
再有髄化におけるGM98(MRF)の調節
本発明は、GM98(また、MRFとしても公知である)の発現および活性を調節することにより、再有髄化を調節し、希突起膠細胞の分化を促進するための組成物および方法を提供する。本明細書ではまた、神経障害を治療し、生物活性剤をスクリーニングするための組成物および方法も提供される。本発明は、MRFの発現および/または活性を調節する方法および組成物を提供する。本明細書では、MRFにより調節される遺伝子の発現および活性もまた説明される。
【発明の詳細な説明】
【技術分野】
【0001】
(相互参照)
本出願は、2008年7月16日に出願された米国仮特許出願第61/081,279号および2008年12月5日に出願された同第61/120,307号の利益を主張し、これらの両出願は本明細書においてその全体が援用される。
【0002】
本出願は、the National Eye InstituteからのRO1 EY10257の下、政府の支援によってなされた。政府は、本発明において一定の権利を有する。
【背景技術】
【0003】
多発性硬化症(MS)とは、発現の再発−寛解パターンから慢性−進行性パターンまでの範囲にわたる臨床的障害を伴う、中枢神経系(CNS)の炎症性脱髄疾患である。MSの病因は未知であるが、自己反応性のCD4+ T細胞反応がミエリンおよび希突起膠細胞に対する炎症性損傷を媒介する(非特許文献1)。CNS病変は、ミエリン損傷の巣状領域を有し、また、軸索の病態、神経の損傷、および星状膠細胞における瘢痕形成とも関連する(非特許文献2)。臨床症状には、失明、麻痺、感覚および協調運動の喪失、ならびに認知障害を含めた、各種の神経学的機能不全が含まれる。
【0004】
ミエリンへの損傷または傷害は、導電速度、および軸索の破壊に対するニューロンの脆弱性に対して重大な影響を及ぼす。軸索の喪失と、進行性の臨床障害との間には相関があり、軸索の完全性を維持するには、ミエリンが無傷であることが重要である(非特許文献3)。ヒトMSの初期には、自発的な再有髄化が生じるが、該疾患の後期段階におけるCNS炎症の遷延、およびミエリン修復の失敗は、最終的に恒常的な衰弱をもたらす。
【0005】
再有髄化をもたらすのは、成熟希突起膠細胞(OL)である。したがって、再有髄化の失敗は、成熟希突起膠細胞の生成、それらが有髄化をもたらす能力の欠損、および/または有髄化に対するニューロンの非受容性と関連することが典型的である。多発性硬化症などの脱髄性疾患では、生存するOLおよびそれらの前駆細胞(希突起膠細胞の前駆細胞、またはOPC)が、脱髄領域の内部および周囲に見出されることが多い。軸索近傍において、これらの生存細胞が再有髄化に失敗することは、OPCが分化できないこと、または有糸分裂後における希突起膠細胞が、有髄化に必要とされる一連の遺伝子発現を再開できないことを反映しうる。有髄化は、希突起膠細胞およびそれらの前駆細胞を正確に局在化させ(非特許文献4;非特許文献5)、適切な細胞数を調節し(非特許文献6; 非特許文献7)、また、希突起膠細胞とそれらの標的である軸索との相互作用を媒介する(非特許文献8)シグナルを含めた、多数のシグナルの調整に依拠し、したがって、これらの過程のいずれかに欠損があれば、再有髄化の失敗がもたらされうる。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Bruckら、J. Neurol. Sci.、206巻、181〜185頁(2003年)
【非特許文献2】Compstonら、Lancet、359巻、1221〜1231頁(2002年)
【非特許文献3】Dubois−Dalcqら、Neuron、48巻、9〜12頁(2005年)
【非特許文献4】Tsaiら、Cell、110巻、373〜383頁(2002年)
【非特許文献5】Tsaiら、J. Neurosci.、26巻、1913〜22頁(2006年)
【非特許文献6】Barresら、Cell、70巻、31〜46頁(1992年)
【非特許文献7】Calverら、Neuron、20巻、869〜882頁(1998年)
【非特許文献8】ShermanおよびBrophy、Nat Rev Neurosci.、6巻、683〜690頁(2005年)
【発明の概要】
【発明が解決しようとする課題】
【0007】
有髄化または再有髄化を増強および促進する効果的な方法を開発する必要がある。OPC、前駆細胞プールの分化、または既存の有糸分裂以後における希突起膠細胞による有髄化の再開を促進する戦略は、再有髄化の確立に有益でありうる。本発明は、再有髄化の促進を対象とする組成物および方法を提供する。本明細書で開示される知見は、ミエリン生成に関与する遺伝子の発現が、MRF(ミエリン遺伝子調節因子)としても公知のGM98(遺伝子モデル98)、および有髄化により影響されることを裏付ける。
【課題を解決するための手段】
【0008】
本発明は、MRFの発現および/または活性を調節する方法および組成物を提供する。本明細書では、MRFにより調節される遺伝子の発現および活性もまた説明される。該方法および組成物を用いて、再有髄化を促進し、MRFの発現/活性、またはMRFにより調節される遺伝子を調節する生物活性剤をスクリーニングするほか、神経障害を治療することもできる。
【0009】
本発明の一態様は、MRFまたはその機能的な変異体をコードする核酸配列に作動的に連結された細胞型特異的な発現調節エレメントを含む、単離核酸分子である。さらに、該細胞型特異的な調節エレメントは、誘導的プロモーターまたは構成的プロモーターである。本発明はまた、本明細書に記載の核酸を含むベクターも提供し、宿主細胞型は、本発明のベクターおよび核酸を含む。
【0010】
本明細書ではまた、MRF導入遺伝子を含むトランスジェニック動物も提供される。導入遺伝子は、本明細書に記載の核酸配列を含みうる。さらに、導入遺伝子は、突然変異、およびエキソンの欠失などの欠失を含みうる。エキソンは、エキソン8など、推定DNA結合ドメイン内のエキソンでありうる。エキソンはまた、CreまたはFlpに対応する部位などのリコンビナーゼ部位にも隣接する。トランスジェニック動物はまた、CreリコンビナーゼまたはFlpなど、リコンビナーゼ導入遺伝子も含みうる。該導入遺伝子はまた、細胞型特異的な発現調節エレメントに作動的に連結されうる。該トランスジェニック動物は、マウスまたはラットなどの哺乳動物でありうる。
【0011】
核酸または導入遺伝子の特異的な発現は、神経膠細胞などの神経細胞内でありうる。神経膠細胞は、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞でありうる。細胞型特異的な調節エレメントは、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来しうる。
【0012】
トランスジェニック動物はまた、再有髄化/有髄化を促進するのに有効な候補生物活性剤をスクリーニングするのに用いることもできる。該方法は、候補生物活性剤を該動物に投与するステップ;対照動物と比較した、表1中の少なくとも1つの遺伝子の発現レベルの上昇についてアッセイし、該上昇が、該動物における再有髄化を促進する前記生物活性剤を示すステップ;および/または対照動物と比較した、該動物における有髄化の変化を観察するステップを含みうる。生物活性剤は、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子でありうる。本発明の別の態様では、動物における再有髄化/有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法であって、候補生物活性剤を動物に投与するステップと、対照動物と比較した、MRFの発現レベルの上昇についてアッセイし、該上昇が、該動物における有髄化を促進する該生物活性剤を示すステップとを含む方法が提供される。本発明はまた、MRF活性を調節するのに有効な候補生物活性剤をスクリーニングする方法も提供する。一態様において、該方法は、被験細胞を候補生物活性剤と接触させるステップと、対照細胞と比較したMRFの発現レベルの変化についてアッセイするステップとを含む。
【0013】
本発明のさらに別の態様では、被験体における神経障害を治療するための組成物であって、前記被験体におけるMRF活性を調節する生物活性剤を含む組成物が提供される。組成物は、被験体における再有髄化/有髄化を促進しうる。組成物は、幹細胞または胚性幹細胞が希突起膠細胞へと分化することを促進しうる。組成物は、MRF、またはMRFの活性もしくは発現を調節する薬剤など、第1の生物活性剤と、希突起膠細胞の分化を誘導する第2の生物活性剤とを含みうる。例えば、第2の生物活性剤は、Sox10、Nkx2.2、Olig1、および/またはOlig2を促進しうる。第2の生物活性剤は、Sox10、Nkx2.2、Olig1、またはOlig2でありうる。組成物は、MRF、Sox10、Nxk2.2、Olig1、Olig2、またはこれらの組合せを含みうる。組成物は、多発性硬化症などの脱髄状態など、神経障害を治療するのに用いることができる。本明細書で提供される組成物は、被験体における神経障害を治療する方法であって、該被験体に、治療有効量の、MRF活性を調節する生物活性剤を投与するステップを含む方法において用いることができる。方法は、被験体における再有髄化/有髄化を促進しうる。投与は、MRF、またはMRFの活性もしくは発現を調節する生物活性剤など、第1の生物活性剤と、また、希突起膠細胞の分化も誘導する第2の生物活性剤とを投与するステップとを含みうる。第2の生物活性剤を投与するステップは、第1の生物活性剤を投与するステップの前の場合もあり、これと同時の場合もあり、この後の場合もある。第2の生物活性剤は、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進しうる。第2の生物活性剤は、相乗効果を有しうる。
【0014】
本発明の新規の特徴は、添付される特許請求の範囲において具体的に記載される。本発明の特徴および利点についてのよりよい理解は、本発明の原理が用いられる例示的な実施形態を記載する以下の詳細な説明、およびそれに付随する図面を参照することにより得られる。
【図面の簡単な説明】
【0015】
【図1】図1は、CNS内におけるOL特異的な転写物であるGM98/MRFの同定を示す図である。A)図1Aは、CNSから急性摘出された細胞におけるGM98/MRFの発現レベルを、Affymetrix解析(プローブセット1439506_at)により決定したことを示す図である。B)図1Bは、ノーザンブロットが、MRFの発現を、P20における脳および培養希突起膠細胞(OL)内では、約5.5Kbの単一の転写物として確認するが、心臓組織内ではこれを確認せず、これは、確立されたOLマーカーであるCNP1の発現パターンを反映することを示す図である。GAPDHは、mRNAレベルの対照として示される。C)図1Cは、P16における脳の矢状断面内における、確立されたOLマーカーであるPLPおよびMRFについてのin situハイブリダイゼーションを示す図である。いずれの遺伝子も、明白な白質の発現パターンを示す。D)図1Dは、標識化された細胞の同一の分布を示す、脳梁側部内におけるPLPおよびMRFの発現についての高倍率画像である。
【図2】図2は、MRFのペプチド構造および細胞内局在化を示す図である。A)図2Aは、マウスMRF遺伝子およびヒトC11Orf9遺伝子のペプチド配列の比較を示す図である。B)図2Bは、マウスMRFタンパク質、およびヒトC11Orf9タンパク質の構造領域、ならびにこれらの領域内におけるそれらの相同性を示すダイアグラムである。C〜E)図2C〜Eは、HEK細胞内におけるmycタグを付したMRFの細胞内局在化を示す図である。抗mycによる、Myc−MRFをトランスフェクトしたHEK細胞に対する染色(E)は、トランスフェクトされた細胞内における、核内対比染色色素であるDAPIとの本質的に完全な共局在化を伴う、該タンパク質の強い核内局在化を示す。mycタグを付していないMRF(D)、およびmycタグを付したタンパク質をトランスフェクトした細胞は、細胞質内局在化(C)を示し、これにより、染色についての陰性対照、および比較がそれぞれもたらされる。
【図3】図3は、MRFの発現が、OLの成熟にとって重要であることを示す図である。A)図3Aは、24〜96時間にわたる分化時における、siContまたはsiMRFをトランスフェクトしたOLの生存率および形態を示す、生存:死滅アッセイ(生存細胞は、カルセイン(Calcien)AMにより染色されて緑色チャネルで示され、非生存核は、エチジウムホモ二量体により染色されて赤色チャネルで示される)を示す図である。siMRFをトランスフェクトした細胞は、siContトランスフェクト細胞と比べて膜シートの沈着の減少および生存率の低下を示した。B)図3Bは、分化条件へと移した24〜96時間後における、siContトランスフェクト細胞と比べたsiMRFトランスフェクト細胞の生存率を示す図である。siMRFトランスフェクト細胞は、分化の48時間後以降、siContトランスフェクト細胞と比べて著明な生存率の低下を示した。**P<0.01。C)図3Cは、分化の24、48、および96時間後において、siContまたはsiMRFをトランスフェクトしたOLにおける、OPCマーカーであるNG2、および初期OLマーカーであるMBPの発現を示す図である。siMRFトランスフェクト細胞は、siContトランスフェクト細胞と同様の時間プロファイルによりNG2の発現を下方調節するが、MBP発現の遅延およびその誘導の低下を示した。D)図3Dは、分化の24〜96時間後における、MBPを発現するsiContトランスフェクトOLおよびsiMRFトランスフェクトOLの比率についての定量化を示す図である。siMRFトランスフェクト細胞は、すべての時点において、MRF発現率の低下を示した。**P<0.01。E)図3Eは、24〜96時間にわたる分化時の、siContまたはsiMRFをトランスフェクトしたOL内における、後期OLマーカーであるMOGの発現を示す図である。siMRFトランスフェクト細胞は、MOG発現に対する誘導の低下を示した。F)図3Fは、分化の24〜96時間後における、MOGを発現するsiContトランスフェクトOLおよびsiMRFトランスフェクトOLの比率についての定量化を示す図である。siMRFトランスフェクト細胞は、分化の48時間後以降、MRF発現率の低下を示した。**P<0.01。結果は、3つの個別の実験を表わす。
【図4】図4は、MRFノックダウン動物による、OL遺伝子発現の解析を示す図である。A)図4Aは、OPCとしてsiContまたはsiMRFをトランスフェクトされ、次いで、分化条件で48時間にわたり培養された細胞における遺伝子発現についてのノーザンブロット解析を示す図である。脳試料、心臓試料、および培養星状細胞試料に由来するRNAを、それぞれ、陽性対照および陰性対照として与えた。細胞にsiMRFをトランスフェクトしたところ、存在するMRF転写物の量が大幅に減少し、また、結果として、PLPの発現、また、より低度ではあるがCNP1の発現も明白に阻害された。GAPDHのノーザンブロット結果、および18Sリボソームバンドの可視化を、ローディング対照として示す。B)図4Bは、OPC、およびOPCとしてsiContまたはsiMRFをトランスフェクトされ、次いで、分化条件で48時間にわたり培養された細胞における遺伝子発現についてのAffymetrix解析の結果を示す図であり、これにより、選択されたOPCマーカーであるNG2、PDGFRα、およびKi67の発現レベルが示される。これらのOPCマーカーの下方調節は、MRFのノックダウンによる影響を受けなかった。C)図4Cは、対照(siContトランスフェクト細胞の値)に対する比率として表わされた、siMRFをトランスフェクトした細胞における、汎OL系統のマーカーであるSox10、初期OLマーカー(Ugt8、CNP1、PLP1、MBP)、および後期OLマーカー(MAG、トランスフェリン、MOBP、およびMOG)の発現を示す図である。siMRFトランスフェクト細胞において、OL遺伝子の発現は、siContトランスフェクト細胞の場合と比べて強く阻害され、分化後期のOLマーカー(トランスフェリン、MOG、およびMOBP)が、同初期マーカー(CNP1またはUgt8)または同中間段階マーカー(PLP1、MBP)より大きな影響を受けることが典型的であった。結果は、3つの個別の実験の平均であり、siContによる発現レベルの平均百分率±SEMとして表わされる。D)図4Dは、分化により>4倍に誘導された遺伝子と、siMRFのトランスフェクションにより<4分の1に抑制された遺伝子との重複を示すベン図である。siMRFにより阻害された遺伝子の大半(81%)は、OL分化時において通常は上方調節される遺伝子であった;これに対し、OLの分化時において通常は誘導される遺伝子のうち、MRFの発現に依存するのは13%に過ぎなかった。
【図5】図5は、MRF発現の不在下におけるOL遺伝子の発現についての表である。OPCとしてsiContもしくはsiMRFをトランスフェクトされ、次いで、分化条件において48時間にわたり培養された細胞、または遺伝子発現のベースラインを与える培養OPCにおける遺伝子発現についてのAffymetrix解析であり、これにより、MRFの不在下において発現の抑制を示す上位50の遺伝子が示される。siMRFの存在下において下方調節される上位50の遺伝子のうち、47は、OPCとsiCont OLとの間で4倍以上に上方調節される遺伝子であった。1つの遺伝子について複数のAffymetrixプローブセットが使用可能である場合、最も強く発現したものだけを示す。
【図6】図6は、MRFの異所性発現によりOLの分化が誘導されることを示す図である。A〜J)図6A〜Jは、pEGFP、および対照(空)ベクター、pSport6−MRF、またはpSport6−Sox10によるトランスフェクション後、48時間にわたり増殖条件(+PDGF、−T3)において培養したOPCを、NG2、MBP、またはMOGについて染色したことを示す図である。A〜C)図6A〜Cは、対照ベクター(A)、CMV−MRF(B)、またはCMV−Sox10をトランスフェクトした細胞を、NG2について染色したことを示す図である。対照ベクターまたはCMV−Sox10をトランスフェクトされたほとんどすべての細胞(GFPの共発現により同定される)は、NG2陽性を維持した(黄色の矢印)。これに対し、CMV−MRFをトランスフェクトされた多くの細胞が、NG2発現を下方調節する(緑色の矢印)一方、同じ培養物中における非トランスフェクト細胞はNG2発現を維持した(赤色の矢印)。D〜J)図6D〜Jは、対照ベクター(D、H)、pSport6−MRF(E、I)、またはpSport6−Sox10(F、J)をトランスフェクトした細胞を、MBP(D、E、F)、またはMOG(H、I、J)について染色したことを示す図である。対照ベクターまたはpSport6−Sox10をトランスフェクトされたほとんどすべての細胞は、MBP陰性およびMOG陰性を維持した(黄色の矢印)。これに対し、pSport6−MRFをトランスフェクトされた多くの細胞は、MBP発現およびMOG発現について陽性であった(黄色の矢印)。L〜M)図6L〜Mは、対照ベクター、pSport6−MRF、またはpSport6−Sox10をトランスフェクトされ、トランスフェクションの48または120時間後においてMBP(L)またはMOG(M)について陽性であった細胞の平均比率を示すグラフである。pSport6−MRFによるトランスフェクションは、大半の細胞がMBP陰性およびMOG陰性を維持した対照ベクターだけのトランスフェクションと比べ、48時間後および120時間後のいずれにおいても、MBP発現およびMOG発現の強力な誘導をもたらした。pSport6−Sox10による細胞へのトランスフェクションは、トランスフェクションの120時間後において、MBPおよびMOGに対する比較的低度の誘導を結果としてもたらした。**P<0.01。結果は、3つの個別の実験を表わす。
【図7】図7は、発生時のニワトリ脊髄内においてMRFを電気穿孔すると、早発性のMBP発現が引き起こされることを示す図である。E3における胚に、MRFおよびEGFPを含有するpCAGGSプラスミドを共電気穿孔し、P8において回収した。A)図7Aは、電気穿孔した脊髄側方だけにおいてMRF RNAの発現を示す、MRFについてのin situハイブリダイゼーションを示す図である。B)図7Bは、脊髄横断面におけるEGFPおよびMBPについての染色を示す図である。電気穿孔された脊髄側方では、偶発的なMBP+細胞(1断面当たり1または2個だけであることが典型的)が見出されたが、対照の側方では見出されなかった。C)図7Cは、脊髄の縦断面におけるEGFPおよびMBPについての染色が、MBPに対する免疫反応性が電気穿孔された側方に限定されることを示す図である。D)図7Dは、図7BおよびCにおいて示されるMBP+細胞の高倍率画像を示す図である。
【図8】図8は、MRF条件付きノックアウトマウスの解析および作製を示す図である。A)図8Aは、MRFを破壊するための戦略についての概略図である。野生型のMRF遺伝子座、標的化ベクター、および相同組換え後において予測される遺伝子座を示す。標的とされるマウスを、Flperマウスと交配させることにより、ネオマイシン耐性カセットを欠失させ、結果として、MRFのエキソン8をloxPで挟んだマウスを得る。略号:Ex8、エキソン8;プライマー1;P2、プライマー2;P3、プライマー3;FRT、Flpリコンビナーゼ部位;SP、サザンプローブ。B)図8Bは、MRF野生型マウス、MRFをloxPで挟んだヘテロ接合マウス、およびMRFをloxPで挟んだホモ接合マウスに由来するゲノムDNAに対するPCR増幅を示す図である。プライマー1および2は、460bpの野生型バンドを発生させ、また、loxP部位の挿入により、対立遺伝子をloxPで挟んだマウスにおいては、668bpのバンドも発生させた。プライマー1および3は、対立遺伝子をloxPで挟んだマウスに特異的な269bpのバンドを発生させた。プライマー1は、イントロン7〜8の上側鎖である。プライマー2は、エキソン8の下側鎖である。プライマー3は、プラスミド挿入物の下側鎖である。C)図8Cは、MRF遺伝子によりコードされる全長タンパク質、また、DNA結合ドメインおよび後に続くC末端領域を欠く、Creを介する切断後の切断タンパク質についての概略図である。
【図9】図9は、CNSの有髄化不全を示す、MRF条件付きノックアウトマウスを示す図である。A〜B)図9A〜Bは、P13においてMBP、NeuN、およびGFAPにより染色した、対照(MRFwt/fl;Olig2wt/Cre)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスの海馬、脳梁、ならびに皮質上部A)および脊髄B)を表わす画像である。C)図9Cは、P13におけるMRF対照マウスおよびMRF条件付きノックアウトマウスの脊髄における、CNP、MBP、MOG、GFAP、およびニューロフィラメントの発現についてのウェスタンブロット解析を示す図である。D)図9Dは、P13における対照(MRFwt/fl;Olig2wt/Cre)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスにおける脊髄の脊髄根および側方白質に対するフルオロミエリン染色を表わす画像である。E)図9Eは、P13における対照動物および条件付きノックアウト動物の神経を表わす電子顕微鏡画像である。対照動物の神経は、著明量の有髄化が進行中であることを示している。これに対し、条件付きノックアウト動物の神経は、軸索の有髄化を一切示さなかった。
【図10】図10は、視神経および坐骨神経内におけるPLPおよびMRFについてのin situハイブリダイゼーションを示す図である。PLPについてのシグナルは、既に報告されている通り(Puckettら、J. Neurosci. Res.、18巻、511〜518頁(1987年))、坐骨神経では低レベルまでであるが、視神経内および坐骨神経内のいずれにおいても(CNS神経およびPNS神経のいずれにおいても)容易に検出可能であった。これに対し、MRFについてのシグナルが視神経内で検出されたことは、これが、OLによっては発現するが、シュワン細胞によっては発現しない(CNSの神経膠細胞によっては発現するが、PNSの神経膠細胞によっては発現しない)ことを示す。
【図11】A〜B)図11A〜Bは、siCont(A)またはsiMRF(B)をトランスフェクトした後で48時間にわたり分化させた、培養希突起膠細胞についての位相イメージングを示す図である。いずれの場合も、大半の細胞は希突起膠細胞の形態を帯びるが、siMRFをトランスフェクトした細胞は、それほど広範な突起を示さなかった。C〜D)図11C〜Dは、siCont(C)またはsiMRF(D)をトランスフェクトした後で72時間にわたり分化させた培養希突起膠細胞上における、MOGおよびGalCについての表面染色(O1抗体による)を示す図である。siContトランスフェクト細胞およびsiMRFトランスフェクト細胞のいずれも、O1抗体により容易に標識化されたが、MOGについて陽性であったのは、siCont細胞だけであった。
【図12】A)図12Aは、対照のsiRNAプール、またはMRFに対するsiRNAプールをトランスフェクトした培養希突起膠細胞内において、MRFおよびMBPを検出するRT−PCR(30サイクル)を示す図である。siMRFトランスフェクト細胞では、MRF転写物レベルおよびMBP転写物レベルのいずれもが明白な低下を示した。B)図12Bは、対照のsiRNAプール、またはMRFに対する個別のsiRNAをトランスフェクトした培養希突起膠細胞内において、MRFおよびMBPを検出するRT−PCR(30サイクル)を示す図である。4つの個別のsiRNAのうちの複数が、siContトランスフェクト細胞と比べて検出可能な、MRFレベルおよびMBPレベルの低下を引き起こした。C)図12Cは、図12Bと同じトランスフェクションに由来する細胞が、分化条件における4日間にわたる培養後において示す、MOGを発現する生存細胞の比率の高さを示す図である。MRFに対する4つの個別のsiRNAのうちの3つが、siContトランスフェクト細胞と比べて、MOGを発現する細胞の比率を著明に低下させたが、これは、図12Bにおいて観察されたMRFのノックダウンと良好に相関する。
【図13】図13は、MRF条件付きノックアウト動物が、成熟希突起膠細胞の喪失を示すことを明らかにする図である。A)図13Aは、P13における対照(MRFwt/fl;Olig2wt/CreおよびMRFfl/fl;Olig2wt/wt)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスの視神経内における、MBP、CC1、NG2、GFAP、ならびにCC1およびPDGFRαと共染色されたOlig2についての免疫染色を示す図である。スケールバー=50μm。B)図13Bは、視神経内におけるOlig2免疫陽性核の密度についての定量化を示す図である。C)図13Cは、視神経内におけるOlig2+/CC1+二重免疫陽性OLの密度についての定量化を示す図である。D)図13Dは、視神経内におけるOlig2+/PDGFRa+二重免疫陽性細胞の密度についての定量化を示す図である。すべての結果は、平均±SEM、1遺伝子型当たりn=4〜5として表わされる。*P<0.05、**P<0.01。E)図13Eは、CC1についてもまた陽性であるOlig2+細胞(OL)、PDGFR1αについてもまた陽性であるOlig2+細胞(OPC)、またはいずれのマーカーについても陽性でないOlig2+細胞に分類される、視神経内における各遺伝子型のOlig2免疫陽性細胞の密度を示す図である。
【図14】図14は、条件付きノックアウト動物の培養希突起膠細胞および脊髄における遺伝子発現を示す図である。対照(MRFwt/fl;Olig2wt/Cre)動物の脳、および条件付きノックアウト(CKO;MRFfl/fl;Olig2wt/Cre)動物の脳に由来するOL培養物(4日間にわたり分化させた)、ならびにP13における対照(MRFwt/fl;Olig2wt/Cre)マウス、および条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスから急性摘出された脊髄に対するGeneChip解析の結果である。A)図14Aは、CKO動物の培養OLにおいて、対照動物の培養OLと比べて最も大幅に下方調節された20遺伝子を、対照動物のOLおよびCKO動物のOL、ならびに対照動物の脊髄およびCKO動物の脊髄におけるMAS 5.0遺伝子の発現値と共に列挙する。また、培養OL(対照値/CKO値)、脊髄(対照値/CKO値)、およびsiRNA実験(siCont値/siMRF値)についても、抑制倍率の値を示す。B)図14Bは、対照動物のOLおよびCKO動物のOL、ならびに対照動物の脊髄およびCKO動物の脊髄における汎OL系統マーカー、OPCマーカー、初期OLマーカー、後期OLマーカーを表わすプローブセット、およびこれらの発現値を示す図である。「A」と表示した遺伝子を発現しない遺伝子と判定し、「B」と表示した遺伝子を発現した遺伝子と考える。「B」とは、わずかな発現に相当する。アレイ上における所与の遺伝子に対して複数のプローブセットが存在した場合は、最も強く発現したプローブセットだけを示す。
【図15】図15は、MRF欠損OPC/OL培養物が、分化条件では欠損を示すが、増殖条件では欠損を示さないことを明らかにする図である。A)図15Aは、増殖(+PDGF、−T3)条件における、NG2およびKi67についての、対照(MRFwt/fl;Olig2wt/Cre)動物による培養物、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)動物による培養物の免疫染色を示す図である。いずれの場合においても、大半の細胞は、NG2陽性前駆細胞およびKi67陽性前駆細胞(Ki67+核を矢印により示す)として維持された。B)図15Bは、分化(−PDGF、+T3)条件において4日間にわたり増殖させた、対照動物による培養物、およびMRF条件付きノックアウト動物による培養物に対する、Ki67、CNP、およびMBPについての免疫染色を示す図である。いずれの場合においても、大半の細胞は、Ki67の発現を下方調節し、OLに特徴的な多極形態を帯び(CNP染色により可視化される)、GFAPについては陰性であった。しかし、頑健なMBP染色が見られたのは、対照(MRFwt/fl;Olig2wt/Cre)動物による培養物中だけであった。スケールバー=100μm。C)図15Cは、OL系統の特殊化および分化に対する転写制御についての概略図である。Olig2は、OL系統の特殊化に必要である。Sox10、Nkx2.2、Yin Yang 1、およびOlig1を含めた複数の遺伝子が、OLへのOPCの頑健な分化には必要である。MRFは、ミエリン遺伝子の完全な相補体を発現する、成熟OLへの未成熟OLの成熟化に必要であり、その誘導はおそらくYin Yang 1により調節される。
【図16】図16は、CNP−Creを介してMRFが欠失する結果として、MRFfl/fl;Olig2wt/Creマウスと同等の、有髄化不全を引き起こす表現型がもたらされることを示す図である。A)図16Aは、MBPにより染色された、P16における対照(MRFfl/fl;CNPwt/wt)動物の脳、および条件付きノックアウト(MRFfl/fl;CNPwt/Cre)動物の脳を表わす画像であり、これらにより、CNP−Creを介するMRF条件付きノックアウト動物の脳梁(cc)内におけるMBP染色の大幅な喪失が示される。一部の、わずかにMBP+であり、有髄化しないOLが、条件付きノックアウト動物の脳内に存在する(矢印)。スケールバー=200μm。B)図16Bは、フルオロミエリンにより染色された、P16における対照(MRFfl/fl;CNPwt/wt)動物の脊髄、および条件付きノックアウト(MRFfl/fl;CNPwt/Cre)動物の脊髄を表わす画像である。条件付きノックアウト動物では、末梢における有髄化(脊髄根;s.r.)は影響を受けないと考えられるが、CNSにおける有髄化は見られない。
【図17】図17は、条件付きノックアウトマウスが、視神経内において細胞アポトーシスの増大を示すことを明らかにする図である。A)図17Aは、抗MBP抗体および抗活性化カスパーゼ3抗体により染色された、P10における対照動物の神経、および条件付きノックアウト動物の視神経を表わす画像である。スケールバー=100μm。B)図17Bは、P10における対照動物の視神経、および条件付きノックアウト動物の視神経内における活性化カスパーゼ3免疫陽性細胞の密度についての定量化により、条件付きノックアウト動物において、アポトーシス細胞の密度の著明な上昇が明らかとなったことを示す図である(**P<0.01。n=5〜6/遺伝子型)。C)図17Cは、図17Aで条件付きノックアウト動物の神経について枠囲いした領域を高拡大率で示す図であり、これにより、核がフラグメント化し、活性化カスパーゼ3が染色される(矢印)、低度のMBP陽性細胞が示される。
【図18】図18は、条件付きノックアウト動物の遺伝子プロファイリングを示す図である。A)図18Aは、対照(MRFwt/fl;Olig2wt/CreおよびMRFfl/fl;Olig2wt/wt)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスに由来する脊髄または培養OLに由来するRNA中における選択遺伝子についてのRT−PCR(30サイクル)解析を示す図であり、これにより、条件付きノックアウト動物による試料中において、OLマーカーであるMOGおよびMOBPの喪失が示される。該遺伝子のエキソン8によりコードされるRNA配列を認識するプライマーを用いて、条件付きノックアウト動物に由来する脊髄または培養OLからPCR産物を検出することはできず、これにより、このエキソンの欠失が確認された。これに対し、MRFのエキソン8の外側に位置するプライマーは、条件付きノックアウト動物による培養OL中におけるMRF発現を検出したが、脊髄中におけるMRF発現は検出しなかった。
【発明を実施するための形態】
【0016】
参照による組込み
本出願において言及されるすべての刊行物および特許出願は、各個別の刊行物または特許出願が、参照により組み込まれるように具体的かつ個別に指示されたと仮定する場合と同程度に、参照により本明細書に組み込まれる。
発明の詳細な説明
本発明は、有髄化/再有髄化の促進において、ミエリン遺伝子調節因子(MRF)とも称する遺伝子モデル98(GM98)、および/またはそれに関連する遺伝子を調節する方法および組成物を提供する。例えば、MRFを標的とし、MRFを介して直接的または間接的に調節しうる、ミエリン塩基性タンパク質(MBP)、ミエリン希突起膠細胞糖タンパク質(MOG)、ミエリン関連希突起膠細胞塩基性タンパク質(MOBP)、およびプロテオリピドタンパク質(PLP)など、ミエリン生成における主要な成分と考えられることが典型的な多くの遺伝子の発現に対する下方調節を結果としてもたらすsiRNAにより、希突起膠細胞におけるMRFの発現を阻害することができる。逆に、例えば、MRF cDNAを含有する発現プラスミドを用いてMRFを発現させることにより、希突起膠細胞の前駆細胞(OPC)におけるMRFの発現を誘導し、これにより、再有髄化を促進することができる。
【0017】
本明細書ではまた、MRFの発現を調節し、有髄化を促進する生物活性剤をスクリーニングする方法も提供される。例えば、それらを、Olig2プロモーターの下流でCreリコンビナーゼを発現するマウス株と交配させることにより、希突起膠細胞系統の細胞内において、loxPに隣接するMRF遺伝子(DNA結合領域の一部をコードする)のエキソン8を切除しうるマウス。該マウスは、MBP陽性の希突起膠細胞またはCNSミエリンの発現に失敗し、広範にわたるCNSの脱髄により、生後3週間で死亡することが典型的である。これらのマウスを用いて、死亡を遅延させ、有髄化不全を軽減し、かつ/またはMBP陽性OLの発生を促進する生物活性剤をスクリーニングすることができる。
【0018】
本明細書に記載の方法および組成物は、神経障害を治療するのに重要でありうる。本明細書で開示される方法および組成物を用いて、多発性硬化症(MS)、進行性多巣性白質脳症(PML)、脳脊髄炎、橋中央ミエリン溶解(CPM)、抗MAG抗体陽性疾患、白質萎縮症:副腎白質萎縮症(ALD)、アレキサンダー病、カナバン病、クラッベ病、異染性白質萎縮症(MLD)、ペリツェウス−メルツバッハー病、レフサム病、コケイン症候群、ファンデルクナップ症候群、ツェルヴェーガー症候群、ギラン−バレー症候群(GBS)、慢性炎症性脱髄性多発神経障害(CIDP)、多巣性運動神経障害(MMN)、脊髄傷害(例えば、脊髄の外傷または切断)、アルツハイマー病、ハンチントン病、筋萎縮性側索硬化症、パーキンソン病、および視神経炎などであるがこれらに限定されない各種のCNS障害およびPNS障害を治療することができる。
【0019】
一般的な技法
本発明を実施する際には、別段に指示しない限り、当技術分野内にある免疫学、生化学、化学、分子生物学、微生物学、細胞生物学、遺伝学、および組換えDNAの従来の技法が用いられる。Sambrook, Fritsch、およびManiatis、「MOLECULAR CLONING: A LABORATORY MANUAL」、第2版(1989年);「CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubelら編(1987年));「METHODS IN ENZYMOLOGY」のシリーズ(Academic Press社)、「PCR 2: A PRACTICAL APPROACH」(M.J. MacPherson、B.D. Hames、およびG.R. Taylor編(1995年));HarlowおよびLane編(1988年)、「ANTIBODIES, A LABORATORY MANUAL」;ならびに「ANIMAL CELL CULTURE」(R.I. Freshney,編(1987年))を参照されたい。
【0020】
定義
本明細書および特許請求の範囲で用いられる単数形の「ある(a)」、「ある(an)」、および「その(the)」は、文脈により別段に明確な指示がなされない限り、複数形の指示を包含する。例えば、「ある細胞(a cell)」は、複数の細胞を、それらの混合物を含めて包含する。
【0021】
「対照」という用語は、実験において比較の目的で用いられる代替的な被験体、細胞、または試料である。対照は、「陽性」の場合もあり、「陰性」の場合もある。例えば、対照細胞は、所与の対象細胞における遺伝子産物の差次的発現についてアッセイする際に用いることができる。対照細胞による遺伝子産物の発現を、被験細胞、例えば、生物活性剤と接触させた被験細胞と比較することができる。さらに、「対照」はまた、異なる時点について比較する実験における同じ被験体、細胞、または試料も表わしうる。生物活性剤をスクリーニングする文脈において、対照細胞は、被験生物活性剤と接触させていない神経細胞でありうる。
【0022】
「ポリヌクレオチド」、「ヌクレオチド」、「ヌクレオチド配列」、「核酸」、および「オリゴヌクレオチド」という用語は、互換的に用いられる。これらは、デオキシリボヌクレオチドもしくはリボヌクレオチド、またはこれらの類似体である、任意の長さのヌクレオチドのポリマー形態を指す。ポリヌクレオチドは、任意の3次元構造を有することが可能であり、公知のものであれ、未知のものであれ、任意の機能を果たしうる。以下が、ポリヌクレオチドの非限定的な例である:遺伝子または遺伝子フラグメントのコード領域または非コード領域、連鎖解析により定義される(1または複数の)遺伝子座、エキソン、イントロン、メッセンジャーRNA(mRNA)、トランスファーRNA、リボソームRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝鎖ポリヌクレオチド、プラスミド、ベクター、任意の配列を有する単離DNA、任意の配列を有する単離RNA、核酸プローブ、およびプライマー。ポリヌクレオチドは、メチル化されたヌクレオチドおよびヌクレオチド類似体など、修飾ヌクレオチドを含みうる。存在させる場合、ヌクレオチド構造に対する修飾の付与は、ポリマーのアセンブリー前の場合もあり、アセンブリー後の場合もある。ヌクレオチド配列には、ヌクレオチド以外の成分を介在させることができる。ポリヌクレオチドは、標識化成分とのコンジュゲーションなどにより、重合化の後においてさらに修飾することができる。
【0023】
本明細書で用いられる「発現」とは、ポリヌクレオチドがmRNAへと転写される過程、および/または、転写されたmRNA(また、「転写物」とも称する)が、その後、ペプチド、ポリペプチド、またはタンパク質へと翻訳される過程を指す。転写物およびコードされたポリペプチドを、集合的に、「遺伝子産物」と称する。ポリヌクレオチドがゲノムDNAに由来する場合、発現は、真核細胞におけるmRNAのスプライシングを包含しうる。
【0024】
「接触」、「送達」、および「投与」という用語は、薬剤が、被験体、組織、または細胞に入ることを意味するように用いることができる。本明細書の開示全体で用いられる該用語には、特定の用語の文法的な変化形も含まれる。例えば、「送達」には、「送達すること」、「送達された」、「送達する」などが含まれる。当技術分野では、生物活性剤を送達または投与する各種の方法が公知である。例えば、本明細書に記載の1または複数の薬剤は、非経口送達、経口送達、腹腔内送達、静脈内送達、動脈内送達、経皮送達、筋肉内送達、リポソーム送達、カテーテルもしくはステントによる局所送達、皮下送達、脂肪組織内送達、または髄腔内送達することができる。
【0025】
ヌクレオチド配列またはポリペプチド配列に適用される、「差次的発現」という用語は、対照において検出される発現と比較した、その配列の過剰発現または過小発現を指す。過小発現はまた、対照と比較した、被験対象における検出可能な発現の不在を証拠とする、特定の配列の発現の不在も包含する。
【0026】
本明細書において、「ポリペプチド」、「ペプチド」、および「タンパク質」という用語は互換的に用いられ、任意の長さのアミノ酸ポリマーを指す。該ポリマーは、直鎖状の場合もあり、分枝状の場合もあり、修飾アミノ酸を含む場合もあり、また、アミノ酸以外の分子を介在させる場合もある。該用語はまた、修飾されたアミノ酸ポリマー、例えば、ジスルフィド結合の形成、グリコシル化、脂質化、アセチル化、リン酸化、または、標識化成分とのコンジュゲーションなど、他の任意の操作を施されたアミノ酸ポリマーも包含する。本明細書で用いられる「アミノ酸」とは、グリシン、およびD光学異性体またはL光学異性体、ならびにアミノ酸類似体およびペプチド模倣剤を含めた、天然アミノ酸および/または非天然アミノ酸もしくは合成アミノ酸を指す。
【0027】
本明細書において、「被験体」、「個体」、または「患者」は、互換的に用いられ、脊椎動物、好ましくは、哺乳動物、より好ましくはヒトを指す。哺乳動物には、マウス、ラット、イヌ、ブタ、サル(類人猿)、ヒト、農場動物、競技動物、およびペットが含まれるがこれらに限定されない。in vivoにおいて得られたか、またはin vitroにおいて培養された生物学的実体を有する組織、細胞、およびそれらの前駆体もまた包含される。
【0028】
本明細書において、「治療」、または「治療すること」、または「改善すること」は、互換的に用いられる。これらの用語は、臨床的な結果を含めた、また、臨床的結果であることが好ましい、有益または所望の結果を得るための手法を指す。本発明を目的とする場合、有益または所望の臨床的結果には、以下:脱髄病変サイズの縮減(例えば、脱髄障害の関連において)、OPCの増殖および成長もしくは病変部位へのOPCの遊走の促進、希突起膠細胞分化の促進、神経障害発症の遅延化、脱髄障害発生の遅延化、神経障害から生じる症状の軽減、疾患を患う患者の生活の質の向上、疾患を治療するのに必要な他の薬物用量の低減、標的化および/もしくは封入などによる別の薬物の効果の増強、疾患の進行の遅延化、ならびに/または個体の生存の延長のうちの1または複数が含まれるがこれらに限定されない。治療は、疾患を防止すること、すなわち、疾患が誘発される前において、保護的組成物を投与することにより、疾患の臨床症状を発生させないようにすること;疾患を抑制すること、すなわち、疾患の誘発的事象の後ではあるが、その臨床的な発現または再発現の前において、保護的組成物を投与することにより、疾患の臨床症状を発生させないようにすること;疾患を阻害すること、すなわち、それらの最初の発現後において、保護的組成物を投与することにより、臨床症状の発生を停止させること;疾患の再発を防止すること、および/または疾患を緩和すること、すなわち、それらの最初の発現後において、保護的組成物を投与することにより、臨床症状を退縮させることを包含する。
【0029】
「薬剤」、「生物学的に活性な薬剤」、「生物活性剤」、「生物活性化合物」、または「生物学的に活性な化合物」という用語は互換的に用いられ、また、言及される文脈において、複数の指示対象も包含する。本明細書で記載される、本発明の1または複数の組合せ治療方法において用いられるこのような化合物には、単純有機分子もしくは複合有機分子、または単純無機分子もしくは複合無機分子、ペプチド、ペプチド模倣剤、タンパク質(例えば、抗体)、DNA、RNA、およびこれらの類似体を含めた核酸分子、炭水化物含有分子、リン脂質、リポソーム、低分子干渉RNA、またはポリヌクレオチド(例えば、アンチセンス)が含まれるがこれらに限定されない。
【0030】
このような薬剤は、神経細胞の増殖または分化と関連する細胞周期経路の構成要素に対するアゴニストまたはアンタゴニストでありうる。本発明の一部の実施形態では、結合部位において同じ3次元構造を有する化合物は、アンタゴニストとして用いうることが意図される。化学構造の3次元解析を用いて、神経細胞周期に関連するポリペプチドに対する結合部位を含めた、活性部位の構造を決定する。
【0031】
本明細書で用いられる「アンタゴニスト」という用語は、標的ポリペプチドの生物学的機能を阻害する能力を有する分子を指す。したがって、「アンタゴニスト」という用語は、標的ポリペプチドの生物学的な役割の文脈で定義される。本明細書で好ましいアンタゴニストが、標的と特異的に相互作用する(例えば、標的に特異的に結合する)一方で、該標的ポリペプチドがそのメンバーであるシグナル伝達経路の他のメンバーと相互作用することにより、該標的ポリペプチドの生物学的活性を阻害する分子もまた、この定義内に具体的に包含される。アンタゴニストにより阻害されることが好ましい生物学的活性は、OPC増殖の増大、OPC増殖の減少、OL分化の増大、または星状細胞増殖の増大、および/または再有髄化の促進と関連する。例えば、アンタゴニストは、神経細胞周期と関連するペプチドと直接的に相互作用する場合もあり、間接的に相互作用する場合もある。限定なしに述べれば、本明細書で定義されるアンタゴニストには、オリゴヌクレオチドデコイ、アプタマー、抗ケモカイン抗体および抗ケモカイン抗体変異体、ペプチド、ペプチド模倣剤、ペプチド以外の低分子、アンチセンス分子、ならびに有機低分子が含まれる。
【0032】
本明細書で用いられる「アゴニスト」という用語は、標的ポリペプチドの生物学的機能を誘発または増強する能力を有する分子を指す。したがって、「アゴニスト」という用語は、標的ポリペプチドの生物学的な役割の文脈で定義される。本明細書で好ましいアゴニストが、標的と特異的に相互作用する(例えば、標的に特異的に結合する)一方で、該標的ポリペプチドがそのメンバーであるシグナル伝達経路の他のメンバーと相互作用することにより、該標的ポリペプチドの生物学的活性を阻害する分子もまた、この定義内に具体的に包含される。アゴニストにより阻害されることが好ましい生物学的活性は、OPC増殖の増大、OPC増殖の減少、OL分化の増大、または星状細胞増殖の増大、それによる再有髄化の促進と関連する。限定なしに述べれば、本明細書で定義されるアンタゴニストには、オリゴヌクレオチドデコイ、アプタマー、抗ケモカイン抗体および抗ケモカイン抗体変異体、ペプチド、ペプチド模倣剤、ペプチド以外の低分子、アンチセンス分子、ならびに有機低分子が含まれる。
【0033】
