
再ミエリン化および脊髄損傷の治療のためのヒト胚性幹細胞に由来するオリゴデンドロサイト
【課題】オリゴデンドロサイトおよびその前駆細胞等の、グリア細胞のマーカーを有する神経系細胞の集団を提供する。
【解決手段】細胞集団は、所望の表現型または機能的能力を有する細胞の濃縮を促進する条件下で、ヒト胚性幹細胞等の多能性幹細胞を分化させることによって作製される。様々な組み合わせの分化因子およびマイトジェンを用いて、オリゴデンドロサイト前駆細胞のマーカーを有する細胞集団を産生することができる。さらなる分化において、成熟オリゴデンドロサイトに特有の複雑な突起が形成される。細胞は、ミエリン鞘を形成する能力があり、中枢神経系の機能を改善するために治療に用いることができる。
【解決手段】細胞集団は、所望の表現型または機能的能力を有する細胞の濃縮を促進する条件下で、ヒト胚性幹細胞等の多能性幹細胞を分化させることによって作製される。様々な組み合わせの分化因子およびマイトジェンを用いて、オリゴデンドロサイト前駆細胞のマーカーを有する細胞集団を産生することができる。さらなる分化において、成熟オリゴデンドロサイトに特有の複雑な突起が形成される。細胞は、ミエリン鞘を形成する能力があり、中枢神経系の機能を改善するために治療に用いることができる。
【発明の詳細な説明】
【技術分野】
【0001】
技術分野
本発明は、一般に、胚細胞および神経系前駆細胞の細胞生物学の分野に関する。より具体的には、本発明は、生物学的研究、薬剤スクリーニング、およびヒトの治療に使用するのに適した、オリゴデンドロサイトおよびその前駆細胞の濃縮された集団を提供する。
【0002】
関連出願
本出願は、2002年7月11日に出願された米国仮出願第60/395,382号、および2003年4月4日に出願された米国実用出願第10/406,817号の利益を主張する。優先権出願は、本明細書によって完全に参照として本明細書に組み入れられる。
【背景技術】
【0003】
背景
オリゴデンドロサイトは、中枢神経系の支持において重要な生理的役割を担う。ヒトの治療にオリゴデンドロサイトが利用できれば、神経細胞を絶縁するミエリン鞘の欠陥によって生じる障害状態の治癒が促進され得る。
【0004】
多発性硬化症は、脳および脊髄の神経細胞を囲むミエリン鞘のゆっくりとした破壊を伴う、進行性かつ障害性の脱髄疾患である。症状は、しびれ感、視覚障害、および認知変化から麻痺に及ぶ。この疾患は免疫学的および遺伝学的要素を有すると考えられており、20歳から40歳の間で臨床型に現れる場合が多い。米国だけでも約300,000人がこの疾患に冒されている。治療法は、現在のところβインターフェロンまたは副腎皮質ステロイドの過程を伴う。これらの薬剤により発病の間の症状を伴う期間が短縮され得るが、一般に長期の障害は妨げられない。
【0005】
脊髄の外傷により外傷部位の近傍で無傷の軸索の脱髄が起こり、それによって軸索の神経伝達能力が奪われる。米国では、毎年約11,000件の脊髄損傷の新たな症例がある。SCI情報ネットワークでは、任意のレベルでの不完全な運動機能に苦しむ患者にかかる生涯直接費は、減給分および生活の質への影響を含めずに、$400,000〜$2,200,000に及ぶと推定している。
【0006】
中枢神経系の細胞上のミエリンは、軸索をその近傍で覆いミエリン鞘を形成するオリゴデンドロサイトによって正しい位置に整えられている。疾患状態におけるオリゴデンドロサイトおよびその前駆細胞の役割は、KeirsteadおよびBlakemore(Adv. Exp. Med. Biol. 468:183、1999)によって概説されている。O−2A細胞として知られているオリゴデンドロサイト前駆細胞は、健常成人のCNSおよび多発性硬化症の病変に存在し、再ミエリン化に関与する(Scoldingら、Brain 121:2221、1998;およびScoldingら、Neuroscience 89:1、1999)。オリゴデンドロサイトが均整をとって増殖し、損傷が大きい箇所では蓄積された前駆細胞を使い果たしてしまうため、適切な再ミエリン化ができないということが起こり得る。
【0007】
神経機能を回復するための再生医療で用い得る細胞集団を作製することを目的として、かなりの研究が行われてきた(Parkら、J. Neurotrauma 16:675、1999によって概説されている)。Keirsteadら(J. Neuroscience 19:7529、1999)は、出生後のラット脳から、移植後にオリゴデンドロサイトおよびシュワン細胞を生じるCNS前駆細胞を単離した。Svendsenら(J. Neurosci. Meth. 85:141、1998)は、発達過程のヒト皮質から前駆細胞を単離した。Mujtabaら(Dev. Biol. 214:113、1999)は、胚性幹細胞からの神経系前駆細胞の単離について報告している。
【0008】
国際公開公報第97/07200号(スタンフォード大学)は、成体ラット脳から単離したオリゴデンドロサイトの培養を示している。国際公開公報第01/28342号(ワシントン大学)は、あらかじめ馴化したオリゴデンドロサイト培地中で神経細胞を培養する特定の方法を提案している。米国特許第5,753,506号(Johe、CNS Stem Cell Technology)は、ニューロン、アストロサイト、およびオリゴデンドロサイトに分化する能力を有する、神経組織から単離した幹細胞を維持する培養系に関する。米国特許第6,238,922号(Stem Cells Inc.)は、神経組織の、ニューロンおよびグリアに分化する能力を有する細胞への解離を提案している。米国特許第6,235,527号(Raoら、ユタ大学)は、哺乳動物神経管組織から解離し、A2B5細胞表面マーカーに基づいて選択した、哺乳動物CNSグリア限定前駆細胞の集団に関する。
【0009】
米国特許第5,968,829号(Cytotherapeutics)は、ニューロン、アストロサイト、およびオリゴデンドロサイトを産生する能力を有するCNS神経幹細胞を含む培養液を請求している。国際公開公報第97/32608号は、移植を介してCNSに送達するための、遺伝子操作した初代オリゴデンドロサイトに関する。米国特許第5,830,621号(Signal Pharmaceuticals)は、アクセッション番号CRL 11881でATCCに寄託したヒトオリゴデンドロサイト細胞株について記載している。この株は、特徴的なマーカーGFAP、GalC、O4、およびA2B5を本質的に含まない。
【0010】
残念なことに、神経組織から単離した前駆細胞がヒトの臨床治療に必要な細胞数を産生するのに十分な複製能力を有するのかどうかは、いまだ明らかではない。
【0011】
別の供給源は、初期の胚組織から単離される多能性細胞である。胚性幹(ES)細胞は、25年以上前に初めてマウス胚から単離された(G.R. Martin、Proc. Natl. Acad. Sci. U.S.A. 78:7634、1981)。ES細胞は、同じ種の実質的に任意の組織種の子孫を生じ得ると考えられている。
【0012】
Fraichardら(J. Cell Sci. 108:3181、1995)は、マウスES細胞のグリア細胞および機能的なニューロンへのインビトロ分化について報告している。Mujtabaら(Dev. Biol. 214:113、1999)は、マウスES細胞からの神経前駆細胞の単離について報告している。Li、Smithら(Cur. Biol. 8:971、 1998)は、細胞系譜選択によるマウスES細胞からのニューロン前駆細胞の産生について報告している。Brustle、Mckayら(Proc. Natl. Acad. Sci. USA 94:14809、1997;Science 287:754、1999)は、有髄化移植片の潜在的な供給源として、マウスES細胞に由来するグリア前駆細胞について報告している。McDonaldら(Nat. Med 5:1410、1999;Proc. Natl. Acad. Sci. USA 97:6126、2000)は、培養においておよび脊髄移植後に、マウスES細胞がオリゴデンドロサイトを形成し、有髄化することを報告している。
【0013】
ヒトES細胞は、ごく最近になって初めて単離された(Thomsonら、Science 282:114、1998)。ヒトES細胞は、未分化状態で維持するためまたは特定の分化経路に沿って導くために、非常に異なる条件を必要とする(米国特許第6,090,622号および第6,200,806号;国際公開公報第99/20741号および国際公開公報第01/51616号)。このため、ヒトES細胞から分化細胞の比較的均一な集団をいかにして調製するかについては、ほとんどわかっていない。
【0014】
国際公開公報第01/88104号(Carpenter、Geron Corporation)は、ヒトES細胞を分化させることによって得られる神経系前駆細胞集団について記載している。90% NCAM陽性、35%βチューブリン陽性、および75% A2B5陽性を超える集団が得られた。Zhangら(Nature Biotech. 19:1129、2001)は、ヒトES細胞からの移植可能な神経系前駆細胞の分化について報告している。国際特許出願PCT/US02/19477(Carpenterら、Geron Corporation)は、生じた集団内の少なくとも10%のMAP−2陽性細胞がドーパミン作動性ニューロンのマーカーであるチロシン水酸化酵素を発現する、ES細胞由来の神経系細胞集団について記載している。
【0015】
最近になって、Billonら(J. Cell Sci. 115:3657、2002)は、遺伝子操作したマウスES細胞からのオリゴデンドロサイト発生のタイミングについて記載している。Kuoら(Biol. Reprod. Dec 11/02)は、28% GFAP陽性であるサルES由来細胞の集団について記載しており;Xianら(Stem Cells 21:41、2003)は、細胞系譜特異的転写因子Olig2を用いた、マウスES細胞からのオリゴデンドロサイトの作製について報告している。
【0016】
ヒトの健康および疾患の管理におけるpPS細胞の全般的な可能性を理解するため、脱髄状態の治療に有用な細胞の濃縮された集団を作製する新しいパラダイムを開発することが必要である。
【発明の概要】
【0017】
概要
本発明は、研究および薬学的組成物の調製に用いるための、グリア系譜の霊長動物細胞を効率的に産生する系を提供する。
【0018】
本発明の分化細胞集団は単離されまたはインビトロで培養され、グリア細胞または神経組織を有髄化し得る細胞の特徴に関して高度に濃縮されている。この細胞は、オリゴデンドロサイトの形態的特徴、本開示で後に記載する特定の抗体で検出可能なもしくは増幅可能なマーカーの発現、またはさらなる分化においてオリゴデンドロサイトを形成する能力を有し得る。この細胞はまた、同時培養アッセイにおいて神経節を有髄化する能力、インビボにおいて脱髄した軸索に対してミエリンを回復する能力、またはヒトもしくは非ヒト動物において神経機能を改善する能力等の、オリゴデンドロサイトの特定の機能的特徴を有し得る。これらの特徴の1つ、2つ、3つ、またはそれ以上が任意の組み合わせで存在し得る。
【0019】
この細胞集団は、あまり分化していない様々な種類の幹細胞から調製することができる。起源となり得る細胞には、胚盤胞由来の霊長動物多能性幹(pPS)細胞(胚性幹細胞に例証される)または初期胚の胚組織が含まれる。したがって、細胞はその起源の元となった組織の子孫であるという特徴を有することになり、このことは起源細胞と分化細胞が同じゲノムを有することを示すことによって確認され得る。
【0020】
本発明の他の態様は、すでに記載した分化細胞を作製または維持する方法に関する。この方法は、本開示において後に説明するような1つまたは複数の成長因子または分化因子の存在下で、多分化能幹細胞または多分化能性幹細胞を培養する段階を含む。
【0021】
例証として、線維芽細胞成長因子(FGF)等のマイトジェンと共にまたはそれなしで、トリヨードチロニン(T3)、セレン、またはレチノイン酸等の1つまたは複数の分化因子を含む培地中で幹細胞を培養することができる。分化細胞の最初の形成は、オリゴデンドロサイト系譜細胞が比較的均一な球体を形成し得る懸濁培養において起こり得る。他の細胞型は、所望の細胞型に選択的に付着する表面上に培養物をプレーティングする等の適切な分離手段によって、除去することができる。任意で、多くは細胞の最初の誘導に用いたような1つまたは複数の分化因子の存在下で、FGFまたは上皮成長因子等のマイトジェンと共に培養することによる、選択の前または後に分化細胞をさらに増殖させることができる。次に、任意でマイトジェンの非存在下で、または後期分化を亢進するポリ−L−リジン等の表面の存在下で培養することにより、細胞をさらに成熟させることができる。
【0022】
本発明の細胞は、多くの商業上重要な用途に用いることができる。例えば、この細胞は、その存在が細胞の維持、毒性、さらなる分化、またはグリア細胞として機能する能力に関連する化合物を、グリア細胞に及ぼす影響についてスクリーニングするのに適している。この細胞はまた、インビトロおよびインビボにおいて隣接する神経組織の髄鞘形成をもたらすのにも適している。
【0023】
ヒトまたは動物治療用の薬学的組成物の調製において、本発明の細胞を用いることも可能である。次にこの組成物を用いて、軸索の髄鞘形成障害に関連するような病態または脊髄損傷を治療することができる。
本発明のさらなる局面は、以下の記載により明らかになると考えられる。
【図面の簡単な説明】
【0024】
【図1】塩基性FGFおよびオリゴデンドロサイト分化因子を含む培地中、懸濁状態で2日間培養したヒト胚性幹(ES)細胞を示す、倒立顕微鏡を用いて撮影した位相差像である。
【図2】レチノイン酸と共に7日間培養した後の細胞を示す。培養物中の80〜90%を示す、細胞の大きな透明な球体が存在した。
【図3】レチノイン酸を除去し、細胞を低濃度のFGFの存在下で維持した後に何が起こるかを示す。
【図4】bFGFを除去した後に現れる変化を示す。大きな凝集塊が解離し、培養液全体が単一細胞および小さな細胞塊でいっぱいになった。同時に、新たに鮮黄色の球体が観察された(矢印)。
【図5】細胞をFGFの非存在下で上皮成長因子と共に培養した場合に成長する鮮黄色の球体(矢印)を示す。
【図6】マトリゲル(登録商標)上にプレーティングしてから2日または3日後の、球体からのグリアに拘束された神経前駆細胞の遊走および分枝を示す。
【図7】10〜20時間だけ細胞をマトリゲル(登録商標)に接着させることにより、オリゴデンドロサイト系譜細胞が選択されたという結果を示す。非接着細胞を除去したが、これによりオリゴデンドロサイトマーカーをもたない細胞は実質的にすべて除去される。次に接着細胞を再懸濁して、FGF、EGF、およびグリア前駆細胞培地の存在下で7日間拡大した。これは、治療的投与および他の目的に対してより適したより分散した細胞集団を産生する利点を有する。この顕微鏡写真は、ヘマトキシンを用いて細胞核を対比染色した(黒矢印)、ガラクトセレブロシドに対する染色(GalC;白矢印)を示す。この条件下でGalCに染色された細胞の割合は、少なくとも約95%であった。
【図8】より高い倍率のES由来オリゴデンドロサイトを示す。細胞は、成熟オリゴデンドロサイトに特有の形態的特徴(間にミエリン皮膜を有するように見える多数の複雑な突起)を有する。
【図9】分化過程における細胞形態の進行を示す。(A): 未分化hES細胞。(B): レチノイン酸を含む懸濁培養において胚様体から成長する透明な球体。(C): EGFの存在下における、オリゴデンドロサイト前駆細胞を含む黄色球体の拡大。 (D): マトリゲル(登録商標)上にプレーティングすることによりポジティブ選択されたオリゴデンドロサイト系譜細胞。(E、F): 培養におけるオリゴデンドロサイト前駆細胞の隆起の増加。(G、H): 次に続くプレーティングによって、成熟オリゴデンドロサイトへのさらなる分化が起こる。
【図10】成熟オリゴデンドロサイトの免疫細胞化学的解析を示す。プレーティングしてから1週間後、細胞は初期グリア細胞のマーカーNG2に関して陽性である(A)。プレーティングしてから8週間後、細胞は、GalC(C)、O4(D)、およびRIP(E)に関して陽性である。パネルBは、示したマーカーで染色された細胞の数を示す。
【図11】ヒト核タンパク質に特異的な抗体で染色した、ラットの脊髄に投与したオリゴデンドロサイト前駆細胞の組織切片を示す
【図12】移植してから9週間後、細胞が白質に向けて遊走または増殖したことを示す。
【図13】移植したオリゴデンドロサイトが、脊髄の断面積として測定される、挫傷後の二次拡大に起因する損傷を悪化させないことを示す。
【図14】hES細胞が軸索分枝を誘導することを示す。オリゴデンドロサイトを移植した動物の像では(上のパネル)、BDA染色の濃い細い線として新たな軸索が示される。未処置の動物では、分枝は認められなかった。移植した細胞は、再生可塑性を誘導する
【図15】損傷部位(中心点)からのニューロン分枝の定量を示す(ブロック当たり3切片の平均値±SEM)。処置した動物は、有意に高いレベルで、中心点の損傷のすぐ頭側にまで標識軸索を有した。
【図16】hES由来オリゴデンドロサイトを移植した動物における、実質的な再ミエリン化の証拠を示す。上段の電子顕微鏡写真における厚い環は、正常にミエリン化した繊維である。視野内の残りの軸索は、薄い層のミエリンを示す。右上のパネルの軸索は、約5層または6層の外被、および進行中の再ミエリン化の証拠を示す。移植した動物のみが、オリゴデンドロサイト活性に起因する新たなミエリンを示した。これにより、移植動物における動作の改善を説明する手段が提供される。
【図17】未分化hES細胞コロニーで検出されたマーカーを示す(左側:抗体+DAPI;右側:抗体染色のみ)。上列は、多能性細胞のマーカーであるSSEA−4に標識されたコロニーを示す。下列は、中胚葉マーカーBMP4に標識されたコロニーを囲む間質細胞を示す。
【図18】分化過程における転写因子Pax6の一過性の出現を示す。上列は、10日目の細胞塊の中心に向かった染色、周辺に向かってより分化した細胞でのすでにダウンレギュレートされた染色を示す。下列は、分化35日目では実質的に染色されないことを示す。
【図19】分化10日目に存在する初期のオリゴデンドロサイト系譜細胞で検出されるマーカーを示す。上列:転写因子Olig1。中列:転写因子SOX10。下列:オリゴデンドロサイト前駆細胞マーカーA2B5。
【図20A】35日目の移植に適したオリゴ前駆細胞において優位を占めるマーカーを示す。1列目および2列目(20A):NG2(コンドロイチン硫酸プロテオグリカン、オリゴデンドロサイト前駆細胞のマーカー); 3列目(20A):GalC;4列目(20B):O4;5列目(20B):Tuj1(ニューロンのマーカー)。実質的にすべての細胞がオリゴデンドログリアマーカーを有したが、神経細胞、間葉細胞、または未分化hES細胞のマーカーは有さない(20B、下のグラフ)。
【図20B】35日目の移植に適したオリゴ前駆細胞において優位を占めるマーカーを示す。1列目および2列目(20A):NG2(コンドロイチン硫酸プロテオグリカン、オリゴデンドロサイト前駆細胞のマーカー); 3列目(20A):GalC;4列目(20B):O4;5列目(20B):Tuj1(ニューロンのマーカー)。実質的にすべての細胞がオリゴデンドログリアマーカーを有したが、神経細胞、間葉細胞、または未分化hES細胞のマーカーは有さない(20B、下のグラフ)。
【図21】ミエリン塩基性タンパク質を作り出す能力が遺伝的に欠損しているシバラーマウスにオリゴデンドロサイトを投与した結果を示す。超微細構造的に、シバラーマウスの軸索はミエリンを欠いているか、または1もしくは2層の緻密でないミエリンによって囲まれている(上のパネル)。細胞を移植してから6週間後、電子顕微鏡解析により多層の緻密なミエリンが示され、このことから移植集団のミエリン形成能が示唆された(下のパネル)。緻密なミエリンは、投与したオリゴデンドロサイトによって直接産生されている。
【図22】衝撃装置を用いてラットの脊髄を挫傷させ、BBB尺度での定量によりその後の期間の脊髄機能について評価する実験の結果を示す。上のグラフ:200キロダイン挫傷;下のグラフ:250キロダイン挫傷。 (■)損傷から1週間後にES由来オリゴデンドロサイトで処置した動物(n=5);(▲)同じ挫傷を被るが細胞を投与していない対照動物(n=3)。盲検で評価した平均値±SEM。hES由来オリゴデンドロサイトを移植した動物は、治療後5週間を超えて持続する有意に優れた地上歩行を示した。
【発明を実施するための形態】
【0025】
詳細な説明
本発明は、多能性幹細胞からオリゴデンドロサイトおよびその前駆細胞を効率的に産生することにより、それらの細胞の大きな集団を作製する問題を解決する。
【0026】
分化細胞集団は著しく均一である。図7は、オリゴデンドロサイトに特有である形態学的特徴を有するヒト細胞の例示的な調製を示す。集団を、オリゴデンドロサイト系譜の細胞のマーカーであるGalCについて染色した。図8はより高い倍率の視野であり、近傍の任意のニューロンを有髄化しようとしているミエリンのシートの生成を示している。これらの細胞の機能的特徴から、これらの細胞はオリゴデンドロサイトの性質のさらなる特徴付けおよびヒトの治療での使用によく適している。
【0027】
本発明の細胞の理想的な供給源は、様々な種類の霊長動物多能性幹(pPS)細胞である。以下に記載する戦略に従っていくつもの適切な培養条件および補因子の中から選択することにより、pPS細胞をオリゴデンドロサイト経路に導出することができる。未分化pPS細胞のオリゴデンドロサイトへの変換は、最適条件下で25%程度の効率で起こり得ることが明らかになった。
【0028】
本発明の組成物および方法により、これまでに利用可能であった技術を超える重要な利点が提供される。所望のアロタイプのpPS細胞からオリゴデンドロサイトを分化させることにより、任意の組織適合性を有するオリゴデンドロサイトを産生することができる。必要に応じて、移植を亢進する任意の方法で、分化の前または後に細胞を遺伝子改変することができる。細胞は天然細胞株に由来するが継続的な組織解剖を必要としないため、規制認可の品質管理要求事項を満たすのに最適である。
【0029】
重要なことには、各出発幹細胞集団から産生され得る細胞の供給量はほぼ無限である。実施例1に説明するように、ひとたびオリゴデンドロサイト系譜細胞を産生かつ選択すれば、成長因子の存在下で培養することによりその細胞を大幅に拡大することができる。さらに、起源pPS細胞をほぼ無期限に増殖させて、それによってより分化した細胞の供給源を連続的に提供することができる。
【0030】
以下の開示により、本発明の分化細胞をいかにして作製および使用するかのさらなる説明を提供する。集団は著しく均一であり、したがって多くの商業的に重要な用途における使用に適している。
【0031】
本発明の1つの局面により、少なくとも約80%の細胞が、霊長動物多能性幹(pPS)細胞の子孫であり;NG2プロテオグリカン(または他のオリゴデンドロサイトマーカー)に特異的な抗体で染色され;かつNeuN(または神経細胞および他の潜在的混入物の他のマーカー)に関して陰性である、分化細胞集団を提供する。分化細胞集団は、オリゴデンドロサイト系譜細胞を産生するのに適した成分のシステムの一部であってよい。このシステムは、分化細胞が産生される元ととなったpPS細胞(ヒト胚性幹細胞等)の系統をさらに含み得る。任意で、分化細胞集団の少なくとも80%の細胞が、A2B5またはPDGFRαも発現する。細胞の少なくとも20%が、オリゴデンドロサイト前駆細胞に特有である双極性形態を示す可能性がある。
【0032】
ある状況下では、シバラー変異(shiverer mutant)マウスの脊髄に分化細胞集団を移植した後、その集団により神経軸索の周りに緻密なミエリンの沈着が起こる;または挫傷ラットの脊髄内もしくは脊髄の周りに集団を移植した後、地上歩行が改善される。あるいは、本発明のオリゴデンドロサイト前駆細胞集団を(例えば、マイトジェンの非存在下でポリ−L−リジンおよびラミニン上で3日間培養することにより)インビトロでさらに分化させてもよい。これにより、少なくとも10%の細胞が成熟オリゴデンドロサイトに特有である複雑な突起を有し;かつ約80%、90%、95%、またはそれ以上がGalC等の成熟オリゴデンドロサイトマーカーを有する、より成熟した集団が産生され得る。増殖段階中に産生されたオリゴデンドロサイト系譜細胞は、前駆細胞段階で発現しているマーカーばかりでなく、成熟細胞の濃縮された集団を産生する能力またはインビボで所望の機能を遂行する能力によっても特徴付けることができる。
【0033】
以下に説明するように、本発明のいくつかの分化細胞集団は、マイトジェンおよび少なくとも2つのオリゴデンドロサイト分化因子を含む培地中で未分化pPS細胞を培養する過程により得ることができる。例えば、細胞凝集体を形成させるために、塩基性線維芽細胞成長因子(FGF)、トリヨードチロニン(T3)、レチノイン酸、およびおそらくセレンの存在下で、未分化pPS細胞を懸濁状態で培養してもよい。
【0034】
本発明のさらなる説明および例証を以下の項で提供する。
【0035】
定義
オリゴデンドロサイトとは、中枢神経系の外膜構造(神経膠)の一部を形成する外胚葉起源の神経系細胞である。この細胞は、個々の軸索の周りを覆いCNSのミエリン鞘を形成する、不定数のベール様またはシート様突起を有する。この細胞は、本開示で後に説明する形態学的基準、表現型基準、または機能的基準によって同定することができる。
【0036】
「神経系前駆細胞」または「神経上皮幹細胞」とは、神経細胞(ニューロン前駆細胞もしくは成熟ニューロン等)またはグリア細胞のいずれかである子孫を産生し得る細胞である。「グリア細胞」という用語は、成熟オリゴデンドロサイト、アストロサイト、およびこれらの細胞のいずれかまたは両方に拘束された前駆細胞を包含する。
【0037】
「オリゴデンドロサイト前駆細胞」は、ニューロンまたは非神経組織に優先して、成熟オリゴデンドロサイトおよび/またはさらなる前駆細胞を含む子孫を形成するように拘束された神経系細胞である。特記しない限りこの細胞は、必ずしも必要ではないが、アストロサイト等の他の種類のグリア細胞を産生する能力を有してもよい。特記しない限り、本開示での「オリゴデンドロサイト」または「オリゴデンドロサイト系譜細胞」への言及は、オリゴデンドロサイト前駆細胞および成熟細胞の両方を指す。
【0038】
細胞の個体発生との関連では、形容詞「分化した」とは相対語である。「分化細胞」とは、比較する細胞よりも発生経路をさらに下方に進行した細胞のことである。したがって、多能性胚性幹細胞は、上記の様々な種類の前駆細胞等の系譜を限定した前駆細胞に分化することが可能である。これらの細胞は、次に経路のさらに下流にある細胞、または成熟オリゴデンドロサイト等の最終段階の分化細胞にさらに分化することが可能である。
【0039】
本開示で用いる「分化剤」とは、オリゴデンドロサイト系譜の分化細胞(前駆細胞および最終分化細胞を含む)を産生するために、本発明の培養系で用いる一群の化合物の1つを指す。化合物の作用形態に関しては限定しない。例えば、薬剤は、表現型の変化を誘導もしくは補助することにより、特定の表現型を有する細胞の成長を促進することによりもしくは他の細胞の成長を遅らせることにより、または他の薬剤と合わせて未知の機構で作用することにより、分化過程を補助する可能性がある。
【0040】
特記しない限り、本開示における「セレン」への言及は、溶液中で任意の対イオンを伴う亜セレン酸(SeO32−)、セレン酸(SeO42−)、またはセレン化物(Se2−)を含む、Seの任意の酸化型を指す。
【0041】
原型の「霊長動物多能性幹細胞」(pPS細胞)とは、前胚組織、胚組織、または受精後の任意の時期の胎児組織に由来する多能性細胞であり、8〜12週齢のSCIDマウスで奇形腫を形成する能力等の標準技術として公認の試験によると、適切な条件下で3つの胚葉(内胚葉、中胚葉、および外胚葉)すべての派生物である様々な異なった細胞型の子孫を産生し得る特徴を有する。pPS細胞の定義には、ヒト胚性幹(hES)細胞およびヒト胚性生殖(hEG)細胞に例証される様々な種類の胚細胞が含まれる。pPS細胞は、悪性供給源に由来しないことが好ましい。細胞は正倍数体であることが望ましい(が必ずしも必要ではない)。その供給源および培養法に依存して、pPS細胞は、人体のすべての異なる細胞型に発達する能力を有するという意味で、全能性であってもなくてもよい。
【0042】
pPS細胞培養物は、集団中の幹細胞およびその派生物の実質的な割合が、胚起源または成体起源の分化細胞と区別できる未分化細胞の形態学的特徴を示す場合に、「未分化である」と記載される。集団内の未分化細胞のコロニーは、分化した隣接細胞に囲まれていることが多いと理解されている。
【0043】
「フィーダー細胞」または「フィーダー」とは、別の種類の細胞と共培養され、第2の種類の細胞が成長できる環境を提供する1つの種類の細胞を表すのに用いられる用語である。pPS細胞集団は、分割後にpPSの成長を支持するために新鮮なフィーダー細胞を添加せずに少なくとも1回成長した場合、フィーダー細胞を「本質的に含まない」と称される。
【0044】
「胚様体」という用語は、pPS細胞が単層培養において過剰成長する場合、または懸濁培養で維持される場合に現れる、分化および未分化細胞の凝集体を指す。胚様体は、形態学的基準および免疫細胞化学法により検出可能な細胞マーカーによって識別可能ないくつかの胚葉に典型的に由来する、様々な細胞型の混合物である。
【0045】
「成長環境」とは、関心対象の細胞がインビトロにおいて増殖、分化、または成熟する環境のことである。環境の特徴には、細胞を培養する培地、存在し得る任意の成長因子または分化誘導因子、および存在する場合には支持構造(例えば、固体表面上の基層)が含まれる。
【0046】
特記しない限り、個々の細胞および細胞集団における表現型マーカーの判定では、細胞は、アイソタイプ対照と比較して、二次免疫細胞化学染色において特異的抗体を用いて実質的により高い染色を示した場合にマーカーに関して「陽性」であると称される。特記しない限り細胞は、この種の免疫細胞化学的解析によりマーカーが抗体で検出不可能である場合に、「陰性」であると称される。
【0047】
任意の適切な人工的操作手段によりポリヌクレオチドが細胞に導入された場合、または細胞がそのポリヌクレオチドを受け継いだ最初に改変された細胞の子孫である場合、その細胞は「遺伝子改変された」、「トランスフェクションされた」、または「 遺伝的に形質転換された」と称される。ポリヌクレオチドは関心対象のタンパク質をコードする転写可能な配列を含む場合が多く、これにより細胞はこのタンパク質を高いレベルで発現することが可能になる。改変した細胞の子孫が同じ改変を有する場合、遺伝的改変は「遺伝性である」と称される。
【0048】
一般的技術
