecalis] ABO46008 [GI:134034945, 05-NOV-2009, Rhodococcus rhodochrous], AAX18182 [GI:60280369, 02-MAR-2005, Nocardia sp. C-14-1], BAA02127 [GI:216932, 16-FEB-2008, Rhodococcus rhodochrous], Q02068 [GI:417382, 20-JAN-2009, Rhodococcus rhodochrous], CAK02877 [GI:115259785, 13-MAY-2009, Rhizobium leguminosarum bv. viciae 3841], CAF05970 [GI:40882143, 10-OCT-2009, Neurospora crassa], CAK08726 [GI:115257629, 13-MAY-2009, Rhizobium leguminosarum bv. viciae 3841], BAD58116 [GI:54016746, 10-MAY-2008, Nocardia farcinica IFM 10152], および Q5Z1U0 [GI:81603033, 03-NOV-2009, Nocardia farcinica]を含むグループから選ばれてもよい。このリストは「Genbank登録番号[Genbank識別子、データベース・エントリーの日付、種/系統。」のフォーマットを使用した。このグループのニトリラーゼの配列は、Genbank登録番号、Genbank識別子(GI)、Genbankデータベース・エントリーの日付および種および/または系統から得られる配列によって特定することができる。
【0033】
ここに使用されるような、用語「ニトリラーゼ」は、ここに開示されるニトラーゼと、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%あるいは少なくとも99%の同一性を持ち合わせているどんなニトリラーゼも含んでいる。熟練した人は、例えばBLASTまたはPBLASTによって、アミノ酸配列レベルで同一性の程度の決定するためのふさわしい方法を知っている。
【0034】
上に示されるように、活性のあるニトリラーゼはおよそ14のサブユニットからなるホモオリゴマー(homooligomeric)の酵素でもよい。一般に、ニトリラーゼは不活性な二量体として存在する。本発明では、宿主細胞の表面に表示されたニトリラーゼは3つ以上の同一のサブユニットを含むホモマルチマー(homomultimer)でもよい。特にホモマルチマー(homomultimer)は、7、8、9、10、11、12、13、14、15あるいは16の同一のサブユニットのような、7〜16の同一のサブユニットを含んでもよい。いくつかのおよそ14のサブユニット、つまり11、12、13、14、15、あるいは16のサブユニットが好ましい。ホモマルチマー(さらに「ホモオリゴマー」としてここに名付けられた)は、宿主細胞細胞膜に表示されたいくつかの同一のポリペプチド・サブユニットの自発的な会合によって形成されてもよい。
ここに使用されるように、「ニトリル」は少なくとも1つの―CN官能基(ニトリル基)を含む任意の化合物を指します。ニトリルはさらに1つ、2、3、4あるいはより多くのニトリル基を含んでもよい。
【0035】
ニトリル基は、6から14個の炭素原子を、好ましくは6から10個の炭素原子を含むアリール基のような芳香族部分に結合されてもよい。アリール基は1つ、2つのあるいは3つの濃縮された芳香環を含んでもよい。より好ましくは、アリール基はフェニル基でもよい。アリール基は、1、2、3、4、5あるいはより多くの置換基のような1つ以上の置換基によって代用されてもよい。
ニトリル基は、1〜8個の炭素原子、1〜6個の炭素原子、1〜4個の炭素原子あるいは1〜3個の炭素原子を含むアルキル基のような脂肪族の部分につながれてもよい。アルキル基は直鎖でもよいし或いは分岐していてもよい。アルキル基は環状の部分を含んでもよい。アルキル基は、1、2、3、4、5あるいはより多くの置換基のような1つ以上の置換基によって代用されてもよい。
【0036】
ニトリルはアルキル・アリール基を含んでもよく、そこではアルキルとアリールはここに記述されるような意味(meanings)を持っています。少なくとも1つのニトリル基はアルキル基および/またはアリール基につながれてもよい。アルキルおよび/またはアリール部分(moiety)は、1、2、3、4、5あるいはより多くの置換基のような1つ以上の置換基によって代用されるかもしれない。
【0037】
ここに使用された1つ以上の置換基は、−OH、ヨード、ブロモ、クロロ、フルオロ、アリール、アルキル、アルコキシ、などから独立して選ばれてもよく、そこではアルキルとアリールはここに記述されるような意味を持っており、アルコキシが1〜8個の炭素原子、1〜6個の炭素原子、1〜4個の炭素原子あるいは1〜3個の炭素原子を含んでいる。好ましいアルコキシ基はメトキシである。アルコキシ基は直鎖でもよいし或いは分岐していてもよい。アルコキシ基は環状の部分を含んでもよい。
【0038】
アルキル基、芳香族基、および/またはアルコキシ基、少なくとも1つのヘテロ原子が炭素原子と置換されるように、1、2、3、4あるいはそれより多くのヘテロ原子のような少なくとも1つのヘテロ原子が独立して含んでもよい。ヘテロ原子は、N、OおよびSから独立して選ばれてもよい。
【0039】
発明のニトリラーゼの基質として使用される典型的なニトリルは、マンデロニトリル、ベンゾニトリル、フェニルプロピオニトリル(phenylpropionitrile)、フェニルグリシノニトリル(phenylgycinonitrile)、ブロモキシニル、イオキシニル、クロロキシニル(chloroxynile)、アニソニトリル(anisonitrile)、3-ブロモ-4-ヒドロキシベンゾニトリル、3-フルオロ-4-ヒドロキシベンゾニトリル、4-ヒドロキシ-3,5-ジメトベンゾニトリル(dimethobenzonitrile)、およびプルナシンのような芳香性のニトリルであり、またn-ブチロニトリル、n-バレロニトリル、イソ-ブチロニトリルおよびスクシノニトリルのような脂肪族ニトリルである。 発明の特に好ましい基質はマンデロニトリルである。
本発明によれば、Alcaligenes faecalisからのニトリラーゼのための好ましい基質はマンデロニトリルとプルナシンである。Klebsiella pneumoniae からのニトリラーゼのための好ましい基質はブロモキシニル、イオキシニル、クロロキシニル、アニソニトリル、3-ブロモ-4-ヒドロキシベンゾニトリル、3-フルオロ-4-ヒドロキシベンゾニトリル、4-ヒドロキシ-3,5-ジメトベンゾニトリル。Saccharomyces cerevisiaeからのニトリラーゼのための好ましい基質は、ベンゾニトリル、フェニルプロピオニトリル(phenylprobionitrile)、マンデロニトリル、フェニルグリシノニトリル、n-ブチロニトリル、n-バレロニトリル、イソ-ブチロニトリルおよびスクシノニトリルである。
発明の1つの側面では、ニトリラーゼをコード化(encoding)する遺伝子は、Alcaligenes faecalisの全体のDNAからPCRによって増幅されクローン化された。その遺伝子は、PCRプライマーに結合された適切な制限部位(restriction sites)による自動トランスポーターの読み枠と正確に融合できる。融合タンパク質の発現は、標準の研究室的手法で示すことができる。表面上で発現され、その上で動けなくされた、酵素は、加えられた基質(この場合、ラセミのマンデロニトリル)を細胞外で変換することができる。
【0040】
ここに適用される培養条件および発現条件で、約50%、好ましくは約60%、より好ましくは約70%、またさらにより好ましくは約80%の変換率は、5日後に達成されるだろう。非対称のC原子がある化合物は、本発明の方法によってある鏡像体過剰率(enantiomeric)中で生産されてもよい。鏡像体過剰率(%ee)は、少なくとも約50%、60%、70%、80%、90%、95%、96%、97%、98%あるいは99%であってもよく、製品、特にカルボン酸用には、少なくとも約99%以上であることが好ましい。キラルHPLCによって示されたように、特に(R)−マンデル酸では、5日後の約80%の変換率および99%以上の鏡像体過剰率(% ee)が達成された。
【0041】
ここに使用される物質の鏡像体過剰率(ee)は物質の光学的な純度のものさしです。鏡像体過剰率は分析される化合物の溶液中で決定されてもよい。適切な溶媒(疎水性・親水性の溶媒のような)およびそれの混合物が知られている。
【0042】
鏡像体過剰率は次の方程式によって計算することができる:
ee% = 100 * ( [ R ] - [ S ] ) / ( [ R ] + [ S ] )
そこでは、[R]、[S]は、種類の濃度を示し、そして、RとSは、(R)−そして(S)−異性体をそれぞれ指す。したがって、ラセミ混合物中では、つまり[R]=[S]となり、ee%=100* 0/([R]+[S])=0%であり、つまりRがSよりも過剰にはならずあるいは逆も同様である。したがって、鏡像異性的に(enantiomerically)純粋な化合物の鏡像体過剰率は100%であるのに対して、ラセミ混合物の鏡像体過剰率は0%である。(R)−および(S)−異性体の濃度は、混合物の比旋光度の決定、キラル・カラム・クロマトグラフィ(例えば、キラルHPLC)およびNMR分光学のように(しかしこれに限定されない)、非対称のC-原子がある化合物の決定用の技術の中で既知の任意の方法によって決定されてもよい。
【0043】
ここに記述された発明は、単にこの目的の反応の一回分から細胞を遠心分離機にかけるだけなので、産物を精製する容易な方法と結合した、ニトリルの加水分解中の穏やかな反応状態という長所を持つ。
【0044】
ニトリラーゼは、微生物の中で組み換え的に発現され、細胞表面へ輸送されてもよい。微生物は独立して繁殖し、生体触媒の精巧な精製がもはや必要でないように、酵素自体を生産する。そのような微生物用の例は、この例1に記述された大腸菌種である。
【0045】
ここに使用された用語「組み換え個体」は、本来は互いにつながれない、互いに効果的につながれた少なくとも2つの核酸配列で作られていた核酸配列を指す。少なくとも2つの配列は同じ生物から得られてもよい。例えば、組み換えの構成物は、Alcaligenes種のシグナル配列および同じAlcaligenes種のニトリラーゼ(それは前記シグナル配列に通常つながれない)をコードする配列を含んでもよい。少なくとも2つの配列は少なくとも2つの異なる生物から得られてもよい。例えば、大腸菌(E. coli)のシグナル・ペプチド、Alcaligenesからのニトリラーゼ遺伝子、大腸菌からの膜貫通のリンカーおよび大腸菌からの自動トランスポータードメインが、効果的につながれてもよい。別の例は、組み換えのポリペプチドであり、そこでは、シグナル・ペプチドがコレラ毒素βサブユニット(CTB)に由来し、自動トランスポータードメインおよび膜貫通のリンカーは大腸菌に由来し、ニトリラーゼはAlcaligenesに由来する。
【0046】
本発明の好ましい実施形態の中で、用語「組み換え的に発現する」は、ここに使用されるように、用語「組み換え式に発現された」などで同義的に使用され、組み換え核酸配列の発現のことを指す。
【0047】
例えば大腸菌、或いはそれどころかあらゆる生物における核酸配列の発現は、大腸菌シグナル・ペプチドと、組み換えの発現を構成するAlcaligenesニトリラーゼ配列を含んでいる。
【0048】
ニトリラーゼが自動トランスポーターのトランスポータードメインへ融合されることは好ましい。発明による自動トランスポーターのトランスポータードメインは、自動トランスポーターの任意のトランスポータードメインでよく、好ましくはβ−バレル構造を形成することができる。β-バレル構造の詳細な説明およびβ−バレル自動トランスポーターの好ましい例は、WO97/35022に示され、参照されることでここに取り込まれる。ヘンダーソンら (2004)は、適切な自動トランスポータードメインを含む自動トランスポータータンパク質について記述した(要約に関しては、Henderson et al.、2004年のテーブル1を参照)。 ヘンダーソンらの開示 (2004)は、参照によってここに取り込まれる。例えば、自動トランスポーターのトランスポータードメインは、Ssp(P09489(S.marcescens))、Ssp-h1(BAA33455(S.marcescens))、Ssp-h2(BAA11383(S.marcescens))、PspA(BAA36466(P.fluorescens))、PspB(BAA36467(P.fluorescens))、Ssa1(AAA80490(P.haemolytica))、SphB1(CAC44081(B. pertussis))、AspA/NalP(AAN71715、N. meningitidis)、 VacA(Q48247、H.pylori)、AIDA-I(Q03155、大腸菌)、IcsA(AAA26547、S.flexneri)、MisL(AAD16954、S.enterica)、TibA(AAD41751、大腸菌)、Ag43(P39180、大腸菌)、ShdA(AAD25110、S.enterica)、AutA(CAB89117、N. meningitidis)、Tsh(I54632、大腸菌)、SepA(CAC05786、S.flexneri)、 EspC(AAC44731、大腸菌)、EspP(CAA66144、大腸菌)、Pet(AAC26634、大腸菌)、Pic(AAD23953、大腸菌)、SigA(AAF67320、S. flexneri)、Sat(AAG30168、大腸菌)、Vat(AAO21903、大腸菌)、EpeA(AAL18821、大腸菌)、EatA(AAO17297、大腸菌)、EspI(CAC39286、大腸菌)、EaaA(AAF63237、大腸菌)、EaaC(AAF63038、大腸菌)、Pertactin(P14283、B. pertussis)、BrkA(AAA51646、B. pertussis)、Tef(AAQ82668、B. pertussis)、Vag8(AAC31247、B. pertussis)、PmpD(O84818、C. trachomatis)、Pmp20(Q9Z812、 C. pneumoniae)、Pmp21(Q9Z6U5、C. pneumoniae)、IgA1プロテアーゼ(NP_283693、N. meningitidis)、App(CAC14670、N. meningitidis)、IgA1プロテアーゼ (P45386, H. influenzae)、Hap(P45387, H. influenzae)、rOmpA(P15921、R. rickettsii)、rOmpB(Q53047、R. rickettsii)、ApeE (AAC38796、S. enterica)、EstA(AAB61674、P. aeruginosa)、Lip-1 (P40601、X. luminescens)、McaP(AAP97134、M. catarrhalis)、BabA (AAC38081、H. pylori)、SabA(AAD06240、H. pylori)、AlpA(CAB05386、H. pylori)、Aae(AAP21063、A. actinomycetemcomitans)、NanB(AAG35309、P. haemolytica)、およびこれらの自動トランスポーターの変異体(variants)から選ばれてもよい。
【0049】
典型的な自動トランスポータータンパク質のそれぞれに括弧の中に与えられたものは、適切なgenbank登録番号および自動トランスポーターが得られた種の例である。自動トランスポーターのトランスポータードメインが好ましくは、大腸菌のAIDA-Iタンパク質であるか、あるいは例えば、Niewert U.、Frey A.、Voss T.、Le Bouguen C.、Baljer G.、Franke S.、Schmidt MAによって記述された、ようなそれの1つの変異体でもよい。AIDA自動トランスポーター系は、浮腫病および離乳後下痢と診断されたブタから単離された大腸菌の中でF18とStx2eと関係する、Clin. Diagn. Lab. Immunol. 2001 Jan, 8(1):143-149;9。
【0050】
上記の示された自動トランスポーター配列の変異体は、例えば、膜貫通部分の一部ではない、β−バレルのループ構造中のアミノ酸配列の変更により得られるだろう。自由に、表面ループをコードする核酸部分は完全に削除してもよい。さらに、保存されたアミノ交換、つまり、1つの親水性のアミノ酸を別の親水性のアミノ酸へ交換すること、および/または1つの疎水性のアミノ酸を別の疎水性のアミノ酸に交換すること、が両親媒性のβ-シート内で起こってもよい。1つの変異体(variant)は、自動トランスポータードメイン(特に、β−シートの範囲内で)のそれぞれの天然の(native)配列に対してアミノ酸レベルで少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%あるいは少なくとも98%の配列同一性を保持する好ましい。
【0051】
発明の方法のステップ(i)が以下のステップを含むことは好ましい、
(a)発現制御配列と効果的につながれた核酸配列を含む微生物を用意し、前記核酸配列は、
(1)シグナル・ペプチドをコード化(encoding)する部分、
(2)表示される組み換えニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4)膜貫通のリンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、
を含み、
(b)前記(a)の核酸配列が発現され、前記核酸配列の発現産物は微生物の表面に表示される条件の下で微生物を培養する。
【0052】
当業者は、核酸配列が発現される条件を考案することができ、また、これはルーチン実験(例えば様々な温度、培地、細胞密度、および/または発現を引き起こす化学薬品の濃度をテストすること)のみを含んでいる。
【0053】
ステップ(a)によって提供される微生物は、発現制御配列と効果的につながれた核酸配列を用いた形質転換(transformation)によって得られてもよく、そこでは前記核酸は次のものを含む、
(1)シグナル・ペプチドをコード化する部分、
(2)表示される組み換えのニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4)膜貫通のリンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、
そこでは成分(1)から(5)は、以下に記述されるような核酸配列である。
【0054】
形質転換のどんな既知の方法も使用することができる。ここに使用される用語「形質転換」は、好ましくは、異種起源の構成物の細胞(好ましくは細菌細胞)中への導入を指し、それを今後は、形質転換されたという。当業者は形質転換のプロトコルおよび手順(例えばコンピテントセルの電気的あるいは化学的な形質転換)を知っている。
【0055】
別の実施形態では、進歩性がある方法はさらにステップを含む。
【0056】
(c) (b)の微生物から膜標本(membrane preparation)を生産すること。
本発明の中で使用された膜標本は触媒現象的に活性のあるニトリラーゼを含む。 ここに使用された用語「膜標本」は、好ましくは薄膜成分中で産物を豊富にしたもの指す。当業者は、膜標本を得るために使用することができるプロトコルおよび手順に精通している。例えば、細菌細胞は培養から収穫され、例えば、凍結解凍サイクル、超音波処理、溶菌緩衝液中の再懸濁などによって溶菌にさらされ、続いて細胞の細胞膜断片を分離するために分画遠心分離がされる。本発明の好ましい実施形態では、膜標本は、外膜標本であり、つまり他の膜の成分、および、細胞質ゾル、内部の薄膜および周辺質(periplasm)のような区画に対して外膜の成分を豊富にした標本である。当業者は、外膜を分離するために使用することができるプロトコルおよび手順、例えば細菌細胞のリゾチーム処理および続いて起こる遠心分離ステップ、に精通している。本発明の好ましい実施形態では、細胞膜標本は触媒現象的に活性のあるニトリラーゼを含む外膜標本で、その膜標本は、細胞質ゾル中の前記ニトリラーゼを合成し、続いて、外膜にそれを輸送する細胞から作られている。別の実施形態では、前記膜標本は、ニトリラーゼ融合タンパク質のような膜タンパクを人工小胞に組み入れることにより例えば十分に処理される。当業者は、例えば適切な洗剤を使用して、小胞中の膜蛋白を機能的に再構成する方法を持っている。
本発明の方法のステップ(a)は、微生物の準備に言及する。微生物はここに記述された任意の微生物でもよい。ステップ(a)で、宿主細胞、特に、宿主バクテリアは、発現制御配列(つまりプロモーターおよびそれぞれの宿主細胞中の遺伝子発現に必要なさらなる任意の配列)と効果的につながれた核酸配列を含む。ここに使用されるように、「発現制御配列と効果的につながれた核酸配列」は、発現制御配列との間の核酸配列の機能的な関係を指す。熟練した人は、適切なプロモーターおよび発現制御配列を知っている。プロモーターおよび/または発現制御配列は、宿主細胞に対して同種起源(homologous)か或いは異種起源であってもよい。
好ましくは、核酸配列は組み換えのベクター、例えばプラスミドベクター、上に置かれる。
ステップ(a)による核酸配列は、(1)シグナル・ペプチド(好ましくは内部の細胞膜を通って周辺質の中への輸送を許容するグラム陰性のシグナル・ペプチドをコードする部分)をコード化する部分を含む。シグナル・ペプチドは宿主細胞に同種起源のシグナル・ペプチドでもよい。シグナル・ペプチドはさらに宿主細胞に異種起源のシグナル・ペプチドでもよい。
さらに、核酸配列は、(2)表示されるニトリラーゼ(「=乗客ポリペプチド」あるいは「乗客」)をコード化する部分を含む。ここに記述されるどんなニトリラーゼをコード化する核酸を使用してもよい。
【0057】
核酸配列は、任意に、(3)プロテアーゼ認識部位をコード化する部分を含む。ここに使用された用語「プロテアーゼ認識部位」は、プロテアーゼ認識部位によって示されたアミド結合の加水分解のやり方で、ポリペプチドをその後切断する特定のプロテアーゼによって認識される特定のアミノ酸配列を指す。プロテアーゼ認識部位は、固有のプロテアーゼ(つまり宿主細胞に自然に生じるプロテアーゼ)あるいは外部から追加されたプロテアーゼ用の認識部位でもよい。例えば、外部から追加されたプロテアーゼは、IgAプロテアーゼ(EPA-0 254 090参照)、トロンビンあるいは因子X(factor X)でもよい。固有のプロテアーゼは、例えば、OmpT、OmpKあるいはプロテアーゼXをから選択されてもよい。プロテアーゼ認識部位は宿主細胞と同種起源でもよい。 プロテアーゼ認識部位は、さらに宿主細胞に対して異種起源でもよい。
【0058】