神経細胞の増殖/分化に対するアゴニスト、アンタゴニスト、および他の調節物質が、本発明の範囲内に明示的に包含される。特定の実施形態において、アゴニスト、アンタゴニスト、および他の調節物質は、神経細胞周期、すなわち、増殖または分化の調節に関与するポリペプチドに結合する抗体および免疫グロブリンの変異体である。これらのアゴニスト性、アンタゴニスト性の調節化合物は、直鎖状形態で提供することもでき、環状形態で提供することもでき、また、場合によって、一般に天然では見出されない、少なくとも1つのアミノ酸残基、または少なくとも1つのアミドイソステアを含みうる。これらの化合物は、グリコシル化、リン酸化、硫酸化、脂質化、または他の過程により修飾することができる。
【0034】
「有効量」または「治療有効量」という用語は、限定せずに述べれば、脱髄病変サイズの縮減(例えば、脱髄障害の関連において)、OPCの増殖および成長の促進、神経障害発症の遅延化、脱髄障害発生の遅延化、神経障害から生じる症状の軽減、疾患を患う患者の生活の質の向上、疾患を治療するのに必要な他の薬物用量の低減、標的化および/もしくは封入などによる別の薬物の効果の増強、疾患の進行の遅延化、ならびに/または個体の生存の延長などの臨床的結果を含めた、有益または所望の結果をもたらすのに十分な量のアンタゴニストを指す。治療有効量は、治療される被験体および疾患状態、被験体の体重および年齢、疾患状態の重症度、投与方式などに応じて変化するが、これらは、当業者により容易に決定されうる。該用語はまた、本明細書に記載の造影方法のいずれか1つにより検出される画像をもたらす用量にも適用される。具体的な用量は、選択される具体的なアンタゴニスト、従う投与レジメン、それが他の化合物と組み合わせて投与されるかどうか、投与回数、造影される組織、それが移送される物理的な送達系に応じて変化する。
【0035】
本明細書で用いられる「抗体」という用語には、組換え抗体、ヒト化抗体、キメラ抗体、単鎖抗体、ヒト化抗体、融合タンパク質、モノクローナル抗体など、すべての抗体形態が含まれる。本発明はまた、神経細胞周期に関与するポリペプチドに対する結合(例えば、神経細胞の増殖/分化の調節に関与する転写因子またはタンパク質に対する結合)が可能な、抗体の機能的フラグメントにも適用可能である。
【0036】
一実施形態では、比較的低用量の全抗体、裸抗体、または全抗体、裸抗体の組合せが用いられる。一部の実施形態では、抗体フラグメント、したがって、完全抗体未満のフラグメントが用いられる。他の実施形態では、薬物、毒素、または治療用放射性同位体との抗体のコンジュゲートが有用である。複数の抗原に結合するハイブリッド抗体を含め、ケモカイン抗原に結合する二特異性の抗体融合タンパク質を、本発明に従い用いることができる。したがって、抗体は、一特異性の場合もあり、多重特異性の場合もある、裸抗体およびコンジュゲート抗体および抗体フラグメントを包含する。
【0037】
「調節すること」、「調節された」、または「調節」という用語は互換的に用いられ、所与の文脈における直接的または間接的な変化を意味する。例えば、MRF発現の調節は、結果として、神経細胞の分化および/または有髄化を変化させる。別の例において、調節は、それ自体が、MRFに関与する遺伝子の発現を調節しうる遺伝子/遺伝子産物の調節でありうる。
【0038】
生物活性剤に適用される「アプタマー」という用語には、特定の分子に対する特異的な結合特性に基づいて選択されるDNA、RNA、またはペプチドが含まれる。例えば、本明細書で開示される、神経細胞周期に関与する特定の遺伝子または遺伝子産物に対する結合について、(1または複数の)アプタマーを選択することができ、この場合、選択は、当技術分野において公知であり、当業者に周知の方法によりなされる。その後、(1または複数の)前記アプタマーを被験体に投与して、免疫反応を調節または制御することができる。特定のタンパク質、DNA、アミノ酸、およびヌクレオチドに対する親和性を有する一部のアプタマーについては説明されている(K. Y. Wangら、Biochemistry、32巻、1899〜1904頁(1993年); Pitner ら、米国特許第5,691,145号;Goldら、Ann. Rev. Biochem.、64巻、763〜797頁(1995年);Szostakら、米国特許第5,631,146号)。組合せライブラリーからは、高親和性で高特異性の結合アプタマーが誘導されている(前出、Goldら)。アプタマーは、用いられる選択に応じて、平衡解離定数がマイクロモルからナノモル未満の範囲にわたる高親和性を有しうる。アプタマーはまた、高度の選択性も示し、例えば、7−メチルGとGとの間で(HallerおよびSarnow、Proc. Natl. Acad. Sci. USA、94巻、8521〜8526頁(1997年))、またはD−トリプトファンとL−トリプトファンとの間で(前出、Goldら)1000倍の識別力を示す。
【0039】
MRF発現の調節
本発明の各種の態様では、細胞型または組織に特異的な発現調節エレメントが、MRFまたはその機能的な変異体をコードする核酸配列に作動的に連結される。MRFをコードする1または複数の核酸配列に、1または複数の調節エレメントを連結することができる。MRF配列は、ヒトオルトログ(図2Aに示されるC11Orf9)、他のオルトログ、相同体、またはこれらの機能的変異体の配列でありうる。MRF配列は、in vitroでもin vivoでも生物学的効果を及ぼすことが可能であり、したがって、生物活性剤でありうる。例えば、MRFは、OLの成熟および/または有髄化を促進しうる。調節エレメントは、選択的なMRF発現に影響を及ぼすことが可能であり、かつ/またはMRFの誘導的発現もしくは構成的発現を提供しうる。MRF発現はまた、他の生物活性剤によっても調節または制御することができる。
【0040】
対象の方法で用いられる生物活性剤は、MRF、また、図5に示される、MRFにより調節される遺伝子など、それに相関する遺伝子、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、もしくはMogなど、OL分化において発現する他の遺伝子の活性または発現のレベルを調節するのに有効である。代替的に、このような遺伝子のサブセット、例えば、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子もまた、本明細書で提供される。相関する遺伝子は、成熟OLにおいて特異的に上方調節される遺伝子でありうる。生物活性剤は、MRFの発現および/または活性を調節することにより、OLの成熟および有髄化/再有髄化に影響する。調節は、MRFの活性または発現のレベルを上昇または低下させることを伴いうる。例えば、薬剤は、MRFまたはMRFの調節に関与する他の遺伝子産物に対するアゴニストの場合もあり、アンタゴニストの場合もある。このような生物活性剤の非限定的な例示的類型は、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、低分子、調合薬、またはこれらの組合せである。
【0041】
MRFなどを含めた生物活性剤は、このような薬剤が発現して、OLの分化もしくは成熟、および/または有髄化を促進するなど、それらの所望の機能を付与するように、細胞または組織内において発現しうる。遺伝子発現は、構成的プロモーターまたは誘導的プロモーター、細胞型特異的な発現調節エレメント、およびエンハンサーが含まれるがこれらに限定されない特定の調節エレメントの制御下に置かれることが典型的である。このような遺伝子は、調節エレメントに作動的に連結されるという。例えば、構成的プロモーター、誘導的プロモーター、または細胞/組織特異的プロモーターを発現ベクター内に組み込んで、宿主細胞内で発現する遺伝子の発現を調節することができる。したがって、用いられるプロモーターエレメントに応じて、生物活性剤は、MRFの発現またはその活性を遮断、増強、または促進するように、所望に応じて発現させることができる。例えば、MRFの機能を促進する薬剤を細胞内において一時的に発現させる結果、OLの分化を増強させ、この結果、最終的に、有髄化/再有髄化をもたらすことができる。調節配列は、中枢神経系または末梢神経系の特定の細胞型における、生物活性剤の異所性発現を可能とする。例えば、選択的なMRFの調節は、神経膠細胞などの神経細胞などであるがこれらに限定されない細胞内において達成することができる。神経膠細胞には、希突起膠細胞、小膠細胞、シュワン細胞、または星状細胞が含まれうる。
【0042】
調節配列の例示的な発現には、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、ミエリン関連糖タンパク質(MAG)遺伝子、ミエリン希突起膠細胞糖タンパク質(MOG)遺伝子、 希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFR−α遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子、2(VEGFR2)遺伝子、スーパーオキシドディスムターゼ(SOD1)遺伝子、チロシンヒドロキシラーゼ遺伝子、ニューロン特異的エノラーゼ遺伝子、パーキン遺伝子(PARK2)、パーキン共調節遺伝子(PACRG)、ニューロン特異的Tα1 α−チューブリン(Tα1)遺伝子、小胞モノアミントランスポーター(VMAT2)遺伝子、およびα−シヌクレイン(SNCA)遺伝子、PDGFR−β遺伝子,Olig1遺伝子、Olig2遺伝子、またはプロテオリピドタンパク質(PLP)遺伝子が含まれるがこれらに限定されない遺伝子から選択される調節配列が含まれる。
【0043】
当技術分野では、その関連する開示が参照により本明細書に組み込まれる、米国特許出願公開第2003/0110524号;同第2003/0199022号;同第2006/0052327号、同第2006/0193841号、同第2006/0040386号、同第2006/0034767号、同第2006/0030541号;米国特許第6472520号、同第6245330号、同第7022319号、同第7033595号において開示されるなど、神経細胞特異的なプロモーターのさらなる例が公知である。また、ウェブサイト<chinook.uoregon.edu/promoters.html>;または<tiprod.cbi.pku.edu.cn:8080/index>(特定の細胞/組織に特異的な遺伝子のプロモーターを列挙する);およびPattersonら、J. Biol. Chem.(1995年)、270巻、23111〜23118頁も参照されたい。
【0044】
MRFおよびMRFの調節物質の発現はまた、細胞型/組織特異的プロモーターを用いる発現系以外の発現系(例えば、エフェクター分子を局所投与する発現系)を用いることによっても、一時的に調節することができる。したがって、一部の実施形態では、細胞死のメディエータータンパク質をコードする遺伝子を、tet応答性プロモーターなど、制御可能なプロモーターエレメントに作動的に連結することができる。例えば、所望の場所および時点において、誘導性薬剤(例えば、テトラサイクリンまたはその類似体)を細胞または被験体に投与し、細胞/組織特異的な形で(例えば、テトラサイクリンだけが、局所的/限定的な形で送達される)、細胞死のメディエータータンパク質の発現を誘導することができる。このような系は、テトラサイクリンの存在が発現を阻害する「オフスイッチ」系、またはテトラサイクリンの存在が発現を誘導するように、大腸菌TetRの突然変異体が用いられる「可逆性の」Tet系を包含することにより、真核細胞における遺伝子発現の緻密な制御をもたらしうる。これらの系は、例えば、GossenおよびBujard(Proc. Natl. Acad. Sci. U.S.A.(1992年)、89巻、5547頁)およびBujardら、による米国特許第5,464,758号;同第5,650,298号;および同第5,589,362号において開示されている。
【0045】
誘導的プロモーターのさらなる例には、MMTV、熱ショック70プロモーター、GAL1−GAL10プロモーター、メタロチオネイン(metallothien)誘導プロモーター(例えば、銅誘導ACE1;他の金属イオン)、ホルモン応答エレメント(例えば、グルココルチコイド、エストロゲン、プロゲステロン(progestrogen))、ホルボールエステル(TREエレメント)、カルシウムイオノフォア応答エレメント、または脱共役タンパク質3、ヒト葉酸受容体α、乳清酸性タンパク質、プロテアーゼ特異的プロモーターのほか、米国特許第6,313,373号において開示されるプロモーターが含まれるがこれらに限定されず;また、<biobase/de/pages/products/transpor.html>におけるオンライン(遺伝子/活性により分類される15,000を超える異なるプロモーター配列を伴うデータベースを提供する);また、Chenら、Nuc. Acids. Res.、2006年、34巻、データベース版、D104〜107頁を参照されたい。
【0046】
さらに他の誘導的プロモーターには、成長ホルモンプロモーター;アデノウイルスE1Aタンパク質により誘導される、アデノウイルス初期遺伝子プロモーター、もしくはアデノウイルス主要後期プロモーターなど、プロモーターヘルパーウイルスにより誘導されるプロモーター;VP16もしくは1CP4などのヘルペスウイルスタンパク質により誘導されるヘルペスウイルスプロモーター;牛痘ウイルスもしくはポックスウイルスのRNAポリメラーゼにより誘導されるプロモーター;または、T7 RNAポリメラーゼ、T3 RNAポリメラーゼ、またはSP6 RNAポリメラーゼによりそれぞれ誘導される、T7プロモーター、T3プロモーター、およびSP6プロモーターなどのバクテリオファージプロモーターが含まれる。
【0047】
他の実施形態では、構成的プロモーターが所望でありうる。例えば、アデノウイルス主要後期プロモーター、サイトメガロウイルス即初期プロモーター、βアクチンプロモーター、またはβグロビンプロモーターを含め、本発明において用いるのに適する多くの構成的プロモーターが存在する。当技術分野では、他の多くの構成的プロモーターが公知であり、本発明において用いることができる。またさらなる実施形態では、調節配列を変化させるかまたは修飾して、発現を増強する(すなわち、プロモーターの強度を増大させる)ことができる。例えば、エンハンサー機能を含むイントロン配列を用いて、プロモーター機能を増大させることができる。ミエリンプロテオリピドタンパク質(PLP)遺伝子は、エンハンサーエレメントとして機能するイントロン配列を含む。この調節エレメント/領域であるASE(アンチサイレンサー/エンハンサー)は、エキソン1 DNAの約1kb下流に位置し、約100bpを包含する。Mengら、J Neurosci Res.、82巻、346〜356頁(2005年)を参照されたい。
【0048】
さらに、特定の細胞内位置では、MRFまたはその調節物質の発現が望ましい場合があり、当技術分野において広範に実施されている組換えDNA法により、MRFまたはその調節物質をコードする核酸配列を、対応する細胞内局所化配列に作動的に連結することができる。例示的な細胞内局所化配列には、(a)細胞外への遺伝子産物の分泌を方向づけるシグナル配列;(b)細胞の細胞質膜または他の膜性区画へのタンパク質の付着を可能とする膜アンカードメイン;(c)コードされたタンパク質の核への移動を媒介する核内局在化配列;(d)コードされたタンパク質を、主に小胞体へと封じ込める、ER保持配列(例えば、KDEL配列);(e)タンパク質がファルネシル化して細胞膜と会合するようにこれをデザインしうる;または(f)コードされたタンパク質産物の特異的な細胞内分布において役割を果たす他の任意の配列が含まれるがこれらに限定されない。
【0049】
他の実施形態では、外部ガイド配列(EGS)を用いて、MRF阻害剤を標的とすることができる(例えば、US5728521、同6057153を参照されたい)。一態様において、本発明の生物活性剤は、MRFの発現および/または活性を調節する機構として、RNA干渉(RNAi)を用いうる。例えば、RNAiを用いて、MRFの発現および/または活性の阻害剤を標的とし、これにより、OLの成熟化および/または有髄化を促進することができる。RNAiは、二本鎖RNA(dsRNA)またはsiRNAにより誘発される、配列特異的な、転写後における遺伝子サイレンシング過程である。RNAiは、Drosophila、線虫、真菌、および植物など、多数の生物において見られ、抗ウイルス防御、トランスポゾン活性の調節、および遺伝子発現の調節に関与していると考えられる。RNAiにおいて、dsRNAまたはsiRNAは、結果としての、配列特異的な遺伝子発現の阻害により、標的mRNAの分解を誘導する。一部の実施形態において、miRNAは、MRF阻害剤を標的とするのに用いられる。
【0050】
本明細書で用いられる低分子干渉RNA(siRNA)とは、被験体の遺伝子を標的とする、ヌクレオチドのRNA二重鎖である。RNA二重鎖とは、RNA分子の2つの領域間における相補的対合により形成される構造を指す。siRNAが遺伝子を標的とするのは、該siRNAの二重鎖部分のヌクレオチド配列が、標的とされる遺伝子のヌクレオチド配列に相補的であるからである。一部の実施形態において、siRNA二重鎖の長さは、30ヌクレオチド未満である。一部の実施形態において、該二重鎖は、29、28、27、26、25、24、23、22、21、20、19、18、17、16、15、14、13、12、11、または10ヌクレオチドの長さである。一部の実施形態において、該二重鎖の長さは、19〜25ヌクレオチドの長さである。siRNAのRNA二重鎖部分は、ヘアピン構造の一部でありうる。二重鎖部分に加えて、ヘアピン構造は、該二重鎖を形成する2つの配列間に位置するループ部分を含有しうる。ループの長さは変化しうる。一部の実施形態において、ループは、5、6、7、8、9、10、11、12、または13ヌクレオチドの長さである。ヘアピン構造はまた、3’突出部分および/または5’突出部分も含有しうる。一部の実施形態において、突出は、0、1、2、3、4、または5ヌクレオチドの長さの3’突出および/または5’突出である。siRNAは、核酸配列によりコードすることが可能であり、該核酸配列はまた、プロモーターも包含しうる。該核酸配列はまた、ポリアデニル化シグナルも包含しうる。一部の実施形態において、ポリアデニル化シグナルは、最小の合成ポリアデニル化シグナルである。
【0051】
生物活性剤はまた、神経細胞分化に関与する1または複数の遺伝子、例えば、MRF阻害剤を標的とする抗体でもありうる。前出の遺伝子のうちのいずれかによりコードされるポリペプチドに特異的な(または同ポリペプチドの活性部位に特異的な)抗体の作製は、それらの各々が参照により本明細書に組み込まれる、米国特許第6,491,916号;同第6,982,321号;同第5,585,097号;同第5,846,534号;同第6,966,424号;および米国特許出願公開第2005/0054832号;同第2004/0006216号;同第2003/0108548号;同第2006/002921号;および同第2004/0166099号において開示されるなど、当業者に公知である。例えば、マウスに抗原を含む組成物を注射し、血清試料を採取することにより抗体生成の存在を検証し、脾臓を摘出してBリンパ球を得、該Bリンパ球を骨髄腫細胞と融合させて、ハイブリドーマを作製し、該ハイブリドーマをクローニングし、注射された抗原に対する抗体を生成する陽性クローンを選択し、該抗原に対する抗体を生成するクローンを培養し、該ハイブリドーマ培養物から該抗体を単離することにより、モノクローナル抗体を得ることができる。十分に確立された各種の技法により、ハイブリドーマ培養物から、モノクローナル抗体を単離および精製することができる。このような単離法には、プロテインAセファロースによるアフィニティークロマトグラフィー、サイズ除外クロマトグラフィー、およびイオン交換クロマトグラフィーが含まれる。例えば、Coliganら(編)、「CURRENT PROTOCOLS IN IMMUNOLOGY」、2.7.1〜2.7.12頁、および2.9.1〜2.9.3頁(John Wiley & Sons社、1991年)を参照されたい。また、Bainesら、「Purification of Immunoglobulin G (IgG)」、「METHODS IN MOLECULAR BIOLOGY」、第10巻、79〜104頁(The Humana Press社、1992年)も参照されたい。
【0052】
標準的な技法を用いて、抗体を作製するのに適切な量の、十分に特徴づけられた抗原を得ることができる。例として述べると、Tedderら、米国特許第5,484,892号により説明される、被覆抗体を用いて、抗原を、細胞から免疫沈降させることができる。代替的に、このような抗原は、対象の抗原を過剰生成する、トランスフェクトされた培養細胞から得ることもできる。公表されたヌクレオチド配列を用いて、これらのタンパク質の各々をコードするDNA分子を含む発現ベクターを構築することができる。例えば、Wilsonら、J. Exp. Med.、173巻、137〜146頁(1991年);Wilsonら、J. Immunol.、150巻、5013〜5024頁(1993年)を参照されたい。例示として述べると、CD3をコードするDNA分子は、相互にプライミングする長いオリゴヌクレオチドを用いてDNA分子を合成することにより得ることができる。例えば、Ausubelら(編)、「CURRENT PROTOCOLS IN MOLECULAR BIOLOGY」、8.2.8〜8.2.13頁(1990年)を参照されたい。また、Wosnickら、Gene、60巻、115〜127頁(1987年);およびAusubelら(編)、「SHORT PROTOCOLS IN MOLECULAR BIOLOGY」、第3版、8−8〜8−9頁(John Wiley & Sons社、1995年)も参照されたい。ポリメラーゼ連鎖反応を用いる確立された技法は、1.8キロ塩基の長さまでの大きさの遺伝子を合成することを可能とする(Adangら、Plant Molec. Biol.、21巻、1131〜1145頁(1993年);Bambotら、PCR Methods and Applications、2巻、266〜271頁(1993年);Dillonら、「Use of the Polymerase Chain Reaction for the Rapid Construction of Synthetic Genes」、「METHODS IN MOLECULAR BIOLOGY」、第15巻、「PCR PROTOCOLS: CURRENT METHODS AND APPLICATIONS」、White(編)、263〜268頁(Humana Press社、1993年))。変化形として、対象の抗原をコードするcDNAを安定的にトランスフェクトしたマウスプレB細胞系により免疫感作したマウスに由来する脾臓細胞と、骨髄腫細胞を融合することにより、モノクローナル抗体を得ることができる。Tedderら、米国特許第5,484,892号を参照されたい。
【0053】
本発明の生物活性剤はまた、MRFまたはその機能的変異体をコードする核酸配列を含むベクターなど、ベクターの形態でもありうる。in vivoまたはin vitroの方法において用いられるベクターには、SV−40、アデノウイルス、レトロウイルスに由来するDNA配列の誘導体、ならびに哺乳動物の機能性ベクターと、機能性プラスミドおよび機能性ファージDNAとの組合せに由来するシャトルベクターが含まれうる。例えば、参照により本明細書に組み込まれる、SouthernおよびBerg、J. Mol. Appl. Genet.、1巻、327〜341頁(1982年);Subraminiら、Mol. Cell. Biol.、1巻、854〜864頁(1981年);KaufinannおよびSharp、J. Mol. Biol.、159巻、601〜621頁(1982年);Scahillら、Proc. Natl. Acad. Sci. USA、80巻、4654〜4659頁(1983年);ならびにUrlaubおよびChasin、Proc. Natl. Acad. Sci. USA、77巻、4216〜4220頁(1980年)により説明されるベクターなど、真核生物の発現ベクターがよく知られている。本発明の方法において用いられるベクターは、ウイルスベクター、好ましくはレトロウイルスベクターでありうる。複製欠損アデノウイルスが好ましい。参照により本明細書に組み込まれるEglitisおよびAndersen、BioTechniques、6巻、7号、608〜614頁(1988年)により説明される、例えば、モロニーマウス白血病ウイルス(MoMulV)、ハーベイマウス肉腫ウイルス(HaMuSV)、マウス乳癌ウイルス(MuMTV)、およびマウス骨髄増殖性肉腫ウイルス(MuMPSV)、ならびに細網内皮症ウイルス(Rev)およびラウス肉腫ウイルス(RSV)などの鳥類レトロウイルスなど、例えば、長い末端反復中に含有されるウイルスの調節配列の制御下において、レトロウイルスの構造遺伝子を、対象の単一の遺伝子により置換する、「単一遺伝子ベクター」を用いることができる。発現構築物は、ウイルスベクターの場合もあり、非ウイルスベクターの場合もある。本発明の一部と考えられるウイルスベクターには、アデノウイルス、アデノ関連ウイルス、ヘルペスウイルス、レトロウイルス(レンチウイルスを含めた)、ポリオーマウイルス、または牛痘ウイルスが含まれるがこれらに限定されない。
【0054】
複数の遺伝子を導入しうる組換えレトロウイルスベクターもまた、本発明の方法により用いることができる。独立プロモーターの調節下にあるcDNAを含有する、内部プロモーターを伴うベクター、例えば、ヒトアデノシン脱アミノ酵素(hADA)のcDNAがそれ自体の調節配列と共に挿入された選択マーカー(noe.sup.R)を伴う、N2ベクターに由来するSAXベクターは、当技術分野において公知の方法を介して、本発明の方法に従いデザインし、用いることができる。
【0055】
MRFまたは他の生物活性剤をコードする核配列の効率的な翻訳には、特定の開始シグナルもまた必要とされうる。これらのシグナルには、ATG開始コドンおよび隣接する配列が含まれる。それ自体の開始コドンおよび隣接する配列を含めた全遺伝子またはcDNAが、適切な発現ベクター内へと挿入される場合、さらなる翻訳制御シグナルは必要とされない場合がある。しかし、該コード配列の一部だけが挿入される場合、おそらく、ATG開始コドンを含めた外因性翻訳制御シグナルを与える場合がある。さらに、全挿入配列の翻訳を確保するには、開始コドンが、所望のコード配列のリーディングフレームと一致すべきである。これらの外因性の翻訳制御シグナルおよび開始コドンは、天然および合成両方の多様な由来でありうる。発現の効率は、適切な転写エンハンサーエレメント、転写ターミネーターなどを包含することにより増強することができる(例えば、Bittnerら、Methods in Enzymol.、153巻、516〜544頁(1987年)を参照されたい)。
【0056】
本発明の宿主細胞は一般に、前述のベクター内における分子など、前出の核酸分子を用いることにより遺伝子改変することができる。したがって、宿主細胞は、MRFなど、結果として希突起膠細胞の分化をもたらす、異なる発現レベルの遺伝子産物を生成しうる。in vitroまたはin vivoにおける細胞の遺伝子改変またはトランスフェクションは、本明細書の上記で言及した参考文献において説明され、また、米国特許第6,998,118号、同第6,670,147号、または同第6,465,246号などにおいて開示される、当技術分野で公知の方法を用いて実施することができる。薬剤の特徴に応じて、全身送達または局所送達、プラスミドベクター、ウイルスベクター、または非ウイルスベクター系による送達、リポソーム製剤およびミニ細胞を含めた調合薬を含め、当業者に公知の送達方式のいずれかを介して薬剤を送達することができる。例えば、哺乳動物の宿主細胞では、多数のウイルスベースの発現系を用いることができる。発現ベクターとしてアデノウイルスを用いる場合、対象のヌクレオチド配列(例えば、治療能がある薬剤をコードするヌクレオチド配列)を、アデノウイルスの転写制御複合体または翻訳制御複合体、例えば、後期プロモーター配列および三連リーダー配列へとライゲーションすることができる。次いで、このキメラ遺伝子を、in vitroまたはin vivoにおける組換えにより、アデノウイルスゲノム内に挿入することができる。ウイルスゲノムの非本質的領域(例えば、領域E1またはE3)内における挿入は、生存可能であり、また、感染した宿主内において、AQP1遺伝子産物を発現することが可能な組換えウイルスを結果としてもたらす(例えば、Logan & Shenk、Proc. Natl. Acad. Sci. USA、8巻、1号、3655〜3659頁(1984年)を参照されたい)。宿主細胞は、神経膠細胞など、神経細胞でありうる。神経細胞には、OPCまたは成熟OLなど、希突起膠細胞のほか、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)が含まれうる。
【0057】
MRFの活性レベルまたは発現レベルの調節は、各種の方法により確認することができる。例えば、遺伝子発現レベルの変化の検出は、増幅アッセイにおいてリアルタイムで実施することができる。一態様において、増幅産物は、DNA挿入剤およびDNA溝結合剤が含まれるがこれらに限定されない蛍光DNA結合剤により直接的に可視化することができる。二本鎖DNA分子内に組み込まれた挿入剤の量は、増幅されるDNA産物の量に比例することが典型的であるので、当技術分野における従来の光学系を用いて、挿入された色素の蛍光発光を定量化することにより、増幅産物の量を簡便に決定することができる。この適用に適するDNA結合色素には、SYBRグリーン、SYBRブルー、DAPI、ヨウ化プロピジウム、ヘキスト(Hoeste)、SYBRゴールド、臭化エチジウム、アクリジン、プロフラビン、アクリジンオレンジ、アクリフラビン、フルオロクマニン、エリピチシン、ダウノマイシン、クロロキン、ジスタマイシンD、クロモマイシン、ホミジウム、ミトラマイシン、ルテニウムポリピリジル、アントラマイシンなどが含まれる。
【0058】
別の態様では、増幅反応において、配列特異的プローブなど、他の蛍光標識を用いて、増幅産物の検出および定量化を容易にすることができる。プローブベースの定量的増幅は、所望の増幅産物の配列特異的な検出に依拠する。それは、特異性および感度の上昇を結果としてもたらす、標的特異的な蛍光プローブ(例えば、TaqManプローブ)を用いる。プローブベースの定量的増幅を実施する方法は当技術分野で十分に確立されており、米国特許第5,210,015号において教示されている。
【0059】
さらに別の態様では、神経周期関連遺伝子との配列相同性を共有するハイブリダイゼーションプローブを用いる、従来のハイブリダイゼーションアッセイを実施することができる。プローブは、ハイブリダイゼーション反応において、被験対象に由来する生物学的試料中に含有される標的ポリヌクレオチドと安定的な複合体を形成することが可能なことが典型的である。プローブ核酸としてアンチセンスを用いる場合、試料中においてもたらされる標的ポリヌクレオチドは、アンチセンス核酸の配列に対して相補的であるように選択されることが、当業者により理解されるであろう。逆に、ヌクレオチドプローブがセンス核酸である場合、標的ポリヌクレオチドは、該センス核酸の配列に対して相補的であるように選択される。
【0060】
当業者に公知である通り、ハイブリダイゼーションは、様々な厳密性の条件下で実施することができる。本発明を実施するのに適するハイブリダイゼーション条件は、プローブと、標的の神経細胞周期遺伝子との認識相互作用が、十分に特異的であると同時に、十分に安定的でもあるような条件である。ハイブリダイゼーション反応の厳密性を上昇させる条件は当技術分野の広い範囲において公知であり、公表されている(例えば、Sambrookら(1989年)、前出;「Nonradioactive In Situ Hybridization Application Manual」、Boehringer Mannheim社、第2版を参照されたい)。ニトロセルロース、ガラス、シリコンが含まれるがこれらに限定されない任意の固体支持体上に固定化されたプローブ、および各種の遺伝子アレイを用いて、ハイブリダイゼーションアッセイを行うことができる。好ましいハイブリダイゼーションアッセイは、米国特許第5,445,934号において説明される、高密度の遺伝子チップ上において実施される。
【0061】
ハイブリダイゼーションアッセイにおいて形成されるプローブ−標的複合体を簡便に検出するには、ヌクレオチドプローブを検出可能な標識へとコンジュゲートさせる。本発明において用いるのに適する検出可能な標識には、光化学的手段、生化学的手段、分光学的手段、免疫化学的手段、電子的手段、光学的手段、または化学的手段により検出可能な任意の組成物が含まれる。当技術分野では、多種多様の適切な検出可能標識が公知であり、これらには、蛍光標識または化学発光標識、放射性同位体標識、酵素リガンド、または他のリガンドが含まれる。好ましい実施形態では、ジゴキシゲニン、β−ガラクトシダーゼ、ウレアーゼ、アルカリホスファターゼ、またはペルオキシダーゼ、アビジン/ビオチン複合体など、蛍光標識または酵素タグを用いることが所望される可能性が高い。
【0062】
ハイブリダイゼーション強度を検出または定量化するのに用いられる検出法は、上記で選択される標識に依存することが典型的である。例えば、写真フィルムまたはホスホイメージャーを用いて、放射性標識を検出することができる。発光された光を検出する光検出器を用いて、蛍光マーカーを検出および定量化することができる。酵素に基質を供給し、基質上における酵素の作用により生成される反応産物を測定することにより、酵素標識を検出することが典型的であり、また、最後に、単に、着色標識を可視化することにより、比色分析標識を検出する。
【0063】
MRF発現の調節はまた、対応するMRFの遺伝子産物、および/もしくは、図5における遺伝子産物など、それに相関する遺伝子産物、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、もしくはMogを検討することによっても決定することができる。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子など、サブセットの発現を評価することもできる。タンパク質レベルの決定は、a)生物学的試料中に含有されるタンパク質を、MRFタンパク質、またはそれに相関するタンパク質に特異的に結合する検出剤と接触させるステップと、(b)このようにして形成される、任意の検出剤:タンパク質複合体を同定するステップとを伴うことが典型的である。一態様において、該タンパク質に特異的に結合する検出剤は、モノクローナル抗体などの抗体である。
【0064】
スクリーニングアッセイ
本発明はまた、MRF活性を調節するのに有効な、候補生物活性剤をスクリーニングする方法も提供する。該方法は、被験細胞を候補生物活性剤と接触させるステップと、対照細胞と比較した、MRFの発現レベル、またはその活性の変化についてアッセイするステップとを含む。本発明の1または複数の方法においてアッセイされる候補生物活性剤はまた、異なる時点で比較するほか、基準または対照と比較して、生物活性剤に対する応答において全体的な差違が見られるかどうかを決定するためにもアッセイされうる。
【0065】
被験細胞は、神経膠細胞などの神経細胞でありうる。被験細胞は、希突起膠細前駆細胞(OPC)、成熟OL、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)でありうるが、これらに限定されない。被験細胞はまた、幹細胞または胚性幹(ES)細胞でもありうる。候補生物活性剤は、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子でありうる。
【0066】
MRF発現の変化は、被験神経細胞または被験幹細胞の分化を調節するのに有効な候補生物活性剤を示すことが典型的である。これもまた候補生物活性剤の有効性を示しうる他の変化には、図5における遺伝子、または成熟希突起膠細胞内で特異的に上方調節される遺伝子の発現変化が含まれうる。例えば、Sox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、またはMogの発現変化。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子など、このような遺伝子のサブセットの発現変化を評価することもできる。したがって、被験細胞を用いて、候補薬剤をスクリーニングし、このような薬剤がMRFを調節し、これにより、OLの成熟を促進または阻害するかどうかを決定することができる。MRFの発現または活性を上昇させるのに有効な生物活性剤は、OLの成熟および再有髄化を促進するのに有効でありうる。それらはまた、ES細胞などからのOLの分化を促進するのにも有効でありうる。このような候補薬剤は、本明細書に記載の動物モデルなどの動物モデルにおいてさらにアッセイすることができ、また、神経障害を治療するための組成物および方法など、神経細胞の分化を誘導する方法において用いることができる。
【0067】
MRF発現レベルの変化は、上記で説明した方法を含め、当技術分野において公知の方法により実施することができる。例えば、発現レベルの変化は、被験細胞および対照細胞に由来するMRFの遺伝子発現プロファイルを解析または比較することによりアッセイすることができる。図5に列挙した遺伝子など、MRFの発現と相関する他の遺伝子、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、および/もしくはMogなど、成熟希突起膠細胞において特異的に上方調節される遺伝子の変化もアッセイすることができる。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子など、このような遺伝子のサブセットの発現変化をアッセイすることもできる。
【0068】
候補薬剤は、該薬剤が、初期遺伝子または後期遺伝子に影響を及ぼすかどうか、したがって、初期分化細胞または成熟分化細胞に影響を及ぼすかどうかを決定するように、前駆細胞周期の異なる時間段階において送達することができる。例えば、候補薬剤をスクリーニングして、初期または後期に誘導される遺伝子、例えば、OL分化の初期におけるUgt8、CNP1、Plp1、および/もしくはMbp、またはMag、Trf、Mobp、および/もしくはMogなど、幼若OLまたは成熟OLと関連するいずれの遺伝子が影響を受けるのかを決定することができる。OPCを、初期遺伝子または後期遺伝子の誘導/下方調節欠損についてスクリーニングすると、OL分化の初期段階および後期段階と関連することが本明細書において同定される遺伝子の活性を調節する薬剤を選択することにより、OLの分化を促進するより良好な治療的標的化を行うことができる。さらに、このような薬剤を被験体に投与して、未分化OPCまたは未成熟OLの異なる段階から、正常で完全なOLの成熟を促進することもできる。このような薬剤はまた、OL分化の異なる段階から、有髄化するOLを生成させるのに必要な遺伝子プログラムを再構成するのにも用いることができる。
【0069】
一部の態様では、アッセイするステップを、in vitroにおいて実施する。該方法の別の態様では、アッセイするステップを、in vivoにおいて実施する。当技術分野では、in vitroおよびin vivoにおける各種の方法が用いられる。例えば、in vitroアッセイを用いて、細胞培養物中におけるOL分化を促進しうる一方、以下でさらに説明される通り、動物モデルにより、in vivoアッセイを実施することができる。当業者には、遺伝子チップ法またはアレイ法(例えば、遺伝子チップは、Agilent社、Affymetrix社、Nanogen社など、複数の販売元により容易に入手可能である)、免疫ブロット解析、RT−PCR、および他の手段などによる発現プロファイルアッセイが周知であり、また、上記でもさらに説明されている。
【0070】
本発明の一部の態様では、1または複数の候補生物活性剤を、細胞培養物と接触させ、このような接触の前において、これと同時に、またはこの後において、ミエリン修復誘導剤または軸索保護誘導剤など、1または複数の他の生物活性剤もまた細胞へと送達し、生物活性剤およびミエリン修復剤または軸索保護剤のどの組合せにより相乗効果がもたらされるかを決定する。1または複数の生物活性剤は、ES細胞などの幹細胞が、OL(またはOPC)へと分化することを誘導する因子でありうる。例えば、該因子は、Sox10、Nkx2.2、Olig1、またはOlig2などの転写因子でありうる。例えば、前駆細胞型から有髄化する希突起膠細胞への移行を示す低速度撮影顕微鏡法を用いることにより、または本明細書で説明されるOL特異的マーカーの発現をアッセイすることにより、培養物中において相乗効果を観察することができる。さらに、前駆細胞に、増強緑色蛍光タンパク質(EGFP)の膜標的化形態をトランスフェクトし、分化細胞の検出において好都合な蛍光顕微鏡法を容易にすることができる。したがって、各種の実施形態では、米国特許第7,008,634号;同第6,972,195号;同第6,982,168号;同第6,962,980号;同第6,902,881号;同第6,855,504号;または同第6,846,625号において開示される技法など、当技術分野において公知の技法を用いて、細胞を培養および/または遺伝子改変し、標的ポリペプチドを発現させることができる。
【0071】
スクリーニングアッセイにおいて用いられる細胞には、被験体から得、約5、6、7、8、9〜約14日間にわたり培養物中において増殖させたOPCが含まれうる。細胞は、約1、2、3、4、5、6、7、8、または9日間にわたり培養することができる。培養物中において増殖させる際、このような細胞には、1または複数のベクターをトランスフェクトすることができる。
【0072】
本発明の別の態様は、スクリーニングアッセイのためのシステムである。したがって、それを介してスクリーニングアッセイに関するデータを発生させうる論理デバイスを表わす例もまた提供される。候補生物活性剤との接触を伴うかまたは伴わない被験細胞の発現プロファイルなどのデータを受信および保存するコンピュータシステム(またはデジタルデバイス)。コンピュータシステムは、また、生物活性剤と接触させた神経細胞と、生物活性剤と接触させなかった対照細胞との発現プロファイルの比較など、データに対する解析も実施しうる。コンピュータシステムは、媒体、および/または、場合によって、固定化された媒体を有するサーバーに接続されうるネットワークポートからの指示を読み取りうる論理装置として理解することができる。システムには、CPU、ディスクドライブ、ならびに、キーボードおよび/またはマウスなど、任意選択の入力デバイス、ならびに任意選択のモニターを包含することが典型的である。示される通信媒体を介し、ローカルまたは遠隔の位置にあるサーバーへのデータ通信を達成することができる。
【0073】
通信媒体には、データを送信および/または受信する任意の手段が含まれうる。例えば、通信媒体は、ネットワーク接続、無線接続、またはインターネット接続でありうる。このような接続は、ウェブ上の通信を提供しうる。本発明に関連するデータを、このようなネットワークまたは接続により送信し、利用者が受信および/または参照することができる。受信者は、個人でありうるが、これに限定されない。コンピュータにより読み取り可能な媒体には、候補生物活性剤と接触させた神経細胞から得られる発現プロファイルの解析結果の送信に適する媒体が含まれうる。媒体は、生物活性剤が、MRFまたは他の相関する遺伝子の発現を調節するかどうかなど、本明細書に記載の方法を用いて導出された結果を包含しうる。
【0074】
本発明のスクリーニング方法を実施する場合、本明細書において例示される方法を含め、希突起膠細胞の分化を確認するのに適用可能な任意の公知の方法を用いることができる。候補生物活性剤は、それらが、活性を促進する(例えば、MRFの発現レベルを増強する)か、または活性を阻害する(例えば、MRF阻害剤などの標的分子に対する結合を介して、発現レベルを低下させるか、または機能を遮断する)かに基づき、選択することができる。
【0075】
マイクロアレイ
本明細書に記載のスクリーニング方法はまた、マイクロアレイ、またはその上に固定化させた遺伝子チップ、その少なくとも1つがMRFに対応する複数のプローブを用いても実施することができる。これらのマイクロアレイはまた、複数種類のヒト疾患組織、例えば、多発性硬化症病変または希突起膠細胞腫瘍組織において存在する、希突起膠細胞系統細胞の分化状態を評価するのにも用いることができる。したがって、本発明は、そのようなマイクロアレイを含む組成物を提供する。
【0076】
マイクロアレイは、MRFに対応する少なくとも1つのプローブ、また、MRFによって調節される遺伝子に相関する1または複数のプローブを包含しうる。複数のプローブは、MRF、また、図5における少なくとも1つの遺伝子に対応しうる。複数のプローブは、MRF、および図5におけるすべての遺伝子に対応しうる。プローブはまた、MRF、また、Sox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、および/またはMogなど、成熟希突起膠細胞において特異的に発現する遺伝子にも対応しうる。代替的に、マイクロアレイ上における複数のプローブは、MRF、また、OL分化の初期におけるUgt8、CNP1、Plp1、および/もしくはMbp、OL分化の後期におけるMag、Trf、Mobp、および/もしくはMog、またはPlp1、Mbp、もしくはMagなど、OL分化の中間段階において発現する遺伝子など、OL分化の異なる段階で発現する遺伝子など、遺伝子のサブセットも含みうる。
【0077】
プローブとは、ハイブリダイゼーション反応において、それに対応する標的ポリヌクレオチドを検出または同定するのに用いられるポリヌクレオチドを指す。ポリヌクレオチドに対して適用される「ハイブリダイズする」という用語は、該ポリヌクレオチドが、ハイブリダイゼーション反応において、ヌクレオチド残基の塩基間における水素結合により安定化する複合体を形成する能力を指す。1つのポリヌクレオチドが、最終的に別のポリヌクレオチドに由来する場合、異なるポリヌクレオチドは互いに「対応する」という。例えば、センス鎖は、同じ二本鎖配列のアンチセンス鎖に対応する。mRNA(また、遺伝子転写物としても公知である)は、それが転写される元の遺伝子に対応する。cDNAは、逆転写反応、またはRNA配列の知識に基づくDNAの化学合成などにより、それが生成される元のRNAに対応する。cDNAはまた、RNAをコードする遺伝子にも対応する。ポリヌクレオチドは、その対の一方が、他方の一部だけに由来する場合も対応するという。
【0078】
本発明のアレイは、比較を目的とする、陽性または陰性の対照プローブを含みうる。適切な対照プローブの選択は、最初に選択する試料プローブ、および調べているその発現パターンに依存する。任意の種類の対照プローブを、アレイ内の任意の位置、またはアレイ全体の複数の位置において局在化させて、ハイブリダイゼーションアッセイにおける空間的なばらつき、全体の発現レベル、または非特異的な結合についての対照とすることができる。
【0079】
本発明において実施されるポリヌクレオチドプローブは、化学合成、組換えによるクローニング、例えば、PCR、またはこれらの任意の組合せにより得ることができる。ポリヌクレオチドを化学合成する方法は当技術分野において周知であり、本明細書で詳細に説明する必要はない。当業者は、本明細書に記載の配列データを用いて、DNA合成器、PCR機を用いるか、または市販品を取り寄せることにより、所望のポリヌクレオチドを得ることができる。選択されたプローブは、該プローブを、固体支持体表面と安定的に会合させる際に有効である任意の適切な技法により、固体支持体の所定の領域上に固定化させる。「安定的に会合した」とは、ハイブリダイゼーション条件および洗浄条件下において、ポリヌクレオチドが、所定領域に局在化された状態を維持することを意味する。ポリヌクレオチドは、それ自体、支持体表面と共有結合させることもでき、これに非共有結合させることもできる。非共有結合の例には、非特異的な吸着、イオン性相互作用、疎水性相互作用、または水素結合による相互作用の結果としての結合が含まれる。共有結合は、ポリヌクレオチドと、支持体表面上に存在する官能基との化学結合の形成を伴う。官能基は、天然の場合もあり、リンカーとして導入される場合もある。非限定的な官能基には、ヒドロキシル、アミン、チオール、およびアミドが含まれるがこれらに限定されない。ポリヌクレオチドプローブの共有結合による固定化に適用可能な例示的な技法には、UVによる架橋形成または他の光誘導型の化学結合形成、および機械誘導型の結合形成が含まれるが、これらに限定されない(例えば、米国特許第5,837,832号、同第5,143,854号、同第5800992号、WO92/10092、WO93/09668、およびWO97/10365を参照されたい)。好ましい方法は、ポリヌクレオチドプローブの末端のうちの1つを、一重の共有結合を介して支持体表面へと連結することである。プローブがより大きな自由度を有し、相補的な標的との複合体相互作用に用いられるとき、このような立体構造は、高度のハイブリダイゼーション効率を可能とする。