分子遺伝学および遺伝子工学における一般的な方法は、Molecular Cloning: A Laboratory Manual (Sambrookら、Cold Spring Harbor);Gene Transfer Vectors for Mammalian Cells (MillerおよびCalos編);およびCurrent Protocols in Molecular Biology(F.M. Ausubelら編、Wiley & Sons) の現行版に記載されている。分子生物学、タンパク質化学、および抗体技法は、Current Protocols in Protein Science(J.E. Colliganら編、Wiley & Sons);Current Protocols in Cell Biology(J.S. Bonifacinoら、Wiley & Sons)、およびCurrent Protocols in Immunology(J.E. Colliganら編、Wiley & Sons)に見出すことができる。本開示で引用する遺伝子操作用の試薬、クローニングベクター、およびキットは、BioRad、Stratagene、Invitrogen、ClonTech、およびSigma−Aldrich Co.等の市販業者から入手可能である。
【0049】
細胞培養法は、一般に、Culture of Animal Cells: A Manual of Basic Technique(R.I. Freshney編、Wiley & Sons);General Techniques of Cell Culture(M.A. HarrisonおよびI.F. Rae、Cambridge Univ. Press)、およびEmbryonic Stem Cells: Methods and Protocols(K. Turksen編、Humana Press)の現行版に記載されている。組織培養用の供給品および試薬は、Gibco/BRL、Nalgene−Nunc International、Sigma Chemical Co.、およびICN Biomedicals等の市販業者から入手可能である。
【0050】
本開示に関連する専門的な参考文献には、Principles of Neuroscience、第4版、Kandelら編、McGraw−Hill 2000;CNS Regeneration: Basic Science and Clinical Advances、M.H. TuszynskiおよびJ.H. Kordower編、Academic Press、1999;The Neuron: Cell and Molecular Biology、第3版、I.B. LevitanおよびL.K. Kaczmarek、Oxford U. Press、2001;Glial Cells: Their Role in Behaviour、P.R. Lamingら編、Cambridge U. Press、1998;The Functional Roles of Glial Cells in Health and Disease、MatsasおよびTsacopoulos編、Plenum Pub. Corp、1999;Glial Cell Development、JwessenおよびRichardson編、Oxford U. Press 2001;およびMan of Steel、Adrian Havill、1996が含まれる。
【0051】
幹細胞の供給源
本発明は、様々な種類の幹細胞を用いて実施することができる。本発明の使用に特に適しているのは、胚盤胞等の妊娠後に形成される組織または妊娠期間の任意の時期に採取した胎児組織もしくは胚組織等に由来する霊長動物多能性幹(pPS)細胞である。限定されない例は、以下に記載するような胚性幹細胞もしくは胚性生殖細胞の初代培養または樹立株である。本発明の技法を初代胚組織または胎児組織に直接実行し、最初に未分化細胞株を樹立することなく初代胚性細胞から神経系細胞を直接導出することも可能である。
【0052】
実施例の項に提供する例証は、ヒト胚性幹細胞を用いて行った研究の結果である。しかし、特記しない限り、本発明は任意の脊椎動物種の幹細胞を用いて実施することが可能である。これには、ヒト、ならびに非ヒト霊長動物および他の非ヒト哺乳動物の多能性幹細胞が含まれる。
【0053】
胚性幹細胞
胚性幹細胞は、霊長動物組織から単離することができる(米国特許第5,843,780号;Thomsonら、Proc. Natl. Acad. Sci. USA 92:7844、1995)。ヒト胚性幹(hES)細胞は、Thomsonら(米国特許第6,200,806号;Science 282:1145、1998;Curr. Top. Dev. Biol. 38:133ページ以降、1998)およびReubinoffら、Nature Biotech. 18:399、2000の記載する技法により、ヒト割球から調製することが可能である。hES細胞に相当する細胞型には、国際公開公報第01/51610号(Bresagen)に概説されるような、原始外胚葉様(EPL)細胞等のその多能性派生物が含まれる。
【0054】
1つの方法では、プロナーゼ(Sigma)に短時間曝露することにより、発生した胚盤胞から透明帯を除去する。1:50希釈したウサギ抗ヒト脾臓細胞抗血清に胚盤胞を30分間曝露し、次にDMEMで5分間3回洗浄し、1:5希釈したモルモット補体(Gibco)に3分間曝露する免疫手術により、内部細胞塊を単離する(Solterら、Proc. Natl. Acad. Sci. USA 72:5099、1975)。DMEMでさらに2回洗浄した後、穏やかにピペッティングして無傷の内部細胞塊(ICM)から溶解した栄養外胚葉細胞を除去し、ICMをmEFフィーダー層上にプレーティングする。
【0055】
9〜15日後、1 mM EDTAを添加したカルシウムおよびマグネシウムを含まないリン酸緩衝食塩水(PBS)に曝露することにより、ディスパーゼもしくはトリプシンに曝露することにより、またはマイクロピペットで機械的に解離することにより、内部細胞塊に由来する増殖物を凝集塊に解離し;次に新鮮な培地中でmEF上に再度プレーティングする。未分化の形態を有して増殖するコロニーをマイクロピペットで個別に選択し、機械的に凝集塊に解離し、再度プレーティングする。ES様の形態は、細胞質に対して核の比率が明らかに高くかつ顕著な核小体を有する小型のコロニーとして特徴付けられる。次に、得られたES細胞を短時間トリプシン処理してダルベッコPBS(2 mM EDTAを含む)に曝露し、IV型コラゲナーゼ(約200 U/mL;Gibco)に曝露することにより、またはマイクロピペットで個別にコロニーを選択することにより、1〜2週間ごとに日常的に分割する。凝集塊の大きさは、約50〜100細胞が最適である。
【0056】
胚性生殖細胞
ヒト胚性生殖(hEG)細胞は、Shamblottら、Proc. Natl. Acad. Sci. USA 95:13726、1998および米国特許第6,090,622号に記載されているように、始原生殖細胞から調製することができる。
【0057】
簡潔に説明すると、約8〜11週間後に採取した生殖隆起を等張緩衝液ですすぎ、0.05%トリプシン/0.53 mM EDTAナトリウム溶液(BRL)0.1 mL中に入れ、<1 mm3の塊に切断する。脱凝集した後、細胞をEG増殖培地(D−グルコース、NaHCO3;15% ES認定ウシ胎仔血清;2 mMグルタミン;1 mMピルビン酸ナトリウム;1000〜2000 U/mLヒト組換え白血病抑制因子;1〜2 ng/mL ヒト組換えbFGF;および10μMフォルスコリン(10% DMSO中)を含むDMEM)約3.5 mL中、37℃で1時間または一晩インキュベートする。
【0058】
次に細胞をEG増殖培地1〜3 mLに再懸濁し、フィーダー層上にプレーティングする(例えば、5000radのγ−照射で不活性化したSTO細胞、ATCC番号CRL 1503)。7〜10日後に1回目の継代を行い、EG細胞と一致する細胞形態が観察されるまで、毎日培地を交換して培養するが、典型的にこれは7〜30日後または1〜4継代後である。
【0059】
他の幹細胞
オリゴデンドロサイトを作製するための出発材料として有用なpPSまたは胚性幹細胞を産生するために、本発明の実施はヒト胚または胚盤胞を脱凝集することを決して必要としない。hES細胞は、公共の受託所(例えば、WiCell Research Institute、米国、ウィスコンシン州、マディソン、またはAmerican Type Culture Collection、米国、バージニア州、マナッサス)から入手可能な樹立株から得ることができる。米国特許公報第2003−0113910A1号は、胚または胎児組織を用いずに導出された多能性幹細胞について報告している。多能性表現型を誘導する因子を使用することにより、臍帯血または他の前駆細胞をpPS細胞に再プログラミングすることも可能であると考えられる(Chambersら、Cell 113:643、2003;Mitsuiら、Cell 113:631、2003)。適切な条件下において、別の方法でpPSまたはhES細胞の定義に見合う任意の細胞を、本発明によりオリゴデンドロサイト系譜細胞の導出に用いることができる。
【0060】
本開示で提供する技法のいくつかを用いて、外胚葉細胞、および胎児組織もしくは成体組織から得られた神経系細胞、または神系前駆細胞等のより拘束された細胞型の分化を維持または促進することも可能である。そのような細胞を得る方法は、例えば、米国特許第5,852,832号;第5,654,183号;第5,849,553号;および第5,968,829号;ならびに国際公開公報第98/50526号および国際公開公報第99/01159号に記載されている。
【0061】
未分化状態でのpPS細胞の増殖
pPS細胞は、分化を促進せずに増殖を促進する培養条件を用いて培養し、連続して増殖させることが可能である。血清を含む例示的なES培地は、80% DMEM(ノックアウトDMEM、Gibco等)、20%の既知組成ウシ胎仔血清(FBS、Hyclone)または血清代替品(国際公開公報第98/30679号)のいずれか、1%非必須アミノ酸、1 mM L−グルタミン、および0.1 mMβ−メルカプトエタノールから作製する。使用する直前に、ヒトbFGFを4 ng/mLになるように添加する(国際公開公報第99/20741号、Geron Corp.)
【0062】
分化を阻害する環境において培養することによってのみ、pPS細胞を未分化状態で拡大することができる。慣例的に、pPS細胞は、マウスの胚組織または胎児組織に由来するフィーダー細胞層上で培養する。増殖を阻害するがpPS細胞を支持する因子の合成はできるように照射した照射mEFを、培養プレートにウェル当たり375,000個プレーティングし、プレーティングしてから5時間〜4日後に使用する(米国特許第6,200,806号)。最近になって、ヒト胚性幹細胞の増殖を分化させることなく支持するヒトフィーダー細胞が開発された(国際公開公報第01/51616号:米国特許出願第09/888,309号;Geron Corp)。hES細胞を分化させ、所望の活性を有する細胞を選択し、次にテロメラーゼ逆転写酵素を発現するようにトランスフェクションして不死化することにより、細胞が得られる。
【0063】
pPS細胞は、フィーダー細胞なしでも未分化状態で維持することができる。無フィーダー培養の環境には、適切な培養基層、特にマトリゲル(Matrigel)(登録商標)またはラミニン等の細胞外マトリックスが含まれる。pPS細胞は、>15,000細胞cm−2(最適には90,000 cm−2〜170,000 cm−2)でプレーティングする。無フィーダー培養物は、分化させることなく細胞の増殖を支持する因子を含む栄養培地により支持される。そのような因子は、照射した(約4,000rad)初代マウス胚線維芽細胞、テロメル化(telomerize)したマウス線維芽細胞、またはpPS細胞由来のヒトフィーダー細胞等のそのような因子を分泌する細胞と共に培地を培養することにより、培地中に導入され得る。フィーダーを、20% 血清代替品および4〜8 ng/mL bFGFを添加したKO DMEM等の無血清培地で約5〜6 x 104 cm−2の密度でプレーティングすることにより、培地を馴化することができる。1〜2日間馴化した培地にさらにbFGFを添加し、これを用いてpPS細胞培養物を1〜2日間支持する。無フィーダー培養法の特徴は、国際公開公報第99/20741号および国際公開公報第01/51616号;ならびにXuら、Nat.Biotechnol. 19:971、2001においてさらに考察されている。
【0064】
ES細胞は顕微鏡下で、高い核/細胞質比、顕著な核小体、および細胞の接合部が識別困難な小型のコロニー形成を伴って見える。霊長動物ES細胞は典型的に、時期特異的胚抗原(SSEA)3および4、Tra−1−60およびTra−1−81と称する抗体を用いて検出できるマーカー(Thomsonら、Science 282:1145、1998)、ならびにテロメラーゼ活性を発現する。インビトロにおけるpPS細胞の分化により、SSEA−4、Tra−1−60、およびTra−1−81の発現が消失し、未分化hEG細胞においても認められるSSEA−1の発現が増加する。
【0065】
幹細胞からのオリゴデンドロサイトの作製
本発明のオリゴデンドロサイト系譜細胞は、所望の表現型を有する細胞を濃縮および拡大する特殊な成長環境において幹細胞を培養することによって得られる。成長環境によって、オリゴデンドロサイト系譜への分化が特異的に方向づけられ得る、所望の細胞の成長が促進され得る、他の細胞型の成長が阻害され得る、またはこれらの活性の任意の組み合わせが起こり得る。
【0066】
本項は、本発明のオリゴデンドロサイトを得るためにとり得るいくつかのアプローチを当業者に説明する指針である。別に規定する場合を除き、過程の根本的な機構に関する説明は、さらなる詳細を支援するための作業仮説としてのみ提供する。使用者が仮説を理解する必要はなく、また本発明を実施するためにこれを仮説に合わせる必要もない。多能性幹細胞からオリゴデンドロサイトを作製することができ、手順のさらなる適合化を行うことができ、本開示に記載する新規産物を得るために別法を見出すことが可能であることを、本出願者らがこれから実証する。
【0067】
pPS細胞からオリゴデンドロサイトを産生する段階には、a) オリゴデンドロサイトを産生するように拘束された細胞の集団を得る段階;b) オリゴデンドロサイト系譜細胞の拡大;およびc) 細胞の後期オリゴデンドロサイトへのさらなる成熟が含まれ得る。
【0068】
幹細胞のオリゴデンドロサイト系譜への方向付け
オリゴデンドロサイトを産生する過程は、典型的に2つの局面、起源幹細胞集団に分化を引き起こす段階;およびオリゴデンドロサイト系譜細胞を主要細胞型にする段階を含む。これらの事象は、経時的にまたは同時に起こり得る。
【0069】
例えば、ドナーpPS細胞培養物の過剰成長により、または胚様体の形成を可能にする低接着性特性を備えた基層を有する培養容器中でpPS細胞を懸濁状態で培養することにより、pPS細胞に胚様体または凝集体の形成を引き起こすことによって分化の過程を誘導することができる。例示的な方法では、hES細胞のコンフルエントな単層培養物を回収し、非接着性細胞培養プレートにプレーティングし、細胞を懸濁状態に保ち、定期的に栄養培地でフィードする。
【0070】
別法としてまたはさらに加えて、細胞が未分化表現型を維持するのを妨げる特定の因子と共に培養することによって、分化過程を開始することができる。最初の分化因子は分化をオリゴデンドロサイト系譜に限定する必要はないが、分化集団内の細胞型の範囲内にオリゴデンドロサイトまたはその前駆細胞を含めるべきである。この種の例示的な成長因子は、レチノイド受容体に結合する、または細胞外シグナル制御キナーゼ(ERK)経路を活性化するリガンドである。典型的に線維芽細胞成長因子(または次項に記載するマイトジェン)等のマイトジェンを培地に含めて、分化過程の進行中に増殖を促進する。
【0071】
ある段階で、培養物をより特異的にオリゴデンドロサイト系譜に方向づけることができる。これは、オリゴデンドロサイトの成長をより特異的に促進する因子を培養物に含めることによって行うことができる。例示的なオリゴデンドロサイト分化因子は、約40 ng/mlのT3(3,5,3’−トリヨード−L−チロニン)およびT4(L−チロキシン)によって例証される、細胞表面上または核内の甲状腺ホルモン受容体に結合するリガンドおよび抗体である。甲状腺ホルモンは、レチノイン酸受容体の発現を増加させる、さもなければオリゴデンドロサイト系譜の細胞への分化を促進すると考えられている。
【0072】
別のオリゴデンドロサイト分化因子は、分化しているオリゴデンドロサイトにおいてミエリン遺伝子の上方制御に関与すると考えられている抗酸化剤であるセレンである。他の候補分化因子は、ビタミンEのような他の抗酸化剤、ならびにチオレドキシン還元酵素およびヨードチロニン脱ヨウ素酵素のファミリー等のセレンが補因子である酵素の活性を増加させる因子である。セレンは、亜セレン酸(SeO32−)の形で少なくとも20 ng/mLまたは100 ng/mLの比較的高い濃度で培地に含めた場合に、特に効果的である。他の候補分化因子または補因子には、骨形成タンパク質(BMP)、ソニックヘッジホッグ(SHH)、および白血病抑制因子(LIF)が含まれる。神経組織によって産生されるオリゴデンドロサイト分化因子の組み合わせも意図するが、これは、胚性CD−1マウス脳および組織または細胞株から得られる因子含有抽出物によって例証される、選択された解離した神経組織または細胞株と共培養することによって産生され得る。
【0073】
これらの技法をすべて組み合わせることにより:マイトジェン、レチノイン酸等の一般的な分化因子、ならびに甲状腺ホルモンおよび亜セレン酸ナトリウムのような特異的分化因子の存在下で胚様体を形成させることによって分化を開始することにより、驚くほど効率的にオリゴデンドロサイトを産生できることを見出した。実施例の項に提供する限定されない例証には、これらの因子、ならびに栄養補充物、インスリン等の成長因子、および抗生物質等の他の標準的な培養成分が含まれる。分化の開始時に適切な組み合わせの分化因子を含めることによって、培養物はオリゴデンドロサイト系譜により拘束された細胞を迅速に産生することになる。
【0074】
オリゴデンドロサイト系譜細胞の拡大
所望の目的のために細胞を用いる以前の任意の段階として、本発明の使用者は、オリゴデンドロサイト系譜細胞を培養において増殖させることによりこの細胞の数を増やしてもよい。
【0075】
これは、1つまたは複数のマイトジェンの存在下で、細胞を栄養培地中で培養することによって達成できる。FGF−2(塩基性FGF)およびFGF−4等の線維芽細胞成長因子ファミリーのメンバーが例示的である。上皮成長因子(EGF)、機能的相同体、およびEGF受容体に結合する他の因子も例示的である。他の候補成長因子は、血小板由来成長因子(PDGF)、インスリン様成長因子(IGF)、およびフォルスコリン等のサイクリックAMPレベルを上昇させる因子である。
【0076】
そのようなマイトジェンの存在下では、レチノイン酸等の分化因子イニシエーターを取り除くことができる。マイトジェンによって細胞は非特異的に成長することから、オリゴデンドロサイト系譜細胞の優先的な成長を維持するために、培地中にオリゴデンドロサイト特異的分化因子を含め続けることが有利な場合が多い。培養液中のマイトジェンのバランスを調整することによって、オリゴデンドロサイトの優先的な成長を支持することも可能である。例えば、EGFは外胚葉を維持するが線維芽細胞を維持せず、線維芽細胞はFGFが存在しなくなれば死滅し得る。FGFに厳密に依存する細胞が除去されたならば、培養液にFGFを再度添加して所望の細胞型の成長を促進できる。
【0077】
細胞の調製における別の任意の段階として、本発明の使用者は、培養物中に存在し得る他の細胞型からオリゴデンドロサイト系譜細胞を分離してもよい。抗体もしくはレクチンを介した接着または細胞表面マーカーについてのソーティング等の、様々な分離手順を意図する。ポジティブ選択およびネガティブ選択の両方に関する適切な表現型マーカーを以下に記載する。UDP−ガラクトース・セラミド・ガラクトシルトランスフェラーゼ(CGT)、2’,3’−サイクリックヌクレオチド3’−ホスホジエステラーゼ(CNP)、NG2プロテオグリカン、NCAM、ミエリン塩基性タンパク質(MBP)、または様々なミエリン関連タンパク質のプロモーター等の組織特異的プロモーターを、アルカリホスファターゼ、緑色蛍光タンパク質、またはルシフェラーゼ等のレポーター遺伝子に連結して構築したプロモーター−レポータープラスミドを用いた、オリゴデンドロサイト系譜細胞のソーティングも意図する。
【0078】
細胞を適切な基層に接着させることによる単純な様式で、オリゴデンドロサイト分化因子の存在下で増殖する他の細胞からオリゴデンドロサイト系譜細胞を分離することができることを見出した。オリゴデンドロサイトは細胞特異的糖質および細胞表面受容体を有し、複合リガンドに優先的に接着することになる。詳細には、ラミニン、ゼラチン、または室温でゲル化し再構成された基底膜を形成するエンゲルブレス・ホルム・スワム(Engelbreth−Holm−Swarm)腫瘍細胞に由来する市販の細胞外マトリックス調製品マトリゲル(登録商標)等の、ある種の基底膜成分への接着によって、オリゴデンドロサイトを分離することができる(実施例1)。オリゴデンドロサイトがマトリックスに接着したら(数時間〜数日)、他の細胞型を洗浄除去し、例えば短時間トリプシン消化することによって接着細胞を回収することができる。この手順は、所望の細胞型の濃縮に加えて、さらなる操作を容易にするために細胞の大きな凝集塊を解離する利点を有する。
【0079】
分離手順の効果は、以下に記載するマーカーに関して濃縮を測定することによって判定することができる。このような方法でオリゴデンドロサイトを精製したならば、すでに記載したようにマイトジェンの存在下でこの細胞をさらに拡大する、以下の項に記載するように成熟させる、または最終用途に適した方法で調製することができる。
【0080】
さらなる成熟
必要に応じて、本発明の細胞を、複製段階を超えて機能的表現型にさらに成熟させることができる。これを行うことで、前駆細胞の可能性を特徴付ける、または治療目的もしくは薬剤スクリーニング目的のために最終段階の細胞を得ることができる。
【0081】
成熟は、前駆細胞表現型のさらなる増殖を阻害する様式で成長条件を変化させることによって達成される。例えば、成熟表現型の出現を促進するポリ−L−リジン等の基層上に細胞をプレーティングすることができる。別法としてまたはさらに加えて、細胞の拡大に用いた1つまたは複数の成長因子を除去する。繊毛様神経栄養因子(CNTF)およびCNTF受容体の他のアゴニスト等の成熟を亢進する因子を、成熟培地中に含めることができる。細胞が所望の成熟度に達したならば、培養液から細胞を回収し(例えば、トリプシンまたはコラーゲンを用いて)、解析または最終的な使用のために調製することができる。
【0082】
分化細胞の特徴
多くの表現型基準に従って、細胞を特徴付けることができる。基準には、形態学的特徴の顕微鏡観察、発現した細胞マーカーの検出または定量、インビトロで測定可能な機能的基準、および宿主動物に注入した際の反応が含まれるが、これらに限定されない。
【0083】
表現型マーカー
オリゴデンドロサイトに特有の表現型マーカーを発現するかどうかに従って、本発明の分化細胞を特徴付けることができる。細胞集団の成熟度に従って存在し得るオリゴデンドロサイト細胞の古典的な免疫細胞化学的マーカーは、以下の通りである:
・NG2、マクロファージおよびオリゴデンドロサイト前駆細胞によって発現されるコンドロイチン硫酸プロテオグリカン
・ガラクトセレブロシド(GalC)、拘束されたオリゴデンドロサイトのマーカー
・ミエリン塩基性タンパク質(MBP)、成熟ミエリンおよびミエリン産生細胞のマーカー。
【0084】
オリゴデンドロサイト系譜細胞によって発現される他の有用なマーカーには、以下が含まれる:
・PDGFRα、オリゴ前駆細胞、オリゴデンドロサイト、および他の細胞型によって発現されるPDGFの膜受容体
・TRα1、オリゴ前駆細胞、オリゴデンドロサイト、ニューロン、および他の細胞型によって発現される甲状腺ホルモンの核受容体
・ミエリンプロテオリピドタンパク質、オリゴデンドロサイトおよびグリア前駆細胞で発現されるミエリンの成分
・O4抗体によって規定されるエピトープ、オリゴデンドロサイト、アストロサイト、およびそれらの前駆細胞のマーカー
・ビメンチン、アストロサイト前駆細胞を特徴付ける線維芽細胞型フィラメントタンパク質(オリゴデンドロサイトでは陰性である場合が多い)
・神経膠繊維酸性タンパク質(GFAP)、アストロサイトのマーカー(オリゴデンドロサイトでは陰性)
・A2B5、II型アストロサイト、グリア前駆細胞、オリゴデンドロサイト前駆細胞、および膵臓β細胞で発現されるエピトープ
・オリゴデンドロサイトおよびその突起を染色し、脊髄および小脳両方の有髄化軸索と一致する、RIP抗体によって認識されるエピトープ。
【0085】
オリゴデンドロサイト分化の経路における様々な時点で発現される転写因子には、以下が含まれる:
・Olig1、オリゴ前駆細胞、運動ニューロン前駆細胞、および腎細胞によって発現されるヘリックス・ループ・ヘリックス(HLH)ファミリー転写因子
・Olig2、オリゴ前駆細胞、運動ニューロン前駆細胞、および松果体によって発現される別のHLHファミリー転写因子
・Sox10、オリゴ前駆細胞、オリゴデンドロサイト、シュワン細胞、神経堤、蝸牛、前立腺、およびメラノサイトによって発現されるSoxファミリー転写因子
・Nkx2.2、オリゴ前駆細胞、オリゴデンドロサイト、ニューロン前駆細胞、膵臓α細胞および膵臓β細胞によって発現されるHoxファミリー転写因子
・Pax6、オリゴ前駆細胞、ニューロン前駆細胞、膵αおよびβ細胞、水晶体網膜、下垂体、肝臓、および脾臓によって発現されるHLHファミリー転写因子。
【0086】
他の細胞型の有用なマーカーには以下が含まれる:
・ニューロン核抗原(NeuN)、ニューロン成熟のマーカー(通常、オリゴデンドロサイト系譜細胞では陰性)
・クラスIIIβチューブリン(TuJ1)、神経細胞の別のマーカー
・微小管関連タンパク質2(MAP−2)、CNS細胞のマーカー(おそらく陽性)
・SSEA−4、Oct−4、およびテロメラーゼ逆転写酵素(TERT)、未分化pPS細胞のマーカー(オリゴデンドロサイトおよびその前駆細胞では陰性)。
【0087】
組織特異的マーカーは、細胞表面マーカーについてのフロー免疫細胞化学法、または細胞内もしくは細胞表面マーカーについての免疫組織化学法(例えば、固定化細胞または組織切片)等の、任意の適切な免疫学的技法を用いて検出することができる。フローサイトメトリー解析の詳細な方法は、Gallacherら、Blood 96:1740、2000に記載されている。細胞表面抗原の発現は、標準的な免疫細胞化学法またはフローサイトメトリーアッセイ法において、選択的に細胞を固定化した後、および選択的に標識二次抗体または標識を増幅する他のコンジュゲートを用いて、有意に検出可能な量の抗体が抗原に結合した場合に陽性と定義される。
【0088】
組織特異的遺伝子産物の発現は、ノーザンブロット解析法、ドットブロットハイブリダイゼーション解析法により、または標準的な増幅方法において配列特異的プライマーを用いる逆転写酵素ポリメラーゼ連鎖反応法(RT−PCR法)により、mRNAレベルで検出することもできる。さらなる詳細については、米国特許第5,843,780号を参照されたい。特定マーカーの配列データは、GenBank等の公的データベースから入手可能である。例えば、ミエリンガラクトリピド生合成酵素UDP−ガラクトース・セラミド・ガラクトシルトランスフェラーゼ(CGT;GenBankアクセッション番号AH006651)は、GalCの合成の最終段階を触媒する酵素である。
【0089】
研究または治療における使用を容易にするために、集団内のオリゴデンドロサイトまたはその前駆細胞の特徴を有する細胞の割合を最大にすることが有利である場合が多い。以下の実施例の項で説明するように、上記のようなオリゴデンドロサイトまたは成熟細胞に特有の表現型マーカーのいずれかの1つ、2つ、3つ、またはそれ以上に関して陽性であると同定されるそのような細胞(または両方の混合物)が少なくとも約20%から約60%、80%、90%、95%、さらに98%を超えるまでのいずれかの細胞集団を得ることが可能である。
【0090】
神経機能の再構成に関連する治療的応用では、細胞集団が他の細胞型 − 詳細には未分化pPS細胞および非外胚葉系譜の細胞を形成する能力を最小限に抑えることが望ましい場合が多い。用途に応じて、ニューロン系譜およびその拘束前駆細胞、アストロサイト系譜およびその拘束前駆細胞、ならびにグリア細胞またはすべての種類の神経系細胞の共通の前駆細胞の割合を最小限に抑えることが有利である可能性もある。特定の態様では、本発明によるオリゴデンドロサイト集団は、これらの他の細胞型による混入を約1%、0.2%、または0.05%未満しか有さない。
【0091】
当業者は、オリゴデンドロサイト系譜細胞に特有の形態学的特徴をすでに理解していると考えられる。オリゴデンドロサイト前駆細胞は、中心体から反対極に2つの突起を伸長した双極の形状をとる場合がある。この細胞はまた、同様のマーカーおよびオリゴデンドロサイトの他の特徴の多くを有する、比較的扁平な細胞の形態をとる場合もある。この細胞は、比較的遅い成長速度ならびに細胞外マトリックスおよび可溶性因子への依存によって、他の細胞(線維芽細胞のような)から識別することができる。双極細胞と扁平細胞は、扁平細胞の優位性を促進する成長因子(EGF、bFGF等)の有無により、相互変換が可能である。本発明の前駆細胞集団は、少なくとも約20%、40%。60%、80%、またはそれ以上の双極細胞または扁平細胞表現型を含み得る。
【0092】
より成熟したオリゴデンドロサイトへの分化に際して、突起の数および複雑度は典型的に増加する。突起はそれらの間にミエリン皮膜を有するようであり、個々の軸索の周りを覆って軸索に沿った神経伝達を促進するミエリン鞘を形成する。
【0093】
遺伝子型特性
単離したpPS細胞またはpPS細胞の樹立株に由来する場合、本発明のオリゴデンドロサイトは起源細胞または細胞株の子孫であると特徴付けられ得る。したがって、オリゴデンドロサイトはその由来の元となった細胞と同じゲノムを有することになる。このことは、任意の核型変化のほかに、pPS細胞とオリゴデンドロサイトとの間で染色体DNAが90%を超えて同一であることを意味する。導入遺伝子を導入する、または内因性遺伝子をノックアウトするために組換え法により処理したオリゴデンドロサイトは、未操作の遺伝子要素はすべて保存されているため、依然として由来の元となった株と同じゲノムを有すると見なされる。
【0094】
オリゴデンドロサイトおよびpPS細胞は、標準的な遺伝子フィンガープリント技法により同じゲノムを有することが同定され得る。オリゴデンドロサイトが、未分化株から正常な有糸分裂の過程を経て得られる場合にも、同じゲノムを所有することが示され得る。
【0095】
特定の産業用途において、この特徴は本発明のオリゴデンドロサイトの貴重な特性である。詳細には、pPS細胞に増殖を引き起こし、細分し、要求に応じてさらなるオリゴデンドロサイトに分化させることができるため、起源pPS細胞が利用できることによって遺伝的に一致したオリゴデンドロサイトがさらにまた供給される。さらに、pPS細胞を他の治療的に重要な系譜に分化させることもできる。例えば、pPS細胞を免疫寛容細胞集団に分化させることが可能であり、この細胞集団は対象とするレシピエントを対応するオリゴデンドロサイトを用いた移植に対して寛容にするのに役立ち得る(国際出願PCT/US01/43434、Geron Corp.)