更に、核酸配列は、(4)乗客ポリペプチド(つまり、宿主細胞の外膜の外側表面上の組み換えニトリラーゼ(2))の提示(presentation)に必要な膜貫通リンカーをコード化する部分を含む。自動トランスポーターに関して同種起源の膜貫通のリンカー・ドメインが使用されてもよく、つまり、膜貫通のリンカー・ドメインは、自動トランスポータードメインをコード化する核酸配列の5'末端へ直接融合された核酸部分によってコード化される。さらに、自動トランスポーターに関して異種起源の膜貫通のリンカー・ドメインが使用されてもよい。膜貫通のリンカーの長さは好ましくは30-160アミノ酸である。
【0059】
さらに、核酸配列は、(5)自動トランスポーターのトランスポータードメインをコード化する部分を含む。本発明の背景では、自動ディスプレイは、任意のグラム陰性のバクテリア中の自動トランスポーターによるタンパク質或いはポリペプチドの組み換え表面への表示でもよい。トランスポータードメインはここに記述された任意のトランスポータードメインでもよい。
【0060】
本発明の核酸配列中の(1)から(5)の要素は、好ましくは、5'から3'に向けて方向づけられる。ステップ(b)で得られた発現産物では、(1)から(5)の核酸配列によってコード化されたアミノ酸配列は、好ましくはN末端からC末端に向けて配置される。
【0061】
本発明の方法のステップ(b)は、ステップ(a)の核酸配列が発現され、組み換えニトリラーゼを含む表現産物が微生物の表面に表示される、条件の下で微生物を培養することを指します。当業者は適切な培養条件を知っている。発明による方法は、次の組成物の液体培地の使用により、微生物、特に1つの細胞当たり100000まで或いはそれ以上の分子である大腸菌あるいは他のグラム陰性の細菌細胞、の表面の乗客タンパクの効率的な発現を許容する:5g/l〜20g/l、好ましくは約10g/lのトリプトン、2g/lから10g/l、好ましくは約5g/lの酵母エキス(yeast extract)、5g/lから20g/l、特に約10g/lのNaCl、および残りの部分の水。培地は恐らくできるだけ低い量の二価の陽イオンを含んでいるべきであり、好ましくはAqua bidestあるいは高度に精製された水、例えば、ミリポア(Millipore)の水が使用されます。液体培地はさらに好ましくは、2μMから20μMの濃度、特に10μMのEDTAを付加的に含んでいてもよい。さらに、それは好ましくは、2 mMから20 mMの好ましい濃度の、2−メルカプト・エタノール或いはジチオトレイトール或いはジチオエリトリトールのような還元試薬(reducing reagents)を含んでいてもよい。還元試薬は、輸送中のポリペプチドの折り畳まれない構造を好む。液体培地は、追加の炭素供給源(C-sources)、好ましくはグルコース(例えば、分泌を促進(つまり、周囲の培地に乗客に移す)するために10g/l以内の量の中で)、をさらに含んでいてもよい。表面のディスプレイのため、好ましくは追加の炭素供給源は添加されません。大腸菌のようなグラム陰性の細胞用の好ましい培養条件は、例に記述される。
【0062】
ステップ(ii)は好気性のおよび/またはを酸化させる条件の下で行なわれてもよい。好ましい実施形態では、用語「好気性の条件」は、酸素の存在によって特徴づけられた条件を指す。別の好ましい実施形態の中で、用語「好気性の条件」は、既知の温度および液体に関して正常なレベルと比較して、高いレベルの酸素の存在を指す。別の好ましい実施形態の中で、用語「好気性の」条件は、例えば液体(例えば培地)と、酸素を含む周囲のガス環境(例えば空気)との間の高いガス交換によって特徴づけられた条件を指す。当業者は、適切な酸素感受性の電極の使用を通じて例えば酸素レベルを測定することができる。さらに、当業者は、例えば空気または酸素によって緩衝液あるいは培地を洗い流したり(flushing)、酸素を放出する化学薬品を添加したり、あるいは単に酸素をあるいは空気の存在下でそのような緩衝液あるいは培地を活発に振ったりして、好気性の条件にすることができる。対照的に、当業者は、例えばアルゴンまたは窒素によって培地または緩衝液を広範囲に洗い流す(flushing)ことにより、あるいはガス交換を最小限にするために培養容器を密閉することにより、嫌気性の条件をもたらすことができる。
【0063】
好ましい実施形態では、ここに使用された用語「酸化条件」は、培地または緩衝液への追加あるいは酸化剤(oxidants)(つまり他の化学薬品に電子(その中で酵素は酸化する傾向がある)を放出させる化学薬品)の存在を指す。例えば、還元された酸化還元酵素は、酸素の存在下において酸化される(oxidise)傾向がある。対照的に、ここに使用された用語「還元する」或いは「還元条件」は、培地または緩衝液への追加あるいは還元剤(reductants)(つまり他の化学薬品に電子を取り上げさせる化学薬品)の存在を指す。当業者は、例えば、酸素のような酸化剤の追加、および、例えばメルカプトエタノールまたはDTTのようなチオールである還元剤の追加を通じて、例えば選択的な酸化還元反応の環境を設立(set up)することができる。さらに、当業者は、例えば酸化還元電極の使用を通じて溶液の酸化還元電位を測定することができる。
大腸菌のようなグラム陰性のバクテリアの宿主細胞では、組み換えの乗客の転置の後、乗客はβ−バレル(それは外膜内のアンカーとして役立つ)によって外膜の表面へ付けられ続ける。外膜内のβ-バレルのコントロールされた結合によって、β-バレルのC末端部は外膜の内部側に向けられるが、しかし、リンカーのN-末端部(組み換え乗客タンパク質はそれに共有結合で結び付けられる)は、外膜の外部の表面(つまり環境)に向けられる。
任意のステップ(c)は、ステップ(b)の微生物からの膜標本の生産を指す。 膜標本を得る適切な方法は当業者に有名である。
ここに使用されるような膜標本は好ましくは外膜標本である。
【0064】
本発明の方法のステップ(ii)は、ニトリラーゼ活性と適合性をもつ条件の下、微生物および/またはそれの膜標本を、1つ以上のニトリラーゼ基質に接触させることを指す。上に示されるように、好ましい実施形態では、ステップ(ii)は、カルボン酸の構成に帰着するニトリルの転換にニトリラーゼが触媒作用を及ぼす条件の下で、微生物および/またはとそれの膜標本をニトリルと接触させることを指す。技術の中で知られている任意の適切な条件が使用されてもよい。その反応は、特に液体培地の中で行なわれる。液体培地では、微生物は懸濁液を形成してもよい。微生物も動けなくされてもよい。膜標本が使用される場合、膜標本は液体培地の中で分散する粒子を形成してもよい。その反応は、約5.0から約9.0までの範囲中のpHで行なわれ、好ましくは約6.0〜約8.0以上で、より好ましくは約6.5から約7.5まででもよい。
【0065】
本発明の方法のステップ(iii)は、微生物の回収を指し、その微生物はステップ(ii)で使用される。本発明の例によって実証されるように、例えば24時間の反応周期の後に、細菌細胞はまだ著しい活動を示す。従って、細胞はさらなる反応周期で使用することができる。回収は反応混合物の遠心分離によって行なわれる。もし必要であれば、細胞が洗われてもよい。
【0066】
発明の別の側面は、ニトリラーゼによって触媒作用を及ぼされた反応の産物の生産のため、特にカルボン酸の生産のため、その表面にニトリラーゼを発現する微生物および/またはそれの膜標本の使用に関連する。生産されたカルボン酸がマンデル酸(特に(R)-マンデル酸)であることが好ましい。
【0067】
発明の別の側面は組み換えのニトリラーゼを表面上に表示する微生物に関係し、そこでは微生物はここに記述されるような微生物で、また、ニトリラーゼはここに記述されるようなニトリラーゼである。微生物は、シグナル・ペプチド、組み換えのニトリラーゼ、任意のプロテアーゼ認識部位、膜貫通のリンカー、およびその表面上の自動トランスポーターのトランスポータードメインを含む融合ポリペプチドを特に表示し、そこでは融合タンパク質の構成要素はここに記述された通りである。
【0068】
まだ、発明の別の側面はニトリラーゼを含む膜標本に関係がある。膜標本は、ここに記述されるような微生物、例えば本発明の方法のステップ(c)の中で得られてもよい。ニトリラーゼはここに記述されるような任意のニトリラーゼでもよい。
【0069】
しかし、発明の別の側面は、微生物へ導入された核酸配列を含み、その表面上に組み換えのニトリラーゼを表示する微生物を生産する方法であり、前記核酸配列は次のものを含む。
【0070】
(1)シグナル・ペプチドをコード化する部分、
(2)表示される組み換えのニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する任意の部分、
(4)膜貫通のリンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、
そこでは、核酸は、発現制御配列と効果的につながれる。(1)から(5)の要素は、上記に記述されるような核酸配列である。微生物はここに記述されるようなあらゆる微生物、例えばバクテリア、特にグラム陰性のバクテリア、さらに詳細には大腸菌でもよい。ニトリラーゼはここに記述されるような任意のニトリラーゼでもよい。「前記微生物へ核酸を導入する」ことは、ここに記述されるように、形質転換を含んでいる。
【図面の簡単な説明】
【0071】
発明は、次の図および例によってさらに例証され、それらは発明の範囲の制限と見なされるべきでない。
【図1】pAT-NitAfによってコード化された自動トランスポーター融合タンパク質の体系。融合サイトの環境は配列として与えられる。クローニング手順により添加された、N-末端での2つのアミノ酸は、イタリック体で太字で示される。シグナル・ペプチダーゼ切断サイトが示される。SP、シグナル・ペプチダーゼ。
【図2】過剰発現された自動トランスポーター融合タンパク質AFP(1)を含む、大腸菌BL21(DE3)pAT-NitAfの外膜断片(fraction)のSDS-PAGEであり、自動トランスポーターを過剰表現した全体細胞をプロテイナーゼK(2)によって消化したもの、およびマーカー・レーン(M)。
【図3】ニトリラーゼ活性の実証: (R、S)−マンデロニトリルからの(R)−マンデル酸の生産。反応混合物は、全量1ml中で、Na-リン酸緩衝液(50 mM、pH 7.5)、(R、S)−マンデロニトリル(10 mM)、10 OD578に対応する大腸菌BL21(DE3)pAT-NitAfの細胞が含まれる。データポイントは3回の実験の平均であり、標準誤差を表わす棒は小さすぎるので、目に見えることができない。塗り潰された記号は、OD5781での導入を表わし、また、空いた記号はOD5780.5での導入を表わす。Tris-HCI緩衝液pH 7(▼/)、リン酸緩衝液pH 7.5(■/□)。
【図4】(R)−マンデル酸キラルHPLC検出。メタノール中の純粋(R)-マンデル酸(10 mM)(前)、大腸菌BL21 pAT-NitAfを用いたラセミのマンデロニトリル転換からのマンデル酸抽出物(後)。
【図5】R−マンデル酸への(R、S)−マンデロニトリルの転換率のpH依存。その反応は、50mMの酢酸緩衝液(空の棒)あるいは50mMのTris-HCI緩衝液(塗り潰された棒)のいずれかの中で行なわれた。反応時間は37°Cで48時間である。
【図6】-18°Cの保管中の全細胞生体触媒の安定性。細胞は20%のグリセロールを含む培地中で冷凍され、溶かした後に50mMリン酸緩衝液pH 7.5中の大腸菌BL21(DE3)pAT-NitAfによってなされた、10mMマンデロニトリルの転換(マンデル酸生産)が、37°Cで120時間以上で始められた。
【図7】異なる緩衝液および温度条件の大腸菌BL21(DE3)pAT-NitAfの再利用可能性。各周期はそれぞれ24時間の転換期間に対応する。続いて、生体触媒は遠心分離によって除去され(13000rpmで60秒)、新鮮な緩衝液の中で再懸濁され、新鮮な基質(最終の濃度10 mM)と結合し、さらに24時間培養した。塗り潰された記号はTris-HCI緩衝液(50 mM、pH 7) を表わし、空の記号はリン酸ナトリウム緩衝液(50 mM、pH 7.5)、45°C(●/o)、42°C(□/■)、37°C(▲/Δ)、30°C(▼/)を表わす。
【図8】KpnIとXhoIを使用するプラスミドDNAの制限消化。レーン1)1kbのマーカー、2)〜 12) KpnI/XhoIを使用して消化された、形質転換体からのプラスミドDNA、13)1kbのマーカー。
【図9】クローン大腸菌UT 5600(DE3)pAT-NitAfおよびpAT-NitScのトリプシン消化に続く分離された外膜。レーン1)pAT-NitSc、トリプシンで消化、2)pAT-NitSc、消化されていない3)pAT-NitAf、プロテイナーゼKで消化された、4)pAT-NitAf、消化されていな、5)分子量標準(kDa)。
【図10】大腸菌BL21(DE3)pAT-NitAfによるマンデロニトリルの転換に続くHPLC解析。A)ネガティブコントロール。 大腸菌BL21(DE3)、OD578=10、培養時間: 120時間/30°C。1) HPLC、2)マンデロニトリル、3)ベンズアルデヒドの溶媒先端 B) 大腸菌BL21(DE3)pAT-NitAf、OD578=10、120時間/30°Cの培養時間による転換。1) HPLC、2)マンデロニトリル、3)ベンズアルデヒドの溶媒先端。
【図11】組み換えの全細胞生体触媒である大腸菌BL21(DE3)pAT-NitAfを使用して生産されたマンデル酸。
【図12】30°Cおよび37°Cのそれぞれで、リン酸緩衝液pH 7.5中での大腸菌BL21(DE3)pAT-NitAfによるマンデル酸生産。1mMのIPTGを1時間、30°Cで使用して、OD578=1の細胞が導入された。時間t=0の基質濃度は10mM マンデロニトリルであった。
【図13】37°Cでリン酸緩衝液中で、pH 7.5および様々な基質濃度を使用した、大腸菌BL21(DE3)pAT-NitAfによるマンデル酸生産。OD578=1に対して1mMのIPTGを1時間30°Cで使用して、OD578=1細胞が導入された。その後、転換は120時間起こった。
【図14】様々な緩衝液系中の37°C(図の中で示されるような例外がある)での大腸菌BL21(DE3)pAT-NitAfによるマンデル酸生産。30°Cで1時間、1mM IPTGを使用して、OD578=1の細胞を導入した。時間t=0での基質濃度は、10mMマンデロニトリルであり、転換は120時間起こった。
【図15】5dの周期にわたる大腸菌BL21(DE3)pAT-NitAfによる(R)−マンデル酸の生産および同一性の検出。ヘキサンを用いたキラル−OMカラム(company CS-Chromatographie Service):2つのプロパノール:TFA(90:10:0.1)、流量(Flow): 0.5ml/分。 210nmの検出では、A) 酢酸エチルで抽出された大腸菌BL21(DE3)pAT-NitAfの細胞上澄みの分析。B)Aからの転換と結合したラセミのマンデル酸。
【図16】6°C(図16)での異なる保管時間の経過後のマンデル酸生産。50mMリン酸緩衝液pH 7.5中で、10mM マンデロニトリル、37°Cで120時間での、大腸菌BL21(DE3)pAT-NitAfによる転換。
【図17】異なる緩衝液中で、24時間の間で互いに異なる温度で、10mMのマンデロニトリルを1000rpmの熱攪拌機(Thermomixer)中で用いた、大腸菌BL21(DE3)pAT-NitAfによるマンデル酸の生産。
【図18】大腸菌JK321 pAT-NitKpによる3,5-ジブロモ-4-ヒドロキシ-安息香酸の生産の検出。培養時間120時間、OD578=10、基質2.5 mM ブロモキシニル。
【0072】
例
次の例は、本発明者が異なる微生物からの3つのニトリラーゼ遺伝子を同定できたとともに、自動ディスプレイ系へ入る前記遺伝子をクローン化できたことを実証する。
【0073】
例1: Alcaligenes faecalisからのニトリラーゼ
全細胞生体触媒は、特に大量の(bulk)およびまれな化学薬品の生産用の魅力的な技術的なツールで、領域−あるいは鏡像異性選択的な合成が必要である。本研究で、基質としてマンデロニトリルを使用する(R)−マンデル酸を合成するための全細胞生体触媒が構築された。この目的のために、Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750からのニトリラーゼは、自動ディスプレイを使用して、大腸菌(大腸菌)の表面上で発現された。自動ディスプレイ系は、グラム陰性菌のための強力な表面の表示システムで、自動トランスポーター経路に基づく。この技術で、大腸菌の表面上の活性のある形態の複数のホモマー(multihomomeric)のニトリラーゼを発現し、99%以上の鏡像体過剰率を持った鏡像異性的に(enantiomerically)純粋な(R)−マンデル酸を生産することが可能になった。外膜中のニトリラーゼ-自動トランスポーター融合タンパク質の結合性(integration)はSDS-PAGEによってモニターされ、酵素の表面露出はプロテアーゼ接触可能性(accessibility)テストによって確認できる。最適な条件の下では、スモールスケールの生物学的形質転換(biotransformation)(1ml)中のOD57810のバクテリア懸濁を用いて、24時間以内に2.6mMの(R)−マンデル酸を生産することは可能であった。
【0074】
複数のホモマー(multihomomeric)の酵素のためのモデルとして、Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750からのニトリラーゼが使用された。
【0075】
全細胞消化中にプロテアーゼ切断のためニトリラーゼはアクセス可能で、自由な酵素が行ったように、ニトリラーゼはベンズアルデヒド用の同じ基質阻害を示した。
結果
融合タンパク質構築
ニトリラーゼ遺伝子は、公表された塩基配列データから推定されたオリゴヌクレオチドの使用によって、Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750のゲノムDNAからポリメラーゼ連鎖反応(PCR)によって増幅された(実験のセクションを参照)。自動ディスプレイに必要な自動トランスポータードメインの翻訳領域中にニトリラーゼを融合させるために必要なニトリラーゼコード化領域の5`端のXhoIサイトと、3`端のKpnIサイトにPCRプライマーを添加した。
【0076】
ベクターの背骨として、自動トランスポーターの骨組みをコード化する、pET-Adxが上記のように使用された(Jose、2001年)。生じたPCR産物は、XhoI/KpnIで切断されたpET-Adxにつながれ(ligated)、pAT-NitAfを与えるために前の乗客Adxと置き換える。生じる自動トランスポーター融合タンパク質は、シグナル・ペプチド、乗客としてのニトリラーゼ、膜貫通リンカーおよびβ-バレルを含む典型的な構造を示す(図1)。コレラ毒素βサブユニット(CTB)に由来したシグナル・ペプチドは、内部の膜を通る輸送に使用される。外膜を通る乗客の転置のためのβ-バレル(自動トランスポーターのトランスポータードメイン)は、および十分な表面アクセスを達成するための連結地域(膜貫通のリンカー)は、AIDA-I(ホセ、2001年)に由来する。
【0077】
pAT-NitAfによってコード化された、生じた成熟した融合タンパク質は、シグナル・ペプチダーゼによって処理された後に約90 kDaの予想された分子量を持っている。表示されたニトリラーゼの切断を防ぐpAT-NitAfは、大腸菌BL21(DE3)(OmpTネガティブ突然変異体)に形質転換された。この系統は大腸菌BL21(DE3)pAT-NitAfと名付けられた。
【0078】
ニトリラーゼ自動トランスポーター融合タンパク質の発現
人工の融合タンパク質の蛋白質生合成は、実験のセクションに従って1mM IPTGの追加によって始められた。ニトリラーゼが表面または周辺質に完全に露出されたか否かを調査するために、大腸菌BL21(DE3)pAT-NitAfの無傷の細胞を用いた全細胞消化が行なわれた。 大腸菌の外膜がプロテイナーゼK(proteinase K)あるいはトリプシンのようなプロテアーゼには浸透性ではないので、外部的に追加されたプロテアーゼによるタンパク質の分解はその表面の露出になるに違いない。発現と全細胞の消化は、ナトリウム・ドデシルスルファート・ポリアクリルアミドゲル等電点電気泳動(SDS-PAGE)によってモニターされた。続くクマシ・ブリリアント・ブルーによる染色は、プロテイナーゼKの追加が融合タンパク質帯域(図2)の大きな縮小に結びついたことを示した。
分析された細胞の完全性に対して感受性のあるマーカーは、Omp F/CおよびOmpAのタンパク質帯域の強度です。消化されたサンプルおよび未消化のサンプル内の2つのOmp帯域の比率で変更はないに違いない。もしOmpAの帯域が消える場合、外膜は漏れるようになりました。また、OmpAの周辺質内の部分はプロテイナーゼKによって分解された。私たちの実験では、OmpAのサイズおよび量はプロテアーゼで処理された細胞および処理されなかった細胞において同一でした。
【0079】
要約すると、私たちは、ほとんどすべて乗客酵素がバクテリアの表面に露出されたと結論を下した。
【0080】
表面に表示されたニトリラーゼの酵素活性
大腸菌BL21(DE3)の分析のために、pAT-NitAfプラスミドを含む細胞は、1mlの全容量中の50mMリン酸ナトリウム緩衝液pH 7.5において、10の578nm(OD578)の光学濃度に調節された。酵素がヒドロキシニトリル(hydroxynitriles)のための基質特異性を示すと説明されるので、ラセミのマンデロニトリルは最終濃度10 mMで基質として使用された。生物学的変換はHPLCによって分析され、生産されたマンデル酸が計算された。最初の生物学的変換は、0.5のOD578でIPTGによって誘導された細胞で試みられた。これは72時間の生物学的変換の後に2.1mMのマンデル酸の生産に結びついた。120時間までのさらなる培養は、生産されたマンデル酸のさらなる増加に帰着しない。生物学的変換は、1のOD578でIPTGによる誘導によって2つの大きさの中で著しく改善された。この修正で、生物学的変換は72時間で横ばいにはならないが、しかし対照的に120時間の観察期間に合計6.6mMのマンデル酸に増加した(図3)。追加基質の量は、自由なベンズアルデヒドによって引き起こされた酵素阻害の発生により制限されており、それは、マンデロニトリルの分解によって自然に形成される(Yamamoto et al.、1992年)。
【0081】
マンデル酸へのマンデロニトリルの転換
HPLC法は、産物(マンデル酸)から基質(マンデロニトリル)を分けて、かつそれらを検出するために開発された。大腸菌BL21(DE3)pAT-NitAfを用いたマンデロニトリルからマンデル酸への転換が確認された(図10)。HPLC解析は、予想通りに、水性系(aqueous system)中のマンデロニトリルがベンズアルデヒド(+HCN)との平衡の中にあることをさらに明らかにした(図10)。いくつかの反応パラメータの適用は、10のOD578の細胞懸濁が使用された時(図11)、5mMの基質濃度からスタートして120時間後に、2.4 mM(それは1mlのサンプル当たり0.36mgの塊り(masse)に相当する)の産物濃度に帰着した。
鏡像体過剰率