【0080】
手動で、またはより一般的には、一連の操作により、試料を多数の所定領域へとスポットする自動デバイスを用いて、複数のプローブ試料を付着させることにより、各アレイを作製することが典型的である。ポリヌクレオチドアレイの作製には、一般に、各種の自動スポッティングデバイスが用いられる。このようなデバイスには、誘電式デバイスまたはインクジェット式デバイス、自動式マイクロピペット、および市販されるこれらのデバイスのうちのいずれか(例えば、Beckman社製、Biomek2000)が含まれる。
【0081】
動物モデル
本発明の別の態様において、スクリーニングアッセイは、in vivoで実施される。例えば、有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法は、候補生物活性剤を動物に投与するステップと、対照動物と比較したMRFの発現レベルの上昇についてアッセイし、該上昇が、動物における有髄化を促進する前記生物活性剤を示すステップとを含みうる。本発明の1または複数の方法においてアッセイされる候補生物活性剤はまた、異なる時点で比較するほか、基準または対照と比較して、生物活性剤に対する応答において全体的な差違が見られるかどうかを決定するためにもアッセイされうる。
【0082】
動物被験体を用いて、候補生物活性剤をスクリーニングし、このような薬剤がMRFを調節するかどうかを決定し、したがって、MRFを下方調節または上方調節する候補薬剤を同定し、これにより、OLの成熟または分化および再有髄化を促進または阻害する薬剤を同定することができる。被験体のスクリーニング方法に有用な候補生物活性剤は、ペプチド、ポリペプチド、ペプチド模倣剤、抗体、アンチセンス、アプタマー、siRNA、および/または低分子を含みうる。MRF発現/活性、および/または神経細胞の分化を調節または改変する能力を有することが疑われる任意の薬剤を、本明細書で開示されるスクリーニング方法にかけることができる。
【0083】
MRF、また、図5に列挙した遺伝子など、MRFの発現と相関する他の遺伝子、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、および/もしくはMogなど、成熟希突起膠細胞において特異的に上方調節される遺伝子の変化もアッセイすることができる。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階において発現する遺伝子など、このような遺伝子のサブセットの発現変化をアッセイすることもできる。有髄化および発現レベルのアッセイは、当業者に周知であり(例えば、遺伝子チップは、複数の販売元から容易に入手することができる)、本明細書でさらに説明されている。
【0084】
動物は、げっ歯動物種または類人猿動物種など、哺乳動物であることが典型的である。動物は、マウス、ラット、モルモット、またはサルでありうる。動物はまた、1または複数の所望の特徴を有する「ノックアウト」または「ノックイン」である動物などのトランスジェニック動物でもありうる。「ノックアウト」は、好ましくは、標的遺伝子の発現が無視できるかまたは検出不可能となるように、標的遺伝子の機能低下を結果としてもたらす、トランスジェニック配列の導入による標的遺伝子の変化を有する。「ノックイン」は、例えば、標的遺伝子のさらなるコピーを導入することにより、または標的遺伝子の内因性のコピーの発現を増強する調節配列を作動的に挿入することにより、標的遺伝子の発現の増大を結果としてもたらす、宿主細胞ゲノムの変化を有するトランスジェニック動物である。ノックインまたはノックアウトのトランスジェニック動物は、標的遺伝子に関して、ヘテロ接合の場合もあり、ホモ接合の場合もある。ノックアウトおよびノックイン共に、「バイジェニック」でありうる。バイジェニック動物は、少なくとも2つの宿主細胞遺伝子が変化している。
【0085】
胚のマイクロ操作技術の進歩により、今日では、哺乳類の有精卵内に異種DNAを導入することが可能である。例えば、マイクロインジェクション、リン酸カルシウムを介する沈殿、リポソーム融合、レトロウイルス感染、または他の手段により、全能性幹細胞または多能性幹細胞を形質転換することができる。次いで、形質転換細胞を胚内に導入し、次いで、該胚をトランスジェニック動物へと発育させる。好ましい実施形態では、感染した胚から導入遺伝子を発現するトランスジェニック動物を作製しうるように、発育する胚に、所望の導入遺伝子を含有するウイルスベクターを感染させる。別の好ましい実施形態では、所望の導入遺伝子を、好ましくは単一細胞段階において、胚の前核または細胞質内へと共注入し、該胚を、成熟トランスジェニック動物へと発育させる。トランスジェニック動物を作製するためのこれらの方法および他の変法は当技術分野で十分に確立されており、したがって、本明細書では詳述しない。例えば、米国特許第5,175,385号、および同第5,175,384号を参照されたい。
【0086】
本発明は、モノジェニック動物およびバイジェニック動物を提供する。例えば、本明細書では、MRFノックインまたはMRFノックアウトを含む動物が開示される。トランスジェニック動物は、MRF導入遺伝子を含むことが可能であり、導入遺伝子は、該動物のゲノム内へと安定的に組み込まれされ、該トランスジェニック動物の野生型コピーを置換する。代替的に、トランスジェニック動物は、該動物のゲノム内にあるMRFの野生型コピーに加え、MRF導入遺伝子も有しうる。例えば、導入遺伝子は、単一のコピーとして、またはコンカタマー、例えば、ヘッドからヘッドへのタンデム配列またはヘッドからテールへのタンデム配列において組み込むことができる。MRF導入遺伝子は、1または複数の突然変異、例えば、特定のドメインまたはエキソンの欠失を有しうる。欠失は、エキソン8(図8)など、推定DNA結合ドメイン内におけるエキソンの欠失でありうる。欠失は、構成的な場合もあり、条件的な場合もある。例えば、MRF導入遺伝子は、リコンビナーゼ部位に隣接する場合がある。トランスジェニック動物はまた、そのゲノム内に、同類の認識配列であるloxP配列(すなわち、loxP部位)を認識する、Creなどのリコンビナーゼをコードする配列を安定的に組み込みうる。
【0087】
当技術分野では、他のリコンビナーゼ認識配列も公知である。例えば、説明される通り、Creリコンビナーゼの認識配列は、8塩基対のコア配列を挟む、2つの13塩基対による逆行反復配列を含む、34塩基対の配列である、loxPである(Sauer、Curr. Opin. Biotech.、5巻、521〜527頁(1994年)を参照されたい)。認識配列の他の例は、リコンビナーゼ酵素であるλインテグラーゼにより認識されるattB、attP、attL、およびattR配列である。attBは、9塩基対による2つのコア型のInt結合部位と、7塩基対の重複領域とを含有する、約25塩基対の配列である。attPは、コア型のInt結合部位と、アーム型のInt結合部位のほか、補酵素IHF、FIS、およびXisに対する部位も含有する、約240塩基対の配列である。Landy、Curr. Opin. Biotech.、3巻、699〜707頁(1993年)を参照されたい。このような部位はまた、Hartleyら、米国特許出願公開第20060035269号による開示において開示されるような、当技術分野において公知の方法および生成物を用いて組換えを増強するように、本発明に従い操作することもできる。
【0088】
本発明のCreリコンビナーゼは、野生型の場合もあり、野生型の変異体の場合もある。Creリコンビナーゼは、トランスジェニック動物(またはトランスジェニック細胞)において誘導可能でありうる。変異体のCreリコンビナーゼは、組換え部位に対する特異性が拡張されている。具体的に述べると、該変異体は、loxP配列以外の配列と、野生型のCreリコンビナーゼが活性である他のlox部位配列との間の組換えを媒介する。一般に、開示されるCre変異体は、野生型のCreが作用しうるlox部位(野生型のlox部位と称する)間における効率的な組換え、野生型Creにより効率的に用いられない変異体lox部位(変異体lox部位と称する)間における効率的な組換え、および野生型lox部位と変異体lox部位との間における効率的な組換えを媒介する。例えば、Cre変異体は、Creリコンビナーゼ(またはFLPなど、他の類似のリコンビナーゼ)を用いうる任意の方法または技法において用いることができる。加えて、Cre変異体は、野生型lox部位と、変異体lox部位との間のはるかにより効率的な組換えを可能とするので、異なる代替的な組換えの実施も可能とする。このような代替的な組換えの制御を用いて、より洗練された配列組換えを達成し、野生型のCreリコンビナーゼによっては不可能な結果を達成することができる。変異体Creリコンビナーゼは、米国特許第6,890,726号において開示されるなど、当技術分野において公知である。Cre活性の誘導可能性は、Creタンパク質の局在化により制御することができる。例えば、Creタンパク質は、Cre融合体である、CreERt2を結果としてもたらす、エストロゲン受容体の突然変異形とのCreリコンビナーゼの融合体でありうる。リガンドの不在下において、CreERt2は、細胞質に存在する。しかし、合成ステロイドホルモン(タモキシフェン)の投与後において、CreERt2タンパク質は、それが機能的である核内へと移動する(すなわち、タモキシフェン誘導性)。
【0089】
リコンビナーゼはまた、Saccharomyces cerevisiaeの2ミクロンのプラスミドの天然酵素である、FLPリコンビナーゼでもありうる。FLPリコンビナーゼは、FRT(FLPリコンビナーゼ標的)配列と称する、特定の34塩基対のDNA配列において活性である。Creリコンビナーゼと同様、当技術分野において公知である(例えば、米国特許第7371577号、同第7060499号、同第6956146号、同第6774279号において説明される)FlpERなどの変異体も用いることができる。
【0090】
本発明の導入遺伝子はまた、中枢神経系内の細胞など、特定の組織型または細胞型へと選択的に導入し、活性化させることもできる。このような細胞型特異的な活性化に必要とされる調節配列は、対象の特定の細胞型に依存するが、当業者には明らかであろう。例えば、リコンビナーゼおよび/またはMRF導入遺伝子をコードする核酸配列を、細胞型特異的な調節エレメントへと作動的に連結することができる。該調節エレメントは、希突起膠細前駆細胞(OPC)、成熟OL、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)などであるがこれらに限定されない神経細胞に対して特異的でありうる。例えば、神経細胞特異的な調節エレメントは、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来しうる。さらに、細胞型特異的な調節エレメントはまた、誘導的プロモーターの場合もあり、構成的プロモーターの場合もある。
【0091】
動物モデルは、免疫調節、ミエリン修復/再有髄化、および/または軸索保護を促進する薬剤など、再有髄化の促進を対象とする生物活性剤についての、治療的に有効で有益な組合せを決定するスクリーニングアッセイにおいて用いることができる。例えば、トランスジェニック動物を改変して、MRF、Sox10、Nkx2.2、Olig1、および/またはOlig2の発現または活性の低下または上昇など、免疫調節、ミエリン修復/再有髄化、および/または軸索保護を促進する薬剤を発現するか、またはこれを発現するレベルを変化させる(すなわち、上昇または低下させる)ことができる。したがって、このような動物を用いて、これもまた、免疫調節、ミエリン修復/再有髄化、および/または軸索保護を対象とする、異なる複数の生物活性剤をスクリーニングし、ここで、該トランスジェニック動物が、1つの評価項目を対象とする薬剤を含む場合は、該動物に(1または複数の)異なる評価項目を対象とする薬剤を投与し、また、この逆を行い、神経障害または本明細書に記載の関連する状態に対して相乗作用的な治療結果をもたらす治療剤の候補となる組合せを同定することができる。
【0092】
動物モデル系を用いて、ニューロンの再有髄化を促進するか、またはこれに有益な生物活性剤の開発を行うことができる。例えば、MRF活性の上昇により、結果として、例えば、免疫調節、ミエリン修復、または軸索保護をもたらす薬剤を発現するように改変されるトランスジェニック動物は、未知の化合物をスクリーニングして、(1)化合物が、免疫忍容性を増強するか、炎症反応を抑制するか、もしくは再有髄化を促進するかどうか、および/または(2)化合物が、動物モデルにおいて、相乗的な治療効果を結果としてもたらしうるかどうかを決定する方法において用いることができる。代替的に、MRFにおいてエキソン8が欠失するトランスジェニック動物など、MRF活性が損なわれた動物を用いて、CNSにおける有髄化不全を緩和し、該動物のOLの成熟を促進する化合物をスクリーニングすることができる。さらに、本発明のトランスジェニック動物から神経細胞を単離して、ex vivo法を含め、細胞ベースの環境または細胞培養物の環境において、さらなる研究またはアッセイを実施することができる。モデル系を用いて、被験薬剤が、有害作用を付与するか、または再有髄化を低下させる、例えば、脱髄後における損傷をもたらすかどうかを調べることができる。
【0093】
動物モデルはまた、組合せ方式により薬剤をスクリーニングするのにも用いることができる。例えば、候補薬剤は、免疫調節、ミエリン修復/再有髄化、または軸索保護をもたらす第2の薬剤など、別の薬剤と共に投与することができる。例えば、MRF、Nkx2.2、Sox10、Olig1、Olig2、またはこれらの組合せなど、公知の生物活性剤を、候補生物活性剤の前に投与することもでき、これと同時に投与することもでき、またはこの後に投与することもできる。(1または複数の)細胞特異的なマーカー遺伝子の発現を検出および/または定量化し、再有髄化が生じたかどうか、また、それはどのくらいであったか、また、このような再有髄化が、対照と比較して増強されているかどうかを決定することなど、合成組合せ治療の検出および定量化により組合せ効果を決定することができる。このような例においてならば、対照は、疾患モデルが誘導される野生型でもありえようし、候補薬剤が投与されないトランスジェニック動物でもありえよう。
【0094】
本発明の動物モデルはまた、脱髄を受けるように誘導することができ、MRF発現を評価することにより、再有髄化に対する生物活性剤の効果を評価することができる。被験動物において脱髄を誘導する多数の方法が確立されている。例えば、神経の脱髄は、被験動物における病原体または物理的傷害、炎症反応および/または自己免疫反応を誘導する薬剤により与えられる場合がある。好ましい方法では、IFN−γおよびクプリゾン(ビス−シクロヘキサノンオキサルジヒドラゾン)が含まれるがこれらに限定されない脱髄誘導剤が用いられる。クプリゾン誘導型脱髄モデルは、Matsushimaら、Brain Pathol.、11巻、107〜116頁(2001年)において説明されている。この方法では、約1〜約10週間の範囲の数週間にわたり、クプリゾンを含有する食餌を被験動物に摂取させることが典型的である。
【0095】
適切な方法により脱髄状態を誘導した後は、既に脱髄させた病変において、またはその近傍において再有髄化を可能とするのに十分な長さの時間にわたり、動物を回復させる。軸索の再有髄化を発生させるのに必要とされる時間の長さは動物ごとに異なるが、少なくとも約1週間を要することが一般的であり、少なくとも約2〜10週間を要することがよりしばしばであり、約4〜約10週間を要することがさらによりしばしばである。
【0096】
神経系(例えば、中枢神経系または末梢神経系)において有髄化した軸索の増加を観察することにより、またはMRFなど、有髄化細胞のマーカータンパク質のレベルの上昇を検出することにより、本明細書に記載の動物モデルにおける再有髄化を確認することができる。脱髄を検出する同じ方法を用いて、再有髄化が生じたかどうかを決定することができる。例えば、当技術分野で公知の免疫組織化学法またはタンパク質解析により、脱髄/再有髄化現象を観察することができる。例えば、MRFなど、希突起膠細胞マーカーを特異的に認識する抗体により、被験動物の脳の断面を染色することができる。別の態様では、免疫ブロッティング、ハイブリダイゼーション法、および増幅手順、ならびに当技術分野、例えば、それらの各々の開示が、参照により本明細書に組み込まれる、Mukouyamaら、Proc.Natl.Acad.Sci.(2006年)、103巻、1551〜1556頁;Zhangら(2003年)、前出;Girardら、J.Neuroscience.(2005年)、25巻、7924〜7933頁;および米国特許第6,909,031号;同第6,891,081号;同第6,903,244号;同第6,905,823号;同第6,781,029号;および同第6,753,456号において十分に確立された他の任意の方法により、MRFなど、希突起膠細胞マーカーの発現レベルを定量化することができる。
【0097】
治療剤
本明細書に記載の組成物は、治療剤として用いることができる。神経障害を有する被験体は、治療有効量の、MRF活性を調節する生物活性剤により治療することができる。本明細書に記載の組成物および方法を用いて、多発性硬化症(MS)、進行性多巣性白質脳症(PML)、脳脊髄炎、橋中央ミエリン溶解(CPM)、抗MAG抗体陽性疾患、白質萎縮症:副腎白質萎縮症(ALD)、アレキサンダー病、カナバン病、クラッベ病、異染性白質萎縮症(MLD)、ペリツェウス−メルツバッハー病、レフサム病、コケイン症候群、ファンデルクナップ症候群、ツェルヴェーガー症候群、ギラン−バレー症候群(GBS)、慢性炎症性脱髄性多発神経障害(CIDP)、多巣性運動神経障害(MMN)、脊髄傷害(例えば、脊髄の外傷または切断)、アルツハイマー病、ハンチントン病、筋萎縮性側索硬化症、パーキンソン病、および視神経炎などであるが、これらに限定されない各種の神経障害を治療することができる。
【0098】
本明細書に記載の生物活性剤を被験体に投与してMRFの発現および/または活性を調節することにより、成熟希突起膠細胞(OL)への希突起膠細胞の前駆細胞(OPC)の分化を促進することができる。神経障害は、多発性硬化症など、脱髄障害でありうる。治療有効量の、MRF活性を調節する生物活性剤を被験体に投与することにより、被験体における再有髄化を促進することができる。
【0099】
再有髄化
本明細書に記載の生物活性剤を被験体に投与して、神経細胞の分化を増強することができる。生物活性剤は、MRFまたはそれにより調節される遺伝子の発現を調節することにより再有髄化を促進しうる。本明細書に記載の生物活性剤は、MRF自体でありうる。このような生物活性剤の投与は、薬剤自体の外因的投与により達成することもでき、本明細書で説明され、また、当業者に公知である、適切な転写調節エレメントを介して、該薬剤を、構成的、誘導的、または細胞特異的な形でコードおよび発現する核酸ベクターを提供することにより達成することもできる。このようにして発現される生物活性剤は、それ自体、神経細胞分化を促進しうる。このような神経細胞には、OL、OPC、SC、NSC、星状細胞、および小膠細胞が含まれる。
【0100】
したがって、神経細胞の分化および/または再有髄化を促進するように、内因性のMRF発現を誘導する生物活性剤を、細胞/被験体へと投与することができる。一部の実施形態では、生物活性剤を用いて、ES細胞が、OL(またはOPC)へと分化するように誘導することができる。ポリペプチド、またはポリペプチドをコードする核酸を投与することにより、MRF発現を調節して、OLへの分化を増強することができる。
【0101】
相同組換え、組込みにより、または本明細書で記載され、また、当業者に周知の成分および方法を用いるプラスミドもしくはウイルスベクターを用いることにより、所望のポリペプチドをコードする核酸を、標的細胞内へと形質転換することができる。トランスフェクトされうる神経細胞には、OL、OPC、SC、NSC、星状細胞、および小膠細胞が含まれる。一部の実施形態において、このような神経細胞には、同時に、または異なる時点において、複数のベクターをトランスフェクトすることができる。さらに、本明細書で開示されるポリペプチドのうちのいずれかをコードする核酸は、本明細書で開示され、また、当業者により認知される、構成的プロモーター、誘導的プロモーター、または細胞特異的プロモーターへと作動的に連結することができる。
【0102】
生物活性剤を投与して、OLの成熟など、神経細胞の分化を結果としてもたらすMRFの発現を増大させ、再有髄化を促進することができる。投与される生物活性剤は、MRF自体でありうる。投与される生物活性剤はまた、MRF発現の調節物質をコードするか、またはMRF自体をコードする核酸ベクターでもありうる。核酸ベクターは、構成的、誘導的、または細胞特異的な転写調節エレメントを含有し、これにより、所望の生物活性剤の持続的な発現または時点により異なる発現をもたらしうる。例えば、テトラサイクリン抑制系を用いるドキシサイクリンにより、細胞におけるMRF発現を誘導することができる。代替的に、発現ベクターは、本明細書で記載されるか、または当業者に周知の、神経特異的プロモーターを含みうる。したがって、それを必要とする被験体を治療する方法において、OPCの増殖と、OLの分化とを交互に行って再有髄化を増強する必要がある場合、神経細胞に対する生物活性剤の発現を調節することができる。
【0103】
例えば、構成的プロモーター、誘導的プロモーター、または神経細胞特異的プロモーターに作動的に連結され、MRF遺伝子、またはMRFの発現/活性を調節する別の遺伝子をコードする核酸分子を、神経細胞にトランスフェクト(神経細胞を遺伝子改変)することができる。このような細胞を形質転換させて、異なる発現レベルでMRFを発現させ、したがって、神経細胞の増殖を調節することができる。例えば、ポリペプチドは、MRFでありうる。
【0104】
また、細胞または被験体に増殖因子またはホルモンを投与して、神経細胞の分化または増殖を促進することもできる。増殖因子およびホルモンは、それ自体、本明細書で開示される任意の生物活性剤の投与と同時に、この前において、またはこの後において投与することができる。このような増殖因子またはホルモンの例には、甲状腺ホルモンT3、インスリン様増殖因子1、線維芽細胞増殖因子2、血小板由来増殖因子(PDGF)、神経成長因子、ニューロトロフィン、ニューレグリン、またはこれらの組合せが含まれる。このような増殖因子またはホルモンはまた、本明細書で開示される他の任意の生物活性剤と同時に、この前において、またはこの後において施される核酸ベクターによってもコードされうる。
【0105】
さらに、生物活性剤の投与の前において、これと同時に、またはこの後において、OPC、シュワン細胞、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)が含まれるがこれらに限定されない神経細胞もまた投与することができる。1種または複数種の生物活性剤と共に、1種または複数種の神経細胞を投与することができる。明確さを目的として述べると、種類とは、例えば、異なる種類の細胞(例えば、希突起膠細胞および星状細胞)または異なる種類の生物活性剤(例えば、抗体およびアンチセンス)を意味する。
【0106】
異なる生物活性剤を投与することができる、例えば、第2の生物活性剤の投与と同時に、この前において、またはこの後において、第1の生物活性剤を投与することができる。複数の生物活性剤を投与することができる。例えば、第1の生物活性剤は、MRF活性を調節する生物活性剤、またはMRF自体でありうる。Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進する生物活性剤など、第2の生物活性剤は、MRFを調節する薬剤の投与と同時に、この前において、またはこの後において投与することができる。代替的に、MRFは、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの投与と同時に、この前において、またはこの後において投与することもできる。各種の生物活性剤の投与は、再有髄化の促進において相乗効果をもたらしうる。
【0107】
前出はまた、標的細胞をトランスフェクトするのに用いうる核酸ベクターによる製剤にも適用しうることを理解されたい。このようなベクターは、標的細胞をトランスフェクトし、所望のポリペプチドを発現することが可能であるものとして、本明細書で記載され、また、当業者により認知されている。まとめると、このようなベクターはまた、本明細書に記載の医薬製剤または治療剤中においても用いることができる。
【0108】
再有髄化細胞の移植
各種の動物モデルにおいて、CNS軸索の再有髄化が示されている。以来、近年の多くの研究により、脱髄疾患における細胞移植の使用と関連する新規の技法および新規の機構が示されている。成人脳から単離されたヒトOP細胞を、有髄化不全のマウス突然変異体へと移植したところ、無髄軸索を有髄化することができた。成人の前駆細胞を用いれば、倫理的問題が回避されることが重要である。OP細胞が、内因的な再有髄化の一因となるのに対し、NSCは、ミエリンの修復を促進する代替的な細胞供給源である。NSCは、成人CNSにおいて見出され、in vitroにおいて大幅に増殖させることができ、また、OL、星状細胞、またはニューロンを形成するように分化させることができる。実験的な自己免疫性脳脊髄炎(EAE)の再発形態または慢性形態を有するげっ歯動物内へと移植したところ、NSCは、CNSの炎症領域および脱髄領域へと遊走し、神経膠細胞原基を優先的に選択することが示されている。さらに、移植を受けたマウスにおける臨床的疾患の緩和は、脱髄病変の修復および軸索傷害の軽減と関連していた。組織学的解析により、移植されたNSCは、大半が、PDGFR+ OP細胞へと分化することが確認された。
【0109】
本発明の態様において、被験体の生物活性剤は、MRF、またはMRFにより上方調節される遺伝子を過剰発現するように改変され、被験体に投与されると、ミエリンの修復または脱髄した軸索の再有髄化に関与する細胞を含みうる。このような細胞を適切なベクターと共に培養し、また、このような細胞に適切なベクターをトランスフェクトして、細胞の成熟、またはOLの分化を増強するポリペプチドを発現させることができる。また、細胞を改変して、Sox10、Nkx2.2、Olig1、またはOlig2を過剰発現させることもできる。1または複数の細胞型を改変して、MRF、Nkx2.2、Sox10、Olig1、Olig2、またはこれらの組合せを過剰発現させることができる。例えば、細胞を改変してMRF、およびSox10、Nkx2.2、Olig1、またはOlig2を過剰発現させ、これを被験体に投与して神経障害を治療することができる。OPCおよび星状細胞など、異なる細胞型もまた投与することができる。一部の実施形態において、ミエリンを生成する細胞またはこれらの前駆細胞には、胚性OPCまたは成体OPCが含まれるがこれらに限定されない。
【0110】
別の生物活性剤の投与の前において、これと同時に、またはこの後において投与しうる細胞(「細胞型」)は、希突起膠細前駆細胞(OPC)、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)でありうる。一部の実施形態において、細胞は、生物活性剤により、OPCまたはOLへと分化するように誘導されたES細胞などのES細胞でありうる。一部の実施形態では、このような細胞を動物被験体に投与して、OLの成熟など、神経細胞の分化を増強することができる。
【0111】
一実施形態において、細胞は、成熟希突起膠細胞をもたらすそれらの能力に基づき、中枢神経系(CNS)内における希突起膠細胞の前駆細胞(OPC)であると考えられる、NG2プロテオグリカンを発現する神経膠細胞(NG2(+)細胞)である。一部の実施形態では、希突起膠細前駆細胞(OPC)、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)を、ケモカインをコードするベクターと共に培養し、これにより形質転換し、in vitroにおいて増殖させた後で移植する。他の実施形態では、in vitroにおいて細胞にトランスフェクトするか、またはこれらを遺伝子改変し、MRF遺伝子、Sox10、Nkx2.2、Olig1、および/またはOlig2によりコードされるタンパク質を発現させる。
【0112】
一部の実施形態では、当技術分野において公知であるか、または本明細書で開示される方法を用いて、1または複数の所望の生物活性剤の発現を可能とするように、希突起膠細前駆細胞(OPC)、シュワン細胞(SC)、嗅球鞘細胞、および神経幹細胞(NSC)に、1または複数の発現ベクターをトランスフェクトする。このような生物活性剤は、MRF、Nkx2.2、Sox10、Olig1、またはOlig2の発現/活性を調節しうる。
【0113】
侵襲性の手順、手術手順、侵襲性が最小限の手順、および手術以外の手順を含め、当技術分野において公知の方法を用いて移植を実施することが理解される。被験体、標的部位、および送達される(1または複数の)薬剤に応じ、当技術分野において公知の方法を用いて細胞型および細胞数を選択することができる。
【0114】
医薬組成物
本発明はまた、被験体における神経障害を治療するための組成物であって、MRFなど、本明細書に記載の生物活性剤を含む組成物も提供する。組成物はまた、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せもさらに含みうる。意図される医薬組成物には、ペプチド、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、核酸発現ベクター、抗体もしくは抗体フラグメント、低分子、またはこれらの組合せが含まれるが、これらに限定されない。このような組成物は、治療有効量で用いることができる。
【0115】
所望の程度の純度を有するこのような薬剤を、薬学的に許容される任意選択の担体、賦形剤、または安定化剤(「Remington’s Pharmaceutical Sciences」、第16版、Osol,A.編、1980年)と共に、凍結乾燥させた製剤または水溶液の形態で混合することにより、このような薬剤による製剤を保存のために調製する。許容される担体、賦形剤、または安定化剤は、用いられる用量および濃度においてレシピエントに対して非毒性であり、これらには、リン酸緩衝液、クエン酸緩衝液、酢酸緩衝液、および他の有機酸緩衝液などの緩衝液;アスコルビン酸およびメチオニンを含めた抗酸化剤;防腐剤(オクタデシルジメチルベンジルアンモニウムクロリド;塩化ヘキサメトニウム;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチルオルベンジルアルコール;メチルパラベンもしくはプロピルパラベンなどのアルキルパラベン;カテコール;レソルシノール;シクロヘキサノール;3−ペンタノール;およびm−クレゾール);低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチン、もしくは免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、もしくはリシンなどのアミノ酸;単糖、二糖、および、グルコース、マンノース、またはデキストリンを含めた他の炭水化物;EDTAなどのキレート化剤;スクロース、マンニトール、トレハロース、もしくはソルビトールなどの糖;甘味剤および他の芳香剤;微晶質セルロース、ラクトース、トウモロコシ、および他のデンプンなどの充填剤;結合剤;添加剤;着色剤;ナトリウムなどの塩形成対イオン;金属錯体(例えば、Zn−タンパク質錯体);ならびに/またはTWEEN(商標)、PLURONICS(商標)、もしくはポリエチレングリコール(PEG)などの非イオン性界面活性剤が含まれる。
【0116】
好ましい実施形態において、本発明の生物活性剤を含む医薬組成物は、酸添加塩および塩基添加塩の両方が含まれることを意図する、薬学的に許容される塩として存在するなど、水溶性形態である。「薬学的に許容される酸添加塩」とは、遊離塩基の生物学的有効性を保持し、生物学的または他の形で有害でなく、塩酸、臭化水素酸、硫酸、硝酸、リン酸などの無機酸、および酢酸、プロピオン酸、グリコール酸、ピルビン酸、シュウ酸、マレイン酸、マロン酸、コハク酸、フマル酸、酒石酸、クエン酸、安息香酸、ケイ皮酸、マンデリン酸、メタンスルホン酸、エタンスルホン酸、p−トルエンスルホン酸、サリチル酸などの有機酸により形成される塩を指す。「薬学的に許容される塩基添加塩」には、ナトリウム塩、カリウム塩、リチウム塩、アンモニウム塩、カルシウム塩、マグネシウム塩、鉄塩、亜鉛塩、銅塩、マンガン塩、アルミニウム塩など、無機塩基に由来する塩が含まれる。アンモニウム塩、カリウム塩、ナトリウム塩、カルシウム塩、およびマグネシウム塩が特に好ましい。薬学的に許容される非毒性の有機塩基に由来する塩には、第一級アミン、第二級アミン、および第三級アミン、天然の置換アミンを含めた置換アミン、環状アミン、およびイソプロピルアミン、トリメチルアミン、ジエチルアミン、トリエチルアミン、トリプロピルアミン、およびエタノールアミンなど、塩基性のイオン交換樹脂による塩が含まれる。in vivoにおける投与に用いられる製剤は、滅菌であることが好ましい。これは、滅菌濾過膜または当技術分野において公知の他の方法を介する濾過により、容易に達成される。
【0117】
薬剤はまた、イムノリポソームとしても調合することができる。リポソームとは、哺乳動物に治療剤を送達するのに有用な、各種の脂質、リン脂質、および/または界面活性剤を含む小胞体である。生物活性剤を含有するリポソームは、Eppsteinら、Proc. Natl. Acad. Sci. USA、82巻、3688〜3692頁(1985年);Hwangら、Proc. Natl. Acad. Sci. USA、77巻、4030〜4034頁(1990年);米国特許第4,485,045号;同第4,544,545号;およびPCT WO97/38731において説明される方法など、当技術分野において公知の方法により調製される。循環時間が延長されたリポソームは、米国特許第5,013,556号において開示されている。リポソームの成分は一般に、生体膜の脂質配置と同様、二層形成で配置される。特に有用なリポソームは、ホスファチジルコリン、コレステロール、およびPEGにより誘導体化されたホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を伴う逆相蒸発法により作製することができる。リポソームは、所望の直径を有するリポソームをもたらすように、規定の小孔サイズによるフィルターを介して押出し成形される。化学療法剤または他の治療的に活性な薬剤も、場合によって、リポソーム内に含有される(Gabizonら、J. National Cancer Inst、81巻、1484〜1488頁(1989年))。
【実施例】
【0118】
(実施例1)
マウスOPCの単離、培養、およびトランスフェクション
本質的に、既に説明される通り(Cahoyら、J Neurosci、28巻、264〜278頁(2008年))に、マウスOPCを単離した。略述すると、P7 C57/B6マウスの脳を摘出し、賽の目に刻み、酵素的に分解して、単一細胞の懸濁液を作製した。これらの細胞を、BSL1(Vector Laboratories社製、L−1100)でコーティングした4枚のペトリディッシュ上において、各15分間ずつにわたり逐次パニングして小膠細胞を枯渇させ、次いで、抗PDGFRαラットモノクローナル抗体(BD Pharmingen社製、558774型)でコーティングしたペトリディッシュ上において、1時間にわたりパニングしてOPCを陽性選択した。付着しない細胞はDPBSにより洗い落とし、付着するOPCは、トリプシンによりペトリディッシュから採取した。2%のB−27(Invitrogen社製)ヒトトランスフェリン(100mg/ml)、ウシ血清アルブミン(100mg/ml)、プトレシン(16mg/ml)、プロゲステロン(60ng/ml)、亜セレン酸ナトリウム(40ng/ml)、N−アセチル−L−システイン(5mg/ml)、D−ビオチン(10ng/ml)、フォルスコリン(4.2mg/ml)、ウシインスリン(5mg/ml)(すべて、Sigma社製)、グルタミン(2mM)、ピルビン酸ナトリウム(1mM)、ペニシリン−ストレプトマイシン(各100U)(すべて、Invitrogen社製)、微量元素B(1倍濃度;バージニア州、ハーンドン、Mediatech社製)、CNTF(10ng/ml;ニュージャージー州、テリータウン、Regeneron社からの恵与)、ならびにPDGF−AA(10ng/ml)およびNT−3(1ng/ml)(いずれも、ニュージャージー州、ロッキーヒル、PeproTech社製)を含有するDMEM(カリフォルニア州、カールスバッド、Invitrogen社製)中、37℃、10%CO2で、PDLによりコーティングしたT175組織培養フラスコ内において、OPCを培養した。分化実験のため、以下の通りに細胞にトランスフェクトし、これらを、上記の通りであるが、PDGF−AAを伴わず、トリヨードチロニン(T3)(40ng/ml;シグマ社製)を伴う培地へと移した。
【0119】
OPCへのトランスフェクション:EBSSによりフラスコを1回洗浄することにより、マウスOPCの培養物を継代培養し、次いで、EBSS中に1:10のトリプシン:EDTA(Sigma社製)を用いて37℃で5分間にわたり該フラスコを処理した。30%のFCS/DPBSにより細胞を回収し、220RCFで15分間にわたり遠心分離し、増殖培地中に再懸濁させ、さらに15分間にわたり遠心分離して、微量のトリプシンを除去した。細胞ペレットを、OPC Nucleofection液(Amaxa社製)中に細胞50×106個/mlで再懸濁させた。100ul(細胞5×106個)を、発現構築物(約4ug)またはsiRNA(20uMの、MRFに対するsiRNAのプール、もしくは個別のsiRNA、または非標的化siRNAのプールであるsiControlである、それぞれ、Dharmacon社製L−056814−00、LU−056814−00、およびD001206−13を10ulずつ)に添加し、Amaxa社製ヌクレオフェクション装置のプログラム0−17により電気穿孔した。次いで、細胞を、OLマーカーアッセイ用には、24ウェルプレートの分化培地中に細胞50,000個/PDLコーティングしたカバースリップ1枚で、異所性発現/分化アッセイ用には、24ウェルプレートの増殖条件に細胞5,000個/PDLコーティングしたカバースリップ1枚で、RNA単離用には、分化培地中に細胞5×106個/PDLコーティングした10cmのディッシュで播種した。
【0120】
(実施例2)
ニワトリ胚のin ovoにおける電気穿孔
可視化を可能とする0.2%ファーストグリーンを伴うTE緩衝液中に約1ulのDNA溶液(1:4のpCIGおよびpCAGGS−MRF混合物約4ug/ul)を、ハンバーガー−ハミルトン段階にある胚12個の神経管内腔へと注入した。ECM830 ElectroSquare Porator(Genetronics社製)を用いて、胚を隔てた30ボルトで5×50ミリ秒にわたるパルスを用いて、電気穿孔を実施した。電気穿孔の4日後に胚を回収し、4%PFA中において2時間にわたり浸漬固定した後、以下の通りに処理して、in situハイブリダイゼーションまたは免疫組織化学にかけた。
【0121】
(実施例3)
免疫組織化学、in situハイブリダイゼーション、およびTEM顕微鏡法
免疫組織化学:灌流固定したマウスに由来する10μmの凍結断面、またはPDLコーティングしたカバースリップ上で増殖させた細胞を、PBS中に4%のパラホルムアルデヒド中において10分間にわたり固定し、PBS中で3分間ずつ5回にわたり洗浄し、次いで、ブロッキング液(表面抗原用には、PBS中に10%のウシ胎仔血清、細胞内抗原用には、0.3%のトリトンXを添加する)中で30分間にわたりインキュベートした。ブロッキング液(1:500のラット抗MBP抗体、1:500のウサギ抗NG2抗体;Chemicon社製、1:500の抗mycモノクローナル4A6抗体;英国、ロンドン、インペリアルカレッジ、R.Reynolds氏のご恵与による、Upstate社製、1:1,000のウサギ抗活性化カスパーゼ3抗体、BD Pharmingen社製、1:50のマウス抗MOG抗体クローン8−18C5)中において、一次抗体と共に一晩にわたり細胞をインキュベートした。PBS中で3分間ずつ5回にわたりカバースリップを洗浄し、適切なフルオロフォアをコンジュゲートさせた二次抗体(Molecular Probes社製;ブロッキング液中において1:500に希釈した)と共に30分間にわたりインキュベートし、PBS中で3分間ずつ5回にわたり洗浄し、DAPI核内対比染色色素を伴うDAKI蛍光マウント用培地によりマウントした。製造元の指示書に従い、10μmの凍結断面上において、Fluoromyelin(Invitrogen社製)染色を実施した。
【0122】
in situハイブリダイゼーション:プライマーGGTGGGTTTGAGTTTGGAGGTTおよびGGGGAAACGCTCTATGAACAGGを用いて、3’UTRから941bpに対応するin situハイブリダイゼーションプローブを、OL cDNAから増幅し、PCR−II−Topoベクター(Invitrogen社製)内へとサブクローニングした。PLPプローブは、William Richardson教授により恵与された。製造元の指示書に従い、T7ポリメラーゼおよびDIG RNA標識化キット(Roche社製)を用い、アンチセンスDIGにより標識化したリボプローブを合成した。P16における脳神経ならびにP17における視神経および坐骨神経に由来する10μmの断面上において、本質的に説明される(Cahoyら、J Neurosci、28巻、264〜278頁(2008年);Schaeren−Wiemers and Gerfin−Moser、Histochemistry、100巻、431〜440頁(1993年))通りに、in situハイブリダイゼーションを実施した。
【0123】
TEM顕微鏡法:麻酔下にあるP13マウスをPBSにより灌流した後で、カコジル酸ナトリウム緩衝液中に2%のグルタルアルデヒド/4%のパラホルムアルデヒドで処理した。視神経を切開摘出し、4℃で一晩にわたり後固定した。1%四酸化オスミウムおよび1%酢酸ウラニルによる処理後、神経をエポン樹脂内に包埋した。スタンフォード大学の微生物学および免疫学電子顕微鏡施設において断面作製および電子顕微鏡法を実施した。
【0124】
(実施例4)
構築物の作製
pCS6−MRF:プライマーCCCGGGCGCCACCATGGAGGTGGTGGACGAGACおよびCTCGAGGGAGGCAGCTCAGTCACACAGGを用いて、培養したマウスOLのcDNAから、MRFのコード領域を増幅した。結果として得られるXma1/Xho1を連結したPCRフラグメントを、Xma1/Xho1により2回にわたり消化したpCMV−Sport6(pCS6)ベクター(Invitrogen社製)内へとライゲーションし、配列決定により確認した。
【0125】
pCS6−Myc−MRF:プライマーCTCGAGGAGGTGGTGGACGAGACCGAAGおよびCTCGAGGGAGGCAGCTCAGTCACACAGGを用いて、培養したマウスOLのcDNAから、開始コドンを除いたMRFのコード領域を増幅した。結果として得られるXma1/Xho1を連結したPCRフラグメントを、Xma1/Xho1により2回にわたり消化したpCS6ベクター内へとライゲーションした。次いで、Mycペプチド(MEQKLISEEDL)をコードする自己アニール化オリゴヌクレオチドを、結果として得られる構築物のXma1部位内へとライゲーションし、N末端においてmyc−MRF融合構築物をもたらした。
【0126】
pCAGGS−MRF:Xho1およびXba1で挟んだプライマーにより、上記のpCS6−MRFからMRFのコード領域を増幅した。結果として得られるPCR産物を、pCAGGSのXba1部位およびXho1部位内へとライゲーションした。
【0127】
(実施例5)
HEK293細胞へのトランスフェクション
トランスフェクションの1日前に、10%ウシ胎仔血清を伴うダルベッコ改変イーグル培地中に50〜70%のコンフルエンスで、HEK293細胞を播種した。製造元の指示書に従い、Lipofectamine 2000(Invitrogen社製)を用いて、トランスフェクションを実施した。トランスフェクションの48時間後において、免疫蛍光法により細胞を解析した。
【0128】
(実施例6)
ノーザンハイブリダイゼーション
全RNA(5〜15μg)を、変性ホルムアルデヒドゲル上で泳動し、Hybond−N+(Amersham社製)へと転写し、UVにより架橋させた。次いで、100mlのハイブリダイゼーション液(7%SDS、0.5M Na2HPO4 pH 7.2、100ug/mlのニシン精巣DNA)中68℃で約2時間にわたり、膜をプレハイブリダイズさせた。製造元の指示書に従い、α−32P−dCTP(GE Healthcare社製)を伴う、Prime It II random Primerキット(Stratagene社製)を用いて、約1kbのMRF cDNA、CNP cDNA、PLP cDNA、またはGAPDH cDNAを含有するプラスミドから、放射性標識化したDNAプローブを生成させた。400gで2分間にわたり、Probe Quant G−50マイクロカラム(Amersham Biosciences社製)により、プローブをスピンし、組み込まれなかった32d−CTPを除去し、5分間にわたり95℃まで加熱した後、15mlのハイブリダイゼーション液中の膜に添加した。68℃で一晩にわたりハイブリダイゼーションを行い、1倍濃度SSC、0.1%SDS中、室温で2分間ずつ10回にわたり洗浄し、次いで、0.5倍濃度SSC、0.1%SDS中68℃で3分間ずつ10回にわたり洗浄した。次いで、X線フィルム(Amersham社製)を−80℃で膜に露光した後、現像した。沸騰する0.1倍濃度のSSC、0.1%SDS中で3回にわたる洗浄により膜をはがし、一晩にわたりフィルムを静置し、シグナルの不在について点検した後で、再プロービングした。
【0129】
(実施例7)
Affimetrix解析
Qiagen社製のオンカラム用DNアーゼ処理を用いるRNeasyマイクロキット(カリフォルニア州、バレンシア、Qiagen社製)により、細胞から全RNAを単離し、夾雑物であるゲノムDNAを除去した。Agilent 2100 Bioanalyzer(Agilent Technologies社製)を用いてRNAの完全性を評価し、NanoDrop ND−1000分光光度計(DE、ロックランド、NanoDrop社製)を用いて、RNA濃度を決定した。
【0130】
対照中におけるスパイクにより、Affymetrixによる2サイクルの標的標識化アッセイ(カリフォルニア州、サンタクララ、Affymetrix社製、900494型)を用いて、1ugの全RNAから、Affymetrix 3’アレイに対するハイブリダイゼーション用にビオチニル化したcRNAを調製した。製造元のプロトコールに従い、標識化したcRNAをフラグメント化し、マウスゲノム430 2.0アレイ(3’アレイ、Affymetrix社製、900495型)へとハイブリダイズした。
【0131】