【0096】
本明細書に記載する技法により、分化の前または後に細胞を拡大することによって、同じゲノムを共有するオリゴデンドロサイト系譜細胞の大きな集団の産生が可能となる。理論的に、108、1010、または1012個の細胞の集団が可能である。そのような大きな集団は、通常、さらなる培養、薬剤スクリーニング、または治療投与に適した個々の容器に分割する。
【0097】
本発明の特定の態様は、グリア細胞またはオリゴデンドロサイトの特徴を有する分化細胞と組み合わせた起源細胞(未分化pPS細胞株または中間の集団等)を含む。2つの集団は同じ容器中に存在しても、同じ施設内の別の容器に存在しても、または2つの別の場所に存在してもよい。すでに記載したように未分化細胞の培養物にオリゴデンドロサイト系譜細胞への分化を全体として引き起こす場合のように、未分化細胞および分化細胞は同時に存在しても別の時点で存在してもよい。
【0098】
機能特性
品質管理の目的で、本発明の分化細胞の特性を機能的基準によって特徴付けることが望ましい場合が多い。目的とする最終的な用途により、異なる機能が異なる相対的関心となり得る。例えば、組織培養において神経組織を再ミエリン化する能力、インビボで脱髄が誘導された部位を修復する能力、または損傷した対象において神経機能を回復する能力について、細胞を評価することができる。これらの特性を判定するための、以下の限定されない例証を含む多くの実験モデルが存在する。
【0099】
1. 共培養におけるインビトロ髄鞘形成
2〜3ヶ月齢雌ラットから腰椎および頚椎後根神経節(DRG)ニューロンを回収することにより、成体DRG培養物を調製する。細胞を粉砕し、遠心分離し、mL当たり約100,000個の生存ニューロンでDMEM F12培地に再懸濁し、ラミニンコーティングした皿にプレーティングして4週間培養する。約2.5 x 104個のオリゴデンドロサイト系譜細胞を添加し、毎日フィードしながらさらに4週間培養することにより、インビトロ髄鞘形成を行う。ヒトオリゴデンドロサイトによるげっ歯類軸索の髄鞘形成は、TargetおよびBlakemore、Eye 8(part2):238、1994に例証されている。
【0100】
オリゴデンドロサイト系譜細胞がミエリンを形成する能力を判定するため、共培養物を4%パラフォルムアルデヒドで一晩固定し、GalCについて染色する。神経節に隣接した視野を選択し、視野当たりのミエリン部分の数をスコアリングする。ネスチン、βチューブリンIII、GFAP、CNP、GalC、Ki−67抗原、NeuN、または神経フィラメント70等のマーカーについて低密度培養物を染色することによって、分化の潜在能力を判定する。
【0101】
培養におけるDRGの髄鞘形成の証拠は、一般的にオリゴデンドロサイトの生物学的有効性と関連がある。細胞がインビボで生存し髄鞘形成または軸索の再成長を亢進する能力は、適切な動物モデルを用いてより一層得られる。これに反して、共培養における髄鞘形成は培養の条件に依存することになり、否定的な結果によってインビボにおける有効性は排除されない。インビトロ試験の予測値は、例えばIGFまたはNT−3のようなニューロトロフィン等の髄鞘形成を亢進すると考えられている因子を添加することにより、培養条件を調製することによって、改善することができる。
【0102】
2. 慢性脱髄モデル
成体ラット脊柱において、慢性脱髄の領域を誘導することができる(Keirsteadら、J Neurosci. 19:7529、1999)。鉛遮蔽を用いてT9を中心に、2 cmを超える距離で脊髄を40 GのX線照射に曝露するが、これにより曝露細胞のDNAにニックが導入され、細胞分裂している細胞が死滅する。2日後に、T9においてエチジウムブロマイドを脊髄内に直接注入する。エチジウムブロマイドはこれに曝露した細胞を死滅させるインターカレート剤であり、これによって慢性脱髄の無細胞領域ならびに脊柱領域の約60%を通して生存オリゴデンドロサイトおよびアストロサイトがない状態が提供される。
【0103】
3日後、ラットの脱髄部位にオリゴデンドロサイト系譜細胞の移植を行う。任意で、48時間前に培地にブロモデオキシウリジン(BrdU)を添加して細胞を予め標識することができる。第一に、細胞を前駆細胞約30個の凝集塊として調製し、μL当たり約60,000個の細胞密度に濃縮することができる。外径約80 umのガラスマイクロピペットを用いて、約10分を上回る時間をかけて細胞1μLを損傷部位に投与する。約2〜4週間後、組織試料を調製し、1 mmの横軸ブロックになるように樹脂またはクリオスタット切片を作製する。
【0104】
冠状面の切片をトルイジンブルーで染色し、一般病理、再ミエリン化の証拠、および細胞形態について解析する。誘導した部位は無細胞であるため、移植後に存在する細胞は投与した細胞に由来する。クリオスタット切片を、GFAP(アストロサイト)、CNP(オリゴデンドロサイト)、RIP(オリゴデンドロサイト)、またはNeuN(ニューロン)等の関連する細胞型のマーカーについて染色することができる。超薄切片を、ミエリン層板の数および細胞の超微細構造について電子顕微鏡により解析することもできる。脱髄部位にわたる移植細胞の再分布、および成熟有髄化細胞への分化を判定することができる。約25%、50%、または75%のレベルでの脱髄した軸索に占める再ミエリン化が、生物学的有効性の増加の証拠である。
【0105】
オリゴデンドロサイト系譜細胞の治療能力は、脱髄の先天性モデルにおいて試験することも可能である。ミエリン塩基性タンパク質における変異または欠陥を含む樹立モデルには、シバラー変異マウス(Roachら、Cell 42:149、1985)およびロングエバンス・シェーカー(Long Evans shaker)ラット(Kwiecienら、J. Neurocytol. 27:581、1989;Delaneyら、Lab. Anim. Sci. 45:547、1995)が含まれる。脳室内または大槽移植により(Mitomeら、Brain 134:2147、2001)、または脊髄への直接投与により(Liu、McDonaldら、Proc. Natl. Acad. Sci. USA 97:6126、2000)、再構築を試験することができる。MBPにおける変異または欠陥を伴うモデルは通常は適切な髄鞘形成の能力がなく、結果として神経欠損を被るため、これらのモデルでの緻密なミエリンの形成は投与細胞に直接起因する。
【0106】
3. 脊髄損傷モデル
SCIモデルは、挫傷および背側片側切断を含む。挫傷では、適切な脊髄損傷装置を用いて、脊髄層を23ミリ秒を上回る時間をかけて約0.9 mm除去する(中程度の損傷)。片側切断損傷では、定位的なマニピュレーターを用いて先のとがった手術用メスで脊髄の背側の半分を切断する。どちらの手順とも、その後適切な術後処置を行う。
【0107】
移植した細胞の遊走を促進しかつミエリン関連成長抑制物質を除去するため、任意で脊髄もまた脱髄する(Keirsteadら、Exp Neurol. 151:303、1998)。脊柱管の中央かつ軸索損傷部位の後方に2 mmの孔を作製する。露出した脊髄に、リン酸緩衝食塩水に溶解した33%モルモット補体(Harlan SeraLab)で1:2に希釈したポリクローナル抗GalC抗体(Chemicon)約4μLを注入する。
【0108】
損傷から約24時間後に、ガラスマイクロピペットを通して動物にオリゴデンドロサイト系譜細胞を移植する。あるいは、1〜3ヶ月間治療を保留することにより、慢性損傷モデルを作製することができる。細胞を用いて処置した後、ビデオテープに機能的反応を記録し、臨床的改善の証拠を得るために定期的にモニターすることができる。例えば、BBB尺度、関節運動に基づいた21ポイント尺度、体重支持、四肢の協調、および他の特性を用いて、地上歩行を定量化することができる(実施例2)。
【0109】
組織学的精密検査をするため、ビオチン標識コレラ毒素βサブユニット(CTB、坐骨神経の両側に0.1%を4μL注入)またはビオチン化デキストランアミン(BDA、感覚運動皮質に10%を10μL注入)等のトレーサーを動物に予め注射することができる。約2週間後、脱髄モデルでのように組織切片を調製し、移植した細胞の位置および再ミエリン化の証拠について検討するが、再ミエリン化は形態学的基準によって正常な髄鞘形成または部分的脱髄と識別可能である。特徴的な神経突起の成長円錐の証拠を得るために、注射したトレーサーで標識した軸索についても切片を解析する。軸索再生の免疫化学的証拠は、RT97抗原(神経フィラメントのマーカー)、セロトニン(5HT)、ノルエピネフリン(NE)、およびカルシトニン遺伝子関連ペプチド(CGRP)等のニューロンマーカーについて染色することによって評価することができる。
【0110】
オリゴデンドロサイトの遺伝子改変
本発明の特定のオリゴデンドロサイト前駆細胞集団は、実質的な増殖能を有する。必要に応じて、内因性遺伝子からの転写を増加させることによって、または導入遺伝子を導入することによって、細胞内ののテロメラーゼ逆転写酵素(TERT)のレベルを増加させることにより複製能をさらに亢進することができる。特に適切なのは、国際公開公報第98/14592号に提供されているヒトテロメラーゼ(hTERT)の触媒成分である。ヒト細胞におけるテロメラーゼのトランスフェクションおよび発現は、Bodnarら、Science 279:349、1998、およびJiangら、Nat. Genet. 21:111、1999に記載されている。
【0111】
未分化pPS細胞においてテロメラーゼ発現を増加させることができ、その後本発明に従ってオリゴデンドロサイトに分化させることができる。あるいは、pPS細胞をオリゴデンドロサイト前駆細胞に分化させ、その後トランスフェクションしてTERT発現を増加させることもできる。遺伝子改変細胞は、標準的な方法に従って、RT−PCR法、テロメラーゼ活性(TRAPアッセイ法)、TERTの免疫細胞化学染色、または複製能により、TERTの発現について評価することができる。用途によっては、myc、SV40ラージT抗原、またはMOT−2をコードするDNAで細胞を形質転換する等の、細胞を不死化する他の方法もまた意図している(米国特許第5,869,243号、国際公開公報第97/32972号、および国際公開公報第01/23555号)。
【0112】
必要に応じて、本発明の細胞を調製もしくはさらに処理し、インビトロで未分化細胞を除去すること、またはインビボで復帰細胞を防ぐことができる。集団から未分化幹細胞を減少させる1つの方法は、エフェクター遺伝子が、TERTプロモーターまたはOCT−4プロモーター等の未分化細胞において優先的な発現を引き起こすプロモーターの制御下にあるベクターを、集団にトランスフェクションすることである。エフェクター遺伝子は、緑色蛍光タンパク質等のセルソーティング導くレポーターであってよい。エフェクターは、例えば毒素またはカスパーゼ等のアポトーシスの介在物質をコードし、細胞に対して直接溶解性であってもよい(Shinouraら、Cancer Gene Ther. 7:739, 2000)。エフェクター遺伝子は、抗体またはプロドラッグ等の外部薬剤の毒素効果に対して細胞を感受性にする効果を有してもよい。発現する細胞をガンシクロビルに対して感受性にする単純ヘルペスチミジンキナーゼ(tk)遺伝子が例示的である(米国特許第6,576,464 B1号)。あるいは、エフェクターは、未分化表現型に復帰する任意の細胞をインビボで天然抗体に感受性にする外来決定因子を、細胞表面に発現させてもよい(U.S. 2003−0032187 A1)。
【0113】
本発明の細胞はまた、組織再生に関与する能力、または治療する対象に治療遺伝子を送達する能力を増強するために、遺伝子改変することも可能である。所望の遺伝子をコードする既知の配列を用いてベクターを設計し、構成的(CMVプロモーター等)またはオリゴデンドロサイト系譜細胞の細胞において特異的(ミエリン塩基性タンパク質のプロモーター等)に活性のあるプロモーターに機能的に連結する。この戦略に従って、オリゴデンドロサイトの成長を亢進する、再ミエリン化を活性化する、または軸索の再生を促進するような、様々な導入遺伝子を発現させることができる。神経成長因子をコードする遺伝子が例示的である(米国特許第5,885,584号および第6,268,340号)。
【0114】
研究および臨床治療におけるオリゴデンドロサイトの使用
本発明は、様々な重要な研究、開発、および商業目的のために、多数のオリゴデンドロサイトを産生する方法を提供する。
【0115】
本発明の細胞を用いて、他の系譜の細胞で優先的に発現されるcDNAが比較的混入していないcDNAライブラリーを調製することができる。本発明の分化細胞を用いて、標準的な方法に従い、オリゴデンドロサイトおよびその派生物のマーカーに特異的なモノクローナル抗体またはポリクローナル抗体を調製することも可能である。
【0116】
特に関心が持たれるのは、薬剤開発および臨床治療を目的とした本発明の組成物の使用である。
【0117】
薬剤スクリーニング
本発明の細胞を用いて、オリゴデンドロサイト前駆細胞および成熟オリゴデンドロサイト両方の特徴に影響を及ぼす因子(溶媒、小分子薬剤、ペプチド、ポリヌクレオチド等)または環境条件(培養環境または操作等)をスクリーニングすることができる。
【0118】
一例では、pPS細胞(未分化または分化パラダイムに向けて起動した)を用いて、オリゴデンドロサイトへの成熟を促進する因子、または長期培養におけるオリゴデンドロサイトの増殖および維持を促進する因子をスクリーニングする。例えば、別のウェル内の細胞に候補成熟因子または成長因子を添加し、さらなる培養および細胞用途の所望の基準に従って生じる任意の表現型変化を判断することにより、それら因子を試験する。これにより、pPS由来オリゴデンドロサイトばかりでなく初代神経組織から単離したオリゴデンドロサイトおよびその前駆細胞に関しても、導出および培養法の改良がもたらされ得る。
【0119】
本発明の他のスクリーニング法は、オリゴデンドロサイトの成長、発達、または毒性に及ぼす潜在的副作用についての薬学的化合物の試験に関する。この種のスクリーニングは、化合物がオリゴデンドロサイト自体への薬学的効果を有するように設計される場合ばかりでなく、他所での主要な薬学的効果のために設計された化合物のオリゴデンドロサイトに関連する副作用を試験するためにも適している。
【0120】
他のスクリーニング法は、軸索を有髄化する役割におけるオリゴデンドロサイト活性に影響を及ぼす可能性を有する小分子薬剤の効果を測定するための、オリゴデンドロサイトの使用に関する。この目的のため、細胞をインビトロで試験化合物と混合することができ、ミエリンに関連する活性に及ぼす化合物の効果を判定する − 例えば、ミエリン塩基性タンパク質等のミエリン関連成分の発現;組織学的に検出可能なミエリン鞘の形成;および神経細胞との共培養において隣接する軸索を有髄化する能力。
【0121】
薬剤スクリーニングの一般原理に関しては、当業者には、米国特許第5,030,015号および教科書In vitro Methods in Pharmaceutical Research、Academic Press、1997を参照されたい。候補薬学的化合物の活性の評価は、一般に本発明の分化細胞を、単独または他の薬剤との組み合わせのどちらかで、候補化合物と混合することを伴う。研究者は、化合物に起因する細胞の形態、マーカー表現型、または機能的活性の任意の変化を判断し(未処理細胞または不活性化合物で処理した細胞と比較して)、化合物の効果を観察した変化と関連づける。
【0122】
臨床治療におけるオリゴデンドロサイト
本発明は、治療を必要とする患者において神経機能を保持または回復するための、オリゴデンドロサイト前駆細胞およびその派生物の使用を提供する。特に、神経組織を再ミエリン化するため、またはさもなければ神経ネットワークの維持もしくは再生を支持するために、本発明の細胞を投与することができる。いかなる限定も意味することなく、投与した細胞は、すでに所定の位置にあるニューロンの機能を安定化するもしくは改善する効果を有する、またはニューロンが相互にもしくはそれが制御する組織と新たな連結を形成するのを支持する可能性がある。
【0123】
薬学的組成物の性質は、部分的には治療する病態に依存する。ある症例では、優れた複製能および操作に必要な回復力を有する前駆細胞(NP2またはGalCに関して陽性)で組成物を製剤化することが適切である場合がある。別の症例では、より迅速に髄鞘形成能を提供するために、より成熟した細胞(GalCまたはMBPに関して陽性)で組成物を製剤化することが適切である場合がある。
【0124】
懸濁培養において形成される大きな凝集体をマトリゲル(登録商標)等の適切なマトリックスに接着させることにより、pPS由来細胞からオリゴデンドロサイト系譜細胞の比較的分散した集団が容易に得られること見出した。培養した細胞を基層上にプレーティングして数時間〜数日間培養し、非接着細胞を除去する。次に、短時間酵素消化した後に粉砕する等の適切な化学的または機械的手段によって、オリゴデンドロサイト系譜細胞を回収する。細胞操作、保存特性、および投与に際して有利な効果を提供する能力の点から見て、単分散集団または約30個以下の細胞の凝集塊は利点を有すると考えられる。細胞は典型的に洗浄し、その後疾患部位に細胞を保持するために効果的な用量を投与するのに適した濃度で(例えば、μL当たり細胞20,000個〜100,000個、治療領域の量によって調整する)、薬学的に適合する培地中に懸濁する。
【0125】
神経に適応した治療(neurological adaptive therapy)の通常の手順に従って、患者の準備をする。拒絶を防ぐために、シクロスポリンA等の標準的な免疫抑制療法により患者を処置してもよい。別法としてまたはさらに加えて、同じpPS細胞株から調製した造血細胞または未分化細胞を用いて、患者をオリゴデンドロサイトのアロタイプに対して特異的に寛容化してもよい(国際公開公報第02/44343号;国際公開公報第03/050251号)。症例によっては、損傷部位においてニューロンを一次的に脱髄すること、投与細胞の接近を促進すること、または再ミエリン化を阻害し得る因子を除去することが有利である場合がある。これを達成する1つの手段は、GalC、O4、またはエリン関連糖タンパク質等の、ミエリン鞘上の1つまたは複数のエピトープに対する補体結合抗体を局所環境に投与することである。Keirsteadら、Brain Res. Bul. 44:727、1997;およびカナダ特許第2,253,078号を参照されたい。次に、本発明の分化細胞を、髄鞘形成もしくは神経機能の再生を所望する部位またはその周りに、1回または複数回投与する。
【0126】
その後対象に支持的な術後処置を施し、移植片の許容または神経機能の再生についてモニターする。適切であれば、患者をファムプリジン(Fampridine)−SR(4−アミノピリジン)等の、オリゴデンドロサイト機能を回復すると考えられる他の手段を用いて同時に治療してもよい。組織試料を、すでに記載した関連マーカーに関して免疫細胞化学法によって解析し、軸索の再ミエリン化、投与細胞の存在に起因する神経突起の発芽等の機能的事象に関して評価することができる。それらの疾患の典型的な病理学的特性により病態の維持または改善に関して、および例えば障害状態尺度(expanded disability status scale; EDSS)での機能の評価に関して、患者をモニターすることができる。
【0127】
本発明の組成物を用いた治療に適切な病態には、ミエリンを維持するまたは産生する能力が、治癒またはさらなる悪化の防止に役立つ進行性脱髄および中枢神経系の外傷が含まれるが、これらに限定されない。
【0128】
多発性硬化症は、脳および脊髄における脱髄部分の散在を特徴とする緩徐進行性疾患である。オリゴデンドログリアの破壊および血管周囲の炎症を伴う脱髄プラークは、CNSを通じて、主として白質(特に、頚部および背部)、視神経、および周室部に起こる。中脳、脳橋、および小脳の路、ならびに大脳および脊髄の灰白質もまた影響を受ける。
【0129】
急性散在性脳脊髄炎(感染後脳脊髄炎)は血管周囲のCNS脱髄を特徴とし、これは自発的に起こり得るが、通常はウイルス感染またはウイルスワクチン接種の後に起こる。慢性炎症性脱髄性多発根神経炎(CIDP)は、炎症性T細胞およびマクロファージを伴う神経内膜の間質および血管周囲浸潤を特徴とし、末梢神経の分節性脱髄を引き起こす。HTLV関連ミエロパシーは、両脚の痙性衰弱を特徴とする緩徐進行性脊髄疾患である。ギランバレー症候群等のいくつかの末梢神経障害も、脱髄を特徴とする。
【0130】
先天性代謝病(フェニルケトン尿症および他のアミノ酸尿症;テイ・サックス病、ニ−マン・ピック病、およびゴーシェ病;フルラー症候群;クラッベ病および他の白質萎縮症等)は、CNSにおけるミエリン鞘の発達に影響を及ぼし、永久的な広汎性神経欠損を起こす可能性がある。副腎白質萎縮症および副腎脊髄神経障害は、副腎機能不全および神経系の広範な脱髄を特徴とする伴性劣性代謝病である。ペリツェウス・メルツバッハー病は、プロテオリピドタンパク質遺伝子の点突然変異のために患者がミエリンを形成しない病態である。レーバー遺伝性視神経萎縮および関連するミトコンドリア障害は、主に両側の中心視力の喪失を特徴とする。
【0131】
CNSの外傷によって起こる任意の急性または長期異常も、無酸素症および虚血によるミエリンの喪失に関連する病態と同様に、治療が検討され得る。この分類には、脳卒中および外傷性脳損傷に関連する病態が含まれる。
【0132】
対麻痺または任意のレベルの不完全な運動機能を起こす様々な種類の脊髄損傷が、本発明の細胞を用いた治療の第一候補である。性質および到達性次第で、頚椎、腰椎、胸椎、および仙椎の損傷は、すべてその病態の改善または安定化を得る可能性がある。SC機能の完全なまたは不完全な喪失を起こす急性損傷は、減圧手術と同時にまたはその直後に治療することができる。慢性病態は、必要に応じていつでも治療または再治療することができる。針、カニューレ、または他の適切な装置を使用して、手術、内視鏡、または経皮注射により、細胞を損傷部位にまたはその近傍に投与することができる。保証されている場合、誘発電位試験により電気生理学的効果を測定することができる。米国脊髄損傷学会(ASIA)の運動スコアならびにフェランズおよびパワーズの生活の質指標(QLI)等の基準により、患者を運動経路および感覚経路の臨床的改善について評価することもできる。
【0133】
いつもと同様、患者の選択、投与形態、およびリハビリテーションのモニタリングの最終責任は、管理する臨床医の責任である。
【0134】
商品流通できるように、本発明のオリゴデンドロサイトは典型的に等張賦形剤を含む薬学的組成物の形態で提供し、ヒト投与に十分に無菌的な条件下で調製する。本発明はまた、生産、流通、または使用の任意の時点で存在する細胞のセットまたは組み合わせを含む試薬系を含む。細胞セットは、同じゲノムを共有する場合がある未分化pPS細胞または他の分化細胞型と組み合わせた、分化したpPS由来細胞の種類(グリア細胞、オリゴデンドロサイト、それらの前駆細胞およびサブタイプ等)に例証されるがこれらに限定されない、本開示に記載する2つまたはそれ以上の細胞集団の任意の組み合わせを含む。セット中の各細胞型は、同じ事業体もしくは事業関係を共有する別の事業体の管理下で、同時にもしくは別の時点で、一緒にもしくは同じ施設内の別の容器に包装しても、または別の場所で包装してもよい。
【0135】
細胞組成物の医薬品製剤化における一般原則については、当業者には、G. MorstynおよびW. Sheridan編によるCell Therapy: Stem Cell Transplantation, Gene Therapy, and Cellular Immunotherapy、Cambridge University Press、1996、およびCell Transplantation for Neurological Disorders、T.B. Freemanら編、Humana Press 1998を参照されたい。細胞は、任意で細胞の保存または上記の病態を治療するための医薬品としての使用に関する情報を添付して、流通もしくは臨床用途に適した装置もしくは容器に、または他の価値ある目的に適した装置もしくは容器に包装する。
【0136】
本発明の特定の態様の限定されないさらなる例証として、以下の実施例を提供する。
【0137】
実施例
実施例1:hES細胞のオリゴデンドロサイトへの分化
H7株およびH1株のヒト胚性幹(hES)細胞を、初代マウスフィーダー細胞により馴化しbFGFを添加したノックアウト(無血清)培地中、マトリゲル(登録商標)基層上で無フィーダー条件において未分化状態で増殖させた(国際公開公報第99/20741号; 国際公開公報第01/51616号、Geron Corp)。
【0138】
以下の計画に従って、hES細胞をオリゴデンドロサイト系譜の細胞に分化させた:
【0139】
(表1)hES細胞からのオリゴデンドロ前駆体(oligodendroprogenitor)の産生
【表1】

GRMはグリア制限培地である(表2を参照のこと)。
移行培地(TR)は、GRMおよびhES細胞を作製するための馴化培地の1:1混合物として作製する。
【0140】
(表2)グリア制限培地(GRM)の成分
【表2】
【0141】
分化手順は以下の通りに行った:
・1日目:コラゲナーゼIV(Gibco 17101−015)を用いて接着基層からESコロニーを解離し、コロニーを4 ng/mL bFGF(Gibco 13256−029)を有するTR中で低接着性6ウェルプレート(Corning 3471)に入れた。分化手順の最初の3日間のみ、ペニシリン−ストレプトマイシン(Gibco 10378−016)を用いた。
・2〜10日目:細胞をレチノイン酸(RA;オールトランス−レチノイン酸、Sigma 223018)、10μMと共に培養し、合計8日間毎日フィードした。RA保存液は、6 mg/mLの濃度で(約0.02 M)DMSOに溶解して調製した。
・3日目から:培地をGRMに置換し、それ以上はbFGFを添加しなかった。培地は毎日交換した。RAは光感受性であるため、フィード中は照明を最小限に落とした。ES細胞凝集体を含むウェルの培地を、15 mLチューブに回収した。低速で短時間遠心分離した後(800 rpm、1分間)、上清を吸引除去し、新鮮培地を添加した。穏やかに上下にピペッティングし(2〜3回)、6ウェルプレートの各ウェルに4 mLずつ確実に均一に分配した。
・10〜15日目:10日目以後は、毎日フィードする際に、20 ng/mL濃度のEGF(Sigma E9644)および2 ng/mL濃度のbFGFを添加した。
・15〜21日目:EGFは20 ng/mL濃度で添加し続けたが、bFGFは培地から除去した。
・21日目以後:20 ng/mL EGFを有するGRM培地を維持することにより、42日目以降でも神経の特徴を有する新たな凝集体/凝集塊が産生され得た。
・これらの神経球を選択するため、分化手順の28日目に、マトリゲル(登録商標)をコーティングした6ウェルプレート(マトリゲル(登録商標)1:30、BD Bioscience 356231)に、培養物全体を解離手順を行うことなく、12〜20時間(一晩)移した。翌日、培養物を穏やかに振盪すると、神経球のみが接着したままであるので、培養物の残りを新鮮なGRM培地で置換した。
・35日目:ポリ−L−リジン(Sigma P2636)およびラミニンでコーティングした4チャンバー・イメージングスライド(Nalgene−Nunc International 154917)に、神経凝集体をプレーティングした。トリプシン−EDTAで短時間(5分)処理し、細胞の凝集を解離させた。PLL−ラミニン上で細胞を7日間培養し、その間一日おきに培養物にフィードした。
・42日目:培養物をパラフォルムアルデヒドで固定し、免疫組織化学法により細胞を特徴付けした。使用したマーカー:NeuN、ガラクトセレブロシド(GalC)、Map2、O4、ビメンチン、GFAP。
【0142】
結果を添付の位相倒立顕微鏡写真(phase inverted photomicrograph)に示す。
【0143】
図1は、移行培地(50%グリア前駆細胞培地)中、懸濁状態で2日間培養した胚様体を示す。
【0144】
図2は、レチノイン酸中での培養の最終時点における、分化7日目の球体を示す。3日目から、透明な球体が懸濁培養物中に現れ始めた。7日目までに、これらの球体が培養物中の細胞の80〜90%を構成した。