次のステップは生産されたマンデル酸の鏡像体過剰率を確認だった。この目的のために、マンデル酸は、酢酸エチルを用いた溶媒抽出によって水系の緩衝液から抽出された。抽出されたマンデル酸は、メタノールの中で溶解され、キラルHPLC解析に直接使用された。鏡像体過剰率(ee)は37°Cの生物学的変換で、ee>99%であるとわかった(図4および15A)。これは、A.faecalisの表面の表示されたニトリラーゼが、高い鏡像異性的な純度をもった(R)-マンデル酸を生産するのに有能なことを示す。図15Aから明らかなように、マンデル酸の(R)−鏡像異性体のほとんど排他的なピークで特徴的な保持時間が検知できた。計算された鏡像異性的な純度は99%以上だった。図15Bは、図15Aからの転換された純粋(R)-および(S)−鏡像異性体のスパイク波形により決定されるような、(R)−マンデル酸の個性を示す。
pH最適条件
Alcaligenes faecalis ニトリラーゼのpH最適条件はpH 7.5であると以前に公表されていたが、私たちは、表面露出によりpH最適条件に変化があるかどうかを調査した。細胞は、最終で10のOD578になるように酢酸緩衝液あるいはTris-HCl緩衝液(両方とも50 mM)の中で再懸濁された。37°Cで120時間の培養の後、上澄みはHPLCによって分析され、また、変換されたマンデル酸の量が計算された。私たちは5.5mMマンデル酸の転換でpH7がpH最適条件であると決定した。pH 7とpH 8の間で強い活性の損失が検出できた。追加的に調査されたリン酸緩衝液pH 7.5は、その他の点は同じ条件の下で、たった4.4mMのマンデル酸の転換に結びついた(データを示さず)(図5)。
【0082】
反応温度およびpH値の最適化で、24時間以内の2.6mMまでのマンデル酸を変換することが可能であった。マンデルアミドは常に検知できなかった。
【0083】
反応速度
一連の実験では、いくつかのパラメータは、実験室スケールでのマンデル酸生産に対するそれらの影響のために分析された。図12は、異なる温度でのマンデル酸生産の反応速度を表わす。37°Cでは、30°Cでの転換と比較して、産物の濃度が約20%増加したことが検知された。30°Cおよび37°Cの両方の転換の進行は、生産されたマンデル酸の濃度が約144時間の転換の後も単にゆっくり増加することをさらに実証した。
基質濃度
さらに、マンデル酸の生産に対する基質濃度の影響が分析された。水溶液(aqueous solution)では、基質のマンデロニトリルは、ベンズアルデヒドおよびHCNと平衡状態にある。しかしながら、過度の濃度にあるベンズアルデヒドは、細胞に毒作用を示し、それによって図13に示されるように基質濃度の増加を制限する。10 mM未満へのニトリル濃度のさらなる縮小が生産速度のさらなる増加に帰着しなかったことは、最適な基質濃度として10mMマンデロニトリルの使用を示唆する。
【0084】
異なる緩衝液システム
全細胞触媒の大腸菌BL21(DE3)pAT-NitAfを使用するマンデル酸生産の相対的な試験も異なる緩衝液システムを使用して行なわれた。図14は、最良の結果が、使用される他の緩衝液と比較して、pH 6.5のトリスバッファーを用いた溶液を使用して達成されたことを示す。
【0085】
貯蔵安定性
構築された全細胞生体触媒全体の貯蔵安定性を決定するために、大腸菌BL21(DE3)pAT-NitAfの細胞(IPTGで誘導される)は、20%(v v-1)のグリセロールを含み、格納されて−18°Cで保存されて、50mMリン酸ナトリウム緩衝液pH 7.5中の、50のOD578に調節された。グリセロールは、細胞の溶解を防ぐための不凍薬剤として-18°Cでサンプルに添加された。異なる時間間隔で、サンプルは、一度リン酸緩衝液の中で洗われ、10のOD578に調節された。あるいは、IPTG誘導ののち、細胞の部分標本は冷蔵庫中で6°Cで保管され、異なる保管時間の後にニトリラーゼ分析にさらされた。反応は10mMマンデロニトリルの追加によって始められ、また、サンプルは37°Cで120時間培養された。 -18°Cで35日の保管の後、細胞は、まだそれらの初期の活性の72%を保持していた(図6)。1週後の、+6°Cの全細胞触媒の活性は、初期活性のわずか約50%だった(図16)。さらに、これらの実験では、これは観察された細胞の溶解と関連した。ここで、再び、細胞の一部が測定された期間にわたる比較的安定な残余活性を示すことは明白である。
【0086】
全細胞触媒の温度依存性
A.faecalisから精製された酵素の最適温度が40−45°Cであることが説明される。 これらの発見は、30°Cから45°Cまでの範囲の温度テストについて記述する図17によって確認された。反応混合物中のリン酸緩衝液あるいはトリス緩衝液の使用は、特別の影響があるようには見えない。
【0087】
全細胞触媒の再利用性(図7)
24時間5周期の転換反応の全細胞生体触媒の再利用性および安定性を調査することは、異なる緩衝液および温度条件で行なわれた。「周期」は、大腸菌細胞の遠心分離および次の周期のための新鮮な基質の追加に続く24時間の転換反応である。24時間以内の最も高い産出が、2.62mMマンデル酸でリン酸ナトリウム緩衝液(pH 7.5および45°C)中の大腸菌BL21(DE3)pAT-NitAfで達成されたが、しかし、活性は、5つの周期の後に最小で当初の活性のたった10%に速く落ちた。一次的転換レートの33%を超える5つの周期を達成できた後、Tris-HCl緩衝液(pH 7および45°C)を用いて、2.53mMマンデル酸の転換が24時間以内に達成可能だった。Tris-HCl(37°C)の中の細胞の活性は最初の周期の2.24 mMから5つの周期の後には1.25 mMまで落ちたが、これは、55%を超える残余の活性を表わす。最初の周期中では、リン酸ナトリウム緩衝剤あるいはTris-HCl緩衝液の中で再懸濁された細胞間の生産速度に小さな違いだけがあったが、だが対照的にすべて成績においてTris-HCl緩衝液を用いた試験サンプルの方が常によかった。
【0088】
実験のセクション
バクテリア株およびプラスミド
サブクローン化の目的で、Invitrogen (Carlsbad, CA, USA)から提供される大腸菌TOP10 [F- mcrA (mrr-hsd RMS-mcrBC) Φ80lacZ M15 lacΧ74 recA1 araD139 (ara-leu)7697 galU galK rpsL (StrR) endA1 nupG]が使用された。大腸菌BL21 (DE3) [B, F-, dcm, ompT, lon, hsdS (rB- mB-), gal, λ (DE3)]は、自動トランスポーター融合タンパク質の発現に使用された。
【0089】
大腸菌系統は、活発な揺れ(200rpm)とともにアンピシリン(100 μgmL-1)を含んでいるLuria-Bertani (LB)培地で37°Cで慣例的に培養された。固形培地は、培地に寒天(agar) (1.5 %, wv-1)の追加して調製された。
【0090】
Invitrogen (Carlsbad, CA, USA)からのプラスミドpCR4-TOPO(登録商標)はニトリラーゼ遺伝子のサブクローン化のために使用された。AIDA-I自動トランスポーターの骨組みをコード化するプラスミドpET-Adx04の構築は、以前に記述された(Rustler et al.、2008年; 米国特許6,180,359番)。図8は、11の形質転換体から分離されたプラスミドを示し、そこではPCR破片に対応する約1kbの正確なDNA断片は、KpnIとXhoIを使用した続く消化によって、7つのクローン中の正しい自動トランスポータープラスミド(約7kb)内に検知された。
【0091】
組み換えDNA技術
ニトリラーゼをコード化する遺伝子は、Alcaligenes faecalisの亜種のfaecalis ATCC 8750 (LGC Standards GmbH, Wesel, Germany)のゲノムDNAから増幅された。
【0092】
ニトリラーゼ遺伝子の増幅用のプライマーは、A.faecalis ATCC 8750(Kaul et al.、2007年ら)のニトリラーゼ遺伝子のための既に記述されていた配列情報に基づいた。前方のプライマーNit-XhoI (5´- CCGCTCGAGCAGACAAGAAAAATCGTCC)および逆のプライマーNit-KpnI (3´- GGGGTACCGGACGGTTCTTGCACC)は、ニトリラーゼ遺伝子を含む1073bp破片を増幅するために使用された。PCR産物のクローニングを容易にするためにXhoIおよびKpnIの制限部位が添加された(プライマー配列中の強調された部分は、それぞれの酵素用の制限部位を表わす)。PCR反応は、10 μLの体積中において、10 ngのゲノムDNA、Eppendorf (Hamburg, Germany)から提供される適切な量のEppendorf PCR Mastermix、1μL(10μMの原液)の前側および後ろ側プライマー(Sigma-Aldrich Chemical Company, Milwaukee, USA)を含む。
【0093】
PCR反応は、Eppendorf Mastercycler gradient from Eppendorf (Hamburg, Germany)によって実現される。PCRプロトコルは、最初の変性ステップが95°Cで5min、30サイクルの、94°Cの30秒、64°Cの1分、72°Cの1分、が続き、および72°Cで6分の最終の延長ステップで終わる。
【0094】
PCR産物は、メーカーの指示によってpCR4-TOPO Vectorへのサブクローン化のために直接使用された。別記されるように、プラスミドDNAの調製が行なわれた。生じたプラスミドは、XhoI/KpnIで消化され、Qiagenゲル抽出キット(Qiagen, Hilden, Germany)を使用して、アガロース・ゲル電気泳動によって精製された。
【0095】
pAT-NitAfに与えるために、生じた破片は、XhoI/KpnIで切断されたpET-Adx04ベクターに挿入され、そこではAdx04をコード化する遺伝子に交換された。
【0096】
外膜標本およびSDS-PAGE
大腸菌細胞はオーバーナイトで成長され、そして、適切な量の培養は、2−メルカプトエタノール(10 mM)およびアンピシリン(100 μgmL-1)を含むLB培地への接種(inoculation)に使用された。0.5または1の光学濃度(OD578)が到達するまで、細胞は37°C(200rpm)で培養された。誘致可能な発現のため、Roth (Karlsruhe, Germany)から購入されたIPTG(1mMの最終濃度)が添加された。30°Cに60分おいた後、細胞は収穫され、Tris-HCl(0.2M、pH 8.2)で洗われた。プロテアーゼ接触可能性テストのため、細胞は、25μLのプロテイナーゼK-溶液(2%)を含む2mlのTris-HCl (0.2M、pH 6.8)の中で再懸濁され、37°Cのウォーター・バスの中で60分培養された。異なる細胞の分別法は、Hantke(Hantke、1981年)の方法の修正によって行なわれた。
【0097】
SDS-PAGEのため、単離された外膜は、Tris-HCl(100 mM、pH 6.8)、ドデシル硫酸ナトリウム(4%)、ブロモフェノール・ブルー(0.2%)、2−メルカプトエタノール(2%)、ジチオスレイトール(0.2M)およびグリセロール(20%)から成るサンプル緩衝液で、1:2で薄められ、それは5分煮沸され、10%のポリアクリルアミドゲルで分析された。タンパク質はクマシ・ブリリアント・ブルーで染色され、Fermentas(St. Leon-Rot, Germany)から購入されたPageruler不染色タンパク質ラダーは、サイズ決定に使用されました。
【0098】
標準転換を行なうこと
標準的な実験は以下のように実行される。プレカルチャーされた組み換え大腸菌BL21(DE3)pAT-NitAf系統が、1:100の比率でメインカルチャーに接種するために使用され、振盪培養器の中で37°Cおよび200rpmで培養される。1のOD578に達する際、IPTGは1mMの最終濃度で使用され、誘導のために添加される。その後、誘導は30°C、200rpmで1時間起こる。その細胞が50mMリン酸緩衝液pH 7.5で一度洗われた後、また同じ緩衝液を使用して、最終的に10のOD578に調節されました。その後、前記細胞懸濁の1mlの部分標本は、20μlのマンデロニトリル(0.5M)と一緒にされ、1000rpmあるいは200rpmで熱攪拌機の中で異なる温度(特に37°Cの中で)で培養される。サンプルは異なる時間間隔で得られ、遠心分離機にかけられ(6000 x g、5分)、VWR (Darmstadt, Germany)から購入された0.2μmのポリエーテルサルホン(polyethersulfone)薄膜フィルタで滅菌ろ過された。上澄みは、HPLCによって直接分析された。それ以上、生物学的変換の滅菌ろ過の後に検知できなかった。
【0099】
分析手法
ニトリル転換は、Waters (MA, USA)からのダイオード・アレー検波器996と2695分離モジュールを装備した、HPLC(Millenium Chromatography Manager 3.2)によって分析された。 アキラル(achiral)な分析の目的で、210nmで分光測光法で検知された転換産物を識別するために、ヒューレット・パッカード(Boblingen, Germany)から提供される逆相カラム[100mm x 4.6mm(内部直径)、Hypersil ODS 5μm直径粒子で満たされた]が使用された。水の中のメタノール(40% vv-1)およびリン酸(0.1%)は、移動相としてHPLC分離に使用された。マンデル酸鏡像異性体の分離のために、CS-Chromatographie Service (Langerwehe, Germany)から提供される、キラル位相カラム[150mm x 4.6mm(内部直径)、Reprosil Chiral-OM 5μm直径の粒子で満たされた]が使用された。
【0100】
移動相は、ヘキサン(90 % vv-1)、イソプロピルアルコール(10 % vv-1)およびトリフルオロ酢酸(0.1 % vv-1)から構成された。マンデル酸は、ジクロロメタン(dichlormethane)の代わりに酢酸エチルが使用された少しの修正(Yamamoto, 1991)と共に、別記されるように生物学的変換サンプルから抽出された。
【0101】
例2: Klebsiella pneumoniaeからのニトリラーゼ
Klebsiella pneumoniaeからのニトリラーゼ遺伝子は、例1に記述されるように、PCRを使用して増幅され、大腸菌標準ベクターへの中間体クローニングおよび塩基配列分析に続いて、対応する自動ディスプレイ・ベクターに挿入された。K. pneumoniaeニトリラーゼを含む融合タンパク質、自動トランスポータードメイン、膜貫通リンカーおよびシグナル・ペプチドをコード化する自動ディスプレイの構成物は、例1にpAT-NitAfのために記述されたとともに、pAT-NitKpと名付けられた。表面上に融合ポリペプチドを表示する、対応する大腸菌系統は、大腸菌BL21(DE3)pAT-NitKpと名付けられた。
Klebsiella pneumoniaeニトリラーゼの発現および表面の局在化の検出は、Alcaligenes faecalisニトリラーゼ(例1参照)と類似して実行された。
【0102】
それは、前記ニトリラーゼがブロモキシニルのための非常に高い基質特異性を示すと言った文献から有名である。前記有毒化合物は、農業で除草剤として広く使用され、毎年、ドイツで工業規模に適用されます。対応するカルボン酸は、低い毒性と自然界での安定性を示す。
【0103】
大腸菌JK321 pAT-NitKpによるブロモキシニルからの3,5-ジブロモ-4-ヒドロキシ-安息香酸の生産は、図18に示されます。この図は、大腸菌細胞の表面上で発現されたK. pneumoniaeニトリラーゼがニトリルをその対応するカルボン酸に変換することができることを示す。
【0104】
例3:Saccharomyces cerevisiaeからのニトリラーゼ
例1に記述されるように、S.cerevisiaeのニトリラーゼ遺伝子はPCRによって増幅されクローン化された。塩基配列分析は、データベースから得られたS.cerevisiae ニトリラーゼ配列と100%の同一性を示した。例1に記述されるように、ニトリラーゼは、自動トランスポータードメイン、膜貫通リンカーおよびシグナル・ペプチドを含む融合タンパク質として大腸菌中でクローン化された。再び、塩基配列分析はデータベースからの配列と100%の同一性を示した。発現の検出は例1と類似して実行された。
【0105】
S.cerevisiaeニトリラーゼ、自動トランスポータードメイン、膜貫通リンカー、およびシグナル・ペプチド、を含む融合タンパク質をコード化する自動ディスプレイの構成物は、例1にpAT-NitAfのために記述されるとともに、pAT-NitScと名付けられた。表面上に融合ポリペプチドを表示する、対応する大腸菌系統は、大腸菌BL21(DE3)pAT-NitScと名付けられた。
【0106】
IPTGの誘導に続いて、S.cerevisiae ニトリラーゼを含む融合タンパク質は、表面に局在化されると分かった。例1に記述された全細胞のトリプシン消化が適用された。結果は図9に要約された。
参考文献
1. Jose J, Meyer TF (2007)、自動ディスプレイの話、発見からバイオテクノロジー的・バイオ医学的な適用まで。Microbiol Mol Biol Rev 71 (4): 600-19
2. Nagasawa T, Mauger J, Yamada H (1990)、新規なニトリラーゼ、Alcaligenes faecalis JM3のアリールアセトニトリラーゼ(arylacetonitrilase)、精製および特徴の描写。Eur J Biochem 194(3): 765-72
3. Yamamoto K, Oishi K. Fujimatsu I, Komatsu K (1991)、Alcaligenes faecalis ATCC 8750によるマンデロニトリルからのR-(-)-マンデル酸の生産。Appl Environ Microbiol. 57(10): 3028-32
4. Rey P, Rossi JC, Taillades J, Gros G, Nore O (2004)、動けなくされたニトリラーゼを使用するニトリルの加水分解:メチオニン・ヒドロキシ類似誘導体(methionine hydroxy analogue derivatives)の合成への適用。J Agric Food Chem 52 (26): 8155-62
5. Kiziak C, Conradt D, Stolz A, Mattes R, Klein J (2005)、Pseudomonas fluorescens EBC191からのニトリラーゼ: 遺伝子のクローニングおよび異種起源の発現および組み換えの酵素の生化学の特徴の描写(characterization)。Microbiology 151 (Pt 11): 3639-48.
6. Kaul P, Stolz A, Banerjee UC (2007)、鏡像異性選択的なニトリル加水分解のための架橋された無定形のニトリラーゼの集合。Adv Synth Catal 349: 2167-2176
7. Banerjee A, Dubey S, Kaul P, Barse B, Piotrowski M, Banerjee UC (2008) 、Pseudomonas putidaからの鏡像異性選択的なニトリラーゼ: クローニング、異種起源の発現およびバイオリアクター研究。Mol Biotechnol [Epub ahead of print]
8. Luo H, Fan L, Chang Y, Ma J, Yu H, Shen Z (2008)、大腸菌中のRhodococcus rhodochrous tg1-A6からのニトリラーゼの、遺伝子クローニング、過剰発現、および特徴の描写。
Appl Biochem Biotechnol [Epub ahead of print]
9. Rustler S, Motejadded H, Altenbuchner J, Stolz A (2008)、Pseudomonas fluorescensからのアリールアセトンニトリラーゼ(arylacetonitrilase)の同時の発現、そして(S)―マンデル酸合成用のPichia pastorisのManihot esculentaからの(S)−オキシニトリラーゼ。Appl Microbiol Biotechnol 80(1): 87-97
10. ニトリラーゼを使用する2-ヒドロキシ-4-メチル酪酸の調製のための工業規模プロセス。米国特許6180359。
11. Ress-Loschke M. Friedrich T, Hauer B, Mattes R (1998)、ニトリラーゼまたはニトリラーゼの遺伝子を含む微生物を用いてニトリルからの光学活性カルボン酸の製造方法。ドイツ特許出願DE 19848129A1
12. J. Jose, Appl. Microbiol. Biotechnol. 2006, 69, 607-614.
13. J. Jose, R. Bernhardt, F. Hannemann, J. Biotechnol. 2002, 95, 257-268.
14. J. Jose, S. von Schwichow, ChemBioChem 2004, 5, 491-499.
15. K. V. Thimann, S. Mahadevan, Arch. Biochem. Biophys. 1964,105,133-141.
16. A. Banerjee, R. Sharma, U. C. Banerjee, Appl. Microbiol. Biotechnol. 2002, 60, 33-44.
17. R. N. Thuku, D. Brady, M. J. Benedik, B. T. Sewell, J. Appl. Microbiol. 2009, 106, 703-727.
18. K. Yamamoto, I. Fujimatsu, K.-I. Komatsu, J. Ferment. Bioeng. 1992, 73, 425-430.
19. J. Jose, R. Bernhardt, F. Hannemann, ChemBioChem 2001, 2, 695-701.
20. J. Maurer, J. Jose, T. F. Meyer, J. Bacteriol. 1997, 179, 794-804.
21. H. C. Birnboim, J. Doly, Nucleic Acids Res. 1979, 7, 1513-1523.
22. K. Hantke, Mol Gen Genet 1981, 182, 288-292.
23. Henderson I et al., 2004、タイプVタンパク質分泌経路: 自動トランスポーターの話。Microbiology and Molecular Biology Reviews, 68(4), 692-744.
24. Niewert U., Frey A., Voss T., Le Bouguen C., Baljer G., Franke S., Schmidt MA. AIDA自動トランスポーター系は、浮腫病およびポスト離乳食下痢と診断されたブタから単離された大腸菌のF18とStx2eに関係する。Clin. Diagn. Lab. Immunol. 2001 Jan, 8(1):143-149;9.