Affymetrix GCOS 1.3ソフトウェアを用いて生の画像ファイルを処理し、個々のプローブセルの強度データを計算し、CELデータファイルを作製した。GCOSおよびMAS 5.0アルゴリズムを用いて、強度データをチップ当たりのデータに標準化して、標的強度のTGT値を500とし、個々のプローブセットに対する発現データおよび存在コール/不在コールを計算した。MAS 5.0アルゴリズムにより解析されるデータに対応する遺伝子記号および遺伝子名は、Affymetrix Netaffx Mouse430_2注釈ファイル(http://www.affymetrix.com/support/technical/byproduct.affx?product=moe430−20)に拠った。異常について生のDAT画像ファイルを検討することにより品質管理を実施し、各GeneChipアレイのバックグラウンド値が100未満であることを確認し、百分率による存在コールが、細胞型に適切であるかどうかモニタリングし、対照中におけるポリ(A)スパイク、ハウスキーピング遺伝子、およびハイブリダイゼーション対照を点検し、標識化とハイブリダイゼーションとの符合を確認した。
【0132】
(実施例8)
RT−PCR
上記に従い調製したRNAを、製造元の指示書に従い、Invitrogen社製Superscript III逆転写酵素を用いる逆転写にかけた。cDNAを、遺伝子特異的なプライマーにより、25または30サイクルにわたり増幅にかけ、2%アガロースゲル上において泳動した。
【0133】
(実施例9)
MRF条件付きノックアウトマウスの作製
MRFのエキソン8を、Pez−Frt−lox−DT標的化ベクターのSal1部位内にクローニングすることにより、エキソン8をloxPで挟んだマウスを作製した。5kbの5’アームおよび3kbの3’アームを、それぞれ、Not1部位およびXho1部位内へとクローニングし、E14ES胚性幹細胞内への相同組換えに対する標的化を可能とした。HindIIIにより消化したDNAに対するサザンブロット法、および5’loxP部位の挿入に対するPCR検証により、正確に標的化されたネオマイシン耐性クローンを同定した。標的化された細胞を線維芽細胞内へと注入し、キメラマウスを作製し、これらを、C57/B6マウスへと交配させ、ヘテロ接合マウスを作製した。これらのマウスを、FlpER株(Farleyら、Genesis、28巻、106〜110頁(2000年))へと交配させ、ネオマイシンカセットを欠失させ、第2世代のFlpER陰性マウスにおけるPCRによりこれを確認した。ヘテロ接合のMRFflox化マウスを、2世代にわたり、Olig2−Creマウス(Schullerら、近刊)またはCNP−Creマウス(Lappe−Siefkeら、Nat Genet、33巻、366〜374頁(2003年))へと交配させ、MRFfl/fl;Olig2wt/creマウスまたはMRFfl/fl;CNPwt/creマウスを得た。共通の上側プライマー(GGGAGGGGGCTTCAAGGAGTGT)、ならびに野生型の対立遺伝子を同定する下側プライマー(CCCCCAGCATGCCGATGTACAC)、およびflox化対立遺伝子を同定する下側プライマー(CCTTTCGCCAGGGGGATCTTG)を用いるPCRにより、マウスの遺伝子型を解析した。Creリコンビナーゼ陽性マウスは、プライマーGCTAAGTGCCTTCTCTACACCTGCおよびGGAAAATGCTTCTGTCCGTTTGにより同定した。
【0134】
(実施例10)
定量化および統計学
培養細胞に対してカウントを実施する場合(マーカーなどの発現を解析するため)、1条件当たり少なくとも3枚のカバースリップを盲検でカウントし、カバースリップ1枚当たり10ずつの視野(20倍の対物レンズ)をカウントした。各カバースリップの平均から、各条件についての平均およびSEMを計算し、多重比較についてのボンフェローニ補正を用いる、対応のない2方向のt検定により条件を比較した。示される実験は、少なくとも2つの個別の実験を表わす。組織断面から細胞を定量化する場合、1遺伝子型当たり4〜6匹ずつのマウスを用いた。解析(Photoshopにより実施する細胞カウント、およびImageJにより定量化するカウント領域の面積を伴う)用に60〜100umの距離を置いて断面を写真撮影(20倍の対物レンズ)することにより、マウス1匹当たり3枚ずつの断面を解析した。活性カスパーゼ3免疫陽性細胞を定量化する場合、20倍の対物レンズで、マウス1匹当たり3枚ずつ、60umの距離を置いた視神経縦断面について、免疫陽性細胞の数を定量化し、次いで、該神経を低倍率(4倍の対物レンズ)で写真撮影し、ImageJにより、該神経の面積を決定した。1遺伝子型当たり5〜6匹ずつのマウスを用いた。各マウスの平均から、各遺伝子型についての平均およびSEMを計算し、多重比較についてのボンフェローニ補正を用いる、対応のない2方向のt検定により遺伝子型を比較した。
【0135】
(実施例11)
CNS内におけるOL特異的な転写物である、GM98/MRFの同定
イムノパニング/FACSによる細胞精製法を、遺伝子プロファイリングと組み合わせて、マウスCNS内における細胞型特異性を示す遺伝子を同定した。急性精製した星状細胞、ニューロン、希突起膠細胞の前駆細胞(OPC)、新たに分化した希突起膠細胞(OL)、有髄化する成熟OLを転写プロファイルの生成に用いたスクリーンの一部として、遺伝子モデル98/ミエリン遺伝子調節因子を同定した。MRFについてのAffymetrixプローブセット(1439506_at型)は、MBP、PLP、およびMOGなど、典型的なOLマーカーが示したのと同様なレベルのOL濃縮を他の細胞型に対しても示し、ニューロンおよび星状細胞においては本質的に検出不能なレベルの発現であり、OPCにおいては低レベルの発現であり、OL内においては比較的強い発現であった。
【0136】
新たに分化したOL(GalC+、MOG−)におけるMRFの発現が、より成熟したOL(MOG+)と少なくとも同程度に高度であったことは、該遺伝子が、有糸分裂後細胞へと移行すると急速に誘導されることを示す(図1A)。全脳、全心臓、および培養OLから単離したRNAに対する、培養OLから増幅したMRF cDNAとのノーザンハイブリダイゼーションにより、脳内において、約5.5Kbのサイズの明白な転写物が確認されたが、これは、心臓に由来する試料中では不在であり、培養OL中では高度に濃縮されていた。確立されたOLマーカーであるプロテオリピドタンパク質(PLP)およびMRFに対するプローブを用いるin situハイブリダイゼーションにより、該2つの遺伝子について同一の発現パターンが確認され、MRFは脳内におけるすべての白質路内において発現し(図1C)、白質内では、PLPと同じ染色色素の細胞内分布を示した(図1D)。MRFの発現は、坐骨神経では検出されず(図10)、これにより、有髄化細胞内におけるMRFの発現が、CNSに制約されることが示された。ESTのデータベース(Unigene社製)は、MRFが他の組織、中でもとりわけ、膵臓および肺内において発現することを示すが、この発現はおそらく、CNS内で見られる発現よりも大幅に低度である。
【0137】
データベースの検索により、MRFは、ヒト遺伝子であるC11Orf9のマウスオルトログであることが明らかとなった。MRFおよびC11Orf9共に、推定のDNA結合ドメインとしてオンラインデータベースに列挙される酵母転写因子であるNdt80に対して相同的な領域を有する高分子タンパク質をコードする(Montanoら、Proc. Natl. Acad. Sci. U S A、99巻、14041〜14046頁(2002年))。OLから単離されたcDNAの配列決定により、GM98/MRFの転写物は、1139アミノ酸のタンパク質をコードすることが示された(図2A)。このタンパク質は、プロリンに富む複数のドメインを含有するN末端領域である、Ntd80様DNA結合領域と、複数の疎水性領域を含有するC末端領域とを含有する。ヒトC11Orf9とのこのタンパク質のアライメントにより、88.6パーセントの全体的相同性が明らかとなり、DNA結合領域内におけるこの相同性は100%であった(図2B)。C11Orf9は、そのC末端内における2つの疎水性領域が膜貫通ヘリックスとして作用する膜貫通タンパク質でありうることが既に示唆されている(Stohrら、Ctyogenet. Cell Genet.、88巻、211〜216頁(2000年))。MRFの細胞内における局在化を確立するため、HEK細胞内において、N末端にMycタグを付した融合体を発現させた。このMycタグを付したタンパク質は、明白な核内発現を示し(図2C)、培養した希突起膠細胞内において該タグを付したタンパク質が発現すると、核内局在化もまた見られた。N末端のMycタグを指向する抗Myc抗体、および該タンパク質のDNA結合領域に対して生成させた抗体によっても同じ細胞内における核内局在化が見られ、これにより、少なくともこれらの領域が、膜内局在化ではなく、核内局在化を示すことが示唆された。
【0138】
(実施例12)
MRFはin vitroにおける希突起膠細胞によるミエリン遺伝子の発現に必要である。
【0139】
MRFが、OLの転写調節において役割を果たすかどうかを確立するため、MRFのコード領域を標的とするsiRNAのプール(siMRF)、またはこれを標的としないsiRNAのプール(siCont)をトランスフェクトすることにより、OL培養物中におけるMRFの発現を遮断した。ノーザンブロットおよびRT−PCRにより評価したところ、siMRFトランスフェクト細胞は、siContトランスフェクト細胞と比べ、MRF mRNAに対する明白で一貫した下方調節を示し(図4A、図12)、これにより、siRNAのプールは、MRFレベルの低減に有効であることが示された。MRFに対するsiRNAを、増殖条件で培養されたOPCにトランスフェクトしたところ、その影響は見られず、大半のsiMRF細胞およびsiCont細胞がNG2陽性前駆細胞として分裂し続けたことは、急性摘出されたOPC内において見られるMRF発現の欠如と符合する。
【0140】
siRNAトランスフェクト細胞を、分化条件(−PDGF、+T3)へと移したところ、siContトランスフェクト細胞と、siMRFトランスフェクト細胞との間で差違が生じ始めた。分化条件への移動から24時間以内に、siMRFトランスフェクト細胞およびsiContトランスフェクト細胞共に、分裂を停止し、突起を拡大させ始めたのに対し、48時間以内に、siMRFトランスフェクト細胞は、siContトランスフェクト細胞より、膜シートの沈着範囲を狭め、また、カルセイン(Calcien)AM/EthHD1生存/死亡アッセイにより評価される生存率も、さほどではないが有意に低下させた(図3A、B)。分化の誘導から48時間以内に85%超が強くMBP陽性であったsiContトランスフェクト細胞とは対照的に、siMRFトランスフェクト細胞は、MBP発現の明白な遅延および低下を示し、分化の48時間後において、MBPについて陽性の細胞は23.6%に過ぎず、分化の96時間後においても、64.4%の細胞で、発現は低度に過ぎなかった(図3A、C)。
【0141】
これに反し、また、それらが単純な前駆細胞様形態から移行することとも符合するが、siMRFトランスフェクト細胞がやはり、siContトランスフェクト細胞に対して同様の割合で、OPCマーカーであるNG2の発現を下方調節したことは、有糸分裂後OLへの初期分化が、MRFのノックダウンによる影響を受けなかったことを強く示唆する。MRFの不在下において、後期OL遺伝子であるMOGの発現は、さらにより強く阻害され、分化の96時間後において、MOGを発現するsiMRFトランスフェクト細胞が5%未満であったの対し、この時点において、MOGについて陽性のsiCont細胞は81%であった(図3A、D)。48時間後および96時間後における、siMRFによる、マーカーMBPおよびMOGを発現する細胞の低減は、生存率の低下よりはるかに大幅であり(例えば、48時間の分化後において、siMRFトランスフェクト細胞が示す生存率は、siContを発現する細胞と比べて20.3%低下したに過ぎなかったが、MBPを発現する細胞の比率は、siCont発現細胞と比べて62.5%低下した)、siMRFトランスフェクト細胞において観察される細胞死の増大が、MBPおよびMOGの発現の喪失を説明するのに十分ではないことを強く示唆した。
【0142】
興味深いことに、位相顕微鏡法下において、siMRFトランスフェクト細胞はほとんど常に、OLの一般的な形態を帯びたが、突起および膜シートの増殖が、siContトランスフェクト細胞より大幅に阻害されることが典型的であり、また一方、MOG陰性細胞が、OLマーカーであるGalCを明白に発現した(図11)ことにより、細胞原基の特定化の変化が、表現型の原因ではないことが裏付けられた。
【0143】
(実施例13)
MRFの下流にある遺伝子の同定
MRFの不在下にあるOLの分化において見られる転写欠損をさらに特徴づけるため、また、MRFの下流にある遺伝子を同定するため、siMRFプールまたはsiContプールによるトランスフェクション後の48時間にわたり分化させたOLに由来するRNAのほか、分化前における遺伝子発現のベースラインを与える、培養OPCに由来するRNAも単離した。48時間後という時点を選択したのは、OL/ミエリン遺伝子の大半が、この段階までにある程度のレベルの誘導を示す(Dugasら、J. Neurosci.、26巻、10967〜10983頁(2006年))一方、siMRFトランスフェクト細胞が、siContトランスフェクト細胞と比べ、やはりほぼ同等レベルの生存を示す時点に依然としてあり、これにより、結果に対する細胞死の交絡による影響の可能性{Dugas、2006年、#9}が制限されるためである。全RNAは、mRNAの3’端を増幅するポリAプライマーを伴う2段階の直鎖状増幅プロトコールを用いて、標識化されたcRNAを作製するのに用いた。この標識化したcRNAを、20,832の固有の遺伝子を表わす45,037のオリゴヌクレオチドプローブセットによる、mRNAの3’端に相補的なオリゴヌクレオチドのプローブセット(3’アレイ)を含有する、Affymetrix Mouse 430 2.0マイクロアレイへとハイブリダイズさせた。また、ノーザンブロット解析も用いて、この時点におけるMRFおよびミエリン遺伝子(PLPおよびCNP)の下方調節も確認した(図4A)。
【0144】
公知のOPC発現遺伝子であるNG2およびPDGFRαの解析により、細胞を分化条件へと移すと、siRNAによりMRFの発現が遮断されていたかどうかに関わらず、これらの遺伝子は、OPCと比べ、本質的に検出不可能なレベルまで下方調節され(図4B)、これにより、MRFが、これらの条件におけるOPCからOLへの移行には必要でないことが裏付けられた。MRFのノックダウンによる、CNP1およびUgt8aなど、極めて初期のOL遺伝子の発現低下がごくわずかであったこともおそらく同様であり、これにより、MRFの不在下においても、OPCはOLへの移行を開始することが可能であり、汎OL系統のマーカーであるSox10は、siMRFのトランスフェクションによりまったく影響を受けなかったことが示唆される。これに対し、OLマーカーであるPLP1、MBP、およびMAGでは、MRFのノックダウンにより、siContトランスフェクト細胞と比べて極めて明白な低下(約80%の)が見られ、後期OL遺伝子であるトランスフェリン、MOBP、およびMOGについては、ほぼ完全な低下(>90%)が見られた(図4C)。培養物中において2型星状細胞の原基を帯びるOPCの比率が小さい(>5%)ため、血清の不在下においても、分化条件へと移すことによる誘導は中程度であったが、MRFのノックダウンにより、おそらくBMPによるシグナル伝達の下流にある星状膠細胞遺伝子であるGFAP、S100b、およびアクアポリン4だけがわずかながらの上昇を示した(siCont値のそれぞれ147%、255%、および153%)ことにより、大部分の細胞は、MRF発現の不在下においてGFAP陰性を維持し、星状細胞系統への分化が、これらの細胞によるOL/ミエリン遺伝子の発現の喪失を説明するわけではないというかつての観察が裏付けられる。
【0145】
MRFの下流にある遺伝子を同定するため、siContとsiMRRとの間で、抑制レベルにより遺伝子を序列化した。MRFのノックダウンにより、104の遺伝子を表わす128のプローブセットが、4分の1<に抑制された。MRFのノックダウンによる抑制レベルが最も大きな50の遺伝子を、図5に示す(図14もまた参照されたい)。したがって、siMRFにより阻害されることが示される、128のプローブセットのうちの104はまた、OPC試料と、2日間にわたり分化させたOL試料との間で>4倍に上方調節されることが示されるプローブセットでもあり、これにより、MRFに依存する遺伝子の大半は通常、OPCからOLへの移行時において調節される遺伝子であることが示された。しかし、逆に、分化により>4倍に誘導される793のプローブセットのうち、siMRFにより強く阻害されたのは104だけであったことにより、OLの発生時において通常調節される遺伝子の大部分が、MRF発現には依存しないことが示唆される(図4D)。
【0146】
(実施例14)
MRFの強制発現はin vitroにおけるOLの分化を誘導する
MRFが、OL/ミエリン遺伝子の発現を誘導するのに十分であるかどうかを評価するため、OPCに、MRF発現構築物(pCMV−Sport6−MRF)、またはEGFPをコードする対照のGFPプラスミドをトランスフェクトして、大半の細胞が通常、分裂するOPCであることを維持し、自発的分化は低レベルにとどまる増殖条件(+PDGF、−T3)へと播種した。トランスフェクションの2日後において、対照のトランスフェクト細胞が示した分化は低レベルであり、MBP陽性およびMOG陽性とカウントされた生細胞は、それぞれ、1.4パーセントおよび0.4パーセントに過ぎなかった。これに対し、MRF発現構築物をトランスフェクトした細胞が、MBPおよびMOGについて、それぞれ、32.8パーセントおよび41.0パーセントの陽性であったことは、分化率が、対照ベクターをトランスフェクトした細胞と比べて約30〜40倍に上昇した(p<0.001)ことを示す。MBPおよびMOGのこの誘導は、MRFトランスフェクト細胞が、OPCマーカーであるNG2を下方調節することにより反映された(図6)。トランスフェクションの5日後までに、MRFトランスフェクト細胞の大半がMBP陽性およびMOG陽性(それぞれ、81.9および77.2%)となったのに対し、対照トランスフェクト細胞の大半は、依然としてMBP/MOG陰性OPCであった(図6)。MRFが異所性発現したMBP陽性細胞およびMOG陽性細胞が、ほとんど均一に成熟OLの一般的形状(突起の高度な分岐および膜シート)を示したことにより、OPC内におけるOL/ミエリン遺伝子の異所性発現ではなく、MRFの異所性発現が、OLへの該細胞の分化を引き起こしたことを強く示唆する。
【0147】
(実施例15)
MRFの強制発現はin vivoにおけるMBPの発現を誘導する
MRFの発現が、in vivoにおけるミエリン遺伝子の発現を駆動するのに十分でありうるかどうかを評価するため、MRF発現構築物(pCAGGS−MRF)を、E3において、ニワトリ脊髄内へと電気穿孔し、ニワトリの脊髄内には通常検出可能なMBPの発現または希突起膠細胞の分化が見られない発生時点であるE8において該胚を屠殺した。MRF発現構築物(共電気穿孔されたEGFP発現プラスミドからのEGFPの発現により同定される)を電気穿孔した脊髄側方内では、偶発的なMBP陽性細胞を見出すことができたが、強いMBP陽性細胞が見出されるのは、断面1枚当たり1〜3個だけであることが典型的であった(図7A、B)。電気穿孔されなかった、対照の脊髄側方において、MBP陽性細胞は観察されなかった。しかし、電気穿孔された脊髄側方におけるMBP陽性細胞の数が、EGFPまたはMFR導入遺伝子を発現する細胞の数よりはるかに少なかった(図7A)ことは、MRFが希突起膠細胞の分化を促進する能力には、他の陽性または陰性の調節因子が影響している可能性が高いことを示すことに注目すべきである。
【0148】
(実施例16)
MRFはマウスにおけるCNSの有髄化に必要である
CNSの有髄化におけるMRFの必要性を評価するため、MRF遺伝子のエキソン8をloxP部位で挟んだマウスを作製した(図8)。エキソン8は、MRFのDNA結合領域の一部をコードする。このエキソンの欠失は、フレームシフトに起因する結果として、DNA結合領域および下流のタンパク質の喪失をもたらすことが予測された(図8C)。エキソン8の細胞特異的欠失または組織特異的欠失は、loxPフランキングマウス(MRFfl/fl)を、対象の細胞型においてCreリコンビナーゼを発現するマウスと交配させることにより実施することができる。MRFfl/flマウスを、Olig2プロモーターの下流でCreを発現するマウス株へと交配させた(結果として、希突起膠細胞および下肢運動ニューロン前駆細胞、OPCおよび成熟OLにおけるCre発現がもたらされた)ところ、結果として得られるMRF条件付きノックアウトマウス(MRFfl/fl;Olig2wt/Cre)は、メンデル頻度で生誕し、生後10日間は、それらの対照同腹子(MRFwt/fl;Olig2wt/cre、およびMRFfl/fl;Olig2wt/wt)から明確には識別できなかった。しかし、P11から、MRF条件付きノックアウト動物は、振戦および発作を発生させたため、それらの同腹子から識別されるようになった。その後数日間にわたり、条件付きノックアウト動物は発作を発生させ、生後3週目のP13〜P17において例外なく死亡した。この表現型は、CNSのミエリンに影響のある他の突然変異体の表現型と符合する。
【0149】
P13における条件付きノックアウトマウスの脳および脊髄に対する免疫組織化学解析は、NeuNおよびGFAPの染色は正常だと考えられ、CNSの全体的構造には影響がないことを示した。これに対し、脳内のMBP染色には大幅な喪失が見られ、対照マウスの白質路内におけるほぼ遍在的なMBP染色と比較して、白質路内で見られるMBP+希突起膠細胞は、偶発的なものに過ぎなかった(図9A)。これらのまれに残存するMBP+ OLは、Creを介する組換えを受けなかった、偶発的なOlig2非依存性細胞を表わす可能性がある。その脊髄根(シュワン細胞により有髄化される)は依然として強くMBP染色されていたが、条件付きノックアウト動物の脊髄内におけるこのMBP染色の喪失は本質的に完全であった(図9B)。このMBP発現の喪失は、脊髄溶解物に対するウェスタンブロットにより確認され、また、これにより、著明に低度のCNP発現、およびMOG発現の完全な喪失も示された(図9C)。条件付きノックアウト動物における星状細胞およびニューロンのタンパク質(それぞれ、GFAPおよびニューロフィラメント)には影響がなかった。白質路内におけるフルオロミエリン染色により、本質的に完全な再有髄化の喪失が明白に立証された(脊髄側柱の白質について示された、図9D)。これに対し、末梢神経は、予測される通り、条件付きノックアウト動物および対照動物のいずれにおいても同等に有髄化されたが、これは、シュワン細胞が、MRFもOlig2も発現しないからである。条件付きマウスの視神経では、電子顕微鏡法により、有髄化の喪失が確認された;対照マウスが、P13までに、視神経における有髄化の明白な証拠を示しつつあったのに対し、条件付きマウスにおいては、神経の有髄化が観察されなかった(図9E)。MRFflマウスを、CNPプロモーターの下流でCreリコンビナーゼを発現するマウスと交配させたところ、同等の表現型(振戦、発作、ならびにミエリン遺伝子の発現および有髄化の喪失)が見られたことにより、条件付きノックアウト動物の表現型は、Olig2−cre対立遺伝子の挿入により引き起こされるOlig2ヘテロ接合によるバックグラウンドには依存しないことが確認された(図16)。これらの知見は、MRFが、in vivoにおいて希突起膠細胞が発生してCNSが有髄化する正常な過程に必要であることを裏付ける。
【0150】
(実施例17)
OLはMRF条件付きノックアウトマウスにおいて分化するが、その後、成熟するとアポトーシスを受ける
MRF条件付きノックアウトマウスにおけるミエリンの欠如が、OL生成の欠如により引き起こされたのか、そうではなくて、有髄化が不可能なOLにより引き起こされたのかを見極めるため、OL系統の各段階の発生に対するMRF欠失の効果を評価した。MBP、成熟OLマーカーであるCC1、OPCマーカーであるNG2およびPDGFRα、および汎OL/星状細胞マーカーであるOlig2を含めた各種のマーカーについて免疫染色した、条件付きノックアウトマウスおよび対照マウスに由来する、P13の視神経断面におけるOPCおよびOLの密度を評価した(図13A)。視神経内における、Olig2免疫陽性核密度のカウントにより、条件付きノックアウト動物の神経における陽性細胞密度の著明な低下(約45%の)が示された;この低下は、条件付きノックアウトマウスにおいて見られるCC1免疫陽性細胞の完全な喪失と本質的に符合した(図13B、C、E)。これに対し、条件付きノックアウトマウスにおいても、PDGFRαに対して免疫陽性であるOlig2+細胞の密度は、わずかな影響を受けただけであり(P<0.05のt検定において、MRFfl/fl;Olig2wt/wtマウスと比べ、条件付きノックアウト動物では18.7%の低下であったが、MRFwt/fl;Olig2wt/Cre対照との有意差は見られなかった;図13D、E)、これにより、条件付きノックアウト動物においてもOPC段階が受ける影響は最小限であることが示され、この観察は、見かけ上正常なNG2染色により裏付けられた。同様に、CC1およびPDGFRαのいずれについても陰性のOlig2+細胞(おそらく、Olig2を発現する星状細胞)の密度は、遺伝子型間において同様であった(図9E)。条件付きノックアウトマウスの視神経におけるMBP染色は、対照マウスと比べて大幅に低下したが、にもかかわらず、低度のMBP+(CC1−であるが)細胞は少数ながら存在したことは興味深く、これにより、条件付きノックアウトマウスにおいて、ごく最小限であれ、少数の有糸分裂後OLが生成されたことが示唆される。これらの細胞の密度は極めて低く(細胞35±4個/mm2);対照マウスにおけるCC1染色により測定される通り、成熟OL密度の約4%に過ぎなかった。条件付きノックアウトマウスにおけるMBP+ OLおよびCC1+ OLの喪失は、これらのマーカーの喪失が、上記のsiRNA実験においてMRF発現の下流には存在しないことが判明しているマーカーであるOlig2の陽性細胞密度の大幅な低下と符合したことを踏まえるなら、MRF欠損OLが、これらのマーカーを発現できないことに加えて、成熟OLの喪失も反映する可能性が高い。まとめると、これらのデータにより、MRF条件付きノックアウト動物では、OPC集団が大部分影響を受けない一方、成熟OL集団は大幅かつ選択的に喪失されるという証拠がもたらされる。
【0151】
条件付きノックアウトマウスにおいて見られる有糸分裂後OLが極めて少数であることは、OPCの分化に対する遮断に起因する可能性もあり、代替的に、それらの分化直後におけるOLの喪失に起因する可能性もある。MRFを欠くOPCがマイトジェンを引き込むと、ミエリン遺伝子を発現することはないが、典型的なOL形状を有するGC+有糸分裂後細胞へと分化しうることを示す、上記のin vitroにおけるsiRNA実験を踏まえるなら、前者の可能性は低いと考えられる。OL死滅の可能性は、その一部がアポトーシス細胞に特徴的な、ブレブ形成し、凝縮またはフラグメント化した核を示す、低度のMBP陽性細胞が、条件付きノックアウト動物の視神経において存在する(図13A、矢印)ことにより示唆された。P13において観察される有糸分裂後OLの欠如が、それらが発生した後におけるOLのアポトーシスの増大により説明しうるかどうかを確立するため、P10の条件付きノックアウトマウスおよび対照マウスを、アポトーシスマーカーである活性化カスパーゼ3に対する抗体により染色した。すべての遺伝子型の視神経において、核が凝縮またはフラグメント化されることが多い、活性化カスパーゼ3免疫陽性細胞が存在したことは、視神経において新たに発生したOLの約半数による発生喪失についての先行報告と符合する。条件付きノックアウトマウスは、それらの多くが、条件付きノックアウト動物の神経において観察される低度のMBP+細胞に対応する、活性化カスパーゼ3陽性細胞である、これらのアポトーシス細胞の密度において、統計学的に有意な約2倍の増大を示した(図17)。これらのデータにより、MRF条件付きノックアウトマウスにおいて、OLは発生するが、その後速やかにアポトーシスを受けることが示される。
【0152】
(実施例18)
MRF条件付きノックアウト動物における有髄化不全は細胞自律的であり、細胞死だけに起因するわけではない
条件付きノックアウトマウスにおけるOLのアポトーシスは、これらのマウスにおいて見られる有髄化不全が、OLの喪失に起因したのか、ミエリン遺伝子を発現できないことに起因したのか、またはこれらの両方であったのかを評価することを困難にした。この問題を明らかにするため、P7における対照マウスおよび条件付きマウスから高度に精製したOPC培養物を作製した。イムノパニングを用いて、対照動物および条件付きノックアウト動物の脳から、同等数のPDGFRα+ OPC(約100万個/脳)を単離することができた。増殖条件において、対照マウスに由来するOPC、および条件付きノックアウトマウスに由来するOPCのいずれも、NG2陽性細胞およびKi67陽性細胞として増殖したことは、MRFが、OL系統のOPC期に必要でないことを裏付けた(図15A)。
【0153】
マイトジェンを引き込むと、対照細胞および条件付きノックアウト細胞のいずれもが、Ki67を下方調節し、CNPに対する抗体により染色したところ、OLの形状を帯び(図15B)、ノックアウト細胞も、それらのin vivoにおける対応物とは異なり、分化の4日後において細胞死をほとんど示さなかった。しかし、ノックアウト細胞には、対照細胞と比較して、基質上における膜シートの拡大をはるかに狭める傾向があった。培養物中では高度の生存率を有するにもかかわらず、条件付きノックアウト動物による培養物が、MBPに対する抗体による染色を示さなかった(図15B)ことは、ミエリン遺伝子発現の喪失が、単に細胞死に後続するのではないことを示した。対照動物および条件付きノックアウト動物の脊髄およびOL培養物からRNAを単離し、RT−PCRおよび遺伝子アレイにより解析したところ、MAGおよびMOBPなど、多くのミエリン遺伝子の発現が、条件付きノックアウト動物の脊髄およびOL培養物のいずれにおいても消失したことは、これらの遺伝子の発現に対するMRFの必要性を示す(図14、図18)。予測される通り、MRFの欠損は、in vivoにおけるOL死滅の誘導に影響を及ぼすため、MRFヌルの脊髄では、MRF依存性OL遺伝子およびMRF非依存性OL遺伝子すべての発現の喪失が観察された。in vitroにおけるOL遺伝子の発現に対するMRF欠損の効果は、siMRFノックダウンについて観察される効果(図5)と極めて類似し、予測される通り、大半のミエリン遺伝子および多くのOL成熟遺伝子が強く下方調節され、またこれには、Ugt8およびCldn11など、複数の初期OL遺伝子に対するMRF欠損の効果も含まれた。その発現が通常、新たに形成された希突起膠細胞による一過性発現に限定される、p57/cdkn1cなど、複数の遺伝子が、in vitroのMRF欠損OLにおいて著明に上方調節されたことは、MRF欠損OLが、依然として大半のミエリン遺伝子を発現する前の成熟の初期段階にある、分化の直前に失速する可能性が高いことを示す。
【0154】
(実施例19)
MRFの発現による脱髄の阻害
MRF発現が誘導可能なトランスジェニックマウスを用いて、該動物におけるMRFの発現または活性の上昇により、脱髄が阻害されることを示す。PDGFRα−CreERTマウスを、CMV−lox−stop−lox−MRFマウスと交配する結果として、PDGFRα−CreERT/CMV−lox−stop−lox−MRFマウスをもたらすことにより、誘導可能なMRF発現を含むトランスジェニックマウスを作製した。PDGFRα−CreERT/CMV−lox−stop−lox−MRFマウスは、lox部位により隣接する上流の終止コドンのため、また、Creリコンビナーゼ変異体であるCreERTが、flox化終止コドンに対して作用できないため、導入遺伝子からMRFを発現することがない。しかし、タモキシフェンの投与によりCreERTの活性を誘導すると、したがって、該マウスにタモキシフェンを投与すると、該終止コドンが切断され、MRFが発現される。
【0155】
この実験では、被験群である1つの群にはタモキシフェンを投与し、対照群である第2の群にはタモキシフェンを投与しない、PDGFRα−CreERT/CMV−lox−stop−lox−MRF動物の2つの群を用いた。いずれの群にも、クプリゾンを施して、脱髄を誘導した。ミエリン特異的遺伝子についての免疫組織化学による遺伝子発現の検出、または軸索についての電子顕微鏡法などにより、両方の群を脱髄について解析した。
【0156】
タモキシフェンを投与された被験群のマウスは、対照群と比較して、より低い程度または度合の脱髄を示す。
【0157】
(実施例20)
MRF発現による再有髄化の促進
この実験では、PDGFRα−CreERT/CMV−lox−stop−lox−MRF動物の2つの群を用いて、MRFの発現または活性の上昇による再有髄化の促進を示す。いずれの群にも、クプリゾンを施して、脱髄を誘導した。被験群である1つの群にはタモキシフェンを投与する一方、対照群である第2の群にはタモキシフェンを投与しない。ミエリン特異的遺伝子についての免疫組織化学による遺伝子発現の検出、または軸索についての電子顕微鏡法などにより、両方の群を脱髄について解析した。タモキシフェンを投与された被験群のマウスは、対照マウスより迅速に、またはより頑健に再有髄化することが可能である。
【0158】
(実施例21)
in vitroにおける生物活性剤のスクリーニング
神経膠細胞株を用いて、再有髄化を促進するか、または脱髄を阻害する生物活性剤をスクリーニングする。マウスからOPCを得、候補生物活性剤により処理する。OPCを、MRFの発現について解析し、候補生物活性剤を投与されなかったOPCと比較した。候補生物活性剤を投与されたOPCが、これを投与されなかったOPCと比較して、より高レベルのMRF発現を示す場合は、該候補生物活性剤を、実施例22に記載の動物モデルにおける試験など、さらなる解析用に選択する。
【0159】
また、ミエリン特異的遺伝子についての免疫組織化学または遺伝子発現の検出によって、細胞を解析することもできる。候補生物活性剤を投与された細胞が、該候補生物活性剤を投与されなかった細胞と比較して、ミエリン特異的遺伝子の発現または軸索の有髄化の増大を示す場合は、該候補生物活性剤を、実施例22に記載の動物モデルにおける試験など、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。
【0160】
(実施例22)
in vivoにおける生物活性剤のスクリーニング
実施例16に記載のMRF条件付きノックアウトマウス(MRFfl/fl;Olig2wt/Cre)を用いて、生物活性剤をスクリーニングする。MRF条件付きノックアウトマウス(MRFfl/fl;Olig2wt/Cre)は、P11において、振戦および発作を発生させる。このスクリーニングでは、P11以前に、MRF条件付きノックアウトマウスに候補生物活性剤を投与する。該動物が、振戦もしくは発作を発生させないか、または発生させる振戦もしくは発作が、該候補生物活性剤を投与されなかったMRF条件付きノックアウトマウスと比較して、それほど重度でない場合は、該候補生物活性剤を、再有髄化を促進するか、または脱髄を阻害する治療剤としてのさらなる開発用に選択する。また、ミエリン特異的遺伝子についての免疫組織化学もしくは遺伝子発現の検出、または軸索についての電子顕微鏡法などによっても、動物を解析し、候補生物活性剤を投与された動物が、該候補生物活性剤を投与されなかった動物と比較して、ミエリン特異的遺伝子の発現または軸索の有髄化の増大を示す場合は、該候補生物活性剤を、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。
【0161】
第2のスクリーニングでは、MRF条件付きノックアウトマウスが、振戦または発作を示した後において、すなわち、約P11以後において、該動物に候補生物活性剤を投与する。該候補生物活性剤を投与されなかったMRF条件付きノックアウトマウスと比較して、振戦または発作の程度が低下するなど、該動物の状態が改善される場合は、該候補生物活性剤を、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。また、ミエリン特異的遺伝子についての免疫組織化学もしくは遺伝子発現の検出、または軸索についての電子顕微鏡法などによっても、動物を解析し、候補生物活性剤を投与された動物が、該候補生物活性剤を投与されなかった動物と比較して、ミエリン特異的遺伝子の発現または軸索の有髄化の増大を示す場合は、該候補生物活性剤を、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。
【0162】
本明細書では、本発明の好ましい実施形態を示し、説明してきたが、このような実施形態が例示だけを目的とするものであることは、当業者には明らかであろう。本発明から逸脱しない限りにおいて、今や、多くの変更、変化、および置換が当業者に生じるであろう。本発明を実施する際に、本明細書で記載される本発明の実施形態に対する各種の代替物を用いうることを理解されたい。本明細書における特許請求の範囲が、本発明の範囲を規定し、これらの特許請求の範囲が、これらおよびこれらの同等物の範囲内にある方法および構造を対象とすることが意図される。
【技術分野】
【0001】
(相互参照)
本出願は、2008年7月16日に出願された米国仮特許出願第61/081,279号および2008年12月5日に出願された同第61/120,307号の利益を主張し、これらの両出願は本明細書においてその全体が援用される。
【0002】
本出願は、the National Eye InstituteからのRO1 EY10257の下、政府の支援によってなされた。政府は、本発明において一定の権利を有する。
【背景技術】
【0003】
多発性硬化症(MS)とは、発現の再発−寛解パターンから慢性−進行性パターンまでの範囲にわたる臨床的障害を伴う、中枢神経系(CNS)の炎症性脱髄疾患である。MSの病因は未知であるが、自己反応性のCD4+ T細胞反応がミエリンおよび希突起膠細胞に対する炎症性損傷を媒介する(非特許文献1)。CNS病変は、ミエリン損傷の巣状領域を有し、また、軸索の病態、神経の損傷、および星状膠細胞における瘢痕形成とも関連する(非特許文献2)。臨床症状には、失明、麻痺、感覚および協調運動の喪失、ならびに認知障害を含めた、各種の神経学的機能不全が含まれる。
【0004】
ミエリンへの損傷または傷害は、導電速度、および軸索の破壊に対するニューロンの脆弱性に対して重大な影響を及ぼす。軸索の喪失と、進行性の臨床障害との間には相関があり、軸索の完全性を維持するには、ミエリンが無傷であることが重要である(非特許文献3)。ヒトMSの初期には、自発的な再有髄化が生じるが、該疾患の後期段階におけるCNS炎症の遷延、およびミエリン修復の失敗は、最終的に恒常的な衰弱をもたらす。
【0005】
再有髄化をもたらすのは、成熟希突起膠細胞(OL)である。したがって、再有髄化の失敗は、成熟希突起膠細胞の生成、それらが有髄化をもたらす能力の欠損、および/または有髄化に対するニューロンの非受容性と関連することが典型的である。多発性硬化症などの脱髄性疾患では、生存するOLおよびそれらの前駆細胞(希突起膠細胞の前駆細胞、またはOPC)が、脱髄領域の内部および周囲に見出されることが多い。軸索近傍において、これらの生存細胞が再有髄化に失敗することは、OPCが分化できないこと、または有糸分裂後における希突起膠細胞が、有髄化に必要とされる一連の遺伝子発現を再開できないことを反映しうる。有髄化は、希突起膠細胞およびそれらの前駆細胞を正確に局在化させ(非特許文献4;非特許文献5)、適切な細胞数を調節し(非特許文献6; 非特許文献7)、また、希突起膠細胞とそれらの標的である軸索との相互作用を媒介する(非特許文献8)シグナルを含めた、多数のシグナルの調整に依拠し、したがって、これらの過程のいずれかに欠損があれば、再有髄化の失敗がもたらされうる。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Bruckら、J. Neurol. Sci.、206巻、181〜185頁(2003年)
【非特許文献2】Compstonら、Lancet、359巻、1221〜1231頁(2002年)
【非特許文献3】Dubois−Dalcqら、Neuron、48巻、9〜12頁(2005年)
【非特許文献4】Tsaiら、Cell、110巻、373〜383頁(2002年)
【非特許文献5】Tsaiら、J. Neurosci.、26巻、1913〜22頁(2006年)
【非特許文献6】Barresら、Cell、70巻、31〜46頁(1992年)
【非特許文献7】Calverら、Neuron、20巻、869〜882頁(1998年)
【非特許文献8】ShermanおよびBrophy、Nat Rev Neurosci.、6巻、683〜690頁(2005年)
【発明の概要】
【発明が解決しようとする課題】
【0007】
有髄化または再有髄化を増強および促進する効果的な方法を開発する必要がある。OPC、前駆細胞プールの分化、または既存の有糸分裂以後における希突起膠細胞による有髄化の再開を促進する戦略は、再有髄化の確立に有益でありうる。本発明は、再有髄化の促進を対象とする組成物および方法を提供する。本明細書で開示される知見は、ミエリン生成に関与する遺伝子の発現が、MRF(ミエリン遺伝子調節因子)としても公知のGM98(遺伝子モデル98)、および有髄化により影響されることを裏付ける。
【課題を解決するための手段】
【0008】
本発明は、MRFの発現および/または活性を調節する方法および組成物を提供する。本明細書では、MRFにより調節される遺伝子の発現および活性もまた説明される。該方法および組成物を用いて、再有髄化を促進し、MRFの発現/活性、またはMRFにより調節される遺伝子を調節する生物活性剤をスクリーニングするほか、神経障害を治療することもできる。
【0009】
本発明の一態様は、MRFまたはその機能的な変異体をコードする核酸配列に作動的に連結された細胞型特異的な発現調節エレメントを含む、単離核酸分子である。さらに、該細胞型特異的な調節エレメントは、誘導的プロモーターまたは構成的プロモーターである。本発明はまた、本明細書に記載の核酸を含むベクターも提供し、宿主細胞型は、本発明のベクターおよび核酸を含む。
【0010】
本明細書ではまた、MRF導入遺伝子を含むトランスジェニック動物も提供される。導入遺伝子は、本明細書に記載の核酸配列を含みうる。さらに、導入遺伝子は、突然変異、およびエキソンの欠失などの欠失を含みうる。エキソンは、エキソン8など、推定DNA結合ドメイン内のエキソンでありうる。エキソンはまた、CreまたはFlpに対応する部位などのリコンビナーゼ部位にも隣接する。トランスジェニック動物はまた、CreリコンビナーゼまたはFlpなど、リコンビナーゼ導入遺伝子も含みうる。該導入遺伝子はまた、細胞型特異的な発現調節エレメントに作動的に連結されうる。該トランスジェニック動物は、マウスまたはラットなどの哺乳動物でありうる。
【0011】
核酸または導入遺伝子の特異的な発現は、神経膠細胞などの神経細胞内でありうる。神経膠細胞は、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞でありうる。細胞型特異的な調節エレメントは、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来しうる。
【0012】
トランスジェニック動物はまた、再有髄化/有髄化を促進するのに有効な候補生物活性剤をスクリーニングするのに用いることもできる。該方法は、候補生物活性剤を該動物に投与するステップ;対照動物と比較した、表1中の少なくとも1つの遺伝子の発現レベルの上昇についてアッセイし、該上昇が、該動物における再有髄化を促進する前記生物活性剤を示すステップ;および/または対照動物と比較した、該動物における有髄化の変化を観察するステップを含みうる。生物活性剤は、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子でありうる。本発明の別の態様では、動物における再有髄化/有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法であって、候補生物活性剤を動物に投与するステップと、対照動物と比較した、MRFの発現レベルの上昇についてアッセイし、該上昇が、該動物における有髄化を促進する該生物活性剤を示すステップとを含む方法が提供される。本発明はまた、MRF活性を調節するのに有効な候補生物活性剤をスクリーニングする方法も提供する。一態様において、該方法は、被験細胞を候補生物活性剤と接触させるステップと、対照細胞と比較したMRFの発現レベルの変化についてアッセイするステップとを含む。
【0013】
本発明のさらに別の態様では、被験体における神経障害を治療するための組成物であって、前記被験体におけるMRF活性を調節する生物活性剤を含む組成物が提供される。組成物は、被験体における再有髄化/有髄化を促進しうる。組成物は、幹細胞または胚性幹細胞が希突起膠細胞へと分化することを促進しうる。組成物は、MRF、またはMRFの活性もしくは発現を調節する薬剤など、第1の生物活性剤と、希突起膠細胞の分化を誘導する第2の生物活性剤とを含みうる。例えば、第2の生物活性剤は、Sox10、Nkx2.2、Olig1、および/またはOlig2を促進しうる。第2の生物活性剤は、Sox10、Nkx2.2、Olig1、またはOlig2でありうる。組成物は、MRF、Sox10、Nxk2.2、Olig1、Olig2、またはこれらの組合せを含みうる。組成物は、多発性硬化症などの脱髄状態など、神経障害を治療するのに用いることができる。本明細書で提供される組成物は、被験体における神経障害を治療する方法であって、該被験体に、治療有効量の、MRF活性を調節する生物活性剤を投与するステップを含む方法において用いることができる。方法は、被験体における再有髄化/有髄化を促進しうる。投与は、MRF、またはMRFの活性もしくは発現を調節する生物活性剤など、第1の生物活性剤と、また、希突起膠細胞の分化も誘導する第2の生物活性剤とを投与するステップとを含みうる。第2の生物活性剤を投与するステップは、第1の生物活性剤を投与するステップの前の場合もあり、これと同時の場合もあり、この後の場合もある。第2の生物活性剤は、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進しうる。第2の生物活性剤は、相乗効果を有しうる。
【0014】
本発明の新規の特徴は、添付される特許請求の範囲において具体的に記載される。本発明の特徴および利点についてのよりよい理解は、本発明の原理が用いられる例示的な実施形態を記載する以下の詳細な説明、およびそれに付随する図面を参照することにより得られる。
【図面の簡単な説明】
【0015】
【図1】図1は、CNS内におけるOL特異的な転写物であるGM98/MRFの同定を示す図である。A)図1Aは、CNSから急性摘出された細胞におけるGM98/MRFの発現レベルを、Affymetrix解析(プローブセット1439506_at)により決定したことを示す図である。B)図1Bは、ノーザンブロットが、MRFの発現を、P20における脳および培養希突起膠細胞(OL)内では、約5.5Kbの単一の転写物として確認するが、心臓組織内ではこれを確認せず、これは、確立されたOLマーカーであるCNP1の発現パターンを反映することを示す図である。GAPDHは、mRNAレベルの対照として示される。C)図1Cは、P16における脳の矢状断面内における、確立されたOLマーカーであるPLPおよびMRFについてのin situハイブリダイゼーションを示す図である。いずれの遺伝子も、明白な白質の発現パターンを示す。D)図1Dは、標識化された細胞の同一の分布を示す、脳梁側部内におけるPLPおよびMRFの発現についての高倍率画像である。