【0145】
図3は、RA処理の抑制後の球体の崩壊を示す。いくつかの小胞は、細胞を球体の内部および外部に添加することにより成長し続けた。細胞集塊は濃い陰影の球状の形をとっていた。球体は、20 ng EGFのみの存在下では崩壊した。しかし、低濃度のbFGFを添加することにより、球体は維持され得た。いくつかの球体は新たな細胞を添加することによって成長し続け、球状の濃い陰影の細胞集塊を生じた。厚い壁を有するいくつかの小さな球体は成長し続けたが、数日後に完全に消失した。従って、bFGFを低濃度(2 ng/mL)で培養物中に5日間保持し続け、それにより球体は層の添加で成長し続けた。
【0146】
図4は、bFGFを除去した後に現れる変化を示す。細胞集塊のほとんどが解離し始め、培養液全体が単一細胞および小さな細胞塊でいっぱいになった。同時に、新たな鮮黄色の球体が観察された(矢印)。
【0147】
図5は、鮮黄色の球体(矢印)が培養を続けながら選択的に維持されたことを示す。あるものは他の球体から、あるものは単離した細胞集団から、あるものは濃い細胞集塊から発芽し、新たな球体が毎日出現した。
【0148】
28日目に、培養物全体をマトリゲル(登録商標)基層上に一晩接着させた。神経に拘束された球体のみが基質に接着し、その他の培養物は浮遊したままであった。
【0149】
図6は、マトリゲル(登録商標)上にプレーティングしてから2、3日後の、球体からのグリアに拘束された神経前駆細胞の遊走および分枝を示す。より小さいコロニーは個々の細胞の縦裂により環を形成したのに対し、大きなコロニーは長い突起を有する星型の遊走を示した。マトリゲル(登録商標)で培養してから数日後に、より多くの遊走細胞が出現した。
【0150】
一週間後、トリプシンを用いてマトリゲル(登録商標)から培養物を解離し、さらなる特徴付けを行うために、ポリ−L−リジンおよびラミニン(Gibco 12163−010)でコーティングしたNuncイメージングチャンバーに低密度でプレーティングした。プレーティングした細胞は、ラミニン基層上で成長してから3〜4日後に、特徴としてオリゴデンドロサイト形状をとった。
【0151】
次に、マトリゲル(登録商標)選択段階が10〜20時間に短縮できることを見出した。非接着細胞を除去し、次に接着細胞を再懸濁して、FGF、EGF、およびグリア前駆細胞培地の存在下で拡大した。これは、治療投与および他の目的に対してより適した、より分散した細胞集団を産生する利点を有する。
【0152】
図7はこの技法の結果を示す。細胞をマトリゲル(登録商標)に接着させた後、7日間拡大し、ポリ−L−リジン・ラミニン上にプレーティングし、マイトジェンの非存在下で培養した。4%パラフォルムアルデヒドで細胞を固定し、3%ヤギ血清および0.3% Triton−X 100(登録商標)界面活性剤中でブロッキングした。ガラクトセレブロシドに対する抗体(GalC、Chemicom)、次にペルオキシダーゼ標識した抗免疫グロブリンを用いて、免疫細胞化学法を行った。ヘマトキシンにより核を対比染色した。(第1パネル倍率20 X;第2パネル倍率40 X)。
【0153】
視野内の少なくとも約95%の細胞が、成熟オリゴデンドロサイト系譜細胞のマーカーであるGalCの染色を示す。
【0154】
図8は、GalCに関して染色された、高倍率のES由来オリゴデンドロサイトを示す(倍率60 X)。少なくとも約10%〜20%の細胞がオリゴデンドロサイトに特有の形態(具体的には、ミエリンシートを示唆する、間に皮膜を有する多数の突起)を示す。
【0155】
実施例2:hES由来オリゴデンドロサイトはインビボにおいて再ミエリン化および神経発芽を起こす
本実施例では、実施例1と同様の戦略にいくつかの改良点を加え、H7株由来のhES細胞を移植用のオリゴデンドロサイト前駆細胞に分化させた。GRMの処方は、プロゲステロン濃度を65 ng/mLに再検討する点以外は同様とした。EGFを初めから添加し、2日後にbFGFを除いた。
【0156】
(表3)hES細胞からのオリゴデンドロ前駆体の産生
【表3】
【0157】
最初の週は、低速で4〜5分間遠心分離してからフィードした。球体が成長を開始した後は、15〜50 mL円錐管内に入れインキュベーター中で5〜10分間培養物を沈降させてフィードした。接着因子を発現する細胞は凝集体(球体)を形成し、単一細胞懸濁液中では非接着細胞よりも早く沈降する。これにより、細胞間接着特性を有する培養物の精度が増す。
【0158】
図9は、分化計画過程における細胞形態の進行を示す。(A): mEF馴化培地中、マトリゲル(登録商標)上で無フィーダー培養にて成長する未分化hES細胞。(B): 3日目までに、レチノイン酸を含む培地での懸濁培養で胚様体から透明な球体が成長し、迅速な細胞の分散を示す。(C): 黄色い球体が大きな塊状で蓄積することから、レチノイン酸による神経系譜細胞の誘導(段階3)、その後のEGFの存在下でのオリゴデンドロサイト前駆細胞の拘束および拡大(段階4)が明白であった。(D): 次に、所望の細胞型のポジティブセレクションのため細胞をマトリゲル(登録商標)上にプレーティングした。(E、F): 拡大段階が進行するにつれて、球体の大きさおよび培養物の割合が増加した。優先的に選択した後、最大で8週までグリア拘束前駆細胞を維持し増幅することができた。(G、H): 次にマイトジェンの非存在下でポリ−L−リジンおよびラミニン上にオリゴデンドロサイト前駆細胞をプレーティングすると、細胞は1週間以内に成熟オリゴデンドロサイトの形態をとった。
【0159】
図10は、成熟オリゴデンドロサイトの免疫細胞化学的解析を示す。プレーティングしてから1週間後、94%を超える細胞が初期グリア細胞のマーカーであるNG2に対する抗体で標識された(A)。プレーティングしてから8週間後、95%を超える細胞がGalC(C)、O4(D)、およびRIP(E)について陽性に染色された。細胞を抗GFAPで染色し、かつDAPIで対比染色したところ、オリゴデンドログリアマーカーで標識されなかった細胞の実質的にすべてが、GFAP陽性であった(B)。
【0160】
次に、Infinite Horizon(登録商標)衝撃装置(Precision Systems and Instrumentation, LLC)を用いて中程度の脊椎挫傷を誘導した成体スプラーグ・ド−リー(Sprague Dawley)ラットを麻酔し、低位胸髄にオリゴデンドロサイト前駆細胞を投与した。
【0161】
図11は、細胞を投与した後に得られた、ヒト核タンパク質に特異的な抗体で染色した組織切片を示す。濃く染色されたことから、移植後にヒト細胞が生存していることが確認される。
【0162】
図12は、移植してから9週間後の、ヒト核タンパク質について染色した脊髄の周囲領域の横断切片を示す。拒絶を防ぐために、シクロスポリンAで動物を処置しておいた。細胞は白質に向けて遊走または増殖した。
【0163】
図13は、移植細胞が投与部位に副作用をもたらすかどうかを判定する詳細な解析の結果を示す。未処置のラットおよび移植したラットの損傷部位にわたり、脊髄の長さに沿って連続切片を得た。各棒グラフは、5枚の切片で測定した平均断面積を示す(平均値±SEM)。挫傷近傍の小さな断面積は、損傷後の二次拡大の結果である。オリゴデンドロサイトは救済(sparing)を誘導しないが、損傷を悪化させることもない。このことから、hES由来オリゴデンドロサイトは脊髄損傷の1 mm以内に投与しても安全であることが示される。
【0164】
図14は、ニューロン分枝を測定する実験から得られたものである。損傷から8週間後、運動皮質の順行性トレーサーとして、動物の皮質脊髄路にBDAを注射した。これらの切片は2週間後に採取したものである。hES由来オリゴデンドロサイトで処置した動物においてのみ、軸索分枝の証拠が存在した(上の2枚のパネルにおいて、濃く染色された細い線として示される)。分枝は、損傷部位の真上の脊髄の領域で観察された。
【0165】
図15は、損傷および細胞投与部位(中心点)から1 mmの間隔で測定したニューロン分枝の定量を示す(ブロック当たり3切片の平均値±SEM)。処置を受けなかった損傷動物では、部位の3 mm上で標識軸索を計測した。処置した動物は、有意に高いレベルで、中心点の損傷のすぐ頭側にまで標識軸索を有した。このことから、移植細胞が再生可塑性を誘導することが確認される。
【0166】
図16は、移植した動物から採取した、実質的な再ミエリン化の証拠を示す切片の一連の電子顕微鏡写真である。左上のパネルの厚い環は、正常にミエリン化した繊維である。視野内の残りの軸索は、薄い層のミエリンを示す。軸索直径に対する鞘の比率から、これらの軸索が新たにミエリン化されたことが示唆される。右上のパネルは、新たにミエリン化した軸索のより解像度の高い像である。約5または6層の外被、および上部にはオリゴデンドロサイトの先端を示す軸索の舌状の縁が存在する。これは、この軸索のミエリン化が進行中であることを示す。下のパネルは、シュワン細胞によって沈着した厚いミエリン鞘を示す。これは移植にかかわらず生じ、未処置動物において限られた程度の回復が提供される。しかし、hES由来オリゴデンドロサイトを移植した動物のみが、オリゴデンドロサイトに特有の、上のパネルで観察される性質のミエリン化の証拠を示した。
【0167】
実施例3:オリゴデンドロサイト系譜細胞の表現型特徴付け
本実施例では、表4に示す手順に従ったhES株H7からのオリゴデンドロサイトの産生過程におけるマーカー発現を追跡する。
【0168】
(表4)hES細胞からのオリゴデンドロ前駆体の産生
【表4】
【0169】
本手順の一般的な適用では、固相選択段階後(約23日目以後の任意の時点)に移植に適したオリゴデンドロサイト前駆細胞が得られる。トリプシンを用いて週に一度継代することにより、最終段階の培養を行い必要に応じて細胞を拡大することができる(少なくとも8週間)。
【0170】
本実施例での表現型解析のために、5%ノックアウト血清代替物を含むGRMを用いて細胞を低密度培養に適合化した。21日目に5% KO−SRを存在させ始めることによって、マトリゲル(登録商標)上にプレーティングした際に双極細胞の増殖が改善された。細胞を低密度でラミニン(ポリ−L−リジン上で15μg/mL)上にプレーティングした場合には、KO−SRにより細胞の生存度および成長が改善された。手順の早い時期にKO−SRを使用しても、成長速度および黄色球体の比率から見て分化の過程に影響を及ぼさなかった。
【0171】
図17は、未分化hES細胞コロニーで検出されたマーカーを示す。左側のパネルは、視野内のすべての細胞核の位置を表すDAPI染色の青蛍光の上に重ね合わせた、各マーカーに特異的な抗体による赤蛍光を白黒で示す。右側のパネルは、マーカー染色のみを示す。2つの像を比較することにより、問題のマーカーを発現する、視野内の全細胞の比率の推定が可能となる。
【0172】
上列は、多能性細胞のマーカーであるSSEA−4に陽性標識されたコロニーを示す。下列は、中胚葉マーカーBMP4に陽性標識されたコロニーを囲む間質細胞を示す。各コロニー内の未分化細胞はBMP4陰性である。
【0173】
図18は、分化過程における転写因子Pax6の一過性の出現を示す(左側:抗体+DAPI;右側:抗体染色のみ)。上列は、10日目の細胞塊の中心に向かった染色、周辺に向かってより分化した細胞でのすでにダウンレギュレートされた染色を示す。下列は、35日目に採取した細胞では実質的に染色されないことを示す。
【0174】
図19は、10日目にレチノイン酸を除去した直後に存在する、初期のオリゴデンドロサイト系譜細胞で検出されるマーカーを示す(左側:抗体+DAPI;右側:抗体染色のみ)。上列:転写因子Olig1(83%±7%)。中列:転写因子SOX10(72%±12%)。下列:非特異的オリゴデンドロサイト前駆細胞マーカーA2B5(97%±3%)。
【0175】
図20A〜20Bは、35日目の完全に分化したオリゴ前駆細胞において優位を占めるマーカーを示す(左側:抗体+DAPI;右側:抗体染色のみ)。1列目および2列目(20A):NG2(コンドロイチン硫酸プロテオグリカン、オリゴデンドロサイト前駆細胞のマーカー); 3列目(20A):GalC;4列目(20B):O4;5列目(20B):Tuj1(ニューロンのマーカー)。
【0176】
結果から、少なくとも約80%の細胞がオリゴデンドログリアマーカーNG2、GalC、およびO4に陽性であることが示される。オリゴデンドログリアマーカーで標識されなかった細胞は、主にニューロンマーカーGFAPまたはTuj1(20B、下列)に陽性であった。二重免疫細胞化学法により、GFAPまたはTuj1陽性細胞はオリゴデンドログリアマーカーを同時に発現しないことが示される。さらに、BMP4またはSSEA−4は検出されず、このことからこれらの培養物が未分化細胞または中胚葉系譜細胞を欠いていることが示唆される。この細胞集団は、少なくとも8回の連続した継代を通して増殖させることが可能であると考えられる。
【0177】
実施例4:hES由来オリゴデンドロサイトは、ミエリン欠損動物において軸索
を再ミエリン化する
これらの細胞によってインビボで起こった髄鞘形成が(内因性オリゴデンドロサイトの誘導によるのではなく)直接的な効果であることを実証するため、分化手順の28日目の細胞を脱髄のシバラーマウスモデルに移植した。シバラーマウスは、染色体18に位置するミエリン塩基性タンパク質遺伝子の変異に関してホモ接合性である(Mbpshi/Mbpshi)。この遺伝子は重複しており、重複遺伝子の大部分が反転し、アンチセンスRNAの形成を引き起こす。これにより、CNSを通じて重篤なミエリン欠損が生じる。
【0178】
培養物からオリゴデンドロサイト前駆細胞を回収し、DMEMで洗浄し、濃縮し、シリコンコーティングしたハミルトンシリンジに装填する。すべての実験と同様に、ここでも10 mg/kg シクロスポリンAにより動物を免疫抑制した。6週間後、動物を屠殺しEM解析のために切片を作製した。
【0179】
図21はその結果を示す。超微細構造的に、シバラーマウスの軸索はミエリンを欠いているか、または1もしくは2層の緻密でないミエリンによって囲まれている(上のパネル)。細胞を移植してから6週間後、電子顕微鏡解析により多層の緻密なミエリンが示され、このことから移植集団のミエリン形成能が示唆された(下のパネル)。
【0180】
マウスはミエリン塩基性タンパク質を産生する能力を欠いているため、緻密なミエリンは栄養作用ではなく、投与したオリゴデンドロサイトによって直接産生されるに違いない。
【0181】
実施例5:ES由来オリゴデンドロサイトによる脊髄機能の回復
ES由来オリゴデンドロサイト系譜細胞が神経活性を回復する能力を、脊髄損傷のラット挫傷モデルで判定した。
【0182】
衝撃装置を用いて、麻酔した成体スプラーグ・ド−リーラットの低位胸髄に中程度の挫傷を誘導した。損傷から1週間後、4μL培地中の細胞2.5 x 105個を挫傷部位に移植した。分化計画(実施例1)の35日目の細胞を、トリプシンおよびEDTAに3分間曝露して細胞を非接着性にすることにより、移植細胞集団を調製した。次に細胞を基礎培地ですすぎ、μl当たり細胞6 x 104個に濃縮した。
【0183】
損傷から6週間後、順行性トレーサーBDA(ビオチン化デキストランアミン)を動物の運動皮質に注射した。腰椎にエメラルドグリーンを注射し、腰椎路を標識した。8週目に実験を終了した時点で、これらのマーカーの位置を判定した。
【0184】
図22は、本実験の2群それぞれについての地上歩行スコアを示す。上のグラフは、200キロダイン挫傷を受けた動物の結果を示す。下のグラフは、250キロダイン挫傷の結果を示す。BBB尺度は、関節運動、体重支持、四肢の協調、足の配置、および歩行安定性に基づく正常な地上歩行を評価する21ポイント尺度である(Basso、Beattie、およびBresnahan、J. Neurotrauma、12:1、1995)。(■)損傷から1週間後にES由来オリゴデンドロサイトで処置した動物(n=5);(▲)同じ挫傷を被るが細胞を投与していない対照動物(n=3)。盲検で評価した平均値±SEM。
【0185】
hES由来オリゴデンドロサイトを移植した動物は、処置後5週間を超えて持続する有意に優れた機能を示した(*≡p<0.05)。
【0186】
これらの結果から、ES由来オリゴデンドロサイトが脊髄損傷の動物モデルにおいて機能の回復を助けることが実証される。先の実施例から、移植したオリゴデンドロサイトにより神経発芽およびミエリン欠損軸索の再ミエリン化が起こることが示される。これらの効果のいずれかまたは両方が、ここで認められた動作の改善に寄与する可能性がある。
【0187】
本発明のある種の適応は日常的な最適化の問題であり、本発明の精神または添付の特許請求の範囲から逸脱することなく実施可能であることは自明である。
【技術分野】
【0001】
技術分野
本発明は、一般に、胚細胞および神経系前駆細胞の細胞生物学の分野に関する。より具体的には、本発明は、生物学的研究、薬剤スクリーニング、およびヒトの治療に使用するのに適した、オリゴデンドロサイトおよびその前駆細胞の濃縮された集団を提供する。
【0002】
関連出願
本出願は、2002年7月11日に出願された米国仮出願第60/395,382号、および2003年4月4日に出願された米国実用出願第10/406,817号の利益を主張する。優先権出願は、本明細書によって完全に参照として本明細書に組み入れられる。
【背景技術】
【0003】
背景
オリゴデンドロサイトは、中枢神経系の支持において重要な生理的役割を担う。ヒトの治療にオリゴデンドロサイトが利用できれば、神経細胞を絶縁するミエリン鞘の欠陥によって生じる障害状態の治癒が促進され得る。
【0004】
多発性硬化症は、脳および脊髄の神経細胞を囲むミエリン鞘のゆっくりとした破壊を伴う、進行性かつ障害性の脱髄疾患である。症状は、しびれ感、視覚障害、および認知変化から麻痺に及ぶ。この疾患は免疫学的および遺伝学的要素を有すると考えられており、20歳から40歳の間で臨床型に現れる場合が多い。米国だけでも約300,000人がこの疾患に冒されている。治療法は、現在のところβインターフェロンまたは副腎皮質ステロイドの過程を伴う。これらの薬剤により発病の間の症状を伴う期間が短縮され得るが、一般に長期の障害は妨げられない。
【0005】
脊髄の外傷により外傷部位の近傍で無傷の軸索の脱髄が起こり、それによって軸索の神経伝達能力が奪われる。米国では、毎年約11,000件の脊髄損傷の新たな症例がある。SCI情報ネットワークでは、任意のレベルでの不完全な運動機能に苦しむ患者にかかる生涯直接費は、減給分および生活の質への影響を含めずに、$400,000〜$2,200,000に及ぶと推定している。
【0006】
中枢神経系の細胞上のミエリンは、軸索をその近傍で覆いミエリン鞘を形成するオリゴデンドロサイトによって正しい位置に整えられている。疾患状態におけるオリゴデンドロサイトおよびその前駆細胞の役割は、KeirsteadおよびBlakemore(Adv. Exp. Med. Biol. 468:183、1999)によって概説されている。O−2A細胞として知られているオリゴデンドロサイト前駆細胞は、健常成人のCNSおよび多発性硬化症の病変に存在し、再ミエリン化に関与する(Scoldingら、Brain 121:2221、1998;およびScoldingら、Neuroscience 89:1、1999)。オリゴデンドロサイトが均整をとって増殖し、損傷が大きい箇所では蓄積された前駆細胞を使い果たしてしまうため、適切な再ミエリン化ができないということが起こり得る。
【0007】
神経機能を回復するための再生医療で用い得る細胞集団を作製することを目的として、かなりの研究が行われてきた(Parkら、J. Neurotrauma 16:675、1999によって概説されている)。Keirsteadら(J. Neuroscience 19:7529、1999)は、出生後のラット脳から、移植後にオリゴデンドロサイトおよびシュワン細胞を生じるCNS前駆細胞を単離した。Svendsenら(J. Neurosci. Meth. 85:141、1998)は、発達過程のヒト皮質から前駆細胞を単離した。Mujtabaら(Dev. Biol. 214:113、1999)は、胚性幹細胞からの神経系前駆細胞の単離について報告している。
【0008】
国際公開公報第97/07200号(スタンフォード大学)は、成体ラット脳から単離したオリゴデンドロサイトの培養を示している。国際公開公報第01/28342号(ワシントン大学)は、あらかじめ馴化したオリゴデンドロサイト培地中で神経細胞を培養する特定の方法を提案している。米国特許第5,753,506号(Johe、CNS Stem Cell Technology)は、ニューロン、アストロサイト、およびオリゴデンドロサイトに分化する能力を有する、神経組織から単離した幹細胞を維持する培養系に関する。米国特許第6,238,922号(Stem Cells Inc.)は、神経組織の、ニューロンおよびグリアに分化する能力を有する細胞への解離を提案している。米国特許第6,235,527号(Raoら、ユタ大学)は、哺乳動物神経管組織から解離し、A2B5細胞表面マーカーに基づいて選択した、哺乳動物CNSグリア限定前駆細胞の集団に関する。
【0009】
米国特許第5,968,829号(Cytotherapeutics)は、ニューロン、アストロサイト、およびオリゴデンドロサイトを産生する能力を有するCNS神経幹細胞を含む培養液を請求している。国際公開公報第97/32608号は、移植を介してCNSに送達するための、遺伝子操作した初代オリゴデンドロサイトに関する。米国特許第5,830,621号(Signal Pharmaceuticals)は、アクセッション番号CRL 11881でATCCに寄託したヒトオリゴデンドロサイト細胞株について記載している。この株は、特徴的なマーカーGFAP、GalC、O4、およびA2B5を本質的に含まない。
【0010】
残念なことに、神経組織から単離した前駆細胞がヒトの臨床治療に必要な細胞数を産生するのに十分な複製能力を有するのかどうかは、いまだ明らかではない。
【0011】
別の供給源は、初期の胚組織から単離される多能性細胞である。胚性幹(ES)細胞は、25年以上前に初めてマウス胚から単離された(G.R. Martin、Proc. Natl. Acad. Sci. U.S.A. 78:7634、1981)。ES細胞は、同じ種の実質的に任意の組織種の子孫を生じ得ると考えられている。
【0012】
Fraichardら(J. Cell Sci. 108:3181、1995)は、マウスES細胞のグリア細胞および機能的なニューロンへのインビトロ分化について報告している。Mujtabaら(Dev. Biol. 214:113、1999)は、マウスES細胞からの神経前駆細胞の単離について報告している。Li、Smithら(Cur. Biol. 8:971、 1998)は、細胞系譜選択によるマウスES細胞からのニューロン前駆細胞の産生について報告している。Brustle、Mckayら(Proc. Natl. Acad. Sci. USA 94:14809、1997;Science 287:754、1999)は、有髄化移植片の潜在的な供給源として、マウスES細胞に由来するグリア前駆細胞について報告している。McDonaldら(Nat. Med 5:1410、1999;Proc. Natl. Acad. Sci. USA 97:6126、2000)は、培養においておよび脊髄移植後に、マウスES細胞がオリゴデンドロサイトを形成し、有髄化することを報告している。
【0013】
ヒトES細胞は、ごく最近になって初めて単離された(Thomsonら、Science 282:114、1998)。ヒトES細胞は、未分化状態で維持するためまたは特定の分化経路に沿って導くために、非常に異なる条件を必要とする(米国特許第6,090,622号および第6,200,806号;国際公開公報第99/20741号および国際公開公報第01/51616号)。このため、ヒトES細胞から分化細胞の比較的均一な集団をいかにして調製するかについては、ほとんどわかっていない。
【0014】
国際公開公報第01/88104号(Carpenter、Geron Corporation)は、ヒトES細胞を分化させることによって得られる神経系前駆細胞集団について記載している。90% NCAM陽性、35%βチューブリン陽性、および75% A2B5陽性を超える集団が得られた。Zhangら(Nature Biotech. 19:1129、2001)は、ヒトES細胞からの移植可能な神経系前駆細胞の分化について報告している。国際特許出願PCT/US02/19477(Carpenterら、Geron Corporation)は、生じた集団内の少なくとも10%のMAP−2陽性細胞がドーパミン作動性ニューロンのマーカーであるチロシン水酸化酵素を発現する、ES細胞由来の神経系細胞集団について記載している。
【0015】
最近になって、Billonら(J. Cell Sci. 115:3657、2002)は、遺伝子操作したマウスES細胞からのオリゴデンドロサイト発生のタイミングについて記載している。Kuoら(Biol. Reprod. Dec 11/02)は、28% GFAP陽性であるサルES由来細胞の集団について記載しており;Xianら(Stem Cells 21:41、2003)は、細胞系譜特異的転写因子Olig2を用いた、マウスES細胞からのオリゴデンドロサイトの作製について報告している。
【0016】
ヒトの健康および疾患の管理におけるpPS細胞の全般的な可能性を理解するため、脱髄状態の治療に有用な細胞の濃縮された集団を作製する新しいパラダイムを開発することが必要である。
【発明の概要】
【0017】
概要
本発明は、研究および薬学的組成物の調製に用いるための、グリア系譜の霊長動物細胞を効率的に産生する系を提供する。
【0018】
本発明の分化細胞集団は単離されまたはインビトロで培養され、グリア細胞または神経組織を有髄化し得る細胞の特徴に関して高度に濃縮されている。この細胞は、オリゴデンドロサイトの形態的特徴、本開示で後に記載する特定の抗体で検出可能なもしくは増幅可能なマーカーの発現、またはさらなる分化においてオリゴデンドロサイトを形成する能力を有し得る。この細胞はまた、同時培養アッセイにおいて神経節を有髄化する能力、インビボにおいて脱髄した軸索に対してミエリンを回復する能力、またはヒトもしくは非ヒト動物において神経機能を改善する能力等の、オリゴデンドロサイトの特定の機能的特徴を有し得る。これらの特徴の1つ、2つ、3つ、またはそれ以上が任意の組み合わせで存在し得る。
【0019】
この細胞集団は、あまり分化していない様々な種類の幹細胞から調製することができる。起源となり得る細胞には、胚盤胞由来の霊長動物多能性幹(pPS)細胞(胚性幹細胞に例証される)または初期胚の胚組織が含まれる。したがって、細胞はその起源の元となった組織の子孫であるという特徴を有することになり、このことは起源細胞と分化細胞が同じゲノムを有することを示すことによって確認され得る。
【0020】
本発明の他の態様は、すでに記載した分化細胞を作製または維持する方法に関する。この方法は、本開示において後に説明するような1つまたは複数の成長因子または分化因子の存在下で、多分化能幹細胞または多分化能性幹細胞を培養する段階を含む。
【0021】
例証として、線維芽細胞成長因子(FGF)等のマイトジェンと共にまたはそれなしで、トリヨードチロニン(T3)、セレン、またはレチノイン酸等の1つまたは複数の分化因子を含む培地中で幹細胞を培養することができる。分化細胞の最初の形成は、オリゴデンドロサイト系譜細胞が比較的均一な球体を形成し得る懸濁培養において起こり得る。他の細胞型は、所望の細胞型に選択的に付着する表面上に培養物をプレーティングする等の適切な分離手段によって、除去することができる。任意で、多くは細胞の最初の誘導に用いたような1つまたは複数の分化因子の存在下で、FGFまたは上皮成長因子等のマイトジェンと共に培養することによる、選択の前または後に分化細胞をさらに増殖させることができる。次に、任意でマイトジェンの非存在下で、または後期分化を亢進するポリ−L−リジン等の表面の存在下で培養することにより、細胞をさらに成熟させることができる。
【0022】