25. Buchholz, Kasche und Bornscheuer (2005)。生体触媒および酵素技術。Enzymes in Organic Chemistry, Wiley-VCH-Verlag, Weinheim.
26. Hartl, F. U., and Hayer-Hartl, M. (2002) Science 295(5561):1852-8
【説明】
【0001】
本発明は、ニトリラーゼ(nitrilase)によって触媒作用を及ぼされた反応の産物を生産する方法に関し、その方法は、(i)その表面に置かれた前記ニトリラーゼを含む微生物、および/または、前記微生物の膜標本(membrane preparation)を用意し、そして(ii)ニトリラーゼ活性と適合性をもつ条件の下で、微生物および/またはそれの膜標本を1つ以上のニトリラーゼ基質と接触させる、ステップを含む。特に、本発明は、(i)その表面上のニトリラーゼを含む微生物および/またはそれの膜標本を用意し、(ii)ニトリラーゼがニトリルのカルボン酸への転換に触媒するように、微生物および/またはそれの膜標本をニトリルと接触させる、のステップを含むカルボン酸を生産する方法に関する。本発明は、さらに、ラセミ(racemic)のマンデロニトリルの処理に使用する、ニトリラーゼを提示する全細胞生体触媒あるいはそれの膜標本を使用して、純粋な(R)-マンデル酸を鏡像異性的に生産する方法に関する。
ニトリルは、様々な産物(例えばカルボン酸、アミド(Banerjee et al.、2002年))を生産するための重要な前駆体分子である。ニトリルの高い安定性により、しかしながら、ニトリルの化学的転換のために高温でそれらを強い塩基性条件あるいは酸性条件にさらすことが必要である(Banerjee et al.、2002年、Nagasawa et al、1990年)。さらに、それぞれの反応の混合物からのこれらの産物を精製することは非常に長時間かかりうんざりするものである。
【0002】
ニトリラーゼ (例えばEC 3.5.5.1)は、ニトリルを一つのステップ(ThimannおよびMahadevan、1964年)で対応するカルボン酸およびアンモニアに変換することができる酵素である。一方では、カルボキシリック(carboxylic)な産物が種々様々の化学の生産工程の中で中間体としてしばしば使用され、他方では、ニトリラーゼが、廃液(waste)の解毒に使用されてもよい(Banerjee et al.、2002年; Thuku et al.、2009年)ので、ニトリラーゼ酵素の使用は本質的に産業的な興味を引きつけた。更に、従来の化学(Banerjee et al.、2002年)において必要な、集中的な(intensive)ブロッキングおよび脱ブロッキングステップおよび触媒を必要としないで、穏やかな条件の下でニトリラーゼは高度に選択的で、鏡像異性的に純粋なカルボン酸を産出する。前記選択性の高さは、従来の方法を使用して達成しなかった、或いは困難とともに達成された産物合成を可能にする。
【0003】
量に関するニトリラーゼの産業適用で最も重要なものは、ラセミのマンデロニトリルを鏡像異性的に純粋な(R)-マンデル酸に転換することである(Rey et al.、2004年)。(R)-マンデル酸は、ジアステレオマー塩を使用して、ラセミ化合物中の鏡像異性体を分離するために使用された。更に、それは、新規の活動的な薬剤の化学合成中での重要なキラル(chiral)中間生成物である。
【0004】
A.faecalisからのニトリラーゼがマンデロニトリルを高い鏡像異性選択性を備えたマンデル酸に変換することができることは文献から有名である。マンデル酸のようなキラルのカルボン酸は、それらが多くの薬学的薬剤か病虫害防除薬剤のための基礎製品として役立つので、有機化学の中で望まれた化合物を表す。したがって、例えば(R)−あるいは(S)−マンデル酸、ラセミのアミンのラセミ溶液のために使用されます。さらに(R)−マンデル酸は中間体として合成に使用されます。
しかしながら、ニトリラーゼは悪評高いほどに不安定な酵素(Buchholz、KascheおよびBornscheuer、2005年)であると見なされており、従って、産業の中で精製された酵素としてめったに使用されないか、全体細胞の標本(preparation)として使用されます。ニトリラーゼの活性中心は、自動酸化(Kobayashi et al.、1992年)に弱く、チオール含んでいる試薬のような多くの化学薬品の方へ反応的である、保存された活性部位のシステイン残基(Kobayashi et al.、1992年)を含む3つのアミノ酸残基(触媒現象のトリオ)の触媒配列を含んでいる。
【0005】
細胞の内部に存在する間、ニトリラーゼはバクテリアの細胞質ゾルの酸化還元反応の環境によって保護され、それは減少され(ChoiおよびLee、2004年)、そして、チオール基(thiol groups)の酸化を防ぐ。しかしながら、ニトリラーゼは酸素へ接触すると活動を失うことが示され、また、この所見は、活性中心(Mateo et al.、2006年)中の重要な酸化還元反応感受性のトリオ・システインの感受性に起因した。
【0006】
いくつかの戦略は不十分な活性および/または安定性の点からニトリラーゼの欠点を克服するために使用された。
【0007】
1つのアプローチでは、精製した酵素製剤ではなくむしろニトリラーゼを発現する全細胞が使用され、それは系統特異的か或いは組み換えの形式で(Kaul et al.、2007年)ニトリラーゼを細胞内で提供される。例えば、研究者は、細胞の内部で置いたニトリラーゼを含む全細胞触媒として、ニトリラーゼを発現するAlcaligenes faecalisの細胞を使用した(Kaul, A、BanerjeeおよU. C. Banerjee、2006年)。97%の鏡像異性体過剰率(enantiomeric)が達成された。このアプローチの基礎となる論理的根拠は、酵素がそれらの自然環境、すなわちバクテリアの細胞質ゾルに残るということであり、そこでは内発的な折り畳み装置類および連続的な転覆(turnover)および蛋白質生合成が、安定で活性のあるタンパク質を一定のレベルの維持する。しかしながら、時間を消費し労力の大きいの手順を使用して、全細胞触媒は調製されるに違いないし、適切なマトリックス系(アルギン酸塩)に付けられるに違いない。さらに、ひっくり返される基質は、選択的な細菌細胞の細胞膜を横切るとともに、選択的な活性を有するとともに基質競合的な酵素の射程範囲内である細胞質ゾル内に一度入るように対向する必要がある。最後に、全細胞触媒は、望ましい産物の調製を汚染するので、したがって、敏感な適用、例えば飲料および食料と同様に医学、化粧品および衛生的製品の調製、のためのそのような製品への使用を除外される。
【0008】
工業的用途用の機能的なニトリラーゼの高いレベルの生産への別のアプローチは、シャペロン(chaperones)の使用に関係する。GroELファミリーのシャペロンは細胞質ゾルにある不可欠なタンパク質で、それは、ATP依存性の結合の多数の輪(rounds)、カプセル化および基質タンパク質の放出を経由した、正確に折り重ねられたタンパク質の生産に必要である(Hartl, F.U.およびHayer-Hartl, M、2002年)。GroELの削除は、結局、機能的な基質タンパク質の低いレベルだけでなく、大量の折り畳まれないで蓄積されたタンパク質に起因する細菌細胞の死に導かれる。シャペロン、より明確に、GroEL系の構成要素は、大腸菌の細胞質ゾル中で組み換えのニトリラーゼとともに共発現(co-expressed)された(米国特許5,629,190番および図9)。このアプローチは、内発的なレベルのシャペロンだけをもつ細胞に比して、より高いレベルの可溶のタンパク質に至った。しかしながら、そのような折り畳み装置が設けられた全細胞の使用、あるいは単独に精製されたシャペロンおよびそれらの共同因子(cofactors)およびニトリラーゼ調製のために追加的な基質の付加は、そのようなアプローチの必要条件である。
米国特許5,629,190番は、ニトリラーゼを組換え的(recombinantly)に発現する全(whole)大腸菌細胞の使用を教示する。しかしながら、たとえGroELのようなタンパク質薬剤、微生物が合成するポリペプチドを折り畳むことを支援するための、例えば、組み換え的に発現されたニトリラーゼ、を組み換え的に発現する場合であっても、細胞の特定の細胞内酵素設備のため、生じる反応は複雑で、それらの結果は予測不能である。米国特許5,629,190番に示されるように、ニトリラーゼが触媒作用するニトリルのカルボン酸への転換は、産物の減少および除去するのが難しい望まれない副産物に結びつく、ニトリルヒドラターゼ(hydratase)が触媒作用する反応(酸アミド形成に帰着する)と競合する。細胞の中にある他の酵素はニトリル基あるいは他の半分(moiety)でもニトリルを修飾するかもしれない。
【0009】
別のアプローチは、および架橋された酵素集合体(つまり可溶の組み換えの酵素、およびグルタルアルデヒドまたはポリ(エチレンイミン)のような反応性の二価性の化学薬品を使用する二次的架橋)の調製および使用に関係する。しかしながらKaul et al. (2007)は、そのような固定が反応速度の著しい減少および一定の特異性の減少に帰着したことを実証した。さらに、化学的架橋は、変更された基質特異性(特定の形態に「固定された」タンパク質分子を容易に産出した架橋プロセスに関連された観察)に結びついた。さらに、それらの分解された産物および残骸と同様に動けなくされた酵素の一群は、望ましい反応生成物から分離するには簡単ではないかもしれない。
【0010】
Yamamoto et al.(1991)は、マンデロニトリルを(R)-(-)-マンデル酸に変換するそれらの能力に関して、様々な分離物(野生種(wildtype strains))をテストした。A.faecalis ATCC 8750は、ラセミのマンデロニトリルからの(R)-(-)-マンデル酸を生産するために最高レベルの活性と鏡像異性選択性とを持つことが発見された(Yamamoto et al.、1991年)。特に、休眠細胞(resting cells)によってマンデロニトリルから作られた(R)-(-)-マンデル酸は、100%の鏡像体過剰率にあった。A.faecalis ATCC 8750は、マンデロニトリル(マンデロニトリル)およびマンデルアミド(mandelamide)、をそれぞれ処理するために使用される1つの(R)−鏡像異性選択的なニトリラーゼおよびアミダーゼを持っている。(R)-(-)-マンデル酸は、91%の収量でのラセミのマンデロニトリルから生まれるが、しかしながら、反応で残る(S)-マンデロニトリルは、一方では(R)―および(S)―異性体のマンデロニトリルの化学平衡、および他方ではベンズアルデヒド/HCNの化学平衡のために、(S)-マンデロニトリルは残されず自発的にラセミ化される。従って、マンデロニトリルはほとんどすべて消費され、(R)-(-)-マンデル酸に転換される。Yamamoto et al. (1991)によれば、(R)―マンデル酸の生産のために使用されたA.faecalis細胞は、特に最適化された培養条件を要することが注目される。
【0011】
バクテリアの起源の異なるニトリラーゼは、クローン化され、大腸菌の中で組み換え的に発現された(Kiziak et al.、2005年; Luo et al.、2008年; Rustler et al.、2008年; 米国特許6,180,359番; Ress-Loschke et al.、1998年)。ニトリラーゼは生化学的な特徴が述べられた(Kiziak et al.、2005年)。さらに、大腸菌のA.faecalis ATCC8750のニトリラーゼ遺伝子を組み換え的に発現させることが可能で、それは、精製後に「架橋した酵素集合体」として、動けなくされた実験の中でその後使用された(リイ、2004年ら)。この方法では、宿主微生物、つまり大腸菌、からニトリラーゼを分離し、浄化しなければならない。このアプローチは全く長時間かかりうんざりするものである。さらに、ニトリラーゼは手順(procedure)の間に非活性化されるかもしれない。
【0012】
Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750からのニトリラーゼは、1つのサブユニット当たりの分子量32 kDを備えたおよそ14のサブユニットから構成されるホモオリゴマー(homooligomeric)の酵素で、よって、天然タンパク質の合計の結合した分子量はおよそ460 kDであるとして、記述された(Yamamoto et al.、1992年)。一般に、ニトリラーゼは不活発な二量体として存在し、自己会合を通じて活発なオリゴマーを形成する。しかしながら、活動に必要とされたサブユニットの数はニトリラーゼごとに異なる(Thuku et al.、2009年)。
要約すると、ニトリラーゼには有機化合物の生産でのニトリラーゼの使用を妨げるいくつかの欠点がある:
ニトリラーゼは、酸化に対して敏感であり、時間とともにニトリラーゼ活動の縮小に帰着する。
【0013】
全細胞生体触媒の形で使用された時、細胞の中にある他の酵素(ニトリルヒドラターゼのような)によって触媒作用を及ぼされた望まれない副反応は、ニトリラーゼによって触媒作用を及ぼされた反応を妨害するかもしれない。
ニトリラーゼは反応混合物から回復(recovered)することができない。
自動表示(Autodisplay)は、組換え型タンパク質のバクテリアの表面の表示用の簡潔なツールを表わす。この表現系は、タイプV分泌系に属するタンパク質の自動トランスポーター・ファミリー(autotransporter family)の分泌メカニズムに基づく。
グラム陰性菌では、自動トランスポーター経路は、細胞表面へのタンパク質の輸送、および細胞外の環境の中へのタンパク質の分泌の両方のために発展した(Jose and Meyer、2007年)。自動トランスポータータンパク質は、細胞表面への輸送のための構造の必要条件をすべて含んでいる前駆体タンパク質として合成される(Jose、2006年)。それらは、内部の薄膜の交差を可能にするSec 経路には典型的なN-末端のシグナル・ペプチドで合成される。一旦、周辺質(periplasm)では、シグナル・ペプチドの切断の後、前駆体のC-終末部がポーリン(porine)状の構造、いわゆるβ-バレルとしての外膜内へ折り畳まれる。この孔によって、N-末端に付属した乗客ドメインは、表面へ移動させられる(Jose、2002年)。そこで、それは切断(自己タンパク分解的に(autoproteolytically)あるいは追加のプロテアーゼによって)されるかもしれないし、あるいはトランスポータードメインによって細胞外被にアンカーされ続けるかもしれない。元の乗客を組換え型タンパク質に取り替えることはその適切な表面の転置(translocation)に帰着する。この目的のために、シグナル・ペプチド、組み換えの乗客、β−バレルおよびリンク地域、それらの中間にある連結位置(linking region)(それは全表面へのアクセスを達成するために必要です)、をからなる、人工の前駆体が遺伝子工学によって構築されなければならない。AIDA-I自動トランスポーターは、様々な乗客ドメインの効率的な表面への表示するため、このように使用されて成功した(Henderson et al.、2004年)。
【0014】
自己ディスプレイ(autodisplay)系では、活性のある酵素へのサブユニットの自己会合は、例えば二量体酵素ソルビトール・デヒドロゲナーゼの中で観察された(Jose (2002年); Jose and von Schwichow、2004年)。
特に、自動表示技術は、大腸菌および他のグラム陰性菌の外膜の表面への前もって定義したタンパク質のための発現方法である。そこでは自動ディスプレイ系は、自動トランスポータータンパク質の自然分泌メカニズムに基づく(Jose and Meyer、2007年)。このプロセスでは、組み換えの乗客タンパク質は、遺伝子工学の標準的方法を使用して、単にシグナル・ペプチドと自動表示ベクターの移動(translocating)ドメインの間で翻訳領域内(in-frame)にそのコード配列を導入することにより輸送されるかもしれない。 シグナル・ペプチドはコレラ毒素サブユニット(CTB)から得られるかもしれないし、人工プロモータと結合されるかもしれない。したがって、自動トランスポータータンパクと呼ばれる別のタンパク質を備えた組み換え融合タンパク質として、外膜を横切る移動が意図された乗客タンパク質が大腸菌(AIDA-I)の外膜で発現される(Jose、2006年)。自動トランスポータータンパク質のC-末端部は大腸菌の外膜内にポーリン(porine)状の構造(β-バレル)を形成する。このポーリン状の構造は、大腸菌の外膜の表面への組み換え乗客タンパク質の移動を促進する(Jose、1995年、2006年、2007年)。
【0015】
本発明の基礎となる問題は、基質分子に容易にアクセス可能な活性状態のニトリラーゼの生産および調製を可能にする方法を提供することである。更に、他の酵素の存在は望まれない副反応を防ぐために回避されるべきである。本発明のさらなる基礎となる問題は、組み換えのニトリラーゼを使用した、ニトリラーゼによって触媒作用を及ぼされた反応の産物(例えばカルボン酸)を生産する方法を提供することであり、そこでは時間とともに活性が低下する問題は少なくとも部分的に解決される。本発明の基礎となる別の問題は方法を提供することであり、そこではニトリラーゼによって触媒作用を及ぼされた反応の産物(例えばカルボン酸)は、容易に取り出されるかもしれない。
驚いたことに、本発明者らは、微生物のような宿主細胞の表面上でニトリラーゼが発現されるかもしれないことを発見し、そこではニトリラーゼは完全に機能的(つまり、カルボン酸の形成に帰着するニトリルの転換に触媒作用を及ぼすことができる。)である。
【0016】
本発明の最初の側面は、ニトリラーゼによって触媒作用を及ぼされた反応の産物を生産する方法で、次のステップを含む、
(i)その表面に置かれた前記ニトリラーゼを含む微生物、および/または前記微生物の膜標本を用意し、
(ii)ニトリラーゼ活性と適合性をもつ条件の下で、前記微生物および/またはそれの膜標本と1つ以上のニトリラーゼ基質とを接触させる。
【0017】
本発明の方法では、ニトリラーゼのどんな既知の基質が使用されてもよい。産物は、ニトリラーゼによって触媒作用を及ぼされた反応の産物であるあらゆる化合物であってもよい。ここに使用されるような「ニトリラーゼ活性と適合性をもつ条件」は、ニトリラーゼが活発な条件を含んでおり、つまり、そこではニトリラーゼは1つ以上の基質を産物に変換することができる。ニトリラーゼが活発になるあらゆる既知の条件が使用されてもよい。
【0018】
本発明の方法の中で使用された1あるいは少なくとも1個のニトリラーゼ基質は、ニトリルであることが好ましい。本発明の方法によって得られた産物がカルボン酸であることはさらに好まれます。「ニトリラーゼ活性と適合性をもつ条件」は、ニトリラーゼがカルボン酸の形成に帰着するニトリルの転換に触媒作用を及ぼすような条件であってよい。本発明の好ましい実施形態はカルボン酸を生産する方法であり、その方法は以下のステップを含む、
(i)その表面に置かれたニトリラーゼを含む微生物および/またはそれの膜標本を用意し、
(ii)カルボン酸の形成に帰着するニトリルの転換に前記ニトリラーゼが触媒作用を及ぼすように、前記微生物および/またはそれの前記膜標本をニトリルに接触させる。
【0019】
任意に、本発明の方法は、ステップ(ii)で使用された微生物を取り出すステップ(iii)をさらに含む。
【0020】
ここに使用されるように、「ステップ」(i)、「ステップ」(ii)あるいは「ステップ」(iii)への言及は、ここに記述されるように、ニトリラーゼによって触媒作用を及ぼされた反応の産物を生産する方法、およびカルボン酸を生産する好ましい方法のそれぞれのステップへの言及である。
【0021】
本発明は、組み換え法によって適切な自動トランスポーター系に融合された時、細菌のニトリラーゼが、それらが数日に対する触媒能力を保持したままで、産業上適切な反応に触媒作用を及ぼすために使用されるように、細菌細胞の表面へ移動させられ、細菌の細胞膜中への活性のある折りたたみ形態に統合されるという驚くべき発見に基づく。
【0022】
細菌細胞の外膜は、細胞質ゾル中のタンパク質に適切なタンパク質折り畳み装置類を欠いているので、これは特に驚くべきことである。更に、本発明者らは、もっとも驚いたことに、還元性のあるバクテリアの細胞質ゾルから、酸素が高いレベルで存在する(非常に酸化しやすい環境を形成する)外膜まで、酸化還元反応に敏感な酵素を転送した後或いは転送中に、酵素が失活しないことを見つけ出した。
【0023】
任意の理論に結合される意図なしで、本発明者らは、ニトリラーゼは、自動ディスプレイ経由で細菌細胞の表面上で動けなくされた時、大量の細菌細胞を育てるために使用されるルーチン方法の一部としての、高い酸素レベルおよび活発な揺れに関連した強い機械的な力にさらされた時さえ、反応性のある触媒アミノ酸残基を非活性化から保護する集合体あるいは3次元構造を形成すると仮定する。
【0024】
ここに記述された全細胞生体触媒は、大腸菌のようなグラム陰性細胞の表面上に活性のあるニトリラーゼを配置することを初めて可能にし、その細胞は、ニトリル転換および対応するカルボン酸の生産に使用されてもよい。特に、ここに記述されたニトリラーゼを表示している全細胞生体触媒は、ニトリルの生物工学的な転換に使用されてもよい。
【0025】
重要なことは、前記反応は、穏やかな条件の下で、よい生産速度で、および有毒な副産物の生成なしで遂行されることである。特に、微生物の表面上で組み換え的に発現されたニトリラーゼは、pH7.0前後のように穏やかな条件の下でニトリルの転換に触媒作用を及ぼすことができる。
【0026】
ここに使用されるように、「全細胞生体触媒」は、有機化合物、特にニトリルに対して化学変化を行なうための、タンパク質酵素、特にニトリラーゼのような天然の触媒を含む、全体の細胞、特に微生物を表し、そこでは天然の触媒は、好ましくは細胞の表面に置かれる。発明によれば、触媒は組み換え的に発現されてもよい。
【0027】
細菌細胞は、疎水性の細胞膜によって互いから分けられたある範囲の区画を含む。グラム陽性の細菌細胞は、細胞質ゾル(細胞の内部)を閉じ込める原形質膜を含む。 原形質膜はペプチドグリカンの層に包まれる。対照的に、グラム陰性菌には原形質膜に加えて、外膜と呼ばれた別の薄膜がある。用語「表面」は、ここに使用されるように、好ましくは環境(例えば液体培養培地)に接している微生物の層を指す。本発明の好ましい実施形態では、表面は、グラム陰性の細菌細胞の液体培養培地に接している側である。ここに使用されるように、「表面に表示され」および「表面上で発現された」の表現は、交換可能に使用され、細胞表面上にニトリラーゼを局在化させることを指す。
【0028】
ここに使用されるように、「微生物」は宿主細胞を指す。発明による宿主細胞は、1つの原核生物あるいは真核生物の微生物、好ましくは原核生物の微生物、より好ましくは細菌細胞、さらに好ましくはグラム陰性の細菌細胞であってよい。最も好ましくは、微生物は大腸菌細胞(E. coli cell)である。「微生物」、「宿主細胞」という用語はここで交換可能に使用される。
ここに使用される「ニトリラーゼ」はニトリラーゼ酵素、触媒現象的に活性のある部分、それの誘導体あるいは類似体を指し、それは、ニトリルのカルボン酸およびアンモニアへの転換に触媒作用を及ぼす。ここに使用されるように、分子の「誘導体」あるいは「類似体」は、その分子から由来された部分或いは分子の変形版(modified version)を指すことが注目されるべきである。本発明では、ニトリラーゼは、カルボン酸の形成に帰着するニトリルの転換に触媒作用を及ぼすことができるニトリラーゼのような任意のニトリラーゼでもよい。好ましいニトリラーゼはEC 3.5.5.1による酵素である(さらに「ニトリルアミノヒドラーゼ(nitrile aminohydrolase)」と呼ばれる)。
【0029】