【図2】図2は、MRFのペプチド構造および細胞内局在化を示す図である。A)図2Aは、マウスMRF遺伝子およびヒトC11Orf9遺伝子のペプチド配列の比較を示す図である。B)図2Bは、マウスMRFタンパク質、およびヒトC11Orf9タンパク質の構造領域、ならびにこれらの領域内におけるそれらの相同性を示すダイアグラムである。C〜E)図2C〜Eは、HEK細胞内におけるmycタグを付したMRFの細胞内局在化を示す図である。抗mycによる、Myc−MRFをトランスフェクトしたHEK細胞に対する染色(E)は、トランスフェクトされた細胞内における、核内対比染色色素であるDAPIとの本質的に完全な共局在化を伴う、該タンパク質の強い核内局在化を示す。mycタグを付していないMRF(D)、およびmycタグを付したタンパク質をトランスフェクトした細胞は、細胞質内局在化(C)を示し、これにより、染色についての陰性対照、および比較がそれぞれもたらされる。
【図3】図3は、MRFの発現が、OLの成熟にとって重要であることを示す図である。A)図3Aは、24〜96時間にわたる分化時における、siContまたはsiMRFをトランスフェクトしたOLの生存率および形態を示す、生存:死滅アッセイ(生存細胞は、カルセイン(Calcien)AMにより染色されて緑色チャネルで示され、非生存核は、エチジウムホモ二量体により染色されて赤色チャネルで示される)を示す図である。siMRFをトランスフェクトした細胞は、siContトランスフェクト細胞と比べて膜シートの沈着の減少および生存率の低下を示した。B)図3Bは、分化条件へと移した24〜96時間後における、siContトランスフェクト細胞と比べたsiMRFトランスフェクト細胞の生存率を示す図である。siMRFトランスフェクト細胞は、分化の48時間後以降、siContトランスフェクト細胞と比べて著明な生存率の低下を示した。**P<0.01。C)図3Cは、分化の24、48、および96時間後において、siContまたはsiMRFをトランスフェクトしたOLにおける、OPCマーカーであるNG2、および初期OLマーカーであるMBPの発現を示す図である。siMRFトランスフェクト細胞は、siContトランスフェクト細胞と同様の時間プロファイルによりNG2の発現を下方調節するが、MBP発現の遅延およびその誘導の低下を示した。D)図3Dは、分化の24〜96時間後における、MBPを発現するsiContトランスフェクトOLおよびsiMRFトランスフェクトOLの比率についての定量化を示す図である。siMRFトランスフェクト細胞は、すべての時点において、MRF発現率の低下を示した。**P<0.01。E)図3Eは、24〜96時間にわたる分化時の、siContまたはsiMRFをトランスフェクトしたOL内における、後期OLマーカーであるMOGの発現を示す図である。siMRFトランスフェクト細胞は、MOG発現に対する誘導の低下を示した。F)図3Fは、分化の24〜96時間後における、MOGを発現するsiContトランスフェクトOLおよびsiMRFトランスフェクトOLの比率についての定量化を示す図である。siMRFトランスフェクト細胞は、分化の48時間後以降、MRF発現率の低下を示した。**P<0.01。結果は、3つの個別の実験を表わす。
【図4】図4は、MRFノックダウン動物による、OL遺伝子発現の解析を示す図である。A)図4Aは、OPCとしてsiContまたはsiMRFをトランスフェクトされ、次いで、分化条件で48時間にわたり培養された細胞における遺伝子発現についてのノーザンブロット解析を示す図である。脳試料、心臓試料、および培養星状細胞試料に由来するRNAを、それぞれ、陽性対照および陰性対照として与えた。細胞にsiMRFをトランスフェクトしたところ、存在するMRF転写物の量が大幅に減少し、また、結果として、PLPの発現、また、より低度ではあるがCNP1の発現も明白に阻害された。GAPDHのノーザンブロット結果、および18Sリボソームバンドの可視化を、ローディング対照として示す。B)図4Bは、OPC、およびOPCとしてsiContまたはsiMRFをトランスフェクトされ、次いで、分化条件で48時間にわたり培養された細胞における遺伝子発現についてのAffymetrix解析の結果を示す図であり、これにより、選択されたOPCマーカーであるNG2、PDGFRα、およびKi67の発現レベルが示される。これらのOPCマーカーの下方調節は、MRFのノックダウンによる影響を受けなかった。C)図4Cは、対照(siContトランスフェクト細胞の値)に対する比率として表わされた、siMRFをトランスフェクトした細胞における、汎OL系統のマーカーであるSox10、初期OLマーカー(Ugt8、CNP1、PLP1、MBP)、および後期OLマーカー(MAG、トランスフェリン、MOBP、およびMOG)の発現を示す図である。siMRFトランスフェクト細胞において、OL遺伝子の発現は、siContトランスフェクト細胞の場合と比べて強く阻害され、分化後期のOLマーカー(トランスフェリン、MOG、およびMOBP)が、同初期マーカー(CNP1またはUgt8)または同中間段階マーカー(PLP1、MBP)より大きな影響を受けることが典型的であった。結果は、3つの個別の実験の平均であり、siContによる発現レベルの平均百分率±SEMとして表わされる。D)図4Dは、分化により>4倍に誘導された遺伝子と、siMRFのトランスフェクションにより<4分の1に抑制された遺伝子との重複を示すベン図である。siMRFにより阻害された遺伝子の大半(81%)は、OL分化時において通常は上方調節される遺伝子であった;これに対し、OLの分化時において通常は誘導される遺伝子のうち、MRFの発現に依存するのは13%に過ぎなかった。
【図5】図5は、MRF発現の不在下におけるOL遺伝子の発現についての表である。OPCとしてsiContもしくはsiMRFをトランスフェクトされ、次いで、分化条件において48時間にわたり培養された細胞、または遺伝子発現のベースラインを与える培養OPCにおける遺伝子発現についてのAffymetrix解析であり、これにより、MRFの不在下において発現の抑制を示す上位50の遺伝子が示される。siMRFの存在下において下方調節される上位50の遺伝子のうち、47は、OPCとsiCont OLとの間で4倍以上に上方調節される遺伝子であった。1つの遺伝子について複数のAffymetrixプローブセットが使用可能である場合、最も強く発現したものだけを示す。
【図6】図6は、MRFの異所性発現によりOLの分化が誘導されることを示す図である。A〜J)図6A〜Jは、pEGFP、および対照(空)ベクター、pSport6−MRF、またはpSport6−Sox10によるトランスフェクション後、48時間にわたり増殖条件(+PDGF、−T3)において培養したOPCを、NG2、MBP、またはMOGについて染色したことを示す図である。A〜C)図6A〜Cは、対照ベクター(A)、CMV−MRF(B)、またはCMV−Sox10をトランスフェクトした細胞を、NG2について染色したことを示す図である。対照ベクターまたはCMV−Sox10をトランスフェクトされたほとんどすべての細胞(GFPの共発現により同定される)は、NG2陽性を維持した(黄色の矢印)。これに対し、CMV−MRFをトランスフェクトされた多くの細胞が、NG2発現を下方調節する(緑色の矢印)一方、同じ培養物中における非トランスフェクト細胞はNG2発現を維持した(赤色の矢印)。D〜J)図6D〜Jは、対照ベクター(D、H)、pSport6−MRF(E、I)、またはpSport6−Sox10(F、J)をトランスフェクトした細胞を、MBP(D、E、F)、またはMOG(H、I、J)について染色したことを示す図である。対照ベクターまたはpSport6−Sox10をトランスフェクトされたほとんどすべての細胞は、MBP陰性およびMOG陰性を維持した(黄色の矢印)。これに対し、pSport6−MRFをトランスフェクトされた多くの細胞は、MBP発現およびMOG発現について陽性であった(黄色の矢印)。L〜M)図6L〜Mは、対照ベクター、pSport6−MRF、またはpSport6−Sox10をトランスフェクトされ、トランスフェクションの48または120時間後においてMBP(L)またはMOG(M)について陽性であった細胞の平均比率を示すグラフである。pSport6−MRFによるトランスフェクションは、大半の細胞がMBP陰性およびMOG陰性を維持した対照ベクターだけのトランスフェクションと比べ、48時間後および120時間後のいずれにおいても、MBP発現およびMOG発現の強力な誘導をもたらした。pSport6−Sox10による細胞へのトランスフェクションは、トランスフェクションの120時間後において、MBPおよびMOGに対する比較的低度の誘導を結果としてもたらした。**P<0.01。結果は、3つの個別の実験を表わす。
【図7】図7は、発生時のニワトリ脊髄内においてMRFを電気穿孔すると、早発性のMBP発現が引き起こされることを示す図である。E3における胚に、MRFおよびEGFPを含有するpCAGGSプラスミドを共電気穿孔し、P8において回収した。A)図7Aは、電気穿孔した脊髄側方だけにおいてMRF RNAの発現を示す、MRFについてのin situハイブリダイゼーションを示す図である。B)図7Bは、脊髄横断面におけるEGFPおよびMBPについての染色を示す図である。電気穿孔された脊髄側方では、偶発的なMBP+細胞(1断面当たり1または2個だけであることが典型的)が見出されたが、対照の側方では見出されなかった。C)図7Cは、脊髄の縦断面におけるEGFPおよびMBPについての染色が、MBPに対する免疫反応性が電気穿孔された側方に限定されることを示す図である。D)図7Dは、図7BおよびCにおいて示されるMBP+細胞の高倍率画像を示す図である。
【図8】図8は、MRF条件付きノックアウトマウスの解析および作製を示す図である。A)図8Aは、MRFを破壊するための戦略についての概略図である。野生型のMRF遺伝子座、標的化ベクター、および相同組換え後において予測される遺伝子座を示す。標的とされるマウスを、Flperマウスと交配させることにより、ネオマイシン耐性カセットを欠失させ、結果として、MRFのエキソン8をloxPで挟んだマウスを得る。略号:Ex8、エキソン8;プライマー1;P2、プライマー2;P3、プライマー3;FRT、Flpリコンビナーゼ部位;SP、サザンプローブ。B)図8Bは、MRF野生型マウス、MRFをloxPで挟んだヘテロ接合マウス、およびMRFをloxPで挟んだホモ接合マウスに由来するゲノムDNAに対するPCR増幅を示す図である。プライマー1および2は、460bpの野生型バンドを発生させ、また、loxP部位の挿入により、対立遺伝子をloxPで挟んだマウスにおいては、668bpのバンドも発生させた。プライマー1および3は、対立遺伝子をloxPで挟んだマウスに特異的な269bpのバンドを発生させた。プライマー1は、イントロン7〜8の上側鎖である。プライマー2は、エキソン8の下側鎖である。プライマー3は、プラスミド挿入物の下側鎖である。C)図8Cは、MRF遺伝子によりコードされる全長タンパク質、また、DNA結合ドメインおよび後に続くC末端領域を欠く、Creを介する切断後の切断タンパク質についての概略図である。
【図9】図9は、CNSの有髄化不全を示す、MRF条件付きノックアウトマウスを示す図である。A〜B)図9A〜Bは、P13においてMBP、NeuN、およびGFAPにより染色した、対照(MRFwt/fl;Olig2wt/Cre)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスの海馬、脳梁、ならびに皮質上部A)および脊髄B)を表わす画像である。C)図9Cは、P13におけるMRF対照マウスおよびMRF条件付きノックアウトマウスの脊髄における、CNP、MBP、MOG、GFAP、およびニューロフィラメントの発現についてのウェスタンブロット解析を示す図である。D)図9Dは、P13における対照(MRFwt/fl;Olig2wt/Cre)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスにおける脊髄の脊髄根および側方白質に対するフルオロミエリン染色を表わす画像である。E)図9Eは、P13における対照動物および条件付きノックアウト動物の神経を表わす電子顕微鏡画像である。対照動物の神経は、著明量の有髄化が進行中であることを示している。これに対し、条件付きノックアウト動物の神経は、軸索の有髄化を一切示さなかった。
【図10】図10は、視神経および坐骨神経内におけるPLPおよびMRFについてのin situハイブリダイゼーションを示す図である。PLPについてのシグナルは、既に報告されている通り(Puckettら、J. Neurosci. Res.、18巻、511〜518頁(1987年))、坐骨神経では低レベルまでであるが、視神経内および坐骨神経内のいずれにおいても(CNS神経およびPNS神経のいずれにおいても)容易に検出可能であった。これに対し、MRFについてのシグナルが視神経内で検出されたことは、これが、OLによっては発現するが、シュワン細胞によっては発現しない(CNSの神経膠細胞によっては発現するが、PNSの神経膠細胞によっては発現しない)ことを示す。
【図11】A〜B)図11A〜Bは、siCont(A)またはsiMRF(B)をトランスフェクトした後で48時間にわたり分化させた、培養希突起膠細胞についての位相イメージングを示す図である。いずれの場合も、大半の細胞は希突起膠細胞の形態を帯びるが、siMRFをトランスフェクトした細胞は、それほど広範な突起を示さなかった。C〜D)図11C〜Dは、siCont(C)またはsiMRF(D)をトランスフェクトした後で72時間にわたり分化させた培養希突起膠細胞上における、MOGおよびGalCについての表面染色(O1抗体による)を示す図である。siContトランスフェクト細胞およびsiMRFトランスフェクト細胞のいずれも、O1抗体により容易に標識化されたが、MOGについて陽性であったのは、siCont細胞だけであった。
【図12】A)図12Aは、対照のsiRNAプール、またはMRFに対するsiRNAプールをトランスフェクトした培養希突起膠細胞内において、MRFおよびMBPを検出するRT−PCR(30サイクル)を示す図である。siMRFトランスフェクト細胞では、MRF転写物レベルおよびMBP転写物レベルのいずれもが明白な低下を示した。B)図12Bは、対照のsiRNAプール、またはMRFに対する個別のsiRNAをトランスフェクトした培養希突起膠細胞内において、MRFおよびMBPを検出するRT−PCR(30サイクル)を示す図である。4つの個別のsiRNAのうちの複数が、siContトランスフェクト細胞と比べて検出可能な、MRFレベルおよびMBPレベルの低下を引き起こした。C)図12Cは、図12Bと同じトランスフェクションに由来する細胞が、分化条件における4日間にわたる培養後において示す、MOGを発現する生存細胞の比率の高さを示す図である。MRFに対する4つの個別のsiRNAのうちの3つが、siContトランスフェクト細胞と比べて、MOGを発現する細胞の比率を著明に低下させたが、これは、図12Bにおいて観察されたMRFのノックダウンと良好に相関する。
【図13】図13は、MRF条件付きノックアウト動物が、成熟希突起膠細胞の喪失を示すことを明らかにする図である。A)図13Aは、P13における対照(MRFwt/fl;Olig2wt/CreおよびMRFfl/fl;Olig2wt/wt)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスの視神経内における、MBP、CC1、NG2、GFAP、ならびにCC1およびPDGFRαと共染色されたOlig2についての免疫染色を示す図である。スケールバー=50μm。B)図13Bは、視神経内におけるOlig2免疫陽性核の密度についての定量化を示す図である。C)図13Cは、視神経内におけるOlig2+/CC1+二重免疫陽性OLの密度についての定量化を示す図である。D)図13Dは、視神経内におけるOlig2+/PDGFRa+二重免疫陽性細胞の密度についての定量化を示す図である。すべての結果は、平均±SEM、1遺伝子型当たりn=4〜5として表わされる。*P<0.05、**P<0.01。E)図13Eは、CC1についてもまた陽性であるOlig2+細胞(OL)、PDGFR1αについてもまた陽性であるOlig2+細胞(OPC)、またはいずれのマーカーについても陽性でないOlig2+細胞に分類される、視神経内における各遺伝子型のOlig2免疫陽性細胞の密度を示す図である。
【図14】図14は、条件付きノックアウト動物の培養希突起膠細胞および脊髄における遺伝子発現を示す図である。対照(MRFwt/fl;Olig2wt/Cre)動物の脳、および条件付きノックアウト(CKO;MRFfl/fl;Olig2wt/Cre)動物の脳に由来するOL培養物(4日間にわたり分化させた)、ならびにP13における対照(MRFwt/fl;Olig2wt/Cre)マウス、および条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスから急性摘出された脊髄に対するGeneChip解析の結果である。A)図14Aは、CKO動物の培養OLにおいて、対照動物の培養OLと比べて最も大幅に下方調節された20遺伝子を、対照動物のOLおよびCKO動物のOL、ならびに対照動物の脊髄およびCKO動物の脊髄におけるMAS 5.0遺伝子の発現値と共に列挙する。また、培養OL(対照値/CKO値)、脊髄(対照値/CKO値)、およびsiRNA実験(siCont値/siMRF値)についても、抑制倍率の値を示す。B)図14Bは、対照動物のOLおよびCKO動物のOL、ならびに対照動物の脊髄およびCKO動物の脊髄における汎OL系統マーカー、OPCマーカー、初期OLマーカー、後期OLマーカーを表わすプローブセット、およびこれらの発現値を示す図である。「A」と表示した遺伝子を発現しない遺伝子と判定し、「B」と表示した遺伝子を発現した遺伝子と考える。「B」とは、わずかな発現に相当する。アレイ上における所与の遺伝子に対して複数のプローブセットが存在した場合は、最も強く発現したプローブセットだけを示す。
【図15】図15は、MRF欠損OPC/OL培養物が、分化条件では欠損を示すが、増殖条件では欠損を示さないことを明らかにする図である。A)図15Aは、増殖(+PDGF、−T3)条件における、NG2およびKi67についての、対照(MRFwt/fl;Olig2wt/Cre)動物による培養物、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)動物による培養物の免疫染色を示す図である。いずれの場合においても、大半の細胞は、NG2陽性前駆細胞およびKi67陽性前駆細胞(Ki67+核を矢印により示す)として維持された。B)図15Bは、分化(−PDGF、+T3)条件において4日間にわたり増殖させた、対照動物による培養物、およびMRF条件付きノックアウト動物による培養物に対する、Ki67、CNP、およびMBPについての免疫染色を示す図である。いずれの場合においても、大半の細胞は、Ki67の発現を下方調節し、OLに特徴的な多極形態を帯び(CNP染色により可視化される)、GFAPについては陰性であった。しかし、頑健なMBP染色が見られたのは、対照(MRFwt/fl;Olig2wt/Cre)動物による培養物中だけであった。スケールバー=100μm。C)図15Cは、OL系統の特殊化および分化に対する転写制御についての概略図である。Olig2は、OL系統の特殊化に必要である。Sox10、Nkx2.2、Yin Yang 1、およびOlig1を含めた複数の遺伝子が、OLへのOPCの頑健な分化には必要である。MRFは、ミエリン遺伝子の完全な相補体を発現する、成熟OLへの未成熟OLの成熟化に必要であり、その誘導はおそらくYin Yang 1により調節される。
【図16】図16は、CNP−Creを介してMRFが欠失する結果として、MRFfl/fl;Olig2wt/Creマウスと同等の、有髄化不全を引き起こす表現型がもたらされることを示す図である。A)図16Aは、MBPにより染色された、P16における対照(MRFfl/fl;CNPwt/wt)動物の脳、および条件付きノックアウト(MRFfl/fl;CNPwt/Cre)動物の脳を表わす画像であり、これらにより、CNP−Creを介するMRF条件付きノックアウト動物の脳梁(cc)内におけるMBP染色の大幅な喪失が示される。一部の、わずかにMBP+であり、有髄化しないOLが、条件付きノックアウト動物の脳内に存在する(矢印)。スケールバー=200μm。B)図16Bは、フルオロミエリンにより染色された、P16における対照(MRFfl/fl;CNPwt/wt)動物の脊髄、および条件付きノックアウト(MRFfl/fl;CNPwt/Cre)動物の脊髄を表わす画像である。条件付きノックアウト動物では、末梢における有髄化(脊髄根;s.r.)は影響を受けないと考えられるが、CNSにおける有髄化は見られない。
【図17】図17は、条件付きノックアウトマウスが、視神経内において細胞アポトーシスの増大を示すことを明らかにする図である。A)図17Aは、抗MBP抗体および抗活性化カスパーゼ3抗体により染色された、P10における対照動物の神経、および条件付きノックアウト動物の視神経を表わす画像である。スケールバー=100μm。B)図17Bは、P10における対照動物の視神経、および条件付きノックアウト動物の視神経内における活性化カスパーゼ3免疫陽性細胞の密度についての定量化により、条件付きノックアウト動物において、アポトーシス細胞の密度の著明な上昇が明らかとなったことを示す図である(**P<0.01。n=5〜6/遺伝子型)。C)図17Cは、図17Aで条件付きノックアウト動物の神経について枠囲いした領域を高拡大率で示す図であり、これにより、核がフラグメント化し、活性化カスパーゼ3が染色される(矢印)、低度のMBP陽性細胞が示される。
【図18】図18は、条件付きノックアウト動物の遺伝子プロファイリングを示す図である。A)図18Aは、対照(MRFwt/fl;Olig2wt/CreおよびMRFfl/fl;Olig2wt/wt)マウス、およびMRF条件付きノックアウト(MRFfl/fl;Olig2wt/Cre)マウスに由来する脊髄または培養OLに由来するRNA中における選択遺伝子についてのRT−PCR(30サイクル)解析を示す図であり、これにより、条件付きノックアウト動物による試料中において、OLマーカーであるMOGおよびMOBPの喪失が示される。該遺伝子のエキソン8によりコードされるRNA配列を認識するプライマーを用いて、条件付きノックアウト動物に由来する脊髄または培養OLからPCR産物を検出することはできず、これにより、このエキソンの欠失が確認された。これに対し、MRFのエキソン8の外側に位置するプライマーは、条件付きノックアウト動物による培養OL中におけるMRF発現を検出したが、脊髄中におけるMRF発現は検出しなかった。
【発明を実施するための形態】
【0016】
参照による組込み
本出願において言及されるすべての刊行物および特許出願は、各個別の刊行物または特許出願が、参照により組み込まれるように具体的かつ個別に指示されたと仮定する場合と同程度に、参照により本明細書に組み込まれる。
発明の詳細な説明
本発明は、有髄化/再有髄化の促進において、ミエリン遺伝子調節因子(MRF)とも称する遺伝子モデル98(GM98)、および/またはそれに関連する遺伝子を調節する方法および組成物を提供する。例えば、MRFを標的とし、MRFを介して直接的または間接的に調節しうる、ミエリン塩基性タンパク質(MBP)、ミエリン希突起膠細胞糖タンパク質(MOG)、ミエリン関連希突起膠細胞塩基性タンパク質(MOBP)、およびプロテオリピドタンパク質(PLP)など、ミエリン生成における主要な成分と考えられることが典型的な多くの遺伝子の発現に対する下方調節を結果としてもたらすsiRNAにより、希突起膠細胞におけるMRFの発現を阻害することができる。逆に、例えば、MRF cDNAを含有する発現プラスミドを用いてMRFを発現させることにより、希突起膠細胞の前駆細胞(OPC)におけるMRFの発現を誘導し、これにより、再有髄化を促進することができる。
【0017】
本明細書ではまた、MRFの発現を調節し、有髄化を促進する生物活性剤をスクリーニングする方法も提供される。例えば、それらを、Olig2プロモーターの下流でCreリコンビナーゼを発現するマウス株と交配させることにより、希突起膠細胞系統の細胞内において、loxPに隣接するMRF遺伝子(DNA結合領域の一部をコードする)のエキソン8を切除しうるマウス。該マウスは、MBP陽性の希突起膠細胞またはCNSミエリンの発現に失敗し、広範にわたるCNSの脱髄により、生後3週間で死亡することが典型的である。これらのマウスを用いて、死亡を遅延させ、有髄化不全を軽減し、かつ/またはMBP陽性OLの発生を促進する生物活性剤をスクリーニングすることができる。
【0018】
本明細書に記載の方法および組成物は、神経障害を治療するのに重要でありうる。本明細書で開示される方法および組成物を用いて、多発性硬化症(MS)、進行性多巣性白質脳症(PML)、脳脊髄炎、橋中央ミエリン溶解(CPM)、抗MAG抗体陽性疾患、白質萎縮症:副腎白質萎縮症(ALD)、アレキサンダー病、カナバン病、クラッベ病、異染性白質萎縮症(MLD)、ペリツェウス−メルツバッハー病、レフサム病、コケイン症候群、ファンデルクナップ症候群、ツェルヴェーガー症候群、ギラン−バレー症候群(GBS)、慢性炎症性脱髄性多発神経障害(CIDP)、多巣性運動神経障害(MMN)、脊髄傷害(例えば、脊髄の外傷または切断)、アルツハイマー病、ハンチントン病、筋萎縮性側索硬化症、パーキンソン病、および視神経炎などであるがこれらに限定されない各種のCNS障害およびPNS障害を治療することができる。
【0019】
一般的な技法
本発明を実施する際には、別段に指示しない限り、当技術分野内にある免疫学、生化学、化学、分子生物学、微生物学、細胞生物学、遺伝学、および組換えDNAの従来の技法が用いられる。Sambrook, Fritsch、およびManiatis、「MOLECULAR CLONING: A LABORATORY MANUAL」、第2版(1989年);「CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubelら編(1987年));「METHODS IN ENZYMOLOGY」のシリーズ(Academic Press社)、「PCR 2: A PRACTICAL APPROACH」(M.J. MacPherson、B.D. Hames、およびG.R. Taylor編(1995年));HarlowおよびLane編(1988年)、「ANTIBODIES, A LABORATORY MANUAL」;ならびに「ANIMAL CELL CULTURE」(R.I. Freshney,編(1987年))を参照されたい。
【0020】
定義
本明細書および特許請求の範囲で用いられる単数形の「ある(a)」、「ある(an)」、および「その(the)」は、文脈により別段に明確な指示がなされない限り、複数形の指示を包含する。例えば、「ある細胞(a cell)」は、複数の細胞を、それらの混合物を含めて包含する。
【0021】
「対照」という用語は、実験において比較の目的で用いられる代替的な被験体、細胞、または試料である。対照は、「陽性」の場合もあり、「陰性」の場合もある。例えば、対照細胞は、所与の対象細胞における遺伝子産物の差次的発現についてアッセイする際に用いることができる。対照細胞による遺伝子産物の発現を、被験細胞、例えば、生物活性剤と接触させた被験細胞と比較することができる。さらに、「対照」はまた、異なる時点について比較する実験における同じ被験体、細胞、または試料も表わしうる。生物活性剤をスクリーニングする文脈において、対照細胞は、被験生物活性剤と接触させていない神経細胞でありうる。
【0022】
「ポリヌクレオチド」、「ヌクレオチド」、「ヌクレオチド配列」、「核酸」、および「オリゴヌクレオチド」という用語は、互換的に用いられる。これらは、デオキシリボヌクレオチドもしくはリボヌクレオチド、またはこれらの類似体である、任意の長さのヌクレオチドのポリマー形態を指す。ポリヌクレオチドは、任意の3次元構造を有することが可能であり、公知のものであれ、未知のものであれ、任意の機能を果たしうる。以下が、ポリヌクレオチドの非限定的な例である:遺伝子または遺伝子フラグメントのコード領域または非コード領域、連鎖解析により定義される(1または複数の)遺伝子座、エキソン、イントロン、メッセンジャーRNA(mRNA)、トランスファーRNA、リボソームRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝鎖ポリヌクレオチド、プラスミド、ベクター、任意の配列を有する単離DNA、任意の配列を有する単離RNA、核酸プローブ、およびプライマー。ポリヌクレオチドは、メチル化されたヌクレオチドおよびヌクレオチド類似体など、修飾ヌクレオチドを含みうる。存在させる場合、ヌクレオチド構造に対する修飾の付与は、ポリマーのアセンブリー前の場合もあり、アセンブリー後の場合もある。ヌクレオチド配列には、ヌクレオチド以外の成分を介在させることができる。ポリヌクレオチドは、標識化成分とのコンジュゲーションなどにより、重合化の後においてさらに修飾することができる。
【0023】
本明細書で用いられる「発現」とは、ポリヌクレオチドがmRNAへと転写される過程、および/または、転写されたmRNA(また、「転写物」とも称する)が、その後、ペプチド、ポリペプチド、またはタンパク質へと翻訳される過程を指す。転写物およびコードされたポリペプチドを、集合的に、「遺伝子産物」と称する。ポリヌクレオチドがゲノムDNAに由来する場合、発現は、真核細胞におけるmRNAのスプライシングを包含しうる。
【0024】
「接触」、「送達」、および「投与」という用語は、薬剤が、被験体、組織、または細胞に入ることを意味するように用いることができる。本明細書の開示全体で用いられる該用語には、特定の用語の文法的な変化形も含まれる。例えば、「送達」には、「送達すること」、「送達された」、「送達する」などが含まれる。当技術分野では、生物活性剤を送達または投与する各種の方法が公知である。例えば、本明細書に記載の1または複数の薬剤は、非経口送達、経口送達、腹腔内送達、静脈内送達、動脈内送達、経皮送達、筋肉内送達、リポソーム送達、カテーテルもしくはステントによる局所送達、皮下送達、脂肪組織内送達、または髄腔内送達することができる。
【0025】
ヌクレオチド配列またはポリペプチド配列に適用される、「差次的発現」という用語は、対照において検出される発現と比較した、その配列の過剰発現または過小発現を指す。過小発現はまた、対照と比較した、被験対象における検出可能な発現の不在を証拠とする、特定の配列の発現の不在も包含する。
【0026】
本明細書において、「ポリペプチド」、「ペプチド」、および「タンパク質」という用語は互換的に用いられ、任意の長さのアミノ酸ポリマーを指す。該ポリマーは、直鎖状の場合もあり、分枝状の場合もあり、修飾アミノ酸を含む場合もあり、また、アミノ酸以外の分子を介在させる場合もある。該用語はまた、修飾されたアミノ酸ポリマー、例えば、ジスルフィド結合の形成、グリコシル化、脂質化、アセチル化、リン酸化、または、標識化成分とのコンジュゲーションなど、他の任意の操作を施されたアミノ酸ポリマーも包含する。本明細書で用いられる「アミノ酸」とは、グリシン、およびD光学異性体またはL光学異性体、ならびにアミノ酸類似体およびペプチド模倣剤を含めた、天然アミノ酸および/または非天然アミノ酸もしくは合成アミノ酸を指す。
【0027】
本明細書において、「被験体」、「個体」、または「患者」は、互換的に用いられ、脊椎動物、好ましくは、哺乳動物、より好ましくはヒトを指す。哺乳動物には、マウス、ラット、イヌ、ブタ、サル(類人猿)、ヒト、農場動物、競技動物、およびペットが含まれるがこれらに限定されない。in vivoにおいて得られたか、またはin vitroにおいて培養された生物学的実体を有する組織、細胞、およびそれらの前駆体もまた包含される。
【0028】
本明細書において、「治療」、または「治療すること」、または「改善すること」は、互換的に用いられる。これらの用語は、臨床的な結果を含めた、また、臨床的結果であることが好ましい、有益または所望の結果を得るための手法を指す。本発明を目的とする場合、有益または所望の臨床的結果には、以下:脱髄病変サイズの縮減(例えば、脱髄障害の関連において)、OPCの増殖および成長もしくは病変部位へのOPCの遊走の促進、希突起膠細胞分化の促進、神経障害発症の遅延化、脱髄障害発生の遅延化、神経障害から生じる症状の軽減、疾患を患う患者の生活の質の向上、疾患を治療するのに必要な他の薬物用量の低減、標的化および/もしくは封入などによる別の薬物の効果の増強、疾患の進行の遅延化、ならびに/または個体の生存の延長のうちの1または複数が含まれるがこれらに限定されない。治療は、疾患を防止すること、すなわち、疾患が誘発される前において、保護的組成物を投与することにより、疾患の臨床症状を発生させないようにすること;疾患を抑制すること、すなわち、疾患の誘発的事象の後ではあるが、その臨床的な発現または再発現の前において、保護的組成物を投与することにより、疾患の臨床症状を発生させないようにすること;疾患を阻害すること、すなわち、それらの最初の発現後において、保護的組成物を投与することにより、臨床症状の発生を停止させること;疾患の再発を防止すること、および/または疾患を緩和すること、すなわち、それらの最初の発現後において、保護的組成物を投与することにより、臨床症状を退縮させることを包含する。
【0029】
「薬剤」、「生物学的に活性な薬剤」、「生物活性剤」、「生物活性化合物」、または「生物学的に活性な化合物」という用語は互換的に用いられ、また、言及される文脈において、複数の指示対象も包含する。本明細書で記載される、本発明の1または複数の組合せ治療方法において用いられるこのような化合物には、単純有機分子もしくは複合有機分子、または単純無機分子もしくは複合無機分子、ペプチド、ペプチド模倣剤、タンパク質(例えば、抗体)、DNA、RNA、およびこれらの類似体を含めた核酸分子、炭水化物含有分子、リン脂質、リポソーム、低分子干渉RNA、またはポリヌクレオチド(例えば、アンチセンス)が含まれるがこれらに限定されない。
【0030】
このような薬剤は、神経細胞の増殖または分化と関連する細胞周期経路の構成要素に対するアゴニストまたはアンタゴニストでありうる。本発明の一部の実施形態では、結合部位において同じ3次元構造を有する化合物は、アンタゴニストとして用いうることが意図される。化学構造の3次元解析を用いて、神経細胞周期に関連するポリペプチドに対する結合部位を含めた、活性部位の構造を決定する。
【0031】
本明細書で用いられる「アンタゴニスト」という用語は、標的ポリペプチドの生物学的機能を阻害する能力を有する分子を指す。したがって、「アンタゴニスト」という用語は、標的ポリペプチドの生物学的な役割の文脈で定義される。本明細書で好ましいアンタゴニストが、標的と特異的に相互作用する(例えば、標的に特異的に結合する)一方で、該標的ポリペプチドがそのメンバーであるシグナル伝達経路の他のメンバーと相互作用することにより、該標的ポリペプチドの生物学的活性を阻害する分子もまた、この定義内に具体的に包含される。アンタゴニストにより阻害されることが好ましい生物学的活性は、OPC増殖の増大、OPC増殖の減少、OL分化の増大、または星状細胞増殖の増大、および/または再有髄化の促進と関連する。例えば、アンタゴニストは、神経細胞周期と関連するペプチドと直接的に相互作用する場合もあり、間接的に相互作用する場合もある。限定なしに述べれば、本明細書で定義されるアンタゴニストには、オリゴヌクレオチドデコイ、アプタマー、抗ケモカイン抗体および抗ケモカイン抗体変異体、ペプチド、ペプチド模倣剤、ペプチド以外の低分子、アンチセンス分子、ならびに有機低分子が含まれる。
【0032】
本明細書で用いられる「アゴニスト」という用語は、標的ポリペプチドの生物学的機能を誘発または増強する能力を有する分子を指す。したがって、「アゴニスト」という用語は、標的ポリペプチドの生物学的な役割の文脈で定義される。本明細書で好ましいアゴニストが、標的と特異的に相互作用する(例えば、標的に特異的に結合する)一方で、該標的ポリペプチドがそのメンバーであるシグナル伝達経路の他のメンバーと相互作用することにより、該標的ポリペプチドの生物学的活性を阻害する分子もまた、この定義内に具体的に包含される。アゴニストにより阻害されることが好ましい生物学的活性は、OPC増殖の増大、OPC増殖の減少、OL分化の増大、または星状細胞増殖の増大、それによる再有髄化の促進と関連する。限定なしに述べれば、本明細書で定義されるアンタゴニストには、オリゴヌクレオチドデコイ、アプタマー、抗ケモカイン抗体および抗ケモカイン抗体変異体、ペプチド、ペプチド模倣剤、ペプチド以外の低分子、アンチセンス分子、ならびに有機低分子が含まれる。
【0033】
神経細胞の増殖/分化に対するアゴニスト、アンタゴニスト、および他の調節物質が、本発明の範囲内に明示的に包含される。特定の実施形態において、アゴニスト、アンタゴニスト、および他の調節物質は、神経細胞周期、すなわち、増殖または分化の調節に関与するポリペプチドに結合する抗体および免疫グロブリンの変異体である。これらのアゴニスト性、アンタゴニスト性の調節化合物は、直鎖状形態で提供することもでき、環状形態で提供することもでき、また、場合によって、一般に天然では見出されない、少なくとも1つのアミノ酸残基、または少なくとも1つのアミドイソステアを含みうる。これらの化合物は、グリコシル化、リン酸化、硫酸化、脂質化、または他の過程により修飾することができる。
【0034】
「有効量」または「治療有効量」という用語は、限定せずに述べれば、脱髄病変サイズの縮減(例えば、脱髄障害の関連において)、OPCの増殖および成長の促進、神経障害発症の遅延化、脱髄障害発生の遅延化、神経障害から生じる症状の軽減、疾患を患う患者の生活の質の向上、疾患を治療するのに必要な他の薬物用量の低減、標的化および/もしくは封入などによる別の薬物の効果の増強、疾患の進行の遅延化、ならびに/または個体の生存の延長などの臨床的結果を含めた、有益または所望の結果をもたらすのに十分な量のアンタゴニストを指す。治療有効量は、治療される被験体および疾患状態、被験体の体重および年齢、疾患状態の重症度、投与方式などに応じて変化するが、これらは、当業者により容易に決定されうる。該用語はまた、本明細書に記載の造影方法のいずれか1つにより検出される画像をもたらす用量にも適用される。具体的な用量は、選択される具体的なアンタゴニスト、従う投与レジメン、それが他の化合物と組み合わせて投与されるかどうか、投与回数、造影される組織、それが移送される物理的な送達系に応じて変化する。
【0035】
本明細書で用いられる「抗体」という用語には、組換え抗体、ヒト化抗体、キメラ抗体、単鎖抗体、ヒト化抗体、融合タンパク質、モノクローナル抗体など、すべての抗体形態が含まれる。本発明はまた、神経細胞周期に関与するポリペプチドに対する結合(例えば、神経細胞の増殖/分化の調節に関与する転写因子またはタンパク質に対する結合)が可能な、抗体の機能的フラグメントにも適用可能である。
【0036】
一実施形態では、比較的低用量の全抗体、裸抗体、または全抗体、裸抗体の組合せが用いられる。一部の実施形態では、抗体フラグメント、したがって、完全抗体未満のフラグメントが用いられる。他の実施形態では、薬物、毒素、または治療用放射性同位体との抗体のコンジュゲートが有用である。複数の抗原に結合するハイブリッド抗体を含め、ケモカイン抗原に結合する二特異性の抗体融合タンパク質を、本発明に従い用いることができる。したがって、抗体は、一特異性の場合もあり、多重特異性の場合もある、裸抗体およびコンジュゲート抗体および抗体フラグメントを包含する。
【0037】
「調節すること」、「調節された」、または「調節」という用語は互換的に用いられ、所与の文脈における直接的または間接的な変化を意味する。例えば、MRF発現の調節は、結果として、神経細胞の分化および/または有髄化を変化させる。別の例において、調節は、それ自体が、MRFに関与する遺伝子の発現を調節しうる遺伝子/遺伝子産物の調節でありうる。
【0038】
生物活性剤に適用される「アプタマー」という用語には、特定の分子に対する特異的な結合特性に基づいて選択されるDNA、RNA、またはペプチドが含まれる。例えば、本明細書で開示される、神経細胞周期に関与する特定の遺伝子または遺伝子産物に対する結合について、(1または複数の)アプタマーを選択することができ、この場合、選択は、当技術分野において公知であり、当業者に周知の方法によりなされる。その後、(1または複数の)前記アプタマーを被験体に投与して、免疫反応を調節または制御することができる。特定のタンパク質、DNA、アミノ酸、およびヌクレオチドに対する親和性を有する一部のアプタマーについては説明されている(K. Y. Wangら、Biochemistry、32巻、1899〜1904頁(1993年); Pitner ら、米国特許第5,691,145号;Goldら、Ann. Rev. Biochem.、64巻、763〜797頁(1995年);Szostakら、米国特許第5,631,146号)。組合せライブラリーからは、高親和性で高特異性の結合アプタマーが誘導されている(前出、Goldら)。アプタマーは、用いられる選択に応じて、平衡解離定数がマイクロモルからナノモル未満の範囲にわたる高親和性を有しうる。アプタマーはまた、高度の選択性も示し、例えば、7−メチルGとGとの間で(HallerおよびSarnow、Proc. Natl. Acad. Sci. USA、94巻、8521〜8526頁(1997年))、またはD−トリプトファンとL−トリプトファンとの間で(前出、Goldら)1000倍の識別力を示す。
【0039】
MRF発現の調節
本発明の各種の態様では、細胞型または組織に特異的な発現調節エレメントが、MRFまたはその機能的な変異体をコードする核酸配列に作動的に連結される。MRFをコードする1または複数の核酸配列に、1または複数の調節エレメントを連結することができる。MRF配列は、ヒトオルトログ(図2Aに示されるC11Orf9)、他のオルトログ、相同体、またはこれらの機能的変異体の配列でありうる。MRF配列は、in vitroでもin vivoでも生物学的効果を及ぼすことが可能であり、したがって、生物活性剤でありうる。例えば、MRFは、OLの成熟および/または有髄化を促進しうる。調節エレメントは、選択的なMRF発現に影響を及ぼすことが可能であり、かつ/またはMRFの誘導的発現もしくは構成的発現を提供しうる。MRF発現はまた、他の生物活性剤によっても調節または制御することができる。
【0040】
対象の方法で用いられる生物活性剤は、MRF、また、図5に示される、MRFにより調節される遺伝子など、それに相関する遺伝子、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、もしくはMogなど、OL分化において発現する他の遺伝子の活性または発現のレベルを調節するのに有効である。代替的に、このような遺伝子のサブセット、例えば、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子もまた、本明細書で提供される。相関する遺伝子は、成熟OLにおいて特異的に上方調節される遺伝子でありうる。生物活性剤は、MRFの発現および/または活性を調節することにより、OLの成熟および有髄化/再有髄化に影響する。調節は、MRFの活性または発現のレベルを上昇または低下させることを伴いうる。例えば、薬剤は、MRFまたはMRFの調節に関与する他の遺伝子産物に対するアゴニストの場合もあり、アンタゴニストの場合もある。このような生物活性剤の非限定的な例示的類型は、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、低分子、調合薬、またはこれらの組合せである。
【0041】
MRFなどを含めた生物活性剤は、このような薬剤が発現して、OLの分化もしくは成熟、および/または有髄化を促進するなど、それらの所望の機能を付与するように、細胞または組織内において発現しうる。遺伝子発現は、構成的プロモーターまたは誘導的プロモーター、細胞型特異的な発現調節エレメント、およびエンハンサーが含まれるがこれらに限定されない特定の調節エレメントの制御下に置かれることが典型的である。このような遺伝子は、調節エレメントに作動的に連結されるという。例えば、構成的プロモーター、誘導的プロモーター、または細胞/組織特異的プロモーターを発現ベクター内に組み込んで、宿主細胞内で発現する遺伝子の発現を調節することができる。したがって、用いられるプロモーターエレメントに応じて、生物活性剤は、MRFの発現またはその活性を遮断、増強、または促進するように、所望に応じて発現させることができる。例えば、MRFの機能を促進する薬剤を細胞内において一時的に発現させる結果、OLの分化を増強させ、この結果、最終的に、有髄化/再有髄化をもたらすことができる。調節配列は、中枢神経系または末梢神経系の特定の細胞型における、生物活性剤の異所性発現を可能とする。例えば、選択的なMRFの調節は、神経膠細胞などの神経細胞などであるがこれらに限定されない細胞内において達成することができる。神経膠細胞には、希突起膠細胞、小膠細胞、シュワン細胞、または星状細胞が含まれうる。
【0042】
調節配列の例示的な発現には、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、ミエリン関連糖タンパク質(MAG)遺伝子、ミエリン希突起膠細胞糖タンパク質(MOG)遺伝子、 希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFR−α遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子、2(VEGFR2)遺伝子、スーパーオキシドディスムターゼ(SOD1)遺伝子、チロシンヒドロキシラーゼ遺伝子、ニューロン特異的エノラーゼ遺伝子、パーキン遺伝子(PARK2)、パーキン共調節遺伝子(PACRG)、ニューロン特異的Tα1 α−チューブリン(Tα1)遺伝子、小胞モノアミントランスポーター(VMAT2)遺伝子、およびα−シヌクレイン(SNCA)遺伝子、PDGFR−β遺伝子,Olig1遺伝子、Olig2遺伝子、またはプロテオリピドタンパク質(PLP)遺伝子が含まれるがこれらに限定されない遺伝子から選択される調節配列が含まれる。
【0043】
当技術分野では、その関連する開示が参照により本明細書に組み込まれる、米国特許出願公開第2003/0110524号;同第2003/0199022号;同第2006/0052327号、同第2006/0193841号、同第2006/0040386号、同第2006/0034767号、同第2006/0030541号;米国特許第6472520号、同第6245330号、同第7022319号、同第7033595号において開示されるなど、神経細胞特異的なプロモーターのさらなる例が公知である。また、ウェブサイト<chinook.uoregon.edu/promoters.html>;または<tiprod.cbi.pku.edu.cn:8080/index>(特定の細胞/組織に特異的な遺伝子のプロモーターを列挙する);およびPattersonら、J. Biol. Chem.(1995年)、270巻、23111〜23118頁も参照されたい。
【0044】
MRFおよびMRFの調節物質の発現はまた、細胞型/組織特異的プロモーターを用いる発現系以外の発現系(例えば、エフェクター分子を局所投与する発現系)を用いることによっても、一時的に調節することができる。したがって、一部の実施形態では、細胞死のメディエータータンパク質をコードする遺伝子を、tet応答性プロモーターなど、制御可能なプロモーターエレメントに作動的に連結することができる。例えば、所望の場所および時点において、誘導性薬剤(例えば、テトラサイクリンまたはその類似体)を細胞または被験体に投与し、細胞/組織特異的な形で(例えば、テトラサイクリンだけが、局所的/限定的な形で送達される)、細胞死のメディエータータンパク質の発現を誘導することができる。このような系は、テトラサイクリンの存在が発現を阻害する「オフスイッチ」系、またはテトラサイクリンの存在が発現を誘導するように、大腸菌TetRの突然変異体が用いられる「可逆性の」Tet系を包含することにより、真核細胞における遺伝子発現の緻密な制御をもたらしうる。これらの系は、例えば、GossenおよびBujard(Proc. Natl. Acad. Sci. U.S.A.(1992年)、89巻、5547頁)およびBujardら、による米国特許第5,464,758号;同第5,650,298号;および同第5,589,362号において開示されている。
【0045】