本発明の細胞は、多くの商業上重要な用途に用いることができる。例えば、この細胞は、その存在が細胞の維持、毒性、さらなる分化、またはグリア細胞として機能する能力に関連する化合物を、グリア細胞に及ぼす影響についてスクリーニングするのに適している。この細胞はまた、インビトロおよびインビボにおいて隣接する神経組織の髄鞘形成をもたらすのにも適している。
【0023】
ヒトまたは動物治療用の薬学的組成物の調製において、本発明の細胞を用いることも可能である。次にこの組成物を用いて、軸索の髄鞘形成障害に関連するような病態または脊髄損傷を治療することができる。
本発明のさらなる局面は、以下の記載により明らかになると考えられる。
【図面の簡単な説明】
【0024】
【図1】塩基性FGFおよびオリゴデンドロサイト分化因子を含む培地中、懸濁状態で2日間培養したヒト胚性幹(ES)細胞を示す、倒立顕微鏡を用いて撮影した位相差像である。
【図2】レチノイン酸と共に7日間培養した後の細胞を示す。培養物中の80〜90%を示す、細胞の大きな透明な球体が存在した。
【図3】レチノイン酸を除去し、細胞を低濃度のFGFの存在下で維持した後に何が起こるかを示す。
【図4】bFGFを除去した後に現れる変化を示す。大きな凝集塊が解離し、培養液全体が単一細胞および小さな細胞塊でいっぱいになった。同時に、新たに鮮黄色の球体が観察された(矢印)。
【図5】細胞をFGFの非存在下で上皮成長因子と共に培養した場合に成長する鮮黄色の球体(矢印)を示す。
【図6】マトリゲル(登録商標)上にプレーティングしてから2日または3日後の、球体からのグリアに拘束された神経前駆細胞の遊走および分枝を示す。
【図7】10〜20時間だけ細胞をマトリゲル(登録商標)に接着させることにより、オリゴデンドロサイト系譜細胞が選択されたという結果を示す。非接着細胞を除去したが、これによりオリゴデンドロサイトマーカーをもたない細胞は実質的にすべて除去される。次に接着細胞を再懸濁して、FGF、EGF、およびグリア前駆細胞培地の存在下で7日間拡大した。これは、治療的投与および他の目的に対してより適したより分散した細胞集団を産生する利点を有する。この顕微鏡写真は、ヘマトキシンを用いて細胞核を対比染色した(黒矢印)、ガラクトセレブロシドに対する染色(GalC;白矢印)を示す。この条件下でGalCに染色された細胞の割合は、少なくとも約95%であった。
【図8】より高い倍率のES由来オリゴデンドロサイトを示す。細胞は、成熟オリゴデンドロサイトに特有の形態的特徴(間にミエリン皮膜を有するように見える多数の複雑な突起)を有する。
【図9】分化過程における細胞形態の進行を示す。(A): 未分化hES細胞。(B): レチノイン酸を含む懸濁培養において胚様体から成長する透明な球体。(C): EGFの存在下における、オリゴデンドロサイト前駆細胞を含む黄色球体の拡大。 (D): マトリゲル(登録商標)上にプレーティングすることによりポジティブ選択されたオリゴデンドロサイト系譜細胞。(E、F): 培養におけるオリゴデンドロサイト前駆細胞の隆起の増加。(G、H): 次に続くプレーティングによって、成熟オリゴデンドロサイトへのさらなる分化が起こる。
【図10】成熟オリゴデンドロサイトの免疫細胞化学的解析を示す。プレーティングしてから1週間後、細胞は初期グリア細胞のマーカーNG2に関して陽性である(A)。プレーティングしてから8週間後、細胞は、GalC(C)、O4(D)、およびRIP(E)に関して陽性である。パネルBは、示したマーカーで染色された細胞の数を示す。
【図11】ヒト核タンパク質に特異的な抗体で染色した、ラットの脊髄に投与したオリゴデンドロサイト前駆細胞の組織切片を示す
【図12】移植してから9週間後、細胞が白質に向けて遊走または増殖したことを示す。
【図13】移植したオリゴデンドロサイトが、脊髄の断面積として測定される、挫傷後の二次拡大に起因する損傷を悪化させないことを示す。
【図14】hES細胞が軸索分枝を誘導することを示す。オリゴデンドロサイトを移植した動物の像では(上のパネル)、BDA染色の濃い細い線として新たな軸索が示される。未処置の動物では、分枝は認められなかった。移植した細胞は、再生可塑性を誘導する
【図15】損傷部位(中心点)からのニューロン分枝の定量を示す(ブロック当たり3切片の平均値±SEM)。処置した動物は、有意に高いレベルで、中心点の損傷のすぐ頭側にまで標識軸索を有した。
【図16】hES由来オリゴデンドロサイトを移植した動物における、実質的な再ミエリン化の証拠を示す。上段の電子顕微鏡写真における厚い環は、正常にミエリン化した繊維である。視野内の残りの軸索は、薄い層のミエリンを示す。右上のパネルの軸索は、約5層または6層の外被、および進行中の再ミエリン化の証拠を示す。移植した動物のみが、オリゴデンドロサイト活性に起因する新たなミエリンを示した。これにより、移植動物における動作の改善を説明する手段が提供される。
【図17】未分化hES細胞コロニーで検出されたマーカーを示す(左側:抗体+DAPI;右側:抗体染色のみ)。上列は、多能性細胞のマーカーであるSSEA−4に標識されたコロニーを示す。下列は、中胚葉マーカーBMP4に標識されたコロニーを囲む間質細胞を示す。
【図18】分化過程における転写因子Pax6の一過性の出現を示す。上列は、10日目の細胞塊の中心に向かった染色、周辺に向かってより分化した細胞でのすでにダウンレギュレートされた染色を示す。下列は、分化35日目では実質的に染色されないことを示す。
【図19】分化10日目に存在する初期のオリゴデンドロサイト系譜細胞で検出されるマーカーを示す。上列:転写因子Olig1。中列:転写因子SOX10。下列:オリゴデンドロサイト前駆細胞マーカーA2B5。
【図20A】35日目の移植に適したオリゴ前駆細胞において優位を占めるマーカーを示す。1列目および2列目(20A):NG2(コンドロイチン硫酸プロテオグリカン、オリゴデンドロサイト前駆細胞のマーカー); 3列目(20A):GalC;4列目(20B):O4;5列目(20B):Tuj1(ニューロンのマーカー)。実質的にすべての細胞がオリゴデンドログリアマーカーを有したが、神経細胞、間葉細胞、または未分化hES細胞のマーカーは有さない(20B、下のグラフ)。
【図20B】35日目の移植に適したオリゴ前駆細胞において優位を占めるマーカーを示す。1列目および2列目(20A):NG2(コンドロイチン硫酸プロテオグリカン、オリゴデンドロサイト前駆細胞のマーカー); 3列目(20A):GalC;4列目(20B):O4;5列目(20B):Tuj1(ニューロンのマーカー)。実質的にすべての細胞がオリゴデンドログリアマーカーを有したが、神経細胞、間葉細胞、または未分化hES細胞のマーカーは有さない(20B、下のグラフ)。
【図21】ミエリン塩基性タンパク質を作り出す能力が遺伝的に欠損しているシバラーマウスにオリゴデンドロサイトを投与した結果を示す。超微細構造的に、シバラーマウスの軸索はミエリンを欠いているか、または1もしくは2層の緻密でないミエリンによって囲まれている(上のパネル)。細胞を移植してから6週間後、電子顕微鏡解析により多層の緻密なミエリンが示され、このことから移植集団のミエリン形成能が示唆された(下のパネル)。緻密なミエリンは、投与したオリゴデンドロサイトによって直接産生されている。
【図22】衝撃装置を用いてラットの脊髄を挫傷させ、BBB尺度での定量によりその後の期間の脊髄機能について評価する実験の結果を示す。上のグラフ:200キロダイン挫傷;下のグラフ:250キロダイン挫傷。 (■)損傷から1週間後にES由来オリゴデンドロサイトで処置した動物(n=5);(▲)同じ挫傷を被るが細胞を投与していない対照動物(n=3)。盲検で評価した平均値±SEM。hES由来オリゴデンドロサイトを移植した動物は、治療後5週間を超えて持続する有意に優れた地上歩行を示した。
【発明を実施するための形態】
【0025】
詳細な説明
本発明は、多能性幹細胞からオリゴデンドロサイトおよびその前駆細胞を効率的に産生することにより、それらの細胞の大きな集団を作製する問題を解決する。
【0026】
分化細胞集団は著しく均一である。図7は、オリゴデンドロサイトに特有である形態学的特徴を有するヒト細胞の例示的な調製を示す。集団を、オリゴデンドロサイト系譜の細胞のマーカーであるGalCについて染色した。図8はより高い倍率の視野であり、近傍の任意のニューロンを有髄化しようとしているミエリンのシートの生成を示している。これらの細胞の機能的特徴から、これらの細胞はオリゴデンドロサイトの性質のさらなる特徴付けおよびヒトの治療での使用によく適している。
【0027】
本発明の細胞の理想的な供給源は、様々な種類の霊長動物多能性幹(pPS)細胞である。以下に記載する戦略に従っていくつもの適切な培養条件および補因子の中から選択することにより、pPS細胞をオリゴデンドロサイト経路に導出することができる。未分化pPS細胞のオリゴデンドロサイトへの変換は、最適条件下で25%程度の効率で起こり得ることが明らかになった。
【0028】
本発明の組成物および方法により、これまでに利用可能であった技術を超える重要な利点が提供される。所望のアロタイプのpPS細胞からオリゴデンドロサイトを分化させることにより、任意の組織適合性を有するオリゴデンドロサイトを産生することができる。必要に応じて、移植を亢進する任意の方法で、分化の前または後に細胞を遺伝子改変することができる。細胞は天然細胞株に由来するが継続的な組織解剖を必要としないため、規制認可の品質管理要求事項を満たすのに最適である。
【0029】
重要なことには、各出発幹細胞集団から産生され得る細胞の供給量はほぼ無限である。実施例1に説明するように、ひとたびオリゴデンドロサイト系譜細胞を産生かつ選択すれば、成長因子の存在下で培養することによりその細胞を大幅に拡大することができる。さらに、起源pPS細胞をほぼ無期限に増殖させて、それによってより分化した細胞の供給源を連続的に提供することができる。
【0030】
以下の開示により、本発明の分化細胞をいかにして作製および使用するかのさらなる説明を提供する。集団は著しく均一であり、したがって多くの商業的に重要な用途における使用に適している。
【0031】
本発明の1つの局面により、少なくとも約80%の細胞が、霊長動物多能性幹(pPS)細胞の子孫であり;NG2プロテオグリカン(または他のオリゴデンドロサイトマーカー)に特異的な抗体で染色され;かつNeuN(または神経細胞および他の潜在的混入物の他のマーカー)に関して陰性である、分化細胞集団を提供する。分化細胞集団は、オリゴデンドロサイト系譜細胞を産生するのに適した成分のシステムの一部であってよい。このシステムは、分化細胞が産生される元ととなったpPS細胞(ヒト胚性幹細胞等)の系統をさらに含み得る。任意で、分化細胞集団の少なくとも80%の細胞が、A2B5またはPDGFRαも発現する。細胞の少なくとも20%が、オリゴデンドロサイト前駆細胞に特有である双極性形態を示す可能性がある。
【0032】
ある状況下では、シバラー変異(shiverer mutant)マウスの脊髄に分化細胞集団を移植した後、その集団により神経軸索の周りに緻密なミエリンの沈着が起こる;または挫傷ラットの脊髄内もしくは脊髄の周りに集団を移植した後、地上歩行が改善される。あるいは、本発明のオリゴデンドロサイト前駆細胞集団を(例えば、マイトジェンの非存在下でポリ−L−リジンおよびラミニン上で3日間培養することにより)インビトロでさらに分化させてもよい。これにより、少なくとも10%の細胞が成熟オリゴデンドロサイトに特有である複雑な突起を有し;かつ約80%、90%、95%、またはそれ以上がGalC等の成熟オリゴデンドロサイトマーカーを有する、より成熟した集団が産生され得る。増殖段階中に産生されたオリゴデンドロサイト系譜細胞は、前駆細胞段階で発現しているマーカーばかりでなく、成熟細胞の濃縮された集団を産生する能力またはインビボで所望の機能を遂行する能力によっても特徴付けることができる。
【0033】
以下に説明するように、本発明のいくつかの分化細胞集団は、マイトジェンおよび少なくとも2つのオリゴデンドロサイト分化因子を含む培地中で未分化pPS細胞を培養する過程により得ることができる。例えば、細胞凝集体を形成させるために、塩基性線維芽細胞成長因子(FGF)、トリヨードチロニン(T3)、レチノイン酸、およびおそらくセレンの存在下で、未分化pPS細胞を懸濁状態で培養してもよい。
【0034】
本発明のさらなる説明および例証を以下の項で提供する。
【0035】
定義
オリゴデンドロサイトとは、中枢神経系の外膜構造(神経膠)の一部を形成する外胚葉起源の神経系細胞である。この細胞は、個々の軸索の周りを覆いCNSのミエリン鞘を形成する、不定数のベール様またはシート様突起を有する。この細胞は、本開示で後に説明する形態学的基準、表現型基準、または機能的基準によって同定することができる。
【0036】
「神経系前駆細胞」または「神経上皮幹細胞」とは、神経細胞(ニューロン前駆細胞もしくは成熟ニューロン等)またはグリア細胞のいずれかである子孫を産生し得る細胞である。「グリア細胞」という用語は、成熟オリゴデンドロサイト、アストロサイト、およびこれらの細胞のいずれかまたは両方に拘束された前駆細胞を包含する。
【0037】
「オリゴデンドロサイト前駆細胞」は、ニューロンまたは非神経組織に優先して、成熟オリゴデンドロサイトおよび/またはさらなる前駆細胞を含む子孫を形成するように拘束された神経系細胞である。特記しない限りこの細胞は、必ずしも必要ではないが、アストロサイト等の他の種類のグリア細胞を産生する能力を有してもよい。特記しない限り、本開示での「オリゴデンドロサイト」または「オリゴデンドロサイト系譜細胞」への言及は、オリゴデンドロサイト前駆細胞および成熟細胞の両方を指す。
【0038】
細胞の個体発生との関連では、形容詞「分化した」とは相対語である。「分化細胞」とは、比較する細胞よりも発生経路をさらに下方に進行した細胞のことである。したがって、多能性胚性幹細胞は、上記の様々な種類の前駆細胞等の系譜を限定した前駆細胞に分化することが可能である。これらの細胞は、次に経路のさらに下流にある細胞、または成熟オリゴデンドロサイト等の最終段階の分化細胞にさらに分化することが可能である。
【0039】
本開示で用いる「分化剤」とは、オリゴデンドロサイト系譜の分化細胞(前駆細胞および最終分化細胞を含む)を産生するために、本発明の培養系で用いる一群の化合物の1つを指す。化合物の作用形態に関しては限定しない。例えば、薬剤は、表現型の変化を誘導もしくは補助することにより、特定の表現型を有する細胞の成長を促進することによりもしくは他の細胞の成長を遅らせることにより、または他の薬剤と合わせて未知の機構で作用することにより、分化過程を補助する可能性がある。
【0040】
特記しない限り、本開示における「セレン」への言及は、溶液中で任意の対イオンを伴う亜セレン酸(SeO32−)、セレン酸(SeO42−)、またはセレン化物(Se2−)を含む、Seの任意の酸化型を指す。
【0041】
原型の「霊長動物多能性幹細胞」(pPS細胞)とは、前胚組織、胚組織、または受精後の任意の時期の胎児組織に由来する多能性細胞であり、8〜12週齢のSCIDマウスで奇形腫を形成する能力等の標準技術として公認の試験によると、適切な条件下で3つの胚葉(内胚葉、中胚葉、および外胚葉)すべての派生物である様々な異なった細胞型の子孫を産生し得る特徴を有する。pPS細胞の定義には、ヒト胚性幹(hES)細胞およびヒト胚性生殖(hEG)細胞に例証される様々な種類の胚細胞が含まれる。pPS細胞は、悪性供給源に由来しないことが好ましい。細胞は正倍数体であることが望ましい(が必ずしも必要ではない)。その供給源および培養法に依存して、pPS細胞は、人体のすべての異なる細胞型に発達する能力を有するという意味で、全能性であってもなくてもよい。
【0042】
pPS細胞培養物は、集団中の幹細胞およびその派生物の実質的な割合が、胚起源または成体起源の分化細胞と区別できる未分化細胞の形態学的特徴を示す場合に、「未分化である」と記載される。集団内の未分化細胞のコロニーは、分化した隣接細胞に囲まれていることが多いと理解されている。
【0043】
「フィーダー細胞」または「フィーダー」とは、別の種類の細胞と共培養され、第2の種類の細胞が成長できる環境を提供する1つの種類の細胞を表すのに用いられる用語である。pPS細胞集団は、分割後にpPSの成長を支持するために新鮮なフィーダー細胞を添加せずに少なくとも1回成長した場合、フィーダー細胞を「本質的に含まない」と称される。
【0044】
「胚様体」という用語は、pPS細胞が単層培養において過剰成長する場合、または懸濁培養で維持される場合に現れる、分化および未分化細胞の凝集体を指す。胚様体は、形態学的基準および免疫細胞化学法により検出可能な細胞マーカーによって識別可能ないくつかの胚葉に典型的に由来する、様々な細胞型の混合物である。
【0045】
「成長環境」とは、関心対象の細胞がインビトロにおいて増殖、分化、または成熟する環境のことである。環境の特徴には、細胞を培養する培地、存在し得る任意の成長因子または分化誘導因子、および存在する場合には支持構造(例えば、固体表面上の基層)が含まれる。
【0046】
特記しない限り、個々の細胞および細胞集団における表現型マーカーの判定では、細胞は、アイソタイプ対照と比較して、二次免疫細胞化学染色において特異的抗体を用いて実質的により高い染色を示した場合にマーカーに関して「陽性」であると称される。特記しない限り細胞は、この種の免疫細胞化学的解析によりマーカーが抗体で検出不可能である場合に、「陰性」であると称される。
【0047】
任意の適切な人工的操作手段によりポリヌクレオチドが細胞に導入された場合、または細胞がそのポリヌクレオチドを受け継いだ最初に改変された細胞の子孫である場合、その細胞は「遺伝子改変された」、「トランスフェクションされた」、または「 遺伝的に形質転換された」と称される。ポリヌクレオチドは関心対象のタンパク質をコードする転写可能な配列を含む場合が多く、これにより細胞はこのタンパク質を高いレベルで発現することが可能になる。改変した細胞の子孫が同じ改変を有する場合、遺伝的改変は「遺伝性である」と称される。
【0048】
一般的技術
分子遺伝学および遺伝子工学における一般的な方法は、Molecular Cloning: A Laboratory Manual (Sambrookら、Cold Spring Harbor);Gene Transfer Vectors for Mammalian Cells (MillerおよびCalos編);およびCurrent Protocols in Molecular Biology(F.M. Ausubelら編、Wiley & Sons) の現行版に記載されている。分子生物学、タンパク質化学、および抗体技法は、Current Protocols in Protein Science(J.E. Colliganら編、Wiley & Sons);Current Protocols in Cell Biology(J.S. Bonifacinoら、Wiley & Sons)、およびCurrent Protocols in Immunology(J.E. Colliganら編、Wiley & Sons)に見出すことができる。本開示で引用する遺伝子操作用の試薬、クローニングベクター、およびキットは、BioRad、Stratagene、Invitrogen、ClonTech、およびSigma−Aldrich Co.等の市販業者から入手可能である。
【0049】
細胞培養法は、一般に、Culture of Animal Cells: A Manual of Basic Technique(R.I. Freshney編、Wiley & Sons);General Techniques of Cell Culture(M.A. HarrisonおよびI.F. Rae、Cambridge Univ. Press)、およびEmbryonic Stem Cells: Methods and Protocols(K. Turksen編、Humana Press)の現行版に記載されている。組織培養用の供給品および試薬は、Gibco/BRL、Nalgene−Nunc International、Sigma Chemical Co.、およびICN Biomedicals等の市販業者から入手可能である。
【0050】
本開示に関連する専門的な参考文献には、Principles of Neuroscience、第4版、Kandelら編、McGraw−Hill 2000;CNS Regeneration: Basic Science and Clinical Advances、M.H. TuszynskiおよびJ.H. Kordower編、Academic Press、1999;The Neuron: Cell and Molecular Biology、第3版、I.B. LevitanおよびL.K. Kaczmarek、Oxford U. Press、2001;Glial Cells: Their Role in Behaviour、P.R. Lamingら編、Cambridge U. Press、1998;The Functional Roles of Glial Cells in Health and Disease、MatsasおよびTsacopoulos編、Plenum Pub. Corp、1999;Glial Cell Development、JwessenおよびRichardson編、Oxford U. Press 2001;およびMan of Steel、Adrian Havill、1996が含まれる。
【0051】
幹細胞の供給源
本発明は、様々な種類の幹細胞を用いて実施することができる。本発明の使用に特に適しているのは、胚盤胞等の妊娠後に形成される組織または妊娠期間の任意の時期に採取した胎児組織もしくは胚組織等に由来する霊長動物多能性幹(pPS)細胞である。限定されない例は、以下に記載するような胚性幹細胞もしくは胚性生殖細胞の初代培養または樹立株である。本発明の技法を初代胚組織または胎児組織に直接実行し、最初に未分化細胞株を樹立することなく初代胚性細胞から神経系細胞を直接導出することも可能である。
【0052】
実施例の項に提供する例証は、ヒト胚性幹細胞を用いて行った研究の結果である。しかし、特記しない限り、本発明は任意の脊椎動物種の幹細胞を用いて実施することが可能である。これには、ヒト、ならびに非ヒト霊長動物および他の非ヒト哺乳動物の多能性幹細胞が含まれる。
【0053】
胚性幹細胞
胚性幹細胞は、霊長動物組織から単離することができる(米国特許第5,843,780号;Thomsonら、Proc. Natl. Acad. Sci. USA 92:7844、1995)。ヒト胚性幹(hES)細胞は、Thomsonら(米国特許第6,200,806号;Science 282:1145、1998;Curr. Top. Dev. Biol. 38:133ページ以降、1998)およびReubinoffら、Nature Biotech. 18:399、2000の記載する技法により、ヒト割球から調製することが可能である。hES細胞に相当する細胞型には、国際公開公報第01/51610号(Bresagen)に概説されるような、原始外胚葉様(EPL)細胞等のその多能性派生物が含まれる。
【0054】
1つの方法では、プロナーゼ(Sigma)に短時間曝露することにより、発生した胚盤胞から透明帯を除去する。1:50希釈したウサギ抗ヒト脾臓細胞抗血清に胚盤胞を30分間曝露し、次にDMEMで5分間3回洗浄し、1:5希釈したモルモット補体(Gibco)に3分間曝露する免疫手術により、内部細胞塊を単離する(Solterら、Proc. Natl. Acad. Sci. USA 72:5099、1975)。DMEMでさらに2回洗浄した後、穏やかにピペッティングして無傷の内部細胞塊(ICM)から溶解した栄養外胚葉細胞を除去し、ICMをmEFフィーダー層上にプレーティングする。
【0055】
9〜15日後、1 mM EDTAを添加したカルシウムおよびマグネシウムを含まないリン酸緩衝食塩水(PBS)に曝露することにより、ディスパーゼもしくはトリプシンに曝露することにより、またはマイクロピペットで機械的に解離することにより、内部細胞塊に由来する増殖物を凝集塊に解離し;次に新鮮な培地中でmEF上に再度プレーティングする。未分化の形態を有して増殖するコロニーをマイクロピペットで個別に選択し、機械的に凝集塊に解離し、再度プレーティングする。ES様の形態は、細胞質に対して核の比率が明らかに高くかつ顕著な核小体を有する小型のコロニーとして特徴付けられる。次に、得られたES細胞を短時間トリプシン処理してダルベッコPBS(2 mM EDTAを含む)に曝露し、IV型コラゲナーゼ(約200 U/mL;Gibco)に曝露することにより、またはマイクロピペットで個別にコロニーを選択することにより、1〜2週間ごとに日常的に分割する。凝集塊の大きさは、約50〜100細胞が最適である。
【0056】
胚性生殖細胞
ヒト胚性生殖(hEG)細胞は、Shamblottら、Proc. Natl. Acad. Sci. USA 95:13726、1998および米国特許第6,090,622号に記載されているように、始原生殖細胞から調製することができる。
【0057】
簡潔に説明すると、約8〜11週間後に採取した生殖隆起を等張緩衝液ですすぎ、0.05%トリプシン/0.53 mM EDTAナトリウム溶液(BRL)0.1 mL中に入れ、<1 mm3の塊に切断する。脱凝集した後、細胞をEG増殖培地(D−グルコース、NaHCO3;15% ES認定ウシ胎仔血清;2 mMグルタミン;1 mMピルビン酸ナトリウム;1000〜2000 U/mLヒト組換え白血病抑制因子;1〜2 ng/mL ヒト組換えbFGF;および10μMフォルスコリン(10% DMSO中)を含むDMEM)約3.5 mL中、37℃で1時間または一晩インキュベートする。
【0058】
次に細胞をEG増殖培地1〜3 mLに再懸濁し、フィーダー層上にプレーティングする(例えば、5000radのγ−照射で不活性化したSTO細胞、ATCC番号CRL 1503)。7〜10日後に1回目の継代を行い、EG細胞と一致する細胞形態が観察されるまで、毎日培地を交換して培養するが、典型的にこれは7〜30日後または1〜4継代後である。
【0059】
他の幹細胞
オリゴデンドロサイトを作製するための出発材料として有用なpPSまたは胚性幹細胞を産生するために、本発明の実施はヒト胚または胚盤胞を脱凝集することを決して必要としない。hES細胞は、公共の受託所(例えば、WiCell Research Institute、米国、ウィスコンシン州、マディソン、またはAmerican Type Culture Collection、米国、バージニア州、マナッサス)から入手可能な樹立株から得ることができる。米国特許公報第2003−0113910A1号は、胚または胎児組織を用いずに導出された多能性幹細胞について報告している。多能性表現型を誘導する因子を使用することにより、臍帯血または他の前駆細胞をpPS細胞に再プログラミングすることも可能であると考えられる(Chambersら、Cell 113:643、2003;Mitsuiら、Cell 113:631、2003)。適切な条件下において、別の方法でpPSまたはhES細胞の定義に見合う任意の細胞を、本発明によりオリゴデンドロサイト系譜細胞の導出に用いることができる。
【0060】
本開示で提供する技法のいくつかを用いて、外胚葉細胞、および胎児組織もしくは成体組織から得られた神経系細胞、または神系前駆細胞等のより拘束された細胞型の分化を維持または促進することも可能である。そのような細胞を得る方法は、例えば、米国特許第5,852,832号;第5,654,183号;第5,849,553号;および第5,968,829号;ならびに国際公開公報第98/50526号および国際公開公報第99/01159号に記載されている。
【0061】
未分化状態でのpPS細胞の増殖
pPS細胞は、分化を促進せずに増殖を促進する培養条件を用いて培養し、連続して増殖させることが可能である。血清を含む例示的なES培地は、80% DMEM(ノックアウトDMEM、Gibco等)、20%の既知組成ウシ胎仔血清(FBS、Hyclone)または血清代替品(国際公開公報第98/30679号)のいずれか、1%非必須アミノ酸、1 mM L−グルタミン、および0.1 mMβ−メルカプトエタノールから作製する。使用する直前に、ヒトbFGFを4 ng/mLになるように添加する(国際公開公報第99/20741号、Geron Corp.)