ニトリラーゼはニトリラーゼ活性のみを表示してもよい。ニトリラーゼはさらに多機能酵素でもよく、それは、他の活性に加えて、ニトリラーゼ活性を示す。
【0030】
熟練した人は、ニトリラーゼが設けられた多くの種を知っており、その種は原核生物と真核生物を含む。例えば、本発明の中で使用されたニトリラーゼは、Bordetella, Klebsiella, Aspergillus, Alcaligenes, Saccharomyces, Burkholderia, Neurospora, Lachancea, Debaryomyces, Yarrowia, Candida, Kluyveromyces, Rhodococcus, Nocardia, and/or Rhizobium.および/またはRhizobiumから得られるものでもよい。本発明の中で使用することができるニトリラーゼを有した種の例は、Bordetella bronchiseptica, Klebsiella pneumoniae, Aspergillus niger, Alcaligenes faecalis, Saccharomyces cerevisiae, Burkholderia multivorans, Aspergillus fumigatus, Neurospora crassa, Lachancea thermotolerans, Debaryomyces hansenii, Yarrowia lipolytica, Candida glabrata, Kluyveromyces lactis, Rhodococcus rhodochrous, Nocardia sp., Rhizobium leguminosarum, および/またはNocardia farcinicaである。
【0031】
本発明のニトリラーゼは、好ましくはAlcaligenes, Klebsiella および/または Saccharomycesから得られる。より好ましいニトリラーゼは、Alcaligenes faecalis, Klebsiella pneumoniae および Saccharomyces cerevisiaeから得られたニトリラーゼから選ばれる。Alcaligenes faecalisは、Alcaligenes faecalisの亜種のfaecalisでもよい。Klebsiella pneumoniaeは、Klebsiella pneumoniaeの亜種ozaenaeでもよい。より好ましいニトリラーゼは、Alcaligenes faecalis のニトリラーゼである。
【0032】
本発明のニトリラーゼは、ニトリラーゼ P_887662 [GI:33600102, 01-MAY-2009, Bordetella bronchiseptica RB50], AAA25057 [GI:149175, 26-APR-1993, Klebsiella pneumoniae], NP_943299 [GI:38639530, 30-APR-2009, Klebsiella pneumoniae], P10045 [GI:115192, 20-JAN-2009, Klebsiella pneumoniae subsp. ozaenae], XP_001389617 [GI:145230706, 28-FEB-2008, Aspergillus niger], ACS13754 [GI:239738518, 15-JUN-2009, Alcaligenes sp. ECU0401], BAA02684 [GI:216203, 16-FEB-2008, Alcaligenes faecalis], CAK46957 [GI:134083480, 24-MAR-2007, Aspergillus niger], EDV09642 [GI:190406375, 16-JUN-2008, Saccharomyces cerevisiae RM11-1a], YP_001945058 [GI:189349430, 07-MAY-2009, Burkholderia multivorans ATCC 17616], NP_012102 [GI:6322027, 16-JUN-2008, Saccharomyces cerevisiae RM11-1a], 1F89B [GI:16975400, 19-NOV-2001, Saccharomyces cerevisiae], NP_012409 [GI:6322335, 05-NOV-2009, Saccharomyces cerevisiae], EDN63257 [GI:151945002, 13-JUL-2007, Saccharomyces cerevisiae YJM789], EDN59257 [GI:151940875, 13-JUL-2007, Saccharomyces cerevisiae YJM789], CAY80342 [GI:259147089, 23-SEP-2009, Saccharomyces cerevisiae EC1118], EEU05439 [GI:256270219, 20-AUG-2009, Saccharomyces cerevisiae JAY291] P40447 [GI:731891, 03-NOV-2009, Saccharomyces cerevisiae], XP_751200 [GI:70992703, 27-FEB-2008, Aspergillus fumigatus Af293], CAD71250 [GI:28950282, 14-NOV-2006, Neurospora crassa], CAK48039 [GI:134075478, 24-MAR-2007, Aspergillus niger], CAR23067 [GI:238934886, 08-OCT-2009, Lachancea thermotolerans], CAG86637 [GI:199431326, 10-SEP-2008, Debaryomyces hansenii], XP_500602 [GI:50546150, 29-OCT-2008, Yarrowia lipolytica], CAG78819 [GI:49651877, 23-OCT-2008, Yarrowia lipolytica], CAG59341 [GI:49525722, 16-DEC-2008, Candida glabrata], XP_454637 [GI:50309261, 18-APR-2008, Kluyveromyces lactis], P20960 [GI:417386, 05-MAY-2009, Alcaligenes faecalis] ABO46008 [GI:134034945, 05-NOV-2009, Rhodococcus rhodochrous], AAX18182 [GI:60280369, 02-MAR-2005, Nocardia sp. C-14-1], BAA02127 [GI:216932, 16-FEB-2008, Rhodococcus rhodochrous], Q02068 [GI:417382, 20-JAN-2009, Rhodococcus rhodochrous], CAK02877 [GI:115259785, 13-MAY-2009, Rhizobium leguminosarum bv. viciae 3841], CAF05970 [GI:40882143, 10-OCT-2009, Neurospora crassa], CAK08726 [GI:115257629, 13-MAY-2009, Rhizobium leguminosarum bv. viciae 3841], BAD58116 [GI:54016746, 10-MAY-2008, Nocardia farcinica IFM 10152], および Q5Z1U0 [GI:81603033, 03-NOV-2009, Nocardia farcinica]を含むグループから選ばれてもよい。このリストは「Genbank登録番号[Genbank識別子、データベース・エントリーの日付、種/系統。」のフォーマットを使用した。このグループのニトリラーゼの配列は、Genbank登録番号、Genbank識別子(GI)、Genbankデータベース・エントリーの日付および種および/または系統から得られる配列によって特定することができる。
【0033】
ここに使用されるような、用語「ニトリラーゼ」は、ここに開示されるニトラーゼと、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%あるいは少なくとも99%の同一性を持ち合わせているどんなニトリラーゼも含んでいる。熟練した人は、例えばBLASTまたはPBLASTによって、アミノ酸配列レベルで同一性の程度の決定するためのふさわしい方法を知っている。
【0034】
上に示されるように、活性のあるニトリラーゼはおよそ14のサブユニットからなるホモオリゴマー(homooligomeric)の酵素でもよい。一般に、ニトリラーゼは不活性な二量体として存在する。本発明では、宿主細胞の表面に表示されたニトリラーゼは3つ以上の同一のサブユニットを含むホモマルチマー(homomultimer)でもよい。特にホモマルチマー(homomultimer)は、7、8、9、10、11、12、13、14、15あるいは16の同一のサブユニットのような、7〜16の同一のサブユニットを含んでもよい。いくつかのおよそ14のサブユニット、つまり11、12、13、14、15、あるいは16のサブユニットが好ましい。ホモマルチマー(さらに「ホモオリゴマー」としてここに名付けられた)は、宿主細胞細胞膜に表示されたいくつかの同一のポリペプチド・サブユニットの自発的な会合によって形成されてもよい。
ここに使用されるように、「ニトリル」は少なくとも1つの―CN官能基(ニトリル基)を含む任意の化合物を指します。ニトリルはさらに1つ、2、3、4あるいはより多くのニトリル基を含んでもよい。
【0035】
ニトリル基は、6から14個の炭素原子を、好ましくは6から10個の炭素原子を含むアリール基のような芳香族部分に結合されてもよい。アリール基は1つ、2つのあるいは3つの濃縮された芳香環を含んでもよい。より好ましくは、アリール基はフェニル基でもよい。アリール基は、1、2、3、4、5あるいはより多くの置換基のような1つ以上の置換基によって代用されてもよい。
ニトリル基は、1〜8個の炭素原子、1〜6個の炭素原子、1〜4個の炭素原子あるいは1〜3個の炭素原子を含むアルキル基のような脂肪族の部分につながれてもよい。アルキル基は直鎖でもよいし或いは分岐していてもよい。アルキル基は環状の部分を含んでもよい。アルキル基は、1、2、3、4、5あるいはより多くの置換基のような1つ以上の置換基によって代用されてもよい。
【0036】
ニトリルはアルキル・アリール基を含んでもよく、そこではアルキルとアリールはここに記述されるような意味(meanings)を持っています。少なくとも1つのニトリル基はアルキル基および/またはアリール基につながれてもよい。アルキルおよび/またはアリール部分(moiety)は、1、2、3、4、5あるいはより多くの置換基のような1つ以上の置換基によって代用されるかもしれない。
【0037】
ここに使用された1つ以上の置換基は、−OH、ヨード、ブロモ、クロロ、フルオロ、アリール、アルキル、アルコキシ、などから独立して選ばれてもよく、そこではアルキルとアリールはここに記述されるような意味を持っており、アルコキシが1〜8個の炭素原子、1〜6個の炭素原子、1〜4個の炭素原子あるいは1〜3個の炭素原子を含んでいる。好ましいアルコキシ基はメトキシである。アルコキシ基は直鎖でもよいし或いは分岐していてもよい。アルコキシ基は環状の部分を含んでもよい。
【0038】
アルキル基、芳香族基、および/またはアルコキシ基、少なくとも1つのヘテロ原子が炭素原子と置換されるように、1、2、3、4あるいはそれより多くのヘテロ原子のような少なくとも1つのヘテロ原子が独立して含んでもよい。ヘテロ原子は、N、OおよびSから独立して選ばれてもよい。
【0039】
発明のニトリラーゼの基質として使用される典型的なニトリルは、マンデロニトリル、ベンゾニトリル、フェニルプロピオニトリル(phenylpropionitrile)、フェニルグリシノニトリル(phenylgycinonitrile)、ブロモキシニル、イオキシニル、クロロキシニル(chloroxynile)、アニソニトリル(anisonitrile)、3-ブロモ-4-ヒドロキシベンゾニトリル、3-フルオロ-4-ヒドロキシベンゾニトリル、4-ヒドロキシ-3,5-ジメトベンゾニトリル(dimethobenzonitrile)、およびプルナシンのような芳香性のニトリルであり、またn-ブチロニトリル、n-バレロニトリル、イソ-ブチロニトリルおよびスクシノニトリルのような脂肪族ニトリルである。 発明の特に好ましい基質はマンデロニトリルである。
本発明によれば、Alcaligenes faecalisからのニトリラーゼのための好ましい基質はマンデロニトリルとプルナシンである。Klebsiella pneumoniae からのニトリラーゼのための好ましい基質はブロモキシニル、イオキシニル、クロロキシニル、アニソニトリル、3-ブロモ-4-ヒドロキシベンゾニトリル、3-フルオロ-4-ヒドロキシベンゾニトリル、4-ヒドロキシ-3,5-ジメトベンゾニトリル。Saccharomyces cerevisiaeからのニトリラーゼのための好ましい基質は、ベンゾニトリル、フェニルプロピオニトリル(phenylprobionitrile)、マンデロニトリル、フェニルグリシノニトリル、n-ブチロニトリル、n-バレロニトリル、イソ-ブチロニトリルおよびスクシノニトリルである。
発明の1つの側面では、ニトリラーゼをコード化(encoding)する遺伝子は、Alcaligenes faecalisの全体のDNAからPCRによって増幅されクローン化された。その遺伝子は、PCRプライマーに結合された適切な制限部位(restriction sites)による自動トランスポーターの読み枠と正確に融合できる。融合タンパク質の発現は、標準の研究室的手法で示すことができる。表面上で発現され、その上で動けなくされた、酵素は、加えられた基質(この場合、ラセミのマンデロニトリル)を細胞外で変換することができる。
【0040】
ここに適用される培養条件および発現条件で、約50%、好ましくは約60%、より好ましくは約70%、またさらにより好ましくは約80%の変換率は、5日後に達成されるだろう。非対称のC原子がある化合物は、本発明の方法によってある鏡像体過剰率(enantiomeric)中で生産されてもよい。鏡像体過剰率(%ee)は、少なくとも約50%、60%、70%、80%、90%、95%、96%、97%、98%あるいは99%であってもよく、製品、特にカルボン酸用には、少なくとも約99%以上であることが好ましい。キラルHPLCによって示されたように、特に(R)−マンデル酸では、5日後の約80%の変換率および99%以上の鏡像体過剰率(% ee)が達成された。
【0041】
ここに使用される物質の鏡像体過剰率(ee)は物質の光学的な純度のものさしです。鏡像体過剰率は分析される化合物の溶液中で決定されてもよい。適切な溶媒(疎水性・親水性の溶媒のような)およびそれの混合物が知られている。
【0042】
鏡像体過剰率は次の方程式によって計算することができる:
ee% = 100 * ( [ R ] - [ S ] ) / ( [ R ] + [ S ] )
そこでは、[R]、[S]は、種類の濃度を示し、そして、RとSは、(R)−そして(S)−異性体をそれぞれ指す。したがって、ラセミ混合物中では、つまり[R]=[S]となり、ee%=100* 0/([R]+[S])=0%であり、つまりRがSよりも過剰にはならずあるいは逆も同様である。したがって、鏡像異性的に(enantiomerically)純粋な化合物の鏡像体過剰率は100%であるのに対して、ラセミ混合物の鏡像体過剰率は0%である。(R)−および(S)−異性体の濃度は、混合物の比旋光度の決定、キラル・カラム・クロマトグラフィ(例えば、キラルHPLC)およびNMR分光学のように(しかしこれに限定されない)、非対称のC-原子がある化合物の決定用の技術の中で既知の任意の方法によって決定されてもよい。
【0043】
ここに記述された発明は、単にこの目的の反応の一回分から細胞を遠心分離機にかけるだけなので、産物を精製する容易な方法と結合した、ニトリルの加水分解中の穏やかな反応状態という長所を持つ。
【0044】
ニトリラーゼは、微生物の中で組み換え的に発現され、細胞表面へ輸送されてもよい。微生物は独立して繁殖し、生体触媒の精巧な精製がもはや必要でないように、酵素自体を生産する。そのような微生物用の例は、この例1に記述された大腸菌種である。
【0045】
ここに使用された用語「組み換え個体」は、本来は互いにつながれない、互いに効果的につながれた少なくとも2つの核酸配列で作られていた核酸配列を指す。少なくとも2つの配列は同じ生物から得られてもよい。例えば、組み換えの構成物は、Alcaligenes種のシグナル配列および同じAlcaligenes種のニトリラーゼ(それは前記シグナル配列に通常つながれない)をコードする配列を含んでもよい。少なくとも2つの配列は少なくとも2つの異なる生物から得られてもよい。例えば、大腸菌(E. coli)のシグナル・ペプチド、Alcaligenesからのニトリラーゼ遺伝子、大腸菌からの膜貫通のリンカーおよび大腸菌からの自動トランスポータードメインが、効果的につながれてもよい。別の例は、組み換えのポリペプチドであり、そこでは、シグナル・ペプチドがコレラ毒素βサブユニット(CTB)に由来し、自動トランスポータードメインおよび膜貫通のリンカーは大腸菌に由来し、ニトリラーゼはAlcaligenesに由来する。
【0046】
本発明の好ましい実施形態の中で、用語「組み換え的に発現する」は、ここに使用されるように、用語「組み換え式に発現された」などで同義的に使用され、組み換え核酸配列の発現のことを指す。
【0047】
例えば大腸菌、或いはそれどころかあらゆる生物における核酸配列の発現は、大腸菌シグナル・ペプチドと、組み換えの発現を構成するAlcaligenesニトリラーゼ配列を含んでいる。
【0048】
ニトリラーゼが自動トランスポーターのトランスポータードメインへ融合されることは好ましい。発明による自動トランスポーターのトランスポータードメインは、自動トランスポーターの任意のトランスポータードメインでよく、好ましくはβ−バレル構造を形成することができる。β-バレル構造の詳細な説明およびβ−バレル自動トランスポーターの好ましい例は、WO97/35022に示され、参照されることでここに取り込まれる。ヘンダーソンら (2004)は、適切な自動トランスポータードメインを含む自動トランスポータータンパク質について記述した(要約に関しては、Henderson et al.、2004年のテーブル1を参照)。 ヘンダーソンらの開示 (2004)は、参照によってここに取り込まれる。例えば、自動トランスポーターのトランスポータードメインは、Ssp(P09489(S.marcescens))、Ssp-h1(BAA33455(S.marcescens))、Ssp-h2(BAA11383(S.marcescens))、PspA(BAA36466(P.fluorescens))、PspB(BAA36467(P.fluorescens))、Ssa1(AAA80490(P.haemolytica))、SphB1(CAC44081(B. pertussis))、AspA/NalP(AAN71715、N. meningitidis)、 VacA(Q48247、H.pylori)、AIDA-I(Q03155、大腸菌)、IcsA(AAA26547、S.flexneri)、MisL(AAD16954、S.enterica)、TibA(AAD41751、大腸菌)、Ag43(P39180、大腸菌)、ShdA(AAD25110、S.enterica)、AutA(CAB89117、N. meningitidis)、Tsh(I54632、大腸菌)、SepA(CAC05786、S.flexneri)、 EspC(AAC44731、大腸菌)、EspP(CAA66144、大腸菌)、Pet(AAC26634、大腸菌)、Pic(AAD23953、大腸菌)、SigA(AAF67320、S. flexneri)、Sat(AAG30168、大腸菌)、Vat(AAO21903、大腸菌)、EpeA(AAL18821、大腸菌)、EatA(AAO17297、大腸菌)、EspI(CAC39286、大腸菌)、EaaA(AAF63237、大腸菌)、EaaC(AAF63038、大腸菌)、Pertactin(P14283、B. pertussis)、BrkA(AAA51646、B. pertussis)、Tef(AAQ82668、B. pertussis)、Vag8(AAC31247、B. pertussis)、PmpD(O84818、C. trachomatis)、Pmp20(Q9Z812、 C. pneumoniae)、Pmp21(Q9Z6U5、C. pneumoniae)、IgA1プロテアーゼ(NP_283693、N. meningitidis)、App(CAC14670、N. meningitidis)、IgA1プロテアーゼ (P45386, H. influenzae)、Hap(P45387, H. influenzae)、rOmpA(P15921、R. rickettsii)、rOmpB(Q53047、R. rickettsii)、ApeE (AAC38796、S. enterica)、EstA(AAB61674、P. aeruginosa)、Lip-1 (P40601、X. luminescens)、McaP(AAP97134、M. catarrhalis)、BabA (AAC38081、H. pylori)、SabA(AAD06240、H. pylori)、AlpA(CAB05386、H. pylori)、Aae(AAP21063、A. actinomycetemcomitans)、NanB(AAG35309、P. haemolytica)、およびこれらの自動トランスポーターの変異体(variants)から選ばれてもよい。
【0049】
典型的な自動トランスポータータンパク質のそれぞれに括弧の中に与えられたものは、適切なgenbank登録番号および自動トランスポーターが得られた種の例である。自動トランスポーターのトランスポータードメインが好ましくは、大腸菌のAIDA-Iタンパク質であるか、あるいは例えば、Niewert U.、Frey A.、Voss T.、Le Bouguen C.、Baljer G.、Franke S.、Schmidt MAによって記述された、ようなそれの1つの変異体でもよい。AIDA自動トランスポーター系は、浮腫病および離乳後下痢と診断されたブタから単離された大腸菌の中でF18とStx2eと関係する、Clin. Diagn. Lab. Immunol. 2001 Jan, 8(1):143-149;9。
【0050】
上記の示された自動トランスポーター配列の変異体は、例えば、膜貫通部分の一部ではない、β−バレルのループ構造中のアミノ酸配列の変更により得られるだろう。自由に、表面ループをコードする核酸部分は完全に削除してもよい。さらに、保存されたアミノ交換、つまり、1つの親水性のアミノ酸を別の親水性のアミノ酸へ交換すること、および/または1つの疎水性のアミノ酸を別の疎水性のアミノ酸に交換すること、が両親媒性のβ-シート内で起こってもよい。1つの変異体(variant)は、自動トランスポータードメイン(特に、β−シートの範囲内で)のそれぞれの天然の(native)配列に対してアミノ酸レベルで少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%あるいは少なくとも98%の配列同一性を保持する好ましい。
【0051】
発明の方法のステップ(i)が以下のステップを含むことは好ましい、
(a)発現制御配列と効果的につながれた核酸配列を含む微生物を用意し、前記核酸配列は、