誘導的プロモーターのさらなる例には、MMTV、熱ショック70プロモーター、GAL1−GAL10プロモーター、メタロチオネイン(metallothien)誘導プロモーター(例えば、銅誘導ACE1;他の金属イオン)、ホルモン応答エレメント(例えば、グルココルチコイド、エストロゲン、プロゲステロン(progestrogen))、ホルボールエステル(TREエレメント)、カルシウムイオノフォア応答エレメント、または脱共役タンパク質3、ヒト葉酸受容体α、乳清酸性タンパク質、プロテアーゼ特異的プロモーターのほか、米国特許第6,313,373号において開示されるプロモーターが含まれるがこれらに限定されず;また、<biobase/de/pages/products/transpor.html>におけるオンライン(遺伝子/活性により分類される15,000を超える異なるプロモーター配列を伴うデータベースを提供する);また、Chenら、Nuc. Acids. Res.、2006年、34巻、データベース版、D104〜107頁を参照されたい。
【0046】
さらに他の誘導的プロモーターには、成長ホルモンプロモーター;アデノウイルスE1Aタンパク質により誘導される、アデノウイルス初期遺伝子プロモーター、もしくはアデノウイルス主要後期プロモーターなど、プロモーターヘルパーウイルスにより誘導されるプロモーター;VP16もしくは1CP4などのヘルペスウイルスタンパク質により誘導されるヘルペスウイルスプロモーター;牛痘ウイルスもしくはポックスウイルスのRNAポリメラーゼにより誘導されるプロモーター;または、T7 RNAポリメラーゼ、T3 RNAポリメラーゼ、またはSP6 RNAポリメラーゼによりそれぞれ誘導される、T7プロモーター、T3プロモーター、およびSP6プロモーターなどのバクテリオファージプロモーターが含まれる。
【0047】
他の実施形態では、構成的プロモーターが所望でありうる。例えば、アデノウイルス主要後期プロモーター、サイトメガロウイルス即初期プロモーター、βアクチンプロモーター、またはβグロビンプロモーターを含め、本発明において用いるのに適する多くの構成的プロモーターが存在する。当技術分野では、他の多くの構成的プロモーターが公知であり、本発明において用いることができる。またさらなる実施形態では、調節配列を変化させるかまたは修飾して、発現を増強する(すなわち、プロモーターの強度を増大させる)ことができる。例えば、エンハンサー機能を含むイントロン配列を用いて、プロモーター機能を増大させることができる。ミエリンプロテオリピドタンパク質(PLP)遺伝子は、エンハンサーエレメントとして機能するイントロン配列を含む。この調節エレメント/領域であるASE(アンチサイレンサー/エンハンサー)は、エキソン1 DNAの約1kb下流に位置し、約100bpを包含する。Mengら、J Neurosci Res.、82巻、346〜356頁(2005年)を参照されたい。
【0048】
さらに、特定の細胞内位置では、MRFまたはその調節物質の発現が望ましい場合があり、当技術分野において広範に実施されている組換えDNA法により、MRFまたはその調節物質をコードする核酸配列を、対応する細胞内局所化配列に作動的に連結することができる。例示的な細胞内局所化配列には、(a)細胞外への遺伝子産物の分泌を方向づけるシグナル配列;(b)細胞の細胞質膜または他の膜性区画へのタンパク質の付着を可能とする膜アンカードメイン;(c)コードされたタンパク質の核への移動を媒介する核内局在化配列;(d)コードされたタンパク質を、主に小胞体へと封じ込める、ER保持配列(例えば、KDEL配列);(e)タンパク質がファルネシル化して細胞膜と会合するようにこれをデザインしうる;または(f)コードされたタンパク質産物の特異的な細胞内分布において役割を果たす他の任意の配列が含まれるがこれらに限定されない。
【0049】
他の実施形態では、外部ガイド配列(EGS)を用いて、MRF阻害剤を標的とすることができる(例えば、US5728521、同6057153を参照されたい)。一態様において、本発明の生物活性剤は、MRFの発現および/または活性を調節する機構として、RNA干渉(RNAi)を用いうる。例えば、RNAiを用いて、MRFの発現および/または活性の阻害剤を標的とし、これにより、OLの成熟化および/または有髄化を促進することができる。RNAiは、二本鎖RNA(dsRNA)またはsiRNAにより誘発される、配列特異的な、転写後における遺伝子サイレンシング過程である。RNAiは、Drosophila、線虫、真菌、および植物など、多数の生物において見られ、抗ウイルス防御、トランスポゾン活性の調節、および遺伝子発現の調節に関与していると考えられる。RNAiにおいて、dsRNAまたはsiRNAは、結果としての、配列特異的な遺伝子発現の阻害により、標的mRNAの分解を誘導する。一部の実施形態において、miRNAは、MRF阻害剤を標的とするのに用いられる。
【0050】
本明細書で用いられる低分子干渉RNA(siRNA)とは、被験体の遺伝子を標的とする、ヌクレオチドのRNA二重鎖である。RNA二重鎖とは、RNA分子の2つの領域間における相補的対合により形成される構造を指す。siRNAが遺伝子を標的とするのは、該siRNAの二重鎖部分のヌクレオチド配列が、標的とされる遺伝子のヌクレオチド配列に相補的であるからである。一部の実施形態において、siRNA二重鎖の長さは、30ヌクレオチド未満である。一部の実施形態において、該二重鎖は、29、28、27、26、25、24、23、22、21、20、19、18、17、16、15、14、13、12、11、または10ヌクレオチドの長さである。一部の実施形態において、該二重鎖の長さは、19〜25ヌクレオチドの長さである。siRNAのRNA二重鎖部分は、ヘアピン構造の一部でありうる。二重鎖部分に加えて、ヘアピン構造は、該二重鎖を形成する2つの配列間に位置するループ部分を含有しうる。ループの長さは変化しうる。一部の実施形態において、ループは、5、6、7、8、9、10、11、12、または13ヌクレオチドの長さである。ヘアピン構造はまた、3’突出部分および/または5’突出部分も含有しうる。一部の実施形態において、突出は、0、1、2、3、4、または5ヌクレオチドの長さの3’突出および/または5’突出である。siRNAは、核酸配列によりコードすることが可能であり、該核酸配列はまた、プロモーターも包含しうる。該核酸配列はまた、ポリアデニル化シグナルも包含しうる。一部の実施形態において、ポリアデニル化シグナルは、最小の合成ポリアデニル化シグナルである。
【0051】
生物活性剤はまた、神経細胞分化に関与する1または複数の遺伝子、例えば、MRF阻害剤を標的とする抗体でもありうる。前出の遺伝子のうちのいずれかによりコードされるポリペプチドに特異的な(または同ポリペプチドの活性部位に特異的な)抗体の作製は、それらの各々が参照により本明細書に組み込まれる、米国特許第6,491,916号;同第6,982,321号;同第5,585,097号;同第5,846,534号;同第6,966,424号;および米国特許出願公開第2005/0054832号;同第2004/0006216号;同第2003/0108548号;同第2006/002921号;および同第2004/0166099号において開示されるなど、当業者に公知である。例えば、マウスに抗原を含む組成物を注射し、血清試料を採取することにより抗体生成の存在を検証し、脾臓を摘出してBリンパ球を得、該Bリンパ球を骨髄腫細胞と融合させて、ハイブリドーマを作製し、該ハイブリドーマをクローニングし、注射された抗原に対する抗体を生成する陽性クローンを選択し、該抗原に対する抗体を生成するクローンを培養し、該ハイブリドーマ培養物から該抗体を単離することにより、モノクローナル抗体を得ることができる。十分に確立された各種の技法により、ハイブリドーマ培養物から、モノクローナル抗体を単離および精製することができる。このような単離法には、プロテインAセファロースによるアフィニティークロマトグラフィー、サイズ除外クロマトグラフィー、およびイオン交換クロマトグラフィーが含まれる。例えば、Coliganら(編)、「CURRENT PROTOCOLS IN IMMUNOLOGY」、2.7.1〜2.7.12頁、および2.9.1〜2.9.3頁(John Wiley & Sons社、1991年)を参照されたい。また、Bainesら、「Purification of Immunoglobulin G (IgG)」、「METHODS IN MOLECULAR BIOLOGY」、第10巻、79〜104頁(The Humana Press社、1992年)も参照されたい。
【0052】
標準的な技法を用いて、抗体を作製するのに適切な量の、十分に特徴づけられた抗原を得ることができる。例として述べると、Tedderら、米国特許第5,484,892号により説明される、被覆抗体を用いて、抗原を、細胞から免疫沈降させることができる。代替的に、このような抗原は、対象の抗原を過剰生成する、トランスフェクトされた培養細胞から得ることもできる。公表されたヌクレオチド配列を用いて、これらのタンパク質の各々をコードするDNA分子を含む発現ベクターを構築することができる。例えば、Wilsonら、J. Exp. Med.、173巻、137〜146頁(1991年);Wilsonら、J. Immunol.、150巻、5013〜5024頁(1993年)を参照されたい。例示として述べると、CD3をコードするDNA分子は、相互にプライミングする長いオリゴヌクレオチドを用いてDNA分子を合成することにより得ることができる。例えば、Ausubelら(編)、「CURRENT PROTOCOLS IN MOLECULAR BIOLOGY」、8.2.8〜8.2.13頁(1990年)を参照されたい。また、Wosnickら、Gene、60巻、115〜127頁(1987年);およびAusubelら(編)、「SHORT PROTOCOLS IN MOLECULAR BIOLOGY」、第3版、8−8〜8−9頁(John Wiley & Sons社、1995年)も参照されたい。ポリメラーゼ連鎖反応を用いる確立された技法は、1.8キロ塩基の長さまでの大きさの遺伝子を合成することを可能とする(Adangら、Plant Molec. Biol.、21巻、1131〜1145頁(1993年);Bambotら、PCR Methods and Applications、2巻、266〜271頁(1993年);Dillonら、「Use of the Polymerase Chain Reaction for the Rapid Construction of Synthetic Genes」、「METHODS IN MOLECULAR BIOLOGY」、第15巻、「PCR PROTOCOLS: CURRENT METHODS AND APPLICATIONS」、White(編)、263〜268頁(Humana Press社、1993年))。変化形として、対象の抗原をコードするcDNAを安定的にトランスフェクトしたマウスプレB細胞系により免疫感作したマウスに由来する脾臓細胞と、骨髄腫細胞を融合することにより、モノクローナル抗体を得ることができる。Tedderら、米国特許第5,484,892号を参照されたい。
【0053】
本発明の生物活性剤はまた、MRFまたはその機能的変異体をコードする核酸配列を含むベクターなど、ベクターの形態でもありうる。in vivoまたはin vitroの方法において用いられるベクターには、SV−40、アデノウイルス、レトロウイルスに由来するDNA配列の誘導体、ならびに哺乳動物の機能性ベクターと、機能性プラスミドおよび機能性ファージDNAとの組合せに由来するシャトルベクターが含まれうる。例えば、参照により本明細書に組み込まれる、SouthernおよびBerg、J. Mol. Appl. Genet.、1巻、327〜341頁(1982年);Subraminiら、Mol. Cell. Biol.、1巻、854〜864頁(1981年);KaufinannおよびSharp、J. Mol. Biol.、159巻、601〜621頁(1982年);Scahillら、Proc. Natl. Acad. Sci. USA、80巻、4654〜4659頁(1983年);ならびにUrlaubおよびChasin、Proc. Natl. Acad. Sci. USA、77巻、4216〜4220頁(1980年)により説明されるベクターなど、真核生物の発現ベクターがよく知られている。本発明の方法において用いられるベクターは、ウイルスベクター、好ましくはレトロウイルスベクターでありうる。複製欠損アデノウイルスが好ましい。参照により本明細書に組み込まれるEglitisおよびAndersen、BioTechniques、6巻、7号、608〜614頁(1988年)により説明される、例えば、モロニーマウス白血病ウイルス(MoMulV)、ハーベイマウス肉腫ウイルス(HaMuSV)、マウス乳癌ウイルス(MuMTV)、およびマウス骨髄増殖性肉腫ウイルス(MuMPSV)、ならびに細網内皮症ウイルス(Rev)およびラウス肉腫ウイルス(RSV)などの鳥類レトロウイルスなど、例えば、長い末端反復中に含有されるウイルスの調節配列の制御下において、レトロウイルスの構造遺伝子を、対象の単一の遺伝子により置換する、「単一遺伝子ベクター」を用いることができる。発現構築物は、ウイルスベクターの場合もあり、非ウイルスベクターの場合もある。本発明の一部と考えられるウイルスベクターには、アデノウイルス、アデノ関連ウイルス、ヘルペスウイルス、レトロウイルス(レンチウイルスを含めた)、ポリオーマウイルス、または牛痘ウイルスが含まれるがこれらに限定されない。
【0054】
複数の遺伝子を導入しうる組換えレトロウイルスベクターもまた、本発明の方法により用いることができる。独立プロモーターの調節下にあるcDNAを含有する、内部プロモーターを伴うベクター、例えば、ヒトアデノシン脱アミノ酵素(hADA)のcDNAがそれ自体の調節配列と共に挿入された選択マーカー(noe.sup.R)を伴う、N2ベクターに由来するSAXベクターは、当技術分野において公知の方法を介して、本発明の方法に従いデザインし、用いることができる。
【0055】
MRFまたは他の生物活性剤をコードする核配列の効率的な翻訳には、特定の開始シグナルもまた必要とされうる。これらのシグナルには、ATG開始コドンおよび隣接する配列が含まれる。それ自体の開始コドンおよび隣接する配列を含めた全遺伝子またはcDNAが、適切な発現ベクター内へと挿入される場合、さらなる翻訳制御シグナルは必要とされない場合がある。しかし、該コード配列の一部だけが挿入される場合、おそらく、ATG開始コドンを含めた外因性翻訳制御シグナルを与える場合がある。さらに、全挿入配列の翻訳を確保するには、開始コドンが、所望のコード配列のリーディングフレームと一致すべきである。これらの外因性の翻訳制御シグナルおよび開始コドンは、天然および合成両方の多様な由来でありうる。発現の効率は、適切な転写エンハンサーエレメント、転写ターミネーターなどを包含することにより増強することができる(例えば、Bittnerら、Methods in Enzymol.、153巻、516〜544頁(1987年)を参照されたい)。
【0056】
本発明の宿主細胞は一般に、前述のベクター内における分子など、前出の核酸分子を用いることにより遺伝子改変することができる。したがって、宿主細胞は、MRFなど、結果として希突起膠細胞の分化をもたらす、異なる発現レベルの遺伝子産物を生成しうる。in vitroまたはin vivoにおける細胞の遺伝子改変またはトランスフェクションは、本明細書の上記で言及した参考文献において説明され、また、米国特許第6,998,118号、同第6,670,147号、または同第6,465,246号などにおいて開示される、当技術分野で公知の方法を用いて実施することができる。薬剤の特徴に応じて、全身送達または局所送達、プラスミドベクター、ウイルスベクター、または非ウイルスベクター系による送達、リポソーム製剤およびミニ細胞を含めた調合薬を含め、当業者に公知の送達方式のいずれかを介して薬剤を送達することができる。例えば、哺乳動物の宿主細胞では、多数のウイルスベースの発現系を用いることができる。発現ベクターとしてアデノウイルスを用いる場合、対象のヌクレオチド配列(例えば、治療能がある薬剤をコードするヌクレオチド配列)を、アデノウイルスの転写制御複合体または翻訳制御複合体、例えば、後期プロモーター配列および三連リーダー配列へとライゲーションすることができる。次いで、このキメラ遺伝子を、in vitroまたはin vivoにおける組換えにより、アデノウイルスゲノム内に挿入することができる。ウイルスゲノムの非本質的領域(例えば、領域E1またはE3)内における挿入は、生存可能であり、また、感染した宿主内において、AQP1遺伝子産物を発現することが可能な組換えウイルスを結果としてもたらす(例えば、Logan & Shenk、Proc. Natl. Acad. Sci. USA、8巻、1号、3655〜3659頁(1984年)を参照されたい)。宿主細胞は、神経膠細胞など、神経細胞でありうる。神経細胞には、OPCまたは成熟OLなど、希突起膠細胞のほか、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)が含まれうる。
【0057】
MRFの活性レベルまたは発現レベルの調節は、各種の方法により確認することができる。例えば、遺伝子発現レベルの変化の検出は、増幅アッセイにおいてリアルタイムで実施することができる。一態様において、増幅産物は、DNA挿入剤およびDNA溝結合剤が含まれるがこれらに限定されない蛍光DNA結合剤により直接的に可視化することができる。二本鎖DNA分子内に組み込まれた挿入剤の量は、増幅されるDNA産物の量に比例することが典型的であるので、当技術分野における従来の光学系を用いて、挿入された色素の蛍光発光を定量化することにより、増幅産物の量を簡便に決定することができる。この適用に適するDNA結合色素には、SYBRグリーン、SYBRブルー、DAPI、ヨウ化プロピジウム、ヘキスト(Hoeste)、SYBRゴールド、臭化エチジウム、アクリジン、プロフラビン、アクリジンオレンジ、アクリフラビン、フルオロクマニン、エリピチシン、ダウノマイシン、クロロキン、ジスタマイシンD、クロモマイシン、ホミジウム、ミトラマイシン、ルテニウムポリピリジル、アントラマイシンなどが含まれる。
【0058】
別の態様では、増幅反応において、配列特異的プローブなど、他の蛍光標識を用いて、増幅産物の検出および定量化を容易にすることができる。プローブベースの定量的増幅は、所望の増幅産物の配列特異的な検出に依拠する。それは、特異性および感度の上昇を結果としてもたらす、標的特異的な蛍光プローブ(例えば、TaqManプローブ)を用いる。プローブベースの定量的増幅を実施する方法は当技術分野で十分に確立されており、米国特許第5,210,015号において教示されている。
【0059】
さらに別の態様では、神経周期関連遺伝子との配列相同性を共有するハイブリダイゼーションプローブを用いる、従来のハイブリダイゼーションアッセイを実施することができる。プローブは、ハイブリダイゼーション反応において、被験対象に由来する生物学的試料中に含有される標的ポリヌクレオチドと安定的な複合体を形成することが可能なことが典型的である。プローブ核酸としてアンチセンスを用いる場合、試料中においてもたらされる標的ポリヌクレオチドは、アンチセンス核酸の配列に対して相補的であるように選択されることが、当業者により理解されるであろう。逆に、ヌクレオチドプローブがセンス核酸である場合、標的ポリヌクレオチドは、該センス核酸の配列に対して相補的であるように選択される。
【0060】
当業者に公知である通り、ハイブリダイゼーションは、様々な厳密性の条件下で実施することができる。本発明を実施するのに適するハイブリダイゼーション条件は、プローブと、標的の神経細胞周期遺伝子との認識相互作用が、十分に特異的であると同時に、十分に安定的でもあるような条件である。ハイブリダイゼーション反応の厳密性を上昇させる条件は当技術分野の広い範囲において公知であり、公表されている(例えば、Sambrookら(1989年)、前出;「Nonradioactive In Situ Hybridization Application Manual」、Boehringer Mannheim社、第2版を参照されたい)。ニトロセルロース、ガラス、シリコンが含まれるがこれらに限定されない任意の固体支持体上に固定化されたプローブ、および各種の遺伝子アレイを用いて、ハイブリダイゼーションアッセイを行うことができる。好ましいハイブリダイゼーションアッセイは、米国特許第5,445,934号において説明される、高密度の遺伝子チップ上において実施される。
【0061】
ハイブリダイゼーションアッセイにおいて形成されるプローブ−標的複合体を簡便に検出するには、ヌクレオチドプローブを検出可能な標識へとコンジュゲートさせる。本発明において用いるのに適する検出可能な標識には、光化学的手段、生化学的手段、分光学的手段、免疫化学的手段、電子的手段、光学的手段、または化学的手段により検出可能な任意の組成物が含まれる。当技術分野では、多種多様の適切な検出可能標識が公知であり、これらには、蛍光標識または化学発光標識、放射性同位体標識、酵素リガンド、または他のリガンドが含まれる。好ましい実施形態では、ジゴキシゲニン、β−ガラクトシダーゼ、ウレアーゼ、アルカリホスファターゼ、またはペルオキシダーゼ、アビジン/ビオチン複合体など、蛍光標識または酵素タグを用いることが所望される可能性が高い。
【0062】
ハイブリダイゼーション強度を検出または定量化するのに用いられる検出法は、上記で選択される標識に依存することが典型的である。例えば、写真フィルムまたはホスホイメージャーを用いて、放射性標識を検出することができる。発光された光を検出する光検出器を用いて、蛍光マーカーを検出および定量化することができる。酵素に基質を供給し、基質上における酵素の作用により生成される反応産物を測定することにより、酵素標識を検出することが典型的であり、また、最後に、単に、着色標識を可視化することにより、比色分析標識を検出する。
【0063】
MRF発現の調節はまた、対応するMRFの遺伝子産物、および/もしくは、図5における遺伝子産物など、それに相関する遺伝子産物、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、もしくはMogを検討することによっても決定することができる。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子など、サブセットの発現を評価することもできる。タンパク質レベルの決定は、a)生物学的試料中に含有されるタンパク質を、MRFタンパク質、またはそれに相関するタンパク質に特異的に結合する検出剤と接触させるステップと、(b)このようにして形成される、任意の検出剤:タンパク質複合体を同定するステップとを伴うことが典型的である。一態様において、該タンパク質に特異的に結合する検出剤は、モノクローナル抗体などの抗体である。
【0064】
スクリーニングアッセイ
本発明はまた、MRF活性を調節するのに有効な、候補生物活性剤をスクリーニングする方法も提供する。該方法は、被験細胞を候補生物活性剤と接触させるステップと、対照細胞と比較した、MRFの発現レベル、またはその活性の変化についてアッセイするステップとを含む。本発明の1または複数の方法においてアッセイされる候補生物活性剤はまた、異なる時点で比較するほか、基準または対照と比較して、生物活性剤に対する応答において全体的な差違が見られるかどうかを決定するためにもアッセイされうる。
【0065】
被験細胞は、神経膠細胞などの神経細胞でありうる。被験細胞は、希突起膠細前駆細胞(OPC)、成熟OL、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)でありうるが、これらに限定されない。被験細胞はまた、幹細胞または胚性幹(ES)細胞でもありうる。候補生物活性剤は、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子でありうる。
【0066】
MRF発現の変化は、被験神経細胞または被験幹細胞の分化を調節するのに有効な候補生物活性剤を示すことが典型的である。これもまた候補生物活性剤の有効性を示しうる他の変化には、図5における遺伝子、または成熟希突起膠細胞内で特異的に上方調節される遺伝子の発現変化が含まれうる。例えば、Sox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、またはMogの発現変化。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子など、このような遺伝子のサブセットの発現変化を評価することもできる。したがって、被験細胞を用いて、候補薬剤をスクリーニングし、このような薬剤がMRFを調節し、これにより、OLの成熟を促進または阻害するかどうかを決定することができる。MRFの発現または活性を上昇させるのに有効な生物活性剤は、OLの成熟および再有髄化を促進するのに有効でありうる。それらはまた、ES細胞などからのOLの分化を促進するのにも有効でありうる。このような候補薬剤は、本明細書に記載の動物モデルなどの動物モデルにおいてさらにアッセイすることができ、また、神経障害を治療するための組成物および方法など、神経細胞の分化を誘導する方法において用いることができる。
【0067】
MRF発現レベルの変化は、上記で説明した方法を含め、当技術分野において公知の方法により実施することができる。例えば、発現レベルの変化は、被験細胞および対照細胞に由来するMRFの遺伝子発現プロファイルを解析または比較することによりアッセイすることができる。図5に列挙した遺伝子など、MRFの発現と相関する他の遺伝子、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、および/もしくはMogなど、成熟希突起膠細胞において特異的に上方調節される遺伝子の変化もアッセイすることができる。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階おいて発現する遺伝子など、このような遺伝子のサブセットの発現変化をアッセイすることもできる。
【0068】
候補薬剤は、該薬剤が、初期遺伝子または後期遺伝子に影響を及ぼすかどうか、したがって、初期分化細胞または成熟分化細胞に影響を及ぼすかどうかを決定するように、前駆細胞周期の異なる時間段階において送達することができる。例えば、候補薬剤をスクリーニングして、初期または後期に誘導される遺伝子、例えば、OL分化の初期におけるUgt8、CNP1、Plp1、および/もしくはMbp、またはMag、Trf、Mobp、および/もしくはMogなど、幼若OLまたは成熟OLと関連するいずれの遺伝子が影響を受けるのかを決定することができる。OPCを、初期遺伝子または後期遺伝子の誘導/下方調節欠損についてスクリーニングすると、OL分化の初期段階および後期段階と関連することが本明細書において同定される遺伝子の活性を調節する薬剤を選択することにより、OLの分化を促進するより良好な治療的標的化を行うことができる。さらに、このような薬剤を被験体に投与して、未分化OPCまたは未成熟OLの異なる段階から、正常で完全なOLの成熟を促進することもできる。このような薬剤はまた、OL分化の異なる段階から、有髄化するOLを生成させるのに必要な遺伝子プログラムを再構成するのにも用いることができる。
【0069】
一部の態様では、アッセイするステップを、in vitroにおいて実施する。該方法の別の態様では、アッセイするステップを、in vivoにおいて実施する。当技術分野では、in vitroおよびin vivoにおける各種の方法が用いられる。例えば、in vitroアッセイを用いて、細胞培養物中におけるOL分化を促進しうる一方、以下でさらに説明される通り、動物モデルにより、in vivoアッセイを実施することができる。当業者には、遺伝子チップ法またはアレイ法(例えば、遺伝子チップは、Agilent社、Affymetrix社、Nanogen社など、複数の販売元により容易に入手可能である)、免疫ブロット解析、RT−PCR、および他の手段などによる発現プロファイルアッセイが周知であり、また、上記でもさらに説明されている。
【0070】
本発明の一部の態様では、1または複数の候補生物活性剤を、細胞培養物と接触させ、このような接触の前において、これと同時に、またはこの後において、ミエリン修復誘導剤または軸索保護誘導剤など、1または複数の他の生物活性剤もまた細胞へと送達し、生物活性剤およびミエリン修復剤または軸索保護剤のどの組合せにより相乗効果がもたらされるかを決定する。1または複数の生物活性剤は、ES細胞などの幹細胞が、OL(またはOPC)へと分化することを誘導する因子でありうる。例えば、該因子は、Sox10、Nkx2.2、Olig1、またはOlig2などの転写因子でありうる。例えば、前駆細胞型から有髄化する希突起膠細胞への移行を示す低速度撮影顕微鏡法を用いることにより、または本明細書で説明されるOL特異的マーカーの発現をアッセイすることにより、培養物中において相乗効果を観察することができる。さらに、前駆細胞に、増強緑色蛍光タンパク質(EGFP)の膜標的化形態をトランスフェクトし、分化細胞の検出において好都合な蛍光顕微鏡法を容易にすることができる。したがって、各種の実施形態では、米国特許第7,008,634号;同第6,972,195号;同第6,982,168号;同第6,962,980号;同第6,902,881号;同第6,855,504号;または同第6,846,625号において開示される技法など、当技術分野において公知の技法を用いて、細胞を培養および/または遺伝子改変し、標的ポリペプチドを発現させることができる。
【0071】
スクリーニングアッセイにおいて用いられる細胞には、被験体から得、約5、6、7、8、9〜約14日間にわたり培養物中において増殖させたOPCが含まれうる。細胞は、約1、2、3、4、5、6、7、8、または9日間にわたり培養することができる。培養物中において増殖させる際、このような細胞には、1または複数のベクターをトランスフェクトすることができる。
【0072】
本発明の別の態様は、スクリーニングアッセイのためのシステムである。したがって、それを介してスクリーニングアッセイに関するデータを発生させうる論理デバイスを表わす例もまた提供される。候補生物活性剤との接触を伴うかまたは伴わない被験細胞の発現プロファイルなどのデータを受信および保存するコンピュータシステム(またはデジタルデバイス)。コンピュータシステムは、また、生物活性剤と接触させた神経細胞と、生物活性剤と接触させなかった対照細胞との発現プロファイルの比較など、データに対する解析も実施しうる。コンピュータシステムは、媒体、および/または、場合によって、固定化された媒体を有するサーバーに接続されうるネットワークポートからの指示を読み取りうる論理装置として理解することができる。システムには、CPU、ディスクドライブ、ならびに、キーボードおよび/またはマウスなど、任意選択の入力デバイス、ならびに任意選択のモニターを包含することが典型的である。示される通信媒体を介し、ローカルまたは遠隔の位置にあるサーバーへのデータ通信を達成することができる。
【0073】
通信媒体には、データを送信および/または受信する任意の手段が含まれうる。例えば、通信媒体は、ネットワーク接続、無線接続、またはインターネット接続でありうる。このような接続は、ウェブ上の通信を提供しうる。本発明に関連するデータを、このようなネットワークまたは接続により送信し、利用者が受信および/または参照することができる。受信者は、個人でありうるが、これに限定されない。コンピュータにより読み取り可能な媒体には、候補生物活性剤と接触させた神経細胞から得られる発現プロファイルの解析結果の送信に適する媒体が含まれうる。媒体は、生物活性剤が、MRFまたは他の相関する遺伝子の発現を調節するかどうかなど、本明細書に記載の方法を用いて導出された結果を包含しうる。
【0074】
本発明のスクリーニング方法を実施する場合、本明細書において例示される方法を含め、希突起膠細胞の分化を確認するのに適用可能な任意の公知の方法を用いることができる。候補生物活性剤は、それらが、活性を促進する(例えば、MRFの発現レベルを増強する)か、または活性を阻害する(例えば、MRF阻害剤などの標的分子に対する結合を介して、発現レベルを低下させるか、または機能を遮断する)かに基づき、選択することができる。
【0075】
マイクロアレイ
本明細書に記載のスクリーニング方法はまた、マイクロアレイ、またはその上に固定化させた遺伝子チップ、その少なくとも1つがMRFに対応する複数のプローブを用いても実施することができる。これらのマイクロアレイはまた、複数種類のヒト疾患組織、例えば、多発性硬化症病変または希突起膠細胞腫瘍組織において存在する、希突起膠細胞系統細胞の分化状態を評価するのにも用いることができる。したがって、本発明は、そのようなマイクロアレイを含む組成物を提供する。
【0076】
マイクロアレイは、MRFに対応する少なくとも1つのプローブ、また、MRFによって調節される遺伝子に相関する1または複数のプローブを包含しうる。複数のプローブは、MRF、また、図5における少なくとも1つの遺伝子に対応しうる。複数のプローブは、MRF、および図5におけるすべての遺伝子に対応しうる。プローブはまた、MRF、また、Sox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、および/またはMogなど、成熟希突起膠細胞において特異的に発現する遺伝子にも対応しうる。代替的に、マイクロアレイ上における複数のプローブは、MRF、また、OL分化の初期におけるUgt8、CNP1、Plp1、および/もしくはMbp、OL分化の後期におけるMag、Trf、Mobp、および/もしくはMog、またはPlp1、Mbp、もしくはMagなど、OL分化の中間段階において発現する遺伝子など、OL分化の異なる段階で発現する遺伝子など、遺伝子のサブセットも含みうる。
【0077】
プローブとは、ハイブリダイゼーション反応において、それに対応する標的ポリヌクレオチドを検出または同定するのに用いられるポリヌクレオチドを指す。ポリヌクレオチドに対して適用される「ハイブリダイズする」という用語は、該ポリヌクレオチドが、ハイブリダイゼーション反応において、ヌクレオチド残基の塩基間における水素結合により安定化する複合体を形成する能力を指す。1つのポリヌクレオチドが、最終的に別のポリヌクレオチドに由来する場合、異なるポリヌクレオチドは互いに「対応する」という。例えば、センス鎖は、同じ二本鎖配列のアンチセンス鎖に対応する。mRNA(また、遺伝子転写物としても公知である)は、それが転写される元の遺伝子に対応する。cDNAは、逆転写反応、またはRNA配列の知識に基づくDNAの化学合成などにより、それが生成される元のRNAに対応する。cDNAはまた、RNAをコードする遺伝子にも対応する。ポリヌクレオチドは、その対の一方が、他方の一部だけに由来する場合も対応するという。
【0078】
本発明のアレイは、比較を目的とする、陽性または陰性の対照プローブを含みうる。適切な対照プローブの選択は、最初に選択する試料プローブ、および調べているその発現パターンに依存する。任意の種類の対照プローブを、アレイ内の任意の位置、またはアレイ全体の複数の位置において局在化させて、ハイブリダイゼーションアッセイにおける空間的なばらつき、全体の発現レベル、または非特異的な結合についての対照とすることができる。
【0079】
本発明において実施されるポリヌクレオチドプローブは、化学合成、組換えによるクローニング、例えば、PCR、またはこれらの任意の組合せにより得ることができる。ポリヌクレオチドを化学合成する方法は当技術分野において周知であり、本明細書で詳細に説明する必要はない。当業者は、本明細書に記載の配列データを用いて、DNA合成器、PCR機を用いるか、または市販品を取り寄せることにより、所望のポリヌクレオチドを得ることができる。選択されたプローブは、該プローブを、固体支持体表面と安定的に会合させる際に有効である任意の適切な技法により、固体支持体の所定の領域上に固定化させる。「安定的に会合した」とは、ハイブリダイゼーション条件および洗浄条件下において、ポリヌクレオチドが、所定領域に局在化された状態を維持することを意味する。ポリヌクレオチドは、それ自体、支持体表面と共有結合させることもでき、これに非共有結合させることもできる。非共有結合の例には、非特異的な吸着、イオン性相互作用、疎水性相互作用、または水素結合による相互作用の結果としての結合が含まれる。共有結合は、ポリヌクレオチドと、支持体表面上に存在する官能基との化学結合の形成を伴う。官能基は、天然の場合もあり、リンカーとして導入される場合もある。非限定的な官能基には、ヒドロキシル、アミン、チオール、およびアミドが含まれるがこれらに限定されない。ポリヌクレオチドプローブの共有結合による固定化に適用可能な例示的な技法には、UVによる架橋形成または他の光誘導型の化学結合形成、および機械誘導型の結合形成が含まれるが、これらに限定されない(例えば、米国特許第5,837,832号、同第5,143,854号、同第5800992号、WO92/10092、WO93/09668、およびWO97/10365を参照されたい)。好ましい方法は、ポリヌクレオチドプローブの末端のうちの1つを、一重の共有結合を介して支持体表面へと連結することである。プローブがより大きな自由度を有し、相補的な標的との複合体相互作用に用いられるとき、このような立体構造は、高度のハイブリダイゼーション効率を可能とする。
【0080】
手動で、またはより一般的には、一連の操作により、試料を多数の所定領域へとスポットする自動デバイスを用いて、複数のプローブ試料を付着させることにより、各アレイを作製することが典型的である。ポリヌクレオチドアレイの作製には、一般に、各種の自動スポッティングデバイスが用いられる。このようなデバイスには、誘電式デバイスまたはインクジェット式デバイス、自動式マイクロピペット、および市販されるこれらのデバイスのうちのいずれか(例えば、Beckman社製、Biomek2000)が含まれる。
【0081】
動物モデル
本発明の別の態様において、スクリーニングアッセイは、in vivoで実施される。例えば、有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法は、候補生物活性剤を動物に投与するステップと、対照動物と比較したMRFの発現レベルの上昇についてアッセイし、該上昇が、動物における有髄化を促進する前記生物活性剤を示すステップとを含みうる。本発明の1または複数の方法においてアッセイされる候補生物活性剤はまた、異なる時点で比較するほか、基準または対照と比較して、生物活性剤に対する応答において全体的な差違が見られるかどうかを決定するためにもアッセイされうる。
【0082】
動物被験体を用いて、候補生物活性剤をスクリーニングし、このような薬剤がMRFを調節するかどうかを決定し、したがって、MRFを下方調節または上方調節する候補薬剤を同定し、これにより、OLの成熟または分化および再有髄化を促進または阻害する薬剤を同定することができる。被験体のスクリーニング方法に有用な候補生物活性剤は、ペプチド、ポリペプチド、ペプチド模倣剤、抗体、アンチセンス、アプタマー、siRNA、および/または低分子を含みうる。MRF発現/活性、および/または神経細胞の分化を調節または改変する能力を有することが疑われる任意の薬剤を、本明細書で開示されるスクリーニング方法にかけることができる。
【0083】
MRF、また、図5に列挙した遺伝子など、MRFの発現と相関する他の遺伝子、またはSox10、Ugt8、CNP1、Plp1、Mbp、Mag、Trf、Mobp、および/もしくはMogなど、成熟希突起膠細胞において特異的に上方調節される遺伝子の変化もアッセイすることができる。代替的に、Ugt8、CNP1、Plp1、および/もしくはMbpなど、OL分化の初期において発現する遺伝子;Mag、Trf、Mobp、および/もしくはMogなど、OL分化の後期において発現する遺伝子;またはPlp1、Mbp、もしくはMagなど、中間段階において発現する遺伝子など、このような遺伝子のサブセットの発現変化をアッセイすることもできる。有髄化および発現レベルのアッセイは、当業者に周知であり(例えば、遺伝子チップは、複数の販売元から容易に入手することができる)、本明細書でさらに説明されている。
【0084】
動物は、げっ歯動物種または類人猿動物種など、哺乳動物であることが典型的である。動物は、マウス、ラット、モルモット、またはサルでありうる。動物はまた、1または複数の所望の特徴を有する「ノックアウト」または「ノックイン」である動物などのトランスジェニック動物でもありうる。「ノックアウト」は、好ましくは、標的遺伝子の発現が無視できるかまたは検出不可能となるように、標的遺伝子の機能低下を結果としてもたらす、トランスジェニック配列の導入による標的遺伝子の変化を有する。「ノックイン」は、例えば、標的遺伝子のさらなるコピーを導入することにより、または標的遺伝子の内因性のコピーの発現を増強する調節配列を作動的に挿入することにより、標的遺伝子の発現の増大を結果としてもたらす、宿主細胞ゲノムの変化を有するトランスジェニック動物である。ノックインまたはノックアウトのトランスジェニック動物は、標的遺伝子に関して、ヘテロ接合の場合もあり、ホモ接合の場合もある。ノックアウトおよびノックイン共に、「バイジェニック」でありうる。バイジェニック動物は、少なくとも2つの宿主細胞遺伝子が変化している。
【0085】
胚のマイクロ操作技術の進歩により、今日では、哺乳類の有精卵内に異種DNAを導入することが可能である。例えば、マイクロインジェクション、リン酸カルシウムを介する沈殿、リポソーム融合、レトロウイルス感染、または他の手段により、全能性幹細胞または多能性幹細胞を形質転換することができる。次いで、形質転換細胞を胚内に導入し、次いで、該胚をトランスジェニック動物へと発育させる。好ましい実施形態では、感染した胚から導入遺伝子を発現するトランスジェニック動物を作製しうるように、発育する胚に、所望の導入遺伝子を含有するウイルスベクターを感染させる。別の好ましい実施形態では、所望の導入遺伝子を、好ましくは単一細胞段階において、胚の前核または細胞質内へと共注入し、該胚を、成熟トランスジェニック動物へと発育させる。トランスジェニック動物を作製するためのこれらの方法および他の変法は当技術分野で十分に確立されており、したがって、本明細書では詳述しない。例えば、米国特許第5,175,385号、および同第5,175,384号を参照されたい。
【0086】
本発明は、モノジェニック動物およびバイジェニック動物を提供する。例えば、本明細書では、MRFノックインまたはMRFノックアウトを含む動物が開示される。トランスジェニック動物は、MRF導入遺伝子を含むことが可能であり、導入遺伝子は、該動物のゲノム内へと安定的に組み込まれされ、該トランスジェニック動物の野生型コピーを置換する。代替的に、トランスジェニック動物は、該動物のゲノム内にあるMRFの野生型コピーに加え、MRF導入遺伝子も有しうる。例えば、導入遺伝子は、単一のコピーとして、またはコンカタマー、例えば、ヘッドからヘッドへのタンデム配列またはヘッドからテールへのタンデム配列において組み込むことができる。MRF導入遺伝子は、1または複数の突然変異、例えば、特定のドメインまたはエキソンの欠失を有しうる。欠失は、エキソン8(図8)など、推定DNA結合ドメイン内におけるエキソンの欠失でありうる。欠失は、構成的な場合もあり、条件的な場合もある。例えば、MRF導入遺伝子は、リコンビナーゼ部位に隣接する場合がある。トランスジェニック動物はまた、そのゲノム内に、同類の認識配列であるloxP配列(すなわち、loxP部位)を認識する、Creなどのリコンビナーゼをコードする配列を安定的に組み込みうる。
【0087】
当技術分野では、他のリコンビナーゼ認識配列も公知である。例えば、説明される通り、Creリコンビナーゼの認識配列は、8塩基対のコア配列を挟む、2つの13塩基対による逆行反復配列を含む、34塩基対の配列である、loxPである(Sauer、Curr. Opin. Biotech.、5巻、521〜527頁(1994年)を参照されたい)。認識配列の他の例は、リコンビナーゼ酵素であるλインテグラーゼにより認識されるattB、attP、attL、およびattR配列である。attBは、9塩基対による2つのコア型のInt結合部位と、7塩基対の重複領域とを含有する、約25塩基対の配列である。attPは、コア型のInt結合部位と、アーム型のInt結合部位のほか、補酵素IHF、FIS、およびXisに対する部位も含有する、約240塩基対の配列である。Landy、Curr. Opin. Biotech.、3巻、699〜707頁(1993年)を参照されたい。このような部位はまた、Hartleyら、米国特許出願公開第20060035269号による開示において開示されるような、当技術分野において公知の方法および生成物を用いて組換えを増強するように、本発明に従い操作することもできる。
【0088】
本発明のCreリコンビナーゼは、野生型の場合もあり、野生型の変異体の場合もある。Creリコンビナーゼは、トランスジェニック動物(またはトランスジェニック細胞)において誘導可能でありうる。変異体のCreリコンビナーゼは、組換え部位に対する特異性が拡張されている。具体的に述べると、該変異体は、loxP配列以外の配列と、野生型のCreリコンビナーゼが活性である他のlox部位配列との間の組換えを媒介する。一般に、開示されるCre変異体は、野生型のCreが作用しうるlox部位(野生型のlox部位と称する)間における効率的な組換え、野生型Creにより効率的に用いられない変異体lox部位(変異体lox部位と称する)間における効率的な組換え、および野生型lox部位と変異体lox部位との間における効率的な組換えを媒介する。例えば、Cre変異体は、Creリコンビナーゼ(またはFLPなど、他の類似のリコンビナーゼ)を用いうる任意の方法または技法において用いることができる。加えて、Cre変異体は、野生型lox部位と、変異体lox部位との間のはるかにより効率的な組換えを可能とするので、異なる代替的な組換えの実施も可能とする。このような代替的な組換えの制御を用いて、より洗練された配列組換えを達成し、野生型のCreリコンビナーゼによっては不可能な結果を達成することができる。変異体Creリコンビナーゼは、米国特許第6,890,726号において開示されるなど、当技術分野において公知である。Cre活性の誘導可能性は、Creタンパク質の局在化により制御することができる。例えば、Creタンパク質は、Cre融合体である、CreERt2を結果としてもたらす、エストロゲン受容体の突然変異形とのCreリコンビナーゼの融合体でありうる。リガンドの不在下において、CreERt2は、細胞質に存在する。しかし、合成ステロイドホルモン(タモキシフェン)の投与後において、CreERt2タンパク質は、それが機能的である核内へと移動する(すなわち、タモキシフェン誘導性)。
【0089】
リコンビナーゼはまた、Saccharomyces cerevisiaeの2ミクロンのプラスミドの天然酵素である、FLPリコンビナーゼでもありうる。FLPリコンビナーゼは、FRT(FLPリコンビナーゼ標的)配列と称する、特定の34塩基対のDNA配列において活性である。Creリコンビナーゼと同様、当技術分野において公知である(例えば、米国特許第7371577号、同第7060499号、同第6956146号、同第6774279号において説明される)FlpERなどの変異体も用いることができる。
【0090】
本発明の導入遺伝子はまた、中枢神経系内の細胞など、特定の組織型または細胞型へと選択的に導入し、活性化させることもできる。このような細胞型特異的な活性化に必要とされる調節配列は、対象の特定の細胞型に依存するが、当業者には明らかであろう。例えば、リコンビナーゼおよび/またはMRF導入遺伝子をコードする核酸配列を、細胞型特異的な調節エレメントへと作動的に連結することができる。該調節エレメントは、希突起膠細前駆細胞(OPC)、成熟OL、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)などであるがこれらに限定されない神経細胞に対して特異的でありうる。例えば、神経細胞特異的な調節エレメントは、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来しうる。さらに、細胞型特異的な調節エレメントはまた、誘導的プロモーターの場合もあり、構成的プロモーターの場合もある。