【0062】
分化を阻害する環境において培養することによってのみ、pPS細胞を未分化状態で拡大することができる。慣例的に、pPS細胞は、マウスの胚組織または胎児組織に由来するフィーダー細胞層上で培養する。増殖を阻害するがpPS細胞を支持する因子の合成はできるように照射した照射mEFを、培養プレートにウェル当たり375,000個プレーティングし、プレーティングしてから5時間〜4日後に使用する(米国特許第6,200,806号)。最近になって、ヒト胚性幹細胞の増殖を分化させることなく支持するヒトフィーダー細胞が開発された(国際公開公報第01/51616号:米国特許出願第09/888,309号;Geron Corp)。hES細胞を分化させ、所望の活性を有する細胞を選択し、次にテロメラーゼ逆転写酵素を発現するようにトランスフェクションして不死化することにより、細胞が得られる。
【0063】
pPS細胞は、フィーダー細胞なしでも未分化状態で維持することができる。無フィーダー培養の環境には、適切な培養基層、特にマトリゲル(Matrigel)(登録商標)またはラミニン等の細胞外マトリックスが含まれる。pPS細胞は、>15,000細胞cm−2(最適には90,000 cm−2〜170,000 cm−2)でプレーティングする。無フィーダー培養物は、分化させることなく細胞の増殖を支持する因子を含む栄養培地により支持される。そのような因子は、照射した(約4,000rad)初代マウス胚線維芽細胞、テロメル化(telomerize)したマウス線維芽細胞、またはpPS細胞由来のヒトフィーダー細胞等のそのような因子を分泌する細胞と共に培地を培養することにより、培地中に導入され得る。フィーダーを、20% 血清代替品および4〜8 ng/mL bFGFを添加したKO DMEM等の無血清培地で約5〜6 x 104 cm−2の密度でプレーティングすることにより、培地を馴化することができる。1〜2日間馴化した培地にさらにbFGFを添加し、これを用いてpPS細胞培養物を1〜2日間支持する。無フィーダー培養法の特徴は、国際公開公報第99/20741号および国際公開公報第01/51616号;ならびにXuら、Nat.Biotechnol. 19:971、2001においてさらに考察されている。
【0064】
ES細胞は顕微鏡下で、高い核/細胞質比、顕著な核小体、および細胞の接合部が識別困難な小型のコロニー形成を伴って見える。霊長動物ES細胞は典型的に、時期特異的胚抗原(SSEA)3および4、Tra−1−60およびTra−1−81と称する抗体を用いて検出できるマーカー(Thomsonら、Science 282:1145、1998)、ならびにテロメラーゼ活性を発現する。インビトロにおけるpPS細胞の分化により、SSEA−4、Tra−1−60、およびTra−1−81の発現が消失し、未分化hEG細胞においても認められるSSEA−1の発現が増加する。
【0065】
幹細胞からのオリゴデンドロサイトの作製
本発明のオリゴデンドロサイト系譜細胞は、所望の表現型を有する細胞を濃縮および拡大する特殊な成長環境において幹細胞を培養することによって得られる。成長環境によって、オリゴデンドロサイト系譜への分化が特異的に方向づけられ得る、所望の細胞の成長が促進され得る、他の細胞型の成長が阻害され得る、またはこれらの活性の任意の組み合わせが起こり得る。
【0066】
本項は、本発明のオリゴデンドロサイトを得るためにとり得るいくつかのアプローチを当業者に説明する指針である。別に規定する場合を除き、過程の根本的な機構に関する説明は、さらなる詳細を支援するための作業仮説としてのみ提供する。使用者が仮説を理解する必要はなく、また本発明を実施するためにこれを仮説に合わせる必要もない。多能性幹細胞からオリゴデンドロサイトを作製することができ、手順のさらなる適合化を行うことができ、本開示に記載する新規産物を得るために別法を見出すことが可能であることを、本出願者らがこれから実証する。
【0067】
pPS細胞からオリゴデンドロサイトを産生する段階には、a) オリゴデンドロサイトを産生するように拘束された細胞の集団を得る段階;b) オリゴデンドロサイト系譜細胞の拡大;およびc) 細胞の後期オリゴデンドロサイトへのさらなる成熟が含まれ得る。
【0068】
幹細胞のオリゴデンドロサイト系譜への方向付け
オリゴデンドロサイトを産生する過程は、典型的に2つの局面、起源幹細胞集団に分化を引き起こす段階;およびオリゴデンドロサイト系譜細胞を主要細胞型にする段階を含む。これらの事象は、経時的にまたは同時に起こり得る。
【0069】
例えば、ドナーpPS細胞培養物の過剰成長により、または胚様体の形成を可能にする低接着性特性を備えた基層を有する培養容器中でpPS細胞を懸濁状態で培養することにより、pPS細胞に胚様体または凝集体の形成を引き起こすことによって分化の過程を誘導することができる。例示的な方法では、hES細胞のコンフルエントな単層培養物を回収し、非接着性細胞培養プレートにプレーティングし、細胞を懸濁状態に保ち、定期的に栄養培地でフィードする。
【0070】
別法としてまたはさらに加えて、細胞が未分化表現型を維持するのを妨げる特定の因子と共に培養することによって、分化過程を開始することができる。最初の分化因子は分化をオリゴデンドロサイト系譜に限定する必要はないが、分化集団内の細胞型の範囲内にオリゴデンドロサイトまたはその前駆細胞を含めるべきである。この種の例示的な成長因子は、レチノイド受容体に結合する、または細胞外シグナル制御キナーゼ(ERK)経路を活性化するリガンドである。典型的に線維芽細胞成長因子(または次項に記載するマイトジェン)等のマイトジェンを培地に含めて、分化過程の進行中に増殖を促進する。
【0071】
ある段階で、培養物をより特異的にオリゴデンドロサイト系譜に方向づけることができる。これは、オリゴデンドロサイトの成長をより特異的に促進する因子を培養物に含めることによって行うことができる。例示的なオリゴデンドロサイト分化因子は、約40 ng/mlのT3(3,5,3’−トリヨード−L−チロニン)およびT4(L−チロキシン)によって例証される、細胞表面上または核内の甲状腺ホルモン受容体に結合するリガンドおよび抗体である。甲状腺ホルモンは、レチノイン酸受容体の発現を増加させる、さもなければオリゴデンドロサイト系譜の細胞への分化を促進すると考えられている。
【0072】
別のオリゴデンドロサイト分化因子は、分化しているオリゴデンドロサイトにおいてミエリン遺伝子の上方制御に関与すると考えられている抗酸化剤であるセレンである。他の候補分化因子は、ビタミンEのような他の抗酸化剤、ならびにチオレドキシン還元酵素およびヨードチロニン脱ヨウ素酵素のファミリー等のセレンが補因子である酵素の活性を増加させる因子である。セレンは、亜セレン酸(SeO32−)の形で少なくとも20 ng/mLまたは100 ng/mLの比較的高い濃度で培地に含めた場合に、特に効果的である。他の候補分化因子または補因子には、骨形成タンパク質(BMP)、ソニックヘッジホッグ(SHH)、および白血病抑制因子(LIF)が含まれる。神経組織によって産生されるオリゴデンドロサイト分化因子の組み合わせも意図するが、これは、胚性CD−1マウス脳および組織または細胞株から得られる因子含有抽出物によって例証される、選択された解離した神経組織または細胞株と共培養することによって産生され得る。
【0073】
これらの技法をすべて組み合わせることにより:マイトジェン、レチノイン酸等の一般的な分化因子、ならびに甲状腺ホルモンおよび亜セレン酸ナトリウムのような特異的分化因子の存在下で胚様体を形成させることによって分化を開始することにより、驚くほど効率的にオリゴデンドロサイトを産生できることを見出した。実施例の項に提供する限定されない例証には、これらの因子、ならびに栄養補充物、インスリン等の成長因子、および抗生物質等の他の標準的な培養成分が含まれる。分化の開始時に適切な組み合わせの分化因子を含めることによって、培養物はオリゴデンドロサイト系譜により拘束された細胞を迅速に産生することになる。
【0074】
オリゴデンドロサイト系譜細胞の拡大
所望の目的のために細胞を用いる以前の任意の段階として、本発明の使用者は、オリゴデンドロサイト系譜細胞を培養において増殖させることによりこの細胞の数を増やしてもよい。
【0075】
これは、1つまたは複数のマイトジェンの存在下で、細胞を栄養培地中で培養することによって達成できる。FGF−2(塩基性FGF)およびFGF−4等の線維芽細胞成長因子ファミリーのメンバーが例示的である。上皮成長因子(EGF)、機能的相同体、およびEGF受容体に結合する他の因子も例示的である。他の候補成長因子は、血小板由来成長因子(PDGF)、インスリン様成長因子(IGF)、およびフォルスコリン等のサイクリックAMPレベルを上昇させる因子である。
【0076】
そのようなマイトジェンの存在下では、レチノイン酸等の分化因子イニシエーターを取り除くことができる。マイトジェンによって細胞は非特異的に成長することから、オリゴデンドロサイト系譜細胞の優先的な成長を維持するために、培地中にオリゴデンドロサイト特異的分化因子を含め続けることが有利な場合が多い。培養液中のマイトジェンのバランスを調整することによって、オリゴデンドロサイトの優先的な成長を支持することも可能である。例えば、EGFは外胚葉を維持するが線維芽細胞を維持せず、線維芽細胞はFGFが存在しなくなれば死滅し得る。FGFに厳密に依存する細胞が除去されたならば、培養液にFGFを再度添加して所望の細胞型の成長を促進できる。
【0077】
細胞の調製における別の任意の段階として、本発明の使用者は、培養物中に存在し得る他の細胞型からオリゴデンドロサイト系譜細胞を分離してもよい。抗体もしくはレクチンを介した接着または細胞表面マーカーについてのソーティング等の、様々な分離手順を意図する。ポジティブ選択およびネガティブ選択の両方に関する適切な表現型マーカーを以下に記載する。UDP−ガラクトース・セラミド・ガラクトシルトランスフェラーゼ(CGT)、2’,3’−サイクリックヌクレオチド3’−ホスホジエステラーゼ(CNP)、NG2プロテオグリカン、NCAM、ミエリン塩基性タンパク質(MBP)、または様々なミエリン関連タンパク質のプロモーター等の組織特異的プロモーターを、アルカリホスファターゼ、緑色蛍光タンパク質、またはルシフェラーゼ等のレポーター遺伝子に連結して構築したプロモーター−レポータープラスミドを用いた、オリゴデンドロサイト系譜細胞のソーティングも意図する。
【0078】
細胞を適切な基層に接着させることによる単純な様式で、オリゴデンドロサイト分化因子の存在下で増殖する他の細胞からオリゴデンドロサイト系譜細胞を分離することができることを見出した。オリゴデンドロサイトは細胞特異的糖質および細胞表面受容体を有し、複合リガンドに優先的に接着することになる。詳細には、ラミニン、ゼラチン、または室温でゲル化し再構成された基底膜を形成するエンゲルブレス・ホルム・スワム(Engelbreth−Holm−Swarm)腫瘍細胞に由来する市販の細胞外マトリックス調製品マトリゲル(登録商標)等の、ある種の基底膜成分への接着によって、オリゴデンドロサイトを分離することができる(実施例1)。オリゴデンドロサイトがマトリックスに接着したら(数時間〜数日)、他の細胞型を洗浄除去し、例えば短時間トリプシン消化することによって接着細胞を回収することができる。この手順は、所望の細胞型の濃縮に加えて、さらなる操作を容易にするために細胞の大きな凝集塊を解離する利点を有する。
【0079】
分離手順の効果は、以下に記載するマーカーに関して濃縮を測定することによって判定することができる。このような方法でオリゴデンドロサイトを精製したならば、すでに記載したようにマイトジェンの存在下でこの細胞をさらに拡大する、以下の項に記載するように成熟させる、または最終用途に適した方法で調製することができる。
【0080】
さらなる成熟
必要に応じて、本発明の細胞を、複製段階を超えて機能的表現型にさらに成熟させることができる。これを行うことで、前駆細胞の可能性を特徴付ける、または治療目的もしくは薬剤スクリーニング目的のために最終段階の細胞を得ることができる。
【0081】
成熟は、前駆細胞表現型のさらなる増殖を阻害する様式で成長条件を変化させることによって達成される。例えば、成熟表現型の出現を促進するポリ−L−リジン等の基層上に細胞をプレーティングすることができる。別法としてまたはさらに加えて、細胞の拡大に用いた1つまたは複数の成長因子を除去する。繊毛様神経栄養因子(CNTF)およびCNTF受容体の他のアゴニスト等の成熟を亢進する因子を、成熟培地中に含めることができる。細胞が所望の成熟度に達したならば、培養液から細胞を回収し(例えば、トリプシンまたはコラーゲンを用いて)、解析または最終的な使用のために調製することができる。
【0082】
分化細胞の特徴
多くの表現型基準に従って、細胞を特徴付けることができる。基準には、形態学的特徴の顕微鏡観察、発現した細胞マーカーの検出または定量、インビトロで測定可能な機能的基準、および宿主動物に注入した際の反応が含まれるが、これらに限定されない。
【0083】
表現型マーカー
オリゴデンドロサイトに特有の表現型マーカーを発現するかどうかに従って、本発明の分化細胞を特徴付けることができる。細胞集団の成熟度に従って存在し得るオリゴデンドロサイト細胞の古典的な免疫細胞化学的マーカーは、以下の通りである:
・NG2、マクロファージおよびオリゴデンドロサイト前駆細胞によって発現されるコンドロイチン硫酸プロテオグリカン
・ガラクトセレブロシド(GalC)、拘束されたオリゴデンドロサイトのマーカー
・ミエリン塩基性タンパク質(MBP)、成熟ミエリンおよびミエリン産生細胞のマーカー。
【0084】
オリゴデンドロサイト系譜細胞によって発現される他の有用なマーカーには、以下が含まれる:
・PDGFRα、オリゴ前駆細胞、オリゴデンドロサイト、および他の細胞型によって発現されるPDGFの膜受容体
・TRα1、オリゴ前駆細胞、オリゴデンドロサイト、ニューロン、および他の細胞型によって発現される甲状腺ホルモンの核受容体
・ミエリンプロテオリピドタンパク質、オリゴデンドロサイトおよびグリア前駆細胞で発現されるミエリンの成分
・O4抗体によって規定されるエピトープ、オリゴデンドロサイト、アストロサイト、およびそれらの前駆細胞のマーカー
・ビメンチン、アストロサイト前駆細胞を特徴付ける線維芽細胞型フィラメントタンパク質(オリゴデンドロサイトでは陰性である場合が多い)
・神経膠繊維酸性タンパク質(GFAP)、アストロサイトのマーカー(オリゴデンドロサイトでは陰性)
・A2B5、II型アストロサイト、グリア前駆細胞、オリゴデンドロサイト前駆細胞、および膵臓β細胞で発現されるエピトープ
・オリゴデンドロサイトおよびその突起を染色し、脊髄および小脳両方の有髄化軸索と一致する、RIP抗体によって認識されるエピトープ。
【0085】
オリゴデンドロサイト分化の経路における様々な時点で発現される転写因子には、以下が含まれる:
・Olig1、オリゴ前駆細胞、運動ニューロン前駆細胞、および腎細胞によって発現されるヘリックス・ループ・ヘリックス(HLH)ファミリー転写因子
・Olig2、オリゴ前駆細胞、運動ニューロン前駆細胞、および松果体によって発現される別のHLHファミリー転写因子
・Sox10、オリゴ前駆細胞、オリゴデンドロサイト、シュワン細胞、神経堤、蝸牛、前立腺、およびメラノサイトによって発現されるSoxファミリー転写因子
・Nkx2.2、オリゴ前駆細胞、オリゴデンドロサイト、ニューロン前駆細胞、膵臓α細胞および膵臓β細胞によって発現されるHoxファミリー転写因子
・Pax6、オリゴ前駆細胞、ニューロン前駆細胞、膵αおよびβ細胞、水晶体網膜、下垂体、肝臓、および脾臓によって発現されるHLHファミリー転写因子。
【0086】
他の細胞型の有用なマーカーには以下が含まれる:
・ニューロン核抗原(NeuN)、ニューロン成熟のマーカー(通常、オリゴデンドロサイト系譜細胞では陰性)
・クラスIIIβチューブリン(TuJ1)、神経細胞の別のマーカー
・微小管関連タンパク質2(MAP−2)、CNS細胞のマーカー(おそらく陽性)
・SSEA−4、Oct−4、およびテロメラーゼ逆転写酵素(TERT)、未分化pPS細胞のマーカー(オリゴデンドロサイトおよびその前駆細胞では陰性)。
【0087】
組織特異的マーカーは、細胞表面マーカーについてのフロー免疫細胞化学法、または細胞内もしくは細胞表面マーカーについての免疫組織化学法(例えば、固定化細胞または組織切片)等の、任意の適切な免疫学的技法を用いて検出することができる。フローサイトメトリー解析の詳細な方法は、Gallacherら、Blood 96:1740、2000に記載されている。細胞表面抗原の発現は、標準的な免疫細胞化学法またはフローサイトメトリーアッセイ法において、選択的に細胞を固定化した後、および選択的に標識二次抗体または標識を増幅する他のコンジュゲートを用いて、有意に検出可能な量の抗体が抗原に結合した場合に陽性と定義される。
【0088】
組織特異的遺伝子産物の発現は、ノーザンブロット解析法、ドットブロットハイブリダイゼーション解析法により、または標準的な増幅方法において配列特異的プライマーを用いる逆転写酵素ポリメラーゼ連鎖反応法(RT−PCR法)により、mRNAレベルで検出することもできる。さらなる詳細については、米国特許第5,843,780号を参照されたい。特定マーカーの配列データは、GenBank等の公的データベースから入手可能である。例えば、ミエリンガラクトリピド生合成酵素UDP−ガラクトース・セラミド・ガラクトシルトランスフェラーゼ(CGT;GenBankアクセッション番号AH006651)は、GalCの合成の最終段階を触媒する酵素である。
【0089】
研究または治療における使用を容易にするために、集団内のオリゴデンドロサイトまたはその前駆細胞の特徴を有する細胞の割合を最大にすることが有利である場合が多い。以下の実施例の項で説明するように、上記のようなオリゴデンドロサイトまたは成熟細胞に特有の表現型マーカーのいずれかの1つ、2つ、3つ、またはそれ以上に関して陽性であると同定されるそのような細胞(または両方の混合物)が少なくとも約20%から約60%、80%、90%、95%、さらに98%を超えるまでのいずれかの細胞集団を得ることが可能である。
【0090】
神経機能の再構成に関連する治療的応用では、細胞集団が他の細胞型 − 詳細には未分化pPS細胞および非外胚葉系譜の細胞を形成する能力を最小限に抑えることが望ましい場合が多い。用途に応じて、ニューロン系譜およびその拘束前駆細胞、アストロサイト系譜およびその拘束前駆細胞、ならびにグリア細胞またはすべての種類の神経系細胞の共通の前駆細胞の割合を最小限に抑えることが有利である可能性もある。特定の態様では、本発明によるオリゴデンドロサイト集団は、これらの他の細胞型による混入を約1%、0.2%、または0.05%未満しか有さない。
【0091】
当業者は、オリゴデンドロサイト系譜細胞に特有の形態学的特徴をすでに理解していると考えられる。オリゴデンドロサイト前駆細胞は、中心体から反対極に2つの突起を伸長した双極の形状をとる場合がある。この細胞はまた、同様のマーカーおよびオリゴデンドロサイトの他の特徴の多くを有する、比較的扁平な細胞の形態をとる場合もある。この細胞は、比較的遅い成長速度ならびに細胞外マトリックスおよび可溶性因子への依存によって、他の細胞(線維芽細胞のような)から識別することができる。双極細胞と扁平細胞は、扁平細胞の優位性を促進する成長因子(EGF、bFGF等)の有無により、相互変換が可能である。本発明の前駆細胞集団は、少なくとも約20%、40%。60%、80%、またはそれ以上の双極細胞または扁平細胞表現型を含み得る。
【0092】
より成熟したオリゴデンドロサイトへの分化に際して、突起の数および複雑度は典型的に増加する。突起はそれらの間にミエリン皮膜を有するようであり、個々の軸索の周りを覆って軸索に沿った神経伝達を促進するミエリン鞘を形成する。
【0093】
遺伝子型特性
単離したpPS細胞またはpPS細胞の樹立株に由来する場合、本発明のオリゴデンドロサイトは起源細胞または細胞株の子孫であると特徴付けられ得る。したがって、オリゴデンドロサイトはその由来の元となった細胞と同じゲノムを有することになる。このことは、任意の核型変化のほかに、pPS細胞とオリゴデンドロサイトとの間で染色体DNAが90%を超えて同一であることを意味する。導入遺伝子を導入する、または内因性遺伝子をノックアウトするために組換え法により処理したオリゴデンドロサイトは、未操作の遺伝子要素はすべて保存されているため、依然として由来の元となった株と同じゲノムを有すると見なされる。
【0094】
オリゴデンドロサイトおよびpPS細胞は、標準的な遺伝子フィンガープリント技法により同じゲノムを有することが同定され得る。オリゴデンドロサイトが、未分化株から正常な有糸分裂の過程を経て得られる場合にも、同じゲノムを所有することが示され得る。
【0095】
特定の産業用途において、この特徴は本発明のオリゴデンドロサイトの貴重な特性である。詳細には、pPS細胞に増殖を引き起こし、細分し、要求に応じてさらなるオリゴデンドロサイトに分化させることができるため、起源pPS細胞が利用できることによって遺伝的に一致したオリゴデンドロサイトがさらにまた供給される。さらに、pPS細胞を他の治療的に重要な系譜に分化させることもできる。例えば、pPS細胞を免疫寛容細胞集団に分化させることが可能であり、この細胞集団は対象とするレシピエントを対応するオリゴデンドロサイトを用いた移植に対して寛容にするのに役立ち得る(国際出願PCT/US01/43434、Geron Corp.)