(1)シグナル・ペプチドをコード化(encoding)する部分、
(2)表示される組み換えニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4)膜貫通のリンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、
を含み、
(b)前記(a)の核酸配列が発現され、前記核酸配列の発現産物は微生物の表面に表示される条件の下で微生物を培養する。
【0052】
当業者は、核酸配列が発現される条件を考案することができ、また、これはルーチン実験(例えば様々な温度、培地、細胞密度、および/または発現を引き起こす化学薬品の濃度をテストすること)のみを含んでいる。
【0053】
ステップ(a)によって提供される微生物は、発現制御配列と効果的につながれた核酸配列を用いた形質転換(transformation)によって得られてもよく、そこでは前記核酸は次のものを含む、
(1)シグナル・ペプチドをコード化する部分、
(2)表示される組み換えのニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4)膜貫通のリンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、
そこでは成分(1)から(5)は、以下に記述されるような核酸配列である。
【0054】
形質転換のどんな既知の方法も使用することができる。ここに使用される用語「形質転換」は、好ましくは、異種起源の構成物の細胞(好ましくは細菌細胞)中への導入を指し、それを今後は、形質転換されたという。当業者は形質転換のプロトコルおよび手順(例えばコンピテントセルの電気的あるいは化学的な形質転換)を知っている。
【0055】
別の実施形態では、進歩性がある方法はさらにステップを含む。
【0056】
(c) (b)の微生物から膜標本(membrane preparation)を生産すること。
本発明の中で使用された膜標本は触媒現象的に活性のあるニトリラーゼを含む。 ここに使用された用語「膜標本」は、好ましくは薄膜成分中で産物を豊富にしたもの指す。当業者は、膜標本を得るために使用することができるプロトコルおよび手順に精通している。例えば、細菌細胞は培養から収穫され、例えば、凍結解凍サイクル、超音波処理、溶菌緩衝液中の再懸濁などによって溶菌にさらされ、続いて細胞の細胞膜断片を分離するために分画遠心分離がされる。本発明の好ましい実施形態では、膜標本は、外膜標本であり、つまり他の膜の成分、および、細胞質ゾル、内部の薄膜および周辺質(periplasm)のような区画に対して外膜の成分を豊富にした標本である。当業者は、外膜を分離するために使用することができるプロトコルおよび手順、例えば細菌細胞のリゾチーム処理および続いて起こる遠心分離ステップ、に精通している。本発明の好ましい実施形態では、細胞膜標本は触媒現象的に活性のあるニトリラーゼを含む外膜標本で、その膜標本は、細胞質ゾル中の前記ニトリラーゼを合成し、続いて、外膜にそれを輸送する細胞から作られている。別の実施形態では、前記膜標本は、ニトリラーゼ融合タンパク質のような膜タンパクを人工小胞に組み入れることにより例えば十分に処理される。当業者は、例えば適切な洗剤を使用して、小胞中の膜蛋白を機能的に再構成する方法を持っている。
本発明の方法のステップ(a)は、微生物の準備に言及する。微生物はここに記述された任意の微生物でもよい。ステップ(a)で、宿主細胞、特に、宿主バクテリアは、発現制御配列(つまりプロモーターおよびそれぞれの宿主細胞中の遺伝子発現に必要なさらなる任意の配列)と効果的につながれた核酸配列を含む。ここに使用されるように、「発現制御配列と効果的につながれた核酸配列」は、発現制御配列との間の核酸配列の機能的な関係を指す。熟練した人は、適切なプロモーターおよび発現制御配列を知っている。プロモーターおよび/または発現制御配列は、宿主細胞に対して同種起源(homologous)か或いは異種起源であってもよい。
好ましくは、核酸配列は組み換えのベクター、例えばプラスミドベクター、上に置かれる。
ステップ(a)による核酸配列は、(1)シグナル・ペプチド(好ましくは内部の細胞膜を通って周辺質の中への輸送を許容するグラム陰性のシグナル・ペプチドをコードする部分)をコード化する部分を含む。シグナル・ペプチドは宿主細胞に同種起源のシグナル・ペプチドでもよい。シグナル・ペプチドはさらに宿主細胞に異種起源のシグナル・ペプチドでもよい。
さらに、核酸配列は、(2)表示されるニトリラーゼ(「=乗客ポリペプチド」あるいは「乗客」)をコード化する部分を含む。ここに記述されるどんなニトリラーゼをコード化する核酸を使用してもよい。
【0057】
核酸配列は、任意に、(3)プロテアーゼ認識部位をコード化する部分を含む。ここに使用された用語「プロテアーゼ認識部位」は、プロテアーゼ認識部位によって示されたアミド結合の加水分解のやり方で、ポリペプチドをその後切断する特定のプロテアーゼによって認識される特定のアミノ酸配列を指す。プロテアーゼ認識部位は、固有のプロテアーゼ(つまり宿主細胞に自然に生じるプロテアーゼ)あるいは外部から追加されたプロテアーゼ用の認識部位でもよい。例えば、外部から追加されたプロテアーゼは、IgAプロテアーゼ(EPA-0 254 090参照)、トロンビンあるいは因子X(factor X)でもよい。固有のプロテアーゼは、例えば、OmpT、OmpKあるいはプロテアーゼXをから選択されてもよい。プロテアーゼ認識部位は宿主細胞と同種起源でもよい。 プロテアーゼ認識部位は、さらに宿主細胞に対して異種起源でもよい。
【0058】
更に、核酸配列は、(4)乗客ポリペプチド(つまり、宿主細胞の外膜の外側表面上の組み換えニトリラーゼ(2))の提示(presentation)に必要な膜貫通リンカーをコード化する部分を含む。自動トランスポーターに関して同種起源の膜貫通のリンカー・ドメインが使用されてもよく、つまり、膜貫通のリンカー・ドメインは、自動トランスポータードメインをコード化する核酸配列の5'末端へ直接融合された核酸部分によってコード化される。さらに、自動トランスポーターに関して異種起源の膜貫通のリンカー・ドメインが使用されてもよい。膜貫通のリンカーの長さは好ましくは30-160アミノ酸である。
【0059】
さらに、核酸配列は、(5)自動トランスポーターのトランスポータードメインをコード化する部分を含む。本発明の背景では、自動ディスプレイは、任意のグラム陰性のバクテリア中の自動トランスポーターによるタンパク質或いはポリペプチドの組み換え表面への表示でもよい。トランスポータードメインはここに記述された任意のトランスポータードメインでもよい。
【0060】
本発明の核酸配列中の(1)から(5)の要素は、好ましくは、5'から3'に向けて方向づけられる。ステップ(b)で得られた発現産物では、(1)から(5)の核酸配列によってコード化されたアミノ酸配列は、好ましくはN末端からC末端に向けて配置される。
【0061】
本発明の方法のステップ(b)は、ステップ(a)の核酸配列が発現され、組み換えニトリラーゼを含む表現産物が微生物の表面に表示される、条件の下で微生物を培養することを指します。当業者は適切な培養条件を知っている。発明による方法は、次の組成物の液体培地の使用により、微生物、特に1つの細胞当たり100000まで或いはそれ以上の分子である大腸菌あるいは他のグラム陰性の細菌細胞、の表面の乗客タンパクの効率的な発現を許容する:5g/l〜20g/l、好ましくは約10g/lのトリプトン、2g/lから10g/l、好ましくは約5g/lの酵母エキス(yeast extract)、5g/lから20g/l、特に約10g/lのNaCl、および残りの部分の水。培地は恐らくできるだけ低い量の二価の陽イオンを含んでいるべきであり、好ましくはAqua bidestあるいは高度に精製された水、例えば、ミリポア(Millipore)の水が使用されます。液体培地はさらに好ましくは、2μMから20μMの濃度、特に10μMのEDTAを付加的に含んでいてもよい。さらに、それは好ましくは、2 mMから20 mMの好ましい濃度の、2−メルカプト・エタノール或いはジチオトレイトール或いはジチオエリトリトールのような還元試薬(reducing reagents)を含んでいてもよい。還元試薬は、輸送中のポリペプチドの折り畳まれない構造を好む。液体培地は、追加の炭素供給源(C-sources)、好ましくはグルコース(例えば、分泌を促進(つまり、周囲の培地に乗客に移す)するために10g/l以内の量の中で)、をさらに含んでいてもよい。表面のディスプレイのため、好ましくは追加の炭素供給源は添加されません。大腸菌のようなグラム陰性の細胞用の好ましい培養条件は、例に記述される。
【0062】
ステップ(ii)は好気性のおよび/またはを酸化させる条件の下で行なわれてもよい。好ましい実施形態では、用語「好気性の条件」は、酸素の存在によって特徴づけられた条件を指す。別の好ましい実施形態の中で、用語「好気性の条件」は、既知の温度および液体に関して正常なレベルと比較して、高いレベルの酸素の存在を指す。別の好ましい実施形態の中で、用語「好気性の」条件は、例えば液体(例えば培地)と、酸素を含む周囲のガス環境(例えば空気)との間の高いガス交換によって特徴づけられた条件を指す。当業者は、適切な酸素感受性の電極の使用を通じて例えば酸素レベルを測定することができる。さらに、当業者は、例えば空気または酸素によって緩衝液あるいは培地を洗い流したり(flushing)、酸素を放出する化学薬品を添加したり、あるいは単に酸素をあるいは空気の存在下でそのような緩衝液あるいは培地を活発に振ったりして、好気性の条件にすることができる。対照的に、当業者は、例えばアルゴンまたは窒素によって培地または緩衝液を広範囲に洗い流す(flushing)ことにより、あるいはガス交換を最小限にするために培養容器を密閉することにより、嫌気性の条件をもたらすことができる。
【0063】
好ましい実施形態では、ここに使用された用語「酸化条件」は、培地または緩衝液への追加あるいは酸化剤(oxidants)(つまり他の化学薬品に電子(その中で酵素は酸化する傾向がある)を放出させる化学薬品)の存在を指す。例えば、還元された酸化還元酵素は、酸素の存在下において酸化される(oxidise)傾向がある。対照的に、ここに使用された用語「還元する」或いは「還元条件」は、培地または緩衝液への追加あるいは還元剤(reductants)(つまり他の化学薬品に電子を取り上げさせる化学薬品)の存在を指す。当業者は、例えば、酸素のような酸化剤の追加、および、例えばメルカプトエタノールまたはDTTのようなチオールである還元剤の追加を通じて、例えば選択的な酸化還元反応の環境を設立(set up)することができる。さらに、当業者は、例えば酸化還元電極の使用を通じて溶液の酸化還元電位を測定することができる。
大腸菌のようなグラム陰性のバクテリアの宿主細胞では、組み換えの乗客の転置の後、乗客はβ−バレル(それは外膜内のアンカーとして役立つ)によって外膜の表面へ付けられ続ける。外膜内のβ-バレルのコントロールされた結合によって、β-バレルのC末端部は外膜の内部側に向けられるが、しかし、リンカーのN-末端部(組み換え乗客タンパク質はそれに共有結合で結び付けられる)は、外膜の外部の表面(つまり環境)に向けられる。
任意のステップ(c)は、ステップ(b)の微生物からの膜標本の生産を指す。 膜標本を得る適切な方法は当業者に有名である。
ここに使用されるような膜標本は好ましくは外膜標本である。
【0064】
本発明の方法のステップ(ii)は、ニトリラーゼ活性と適合性をもつ条件の下、微生物および/またはそれの膜標本を、1つ以上のニトリラーゼ基質に接触させることを指す。上に示されるように、好ましい実施形態では、ステップ(ii)は、カルボン酸の構成に帰着するニトリルの転換にニトリラーゼが触媒作用を及ぼす条件の下で、微生物および/またはとそれの膜標本をニトリルと接触させることを指す。技術の中で知られている任意の適切な条件が使用されてもよい。その反応は、特に液体培地の中で行なわれる。液体培地では、微生物は懸濁液を形成してもよい。微生物も動けなくされてもよい。膜標本が使用される場合、膜標本は液体培地の中で分散する粒子を形成してもよい。その反応は、約5.0から約9.0までの範囲中のpHで行なわれ、好ましくは約6.0〜約8.0以上で、より好ましくは約6.5から約7.5まででもよい。
【0065】
本発明の方法のステップ(iii)は、微生物の回収を指し、その微生物はステップ(ii)で使用される。本発明の例によって実証されるように、例えば24時間の反応周期の後に、細菌細胞はまだ著しい活動を示す。従って、細胞はさらなる反応周期で使用することができる。回収は反応混合物の遠心分離によって行なわれる。もし必要であれば、細胞が洗われてもよい。
【0066】
発明の別の側面は、ニトリラーゼによって触媒作用を及ぼされた反応の産物の生産のため、特にカルボン酸の生産のため、その表面にニトリラーゼを発現する微生物および/またはそれの膜標本の使用に関連する。生産されたカルボン酸がマンデル酸(特に(R)-マンデル酸)であることが好ましい。
【0067】
発明の別の側面は組み換えのニトリラーゼを表面上に表示する微生物に関係し、そこでは微生物はここに記述されるような微生物で、また、ニトリラーゼはここに記述されるようなニトリラーゼである。微生物は、シグナル・ペプチド、組み換えのニトリラーゼ、任意のプロテアーゼ認識部位、膜貫通のリンカー、およびその表面上の自動トランスポーターのトランスポータードメインを含む融合ポリペプチドを特に表示し、そこでは融合タンパク質の構成要素はここに記述された通りである。
【0068】
まだ、発明の別の側面はニトリラーゼを含む膜標本に関係がある。膜標本は、ここに記述されるような微生物、例えば本発明の方法のステップ(c)の中で得られてもよい。ニトリラーゼはここに記述されるような任意のニトリラーゼでもよい。
【0069】
しかし、発明の別の側面は、微生物へ導入された核酸配列を含み、その表面上に組み換えのニトリラーゼを表示する微生物を生産する方法であり、前記核酸配列は次のものを含む。
【0070】
(1)シグナル・ペプチドをコード化する部分、
(2)表示される組み換えのニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する任意の部分、
(4)膜貫通のリンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、
そこでは、核酸は、発現制御配列と効果的につながれる。(1)から(5)の要素は、上記に記述されるような核酸配列である。微生物はここに記述されるようなあらゆる微生物、例えばバクテリア、特にグラム陰性のバクテリア、さらに詳細には大腸菌でもよい。ニトリラーゼはここに記述されるような任意のニトリラーゼでもよい。「前記微生物へ核酸を導入する」ことは、ここに記述されるように、形質転換を含んでいる。
【図面の簡単な説明】
【0071】
発明は、次の図および例によってさらに例証され、それらは発明の範囲の制限と見なされるべきでない。
【図1】pAT-NitAfによってコード化された自動トランスポーター融合タンパク質の体系。融合サイトの環境は配列として与えられる。クローニング手順により添加された、N-末端での2つのアミノ酸は、イタリック体で太字で示される。シグナル・ペプチダーゼ切断サイトが示される。SP、シグナル・ペプチダーゼ。
【図2】過剰発現された自動トランスポーター融合タンパク質AFP(1)を含む、大腸菌BL21(DE3)pAT-NitAfの外膜断片(fraction)のSDS-PAGEであり、自動トランスポーターを過剰表現した全体細胞をプロテイナーゼK(2)によって消化したもの、およびマーカー・レーン(M)。
【図3】ニトリラーゼ活性の実証: (R、S)−マンデロニトリルからの(R)−マンデル酸の生産。反応混合物は、全量1ml中で、Na-リン酸緩衝液(50 mM、pH 7.5)、(R、S)−マンデロニトリル(10 mM)、10 OD578に対応する大腸菌BL21(DE3)pAT-NitAfの細胞が含まれる。データポイントは3回の実験の平均であり、標準誤差を表わす棒は小さすぎるので、目に見えることができない。塗り潰された記号は、OD5781での導入を表わし、また、空いた記号はOD5780.5での導入を表わす。Tris-HCI緩衝液pH 7(▼/)、リン酸緩衝液pH 7.5(■/□)。
【図4】(R)−マンデル酸キラルHPLC検出。メタノール中の純粋(R)-マンデル酸(10 mM)(前)、大腸菌BL21 pAT-NitAfを用いたラセミのマンデロニトリル転換からのマンデル酸抽出物(後)。
【図5】R−マンデル酸への(R、S)−マンデロニトリルの転換率のpH依存。その反応は、50mMの酢酸緩衝液(空の棒)あるいは50mMのTris-HCI緩衝液(塗り潰された棒)のいずれかの中で行なわれた。反応時間は37°Cで48時間である。
【図6】-18°Cの保管中の全細胞生体触媒の安定性。細胞は20%のグリセロールを含む培地中で冷凍され、溶かした後に50mMリン酸緩衝液pH 7.5中の大腸菌BL21(DE3)pAT-NitAfによってなされた、10mMマンデロニトリルの転換(マンデル酸生産)が、37°Cで120時間以上で始められた。
【図7】異なる緩衝液および温度条件の大腸菌BL21(DE3)pAT-NitAfの再利用可能性。各周期はそれぞれ24時間の転換期間に対応する。続いて、生体触媒は遠心分離によって除去され(13000rpmで60秒)、新鮮な緩衝液の中で再懸濁され、新鮮な基質(最終の濃度10 mM)と結合し、さらに24時間培養した。塗り潰された記号はTris-HCI緩衝液(50 mM、pH 7) を表わし、空の記号はリン酸ナトリウム緩衝液(50 mM、pH 7.5)、45°C(●/o)、42°C(□/■)、37°C(▲/Δ)、30°C(▼/)を表わす。
【図8】KpnIとXhoIを使用するプラスミドDNAの制限消化。レーン1)1kbのマーカー、2)〜 12) KpnI/XhoIを使用して消化された、形質転換体からのプラスミドDNA、13)1kbのマーカー。
【図9】クローン大腸菌UT 5600(DE3)pAT-NitAfおよびpAT-NitScのトリプシン消化に続く分離された外膜。レーン1)pAT-NitSc、トリプシンで消化、2)pAT-NitSc、消化されていない3)pAT-NitAf、プロテイナーゼKで消化された、4)pAT-NitAf、消化されていな、5)分子量標準(kDa)。
【図10】大腸菌BL21(DE3)pAT-NitAfによるマンデロニトリルの転換に続くHPLC解析。A)ネガティブコントロール。 大腸菌BL21(DE3)、OD578=10、培養時間: 120時間/30°C。1) HPLC、2)マンデロニトリル、3)ベンズアルデヒドの溶媒先端 B) 大腸菌BL21(DE3)pAT-NitAf、OD578=10、120時間/30°Cの培養時間による転換。1) HPLC、2)マンデロニトリル、3)ベンズアルデヒドの溶媒先端。
【図11】組み換えの全細胞生体触媒である大腸菌BL21(DE3)pAT-NitAfを使用して生産されたマンデル酸。
【図12】30°Cおよび37°Cのそれぞれで、リン酸緩衝液pH 7.5中での大腸菌BL21(DE3)pAT-NitAfによるマンデル酸生産。1mMのIPTGを1時間、30°Cで使用して、OD578=1の細胞が導入された。時間t=0の基質濃度は10mM マンデロニトリルであった。
【図13】37°Cでリン酸緩衝液中で、pH 7.5および様々な基質濃度を使用した、大腸菌BL21(DE3)pAT-NitAfによるマンデル酸生産。OD578=1に対して1mMのIPTGを1時間30°Cで使用して、OD578=1細胞が導入された。その後、転換は120時間起こった。
【図14】様々な緩衝液系中の37°C(図の中で示されるような例外がある)での大腸菌BL21(DE3)pAT-NitAfによるマンデル酸生産。30°Cで1時間、1mM IPTGを使用して、OD578=1の細胞を導入した。時間t=0での基質濃度は、10mMマンデロニトリルであり、転換は120時間起こった。
【図15】5dの周期にわたる大腸菌BL21(DE3)pAT-NitAfによる(R)−マンデル酸の生産および同一性の検出。ヘキサンを用いたキラル−OMカラム(company CS-Chromatographie Service):2つのプロパノール:TFA(90:10:0.1)、流量(Flow): 0.5ml/分。 210nmの検出では、A) 酢酸エチルで抽出された大腸菌BL21(DE3)pAT-NitAfの細胞上澄みの分析。B)Aからの転換と結合したラセミのマンデル酸。
【図16】6°C(図16)での異なる保管時間の経過後のマンデル酸生産。50mMリン酸緩衝液pH 7.5中で、10mM マンデロニトリル、37°Cで120時間での、大腸菌BL21(DE3)pAT-NitAfによる転換。
【図17】異なる緩衝液中で、24時間の間で互いに異なる温度で、10mMのマンデロニトリルを1000rpmの熱攪拌機(Thermomixer)中で用いた、大腸菌BL21(DE3)pAT-NitAfによるマンデル酸の生産。
【図18】大腸菌JK321 pAT-NitKpによる3,5-ジブロモ-4-ヒドロキシ-安息香酸の生産の検出。培養時間120時間、OD578=10、基質2.5 mM ブロモキシニル。
【0072】
例
次の例は、本発明者が異なる微生物からの3つのニトリラーゼ遺伝子を同定できたとともに、自動ディスプレイ系へ入る前記遺伝子をクローン化できたことを実証する。
【0073】
例1: Alcaligenes faecalisからのニトリラーゼ
全細胞生体触媒は、特に大量の(bulk)およびまれな化学薬品の生産用の魅力的な技術的なツールで、領域−あるいは鏡像異性選択的な合成が必要である。本研究で、基質としてマンデロニトリルを使用する(R)−マンデル酸を合成するための全細胞生体触媒が構築された。この目的のために、Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750からのニトリラーゼは、自動ディスプレイを使用して、大腸菌(大腸菌)の表面上で発現された。自動ディスプレイ系は、グラム陰性菌のための強力な表面の表示システムで、自動トランスポーター経路に基づく。この技術で、大腸菌の表面上の活性のある形態の複数のホモマー(multihomomeric)のニトリラーゼを発現し、99%以上の鏡像体過剰率を持った鏡像異性的に(enantiomerically)純粋な(R)−マンデル酸を生産することが可能になった。外膜中のニトリラーゼ-自動トランスポーター融合タンパク質の結合性(integration)はSDS-PAGEによってモニターされ、酵素の表面露出はプロテアーゼ接触可能性(accessibility)テストによって確認できる。最適な条件の下では、スモールスケールの生物学的形質転換(biotransformation)(1ml)中のOD57810のバクテリア懸濁を用いて、24時間以内に2.6mMの(R)−マンデル酸を生産することは可能であった。
【0074】
複数のホモマー(multihomomeric)の酵素のためのモデルとして、Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750からのニトリラーゼが使用された。
【0075】
全細胞消化中にプロテアーゼ切断のためニトリラーゼはアクセス可能で、自由な酵素が行ったように、ニトリラーゼはベンズアルデヒド用の同じ基質阻害を示した。
結果
融合タンパク質構築
ニトリラーゼ遺伝子は、公表された塩基配列データから推定されたオリゴヌクレオチドの使用によって、Alcaligenes faecalisの亜種(subsp)であるfaecalis ATCC 8750のゲノムDNAからポリメラーゼ連鎖反応(PCR)によって増幅された(実験のセクションを参照)。自動ディスプレイに必要な自動トランスポータードメインの翻訳領域中にニトリラーゼを融合させるために必要なニトリラーゼコード化領域の5`端のXhoIサイトと、3`端のKpnIサイトにPCRプライマーを添加した。