【0091】
動物モデルは、免疫調節、ミエリン修復/再有髄化、および/または軸索保護を促進する薬剤など、再有髄化の促進を対象とする生物活性剤についての、治療的に有効で有益な組合せを決定するスクリーニングアッセイにおいて用いることができる。例えば、トランスジェニック動物を改変して、MRF、Sox10、Nkx2.2、Olig1、および/またはOlig2の発現または活性の低下または上昇など、免疫調節、ミエリン修復/再有髄化、および/または軸索保護を促進する薬剤を発現するか、またはこれを発現するレベルを変化させる(すなわち、上昇または低下させる)ことができる。したがって、このような動物を用いて、これもまた、免疫調節、ミエリン修復/再有髄化、および/または軸索保護を対象とする、異なる複数の生物活性剤をスクリーニングし、ここで、該トランスジェニック動物が、1つの評価項目を対象とする薬剤を含む場合は、該動物に(1または複数の)異なる評価項目を対象とする薬剤を投与し、また、この逆を行い、神経障害または本明細書に記載の関連する状態に対して相乗作用的な治療結果をもたらす治療剤の候補となる組合せを同定することができる。
【0092】
動物モデル系を用いて、ニューロンの再有髄化を促進するか、またはこれに有益な生物活性剤の開発を行うことができる。例えば、MRF活性の上昇により、結果として、例えば、免疫調節、ミエリン修復、または軸索保護をもたらす薬剤を発現するように改変されるトランスジェニック動物は、未知の化合物をスクリーニングして、(1)化合物が、免疫忍容性を増強するか、炎症反応を抑制するか、もしくは再有髄化を促進するかどうか、および/または(2)化合物が、動物モデルにおいて、相乗的な治療効果を結果としてもたらしうるかどうかを決定する方法において用いることができる。代替的に、MRFにおいてエキソン8が欠失するトランスジェニック動物など、MRF活性が損なわれた動物を用いて、CNSにおける有髄化不全を緩和し、該動物のOLの成熟を促進する化合物をスクリーニングすることができる。さらに、本発明のトランスジェニック動物から神経細胞を単離して、ex vivo法を含め、細胞ベースの環境または細胞培養物の環境において、さらなる研究またはアッセイを実施することができる。モデル系を用いて、被験薬剤が、有害作用を付与するか、または再有髄化を低下させる、例えば、脱髄後における損傷をもたらすかどうかを調べることができる。
【0093】
動物モデルはまた、組合せ方式により薬剤をスクリーニングするのにも用いることができる。例えば、候補薬剤は、免疫調節、ミエリン修復/再有髄化、または軸索保護をもたらす第2の薬剤など、別の薬剤と共に投与することができる。例えば、MRF、Nkx2.2、Sox10、Olig1、Olig2、またはこれらの組合せなど、公知の生物活性剤を、候補生物活性剤の前に投与することもでき、これと同時に投与することもでき、またはこの後に投与することもできる。(1または複数の)細胞特異的なマーカー遺伝子の発現を検出および/または定量化し、再有髄化が生じたかどうか、また、それはどのくらいであったか、また、このような再有髄化が、対照と比較して増強されているかどうかを決定することなど、合成組合せ治療の検出および定量化により組合せ効果を決定することができる。このような例においてならば、対照は、疾患モデルが誘導される野生型でもありえようし、候補薬剤が投与されないトランスジェニック動物でもありえよう。
【0094】
本発明の動物モデルはまた、脱髄を受けるように誘導することができ、MRF発現を評価することにより、再有髄化に対する生物活性剤の効果を評価することができる。被験動物において脱髄を誘導する多数の方法が確立されている。例えば、神経の脱髄は、被験動物における病原体または物理的傷害、炎症反応および/または自己免疫反応を誘導する薬剤により与えられる場合がある。好ましい方法では、IFN−γおよびクプリゾン(ビス−シクロヘキサノンオキサルジヒドラゾン)が含まれるがこれらに限定されない脱髄誘導剤が用いられる。クプリゾン誘導型脱髄モデルは、Matsushimaら、Brain Pathol.、11巻、107〜116頁(2001年)において説明されている。この方法では、約1〜約10週間の範囲の数週間にわたり、クプリゾンを含有する食餌を被験動物に摂取させることが典型的である。
【0095】
適切な方法により脱髄状態を誘導した後は、既に脱髄させた病変において、またはその近傍において再有髄化を可能とするのに十分な長さの時間にわたり、動物を回復させる。軸索の再有髄化を発生させるのに必要とされる時間の長さは動物ごとに異なるが、少なくとも約1週間を要することが一般的であり、少なくとも約2〜10週間を要することがよりしばしばであり、約4〜約10週間を要することがさらによりしばしばである。
【0096】
神経系(例えば、中枢神経系または末梢神経系)において有髄化した軸索の増加を観察することにより、またはMRFなど、有髄化細胞のマーカータンパク質のレベルの上昇を検出することにより、本明細書に記載の動物モデルにおける再有髄化を確認することができる。脱髄を検出する同じ方法を用いて、再有髄化が生じたかどうかを決定することができる。例えば、当技術分野で公知の免疫組織化学法またはタンパク質解析により、脱髄/再有髄化現象を観察することができる。例えば、MRFなど、希突起膠細胞マーカーを特異的に認識する抗体により、被験動物の脳の断面を染色することができる。別の態様では、免疫ブロッティング、ハイブリダイゼーション法、および増幅手順、ならびに当技術分野、例えば、それらの各々の開示が、参照により本明細書に組み込まれる、Mukouyamaら、Proc.Natl.Acad.Sci.(2006年)、103巻、1551〜1556頁;Zhangら(2003年)、前出;Girardら、J.Neuroscience.(2005年)、25巻、7924〜7933頁;および米国特許第6,909,031号;同第6,891,081号;同第6,903,244号;同第6,905,823号;同第6,781,029号;および同第6,753,456号において十分に確立された他の任意の方法により、MRFなど、希突起膠細胞マーカーの発現レベルを定量化することができる。
【0097】
治療剤
本明細書に記載の組成物は、治療剤として用いることができる。神経障害を有する被験体は、治療有効量の、MRF活性を調節する生物活性剤により治療することができる。本明細書に記載の組成物および方法を用いて、多発性硬化症(MS)、進行性多巣性白質脳症(PML)、脳脊髄炎、橋中央ミエリン溶解(CPM)、抗MAG抗体陽性疾患、白質萎縮症:副腎白質萎縮症(ALD)、アレキサンダー病、カナバン病、クラッベ病、異染性白質萎縮症(MLD)、ペリツェウス−メルツバッハー病、レフサム病、コケイン症候群、ファンデルクナップ症候群、ツェルヴェーガー症候群、ギラン−バレー症候群(GBS)、慢性炎症性脱髄性多発神経障害(CIDP)、多巣性運動神経障害(MMN)、脊髄傷害(例えば、脊髄の外傷または切断)、アルツハイマー病、ハンチントン病、筋萎縮性側索硬化症、パーキンソン病、および視神経炎などであるが、これらに限定されない各種の神経障害を治療することができる。
【0098】
本明細書に記載の生物活性剤を被験体に投与してMRFの発現および/または活性を調節することにより、成熟希突起膠細胞(OL)への希突起膠細胞の前駆細胞(OPC)の分化を促進することができる。神経障害は、多発性硬化症など、脱髄障害でありうる。治療有効量の、MRF活性を調節する生物活性剤を被験体に投与することにより、被験体における再有髄化を促進することができる。
【0099】
再有髄化
本明細書に記載の生物活性剤を被験体に投与して、神経細胞の分化を増強することができる。生物活性剤は、MRFまたはそれにより調節される遺伝子の発現を調節することにより再有髄化を促進しうる。本明細書に記載の生物活性剤は、MRF自体でありうる。このような生物活性剤の投与は、薬剤自体の外因的投与により達成することもでき、本明細書で説明され、また、当業者に公知である、適切な転写調節エレメントを介して、該薬剤を、構成的、誘導的、または細胞特異的な形でコードおよび発現する核酸ベクターを提供することにより達成することもできる。このようにして発現される生物活性剤は、それ自体、神経細胞分化を促進しうる。このような神経細胞には、OL、OPC、SC、NSC、星状細胞、および小膠細胞が含まれる。
【0100】
したがって、神経細胞の分化および/または再有髄化を促進するように、内因性のMRF発現を誘導する生物活性剤を、細胞/被験体へと投与することができる。一部の実施形態では、生物活性剤を用いて、ES細胞が、OL(またはOPC)へと分化するように誘導することができる。ポリペプチド、またはポリペプチドをコードする核酸を投与することにより、MRF発現を調節して、OLへの分化を増強することができる。
【0101】
相同組換え、組込みにより、または本明細書で記載され、また、当業者に周知の成分および方法を用いるプラスミドもしくはウイルスベクターを用いることにより、所望のポリペプチドをコードする核酸を、標的細胞内へと形質転換することができる。トランスフェクトされうる神経細胞には、OL、OPC、SC、NSC、星状細胞、および小膠細胞が含まれる。一部の実施形態において、このような神経細胞には、同時に、または異なる時点において、複数のベクターをトランスフェクトすることができる。さらに、本明細書で開示されるポリペプチドのうちのいずれかをコードする核酸は、本明細書で開示され、また、当業者により認知される、構成的プロモーター、誘導的プロモーター、または細胞特異的プロモーターへと作動的に連結することができる。
【0102】
生物活性剤を投与して、OLの成熟など、神経細胞の分化を結果としてもたらすMRFの発現を増大させ、再有髄化を促進することができる。投与される生物活性剤は、MRF自体でありうる。投与される生物活性剤はまた、MRF発現の調節物質をコードするか、またはMRF自体をコードする核酸ベクターでもありうる。核酸ベクターは、構成的、誘導的、または細胞特異的な転写調節エレメントを含有し、これにより、所望の生物活性剤の持続的な発現または時点により異なる発現をもたらしうる。例えば、テトラサイクリン抑制系を用いるドキシサイクリンにより、細胞におけるMRF発現を誘導することができる。代替的に、発現ベクターは、本明細書で記載されるか、または当業者に周知の、神経特異的プロモーターを含みうる。したがって、それを必要とする被験体を治療する方法において、OPCの増殖と、OLの分化とを交互に行って再有髄化を増強する必要がある場合、神経細胞に対する生物活性剤の発現を調節することができる。
【0103】
例えば、構成的プロモーター、誘導的プロモーター、または神経細胞特異的プロモーターに作動的に連結され、MRF遺伝子、またはMRFの発現/活性を調節する別の遺伝子をコードする核酸分子を、神経細胞にトランスフェクト(神経細胞を遺伝子改変)することができる。このような細胞を形質転換させて、異なる発現レベルでMRFを発現させ、したがって、神経細胞の増殖を調節することができる。例えば、ポリペプチドは、MRFでありうる。
【0104】
また、細胞または被験体に増殖因子またはホルモンを投与して、神経細胞の分化または増殖を促進することもできる。増殖因子およびホルモンは、それ自体、本明細書で開示される任意の生物活性剤の投与と同時に、この前において、またはこの後において投与することができる。このような増殖因子またはホルモンの例には、甲状腺ホルモンT3、インスリン様増殖因子1、線維芽細胞増殖因子2、血小板由来増殖因子(PDGF)、神経成長因子、ニューロトロフィン、ニューレグリン、またはこれらの組合せが含まれる。このような増殖因子またはホルモンはまた、本明細書で開示される他の任意の生物活性剤と同時に、この前において、またはこの後において施される核酸ベクターによってもコードされうる。
【0105】
さらに、生物活性剤の投与の前において、これと同時に、またはこの後において、OPC、シュワン細胞、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)が含まれるがこれらに限定されない神経細胞もまた投与することができる。1種または複数種の生物活性剤と共に、1種または複数種の神経細胞を投与することができる。明確さを目的として述べると、種類とは、例えば、異なる種類の細胞(例えば、希突起膠細胞および星状細胞)または異なる種類の生物活性剤(例えば、抗体およびアンチセンス)を意味する。
【0106】
異なる生物活性剤を投与することができる、例えば、第2の生物活性剤の投与と同時に、この前において、またはこの後において、第1の生物活性剤を投与することができる。複数の生物活性剤を投与することができる。例えば、第1の生物活性剤は、MRF活性を調節する生物活性剤、またはMRF自体でありうる。Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進する生物活性剤など、第2の生物活性剤は、MRFを調節する薬剤の投与と同時に、この前において、またはこの後において投与することができる。代替的に、MRFは、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの投与と同時に、この前において、またはこの後において投与することもできる。各種の生物活性剤の投与は、再有髄化の促進において相乗効果をもたらしうる。
【0107】
前出はまた、標的細胞をトランスフェクトするのに用いうる核酸ベクターによる製剤にも適用しうることを理解されたい。このようなベクターは、標的細胞をトランスフェクトし、所望のポリペプチドを発現することが可能であるものとして、本明細書で記載され、また、当業者により認知されている。まとめると、このようなベクターはまた、本明細書に記載の医薬製剤または治療剤中においても用いることができる。
【0108】
再有髄化細胞の移植
各種の動物モデルにおいて、CNS軸索の再有髄化が示されている。以来、近年の多くの研究により、脱髄疾患における細胞移植の使用と関連する新規の技法および新規の機構が示されている。成人脳から単離されたヒトOP細胞を、有髄化不全のマウス突然変異体へと移植したところ、無髄軸索を有髄化することができた。成人の前駆細胞を用いれば、倫理的問題が回避されることが重要である。OP細胞が、内因的な再有髄化の一因となるのに対し、NSCは、ミエリンの修復を促進する代替的な細胞供給源である。NSCは、成人CNSにおいて見出され、in vitroにおいて大幅に増殖させることができ、また、OL、星状細胞、またはニューロンを形成するように分化させることができる。実験的な自己免疫性脳脊髄炎(EAE)の再発形態または慢性形態を有するげっ歯動物内へと移植したところ、NSCは、CNSの炎症領域および脱髄領域へと遊走し、神経膠細胞原基を優先的に選択することが示されている。さらに、移植を受けたマウスにおける臨床的疾患の緩和は、脱髄病変の修復および軸索傷害の軽減と関連していた。組織学的解析により、移植されたNSCは、大半が、PDGFR+ OP細胞へと分化することが確認された。
【0109】
本発明の態様において、被験体の生物活性剤は、MRF、またはMRFにより上方調節される遺伝子を過剰発現するように改変され、被験体に投与されると、ミエリンの修復または脱髄した軸索の再有髄化に関与する細胞を含みうる。このような細胞を適切なベクターと共に培養し、また、このような細胞に適切なベクターをトランスフェクトして、細胞の成熟、またはOLの分化を増強するポリペプチドを発現させることができる。また、細胞を改変して、Sox10、Nkx2.2、Olig1、またはOlig2を過剰発現させることもできる。1または複数の細胞型を改変して、MRF、Nkx2.2、Sox10、Olig1、Olig2、またはこれらの組合せを過剰発現させることができる。例えば、細胞を改変してMRF、およびSox10、Nkx2.2、Olig1、またはOlig2を過剰発現させ、これを被験体に投与して神経障害を治療することができる。OPCおよび星状細胞など、異なる細胞型もまた投与することができる。一部の実施形態において、ミエリンを生成する細胞またはこれらの前駆細胞には、胚性OPCまたは成体OPCが含まれるがこれらに限定されない。
【0110】
別の生物活性剤の投与の前において、これと同時に、またはこの後において投与しうる細胞(「細胞型」)は、希突起膠細前駆細胞(OPC)、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)でありうる。一部の実施形態において、細胞は、生物活性剤により、OPCまたはOLへと分化するように誘導されたES細胞などのES細胞でありうる。一部の実施形態では、このような細胞を動物被験体に投与して、OLの成熟など、神経細胞の分化を増強することができる。
【0111】
一実施形態において、細胞は、成熟希突起膠細胞をもたらすそれらの能力に基づき、中枢神経系(CNS)内における希突起膠細胞の前駆細胞(OPC)であると考えられる、NG2プロテオグリカンを発現する神経膠細胞(NG2(+)細胞)である。一部の実施形態では、希突起膠細前駆細胞(OPC)、シュワン細胞(SC)、嗅球鞘細胞、星状細胞、小膠細胞、および神経幹細胞(NSC)を、ケモカインをコードするベクターと共に培養し、これにより形質転換し、in vitroにおいて増殖させた後で移植する。他の実施形態では、in vitroにおいて細胞にトランスフェクトするか、またはこれらを遺伝子改変し、MRF遺伝子、Sox10、Nkx2.2、Olig1、および/またはOlig2によりコードされるタンパク質を発現させる。
【0112】
一部の実施形態では、当技術分野において公知であるか、または本明細書で開示される方法を用いて、1または複数の所望の生物活性剤の発現を可能とするように、希突起膠細前駆細胞(OPC)、シュワン細胞(SC)、嗅球鞘細胞、および神経幹細胞(NSC)に、1または複数の発現ベクターをトランスフェクトする。このような生物活性剤は、MRF、Nkx2.2、Sox10、Olig1、またはOlig2の発現/活性を調節しうる。
【0113】
侵襲性の手順、手術手順、侵襲性が最小限の手順、および手術以外の手順を含め、当技術分野において公知の方法を用いて移植を実施することが理解される。被験体、標的部位、および送達される(1または複数の)薬剤に応じ、当技術分野において公知の方法を用いて細胞型および細胞数を選択することができる。
【0114】
医薬組成物
本発明はまた、被験体における神経障害を治療するための組成物であって、MRFなど、本明細書に記載の生物活性剤を含む組成物も提供する。組成物はまた、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せもさらに含みうる。意図される医薬組成物には、ペプチド、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、核酸発現ベクター、抗体もしくは抗体フラグメント、低分子、またはこれらの組合せが含まれるが、これらに限定されない。このような組成物は、治療有効量で用いることができる。
【0115】
所望の程度の純度を有するこのような薬剤を、薬学的に許容される任意選択の担体、賦形剤、または安定化剤(「Remington’s Pharmaceutical Sciences」、第16版、Osol,A.編、1980年)と共に、凍結乾燥させた製剤または水溶液の形態で混合することにより、このような薬剤による製剤を保存のために調製する。許容される担体、賦形剤、または安定化剤は、用いられる用量および濃度においてレシピエントに対して非毒性であり、これらには、リン酸緩衝液、クエン酸緩衝液、酢酸緩衝液、および他の有機酸緩衝液などの緩衝液;アスコルビン酸およびメチオニンを含めた抗酸化剤;防腐剤(オクタデシルジメチルベンジルアンモニウムクロリド;塩化ヘキサメトニウム;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチルオルベンジルアルコール;メチルパラベンもしくはプロピルパラベンなどのアルキルパラベン;カテコール;レソルシノール;シクロヘキサノール;3−ペンタノール;およびm−クレゾール);低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチン、もしくは免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、もしくはリシンなどのアミノ酸;単糖、二糖、および、グルコース、マンノース、またはデキストリンを含めた他の炭水化物;EDTAなどのキレート化剤;スクロース、マンニトール、トレハロース、もしくはソルビトールなどの糖;甘味剤および他の芳香剤;微晶質セルロース、ラクトース、トウモロコシ、および他のデンプンなどの充填剤;結合剤;添加剤;着色剤;ナトリウムなどの塩形成対イオン;金属錯体(例えば、Zn−タンパク質錯体);ならびに/またはTWEEN(商標)、PLURONICS(商標)、もしくはポリエチレングリコール(PEG)などの非イオン性界面活性剤が含まれる。
【0116】
好ましい実施形態において、本発明の生物活性剤を含む医薬組成物は、酸添加塩および塩基添加塩の両方が含まれることを意図する、薬学的に許容される塩として存在するなど、水溶性形態である。「薬学的に許容される酸添加塩」とは、遊離塩基の生物学的有効性を保持し、生物学的または他の形で有害でなく、塩酸、臭化水素酸、硫酸、硝酸、リン酸などの無機酸、および酢酸、プロピオン酸、グリコール酸、ピルビン酸、シュウ酸、マレイン酸、マロン酸、コハク酸、フマル酸、酒石酸、クエン酸、安息香酸、ケイ皮酸、マンデリン酸、メタンスルホン酸、エタンスルホン酸、p−トルエンスルホン酸、サリチル酸などの有機酸により形成される塩を指す。「薬学的に許容される塩基添加塩」には、ナトリウム塩、カリウム塩、リチウム塩、アンモニウム塩、カルシウム塩、マグネシウム塩、鉄塩、亜鉛塩、銅塩、マンガン塩、アルミニウム塩など、無機塩基に由来する塩が含まれる。アンモニウム塩、カリウム塩、ナトリウム塩、カルシウム塩、およびマグネシウム塩が特に好ましい。薬学的に許容される非毒性の有機塩基に由来する塩には、第一級アミン、第二級アミン、および第三級アミン、天然の置換アミンを含めた置換アミン、環状アミン、およびイソプロピルアミン、トリメチルアミン、ジエチルアミン、トリエチルアミン、トリプロピルアミン、およびエタノールアミンなど、塩基性のイオン交換樹脂による塩が含まれる。in vivoにおける投与に用いられる製剤は、滅菌であることが好ましい。これは、滅菌濾過膜または当技術分野において公知の他の方法を介する濾過により、容易に達成される。
【0117】
薬剤はまた、イムノリポソームとしても調合することができる。リポソームとは、哺乳動物に治療剤を送達するのに有用な、各種の脂質、リン脂質、および/または界面活性剤を含む小胞体である。生物活性剤を含有するリポソームは、Eppsteinら、Proc. Natl. Acad. Sci. USA、82巻、3688〜3692頁(1985年);Hwangら、Proc. Natl. Acad. Sci. USA、77巻、4030〜4034頁(1990年);米国特許第4,485,045号;同第4,544,545号;およびPCT WO97/38731において説明される方法など、当技術分野において公知の方法により調製される。循環時間が延長されたリポソームは、米国特許第5,013,556号において開示されている。リポソームの成分は一般に、生体膜の脂質配置と同様、二層形成で配置される。特に有用なリポソームは、ホスファチジルコリン、コレステロール、およびPEGにより誘導体化されたホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を伴う逆相蒸発法により作製することができる。リポソームは、所望の直径を有するリポソームをもたらすように、規定の小孔サイズによるフィルターを介して押出し成形される。化学療法剤または他の治療的に活性な薬剤も、場合によって、リポソーム内に含有される(Gabizonら、J. National Cancer Inst、81巻、1484〜1488頁(1989年))。
【実施例】
【0118】
(実施例1)
マウスOPCの単離、培養、およびトランスフェクション
本質的に、既に説明される通り(Cahoyら、J Neurosci、28巻、264〜278頁(2008年))に、マウスOPCを単離した。略述すると、P7 C57/B6マウスの脳を摘出し、賽の目に刻み、酵素的に分解して、単一細胞の懸濁液を作製した。これらの細胞を、BSL1(Vector Laboratories社製、L−1100)でコーティングした4枚のペトリディッシュ上において、各15分間ずつにわたり逐次パニングして小膠細胞を枯渇させ、次いで、抗PDGFRαラットモノクローナル抗体(BD Pharmingen社製、558774型)でコーティングしたペトリディッシュ上において、1時間にわたりパニングしてOPCを陽性選択した。付着しない細胞はDPBSにより洗い落とし、付着するOPCは、トリプシンによりペトリディッシュから採取した。2%のB−27(Invitrogen社製)ヒトトランスフェリン(100mg/ml)、ウシ血清アルブミン(100mg/ml)、プトレシン(16mg/ml)、プロゲステロン(60ng/ml)、亜セレン酸ナトリウム(40ng/ml)、N−アセチル−L−システイン(5mg/ml)、D−ビオチン(10ng/ml)、フォルスコリン(4.2mg/ml)、ウシインスリン(5mg/ml)(すべて、Sigma社製)、グルタミン(2mM)、ピルビン酸ナトリウム(1mM)、ペニシリン−ストレプトマイシン(各100U)(すべて、Invitrogen社製)、微量元素B(1倍濃度;バージニア州、ハーンドン、Mediatech社製)、CNTF(10ng/ml;ニュージャージー州、テリータウン、Regeneron社からの恵与)、ならびにPDGF−AA(10ng/ml)およびNT−3(1ng/ml)(いずれも、ニュージャージー州、ロッキーヒル、PeproTech社製)を含有するDMEM(カリフォルニア州、カールスバッド、Invitrogen社製)中、37℃、10%CO2で、PDLによりコーティングしたT175組織培養フラスコ内において、OPCを培養した。分化実験のため、以下の通りに細胞にトランスフェクトし、これらを、上記の通りであるが、PDGF−AAを伴わず、トリヨードチロニン(T3)(40ng/ml;シグマ社製)を伴う培地へと移した。
【0119】
OPCへのトランスフェクション:EBSSによりフラスコを1回洗浄することにより、マウスOPCの培養物を継代培養し、次いで、EBSS中に1:10のトリプシン:EDTA(Sigma社製)を用いて37℃で5分間にわたり該フラスコを処理した。30%のFCS/DPBSにより細胞を回収し、220RCFで15分間にわたり遠心分離し、増殖培地中に再懸濁させ、さらに15分間にわたり遠心分離して、微量のトリプシンを除去した。細胞ペレットを、OPC Nucleofection液(Amaxa社製)中に細胞50×106個/mlで再懸濁させた。100ul(細胞5×106個)を、発現構築物(約4ug)またはsiRNA(20uMの、MRFに対するsiRNAのプール、もしくは個別のsiRNA、または非標的化siRNAのプールであるsiControlである、それぞれ、Dharmacon社製L−056814−00、LU−056814−00、およびD001206−13を10ulずつ)に添加し、Amaxa社製ヌクレオフェクション装置のプログラム0−17により電気穿孔した。次いで、細胞を、OLマーカーアッセイ用には、24ウェルプレートの分化培地中に細胞50,000個/PDLコーティングしたカバースリップ1枚で、異所性発現/分化アッセイ用には、24ウェルプレートの増殖条件に細胞5,000個/PDLコーティングしたカバースリップ1枚で、RNA単離用には、分化培地中に細胞5×106個/PDLコーティングした10cmのディッシュで播種した。
【0120】
(実施例2)
ニワトリ胚のin ovoにおける電気穿孔
可視化を可能とする0.2%ファーストグリーンを伴うTE緩衝液中に約1ulのDNA溶液(1:4のpCIGおよびpCAGGS−MRF混合物約4ug/ul)を、ハンバーガー−ハミルトン段階にある胚12個の神経管内腔へと注入した。ECM830 ElectroSquare Porator(Genetronics社製)を用いて、胚を隔てた30ボルトで5×50ミリ秒にわたるパルスを用いて、電気穿孔を実施した。電気穿孔の4日後に胚を回収し、4%PFA中において2時間にわたり浸漬固定した後、以下の通りに処理して、in situハイブリダイゼーションまたは免疫組織化学にかけた。
【0121】
(実施例3)
免疫組織化学、in situハイブリダイゼーション、およびTEM顕微鏡法
免疫組織化学:灌流固定したマウスに由来する10μmの凍結断面、またはPDLコーティングしたカバースリップ上で増殖させた細胞を、PBS中に4%のパラホルムアルデヒド中において10分間にわたり固定し、PBS中で3分間ずつ5回にわたり洗浄し、次いで、ブロッキング液(表面抗原用には、PBS中に10%のウシ胎仔血清、細胞内抗原用には、0.3%のトリトンXを添加する)中で30分間にわたりインキュベートした。ブロッキング液(1:500のラット抗MBP抗体、1:500のウサギ抗NG2抗体;Chemicon社製、1:500の抗mycモノクローナル4A6抗体;英国、ロンドン、インペリアルカレッジ、R.Reynolds氏のご恵与による、Upstate社製、1:1,000のウサギ抗活性化カスパーゼ3抗体、BD Pharmingen社製、1:50のマウス抗MOG抗体クローン8−18C5)中において、一次抗体と共に一晩にわたり細胞をインキュベートした。PBS中で3分間ずつ5回にわたりカバースリップを洗浄し、適切なフルオロフォアをコンジュゲートさせた二次抗体(Molecular Probes社製;ブロッキング液中において1:500に希釈した)と共に30分間にわたりインキュベートし、PBS中で3分間ずつ5回にわたり洗浄し、DAPI核内対比染色色素を伴うDAKI蛍光マウント用培地によりマウントした。製造元の指示書に従い、10μmの凍結断面上において、Fluoromyelin(Invitrogen社製)染色を実施した。
【0122】
in situハイブリダイゼーション:プライマーGGTGGGTTTGAGTTTGGAGGTTおよびGGGGAAACGCTCTATGAACAGGを用いて、3’UTRから941bpに対応するin situハイブリダイゼーションプローブを、OL cDNAから増幅し、PCR−II−Topoベクター(Invitrogen社製)内へとサブクローニングした。PLPプローブは、William Richardson教授により恵与された。製造元の指示書に従い、T7ポリメラーゼおよびDIG RNA標識化キット(Roche社製)を用い、アンチセンスDIGにより標識化したリボプローブを合成した。P16における脳神経ならびにP17における視神経および坐骨神経に由来する10μmの断面上において、本質的に説明される(Cahoyら、J Neurosci、28巻、264〜278頁(2008年);Schaeren−Wiemers and Gerfin−Moser、Histochemistry、100巻、431〜440頁(1993年))通りに、in situハイブリダイゼーションを実施した。
【0123】
TEM顕微鏡法:麻酔下にあるP13マウスをPBSにより灌流した後で、カコジル酸ナトリウム緩衝液中に2%のグルタルアルデヒド/4%のパラホルムアルデヒドで処理した。視神経を切開摘出し、4℃で一晩にわたり後固定した。1%四酸化オスミウムおよび1%酢酸ウラニルによる処理後、神経をエポン樹脂内に包埋した。スタンフォード大学の微生物学および免疫学電子顕微鏡施設において断面作製および電子顕微鏡法を実施した。
【0124】
(実施例4)
構築物の作製
pCS6−MRF:プライマーCCCGGGCGCCACCATGGAGGTGGTGGACGAGACおよびCTCGAGGGAGGCAGCTCAGTCACACAGGを用いて、培養したマウスOLのcDNAから、MRFのコード領域を増幅した。結果として得られるXma1/Xho1を連結したPCRフラグメントを、Xma1/Xho1により2回にわたり消化したpCMV−Sport6(pCS6)ベクター(Invitrogen社製)内へとライゲーションし、配列決定により確認した。
【0125】
pCS6−Myc−MRF:プライマーCTCGAGGAGGTGGTGGACGAGACCGAAGおよびCTCGAGGGAGGCAGCTCAGTCACACAGGを用いて、培養したマウスOLのcDNAから、開始コドンを除いたMRFのコード領域を増幅した。結果として得られるXma1/Xho1を連結したPCRフラグメントを、Xma1/Xho1により2回にわたり消化したpCS6ベクター内へとライゲーションした。次いで、Mycペプチド(MEQKLISEEDL)をコードする自己アニール化オリゴヌクレオチドを、結果として得られる構築物のXma1部位内へとライゲーションし、N末端においてmyc−MRF融合構築物をもたらした。
【0126】
pCAGGS−MRF:Xho1およびXba1で挟んだプライマーにより、上記のpCS6−MRFからMRFのコード領域を増幅した。結果として得られるPCR産物を、pCAGGSのXba1部位およびXho1部位内へとライゲーションした。
【0127】
(実施例5)
HEK293細胞へのトランスフェクション
トランスフェクションの1日前に、10%ウシ胎仔血清を伴うダルベッコ改変イーグル培地中に50〜70%のコンフルエンスで、HEK293細胞を播種した。製造元の指示書に従い、Lipofectamine 2000(Invitrogen社製)を用いて、トランスフェクションを実施した。トランスフェクションの48時間後において、免疫蛍光法により細胞を解析した。
【0128】
(実施例6)
ノーザンハイブリダイゼーション
全RNA(5〜15μg)を、変性ホルムアルデヒドゲル上で泳動し、Hybond−N+(Amersham社製)へと転写し、UVにより架橋させた。次いで、100mlのハイブリダイゼーション液(7%SDS、0.5M Na2HPO4 pH 7.2、100ug/mlのニシン精巣DNA)中68℃で約2時間にわたり、膜をプレハイブリダイズさせた。製造元の指示書に従い、α−32P−dCTP(GE Healthcare社製)を伴う、Prime It II random Primerキット(Stratagene社製)を用いて、約1kbのMRF cDNA、CNP cDNA、PLP cDNA、またはGAPDH cDNAを含有するプラスミドから、放射性標識化したDNAプローブを生成させた。400gで2分間にわたり、Probe Quant G−50マイクロカラム(Amersham Biosciences社製)により、プローブをスピンし、組み込まれなかった32d−CTPを除去し、5分間にわたり95℃まで加熱した後、15mlのハイブリダイゼーション液中の膜に添加した。68℃で一晩にわたりハイブリダイゼーションを行い、1倍濃度SSC、0.1%SDS中、室温で2分間ずつ10回にわたり洗浄し、次いで、0.5倍濃度SSC、0.1%SDS中68℃で3分間ずつ10回にわたり洗浄した。次いで、X線フィルム(Amersham社製)を−80℃で膜に露光した後、現像した。沸騰する0.1倍濃度のSSC、0.1%SDS中で3回にわたる洗浄により膜をはがし、一晩にわたりフィルムを静置し、シグナルの不在について点検した後で、再プロービングした。
【0129】
(実施例7)
Affimetrix解析
Qiagen社製のオンカラム用DNアーゼ処理を用いるRNeasyマイクロキット(カリフォルニア州、バレンシア、Qiagen社製)により、細胞から全RNAを単離し、夾雑物であるゲノムDNAを除去した。Agilent 2100 Bioanalyzer(Agilent Technologies社製)を用いてRNAの完全性を評価し、NanoDrop ND−1000分光光度計(DE、ロックランド、NanoDrop社製)を用いて、RNA濃度を決定した。
【0130】
対照中におけるスパイクにより、Affymetrixによる2サイクルの標的標識化アッセイ(カリフォルニア州、サンタクララ、Affymetrix社製、900494型)を用いて、1ugの全RNAから、Affymetrix 3’アレイに対するハイブリダイゼーション用にビオチニル化したcRNAを調製した。製造元のプロトコールに従い、標識化したcRNAをフラグメント化し、マウスゲノム430 2.0アレイ(3’アレイ、Affymetrix社製、900495型)へとハイブリダイズした。
【0131】
Affymetrix GCOS 1.3ソフトウェアを用いて生の画像ファイルを処理し、個々のプローブセルの強度データを計算し、CELデータファイルを作製した。GCOSおよびMAS 5.0アルゴリズムを用いて、強度データをチップ当たりのデータに標準化して、標的強度のTGT値を500とし、個々のプローブセットに対する発現データおよび存在コール/不在コールを計算した。MAS 5.0アルゴリズムにより解析されるデータに対応する遺伝子記号および遺伝子名は、Affymetrix Netaffx Mouse430_2注釈ファイル(http://www.affymetrix.com/support/technical/byproduct.affx?product=moe430−20)に拠った。異常について生のDAT画像ファイルを検討することにより品質管理を実施し、各GeneChipアレイのバックグラウンド値が100未満であることを確認し、百分率による存在コールが、細胞型に適切であるかどうかモニタリングし、対照中におけるポリ(A)スパイク、ハウスキーピング遺伝子、およびハイブリダイゼーション対照を点検し、標識化とハイブリダイゼーションとの符合を確認した。
【0132】
(実施例8)
RT−PCR
上記に従い調製したRNAを、製造元の指示書に従い、Invitrogen社製Superscript III逆転写酵素を用いる逆転写にかけた。cDNAを、遺伝子特異的なプライマーにより、25または30サイクルにわたり増幅にかけ、2%アガロースゲル上において泳動した。
【0133】
(実施例9)
MRF条件付きノックアウトマウスの作製
MRFのエキソン8を、Pez−Frt−lox−DT標的化ベクターのSal1部位内にクローニングすることにより、エキソン8をloxPで挟んだマウスを作製した。5kbの5’アームおよび3kbの3’アームを、それぞれ、Not1部位およびXho1部位内へとクローニングし、E14ES胚性幹細胞内への相同組換えに対する標的化を可能とした。HindIIIにより消化したDNAに対するサザンブロット法、および5’loxP部位の挿入に対するPCR検証により、正確に標的化されたネオマイシン耐性クローンを同定した。標的化された細胞を線維芽細胞内へと注入し、キメラマウスを作製し、これらを、C57/B6マウスへと交配させ、ヘテロ接合マウスを作製した。これらのマウスを、FlpER株(Farleyら、Genesis、28巻、106〜110頁(2000年))へと交配させ、ネオマイシンカセットを欠失させ、第2世代のFlpER陰性マウスにおけるPCRによりこれを確認した。ヘテロ接合のMRFflox化マウスを、2世代にわたり、Olig2−Creマウス(Schullerら、近刊)またはCNP−Creマウス(Lappe−Siefkeら、Nat Genet、33巻、366〜374頁(2003年))へと交配させ、MRFfl/fl;Olig2wt/creマウスまたはMRFfl/fl;CNPwt/creマウスを得た。共通の上側プライマー(GGGAGGGGGCTTCAAGGAGTGT)、ならびに野生型の対立遺伝子を同定する下側プライマー(CCCCCAGCATGCCGATGTACAC)、およびflox化対立遺伝子を同定する下側プライマー(CCTTTCGCCAGGGGGATCTTG)を用いるPCRにより、マウスの遺伝子型を解析した。Creリコンビナーゼ陽性マウスは、プライマーGCTAAGTGCCTTCTCTACACCTGCおよびGGAAAATGCTTCTGTCCGTTTGにより同定した。
【0134】
(実施例10)
定量化および統計学
培養細胞に対してカウントを実施する場合(マーカーなどの発現を解析するため)、1条件当たり少なくとも3枚のカバースリップを盲検でカウントし、カバースリップ1枚当たり10ずつの視野(20倍の対物レンズ)をカウントした。各カバースリップの平均から、各条件についての平均およびSEMを計算し、多重比較についてのボンフェローニ補正を用いる、対応のない2方向のt検定により条件を比較した。示される実験は、少なくとも2つの個別の実験を表わす。組織断面から細胞を定量化する場合、1遺伝子型当たり4〜6匹ずつのマウスを用いた。解析(Photoshopにより実施する細胞カウント、およびImageJにより定量化するカウント領域の面積を伴う)用に60〜100umの距離を置いて断面を写真撮影(20倍の対物レンズ)することにより、マウス1匹当たり3枚ずつの断面を解析した。活性カスパーゼ3免疫陽性細胞を定量化する場合、20倍の対物レンズで、マウス1匹当たり3枚ずつ、60umの距離を置いた視神経縦断面について、免疫陽性細胞の数を定量化し、次いで、該神経を低倍率(4倍の対物レンズ)で写真撮影し、ImageJにより、該神経の面積を決定した。1遺伝子型当たり5〜6匹ずつのマウスを用いた。各マウスの平均から、各遺伝子型についての平均およびSEMを計算し、多重比較についてのボンフェローニ補正を用いる、対応のない2方向のt検定により遺伝子型を比較した。
【0135】
(実施例11)
CNS内におけるOL特異的な転写物である、GM98/MRFの同定
イムノパニング/FACSによる細胞精製法を、遺伝子プロファイリングと組み合わせて、マウスCNS内における細胞型特異性を示す遺伝子を同定した。急性精製した星状細胞、ニューロン、希突起膠細胞の前駆細胞(OPC)、新たに分化した希突起膠細胞(OL)、有髄化する成熟OLを転写プロファイルの生成に用いたスクリーンの一部として、遺伝子モデル98/ミエリン遺伝子調節因子を同定した。MRFについてのAffymetrixプローブセット(1439506_at型)は、MBP、PLP、およびMOGなど、典型的なOLマーカーが示したのと同様なレベルのOL濃縮を他の細胞型に対しても示し、ニューロンおよび星状細胞においては本質的に検出不能なレベルの発現であり、OPCにおいては低レベルの発現であり、OL内においては比較的強い発現であった。
【0136】
新たに分化したOL(GalC+、MOG−)におけるMRFの発現が、より成熟したOL(MOG+)と少なくとも同程度に高度であったことは、該遺伝子が、有糸分裂後細胞へと移行すると急速に誘導されることを示す(図1A)。全脳、全心臓、および培養OLから単離したRNAに対する、培養OLから増幅したMRF cDNAとのノーザンハイブリダイゼーションにより、脳内において、約5.5Kbのサイズの明白な転写物が確認されたが、これは、心臓に由来する試料中では不在であり、培養OL中では高度に濃縮されていた。確立されたOLマーカーであるプロテオリピドタンパク質(PLP)およびMRFに対するプローブを用いるin situハイブリダイゼーションにより、該2つの遺伝子について同一の発現パターンが確認され、MRFは脳内におけるすべての白質路内において発現し(図1C)、白質内では、PLPと同じ染色色素の細胞内分布を示した(図1D)。MRFの発現は、坐骨神経では検出されず(図10)、これにより、有髄化細胞内におけるMRFの発現が、CNSに制約されることが示された。ESTのデータベース(Unigene社製)は、MRFが他の組織、中でもとりわけ、膵臓および肺内において発現することを示すが、この発現はおそらく、CNS内で見られる発現よりも大幅に低度である。
【0137】
データベースの検索により、MRFは、ヒト遺伝子であるC11Orf9のマウスオルトログであることが明らかとなった。MRFおよびC11Orf9共に、推定のDNA結合ドメインとしてオンラインデータベースに列挙される酵母転写因子であるNdt80に対して相同的な領域を有する高分子タンパク質をコードする(Montanoら、Proc. Natl. Acad. Sci. U S A、99巻、14041〜14046頁(2002年))。OLから単離されたcDNAの配列決定により、GM98/MRFの転写物は、1139アミノ酸のタンパク質をコードすることが示された(図2A)。このタンパク質は、プロリンに富む複数のドメインを含有するN末端領域である、Ntd80様DNA結合領域と、複数の疎水性領域を含有するC末端領域とを含有する。ヒトC11Orf9とのこのタンパク質のアライメントにより、88.6パーセントの全体的相同性が明らかとなり、DNA結合領域内におけるこの相同性は100%であった(図2B)。C11Orf9は、そのC末端内における2つの疎水性領域が膜貫通ヘリックスとして作用する膜貫通タンパク質でありうることが既に示唆されている(Stohrら、Ctyogenet. Cell Genet.、88巻、211〜216頁(2000年))。MRFの細胞内における局在化を確立するため、HEK細胞内において、N末端にMycタグを付した融合体を発現させた。このMycタグを付したタンパク質は、明白な核内発現を示し(図2C)、培養した希突起膠細胞内において該タグを付したタンパク質が発現すると、核内局在化もまた見られた。N末端のMycタグを指向する抗Myc抗体、および該タンパク質のDNA結合領域に対して生成させた抗体によっても同じ細胞内における核内局在化が見られ、これにより、少なくともこれらの領域が、膜内局在化ではなく、核内局在化を示すことが示唆された。
【0138】
(実施例12)
MRFはin vitroにおける希突起膠細胞によるミエリン遺伝子の発現に必要である。
【0139】
MRFが、OLの転写調節において役割を果たすかどうかを確立するため、MRFのコード領域を標的とするsiRNAのプール(siMRF)、またはこれを標的としないsiRNAのプール(siCont)をトランスフェクトすることにより、OL培養物中におけるMRFの発現を遮断した。ノーザンブロットおよびRT−PCRにより評価したところ、siMRFトランスフェクト細胞は、siContトランスフェクト細胞と比べ、MRF mRNAに対する明白で一貫した下方調節を示し(図4A、図12)、これにより、siRNAのプールは、MRFレベルの低減に有効であることが示された。MRFに対するsiRNAを、増殖条件で培養されたOPCにトランスフェクトしたところ、その影響は見られず、大半のsiMRF細胞およびsiCont細胞がNG2陽性前駆細胞として分裂し続けたことは、急性摘出されたOPC内において見られるMRF発現の欠如と符合する。
【0140】
siRNAトランスフェクト細胞を、分化条件(−PDGF、+T3)へと移したところ、siContトランスフェクト細胞と、siMRFトランスフェクト細胞との間で差違が生じ始めた。分化条件への移動から24時間以内に、siMRFトランスフェクト細胞およびsiContトランスフェクト細胞共に、分裂を停止し、突起を拡大させ始めたのに対し、48時間以内に、siMRFトランスフェクト細胞は、siContトランスフェクト細胞より、膜シートの沈着範囲を狭め、また、カルセイン(Calcien)AM/EthHD1生存/死亡アッセイにより評価される生存率も、さほどではないが有意に低下させた(図3A、B)。分化の誘導から48時間以内に85%超が強くMBP陽性であったsiContトランスフェクト細胞とは対照的に、siMRFトランスフェクト細胞は、MBP発現の明白な遅延および低下を示し、分化の48時間後において、MBPについて陽性の細胞は23.6%に過ぎず、分化の96時間後においても、64.4%の細胞で、発現は低度に過ぎなかった(図3A、C)。
【0141】
これに反し、また、それらが単純な前駆細胞様形態から移行することとも符合するが、siMRFトランスフェクト細胞がやはり、siContトランスフェクト細胞に対して同様の割合で、OPCマーカーであるNG2の発現を下方調節したことは、有糸分裂後OLへの初期分化が、MRFのノックダウンによる影響を受けなかったことを強く示唆する。MRFの不在下において、後期OL遺伝子であるMOGの発現は、さらにより強く阻害され、分化の96時間後において、MOGを発現するsiMRFトランスフェクト細胞が5%未満であったの対し、この時点において、MOGについて陽性のsiCont細胞は81%であった(図3A、D)。48時間後および96時間後における、siMRFによる、マーカーMBPおよびMOGを発現する細胞の低減は、生存率の低下よりはるかに大幅であり(例えば、48時間の分化後において、siMRFトランスフェクト細胞が示す生存率は、siContを発現する細胞と比べて20.3%低下したに過ぎなかったが、MBPを発現する細胞の比率は、siCont発現細胞と比べて62.5%低下した)、siMRFトランスフェクト細胞において観察される細胞死の増大が、MBPおよびMOGの発現の喪失を説明するのに十分ではないことを強く示唆した。