【0096】
本明細書に記載する技法により、分化の前または後に細胞を拡大することによって、同じゲノムを共有するオリゴデンドロサイト系譜細胞の大きな集団の産生が可能となる。理論的に、108、1010、または1012個の細胞の集団が可能である。そのような大きな集団は、通常、さらなる培養、薬剤スクリーニング、または治療投与に適した個々の容器に分割する。
【0097】
本発明の特定の態様は、グリア細胞またはオリゴデンドロサイトの特徴を有する分化細胞と組み合わせた起源細胞(未分化pPS細胞株または中間の集団等)を含む。2つの集団は同じ容器中に存在しても、同じ施設内の別の容器に存在しても、または2つの別の場所に存在してもよい。すでに記載したように未分化細胞の培養物にオリゴデンドロサイト系譜細胞への分化を全体として引き起こす場合のように、未分化細胞および分化細胞は同時に存在しても別の時点で存在してもよい。
【0098】
機能特性
品質管理の目的で、本発明の分化細胞の特性を機能的基準によって特徴付けることが望ましい場合が多い。目的とする最終的な用途により、異なる機能が異なる相対的関心となり得る。例えば、組織培養において神経組織を再ミエリン化する能力、インビボで脱髄が誘導された部位を修復する能力、または損傷した対象において神経機能を回復する能力について、細胞を評価することができる。これらの特性を判定するための、以下の限定されない例証を含む多くの実験モデルが存在する。
【0099】
1. 共培養におけるインビトロ髄鞘形成
2〜3ヶ月齢雌ラットから腰椎および頚椎後根神経節(DRG)ニューロンを回収することにより、成体DRG培養物を調製する。細胞を粉砕し、遠心分離し、mL当たり約100,000個の生存ニューロンでDMEM F12培地に再懸濁し、ラミニンコーティングした皿にプレーティングして4週間培養する。約2.5 x 104個のオリゴデンドロサイト系譜細胞を添加し、毎日フィードしながらさらに4週間培養することにより、インビトロ髄鞘形成を行う。ヒトオリゴデンドロサイトによるげっ歯類軸索の髄鞘形成は、TargetおよびBlakemore、Eye 8(part2):238、1994に例証されている。
【0100】
オリゴデンドロサイト系譜細胞がミエリンを形成する能力を判定するため、共培養物を4%パラフォルムアルデヒドで一晩固定し、GalCについて染色する。神経節に隣接した視野を選択し、視野当たりのミエリン部分の数をスコアリングする。ネスチン、βチューブリンIII、GFAP、CNP、GalC、Ki−67抗原、NeuN、または神経フィラメント70等のマーカーについて低密度培養物を染色することによって、分化の潜在能力を判定する。
【0101】
培養におけるDRGの髄鞘形成の証拠は、一般的にオリゴデンドロサイトの生物学的有効性と関連がある。細胞がインビボで生存し髄鞘形成または軸索の再成長を亢進する能力は、適切な動物モデルを用いてより一層得られる。これに反して、共培養における髄鞘形成は培養の条件に依存することになり、否定的な結果によってインビボにおける有効性は排除されない。インビトロ試験の予測値は、例えばIGFまたはNT−3のようなニューロトロフィン等の髄鞘形成を亢進すると考えられている因子を添加することにより、培養条件を調製することによって、改善することができる。
【0102】
2. 慢性脱髄モデル
成体ラット脊柱において、慢性脱髄の領域を誘導することができる(Keirsteadら、J Neurosci. 19:7529、1999)。鉛遮蔽を用いてT9を中心に、2 cmを超える距離で脊髄を40 GのX線照射に曝露するが、これにより曝露細胞のDNAにニックが導入され、細胞分裂している細胞が死滅する。2日後に、T9においてエチジウムブロマイドを脊髄内に直接注入する。エチジウムブロマイドはこれに曝露した細胞を死滅させるインターカレート剤であり、これによって慢性脱髄の無細胞領域ならびに脊柱領域の約60%を通して生存オリゴデンドロサイトおよびアストロサイトがない状態が提供される。
【0103】
3日後、ラットの脱髄部位にオリゴデンドロサイト系譜細胞の移植を行う。任意で、48時間前に培地にブロモデオキシウリジン(BrdU)を添加して細胞を予め標識することができる。第一に、細胞を前駆細胞約30個の凝集塊として調製し、μL当たり約60,000個の細胞密度に濃縮することができる。外径約80 umのガラスマイクロピペットを用いて、約10分を上回る時間をかけて細胞1μLを損傷部位に投与する。約2〜4週間後、組織試料を調製し、1 mmの横軸ブロックになるように樹脂またはクリオスタット切片を作製する。
【0104】
冠状面の切片をトルイジンブルーで染色し、一般病理、再ミエリン化の証拠、および細胞形態について解析する。誘導した部位は無細胞であるため、移植後に存在する細胞は投与した細胞に由来する。クリオスタット切片を、GFAP(アストロサイト)、CNP(オリゴデンドロサイト)、RIP(オリゴデンドロサイト)、またはNeuN(ニューロン)等の関連する細胞型のマーカーについて染色することができる。超薄切片を、ミエリン層板の数および細胞の超微細構造について電子顕微鏡により解析することもできる。脱髄部位にわたる移植細胞の再分布、および成熟有髄化細胞への分化を判定することができる。約25%、50%、または75%のレベルでの脱髄した軸索に占める再ミエリン化が、生物学的有効性の増加の証拠である。
【0105】
オリゴデンドロサイト系譜細胞の治療能力は、脱髄の先天性モデルにおいて試験することも可能である。ミエリン塩基性タンパク質における変異または欠陥を含む樹立モデルには、シバラー変異マウス(Roachら、Cell 42:149、1985)およびロングエバンス・シェーカー(Long Evans shaker)ラット(Kwiecienら、J. Neurocytol. 27:581、1989;Delaneyら、Lab. Anim. Sci. 45:547、1995)が含まれる。脳室内または大槽移植により(Mitomeら、Brain 134:2147、2001)、または脊髄への直接投与により(Liu、McDonaldら、Proc. Natl. Acad. Sci. USA 97:6126、2000)、再構築を試験することができる。MBPにおける変異または欠陥を伴うモデルは通常は適切な髄鞘形成の能力がなく、結果として神経欠損を被るため、これらのモデルでの緻密なミエリンの形成は投与細胞に直接起因する。
【0106】
3. 脊髄損傷モデル
SCIモデルは、挫傷および背側片側切断を含む。挫傷では、適切な脊髄損傷装置を用いて、脊髄層を23ミリ秒を上回る時間をかけて約0.9 mm除去する(中程度の損傷)。片側切断損傷では、定位的なマニピュレーターを用いて先のとがった手術用メスで脊髄の背側の半分を切断する。どちらの手順とも、その後適切な術後処置を行う。
【0107】
移植した細胞の遊走を促進しかつミエリン関連成長抑制物質を除去するため、任意で脊髄もまた脱髄する(Keirsteadら、Exp Neurol. 151:303、1998)。脊柱管の中央かつ軸索損傷部位の後方に2 mmの孔を作製する。露出した脊髄に、リン酸緩衝食塩水に溶解した33%モルモット補体(Harlan SeraLab)で1:2に希釈したポリクローナル抗GalC抗体(Chemicon)約4μLを注入する。
【0108】
損傷から約24時間後に、ガラスマイクロピペットを通して動物にオリゴデンドロサイト系譜細胞を移植する。あるいは、1〜3ヶ月間治療を保留することにより、慢性損傷モデルを作製することができる。細胞を用いて処置した後、ビデオテープに機能的反応を記録し、臨床的改善の証拠を得るために定期的にモニターすることができる。例えば、BBB尺度、関節運動に基づいた21ポイント尺度、体重支持、四肢の協調、および他の特性を用いて、地上歩行を定量化することができる(実施例2)。
【0109】
組織学的精密検査をするため、ビオチン標識コレラ毒素βサブユニット(CTB、坐骨神経の両側に0.1%を4μL注入)またはビオチン化デキストランアミン(BDA、感覚運動皮質に10%を10μL注入)等のトレーサーを動物に予め注射することができる。約2週間後、脱髄モデルでのように組織切片を調製し、移植した細胞の位置および再ミエリン化の証拠について検討するが、再ミエリン化は形態学的基準によって正常な髄鞘形成または部分的脱髄と識別可能である。特徴的な神経突起の成長円錐の証拠を得るために、注射したトレーサーで標識した軸索についても切片を解析する。軸索再生の免疫化学的証拠は、RT97抗原(神経フィラメントのマーカー)、セロトニン(5HT)、ノルエピネフリン(NE)、およびカルシトニン遺伝子関連ペプチド(CGRP)等のニューロンマーカーについて染色することによって評価することができる。
【0110】
オリゴデンドロサイトの遺伝子改変
本発明の特定のオリゴデンドロサイト前駆細胞集団は、実質的な増殖能を有する。必要に応じて、内因性遺伝子からの転写を増加させることによって、または導入遺伝子を導入することによって、細胞内ののテロメラーゼ逆転写酵素(TERT)のレベルを増加させることにより複製能をさらに亢進することができる。特に適切なのは、国際公開公報第98/14592号に提供されているヒトテロメラーゼ(hTERT)の触媒成分である。ヒト細胞におけるテロメラーゼのトランスフェクションおよび発現は、Bodnarら、Science 279:349、1998、およびJiangら、Nat. Genet. 21:111、1999に記載されている。
【0111】
未分化pPS細胞においてテロメラーゼ発現を増加させることができ、その後本発明に従ってオリゴデンドロサイトに分化させることができる。あるいは、pPS細胞をオリゴデンドロサイト前駆細胞に分化させ、その後トランスフェクションしてTERT発現を増加させることもできる。遺伝子改変細胞は、標準的な方法に従って、RT−PCR法、テロメラーゼ活性(TRAPアッセイ法)、TERTの免疫細胞化学染色、または複製能により、TERTの発現について評価することができる。用途によっては、myc、SV40ラージT抗原、またはMOT−2をコードするDNAで細胞を形質転換する等の、細胞を不死化する他の方法もまた意図している(米国特許第5,869,243号、国際公開公報第97/32972号、および国際公開公報第01/23555号)。
【0112】
必要に応じて、本発明の細胞を調製もしくはさらに処理し、インビトロで未分化細胞を除去すること、またはインビボで復帰細胞を防ぐことができる。集団から未分化幹細胞を減少させる1つの方法は、エフェクター遺伝子が、TERTプロモーターまたはOCT−4プロモーター等の未分化細胞において優先的な発現を引き起こすプロモーターの制御下にあるベクターを、集団にトランスフェクションすることである。エフェクター遺伝子は、緑色蛍光タンパク質等のセルソーティング導くレポーターであってよい。エフェクターは、例えば毒素またはカスパーゼ等のアポトーシスの介在物質をコードし、細胞に対して直接溶解性であってもよい(Shinouraら、Cancer Gene Ther. 7:739, 2000)。エフェクター遺伝子は、抗体またはプロドラッグ等の外部薬剤の毒素効果に対して細胞を感受性にする効果を有してもよい。発現する細胞をガンシクロビルに対して感受性にする単純ヘルペスチミジンキナーゼ(tk)遺伝子が例示的である(米国特許第6,576,464 B1号)。あるいは、エフェクターは、未分化表現型に復帰する任意の細胞をインビボで天然抗体に感受性にする外来決定因子を、細胞表面に発現させてもよい(U.S. 2003−0032187 A1)。
【0113】
本発明の細胞はまた、組織再生に関与する能力、または治療する対象に治療遺伝子を送達する能力を増強するために、遺伝子改変することも可能である。所望の遺伝子をコードする既知の配列を用いてベクターを設計し、構成的(CMVプロモーター等)またはオリゴデンドロサイト系譜細胞の細胞において特異的(ミエリン塩基性タンパク質のプロモーター等)に活性のあるプロモーターに機能的に連結する。この戦略に従って、オリゴデンドロサイトの成長を亢進する、再ミエリン化を活性化する、または軸索の再生を促進するような、様々な導入遺伝子を発現させることができる。神経成長因子をコードする遺伝子が例示的である(米国特許第5,885,584号および第6,268,340号)。
【0114】
研究および臨床治療におけるオリゴデンドロサイトの使用
本発明は、様々な重要な研究、開発、および商業目的のために、多数のオリゴデンドロサイトを産生する方法を提供する。
【0115】
本発明の細胞を用いて、他の系譜の細胞で優先的に発現されるcDNAが比較的混入していないcDNAライブラリーを調製することができる。本発明の分化細胞を用いて、標準的な方法に従い、オリゴデンドロサイトおよびその派生物のマーカーに特異的なモノクローナル抗体またはポリクローナル抗体を調製することも可能である。
【0116】
特に関心が持たれるのは、薬剤開発および臨床治療を目的とした本発明の組成物の使用である。
【0117】
薬剤スクリーニング
本発明の細胞を用いて、オリゴデンドロサイト前駆細胞および成熟オリゴデンドロサイト両方の特徴に影響を及ぼす因子(溶媒、小分子薬剤、ペプチド、ポリヌクレオチド等)または環境条件(培養環境または操作等)をスクリーニングすることができる。
【0118】
一例では、pPS細胞(未分化または分化パラダイムに向けて起動した)を用いて、オリゴデンドロサイトへの成熟を促進する因子、または長期培養におけるオリゴデンドロサイトの増殖および維持を促進する因子をスクリーニングする。例えば、別のウェル内の細胞に候補成熟因子または成長因子を添加し、さらなる培養および細胞用途の所望の基準に従って生じる任意の表現型変化を判断することにより、それら因子を試験する。これにより、pPS由来オリゴデンドロサイトばかりでなく初代神経組織から単離したオリゴデンドロサイトおよびその前駆細胞に関しても、導出および培養法の改良がもたらされ得る。
【0119】
本発明の他のスクリーニング法は、オリゴデンドロサイトの成長、発達、または毒性に及ぼす潜在的副作用についての薬学的化合物の試験に関する。この種のスクリーニングは、化合物がオリゴデンドロサイト自体への薬学的効果を有するように設計される場合ばかりでなく、他所での主要な薬学的効果のために設計された化合物のオリゴデンドロサイトに関連する副作用を試験するためにも適している。
【0120】
他のスクリーニング法は、軸索を有髄化する役割におけるオリゴデンドロサイト活性に影響を及ぼす可能性を有する小分子薬剤の効果を測定するための、オリゴデンドロサイトの使用に関する。この目的のため、細胞をインビトロで試験化合物と混合することができ、ミエリンに関連する活性に及ぼす化合物の効果を判定する − 例えば、ミエリン塩基性タンパク質等のミエリン関連成分の発現;組織学的に検出可能なミエリン鞘の形成;および神経細胞との共培養において隣接する軸索を有髄化する能力。
【0121】
薬剤スクリーニングの一般原理に関しては、当業者には、米国特許第5,030,015号および教科書In vitro Methods in Pharmaceutical Research、Academic Press、1997を参照されたい。候補薬学的化合物の活性の評価は、一般に本発明の分化細胞を、単独または他の薬剤との組み合わせのどちらかで、候補化合物と混合することを伴う。研究者は、化合物に起因する細胞の形態、マーカー表現型、または機能的活性の任意の変化を判断し(未処理細胞または不活性化合物で処理した細胞と比較して)、化合物の効果を観察した変化と関連づける。
【0122】
臨床治療におけるオリゴデンドロサイト
本発明は、治療を必要とする患者において神経機能を保持または回復するための、オリゴデンドロサイト前駆細胞およびその派生物の使用を提供する。特に、神経組織を再ミエリン化するため、またはさもなければ神経ネットワークの維持もしくは再生を支持するために、本発明の細胞を投与することができる。いかなる限定も意味することなく、投与した細胞は、すでに所定の位置にあるニューロンの機能を安定化するもしくは改善する効果を有する、またはニューロンが相互にもしくはそれが制御する組織と新たな連結を形成するのを支持する可能性がある。
【0123】
薬学的組成物の性質は、部分的には治療する病態に依存する。ある症例では、優れた複製能および操作に必要な回復力を有する前駆細胞(NP2またはGalCに関して陽性)で組成物を製剤化することが適切である場合がある。別の症例では、より迅速に髄鞘形成能を提供するために、より成熟した細胞(GalCまたはMBPに関して陽性)で組成物を製剤化することが適切である場合がある。
【0124】
懸濁培養において形成される大きな凝集体をマトリゲル(登録商標)等の適切なマトリックスに接着させることにより、pPS由来細胞からオリゴデンドロサイト系譜細胞の比較的分散した集団が容易に得られること見出した。培養した細胞を基層上にプレーティングして数時間〜数日間培養し、非接着細胞を除去する。次に、短時間酵素消化した後に粉砕する等の適切な化学的または機械的手段によって、オリゴデンドロサイト系譜細胞を回収する。細胞操作、保存特性、および投与に際して有利な効果を提供する能力の点から見て、単分散集団または約30個以下の細胞の凝集塊は利点を有すると考えられる。細胞は典型的に洗浄し、その後疾患部位に細胞を保持するために効果的な用量を投与するのに適した濃度で(例えば、μL当たり細胞20,000個〜100,000個、治療領域の量によって調整する)、薬学的に適合する培地中に懸濁する。
【0125】
神経に適応した治療(neurological adaptive therapy)の通常の手順に従って、患者の準備をする。拒絶を防ぐために、シクロスポリンA等の標準的な免疫抑制療法により患者を処置してもよい。別法としてまたはさらに加えて、同じpPS細胞株から調製した造血細胞または未分化細胞を用いて、患者をオリゴデンドロサイトのアロタイプに対して特異的に寛容化してもよい(国際公開公報第02/44343号;国際公開公報第03/050251号)。症例によっては、損傷部位においてニューロンを一次的に脱髄すること、投与細胞の接近を促進すること、または再ミエリン化を阻害し得る因子を除去することが有利である場合がある。これを達成する1つの手段は、GalC、O4、またはエリン関連糖タンパク質等の、ミエリン鞘上の1つまたは複数のエピトープに対する補体結合抗体を局所環境に投与することである。Keirsteadら、Brain Res. Bul. 44:727、1997;およびカナダ特許第2,253,078号を参照されたい。次に、本発明の分化細胞を、髄鞘形成もしくは神経機能の再生を所望する部位またはその周りに、1回または複数回投与する。
【0126】
その後対象に支持的な術後処置を施し、移植片の許容または神経機能の再生についてモニターする。適切であれば、患者をファムプリジン(Fampridine)−SR(4−アミノピリジン)等の、オリゴデンドロサイト機能を回復すると考えられる他の手段を用いて同時に治療してもよい。組織試料を、すでに記載した関連マーカーに関して免疫細胞化学法によって解析し、軸索の再ミエリン化、投与細胞の存在に起因する神経突起の発芽等の機能的事象に関して評価することができる。それらの疾患の典型的な病理学的特性により病態の維持または改善に関して、および例えば障害状態尺度(expanded disability status scale; EDSS)での機能の評価に関して、患者をモニターすることができる。
【0127】
本発明の組成物を用いた治療に適切な病態には、ミエリンを維持するまたは産生する能力が、治癒またはさらなる悪化の防止に役立つ進行性脱髄および中枢神経系の外傷が含まれるが、これらに限定されない。
【0128】
多発性硬化症は、脳および脊髄における脱髄部分の散在を特徴とする緩徐進行性疾患である。オリゴデンドログリアの破壊および血管周囲の炎症を伴う脱髄プラークは、CNSを通じて、主として白質(特に、頚部および背部)、視神経、および周室部に起こる。中脳、脳橋、および小脳の路、ならびに大脳および脊髄の灰白質もまた影響を受ける。
【0129】
急性散在性脳脊髄炎(感染後脳脊髄炎)は血管周囲のCNS脱髄を特徴とし、これは自発的に起こり得るが、通常はウイルス感染またはウイルスワクチン接種の後に起こる。慢性炎症性脱髄性多発根神経炎(CIDP)は、炎症性T細胞およびマクロファージを伴う神経内膜の間質および血管周囲浸潤を特徴とし、末梢神経の分節性脱髄を引き起こす。HTLV関連ミエロパシーは、両脚の痙性衰弱を特徴とする緩徐進行性脊髄疾患である。ギランバレー症候群等のいくつかの末梢神経障害も、脱髄を特徴とする。
【0130】
先天性代謝病(フェニルケトン尿症および他のアミノ酸尿症;テイ・サックス病、ニ−マン・ピック病、およびゴーシェ病;フルラー症候群;クラッベ病および他の白質萎縮症等)は、CNSにおけるミエリン鞘の発達に影響を及ぼし、永久的な広汎性神経欠損を起こす可能性がある。副腎白質萎縮症および副腎脊髄神経障害は、副腎機能不全および神経系の広範な脱髄を特徴とする伴性劣性代謝病である。ペリツェウス・メルツバッハー病は、プロテオリピドタンパク質遺伝子の点突然変異のために患者がミエリンを形成しない病態である。レーバー遺伝性視神経萎縮および関連するミトコンドリア障害は、主に両側の中心視力の喪失を特徴とする。
【0131】
CNSの外傷によって起こる任意の急性または長期異常も、無酸素症および虚血によるミエリンの喪失に関連する病態と同様に、治療が検討され得る。この分類には、脳卒中および外傷性脳損傷に関連する病態が含まれる。
【0132】
対麻痺または任意のレベルの不完全な運動機能を起こす様々な種類の脊髄損傷が、本発明の細胞を用いた治療の第一候補である。性質および到達性次第で、頚椎、腰椎、胸椎、および仙椎の損傷は、すべてその病態の改善または安定化を得る可能性がある。SC機能の完全なまたは不完全な喪失を起こす急性損傷は、減圧手術と同時にまたはその直後に治療することができる。慢性病態は、必要に応じていつでも治療または再治療することができる。針、カニューレ、または他の適切な装置を使用して、手術、内視鏡、または経皮注射により、細胞を損傷部位にまたはその近傍に投与することができる。保証されている場合、誘発電位試験により電気生理学的効果を測定することができる。米国脊髄損傷学会(ASIA)の運動スコアならびにフェランズおよびパワーズの生活の質指標(QLI)等の基準により、患者を運動経路および感覚経路の臨床的改善について評価することもできる。
【0133】
いつもと同様、患者の選択、投与形態、およびリハビリテーションのモニタリングの最終責任は、管理する臨床医の責任である。
【0134】
商品流通できるように、本発明のオリゴデンドロサイトは典型的に等張賦形剤を含む薬学的組成物の形態で提供し、ヒト投与に十分に無菌的な条件下で調製する。本発明はまた、生産、流通、または使用の任意の時点で存在する細胞のセットまたは組み合わせを含む試薬系を含む。細胞セットは、同じゲノムを共有する場合がある未分化pPS細胞または他の分化細胞型と組み合わせた、分化したpPS由来細胞の種類(グリア細胞、オリゴデンドロサイト、それらの前駆細胞およびサブタイプ等)に例証されるがこれらに限定されない、本開示に記載する2つまたはそれ以上の細胞集団の任意の組み合わせを含む。セット中の各細胞型は、同じ事業体もしくは事業関係を共有する別の事業体の管理下で、同時にもしくは別の時点で、一緒にもしくは同じ施設内の別の容器に包装しても、または別の場所で包装してもよい。
【0135】
細胞組成物の医薬品製剤化における一般原則については、当業者には、G. MorstynおよびW. Sheridan編によるCell Therapy: Stem Cell Transplantation, Gene Therapy, and Cellular Immunotherapy、Cambridge University Press、1996、およびCell Transplantation for Neurological Disorders、T.B. Freemanら編、Humana Press 1998を参照されたい。細胞は、任意で細胞の保存または上記の病態を治療するための医薬品としての使用に関する情報を添付して、流通もしくは臨床用途に適した装置もしくは容器に、または他の価値ある目的に適した装置もしくは容器に包装する。
【0136】
本発明の特定の態様の限定されないさらなる例証として、以下の実施例を提供する。
【0137】
実施例
実施例1:hES細胞のオリゴデンドロサイトへの分化
H7株およびH1株のヒト胚性幹(hES)細胞を、初代マウスフィーダー細胞により馴化しbFGFを添加したノックアウト(無血清)培地中、マトリゲル(登録商標)基層上で無フィーダー条件において未分化状態で増殖させた(国際公開公報第99/20741号; 国際公開公報第01/51616号、Geron Corp)。
【0138】
以下の計画に従って、hES細胞をオリゴデンドロサイト系譜の細胞に分化させた:
【0139】
(表1)hES細胞からのオリゴデンドロ前駆体(oligodendroprogenitor)の産生
【表1】
GRMはグリア制限培地である(表2を参照のこと)。
移行培地(TR)は、GRMおよびhES細胞を作製するための馴化培地の1:1混合物として作製する。
【0140】
(表2)グリア制限培地(GRM)の成分
【表2】
【0141】
分化手順は以下の通りに行った:
・1日目:コラゲナーゼIV(Gibco 17101−015)を用いて接着基層からESコロニーを解離し、コロニーを4 ng/mL bFGF(Gibco 13256−029)を有するTR中で低接着性6ウェルプレート(Corning 3471)に入れた。分化手順の最初の3日間のみ、ペニシリン−ストレプトマイシン(Gibco 10378−016)を用いた。
・2〜10日目:細胞をレチノイン酸(RA;オールトランス−レチノイン酸、Sigma 223018)、10μMと共に培養し、合計8日間毎日フィードした。RA保存液は、6 mg/mLの濃度で(約0.02 M)DMSOに溶解して調製した。