【0076】
ベクターの背骨として、自動トランスポーターの骨組みをコード化する、pET-Adxが上記のように使用された(Jose、2001年)。生じたPCR産物は、XhoI/KpnIで切断されたpET-Adxにつながれ(ligated)、pAT-NitAfを与えるために前の乗客Adxと置き換える。生じる自動トランスポーター融合タンパク質は、シグナル・ペプチド、乗客としてのニトリラーゼ、膜貫通リンカーおよびβ-バレルを含む典型的な構造を示す(図1)。コレラ毒素βサブユニット(CTB)に由来したシグナル・ペプチドは、内部の膜を通る輸送に使用される。外膜を通る乗客の転置のためのβ-バレル(自動トランスポーターのトランスポータードメイン)は、および十分な表面アクセスを達成するための連結地域(膜貫通のリンカー)は、AIDA-I(ホセ、2001年)に由来する。
【0077】
pAT-NitAfによってコード化された、生じた成熟した融合タンパク質は、シグナル・ペプチダーゼによって処理された後に約90 kDaの予想された分子量を持っている。表示されたニトリラーゼの切断を防ぐpAT-NitAfは、大腸菌BL21(DE3)(OmpTネガティブ突然変異体)に形質転換された。この系統は大腸菌BL21(DE3)pAT-NitAfと名付けられた。
【0078】
ニトリラーゼ自動トランスポーター融合タンパク質の発現
人工の融合タンパク質の蛋白質生合成は、実験のセクションに従って1mM IPTGの追加によって始められた。ニトリラーゼが表面または周辺質に完全に露出されたか否かを調査するために、大腸菌BL21(DE3)pAT-NitAfの無傷の細胞を用いた全細胞消化が行なわれた。 大腸菌の外膜がプロテイナーゼK(proteinase K)あるいはトリプシンのようなプロテアーゼには浸透性ではないので、外部的に追加されたプロテアーゼによるタンパク質の分解はその表面の露出になるに違いない。発現と全細胞の消化は、ナトリウム・ドデシルスルファート・ポリアクリルアミドゲル等電点電気泳動(SDS-PAGE)によってモニターされた。続くクマシ・ブリリアント・ブルーによる染色は、プロテイナーゼKの追加が融合タンパク質帯域(図2)の大きな縮小に結びついたことを示した。
分析された細胞の完全性に対して感受性のあるマーカーは、Omp F/CおよびOmpAのタンパク質帯域の強度です。消化されたサンプルおよび未消化のサンプル内の2つのOmp帯域の比率で変更はないに違いない。もしOmpAの帯域が消える場合、外膜は漏れるようになりました。また、OmpAの周辺質内の部分はプロテイナーゼKによって分解された。私たちの実験では、OmpAのサイズおよび量はプロテアーゼで処理された細胞および処理されなかった細胞において同一でした。
【0079】
要約すると、私たちは、ほとんどすべて乗客酵素がバクテリアの表面に露出されたと結論を下した。
【0080】
表面に表示されたニトリラーゼの酵素活性
大腸菌BL21(DE3)の分析のために、pAT-NitAfプラスミドを含む細胞は、1mlの全容量中の50mMリン酸ナトリウム緩衝液pH 7.5において、10の578nm(OD578)の光学濃度に調節された。酵素がヒドロキシニトリル(hydroxynitriles)のための基質特異性を示すと説明されるので、ラセミのマンデロニトリルは最終濃度10 mMで基質として使用された。生物学的変換はHPLCによって分析され、生産されたマンデル酸が計算された。最初の生物学的変換は、0.5のOD578でIPTGによって誘導された細胞で試みられた。これは72時間の生物学的変換の後に2.1mMのマンデル酸の生産に結びついた。120時間までのさらなる培養は、生産されたマンデル酸のさらなる増加に帰着しない。生物学的変換は、1のOD578でIPTGによる誘導によって2つの大きさの中で著しく改善された。この修正で、生物学的変換は72時間で横ばいにはならないが、しかし対照的に120時間の観察期間に合計6.6mMのマンデル酸に増加した(図3)。追加基質の量は、自由なベンズアルデヒドによって引き起こされた酵素阻害の発生により制限されており、それは、マンデロニトリルの分解によって自然に形成される(Yamamoto et al.、1992年)。
【0081】
マンデル酸へのマンデロニトリルの転換
HPLC法は、産物(マンデル酸)から基質(マンデロニトリル)を分けて、かつそれらを検出するために開発された。大腸菌BL21(DE3)pAT-NitAfを用いたマンデロニトリルからマンデル酸への転換が確認された(図10)。HPLC解析は、予想通りに、水性系(aqueous system)中のマンデロニトリルがベンズアルデヒド(+HCN)との平衡の中にあることをさらに明らかにした(図10)。いくつかの反応パラメータの適用は、10のOD578の細胞懸濁が使用された時(図11)、5mMの基質濃度からスタートして120時間後に、2.4 mM(それは1mlのサンプル当たり0.36mgの塊り(masse)に相当する)の産物濃度に帰着した。
鏡像体過剰率
次のステップは生産されたマンデル酸の鏡像体過剰率を確認だった。この目的のために、マンデル酸は、酢酸エチルを用いた溶媒抽出によって水系の緩衝液から抽出された。抽出されたマンデル酸は、メタノールの中で溶解され、キラルHPLC解析に直接使用された。鏡像体過剰率(ee)は37°Cの生物学的変換で、ee>99%であるとわかった(図4および15A)。これは、A.faecalisの表面の表示されたニトリラーゼが、高い鏡像異性的な純度をもった(R)-マンデル酸を生産するのに有能なことを示す。図15Aから明らかなように、マンデル酸の(R)−鏡像異性体のほとんど排他的なピークで特徴的な保持時間が検知できた。計算された鏡像異性的な純度は99%以上だった。図15Bは、図15Aからの転換された純粋(R)-および(S)−鏡像異性体のスパイク波形により決定されるような、(R)−マンデル酸の個性を示す。
pH最適条件
Alcaligenes faecalis ニトリラーゼのpH最適条件はpH 7.5であると以前に公表されていたが、私たちは、表面露出によりpH最適条件に変化があるかどうかを調査した。細胞は、最終で10のOD578になるように酢酸緩衝液あるいはTris-HCl緩衝液(両方とも50 mM)の中で再懸濁された。37°Cで120時間の培養の後、上澄みはHPLCによって分析され、また、変換されたマンデル酸の量が計算された。私たちは5.5mMマンデル酸の転換でpH7がpH最適条件であると決定した。pH 7とpH 8の間で強い活性の損失が検出できた。追加的に調査されたリン酸緩衝液pH 7.5は、その他の点は同じ条件の下で、たった4.4mMのマンデル酸の転換に結びついた(データを示さず)(図5)。
【0082】
反応温度およびpH値の最適化で、24時間以内の2.6mMまでのマンデル酸を変換することが可能であった。マンデルアミドは常に検知できなかった。
【0083】
反応速度
一連の実験では、いくつかのパラメータは、実験室スケールでのマンデル酸生産に対するそれらの影響のために分析された。図12は、異なる温度でのマンデル酸生産の反応速度を表わす。37°Cでは、30°Cでの転換と比較して、産物の濃度が約20%増加したことが検知された。30°Cおよび37°Cの両方の転換の進行は、生産されたマンデル酸の濃度が約144時間の転換の後も単にゆっくり増加することをさらに実証した。
基質濃度
さらに、マンデル酸の生産に対する基質濃度の影響が分析された。水溶液(aqueous solution)では、基質のマンデロニトリルは、ベンズアルデヒドおよびHCNと平衡状態にある。しかしながら、過度の濃度にあるベンズアルデヒドは、細胞に毒作用を示し、それによって図13に示されるように基質濃度の増加を制限する。10 mM未満へのニトリル濃度のさらなる縮小が生産速度のさらなる増加に帰着しなかったことは、最適な基質濃度として10mMマンデロニトリルの使用を示唆する。
【0084】
異なる緩衝液システム
全細胞触媒の大腸菌BL21(DE3)pAT-NitAfを使用するマンデル酸生産の相対的な試験も異なる緩衝液システムを使用して行なわれた。図14は、最良の結果が、使用される他の緩衝液と比較して、pH 6.5のトリスバッファーを用いた溶液を使用して達成されたことを示す。
【0085】
貯蔵安定性
構築された全細胞生体触媒全体の貯蔵安定性を決定するために、大腸菌BL21(DE3)pAT-NitAfの細胞(IPTGで誘導される)は、20%(v v-1)のグリセロールを含み、格納されて−18°Cで保存されて、50mMリン酸ナトリウム緩衝液pH 7.5中の、50のOD578に調節された。グリセロールは、細胞の溶解を防ぐための不凍薬剤として-18°Cでサンプルに添加された。異なる時間間隔で、サンプルは、一度リン酸緩衝液の中で洗われ、10のOD578に調節された。あるいは、IPTG誘導ののち、細胞の部分標本は冷蔵庫中で6°Cで保管され、異なる保管時間の後にニトリラーゼ分析にさらされた。反応は10mMマンデロニトリルの追加によって始められ、また、サンプルは37°Cで120時間培養された。 -18°Cで35日の保管の後、細胞は、まだそれらの初期の活性の72%を保持していた(図6)。1週後の、+6°Cの全細胞触媒の活性は、初期活性のわずか約50%だった(図16)。さらに、これらの実験では、これは観察された細胞の溶解と関連した。ここで、再び、細胞の一部が測定された期間にわたる比較的安定な残余活性を示すことは明白である。
【0086】
全細胞触媒の温度依存性
A.faecalisから精製された酵素の最適温度が40−45°Cであることが説明される。 これらの発見は、30°Cから45°Cまでの範囲の温度テストについて記述する図17によって確認された。反応混合物中のリン酸緩衝液あるいはトリス緩衝液の使用は、特別の影響があるようには見えない。
【0087】
全細胞触媒の再利用性(図7)
24時間5周期の転換反応の全細胞生体触媒の再利用性および安定性を調査することは、異なる緩衝液および温度条件で行なわれた。「周期」は、大腸菌細胞の遠心分離および次の周期のための新鮮な基質の追加に続く24時間の転換反応である。24時間以内の最も高い産出が、2.62mMマンデル酸でリン酸ナトリウム緩衝液(pH 7.5および45°C)中の大腸菌BL21(DE3)pAT-NitAfで達成されたが、しかし、活性は、5つの周期の後に最小で当初の活性のたった10%に速く落ちた。一次的転換レートの33%を超える5つの周期を達成できた後、Tris-HCl緩衝液(pH 7および45°C)を用いて、2.53mMマンデル酸の転換が24時間以内に達成可能だった。Tris-HCl(37°C)の中の細胞の活性は最初の周期の2.24 mMから5つの周期の後には1.25 mMまで落ちたが、これは、55%を超える残余の活性を表わす。最初の周期中では、リン酸ナトリウム緩衝剤あるいはTris-HCl緩衝液の中で再懸濁された細胞間の生産速度に小さな違いだけがあったが、だが対照的にすべて成績においてTris-HCl緩衝液を用いた試験サンプルの方が常によかった。
【0088】
実験のセクション
バクテリア株およびプラスミド
サブクローン化の目的で、Invitrogen (Carlsbad, CA, USA)から提供される大腸菌TOP10 [F- mcrA (mrr-hsd RMS-mcrBC) Φ80lacZ M15 lacΧ74 recA1 araD139 (ara-leu)7697 galU galK rpsL (StrR) endA1 nupG]が使用された。大腸菌BL21 (DE3) [B, F-, dcm, ompT, lon, hsdS (rB- mB-), gal, λ (DE3)]は、自動トランスポーター融合タンパク質の発現に使用された。
【0089】
大腸菌系統は、活発な揺れ(200rpm)とともにアンピシリン(100 μgmL-1)を含んでいるLuria-Bertani (LB)培地で37°Cで慣例的に培養された。固形培地は、培地に寒天(agar) (1.5 %, wv-1)の追加して調製された。
【0090】
Invitrogen (Carlsbad, CA, USA)からのプラスミドpCR4-TOPO(登録商標)はニトリラーゼ遺伝子のサブクローン化のために使用された。AIDA-I自動トランスポーターの骨組みをコード化するプラスミドpET-Adx04の構築は、以前に記述された(Rustler et al.、2008年; 米国特許6,180,359番)。図8は、11の形質転換体から分離されたプラスミドを示し、そこではPCR破片に対応する約1kbの正確なDNA断片は、KpnIとXhoIを使用した続く消化によって、7つのクローン中の正しい自動トランスポータープラスミド(約7kb)内に検知された。
【0091】
組み換えDNA技術
ニトリラーゼをコード化する遺伝子は、Alcaligenes faecalisの亜種のfaecalis ATCC 8750 (LGC Standards GmbH, Wesel, Germany)のゲノムDNAから増幅された。
【0092】
ニトリラーゼ遺伝子の増幅用のプライマーは、A.faecalis ATCC 8750(Kaul et al.、2007年ら)のニトリラーゼ遺伝子のための既に記述されていた配列情報に基づいた。前方のプライマーNit-XhoI (5´- CCGCTCGAGCAGACAAGAAAAATCGTCC)および逆のプライマーNit-KpnI (3´- GGGGTACCGGACGGTTCTTGCACC)は、ニトリラーゼ遺伝子を含む1073bp破片を増幅するために使用された。PCR産物のクローニングを容易にするためにXhoIおよびKpnIの制限部位が添加された(プライマー配列中の強調された部分は、それぞれの酵素用の制限部位を表わす)。PCR反応は、10 μLの体積中において、10 ngのゲノムDNA、Eppendorf (Hamburg, Germany)から提供される適切な量のEppendorf PCR Mastermix、1μL(10μMの原液)の前側および後ろ側プライマー(Sigma-Aldrich Chemical Company, Milwaukee, USA)を含む。
【0093】
PCR反応は、Eppendorf Mastercycler gradient from Eppendorf (Hamburg, Germany)によって実現される。PCRプロトコルは、最初の変性ステップが95°Cで5min、30サイクルの、94°Cの30秒、64°Cの1分、72°Cの1分、が続き、および72°Cで6分の最終の延長ステップで終わる。
【0094】
PCR産物は、メーカーの指示によってpCR4-TOPO Vectorへのサブクローン化のために直接使用された。別記されるように、プラスミドDNAの調製が行なわれた。生じたプラスミドは、XhoI/KpnIで消化され、Qiagenゲル抽出キット(Qiagen, Hilden, Germany)を使用して、アガロース・ゲル電気泳動によって精製された。
【0095】
pAT-NitAfに与えるために、生じた破片は、XhoI/KpnIで切断されたpET-Adx04ベクターに挿入され、そこではAdx04をコード化する遺伝子に交換された。
【0096】
外膜標本およびSDS-PAGE
大腸菌細胞はオーバーナイトで成長され、そして、適切な量の培養は、2−メルカプトエタノール(10 mM)およびアンピシリン(100 μgmL-1)を含むLB培地への接種(inoculation)に使用された。0.5または1の光学濃度(OD578)が到達するまで、細胞は37°C(200rpm)で培養された。誘致可能な発現のため、Roth (Karlsruhe, Germany)から購入されたIPTG(1mMの最終濃度)が添加された。30°Cに60分おいた後、細胞は収穫され、Tris-HCl(0.2M、pH 8.2)で洗われた。プロテアーゼ接触可能性テストのため、細胞は、25μLのプロテイナーゼK-溶液(2%)を含む2mlのTris-HCl (0.2M、pH 6.8)の中で再懸濁され、37°Cのウォーター・バスの中で60分培養された。異なる細胞の分別法は、Hantke(Hantke、1981年)の方法の修正によって行なわれた。
【0097】
SDS-PAGEのため、単離された外膜は、Tris-HCl(100 mM、pH 6.8)、ドデシル硫酸ナトリウム(4%)、ブロモフェノール・ブルー(0.2%)、2−メルカプトエタノール(2%)、ジチオスレイトール(0.2M)およびグリセロール(20%)から成るサンプル緩衝液で、1:2で薄められ、それは5分煮沸され、10%のポリアクリルアミドゲルで分析された。タンパク質はクマシ・ブリリアント・ブルーで染色され、Fermentas(St. Leon-Rot, Germany)から購入されたPageruler不染色タンパク質ラダーは、サイズ決定に使用されました。
【0098】
標準転換を行なうこと
標準的な実験は以下のように実行される。プレカルチャーされた組み換え大腸菌BL21(DE3)pAT-NitAf系統が、1:100の比率でメインカルチャーに接種するために使用され、振盪培養器の中で37°Cおよび200rpmで培養される。1のOD578に達する際、IPTGは1mMの最終濃度で使用され、誘導のために添加される。その後、誘導は30°C、200rpmで1時間起こる。その細胞が50mMリン酸緩衝液pH 7.5で一度洗われた後、また同じ緩衝液を使用して、最終的に10のOD578に調節されました。その後、前記細胞懸濁の1mlの部分標本は、20μlのマンデロニトリル(0.5M)と一緒にされ、1000rpmあるいは200rpmで熱攪拌機の中で異なる温度(特に37°Cの中で)で培養される。サンプルは異なる時間間隔で得られ、遠心分離機にかけられ(6000 x g、5分)、VWR (Darmstadt, Germany)から購入された0.2μmのポリエーテルサルホン(polyethersulfone)薄膜フィルタで滅菌ろ過された。上澄みは、HPLCによって直接分析された。それ以上、生物学的変換の滅菌ろ過の後に検知できなかった。
【0099】
分析手法
ニトリル転換は、Waters (MA, USA)からのダイオード・アレー検波器996と2695分離モジュールを装備した、HPLC(Millenium Chromatography Manager 3.2)によって分析された。 アキラル(achiral)な分析の目的で、210nmで分光測光法で検知された転換産物を識別するために、ヒューレット・パッカード(Boblingen, Germany)から提供される逆相カラム[100mm x 4.6mm(内部直径)、Hypersil ODS 5μm直径粒子で満たされた]が使用された。水の中のメタノール(40% vv-1)およびリン酸(0.1%)は、移動相としてHPLC分離に使用された。マンデル酸鏡像異性体の分離のために、CS-Chromatographie Service (Langerwehe, Germany)から提供される、キラル位相カラム[150mm x 4.6mm(内部直径)、Reprosil Chiral-OM 5μm直径の粒子で満たされた]が使用された。
【0100】
移動相は、ヘキサン(90 % vv-1)、イソプロピルアルコール(10 % vv-1)およびトリフルオロ酢酸(0.1 % vv-1)から構成された。マンデル酸は、ジクロロメタン(dichlormethane)の代わりに酢酸エチルが使用された少しの修正(Yamamoto, 1991)と共に、別記されるように生物学的変換サンプルから抽出された。
【0101】
例2: Klebsiella pneumoniaeからのニトリラーゼ
Klebsiella pneumoniaeからのニトリラーゼ遺伝子は、例1に記述されるように、PCRを使用して増幅され、大腸菌標準ベクターへの中間体クローニングおよび塩基配列分析に続いて、対応する自動ディスプレイ・ベクターに挿入された。K. pneumoniaeニトリラーゼを含む融合タンパク質、自動トランスポータードメイン、膜貫通リンカーおよびシグナル・ペプチドをコード化する自動ディスプレイの構成物は、例1にpAT-NitAfのために記述されたとともに、pAT-NitKpと名付けられた。表面上に融合ポリペプチドを表示する、対応する大腸菌系統は、大腸菌BL21(DE3)pAT-NitKpと名付けられた。
Klebsiella pneumoniaeニトリラーゼの発現および表面の局在化の検出は、Alcaligenes faecalisニトリラーゼ(例1参照)と類似して実行された。
【0102】
それは、前記ニトリラーゼがブロモキシニルのための非常に高い基質特異性を示すと言った文献から有名である。前記有毒化合物は、農業で除草剤として広く使用され、毎年、ドイツで工業規模に適用されます。対応するカルボン酸は、低い毒性と自然界での安定性を示す。
【0103】
大腸菌JK321 pAT-NitKpによるブロモキシニルからの3,5-ジブロモ-4-ヒドロキシ-安息香酸の生産は、図18に示されます。この図は、大腸菌細胞の表面上で発現されたK. pneumoniaeニトリラーゼがニトリルをその対応するカルボン酸に変換することができることを示す。
【0104】
例3:Saccharomyces cerevisiaeからのニトリラーゼ
例1に記述されるように、S.cerevisiaeのニトリラーゼ遺伝子はPCRによって増幅されクローン化された。塩基配列分析は、データベースから得られたS.cerevisiae ニトリラーゼ配列と100%の同一性を示した。例1に記述されるように、ニトリラーゼは、自動トランスポータードメイン、膜貫通リンカーおよびシグナル・ペプチドを含む融合タンパク質として大腸菌中でクローン化された。再び、塩基配列分析はデータベースからの配列と100%の同一性を示した。発現の検出は例1と類似して実行された。
【0105】
S.cerevisiaeニトリラーゼ、自動トランスポータードメイン、膜貫通リンカー、およびシグナル・ペプチド、を含む融合タンパク質をコード化する自動ディスプレイの構成物は、例1にpAT-NitAfのために記述されるとともに、pAT-NitScと名付けられた。表面上に融合ポリペプチドを表示する、対応する大腸菌系統は、大腸菌BL21(DE3)pAT-NitScと名付けられた。
【0106】
IPTGの誘導に続いて、S.cerevisiae ニトリラーゼを含む融合タンパク質は、表面に局在化されると分かった。例1に記述された全細胞のトリプシン消化が適用された。結果は図9に要約された。
参考文献
1. Jose J, Meyer TF (2007)、自動ディスプレイの話、発見からバイオテクノロジー的・バイオ医学的な適用まで。Microbiol Mol Biol Rev 71 (4): 600-19
2. Nagasawa T, Mauger J, Yamada H (1990)、新規なニトリラーゼ、Alcaligenes faecalis JM3のアリールアセトニトリラーゼ(arylacetonitrilase)、精製および特徴の描写。Eur J Biochem 194(3): 765-72
3. Yamamoto K, Oishi K. Fujimatsu I, Komatsu K (1991)、Alcaligenes faecalis ATCC 8750によるマンデロニトリルからのR-(-)-マンデル酸の生産。Appl Environ Microbiol. 57(10): 3028-32
4. Rey P, Rossi JC, Taillades J, Gros G, Nore O (2004)、動けなくされたニトリラーゼを使用するニトリルの加水分解:メチオニン・ヒドロキシ類似誘導体(methionine hydroxy analogue derivatives)の合成への適用。J Agric Food Chem 52 (26): 8155-62
5. Kiziak C, Conradt D, Stolz A, Mattes R, Klein J (2005)、Pseudomonas fluorescens EBC191からのニトリラーゼ: 遺伝子のクローニングおよび異種起源の発現および組み換えの酵素の生化学の特徴の描写(characterization)。Microbiology 151 (Pt 11): 3639-48.