【0142】
興味深いことに、位相顕微鏡法下において、siMRFトランスフェクト細胞はほとんど常に、OLの一般的な形態を帯びたが、突起および膜シートの増殖が、siContトランスフェクト細胞より大幅に阻害されることが典型的であり、また一方、MOG陰性細胞が、OLマーカーであるGalCを明白に発現した(図11)ことにより、細胞原基の特定化の変化が、表現型の原因ではないことが裏付けられた。
【0143】
(実施例13)
MRFの下流にある遺伝子の同定
MRFの不在下にあるOLの分化において見られる転写欠損をさらに特徴づけるため、また、MRFの下流にある遺伝子を同定するため、siMRFプールまたはsiContプールによるトランスフェクション後の48時間にわたり分化させたOLに由来するRNAのほか、分化前における遺伝子発現のベースラインを与える、培養OPCに由来するRNAも単離した。48時間後という時点を選択したのは、OL/ミエリン遺伝子の大半が、この段階までにある程度のレベルの誘導を示す(Dugasら、J. Neurosci.、26巻、10967〜10983頁(2006年))一方、siMRFトランスフェクト細胞が、siContトランスフェクト細胞と比べ、やはりほぼ同等レベルの生存を示す時点に依然としてあり、これにより、結果に対する細胞死の交絡による影響の可能性{Dugas、2006年、#9}が制限されるためである。全RNAは、mRNAの3’端を増幅するポリAプライマーを伴う2段階の直鎖状増幅プロトコールを用いて、標識化されたcRNAを作製するのに用いた。この標識化したcRNAを、20,832の固有の遺伝子を表わす45,037のオリゴヌクレオチドプローブセットによる、mRNAの3’端に相補的なオリゴヌクレオチドのプローブセット(3’アレイ)を含有する、Affymetrix Mouse 430 2.0マイクロアレイへとハイブリダイズさせた。また、ノーザンブロット解析も用いて、この時点におけるMRFおよびミエリン遺伝子(PLPおよびCNP)の下方調節も確認した(図4A)。
【0144】
公知のOPC発現遺伝子であるNG2およびPDGFRαの解析により、細胞を分化条件へと移すと、siRNAによりMRFの発現が遮断されていたかどうかに関わらず、これらの遺伝子は、OPCと比べ、本質的に検出不可能なレベルまで下方調節され(図4B)、これにより、MRFが、これらの条件におけるOPCからOLへの移行には必要でないことが裏付けられた。MRFのノックダウンによる、CNP1およびUgt8aなど、極めて初期のOL遺伝子の発現低下がごくわずかであったこともおそらく同様であり、これにより、MRFの不在下においても、OPCはOLへの移行を開始することが可能であり、汎OL系統のマーカーであるSox10は、siMRFのトランスフェクションによりまったく影響を受けなかったことが示唆される。これに対し、OLマーカーであるPLP1、MBP、およびMAGでは、MRFのノックダウンにより、siContトランスフェクト細胞と比べて極めて明白な低下(約80%の)が見られ、後期OL遺伝子であるトランスフェリン、MOBP、およびMOGについては、ほぼ完全な低下(>90%)が見られた(図4C)。培養物中において2型星状細胞の原基を帯びるOPCの比率が小さい(>5%)ため、血清の不在下においても、分化条件へと移すことによる誘導は中程度であったが、MRFのノックダウンにより、おそらくBMPによるシグナル伝達の下流にある星状膠細胞遺伝子であるGFAP、S100b、およびアクアポリン4だけがわずかながらの上昇を示した(siCont値のそれぞれ147%、255%、および153%)ことにより、大部分の細胞は、MRF発現の不在下においてGFAP陰性を維持し、星状細胞系統への分化が、これらの細胞によるOL/ミエリン遺伝子の発現の喪失を説明するわけではないというかつての観察が裏付けられる。
【0145】
MRFの下流にある遺伝子を同定するため、siContとsiMRRとの間で、抑制レベルにより遺伝子を序列化した。MRFのノックダウンにより、104の遺伝子を表わす128のプローブセットが、4分の1<に抑制された。MRFのノックダウンによる抑制レベルが最も大きな50の遺伝子を、図5に示す(図14もまた参照されたい)。したがって、siMRFにより阻害されることが示される、128のプローブセットのうちの104はまた、OPC試料と、2日間にわたり分化させたOL試料との間で>4倍に上方調節されることが示されるプローブセットでもあり、これにより、MRFに依存する遺伝子の大半は通常、OPCからOLへの移行時において調節される遺伝子であることが示された。しかし、逆に、分化により>4倍に誘導される793のプローブセットのうち、siMRFにより強く阻害されたのは104だけであったことにより、OLの発生時において通常調節される遺伝子の大部分が、MRF発現には依存しないことが示唆される(図4D)。
【0146】
(実施例14)
MRFの強制発現はin vitroにおけるOLの分化を誘導する
MRFが、OL/ミエリン遺伝子の発現を誘導するのに十分であるかどうかを評価するため、OPCに、MRF発現構築物(pCMV−Sport6−MRF)、またはEGFPをコードする対照のGFPプラスミドをトランスフェクトして、大半の細胞が通常、分裂するOPCであることを維持し、自発的分化は低レベルにとどまる増殖条件(+PDGF、−T3)へと播種した。トランスフェクションの2日後において、対照のトランスフェクト細胞が示した分化は低レベルであり、MBP陽性およびMOG陽性とカウントされた生細胞は、それぞれ、1.4パーセントおよび0.4パーセントに過ぎなかった。これに対し、MRF発現構築物をトランスフェクトした細胞が、MBPおよびMOGについて、それぞれ、32.8パーセントおよび41.0パーセントの陽性であったことは、分化率が、対照ベクターをトランスフェクトした細胞と比べて約30〜40倍に上昇した(p<0.001)ことを示す。MBPおよびMOGのこの誘導は、MRFトランスフェクト細胞が、OPCマーカーであるNG2を下方調節することにより反映された(図6)。トランスフェクションの5日後までに、MRFトランスフェクト細胞の大半がMBP陽性およびMOG陽性(それぞれ、81.9および77.2%)となったのに対し、対照トランスフェクト細胞の大半は、依然としてMBP/MOG陰性OPCであった(図6)。MRFが異所性発現したMBP陽性細胞およびMOG陽性細胞が、ほとんど均一に成熟OLの一般的形状(突起の高度な分岐および膜シート)を示したことにより、OPC内におけるOL/ミエリン遺伝子の異所性発現ではなく、MRFの異所性発現が、OLへの該細胞の分化を引き起こしたことを強く示唆する。
【0147】
(実施例15)
MRFの強制発現はin vivoにおけるMBPの発現を誘導する
MRFの発現が、in vivoにおけるミエリン遺伝子の発現を駆動するのに十分でありうるかどうかを評価するため、MRF発現構築物(pCAGGS−MRF)を、E3において、ニワトリ脊髄内へと電気穿孔し、ニワトリの脊髄内には通常検出可能なMBPの発現または希突起膠細胞の分化が見られない発生時点であるE8において該胚を屠殺した。MRF発現構築物(共電気穿孔されたEGFP発現プラスミドからのEGFPの発現により同定される)を電気穿孔した脊髄側方内では、偶発的なMBP陽性細胞を見出すことができたが、強いMBP陽性細胞が見出されるのは、断面1枚当たり1〜3個だけであることが典型的であった(図7A、B)。電気穿孔されなかった、対照の脊髄側方において、MBP陽性細胞は観察されなかった。しかし、電気穿孔された脊髄側方におけるMBP陽性細胞の数が、EGFPまたはMFR導入遺伝子を発現する細胞の数よりはるかに少なかった(図7A)ことは、MRFが希突起膠細胞の分化を促進する能力には、他の陽性または陰性の調節因子が影響している可能性が高いことを示すことに注目すべきである。
【0148】
(実施例16)
MRFはマウスにおけるCNSの有髄化に必要である
CNSの有髄化におけるMRFの必要性を評価するため、MRF遺伝子のエキソン8をloxP部位で挟んだマウスを作製した(図8)。エキソン8は、MRFのDNA結合領域の一部をコードする。このエキソンの欠失は、フレームシフトに起因する結果として、DNA結合領域および下流のタンパク質の喪失をもたらすことが予測された(図8C)。エキソン8の細胞特異的欠失または組織特異的欠失は、loxPフランキングマウス(MRFfl/fl)を、対象の細胞型においてCreリコンビナーゼを発現するマウスと交配させることにより実施することができる。MRFfl/flマウスを、Olig2プロモーターの下流でCreを発現するマウス株へと交配させた(結果として、希突起膠細胞および下肢運動ニューロン前駆細胞、OPCおよび成熟OLにおけるCre発現がもたらされた)ところ、結果として得られるMRF条件付きノックアウトマウス(MRFfl/fl;Olig2wt/Cre)は、メンデル頻度で生誕し、生後10日間は、それらの対照同腹子(MRFwt/fl;Olig2wt/cre、およびMRFfl/fl;Olig2wt/wt)から明確には識別できなかった。しかし、P11から、MRF条件付きノックアウト動物は、振戦および発作を発生させたため、それらの同腹子から識別されるようになった。その後数日間にわたり、条件付きノックアウト動物は発作を発生させ、生後3週目のP13〜P17において例外なく死亡した。この表現型は、CNSのミエリンに影響のある他の突然変異体の表現型と符合する。
【0149】
P13における条件付きノックアウトマウスの脳および脊髄に対する免疫組織化学解析は、NeuNおよびGFAPの染色は正常だと考えられ、CNSの全体的構造には影響がないことを示した。これに対し、脳内のMBP染色には大幅な喪失が見られ、対照マウスの白質路内におけるほぼ遍在的なMBP染色と比較して、白質路内で見られるMBP+希突起膠細胞は、偶発的なものに過ぎなかった(図9A)。これらのまれに残存するMBP+ OLは、Creを介する組換えを受けなかった、偶発的なOlig2非依存性細胞を表わす可能性がある。その脊髄根(シュワン細胞により有髄化される)は依然として強くMBP染色されていたが、条件付きノックアウト動物の脊髄内におけるこのMBP染色の喪失は本質的に完全であった(図9B)。このMBP発現の喪失は、脊髄溶解物に対するウェスタンブロットにより確認され、また、これにより、著明に低度のCNP発現、およびMOG発現の完全な喪失も示された(図9C)。条件付きノックアウト動物における星状細胞およびニューロンのタンパク質(それぞれ、GFAPおよびニューロフィラメント)には影響がなかった。白質路内におけるフルオロミエリン染色により、本質的に完全な再有髄化の喪失が明白に立証された(脊髄側柱の白質について示された、図9D)。これに対し、末梢神経は、予測される通り、条件付きノックアウト動物および対照動物のいずれにおいても同等に有髄化されたが、これは、シュワン細胞が、MRFもOlig2も発現しないからである。条件付きマウスの視神経では、電子顕微鏡法により、有髄化の喪失が確認された;対照マウスが、P13までに、視神経における有髄化の明白な証拠を示しつつあったのに対し、条件付きマウスにおいては、神経の有髄化が観察されなかった(図9E)。MRFflマウスを、CNPプロモーターの下流でCreリコンビナーゼを発現するマウスと交配させたところ、同等の表現型(振戦、発作、ならびにミエリン遺伝子の発現および有髄化の喪失)が見られたことにより、条件付きノックアウト動物の表現型は、Olig2−cre対立遺伝子の挿入により引き起こされるOlig2ヘテロ接合によるバックグラウンドには依存しないことが確認された(図16)。これらの知見は、MRFが、in vivoにおいて希突起膠細胞が発生してCNSが有髄化する正常な過程に必要であることを裏付ける。
【0150】
(実施例17)
OLはMRF条件付きノックアウトマウスにおいて分化するが、その後、成熟するとアポトーシスを受ける
MRF条件付きノックアウトマウスにおけるミエリンの欠如が、OL生成の欠如により引き起こされたのか、そうではなくて、有髄化が不可能なOLにより引き起こされたのかを見極めるため、OL系統の各段階の発生に対するMRF欠失の効果を評価した。MBP、成熟OLマーカーであるCC1、OPCマーカーであるNG2およびPDGFRα、および汎OL/星状細胞マーカーであるOlig2を含めた各種のマーカーについて免疫染色した、条件付きノックアウトマウスおよび対照マウスに由来する、P13の視神経断面におけるOPCおよびOLの密度を評価した(図13A)。視神経内における、Olig2免疫陽性核密度のカウントにより、条件付きノックアウト動物の神経における陽性細胞密度の著明な低下(約45%の)が示された;この低下は、条件付きノックアウトマウスにおいて見られるCC1免疫陽性細胞の完全な喪失と本質的に符合した(図13B、C、E)。これに対し、条件付きノックアウトマウスにおいても、PDGFRαに対して免疫陽性であるOlig2+細胞の密度は、わずかな影響を受けただけであり(P<0.05のt検定において、MRFfl/fl;Olig2wt/wtマウスと比べ、条件付きノックアウト動物では18.7%の低下であったが、MRFwt/fl;Olig2wt/Cre対照との有意差は見られなかった;図13D、E)、これにより、条件付きノックアウト動物においてもOPC段階が受ける影響は最小限であることが示され、この観察は、見かけ上正常なNG2染色により裏付けられた。同様に、CC1およびPDGFRαのいずれについても陰性のOlig2+細胞(おそらく、Olig2を発現する星状細胞)の密度は、遺伝子型間において同様であった(図9E)。条件付きノックアウトマウスの視神経におけるMBP染色は、対照マウスと比べて大幅に低下したが、にもかかわらず、低度のMBP+(CC1−であるが)細胞は少数ながら存在したことは興味深く、これにより、条件付きノックアウトマウスにおいて、ごく最小限であれ、少数の有糸分裂後OLが生成されたことが示唆される。これらの細胞の密度は極めて低く(細胞35±4個/mm2);対照マウスにおけるCC1染色により測定される通り、成熟OL密度の約4%に過ぎなかった。条件付きノックアウトマウスにおけるMBP+ OLおよびCC1+ OLの喪失は、これらのマーカーの喪失が、上記のsiRNA実験においてMRF発現の下流には存在しないことが判明しているマーカーであるOlig2の陽性細胞密度の大幅な低下と符合したことを踏まえるなら、MRF欠損OLが、これらのマーカーを発現できないことに加えて、成熟OLの喪失も反映する可能性が高い。まとめると、これらのデータにより、MRF条件付きノックアウト動物では、OPC集団が大部分影響を受けない一方、成熟OL集団は大幅かつ選択的に喪失されるという証拠がもたらされる。
【0151】
条件付きノックアウトマウスにおいて見られる有糸分裂後OLが極めて少数であることは、OPCの分化に対する遮断に起因する可能性もあり、代替的に、それらの分化直後におけるOLの喪失に起因する可能性もある。MRFを欠くOPCがマイトジェンを引き込むと、ミエリン遺伝子を発現することはないが、典型的なOL形状を有するGC+有糸分裂後細胞へと分化しうることを示す、上記のin vitroにおけるsiRNA実験を踏まえるなら、前者の可能性は低いと考えられる。OL死滅の可能性は、その一部がアポトーシス細胞に特徴的な、ブレブ形成し、凝縮またはフラグメント化した核を示す、低度のMBP陽性細胞が、条件付きノックアウト動物の視神経において存在する(図13A、矢印)ことにより示唆された。P13において観察される有糸分裂後OLの欠如が、それらが発生した後におけるOLのアポトーシスの増大により説明しうるかどうかを確立するため、P10の条件付きノックアウトマウスおよび対照マウスを、アポトーシスマーカーである活性化カスパーゼ3に対する抗体により染色した。すべての遺伝子型の視神経において、核が凝縮またはフラグメント化されることが多い、活性化カスパーゼ3免疫陽性細胞が存在したことは、視神経において新たに発生したOLの約半数による発生喪失についての先行報告と符合する。条件付きノックアウトマウスは、それらの多くが、条件付きノックアウト動物の神経において観察される低度のMBP+細胞に対応する、活性化カスパーゼ3陽性細胞である、これらのアポトーシス細胞の密度において、統計学的に有意な約2倍の増大を示した(図17)。これらのデータにより、MRF条件付きノックアウトマウスにおいて、OLは発生するが、その後速やかにアポトーシスを受けることが示される。
【0152】
(実施例18)
MRF条件付きノックアウト動物における有髄化不全は細胞自律的であり、細胞死だけに起因するわけではない
条件付きノックアウトマウスにおけるOLのアポトーシスは、これらのマウスにおいて見られる有髄化不全が、OLの喪失に起因したのか、ミエリン遺伝子を発現できないことに起因したのか、またはこれらの両方であったのかを評価することを困難にした。この問題を明らかにするため、P7における対照マウスおよび条件付きマウスから高度に精製したOPC培養物を作製した。イムノパニングを用いて、対照動物および条件付きノックアウト動物の脳から、同等数のPDGFRα+ OPC(約100万個/脳)を単離することができた。増殖条件において、対照マウスに由来するOPC、および条件付きノックアウトマウスに由来するOPCのいずれも、NG2陽性細胞およびKi67陽性細胞として増殖したことは、MRFが、OL系統のOPC期に必要でないことを裏付けた(図15A)。
【0153】
マイトジェンを引き込むと、対照細胞および条件付きノックアウト細胞のいずれもが、Ki67を下方調節し、CNPに対する抗体により染色したところ、OLの形状を帯び(図15B)、ノックアウト細胞も、それらのin vivoにおける対応物とは異なり、分化の4日後において細胞死をほとんど示さなかった。しかし、ノックアウト細胞には、対照細胞と比較して、基質上における膜シートの拡大をはるかに狭める傾向があった。培養物中では高度の生存率を有するにもかかわらず、条件付きノックアウト動物による培養物が、MBPに対する抗体による染色を示さなかった(図15B)ことは、ミエリン遺伝子発現の喪失が、単に細胞死に後続するのではないことを示した。対照動物および条件付きノックアウト動物の脊髄およびOL培養物からRNAを単離し、RT−PCRおよび遺伝子アレイにより解析したところ、MAGおよびMOBPなど、多くのミエリン遺伝子の発現が、条件付きノックアウト動物の脊髄およびOL培養物のいずれにおいても消失したことは、これらの遺伝子の発現に対するMRFの必要性を示す(図14、図18)。予測される通り、MRFの欠損は、in vivoにおけるOL死滅の誘導に影響を及ぼすため、MRFヌルの脊髄では、MRF依存性OL遺伝子およびMRF非依存性OL遺伝子すべての発現の喪失が観察された。in vitroにおけるOL遺伝子の発現に対するMRF欠損の効果は、siMRFノックダウンについて観察される効果(図5)と極めて類似し、予測される通り、大半のミエリン遺伝子および多くのOL成熟遺伝子が強く下方調節され、またこれには、Ugt8およびCldn11など、複数の初期OL遺伝子に対するMRF欠損の効果も含まれた。その発現が通常、新たに形成された希突起膠細胞による一過性発現に限定される、p57/cdkn1cなど、複数の遺伝子が、in vitroのMRF欠損OLにおいて著明に上方調節されたことは、MRF欠損OLが、依然として大半のミエリン遺伝子を発現する前の成熟の初期段階にある、分化の直前に失速する可能性が高いことを示す。
【0154】
(実施例19)
MRFの発現による脱髄の阻害
MRF発現が誘導可能なトランスジェニックマウスを用いて、該動物におけるMRFの発現または活性の上昇により、脱髄が阻害されることを示す。PDGFRα−CreERTマウスを、CMV−lox−stop−lox−MRFマウスと交配する結果として、PDGFRα−CreERT/CMV−lox−stop−lox−MRFマウスをもたらすことにより、誘導可能なMRF発現を含むトランスジェニックマウスを作製した。PDGFRα−CreERT/CMV−lox−stop−lox−MRFマウスは、lox部位により隣接する上流の終止コドンのため、また、Creリコンビナーゼ変異体であるCreERTが、flox化終止コドンに対して作用できないため、導入遺伝子からMRFを発現することがない。しかし、タモキシフェンの投与によりCreERTの活性を誘導すると、したがって、該マウスにタモキシフェンを投与すると、該終止コドンが切断され、MRFが発現される。
【0155】
この実験では、被験群である1つの群にはタモキシフェンを投与し、対照群である第2の群にはタモキシフェンを投与しない、PDGFRα−CreERT/CMV−lox−stop−lox−MRF動物の2つの群を用いた。いずれの群にも、クプリゾンを施して、脱髄を誘導した。ミエリン特異的遺伝子についての免疫組織化学による遺伝子発現の検出、または軸索についての電子顕微鏡法などにより、両方の群を脱髄について解析した。
【0156】
タモキシフェンを投与された被験群のマウスは、対照群と比較して、より低い程度または度合の脱髄を示す。
【0157】
(実施例20)
MRF発現による再有髄化の促進
この実験では、PDGFRα−CreERT/CMV−lox−stop−lox−MRF動物の2つの群を用いて、MRFの発現または活性の上昇による再有髄化の促進を示す。いずれの群にも、クプリゾンを施して、脱髄を誘導した。被験群である1つの群にはタモキシフェンを投与する一方、対照群である第2の群にはタモキシフェンを投与しない。ミエリン特異的遺伝子についての免疫組織化学による遺伝子発現の検出、または軸索についての電子顕微鏡法などにより、両方の群を脱髄について解析した。タモキシフェンを投与された被験群のマウスは、対照マウスより迅速に、またはより頑健に再有髄化することが可能である。
【0158】
(実施例21)
in vitroにおける生物活性剤のスクリーニング
神経膠細胞株を用いて、再有髄化を促進するか、または脱髄を阻害する生物活性剤をスクリーニングする。マウスからOPCを得、候補生物活性剤により処理する。OPCを、MRFの発現について解析し、候補生物活性剤を投与されなかったOPCと比較した。候補生物活性剤を投与されたOPCが、これを投与されなかったOPCと比較して、より高レベルのMRF発現を示す場合は、該候補生物活性剤を、実施例22に記載の動物モデルにおける試験など、さらなる解析用に選択する。
【0159】
また、ミエリン特異的遺伝子についての免疫組織化学または遺伝子発現の検出によって、細胞を解析することもできる。候補生物活性剤を投与された細胞が、該候補生物活性剤を投与されなかった細胞と比較して、ミエリン特異的遺伝子の発現または軸索の有髄化の増大を示す場合は、該候補生物活性剤を、実施例22に記載の動物モデルにおける試験など、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。
【0160】
(実施例22)
in vivoにおける生物活性剤のスクリーニング
実施例16に記載のMRF条件付きノックアウトマウス(MRFfl/fl;Olig2wt/Cre)を用いて、生物活性剤をスクリーニングする。MRF条件付きノックアウトマウス(MRFfl/fl;Olig2wt/Cre)は、P11において、振戦および発作を発生させる。このスクリーニングでは、P11以前に、MRF条件付きノックアウトマウスに候補生物活性剤を投与する。該動物が、振戦もしくは発作を発生させないか、または発生させる振戦もしくは発作が、該候補生物活性剤を投与されなかったMRF条件付きノックアウトマウスと比較して、それほど重度でない場合は、該候補生物活性剤を、再有髄化を促進するか、または脱髄を阻害する治療剤としてのさらなる開発用に選択する。また、ミエリン特異的遺伝子についての免疫組織化学もしくは遺伝子発現の検出、または軸索についての電子顕微鏡法などによっても、動物を解析し、候補生物活性剤を投与された動物が、該候補生物活性剤を投与されなかった動物と比較して、ミエリン特異的遺伝子の発現または軸索の有髄化の増大を示す場合は、該候補生物活性剤を、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。
【0161】
第2のスクリーニングでは、MRF条件付きノックアウトマウスが、振戦または発作を示した後において、すなわち、約P11以後において、該動物に候補生物活性剤を投与する。該候補生物活性剤を投与されなかったMRF条件付きノックアウトマウスと比較して、振戦または発作の程度が低下するなど、該動物の状態が改善される場合は、該候補生物活性剤を、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。また、ミエリン特異的遺伝子についての免疫組織化学もしくは遺伝子発現の検出、または軸索についての電子顕微鏡法などによっても、動物を解析し、候補生物活性剤を投与された動物が、該候補生物活性剤を投与されなかった動物と比較して、ミエリン特異的遺伝子の発現または軸索の有髄化の増大を示す場合は、該候補生物活性剤を、脱髄を阻害するか、または再有髄化を促進する治療剤としてのさらなる開発用に選択する。
【0162】
本明細書では、本発明の好ましい実施形態を示し、説明してきたが、このような実施形態が例示だけを目的とするものであることは、当業者には明らかであろう。本発明から逸脱しない限りにおいて、今や、多くの変更、変化、および置換が当業者に生じるであろう。本発明を実施する際に、本明細書で記載される本発明の実施形態に対する各種の代替物を用いうることを理解されたい。本明細書における特許請求の範囲が、本発明の範囲を規定し、これらの特許請求の範囲が、これらおよびこれらの同等物の範囲内にある方法および構造を対象とすることが意図される。
【特許請求の範囲】
【請求項1】
MRFまたはその機能的な変異体をコードする核酸配列に作動的に連結された細胞型特異的な発現調節エレメントを含む、単離核酸分子。
【請求項2】
前記細胞型が神経細胞である、請求項1に記載の単離核酸分子。
【請求項3】
前記神経細胞が神経膠細胞である、請求項2に記載の単離核酸分子。
【請求項4】
前記神経膠細胞が、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞である、請求項3に記載の単離核酸分子。
【請求項5】
前記神経細胞特異的な調節エレメントが、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来する、請求項1に記載の単離核酸分子。
【請求項6】
前記細胞型特異的な調節エレメントが、誘導的プロモーターまたは構成的プロモーターである、請求項1に記載の単離核酸分子。
【請求項7】
請求項1、2、3、4、5、または6に記載の前記単離核酸分子を含むベクター。
【請求項8】
請求項1、2、3、4、5、または6に記載の前記単離核酸分子を含む宿主細胞。
【請求項9】
MRF導入遺伝子を含むトランスジェニック動物。
【請求項10】
前記MRF導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項9に記載のトランスジェニック動物。
【請求項11】
前記MRF導入遺伝子が、エキソンの欠失を含む、請求項9に記載のトランスジェニック動物。
【請求項12】
前記エキソンが、エキソン8である、請求項11に記載のトランスジェニック動物。
【請求項13】
前記MRF導入遺伝子が、リコンビナーゼ部位に隣接する、請求項11に記載のトランスジェニック動物。
【請求項14】
前記動物が、リコンビナーゼ導入遺伝子を含む、請求項9に記載のトランスジェニック動物。
【請求項15】
前記リコンビナーゼ導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項15に記載のトランスジェニック動物。
【請求項16】
前記リコンビナーゼが、CreリコンビナーゼまたはFlpである、請求項13または14に記載のトランスジェニック動物。
【請求項17】
前記細胞型が神経細胞である、請求項10または15に記載のトランスジェニック動物。
【請求項18】
前記神経細胞が神経膠細胞である、請求項17に記載のトランスジェニック動物。
【請求項19】
前記神経膠細胞が、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞である、請求項18に記載のトランスジェニック動物。
【請求項20】
前記神経細胞特異的な調節エレメントが、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来する、請求項17に記載のトランスジェニック動物。
【請求項21】
前記細胞型特異的な調節エレメントが、誘導的プロモーターまたは構成的プロモーターである、請求項10または15に記載のトランスジェニック動物。
【請求項22】
前記動物が哺乳動物である、請求項9に記載のトランスジェニック動物。
【請求項23】
前記動物がマウスまたはラットである、請求項9に記載のトランスジェニック動物。
【請求項24】
被験体における神経障害を治療するための組成物であって、前記被験体におけるMRF活性を調節する生物活性剤を含む組成物。
【請求項25】
被験体における再有髄化を促進するための組成物であって、前記被験体におけるMRF活性を調節する生物活性剤を含む組成物。
【請求項26】
幹細胞の希突起膠細胞への分化を促進するための組成物であって、前記幹細胞におけるMRF活性を調節する生物活性剤を含む組成物。
【請求項27】
希突起膠細胞の分化を誘導する第2の生物活性剤をさらに含む、請求項24、25、または26に記載の組成物。
【請求項28】
前記第2の生物活性剤が、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進する、請求項27に記載の組成物。
【請求項29】
前記神経障害が脱髄を含む、請求項24に記載の組成物。
【請求項30】
前記神経障害が多発性硬化症を含む、請求項24に記載の組成物。
【請求項31】
前記生物活性剤が、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子である、請求項24、25、または26に記載の組成物。
【請求項32】
被験体における神経障害を治療する方法であって、前記被験体に、治療有効量の、MRF活性を調節する生物活性剤を投与するステップを含む方法。
【請求項33】
被験体における再有髄化を促進する方法であって、前記被験体に、治療有効量の、MRF活性を調節する生物活性剤を投与するステップを含む方法。
【請求項34】
幹細胞の希突起膠細胞への分化を促進する方法であって、MRF活性を調節し、これにより、前記幹細胞の希突起膠細胞への分化を促進する生物活性剤を、前記幹細胞に導入するステップを含む方法。
【請求項35】
希突起膠細胞への分化を誘導する第2の生物活性剤を投与または導入するステップをさらに含む、請求項32、33、または34に記載の方法。
【請求項36】
前記第2の生物活性剤を投与または導入するステップが、MRFの活性を調節する前記生物活性剤を投与するステップの前であるか、これと同時であるか、またはこの後である、請求項35に記載の方法。
【請求項37】
前記第2の生物活性剤が、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進する、請求項35に記載の方法。
【請求項38】
前記第2の生物活性剤を投与するステップが、相乗効果を有する、請求項35に記載の方法。
【請求項39】
前記神経障害が脱髄を含む、請求項32に記載の方法。
【請求項40】
前記神経障害が多発性硬化症である、請求項32に記載の方法。
【請求項41】
前記生物活性剤が、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子である、請求項32、33、または34に記載の方法。
【請求項42】
前記被験体が、マウス、ラット、またはヒトである、請求項32または33に記載の方法。
【請求項43】
MRF活性を調節するのに有効な候補生物活性剤をスクリーニングする方法であって、
(a)被験細胞を、前記候補生物活性剤と接触させるステップと、
(b)対照動物と比較したMRFの発現レベルの変化についてアッセイするステップと
を含む方法。
【請求項44】
動物における有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法であって、
(a)候補生物活性剤を動物に投与するステップと、
(b)対照動物と比較したMRFの発現レベルの上昇についてアッセイするステップであって、ここで、前記上昇は、前記生物活性剤が前記動物における有髄化を促進することを示すステップと
を含む方法。
【請求項45】
動物における再有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法であって、
(a)候補生物活性剤を、MRF導入遺伝子を含む動物に投与するステップ;
(b)対象動物と比較した、表1中の少なくとも1つの遺伝子の発現レベルの上昇についてアッセイするステップであって、ここで、前記上昇は、前記生物活性剤が前記動物における再有髄化を促進することを示すステップ;および/または
(c)対象動物と比較した、前記動物における有髄化の変化を観察するステップ
を含む方法。
【請求項46】
前記MRF導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項44に記載の方法。
【請求項47】
前記MRF導入遺伝子が、エキソンの欠失を含む、請求項44に記載の方法。
【請求項48】
前記エキソンが、エキソン8である、請求項46に記載の方法。
【請求項49】
前記MRF導入遺伝子が、リコンビナーゼ部位に隣接する、請求項44に記載の方法。
【請求項50】
前記リコンビナーゼ部位が、loxPまたはFRTである、請求項48に記載の方法。
【請求項51】
前記動物が、リコンビナーゼ導入遺伝子を含む、請求項44に記載の方法。
【請求項52】
前記リコンビナーゼ導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項50に記載の方法。
【請求項53】
前記リコンビナーゼが、CreリコンビナーゼまたはFlpである、請求項48または50に記載の方法。
【請求項54】
前記細胞型が神経細胞である、請求項45に記載の方法。
【請求項55】
前記神経細胞が神経膠細胞である、請求項53に記載の方法。
【請求項56】
前記神経膠細胞が、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞である、請求項54に記載の方法。
【請求項57】
前記神経細胞特異的な調節エレメントが、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来する、請求項54に記載の方法。
【請求項58】
前記細胞型特異的な調節エレメントが、誘導的プロモーターまたは構成的プロモーターである、請求項45に記載の方法。
【請求項59】
前記動物がマウスである、請求項44または44に記載の方法。
【請求項60】
前記生物活性剤が、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子である、請求項42、43、または44に記載の方法。
【請求項1】
MRFまたはその機能的な変異体をコードする核酸配列に作動的に連結された細胞型特異的な発現調節エレメントを含む、単離核酸分子。
【請求項2】
前記細胞型が神経細胞である、請求項1に記載の単離核酸分子。
【請求項3】
前記神経細胞が神経膠細胞である、請求項2に記載の単離核酸分子。
【請求項4】
前記神経膠細胞が、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞である、請求項3に記載の単離核酸分子。
【請求項5】
前記神経細胞特異的な調節エレメントが、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来する、請求項1に記載の単離核酸分子。
【請求項6】
前記細胞型特異的な調節エレメントが、誘導的プロモーターまたは構成的プロモーターである、請求項1に記載の単離核酸分子。
【請求項7】
請求項1、2、3、4、5、または6に記載の前記単離核酸分子を含むベクター。
【請求項8】
請求項1、2、3、4、5、または6に記載の前記単離核酸分子を含む宿主細胞。
【請求項9】
MRF導入遺伝子を含むトランスジェニック動物。
【請求項10】
前記MRF導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項9に記載のトランスジェニック動物。
【請求項11】
前記MRF導入遺伝子が、エキソンの欠失を含む、請求項9に記載のトランスジェニック動物。
【請求項12】
前記エキソンが、エキソン8である、請求項11に記載のトランスジェニック動物。
【請求項13】
前記MRF導入遺伝子が、リコンビナーゼ部位に隣接する、請求項11に記載のトランスジェニック動物。
【請求項14】
前記動物が、リコンビナーゼ導入遺伝子を含む、請求項9に記載のトランスジェニック動物。
【請求項15】
前記リコンビナーゼ導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項15に記載のトランスジェニック動物。
【請求項16】
前記リコンビナーゼが、CreリコンビナーゼまたはFlpである、請求項13または14に記載のトランスジェニック動物。
【請求項17】
前記細胞型が神経細胞である、請求項10または15に記載のトランスジェニック動物。
【請求項18】
前記神経細胞が神経膠細胞である、請求項17に記載のトランスジェニック動物。
【請求項19】
前記神経膠細胞が、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞である、請求項18に記載のトランスジェニック動物。
【請求項20】
前記神経細胞特異的な調節エレメントが、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来する、請求項17に記載のトランスジェニック動物。
【請求項21】
前記細胞型特異的な調節エレメントが、誘導的プロモーターまたは構成的プロモーターである、請求項10または15に記載のトランスジェニック動物。
【請求項22】
前記動物が哺乳動物である、請求項9に記載のトランスジェニック動物。
【請求項23】
前記動物がマウスまたはラットである、請求項9に記載のトランスジェニック動物。
【請求項24】
被験体における神経障害を治療するための組成物であって、前記被験体におけるMRF活性を調節する生物活性剤を含む組成物。
【請求項25】
被験体における再有髄化を促進するための組成物であって、前記被験体におけるMRF活性を調節する生物活性剤を含む組成物。
【請求項26】
幹細胞の希突起膠細胞への分化を促進するための組成物であって、前記幹細胞におけるMRF活性を調節する生物活性剤を含む組成物。
【請求項27】
希突起膠細胞の分化を誘導する第2の生物活性剤をさらに含む、請求項24、25、または26に記載の組成物。
【請求項28】
前記第2の生物活性剤が、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進する、請求項27に記載の組成物。
【請求項29】
前記神経障害が脱髄を含む、請求項24に記載の組成物。
【請求項30】
前記神経障害が多発性硬化症を含む、請求項24に記載の組成物。
【請求項31】
前記生物活性剤が、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子である、請求項24、25、または26に記載の組成物。
【請求項32】
被験体における神経障害を治療する方法であって、前記被験体に、治療有効量の、MRF活性を調節する生物活性剤を投与するステップを含む方法。
【請求項33】
被験体における再有髄化を促進する方法であって、前記被験体に、治療有効量の、MRF活性を調節する生物活性剤を投与するステップを含む方法。
【請求項34】
幹細胞の希突起膠細胞への分化を促進する方法であって、MRF活性を調節し、これにより、前記幹細胞の希突起膠細胞への分化を促進する生物活性剤を、前記幹細胞に導入するステップを含む方法。
【請求項35】
希突起膠細胞への分化を誘導する第2の生物活性剤を投与または導入するステップをさらに含む、請求項32、33、または34に記載の方法。
【請求項36】
前記第2の生物活性剤を投与または導入するステップが、MRFの活性を調節する前記生物活性剤を投与するステップの前であるか、これと同時であるか、またはこの後である、請求項35に記載の方法。
【請求項37】
前記第2の生物活性剤が、Sox10、Nkx2.2、Olig1、Olig2、またはこれらの組合せの活性を促進する、請求項35に記載の方法。
【請求項38】
前記第2の生物活性剤を投与するステップが、相乗効果を有する、請求項35に記載の方法。
【請求項39】
前記神経障害が脱髄を含む、請求項32に記載の方法。
【請求項40】
前記神経障害が多発性硬化症である、請求項32に記載の方法。
【請求項41】
前記生物活性剤が、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子である、請求項32、33、または34に記載の方法。
【請求項42】
前記被験体が、マウス、ラット、またはヒトである、請求項32または33に記載の方法。
【請求項43】
MRF活性を調節するのに有効な候補生物活性剤をスクリーニングする方法であって、
(a)被験細胞を、前記候補生物活性剤と接触させるステップと、
(b)対照動物と比較したMRFの発現レベルの変化についてアッセイするステップと
を含む方法。
【請求項44】
動物における有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法であって、
(a)候補生物活性剤を動物に投与するステップと、
(b)対照動物と比較したMRFの発現レベルの上昇についてアッセイするステップであって、ここで、前記上昇は、前記生物活性剤が前記動物における有髄化を促進することを示すステップと
を含む方法。
【請求項45】
動物における再有髄化を促進するのに有効な候補生物活性剤をスクリーニングする方法であって、
(a)候補生物活性剤を、MRF導入遺伝子を含む動物に投与するステップ;
(b)対象動物と比較した、表1中の少なくとも1つの遺伝子の発現レベルの上昇についてアッセイするステップであって、ここで、前記上昇は、前記生物活性剤が前記動物における再有髄化を促進することを示すステップ;および/または
(c)対象動物と比較した、前記動物における有髄化の変化を観察するステップ
を含む方法。
【請求項46】
前記MRF導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項44に記載の方法。
【請求項47】
前記MRF導入遺伝子が、エキソンの欠失を含む、請求項44に記載の方法。
【請求項48】
前記エキソンが、エキソン8である、請求項46に記載の方法。
【請求項49】
前記MRF導入遺伝子が、リコンビナーゼ部位に隣接する、請求項44に記載の方法。
【請求項50】
前記リコンビナーゼ部位が、loxPまたはFRTである、請求項48に記載の方法。
【請求項51】
前記動物が、リコンビナーゼ導入遺伝子を含む、請求項44に記載の方法。
【請求項52】
前記リコンビナーゼ導入遺伝子が、細胞型特異的な調節エレメントに作動的に連結される、請求項50に記載の方法。
【請求項53】
前記リコンビナーゼが、CreリコンビナーゼまたはFlpである、請求項48または50に記載の方法。
【請求項54】
前記細胞型が神経細胞である、請求項45に記載の方法。
【請求項55】
前記神経細胞が神経膠細胞である、請求項53に記載の方法。
【請求項56】
前記神経膠細胞が、希突起膠細胞、希突起膠細胞の前駆細胞、シュワン細胞、星状細胞、または小膠細胞である、請求項54に記載の方法。
【請求項57】
前記神経細胞特異的な調節エレメントが、CC1遺伝子、ミエリン塩基性タンパク質(MBP)遺伝子、セラミドガラクトシルトランスフェラーゼ(CGT)遺伝子、希突起膠細胞−ミエリン糖タンパク質(OMG)遺伝子、サイクリックヌクレオチドホスホジエステラーゼ(CNP)遺伝子、NOGO遺伝子、ミエリンタンパク質ゼロ(MPZ)遺伝子、末梢ミエリンタンパク質22(PMP22)遺伝子、プロテイン2(P2)遺伝子、GFAP遺伝子、AQP4遺伝子、PDGFα遺伝子、RG5遺伝子、p糖タンパク質遺伝子、ニュールツリン(NRTN)遺伝子、アルテミン(ARTN)遺伝子、ペルセフィン(PSPN)遺伝子、スルファチド遺伝子もしくはプロテオリピドタンパク質(PLP)遺伝子、Olig1遺伝子、またはOlig2遺伝子に由来する、請求項54に記載の方法。
【請求項58】
前記細胞型特異的な調節エレメントが、誘導的プロモーターまたは構成的プロモーターである、請求項45に記載の方法。
【請求項59】
前記動物がマウスである、請求項44または44に記載の方法。
【請求項60】
前記生物活性剤が、ペプチド、抗体、アプタマー、siRNA、miRNA、EGS、アンチセンス分子、ペプチド模倣剤、または低分子である、請求項42、43、または44に記載の方法。
【図5】


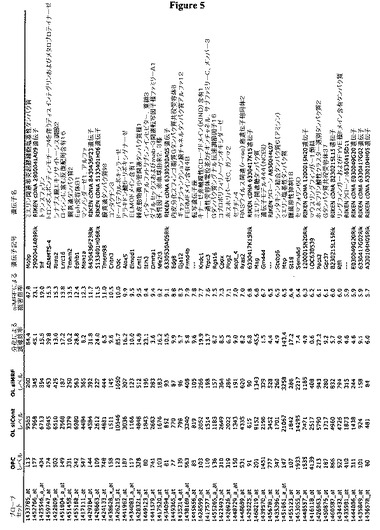
【図11A】

【図11B】

【図11C】

【図11D】

【図14】
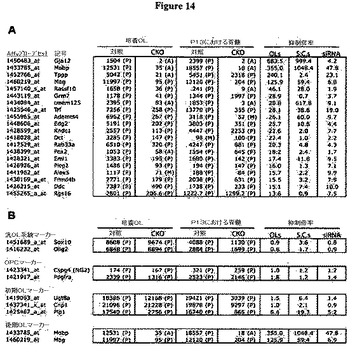
【図16A】
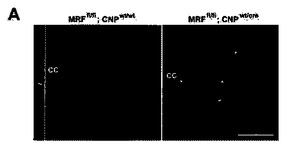
【図16B】
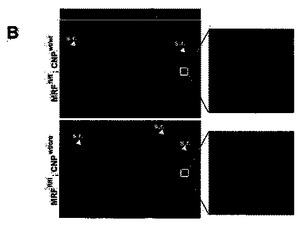
【図1】
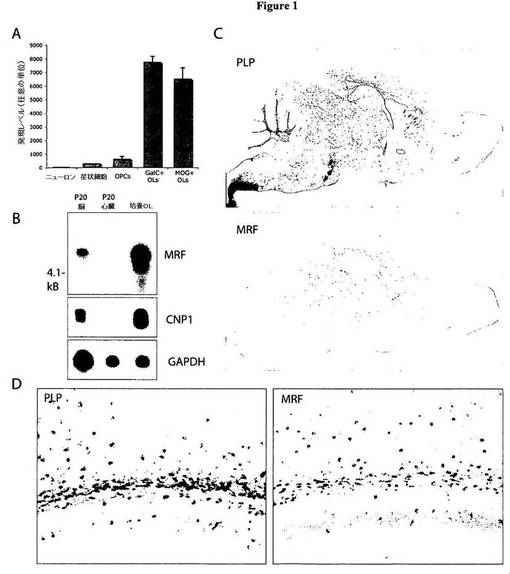
【図2】

【図3】
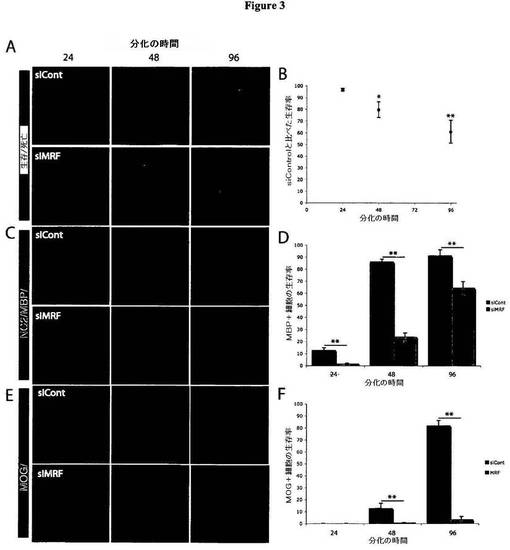
【図4】
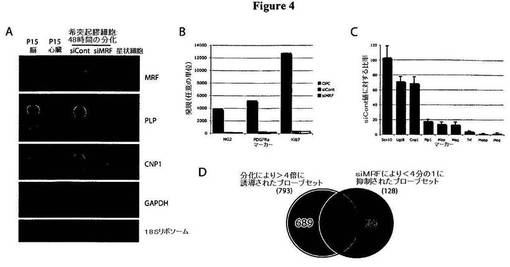
【図6】
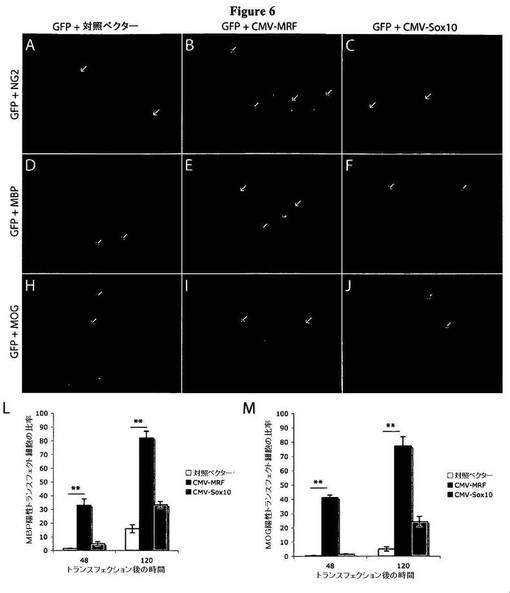
【図7】
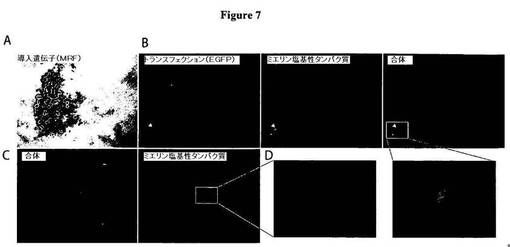
【図8】
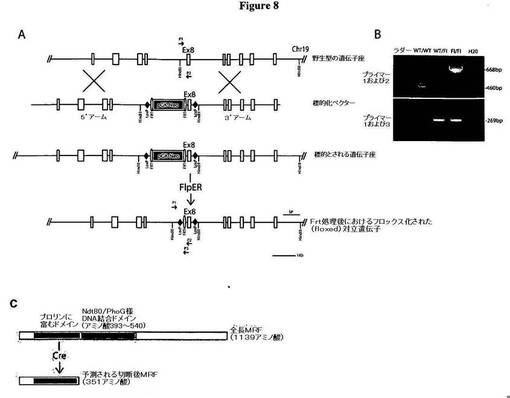
【図9】
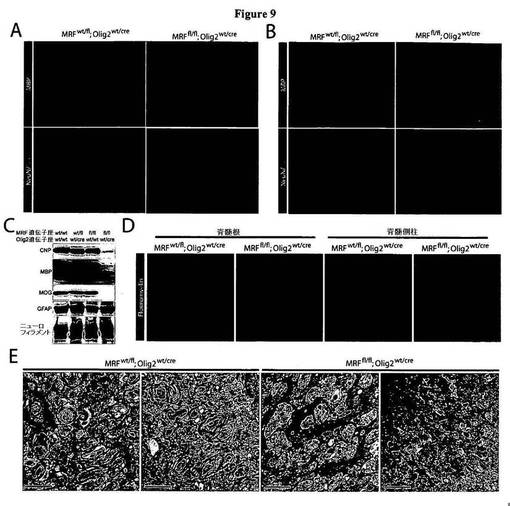
【図10】
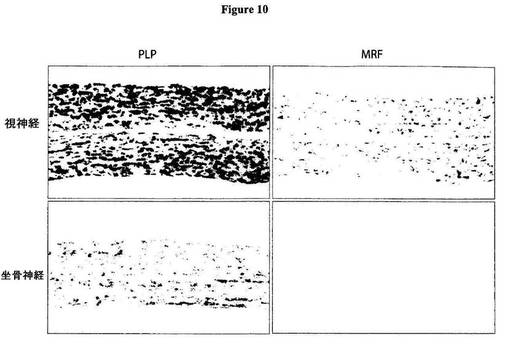
【図12】
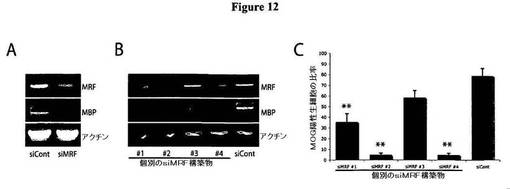
【図13】
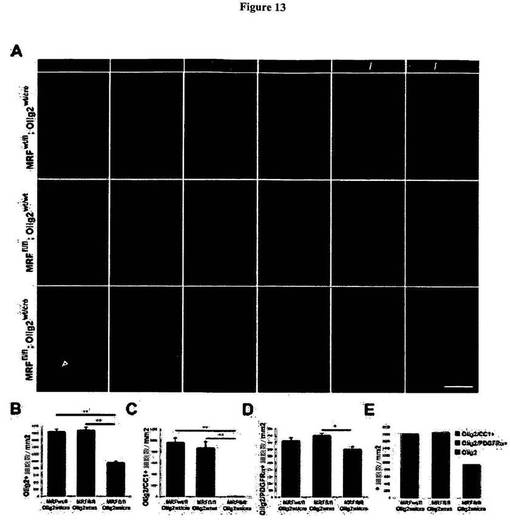
【図15】
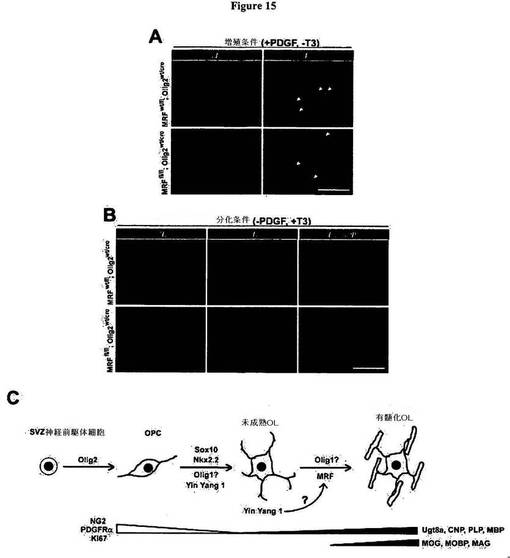
【図17】
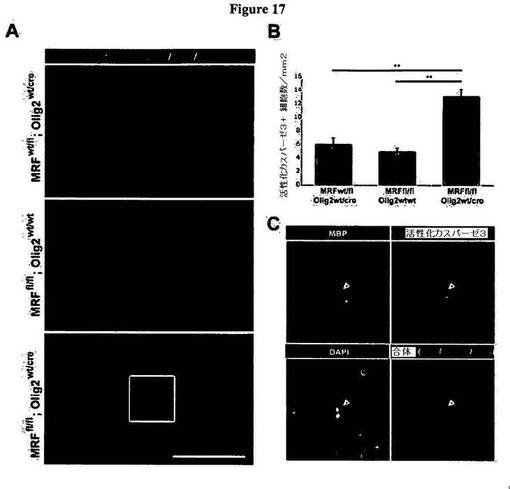
【図11A】
【図11B】
【図11C】
【図11D】
【図14】
【図16A】
【図16B】
【図1】
【図2】
【図3】
【図4】
【図6】
【図7】
【図8】
【図9】
【図10】
【図12】
【図13】
【図15】
【図17】
【公表番号】特表2011−528227(P2011−528227A)
【公表日】平成23年11月17日(2011.11.17)
【国際特許分類】
【出願番号】特願2011−518738(P2011−518738)
【出願日】平成21年7月16日(2009.7.16)
【国際出願番号】PCT/US2009/004155
【国際公開番号】WO2010/008588
【国際公開日】平成22年1月21日(2010.1.21)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
【公表日】平成23年11月17日(2011.11.17)
【国際特許分類】
【出願日】平成21年7月16日(2009.7.16)
【国際出願番号】PCT/US2009/004155
【国際公開番号】WO2010/008588
【国際公開日】平成22年1月21日(2010.1.21)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
[ Back to top ]