・3日目から:培地をGRMに置換し、それ以上はbFGFを添加しなかった。培地は毎日交換した。RAは光感受性であるため、フィード中は照明を最小限に落とした。ES細胞凝集体を含むウェルの培地を、15 mLチューブに回収した。低速で短時間遠心分離した後(800 rpm、1分間)、上清を吸引除去し、新鮮培地を添加した。穏やかに上下にピペッティングし(2〜3回)、6ウェルプレートの各ウェルに4 mLずつ確実に均一に分配した。
・10〜15日目:10日目以後は、毎日フィードする際に、20 ng/mL濃度のEGF(Sigma E9644)および2 ng/mL濃度のbFGFを添加した。
・15〜21日目:EGFは20 ng/mL濃度で添加し続けたが、bFGFは培地から除去した。
・21日目以後:20 ng/mL EGFを有するGRM培地を維持することにより、42日目以降でも神経の特徴を有する新たな凝集体/凝集塊が産生され得た。
・これらの神経球を選択するため、分化手順の28日目に、マトリゲル(登録商標)をコーティングした6ウェルプレート(マトリゲル(登録商標)1:30、BD Bioscience 356231)に、培養物全体を解離手順を行うことなく、12〜20時間(一晩)移した。翌日、培養物を穏やかに振盪すると、神経球のみが接着したままであるので、培養物の残りを新鮮なGRM培地で置換した。
・35日目:ポリ−L−リジン(Sigma P2636)およびラミニンでコーティングした4チャンバー・イメージングスライド(Nalgene−Nunc International 154917)に、神経凝集体をプレーティングした。トリプシン−EDTAで短時間(5分)処理し、細胞の凝集を解離させた。PLL−ラミニン上で細胞を7日間培養し、その間一日おきに培養物にフィードした。
・42日目:培養物をパラフォルムアルデヒドで固定し、免疫組織化学法により細胞を特徴付けした。使用したマーカー:NeuN、ガラクトセレブロシド(GalC)、Map2、O4、ビメンチン、GFAP。
【0142】
結果を添付の位相倒立顕微鏡写真(phase inverted photomicrograph)に示す。
【0143】
図1は、移行培地(50%グリア前駆細胞培地)中、懸濁状態で2日間培養した胚様体を示す。
【0144】
図2は、レチノイン酸中での培養の最終時点における、分化7日目の球体を示す。3日目から、透明な球体が懸濁培養物中に現れ始めた。7日目までに、これらの球体が培養物中の細胞の80〜90%を構成した。
【0145】
図3は、RA処理の抑制後の球体の崩壊を示す。いくつかの小胞は、細胞を球体の内部および外部に添加することにより成長し続けた。細胞集塊は濃い陰影の球状の形をとっていた。球体は、20 ng EGFのみの存在下では崩壊した。しかし、低濃度のbFGFを添加することにより、球体は維持され得た。いくつかの球体は新たな細胞を添加することによって成長し続け、球状の濃い陰影の細胞集塊を生じた。厚い壁を有するいくつかの小さな球体は成長し続けたが、数日後に完全に消失した。従って、bFGFを低濃度(2 ng/mL)で培養物中に5日間保持し続け、それにより球体は層の添加で成長し続けた。
【0146】
図4は、bFGFを除去した後に現れる変化を示す。細胞集塊のほとんどが解離し始め、培養液全体が単一細胞および小さな細胞塊でいっぱいになった。同時に、新たな鮮黄色の球体が観察された(矢印)。
【0147】
図5は、鮮黄色の球体(矢印)が培養を続けながら選択的に維持されたことを示す。あるものは他の球体から、あるものは単離した細胞集団から、あるものは濃い細胞集塊から発芽し、新たな球体が毎日出現した。
【0148】
28日目に、培養物全体をマトリゲル(登録商標)基層上に一晩接着させた。神経に拘束された球体のみが基質に接着し、その他の培養物は浮遊したままであった。
【0149】
図6は、マトリゲル(登録商標)上にプレーティングしてから2、3日後の、球体からのグリアに拘束された神経前駆細胞の遊走および分枝を示す。より小さいコロニーは個々の細胞の縦裂により環を形成したのに対し、大きなコロニーは長い突起を有する星型の遊走を示した。マトリゲル(登録商標)で培養してから数日後に、より多くの遊走細胞が出現した。
【0150】
一週間後、トリプシンを用いてマトリゲル(登録商標)から培養物を解離し、さらなる特徴付けを行うために、ポリ−L−リジンおよびラミニン(Gibco 12163−010)でコーティングしたNuncイメージングチャンバーに低密度でプレーティングした。プレーティングした細胞は、ラミニン基層上で成長してから3〜4日後に、特徴としてオリゴデンドロサイト形状をとった。
【0151】
次に、マトリゲル(登録商標)選択段階が10〜20時間に短縮できることを見出した。非接着細胞を除去し、次に接着細胞を再懸濁して、FGF、EGF、およびグリア前駆細胞培地の存在下で拡大した。これは、治療投与および他の目的に対してより適した、より分散した細胞集団を産生する利点を有する。
【0152】
図7はこの技法の結果を示す。細胞をマトリゲル(登録商標)に接着させた後、7日間拡大し、ポリ−L−リジン・ラミニン上にプレーティングし、マイトジェンの非存在下で培養した。4%パラフォルムアルデヒドで細胞を固定し、3%ヤギ血清および0.3% Triton−X 100(登録商標)界面活性剤中でブロッキングした。ガラクトセレブロシドに対する抗体(GalC、Chemicom)、次にペルオキシダーゼ標識した抗免疫グロブリンを用いて、免疫細胞化学法を行った。ヘマトキシンにより核を対比染色した。(第1パネル倍率20 X;第2パネル倍率40 X)。
【0153】
視野内の少なくとも約95%の細胞が、成熟オリゴデンドロサイト系譜細胞のマーカーであるGalCの染色を示す。
【0154】
図8は、GalCに関して染色された、高倍率のES由来オリゴデンドロサイトを示す(倍率60 X)。少なくとも約10%〜20%の細胞がオリゴデンドロサイトに特有の形態(具体的には、ミエリンシートを示唆する、間に皮膜を有する多数の突起)を示す。
【0155】
実施例2:hES由来オリゴデンドロサイトはインビボにおいて再ミエリン化および神経発芽を起こす
本実施例では、実施例1と同様の戦略にいくつかの改良点を加え、H7株由来のhES細胞を移植用のオリゴデンドロサイト前駆細胞に分化させた。GRMの処方は、プロゲステロン濃度を65 ng/mLに再検討する点以外は同様とした。EGFを初めから添加し、2日後にbFGFを除いた。
【0156】
(表3)hES細胞からのオリゴデンドロ前駆体の産生
【表3】
【0157】
最初の週は、低速で4〜5分間遠心分離してからフィードした。球体が成長を開始した後は、15〜50 mL円錐管内に入れインキュベーター中で5〜10分間培養物を沈降させてフィードした。接着因子を発現する細胞は凝集体(球体)を形成し、単一細胞懸濁液中では非接着細胞よりも早く沈降する。これにより、細胞間接着特性を有する培養物の精度が増す。
【0158】
図9は、分化計画過程における細胞形態の進行を示す。(A): mEF馴化培地中、マトリゲル(登録商標)上で無フィーダー培養にて成長する未分化hES細胞。(B): 3日目までに、レチノイン酸を含む培地での懸濁培養で胚様体から透明な球体が成長し、迅速な細胞の分散を示す。(C): 黄色い球体が大きな塊状で蓄積することから、レチノイン酸による神経系譜細胞の誘導(段階3)、その後のEGFの存在下でのオリゴデンドロサイト前駆細胞の拘束および拡大(段階4)が明白であった。(D): 次に、所望の細胞型のポジティブセレクションのため細胞をマトリゲル(登録商標)上にプレーティングした。(E、F): 拡大段階が進行するにつれて、球体の大きさおよび培養物の割合が増加した。優先的に選択した後、最大で8週までグリア拘束前駆細胞を維持し増幅することができた。(G、H): 次にマイトジェンの非存在下でポリ−L−リジンおよびラミニン上にオリゴデンドロサイト前駆細胞をプレーティングすると、細胞は1週間以内に成熟オリゴデンドロサイトの形態をとった。
【0159】
図10は、成熟オリゴデンドロサイトの免疫細胞化学的解析を示す。プレーティングしてから1週間後、94%を超える細胞が初期グリア細胞のマーカーであるNG2に対する抗体で標識された(A)。プレーティングしてから8週間後、95%を超える細胞がGalC(C)、O4(D)、およびRIP(E)について陽性に染色された。細胞を抗GFAPで染色し、かつDAPIで対比染色したところ、オリゴデンドログリアマーカーで標識されなかった細胞の実質的にすべてが、GFAP陽性であった(B)。
【0160】
次に、Infinite Horizon(登録商標)衝撃装置(Precision Systems and Instrumentation, LLC)を用いて中程度の脊椎挫傷を誘導した成体スプラーグ・ド−リー(Sprague Dawley)ラットを麻酔し、低位胸髄にオリゴデンドロサイト前駆細胞を投与した。
【0161】
図11は、細胞を投与した後に得られた、ヒト核タンパク質に特異的な抗体で染色した組織切片を示す。濃く染色されたことから、移植後にヒト細胞が生存していることが確認される。
【0162】
図12は、移植してから9週間後の、ヒト核タンパク質について染色した脊髄の周囲領域の横断切片を示す。拒絶を防ぐために、シクロスポリンAで動物を処置しておいた。細胞は白質に向けて遊走または増殖した。
【0163】
図13は、移植細胞が投与部位に副作用をもたらすかどうかを判定する詳細な解析の結果を示す。未処置のラットおよび移植したラットの損傷部位にわたり、脊髄の長さに沿って連続切片を得た。各棒グラフは、5枚の切片で測定した平均断面積を示す(平均値±SEM)。挫傷近傍の小さな断面積は、損傷後の二次拡大の結果である。オリゴデンドロサイトは救済(sparing)を誘導しないが、損傷を悪化させることもない。このことから、hES由来オリゴデンドロサイトは脊髄損傷の1 mm以内に投与しても安全であることが示される。
【0164】
図14は、ニューロン分枝を測定する実験から得られたものである。損傷から8週間後、運動皮質の順行性トレーサーとして、動物の皮質脊髄路にBDAを注射した。これらの切片は2週間後に採取したものである。hES由来オリゴデンドロサイトで処置した動物においてのみ、軸索分枝の証拠が存在した(上の2枚のパネルにおいて、濃く染色された細い線として示される)。分枝は、損傷部位の真上の脊髄の領域で観察された。
【0165】
図15は、損傷および細胞投与部位(中心点)から1 mmの間隔で測定したニューロン分枝の定量を示す(ブロック当たり3切片の平均値±SEM)。処置を受けなかった損傷動物では、部位の3 mm上で標識軸索を計測した。処置した動物は、有意に高いレベルで、中心点の損傷のすぐ頭側にまで標識軸索を有した。このことから、移植細胞が再生可塑性を誘導することが確認される。
【0166】
図16は、移植した動物から採取した、実質的な再ミエリン化の証拠を示す切片の一連の電子顕微鏡写真である。左上のパネルの厚い環は、正常にミエリン化した繊維である。視野内の残りの軸索は、薄い層のミエリンを示す。軸索直径に対する鞘の比率から、これらの軸索が新たにミエリン化されたことが示唆される。右上のパネルは、新たにミエリン化した軸索のより解像度の高い像である。約5または6層の外被、および上部にはオリゴデンドロサイトの先端を示す軸索の舌状の縁が存在する。これは、この軸索のミエリン化が進行中であることを示す。下のパネルは、シュワン細胞によって沈着した厚いミエリン鞘を示す。これは移植にかかわらず生じ、未処置動物において限られた程度の回復が提供される。しかし、hES由来オリゴデンドロサイトを移植した動物のみが、オリゴデンドロサイトに特有の、上のパネルで観察される性質のミエリン化の証拠を示した。
【0167】
実施例3:オリゴデンドロサイト系譜細胞の表現型特徴付け
本実施例では、表4に示す手順に従ったhES株H7からのオリゴデンドロサイトの産生過程におけるマーカー発現を追跡する。
【0168】
(表4)hES細胞からのオリゴデンドロ前駆体の産生
【表4】
【0169】
本手順の一般的な適用では、固相選択段階後(約23日目以後の任意の時点)に移植に適したオリゴデンドロサイト前駆細胞が得られる。トリプシンを用いて週に一度継代することにより、最終段階の培養を行い必要に応じて細胞を拡大することができる(少なくとも8週間)。
【0170】
本実施例での表現型解析のために、5%ノックアウト血清代替物を含むGRMを用いて細胞を低密度培養に適合化した。21日目に5% KO−SRを存在させ始めることによって、マトリゲル(登録商標)上にプレーティングした際に双極細胞の増殖が改善された。細胞を低密度でラミニン(ポリ−L−リジン上で15μg/mL)上にプレーティングした場合には、KO−SRにより細胞の生存度および成長が改善された。手順の早い時期にKO−SRを使用しても、成長速度および黄色球体の比率から見て分化の過程に影響を及ぼさなかった。
【0171】
図17は、未分化hES細胞コロニーで検出されたマーカーを示す。左側のパネルは、視野内のすべての細胞核の位置を表すDAPI染色の青蛍光の上に重ね合わせた、各マーカーに特異的な抗体による赤蛍光を白黒で示す。右側のパネルは、マーカー染色のみを示す。2つの像を比較することにより、問題のマーカーを発現する、視野内の全細胞の比率の推定が可能となる。
【0172】
上列は、多能性細胞のマーカーであるSSEA−4に陽性標識されたコロニーを示す。下列は、中胚葉マーカーBMP4に陽性標識されたコロニーを囲む間質細胞を示す。各コロニー内の未分化細胞はBMP4陰性である。
【0173】
図18は、分化過程における転写因子Pax6の一過性の出現を示す(左側:抗体+DAPI;右側:抗体染色のみ)。上列は、10日目の細胞塊の中心に向かった染色、周辺に向かってより分化した細胞でのすでにダウンレギュレートされた染色を示す。下列は、35日目に採取した細胞では実質的に染色されないことを示す。
【0174】
図19は、10日目にレチノイン酸を除去した直後に存在する、初期のオリゴデンドロサイト系譜細胞で検出されるマーカーを示す(左側:抗体+DAPI;右側:抗体染色のみ)。上列:転写因子Olig1(83%±7%)。中列:転写因子SOX10(72%±12%)。下列:非特異的オリゴデンドロサイト前駆細胞マーカーA2B5(97%±3%)。
【0175】
図20A〜20Bは、35日目の完全に分化したオリゴ前駆細胞において優位を占めるマーカーを示す(左側:抗体+DAPI;右側:抗体染色のみ)。1列目および2列目(20A):NG2(コンドロイチン硫酸プロテオグリカン、オリゴデンドロサイト前駆細胞のマーカー); 3列目(20A):GalC;4列目(20B):O4;5列目(20B):Tuj1(ニューロンのマーカー)。
【0176】
結果から、少なくとも約80%の細胞がオリゴデンドログリアマーカーNG2、GalC、およびO4に陽性であることが示される。オリゴデンドログリアマーカーで標識されなかった細胞は、主にニューロンマーカーGFAPまたはTuj1(20B、下列)に陽性であった。二重免疫細胞化学法により、GFAPまたはTuj1陽性細胞はオリゴデンドログリアマーカーを同時に発現しないことが示される。さらに、BMP4またはSSEA−4は検出されず、このことからこれらの培養物が未分化細胞または中胚葉系譜細胞を欠いていることが示唆される。この細胞集団は、少なくとも8回の連続した継代を通して増殖させることが可能であると考えられる。
【0177】
実施例4:hES由来オリゴデンドロサイトは、ミエリン欠損動物において軸索
を再ミエリン化する
これらの細胞によってインビボで起こった髄鞘形成が(内因性オリゴデンドロサイトの誘導によるのではなく)直接的な効果であることを実証するため、分化手順の28日目の細胞を脱髄のシバラーマウスモデルに移植した。シバラーマウスは、染色体18に位置するミエリン塩基性タンパク質遺伝子の変異に関してホモ接合性である(Mbpshi/Mbpshi)。この遺伝子は重複しており、重複遺伝子の大部分が反転し、アンチセンスRNAの形成を引き起こす。これにより、CNSを通じて重篤なミエリン欠損が生じる。
【0178】
培養物からオリゴデンドロサイト前駆細胞を回収し、DMEMで洗浄し、濃縮し、シリコンコーティングしたハミルトンシリンジに装填する。すべての実験と同様に、ここでも10 mg/kg シクロスポリンAにより動物を免疫抑制した。6週間後、動物を屠殺しEM解析のために切片を作製した。
【0179】
図21はその結果を示す。超微細構造的に、シバラーマウスの軸索はミエリンを欠いているか、または1もしくは2層の緻密でないミエリンによって囲まれている(上のパネル)。細胞を移植してから6週間後、電子顕微鏡解析により多層の緻密なミエリンが示され、このことから移植集団のミエリン形成能が示唆された(下のパネル)。
【0180】
マウスはミエリン塩基性タンパク質を産生する能力を欠いているため、緻密なミエリンは栄養作用ではなく、投与したオリゴデンドロサイトによって直接産生されるに違いない。
【0181】
実施例5:ES由来オリゴデンドロサイトによる脊髄機能の回復
ES由来オリゴデンドロサイト系譜細胞が神経活性を回復する能力を、脊髄損傷のラット挫傷モデルで判定した。
【0182】
衝撃装置を用いて、麻酔した成体スプラーグ・ド−リーラットの低位胸髄に中程度の挫傷を誘導した。損傷から1週間後、4μL培地中の細胞2.5 x 105個を挫傷部位に移植した。分化計画(実施例1)の35日目の細胞を、トリプシンおよびEDTAに3分間曝露して細胞を非接着性にすることにより、移植細胞集団を調製した。次に細胞を基礎培地ですすぎ、μl当たり細胞6 x 104個に濃縮した。
【0183】
損傷から6週間後、順行性トレーサーBDA(ビオチン化デキストランアミン)を動物の運動皮質に注射した。腰椎にエメラルドグリーンを注射し、腰椎路を標識した。8週目に実験を終了した時点で、これらのマーカーの位置を判定した。
【0184】
図22は、本実験の2群それぞれについての地上歩行スコアを示す。上のグラフは、200キロダイン挫傷を受けた動物の結果を示す。下のグラフは、250キロダイン挫傷の結果を示す。BBB尺度は、関節運動、体重支持、四肢の協調、足の配置、および歩行安定性に基づく正常な地上歩行を評価する21ポイント尺度である(Basso、Beattie、およびBresnahan、J. Neurotrauma、12:1、1995)。(■)損傷から1週間後にES由来オリゴデンドロサイトで処置した動物(n=5);(▲)同じ挫傷を被るが細胞を投与していない対照動物(n=3)。盲検で評価した平均値±SEM。
【0185】
hES由来オリゴデンドロサイトを移植した動物は、処置後5週間を超えて持続する有意に優れた機能を示した(*≡p<0.05)。
【0186】
これらの結果から、ES由来オリゴデンドロサイトが脊髄損傷の動物モデルにおいて機能の回復を助けることが実証される。先の実施例から、移植したオリゴデンドロサイトにより神経発芽およびミエリン欠損軸索の再ミエリン化が起こることが示される。これらの効果のいずれかまたは両方が、ここで認められた動作の改善に寄与する可能性がある。
【0187】
本発明のある種の適応は日常的な最適化の問題であり、本発明の精神または添付の特許請求の範囲から逸脱することなく実施可能であることは自明である。
【特許請求の範囲】
【請求項1】
未分化ヒト胚性幹(hES)細胞、
および前記未分化細胞から分化した細胞集団であって、細胞表面NG2プロテオグリカンを発現し、かつニューロンマーカーNeuNに対して陰性であるグリア細胞を少なくとも80%含む細胞集団を含むシステムであって、
前記分化した細胞集団は、前記未分化ヒト胚性幹(hES)細胞を、成長因子、甲状腺ホルモン受容体のリガンド、およびレチノイン酸受容体のリガンドを含む培地において培養することにより得られる、グリア細胞を産生するためのシステム。
【請求項2】
前記成長因子が塩基性線維芽細胞成長因子である、請求項1記載のシステム。
【請求項3】
前記甲状腺ホルモン受容体のリガンドが甲状腺ホルモンT3であり、前記レチノイン酸受容体のリガンドがレチノイン酸である、請求項1記載のシステム。
【請求項4】
前記甲状腺ホルモン受容体のリガンドが甲状腺ホルモンT3であり、前記培地がセレンを更に含む、請求項1記載のシステム。
【請求項5】
分化細胞集団が以下の特徴の1つまたは複数を有する、請求項1記載のシステム:
・分化細胞集団内の約94%の細胞が、NG2プロテオグリカンを発現する;
・少なくとも80%の細胞が血小板由来成長因子受容体α(PDGFRα)も発現する;
・少なくとも20%の細胞がオリゴデンドロサイト前駆細胞に特有である双極性形態を示す;または
・マイトジェンの非存在下でポリ−L−リジンおよびラミニン上で細胞を3日間培養した後に、少なくとも10%の細胞が成熟オリゴデンドロサイトに特有である複雑な突起を有する。
【請求項6】
・シバラー変異(shiverer mutant)マウスの脊髄に分化細胞集団を移植することによりニューロン軸索の周りに緻密なミエリンの沈着が起こる;または
・挫傷ラットの脊髄内または脊髄の周りに分化細胞集団を移植することにより地上歩行が改善される、
請求項1から5のいずれか一項記載のシステム。
【請求項7】
未分化ヒト胚性幹(hES)細胞の系統、
成長因子、甲状腺ホルモン受容体のリガンド、およびレチノイン酸受容体のリガンドを含む培地、
および細胞表面NG2プロテオグリカンを発現する少なくとも80%のグリア細胞を含むが、神経細胞を実質的に含まない、それらから分化した細胞集団を含む、グリア細胞を産生するためのシステム。
【請求項1】
未分化ヒト胚性幹(hES)細胞、
および前記未分化細胞から分化した細胞集団であって、細胞表面NG2プロテオグリカンを発現し、かつニューロンマーカーNeuNに対して陰性であるグリア細胞を少なくとも80%含む細胞集団を含むシステムであって、
前記分化した細胞集団は、前記未分化ヒト胚性幹(hES)細胞を、成長因子、甲状腺ホルモン受容体のリガンド、およびレチノイン酸受容体のリガンドを含む培地において培養することにより得られる、グリア細胞を産生するためのシステム。
【請求項2】
前記成長因子が塩基性線維芽細胞成長因子である、請求項1記載のシステム。
【請求項3】
前記甲状腺ホルモン受容体のリガンドが甲状腺ホルモンT3であり、前記レチノイン酸受容体のリガンドがレチノイン酸である、請求項1記載のシステム。
【請求項4】
前記甲状腺ホルモン受容体のリガンドが甲状腺ホルモンT3であり、前記培地がセレンを更に含む、請求項1記載のシステム。
【請求項5】
分化細胞集団が以下の特徴の1つまたは複数を有する、請求項1記載のシステム:
・分化細胞集団内の約94%の細胞が、NG2プロテオグリカンを発現する;
・少なくとも80%の細胞が血小板由来成長因子受容体α(PDGFRα)も発現する;
・少なくとも20%の細胞がオリゴデンドロサイト前駆細胞に特有である双極性形態を示す;または
・マイトジェンの非存在下でポリ−L−リジンおよびラミニン上で細胞を3日間培養した後に、少なくとも10%の細胞が成熟オリゴデンドロサイトに特有である複雑な突起を有する。
【請求項6】
・シバラー変異(shiverer mutant)マウスの脊髄に分化細胞集団を移植することによりニューロン軸索の周りに緻密なミエリンの沈着が起こる;または
・挫傷ラットの脊髄内または脊髄の周りに分化細胞集団を移植することにより地上歩行が改善される、
請求項1から5のいずれか一項記載のシステム。
【請求項7】
未分化ヒト胚性幹(hES)細胞の系統、
成長因子、甲状腺ホルモン受容体のリガンド、およびレチノイン酸受容体のリガンドを含む培地、
および細胞表面NG2プロテオグリカンを発現する少なくとも80%のグリア細胞を含むが、神経細胞を実質的に含まない、それらから分化した細胞集団を含む、グリア細胞を産生するためのシステム。
【図1】


【図2】
【図3】
【図4】

【図5】

【図6】

【図7】
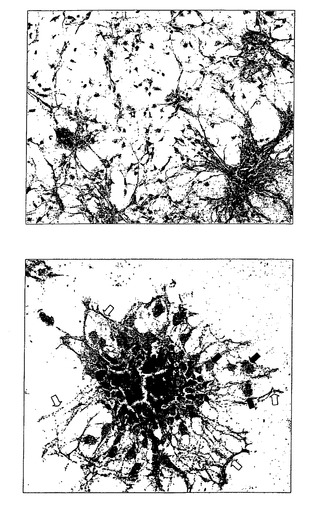
【図8】
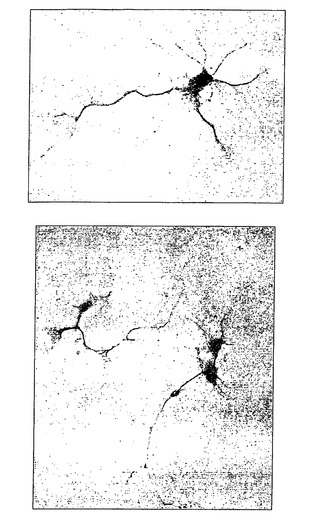
【図9】
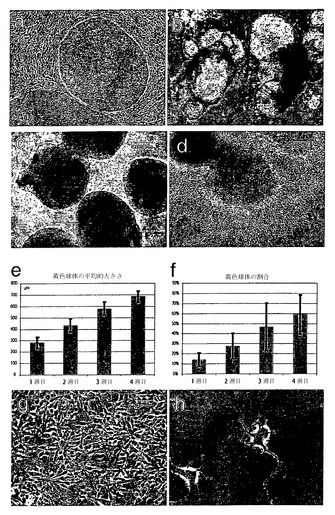
【図10】
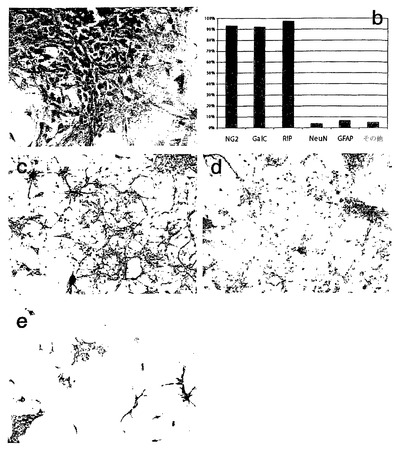
【図11】

【図12】

【図13】
【図14】

【図15】
【図16】

【図17】
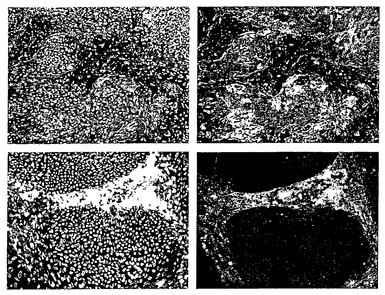
【図18】
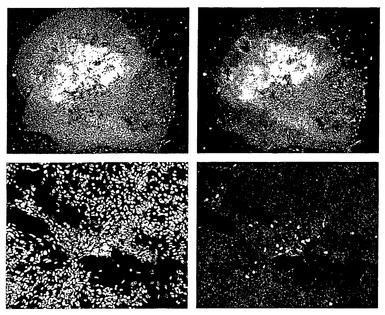
【図19】
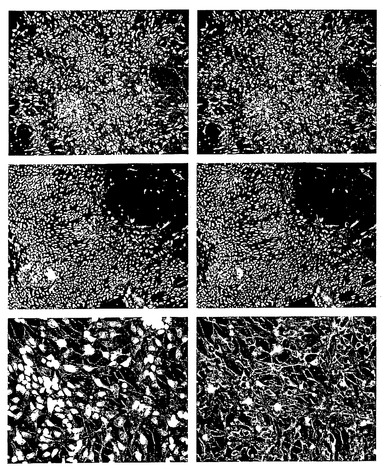
【図20A】

【図20B】
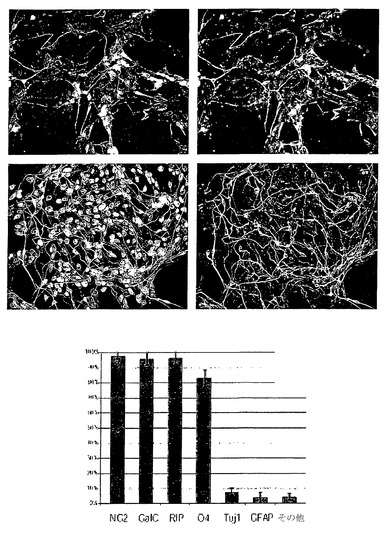
【図21】
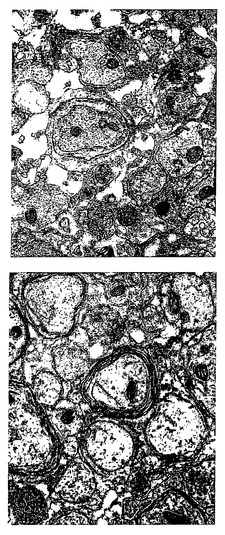
【図22】
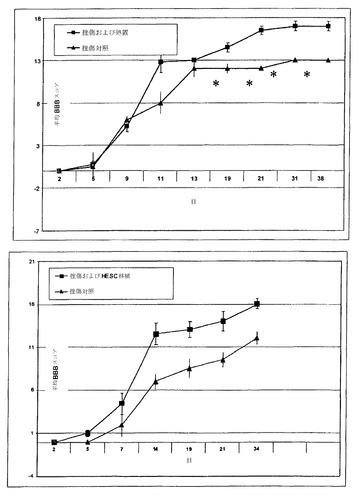
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20A】
【図20B】
【図21】
【図22】
【公開番号】特開2011−155978(P2011−155978A)
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願番号】特願2011−47716(P2011−47716)
【出願日】平成23年3月4日(2011.3.4)
【分割の表示】特願2005−505090(P2005−505090)の分割
【原出願日】平成15年7月11日(2003.7.11)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【Fターム(参考)】
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願日】平成23年3月4日(2011.3.4)
【分割の表示】特願2005−505090(P2005−505090)の分割
【原出願日】平成15年7月11日(2003.7.11)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【Fターム(参考)】
[ Back to top ]