6. Kaul P, Stolz A, Banerjee UC (2007)、鏡像異性選択的なニトリル加水分解のための架橋された無定形のニトリラーゼの集合。Adv Synth Catal 349: 2167-2176
7. Banerjee A, Dubey S, Kaul P, Barse B, Piotrowski M, Banerjee UC (2008) 、Pseudomonas putidaからの鏡像異性選択的なニトリラーゼ: クローニング、異種起源の発現およびバイオリアクター研究。Mol Biotechnol [Epub ahead of print]
8. Luo H, Fan L, Chang Y, Ma J, Yu H, Shen Z (2008)、大腸菌中のRhodococcus rhodochrous tg1-A6からのニトリラーゼの、遺伝子クローニング、過剰発現、および特徴の描写。
Appl Biochem Biotechnol [Epub ahead of print]
9. Rustler S, Motejadded H, Altenbuchner J, Stolz A (2008)、Pseudomonas fluorescensからのアリールアセトンニトリラーゼ(arylacetonitrilase)の同時の発現、そして(S)―マンデル酸合成用のPichia pastorisのManihot esculentaからの(S)−オキシニトリラーゼ。Appl Microbiol Biotechnol 80(1): 87-97
10. ニトリラーゼを使用する2-ヒドロキシ-4-メチル酪酸の調製のための工業規模プロセス。米国特許6180359。
11. Ress-Loschke M. Friedrich T, Hauer B, Mattes R (1998)、ニトリラーゼまたはニトリラーゼの遺伝子を含む微生物を用いてニトリルからの光学活性カルボン酸の製造方法。ドイツ特許出願DE 19848129A1
12. J. Jose, Appl. Microbiol. Biotechnol. 2006, 69, 607-614.
13. J. Jose, R. Bernhardt, F. Hannemann, J. Biotechnol. 2002, 95, 257-268.
14. J. Jose, S. von Schwichow, ChemBioChem 2004, 5, 491-499.
15. K. V. Thimann, S. Mahadevan, Arch. Biochem. Biophys. 1964,105,133-141.
16. A. Banerjee, R. Sharma, U. C. Banerjee, Appl. Microbiol. Biotechnol. 2002, 60, 33-44.
17. R. N. Thuku, D. Brady, M. J. Benedik, B. T. Sewell, J. Appl. Microbiol. 2009, 106, 703-727.
18. K. Yamamoto, I. Fujimatsu, K.-I. Komatsu, J. Ferment. Bioeng. 1992, 73, 425-430.
19. J. Jose, R. Bernhardt, F. Hannemann, ChemBioChem 2001, 2, 695-701.
20. J. Maurer, J. Jose, T. F. Meyer, J. Bacteriol. 1997, 179, 794-804.
21. H. C. Birnboim, J. Doly, Nucleic Acids Res. 1979, 7, 1513-1523.
22. K. Hantke, Mol Gen Genet 1981, 182, 288-292.
23. Henderson I et al., 2004、タイプVタンパク質分泌経路: 自動トランスポーターの話。Microbiology and Molecular Biology Reviews, 68(4), 692-744.
24. Niewert U., Frey A., Voss T., Le Bouguen C., Baljer G., Franke S., Schmidt MA. AIDA自動トランスポーター系は、浮腫病およびポスト離乳食下痢と診断されたブタから単離された大腸菌のF18とStx2eに関係する。Clin. Diagn. Lab. Immunol. 2001 Jan, 8(1):143-149;9.
25. Buchholz, Kasche und Bornscheuer (2005)。生体触媒および酵素技術。Enzymes in Organic Chemistry, Wiley-VCH-Verlag, Weinheim.
26. Hartl, F. U., and Hayer-Hartl, M. (2002) Science 295(5561):1852-8
【特許請求の範囲】
【請求項1】
以下のステップを含む、ニトリラーゼによって触媒される反応の産物を生産する方法、
(i) その表面に置かれた前記ニトリラーゼを含む微生物、および/または、前記微生物の膜標本を用意し、
(ii) ニトリラーゼ活性と適合性をもつ条件の下、微生物および/またはそれの膜標本を1つ以上のニトリラーゼ基質と接触させる。
【請求項2】
ニトリラーゼによって触媒された反応の産物はカルボン酸であり、少なくとも1つのニトリラーゼの基質はニトリルである請求項1に記載の方法。
【請求項3】
前記ニトリラーゼは、前記微生物中で組み換え的に発現され、その表面へ輸送される請求項1または請求項2に記載の方法。
【請求項4】
前記ニトリラーゼは、自動トランスポーターのトランスポータードメインに融合される請求項1から請求項3のいずれか1項に記載の方法。
【請求項5】
ステップ(i)は以下を含む請求項1から請求項4のいずれか1項に記載の方法、
(a) 発現制御配列と効果的につながれた核酸配列を含む微生物を用意し、前記核酸配列は、以下を含む、
(1) シグナル・ペプチドをコード化する部分、
(2) 表示される組み換えのニトリラーゼをコード化する部分、
(3) プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4) 膜貫通リンカーをコード化する部分、
(5) 自動トランスポーターのトランスポータードメインをコード化する部分、
(b) (a)の前記核酸配列が発現されるとともに前記核酸配列の発現産物が前記微生物の表面に表示されるような条件の下で前記微生物を培養する。
【請求項6】
以下のステップを含む請求項5に記載の方法、
(c) (b)の前記微生物から膜標本を作る。
【請求項7】
前記自動トランスポーターの前記トランスポータードメインは、Ssp、Ssp-h1、Ssp-h2、PspA、PspB、Ssa1、SphB1、AspA/NalP、VacA、AIDA-I、IcsA、MisL、TibA、Ag43、ShdA、AutA、Tsh、SepA、EspC、EspP、Pet、Pic、SigA、Sat、Vat、EpeA、EatA、EspI、EaaA、EaaC、Pertactin、BrkA、Tef、Vag8、PmpD、Pmp20、Pmp21、IgA1 protease、App、Hap、rOmpA、rOmpB、ApeE、EstA、Lip-1、McaP、BabA、SabA、AlpA、Aae、NanBおよびそれの変異体からなるグループから選択される請求項4から請求項6のいずれか1項に記載の方法。
【請求項8】
前記ニトリラーゼは、Alcaligenesニトリラーゼ、Klebsiellaニトリラーゼ、およびSaccharomycesニトリラーゼからなるグループから選択される先行する請求項のいずれか1項に記載の方法。
【請求項9】
前記微生物は、グラム陰性のバクテリアであり、特に大腸菌である先行する請求項のいずれか1項に記載の方法。
【請求項10】
ステップ(ii)は、約6.5から約7.5までの範囲中のpHで行なわれる先行する請求項のいずれか1項に記載の方法。
【請求項11】
ステップ(ii)は、好気性のおよび/またはを酸化させる条件の下で行なわれる先行する請求項のいずれか1項に記載の方法。
【請求項12】
前記ニトリルは、マンデロニトリル、プルナシン、ブロモキシニル、イオキシニル、クロロキシニル(chloroxynile)、アニソニトリル(anisonitrile)、3-ブロモ-4-ヒドロキシベンゾニトリル、3-フルオロ-4-ヒドロキシベンゾニトリル、4-ヒドロキシ-3,5-ジメトベンゾニトリル(dimethobenzonitrile)、ベンゾニトリル、フェニルプロピオニトリル(phenylprobionitrile)、フェニルグリシノニトリル、n-ブチロニトリル(butyronitrile)、n-バレロニトリル(valeronitrile)、イソブチロニトリルおよびスクシノニトリルからなるグループから選ばれる請求項2から請求項11のいずれか1項に記載の方法。
【請求項13】
ニトリラーゼによって触媒された反応の産物はカルボン酸であり、前記カルボン酸が少なくとも約70%の鏡像体過剰率で生産される先行する請求項のいずれか1項に記載の方法。
【請求項14】
さらに以下のステップを含む、先行する請求項のいずれか1項に記載の方法、
(iii) ステップ(ii)で使用された微生物を回収する。
【請求項15】
その表面上に組み換えニトリラーゼを表示する微生物。
【請求項16】
請求項15の前記微生物から得られた組み換えニトリラーゼを備えた膜標本。
【請求項17】
ニトリラーゼによって触媒された反応の産物の生産のための請求項15の前記微生物および/または請求項16の膜標本の使用。
【請求項18】
微生物に核酸配列を導入することを含む、その表面上に組み換えニトリラーゼを表示する微生物を生産する方法、前記核酸配列は以下を含む、
(1)シグナル・ペプチドをコード化する部分、
(2)表示される組み換えのニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4)膜貫通リンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、そこでは前記核酸配列が発現制御配列と効果的につながれる。
【請求項1】
以下のステップを含む、ニトリラーゼによって触媒される反応の産物を生産する方法、
(i) その表面に置かれた前記ニトリラーゼを含む微生物、および/または、前記微生物の膜標本を用意し、
(ii) ニトリラーゼ活性と適合性をもつ条件の下、微生物および/またはそれの膜標本を1つ以上のニトリラーゼ基質と接触させる。
【請求項2】
ニトリラーゼによって触媒された反応の産物はカルボン酸であり、少なくとも1つのニトリラーゼの基質はニトリルである請求項1に記載の方法。
【請求項3】
前記ニトリラーゼは、前記微生物中で組み換え的に発現され、その表面へ輸送される請求項1または請求項2に記載の方法。
【請求項4】
前記ニトリラーゼは、自動トランスポーターのトランスポータードメインに融合される請求項1から請求項3のいずれか1項に記載の方法。
【請求項5】
ステップ(i)は以下を含む請求項1から請求項4のいずれか1項に記載の方法、
(a) 発現制御配列と効果的につながれた核酸配列を含む微生物を用意し、前記核酸配列は、以下を含む、
(1) シグナル・ペプチドをコード化する部分、
(2) 表示される組み換えのニトリラーゼをコード化する部分、
(3) プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4) 膜貫通リンカーをコード化する部分、
(5) 自動トランスポーターのトランスポータードメインをコード化する部分、
(b) (a)の前記核酸配列が発現されるとともに前記核酸配列の発現産物が前記微生物の表面に表示されるような条件の下で前記微生物を培養する。
【請求項6】
以下のステップを含む請求項5に記載の方法、
(c) (b)の前記微生物から膜標本を作る。
【請求項7】
前記自動トランスポーターの前記トランスポータードメインは、Ssp、Ssp-h1、Ssp-h2、PspA、PspB、Ssa1、SphB1、AspA/NalP、VacA、AIDA-I、IcsA、MisL、TibA、Ag43、ShdA、AutA、Tsh、SepA、EspC、EspP、Pet、Pic、SigA、Sat、Vat、EpeA、EatA、EspI、EaaA、EaaC、Pertactin、BrkA、Tef、Vag8、PmpD、Pmp20、Pmp21、IgA1 protease、App、Hap、rOmpA、rOmpB、ApeE、EstA、Lip-1、McaP、BabA、SabA、AlpA、Aae、NanBおよびそれの変異体からなるグループから選択される請求項4から請求項6のいずれか1項に記載の方法。
【請求項8】
前記ニトリラーゼは、Alcaligenesニトリラーゼ、Klebsiellaニトリラーゼ、およびSaccharomycesニトリラーゼからなるグループから選択される先行する請求項のいずれか1項に記載の方法。
【請求項9】
前記微生物は、グラム陰性のバクテリアであり、特に大腸菌である先行する請求項のいずれか1項に記載の方法。
【請求項10】
ステップ(ii)は、約6.5から約7.5までの範囲中のpHで行なわれる先行する請求項のいずれか1項に記載の方法。
【請求項11】
ステップ(ii)は、好気性のおよび/またはを酸化させる条件の下で行なわれる先行する請求項のいずれか1項に記載の方法。
【請求項12】
前記ニトリルは、マンデロニトリル、プルナシン、ブロモキシニル、イオキシニル、クロロキシニル(chloroxynile)、アニソニトリル(anisonitrile)、3-ブロモ-4-ヒドロキシベンゾニトリル、3-フルオロ-4-ヒドロキシベンゾニトリル、4-ヒドロキシ-3,5-ジメトベンゾニトリル(dimethobenzonitrile)、ベンゾニトリル、フェニルプロピオニトリル(phenylprobionitrile)、フェニルグリシノニトリル、n-ブチロニトリル(butyronitrile)、n-バレロニトリル(valeronitrile)、イソブチロニトリルおよびスクシノニトリルからなるグループから選ばれる請求項2から請求項11のいずれか1項に記載の方法。
【請求項13】
ニトリラーゼによって触媒された反応の産物はカルボン酸であり、前記カルボン酸が少なくとも約70%の鏡像体過剰率で生産される先行する請求項のいずれか1項に記載の方法。
【請求項14】
さらに以下のステップを含む、先行する請求項のいずれか1項に記載の方法、
(iii) ステップ(ii)で使用された微生物を回収する。
【請求項15】
その表面上に組み換えニトリラーゼを表示する微生物。
【請求項16】
請求項15の前記微生物から得られた組み換えニトリラーゼを備えた膜標本。
【請求項17】
ニトリラーゼによって触媒された反応の産物の生産のための請求項15の前記微生物および/または請求項16の膜標本の使用。
【請求項18】
微生物に核酸配列を導入することを含む、その表面上に組み換えニトリラーゼを表示する微生物を生産する方法、前記核酸配列は以下を含む、
(1)シグナル・ペプチドをコード化する部分、
(2)表示される組み換えのニトリラーゼをコード化する部分、
(3)プロテアーゼ認識部位をコード化する、任意に設けられる部分、
(4)膜貫通リンカーをコード化する部分、
(5)自動トランスポーターのトランスポータードメインをコード化する部分、そこでは前記核酸配列が発現制御配列と効果的につながれる。
【図1】

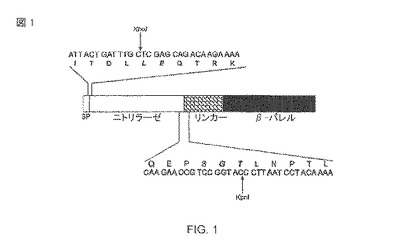
【図2】
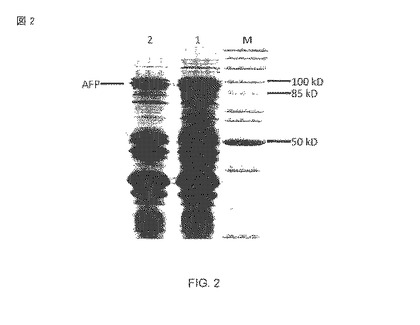
【図3】
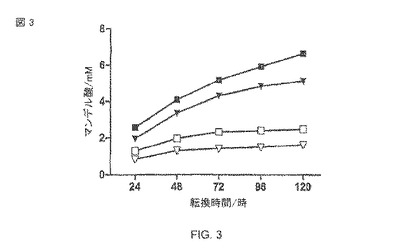
【図4】
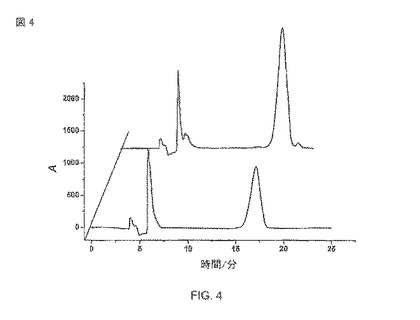
【図5】
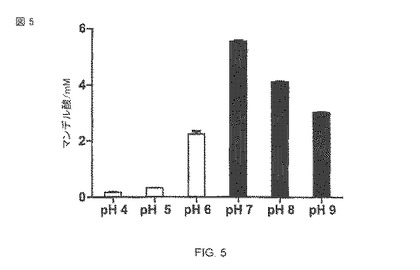
【図6】
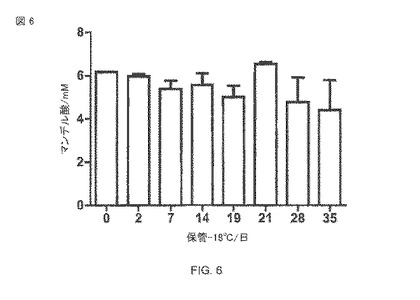
【図7】
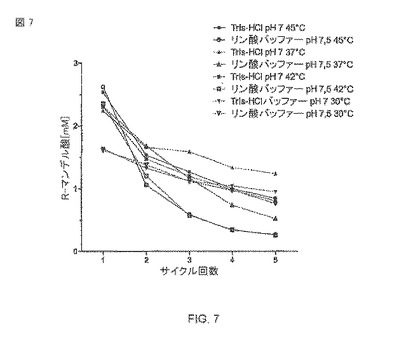
【図8】
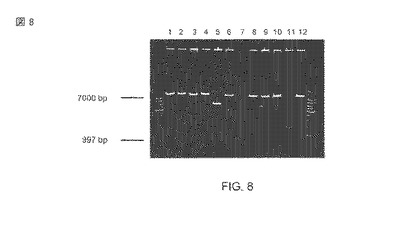
【図9】
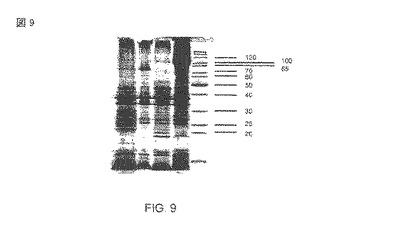
【図10】
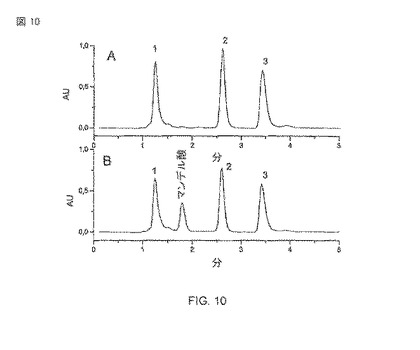
【図11】
【図12】
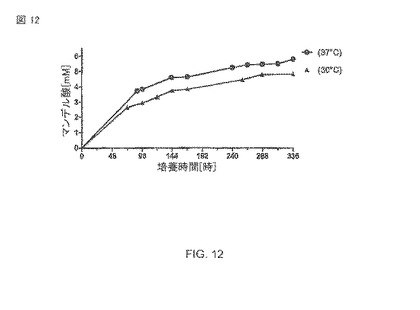
【図13】
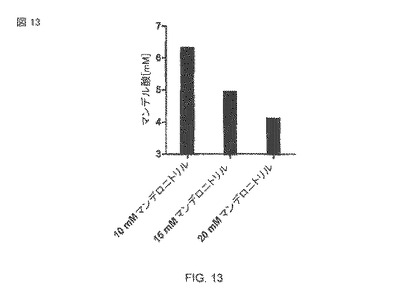
【図14】
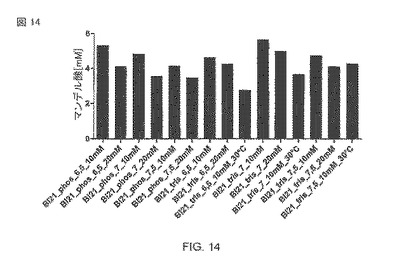
【図15】
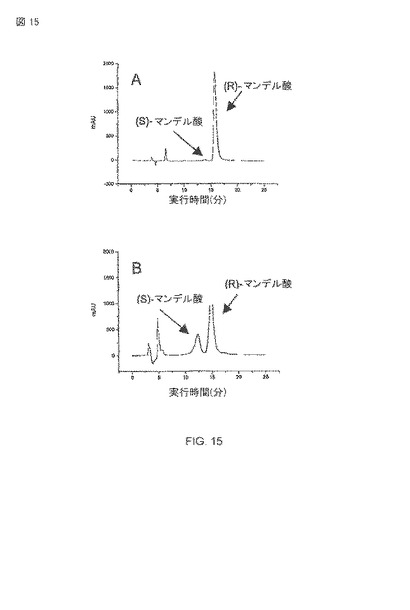
【図16】
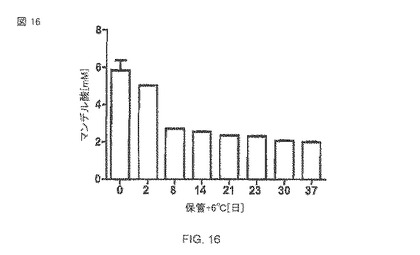
【図17】
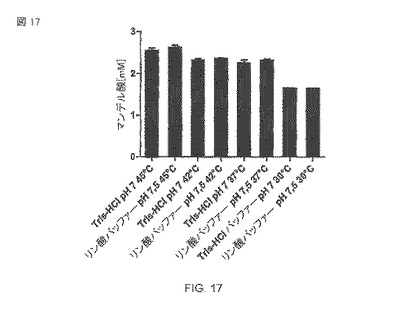
【図18】
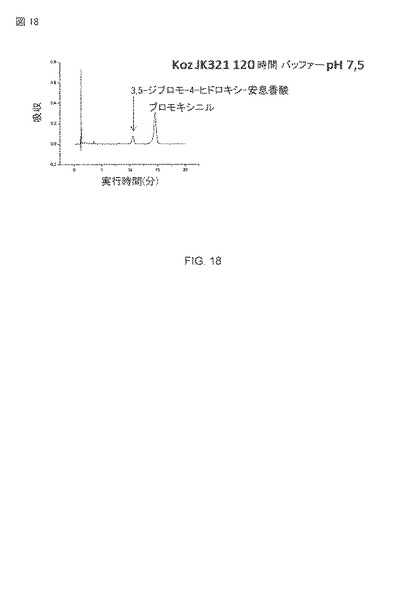
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2013−510587(P2013−510587A)
【公表日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願番号】特願2012−539221(P2012−539221)
【出願日】平成22年11月16日(2010.11.16)
【国際出願番号】PCT/EP2010/006951
【国際公開番号】WO2011/057820
【国際公開日】平成23年5月19日(2011.5.19)
【出願人】(512128553)ジルス・ベタイリグングスゲゼルシャフト・エムベーハー・ウント・コンパニー・パテンテ・ザ・ファースト・カーゲー (1)
【氏名又は名称原語表記】ZYRUS Beteiligungsgesellschaft mbH & Co. Patente I KG
【住所又は居所原語表記】Berliner Str. 1, 12529 Schoenefeld/Waltersdorf, Germany
【Fターム(参考)】
【公表日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願日】平成22年11月16日(2010.11.16)
【国際出願番号】PCT/EP2010/006951
【国際公開番号】WO2011/057820
【国際公開日】平成23年5月19日(2011.5.19)
【出願人】(512128553)ジルス・ベタイリグングスゲゼルシャフト・エムベーハー・ウント・コンパニー・パテンテ・ザ・ファースト・カーゲー (1)
【氏名又は名称原語表記】ZYRUS Beteiligungsgesellschaft mbH & Co. Patente I KG
【住所又は居所原語表記】Berliner Str. 1, 12529 Schoenefeld/Waltersdorf, Germany
【Fターム(参考)】
[ Back to top ]